/






























































































































































































































































Text
ФИЗИОЛОГИЯ ЧЕЛОВЕКА
ФИЗКУЛЬТУРА И СПОРТ-1975
ФИЗИОЛОГИЯ ЧЕЛОВЕКА
5-е издание
Под общей редакцией профессора
Н. В. ЗИМКИНА
Допущено
Комитетом по физической культуре и спорту при Совете Министров СССР в качестве учебника для институтов физической культуры
V
МОСКВА ФИЗКУЛЬТУРА И СПОРТ 1975
7А.06 Ф 50
Физиология человека. (Учебник для институтов Ф50 физической культуры. Изд. 5-е ). Под ред. Н. В. Зим-кина. М., «Физкультура и спорт», 1975.
496 с. с ил.
Учебник написан в соответствии с новой (утвержденной в 1974 г.) программой по физиологии для институтов физической культуры. В первой его части излагаются вопросы общей физиологии человека, во второй — физиологические основы физического воспитания и спорта.
Настоящее (пятое) издание учебника почти полностью переработано, авторский коллектив увеличен. Изменено расположение глав. Ряд материалов выделен в отдельные главы (1, 16, 21); по новому плану написаны главы по физиологии различных видов спорта.
ф 60901—152
009(01)—75
7А.06
ФИЗИОЛОГИЯ ЧЕЛОВЕКА
5-е издание
Под общей редакцией профессора Н. В. Зимкина
Заведующая редакцией А. К. Гринкевич. Редактор Г. Б. Хот янова. Художественный редактор Ю. В. Архангельский. Технический редактор Н, И. Бурова. Корректор Л. А. Кирюхина. А 03892. Сдано в производство 13/11 1975 г. Подписано к печати 14/VII 1975 г. Формат 60X90/16* Бумага тип. № 2. Печ. л. 31. Уч.-изд. л. 35,9. Тираж 90 000 экз. Издат. № 5139. Цена 1 р. 36 к. Зак. 113. Ордена «Знак Почета» издательство «Физкультура и спорт» Государственного комитета Совета Министров СССР по делам издательств, полиграфии и книжной торговли. 103006. Москва, К-6, Каляевская ул., 27. Ярославский полиграфкомбинат «Со-юзполиграфпрома» при Государственном комитете Совета Министров СССР по делам издательств, полиграфии н книжной торговли. 150014, Ярославль, ул. Свободы, 97.
(6) Издательство «Физкультура и спорт», 1975 г.
ВВЕДЕНИЕ
§ 1. Физиология и ее содержание
Физиология — наука, изучающая функции и механизмы деятельности отдельных клеток, органов и систем органов, а также организма в целом.
Являясь ветвью биологии, физиология опирается на данные многих других биологических наук. Без знания морфологического строения клеток, тканей органов и систем органов изучение их функций не может быть полноценным. Важнейшее значение имеет изучение химии и физики,' так как все явления, происходящие в организме, обусловлены материальными процессами и в основе этих явлений лежат законы физико-химических наук. Как указывает Ф. Энгельс, «физиология есть, разумеется, физика и в особенности химия живого тела, но вместе с тем она перестает быть специальной химией: с одной стороны, сфера ее действия здесь ограничивается, но с другой стороны, она... поднимается на более высокую ступень» *.
Функциональные изменения в живых организмах и механизмы их регуляции физиология изучает вместе с такими сопредельными науками, как биохимия и биофизика. Некоторые разделы физиологии, в особенности физиология нервной системы и анализаторов, тесно соприкасаются с психологией.
Знание физиологических механизмов, обеспечивающих функции организма, имеет большое значение для формирования диалектикоматериалистического мировоззрения. В то же время данные о функциональных свойствах организма получают наиболее правильное освещение только при трактовке их с позиций общетеоретических и методологических основ диалектического материализма.
Важными вопросами, вокруг которых происходит борьба между материализмом и идеализмом, являются развитие живого вещества из неживого и природа сознания человека. Результаты физиологических исследований в сочетании с данными общей биологии, морфологии и других наук объясняют причины перехода простейших форм жизни к более сложным и постепенное развитие функций организма. Тем самым физиология содействует формированию научных представлений об эволюции животного мира, позволяет более четко представить процесс развития, благодаря которому человек
Ф. Энгельс. Диалектика природы. Госполитиздат, 1964, стр. 222.
И. М. Сеченов
И. П. Павлов
завял совершенно особое положение в ряду других представителей живых организмов.
Важное значение в борьбе диалектико-материалистического мировоззрения с идеализмом, в частности с витализмом, и религиозными предрассудками, имеет учение И. М. Сеченова и И. П. Павлова об условнорефлекторных, или индивидуально приобретенных, формах поведения животных и человека. Это учение раскрывает материальную природу сложнейших форм приспособления организма к постоянно изменяющимся условиям внешней среды. Экспериментальные исследования условнорефлекторной деятельности, проведенные И. П. Павловым и его школой, позволили получить ценнейшие данные для естественнонаучного материалистического обоснования природы сознания и познаваемости всех явлений, протекающих в организме человека. Эти данные явились новым доказательством диалектико-материалистического положения о первичности материи и вторичности сознания.
Для специалистов в области физического воспитания, спорта изучение физиологии имеет важное теоретическое и практическое значение. Работа их не может быть полноценной, если они не будут хорошо знать физиологических закономерностей деятельности организма и воздействия на него физических упражнений.
В физиологии применяются различные экспериментальные методы исследования, дополняемые наблюдениями. С усовершенствованием методов исследования острые опыты (при которых наркоз, боль и другие факторы извращают нормальное течение функций организма) все больше уступают место хроническим экспериментам. Благодаря этому стало возможным изучение функций организ-
4
ма в условиях его нормального взаимодействия с окружающей средой.
В опытах на животных широко используются хирургические методы, связанные с экстирпациями (удалением) и пересадками органов и тканей, с созданием фистул для получения через них пищеварительных соков, мочи, крови и лимфы, с вживлением электродов для раздражения различных органов и тканей и отведения биотоков и т. д.
В физиологических исследованиях, в частности тех, которые проводятся на человеке, все большее значение приобретают биоэлектрические, тензометрические, фотометрические, телеметрические и другие физические методы исследования. Особенно ценным для изучения организма является использование современной электронной и вычислительной аппаратуры. Это позволяет изучать функции не только периферических, но и глубоко расположенных органов и тканей, регистрировать различные показатели функций организма как контактным, так и радиотелеметрическим путем и подвергать полученные материалы обработке и математическому анализу с использованием вычислительных машин.
§ 2. Краткие сведения о развитии физиологии
Первоначальные представления о функциях организма сложились еще у врачей и ученых Древней Греции (Гиппократ — 460— 377 гг. до н. э., Аристотель — 384—322 гг. до н. э.), Древнего Рима (Гален — 201—131 гг. до н. э.), Древнего Китая, Древней Индии и других стран. Изучение ими строения тела проводилось одновре-местно с исследованием функций организма. Самостоятельной научной дисциплиной физиология стала лишь в XVII веке, когда наряду с наблюдениями, проводившимися над людьми и животными, началась разработка экспериментальных методов исследования. Важнейшее значение для формирования физиологии как отдельной науки имели результаты исследований ученых XVII века: открытие В. Гарвеем (1578—1657) кругов кровообращения, формулирование Р. Декартом (1596—1650) понятия о рефлексе и разработка Д. Бо-релли (1608—1679) механики движений. Значительного расцвета физиология как наука достигла в XIX и особенно в XX веке.
Большое влияние на развитие физиологии оказали французские исследователи — Ф. Мажанди (1783—1855), К. Бернар (1813— 1878), Л. Лапик (1866—1952), немецкие — И. Мюллер (1801— 1858)., К. Людвиг (1816—1895), Э. Дюбуа-Реймон (1818—1896), Г. Гельмгольц (1821—1894), Э. Пфлюгер (1828—1910), английские— Ч. Белл (1774—1842), Дж. Ленгли (1852—1925), Ч. Шеррингтон (1859—1952), Д. Холден (1860—1936), Э. Старлинг (1866—1927), Д. Баркрофт (1872—1947), итальянские — Л. Гальвани (1737—1798), А. Mocco (1846—1910), Л. Лючиани (1840— 1919), голландские—В. Эйнтговен (1860—1927), Р. Магнус (1872— 1927), американский исследователь В. Кэннон (1871 —1945),
5
датский — А. Крот (1874—1949), чешский — Ян Пуркеиье (1787— 1869) и многие другие.
Большой вклад в развитие физиологии внесли отечественные ученые. Важное научное значение имели представления гениального русского ученого М. В. Ломоносова (1711—1765) о сохранении материи и о трехкомпонентности цветового зрения. Ценными были работы А. М. Филомафитского (1807—1849) о действии наркоза и В. А. Басова (1812—1879) об изучении функций желудка с помощью наложения фистул.
Для создания новых оригинальных путей развития русской физиологии огромное значение имели материалистические идеи, содержавшиеся в работах великих русских революционных демократов: В. Г. Белинского, А. И. Герцена, Н. Г. Чернышевского, Н. А. Добролюбова, Д. И. Писарева. Под влиянием этих идей сложилось мировоззрение «отца русской физиологии» И. М. Сеченова (1829—1905), развившего научные представления о физиологии нервной системы, дыхания, утомления и т. д. В работе «Рефлексы головного мозга» (1863) И. М. Сеченов изложил свои материалистические взгляды на природу произвольных движений и психических явлений.
Большая роль в развитии современной физиологии принадлежит И. П. Павлову (1849—1936). Разработанные им новые подходы к исследованию физиологических функций позволили значительно расширить возможности физиологической науки, включив в сферу изучения не только отдельные органы и системы органов, но и жизнедеятельность целостного организма, непрерывно взаимодействующего с внешней средой. Используя новые методы, И. П. Павлов и его сотрудники установили основные закономерности деятельности пищеварительных желез. Большую ценность представляют его исследования по кровообращению и нервной трофике. Изучая условные рефлексы, И. П. Павлов создал совершенно новый раздел физиологии— физиологию высшей нервной деятельности.
Работы И. П. Павлова имеют важнейшее значение и для формирования диалектико-материалистического мировоззрения. Изучение высшей нервной деятельности и закономерностей приспособления организма к воздействиям внутренней и внешней среды (не только природной, но и социальной), эспериментально подтверждает правильность основного положения марксизма-ленинизма о первичности материи и вторичности сознания.
И. П. Павлов был учителем многих видных советских физиологов. Среди них прежде всего следует назвать Л. А. Орбели (1882— 1958)—одного из создателей нового направления в физиологической науке — эволюционной физиологии. Он внес важный вклад почти во все разделы физиологической науки, в особенности в физиологию мышечной деятельности, симпатической нервной системы, мозжечка и органов чувств. Велика его роль в развитии физиологических основ физической культуры, спорта. Им впервые организована кафедра физиологии в физкультурном вузе (ныне Ленинградский институт физической культуры им. П. Ф. Лесгафта), которой
6
Н. Е. Введенский
А. А. Ухтомский
он заведовал с 1919 по 1927 г. Вместе с многочисленными учениками, в том числе такими известными физиологами, как А. Г. Гине-цинский (1895—1962) и А. В. Лебединский (1902—1965), Л. А. Ор-бели исследовал многие вопросы, связанные с двигательной деятельностью: механизмы координации движений, взаимодействие афферентных систем, адаптационно-трофические влияния симпатической нервной системы, функции мозжечка. Многие его ученики стали ведущими специалистами в области физиологии спорта.
Учениками И. П. Павлова были также К. М. Быков (1886-— 1959), в лаборатории которого исследовалась условнорефлекторная регуляция деятельности внутренних органов; Г. В. Фольборт (1885—1960), внесший большой вклад в изучение пищеварительных процессов и вопросов утомления и восстановления; П. С. Купалов (1888—1964), изучавший различные закономерности условиореф-лекторных реакций; П. К. Анохин (1898—1974), разрабатывавший многие актуальные вопросы нейрофизиологии и высшей нервной деятельности и выдвинувший концепцию о системной деятельности нервной системы.
Крупнейшими представителями отечественной физиологии являются Н. Е. Введенский (1852—1922) и А. А. Ухтомский (1875— 1942). Их труды по физиологии нервной системы считаются классическими. Н. Е. Введенский первым обнаружил в мышцах явления оптимума и пессимума и частые колебания биотоков. Он разработал оригинальную теорию о физиологической природе возбуждения и торможения, сформулировал понятие о лабильности нервов и мышц и создал учение о парабиозе. Ценнейшими являются также его работы о координации деятельности мышц. Дальнейшее развитие идеи Н. Е. Введенского получили в лабораториях его ученика
7
A., А. Ухтомского, открывшего закон доминанты в деятельности нервной системы. Его работы были важным этапом в разработке идей Н. Е. Введенского, касавшихся природы явлений возбуждения, торможения, парабиоза, лабильности и др. Многие работы А. А. Ухтомского, связанные с физиологией двигательного аппарата, легли в основу вновь возникшего раздела физиологии — физиологии труда.
Большое значение для развития общей физиологии имели труды и других отечественных ученых. Ф. В. Овсянниковым (1827—1906) выявлена локализация сосудодвигательного центра в продолговатом мозгу; И. Ф. Ционом (1842—1912) обнаружены в дуге аорты рецепторы, раздражаемые изменением кровяного давления. И. Ф. Тарханов (1946—1908) внес крупный вклад в физиологию нервной системы. В. Я- Данилевским (1852—1939) разработан ряд вопросов в области биоэлектрических явлений и термодинамики мышечной ткани; Н. А. Миславским (1854—1928) уточнена локализация дыхательного центра в продолговатом мозгу. А. Ф. Самойлов (1867—1930) явился одним из основателей учения об электрокардиографии. П. П. Лазарев (1878—1942), создатель советской биофизики, был крупным специалистом в области общей физиологии органов чувств. В. В. Парин (1903—1971) выполнил ценные работы в области кровообращения и содействовал внедрению кибернетики в физиологию. И. С. Бериташвили (1885—1974) внес большой научный вклад в разработку механизмов координации движений, условных рефлексов и памяти.
Из работающих в настоящее время отечественных физиологов большой вклад в разработку физиологии нервной системы, анализаторов, высшей нервной деятельности, двигательного аппарата, кровообращения, дыхания, обмена веществ и энергии, пищеварения, выделительных процессов, терморегуляции и желез внутренней секреции внесли Э. А. Астратян, Г. В. Гершуни, Н. В. Голиков, А. Б. Коган, П. Г. Костюк, М. Н. Ливанов, М. Е. Маршак, С. П. На-рикашвили, А. Н. Ройтбак, М. В. Сергиевский, В. Н. Черниговский, А. М. Уголев, Ю. М. Уфлянд и др.
Бурное развитие физкультурного движения и прогресс науки в Советском Союзе обусловили появление в 30-х годах нового раздела физиологии — физиологии спорта. Теоретические предпосылки для возникновения и развития физиологии спорта можно найти в многочисленных работах И. М. Сеченова, И. П. Павлова, Н. Е. Введенского, А. А. Ухтомского, Л. А. Орбели, И. С. Бериташвили, К. М. Быкова и других отечественных физиологов. В этих работах рассматриваются многие вопросы, имеющие важное значение для характеристики двигательной деятельности: 1) механизмы мышечной деятельности, в частности координации движений; 2) формирование двигательных условных рефлексов; 3) функции анализаторов; 4) особенности вегетативных функций (кровообращения, дыхания, обмена веществ и др.) при мышечных напряжениях; 5) природа утомления.
Отдельные работы, посвященные изучению физиологических
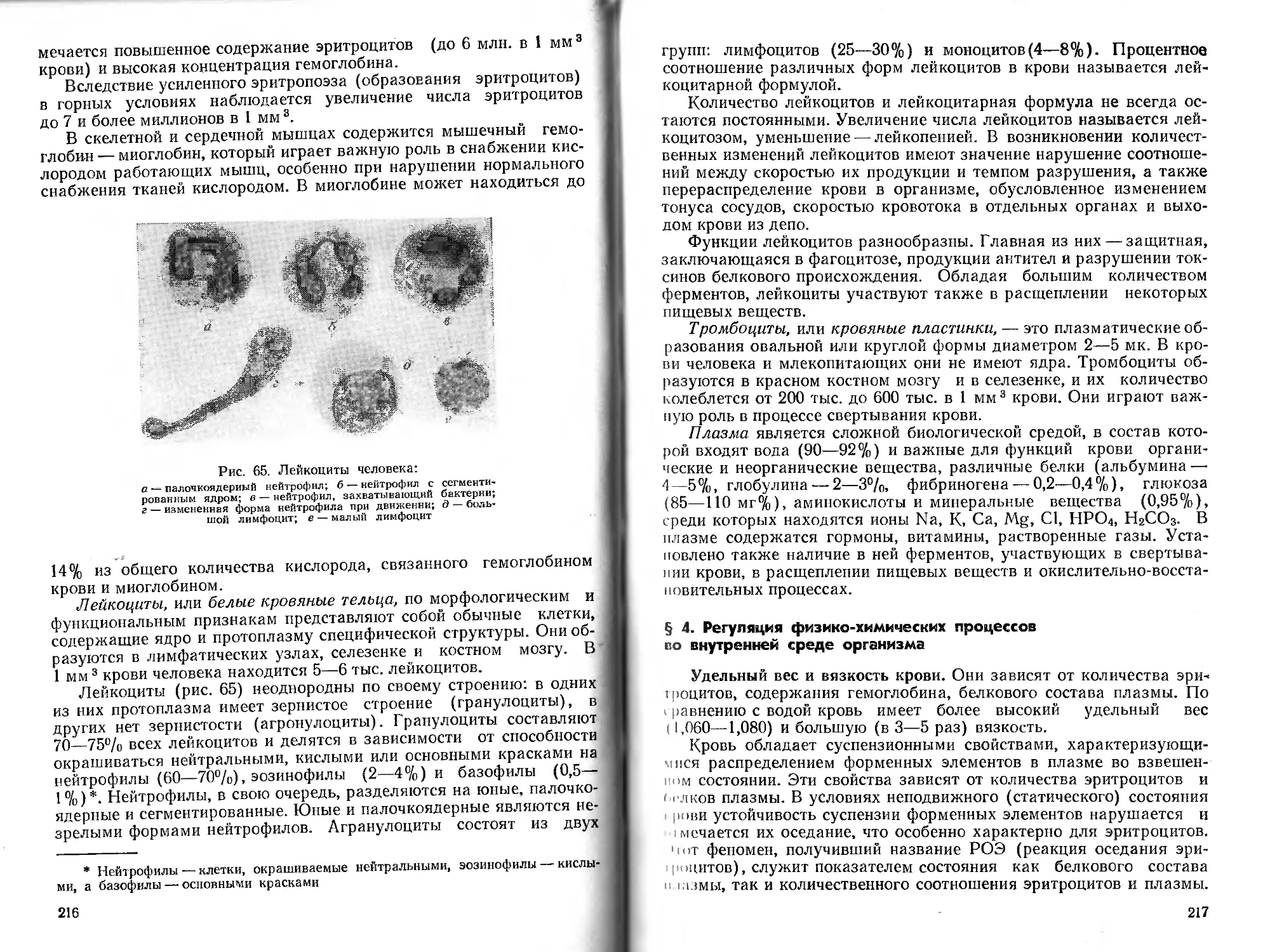
8
Л. А. Орбели А. Н. Крестовников
функций организма при физических упражнениях, были опубликованы еще в конце прошлого века (И. О. Розанов, С. С. Груздев, Ю. В. Блажсвич, П. К. Горбачев и др.). Однако систематическое изучение физиологических основ физического воспитания началось значительно позже — лишь после Октябрьской революции. В создании этого раздела физиологии особенно большая заслуга принадлежит ученику И. П. Павлова и Л. А. Орбели — А. Н. Крестовникову (1885—1955). Вместе со своими сотрудниками он участвовал в разработке важнейших разделов физиологии спорта. А. Н. Крестовников был первым автором отечественной монографии по физиологии спорта и учебника по физиологии для физкультурных вузов.
В настоящее время изучение различных проблем физиологии спорта значительно расширилось и углубилось. Исследования ведутся во всех институтах физической культуры, во многих педагогических и медицинских институтах нашей страны. В целом ряде работ освещена разработка вопросов, касающихся функций анализаторов и отражения мышечной деятельности в биоэлектрической активности мозга (Е. В. Сологуб, В. С. Фарфель, А. И. Яроцкий и др.), физических качеств и физиологической характеристики двигательного навыка (Н. В. Зимкин, Я. М. Коц, А. В. Коробков, В. С. Фарфель, Я. А. Эголинский и др.), функций сердечно-сосудистой системы и дыхания (В. В. Васильева, В. Л. Карпман, В. В. Михайлов, К. М. Смирнов и др.), эндокринных функций (А. А. Виру, Н.Н. Яковлев и др.), стартового состояния, врабатывания и утомления (Ю. И. Данько, В. В. Розенблат и др.). Большая работа по физиологии спорта проводится и за рубежом. В результате всех этих исследований получены данные, имеющие важное значение для физического воспитания и спорта.
б
Часть первая
ОБЩАЯ ФИЗИОЛОГИЯ
Глава I
ОБЩИЕ ФИЗИОЛОГИЧЕСКИЕ СВОЙСТВА
И ЗАКОНОМЕРНОСТИ ОСНОВНЫХ ЖИЗНЕННЫХ ПРОЦЕССОВ
§ 1. Обмен веществ. Организм и внешняя среда. Гомеостаз
Обязательным процессом, без которого немыслима жизнь, является обмен веществ. Он возможен только при условии затрат свободной энергии, т. е. при совершении работы. По Ф. Энгельсу, под работой, в широком смысле слова, следует понимать изменение форм движения материи, рассматриваемое с количественной стороны.
Обмен веществ (метаболизм) —это одновременные, но не всегда одинаково протекающие по своей интенсивности процессы ассимиляции (анаболизма) и диссимиляции (катаболизма).
За счет ассимиляции происходит накопление пластических веществ, идущих на формирование различных тканей организма (массы тела), и энергетических веществ, необходимых для осуществления всех процессов жизнедеятельности, в том числе движения.
За счет диссимиляции совершаются распад химических веществ, разрушение тканевых элементов тела (старых, отмерших и поврежденных), и освобождение энергии из энергетических веществ, накопленных в процессе ассимиляции.
Оба процесса осуществляются при условии поступления, переработки и усвоения пластических и энергетических веществ (белки, жиры и углеводы), витаминов, минеральных веществ и микроэлементов из внешней среды в виде продуктов питания, а также удаления из организма продуктов распада. То или иное течение обмена веществ зависит от складывающихся в каждый отдельный момент взаимоотношений организма с окружающей его средой.
Влияние среды на организм многогранно. Она является поставщиком всех необходимых для его жизнедеятельности и развития веществ, она же служит источником постоянного и бесчисленного потока возмущающих воздействий (раздражений). Существование организма в этих условиях возможно только в том случае, если он своевременно реагирует на все воздействия соответствующими приспособительными реакциями. Эти реакции не должны сопровождаться изменениями функций, выходящими за рамки пределов физиологических колебаний. В противном случае может нарушиться нормальная жизнедеятельность организма, что вызовет заболевание, а в некоторых случаях даже смерть. Поэтому всеми животными и растительными организмами в процессе формирования взаимоот
10
ношений со средой выработана способность не только приобретать новые, наследственно закрепляемые качества, но и сохранять имеющееся относительное постоянство химического состава и функций своего организма, т. е. гомеостаз (от греч. homoios — одинаковый, подобный, stasis — состояние).
Несмотря на то что животные и растительные организмы по своему химическому составу (концентрация веществ), в том числе по составу циркулирующих в нем жидкостей (кровь, лимфа, тканевая жидкость), и по температуре отличаются от концентрации веществ и температурного режима окружающей их среды, все эти организмы, вопреки второму закону термодинамики, требующему уравновешивания, сохраняют свое динамическое неравновесное состояние. Живые организмы подчиняются всеобщему закону биологии: «Живые системы никогда не бывают в равновесии и исполняют за счет своей свободной энергии постоянную работу против равновесия, требуемого законами физики и химии при существующих внешних условиях» (Э. Бауэр). Основным выражением этого закона и является способность живых объектов сохранять свой гомеостаз путем использования различных по степени своей активности механизмов приспособления.
Так, для одних представителей живых существ средством сохранения гомеостаза является пассивный способ приспособления к окружающей среде. Они приобрели способность переходить на максимально допустимый низкий уровень функциональной активности вплоть до анабиоза (от греч. ana — обратно, bios — жизнь), точнее, до парабиоза (от греч. рага — около) или гипобиоза (от греч. hypo — уменьшение, ослабление). Однако подобный способ приспособления (адаптации) к изменяющимся условиям существования в эволюционном отношении недостаточно надежен, так как неблагоприятные условия могут оставаться без изменения более длительное время, чем то, в течение которого организм в состоянии сохранить в условиях анабиоза свою жизнеспособность. Поэтому обстоятельства могут сложиться так, что восстановление благоприятных жизненных условий произойдет после утраты организмом способности вернуться из этого состояния к активной жизнедеятельности.
У других представителей живой природы сформировались активные формы приспособления, позволяющие вести поиск более подходящих условий существования, обеспечивающих сохранение гомеостаза. Естественно, активный поиск возможен только в том случае, если живое существо способно перемещаться в окружающем пространстве, т. е. двигаться.
У третьих представителей живого преимущественно развились механизмы активного вмешательства в окружающий их внешний мир. Высшего совершенства подобная форма адаптации достигла у человека в виде трудовой деятельности. Эта адаптация является прямым ее следствием и причиной дальнейшего совершенствования двигательной активности. Движение выступает здесь не просто как средство перемещения в пространстве, а как тончайший механизм осуществления всех форм трудовой, творческой преобразующей
11
деятельности. Таким образом, движение во всем многообразии своего выражения — наиболее совершенный способ приспособления к окружающей среде и активного воздействия на нее. Это — способ активного преобразования.
Организму свойствен принцип целостности, характеризующийся теснейшей взаимосвязью всех его органов и систем органов. Полноценное движение, обусловливающее приспособление организма к новым условиям среды, возможно лишь при обеспечении двигательного аппарата, в первую очередь мускулатуры, необходимыми для его деятельности продуктами питания, кислородом и при выведении продуктов распада. Это требует координированной деятельности органов кровообращения, дыхания, пищеварения, выделения и других, регулируемых нервной системой. Рационально используемые физическая культура, спорт способствуют поддержанию у человека гомеостаза, восполняя ограничение двигательной активности, возникшее как следствие научно-технического прогресса.
У живых существ уже на самых ранних уровнях эволюционного развития эти три способа приспособления к окружающей среде не исключают друг друга, а существуют во взаимосвязи, имея лишь различную степень выраженности.
§ 2. Раздражители и возбудимость
Любая приспособительная реакция возможна только в том случае, если для ее возникновения есть побудительные причины. Ими являются различные факторы внешней и внутренней среды. В физиологии они получили название раздражитель, стимул, возмущающее воздействие. Процесс воздействия раздражителя на живое образование называется раздражением.
По своей энергетической природе раздражители могут быть химическими, электрическими, механическими, температурными, радиационными и др. По биологическому значению — адекватными (от лат. adaequatus— соответствующий, специальный) и неадекватными (общими).
Адекватные раздражители — это раздражители, которые при минимальной энергии раздражения вызывают возбуждение в рецепторах и клетках, обладающих специальной способностью реагировать на данный раздражитель. Например, для чувствительных клеток сетчатки глаза адекватным раздражителем является часть спектра электромагнитных волн (световые излучения), для органов слуха — частота воздушных колебаний в пределах от 16 до 20 000 герц, для мышцы — нервный импульс. Все остальные раздражители для перечисленных образований будут неадекватными. Неадекватные, или общие, раздражители — это те виды воздействий, которые могут восприниматься многими возбудимыми образованиями и вызыватЕ> ответные реакции (как возбуждение, так и торможение) только при относительно значительной силе и длительности своего воздействия. Примером может служить ощущение вспышки
12
света при механическом раздражении сетчатки глаза в результате сдавливания глазного яблока.
Следовательно, многие (как адекватные, так и неадекватные) воздействия внешней среды на организм и сдвиги во внутренней его среде могут выполнять роль раздражителей, вызывающих ответную реакцию. Однако реакция на адекватный раздражитель возникает легче и имеет более совершенный приспособительный характер, чем реакция на неадекватный раздражитель.
Способность живого образования отвечать на действия раздражителя изменениями своих физиологических свойств и возникновением состояния возбуждения называется возбудимостью.
§ 3. Функциональная активность
В живых объектах следует различать три уровня состояния функциональной активности: физиологический покой, возбуждение, торможение.
Состояние физилогического покоя характеризуется отсутствием внешне выраженных проявлений специфической активности, например движения, секреции. Вместе с тем это не бездеятельное состояние. Оно служит основой, без которой специфическая активность не может проявиться. В условиях физиологического покоя протекают процессы обмена веществ на уровне, необходимом для поддержания жизнедеятельности организма, обеспечения его готовности реагировать на воздействия, требующие проявления специфической активности. Поэтому правильнее говорить не просто о состоянии физиологического покоя, а о состоянии относительного физиологического покоя.
Возбуждение — это процесс, возникающий в результате того или иного воздействия на живой объект как проявление свойства возбудимости. Различают местное и распространяющееся возбуждения. Первое охватывает сравнительно ограниченный участок вблизи от места нанесения раздражения, второе распространяется на более значительные расстояния от него, например по всему нерву и далее по мышечному волокну.
Проявления возбуждения могут быть неспецифическими, общими для всех или многих живых объектов (например, изменения процесса обмена веществ и температуры, связанные с освобождением энергии и возникновением местного потенциала и потенциала действия— см. ниже), и специфическими, выражающимися в осуществлении физиологической функции, свойственной только данному органу или ткани (например, сокращение мышцы, секреция железы).
Торможение представляет собой активный процесс, проявляющийся в ослаблении или прекращении специфических функциональных отправлений. Оно осуществляется за счет усиления или ослабления обмена веществ. В некоторых случаях торможение может проявляться и в виде снижения энергетического уровня при состоянии физиологического покоя.
13
§ 4. Биологические структуры и процессы обмена веществ
Все формы приспособительного поведения возможны только на базе обмена веществ (метаболизма), свойственного живым организмам. Оптимальное в каждом отдельном случае течение обмена веществ определяется строгой упорядоченностью химических процессов в живом организме. Эта упорядоченность зависит как от соотношения скоростей, регулируемых ферментами химических реакций, так и от наличия высокой степени структурной организованности всех частей живого организма.
Рассмотреть это можно на примере такой составной части организма, как клетка. Она представляет собой самостоятельную структурную единицу. Каждая клетка пространственно отделена от других клеток клеточной мембраной, окружающей ее со всех сторон. Существование клетки и входящих в ее состав элементов (ядро, митохондрии и др.) возможно только при наличии отделяющих их друг от друга мембран. Разрушение мембраны автоматически приводит к разрушению структурной единицы. Это обусловлено тем, что клеточные мембраны являются не только механическими барьерами, но и регуляторами обмена веществ между клеткой и окружающей ее средой, так как через мембрану осуществляется избирательный транспорт (перенос) веществ.
Различают «пассивный» и «активный» транспорт веществ через мембраны.
Пассивный транспорт через мембрану в клетку или из нее в окружающую среду практически происходит без существенных затрат энергии обмена веществ, протекающего в клетке. Перенос веществ осуществляется по градиенту* — от большей величины к меньшей (например, осмотический градиент возникает при разности осмотического давления внутри клетки и вне ее). Градиент —это только условие, необходимое для переноса веществ через мембрану. Основным механизмом, обеспечивающим проникновение их в клетку, является диффузия.
Проникновение веществ в клетку происходит главным образом двумя путями: 1) через поры клеточной мембраны (уже существующие в ней или образующиеся под влиянием тех или иных воздействий) и 2) за счет растворения проникающих веществ в липидах клеточной мембраны. Первый способ проникновения характерен главным образом для водорастворимых веществ, а также ионов, а второй— для органических соединений, не растворимых в воде.
При изучении проницаемости клеточных мембран было замечено, что многие вещества проходят через них значительно быстрее, чем это может обеспечить диффузия (концентрационный градиент), а во многих случаях они проходят через мембрану в клетку и из клетки в окружающую ее среду против градиента, Т. е. направлены от меньшей концентрации к большей. Естественно, при переносе ве
* Градиент — мера возрастания или убывания на единицу измерителя (концентрация, вес, длина, давление и др.).
14
щества от меньшей концентрации к большей клетка должна совершать работу, т. е. затрачивать энергию. Поэтому транспорт вещества через клеточную мембрану с затратой энергии обмена вещества клетки получил название активного транспорта. Активный перенос всегда является избирательным. С его помощью обеспечивается доставка в клетку необходимых для ее жизнедеятельности веществ, а также удаление ненужных продуктов обмена веществ.
Например, одним из механизмов активного переноса веществ через мембрану являются так называемые «ионные насосы». Известно, в частности, что в условиях физиологического покоя и нормальной температуры внеклеточная среда богата содержанием ионов натрия и бедна содержанием ионов калия. Так, в эритроцитах, мышечных и нервных волокнах ионов калия в 30—50 раз больше, чем в плазме крови, лимфе и межклеточной жидкости, а ионов натрия в цитоплазме этих клеток в 8—10 раз меньше. Объяснить подобное резко ассиметричное распределение ионов натрия и калия по обе стороны клеточной мембраны механизмом только пассивного переноса (диффузии) нельзя. Предполагают, что в мембране существует специальный «натрий-калиевый насос», участвующий в поддержании ионной асимметрии. В условиях физиологического покоя он обеспечивает «откачивание» натрия из клетки и «подсасывание» калия внутрь ее. Осуществляется это при помощи особых химических «переносчиков», образующихся в мембране в процессе обмена веществ. Они захватывают у внутренней поверхности мембраны ионы натрия и переносят их на наружную поверхность, где ионы отщепляются и таким образом выводятся наружу из клетки. На наружной же поверхности мембраны к переносчику присоединяются ионы калия, которые и транспортируются на внутреннюю поверхность мембраны, т. е. внутрь клетки.
Работа эта осуществляется с затратой энергии, источником которой является расщепление АТФ и фосфагена при участии фермента аденозин-трифосфатазы.
Известны подобные «насосы» и для переноса других ионов.
Более сложны и менее изучены механизмы активного переноса сахара, аминокислот и других веществ. Во всех случаях активный перенос в отличие от пассивного, имеющего преимущественно физико-химическую природу, является чисто биохимическим процессом, осуществляющимся в мембране. Таким образом, мембрана — это не столько механический барьер, отделяющий одну клетку от другой, сколько сложная функциональная структура, внутри и на поверхности которой совершаются важнейшие ферментативные, окислительно-восстановительные метаболические реакции.
§ 5. Мембранный потенциал
Благодаря механизмам пассивного и активного избирательного переноса через клеточную мембрану органических и неорганических катионов (положительно заряженных ионов) и анионов (отрицательно заряженных ионов) образуется по одну сторону клеточной
15
мембраны избыток катионов, а по другую — избыток анионов. В результате этого между внешней и внутренней поверхностями мембраны возникает разность потенциалов. Эта разность называется мембранным потенциалом. Существуют три отличающихся друг от друга проявления мембранного потенциала — потенциал покоя, местный потенциал и потенциал действия.
Потенциал покоя. В 1838 г. Ш. Маттеучи показал, что с наружной стороны мышца заряжена положительно, а внутри — отрицательно. Позже было установлено, что это характерно для большинства клеток животных и растений. Разность потенциалов, существующая в условиях физиологического покоя, получила название мембранного потенциала покоя. Из имеющихся теорий, объясняющих природу его возникновения и поддержания, наибольшее признание получила мембранная теория. Сущность ее состоит в следующем. Избирательно проницаемая (полупроницаемая) мембрана разделяет два раствора — цитоплазму, находящуюся внутри клетки, и межклеточною жидкость. При этом внутренняя поверхность мембраны соприкасается с цитоплазмой, наружная — с межклеточной жидкостью. Как уже указывалось, содержание положительно заряженных ионов калия (К+) в цитоплазме в несколько раз превосходит содержание их вне клетки, а содержание ионов натрия (Na+), напротив, вне клетки больше, чем в цитоплазме.
Известно, что если в обычном растворе находится какое-то количество катионов, то и анионов в нем должно быть столько же. Баланс ионов, необходимый для электронейтральности, восполняется неорганическими анионами (С1", НРО43-, НСО3~), а также крупными (белки, аминокислоты и др.) органическими анионами (А-), накапливающимися в цитоплазме в процессе внутриклеточного обмена веществ. Электронейтральность по обе стороны клеточной мембраны не сохраняется. Одна из основных причин этого та, что проницаемость клеточной мембраны в условиях покоя для ионов калия достаточно высокая, а для ионов натрия — низкая. В связи с существующим концентрационным градиентом для калия какое-то количество его ионов выходит из клетки через поры мембраны на ее наружную поверхность, создавая на ней избыток положительно заряженных частиц. Крупные же молекулы органических ионов (А-), находящиеся внутри клетки, не могут пройти через мелкие поры мембраны на ее наружную поверхность. Оставшись без нейтрализующих их положительно заряженных ионов калия, они создают избыток у внутренней поверхности мембраны отрицательно заряженных частиц. В результате возникает разность потенциалов — потенциал покоя.
Возникновение и поддержание потенциала покоя представляет собою активный саморегулирующийся процесс, для обеспечения которого требуются определенные затраты энергии. При этом чем больше поляризована мембрана, тем выше этот потенциал.
У различных представителей животного мира в клетках одних и тех же органов потенциал покоя может быть неодинаковым. Он специфичен также для клеток различных тканей одного и того же
16
организма. Например, в клетках скелетных мышц мембранный потенциал покоя колеблется в пределах 80—90 мв, а в клетках эпителиальной ткани — 20—30 мв.
В одной и той же клетке потенциал покоя изменяется незначительно. Сохранение относительно устойчивого значения потенциала покоя, свойственного данной клетке, — один из важных показателей ее физиологического состояния. Резкое снижение этого потенциа
ла— показатель ухудшения состояния клетки, возникновения патологических изменений. Сни-
МП, МВ
жение потенциала покоя до нуля свидетельствует о полном нарушении избирательной проницаемости клеточной мембраны для различных ионов и, следовательно, о гибели клетки. __
Местный потенциал. При действии на клетку различных раздражителей мембранный потенциал начинает уменьшаться, т. е. происходит процесс деполяризации. По мере роста интенсивности раздражения (до определенного предела) мембранный
Рис. 1. Изменение мембранного потенциала в зависимости от силы наносимого раздражения длительностью 2 мсек.:
1—5— изменение мембранного потенциала (МП). Внизу — относительная сила наносимого раздражения (электрический ток). Горизонтальные линии: сплошная — уровень потенциала покоя; штрих-пунктирная — критический уровень местного потенциала, т. е. момент возникновения потенциала действия; штриховая — уровень перехода деполяризации в инверсию знака потенциала действия (по Ба Катцу и Е. К. Жукову)
потенциал в месте нанесения раздражения изменяется градуально (рис. 1).
Снижение величины потенциала покоя происходит в связи с уменьшением поляризации поверхностей клеточной мембраны и увеличением ее проницаемости для потенциалообразующих ио-
нов, в первую очередь ионов
Na+. Проникающие внутрь клетки катионы, понижая величину положительного заряда у наружной поверхности мембраны, одновременно увеличивают количество ионов с положительными зарядами у внутренней ее поверхности. В результате уменьшается величина отрицательного заряда у внутренней поверхности мембраны и в целом величина потенциала покоя. С прекращением раздражения восстанавливается исходная величина этого потенциала.
Эти изменения потенциала покоя распространяются от места своего возникновения, постепенно ослабевая (с декрементом) вплоть до исчезновения. В мышечной и нервных тканях изменение потенциала ограничено небольшим участком в месте нанесения раздражения. Такое изменение потенциала получило название местно
го потенциала.
2—113
17
Местный потенциал становится роваться при силе раздражения в
порогового раздражения, вызывающая потенциал действия.
При действии некоторых раздражителей, например тормозных медиаторов (см. § 6 в этой главе), местные изменения потенциала
Рис. 2. Ионный механизм возникновения мембранного потенциала (МП) действия и восстановления потенциала покоя (на примере нервного волокна).
Сплошная горизонтальная линия — уровень потенциала покоя, а—потенциал действия; б — направление движения ионов Na+ и К+ в различные фазы развития потенциала действия; в — изменение знака электрического заряда на поверхностях мембраны во время развития потенциала действия; 1—2 — возникновение потенциала действия (восходящее колено): / — фаза деполяризации; 2 — фаза инверсии; 3—4 — восстановление потенциала покоя (нисходящее колено потенциала действия): 3 — фаза реверсии, 4 — фаза репо-лярнзации. Размеры кружков вокруг символов ионов Na+ и К+ указывают на интенсивность переноса ионов через мембрану. Стрелками показано направление переноса (по А. Ходжкину)
личины (см. рис. 1). Оказалось, что, как только местный потенциал достигает некоторой критической величины (рис. 2), проницаемость клеточной мембраны для ионов Na+ лавинообразно возрастает.
заметным и начинает регистри-1,5—2 раза меньшей, чем сила
приводят не к деполяризации, а к гиперполяризации, т. е. к снижению возбудимости.
Для клеток соединительной и эпителиальной тканей, т. е. тканей, из которых в основном образованы почти все внутренние органы (печень, железы, органы пищеварения, дыхания и др.), практически не имеющих периода физиологического покоя, единственной формой электрического ответа на раздражение являются градуальные изменения потенциала покоя. Когда эти изменения достигают критического уровня, возникает специфическая функциональная активность данной клетки (например, для железы— секреция).Местная реакция на раздражение, как уже упоминалось выше, получила название местного возбуждения. Местный потенциал есть одно из проявлений этого возбуждения, а его величина — в определенной степени мера состояния возбуждения, т. е. функциональной активности.
Потенциал действия. При изучении электрических свойств мышечной и нервной тканей (клеток) было замечено, что зависимость величины местного потенциала от силы действующего раздражения нарушается при достижении этим потенциалом определенной ве-
Стремительно проникающий в клетку поток их способствует процессу деполяризации мембраны. Потенциал внутри клетки стано-
18
вится равным нулю, продолжающееся же далее поступление ионов Na+ не только «гасит» избыток имеющихся там анионов (А~), но и приводит к накоплению ионов Na+ у внутренней поверхности мембраны и недостатку их на наружной. В результате в месте нанесения раздражения у наружной поверхности мембраны положительный заряд становится отрицательным, а у внутренней — положительным, т. е. происходит изменение знака потенциала мембраны на обратный (инверсия) и возникает потенциал действия. Величина его находится в прямой зависимости от количества ионов Na+, перешедших с наружной поверхности мембраны на внутреннюю.
Как только количество ионов Na+, проникших в клетку, становится равным количеству оставшихся вне клетки, т. е. как только наступает равновесие, поступление ионов Na+ в клетку прекращается. Это происходит через 0,5—1 мсек, от начала возникновения потенциала действия. К этому моменту проницаемость мембраны существенно возрастает для ионов К+, и они в большом количестве выходят из клетки на наружную поверхность мембраны. В результате этого у наружной поверхности мембраны вновь создается положительный заряд, а у внутренней — отрицательный. Все это происходит преимущественно за счет пассивного переноса ионов по градиенту концентрации. Однако если бы в процессе возникновения и тем более «затухания» потенциала действия осуществлялся только пассивный перенос ионов Na+ и К+ по градиенту концентрации, то очень скоро установилось бы стойкое равенство их содержания у внутренней и наружной поверхностей мембраны. В действительности этого не происходит, так как включается, особенно в фазе восстановления (см. рис. 2), механизм активного переноса — «калийнатриевый насос». С его помощью из клетки «откачивается» натрий и «подкачивается» в нее калий. В результате восстанавливается потенциал покоя с ассимметричной исходной концентрацией Na+ и К+ у внешней и внутренней поверхностей мембраны.
Для возникновения потенциала действия и местного потенциала обязательно присутствие катионов кальция во внеклеточной среде.
Процесс деполяризации и инверсии знака (см. рис. 2) при возникновении потенциала действия в нервных и мышечных клетках (восходящее колено) развивается очень быстро (0,1—0,5 мсек.). Реполяризация (от лат. ге — приставка, обозначающая назад, обратно) — восстановление потенциала покоя (нисходящее колено) — протекает значительно медленнее (0,5—5,5 мсек.). Общая продолжительность потенциала действия— 1—5 мсек.
Потенциал действия отличается от местного потенциала тем, что он возникает при достижении критического уровня деполяризации мембраны под воздействием строго определенной в каждом случае силы раздражения, называемой пороговой. Величина потенциала действия при раздражениях, сила которых выше пороговой, не изменяется, т. е. он градуально независим от силы надпороговых раздражений. Потенциал действия возникает только через какое-то время (латентный период) от начала нанесения раздражения. Это время равно длительности достижения местным потенциалом кри
19
тической величины. Возбуждение, сопровождающееся возникновением потенциала действия, в отличие от возбуждения, сопровождающегося местным потенциалом, способно распространяться, например по нерву, без ослабления (без декремента) на большие расстояния от места своего возникновения.
Эти свойства потенциала действия определяют и биологический смысл распространения волны возбуждения — включение систем, обеспечивающих проявление и регулирование моторных и вегетативных реакций. Включение их происходит тогда, когда факторы внешней среды достигают значений, вызывающих существенные отклонения в жизнедеятельности организма. При отсутствии подобной необходимости эти системы могут находиться в состоянии физиологического покоя или близком к нему. Это имеет отношение в первую очередь к аппарату произвольных движений — скелетной мускулатуре.
§ 6. Распространение (проведение) возбуждения
Для поддержания гомеостатического единства все элементы, из которых состоит организм (клетки, органы, системы органов), должны иметь возможность общаться друг с другом, обмениваться информацией, продуктами своей жизнедеятельности, т. е. пространственно взаимодействовать.
Существует несколько способов обеспечения этого пространственного взаимодействия. Один из основных — распространение возбуждения от места его возникновения до исполнительных приборов. Механизм подобного распространения волны возбуждения легче всего представить на примере продвижения потенциала действия вдоль нервного волокна, а также на примере передачи возбуждения через синапсы с одной нервной клетки на другую или с нервной клетки на мышечное волокно (см. далее рис. 3 и 4).
Возникший в месте нанесения раздражения потенциал действия является не только показателем состояния возбуждения в этом месте, но и источником раздражения соседнего, невозбужденного участка нервного или мышечного волокна. По своей силе потенциал действия в 5—10 раз превышает силу раздражения, необходимую для возбуждения расположенного рядом участка, находящегося в состоянии покоя. Благодаря этому последовательно возникающие потенциалы действия создают как бы ток действия (рис. 3). В отличие от местного потенциала, он распространяется по всей длине нервного волокна. В безмякотных нервных волокнах, не имеющих миелиновой оболочки, возбуждение проводится с некоторым затуханием (декрементом), а в волокнах с миелиновой оболочкой — без него.
Проведение возбуждения, как и его возникновение, не сводится к чисто физическим процессам, в частности к току действия, а сопровождается изменением обмена веществ с потреблением энергии.
Структурное образование, обеспечивающее передачу возбуждения с одной нервной клетки на другую или с нервного волокна на
20
иннервированную им клетку — мышечную, железистую и др., называется синапсом (от греч. synapsis — соединение, связь) (рис. 4). Он состоит из синаптической пуговки (бляшки) —утолщения нерв-
ного окончания, покрытого мембраной, получившей название пресинаптической. На прилежащей к ней другой нервной или иной клетке также находится мембрана. Она называется постсинаптической. Между этими двумя мембранами находится узкое пространство — синаптическая щель. В синаптических пуговках синапсов есть мельчайшие пузырьки, содержащие в себе особые химические передатчики нервного возбуждения — медиаторы (от лат. mediator — посредник). К таким медиаторам относятся: для возбуждающих синапсов — ацетилхолин и норадреналин. В связи с этим различают холинэргические и адренэр
Б
гические синапсы.
Установлено, что как только волна возбуждения (ток действия) достигает синаптической пуговки, происходит массовое перемещение пузырьков, содержащих медиаторы, к пресинаптической мембране. Здесь они располагаются у ее пор, лопаются и выбрасывают через
Рис. 3. Схема, иллюстрирующая мембранную теорию распространения возбуждения с помощью локальных потенциалов действия (на примере нервного волокна):
А — состояние физиологического покоя. Мембрана поляризована. С наружной ее стороны заряд положительный, с внутренней — отрицательный; Б, В. Г — последовательные стадии распространения возбуждения (слева направо): сплошная линия —- волна деполяризации и сопровождающий ее потенциал действия, распространяющийся по мембране. Вертикальные стрелки указывают направление перемещения ионов Na+ и К+ через мембрану, горизонтальные стрелки — направление распространения возбуждения (по К. Вилли и В. Детье)
поры медиатор в синаптиче-скую щель. Попав в щель, медиатор образует соединение с химическим веществом (рецептором), находящимся в постсинаптической мембране. Под влиянием этого соединения проницаемость постсинаптической мембраны увеличивается, в результате чего (по указанному выше механизму) на ней возникает местный постсинаптический потенциал. При достижении критической величины он возбуждает клетку, на которой расположена постсинаптическая мембрана. Этот потенциал носит название возбудительный постсинаптический потенциал. Под его влиянием в клетке возникает распространяющееся возбуждение, а следовательно, и потенциал действия, приводя ее в рабочее состоя-
21
Синаптическая щель
Рис. 4. Схема строения синапса
Пресинаптическая мембрана
Аксон Митохондрий ’ уэырькй медиатора
ние (для нервной клетки это дальнейшая передача возбуждения по цепи нервных клеток, для мышечной — сокращение, для железистой — секреция и т. д.). В постсинаптической щели избыток медиатора разрушается (инактивируется) специальными ферментами. При недостаточности медиатора не обеспечивается передача возбуждения, но создаются более благоприятные условия для его возникновения при поступлении следующего импульса. Поэтому в синаптической щели всегда находится некоторое количество медиатора, поддерживающего состояние готовности постсинаптической мембраны к передаче возбуждения.
Есть синапсы, в которых под влиянием возбуждения в синаптическую щель поступают тормозные медиаторы (например, гамма-аминомасляная кислота), вызывающие не деполяризацию, а, напротив, увеличение поляризации —гиперополяризацию постсинаптической мембраны. Естественно, что в этих условиях возбудимость постсинаптической мембраны снижается. Это служит препятствием для продвижения возбуждения. Такая гиперполяризация получила название тормозный
постсинаптический потенциал. Синапсы, работающие по этому принципу, называются тормозящими (тормозными) синапсами.
Благодаря тому что медиаторы выделяются из синаптической пуговки и местом их действия является постсинаптическая мембрана, синапсы передают возбуждение или создают торможение всегда только в одном направлении: пресинаптическая мембрана—> постсинаптическая мембрана.
При проведении возбуждения в одних образованиях срабатывают возбуждающие синапсы, в других — тормозящие. Это приводит к усилению деятельности одних органов и систем и торможению деятельности других. В результате организм может в зависимости от обстоятельств избирательно использовать различные мышцы и вегетативные органы при осуществлении приспособительных реакций в ответ на то или иное воздействие.
Постсинаптмческая мембрана
Дендрит
§ 7. Раздражение и основные свойства и параметры возбудимости
Для лучшего понимания сущности и механизмов осуществления приспособительных реакций организма со средой, а также произвольного управления ими важное значение имеет количественная оценка основных свойств и закономерностей проявления жизнедеятельности организма.
Значение силы раздражения. Для перехода той или иной биологической системы из состояния физиологического покоя в состояние возбуждения, проявления специфической для нее активности
22
Рис. 5. Схема зависимости ответной реакции мышцы, волокна которой имеют различные пороги, от силы раздражения: А — подпороговая сила раздражения; Б— пороговая сила; В — субмаксимальная сила; Г — максимальная сила; Д — супермаксимальная сила. Штриховая линия — величина суммарных энергетических сдвигов; сплошные вертикальные линии — величина функционального ответа. По оси абсцисс— сила раздражения, по оси ординат — величина ответа (в усл. ед.)
(по Л. В. Латманизовой)
(например, для мышцы — сокращение, для нерва — возникновение распространяющегося потенциала), необходимо, чтобы сила действующего раздражителя достигла критической, т. е. пороговой, величины. Порогом возбуждения называется минимальная сила раздражения, при воздействии которой на биологический объект возникают возбуждение в виде местного потенциала и потенциала действия, сокращения мышцы, секреции железы и т. п. Одним из показателей порога возбуждения в органах, образованных из эпителиальных тканей (железы и др.), может служить возникновение критического местного потенциала, а для нервной и мышечной тканей — возникновение потенциала действия.
Сила раздражения ниже пороговой называется подпороговой (в ответ на нее может возникать только местное возбуждение — местный потенциал, — не сопровождающееся специфической реакцией), сила раздражения выше пороговой — надпороговой или сверхпороговой. В частности, в целой мышце или в целом нервном стволе имеются волокна с разными порогами. Сначала возбуждаются волокна с низкими порогами. При последующем увеличении силы раздражения каждый раз вовлекаются все новые и новые волокна, число их увеличивается и вследствие этого становится более выраженной ответная реакция. С достижением такой силы раздражения, когда возбуждаются волокна не только с низкими и средними, но и с самыми высокими порогами, ответная реакция достигает наибольшей величины. Дальше повышение интенсивности раздражения уже не вызывает увеличения ответной реакции.
Сила раздражения для органа с разными порогами возбуждения составляющих его элементов, вызывающая наиболее выраженную ответную реакцию, называется максимальной, сила же выше максимальной, при которой ответ уже не увеличивается, — супермаксимальной. Все степени силы раздражителя, находящиеся между пороговой и максимальной, называются субмаксимальными (рис. 5).
При исследованиях обычно определяют минимальный порог, т. е. порог для наиболее возбудимых элементов, например мышцы или нерва. Чем ниже порог силы, тем выше возбудимость этих элементов мышц и нервов, т. е. способность реагировать на минимальные по силе раздражения (см. рис. 5).
Значение длительности раздражения. Для возникновения возбуждения помимо порога силы имеет значение и порог времени (рис. 6).
23
На основании изучения соотношения между силой и длительностью раздражения, которые необходимы для возникновения возбуждения, Л. Гоорвегом, Г. Вейсом и Л. Лапиком в опытах с раздражением электрическим током была построена кривая силы — длительности. По закону обратной зависимости между силой и продолжительностью раздражения чем больше сила раздражения, тем меньше требуется времени для того, чтобы возникло возбуждение. Таким образом, для каждой силы раздражения (до определенного предела) существует свой порог времени возбуждения.
Анализ кривой силы — длительности раздражения (рис. 7) пока
Рис. 6. Изменение мембранного потенциала до критического уровня деполяризации мембраны при действии раздражения (электрический ток) разной силы и длительности.
Критический (пороговый) уровень показан штриховой линией. Внизу — раздражающие стимулы различной силы и длительности, при воздействии которых получены ответы (потенциалы действия) — А, Б, В (по Е. Б. Бабскому)
Рис. 7. Кривая силы — длительности раздражения:
ОБ — реобаза; ОА (Хр) — хронаксия; Р реобаза
зывает, что при увеличении длительности раздражения отрезок кривой начиная с определенного момента идет параллельно оси абсцисс, т. е. дальнейшее увеличение продолжительности раздражения не сопровождается изменением пороговой силы раздражения. Минимальная сила раздражения, вызывающая при практически неорга-ниченной длительности действия ответную реакцию, называется реобазой. Наименьшая длительность действия раздражения силой в одну реобазу, достаточная для возникновения ответной реакции, называется полезным временем.
Поскольку на кривой силы — длительности точка полезного времени находится в той части, которая идет почти параллельно оси абсцисс, точно определить его трудно: малейшая ошибка при определении реобазы значительно сдвигает порог времени. В связи с этим для повышения точности определения временного порога Л. Лапиком было предложено измерять не полезное время, а условную величину названную им хронаксией (от греч. chronos — время, франц, action — действие).
24
Рис. 8. Влияние крутизны нарастания силы раздражения на возникновение потенциала действия (распространяющегося возбуждения) и его величину (по Е. Б. Бабскому) .
Ось ординат — сила раздражения (ив), ось абсцисс — время (мсек.). / — раздражение прямоугольным («мгновенным») толчком тока; 2, 3, 4, 5 — линейно нарастающая сила раздражений. Стрелки — отметка критического (порогового) уровня деполяризации мембраны при данной крутизне раздражения. При уменьшении крутизны нарастания силы раздражения ниже градиента (кривая 5) возникает только местный потенциал (местное возбуждение)
Хронаксия— это наименьшее время, необходимое для возникновения возбуждения при удвоенной реобазе. Как видно из кривой силы — длительности (см. рис. 7), хронаксии соответствует отрезок ОА на абсциссе, а точка А находится на самой крутой части изгиба кривой и поэтому может быть определена более точно, чем полезное время. В практике для определения хронаксии удобнее всего применять легко дозируемый по силе и величине действия электрический ток.
Значение градиента крутизны (нарастания) раздражения. В сложных взаимоотношениях со средой организму приходится сталкиваться не только с изменяющейся силой и длительностью действия на него факторов окружающей среды (раздражений), но и с быстротой изменения силы этих раздражений (воздействий). Степень крутизны («перепада»), т. е. величина прироста силы раздражения в единицу времени, представляет собой градиент раздражения.
Установлено, что реакция (возникновение возбуждения) зависит от градиента раздражения, т. е. крутизны нарастания его во времени (рис. 8). Чем выше градиент, тем быстрее (до известных пределов) возникает реакция возбуждения. При медленном нарастании силы раздражения (даже при достижении ею чрезвычайных значений) специфическая ответная реакция не возникает. Например, мышечное волокно отвечает сокращением на быстро возросшую от нуля до порогового значения силу раздражения. Сила же, во много раз превосходящая пороговую, но нарастающая во времени очень медленно, не вызывает сокращения. Сдвиги в мышце ограничиваются развитием местного возбуждения — местного потенциала.
Разностный (дифференциальный) порог. Возможность установления оптимальных отношений организма со средой зависит не только от его способности быстро реагировать на минимальные по силе раздражения или адаптироваться к медленно нарастающим раздражениям большой силы. Очень часто на организм действует серия одновременно или последовательно поступающих раздражений, каждое из которых требует от него соответствующего ответа. Для осуществления подобных реакций живой объект, на который действуют подобные раздражения, должен «различать» их не толь-
25
ки по качественным показателям, но и по силе и длительности действия. Поэтому существует еще одна мера оценки функциональных возможностей объектов — разностный порог, или порог разности. Это та минимальная разность силы или длительности действия между двумя раздражениями, достаточная для возникновения нового уровня активности ткани (органов, систем органов и всего организма) . Чем меньше эта разность, тем точнее осуществляется переход на необходимый новый уровень активности. Определение разностных порогов имеет важное значение для успешного выполнения физических упражнений.
§ 8. Изменение возбудимости при протекании одиночного цикла возбуждения
Возбудимость, т. е. способность реагировать на поступающее раздражение в различные фазы развития одного цикла возбуждения, является переменной величиной. Если принять состояние воз-
Рис. 9. Изменение возбудимости при протекании одиночного цикла возбуждения (схема): А—электрическая реакция (потенциал действия); Б — изменение возбудимости (уровень состояния покоя принят за 100%). 1 — начальное повышение возбудимости (развитие местного потенциала); 2—фаза абсолютной рефрактерности (пик потенциала действия — деполяризация и инверсия — восходящее колено); 3—фаза относительной рефрактерности (восстановление потенциала покоя — реполяризацня быстрая и часть медленной, начальная часть отрицательного следового потенциала); 4—фаза экзальтации (продолжение восстановления потенциала покоя — отрицательный следовой потенциал); 5 — фаза субнормальной возбудимости (положительный следовой потенциал)
будимости в условиях физиологического покоя за 100%, то в ходе развития одного цикла возбуждения возбудимость изменяется в сторону как повышения, так и понижения (рис. 9). Повышение возбудимости называется экзальтацией (от лат. exal-t at io — очень возбужденное), понижение — ре-фрактерностью (от лат. refracta — невосприимчивый) .
В изменении возбудимости от момента нанесения раздражения до завершения одиночного цикла возбуждения отмечается несколько периодов (фаз). В период развития местного возбуждения (местного потенциала) наблюдается некоторое повышение возбудимости. Об этом свидетельствует то, что каждое нанесенное
в это время дополнитель-
ное раздражение, по силе даже ниже порогового, ускоряет развитие местного потенциала. Как только он достигает критической величины и переходит в высоковольтный потенциал действия — пик
26
(восходящее колено — деполяризация и инверсия), возбудимость быстро снижается и практически становится равной нулю (особенно в верхней точке пика потенциала). Время, в течение которого происходит это снижение возбудимости, называется абсолютной рефрактерной фазой (периодом), а само снижение возбудимости — абсолютной рефрактерностъю. Раздражение любой сверхпороговой силы, нанесенное в этот период, практически не может повлиять на развитие текущего возбуждения (потенциала действия).
В фазе восстановления потенциала покоя (нисходящее колено — реверсия и реполяризация) возбудимость последовательно восстанавливается. Время восстановления ее до исходного уровня называется относительной рефрактерной фазой (периодом), а состояние, в котором находится живой объект — относительной рефрактер-ностыо (невозбудимостыо). Раздражение, нанесенное в этот период, может вызвать усиление возбуждения только в том случае, если по сравнению с исходной величиной оно по силе будет надпороговым. Длительность относительной рефрактерной фазы значительно больше, чем абсолютной. Она захватывает начальную часть отрицательного следового потенциала действия.
Вслед за периодом относительной невозбудимости наступает фаза экзальтации (повышенной возбудимости). В этой фазе может возникнуть повторная волна возбуждения даже на раздражения, которые в исходном состоянии, т. е. в состоянии физиологического покоя, были по своей силе допороговыми. Фаза экзальтации совпадает по времени с периодом окончания отрицательного и началом развития положительного следового потенциала действия.
Биологический смысл фазового изменения возбудимости в ходе развития одиночной волны возбуждения заключается в следующем.
Начальная фаза повышения возбудимости обеспечивает условие, при котором каждый дополнительный раздражитель ускоряет процесс подготовки (местное возбуждение) к специфической (для данной ткани) приспособительной реакции.
Состояние абсолютной рефрактерности позволяет данной ткани «без помех» осуществлять текущую приспособительную реакцию. Если бы в этих условиях возбудимость была нормальной, то дополнительное раздражение, вызвав дополнительное возбуждение, могло бы исказить эту реакцию, превратив ее в чрезмерную или в недостаточную для данных условий. Абсолютная рефрактерность защищает ткань от чрезмерных энергетических трат в процессе осуществления текущей приспособительной реакции. Сходную роль играет и относительная рефрактерность, с той разницей, что в данном случае живое образование в состоянии реагировать на раздражения, требующие срочного ответа. Именно поэтому для большинства тканей и органов, работающих непрерывно и не имеющих длительных периодов физиологического покоя (например, сердце), характерна более длительная по сравнению со скелетной мускулатурой рефрактерность.
Состояние экзальтации создает условия готовности ткани к от
27
вету на повторное раздражение не только прежней силы, но и более слабой, являвшейся ранее подпороговой. Раздражение, поступившее в эту фазу, вызывает ответную реакцию с большей легкостью.
§ 9. Функциональная подвижность (лабильность)
Чем короче в возбудимых образованиях длительность рефрактерной фазы, тем с большей частотой они могут отвечать на повторные раздражения, и наоборот. Скорость протекания процессов возбуждения была названа Н. Е. Введенским функциональной подвижностью или лабильностью. Мерой функциональной подвижности является максимально доступное количество циклов возбуждения, воспроизводимых в единицу времени (секунду). Таким образом, зная величину лабильности, можно определить полную длительность одного цикла возбуждения.
Функциональная подвижность непостоянна и может как увеличиваться, так и уменьшаться. Особенно четко выражена эта изменчивость у малолабильных образований, например у мышцы сердца. В процессе деятельности может происходить сонастраивание, т. е. «усвоение» ритма (А.А. Ухтомский). Органы и ткани, изменяя свою лабильность, в состоянии усваивать как более высокие, так и более низкие ритмы. Усвоение ритма различными системами организма может наблюдаться в процессе врабатывания при занятиях физическими упражнениями. Для каждой ткани характерен некоторый диапазон (в пределах шкалы функциональной подвижности) рабочего ритма, в котором ее функция оказывается наиболее эффективной по производительности и энергетической экономичности и может продолжаться более длительное время.
§ 10. Оптимум и пессимум частоты
Н. Е. Введенским в опытах на нервно-мышечном препарате лягушки было обнаружено, что если наносить раздражения одной и той же силы, последовательно увеличивая в единицу времени (секунду) частоту подаваемых сигналов, то ответная реакция также последовательно нарастает. Достигнув своей максимальной величины, она, несмотря на продолжающийся рост частоты наносимых раздражений, не только не увеличивается, но постепенно уменьшается и прекращается совсем.
Частота раздражений, вызывающая максимальную по интенсивности функциональную реакцию, была названа Н. Е. Введенским оптимальной (от лат. optimum — наилучший), а частота, ведущая к ослаблению реакций,— пессимальной (от лат. pessimum — наихудший). Здесь максимальная ответная реакция характеризуется тем, что каждое следующее раздражение попадает в фазу экзальтации, т. е. наилучшей готовности ткани к ответу. При последовательном повышении частоты каждое следующее раздражение начинает попадать сперва в относительную, а затем и в абсолютную рефрактер-
28
ную фазу. Естественно, ответная реакция в этих условиях будет прогрессивно снижаться вплоть до нуля, так как в фазах рефрак-терности возбудимость, т. е. способность ткани реагировать на раздражение, также прогрессивно снижается в направлении от относительной рефрактерности к абсолютной.
Природа возникновения пессимума может быть обусловлена также угнетением интенсивности обмена веществ. Опыты показали,
что если в условиях развивающегося пессимума уменьшить частоту
раздражения до оптимальных значений, то ответная реакция тотчас
же повышается (рис. 10). Следовательно, пессимум — особая форма активной реакции ткани, направленной на защиту ее от чрезмерных по частоте раздражений, т. е. пессимум — одна из форм проявления торможения. Это, по существу, «обратная» сторона возбуждения, достигшего своего критического уровня и превратившегося в свою функциональную противоположность.
Сопоставляя два вида торможения — синаптическое (см.§ 6) и пес-симальное, можно видеть, что оба они — проявление активной деятельности тканей и органов. В первом случае торможение возникает вследствие возбуждения тормозящих си-
Рис. 10. Оптимум и пессимум частоты раздражения.
На рисунке видно, что частоты выше 100 раздражений в 1 сек. вызывают пессимальный эффект — снижение ответной реакции. Оптимальная частота раздражений (100) в условиях изменяющегося функционального состояния мышцы (утомление) во всех случаях дает больший эффект, чем пессималь-ная (150—250) (по Н. Е. Введенскому)
напсов и выделения ими тормозящих медиатров. Это торможение называется первичным. Во втором наблюдается переход возбужде-
ния в торможение без участия особых тормозящих структур. Такое торможение называется вторичным. Самым важным здесь является то, что возбуждение и торможение — это не просто состояния, сменяющие друг друга, а саморегулирующиеся процессы, обеспе-
чивающие установление оптимального уровня отношений организма со средой.
§11. Парабиоз
Своеобразие взаимосвязи состояний возбуждения и торможения и их взаимоперехода на примере оптимума и пессимума побудили Н. Е. Введенского специально заняться выяснением этого вопроса. Им было, в частности, обнаружено, что на различные воздействия нерв отвечает однотипной местной реакцией, в которой можно различить несколько последовательно протекающих стадий. Основной опыт для выявления этих стадий сводился к следующему. На нерв нервно-мышечного препарата лягушки накладывались на разных расстояниях от мышцы электроды (рис. 11, а, б). Нанесение раздражений нарастающей частоты или силы (от пороговых и выше-условно «слабый», «средний» и «большой») на месте приложения
29
Рис. 11. Схема постановки опытов Введенского по изучению парабиоза: 1 — раздражение током малой частоты; 2 — раздражение током средней частоты; 3 — раздражение током высокой частоты. *•* Столбики — величина ответной реакции нерва на различные стадии парабиоза: А —* норма; Б — уравнительная стадия;
Б — парадоксальная стадия; Г — схема нервно-мышечного препарата; а, б — места наложения электродов на нерв. Участок нерва, подвергающийся воздействию (химическому, температурному и др.), заштрихован; в — икроножная мышца (по Л. В. Латмаиизовой).
каждого из этих электродов вызывало сокращение мышцы в соответствии с законом силы раздражения (рис. 11, 4). Если же на небольшой участок нерва между электродами а и б воздействовали каким-либо химическим (эфир, хлороформ и др.) или физическим (температурным и др.) раздражителем, изменяющим функциональное состояние нерва, то на повторные раздражения на месте приложения электрода б ответная реакция не изменялась, а на раздражения на месте приложения электрода а изменялась, так как возбуждение, возникшее в точке а, должно было пройти через функционально измененный участок нерва. Вначале это проявляется в виде уравнительной стадии, когда на все три силы интенсивности раздражения возникает равная по величине ответная реакция мышцы (см. рис. 11, Б). Несколько позже эффект раздражения с точки а становится
еще более необычным: возникает парадоксальная стадия, когда слабые раздражения вызывают более сильный эффект, чем средние, а средние — больший по сравнению с сильными раздражениями (см. рис. И, В). Эта стадия прежде всего является результатом снижения лабильности в участке нерва, подвергавшегося воздействию тех или иных факторов, а следовательно, результатом снижения его пропускной способности для сильных высокочастотных раздражений. Через некоторое время, когда под влиянием химического или физического воздействия возбудимость и лабильность этого участка нерва снизятся еще более, наступает тормозящая стадия. В этом случае при раздражении нерва в точке а мышца не реагирует на раздражения любой силы. Возбуждение, возникающее в этой точке, доходит до зоны с измененной функциональной активностью нерва, через которую пройти не может, и далее не распространяется. Таким образом, состояние возбуждения последовательно может перейти в состояние торможения. Тормозящая стадия обратима. В случае же ее дальнейшего углубления возникает необратимый процесс и наступает смерть. Именно поэтому обнаруженное явление было названо парабиозом, т. е. «на грани жизни». Последовательное развитие стадий парабиоза получило название парабиотического процесса. Подобное развитие стадий ответной приспособительной реакции на любое воздействие присуще всем живым клеткам, тканям и органам.
30
§12. Механизмы регулирования функций организма
Различают два основных механизма регуляции функций — гуморальный (от лат. humor — жидкость) и нервный.
Гуморальный механизм. Он осуществляется за счет химических веществ, находящихся в циркулирующих в организме жидкостях (крови, лимфе, тканевой жидкости). Химическими регуляторами функций могут быть: вещества, поступающие извне вместе с продуктами питания, при дыхании, через кожу; неспецифические продукты обмена веществ органов и тканей — углекислота (например, накопление ее в крови возбуждает центр дыхания), мочевина и др.; некоторые специфические продукты обмена веществ клеток, в том числе медиаторы — передатчики возбуждения в синапсах (ацетилхолин, норадреналин и др.); гормоны — физиологически активные вещества, вырабатываемые железами внутренней секреции.
Гормоны (от греч. hormon — движущий) можно назвать важнейшими специализированными химическими регуляторами жизнедея-гелыюсти организма. Они оказывают влияние на течение обмена веществ, формирование органов и тканей, могут «запускать» деятельность различных органов и, наконец, корригировать интенсивность функций организма или его органов и др.
Механизмы действия гормонов могут быть различными. Существует прямой механизм, осуществляемый путем непосредственного влияния гормонов на обмен веществ органов и другие процессы через ферментные системы, а также непрямой, более сложный — через нервную систему. Во втором случае гормоны оказывают влияние, изменяя функциональное состояние нервных центров или воздействуя сперва на ее воспринимающие аппараты — рецепторы.
Отличительной чертой гуморальной регуляции является то, что химический регулятор, попадая в кровоток, поступает ко всем органам и тканям, независимо от того, участвует он в регуляции функций или нет. Только избирательное отношение тканей к этому веществу обусловливает включение гормонов в процесс регуляции. Скорость распространения химического регулятора, например гормона, от места образования до места приложения относительно небольшая — соответствует скорости кровотока.
Между различными химическими регуляторами, особенно гормонами, очень четко проявляется принцип саморегуляции (см. § 13 лой главы). Так, если гормон поджелудочной железы инсулин способствует снижению уровня сахара в крови, то гормон мозгового слоя надпочечников адреналин — его увеличению. Если продукция инсулина становится избыточной, тотчас же усиливается продукция . феналина, и наоборот. То же самое характерно и для других желез.
Нервный механизм. Этот механизм регуляции эволюционно более молодой. Он отличается от гуморального тем, что нервные им-пульсы распространяются по нервным путям с достаточно большой скоростью (в зависимости от характера нерва — от 0,5 до 80— Г. 0 м/сек) и идут по определенным нервным волокнам к строго
31
определенным органам или частям организма, которые эти нервы иннервируют.
Основным нервным механизмом регуляции функций является рефлекс — ответная реакция организма, осуществляемая через клетки нервной системы. Он реализуется по рефлекторной дуге — пути, по которому пробегает возбуждение от рецепторов до исполнительных органов (мышц, желез), осуществляющих ответную реакцию на раздражение.
В состав рефлекторной дуги входят: 1) воспринимающие раздражение рецепторы (от лат. receptor — воспринимающее образование) ; 2) афферентные нервные волокна — отростки чувствительных
Рис. 12. Схема двухиейронной (вверху) и трехнейронной (внизу) дуги спинномозгового рефлекса.
ЧН — чувствительный нейрон; ВН — вставочный нейрон, МН — мотонейрон
нейронов, несущие возбуждение к клеткам центральной нервной системы; 3) передаточные нейроны и синапсы, проводящие возбуждение к эффекторным нейронам; 4) эфферентные нервные волокна, передающие возбуждение к исполнительному органу.
Простейшая рефлекторная дуга состоит минимум из двух нейронов— афферентного (от лат. afferens — приносящий) и эфферентного (от лат. effero — выносить) (рис. 12). В большинстве случаев рефлекторные дуги многонейронные.
В механизме нервной регуляции функций различают два вида рефлексов: безусловные — врожденные, наследственно передающиеся реакции организма, и условные — приобретенные организмом в течение жизни.
Безусловные рефлексы являются видовыми, т. е. свойственными всем представителям данного вида, условные — индивидуальными (у одних представителей данного вида они могут быть, у других — нет).
32
Важный признак условных рефлексов — их сигнальный характер. Они развиваются как «предупредительная» (И. П. Павлов) деятельность, т, е. деятельность, предвосхищающая (прогнозирующая) ход последовательно развивающихся событий. Например, звук звонка не имеет для собаки пищевого значения, но многократное подкрепление этого звука пищей приводит к тому, что этот раздражитель начинает вызывать у собаки рефлексы пищевого поведения, т. е. звонок приобретает для этого рефлекса сигнальное значение, «предупреждает» животных о возможности получения пищи, к которому следует соответствующим образом подготовиться.
Нервная регуляция функций складывается из сложнейших взаимоотношений безусловных и условных рефлексов.
Единство нейрогуморальной регуляции функций. Нервная и гуморальная регуляции функций взаимосвязаны и образуют единую нейрогуморальную регуляцию. Так, передатчиком нервного возбуждения является гуморальный компонент (медиатор). В то же время деятельность многих желез внутренней секреции «запускается» нервным импульсом. Следовательно, органической частью гуморальной регуляции при действии гормонов или метаболитов (углекислота и др.) является нервный компонент. В качестве эффектора при нанесении раздражения на какой-либо рецептор могут принимать участие многие органы: мышцы, сердце, аппарат пищеварения, железы внутренней секреции и др. Так, при двигательной деятельности сокращаются мышцы, изменяет свою работу сердце, железы внутренней секреции выделяют в кровь гормоны, которые по принципу обратной связи (см. § 13) могут оказать и на мышцу, и на сердце, и на нервную систему, и на другие органы в одних случаях усиливающее, в других ослабляющее воздействие. Таким образом, рефлекторная реакция сопровождается гуморальными сдвигами, равно как и гуморальный сдвиг сопровождается изменением рефлекторной регуляции.
§13. Саморегулирование
Саморегулирование — это основное свойство живых систем. Оно направлено на оптимальное взаимодействие между элементами, составляющими организм, и процессами, обеспечивающими поддержание его целостности. Можно выделить четыре основных принципа управления и регулирования; принцип неравновесное™, градиента; принцип замкнутого контура регулирования; принцип прогнозирования; принцип целостности (см. § 1).
Принцип неравновесное™, градиента. Биологическая сущность жизни заключается в способности живого сохранять свой гомеостаз, т. е. поддерживать динамическое неравновесное, асимметричное состояние относительно окружающей среды. Следовательно, для обеспечения жизни необходимы процессы регулирования, обеспечивающие при наличии соответствующих механизмов нужный для сохранения жизни уровень неравновесное™, асимметричности. Примером может быть механизм возникновения и поддержания потен
2-113
33
циала покоя, в основе которого лежит асимметричное распределение ионов Na+ и К+ на поверхностях мембраны (см. § 5). Другим механизмом осуществления этого принципа регулирования является то, что все события совершаются во времени. Это значит, что с того момента, когда на вход системы (рис. 13) поступает какой-то сигнал, требующий ответа, проходит какое-то время до момента его возникновения на выходе. Следовательно, ответ на поступающий сигнал всегда возникает с некоторым опозданием, т. е. существует рассогласование во времени между поступлением в спе-
Рис. 13. Схема замкнутого контура регулирования (по А. Б. Когану)
циальные образования (рецепторы) раздражения и возникновением ответа.
Принцип замкнутого контура регулирования. Живому объекту недостаточно просто ответить на поступившее раздражение той или иной реакцией. Необходимо еще оценить, насколько эта реакция на выходе соответствует требованиям раздражения на входе системы и насколько она адекватна сохранению наиболее оптимальных жизненных свойств организма в данных условиях (см.' рис. 13).
Общая схема регулирования по этому принципу может быть рассмотрена на примере работы скелетных мышц:
1) на вход (например, воспринимающие приборы-рецепторы) объекта управления регулирования (например, скелетную мышцу) поступает возмущающее воздействие (химическое раздражение — молочная кислота, в избытке образовавшаяся в результате длительной работы этой мышцы и вызывающая болевое ощущение);
2) на выходе системы (мышцы) возникает реакция управляемой величины (сокращение);
3) в описываемых условиях сильное сокращение сопровождается усилением болевых ощущений и вызывает защитную реакцию. Поэтому сигнал с сокращающейся мышцы по обратной связи поступает в управляющее устройство (функции его выполняют соответствующие отделы центральной нервной системы). Здесь оценивается качество поступившего сигнала, его биологическое значение. Вследствие этого управляющее устройство оказывает на объект управления (в нашем примере скелетную мышцу) тормозящее воздействие, используя существующие способы торможения.
24
В результате управляемая величина (сокращение) на выходе уменьшается, благодаря чему снижается болевое ощущение, о чем но обратной связи информируется управляющее устройство. Саморегуляция осуществляется по принципу работы замкнутого контура с использованием обратных связей. В данном случае описана работа отрицательной обратной связи. Она называется так потому, что вызывает ослабление управляемой величины (работы органа).
Под обратной связью следует понимать поступающую через соответствующий канал на вход управляющего устройства информацию о событиях, происходящих на выходе системы. Функцию каналов обратной связи могут выполнять рецепторы, нервные проводники и нервные клетки, циркулирующие в организме жидкости и др.
Если обратная связь вызывает усиление работы того или иного органа, ее называют положительной обратной связью, если она вызывает ослабление работы, — отрицательной обратной связью.
Помимо обратных, есть и прямые связи. По ним управляющее устройство включает объекты управления, обусловливающие ту или иную форму активности.
Принцип прогнозирования. Этот принцип заключается в прогнозировании биосистемой своего поведения в будущем на основе оценки вероятности повторения прошлого опыта. Примером может служить прогнозирующая сигнальная функция условного рефлекса.
Принципиальный прогноз обеспечивает основу предупредительной регуляции — настройку на встречу с ожидаемым явлением, а встреча с ним включает механизмы корригирующей регуляции. По-пому при обучении двигательным навыкам формирование и сохранение в памяти идеальной (основной, принципиальной) модели, а также наличие некоторого набора моделей движений для обеспечения оптимального выбора их в возникающих конкретных условиях— одно из главных условий эффективной двигательной активности, в том числе в спорте.
Глава II
СЕНСОРНЫЕ (АФФЕРЕНТНЫЕ) СИСТЕМЫ
§ 1. Общая характеристика деятельности сенсорных систем
Организм животных и человека функционирует, постоянно получая информацию как об особенностях внешней среды, в которой он находится, так и о состоянии всех частей тела: внутренних органов, мышц, кожи и т. д. Физиологические аппараты, воспринимающие ну информацию, получили название органы чувств.
В прошлом веке из-за отсутствия достаточных знаний о механизмах восприятия различных раздражений из внешней и внутренней < реды внимание исследователей было обращено в основном на периферические звенья сенсорных (от лат. sensus — чувство, ощущение) аппаратов — рецепторы и особые приспособления, благодаря юиорым энергия внешнего раздражения воздействует на рецепторы
т
35
в сетчатке (светопреломляющий аппарат глаз), внутреннем ухе (звукопроводящий аппарат уха) и др. Между тем эти образования представляют собой лишь часть того сложного физиологического аппарата, который обусловливает восприятие раздражений внешней и внутренней среды и обязательно включает в себя комплекс центростремительных нейронов в различных отделах нервной системы, в том числе проекционные зоны зрения, слуха и других видов чувствительности в коре больших полушарий. Поэтому И. П. Павлов, подчеркивая необходимость учета всех звеньев тех приборов, которые воспринимают раздражения со стороны внешней и внутренней среды, предложил назвать органы чувств анализаторами. Но и это название лишь частично отражает выполняемые ими функции, так как в аппарате восприятия внешних и внутренних раздражений наряду с анализом осуществляются сложные процессы синтеза. Вследствие этого в физиологической литературе в настоящее время все большее распространение получает термин «сенсорные системы», более соответствующий сущности работы воспринимающих приборов.
Однако физиологи и представители других медико-биологических, а также психологических дисциплин вкладывают в содержание терминов «органы чувств» и «анализатор» то же содержание, что и в термин «сенсорные системы». Поэтому в литературе все эти три термина используются как синонимы и часто заменяют друг друга.
Структура и деятельность сенсорных систем весьма сложные. Возбуждение, возникшее в каком-либо рецепторе, проводится в высшие отделы центральной нервной системы несколькими путями. Во-первых, через так называемый специфический путь, который включает в себя: 1) рецептор, 2) первый чувствительный нейрон, расположенный всегда вне центральной нервной системы — в межпозвоночных спинномозговых ганглиях (от греч. ganglion — нервный узел, скопление нервных клеток), в полулунном, или Гассеровом, яремном, спиральном и других ганглиях черепномозговых нервов; 3) второй нейрон — в спинном, продолговатом или среднем мозге; 4) третий нейрон — в зрительных буграх, 5) четвертый нейрон — в проекционной зоне данного анализатора коры больших полушарий. Кроме этого, в среднем, спинном и продолговатом мозге происходит переключение на пути, ведущие в другие отделы головного мозга, в том числе и мозжечок, ретикулярную формацию и т. д. Из ретикулярной же формации возбуждение может направляться по так называемым неспецифическим путям во все отделы коры больших полушарий.
Таким образом, возбуждение, возникшее в каком-либо одном рецепторе, распространяющееся по нервному волокну первого нейрона, в дальнейшем, в разных отделах нервной системы, переключается на целый ряд нейронов и приходит в высшие отделы, в том числе в кору больших полушарий, по огромному числу различных нервных путей. Запись биотоков показывает, что при раздражении какого-либо рецептора возбуждение вначале регистрируется в проекционной зоне данного вида чувствительности (первичный ответ),
36
а спустя несколько миллисекунд может наблюдаться и в других зонах коры (вторичный ответ). Поэтому различение раздражений разных рецепторов (даже в одном и том же анализаторе) обусловлено возникновением в коре больших полушарий различных сложнейших мозаик возбужденных и заторможенных пунктов, охватывающих разные части коры больших полушарий и других отделов мозга.
Анализ раздражителей происходит во всех звеньях анализатора. Первичный анализ осуществляется уже в рецепторах, которые реагируют только на строго определенные раздражители среды: рецепторы сетчатки — на свет, рецепторы внутреннего уха — на звуковые колебания и т. д. Более сложный анализ происходит в спинном мозгу, благодаря чему на тактильные или другие раздражители у спинального животного можно получить локальные ответные реакции. Наиболее сложный анализ осуществляется в корковых концах сенсорных систем, в различных проекционных зонах коры больших полушарий.
Импульсы, поступающие через анализаторы в центральную нервную систему, поддерживают ее тонус на высоком уровне. При выключении у животных хирургическим путем нескольких сенсорных систем (зрительной, слуховой, вестибулярной, обонятельной) тонус нервной системы снижается и такие животные почти все время спят.
Как отмечалось выше, организм является саморегулирующейся системой. Кибернетика, представляющая собой науку об общих основах управления, придает огромное значение в саморегулирующихся системах восприятию, хранению и переработке информации. В живом организме, в том числе организме человека, в осуществлении информации важнейшая роль принадлежит сенсорным системам (особенно в процессах тонкой регуляции двигательных функций). Информация, получаемая различными органами и тканями гуморальным путем, имеет менынее значение.
Сигналы, поступающие в центральную нервную систему через анализаторы, используются для регуляции функций в организме, в одних случаях вызывая новые реакции, в других — корригируя течение происходящей в данный момент деятельности.
Как уже указывалось, сигнализацию, исходящую из различных органов во время их деятельности, в настоящее время часто обозначают термином, взятым из кибернетики, — «обратные связи». Благодаря обратным связям, осуществляемым через сенсорные системы, центральная нервная система непрерывно оповещается о характере выполняемой двигательной или вегетативной функции, что используется как для коррекции продолжающейся, так и для улучшения последующей деятельности. Искусственное выключение важных для данной деятельности обратных связей, например осуществляемых при раздражении рецепторов периферии или центра < етчатки глаза, резко ухудшает или даже исключает возможность выполнения многих действий, в том числе спортивных упражнений (А. Н. Крестовников).
37
Необходимая для регуляции той или иной деятельности информация, поступающая через сенсорные системы обычным путем — через органы чувств и центральную нервную систему, не всегда является достаточной. Эта недостаточность в целом ряде случаев компенсируется различного рода приборами. Например, микроскоп позволяет видеть объекты, которые в тысячи раз меньше различаемых простым глазом, тензометр — определять ничтожные величины давления, рапидная киносъемка — выявлять особенности последовательности движений в микроинтервалах времени.
В спорте дополнительная информация о параметрах движения, получаемая благодаря использованию специальных приборов (сигнализация о переходе от спортивной ходьбы к бегу, о соответствии бега заданной скорости, о продолжительности различных элементов движения при плавании, о параметрах маховых движений в гимнастике и т. д.), служит критерием, необходимым для улучшения техники спортивных движений. При этом важно, чтобы информация воспринималась спортсменом в момент самого движения или же сообщалась ему сразу после выполнения упражнения (В. С. Фарфель). Такая информация, получаемая при спортивной деятельности, получила название срочная инструментальная информация.
Классификация сенсорных систем. В зависимости от характера раздражителей можно условно разделить все сенсорные системы на несколько групп, реагирующих на следующие виды раздражений: 1) механические (тактильный, болевой, проприоцептивный, или двигательный, вестибулярный анализаторы, барорецептивный сосудистый отдел висцерального, или интероцептивного, анализатора), 2) химические (вкусовой, обонятельный анализаторы, хеморе-цептивный отдел висцерального анализатора в сосудах, в пищеварительном тракте и в других органах), 3) световые (зрительный анализатор), 4) звуковые (слуховой анализатор) и 5) температурные (температурный анализатор).
По среде, из которой воспринимаются раздражения, сенсорные системы делятся на две главные группы: 1) внешние и 2) внутренние (воспринимающие раздражения со стороны внутренней среды организма).
К внешним сенсорным системам принадлежат зрительная, слуховая, обонятельная, вкусовая и тактильная (осязательная), к внутренним — химическая (реагирующая на изменения химического состава крови и ткани), баростезическая (от греч. Ьаго — тяжесть, aisthesis — ощущение, чувство), реагирующая на изменения давления, например, в кровеносных сосудах. Температурная, болевая, вестибулярная и двигательная сенсорные системы могут возбуждаться при действии раздражителей как внешней, так и внутренней среды. Так, наряду с температурными рецепторами кожи, реагирующими преимущественно на изменение температуры внешней среды, имеются температурные рецепторы внутри тканей и органов, функционирующие в связи с изменениями температуры внутри организма; рецепторы вестибулярной и двигательной сен-
38
Рис. 14. Схема передачи возбуждения с рецептора на окончание чувствительного
сорных систем могут возбуждаться как при действии внешних сил, так и во время перемещения частей или всего тела, обусловленного сокращениями различных мышечных групп.
Пороги раздражения. Важнейшей особенностью рецепторов всех анализаторов является высокая их чувствительность при адекватном раздражении. Адекватные раздражители вызывают возбуждение в рецепторах при минимальной энергии соответствующего агента. Например, в зрительной сенсорной системе возбуждение может возникнуть при действии световой энергии равной 2,5-10~10 эрг/сек. Чтобы 1 мл воды нагреть на 1°, нужно эту энергию накапливать в течение шестидесяти тысяч лет. В звуковой системе порог раздражения может быть еще меньше — 1,6-10-11 эрг/сек. Некоторые химические вещества при действии на обонятельную сенсорную систему также распознаются при весьма малых концентрациях: например, ацетон — 0,4-10-8 г/мл, камфора — 1,6-10-11 г/мл.
Высокая возбудимость ряда рецепторов обусловлена наличием в них особых мембран, чувствительных к стимуляции именно адекватными раздражителями. При действии соответствующих стимулов повышается проницаемость поверхностной мембраны в рецепторах, что вызывает ее деполяризацию. Возникающий при этом рецепторный потенциал, достигая определенной величины, воздействует на окончания нервных волокон, деполяризует их и приводит к последующей передаче возбуждения по этим волокнам в центральную нервную систему (рис. 14). В других рецепторах выделяется медиатор, действующий на разветвления нервных волокон.
Пороги раздражения не являются постоянной величиной, так как и возбудимость рецепторов и состояние нервных клеток сенсорной системы в различных частях нервной системы могут значительно колебаться как в сторону улучшения, так и в сторону ухудшения. Это обусловлено, во-первых, тем, что высшие нервные центры.могут регулировать возбудимость афферентных нейронов на всех уровнях нервной системы. Например, при заторможенности нервной системы, наблюдаемой сразу после сна или в состоянии значительного утомления после работы, пороги раздражения увеличиваются. Наоборот, если нервные центры сенсорной системы находятся в состоянии повышенной возбудимости, пороги раздражения уменьшаются.
нерва.
Токи действия, возникающие при раздражении рецептора (/), воздействуют на окончания (2) чувствительного нервного волокна (3), вызывая в нем деполяризацию с последующим распространением волны возбуждения через перехваты Ранвье (4) в направлении к нервным центрам.
39
Во-вторых, возбудимость самих рецепторов может изменяться в еще большей степени (например, в зрительной системе — в десятки ты сяч раз).
Наряду с абсолютными порогами, характеризующимися минимальной энергией, при которой возникает возбуждение, в физиологии сенсорных систем, в том числе в физиологии спорта, часто исследуется разностный (дифференциальный) порог, т. е. разница между двумя интенсивностями раздражения, которая еще воспринимается организмом (см. главу I). Такими порогами являются минимальная разница между различными интенсивностями (давления, растяжения, яркости света, цветовых оттенков, величиной объектов, высотой звуков, углами движений в суставе, скоростью передвижений и пр.) или длительностью действия раздражителя.
Адаптация. Характерным свойством деятельности анализаторов является их адаптация (от лат. adaptatio — приспособление), т. е. свойство приспосабливаться к интенсивности действия раздражителя. При действии раздражителей значительной интенсивности возбудимость анализаторов уменьшается (т. е. пороги раздражения увеличиваются), при воздействиях малой интенсивности возбудимость увеличивается (т. е. пороги уменьшаются).
Изменение порогов раздражения в процессе адаптации наблюдается в условиях действия как сильных, так и слабых раздражителей. Так, снижение чувствительности при раздражении глаза сильным светом говорит о световой адаптации, характеризующейся увеличением порогов раздражения. Темновая адаптация, проявляющаяся в снижении порогов раздражения, наблюдается при действии слабого света. В одних случаях процесс адаптации происходит в течение десятков минут (темновая адаптация при переходе от яркого света к темноте), в других — на протяжении десятков секунд (световая адаптация при переходе от темноты к дневному свету).
В некоторых случаях в результате адаптации раздражители полностью перестают восприниматься. Например, после длительного пребывания в комнате перестают субъективно восприниматься запахи, хорошо различавшиеся при входе в нее.
Адаптация характеризуется известной степенью специализации. Так, при адаптации к действию сильного звука очень высокого топа сохраняется достаточно хорошая чувствительность к действию низких тонов.
Физиологическое значение адаптации во всех анализаторах заключается в установлении оптимального количества сигналов, поступающих в центральную нервную систему. Например, в темноте при действии слабых световых раздражений видимость предметов становится возможной только благодаря повышению чувствительности рецепторов зрительного анализатора. Наоборот, при ярком освещении чувствительность зрительных рецепторов резко снижается, что предупреждает избыточное поступление от них в центральную нервную систему информации, приводящее к ухудшению различения видимых объектов.
40
Следует отличать пороги ощущения от порогов, при которых возникают физиологические реакции, субъективно не воспринимаемые человеком. Неощущаемые пороги реакций особенно выражены в восприятии ряда вегетативных функций, связанных с регуляцией кровообращения, дыхания, выделительных процессов, пищевари
тельных и др.
Иррадиация и индукция. Возбуждение, возникающее в отдельных нервных клетках сенсорной системы, может иррадиировать (от лат. irradiare — сиять), т. е. распространяться на другие нерв
ные клетки того же анализатора.
Иррадиация свойственна всем анализаторам. Например, в зрительной системе она обнаруживается при наблюдении за величиной и формой солнца. Если смотреть на солнце через сильно закопченное стекло, оно кажется круглым пятном определенного размера с рельефно очерченными краями. При постепенном уменьшении степени
Рис. 15. Иррадиация возбуждения при восприятии белого и черного квадратов одинаковой величины: белый квадрат кажется больше, чем черный.
ние. 16. Схема одновременной индукции— отрицательной (I) и положительной (II)
законченности стекла солнце утрачивает свою правильную круглую форму, причем кажется, что размеры его сильно увеличиваются. Иррадиация четко проявляется, например, при рассматривании белого квадрата на черном фоне (рис. 15).
Одновременная индукция (боковое торможение) является процессом, противоположным иррадиации. Если иррадиация содействует распространению на соседние нервные клетки того же самого процесса (возбуждения или торможения), то одновременная индукция вызывает в них процесс с обратным знаком (рис. 16).
Сущность одновременной индукции (от лат. inductio — наведение) в функциях анализаторов заключается в том, что возбуждение нервных клеток каких-либо одних функциональных элементов анализатора одновременно вызывает торможение соседних или взаимосвязанных нервных клеток других функциональных элементов того же анализатора (см. рис. 16).
Последовательная индукция состоит в том, что после прекраще
41
ния возбуждения в нервных центрах развивается процесс торможения, а после прекращения торможения — процесс возбуждения.
Последовательную индукцию можно наблюдать, например, при деятельности зрительного анализатора. Если в течение 10—15 сек. смотреть на черный квадрат на белом фоне, затем перевести взор и фиксировать другую точку на этом же белом фоне, то спустя 1— 3 сек. (скрытый, или латентный, период) на его месте будет виден в течение некоторого времени (обычно 5—15 сек.) белый квадрат, кажущийся значительно светлее, чем фон (рис. 17). При демонст-
Рис. 17. Последовательная индукция после фиксации взора на центре черного квадрата в течение 10—15 сек. и последующего перевода точки фиксации взора на черную точку справа
рации белого квадрата на черном фоне последовательная индукция проявляется в возникновении на черном фоне еще более темного квадрата.
Таким образом, в основе явлений контраста, наблюдаемого при деятельности различных анализаторов, лежат процессы одновременной и последовательной индукции.
Следовые процессы в анализаторах. Физиологические процессы, протекающие в анализаторах, не заканчиваются с прекращением раздражения, а продолжаются еще некоторое время в виде положительных и отрицательных следовых явлений. Положительные следовые процессы имеют большое практическое значение. Например, наличие их при раздражении зрительного анализатора обеспечивает слитное восприятие раздельных кадров в кинофильмах.
По своему характеру положительные следовые процессы аналогичны процессам при действии раздражителя и являются непосредственным их продолжением. Обычно они длятся короткое время. Их сменяют отрицательные следовые явления, характеризующиеся нервными процессами, противоположными тем, которые имеют место при действии непосредственного раздражителя. Отрицательные следовые явления возникают спустя некоторое время после прекращения действия раздражителя и могут быть более длительными, чем положительные.
Взаимодействие сенсорных систем. Все анализаторы функционируют не изолированно, а в непрерывном взаимодействии друг сдру-
42
гом. Явления иррадиации, индукции, генерализации и дефференци-рования, без которых невозможна деятельность ни одной из сенсорных систем, — один из многочисленных примеров этого взаимодействия.
Раздражения, впервые воздействующие на организм, вначале воспринимаются в генерализованной форме. Например, при отсутствии специальной тренировки человек различает прикосновение к коже спины двух ножек циркуля только в том случае, если расстояние между ними равно 60—100 мм. Это обусловлено иррадиацией процесса возбуждения. После тренировки, когда уменьшаются явления иррадиации и усиливаются процессы одновременной индукции, точность дифференциации раздражителей по месту повышается и это расстояние уменьшается.
Любое различение по месту раздражения в тактильной, температурной, болевой, зрительной и других сенсорных системах происходит при условии взаимодействия отдельных функциональных элементов в этих системах. На взаимодействии различных функциональных элементов одного и того же анализатора основаны различение формы, цвета и расположения предметов в пространстве — в зрительной системе, различение направления к источнику звука — в слуховой и т. д.
При функционировании органов чувств взаимодействуют различные сенсорные системы: воздействия внешней среды на организм, как правило, воспринимаются не одним, а несколькими анализаторами, которые взаимодействуют. Например, при введении пищи в рот одновременно раздражаются вкусовая, тактильная и температурная, а иногда и обонятельная сенсорные системы; при прикосновении какого-либо предмета к коже одновременно раздражаются тактильная и температурная системы, причем раздражитель часто бывает виден, т. е. при этом участвует зрение.
Ленинская теория отражения и деятельность сенсорных систем. Изучение сенсорных систем стоит в теснейшей связи с вопросами марксистско-ленинской методологии. Органы чувств (сенсорные системы) являются тем материальным субстратом, который совершенно необходим для возникновения ощущений. В. И. Ленин указывал, что ощущения, т. е. образы внешнего мира, существуют в нас, порождаемые действием вещей на наши органы чувств, и что они представляют собой отражение вещей внешнего мира.
Ощущения, по В. И. Ленину, — это не знаки, символы или иероглифы, а именно копии отображаемых предметов. Знаки, символы, иероглифы, являясь условными обозначениями, не предполагают необходимость соответствия реальному объекту. Копии же всегда характеризуют обязательное сходство с изображаемым объектом, г. е. с объективной, существующей вне нас, реальностью. Критерий истины — практика — подтверждает это.
Однако у человека процесс познания объектов внешнего мира не ограничивается лишь просто ощущениями. Последние находятся в неразрывной связи с мышлением. Познание, по В. И. Ленину, есть отражение человеком природы. Но это не простое, не непосрёдствен-
43
ное, не цельное отражение, а процесс ряда абстракций, формирования, образования понятий, законов.
Таким образом, путь познания человеком истины, как указывает В. И. Ленин, идет от чувственного созерцания к абстрактному мышлению и от мышления к практике. Практика, непрерывно проверяющая правильность отражения в нашем сознании объективной реальности, приобретает еще большее значение в процессе преднамеренного воздействия на внешнюю природу, имеющего место лишь у человека. При этом человек использует не только личную практику, полученную в индивидуальном опыте, но и всю ту практику, которая обусловлена общественно-историческим развитием человечества. Поэтому органы чувств (сенсорные системы) человека отражают внешний мир глубже и полнее, чем органы чувств животных.
§ 2. Зрительная сенсорная система
Зрительная сенсорная система — одна из важнейших систем, воспринимающих воздействия со стороны находящихся на различных расстояниях объектов внешней среды.
Общие сведения о строении глаза. Глазное яблоко (рис. 18) расположено в полости глазницы. Оно имеет шаровидную форму. Стен-
ки его состоят из трех оболочек: наружной, сосудистой и внутренней. Наружная оболочка спереди состоит из роговицы, переходящей в склеру, или белочную оболочку. Сосудистая оболочка, выстилающая глазное яблоко, переходит в ресничное тело и радужную оболочку. Внутренней оболочкой является сетчатая, в которой расположены рецепторы зрительного анализатора — палочки и колбочки.
Светопроводящие среды глаза характеризуются прозрачностью. Они состоят из роговицы, влаги передней камеры, хрусталика и стекловидно-
Рис. 18. Схема строения глаза человека го Тела-
Между передней камерой и хрусталиком расположена радужка (радужная оболочка), окрашенная у разных людей в разные цвета. Цвет глаз обусловлен окраской радужки, просвечивающей через бесцветную, совершенно прозрачную роговицу. В середине радужки имеется отверстие — зрачок, через которое световые лучи падают на хрусталик.
44
Рис. 19. Глазные мышцы:
/ — верхняя косая; 2 — мышца, поднимающая веко; 3 — верхняя прямая; 4 — наружная прямая; 5 — нижняя прямая; 6 — зрительный нерв.
Хрусталик со всех сторон окружен связкой, прикрепляющейся к ресничному телу. Основную массу ресничного тела составляют мышечные волокна (ресничная мышца), при сокращении которых ослабляется натяжение ресничной (цинновой, цилиарной) связки.
Вблизи заднего полюса глазного яблока, несколько внутрь от него, находится место входа зрительного нерва, лишенное светочувствительных элементов, — слепое пятно. У самого заднего полюса расположен слегка углубленный участок — желтое пятно, в середине которого находится центральная ямка.
К глазному яблоку прикрепляются наружные глазные мышцы (рис. 19): верхняя прямая, нижняя прямая, внутренняя прямая, наружная прямая, верхняя косая (блоковая) и нижняя косая. Различные комбинации сокращений этих мышц обеспечивают все разнообразие движений глазного яблока.
Роговица и передняя часть белочной оболочки покрыты тонкой прозрачной слизистой оболоч
кой — конъюнктивой, переходящей на веки. Веки служат защитными приспособлениями глаза, предохраняя конъюнктиву от повреждения при соприкосновении с различными внешними объектами.
Преломляющий аппарат глаза. При переходе световых лучей из одной среды в другую происходит их преломление — рефракция. Преломляющие среды глаза — это сложная оптическая система, позволяющая получать на сетчатке глаза четкое изображение (в уменьшенном виде) предметов окружающего мира. Лучи, падающие перпендикулярно к поверхности преломляющей среды, распространяются в ней без преломления.
Если на правильную сферическую поверхность пада
ет пучок параллельных световых лучей, то лучи, падающие перпендикулярно, не преломляются, а все остальные преломляются и сходятся в одной точке. Эта точка, в которой сходятся параллельные световые лучи, называется фокусной точкой или фокусом (рис. 20).
Рис. 20. Схема преломления лучей в дальнозорком (А), нормальном (Б) и близоруком (В) глазах
45
Преломляющая сила оптических линз измеряется диоптриями (Д) и рассчитывается по формуле Д ь=—, где f (фокусное расстояние) выражено в метрах. В человеческом глазу фокусное расстояние равно примерно 1,7 см, т. е. 0,017 м. Следовательно, для получения фокусного изображения на сетчатке пои падении ( ! \ параллельных лучей требуется преломляющая сила около 58,7 диоптрий! -1.
Основными преломляющими средами глаза являются роговица и хрусталик. При восприятии объектов, находящихся на расстоянии свыше 5 м, нормальным глазом примерно 2/з преломляющей его силы принадлежит роговице и '/з — хрусталику.
Рефракция глаза. Для того чтобы при преломлении параллельных световых лучей они сходились в фокусе на сетчатке, необходимо соответствие суммарной преломляющей силы всех сред глаза его длиннику.
Нормальная (эмметропическая) рефракция характеризуется таким преломлением параллельных лучей, когда длинник глаза полностью соответствует рефракции и фокус оказывается именно на сетчатке (см. рис. 20, Б). Это обеспечивает ясное изображение на ней видимых объектов.
Близорукая (миопическая) рефракция возникает обычно в результате врожденного удлинения глаза в передне-заднем направлении. При этом из-за большей преломляющей силы, чем это требуется для данной длины глаза, фокусное изображение при преломлении параллельных лучей получается перед сетчаткой (см. рис. 20, В) и на ней вместо точечного изображения возникает круг светорассеяния, тем больший, чем сильнее близорукость.
Дальнозоркая (гиперметропическая) рефракция наблюдается, как правило, при укорочении длинника глаза. Она обусловлена недостаточностью преломляющей силы для данной длины глаза и характеризуется схождением параллельных лучей в фокусе за сетчаткой (см. рис. 20, Д). Вместо точечного изображения на сетчатке получается, как и при близорукости, круг светорассеяния.
При неодинаковом преломлении в различных меридианах одного и того же глаза, которое называется астигматизмом, круги светорассеяния получаются на сетчатке в виде эллипсов или линий.
Аккомодация и зрачковый рефлекс. Приспособление глаза к получению на сетчатке фокусного изображения видимых объектов путем изменения преломляющей силы хрусталика называется аккомодацией.
При нормальной (эмметропической) рефракции световые лучи, идущие к глазу от какой-либо точки, сходятся на сетчатке лишь в том случае, если они параллельные (см. рис. 20, Б). Все расходящиеся лучи, например от точек близко расположенных объектов, сходятся, согласно законам физики, за пределами сетчатки. Вследствие этого человек с нормальной рефракцией должен был бы все объекты, находящиеся на относительно близком расстоянии, видеть в кругах светорассеяния с фокусным изображением их за пределами сетчатки. В действительности же возможно при наличии акко-
46
модации отчетливое видение как удаленных, так и близких объектов.
Физиологический механизм аккомодации состоит в следующем (рис. 21). К хрусталику, представляющему собой эластическое те-
ло, заключенное в прозрачную капсулу, прикреплена ресничная (циннова) связка. Эта связка, опоясывающая хрусталик, растягивает капсулу, придавая хрусталику чечевицеобразную форму. Однако при уменьшении натяжения ресничной связки хрусталик в силу своей эластичности становится более выпуклым. В результате радиус его кривизны уменьшается и преломляющая сила увеличивается.
Из-за ослабления натяжения ресничной связки при сокращении ресничной мышцы уменьшается растяжение капсулы хрусталика, и он становится более выпуклым.
Практически степень сокращения аккомодационной (рес-
Рис. 21. Схема аккомодации:
I — преломление хрусталиком параллельных световых лучей без аккомодации; II — преломление расходящихся световых лучей при рассматривании близкого объекта, т. е. при аккомодации, приводящей к увеличению кривизны хрусталика и его преломляющих свойств, 1 — роговица; 2 — радужная оболочка; 3 — хрусталик (вверху — в состоянии покоя, внизу — при аккомодации); 4 — ресничная мышца; 5 — Циннова связка, растягивающая хрусталик.
Рис. 22. Уменьшение аккомодации глаза (шкала слева в диоптриях) и удаление ближайшей точки ясного видения (шкала справа в см) в связи с возрастом
ничной) мышцы, а следовательно, и увеличение кривизны хрусталика соответствуют расстоянию, на котором расположен рассматриваемый предмет. Поэтому человек может ясно видеть предметы, находящиеся как на далеком, так и на близком расстоянии.
Аккомодация уменьшается с возрастом. Это объясняется тем, что хрусталик, как и другие эластические образования, постепенно утрачивает v свою эластичность. Поэтому если в молодом возрасте изменения его кривизны могут увеличивать преломляющую силу на 10—/5 диоптрий, то после 40 лет преломляющая сила
хрусталика при аккомодации
уменьшается (рис. 22) и необходимо пользоваться очками при чтении и работе с мелкими предметами на близком расстоянии.
Невозможность различения на близком расстоянии мелких объ
47
ектов, наступающая в результате уменьшения эластичности хрусталика (обычно после 40 лет), называется пресбиопией или старческой дальнозоркостью.
Аккомодация глаза является рефлекторным процессом. Цент-тробежными нервами, участвующими в ее осуществлении, являются волокна глазодвигательного нерва, иннервирующие ресничную
ю
Рис. 23. Строение сетчатки:
I, II, /П — первый, второй и' третий нейроны; I — пигментный слой, прилежащий к сетчатке; 2 — слой палочек и колбочек; 3 — наружная пограничная перепонка; 4 — внешний зернистый слой; 5 — внешний межзернистый слой; 6 — внутревиий зернистый слой; 7 — внутренний межзернистый слой; 3 — ганглиозные клетки зрительного нерва; 9—волокна зрительного нерва; -10 — внутренняя пограничная перепонка
мышцу.
Зрачок представляет собою отверстие в радужной оболочке (радужке). В ее толще заложены гладкие мышечные воло'К-на двух родов: круговые (циркулярные) и радиальные (см. рис. 21). При сокращении циркулярных мышечных волокон, иннервируемых парасимпатическими нервными волокнами глазодвигательного нерва, зрачок суживается. При сокращении радиальных мышечных волокон, иннервируемых симпатическими нервами, происходит расширение зрачка. Величина зрачка регулируется рефлекторно. Свет, действуя на ре
цепторы зрительного анализатора, вызывает сужение зрачка, в темноте он расширяется.
Составные элементы зрительной сенсорной системы и их локализация. Рецепторы зрительной сенсорной системы — па
лочки и колбочки, соединенные с волокнами первых нейронов этой
системы, находятся в сетчатке. В ней же расположены нервные клетки — первый и второй нейроны зрительного анализатора (рис. 23).
Нервные волокна второго нейрона в составе зрительного нерва выходят из глазного яблока, направляясь к зрительным буграм промежуточного мозга, где расположен третий нейрон этого анализатора. Нервное волокно третьего нейрона оканчивается в коре больших полушарий, в затылочной доле которой находится четвертый, последний, нейрон зрительной сенсорной системы.
Палочки и колбочки распределены в сетчатке неравномерно.
4S
Основная масса колбочек сосредоточена в желтом пятне, причем в центральной его ямке имеются только колбочки. Чем дальше от центральной ямки находится участок сетчатки, тем меньше колбочек он содержит. Палочки занимают всю основную часть сетчатки, за исключением центральной ямки.
В месте входа зрительного нерва в сетчатку нет светочувствительных элементов — ни палочек, ни колбочек. Проецируемое здесь изображение какого-либо объекта не может быть воспринято. В связи с этим это место названо слепым пятном.
Адаптация к свету и темноте. С уменьшением освещенности чувствительность зрительного анализатора возрастает, что называется темновой адаптацией. С увеличением освещенности чувствительность этого анализатора снижается, что носит название световая адаптация. При переходе из условий высокой освещенности в полную темноту чувствительность зрительного анализатора увеличивается в десятки и даже сотни тысяч раз, причем нарастает в первые 2—3 сек. незначительно, затем с 3—4 сек. до 15—20 сек. быстро, далее вновь постепенно, приближаясь к максимуму только после пребывания в темноте в течение 30—40 мин. Поэтому человек, попадая, например, из ярко освещенного в слабо освещенное помещение (из фойе в кинозал, в котором демонстрируется фильм), вначале совершенно не различает окружающие предметы, а спустя несколько десятков секунд или минут они становятся хорошо различимыми.
При изменении освещенности в обратном направлении — от темноты к свету — чувствительность снижается значительно быстрее, и В условиях высокой освещенности (например, естественный дневной свет) процесс этот заканчивается в течение 1—2 мин.
Чувствительность зрительных рецепторов к действию света обусловлена наличием в . них особых светочувствительных веществ — пигментов. Так, в палочках сетчатки содержится родопсин (зрительный пурпур). При действии света он разлагается, что вызывает резкое снижение их чувствительности. В темноте происходит восстановление ( ресинтез) родопсина. В процессах ресинтеза важная роль принадлежит витамину А.
При некоторых заболеваниях и при отсутствии или недостатке в пище витамина А у человека ухудшается возбудимость палочек сетчатки. Вследствие этого у лиц, страдающих авитаминозом или гиповитаминозом А, наблюдается ухудшение способности видеть в условиях слабой (например, ночью) освещенности — куриная слепота.
Поле зрения и острота зрения. Границы видимого пространства при неподвижном состоянии глазного яблока называются полем зрения. Величина поля зрения для белого цвета в обычных условиях ограничивается стенками глазницы, затрудняющими доступ световым лучам, падающим на зрачок с разных сторон окружающего пространства. Размеры поля зрения для белого цвета в среднем следующие: с наружной стороны — 90°, с внутренней — 65°, кверху — 65°, книзу — 75°. Уменьшение поля зрения с внутренней и
49
верхней сторон объясняется ограниченным (из-за носа и верхней стенки орбиты глаза) доступом световых лучей.
Поле зрения для цветных объектов значительно меньше, чем для объектов белого цвета. Это обусловлено тем, что цветные объекты воспринимаются только колбочками, которые на периферии сетчатки отсутствуют.
Минимальный угол, при котором еще можно видеть раздельно две точки, называется остротой зрения. У большинства людей она достигает 30—40 угловых секунд. Границей нормальной остроты зрения является 1 угловая минута. В специальных таблицах, применяемых для исследования, острота зрения, условно оцениваемая цифрой 1, обозначает остроту зрения в 1 угловую минуту. Острота зрения 0,5 соответствует 2 угловым минутам, а 0,1—10 угловым минутам.
Чем больше удален какой-либо участок сетчатки от середины центральной ямки, тем хуже острота зрения (на крайних точках сетчатки — в сотни раз). Палочки, расположенные на периферии сетчатки, сигнализируют об исчезновении ранее видимых объектов или о появлении в поле зрения новых. Благодаря движению головы и глаз соответствующие объекты начинают проецироваться в центральной ямке, что обеспечивает максимальное различение. Поэтому элементы анализатора, связанные с рецепторами в центральной ямке, образно называют анализатором различения, а элементы, связанные с периферией сетчатки, — анализатором движения. Некоторые физические упражнения, например упражнения с мячом в спортивных играх, практически оказываются невыполнимыми, если выключается периферическое зрение, так как объект непрерывно исчезает из поля зрения и виден только в отдельные моменты. Резко затрудняются при выключении периферического зрения движения в слаломе и ряде других физических упражнений (А. Н. Крестовников) .
Когда световые лучи попадают в глаз не из воздушной, а непосредственно из водной среды, то роговица, коэффициент преломления которой (1,376) близок к коэффициенту преломления воды (1,333), почти совсем теряет свою преломляющую силу. Поэтому нормальный глаз видит под водой, подобно глазу с огромной гиперметропической рефракцией. Так, резко уменьшается (в 200 раз), например, острота зрения в воде при нырянии: человек хорошо видит при нырянии свои пальцы, так как угловая величина (толщина) их более чем в тысячу раз превосходит остроту зрения, и не различает складки на сгибе пальца, угловая величина которых превышает остроту зрения в воздушной среде менее чем в 200 раз.
Цветоразличение. Восприятие цветности обеспечивается колбочковым аппаратом сетчатки. При функционировании одних палочек (например, в темноте) цвета не различаются человеком.
Рецепторы глаза человека могут возбуждаться в обычных условиях при действии световых лучей с длиной волны примерно от 400 до 750 миллимикрон. Лучи с длиной волны менее 400 милли-
50
микрон (ультрафиолетовые) и с длиной волны более 800 миллимикрон (инфракрасные) не воспринимаются рецепторами зритель-
ного анализатора.
Каждый цвет характеризуется цветностью, насыщенностью и яркостью. Цветностью называется то, что отличает цветной объект (красный, синий, голубой, зеленый и др.) от белого. Насыщенность цвета определяется степенью примеси к нему белого цвета. Яркость — интенсивность степени излучения. Спектральные цвета далеко не выявляют всего богатства цветовых оттенков, наблюдаемых в природе. Изменяя насыщенность и яркость одного и того же спектрального цвета, можно получить свыше тысячи различных цветовых оттенков. Смешивая два или несколько различных цветов, можно получить новый, отличающийся от смешиваемых компонентов. Все пары цветов, дающие при смешении белый цвет (например, желтый и синий, красный и голубовато-зеленый, зеленый и пурпурный), называются дополнительными.
Относительно природы цветового зрения еще в 1757 г. М. В. Ломоносовым было высказано предположение, что различение цветов
основано на наличии в глазу трех родов приемников, неодинаково реагирующих на различный состав света. В дальнейшем трехкомпонентная теория зрения получила обоснование в исследованиях Г. Гельмгольца.
Сущность трехкомпонентной теории цветового зрения заключается в следующем. В сетчатке имеется три рода приемников (рецепторов) , из которых
Рис. 24. Схема возбуждения трех родов приемников света при раздражении различными длинами волн, характеризующих восприятие разного цвета.
Буквы под линиями осей абсцисс обозначают цвета спектра (К — красный, О — оранжевый и т. д.). А, Б, В — раздельные схемы возбуждения трех родов приемников света; Г — совместная схема для всех приемников
красновоспринимающие больше всего возбуждаются длинноволновой частью видимого спектра, зеленовосприни-мающие — средневолновой частью, фиолетововоспринимающие — коротковолновой частью спектра (рис. 24). При изолированном действии волн разной длины получаются различные степени возбуждения этих трех родов приемников. Вследствие этого раздражение каждой длиной волны воспринимается как особый цвет.
Если одновременно и в одинаковой степени возбуждаются все три рода приемников, получается ощущение белого цвета. Это имеет место при действии световых лучей, содержащих все длины волн видимого спектра (например, солнечный свет, свет от электрической лампочки, свет, отражаемый белой бумагой), а также при одновременном действии дополнительных цветов. Для дополнительных цветов характерно то, что при смешении^эни возбуждают в одинаковой степени приемники всех трех родов.
61
При отсутствии у человека одного из трех видов приемников света (чаще всего красновоспринимающего или зеленовосприни-мающего) восприятие цвета нарушается и возникает цветослепота. Такой человек не может отличить красный цвет от зеленого.
Восприятие пространства. Зрительное восприятие человеком пространства основано на ряде физиологических механизмов. Важная роль в восприятии пространства принадлежит глубинному зрению, которое может осуществляться как монокулярно (от греч. monos — один, от лат. oculus — глаз), так и бинокулярно (от греч. bi, bin — приставка, указывающий на наличие двух объектов).
Благодаря глубинному зрению человек с большой точностью может определить расположение в видимом пространстве различных предметов, неодинаково удаленных от глаза.
При монокулярном зрении глубина (удаление) находящихся в поле зрения объектов обусловлена следующими факторами: 1) объекты, находящиеся ближе, заслоняют (прикрывают) собою более удаленные предметы; 2) основания близких предметов проецируются в поле зрения ниже, чем основания дальних; 3) проекция ближе расположенных предметов одинаковой величины на сетчатке больше, чем предметов, расположенных дальше; 4) далеко расположенные предметы видны менее отчетливо («в дымке»), чем близко расположенные; 5) цвета далеко расположенных объектов теряют по мере удаления свою насыщенность, все больше приближаясь к серому цвету; 6) если наблюдатель или видимые объекты передвигаются, то угловая скорость перемещения на сетчатке изображений дальних предметов меньше, чем изображений близких предметов. Например, при наблюдении из окна движущегося вагона изображение на сетчатке расположенных вдоль полотна телеграфных столбов смещается со значительно большей быстротой, чем изображение находящегося на расстоянии 3—4 км высокого здания. Аналогичное перемещение на сетчатке изображения близких и далеких объектов происходит при движениях головы или глаз.
При рассматривании объектов, расположенных на расстоянии свыше 100—200 м, точность определения глубины находящихся в поле зрения предметов одинакова при монокулярном и бинокулярном зрении. При меньших дистанциях, особенно до 1 м, бинокулярное зрение позволяет точнее определять глубину, чем монокулярное. Убедительный пример — опыт с вдеванием нитки в ушко иглы при бинокулярном и монокулярном зрении: при видении одним глазом человек вдевает нитку с большим трудом, чем при бинокулярном зрении.
Сущность бинокулярного зрения состоит в различении степени смещения на сетчатке правого и левого глаз изображения разноудаленных предметов от изображений точки фиксации.
При утомлении зрительного анализатора может нарушиться мышечный баланс глазной мускулатуры, обусловливающий соответственно взаимосвязанное положение зрительных осей при наблюдении далеко и близко расположенных объектов. Это приводит к некоторому ухудшению бинокулярного зрения. Исследования, осу
52
ществленные А. Н. Крестовниковым и его сотрудниками, показали, что после интенсивных физических упражнений мышечный баланс может нарушиться. Особенно выражены его нарушения у представителей спортивных игр, однако значительно меньше у мастеров спорта.
С состоянием бинокулярного зрения в известной мере связан глазомер -— способность оценивать расстояния невооруженным глазом. Бинокулярное зрение улучшает глазомер только на близких расстояниях, особенно до 1—5 м. На дистанциях свыше 100—200 м бинокулярное зрение без использования специальных приборов (дальномеров) не имеет преимуществ перед монокулярным.
Определение длины линий и размеров плоскостных и объемных объектов глазомерным путем может быть весьма точным. Например, разностный порог при оценке длины линий приближается к 1/40—1/60. При оценке площади диска порог различения также может доходить до 1/50—1/60, различение же высот треугольников возможно даже при разнице в 1/60—1/154.
Восприятие скорости движения предметов в пространстве связано со скоростью перемещения изображения на сетчатке и импуль-сацией, поступающей в центральную нервную систему из мышц гла-га при его движениях.
§ 3. Слуховая сенсорная система
Слуховая сенсорная система, имеющая важнейшее значение в речевой деятельности, воспринимает звуковые колебания внешней среды. Звуковоспринимающая часть этой системы тесно связана с функциями звукопроводящего аппарата. Для ясного представления о работе слухового анализатора необходимо хорошо знать устройство этого аппарата.
Наружное, среднее и внутреннее ухо. Ухо разделяется на три части: наружное ухо, среднее и внутреннее (рис. 25).
Наружное ухо состоит из ушной раковины и наружного слухового прохода. Оно отделяется барабанной перепонкой от среднего уха, представляющего собой небольшую воздушную (барабанную) полость с тремя слуховыми косточками: молоточком, наковальней и стремячком (см. рис. 25). Среднее ухо соединяется с полостью носоглотки через Евстахиеву (слуховую) трубу. Стремячко примыкает к закрытому мембраной (перепонкой) овальному окну, через которое звуковые колебания передаются во внутреннее ухо.
Внутреннее ухо находится в улитке, расположенной в пирамидке височной кости. Улитка содержит несколько спиральных завитков (у человека 2*/4—23/4), имеющих внутренние перепончатые и наружные костные стенки. Полость костного и перепончатых каналов заполнена жидкостью. В перепончатых каналах находится жидкость, называемая эндолимфой, а в костном канале, т. е. между перепончатой и костной стенками улитки, — жидкость, называемая перилимфой. К внутренней стенке костного канала, начиная от основания улитки до ее верхушки, прикреплена тонкая костная
53
Рис. 25. Схема строения слухового аппарата. Слева — звукопроводящий аппарат уха, справа — схема строения улитки (в разрезе): 1 — рейсиерова мембрана; 2 — покровная мембрана; 3 — кортиев орган; •4—основная мембрана; 5 —волокна слухового нерва; 6 — костная спиральная пластинка
пластинка, следующая по всему ходу спирали. Эта пластинка в полости канала улитки продолжается до ее наружной стенки (см. рис. 25) в виде так называемой основной мембраны (перепонки). Вследствие этого полость перепончатого канала делится на две части — верхнюю и нижнюю лестницы (см. рис. 25). Они соединяются друг с другом у вершины улитки через отверстие (гелико-трему) диаметром около 0,03 мм. Верхняя лестница делится тонкой перепонкой (рейснеровой мембраной) на две части, образуя среднюю лестницу. Над основной мембраной в средней лестнице имеется еще покровная мембрана, расположенная над внутренней частью основной мембраны.
Костная пластинка в улитке, к которой прикреплена основная мембрана, у основания более широкая и по направлению к вершине улитки постепенно суживается. Поэтому волокна основной мембраны наиболее длинные у вершины и укорачиваются по мере приближения к основанию улитки.
На основной мембране в полости средней лестницы расположен кортиев орган, в котором имеются опорные и воспринимающие волосковые клетки. От этих клеток, являющихся рецепторами звуковых колебаний, начинаются чувствительные слуховые нервные волокна.
Механизм передачи звуковых волн. Колебания воздушной среды, возникающие при действии различных генераторов звуковых волн, передаются через ушную раковину и наружный слуховой проход барабанной перепонке. Колебания барабанной перепонки воздействуют через слуховые косточки (молоточек, наковальню и стре-мячко) на мембрану овального окна.
Слуховые косточки, действуя как рычаги, уменьшают амплитуду колебаний мембраны овального окна и вместе с тем увеличивают (до 50 раз) силу этих колебаний. При воздействии звуков большой силы специальные мышцы (мышца, напрягающая барабанную пе
54
репонку, и мышца стремени) усиливают натяжение барабанной перепонки и мембраны овального окна. Это приводит к уменьшению колебаний перепонки, что предохраняет ее от разрыва.
Колебания мембраны овального окна передаются перилимфе нижней (барабанной) лестницы, а следовательно, и основной мембране. При колебаниях основной мембраны возникает возбуждение в расположенных на ней волосковых клетках, являющихся слуховыми рецепторами. В них возникает возбуждение, если они касаются покровной мембраны. Это возбуждение передается в центральную нервную систему.
Помимо описанного механизма проведения звуковых колебаний путем воздушной проводимости существует еще и костная проводимость — звуковые волны вызывают колебания в костях черепа, которые, минуя наружное и среднее ухо, передаются непосредственно улитке.
В воздушной среде у человека преобладает воздушная проводимость, так как звуки средней и в особенности низкой высоты плохо передаются через кости. Очень высокие звуки, например 10 000 гц и выше, передаваемые путем костной проводимости, воспринимаются почти так же хорошо, как и передаваемые путем воздушной.
Под водой (ныряние, подводное плавание) хорошо воспринимаются благодаря костной проводимости не только высокие, но и относительно низкие звуки, например речь человека.
В нервной системе передача возбуждения от слуховых рецепторов осуществляется через ряд нейронов. Клетки первых нейронов слухового анализатора расположены в спиральном узле, клетки вторых — в продолговатом мозгу, третьих — в зрительных буграх и четвертых — в височной доле коры больших полушарий головного мозга.
Физиологический механизм восприятия звука. Восприятие звука основано на резонансе. По теории Г. Гельмгольца, звуковые колебания различной частоты, передаваемые в улитку через звукопроводящий аппарат, вызывают в результате резонанса колебания различных волокон основной мембраны. При этом высокие звуки приводят в состояние колебания более короткие волокна мембраны у основания, а низкие — более длинные волокна у верхушки улитки. Считалось также, что каждой частоте соответствует свой ограниченный участок основной мембраны.
В настоящее время установлено, что резонируют не волокна основной мембраны, а находящийся в улитке столб жидкости. Высокие тоны вызывают колебания жидкости только у основания улитки. При меньшей частоте возникают колебания и в средней части, а при низких тонах — во всей улитке. Эти колебания передаются волокнами основной мембраны. Резонанс при действии звуков разной высоты зависит не только от длины столба колеблющейся жидкости, но и от места, где отмечается максимальная амплитуда колебаний. Чем ниже звук, тем дальше от основания улитки находится участок жидкости с максимальной амплитудой колебаний.
Таким образом, каждой высоте звука соответствует особый ха
55
рактер колебаний жидкости в улитке. Это вызывает разную интенсивность колебаний основной мембраны и, следовательно, возбуждение неодинаковых ансамблей рецепторов.
Характеристика некоторых показателей функций звуковой сенсорной системы. Человек воспринимает звуки с частотой колебаний от 15—20 до 20000 гц. Звуки, используемые в речи, содержат частоты звуковых колебаний от 150 до 2500 гц. В старости ухудшается в первую очередь восприятие звуков высокой частоты.
Если выслушивается звук одной частоты, он воспринимается как чистый тон. К нему могут примешиваться обертоны, колебания которых в кратное число раз больше, чем колебания основного тона. По своей силе обертоны слабее основного тона. Различные сочетания основного тона с его обертонами характеризуют тембр звука. Беспорядочные, непериодические колебания вызывают ощущение шума.
Громкость звука определяется в белах (логарифмах отношения мощности данного звука к пороговой, принятой за единицу). В практике, однако, расчеты ведутся в децибелах (0,1 бела). Диапазон громкости звуков, воспринимаемых человеком, весьма велик — от 1 до 140 децибелов (дб), т. е. в пределах от 1 до 1014 раз. Например, громкость звуков, воспринимаемых человеком в обычной жизни, следующая: тихий шепот на расстоянии 1,5 м — 10 дб, тиканье часов — 20 дб, тихий разговор — 40 дб, громкий разговор — 60 дб, крик — 80 дб, шум трамвая — 70—75 дб, шум пропеллера самолета на расстоянии 5 м — НО—120 дб, гром —- 120 дб, шум при клепке в котельных — 130—140 дб. Звуки свыше 100 и в особенности 120—140 дб при длительном хроническом действии приводят к травме слухового аппарата человека и снижению слуха.
Пороги слышимости для различных звуковых частот неодинаковы, они наименьшие в диапазоне частот от 1000 до 5000 гц.
При действии звуков разной громкости слуховой анализатор адаптируется к ним: чувствительность его повышается при действии тихих и снижается при действии громких звуков. При адаптации пороги могут изменяться в сотни тысяч и даже миллионы раз.
Человек, слыша звук, способен определять направление к его источнику. Точность определения направления большая (до 1—2°) при бинауральном слухе, т. е. слушании двумя ушами, и хуже (5—10°) при моноуральном слухе, т. е. слушании одним ухом. Бинауральный слух основан на разности фаз звуковых волн и разнице в громкости и времени достижения звуковыми волнами правого уха и левого в том случае, если источник звука находится не в плоскости симметрии головы, а где-то справа или слева.
§ 4. Вестибулярная сенсорная система
Функции вестибулярной сенсорной системы связаны с влиянием на организм механических факторов: действия силы тяжести и ускорений прямолинейного и центробежного характера при перемещениях головы или всего тела.
56
Вестибулярный аппарат. Рецепторы вестибулярного анализатора находятся в различных отделах лабиринта, расположенного в пирамидке височной кости (рис. 26). Лабиринт имеет костные и перепончатые стенки. Полость между костными и перепончатыми стенками заполнена жидкостью — перилимфой. Жидкость, находящаяся в полости перепончатого лабиринта, называется эндолимфой.
Рис. 26. Схема вестибулярного аппарата:
/, 2, 3 — корешки слухового нерва (1 и 2 — вестибулярные нервы);
4— улитковый ход; 5 — передняя ампула; 6 — задняя ампула; 7 — горизонтальная ампула; 8 — общее колено; 9 — отолитовый прибор
Составными частями лабиринта являются преддверие, полукружные каналы и улитка.
Улитка, в которой расположены рецепторная часть слуховой сенсорной системы, не относится к собственно вестибулярному аппарату.
Преддверие представляет собою полость, разделенную костным гребешком на две части — переднюю, называемую мешочком (от лат. sacculus), и заднюю, носящую название маточка (от лат. ut-riculus).
На стенках преддверия имеется 5 отверстий, от которых в виде дугообразных ходов начинаются 3 полукружных канала: боковой, верхний и задний. Они расположены в трех плоскостях: боковой — н горизонтальной, верхний — во фронтальной и задний — в сигит-।альной. Около места соединения с преддверием один конец каждого канала образует расширение — ампулу.
57
В аппарате преддверия на дне маточки и на внутренней стенке мешочка имеются так называемые статические холмики, в области
Схема отолитового прибора: 2—чувствительные клетки; 3—опор-4 — вестибулярный нерв; 5 — студе-i; 6 — волоски опорных клеток; 7 —
Рис. 27.
1 — отолиты; . ные клетки; нистая масса;
перепончатая стенка; 8 — отолитовая мембрана
которых расположены рецепторы чувствительных клеток, переме-мающихся с опорными клетками (рис. 27). Важной частью воспринимающего аппарата преддверия является отолитовый прибор. Волоски опорных клеток переплетаются вместе и, склеиваясь, образуют плотную пластинку — отолитовую мембрану, на поверхности которой находятся отолитовые кристаллы углекислого и фосфорнокислого кальция.
Под действием силы тяжести при перемещениях тела или движениях головы отолитовая мембрана может свободно перемещаться в эндолимфе мешочка или маточки и оказывать различное давление на чувствительность клетки в разных частях статического холмика. Тем самым изменяется характер сигнализации от аппарата преддверия в центральную нервную систему.
В маточке отолитовая мембрана расположена в горизонтальной плоскости. В мешочке мембрана несколько отклонена кнаружи, вследствие чего может воспринимать изменения силы тяжести в сагиттальной и фронтальной плоскостях. В целом же аппа
рат преддверия воспринимает раздражения при изменении силы тяжести во всех трех плоскостях.
В полукружных каналах также имеются особые образования — ампулярные гребешки, в которых расположены чувствительные и опорные клетки. Волоски чувствительных клеток окружены студенистым веществом, образующим над поверхностью ампулы особую каемку -— ампулярную купулу, способную колебаться при изменениях давления эндолимфы полукружных каналов. При действии углового ускорения, например при вращениях, в ампулах изменяется давление эндолимфы. В начале движения или при увеличении скорости эндолимфа полукружных каналов отстает от движения костной части, вследствие чего давление в ампуле увеличивается. При уменьшении скорости вращения или прекращении движения эндолимфа в силу инерции продолжает поступательное движение, в связи с чем давление в ампуле снижается.
Поскольку полукружные каналы расположены во всех трех взаимноперпендикулярных плоскостях, при любом угловом ускорении происходит изменение давления в тех или иных ампулах, воспринимаемое соответствующими рецепторами..
Первые нейроны вестибулярного анализатора, аксоны которых соединены с рецепторами, расположены в вестибулярном ганглии.
58
Вестибулярный нерв присоединяется к слуховому, и в его составе входит в продолговатый мозг. Отсюда начинаются аксоны вторых нейронов, направляющихся в зрительные бугры и далее в височную долю больших полушарий. Вестибулярный анализатор тесно связан с мозжечком, регулирующим его функции.
Вестибулярные рефлексы. Рефлексы, вызываемые при нормальном раздражении вестибулярного анализатора, имеют важное значение как для анализа положения и перемещений головы в пространстве, так и для активации тонуса мышц и поддержания равновесия тела. Существенное влияние вестибулярные рефлексы оказывают (в частности, при значительных раздражениях вестибулярного аппарата) на вегетативные функции.
Адекватными раздражителями для аппарата преддверия являются сила земного притяжения, ускорения при прямолинейных движениях и центробежная сила.
При увеличении давления отолитовой мембраны маточки на чувствительные клетки рефлекторно повышается тонус сгибателей конечностей, туловища и шеи и снижается тонус их разгибателей. Это наблюдается, например, при быстром подъеме и спуске в виде так называемых лифтных рефлексов. В начале подъема и при окончании спуска происходит сгибание конечностей, шеи и туловища, при окончании подъема и в начале спуска — их разгибание. Лифт-ные рефлексы в спорте встречаются при различных прыжках.
С увеличением давления на рецепторы мешочка на той же стороне тела рефлекторно повышается тонус отводящих мышц конечностей и боковых мышц шеи и туловища, при уменьшении давления — тонус их понижается. Эти рефлексы играют важную роль в сохранении равновесия тела при перемещениях во фронтальной плоскости в борьбе, спортивных играх и др.
Для полукружных каналов адекватными раздражениями являются угловые ускорения и ускорение Кориолиса (добавочное ускорение, достигаемое, например, путем наклона головы вниз при вращении человека вокруг вертикальной оси).
Угловое и добавочное ускорения вызывают у человека нистагм, т. е. вынужденные ритмические движения глазных яблок (нистагм глаз) и головы (нистагм головы). Нистагм характеризуется движением, состоящим из двух компонентов: медленного, противоположного стороне вращения, и быстрого, направленного в сторону вращения. Общее направление нистагма определяют по быстрому -компоненту.
По прекращении вращения нистагм продолжается еще некоторое время, но уже в направлении против вращения. Это изменение направления нистагма объясняется тем, что после остановки движения ток эндолимфы в полукружных каналах в силу инерции приобретает противоположное действие.
Кроме нистагма во время и после вращения наблюдается изменение тонуса мышц. Вследствие этого после вращения человек не всегда может идти прямолинейно и отклоняется в ту сторону, на которой уменьшился тонус мышц.
59
Перераздражение вестибулярного аппарата. Вестибулярные рефлексы, возникающие при сильном или среднем, но длительном раздражении рецепторов преддверия и полукружных каналов, метут способствовать появлению рефлексов, ухудшающих состояние организма, вызывающих состояние укачивания, называемое также «морской» или «воздушной» болезнью.
При перераздражении вестибулярного прибора нарушается нормальное протекание целого ряда рефлексов. При этом наблюдаются бледность, потливость, головокружение, слабость, тошнота, рвота, нарушение нормального тонуса мышц, понижение работоспособности.
Роль функций вестибулярной сенсорной системы в спорте. Естественным раздражителем вестибулярной системы является важнейший компонент физических упражнений — движение. Поэтому тренировка функций этого анализатора имеет в спорте очень большое значение.
При выполнении многих движений в спорте, особенно связанных с сохранением равновесия, сигнализация со стороны вестибулярного анализатора обеспечивает информацию о различных перемещениях тела человека. Эта информация, поступая в центральную нервную систему, улучшает программирование движений и тем самым их координацию.
В результате тренировки в гимнастических упражнениях, прыжках на батуте, прыжках в воду и др. снижаются пороги вестибулярной чувствительности. Это позволяет определять даже малейшие перемещения тела, при которых происходит и смещение головы.
Устойчивость к сильному или длительному раздражению вестибулярного аппарата варьирует у различных лиц в широких пределах. Одни из них укачиваются даже при езде в трамвае или в автомобиле, другие же сохраняют нормальное состояние и при значительном раздражении этого анализатора, например при выполнении фигур высшего пилотажа в авиации, при сильной качке на кораблях.
Повышению устойчивости организма к укачиванию особенно способствуют физические упражнения, связанные с различными вращениями и быстрыми поворотами (круговые движения туловища, кувырки, обороты на перекладине, вращение на кольцах, фигурное катание на коньках, прыжки на батуте, на лыжах с трамплина и др.). Весьма эффективны предложенные А. И. Яроцким упражнения с быстрыми движениями головой: 1) повороты головы вниз-вверх, 2) наклоны головы вправо-влево, 3) повороты головы вправо-влево, 4) вращение головы справа налево и 5) то же слева направо.
Выполнение комплекса этих упражнений (по 1 мин. каждое) 2—3 раза в неделю значительно улучшает устойчивость вестибулярного аппарата у лиц, подверженных укачиванию.
Организм плохо переносит не только перераздражение, но и отсутствие раздражения вестибулярного аппарата (например, в космическом полете вследствие отсутствия силы тяжести вестибуляр
60
ный аппарат не раздражается/. Если нет соответствующей тренировки, это приводит к расстройству нормального течения физиологических функций и резкому ухудшению самочувствия.
§ 5. Двигательная сенсорная система
Роль двигательной сенсорной системы в функциях организма. Двигательная сенсорная система (называемая также проприоцептивной или суставно-мышечной чувствительностью) связана с деятельностью различных звеньев двигательного аппарата. При сохранении какого-либо положения тела и при движениях этот анализатор осуществляет обратные связи, информируя центральную нервную систему о степени сокращения мышц, о натяжении сухожилий и связок, о положении суставов.
Импульсация, поступающая через двигательный анализатор, необходима также для поддержания тонуса мышц. При перерыве чувствительных путей двигательной сенсорной системы (проприоцептивной чувствительности) в соответствующих мышцах исчезает тонус.
При движениях отсутствие обратных связей через двигательную сенсорную систему резко нарушает координацию движений. Люди, у которых в результате заболевания спинной сухоткой перерождаются нервные волокна проприоцептивной чувствительности, не способны в темноте ни стоять, ни двигаться, так как их центральная нервная система не получает информации о состоянии двигательного аппарата. На свету такая информация поступает через зрительный анализатор, и стояние и ходьба возможны, но в весьма несовершенной форме.
Высокое совершенство выполнения спортивных упражнений всегда связано с непрерывным получением из органов движения через двигательный анализатор точной информации о деятельности мышц и состоянии сухожилий, связок и суставов.
И. М. Сеченов еще в 1863 г. обратил внимание на роль мышечной чувствительности — «темного мышечного чувства» в координации движений, отметив также его значение для пространственного восприятия внешнего мира.
Рецепторы и нервные центры. Механорецепторы двигательной сенсорной системы (проприорецепторы, т. е. собственные рецепторы двигательного аппарата) расположены в мышцах, сухожилиях и суставно-связочном аппарате. Проприорецепторы разделяются на 3 основных типа. Первый тип — тельца Гольджи. Они составляют простые разветвления окончаний афферентного нерва, свободно лежащие или оплетающие сухожильные и отчасти мышечные волокна (рис. 28). Второй тип — тельца Паччини. Они расположены в фасциях, суставах и сухожилиях. Третий тип представлен более сложными, покрытыми капсулой образованиями удлиненной формы — особыми мышечными веретенами. Это интрафузальные волокна, отличающиеся от обычных — экстрафузальных волокон — менее выраженной поперечной исчерченностью.
61
Рис. 28. Механорецепторы мышцы: а — свободные окончания афферентного волокна, оплетающие конец мышечного волокна; б — мышечное веретено; 1 — капсула; 2 — внутренние мышечные волокна; 3 — афферентное волокно; 4 — его окончание; 5 — эфферентные волокна; 6 — нх окончания
Рецепторы первого и второго типов возбуждаются при сокращении мышцы, рецепторы же третьего типа (расположенные в интра-фузальных веретенах) — преимущественно при расслаблении. Таким образом, поток афферентных импульсов поступает в центральную нервную систему при любом положении мышцы, т. е. обратные связи непрерывно сигнализируют о состоянии двигательного аппарата, о всех изменениях, даже самых тонких, возникающих в нем.
Явления адаптации в волокнах двигательного аппарата выражены очень слабо. Это имеет важное биологическое значение, так как ряд мышц непрерывно работает на протяжении многих десятков минут и даже часов. Если бы двигательный анализатор быстро адаптировался, то снижение порогов чувствительности нарушило бы нормальное осуществление обратных связей и координацию движений. Однако имеется разница между мышцами, осуществляющими преимущественно фазическую или тоническую деятельность. В первых адаптация рецепторов почти отсутствует, во вторых она имеет место.
Чувствительность рецепторов в веретенах регулируется центральной нервной системой. К мышцам подходят два рода афферентных нервных волокон: альфа-волокна (70%) и гамма-волокна (30%)- Через альфа-волокна, имеющие диаметр от 9 до 13 мк, поступают импульсы, вызывающие сокращения мышцы. Гамма-волокна, диаметр которых от 3 до 6 мк, не вызывают общего мышечного сокращения. Они сокращают только интрафузальные мы
шечные веретена, угнетая возбудимость рецепторов двигательного анализатора.
Возбуждение, возникшее в рецепторах двигательной сенсорной системы, идет по волокнам первых нейронов проприоцептивной чувствительности (расположенных в спинальных ганглиях) до вторых нейронов в продолговатом мозгу. Третьи нейроны находятся в зри
62
тельных буграх. В высшем отделе центральной нервной системы — в передней центральной извилине коры больших полушарий — локализуются четвертые нейроны двигательного анализатора.
Значение двигательной сенсорной системы для функций вегетативных органов. Мышечная деятельность является одним из важнейших стимуляторов функций вегетативных органов. Во время самой мышечной работы усиливаются функции сердечно-сосудистой, дыхательной, выделительной и других систем. После ее окончания повышается потребность в пище и усиливается деятельность пищеварительных органов. Таким образом, импульсация, поступающая в центральную нервную систему через двигательную сенсорную систему, влияет на работу различных вегетативных органов. Это влияние, т. е. моторно-висцеральные реакции, осуществляется как безусловно, так и условнорефлекторным путем.
Работа внутренних органов, в свою очередь, может вызывать двигательные реакции — висцеромоторные рефлексы (М. Р. Моген-дович, В. Н. Черниговский).
Влияние спортивной тренировки на двигательную (проприоцептивную) сенсорную систему. Спортивная тренировка оказывает большое влияние на проприоцептивную чувствительность. Специальный анализ показал, что человек воспринимает различия в пространственных перемещениях звеньев тела в суставах точнее, чем различия в весе или в степени развиваемого произвольного напряжения мышц. Вследствие этого ошибки в воспроизведении спортсменами заданного усилия при статических напряжениях выражены значительно больше, чем при движении, т. е. в условиях перемещения звеньев тела в суставах. Это объясняется тем, что раздражение рецепторов суставов лучше отражается в сознании человека, чем раздражение рецепторов мышц и сухожилий (В. С. Фарфель).
В результате тренировки у спортсменов повышается чувствительность, т. е. снижаются абсолютные пороги раздражения. Это проявляется, в частности, в восприятии человеком меньших минимальных углов смещения звеньев конечностей в суставах. Улучшаются также дифференциальные пороги: снижается минимальная разность в величинах углов перемещения звеньев тела, веса удерживаемых грузов и временных интервалов между последующими раздражениями.
Важная роль в улучшении эффективности функций двигательной сенсорной системы при спортивной тренировке принадлежит инструментальной срочной информации о пространственных и временных особенностях структуры движений. Благодаря ей компенсируется недостаточная осознаваемость информации, поступающей в центральную нервную систему через этот анализатор.
§ 6. Висцеральная, болевая, температурная, тактильная, обонятельная и вкусовая сенсорные системы
Висцеральная (интероцептивная, внутренняя) сенсорная система. Полноценная работа внутренних органов может быть обеспечена центральной нервной системой только при наличии обратных
63
связей, т. е. сигнализации о состоянии и изменениях в их деятель-
ности.
Все внутренние органы богато снабжены рецепторами висцеральной (от лат. viscera — внутренние органы) сенсорной системы. Эти рецепторы многообразны и реагируют на различные раздражители: химические (хеморецепторы), осмотические (осморецепторы),
механические, связанные с изменением давления и растяжения (ба-
рорецепторы), температурные (терморецепторы) и болевые.
Возбуждение при раздражении рецепторов висцеральной сенсорной системы поступает в центральную нервную систему через различные нервы. Третий нейрон этой системы расположен в зрительных буграх. Четвертый нейрон локализуется в зависимости от характера раздражителей в разных частях коры больших полушарий.
Сдвиги в состоянии внутренних органов, связанные с изменениями химизма, температуры и осмотического и механического давлений, вызывают перестройку характера сигнализации в центральную нервную систему. В ответ на это изменяется нервная и нейрогуморальная регуляция деятельности внутренних органов.
Висцеральная сенсорная система играет важную роль при мышечной работе, вызывающей существенные сдвиги в деятельности всех внутренних органов. Сигнализация, осуществляемая этой системой, обеспечивает регуляцию обмена веществ, работы сердца, кровяного давления и т. д.
Характерной особенностью висцеральной сенсорной системы является то, что ее сигнализация, как правило, не ощущается человеком.
Болевая сенсорная система. Рецепторы болевой сенсорной системы в виде свободных окончаний (рис. 29) чувствительных нервов находятся как на поверхности (коже), так и внутри тела.
Первый нейрон этой системы расположен в межпозвоночных спинальных ганглиях, второй — в спинном мозгу, третий — в зрительных буграх. Локализация болевой чувствительности в коре больших полушарий точно неизвестна.
Рис. 29. Рецепторы кожи:
а — паччиниево тельце; б — мейснерово тельце; в — нервное сплетение вокруг волосяной луковицы; е — колба Краузе; д — свободное нервное окончание
64
Роль болевой чувствительности заключается в сигнализации о повреждающих воздействиях на организм и о необходимости их устранения. При систематических воздействиях на болевую сенсорную систему чувствительность ее снижается (например, у боксеров) .
Температурная сенсорная система. Изменения температуры воспринимаются двумя главными видами рецепторов (см. рис. 29), из которых одни реагируют на повышение температуры (клетки Руффини и Гольджи — Маццони), другие — на ее снижение (концевые колбы Краузе). Для температурных рецепторов кожи нейтральная точка, при которой человек не ощущает ни тепла, ни холода, находится в пределах от +29 до +32°.
Рецепторы температурной чувствительности расположены в коже и слизистых оболочках носа, рта, гортани и во внутренних органах.
Тактильная сенсорная система. Рецепторы тактильной сенсорной системы, реагирующие на прикосновение и давление, расположены в коже в виде телец Паччини, Мейснера и др. (см. рис. 29). Наибольшее количество этих рецепторов находится на губах, кончике языка и пальцах, наименьшее — на коже туловища. Вследствие этого прикосновение двух ножек циркуля различается на следующих расстояниях: кончик языка — 1,1 мм, ладонная поверхность пальцев — 2,2 мм, кончик носа — 6,8 мм, середина ладони — 8,9 мм, средняя линия спины — 67 мм.
Первый нейрон тактильной сенсорной системы находится в межпозвоночных спинальных ганглиях, второй — в спинном мозгу, третий— в зрительных буграх, четвертый — в задней центральной из-?вилине коры больших полушарий.
В спортивной деятельности тактильная чувствительность имеет значение при выполнении упражнений, связанных с осуществлением сложных координаций. При этом тактильные рецепторы возбуждаются при прикосновениях к снарядам и телу противника, при растягивании кожи во время движений.
Обонятельная сенсорная система. Рецепторы обонятельной сенсорной системы расположены между эпителиальными клетками оболочки верхней носовой раковины. Они обладают высокой чувствительностью и реагируют на присутствие в воздухе ничтожного количества некоторых веществ. Например, меркаптан ощущается в концентрации 0,006 мл на 1 л воздуха, уксусный этил —0,686 мл, этиловый эфир — 5,833 мл. Чувствительность обонятельного анализатора повышается при быстрых дыхательных движениях при дыхании через нос, способствующих проникновению воздуха к верхней носовой раковине. Набухание слизистой оболочки носа (например, при насморке) снижает чувствительность анализатора.
Корковый отдел обонятельной сенсорной системы локализуется в обонятельном мозгу, крючке и в области извилины морского коня.
Вкусовая сенсорная система. На кончике, боковых поверхностях и корне языка, а также на задней стенке глотки и поверхности мягкого нёба находятся рецепторы вкусовой сенсорной системы. Одни
3—113
65
из них реагируют на сладкое, другие — на горькое, третьи — на кислое, четвертые — на соленое.
Рецепторы вкусового анализатора связаны с чувствительными волокнами язычного, языкоглоточного и блуждающего нервов. Корковый центр вкусовой сенсорной системы расположен в области извилины морского коня.
Вкусовые ощущения, получаемые от пищи, обусловлены одновременным раздражением рецепторов не только вкусовой, но и других сенсорных систем: тактильной, температурной и обонятельной.
§ 7. Роль различных сенсорных систем при занятиях физическими упражнениями
Как и при любой деятельности, значение функций большинства сенсорных систем в процессе занятий физическими упражнениями весьма велико. Выполнение многих упражнений невозможно при значительном расстройстве двигательной и висцеральной сенсорных систем и резко затруднено при выключении функций зрительного, слухового, вестибулярного и тактильного анализаторов.
При выполнении физических упражнений всегда одновременно функционирует несколько сенсорных систем. В результате между ними по механизму условного рефлекса складываются определенные взаимоотношения, имеющие специфический характер при каждой новой комбинации комплекса раздражителей. Субъективно это воспринимается в виде особых ощущений, имеющих специализированный характер. Так, у спортсменов формируются ощущения, образно называемые «чувством снега» (у лыжников), «чувством воды» (у пловцов), «чувством льда» (у конькобежцев) и т. д. (А. Н. Крестовников). Эти ощущения позволяют спортсменам лучше координировать двигательную деятельность с учетом условий внешней среды.
В результате систематической физической тренировки функции многих анализаторов улучшаются. Это выражается, например, в снижении порогов проприоцептивной чувствительности. Так, у тяжелоатлетов и боксеров наблюдается высокая чувствительность двигательного анализатора при движениях в локтевом и плечевом суставах, у лыжников, прыгунов и слаломистов — при движениях в голеностопных суставах. Совершенствование функций зрительного аппарата (увеличение поля зрения, улучшение баланса глазных мышц и глубинного зрения) отмечается у представителей спортивных игр. Функции вестибулярной сенсорной системы, в частности связанные с устойчивостью к укачиванию, улучшаются в результате тренировки в гимнастических упражнениях, плавании и др.
В некоторых случаях происходит и снижение чувствительности. Например, у боксеров понижается болевая и тактильная чувствительность в тех областях тела, которые часто подвергаются ударам.
Данные о состоянии анализаторов используются для характеристики уровня тренированности спортсменов, определения степени утомления и т. д.
66
Глава III
ФИЗИОЛОГИЯ НЕРВНО-МЫШЕЧНОГО АППАРАТА
§ 1. Строение нервно-мышечного аппарата
Сокращение скелетных мышц возникает в ответ на нервные
импульсы, идущие от специальных нервных клеток — мотонейронов. Мышцы и иннервирующие их мотонейроны составляют нервно-мы-
шечный аппарат человека.
Связь мотонейронов с мышцами осуществляется через ак-<оны. Это длинные отростки, которые отходят от тел мотонейронов и в составе периферических нервов достигают мышцы. Внутри нее каждый аксон многократно ветвится, образуя концевые веточки. Каждая веточка оканчивается на одном мышечном волокне, образуя нервно-мышечный синапс, или концевую пластинку. Следовательно, один мотонейрон иннервирует столько мышечных волокон, сколько концевых веточек имеет его аксон. Мотонейрон, его аксон и мышечные волокна, иннервируемые этим аксоном, составляют двигательную единицу (рис. 30, Д). Она представляет собой основной функционально-структурный элемент нервно-мышечного аппарата. Таким образом, нервно-мышечный аппарат можно рассматривать как совокупность двигательных единиц.
По своему строению, как и ио функциональным особенностям, двигательные единицы неодинаковы. Они отличаются друг от друга размерами тела мотонейрона, толщиной его аксона и числом мышечных волокон, входящих в состав двигательной единицы.
Так, малая двигательная единица включает относитель-
Рис. 30. Двигательная единица (А) и нервно-мышечный синапс (Ь):
А — двигательная единица: / — тело мотонейрона; 2 — ядро; 3 — дендриты; 4 — аксон; 5 — миелиновые оболочки аксона; б — концевые веточки аксона; 7 — нервно-мышечные синапсы (показанные крупно на Б).
Б — нервно-мышечный синапс: I — концевая веточка аксона, II — мышечное волокно. 1 — пресинаптнческая мембрана, покрывающая концевую веточку в зоне нервно-мышечного синапса; 2 — пузырьки, содержащие ацетилхолин; 3 — митохондрии; 4 — синаптическая щель; 5 — постсннаптнческая мембрана, покрывающая мышечное волокно в зоне нервно-мышечного синапса; 6 — ядро мышечной клетки; 7 — миофибрилла
3'
67
Рис. 31. Схематическое изображение мышцы. Мышца (а) состоит из мышечных волокон (б), каждое из них — из миофибрнлл (в). Миофиб-рнлла (г) составлена из толстых и тонких мно-филаментов (д).
На рисунке показан один саркомер, ограниченный с двух сторон Z-линиями. / — изотропный диск, А — анизотропный диск, Н — участок с меньшей анизотропностью. Поперечный срез миофибриллы (е) дает представление о гексагональном распределении толстых и тонких миофиламентов.
но маленький мотонейрон стойким аксоном, который имеет небольшое число концевых веточек и соответственно иннервирует небольшое число мышечных волокон (самая малая— до нескольких десятков). Малые двигательные единицы входят в состав всех мелких мышц лицевой мускулатуры, пальцев рук и ног, кистей и частично в состав больших мышц туловища и конечностей. Большая двигательная единица включает крупный мотонейрон с относительно толстым аксоном, который образует большое число концевых веточек в мышце и соответственно иннервирует большое число (до нескольких тысяч) мышечных волокон. Таким образом, чем крупнее тело мотонейрона, тем толще его аксон и тем больше мышечных волокон иннервируется этим мотонейроном. Большие двигательные единицы входят преимущественно в состав больших мышц туловища и конечностей. Следовательно, каждая мышца составлена из самых разных по размеру двигательных единиц — от малых до больших. Вместе с тем самая малая двигательная единица той или иной мышцы может быть значительно больше, чем самая большая двигательная единица другой мышцы (например, малая двигательная единица мышцы туловища превосходит по размеру большую двигательную единицу мышцы, осуществляющей движение глаза).
Скелетная мышца (рис. 31) состоит из группы мышечных пучков. Каждый из них состоит из тысяч мышечных волокон.
68
У человека количество этих волокон в мышце устанавливается через 4—5 месяцев после рождения и затем практически не изменяется. При рождении толщина (диаметр) их составляет примерно '/ь толщины волокон у взрослых людей. Диаметр мышечных волокон может значительно изменяться под воздействием тренировки (см. ниже).
Мышечное волокно представляет собой клетку цилиндрической формы. В мышце с параллельным ходом волокон они обычно крепятся к обоим сухожилиям, но в очень длинных мышцах (например, портняжной) большое число волокон короче всей мышцы. Такие
мышечные волокна крепятся одним концом к сухожилию, а другим — к соединительнотканным перемычкам внутри мышцы. Мышечное волокно покрыто тонкой эластичной мембраной—сарколеммой. Ее структура подобна структуре мембран других клеток, в частности нервных. Мембрана мышечных клеток играет важную роль в возникновении и проведении возбуждения.
Внутреннее содержимое мышеч
Рис. 32. Схема основных элементов саркоплазматического ретикулума (продольный разрез мышечного волокна):
1 — поверхностная мембрана мышечного волокна; 2 — поперечная трубочка; 3 — продольная трубочка саркоплазматического ретикулума;
4 — концевое утолщение продольной трубочки (цистерна), 5 — саркоплазма; 6 — миофиламент. Остальные обозначения пояснены на рнс. 31.
ного волокна называется саркоплазмой. Она состоит из двух частей. Первая — саркоплазматический матрикс — представляет собой жидкость, в которую погружены сократительные элементы мышечного волокна — миофибриллы (см. рис. 31). В этой жидкости находятся растворимые белки (например, миоглобин), гранулы гликогена, капельки жира, фосфатсодержащие вещества
и другие малые молекулы и ионы. Вторая часть саркоплазмы — саркоплазматический ретикулум (рис. 32). Так обозначается систе-
ма сложно связанных между собой элементов в виде вытянутых мешочков и продольных трубочек, расположенных между миофибриллами параллельно им. Мышечное волокно внутри пересекают поперечные трубочки. Выстилающие их мембраны по своей структуре сходны с сарколеммой. Поперечные трубочки соединяются с
поверхностной мембраной мышечного волокна, связывая его внутренние части с межклеточным пространством. Продольные трубочки примыкают к поперечным, образуя так называемые цистерны в зоне контактов. Эти цистерны отделены от поперечных трубочек очень узкой щелью. На продольном разрезе волокна видна харак-
терная структура — триада, включающая поперечную трубочку с прилегающими к ней с двух сторон цистернами. Ретикулярные триады фиксированы так, что их центр находится вблизи границы Till /-дисков (см. ниже). Саркоплазматический ретикулум играет
69
важную роль в передаче возбуждения от поверхностной мембраны волокна вглубь к миофибриллам и в акте сокращения. Через саркоплазматический ретикулум и поперечные трубочки может также происходить выделение продуктов обмена (в частности, молочной кислоты) из мышечной клетки в межклеточное пространство и далее в кровь.
В мышечном волокне содержится до 1000 и более миофибрилл (см. рис. 31, в). Каждая из них имеет диаметр 1—3 мк и состоит из параллельно лежащих нитей двух типов — толстых и тонких миофиламентов. Толстые нити состоят из миозина, а тонкие из актина, представляющих два основных типа сократительных белков. На всем протяжении миофибриллы расположение толстых миозиновых и тонких актиновых миофиламентов строго упорядочено — группа параллельных толстых миофиламентов образует так называемый темный A-диск, который чередуется со светлым /-диском, образованным группой тонких актиновых миофиламентов. Светлые /-диски делятся пополам поперечной Z-линией, с обеих сторон которой крепятся тонкие актиновые миофиламенты. Участок между двумя соседними Z-линиями называется саркомером. Он представляет собой основную повторяющуюся структуру мышцы. В центральной области А-диска — Н-зоне тонкие и толстые миофиламенты не перекрывают друг друга. Если поперечное сечение проходит через Н-зону, то на срезе видны только толстые миофиламенты, которые располагаются в правильной гексагональной (шестиугольной) форме. На поперечном срезе A-диска вне центральной области каждый толстый миофиламент окружен шестью тонкими нитями, а каждый тонкий миофиламент — тремя толстыми. На поперечном срезе /-диска видны только тонкие миофиламенты.
Толстый миофиламент образован удлиненными молекулами миозина, длинные «хвосты» которых уложены в нить, а «головы» образуют поперечные отростки, подходящие к тонким миофиламентам. «Голова» миозиновой молекулы состоит из тяжелого меромиозина, который обладает ферментативной АТФ-азной активностью (катализирует гидролиз АТФ с образованием АДФ и фосфора). Средняя часть толстого миофиламента не имеет поперечных отростков. В двух половинах толстого миофиламента «головы» молекул миозина имеют противоположное друг другу направление (см. рис. 31, д). Тонкий миофиламент представляет собой две скрученные, обвитые одна вокруг другой в виде двойной спирали актиновые нити. Тонкие миофиламенты по одну сторону Z-линии ориентированы в одном направлении, по другую сторону — в противоположном. Благодаря такому расположению актиновых и миозиновых молекул в двух половинах A-диска актиновые филаменты движутся с двух сторон саркомера в направлении друг к другу, т. е к середине саркомера.
Нервно-мышечный синапс (см. рис. 30, Б), с помощью которого мотонейрон связан с мышечным волокном, имеет две основные части — нервную (пресинаптическую) и мышечную (постсинаптическую). Пресинаптическая часть представлена концевой веточкой
70
аксона, погруженной в углубление на поверхности мышечного волокна. Поверхностная мембрана концевой веточки носит название пресинаптическая мембрана. Нервное окончание содержит более миллиона пузырьков ацетилхолина (АХ)—медиатора нервно-мышечного синапса. Мембрана, покрывающая мышечное волокно в области нервно-мышечного синапса, носит название постсинаптическая мембрана (или концевая пластинка). Она образует многочисленные складки, уходящие в глубь волокна, благодаря чему увеличивается ее поверхность. Постсинаптическая мембрана имеет так называемые холинорецепторные участки и содержит фермент аце-тилхолинэстеразу (АХЭ). Пре- и постсинаптические мембраны разделены узкой синаптической щелью, открывающейся во внеклеточное пространство.
§ 2. Нервно-мышечная передача
Проведение возбуждения с нервного волокна на мышечное можно представить схематически следующим образом:
1) при достижении нервным импульсом концевой веточки аксона из нее выделяется некоторое количество АХ;
2) выделившийся АХ диффундирует через малое пространство между нервным окончанием и концевой пластинкой (синаптическую щель) и вступает в реакцию с имеющимися на постсинаптической мембране мышечного волокна холинорецепторами;
3) при образовании комплекса «АХ-рецептор» увеличивается ионная проницаемость мембраны концевой пластинки, что приводит к возникновению локального электрического потенциала — потенциала концевой пластинки (ПКП);
4) ПКП. достигнув пороговой величины, электрически возбуждает соседнюю (внесинаптическую) мембрану мышечного волокна. Поэтому вначале потенциал действия мышечного волокна возникает вблизи синапса, а затем распространяется по всем направлениям от концевой пластинки, охватывая возбуждением все мышечное волокно;
5) комплекс «АХ-рецептор» быстро разрушается специальным ферментом — АХЭ, которая, как уже отмечалось, в большой концентрации содержится в области мембраны концевой пластинки. Разрушение комплекса сопровождается восстановлением исходной проницаемости постсинаптической мембраны и затуханием ПКП.
Даже в том случае, когда к нервно-мышечному синапсу не поступают нервные импульсы, он находится в состоянии непрерывной активности. Это проявляется в том, что на постсинаптической мембране постоянно образуются небольшие локальные электрические потенциалы — миниатюрные ПКП. Эти потенциалы возникают в результате действия небольших порций (или квантов) АХ, которые постоянно выделяются из нервного окончания. Согласно расчетам, квант содержит постоянное число молекул АХ (около 10 000). Выделение кванта АХ в синаптическую щель происходит благодаря опорожнению одного синаптического пузырька в концевой веточке
71
аксона. Пузырьки, содержащие АХ, находятся внутри концевой веточки в состоянии броуновского движения. Тот или иной пузырек, оказавшись вблизи мембраны нервного окончания, подпадает под действие сил, притягивающих его и разрушающих оболочку, и содержимое пузырька переходит в синаптическую щель. В покое частота освобождения квантов АХ варьирует в широких пределах, но относительно невелика.
С приходом нервного импульса происходит кратковременная деполяризация мембраны нервного окончания. Это сопровождается резким увеличением частоты выделения квантов АХ, что вызывает большой суммированный ПКП. В механизме выделения АХ важная роль принадлежит ионам кальция. При деполяризации пресинапти-ческой мембраны ее проницаемость для этих ионов увеличивается и они входят внутрь нервного окончания. Здесь кальций способствует освобождению АХ, после чего этот медиатор выделяется в синаптическую щель.
Для проведения возбуждения через синапс необходимо около 0,5 мсек. Это время называется синаптической задержкой. Оно складывается из: а) времени, необходимого для освобождения АХ из нервного окончания, б) времени диффузии АХ от пресинапти-ческой мембраны к постсинаптической и в) времени, требующегося для взаимодействия АХ с рецепторами концевой пластинки.
Проведение одиночного импульса сопровождается расходом лишь незначительной части АХ, содержащегося в концевой веточке аксона. Запасов АХ в нервном окончании достаточно для проведения около 10 тыс. импульсов. Кроме того, запасы АХ непрерывно восполняются путем синтеза этого вещества из глюкозы и холина, происходящего под влиянием холинацетилазы. Синтез АХ осуществляется в самом нервном окончании. Количество образующегося АХ возрастает с учащением импульсации мотонейрона. Как уже указывалось, в синаптической щели АХ подвергается гидролизу под действием АХЭ. В результате образуются уксусная кислота и холин. Последний переходит затем обратно в нервное окончание, где используется для синтеза АХ.
После выделения АХ в синаптическую щель он присоединяется к холинорецептору, в связи с чем увеличивается проницаемость постсинаптической мембраны мышечного волокна для ионов натрия и калия. Это приводит к возникновению ПКП. Достигнув критического, порогового, уровня (около 30 мв), ПКП вызывает генерацию распространяющегося потенциала действия мышечного волокна. Механизм генерации этого потенциала аналогичен тому, который наблюдается при проведении возбуждения по нервному волокну. Потенциал действия возникает в той зоне мышечного волокна, которая граничит с постсинаптической мембраной. По мере развития ПКП возникает разность потенциалов между деполяризованной постсинаптической мембраной и недеполяризованной мембраной, окружающей синапс, и появляются местные электрические токи. Эти токи входят в волокно через постсинаптическую мембрану и выходят через мембрану прилежащих внесинаптических участков.
72
Когда токи достигают пороговой величины, они возбуждают мембрану этих участков. Возникает мышечный потенциал действия, который затем последовательно распространяется по поверхности мышечного волокна. Скорость проведения по мембране мышечного волокна ниже, чем в нервных волокнах, — около 5 м/сек.
Потенциал действия распространяется не только по поверхностной мембране мышечного волокна, но и по мембранам, выстилающим поперечные трубочки саркоплазматического ретикулума. Такой процесс является составной частью механизма, с помощью которого осуществляется связь между возбуждением и сокращением мышечного волокна. Этот механизм носит разные названия: связь «возбуждение — сокращение», электромеханическая связь (ЭМС) или мембранно-фибриллярная связь. Распространение волны деполяризации по поперечным трубочкам вызывает деполяризацию мембран цистерн продольных трубочек саркоплазматического ретикулума. Эта деполяризация сопровождается выходом ионов кальция в межфибриллярное пространство, что запускает процесс сокращения мышечного волокна.
Механизм стимулирующего действия ионов кальция на сократительный аппарат мышцы еще неизвестен. Есть основания предполагать, что эти ионы необходимы для возникновения механического взаимодействия между актиновыми и миозиновыми нитями (см. ниже). Кроме того, ионы кальция стимулируют АТФ-азную активность миозина, способствуя тем самым освобождению энергии, необходимой для сокращения. Повышенная концентрация ионов кальция в межфибриллярном пространстве сохраняется всего несколько миллисекунд, а затем они «перекачиваются» обратно в цистерны с помощью специального кальциевого насоса. Для этой работы, которая совершается против диффузионного градиента, нужна затрата определенной энергии. Ее источником является расщепление АТФ. Подсчитано, что на возврат двух ионов кальция из межфибриллярного пространства в полость цистерн расходуется одна молекула АТФ. Снижение концентрации ионов кальция в межфибриллярном пространстве приводит к расслаблению мышечного волокна. Таким образом, в саркоплазматическом ретикулуме ионы кальция играют роль своеобразного медиатора в процессе преобразования возбуждения в сокращение.
§ 3. Механизм и энергетика мышечного сокращения
Изменение механического состояния миофибриллярного сократительного аппарата мышечных волокон в результате действия нервных импульсов называется сокращением. Внешне сокращение проявляется в изменении или напряжения, или длины мышцы, или того и другого. При этом потенциальная химическая энергия превращается в механическую и может совершаться механическая работа. Период химико-механических изменений в сократительном аппарате, в течение которого возникает механическая тяга, определяющая акт сокращения, носит название период активного состояния.
73
По теории скольжения, в основе сокращения лежит механическое взаимодействие между миозиновыми и актиновыми миофиламентами благодаря образованию между ними в период активности поперечных мостиков. При этом косо расположенные поперечные мостики осуществляют тягу, благодаря которой происходит «втягивание» тонких актиновых миофиламентов в промежутки между толстыми миозиновыми миофиламентами. В образовании поперечных мостиков между толстыми и тонкими миофиламентами участвует АТФ. Согласно одной из моделей сокращения, при расслабленном состоянии мышцы АТФ прикреплена только с одной стороны мостика — на «голове» миозиновой молекулы. Здесь АТФ находится вблизи АТФ-азы, что предотвращает соединение актина с миозином. В результате толстые и тонкие миофиламенты могут свободно скользить друг относительно друга при пассивном растяжении или укорочении мышцы. С выделением ионов кальция из цистерн саркоплазматического ретикулума они присоединяются с одной стороны к АТФ на «голове» миозина, а с другой стороны — к специальным местам на тонком актиновом миофиламенте, образуя поперечные мостики между актиновыми и миозиновыми миофиламентами. Продвижение тонких миофиламентов относительно толстых сопровождается приближением поперечных мостиков к зоне расположения АТФ-азы, что вызывает гидролиз АТФ и соответственно разрушение мостиков. Далее АТФ восстанавливается, и в следующем цикле формируются новые поперечные мостики, вследствие чего образуется связь толстых миофиламентов со следующими участками тонких миофиламентов. Этот процесс происходит повторно и во многих участках миофиламентов. В результате сокращение носит непрерывный и плавный характер.
Если укорочения мышцы не происходит (см. ниже), усилие может создаваться за счет повторного взаимодействия толстых и тонких миофиламентов на одних и тех же фиксированных участках. В этом случае никакой внешней работы не совершается, а энергия расходуется на поддержание напряжения мышцы. Напряжение, развиваемое сократительными элементами в миофибриллах, передается сарколемме и через нее — сухожилию мышцы.
Непосредственным источником энергии для мышечного сокращения является расщепление высокоэнергетического вещества АТФ. В мышце происходит также промежуточная реакция, вовлекающая второе высокоэнергетическое вещество — креатинфосфат (КФ). Оно не может действовать как непосредственный источник энергии, поскольку его расщепление не оказывает влияния на сократительные белки мышцы. КФ обеспечивает энергией ресинтез АТФ. В свою очередь, энергия для ресинтеза КФ обеспечивается окислением. Суммируем эти реакции:
1) АТФ -> АДФ + Р + энергия для сокращения
2) КФ + АДФ -> К + АТФ
3) К + Р + энергия от окисления -> КФ
74
Таким образом, освобождение энергии, которое обеспечивает сокращение мышцы, не требует наличия кислорода (т. е. является анаэробной реакцией). Это отчасти объясняет способность мышцы сокращаться в условиям неадекватного снабжения кислородом, как это имеет место в начальной стадии работы и во время тяжелой работы. Наличное содержание АТФ в мышце достаточно для интенсивной работу в течение лишь нескольких десятых долей секунды. Поэтому мышца должна обладать анаэробным механизмом для быстрой регенерации АТФ, что и обеспечивается расщеплением КФ-Однако содержание КФ также ограничено, и его полное расщепление может ресинтезировать АТФ в количестве, достаточном лишь для интенсивной активности в течение всего нескольких секунд. Значит, для восстановления КФ и, следовательно, восстановления АТФ во время напряженной мышечной работы необходимо участие других анаэробных механизмов. Таким механизмом является расщепление гликогена до молочной кислоты. Этот процесс может происходить в анаэробных условиях до тех пор, пока мышечная активность не заканчивается из-за истощения мышечного гликогена или повышения кислотности (в результате накопления молочной кислоты). Это очень расточительный процесс, так как превращение гликогена в молочную кислоту, позволяя мышце сокращаться, несмотря на недостаточное снабжение кислородом, освобождает значительно меньше энергии, чем ее полное окисление до СОг и Н2О.
§ 4. Формы мышечного сокращения
В результате сокращения в мышечных волокнах возникает напряжение. Эта способность развивать напряжение является основным свойством мышцы. Даже при спокойной ходьбе трехглавая мышца голени может развивать напряжение, которое почти в 4 раза больше веса идущего человека, а при беге икроножная мышца может развить напряжение, в 6 раз превышающее вес тела. Если бы все мышцы, содержащие примерно около 300 млн. волокон, возбуждались одновременно и максимально и осуществляли тягу в одном направлении, они смогли бы развить силу, по крайней мере, в 25 тонн.
Напряжение, развиваемое мышцами при сокращении, реализуется по-разному. Если внешняя нагрузка меньше, чем напряжение сокращающейся мышцы, то мышца укорачивается и вызывает движение. Это концентрический тип сокращения (рис. 33, внизу). Поскольку в экспериментальных условиях при электрическом раздражении укорочение мышцы происходит при постоянном напряжении, равном внешней нагрузке (см. рис. 33, Б). Этот тип сокращения называют также изотоническим (равное напряжение). Другое название концентрического сокращения — миометрическое. Если внешняя нагрузка больше, чем напряжение, развиваемое мышцей во время сокращения, то такая мышца растягивается (удлиняется) при сокращении. Это эксцентрический, или плиометрический, тип сокращения.
75
Концентрический и эксцентрический типы сокращения, т. е. сокращения, при которых мышца изменяет свою длину, относятся к динамической форме сокращения.
Сокращение мышцы, при котором она развивает напряжение, но не изменяет своей длины, называется изометрическим (равная дли-
А
Рис. 33. Схемы, поясняющие механику изометрического и изотонического сокращений.
Вверху: А — механическая модель мышечного волокна, включающего сократительный компонент (СК) и пассивные упругие компоненты — последовательный упругий компонент (ПОУК) н параллельный упругий компонент (ПУК); Ь — схематическое изображение модели изометрического сокращения, (l •— общая длина мышцы, F' — напряжение покоя, F" — напряжение при сокращении);/? — график изменения напряжения во время изометрического сокращения.
Внизу: А — схематическое изображение модели изотонического сокращения, Б — график изменения напряжения, В — длины мышцы при изотоническом сокращении. Видно, что в начале изотонического сокращения увеличивается «внутреннее напряжение»; когда оно достигает максимального значения, начинается укорочение мышцы. Во время укорочения напряжение (/•') остается неизменным и равным внешней нагрузке (Р)
на). Это статическая форма сокращения (см. рис. 33, наверху). Она возникает в двух случаях: либо когда внешняя нагрузка равна напряжению, развиваемому мышцей яри сокращении, либо когда внешняя нагрузка превышает напряжение мышцы, но отсутствуют
76
условия для растяжения мышцы под влиянием этой внешней нагрузки.
При динамических формах сокращения производится внешняя работа — при концентрическом сокращении положительная, при эксцентрическом отрицательная. Величина работы в обоих случаях может быть определена как произведение внешней нагрузки (поднятого веса) на пройденное расстояние. При изометрическом сокращении «расстояние» равно нулю, и, согласно физическому закону, в этом случае мышца не производит никакой работы. Однако с физиологической точки зрения ясно, что изометрическое сокращение требует энергии и может быть очень утомительным. В этом случае «работа» может быть определена как произведение напряжения мышцы на время ее сокращения (т. е. эквивалентно импульсу силы в физике).
Активация мышечных волокон (их сокращение) всегда связана с увеличенной продукцией тепла. Часть этой теплопродукции предшествует подъему напряжения во время изометрического сокращения или укорочению мышцы при ее изотоническом сокращении. Это так называемое тепло активации. Во время изотонического сокращения тепло также продуцируется, причем пропорционально степени укорочения, которому подвергается мышца (тепло укорочения). Некоторое количество тепла освобождается при расслаблении мышцы. Начальная продукция тепла, связанная с сокращением мышцы, не зависит от снабжения ее кислородом. После прекращения механических явлений в условиях адекватного снабжения мышцы кислородом имеет место теплопродукция, примерно равная начальному теплообразованию (тепло восстановления, или задержанное тепло). Во время изометрического сокращения в тепло превращается вся выделяемая мышцей энергия, а при динамическом сокращении — не менее 75%• В табл. 1 приведены основные характеристики различных форм и типов сокращений.
Таблица 1
Формы и типы мышечного сокращения
Форма сокращения Тип сокращения Функция Внешняя нагрузка на мышцу Внешняя работа мышцы
Динамическая Концентрический, или миометри-ческнй (изотонический) Ускорение Меньше, чем напряжение мышцы Позитивная
Эксцентри чески й, или плиометри-ческнй Замедление Больше, чем напряжение мышцы Негативная
Статическая Изометрический Фиксация Такая же, как напряжение мышцы Равна нулю
77
В реальных условиях деятельности мышц практически не встречается чисто изометрическое или чисто изотоническое сокращение. В частности, при осуществлении движений внешняя нагрузка на сокращающиеся мышцы изменяется хотя бы уж потому, что изменяются механические условия их действия (плечи действия сил и угол их приложения). Следовательно, такое сокращение мышц уже не является чисто изотоническим, т. е. с равной нагрузкой. Чисто изометрическое сокращение мышцы также вряд ли может быть, так как благодаря наличию упругих образований, через которые передается ее напряжение, возможно некоторое их растяжение, а следовательно, укорочение мышцы. Даже в экспериментальных условиях, когда сухожилия мышцы фиксированы, напряжение собственных сократительных элементов передается упругим (растяжимым) пассивным элементам мышцы, вызывая некоторое их растяжение, в результате чего происходит укорочение собственно сократительных ее компонентов. Смешанную форму сокращения, при которой изменяется и длина и напряжение мышцы, называют ауксотонической.
§ 5. Режимы сокращения
В ответ на импульс, приходящий к мышечным волокнам от мотонейрона (а в условиях опыта — в ответ на однократное электрическое раздражение мышцы или ее нерва), возникает быстрая сократительная реакция этих волокон. Такой процесс называется одиночным сокращением (рис. 34, Л). Сущность этого процесса заключается в активации сократительных элементов, вызывающей подъем напряжения в них и последующее укорочение. При изометрическом сокращении сократительные элементы мышечных волокон укорачиваются за счет растяжения последовательных упругих элементов мышцы и сухожилия, передавая напряжение на регистрирующее устройство, а в условиях обычной деятельности мышцы — на костные рычаги. При изотоническом сокращении активация сократительных элементов приводит к подъему внутреннего напряжения, что вызывает укорочение мышцы. Таким образом, кривые изометрического или изотонического одиночного сокращения служат внешним проявлением активации сократительного аппарата.
В кривой одиночного сокращения можно выделить две фазы (см. рис. 34, Л) —фазу подъема напряжения и фазу расслабления (при изометрическом сокращении) или фазу укорочения и фазу удлинения (при изотоническом сокращении). Длительность фазы напряжения (укорочения) примерно вдвое короче, чем длительность фазы расслабления (удлинения). Длительности этих фаз различны у разных мышечных волокон.
Обычно по длительности фазы напряжения при изометрическом сокращении судят о скорости сокращения мышечных волокон: чем короче фаза напряжения, тем выше скорость сокращения. У наиболее быстросокрашающихся волокон глазных мышц время одиночного сокращения (точнее, фазы подъема напряжения) состав
78
ляет около 7,5 мсек., у наиболее медленных мышечных волокон, например волокон камбаловидной мышцы,— до 150 мсек. Время расслабления хорошо коррелирует с временем напряжения: чем короче фаза напряжения, тем короче и фаза расслабления.
Внешняя кривая одиночного сокращения отличается по форме и времени развития от «внутреннего» сократительного процесса.
Механограмма
Потенциалы действия
25имп/сек ^^пЛек
Режим одиночных сокращений
4- W-H ншншшжн ми
Тетанические сокращения
Суммарный эффект
Рис. 34. Режимы сокращений двигательной единицы (Л) и суммация эффектов сокращения четырех двигательных единиц (Б)
Потенциал действия мышечного волокна развивается почти полностью еще до начала сокращения. Через 1—2 мсек, после начала потенциала действия возникает состояние активности в сократительном аппарате. Эта временная задержка необходима для деполяризации поперечных трубочек саркоплазматического ретикулума, освобождения ионов кальция и их продвижения к нитям миозина и актина, взаимодействия этих ионов с активными участками (поперечными мостиками). Однако и после развития активного состояния напряжение мышечных волокон повышается не сразу. Вначале сократительные элементы вызывают растяжение расположен
79
ных последовательно с ними упругих компонентов — пассивных несократи4ёльных элементов Мышцы. В результате напряжение целой мышцы (сухожилия) повышается позднее и медленнее, чем напряжение в ее контрактильных элементах.
Активное состояние возбужденных мышечных волокон длится всего несколько миллисекунд. При этом чем короче длительность активного состояния, тем меньше длительность одиночного сокращения. Иначе говоря, более быстро сокращающиеся мышечные волокна имеют более короткий период активного состояния. Таким образом, сократительные элементы начинают расслабляться еще до того, как напряжение в сухожилии мышцы достигает максимума. В результате в процессе одиночного сокращения мышечные волокна не успевают проявить свое максимально возможное напряжение. Поэтому напряжение при одиночном сокращении обычно в несколько раз меньше, чем максимально возможное напряжение данных мышечных волокон.
В естественных условиях мышечной деятельности мотонейроны посылают не одиночные импульсы, а серию их. Частота их (или обратная величина — интервалы между импульсами) в разных условиях может быть различной. В зависимости от частоты импуль-сации мотонейрона мышечные волокна двигательной единицы могут работать в двух режимах — режиме одиночных сокращений и режиме тетанического сокращения (см. рис. 34, А).
Режим одиночных сокращений. Мышечные волокна работают в таком режиме при относительно низкой частоте импульсации их мотонейронов. В этих случаях интервалы между соседними импульсами мотонейрона равны или больше длительности одиночного сокращения иннервируемых им мышечных волокон. Следовательно, еще до прихода следующего импульса от мотонейрона мышечные волокна успевают полностью расслабиться. Поэтому каждое новое сокращение возникает после полного их расслабления. Это обусловливает характерную черту режима одиночных сокращений — практическую неутомляемость мышечных волокон, т. е. способность работать в таком режиме очень длительно. Вместе с тем, работая в таком режиме, мышечные волокна развивают относительно небольшое напряжение, не успевая из-за малой длительности активного состояния проявить свои максимальные силовые возможности.
Частота импульсации мотонейронов, при которой их мышечные волокна работают в режиме одиночных сокращений, неодинакова у разных двигательных единиц. Например, для медленной двигательной единицы, мышечные волокна которой имеют длительность одиночного сокращения 100 мсек., режим одиночных сокращений наблюдается при частоте импульсации мотонейрона, не превышающей 10 имп/сек. Для быстрой двигательной единицы, мышечные волокна которой имеют длительность одиночного сокращения, например, 20 мсек. (0,02 сек.), режим одиночных сокращений обеспечивается частотой импульсации мотонейрона менее 50 имп/сек. Таким образом, чем медленнее двигательная единица, тем реже «о
частота импульсации мотонейрона, при которой его мышечные волокна работают в режиме одиночных сокращений.
Режим тетанического сокращения, (см. рис. 34, А). Такой режим работы мышечных волокон возникает при относительно высокой частоте импульсации мотонейронов. В этих случаях интервалы между соседними импульсами мотонейрона короче, чем длительность одиночного сокращения иннервируемых им мышечных волокон. Если второй импульс от мотонейрона приходит до того, как закончился первый цикл сокращения, то второй цикл накладывается на предыдущий и суммарный ответ мышечных волокон становится больше, чем при одиночном сокращении. Это превышение в силе изометрического напряжения (или в величине укорочения при изотоническом сокращении) зависит от интервалов между импульсами, иначе говоря, от частоты импульсации мотонейрона.
Если к мышечным волокнам приходит серия импульсов с большой частотой, то сокращение, вызванное вторым импульсом, накладывается на первое сокращение, на второе сокращение накладывается механический ответ на третий импульс и т. д., т. е. происходит суперпозиция (наложение отдельных ответов друг на друга). При этом ответ (прибавка к напряжению или укорочению) на каждый последующий импульс меньше, чем на предыдущий. После нескольких первых импульсов последующие ничего не добавляют к достигнутому напряжению (укорочению), но поддерживают это напряжение (или укорочение при изотоническом сокращении). Такой режим сокращения мышечных волокон называется полным или гладким тетанусом
Он возникает тогда, когда все сократительные элементы максимально активированы (благодаря суммации активных состояний), а эластические элементы достигают фиксированной длины. Частота импульсации мотонейрона, при которой его мышечные волокна развивают полный тетанус, называется частотой слияния или частотой полного, гладкого, тетануса. Повышение частоты сверх той, при которой достигается полный тетанус, увеличивает напряжение мышечных волокон очень мало. Поэтому иногда частоту импульсации мотонейрона для полного тетануса называют максимальной.
Если увеличение частоты импульсации мотонейронов сверх максимальной (при которой возникает полный тетанус) почти не вызывает изменения в максимальном напряжении мышечных волокон, то для повышения скорости нарастания их напряжения («градиента силы») это увеличение играет важную роль.
Напряжение, развиваемое мышечными волокнами при полном тетанусе, обычно в 2—4 раза больше, чем при одиночном сокращении. Отношение максимального напряжения, достигаемого мышечными волокнами при одиночном сокращении, к напряжению при полном тетанусе называется тетаническим индексом. Обычно быстрые двигательные единицы имеют более низкий тетанический индекс, чем медленные двигательные единицы. Это связано, по-видимому, с более коротким периодом активного состояния у быстрых мышечных волокон по сравнению с длительностью активного состоя
4—113
81
ния у медленных мышечных волокон. Поэтому первые успевают развить во время одиночного цикла сокращения лишь слабое напряжение.
Если частота импульсации мотонейронов выше частоты, характерной для режима одиночных сокращений, но ниже частоты полного тетануса, то происходит суперпозиция соседних циклов сокращения. Однако их полного механического слияния (как при полном тетанусе) в этом случае не возникает, а наблюдается волнообразный ответ мышцы, обозначаемый как неполный, или зубчатый, тетанус (см. рис. 34). При неполном тетанусе величина напряжения (укорочения) меньше, чем при полном тетанусе и, кроме того, она колеблется на протяжении всего сокращения.
Режим тетанического сокращения мышечных волокон в отличие от режима одиночных сокращений утомителен и не может поддерживаться длительное время. Это объясняется тем, что из-за укорочения (или полного отсутствия при полном тетанусе) фазы расслабления мышечные волокна не успевают восстановить энергетический потенциал, израсходованный в фазы напряжения (укорочения). В результате сокращение мышечных волокон при тетаническом режиме происходит в «долг». Особенно утомителен режим полного тетануса, который может поддерживаться считанные секунды или даже доли секунды.
Частота импульсации мотонейронов, при которой возникает режим тетанического сокращения, выше для быстрых мышечных волокон с их относительно коротким периодом одиночного сокращения и ниже для медленных мышечных волокон с их более длительным периодом одиночного сокращения. Так, полный тетанус внутренней прямой мышцы глаза возникает при частоте импульсации мотонейронов около 350 имп/сек, а для медленных двигательных единиц камбаловидной мышцы для этого достаточна частота 30 имп/сек.
§ 6. Регуляция напряжения мышцы
Управление движениями и сохранение определенного положения тела и его звеньев в поле тяжести Земли связано с сокращением мышц. Помимо выбора нужных мышц и моментов их включения центральная нервная система при управлении движениями и сохранении позы должна регулировать степень напряжения (укорочения этих мышц). Для решения последней задачи она использует три механизма: 1) регуляцию числа активных двигательных единиц (мотонейронов) данной мышцы, 2) выбор режима их работы, 3) определение характера временной связи активности двигательных единиц. Рассмотрим последовательно эти механизмы.
1. Число активных двигательных единиц. Активная двигательная единица — это такая единица, мотонейроны которой посылают импульсы, а соответствующие мышечные волокна отвечают на них сокращением. Чем больше активных двигательных единиц у данной мышцы, тем большее напряжение она развивает.
82
Число активных двигательных единиц определяется интенсивностью возбуждающих влияний, которым подвергаются мотонейроны данной мышцы со стороны нейронов более высоких моторных уровней (моторной коры, подкорковых моторных центров), промежуточных нейронов спинного мозга и рецепторов. Если для выполнения двигательной задачи мышца должна развить небольшое напряжение, то ее мотонейроны подвергаются относительно слабым возбуждающим влияниям. Поскольку любая мышца иннервируется мотонейронами, имеющими неодинаковые размеры (см. выше), то реакция их на эти возбуждающие влияния будет различной. Известно, что чем меньше размер тела мотонейрона, тем ниже порог возбуждения. Поэтому при относительно слабых возбуждающих влияниях потенциалы действия возникают лишь у наименьших из мотонейронов данной мышцы. В результате слабые напряжения мышцы обеспечиваются в основном активностью низкопороговых, малых, двигательных единиц. Большее напряжение мышцы требует большей интенсивности возбуждающих влияний на ее мотонейроны. При этом помимо малых двигательных единиц активными становятся все большие по размеру двигательные единицы. Таким образом, значительные напряжения мышцы обеспечиваются активностью многих ее двигательных единиц, начиная от низкопороговых (малых) и кончая высокопороговыми (большими). Этот механизм включения двигательных единиц в соответствии с их размерами носит название «правило размера». Согласно этому правилу, самые малые двигательные единицы мышцы активны при любом ее напряжении, тогда как большие двигательные единицы, входящие в состав данной мышцы, активны лишь при больших ее напряжениях. 11оэтому у одной и той же мышцы степень использования больших двигательных единиц по сравнению с малыми единицами ниже. Иначе говоря, в условиях обычной деятельности мышечные волокна больших двигательных единиц тренируются относительно мало.
2. Режим активности двигательных единиц. Как уже отмечалось, чем выше (в определенных границах) частота импульсации мотонейрона, тем большее напряжение развивает двигательная единица и тем, следовательно, значительнее ее вклад в общее напряжение мышцы. Поэтому регуляция частоты импульса-IIпи мотонейронов является важным механизмом, определяющим напряжение мышцы.
Частота импульсации мотонейронов зависит от интенсивности возбуждающих влияний, которым они подвергаются. Если интенсивность небольшая , то работают низкопороговые, малые мотонейроны и частота их импульсации относительно невелика. В этом случае малые двигательные единицы работают в режиме одиночных сокращений. Такая активность двигательных единиц обеспечи-нает лишь слабое, но зато мало утомительное сокращение мышцы. I ю достаточно, например, для сохранения вертикальной позы нла.
В связи с этим понятно, почему позная активность мышц может поддерживаться без утомления много часов подряд.
г
83
Увеличение напряжения мышцы возникает благодаря усилению возбуждающих влияний на ее мотонейроны. Это усиление приводит не только к включению новых, более высокопороговых мотонейронов, но и к повышению частоты импульсации относительно низкопороговых мотонейронов. При этом интенсивность возбуждающих влияний, которым подвержены наиболее высокопороговые из активных мотонейронов, еще недостаточна, чтобы вызвать их высокочастотный разряд. Поэтому из активных двигательных единиц более низкопороговые работают с относительно большой для них частотой и, следовательно, в режиме тетанического сокращения, а наиболее высокопороговые — в режиме одиночных сокращений.
При очень больших напряжениях мышцы большинство двигательных единиц работают в тетаническом режиме (из-за высокой частоты импульсации их мотонейронов), и потому большие напряжения мышцы могут поддерживаться недолго (см. ниже).
3. Связь во времени активности разных двигательных единиц. Напряжение мышцы в определенной мере зависит от того, как связаны во времени импульсы, посылаемые разными мотонейронами данной мышцы. Для пояснения рассмотрим упрощенный пример активности четырех двигательных единиц одной мышцы, работающих в режиме одиночных сокращений В одном случае все четыре двигательные единицы сокращаются одновременно (синхронно), так как их мотонейроны посылают импульсы одновременно. В другом случае двигательные единицы работают с разной частотой и неодновременно (асинхронно) и, следовательно, фазы сокращений их мышечных волокон не совпадают (см. рис. 34, Б). В первом случае общее напряжение мышцы больше, чем во втором, но колебания напряжения при этом очень большие; во втором случае общее напряжение мышцы меньше, чем в первом, но колебания напряжения значительно меньше. Из этого примера ясно, что если двигательные единицы работают в режиме одиночных сокращений, но асинхронно, то общее напряжение всей мышцы колеблется незначительно. Чем больше число асинхронно сокращающихся двигательных единиц, тем меньше колебания в напряжении мышц. Соответственно более плавно совершается движение или точнее удерживается необходимая поза (меньше амплитуда физиологического тремора). В нормальных условиях большинство двигательных единиц одной мышцы работают асинхронно, независимо друг от друга, что и обеспечивает нормальную плавность ее сокращения. При утомлении, вызванном большой и длительной мышечной работой, нарушается нормальная деятельность двигательных единиц, и они начинают возбуждаться одновременно (синхронно). В результате движение теряет плавность, нарушается его точность, возникает тремор утомления — большие размашистые дрожательные движения с частотой около 6 колебаний в 1 сек.
Если двигательные единицы работают в режиме полного (или почти полного (тетануса, то характер временной связи их активности почти не влияет на величину максимального напряжения, развиваемого мышцей в целом. Дело в том, что при полном тетану-
84
< г уровень напряжения каждой из работающих двигательных единиц поддерживается почти постоянным (гладкий тетанус). Поэтому при относительно длительных и сильных сокращениях мышцы характер связи во времени импульсной активности мотонейронов практически не отражается на максимальном напряжении мышцы.
Наоборот, при кратковременных сокращениях или в начале любого сокращения мышц синхронизация импульсной активности мотонейронов играет важную роль, влияя на скорость развития напряжения, т. е. на величину «градиента силы». Ясно, что чем больше совпадений в сократительных циклах разных двигательных единиц в начале развития напряжения мышцы, тем быстрее оно нарастает. Такая синхронизация происходит особенно часто в начале выполнения быстрых движений, совершаемых против большой внешней нагрузки. Это в значительной степени связано с тем, что в начале разряда частота импульсации мотонейронов выше, чем в дальнейшем. Благодаря высокой начальной частоте импульсации и активности большого числа мотонейронов вероятность совпадения сократительных циклов многих двигательных единиц (синхронизации) в начале движения очень велика. Таким образом, скорость нарастания напряжения мышцы («градиент силы», или «взрывная сила») зависит как от числа активируемых двигательных единиц, так и от начальной частоты и степени синхронизации импульсации мотонейронов данной мышцы.
§ 7. Характеристические кривые мышцы
Мышца является упругим телом: упругостью обладают как сократительные, так и несократительные, «пассивные», ее элементы. Выделяют две группы упругих элементов мышцы — последовательные и параллельные. К последовательным упругим элементам относятся участки миозиновых поперечных мостиков, участки перехода миофибрилл в соединительнотканные образования и сухожилия мышцы. Кроме того, в мышце имеются соединительнотканные образования, которые формируют параллельный упругий компонент мышцы. К ним относятся оболочки, покрывающие пучки мышечных волокон (эпимизий), оболочки, отделяющие эти пучки друг от друга внутри мышцы (перимизий), и оболочки мышечных волокон (эндо-мизий).
Упругие свойства покоящейся (пассивной) мышцы проявляются в том, что при растяжении в ней развивается напряжение. Изолированная мышца имеет равновесную длину, при которой ее упругое напряжение равно нулю. При растяжении пассивной мышцы ее упругое напряжение прогрессивно увеличивается в пределах растяжения мышцы вдвое от ее равновесной длины (рис. 35). В теле человека даже в условиях покоя скелетные мышцы имеют слабое напряжение, так как они несколько растянуты. Большинство мышц при крайне возможных степенях их растяжения (при максимальном суставном угле) имеют длину, которая примерно на 20% больше равновесной длины (у отдельных мышц —до 40%).
85
Напряжение, которое развивает мышца при сокращении, вызванном импульсацией от мотонейронов (а в условиях опыта —в результате электрического раздражения), также зависит от длины мышцы. Эта зависимость выражается в форме характеристической кривой «длина — напряжение». Напряжение сокращающейся мыш-
цы максимально, если во время электрической стимуляции или в
условиях натуральной активации длина мышцы примерно на 20%
Изменение в длине.см
Рис. 35. Кривые «длина — напряжение», полученные в исследованиях у человека на трехглавой мышце плеча:-/ — кривая, полученная при регистрации максимального произвольного (общего) напряжения мышцы при разной ее длине;
2 — кривая пассивного напряжения мышцы в состоянии ее расслабления; 3 — кривая «чистого» (активного) произвольного
больше равновесной.
Длина мышцы, при которой она развивает максимальное напряжение при сокращении, называется длиной покоя. Как уже отмечалось, большинство мышц в естественных условиях достигают длины покоя при максимально возможном или близком к нему растяжении. Соответственно у большинства мышц максимальное напряжение развивается при максимально возможном их растяжении, т. е. при крайнем удалении друг от друга костных рычагов, к Ко-
торым крепится данная мышца.
Рис. 35 показывает связь между напряжением, развиваемым во время максимального произвольного усилия, и изменением в длине трехглавой мышцы плеча у человека. Кривая «чистого», или активного.
напряжения мышцы, полученная путем вычитания кривой 2 из кривой •/. Вертикальная штриховая линия пересекает ось абсцисс при длине покоя данной мышцы. За ноль на оси абсцисс принята длина, при которой общее напряжение мышцы (кривая 1) равно нулю. Заштрихованные области соответствуют разбросу данных у разных испытуемых и в разных опытах
произвольного напряжения определяется путем измерения пассивного напряжения, возникающего при рас-тяжении расслабленной мышцы до данной длины, и затем вычитания этой величины «пассивного» напряжения из общего напряжения, развиваемого сокращающейся мышцей
при той же длине. Как видно на графике, «чистое» напряжение повышается более или менее линейно с увеличением начальной длины мышцы вплоть до достижения максимума. Если исходная дли-
на больше, чем этот максимум, то развивается меньшее напряжение. Таким образом, увеличение или уменьшение длины мышцы
сверх длины покоя приводит к падению активного напряжения.
Особенности характеристической кривой «длина — напряжение» становятся понятными при рассмотрении условий взаимодействия актиновых и миозиновых нитей в процессе сокращения (рис. 36). Дело в том, что напряжение, которое могут развивать миофибриллы, зависит от числа поперечных мостиков, образующихся в процессе
сокращения между миозиновыми и актиновыми нитями, так как мостики являются местом взаимодействия и развития усилия между
86
двумя типами нитей. Наибольшее число поперечных мостиков может образоваться при длине покоя мышцы (соответственно длине покоя саркомеров миофибрилл). При меньшей длине актиновые нити входят в Н-зону, в которой миозиновые нити лишены поперечных мо-
стиков, при сильном укорочении мышцы концы противостоящих актиновых нитей перекрываются. При этом часть центральных мостиков становится неактивной. Наконец, при значительном укорочении мышцы толстые миозиновые нити упираются в Z-линии и даже могут при этом несколько изгибаться. Все это ведет к уменьшению числа образующихся поперечных мостиков и в результате к снижению силы активного сокращения мышцы.
При значительном растяжении мышцы сверх длины покоя актиновые и миозиновые нити перестают перекрываться, поэтому между ними уже не возникают поперечные
Рис. 36. Кривая «длина — напряжение» для отдельного саркомера.
Внизу схематически показаны взаимоотношения толстых и тонких нитей при разной длине саркомера (в микронах). Каждой схеме соответствует на кривой определенная точка (указано вертикальными стрелками)
связи. Соответственно отсутствует скольжение нитей относительно
друг друга и не развивается напряжение.
В экспериментальных условиях мышца может быть растянута до длины, при которой в ней вообще отсутствует активное напряжение, т. е. в ответ на ее раздражение она не отвечает подъемом напряжения. Как уже отмечалось, только для некоторых мышц в условиях нормального функционирования значительное растяжение их может вести к некоторому снижению активного напряжения по сравнению с максимальным напряжением при длине покоя. Наименьшая длина мышц в естественных условиях составляет у большинства из них примерно 0,7 от равновесной длины.
Другая характеристическая кривая мышцы определяет зависимость скорости укорочения мышцы во время сокращения от внешней нагрузки на нее (груза). Такая характеристическая кривая -сила (груз) — скорость» может быть получена как на изолированной мышце, так и в условиях произвольной активации мышц.
Для этого стимулируют мышцу (или группу мышц-синергистов), вызывая ее максимальное сокращение. Во время стимуляции концы мышцы фиксированы
87
или груз закреплен так, чтобы препятствовать ее укорочению. Затем фиксаторы снимают, и мышца укорачивается, поднимая груз. Вначале скорость укорочения мышцы наибольшая, а затем начинает падать и достигает нуля при максимальном укорочении, мышцы. Начальная скорость укорочения определяется как скорость укорочения. Скорость укорочения измеряется при определенной длине мышцы, поскольку напряжение, развиваемое ею, зависит от длины.
Как следует из кривой «груз — скорость» (рис. 37), при большом грузе мышца не укорачивается. Напряжение, которое она при этом развивает, соответствует ее максимальному изометрическому напряжению при данной длине. Если величина груза меньше, чем максимальное изометрическое напряжение, то мышца укорачивается.
Рис. 37. Кривая «груз — скорость» для грудных мышц человека.
Мышцы предварительно были слегка растянуты небольшим грузом до длины, несколько превышающей равновесную. Последовательно подвешивались разные грузы через блок, но стопор предотвращал движение груза. Испытуемый должен был максимально произвольно напрячь грудные мышцы. Затем стопор снимался и регистрировалась скорость движения груза.
большого усилия, как при
При этом скорость укорочения тем больше, чем меньше величина груза. Произведение действующего на мышцу груза (плюс некоторая константа) на скорость укорочения (также плюс некоторая константа) — постоянная для данной мышцы величина.
Общая форма кривой «груз — скорость» хорошо объясняется теорией сокращения. Когда мышца укорачивается быстро, взаимодействие скользящих друг относительно друга актиновых и миозиновых нитей очень кратковременно. В результате число взаимодействующих участков поперечных мостиков и развиваемое напряжение меньше, чем при медленном укорочении мышцы.
В случае изотонического (динамического) сокращения при любых скоростях укорочения мышца никогда не развивает столь 'рнческом сокращении (рис. 38).
Рассмотрим пример определения силы при изометрическом и динамическом (концентрическом и эксцентрическом) сокращениях мышц-сгибателей локтевого сустава. В этих опытах испытуемые должны были с максимальным усилием тянуть за кольцо в направлении к туловищу. Изометрическая сила (см. рис. 38) измерялась при 10 разных положениях руки: исходное положение кольца — на расстоянии прямой вытянутой вперед руки (0 на оси абсцисс), остальные поло; Кения— по мере приближения его к туловищу (ось абсцисс). Для измерения силы во время динамического (изотонического) концентрического сокращения мышц испытуемый осуществлял тягу за кольцо против сопротивления, создаваемого мотором. Варьируя силу сопротивления мотора, можно было изменять скорость движения (последняя тем меньше, чем больше сопротивление—-см. зависимость «груз — скорость»), В случае динамического эксцентрического сокращения испытуемый тянул за кольцо, сопротивляясь его движению в противоположную сторону. Это движение кольца осуществлялось мотором с разной силой тяги. Соответственно скорость вынужденного разгибательного движения в локтевом суставе оказывалась тем больше, чем сильнее тяга мотора. Сила динамичес-
Ь8
кого сокращения измерялась при тех же положениях руки, что и при измерении изометрической силы.
Как следует из приведенных данных, динамическая сила, развиваемая во время укорочения мышцы (динамическое концентрическое сокращение), всегда меньше, чем изометрическая сила при тех же положениях конечности (т. е. при тех же длинах мышц). Как изометрическая, так и динамическая сила сокращения
мышц уменьшается с их укорочением.
Кривые на рис. 38 показывают также, что при максимальном концентрическом сокращении развиваемая сила тем меньше, чем быстрее движение (три нижние кривые), и, как уже отмечалось, при любой скорости укорочения динамическая сила меньше, чем изометрическая. При динамическом эксцентрическом сокращении, наоборот, максимальная сила во время удлинения сокращающихся мышц всегда больше, чем изометрическая (см. три верхних графика). Это превышение возрастает с увеличением скорости движения (удлинения мышц). При данной скорости имеется высокая корреляция между изометрической силой и динамической. Поэтому максимальная динамическая сила может быть с высокой вероятностью предсказана по результатам более
Рпс. 38. Зависимость статической и динамической сил от длины мышц (положения конечности):
простого измерения максимальной изометрической силы тех же мышц. Однако у спортсме-
А — максимальная горизонтальная тяга при разных положениях руки. Верхний ряд чисел под абсциссой — степень приближения кольца к туловищу (слева направо — укорочение сгибателей), нижний ряд чисел — приближение в % от общей
нов используется в тренировках преимущественно один из видов упражнений — изометри-ческие или динамические. Поэтому показатели максимальной изометрической и динамической силы у них могут не коррелировать между собой.
длины руки (динамическая сила измерялась при движении со скоростью, при которой за 1 сек. рука проходит расстояние, составляющее 15% от ее общей длины).
Б — максимальная тяга при движениях руки с разной скоростью. Сила тяги выражена в % от максимальной произвольной изометрической силы, принятой за 100% (ось ординат). Три нижние кривые —• при концентрических, три верхние — при эксцентрических сокращениях. Для определения скорости движения пройденное рас-стояие выражали в % от длины руки испытуемого (по Э. Асмуссену и др.)
§ 8. Работа мышц и ее механическая эффективность
Мышцы — это машины, преобразующие химическую энергию в механическую работу или напряжение. При укорочении мышцы во время сокращения против груза или сопротивления (изотониче-
89
ское, или динамическое, сокращение) производится работа. При развитии же напряжения без укорочения мышцы (изометрическое сокращение) внешняя работа не производится. Оценивая деятельность мышц, обычно учитывают только производимую ими внешнюю работу. В наиболее простом случае работа мышцы (IT) по подъему груза на некоторую высоту может быть рассчитана как произведение веса груза (Р) на высоту подьема (Л) и выражена в килограммометрах: W=P-h кгм.
Величина работы, производимой мышцей, зависит от внешней нагрузки на нее. В качестве иллюстрации ниже приводятся данные одного из опытов на изолированной мышце лягушки. Сокращаясь под действием электрического раздражения, эта мышца поднимала различные грузы:
Груз (г) О
Укорочение (мм) 14
Механическая работа (г-мм) О
50 100 150 200 250
9 7 5 2 0
450 700 750 400 0
Видно, что с увеличением груза степень сокращения (укорочения) уменьшается и при грузе 250 г укорочения мышцы уже не происходит (максимальная изометрическая сила данной мышцы). Внешняя механическая работа мышцы по мере возрастания груза вначале увеличивается, а затем уменьшается. Наибольшую внешнюю работу мышца производит при средних для нее нагрузках. Это явление носит название закон средних нагрузок.
Внешняя механическая работа зависит также от скорости сокращения мышцы. Наибольшая внешняя работа выполняется ею при средних скоростях укорочения. При высоких скоростях укорочения мышцы значительная часть ее энергии расходуется на преодоление внутреннего трения (вязкости). При этом чем выше скорость укорочения, тем больше внутреннее трение. При слишком медленном укорочении мышцы часть энергии идет не на укорочение, а на поддержание достигнутой степени укорочения мышцы.
Закон средних нагрузок и средних скоростей сокращения в значительной степени определяет механическую эффективность или производительность (R) мышц. При выполнении любой работы лишь часть потенциальной химической энергии превращается в механическую работу, большая же часть неизбежно переходит в тепло. Поэтому общий энергетический расход (Е) есть сумма расхода энергии на механическую работу (U7) и расхода энергии на образование тепла (Н), т. е. Е = W+H. Механическая эффективность, или производительность (R), мышечной работы (иначе —коэффициент полезного действия) представляет собой отношение (%) внешне выполненной механической работы, представленной в калориях как механическая энергия (W7), к общей внешней энергопродукции (Е):
7?=-^- 100%.
Если исследование ведется на изолированной мышце, то механическая эффективность может быть рассчитана на основании данных
90
измерения совершенной работы (IV') и энергии, проявляющейся в форме тепла. Работа и тепло в этом случае единственные формы проявления энергии. Расчет механической эффективности производится по следующей формуле:
H+W
Если исследование ведется на мышце с нормальным кровоснабжением, то расходуемая ею энергия (Ej рассчитывается по величине потребления кислорода. У изолированной мышцы механическая эффективность зависит от внешней нагрузки и скорости укорочения мышцы и может достигать 45—50% (рис. 39, А). Наиболее высокая
Рис. 39. Зависимость механической эффективности мышечной работы от силы сокращения при изометрическом режиме (4) и от мощности работы при динамическом режиме (Б, В, Г)
А — механическая эффективность (ось ординат) изолированной мышцы, подвергавшейся электрической тетанической стимуляции и удерживающей различные грузы (ось абсцисс); Б — механическая эффективность при работе на велоэргометре с разной мощностью (Г. Галлетти); В — механическая эффективность при работе на велоэргометре с разной длительностью цикла, т. е. с разной частотой вращения педалей (Д. Дикинсон); Г — энергетические траты при ходьбе с разной скоростью без обуви и в обуви разного веса. Наименьшие величины энерготрат отмечаются при скорости ходьбы 60—80 м/мин, т, е. 3,6—4,8 км/час (Т. Хеттингер и Э. Мюллер)
механическая эффективность изолированной мышцы обнаруживается при внешней нагрузке, составляющей около 50% от максимальной изометрической силы данной мышцы и при скорости укорочения около 30% от максимальной.
Для расчета общей производительности мышечной работы у человека количество израсходованной энергии (Е) определяют по
91
объему кислорода, потребленного во время выполнения работы и в период восстановления (см. ниже). В этом случае производительность рассчитывается по следующей формуле:
R% = 0,49 • 100.
В этой формуле 0,49 — коэффициент эквивалентности между механической работой и объемом потребленного кислорода, т. е. при 100 %-ной производительности для осуществления 1 кгм работы необходимо 0,49 мл О2. Например, для выполнения работы в 120 кгм потребовалась затрата 300 мл О2 (К2), отсюда производительность составила:
R = 0,49 • — • 100 = 19,6%.
300
В данном случае определяется общая производительность, так как часть кислорода, поглощаемого сверх уровня покоя, используется не только скелетными мышцами, но и другими органами и тканями, участвующими в мышечной деятельности организма. Поэтому реальная механическая эффективность мышц в этом случае будет выше. Наиболее высокие показатели производительности у человека (20—25%) обнаруживаются во время работы с участием большого числа мышечных групп (например, работы на велоэргометре). Производительность во время локальной мышечной работы, т. е. работы с участием небольшого числа мышц (например, сгибание — разгибание в локтевом суставе), обычно ниже, чем при работе с участием большого числа мышц (регионарная или глобальная работа).
Производительность варьирует в зависимости от условий выполнения мышечной работы. Как видно на рис. 39, Б, с увеличением мощности работы на велоэргометре производительность падает, так что при мощности 40 вт она составляет 25%, а при мощности 200 вт—менее 23%. При локальной динамической работе производительность примерно одинакова при разных, но относительно небольших мощностях работы (ниже критического уровня — см. дальше). При мощности выше этого уровня производительность снижается, что связано с включением «ненужных» мышечных групп.
В случаях работы с постоянной мощностью самая большая производительность наблюдается при средних значениях внешней нагрузки (груза), амплитуды и частоты движений. При увеличении или уменьшении этих характеристик производительность снижается. Дело в том, что при движениях с небольшой амплитудой или при малой внешней нагрузке поддержание постоянной мощности работы обеспечивается за счет более высокой частоты движений. При этом производительность падает, так как возрастает число мышечных сокращений, уменьшаются паузы между ними и увеличивается скорость укорочения мышц. Так, при работе на велоэргометре производительность наибольшая при длительности одного цикла педалирования около 1 сек. (см. рис. 39, В.). Наоборот, при большой
92
внешней нагрузке скорость укорочения снижается, что также ведет к падению производительности. При ходьбе наименьшие энергетические затраты на работу по вертикальному перемещению веса тела, т. е. наибольшая производительность, обнаруживаются при ходьбе со скоростью 3,6—4,8 км/час. Энергетически оптимальная скорость ходьбы снижается с увеличением веса обуви, т. е. внешней нагрузки (см. рис. 39, Г).
Производительность изменяется в зависимости от уровня тренированности спортсмена. С повышением тренированности отмечается уменьшение энергетических затрат (потребления кислорода) во время выполнения одной и той же внешней работы. Это повышение производительности определяется тремя главными факторами. Во-первых, улучшается деятельность сердечно-сосудистой и дыхательной систем, обеспечивающих работающие мышцы кислородом и другими энергетическими веществами. Во-вторых, улучшается координация движений, происходит совершенствование межмышечных координаций, исчезает активность «ненужных» мышечных групп. В-третьих, повышается сила тренированных мышц, которые могут теперь выполнять более интенсивную в абсолютном значении работу, так что работа той же мощности требует относительно меньших усилий активных мышечных групп.
Важно подчеркнуть, что повышение производительности проявляется лишь у тренируемых мышц. Так, общая мышечная тренировка мало изменяет производительность при локальной мышечной работе. Иначе говоря, повышение производительности мышц в результате тренировки чрезвычайно специфично. Используя один вид упражнения, можно рассчитывать на существенное повышение производительности при выполнении только этого вида упражнения.
§ 9. Мышечная сила
Сила мышцы может быть определена по максимальному напряжению, которое она развивает в условиях изометрического сокращения. Как уже отмечалось, сила сокращения мышцы зависит от ее длины. Определяя силу мышцы, важно учитывать длину, при которой она сокращается. Изометрически сокращающаяся мышца развивает максимально возможное для нее напряжение при соблюдении следующих трех условий: первое — активация всех двигательных единиц (мышечных волокон) данной мышцы; второе — режим полного тетануса у всех двигательных единиц; третье — сокращение мышцы при длине покоя. В этом случае изометрическое напряжение мышцы соответствует ее максимальной силе.
Максимальная сила мышцы зависит от числа мышечных волокон, составляющих данную мышцу, и от толщины этих волокон. Число и толщина их определяют толщину мышцы в целом, или, иначе, площадь поперечного сечения мышцы (анатомический поперечник). Отношение максимальной силы мышцы к ее анатомическому поперечнику называется относительной силой мышцы. Она измеряется в кг/см2. Анатомический поперечник определяется как площадь
93
поперечного разреза мышцы, проведенного перпендикулярно ее длине. При этом для мышц с ходом волокон, совпадающим с длинной продольной осью мышцы (мышцы с параллельным ходом волокон), такой поперечный разрез проходит перпендикулярно ходу волокон. Для точного определения поперечника мышц с косым расположением волокон разрез должен проходить также перпендикулярно ходу волокон и потому расположен под углом к длиннику мышцы. Поперечный разрез мышцы, перпендикулярный ходу ее волокон, позволяет получить физиологический поперечник мышцы. Для мышц с параллельным ходом волокон физиологический поперечник совпадает с анатомическим. Отношение максимальной силы мышцы к ее физиологическому поперечнику называется абсолютной силой мыищы. Она колеблется в пределах 4—8 кг/см2.
Поскольку сила мышцы зависит от ее поперечника, увеличение последнего сопровождается ростом силы данной мышцы. Увеличение мышечного поперечника в результате мышечной тренировки называется рабочей гипертрофией мышцы (от лат. hypertrophia— усиленное питание). Мышечные волокна, являющиеся высокоспециализированными дифференцированными клетками, по-видимому, не способны к делению с образованием новых волокон. (Как известно, нервные клетки также не способны к делению.) Во всяком случае, если деление мышечных клеток и имеет место, то только в особых случаях и в очень небольшом количестве. Рабочая гипертрофия мышцы происходит отчасти за счет продольного расщепления, а главным образом за счет утолщения (увеличения объема) мышечных волокон.
Можно выделить два основных типа рабочей гипертрофии мышечных волокон. Первый тип (саркоплазматический)—утолщение мышечных волокон за счет преимущественного увеличения объема саркоплазмы, т. е. несократительной части мышечных волокон. Этот тип гипертрофии приводит к повышению метаболических резервов мышцы: гликогена, безазотистых веществ, креатинфосфата, миоглобина и др. Значительное увеличение числа капилляров в результате тренировки также может в какой-то мере вызывать некоторое утолщение мышцы.
В мышцах тренирующихся животных содержание воды несколько больше, чем в мышцах нетренирующихся животных. После приема гормона питуитрина, который способствует задержке воды в организме, четырехглавая мышца бедра у тренированных крыс становится на 20—30% тяжелее и поперечное сечеипе ее на 6—12% больше, чем у нетренированных. Однако никаких функциональных преимуществ такие «гипертрофированные» мышцы не имеют. Действительно, мышцы у животных, получающих питуитрин, развивают меньшее напряжение на 1 г своего веса, чем у контрольных животных.
Первый тип рабочей гипертрофии мало влияет на рост силы мышц, но зато значительно повышает способность их к продолжительной работе, т. е. выносливость.
Второй тип рабочей гипертрофии (миофибриллярный) связан с увеличением объема миофибрилл, т. е. собственно сократительного аппарата мышечных волокон. При этом мышечный поперечник мо
94
жет увеличиваться не очень значительно, так как в основном возрастает плотность укладки миофибрилл в мышечном волокне. Второй тип рабочей гипертрофии ведет к значительному росту максимальной силы мышцы. Существенно увеличивается и абсолютная сила мышцы, тогда как при первом типе рабочей гипертрофии опа или совсем не изменяется или даже несколько уменьшается.
Преимущественное развитие первого или второго типа рабочей гипертрофии определяется характером мышечной тренировки. Вероятно, длительные динамические упражнения с относительно небольшой нагрузкой вызывают рабочую гипертрофию главным образом первого типа (преимущественное увеличение объема саркоплазмы, а не миофибрилл). Изометрические упражнения с применением больших мышечных напряжений (более 2/3 от максимальной произвольной силы тренируемых мышечных групп), наоборот, способствуют развитию рабочей гипертрофии второго типа (миофибрил-лярной гипертрофии).
В основе рабочей гипертрофии лежит интенсивный синтез мышечных белков. Усиленная мышечная активность каким-то образом воздействует на генетический аппарат мышечных клеток, ответственный за регуляцию синтеза белков. Соответственно концентрация ДНК и РНК в гипертрофированной мышце больше, чем в нормальной.
Очень важную роль в регуляции объема мышечной массы, в частности в развитии рабочей гипертрофии мышцы, играют особые гормоны — андрогены (см. главу XIII).
У мужчин эти гормоны вырабатываются половыми железами (семенниками) и в коре надпочечников, а у женщин — только в коре надпочечников. Соответственно у мужчин количество андрогенов в организме больше, чем у женщин. Роль андрогенов в увеличении мышечной массы проявляется в следующем.
Во-первых, возрастное развитие мышечной массы в организме идет параллельно с увеличением продукции андрогенных гормонов. Первое заметное утолщение мышечных волокон наблюдается в 6—7-летнем возрасте, когда усиливается • образование андрогенов. С наступлением полового созревания (в возрасте 11— 15 лет) начинается интенсивный прирост мышечной массы у юношей, который продолжается и после периода полового созревания. У девушек развитие мышечной массы в основном заканчивается с периодом полового созревания. Соответствующий характер имеет и рост мышечной силы в школьном возрасте. Датский физиолог Э. Асмуссен установил, что связь между показателями мышечной силы и ростом у детей 7—17 лет на двойной логарифмической шкале выражена прямой линией (рис. 40, А). Достоверных различий в силе мышц ног у мальчиков и девочек одинакового роста нет, но сила мышц туловища и рук во все возрастные периоды (начиная с 7 лет) у мальчиков значительно больше, чем у девочек. После того как рост мальчиков достигает 150 см и более (что наблюдается в 13 лет, когда обычно начинается половое созревание), у них отмечается значительный прирост силы некоторых мышечных групп, особенно мышц рук.
Во-вторых, роль андрогенов проявляется в различиях мышечной силы у мужчин и женщин (см. рис. 40, Б). Большие показатели ее у мужчин, чем у женщин, отчасти объясняются половыми различиями в линейных размерах тела. Поскольку сила пропорциональна площади поперечного сечения мышц, мышечная сила должна зависеть от длины тела (Н) как №. Однако, скорректировав показатели силы с размерами тела, Асмуссен нашел, что силовые показатели у взрослых женщин в среднем составляют только около 80% от показателей мышечной силы у мужчин (см. рис. 40, Б — правый график). Вместе с тем если у женщин в результате некоторых заболеваний усиливается секреция андрогенов
96
надпочечниками, то происходит интенсивное увеличение мышечной массы, появляется хорошо развитый мышечный рельеф, возрастает мышечная сила.
В экспериментах на животных было установлено, что введение препаратов андрогенных гормонов (анаболиков) вызывает значительную интенсификацию синтеза мышечных белков, в результате чего увеличивается масса тренируемых мышц и как результат — их сила.
Рис. 40. Возрастно-половые изменения максимальной призвольной силы. А — показатели роста (ось абсцисс) и силы (ось ординат) сгибателей туловища (слева) и мышц рук (справа), отложенные на логарифмической шкале (Э. Асмуссен и др.); В — изменение мышечной силы у мужчин и женщин с возрастом. На правой диаграмме кривая б получена из кривой а с учетом различий в росте у мужчин и женщин. Средние показатели силы у мужчин в возрасте 22 лет приняты за 100%.
Измерение мышечной силы у человека осуществляется при произвольном напряжении мышц. Поэтому когда говорят о мышечной силе у человека, речь идет q максимальной произвольной мышечной силе, т. е. о суммарной величине изометрического напряжения (точнее — о суммарном моменте) группы мышц при максимальном произвольном усилии испытуемого *. Максимальная произвольная мышечная сила зависит от двух групп факторов, которые можно обозначить как мышечные (периферические) факторы и координационные (нервные) факторы.
К мышечным (периферическим) факторам относятся: а) механические условия действия мышечной тяги — плечо рычага дейст
* В спортивной педагогике этому понятию эквивалентно понятию «абсолют-। на я сила мышц».
S6
вия мышечной силы и угол приложения этой силы к костным рычагам; б) длина мышц, так как напряжение мышцы зависит от ее длины; в) поперечник активируемых мышц, так как при прочих равных условиях проявляемая мышечная сила тем больше, чем больше суммарный поперечник произвольно сокращающихся мышц.
К координационным (нервным) факторам относится совокупность центральнонервных координационных механизмов управления мышечным аппаратом. Их можно разделить на две группы — механизмы внутримышечной и межмышечной координации. О механизмах внутримышечной координации (числе активных двигательных единиц данной мышцы .частоте импульсации ее мотонейронов и связи их импульсации во времени) написано выше (см. § 6). Эти три механизма центральной регуляции напряжения мышцы определяют, какой вклад в общий силовой показатель вносит каждая из них.
Поскольку при измерении силы сокращается много мышц — синергисты и антагонисты нескольких суставов—показатель максимальной произвольной силы зависит также от координации активности всех этих мышц (межмышечная координация). В частности, совершенство межмышечной координации проявляется в правильном выборе активируемых мышц-синергистов, в адекватном ограничении активности мышц-антагонистов данного сустава и усилении активности мышц-антагонистов, обеспечивающих фиксацию смежных суставов и т. п.
Все сказанное позволяет сделать следующее заключение. Управление мышцами в случае, когда требуется проявить максимальную произвольную силу, является сложной задачей для центральной нервной системы. Отсюда понятно, почему в обычных условиях максимальная произвольная сила тех или иных групп мышц меньше, чем их максимальная сила. Разница между максимальной силой мышц и их силой, проявляемой при максимальном произвольном усилии, называется силовым дефицитом.
Определение силового дефицита у человека производится следующим образом. С помощью специальной динамометрической установки измеряют максимальную произвольную силу выбранной группы мышц, затем — ее максимальную силу. Для этого раздражают электрическим током нерв, иннервирующий данную мышечную группу, импульсами такой силы, которая возбуждает все моторные волокна (аксоны мотонейронов) этой мышечной группы. При этом применяют частоту раздражения, достаточную для возникновения полного тетануса мышечных волокон (обычно 50—100 пмп/сек). Таким образом сокращаются все мышечные волокна данной мышечной группы, развивая максимально возможное для них напряжение.
Различие между максимальной силой и произвольной максимальной силой данной мышечной группы (силовой дефицит) тем меньше, чем совершеннее центральное управление мышечным аппаратом.
Величина силового дефицита зависит от трех факторов: психологического состояния испытуемого, количества одновременно активируемых мышечных групп и степени совершенства произвольного управления ими.
97
Первый фактор. Известно, что при некоторых эмоциональных состояниях человек может проявлять такую силу, которая намного превышает его максимальные силовые возможности в обычных условиях. К таким эмоциональным (стрессовым) состояниям относится, в частности, состояние спортсмена во время соревнования. В экспериментальных условиях значительное повышение показателей максимальной произвольной силы (т. е. уменьшение силового дефицита) обнаруживается в ситуациях, вызывающих сильную эмоциональную реакцию испытуемого, например после неожиданного выстрела. То же отмечается при гипнозе, приеме некоторых лекарственных препаратов. При этом положительный эффект (уменьшение силового дефицита) более выражен у нетренированных испытуемых и слабее или совсем отсутствует у хорошо тренированных спортсменов, например тяжелоатлетов (М. Икай и А. Штейнхауз, 1961).
Второй фактор. При одинаковых условиях измерения величина силового дефицита, по-видимому, тем больше, чем больше число одновременно сокращающихся мышечных групп. Например, когда измеряется максимальная произвольная сила только приводящих мышц большого пальца руки, силовой дефицит составляет у разных испытуемых 5—15% от максимальной силы этих мышц. При определении же произвольной силы двух групп мышц, приводящих этот палец и сгибающих его концевую фалангу, силовой дефицит возрастает до 20%.
Третий фактор. Значение этого фактора (степень совершенства произвольного управления мышцами) для проявляемой человеком максимальной произвольной силы доказывается различными экспериментами. Показано, например, что изометрическая тренировка, проводимая при определенном положении конечности, может вызвать значительное повышение максимальной произвольной силы, измеряемой в том же положении. Если измерения силы проводятся при другие положениях конечности, то прирост мышечной силы оказывается незначительным или отсутствует совсем. Если бы увеличение силы зависело лишь от прироста поперечника тренируемых мышц, то оно должно бы обнаружиться при измерениях в любом положении конечности. Однако увеличение произвольной мышечной силы выявляется в основном при измерениях в определенной (тренируемой) позе. Это означает, что в данном случае прирост силы обусловлен более совершенным, чем до тренировки, центральным управлением мышцами, т. е. совершенствованием координационных (нервных) механизмов.
Роль третьего фактора выявляется также при изучении показателя относительной произвольной силы *, которая определяется пу
* Так как у живого человека удается измерять только анатомический поперечник, для большинства мышц определяется не абсолютная произвольная их сила (отношение максимальной произвольной силы к физиологическому поперечнику), а относительная произвольная сила. Таким образом, последнее понятие шире и потому принято в нашем учебнике. В спортивной педагогике понятием «относительная сила» обозначают отношение максимальной произвольной силы к весу спортсмена.
98
тем деления максимальной произвольной силы на величину мышечного поперечника.
Как уже отмечалось выше, в период от 12 до 20 лет максимальная произвольная сила у юношей больше, чем у девушек (см. рис. 40, Л). В пределах этого возраста увеличивается и мышечная сила и мышечная масса. В результате относительная произвольная сила в среднем одинакова у мужчин и женщин и составляет около 6 кг/см2. Вместе с тем имеются значительные различия в относительной произвольной силе у разных людей. Таким образом, хотя степень использования силовых возможностей мышечного аппарата в среднем одинакова у мужчин и женщин разного возраста, она существенно отличается у разных людей. Эти различия могут быть отчасти следствием тренировки.
Так, по данным М. Икай, после 100-дневной тренировки с использованием изометрических упражнений максимальная произвольная сила выросла у не тренирующихся до этого людей на 92%, а площадь поперечного сечения тренируемых мышц только на 23%. В результате относительная произвольная сила увеличилась в среднем с 6,3 до 10 кг/см2. Следовательно, систематическая тренировка может повышать степень совершенства произвольного управления мышцами и таким образом уменьшать силовой дефицит.
Степень совершенства произвольного управления мышцами определяется эффективностью действия двух координационных механизмов— межмышечных и внутримышечных (см. выше). В частности, максимальная произвольная сила может быть снижена из-за «ненужного» напряжения мышц-антагонистов, которое уменьшает суммарное напряжение, развиваемое мышцами-агонистами (дефект механизмов межмышечной координации). В результате «дефектов» внутримышечной координации часть наиболее высокопороговых двигательных единиц не вовлекается в произвольную активность и не все активные двигательные единицы работают в режиме полного тетануса. Обычно силовой дефицит возникает как следствие обеих этих причин. Во всех таких случаях максимальная произвольная сила мышцы меньше, чем ее максимальная сила.
Как указывалось выше (см. § 6), наиболее высокопороговыми являются большие двигательные единицы мышцы. Вместе с тем их вклад в общее напряжение мышцы особенно велик, так как каждая из них содержит большое число мышечных волокон. Поэтому когда перед спортсменом стоит задача развивать мышечную силу, он должен систематически применять на тренировках упражнения, которые требуют проявления большой мышечной силы, составляющей не менее 2/з от его максимальной произвольной силы. В этом случае совершенствуется произвольное управление мышцами, и в частности механизмы внутримышечной координации, обеспечивающие включение как можно большего числа двигательных единиц основных мышц, в том числе наиболее высокопороговых, больших, двигательных единиц.
До сих пор речь шла об изометрической (статической) мышечной силе. Мышечная сила, измеряемая в условиях дмнамическо-
99
го режима работы мышц (концентрического или эксцентрического сокращения), обозначается как динамическая сила (F). Она определяется по ускорению (а), сообщаемому внешней массе (т) при концентрическом сокращении, или по замедлению (ускорению с обратным знаком) движения массы при эксцентрическом сокращении мышц. Такое определение основано на физическом законе, согласно которому
F = т • а.
При этом проявляемая мышечная сила зависит от величины перемещаемой массы: в некоторых пределах с увеличением веса перемещаемого тела показатели силы растут, а далее увеличение веса не сопровождается приростом показателя динамической силы.
При измерении динамической силы испытуемый выполняет движение, которое требует сложных внемышечной и внутримышечной координаций. Поэтому показатели динамической силы значительно отличаются у разных людей и при повторных измерениях у одного и того же человека колеблются больше, чем показатели изометрической (статической) силы.
Динамическая сила, измеряемая при концентрическом сокращении мышц, меньше, чем статическая (см. рис. 38). Конечно, такое сравнение проводится при максимальных усилиях испытуемого в обоих случаях и одинаковом суставном угле.
Как было отмечено выше (§ 4), в режиме эксцентрических сокращений (уступающий режим) мышцы способны проявлять динамическую силу, значительно превышающую максимальную изометрическую. Чем больше скорость движения, тем больше проявляемая динамическая сила при уступающем режиме сокращения мышц (см. рис. 38, Б).
У одних и тех же испытуемых при определенных условиях измерения обнаруживается умеренная корреляция между показателями статической и динамической силы (коэффициенты корреляции в пределах 0,6—0,8).
Увеличение динамической силы в результате тренировки может не вызывать повышения статической силы и наоборот. Изометрические упражнения увеличивают динамическую силу, но значительно меньше, чем статическую. Все это указывает на чрезвычайную специфичность тренировочных эффектов: использование определенного вида упражнений вызывает наиболее значительное повышение результата именно в этом виде упражнений. Такая специфичность тренировочного эффекта обнаруживается и в отношении многих других физиологических процессов (см. ниже).
К одной из разновидностей мышечной силы относят так называемую взрывную силу, которая характеризует способность к быстрому проявлению мышечной силы. Она в значительной мере определяет, например, высоту вертикального прыжка или прыжка в длину с места, переместительную скорость на коротких отрезках бега с максимально возможной скоростью и др. В качестве показателей взрывной силы используют отношение максимальной прояв
100
Ляемой силы к времени ее достижения или времени достижения половины этой силы. Показатели взрывной силы мало зависят от максимальной произвольной изометрической силы соответствующих мышечных групп. Так, изометрические упражнения, увеличивая статическую силу, незначительно изменяют показатели прыгучести (вертикального прыжка или прыжка с места в длину). Следовательно, физиологические механизмы, ответственные за взрывную силу, отличны от механизмов, определяющих статическую силу. Среди координационных факторов важную роль в проявлении взрывной силы играет характер импульсации мотонейронов активных мышц — частота их импульсации в начале разряда и синхронизация импульсации разных мотонейронов (см. § 6). Среди «мышечных» факторов определенное значение, видимо, имеют скоростные сократительные свойства мышечных волокон (см. § 10).
§10. Скоростные свойства мышц
Мышцы человека состоят из мышечных волокон, обладающих разной скоростью сокращения (см. § 5), причем мышечные волокна, входящие в состав одной двигательной единицы, имеют одинаковую скорость сокращения. Характеристика скоростных сократительных свойств мышцы в целом определяется количественным соотношением двигательных единиц с разными скоростями сокращения. Большинство мышц человека являются смешанными: они включают двигательные единицы с разными скоростями сокращения. Двигательные единицы части мышц у человека можно разделить на две группы: медленные двигательные единицы (со средним временем одиночного сокращения около 120 мсек.) и быстрые (со средним временем одиночного сокращения около 60 мсек.). Соотношение быстрых и медленных двигательных единиц в разных мышцах может заметно отличаться. Так, в трехглавой мышце плеча человека только 2% волокон имеют время одиночного сокращения, превышающие 60 мсек., тогда как в двуглавой мышце плеча таких медленных волокон более 30%. Соответственно «среднее» время одиночного сокращения у двуглавой мышцы больше, чем у трехглавой.
При рождении все мышечные волокна являются медленными. В первые недели жизни начинает происходить дифференцировка двигательных единиц. Скорость сокращения мышечных волокон одного типа двигательных единиц постепенно увеличивается, и они превращаются в быстрые. Мышечные волокна другого типа двигательных единиц остаются медленными, составляя медленные двигательные единицы мышц. Опыты на животных показали, что превращение мышечных волокон в медленные или быстрые регулируется специальными нервными влияниями, которым онн подвергаются со стороны своих мотонейронов. Быстрыми становятся мышечные волокна, иннервируемые «быстрыми» мотонейронами, для которых характерна относительно высокая частота импульсации. Медленные мышечные волокна связаны с «медленными» мотонейронами, которые имеют относительно низкую частоту импульсации. Если у животного произвести операцию с перекрестным перешиванием нервов — нерв, связанный с быстрой мышцей (состоящей в основном из быстрых мышечных волокон), подшить к медленной мышце, а нерв медленной мышцы подшить к быстрой мышце, — то через некэто-
101
рое время быстрая мышца превращается в медленную, а медленная — в быструю. Помимо изменения скоростных сократительных свойств мышечных волокон в них происходят и значительные биохимические сдвиги, изменения в характере кровоснабжения и др.
Тип нервных влияний, который регулирует сократительные, биохимические и другие свойства мышечных волокон, относится к трофическим влияниям, (от греч. trophos — витание) нервной системы. Этим типом влияний обусловлены изменения, возникающие в мышечном аппарате в процессе систематической тренировки. Предполагается, что непрерывное выделение АХ из нервных моторных окончаний даже в отсутствие нервных импульсов от мотонейронов (см. § 2) есть одно из проявлений этого нервнотрофического контроля за мышцей.
Отмеченная выше корреляция между частотным диапазоном импульсной активности мотонейронов и скоростными сократительными свойствами их мышечных волокон показывает, что частота импульсации мотонейронов каким-то (пока неизвестным) образом влияет на эти свойства. Действительно, если систематически раздражать быструю мышцу кошки импульсами тока с редкой частотой (до 10 имп/сек), то мышца становится медленной. Стимуляция той же мышцы с частотой 50 имп/сек не снижает скорости ее сокращения.
Скоростные сократительные свойства мышц, по-видимому, играют определенную роль в ряде характеристик мышечной деятельности. В частности, как отмечалось выше, градиент силы, или взрывная сила, зависит от скоростных сократительных свойств мышц. Чем быстрее сокращаются мышечные волокна, тем при прочих равных условиях выше скорость движения. Во всяком случае у спортсменов-легкоатлетов высших разрядов прослеживается четкая зависимость скоростных сократительных свойств мышц от специализации, требующей высокой переместительной скорости. Наиболее быстрыми мышцами-разгибателями голени обладают бегуны на 100 и 200 м. Прыгуны в длину и бегуны на 400 м имеют более медленные мышцы, чем спринтеры.
Некоторые виды тренировки могут сопровождаться замедлением скорости сокращения тренируемых мышц. Так, наиболее загружаемые мышцы толчковой ноги у прыгунов характеризуются некоторой замедленностью сокращения по сравнению с мышцами маховой ноги. Известно также, что изометрическая тренировка, вызывая увеличение силы тренируемых мышц, может вести к снижению скорости их динамического сокращения.
§ 11. Мышечная выносливость
Мышечную выносливость можно оценивать по предельному времени выполнения заданной статической или динамической работы. При статической работе она определяется по времени, в течение которого поддерживается постоянная сила давления или удерживается в постоянном положении некоторый груз. Выполненная при этом
102
«работа» может быть определена как произведение силы давления (F) или груза на время работы (<), т. е. как импульс силы: F»t. Для того чтобы сравнивать статическую работу, выполненную различными людьми или производимую разными мышцами, необходимо
Рис. 41. Показатели статической (Л) и динамической (Б, В, Г) выносливости:
А—связь между силой сокращения (в % от максимальной! и длительностью его удержания.
Б — соотношение между предельным временем работы (ось абсцисс) и предельной работой (ось ординат). Основная работающая мышца — трехглавая мышца плеча. Работа состояла в подъеме груза 6 кг (сплошные линии) и 10 кг (прерывистые линии) на постоянную высоту с разной частотой движений (от 9 до 25 в 1 мин.).
В — зависимость предельного времени (сплошные линии) и предельной динамической работы (прерывистые линии) от мощности работы, т. е. частоты движений (ось абсцисс). Работа состояла в подъеме груза 6 кг с разной частотой. Основная работающая мышца — трехглавая мышца плеча.
Г — динамическая работа критической мощности с разными частотой (ось абсцисс) и грузом (ось ординат).
либо вычислять (или прямо измерять) силу напряжения этих мышц (если известны величины длины плеч рычагов и углы приложения силы сокращающихся мышц), либо выражать проявляемую силу сокращения в долях (процентах) от максимальной произвольной силы этой же мышечной группы.
103
Предельное время статической работы (статическая выносливость) находится в обратной зависимости от F. На графике рис. 41, Л кривая «сила давления=предельное время работы» в левой части идет почти параллельно прерывистой вертикальной линии, которая пересекает ось абсцисс в точке, соответствующей примерно 20% от максимальной силы. Эта линия называется асимптотой кривой «сила давления (груз) — предельное время работы». Она характеризует критический уровень статической работы. На рис. 41, Л видно, что, когда требуемая сила давления (груз) составляет менее 20% от максимальной силы, т. е. меньше критического уровня, статическая работа может выполняться в течение очень длительного времени («бесконечно долго»). Если сила давления (груз) превышает критический уровень, то предельное время статической работы непродолжительно. В диапазоне силы давления (груза) 20—80% от максимальной силы предельное время статической работы уменьшается с увеличением силы давления (груза) согласно следующей зависимости:
t - к ”РМ (F/Z-MaKc)"’
где К— константа, п — показатель степени, равный примерно 2,5. Эта формула показывает, что даже небольшое снижение силы статического сокращения приводит к значительному удлинению времени, в течение которого возможно поддержание этого сокращения. Максимальное же произвольное сокращение мышц может поддерживаться лишь несколько секунд.
Если статическая работа периодически прерывается фазами отдыха, то общее ее количество, которое может быть выполнено, возрастает. В этих случаях кривая «сила давления (груз) —предельное время работы» (см. рис. 41, Л) смещается вправо. Этот сдвиг усиливается с увеличением длительности фаз отдыха.
Для измерения динамической работы можно использовать мощность выполняемой работы, т. е. отношение работы (выраженной в килограммометрах или джоулях) к времени, затраченному на ее выполнение. Выделяют два вида мощности — пиковую и критическую.
Пиковая мощность представляет собой максимальную мощность, которая может быть достигнута в какой-то момент выполнения динамической работы.
Пиковая мощность измеряется во время максимального произвольного динамического сокращения мышц при подъеме груза или сообщении ускорения определенной массе. В опытах на изолированных мышцах для измерения пиковой мощности они подвергаются тетанической электрической стимуляции. Укорачиваясь при сокращении, мышца поднимает груз. Пиковая мощность достигается при движении (укорочении) с оптимальной скоростью, равной менее половины от максимальной, и с оптимальной внешней нагрузкой. Последнюю определяют, используя соотношение «груз — скорость» (см. § 7).
Критическая мощность динамической работы — это такая мощность, которая может поддерживаться в течение длительного времени
ни (на протяжении многих часов). Она аналогична критическому уровню для статической работы. Если мощность динамической работы превышает критическую, то продолжительность такой работы ограничена.
Выносливость при динамической работе (динамическая выносливость) может быть оценена по предельному времени (/пред.) и предельной работе (ГпрСд.) заданной мощности. При выполнении динамической работы, мощность которой выше критической, время от начала до момента отказа от нее есть предельное время работы с данной мощностью (/пРед.). а общее количество работы, выполненной за этот отрезок времени, представляет собой предельную работу (Тпред.)- Оба эти показателя снижаются с увеличением и повышаются с уменьшением мощности выполняемой динамической работы. Между предельной работой (/’пред:) и предельным временем работы (/пред.) заданной мощности имеется линейная зависимость (см. рис. 41, Б):
^пред в Ь • /ПрСд_, |1|
где а и Ъ — константы, которые имеют размерность мощности работы (кгм/мин). Несколько схематизируя, эту формулу интерпретируют следующим образом. Предельная работа определяется суммой энергетического резерва (а) и энергии восстановления (b-t). Константа b отражает максимальную скорость восстановления энергетического потенциала сокращающимися мышцами. Если мощность выполняемой динамической работы ниже этой константы или равна ей, то такая работа может выполняться длительное время ( «бесконечно долго»). Если выполняемая работа имеет более высокую мощность, чем Ь, то предельное время такой работы ограничено. Таким образом, критическая мощность работы равна значению фактора Ь.
Как уже указывалось, предельное время динамической работы зависит от ее мощности. Мощность выполняемой работы может быть определена как отношение предельной работы к предельному времени ее выполнения:
р '/'пред, /пред.
После преобразований получается, что
'пред р_Ь
Это уравнение показывает, что при Р = Ь предельное время динамической работы не определяется (критическая мощность). В случаях, когда Р>Ь, предельное время является обратной функцией от мощности выполняемой работы. На графике (см. рис. 41, В) кривая мощность = предельное время» динамической работы имеет такой же характер (гипербола), как и кривая зависимости предельной длительности статической работы от силы статического сокра-
105
тения мышц. Критическая мощность динамической работы составляет менее '/ю от пиковой мощности.
Выносливость при статической и динамической работе зависит в. большой мерс от различных физиологических механизмов. Поэтому между этими двумя видами выносливости существует слабая связь. Соответственно систематическое применение упражнений, требующих статической выносливости, позволяет намного повышать ее, но мало изменяет динамическую выносливость. Длительные же динамические упражнения вызывают значительное повышение динамической выносливости без заметного изменения статической.
Между показателями мышечной силы и выносливости существует сложная связь. Между максимальной произвольной силой и статической выносливостью одной и той же мышечной группы имеется прямая зависимость: чем больше сила мышечной группы, тем длительнее удержание выбранного усилия (больше «абсолютная выносливость»). Иная связь между силой и выносливостью обнаруживается в опытах, в которых разные испутыемые удерживают одинаковые относительные усилия, например 60% от их силового максимума (при этом чем сильнее испытуемый, тем большее по абсолютной величине мышечное усилие он развивает). В этих случаях среднее предельное время работы («относительная выносливость») не отличается у людей с разной мышечной силой.
При определении силы и динамической выносливости мышц-сгибателей плеча и разгибателей бедра у неспортсменов и спортсменов различных специализаций не было обнаружено связи между этими показателями. В опытах, результаты которых проведены в табл. 2, испытуемые должны были поднимать груз в */з от максимальной силы данной мышечной группы в ритме одно движение в 1 сек. При этом средние показатели мышечной силы и динамической выносливости обнаруживают специфическую связь со спортивной специализацией. Например, как у мужчин, так и у женщин наиболее сильными мышцами ног обладают дискоболы, но у них самые низкие показатели динамической выносливости. Бегуны на средние и длинные дистанции по силе мышц ног не отличаются от неспортсменов, но у первых выявляется чрезвычайная большая динамическая выносливость. В то же время при работе руками у этих же бегунов не выявлено повышенной динамической выносливости. Все это еще раз свидетельствует о высокой специфичности тренировочных эффектов: больше всего повышаются функциональные показатели в тех упражнениях, которые являются основными в тренировке спортсмена.
Рассматривая физиологические механизмы, определяющие такие различия, следует иметь в виду, что изменения в структурных, биохимических и физиологических особенностях тренируемых мышц и в характере их нервной регуляции специфически связаны с используемыми в тренировках видами упражнений и режимом их применения. Поэтому тренировка, направленная преимущественно на развитие мышечной силы, совершенствует механизмы, способствующие улучшению этого качества, значительно меньше влияя на мышечную выносливость, и наоборот.
106
§12. Гладкие мышцы
К гладким мышцам относятся мышцы внутренних органов и стенок сосудов. Эти мышцы по ряду анатомических и физиологических характеристик отличаются от скелетных мышц, осуществляющих движения звеньев тела. Сократительный аппарат гладких мышц, как и скелетных, включает тонкие (актиновые) и толстые (миозиновые) нити. Эти нити в гладких мышечных клетках не имеют такого упорядоченного расположения, как в скелетных (поперечнополосатых). Поэтому под микроскопом у гладких мышц не видно поперечных полос. Отсюда название «гладкие».
Клетки гладких мышц имеют ветвления на обоих концах и плотно прилегают друг к другу, образуя длинные тонкие цилиндрические пучки диаметром 0,05—0,01 мм. Эти пучки ветвятся и через несколько миллиметров соединяются с другими пучками. Их сеть образует во внутренних органах либо слои (пласты), либо еще более толстые пучки. Соседние гладкомышечные клетки имеют два вида контактов друг с другом: первый — тесное соприкосновение мембран контактирующих клеток на относительно большом протяжении и второй—соединения в виде межклеточных плазматических мостиков. Оба вида контактов обеспечивают распространение возбуждения от одной гладкомышечной клетки к другой. Поэтому сеть таких клеток действует как единое целое: возбуждение, возникшее в группе клеток, распространяется на другие клетки сети.
Таблица 2
Мышечная сила и динамическая выносливость (по М. Икай, 1964)
Испытуемые Число испытуемых Разгибатели коленного сустава Сгибатели локтевого сустава
сила (кг) ВЫНОСЛИВОСТЬ (колич. движений) сила (кг) выносливость (колич. движений)
Мужчины Неспортсмены Спринтеры Бегуны на средние и длинные дистанции Барьеристы Прыгуны Дискоболы Женщины Неспортсменки Спринтеры Бегуньи на средние и длинные дистанции Барьеристки Прыгуньи Дискоболки 15 7 6 3 8 8 10 3 4 5 2 1 55 71 55 61 68 88 48 53 48 56 68 48 52 399 67 49 38 71 68 67 57 43 17 19 19 19 21 26 9 12 10 10 12 17 75 65 48 46 45 51 70 71 68 67 58 43
107
По функциональным особенностям гладкие мышцы делят на два типа — висцеральные (т. е. внутренние) гладкие мышцы, выстилающие стенки желудочно-кишечного тракта и мочевыделительного тракта, и унитарные гладкие мышцы, расположенные в стенках сосудов, в зрачке и хрусталике глаза и у корней волос кожного покрова (мышцы, взъерошивающие шерсть у животных).
Нервная регуляция гладких мышц осуществляется через симпатические и парасимпатические волокна вегетативной нервной системы.
Особенность висцеральных гладкомышечных клеток состоит в том, что они способны сокращаться и при отсутствии прямых нервных влияний. Так, гладкие мышцы кишечного тракта даже после перерезки нервных волокон продолжают ритмически сокращаться, обеспечивая перистальтику кишечника. Постоянный потенциал покоя в висцеральных гладкомышечных клетках отсутствует. Мембранный потенциал этих клеток медленно уменьшается («дрейфует»). В связи с тем, что такие изменения мембранного потенциала не зависят от нервных влияний, они получили название спонтанных изменений. Когда мембранный потенциал в результате «дрейфа» достигает критического уровня (примерно 30 мв), возникает потенциал действия, сопровождающийся волной сокращения мышечной клетки. Продолжительность потенциала действия очень большая — до нескольких секунд (папомним, что длительность потенциала действия у мышечных волокон скелетных мышц — несколько миллисекунд). Несколько секунд длится и одиночное сокращение гладкомышечных клеток. Таким образом, гладкие мышцы —это очень медленно сокращающиеся мышцы.
При возникновении потенциала действия в какой-нибудь группе мышечных клеток возбуждение распространяется от них к соседним, что приводит к их возбуждению и сокращению. В результате последовательно сокращается один участок мышечного слоя за другим. Скорость проведения возбуждения по сети гладкомышечных клеток не превышает 3—5 см/сек.
Спонтанная активность гладкомышечных клеток отчасти связана со способностью этих клеток возбуждаться при растяжении. Оно вызывает деполяризацию мембраны и возникновение потенциалов действия с последующим сокращением клеток. Частота потенциалов действия (и соответственно частота сокращений) тем больше, чем больше и быстрее растягивается гладкая мышца. Благодаря такому механизму обеспечивается, в частности, продвижение пищевого комка по пищеварительному тракту. Растянутая комком пищи мышечная стенка кишки отвечает сокращением и таким образом проталкивает комок в следующий участок кишки. Растяжение последнего приводит к сокращению его мышечного аппарата, что обеспечивает дальнейшее продвижение пищи.
Как уже отмечалось выше, волокна скелетных мышц сокращаются только тогда, когда к ним приходят импульсы от мотонейронов. Следовательно, нервная система осуществляет в данном случае пусковую роль. Висцеральные гладкие мышцы контролируются ве
108
гетативной нервной системой, которая осуществляет в отношении этих мышц не пусковую, а настроечную, регулирующую, функцию. Это означает, что сама активность висцеральных гладких мышц возникает спонтанно, без нервных влияний, но уровень этой активности (сила и частота сокращений) изменяется под влиянием вегетативной нервной системы. В частности, изменяя скорость «дрейфа» мембранного потенциала, нервные импульсы вегетативных волокон воздействуют на частоту сокращений висцеральных гладкомышечных волокон.
Унитарные гладкие мышцы также могут быть спонтанно активными, но они сокращаются в основном под влиянием нервных импульсов вегетативных волокон. Особенность унитарных гладкомышечных волокон состоит в том, что приходящий к ним отдельный нервный импульс не способен вызвать их сокращение. В ответ на такой импульс возникает лишь временная подпороговая деполяризация мембраны мышечной клетки, которая длится несколько десятых долей секунды. Когда по вегетативному нервному волокну следует серия импульсов с частотой около 1 импульса в 1 сек. и более, этого уже достаточно для суммации подпороговых сдвигов мембранного потенциала, развития потенциала действия мышечного волокна и его сокращений. В унитарной гладкой мышце, как и в висцеральных гладких мышцах, возбужденные мышечные клетки оказывают влияние на соседние клетки. В результате возбуждение захватывает много клеток (отсюда название этих мышц — унитарные, т. е. состоящие из унит — «единиц» с большим числом мышечных волокон в каждой из них).
В нервной регуляции сокращения гладких мышц участвуют два медиатора — ацетилхолин (АХ) и адреналин (норадреналин). Способ действия АХ в гладких мышцах такой же, каки в скелетных: АХ увеличивает ионную проницаемость мембраны, вызывая ее деполяризацию. Механизм действия адреналина неизвестен. Скелетные мышечные волокна реагируют на действие медиатора только в области концевой пластинки (нервно-мышечного синапса), тогда как гладкомышечные волокна отвечают на действие медиатора независимо от места его приложения. Поэтому на гладкие мышцы могут влиять медиаторы, содержащиеся в крови (например, адреналин, оказывающий длительное влияние на гладкие мышцы, вызывает их сокращение).
Характерной особенностью гладких мышц является то, что их сокращение не требует больших энергетических затрат,
Г л г в г IV
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Центральную нервную систему составляют спинной и головной мозг. Основными функциями центральной нервной системы являются: 1) регуляция деятельности всех тканей и органов и объединение
109
их в единое целое; 2) обеспечение приспособления организма к условиям внешней среды (организация адекватного поведения соответственно потребностям организма).
Управление различными функциями осуществляется и гуморальным путем (через кровь, лимфу, тканевую жидкость), однако нервная система играет главенствующую роль. У высших животных и человека ведущим отделом центральной нервной системы является кора больших полушарий, которая управляет также наиболее сложными функциями в жизнедеятельности человека — психическими процессами (сознание, мышление, память и др.).
§ 1. Физиология нервной клетки
Основным структурным элементом нервной системы является нервная клетка, или нейрон. Через нейроны осуществляется пере
дача информации от одного участка нервной системы к другому, обмен информацией между нервной системой и различными участ-
Рис. 42. Клетки центральной нервной системы.
Основные формы клеток коры больших полушарий — пирамидная, звездчатая и клетки глии.
ками тела. В нейронах происходят сложнейшие процессы обработки информации. С их помощью формируются ответные реакции организма (рефлексы) на внешние и внутренние раздражения.
Нейроны разделяются на три основных типа: афферентные, эфферентные и промежуточные нейроны. Афферентные нейроны (чувствительные, или центростремительные) передают информацию от рецепторов в цент
ральную нервную систему. Тела этих нейронов расположены вне центральной нервной системы — в спинномоз
говых ганглиях и в ганглиях
черепно-мозговых нервов.
Афферентный нейрон имеет ложноуниполярную форму, т. е. оба его отростка выходят из одного полюса клетки. Далее нейрон разделяется на длинный дендрит, образующий на периферии воспринимающее образование — рецептор, и аксон, входящий через задние рога в спинной мозг. К афферентным нейронам относят также нервные клетки, аксоны которых составляют восходящие пути спинного и головного мозга. Эфферентные нейроны (центробежные) связаны с передачей нисходящих влияний от вышележащих этажей нервной системы к нижележащим (например, пирамидные нейроны коры больших полушарий — рис. 42) или из центральной нервной системы к рабочим органам (например, в передних
НО
, более мелкие различными (в частности,
Яд.
MB
Рис. 43. Строение нейрона: Я — ядро, Яд. — ядрышко, М — митохондрии, Р — отдельные рибосомы и их скопления, Д — дендриты, С — синапсы, X — аксонный холмик, НС — начальный сегмент, А — аксон, Ш —-Шванновская клетка, составляющая миелиновую оболочку аксона, и ее ядро, П — перехват Ранвье. НМ — нервно-мышечное окончание, МВ — мышечное волокно..
рогах спинного мозга расположены тела двигательных нейронов, или мотонейронов, от которых идут волокна к скелетным мышцам; в боковых рогах спинного мозга находятся клетки вегетативной нервной системы, от которых идут пути к внутренним органам). Для эфферентных нейронов характерны разветвленная сеть дендритов и один длинный отросток — аксон. Промежуточные нейроны (интернейроны, или вставочные) — это, как правило, клетки, осуществляющие связь между афферентными и эфферентными) нейронами. Они передают нервные влияния в горизонтальном направлении (например, в пределах одного сегмента спинного мозга) и в вертикальном (например, из одного сегмента спинного мозга в другие — выше или нижележащие сегменты). Благодаря многочисленным разветвлениям аксона промежуточные нейроны могут одновременно возбуждать большое число других нейронов (например, звездчатые клетки коры — см. рис. 42).
Функциональное значение различных структурных элементов нервной клетки. Различные структурные элементы нейрона имеют свои функциональные особенности и разное физиологическое значение. Нервная клетка состоит из тела, или сомы (рис. 43), и различных отростков. Многочисленные древовидно разветвленные отростки дендриты (от греч. dendron — дерево) служат входами нейрона, через которые сигналы поступают в нервную клетку. Выходом нейрона является отходящий от тела клетки отросток аксон (от греч. axis — ось), который передает нервные импульсы дальше — другой нервной клетке или рабочему органу (мышце, железе). Форма нервной клетки, длина и расположение отростков чрезвычайно разнообразны и зависят от функционального назначения нейрона.
Среди нейронов встречаются самые крупные клеточные элементы организма. Размеры их поперечника колеблются от 6—7 мк (мелкие зернистые клетки мозжечка) до 70 мк (моторные нейроны головного и спинного мозга). Плотность их расположения в некоторых отделах центральной нервной системы очень велика. Например, в коре больших полушарий человека на 1 мм3 приходится почти 40 тыс. нейронов. Тела и дендриты нейронов коры занимают в целом примерно половину объема коры.
111
В крупных нейронах почти */з—'Д величины их тела составляет ядро. Оно содержит довольно постоянное количество дезоксирибонуклеиновой кислоты (ДНК). Входящие в его состав ядрышки участвуют в снабжении клетки рибонуклеиновыми кислотами (РНК) и протеинами. В моторных клетках при двигательной деятельности ядрышки заметно увеличиваются в размерах. Нервная клетка покрыта плазматической мембраной — полупроницаемой клеточной оболочкой, которая обеспечивает регуляцию концентрации ионов внутри клетки и ее обмен с окружающей средой. При возбуждении проницаемость клеточной мембраны изменяется, что играет важнейшую роль в возникновении потенциала действия и передаче нервных импульсов. Аксоны многих нейронов покрыты миелиновой оболочкой, образованной Шванновскими клетками, многократно «обернутыми» вокруг ствола аксона. Однако начальная часть аксона и расширение в месте его выхода из тела клетки — аксонный холмик лишены такой оболочки. Мембрана этой немиелинизированной части нейрона — так называемого начального сегмента-—обладает высокой возбудимостью.
Внутренняя часть клетки заполнена цитоплазмой, в которой расположены ядро и различные органоиды. Цитоплазма очень богата ферментными системами (в частности, обеспечивающими гликолиз) и белком. Ее пронизывает сеть трубочек и пузырьков — эндоплазматический ретикулюм. В цитоплазме имеются также отдельные зернышки — рибосомы и скопления этих зернышек — тельца Ниссли, представляющие собой белковые образования, содержащие до 50% РНК. Это белковые депо нейронов, где также происходит синтез белков и РНК. При чрезмерно длительном возбуждении нервной клетки, вирусных поражениях центральной нервной системы и других неблагоприятных воздействиях величина этих рибосомных зернышек резко уменьшается.
В специальных аппаратах нервных клеток — митохондриях совершаются окислительные процессы с образованием богатых энергией соединений (макроэргических связей АТФ). Это энергетические станции нейрона. В них происходит трансформация энергии химических связей в такую форму, которая может быть использована нервной клеткой. Митохондрии концентрируются в наиболее активных частях клетки. Их дыхательная функция усиливается при мышечной тренировке. Интенсивность окислительных процессов нарастает в нейронах более высоких отделов центральной нервной системы, особенно в коре больших полушарий. Резкие изменения митохондрий вплоть до разрушения, а следовательно, и угнетение деятельности нейронов отмечаются при различных неблагоприятных воздействиях (длительном торможении в центральной нервной системе, при интенсивном рентгеновском облучении, кислородном голодании и гипотермии).
Обмен веществ в нейроне. Основной особенностью обмена веществ в нейроне является высокая скорость обмена и преобладание аэробных процессов. Потребность мозга в кислороде очень велика (в состоянии покоя поглощается около 46 мл,/мин кислорода). Хотя 1)2
вес мозга по отношению к весу тела составляет всего 2%, потребление кислорода мозгом достигает в состоянии покоя у взрослых людей 25% от общего его потребления организмом, а у маленьких детей — 50%. Даже кратковременное нарушение доставки кислорода кровью может вызвать необратимые изменения в деятельности нервных клеток: в спинном мозгу — через 20—30 мин., в стволе головного мозга — через 15—20 мин., а в коре больших полушарий — уже через 5—6 мин.
Энерготраты мозга составляют */6 — 7в суточных затрат организма человека. Основным источником энергии для мозговой ткани является глюкоза. Мозг человека требует для обмена около 115 г глюкозы в сутки. Содержание ее в клетках мозга очень мало, и она постоянно черпается из крови.
Деятельное состояние нейронов сопровождается трофическими процессами — усилением в них синтеза белков. При различных воздействиях, вызывающих возбуждение нервных клеток, в том числе при мышечной тренировке, в их ткани значительно возрастает количество белка и РНК, при тормозных же состояниях и утомлении нейронов содержание этих веществ уменьшается. В процессе восстановления оно возвращается к исходному уровню или превышает его. Часть синтезированного в нейроне белка компенсирует его расходы в теле клетки во время деятельности, а другая часть перемещается вдоль по аксону (со скоростью около 1—3 мм в сутки) и, вероятно, участвует в биохимических процессах в синапсах.
Кровоснабжение нервных клеток. Высокая потребность нейронов в кислороде и глюкозе обеспечивается интенсивным кровотоком.
Кровь протекает через мозг в 5—7 раз скорее, чем через покоящиеся мышцы. Мозговая ткань обильно снабжена кровеносными сосудами. Наиболее густая сеть их находится в коре больших полушарий (занимает около 10% объема коры). В отдельных слоях ее средняя длина капиллярной сети достигает у человека 1 м на 1 мм3 ткани. Каждый крупный нейрон имеет несколько собственных капилляров у основания тела клетки, а группы мелких клеток окутаны общей капиллярной сетью. При активном состоянии нервной клетки она нуждается в усиленном поступлении через кровь кислорода и питательных веществ. Вместе с тем жесткий каркас черепа и малая сжимаемость нервной ткани препятствуют резкому увеличению кровоснабжения мозга при работе. Однако это компенсируется выраженными в мозгу процессами перераспределения крови, в результате которых активный участок нервной ткани получает значительно больше крови, чем находящийся в покое. Возможность перераспределения крови в мозгу обеспечена наличием в основаниях артериальных ветвей крупных пучков гладких мышечных волокон— сфинктерных валиков. Эти валики могут уменьшать или увеличивать диаметр сосудов и тем самым производить раздельную регуляцию кровоснабжения разных участков мозга.
Мышечная работа вызывает снижение тонуса стенок мозговых артерий. При развитии физического или умственного утомления то
5—113
113
нус артериальных сосудов повышается, что ведет к уменьшению кровотока через нервную ткань.
В головном мозгу имеется богато развитая система анастомозов между различными артериями, между венозными сосудами и между артериями и венами. Эта система уменьшает пульсацию внутричерепного кровотока, обусловленную ритмическими сокращениями сердца и дыхательными движениями грудной клетки. Уменьшение пульсовых колебаний способствует улучшению тканевого кровотока. Благодаря наличию артерио-венозных анастомозов пульсовые колебания кровотока передаются с артерий мозга на вены, минуя капилляры. Анастомоз между системами сонных и позвоночных артерий (Виллизиев круг) гарантирует постоянство кровотока в различных отделах головного мозга при любом положении головы по отношению к туловищу и направлению силы тяжести, связанном с изменением положения тела в пространстве.
Клетки глии. В процессах питания нервных клеток и их обмене веществ участвуют также окружающие нейрон клетки глии (глиальные клетки, или нейроглия). Эти клетки заполняют в мозгу все пространство между нейронами. В коре больших полушарий их примерно в 5 раз больше, чем нервных клеток. Капилляры в центральной нервной системе плотно окружены клетками глии, которые покрывают сосуд полностью или оставляют небольшую часть (15%) свободной. Выросты некоторых глиальных клеток расположены частично на кровеносных сосудах и частично на нейроне (см. рис. 42). Полагают, что расположение этих клеток между сосудом и нейроном указывает на их роль в снабжении нервных клеток питательными веществами из крови. Глиальные клетки активно участвуют в функционировании нейрона. Показано, что при длительном возбуждении в нейроне высокое содержание белка и нуклеиновых кислот поддерживается за счет клеток глии, в которых их количество соответственно уменьшается. В процессе восстановления после работы запасы белка и нуклеиновых кислот сначала нарастают в клетках глии, а затем и в цитоплазме нейрона. Глиальные клетки обладают способностью перемещаться в пространстве по направлению к наиболее активным нейронам. Это наблюдается при различных афферентных раздражениях и при мышечной нагрузке. Например, уже через 20 мин. плавания у крыс было обнаружено увеличение числа глиальных клеток вокруг мотонейронов переднего рога спинного мозга.
Возможно, клетки глии участвуют в условнорефлекторной деятельности мозга и в процессах памяти.
Основные функции нервной клетки. Основными функциями нервной клетки являются восприятие внешних раздражений (рецепторная функция), их переработка (интегративная функция) и передача нервных влияний на другие нейроны или различные рабочие органы (эффекторная функция).
Особенности осуществления этих функций позволяют разделить все нейроны центральной нервной системы на 2 большие группы: 1) клетки, передающие информацию на большие расстояния (из
114
одного отдела центральной нервной системы в другой, от периферии к центру, от центров к исполнительному органу). Это крупные, афферентные и эфферентные нейроны, имеющие на своем теле и отростках большое количество синапсов, как возбуждающих, так и
тормозящих, и способные к сложным процессам переработки посту-
пающих через них влияний; 2) клетки, обеспечивающие межнейроальныс связи в пределах ограниченных нервных структур (промежуточные нейроны спинного мозга, коры больших полушарий и др.). Это мелкие клетки, воспринимающие нервные влияния только через возбуждающие синапсы. Эти клетки не способны к сложным процессам интеграции локальных синаптических влияний потенциалов, они служат передатчиками возбуждающих или тормозя
Рис. 44. Межнейронные синапсы:
А — синапсы на теле и отростках гигантских пирамидных клеток Беца (по М. В. Моториной); И — синапс на шипике дендрита пирамидной клетки коры больших полушарий (по Дж. Экклсу)
Рис. 45. Схема проведения возбуждения в межнейронном синапсе (по Дж. Экклсу). Стрелка — одностороннее направление проведения через синапс,. Остальные пояснения даны в тексте
щих влияний на другие нерв-
ные клетки.
Воспринимающая функция нейрона. Все раздражения, поступающие в нервную систему, передаются на нейрон через определенные участки его мембраны, находящиеся в области синаптических контактов. В большинстве нервных клеток эта передача осуществляется химическим путем с помощью медиаторов. Ответом нейронов на внешнее раздражение является изменение величины мембранного потенциала.
Чем больше синапсов на
нервной клетке, тем больше воспринимается различных раздражений и, следовательно, шире сфера влияний на ее деятельность и возможность участия нервной клетки в разнообразных реакциях организма. На телах крупных мотонейронов спинного мозга насчитывают до 15000—20000 синапсов. Разветвления аксонов могут образовывать синапсы иа дендритах (аксодендритические синапсы) и на соме (теле) нервных клеток (аксосоматические синапсы), в ряде случаев — на аксоне (аксоаксональные синапсы). Наибольшее чис-
115
ло (до 50%) синапсов находится на дендритах. Особенно густо они покрывают средние части и окончания дендритных отростков, причем многие контакты расположены на специальных шипиковидных выростах, или шипиках (рис. 44), которые еще больше увеличивают воспринимающую поверхность нейрона. В мотонейронах спинного мозга и пирамидных клетках коры поверхность дендритов в 10—20 раз больше поверхности тела клетки.
Чем сложнее интегративная функция нейрона, тем большее развитие имеют аксодендритические синапсы (в первую очередь те, которые расположены на шипиках). Особенно они характерны для нейрональных связей пирамидных клеток в коре больших полушарий.
Промежуточные нейроны (например, звездчатые клетки коры) таких шипиков лишены.
Приходящие в пресинаптическую часть контакта нервные импульсы вызывают опорожнение синаптических пузырьков с выведением медиатора в синаптическую щель (рис. 45). Веществами, передающими нервные влияния в синапсах нервных клеток, или медиаторами, могут быть ацетилхолин (в некоторых клетках спинного мозга, в вегетативных ганглиях), норадреналин (в окончаниях симпатических нервных волокон, в гипоталамусе), некоторые аминокислоты и др. Диаметр пузырьков примерно равен ширине синаптической щели. В клетках передней центральной извилины коры больших полушарий у людей 18—30 лет синаптические пузырьки имеют диаметр 250—300 А при ширине синаптической щели 200— 300 А. Выделение медиатора облегчается тем, что синаптические пузырьки скапливаются вблизи от синаптической щели— в так называемых активных, или оперативных, зонах. Чем больше нервных импульсов проходит через синапс, тем больше пузырьков перемещается в эту зону и прикрепляется к пресинаптической мембране. В результате облегчается выделение медиатора последующими нервными импульсами.
Эффекты, возникающие при активации синапса, могут быть возбуждающими или тормозящими. Это зависит от качества медиатора и свойств постсинаптической мембраны. Возбуждающие нейроны выделяют возбуждающий медиатор, а тормозные — тормозной. Кроме того, один и тот же медиатор может оказывать различное воздействие в разных органах (например ацетилхолин возбуждает скелетные мышечные волокна и тормозит сердечные).
В состоянии покоя мембрана поляризована: снаружи регистрируется положительный заряд, а внутри — отрицательный. В нервной клетке мембранный потенциал покоя равен примерно 70 мв.
При возбуждающих влияниях увеличение проницаемости мембраны обусловливает вхождение положительно заряженных ионов натрия в клетку и, следовательно, уменьшение разности потенциалов по обе стороны мембраны, т. е. ее деполяризацию. В постсинаптической мембране данного участка клетки при этом регистрируется небольшое отрицательное колебание мембранного потенциала с амплитудой около 10 мв, или возбуждающий постсинаптический по-
ив
тенциал (сокращенно ВПСП), нарастающий примерно за 1, 2 мсек, до своего максимума и затем убывающий.
При тормозных воздействиях проницаемость мембраны увеличивается незначительно — главным образом для ионов калия (диаметр гидратированного иона калия меньше, чем диаметр иона натрия). Поскольку ионов калия больше внутри клетки, они выходят из нее наружу, увеличивая поляризацию мембраны, т. е. вызывают ее гиперполяризацию. При этом регистрируется положительное колебание с амплитудой около 5 мв — тормозный постсинаптический потенциал (сокращенно ТПСП). Обычно ВПСП и ТПСП длятся всего несколько миллисекунд. Однако в некоторых нейронах головного мозга постсинаптические потенциалы могут иметь значительно большую длительность: ВПСП — до 80 мсек., ТПСП — более 100 мсек.
Интегративная функция нейрона. Общее изменение мембранного потенциала нейрона является результатом сложного взаимодействия (интеграции) местных ВПСП и ТПСП всех многочисленных активированных синапсов на теле и дендритах клетки. На мембране нейрона происходит процесс алгебраического суммирования положительных и отрицательных колебаний потенциала. При одновременной активации нескольких возбуждающих синапсов общий ВПСП нейрона представляет собой сумму отдельных местных ВПСП каждого синапса. При одновременном возникновении двух различных синаптических влияний— ВПСП и ТПСП — происходит взаимное вычитание их эффектов. В конечном итоге реакция нервной клетки определяется суммой всех синаптических влияний. Преобладание тормозных синаптических воздействий приводит к гиперполяризации мембраны и торможению деятельности клетки. При сдвиге мембранного потенциала в сторону деполяризации повышается возбудимость клетки. Ответный разряд нейрона возникает лишь тогда, когда изменения мембранного потенциала достигают порогового значения — критического уровня деполяризации. Для этого величина ВПСП клетки должна составлять примерно 10 мв.
В крупных (афферентных и эфферентных) нейронах возбудимость различных участков мембраны неодинакова. В области начального сегмента нервной клетки (аксонного холмика и начальной немиелинизированной части аксона) имеется низкопороговая зона, мембрана которой обладает в несколько раз более высокой возбудимостью, чем на других участках клетки (порог возбудимости мембраны начального сегмента равен 10 мв, а порог возбудимости соматодендритической мембраны — 20—30 мв). В этой зоне с момента достижения критического уровня деполяризации начинается лавинообразное вхождение натрия в клетку и регистрируется потенциал действия (ПД). В ПД различают кратковременную высоковольтную часть, или спайк (пик), и длительные низкоамплитудные колебания — следовые потенциалы. ПД мотонейронов имеют амплитуду пика около 80—100 мв и длительность его около 1,5 мсек.
Эффекторная функция нейрона. С появлением ПД, который в отличие от местных изменений мембранного потенциала
117
(ВПСП и ТПСП) является распространяющимся процессом, нервный импульс начинает проводиться от тела нервной клетки вдоль по аксону к другой нервной клетке или рабочему органу, т. е. осуществляется эффекторная функция нейрона. Синапсы, расположенные ближе к возбудимой низкопороговой зоне на теле клетки (аксосо-матические), оказывают большее влияние на возникновение потенциала действия, чем более удаленные, расположенные на окончаниях дендритов (аксодендритические). Импульсы, приходящие через аксосоматический синапс, как правило, вызывают ответный разряд нейрона, а импульсы, действующие на аксодендритический синапс,— лишь подпороговое изменение его возбудимости. Так, разряды мотонейронов спинного мозга и пирамидных нейронов коры, вызывающие двигательные реакции организма, являются ответом на специфические аксосоматические влияния. Но возникнет ли этот ответ или нет, определяется характером воздействий, поступающих через аксодендритические синапсы от других нервных путей. Так складываются адекватные реакции, зависящие от многих раздражений, действующих на организм в данный момент времени, и осуществляется тонкое приспособление поведения к меняющимся условиям внешней среды.
Процессы, происходящие в активном нейроне, можно представить в виде следующей цепи: потенциал действия в пресинаптиче-ском окончании предыдущего нейрона—^-выделение медиатора в синаптическую щель—^увеличение проницаемости постсинаптической мембраны—>ее деполяризация (ВПСП) или гиперполяризация (ТПСП)—^-взаимодействие ВПСП и ТПСП на мембране сомы и дендритов нейрона—->сдвиг мембранного потенциала в случае преобладания возбуждающих влияний—^достижение критического уровня деполяризации—^-возникновение потенциала действия в низкопороговой зоне (мембране начального сегмента) нейрона—> распространение потенциала действия вдоль по аксону (процесс проведения нервного импульса)—^-выделение медиатора в окончаниях аксона (передача нервного процесса на следующий нейрон или на рабочий орган).
Таким образом, передача информации в нервной системе происходит с помощью двух механизмов — электрического (ВПСП, ТПСП, потенциалы действия) и химического (медиаторы).
§ 2. Физиологические показатели функционального состояния нейрона
Величина мембранного потенциала является основным параметром, который определяет значения важнейших показателей функционального состояния нейрона — его возбудимость и лабильность.
Возбудимость нейрона. Возбудимость нейрона — это его способность отвечать на синаптическое воздействие потенциалом действия. Она зависит от соотношения двух параметров — мембранного потенциала и критического уровня деполяризации (рис. 46). В нормальных условиях деятельности критический уровень деполяриза-118
ции нейрона относительно постоянный, поэтому возбудимость ней-
рона определяется в основном величиной мембранного потенциала.
Подпороговые изменения мембранного потенциала. Мембранный потенциал может иметь нормальную величину (для нервной клетки — около 70 мв), уменьшенную (явление деполяризации) или большую (явление гиперполяризации). Для по-
явления ПД при нормальном уровне мембранного потенциала требуется его сдвиг примерно на 10 мв, т. е. амплитуда порогового
ВПСП должна быть около 10 мв для достижения критического уровня деполяризации. Эта величина и называется порогом возбудимости нейрона (см. рис. 46, /1).
При активной деятельности нейрона в нем может некоторое время сохраняться состояние подпороговой деполяризации. Такое же состояние возникает при слабых синаптических воздействиях, когда появляются под-подпороговые ВПСП. При этом разница между уровнем мембранного потенциала и критическим уровнем деполяризации уменьшается (см.
Рис. 46. Возбудимость нейрона:
А — нормальная возбудимость; Б — повышенная возбудимость (исходная деполяризация мембраны и уменьшение пороговой величины ВПСП); В — пониженная возбудимость (исходная гиперполяризация мембраны и увеличение пороговой величины ВПСП); Г — сильное возбуждение нейрона (сверхпороговая деполяризация и возникновение не одного, а серии потенциалов действия).
Стрелки — пороговая величина возбуждающего постсииаптического потенциала (ВПСП), необходимая для возникновения потенциала действия. Исх. — исходный уровень поляризации мембраны; Кр — критический уровень. Ось ординат — мембранный потенциал (мв)
рис. 46, Б), т. е. требуется
уже меньшая сила раздражения (меньшая амплитуда порогового ВПСП) для появления ответного ПД. Возбудимость нейрона в
этом случае повышена.
При понижении возбудимости нервной клетки разница между уровнем мембранного потенциала и критическим уровнем деполяризации увеличена и, следовательно, требуется значительно большее, чем в норме, раздражение (большая амплитуда ВПСП) для появления ПД (см. рис. 46, В).
Сверхпороговое изменен не мембранного потенциала. При сильном возбуждении в нервной клетке могут возникать ВПСП очень большой амплитуды, которая значительно превосходит критический уровень деполяризации и длительное время сохраняется на этом сверхпороговом уровне. В результате в течение всего периода сверхпороговой деполяризации нейрон генерирует потенциалы действия (см. рис. 46, Г). Такие резкие сдвиги возбудимости нейронов возникают, например, под влиянием вышележащих отделов центральной нервной системы. Импульсы, приходящие к мотонейронам спинного мозга из головного мозга, способны вызывать мощные ВПСП с амплитудой 20—25 мв, которые гораздо выше пороговой величины (10 мв). При этом мотонейрон отвечает на одно раздражение залпом нескольких ПД. Этим дости
119
гается значительное усиление двигательного ответа при рефлекторных реакциях.
Величина деполяризации нервных клеток находится в линейной зависимости от частоты раздражающих импульсов. Высшие отделы мозга, посылая импульсы различной частоты к нейронам нижележащих отделов, регулируют их возбудимость, осуществляя контроль за ответными реакциями организма.
Изменения возбудимости нейрона при его деятельности. Протекание каждого ПД сопровождается закономерными изменениями возбудимости нейрона. В момент возникновения высоковольтной части —спайка нервная клетка не может ответить на раздражение новым ПД, т. е. является абсолютно невозбудимой. Эта фаза (абсолютная рефрактерная фаза) длится примерно 0,5 мсек. Затем, в период постепенного снижения деполяризации мембраны, возбудимость нейрона восстанавливается до исходного уровня (фаза пониженной возбудимости, или относительная рефрактерная фаза) и даже некоторое время может его превышать (супернормальная, или экзальтационная, фаза). После этого может наблюдаться еще некоторое понижение возбудимости во время развития следовой гиперполяризации (субнормальная фаза). Длительность описанных фаз изменения возбудимости определяет протяженность периодов повышенной активности (нормальная и супернормальная возбудимость) и пониженной активности (абсолютная и относительная рефрактерные фазы, субнормальная фаза) нейронов. Так, большинство нейронов промежуточного мозга и коры больших полушарий после очередного разряда импульсов во время протекания абсолютной и относительной рефрактерной фаз неактивны в течение примерно 100 мсек. Следовательно, наиболее удобным ритмом их спонтанной активности может быть ритм около 10 разрядов в 1 сек., который и является одним из факторов, определяющих частоту электрических колебаний на поверхности мозга в состоянии покоя.
Лабильность нейрона. Лабильность нейрона-—это скорость протекания элементарных реакций, лежащих в основе его возбуждения. Наиболее удобная мера лабильности нейрона — максимальная скорость протекания ПД. В качестве меры лабильности используют также максимальную частоту импульсов, воспроизводимых нейроном без трансформации в единицу времени, или оптимальную, наиболее устойчивую, частоту импульсов (текущая лабильность). Разные по функциям и величине нервные клетки обладают различными величинами лабильности. Даже в пределах одного нейрона лабильность разных его структур (дендритов, сомы, начального сегмента и аксона) резко различается.
Величина лабильности нейрона определяется уровнем его мембранного потенциала.
Лишь при определенной величине мембранного потенциала достигается оптимальный уровень возбудимости и лабильности нервной клетки, а также наиболее высокий уровень ее ритмической активности, что является важным условием для передачи информации
120
в нервной системе и осуществления целесообразных реакций. Например, наибольший максимальный ритм мотонейронов спинного мозга (до 100 импульсов в 1 сек.) отмечается при средней, оптимальной, величине мембранного потенциала — 55 мв (т. е. при повышенной возбудимости мотонейрона).
В естественных условиях деятельности частота разрядов мотонейрона обычно не превышает 50—60 импульсов в 1 сек. Лишь при мощных влияниях вышележащих отделов нервной системы резко изменяется мембранный потенциал мотонейронов и максимальная частота их разрядов может повыситься до 100 и даже до 300 импульсов в 1 сек. Однако разряды с такой частотой очень кратковременны. Они сменяются более медленным и устойчивым ритмом активности. В мелких промежуточных нейронах спинного мозга частота разрядов в начальные моменты активации достигает 600—• 800 импульсов в 1 сек., а иногда превышает 1000 импульсов в 1 сек. Такая частота необходима для резкого и быстрого воздействия этих клеток на мотонейроны; через некоторое время она резко замедляется.
Изменения в нейронах при активном и бездеятельном состояниях. Афферентные раздражения, возникающие при различных влияниях внешней и внутренней среды, в том числе при мышечной работе (с рецепторов мышц и сухожилий), повышают возбудимость и лабильность нейронов, их способность к стабильной ритмической активности. Бездеятельное же состояние снижает эти показатели, ухудшая функциональное состояние нервной клетки.
Активная деятельность клетки вызывает не только функциональные изменения, но и значительные структурные и биохимические перестройки различных ее элементов. Прогрессивные изменения наблюдаются в синаптическом аппарате нейронов. Эксперименты с тренировкой животных (тренировка задних конечностей собаки путем массажа, электрического раздражения и пассивной гимнастики, тренировка крыс с помощью плавания) показали, что средняя величина (диаметр) синапсов на мотонейронах поясничного отдела спинного мозга достоверно возрастает (примерно на 35%)- С увеличением размеров синапсов улучшается проведение нервных импульсов по рефлекторным путям.
Активное участие корковых нейронов в условнорефлекторной деятельности вызывает усиленное развитие шипикового аппарата на дендритах. Это способствует улучшению межнейронных взаимосвязей в коре больших полушарий. Подавление условнорефлектор-пой деятельности различными фармакологическими воздействиями приводит к ухудшению рецепторной функции нейронов — нарушению синаптических контактов (исчезновению шипиков), а затем и к чеформации дендритов.
При усилении афферентной импульсации, поступающей к нейрону, в нейрональных синапсах происходят увеличение числа синап-шческих пузырьков, их усиленное новообразование и интенсивное перемещение в оперативную зону пресинаптической мембраны. Кроме того, в пресинаптических образованиях отмечается еще боль
121
шее усиление (в 7 раз по сравнению с покоем) синтеза медиатора (ацетилхолина) и выраженное увеличение его выделения (в 70 раз). В нейронах повышается уровень окислительных процессов, возрастает количество белков и РНК в телах нервных клеток и в клетках глии.
При усиленной двигательной активности увеличиваются размеры эффекторных нейронов — пирамидных клеток коры и мотонейронов спинного мозга.
Все прогрессивные структурные, биохимические и функциональные изменения улучшают осуществление нервными клетками их сложных функций. Это лежит в основе совершенствования различных поведенческих реакций целого организма, в том числе двигательной деятельности и психических процессов у человека.
§ 3. Системы нервных клеток
Диффузная нервная сеть и процесс централизации. У простейших одноклеточных животных одна клетка осуществляет разнообразные функции. Передвижение этих животных обеспечивается либо выпячиванием части клетки (ложноножки) и переливанием в нее содержимого клетки (амебоидное движение, характерное и для белых клеток крови человека), либо с помощью специальных образований — ресничек или жгутиков.
Усложнение деятельности организма привело к разделению функций различных клеток —их специализации. Одни из них приобрели способность к сокращению (мышечные клетки), другие — к восприятию внешних и внутренних раздражений, переработке поступающей информации и передаче управляющих сигналов на органы движения и другие органы тела (нервные клетки). Наиболее простая форма организации нервной системы — диффузная нервная сеть низших беспозвоночных животных (губок, гидр, актиний и медуз). Нейроны такой сети имеют многочисленные взаимные связи, по которым возбуждение распространяется диффузно, по всем направлениям.
Более сложной формой является сегментарная организация нервной системы у высших беспозвоночных животных (червей, насекомых). Тело их состоит из имеющих одинаковое строение участков — сегментов, иннервация которых осуществляется расположенными в этих же сегментах нервными клетками.
С усложнением поведенческих реакций происходит развитие у позвоночных животных головного конца нервной системы — энце-фалона. В нем сосредоточиваются группы нервных клеток, управляющих важнейшими функциями, — нервные центры. При этом утрачивается автономность отдельных сегментов и все большая часть функций передается вышележащим отделам нервной системы. Этот процесс получил название энцефализации (централизация) функций.
С формированием коры больших полушарий происходит подчи
122
нение ей всех других отделов нервной системы, т. е. процесс корти-кализации.
Строение и функции центральной нервной системы у человека отражают этот эволюционный процесс. Более древние и менее сложно организованные ее отделы имеют более выраженную, сегментарную организацию. Это спинной мозг, продолговатый и средний, многие центры которых иннервируют расположенные на том же уровне участки тела (рис. 47). Находящиеся над ними более молодые отделы, так называемые надсегментарные — промежуточный
Рис. 47. Сегментарное распределение волокон задних корешков спинного мозга в коже.
Буквы Ct L и S обозначают отделы спинного мозга, цифры — номера сегментов.
мозг, мозжечок, кора больших полушарий, — не имеют непосредственной связи с органами тела, а управляют их деятельностью через сегментарные центры. Надсегментарные отделы приобретают по мере совершенствования все более важную роль в жизнедеятельности органов. Они не только подчиняют, но и значительно перестраивают функции нижележащих структур, оставляя им лишь самые элементарные рефлексы. Так, отделение спинного мозга от головного вызывает угнетение всех его рефлексов — спинальный шок, который выражен гораздо слабее и длится меньше (всего несколько минут) у рыб и земноводных и более глубок и длителен (несколько недель) у млекопитающих. После исчезновения явлений шока спинальная лягушка (лишенная головного мозга) может осуществлять довольно сложные координированные движения (например, удаление пальцами задней лапки кислотного раздражителя с бедра), у спинальных кошек и собак могут наблюдаться сгибание, разгибание, маятникообразные движения конечностей, а у человека при перерыве связи головного мозга со спинным — практически только сгибательные рефлексы (разгибательные движения у него ниже места перерыва спинного мозга почти не наблюдаются, перекрестный разгибательный рефлекс выражен намного слабее, чем у спинальной собаки).
Соматотопическое представительство функций. Системы нейронов, образующих восходящие пути от рецепторов или нисходящие пути к рабочим органам, расположены в определенном порядке —
123
по типу проекции «точку в точку». Так, каждый участок воспринимающей поверхности глаза (сетчатой оболочки с фоторецепторами) передает своп сигналы определенному участку в зрительной области коры больших полушарий. Эти чувствительные корковые центры расположены таким образом, что они как бы образуют экран, отражающий расположение фоторецепторов на периферии. Следовательно, наш мозг получает объективную информацию о пространственных особенностях внешнего мира. Определенная пространственная организация в нервной системе сигналов от различных органов чувств (от зрительных, слуховых, мышечных, тактильных и других рецепторов) позволяет человеку иметь необходимое представление о схеме пространства.
Таким же образом в мозгу обеспечивается представление о схеме тела. Порядок размещения афферентных волокон в проводящих путях и локализация нервных центров в проекционных областях разных этажей нервной системы соответствуют порядку размещения участков кожи и скелетных мышц в теле. Этому принципу отвечает представительство моторных функций в различных этажах нервной системы. Так, например, в самой верхней части передней центральной извилины коры больших полушарий находятся центры, посылающие импульсы к мышцам нижних конечностей, несколько ниже — моторные центры мышц туловища, еще ниже — моторное представительство мышц верхних конечностей и, наконец, центры мышц шеи и головы. Таков же порядок расположения этих центров в моторных ядрах нижележащих этажей нервной системы (например, в красном ядре среднего мозга).
Такая организация связей в центральной нервной системе, при которой пространственное расположение нейронов, связанных с определенными рецепторами или рабочими органами на периферии, отражает специфику их пространственного расположения в организме, называется соматопическим (от лат. somaticus — телесный и topicus — местный) или экранным представительством функций. В соматопической организации нервной системы важное значение имеют: 1) функциональная значимость отдельного органа тела и 2) роль данного участка мозга в его иннервации. В связи с этим возникает несоответствие размеров органа тела и размеров территории мозга, которая управляет его деятельностью. Например, мышцы пальцев руки выполняют чрезвычайно сложные функции в произвольных движениях человека. В их деятельности основную роль играют импульсы, идущие от коры больших полушарий. Этим объясняется то, что представительство (как сенсорное, так и в особенности моторное) этих мышц в коре занимает значительно большую территорию, чем представительство всех остальных мышц тела.
Механизмы взаимодействия нервных клеток. Нервные клетки функционируют в тесном взаимодействии друг с другом.
Значение нервных импульсов. Все взаимодействия между нервными клетками осуществляются благодаря двум механизмам: 1) влияниям электрических полей нервных клеток (элект-ротоническим влияниям) и 2) влияниям нервных импульсов.
124
Первые распространяются на очень небольшие территории мозга. Электрический заряд нервной клетки создает вокруг нее электрическое поле, колебания которого вызывают изменения электрических полей лежащих рядом нейронов, что приводит к изменениям их возбудимости, лабильности и проводимости. Электрическое поле нейрона имеет сравнительно небольшую протяженность — около 100 мк, оно быстро затухает по мере удаления от клетки и может оказывать воздействие лишь на соседние нейроны.
Второй механизм обеспечивает не только ближайшие взаимодействия, но и передачу нервных влияний на большие расстояния. Именно с помощью нервных импульсов происходит объединение отдаленных и изолированных участков мозга в общую, синхронно работающую систему, что необходимо для протекания сложных форм деятельности организма. Нервный импульс, следовательно, является основным средством связи между нейронами. Высокая скорость распространения импульсов и локальное их воздействие на избранную точку мозга способствуют быстрой и точной передаче информации в нервной системе. В межнейронных взаимодействиях используется частотный код, т. е. изменения функционального состояния и характера ответных реакций одной нервной клетки кодируются изменением частоты импульсов (потенциалов действия), которые она посылает к другой нервной клетке. Общее количество импульсов, отправляемых нервной клеткой в единицу времени, или ее суммарная импульсная активность, — важный физиологический показатель деятельности нейрона.
Значение синапсов. В передаче нервных импульсов от одного нейрона к другому большое значение имеют синапсы. Особенно много их в высших отделах нервной системы и у нейронов с наиболее сложными функциями. С совершенствованием условнорефлекторной деятельности в процессе эволюции животных организмов увеличивается число синаптических контактов на пирамидных нейронах коры больших полушарий, в частности за счет увеличения числа шипиков на их дендритах. Состояние синапсов на этих шипи-ках коррелирует с состоянием высшей нервной деятельности. При подавлении условных рефлексов некоторыми веществами (например, аминазином) контакты на шипиках разрушаются, исчезают сами шипики, а затем деформируются и дендритные отростки нейрона.
Способность синапсов передавать нервные импульсы непостоянна. Она повышается после активной деятельности синапса и снижается при отсутствии активности. Понижение функциональных возможностей синапсов (гипосинапсия) ведет к ухудшению проведения через них нервных импульсов, а их полное нарушение (асинапсия) вызывает окончательное разобщение нервных клеток.
При отсутствии активации синапса в течение нескольких дней уже наблюдаются уменьшение размера синаптических пузырьков, запасов медиатора, нарастание длительности синаптической задержки и т. п. Бездействие синаптических контактов на мотонейронах спинного мозга, вызванное перерезкой чувствительных путей от
125
соответствующих мышечных волокон, через 2—4 недели уменьшает их возбуждающее влияние на мембрану мотонейрона вдвое по сравнению с влияниями других афферентных путей, оканчивающихся на том же мотонейроне. У кролика через несколько дней содержания в абсолютной темноте, т. е. после полного бездействия рецепторных клеток глаза, уменьшаются синаптические пузырьки в окончаниях их аксонов.
Активная деятельность нервных клеток способствует улучшению синаптического взаимодействия между ними-, росту площади синаптических контактов, ускорению проведения через синапсы и т. д. Это наблюдается, например, при двигательной деятельности. Как показали эксперименты на животных, ежедневная принудительная ходьба их по 1 часу приводит к значительной активации синапсов в соответствующих рефлекторных дугах спинного мозга.
Синапсы являются самым чувствительным местом нервной системы. Именно синаптические контакты между нейронами первыми реагируют на различные повреждающие воздействия. При травмах нервной ткани, отеках, нарушениях кровообращения, рентгеновских облучениях, отравлениях и других неблагоприятных воздействиях в синапсах сначала возникают обратимые изменения — сильное набухание (увеличение их размеров в несколько раз) и деформация, а затем происходят необратимые изменения синапсов — их разрушение, в результате чего полностью нарушается нервная деятельность.
§ 4. Свойства нервных центров
Понятие о нервном центре. В сложных многоклеточных организмах животных и человека отдельная нервная клетка не в состоянии регулировать какие-либо функции. Все основные формы деятельности нервной системы связаны с участием в ее функциях определенных групп нервных клеток — нервных центров.
Нервным центром называют совокупность нервных клеток, необходимых для осуществления какой-либо функции. Эти центры отвечают соответствующими рефлекторными реакциями на внешнее раздражение, поступившее от связанных с ними рецепторов. Клетки нервных центров реагируют и на непосредственное их раздражение веществами, находящимися в протекающей через них крови (гуморальные влияния).
Для выявления функций нервного центра используют ряд методов: метод раздражения (усиление его деятельности), метод разрушения или удаления — экстирпации (нарушение регулируемой им функции), электрофизиологический метод — регистрация электрических явлений в нервном центре и др. Так, функциональное значение, а также локализация дыхательного центра в продолговатом мозгу были выявлены путем последовательных перерезок головного мозга. При перерезках выше области его расположения (выше продолговатого мозга) дыхание сохранялось, а при перерезках ниже продолговатого мозга дыхательные движения исчезали.
126
Сложные реакпии в целостном организме обычно связаны с участием многих нервных центров, расположенных в различных этажах центральной нервной системы. Например, произвольное изменение дыхания осуществляется человеком при участии центров коры больших полушарий, особого центра в среднем мозгу, дыхательного центра продолговатого мозга и центров спинного мозга, иннервирующих дыхательные мышцы.
Физиологические свойства нервных центров определяют характер ответных реакций. Эти свойства в значительной мере связаны с особенностями проведения нервных импульсов через синапсы, соединяющие различные нервные клетки.
Одностороннее проведение возбуждения. В отличие от нервного волокна, в котором возможно двустороннее проведение волны возбуждения от места нанесения раздражения, в нервных центрах вло-на возбуждения проводится только в одном направлении: раздражение задних (чувствительных) корешков спинного мозга вызывает возникновение нервного импульса в передних двигательных корешках, однако при раздражении передних корешков волна возбуждения в задних корешках не появляется. Это обусловлено особенностями синаптической передачи возбуждения в отдельных нервных клетках.
Поскольку проведение волны возбуждения от одного нейрона к другому через синапс происходит химическим путем — с помощью медиатора, а медиатор содержится лишь в пресинаптической части синапса и отсутствует в постсинаптической мембране, — проведение нервных влияний через синапс возможно только от пресинаптической мембраны к постсинаптической и невозможно в обратном направлении (см. рис. 45). В связи с этим поток нервных импульсов в рефлекторной дуге имеет определенное направление: от афферентных нейронов к вставочным и затем к эффекторным — мотонейронам и вегетативным нейронам.
Замедленное проведение возбуждения. В основе проведения нервных импульсов по цепочке нейронов лежат два различных механизма: электрический (проведение потенциала действия — ПД — по нервным волокнам) и химический (передача через синапс с помощью медиатора). Первый осуществляется с большой скоростью (до 100—140 м/сек), второй — в тысячу раз медленнее. Замедление проведения связано с затратой времени на процессы, происходящие от момента прихода пресинаптического импульса в синапс до появления в постсинаптической мембране возбуждающих или тормозных потенциалов. Этот интервал называется синаптической задержкой и составляет в мотонейронах и большинстве других клеток центральной нервной системы примерно 0,3 мсек. В вегетативной нервной системе длительность синаптической задержки больше — не менее 10 мсек. За это время пресинаптический импульс вызывает опорожнение синаптических пузырьков, происходит диффузия медиатора через синаптическую щель, увеличение под его влиянием ионной проницаемости постсинаптической мембраны и возникает постсинаптический потенциал. После этого требуется еще около
127
1,2 мсек, на развитие возбуждающего постсинаптического потенциала (ВПСП) до максимума, чтобы возник (в случае достижения критического уровня деполяризации) ПД. Таким образом, весь процесс передачи нервного импульса (от ПД одной клетки до ПД следующей клетки) через один синапс занимает примерно 1,5 мсек. При утомлении, охлаждении и ряде других воздействий длительность синаптической задержки возрастает. Если же для осуществления какой-либо реакции требуется участие большого числа нейронов (многих сотен и даже тысяч), то суммарная величина задержки проведения по нервным центрам — так называемое центральное время проведения — может составить десятые доли секунды и даже целые секунды. Поскольку проведение импульсов по нервным волокнам от периферических рецепторов в нервные центры и от нервных центров к исполнительным органам занимает сравнительно небольшое время, общее время от момента нанесения внешнего раздражения до появления ответной реакции организма (латентный период рефлекса) определяется центральным временем проведения. Величина латентного периода рефлекса служит важным показателем функционального состояния нервных центров и широко используется в практике. Латентный период двигательной реакции измеряется от момента подачи сигнала до момента начала движения (или до появления электрической активности соответствующей мышцы). При осуществлении человеком простых движений в ответ на внешние сигналы, например нажимание на кнопку при световом сигнале (рис. 48, Л), длительность латентного периода двигательной реакции составляет около 120—220 мсек., а при утомлении, недостатке кислорода и пр. она может увеличиваться до 300—500 и более миллисекунд. При более сложных движениях, например выполнении фехтовальщиком укола в мишень с выпадом в ответ на световой сигнал, этот период значительно больше — около 300— 400 мсек, и более (см. рис. 48, Б).
Укорочение латентного периода одной и той же двигательной реакции человека в процессе спортивной тренировки может отражать повышение функционального состояния нервных центров (ускорение проведения) и перестройку нервных цепей, участвующих в передаче нервных импульсов (укорочение проводящего пути).
Суммация. В ответ на одиночную афферентную волну, идущую от мышц к мотонейронам, в пресинаптической части синапса освобождается 1 квант медиатора. При этом в пресинаптической мембране мотонейрона обычно возникает ВПСП подпороговой величины— 0,1—0,2 мв, недостаточной для вызова ответной реакции клетки. Чтобы величина ВПСП достигла критического уровня (порога возникновения ПД — примерно 10 мв-), требуется суммация на мембране клетки многих подпороговых ВПСП.
Различают пространственную и временную суммацию. Пространственная суммация наблюдается в случае одновременного поступления нескольких импульсов в один и тот же нейрон по разным пресинаптическим волокнам. Одномоментное возбуждение синапсов в различных участках мембраны мотонейрона повышает амплитуду
128
суммарного ВПСП до пороговой величины, возникает ответный импульс мотонейрона и рефлекторно осуществляется двигательная реакция. Так, при синхронном раздражении трех афферентных нервных волокон — от короткой малоберцовой мышцы, длинного разгибателя пальцев и длинной малоберцовой мышцы — в мотонейроне длинной малоберцовой мышцы регистрируется ВПСП, равный сумме отдельных ВПСП, получаемых при изолированном раздражении каждого волокна (рис. 49, Б). Обычно требуется одновременная ак-
150 мсек.
1000 мсек.'
Рис. 48. Латентный период двигательной реакции человека:
А — нажимание на кнопку (/) в ответ на световой сигнал (2); Б — выполнение фехтовальщиком укола в мишень с выпадом по световому сигналу: 1 — отметка светового сигнала и электромиограмма трехглавой мышцы плеча вооруженной руки (справа — калибровка ЭМГ); 2— отметка момента нанесения укола.
150 мсек, и 450 мсек. — величины латентного периода двигательной реакции, 1000 мсек. — общая длительность двигательной реакции (латентный и моторный компоненты)
Рис. 49. Временная (Л) и пространственная (Б) суммация возбуждения в нервных центрах.
На А: ПД — потенциал действия, Кр.— критический уровень деполяризации, ВПСП — возбуждающие постси иапти-ческие потенциалы, МГ — механограмма мышечного сокращения, Р — после-, довательные подпороговые раздражения (1—6) одного и того же нервного' волокна.
На Б: ВПСП в мотонейроне длинной малоберцовой мышцы в ответ на раздражение (Р) нервных волокон, идущих от короткой малоберцовой мышцы (/), длиного разгибателя пальцев (2), длинной малоберцовой мышцы (3), и на одновременное раздражение всех трех нервов вместе (1+2+3) (по Дж. Экклсу)
тивация 50—100 афферентных волокон для получения ответа двигательной клетки спинного мозга. Временная суммация происходит при активации одного и того же афферентного пути серией последовательных раздражений. Если интервалы между поступающими импульсами достаточно коротки и ВПСП мотонейрона от предыдущего раздражения не успевают затухать, то последующие ВПСП накладываются друг на друга, пока деполяризация мембраны не достигнет критического уровня для возникновения ПД (см. рис. 49, Л).
Явления суммации могут наблюдаться при осуществлении отдельных рефлексов (например, рефлексов чихания и кашля в ответ
129
одиночные импульсы, В — пачка импульсов
на слабые раздражения слизистой оболочки дыхательных путей). Они лежат в основе формирования нейронных комплексов, определяющих поведение целого организма, в основе развития условных рефлексов.
Фоновая активность. Многие нервные клетки обладают способностью к постоянной импульсной активности. Их мембранный потенциал периодически колеблется, то увеличиваясь, то уменьшаясь. Каждый раз, когда его увеличение превышает критический уровень деполяризации (порог возбудимости клетки), возникает импульсный разряд. Такая автоматическая деятельность называется фоновой активностью нейрона (рис. 50). Ее могут вызывать случайные импульсы, циркулирующие в нервной системе, но она может проявляться и при отсутствии приходящих к нейрону раздражений — в результате изменений ионных и обменных процессов, колебаний температуры и других причин. В последнем случае ее называют спонтанной.
Фоновая активность играет важную роль в деятельности целого организма. Во-первых, импульсы, приходящие по афферентным путям, легче возбуждают клетки со спонтанной активностью, чем бездеятельные. Это облегчает протекание многих рефлекторных реакций. Во-вторых, клетки, обладающие постоянным ритмом фоновой активности, могут иметь значение водителя ритма (пейсме-кера), т. е. задавать этот ритм другим нервным клеткам. При этом возникают большие комплексы синхронно работающих клеток, в которых благодаря общему ритму особенно хорошо выражено взаимодействие между отдельными нейронами. В-третьих, некоторые такие клетки выполняют роль счетчиков времени в центральной нервной системе. Их деятельность помогает поддерживать определенный ритм движений, субъективно оценивать интервалы времени, через определенные сроки включать необходимые функции и т. п.
Автоматическая деятельность присуща клеткам дыхательного центра. Их спонтанные разряды позволяют поддерживать ритмические дыхательные движения не только при бодрствовании, но и во время сна человека, в условиях наркоза; периодические разряды дыхательного центра регистрируются даже при полной изоляции продолговатого мозга от других отделов центральной нервной системы.
Вызванная активность. Импульсные разряды нейрона, возникающие в ответ на внешнее раздражение, называются вызванной ак-
130
лишь на новые раздражители,
ззг
А
Б
HllIlllilJilHlinillllliWUL р
Рис. 51. Усвоение ритма в коре больших полушарий (Л) и в дыхательном центре продолговатого мозга (Б). На А: ЭЭГ — запись электрической актив-иости затылочной области коры у человека (зрительная проекция), Р — ритм световых мельканий.
На Б: ПГ — запись дыхательных движений (пеймограмма) кролика, Р — ритмическое раздражение блуждающего нерва^ Стрелки — начало усвоения ритма
тивностью. Ответ нервной клетки может возникнуть в форме одиночного ПД, серии импульсов с затухающей частотой, а также в виде пачек импульсов, появляющихся через определенные интервалы. В различных нейронах наблюдаются разные типы вызванной активности. Одни клетки дают разряды только в момент включения раздражителя (эффект включения) —это наиболее частая реакция клеток, другие — только в момент его выключения (эффект выключения), третьи — в обоих случаях. В центральной нервной системе имеются клетки, дающие разряды что обеспечивает ориентировочную реакцию. Известны также клетки, в которых отмечается вызванная активность при неправильных реакциях организма, т. е. сигнализирующие об ошибках, что позволяет вносить коррекции в действия человека.
Усвоение и трансформация ритма. При ритмических раздражениях вызванная активность нейрона может настроиться на ритм приходящих импульсов (явление усвоения ритма— рис. 51). Эта настройка деятельности нейрона на заданный извне ритм (реакция усвоения, перестройки или на-
вязывания ритма) имеет большое значение для организации взаимодействия между различными нервными центрами и различными этажами нервной системы, в частности для организации ритмических движений у человека.
Активность нейрона может перестроиться как на более высокий, так и на более низкий ритм. В результате такой способности многие нервные клетки получают возможность сонастраиваться, т. е. работать сообща, в одном комплексе, не мешая друг другу различной по темпу импульсацией.
Впервые вопрос о значении усвоения ритма для протекания реакций организма был поставлен А. А. Ухтомским и его учениками (Н. В. Голиковым, Е. К. Жуковым и др.). Особенно важно, что в процессе ритмического раздражения рефлекторной дуги нервные центры способны усвоить ритм активности, превышающий их первоначальную лабильность. Это явление лежит в основе повышения работоспособности отдельных рефлекторных дуг, а также целостного организма (в частности, это имеет место в спортивной тренировке).
Очень часто вызванная активность нейрона не соответствует ритму приходящей к данной клетке импульсации. Нервные клетки обладают свойством изменять частоту передающихся импульсов, т. е. свойством трансформации ритма. Характер ответного разряда
131
зависит, во-первых, от свойств раздражителя и, во-вторых, от функционального состояния нейрона (его мембранного заряда, возбудимости, лабильности). В обычных условиях чем сильнее раздражение, тем чаще проявляется ответная импульсация.
При высокой возбудимости нейрона может возникать учащение импульсации (мультипликация ритма). В этом случае афферентная волна вызывает длительные надпороговые ВПСП в клетке, и на каждое одиночное раздражение клетка отвечает залпом из нескольких ПД (пока ВПСП превышает пороговый уровень). При низком уровне возбудимости и лабильности нейрона чаще происходит уре-жение ритма, так как несколько приходящих импульсов должны суммироваться, чтобы наконец достичь порога возникновения ПД. В результате на несколько приходящих импульсов нейрон отвечает лишь одним импульсом.
В некоторых крупных клетках, например мотонейронах спинного мозга, имеются специальные механизмы, регулирующие ритм ответных реакций. После каждого ПД в этих клетках наблюдаются выраженные' следовые процессы — следовая гиперполяризация мембраны. В этот момент возбудимость клетки понижена. Следующий разряд возникает в клетке лишь тогда, когда заканчивается этот следовой потенциал. Поэтому ритм разрядов мотонейрона, направляющихся к скелетной мышце, обычно не превышает 60 импульсов в 1 сек. Мелкие клетки, например вставочные нейроны, такого механизма не имеют и могут давать в начальные моменты ответа очень высокую частоту разрядов — до 1600 импульсов в 1 сек., которая, однако, быстро затухает.
Следовые процессы. После окончания действия раздражителя активное состояние нервной клетки или нервного центра обычно продолжается еще некоторое время. Длительность следовых процессов различна: небольшая в спинном мозгу (несколько секунд или минут), значительно больше в центрах головного мозга (десятки минут, часы и даже дни) и очень велика в коре больших полушарий (до нескольких десятков лет). Различают явные и скрытые следы прежних раздражений. В явной форме сохраняются лишь кратковременные следовые процессы или последствия. Они могут быть одноименными — продолжение возникшего в клетке состояния (тормозное или экзальтационное последействие) или контрастными (возбуждение вслед за торможением, торможение вслед за возбуждением) .
Поддерживать состояние возбуждения или торможения в нервном центре могут импульсы, блуждающие в нервных цепях — задержавшиеся на длинных путях передачи или возвращающиеся повторно к нейрону по замкнутым цепям нейронов.
Значительно сложнее по природе скрытые следы, характеризующиеся отсутствием импульсации в нервных центрах. Предполагают, что длительное сохранение в нервной клетке скрытых следов со всеми характерными свойствами раздражителя основано на изменении структуры составляющих клетку белков (возможно, также и белков глиальных клеток).
132
Согласно биохимической теории памяти (X. Хиден), в процессе запоминания происходят структурные изменения в молекулах рибонуклеиновой кислоты (РНК), от которой зависит образование белков клетки. В результате этого в нервной клетке начинают синтезироваться «измененные» белки с отпечатками прежних раздражений, составляющие биохимическую основу памяти.
Эксперименты на крысах и других животных показали, что способность их запоминать следы прежних раздражений или способность к обучению коррелирует с изменением содержания рибонуклеиновой кислоты в нервной ткани. Интересные опыты были проведены на червях. У них вырабатывали условные рефлексы — включение света сопровождалось электрическим ударом, вызывающим у червя реакцию сжимания. После ряда сочетаний этих раздражений одно включение света заставляло червей сжиматься. Эту способность реагировать на включение света двигательной реакцией приобретали также и «необученные» черви после того, как их кормили кашицей из растолченных «обученных» червей или вводили им рибонуклеиновую кислоту, полученную из тканей «обученных» червей. Следовательно, приобретенный опыт может передаваться от одного организма к другому с химическими веществами.
При формировании новых двигательных навыков увеличивается содержание измененной рибонуклеиновой кислоты в соответствующих нервных центрах. Такие изменения, сохраняющиеся в двигательных нейронах, могут составлять основу так называемой «мышечной памяти». Известно, что человек, забывший в результате ранения одного из участков мозга начертания букв алфавита, может писать слова автоматически, пользуясь сохранившейся моторной памятью. Мышечная память позволяет человеку вспомнить заученные движения, которые он забыл после длительного перерыва.
Непродолжительные последействия (длительностью до 1 часа) лежат в основе так называемой кратковременной памяти, а длительные следы, связанные с биохимическими перестройками в клетках, — в основе долгосрочной памяти. Кратковременная память служит человеку недолго (например, спросив по телефону справочной необходимый номер телефона, можно набрать его и сразу забыть). Долгосрочная память обеспечивает длительное сохранение следов. Она имеет важнейшее значение в обучении человека (например, для усвоения определенного объема теоретических знаний, фиксирования в мозгу выработанных двигательных навыков).
§ 5. Координация деятельности центральной нервной системы
Для деятельности центральной нервной системы характерна определенная упорядоченность и согласованность рефлекторных ре акций, т. е. их координация. Взаимодействие двух нервных процессов — возбуждения и торможения, лежащих в основе всех сложных регуляторных функций организма, закономерности их одновремен
133
ного протекания в различных нервный центрах, а также последовательная смена во времени определяют точность и своевременность ответных реакций организма на внешние и внутренние воздействия.
Иррадиация и концентрация нервных процессов. Проведение афферентной волны по рефлекторной дуге вызывает в ее нервных центрах состояние возбуждения или торможения. Эти процессы при определенных условиях могут охватывать и другие рефлекторные центры. Распространение процесса возбуждения на другие нервные центры называют иррадиацией. Она осуществляется благодаря многочисленным взаимосвязям нейронов одной рефлекторной дуги с нейронами других рефлекторных дуг, так что при раздражении одного рецептора возбуждение в принципе может распространяться в центральной нервной системе в любом направлении и на любую нервную клетку.
Чем сильнее афферентное раздражение и чем выше возбудимость окружающих нейронов, тем больше нейронов охватывает процесс иррадиации. Это явление можно наблюдать на спинальной лягушке. Слабое давление на пальцы задней лапки вызывает ответный рефлекс сгибания этой же лапки. Небольшое усиление давления приводит к сгибанию другой задней лапки, хотя рецепторы последней не раздражаются. Этот ответ возникает в результате того, что в сферу возбуждения помимо нервных центров одноименной половины спинного мозга вовлекаются центры другой его половины. При еще более сильном раздражении волна возбуждения охватывает вышележащие и нижележащие нервные центры и вызывает движения- верхних конечностей (сначала на стороне тела, подвергшейся раздражению, а затем на противоположной).
Аналогичное явление иррадиации возбуждения можно наблюдать при действии различных раздражений в коре больших полушарий.
Иррадиация через некоторое время сменяется явлением концентрации процессов возбуждения в том же исходном пункте центральной нервной системы. Концентрация происходит в несколько раз медленнее, чем иррадиация нервных процессов.
Процесс иррадиации играет положительную роль при формировании новых реакций организма (ориентировочных реакций, условных рефлексов). Активация большого количества различных нервных центров позволяет отобрать из их числа наиболее нужные для последующей деятельности, т. е. совершенствовать ответные действия организма. Благодаря иррадиации возбуждения между различными нервными центрами возникают новые функциональные связи— условные рефлексы. На этой основе возможно, например, формирование новых двигательных навыков.
Вместе с тем иррадиация возбуждения может оказать и отрицательное воздействие на состояние и поведение организма. Так, иррадиация возбуждения в центральной нервной системе нарушает тонкие взаимоотношения, сложившиеся между процессами возбуждения и торможения в нервных центрах, и приводит к расстройству двигательной деятельности.
134
Торможение в центральной нервной системе. Явление торможения в нервных центрах было впервые открыто И. М. Сеченовым в 1862 г. Значение этого процесса было рассмотрено им в книге «Рефлексы головного мозга».
Опуская лапку лягушки в кислоту и одновременно раздражая некоторые участки головного мозга (например, накладывая кристаллик поваренной соли на область промежуточного мозга), И. М. Сеченов наблюдал резкую задержку и даже полное отсутствие «кислотного» рефлекса спинного мозга (отдергивания лапки). Отсюда он сделал заключение, что одни нервные центры могут существенно изменять рефлекторную деятельность в других центрах, в частности вышележащие нервные центры могут тормозить деятельность нижележащих. Описанный опыт вошел в историю физиологии под названием «Сеченовского торможения».
Тормозные процессы — необходимый компонент в координации нервной деятельности. Во-первых, процесс торможения ограничивает иррадиацию возбуждения, чем способствует его концентрации в необходимых участках нервной системы. Во-вторых, возникая в одних нервных центрах параллельно с возбуждением других нервных центров, процесс торможения тем самым выключает деятельность ненужных в данный момент органов, осуществляя координационную функцию. В-третьих, развитие торможения в нервных центрах предохраняет их от чрезмерного перенапряжения при работе, т. е. играет охранительную роль.
По месту возникновения различают пресинаптическое торможение и постсинаптическое.
Постсинаптическое торможение. Оно возникает в постсинаптической мембране нейрона в результате действия тормозного медиатора и связано с наличием в центральной нервной системе специальных тормозных нейронов. Это особый тип вставочных нейронов, у которых окончания аксонов выделяют тормозной медиатор. Природа тормозного медиатора в настоящее время точно не установлена. Нервные импульсы, подходя к тормозным нейронам, вызывают в них такой же процесс возбуждения, как и в других нервных клетках. В ответ по аксону тормозной клетки распространяется обычный ПД, по в отличие от других нейронов окончания аксона при этом выделяют не возбуждающий, а тормозной медиатор. Под влиянием этого медиатора возникает кратковременная гиперполяризация постсинаптической мембраны следующего нейрона и регистрируется тормозной постсинаптический потенциал (ТПСП). В результате тормозные клетки не возбуждают, а тормозят те нейроны, на которых оканчиваются их аксоны. Такой вид торможения называют прямым, так как оно возникает сразу, без предварительного возбуждения.
Специальные тормозные нейроны — это клетки Рэншоу в спинном мозгу и корзинчатые клетки в промежуточном мозгу. Клеткам Рэншоу принадлежит важная роль в координации деятельности спинного мозга. Большое значение, например, эти клетки имеют при регуляции деятельности мышц-антагонистов. Они обеспечивают
135
Пр.
Рис. 52. Торможение в центральной нервной системе (по Дж. Экклсу):
А — пресииаптическое торможение: Пр — преси-наптическое волокно, Т — тормозная клетка, Н — нейрон;
Б — возвратное (постсинаптическое) торможение: МН — мотонейрон, Т — тормозная клетка Рэншоу, М — мышца;
В — участие клеток Рэншоу (Т) в регуляции деятельности мышц-анта гон истов: торможение (—) мотонейрона мышцы-разгибателя (МР) при возбуждении (+) мотонейрона мышцы-сгибателя (МС), Р — мышца-разгибатель, С — мышца-сгибатель; Т — торможение нейронов промежуточного мозга (Тал.— таламус) с участием тормозной корзинчатой клетки (Т)
развитие торможения в мотонейронах мышц-антагонистов (рис. 52, В), что облегчает осуществление сокращения этих мышц. Клетки Рэншоу участвуют в регуляции уровня активности отдельных мотонейронов, ограничивая (тормозя) чрезмерное их возбуждение.
Корзинчатые клетки играют важную роль в регуляции деятельности высших отделов мозга — промежуточного мозга и коры больших полушарий. Они являются как бы воротами, которые пропускают или не пропускают импульсы, идущие в кору больших полушарий.
Это обусловлено тем, что эти клетки вызывают синхронное торможение большого числа связанных с ними нейронов промежуточного мозга, регулируя тем самым прохождение восходящих импульсов через эти нейроны, время поступления их в кору больших полушарий и ритм корковой активности (см. рис. 52, Г).
При возбуждении мотонейрона спинного мозга импульсы поступают по его аксону к мышечным волокнам и одновременно по коллатералям аксона — к тормозной клетке Рэншоу. Аксоны последней «возвращаются» к этому же мотонейрону, вызывая его торможение (см.
рис. 52, Б). Следовательно, через клетку Рэншоу мотонейрон может сам себя затормаживать. Чем больше возбуждающих импульсов посылает мотонейрон на периферию (а значит, и к тормозной клетке), тем сильнее это возвратное торможение (разновидность постсинаптического торможения). Такая замкнутая система действует как механизм саморегуляции нейрона, ограничивая степень его возбуждения и предохраняя от чрезмерной активности.
Пресииаптическое торможение. Оно возникает перед синаптическим контактом — в пресинаптической области. Окончания аксонов одной нервной клетки образуют аксоаксональный синапс на окончании аксона другой нервной клетки и блокируют передачу возбуждения в последнем (см. рис. 52, А). В области такого пресинаптического контакта развивается чрезмерно сильная деполяризация мембраны аксона, которая приводит к состоянию парабиоза (пессимального торможения, по Н. Е. Введенскому). Заторможенный концевой участок аксона прекращает проведение нервных импульсов к синапсу и, значит, к следующей нервной клетке. Этот вид торможения обусловливает ограничение притока афферентных импульсов к нервным центрам.
136
Доминанта. Активность нервных центров непостоянна, и преобладание активности одних из них над активностью других вызывает заметные перестройки в процессах координации рефлекторных реакций.
Исследуя особенности межцентральных отношений, А. Л. Ухтомский обнаружил, что если в организме животного осуществляется сложная рефлекторная реакция, например повторяющиеся акты глотания, то электрические раздражения моторных центров коры не только перестают вызывать в этот момент движения конечностей, но и усиливают и ускоряют протекание начавшейся цепной реакции глотания, оказавшейся главенствующей. Аналогичное явление наблюдалось при отравлении фенолом передних отделов спинного мозга лягушки. Повышение возбудимости мотонейронов приводило к тому, что отравленная лапка отвечала рефлексом потирания (отряхивания) не только на непосредственное раздражение ее кожи кислотой, но и на самые разнообразные посторонние раздражители: поднимание животного со стола в воздух, удар по столу, где оно сидит, прикосновение к передней лапке животного и др.
Подобные эффекты, когда разнообразные поводы вызывают не адекватный им ответ, а реакцию, уже подготовленную в организме, постоянно встречаются и в поведении человека (смысл этого точно передают, например, такие пословицы, как «у кого что болит, тот о том и говорит», «голодной куме все пирог на уме»).
В 1923 г. А. А. Ухтомский сформулировал принцип доминанты как рабочий принцип деятельности нервных центров.
Термином доминанта был обозначен господствующий очаг возбуждения в центральной нервной системе, определяющий текущую деятельность организма.
Основные черты доминанты следующие: 1) повышенная возбудимость нервных центров, 2) стойкость возбуждения во времени, 3) способность к суммации посторонних раздражений и 4) инерция доминанты. Доминирующий (господствующий) очаг может возникнуть лишь при определенном функциональном состоянии нервных центров. Одним из условий его образования является повышенный уровень возбудимости нервных клеток, который обусловливается различными гуморальными и нервными влияниями (длительными афферентными импульсациями, гормональными перестройками в организме, воздействиями фармакологических веществ, сознательным управлением нервной деятельностью у человека и пр.).
Установившаяся доминанта может быть длительным состоянием, которое определяет поведение организма на тот или иной срок. Способность стойко поддерживать возбуждение во времени — характерная черта доминанты. Однако далеко не всякий очаг возбуждения становится доминантным. Повышение возбудимости нервных клеток и их функционального значения определяется способностью суммировать возбуждение при поступлении любого случайного импульса.
Восходящие нервные импульсы могут направляться не только по прямому специфическому пути — в соответствующие проекционные
137
зоны мозга, но и через боковые ответвления — в любые зоны центральной нервной системы (см. § 6 этой главы). В связи с этим при наличии в каком-либо участке нервной системы очага с оптимальным уровнем возбудимости этот очаг приобретает возможность повышать свою возбудимость за счет суммирования не только собственных афферентных раздражений, но и посторонних, адресованных другим центрам. Не сила возбуждения, а способность накапливать и суммировать его превращает нервный центр в доминирующий. Явления суммации лучше всего выражены лишь при умеренном, оптимальном, повышении возбудимости нейронов. Это выражается в том, что доминанта легче всего подкрепляется слабыми раздражениями и гасится сильными.
Чем больше нейронов вовлечено в данный очаг возбуждения, тем прочнее доминанта и тем больше она подавляет деятельность других отделов мозга, вызывая в них так называемое сопряженное торможение. Нервные клетки, входящие в доминирующий очаг, расположены не обязательно в одном участке нервной системы. Чаще всего они составляют определенную систему клеток (по А. А. Ухтомскому, «созвездие», или констелляцию, нейронов), расположенных в разных этажах головного и спинного мозга. Такими сложными являются, например, доминанты, обеспечивающие выполнение мышечной работы. Внешним их выражением могут быть стационарно поддерживаемые движение и рабочая поза, а также исключение в этот момент других движений и поз. В эти доминанты включаются клетки разнообразных областей коры больших полушарий и подкорковых отделов, связанных с организацией двигательной деятельности, а также клетки различных эмоциональных и вегетативных центров (дыхательного, сердечно-сосудистого, терморегуляционного и др.).
Объединение большого числа нейронов в одну рабочую систему происходит путем взаимного сонастраивания на общий темп активности, т. е. путем усвоения ритма. Одни нервные клетки снижают свой более высокий темп деятельности, другие повышают низкий темп до какого-то среднего, оптимального, ритма. Доминирующая группа нервных центров, работающих в общем ритме, тормозит центры с иными ритмами активности. Значение явления усвоения ритма как механизма формирования доминирующего очага и механизма его функционального выделения из общей массы нервных клеток подтверждено в последнее время электрофизиологическими исследованиями на животных и человеке.
Важным свойством доминанты является инерция. Однажды возникшая доминанта может длительное время поддерживаться и после удаления первоначального стимула, например при осуществлении цепных двигательных рефлексов. Инерция выражается также в том, что доминанта может надолго сохраняться как следовое состояние (потенциальная доминанта). При возобновлении прежнего состояния или прежней внешней ситуации доминанта может возникнуть снова. Такое воспроизведение доминанты происходит в организме спортсмена условнорефлекторно в предстартовом состоя-
138
нин. когда в известной мере активизируются все те нервные центры, которые входили в рабочую систему во время предыдущих тренировок. Это проявляется в усилении всего комплекса функций, связанных с мышечной работой: центральных, мышечных, выделительных, сосудистых и др. Мысленное выполнение физических упражнений также воспроизводит (актуализирует) доминирующую систему центров, что обеспечивает тренирующий эффект представления движений и является основой так называемой идеомоторной тренировки.
В норме в нервной системе редко отсутствуют какие-либо доминанты. Бездоминантное состояние — это очень слабое возбуждение, разлитое более или менее равномерно по различным нервным центрам. Сходное состояние возникает у спортсменов в процессе полного расслабления, при аутогенной тренировке. Путем такого расслабления добиваются устранения мощных рабочих доминант и восстановления работоспособности нервных центров.
Как фактор поведения доминанта связана с высшей нервной деятельностью, с психологией человека. Доминанта является физиологической основой акта внимания. Она определяет характер восприятия раздражений из внешней среды, делая его односторонним, но зато более целеустремленным. При наличии доминанты многие влияния внешней среды остаются вне внимания, но зато более интенсивно улавливаются и анализируются те, которые особенно интересуют человека. Доминанта — мощный фактор отбора биологически и социально наиболее значимых раздражений.
Возникновение в коре больших полушарий доминантных состояний наблюдается в начале образования временных связей. Условный рефлекс образуется, когда доминантный очаг возбуждения начинает отвечать не на любое афферентное раздражение, а лишь на специфическое раздражение, ставшее сигнальным.
Поскольку доминанта связана с определенной реакцией, она определяет одностороннее выражение поведения. Чем выраженнее доминанта, тем больше она тормозит другие текущие рефлексы. Таким образом, из многих степеней свободы выбирается одна — при наличии в определенных двигательных центрах доминанты работает интенсивно лишь та часть мускулатуры, которая управляется этими центрами, а остальная выключена из сферы деятельности в результате сопряженного торможения. При этом заторможены также многие вегетативные центры. В начальный момент интенсивной мышечной работы почти полностью могут исчезать условные рефлексы: слюноотделительные, мигательные и др. Это обеспечивает целесообразность движений и экономичность энерготрат. Мощная двигательная доминанта при статических усилиях за счет сопряженного торможения приводит к задержке дыхания и угнетению деятельности сердечно-сосудистой системы.
По мере формирования двигательного навыка система доминирующих нервных центров совершенствуется. Из нее исключаются все лишние нервные центры, остаются лишь те, которые необходимы и достаточны для осуществления двигательной задачи.
139
Взаимодействие процессов возбуждения и торможения в центральной нервной системе. Важнейшей характеристикой деятельности нервных центров является постоянное взаимодействие процессов возбуждения и торможения как между разными центрами, так и в пределах каждого из них.
Взаимосочетанная (реципрокная) иннервация мышц-антагонистов. Для сгибательного движения в суставе необходимо не только сокращение мышц-сгибателей, но и одновременное расслабление мышц-разгибателей. При этом в мотонейронах мышц-сгибателей возникает процесс возбуждения, а в мотонейронах мышц-разгибателей — процесс торможения. При возбуждении же центров разгибателей, наоборот, тормозятся центры сгибателей. Такие координационные взаимоотношения между моторными центрами спинного мозга были названы взаимосочетанной или реципрокной иннервацией мышц-антагонистов.
Появление и усиление в нервных центрах процесса торможения при одновременном возбуждении других центров получило по аналогии с физическими процессами название индукции (в данном случае это одновременная индукция). В настоящее время выяснены механизмы проявления реципрокной иннервации. Афферентное раздражение (например, болевое раздражение рецепторов кожи) направляется не только по собственному рефлекторному пути к мотонейронам мышц-сгибателей, но и одновременно через коллатерали активирует тормозные клетки Рэншоу. Окончания этих клеток образуют тормозные синапсы на мотонейронах мышц-разгибателей, вызывая в них торможение.
Реципрокные отношения между центрами мышц-антагонистов не являются постоянными и единственно возможными. В необходимых ситуациях (например, при фиксации суставов, при точностных движениях) они сменяются одновременным их возбуждением. В этом проявляется большая гибкость, целесообразность сложившихся в организме координаций.
Показано, что у человека во время ходьбы и бега основной формой координации являются реципрокные отношения, но помимо них имеются фазы одновременной активности мышц-антагонистов голеностопного и, особенно коленного и тазобедренного суставов. Длительность фаз одновременной активности увеличивается с повышением скорости перемещения.
Взаимосочетанные (реципрокные) отношения характерны не только для моторных центров спинного мозга, но и для других центров. Еще в 1896 г. Н. Е. Введенский в опытах на животных наблюдал при раздражении двигательной зоны одного полушария коры головного мозга реципрокное торможение моторных центров другого полушария. При этом сокращение мышц одной половины тела сопровождалось расслаблением одноименных мышц другой.
Реципрокные отношения формируются также при возникновении доминанты, когда при возбуждении одних центров с помощью сопряженного торможения выключается деятельность других, посторонних, нервных центров.
140
Последовательная смена процессов возбуждения и торможения. Взаимоотношения процессов возбуждения и торможения в центральной нервной системе могут проявляться во времени в виде последовательной смены возбуждения и торможения в одних и тех же нервных центрах.
«Возбуждение вслед за торможением» впервые наблюдал И. М. Сеченов. Он описал резкое усиление у лягушки рефлекторной деятельности после ее торможения сильным раздражителем: резкий прыжок с голосовой реакцией и восстановлением кожной чувствительности («рефлекс Сеченова»). Позднее было обнаружено «торможение вслед за возбуждением» (А. А. Ухтомский). После сильных ритмических раздражений лапки лягушки выключение раздражителя приводит к мгновенному расслаблению лапки—поднятая конечность падает, как плеть. Описанные взаимоотношения процессов возбуждения и торможения часто встречаются в коре больших полушарий при условнорефлекторной деятельности.
Контрастное усиление одного процесса после другого в одном и том же нервном центре получило название последовательной индукции. Оно имеет большое значение при организации ритмической двигательной деятельности, обеспечивая попеременное сокращение и расслабление мышц.
Принцип конвергенции. К одной и той же нервной клетке благодаря многочисленным побочным взаимосвязям рефлекторных дуг могут поступать импульсы от различных рецепторов тела, т. е. сигналы о самых разнообразных раздражениях. Схождение импульсов, поступивших по различным афферентным путям, в каком-либо одном центральном нейроне или нервном центре называется конвергенцией.
В низших отделах нервной системы — спинном и продолговатом мозгу — конвергенция выражена гораздо меньше. Нейроны этих отделов получают информацию от рецепторов сравнительно небольших участков тела — рецептивных полей одного и того же рефлекса. В надсегментарных отделах, особенно в коре больших полушарий, происходит конвергенция импульсов различного происхождения от разных рефлекторных путей. Нейроны надсегментарных отделов могут получать сигналы о световых, звуковых, проприоцептивных и прочих раздражениях, т. е. сигналы разной модальности. На теле нейронов постоянно изменяются «конвергентные узоры» — возбужденные и заторможенные участки. Подсчитано, что размеры рецептивных полей корковых нейронов, т. е. участков тела, от которых к ним могут поступать афферентные раздражения, в 16— 100 раз больше, чем размеры тех же полей для афферентных клеток спинальных рефлекторных дуг. Благодаря такому разнообразию поступающей информации в нейронах вышележащих отделов головного мозга может происходить ее широкое взаимодействие, сопоставление, отбор, выработка адекватных реакций и установление новых связей между рефлексами.
Принцип общего конечного пути. Афферентных нейронов в центральной нервной системе в несколько раз больше, чем эфферентных.
141
В связи с этим многие афферентные влияния поступают к одним и тем же вставочным и эфферентным нейронам, которые являются для них общими конечными путями к рабочим органам. Система реагирующих нейронов образует таким образом как бы воронку («воронка Шеррингтона»), Множество разнообразных раздражений может возбудить одни и те же мотонейроны спинного мозга и вызвать одну и ту же двигательную реакцию (например, сокращение мышц-сгибателей верхней конечности). Английский физиолог Ч. Шеррингтон, установивший принцип общего конечного пути, предложил различать союзные (аллиированные) и антагонистические рефлексы. Встречаясь на общих конечных путях, союзные рефлексы взаимно усиливают друг друга, а антагонистические — тормозят. В первом случае в нейронах общего конечного пути имеет место пространственная суммация (например, сгибательный рефлекс усиливается при одновременном раздражении нескольких участков кожи). Во втором случае происходит борьба конкурирующих влияний за обладание общим конечным путем, в результате чего один рефлекс осуществляется, а другие затормаживаются. При этом освоенные движения выполняются с меньшим трудом, так как в их основе лежат упорядоченные во времени синхронизированные потоки импульсов, которые проходят через конечные пути легче, чем импульсы, поступающие в случайном порядке.
Преобладание на конечных путях той или иной рефлекторной реакции обусловлено ее значением для жизнедеятельности организма в данный момент.
В таком отборе важную роль играет наличие в центральной нервной системе доминанты. Она обеспечивает протекание главной реакции. Например, ритмический шагательный рефлекс и одиночный, непрерывный рефлекс сгибания при болевом раздражении являются антагонистическими. Однако спортсмен, внезапно получивший травму, может продолжать бег к финишу, т. е. осуществлять ритмический рефлекс и подавлять болевые раздражения, которые, поступая к мотонейронам сгибательных мышц, препятствуют попеременному сгибанию и разгибанию ноги.
§ 6. Специфические и неспецифические системы
В нервной системе в настоящее время различают два отдела — специфическую и неспецифическую системы. Специфическая система расположена в наружных и боковых частях центральной нервной системы, а неспецифическая занимает срединную ее часть (рис. 53). Эти системы различаются по строению и функциональным особенностям.
К специфической системе относятся все пути и нервные центры, проводящие афферентную импульсацию от различных рецепторов тела и эфферентную импульсацию к рабочим органам. Это пути сенсорных систем и нисходящие пути.
Афферентные пути этой системы проводят сигналы какой-либо специфической чувствительности (например, мышечно-суставной.
142
тактильной, слуховой, зрительной и других) от рецепторов, воспринимающих- раздражения определенного качества, к соответствующим нервным центрам, где происходит анализ этих сигналов и воз
Рис. 53. Специфические и неспецифические отделы центральной нервной системы (по Р. Галамбосу)
никают ощущения и восприятия.
Эфферентные импульсы от нервных центров к исполнительным органам тела — мышцам и железам — направляются по специфическим нисходящим путям, образующим проводящие пути, или ^тракты (например, пирамидный тракт), и служат для управления определенными функциями на периферии (например, движениями скелетных мышц, изменением просвета сосудов и т. д.). Это эфферентные отделы, обеспечивающие различные рефлекторные реакции, описание которых дано в соответствующих разделах учебника.
Следовательно, деятельность специфической системы связана с анализом различных раздражителей и определенным характером ответных реакций организма. Б этой деятельности активное участие принимает также неспецифическая система организма, регулирующая и изменяющая как восприятие организмом специфических раздражений, так и его эфферентную деятельность.
Неспецифическая система не
связана с анализом какой-либо специфической чувствительности или с выполнением определенных рефлекторных реакций. Им пульсация в эту систему поступает через боковые ответвления от специфических путей, передающих проприоцептивные, слуховые и другие специфические сигналы. К одному и тому же нейрону могут приходить импульсы различного происхождения и от разных рецепторов тела. В результате такого схождения (конвергенции) импульсов создаются широкие возможности для их взаимодействия. Вследствие этого неспецифическая система играет большую роль в процессах интеграции функций в организме.
Характерной особенностью нейронов неспецифической системы являются также обилие и разнообразие их отростков. Они обеспечивают широкую циркуляцию импульсов в неспецифической системе. Благодаря этому здесь возможно взаимодействие одной клетки почти с 30 000 других нейронов. По характеру расположения нейронов и обилию их связей неспецифические отделы ствола головного мозга получили название сетевидного образования (или ретикулярной формации).
Функции ретикулярной формации. Различают два основных типа влияния неспецифической системы на работу других нервных цент
143
ров — активирующее и тормозящее влияния. Оба они могут адресоваться как вышележащим центрам (восходящие влияния), так и нижележащим (нисходящие влияния).
Восходящие влияния. В опытах на животных было показано, что из сетевидного образования среднего мозга исходит мощное активирующее влияние на кору больших полушарий. Электрические раздражения этих отделов неспецифической системы через вживленные электроды вызывали пробуждение спящего животного. У бодрствующего животного подобное раздражение повышало уровень корковой активности, усиливало внимание к внешним сигналам и улучшало их восприятие. Эти опыты свидетельствуют о большом значении импульсов среднемозговых отделов неспецифической системы в повышении функционального состояния нейронов коры больших полушарий. Восходящие влияния из этих областей охватывают диффузно всю кору, вызывая общее изменение ее функционального состояния. Они особенно усиливаются при действии новых раздражений, заранее подготавливая организм к реакции на неожиданную ситуацию. Это всегда имеет место при ориентировочных реакциях и в начальных стадиях выработки условного рефлекса. В результате активирующих воздействий во всех областях коры больших полушарий повышается возбудимость и лабильность корковых нейронов, быстрее возникают ответы на внешние раздражения и легче происходит вовлечение новых нервных клеток в протекающую реакцию. Увеличение числа активированных нейронов позволяет в процессе обучения, тренировки «выбрать» из них клетки, наиболее важные для осуществления ответных действий организма.
Активирующие влияния, исходящие из промежуточного мозга (от неспецифических ядер таламуса), в отличие от среднемозговых, воздействуют лишь на ограниченные области коры. Считают, что такое избирательное повышение активности небольшой территории коры имеет значение в организации внимания, выделяя на общем фоне работу небольшого числа корковых клеток. Обычно активируются нейроны именно тех отделов коры, которые заняты в данный момент наиболее важной для организма деятельностью. Это в значительной мере способствует протеканию специфических функций мозга — восприятию и переработке наиболее нужных сигналов, осуществлению первоочередных двигательных актов.
Неспецифическая система способна оказывать на кору помимо активирующих и тормозящие влияния. Эти влияния различны по характеру: локальные — из области промежуточного мозга и общие (диффузные) — из задних отделов мозга. Первые участвуют в создании «рабочей мозаики» в коре, разделяя ее на более активные и менее активные участки. Вторые обусловливают диффузное угнетение корковых функций. Широко распространенное по всей коре торможение деятельности корковых нейронов возникает с участием заднёстволовых отделов ретикулярной формации — неспецифических нейронов задних частей среднего и продолговатого мозга. Оно имеет место в случае длительной и монотонной работы (например,
144
в производственных условиях при работе на конвейере или в спорте при прохождении длинных и сверхдлинных дистанций).
В электрических проявлениях деятельности мозга активирующие влияния проявляются в виде возникновения частой асинхронной активности (десинхронизация), а тормозящие влияния-—в виде медленных ритмичных колебаний (синхронизация).
Нисходящие влияния. Все отделы неспецифической системы оказывают помимо восходящих значительные нисходящие влияния. Отделы ствола мозга регулируют (активируют или угнетают) активность нейронов спинного мозга и проприорецепторов мышц (мышечных веретен). Эти влияния совместно с воздействиями из экстрапирамидной системы и мозжечка играют большую роль в регуляции тонуса мышц и обеспечении позы человека. Непосредственные команды к осуществлению движений и влияния, формирующие перестройки тонуса мышц, передаются по специфическим путям. Однако неспецифические влияния позволяют существенно изменить протекание этих реакций. При усилении активирующих воздействий из ретикулярной формации среднего мозга на нейроны спинного мозга увеличивается амплитуда производимых движений и повышается тонус скелетных мышц. Включение этих влияний при некоторых эмоциональных состояниях помогает повысить эффективность двигательной деятельности человека и выполнить значительно большую работу, чем в обычных условиях.
Таким образом, в формировании целостных актов организма участвуют обе системы головного мозга — специфическая и неспецифическая. Проведение и обработку специфической информации, а также управление ответными реакциями осуществляет специфическая система. На эти процессы значительно влияет неспецифическая система. В свою очередь, деятельность этого неспецнфического отдела контролируется специфическими системами мозга. В объединении функций обеих систем и общем контроле за их активностью ведущая роль принадлежит коре больших полушарий.
§ 7. Основные рефлекторные центры нервной системы
Спинной мозг. Спинной мозг является низшим и наиболее древним отделом центральной нервной системы. Он имеет значительно меньшую самостоятельность у человека по сравнению с животными. У человека его вес по отношению к головному мозгу составляет всего 2% (у кошек — 25%, у кролика — 45%, у черепах— 120%).
Надежность сегментарных функций спинного мозга обеспечена множественностью его связей с периферией: каждый сегмент спинного мозга иннервирует 3 метамера (участка) тела — собственный, половину вышележащего и половину нижележащего, а каждый метамер тела получает иннервацию от 3 сегментов спинного мозга. Такое устройство гарантирует осуществление функций спинного мозга при возможных его перерывах и других поражениях.
Распределение функций входящих и выходящих волокон спинного мозга подчиняется определенному закону: все чувствительные
6—113
145
(афферентные) волокна входят в спинной мозг через его задние корешки, а двигательные и вегетативные (эфферентные) выходят через передние корешки. В задних корешках волокон гораздо больше, чем в передних (их соотношение у человека примерно 5:1). т. е. при большом разнообразии поступающей информации организм использует незначительное количество исполнительных приборов. Основную часть волокон в спинномозговых корешках составляют мякотные волокна. По задним корешкам в спинной мозг поступают импульсы от рецепторов скелетных мышц, сухожилий, кожи, сосудов, внутренних органов. Передние корешки содержат волокна к скелетным мышцам и вегетативным ганглиям.
Задние корешки образованы волокнами одного из отростков афферентных нейронов, тела которых расположены вне центральной нервной системы — в межпозвоночных ганглиях, а волокна другого отростка связаны с рецептором. Общее число афферентных волокон у человека достигает примерно 1 млн. Они различаются по диаметру. Наиболее толстые идут от рецепторов мышц и сухожилий, средние по толщине — от тактильных рецепторов кожи, от части мышечных рецепторов и от рецепторов внутренних органов (мочевого пузыря, желудка, кишечника и др.), наиболее тонкие миелинизированные и немиелинизированные волокна — от болевых рецепторов и терморецепторов. Одна часть афферентных волокон заканчивается на нейронах спинного мозга, другая часть направляется к нейронам продолговатого мозга, образуя спинно-бульбарный путь.
Передние корешки состоят из отростков мотонейронов передних рогов спинного мозга и нейронов боковых рогов. Волокна первых направляются к скелетной мускулатуре, а волокна вторых переключаются в вегетативных ганглиях на другие нейроны и иннервируют внутренние органы.
В составе серого вещества спинного мозга человека насчитывают около 13,5 млн. нервных клеток. Из них двигательные клетки — мотонейроны— составляют всего 3%, а 97% представляют промежуточные клетки (вставочные, или интернейроны). Среди мотонейронов спинного мозга различают крупные клетки — альфа-мотонейроны и мелкие клетки — гамма-мотонейроны. От альфа-мотонейронов отходят наиболее толстые и быстропроводящие волокна двигательных нервов, вызывающие сокращение скелетных мышечных волокон. Тонкие волокна гамма-мотонейронов не вызывают сокращения мышц. Они подходят к проприорецепторам — мышечным веретенам и вызывают сокращение их внутренних (интрафузальных) мышечных волокон. При этом сокращении растягиваются рецепторы веретен, повышается их чувствительность, усиливается поток афферентных импульсов от скелетных мышц к нервным центрам. Таким образом, альфа-мотонейроны вызывают двигательные акты, а гамма-мотонейроны регулируют чувствительность мышечных рецепторов, информирующих мозг о выполнении этих движений.
Группу альфа-мотонейронов, иннервирующих отдельную скелетную мышцу, называют ее моторным ядром. Ядра крупных скелетных мышц состоят из мотонейронов, расположенных в 2—3 сег
146
ментах спинного мозга. Отростки этих клеток выходят из спинного мозга в составе 2—3 передних корешков. Мелкие же мышцы иннервируются мотонейронами одного сегмента, волокна которого идут в составе одного переднего корешка.
Особое место в деятельности спинного мозга занимают его промежуточные нейроны, или интернейроны. Это в основном мелкие клетки, через которые осуществляются межнейронные взаимодействия в спинном мозгу и координация деятельности мотонейронов. К промежуточным нейронам относятся и тормозные клетки Рэншоу, с помощью которых осуществляются возвратное торможение альфа-мотонейронов и реципрокное торможение центров мышц-антагонистов.
Огромное значение в сложных процессах координации имеют межнейронные взаимодействия на уровне спинного мозга. Это может быть продемонстрировано следующими данными: из огромного количества межнейронных синапсов лишь 10% образовано волокнами, приходящими из головного мозга, и всего около 1 % — афферентными волокнами, т. е. почти 90% остальных синаптических контактов на спинальных клетках образовано волокнами, которые начинаются и кончаются в самом спинном мозгу. Это указывает на существенную роль собственной интегративной деятельности спинного мозга. Благодаря такому множеству существующих связей имеются широкие возможности комбинаций различных нервных клеток для организации любой целесообразной ответной реакции организма.
У человека процессы координации на уровне спинного мозга в значительно большей мере подчинены регулирующим влияниям головного мозга, чем у животных. Нарушение связей спинного мозга с головным приводит к выраженному расстройству протекания спинно-мозговых рефлексов (спинальный шок). На вставочных и моторных нейронах импульсы, приходящие в спинной мозг из головного, взаимодействуют с сегментарными афферентными влияниями. Приказы вышележащих этажей нервной системы увязываются таким образом с текущим состоянием двигательного аппарата.
Рефлексы спинного мозга можно подразделить на двигательные, осуществляемые альфа-мотонейронами передних рогов, и вегетативные, осуществляемые эфферентными клетками боковых рогов, Мотонейроны спинного мозга иннервируют все скелетные мышцы (за исключением мышц лица). Спинной мозг осуществляет элементарные двигательные рефлексы — сгибательные и разгибательные, возникающие при раздражении рецепторов кожи или проприореиепто-ров мышц и сухожилий, а также посылает постоянную импульсацию к мышцам, поддерживая их напряжение — мышечный тонус.
Мышечный тонус возникает в результате раздражения проприо-рецепторов мышц и сухожилий при их растяжении во время движений человека или при воздействии силы тяжести. Импульсы от про-нриорецепторов поступают к мотонейронам спинного мозга, а импульсы от мотонейронов направляются к мышцам, обеспечивая поддержание их тонуса. При разрушении нервных центров спинного мозга или при перерезке нервных волокон, идущих от мотонейронов
I
147
к мышцам, исчезаег тонус скелетных мышц. Участие спинного мозга в двигательной деятельности проявляется не только в поддержании тонуса, но и в организации элементарных двигательных актов и сложной координации деятельности различных мышц (например, согласованной деятельности мышц-антагонистов). Это возможно благодаря мощному развитию системы вставочных нейронов и их богатым взаимосвязям внутри спинного мозга.
Специальные мотонейроны иннервируют дыхательную мускулатуру — межреберные мышцы и диафрагму и обеспечивают дыхательные движения. Вегетативные нейроны иннервируют все внутренние органы (сердце, сосуды, железы внутренней секреции, пищеварительный тракт и др.) и осуществляют рефлексы, регулирующие их деятельность.
П роводниковая функция спинного мозга связана с передачей в вышележащие отделы нервной системы получаемого с периферии потока информации и с проведением импульсов, идущих из головного мозга в спинной. Наиболее важными восходящими путями спинного мозга являются: 1) путь в продолговатый мозг — спиннобульбарный; 2) в мозжечок — спинно-мозжечковый, несущие импульсы от проприорецепторов мышц, суставов и сухожилий, частично от рецепторов кожи; 3) в промежуточный мозг — спинно-таламический путь (от тактильных, болевых и терморецепторов). По различным восходящим путям передаются в головной мозг сигналы от интерорецепторов внутренних органов
Продолговатый мозг и варолиев мост. Продолговатый мозг и варолиев мост относят к заднему мозгу. Он является частью ствола мозга. Задний мозг осуществляет сложную рефлекторную деятельность и служит для соединения спинного мозга с вышележащими отделами головного мозга. В срединной его области расположены задние отделы ретикулярной формации, оказывающие неспецифические тормозные влияния на спинной и головной мозг.
Через продолговатый мозг проходят восходящие пути от рецепторов слуховой и вестибулярной чувствительности. Функции нейронов вестибулярных ядер продолговатого мозга разнообразны. Одна часть их реагирует на перемещение тела (например, при горизонтальных ускорениях в одну сторону они увеличивают частоту разрядов, а при ускорениях в другую сторону уменьшают их). Другая часть предназначена для связи с моторными системами. Эти вестибулярные нейроны, повышая возбудимость мотонейронов спинного мозга и нейронов двигательной зоны коры больших полушарий, позволяют регулировать двигательные акты в соответствии с вестибулярными влияниями.
В продолговатом мозгу оканчиваются афферентные нервы, несущие информацию от рецепторов кожи и мышечных рецепторов. Здесь они переключаются на другие нейроны, образуя путь в таламус и далее в кору больших полушарий. Восходящие пути кожномышечной чувствительности (как и большая часть нисходящих кортико-спинальных волокон) перекрещиваются на уровне продолговатого мозга.
148
В продолговатом мозгу и варолиевом мосту находится большая группа черепно-мозговых ядер (от V до XII пары), иннервирующих кожу, слизистые оболочки, мускулатуру головы и ряд внутренних органов (сердце, легкие, печень). Совершенство этих рефлексов обусловлено наличием большого количества нейронов, образующих ядра, и соответственно большого числа нервных волокон. Так, только в одном нисходящем корешке тройничного нерва, проводящем болевую, температурную и тактильную чувствительность от головы, содержится во много раз больше волокон, чем в спинно-таламическом пути, содержащем волокна, идущие от болевых и температурных рецепторов остальной части тела.
На дне IV желудочка в продолговатом мозгу находится жизненно важный дыхательный центр, состоящий из центров вдоха и выдоха. Его составляют мелкие клетки, посылающие импульсы к дыхательным мышцам через мотонейроны спинного мозга. В непосредственной близости расположен сердечно-сосудистый центр. Его крупные клетки регулируют деятельность сердца и состояние сосудов. Функции этих центров взаимосвязаны. Ритмические разряды дыхательного центра изменяют частоту сердечных сокращений, вызывая дыхательную аритмию — учащение сердцебиений на вдохе и замедление их на выдохе.
В продолговатом мозгу находится ряд рефлекторных центров, связанных с процессами пищеварения. Это группа центров моторных рефлексов (жевания, глотания, движений желудка и части кишечника), а также секреторных (слюноотделение, выделение пищеварительных соков желудка, поджелудочной железы и др.). Кроме того здесь находятся центры некоторых защитных рефлексов: чихания, кашля, мигания, слезоотделения, рвоты.
Продолговатый мозг играет важную роль в осуществлении двигательных актов и в регуляции тонуса скелетных мышц (см. ниже). Влияния, исходящие из вестибулярных ядер продолговатого мозга, усиливают тонус мышц-разгибателей, что важно для организации позы.
Неспецифические отделы продолговатого мозга, наоборот, оказывают угнетающее влияние на тонус скелетных мышц, снижая его и в мышцах-разгибателях. Продолговатый мозг участвует в осуществлении рефлексов поддержания и восстановления позы тела, так называемых установочных рефлексов (см. ниже).
Средний мозг. Через средний мозг, являющийся продолжением ствола мозга, проходят восходящие пути от спинного и продолговатого мозга к таламусу, коре больших полушарий и мозжечку.
В состав среднего мозга входят четверохолмия, черная субстанция и красные ядра. Срединную его часть занимает ретикулярная формация (см. § 6 этой главы), нейроны которой оказывают мощное активирующее влияние на всю кору больших полушарий, а также на спинной мозг.
Передние бугры четверохолмия представляют собой первичные фительные центры, а задние бугры — первичные слуховые центры. Они осуществляют также ряд реакций, являющихся компонентами
149
ориентировочного рефлекса при появлении неожиданных раздражителей. В ответ на внезапное раздражение происходит поворот головы и глаз в сторону раздражителя, а у животных — настораживание ушей. Этот рефлекс (по Й. П. Павлову, рефлекс «Что такое?») необходим для подготовки организма к своевременной реакции на любое новое воздействие. Он сопровождается усилением тонуса мышц-сгибателей (подготовка к двигательной реакции) и изменениями вегетативных функций (дыхание, сердцебиения).
Средний мозг играет важную роль в регуляции движений глаз. Управление глазодвигательным аппаратом осуществляют расположенные в среднем мозгу ядра блокового (IV) нерва, иннервирующего верхнюю косую мышцу глаза, и глазодвигательного (III) нерва, иннервирующего верхнюю, нижнюю и внутреннюю прямые мышцы, нижнюю косую мышцу и мышцу, поднимающую веко, а также расположенное в заднем мозгу ядро отводящего (VI) нерва, иннервирующего наружную прямую мышцу глаза. С участием этих ядер осуществляются поворот глаза в любом направлении, аккомодация глаза, фиксация взгляда на близких предметах путем сведения зрительных осей, зрачковый рефлекс (расширение зрачков в темноте и сужение их на свету).
У человека при ориентации во внешней среде ведущим является зрительный анализатор, поэтому особое развитие получили передние бугры четверохолмия (зрительные подкорковые центры). У животных с преобладанием слуховой ориентации (собака, летучая мышь), наоборот, в большей степени развиты задние бугры (слуховые подкорковые центры).
Черная субстанция среднего мозга имеет отношение к рефлексам жевания и глотания, участвует в регуляции тонуса мышц (особенно при выполнении мелких движений пальцами рук).
В среднем мозгу важные функции осуществляет красное ядро. О возрастании роли этого ядра в процессе эволюции свидетельствует резкое увеличение его размеров по отношению к остальному объему среднего мозга. Красное ядро тесно связано с корой больших полушарий, ретикулярной формацией ствола, мозжечком и спинным мозгом.
От красного ядра начинается руброспинальный путь к мотонейронам спинного мозга. С его помощью осуществляется регуляция тонуса скелетных мышц, происходит усиление тонуса мышц-сгибателей. Это имеет большое значение как при поддержании позы в состоянии покоя, так и при осуществлении движений. Импульсы, приходящие в средний мозг от рецепторов сетчатки глаза и от про-приорецепторов глазодвигательного аппарата, участвуют в осуществлении глазодвигательных реакций, необходимых для ориентации в пространстве, выполнении точностных движений.
Промежуточный мозг. В состав промежуточного мозга, который является передним концом ствола мозга, входят зрительные бугры — таламус и подбугровая область — гипоталамус.
Таламус представляет собой важнейшую «станцию» на пути афферентных импульсов в кору больших полушарий.
150
Ядра таламуса подразделяют на специфические и неспецифические.
К специфическим относят переключательные (релейные) ядра и ассоциативные. Через переключательные ядра таламуса передаются афферентные влияния от всех рецепторов тела. Это так называемые специфические восходящие пути. Они характеризуются со-матотопической организацией. Особенно большое представительство в таламусе имеют эфферентные влияния, поступающие от рецепторов лица и пальцев рук. От таламических нейронов начинается путь, к соответствующим воспринимающим областям коры — слуховым, зрительным и др. Ассоциативные ядра непосредственно не связаны с периферией. Они получают импульсы от переключающих ядер и обеспечивают их взаимодействие на уровне таламуса, т. е. осуществляют подкорковую интеграцию специфических влияний. Импульсы от ассоциативных ядер таламуса поступают в ассоциативные области коры больших полушарий, где участвуют в процессах высшего афферентного синтеза.
Помимо этих ядер, в таламусе имеются неспецифические ядра, которые могут оказывать как активирующее, так и тормозящее влияние на кору (см. § 6 этой главы).
Благодаря обширным связям таламус играет важнейшую роль в жизнедеятельности организма. Импульсы, идущие от таламуса в кору, изменяют состояние корковых нейронов и регулируют ритм корковой активности. Между корой и таламусом существуют кольцевые кортико-таламические взаимосвязи, лежащие в основе образования условных рефлексов. С непосредственным участием таламуса происходит формирование эмоций человека. Таламусу принадлежит большая роль в возникновении ощущений, в частности ощущения боли.
Подбугровая. область расположена под зрительными буграми и имеет тесные нервные и сосудистые связи с прилежащей железой внутренней секреции — гипофизом. Здесь расположены важные вегетативные нервные центры, регулируюьцие обмен веществ в организме, обеспечивающие поддержание постоянства температуры тела (у теплокровных) и другие вегетативные функции.
Участвуя в выработке условных рефлексов и регулируя вегета-। явные реакции организма, промежуточный мозг играет очень важную роль в двигательной деятельности, особенно при формировании новых двигательных актов и выработке двигательных навыков.
Подкорковые узлы. Подкорковыми узлами называют группу ядер серого вещества, расположенных непосредственно под полушариями большого мозга. К ним относятся парные образования: хвоста-н к' тело и скорлупа, составляющие вместе полосатое тело (стриа-।\м), и бледное ядро (паллидум). Подкорковые ядра получают ш палы от рецепторов тела через зрительные бугры. Эфферентные импульсы подкорковых ядер направляются к нижележащим центам экстрапирамидной системы. Подкорковые узлы функционируют । < ишстве с корой больших полушарий, промежуточным мозгом
151
и другими отделами мозга. Это обусловлено наличием кольцевых связей между ними. Через подкорковые ядра могут соединяться между собою разные отделы коры больших полушарий, что имеет большое значение при образовании условных рефлексов. Совместно с промежуточным мозгом подкорковые ядра участвуют в осуществлении сложных безусловных рефлексов: оборонительных, пищевых и др.
Представляя собой высший отдел мозгового ствола, подкорковые узлы объединяют деятельность нижележащих образований, регулируя мышечный тонус и обеспечивая необходимое положение тела во время физической работы. Бледное ядро выполняет моторную функцию. Оно обеспечивает проявление древних автоматизмов — ритмических рефлексов. С его деятельностью связано также выполнение содружественных (например, движения туловища и рук при ходьбе), мимических и других движений.
Полосатое тело оказывает на двигательную деятельность тормозящее, регулирующее влияние, угнетая функции бледного ядра, а также моторной области коры больших полушарий. При заболевании полосатого тела возникают непроизвольные беспорядочные сокращения мышц (гиперкинезы). Они обусловливают некоординированные толчкообразные движения головы, рук и ног. Нарушения возникают также в чувствительной сфере — понижается болевая чувствительность, расстраиваются внимание и восприятие.
В настоящее время выявлено значение хвостатого тела в самооценке поведения человека. При неправильных движениях или умственных операциях из хвостатого ядра в кору больших полушарий поступают импульсы, сигнализирующие об ошибке.
Мозжечок. Это — надсегментарное образование, не имеющее непосредственной связи с исполнительными аппаратами. Мозжечок входит в состав экстрапирамидной системы. Он состоит из двух полушарий и червя, находящегося между ними. Наружные поверхности полушарий покрыты серым веществом — корой мозжечка, а скопления серого вещества в белом веществе образуют ядра мозжечка.
Мозжечок получает импульсы от рецепторов кожи, мышц и сухожилий через спинно-мозжечковые пути и через ядра продолговатого мозга (от спинно-бульбарного пути). Из продолговатого мозга в мозжечок поступают также вестибулярные влияния, а из среднего мозга — зрительные и слуховые. Корково-мосто-мозжечковый путь связывает мозжечок с корой больших полушарий. В коре мозжечка представительство различных периферических рецепторов имеет со-матотопическую организацию. Кроме того, наблюдается упорядоченность связей этих зон с соответствующими воспринимающими областями коры. Так, зрительная зона мозжечка связана со зрительной зоной коры, представительство каждой группы мышц в мозжечке — с представительством одноименных мышц в коре и т. д. Такое соответствие облегчает совместную деятельность мозжечка и коры в управлении различными функциями организма.
Эфферентные импульсы от мозжечка поступают к красным яд
152
рам, ретикулярной формации, продолговатому мозгу, таламусу, коре и подкорковым ядрам.
Мозжечок участвует в регуляции двигательной деятельности. Электрические раздражения поверхности мозжечка вызывают движения глаз, головы и конечностей, которые отличаются от корковых моторных эффектов тоническим характером и большой длительностью. Мозжечок регулирует изменение и перераспределение тонуса скелетных мышц, что необходимо для организации нормальной позы и двигательных актов.
Функции мозжечка изучались в клинике при его поражениях у человека, а также у животных путем удаления (экстирпации мозжечка) (Л. Лючиани, Л. А. Орбели). В результате выпадения функций мозжечка возникают двигательные расстройства: атония — резкое падение и неправильное распределение тонуса мышц, астазия— невозможность сохранения неподвижного положения, непрерывные качательные движения, дрожание головы, туловища и конечностей, астенця — повышенная утомляемость мышц, атаксия — нарушение координированных движений, походки и др.
Мозжечок оказывает влияние также на ряд вегетативных функций, например желудочно-кишечного тракта, на уровень кровяного давления, на состав крови.
Таким образом, в мозжечке происходит интеграция самых различных сенсорных влияний, в первую очередь проприоцептивных и вестибулярных. Мозжечок даже ранее считали центром равновесия и регуляции мышечного тонуса. Однако его функции, как оказалось, гораздо обширнее — охватывают также регуляцию деятельности вегетативных органов. Деятельность мозжечка протекает в непо-средственой связи с корой больших полушарий, под ее контролем.
§ 8. Кора больших полушарий головного мозга
Общий план организации коры. Кора больших полушарий является высшим отделом центральной нервной системы, который в процессе филогенетического развития появляется позже всего и формируется в ходе индивидуального (онтогенетического) развития позже других отделов мозга. Кора представляет собой слой серого вещества толщиной 2—3 мм, содержащий в среднем около 14 млрд, (от 10 до 18 млрд.) нервных клеток, нервные волокна и межуточную ткань (нейроглию). На поперечном ее срезе по расположению нейронов и их связей различают 6 горизонтальных слоев. Благодаря многочисленным извилинам и бороздам площадь поверхности коры достигает 0,2 м2. Непосредственно под корой находится белое вещество, состоящее из нервных волокон, которые передают возбуждение в кору и из нее, а также от одних участков коры дру-। нм.
Корковые нейроны и их связи. Несмотря на огромное число нейринов в коре, известно очень немного их разновидностей. Основными типами их являются пирамидные и звездчатые нейроны.
153
В афферентной функции коры и в процессах переключения возбуждения на соседние нейроны основная роль принадлежит звездчатым нейронам. Они составляют у человека более половины всех клеток коры. Эти клетки имеют короткие ветвящиеся аксоны, не выходящие за пределы серого вещества коры, и короткие ветвящиеся дендриты. Звездчатые нейроны участвуют в процессах восприятия раздражений и объединении деятельности различных пирамидных нейронов.
Пирамидные нейроны осуществляют эфферентную функцию коры и внутрикорковые процессы взаимодействия между удаленными друг от друга нейронами. Они делятся на крупные пирамиды, от которых начинаются проекционные, или эфферентные, пути к подкорковым образованиям, и мелкие пирамиды, образующие ассоциативные пути к другим отделам коры. Наиболее крупные пирамидные клетки — гигантские пирамиды Беца — находятся в передней центральной извилине, в так называемой моторной зоне коры. Характерная особенность крупных пирамид — их вертикальная ориентация в толще коры. От тела клетки вертикально вверх к поверхности коры направлен наиболее толстый (верхушечный) дендрит, через который в клетку поступают различные афферентные влияния от других нейронов, а вертикально вниз отходит эфферентный отросток — аксон.
Многочисленность контактов (например, только на дендритах крупной пирамиды их насчитывают от 2 до 5 тыс.) обеспечивает возможность широкой регуляции деятельности пирамидных клеток со стороны множества других нейронов. Это позволяет координировать ответные реакции коры (в первую очередь ее моторную функцию) с разнообразными воздействиями из внешней среды и внутренней среды организма.
Для коры больших полушарий характерно обилие межнейронных связей. По мере развития мозга человека после его рождения увеличивается число межцентральных взаимосвязей, особенно интенсивно до 18 лет.
Функциональной единицей коры является вертикальная колонка взаимосвязанных нейронов. Вытянутые по вертикали крупные пирамидные клетки с расположенными над ними и под ними ней-"ронами образуют функциональные объединения нейронов. Все нейроны вертикальной колонки отвечают на одно и то же афферентное раздражение (от одного и того же рецептора) одинаковой реакцией и совместно формируют эфферентные ответы пирамидных нейронов.
Распространение возбуждения в поперечном направлении — от одной вертикальной колонки к другой — ограничено процессами торможения. Возникновение активности в вертикальной колонке приводит к возбуждению спинальных мотонейронов и сокращению связанных с ними мышц. Этот путь используется, в частности, при произвольном управлении движениями конечностей.
Первичные, вторичные и третичные поля коры. Особенности строения и функционального значения отдельных участков коры позволяют выделить отдельные корковые поля.
154
Рис. 54. Первичные (проекционные) поля коры больших полушарий (по В. Пен-фильду, Л. Робертсу)
информации. Отдельные
Различают три основные группы полей в коре: первичные, вторичные и третичные поля.
Первичные поля связаны с органами чувств и органами движения на периферии, они раньше других созревают в онтогенезе, имеют наиболее крупные клетки. Это так называемые ядерные зоны анализаторов, по И. П. Павлову (например, поле болевой, температурной, тактильной и мышечно-суставной чувствительности в задней центральной извилине коры, зрительное поле в затылочной облас-сти, слуховое поле в височной области и двигательное поле в передней центральной извилине коры) (рис. 54). Эти поля осуществляют анализ отдельных раздражений, поступающих в кору от соответствующих рецепторов. При разрушении первичных полей возникают так называемая корковая слепота, корковая глухота и т. п. Рядом расположены вторичные поля, или периферические зоны анализаторов, которые связаны с отдельными органами только через первичные поля. Они служат для обобщения и дальнейшей обработки поступав
щения синтезируются в них в комплексы, обусловливающие процессы восприятия. При поражении вторичных полей сохраняется способность видеть предметы, слышать звуки, но человек их не узнает, не помнит их значения. Первичные и вторичные поля имеются и у человека, и у животных.
Наиболее далеки от непосредственных связей с периферией третичные поля, или зоны перекрытия анализаторов. Эти поля есть только у человека. Они занимают почти половину территории коры п имеют обширные связи с другими отделами коры и с неспецифическими системами мозга. В этих полях преобладают наиболее мелкие и разнообразные клетки. Основным клеточным элементом здесь являются звездчатые нейроны. Третичные поля находятся в задней половине коры -— на границах теменных, височных и затылочных ее областей и в передней половине — в передних частях лобных областей. В этих зонах оканчивается наибольшее число нервных волокон, соединяющих левое и правое полушария, поэтому роль их особенно велика в организации согласованной работы обоих полушарий. Третичные поля созревают у человека позже других корковых полей, они осуществляют наиболее сложные функции коры. Оесь происходят процессы высшего анализа и синтеза. В третичных полях на основе синтеза всех афферентных раздражений и с учетом следов прежних раздражений вырабатываются цели и за-Личи поведения. Согласно им происходит программирование двига
155
тельной деятельности. Развитие третичных полей у человека связывают с функцией речи. Мышление (внутренняя речь) возможно только при совместной деятельности анализаторов, объединение информации от которых происходит в третичных полях.
При врожденном недоразвитии третичных полей человек не в состоянии овладеть речью (произносит лишь бессмысленные звуки) и даже простейшими двигательными навыками (не может одеваться, пользоваться орудиями труда и т. п.).
Воспринимая и оценивая все сигналы из внутренней и внешней среды, кора больших полушарий осуществляет высшую регуляцию всех двигательных и эмоционально-вегетативных реакций.
Функции коры больших полушарий. Кора больших полушарий выполняет наиболее сложные функции организации приспособительного поведения организма во внешней среде. Это прежде всего функция высшего анализа и синтеза всех афферентных раздражений.
Афферентные сигналы поступают в кору по разным каналам, в разные ядерные зоны анализаторов (первичные поля), а затем синтезируются во вторичных и третичных полях, благодаря деятельности которых создается целостное восприятие внешнего мира. Этот синтез лежит в основе сложных психических процессов восприятия, представления, мышления. Кора больших полушарий представляет собою орган, тесно связанный с возникновением у человека сознания и регуляцией его общественного поведения. Важной стороной деятельности коры больших полушарий является замыкательная функция — образование новых рефлексов и их систем (условные рефлексы, динамические стереотипы — см. главу XV).
Благодаря необычайно большой продолжительности сохранения в коре следов прежних раздражений (памяти) в ней накапливается огромный объем информации. Это имеет большое значение для сохранения индивидуального опыта, который используется по мере необходимости.
Электрическая активность коры больших полушарий. Изменения функционального состояния коры отражаются на характере ее биопотенциалов. Регистрация электроэнцефалограммы (ЭЭГ), т. е. электрической активности коры, производится непосредственно с ее обнаженной поверхности (в опытах на животных и при операциях на человеке) или через неповрежденные покровы головы (в естественных условиях на животных и человеке). Современные элекгроэнцефалографы усиливают эти потенциалы в 2—3 млн. раз и дают возможность исследовать ЭЭГ от многих точек коры одновременно.
В ЭЭГ различают определенные диапазоны частот, называемые ритмами ЭЭГ (рис. 55). В состоянии относительного покоя чаще всего регистрируется альфа-ритм (8—12 колебаний в 1 сек.), в состоянии активного внимания — бета-ритм (выше 13 колебаний в 1 сек.), при засыпании, некоторых эмоциональных состояниях — тета-ритм (4—-7 колебаний в 1 сек.), при глубоком сне, потере сознания, наркозе — дельта-ритм (1—3 колебания в 1 сек.).
156
В ЭЭГ отражаются особенности взаимодействия корковых нейронов при умственной и физической работе. Отсутствие налаженной координации при выполнении непривычной или тяжелой работы приводит к так называемой десинхронизации ЭЭГ — быстрой асинхронной активности (см. рис. 55). По мере формирования двигательного навыка происходит сонастраивание активности отдельных связанных с данным движением нейронов и отключение посторонних.
В ЭЭГ при этом возникают различные формы синхронизации (см. рис. 55, ж, з). Выполнение освоенного и автоматизированного
движения может протекать при незначительной активности очень небольшого числа корковых нейронов, находящихся в ограниченных областях коры. При этом почти на всей остальной поверхности коры восстанавливается исходный ритм колебаний— альфа-ритм (см. рис. 55, з).
В процессе спортивной тренировки происходит перестройка и совершенствование функций коры больших полушарий. С ростом спортивного мастерства увеличиваются амплитуда и регулярность проявления фоновой активности — альфа-ритма в состоянии покоя. При развитии качества быстроты (например, у баскетболистов) повышается частота волн альфа-ритма, что способствует ускорению произвольных движений.
В процессе мышечной работы
значительно усиливается по сравнению с состоянием относительного покоя взаимосвязанность (синхронность и синфазность) электрической активности различных областей коры. Это облегчает функциональные взаимодействия между различными корковыми центрами. Процесс формирования двигательного навыка сопровождается концентрацией взаи-
I сек. 50 мкв
i сек.
Рис. 55. Электроэнцефалограмма затылочной (а. б, в, г, о) и моторной (е, ж, з) областей коры больших полушарий человека при различных состояних и во время мышечной работы:
а — при открытых глазах (видны по преимуществу бета-волны); б — при закрытых глазах в покое (видны альфа-волны); в — при дремотном состоянии; г — при засыпании; д — при глубоком сне; е — асинхронная частая активность при выполнении непривычной или тяжелой работы (явление десии-хронизации); ас, з — различные формы синхронизации: ж — медленные потенциалы в темпе движения при выполнении циклических движений; з — появление альфа-ритма при выполнении освоенного движения
несвязанной активности в ограниченных зонах коры, наиболее важных для текущей деятельности. Между этими зонами устанавливается общий ритм активности. В такие характерные системы взаимо-
действующих корковых зон включаются не только первичные поля (моторные, зрительные и др.), но и вторичные (например, премо-горные и др.) и особенно третичные поля: передние — программирующие лобные области и задние — зоны афферентного синтеза (нпжнетеменные и др.).
157
§ 9. Нисходящие влияния головного мозгв на двигательную деятельность
Основные нисходящие системы головного мозга. Высшие отделы головного мозга осуществляют свои влияния на деятельность нижележащих отделов, в том числе спинного мозга, через нисходящие пути.
У млекопитающих животных и человека контроль за рефлекторной деятельностью спинного мозга осуществляется главным образом тремя нисходящими системами: 1) кортико-спинальной, 2) кор-тико-рубро-спинальной и 3) кортико-ретикуло-спинальной. Пути этих систем идут в спинной мозг непосредственно от нейронов коры больших полушарий, а также через нейроны красного ядра среднего мозга — рубро-спинальный путь, ядра ретикулярной формации продолговатого и среднего мозга — ретикуло-спинальный путь и вестибулярные ядра продолговатого мозга — вестибуло-спинальный путь. Все эти системы функционируют в тесном взаимодействии и под контролем коры больших полушарий. Окончания волокон образуют, как правило, контакты с дендритами спинальных клеток (аксодендритные синапсы).
Нисходящие пути головного мозга группируют обычно в 2 основные нисходящие системы — пирамидную и экстрапирамидную. Под пирамидной системой, или пирамидным трактом, понимают прямые пути от корковых пирамидных нейронов к нейронам спинного мозга (кортико-спинальный путь) и ядрам черепно-мозговых нервов ствола мозга (кортико-бульбарный путь). Экстрапирамидную систему составляют многоступенчатые связи коры больших полушарий со спинным мозгом. К ней относят подкорковые узлы, люисово тело промежуточного мозга, красные ядра и черную субстанцию среднего мозга, мозжечок, ретикулярную формацию ствола мозга и вестибулярные ядра продолговатого мозга. Важнейшими путями этой системы являются кортико-рубро-спинальная и кортико-рети-куло-спинальная системы. Группировка путей на указанные 2 системы в значительной мере условна, так как не соответствует особенностям их морфологии и функционирования.
В настоящее время предлагают подразделять основные нисходящие пути, исходя из расположения проводящих волокон в белом веществе спинного мозга, на следующие 2 системы: 1) латеральную, волокна которой расположены в боковых (латеральных) частях спинного мозга (сюда относят кортико-спинальную и рубро-спи-гщльную системы), и 2) медиальную, волокна которой идут во внутренней (медиальной) части белого вещества (состоящую из вести-було-спинальной и ретикуло-спинальной систем).
Волокна латеральной системы оканчиваются преимущественно в задних рогах спинного мозга. Они связаны со специальной группой вставочных нейронов, передающих их влияния на мотонейроны в передние рога спинного мозга. Только у обезьян и человека имеются волокна, непосредственно оканчивающиеся на мотонейронах. Латеральные системы — это самые молодые в эволюционном отно-
158
шепни системы, характерные лишь для млекопитающих. Больше всего они развиты у наиболее высокоорганизованных животных и особенно у человека.
Волокна медиальной системы направляются к сегментарным вставочным нейронам и отчасти мотонейронам медиальной (внутренней) части передних рогов спинного мозга. Это древние системы, которые характерны для всех классов позвоночных животных.
Кортико-спинальная система. Ей принадлежит важнейшая роль в осуществлении произвольных движений у человека. Начало этой системе дают крупные и мелкие пирамидные клетки коры больших полушарий. У человека около 30% волокон пирамидного пути начинается в передней центральной извилине, примерно 20% — в задней центральной извилине, остальные волокна — в других отделах коры и в подкорковых образованиях головного мозга. Большая часть (80%) волокон пирамидного пути перекрещивается на уровне продолговатого мозга, меньшая часть — на соответствующем сегменте спинного мозга. В результате этого корковые клетки левого полушария управляют преимущественно деятельностью мышц правой половины тела, а корковые клетки правого полушария — левой половины тела. У человека количество перекрещивающихся волокон может варьировать, по современным данным, от 0 до 100%, и многие волокна могут идти от корковых клеток к спинному мозгу без перекреста. Кроме того, в составе пирамидного пути имеются волокна с двойным перекрестом — на уровне межполушарных волокон (мозолистого тела) и в продолговатом мозгу. Эти морфологические особенности создают возможность корковой иннервации двигательного аппарата своей стороны тела. Такое дублирование управления мышцами со стороны левого и правого полушарий позволяет компенсировать двигательные функции при односторонних поражениях мозга, а также обеспечивает взаимозамещение деятельности полушарий в процессе длительной мышечной работы.
Волокна пирамидного тракта различаются по диаметру и скорости проведения. У человека в каждом пирамидном пучке на уровне продолговатого мозга содержится около 1 млн. нервных волокон — около ’/з мякотных и 2/3 безмякотных. В составе пирамидного тракта находятся неоднородные волокна, диаметр которых колеблется от 1 до 25 мк. Большей частью это тонкие волокна. Лишь около 2— 4% волокон имеют диаметр более 10 мк. Это волокна, характеризующиеся наибольшей скоростью проведения — до 120—140 м/сек. Они начинаются от гигантских пирамидных клеток Беца в передней центральной извилине коры больших полушарий. Примерно такие скорости были обнаружены у человека во время нейрохирургических операций.
Пирамидная система выполняет 3 основные функции: 1) посылает мотонейронам спинного мозга импульсы-команды к движениям (пусковые влияния); 2) изменяет проведение нервных импульсов во вставочных нейронах (влияния, регулирующие протекание спинномозговых рефлексов) и 3) осуществляет контроль за потоком афферентных сигналов в нервные центры (обеспечение обратных свя-
159
зей от работающих органов). Первая функция — двигательная функция кортико-спинальной системы — заключается в преимущественном возбуждении мотонейронов мышц-сгибателей. При этом импульсы, поступающие в спинной мозг, вызывают двоякий эффект — не только возбуждают мотонейроны мышц-сгибателей, но и одновременно затормаживают мотонейроны мышц-разгибателей, что обеспечивает беспрепятственное сгибание в соответствующем суставе.
Влияния пирамидной системы больше выражены по отношению к мышцам верхней части туловища и верхних конечностей, чем к мышцам нижних конечностей. Эта неравномерность воздействий отражена в самом устройстве пирамидного пути: значительно большее количество кортико-спинальных волокон проходит в шейной и грудной областях спинного мозга по сравнению с пояснично-крестцовой областью (табл. 3). Двигательные сигналы адресуются преимущественно мотонейронам дистальных мышц конечностей. Этим обеспечивается более тонкое влияние на отдельные мышцы и даже части мышц верхней конечности, особенно на мышцы фаланг пальцев руки.
Развитие специализированного управления дистальными мышцами верхней конечности со стороны коры больших полушарий связано не только с общим совершенствованием движений человека, но главным образом с особой ролью руки — участием ее в сложнейших двигательных операциях, требующих большой точности и быстроты.
Таблица 3
Распределение волокон пирамидного тракта в различных сегментах спинного мозга у человека (по А.Вейль и А.Лассек, 1929; А.Лассек, Л.Дауд и ______________________________А .Вейль, 1930)
Сегменты спинного мозга Среднее (для 10 взрослых людей) количество (%) волокон Колич. волокон на 1 г мышцы
З-й шейный 100 95
4—7-й шейные 81 24
З-й грудной 55 )
6-й грудной 46 > 62
12-грудной 35 J
4-й поясничный 20 13
В процессе эволюционного развития у обезьян и человека появляются прямые пути (без вставочных нейронов) от пирамидных клеток коры к мотонейронам спинного мозга. Чем больше развиты у них дифференцированные движения пальцев верхней конечности, особенно хватательные, тем большее количество корковых волокон оканчивается непосредственно на спинальных мотонейронах. Так, у макаки их число составляет около 2% всех волокон пирамидного пути, у шимпанзе — 5%, а у человека — 8%. Наличие в этих путях всего одного синапса (моносинаптические пути) ускоряет проведение корковых импульсов к двигательному аппарату.
160
Считают, что эти пути передают влияния от мозговых механизмов, обеспечивающих принятие быстрого решения при произвольных движениях человека.
В процессе филогенетического развития произошло разделение пирамидной системы на 2 подсистемы с различным функциональным значением — «быструю» и «медленную». К «быстрой» подсистеме относятся крупные клетки моторной области коры, аксоны которые обладают высокой скоростью проведения — от 80 до 25 м/сек (средняя скорость — 50 м/сек). К «медленной» подсистеме — средние и мелкие клетки моторной и других областей коры, обладающие меньшей скоростью проведения в аксонах-—от 25 до 7 м/сек (средняя скорость 14 м/сек). В функциях данных нейронов имеются существенные различия. «Быстрые» пирамидные нейроны значительно быстрее отвечают на афферентное раздражение (латентный период их активации меньше), процесс возбуждения в них короче (меньше длительность пиков потенциала действия и следовых потенциалов). Импульсы, идущие по быстрым волокнам, подходят к клеткам спинного мозга значительно раньше, чем импульсы, передающиеся по медленным волокнам. В состоянии мышечного покоя крупные пирамидные нейроны с быстропроводящими аксонами имеют слабую импульсную активность, а при возникновении движения частота их разрядов резко повышается. Полагают, что «быстрые» нейроны обеспечивают срочную передачу информации от коры к спинному мозгу и контролируют фазные рефлекторные движения. Их называют нейронами движения.
В отличие от них «медленные» нейроны характеризуются постоянной ритмической импульсацией в состоянии покоя. При движениях их активность изменяется медленно и незначительно. Считают, что эти нейроны обеспечивают регуляцию позы тела — ее организацию и приспособление к условиям внешней среды. Соответственно их называют нейронами положения.
Функции «медленной» и «быстрой» подсистем кортико-спинального пути взаимосвязаны. Усиление активности «медленных» нейронов облегчает появление разрядов «быстрых» нейронов.
Вторая функция пирамидной системы заключается в активации вставочных нейронов спинного мозга, передающих импульсы от чувствительных нейронов к двигательным. Передача возбуждения в многонейронных (полисинаптических) рефлекторных дугах спинного мозга при этом облегчается. Важно, что повышение возбудимости вставочных нейронов наступает под влиянием коры до начала движения и сохраняется в течение всего периода работы мышц, облегчая ее протекание. Однако корковые влияния носят избира-гельный характер — проведение улучшается не во всех рефлекторных дугах, а лишь в наиболее нужных для данного момента.
Таким образом, спинной мозг самостоятельно осуществляет присущую ему рефлекторную деятельность со всеми особенностями процессов координации (сгибательные реакции сопровождаются <i (повременным торможением центров мышц-антагонистов, возникают перекрестные рефлексы и т. п.). Роль коры в этом случае про-
161
является в регуляции этой деятельности (усилении или ослаблении отдельных рефлексов, выборе и запуске адекватных двигательных реакций).
Третья функция пирамидного тракта связана с выключением излишних афферентных воздействий на нервные центры. Аксоны кортико-спинальной системы образуют окончания на аксонах чувствительных нервных клеток, передающих импульсы от рецепторов на периферии в спинной мозг (аксоаксональные синапсы). В месте их контакта развивается под влиянием корковых импульсов пре-синаптическое торможение (сильная деполяризация чувствительных волокон), которое блокирует проведение афферентных импульсов к нервным центрам спинного и головного мозга. Главной особенностью этого торможения является то, что, подавляя все виды афферентных импульсов, оно не распространяется на импульсы, идущие от проприорецепторов мышц. В результате этого в процессе движения особенно усиливается роль сигналов от работающих мышц, анализ которых необходим для совершенного управления двигательными актами. Одновременно ослабевает импульсация от других рецепторов тела, посторонних для выполняемой работы.
Поражения пирамидного тракта у человека (кровоизлияния, травмы и др.) приводят к невозможности выполнять произвольные движения. Однако через некоторое время эти движения могут восстановиться за счет деятельности других нисходящих систем, связывающих кору головного мозга со спинным мозгом и способных дублировать функции пирамидной системы. Частично утраченными остаются лишь тонкие движения пальцев. При разрушении патологическим очагом пирамидного тракта на одной стороне тела возможна также компенсация двигательных функций за счет гипертрофии (увеличения диаметра) пирамидных волокон на другой стороне тела.
Кортико-рубро-спинальная система. Корковые нейроны образуют прямые связи с клетками красного ядра среднего мозга, от которых начинаются волокна рубро-спинальной системы. Окончания этих волокон находятся в латеральных частях серого вещества спинного мозга — в основании его задних рогов. У человека этот путь развит в меньшей степени, чем у других млекопитающих. Он образован преимущественно тонкими волокнами, идущими от мелких клеток красного ядра, которые связаны с мотонейронами спинного мозга лишь через специальную систему вставочных нейронов. Основная часть волокон рубро-спинальной системы заканчивается в шейных отделах спинного мозга. В состав кортико-рубро-спиналь-ного пути входят волокна, скорость проведения которых значительно выше, чем в большинстве волокон пирамидного пути,— до 105— 124 м/сек (средняя скорость — около 70 м/сек). Эти волокна могут пропускать импульсы с очень высокой частотой —свыше 500 в 1 сек.
Афферентные импульсы поступают к красным ядрам от рецепторов кожи, суставных сумок и болевых рецепторов. Красные ядра являются важным промежуточным этапом между корой и спинным мозгом, где происходит подключение регулирующих влияний от 162
мозжечка, промежуточного мозга и подкорковых ядер. В красном ядре хорошо выражено соматотопическое представительство функций: эфферентные клетки рубро-спинальных путей активируются лишь раздражениями, поступающими от того участка тела, с двигательным аппаратом которого они связаны.
Функции кортико-рубро-спинальной системы подобны функциям кортико-спинальной (пирамидной) системы. Это — преимущественное возбуждение мотонейронов мышц-сгибателей, повышение активности вставочных нейронов и контроль за афферентной импуль-сацией, идущей по восходящим путям. Однако, в отличие от пирамидной системы, здесь нет подразделения на функционально различные подсистемы.
Деятельность кортико-рубро-спинальной системы ограничивается лишь тоническим контролем. По рубро-спинальным волокнам в клетки спинного мозга постоянно поступает тоническая импульса-ция с частотой около 40—60 имп/сек. Во время движения головы или конечностей частота ее возрастает (до 100—200 имп/сек). Таким образом, функционально кортико-руброспинальный путь представляет собой единое целое с «медленной» подсистемой кортикоспинального пути, регулируя уровень активности спинальных клеток и тонус соответствующих скелетных мышц. Эта система, обобщая влияния от коры, мозжечка, промежуточного мозга и подкорковых ядер, играет большую роль в организации позно-тонических. реакций организма. Формирование этих реакций важно не только в состоянии покоя, но и при выполнении произвольных движений. Еще до начала двигательного акта должна начаться подготовка к нему, в частности в виде изменения позы тела. В связи с этим наличие быстропроводящих волокон в кортико-рубро-спинальной системе может обеспечить своевременное перераспределение тонуса соответствующих мышц.
Медиальная система (ретикуло-спинальный и вестибуло-спи-нальный компоненты). Медиальная, более древняя, система служит главным образом для тонических воздействий на нейроны спинного мозга. Она подразделяется на ретикуло-спинальную и вестибулоспинальную. Обе системы контролируются корой больших полушарий. Нисходящие влияния, передаваемые ими, отчетливо различаются по своему влиянию на мотонейроны мышц сгибателей и разгибателей.
Кортико-ретикулярные волокна идут в составе пирамидного |ракта от клеток сенсомоторной и других областей коры к ядрам варолиева моста и отчасти продолговатого мозга. От этих ядер начинаются волокна ретикуло-спинального пути к вставочным и дви-1 ательным нейронам спинного мозга (в медиальной части передне-|(> рога). Количество волокон в этой системе значительно меньше, чем в пирамидной (около 3000—4000). Они не перекрещиваются и иннервируют нейроны спинного мозга на своей стороне. Связи них волокон с мотонейронами большей частью полисинаптичес-кис— через множество вставочных клеток. Никакой соматотопичес-||>п организации этого пути не обнаружено. Основная функция
163
кортико-ретикулярного пути — тоническое повышение возбудимости мотонейронов мышц-сгибателей, что приводит к усилению тонуса этих мышц.
Вестибуло-спинальный компонент медиальной системы перелает импульсы от вестибулярных ядер продолговатого мозга (например, от ядра Дейтерса) к вставочным и двигательным нейронам переднего рога спинного мозга. Основная функция этого пути — тоническое повышение возбудимости мотонейронов мышц-разгибателей, что обеспечивает усиление тонуса этих мышц.
Таким образом, среди нисходящих систем, осуществляющих функцию контроля активности спинного мозга, три системы обеспечивают повышение возбудимости мотонейронов мышц-сгибателей: кортико-спинальная, кортико-рубро-спинальная и кортико-ретику-ло-спинальная, а вестибуло-спинальная система тормозит эти мотонейроны. Лишь одна часть кортико-спинальной системы обусловливает фазную двигательную деятельность. Остальные же нисходящие системы обеспечивают регуляцию тонуса мышц и поз-ных реакций организма.
Роль спинного и головного мозга. Несмотря на совершенство процессов координации в спинном мозгу, они находятся под постоянным контролем головного мозга, в первую очередь коры больших полушарий.
В организме имеются специальные механизмы, обусловливающие преимущественное воздействие коры больших полушарий на общие конечные пути к мышцам — спинальные мотонейроны. Большая эффективность кортико-спинальных влияний по сравнению с сегментарными афферентными влияниями обеспечивается, во-первых, наличием прямых путей из коры к мотонейронам спинного мозга и, во-вторых, возможностью особенно быстрой их активации корковыми импульсами. Электрофизиологическими исследованиями показано, что ритмические воздействия из двигательной области коры вызывают чрезвычайно резкое нарастание суммарной амплитуды возбуждающих постсинаптических потенциалов спинальных мотонейронов. Амплитуда каждого последующего возбуждающего постсинаптического потенциала увеличивается примерно в 6 раз больше, чем при поступлении к тем же мотонейронам импульсов от проприорецепторов по афферентным путям. Таким образом, достаточно уже 2—3 импульсов, идущих от коры, чтобы деполяризация в мотонейроне достигла порогового уровня, необходимого для возникновения ответного разряда в скелетную мышцу. В результате кора больших полушарий может вызывать двигательные действия быстрее, чем периферические раздражения, и часто даже вопреки им.
§ 10. Нервная регуляция эмоций и вегетативных функций
Значение эмоций и их нервная регуляция. Возникновение эмоций связывают с деятельностью лимбической системы, в которую входят некоторые подкорковые образования и участки коры. Кор-164
новые отделы лимбической системы, представляющие ее высший отдел, находятся на нижних и внутренних поверхностях больших полушарий (поясная извилина, гиппокамп и др.). К подкорковым структурам лимбической системы относят гипоталамус, некоторые ядра таламуса, среднего мозга и ретикулярной формации. Между всеми этими образованиями имеются тесные прямые и обратные связи, образующие «лимбическое кольцо».
Лимбическая система участвует в самых разнообразных проявлениях деятельности организма. Она формирует положительные и отрицательные эмоции со всеми двигательными, вегетативными и эндокринными их компонентами (изменением дыхания, сердцебиения, кровяного давления, деятельности желез внутренней секреции, скелетных и мимических мышц и др.). От нее зависит эмоциональная окраска психических процессов и изменения двигательной активности. Она создает мотивацию поведения (определенную предрасположенность). Возникновение эмоций имеет «оценочное влияние» на деятельность специфических систем, так как, подкрепляя определенные способы действий, пути решения поставленных задач, они обеспечивают избирательный характер поведения в ситуациях со многими выборами.
Лимбическая система участвует в формировании ориентировочных и условных рефлексов. Благодаря центрам лимбической системы могут вырабатываться даже без участия других отделов коры оборонительные и пищевые условные рефлексы. При поражениях этой системы затрудняется упрочение условных рефлексов, нарушаются процессы памяти, теряется избирательность реакций и отмечается неумеренное их усиление (чрезмерно повышенная двигательная активность и т. д.). Известно, что так называемые психотропные вещества, изменяющие нормальную психическую деятельность человека, действуют именно на структуры лимбической системы.
Электрические раздражения различных участков лимбической системы через вживленные электроды (в эксперименте на животных и в клинике в процессе лечения больных) выявили наличие центров удовольствия, формирующих положительные эмоции, и центров неудовольствия, формирующих отрицательные эмоции. Изолированное раздражение таких точек в глубоких структурах мозга человека вызывало появление чувства «беспричинной радости», «беспредметной тоски», «безотчетного страха».
В специальных опытах с самораздражением на крысах животное приучали нажимом лапы на педаль замыкать цепь и производить электрическое раздражение собственного мозга через вживленные электроды. При локализации электродов в центрах отрицательных эмоций (некоторые области таламуса) животное старалось избегать замыкания цепи, а при их расположении в центрах положительных эмоций (гипоталамус, средний мозг) нажимы лапой на педаль следовали почти непрерывно, доходя до 8 тыс. раздражений в 1 час.
Велика роль эмоциональных реакций в спорте (положительные
165
эмоции при выполнении физических упражнений — «мышечная радость», радость победы и отрицательные — неудовлетворенность спортивным результатом и др.). Положительные эмоции могут значительно повышать, а отрицательные — понижать работоспособность человека. Большие напряжения, сопровождающие спортивную деятельность, особенно во время соревнований, создают и эмоциональное напряжение — так называемый эмоциональный стресс. От характера протекания в организме реакций эмоционального стресса зависит успешность двигательной деятельности спортсмена.
Вегетативная нервная система. Регуляция деятельности внутренних органов осуществляется нервной системой через специальный ее отдел — вегетативную нервную систему.
Особенности строения вегетативной нервной системы. Все функции организма можно разделить на соматические, или анимальные (от лат. anirnai — животное), связанные с деятельностью скелетных мышц, — организация позы и перемещение в пространстве, и вегетативные (от лат. vegetativus — растительный), связанные с деятельностью внутренних органов, — процессы дыхания, кровообращения, пищеварения, выделения, обмена веществ, роста и размножения. Деление это условно, так как вегетативные процессы присущи также и двигательному аппарату (например, обмен веществ и др.); двигательная деятельность неразрывно связана с изменением дыхания, кровообращения и пр.
Раздражения различных рецепторов тела и рефлекторные ответы нервных центров могут вызывать изменения как соматических, гак и вегетативных функций, т. е. афферентные и центральные отделы этих рефлекторных дуг общие. Различны лишь их эфферентные отделы.
Совокупность эфферентных нервных клеток спинного и головного мозга, а также клеток особых узлов (ганглиев), иннервирующих внутренние органы, называют вегетативной нервной системой. Следовательно, эта система представляет собой эфферентный отдел нервной системы, через который центральная нервная система управляет деятельностью внутренних органов.
Характерной особенностью эфферентных путей, входящих в рефлекторные дуги вегетативных рефлексов, является их двухней-оонное строение. От тела первого эфферентного нейрона, который находится в центральной нервной системе (в спинном, продолговатом или среднем мозгу), отходит длинный аксон, образующий предузловое (или преганглионарное) волокно. В вегетативных ганглиях— скоплениях клеточных тел вне центральной нервной системы — возбуждение переключается на второй эфферентный нейрон, от которого отходит послеузловое (или постганглионарное) волокно к иннервируемому органу (рис. 56).
Вегетативная нервная система подразделяется на 2 отдела — симпатический и парасимпатический. Эфферентные пути симпатической нервной системы начинаются в грудном и поясничном отделах спинного мозга от нейронов его боковых рогов. Передача возбуждения с предузловых симпатических волокон на послеузловые
166
происходит в ганглиях пограничных симпатических стволов с участием медиатора ацетилхолина, а передача возбуждения с послеуз-ловых волокон на иннервируемые органы — с участием медиатора норадреналина, или симпатина. Эфферентные пути парасимпатической нервной системы начинаются в головном мозгу от некоторых ядер среднего и продолговатого мозга и от нейронов крестцового отдела спинного мозга. Парасимпатические ганглии расположены в непосредственной близости от иннервируемых органов или внутри
Рис. 56. Вегетативная нервная система.
Слева —• области выхода волокон: парасимпатической (черный цвет) и симпатической (заштриховано) систем.
Справа — строение эфферентной части рефлекторной дуги вегетативных рефлексов. Слева — схема среднего продолговатого и спинного мозга. Арабские цифры — номера грудных сегментов, римские — номера поясничных сегментов
их. Проведение возбуждения в синапсах парасимпатического пути происходит с участием медиатора ацетилхолина.
Роль вегетативной нервной системы в орган из-м е. Вегетативная нервная система, регулируя деятельность внутренних органов, повышая обмен веществ скелетных мышц, улучшая их кровоснабжение, повышая функциональное состояние нервных центров и т. д., способствует осуществлению функций соматической и нервной системы, которая обеспечивает активную приспособительную деятельность организма во внешней среде (прием внешних сигналов, их обработку, двигательную деятельность, направленную па защиту организма, на поиски пищи, у человека — двигательные акты, связанные с бытовой, трудовой, спортивной деятельностью и пр.). Передача нервных влияний в соматической нервной системе осуществляется с большой скоростью (толстые соматические волокна имеют высокую возбудимость и скорость проведения 50— 140 м/сек). Соматические воздействия на отдельные части двигательного аппарата характеризуются высокой избирательностью. Вегетативная нервная система участвует в этих приспособительных реакциях организма, особенно при чрезвычайных напряжениях (стресс).
167
Другой существенной стороной деятельности вегетативной нервной системы является ее огромная роль в поддержании постоянства внутренней среды организма.
Постоянство физиологических показателей может обеспечиваться различными путями. Например, постоянство уровня кровяного давления поддерживается изменениями деятельности сердца, просвета сосудов, количества циркулирующей крови, ее перераспределением в организме и т. п. В гомеостатических реакциях наряду с нервными влияниями, передающимися по вегетативным волокнам, имеют значение гуморальные влияния. Все эти влияния в отличие от соматических передаются в организме значительно медленнее и более диффузно. Тонкие вегетативные нервные волокна отличаются низкой возбудимостью и малой скоростью проведения возбуждения (в предузловых волокнах скорость проведения составляет 3— 20 м/сек, а в послеузловых — 0,5—3 м/сек).
Согласно представлениям И. П. Павлова и Л. А. Орбели, все нервные влияния делятся на пусковые, включающие деятельность органа, и трофические, изменяющие его обмен веществ и функциональное состояние. Многие влияния вегетативной нервной системы можно рассматривать как трофические.
Функции симпатического отдела вегетативной нервной системы. С участием этого отдела протекают многие важные рефлексы в организме, направленные на обеспечение его деятельного состояния, в том числе двигательной деятельности. К ним относятся рефлексы расширения бронхов, учащения и усиления сердечных сокращений, расширения сосудов сердца и легких при одновременном сужении сосудов кожи и органов брюшной полости (обеспечение перераспределения крови), выброс депонированной крови из печени и селезенки, расщепление гликогена до глюкозы в печени (мобилизация углеводных источников энергии), усиление деятельности желез внутренней секреции потовых желез. Симпатический отдел нервной системы снижает деятельность ряда внутренних органов: в результате сужения сосудов в почках уменьшаются процессы мочеобразования, угнетается секреторная и моторная деятельность органов желудочно-кишечного тракта, предотвращается акт мочеиспускания (расслабляется мышца стенки мочевого пузыря и сокращается его сфинктер). Повышенная активность организма сопровождается симпатическим рефлексом расширения зрачка.
Огромное значение для двигательной деятельности организма имеет трофическое влияние симпатических нервов на скелетные мышцы. Раздражение этих нервов не вызывает сокращения мышц. Однако сниженная амплитуда сокращений утомленной мышцы может снова увеличиться при возбуждении симпатической нервной системы — эффект Орбели—Гинецинского. Усиление сокращений можно наблюдать и на неутомленной мышце, присоединяя к раздражениям двигательных нервов раздражения симпатических волокон (Я. Б. Лехтман). Более того, симпатические влияния на скелетные мышцы в целостном организме возникают раньше, чем пуско-
168
вне влияния двигательных нервов, заранее подготавливая мышцы к работе. Л. А. Орбели подчеркивал важнейшее значение симпатических воздействий для приспособления (адаптации) организма к работе, к различным условиям внешней среды, что отражено в его учении об адаптационно-трофической роли симпатической нервной системы.
Функции парасимпатического отдела вегетативной нервной системы. Этот отдел нервной системы принимает активное участие в регуляции деятельности внутренних органов, в процессах восстановления организма после деятельного состояния.
Парасимпатическая нервная система осуществляет сужение бронхов, замедление и ослабление сердечных сокращений; сужение сосудов сердца; пополнение энергоресурсов (синтез гликогена в печени и усиление процессов пищеварения); усиление процессов мочеобразования в почках и обеспечение акта мочеиспускания (сокращение мышц мочевого пузыря и расслабление его сфинктера) и др.
Парасимпатическая нервная система в противоположность симпатической преимущественно оказывает пусковые влияния: сужение зрачка, включение деятельности пищеварительных желез и т. д.
§ 11. Общие принципы регуляции двигательной деятельности
Теория функциональной системы. Современные представления об организации целостных актов поведения живых организмов отражены в теории функциональной системы (П. К. Анохин). Для удовлетворения своих потребностей в условиях постоянного изменения внешней среды организму необходимо ставить перед собой определенные задачи и в своей поведенческой деятельности добиваться достижения намеченного результата.
Согласно теории П. К. Анохина, именно полезный результат является решающим фактором поведения и для его достижения в нервной системе формируется группа взаимосвязанных нейронов — так называемая функциональная система. Сколько нервных клеток из огромного их числа будет включено в эту систему, какой уровень активности этих клеток необходим в данный момент, какие взаимосвязи должны возникнуть между ними и какие возможные взаимосвязи должны быть исключены — все это определяется намечаемым результатом. Следовательно, важная задача деятельности системы — выявление и оценка результата действия.
Организм в своем поведении использует далеко не все структурные и функциональные свои возможности (степени свободы), а оставляет лишь необходимые. Без этого деятельность его была бы хаотичной, нецелесообразной. Создание функциональной системы необходимо для того, чтобы отдельные ее элементы действовали не самостоятельно и независимо друг от друга, а подчинялись общей задаче— получить искомый результат.
169
Деятельность функциональной системы можно разбить на отдельные последовательные этапы: 1) обработка всех сигналов, поступающих из внешней и внутренней среды организма — так называемый афферентный синтез; 2) принятие решения; 3) создание (па основе принятого решения) представления об ожидаемом результате действия и формирование конкретной программы действий для достижения этого результата; 4) анализ полученного результата и уточнение программы действий.
На первом этапе (афферентный синтез) в нервных центрах одновременно взаимодействуют 4 типа поступающих к ним раздражений: 1) пусковая афферентация—сигналы, вызывающие действие; 2) обстановочная афферентация— все остальные внешние раздражения, создающие афферент1Шй фон; 3) мотивация — собственные потребности организма, доминирующие в данный момент: биологические мотивации — жажда, голод, страх и другие, а также высшие мотивации, формирующиеся у человека под влиянием социальных факторов (желание добиться какой-либо цели в жизни, достичь определенного результата в спорте и др.); 4) память — имеющиеся в нервной системе следы прежних раздражений, накопленный опыт.
Все эти виды сигналов должны одновременно обрабатываться в нервных клетках и сопоставляться друг с другом. При оценке текущих афферентных раздражений производится их сопоставление со следами прежних раздражений, хранящихся в памяти. Большую роль в отборе сигналов играют доминирующие мотивации, так как они повышают восприимчивость организма к наиболее важным в данный момент сигналам.
На основании афферентного синтеза принимается решение («что надо делать») и вырабатывается программа действий («как надо делать»). При этом производится оценка всех возможных путей решения возникших перед организмом задач и ответных реакций организма и выбирается лишь одно необходимое решение. Одновременно в мозгу создается представление о том, какой результат должен быть получен.
Динамическая функциональная система — это организованная система, результат действия которой сообщается в центральную нервную систему.
Без постоянной оценки результата действия невозможно выполнить задуманную программу, вносить в нее поправки для улучшения результата. Например, совершенствование техники выполнения упражнения невозможно без постоянной информации о его результатах.
Представление о функциональной системе П. К. Анохина имеет универсальное значение, так как помогает понять различные стороны деятельности организма (механизмы его поведения, закономерности формирования различных функций в процессе индивидуального развития, особенности их нарушения при патологии и возможные пути восстановления).
Циклическая многоуровневая система регуляции движений. Выполнение двигательных актов осуществляется обширным комплек
170
сом нейронов, расположенных в различных отделах центральной нервной системы. Каждый из нервных центров, входящих в данный комплекс, может воздействовать как на нижележащие, так и на вышележащие центры. При этом между ними могут возникать циклы взаимных влияний от высших центров к низшим и обратно. Например, очень часто наблюдается циркуляция импульсов между корой и промежуточным мозгом: импульсы из коры больших полушарий могут влиять на активность нейронов промежуточного мозга, а те, в свою очередь, могут воздействовать на корковые нейроны. В каждом подобном цикле роль задающих звеньев принадлежит преимущественно вышележащим центрам, а активность нижележащих центров является регулируемым звеном. Эта активность поддерживается на заданном уровне в зависимости от потребностей организма.
Наличие циклических взаимосвязей между различными отделами нервной системы позволяет говорить о замкнутой системе регулирования, или замкнутом цикле регулирования.
В процессе двигательной деятельности одновременно возникает множество замкнутых циклов регулирования (например, между корой и ретикулярной формацией, между корой и мозжечком, между мотонейронами спинного мозга и мышцей и др.).
Таким образом, система регуляции движения является многоэтажной (многоуровневой) системой. Важную роль в этой сложной функциональной системе играет кора больших полушарий, которая управляет процессами, протекающими в этих циклах регулирования, и направляет их на решение общей задачи — достижение рабочего эффекта, или цели движения.
Программирование. Для решения поставленных задач в центральной нервной системе вырабатываются программы двигательных действий. Они определяют набор и последовательность включения двигательных актов. В соответствии с этой программой включаются команды к эфферентным нейронам и далее — команды к мышцам.
Программы двигательных действий у человека разнообразны. Они обеспечивают бытовую, производственную и спортивную его деятельность. При этом команды к действию (например, выстрел стартового пистолета, команда «марш!») являются лишь сигналами о начале работы, сама же мышечная работа развертывается по имеющейся программе. В ряде случаев программирование протекает в условиях крайнего дефицита времени, (например при спортивных играх, фехтовании, борьбе и др.). В этих случаях требуется высокая скорость переработки поступающей информации и выполнения ответных действий.
Программы двигательных действий могут быть очень сложными, особенно при нестандартной работе, так как для достижения одного н того же результата в изменяющихся условиях (например, для успешного выполнения броска в баскетбольную корзину при разных шровых ситуациях) могут потребоваться включение различных мы-печных групп, различная тактика поведения.
171
Рис. 57. Схема регуляции движений
Обратные связи. Для функциональной системы регуляции движений недостаточно лишь наличия программирующих и управляющих нервных центров. Характерной особенностью подобных систем регуляции является наличие обратной афферентации, или так называемой обратной связи, от рабочего органа к регулирующим нервным центрам (рис. 57). Осведомление центральной нервной системы о деятельности, вызванной на периферии ее управляющими импульсами, происходит при помощи афферентных сигналов, отправляемых органами зрения, . проприорецепторами мышечносуставного аппарата и другими рецепторами.
Таким образом, с помощью обратных связей производится оценка выполненных движений, определяется их эффективность.
Аппараты сравнения. Эти нервные аппараты, получая обратную сигнализацию, сличают заданную программу с реальными результатами ее выполнения (П. К. Анохин, Н. А. Бернштейн и др.). Имеются данные, что та
кие аппараты находятся в лобных долях, подкорковых ядрах (хвостатые тела) и других образованиях.
Если есть расхождения фактического и требуемого действия, то в аппаратах сравнения возникают сигналы об ошибках — так называемые импульсы рассогласования. Соответственно этим импульсам в систему, управляющую движением, вносятся необходимые поправки — коррекции. В результате этого в процессе выполнения движения можно сохранить его основные черты, целесообразность, несмотря на изменение условий его осуществления. Так, сохраняется, например, индивидуальный характер походки человека при различиях в обуви, грунте, скорости движения, положении центра тя-. жести и т. д.
§ 12. Позно-тонические реакции
Общее представление о двигательной деятельности человека. Двигательная деятельность включает в себя процессы осуществления двигательных актов и процессы поддержания позы.
В основе двигательной деятельности человека лежат 2 формы механической реакции мышечных волокон: 1) их фазное (тетаническое) сокращение и 2) длительное тоническое напряжение. Фазная деятельность обеспечивает быстрые и сильные сокращения мышц
172
при выполнении движений, а также коррекции позы. Эти сокращения осуществляются быстрыми двигательными единицами. Тоническое напряжение отличается от фазного медленным развитием, большей экономичностью, преобладанием изометрического режима и преимущественным участием медленных двигательных единиц. Оно лежит в основе поддержания определенной позы тела. Если при поддержании позы требуется значительное усилие мышц (например, при выполнении угла в упоре), то включаются фазные сокращения мышц.
Процессы поддержания позы и двигательные акты скелетных мышц тесно связаны между собой. С одной стороны, движения всегда происходят на фоне определенной позы тела; с другой стороны, процессы поддержания позы обеспечиваются не только тоническим напряжением мышц, но и их фазной деятельностью.
В двигательной деятельности человека различают произвольные и непроизвольные движения и позы. Произвольные движения вырабатываются с участием сознания, а по мере их освоения могут происходить и бессознательно. Ведущую роль в осуществлении произвольных движений играют высшие интегративные области коры больших полушарий (лобные доли и др.). Еще до их выполнения в мозгу создается модель предстоящего движения, его программа. Человек сознает цель этого движения или необходимость принятия определенной позы. В процессе выполнения двигательного акта он, внося коррекции в его выполнение, добивается запрограммированного результата действия.
Непроизвольные движения и позы, выполняемые бессознательно, являются преимущественно безусловными рефлексами (ориентировочными. защитными и др.). Непроизвольными также становятся те произвольные движения и позы, которые автоматизируются при многократном их повторении. Непроизвольная двигательная деятельность может осуществляться различными отделами нервной системы.
Рефлекторная природа мышечного тонуса. В поддержании тонического напряжения мышц участвует сложная многоуровневая система нервных центров, находящаяся под контролем коры больших полушарий. Многочисленные опыты па животных, а также наблюдения за нарушениями мышечной деятельности у человека в клинике позволили выявить значение отдельных участков мозга в осуществлении и регуляции мышечного тонуса.
По своей природе мышечный тонус является рефлекторным актом.
Для возникновения тонуса в мышцах достаточна уже рефлекторная деятельность спинного мозга. Однако тонкое перераспределение тонуса между различными мышцами, необходимое для протекания реакций целостного организма, осуществляется более высокими этажами центральной нервной системы, а его произвольная регуляция — корой больших полушарий.
В наиболее простом случае появление тонуса обусловлено сокращением мышцы в ответ на раздражение проприорецепторов при
173
ее растяжении. Такой рефлекс, осуществляемый спинным мозгом, называется миотатическим рефлексом на растяжение. Если это растяжение длительно, то рефлекторное сокращение носит тоже длительный тонический характер. При растяжении мышцы возбужда
ются чувствительные нервные окончания в мышечных веретенах. Импульсы ог них направляются по толстым афферентным волокнам
РФ
Рис. 58. Гамма-регуляция деятельности мышц:
РФ — неспецифические влияния ретикулярной формации ствола головного мозга, регулирующие активность гамма-мотоиейро-иов; а — толстые альфа-волокна от крупных альфа-мотонейро-иов спинного мозга, вызывающие сокращения волокон скелетных мышц (э); у — тонкие гамма-волокна от мелких гамма-мотонейронов спинного мозга, изменяющие натяжение
внутр иверетенных (интрафу-
зальных — и) мышечных волокон и, следовательно, чувствительность проприорецепторов
(р); а — афферентное волокно, проводящее проприоцептивную импульсацию от рецепторов мышечного веретена, влияющую на деятельность альфа-мотонейронов
в передние рога спинного мозга, где не-посредственно (без участия вставочных нейронов) передаются на альфа-мотонейроны и вызывают сокращение мышц. Такая же двухнейронная рефлекторная дуга лежит в основе аналогичного сухожильного рефлекса — сокращения мышцы при ударе по ее сухожилию. К сухожильным рефлексам относят рефлекс ахиллова сухожилия, коленный рефлекс и др.
В естественных условиях основным раздражителем рецепторов мышечных и сухожильных веретен является сила тяжести, растягивающая скелетные мышцы, особенно мышцы-разгибатели. Поражение спинного мозга приводит к исчезновению тонуса мышц.
Гамма-регуляция мышечного тонуса. Степень тонического напряжения мышцы зависит от частоты импульсов, посылаемых к ней тоническими альфа-мотонейронами. Частота разрядов альфа-мотонейронов, в свою очередь, регулируется импульсами от проприорецепторов той же самой мышцы. Получается замкнутое кольцо между мышцей и иннервирующими ее мотонейронами. Однако потоки импульсов в этом кольце могут регулироваться вышележащими этажами нервной системы с помощью гамма-мотонейронов спинного мозга (рис. 58). Разряд гамма-мотонейронов повышает чувствительность мышечных веретен. В результате увеличивается поток импульсов от рецепторов к альфа-мотонейронам и от альфа-мото-нейронов к мышце. Тем самым повышается мышечный тонус.
Изменение напряжения мышц под влиянием деятельности гдм-ма-мотонейронов называется гамма-регуляцией. Активность гамма-мотонейронов находится под контролем ретикулярной формации ствола мозга. В регуляции их деятельности, а следовательно, и в ре-
гуляции тонуса скелетных мышц участвуют мозжечок, подкорковые
174
ядра и другие структуры экстрапирамидной системы. Высший контроль осуществляется корой больших полушарий.
Значение различных отделов головного мозга в регуляции тонуса скелетных мышц. Различные отделы головного мозга, посылая импульсы к вставочным нейронам и мотонейронам спинного мозга, могут регулировать тонус скелетных мышц, изменяя тем самым поз-ные и двигательные реакции организма. Влияния на мышечный тонус оказывают обе системы головного мозга — как специфическая, так и неспецифическая.
Неспецифическая система вызывает общее изменение тонуса различных мышц: усиление тонуса осуществляет активирующий отдел среднего мозга, а угнетение — тормозящий отдел продолговатого мозга.
Специфические системы действуют избирательно — на отдельные группы мышц. Усиление тонуса мышц-сгибателей вызывают кортико-спинальная, кортико-рубро-спинальная и частично кор-тико-ретикуло-спинальная системы (последняя оказывает также неспецифическое диффузное влияние). Одновременно эти системы снижают тонус мышц-разгибателей. В противоположность им вес-тибуло-спинальная система повышает тонус мышц-разгибателей и тормозит тоническое напряжение мышц-сгибателей. Изменения тонуса могут происходить очень быстро, нередко опережая двигательные акты. Это позволяет организму подготовиться к необходимому движению, заранее изменив позу тела.
В опытах на животных было показано, что при перерезке головного мозга между продолговатым и средним мозгом резко повышается тонус мышц-разгибателей. Это явление было названо децеребрационной ригидностью (децеребрация — лишение животного всех отделов головного мозга выше продолговатого, а ригидность — это твердость). После децеребрации у кошки напряжены мышцы спины, голова запрокинута назад, а хвост и конечности вытянуты. Этого не происходит в целостном организме, так как влияния из вестибулярных ядер продолговатого мозга, усиливающие тонус мышц-разгиба-телей, уравновешиваются в необходимой степени воздействиями других нисходящих систем, усиливающих тонус мышц-сгибателей. Правильное распределение тонуса различных мышц, которое лежит в основе организации определенной позы, обеспечивается деятельностью мозжечка, подкорковых ядер (полосатого тела и бледного ядра) и коры больших полушарий.
Мозжечок не имеет прямых связей со спинным мозгом, но через । расные ядра среднего мозга повышает тонус мышц-сгибателей, а через вестибулярные ядра продолговатого мозга усиливает тонус мышц-разгибателей. Поражения мозжечка нарушают правильное перераспределение тонуса скелетных мышц, что изменяет не только позные, но и двигательные реакции организма. При мозжечковых расстройствах вследствие снижения и ненормального распределения тонуса мышц конечностей нарушается походка (атаксия), бледное ядро угнетает тонус мышц, а при его поражениях тонус v вливается. Полосатое тело, регулируя деятельность бледного яд-
175
Рис 59. Деятельность мышц при поддержании позы (по А. Лун-дервольду):
А — поза сидя с выпрямленной спиной (неудобная поза), Б — естественная поза сидя (удобная поза). /—4 — электромиограммы: правой трапециевидной мышцы (7), правой широчайшей мышцы спины (2), правой крестцово-спинальной (3) и левой крестцово-спинальной (7) мышц
ра, снижает его угнетающее действие. Высший контроль тонической активности мышц осуществляет кора больших полушарий, в частности ее моторные, премоторные и лобные области. С ее участием происходит выбор наиболее целесообразной для данного момента позы организма, обеспечивается ее соответствие двигательным задачам.
Непосредственное отношение к регуляции тонуса имеют медленные пирамидные нейроны коры — так называемые нейроны положения, образующие медленную часть кортико-спинального пути (пирамидного тракта). По этому пути передаются прямые корковые воздействия на спинной мозг. Кроме того, изменяя активность ретикулярной формации ствола мозга, подкорковых ядер, мозжечка, ядер среднего и продолговатого мозга, кора больших полушарий оказывает через них параллельное влияние на тонус мышц.
Позы. Позой называется закрепление частей скелета в определенном положении.
Значение поз в двигательной деятельности человека. Организация позы необходима для преодоления силы земного притяжения. Поза человека служит для сохранения равновесия тела в состоянии двигательного покоя и при выполнении статической или динамической работы, а также для фиксации ряда суставов, необходимой для осуществления движения в других суставах.
При многих видах трудовой и спортивной деятельности человеку приходится принимать новые необычные позы, без которых невозможно движение. Поза, принимаемая при работе, называется рабочей позой. Правильная ее организация способствует повышению работоспособности человека. Наконец, большое значение имеют предрабочие изменения позы, которые заранее компенсируют возможные изменения центра тяжести тела и предотвращают его падение, а также препятствуют действию реактивных сил. Реакции поддержания позы обеспечивают
ся чаще всего тоническим напряжением мышц, а при больших нагрузках и при коррекциях, исправляющих позу,— фазными сокращениями мышц.
176
Во время отдыха или выполнения работы человек выбирает наиболее удобную позу, при которой возможно минимальное напряжение скелетных мышц (рис. 59). Организация такой оптимальной пони часто требует длительной тренировки. Например, при стрельбе из положения стоя только у высококвалифицированных стрелков топическая активность мышц, обеспечивающих стояние, минимальна (на электромиограмме регистрируется наименьшая электрическая активность мышц). У спортсменов низкой квалификации и новичков наблюдается значительное напряжение этих мышц.
Частным случаем позных реакций является поддержание заданного усилия мышцы и заданного суставного угла. Усилия мышц не сохраняются постоянными. Каждый раз, когда мышца расслабляется и возникают отклонения от заданного усилия или от определенного положения костей, появляются небольшие фазные сокращения работающих мышц, исправляющие эти отклонения. Нарастание усилия мышц достигается благодаря увеличению числа активных альфа-мотонейронов спинного мозга. Таким образом, сохранение постоянного усилия и суставного угла поддерживается как тоническими, так и фазными сокращениями мышц.
Известно, что человек может (особенно в результате мышечной тренировки) очень точно дозировать усилия своих мышц и определять положение суставов. Информацию о положении конечностей сообщают в кору больших полушарий многие рецепторы — мышечные и сухожильные, зрительные, кожные, но особенно большое значение имеют рецепторы суставных сумок.
Позно-тонические реакции мышц часто бывают кратковременными, так как возникают только в определенные фазы движения. Они способствуют, в частности, фиксации суставов при отталкивании от опоры в процессе ходьбы. Электромиографические исследования ходьбы показали, что наряду с работой мышц-антагонистов, вызывающих сгибания и разгибания нижних конечностей (например, передней большеберцовой мышцы и икроножной мышцы ноги), в фазу отталкивания носком от опоры происходит тоническое напряжение мышц стопы, фиксирующих ее свод.
Установочные рефлексы. Эти рефлексы способствуют сохранению позы. К ним относятся статические и статокинетические рефлексы, в осуществлении которых большое значение имеет продолговатый и средний мозг.
Статические рефлексы возникают при изменении положения тела или его частей в пространстве: 1) при изменении положения го-ювы в пространстве — это так называемые лабиринтные рефлексы, возникающие в результате раздражения рецепторов вестибулярно-io аппарата; 2) при изменении положения головы по отношению к |уловищу — шейные рефлексы с проприорецепторов мышц шеи и <) при нарушении нормальной позы тела — выпрямительные реф-и'ксы с рецепторов кожи, вестибулярного аппарата и сетчатки глаз. Например, при отклонении головы назад повышается тонус мышц-Р । иибателей спины, а при наклоне вперед — тонус сгибателей (лабиринтный рефлекс). Подобные реакции имеют место при вы-
I из
177
полпенни многих физических упражнений. Так, выполнение гимнастом стойки на кистях облегчается при отклонении головы назад, выполнение группировки во время прыжков в воду или акробатических прыжков — при наклоне головы вперед. Выпрямительные рефлексы — это последовательные сокращения мышц шеи и туловища, которые обеспечивают возвращение тела в вертикальное положение теменем кверху. У. человека они проявляются, например, во время ныряния.
Статокинетические рефлексы компенсируют отклонения тела при ускорении или замедлении прямолинейного движения, а также при вращениях. Например, при быстром подъеме усиливается тонус сгибателей, и человек приседает, а при быстром спуске усиливается тонус разгибателей, и человек выпрямляется — это так называемый лифтный рефлекс. При вращении тела реакции противовращения проявляются в отклонении головы, тела и глаз в сторону, противоположную движению. Движение глаз со скоростью вращения тела, но в противоположную сторону и быстрое возвращение в исходное положение — нистагм глаз — обеспечивают сохранение изображения внешнего мира на сетчатке глаз и тем самым зрительную ориентацию.
В двигательной деятельности человека часто возникают ситуации, когда требуется подавить эти установочные рефлексы. Произвольное подавление врожденных установочных рефлексов продолговатого и среднего мозга обеспечивается тормозящими влияниями из коры полушарий. Например, для спринтера невыгодно раннее выпрямление туловища при стартовом разбеге, поэтому корой больших полушарий тормозится выпрямительный рефлекс.
§13. Двигательная деятельность
Движения человека являются комплексом различных двигательных актов, которые обеспечиваются как сравнительно простыми двигательными рефлексами, так и сложнейшими их комбинациями.
Двигательные акты различной сложности. Рассматривая различные двигательные акты человека, можно выделить элементарные двигательные рефлексы, более сложные—ритмические рефлексы и, наконец, особенно сложные формы двигательной деятельности, обеспечивающие поведение человека.
Элементарные двигательные рефлексы. При сложных движениях человека используются элементарные двигательные рефлексы, осуществляемые спинным мозгом. К простым безусловным двигательным рефлексам спинного мозга относятся: 1) рефлексы на растяжение (миотатические и сухожильные рефлексы— рис. 60, Д), 2) сгибательные рефлексы на раздражение кожных рецепторов (рефлексы удаления от раздражителя — см. рис. 60, Б) и 3) рефлексы отталкивания (рефлексы сближения с опорой — см. рис. 60, В).
178
Миотатические и сухожильные рефлексы возникают при растяжении рецепторов мышц и сухожилий. Быстрое растягивание этих рецепторов рефлекторно вызывает фазное сокращение скелетных мышц. Это рефлекс активного противодействия мышцы ее растяжению. Рефлекторная дуга рефлекса на растяжение — моносинап-тическая. Она состоит всего из двух нейронов — афферентного и эфферентного, между которыми имеется лишь один синапс. Простота рефлекторной дуги обеспечивает быструю и точную реакцию мышцы, меньшую утомляемость рефлекторных ответов. Рефлексы
Рис. 60. Элементарные двигательные рефлексы спинного мозга:
А — сухожильный рефлекс на растяжение (сгибательный коленный рефлекс), Б — кожный сгибательный рефлекс (защитный), В — рефлекс отталкивания (давления на опору)
на растяжение имеют большое значение при статической деятельности мышц для поддержания определенной позы, а также при осуществлении локомоторных актов (ходьбы, бега и др.). В произвольной двигательной деятельности человека иногда необходимо подавлять эти рефлексы (например, выполнение гимнастического шпагата требует подавления рефлекса на растяжение).
При раздражении рецепторов кожи (болевых, температурных и др.) возникает сокращение мышц-сгибателей, которое позволяет отдернуть конечность от повреждающего раздражителя. Этот сгибательный кожный рефлекс носит защитный характер. В его основе лежит иолисинаптическая рефлекторная дуга. В отличие от моно-гинаптического рефлекса на растяжение при сгибательном рефлексе быстрее наступает утомление; время этого рефлекса в связи с наличием вставочных нейронов значительно больше.
Одна из разновидностей элементарных рефлексов спинного мозга — рефлекс отталкивания, описанный Ч. Шеррингтоном. Этот рефлекс возникает при раздражении кожи стопы опорой. Однако, в шличие от сгибательного рефлекса, он приводит не к отдергиванию конечности от раздражителя, а к сближению с раздражителем. В первом случае мы имеем защитный рефлекс, предохраняющий орга-
179
низм от разрушения. Во втором случае — биологически целесообразный акт, обеспечивающий контакт с опорой, при стоянии и отталкивание от нее при передвижениях. Этот рефлекс лежит в основе сложных локомоций: ходьбы, бега, прыжков в длину или высоту и др.
Ритмические рефлексы. Составной частью различных сложных двигательных действий, как произвольных, так и непроизвольных, часто являются ритмические рефлексы. Они особенно выражены при выполнении циклических движений. Их возникновение и протекание связаны с проявлением механизмов взаимосоче-танной (реципрокной) иннервации мышц-антагонистов.
Эта форма рефлексов — одна из древних и относительно простых форм — имеет большое значение в организации многих сложных движений. Например, включение ритмических циклоидных движений в акт письма позволяет человеку перейти от отдельного начертания букв к обычной письменной скорописи; то же самое происходит при освоении акта ходьбы — переход от отдельных шагов к ритмической походке (Н. А. Бернштейн).
Наиболее простая форма ритмического рефлекса — чесательный рефлекс животных, который обеспечивается попеременным сокращением и расслаблением одних и тех же мышц одной конечности. Возбуждение центров-сгибателей сопровождается одновременным торможением центров мышц-разгибателей, в чем проявляется реципрокная их иннервация. В последующей фазе движения в этих центрах возникают противоположные процессы — возбуждение вслед за торможением и торможение вслед за возбуждением. При этом проявляется так называемый рефлекс отдачи: после акта сокращения наступает расслабление, а затем снова акт сокращения. Рефлекс отдачи — это рефлекс на растяжение с проприорецепторов мышц и сухожилий, в результате которого возникает сокращение растянутого разгибателя. Такая простая форма ритмического рефлекса часто встречается в двигательной деятельности человека (например, при забивании молотком гвоздей).
Более сложная форма ритмического рефлекса — шагательный рефлекс, лежащий в основе ходьбы, бега и других локомоций. Для осуществления шагательных движений требуется участие уже не одной, а двух симметричных конечностей. Подключение второй конечности к координированному двигательному акту происходит в результате осуществления перекрестного рефлекса- Он хорошо виден на спинальном животном. Раздражение рецепторов кожи вызывает сгибание раздражаемой конечности и одновременное разгибание одноименной конечности на противоположной стороне тела. Этот рефлекс осуществляется с участием вставочных тормозных нейронов, вызывающих торможение мотонейронов на противоположной стороне спинного мозга. В целостном организме человека сгибание одной ноги вызывает перекрестный разгибательный рефлекс другой ноги, принимающей на себя тяжесть тела при стоянии, ходьбе и пр.
Следовательно, механизмы шагательных движений заложены уже на уровне спинного мозга. У спинального животного в первой 180
фазе движения болевое раздражение рецепторов кожи вызывает сгибательный рефлекс раздражаемой конечности (табл. 4). Одновременно при участии тормозных клеток Рэншоу происходит торможение центров мышц-разгибателей этой же конечности и торможение центров мышц-сгибателей симметричной конечности. Во вторую фазу движения возникает рефлекс отдачи с проприорецепторов растянутых мышц-разгибателей раздражаемой конечности и мышц-сгибателей другой стороны тела. Импульсы от этих рецепторов вызывают рефлекторное сокращение мышц-разгибателей и одновременное торможение центров мышц-сгибателей той же конечности и мышц разгибателей симметричной конечности. Последовательная смена этих двух фаз движения и составляет ритмический акт шагания.
Включение этого спинального механизма производится с более высоких этажей нервной системы. В так называемой «локомоторной области» среднего мозга имеются центры, электрическое раздражение которых вызывает шагательные движения конечностей.
В осуществлении шагательного рефлекса принимает участие мозжечок. Удаление одного его полушария у животных приводит к искажению движений (особенно передней конечности). Движения сильно варьируют по амплитуде, заметно искажаются при малейших посторонних воздействиях, происходит чрезмерное сгибание локтевого сустава в фазе переноса конечности.
Таблица 4
Процессы возбуждения (-(-) и торможения (—) в нервных центрах спинного мозга при осуществлении шагательного рефлекса
Рецепторы, вызывающие рефлекс Правая лапа Левая лапа
сгибатели разгибатели сгибатели разгиба тели
Нолевые рецепторы кожи правой лапы + +
11роприорецепторы разгибателей правой лапы и сгибателей левой + +
11ропрнорецепторы сгибателей правой i.iiibi и разгибателей левой + — — +
Протекание ритмических рефлексов, в том числе шагательного, i вязано с деятельностью различных отделов неспецифической сис-1смы. При активирующих влияниях происходит нарастание ампли-|\ды движений, а при угнетающих влияниях — ее снижение.
Важное значение в регуляции ритмических рефлексов имеют подкорковые ядра — бледное ядро и полосатое тело, обеспечивающие их автоматическое протекание, содружественные движения конечностей, вспомогательные реакции (фиксацию суставов и др.).
Высшим регулятором рефлексов является кора больших полушарий, особенно ее премоторная область. Благодаря коре ритмиче-кие движения (например, простой акт ходьбы) приобретают опре-
181
деленное смысловое значение, включаются как составной элемент в сложные акты поведения.
Об участии различных корковых областей в регуляции ритмических рефлексов свидетельствует появление на электроэнцефалограмме медленных потенциалов, синхронных по частоте с темпом движений. Протекание ритмических рефлексов облегчается тем, что
Рис. 61. Деятельность мышц стопы и голени при ходьбе (по Р. Манн, В. Инмани).
Цифры наверху — время в % от длительности одного цикла (шага). Цифры слева: /—6 — электромиограммы мышц-синергис-тов и мышц-антагонистов стопы, одновременно сокращающихся для фиксации стопы в фазу отталкивания; 7—S —электромиограммы мышц-антагонистов голени: передней большеберцовой (7) и икроножной (8), работающих попеременно. Стрелка внизу и пунктирная линия момент отделения стопы от опоры
между нервными центрами и ра-бодающими мышцами возникают согласованные во времени потоки нервных импульсов. Прохождение импульсов от нервных центров на периферию к рабочим органам и от мышц обратно к нервным центрам осуществляется в определенном темпе — в темпе рабочих движений. Налаживание этого темпа и стабильное его поддержание, включение древних ритмических автоматизмов в систему регуляции движений значительно упрощают управление ритмическими рефлексами, делают эту двигательную деятельность более экономичной.
Сложные формы двигательной деятельности. В целостном поведении простые рефлексы, сочетаясь, обусловливают сложные двигательные действия. Социальные условия жизни человека намного усложняют его двигательную деятельность, приводя к появлению специально человеческих форм движений: бытовых, производственных, спортивных. Простые и сложные ритмические рефлексы лежат в основе циклической двигательной деятельности человека: ходьбы, бега, плавания, гребли, ходьбы на лыжах, езды на велосипеде и пр. Большое значение в их осущест
влении имеют механизмы реципрокной иннервации мышц-антагонистов (рис. 61).
Однако в целом ряде движений требуется одноименная работа мыищ-антагонистов — в одной конечности или даже ц обеих. В этих
случаях реципрокные отношения, характерные для спинного мозга, подавляются центрами головного мозга. Особенно часто встречаются симметричные (а не реципрокно-перекрестные) движения в дея-
182
зольности верхних конечностей человека, т. е. происходит одновременное сокращение одноименных мышц левой и правой рук.
Наиболее сложные формы движений представляют собой многофазную цепь рефлексов, которая основана уже не на элементарных реципрокных отношениях, а является целостным двигательным навыком, образующимся в процессе обучения по механизму условных рефлексов.
Значение коры больших полушарий в регуляции произвольных движений. Ведущую роль в регуляции произвольных движений играет кора больших полушарий, особенно ее высшие отделы—зоны перекрытия анализаторов, или третичные поля. В коре больших полушарий происходит выработка цели и задачи движений, соответственно этому строится и программа конкретных действий, которые нужны человеку для осуществления цели. В сложные поведенческие акты включаются не только моторные компоненты, но и необходимые вегетативные компоненты. Еще до начала движений кора больших полушарий повышает активность тех вставочных и моторных нейронов спинного мозга, которым предстоит участвовать в движении. В предстартовый период перед началом циклических движений в электрической активности коры происходит настройка на темп предстоящих движений. В тот момент, когда производится движение, кора тормозит деятельность всех посторонних афферентных путей и оказывается особенно восприимчивой к сигналам от рецепторов мышц, сухожилий и суставных сумок.
В организации двигательного акта участвуют самые различные отделы коры больших полушарий. Моторная зона коры (поле 4) посылает импульсы к отдельным мышцам, преимущественно к дистальным мышцам конечностей. Объединение отдельных элементов движения в целостный акт осуществляют вторичные поля (б-е и е) премоторной области. Они определяют последовательность двигательных актов, формируют ритмические серии движений, регу-. лируют тонус мышц. Задняя центральная извилина коры — общечувствительная область — обеспечивает субъективное ощущение движения. Здесь имеются нейроны, сигнализирующие только о возникновении движений в суставе, и нейроны, постоянно информирующие мозг о положении конечности (нейроны движения и нейроны положения).
К пространственной организации движений прямое отношение имеют задние третичные поля — нижнетеменные и теменно-затылочно-височные области коры. С их участием производится оценка удаленности и расположения предметов, оценка расположения отдельных частей собственного тела в пространстве и др. При поражении них областей у человека теряется представление о «схеме тела» (о к'М, где находится нос, глаз, ухо, предплечье, спина, как опустить, например, «руки по швам»). Нарушается также представление о * схеме пространства», пространственная ориентация движений (Л. Р. Лурия). Трудности возникают при выполнении самых прос-ti.ix актов: человек видит стул и узнает его, но садится мимо него;
1,83
он не понимает, откуда идет звук, что означает «влево», «вправо», «вперед», «назад», не может правильно есть (например, ложка с супом попадает мимо рта) и т. д. Становится невозможным использование каких-либо орудий для трудовой или спортивной деятельности.
Области коры, относящиеся к лимбической системе (нижние и внутренние части коры), обеспечивают эмоциональную окраску движений и управляют вегетативными реакциями организма при работе.
В высшей регуляции произвольных движений важнейшая роль принадлежит лобным долям. В третичных полях лобной коры происходит сознательное программирование произвольных движений, определение цели поведения, двигательных задач и необходимых для их выполнения двигательных актов, а также сопоставление намеченной программы с результатами ее реализации. При регуляции лобными долями движений используется вторая сигнальная система. Движения программируются в ответ па поступающие извне словесные сигналы (словесные указания тренера, спортивные команды и пр.), а также благодаря участию внешней и внутренней речи (мышления) самого человека.
При поражениях лобных долей человек утрачивает способность ставить перед собой задачи, находить пути их решения, сличать выполнение действия с замыслом. При этом сохраняется способность осуществлять любой моторный акт, непосредственно повторяя или копируя его при движениях других людей. Однако сознательно использовать эти движения для достижения какой-либо определенной цели уже невозможно. Появляется стереотипность движений: происходит автоматическое и монотонное повторение одних и тех же же однородных двигательных актов, не осознается ошибочность действий, сильно затрудняются реакции выбора.
Таким образом, произвольные движения человека — результат объединенной деятельности самых различных отделов центральной нервной системы. В регуляции таких действий участвует многоэтажная и многозвенная функциональная система, состоящая из многих сотен, тысяч и миллионов нейронов. Работа этой системы сводится к определению оптимальных способов решения двигательных задач: удачного момента для начала движения, наиболее подходящей его структуры (сочетание мышц, степень и скорость их напряжения, порядок включения и т. п ), оптимального уровня функционирования вегетативных систем, постоянной и возможно более эффективной коррекции движений по ходу его выполнения и др.
Речевая регуляция движений. Спецификой регуляции движений у человека является то, чго они подчинены речевым воздействиям (как внешним — звуковая и письменная речь, так и внутренним — собственная внутренняя речь). Речевая регуляция движений связана в первую очередь с функцией лобных долей. Исследования покали, что на первых этапах развития — у ребенка 2—3 лет — регуляция произвольных двигательных реакций соответственно речевым инструкциям оказывается недостаточной. Он не может управлять дви
184
жением в полном соответствии с указаниями другого лица и еще не в состоянии полностью подчинить движения собственным речевым командам. К 3—4 годам ребенок осваивает речевые навыки и сам произносит «надо» или «не надо» делать движения. Эти речевые отчеты помогают ему в регуляции действий. Афферентная импуль-сация, идущая от речевой мускулатуры, служит важным ориентиром, дополняющим проприоцептивные сигналы от работающих мышц. Речевые самоотчеты помогают осмысливать и направлять движения. Постепенно простые речевые реакции усложняются и образуют сложную систему саморегуляции. Внешняя речь ребенка (произносимые им слова) сначала заменяется шепотом, а затем переходит во внутреннюю речь.
Внутренняя речь (процессы мышления) становится важным регулятором действий взрослого человека. При поражении лобной доли у него может сохраняться речь, но нарушается ее регулирующая роль в двигательных действиях. Больной не может выполнять словесные инструкции, исходящие от другого лица, или производить движения согласно собственной речи (например, под собственную команду «раз», «два»). Нарушается сличение результата движения и задуманной программы его выполнения. Движения становятся стереотипными, инертными, теряют свой смысл.
Глава V
ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ [УСЛОВНЫЕ РЕФЛЕКСЫ)
§ 1. Роль высшей нервной деятельности в адаптации организма к изменяющимся условиям среды
Высшая нервная деятельность обеспечивает индивидуальное приспособление организма к изменяющимся условиям внешней и внутренней среды. Она детерминирована (от лат. determinare — определять) совокупным действием многих факторов. К ним относятся, с одной стороны, афферентная импульсация, которая поступает в центральную нервную систему от рецепторов, воспринимающих эндогенные (от греч. endon — внутри, genos — род, происхождение) и экзогенные (от греч. ехо — снаружи) раздражения, т. е. раздражения из внутренней и внешней среды, с другой стороны — следовые явления от прежней деятельности нервной системы, т. е. память. Важная роль в афферентной импульсации принадлежит сенсорным коррекциям (обратным связям), передающим через внутренние и внешние анализаторы сигналы о характере и эффективности реакций организма (в спорте, например, о перемещениях звеньев тела, траектории перемещающихся снарядов при метаниях).
На основе анализа и синтеза афферентной импульсации (в том числе сенсорных коррекций) и следовых процессов формируются новые рефлекторные акты и целостное поведение организма.
185
Высшая нервная деятельность имеет важнейшее значение в процессе приобретения новых двигательных навыков и адаптации к различным физическим упражнениям.
Диалектико-материалистические представления о природе и детерминированности высших проявлений жизни, развитые К. Марксом, Ф. Энгельсом и В. И. Лениным, получили физиологическое обоснование в трудах И. М. Сеченова и И. П. Павлова. И. М. Сеченов впервые высказал (в книге «Рефлексы головного мозга») мысль о том, что произвольные движения, психические процессы и мышление человека по своей природе являются отраженными (рефлекторными). И. П. Павлов в учении об условных рефлексах не только экспериментально подтвердил правильность материалистических идей И. М. Сеченова, но и внес огромный вклад в дальнейшее развитие этих идей.
Учение И. П. Павлова о высшей нервной деятельности, экспериментально обосновывающее первичность материи и вторичность сознания, является одной из важных естественнонаучных сторон марксистско-ленинской философии. Рефлекторная теория И. П. Павлова, основанная на конкретных научных исследованиях, вскрыла многие закономерности нервного процесса как процесса отражения и его пластичный характер. Она содержит экспериментальные доказательства философских положений о познаваемости материальных процессов, лежащих в основе психической деятельности, о причинной обусловленности произвольных движений и поступков человека материальными процессами в нервной системе, вызванными раздражениями из внешней или внутренней среды и следами от всей предшествующей деятельности.
Механизмы высшей нервной деятельности у высших животных и человека связаны с деятельностью ряда отделов головного мозга. Основная роль в этих механизмах принадлежит коре больших полушарий (И. П. Павлов). Экспериментально показано, что у высших представителей животного мира после полного оперативного удаления коры высшая нервная деятельность резко ухудшается. Они теряют способность тонко приспосабливаться к внешней среде и самостоятельно существовать в ней.
Отсутствие активных пищедобывательных рефлексов и защитных реакций на дистантные (от лат. distantia — расстояние) раздражители может привести к гибели от голода или биологических врагов.
У человека кора больших полушарий головного мозга выполняет роль «распорядителя и распределителя» всех жизненных функций (И. П. Павлов). Это обусловлено тем, что в ходе филогенетического развития происходит процесс кортикализации функций. Он выражается во все большем подчинении соматических и вегетативных отправлений организма регуляторным влияниям коры мозга. В случае гибели нервных клеток в значительной части коры головного мозга человек оказывается нежизнеспособным и быстро погибает при заметном нарушении гомеостаза важнейших вегетативных функций.
186
& 2. Безусловные и условные рефлексы
И. П. Павлов разделил все рефлексы на 2 группы — безусловные и условные рефлексы. Они представляют собой сложные многокомпонентные реакции, включающие деятельность ряда как соматических, так и вегетативных органов. Среди компонентов рефлекса можно выделить главные и дополнительные. Например, в оборонительном рефлексе главным является двигательный компонент, в пищевом — двигательный и секреторный.
Безусловными рефлексами называются врожденные рефлексы. К моменту рождения у животных и человека закладывается основной наследственный фонд таких рефлексов. Но некоторые из них, в частности половые, формируются после рождения, по мере соответствующего морфологического и функционального созревания нервной, эндокринной и других систем.
Безусловные рефлексы обеспечивают первое, грубое приспособление организма к изменениям внешней и внутренней среды. Так, организм новорожденного адаптируется к среде за счет безусловных рефлексов дыхания, сосания, глотания и др.
Безусловные рефлексы отличаются стабильностью, которая обусловливается наличием в центральной нервной системе готовых стойких нервных связей для проведения рефлекторного возбуждения. Эти рефлексы носят видовой характер. Представители одного п того же вида животных имеют примерно одинаковый фонд безусловных рефлексов. Каждый из них проявляется при раздражении определенного рецептивного поля (рефлексогенной зоны). Например, глоточный рефлекс возникает при раздражении задней стенки глотки, рефлекс слюноотделения — при раздражении рецепторов полости рта, коленный, ахиллов, локтевой рефлексы — при раздражении рецепторов сухожилий определенных мышц, зрачковый — при действии на сетчатку резкого изменения освещенности и т. д. При раздражении других рецептивных полей эти реакции не вызываются.
Большинство безусловных рефлексов могут возникать без учас-i ня коры больших полушарий и подкорковых узлов. Вместе с тем центры безусловных рефлексов находятся под контролем коры больших полушарий и подкорковых узлов, которые оказывают субординационное (от лат. sub — подчинение, ordinatio — приведение г. порядок) влияние.
К сложным безусловным рефлексам относятся инстинкты. Животным присуши четыре основных вида инстинктов: пищевой, половой, родительский, оборонительный. В отличие от простых безусловных рефлексов, инстинкты обычно регулируются стимулами из внутренней среды организма, в том числе гормональными: голодом, жаждой, болью и др. Такого рода эндогенные мотивации (от Франц, motif — побудительная причина, повод к действию) стиму- пруют поисковую и другие виды деятельности животных в среде в х обитания и направлены на удовлетворение возникающих по-буждений.
187
Инстинкты весьма прочны. Сложные формы поведения, связанные с их проявлением, нередко свидетельствуют об их высоком приспособительном значении. Например, у щенков из породы охотничьих собак без предварительного обучения в условиях охоты проявляются в поведении многие нюансы, характерные для тренированных животных.
При росте и развитии организма система безусловнорефлекторных связей все же оказывается ограниченной, инертной, неспособной обеспечить достаточно подвижные адаптационные реакции, соответствующие колебаниям внешней и внутренней среды. Более совершенная адаптация организма к постоянно изменяющимся условиям существования происходит благодаря условнорефлекторным, т. е. индивидуально приобретенным, реакциям. Условнорефлекторные механизмы головного мозга имеют отношение ко всем видам деятельности организма (к соматическим и вегетативным функциям, к поведению), обеспечивая приспособительные реакции, направленные на сохранение целостности и стабильности системы «организм — среда». И. П. Павлов назвал условный рефлекс временной связью раздражителя с ответной деятельностью, образующейся в организме при определенных условиях. Поэтому в литературе вместо термина «условный рефлекс» часто используется термин «временная связь», который включает и более сложные проявления деятельности животных и человека, представляющие собою целые системы рефлексов и поведенческие акты.
Условные рефлексы не являются врожденными и приобретаются в процессе жизни в результате постоянного общения организма с внешней средой. Они не отличаются столь выраженной стабильностью, как безусловные рефлексы, и исчезают при отсутствии подкрепления. При этих рефлексах ответные реакции могут быть связаны с раздражением самых различных рецептивных полей (рефлексогенных зон). Так, условный пищевой секреторный рефлекс можно выработать и воспроизвести при раздражении разных органов чувств (зрение, слух, обоняние и др.).
§ 3. Механизм и условия образования условных рефлексов
Механизм образования условных рефлексов. Если интенсивность раздражения рецепторов достигает в различных рефлексогенных зонах пороговой или надпороговой силы, в них возникает возбуждение, которое, распространяясь по чувствительным нервам, приходит в центральную нервную систему и вызывает ответную рефлекторную реакцию.
Рефлекторное возбуждение, возникающее в какой-либо рефлексогенной зоне, адресуется из центров чувствительных нервов не всем, а строго определенным эффекторным (двигательным или секреторным) нервным центрам. В безусловных рефлексах эта связь между чувствительными и эффекторными центрами является врожденной.
188
Условные рефлексы характеризуются тем, что каждый раздражитель (световой, звуковой, проприоцептивный и др.) может при известных условиях приобрести сигнальное значение и стать раздражителем, вызывающим специальную ответную реакцию организма: двигательную, секреторную, пищевую, оборонительную и т. д. Например, если индифферентный раздражитель — звук звонка сочетается с действием пищи на рефлексогенную зону безусловного слюноотделительного рефлекса, то после нескольких сочетаний возбуждение, возникающее в слуховом чувствительном центре, будет распространяться на эфферентные центры слюноотделения (рис. 62). Обстановка спортивных состязаний, многократно сочетае-
Рис. 62. Схема исследования слюноотделительных условных рефлексов у собаки
мая с выполнением физических упражнений, также может стать условным раздражителем, т. е. сигналом, вызывающим изменение частоты сердечных сокращений, кровяного давления, интенсивности обмена веществ и т. д. Изменения физиологических функций в организме, наблюдаемые при стартовом состоянии, представляют собой примеры условных рефлексов, образовавшихся при сочетании воздействия обстановки стадиона с выполнением физических упражнений в условиях состязаний.
И. И. Павлов объясняет механизм образования условных рефлексов следующим образом. Если в центральной нервной системе возникают два очага возбуждения, то более сильный из них «при-|ягивает» к себе возбуждение из менее сильного. Если такого рода взаимодействие сильного и слабого очагов возбуждения сочетать повторно несколько раз, может образоваться условный рефлекс. Гак, после нескольких сочетаний относительно слабого очага возбуждения в зрительном центре (при действии света) с более сильным очагом возбуждения в пищевом центре (при кормлении) возбуждение из зрительного центра будет распространяться на пищевой
189
центр. В результате действие светового раздражителя будет вызывать пищевую реакцию в виде слюноотделения даже без принятия пищи, т. е. образуется условный рефлекс.
Электрофизиологический анализ формирования условных рефлексов показал, что для механизмов замыкания имеет значение пространственная синхронизация. Она заключается в сходстве просекания (по времени и фазе) биопотенциалов в различных пространственно удаленных друг от друга констелляциях (от лат. constellation — плеяда, совокупность) нервных клеток (М. Н. Ливанов, Е. Б. Сологуб и др.).
Образование условных рефлексов тесно связано с нервными механизмами памяти.
Как уже отмечалось в главе IV, существует 2 основных вида памяти: кратковременная (краткосрочная) и долговременная (долгосрочная). Информация, поступающая в нервную систему, сначала фиксируется в кратковременной памяти. В дальнейшем она или быстро утрачивается или «передается на хранение» в долговременную память.
Информация, фиксированная в долговременной памяти, может сохраняться дни, недели, месяцы и даже многие годы. Но такая передача информации в долговременную память происходит не всегда. Например, если человеку сообщить незнакомый семизначный номер телефона, то в большинстве случаев он может правильно повторить его. Но через несколько минут (особенно несколько десятков минут) этот помер забывается. Передачу информации из кратковременной памяти в долговременную можно искусственно прекратить путем соответствующих воздействий на центральную нервную систему. Так, если при начальном формировании условного рефлекса сразу после его выработки дать наркоз, то образовавшийся ранее условный рефлекс исчезает после снятия наркоза. Если же такой наркоз дать через несколько часов или дней после образования условнорефлекторной связи, то рефлекс сохраняется.
Считается, что фиксация информации связана с длительной (на протяжении десятков минут) реверберацией (от лат. reverberare — отражать), т. е. с циркуляцией возбуждения по нейронным цепям. Наркоз прекращает реверберацию. Поэтому если через 5—20 мин. после выработки условного рефлекса дать наркоз, то длительность реверберации оказывается меньше необходимой. Это препятствует фиксации информации в структурах долговременной памяти. Видимо, структуры, связанные с фиксацией кратковременной и долговременной памяти, различны.
В процессах фиксации информации большая роль отводится рибонуклеиновой кислоте (РНК). В опытах на крысах было показано, что применение препаратов, вызывающих снижение содержания РНК в нервной ткани, приводит к значительному снижению эффективности обучения, т. е. выработки условных рефлексов.
У низших животных образование условных рефлексов может осуществляться за счет подкорковых отделов головного мозга — мозжечка (рыбы), полосатого тела (птицы) и пр.
190
У млекопитающих и у человека в формировании условнорефлек-горных связей важнейшая роль принадлежит коре больших полушарий. Но при этом связи между условнорефлекторными центрами осуществляются как за счет межкортикальных проводящих путей (т. е. между разными зонами коры больших полушарий головного мозга), так и за счет путей, соединяющих кору с различными подкорковыми образованиями (ретикулярной формацией и др.). Так, после удаления больших полушарий у собак сохраняются и могут образовываться только самые простые условные рефлексы. Они вырабатываются очень медленно, отличаются непрочностью и отсутствием целенаправленности. Последнее проявляется, например, в развитии в ответ на условный сигнал беспорядочной двигательной активности (Э. С. Асратян, М. М. Хананашвили, Н. Ю. Беленков и др.).
Основные условия образования условных рефлексов. Условные рефлексы хорошо образуются только при определенных условиях. Главнейшими из них являются: 1) повторное сочетание действия ранее индифферентного условного раздражителя с действием подкрепляющего безусловного или ранее хорошо выработанного условного раздражителя; 2) некоторое предшествование во времени действия индифферентного агента действию подкрепляющего раздражителя; 3) бодрое состояние организма; 4) отсутствие других видов активной деятельности; 5) достаточная степень возбудимости безусловного или хорошо закрепленного условного подкрепляющего раздражителя; 6) надпороговая интенсивность условного раздражителя.
Совпадение действия индифферентного раздражителя с действием подкрепляющего раздражителя (безусловного или хорошо ранее закрепленного условного раздражителя), как правило, должно повторяться несколько раз. При образовании новых условных рефлексов в одной и той же обстановке процесс формирования этих рефлексов ускоряется. Например, у собаки в опытах в камере первый условный рефлекс образуется после 10—20 сочетаний, последующие же — значительно быстрее. У человека многие условные рефлексы, в особенности на словесные раздражители, могут образовываться после одного сочетания.
Длительность времени предшествования действия нового условного раздражителя действию подкрепляющего не должно быть значительным. Так, у собак рефлексы особенно хорошо вырабатываются при длительности этого времени предшествования 5—10 сек. При сочетании в обратном порядке, когда подкрепляющий раздражитель начинает действовать ранее индифферентного, условный рефлекс не вырабатывается.
Образование условнорефлекторных связей, легко протекающее в условиях бодрого состояния организма, затрудняется при его заторможенности. Так, у животных, находящихся в сонливом состоянии, условные рефлексы или не образуются совсем, или же образуются медленно, с трудом. Заторможенное состояние затрудняет формирование условных рефлексов и у человека.
191
При доминировании в. Антральной нервной системе центров, не связанных с образованием данных условных рефлексов, формирование этих рефлексов затрудняется. Так, если у собаки возникло резкое возбуждение, например при виде кошки, то в этих условиях образование пищевого слюноотделительного рефлекса на звук звонка или свет лампочки не происходит. У человека, поглощенного каким-либо делом, образование условных рефлексов на другие виды деятельности в это время также резко затрудняется.
Условные рефлексы образуются только при наличии достаточной возбудимости центров этих подкрепляющих рефлексов. Например, при выработке у собак пищевых условных рефлексов опыты ставятся при условии высокой возбудимости пищевого центра (голодном состоянии животного).
Возникновение и закрепление условнорефлекторной связи происходит при определенном уровне возбуждения нервных центров. В связи с этим сила условного сигнала должна быть достаточной — выше пороговой, но не чрезмерной. На слабые раздражители условные рефлексы не вырабатываются совсем или формируются медленно и отличаются неустойчивостью. Чрезмерно сильные раздражители вызывают развитие в нервных клетках охранительного (запредельного) торможения, что также затрудняет или исключает возможность образования условных рефлексов.
Явления генерализации (обобщения) при образовании условных рефлексов. Характерной особенностью вновь образованных условных рефлексов является их генерализация. Ее сущность заключается в том, что условные раздражители обобщаются и условный рефлекс, образовавшийся на определенный раздражитель, воспроизводится и при действии других, сходных с ним, раздражителей. Процесс возбуждения, возникающий при действии какого-либо одного раздражителя, в результате иррадиации переходит и на корковые центры других, близких по характеру, раздражителей. Например, при образовании условного пищевого слюноотделительного рефлекса на тон 1000 гц условнорефлекторное слюноотделение в первое время после установления временной связи будет вызываться и при действии других тонов: 200 гц, 5000 гц и т. д. В этом случае генерализация выражена при действии близких по частоте тонов больше, чем при действии тонов более отдаленных. Если тоны 700—800 гц дают такой же эффект, как и условный раздражитель, на который вырабатывается рефлекс (1000 гц), то тон 200 гц вызывает значительно меньшее условнорефлекторное слюноотделение.
В двигательных условных рефлексах, например при образовании двигательных навыков, феномен генерализации проявляется в участии в двигательном акте значительного числа мышц, сокращение которых не является необходимым.
Электрофизиологически иррадиация возбуждения выражается в десинхронизации электрокортикограммы (П. К- Анохин) и распространении первичных вызванных ответов по горизонтальным связям коры мозга (А. Б. Коган, Н. Н. Ткаченко).
192
Явления концентрации при упрочении условных рефлексов. Упрочившиеся условные рефлексы генерализованы в меньшей степени, чем только что образованные. Это объясняется тем, что процесс возбуждения при многократном повторении условных раздражений концентрируется и меньше иррадирует на другие центры. При этом чем больше упрочивается условный рефлекс, тем меньше становится выраженность явлений генерализации. Вследствие этого при упрочившемся условном рефлексе генерализация сказывается лишь на отношении раздражителей, весьма близких друг другу. Например, у собаки, имеющей упроченный условный рефлекс на тон в 1000 гц, генерализация может проявиться при раздражении тоном 900 гц, но ее не будет при раздражении тоном 200—500 гц.
При образовании условных рефлексов у человека явления генерализации выражены несравненно меньше, чем у животных. Процессы концентрации, проявляющиеся у человека в значительно большей степени, чем у животных, дают возможность сразу же устранить условные рефлексы на воздействия, существенно отличающиеся от раздражителя, на который вырабатывается данный условный рефлекс.
§ 4. Сенсорные и оперантные условные рефлексы
Каждый рефлекс содержит афферетные (сенсорные) и эфферентные (исполнительные) компоненты (звенья). В одних случаях формирование новых условных рефлексов относится к образованию только новых сенсорных компонентов, в других же — к образованию и эфферентных, т. е. исполнительных, компонентов рефлекса. Вследствие этого условные рефлексы могут быть двух видов — сенсорные и оперантные (эффекторные).
В сенсорных условных реакциях (названных Ю. Конорским условными рефлексами 1-го рода) ответные акты являются или унаследованными (пищевыми, оборонительными, ориентировочными, половыми и другими безусловными рефлексами), или ранее хорошо закрепленными условными рефлексами (условные рефлексы высших порядков). Следовательно, они характеризуются формированием только афферентной части рефлекса, при которой индифферентный раздражитель превращается в активный. Ответная же реакция на условный раздражитель остается той же, что и при безусловном или ранее хорошо выработанном условном раздражителе. Например, при выработке оборонительного условного рефлекса па uinax в центральной нервной системе устанавливаются связи между афферентными клетками, воспринимающими раздражение обонятельного анализатора, и болевым центром. При этом характер ответной условной и безусловной реакций совпадает. В обоих случаях начинается слюноотделение. Аналогично вырабатываются и проявляются другие сенсорные условные реакции (в частности, оборони-и-льные двигательные условные рефлексы в виде отдергивания руки, подкрепляемые болевым раздражением, сухожильные, зрачковые, мигательные рефлексы).
193
Условные рефлексы этого рода не всегда могут в достаточной мере изменить взаимоотношения организма с окружающей средой и полноценно обеспечить адекватное приспособление, так как при этом не организуются новые формы самих ответных реакций. Более адекватное приспособление обеспечивается тем, что животные и человек способны кардинально изменять характер своих эффекторных реакций при взаимоотношениях с окружающей средой.
Оперантные условные рефлексы (по классификации Ю. Конор-ского, рефлексы 2-го рода) характеризуются новой (не унаследованной от предков или не имевшейся в ранее индивидуально приобретенном фонде) формой ответной реакции. Эти рефлексы называются также «инструментальными», так как при их осуществлении используются разные предметы (инструменты). Например, животное конечностью открывает задвижку на дверце и достает находящуюся за ней пищу. Поскольку при формировании таких рефлексов создается сложный комплекс вновь образованных движений, эти рефлексы называют также «манипуляционными».
В образовании соответствующих условных оперантных рефлексов важнейшая роль принадлежит импульсам, поступающим из двигательного аппарата. Элементарная модель условного оперант-ного рефлекса наблюдалась при выработке классического пищевого условного рефлекса на пассивное сгибание лапы у собаки (Ю. Ко-норский). У животного было выявлено два типа условных реакций— условнорефлекторное слюноотделение в ответ на пассивное сгибание лапы (сенсорный условный рефлекс, или рефлекс 1-го рода) и многократное активное сгибание конечности, являвшееся не только сигналом, но и способом получения пищи (оперантный условный рефлекс, или рефлекс 2-го рода).
Одним из видов оперантных рефлексов являются опыты с само-раздражением мозговых структур, при котором получается «чувство удовлетворения потребностей» (см. главу IV, § 10).
При формировании оперантных условных реакций важнейшая роль принадлежит обратной связи между клетками в нервных центрах безусловных или ранее хорошо выработанных условных рефлексов и клетками центров двигательного анализатора. Этому способствует высокий уровень возбудимости двигательных центров, обусловленный потоком афферентных импульсов от проприорецеп-торов сокращающихся мышц.
Таким образом, одним из условий выработки оперантных моторных условных рефлексов является обязательное включение в систему раздражителей импульсации, возникшей спонтанно или вызванной активным или пассивным движением. Образование этих рефлексов зависит от характера подкрепляющего агента. Оперантные условные рефлексы составляют основу двигательных навыков. Закреплению их способствуют обратные связи, осуществляемые через проприорецепторы мышц, выполняющих движение, и через рецепторы ряда других анализаторов. Благодаря этой афферента-ции центральная нервная система сигнализирует о результатах выполнения движения.
194
Формирование новых движений, т. е. не унаследованных от предков, наблюдается не только у человека, но и у животных. Но для человека этот процесс имеет особенно большое значение, так как почти все сложные двигательные акты (в частности, физические упражнения в различных видах спорта) формируются именно в результате обучения.
§ 5. Виды условных рефлексов
В зависимости от особенностей ответных реакций, характера раздражителей, условий их применения и подкрепления и т. д. выделяют различные виды условных рефлексов. Эти виды классифицируются, исходя из различных критериев, в соответствии с поставленными задачами. Некоторые из этих классификаций имеют большое значение как в теоретическом, так и в практическом отношении, в том числе при спортивной деятельности.
Натуральные (естественные) и искусственные условные рефлексы. Условные рефлексы, образующиеся на действие сигналов, характеризующих постоянные свойства безусловных раздражителей (например, запах или вид пищи), называются натуральными условными рефлексами.
Иллюстрацией закономерностей образования натуральных условных рефлексов являются опыты И. С. Цитовича. В этих -опытах щенки одного и того же помета содержались на различном пищевом рационе: одних кормили только мясом, других — только молоком. У животных, которых кормили мясом вид и запах его уже на расстоянии вызывали условную пищевую реакцию с ярко выраженными моторным и секреторным компонентами. Щенки, получавшие лишь молоко, первый раз реагировали на мясо только ориентировочной реакцией (т. е., по образному выражению И. П. Павлова, рефлексом «Что такое?») —обнюхивали его и отворачивались. Однако уже однократное сочетание вида и запаха мяса с едой полностью устраняло это «безразличие». У щенков выработался натуральный пищевой условный рефлекс. Образование натуральных (естественных) условных рефлексов на вид, запах пищи и свойства 1 ругих безусловных раздражителей характерно и для человека..
Натуральные условные рефлексы характеризуются быстрой выработкой и большой стойкостью. Они могут удерживаться всю жизнь при отсутствии последующих подкреплений. Это объясняется п’М, что натуральные условные рефлексы имеют большое биологическое значение, особенно на ранних этапах приспособления организма к окружающей среде. Именно свойства самого безусловного раздражителя (например, вид и запах пищи) являются теми первыми сигналами, которые действуют на организм после рождения.
Но условные рефлексы можно выработать также на различные 'ндифферентные сигналы (свет, звук, запах, изменения температуры и др.), не обладающие в естественных условиях свойствами раз-1р.1жителя, вызывающего безусловный рефлекс. Такого рода реакции, в отличие от натуральных, называются искусственными услов-
195
ними рефлексами. Например, запах мяты не присущ мясу. Однако если этот запах несколько раз сочетать с подкармливанием мясом, то образуется условный рефлекс: запах мяты становится условным сигналом еды и начинает вызывать слюноотделительную реакцию без подкрепления. Искусственные условные рефлексы медленнее вырабатываются и быстрее угасают при неподкреплении.
Примером выработки условных рефлексов на искусственные раздражители может быть образование у человека секреторных и двигательных условных рефлексов на сигналы в виде звучания звонка, ударов метронома, усиления или ослабления освещения, прикосновения к коже и т. д.
Условные рефлексы первого и высших порядков. Реакции, образуемые на основе безусловных рефлексов, называются условными рефлексами первого порядка, а реакции, вырабатываемые на основе ранее приобретенных условных рефлексов, — условными рефлексами высших порядков (второго, третьего и т. д.). При выработке условных рефлексов высших порядков индифферентный сигнал подкрепляется хорошо упроченными условными раздражителями. Если, например, раздражение в виде звонка подкреплять едой (безусловная реакция), то вырабатывается условный рефлекс первого порядка. После упрочения условного рефлекса первого порядка можно выработать на его основе условный рефлекс второго порядка, в частности на свет. На базе условного рефлекса второго порядка можно образовать условный рефлекс третьего порядка, на базе рефлекса третьего порядка — рефлекс четвертого порядка и т. д.
Образование условных рефлексов высших порядков зависит от совершенства организации нервной системы, ее функциональных свойств и биологической значимости безусловного рефлекса, на базе которого выработан условный рефлекс первого порядка. Например, у собак в искусственных условиях па фоне повышенной пищевой возбудимости можно выработать слюноотделительный условный рефлекс третьего порядка. В случае двигательно-оборонительной реакции у этих же животных возможно образование условных рефлексов четвертого порядка. У обезьян, стоящих на более высокой ступени филогенетической лестницы, условные рефлексы высших порядков образуются легче, чем у собак. Для человека процесс образования условных рефлексов высших порядков оказывается наиболее адекватным. При наличии повышенной возбудимости центральной нервной системы даже у детей в возрасте до года вырабатываются условные рефлексы пятого и шестого порядка (Н. И. Красногорский). При развитии функции речи порядковый диапазон этих реакций значительно расширяется. Так, подавляющее большинство двигательных условных рефлексов у человека образуется путем подкрепления не каким-либо безусловным раздражителем, а различными условными сигналами в виде словесных инструкций, объяснений и др.
Биологическое значение условных рефлексов высших порядков состоит в том, что они обеспечивают сигнализацию о предстоящей деятельности при подкреплении не только безусловными, но и ус-
196
ловными раздражителями. В связи с этим более быстро и полно происходит развертывание адаптационных реакций организма.
Положительные и отрицательные условные рефлексы. Условные рефлексы, в динамике которых проявляется активность организма в виде двигательных или секреторных реакций, называются положительными. Условные реакции, не сопровождающиеся внешним двигательным и секреторным эффектами в связи с их угнетением,
относят к отрицательным, или тормозным, рефлексам. В процессе приспособления организма к изменяюшимся условиям среды оба вида рефлексов имеют большое значение. Они тесно
взаимосвязаны, так как проявление одного вида деятельности сочетается с угнетением других видов. Например, при оборонительных двигательных условных рефлексах тормозятся условные пищевые реакции и наоборот. При условном раздражителе в виде команды «Смирно!» вызывается деятельность мышц, обусловливающих стояние в определенном положении и торможение других условных двигательных реакций, которые осуществлялись до этой команды (например, ходьба, бег).
Такое важное качество, как дисциплинированность, всегда связано с одно-
o' 20 40 60 80 100' 120 140 Время,сек.
Рис. 63. Схема сочетания во времени условных раздражителей с подкреплением безусловным рефлексом при наличных
временным сочетанием положительных и отрицательных (тормозных) условных рефлексов. Например, при выполнении некоторых физических упражнений (прыжки в воду с вышки, гимнастическое сальто и др.) для подавления реакций самосохранения и чувства страха требуется торможение сильнейших отрицательных оборонительных
и следовых раздражениях. Белые прямоугольники — время действия условного раздражителя, черные прямоугольники — время действия безусловного раздражителя. При наличных рефлексах условный н безусловный раздражители в какой-то период времени действуют одновременно (А, Б, В). При следовых рефлексах между концом действия условного раздражителя и началом действия безусловного имеется перерыв (Г, Д, Е)
условных рефлексов.
Наличные и следовые рефлексы. Условные рефлексы, в которых условный сигнал предшествует безусловному раздражителю, дей-
ствует совместно с ним и заканчивается одновременно или на несколько секунд раньше или позднее прекращения безусловного раздражения, называются наличными (рис. 63). Как уже отмечалось, для образования условного рефлекса необходимо, чтобы условный сигнал начал действовать раньше начала действия подкрепляющего раздражителя. Интервал между ними, т. е. степень отставления подкрепляющего раздражителя от условного сигнала, может быть различным. В зависимости от длительности отставления безусловного подкрепления от начала действия условного сигнала наличные условные рефлексы у животных, например пищевые, классифици-
197
(см. рис. 63)
руются как совпадающие (0,5—1 сек.), короткоотставленные (3— 5 сек.), нормальные (10—30 сек.) и запаздывающие (1—5 мин. и более).
При следовых условных рефлексах условный раздражитель подкрепляется после прекращения его действия
Между затухающим очагом возбуждения в коре от индифферентного агента и очагом возбуждения в корковом представительстве подкрепляющего безусловного или ранее хорошо выработанного рефлекса образуется временная связь. Следовые условные рефлексы формируются при коротком (10—20 сек.) и длинном (позднем) отставлениях (1—2 мин. и более). К группе следовых условных рефлексов относится, в частности, рефлекс на время, выполняющий роль так называемых «биологических часов».
Наличные и следовые условные рефлексы с большим отставлени-ем являются сложными формами проявления высшей нервной деятельности и доступны лишь животным с достаточно развитой корой головного мозга. Выработка подобных рефлексов у собак связана с большими трудное!ями. У человека следовые условные рефлексы образуются легко.
Следовые условные реакции имеют большое значение при физических упражнениях. Например, в гимнастической комбинации, состоящей из нескольких элементов, следовое возбуждение в коре мозга, вызванное действием первой фазы движения, служит раздражителем для программирования цепи всех последующих. Внутри цепной реакции каждый из элементов является условным сигналом для перехода к следующей фазе движения.
Экстероцептивные, проприоцептивные и интероцептивные рефлексы. В зависимости от анализатора, на основе которого выработаны условные рефлексы, последние делятся на три вида. Реакции выработанные при раздражении внешних анализаторов (зрительного, слухового и др.), называются экстероцептивными, а выработанные при раздражении рецепторов мышц — проприоцептивными, ре цепторов внутренних органов — интероцептивными.
Главным средством связи организма с внешней средой являютсг экстеро- и проприоцептивные условные рефлексы. Реакции, имеющие большую биологическую значимость, вырабатываются быстрее и лучше дифференцируются. В то же время они достаточно дина мичны и могут угасать при изменении сигнального значения раз дражителей и их неподкреплении.
Интероцептивные условные рефлексы вырабатываются и диф ференцируются значительно медленнее, отличаются большой инертностью, не угасая при неподкреплениях в течение длительного вре мепи. Афферентная импульсация от интерорецепторов может много кратно совпадать по времени с осуществлением ответные соматических и вегетативных реакций, возникающих при возденет вии на организм определенных сигналов внешней среды. При этом интероцептивные раздражители приобретают для соответствующих реакций сигнальное значение. В целом интероцептивные раздражения стимулируют координирующее влияние нервных центров, в осо
188
бенности коры больших полушарий, на взаимодействие факторов внутренней и внешней среды, в результате чего развиваются тонкие условнорефлекторные приспособительные реакции. При мышечной деятельности интенсивность проявления вегетативных функций повышается (кровообращение, дыхание и др.). Импульсация со стороны интерорецепторов в центральную нервную систему заметно усиливается, в связи с чем создаются более благоприятные условия для образования интероцептивных условных рефлексов. Определенный характер изменения вегетативных функций в процессе выполнения спортивной работы может сочетаться по механизму условных рефлексов с конкретной двигательной деятельностью и тем самым способствовать более эффективному ее выполнению.
Условные рефлексы на комплексные раздражители. Образование условных рефлексов может происходить при действии не только одиночных, но и комплексных раздражителей, относящихся к одной или разным сенсорным системам. Комплексные раздражители могут действовать одновременно и последовательно. При комплексе одновременно действующих раздражителей поступают сигналы от нескольких раздражителей. Например, условный пищевой рефлекс может вызываться одновременным воздействием запаха, формы и цвета раздражителя. При комплексе последовательно действующих раздражителей первый из них, например световой, сменяется вторым, например звуковым (в виде высокого тона), затем третьим, например звуком метронома. Подкрепление же следует только после действия всего этого комплекса.
§ 6. Первая и вторая сигнальные системы
Взаимоотношения организма со средой осуществляются на основе сигналов, поступающих в нервную систему в результате непосредственного воздействия предметов и явлений внешнего мира на рецепторы. Этот тип сигнализации И. П. Павлов назвал первой сигнальной системой. В животном мире первая сигнальная система является единственным каналом информации организма о состоянии среды. Различные предметы внешнего мира, их физические и химические свойства (звук, цвет, форма, химический состав и др.) приобретают значение условных сигналов, оповещают организм о наступающих вслед за ними явлениях, вызывая тем самым приспособи-1ельные реакции. Например, дремлющее травоядное животное при шуке шагов или запахе хищника спасается бегством, так как эти раздражители сигнализируют об опасности.
Первая сигнальная система высших животных обеспечивает допаточно совершенное отражение внешнего мира и в связи с этим i никое и точное приспособление их к среде. И. П. Павлов рассматривал первую сигнальную систему как систему восприятий, впечатлений от всех воздействий внешнего и внутреннего мира, сигнали-шрующих о биологически полезных или вредных для организма раздражителях. Он писал: «Для животного действительность сигнализируется почти исключительно только раздражениями и следа
199
ми их в больших полушариях, непосредственно приходящими в специальные клетки зрительных, слуховых и других рецепторов организма. Это то, что мы имеем в себе как впечатления, ощущения и представления от окружающей внешней среды, как обшейриродной, так и нашей социальной, исключая слово, слышимое и видимое. Это первая сигнальная система действительности, общая у нас с животными» *.
Сигналы первой сигнальной системы являются конкретными и относятся к определенному предмету.
Образование условных рефлексов через первую сигнальную систему составляет у высших животных физиологическую основу их злементарного конкретного, или предметного, мышления. Первая сигнальная система — одинаковая у человека и животных. В условиях обычной жизни у человека она изолированно функционирует только в первые шесть месяцев жизни.
При воспитании человека развивается вторая сигнальная система, характерная только для человека. Это переводит высшую нервную деятельность человека на более высокую ступень. Она приобретает новые качества, обусловливающие расширение возможностей общения с внешним миром и многогранность его проявлений. И. П. Павлов назвал вторую сигнальную систему «чрезвычайной прибавкой» к механизмам высшей нервной деятельности человека. Вторая сигнальная система — это речь, слово, видимое, слышимое, произносимое мысленно. Это высшая система сигнализации окружающего мира. Она состоит в словесном обозначении всех его сигналов и в речевом общении. Вторая сигнальная система развилась у человека под влиянием социальной среды в процессе труда. Большую роль в этом сыграли кинестезические раздражения мозга, возникающие в результате трудовых процессов. Слово для человека служит таким же физиологическим раздражителем, как предметы и явления окружающего мира.
Словесные сигналы обобщают раздражители первой сигнальной системы. Одно и то же слово «стол» сигнализирует не только об определенном столе, но и о многих других столах, разных по величине, форме, цвету и др. В этом факте выражено не только обобщение, но и отвлечение от конкретных предметов действительности, т. е. переход человека от предметного мышления к абстрактному. Для того чтобы слово «стол» указывало на конкретный стол, необходимо уточнение — «этот стол». В пределах второй сигнальной системы обобщаются раздражители не только первой, но и самой второй сигнальной системы. Например, слово узкого значения «осина» обобщает конкретные раздражители первой сигнальной системы, а слово более широкого значения «дерево» — раздражители второй сигнальной системы.
Таким образом, вторая сигнальная система является всеобъемлющей, способной заменить, абстрагировать и обобщить все раздра
* И. П. Павлов. Двадцатилетии?! опыт объективного изучения высшей нервной деятельности. М., Медгиз, 1951, стр. 467.
200
жители первой сигнальной системы. Благодаря всей предшествующей жизни взрослого человека слово связано со всеми внешними и внутренними раздражениями, приходящими в нервную систему, обо всех них сигнализирует и все их заменяет, вызывая такие же действия, как они.
Другое исключительно важное значение второй сигнальной системы заключается в том, что она резко увеличивает объем информации— за счет использования не только индивидуального, но и коллективного опыта всего человечества. Получаемая человеком от других лиц словесная информация — устная и в особенности письменная— имеет чрезвычайно широкий диапазон (это может быть информация не только от ныне живущих лиц, но и от многих предшествующих поколений). Так, совершенствование спортсмена лишь частично происходит за счет личного его опыта, через словесную информацию он широко использует опыт своего тренера и огромного числа других лиц, изложенный в методических пособиях, учебниках, статьях и т. д.
Первая и вторая сигнальные системы функционально взаимосвязаны. Сигналы первой сигнальной системы, поступающие из различных частей тела и окружающей среды, непрерывно взаимодействуют с сигналами второй сигнальной системы. При этом образуются условные рефлексы второго и высших порядков, функционально связывающие сигнальные системы в единое целое. Кроме того, связь между двумя сигнальными системами, основанная на элективной (избирательной) иррадиации возбуждения, позволяет воспроизводить через вторую сигнальную систему условные рефлексы, выработанные на основе первой сигнальной системы (А. Г. Иванов-Смоленский).
Вторая сигнальная система составляет физиологическую основу абстрактного речевого мышления, присущего только человеку. Афферентные сигналы, поступающие в центральную нервную систему от речевых органов, через слуховой и зрительный анализаторы, формируют у человека сложные рефлексы, обусловливающие звуковую и письменную речь.
Локализация функций второй сигнальной системы в коре головного мозга выяснена еще не полностью. В ее осуществлении участвуют структуры правого и левого полушарий. Доминирующая роль при этом у большинства людей (правшей) принадлежит левому полушарию. Сравнительно обширные его участки осуществляют сложные функции, связанные с пониманием смысла слов, координацией речедвигательного аппарата при их произнесении и другими процессами.
§ 7. Значение торможения в условнорефлекторной деятельности и его виды
Значение торможения в условнорефлекторной деятельности. Наряду с процессами возбуждения существенная роль (как при безусловнорефлекторной, так и при условнорефлекторной деятельно
201
сти) принадлежит процессам торможения, т. е. угнетения, подавления деятельного состояния. Возбуждение и торможение, непрерывно взаимодействуя, обусловливают возможность осуществления одних реакций и задержки других (нарушающих координацию безусловно- или условнорефлёкторной ответной реакции или не являющихся необходимыми). Следовательно, образование любого условного рефлекса в виде координированного ответного акта требует возбуждения одних корковых нервных центров и торможения других.
Виды торможения. Наблюдаемые при условнорефлекторной деятельности виды торможения можно разделить на две основные группы — внешнее торможение и внутреннее.
Внешнее торможение условных рефлексов. Внешнее торможение по своей природе является врожденным видом торможения, присущим всем отделам центральной нервной системы и свойственным как безусловным, так и условным рефлексам. Оно возникает «с места»— без предварительной выработки. К этому виду торможения относятся внешнее торможение и запредельное. Первое из них основано на взаимодействии центров условного рефлекса и постороннего раздражителя. Например, если в момент действия условного сигнала начинает действовать сильный посторонний раздражитель, то условная реакция тормозится. Сильное возбуждение, возникающее в центре постороннего раздражителя, по закону индукции, резко снижает возбудимость корковых центров данного условного рефлекса.
Внешнее торможение постоянно встречается в практике физической культуры и спорта. Например, условнорефлекторные связи, обеспечивающие в обычной обстановке на тренировочном занятии отличное выполнение гимнастических комбинаций, могут нарушаться под влиянием сильных раздражителей (соревнование, присутствие посторонних лиц, резкие замечания тренера и др.), действующих как внешний тормоз. При повторных воздействиях внешнее торможение угасает или проявляется незначительно. Из практики спорта известно, что прочно закрепившиеся условные рефлексы меньше нарушаются под влиянием внешнего торможения. Степень выраженности внешнего торможения определяется функциональным состоянием и типом нервной системы. У лиц с уравновешенным типом нервной системы внешнее торможение при одних и тех же условиях выражено слабее, чем у представителей неуравновешенного типа или у имеющих функциональные расстройства нервной деятельности.
Запредельное (охранительное) торможение, развивающееся при действии сверхсильных или обычных, но длительно продолжающихся раздражений, представляет собой защитный механизм, предотвращающий нервные клетки от истощения и способствующий восстановлению их химического состава и нормального функционального состояния. По своей физиологической природе охранительное торможение может вызываться факторами, связанными как с безусловно- так и с условнорефлекторной деятельностью. Травмы, резкие и длительные раздражения анализаторов и др. способны вызывать охранительное торможение.
202
Необходимо указать, что внешнее торможение значительно сильнее сказывается на относительно мало закрепленных условных рефлексах. Из спортивной практики известно, что чем прочнее в процессе тренировки закреплены двигательные условные рефлексы, тем меньше они нарушаются при наличии внешнего торможения.
Внутреннее торможение условных рефлексов. Если торможение условных рефлексов первично возникает в их центрах, то такого рода торможение называют внутренним. Существует несколько видов внутреннего торможения, к которым относятся: угасательное, дифференцированное и торможение при запаздывании подкрепления условного раздражения. Внутреннее торможение, постепенно захватывающее всю кору больших полушарий, приводит к развитию сонного состояния.
Угасательное торможение. Если раздражители, вызывающие условный рефлекс, не подкреплять действием безусловных рефлексов, то они постепенно угасают (табл. 5).
Таблица 5
Угашение у собаки условного рефлекса на вид мясного порошка при отсутствии подкрепления едой (по И П Павлову)
Время Род раздражителя, щего f течение действую 1 мин. Колич. выделившейся слюны на условный раздражитель (мл)
11 час. 48 мин Вид мясного порошка 0,8
11 час. 51 МИН. ...... 0,7
11 час. 54 мин. » » » 0,5
11 час. 52 мин » » 0,3
12 час. 00 мин » » 0,2
12 час. 03 мин » » 0,1
12 час. 06 мин » » » 0,0
12 час. 09 мин. 0,0
Угашение условных рефлексов является не полным разрывом установившихся временных связей, а лишь их торможением. Во-первых, после перерыва (например, на следующий день) угашенный рефлекс самостоятельно постепенно восстанавливается. Лишь при многократном повторном неподкреплении условного раздражителя безусловным рефлексом угашение становится стойким. Во-вторых, угашенный рефлекс сразу же восстанавливается, если во время действия угашенного условного раздражителя внезапно воздействовать каким-либо сильным посторонним раздражителем. Это явление, связанное с устранением торможения условных рефлексов (например, в результате угашения), называется растормаживанием.
Если после угашения восстанавливать временные связи, подкрепляя условные раздражители безусловным рефлексом, то при повторных опытах без подкрепления условные рефлексы будут угашаться все быстрее и быстрее.
203
У человека при воздействии через вторую сигнальную систему угашение ряда условных рефлексов или их компонентов может происходить очень быстро, даже после однократной инструкции тренера. Однако это не относится к прочно зафиксированным двигательным условным рефлексам. В таких случаях для получения желаемого эффекта при переучивании, связанном с угашением старых и выработкой новых условных рефлексов, требуется длительная тренировка.
Угашение условных рефлексов при неподкреплении соответствующими безусловными или условными (при рефлексах высших порядков) раздражителями имеет большое биологическое значение, так как при этом устраняются именно те условные раздражители, которые потеряли свое сигнальное значение для приспособления к окружающей среде.
Дифференцированное торможение. Приспособление организма к условиям окружающей среды характеризуется тончайшим соответствием ответных реакций особенностям раздражения. Это соответствие приобретается благодаря возможности осуществлять высокую степень дифференцирования (различения) условных раздражителей внешней среды.
Условные рефлексы вначале образуются, как уже указывалось, в генерализованной форме. В дальнейшем происходит процесс дифференцирования, т. е. специализирования, условных раздражителей, который связан с торможением реакции на раздражители, не подкрепляемые безусловными или ранее выработанными условными рефлексами. Например, при образовании условного рефлекса на тон 1000 гц при первой пробе рефлекс будет обнаруживаться и на высокие (2500 гц) и более низкие (500 гц) тоны. Но если тон 2500 и 500 гц не подкреплять, рефлекс на них постепенно угаснет.
Дифференцирование простых раздражителей у животных может быть не только не хуже, но даже лучше, чем у человека. Например, собаки прекрасно различают запахи, тонкие оттенки серого цвета, разницу в высоте различных тонов и пр., которые совершенно недоступны человеку. Однако по способности к образованию дифференцировок при сложных комплексных раздражениях человек превосходит животных. Специальные опыты показали, что если, например, для человека дифференцирование таких двух комплексов, как «шипение — высокий тон — низкий тон — звонок» и «шипение — низкий тон — высокий тон — звонок» не представляет никаких трудностей (различение наблюдается после двух-пяти сочетаний), то для собак такая дифференцировка является уже предельной и для образования ее требуются многие месяцы и даже годы работы.
Сложнейшие дифференцировки образуются у человека при действии сигналов второй сигнальной системы. Он неодинаково реагирует не только на различные слова, но и на интонации, с которыми эти слова сказаны, на место того или иного слова в предложении и др.
Сложные процессы дифференцирования происходят при образовании двигательных условных рефлексов. Например, при обучении
204
различным физическим упражнениям руководитель подкрепляет правильное выполнение двигательного акта и не подкрепляет неправильное. Такого рода словесные подкрепления способствуют дифференцированию правильного выполнения упражнения и неправильного.
Скорость образования дифференцировок зависит от степени филогенетического и онтогенетического развития организма, индивидуальных его особенностей, функционального значения раздражений, которые необходимо дифференцировать, и от других факторов. У животных, стоящих на низком уровне филогенетического развития, сложные дифференцировки образуются, как правило, с большим трудом, чем у животных с более высокой организацией нервной системы. У человека в раннем детском возрасте дифференцировки также вырабатываются труднее, чем в более позднем. Большое значение имеет и функциональное состояние нервной системы. Например, при резком утомлении, при заболевании дифференцировки вырабатываются у человека труднее, ранее же выработанные могут несколько нарушаться.
Раздражители, действующие на различные анализаторы (например, свет и звук, запах и прикосновение), дифференцируются «с места», т. е. сразу же при первом применении. Раздражители того же анализатора, но резко отличающиеся друг от друга по своему характеру (например, тон 3000 гц и шум), также дифференцируются «с места». Раздражители, сходные по характеру (например, тон 1000 и 2000 гц), требуют для дифференцирования нескольких сочетаний, более же близкие по характеру (например, тон 1000 и 1100 гц) будут различаться только после значительного числа сочетаний.
В отдельных случаях дифференцирование сложных и вместе с тем близких по характеру раздражений требует огромного труда, особенно в процессе выработки дифференцировок при образовании двигательных рефлексов. Например, для образования дифференцировок в речедвигательных условных рефлексах, позволяющих говорить без акцента на иностранном языке, нужна большая практика. Она необходима и при обучении многим видам физических упражнений, когда определенный характер движений, необходимый для достижения максимального результата, нужно дифференцировать с другими (близкими по форме, но менее эффективными) движениями.
При наличии внешнего торможения дифференцирование условных раздражителей ухудшается. Поэтому, например, нарушение правильности движений при выполнении физических упражнений в момент действия внешнего торможения объясняется расстройством не только положительных условных рефлексов, но и дифференцировок.
Особым видом дифференцировочного торможения является так называемый условный тормоз. Если новый раздражитель, действующий непосредственно перед хорошо выработанным условным раздражителем, не будет подкрепляться безусловным рефлексом, то он
205
превращается в условный тормоз. Например, после выработки слюноотделительного условного рефлекса на звук метронома к нему присоединяют какой-либо новый раздражитель — свет, непосредственно после которого начинает звучать метроном. Если действие метронома всегда подкреплять введением в рот кислоты, а при действии последовательной комбинации «свет—метроном» не давать такого подкрепления, то свет станет условным тормозом. В этом случае действие метронома станет вызывать условный рефлекс. Если же перед этим будет действовать свет («свет+метроном»), то рефлекс окажется заторможенным.
Условное торможение часто проявляется при выполнении физических упражнений, например в спортивных играх (запрещение некоторых действий в определенных зонах игры).
Торможение при запаздывании подкрепления условного раздражения. Торможение может развиваться при выработанных условных рефлексах с большим отставлением от начала действия условного раздражителя до подкрепления безусловным рефлексом. Удлинение скрытого периода условного рефлекса при длительном оставлении подкрепления безусловным или ранее хорошо выработанным условным раздражителем основано на развитии торможения в первый период действия условного раздражителя.
Торможение при запаздывании подкрепления условного рефлекса возникает не сразу, а вырабатывается постепенно. При этом чем сильнее условный раздражитель, тем с большим трудом вырабатывается этот вид торможения. Существенное значение имеет и сила подкрепляющего раздражителя. Например, при повышении после голодания возбудимости пищевого центра происходит растормаживание и скрытый период условных рефлексов резко уменьшается.
У человека многие условные рефлексы имеют запаздывающий характер. Кратковременные явления запаздывания наблюдаются, например, при строевых командах. Подготовительная часть команд служит условным раздражителем, связанным с развитием запаздывания, а исполнительная часть — условным раздражителем, требующим немедленного исполнения.
§ 8. Динамический стереотип
Различные условные рефлексы постоянно взаимодействуют друг с другом. Если раздражители повторяются в определенном порядке, то между ними формируется взаимосвязь, характеризующаяся стереотипной последовательностью возникновения ответных реакций. При этом рефлексы соответствуют не столько данному раздражителю, сколько месту раздражителя в последовательной цепи их.
Стереотип внешних проявлений реакций в виде секреции или движения был назван И. П. Павловым динамическим стереотипом или функциональной системностью. Термин «динамический» подчеркивает функциональный характер этого стереотипа (формирование и закрепление его только после соответствующих упражнений, воз
206
можность его переделки, угасание при длительных перерывах, ухудшение при утомлении, сильных эмоциях, заболеваниях и пр.).
Динамический стереотип в отношении сенсорных условных рефлексов ярко проявляется при пищевых рефлексах. Например, если в опытах на собаке применять в течение длительного времени систему положительных и отрицательных условных раздражителей, чередуя их через определенные интервалы времени в строгой последовательности, то это фиксируется нервной системой. Каждому раздражителю, примененному на строго фиксированном для него в стереотипе месте, в соответствии с законом силы раздражителя, соответствует определенная величина ответной реакции (Э. А. Асратян, П. С. Купалов и др.). Стереотип может выявляться при замене всех условных раздражителей одним из них, обычно средней силы. Этот раздражитель, примененный в стереотипе вместо других условных раздражителей этого стереотипа, вызывает ответы, величина которых соответствует ответам на замененные раздражители. На месте же отрицательного раздражителя положительный сигнал вызывает очень слабую условную (в частности, слюноотделительную) реакцию.
В двигательной деятельности спортсмена стереотип проявляется, например, в последовательности фаз сложных гимнастических, тяжелоатлетических и других стандартно выполняемых движений.
Переделка стереотипа в некоторых случаях представляет для нервной системы трудный процесс. Чтобы выработать новый стереотип, необходимо сначала угасить старый. По хорошо закрепленный стереотип трудно подается угашению и может проявляться вновь при возникновении условий, которым он соответствовал.
Динамический стереотип может быть связан не только с отдельными вегетативными или двигательными функциями, но и с целостной деятельностью организма, режимом жизни человека. Формирование таких динамических стереотипов имеет большое значение для человека. Социальная среда, воздействующая на него,— быт, учеба, работа, как правило, в течение более или менее длительного времени остается относительно постоянной (домашний и рабочий режимы, их темп и др.). За счет следового возбуждения клеток в нервных центрах стереотип запечатлевается в них в виде сложной функциональной системы, в которой все воздействующие компоненты среды сливаются в единый синтетический комплекс. Таким образом, стереотип можно характеризовать как систему условных рефлексов па совокупность раздражителей естественной среды. Системность облегчает деятельность. Человек, привыкший изо дня в день делать одну и ту же работу, обычно выполняет ее с большей легкостью.
Однако образование прочного динамического стереотипа может наряду с положительным значением иметь и отрицательное. Привычка действовать по определенному стандарту затрудняет приспособление к новым условиям выполнения работы, к новому режиму жизни. В некоторых случаях при изменении ситуации прочный динамический стереотип задерживает приспособление организма к реакциям, более соответствующим новым условиям труда и быта. Изме
207
нение привычных форм работы, режима жизни переживается тяжело и может привести к нарушениям некоторых функций организма, особенно у лиц пожилого возраста. Поэтому, как указывал И. П. Павлов, установление динамического стереотипа является положительным при стандартных условиях деятельности и отрицательным при варьировании этих условий и резком их изменении. Это относится и к физическим упражнениям. Для стандартно выполняемых движений установление стереотипа полезно, для изменяющихся (спортивные игры, единоборство) — нежелательно.
§ 9. Экстраполяция в высшей нервной деятельности
В организме в течение жизни формируется огромное количество различных сенсорных и оперантных условных рефлексов, образующихся в результате действия многочисленных конкретных раздражителей и их комплексов. Вместе с тем число их все же весьма мало по сравнению с огромным разнообразием тех стимулов, которые в процессе жизни воздействуют на организм и требуют экстренных соответствующих обстановке ответных реакций.
Адекватное реагирование организма на возникающие новые раздражения «с места», т. е. без предварительной выработки данного конкретного условного рефлекса, обеспечивается способностью нервной системы к экстраполяции. Она обусловлена способностью-нервной системы в новых, в том числе ранее не встречавшихся, условиях правильно реагировать на основании предшествующего наследственного или индивидуально приобретенного опыта и эффективно решать возникающие задачи.
Экстраполяция наблюдается как при условнорефлекторной, так и при безусловнорефлекторной деятельности, причем в некоторых случаях даже за счет функций низших отделов центральной нервной системы.
В безусловнорефлекторной деятельности экстраполяция четко проявляется в следующем опыте. Обезглавленная (спинальная) лягушка лапкой рефлекторно снимает с кожи спины бумажку, смоченную кислотой. Этот рефлекс всегда осуществляется движением задней лапки той же стороны. Если ее привязать, то для снятия бумажки используется новый рефлекс — в виде движения задней лапки противоположной стороны. При привязывании обеих задних лапок возникают рефлекторные попытки снять раздражающий агент передними лапками.
Экстраполяция достигает особенно высоких форм при наличии значительного фонда условных рефлексов и связана как с сенсорной, так и с оперантной стороной высшей нервной деятельности.
Одним из проявлений экстрополяции в сенсорных компонентах условных рефлексов можно считать обобщение близких по характеру раздражителей. Если, например, на тон в 1000 гц вырабатывается условный рефлекс, то соответствующая реакция, как известно, возникает и при звучании более высоких и низких тонов. Чтобы отдифференцировать от них тон в 1000 гц, требуется специальное угашение действия более высоких или низких тонов.
208
Рис. 64. Экстраполяция в двигательной деятельности с учетом пространства и времени (по Л. В. Крушпнскому):
А — исходное место расположения зверя; В — исходное место расположения собаки в момент, когда она увидела зверя. AAi— линия движения зверя, ВВ\ — линия движения собаки
В результате образования различного рода оперантных условных рефлексов не только человек, но и животные могут правильно реагировать на создавшиеся новые конкретные условия. Например, собака, заметив на охоте бегущего в определенном направлении зверя, экстраполирует скорость и направление передвижения, чтобы бежать к месту встречи по кратчайшему расстоянию. Вследствие этого она направляется не к тому месту, где был замечен зверь, а к месту предстоящего пересечения путей (рис. 64).
Своих наивысших форм экстраполяция достигает у человека. Она постоянно проявляется и при умственной, и при мышечной работе, в частности при спортивной деятельности.
На основании полученного опыта, т. е. приобретения ряда конкретных связей, человек пу
тем экстраполяции правильно решает новые задачи, родственные в определенной мере ранее усвоенным (подробнее об экстраполяции в двигательных навыках см. главу XVI, § 9).
§10. Типы высшей нервной деятельности
На основании изучения условных рефлексов и сопоставления их с динамикой внешнего поведения И. П. Павлов выделил четыре основных типа нервной системы, совпадающие с классификацией темпераментов, предложенной более 2 тысяч лет назад Гиппократом. В основу классификации положено три функциональных показателя свойства процессов возбуждения и торможения: сила, уравновешенность и подвижность (табл. 6).
Сильный неуравновешенный (безудержанный) тип характеризуется сильными нервными процессами с резко выраженным преобладанием возбуждения. Представители этого типа очень активны. У них быстро вырабатываются положительные и медленнее тормозные условные рефлексы. Последние являются недостаточно стойкими. У представителей сильного уравновешенного подвижного и сильного уравновешенного инертного типов быстро вырабатываются и стабилизируются положительные и тормозные условные рефлексы. Различие между этими типами обнаруживается при изменении сигнального значения положительных и тормозных раздражителей, когда положительные перестают подкрепляться, а тормозные подкрепляются безусловными или хорошо выработанными условными рефлексами. Представители сильного уравновешенного подвижного
к—113
209
Таблица 6
Схема четырех типов нервной системы (по И. П. Павлову)
Гии нервной системы Характеристика по показателям свойств нервной системы Соответствие темпераментам по Гиппократу
по силе по уравновешенности по подвиж и ости
Сильный неуравновешенны й (безудержный) Сильный Неуравновешенный Преобладание возбуждения над торможением — Холерик
Сильный уравновешенный подвижный » У рав новешенный Подвижный Сангвиник
Сильный уравновешенный инертный » Инертный Флегматик
Слабый Слабый Неуравновешенный. Преобладание торможения над возбуждением Меланхолик
типа быстро приспосабливаются к изменившемуся значению раздражителей и вырабатывают соответственно изменившимся условиям новые условные рефлексы. Для представителей сильного уравновешенного инертного типа перестройка является более сложным процессом и осуществляется медленнее.
Слабый тип характеризуется легкой тормозимостью даже при самых незначительных внешних влияниях. Представителей этого типа отличает замедленная выработка и неустойчивость положительных условных рефлексов. Тормозные рефлексы вырабатываются у них быстро и характеризуются стабильностью. Животные со слабым типом нервной системы даже при многолетнем пребывании в лаборатории проявляют пассивно-оборонительную реакцию.
Классификация типов высшей нервной деятельности у детей (Н. И. Красногорский, А. Г. Иванов-Смоленский) укладывается в схему четырех типов нервной системы, описанных И. П. Павловым. Критерии оценки типологических особенностей в этих классификациях не выходят за рамки силы, уравновешенности и подвижности нервных процессов. Признаки одного из четырех типов нервной системы наблюдаются у большинства взрослых людей, обследуемых по методике двигательных условных рефлексов на речевое подкрепление.
Однако в резко выраженном виде эти четыре типа встречаются относительно редко. Большинство людей относится к промежуточным формам, число вариантов которых весьма велико. Существенное значение при этом принадлежит воспитанию.
И. П. Павлов рассматривал тип нервной системы как сплав ге-
210
нотипа, т. е. наследственной основы нервной деятельности (обусловливающей конституциональные особенности организма, в том числе и темперамент), с фенотипом, т. е. приобретенными в результате воспитания свойствами.
Таким образом, в процессе воспитания к врожденным свойствам нервной системы добавляются новые, связанные с многообразным воздействием социальной среды. Свойства нервной системы, приобретенные под влиянием внешней среды, могут доминировать и подавлять генетические задатки. Это проявляется даже в опытах иа животных. Так, внешне трусливые, проявляющие пассивно-оборонительную реакцию животные нередко обнаруживают в экспериментальных условиях все черты сильного типа, а внешне активные — черты слабого типа.
У человека, для которого социальная среда служит решающим фактором, определяющим характер высшей нервной деятельности, исключительно важное значение в формировании типа нервной системы имеет воспитание.
§ 11. Фазовые состояния
Здоровая нервная система в бодром состоянии характеризуется нормальным взаимодействием (балансом) между процессами возбуждения и торможения, благодаря чему обеспечивается правильное соотношение величины ответных реакций со значением и физиологической силой раздражителей. Но в некоторых случаях, когда организм оказывается в трудных условиях (например, при переутомлении, заболевании, решении ответственных или особенно трудных задач), баланс между возбуждением и торможением нарушается и в нервной системе возникают фазовые состояния. Признаками развития этих состояний являются неустойчивость структуры и нарушение закона силы в условнорефлекторных реакциях.
Закон силы (см. главу I) заключается в том, что реакции организма на раздражители средней величины оказываются больше, чем па слабые раздражители, а реакции на сильные больше, чем на средние. Например, на слабый световой раздражитель реакция является наименьшей, на средний — несколько большей, и на сильный— самой большой. Эта разница реакции при разной физической интенсивности раздражителей, часто наблюдаемая в опытах на животных, не всегда имеет место в человеческой практике. У людей интенсивность реакции в основном связана со смысловым значением раздражителей, в особенности при словесных сигналах. Тихо ска-шпное слово, например об опасности, имеет значительно большее шачение, чем громко сказанная индифферентная фраза. Поэтому по взаимоотношениях людей физическая сила раздражителя только в некоторых случаях соответствует интенсивности ответной реакции.
Значительно чаще эти ответные реакции, т. е. проявление закона силы, обусловлены смысловой значимостью раздражителей Нарушение закона силы в условнорефлекторных реакциях протекает поти-н* 211
Ч
пу парабиоза Н. Е. Введенского и проявляется в возникновении фазовых состояний (см.главу I).
Первая фаза — уравнительная. Она характеризуется тем, что сильные, средние и слабые раздражители вызывают реакции одинаковой величины.
Вторая фаза — парадоксальная-, слабые условные сигналы по величине реакции превосходят сильные.
Третья фаза — ультрапарадоксальная: при отсутствии условных реакций на положительные сигналы развивается положительный эффект на тормозные раздражители.
Четвертая фаза — тормозная. Она характеризуется полным торможением условнорефлекторных реакций.
Сравнительно недавно описана наркотическая фаза, характеризующаяся равномерным уменьшением величины условнорефлекторных ответов на слабые и сильные раздражители до полного их выпадения.
Четкая последовательность в развитии фазовых состояний иногда может нарушаться вследствие выпадения той или другой фазы. При переходе от парабиотического состояния к нормальному фазы следуют в обратном порядке.
Пограничные фазовые состояния особенно хорошо выражены при медленном погружении в гипнотический сон. Последний представляет собой частичное торможение коры, захватывающее относительно большие участки, особенно сенсомоторную область. При этом ряд центров может находиться в состояии парадоксальной фазы, когда словесный раздражитель действует сильнее, чем сам предмет. В связи с этим предметы воспринимаются в соответствии со словесным раздражением.
Во время гипноза и сна «дежурные» пункты коры, настроенные на восприятие определенных сигналов внешней среды, свободны от фазовых явлениях и обладают высокой степенью возбудимости. При гипнозе эти корковые, центры, связанные с реагированием на голос гипнотизера, обеспечивают речевую связь (так называемый «рапорт»). Во сне эти центры воспринимают раздражители, имеющие для организма специальное значение (например, для матери — плач ребенка).
Нарушение нормального баланса нервных процессов при неврозах, перетренированности и др. может вызвать срыв высшей нервной деятельности и фазовые состояния. При преобладании процессов возбуждения нарушаются все условнорефлекторные реакции, связанные с торможением: дифференцировки, запаздывание и др. При преобладании процессов торможения исчезают многие из ранее выработанных условных рефлексов. Особенно легко срыв высшей нервной деятельности происходит у лиц с сильным неуравновешенным и у лиц со слабым типом нервной системы.
Для восстановления нормального состояния нервных процессов применяется ряд средств, важнейшими из которых являются отдых, облегчение условий работы, перемена обстановки, обеспечение нормального сна и питания, фармакологические средства.
212
§12. Особенности высшей нервной деятельности человека
Человек является существом социальным. Такой социальный фактор, как активная трудовая деятельность, способствовал прогрессивному развитию больших полушарий головного мозга, в том числе корковых структур, являющихся у человека главным материальным субстратом высшей нервной деятельности. Физиологически определенную роль в зтом сыграли кинестезические раздражения, возникающие при двигательной деятельности в проприорецепторах работающих мышц. Качественные особенности высшей нервной деятельности человека связаны главным образом с механизмами второй сигнальной системы. Составляя основу абстрактного речевого мышления, вторая сигнальная система обеспечила человеку возможность словесного мышления, замену словами конкретных предметов и явлений окружающего мира. Результат абстрактно обобщенного отражения мозгом внешнего мира — понятия и умозаключения — стимулирует произвольную деятельность человека, направляя ее на активное преобразование среды, в которой он живет. Тесная функциональная связь двух сигнальных систем, основанная па взаимном подкреплении пергосигнальных и второсигнальных реакций, обеспечивает высокий, качественно новый уровень аналитической и синтетической деятельности мозга человека.
Специфической особенностью высшей нервной деятельности человека, связанной с развитием второй сигнальной системы, является формирование временных связей на мысленное слово. Оно по механизму условного рефлекса стимулирует определенные действия, поступки, способствуя тем самым формированию общего поведения человека в социальной среде.
Высшие формы анализа и синтеза, присущие человеческому мозгу, компенсируют меньшие пороги чувствительности некоторых анализаторов по сравнению с животными. Тонкость анализа внешнего мира человеком отмечена еще Ф. Энгельсом: «Орел видит значительно дальше, чем человек, но человеческий глаз замечает в вещах значительно больше, чем глаз орла *.
Специфические особенности высшей нервной деятельности чело-г.ека, обусловливающие абстрактное мышление, позволили человечеству достичь великих свершений в преобразовании природы и общества и стать, по выражению И. П. Павлова, «хозяином действи-। ельности».
Исключительно важное значение условных рефлексов заключа-гея в том, что еще до каких-либо непосредственных воздействий на организм, при возникновении лишь сигналов возможности этих шндействий, нервная система организма правильно прогнозирует предстоящие события и заранее подготовляет программы своих предстоящих адекватных ответных действий. При этом у человека i.iKoe прогнозирование может производиться не только непосредст
• К. Маркс и Ф. Энгельс. Соч., 2-е изд., т. 20. Госполитиздат, 1961, п 490.
213
венно перед предстоящим действием, но и за часы, дни, недели и даже месяцы и годы.
Физиология высшей нервной деятельности имеет важное значение для естественнонаучного обоснования физического воспитания. Формирование навыков и процесс тренировки могут быть полноценными лишь в том случае, если учитываются закономерности образования временных связей, регулирующих деятельность мышц и вегетативных органов.
Глава Vi
КРОВЬ
§ 1. Кровь как внутренняя среда организма
Кровь вместе с лимфой и межтканевой жидкостью составляет внутреннюю среду организма, в которой протекает жизнедеятельность всех клеток и тканей. Она представляет собой своеобразную форму ткани и характеризуется следующими особенностями-. 1) является жидкой средой, содержащей форменные элементы; 2) находится в постоянном движении; 3) составные части крови в основном образуются и разрушаются вне ее.
Кровь вместе с кроветворными и кроверазрушающими органами (костным мозгом, селезенкой, печенью и лимфатическими узлами) составляет целостную систему крови (Г. Ф. Ланг). Деятельность этой системы регулируется нейрогуморальным и рефлекторным путем.
Благодаря циркуляции в сосудах кровь выполняет в организме следующие важнейшие функции: 1) транспортную, 2) регуляторную, 3) защитную и 4) дыхательную.
Кровь транспортирует питательные вещества (глюкозу, аминокислоты, жиры и др.) к клеткам, а конечные продукты обмена веществ ( аммиак, мочевину, мочевую кислоту и др.) —от них к органам выделения. Осуществляя перенос гормонов и других физиологических активных веществ, воздействующих на различные органы и ткани, она выполняет регуляторную функцию. С функциями крови тесно связана регуляция постоянства температуры тела. Кровь переносит тепло от органов с интенсивным его образованием к органам с менее интенсивной теплопродукцией и к местам, где она охлаждается (поверхность кожи). Защитную функцию кровь выполняет благодаря способности лейкоцитов к фагоцитозу и наличию в ней иммунных тел, обезвреживающих микроорганизмы и их яды, разрушающих чужеродные белки. Доставляя кислород от легких к тканям, принося к легким углекислый газ, кровь осуществляет дыхательную функцию.
214
§ 2. Количество крови в организме
У взрослого человека общее количество крови составляет 5— 8% веса тела, что соответствует 5—6 л. Объем крови принято обозначать по отношению к весу тела ( мл/кг). В среднем он равен у мужчин 61,5 мл/кг, у женщин — 58,9 мл/кг.
В кровеносных сосудах в состоянии покоя циркулирует не вся кровь. Около 40—50% ее находится в кровяных депо (селезенке, печени, сосудах кожи и легких). Депонированная кровь в единице объема содержит больше форменных элементов по сравнению с кровью, циркулирующей в сосудах. Выход крови из депо в общий кровоток обусловливается многими факторами. Наиболее важным из них является дефицит кислорода в организме, возникающий под влиянием различных причин (мышечная деятельность, кровопотери, пониженное атмосферное давление и др.). Уменьшение количества крови более чем на 7з опасно для жизни. Потеря крови в небольших количествах (200—400 мл) для здоровых людей не только не представляет вреда, но даже стимулирует кроветворение.
§ 3. Состав крови
Кровь состоит из форменных элементов (55—58%) —эритроцитов, лейкоцитов и тромбоцитов — и жидкой части — плазмы (42— 45%).
Эритроциты, или красные кровяные тельца, у человека и млекопитающих представляют специализированные безъядерные клетки диаметром 7—8 мк. Они образуются в красном костном мозгу, разрушаются в печени и селезенке. В 1 мм3 крови содержится в среднем 5 млн. эритроцитов у мужчин и 4,5 у женшин. Строение и состав эритроцитов обусловлены выполняемой ими функцией — транспорт газов. Форма эритроцитов в виде двояковогнутого диска увеличивает соприкосновение с окружающей средой, способствуя этим ускорению процессов газообмена.
В начальных фазах своего развития эритроциты имеют ядро и называются ретикулоцитами. По мере созревания ядро замещается дыхательным пигментом — гемоглобином (Нв), составляющим 90% сухого вещества эритроцитов.
Гемоглобин обладает свойством легко связывать и отщеплять кислород. Присоединяя его, он становится оксигемоглобином (НвОг). Отдавая кислород в местах с малым его содержанием, он превращается в восстановленный (редуцированный) гемоглобин. В крови взрослых людей находится 14—15% гемоглобина. Общее I о содержание равно примерно 700 г.
Количество эритроцитов и гемоглобина в крови может изменя-о.ся под влиянием факторов внешней среды (атмосферное давление, сезоны года, особенности климата и др.), физиологической перестройки организма в определенные периоды жизни, под влиянием । и< тематических занятий спортом и т. д. У спортсменов нередко от
215
мечается повышенное содержание эритроцитов (до 6 млн. в 1 мм 3 крови) и высокая концентрация гемоглобина.
Вследствие усиленного эритропоэза (образования эритроцитов) в горных условиях наблюдается увеличение числа эритроцитов до 7 и более миллионов в 1 мм3.
В скелетной и сердечной мышцах содержится мышечный гемоглобин — миоглобин, который играет важную роль в снабжении кислородом работающих мышц, особенно при нарушении нормального снабжения тканей кислородом. В миоглобине может находиться до
Рис. 65. Лейкоциты человека:
а —* палочкоядериый нейтрофил; б — нейтрофил с сегментированным ядром; в — нейтрофил, захватывающий бактерии; г — измененная форма нейтрофила при движении; д — большой лимфоцит; е — малый лимфоцит
14% из общего количества кислорода, связанного гемоглобином крови и миоглобином.
Лейкоциты, или белые кровяные тельца, по морфологическим и
функциональным признакам представляют собой обычные клетки, содержащие ядро и протоплазму специфической структуры. Они образуются в лимфатических узлах, селезенке и костном мозгу. В 1 мм3 крови человека находится 5—6 тыс. лейкоцитов.
Лейкоциты (рис. 65) неоднородны по своему строению: в одних
из них протоплазма имеет зернистое строение (гранулоциты), в других нет зернистости (агронулоциты). Гранулоциты составляют 70—75% всех лейкоцитов и делятся в зависимости от способности окрашиваться нейтральными, кислыми или основными красками на нейтрофилы (60—70%), эозинофилы (2—4%) и базофилы (0,5— 1%)*. Нейтрофилы, в свою очередь, разделяются на юные, палочкоядерные и сегментированные. Юные и палочкоядерные являются незрелыми формами нейтрофилов. Лгранулоциты состоят из двух
* Нейтрофилы — клетки, окрашиваемые нейтральными, эозинофилы — кислыми, а базофилы — основными красками
216
групп: лимфоцитов (25—30%) и моноцитов(4—8%). Процентное соотношение различных форм лейкоцитов в крови называется лейкоцитарной формулой.
Количество лейкоцитов и лейкоцитарная формула не всегда остаются постоянными. Увеличение числа лейкоцитов называется лейкоцитозом, уменьшение — лейкопенией. В возникновении количественных изменений лейкоцитов имеют значение нарушение соотношений между скоростью их продукции и темпом разрушения, а также перераспределение крови в организме, обусловленное изменением тонуса сосудов, скоростью кровотока в отдельных органах и выходом крови из депо.
Функции лейкоцитов разнообразны. Главная из них — защитная, заключающаяся в фагоцитозе, продукции антител и разрушении токсинов белкового происхождения. Обладая большим количеством ферментов, лейкоциты участвуют также в расщеплении некоторых пищевых веществ.
Тромбоциты, или кровяные пластинки, — это плазматические образования овальной или круглой формы диаметром 2—5 мк. В крови человека и млекопитающих они не имеют ядра. Тромбоциты образуются в красном костном мозгу и в селезенке, и их количество колеблется от 200 тыс. до 600 тыс. в 1 мм3 крови. Они играют важную роль в процессе свертывания крови.
Плазма является сложной биологической средой, в состав которой входят вода (90—92%) и важные для функций крови органические и неорганические вещества, различные белки (альбумина — 4—5%, глобулина — 2—3%, фибриногена — 0,2—0,4%), глюкоза (85—ПО мг%), аминокислоты и минеральные вещества (0,95%), среди которых находятся ионы Na, К, Са, Mg, Cl, НРО4, Н2СО3. В плазме содержатся гормоны, витамины, растворенные газы. Установлено также наличие в ней ферментов, участвующих в свертывании крови, в расщеплении пищевых веществ и окислительно-восстановительных процессах.
§ 4. Регуляция физико-химических процессов во внутренней среде организма
Удельный вес и вязкость крови. Они зависят от количества эритроцитов, содержания гемоглобина, белкового состава плазмы. По сравнению с водой кровь имеет более высокий удельный вес (1,060—1,080) и большую (в 3—5 раз) вязкость.
Кровь обладает суспензионными свойствами, характеризующимися распределением форменных элементов в плазме во взвешенном состоянии. Эти свойства зависят от количества эритроцитов и («•яков плазмы. В условиях неподвижного (статического) состояния । рови устойчивость суспензии форменных элементов нарушается и । мечается их оседание, что особенно характерно для эритроцитов, нот феномен, получивший название РОЭ (реакция оседания эри-। роцитов), служит показателем состояния как белкового состава и 1.1ЗМЫ, так и количественного соотношения эритроцитов и плазмы.
217
РОЭ в норме находится у мужчин в пределах 4—6 мм/час, у женщин — 6—10 мм/час.
Для нормальной жизнедеятельности клеток организма и скорости и направленности химических процессов в них весьма важно постоянство физико-химического состава крови (в частности, постоянство осмотического давления, кислотно-щелочного равновесия, уровня водно-солевого и белкового состава крови), которое достигается благодаря действию сложных регуляторных механизмов.
Осмотическое давление крови. В плазме и клетках крови находятся различные растворенные вещества, среди которых наибольший удельный вес имеют минеральные соли. В силу законов осмоса растворитель способен проникать через оболочки-мембраны — в сторону большей концентрации растворенных веществ. Сила, которая приводит в движение растворитель, обеспечивая его проникновение через мембрану, называется осмотическим давлением. У человека и млекопитающих осмотическое давление крови равно 7,6 — 8,1 атм. Поддержание его постоянства особенно важно для состояния эритроцитов, так как они реагируют на колебания осмотического давления изменением своего объема. Если из плазмы крови теряется много воды и концентрация солей в ней повышается, то в силу законов осмоса вода из эритроцитов начинает поступать в плазму через их полупроницаемую оболочку (мембрану), что приводит к сморщиванию эритроцитов. При понижении концентрации солей в плазме вода из нее поступает в эритроциты, которые набухают, теряя свои физиологические свойства. При очень низком осмотическом давлении эритроциты могут лопаться — происходит их гемолиз (растворение) .
Способность эритроцитов сохранять целость своей структуры при изменениях осмотического давления называется осмотической стойкостью или резистентностью. Осмотическое давление может колебаться под влиянием изменений температуры, потребления пищевых веществ, различных условий жизни. Его постоянство регулируется путем изменения выделительных функций почек и потовых желез.
Особое значение для поддержания осмотического давления имеет постоянство белкового состава плазмы. Осмотическое давление определяется не только растворимыми кристаллоидами, но и белковыми соединениями. Осмотическое давление, зависящее от содержания белков в плазме, называется онкотическим. Из общего осмотического давления (7—8 атм.) на долю белков приходится только 1/20—*/зо атм. (30—35 мм рт. ст.). Хотя это небольшая величина, но она весьма важна для процессов фильтрации и распределения воды между кровью и тканями организма. Белки представляют собой поглощающие воду коллоиды, обладающие большим молекулярным весом. Поэтому белки плазмы оказывают некоторое противодействие процессам фильтрации, удерживая воду внутри сосудистой системы.
Поддержание постоянства активной реакции (pH) крови. Кровь имеет слабощелочную реакцию. Активная ее реакция определяется
218
количественным соотношением Н+ и ОН- ионов. Показатель активной реакции (pH) артериальной крови равен 7,4; pH венозной крови— 7,35, что объясняется большим содержанием в ней углекислоты. Физиологические процессы оптимально протекают при определенной активной реакции крови. От величины этой реакции зависят процессы окисления и восстановления в клетках, процессы расщепления и синтеза белков, гликолиза, окисления углеводов и жиров, способность гемоглобина отдавать тканям кислород. Поэтому значительные сдвиги от нормальной концентрации pH (7,35—7,40) могут приводить к нарушению многих физиологических процессов, а при чрезвычайных сдвигах за известные пределы — и к гибели организма.
В процессе обмена в организме постоянно освобождаются вещества кислой реакции. Поэтому всегда имеется тенденция к сдвигу реакции крови в кислую сторону. Несмотря на это, для крови человека характерна высокая устойчивость реакции. Регуляция концентрации Н+ и ОН- крови является одной из весьма совершенных в организме. Тонкая регуляция pH крови обеспечивается буферными свойствами и деятельностью органов выделения.
Буферные свойства крови заключаются в способности препятствовать сдвигу активной реакции крови. Эта способность обусловливается буферными системами, которые состоят из смеси слабых кислот с солями этих кислот и сильных оснований. К ним относится: 1) угольная кислота — двууглекислый натрий (карбонатная буферная система); 2) одноосновный — двуосновный фосфорнокислый натрий (фосфатная буферная система); 3) белки плазмы (буферная система белков плазмы); 4) гемоглобин — калийная соль гемоглобина (буферная система гемоглобина). Буферные свойства крови на 75% зависят от содержания в ней гемоглобина и его солей.
Буферные системы особенно препятствуют сдвигу реакции крови в кислую сторону. Это объясняется способностью сильных кислот (например, молочной кислоты), поступающих в кровь, вытеснять слабые кислоты из их соединения с основаниями. При этом образуются соли сильных кислот, что сдерживает сдвиг реакции крови в кислую сторону. Все буферные системы крови создают в крови щелочной резерв, который в организме относительно постоянен.
Кислотно-щелочное равновесие крови выражает малосдвигаемое и нормальных условиях соотношение кислотных и щелочных эквивалентов. Нервно-гуморальные механизмы, регулирующие деятельность почек, потовых желез, дыхательной и пищеварительной систем, обеспечивают удаление из организма продуктов обмена и этим сохраняют постоянство pH и кислотно-щелочного равновесия.
§ 5. Защитные реакции крови
Повышение сопротивляемости инфекции. Основная функция лейкоцитов связана с процессами иммунногенеза — способностью син-।г hiровать антитела или иммунные тела, которые обезвреживают микробы и продукты их жизнедеятельности. Этим обеспечивается
219
сопротивляемость организма определенным заболеваниям. Иммунные тела особенно активно образуются при некоторых инфекционных болезнях (корь, оспа и др.), сохраняясь на многие годы в активном состоянии, инактивируя токсины различного происхождения и предупреждая этим повторные инфекционные заболевания. В результате создается невосприимчивость организма к вреднодействующим факторам среды (иммунитет). Лейкоциты, обладая способностью к амебовидным движениям, адсорбируют циркулирующие в крови антитела и, проникая через стенки сосудов, доставляют их в ткани к очагам воспаления. Нейтрофилы, содержащие большое количество ферментов, обладают способностью к захватыванию и перевариванию болезнетворных микробов. Это явление, открытое И. И. Мечниковым, было названо фагоцитозом ( от греч. phagos — пожирающий). Перевариванию подвергаются и клетки организма, дегенерирующие в очагах воспаления.
Лейкоциты участвуют также в восстановительных процессах после воспаления тканей.
Защита организма от кровотечений. Эта функция осуществляется благодаря способности крови к свертыванию (гемокоагуляции). Сущность свертывания крови заключается в переходе растворенного в плазме белка фибриногена в нерастворенный белок — фибрин, который образует нити, склеенные с краями раны. Сгусток крови (тромб) преграждает дальнейшее кровотечение, предохраняя организм от кровопотерь.
Превращение фиброногена в фибрин осуществляется при воздействии фермента тромбина, который образуется из белка протромбина под влиянием тромбопластина, появляющегося в крови при разрушении тромбоцитов. Образование тромбопластина и превращение протромбина в тромбин протекают при участии ионов кальция.
Переливание крови. Оно имеет исключительное значение при больших кровепотерях и некоторых заболеваниях. При этом необходим строгий учет совместимости крови лиц, дающих кровь (доноров), с кровью реципиентов — лиц, которым переливают кровь. Несовместимость возникает тогда, когда в эритроцитах переливаемой крови содержится агглютиноген — вещество, способное вызывать склеивание эритроцитов, а в плазме крови реципиента — соответствующий агглютинин, являющийся веществом, запускающим процесс склеивания эритроцитов (агглютинацию).
§ 6. Изменения б крови при двигательной деятельности
Мышечная деятельность вызывает значительные изменения в системе крови. Повышенное образование кислых продуктов обмена в результате кислородного дефицита, возникающего при интенсивной мышечной работе, может привести к нарушению кислотно-щелочного равновесия крови в сторону метаболического ацидоза. Наблюдающееся в этом случае снижение щелочного резерва может происходить без изменения величины pH (компенсированный аци
220
доз), а иногда при его снижении до 6,95 (декомпенсированный ацидоз). Особая роль при этом принадлежит молочной кислоте, концентрация которой иногда может возрастать с 9—12 мг % в покое до 250 мг % •
Величина сдвигов в параметрах кислотно-щелочного равновесия под влиянием нагрузки отражает степень тренированности спортсмена. По сравнению с нетренированными лицами у спортсменов, тренированных в длительных циклических упражнениях, при стандартной работе сдвиги кислотно-щелочного равновесия выражены меньше; при продолжительной работе метаболический ацидоз у них развивается значительно позже, что обеспечивается большей буферной емкостью крови. Под влиянием длительной мышечной работы вязкость крови может повыситься на 70%. Это обусловлено увеличением количества форменных элементов крови, изменением коллоидных свойств, потерей большого количества воды за счет усиленного потения и другими факторами. Повышение вязкости крови, изменяя периферическое сопротивление, влияет на гемодинамику, затрудняя работу сердечно-сосудистой системы.
Мышечная активность вызывает увеличение количества лейкоцитов (миогенный лейкоцитоз) со сдвигами в лейкоцитарной формуле. Степень изменений картины белой крови зависит от объема выполненной физической работы и ее интенсивности. Причиной общего лейкоцитоза является выход крови из кроветворных органов и кровяных депо, где содержится больше клеточных элементов по сравнению с кровью периферических сосудов.
В развитии мышечного лейкоцитоза выделены 3 фазы (А. П. Егоров): лимфоцитарная, первая нейтрофильная и вторая нейтрофильная.
/ фаза — лимфоцитарная — характеризуется незначительным лейкоцитозом (от 10 тыс. до 12 тыс. в 1 мм3) за счет увеличения количества лимфоцитов. Эта фаза наблюдается через 10 мин. после начала мышечной работы. // фаза — первая нейтрофильная — характеризуется увеличением количества лейкоцитов до 16—18 тыс. в 1 мм3 при значительном сдвиге формулы крови за счет увеличения числа нейтрофилов, среди которых появляются юные формы. Возрастает количество юных клеток (до 2—8%), палочкоядерных (до 10—12%), сегментированных (до 65—75%). Одновременно уменьшается количество эозинофилов (до 1—2%) и лимфоцитов (до 10— 12%) Эта фаза резко выражена через 1—2 часа после начала длительной интенсивной работы и является следствием усиленной деятельности кроветворных органов. Ill фаза — вторая нейтрофильная— выражается в резком лейкоцитозе, доходящем до 30—50 тыс. лейкоцитов в 1 мм3 и значительно выраженных сдвигах в формуле крови Количество юных нейтрофилов может возрастать до 6—8%, налочкоядерных — до 20—25%, количество лимфоцитов снижается до 5—10%. Эозинофильные клетки исчезают Эта фаза наблюдается после весьма длительной и интенсивной мышечной деятельности. Вторая нейтрофильная фаза в отличие от первой нейтрофильной служит одним из признаков развивающегося переутомления.
221
Фазные изменения наблюдаются также в картине красной крови, химизме и энергетике клеток (Л. Я. Евгеньева, О. Р. Немирович-Данченко). Выделено 3 основных типа реакции красной крови на физическую нагрузку.
При первом типе реакции отмечается повышение количества эритроцитов (миогенный эритроцитоз) в 1 мм3 до 5,5—6,0 млн. и процента гемоглобина при неизменном цветовом показателе (ЦП*). Активность ферментативных систем крови, как и количество ретикулоцитов, при этом не изменяется. К исходному уровню состояние этих параметров приходит через несколько часов. Такой тип реакции наблюдается при кратковременной и интенсивной работе. Изменения в картине крови наступают вследствие выхода крови из депо, подтверждением чего является опустошение синусов и увеличение количества фолликулов в лимфатических узлах и селезенке (А. Н. Лапутин).
Второй тип реакции связан со значительным усилением функций кроветворных органов, о чем свидетельствует повышение в крови количества незрелых форм эритроцитов — ретикулоцитов. Наблюдается незначительное снижение количества эритроцитов при большей степени падения процента гемоглобина, уменьшение значения цветного показателя, повышение активности ферментативных систем крови. Восстановительный период может продолжаться до 2 дней. Этот тип реакции в основном отмечается при длительной и интенсивной работе.
Третий тип реакции выявляется при некоторых видах мышечной деятельности (3-дневные соревнования в лыжном спорте, многодневные велогонки). Он связан с угнетением кроветворной функции. Количество эритроцитов при этом уменьшается и значительно понижается (иногда ниже 11 гм % по Сали) содержание гемоглобина с резким (до 0,5) падением цветового показателя. Отмечается угнетение активности окислительно-восстановительных ферментов крови. Восстановительный период может продолжаться до 6 дней. Этот тип реакции свидетельствует о развитии чрезмерно выраженного утомления.
При мышечной работе наблюдается также миогенный тромбоцитоз с возрастанием в 2 и более раза количества кровяных пластинок (А. А. Маркосян). Это явление отмечается в течение нескольких часов после работы. С повышением числа тромбоцитов под влиянием физических нагрузок ускоряется свертываемость крови, что наряду с миогенным лейкоцитозом представляет собой защитную реакцию. Миогенный тромбоцитоз имеет большое биологическое значение при мышечной работе, которая может быть связана с опасностью кровотечений.
КГ -100 КЭ 100
16-6 5000000
где КГ—концентрация гемоглобина; 16,6 — нормальная концентрация гемоглобина; КЭ — количество эритроцитов в 1 мм3 в норме. У здорового человека в покое ЦП приближается к единице.
222
Глава VII
КРОВООБРАЩЕНИЕ
Кровообращением называется непрерывное движение крови в организме. К системе органов кровообращения относятся сердце — источник энергии, обеспечивающей движение крови, и сосуды, выполняющие транспортную и перераспределительную функции.
В целостном организме деятельность органов кровообращения регулируется центральной нервной системой и гуморальными факторами. Благодаря этим воздействиям в каждый отдельный момент жизнедеятельности организма устанавливается необходимое соответствие между величиной кровотока и потребностью тканей в кислороде и питательных веществах.
В XVII в. В. Гарвей доказал, что кровь в организме движется по замкнутым кругам. Движение ее осуществляется за счет разности давлений, создаваемой сердцем. Существуют два круга кровообращения: большой и малый. Большой круг начинается от левого желудочка сердца. Он включает аорту, артерии разного калибра, артериолы, капилляры, венулы и вены. Заканчивается большой круг полыми венами, впадающими в правое предсердие. Через стенки капилляров происходит обмен веществ между кровью и тканями; артериальная кровь отдает кислород и, насыщаясь углекислотой, превращается в венозную. Малый круг кровообращения, начинающийся от правого желудочка сердца, включает легочные артерии, артериолы, капилляры, венулы и вены. Он заканчивается легочными венами, впадающими в левое предсердие. В капиллярах венозная кровь, обогащаясь кислородом и освобождаясь от углекислоты, превращается в артериальную.
Сердце
§ 1. Сердце и его физиологические свойства
Общая характеристика. Сердце представляет собой полый мышечный орган, разделенный продольной перегородкой на изолированные друг от друга правую и левую половины. Каждая из них состоит из предсердия и желудочка, разделенных фиброзными перегородками. Односторонний ток крови из предсердий в желудочки и оттуда в аорту и легочную артерию обеспечивается клапанами, находящимися у входного и выходного отверстий желудочков. Открытие и закрытие клапанов зависят от величины давлений по обе их с тороны.
Мышечные волокна сердца содержат миофибриллы, имеющие поперечную исчерченность. Диаметр мышечных волокон составляет 12—24 мк, длина может достигать 50 мк.
Толщина стенок разных отделов сердца неодинакова. Это обусловлено различиями в мощности производимой работы. Наиболь-
223
шая работа выполняется мышцами левого желудочка, толщина стенки которого достигает 10—15 мм. Стенки правого желудочка несколько тоньше (5—8 мм), еще тоньше стенки предсердий (2—Змм)< Размеры сердца обусловлены объемом его полостей и толщиной стенок. Эти величины зависят от размеров тела, возраста, пола и двигательной активности человека. Размеры сердца определяют путем рентгенографии, объемы полостей — при помощи радиокардиографии (введение в кровь радиоактивных веществ и регистрация проходящей через сердце крови при помощи счетчиков Гейгера-Мюллера). У здоровых взрослых мужчин среднего роста и веса длинник сердца равен в среднем 14 см, поперечник 12 см, объем полостей желудочков 250—350 мл. У женщин эти величины несколько меньше.
Общий объем сердца определяют при помощи специального метода — биплановой телерентгенографии. Снимки сердца при этом делаются в двух проекциях. На основании полученных величин вычисляют объем сердца. В среднем он составляет у мужчин 700—900 мл, у женщин 500—600 мл. Тяжелый физический труд и занятия спортом способствуют развитию гипертрофии миокарда и ведут к увеличению объема по-
Рис. 66. Схема проводящей систе- лосТ>еЙ сердца.
мы сердца: Сердце снабжается кровью через
Z — синоатриальный узел Кис —Фляка; ВСНеЧНЫе артерии, Начинающиеся у Т^аа^Г“-ТЖаяТ™аУ™Ш^ МеСта ВЫХОДЯ аОрТЫ. КрОВЬ ПОСТуПЭ-4—5 —правая и левая ножки пучка ет в венечные артерии во время рас-гнса; 6-^н^е разорения про- слабления сердца. При сокращении желудочков вход в венечные артерии прикрывается полулунными клапанами, а сами артерии сжимаются сократившейся мышцей сердца. Поэтому кровоснабжение сердца при его сокращении уменьшается. В венечные артерии поступает около 200—250 мл крови в 1 мин. При физической работе кровоснабжение сердца увеличивается.Объ-ем притекающей к нему крови зависит от мощности выполняемой работы. При очень напряженной работе кровоснабжение сердца может возрастать до 1000 мл.
Физиологические свойства сердечной мышцы. Сердечная мышца обладает способностью к автоматии, возбудимостью, проводимостью и сократимостью.
Автоматия сердца. Способность сердца ритмически сокращаться без внешних раздражений, под влиянием импульсов, возникающих в нем самом, называется автоматией сердца. Возбуждение в нем возникает в месте впадения полых вен в правое пред
224
сердие. Здесь находится скопление атипической мышечной ткани, называемое синоатриальным узлом или узлом Кис-Фляка (рис. 66). Атипическая мышечная ткань по своему строению отличается от основной массы миокарда. Клетки этой ткани богаты протоплазмой, поперечная же исчерченность в них выражена менее четко.
Возникающее в синоатриальном узле — главном водителе ритма сердца — возбуждение распространяется до атриовентрикулярного узла, расположенного в правом предсердии в межпредсердной перегородке (см. рис. 66). От этого узла отходит пучок Гиса, он делится на две ножки, разветвления которых, называемые волокнами Пуркине, проводят возбуждение к мускулатуре желудочков.
Синоатриальный узел обладает наиболее выраженной автома-тией. В нормальных условиях импульсы из этого отдела сердца обеспечивают деятельность всех остальных. Автоматия других участков миокарда, в частности атриовентрикулярного узла, выражена слабее. Она подавляется импульсами от главного водителя ритма сердца.
Если, например, у лягушки изолировать синоатриальный узел (путем перерезки или охлаждения соответствующих участков сердца), то деятельность сердца временно прекращается. Затем сокращения его возникают вновь, но ритм их будет менее частым, чем был до изоляции главного водителя ритма. Этот опыт, впервые проведенный Станниусом, доказывает ведущую роль синоатриального узла для нормальной работы сердца.
Автоматия водителей ритма сердца обусловлена периодическим изменением мембранных потенциалов в их клетках. Во время диастолы происходит постепенная деполяризация мембраны. В тот момент, когда ее потенциал оказывается значительно сниженным, возникает возбуждение, распространяющееся по всем волокнам миокарда. Периодически наступающая деполяризация клеточных мембран обусловлена изменением их проницаемости. По одним данным, во время диастолы уменьшается выход ионов калия из клеток, по другим, наоборот, увеличивается поступление туда ионов натрия. В результате концентрация ионов натрия и калия по обе стороны мембраны начинает изменяться, что ведет к ее деполяризации. Значение ионов натрия для возникновения процессов возбуждения в клетках — водителях ритма подтверждается более высоким содержанием здесь натрия по сравнению с другими участками миокарда.
Возбудимость сердца. Она проявляется в возникновении возбуждения при действии разных раздражителей. Сила раздражителя при этом должна быть не менее пороговой. При некоторых условиях пороговые раздражители вызывают сокращения максимальной силы. Эта особенность возникновения возбуждения в сердце получила название закона «все или ничего». Однако закон этот проявляется не всегда. Степень сокращения сердечной мышцы за-нисит не только от силы раздражителя, но и от величины ее предварительного растяжения, а также от температуры и состава питающей ее крови.
225
Возбудимость сердечной мышцы непостоянна. Она изменяется по ходу возбуждения. В начальном его периоде сердечная мышца невосприимчива (рефрактерна) к повторным раздражениям. Этот период называется фазой абсолютной рефрактерности. У человека она длится 0,2—0,3 сек., т. е. совпадает с временем сокращения сердца. По окончании фазы абсолютной рефрактерности возбудимость сердечной мышцы постепенно восстанавливается и на очень
выше исходной (рис. 67).
короткое время становится
Время, мсек.
Рис. 67. Электрический потенциал (а), механическая реакция сердечной мышцы (б) и изменение ее возбудимости:
Из-за длительного периода абсолютной рефрактерности сердечная мышца в обычных условиях не может сокращаться по типу тетануса, что очень важно для координации работы предсердий и желудочков.
При действии частых раздражителей сердечная мышца не отвечает на те из них, которые поступают в фазе абсолютной рефрактерности. Если же дополнительный внеочередной импульс действует на сердце в тот момент, когда его возбудимость уже вос-
/ — период абсолютной рефрактерности: становилась, ТО Возникает ДОПОЛ-2—период относительной рефрактерности; нИТСЛЬНОе сокращение сердца, НЭ-3 — период повышенной (супериормаль- ‘
ной) возбудимости зываемое экстрасистолой. Следу-
ющий очередной импульс при этом попадает к сердцу в фазе его рефрактерности. Сердце на него не реагирует, и поэтому после экстрасистолы наблюдается удлинен-
ная (компенсаторная) пауза.
Проводимость сердца. Она обеспечивает распространение возбуждения от клеток водителей ритма по всему миокарду.
Распространение возбуждения по сердцу осуществляется электрическим путем. Потенциал действия, возникший в одной мышечной клетке, является раздражителем для других. Способность к проведению возбуждения зависит от структурных особенностей мышечных волокон сердца и от многих других факторов. Например, она
увеличивается при повышении температуры и снижается при недостатке кислорода. Разные отделы сердца имеют разную проводимость. Это зависит от содержания в них гликогена и от длительности рефрактерных фаз. Периферические разветвления проводящей системы сердца расположены непосредственно под эндокардом. Поэтому возбуждение охватывает прежде всего внутренние слои сердца и затем распространяется кнаружи. Вследствие этого скорость распространения возбуждения по сердцу зависит не только от особенностей проводящей системы, но и от толщины мышечных
стенок.
Наибольшей проводимостью обладают клетки проводящей системы сердца, и особенно волокна Пуркине. Скорость же проведения
226
возбуждения от мышечных волокон предсердий к атровентрикуляр-ному узлу невысока. Происходящая здесь задержка распространения возбудительного процесса обеспечивает последовательность в работе предсердий и желудочков.
Сократимость сердечной м ы ш ц ы. Она обусловливает увеличение напряжения или укорочения ее мышечных волокон при возбуждении. Сокращение сердечной мышцы, вызванное одним стимулом, длится дольше, чем одиночное сокращение скелетной мышцы. Это зависит от относительно меньшей лабильности сердечной мышцы. В физиологических условиях каждая волна возбуждения в сердце сопровождается его сокращением. В искусственных условиях эта закономерность может нарушаться. Например, при отсутствии кальция в растворе, питающем сердце, возбуждение не сопровождается его сокращением.
Поставщиком энергии для сокращения сердечной мышцы служат макроэргнческие фосфорсодержащие вещества. Восстановление их происходит за счет энергии, освобождающейся при дыхательном и гликолитическом фосфорилировании. При этом преобладающими являются аэробные реакции.
§ 2. Электрические процессы в сердце
Биопотенциалы, возникающие в сердце при прохождении волны возбуждения, распространяются по всему телу и могут быть зарегистрированы с помощью электродов, присоединенных к разным участкам тела человека (конечности, грудная клетка). Электроды соединяются с прибором, обеспечивающим усиление и регистрацию потенциалов сердца (электрокардиограф). Запись биотоков сердца на пленке или бумаге называется электрокардиограммой (ЭКГ).
Электроды (металлические пластинки, под которые помещается марлевая прокладка, смоченная раствором электролита) размещают на конечностях (стандартные отведения: первое — правая и левая рука, второе — правая рука и левая нога, третье — левая нога и левая рука) и на определенных участках грудной клетки (рис.68). Запись ЭКГ можно производить в покое (в положении лежа, сидя, стоя) и непосредственно при мышечной деятельности (работа на велоэргометре, бег на третбане и др.). Для регистрации 'КГ на расстоянии применяют специальную портативную (весом около 150 г) телеметрическую аппаратуру: транзисторный передатчик, соединенный с электродами, прикрепленными к коже в области । рудины. Запись ЭКГ с помощью приемника и регистрирующей аппаратуры может производиться в радиусе до 500 и более метров.
При анализе ЭКГ определяют величину зубцов в милливольтах и продолжительность интервалов между ними в долях секунды. В каждом сердечном цикле различают зубцы Р, Q, R, S и Т ipnc. 69). Зубец Р отражает процесс возбуждения в предсердиях. Интервал PQ — время, в течение которого возбуждение распространи гея от предсердий до желудочков. В среднем оно равно 0,12— <|.!8 сек. Комплекс зубцов QRS возникает при возбуждении желу
227
дочков. Его продолжительность в покое равна 0,06—0,08 сек. Величина самого большого положительного зубца R в среднем составляет 0,2—2,0 мв. Широкий зубец Т связан с восстановительными процессами в миокарде после его возбуждения (т. е. с реполяризацией) .
Интервал QRST, или сокращенно Q — Т, называется электрической систолой. Это — время, в течение которого миокард охвачен вол-
А.
Б
Рис. 68. Размещение электродов и электрокардиограммы, записанные в трех стандартных (А) и шести грудных (Б) отведениях.
Л 2, 3, 4, 5, 6 — места прикрепления электродов на грудной клетке
ной возбуждения. Оно зависит от продолжительности сердечного цикла, которую удобнее всего определять по интервалу R — R. Зная среднюю продолжительность сердечного цикла и скорость движения ленты в электрокардиографе, нетрудно рассчитать частоту сердцебиений. Например, если лента движется со скоростью 3000 мм/мин., а длительность сердечного цикла составляет в среднем 20 мм, то частота сердцебиений в данном случае равна 150 ударам в 1 мин. (3000:20=150).
По показателям ЭКГ можно судить об автоматии, проводимости и возбудимости сердечной мышцы. Особенности автоматии сердца проявляются в изменениях частоты и ритма комплексов зубцов ЭКГ, особенности изменения проводимости — в продолжительности интервалов, а возбудимости — в динамике ритма и вольтажа зубцов.
У здоровых людей может наблюдаться синусовая аритмия, при которой разница в продолжительности сердечных циклов в покое составляет 0,1—0,3 сек. и более. Иногда аритмия связана с фазами дыхания (дыхательная аритмия). В этих случаях сердцебиения уча
228
щаются при вдохе и урежаются при выдохе. Нарушение ритма сердечных сокращений нередко проявляется в виде экстрасистолии (возникновение дополнительных внеочередных систол). Экстрасистолы могут возникать в синусовом узле, в других отделах проводящей системы и даже в клетках сократительного миокарда.
Под влиянием физических нагрузок происходит укорочение сердечного цикла и большинства интервалов ЭКГ- Зубец Р при этом увеличивается, а вольтаж комплекса зубцов QRS несколько снижается. Нередко наблюдается смещение интервалов Р — Q и S — Т ниже изолинии (Л. А. Бутченко). Изменение показателей ЭКГ при работе обусловлено усилением влияния симпатических нервов на сердце, активацией энергетических процессов в миокарде, а в некоторых случаях и возникновением кислородной недостаточности.
§ 3. Частота сердечных сокращений в покое и при мышечной деятельности
Ритм возбуждения в синоатриальном узле (синусовый ритм) у здоровых людей определяет частоту сердцебиения. В целостном организме на автоматию водителя ритма оказывают влияние импульсы, поступающие из центральной нервной системы, и гуморальные факторы.
Сердечные сокращения подсчитываются по артериальному пульсу или по числу толчков сердца в грудную клетку (в 5-м межре-
Рис. 69. Схема электрокардиограммы здорового человека, зарегистрированной в первом отведении:
1 — интервал PQ; 2 ** интервал QPS; 3 — интервал ST', 4 — интервал Qf (электрическая систола сердца)
Рис. 70. Определение частоты сердцебиений у конькобежцев при помощи радио-телеметрической аппаратуры.
1 — устройство, регистрирующее биотоки сердца и передающее сигналы; 2 — телефон, информирующий спортсмена о частоте сердцебиений
перье слева). В спорте широко распространен также метод опреде-цчшя частоты сердцебиений путем регистрации биопотенциалов грдца, либо при помощи проводной связи, либо с использованием радиоаппаратуры (рис. 70).
229
Частота сердцебиений в состоянии покоя зависит oi возраста, пола, размеров тела, образа жизни человека. У большинства взрослых здоровых людей эта величина составляет 60—70 ударов в 1 мин. У детей частота сердцебиений больше, чем у взрослых, у женщин обычно больше, чем у мужчин. У мелких животных сердце сокращается чаще, чем у крупных. Например, у некоторых птиц частота сердцебиений равна 300—400 ударам в 1 мин.
У физически активных людей в состоянии покоя сердечный
ритм, как правило, реже, чем
у ведущих малоподвижный образ
Рис. 71. Частота сокращений (ЧСС) при работе разной
сердечных у гребпоз мощности (по В. С. Фарфелю).
Ось абсцисс — скорость движущейся лодки (м/сек)
Рис. 72. Частота сердечных сокращений (ЧСС) при беге на 5000 м
жизни. Частота сердцебиений менее 60 ударов в 1 мин. называется брадикардией (от греч. bradys — медленный, cardia — сердце). У спортсменов это явление может быть выражено очень резко. Например, у лыжников и бегунов-стайеров частота сердцебиений в покое нередко составляет 40 и менее ударов в 1мин. Частота сердечных сокращений, превышающая 90 ударов в 1 мин., называется тахикардией (от греч. tachys — быстрый).
Частота сердцебиений зависит от положения тела: при положении стоя она больше, чем при положениях сидя и лежа.
На сердечный ритм оказывают влияние психические факторы. Он учащается при эмоциональных возбуждениях человека.
Мышечная работа вызывает учащение сердцебиений. Изменение сердечного ритма в этих условиях зависит от пола и возраста. При выполнении одинаковой работы частота сердцебиений у женщин больше, чем у мужчин, у детей и подростков — больше, чем у взрослых.
Учащение сердцебиений при стандартной работе зависит от ее мощности (рис. 71). При спортивной деятельности эта зависимость проявляется не столь резко, так как в этих условиях на сердечный ритм влияют психические, метеорологические и многие другие фак
230
торы. В большинстве случаев при физических упражнениях сердечный ритм повышается до 160—180 ударов в Imiih. (рис. 72), но иногда может быть и больше (до 220 ударов).
§ 4. Фазы сердечного цикла
Деятельность сердца характеризуется непрерывной сменой сокращений и расслаблений. Сокращение сердца называется систолой, расслабление — диастолой. Сердечный цикл состоит из трех фаз: 1 — систола предсердий (в этой фазе желудочки рас- —--------------------<Л---------а
слаблены и наполняются кровью) ; 2 — систола желудочков (кровь под большим давлением выбрасывается правым желудочком в легочную артерию, левым — в аорту); 3 — общая диастола сердца (мускулатура предсердий и желудочков расслаблена). Длительность сердечного цикла зависит от частоты сердцебиений. При сердечном ритме 75 ударов в 1 мин. она составляет 0,8 сек. Систола предсердий при этом равна 0,10 сек., систола желудочков — 0,33 сек., диастола желудочков — 0,47 сек. Общая
Рис. 73. Поликардиограмма: а — электрокардиограмма; б — сфигмограмма (пульсация сонной артерии); в — фонокардиограмма (звуковые явления при сокращении сердца); г — отметка времени
диастола сердца, продолжаясь 0,37 сек., занимает несколько меньше половины времени сердечного цикла.
Более подробно фазовая структура сердечного цикла изучается при помощи поликардиографии, которая обеспечивает одновременную регистрацию электрических, звуковых и механических явлений, возникающих при деятельности сердца (рис. 73). Несколько упрощенная схема отдельных <|>аз сердечного цикла представлена на рис. 74.
Начальный период систолы
Рис. 74. Схема сердечного цикла при частоте сердечных сокращений 70 раз в 1 мин.
Систола (площадь с точками) и диастола (белая площадь) предсердий (Д) и желудочков (Ь). / — асинхронная фаза сокращения; 2 — изометрическое сокращение; 3 — фаза изгнания; 4 — протодиастолический период; 5 — изометрическое расслабление; 6 — фаза наполнения желудочков
желудочков, когда еще не все
мышечные волокна его охвачены возбуждением, называется фазой асинхронного сокращения. В покое она продолжается 0,05—0,06 сек. Давление в желудочках в это время постепенно нарастает, что ве-
231
дет к закрытию атриовентрикулярных клапанов. Следующая фаза систолы желудочков, при которой их полости полностью изолированы от предсердий и артериальных сосудов, называется фазой изометрического сокращения. Она длится около 0,03—0,05 сек. В этой фазе давление внутри желудочков резко нарастает, что ведет к раскрытию полулунных клапанов. За изометрической фазой следует период систолы сердца, называемый фазой изгнания крови из желудочков. Она длится около 0,25 сек. Диастола желудочков начинается протодиастолическим периодом. В это время их мускулатура постепенно расслабляется, но полулунные клапаны остаются еще открытыми. Дальнейшее расслабление мускулатуры желудочков и уменьшение в них давления ведут к закрытию полулунных клапанов. Период расслабления желудочков при закрытых клапанах и изоляции их полостей от предсердий называется фазой изометрического расслабления. Она длится 0,08 сек. Затем атриовентрикулярные клапаны раскрываются, и желудочки наполняются кровью из предсердий. Фаза наполнения желудочков длится 0,35 сек., в конце ее происходит сокращение предсердий, продолжающееся 0,1 сек.
При физической работе наряду с учащением сердцебиений изменяется фазовая структура сердечного цикла (табл. 7).
Таблица 7
Частота сердцебиений, длительность сердечного цикла и его фазовая структура при физической работе (по Л. В. Карпману)
Частота сердцебиений (колич. ударов 5 1 мин.)...........................
Сердечный цикл (сек.)...............
Асинхронное сокращение (сек.) .... Изометрическое сокращение (сек.) . . . Фаза изгнания крови (сек.)..........
Диастола (сек.).....................
При работе
а а а Я и
ё £ £ £ S
аК -я
Cl со LO
О
Ч а а а И а
66 140 145 152 155 157
0,906 0,428 0,411 0,394 0,385 0,380
0,069 0,061 0,059 0,060 0,058 0,057
0,053 0,010 0,005 0,005 0,004 0,003
0,234 0,183 0,172 0,167 0,160 0,159
0,550 0,174 0,175 0,162 0,164 0,161
Все систолические фазы при этом укорачиваются. При напряженной мышечной деятельности фаза изометрического сокращения практически приближается к 0 (см. табл. 7). Ее укорочение обусловлено увеличением скорости повышения внутрижелудочкового давления. Длительность периода изгнания может уменьшаться почти вдвое — до 0,12—0,15 сек. (см. рис. 83). Особенно резко укорачивается при работе диастола. Например, при работе, сопровождающейся учащением сердечного ритма до 200 ударов в 1 мин., диастола
232
укорачивается до 0,10—0,13 сек. На наполнение желудочков при таком высоком ритме затрачивается всего 0,05—0,08 сек. После окончания работы фазовая структура сердечного цикла постепенно восстанавливается.
§ 5. Работа сердца как нагнетательного насоса
На протяжении сердечного цикла в результате сокращений и расслаблений миокарда давление в полостях сердца периодически то нарастает, то понижается. Это давление может быть измерено у людей и животных прямым способом. Для этого катетер (тонкий эластичный полый зонд) вводят в вену руки и продвигают по ней в правое предсердие, правый желудочек и легочную артерию. В аорту и левый желудочек катетер вводят через плечевую артерию. Катетеры соединяются с манометрами, регистрирующими величины давления в полостях сердца.
Давление в предсердиях при систоле повышается до 5—8 мм рт. ст., при диастоле снижается до 0, т. е. становится равным атмосферному давлению. При сокращении предсердий кровь перемещается в расслабленные желудочки. При повышении давления в предсердиях кровь могла бы поступать обратно в вены. Однако этого не происходит, так как сокращающаяся мускулатура предсердий сжимает устья вен.
При систоле желудочков давление в них возрастает: в правом — до 25—30 мм рт. ст., в левом — до 115—125. Разница в давлении в обоих желудочках обусловлена более мощным развитием миокарда левого желудочка, которому при изгнании крови в аорту приходится преодолевать значительное сопротивление. При увеличении давления в правом желудочке до 5—12 мм рт. ст., а в левом до 65— 75 полулунные клапаны раскрываются, и кровь устремляется в легочную артерию и аорту. При систоле желудочков кровь не может возвратиться обратно в предсердия, так как атриовентрикулярные клапаны в этот момент закрыты. Давление в легочной артерии и аорте при систоле желудочков почти одинаково с давлением в их полостях. При расслаблении желудочков давление в них становится ниже, чем в артериальных сосудах. Это ведет к захлопыванию полулунных клапанов. По мере расслабления мускулатуры желудочков давление в них продолжает снижаться и в конце диастолы надает до 0. Как только давление в желудочках становится ниже, чем в предсердиях, раскрываются атриовентрикулярные клапаны и начинается наполнение желудочков новой порцией крови. Таким образом, сердечный насос периодически нагнетает кровь в артериальные стволы, поддерживает в них высокое давление, тем самым обеспечивая течение крови.
Наполнению сердца кровью во время диастолы способствует присасывающее действие грудной клетки, которое усиливается во время вдоха, когда давление в полых венах и предсердиях резко снижается.
233
При физической работе возрастает сила сокращения сердца. Это сопровождается увеличением скорости повышения и снижения внутрижелудочкового давления, что играет большую роль в обеспечении нарастающего при работе сердечного выброса крови.
§ 6. Механические и звуковые явления при сокращении сердца
Деятельность сердца сопровождается возникновением механических (сердечный толчок) и звуковых (тоны сердца) явлений.
Сердечный толчок. В момент сокращения сердца у большинства людей в 5-м межреберье слева внутри от срединно-ключичной линии прощупывается сердечный толчок. Он возникает в результате изменения формы сердца при сокращении, увеличения плотности его стенки и подъема верхушки. Колебания грудной клетки, вызванные сердечным толчком, могут быть записаны на кардиографе.
Изгнание крови из желудочков и ее движение по крупным сосудам вследствие реактивной отдачи вызывают колебания всего тела. Регистрация этих колебаний при помощи специальной аппаратуры называется баллистокардиографией. Этот метод позволяет изучать силу, регулярность и скорость выбрасывания крови в большие артериальные стволы, а также особенности наполнения сердца во время диастолы.
Тоны сердца. При работе сердца возникают звуковые явления, называемые сердечными тонами. Чаще они изучаются путем аускультации (выслушивания). Более точную информацию дает запись звуковых явлений — фонокардиография. Этот метод позволяет исследовать звуки сердца в диапазоне частот, не доступных слуховому восприятию, и определять форму тона, его продолжительность и его место в сердечном цикле.
При записи фонокардиограмм используется приемник звуков — микрофон, который преобразует звуковые колебания в электрические. Последние усиливаются электронным усилителем и регистрируются на фотопленке.
При выслушивании сердца определяются два тона: первый, или систолический, тон — низкий и протяжный и второй, или диастолический, — более высокий и отрывистый. Первый тон возникает в результате колебаний атриовентрикулярных клапанов и прикрепленных к ним сухожильных нитей. В происхождении этого тона имеют значение и звуковые явления, возникающие при сокращении волокон миокарда. Второй тон обусловлен захлопыванием полулунных клапанов.
Фонокардиографические исследования показали, что при деятельности сердца возникают кроме двух основных еще третий и четвертый тоньп Третий тон обусловлен колебаниями стенки сердца в момент наполнения кровью желудочков. Четвертый тон предшествует первому. Он возникает в результате усиления колебаний стенки желудочков при систоле предсердий.
234
При неполноценности клапанов сердца и других нарушениях его деятельности выслушиваются шумы. Особенности этих шумов и место их наилучшего выслушивания играют важную роль в диагностике заболеваний сердца.
§ 7. Систолический и минутный объемы крови
Систолическим, или ударным, объемом называется количество место их наилучшего выслушивания играют важную роль в диаг-щении. Эта величина зависит от количества крови, притекающей к сердцу, и от силы сердечных сокращений.
В состоянии покоя у взрослых здоровых людей систолический объем крови равен в среднем 60—80 мл. При систоле желудочков выбрасывается не вся кровь, содержащаяся в них, а лишь около половины. Остающаяся в желудочках кровь называется резервным объемом. Благодаря наличию резервного объема крови систолический объем может резко увеличиваться уже при первых сокращениях сердца после начала работы. Кроме резервного объема в желудочках сердца имеется еще остаточный объем крови, который не выбрасывается даже при самом сильном сокращении.
При мышечной деятельности систолический объем крови возрастает до 100—150 мл. В отдельных случаях он может повышаться до 180—200 мл. Увеличение систолического выброса является одним из важнейших факторов, обеспечивающих усиление кровоснабжения работающих органов. Систолический объем крови достигает наибольших величин при относительно легкой работе, потребность в кислороде при которой не превышает 40% аэробных возможностей организма. При повышении мощности работы систолический объем крови либо стабилизируется на достигнутом уровне, либо начинает снижаться. Механизм этого явления не вполне ясен. Возможно, оно связано с учащением сердцебиений и укорочением наполнения кровью полостей сердца.
Количество крови, выбрасываемое сердцем в течение 1 мин., называется минутным объемом крови. Этот показатель зависит от частоты сердцебиений и систолического объема крови. В состоянии покоя у взрослых людей минутный объем крови составляет в среднем 5—6 л.
На минутный объем крови влияет величина поверхности тела. 11оэтому для сравнительной его оценки определяют так называемый сердечный индекс. Он вычисляется путем деления минутного объема крови на площадь поверхности тела и равен у взрослых людей и среднем 2,5—3,5 л/мин/м2.
При мышечной деятельности минутный объем крови увеличивается, что необходимо для обеспечения кровоснабжения работающих органов.
При легкой работе минутный объем крови нарастает до 10— I > л. У тренированных людей при очень напряженной мышечной деятельности он может достигать 30—35 и более литров.
235
При легкой работе минутный объем крови возрастает за счет увеличения систолического объема и учащения сердечного ритма. При повышении мощности работы дальнейшее увеличение минутного объема крови обусловлено главным образом учащением сердцебиений.
Минутный объем крови является важнейшим показателем производительности работы сердца.
Величина минутного объема крови зависит от кислородной потребности, которая, в свою очередь, обусловлена мощностью выполняемой работы. Поэтому между величиной минутного объема и мощностью работы должна быть прямая зависимость. Однако эта зависимость наблюдается не всегда. Усиление кровоснабжения работающих органов происходит не только за счет абсолютного увеличения минутного объема крови, но и за счет его перераспределения. При этом чем рациональнее распределение циркулирующей крови, тем при прочих равных условиях меньше нарастает минутный объем крови.
Определение систолического и минутного объемов крови у человека производят при помощи разных методов. Наиболее точным из них является прямой метод Фика. Он основан на определении количества потребляемого кислорода и артерио-венозной разности. Последняя определяется путем катетеризации правого предсердия и одной из крупных артерий. Этот метод применяется главным образом в клинике. В настоящее время он используется иногда и для определения минутного объема крови у здоровых людей.
Величины, наиболее близкие к полученным прямым методом, получаются при определении минутного объема путем разведения краски или радиоактивных веществ в крови. Такое вещество, введенное в одну из периферических вен, проходит через правую половину сердца, легкие, левую половину сердца и поступает в артерии большого круга, где и определяется его концентрация. На основании данных о количестве введенного вещества в вену, о концентрации его в артериальной крови и о времени, которое потребовалось на прохождение введенного вещества через всю систему кровообращения, вычисляют величину минутного объема крови.
Кровеносные сосуды
§ 8. Основные сведения о гемодинамике
Движение крови по кровеносным сосудам обусловлено разностью давления в артериях и венах. Оно подчинено законам гидродинамики *. Движущаяся кровь на всех участках своего пути испытывает сопротивление, которое зависит от ее вязкости, а также от длины и диаметра кровеносного сосуда.
* Гидродинамика — учение о движении жидкости. Учение о движении крови з сосудистой системе называется гемодинамикой.
236
Сопротивление кровотоку в разных отделах сосудистой системы неодинаково. Эта величина прежде всего зависит от диаметра сосуда. При уменьшении диаметра в 2 раза сопротивление току крови возрастает в 16 раз. Наибольшее сопротивление кровотоку создается в артериолах и капиллярях, диаметр которых очень мал. Давление крови, обеспечиваемое деятельностью сердца, постепенно снижается по ходу кровеносной системы. При этом в артерии оно уменьшается на 10%, в артериолах и капиллярах — на 85%. Таким образом, наибольшая часть энергии, освобождаемой сердцем при сокращении, затрачивается на продвижение крови по артериям и капиллярам.
Количество крови (Q), проходящее в единицу времени через всю кровеносную систему, тем больше, чем больше разность давлений в ее артериальном (Ра) и венозном (Рв) концах и чем меньше сопротивление току крови (Р):
Это отношение отражает основной гидродинамический закон и определяет как величину общего кровотока в организме, так и величину кровоснабжения отдельных органов. При изучении величины кровотока различают его объемную и линейную скорости.
Объемной скоростью кровотока называют количество крови, просекающее через всю кровеносную систему в единицу времени. Эта величина аналогична минутному объему крови (см. § 7). Она измеряется в мл/мин и мл/сек. Различают общую объемную скорость кровотока и местную, относящуюся к отдельному органу. Как общая, так и местная скорости кровотока непостоянны. При физической работе нарастают и общая объемная скорость и местная в активных органах. В неактивных же органах объемная скорость кровотока при физической работе уменьшается (табл. 8).
Таблица 8
объемная скорость кровотока (общая и в разных органах) у человека в покое и при физической работе (по Вейду и Бишопу)
Показатели Объемная скорость кровотока (мл/мин)
состояние покоя физическая работа
легкая средняя тяжелая
• |б1цая объемная скорость кровотока . . 5800 9500 17 500 25 000
। келстные мышцы 1200 4500 12 500 22 000
Moir 750 750 750 750
' рдце 250 350 750 1 000
1 'рганы брюшной полости (без почек) . 1400 1100 600 300
Почки 1100 900 600 250
hi »ка „ 700 1500 1 900 600
1р\гис органы 600 400 400 100
237
Рис. 75. Линейная скорость кровотока в разных отделах сосудистой системы:
I — аорта, // — артерии, III — артериолы. IV — капилляры, V — вены
сокая в аорте и артериях, она
Данные, приведенные в табл. 8, показывают, что при физической работе наряду с увеличением общей объемной скорости кровотока резко повышается кровоснабжение скелетных мышц и сердца. Кровоток в мозгу при этом остается неизменным, кровоснабжение же внутренних органов резко уменьшается.
Линейной скоростью кровотока называют скорость движения частиц крови вдоль сосуда. Эта величина, измеренная в см/сек, прямо пропорциональна объемной скорости кровотока и обратно пропорциональна площади сечения кровеносного русла. Линейная скорость неодинакова для всех частиц передвигающейся крови: она больше в центре сосуда и меньше около его стенок.
Линейная скорость кровотока неодинакова в разных отделах сосудистой системы (рис. 75). Вы-гзко снижается в капиллярах, об
щая площадь сечения которых в 500—600 раз больше площади сечения аорты. В венах линейная скорость кровотока вновь повышается, что объясняется их меньшим общим кровотоком по сравнению с капиллярами.
О средней линейной скорости кровотока можно судить по времени полного кругооборота крови. При состоянии мышечного покоя он осуществляется в течение 21—23 сек. При физической работе это время уменьшается. Ускорение кровотока зависит от мощности работы. При легкой физической работе полный кругооборот крови происходит в течение 15 сек., при тяжелой — в течение 8—-9 сек. Линейная скорость кровотока по артериям может возрастать при работе более чем в 5 раз.
§ 9. Общее периферическое сопротивление
Общее сопротивление сосудов кровотоку, называемое периферическим сопротивлением, прямо пропорционально разности давлений в артериальном и венозном концах сосудистой системы и обратно пропорционально линейному объему крови. Общее перифериче-р ___________________________________________________р
ское сопротивление вычисляется по формуле: R — —~—-. При
этих расчетах о величине Ра—Рв судят по среднему артериальному давлению, отражающему энергию движения крови. Величина Q показывает количество крови, протекающее по сосудам в течение 1 сек. Величина общего периферического сопротивления выражается в дин-сек1см~ъ. У взрослого человека в состоянии покоя в среднем оно равно 1400—2500 дин-сек1смг-ъ. В сосудах малого круга периферическое сопротнзлсппе меньше, чем в сосудах большого круга.
238
Это объясняется большим диаметром артериол в легких. При физической работе в связи с расширением артериол и капилляров в активных мышнах периферическое сопротивление снижается. При этом чем больше увеличивается минутный объем крови, тем резче снижается общее периферическое сопротивление. Изменение его зависит от мощности выполняемой работы. При легкой работе оно снижается в среднем до 1000 дин-сек!см~ъ, при тяжелой — до 800 дин- сек!см~5, при очень тяжелой — до 600—800 дин -сек! см~ъ.
Общее периферическое сопротивление следует рассматривать как суммарную величину сопротивления всех артериол и капилляров тела. Оно не является показателем кровоснабжения отдельных органов. Однако направление его сдвигов (увеличение или уменьшение), отражая общий уровень кровоснабжения организма, указывает на соотношение сосудосуживающих и сосудорасширяющих реакций. Например, уменьшение периферического сопротивления ври физической работе обусловлено расширением артериол и капилляров в скелетных мышцах.
§10. Кровяное давление
При каждом сокращении сердца в артерии выбрасывается под большим давлением некоторое количество крови. Ее свободному передвижению препятствует сопротивление периферических сосудов. В итоге в кровеносных сосудах создается давление, называемое кровяным давлением. Оно не
одинаково в различных отделах сосудистой системы. Являясь наибольшим в аорте и крупных артериях, кровяное давление снижается в мелких артериях, артериолах, капиллярах, венах и становится ниже атмосферного в полых венах (рис. 76).
Величина артериального давления зависит от количества крови, поступающей в единицу времени из сердца в аорту, от интенсивности оттока
мм
рт.ст. 120
100
80
60
40
20'
Рнс. 76. Кровяное давление в разных отделах сосудистой системы:
а — систолическое давление; б — среднее давление; в — диастолическое давление;
1 — аорта; 2 — крупные артерии; 3 — мелкие артерии; 4 — артериолы; 5 — капилляры; 6 — венулы; 7 — вены; 8 — полые вены
крови из центральных сосудов на периферию, от емкости сосу-
дистого русла, от упругого сопротивления артериальных стенок и от вязкости крови. Поступление крови в артерии, т. е.
истолический объем крови, за-
висит от силы сокращения сердца. Отток крови из артерий зависит
<>г сопротивления в периферических сосудах. Поэтому давление в
239
артериях будет тем выше, чем сильнее сокращения сердца и чем больше периферическое сопротивление.
Давление в артериях неодинаково на протяжении сердечного цикла. Оно больше в момент систолы и меньше при диастоле. Наибольшее давление в артериях называется систолическим или максимальным, наименьшее — диастолическим или минимальным. Давление в артериях во время диастолы желудочков не падает до 0. Оно поддерживается благодаря упругости артериальных стенок, растянутых во время систолы. При систоле желудочков артерии наполняются кровью. Та кровь, которая не успевает пройти далее в периферические сосуды, растягивает стенки крупных артерий. Во время диастолы кровь в артериях не испытывает давления со стороны сердца. В это время на нее оказывают давление лишь артериальные стенки, растянутые при систоле сердца и возвращающиеся благодаря своей упругости в исходное состояние.
Колебания кровяного давления при систоле и диастоле сердца происходят лишь в аорте и артериях. В артериолах же, капиллярах и венах давление крови постоянно на всем протяжении сердечного цикла (см. рис. 76). Давление в артериолах отражает уровень так называемого среднего давления в артериях. Среднее артериальное давление есть величина, результирующая все переменные значения давления на протяжении сердечного цикла. Оно представляет собой ту среднюю величину давления, которое могло бы обеспечить течение крови в артериях без колебаний давления при систоле и диастоле. Это означает, что среднее давление выражает энергию непрерывного движения крови. Величина среднего давления ближе к диастолическому давлению. Это объясняется более продолжительным снижением давления при диастоле по сравнению с его повышением при систоле.
Методы определения артериального давления. Артериальное давление можно измерять прямым или косвенным методом.
При прямом методе в артерию вводится полая игла, соединенная с манометром. Это наиболее точный метод определения артериального давления. Однако он требует хирургического вмешательства и поэтому мало пригоден для массовых исследований.
Более распространенным является косвенный (манжеточный) метод определения величин кровяного давления, предложенный Рива-Роччи в 1896 г. Он основан на измерении той величины давления, которая необходима для полного сжатия артерии и прекращения в ней тока крови. Для этого на конечность испытуемого накладывают манжету, соединенную с резиновой грушей (служащей для нагнетания воздуха) и ртутным или механическим манометром. При нагнетании воздуха в манжету происходит сдавливание конечности и проходящей в ней артерии. В тот момент, когда давление в манжете становится выше систолического, пульсация на периферическом конце артерии прекращается. Появление первого пульсового толчка при уменьшении давления в манжете соответствует величине систолического давления в артерии. Этим способом можно определить лишь величину систолического давления.
240
Для определения систолического и диастолического давлений применяется звуковой, или аускультативный, метод (от лат. ayscul-tatio — выслушивание), предложенный Н. С. Коротковым в 1905 г. При этом методе используются те же манжета и манометр, что и при способе Рива-Роччи. Но о величине кровяного давления в артерии судят не по появлению пульса, а по возникновению и исчезновению звуков, которые выслушиваются на артерии ниже места наложения манжеты. Эти звуки возникают лишь тогда, когда кровь течет по сжатой артерии. При определении величин кровяного давления аускультативным способом воздух нагнетается в манжету до полного сжатия плечевой артерии и прекращения звуков. После этого давление в манжете медленно снижается. Когда оно понижается настолько, что давление крови при систоле оказывается выше, чем давление в манжете, то на артерии начинают выслушиваться звуки, обусловленные током крови под манжетой при систоле. Показание манометра, соответствующее появлению этих звуков, характеризует систолическое давление. При дальнейшем снижении давления в манжете звуки сначала усиливаются, а затем исчезают. В этот момент давление в манжете становится равным давлению в артерии при расслаблении сердца. Показание манометра при этом соответствует величине диастолического давления.
Аускультативный способ дает точные данные при измерении кровяного давления в состоянии покоя. Непосредственно при выполнении физической работы он почти не применим. В периоде восстановления после работы этот метод дает лишь ориентировочные данные. Особенно часто в этих условиях возникают ошибки при определении диастолического давления. Способы Рива-Роччи и Короткова не позволяют определять величину среднего давления. Его измерение производится либо при помощи специального манометра И. М. Сеченова, либо путем планиметрии кривых артериального давления, зарегистрированных оптическим манометром. О величине среднего давления можно судить также по изменению колебаний артериальной стенки при сжатии ее манжетой. Этот способ определения артериального давления называется артериальной осциллографией или тахоосциллографией.
Показатели кровяного давления. У взрослых здоровых людей систолическое давление в плечевой артерии чаще всего находится в пределах от ПО до 125 мм рт. ст. По данным Всемирной организации здравоохранения, у лиц 20—60 лет систолическое давление но 140 мм рт. ст. является нормотоническим, выше 140 мм рт. ст. — гипертоническим, ниже 100 мм рт. ст. — гипотоническим*. Среднее давление у здоровых людей находится в пределах 80—90 мм рт. ст. Диастолическое давление обычно на 10 мм рт. ст. превышает половину систолического давления, т. е. составляет 60—80 мм рт. ст. Разница между систолическим и диастолическим давлениями называется пульсовым давлением или пульсовой амплитудой. Ее величина равна в среднем 40 мм рт. ст. При повышении или понижении
* Гипертония — повышение тонуса, гипотония — понижение тонуса.
''—ИЗ
241
систолического давления соответственно изменяются и среднее, и диастолическое.
У людей пожилого возраста кровяное давление в связи с увеличением жесткости артериальных стенок выше, чем у людей молодого возраста. У детей кровяное давление ниже, чем у взрослых.
Кровяное давление в разных артериях неодинаково. Оно может быть различным даже в артериях одного калибра, например в правой и левой плечевых артериях. Еще чаще разность давлений отмечается в артериях верхних и нижних конечностей. Впервые этот факт был отмечен И. М. Сеченовым (1851). У большинства людей кровяное давление выше в артериях нижних конечностей. Различие в величинах давления в артериях одинакового калибра обусловлено неодинаковым функциональным состоянием их стенок.
Кровяное давление изменяется под воздействием различных факторов. Например, оно повышается при эмоциональном возбуждении (страх, гнев и др.). Это происходит в результате усиления деятельности сердца и сужения сосудистого русла.
Физическая работа также ведет к изменению кровяного давления. Аускультативное определение артериального давления (по Короткову), обычно проводимое уже после окончания работы (в периоде восстановления), показывает, что систолическое давление в это время может быть повышенным до 150—200 мм рт. ст. Степень его увеличения зависит от мощности работы и индивидуальных особенностей человека. Среднее давление и диастолическое под влиянием работы изменяются меньше, чем систолическое. После длительной напряженной мышечной деятельности диастолическое давление, а в некоторых случаях и систолическое оказываются ниже исходных величин, что обусловлено значительным расширением сосудистого русла в работавших мышцах.
Измерения артериального давления прямым методом при работе подтвердили данные, полученные ранее аускультативным способом. Однако сдвиги среднего и диастолического давлений в этих случаях были больше. Кроме того, прямыми исследованиями выявлено, что систолическое давление при работе повышается не сразу, а волнообразно. Это обусловлено запаздыванием расширения периферического сосудистого русла по отношению к увеличению сердечного выброса крови.
Радиотелеметрические измерения кровяного давления, проведенные в последние годы у людей в процессе их разнообразной деятельности, показали, что оно изменяется более значительно, чем это было установлено ранее (табл. 9).
Под влиянием работы кровяное давление в разных сосудистых областях тела изменяется неодинаково. В артериях неработающих конечностей оно возрастает значительнее, чем в работающих |(рис. 77). Это объясняется суммированием центральной волны повышенного давления с волной, отраженной от периферии. Последняя возникает в результате сужения артериол в неактивных мышцах при работе других.
242
таблиц а 9
Изменение артериального давления (мм рт. ст.) при физических нагрузках по данным радиотелеметрии (средние величины) (по Б. Бахману и Р. Церцави)
Артериальное давление
Условия
Артериальное давление
Условия
К
eg ss S CD
Состояние покоя (положение си-Дя) То же
» »
» »
» »
135
93
78
Бег
262
167
100
140
120
116
127
95
95
92
95
80
77
76
80
Плавание
Бег на лыжах
Подъем в гору на лыжах
Спуск с горы на лыжах
221
158
159
185
174
113
114
132
136
80
82
101
§ 11. Артериальный пульс
Артериальным пульсом называют ритмические колебания артериальной стенки, обусловленные систолическим повышением давления в артерии. Пульсация артерий определяется путем легкого прижатия ее к подлежащей кости. По частоте пульса у здоровых людей можно точно судить о ритме сердечных сокращений. При некоторых заболеваниях сердца артериальный пульс не соответствует частоте сердцебиений. В этих случаях подсчитывается число сердечных толчков.
Рис. 77. Систолическое давление в плечевой (И) и бедренной (Б) артериях прн тяжелой работе на ручном (1) и ножном (2) велоэргометрах (по П. О. Астранду)
Рис. 78. Сфигмограмма лучевой артерии:
/ — анакротический подъем; 2 — катакротический спуск; 3 — дикротический подъем
243
Пульсовая волна возникает в аорте в момент изгнания крови из желудочка, когда давление в аорте повышается и стенка ее растягивается. Волна повышенного давления и вызванного этим колебания артериальной стенки распространяется с определенной скоростью от аорты до артериол и капилляров.
Зарегистрированная на бумажной ленте или фотопленке пульсовая кривая называется сфигмограммой. На сфигмограммах аорты и крупных артерий различают анакротический подъем, катакротиче-ский спуск пульсовой кривой и вторичный, или дикротический, подъем (рис. 78). Анакротический подъем обусловлен систолическим повышением давления и вызванным этим растяжением артериальной стенки. В конце систолы левого желудочка давление в нем начинает падать, в связи с чем наступает катокротический спуск пульсовой кривой. В тот момент, когда желудочек начинает расслабляться, давление в нем уменьшается столь резко, что кровь, выброшенная в аорту, устремляется назад к желудочку. Но на своем пути она захлопывает полулунные клапаны, которые препятствуют обратному току крови в левый желудочек. Возникающая при этом волна давления сопровождается его вторичным повышением в артериях, называемым дикротическим подъемом пульсовой кривой.
§12 . Скорость распространения пульсовой волны по артериям
Функциональное состояние артериальных стенок определяется их жесткостью. Последняя зависит от геометрических параметров сосуда, особенностей его соединительнотканной основы, степени напряжения гладких мышц сосудистой стенки и величины кровяного давления. Чем больше жесткость артериальных стенок, тем относительно меньше их растяжимость.
О жесткости артериальных стенок судят по скорости распространения пульсовой волны. Между этими величинами имеется прямая зависимость. Скорость распространения пульсовой волны определяется по времени возникновения пульсации на центральном и периферическохм участках артерии. Для этого регистрируются сфигмограммы либо с двух разных артерий, либо с двух точек одной. Зная расстояние между этими точками и время запаздывания пульсации, можно рассчитать скорость распространения пульсовой волны. У взрослых здоровых людей скорость распространения пульсовой волны по аорте равна в среднем 5—7 м/сек, по магистральным артериям руки — 6—8 м/сек, по магистральным артериям ноги — 8—10 м/сек. С возрастом скорость распространения пульсовой волны по артериям и особенно по аорте увеличивается. Это объясняется повышением жесткости их стенок (в связи со склеротическими изменениями).
Под влиянием физических нагрузок скорость распространения пульсовой волны по аорте и артериям неработающей конечности возрастает. Это происходит благодаря повышению кровяного давления в сосуде и сокращению гладких мышц его стенок. Скорость
244
распространения пульсовой волны по артериям работающих конечностей повышается в меньшей степени или даже снижается (рис. 79).
Уменьшение жесткости артериальных стенок способствует увеличению притока крови к органам, питаемым данной артерией.
§13 . Кровообращение в артериолах и капиллярах
В капиллярах происходит обмен веществ между кровью и тканями. Общее количество капилляров очень велико. Поэтому кровяное русло в области капилляров в 600—800 раз превышает сечение аорты. Скорость кровотока в капиллярах небольшая—всего 0,3—
1,0 мм/сек. Число капилляров в разных тканях неодинаково. Их больше в тех тканях, где интенсивнее обмен веществ. Например, на 1 мм2 сердечной мышцы приходится капилляров в 2 раза больше, чем на 1 мм2 скелетной. Кровяное давление в разных капиллярах колеблется от 8 до 40 мм рт. ст. Неодинаково давление и в одном и том же капилляре. Оно несколько больше в его артери-
альном конце и меньше в веноз-
ном. д5
При покое органа в нем функ-
ционирует лишь небольшое число Г*ис- 79. Изменение скорости распро-капилляров, остальные находятся стРанения пульсовой волны (в % к r п исходным величинам) по артериям
и спавшемся состоянии. При ра- верхних (черные столбики) н нижних боте органа капилляры раскрыва- (заштрихованные столбики) конечно-ются. Это ведет к усилению мест- стей под воздействием плавания на иого кровотока, т. е. к рабочей ги- 100 м при, п°мо,ни РУК (О и ног (2) перемни (от лат. hyperhainua — полнокровие). Изменение числа действующих капилляров в работающем органе было обнаружено Крогом. Он подсчитал, что на 1 мм2 скелетной мышцы в покое приходится 35—85 действующих капилляров, при работе же число их возрастает до 2500—3000. Соответственно увеличивается и общая поверхность капилляров: в покое она равна 3—8 см2 на 1 см2 мышечной ткани, при работе — 360—370 см2. Объем крови по отношению к объему мышцы возрастает при этом с 0,02—0,06 до 5,5— 15%.
Раскрытие спавшихся, так называемых «резервных», капил-.oipoB обусловлено повышением давления в артериолах. Последние играют роль «кранов», регулирующих просвет капилляров и наполнение их кровью.
245
§ 14. Кровообращение в венах
В начале венозной системы (венозное колено капилляров) давление у человека равно в среднем 15 мм рт. ст., в венах конечностей оно составляет 5—9 мм рт. ст. В крупных венах, находящихся вблизи грудной полости, давление почти равно атмосферному и изменяется в связи с дыханием. При вдохе, когда грудная клетка расширена, давление в полых венах становится ниже атмосферного. При выдохе оно повышается до 2—5 мм рт. ст. При форсированном выдохе и натуживании оно возрастает еще больше. Разность давлений в различных венах способствует перемещению крови по венозной системе.
Стенки вен тоньше и более растяжимы, чем стенки артерий. Поэтому емкость венозного сосудистого русла может резко увеличиваться (в 5—6 раз) даже при небольшом повышении давления в крупных венах. В результате в венах может депонироваться значительная часть крови, в связи с чем их называют «емкостными» сосудами в отличие от артерий, обозначаемых термином «резистивные» (от англ, resistance — сопротивление) сосуды.
Основная масса крови из артерий попадает в вены через капилляры. Однако между артериями и венами имеется и более короткий путь — через артерио-венозные анастомозы. Они играют роль шунтов, регулирующих капиллярное кровообращение. Например, в коже артерио-венозные анастомозы открываются при очень высокой или очень низкой температуре окружающей среды. В капилляры при этом поступает меньше крови, что предотвращает излишнее перегревание или охлаждение тела.
Линейная скорость кровотока возрастает в венозной системе от периферии к сердцу. В периферических венах среднего калибра она равна 6—14 см/сек, в полых венах — до 30 см/сек, что значительно меньше линейной скорости кровотока в аорте. Объемная скорость кровотока сохраняется одинаковой во всех отделах венозной системы.
Движение крови в нижней полой вене и в венах нижних конечностей направлено против действия силы тяжести. Этому движению способствуют: во-первых, венозные клапаны, обеспечивающие однонаправленный ток крови — от периферии к сердцу; во-вторых, изменение давления в плевральной полости (оно повышается при выдохе, что способствует накоплению крови в крупных венах, и снижается при вдохе, увеличивая поступление крови из вен к сердцу); в третьих, сокращения скелетных мышц при работе, которые, сдавливая вены, обеспечивают выжиманию из них крови по направлению к сердцу.
Воздействие дыхательных движений на венозное кровообращение образно называется «дыхательным насосом», влияние деятельности скелетных мышц — «мышечным насосом».
При динамической циклической работе движению крови в венах способствуют и дыхательный и мышечный насосы. При статических усилиях и особенно при силовой работе, характеризующей
246
ся натуживанием, внутригрудное давление и давление в полых венах повышаются. Приток крови к сердцу при этом снижается. В результате уменьшается сердечный выброс, снижается артериальное давление, ухудшается кровоснабжение головного мозга, что может привести к обморочному состоянию.
§15 . Кровообращение в малом кругу
Особенности гемодинамики в легких зависят от морфологических и функциональных особенностей сосудов малого круга кровообращения. Легочное русло, пропускающее за 1 мин. то же количество крови, что и большой круг, имеет меньшую протяженность. Крупные легочные артерии более растяжимы, чем артерии большого круга. Поэтому они могут вмещать относительно больше крови без существенных изменений давления. В легких хорошо развиты артерио-венозные анастомозы, благодаря которым в случае необходимости (например, при усилении кровообращения во время мышечной деятельности) укорачивается путь крови. Обширная капиллярная сеть легких не оказывает большого сопротивления кровотоку. Растяжимости легочных сосудов способствует малое противодавление окружающей ткани (оно ниже, чем в других органах). Все это создает условия для малого сопротивления кровотоку в легких. Поэтому правый желудочек выполняет относительно меньшую работу, чем левый.
Непосредственная связь стенок мелких сосудов с тканью легких ставит легочное кровообращение в зависимость от дыхательных движений. Однако наличие мышц в сосудистых стенках и их активные сокращения и расслабления снижают зависимость распределения крови в легких от дыхательных движений, а также от объема притекающей и оттекающей крови. Сосуды легких характеризуются меньшим сопротивлением, чем сосуды большого круга. Это сопротивление может преодолеваться при относительно небольшом давлении, создаваемом работой правого желудочка. В легочной артерии человека систолическое давление составляет 25—30 мм рт. ст., среднее — около 14—15 мм, диастолическое — 5—10 мм. Таким образом, давление в артериях легких во много раз ниже, чем в большом кругу. В легочных венах оно равно в среднем 6—12 мм рг. ст.
Сосуды легких могут депонировать кровь, т. е. вмещать ее избыточный объем, не используемый самим органом. Накопление крови в депо не вызывает существенного подъема давления в его сосудах. Емкость легочных сосудов непостоянна. При вдохе она \ величивается, при выдохе — уменьшается. Легочные сосуды могут вмещать от 10 до 25% всего объема крови.
При физической работе приток крови к легким увеличивается. Давление же при этом в легочных сосудах изменяется мало. Оно остается постоянным даже при увеличении кровотока в 2—3 и более I аз. Лишь в самом начале работы давление в легочной артерии мо-я ci повышаться на 4—5 мм рт. ст., что обусловлено запаздыванием
24/
раскрытия резервных капилляров. При дальнейшей работе давление в сосудах легких, как правило, нормализуется. Некоторые исследователи наблюдали при физической работе понижение артериального давления в легких, обусловленное значительным расширением сосудистого русла в связи с углублением дыхания.
Регуляция кровообращения
§16 . Регуляция деятельности сердца
Кровообращение в организме осуществляется по системе параллельно включенных местных кругов. Объем кровотока в разных областях тела неодинаков. Он больше в активных органах и меньше в неактивных. В зависимости от состояния организма общий объем кровотока также постоянно изменяется. Между работой сердца, просветом и емкостью сосудистого русла и количеством циркулирующей крови имеются определенные соотношения. Они регулируются нервными и гуморальными механизмами.
Работа сердца усиливается при увеличении венозного притока. Мышца сердца при этом сильнее растягивается во время диастолы, что способствует ее более мощному сокращению. Однако эта закономерность, впервые описанная Э. Старлингом, проявляется лишь в определенных условиях. Главную же роль в регуляции деятельности сердца играют нервная система и ряд гуморальных факторов.
Иннервация сердца. Сердце иннервируется эфферентными ветвями блуждающего и симпатического нервов. Эти нервы не являются пусковыми, они лишь регулируют возбудимость и проводимость сердечной мышцы, силу и частоту сердцебиений. Эфферентные волокна, находящиеся в ветвях блуждающего нерва, проводят имт пульсы от рецепторов сердца в нервные центры. Эфферентные волокна блуждающих нервов проводят импульсы, тормозящие деятельность сердца. Этот факт был обнаружен в 1845 г. братьями Вебер. Позднее И. П. Павлов показал, что раздражение одних волокон блуждающего нерва вызывает урежение сердцебиений, а раздражение других — их ослабление. Тормозящий эффект при раздра-
Рис. 80. Изменение деятельности сердца под влиянием раздражения блуждающего (Л) и симпатического (Б) нервов:
1 — запись сокращений сердца, 2 — отметка раздражения нервов
248
из-под значе-жизни
КН
пи
АФС
Рис. 81. Схема иннервации сердца: С — сердце; ПМ — продолговатый мозг; Ц — центры, регулирующие деятельность сердца; СП — спинной мозг; БЫ ~ блуждающий нерв; СН — симпатический нерв; НД — нерв-депрессор; КН — каротидный нерв; АФ — афферентные волокна от правого предсердия и полой вены; АФС — афферентные волокна от сердца, несущие информацию в спинной мозг
нд
женин блуждающего нерва проявляется также в понижении возбудимости и проводимости миокарда. При слабом раздражении блуждающих нервов ритм сердечных сокращений урежается, при более сильном происходит остановка сердца (рис. 80, Л). При длительном раздражении блуждающего нерва прекратившиеся вначале сокращения сердца возобновляются, несмотря на продолжающееся раздражение. Это явление, называемое «ускользанием» сердца влияния блуждающего нерва, имеет большое биологическое ние. Благодаря ему обеспечивается возможность сохранения при длительном раздражении блуждающих нервов, которое могло бы вызвать полную остановку сердца и гибель организма.
Симпатические нервы усиливают деятельность сердца. И. Ф. Цнон (1867) обнаружил, что раздражение этих нервов ведет к учащению и усилению сердцебиений (см. рис.80,Б). И. П. Павлов (1883) установил, что отдельные волокна симпатического нерва, как и волокна блуждающего, по-разному влияют на работу сердца. Одни из них учащают ритм сердечных сокращений, другие усиливают их. Усиливающие нервные волокна являются трофическими, т. е. действующими на сердце путем усиления обмена веществ в миокарде. Раздражение симпатических нервов улучшает проведение возбуждения в сердце и повышает его возбудимость.
Центры сердечных нервов. Центры блуждающих нервов расположены в продолговатом мозгу (рис. 81). Вторые нейроны блуждающего нерва нервных узлах сердца (интрамуральные ганглии*). Отростки этих нейронов иннервируют синоатриальный узел мышцы предсердий и нгриовентрикулярный узел. Миокард желудочков сердца не иннер-нируется блуждающим нервом.
Нейроны симпатических нервов расположены в верхних сегмен-ых грудного отдела спинного мозга (см. рис. 81), отсюда возбуждение передается в шейные и верхние грудные симпатические узлы и лнлее к сердцу. Импульсы с нервных окончаний на сердце переда-К’ия посредством медиаторов. Для блуждающих нервов медиатором служит ацетилхолин, для симпатических — норадреналин.
находятся непосредственно в
• Интрамуральные (от лат. mursis — стена) ганглии — скопления нервных •лггок внутри миокарда.
249
от разных рецепторов те влияют и химически!
Центры блуждающего нерва постоянно находятся в состог некоторого возбуждения (тонуса), степень которого изменяется влиянием центростремительных импульсов ла. Корме того, на состояние этих центров раздражители (адреналин, ионы кальция, угольная кислота и др.)
При стойком повышении тонуса центров блуждающих нервов серд цебиения урежаются, возникает синусовая брадикардия. При пере резке блуждающих нервов ритм сокращений сердца учащается i связи с прекращением поступления к нему тормозящих импульсов
Тонус центров симпатических нервов выражен слабее, чем блуж дающих. Возбуждение в этих центрах усиливается при эмоциях i при мышечной деятельности. Это ведет к учащению и усилен иг сердечных сокращений.
Нервы сердца обеспечивают приспособление его деятельност, к разным условиям. При мышечном покое преобладает влияни блуждающих нервов, при работе — влияние симпатических нервов
Рефлекторная регуляция деятельности сердца. В этом сложнор процессе участвуют центры продолговатого и спинного мозга, гипс таламуса, мозжечка и коры больших полушарий. В рефлекторной регуляции деятельности сердца имеет значение раздражение мне гих рецепторов. Главную роль при этом играют баро- или прессоре цепторы и хеморецепторы. Области сосудистой системы, где сосре доточены эти рецепторы, называются рефлексогенными зонам! К ним относятся дуга аорты, область разветвления сонных артерии и др. (см. рис. 81). Такие же рецепторы имеются и в самом сердце
Баро- или прессорецепторы возбуждаются при повышении дав ления в сосудах и растяжении их стенок. Возникающий при это! поток афферентных импульсов усиливает возбуждение в центрах блуждающих нервов, что ведет к урежению и ослаблению сердеч ных сокращений. В месте впадения полых вен в правое предсердие также расположены рецепторы (см. рис. 81), возбуждающиеся пр повышении давления в этой области. Импульсы от этих рецепторов тормозят центры блуждающих нервов и возбуждают центры сил патических, в результате чего деятельность сердца усиливается) (рефлекс Бейнбриджа). Рефлекторные изменения деятельност сердца происходят также при раздражении рецепторов других об ластей сосудистого русла (легочные артерии, сосуды внутренних ор ганов). Например, рефлекторная остановка сердца может быть вы звана ударом по передней стенке живота. Возникающее при это] раздражение рецепторов кишечника передается на блуждающий] нерв, что и ведет к остановке сердца (рефлекс Гольца).
Хеморецепторы сосудистых рефлексогенных зон возбуждаютс при изменении состава омывающей их крови (увеличение концент рации углекислоты, уменьшение содержания кислорода и др.).
В регуляции деятельности сердца имеют значение и сигналы о проприорецепторов двигательного аппарата. При физической рабе те поток проприоцептивных импульсов усиливается, что тормози! центры блуждающих нервов и ведет к учащению сердцебиений.
Ритм сердечных сокращений может изменяться при изменени
250
температуры. При повышении температуры тела (например, при физической работе в условиях высокой температуры окружающей среды) сердцебиения учащаются более резко, чем при такой же работе, но в условиях нормальной температуры. При охлаждении тела, например при купании в прохладной воде, сердцебиения уре-жаются. Повышение или понижение температуры тела на I ° изменяет частоту сердечных сокращений в среднем на 10 ударов 1 мин. Частота сердцебиений изменяется при болевых раздражениях и различных эмоциях (гнев, страх и т. п.).
В регуляции деятельности сердца принимает участие кора больших полушарий. Это доказано путем выработки условных рефлексов, изменяющих деятельность сердца. Один из примеров кортикальной регуляции сердечного ритма — предстартовое учащение сердцебиений.
Гуморальная регуляция деятельности сердца. На деятельность сердца влияют некоторые гормоны и электролиты.
Гормоны надпочечника адреналин и норадреналин вызывают учащение и усиление сердечных сокращений. Их действие подобно действию симпатического нерва.
Гормон щитовидной железы тироксин усиливает восприимчивость сердца к импульсам, поступающим по блуждающим и симпа-шческим нервам. При избытке тироксина в крови в состоянии покоя. когда преобладает влияние блуждающих нервов, деятельность сердца может быть резко приторможенной. При возбуждении же симпатического нерва сокращения сердца чрезмерно учащаются.
Электролиты имеют большое значение для нормальной деятельности сердца. Изменение концентрации в крови ионов калия и кальция влияет на автоматию сердца и его сократительные свойства. При избытке ионов калия урежается ритм и уменьшается сила сокращений сердца, снижаются его возбудимость и проводимость. При промывании изолированного сердца животных концентрированным раствором калия происходит расслабление миокарда и ос-1ановка сердца в диастоле. Однако не только избыток, но и понижение концентрации калия в растворе, промывающем сердце, вызывает нарушение его деятельности. Йоны кальция учащают ритм и \( вливают сердечные сокращения, повышают возбудимость и проводимость миокарда. При избытке кальция сердце останавливается и систоле. При недостатке кальция происходит ослабление сердечных сокращений.
6 17. Регуляция функционального состояния сосудов
Нервные влияния па сосуды. Сосуды имеют сосудосуживающие и, рвы (вазоконстрикторы). Они относятся к симпатической нервной • иг геме.
А. П. Вальтер (1842), раздражая седалищный нерв лягушки, впервые наблюдал сужение сосудов. Позднее Бернар отметил, что • |.in у кролика на шее перерезать с одной стороны симпатический •и рн, то ухо на этой стороне делается краснее и теплее другого и
251
на свет видно, что сосуды его расширены. Это явление обусловлено тем, что по симпатическому нерву к кровеносным сосудам поступают нервные импульсы, которые поддерживают их стенки в состоянии некоторого напряжения (тонуса). Когда симпатические нервы перерезаны, то поток импульсов прекращается и сосуды расширяются. Правильность этого предположения доказывается тем, что раздражение периферического конца симпатического нерва вызывает сужение сосудов уха.
Главные сосудодвигательные центры расположены в продолговатом мозгу (В. Ф. Овсянников, 1871). В этих центрах имеются сосудосуживающие (прессорные) и сосудо-мм рт.ст. расширяющие (депрессорные) отделы.
г 200
-180
- SOO
-80
- 60
*-40
J40
-120
' ’.О.МЛМ.Ч »АЛЛД J
Время,сек.'
Рис. 82. Понижение артериального давления у собак при раздражении прессорецепторов каротидного синуса:
а — артериальное давление (мм рт. ст,), б — раздражение прессо-рецепторов
Основную роль в регуляции просвета I сосудов играют прессорные центры. При их возбуждении сосуды суживаются, при торможении — расширяются. Депрессорный отдел сосудодвигательного центра ( способствует расширению сосудов путем ( торможения сосудосуживающих центров.] Значение продолговатого мозга в регуляции тонуса сосудов доказано опытами] с перерезкой мозга. Если произвести пере-] резку между продолговатым мозгом и спинным, то расширяются сосуды и понижается кровяное давление. Однако через] некоторое время оно вновь несколько по-1 вышается. Теперь это обусловлено им-1 пульсами из сосудодвигательных центров] спинного мозга, которые в нормальных! условиях подчинены центрам продолго-1 ватого мозга.
Возбуждение и торможение сосудосу4 кивающих центров возникают в результате нервных и гуморальных воздействий^ К нервным воздействиям относятся афферентные импульсы от рецепторов разных] областей тела (стенки кровеносных сосу! дов, органы брюшной полости, кожа) J Наибольшее значение при этом имеют импульсы от прессе- и хемо! рецепторов, находящихся в сосудистых рефлексогенных зонах (дуга аорты, область разветвления сонных артерий). Афферентные импульсы от рефлексогенных зон поступают в сосудодвигательныЯ центр по депрессорному и синокаротидному нервам.
Раздражение прессорецепторов происходит при повышении кровяного давления. Их возбуждение вызывает торможение сосудосуживающего центра и, как следствие этого, расширение сосудов и понижение кровяного давления (рис. 82). Перерезка депрессорных и синокаротидных нервов у животных ведет к резкому и стойкому повышению кровяного давления.
252
Хеморецепторы возбуждаются при изменении состава крови. Накопление в крови углекислоты и уменьшение содержания кислорода, раздражая хеморецепторы, увеличивают тонус сосудодвигательного центра и рефлекторно повышают артериальное давление. Таким образом, раздражение прессо- и хеморецепторов рефлексогенных зон вызывает разнонаправленные реакции. При раздражении прессорецепторов происходит расширение сосудов, при раздражении хеморецепторов — сужение.
Прессорецепторы имеются не только в аорте и сонных артериях, но и в сосудах других областей тела (легкие, кишечник, селезенка и др.).
Хеморецепторы находятся в сосудах селезенки, надпочеченика, почек, костного мозга. Эти хеморецепторы воспринимают изменение не только содержания в крови углекислоты и кислорода, но и концентрации адреналина и некоторых других веществ.
Кроме сосудодвигательных центров продолговатого и спинного мозга в регуляции просвета сосудов участвуют центры промежуточного мозга (гипоталамус) и больших полушарий. Раздражение некоторых ядер гипоталамуса сопровождается сужением сосудов и повышением кровяного давления.
Участие коры больших полушарий в управлении сосудистым тонусом подтверждается наличием нервных вазомоторных (сосудодвигательных) путей, соединяющих этот отдел мозга с гипоталамусом, стволовой частью мозга и спинным мозгом. Раздражение и разрушение некоторых отделов больших полушарий вызывают изменения в состоянии сосудов.
Степень и значимость влияний коры больших полушарий на разные (по механизму и физиологической роли) вазомоторные реакции неодинаковы. Особенно ярко регулирующее воздействие коры проявляется при выработке сосудистых условных реакций, являющихся компонентами сложных безусловных рефлексов (ориентировочных, оборонительных, пищевых, половых и др.). Условнорефлекторное изменение сосудистого тонуса наблюдается и в тех случаях, когда условные раздражители подкрепляются безусловными рефлексами, имеющими свое «представительство» в коре больших полушарий. Влияние же коры на местные вазомоторные реакции (реактивная гиперемия, реакции на воздействие тепла и холода) в настоящее время ставится под сомнение (В. В. Орлов, 1971, Г. П. Конради, 1973).
На сосудистые реакции у человека оказывают влияние различные эмоции. Это проявляется в изменении окраски кожи (побледнение или покраснение) и повышении кровяного давления.
Гуморальные влияния на сосуды. Некоторые химические вещества (гормоны, медиаторы), действуя непосредственно на стенки сосудов, могут вызывать их сужение и расширение. Например, сосудосуживающий эффект оказывают гормоны надпочечника — норадреналин и адреналин и гормон задней доли гипофиза — вазопрессин-Адреналин и норадреналин суживают артерии и артериолы органов брюшной полости, кожи и легких. Увеличение концентрации этих
253
гормонов в крови ведет к резкому повышению давления. Вазопрессин суживает артериолы и капилляры.
Значительный сосудосуживающий эффект вызывает образую* щееся в почках вещество ренин. Выделение его увеличивается при недостаточном кровоснабжении почек и при снижении артериального давления. Ренин активирует находящееся в плазме вещество — гипертензиноген. Превращаясь в гипертензин, оно вызывает сужение сосудов и повышение артериального давления. К сосудосуживающим веществам относится также серотонин. Он образуется в слизистой оболочке кишечника, в мозгу, при распаде кровяных пластинок. Серотонин суживает сосуды при кровотечениях и предохраняет этим организм от потери крови.
К сосудорасширяющим веществам относятся ацетилхолин, гистамин, медуллин, простагландины, брадикинин, аденозинтрифосфор-ная, молочная и угольная кислоты.
Ацетилхолин образуется в окончаниях парасимпатических нервов. Он расширяет мелкие артерии Быстро разрушаясь в крови, ацетилхолин оказывает лишь местный эффект, т. е. расширяет сосуды, находящиеся около нервных окончаний. Гистамин образуется в стенках желудка и кишечника, в коже, в скелетных мышцах (при их работе). Он расширяет артериолы и увеличивает кровенаполнение капилляров, действуя главным образом на сосуды брюшной полости. Введение 1—2 мг гистамина в вену кошки вызывает резкое расширение капиллярной сети, понижение кровяного давления и гибель животного. Медуллин образуется в мозговом слое почки. Простагландины (называются они так потому, что впервые были обнаружены в секрете предстательной железы) образуются в разных тканях. Брадикинин (очень активное сосудорасширяющее вещество) расширяет артериолы и понижает кровяное давление. Он образуется в слюнных и поджелудочной железах, в легких и в коже, особенно при ее нагревании.
§ 18. Рабочая гиперемия
Расширение сосудов в работающем органе и увеличение в нем кровотока называется рабочей или функциональной гиперемией. Это явление было впервые описано Задлером (1875). Гаскелл (1877) объяснил его накоплением продуктов обмена в работающей мышце. Этой точки зрения придерживается большинство исследователей и в настоящее время. При работе мышц в них образуются гистамин, адениловая, молочная, угольная кислоты, повышается концентрация ионов калия. Все эти вещества, действуя на стенки артериол, вызывают расслабление их гладких мышц и расширение просвета. Это ведет к увеличению объема крови, поступающей в капилляры. Число раскрытых капилляров нарастает, что и обеспечивает возникновение рабочей гиперемии. Вещества, образующиеся в сокращающейся мышце (метаболиты), действуют не только на гладкие мышцы артериол, но и на тканевые рецепторы. Возникающее здесь возбуждение передается в сосудосуживающий центр и
254
вызывает рефлекторное сужение сосудов в неактивных областях тела. Этот процесс, называемый компенсаторной вазоконстрикцией, обеспечивает усиление кровотока в работающих органах.
В настоящее время на основе новых электрофизиологических данных разработана гистомеханическая гипотеза, объясняющая возникновение рабочей гиперемии (В. М. Хаютин и др.). Ее авторы предполагают, что расслабление артериальных гладких мышц происходит в результате механических изменений формы артериол при сокращениях скелетной мышцы. Тонус мышц сосудистых стенок регулируется водителями ритма, роль которых выполняют мышечные клетки сосудистой стенки, обладающие высокой автоматией. Импульсы от этих клеток поддерживают тонус гладких мышц артериолярных стенок. При сокращении скелетной мышцы происходит деформация стенок артериол, что нарушает контакт между водителями ритма и не обладающими автоматией гладкими мышцами сосудистой стенки. При этом гладкие мышцы расслабляются, приводя к расширению артериол и возникновению рабочей гиперемии. Нервные импульсы, поступающие по двигательным нервам к мышце и вызывающие ее сокращение, одновременно обеспечивают усиление ее кровоснабжения.
Расширение сосудов в мышце происходит сразу же в начале работы. Даже небольшая начальная частота двигательных импульсов, поступающих к скелетной мышце, уже вызывает гиперемию. Очень частое раздражение мышцы приводит к нарастанию кровотока не при сокращении мышцы, а при последующем ее расслаблении.
Таким образом, рабочая гиперемия обусловлена сложным комплексом химических, нервных и механических факторов.
Перераспределение крови, происходящее при мышечной деятельности, необходимо для обеспечения высокой работоспособности организма. Без повышенной доставки крови нарушаются химические процессы в мышце, освобождающие энергию, которая требуется для ее сокращения.
§ 19. Кровообращение при физической работе
Энергия, необходимая для мышечной деятельности, освобождается главным образом в результате окисления органических веществ в мышцах. Поэтому потребность в кислороде при физической работе значительно возрастает. Поскольку кислород доставляется кровью, то кровоснабжение активных органов, в особенности скелетных мышц и сердца, должно быть при работе увеличенным. Приспособление деятельности сердечно-сосудистой системы к повышенным требованиям достигается посредством нервной и гуморальной регуляций.
Изменение кровотока начинается еще в предрабочем (предстартовом) периоде. Одно лишь представление о работе может вызы-пать подготовительную перестройку деятельности сердца и функционального состояния сосудов. Эта перестройка осуществляется по механизму сложных условно-безусловных рефлексов.
255
Возникающее при подготовке к работе возбуждение центров симпатической инвервации и‘увеличенное образование катехоламинов* ведут к усилению деятельности сердца, перераспределению крови и повышению артериального давления. Все это уже до работы увеличивает кровоток к скелетным мышцам, сердцу и легким.
В начале работы в систему регуляции кровообращения включаются дополнительные рефлекторные, гуморальные и физические факторы. Кора больших полушарий, посылая эфферентные импульсы к двигательному аппарату, одновременно влияет на центры вегетативной нервной системы. При этом наблюдается дальнейшее повышение возбуждения в центрах симпатических нервов и угнетение парасимпатических. В работающих мышцах происходит расслабление гладких мышц артериол и прекапиллярных сфинктеров, расширение и раскрытие нефункционирующих в состоянии покоя капилляров. Все это обеспечивает увеличение местного кровотока. Расширение кровеносных сосудов в работающих мышцах сочетается с сужением сосудов брюшных органов и других неактивных при данной мышечной деятельности областей тела.
Изменения деятельности сердца и состояния сосудов при работе рефлекторно усиливаются импульсами от проприорецепторов двигательного аппарата и от сосудистых хеморецепторов, сигнализирующих о повышении концентрации кислых продуктов в крови.
Работа скелетных мышц, особено динамическая, увеличивая возврат венозной крови к сердцу, способствует усилению его сокращений. Однако наибольшее значение для увеличения мощности сердечного сокращения имеют не механические условия работы сердца, а стимулирующие воздействия симпатических нервов. В результате повышения сократимости сердечной мышцы при работе увеличивается систолический выброс крови. Это сочетается с учащением сердечных сокращений и ведет к значительному повышению производительности сердца. При напряженной мышечной деятельности систолический объем крови может возрастать до 150—180 мл и более, чистота сердцебиений—до 200 и более ударов в 1 мин., минутный объем крови — до 25—35 и более литров. Кровоток к скелетным мышцам увеличивается при этом до 22—25 и более литров.
Обмен веществ между кровью и тканями тела
§ 20. Тканевая жидкость и обмен веществ через капиллярную стенку
Посредником между кровью и клетками органов в большинстве случаев является соединительная ткань, которая заполняет пространство между кровеносными сосудами и клетками органа. Раство
* К катехоламинам относятся адреналин, норадреналин и другие вещества, вырабатываемые мозговым слоем надпочечника,
256
ры газов, органических веществ и минеральных солей, покидающие кровяное русло в начальных участках капилляров, как и растворы веществ, образующихся в процессе тканевого метаболизма, распространяются по поверхности соединительнотканных волокон (в силу закона капиллярности), а также связываются с коллоидами основного вещества соединительной ткани. Эти растворы, непрерывно поступающие в соединительную ткань и как бы пропитывающие ее, образуют тканевую жидкость. Через всю эту сложную систему осуществляется диффузия веществ из крови в клетки тела и из клеток тела в кровь.
Вода, вышедшая из кровеносных капилляров, а также образовавшаяся в самих тканях возвращается вместе с растворенными в ней веществами обратно в кровь. Этот возврат совершается двумя путями: одна часть тканевой жидкости всасывается обратно в кровеносные капилляры, другая в виде лимфы возвращается в кровеносное русло через разветвленную лимфатическую систему.
Капилляры представляют собой микроскопические трубочки, находящиеся между артериолами и венулами. Средний диаметр капилляров равен 5—10 мк, средняя длина — 0,3 мм. Стенка капилляра состоит всего лишь из одного слоя эндотелиальных клеток. Толщина этой стенки— около 1 мк. Общая поверхность капилляров очень велика. У человека она составляет более 6000 м2. Эта поверхность постоянно омывается несколькими литрами крови, которая распределяется в виде тонкого слоя, соответствующего диаметру капилляров. Ясно, насколько может быть велик обмен веществ через такую огромную поверхность. Величина капиллярного ложа не является постоянной, она может увеличиваться или уменьшаться путем изменения просвета и количества действующих капилляров.
Стенка кровеносных капилляров является своеобразным живым фильтром. Вода и растворенные в ней кислород, углекислота и различные кристаллоиды —соли, глюкоза, аминокислоты, мочевина и другие вещества низкого молекулярного веса могут очень быстро проходить сквозь стенку капилляров. Коллоиды же, в том числе белки плазмы, в обычных условиях не проходят через капиллярную стенку из-за большого размера молекул. Проницаемость капиллярной стенки изменчива. Она повышается при недостатке кислорода и увеличении концентрации водородных ионов.
Движение веществ сквозь стенку капилляров представляет собой сложное сочетание процессов диффузии, фильтрации и осмоса. Те вешетва, концентрация которых в крови выше, чем в клетках тела (кислород, глюкоза, аминокислоты и др.), проходят через капиллярную стенку в клетки. Вещества же, концентрация которых выше в клетках (молочная кислота, мочевина и др.), перемещаются в кровь.
Фильтрация воды и растворенных в ней веществ через сосудистую стенку обусловлена гидростатическим давлением крови в капиллярах. Чем выше это давление, тем интенсивнее фильтрация.
Осмотическое давление зависит от разности концентраций веществ, неспособных пройти через полупроницаемую перепонку (бел
257
ки и др.). Поскольку концентрация белков в крови выше, чем в тканевой жидкости, осмотическое давление в крови также выше. Осмотическое давление белков и других коллоидов, называемое также онкотическим, удерживает воду в крови. Таким образом, процессы осмоса противодействуют фильтрации. Чем больше концентрация белков в плазме и чем меньше их концентрация в тканевой жидкости, тем это противодействие сильнее.
Фильтрация жидкой части крови происходит главным образом в артериальном конце капилляра, где гидростатическое давление крови превышает онкотическое. В венозном же конце капилляра гидростатическое давление в результате резкого падения (до 10 мм рт. ст.) становится ниже онкотического, в связи с чем происходит обратное проникновение тканевой жидкости в кровь.
Если онкотическое давление крови очень низкое, что наблюдается, например, при длительном белковом голодании, то обратное проникновение тканевой жидкости в капилляры уменьшается и в тканях могут возникать отеки.
§ 21. Лимфа и ее течение
Тканевая жидкость, всосавшаяся в лимфатические сосуды, называется лимфой. По своему химическому составу она похожа на плазму крови, отличаясь от нее меньшим содержанием белков (4—5%). В связи с этим удельный вес и вязкость лимфы меньше по сравнению с плазмой.
В лимфе имеются форменные элементы. К числу их относятся белые кровяные тельца, главным образом лимфоциты, которые образуются в лимфатических узлах. Эритроциты в лимфе отсутствуют, поэтому она, в отличие от крови, бесцветна.
В норме скорость образования лимфы и скорость ее оттока от тканей одинаковы.
Лимфатическая система начинается сетью анастомозирующих между собой, пронизывающих все ткани капилляров, в которые поступает тканевая жидкость. Эти лимфатические капилляры образуют лимфатические сосуды, которые впадают либо в грудной проток, либо в правый шейный, соединяющиеся с подключичными венами. Через эти вены жидкость, вышедшая из крови в ткани, возвращается в кровяное русло.
У человека за сутки в грудной проток поступает около 1200— 1600 мл лимфы. В состоянии покоя из грудного протока в вену вливается около 1 мл лимфы в 1 мин.
По ходу лимфатических сосудов находится большое количество лимфатических узлов, которые являются местом образования лимфоцитов и играют важную роль в защите организма от болезнетворных микробов.
В передвижении лимфы большую роль играет отрицательное давление в грудной полости. Оно способствует расширению грудного лимфатического протока и присасыванию в него лимфы из периферических лимфатических сосудов. Кроме того, перемещению лим
258
фы способствуют ритмические сокращения стенок лимфатических сосудов. Наличие в них клапанов обеспечивает одностороннее течение лимфы. Некоторое значение для движения лимфы имеет ритмическая пульсация артерий, рядом с которыми расположены лимфатические сосуды. При мышечной работе движение лимфы ускоряется. Сокращающиеся скелетные мышцы сдавливают лимфатические сосуды и способствуют этим оттоку лимфы из конечностей.
Скорость течения лимфы значительно меньше скорости кровотока. В крупных лимфатических сосудах она равна 250—300 мм в I мин.
После приема пищи и во время физической работы скорость лимфотока возрастает. Сокращающиеся скелетные мышцы сдавливают проходящие в них лимфатические сосуды. Это способствует перемещению лимфы по направлению к венам. Массаж также ведет к усилению лимфообращения, что повышает интенсивность восстановительных процессов в тканях.
Лимфатическая система снабжена веточками симпатических нервов, раздражение которых вызывает сокращение стенок крупных лимфатических сосудов. Скорость течения лимфы может изменяться рефлекторно под влиянием болевых раздражений, а также при раздражении рецепторов каротидных синусов и других внутренних органов.
Глава VIII
ДЫХАНИЕ
Дыханием называется комплекс физиологических процессов, обеспечивающих потребление кислорода и выделение углекислого газа тканями живого организма. В основе дыхательной функции лежат тканевые окислительно-восстановительные биохимические процессы, обеспечивающие обмен энергии в организме человека, животных и растений.
Обмен газов, или потребление кислорода и выделение углекислого газа, между организмом и окружающей газовой средой осуществляется путем сложного взаимодействия систем дыхания, кровообращения и крови. Утилизация кислорода и выделение углекислого газа тканями зависят от окислительно-восстановительного потенциала клеток, а также от физических свойств тканевых струк-। у р, определяющих скорость диффузии кислорода и углекислого । аза через клеточные мембраны.
Газообмен является многозвеньевым процессом. Поступающий в кткие кислород переходит в кровь, доставляется к тканям, перехо-1и г через стенки капилляров в межтканевую жидкость и утилизиру-< гея клетками. Углекислый газ из тканей поступает в кровь, доставляется к легким и переходит в альвеолярный воздух, состав которо-н> поддерживается на относительно определенном уровне за счет вентиляции легких. Такой взаимосвязанный обмен газов в организ
259
ме может быть схематически представлен в следующей последовательности: 1) внешнее (легочное) дыхание, или обмен газов между внешней средой и альвеолами легких; 2) обмен газов между альвеолярным воздухом и кровью капилляров легких; 3) перенос кровью кислорода и углекислого газа; 4) обмен газов между кровью капилляров и тканями организма; 5) внутриклетечное, или тканевое, дыхание (подробно рассматривается в курсе биохимии).
§ 1. Внешнее дыхание
Общие положения. Атмосферный воздух попадает через нос и рот в трахею, переходит в правый и левый бронхи, которые древовидно разветвляются. Из мелких бронхов воздух через бронхиолы заполняет легочные пузырьки — альвеолы, стенки которых состоят из эпителиальных клеток и опорной соединительной ткани. Сквозь альвеолярную мембрану происходит обмен газов между альвеолярным воздухом и кровью, протекающей по капиллярам, оплетающим легочные пузырьки.
Обновление воздуха в альвеолах происходит благодаря изменениям объема грудной клетки в результате сокращения межреберных дыхательных мышц и диафрагмы. Дыхательные движения оказывают механическое влияние на органы средостения: сердце с крупными кровеносными сосудами, грудной проток лимфатической системы и пищевод Важное значение для осуществления вдоха и выдоха имеет герметически замкнутая плевральная полость, точнее, плевральная щель. Она образована висцеральным (покрывающим легкое) и париетальным (выстилающим изнутри грудную клетку) листками плевры и заполнена небольшим количеством жидкости.
Механизмы вдоха и выдоха. Дыхательные движения—вдох и выдох — периодически изменяют объем грудной клетки.
При вдохе объем грудной клетки благодаря сокращению наружных межреберных мышц диафрагмы увеличивается. Сокращение наружных межреберных мышц вызывает приподнимание ребер с грудной костью и, следовательно, увеличение грудной полости в передне-заднем и боковых направлениях. Сокращение мышечных во-\ локон диафрагмы вызывает уплощение ее купола. Перегородка между грудной и брюшной полостями опускается, что также приводит к увеличению размера грудной клетки в вертикальном направлении. При этом происходит некоторое сдавливание органов брюшной полости.
После окончания сокращения наружных межреберных и ди-| афрагмальной мышц начинается выдох. В условиях спокойного дыхания опускание ребер при выдохе происходит пассивно, за счет эластической тяги растянутого при вдохе связочного аппарата. При более глубоком дыхании опускание ребер происходит активно, в результате сокращения внутренних межреберных мышц. Органы брюшной полости, смещенные вследствие сокращения диафрагмальной мышцы после ее расслабления, вдавливают купол диафрагмы в грудную полость, уменьшая ее размеры в вертикальном направле
260
нии. Таким образом, выдох характеризуется уменьшением всех размеров грудной полости.
В конце выдоха ребра вместе с грудной костью опущены (рис. 83), купол диафрагмы глубоко вдается в грудную полость. Дыхательные мышцы расслаблены.
Причины перемещения воздуха при дыхательных движениях.
При вдохе легкие следуют за движением грудной клетки, хотя и не
Рис. 83. Изменение положения положения диафрагмы при
грудной клетки при выдохе (А), вдохе (Б) и выдохе (а), обычном вдохе (б) и глубоком вдохе (в)
сращены с ее стенкой. Это происходит потому, что щелевидное плевральное пространство, отделяющее стенку легкого от стенки грудной клетки, не увеличивается, так как не имеет сообщения с воздухом. В результате с увеличением размеров грудной полости легкие расширяются и давление в них становится ниже атмосферного. Между полостью легких и наружным воздухом возникает разность давлений, которая определяет закономерный переход воздуха из атмосферы внутрь легких, в которых давление ниже атмосферного. Таким образом, причиной поступления воздуха в легкие является понижение в них давления вследствие увеличения объема грудной полости.
Во время выдоха объем грудной полости уменьшается и воздух в легких сжимается, чему способствует укорочение эластической ткани легких, растянутой во время вдоха. В силу того, что давление в полости легких становится выше атмосферного, воздух выходит в атмосферу. Следовательно, причиной выхода воздуха из легких является повышение давления в них, которое происходит в результате уменьшения размеров грудной полости. Если дышать через короткую трубку, боковой отросток которой соединен с манометром, то легко убедиться в закономерном снижении давления в легких на вдохе и повышении его на выдохе. Разность давлений (в покое примерно 2 мм рт. ст.) имеет место только во время дыхательных движений. Как только изменение объема грудной полости прекращается, давление в ней становится равным атмосферному и вдыхание или выдыхание воздуха прекращается. Если сузить просвет дыха
261
тельной трубки и тем самым затруднить движение воздуха, то изменение внутрилегочного давления при вдохе и выдохе будет более выраженным. В случае полного замыкания дыхательного отверстия при выдохе происходит натуживание, во время которого давление внутри легких увеличивается и при максимальном выдыхательном усилии может повысить атмосферное до 100 мм рт. ст. и более. Во время максимального вдоха (без поступления воздуха в дыхательные пути) давление в легких по сравнению с атмосферным может снизиться на 70 мм рт. ст. и более. Показатели силы вдоха и выдоха могут характеризовать силу дыхательных мышц.
Эластическая тяга легких. В процессе вдоха и выдоха большое значение имеет растяжимость легочной ткани. В результате возрастного роста и развития легких и грудной клетки возникает постоянная тяга легких, стремящихся спасться в силу высокой эластичности своих тканей. Поэтому в плевральной щели, в которой висцеральный листок плевры (покрывающий легкое) и париетальный листок плевры (выстилающий стенки грудной полости) плотно прижаты друг к другу, всегда поддерживается отрицательное давление. Если в опыте на животном ввести в плевральную щель трубочку и таким образом соединить ее с водухом, эластические легкие спадутся, а плевральная щель заполнится воздухом. В таких условиях вдох и выдох станут невозможными, так как работа дыхательных мышц не вызовет изменения внутрилегочного давления.
Как известно, зависимость объемов и давлений для идеально упругого тела является прямолинейной. Для легочной ткани эта зависимость иная. Если давление в легких увеличить, начиная от уровня выдоха при атмосферном давлении (путем вдувания воздуха через трубочку, вставленную в трахею), вначале объем увеличивается в небольшой степени, а затем быстро возрастает. При приближении к предельному объему увеличение его вновь становится незначительным. Следовательно, между объемом и давлением в легких нет прямолинейной связи, эластические же свойства легких изменяются в зависимости от их предшествующего состояния.
Роль поверхностного нагяжения. В настоящее время доказана существенная роль поверхностного натяжения пленки (внутриаль-веолярой жидкости), выстилающей альвеолы.
Расчеты показали, что величина поверхностного натяжения в альвеолах должна быть столь высокой, что для их расправления при дыхании необходимо давление, в несколько раз превышающее действительное. Если исключить роль поверхностного натяжения, заполнив легкие физиологическим раствором, то можно убедиться, что их растяжимость значительно повышается, взаимосвязь давления и объема приближается к прямолинейной и почти полностью устраняется различие в изменении давления и объема при исходных уровнях спавшегося и растянутого воздухом легкого. Вместе с тем роль поверхностного натяжения в альвеолах оказалась значительно меньшей по сравнению с расчетными данными. Это связано со специальным активно действующим на поверхностное натяжение веществом. Оно образуется в митохондриях клеток альвеолярного эпи
262
телия и условно названо сурфактантом (от англ, surface — поверхность, act — действие). Это вещество значительно снижает величину поверхностного натяжения. Так, расправленные альвеолы при давлении в 20 см водн. ст. в случае введения вещества, подавляющего действие сурфактанта, спадаются уже при снижении давления до 18 см водн. ст. При нормальном действии сурфактанта снижение давления даже до 5 см водн. ст. вызывает снижение объема альвеол всего лишь наполовину.
Кроме эластического сопротивления дыханию тканей легких следует учитывать и неэластическое сопротивление, зависящее от трения при дыхательном перемещении тканей легких. Такое перемещение тканей вызывается разностью давлений в легких и атмосфере, создаваемой работой мышц, взаимодействующих с эластическими силами и силой тяжести тканей грудной клетки. Заполнение легких воздухом происходит весьма неравномерно и зависит от положений грудной клетки и работы дыхательных мышц.
Общая емкость легких. Суммарная величина воздуха, которую могут вместить легкие при максимальном вдохе, называется общей емкостью легких.
В общей емкости легких можно выделить четыре составляющих ее компонента: дыхательный объем, резервный объем вдоха, резервный объем выдоха, остаточный объем.
Дыхательным объемом называется количество воздуха, проходящего через легкие при одном вдохе (выдохе). В покое он равен примерно 350—800 мл, при мышечной работе может достигать 1— 2 л и больше. Резервный объем вдоха составляет воздух, который дополнительно можно вдохнуть после обычного вдоха. Резервным объемом выдоха называется объем воздуха, который еще можно выдохнуть после обычного выдоха. Остаточным называется объем воздуха, который остается в легких после максимального выдоха.
Сумма дыхательного воздуха, резервного объема вдоха и резервного объема выдоха составляет жизненную емкость легких (ЖЕЛ). Прямым путем при помощи спирометра регистрируют ЖЕЛ и три составляющие ее объема. Полученные индивидуальные величины ЖЕЛ сопоставляют с так называемыми должными величинами, т. е. со средними для людей определенного пола, роста, веса и возраста. Должные величины можно определить по номограмме или по специальным таблицам. Отклонения ЖЕЛ в пределах ±20% от должной не являются существенными.
Величина ЖЕЛ зависит от роста, веса, положения тела и многих других факторов и колеблется в широких пределах — от 1500 до 7500 мл. Она несколько больше в положении стоя по сравнению с положениями сидя и лежа.
При спортивных позах величина ЖЕЛ может значительно изменяться. Например, при положении стойка на лопатках согнувшись ЖЕЛ существенно уменьшается, и главным образом из-за резкого уменьшения резервных объемов выдоха и вдоха.
В состав дыхательного воздуха входит объем так называемого мертвого, или вредного, пространства, образуемого воздухоносными
263
путями, заполненными не участвующим в газообмене воздухом. Слово «.вредное» является условным, поскольку заполняющий это пространство воздух играет положительную роль в поддержании оптимальной влажности и температуры альвеолярного воздуха. Различают понятия анатомическое «мертвое пространство» (объемом примерно 140 мл) и функциональное вредное пространство, которое при мышечной работе может увеличиваться в 2 раза и более из-за несоответствия вентиляции и кровотока в значительных участках легочной ткани.
§ 2. Вентиляция легких
Для газообмена между организмом и атмосферным воздухом большое значение имеет вентиляция легких, способствующая обновлению состава альвеолярного воздуха. Количественным показателем легочной вентиляции является минутный объем дыхания (МОД). Он равен произведению дыхательного объема на число дыханий в 1 мин.
Вентиляцию легких определяют путем забора выдыхаемого воздуха в резиновые емкости (мешки Дугласа) с последующим пропусканием воздуха через газовые счетчики. В настоящее время используются и миниатюрные счетчики (турбинки), вмонтированные в дыхательную маску. Для сопоставления величин объемов вентилируемого воздуха их приводят к строго определенным условиям температуры, атмосферного давления и насыщенности парами воды (поправка BTPS). При расчете объемов кислорода и углекислого газов легочная вентиляция приводится к температуре 0°, 760 мм рт. ст. и сухому газу (поправка STPD). Это позволяет точно рассчитать объемы газов в единых условиях.
Из воздуха альвеол кислород переходит в кровь, и в него поступает углекислый газ. Поэтому альвеолярный воздух содержит меньше кислорода и больше углекислого газа по сравнению с воздухом, вдыхаемым из атмосферы. Ниже приводятся данные состава вдыхаемого, выдыхаемого и альвеолярного воздуха при спокойном дыхании (в %).
Воздух Кислород Углекислый газ Азот
Вдыхаемый 20,94 0,03 79,03
Выдыхаемый 16,30 4,00 79,70
Альвеолярный 14,40 5,60 80,00
Выдыхаемый воздух состоит из смеси альвеолярного и воздуха вредного пространства, по составу не отличающегося от атмосферного. Поэтому выдыхаемый воздух содержит больше кислорода и меньше углекислого газа по сравнению с альвеолярным. Назначение легочной вентиляции состоит в поддержании относительно постоянного уровня парциального давления * кислорода и углекислого
* Парциальное давление газа — часть общего давления газовой смеси, приходящаяся на долю того или иного газа в газовой смеси.
264
газа в альвеолярном воздухе. При барометрическом давлении 760 мм рт. ст. парциальное давление кислорода в нем равно 159 мм рт. ст. В альвеолярном воздухе парциальное давление водяного пара равно 47 мм рт. ст., и, следовательно, на другие газы приходится 760—47=713 мм рт. ст. При содержании в альвеолярном воздухе 14,3% кислорода его парциальное давление будет равно 102 мм рт. ст., а 5,6% углекислого газа — 40 мм рт. ст., на долю 80% азота — 571 мм рт. ст.
Следует различать понятия минутный объем вентиляции и минутный объем альвеолярной вентиляции. Для характеристики последней необходимо из величин МОД вычесть суммарную величину вентиляции мертвого пространства, т. е. того объема воздуха, который не достиг альвеол. К воздуху мертвого пространства относится и воздух тех альвеол, в которых газообмен происходит не полностью («параллельное» мертвое пространство). Даже в состоянии покоя имеет место неравномерность вентиляции различных участков легкого. Для оценки эффективности альвеолярной вентиляции важное значение имеет соответствие вентиляции кровотоку в кровеносных капиллярах каждой альвеолы и в их группах. При некоторых состояних организма парциальное давление газов в альвеолярном воздухе и в артериальной крови неодинаково.
Работа дыхательных мышц. Вентиляция легких осуществляется благодаря работе мышц, участвующих в дыхательных движениях. Эта работа связана с преодолением эластического сопротивления и сопротивления дыхательному потоку воздуха (неэластическое сопротивление).
Для регистрации величины работы при дыхании применяется специальный прибор (пневмотахограф). Внутриплевральное давление регистрируют путем записи внутрипищеводного давления, которое равно внутриплевральному. Таким образом, работу дыхательных мышц можно вычислить по следующей формуле: W = (pdv, где W — работа, р — общее давление, a dv — изменение объема системы. Отсюда ясно, что увеличение МОД связано с увеличением работы дыхательных мышц и увеличением стоимости легочной вентиляции. При малых величинах МОД, превышающих величины покоя всего в 2 раза (в пределах 20 л), на работу дыхательных мышц расходуется всего 5—20 мл/мин кислорода, а при МОД в 100 л (десятикратное превышение уровня покоя) кислородная стоимость дыхания достигает 200 мл/мин, т. е. увеличивается в 20 раз. При предельных сдвигах МОД до 150—200 л потребление кислорода достигает стократного увеличения по сравнению с уровнем покоя. В таких условиях не весь поступающий в организм дополнительный кислород может использоваться для полезной работы.
Возможности и ограничения вентиляции легких. Степень обновления альвеолярного воздуха благодаря легочной вентиляции может изменяться в широких пределах. Например, при мышечной работе легочная вентиляция может увеличиваться по сравнению с уровнем покоя до 25—30 раз. Однако имеется ряд существенных ограничений, затрудняющих использование вентиляции легких для
265
эффективной альвеолярной вентиляции. К ним относится биомеханический фактор, определяющий различное сочетание объемов и емкостей общей емкости легких. Так, жизненная емкость легких (ЖЕЛ) у штангистов при выполнении толчка составляет примерно 65% по сравнению с ЖЕЛ в положении стоя; максимальная вентиляция легких у борцов в положении моста равна 80% от максимальной вентиляции легких в положении стоя. Пределы вентиляции легких лимитирует и большая механическая работа мышц и высокая кислородная стоимость дыхания. В случаях высокого уровня МОД или большой длительности работы возможно значительное снижение коэффициента полезного действия при двигательной деятельности.
Эффективность вентиляции легких может резко снижаться при несоответствии ее кровотоку в легких, неравномерности вентиляции и увеличении функционального вредного пространства.
§ 3. Обмен газов между альвеолярным воздухом и кровью капилляров легких
Диффузия газов через альвеолярную мембрану. Между парциальным давлением кислорода альвеолярного воздуха и напряжением * его в венозной крови существует разность: если парциальное
Атмосферный воздух
давление кислорода в альвеолярном воздухе равно примерно 102 мм рт. ст., то в крови, протекающей в капиллярах, оплетающих альвеолярную стенку, оно составляет только 40 мм рт. ст. Причиной перехода СОг из крови в альвеолярный воздух является то, что напряжение газа в венозной крови капилляров (примерно 47 мм рт. ст.) выше парциального давления в алвеолярном воздухе (40 мм рт. ст.).
На рис. 84 представлена схема условий, которые существуют по обе стороны дыхательной поверхности легкого. Как видно из рисунка, парциальное давление
Рис. 84. Обмен газов через кислорода (рО2) в альвеолярном воздухе стенки альвеолы примерно на 62 мм рт. ст. выше, чем в
крови, притекающей к легким, что определяет диффузию О2 в кровь. В оттекающей от легких крови рО2 приближается к 100 мм рт. ст. В связи с тем, что парциальное напряжение углекислого газа (рСО2) в притекающей к легким крови
примерно на 7 мм рт. ст. выше, чем в альвеолярном воздухе, углекислый газ переходит в альвеолярный воздух (см. рис. 84).
Сопротивление диффузии газов. В последние годы придается существенное значение факторам, замедляющим диффузионные
* Термин «напряжение газа» применяется для характеристики растворов газа в жидкости и аналогичен термину «давление газа» для газовых смесей.
266
процессы, особенно диффузию кислорода. Диффузия зависит как от свойств альвеолярной мембраны, так и от условий кровоснабжения легочной ткани. Изменения диффузии наблюдается в условиях сильных воздействий на организм, при мышечной нагрузке, изменениях положений тела и др.
Альвеолярная и капиллярная мембраны являются сложным неоднородным образованием. Внутренняя стенка альвеол выстлана жидкой пленкой, предохраняющей их ткань от высыхания и содержащей важные вещества (сурфактанты), определяющие необходимую способность легочной ткани к растяжению. Растворимость углекислоты в тканях легочной мембраны выше растворимости в ней кислорода более чем в 20 раз. Поэтому выведение СО2 из крови осуществляется, как правило, без существенных трудностей.
Определенное значение для диффузии кислорода может иметь общее сопротивление газообмену, зависящее от величины «мембранного сопротивления» и «внутрикапиллярного сопротивления». Обычно в легких имеется такое соотношение скорости кровотока с емкостью капилляров, которое обеспечивает оптимальные условия для газообмена. Однако в некоторых сложных условиях деятельности кровоток в капиллярах может значительно ускоряться. Вследствие этого время контакта альвеолярного воздуха с кровью, необходимое для диффузии кислорода в кровь через альвеолярную мембрану, оказывается недостаточным. В этом случае кровь вытекает из легочных капилляров с уменьшенным парциальным напряжением кислорода. Неравномерность кровоснабжения и вентиляции альвеол может вызывать нарушение диффузионных возможностей в легких и снижение насыщения крови кислородом.
§ 4. Перенос газов кровью
Перенос кислорода. Дыхательная функция крови прежде всего характеризуется цоставкой к тканям тела необходимого им количества кислорода. Химическим переносчиком кислорода служит пигмент крови гемоглобин (НЬ). Он обладает способностью вступать с кислородом в непрочное соединение — оксигемоглобин (НЬО2), которое легко распадается с освобождением кислорода.
Молекула гемоглабина включает четыре частицы гема железосодержащего вещества, имеющего важнейшее значение для возникновения обратимой связи с кислородом. Белковая часть молеку-1Ы гемоглобина глобин составляет 96% общей массы, а 4% приходится на долю гема. Поскольку молекула гемоглобина содержит 1 частицы гема, она может одновременно или последовательно свя-<ать 4 молекулы кислорода: НЬ4+4О2—*’(НЬО2)4. Кислородная емкость гемоглобина определяется в условиях, когда все молекулы гемоглобина связаны с кислородом. Количество кислорода, связанного гемоглобином в 100 мл крови, носит название кислородная емкость крови. Известно, что каждый грамм гемоглобина связывает 1,33—1,36 мл О2. Следовательно, кислородная емкость крови здоро-ного взрослого мужчины, у которого в 100 мл крови содержится
267
15 г Hb, составляет 20,4 объемных процента (1,36X15=20,4 об%). Величина кислородной емкости крови колеблется в ши-
роких пределах у разных людей и в разных условиях (от 17,42 до 24,12 об %) . Имеются данные о некотором увеличении (на 10—15%) кислородной емкости циркулирующей крови при физических нагрузках за счет выхода богатой эритроцитами крови из депо (селезенки, печени). Кислородная емкость всей крови человека,содержащей примерно 750 г гемоглобина, сос-
Рис. 85. Кривая диссоциации оксигемоглобина в крови человека в покое:
А содержание НсОг в артериальной крови, В — то же в венозной крови
тавляет около 1000 мл О2, что достаточно для кислородного потребления в покое в течение 5 — 6 мин. В реальных условиях используется лишь часть кислородного запаса крови. В трудных условиях мышечной работы кислородная емкость крови может оказаться существенным энергетическим резервом организма. Интересно, что животные, обладающие очень высокой способ-
ностью к осуществлению мощных движений в трудных условиях (например, в воде), имеют высокую кислородную емкость крови. Так, кислородная емкость крови кенгуру равна 27 об %, а тюленя — 29 об %.
Насыщение крови кислородом зависит от рО2 в крови, в свою очередь зависящего от рО2 атмосферного воздуха. По мере увеличения рО2 крови процент насыщения ее кислородом возрастает, а при понижении рО2 уменьшается. Однако в соотношениях рО2 в крови и насыщения крови кислородом нет прямо пропорциональной зависимости: в зоне малых значений рО2 процент НЬО2 повышается значительно быстрее, чем в зоне больших величин рО2. Эта важная функциональная связь отчетливо видна при графическом изображении кривой диссоциации (расщепления) оксигемоглобина (рис. 85). По оси абсцисс на рис. 85 отложены величины рО2, а по оси ординат— соответствующие им величины НЬО2 (в %). При соединении нанесенных на график точек получается кривая, круто поднимающаяся в зоне малых величин рО2 и очень незначительно повышающаяся в зоне больших значений рО2.
Кривая диссоциации оксигемоглобина наглядно показывает условия, определяющие перенос кислорода кровью. В притекающей к легким крови (венозной) рО2 равно 40 мм рт. ст. По мере обогащения в капиллярах легкого кислородом рО2 крови достигает 100 мм рт. ст. и почти все молекулы гемоглобина (96%) переходят в окисленную форму — оксигемоглобин. Близкое к горизонтальному направление кривой диссоциации оксигемоглобина в зоне больших значений рО2 означает, что небольшие колебания рО2 окружающего
268
воздуха или альвеолярного воздуха в этом случае не будут существенно сказываться на связывании кислорода гемоглобином. Даже при подъеме на высоту 4000 м, где рО2 в альвеолах падает со 102 до 60 мм рт. ст. (на 40%), насыщение крови кислородом в капиллярах альвеол уменьшается всего на 11% (с 96 до 85%).
При снижении рО2 в крови тканевых капилляров от 60 до 30 мм рт. ст. насыщение крови кислородом падает уже на 20%, т. е. до 40% НЬО2. Такой процесс диссоциации оксигемоглобина совершается по мере продвижения крови через ткани. Поскольку в диапазоне низких значений рО2 его уменьшение ведет к быстрому расщеплению оксигемоглобина, высокое усиление окислительных процессов в тканях резко повышает скорость снижения насыщения крови кислородом, т. е. увеличивает переход О2 из крови в ткань.
В различных условиях деятельности может возникать острое снижение насыщения крови кислородом — гипоксемия. Степень гипоксемии определяют путем взятия порций крови из артериального или венозного кровеносного сосуда или из полостей сердца с последующим извлечением газов на приборе Ван Слайка и их количественным анализом. Извлечение газов из порции крови осуществляется на основе принципа ртутного насоса, «отсасывающего» из крови весь кислород и углекислый газ, разработанного еще И. М. Сеченовым.
Причины гипоксемии весьма разнообразны. Она может возникать в крови легких в связи со снижением рО2 в альвеолярном воздухе (например, при произвольной задержке дыхания), а также при неравномерности вентиляции в различных частях легких.
Гипоксемия наблюдается даже в состоянии покоя (рО2 снижается со 100% до 96)—физиологическая гипоксемия. Она может усиливаться при двигательной деятельности вследствие нарастания неравномерности вентиляции легких, увеличения скорости кровотока в связи с малым временем контакта воздуха и крови, а также в случаях увеличения толщины легочной мембраны. Гипоксемия может быть связана с перебросом крови через артерио-венозные анастомозы в малом круге кровообращения. В этом случае кровь, бедная кислородом, минует дыхательные поверхности альвеол и примешивается к артериализованной крови, оттекающей от легких, что и создает ту или иную степень гипоксемии. Гипоксемия артериальной крови, протекающей в тканях, связана с недостатком О2 в них н усиленной мобилизацией кислорода из оксигемоглобина крови, быстро наступающие гипоксемические изменения возникают и в венозной крови (венозная гипоксемия), как результат снижения насыщения кислородом крови, оттекающей от тканей с выраженной активностью окислительных процессов (например, от работающих мышц).
Перенос углекислого газа. Перенос кровью углекислоты от тканей к легким важен прежде всего для освобождения организма от избытка углекислого газа. Попадая в кровь, углекислый газ участвует в выравнивании кислотно-щелочного равновесия, и его напря-/кение сказывается на кривой диссоциации оксигемоглобина.
269
Углекислый газ диффундирует из тканей, где его напряжение велико, в кровь с меньшим напряжением СОг. В капиллярах тканей рСО2 относительно большое — 47 мм рт. ст. Поэтому СО2 связывается с кровью, превращаясь в соли угольной кислоты. Всего в венозной крови содержится 55—58 об % углекислого газа. Большая его часть в виде солей угольной кислоты находится в плазме и эритроцитах. В плазме растворено 2,5 об % СО2, 4—5 об % его находится в эритроцитах в соединении с гемоглобином в виде карбогемоглобина.
Рис. 86. Кривые диссоциации оксигемоглобина в зависимости от содержания углекислого газа
Важный процесс образования угольной кислоты из углекислого газа и воды в крови тканей происходит в эритроцитах, где содержится фермент карбоангидраза. Он ускоряет реакцию СО2 + Н2О ч* Н2СО3 как слева направо, так и справа налево. Последнее имеет место в крови легких, где напряжение СО2 ниже. Путем нейтрализации Н2СО3 белками крови связывается до 90% всей образующейся углекислоты. Остальные 10% переносятся в виде системы СО2—Н2СО3.
В связывании СО2 кровью существенную роль играет гемоглобин. Вместе с тем и повышение напряжения углекислого газа повышает диссоциацию оксигемоглобина с ускорением образования восстановленного гемоглобина. Это можно видеть на рис. 86, на котором показано изменение кривой диссоциации оксигемоглобина ,в зависимости от напряжения углекислого газа в крови.
§ 5. Обмен газов между кровью и тканями
Переход кислорода из крови в ткани. В клетках тканей рО2 постоянно стремится к снижению, а в функционирующих мышцах может снизиться до нуля. Поэтому из притекающей к тканям артериальной крови, где исходная величина рО2 большая (около 100 мм рт. ст.), кислород диффундирует в ткани, и рО2 крови снижается все больше и больше по мере протекания ее через ткани и становится равным 40 мм рт. ст. и меньше.
Кровь капилляров большого круга кровообращения отдает не весь кислород. Если в артериях имеется в среднем 19 об% О2, то в оттекающей от тканей венозной крови — около 11 об % О2. Следовательно, ткани утилизировали 8 об % кислорода. Разность между об % О2 в притекающей к тканям артериальной крови и оттекающей от них венозной называется артерио-венозной разностью. Эта величина служит важной характеристикой дыхательной функции крови, показывая, какое количество кислорода доставляют тканям каждые 100 мл крови. Для того чтобы выяснить, какая часть приносимого
270
кровью кислорода переходит в ткани, вычисляют коэффициент утилизации кислорода. Его определяют путем деления величины артерио-венозной разности (по кислороду) на содержание кислорода в венозной крови и умножения на 100. В покое коэффициент утилизации О2 обычно равен 30—40%. При напряженной мышечной работе, когда в оттекающей от мышц венозной крови содержание О2 уменьшается примерно до 8 об % и более (вместо 11 об % в покое), утилизация кислорода участвующими в работе тканями может возрасти до 50—60 об % и более.
В снабжении мышц кислородом в трудных условиях работы может иметь значение и внутримышечный пигмент миоглобин, который связывает дополнительно 1,0—1,5 л О2.
Связь кислорода с миоглобином более прочная, чем с гемоглобином. Оксимиоглобин отдает кислород только при выраженной гипоксемии. При этом существенное значение имеют ферментативные внутриклеточные процессы.
Переход углекислого газа из тканей в кровь. Поскольку рСО2 в тканях достигает значительных величин (50—60 мм рт. ст. и выше), углекислый газ переходит в межтканевую жидкость, где рСО2 равно в среднем 46 мм рт. ст., и в кровь, превращая ее в венозную (рСО2 — около 40 мм рт. ст.). Повышение напряжения углекислоты в крови, а также увеличение сдвига pH в кислую сторону, например при мышечной работе, способствует отдаче кислорода кровью для окислительных процессов в тканях.
§ 6. Регуляция дыхания
Общие сведения. Физиологические механизмы регуляции дыхания многогранны.
Изменение дыхания в соответствии с потребностями организма достигается посредством сложной системы нервно-гуморальных воздействий на дыхательный центр, который расположен в продолговатом мозгу. Вентиляция легких может увеличиваться или уменьшаться в зависимости от афферентных сигналов, приходящих к дыхательному центру из различных рецепторов, т. е. в порядке безусловного рефлекса. Кроме того, к дыхательному центру постоянно поступают импульсы от коры больших полушарий. С участием коры образуются условнорефлекторные связи по механизму условных рефлексов, регулирующие дыхание. Им принадлежит большая роль в формировании дыхания у спортсменов при выполнении специальных движений.
В единстве с нервной регуляцией дыхания осуществляется регуляция гуморальным путем: изменения дыхания происходят в ре-хльтате изменений химического состава крови, влияющего на разнообразные рецепторы в тканях тела и на нервные клетки дыха-пльного центра.
Дыхательный центр. Еще в 1912 г. Легаллуа показал, что если ।слать укол в определенное место продолговатого мозга, то дыхание полностью прекращается. Ф. Флуранс, обнаруживший мгновен
271
ную остановку дыхательных движений при таком уколе, назвал этот участок продолговатого мозга «жизненным узлом». Исследования Н. А. Миславского показали, что располагающийся на дне IV желудочка в сетевидном образовании продолговатого мозга дыхательный центр является парным образованием. По Н. А. Мислав-скому, он делится на центр вдоха (инспираторный) и центр выдоха (экспираторный). Современные электрофизиологические исследования с применением микроэлектродной техники подтвердили нали- . чие различных нейронов, раздражения которых вызывает либо вдох, либо выдох. Наряду с воспринимающими сигналы нервными инспираторными и экспираторными клетками дыхательного центра имеются группы исполнительных нервных клеток, тесно связанных! с межреберными мышцами и диафрагмой. В организации нормального чередования дыхательных движений участвуют также специ-1 альные группы клеток в варолиевом мосту, регулирующие нормальную смену вдоха и выдоха (пневмотаксический центр). Дыхательный центр обладает способностью к автоматической деятельнос-1 ти. Это можно наблюдать на изолированном мозге животного. Авто-1 матизм деятельности дыхательного центра объясняют ритмической динамикой обменных процессов внутри дыхательного центра, в связи с чем в его структурных элементах возникают круговые ритмы возбуждений (М. В. Сергиевский). Автоматия дыхательного центра зависит от сдвигов окружающей его среды и от приходящих к нем) импульсов при раздражении различных рецепторов тела.
Иннервация дыхательных мышц. Проводящие эфферентные пути, несущие импульсы от дыхательного центра, спускаются в спинной мозг и заканчиваются около мотоневронов диафрагмальных и межреберных нервов. Импульсы, поступающие в дыхательны! центр по афферентным путям, возбуждают его невроны, и они, в свою очередь, проводят импульсы к дыхательным мышцам. Таким образом, соответственно периодическому возбуждению дыхательного центра, корригированному афферентными импульсами, происходят периодические сокращения дыхательных мышц.
Гуморальная регуляция дыхания. Изменения дыхания происходят под влиянием сдвигов парциального напряжения углекислого газа и кислорода в крови, т. е. гуморальным путем. Это положение подтверждается в опыте с перекрестным кровообращением мозга у| животных (опыт Фридерика).
Опыт заключался в том, что сонные артерии, снабжающие мозг кровью, при помощи резиновых трубок соединяют друг с другом крест-накрест, а другие сонные артерии перевязывают. В ре! зультате голова собаки I снабжается кровью, притекающей от собаки II, а голова собаки II питается кровью собаки I. Если вызвать снижение насыщения крови кислородом (гипоксемию) и повышение в крови напряжения углекислого газа (гиперкапнию) у одной из собак, то одышка возникает у другой собаки, мозг которой питается кровью собаки, подвергшейся задушению. Поскольку связь через нервную систему между собаками отсутствует, совершенно ясно, что раздражающее действие недостатка кислорода и избытка углекис
272
лоты передается кровью собаки, подвергшейся задушению, на центральную нервную систему собаки со свободным дыханием. В свою очередь усиленное дыхание собаки, у которой трахея не пережата, приводит к вымыванию углекислого газа из ее крови, снабжающей мозг собаки с пережатой трахеей. Действие крови с нормальным напряжением Ог и СО2 на мозг собаки, у которой пережата трахея, приводит к парадоксальному ослаблению у нее активности дыхания, несмотря на выраженную гипоксемию и гиперкапнию. Дыхательный центр особенно чувствителен к увеличению напряжения СО2 в крови и относительно менее чувствителен к изменению содержания кислорода. Уже небольшое повышение об % СО2 в крови, как при усилении обмена веществ, так и при увеличении содержания СО2 во вдыхаемом воздухе, значительно усиливает функцию дыхательного центра. Подобно углекислоте действует на дыхательный центр молочная кислота и вообще любой сдвиг реакции крови в кислую сторону. Например, при уменьшении pH крови на 0,02 вентиляция легких может возрасти на 10 л в 1 мин.
В настоящее время установлено, что избыток СО2 в крови и сдвиги в кислую сторону прежде всего оказывают влияние на хеморецепторы, находящиеся вблизи клеток дыхательного центра. Отсюда возникают рефлекторные воздействия на дыхательный центр. На дыхание влияет также раздражение хеморецепторов клубочков сонных артерий пониженным содержанием в крови кислорода и избытком углекислоты.
Следовательно, опыт с перекрестным кровообращением мозга двух животных следует трактовать как проявление не только гуморальной регуляции, но и рефлекторной. Он подчеркивает неразрывную связь нервной и гуморальной регуляций дыхания.
Значение сдвигов парциального напряжения О2 и СО2 в крови неодинаково. Если недостаток кислорода в крови сказывается прежде всего на учащении дыхания, то избыток СО2 приводит к заметному углублению дыхания.
Рефлексы при раздражении рецепторов легких. Рефлекторное влияние при раздражении механорецепторов легких было описано н 1868 г. Герингом и Брейером. Оно заключается в том, что при вдохе от рецепторов легких в дыхательный центр по веточкам блуждающего нерва поступают импульсы, тормозящие вдох, а при выдохе— импульсы, тормозящие выдох. Если раздувать легкие индифферентным газом (например, азотом), мышцы вдоха перепоют сокращаться, а при искусственном отсасывании воздуха из легких наступает сокращение диафрагмы. Последнее имеет место . ишь при большом спадении тканей легких. При обычном дыхании штудируется переход от вдоха к выдоху.
Рефлексы с рецепторов скелетных мышц. Афферентные импуль-<м, посылаемые работающими мышцами, повышают активность ыхательных движений рефлекторным путем. Это демонстративно проявляется в опыте с пережатием бедренных артерий у человека при работе ног. При таком выключении прямого гуморального влия-•ия химических изменений в мышцах легочная вентиляция поддер
113
273
живалась на таком же уровне, как и до пережатия артерий (опыт Е. Асмуссена, Е. Христенсена и М. Нильсена). Кислородная недостаточность в работающих мышцах доказывалась тем, что общий газообмен снижался на 20—50%, а величина совершаемой работы была такой же, как до пережатия артерий. Таким образом, сигнализация с работающих мышц в нервные центры могла подаваться только по нервным проводникам. Следовательно, поддержание вентиляционной активности дыхания обеспечивается только рефлекторным путем. Очевидно, возбуждение дыхательного центра в этом-опыте является результатом распространения возбуждения по различным сегментам центральной нервной системы, и прежде всего от высшего отдела — коры больших полушарий.
Наибольшее влияние на дыхательный центр оказывают афферентные импульсы при растяжении межреберных и брюшных мышц, наиболее тесно связанных с дыхательными движениями. (
В специальных опытах на животных было показано, что раздражение рецепторов растяжения межреберных мышц и мышц живота может тормозить инспираторные клетки дыхательного центра и усиливать возбуждение в экспираторных клетках.
Роль высших отделов мозга в регуляции дыхания. Приспособле-
ние дыхания к изменяющимся условиям среды и деятельности организма тесно связано с функциями коры больших полушарий. Так, у собак с удаленной корой больших полушарий дыхание в покое и| в стандартных условиях осуществлялось без видимых отклонений!
но при выполнении даже нескольких шагов у них возникла резко выраженная и длительная одышка (Э. А. Асратян). При повторном совпадении усиления дыхания под влиянием специфических раз] дражений хеморецепторов сосудистых рефлексогенных зон с любым «посторонним» раздражителем, например звуковым или световым] возникает условный дыхательный рефлекс: действие одного условного сигнала вызывает изменения, наблюдаемые под влиянием специфического безусловно-рефлекторного воздействия. Исследования, осуществленные К. Л4. Быковым, А. Н. Крестовниковым и их со-
трудниками, произведенные на людях, показали, что условные
рефлексы (например, на сигнальные раздражители, связанные обстановкой, в которой осуществлялась мышечная работа) являю! ся весьма важными для полноценной «настройки» системы дыхани
на эффективную мышечную работу.
Важное значение в произвольной регуляции
дыхания имев
сложная система информации высших центров об изменениях па]
циального напряжения кислорода и углекислого газа в крови в ра| нообразных условиях двигательной деятельности (например, рабо та с дыханием газовой смесью с пониженным парциальным давле нием Ог и др.).
Острое кислородное голодание может приводить к потере созна пня без предваряющих неприятных ощущений (Дж. Холдэн Дж. Пристли). Это наблюдалось, например, у летчиков и альпинио тов, у которых переход в бессознательное состояние иногда сопровождался эйфорией (ложным чувством благополучия).
274
Однако способность человека различать степень кислородного голодания и избытка СО2 в крови, несомненно, существует и может совершенствоваться. В специальных опытах было показано, что люди отличают от воздуха газовые смеси, которые вызывают в организме гипоксические или гиперкапнические сдвиги (И. С. Бреслав). Такая способность различать газовые среды может быть значительно развита путем тренировки, особенно в случае сочетания гипоксемии с гиперкапнией. Так, у выдающихся спортсменов с развитой способностью эффективно распределять мышечную работу на протяжении длительного времени, была установлена поразительная способность тонко дифференцировать состояния гипоксемии и гиперкапнии, возникающие в условиях опыта с острым нарастанием этих состояний Такая способность имеет большое значение для саморегуляции поведения в сложных условиях деятельности, например при нырянии, подводном плавании, при работе в горах (А. Б. Ган-дельсман).
§ 7. Дыхание при физической работе
Общая характеристика. Мышечная работа всегда связана с увеличением газообмена, поскольку энергия черпается в процессе окисления органических веществ. Изменения дыхания четко выражены даже при физических нагрузках, выполняемых малыми мышечными группами. При легкой работе обмен газов может повыситься в 2—3 раза, а при тяжелой — в 20—30 раз по сравнению с уровнем покоя. Исключительно большой удельный вес потребления кислорода при работе зависит не только от его потребления мышцами, непосредственно участвующими в выполнении движений, но и от потребления кислорода мышцами, обеспечивающими высокую легочную вентиляцию, а также мышцей сердца и другими тканями тела.
Согласование дыхания с движениями осуществляется благодаря сложной системе приспособительных изменений в организме, прежде всего биохимическим изменениям в мышечном аппарате и изменениям биомеханических условий при различных движениях.
Активность окислительных процессов тем больше, чем больше мощность выполняемой работы. Это прослеживается в строгих лабораторных условиях, в которых увеличение мощностей сопровож-ыется почти линейным повышением потребления кислорода. Однако в естественных условиях часто не наблюдается строгой пропорциональности между мощностью работы и величиной потребления кислорода. Это может быть связано с различным режимом выполненной работы, когда одна и та же мощность достигается либо пути учащения темпа при малом отягощении, либо путем увеличения перемещаемого груза при редком темпе работы. Исследования показали, что больший удельный вес темпа при одной мощности работы вызывает достоверное усиление дыхания по сравнению с работой такой же мощности при большем удельном весе отягощения. В ряде случаев оказывается, что при одной и той же мощности
io*
27Ь
работы потребление кислорода возрастает при нарушении координации движений или недостаточном использовании инерционных сил (например, при беге на коньках). Это объясняется вовлечением в двигательную деятельность мышц, не имеющих непосредственного отношения к данной работе, а также дополнительным усилением деятельности дыхательной мускулатуры и мышцы сердца, что приводит к повышению потребления кислорода и к понижению коэффициента полезного действия при выполнении работы. Возможны соотношения и
обратного порядка, когда
повышение
мощности ра
4
3
2
I
Рис. 87. Средние данные о структуре общей емкости легких при различных спортивных позах.
Обозначения поз: I— поза велосипедиста; II— борцовский мост; III— поза конькобежца (низкая посадка); IV — стойка на голове и предплечьях; V — стойка на локтях согнувшись. Объемы: / — остаточный воздух; 2 — резервный объем выдоха; 3 — дыхательный воздух; 4 — резервный объем вдоха. Светлые столбики — исходная величина при положении стоя, заштрихованные столбики — величины при спортивных позах
боты достигается без увеличения потребления кислорода
при понижении
кислородной стоимости движений —за счет
или даже
более
экономной координации движений, лучшего использования инерци
онных сил, более экономной работы дыхательных и сердечных мыш1
Такое изменение экономичности работы происходит в процессе по
вышения тренированности спортсменов.
Дыхание при мышечной работе тесно связано с биомеханически ми особенностями позы и механикой самих движений.
При различных позах тела по-разному сочетаются объемы и ем кости общей емкости легких (рис. 87), что может снижать равно мерность вентиляции легких и уменьшать общую площадь вентили руемой части альвеол. Например, как видно на рис. 87, при пол «гимнастический мост» функциональная остаточная емкость легких определяющая общую их поверхность, участвующая в диффузш газов, отчетливо уменьшилась по сравнению с исходным положени ем стоя. При этом же положении тела резко снизилась максималь
276
ная вентиляция легких — до 98,4 со 121,3 л в положении стоя. Подобные изменения наблюдались и при исследовании поз велосипедиста и конькобежца.
Спирографические исследования (запись легочных объемов при дыхательных движениях) показали, что при ряде движений, особенно при напряженной мышечной работе, может наблюдаться существенное лимитирование мышечной работы вследствие биомеханического ограничения внешнего дыхания. Имеет значение и большой удельный вес потребления кислорода, приходящегося на долю мышц, участвующих в усиленном «рабочем» дыхании. Таким образом, особенности внешнего дыхания зависят от целого ряда характеристик движений (особенностей циклических, ациклических, силовых и других видов движений, от их мощности, темпа сопротивления и пр.).
Адаптация дыхания к движениям происходит путем различной интеграции объемов и емкостей общей емкости легких, легочной вентиляции, изменения степени равномерности вентиляции и диффузионной способности альвеолярной мембраны. Важное значение для эффективности дыхания при этом имеет координация функций дыхания и кровообращения (М. Е. Маршак). Обучение движениям приводит к специализированной интеграции дыхательной функции. Чем прочнее связь дыхания и движений, тем легче при прочих равных условиях выполняются движения. При этом сами дыхательные движения становятся как бы компонентами выученных движений.
Вентиляция легких. При выполнении небольших по величине циклических лабораторных нагрузок отмечается линейная зависимость минутного объема дыхания и потребления кислорода. Соответственно увеличивается и выведение углекислого газа, а дыхательный коэффициент (отношение объема СО2 к объему О2) не превышает единицы. У взрослых лиц МОД составляет в покое около Г) —8 л/мин, а при работе нередко достигает 80—100 и даже 150 и более литров в 1мин. Легочная вентиляция может возрастать за гнет углубления дыхания, что повышает эффективность альвеолярной вентиляции. Но это достигается путем большой работы дыха-и-льных мышц, что снижает экономичность доставки кислорода к работающей скелетной мускулатуре. Поэтому углубление дыхания играет благоприятную биологическую роль только до такой степени, при которой не снижается коэффициент полезного действия выполняемой работы и сохраняется синхронность движений тела с нюхательными движениями.
Во время работы нередко отмечается повышение эффективнос-1И использования кислорода из вентилируемого воздуха до 4— ",(> по сравнению с 3—4% в покое. В зависимости от условий рабо-и.| может возникать повышенная вентиляционная активность, при мчорой возрастает об % О2 и уменьшается об % СО2 в альвеолярном воздухе, что характеризует относительную гипервентиляцию. <> шако это не приводит к повышенной диффузии О2 в кровь. «Вымывание» же СО2 из венозной крови может понизить эффективность
277
выполняемой работы в связи с понижением возбудимости дыхательного центра и снижением диссоциации оксигемоглобина. При относительном снижении уровня вентиляции (относительной гиповентиляции) об % О2 в альвеолярном воздухе снижается. Только значительное снижение об % Ог в альвеолярном воздухе, например при произвольной задержке дыхания в процессе работы, может затруднить диффузию О2 в кровь.
При прекращении вентиляции — непрерывной произвольной задержке дыхания — вначале происходит пополнение крови кислородом из воздуха легких, что определяет большую длительность задержки дыхания на вдохе по сравнению с задержкой дыхания на выдохе. Затем может в известной мере использоваться кислород крови до падения напряжения О2 в крови ниже 70% НЬО2 и внутримышечный кислород, содержащийся в митохондриях мышц в соединении с миоглобином в виде оксимиоглобина.
Длительность задержки дыхания в покое на вдохе у человека составляет в среднем 1—2 мин. У спортсменов, особенно у использующих преимущественно упражнения на выносливость, достигает 2,5—3,5 мин. При работе длительность задержки дыхания всегда понижается в зависимости от интенсивности работы и не превышает при трудной работе 15—20 сек. Относительная гиповентиляция, и ее крайняя степень — произвольная задержка дыхания, составляет весьма частый компонент дыхания, повышающий точность выполняемых движений там, где необходима фиксация грудной клетки. Из общего резерва кислорода (в пределах 2 л) около половины может быть использовано для работы, сопровождающейся задержкой дыхания.
Доставка кровью кислорода тканям н углекислого газа к легким. При работе в несколько раз увеличивается объемная скорость кровотока, что обеспечивает доставку нужного количества кислорода к работающим мышцам и транспорт углекислого газа к альвеолярным капиллярам. При этом может возникать венозная гипоксемия (до 20—30% НЬО2 вместо 60% НЬО2 в покое). Если же усиливается i неравномерность вентиляции и возникает недостаточная координация дыхания и кровотока в малом круге кровообращения, то воз-, пикает артериальная гипоксемия, достигающая при тяжелой и длительной работе 80% НЬО2 и ниже. Эти явления отражают приспособительные механизмы повышения утилизации кислорода из крови для удовлетворения окислительных процессов в напряженно работающих мышцах.
Кислородный запрос и его удовлетворение. Количество кислорода, необходимое для окислительных процессов, обеспечивающих ту или иную работу, называется кислородным запросом. Различают суммарный, или общий, кислородный запрос, т. е. количество кислорода, необходимое для выполнения всей работы, и минутный кислородный запрос, т. е. количество кислорода, потребляемое при дан ной работе в течение 1 мин.
Суммарный кислородный запрос вычисляется путем суммирова пня потребления кислорода во время работы и в восстановительном
278
периоде за вычетом всего количества кислорода, необходимого для поддержания уровня покоя.
В зависимости от особенностей мышечной работы устанавливается оптимальный для нее уровень потребления кислорода (рис. 88). Как видно из данных этого рисунка, уровень потребления кислорода при разных видах работы весьма различен: он относительно низок при тяжелоатлетических упражнениях, прыжках на батуте,
при гимнастических упражнениях на снарядах, произвольных упражнениях фигуристов и при спринтерском беге и значительно выше при скоростном беге на коньках (5000 м), велосипедный гонках на треке и особенно высок при беге на длинные дистанции и при скоростном плавании под водою с аквалангом. Эти данные, полученные на высококвалифицированных спортсменах, следует рассматривать как показатели оптимизации дыхания в соответствии со специализацией двигательной деятельности, поскольку повышение спортивного мастерства не сопровождается повышением уровня потребления кислорода в ходе работы и не характеризуется увеличением общего кис-
лородного запроса. Обычно развитие тренированности характеризуется процессом экономизации минутного и общего кислородного запроса от 7—8% ДО 30% и более к исходному уровню. Это происходит при увеличении эффективности выполняемой работы.
При относительно равномерной работе (с колебаниями мощности в пределах ±3%), если она
Рис. 88. Схема потребления кислорода при различных физических нагрузках:
I— тяжелоатлетические упражнения; II— упражнения на батуте; III — гимнастические упражнения на снарядах; IV — произвольные упражнения в фигурном катании; V — бег на 100 м; 17 — бег на коньках (5000 м); VII— велосипедные гонки (1500 м трек); VIII - бег на длинные дистанции; IX — скоростное плавание под водой с аквалангом. Цифры справа — потребление кислорода (л) па 30-й сек. восстановления
легко выполняется спортсменом, может наступить вскоре после начала работы равновесие между кислородным запросом и его удовлетворением, или истинное устойчивое состояние. При напряженной циклической работе, когда минутный кислородный запрос \ довлетворяется не в полной мере (поскольку функции кровообращения и дыхания характеризуются предельно возможным уровнем |ранспорта кислорода), отмечается устойчивая величина потребления кислорода. Однако такая устойчивость потребления кислорода
к'пряжена с нарастанием кислородного долга, не проявляющегося н<> ходу работы. В этих случаях говорят о кажущемся устойчивом
27Э
состоянии. Таким образом, истинное устойчивое состояние может поддерживаться длительное время, кажущееся же устойчивое состояние ограничено временем достижения предельно переносимой величины кислородного долга.
Максимальное потребление кислорода. Оценивая роль дыхания и кровообращения в удовлетворении потребности в кислороде при мышечной работе, необходимо учитывать следующее: 1) усиление дыхания и кровообращения происходит постепенно, и полная мобилизация доставки О2 тканями достигается лишь через несколько минут; 2) производительность дыхательной и сердечно-сосудистой систем имеет индивидуальные пределы («кислородный потолок»), выше которых уже не может увеличиваться количество поглощенного кислорода при нарастающей мощности работы. В этом отношении показатель максимального потребления кислорода (МПК) является важной объективной характеристикой возможностей обеспечения дыхательной и сердечно-сосудистой системами доставки тканям предельных величин кислорода.
Определение МПК осуществляется прямым и косвенным путями. Принцип прямого определения заключается в постепенном (ступенчатом) увеличении физической нагрузки (обычно на велоэргометре) до предельно возможной. Кислородный потолок достигается примерно на 5-й мин. работы — раньше, чем предел мощности. МПК выражает потребление кислорода за 1 мин. Эта величина обозначается литрами, если потребление Ог рассчитывается на весь организм, и миллилитрами при рассчете на 1 кг веса тела. У неспортсменов МПК колеблется в пределах 2,5—3,5 л/мин, а у спортсменов достигает 6 л/мин и более, а при пересчете на 1 кг веса тела соответственно 40—50 мл/кг и до 70—90 мл/кг.
МПК является одним из существенных показателей функционального состояния дыхательной и сердечно-сосудистой систем. Оно отражает преимущественно неспецифическую устойчивость вегетативных систем к предельным напряжениям. Однако в этом показателе могут отражаться и специфические особенности двигательной деятельности. Так, наиболее тесные связи обнаруживаются между
группе спортсменов,
спортивными показателями
специализирую-
в
щихся в длительных циклических и напряженных нагрузках (скоростное плавание под водой, стайерский бег, марафонский бег, велосипедные гонки).
Величины МПК оказываются несколько большими при выполнении нагрузок, близких к привычным. Например, у велосипедис-
тов— при выполнении нагрузок на велоэргометре, у пловцов — при проплывании с предельной скоростью отрезков дистанции «своим» способом и т. д.
Кислородный долг. Количество кислорода, которое поглощается по окончании физической работы сверх уровня покоя, называется кислородным долгом. Его величина после физических нагрузок, выполняемых в условиях истинного устойчивого состояния, примерно соответствует дефициту кислородного потребления в начале рабо ты. В таких условиях абсолютный кислородный долг невелик — 3—
280
5 л. После напряженной циклической работы длительностью до 5— 6 мин. величина кислородного долга может достигать 20—22 л.
Кислородный долг отражает прежде всего процесс расщепления энергетических веществ, не восстанавливающихся во время работы. Это в сновном окислительные реакции, направленные на ресинтез АТФ и синтез углеводов. Часть кислородного долга идет на пополнение кислородного резерва воздуха легких, крови и мышц, составляющего около 2 л. Таким образом, величина кислородного долга отражает не только дефицит кислорода при анаэробных биохимических процессах, но и восполнение кислородного резерва организма, использованного при работе.
Непосредственно после работы наблюдается наиболее интенсивное погашение кислородного долга, связанное с компенсацией анаэробных процессов и восполнением кислородного резерва организма. Затем погашение долга переходит в менее интенсивную фазу (лактатную фазу кислородного долга), отражающую процессы ресинтеза органических соединений, распавшихся при работе.
Глава IX
ПИЩЕВАРЕНИЕ
§ 1. Общая характеристика пищеварительных процессов
Пищеварением называется процесс физической и химической обработки пищи, в результате чего она превращается в такие вещества, которые могут всасываться в кровь и усваиваться. Таким образом, пищеварение является начальным этапом обмена веществ в организме.
Физическая обработка пищи заключается в ее размельчении, перемешивании и растворении. Химическая ее обработка происходит под влиянием гидролитических ферментов, содержащихся в пищеварительных соках. В результате этих процессов сложные вещества, входящие в состав пищи, расщепляются на более простые, которые всасываются в кровь и усваиваются организмом. Белки всасываются в виде аминокислот и низкомолекулярных компонентов, жиры — в виде глицерина и солей жирных кислот, углеводы — в виде моносахаридов. Лишь вода, минеральные соли и небольшое количество низкомолекулярных органических соединений могут всасываться в кровь без предварительной обработки.
Деятельность органов пищеварения регулируется нервными и гуморальными механизмами.
В изучении физиологии пищеварения большая роль принадлежит И. П. Павлову и его ученикам. Они разработали методы исследования пищеварительных процессов у животных. Это позволило подробно характеризовать функции пищеварительных органов и раскрыть сложные механизмы их деятельности. Однако методы, разработанные на животных, не могли быть широко использованы
281
при изучении особенностей пищеварительных процессов у человека. Длительное время изучение двигательных и секреторных функций органов пищеварения у него ограничивалось главным образом зондированием желудка и двенадцатиперстной кишки и применением рентгенологических методов. В настоящее время разработан ряд новых приемов, позволяющих изучать процессы пищеварения у человека. Например, двигательная деятельность желудка исследуется путем регистрации биопотенциалов гладких мышц, находящихся в его стенках. Широко используются и радиотелеметрические методы. Сущность их заключается в проглатывании маленького радиопередатчика (радиопилюля), который свободно проходит по пищеварительному тракту и дает информацию о кислотности содержимого желудка и кишечника, о давлении и температуре в этих органах.
§ 2. Пищевареьие в полости рта
В ротовой полости у человека пища задерживается в течение 15—18 сек. Здесь начинается ее физическая (смачивание слюной и измельчение путем жевания) и химическая (воздействие ферментов) обработка. Находясь в ротовой полости, пища раздражает вкусовые, тактильные и температурные рецепторы слизистой оболочки.
У человека имеются три пары крупных слюнных желез (околоушные, подъязычные и подчелюстные) и много мелких, рассеянных в слизистой оболочке рта.
Слюна — первый пищеварительный сок, действующий на пищу при ее поступлении в организм. В слюне человека содержатся ферменты, вызывающие гидролитическое расщепление сложных углеводов до дисахаридов и частично до глюкозы. Фермент амилаза (птиалин) превращает крахмал в дисахариды, фермент мальтаза расщепляет дисахариды до моносахаридов.
Ферменты слюны наиболее активны в нейтральной среде. Попадая с пищей в желудок (где среда резко кислая), они теряют свою активность. Лишь внутри пищевого комка, где некоторое время сохраняется нейтральная реакция, они продолжают расщеплять углеводы.
Кроме ферментов в состав слюны входят белковые вещества муцин и глобулин, аминокислоты, креатин, мочевая кислота, мочевина и неорганические соли. Муцин слюны делает пищевой комок скользким, что облегчает глотание пищи и продвижение ее по пищеводу.
И. П. Павлов разработал операции, позволившие изучить механизм слюноотделения. У собаки в полости рта отпрепаровывается кусочек слизистой оболочки в том месте, где открывается проток слюнной железы. Вырезанный кусок слизистой оболочки вместе с отверстием протока выводится наружу и вживляется в кожу щеки. В результате слюна, выделяемая железой, поступает не в ротовую
282
полость, а в пробирку, прикрепляемую к специальной капсуле, наклеенной на кожу. У человека деятельность слюнных желез изучается при помощи маленьких резиновых капсул, прикрепляемых к слизистой оболочке ротовой полости в том месте, где находится отверстие протока слюнной железы. Слюна попадает из железы в капсулу и по резиновой трубке, прикрепленной к ней, стекает в пробирку.
При помощи этих операций было установлено, что деятельность слюнных желез регулируется рефлекторно. При поступлении в рот пищи или отвергаемых веществ раздражаются рецепторы слизистой оболочки рта. Слюноотделение при этом происходит по механизму безусловных рефлексов.
Выделение слюны при виде и запахе пищи, а также при действии индифферентных раздражителей (зрительных, звуковых, обонятельных и др.), ранее сочетавшихся с приемом пищи, происходит по механизму условных рефлексов.
Таким образом, деятельность слюнных желез регулируется сложным рефлекторным путем. Центры слюноотделения, как и центры других пищеварительных процессов, находятся в продолговатом мозгу. Центростремительными нервами при безусловном рефлекторном выделении слюны являются веточки тройничного и языкоглоточного нервов. Центробежные функции выполняют парасимпатические и симпатические нервы. При раздражении первых происходит обильное выделение жидкой слюны, при раздражении вторых выделяется густая слюна, содержащая много муцина. Условнорефлекторное выделение слюны может возникать при раздражении разных рецепторов (зрительных, слуховых, обонятельных и др.). Информация при этом поступает в высшие отделы мозга, а импульсы, идущие оттуда, возбуждают центры слюноотделения продолговатого мозга.
Секреция жидкой слюны при раздражении парасимпатических нервов обусловлена усилением кровоснабжения слюнных желез. Выделение же слюны при раздражении симпатических нервов может происходить даже при уменьшении местного кровотока. Эти данные показывают, что образование слюны обусловлено не фильтрацией, а секреторной деятельностью железистых клеток.
Состав слюны зависит от характера веществ, поступающих в ротовую полость. При попадании в рот отвергаемых веществ (камни, песок, кислоты) выделяется много жидкой (отмывной) слюны. При поступлении в рот пищи вырабатывается более густая слюна, богатая ферментами и муцином. При этом на сухую пищу выделяется больше слюны, чем на увлажненную. Опытами И. П. Павлова было установлено, что, например, на мясосухарный порошок выделяется у животных в 6 раз больше слюны, чем на мясо.
Глотание происходит рефлекторно. Пережеванная и смоченная с.поной пища перемещается на корень языка и далее движениями его кзади и кверху проталкивается в глотку.
При сокращениях мышц глотки пищевой комок перемещается в пищевод. Дальнейшее передвижение пищи по пищеводу обеспечи
283
вается перистальтической волной сокращения мышц, находящихся в его стенке. Путь от ротовой полости до желудка твердая пища проходит за 8—9 сек., жидкая — за 1—2 сек.
§ 3. Пищеварение в желудке
Пища подвергается в желудке физической и химической обра- 1 ботке. Перемешивание, перетирание и разминание пищи происходит ' благодаря деятельности гладких мышц желудочной стенки.
Химическая обработка пищи осуществляется желудочным соком, выделяемым железами его слизистой оболочки. Железы же- I лудка состоят из главных, добавочных и обкладочных клеток. Главные клетки образуют ферменты, добавочные — слизь, обкладоч- I ные — соляную кислоту.
Состав желудочного сока. Желудочный сок содержит ферменты протеазы и липазу. К протеазам относятся пепсины, желатиназа и ' химозин. Пепсины выделяются железами желудка в виде неактивных пепсиногенов. Активирует их соляная кислота. При недостатке i соляной кислоты переваривание пищи (особенно белковой) нарушается. Пепсины расщепляют белки до промежуточной стадии (поли- I пептиды). Окончательное расщепление белков (до аминокислот) происходит в кишечнике. Желатиназа способствует перевариванию i белков соединительной ткани. Химозин створаживает молоко, что имеет значение для его переваривания. Липаза расщепляет жиры на глицерин и жирные кислоты. В желудке она действует только I на эмульгированные жиры (молоко).
Желудочный сок имеет кислую реакцию (pH равен 1,5—2,5), что I обусловлено наличием в нем соляной кислоты (0,4—0,5%). Для нейтрализации 100 мл желудочного сока здорового человека тре- I буется 40—60 мл децинормального раствора щелочи. Этот показа- ' тель называется кислотностью желудочного сока.
Соляная кислота желудочного сока играет важную роль в пи- * щеварении. Она активирует пепсиногены, вызывает денатурацию и набухание белков (что облегчает их переваривание), способствует а створаживанию молока, активирует гормон гастрин, образующий- | ся в слизистой оболочке привратника и стимулирующий желудоч-» ную секрецию. Поступая в двенадцатиперстную кишку, соляная кислота действует на ее слизистую оболочку, в которой образуются! гормоны, регулирующие деятельность желудка, поджелудочной железы и печени. Кроме того, соляная кислота задерживает развитие гнилостных процессов в желудке и усиливает его двигательную деятельность. Соляная кислота участвует также в сложном процессе перемещения пищевых масс из желудка в кишечник.
Слизь желудочного сока предохраняет внутреннюю оболочку желудка от вредных механических и химических воздействий. Кроме того, она содержит вещества, усиливающие секрецию желудочных желез, и, адсорбируя витамины, предохраняет их от разрушающего действия желудочного сока.
284
Выделение желудочного сока. И. П. Павлов разработал методы, позволившие детально изучить деятельность желудочных желез. Первый метод заключается в том, что в желудке собаки делают отверстие, в которое вставляют трубку (фистулу). Затем у животного перерезают шейную часть пищевода и концы его вживляют в кожную рану. У оперированной таким образом собаки пища при кормлении выпадает из искусственного отверстия пищевода. Происходит так называемое мнимое кормление (рис. 89), сопровож
дающееся выделением слюны и желудочного сока. После окончания опыта пережеванная собакой пища вкладывается через фистулу в ее желудок, благодаря чему питание животного не нарушается. Опыты с мнимым кормлением показали, что сокоотделение в желудке может начинаться еще задолго до поступле-
ния туда пищи.
Второй метод исследования деятельности желудка состоит в том, что из желудка собаки выкраивается лоскут, из которого создается маленький изолированный желудочек. Его полость полностью отделена от полости большого желудка (см. рис. 89). На изолированный желудочек накладывается постоянная фистула, что позволяет собирать выделяющийся там желудочный сок. При этой операции нервы изолированного желудочка полностью сохраняются. Поэтому его деятельность является как бы «зеркалом» работы боль-.
Рис. 89. Собака с перерезанным пищеводом и фистулой желудка (А) и собака с «маленьким желудочком» (Б)
шого желудка. На животном с изолированным желудочком можно
изучать работу желудочных желез во все периоды пищеварения.
В секреторной деятельности желудка наблюдается три фазы. Первая называется нервной (сложнорефлекторной), вторая — желудочной, третья — кишечной.
В первой фазе желудочный сок выделяется при действии условных раздражителей (вид, запах пищи, индифферентные раздражители, совпадавшие ранее с приемом пищи) и безусловных (механическое и химическое раздражение пищей слизистой оболочки рта
и глотки). Возбуждение, возникающее в рецепторах, передается в пищевые центры, оттуда импульсы по центробежным волокнам блуждающего нерва поступают к железам желудка. При перерезке этих нервов рефлекторное выделение желудочного сока прекращается. Электрическое раздражение периферических концов предварительно перерезанного блуждающего нерва вызывает отделение желудочного сока с высокой ферментативной активностью. Раз-
285
дражение симпатического нерва, наоборот, тормозит желудочную секрецию. ,
Условнорефлекторное выделение желудочного сока ^ожет начинаться задолго до приема пищи. Этот сок называете^' аппетитным. Его выделение обеспечивает готовность желудка к, приему пищи.
Аппетит, т. е. ощущение потребности введения пищи в организм, способствует желудочной секреции. Возникновение аппетита связано с действием условных и безусловных раздражителей. Поэтому обстановка, в которой происходит прием пищи, и ее вкусовые качества имеют значение для процессов пищеварения. Однако, несмотря на большую роль условнорефлекторных раздражителен в возникновении аппетита, главным возбудителем пищевого центра является изменение состава крови, которая в перерывах между приемами пищи, по выражению И. П. Павлова, постепенно становится «голодной».
Вторая фаза секреторной деятельности желудка начинается после поступления пищи в желудок, т. е. несколько позднее сложнорефлекторной. Продолжается она до тех пор, пока все пищевые массы не будут эвакуированы из желудка в кишечник (6—8 час. и более). Вторая фаза обусловлена действием механических и химических (гуморальных) раздражителей. Механические раздражители действуют на механорецепторы слизистой оболочки желудка и рефлекторно вызывают секрецию. Химические воздействия осуществляются через кровь, которая омывает железы желудка. При переваривании пищи в желудке образуются вещества (продукты расщепления белка, экстрактивные вещества, содержащиеся в отварах мяса и овощей), которые всасываются в кровь и стимулируют деятельность желудка.
Большую роль в усилении желудочной секреции играет гормон гастрин, выделяемый слизистой оболочкой привратника. При удалении этой части желудка секреция его резко уменьшается. Введение же гастрина стимулирует деятельность желудка, поджелудочной железы и выделение желчи. Кроме гастрина на желудочную секрецию влияет гистамин, образующийся в слизистой оболочке желудка и поступающий в кровь. Он действует главным образом на обкладочные клетки желудочных желез и стимулирует образование соляной кислоты.
Третья фаза выделения желудочного сока обусловлена химическими воздействиями на железы желудка через кровь. Различные вещества, образующиеся при переваривании пищи в кишечнике (продукты расщепления белков, экстрактивные вещества), всасываются в кровь и усиливают желудочную секрецию. Кроме того, деятельность желудочных желез стимулируется гормоном энтерогастрином, выделяемым слизистой оболочкой двенадцатиперстной кишки.
Производя операции на пищеводе и желудке, И. П. Павлов раскрыл сложный механизм работы желудочных желез у животных. Таким же образом осуществляется желудочная секреция у человека. Например’, после операции с перерезкой пищевода и наложени
ем фистулы желудка у человека можно наблюдать обильное выделение сока по механизму условных и безусловных рефлексов (мнимое кормление). Относительно меньше выделяется сока при изолированном раздражении механорецепторов желудка. Наиболее интенсивна желудочная секреция при мнимом кормлении, сочетающемся с механическим раздражением слизистой оболочки желудка (рис. 90).
Влияние состава пищи на желудочную секрецию. Деятельность желудка зависит от состава и количества поступающей в организм пищи. Например, экстрактивные вещества, содержащиеся в мясе, являются сильным раздражителем желудочных желез. При перева
ривании мяса желудочная секреция нарастает до максимума уже в первые часы пищеварения. Продукты переваривания мяса, всасываясь в кровь, стимулируют деятельность желез желудка в течение многих часов. При углеводной пище максимальное отделение желудочного сока большой переваривающей силы происходит в сложнорефлекторной фазе. Затем секреция уменьшается и продолжается на сниженном уровне 8—9 часов.
Помимо пищевых веществ, стимулирующих деятельность желудочных желез, есть и та-
мл
60 120 0 60 120 0 60 120 мин.
Рис. 90. Секреция желудочных желез (мл) у человека с фистулой желудка после операции с перерезкой пищевода (по Т. И. Курци.чу).
Выделение сока при мнимом кормлении (а), при механическом раздражении слизистой оболочки желудка (б), при мнимом кормлении и механическом раздражении слизистой оболочки желудка (в)
кие, которые оказывают на нее тормозящее воздействие (жир, концентрированные растворы NaCl и др.)- Для лучшего переваривания жир должен содержаться в пище в смеси с веществами, вызывающими обильную желудочную секрецию (овощи, овощные отвары, мясной бульон и т. п.).
Движения желудка. В стенке желудка находятся гладкие мышечные. волокна, обеспечивающие его двигательную деятельность. Последняя способствует перемешиванию пищевых масс и продвижению их из желудка в кишечник. Движения желудка стимулируются нервными импульсами, поступающими по блуждающим нервам. Их перерезка ведет к временному прекращению двигательной активности желудка. Через некоторое время движения восстанавливаются, что обусловлено автоматией гладких мышц желудочных стенок. При возбуждении симпатических (чревных) нервов движения желудка в большинстве случаев тормозятся.
Двигательная активность желудка усиливается при механическом и химическом раздражениях его слизистой оболочки.
Регуляция движений желудка может осуществляться и гуморальным путем, через кровь. Усиливают движения желудка гас-
ipun, гистамин и холин, тормозят — адреналин и норадреналин.
28F
Перемещение пищевых масс из желудка в двенадцатиперстную кишку. Переваривание пищи в желудке обычно продолжается в течение 6—8 часов. Длительность этого процесса зависим от состава пищи, ее объема и консистенции, а также от количества выделившегося желудочного сока. Особенно долго задерживается в желудке жирная пища (до 8—10 час. и более). /
Эвакуация пищевых масс из желудка происходит периодически, отдельными порциями. Это способствует их лучшему перевариванию в кишечнике, ферментативная активность соков которого снижается в присутствии соляной кислоты. Периодическое перемещение пищевых масс из желудка в двенадцатиперстную кишку обеспечивается периодическими сокращениями мускулатуры всего желудка, и в особенности сильными сокращениями мышечного жома (сфинктера) привратника. Мышцы привратника рефлекторно сокращаются при действии соляной кислоты на рецепторы слизистой оболочки двенадцатиперстной кишки. Привратник остается закрытым до тех пор, пока не нейтрализуются кислые пищевые массы.
Рефлекторные сокращения и расслабления мышц привратника могут быть вызваны не только соляной кислотой, но и другими веществами, находящимися в желудке и кишечнике. Например, жир, поступая в двенадцатиперстную кишку, способствует сокращению этих мышц. Кроме этих местных рефлекторных воздействий на эвакуацию пищевых масс влияют центрально-нервные и гуморальные факторы. Прекращение импульсов, поступающих к желудку по блу-. ждающим нервам, задерживает переход пищевых масс в кишечник. Такой же эффект наблюдается при введении в кровь некоторых веществ, образующихся в слизистой оболочке двенадцатиперстной кишки.
§ 4. Пищеварение в кишечнике
Пищеварение в двенадцатиперстной кишке. Пищевые массы подвергаются в двенадцатиперстной кишке воздействию сока кишечных желез, сока поджелудочной железы и желчи. Длина двенадцатиперстной кишки невелика, пищевые массы в ней не задерживаются, и изливающиеся здесь соки расщепляют питательные вещества в нижележащих отделах кишечника. В слизистой оболочке двенадцатиперстной кишки находятся бруннеровы и либеркюновы железы, сок которых содержит ферменты, расщепляющие белки, жиры и углеводы. В кишечном соке находится также фермент энтерокиназа — активатор трипсиногена поджелудочного сока.
Поджелудочная железа и ее роль в пищеварительном процессе. Поджелудочная железа состоит из двух родов клеток. Одни из них сгруппированы в виде островков и вырабатывают гормон инсулин. Другие выделяют пищеварительный поджелудочный сок, который выводится из железы через проток в полость двенадцатиперстной кишки. Поджелудочный сок имеет щелочную реакцию, pH его равен 7,8—8,4. Он содержит ферменты, расщепляющие белки, а также высокомолекулярные полипептиды, углеводы и жиры. Белковый фер
28S
мент трипсин выделяется железой в недеятельном состоянии. Он активизируется энтерокиназой кишечного сока. Действие фермента липазы, расщепляющей жиры, усиливается желчью.
Изучение деятельности поджелудочной железы способствовала операция, разработанная И. П. Павловым. При этой операции кусочек слизистой оболочки двенадцатиперстной кишки вместе с протоком железы выводят у животного наружу и вживляют в кожу. Пищеварение в этом случае не нарушается, так как поджелудочный сок поступает в кишечник по второму протоку.
Секреция поджелудочного сока происходит под влиянием нервных и гуморальных факторов. Она возникает при действии условных и безусловных раздражителей. Условнорефлекторное выделение поджелудочного сока начинается при виде и запахе пищи, а у человека даже при разговоре о ней. При акте еды происходит механическое раздражение рецепторов ротовой полости и глотки. Сигналы отсюда, поступая в продолговатый мозг, вызывают выделение поджелудочного сока по механизму безусловных рефлексов. Секреторными нервами поджелудочной железы служат волокна блуждающего нерва.
Химическими возбудителями поджелудочной железы являются гормоны, вырабатываемые слизистой оболочкой двенадцатиперстной кишки. Главный из них — секретин. Он выделяется в неактивной форме, активируется соляной кислотой и, поступая в кровь, стимулирует секрецию поджелудочной железы.
Секреция поджелудочного сока начинается через 2—3 мин. после приема пищи и продолжается 6—14 часов. Количество выделяемого сока и его ферментный состав зависят от количества и состава поступающей пищи. При употреблении хлеба наибольшая секреция поджелудочной железы наблюдается на первом часу пищеварения, при употреблении мяса — на втором, молока — на третьем. Жирная пища вызывает относительно небольшое сокоотделение.
Роль печени в пищеварении. Клетки печени непрерывно выделяют желчь, которая является одним из важнейших пищеварительных соков. В перерывах между приемами пищи желчь накапливается в желчном пузыре. Здесь происходит обратное всасывание ее жидкой части. Поэтому желчь пузыря гуще по консистенции и темнее по окраске, чем желчь, выделяемая непосредственно из печени.
В состав желчи входят желчные кислоты, желчные пигменты и другие органические и неорганические вещества. Желчные кислоты имеют большое значение в процессе переваривания жира. Желчный пигмент билирубин образуется из гемоглобина, который освобождается при разрушении в печени эритроцитов. Темный цвет желчи обусловлен наличием в ней этого пигмента.
Желчь активирует ферменты поджелудочного и кишечного соков, в особенности липазу. Значение желчи для переваривания жира очень велико. Она эмульгирует жиры и повышает растворимость жирных кислот, что облегчает их всасывание. Усиливая щелочную реакцию в кишечнике, желчь препятствует разрушению трипсина пепсином. Кроме того, она стимулирует движения кишок и, обладая
289
бактерицидными свойствами, задерживает гнилостные пррДессы в кишечнике. В сутки у человека образуется около 500—700Мл желчи.
Усиление желчеобразования при пищеварении и выделение желчи из пузыря в кишку происходят под влиянием нервных и гуморальных воздействий. Вид и запах пищи, акт еды, раздражение пищевыми массами рецепторов желудка и двенадцатиперстной кишки усиливают желчеобразование и вызывают выход желчи в кишку по механизму условных и безусловных рефлексов. Секреторным нервом печени служит блуждающий нерв. Симпатический нерв вызы- , вает угнетение желчеобразования и прекращение эвакуации желчи из пузыря.
Химическими раздражителями печени являются пищеварительные гормоны (секретин и др.), продукты переваривания белка и экстрактивные вещества, всасывающиеся в кровь. Сильным желчегонным свойством обладает сама желчь. Всасываясь в кровь, она стимулирует деятельность печеночных клеток.
Выделение желчи в двенадцатиперстную кишку обусловлено согласованной деятельностью мышц стенок желчного пузыря и его сфинктера. Сокращение мышц пузыря и расслабление сфинктера обеспечивают выход желчи в кишку. Расслабление мышц пузыря и сокращение сфинктера, наоборот, способствуют накоплению желчи в пузыре. Усиление сократительной деятельности пузыря происходит также под влиянием холецистокинина, образующегося в слизис- | той оболочке двенадцатиперстной кишки под влиянием продуктов переваривания белков и жиров.
Желчевыделение после приема пищи продолжается несколько часов. Количество желчи и продолжительность ее выделения зави- I сят от объема и состава принятой пищи. Особенно много желчи вы- ' деляется при употреблении яичных желтков, молока, мяса и жиров. I Первые порции желчи, выделяемые при пищеварении, отличаются темным цветом. Это указывает на выход ее из желчного пузыря. । В дальнейшем желчь начинает поступать в кишку непосредственно < из печени. Окраска этих порций желчи менее темная.
Печень, образуя желчь, выполняет не только секреторную, но и 1 экскректорную (выделительную) функцию. К экскректам печени от- I носятся желчные пигменты, кальций, железо и некоторые другие ве- I шества. Попадая с желчью в кишечник, они выбрасываются из организма.
Пищеварение в тонких кишках. Переработанные в желудке и двенадцатиперстной кишке пищевые массы, называемые химусом, | перемещаются в тонкие кишки, где продолжается их переваривание. 1
Поверхность тонких кишок покрыта ворсинками, между которы- 1 мн открываются протоки трубчатых либеркюновых желез.
Эти железы выделяют кишечный сок—бесцветную жидкость I щелочной реакции, мутную от примеси слизи, спущенных эпителиальных клеток и кристаллов холестерина. В слизистой оболочке не-прерывно происходит смена поверхностного слоя эпителия. Образование новых эпителиальных клеток сочетается с отторжением отмирающих. Последние образуют слизистые комочки, поступающие в 290
просвет кишки и играющие значительную роль в предохранении ее внутренней оболочки от механических повреждений.
Сок тонких кишок содержит ферменты, расщепляющие углеводы, жиры и продукты переваривания белка — полипептиды. Последние расщепляются здесь до аминокислот. В кишечном соке содержатся также ферменты энтерокиназа, активирующая трипсиноген поджелудочного сока, и нуклеаза, действующая на нуклеиновые кислоты.
Для исследования деятельности кишок производят операцию, предложенную Тири—Велла. При этой операции два конца изолированной кишечной петли выводятся наружу и вживляются в кожу. Брыжейка кишки, а следовательно, и ее нервы и сосуды при этом сохраняются. Проходимость кишечника восстанавливается путем сшивания верхнего и нижнего концов отрезка перерезанной кишки.
Выделение кишечного сока происходит под влиянием механических и химических раздражений стенок кишечника. Из химических раздражителей большую роль играют продукты переваривания белка, пряности, молочный сахар и др. Секреция кишечных желез при механическом и химическом раздражениях слизистой оболочки осуществляется рефлекторно. Центры этих периферических рефлексов расположены в нервных сплетениях непосредственно в кишечной стенке.
Влияние центральной нервной системы на секрецию кишечных желез не установлено.
В тонких кишках расщепление питательных веществ наиболее интенсивно происходит непосредственно около их стенок. Эти процессы называются пристеночным пищеварением. Их высокая интенсивность обусловлена адсорбцией ферментов пористой поверхностью кишечной стенки и усилением их активности. Таким образом, питательные вещества наиболее эффективно расщепляются в том месте, где происходит их всасывание.
Пищеварение в толстых кишках. Переваривание пищи заканчивается в основном в тонком кишечнике. Железы слизистой оболочки толстых кишок под влиянием механических и химических раздражителей выделяют пищеварительный сок. Однако ферменты его малоактивны. В толстых кишках может перевариваться растительная клетчатка под воздействием находящихся здесь бактерий, вызывающих сбраживание углеводов и гниение белков. Содержимое растительных клеток при этом освобождается, подвергается ферментативному расщеплению и всасыванию.
Под влиянием бактерий в толстых кишках происходит расщепление невсосавшихся продуктов переваривания белка. При этом образуются вещества, ядовитые для организма. Они всасываются в кровь и по воротной вене транспортируются в печень, где происходит их обезвреживание.
Формирующийся в нижних отделах толстых кишок кал состоит из остатков непереваренной пищи, слизи, желчных пигментов и бактерий. Последние составляют около 30% его сухого веса. Опорож-
291
I
нение прямой кишки (дефекация) происходит рефлекторно. Центр этого рефлекса находится в спинном мозгу, деятельность которого регулируется вышележащими отделами центральной нервной системы.
Движения кишок. Большое значение для переваривания пищи имеют движения кишок, способствующие ее перемешиванию и продвижению вдоль кишечника. Различают маятникообразные и перистальтические движения кишок.
Маятникообразные движения происходят при попеременном ритмическом сокращении ее кольцевых и продольных мышц. При этом на коротком участке кишка то сокращается, то расслабляется, и ее содержимое передвигается то в одном, то в другом направлении. В результате неодновременности сокращений разных участков кишки происходит ритмическая сегментация ее содержимого — оно то разделяется, то вновь соединяется вместе. Маятникообразные движения кишки обеспечивают перемешивание кишечного содержимого с пищеварительными соками.
Перистальтические движения заключаются в том, что сокращение кольцевых мышц и вызываемое этим сужение кишки на одном участке сочетается с расслаблением мышц и расширением соседнего нижележащего участка. В результате содержимое кишки из суженного участка перемещается в нижний расширенный. В следующий момент мышцы того участка кишки, где они были расслаблены, сокращаются, а в соседнем нижележащем отрезке расслабляются. Перистальтические движения обеспечивают передвижение кишечного содержимого вдоль кишечника по направлению к прямой I кишке.
Гладкие мышцы стенки кишечника обладают автоматией. При механическом и химическом раздражении движения кишок усиливаются. Если пища не содержит грубых, плохо перевариваемых веществ, то движения кишок уменьшаются. К химическим раздражителям слизистой оболочки кишки относятся желудочный сок, слабые растворы кислот и щелочей, некоторые продукты переваривания пищи.
Регуляция движения кишок происходит гуморальным и нервным путями. Химическое воздействие на движение кишок оказывают) ацетилхолин и норадреналин. Ацетилхолин образуется при раздражении блуждающего нерва, он усиливает движение кишок. Норад-1 реналин, выделяющийся при раздражении симпатического нерва, оказывает тормозящее действие. Импульсы из центральной нервной системы поступают к кишечнику по блуждающим и симпатическим нервам. Первый из них усиливает движение кишок, второй — тормозит.
Движения толстых кишок очень медленные. Поэтому около половины времени, затрачиваемого на пищеварительный процесс (около 1—2 суток), идет на передвижение остатков пищи по толстым кишкам. Автоматия толстых кишок выражена слабее, чем тонких. Движения их возникают главным образом под влиянием местных механических раздражений.
292
§ 5. Всасывание продуктов переваривания пищи
Всасывание питательных веществ в кровь и лимфу происходит главным образом в тонких кишках. В желудке в небольших количествах всасываются лишь вода, минеральные соли и моносахариды. В толстых кишках обычно всасывается только вода.
Интенсивное всасывание в тонких кишках обеспечивается их большой поверхностью, во много раз превышающей поверхность тела. Поверхность кишечника увеличивается наличием ворсинок, являющихся выростами слизистой оболочки кишки. Внутри каждой ворсинки находятся гладкие мышечные волокна и хорошо развитая кровеносная и лимфатическая сеть.
Всасывание представляет собой сложный процесс, включающий диффузию, фильтрацию и осмос. Кроме того, эпителий кишки обладает способностью избирательно всасывать одни вещества и ограничивать всасывание других. Большое значение для всасывания имеют движения ворсинок, которые стимулируются химическими веществами, находящимися в кишечнике: белки, экстрактивные вещества, желчные кислоты, гормон вилликинин, вырабатываемый слизистой оболочкой двенадцатиперстной кишки, и др.
Белки всасываются в виде аминокислот и в небольшом количестве в виде полипептидов. Аминокислоты после всасывания поступают по воротной вене в печень, где происходит их дезаминирование и переаминирование.
Углеводы всасываются в кровь главным образом в виде глюкозы. При прохождении через стенку кишки она фосфорилируется, что ускоряет всасывание. Этот процесс стимулируется гормоном поджелудочной железы инсулином.
Жиры всасываются в виде жирных кислот и глицерина. Жирные кислоты нерастворимы в воде. Всасывание их, а также холестерина и других липоидов происходит лишь при наличии желчи. Только эмульгированные жиры могут частично всасываться без предварительного расщепления до глицерина и жирных кислот. Большая часть жира всасывается в лимфу. Затем через грудной проток он поступает в кровь. В кишечнике человека за сутки всасывается не более 150—160 г жира.
Вода преимущественно всасывается в тонких и толстых кишках. Ее всасывание происходит главным образом путем осмоса. Если осмотическое давление в кишечнике становится выше, чем в крови, то интенсивность всасывания снижается.
Растворенные в воде соли натрия, калия и кальция всасываются преимущественно в тонком кишечнике.
§ 6. Печень и ее функции
Функции печени многообразны и жизненно необходимы. Кроме образования желчи и участия в пищеварительных процессах, печень играет большую роль в обмене веществ. Продукты переваривания
293
пиши переносятся кровью в печень, и здесь происходит их дальнейшая переработка.
Ядовитые вещества, поступившие в кровь из кишечника, в печени обезвреживаются. Задерживаются здесь и чужеродные для организма белки. Эта важная функция печени называется барьерной. Печень является как бы барьером, задерживающим поступление в организм вредных для него веществ.
Барьерная функция печени изучена при помощи операции Н. В. Экка. У животных при этом производят искусственное соединение воротной и нижней полой вен. Воротная вена перевязывается выше этого соединения, вследствие чего ток крови, минуя печень, направляется в общий круг кровообращения. Оперированные таким образом животные при питании мясом быстро погибают в результате отравления организма продуктами расщепления белка, образующимися в кишечнике и поступающими в кровь. При питании растительной пищей жизнь сохраняется более длительное время.
Барьерная функция печени четко проявляется при введении животному продуктов переваривания пищи в периферические кровеносные сосуды, что сопровождается явлениями тяжелого отравления. Введение же этих веществ в воротную вену не вызывает отрицательных реакций.
Кроме барьерной функции, печень принимает большое участие в обмене органических и неорганических веществ. В ней происходит синтез некоторых белков (фибриногена, альбумина и др.). Здесь же из аммиака синтезируется мочевина. В печени депонируется гликоген. Он образуется здесь не только из углеводов, но также из жиров и аминокислот после дезаминирования последних. Расщепление гликогена в печени до глюкозы и поступление ее в кровь обеспечивают постоянную концентрацию сахара в крови.
Жиры и липоиды в небольших количествах тоже могут депонироваться в печени. Здесь же происходит синтез нейтральных жиров из углеводов и безазотистых компонентов аминокислот. В печени синтезируются и липоиды, например эфиры холестерина.
Большую роль играет печень в обмене витаминов, особенно группы А.
§ 7. Влияние мышечной работы на деятельность пищеварительных органов
Мышечная деятельность, повышая обмен веществ и энергии, увеличивает потребность организма в питательных веществах и тем самым стимулирует желудочную и кишечную секреции, что благоприятно влияет на пищеварительные процессы.
Однако положительное влияние физической работы на пищеварение наблюдается не всегда. Например, физическая работа, выполняемая сразу после приема пищи, не усиливает, а задерживает пищеварительные процессы. Сильнее всего при мышечной деятельности тормозится рефлекторное выделение пищеварительных соков.
294
В лаборатории Н. К. Верещагина установлено, что желудочная секреция у собак уменьшается не только при удержании тяжелого груза, но и при воздействии условных раздражителей, действие которых обычно совпадает с выполнением работы.
Угнетение пищеварительных функций при напряженной мышечной деятельности обусловлено торможением пищевых центров в результате отрицательной индукции с возбужденных двигательных центров. Тормозящее влияние мышечной деятельности на пищеварение усиливается в результате перераспределения крови. Кровоснабжение пищеварительных желез при этом уменьшается, что и ведет к уменьшению секреции.
В связи с угнетением пищеварительных процессов во время мышечной деятельности не рекомендуется сразу после еды приступать к физической работе. При занятиях спортом следует иметь в виду, что не только мышечная работа тормозит пищеварительные процессы, но и переваривание пищи отрицательно влияет на двигательную деятельность. Возбуждение пищевых центров и отток крови от мышц к органам брюшной полости снижают эффективность физической работы. Кроме того, наполненный желудок приподнимает купол диафрагмы, что неблагоприятно сказывается на деятельности органов дыхания и кровообращения. В связи с этим физические упражнения рекомендуется выполнять не ранее чем через 2—2,5 часа после приема пищи.
В том случае, когда человек вынужден сразу же после приема пищи выполнять значительные физические нагрузки, иногда можно для адаптации пищеварительных органов к деятельности в таких условиях принимать пищу и непосредственно перед работой. Это следует делать лишь на тренировочных занятиях, но не на соревнованиях, требующих оптимальных условий для работы скелетных мышц, сердца и органов дыхания.
Глава X
ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ
§ 1. Сущность обмена веществ и энергии
Отличительным признаком живых организмов являются энергетические траты и постоянный обмен веществ с окружающей их внешней средой. Его сущность состоит в том, что из внешней среды в организм поступают разнообразные, богатые потенциальной химической энергией вещества; в организме они расщепляются на более простые; освобождающаяся при этом энергия обеспечивает протекание физиологических процессов и выполнение внешней работы. Кроме того, поступающие в организм вещества используются для восстановления изнашиваемых и построения новых клеток и тка-н -й и для образования гормонов и ферментов. Некоторые органические вещества при избыточном поступлении могут депонировать
295
ся, т. е. откладываться в организме в виде запасов. Образующиеся в процессе обмена продукты распада удаляются из организма во внешнюю среду органами выделения.
Питательными веществами, снабжающими организм энергией и строительным (пластическим) материалом, являются белки, жиры и углеводы. Кроме того, для нормального протекания обмена веществ в организме необходимо поступление витаминов, воды и минеральных солей.
Обмен веществ в организме является сложной системой связанных друг с другом реакций расщепления (диссимиляции) и синтеза (ассимиляции) органических веществ. При реакциях диссимиляции происходит освобождение потенциальной химической энергии, которая обеспечивает деятельность всех органов и выполнение внешней работы. Реакции синтеза требуют для своего осуществления притока энергии извне. Затрачиваемая при этом энергия превращается в потенциальную химическую энергию сложных молекул.
Все химические реакции в организме, в том числе переваривание пищи, окислительно-восстановительные и другие процессы осуществляются при участии биологических катализаторов (фермен тов).
Обмен веществ и энергии в организме регулируется нервным f гуморальным путями. Регуляция, осуществляемая нервной системой, которой принадлежит особенно большая роль в этом процессе происходит путем непосредственной посылки эфферентных нерв ных импульсов к тканям и через железы внутренней секреции, гормоны которых влияют на уровень обмена веществ и энергии. В из менении обмена веществ, обеспечивающем приспособление орга низма к различным условиям жизни, важное значение имею', условные рефлексы. Индифферентные раздражения после повторно го сочетания с факторами, влияющими на обмен веществ и энергии могут условнорефлекторным путем изменять его в нужном направ лении.
§ 2. Обмен белков и его регуляция
Белки построены из аминокислот. Они являются основным пла стическим материалом, из которого построены клетки и ткани орга низма. Например, в составе скелетных мышц находится около 20^ белка. Наличием белка обусловлена сократительная функция мы шечной ткани. Из белков состоят многие ферменты и гормоны. Бе лок крови гемоглобин участвует в транспорте фибриноген необходим для свертывания крови, белки — нуклеопротеиды имеют значение для венных свойств.
Белки могут использоваться организмом и гии. При расщеплении белка из безазотистой образуются углеводы, дальнейшее превращение которых обеспечи
кислорода. Белок Некоторые сложны» передачи наследст
как источники энер части его молекулы
296
вает освобождение энергии. Окисление 1 г белка освобождает 5,3 ккал. Но азотистые продукты расщепления белка (мочевина, аммиак и др.) не подвергаются в организме дальнейшему окислению. Поэтому при окислении 1 г белка в организме освобождается столько же энергии, сколько и при окислении 1 г углеводов, т. е. 4,1 ккал.
Поступая в организм с пищей, белки расщепляются в кишечнике до аминокислот и в таком виде всасываются в кровь и транспортируются в печень. Часть поступивших в печень аминокислот подвергается дезаминированию и переаминированию. Эти процессы обеспечивают синтез некоторых аминокислот и белков. Из печени аминокислоты поступают в ткани и используются для синтеза белков. Избыточные белковые вещества, поступающие с пищей, превращаются в организме в углеводы и жиры.
Конечными продуктами расщепления белков в тканях являются аммиак, мочевина, мочевая кислота, креатинин и некоторые другие. Они выводятся из организма почками и частично потовыми железами.
О белковом обмене можно судить по азоту, так как он почти целиком поступает в организм с белковой пищей. Поэтому для характеристики белкового обмена в организме пользуются определением азотистого баланса, т. е. разницы между количеством азота, поступившего в организм, и количеством азота, выведенного из организма. У лиц, выполняющих среднюю по тяжести физическую работу, при потреблении 100—ПО г белка в сутки наблюдается азотистое равновесие, т. е. выведение азота из организма равно его усвоению. Состояние, при котором усвоение азота превышает его выведение, называется положительным азотистым балансом. Оно характерно для растущего организма (за счет белков пищи происходит рост и увеличение массы тела). Положительный азотистый баланс может быть и у взрослых после голодания и заболеваний, вызвавших усиленный распад тканевых белков без достаточного их восстановления, а также у спортсменов, особенно в подготовительном периоде тренировки, одной из задач которого является развитие скелетной мускулатуры. При полном и частичном (белковом) голодании и при некоторых заболеваниях усваивается меньше азота, чем выделяется. Такое состояние называется отрицательным азотистым балансом.
Нормальная жизнедеятельность организма возможна лишь при азотистом равновесии, или положительном азотистом балансе.
При поступлении в организм избыточного пищевого белка у взрослого человека азотистое равновесие не нарушается. Чем больше при этом содержится азота в пище, тем больше его выводится с мочой. В виде запасов белок в организме не откладывается. При голодании белки одних органов могут использоваться для поддержания жизнедеятельности других, более важных. При этом тратятся в первую очередь белки печени и скелетных мышц, и вес этих органов снижается. Вес же мозга и сердца и содержание в них белков остаются почти без изменения.
297
Обмен белков в организме регулируется нервными центрами, расположенными в подбугровой области промежуточного мозга. При экспериментальном повреждении у животных некоторых ядер этого отдела мозга усиливается белковый обмен, его баланс ста- ’ новится отрицательным, вследствие чего наступает резкое истощение. Нервная система влияет на белковый обмен через гормоны щитовидной железы, передней доли гипофиза (соматотропный гормон) и других желез внутренней секреции.
§ 3. Обмен углеводов и его регуляция
Углеводы поступают в организм с растительной и в меньшем количестве с животной пищей. Кроме того, они синтезируются в нем из продуктов расщепления аминокислот и жиров.
Углеводы — важная составная часть живого организма, хотя количество их в организме значительно меньше, чем белков и жиров, — всего около 2% сухого вещества тела.
Углеводы служат в организме основным источником энергии. При окислении 1 г углеводов освобождается 4,1 ккал энергии. Для окисления углеводов требуется значительно меньше кислорода, чем для окисления жиров. Это особенно повышает роль углеводов при мышечной деятельности. Значение их как источника энергии под- i тверждается тем, что при уменьшении концентрации глюкозы в крови резко снижается физическая работоспособность. Большое значение углеводы имеют для нормальной деятельности нервной i системы.
Пища содержит главным образом сложные углеводы, которые расщепляются в кишечнике и всасываются в кровь, преимущественно в виде глюкозы. В небольших количествах глюкоза содержится во всех тканях. Концентрация ее в крови колеблется от 0,08 до 0,12%. Поступая в печень и мышцы, глюкоза используется там для окислительных процессов, а также превращается в гликоген и от-, кладывается в виде запасов.
При голодании запасы гликогена в печени и концентрация глюкозы в крови уменьшаются. То же самое происходит при длитель-1 ной и напряженной физической работе без дополнительного прие-Ч ма углеводов. Уменьшение концентрации глюкозы в крови ниже 0,07% называется гипогликемией, а увеличение выше 0,12% —гипергликемией.
При гипогликемии появляется мышечная слабость, чувство голода, падает температура тела. Нарушение деятельности нервной системы проявляется при этом в возникновении судорог, помрачении и потере сознания.
Гипергликемия может возникать после приема пищи, богатой легкоусваиваемыми углеводами, при эмоциональном возбуждении, а также при заболеваниях поджелудочной железы или при ее удалении у животных с экспериментальной целью. Избыток глюкозы выводится из крови почками (гликозурия). У здорового человека это можно наблюдать после приема натощак 150—200 г сахара. I
298
В печени содержится около 10% гликогена, в скелетных мышцах—не более 2%. Общие запасы его в организме составляют в среднем 350 г. При уменьшении концентрации глюкозы в крови происходит интенсивное расщепление гликогена печени и выход глюкозы в кровь. Благодаря этому поддерживается постоянный уровень глюкозы в крови и удовлетворяется потребность в ней других органов.
В организме происходит постоянный обмен глюкозой между печенью, кровью, мышцами, мозгом и другими органами. Главный потребитель глюкозы — скелетные мышцы. Расщепление в них углеводов осуществляется по типу анаэробных и аэробных реакций. Одним из продуктов расщепления углеводов является молочная кислота.
Запасы углеводов особенно интенсивно используются при физической работе. Однако полностью они никогда не исчерпываются. При уменьшении запасов гликогена в печени его дальнейшее расщепление прекращается, что ведет к снижению концентрации глюкозы в крови до 0,05—0,06%, а в некоторых случаях до 0,04— 0,038%. В последнем случае мышечная деятельность продолжаться не может. Таким образом, уменьшение содержания глюкозы в крови—один из факторов, снижающих работоспособность организма при длительной и напряженной мышечной деятельности. При такой работе необходимо пополнять углеводные запасы в организме, что достигается увеличением углеводов в пищевом рационе, дополнительным введением их перед началом работы и непосредственно при ее выполнении. Насыщение организма углеводами способствует сохранению постоянной концентрации глюкозы в крови, что необходимо для поддержания высокой работоспособности человека.
Влияние приема углеводов на работоспособность установлено лабораторными экспериментами и наблюдениями при спортивной деятельности. Эффект от принимаемых до работы углеводов при прочих равных условиях зависит от их количества и времени приема.
Углеводный обмен в организме регулируется нервной системой. Это было установлено Клодом Бернаром, который после укола иглой в дно IV желудочка мозга («сахарный укол») наблюдал усиленный выход углеводов из печени с последующими гипергликемией и гликозурией. Эти наблюдения свидетельствуют о наличии в продолговатом мозгу центров, регулирующих углеводный обмен. Позднее было установлено, что высшие центры, регулирующие обмен углеводов, находятся в подбугровой области промежуточного мозга. При раздражении этих центров наблюдаются такие же явления, как и при уколе в дно IV желудочка. Большое значение в регуляции углеводного обмена имеют условнорефлекторные раздражители. Одним из доказательств этого служит увеличение концентрации глюкозы в крови при возникновении эмоций (например, у спортсменов перед ответственными стартами).
Влияние центральной нервной системы на углеводный обмен осуществляется главным образом посредством симпатической ин
299
нервации. Раздражение симпатических нервов усиливает образование адреналина в надпочечниках. Он вызывает расщепление гликогена в печени и скелетных мышцах и повышение в связи с этим концентрации глюкозы в крови. Гормон поджелудочной железы глюкагон также стимулирует эти процессы. Гормон поджелудочной железы инсулин является антагонистом адреналина и глюкогена. Он непосредственно влияет на углеводный обмен печеночных клеток, активирует синтез гликогена и тем самым способствует его депонированию. В регуляции углеводного обмена участвуют гормоны надпочечников, щитовидной железы и гипофиза.
§ 4. Обмен липидов и его регуляция
К липидам относятся нейтральные жиры, фосфатиды и стерины. Нейтральные жиры расщепляются в кишечнике до глицерина и жирных кислот. Эти вещества, проходя через стенку кишки, вновь превращаются в жир, который всасывается в лимфу и в небольшом количестве в кровь. Кровь транспортирует жиры в ткани, где они используются для пластического синтеза и в качестве энергетического материала. Липиды входят в состав клеточных структур. Они необходимы для новообразования клеток.
Жиры могут откладываться в организме в виде запасов, которые в первую очередь используются при голодании организма. Общее количество жира у человека составляет в среднем 10—12% веса тела, при ожирении оно может достигать 40—50%.
Жировые депо в организме непрерывно обновляются. В условиях нормальной жизнедеятельности выход жира из депо сочетается с его депонированием. При полном энергетическом равновесии интенсивность этих процессов в организме одинакова. Если процессы депонирования превышают использование жира, он накапливается в организме; если они недостаточны, жировые запасы уменьшаются.
Жиры, поступающие в ткани из кишечника и из жировых депо, путем сложных превращений окисляются и используются как источник энергии. При окислении 1 г жира освобождается 9,3 ккал энергии. В связи с тем, что в молекуле жира содержится относительно мало кислорода, последнего требуется для окисления жира больше, чем для окисления углеводов. Как энергетический материал жир используется главным образом в состоянии покоя и при выполнении длительной малоинтенсивной физической работы. В начале более напряженной мышечной деятельности используются преимущественно углеводы. Но через некоторое время, в связи с уменьшением запасов гликогена, и при работе начинают окисляться жиры и продукты их расщепления. Процесс замещения углеводов жирами может быть настолько интенсивным, что 80% всей необходимой в этих условиях энергии освобождается в результате расщепления жира.
Жировая ткань, покрывающая различные органы, предохраняет их от механических воздействий. Скопление жира в брюшной поло
300
сти обеспечивает фиксацию внутренних органов. Подкожная жировая клетчатка, являясь плохим проводником тепла, защищает тело от излишних теплопотерь. Жир входит в состав секрета сальных желез, который предохраняет кожу от высыхания и излишнего смачивания при соприкосновении с водой.
Обмен липидов тесно связан с обменом белков и углеводов. Поступающие в организм в избытке углеводы и белки превращаются в жир. Наоборот, при голодании жиры, расщепляясь, служат источником углеводов.
Обмен липидов в организме регулируется центральной нервной системой. При повреждении некоторых ядер подбугровой области промежуточного мозга жировой обмен нарушается. При этом происходит либо ожирение организма, либо его истощение.
Нервная система может оказывать непосредственное воздействие на жировой обмен. Если у животного перерезать нервы, иннервирующие жировую ткань с одной стороны тела, то при голодании жировые запасы на этой стороне не уменьшаются.
Нервная система регулирует жировой обмен и путем воздействия на железы внутренней секреции. Недостаточная функция передней доли гипофиза, щитовидной железы и половых желез ведет к ожирению. Гормон поджелудочной железы инсулин стимулирует образование жиров из углеводов.
Увеличение жировых запасов в здоровом организме происходит главным образом в связи с потреблением пищи, превышающим потребность.
§ 5. Водно-солевой обмен
Вода в организме находится в виде солевых растворов. Это обусловливает тесную связь водного обмена с обменом минеральных веществ, находящихся в организме в виде солей и их ионов.
Тело взрослого человека на 50—60% состоит из воды. Вода составляет около 92% крови, 70% общего веса скелетных мышц, 84% серого вещества мозга, 22% костной ткани.
Входя в состав тканей, вода является одним из структурных компонентов тела. Она служит растворителем многих химических веществ в организме и активно участвует в процессах обмена. Если прекращается поступление воды в организм, то он погибает. Полное голодание, но при условии приема воды переносится человеком в течение 40—45 дней. Потеря веса тела при этом может достигать 40%. При лишении же воды потеря 10% веса уже ведет к тяжелым поражениям, а потеря 20—22% —к смерти.
Поступление воды в организм вызывает незначительное и кратковременное повышение ее содержания в крови. Она быстро переходит в ткани и частично депонируется в печени. Избыток ее выводится из организма почками.
Минеральные вещества входят в состав всех живых тканей. Нормальное функционирование тканей обеспечивается не только наличием в них тех или иных солей, но и строго определенными их
301
количественными соотношениями. Неорганические вещества поддерживают необходимое осмотическое давление в клетках и биологических жидкостях и наряду с белками обеспечивают постоянство pH тканей. При полном отсутствии минеральных солей в пище животные резко теряют в весе и погибают. При избыточном поступлении минеральных солей в организм они могут откладываться в виде запасов. Натрий и хлор депонируются в подкожной клетчатке, калий — в скелетных мышцах, кальций и фосфор — в костях.
Осмотическое давление внутренней среды организма поддерживается путем регуляции поступления воды и солей и их выделения. При повышении осмотического давления возникает чувство жажды. При поступлении воды в организм осмотическое давление снижается.
Выведение воды и солей из организма происходит благодаря нервным и гуморальным влияниям на почки и потовые железы. Нервные центры, регулирующие водно-солевой обмен, расположены в подбугровой области промежуточного мозга. Здесь же находятся осморецепторные клетки, воспринимающие изменения осмотического давления в крови. Воздействие осморецепторов * рефлекторно влияет на водно-солевой обмен, обеспечивая постоянство осмотического давления во внутренней среде организма. Большую роль в регуляции водно-солевого обмена играют антидиуретический гормон гипофиза и гормоны коры надпочечников минералокортикоиды. Гормон гипофиза стимулирует обратное всасывание воды в почке и уменьшает этим мочеобразование. Минералокортикоиды (альдостерон и др.) действуют на эпителий почечных канальцев и повышают обратное всасывание в кровь натрия. На деятельность почек влияют также гормоны щитовидной и паращитовидной желез. Первый увеличивает мочеобразование, второй способствует выведению из организма солей кальция и фосфора.
§ 6. Витамины
Витамины необходимы для нормальной жизнедеятельности организма. Большинство из них человек получает с пищей. Некоторые витамины синтезируются бактериями в кишечнике. Поэтому отсут- ' •ствие их в пище не влияет на состояние организма. Витамины обладают высокой биологической активностью. Находясь в ничтожно малых концентрациях, они влияют на обмен веществ в организме. •Одни витамины входят в состав ферментов, обеспечивающих протекание различных биологических реакций, другие имеют тесные связи с деятельностью желез внутренней секреции. Витамины обеспечивают работоспособность организма и повышают его сопротивляемость различным заболеваниям.
Потребность в витаминах повышается при изменениях барометрического давления и температуры окружающей среды, при на-
* Осморецепторы — рецепторы, возбуждающиеся при изменениях осмотического давления. Они находятся в промежуточном мозгу и в сосудах внутренних органов.
•302
пряженной мышечной деятельности и при некоторых заболеваниях. У молодых, растущих организмов потребность в витаминах больше, чем у взрослых.
Отсутствие отдельных витаминов в пище вызывает нарушения функций организма, называемые авитаминозами. Недостаточное поступление витаминов сопровождается менее резкими изменениями — гиповитаминозами.
§ 7. Обмен энергии
В процессе жизнедеятельности организма его энергетические запасы непрерывно уменьшаются. Энергия расходуется на осуществление различных физиологических функций, на внешнюю работу, поддержание температуры тела и т. п. Продолжение жизни возможно лишь при постоянном пополнении запасов энергии, что и происходит благодаря приему пищи.
Соотношение количества энергии, поступающей с пищей, и энергии, расходуемой организмом, называется энергетическим балансом. При избыточном питании происходит накопление энергетических запасов. В условиях недостаточного питания они уменьшаются.
Методы определения расхода энергии. Интенсивность энергетического обмена в организме определяется при помощи калориметрии. Определение энергетического обмена может производиться тремя методами: 1) измерение тепла, выделяемого организмом,— прямая калориметрия; 2) измерение объема поглощаемого организмом кислорода и выделяемой углекислоты — непрямая респираторная калориметрия; 3) определение калорийности всасываемых питательных веществ, поступающих в организм с пищей, — непрямая алиментарная (от лат. alimentum — пища, питание) калориметрия.
Метод прямой калориметрии применяется в опытах на животных с конца XVIII в. Калориметр для определения энергетического обмена у человека был впервые сконструирован в конце XIX в. Он представляет собой герметически закрытую камеру с двойными стенками, обеспечивающими ее теплонепроницаемость. В камере расположены трубки, по которым циркулирует вода. Тепло, выделяемое человеком или животным, находящимся в калориметре, нагревает эту воду. Зная количество воды, протекающей через камеру, и степень ее нагревания, можно определить количество тепла, освобождаемого организмом.
Прямая калориметрия — наиболее точный метод исследования энергетических трат. Но он требует относительно длительных наблюдений и не позволяет проводить исследования при многих формах профессиональной и спортивной деятельности человека. Использование этого метода позволило сопоставить величину энергии, освобождаемой в виде тепла, с количеством поглощаемого кислорода и выделяемой углекислоты. Результаты этих сопоставлений дали возможность обосновать принципы непрямой респираторной калориметрии — метода, более удобного для оценки энерготрат у человека. Этот метод впервые был применен Лавуазье на животных в
303
конце XVIII в. В дальнейшем он был несколько изменен и в настоящее время широко используется при исследованиях энергозатрат у человека в условиях профессиональной и спортивной деятельности. В разработке метода респираторной калориметрии принимали участие И. М. Сеченов, М. Н. Шатерников, Д. С. Холден, Д. Г. Дуглас и др. Известно, что чем больше энергии расходует организм, тем интенсивнее протекают окислительные процессы в его тканях и тем относительно больше поглощается кислорода и образуется углекислоты. В связи с этим о величине энергетического расхода можно судить не только по количеству освобождаемого тепла, как это делают при прямой калориметрии, но и по количеству поглощаемого кислорода и образующейся углекислоты.
Один литр кислорода окисляет в организме определенные количества тех или иных веществ. При этом в зависимости от природы окисляемого вещества выделяется большее или меньшее количество энергии. При окислении 1 г углеводов требуется 0,8 л кислорода и освобождается 4,1 ккал, тепла. Следовательно, 1 л кислорода может полностью окислить 1,26 г углеводов, при этом освободится 5,05 ккал, энергии. При окислении жира эти соотношения будут иными в связи с малым содержанием кислорода в его молекуле. Один литр кислорода может окислить всего 0,5 г жира, при этом освободится 4,7 ккал, тепла.
Количество энергии, освобожденное при использовании 1 л кислорода, называется его калорическим эквивалентом. При окислении углеводов калорический эквивалент, как это указано выше, равен 5,05 ккал., при окислении жира — 4,7 ккал., белков — 4,85 ккал. В организме обычно окисляется смесь питательных веществ. Поэтому калорический эквивалент кислорода колеблется от 4,7 до 5,05 ккал. С увеличением в окисляемой смеси углеводов калорический эквивалент кислорода повышается, а с увеличением жира снижается.
Дыхательный коэффициент. О величине калорического эквивалента кислорода узнают по величине дыхательного коэффициента— отношению объема выдыхаемой углекислоты к объему поглощаемого кислорода ("от)’ Величина Дыхательного коэффициента зависит от состава окисляемых веществ. При окислении углеводов объем поглощаемого кислорода равен объему выделяемой углекислоты. Следовательно, дыхательный коэффициент при этом равен 1. При окислении жира, в молекуле которого мало кислорода, дыхательный коэффициент очень низок (0,7). При окислении смеси питательных веществ величина его колеблется от 0,7 до 1. Чем больше в этой смеси содержится углеводов, тем он выше. Рассчитав величину дыхательного коэффициента, по специальной таблице определяют калорический эквивалент кислорода (табл. 10).
При расчете энергетического расхода путем непрямой респираторной калориметрии необходимо узнать количество поглощаемого кислорода и выделенной углекислоты. Для этого измеряется объем легочной вентиляции и определяется состав выдыхаемого воздуха.
304
Таблица 10
Калорический эквивалент кислорода при разной величине дыхательного коэффициента
Показатели Величины
Дыхательный коэффициент Калорический эквивалент 0,70 4,70 0,75 4,74 0,80 4,80 0,90 4,92 0,95 4,98 1,00 5,05
Выдыхаемый воздух собирается в прорезиненный мешок, для чего используется специальная газовая маска. Объем его измеряется при помощи газовых часов, а химический анализ производится в специальном приборе. Установив величину легочной вентиляции и состав выдыхаемого воздуха, вычисляют объемы поглощенного кислорода и выделенной углекислоты. Затем определяют дыхательный коэффициент, находят в таблице (см. табл. 10) калорический эквивалент кислорода и вычисляют энерготраты в килокалориях. Поясним это на примере. Предположим, что при выполнении физической работы легочная вентиляция была равна 70 л в 1 мин. В выдыхаемом воздухе содержалось 16,31% кислорода и 4,35% углекислоты. Следовательно, организм потреблял 4,59% кислорода (20,9— —16,31=4,59) и выделял 4,32% углекислоты (4,35—0,03 = 4,32). При переводе в миллилитры эти величины соответственно составляют 3200 и 3020. Дыхательный коэффициент при этом равен 0,95 (3020:3200). В табл. 10 находим калорический эквивалент — он равен 4,98. Значит, в данном случае за 1 мин. работы энерготраты составляют около 16 ккал. Зная продолжительность работы, нетрудно определить суммарный расход энергии, необходимый для ее выполнения.
Непрямая респираторная калориметрия широко применяется в физиологии труда и спорта. Она позволяет определять расход энергии при мышечной деятельности в самых разнообразных условиях. С помощью этого метода определены энергетические затраты при беге на лыжах, при плавании, восхождениях на горы и при других видах физических упражнений. Однако этот метод не лишен недостатков. Необходимость использования газовых масок и мешков для забора выдыхаемого воздуха несколько затрудняет дыхание и делает метод газообмена неприемлемым для определения расхода энергии у спортсменов в условиях соревнований. В настоящее время для исследования газообмена сконструированы точные электрофизические приборы. Дальнейшее их усовершенствование и приспособление к проведению исследований при напряженной мышечной деятельности позволят более широко изучать энергозатраты при занятиях спортом.
При непрямой алиментарной калориметрии учитывают калорийность принимаемой пищи и ведут наблюдения за весом тела. Постоянство веса свидетельствует о балансе между поступлением энергетических ресурсов в организм и их использованием. В таких
11—113
305
тивности
Рис. 91. Изменение основного обмена с возрастом (по А. Флейшу)
условиях количество поступившей пищи может быть показателем энергозатрат. Однако в организме поступление питательных веществ и расход энергии не всегда сбалансированы, что ведет к ошибкам при использовании метода алиментарной калориметрии. Неудобство этого метода обусловлено еще и тем, что он не позволяет определять энерготраты за короткие отрезки времени.
Уровни энергетических трат в организме. В зависимости от ак-;го внешней среды различают три уровня энергетического обмена 1 — основной обмен, 2 — обмен в состоянии относительного покоя, 3 — энерготраты при физической работе.
Основным обменом называют величину энерготрат в состоянии полного мышечного покоя, натощак (спустя 12 час. после приема пищи), при температуре окружающей среды 20—22°.
У взрослого человека весом 70 кг основной об-
мен в сутки оставляет около 1700 ккал. Из этой величины 25% идет на работ)/ вегетативных систем (сердце, почки и др.), 75% —на деятельность всех остальных клеток и тканей организма. У здоровых людей основной обмен может колебаться в пределах ±15%. У женщин он на 5% ниже, чем у мужчин. Основной обмен зависит главным образом от величины поверхности тела и возраста. Чем относительно больше поверхность тела, тем больше основной обмен. Поэтому его величину принято выражать в ккал на 1 м2 поверхности тела в 1 час.
С возрастом (рис. 91) в связи с понижением интенсивности внутриклеточных окислительных процессов основной обмен понижается. Дети до 5 лет расходуют в состоянии основного обмена 50—55 ккал, на 1 м2 в 1 час, подростки — 42 ккал., взрослые — около 37 ккал, лица пожилого возраста — около 34 ккал.
На величину основного обмена влияют уровень двигательной активности организма и особенности питания. Например, при одинаковой поверхности тела обмен веществ у диких животных выше, чем у домашних. Если животное помещено в маленькую клетку, ограничивающую его движения, основной обмен понижается. Длительное голодание организма также ведет к снижению основного обмена.
Таким образом, основной обмен поддерживается на одном уровне только при постоянных условиях жизнедеятельности. При изменении этих условий он становится иным.
£06
Основной обмен в организме регулируется нервной системой и гормонами. Роль нервной системы подтверждена опытами с введением животному кураре. Этот яд парализует окончания двигательных нервов в мышцах. В результате в них уменьшается интенсивность окислительных процессов, что ведет к снижению основного обмена.
На основной обмен влияют гормоны щитовидной железы и гипофиза. При усилении функции щитовидной железы величина его повышается, при ослаблении понижается.
Спортивная тренировка, экономизируя окислительные процессы в организме, в большинстве случаев ведет к снижению основного обмена. Наиболее ярко это проявляется у спортсменов-стайеров.
Энерготраты в состоянии относительного покоя превышают величину основного обмена. Это обусловлено влиянием на обмен пищеварительных процессов, терморегуляцией и энерготратами на поддержание позы тела.
Увеличение расхода энергии после приема пищи обусловлено работой гладких мышц пищеварительных органов и деятельностью пищеварительных желез. Процессы всасывания питательных веществ и их усвоение также требуют дополнительных энергетических трат. Повышение обмена энергии под влиянием приема пищи зависит от ее состава и количества. Наиболее резкие сдвиги вызывает переваривание белковой пищи. Расход энергии при этом возрастает на 20—30%. Переваривание жира сопровождается увеличением энерготрат на 7%, углеводов — на 4—5%. Повышение энергетического обмена под влиянием приема пищи может продолжаться 5—6 час. и более.
Поддержание постоянства температуры тела (вне зоны комфорта) требует значительных энерготрат. Например, при низкой температуре окружающей среды окислительные процессы могут в 3— 4 раза превышать уровень основного обмена.
Поддержание позы тела обусловлено повышением тонуса определенных мышечных групп. Это ведет к увеличению энерготрат. Они особенно нарастают при необходимости сохранять неудобную или непривычную для человека позу.
Энерготраты в состоянии относительного покоя могут быть резко увеличенными в связи с восстановительными процессами после выполнения работы. Это обусловлено химическими реакциями в мышцах, главным образом окислением молочной кислоты.
Уровень энергетического обмена в состоянии относительного покоя может изменяться под влиянием условнорефлекторных раздражителей. Любой из них после нескольких счетаний с безусловнорефлекторными изменениями обмена веществ может стать сигналом, вызывающим такие же сдвиги обмена. Например, производственный шум вызывает увеличение расхода энергии даже у рабочих, находящихся в другом помещении и не занятых работой. Расход энергии еще больше увеличивается в помещении цеха при наблюдении за работой своей бригады. У спортсменов расход энергии повышается в предстартовом периоде. Такое условнорефлекторное no
li*
307
вышение обмена веществ имеет большое значение для подготовки к предстоящей деятельности.
Энерготраты при физической работе резко увеличиваются. Например, при ходьбе расходуется энергии на 80—100% больше по сравнению с покоем, при беге — на 400% и более.
Суточный расход энергии человека включает величину основного обмена и энергию, необходимую для выполнения профессионального труда, спортивной и других видов мышечной деятельности. Умственный труд требует небольших энергетических затрат.
По характеру выполняемой производственной деятельности и величине энерготрат взрослое население может быть разделено на 4 группы. К первой группе относятся лица, занимающиеся умственным трудом, не требующим мышечных напряжений. Суточный расход у них составляет в среднем 2200—3000 ккал. У лиц, выполняющих механизированную работу, расход энергии повышен до 2350— 3200 ккал. При частично механизированном труде суточный расход энергии достигает 2500—3400 ккал. Очень тяжелый, немеханизированный физический труд вызывает расход энергии равный 2900— 3990 ккал. В отдельных случаях при выполнении длительной и тяжелой работы суточный расход достигает еще больших величин.
Спортивная деятельность сопровождается значительным увеличением суточного расхода энергии — до 4500—5000 ккал. В дни тренировок с повышенными нагрузками и в дни соревнований в некоторых видах спорта (лыжные гонки, бег на длинные дистанции и др.) эти величины могут быть еще больше.
Энерготраты при работе, которые рассчитывают на единицу времени или на единицу пути, прямо пропорциональны ее мощности. Суммарный же расход энергии зависит не только от мощности работы, но и от ее длительности. Например, при легкоатлетическом беге энерготраты, рассчитанные на 1 м пути, будут наибольшими при беге на 100 м, суммарный же расход энергии — наибольшим при марафонском беге.
На уровень энерготрат влияют температура и влажность воздуха, барометрическое давление, сила ветра, особенности грунта при беге, условия скольжения при беге па коньках и лыжах. Например, при одинаковой скорости бега на лыжах энерготраты при плохом скольжении будут значительно больше, чем при хорошем. На расход энергии влияют также эмоции, возникающие во время работы и особенно при спортивной деятельности. Эмоции могут усиливать или, наоборот, снижать обмен веществ и энергии в организме.
При выполнении человеком механической работы коэффициент полезного действия может достигать 20—25%. Остальная освобождаемая в организме энергия превращается в тепло. Коэффициент полезного действия при мышечной деятельности зависит от структуры и темпа движений, от количества вовлекаемых в работу мышц и степени тренированности человека, выполняющего работу.
Оптимальный ритм движений, при котором энерготраты минимальны, различен при работе разной мощности. Он зависит также от тренированности человека.
308
По величине энерготрат нельзя судить об утомительности работы, даже физической. Например, весьма утомительные статические усилия требуют для своего выполнения меньше энергии, чем кажущаяся менее тяжелой динамическая работа.
Мышечная работа необходима для нормальной жизнедеятельности организма. Количество энергии, затрачиваемое непосредственно на мышечную деятельность, должно быть не менее 1200— 1300 ккал, в сутки. В связи с этим для лиц, не занимающихся физическим трудом, у которых, следовательно, на мышечную деятельность расходуется меньшее количство энергии, физические упражнения особенно необходимы.
Глава X!
ВЫДЕЛИТЕЛЬНЫЕ ФУНКЦИИ
§ 1. Общая характеристика выделительных процессов
Поддержание оптимальных отношений, организма со средой и сохранение гомеостаза обеспечиваются поступлением различных жизненно необходимых веществ извне и выделением конечных продуктов обмена веществ, а также чужеродных (лекарственных и др.) веществ, поступивших из окружающей среды.
Выделительные функции осуществляются многими системами организма: желудочно-кишечным трактом, органами внешнего дыхания, почками, потовыми, сальными, слезными, молочными и некоторыми другими железами (слизистая носа и др.).
Желудочно-кишечный тракт выводит из организма остатки пищеварительных соков, которые удаляются вместе с неусвоенными остатками пищевых веществ. Через дыхательные пути удаляются газообразные продукты: углекислота, лекарственные вещества и др.
Среди органов выделения особая роль принадлежит молочным, сальным и слезным железам. Первые выделяют молоко, необходимое для вскармливания потомства, вторые — кожное сало, образующее защитный слой на поверхности тела и др., третьи — влагу, смачивающую слизистые оболочки глазного яблока.
Основное значение для полноценного освобождения организма от конечных продуктов обмена и чужеродных веществ имеют выделительные функции почек и потовых желез.
§ 2. Основные функции почек и структура мочеобразовательных единиц
Основные функции почек. К ним относятся: 1) поддержание нормальной концентрации в организме воды, солей и ряда других кристаллоидов (например, глюкозы); 2) регулирование pH крови, кислотно-щелочного равновесия и осмотического давления в тканях 1сла; 3) удаление из организма конечных продуктов белкового об-
309
Рис. 92. Схема строения нефрона и его кровеносных сосудов (А и Б) и получение первичной мочи из капсулы с помощью микропипетки (В).
А: / — сосудистый (мальпигиев) клубочек, окруженный капсулой Боумена; 2 — извитой каналец, 3 — собирательная трубка;
В: 1 — приносящий сосуд; 2 — выносящий сосуд; 3 —* капиллярная сеть клубочка; 4 — полость капсулы; 5 — начало извитого канальца; 6—-наружная оболочка капсулы;
В: 1 — стеклянная палочка; 2 — каналец; 3 — пипетка; 4 — ртуть; 5 — клубочковая жидкость; 6 — артериолы
мена и чужеродных, в том числе лекарственных, веществ; 4) секреция гормона ренина, влияющего на тонус кровеносных сосудов. Все эти функции осуществляются в результате образования мочи в нефронах.
Структура нефрона. В почках человека имеется больше двух миллионов нефронов. Каждый нефрон представляет собой особую функциональную единицу (рис. 92), включающую мальпигиево тельце и мочевые канальцы.
Мальпигиево тельце состоит из капсулы Шу-млянского — Боумена, внутри которой находится сосудистый клубочек (см. рис. 92).
Капсула Шумлянского — Боумена образована вдавле-нием слепого расширенного конца мочевого канальца в его просвет, а потому имеет двойную стенку. Внутренняя стенка капсулы тесно соприкасается со стенками капилляров сосудистого клубочка, образуя базальную фильтрующую мембрану. Между ней и наружной стенкой капсулы находится щелевидная полость, в которую поступает плазма, фильтрующаяся через базальную мембрану из капилляров клубочка.
Клубочек состоит из приносящей артериолы, сложной сети артериальных капилляров и выносящей артериолы. Диаметр выносящей артериаолы меньше по сравнению с приносящей, что способствует поддержанию в капиллярах клубочков относительно высокого кровяного давления.
Мочевые канальцы начинаются от щелевидной полости капсулы, которая непосредственно переходит в проксимальный (каналец первого порядка) извитой каналец. В некотором отдалении от капсулы проксимальный извитой каналец выпрямляется и образует петлю Генле, переходящую, в свою очередь, в дистальный (каналец второго порядка) извитой каналец, открывающийся в собирательную трубку.
Одним из важных факторов, определяющих мочеобразование в нефронах, являются особенности строения их кровеносных сосудов. Почечная артерия, разветвляясь, образущ
310
мелкие так называемые приносящие артериолы, которые входят в состав мальпигиева тельца. Здесь приносящие артериолы переходят в капиллярную сеть — плотный сосудистый клубочек. Сливаясь, капилляры вновь образуют выносящую артериолу, имеющую меньший диаметр, чем приносящая. Разница в диаметрах приносящей и выносящей артериол — важный фактор, обеспечивающий в сосудистом клубочке наличие высокого капиллярного давления крови. В капиллярах клубочка кислород практически не используется, и вследствие этого оттекающая из них кровь является артериальной. Артериола, выйдя из капсулы Шумлянского — Боумена, вновь разветвляется на множество капилляров. Они оплетают извитые канальцы нефрона, питая их ткань.
Кроме описанных выше нефронов, получивших название корковых, в почке имеются еще и другие, отличающиеся по местоположению и кровоснабжению, — ю к ст а м е д у л л я р и ы е нефроны. Они находятся в мозговом слое почек, диаметр выносящей артериолы в них такой же, как и у приносящей. Юкстамедулляр-ные нефроны отличаются также некоторыми другими деталями строения. У места вхождения артериолы в клубочек располагаются специализированные клетки, выполняющие внутрисекреторную функцию. Они вырабатывают особое вещество ренин, участвующее в регуляции кровяного давления и почечного кровотока.
§ 3. Механизм мочеобразования в почке
Мочеобразование представляет собой весьма сложный процесс. В соответствии с современной фильтрационно-реабсорбционной теорией мочеобразования в этом процессе можно выделить две фазы — фильтрацию и реабсорбцию.
Фильтрация. Благодаря высокому давлению крови в сосудистом клубочке (60—70 мм рт. ст.) через стенки капилляров фильтруется в полость капсулы Шумлянского — Боумена плазма крови со всеми растворенными в ней кристаллоидами (соли, глюкоза и др.). Этот фильтрат, поступающий в Боуменовскую капсулу и переходящий затем в мочевые канальцы, называют первичной мочой. Но крупные коллоидные частицы (белки), как и форменные элементы крови, не фильтруются через стенки капилляров клубочков. Поэтому первичную мочу образно называют безбелковой плазмой.
В сутки у человека образуется до 150—180 л первичной мочи. Такая интенсивная фильтрация возможна только в условиях обильного кровоснабжения почки и при особом строении фильтрационной поверхности капилляров клубочка, в которых поддерживается высокое давление крови. При снижении этого давления до 30— 35 мм рт. ст. фильтрация полностью прекращается, так как оно становится равным осмотическому давлению коллоидов крови. В сутки через почки протекает до 1700—1800 л крови, и из каждых 10 л проходящей крови образуется примерно 1 л первичной мочи.
Реабсорбция в мочевых канальцах. Вся образующаяся первичная моча поступает в извитые канальцы и петлю Генле, где происходит обратное всасывание — реабсорбция (от лат. ге — приставка,
311
обозначающая обратное или противоположное действие, absorb-tio — всасывание). Из 150—180 л первичной мочи реабсорбируется около 148—178,5 л воды. Вследствие этого количество конечной мочи, т. е. той, которая из мочевых канальцев и собирательных трубок переходит в почечные лоханки и затем поступает в мочевой пузырь, составляет всего около 1,5 л в сутки. Такое значительное обратное всасывание объясняется тем, что общая суммарная поверхность канальцев почек человека равна 40—50 м2.
Обратному всасыванию подвергается также много других органических и неорганических веществ первичной мочи. Некоторые вещества не реабсорбируются и целиком выводятся из организма с мочой.
Все вещества, всасывающиеся в кровь из первичной мочи, называются пороговыми. К ним относится ряд органических (глюкоза, и др.) и неорганических (соли Na, К- Са, фосфаты и др.) веществ. Они выводятся из организма с конечной мочой только при относительно высокой концентрации их в крови.
Величина концентрации вещества в крови, при которой оно реабсорбируется не полностью и начинает выводиться из организма с конечной мочой, называется порогом выведения. Таким образом, обратное всасывание веществ из первичной мочи находится в определенной зависимости от концентрации их в крови. Например, если концентрация глюкозы в плазме крови не превышает 150—180 мг%, то она в мочевых канальцах полностью подвергается обратному всасыванию. При более высоком содержании глюкозы в крови часть ее не реабсорбируется и поступает в конечную мочу. Такое явление получило название глюкозурии, т. е. выделения глюкозы с мочой. Глюкозурия может возникать при чрезмерном поступлении глюкозы с пищей, при возникновении эмоциональных состояний, в том числе в процессе выполнения физической работы. То же самое наблюдается и в отношении ряда других веществ. Так, при недостатке в организме поваренной соли она полностью реабсорбируется. Если же в крови находится избыточное количество этой соли, то часть ее выводится с конечной мочой.
Непороговые вещества. Вещества, которые не подвергаются обратному всасыванию и полностью выводятся с конечной мочой, называются непороговыми. К ним относятся конечные продукты белкового обмена веществ (мочевина, креатинин), сульфаты, некоторые лекарственные вещества и др.
Реабсорбция — процесс очень сложный, осуществляющийся за счет активного и пассивного переноса веществ через мембрану (см. гл. 1, § 4) извитых канальцев и петли Генле.
Нисходящее и восходящее колена петли Генле (рис. 93) образуют так называемую поворотно-противоточную систему. Тесно соприкасаясь друг с другом, нисходящее и восходящее колена функционируют как единый механизм. Сущность такой совместной работы (см. рис. 93) заключается в том, что из полости нисходящего колена в тканевую жидкость почки обильно поступает вода. Это приводит в данном колене к «сгущению», т. е к повышению концентра
312
ции различных веществ мочи. Из восходящего же колена в тканевую жидкость активно выводятся ионы Na, но не выводится вода. Повышение концентрации ионов Na в тканевой жидкости способствует повышению ее осмотического давления, а следовательно, и усилению «отсасывания» воды из нисходящего колена. Это вызывает еще большее «сгущение» мочи в петле Генле. Здесь, как и везде в живых системах, вновь
проявляет себя феномен саморегуля-
Корковое вещество
Мозговое вещество
з к
12
Тканевая жидкость
Тканевая петля Генле жидкость
12
Петля капилляра
Na+
Собирательная трубочка
Рис. 93. Схема поворотно-противоточного механизма нефрона. Пунктирными стрелками показано общее направление движения жидкости, черными стрелками — пассивный перенос натрия, белыми стрелками — передвижение воды. Цифры — относительные концентрации осмотически активных растворенных веществ (по К. Вилли и В. Детье)
ции. Выход воды из нисходящего колена способствует выходу из восходящего ионов Na, a Na, в свою очередь, обусловливает выход воды. Таким образом, петля Генле работает как концентрационный механизм. «Сгущение» мочи продолжается и далее в собирательных трубочках.
Клетки канальцев способны также выводить из организма многие вещества путем секреции. В левую очередь это имеет отношение к веществам, которые слабо фильтруются или совсем не переходят в первичную мочу (некоторые коллоиды, органические кислоты, аминокислоты и др.). Ряд поступающих в мочу веществ почки синтезируют сами (мочевину, мочевую кислоту, уробилин и др.).
В результате столь сложной фильтрационно-реабсорбционной, секреторной и синтезирующей работы нефрона образуется конечная моча. По своему составу она существенно отличается от первичной мочи (табл. 11).
313
Таблица 11
Содержание в плазме крови н первичной моче различных веществ
Вещества Содержание веществ (%) Отличие содержания вещества в моче от содержания его в плазме крови
в плазме крови и первичной моче в моче, выделяемой в мочеточники
Мочевина Мочевая кислота Глюкоза Калий Натрий Фосфаты Сульфаты 0,03 0,004 0,1—0,15 0,02 0,32 0,009 0,002 2,0 0,05 0,15 0,35 0,15 0,18 В 67 раз больше В 12 раз больше Отсутствует в моче В 7 раз больше Приблизительно столько же В 16 раз больше В 90 раз больше
§ 4. Регуляция мочгобразования
Регуляция работы почек обеспечивается нейрогуморйльным механизмом. Основным гуморальным компонентом этой регуляции является гормон задней доли гипофиза — антидиуретический гормон. Он усиливает реабсорбцию воды и тем самым уменьшает образование и выделение (диурез) мочи. При сокращении поступления этого гормона в кровь снижается обратное всасывание воды из канальцев. Диурез при этом значительно возрастает (иногда до 20— 25 л в сутки). Гормон надпочечников адреналин, суживая выносящую артериолу сосудистого клубочка, повышает в нем давление. Вследствие этого происходит усиление фильтрации и образования мочи. Но при более значительных концентрациях адреналин оказывает противоположный эффект. Он суживает также и приносящие сосуды и тем самым снижает давление в капиллярах сосудистого клубочка. Это приводит к уменьшению в нем фильтрации, а следовательно, и образования первичной мочи. Такого рода снижение мо-чеобразования у спортсменов наблюдается при сильных эмоциях, вызывающих увеличение выделения адреналина в кровь. На деятельность почек оказывают влияние и некоторые пищевые продукты, а также специальные мочегонные средства.
Прямая нервная регуляция работы почек выражена слабее, чем гуморальная. В основном она осуществляется через рефлекторные изменения просвета почечных сосудов под влиянием тех или иных воздействий. Это ведет к изменению почечного кровотока и, следовательно, процесса мочеобразования.
Нервная система может влиять на работу почек как безусловнорефлекторным, так и условнорефлекторным путем. Эти влияния могут изменять функциональное состояние гипоталамической области мозга. Гипоталамус же, в свою очередь, оказывает воздействие на секрецию антидиуретического гормона гипофиза.
314
§ 5. Мочевыведение
Конечная моча по собирательным трубочкам и более крупным коллекторам — белиниевым трубкам — поступает в почечные лоханки, мочеточники и мочевой пузырь. Когда в мочевом пузыре человека собирается 250—300 мл мочи, давление в нем становится равным 15—16 см водн. ст. Оно достаточно для раздражения рецеп торов, расположенных в стенках пузыря. Возбуждение, возникшее в них, поступает по афферентным нервам в крестцовую часть спинного мозга, где располагается центр непроизвольно-рефлекторного мочеиспускания. Эфферентная иннервация мочевого пузыря осуществляется по симпатическим и парасимпатиечским волокнам. Импульсы, поступающие по симпатическим волокнам, расслабляют мышцу пузыря и повышают тонус его жома. Это способствует заполнению пузыря мочой и ее удержанию в нем. Противоположное действие оказывают импульсы, поступающие по парасимпатическим волокнам. В результате моча выжимается из пузыря и по мочеиспускательному каналу изливается наружу. Во время мочеиспускания напрягаются также мышцы брюшного пресса, изменяется дыхание и работа сердца и др.
Мочеиспускание, как и все процессы, протекающие в организме, является следствием целостной реакции его на раздражение. Крестцовый центр мочеиспускания находится под контролирующим влиянием центров, расположенных в продолговатом и среднем мозгу, а также коры больших полушарий головного мозга. Поэтому акт мочеиспускания до определенных пределов наполнения мочевого пузыря (давление не выше 16—18 см водн. ст.) произволен и только при дальнейшем повышении давления становится непроизвольным. Это является выражением защитной реакции организма.
§ 6. Выделительная функция потовых желез
Выделительные функции почек дополняются выделительными функциями потовых желез. Работа их взаимосвязана и частично взаимозаменяема.
Замена выделительной функции почек деятельностью потовых желез происходит не только в случае недостаточности почек при их заболевании, но и при выполнении физической работы.
Различают два вида потоотделения — термическое потоотделение и психическое, или эмоциональное.
Термическое потоотделение происходит по всей поверхности тела, за исключением ладоней и подошвенной стороны стоп, психическое— на ладонях, подошвенной стороне стоп, в подмышечных впадинах и реже на других участках тела.
Причиной термического потоотделения является повышение температуры окружающей среды. Его интенсивность и быстрота наступления находятся в прямой зависимости от интенсивности и величины повышения температуры внешней среды. Однако, достигнув
315
максимальной величины, потоотделение ослабляется даже при высокой температуре.
Психическое потоотделение возникает в результате тех или иных эмоциональных переживаний (страх, радость и др.), умственного напряжения и др., т. е. факторов, не оказывающих существенного влияния на температуру тела. По своим свойствам психическое потоотделение резко отличается от термического. Оно имеет очень короткий латентный период и начинается сразу после возбуждающего воздействия, быстро достигает максимума, соответствующего силе раздражителя, и столь же быстро прекращается с прекращением раздражения.
Потоотделение, вызываемое мышечной деятельностью, представляет собой сочетание двух видов потоотделения — термического (вследствие повышения теплопродукции при мышечной работе) и психического (вследствие эмоциональной настроенности, вызванной этой работой). Таким образом, интенсивность потоотделения при мышечной работе зависит как от ее характера, так и от эмоционального фона.
Потоотделение — процесс постоянный. В условиях физического и эмоционального покоя, при комфортной температуре окружающей среды он совершается со значительно меньшей интенсивностью, чем во время физической работы и эмоционального возбуждения. Количество пота, выделяемое в этих условиях, достигает в сутки 500— 1000 мл. Регуляция деятельности потовых желез осуществляется нервным и гуморальным путями. Секреторными нервами потовых желез служат симпатические нервы. Отличительной особенностью волокон симпатических нервов является то, что они выделяют в качестве медиатора не симпатии, а ацетилхолин.
§ 7. Влияние мышечной работы на функции выделения
Во время длительной работы даже умеренной мощности (бег на длинные дистанции и др.) всегда наблюдается перераспределение крови. Увеличивается приток ее к мышцам и значительно сокращается приток к внутренним органам. Это может привести к резкому снижению давления в капиллярах сосудистых клубочков почек, к уменьшению и даже прекращению мочеобразования. Однако временно возникшая недостаточность почек компенсируется усилением работы потовых желез. Кроме того, усиление потоотделения увеличивает теплоотдачу, что очень важно в условиях избыточной продукции тепла во время выполнения физических упражнений.
При работе субмаксимальной мощности, т. е. сопровождающейся преимущественно анаэробным расщеплением углеводов, концентрация молочной кислоты в моче достигает 0,22—0,24%, тогда как при работе умеренной мощности содержание ее не превышает 0,05— 0,06%. Выведению избытка молочной кислоты, а также мочевины, креатинина и аммиака при мышечной работе умеренной мощности способствует усиленная работа потовых желез.
316
Важным показателем состояния эмоционального возбуждения спортсмена является появление глюкозы в моче. Это происходит в результате активации процесса гликогенолиза под влиянием усиленного выделения адреналина надпочечниками, что приводит к повышению концентрации глюкозы в крови выше пороговой.
Иногда при мышечных и эмоциональных напряжениях может быть обнаружен белок в моче. Исчезновение его через сутки свидетельствует не о патологии, а о физической реакции организма на большие нагрузки.
Количество пота, выделяемое во время работы, может быть весьма значительным. Так, у мужчин на марше со скоростью 5 км/час с грузом 10 кг при температуре воздуха 4-32,6—34,7° за 1 час выделялось 1056—1787 г пота. У рабочих горячих цехов и солдат, несущих службу в тропиках, количество выделяемого пота может достигать до 10—15 кг в день.
Глава Xi!
ТЕПЛООБМЕН
Организм человека находится под воздействием постоянных колебаний внешней температуры. Кроме того, изменяется в зависимости от интенсивности обменных процессов и количество тепла, вырабатываемого внутри организма. Все это могло бы приводить к колебаниям температуры тела человека. Однако этого не происходит: специальная система терморегуляции поддерживает температуру тела человека на определенном уровне, наиболее выгодном для процессов жизнедеятельности.
§ К Пойкилотермия и гомсйотермия
В зависимости от особенностей теплообмена животные организмы принято делить на холоднокровные — пойкилотермные (от греч. poikilos — разнообразный) и теплокровные — гомойотермные (от греч. homos — одинаковый, равный). У первых температура тела изменяется соответственно колебаниям температуры внешней среды, у вторых, несмотря на колебания температуры внешней среды, сохраняется стабильная температура тела. И по числу видов, и по числу особей гомойотермные организмы составляют меньшую часть животного мира: к ним относятся лишь млекопитающие и птицы. Все прочие организмы, от простейших до пресмыкающихся, — пойкилотермные.
Стабильная температура тела — одна из важнейших биологических констант, сформировавшихся в процессе эволюции у высших представителей животного мира. Температура гомойотермных, значительно превышающая обычную температуру внешней среды, обеспечивает высокую скорость химических реакций внутри организма
317
и, следовательно, высокую интенсивность всех процессов жизнедеятельности.
Способность гомойотермных противостоять воздействию холода и тепла, сохраняя стабильную температуру тела, имеет известные пределы. Особо неблагоприятные условия внешней среды (чрезмерно низкая или высокая температура воздуха, сочетающаяся с его повышенной влажностью и большой скоростью движения) могут привести к нарушению гомойотермии. При чрезмерно низкой или чрезмерно высокой температуре среды защитные терморегуляторные механизмы оказываются уже недостаточными, температура гомойотермного организма соответственно начинает падать или повышаться. В первом случае развивается состояние гипотермии, во втором — состояние гипертермии.
§ 2. Температурные «оболочка» и «ядро» тела
36'
32“
31“
Рис. 94. Схема температурных уровней в теле обнаженного человека при температуре воздуха 20°.
Область тела, обозначенная точками,— ядро
В гомойотермном организме принято различать две темпера-турные зоны: наружную и внутреннюю, образно называемые «оболочкой» и «ядром». Только «ядро» характеризуется стабильной температурой. К нему относятся мозг, органы грудной клетки, брюшной полости и малого таза. Органы и ткани, расположенные на периферии те-37" ла (кожа, большая часть скелетной мускулатуры .. и костной системы), составляют «оболочку».Температура ее в определенной степени возрастает при повышении температуры внешней среды и наоборот (рис. 94). Колебания температуры «оболочки» направлены на поддержание гомойотермии «ядра». Так, при опасности перегревания подъем температуры «оболочки» способствует отдаче тепла во внешнюю среду; при опасности охлаждения снижение температуры «оболочки» ограничивает теплоотдачу.
Широко используемый термин «температура тела», как правило, относится к температуре внутренних областей тела, т. е. «ядра». При этом разные участки «ядра» тоже не имеют строго идентичной температуры. Например, при измерении у человека температуры в венозных сосудах, собирающих кровь от отдельных внутренних органов, выявляются различия в пределах нескольких десятых градуса: обычно более высокие цифры регистрируются в крови внутренней яремной вены (т. е. в крови, оттекающей от мозга), на втором месте стоит кровь печеночной вены, далее — кровь коронарного синуса, почечной вены, правого предсердия. Эти различия объясняются неодинаковой интенсивностью энергетического обмена отдельных органов: чем он выше, тем больше при прочих равных условиях
318
и местная температура. Кроме того, имеет значение интенсивность местного кровотока: температура органа, превышающая температуру артериальной крови, будет снижаться по мере усиления кровотока и наоборот. В обычных условиях кровь, проходя по сосудам «ядра», забирает тепло, а проходя по сосудам «оболочки», отдает его тканям.
Наиболее точное представление о средней температуре тела можно получить, измеряя ее в полости сердца (где как бы происходит усреднение температур, характеризующих отдельные участки тела). Но технически осуществить это сложно. На практике обычно пользуются измерением подмышечной температуры или ректальной (в прямой кишке). Реже измеряют температуру в полости рта, пищевода или в глубине слухового прохода.
У человека подмышечная температура в норме варьирует от 36 до 37°, ректальная — от 36,2 до 37,6°, причем имеются небольшие суточные колебания ее в этих пределах: минимальная температура отмечается во время ночного сна, максимальная — во второй половине дня.
§ 3. Температурные зоны внешней среды
Каждый вид гомойотермных характеризуется определенной термонейтральной зоной среды, в пределах которой организм в покое не ощущает ни холода, ни жары. Ниже и выше этой зоны располагаются соответственно зоны умеренного охлаждения и умеренного перегревания, а далее — температурные зоны внешней среды, обусловливающие развитие гипотермии и гипертермии.
В пределах термонейтральной зоны энергетический обмен организма устанавливается на минимальном уровне. При смещении температуры среды ниже или выше этой зоны обмен возрастает. В первом случае очевидна целесообразность возрастания обмена: увеличивая на холоде энергетический обмен и, значит, теплопродукцию, организм стремится компенсировать повышенные потери тепла. Во втором случае, при перегревании, некоторое увеличение энергетического обмена происходит вынужденно, вследствие компенсаторной активизации механизмов теплоотдачи: усиления работы дыхательной мускулатуры в связи с «тепловой одышкой»; усиления работы сердца, которому приходится перекачивать через сосуды «оболочки» возросшее количество крови; активизации потовых желез. Дополнительное значение при этом может иметь и подъем температуры «оболочки» (а в случае развития гипертермии — также «ядра») с соответственным повышением скорости всех химических реакций.
Для человека в обычной домашней одежде нейтральная температура воздуха равна 19—22°, для обнаженного человека — 28—31°. Нейтральная температура воды, теплоемкость и теплопроводность которой выше, чем воздуха, равна для человека, погруженного в воду до подбородка, примерно 35°.
319
§ 4. Механизмы теплопродукции
Обмен веществ в организме всегда сопровождается образованием тепла. Основной источник мобилизации энергии в организме — окисление; дополнительную роль играет гликолиз. Эти химические процессы носят экзотермический (от греч. ехо — снаружи) характер: их энергия лишь частично аккумулируется в молекулах АТФ, значительная же доля переходит в тепло. В виде такого «первичного» тепла в тканях рассеивается около 60% энергии. Остальные 40% после расщепления АТФ обеспечивают работу мышц, многообразные процессы синтеза и секреции, работу «ионного насоса». Но и при этом та или иная часть энергии переходит затем в тепло.
Такое «вторичное» тепло образуется в результате химических реакций, имеющих экзотермический характер, а при сокращении мышечных волокон — в результате их трения. В конечном итоге переходит в тепло или вся энергия, мобилизуемая в организме, или (при наличии внешней работы, к. п. д. которой, как известно, не превышает 25—30%) большая ее часть.
Существует несколько физиологических реакций, обеспечивающих компенсаторное увеличение теплопродукции при действии на организм холода. Один из эффективных способов увеличения теплопродукции в этих условиях — произвольная мышечная деятельность. За счет такой деятельности энергетический обмен организма может возрасти в 10 и более раз. Однако часть мобилизуемой при этом энергии расходуется организмом на внешнюю работу. Кроме того, при любых движениях усиливаются потери тепла с поверхности тела.
Другая реакция компенсаторного повышения теплопродукции — холодовая дрожь. Она более экономна. При такой непроизвольной сократительной деятельности скелетных мышц энергетический обмен может возрасти лишь в 2—4 раза, но зато вся мобилизуемая энергия, перейдя в тепло, сохраняется внутри организма.
Наконец, третья реакция повышения теплопродукции на холоде связана с выделением в кровь норадреналина. Этот гормон, действуя, на скелетные мышцы, а также, по-видимому, на печень и жировую ткань, обусловливает перераспределение энергии, образующейся в них при окислительных процессах: доля энергии, аккумулируемая при этом в молекулах АТФ, уменьшается, а доля, переходящая непосредственно в «первичное» тепло, увеличивается. Особенно характерна эта реакция для акклиматизированного к холоду организма, в котором возрастает и выработка норадреналина, и чувствительность к последнему тканей. В таком организме описанная теплопродукция не уступает по мощности холодовой дрожи и может почти целиком замещать ее. Однако этот способ повышения теплопродукции невыгоден при необходимости выполнения произвольной мышечной работы: неизбежно падает ее КПД, поскольку в акклиматизированном к холоду организме уменьшается доля энергии, аккумулируемой в молекулах АТФ.
820
§ 5. Механизмы теплоотдачи
Существуют следующие физические механизмы теплоотдачи: 1) теплопроведение, 2) теплоизлучение, 3) испарение.
Теплопроведение может осуществляться путем кондукции и конвекции. Кондукция— прямая передача тепла от более нагретого тела к менее нагретому, неподвижно с ним соприкасающемуся (например, от ступней к полу, на котором стоит человек, или с поверхности тела в неподвижный воздух). Конвекция— прямая передача тепла в условиях, когда тело и среда перемещаются относительно друг друга (например, при обдувании тела ветром или при беге даже в условиях полного безветрия). При одних и тех же физических свойствах среды (теплоемкости и теплопроводности) конвекция представляет значительно более мощный механизм теплоотдачи, чем кондукция.
Теплоизлучение характеризуется отдачей тепла лучистым путем, в основном за счет инфракрасной части спектра.,
Испарение сопровождается потерями тепловой энергии, которую уносят молекулы, отрывающиеся от поверхности воды. При переходе в воздух 1 г воды теряется около 0,58 ккал тепла.
Выраженность различных механизмов теплоотдачи определяется рядом условий. Лучистая передача тепла от более нагретого тела к менее нагретому прямо зависит от разницы температур между ними. Кондукция обусловливается помимо такой температурной разницы еще и теплоемкостью и теплопроводностью среды. На конвекцию влияют как перечисленные факторы, так и скорость перемещения тел относительно друг друга. Наконец, теплоотдача испарением (с влажной поверхности в воздух) при прочих равных условиях находится в прямой зависимости от скорости движения воздуха и в обратной — от его относительной влажности.
Процессы теплоотдачи происходят в основном на поверхности кожи и слизистых оболочек дыхательных путей. При теплоотдаче же с калом и мочой соответственно убывает масса тела, а значит, не изменяется его тепловой баланс; вместе с тем некоторое влияние на этот баланс может оказывать прием холодной или горячей пищи, воды.
У многих видов организмов известны определенные области тела, поверхность которых наиболее активно участвует в регуляции теплоотдачи. Например, у кролика — это уши, у мышей — хвост (лишенные хвоста мыши при перегревании погибают намного быстрее). У человека важная роль в регуляции теплоотдачи принадлежит конечностям.
Перечисленные выше физические механизмы теплоотдачи лежат в основе тех или иных физиологических реакций. Одна из таких реакций — сдвиг кровотока в «оболочке». Например, при жаркой погоде кровь приливает к коже, и она становится горячей. В результате увеличивается перепад температур между поверхностью тела и внешней средой, а это усиливает кондукционную, конвекционную и лучистую отдачу тепла.
321
Однако в тех случаях, когда внешняя температура превышает температуру поверхности тела, кондукционный, конвекционный и лучистый перенос тепла приобретает уже противоположную направленность: извне — внутрь организма. В этих условиях организм может противостять перегреванию лишь благодаря таким реакциям теплоотдачи, как «тепловая одышка» и потоотделение. Если, например, в термонейтральной зоне у человека на долю испарения приходится около 25% общей теплоотдачи, то уже при температуре воздуха 34,5°—100%. У человека потооделение — самая эффективная реакция теплоотдачи. Оно может достигать 2 л за 1 час. При испарении такого количества пота во внешнюю среду отдается около 1150 ккал., т. е. количество тепла, примерно в 20 раз превышающее уровень основного обмена.
§ 6. Центральная регуляция теплообмена
Поддержание стабильной температуры тела сводится в принципе к согласованию двух противоположно направленных процессов: теплопродукции и теплоотдачи. Если, например, теплоотдача становится больше теплопродукции, температура тела начинает снижаться, и наоборот. Чтобы избежать этого, организм при вынужденном ускорении одного из названных процессов стремится компенсаторно активизировать во столько же раз и второй процесс.
Включение компенсаторных реакций теплопродукции и теплоотдачи происходит рефлекторным путем. Первоначально, как правило, возбуждаются терморецепторы; одни из них реагируют на холод, другие — на тепло. Лучше всего изучены терморецепторы кожи (особенно много их на коже лица и туловища). Кроме того, известны терморецепторы в слизистой оболочке языка и желудка, трахеи и бронхов, в стенке различных венозных сосудов. Наконец, отдельные нервные клетки, специализировавшиеся на восприятии местных колебаний температуры, обнаружены в межуточном и в спинном мозге. Таким образом, разветвленная сеть чувствительных элементов обеспечивает подробную информацию о температурных сдвигах во внешней и внутренней среде организма.
Эта информация поступает в высшие центры теплообмена, расположенные в гипоталамусе. Так, при эксперименте на животных раздражение электрическим током передней части гипоталамуса (даже при нейтральной температуре среды) включает реакции теплоотдачи: расширение сосудов кожи, «тепловую одышку», потоотделение. После разрушения переднего гипоталамуса животные теряют способность реагировать на перегревание усилением теплоотдачи.
Если центры переднего гипоталамуса противодействуют повышению температуры тела, то центры заднего гипоталамуса регулируют реакции, препятствующие ее понижению. Электрораздражение заднего гипоталамуса вызывает дрожь, а при его разрушении животные теряют способность отвечать на охлаждение дрожью и сужением сосудов кожи.
322
Терморегуляторные центры гипоталамуса вовлекают в свою деятельность и некоторые другие отделы мозга. При этом в соответствующих реакциях участвуют: сосудодвигательный центр продолговатого мозга (при терморегуляторных сдвигах кровотока в «оболочке»), дыхательный центр продолговатого мозга (при «тепловой одышке»), мотонейроны спинного мозга (при холодовой дрожи), симпатические нейроны спинного мозга (при потоотделении, при выделении в кровь норадреналина). В свою очередь, на терморегуляторные центры гипоталамуса оказывает влияние кора больших полушарий. Например, хорошо известен факт подавления дрожи во время выполнения произвольных движений. Условнорефлекторным путем (т. е. при участии коры больших полушарий) у экспериментальных животных можно вызывать «тепловую одышку», терморегуляторные сдвиги кровотока в «оболочке», холодовую дрожь.
§ 7. Особенности теплообмена при мышечной работе
При работе в мышцах образуется большое количество тепла. Это приводит к подъему их температуры. Из работающих мышц тепло разносится током крови по всему телу, вызывая общий подъем его температуры. В обычных условиях, как сказано выше, организм сопротивляется любому смещению температуры «ядра». Напротив, при мышечной работе такое смещение осуществляется сравнительно легко: в этом случае происходит функциональная перестройка самих центров теплообмена.
Рис. 95. Прирост температуры тела в процессе работы на велоэргометре (по В. А. Бернштейну, М. М. Синайскому и В. Г. Федотовой).
Мощность работы соответствует 30, 50 и 70% максимального потребления кислорода (МПК). Ось абсцисс — время работы (мин.), ось орди-нат — прирост температуры тела (градусы)
При работе умеренной мощности после первоначального подъема температура тела стабилизируется на новом уровне; он тем выше, чем интенсивнее работа (рис. 95). Такой подъем температуры
.323
тела имеет регулируемый характер, его выраженность не зависит от колебаний (в широких пределах) температуры внешней среды. Подобные колебания обусловливают лишь компенсаторное (с целью сохранить новый уровень температуры тела) усиление или ослабление теплоотдачи. Среди реакций теплоотдачи наиболее важную роль при мышечной работе играет потоотделение.
Некоторый подъем температуры тела выгоден при работе: возрастает возбудимость, проводимость, лабильность нервных центров, снижается вязкость мышц, в протекающей через них крови улучшаются условия отщепления кислорода от гемоглобина. Поэтому повышение температуры тела перед началом работы — одна из главных задач спортивной разминки. Небольшое повышение температуры можно зарегистрировать в предстартовом состоянии и без разминки (оно возникает условнорефлекторно).
Не выяснен окончательно вопрос о том, до какого уровня подъем температуры тела при работе следует считать благоприятным. По-видимому, этот уровень должен быть тем выше, чем больше мощность работы. Установлено, что при работе в пределах зоны умеренной мощности регулируемый центрами теплообмена подъем температуры тела может составлять 1,5—2°.
Наряду с регулируемым подъемом при мышечной работе может наблюдаться также дополнительный, вынужденный подъем температуры тела. Он происходит при чрезмерно высокой температуре и влажности воздуха, при чрезмерной теплоизоляции работающего. В этих условиях механизмы теплоотдачи уже не справляются с выведением из организма избыточного тепла и температура тела прогрессивно нарастает, что может привести к тепловому удару.
Глава Xh!
ВНУТРЕННЯЯ СЕКРЕЦИЯ
§ 1. Общая характеристика желез внутренней секреции
В регуляции жизнедеятельности организма важное значение имеют вещества высокой биологической активности, выделяемые специальными органами в кровоток и способными, несмотря на их чрезвычайно малые концентрации в крови, вызывать значительные изменения в состоянии организма, в частности обмена веществ в нем. Эти вещества называются гормонами, а выделяющие их органы — эндокринными железами ( от греч. endon — внутрь, krino — выделять) или железами внутренней секреции.
Эндокринные железы (в отличие от желез внешней секреции, например пищеварительного тракта) не имеют протоков. Образованные ими гормоны всасываются непосредственно в кровь протекающую через железу.
Гормоны обладают дистанционным действием, т. е., поступая в кровяное русло, оказывают влияние на органы и ткани, расположен-324
ные вдали от той железы, где они синтезируются. Действие гормонов характеризуется специфичностью. Она выражается в двух формах: одни гормоны (например, половые) влияют только на некото-
рые органы и ткани, другие управляют лишь определенными изменениями в цепи обменных процессов и в активности регулирующих, их ферментов.
Гормоны сравнительно быстро разрушаются. Для поддержания достаточного количества гормона в крови необходимо постоянное выделение его соответствующей железой. Если же в результате патологических процессов деятельность какой-либо эндокринной железы нарушается, что выражается в недостаточной или чрезмерной продукции гормонов, то возникают существенные функциональные расстройства, а в некоторых случаях может наступить даже смерть. Почти все расстройства деятельности эндокринных желез вызывают понижение общей работоспособности.
Важным фактором, регулирующим интенсивность образования и секреции гормонов, является характер регулируемых ими процессов. Как только изменения, вызываемые каким-либо гормоном, достигают определенной величины, образование и выделение этого гормона уменьшаются. В ряде случаев увеличивается продукция другого гормона, дей-
Рис. 96. Схема передачи органам-исполнителям через эндокринные железы влияний нервной системы. 1 — головной мозг, 2 — нейросекреция, 3 — гипофиз, 4 •— тропные гормоны аденогипофиза; 5 — нервный ствол; 6 — железы внутренней секреции; 7 — нервные волокна, через которые поступают нервные импульсы в железы внутренней секреции; 8 — гормоны; 9 — орган-исполнитель; 10 кровеносный сосуд
ствующего противоположно на
данный процесс. Таким образом, высокое содержание в крови продуктов обменных процессов, образующихся под влиянием гормона, нередко подавляет активность соответствующей железы, а низкое содержание этих продуктов, наоборот, может быть стимулятором ее деятельности.
Функции эндокринных желез регулируются центральной нервной системой, которая контролирует выделение всех гормонов (рис. 96). Таким образом, нервное и гуморальное воздействия на различные органы и ткани представляют собой проявление единой системы иейрогуморальной регуляции функций организма.
Классическими методами изучения эндокринных функций являются удаление v подопытных животных какой-либо эндокринной железы, введение в организм экстрактов, в том числе гормонов, меченных радиоактивными изотопами, и ис
325
следование больных с недостаточностью или избыточностью функций той или иной железы. Наблюдаемые при этом изменения в деятельности организма свидетельствуют о роли соответствующей железы.
§ 2. Надпочечники
Надпочечники расположены над верхними полюсами почек. Каждый из них весит 3—5 г и состоит из мозгового и коркового слоев, являющихся, по существу, двумя разными железами.
Мозговой слой надпочечников. Он образует сероватую «сердцевинку», составляя 10% веса всей железы. Основным гормоном, образующимся в ней, является адреналин. Наряду с ним из мозгового слоя поступает в кровоток норадреналин, отличающийся от адреналина отсутствием одной метильной группы.
В биосинтезе адреналина образование норадреналина непосредственно предшествует образованию самого адреналина. В более значительных количествах норадреналин образуется в нервных окончаниях симпатической нервной системы (где он выполняет роль медиаторного вещества) и отсюда в основном поступает в кровь.
Адреналин, как и норадреналин, сразу после образования связывается в виде гранул с эндоплазматической сетью клеток железы. Под влиянием симпатических нервных импульсов, приходящих к железе по чревному нерву, гранулы освобождаются и гормоны поступают в кровоток.
Адреналин и норадреналин очень сходны по своим свойствам и физиологическому влиянию, но пороги их действия различны. Норадреналин сильнее действует на кровеносные сосуды, и ему принадлежит основная роль в сосудодвигательных реакциях. Но в регуляции обменных процессов адреналин в 4—8 раз активнее норадреналина. Адреналин наряду с сосудосуживающим действием на кровеносные сосуды кожи, почки, селезенки и органов пищеварительного тракта обладает также и сосудорасширяющим влиянием на сосуды мозга и мышц.
Под влиянием адреналина ускоряется и усиливается деятельность сердца, повышается его возбудимость и увеличивается скорость проведения импульсов по сердечной мышце. Важное значение имеет (в особенности при мышечных напряжениях) сокращение под влиянием адреналина мышц стенок сосудов в органах, являющихся депо крови, а также расслабление гладких мышц бронхов, что способствует уменьшению сопротивления движения воздуха при дыхании. Таким образом, в результате воздействия адреналина усиливается транспорт кислорода к тканям, в частности к мышцам
Роль адреналина в мобилизации энергетических ресурсов орга низма заключается в том, что под его влиянием расщепляется гли коген печени. В результате этого усиливается поступление глюкозы из печени в кровь и возрастает ее содержание в крови. Анаэробный распад гликогена в мышцах также происходит при действии адреналина на соответствующую ферментативную систему. В связи с этим адреналин имеет важное значение в мобилизации анаэробной работоспособности организма.
326
Другая сторона роли адреналина в мобилизации энергетических ресурсов организма заключается в его липолитическом действии. Оно выражается в ускорении распада жиров, вследствие чего содержание свободных жирных кислот (важного субстрата окислительных процессов) увеличивается как в самой жировой ткани, так и в крови.
С действием адреналина на обмен веществ связан известный факт повышения работоспособности утомленных мышц при его введении. Воздействуя на ретикулярную формацию мозга, адреналин способствует повышению возбудимости центральной нервной системы.
Центральная нервная система может через симпатические нервы быстро увеличить выделение адреналина в кровоток, например, при физической нагрузке, а также при сильных переживаниях. Это способствует мобилизации функций и ресурсов организма для эффективного выполнения необходимых двигательных актов.
Кора надпочечников. Она состоит из трех зон: наружной — клубочковой, средней — пучковой и внутренней — сетчатой. Эти зоны различаются между собой не только по гистоструктуре, но и по функциональной активности и гормонам, образующимся в них.
Кора надпочечников является жизненно важной железой внутренней секреции. Удаление ее быстро приводит к смерти, что не наблюдается у животных при разрушении у них мозгового слоя надпочечников. Патологическое изменение коры надпочечников у людей, известное как болезнь Аддисона, сравнительно быстро приводит при отсутствии лечения к смерти. В обоих случаях типичными симптомами недостаточности гормонов коры надпочечников являются мышечная слабость и быстрая утомляемость. После введения гормонов коры надпочечников работоспособность нормализуется.
Гормоны коры надпочечников являются стероидами и носят общее название кортикостероиды или кортикоиды. Они делятся на три группы:
1) минералокортикоиды — выделяемые в клубочковой зоне и регулирующие минеральный обмен в основном на уровне почек; 2) глюкокортикоиды — выделяемые в пучковой зоне и оказывающие различные регуляторные воздействия в широком диапазоне; 3) аналоги половых гормонов — выделяемые в сетчатой зоне.
Основным и наиболее активным минералокортикоидом является альдостерон. Он увеличивает реабсорбцию натрия в канальцах почек и поддерживает на должном уровне его содержание в плазме крови, лимфе и тканевой жидкости. Это приводит к задержке воды в организме и способствует повышению артериального давления, снижая реабсорбцию калия в канальцах почек, альдостерон уменьшает содержание его в организме. При недостатке минералокортикоидов организм теряет такое количество натрия, что могут возникать изменения внутренней среды, приводящие к смерти.
Во время выполнения физических нагрузок, сопровождающихся \спленным потоотделением, а также в некоторых других условиях, вызывающих значительные потери жидкости (например, при пере
327
гревании), продукция альдостерона усиливается. В результате этого задерживается и даже полностью прекращается выведение с мочой натрия, чем компенсируются значительные его потери, вызванные потоотделением. С потом же теряется и значительное количество калия. Однако во время мышечной работы распад гликогена и тканевых белков ведет к освобождению в больших размерах ионов калия. В этих условиях усиление выведения калия через почки и пот являются более благоприятной реакцией, чем задержка его. Усиление секреции альдостерона предохраняет организм от существенных изменений содержания натрия и калия в плазме крови. Это имеет важное значение при длительных физических упражнениях, например при марафонском беге.
Клубочковая зона коры надпочечников выделяет в небольших количествах дезоксикортикостерон.
Глюкокортикоидами, продуцируемыми корой надпочечников, являются кортизон и кортикостерон. У человека значительно преобладает продукция более активного из них — кортизона. Диапазон воздействия этих гормонов весьма широкий. Они участвуют в регуляции углеводного, белкового, жирового и водно-электролитного обмена, оказывают влияние на функции сердечно-сосудистой и центральной нервной систем, способствуют повышению сопротивляемости организма к различным вредным воздействиям.
Перечисленные физиологические влияния глюкокортикоидов свидетельствуют о важном значении гормонов в процессах приспособления организма к изменениям внешней среды. Поэтому их называют также адаптивными гормонами.
Глюкокортикоиды играют важную роль в приспособлении организма к мышечной работе. Если выполняемая физическая нагрузка достаточно интенсивна, то отмечается повышенная активность коры надпочечников. В результате содержание кортизола и кортикосте-1 рона в крови увеличивается. Благодаря этому мобилизируются бел-] ковые и жировые ресурсы организма; усиливается новообразование гликогена в печени и обеспечивается эффективное перемещение ионов через клеточные мембраны и удаление из клеток воды, образующейся в результате усиления окислительных процессов; тонизируются очень многие приспособительные реакции, в том числе реакции сердечно-сосудистой системы. Однако при длительных утомительных нагрузках вслед за первоначальным усилением наблюдается угнетение продукции глюкокортикоидов. Эту реакцию можно рассматривать как защитную, направленную на предотвращение чрезмерных затрат ресурсов организма.
На уровень секреции глюкокортикоидов влияет поступление в кровь гормона аденогипофиза — адренокортикотропного гормона (А1\ТГ). В свою очередь, секреция АКТГ зависит от влияния продуктов нейросекреторных клеток промежуточного мозга. Так обеспечивается участие центральной нервной системы в управлении секрецией глюкокортикоидов, а также быстрое изменение функциональной активности железы в соответствии с внешними условиями и характером деятельности организма.
328
Общий адаптационный синдром. При действии факторов, веду, щих к изменениям внутренней среды, сохранение жизни в высокоорганизованном организме возможно только при возникновении специальных защитных реакций, обеспечивающих восстановление необходимого постоянства внутренней среды организма. Эти специальные защитные меры и компенсаторные изменения являются специфическими, т. е. они защищают организм только от данного фактора.
Наряду со специфическими существуют и неспецифические защитные реакции, т. е. предохраняющие организм от вредных для него воздействий не одного, а многих факторов. Как показал Г. Селье, особо важное место в общей, неспецифической, приспособительной реакции принадлежит усиленной деятельности коры надпочечников, в частности усиленной продукции глюкокортикоидов. Состояние организма, характеризующееся развертыванием неспецифических приспособительных механизмов, названо Г. Селье «состоянием стресса» (от англ, stress — напряжение), а обусловливающий его фактор — стрессором.
Стрессовая реакция представляет собой нормальное приспособление организма к сильному действию неблагоприятных факторов. По со стрессовой реакцией могут быть связаны и некоторые патологические изменения. Они возникают или при чрезмерной стрессовой реакции, или при недостаточной ее выраженности. У животных, у которых предварительно удалены надпочечники, стрессовая реакция отсутствует. Эти животные исключительно чувствительны к воздействию стрессоров (оно может приводить даже к смертельному исходу) .
Стрессовая реакция представляет собой общую мобилизацию защитных сил организма. В связи с этим начальный период воздействия стрессора на организм носит название стадия тревоги. После повторных воздействий стрессора эта стадия переходит во вторую — стадию резистентности (устойчивости). Она характеризуется тем, что степень активации коры надпочечников постепенно понижается, вплоть до отсутствия заметных изменений функций коры надпочечников. В то же время развиваются высокие резервные возможности коры надпочечников. Сопротивляемость организма стрессору повышается, что обеспечивается уже не благодаря усиленной продукции глюкокортикоидов, а вследствие повышения тканевой устойчивости. Всякое приспособление, однако, имеет свои границы. При длительном или слишком частом повторении воздействия стрессора или при одновременном воздействии на организм нескольких стрессоров фаза резистентности переходит в третью стадию — стадию истощения. Она характеризуется резким снижением сопротивляемости организма по отношению ко всяким стрессорам.
Продуцирование половых гормонов корой надпочечников. В сетчатой зоне коры надпочечников образуются стероидные гормоны, которые сходны со стероидными гормонами, продуцируемыми половыми железами. Так, в коре надпочечников образуются соединения, близкие по химическому составу и физиологическому воздей
328
ствию мужским половым гормонам. Поэтому они носят название андрогены (от греч. andros — мужчина), в также женские половые гормоны — эстрогены. Соответственно в организме мужчин находятся женские, а в организме женщин мужские половые гормоны.
§ 3. Половые железы
Половые железы (гонады) осуществляют две функции. Первая заключается в образовании половых клеток (в семенниках — сперматозоидов, в яичниках — яйцеклеток). Таким образом, гонады обеспечивают размножение. Вторая функция состоит в выделении гормонов.
Мужской половой гормон тестостерон получил свое название в связи с местом его образования (семенники по-латински — testis) и химической характеристикой (является стероидом). Он стимулирует развитие мужских половых органов и формирование вторичных половых признаков (характер оволосения, тембр голоса, распределение жира). От его содержания в крови зависит половая активность мужчин. Очень важна и вторая сторона физиологического действия тестостерона (а также андрогенов надпочечникового происхождения) — влияние на белковый обмен. Тестостерон и другие андрогены усиливают синтез белков и способствуют тем самым развитию гипертрофии скелетных мышц (см. главу III).
Образование женских половых гормонов и общая активность половых желез у женщин характеризуются цикличностью. Половой цикл (овариально-менструальный) длится 27—28 дней. Его разделяют на четыре периода: 1) предовуляционный, 2) овуляционный, 3) послеовуляционный, 4) период покоя (рис. 97). Предовуляционный период характеризуется увеличением фолликулов в яичнике. Один из них выступает над поверхностью яичника. Он содержит в себе яйцеклетку и в то же время выделяет гормоны — эстрогены (эстрогены образуются также в ткани яичника и в плаценте). Они активируют развитие женских половых органов и вторичных половых признаков (в том числе развитие молочных желез); влияют на половое поведение женщин, вызывают координированные сокращения труб и матки; усиливают также синтез белков, в первую очередь в матке.
По окончании стадии созревания фолликула происходит разрыв его оболочки и яйцеклетка поступает в отверстие яйцевода. Данный процесс носит название овуляции (в норме это проходит через 14 дней от начала предыдущей менструации, когда содержание эстрогенов в крови самое высокое). Остатки фолликула превращаются н желтое тело. Оно быстро увеличивается в размерах и становится органом внутренней секреции. Его гормоном является прогестерон. Он подготовляет слизистую оболочку матки к имплантации оплодотворенного яйца и способствует дальнейшему развитих плода и нормальному течению беременности, тормозя в период беременности овуляцию.
330
Если оплодотворения яйцеклетки не произошло, наступает пос-леовуляционный период. Желтое тело дегенерирует. Оно отторгается и удаляется из организма с кровотечением (менструация). По окончании этого происходит быстрая регенерация слизистой оболочки матки.
После завершения послеовуляционного периода наступает период покоя, а за ним — новый предовуляционный период.
Рис. 97. Схема полового цикла женщины на протяжении месяца.
Вверху: влияние гонадотропных гормонов аденогипофиза. ФСГ — фолликулостимулирующий гормон; ЛГ — лютенизирующий гормон; Л — лютенотропный гормон. Римские цифры — состояние фолликулов и желтого тела (/—// — дегенерирующее желтое тело, III—IV — созревание фолликула, V — овуляция, V/— максимальное развитие желтого тела). Арабские цифры — дни месяца, А — Б — степень развития слизистой матки.
Внизу: Э — уровень эстрогенов в крови, II—уровень прогестерона в крови
Активность половых желез регулируется тремя гонадотропными гормонами передней доли гипофиза — аденогипофиза (см. рис. 97); фолликулостимулирующий, лютенизирующий гормоны и пролактин.
Фолликулостимулирующий гормон активирует у женщин развитие и рост фолликулов, а также образование эстрогенов, у мужчин— сперматогенез в семенниках. Лютенизирующий гормон опре
331
деляет у женщин наступление овуляции и образование желтого тела, у мужчин стимулирует продукцию тестостерона. Пролактин и лютенотропный гормон стимулируют образование прогестерона. Они вызывают также лактацию и способствуют пробуждению материнского инстинкта.
§ 4. Поджелудочная железа
Поджелудочная железа (рапкгеаз) является и пищеварительной и эндокринной железой. Эндокринная ткань составляет лишь 1% от всего органа. Она находится в железе в виде островков Лангерганса, содержащих разные клетки. Бета-клетки выделяют гормон инсулин (от лат. insula — остров), альфа-клетки образуют гормон глюкагон.
Инсулин повышает проницаемость мембраны мышечных и жировых клеток для глюкозы. Способствуя транспорту глюкозы внутрь клетки, он обеспечивает осуществление процессов ее утилизации. Инсулин имеет также важное значение в отложении запасов углеводов в печени в виде гликогена. Недостаточность инсулина в организме приводит к тяжелому патологическому состоянию (сахарная болезнь, или диабет), которое характеризуется повышением содержания сахара в крови со 100—120 мг% в норме до 300—400 мг°/о. Инсулин в организме стимулирует образование жира, угнетает мобилизацию его из жировых депо и участвует в синтезе белков.
В начале мышечной работы содержание инсулина в крови увеличивается, при длительных физических нагрузках снижается. Снижение уровня инсулина в крови способствует во время мышечной работы переключению окисления углеводов на окисление жиров.
Глюкагон является антагонистом инсулина. Он стимулирует расщепление гликогена в печени, а также жира в жировой ткани. Во время мышечной работы его уровень в крови повышается.
Секреция инсулина и глюкагона зависит от содержания глюкозы в крови. Увеличение концентрации глюкозы в крови повышает секрецию глюкагона. Секреция инсулина регулируется также вегетативной нервной системой. Раздражение блуждающего нерва усиливает выделение инсулина. Раздражение симпатических волокон уменьшает его.
§ 5. Щитовидная железа
Щитовидная железа самая крупная из эндокринных желез. Ее вес у взрослого мужчины составляет около 20 г. Гормонами щитовидной железы являются тираксин и трийодтиронин (выделяется в небольших количествах).
В молодом организме тироксин имеет большое значение для общего его развития и роста. Недостаточность функции щитовидной железы в детском возрасте приводит к развитию так называемого кретинизма (рис. 98). Характерным признаком этого заболевания является задержка роста и нарушение пропорций тела, задержка
332
полового развития, умственная отсталость. У взрослого человека важнейшее значение гормонов щитовидной железы заключается в регуляции интенсивности окислительных процессов, усиливающихся под влиянием тироксина. Поэтому активность щитовидной железы отражается на уровне основного обмена, который при недостаточ-_шости функций щитовидной железы уменьшается, а при гиперфункции (например, при Базедовой болезни) увеличивается. Повышение секреции тироксина способствует образованию тепла в организме и, следовательно, приспособлению его к воздействию холода.
Тироксин усиливает влияние адреналина и симпатической нервной системы. Он повышает возбудимость центральной нервной системы и увеличивает также синтез белков.
Активность щитовидной железы определяется содержанием в крови тиреотропного гормона (ТТГ) аденогипофиза. В свою очередь, повышенный уровень тироксина в крови угнетает продукцию этих стимуляторов щитовидной железы.
Источником для синтеза тироксина и трийодтиронина служат аминокислота, тирозин и йод. Полноценная функция i щитовидной железы возможна только при достаточном содержании йода в пище. В случае недостатка его количество тироксина в крови снижается. Это обусловливает усиленную секрецию ТТГ. Вслед-
ствие этого щитовидная железа гипертро- Рис- 98- Случай кретинизма фируется, хотя общая продукция гормона (девушка лет) снижена. Это заболевание называется эндемическим зобом.
Влияние физических нагрузок на функцию щитовидной железы выяснено недостаточно. Можно предполагать, что при физических нагрузках происходит активация деятельности щитовидной железы и усиление тканевого расщепления тиреоидных гормонов.
Новыми исследованиями установлено, что в щитовидной железе вырабатывается еще один гормон — тирокальциотонин. Он уменьшает содержание кальция в крови, усиливая переход его из крови в костную ткань.
§ 6. Околощитовидные железы. Вилочковая железа. Эпифиз
Околощитовидные (паращитовидные) железы. Они у человека (обычно их четыре) непосредственно прилегают к задней поверхности щитовидной железы. Гормоном околощитовидных желез являйся паратгормон. Он увеличивает содержание кальция в крови (за счет вымывания его из костей скелета), воздействуя тем самым
333
на возбудимость и лабильность нервно-мышечного аппарата и центральную нервную систему. Удаление паращитовидных желез приводит к клоническим судорогам, тетании и смерти. Увеличение содержания кальция в крови позволяет предполагать активацию околощитовидной железы во время мышечной работы.
Паратгормон и тирокальциотонин регулируют содержание кальция в крови, которое изменяется в узких границах. Падение уровня кальция в крови усиливает активность околощитовидной железы и в результате увеличивает его содержание. Повышение содержания кальция в крови, в свою очередь, приводит к угнетению продукции паратгормона и обусловливает усиленное образование тирокальцио-тонина.
Вилочковая железа (тимус). Эндокринная функция этой железы изучена относительно мало. Известно, что гормон ее влияет на реализацию воздействия различных гормонов (соматотропного гормона, кортизола и инсулина). Имеются данные, указывающие на большое значение вилочковой железы в процессах развития организма и в приспособлении к вредным воздействиям.
Под влиянием различных сильнодействующих факторов и воз. действия физических нагрузок вес вилочковой железы уменьшается. Однако это не связано с угнетением ее эндокринной активности. Предполагается, что, несмотря на уменьшение веса, активность тимуса увеличивается в этих случаях.
Эпифиз. В нем образуется гормон мелатонин. Он угнетает развитие половых желез, предотвращая преждевременное половое развитие, участвует в регуляции электролитного и углеводного обмена.
Секреция эпифиза зависит от освещенности. Под влиянием света продукция мелатонина уменьшается. Считают, что цикличность активности эпифиза, соответствующая смене периодов дня и ночи, представляет собой своеобразные «биологические часы» организма.
§ 7. Гипофиз
Гипофиз содержит 3 доли — переднюю, промежуточную и заднюю.
Передняя доля гипофиза, или аденогипофиз, выполняет в эндокринном оркестре как бы роль дирижера. Как указывалось выше, такую роль он осуществляет с помощью тропных гормонов, регулирующих активность коры надпочечников (адренокортикотропный гормон), половых желез (гонадтропные гормоны) и щитовидной же лезы (тиреотропный гормон). Кроме того, адреногипофиз выделяет соматотропный гормон, или гормон роста.
Под влиянием соматотропного гормона у детей происходит ускорение роста. При недостаточной выработке этого гормона в раннем возрасте человек остается карликом. При избыточной продукции этого гормона в детском возрасте развивается гигантизм (рост человека достигает иногда даже 240—250 см —рис. 99). Избыточ ная продукция соматотропного гормона у взрослых лиц может вес
334
ти к заболеванию, называемому акромегалией. Она проявляется в непропорциональном увеличении размеров дистальных частей тела (пальцев, кистей, стоп, носа, нижней челюсти).
Соматотропный гормон повышает синтез белков, способствует оптимальному использованию клетками питательных веществ, уси. ливает освобождение жирных кислот из жировой ткани и в определенных условиях угнетает использование углеводов тканями.
Факторами, стимулирующими продукцию и выделение тропных гормонов передней доли гипофиза, являются специфические нейрогормоны (освобождающие факторы), выделяемые нейросекреторными клетками гипоталамуса. Продукция тропных гормонов регулируется также содержанием регулируемого гормона, образующегося под их влиянием. Это типичный пример регуляции по принципу отрицательной обратной связи: увеличение содержания регулируемого гормона угнетает, уменьшение его увеличивает продукцию регулятора — тропного гормона.
Физическими нагрузками вызывается усиленная продукция адренокортикотропных и соматотропного гормонов. Вероятно, усиливается также выделение тиреотропного гормона, но угнетается секреция гонадотропного гормона. В состоянии утомления секреция адренокортикотропного гормона угнетена. При работе наблюдается новое усиление секреции это-
Рис. 99. Изменение роста при геперфункции и гипофункции гипофиза.
Случай гигантизма (справа человек нормального роста)
го гормона.
Промежуточная доля гипофиза у человека развита слабо. Ее продукт — меланофорный гормон способствует образованию коричневого пигмента, который обусловливает потемнение окраски кожи.
Задняя доля гипофиза, или нейрогипофиз, выделяет два нейрогормона, местом образования которых являются нейросекреторные клетки. Один из них — антидиуретический гормон — регулирует содержание воды в организме и объем жидкости в кровеносных сосудах. При обеднении организма водой (повышении осмотического давления крови) секреция этого гормона усиливается. Основное действие антидиуретического гормона связано с канальцами почек, в которых усиливается реабсорбция воды. Вследствие этого диурез уменьшается. Физические нагрузки (в частности, те, которые обусловливают увеличение потоотделения) усиливают секрецию антидиуретического гормона.
Другой гормон задней доли гипофиза — окситоцин — способствует сокращению мускулатуры беременной матки во время родов. Во время лактации он способствует отделению молока.
335
Часть вторая
ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ ФИЗИЧЕСКОГО
ВОСПИТАНИЯ И СПОРТА
Глава XIV
ФИЗИОЛОГИЧЕСКАЯ КЛАССИФИКАЦИЯ ПОЗ И РАЗНЫХ
ВИДОВ МЫШЕЧНОЙ ДЕЯТЕЛЬНОСТИ
Мышечная деятельность человека отличается чрезвычайным разнообразием. Однако возможна ее классификация на отдельные группы, объединяемые по тем или иным физиологическим признакам.
§ 1. Статическая и динамическая работа
Работу мышц принято делить по характеру мышечного сокращения на статическую и динамическую. Деятельность мышц в условиях сохранения неподвижного положения тела или его звеньев, а также удерживания какого-либо груза обозначается как статическая работа (статическое усилие). Внешняя работа при этом отсутствует, так как в механике работа измеряется произведением силы на путь, а при статической работе, происходящей в изометрическом режиме деятельности мышц, путь равен нулю. Но и при статической работе мышца расходует энергию, которая тратится на поддержание напряжения мышцы. Между тем для большинства видов мышечной деятельности характерен ауксотонический режим работы, в которой сочетаются и сокращение и напряжение. Именно в ауксото-ническом режиме мышцы выполняют динамическую работу, т. е. работу, при которой мышечная сила приводит в движение части человеческого тела. Статическая работа мышц совершается при поддержании положения тела, динамическая — при его движениях.
§ 2. Позы тела и мышечная деятельность
Поддержание какой-либо позы требует от мышц напряжения, величина которого может широко варьировать. Максимальное напряжение мышцы характеризует е»е максимальную силу, которую принято измерять динамометрами в изометрических условиях. Такое напряжение мышцы длится, как правило, не более 1 сек. Чем меньше величина напряжения мышц, тем дольше оно может поддерживаться (рис. 100). Длительное напряжение, которое обычно поддерживается непроизвольно, характеризует тонус мышц.
Статически напряженная мышца может создавать препятствие своему же собственному кровообращению. Измерения показали, что при большом напряжении в ней развивается значительное внутреннее давление, которое сжимает кровеносные сосуды, уменьшая движение крови. Это ограничение кровотока в мышцах при статичес
336
ких усилиях является одной из причин возникновения ее утомления.
Большинство поз, поддерживаемых активно, представляет собой противодействие мышц силе земного притяжения. Такие позы именуются антигравитационными (от лат. rgavitas — тяжесть). Сложность управления ими зависит от размеров опорной поверхности, положения центра тяжести тела относительно опоры, степени мышеч-
ного напряжения, использования простейших позно-то-нических рефлексов.
Лежание. Самой простой позой является лежание. Как показали опыты, полное расслабление всех мышц тела возможно лишь при лежании на боку с несколько согнутыми звеньями тела. Именно в этих условиях, а также в условиях невесомости сгибатели и разгибатели имеют одинаковый и минимальный мышечный тонус. Если же лежание происходит при выпрямленном теле, то сгибатели оказываются растянутыми, а разгибатели
Рис. 100. Зависимость длительности статического усилия от его величины (по К. Ваххольдеру)
сокращенными и напряжен-
ными. Поэтому лежание на воде при плавании не связано с полным расслаблением мышц. При этой позе требуется активное напряжение разгибателей, противодействующее упругому сопротивлению растянутых сгибателей.
Сидение. Эта поза обусловлена небольшим напряжением разгибателей туловища и шеи при относительно расслабленной мускулатуре ног. В спорте поза сидения встречается в гребле, езде на велосипеде, на мотоцикле, на коне, а также в гимнастике. Однако вез-
де здесь сидение отягощено дополнительными, подчас весьма значи-
тельными, напряжениями мышц не только туловища, но и конечностей.
Стояние. Данная поза требует усилия разгибателей, причем не только туловища и шеи, но и ног, преодолевающих упругие силы растянутых сгибателей и вес выпрямленного тела. Трудности стояния осложнены положением неустойчивого равновесия — центр тяжести тела расположен выше площади опоры.
В спорте стояние — наиболее распространенная поза. В зависимости от размеров опорной площади можно представить различные виды спортивного стояния в такой последовательности по степени возрастающей его сложности: ноги широко расставлены (поза фехтовальщика, стрелка, борца), стойка «смирно», стояние на коньках, стояние на носках, стояние на бревне (ноги на одной линии), стояние на одной ноге, стояние на носке. Стояние может быть осложнено
г>—из
337
изменением положения туловища в пространстве (например, горизонтальное положение туловища и одной ноги в позе «ласточка»).
Вис и упор. Значительную сложность по сравнению со стоянием представляют позы, связанные с опорой на руки. Из них наиболее простой является поза «.вис». Значительно сложнее поза «упор», так как положение центра тяжести здесь более высокое и требуется значительное напряжение мышц плечевого пояса.
Стойка на кистях. Наибольшей сложностью характеризуется поза «стойка на кистях». Малая опорная поверхность кистей, высокое положение центра тяжести, необычное положение головой вниз, необходимость подавления рефлексов вертикального стояния, а также относительная слабость мышц рук по сравнению с мышцами ног — все это делает данную позу весьма сложной, для освоения которой требуется длительное обучение.
§ 3. Классификация физических упражнений
Все виды физических упражнений можно разбить в зависимости от различных физиологических характеристик совершаемых движений на определенные группы (см. схему).
Большая группа видов физических упражнений выполняется в строго постоянных условиях и характеризуется строгой постоянностью движения, доведенной до стандартности. Такие физические упражнения формируются по принципу двигательного динамического стереотипа. В ответ на стереотипную систему раздражителей возникает определенная система нервных процессов, управляющих движениями, что обусловливает стереотипный характер совершаемых движений. К такого рода упражнениям относятся упражнения в легкой атлетике, тяжелой атлетике, плавании, конькобежном и велосипедном спорте, гимнастике, гребле и др. Во всех этих видах спорта формируется определенная последовательность выполнения движений. Эти движения на тренировках и соревнованиях совершаются в строго определенных, заранее известных условиях, предусмотренных спортивными правилами. В рамках такой стандартности движений и постоянства условий их выполнения спортсмен стремится показать наилучший результат в технике выполнения движений, в мышечной силе, в быстроте движений, в проявлении выносливости.
Другая большая группа видов физических упражнений характеризуется нестандартностью, непостоянством условий, отсутствием жесткой стереотипности в совершаемых движениях. Это — единоборства (борьба, бокс, фехтование) и спортивные игры. Здесь характер движений спортсмена, который взаимодействует с противником или с участниками команд, не определен заранее и изменяется в соответствии с действиями противников и партнеров. Конечно, известная стереотипность движений имеется и в этих условиях, например при проведении некоторых приемов защиты и нападения в единоборствах. Однако в основе движений в этой группе видов спорта лежит реагирование на изменяющиеся условия, на переменные си-
338
Движения Позы
Физиологическая классификация движений в спорте
Лежание (плавание, стрельба)
Сидение (на коне, велосипеде, мотоцикле, в лодке)
Стояние: ноги расставлены (стрельба, фехтование, поднимание штанги,
борьба), ноги вместе (стойка «смирно»), ноги на одной линий (на бревне), на одной ноге, на носке (гимнастика), на коньке (фигурное катание)
С опорой
на руки: вис, упор, стойки на предплечьях, на кистях, на одной руке
I. Стереотипные (стандартные) движения
1. Движения количественного значения (оцениваемые системой CCS)
Циклические
По мощности По видам локомоций
осуществляемые осуществляемые
Максимальная Субмаксималь- Естественные Со скольжением ногами Ходьба, бег Бег на коньках с помощью рук Ходьба на лыжах
ная Большая С рычажными передачами Езда на велосипеде Гребля
Умеренная В водной среде Плавание
Ациклические
Скоростно-силовые Собственно-силовые Прицельные
Прыжки Поднимание штанги Стрельба
Метания Подачи и штрафные броски мяча
2. Движения качественного значения (оцениваемые в баллах)
По видам спорта
Спортивная и художественная гимнастика
Акробатика
Фигурное катание
Прыжки в воду и на батуте
По характеристикам движений
Сила
Быстрота
Координация
Ориентировка в пространстве н времени
Равновесие
Гибкость
Безопорность
Выразител ьность
II. Ситуационные (нестандартные) движения
Единоборства
Борьба
Бокс
Фехтование
Спортивные игры
Бадминтон
Теннис Волейбол
Водное поло Ручной мяч Баскетбол Футбол
Хоккей с мячом Хоккей с шайбой
Прессы
Бег
Лыжные гонки на пересеченной местности
Вело- и мотокроссы
Горнолыжный спорт
339
12*
туации. Действия спортсмена постоянно связаны с решением задачи, как ему поступить в данный момент, какое действие целесообразнее всего совершить в соответствии с возникшей в данный момент ситуацией.
Каждая из этих больших групп видов спорта со стандартными и нестандартными движениями, в свою очередь, делится на более мелкие группы.
Виды спорта со стандартным характером движений делятся на две группы. Одна характеризуется движениями, направленными на развитие силы, быстроты и выносливости, показатели которых измеряются в единицах пространства, силы и времени (система CGS — сантиметр, грамм, секунда). Эта группа подразделяется на две подгруппы. Одна из них включает циклические движения, другая — ациклические. В другую группу входят движения хотя и требующие определенной силы, быстроты и выносливости, но не оцениваемые в точных мерах пространства и времени. Основная задача спортсмена при их выполнении — показать способность управлять своими движениями, проявить умение сочетать их в координированные акты разной степени сложности, использовать для движений различные мышечные группы и т. п. Достижения в видах спорта, для которых характерны эти движения, оцениваются в условных единицах — баллах.
§ 4. Циклические движения
К видам спорта с циклическим характером движений относятся ходьба, бег, плавание, бег на коньках и на лыжах, гребля, езда на велосипеде.
Движения во всех этих видах спорта названы циклическими потому, что в основе каждого из них лежит повторение одного и того же цикла — круга движений (от греч. kiklos — кругооборот, круг). Все элементы движений, составляющих один цикл, обязательно присутствуют в одной и той же последовательности во всех циклах. Каждый цикл движений тесно связан с предыдущим и последующим. Эта связь носит рефлекторный характер, подобно цепному рефлексу: один рефлекс, завершаясь, вызывает следующий. Такой цепной характер свойствен ритмическому двигательному рефлексу, который и является физиологической основой циклических движений.
Ритмический двигательный рефлекс лежит в основе шагателыю-го рефлекса, который, в свою очередь, является основой циклических локомоций, т. е. передвижений тела в пространстве посредством повторяющихся движений.
Между отдельными циклическими движениями, используемыми в спорте, имеются и физиологические различия. Они определяются главным образом тем, в какой мере в данном виде локомоции используются элементарные механизмы ритмического двигательного рефлекса, или, наоборот, в какой мере данный вид локомоций отличается от простейшего шагания, какие усложнения возникают в дан-
340
нои локомоции, какова степень трудности овладения этим видом циклического движения и обучения ему.
Простейшим видом циклических движений является ходьба, в которой в наибольшей мере выражен механизм шагательного реф’ лекса. Сложнее по сравнению с ходьбой, бег. Для него характерно чередование периода одиночной опоры с периодом полета.
Более трудными, чем элементарные локомоции — ходьба и бег (в которых отталкивание от земли совершается на основе трения между ногой и грунтом), являются локомоции на скользкой опоре, к которым относятся движения в конькобежном и лыжном спорте. Бег на коньках и лыжах является, по существу, аналогом ходьбы, потому что в нем отсутствует период полета и имеется характерное для ходьбы чередование одиночной и двойной опоры. Особенно близок к ходьбе бег на лыжах, в котором, как и при обыкновенной ходьбе, в момент отталкивания от опоры происходит остановка ноги относительно опоры. При перемещении на коньках толчковая нога продолжает свое движение относительно опоры. Вместе с тем передвижение на лыжах характеризуется дополнительной сложностью, связанной с координацией движений не только ног, но и рук, совершающих отталкивание палками.
От рассмотренных видов локомоций, в которых передвижение тела происходит вследствие отталкивания ногами от почвы, сильно отличается езда на велосипеде. Хотя и здесь передвижение обусловлено работой ног, однако отсутствует опора о почву, сопровождающаяся отталкиванием и периодом переноса. Передвижение па велосипеде совершается с помощью рычажных передач, а движения ногами являются круговыми, не встречающимися в естественных видах локомоций. Основная координационная сложность езды на велосипеде заключается в крайне неустойчивом равновесии, исключающем стояние на месте.
Указанные выше основные виды локомоций представляют собой передвижение в пространстве посредством ног. Принципиально отличными от них являются локомоции, совершаемые с участием рук. К ним относятся ходьба па лыжах, гребля и плавание. В гребле, как и при езде на велосипеде, усилия передаются на опору посредством рычажных передач.
Плавание отличается от всех других видов локомоций не только тем, что происходит при горизонтальном положении тела и с участием как рук, так и ног, по главным образом тем, что опо совершается в непривычной, водной, среде. Обучение плаванию связано с торможением ряда двигательных рефлексов, выработавшихся в условиях наземного передвижения. Отсутствие твердой опоры и необходимость придания телу горизонтального положения требуют подавления выпрямительных рефлексов. Уменьшение веса тела пловца, находящегося в воде, обеспечивает относительную невесомость, что исключает необходимость антигравитационных рефлексов. Большая плотность воды по сравнению с воздухом резко уменьшает инерционность движений, что также требует перестройки характера той нервной импульсации, которая рассчитана на исполь
341
зование инерционности движений в воздушной среде. Реактивные силы, которые при движениях на земле вызывали ответные опорные усилия, требуют при движениях в воде для сохранения нужного положения тела уже совсем иных ответных действий. Поэтому движения при плавании имеют мало общего с передвижениями на земле и требуют образования совершенно новых навыков.
Общей чертой для всех циклических движений является то, что выполняемая работа может характеризоваться разными мощностью
и длительностью.
Мощность, т. е. количество работы в единицу времени, зависит от силы мышечных сокращений, их частоты и амплитуды. Поэтому бег в зависимости от силы отталкивания от земли при каждом шаге, от длины шагов, их частоты, передвижения по горизонтали, в гору, под гору может представлять собой работу различной мощности. В легкоатлетиче ском гладком беге главной величиной, от которой зависит
Рис. 101. Общая кривая гависимости рекордного времени от скорости и бега (Б), плавания (П), бега на коньках (К)
мощность, является скорость бега, представляющая собой произведение длины шагов на их частоту. Сказанное относится к любому циклическому движению: мощность везде линейно связана со скоростью передвижения. При дви-
жениях человека определить мощность не всегда представляется возможным. Поэтому в литературе, в частности в последующих главах, вместо термина «мощность» используется также термин «интенсивность». Кроме того, у разных лиц предельные мощности работы могут значительно отличаться друг от друга. Вследствие этого максимальная мощность для одного человека может соответствовать лишь умеренной мощности для другого.
Время, в течение которого может совершаться та или иная работа, т. е. ее предельная длительность, зависит от мощности. Если она велика, то длительность выполнения работы мала вследствие быстро наступающего утомления. Если мощность работы мала, то ее длительность может быть велика, потому что утомление наступает не скоро. Следовательно, между предельной длительностью работы, т. е. временем наступления утомления, и мощностью существует обратная зависимость. Она выражается уравнением Nf-PR, где N — мощность, t — время, р и К—коэффициенты. Соответственно зависимость предельного времени работы от скорости бега или плавания или другого вида локомоций выражается формулой: vt~p = K, где v — скорость передвижения. Графически эта зависимость представлена кривой на рис. 101. Эта кривая выведена на основании цифр спортивных рекордов в различных циклических видах спорта: бега, плавания, бега на коньках. Такой же кривой выражается за-
242
кривая
Рис. 102. Логарифмическая рекордов в беге и зоны относительной мощности
висимость между рекордным временем и скоростью передвижения в любых иных циклических видах спорта.
Кривая зависимости времени от скорости выглядит на рис. 101 однородной на всем ее протяжении. Однако если отложить на осях координат не абсолютные значения времени и скорости, а их логарифмы, указанная зависимость изображается в виде ломаной линии. Она показана на рис. 102. Примечательно, что места перелома прямых линий приходятся на определенные участки времени. Первый перелом происходит в районе 20 сек., второй — в районе 3—5 мин. и третий— в районе 20—30 мин. Указанные три района времени делят всю зависимость времени от скорости на четыре отрезка, названные зонами относительной мощности (В. С. Фар-фель). Эти зоны являются общими для всех циклических движений. Каждая из этих зон имеет свою физиологическую характеристику, отличающуюся от физиологических характеристик других зон относительной мощности.
Первая зона — работа максимальной мощности. Продолжительность этой работы обычно не превышает 20 сек. При такой максимальной работе явления утомления наступают уже через 10—15 сек., что
проявляется в некотором снижении мощности. Основная характерная черта работы максимальной мощности состоит в том, что работа совершается в анаэробных условиях. Мощность работы настолько велика, что организм не в состоянии обеспечить ее за счет аэробных процессов. Если бы такая работа осуществлялась за счет аэробных реакций, то органы дыхания и кровообращения должны были бы обеспечить доставку к мышцам свыше 40 л кислорода в 1 мин. Между тем даже у высокотренированного спортсмена при полном усилении функций дыхания и кровообращения потребление кислорода составляет не более 5—6 л в 1 мин. В течение же первых 10— 20 сек. работы потребление кислорода не превышает даже четверти литра. Поэтому кислородный запрос организма при работе максимальной мощности удовлетворяется лишь по ее окончании. Во время же самой работы, которая происходит в анаэробных условиях, образуется значительный кислородный долг.
Главная масса энергии, освобождающейся при максимальной мощности работы, образуется за счет энергии распада АТФ и КФ. Потребляемый кислород после работы идет на окислительный ресинтез продуктов алактатного происхождения, т. е продуктов распада АТФ и КФ. Вследствие малой продолжительности работы
343
(например, бег на 100 м) абсолютная величина кислородного долга сравнительно невелика — 7—8 л. Эти цифры характеризуют абсолютную величину кислородного запроса спринтерского бега.
Процессы дыхания и кровообращения при максимальной мощности работы усилены незначительно. Практически во время спринтерского бега осуществляется лишь несколько поверхностных дыхательных движений. Сердце за этот малый отрезок времени несколько увеличивает частоту своих сокращений, но систолический объем возрастает незначительно, чему соответствует сравнительно малое увеличение минутного объема кровообращения.
Вследствие небольшой продолжительности работы невелико поступление в кровь образовавшихся в мышцах продуктов анаэробного распада. Существенных изменений в морфологическом составе крови также не происходит.
Клетки центральной нервной системы, посылающие свои импульсы к мышцам, работающим с максимальной мощностью, функционируют, естественно, тоже на пределе своих возможностей. Рецепторы двигательного аппарата, раздражаемые при движениях, совершаемых с максимальной быстротой и силой, посылают в нервные центры предельно частые импульсы. Следовательно, как эфферентный, так и афферентный отделы нервной системы, связанные с данной двигательной функцией, работают в этом случае с максимальным напряжением. Вероятно, это обстоятельство также обусловливает быстрое утомление при работе максимальной мощности. Причиной же утомления мышц является в данном случае максимальное накопление в единицу времени продуктов аэробного распада и уменьшение АТФ, КФ.
Вторая зона — работа субмаксимальной мощности. Предельная ее продолжительность — не менее 20—30 сек., но не более 3—5 мин. При такой работе наряду с распадом АТФ и КФ происходит распад гексозофосфата, энергия которого обеспечивает ресинтез АТФ и КФ. П, результате гликолиза образуются значительные количества молочной кислоты, которая успевает диффундировать в кровь. По этой причине при работе субмаксимальной мощности в крови обнаруживаются высокий уровень молочной кислоты — свыше 200 мг%. Это вызывает подкисление крови, pH которой может снизиться до 7,0.
Помимо анаэробных процессов, интенсивно развертывающихся при работах субмаксимальной мощности, включаются также и аэробные процессы. Резко усиливаются дыхание и кровообращение. Это обеспечивает увеличение количества кислорода, притекающего с кровью к мышцам. Потребление кислорода непрерывно возрастает, но максимальных величин оно достигает обычно почти в конце работы. Образующийся кислородный долг очень велик — он значительно больше, чем после работы максимальной мощности, что объясняется продолжительностью работы. Потребляемый после работы кислород идет на окислительный ресинтез как АТФ и КФ, так и углеводов. Величина кислородного долга может достигать 20 л.
Работа субмаксимальной мощности вызывает максимальные физиологические сдвиги в организме. Это отражается на характере
344
утомления, проявляющегося к концу дистанции. Спортсмен финиширует в условиях максимального накопления молочной кислоты в мышцах и крови и усиления функций дыхания и кровообращения при изменении pH в кислую сторону, возрастании напряжения СОг уменьшении напряжения О2 в крови. Кроме того, в крови в этот момент повышается осмотическое давление вследствие перехода воды из плазмы в мышцы и начавшегося интенсивного потоотделения. Поэтому работоспособность клеток центральной нервной системы, омываемых кровью, резко изменившей свой химический состав, снижается.
Третья зона — работа большой мощности. Она характеризуется длительностью не менее 3—5 мин. и не более 20—30 мин. Здесь уже вполне достаточно времени для того, чтобы дыхание и кровообращение могли усилиться в полной мере. Поэтому работа, выполняемая через несколько минут после старта, происходит при потреблении кислорода, близком к максимально возможному. Вместе с тем кислородный запрос при такой работе больше, чем возможное потребление кислорода. Интенсивность анаэробных процессов превышает интенсивность аэробных реакций. В связи с этим в мышцах накапливаются продукты анаэробного распада и происходит образование кислородного долга. Кислород используется теперь главным образом на ресинтез углеводов.
Во время работы большой мощности заметную роль играют выделительные процессы. Потоотделение, усиливавшееся в первые минуты работы, полностью включается в терморегуляторную функцию, предохраняя организм от перегревания. Помимо этого, с потом удаляется часть молочной кислоты и других продуктов обмена, поступивших из мышц в кровь. Значительная часть молочной кислоты удаляется почками. Поэтому изменения в химическом составе крови при работе большой мощности и отрицательные воздействия на работоспособность нервных центров со стороны крови могут быть выражены несколько меньше, чем при работе субмаксимальной мощности. Однако такая работа предъявляет более высокие требования к сердцу. Оно не только резко усиливает свою деятельность в первые минуты работы, но должно функционировать почти на пределе своих возможностей в течение десятков минут. Это предъявляет особо высокие требования к выносливости сердца.
Четвертая зона — работа умеренной мощности. Она может продолжаться свыше 20—30 мин. Особенностью, отличающей зону умеренной мощности от всех трех вышеперечисленных зон, является наличие устойчивого состояния, впервые описанного А. Хиллом. Под устойчивым состоянием понимается равенство величин кислородного запроса и потребления кислорода в единицу времени. Лишь в начале работы кислородный запрос превышает потребление кислорода. Однако уже через несколько минут потребление кислорода достигает уровня кислородного запроса. Кислород, потребляемый мышцами во время работы, используется двояко: одна часть идет на окислительный ресинтез АТФ, КФ и углеводов, а другая — на непосредственное окисление жиров и углеводов. Накопление молоч-
345
ной кислоты при истинном устойчивом состоянии отсутствует или же невелико. Вследствие этого при работах умеренной мощности содержание молочной кислоты в крови практически почти не увеличивается. Кислотность крови и ее газовый состав остаются в нор* ме. Функции дыхания и кровообращения при спортивных напряжениях умеренной мощности увеличены сильно, однако не максимально. Уровень потребления кислорода может достигать примерно85% от максимального. х
При многочасовой работе умеренной мощности может наступить частичное истощение углеводных запасов в связи с большим расходом углеводов в мышцах. Начавшееся истощение гликогенных депо печени приводит к уменьшению поступления глюкозы в кровь. Уровень сахара в крови, обычно равный 80-—НО мг%, снижается до 50 и даже 40 мг%. Такой выраженный недостаток сахара в крови (гипогликемия) отрицательно сказывается на состоянии нервных центров и может привести к «отказу» от работы и обморочному состоянию. Поэтому при длительных спортивных напряжениях умеренной мощности (марафонский бег, марафонское плавание, лыжные гонки на 50 км, многочасовые велогонки и т. п.) должен предусматриваться прием спортсменом на дистанции раствора сахара или глюкозы.
Длительная работа умеренной мощности, сопровождающаяся сильным потоотделением, вызывает большие потери воды из организма, а следовательно, и потерю веса (до 0,8—1 кг в 1 час). Длительное и обильное потоотделение приводит к нарушению водно-солевого равновесия в организме и повышению осмотического давления крови. Это, однако, не означает, что восполнение убыли воды в организме и уменьшение концентрации солей в крови должны обеспечиваться только питьем большого количества пресной воды. С потом может выводиться до 3—5 г соли в 1 час, и вследствие этого многочасовая работа при высокой внешней температуре может привести к значительному уменьшению количества солей в организме. Поэтому по окончании работы, сопряженной с сильным потением, спортсмену рекомендуется помимо питья воды принимать с пищей соль.
Интенсивная мышечная работа вызывает усиление функции ряда желез внутренней секреции, в особенности надпочечников. При работе, в особенности связанной с эмоциональным напряжением, происходит усиленное образование катехоламинов и гормонов коры надпочечников. Однако при очень длительной, эмоционально напряженной и утомительной работе, которая характерна для марафонских дистанций, образование гормонов коры надпочечников резко уменьшается. Одним из признаков этого служит уменьшение или даже исчезновение из крови эозинофилов. Таким образом, нарушение функции желез внутренней секреции является одним из факторов утомления при многочасовой работе умеренной мощности.
Общая характеристика физиологических изменений, присущих рассмотренным четырем зонам мощности работы, представлена в табл. 12.
346
Характеристика работы разной относительной мощности
умеренная а а г с Й Е 1 До 0,3 о о о о о CI Ниже максимального Равно 1 О Небольшое Ниже максимального Ниже максимального
Зоны мощности большая я s Е So ’7 <м 5S До 0,5—0,4 До 900 Близко к максимальному Около 5/6 До 12 Большое Максимальное Близко к максимальному
субмаксимальная От 20—30 сек. до 3—5 мин. До 1,5—0,6 До 450 Повышается до максимума Около 1/3 До 20 Максимальное Нарастает до максимума Нарастает до максимума
максимальная С с с До 4 О оо о £3 ф 2 2 ф &• то я то ф X Ниже 1/10 До 8 Небольшое Незначительное Небольшое
Показатели Предельная продолжительность работы ф ТО м S S сд ф Д <т> о X то о. Общий расход энергии (ккал) Потребление кислорода в единицу времени при работе Отношение потребления кислорода при работе к кислородному запросу Кислородный долг (л) Повышение уровня молочной кислоты в крови Усиление дыхания Усиление работы сердца
347
§ 5. Ациклические движения
Ациклические движения не обладают слитной повторяемостью циклов и представляют собой стереотипно следующие фазы движений, имеющие четкое завершение. Наиболее типичными ациклическими движениями являются однократные движения в виде прыжка, метания, поднимания тяжестей. Все они отличаются определенной последовательностью фаз или элементов движений, характеризующихся максимальной силой или быстротой.
Для достижения наилучшего спортивного результата при выполнении силовых и скоростно-силовых упражнений мышцы должны развивать максимальную силу сокращения. Несмотря на эту общую черту, все они условно делятся в зависимости от условий выполнения упражнения на две группы. В одной из них, куда входит поднимание тяжестей, основной переменной величиной, определяющей максимум силы мышц, является преодолеваемое сопротивление — вес штанги. Напряжение, развиваемое мышцей, соответствует весу поднимаемой штанги. В то же время скорость мышечного сокращения при данном способе поднимания штанги изменяется незначительно. Движения такого типа получили название собственно-силовых. К другой группе относятся прыжки и метания, где преодолеваемое сопротивление постоянно и равно весу тела спортсмена или весу метаемого снаряда. Основной переменной величиной, в которой реализуется сила мышц, является скорость мышечного сокращения, обусловливающая дальность прыжка или полета снаряда. Движения такого типа получили название скоростно-силовых.
Помимо общих физиологических черт каждое из скоростно-силовых упражнений имеет свои особенности, определяемые характером двигательных координаций.
§ 6. Прицельные движения
К группе ациклических однократных движений относится, как отмечалось выше, и стрельба во всех ее разновидностях. Ее отличительной чертой является меткость попадания в мишень пулей или стрелой.
Специфической особенностью прицельных движений, относящихся по своей структуре к однократным ациклическим движениям, является то, что результат в них оценивается не в точных количественных мерах. Точность в стрельбе измеряется расстоянием от центра мишени, оцениваемым, однако, не в абсолютных мерах пространства, а в условных единицах — очках. В этом отношении прицельные действия приближаются к описываемой ниже группе упражнений, оцениваемых по качеству выполнения.
§ 7. Упражнения, оцениваемые по качеству выполнения
Как указывалось выше, к стереотипным движениям относятся также сложнокоординированные упражнения, оцениваемые не в количественных мерах, а по качеству выполнения — в баллах. В эту
348
группу входят гимнастика спортивная и художественная, акробатика, прыжки в воду и на батуте, фигурное катание. В этих видах спорта ярко проявляется способность управлять своими движениями.
В разных физических упражнениях демонстрируется совершенство управления силой и быстротой движений, ориентировки в пространстве и во времени. Высокий уровень гибкости в суставах конечностей и позвоночника требуется во всех упражнениях этой группы. Многие из них содержат безопорные положения, создающие особые трудности для управления. В этих видах спорта от исполнителей требуется выразительность движений, художественная, эстетическая их сторона.
§ 8. Нестандартные движения
При нестандартных движениях характер их выполнения зависит целиком от возникших в данный момент условий, в которых они должны быть выполнены. Физиологическая классификация этих движений затруднительна именно из-за их нестандартности. Их можно рассматривать с точки зрения относительной сложности тех ситуаций, в которых приходится действовать спортсмену. Эта сложность характеризуется степенью неопределенности выбора нужного движения. Чем больше число меняющихся переменных факторов, тем больше неопределенность предстоящего действия. Поэтому одним из главных факторов, определяющих сложность выбора нужного действия, является число участников. Другой важный фактор — быстрота, с которой спортсмен должен реагировать на создавшуюся ситуацию и часто возникающий при этом дефицит времени. Чем меньше времени для выбора нужного действия, тем, очевидно, труднее для спортсмена условия решения двигательной задачи. Наконец, фактором физиологической классификации этой группы движений может являться интенсивность (относительная мощность) данного вида спорта. Нестандартные движения делятся на две группы, соответствующие традиционно сложившемуся делению в спорте, на единоборство и спортивные игры.
В единоборствах сложность выбора нужного движения определяется действиями противника, с которым спортсмен находится в условиях непосредственного контакта.
Спортивные игры отличаются от единоборства наличием посредника между противниками в виде мяча или шайбы. Спортсмен реагирует в спортивных играх на действия противника или своих партнеров по команде, но его непосредственные двигательные действия связаны с шайбой или мячом, которые в силу своих упругих свойств приносят дополнительную неопределенность в возникающие двигательные задачи.
Степень сложности действий в спортивных играх определяется также числом участников, размерами площадки, быстротой передвижения, продолжительностью игры, ее правилами.
349
Глава XV
ФИЗИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА СОСТОЯНИЙ
ОРГАНИЗМА ПРИ СПОРТИВНОЙ ДЕЯТЕЛЬНОСТИ
Связанные с работой мышц изменения многих функций организма (увеличение легочной вентиляции, потребления кислорода, частоты сердцебиений, систолического и минутного объемов крови, повышение уровня обмена веществ и энергии) могут наблюдаться еще до начала мышечной деятельности, в результате возникновения предстартового и стартового состояний.
Перед выполнением работы проводят для более полной мобилизации возможностей организма подготовительные упражнения, называемые разминкой- Однако она не может сразу же после начала работы полностью обеспечить мобилизацию всех необходимых функций организма и увеличить работоспособность до требуемого уровня. В начале работы происходит постепенное повышение работоспособности организма, обусловленное сонастраиванием деятельности отдельных физиологических систем. Этот начальный период работы называется периодом врабатывания. После окончания врабатывания при длительной работе наступает устойчивое состояние.
Физическая работа обычно сопровождается утомлением. Оно характеризуется снижением функциональных возможностей организма.
После работы происходит пополнение энергетических запасов в организме и восстановление его функционального состояния. После-рабочий период называется восстановительным.
Предстартовые и стартовые реакции, врабатывание, утомление, процессы восстановления можно наблюдать при любой физической работе. Но наиболее ярко они выражены при спортивной деятельности, характеризующейся большим физическим и психическим напряжением.
§ 1. Предстартовое и стартовое состояния
Предстартовое состояние возникает за много часов и даже суток до начала соревнований, а собственно стартовое является продолжением и в большинстве случаев усилением предстартовых реакций.
В предстартовом и особенно в стартовом состояниях увеличиваются легочная вентиляция и газообмен, повышаются температура тела, частота сердцебиений, изменяется функциональное состояние двигательного аппарата.
Предстартовые и стартовые реакции обусловлены эмоциями, возникающими перед работой. Наиболее ярко они проявляются перед крупными соревнованиями.
По механизму возникновения предстартовые и стартовые реакции являются условными рефлексами. Они могут быть специфическими
250
I
и неспецифическими. Первые обусловлены особенностями предстоящей мышечной деятельности (чем больше мощность работы, тем резче выражены предстартовые сдвиги), вторые — не характером предстоящей работы, а значимостью данного соревнования для спортсмена.
В каждом отдельном случае в предстартовом периоде могут преобладать или специфические или неспецифические реакции. Если более выражены специфические, то степень предстартовых сдвигов соответствует трудности предстоящей работы. Например, на тренировочных занятиях у боксеров частота сердцебиений, кровяное давление и газообмен значительнее увеличиваются перед работой с большой грушей, чем перед работой с малой.
В случае преобладания неспецифических реакций предстартовые и стартовые сдвиги могут не соответствовать тем вегетативным реакциям, которые возникают непосредственно при работе. Например, перед стартом у бегунов и метателей частота сердцебиений может быть одинаковой, хотя характер мышечной деятельности при этих физических упражнениях различен. В то же время предстартовые сдвиги могут быть разными перед выполнением одинаковой работы. Эти факты объясняются тем, что при спортивной деятельности большее значение имеют неспецифические предстартовые реакции.
Таким образом, предстартовые и стартовые реакции обусловлены не только мощностью предстоящей работы, но и другими факторами. к ним относятся значимость соревнований, особенности Их проведения, уровень подготовленности спортсмена и тип его нервной деятельности. При благоприятном соотношении этих факторов предстартовые реакции протекают на оптимальном уровне, способствующем повышению работоспособности организма. При неблагоприятном стечении обстоятельств может возникать либо чрезмерное возбуждение организма, либо торможение его физиологических функций, что ведет к понижению работоспособности.
В психологии рассматриваются три рзновидности предстартовых и стартовых реакций: состояние боевой готовности предстартовая лихорадка и предстартовая апатия. Физиологические исследования подтверждают наличие этих разновидностей предстартового состояния.
Боевая готовность характеризуется оптимальными изменениями в функциональном состоянии центральной нервной системы спортсмена. Это сопровождается умеренными соматическими и вегетативными реакциями: повышается возбудимость и лабильность двигательного аппарата, усиливаются деятельность органов дыхания и кровообращения.
Предстартовая лихорадка характеризуется резко выраженными процессами возбуждения в центральной нервной системе, что снижает способность спортсмена к дифференцированию раздражителей и может вести к тактическим ошибкам, ухудшающим спортивный результат. Чаще такие ошибки возникают при спортивных играх, но иногда бывают и при работе циклического характера. Вегетативные сдвиги при предстартовой лихорадке очень велики. Уча-
351
щение сердцебиений, повышение газообмена и температуры тела могут быть очень значительными.
Предстартовая апатия характеризуется преобладанием тормозных процессов в центральной нервной системе. Изменения соматических и вегетативных функций выражены мало. Предстартовая апатия часто возникает при недостаточной тренированности спортсмена. Иногда она наблюдается при ожидании встречи с очень сильным противником или при неожиданном перенесении старта на более позднее время.
Предстартовая апатия обычно отрицательно влияет на результаты спортивных выступлений.
Разновидность предстартового состояния и выраженность возникающих при этом реакций зависят от уровня тренированности спортсмена. У тренированных вегетативные сдвиги могут быть даже более выраженными, чем у нетренированных, но эти сдвиги сочетаются у первых с лучшей уравновешенностью нервных процессов, что играет большую роль в поддержании высокой работоспособности.
Предстартовые реакции можно регулировать. В процессе тренировки спортсмена следует обучать его управлению эмоциями. Кроме того, необходима правильная организация отдыха в дни и часы, предшествующие спортивному соревнованию. Длительное пребывание перед стартом в обстановке соревнований (непосредственно в зале, на стадионе) может оказывать отрицательное воздействие. В этих случаях для поддержания высокой работоспособности следует переключать внимание спортсмена на иную деятельность.
Одним из важных приемов, регулирующих предстартовые реакции, является разминка. При подборе упражнений, выполняемых при разминке, необходимо учитывать особенности предстартовых реакций. Эффект разминки обусловлен следующим. Если в предстартовом состоянии преобладают тормозные процессы, то разминка может уменьшить или совсем снять это торможение. При преобладании же возбудительных процессов разминка, усиливая возбуждение в двигательном анализаторе, способствует снижению его в других центрах. Таким образом, благоприятное воздействие ее при всех формах предстартовых реакций связано с установлением оптимальных соотношений между возбудительными и тормозными процессами в центральной нервной системе.
Регуляция предстартовых реакций может осуществляться путем словесных воздействий на вторую сигнальную систему спортсмена. Анализ предстоящей борьбы, оценка своих возможностей и сил соперников способствуют оптимизации возбудительных процессов в предстартовом состоянии.
Некоторое значение для регуляции предстартовых реакций имеет массаж незадолго до старта. Усиливая поток афферентных импульсов от двигательного аппарата и кожи, он действует аналогично разминке. Примерно таков же и механизм повторных глубоких дыханий, которые рекомендуется производить перед выходом на старт.
352
§ 2. Изменение функционального состояния организма при разминке
Разминка состоит из общей и специальной частей. Первая способствует созданию оптимальной возбудимости центральной нервной системы и двигательного аппарата, повышению обмена веществ и температуры тела, деятельности органов кровообращения и дыхания. Вторая часть направлена на повышение работоспособности тех звеньев двигательного аппарата, которые будут участвовать в предстоящей деятельности.
Общая часть разминки может быть почти одинаковой во всех видах спорта; специальная же часть должна быть тесно связана со специализацией спортсмена. Например, при игре в баскетбол специальная часть разминки должна отражать специфические особенности игровой деятельности. При этом для повышения эффективности разминки отдельные приемы следует выполнять в условиях, по возможности приближенных к условиям сложной игровой обстановки.
Правильно организованная разминка способствует повышению возбудимости и лабильности нервных центров, что создает оптимальные условия как для возникновения новых временных связей в процессе обучения, так и для осуществления уже усвоенных двигательных навыков в сложных условиях спортивной деятельности. Благодаря этому после разминки укорачивается время между стартовым сигналом и началом движений, улучшается ориентировка в окружающей обстановке, уменьшается количество неправильных реакций при ее изменениях.
Под влиянием разминки повышаются активность ферментов и скорость протекания биохимических реакций в мышцах, возбудимость и лабильность мышц.
Особенно велико значение разминки при физических упражне-нениях, требующих значительного повышения потребления кислорода. Это обусловлено усилением деятельности органов дыхания и сердца, выходом крови из депо, перераспределением крови между работающими и неработающими органами, повышением температуры тела. Последнее способствует более интенсивной диссоциации оксигемоглобина в тканях. Кроме того, при повышении температуры снижается вязкость мышц, что предохраняет спортсмена от травм.
Физиологические сдвиги, вызванные разминкой, не исчезают сразу после ее прекращения. Они оставляют следы, которые обеспечивают улучшение работоспособности при последующей деятельности. Например, при повторении работы легочная вентиляция, несмотря на ее восстановление в интервале отдыха, будет больше, чем при предшествующей работе. Это объясняется следовыми явлениями в нервных центрах, способствующими более быстрой и полной мобилизации дыхания при повторной работе. Следовые явления после работы зависят от ее длительности и от интервала отдыха между предварительной работой и последующей. Например, предва
353
рительная 10-минутная работа вызывает большее увеличение легочной вентиляции, чем 5-минутная, а 10-минутный интервал отдыха при этом более эффективен, чем 15-минутный (табл. 13).
Т
Влияние предыдущей работы на легочную вентиляцию при последующей работе (по Б. С. Гиппенрейтеру)
Продолжительность (мин.) Легочная вентиляция (л) за 5 мин. Легочная вентиляция (л) за 1 мин. перед началом
1-я работа перерыва 1-я работа 2-я работа разница 1-я работа 2-я работа
10 10 251,0 308,9 57,9 9,2 9,0
10 15 220,9 265,1 44,2 7,1 7,3
5 10 238,3 258,8 20,5 7,7 7,6
5 15 253,5 265,1 11,6 8,2 8,3
Предварительная работа укорачивает время до начала потоотделения при последующей работе. Это обусловлено сохранением повышенной температуры тела в течение 20—30 мин. после выполнения первой работы.
Таким образом, благоприятное воздействие разминки заключается не только в возникновении кратковременных физиологических сдвигов, но и в сохранении относительно длительных следовых явлений, обеспечивающих повышение работоспособности.
Оптимальные величины длительности разминки и длительности интервала между ее окончанием и началом работы определяются видом спортивной деятельности, степенью тренированности спортсменов, метеорологическими условиями и другими факторами. В среднем разминка должна продолжаться 10—30 мин. Необходимо, чтобы уже при разминке началось потоотделение, свидетельст-> вующее о готовности теплорегуляционных механизмов к повышенным требованиям во время работы.
Лабораторные исследования показали, что оптимальным для отдыха между разминкой и началом основной работы является 3-минутный интервал. В практике спортивной деятельности эти интервалы обычно более продолжительны. Поэтому непосредственно перед стартом рекомендуется дополнительно выполнить несколько упражнений, соответствующих предстоящей работе.
Разминка не должна вызывать утомления. Количество работы, выполненное при разминке, должно быть строго индивидуализировано. Кроме того, для предупреждения утомления мышц при разминке целесообразно нагружать не только те мышцы, которым предстоит основная работа, но и те, которые не будут участвовать в ее выполнении. Хронаксиметрическими исследованиями мышц после напряженной работы было обнаружено, что хронаксия не участвовавших в работе мышц снижается. Это косвенно свидетельствует о
354
повышении их лабильности, а следовательно, и работоспособности. При выборе упражнений для разминки надо учитывать эту закономерность.
§ 3. Врабатывание
Постепенное повышение работоспособности организма при работе называется врабатыванием. Оно обусловлено усилением деятельности физиологических систем, принимающих участие в работе.
Чем быстрее заканчивается врабатывание, тем при прочих равных условиях выше производительность работы.
Врабатывание следует рассматривать как адаптацию организма к более высокому уровню деятельности. Известно, что работоспособность спортсмена при выполнении физических упражнений нарастает постепенно. Например, при метаниях и прыжках более высокие результаты чаще достигаются при повторных попытках. Эта закономерность менее ярко проявляется при длительной работе умеренной мощности. Однако и здесь, если это не идет вразрез с тактическим замыслом, скорость передвижения в начале работы обычно несколько меньше, чем на последующих километрах.
Большое значение для врабатывания имеет возникновение специфической для данной деятельности доминанты, которая обеспечивает необходимую координацию всех физиологических процессов в организме.
Важную роль при врабатывании играет также автоматически действующая регуляция функций. При этом чем интенсивнее работают мышцы, тем резче выражены изменения во внутренней среде и тем сильнее проприоцептивная и интероцептивная афферентация, рефлекторно регулирующая деятельность организма. Усиление афферентных импульсов изменяет функциональное состояние соответствующих центров и обеспечивает более полную мобилизацию всех возможностей организма.
Адаптация всех систем организма к деятельности на высоком уровне, необходимом при физической работе, происходит преимущественно условнорефлекторным путем. При этом следует различать условнорефлекторные пусковые воздействия, обеспечивающие переход от состояния покоя к работе, и воздействия, регулирующие деятельность организма непосредственно при работе. О значении условнорефлекторных раздражителей для процессов врабатывания свидетельствуют многие факты. Например, уведомление человека о предстоящей тяжелой и длительной работе удлиняет период врабатывания.
При врабатывании происходит изменение функционального состояния многих нервных центров, а следовательно, и межцентральных отношений. Исследования показали, что в начале напряженной работы в центрах некоторых вегетативных и соматических функций возникает торможение (Ю. И. Данько). Это происходит по механизму отрицательной индукции в связи с сильным возбуждением дви-1 ательных центров. Тормозная фаза, характерная для начала вра-
355
батывапия, сменяется возбуждением, обеспечивающим повышение работоспособности. Торможение в начале работы выражено тем резче, чем труднее и непривычнее работа. С развитием тренированности тормозная фаза укорачивается. О постепенном уравновешивании возбудительно-тормозных нервных процессов при врабатывании свидетельствует происходящее в это время изменение условно-рефлекторной деятельности. В начале работы удлиняется скрытый период условных реакций, интенсивность же их снижается. По мере врабатывания условнорефлекторная деятельность нормализуется.
Регистрация биопотенциалов мозга позволяет наблюдать изменения в деятельности нервных центров при врабатывании. Происходит снижение вариативности амплитудных и частотных характеристик электроэнцефалограммы, а также увеличение интенсивности периодических процессов по сравнению со случайными (Е. Б. Сологуб).
Различные системы организма настраиваются на рабочий уровень не одновременно (гетерохронно). Двигательный аппарат, обладающий относительно высокой возбудимостью и лабильностью, настраивается быстрее, чем вегетативные системы. Однако и для повышения работоспособности скелетных мышц требуется определенное время. Например, скорость бега па 100 м на 1-й сек. составляет в среднем 55% от максимальной, па 2-й — 76% и лишь на 5—6-й сек. достигает максимума. Наблюдения за лучшими бегунами-спринтерами показали, что наивысшей скорости при беге на 100 м они достигают не ранее чем на 35—40-м метре дистанции. Такое относительно медленное повышение скорости бега обусловлено биомеханическими особенностями движений на старте и постепенным врабатыванием двигательного аппарата.
Значительно позже двигательных функций достигает необходимого уровня работа отдельных внутренних органов. При этом повышение различных вегетативных показателей происходит не одновременно. Сердечный ритм учащается уже на первых секундах работы. В большинстве случаев к концу 1-й мин. работы этот показатель достигает почти максимального уровня. В одних случаях учащение сердцебиений происходит плавно, в других — скачкообразно. Иногда при этом возникают экстрасистолы, называемые «экстрасистолами включения» Укорочение сердечных циклов в начале работы обычно сочетается с уравниванием их длительности. В некоторых случаях при врабатывании происходит усиление синусовой аритмии, что можно объяснить «поиском» оптимального для данных условий ритма. Укорочение и уравнивание сердечных циклов при врабатывании обусловлено усилением влияния на сердце симпатических нервов и торможением блуждающих. Сердечный ритм имеет тесные связи с дыхательными движениями. При этом чем выше частота дыханий, тем стабильнее длительность сердечных циклов.
Увеличение систолического объема крови при врабатывании происходит одновременно с учащением сердечного ритма. Уже с первым сокращением сердца при работе желудочки выбрасывают крови больше, чем в условиях мышечного покоя.
356
Артериальное давление начинает повышаться в самом начале работы. Систолическое давление при этом изменяется волнообразно, что обусловлено некоторым несоответствием между увеличением объема циркулирующей крови и расширением периферических сосудов.
Врабатывание дыхательных функций происходит в течение нескольких минут. Поэтому в начальном периоде работы потребность организма в кислороде не удовлетворяется, в связи с чем образуется кислородный долг.
При спортивной деятельности период врабатывания может иметь разную длительность. Это зависит от характера выполняемой работы, степени тренированности спортсмена, его индивидуальных особенностей и функционального состояния в день работы. При скоростной работе врабатывание заканчивается через несколько секунд, при более длительной — через несколько минут. Есть данные, показывающие, что врабатывание происходит медленнее при сложных по координации движениях. У более тренированных при прочих равных условиях этот процесс осуществляется быстрее, чем у нетренированных. Укорочению его способствует правильно организованная разминка.
При увеличении мощности работы в процессе ее выполнения также могут возникать явления, сходные с врабатыванием. Но они заканчиваются быстрее, так как происходят на фоне уже повышенной деятельности всего организма.
§ 4. Устойчивое состояние
Устойчивое состояние, возникающее после окончания врабатывания, наблюдается при работе, продолжающейся не менее 4— 6 мин. Потребление кислорода при этом стабилизируется. Деятельность других органов и систем также устанавливается на относительно постоянном уровне.
Различают истинное и кажущееся устойчивое состояние. Первое возникает при работе умеренной мощности, второе — при работе большой мощности.
Истинное устойчивое состояние характеризуется высокой согласованностью функций двигательных и вегетативных систем. Внутренняя среда организма при этом не изменяется. Многие из биохимических поставщиков энергии целиком восстанавливаются непосредственно в процессе работы.
Для поддержания устойчивого состояния при длительной работе необходима мобилизация всех систем организма. Минутный объем крови, легочная вентиляция и потребление кислорода достигают величин, необходимых для данной работы, и удерживаются на этом уровне (рис. 103).
При длительной работе в скелетных мышцах преобладают аэробные реакции, что обеспечивает почти полный ресинтез гликогена. Молочная кислота накапливается в мышцах в минимальном количестве и почти не диффундирует в кровь, что обеспечивает сохранение кислотно-щелочного равновесия.
357
Для поддержания устойчивого состояния большое значение имеет ресинтез макроэргических фосфорных соединений. Существенную роль при длительной работе играет переключение с гликолитического ресинтеза богатых энергией веществ на окислительный. Опыты на животных показали, что искусственное уменьшение окис-
Рис. 103. Потребление кислорода (заштрихованная площадь) и кислородный долг при работе, характеризующейся истинным (А) и кажущимся (Б) устойчивым состоянием
лительного фосфорилирования затрудняет возникновение устойчивого состояния (Н. Н. Яковлев).
Механизмы возникновения и поддержания устойчивого состояния у разных людей могут быть различными. У одних повышенную доставку кислорода обеспечивает главным образом резко нарастающая легочная вентиляция. У других при относительно небольшом увеличении этого показателя устойчивое состояние обеспечивается более эффективным использованием кислорода из вдыхаемого воздуха. Наконец, в некоторых случаях при относительно умеренном увеличении функции дыхательного аппарата устойчивое состояние обусловлено главным образом усилением деятельности сердца.
При кажущемся устойчивом состоянии деятельность дыхательного аппарата и сердца приближается к уровню, необходимому для обеспечения выполняемой работы. Однако, несмотря на это, кислородная потребность полностью не удовлетворяется, и в каждый момент работы образуется и постепенно нарастает кислородный долг (см. рис. 103). Необходимость усилить доставку кислорода к тканям резко повышает требования к органам кровообращения. Частота сердечных сокращений и минутный объем крови при кажущемся устойчивом состоянии близки к предельным величинам. Кислородная недостаточность ведет к усилению анаэробных процессов. В результате в мышцах, а затем и в крови нарастает концентрация
358
молочной кислоты и происходят значительные сдвиги pH в кислую сторону.
При кажущемся устойчивом состоянии внутренние органы, работая в режиме близком к пределу, не могут полностью обеспечить кислородную потребность. О наличии устойчивого состояния в этих случаях говорят лишь потому, что потребление кислорода, постепенно нарастая в периоде врабатывания, достигает определенного уровня, который сохраняется в течение относительно длительного времени (до 20—30 мин.).
Работа при кажущемся устойчивом состоянии требует большого напряжения функций как двигательного аппарата, так и всех систем, обеспечивающих его деятельность.
Стабилизация физиологических процессов, наступающая при повторной работе (например, при многократном пробегании отрезков дистанции на тренировочном занятии), также является своеобразным «устойчивым состоянием». В этих условиях частота сердцебиений, легочная вентиляция, потребление кислорода и другие физиологические показатели сначала нарастают при каждой последующей работе, затем период врабатывания заканчивается, и дальнейшее повторение работы осуществляется при относительном постоянстве физиологических функций.
§ 5. «Мертвая точка» и «второе дыхание»
Напряженная работа не может продолжаться длительное время. Через несколько минут — а при работе максимальной мощности даже через несколько секунд— в организме наступают изменения, вынуждающие его прекратить мышечную деятельность. Эти изменения обусловлены несоответствием между интенсивной деятельностью двигательного аппарата и функциональными возможностями вегетативных систем организма, обеспечивающих доставку кислорода к работающим мышцам. Вследствие этого нарушается внутриклеточный обмен в центральной нервной системе, в сердце и мышцах и происходят значительные сдвиги во внутренней среде организма.
При работе меныпей мощности, характеризующейся устойчивым состоянием, также может возникать несоответствие между деятельностью двигательного аппарата и внутренних органов. Но здесь это несоответствие выражено менее резко, в связи с чем его можно преодолеть и в известной мере восстановить работоспособность. Такое временное снижение работоспособности называют «мертвой точкой», а состояние, возникающее после ее преодоления, — «вторым дыханием».
«Мертвая точка» и «второе дыхание» характерны для работы циклического характера большой и умеренной мощности. Эти состояния впервые были обнаружены в конце XIX в. у бегунов на длинные дистанции. При «мертвой точке» резко учащается дыхание, увеличиваются легочная вентиляция, поглощение кислорода и выведение углекислоты. Несмотря на увеличение легочной вентиляции,
359
напряжение углекислоты в крови и в альвеолярном воздухе нарастает. Частота сердцебиений при этом резко увеличена, кровяное давление повышено, pH крови снижено. Артерио-венозная разность по кислороду резко увеличена.
При выходе из «мертвой точки» поглощение кислорода и артериовенозная разность понижаются, легочная же вентиляция некоторое время еще повышена, что объясняется необходимостью освободить организм от накопившейся углекислоты. При «втором дыхании» постепенно происходит относительное восстановление кислотно-щелочного равновесия.
При «мертвой точке» начинается потоотделение, усиливающееся при «втором дыхании». Это свидетельствует о «настройке» теплорегуляционных механизмов на необходимый уровень, что играет большую роль в поддержании работоспособности. Усиление потоотделения при «втором дыхании» происходит почти всегда. Поэтому некоторые исследователи объясняли выход из «мертвой точки» освобождением организма от избытка молочной кислоты с потом. Однако в дальнейшем было доказано, что «второе дыхание» может опережать возникновение усиленного потоотделения.
Механизм возникновения «мертвой точки» не вполне ясен. По-видимому, она обусловлена временным нарушением соответствия в деятельности скелетных мышц и органов, обеспечивающих доставку кислорода. Возникающие в этот момент неблагоприятные изменения в состоянии нервных центров усиливают разлад в работе физиологических систем. Выходу из «мертвой точки» способствует восстановление нормальных соотношений между возбудительными и тормозными процессами в центральной нервной системе.
Возникающее при «мертвой точке» несоответствие в деятельности отдельных систем организма может быть преодолено не при всякой работе. Например, при работе максимальной и субмаксимальной мощности «второе дыхание» не наступает и работа продолжается при нарастающем утомлении.
При разной по длительности и мощности работе «мертвая точка» возникает в разные сроки. Например, при беге на 5 и 10 км ее можно наблюдать через 5—6 мин. после начала работы. При беге на более длинные дистанции состояние «мертвой точки» наступает позднее. Иногда при такой работе она может наблюдаться повторно.
Время возникновения, продолжительность и степень проявления «мертвой точки» зависят от многих факторов. Главные из них — степень тренированности спортсмена и мощность выполняемой работы. У более тренированных может и не быть «мертвой точки» или она наступает позднее и протекает легче, чем у нетренированных. Быстрое включение организма в интенсивную деятельность ускоряет момент появления «мертвой точки».
Разминка ослабляет проявление «мертвой точки» и способствует более быстрому возникновению «второго дыхания».
Преодоление «мертвой точки» требует значительного волевого напряжения. В процессе тренировочных занятий спортсмен должен быть приучен к неприятным ощущениям, появляющимся при кисло
360
родной недостаточности и накоплении в организме недоокисленных веществ (молочная кислота и др.).
Наступлению «второго дыхания» способствует произвольное увеличение легочной вентиляции. Особенно эффективны при этом глубокие выдохи, усиливающие выведение углекислоты из организма, благодаря чему восстанавливается кислотно-щелочное равновесие. Преодоление «мертвой точки» может быть ускорено снижением мощности работы. Но это нельзя рекомендовать в условиях соревнований. Нецелесообразно снижать мощность работы при возникновении «мертвой точки» также и на тренировочных занятиях. В процессе тренировки адаптация человека к напряженной мышечной деятельности происходит лишь при значительном изменении внутренней среды организма.
§ 6. Утомление
При мышечной деятельности может возникать утомление, характеризующееся комплексом изменений в состоянии различных функций организма. Степень выраженности этих изменений, в том числе чувства усталости, тем больше, чем интенсивнее и продолжительнее была проделанная работа.
Определение и основные показатели утомления. Утомлением называется особое состояние, возникающее как следствие работы и проявляющееся в ухудшении двигательных и вегетативных функций и их координации, понижении работоспособности и появлении чувства усталости. Это состояние имеет временный характер и исчезает через некоторое время после прекращения работы, т. е. во время отдыха.
Внешние проявления мышечного утомления разнообразны. Они зависят от характера выполняемых физических упражнений, особенностей внешней среды и личных индивидуальных особенностей спортсменов. К внешним проявлениям утомления, часто встречающимся в спорте, относятся: нарушение координации движений, падение производительности работы, одышка, чрезмерная потливость, покраснение кожных покровов и др.
Эти внешние проявления обусловлены как ухудшением работы периферических органов, так и расстройством координации их деятельности нервной системой.
Изменение координации функций периферических органов, возникающее через некоторое время после начала работы происходит в одних случаях еще до снижения работоспособности исполнительных аппаратов и представляют собою как бы профилактическое мероприятие, позволяющее более длительно сохранить высокую эффективность работы. В других случаях оно наступает вследствие расстройства функций нервной системы, которое бывает при сильном утомлении.
Ухудшение функций периферических органов при работе, возникающее в результате неполноценной нервной регуляции, может проявляться в различных формах. Во-первых, могут снижаться по
361
казатели деятельности различных органов и систем органов (например, уменьшаются минутный объем легочной вентиляции, минутный объем крови, потребление кислорода). Во-вторых, вследствие нарушения координации может наблюдаться более высокая, чем необходимо, степень мобилизации функций периферических органов. Это выражается в меньшей экономичности работы различных систем организма, в особенности при перерасчете затраченной энергии на 1 кг веса тела, на 1 м пройденной дистанции или на 1 единицу времени.
В целях сохранения работоспособности периферических исполнительных аппаратов нервная система может изменять формы координации их деятельности: заменять работу одних мышечных элементов другими, уменьшать глубину дыхательных движений и т. д.
Сочетание разных показателей утомления. В зависимости от вида работы и состояния различных функций организма наблюдается разное сочетание показателей утомления. В одних случаях отмечаются и снижение функций периферических органов, и ухудшение координации нервной системой их деятельности, и понижение производительности работы, и ощущение усталости. В других случаях могут быть только один или два из этих общих показателей утомления. Например, если выполняется интересная работа, то при наличии ряда перечисленных показателей может отсутствовать субъективная сторона утомления — чувство усталости, которое во многих случаях появляется только в самом конце работы или даже после нее. При неинтересной работе, наоборот, ощущение усталости может обнаруживаться значительно раньше возникновения объективных признаков ухудшения вегетативных и двигательных функций организма и снижения работоспособности.
При циклической длительной работе с предельной или со значительной интенсивностью ухудшается в результате развития утомления состояние ряда периферических органов, менее совершенной становится деятельность нервной ситемы, что приводит к нарушению нормальной координации двигательных и вегетативных функций, снижается производительностъ работы, возникает чувство усталости. При аналогичной же длительной работе малой мощности производительность работы может остаться той же, хотя при этом нарушаются некоторые показатели структуры движений, изменяется деятельность отдельных вегетативных органов и возникает ощущение усталости. Это наблюдается, например, при многочасовых передвижениях с небольшой скоростью.
Расхождение между физиологическими показателями утомления и производительностью работы наблюдается часто прн финишных ускорениях. Спортсмен на финише может на короткое время существенно увеличить скорость передвижения даже при наличии ряда субъективных и объективных признаков утомления.
Утомление и состояние энергетических ресурсов. Несмотря на то что утомление приводит к временному снижению работоспособности, оно имеет важное биологическое значение, являясь сигналом о частичном истощении ресурсов.
362
Снижение или прекращение деятельности скелетных мышц, сердца, желез внутренней секреции и других органов происходит всегда при наличии еще некоторого остаточного запаса энергетических и других веществ. Это связано с тем, что как полное, так и частичное, но резкое снижение содержания этих веществ может вызвать перерождение, а в некоторых случаях даже гибель определенных клеток организма. Утомление при работе возникает еще при наличии значительных резервов, приводя к снижению или прекращению деятельности. Эти резервы частично используются человеком в экстренных случаях, например при спуртах, финишном ускорении.
Роль эмоционального возбуждения при утомлении. При возникновении эмоциональных состояний существенно изменяются воздействия центральной нервной системы на органы и ткани. При положительных эмоциях усиливаются влияния через симпатические нервы. При этом увеличивается секреция катехоламинов—адреналина, норадреналина Повышение деятельности симпато-адреналовой системы способствует увеличению степени мобилизации энергетических ресурсов в работающих органах и улучшает деятельность мышц. При отрицательных эмоциях может наблюдаться ухудшение ряда функций организма и снижение работоспособности.
Эмоциональные факторы играют существенную роль при спуртах и на финише: несмотря на выраженные симптомы утомления, спортсмен может увеличить скорость передвижения.
Теории утомления. По вопросу о физиологических механизмах мышечного утомления было предложено много различных теорий. По одним из этих теорий, причины возникновения утомления непосредственно локализовались в работающих мышцах— теория истощения энергетических ресурсов, теория «задушения» в результате нарастающего недостатка в кислороде, теория «засорения» мышцы продуктами распада энергетических веществ, теория «отравления» в результате накопления «кенотоксинов» (т. е. мышечных ядов) в мышце при работе. Другие теории связывали возникновение утомления только с деятельностью нервной системы, в частности коры больших полушарий.
В настоящее время получены экспериментальные данные, не позволяющие локализовать причины утомления в каком-либо одном органе или системе органов, в том числе в нервной системе. Мышечная деятельность связана с вовлечением в деятельность многих органов. В многозвенной же системе, обеспечивающей производительность мышечной работы, снижение работоспособности может быть вызвано недостаточностью работы не только нервной системы, но и различных других рабочих звеньев: скелетных мышц, органов дыхания, сердца, желез внутренней секреции и др. Согласно современным представлениям о мышечном утомлении, оно связано, во-первых, с наличием «многосистемных» изменений функций, т. е. изменений не в каком-либо одном органе или системе органов, а во многих из них, и, во-вторых, с различным сочетанием деятельности органов, ухудшение функций которых наблюдается при том или ином виде физических упражнений.
363
Первичные и вторичные проявления утомления. Поскольку в организме все проявления деятельности обусловлены одновременным вовлечением в работу целого ряда органов и систем органов, т. е. являются системными, нарушение тех или других функций при развитии утомления наблюдается в показателях работы многих органов.
Ухудшение функции того или другого органа может быть как первичным, так и вторичным. Первичное снижение функций характеризуется тем, что орган начинает работать менее интенсивно вследствие изменений, возникающих непосредственно в нем самом. Например, некоторые мышечные волокна не способны длительно поддерживать сокращение и расслабляются даже в том случае, когда в них продолжают поступать импульсы из центральной нервной системы. При вторичном снижении функций та же мышца, даже если она полностью сохранила свои свойства, начинает плохо сокращаться в результате ухудшения регуляции ее деятельности центральной нервной системой.
Начало утомления обусловлено большей частью первично возникающими изменениями в состоянии некоторых органов. При продолжении работы снижение функций многих систем органов вызывается уже вторичным путем. Например, при длительной мышечной работе таким вторичным путем может ухудшаться как состояние самих мышц и вегетативных систем органов, так и нервная регуляция их деятельности.
Поскольку при мышечной работе в деятельность вовлекается большинство систем органов, то развитие утомления характеризуется причудливым сочетанием и первично и вторично вызванных изменений деятельности разных органов. Вследствие этого в ряде случаев очень сложно отличить первично возникающие изменения от вторичных. Иногда такая дифференциация возможна. Так, в горах первичной причиной, вызывающей возникновение у спортсменов утомления, является гипоксия, которая ухудшает возможность доставки кровью кислорода тканям организма. При низком уровне тренированности утомление первично проявляется в виде недостаточной деятельности сердца и неполноценного перераспределения крови и т. д.
Ярким примером значения первичных изменений вегетативных процессов для вторичного вызова ухудшения деятельности нервной и других систем является изменение содержания глюкозы в крови. Большое потребление во время длительной интенсивной мышечной работы (бег, лыжные гонки и др.) глюкозы через некоторое время приводит к снижению содержания ее в крови. Это снижение вторичным путем ведет к ухудшению состояния мозга, весьма чувствительного к понижению сахара в крови. В свою очередь, ухудшение состояния мозга может нарушить координацию самых различных двигательных и вегетативных функций, снижая тем самым эффективность выполнения физических упражнений. Таким образом, первичные изменения какой-либо одной вегетативной функции приводят к нарушению функций целого ряда органов и систем органов.
364
Во многих случаях, в частности при значительных по мощности усилиях, первичное утомление возникает в результате ухудшения функционального состояния самих мышц. Некоторым функциональным единицам в мышце свойственна высокая степень утомляемости, и они могут выключаться из деятельности спустя весьма короткое время после начала напряжения. Вследствие этого сила данной мышцы может быстро снижаться, иногда даже на протяжении нескольких секунд. Так, при статических усилиях с большим отягощением деятельность некоторых функциональных единиц прекращается через относительно короткое время. Поэтому спустя несколько секунд поддержание усилия становится невозможным (например, удержание штанги, угла на кольцах).
При возникновении утомления снижаются сила и скорость сокращения мышц, затягивается фаза расслабления, повышаются пороги возбудимости, снижается коэффициент полезного действия мышечного сокращения. При сильной же степени утомления после прекращения сокращения может наблюдаться неполное расслабление мышечных волокон, так называемая остаточная контрактура.
Важнейшая роль при утомлении принадлежит процессам, протекающим в центральной нервной системе. Деятельность нервной системы, как и других систем организма, может ухудшаться как первично, так и вторично.
При умственной работе, при трудных для спортсмена ситуациях, при боязни соперников и в некоторых других случаях изменения деятельности нервной системы являются первичным фактором, вызывающим утомление. Вторично деятельность нервной системы угнетается при увеличении в крови содержания недоокисленных продуктов обмена веществ, гипоксии, гипогликемии, изменений содержания гормонов в крови и т. д. В результате ухудшаются программирование и координация двигательных и вегетативных функций, что приводит к снижению производительности работы спортсмена и возникновению чувства усталости.
Каждая клетка, каждый орган обладают способностью в известной мере противостоять утомлению. Но особая роль в этом отношении принадлежит нервной системе, которая, программируя и координируя все двигательные и вегетативные функции, может варьировать формы координации, заменяя в работе утомленные клетки и органы неутомленными или менее утомленными, повышать степень мобилизации ресурсов в работающих органах и тканях и т. д. Вследствие этого значительно отдаляется момент возникновения утомления и снижения работоспособности.
Преодолеваемое (скрытое) утомление. Смена форм координации движений ярко проявляется в ряде циклических упражнений: беге на коньках, плавании и др. В этих упражнениях, например, через некоторое время после начала отмечается даже при неизменной скорости движения учащение темпа и уменьшение длины преодолеваемого расстояния при каждом шаге, т. е. возникает новая форма координации (В. С. Фарфель). В результате уменьшение длины шагов компенсируется учащением их и общая скорость передвижения
365
остается прежней. Снижение же скорости передвижения начинается лишь тогда, когда учащение темпа уже не компенсирует уменьшение шага или когда темп также начинает урежаться. Примечательно, что учащение темпа и уменьшение шага возникают еще задолго до того времени, когда для спортсмена становится невозможным сохранять исходные величины этих показателей. Следовательно, такое рано возникающее изменение координации движений имеет профилактический характер, направлено на предупреждение или задержку развития явлений утомления.
Изменение координации двигательных и вегетативных функций, обусловливающее задержку развития утомления и протекающее без снижения эффективности работы (например, бег, гребля, плавание) квалифицируется как проявление преодолеваемого (скрытого) утомления.
В физиологическом механизме возникновения преодолеваемого утомления важная роль принадлежит условным рефлексам и развитию экстраполяции. Благодаря им хорошо тренированный человек значительно лучше использует возможности организма в отношении смены форм координации двигательных и вегетативных функций для предотвращения или отсрочки развития утомления.
Особенности утомления при разных видах физических упражнений. Специфика физилогических процесов при разных видах физических упражнений обусловливает различную природу возникновения утомления. Существенное значение при этом имеют характер упражнений, их длительность, мощность, сложность выполнения и др.
При циклической работе максимальной мощности основной причиной снижения работоспособности являются развитие торможения и уменьшение подвижности нервных процессов. Это торможение развивается в результате утомления, возникающего в центральной нервной системе под влиянием потока афферентных импульсов, посылаемых работающими мышцами. Немаловажное значение при этом имеет изменение функционального состояния самих мышц, снижение их возбудимости, лабильности и скорости расслабления.
Многообразны физиологические причины утомления при циклической работе субмаксимальной мощности. Так же как и при работе максимальной мощности, афферентная импульсация постепенно приводит к угнетению деятельности нервных центров. Этому способствует резкий недостаток кислорода. В связи с анаэробным характером внутриклеточного метаболизма в мышцах происходит накопление продуктов обмена. В частности, содержание молочной кислоты в крови может увеличиваться в 15—25 раз. Недоокислен-ные продукты обмена веществ, всасываясь в кровь, ухудшают деятельность нервных клеток.
При циклической работе большой мощности, т. е. при работе на уровне кажущегося устойчивого состояния, важная роль в развитии утомления принадлежит недостаточности кардио-респираторных функций, необходимости на протяжении длительного времени 366
поддерживать весьма напряженную работу сердца и дыхательного аппарата для обеспечения интенсивно работающего организма нужным количеством кислорода. Как известно, при этой работе кислородный запрос несколько превышает потребление кислорода. Кислородный долг при работе данной мощности не так велик, как при работе субмаксимальной мощности, но все же значителен и, кроме того, действует длительно — на протяжении многих минут и даже десятков минут. Происходит также снижение в крови гормонов некоторых желез внутренней секреции, в частности коры надпочечников. Известную роль в угнетении функций нервных центров при преодолении спортсменом длинных дистанций может играть монотонное действие на нервные клетки афферентных импульсов, периодически поступающих из работающих мышц.
При циклической работе умеренной мощности основной причиной развития утомления является трудность длительного поддержания на высоком уровне функций кардио-респираторной системы. Кроме того, в нервных центрах может возникать торможение под влиянием многократного однообразного раздражения афферентными импульсами. Угнетение же деятельности этих центров приводит к расстройству координации движений. При работе этой мощности продолжительностью более 40—60 мин. уменьшается в связи с длительным расходованием запасов углеводов содержание сахара в крови. В результате расстраивается деятельность центральной нервной системы. Значительная потеря хлоридов и изменение количественного соотношения ионов натрия, калия и кальция, хлора и фосфора в крови и тканях тела при таких упражнениях, происходящие вследствие обильного потоотделения во время длительных спортивных упражнений, также ведут к понижению работоспособности у спортсмена.
Появлению утомления способствует перегревание, наблюдаемое при длительной работе умеренной мощности, особенно в условиях высокой температуры и большой влажности окружающей среды. При перегревании нарушение нормальной деятельности центральной нервной системы может приводить к «тепловому удару» (головная боль, помутнение, а в тяжелых случаях и потеря сознания). Охлаждение организма также может быть фактором, способствующим развитию утомления.
При ациклических видах физических упражнений отмечаются различные формы утомления.
Во всех спортивных играх в результате необходимости постоянного нового программирования игроками своих действий при решении сложных двигательных задач наблюдается утомление высших отделов мозга. Оно приводит к снижению скорости и координированности движений и ухудшению функций некоторых анализаторов. В таких видах спорта, как хоккей, существенная роль принадлежит (как и при циклических упражнениях субмаксимальной мощности) недостаточности кислородного обеспечения и накоплению кислородного долга.
При гимнастических и тяжелоатлетических упражнениях утом
367
ление может сказываться на функциональном состоянии мышц. Падает их возбудимость, уменьшается сила, изменяются твердость, вязкость и скорость сокращения и расслабления мышц.
При статических усилиях со значительными напряжениями одной из причин возникновения утомления является снижение силы вследствие выключения деятельности некоторых наименее устойчивых мышечных волокон.
Значение утомления в развитии состояния тренированности. Утомление в процессе мышечной или умственной деятельности, не переходящее определенных пределов, — физиологическое, а не патологическое явление и полезно для организма.
Работа до утомления представляет собою важный фактор роста тренированности, в особенности тогда, когда она связана с развитием выносливости. Физиологический смысл этого явления заключается в том, что, тренируясь до наступления утомления,спортсмены адаптируются к повышенным нагрузкам. В случаях же, когда тренировочные упражнения прекращаются до начала возникновения утомления, развитие тренированности приостанавливается. То же происходит и в том случае, если тренировочные занятия приводят к резко выраженной степени утомления. При этом может возникнуть состояние перетренированности. Как ясно из сказанного выше, в спорте следует избегать не утомления «вообще», а лишь чрезмерного его развития. При этом пределы чрезмерности связаны не только с характером выполняемых упражнений, но и с их длительностью.
§ 7. Восстановительные процессы
Мышечная деятельность, как правило, сопровождается времен- | ным снижением работоспособности. После окончания работы, в периоде восстановления, нормализуется внутренняя среда организма, восстанавливаются энергетические запасы, различные функции приходят в состояние рабочей готовности. Все эти процессы не только обеспечивают восстановление работоспособности организма, но и способствуют ее временному увеличению.
Повышение работоспособности в процессе тренировки зависит не только от объема и интенсивности нагрузок, но и от продолжительности интервалов отдыха между выполнением упражнений. В связи с этим при планировании тренировочных занятий необходимо учитывать особенности восстановительных процессов.
Восстановительные процессы частично протекают непосредственно во время мышечной деятельности. Примером этого являются окислительные реакции, обеспечивающие ресинтез богатых энергией химических веществ. Однако при работе процессы диссимиляции преобладают над процессами ассимиляции. Лишь при длительной мышечной деятельности, характеризующейся истинным устойчивым состоянием, устанавливается динамическое равновесие между расщеплением химических веществ и их ресинтезом. Нарушение баланса между этими реакциями выражено при работе тем резче, чем
368
больше оказывается ее мощность и чем меньше подготовлен к ней человек.
В восстановительном периоде преобладают процессы ассимиляции. Это обеспечивает пополнение израсходованных при работе энергетических запасов. Сначала они восстанавливаются до исходного уровня, затем на некоторое время становятся выше его (фаза суперкомпенсации) и далее вновь понижаются (рис. 104).
Рис. 10-1. Схема процессов расхода и восстановления энергетических запасов организма
Фазы восстановления. Различают ранние и поздние фазы восстановления. После легкой работы ранние фазы заканчиваются в течение нескольких минут, после напряженной работы —в течение нескольких часов. Поздние фазы восстановления после длительной и напряженной мышечной деятельности затягиваются на несколько суток.
По уровню работоспособности организма в периоде восстановления различают фазы пониженной и повышенной работоспособности. Первая наблюдается сразу после окончания мышечной деятельности. В дальнейшем работоспособность восстанавливается и, продолжая возрастать, становится выше исходной. Этот период называется фазой повышенной работоспособности. Через некоторое время после окончания мышечной деятельности работоспособность вновь снижается до исходного уровня. Фазовые изменения работоспособности в периоде восстановления установлены экспериментально. Например, у тяжелоатлетов через I мин. после жима штанги двумя руками «до отказа» работоспособность оказалась сниженной в среднем на 60% по сравнению с исходной величиной. На 7-й мин. восстановления опа была ниже па 10%. К 12-й мин. восстановления превысила исходный уровень и оставалась повышенной до 25-й мин. (рис. 105, 106).
Продолжительность отдельных фаз восстановления зависит от особенностей выполненной работы (мощность, длительность, структура движений) и от степени тренированности человека.
Повторные нагрузки целесообразно выполнять в фазу повышенной работоспособности: в этих условиях тренированность организма
13—113
369
развивается наиболее интенсивно. Однако в ряде случаев повторные нагрузки следует назначать ранее этого срока. Работа при неполном восстановлении адаптирует организм к деятельности в условиях измененной внутренней среды. Слишком длинные интервалы отдыха между повторными нагрузками снижают эффективность тренировки. Повторная деятельность при этом выполняется при уже снизившейся работоспособности, что не стимулирует ее дальнейшего раз
Рис. 105. Изменение работоспособности в процессе восстановления после жима штанги двумя руками «до отказа» (по Б. С. Гиппенрейтеру).
Горизонтальная пунктирная линия — исходный уровень работоспособности (100%); столбики — работоспособность при повторных жимах штанги «до отказа» на разных минутах восстановления
Рис. 106. Сила мышц кисти на ЗО Й сек., 2- и 5-й мин. и длительность статического усилия на 6-й мин. восстановления после утомительной работы (по В. М. Волкову и А. В. Ромашеву).
Горизонтальная пунктирная линия — исходные величины силы кисти н длительности поддержания статического усилия, взятые за 100%; заштрихован* ные столбики — сила кисти; черный столбик — длительность поддержания статического усилия
вития. Повышение работоспособности, вызванной мышечной деятельностью, должно подкрепляться последующей работой. Если этого не происходит, то работоспособность снижается до исходного уровня и дальнейшие прогрессивные изменения в организме могут приостанавливаться.
Показатели восстановления работоспособности. При определении оптимальных интервалов отдыха необходимо учитывать интенсивность восстановительных процессов. Наиболее точным показателем этого является уровень работоспособности, объем повторной работы, который может выполнить человек в данных условиях. Однако такой способ связан с выполнением дополнительной напряженной работы и поэтому не может быть рекомендован для спортивной практики. Более удобным и достаточно информативным способом оценки работоспособности является изучение особенностей реакций организма на различные тесты, выполняемые до тренировоч-
370
кого занятия и в периоде восстановления. К таким тестам относятся косвенное определение максимального потребления кислорода (МПК), проба PWCno, исследование оксигенации крови при задержке дыхания, определение способности скелетных мышц к развитию напряжения и к расслаблению и др.
После напряженной мышечной деятельности величины МПК и PWCno обычно оказываются сниженными. Затем они постепенно восстанавливаются и через некоторое время после окончания мышечной деятельности становятся выше исходных. Например, у квалифицированных велосипедистов через 1 час после окончания тренировочного режима в подготовительном периоде величина PWC170 оказалась сниженной в среднем с 1701 кгм/мин до 1573 кгм/мин, через 24 часа этот показатель стал выше, чем был до работы. Такая же динамика работоспособности наблюдалась и у велосипедистов менее высокой квалификации.
Изменения величин МПК и PWCno после мышечной деятельности отражают динамику работоспособности организма в восстановительном периоде. Однако ни одна из этих величин не может считаться достоверным критерием готовности всех систем организма к повторным нагрузкам. Это объясняется разновременным их восстановлением. Например, исследования квалифицированных бегунов показали, что полное восстановление величин МПК после напряженной тренировочной нагрузки нередко сочетается с недостаточным завершением восстановительных процессов в миокарде. Изменения некоторых показателей электрокардиограммы становятся в это время даже более выраженными по сравнению с данными, полученными сразу же после работы.
Тесты с определением оксигенации крови выявили, что в периоде восстановления содержание НвО2 при задержке дыхания снижается быстрее и более значительно (в связи с повышением интенсивности окислительных процессов в тканях), чем до работы. Градиент падения оксигенации крови
Снижение %НЬО2
Длительность задержки дыхания остается при этом увеличенным в течение нескольких суток.
Об изменениях внутриклеточного метаболизма в процессе восстановления можно судить по длительности удержания стабильного уровня оксигенации крови при задержке дыхания (фаза А — Б оксигемограммы). Уменьшаясь под влиянием напряженной мышечной деятельности, этот показатель вновь удлиняется в восстановительном периоде, что свидетельствует о нормализации химических процессов в работавших органах.
О восстановлении работоспособности двигательного аппарата можно судить по особенностям напряжения и расслабления мышц. Снижаясь при утомительной работе, эти функции мышц повышаются в периоде восстановления. По их динамике можно определить степень готовности органов движения к повторной работе.
В спортивной и врачебной практике о восстановлении работоспособности организма нередко судят по восстановлению одной из
13*
371
его функций. Однако это дает лишь ориентировочные сведения. Отдельные функции восстанавливаются в разное время, что обусловлено характером мышечной деятельности и индивидуальными особенностями человека.
Интенсивность восстановительных процессов можно оценивать по динамике частоты сердцебиений. Этот показатель определяется сразу же после работы и затем повторно через строго определенные промежутки времени. Снижение этого показателя по отношению к величине, установленной сразу же после работы, в известной степени позволяет судить о интенсивности восстановления, а следовательно, и о готовности организма к повторной работе. Например, при тренировке бегунов на средние дистанции повторное прохождение отрезков дистанции рекомендуется после снижения частоты сердечных сокращений на 30% по отношению к величине, зарегистрированной сразу же после окончания бега на предыдущем отрезке.
Восстановление других показателей функционального состояния органов кровообращения весьма вариативно. Вследствие этого они менее точно определяют готовность организма к повторной работе. Например, восстановление артериального давления в одних случаях происходит в течение нескольких минут, в других задерживается на длительное время. После длительной и напряженной работы отдельные показатели артериального давления часто становятся ниже исходных величин, что обусловлено гиперемией в работавших мышцах. На длительную послерабочую гиперемию указывает также снижение жесткости артериальных стенок в активных областях тела. По ходу восстановления жесткость артериальных стенок претерпевает фазовые изменения. При этом у более тренированных людей наблюдаются наиболее резко выраженные дифференцированные сосудистые реакции.
Большое значение для восстановления работоспособности организма имеет нормализация его внутренней среды. Продолжительность восстановления pH крови и ее щелочных резервов зависит от мощности и длительности работы. Например, по некоторым данным, эти показатели восстанавливаются быстрее после бега на 200 и 10000 м и медленнее после бега на 400 и 5000 м.
Восстановление форменных элементов крови происходит очень медленно. Эритроциты и гемоглобин могут в зависимости от особенностей мышечной деятельности и степени тренированности человека, восстанавливаться в течение нескольких часов или суток. Если под влиянием работы содержание эритроцитов и гемоглобина резко понижается, то его восстановление до исходного уровня иногда задерживается до 7 и более суток (Л. Я. Евгеньева). Содержание в крови лейкоцитов и тромбоцитов, а также лейкоцитарная формула восстанавливаются после длительной и напряженной работы в течение нескольких суток.
При очень напряженной работе, характеризующейся эмоциональным возбуждением, количество эозинофилов в крови резко уменьшается. Восстановление численности этих клеток происходит в течение 1—2 суток.
372
После выполнения динамической работы максимальной мощности наибольший интерес представляют сдвиги в функциональном состоянии двигательного аппарата. Восстановление работоспособности после скоростной работы тесно коррелирует также с погашением кислородного долга и функциональным состоянием центральной нервной системы. Оптимальным интервалом отдыха в этих случаях будет такой, при котором возбудимость двигательных центров остается еще высокой, а относительно небольшой кислородный долг
моо
1200
1000
900
Рис. 107. Энергетические траты в условиях основного обмена у лыжников в соревновательном периоде (по В. В. Михайлову).
Ось ординат — величина энергетических трат (ккал.); ось абсцисс —дни исследований. Стрелки-величины обмена после дня отдыха; полоса с точками — стандартные величины по Бенедикту и Гаррису
оказывается уже почти ликвидированным.
После работы субмаксимальной мощности для восстановления работоспособности большую роль играет погашение кислородного долга и нормализация внутренней среды организма. Кислородный долг состоит из двух частей (фракций). Первая — алактатная — обусловлена ресинтезом фосфорсодержащих соединений (АТФ и др.), вторая — лак-татная — связана с окислением молочной кислоты. В процессе восстановления происходит сначала быстрая ликвидация кислородного долга, что связано с интенсивными окислительными реакциями в мышцах. В дальнейшем этот процесс протекает менее быстро. Он обусловлен ресинтезом молочной кислоты
При работе субмаксимальной мощности кислородный долг у тренированных спортсменов может достигать 20 л и более. Его ликвидация после работы такой мощности обычно заканчивается в течение 1,5—2 часов.
Выполнение длительной работы большой и умеренной мощности характеризуется медленным восстановлением дыхательных функций и энергетики. Даже у квалифицированных спортсменов энергетические траты снижаются до исходных величин очень долго — в течение нескольких суток. Например, у тренированных лыжников-гонщиков это продолжается 2—3 дня (рис. 107).
Казалось бы, о восстановлении работоспособности организма можно судить по восстановлению его основных двигательных качеств: быстроты, силы и выносливости. Однако экспериментальные данные показывают, что восстановление их также весьма вариа
<73
тивно и протекает гетерохронно. Даже такие взаимосвязанные показатели, как сила и силовая выносливость, восстанавливаются, в разное время. Например, сила кисти после статической работы к 5-й мин. восстановления уже достигает 90% исходной величины. Силовая же выносливость, от которой зависит объем повторной работы, на 6-й мин. восстановления еще на 40% ниже исходного уровня.
Длительность интервалов отдыха между отдельными упражнениями на тренировочных занятиях, между ними и повторными вы-
ступлениями на соревнованиях должна планироваться с учетом того, что эффективность последующей работы будет больше тогда,
когда утомление от предыдущей деятельности почти ликвидировано,
а положительное последействие этой работы еще сохранено.
Рис. 108. Влияние активного
отдыха на работоспособность мышц (опыт И. М. Сеченова).
Вверху: ГЦ — работа правой руки; 1Ъ — повторная работа той же рукой после 10 мин. пассивного отдыха.
Внизу: П( —- работа правой рукой;
Л — работа левой рукой; П2 — повторная работа правой рукой
На тренировочных занятиях оптимальная длительность интервалов отдыха зависит от объема и мощности выполняемых нагрузок, от уровня тренированности спортсменов, от метеорологических условий и др. В среднем она колеблется от 1 до 20 мин.
Оптимальные интервалы отдыха между тренировочными занятиями могут быть разными. Однако продолжительность их не должна быть больше 48 часов. Для достижения высоких спортивных результаов необходимо тренироваться с меньшими
интервалами отдыха (5—6 и более раз в неделю). На тренировочных сборах нагрузки могут выполняться даже 2—3 раза в день. Неполное восстановление в этих условиях не является препятствием для повторной работы.
Средства, ускоряющие процессы восстановления. В спортивной
практике применяются различные средства, ускоряющие восстановительные процессы.
Одним из средств, ускоряющих восстановление после мышечной работы, является активный отд ы х, т. е. переключение на другой вид деятельности. Его значение впервые было установлено И. М. Сеченовым. Он показал, что более быстрое восстановление работоспособности утомленной конечности происходит не при полном покое, а при работе другой конечности. Данные одного из опытов И. М. Сеченова приведены на рис. 108. В этом опыте производилась работа правой рукой на эргографе в Течение 25 мин. Затем ей давался полный 10-минутный отдых, в результате которого работоспособность ее несколько восстанавливалась, но все же оставалась ниже исходной. Последующая работа, выполненная левой рукой, повышала работоспособность правой.
Факты, обнаруженные И. М. Сеченовым, объясняются особенно
стями восстановительных процессов, протекающих в нервных цент-
374'
pax, и межцентральными отношениями, возникающими при активном отдыхе. После утомительной работы правой руки центры, иннервирующие ее мускулатуру, приторможены. При последующей работе левой руки возбуждение в центрах ее мышц усиливает торможение в центрах правой конечности (по механизму отрицательной индукции). Это способствует восстановлению работоспособности мышц правой руки.
Однако такие благоприятные межцентральные отношения возникают при активном отдыхе не всегда. В некоторых случаях возбуждение центров работающих мышц иррадиирует в «отдыхающие» центры, задерживая протекающие там восстановительные процессы. Особенности межцентральных отношений при активном отдыхе зависят от уровня подготовленности человека к выполняемой работе. Например, при непривычной работе правой рукой работа левой в целях активного отдыха не дает положительного эффекта. По мере же развития тренированности преимущество активного отдыха перед пассивным становится более выраженным. Это объясняется тем, что при непривычной работе активный отдых вызывает иррадиирование процессов возбуждения с рабочих центров на отдыхающие. Когда же работа становится привычной, в центрах начинают преобладать индукционные отношения. Работа левой руки в этих условиях приводит к торможению в центрах правой с последующим (после прекращения работы левой руки) усилением в них возбуждения.
При подборе упражнений, применяемых с целью активного отдыха, следует учитывать особенности выполняемой работы и степень подготовленности человека к ней. Чаще для активного отдыха утомленных предшествующей работой мышц производят работу другими мышцами. В некоторых случаях процессы восстановления можно ускорить продолжением той же работы, снизив ее интенсивность. Например, в интервалах между скоростным бегом рекомендуется медленный бег или ходьба. Некоторые исследователи считают, что при активном отдыхе эффективны упражнения с расслаблением утомленных мышц.
Активный отдых дает наибольший эффект при работе средней тяжести. После легкой и кратковременной работы он не нужен, а после длительной и истощающей нецелесообразен. Длительность активного отдыха, структура выполненных при этом движений и время его проведения также зависят от особенностей основной работы. Правильно организованный активный отдых не только укорачивает восстановительный период, но и облегчает врабатывание при последующей деятельности.
Кроме активного отдыха, для более быстрого восстановления работоспособности применяют вдыхание богатых кислородом газовых смесей, водные процедуры, массаж и другие раздражители умеренной силы.
Вдыхание увлажненного воздуха, содержащего .65—75% кислорода, ускоряет ликвидацию кислородного долга, в связи с чем повышает интенсивность восстановления работоспособ-
375
г
ности. Это имеет большое значение в интервалах между забегами, во время перерывов в спортивных играх и боксе и в других видах спорта. Иногда рекомендуется пребывание спортсменов в специальных палатках, воздух которых обогащен кислородом. Кроме этих приемов, повышенная доставка кислорода в процессе восстановления может быть обеспечена питьем так называемых «кислородных коктейлей». В этих случаях поступление кислорода в организм происходит путем его диффузии в кровь из желудочно-кишечного тракта.
Водные процедуры благоприятно воздействуют на центральную нервную систему. Это объясняется тем, что афферентные импульсы от рецепторов кожи вызывают новые очаги возбуждения в определенных отделах мозга, способствуя установлению оптимальных межцентральных отношений.
Механизм воздействия массажа такой же, как и водных процедур. Афферентные импульсы от кожи и мышц изменяют функциональное состояние центральной нервной системы. Особенно эффективны вибрационный и гидромассаж.
Большую роль в повышении интенсивности восстановительных процессов играет питан не. Ойо должно быть достаточно калорийным и содержать все необходимые органические и неорганические вещества. Исключительно важна при этом витаминизация организма.
В настоящее время в спортивной практике применяются специальные напитки и печенье, в состав которых входят сахар, витамины. соли, белковые и другие вещества.
Восстановительные процессы протекают у человека интенсивнее при наличии положительных эмоций. Однако чрезмерное возбуждение после работы отрицательно влияет на восстановление. В связи с этим в некоторых случаях после участия в соревнованиях, вызывающих чрезмерное эмоциональное возбуждение, рекомендуется для усиления восстановительных процессов прием средств, способствующих развитию торможения в центральной нервной системе.
I
Глава XV!
ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ ДВИГАТЕЛЬНЫХ
НАВЫКОВ
§ 1. Временные связи как физиологическая основа формирования двигательных навыков
Двигательные акты, с которыми рождается человек, весьма ограничены как по своему числу, так и в особенности по сложности их координации. Морфологическое и функциональное созревание многих нервных центров и их связей с мышечкой системой происходит и в постнатальном периоде, т. е. после рождения. Поэтому ряд уна-376
следованных относительно простых движений возникает у ребенка спустя некоторое время после появления на свет. Весь же основной двигательный фонд человека, все более или менее сложные двигательные акты приобретаются в результате обучения. Вследствие этого в сложном сплаве из врожденных и индивидуально приобретенных компонентов двигательной деятельности взрослого человека безусловнорефлекторные двигательные рефлексы играют лишь подчиненную роль. Они проявляются только в виде простейших кожных, сухожильных, болевых, вестибулярных, зрительных и некоторых других рефлексов. Все же более или менее сложные движения принадлежат, как указывал еще И. М. Сеченов, не к унаследованным, а к заученным. Они возникают в результате опыта, приобретаемого на протяжении индивидуальной жизни. Следовательно, двигательные навыки, являющиеся индивидуально приобретенными двигательными актами, формируются на основе механизма временных связей.
Индивидуально приобретенные виды двигательной деятельности характеризуются различной сложностью, разными формами вовлечения афферентных и эфферентных функций в условнорефлекторные связи. Работами И. П. Павлова и его многочисленных учеников и последователей (П. К. Анохин, Э. А. Асратян, М. И. Алексеев. 3. И. Бирюкова-Коларова, Л. Г. Воронин, Н. В. Зимкин, Ю. Конор-ский, А. Н. Крестовников, Л. В. Крушинский, П. С. Купалсв и др.) был выявлен целый ряд закономерностей, характеризующих особенности формирования двигательных навыков на основе образования новых временных связей.
§ 2. Сенсорные и эффекторные компоненты двигательного навыка
Двигательные навыки человека по особенностям физиологических механизмов их формирования характеризуются большим разнообразием.
Классические условные реакции в опытах с выделением слюны характеризовались образованием временной связи между индифферентным сигналом и безусловным рефлексом (условные рефлексы первого порядка) или ранее образованной устойчивой условнорефлекторной реакцией (условные рефлексы высших порядков). Это сенсорные, или, по классификации Ю. Конорского, условные рефлексы первого рода (см. главу XII, § 4). В них ответная реакция, например слюноотделение, отдергивание руки при нанесении болевого раздражения, является или безусловнорефлекторной или ранее приобретенной условной реакцией. В этих реакциях только сигнал приобретает новые (условнорефлекторные) свойства.
Но когда речь идет о двигательных навыках, то всегда подразумевается не простое повторение по условному сигналу ранее имевшейся реакции, а образование оперантных условных рефлексов, т. е., по классификации Ю. Конорского, рефлексов второго рода. Эти рефлексы характеризуются новой формой движения или обра
377
зованием комбинации из уже ранее известных элементов нового сложного двигательного акта, до этого не имевшегося у данного ор-ганизма. Следовательно, в этом случае условнорефлекторные процессы связаны не только с афферентным (чувствительным), но и с эфферентным (эффекторным) звеном рефлексов.
Двигательные навыки человека характеризуются тем, что в них сочетаются оба рода условных реакций. С одной стороны, через первую и вторую сигнальные системы устанавливаются временные связи между ранее индифферентными для спортсмена раздражителями и последующей деятельностью (сенсорные условные реакции), а с другой — вырабатываются новые (до этого времени не имевшиеся в двигательном фонде человека) ответные двигательные реакции (оперантные условные реакции) с соответствующим характером протекания не только двигательных, но и вегетативных функций.
При образовании спортивных и других двигательных навыков у человека особенно большое значение имеют временные связи высших порядков, формирующиеся при воздействиях не только через первую, но и через вторую сигнальную систему (обучение различным навыкам всегда производится путем как показа, так и словесного объяснения).
§ 3. Значение для формирования двигательных навыков ранее выработанных координаций
Формирование двигательных навыков всегда происходит на базе ранее выработанных организмом координаций. Например, навык стояния формируется у ребенка на базе навыка сидения, при котором приобретается способность удерживать в вертикальном положении голову и туловище. Навык ходьбы образуется на базе навыка стояния. При формировании различных спортивных навыков многие компоненты двигательного акта также не являются полностью новыми. В этих навыках в качестве составных частей всегда имеются элементы ранее приобретенных навыков.
В тех случаях, когда необходимо усвоить сложный навык, компоненты которого в значительной своей части являются новыми, обычно используются подготовительные упражнения й обучение по элементам. Тем самым на базе освоенных более простых координаций постепенно происходит усложнение навыка.
Наличие прочно закрепившихся навыков в некоторых случаях не только не содействуют, но даже препятствуют формированию нового навыка. Это наблюдается, в частности, тогда, когда структура нового движения связана с переделкой прочно закрепленного старого навыка. В этих случаях чем прочнее закреплен старый навык, тем труднее образовать полноценный новый. При обучении на выкам важно сразу же формировать правильные'движения, так как переделка прочно закрепленных неполноценных двигательных актов-на новые может потребовать длительных упражнений.
378
§ 4. Характеристика формирования двигательного навыка по данным электромиографии
Использование метода электромиографии для изучения особенностей деятельности мышц при формировании двигательного навыка позволяет определить, какие мышцы вовлекаются в деятельность, степень их активности, моменты включения и выключения,
длительность импульсации и т. п.
Принципиально важные факты, подтверждающие условнорефлекторную природу двигательного навыка, были выявлены в работах Ю. М. Уфлянда, В. Гурфинкеля, П. И. Гуменера и др. В их ис-
следованиях, проведенных на человеке в клинике, показана перестройка характера биопотенциалов при пересадке одной из головок мышцы к месту прикрепления ее антагониста. Спустя 8 недель — а после специального обучения через 2—3 недели — деятельность пересаженной головки мышцы, по данным электромиографии, постепенно изменялась (рис. 109). Если до пересадки биотоки возникали в мышце только при движении, которое она ранее осуществляла, то после пересадки биопотенциалы, наоборот, наблюдались при противоположном движении.
В начальных стадиях развития двигательного навыка биопотенциалы регистрируются не только в тех мышцах, которые необходи-
Б
Рис. 109. Электромиограмма сгибателя бедра (полусухожильной мышцы) до и после пересадки сухожилия на надколенник при сгибании и разгибании ноги в коленном суставе (по Ю. М. Уфлянду):
А — при сгибании до пересадки; Б — то же при разгибании; В — при сгибании после пересадки; Г — то же при разгибании. Отметка времени — 0,01 сек.
мы. для осуществления данного двигательного акта, но и в ряде «посторонних» мышц. Это связано с явлениями иррадиации в нервных центрах. По мере укрепления навыка происходит ограничение иррадиации, и при полностью сформированном навыке она наблюдается только в необычных условиях, например при действии сильных посторонних раздражителей, при утомлении (Е. К. Жуков, Ю. 3. Захарьянц и др.).
В результате совершенствования навыка при циклических движениях изменяется длительность периодов импульсации. В начальных стадиях формирования навыка в соответствующих мышцах импульсации наблюдается не только во время активных фаз движения, но и в интервалах между ними. В дальнейшем биотоки концентрируются, их залпы становятся короткими и при относительно медленных циклических движениях во время- пауз уменьшаются вплоть до полного исчезновения (М. А. Киселев и М. Е. Маршак, С. А. Косилов, Р. С. Персон и др.).
379
А 123 456789 10
100- 200 300 400 мсек.
В процессе формирования навыка происходит изменение взаимоотношений между мышцами-антагонистами. В начале обучения может наблюдаться одновременная биоэлектрическая активность в этих мышцах. В дальнейшем, по мере овладения навыком, при относительно медленных движениях обнаруживается реципрокность между ними и биоэлектрическая активность начинает возникать поочередно. Однако даже при сформированном навыке реципрокность может быть выражена не полностью, проявляясь лишь в снижении активности во время сокращения антагониста. При этом чем быстрее темп движений, тем больше биоэлектрическая активность агониста сочетается с одновременной аналогичной активностью антагониста. Это наблюдается даже при таком хорошо усвоенном навыке человека, как бег (рис. ПО). Все это позволяет считать, что одновременная биоэлектрическая активность мышц-антагонистов, особенно при сложных движениях, не всегда является признаком иррадиации, т. е. несовершенства координации. В ряде случаев некоторые формы одновременной деятельности мышц-антагонистов представляют собой выражение специальной формы координации, отмечающейся при высокой степени совершенства данного двигательного навыка.
При выполнении одного и того же сложного двигательного акта картина' биоэлектрической активности в мышцах по мере развития навыка претерпевает существенные изменения. Например, при значительном растяжении мышц, в частности при выполнении новичком упражнения «шпагат», в растяутых мышцах бедра отмечается значительная биоэлектрическая активность. В дальнейшем же, по мере упрочения навыка, она резко снижается (Ю. 3. Захарьянц).
Изменения биоэлектрической активности мышц при формировании навыка у разных лиц протекают неодинаково. В связи с этим
Рис. ПО. Схема движения и величины электрической активности мышц ноги при беге (но И. М. Козлову):
Л — при беге в максимальном темпе; Б — при бе-, ге в темпе 200 шагов в 1 мин. Горизонтальные ряды цифр — последовательные (с интервалом
50 мсек.) положения правой ноги при беге.
Черные и заштрихованные участки — изменяющиеся величины интегрированной электрической активности мышц: черные участки — при преодолевающей работе, участки с косой штриховкой — при уступающей работе, участки с вертикальной штриховкой — при фиксирующей работе (изометрическое напряжение). Римские цифры — мышцы: /—'Ягодичная, // — прямая мышца бедра, III — лвуглавая мышца плеча, IV — большеберцовая,
V — камбаловидная
380
в картине биоэлектрической активности у спортсменов одинаковой квалификации наряду с общими чертами выявляются и существенные различия.
§ 5. Вегетативные компоненты двигательного навыка
Вегетативные функции мобилизуются при двигательной деятельности главным образом по механизму безусловных рефлексов. Например, при мышечной работе безусловнорефлекторным путем происходят резкое усиление функций кровообращения и дыхания, уменьшение доставки крови к неработающим органам, торможение функций пищеварительного аппарата и почек и т. п. Наряду с этим при образовании двигательного навыка может происходить изменение характера протекания вегетативных безусловных рефлексов, приспособление их не вообще к мышечной работе, а именно к данному виду двигательной деятельности. Эти особенности функций вегетативных органов, приобретенные в процессе формирования навыков, и составляют условнорефлекторные дыхательные (М. Е. Маршак, К. М. Смирнов и др.), сердечно-сосудистые (П. Аст-ранд, В. В. Васильева, В. И. Георгиев и др.) и другие вегетативные компоненты произвольного двигательного акта.
Двигательные и вегетативные компоненты двигательного навыка формируются не одновременно. В навыках с относительно простыми движениями раньше заканчивается формирование двигательных компонентов, а в навыках со сложными движениями — формирование вегетативных компонентов.
Характерно, что после образования навыка вегетативные компоненты становятся более инертными, чем двигательные. Например, при изменении привычной формы деятельности — переходе с непрерывной работы на работу с переменной интенсивностью двигательные функции изменяются сразу же. В то же время вегетативные органы еще длительное время функционируют в соответствии с ранее сформировавшимся стереотипом.
§ 6. Афферентный синтез в двигательных навыках
Все реакции организма (в том числе движения при выполнении физических упражнений) являются, как уже отмечалось выше, системными, т. е. обусловленными совместной деятельностью многих отделов нервной системы и периферических органов, направленной на достижение определенной цели. Системный подход к изучению различных функций организма требует при трактовке проявлений двигательного навыка учета как регулирующих функций нервной системы, так и состояния исполнительных периферических органов.
В нервной структуре двигательных навыков, являющихся поведенческими актами, условно можно выделить афферентные (приносящие, чувствительные), программирующие и эфферентные (выносящие, исполнительные) компоненты.
Афферентные, или анализаторные, компоненты, как известно, состоят из рецепторов, чувствительных нейронов и популяций (сово
381
купностей) афферентных нервных клеток в центральной нервной системе. Все эти элементы нервной системы воспринимают раздражение из внешней среды и различных частей самого организма и участвуют в осуществлении так называемого афферентного синтеза.
Значение мотивации, памяти, обстановочной и пусковой информации в афферентном синтезе. По П. К. Анохину, афферентный синтез происходит при взаимодействии четырех основных факторов: 1) мотивации, 2) памяти, 3) обстановочной информации и 4) пусковой информации.
У животных мотивация связана преимущественно с удовлетворением таких потребностей, как голод, влечение к другому полу, стремление освободиться от неблагоприятно действующих на организм факторов (оборонительные и другие защитные рефлексы) и др. Эти мотивации свойственны и человеку. Однако в его трудовой и спортивной деятельности особенно большое значение имеют не эти, а различного рода социально обусловленные виды мотивации.
Как уже указывалось выше, важная роль в афферентном синтезе принадлежит кратковременной и долговременной памяти, т. е. следовым явлениям, обусловленным ранее приобретенным опытом. Значение следовых явлений, как кратковременных, так и долговременных, особенно велико у человека, поскольку все стороны его жизни обусловлены накоплением огромного личного и социального опыта. В виде следов в нервной системе (памяти), имеющихся у человека, предшествующий опыт оказывает сильнейшее влияние на оценку любых событий и ситуаций.
Выполнение физических упражнений проводится в определенной обстановке (стадион, спортивная площадка, спортивный зал и т. п.). Поэтому обстановочная информация, поступающая из окружающей среды, является, несомненно, весьма существенным компонентом для правильного афферентного синтеза.
Наконец, важная роль принадлежит непосредственным пусковым сигналам, какими в спорте являются выстрел, звук свистка, движение флажка, голосовая команда и др.
При многих видах физических упражнений раздражители, требующие ответных двигательных актов, весьма сложны. В этом случае пусковая афферентация представляет собой не единичный сигнал, а ситуацию определенного характера, т. е. здесь пусковой сигнал и обстановочная информация сливаются. Это всегда сильно осложняет афферентный синтез. Например, в разных видах единоборства и спортивных игр многократно нужно начинать новые действия. При этом, особенно в коллективных спортивных играх, начало и характер ответных движений определяются не каким-либо отдельным сигналом, а всей создавшейся ситуацией, т. е. совокупностью многих (в ряде случаев десятков и даже сотен) раздражителей. Так, в спортивных играх для направления своих действий игрок должен учитывать не просто непрерывно изменяющееся расположение всех других партнеров, но и специализацию и мастерство каждого из них. При этом синтез осуществляется при резком
382
дефиците времени, в некоторых случаях за секунды и даже доли секунд.
Таким образом, в спорте в одних случаях (например, у гимнаста) афферентный синтез для принятия решения о начале своих движений относительно прост, в других же (в частности, при единоборстве и в особенности при коллективных спортивных играх) весьма сложен.
Роль сенсорных коррекций (обратных связей) в двигательном навыке. Афферентный синтез происходит не только перед запуском двигательной деятельности, но и при выполнении самого движения. В этом синтезе в процессе движения важнейшая роль принадлежит сенсорным коррекциям, которые осуществляются благодаря информации, поступающей из мышц и внутренних органов. При этом им-иульсация из мышц является совершенно необходимым условием для высококоординированных движений. Такая информация имеет сигнальное значение и для последующих движений. В частности, импульсы, поступающие в центральную нервную систему при циклическом движении или при ациклической сложной многофазной комбинации (гимнастика, фигурное катание и др.), являются своего рода связками между циклами или фазами, сигналами для возникновения последующих фаз или циклов двигательного акта. Внешние сенсорные системы, сигнализируя о положении частей тела, также обеспечивают обратные связи и тем самым вместе с внутренними сенсорными системами содействуют созданию в нервной системе соответствующих программ осуществления и коррекции движений. Внешние анализаторы имеют особенно большое значение для информации об изменениях в окружающей обстановке. Они сигнализируют как о необходимости начать физические упражнения, так и о необходимости их изменения или прекращения.
Таким образом, сенсорные коррекции при физических упражнениях имеют специфические особенности, обусловленные характером движения. При медленном выполнении двигательных актов обратные связи корригируют данное движение или какую-либо его фазу. При сложных быстро осуществляющихся многофазных движениях (например, гимнастических) обратные связи в результате недостатка времени могут корригировать только последующие фазы. Наконец, при кратковременных движениях (в частности, баллистических — метаниях, бросках) обратные связи корригируют двигательный акт только при последующих его повторениях.
§ 7. Программирование в двигательном навыке и учет состояния исполнительных приборов
Афферентный синтез является основой для принятия решения и программирования последующих действий как перед началом физического упражнения, так и в процессе его выполнения.
Программирование движений, т. е. центральный компонент навыка, характеризуется' различной степенью трудности своего осуществления. Это связано, с одной стороны, со степенью сложности
383
двигательного акта, с дургой же —со степенью новизны. Если движение совершалось ранее многократно и навык уже хорошо освоен, то многократно повторяющееся программирование даже сложных стереотипных двигательных актов, например в гимнастике, при метаниях, совершается относительно легко. При новых же движениях, например спортивных играх и единоборстве, процесс программирования является более трудным. Это обусловлено, во-первых, необходимостью вследствие непрерывного изменения обстановки осуществлять программирование, как и афферентный синтез, в течение
Рис. 111. Изменения тетанического сокращения икроножной ньшцы человека до и после статических усилий:
I — до статистических усилий, II — после поддержания 1-го статического усилия «до отказа». III — после повторного поддержания статического усилия «до отказа»
весьма короткого времени и, во-вторых, необходимостью осуществлять программирование движений каждый раз в каком-то новом варианте, поскольку движения в этих видах спорта, как правило, не являются стандартными и выполняются при самых различных положениях собственного тела.
Эффекторный компонент в движениях в спорте- требует соответствия программы выполнения двигательного акта функциональным возможностям мышц и обеспечивающих их работу вегетативных органов. Рассогласование между необходимой программой и фактическим выполнением движения особенно усиливается при из-исполнительного прибора
менении состояния периферического
(мышц, кардио-респираторной и других систем организма).
Функциональные возможности периферических органов, в частности мышц, постоянно изменяются. В результате тренировки мышцы становятся сильнее, при отсутствии ее — слабее. После неболь-
шой работы мышечные сокращения усиливаются, после значительной — ослабляются (рис. 111).
Изменчивость состояния исполнительных приборов требует своевременного поступления соответствующей информации в нервные центры. Лишь- в этом случае нервная система может создать полноценную программу, обеспечивающую эффективное выполнение дви-
гательных задач.
Хотя при нормальных условиях тренировки у спортсменов мышцы не подвергаются более или менее значительной атрофии, все же незначительные изменения их состояния могут происходить даже при коротких (1—2 дня) перерывах в ней. и это может сказываться на спортивных результатах. Ведь на соревнованиях результаты выступлений спортсменов могут отличаться друг от друга очень незначительно: различие может составлять ничтожную величину (например, в спринте — десятые и сотые доли секунды, в прыжках и метаниях — сантиметры, в гимнастике, фигурном катании — десятые и
384
сотые доли балла). Такого рода разница может в отдельных случаях обусловливаться несоответствием программирования (в центральной нервной системе) состоянию периферических аппаратов, в том числе мышц.
При выполнении таких упражнений, как прыжки в высоту, прыжки с шестом, поднятие тяжестей, лучшие результаты достигаются, как правило, не при первом, а при повторных выполнениях. Это частично связано с тем, что во время решения начальных, более легких задач (меньшая высота, меньший вес) нервная система получает точную информацию о фактическом состоянии периферического мышечного аппарата. Поэтому специальная разминка у спортсменов перед выполнением любых сложнокоординированных упражнений содействует наряду с решением многих задач информации нервных центров о состоянии мышц в данный момент.
§ 8. Стереотипность и изменчивость двигательного навыка
Двигательный навык, как правило, представляет собою не элементарный, а комплексный двигательный акт, состоящий из нескольких элементов (фаз), сочетанных друг с другом в единый целостный двигательный акт. Например, при ациклических движениях (поднятие тяжестей, метания, гимнастические упражнения и др.) отдельные фазы в определенном порядке последовательно сменяют друг друга; при циклических движениях (ходьба, бег и др.) также имеется закономерная связь фаз движения в каждом цикле.
Анализ становления двигательного навыка показывает, что в процессе его формирования отдельные фазы движения, представляющие собой как бы различно протекающие двигательные рефлексы, складываются в своеобразную цепь реакций, осуществляющихся в виде определенного динамического стереотипа (см. гл. V, §8).
После закрепления динамического стереотипа выполнение навыка протекает стандартно только по биомеханической структуре, т. е. по общему внешнему характеру осуществления движения. Во внутренней структуре его всегда имеются различия, касающиеся ряда деталей в работе участвующих в движении мышц: длительности скрытых периодов, последовательности включения в деятельность отдельных мышц, продолжительности периода импульсации в них, величины средней и максимальной амплитуды биотоков и т. д. Это объясняется тем, что при наличии в организме большого числа исполнительных приборов (сотни мышц и сотни тысяч и даже миллионы двигательных единиц в них) центральная нервная система имеет возможность достигать одного и того же внешнего эффекта за счет многих вариаций тонкой внутренней структуры движения.
Динамический стереотип является характерным для внешней структуры только тех навыков, в которых последовательность фаз движения может протекать по определенному стандарту. Но существуют и другие виды навыков, в которых необходимо в связи с
385
частыми изменениями ситуации реагировать каждый раз новым движением. К такого рода навыкам относятся навыки в единобор-стве (бокс, борьба, фехтование) и спортивных играх (футбол, хоккей, баскетбол и др.). В этих навыках динамический стереотип в виде стабильной целостной системы движений, как правило, не образуется. Если же он образуется, то это относится не к проявлениям сложных двигательных комбинаций, а лишь к отдельным составным элементам, например к штрафным броскам мяча в баскетболе.
§ 9. Экстраполяция в двигательных навыках
Поскольку двигательная деятельность человека характеризуется большой вариативностью, значительная часть моторных актов новой структуры осуществляется путем экстраполяции (см. главу V, § 9), обеспечивающей так называемый перенос'навыков. Экстраполяция при регуляции моторных функций организма является способностью нервной системы на основании имеющегося опыта адекватно решать вновь возникающие двигательные задачи.
Формы экстраполяции весьма разнообразны. Они имеют отношение к самым разным сторонам двигательной деятельности, в том числе к тому, что связано с правильной оценкой создавшейся ситуации, программированием характера и формы предстоящих движений и пр.
Экстраполяция широко осуществляется нервной системой не только при выполнении совершенно новых двигательных актов. Она постоянно происходит при привычных движениях (например, таких, как ходьба). Человек при ходьбе использует огромное количество различных вариантов комбинаций деятельности мышц, необходимых каждый раз для адекватного приспособления к данным условиям. Любой наклон тела или поворот головы, изменение высоты и длины шага, увеличение или снижение веса переносимого груза, особенности грунта и др. всегда сопровождаются изменениями электромиографической картины деятельности мышц, т. е. изменениями взаимосочетания в работе мышц. Естественно, практически невозможно обучать человека неограниченному числу встречающихся в жизни вариантов ходьбы. Но при овладении даже ограни- ' ченным количеством вариантов этого двигательного акта он оказывается способным путем экстраполяции осуществлять ходьбу в самых различных условиях.
Еще большее значение экстраполяция имеет при движениях, производимых со значительными вариациями характера двигательного акта. Например, футболист может произвести удар по мячу разными частями правой и левой ног, с неодинаковой силой и при различном исходном положении собственного тела. Такого рода разнообразные двигательные задачи после обучения относительно ограниченному числу приемов решаются путем экстраполяции.
Таким образом, благодаря экстраполяции организм, осваивая определенное число вариантов навыков, приобретает способность пра
386
вильно выполнять огромное число других вариантов. При этом по мере усвоения новых вариантов происходит все большее и большее расширение экстраполяции.
Возможности человека к экстраполяции при координации двигательных актов лишь в небольшой степени обусловлены наследственной информацией. Основное значение имеет при этом формирование временных связей, облегчающих экстраполяцию. При однообразном выполнении двигательных актов возможности к экстраполяции суживаются, при разнообразном их выполнении — расширяются.
Диапазон экстраполяции всегда несколько ограничен. Так, навыки, которыми обладает футболист, недостаточны для выполнения путем экстраполяции приемов борца или боксера и наоборот.
В практике физического воспитания развитие экстраполяции в двигательной деятельности имеет весьма большое значение. Экстраполяция должна учитываться при подборе комплекса вспомогательных подготовительных упражнений. Этот комплекс должен включать в первую очередь такие вспомогательные упражнения, которые путем экстраполяции оказывают значительное влияние на освоение основного упражнения. Кроме того, если несколько вспомогательных упражнений дают по механизму экстраполяции один и тот же эффект, то количество их можно уменьшить. При включении каких-либо упражнений в общую физическую подготовку также необходимо всегда учитывать тот эффект, который путем экстраполяции сказывается на выполнении двигательных компонентов основных для специализации спортсмена упражнений и на развитии вегетативных функций (кровообращение, дыхание и т. п.), обеспечивающих двигательную деятельность.
§10. Автоматизация движений
Многие компоненты двигательных навыков, а в ряде случаев все частные особенности навыка автоматизируются, т. е. выполняются без их осознавания.
В организме осуществляется большое число неосознаваемых рефлекторных актов, протекающих всегда непроизвольно. Это так называемые первичные автоматизмы, связанные с различными безусловнорефлекторными реакциями, регулирующими вегетативные и некоторые двигательные функции. Наряду с этим имеются и вторичные автоматизмы, т. е. реакции, которые ранее протекали с осо-знованием их и лишь потом получили возможность осуществляться автоматически. К ним относятся, в частности, двигательные навыки.
По представлениям И. П. Павлова, автоматизация движений связана с состоянием возбудимости нервных центров. Условные рефлексы и дифференцировки хорошо проявляются в том случае, если соответствующие нервные центры, представляющие собою сложную совокупность из возбужденных и заторможенных нервных
387
клеток, находятся в состоянии оптимальной возбудимости. В этих центрах участки с оптимальной возбудимостью не являются закрепленными и постоянно перемещаются в больших полушариях под влиянием следовых процессов и раздражений из внешней и внутренней среды. Если же нервные центры находятся в состоянии известной степени торможения, т. е. пониженной возбудимости, то их творческая деятельность, проявляющаяся в образовании новых условных рефлексов и дифференцировок, затруднена или даже полностью исключена. Вместе с тем нервные центры, находясь в этом состоянии, еще способны осуществлять хорошо закрепленные рефлексы.
Сформировавшиеся двигательные навыки характеризуются именно такими хорошо закрепленными временными связями. Вследствие этого в тех случаях, когда навыки осуществляются в условиях оптимального возбуждения соответствующих нервных центров, имеет место осознавание их; если же они осуществляются в условиях несколько пониженного возбуждения нервных центров, то выполнение их происходит автоматизированно.
Следовые процессы даже после автоматизированно выполненных движений могут оживляться и осознаваться. Например, действия вратаря, борца при внезапно наступившей опасной ситуации, выполненные автоматизированно, могут полностью ими осознаваться после окончания соответствующих действий.
Рассматривая явления автоматизации навыка, следует разграничивать осознавание общих задач движения, связанных с перемещением крупных звеньев тела, и частных сторон двигательного акта, касающихся положения мелких структурных элементов, работы отдельных мышц и двигательных единиц, участвующих в движении. Деятельность мелких мышечных структур, как и отдельных функциональных моторных единиц или их небольших групп, не осознается человеком. Без специальной тренировки не отражается в сфере сознания и деятельность многих отдельных мышц. Хорошо осознаются движения различных крупных звеньев тела и всего тела в целом. Весьма слабо отражаются в сознании вегетативные компоненты навыков.
В нервной системе процессы, связанные с управлением автоматизированными и неавтоматизированными компонентами движения, тесно сочетаются друг с другом. При обучении и тренировке сознательный контроль за движениями имеет весьма важное значение (А. Ц. Пуни и др.). В обучении двигательным навыкам важнейшая роль принадлежит сознательному формулированию стоящих перед спортсменом задач, в частности связанных с общей структурой движений, Это положительно воздействует и на многие из тех процессов в нервных центрах, мышцах и вегетативных органах, которые совершенно не осознаются человеком. На доведении до сознания особенностей выполнения физических упражнений (например, характера совершенных спортсменом ошибок) основано значение срочной информации, получаемой инструментальным путем (В. С. Фарфель).
388
§ 11. Стадии (фазы) формирования двигательного акта как навыка
Становление двигательного акта как навыка проходит через несколько стадий, или фаз. В первой стадии отмечаются иррадиация нервных процессов с генерализацией ответных реакций и вовлечением в работу лишних мышц, объединение отдельных частных действий в целостный акт, во второй — концентрация возбуждения, улучшение координации, устранение излишнего мышечного напряжения и высокая степень стереотипности движений, в третьей — стабилизация, высокая степень координации и автоматизации движений.
В ряде случаев некоторые из фаз могут отсутствовать. Это связано с многими факторами: степенью сложности и мощностью мышечной работы, исходным состоянием двигательного аппарата, квалификацией спортсмена и др. Известно, что новые сложные движения всегда формируются на фоне прежде сложившихся координаций (И. М. Сеченов). Вследствие этого развитие, например, новых навыков выполнения гимнастических упражнений будет происходить совершенно различно у новичков, у спортсменов средней квалификации и у мастеров спорта. Неодинаковыми по целому ряду показателей будут стадии развития навыка при обучении фехтовальщика, теннисиста и бегуна. Совершенно различно будет формироваться навык, требующий значительных мышечных усилий (например, гимнастические упражнения, подъем тяжестей) у лиц с различным развитием опорно-двигательного аппарата. Если этот аппарат не подготовлен к работе со значительными мышечными усилиями, навык будет развиваться сопряженно с укреплением суставно-связочного аппарата и увеличением мышечной силы. Поэтому все схемы стадий развития навыка носят лишь ориентировочный характер.
§12. Устойчивость навыков при разных состояниях организма и длительность сохранения их после прекращения тренировки
Двигательные навыки, как и другие условные рефлексы, недостаточно постоянны в начале образования, в дальнейшем же становятся все более и более стойкими. При этом чем они проще по своей структуре, тем прочнее. Сложнейшие координационные отношения, которыми характеризуется выполнение навыка на особенно высоком уровне, менее стойки. Вследствие этого трудно при повторениях каждый раз показывать свои лучшие результаты. Если даже один какой-либо фактор, влияющий на выполнение упражнения, становится менее полноценным, то результат снижается. К такого рода факторам относятся ухудшение общего состояния нервной системы (например, при утомлении), развитие в организме гипоксии, недостаточная адаптация при значительном изменении поясного времени, неуверенность в результатах при наличии сильных противников
389
и др. Существенное значение имеют тип нервной системы, его устойчивость при действии отрицательных факторов.
После прекращения систематической тренировки навык начинает ухудшаться. Но это ухудшение выражено различно для разных его компонентов. Наиболее сложные двигательные компоненты могут ухудшаться даже при перерывах в несколько дней. Еще больше они страдают при длительных перерывах (недели, месяцы). Поэтому для достижения высоких результатов тренировка должна быть систематической, без длительных интервалов. Вместе с тем более грубые компоненты навыка могут сохраняться месяцами, годами и десятилетиями. Например, человек, научившийся плавать, кататься на коньках или ездить на велосипеде, сохраняет эти навыки в упрощенном виде даже после весьма больших перерывов.
Вегетативные компоненты навыков, связанные с регуляцией функции кровообращения, дыхания и т. д., имеют ряд отличий от двигательных. При кратковременной смене одного вида деятельности другим вегетативные компоненты перестраиваются медленнее, чем двигательные. При длительных же перерывах (месяцы и в особенности годы) вегетативные компоненты навыка, в отличие от двигательных компонентов, угасают полностью.
Глава XVII
СПОРТИВНАЯ ТРЕНИРОВКА И РАЗВИТИЕ ТРЕНИРОВАННОСТИ
§ 1. Спортивная тренировка. Тренированность. Перетренированность.
Спортивная тренировка. Это — специализированный педагогический процесс, направленный на повышение как общей физической работоспособности человека, так и специальной, обеспечивающей высокие достижения в избранном виде спорта.
Тренировка строится на основе общих, педагогических и специфических принципов. К первым относятся активность, сознательность, наглядность, систематичность, последовательность, доступность и прочность. Ко вторым — единство общей и специальной физической подготовки, непрерывность и цикличность тренировочного процесса, постепенное и максимальное повышение тренировочных нагрузок. Принципы систематичности и прочности принадлежат к числу главных при планировании спортивной тренировки. При правильной их реализации создаются оптимальные условия для совер шенствования двигательных навыков и повышения функциональных возможностей организма. Прочность двигательных навыков обеспечивается повторением изучаемых упражнений. Длительные перерывы в тренировке ведут к угасанию временных связей, лежащих в основе спортивной техники, и снижению функциональной подготок ленности спортсмена. Важнейшим специфическим принципом спор'
390
тивной тренировки является принцип единства общей и специальной физической подготовки спортсмена. Общая физическая подготовка служит основой для развития специальной тренированности в избранном виде спорта.
Л. А. Орбели считал, что разностороннее физическое воспитание обеспечивает возможность использования врожденных задатков организма при определенных жизненных ситуациях и для этого необходимо своевременно развивать эти задатки. Он говорил, что одной из важнейших задач физического воспитания является «тренировка тренируемости», т. е. повышение способности организма к овладению различными формами двигательной активности.
Повышение физической подготовленности обеспечивает успех не только в тех видах физических упражнений, которые систематически выполнялись, но и в других видах. Это явление объясняется так называемым «положительным переносом» двигательных навыков и физических качеств. Но некоторые виды физических упражнений являются как бы «несовместимыми». Параллельное занятие ими задерживает спортивное совершенствование, что называют «отрицательным переносом» двигательных навыков и физических качеств. «Положительные» и «отрицательные переносы» обусловлены сходством или различием временных связей, обеспечивающих выполнение движений, а также сходством или различием происходящих при этом морфофункциональных изменений в организме.
Общая физическая подготовка должна предшествовать специальной подготовке и в дальнейшем органически с ней сочетаться. При этом чем выше тренированность спортсмена, тем большее значение приобретают специальные упражнения, близкие по структуре и по физиологическому эффекту к упражнениям, характерным для избранного вида спорта.
Длительные перерывы в тренировке ведут к угасанию временных связей, лежащих в основе техники выполнения движений. При этом раньше других угасают наиболее тонко специализированные и наименее закрепленные связи.
Разнообразные сдвиги в организме, возникающие при мышечной деятельности, сохраняются лишь некоторое время. Поэтому для развития тренированности необходимо, чтобы интервалы отдыха между упражнениями не были излишне продолжительными.
Тренировочный процесс, сохраняя непрерывность, разносторонность и специальную направленность, не остается постоянным на всем своем протяжении. Содержание его и структура должны изменяться в связи с принципом цикличности. На этом основана периодизация спортивной тренировки. В подготовительном периоде тренировка направлена на развитие необходимых физических качеств и совершенствование техники движений. Объем и интенсивность тренировочных нагрузок в это время постепенно повышаются. Соревновательный период характеризуется участием в основных соревнованиях. Весь режим тренировочной работы и отдыха в это время направлен на сохранение или даже повышение достигнутого уровня тренированности. В переходном периоде участие в соревно
391
ваниях прекращается. Тренировочные нагрузки снижаются как по объему, так и по интенсивности. Длительность этого периода обычно не превышает 4—6 недель.
Таким образом, на протяжении круглогодичной тренировки объем и интенсивность нагрузок непостоянны. В связи с этим изменяется и уровень общей и специальной работоспособности спортсмена, что необходимо учитывать при планировании тренировочного процесса. Объем тренировочных нагрузок определяется количеством выполненных упражнений или длиной пройденных отрезков дистанции. Интенсивность тренировочных нагрузок в настоящее время принято оценивать по степени учащения сердечного ритма.
В зависимости от особенностей вида спорта в тренировке преобладают нагрузки той или иной длительности и интенсивности. Например, при подготовке лыжников-гонщиков применяются очень большие по объему нагрузки, интенсивность же их обычно не превышает 70—90% от максимальной.
Спортивная тренировка обеспечивает повышение технической, физической, психической и тактической подготовленности спортсмена в избранном виде спорта. Физиология спорта изучает механизмы повышения технической и физической подготовленности.
Техническая подготовка направлена на формирование и совершенствование двигательных навыков (см. гл. XVI). Физическая подготовка способствует развитию физических качеств человека: быстроты, силы, выносливости и ловкости. Повышение каждого из этих качеств обусловлено биохимическими, морфологическими и функциональными изменениями во всех системах организма, и в первую очередь в двигательном аппарате.
Тренированность. Уровень специальной работоспособности спортсмена называют тренированностью. Она может быть высокой и низкой.
Тренированность повышается при систематической тренировке и резко снижается при ее прекращении. Она изменяется также в процессе круглогодичной тренировки. Постепенно повышаясь в подготовительном периоде, достигает максимального уровня в соревновательном периоде и вновь несколько снижается в переходном. Наивысший для спортсмена уровень тренированности не может поддерживаться постоянно. Длительность его зависит от вида спорта, специфики тренировочного процесса и индивидуальных особенностей человека.
В процессе тренировки повышается резистентность (устойчивость) клеток и органов к происходящим при работе изменениям внутренней среды. Благодаря этому тренированный организм может продолжать работу при значительном кислородном долге, изменении pH, уменьшении концентрации глюкозы в крови и пр. Нетренированный же при этих изменениях обычно вынужден прекратить работу.
Тренированность повышается в результате многократно повторяемой работы и является естественным ее следствием. Еще в начале XIX столетия выдающийся французский естествоиспытатель
392
Ламарк высказал мысль о том, что «работа строит орган». Это важное биологическое явление обусловлено тем, что во время работы происходят не только процессы диссимиляции, но и процессы ассимиляции. После же окончания работы ассимиляция, т. е. восстановление ресурсов организма, может идти «через край», что обеспечивает не только компенсацию расхода, но и увеличение энергетических запасов по сравнению с исходным уровнем (см. рис. 104 в гл. XV). Это необходимо для повышения работоспособности.
При тренировке каждая последующая работа, начинаясь па фоне повышенной работоспособности, ведет к дальнейшему ее росту. Интенсивность восстановительных процессов в тканях усиливается при накоплении продуктов внутриклеточного обмена. Например, ресинтез АТФ, креатинофосфата и углеводов стимулируется продуктами расщепления этих веществ. Большое значение для восстановления химизма тканей имеют импульсы из центральной нервной системы. К числу таких влияний относится адаптационно-трофический эффект при раздражении симпатических нервов (см. гл. IV, § 10). Важную роль в адаптации организма к мышечной деятельности играют гормоны (см. гл. ХШ).
Мышечная деятельность оставляет длительные следы в состоянии различных систем организма. Повторные нагрузки углубляют эти следы и способствуют более выраженным биохимическим, морфологическим и функциональным перестройкам в организме, обеспечивающим повышение его работоспособности.
Перетренированность. Если тренировочный процесс организован неправильно, не соблюдается постепенность в повышении объема и интенсивности нагрузок, чрезмерно часто применяются максимальные нагрузки и отсутствует достаточный отдых между ними, то работоспособность организма снижается. Такое состояние спортсмена называется перетренированностью.
Перетренированность чаще наблюдается у квалифицированных спортсменов, стремящихся форсированной тренировкой повысить свои результаты.
Перетренированность нельзя смешивать с острым перенапряже-, нием. которое возникает при выполнении однократной очень напряженной работы, превышающей функциональные возможности спортсмена. Это состояние обычно проявляется в нарушении деятельности органов кровообращения.
Состояние перетренированное™ может быть выражено в разной' степени. Раньше всего при этом возникают изменения нервной деятельности, что проявляется в ухудшении координации движений, нарушении сна, отсутствии желания продолжать тренировку. Понижение координационных возможностей отрицательно отражается на технике движений и ведет к снижению спортивных результатов.
При перетренированное™ нередко наблюдаются изменения сердечного ритма, резкое увеличение размеров сердца, повышение кровяного давления. Иногда происходит уменьшение показателей максимальной вентиляции и жизненной емкости легких. Отмечается ухудшение аппетита, что может вести к уменьшению веса тела.
393
Нарушение функционального состояния организма при перетренированное™ наиболее ярко проявляется в изменении его реакций на нагрузки. При стандартных нагрузках у перетренированных более резко учащается пульс, нарастают артериальное давление, легочная вентиляция, потребление кислорода и кислородный долг. При более напряженной работе у них нередко возникают нарушение сердечного ритма и другие неблагоприятные изменения деятельности сердца. Восстановление после работы у перетренированных происходит замедленно. Все это свидетельствует об ухудшении регуляции вегетативных процессов, о снижении адаптации организма к физическим нагрузкам.
В случае возникновения перетренированности необходимо сни- । зить объем и интенсивность тренировочных нагрузок. При легкой форме перетренированности этого достаточно для ее ликвидации. При более тяжелых формах спортсмену должен быть предоставлен активный отдых (переключение на другой вид спортивной деятель- 1 ности). При очень тяжелых формах тренировка должна быть временно полностью прекращена.
Легкие формы перетренированности ликвидируются, если приняты соответствующие меры, обычно в течение 15—30 дней. При более тяжелых формах требуется отдых 1—2 месяца с полным прекращением тренировки на 2—3 недели.
§ 2. Физиологические показатели тренированности
. Особенности морфофункционального состояния разных систем I организма, возникающие при спортивной тренировке, называются физиологическими показателями тренированности. Они изучаются у человека в состоянии мышечного покоя, при выполнении стандартных (тестирующих) нагрузок и нагрузок предельной мощности.
Некоторые физиологические показатели тренированности, достигнув определенного уровня уже в первые годы тренировки, в дальнейшем почти не изменяются. Они не снижаются, например, при уменьшении объема и интенсивности нагрузок в переходном периоде тренировки. Обеспечивая информацию о степени физической подготовленности человека, эти показатели не позволяют судить о динамике тренированности в процессе круглогодичной тренировки. К таким показателям относятся, например, морфологические изменения в костной ткани (увеличение диафизов трубчатых костей, утолщение их коркового слоя, увеличение костных выступов, шероховатостей, гребешков). У квалифицированных спортсменов мало изменяются на протяжении годичной тренировки жизненная емкость легких и частота сердечных сокращений. Поэтому эти показатели не дают достоверных сведений о динамике тренированности.
Другие показатели тренированности более лабильны. Они повышаются при нарастании тренированности и достоверно снижаются в переходном периоде, когда уровень тренированности невысок.
394
К таким показателям относятся, например, объем сердца, длительность изометрического периода систолы сердца и др.
Физиологические показатели тренированности зависят от спортивной специализации. Например, у спортсменов-стайеров морфофункциональные изменения органов дыхания и кровообращения выражены более ярко, чем у спринтеров. Различны у них и реакции на одинаковую работу. Так, при одинаковой скоростной работе у стайеров легочная вентиляция увеличивается меньше, а коэффициент использования кислорода нарастает больше, чем у спринтеров (Г. М. Попова).
Кроме того, физиологические показатели тренированности зависят от индивидуальных особенностей человека. У спортсменов, даже специализирующихся в одном виде спорта, одни и те же показатели могут быть выражены по-разному. Например, такой постоянный показатель развития выносливости, как брадикардия, иногда отсутствует у квалифицированных бегунов-стайеров и лыжников.
Адаптация к нагрузкам у разных людей также происходит неодинаково. Например, у одних спортсменов обеспечение кислородной потребности при работе происходит за счет повышения функций главным образом органов дыхания, у других — кровообращения. В связи с этим по динамике одного из физиологических показателей нельзя судить об изменении тренированности спортсмена. Более ценную информацию дают комплексные исследования морфофункционального состояния важнейших физиологических систем. Однако и в этом случае оценка тренированности может быть достоверной лишь при сочетании физиологических исследований с данными педагогических и врачебных наблюдений.
Показатели тренированности в состоянии покоя. По морфофункциональным показателям некоторых систем организма в состоянии покоя можно судить о динамике тренированности спортсмена.
Центральная нервная система. Мозг животных, подвергаемых экспериментальной тренировке, отличается большим весом по сравнению с мозгом нетренированных животных и некоторыми биохимическими особенностями. У тренированных животных увеличено число разветвлений дендритов нервных клеток больших полушарий. Систематическая мышечная деятельность сопровождается повышением буферных свойств нервной ткани и повышением активности окислительных и некоторых других ферментов.
Электроэнцефалограмма тренированного человека характеризуется относительно более частым альфа-ритмом и более выраженной амплитудой его колебаний. Тренированный может быстрее усваивать ритм раздражений и воспроизводить более высокую частоту его, что свидетельствует о повышенной лабильности нервных клеток.
У тренированных, особенно к скоростной работе, увеличена подвижность нервных процессов. Это проявляется в укорочении скрытого периода двигательных реакций, уточнении дифференцировок и повышении скорости переработки поступающей от анализаторов
395
информации. Для спортсменов-стайеров характерна высокая уравновешенность нервных процессов.
Двигательный аппарат. При тренировке происходят значительные морфологические и функциональные изменения во всех звеньях двигательного аппарата (мышцы, кости, сухожилия, связки). Влияние мышечной деятельности на развитие скелета было отмечено П. Ф. Лесгафтом. Он обнаружил, что развитие скелетных мышц сопровождается утолщением костей и увеличением их прочности. У тренированных людей, а также у животных, подвергаемых экспериментальной тренировке, поперечные размеры костей увеличены, корковый слой утолщен. На поверхности костей образуются выступы и шероховатости. Наряду с макроскопическими изменениями костей происходят и гистологические сдвиги в структуре и расположении костных балок, увеличении числа остеонов и др. Все это способствует увеличению механической прочности костей.
При тренировке увеличиваются масса и объем скелетных мышц. Больше всего гипертрофируются мышцы, выполняющие силовые и статические напряжения. Динамическая же работа вызывает меньшие морфологические изменения мышц.
Рабочая гипертрофия мышц происходит за счет увеличения объема отдельных мышечных волокон. При этом утолщается их сарколемма, увеличиваются объем саркоплазмы, число миофибрилл и других структурных внутриклеточных элементов.
Развитие скелетных мышц ведет к увеличению удельного веса тела. Этому способствуют потери воды и жира при физических нагрузках. Прекращение же тренировки вызывает уменьшение мышечной массы.
Гипертрофия скелетных мышц сопровождается улучшением их кровоснабжения. У животных при экспериментальной тренировке увеличивается количество капилляров в скелетных мышцах. Так, в опытах, в которых тренировке подвергались мышцы лишь одной стороны тела, в нетренированных мышцах обнаружено в среднем 46 капилляров в 100 мышечных волокнах, а в тренированных — 98.
В мышечной ткани имеются артерио-венозные анастомозы. При покое в нетренированной мышце часть из них открыта, в тренированной же они закрыты. Это обеспечивает в тренированной мышце тесный контакт всей протекающей крови с мышечной тканью, что снижает потребность последней в кровоснабжении. Поэтому объем кровотока в тренированных мышцах относительно меньше, чем в нетренированных. Этому способствует, в частности, повышение активности окислительных ферментов в тренированных мышцах.
При рабочей гипертрофии происходит развитие иннервацион-ных приборов мышцы. По мере утолщения мышечного волокна в нем разрастаются нервные окончания. В них становится больше количество концевых веточек и ядер шванновской глии. Изменения в иннервациоином приборе мышцы обеспечивают увеличение контактирующей! поверхности между мышечным волокном и нервом.
396
Одновременно с гипертрофией мышечных волокон в них происходят и биохимические изменения. Опыты на животных показали, что в результате экспериментальной тренировки в мышцах повышается содержание белков саркоплазмы и сократительного белка миофибрилл — миозина.
В тренированных мышцах увеличивается количество миоглобина, что повышает кислородную емкость мышц и способствует интенсификации окислительных процессов. Установлено, что у животных, ведущих более подвижный образ жизни, относительно больше миоглобина в мышцах. Например, мышцы комнатной собаки содержат 400 мг% миоглобина, а мышцы охотничьей собаки — 1000 мг%.
Наступающие в процессе тренировки биохимические и морфологические изменения в скелетных мышцах ведут к функциональным сдвигам. У тренированных, особенно к скоростной работе, повышены возбудимость и лабильность мышц. У лиц, не занимающихся физическими упражнениями, возбудимость и лабильность мышц-антагонистов, измеряемые по величине реобазы и хронаксии, как правило, неодинаковы. В процессе тренировки величины их сближаются, что способствует совершенствованию координации двигательной деятельности.
Одним из основных функциональных свойств скелетных мышц является их сила. Повышение ее в процессе тренировки обусловлено гипертрофией мышечных волокон и увеличением возможности одновременно включать в работу максимальное число двигательных единиц.
Наибольшее напряжение, которое мышца может развить в изометрических условиях, называется ее абсолютной силой. Эта величина обычно больше, чем сила мышцы при произвольном напряжении. Специальная силовая подготовка ведет к сближению этих показателей.
Значительная мышечная сила требуется для достижения высоких результатов во многих видах спорта. Наиболее эффективно сила развивается при выполнении физических упражнений, где она является определяющим компонентом (подъем штанги, выполнение висов и упоров в гимнастике и т. п.).
Большое значение, для работоспособности и для координации движений имеет скорость расслабления мышц. По данным Ю. В. Вы-сочина, способность мышц к быстрому расслаблению в процессе тренировки улучшается в большей степени, чем многие другие функциональные параметры (укорочение латентного периода, скорость нарастания усилия и пр.).
Способность мышц к максимальным произвольному напряжению и расслаблению определяют при помощи специального прибора — миотонометра. Он позволяет оценивать твердость мышц при этих состояниях. У тренированных твердость мышцы при произвольном напряжении больше, а при расслаблении меньше, чем у нетренированных. Разница между этими показателями нарастает при повышении тренированности.
397
Обмен веществ и энергии. При нормальном питании у спортсменов обычно наблюдается азотистое равновесие. Лишь в подготовительном периоде тренировки, когда работа направлена на развитие скелетной мускулатуры, азотистый баланс может быть положительным.
В тренированном организме увеличены запасы углеводов, что очень важно для повышения работоспособности (особенно при выполнении длительной и напряженной мышечной деятельности). Запасы жира относительно уменьшены. Основной обмен находится в пределах стандартных величин или несколько понижен. При частых тренировочных занятиях и особенно выступлениях на соревнованиях он может быть значительно выше стандартных величин: В дни после напряженных нагрузок основной обмен повышен, снижаясь в последующие дни отдыха (см. рис. 107). В связи с этим повышение основного обмена у спортсменов надо рассматривать как следствие неполного восстановления после тяжелой работы.
Дыхательная система. В процессе тренировки происходят морфофункциональные изменения органов дыхания. Это является одним из важнейших условий, обеспечивающих увеличенное потребление кислорода при работе. Эти изменения выражаются в развитии дыхательных мышц, о чем можно судить по увеличению жизненной емкости легких (ЖЕЛ) и максимальной вентиляции легких (МВЛ).
Величина ЖЕЛ зависит от специализации спортсмена и его спортивного стажа. У мужчин, занимающихся циклическими видами спорта, ЖЕЛ составляет 5000—7000 мл и более, у женщин — 3500—5000 мл. Наиболее велика ЖЕЛ у пловцов, что обусловлено особенностями дыхания при плавании. Преодоление сопротивления воды при вдохе и выдохе способствует развитию дыхательных мышц.
Для характеристики влияния физических упражнений на дыхательный аппарат нужно учитывать не только абсолютную величину ЖЕЛ, но и соотношение составляющих ее объемов. У более тренированных дополнительный объем вдоха больше дополнительного объема выдоха.
ЖЕЛ зависит от веса тела. Поэтому для оценки дыхательных функций более информативен так называемый жизненный показатель, т. е. отношение ЖЕЛ (в мл) к весу тела (в кг). Он особенно велик у бегунов на длинные и сверхдлинные дистанции.
О выносливости дыхательных мышц судят по динамике величины ЖЕЛ при повторных ее определениях через короткие интервалы отдыха (проба Розенталя). При достаточной выносливости дыхательных мышц ЖЕЛ при повторных определениях не снижается, при недостаточной выносливости снижается вследствие утомления дыхательных мышц.
ЖЕЛ является важным критерием развития дыхательного ап- 4 парата. Однако у квалифицированных спортсменов после достижения определенной величины она остается стабильной и не отражает изменений их работоспособности в разные периоды круглогодич
398
ной тренировки. Поэтому величина ЖЕЛ не может быть достаточно информативным показателем уровня тренированности.
Величина МВЛ более изменчива, чем показатель ЖЕЛ. Наиболее высокого уровня МВЛ достигает в соревновательном периоде, когда спортсмен находится в спортивной форме. В периоды снижения объема и интенсивности тренировочных нагрузок МВЛ несколько уменьшается. Эта величина, как и ЖЕЛ, зависит от пола, веса тела и спортивной специализации. У мужчин, занимающихся циклическими видами спорта, МВЛ составляет 100—150 л/мин и более, у женщин эта величина несколько меньше.
, Разница в объеме грудной клетки при вдохе и выдохе (экскурсия грудной клетки) у тренированных больше, что имеет важное значение для увеличения вентиляции легких при мышечной деятельности.
Частота дыханий у тренированных в состоянии покоя меньше, чем у нетренированных. У первых она не превышает в среднем 10— 14 дыханий в 1 мин. Глубина дыханий при этом оказывается увеличенной до 700—800 мл. Большая глубина дыхания обеспечивает увеличение дыхательной поверхности легких, что создает лучшие условия для обмена газов между альвеолярным воздухом и кровью.
Минутный объем дыхания (легочная вентиляция) при мышечном покое у тренированных почти такой же, как и у нетренированных. Соотношение же компонентов, определяющих уровень легочной вентиляции, в процессе тренировки существенно изменяется. Происходящие при этом урежение дыхания и увеличение его глубины экономизируют дыхательный акт.
Потребление кислорода в состоянии покоя в процессе тренировки, как правило, почти не изменяется. Но в некоторых случаях, особенно когда тренировка ведет к резко выраженной гипертрофии мышц, потребление кислорода в покое несколько увеличивается. В случаях, когда адаптация к работе характеризуется главным образом экономизацией окислительных процессов, потребление кислорода в покое может уменьшаться. По некоторым данным, у тренированных увеличен коэффициент использования кислорода из вдыхаемого воздуха. Легочная вентиляция при этом оказывается сниженной, что ведет к экономизации внешнего дыхания.
Содержание углекислоты в выдыхаемом воздухе у спортсменов, особенно у тренирующихся к длительной циклической работе, несколько повышено. Это обусловлено увеличением ее концентрации в крови в связи с нарастанием щелочных резервов.
Об интенсивности тканевого дыхания и о степени возбудимости дыхательного центра можно судить по времени произвольной задержки дыхания. Обычно это время больше у более тренированных. Оксигемометрические исследования показали, что у них при задержке дыхания продолжительнее как устойчивая, так и особенно гипоксемическая фазы. Восстановление оксигенации крови после окончания задержки дыхания у тренированных происходит быстрее.
S99
Сердечно-сосудистая система. Систематическая тренировка, особенно к длительной циклической работе, сопровождается биохимическими, морфологическими и функциональными изменениями сердца и сосудов. Главную роль в этом играет усиление парасимпатических (холинергических) и угнетение адренергических влияний на органы кровообращения (В. Рааб и др.).
В XIX в. было обнаружено, что занятия спортом ведут к увеличению размеров сердца. Тогда это рассматривалось как неблагоприятное явление. Позднее же было установлено, что систематическая мышечная деятельность, как правило, ведет к увеличению полостей сердца (тоиогенная дилатация) и к умеренной гипертрофии миокарда. В сердечной мышце при этом увеличивается содержание гликогена и белковых соединений, в частности миоглобина. Последнее предохраняет ее от развития гипоксии.
Гипертрофия миокарда сопровождается развитием капиллярной сети, увеличением диаметра капилляров и возникновением между ними поперечных анастомозов.
При экспериментальной тренировке животных вес правого желудочка сердца может возрастать на 10—16%, вес левого — на 10— 13%. Отношение веса сердца к весу тела при этом несколько увеличивается.
Вес сердца у лиц, не занимающихся спортом, обычно находится в прямой зависимости от веса тела. У спортсменов в связи с гипертрофией миокарда эта зависимость выражена в меньшей степени.
С. П. Летунов и Р. Е. Мотылянская путем рентгенологических исследований обнаружили у спортсменов увеличенные размеры сердца в 74% случаев. Наиболее часто это наблюдалось у тренирующихся к длительной циклической работе. Например, у лыжников гипертрофия сердца отмечалась в 84%. У них чаще, чем у специализирующихся в других видах спорта, наблюдалась гипертрофия обоих желудочков сердца.
Степень гипертрофии сердца зависит от особенностей тренировки, а также от возраста, в котором начаты занятия спортом.
В настоящее время установлено, что более эффективной является умеренная гипертрофия миокарда. Резко же выраженная гипертрофия нередко сопровождается понижением сократительных свойств миокарда, что отрицательно сказывается на производительности работы сердца.
Одновременно с гипертрофией стенок сердца увеличивается объем его полостей. Это ведет к увеличению общего объема сердца. По данным Рейнделла, он составляет у спортсменов в среднем около 1000 см3, у не занимающихся спортом на 30—40% меньше. Индивидуальные колебания этого показателя очень велики. Например, у участников Олимпиады 1960 г. объем сердца варьировал в пределах от 505 до 1300 см3.
Объем сердца в определенной мере зависит от веса тела и роста. При расчете на 1 кг веса он составляет v тренированных мужчин 10—18,5 см3 кг, у женщин — 7—14,5 см3 кг.
400
16 см3/кг)
МЛ 1150
1100
Относительную величину объема сердца с учетом веса и роста Объем сердца (см3) п следует определять так: -----------1-- Полученная величина,
вес (кг) рост (см)
выраженная в условных единицах, у тренированных равна 40—152, у нетренированных — 25—80 (В. Л. Карпман).
Увеличенный объем сердца обеспечивает возможность значительного нарастания систолического и минутного объемов крови при мышечной деятельности. Однако при чрезмерном нии (свыше 1200 сма, или соответственно ные свойства миокарда и резервные возможности сердца могут снижаться.
Увеличение объема сердца у спортсменов зависит от их специализации. Оно наиболее выражено у тренирующихся к длительной циклической работе.
Объем сердца изменяется на протяжении круглогодичной тренировки, поэтому он может служить одним из показателей тренированности. В те периоды, когда тренировочные и соревновательные нагрузки очень высоки, объем сердца у спортсменов достигает наибольших величин. В периоды снижения нагрузок он уменьшается. Например, наблюдения за группой квалифицированных велосипедистов-шоссейников по-
его увеличе-сократитель-
1050
1000
950
Рис. 112. (см3) у 1
Периоды тренировки
. Изменение объема сердца велосипедистов в разные периоды тренировки (по С. В. Хрущеву): 1 и 2 —* переходный период, 3 и 4 — подготовительный, 5 — соревновательный
казали, что в конце соревновательного периода объем сердца у них составлял в среднем 1100 см3. В течение переходного периода он снизился до 1000 см 3. Повышение тренировочных нагрузок в подготовительном периоде вновь способствовало увеличению объема сердца в среднем до 1150 см3 (рис. 112).
Частота сердцебиений у тренированных, как правило, реже, чем у лиц, не занимающихся спортом. У спортсменов-мужчин ритм сердечных сокращений составляет в среднем 55 ударов в 1 мин., у женщин — 59, у не занимающихся спортом — 70. Брадикардия у спортсменов обусловлена усилением воздействий блуждающего нерва на сердце и некоторыми биохимическими изменениями в синоатриальном узле. Эти явления больше выражены у тех спортсменов, за-
нимающихся циклическими упражнениями, в тренировке которых преобладает работа над развитием общей выносливости. По средним данным, у бегунов-стайеров сердечный ритм не превышает 52 ударов в 1 мин., у спринтеров же он равен 60. Частота сердцебиений меньше 50 ударов в 1 мин. у бегунов-стайеров наблюдается в 30% случаев, у спринтеров — лишь в 9%. У квалифицированных лыжников-гонщиков сердечный ритм в покое равен в среднем 44 ударам в 1 мин. В отдельных случаях он не превышает 30.
14—113
401
У лыжниц-гонщиц частота сердцебиений несколько больше. У футболистов она составляет в среднем 56 ударов в 1 мин., у волейболистов — 60, у борцов и штангистов — 59. Брадикардия может наблюдаться и у некоторых спортсменов, в деятельности которых преобладает относительно кратковременная силовая и скоростная работа. Это объясняется индивидуальными особенностями человека и воздействием обшей физической подготовки, одной из задач которой является развитие выносливости.
Брадикардия у спортсменов развивается главным образом в первые 2—3 года занятий спортом. В дальнейшем этот показатель стабилизируется и мало изменяется на протяжении круглогодичной тренировки.
В некоторых случаях может развиваться резко выраженная брадикардия при чрезмерных тренировочных нагрузках, превышающих функциональные возможности организма.
У тренированных спортсменов брадикардия нередко сочетается с синусовой аритмией. При умеренной аритмии разница в длительности отдельных сердечных циклов колеблется в пределах 0,10— 0,15 сек., при выраженной аритмии превышает 0,30 сек.
Синусовая аритмия нередко наблюдается у молодых людей, не занимающихся спортом. В этих случаях она связана с фазами дыхания и поэтому называется дыхательной. У спортсменов синусовая аритмия не является дыхательной, она одинаково проявляется во время вдоха и выдоха и обусловлена, как и брадикардия, усилением влияния на сердце блуждающих нервов. Это явление возникает и усиливается в процессе спортивной тренировки. Наличие синусовой аритмии свидетельствует о способности сердца быстро адаптироваться к изменяющимся условиям деятельности.
Происходящие в процессе тренировки изменения в морфофункциональном состоянии сердца отражаются на показателях электрокардиограммы (ЭКГ). По данным Л. А. Бутченко, ЭКГ спортсменов, тренировка которых направлена преимущественно на развитие выносливости, характеризуется наличием синусовой брадикардии и синусовой аритмии, низкими зубцами Р, высокими зубцами Т, смещением сегмента ST выше изолинии и высоким вольтажем зубцов комплекса QHS.
Под влиянием спортивной тренировки значительно изменяется фазовая структура сердечного цикла. Длительность его при этом увеличивается в основном за счет удлинения диастолы. Наряду с этим происходят изменения и в фазовой структуре систолы (табл. 14). Особенно резко удлиняется фаза изометрического сокращения, в связи с чем становится больше и период напряжения миокарда.
Удлинение фазы изометрического сокращения у спортсменов наблюдается в 20% случаев. Иногда (чаще у стайеров) длительность этой фазы превышает границы нормы.
Период изгнания крови из желудочков может изменяться по-разному. В связи с этим по-разному изменяется и длительность механической систолы сердца, т. е. время, затрачиваемое на асин- 1
402
Таблица 14
Средняя длительность фаз сердечного цикла (сек.) у тренированных спортсменов и у людей, не занимающихся спортом (по В.Л. Карпману)
Фазы сердечного цикла Не занимающиеся спортом Спортсмены
Фаза асинхронного сокращения .... 0,051 0,061
Фаза изометрического сокращения . . . 0,031 0,047
Период напряжения 0,082 0,108
Период изгнания крови 0,256 0,258
Механическая систола 0,287 0,305
Сердечный цикл 0,850 0,948
Рис. 113. Объем левого желудочка в состоянии покоя и при работе у не занимающихся спортом (А) и у тренированных спортсменов (Б).
1 — систолический объем крови; 2 — резервный объем крови; 3 — остаточный объем крови.
хронное, изометрическое сокращение и изгнание крови из желудочка.
Величина внутрасиста лического показателя (соотношение длительности периода изгнания крови и длительности механической систолы) при повышении тренированности уменьшается.
Все изменения в фазовой структуре сердечного цикла наиболее ярко выражены у квалифицированных спортсменов, тренирующихся к длительной циклической работе. Изучение этих показателей в процессе круглогодичной тренировки дает информацию о степени адаптации сердца к физическим нагрузкам. Однако в некоторых случаях изменения фазовой структуры сердечного цикла у спортсменов могут быть обусловлены перенапряжением миокарда. Поэтому для правильной оценки наблюдаемых сдвигов показатели фазовой структуры сердечного цикла должны рассматриваться в сочетании с данными электрокардиографии.
Удлинение изометрического сокращения и некоторые другие
изменения в фазовой структуре средечного цикла у тренированных спортсменов указывают на гиподинамию миокарда. Это явление, обусловленное усилением воздействий блуждающего нерва на сердце, означает, что при мышечном покое оно сокращается с меньшей силой. Гиподинамия миокарда свидетельствует о наличии значительных функциональных резервов сердца. Состояние гиподина-
14*
403
мии регулируется путем нервных и гуморальных влияний на сердце и при мышечной деятельности сменяется гипердинамией.
Одним из показателей гиподинамии миокарда у спортсменов может служить увеличенный резервный объем крови (рис. 113). Он обеспечивает повышение сердечного выброса при мышечной деятельности.
Систолический объем крови с ростом тренированности несколько уменьшается, что указывает на экономизацию деятельности сердца при мышечном покое.
Экономизация деятельности сердца у спортсменов проявляется также в снижении мощности сердечных сокращений. Например, у гребцов на байдарке мощность сокращений левого желудочка, рассчитанная по специальной формуле, в подготовительном периоде тренировки составляла в среднем 4,02 вт, в соревновательном — 2,76 вт.
Минутный объем крови при повышении уровня тренированности уменьшается, что обусловлено лучшим использованием кислорода тканями и вследствие этого понижением их потребности в кровоснабжении. Особенно четко это выражено при пересчете минутного объема крови на единицу поверхности тела (сердечный индекс). Например, у квалифицированных лыжников этот показатель в подготовительном периоде тренировки составлял в среднем 2,54 л/мин/м2, в соревновательном же снизился до 1,64 л/мин/м2.
Систолический и минутный объем крови, уменьшаясь в соревновательном периоде тренировки, вновь увеличивается при снижении тренировочных нагрузок в переходном периоде. Следовательно, по изменениям систолического и минутного объема крови при мышечном покое можно судить о динамике тренированности спортсменов.
Показатели артериального давления у спортсменов находятся в пределах возрастных норм. Спортивная специализация не влияет на эти величины. При развитии тренированности наблюдается тенденция к повышению артериального давления. Несколько больше при этом нарастает диастолическое давление, что обусловлено уменьшением потребности тканей в кровоснабжении. Мелкие артерии в связи с этим суживаются, просвет их уменьшается. Сопротивление оттоку крови на периферию во время диастолы возрастает, что и ведет к повышению диастолического давления.
Если в состоянии покоя систолическое давление у спортсменов выше 140 мм рт. ст. или ниже 100 мм рт. ст., то в большинстве случаев это обусловлено нарушением регуляции функционального состояния сердца и сосудов.
О функциональном состоянии артериальных стенок, которое играет большую роль в обеспечении местного (регионарного) кровотока, судят по степени их жесткости, определяемой по скорости распространения пульсовой волны. На начальных этапах тренировки жесткость артериальных стенок у спортсменов в состоянии покоя существенно не отличается от возрастных стандартов. В дальнейшем при развитии тренированности эти показатели изменяются, что обусловлено главным образом изменением тонуса гладких мышц ар-
404
термальных стенок. Характер этих изменений зависит от особенностей выполняемых движений. Изменения более выражены при заня-
тиях теми видами спорта, в которых нагрузка на отдельные мышечные группы распределяется неравномерно. Например, при беге, особенно на длинные дистанции, наиболее активны мышцы нижних
конечностей. Поэтому у бегунов в наибольшие изменения жесткости При изменении структуры выполняемых движений меняется и жесткость артериальных стенок в работающих звеньях тела. Например, если у лыжников-гонщиков в подготовительном периоде тренировки преобладают беговые нагрузки, то у них в состоянии покоя повышается жесткость стенок артериальных сосудов ног (рис. 114). При переходе же к тренировкам на снегу, где нагрузки на разные мышечные группы распределяются более равномерно, жесткость стенок артерий ног и рук становится почти одинаковой.
Изменение жесткости артериальных стенок приразвитиитрени-рованности может*’’ происходить по-разному. Например, у бегунов-новичков на первом этапе тренировки она снижается. Это обус-
процессе тренировки происходят стенок артерий ног.
Рис. 114. Скорость распространения пульсовой волны (цифры над столбиками, см/сек) по артериям верхних (в) и нижних (н) конечностей у лыжников в подготовительном (Л) и соревновательном (Б) периодах тренировки (по В. Д. Евстратову и В. В. Трунину)
ловлено длительным периодом восстановления после тренировочных нагрузок. Кровоснабжение работавших мышц в это время должно быть усиленным. Поэтому капилляры в мышцах остаются расширенными, гладкие мышцы артериол и артерий расслабленными. Последнее и ведет к понижению жесткости артериальных стенок. При развитии тренированности жесткость артериальных стенок в состоянии покоя, особенно в активных областях тела, начинает повышаться. Это объясняется уменьшением потребности тренированных мышц в кровоснабжении, что ведет к сужению артериол, повышению периферического сопротивления и увеличению жесткости стенок крупных артерий.
Система крови. Общее количество крови в организме при развитии тренированности несколько увеличивается. Содержание в ней эритроцитов и гемоглобина повышается. Это увеличивает ды-
хательную поверхность крови и ее кислородную емкость.
Для высоких результатов в спорте, особенно в циклических видах, требуется наличие в 1 мм3 крови не менее 4,7 млн. эритроцитов и не менее 14,5% гемоглобина. В отдельных случаях у спортсменов после тренировки в горной местности число эритроцитов может превышать 6 млн. в 1 мм3, соответственно при этом увели
405
чивается и количество гемоглобина. Эти показатели особенно важны для спортсменов, тренирующихся к длительной работе большой и умеренной мощности.
Лейкоцитарная формула у тренированных, особенно у стайеров, изменена в сторону увеличения количества лимфоцитов.
Изменения, наступающие в плазме крови в процессе тренировки, обусловлены главным образом повышением мощности буферных систем, предохраняющих pH от резких сдвигов в кислую сторону. Увеличение их мощности обусловлено повышением активности некоторых ферментов крови.
Щелочной резерв крови у спортсменов увеличен. У тренированных этот показатель составляет в среднем около 70 объемных процентов, у не занимающихся спортом — 65.
Приведенные данные свидетельствуют о наличии глубоких и разносторонних морфофункциональных особенностей у тренированных спортсменов, находящихся в состоянии мышечного покоя, отличаю щих их от людей, не занимающихся спортом. Этими особенностями обусловлена экономичность физиологических функций в состоянии покоя и готовность организма выполнять напряженную мышечную деятельность.
§ 3. Реакции тренированного и нетренированного организма на стандартные (тестирующие) нагрузки
Стандартные нагрузки, применяемые для оценки тренированности, должны иметь строго определенную мощность и длительность и быть доступными как для тренированных, так и для нетренирован-
ных лиц.
Реакции на тестирующие нагрузки у тренированных характеризуются следующими особенностями: 1) все функции в начале работы (по сравнению с не-
Рис. 115. Схема физиологических реакций на стандартную нагрузку у тренированных (сплошная линия) и нетренированных (прерывистая линия)
тренированными лицами) повышаются у них быстрее-, 2) в процессе работы уровень физио логических процессов у них менее высок; 3) восстановление заканчивается относительно быстрее (рис. 115).
Методы определения общей работоспособности. Для оценки общей работоспособности человека применяют разные тесты: пробу PWCno*,
* PWC (англ.) —первые буквы слов physical ская работоспособность.
working capacity, т. е. физиче-
406
Гарвадский тест и др. Для определения аэробных возможностей используют тест и номограмму П. О. Астранда. Для оценки специальной работоспособности рекомендуются тесты, структура движений и мощность работы при которых адекватны спортивной специализации.
Мощность тестирующих нагрузок наиболее точно дозируется при работе на велоэргометре. Она регулируется при этом величиной сопротивления и темпом вращения педалей. При отсутствии велоэргометра применяют степ-тест *. Мощность работы при этом определяется высотой ступенек и темпом подъемов.
Проба PWCno. Она позволяет определять мощность работы, при которой сердечный ритм достигает 170 ударов в 1 мин. Чем больше мощность работы при таком учащении сердцебиений, тем относительно выше общая физическая работоспособность человека. При пробе PWCno выполняются две 5-минутные нагрузки с 3-минутным отдыхом между ними. Мощность работы при первой нагрузке меньше, чем при второй. Обе они не должны быть предельными для испытуемого и выбираются с учетом пола и физической подготовленности человека (табл. 15).
Таблица 15
Мощность нагрузок (кгм/мин), рекомендуемых для определения физической работоспособности (по В. Л. Карпману)
Обследуемые Женщины Мужчины
1-я нагрузка 2-я нагрузка 1-я нагрузка 2-я нагрузка
Спортсмены Не занимающиеся спортом . 300 150 600 300 600 300 1500 600
В конце каждой нагрузки подсчитывается частота сердцебиений (за 15 сек.). На основании данных о мощности выполненной работы и частоте сердечных сокращений рассчитывается величина PWCno в кгм/мин по следующей формуле:
PWC170 = Mt +
(М2-М,)-
(170—4C.fl
ЧС2 — ЧС, ]•
где Mi — мощность первой работы, М2 — мощность второй работы; ЧС1 — частота сердцебиений в конце первой работы; ЧС2 — то же в конце второй работы.
У спортсменов, особенно у специализирующихся в циклических видах спорта, величина PWCno больше, чем у не занимающихся спортом, причем как по абсолютной величине, так и при расчете на 1 кг веса тела (табл. 16).
* Степ-тест — от англ, step — шаг, test — испытание, проба, где нагрузкой служит восхождение на ступеньку.
407
Таблица 16
Беличика PWCi70 у спортсменов, специализирующихся в разных видах спорта, и неспортсмеиов (по В. Л. Карпману)
Показатели PWC170 Лыжный спорт Конькобежный спорт Велосипедный спорт Ходьба Футбол Хоккей Баскетбол Борьба 1 Не занимающиеся спортом
кгм/мин кгм/мин на 1 кг веса те- 1760 1710 1670 1548 1523 1428 1625 1370 1044 1027
ла .... 25,7 24,0 22,6 22,5 21,7 20,1 18,7 16,6 16,5 15,5
Величина PWCi7o, являясь показателем общей физической работоспособности спортсмена, изменяется в разные периоды тренировки. В циклических видах спорта она достигает максимальной величины в состоянии спортивной формы. Например, у квалифицированных велосипедистов величина PWC]70 составляет в среднем (по В. В. Васильевой с соавт.) в переходном периоде тренировки 1412 (±82) кгм/мин, в подготовительном — 1823 (±64), в соревновательном — 2063 (±74).
Величина PWCno зависит от объема сердца. Чем он больше, тем большую по мощности работу можно выполнить при частоте сердцебиения 170 ударов в 1 мин.
Максимальное потребление кислорода. Очень важным фактором, определяющим общую работоспособность организма, являются его аэробные возможности. Они оцениваются по величине МПК. Для ориентировочного определения МПК разработаны косвенные методы с применением тестирующих нагрузок. В одном из них такой нагрузкой служит восхождение на ступеньку определенной высоты в темпе 90 движений в 1 мин. в течение 5 мин. На последних 15 сек. работы подсчитывается частота сердцебиений и определяется по номограмме, разработанной П. О. Астрандом, величина МПК- Этот метод не очень точен, но он позволяет оценивать динамику общей работоспособности спортсмена в процессе тренировки. Аэробные возможности можно определять и по величине PWC170. На основании этого предложены формулы, позволяющие вычислять максимальное потребление кислорода — МПК (В. Л. Карпман). Для тренированных применяется формула: МПК = 2,2 PWCi70+ ±1070, для нетренированных — формула: МПК= l,7PWCi7l)+1240.
Гарвардский тест. В этом тесте, который разработан в Гарвардском университете, тестирующая нагрузка заключается в подъемах ла ступеньку определенной высоты в темпе 120 движений в 1 мин. Работа продолжается до утомления, но не более 5 мин. В этом тесте индекс работоспособности вычисляется по формуле, в которой учтены длительность работы и количество сердцебиений за первую 408
половину 2-й минуты восстановления. Индекс ниже 50 соответствует низкому уровню работоспособности, индекс 50—80 — среднему, индекс больше 80 — высокому.
Другие тесты работоспособности. Эти тесты, как и описанные выше, различаются по изучаемым функциям и по особенностям тестирующих нагрузок. Однако для всех них является общим требование строгой стандартности нагрузок и доступности по их мощности.
У более тренированных лиц стандартные нагрузки относительно небольшой мощности вызывают менее выраженные изменения во всех системах организма. Это характеризует экономизацию функций, происходящую в процессе тренировки. Величина энергетических затрат при работе тесно связана с экономичностью внутриклеточного обмена веществ, уменьшение которого сопровождается снижением интенсивности функций дыхания и кровообращения.
§ 4. Реакции отдельных систем организма на стандартные
(тестирующие) нагрузки
Центральная нервная система. После выполнения
стандартной работы тренированными лицами скрытый период эле-
ментарных двигательных реакций оказывается у них укороченным,
способность к дифференцировкам повышенной, явления последовательного торможения уменьшенными. У нетренированных же все эти показатели могут изменяться в противоположном направлении в связи с утомлением и развитием охранительного торможения.
Двигательный аппарат. При стандартной работе электри-
ческая активность мышц у трени- в
рованных меньше, чем у нетренированных. Электрические потенциалы при этом сконцентрированы во времени. Они возникают в активных фазах движения и почти полностью исчезают в фазах относительного покоя (рис. 116).
Рис. 116. Электромиограмма трехглавой мышцы плеча при выполнении работы до (а) и после (б) тренировки (по Р. С. Персон), в — отметка времени
Регистрация электрических потенциалов ^отдельных двигательных единиц разных мышц (при помощи игольчатых электродов) показала, что в процессе тренировки при выполнении одинаковой
работы уменьшается частота потенциалов и увеличивается длительность интервалов между электрическими разрядами. Амплитуда
электрических потенциалов ио мере развития тренированности снижается. Особенности электрической активности мышц у тренированных свидетельствуют о лучшей координации деятельности цент-
409
ров и о большей концентрации нервных процессов в двигательных нервных центрах.
Возбудимость и лабильность мышц под влиянием тестирующих нагрузок у тренированных либо не изменяются, либо повышаются. У нетренированных же эти показатели снижаются.
Расход энергии. При стандартной работе расход энергии у тренированных меньше, чем у нетренированных. Например, квалифицированные лыжники расходуют энергии на 18—25% меньше, чем спортсмены менее высоких разрядов при одинаковой скорости передвижения на лыжах.
Дыхательная система. У тренированных дыхание более согласовано с движениями, чем у нетренированных. В процессе тренировки формируется тип дыхания, наиболее эффективный при данной двигательной деятельности.
Легочная вентиляция, кислородный запрос и кислородный долг при стандартной работе у тренированных меньше, чем у нетренированных, а коэффициент использования кислорода из вдыхаемого воздуха, наоборот, больше.
Сердечно-сосудистая система. Относительно меньший кислородный запрос и лучшая утилизация кислорода тканями у тренированных обеспечивают меньшие требования к органам кровообращения.
При выполнении стандартной работы у тренированных частота сердцебиений в абсолютных величинах меньше, а в процентном отношении к исходному уровню может быть больше, чем у нетренированных.
Изменение сердечного ритма в процессе работы неодинаково у лиц разной степени тренированности. В периоде врабатывания он интенсивнее нарастает у тренированных, при дальнейшем же продолжении работы больше учащается у нетренированных. Восстановление этого показателя после работы происходит быстрее у более тренированных. Эти различия столь закономерны, что по скорости восстановления сердечного ритма можно судить о степени адаптации органов кровообращения к физическим нагрузкам.
Синусовая аритмия, часто наблюдающаяся у спортсменов в состоянии покоя, уменьшается и даже исчезает при мышечной деятельности. У тренированных это происходит позднее, чем у нетренированных.
Систолический и минутный объем крови нарастает меньше у более тренированных.
Кровяное давление в плечевой артерии у тренированных повышается меньше, но иногда может быть и больше, чем у нетрениро- I ванных. Это зависит от особенностей работы. В активных областях тела артериальное давление повышается у тренированных меньше, чём у нетренированных, а в неактивных несколько больше. Следо- ,Л вательно, при выполнении степ-теста или работы на велоэргометре давление в плечевой артерии у тренированных может нарастать больше, чем у нетренированных. Жесткость артериальных стенок, ] при стандартных нагрузках в активных областях тела у трениро
410
вануых увеличивается меньше, а в неактивных больше, чем у нетренированных. Различия в изменениях кровяного давления и жесткости артериальных стенок в разных областях тела служат одним из показателей адаптации сосудистой системы к мышечной деятельности. Эти изменения способствуют соответствующему перераспределению крови, что необходимо для поддержания рабочей гиперемии.
Система крови. Изменения во внутренней среде у тренированных выражены при стандартной работе меньше, чем у нетренированных. У тренированных менее выражены снижение pH, нарастание концентрации молочной кислоты в крови и сдвиги в составе форменных элементов.
Таким образом, изучение реакций на стандартные нагрузки позволяют сделать вывод, что у тренированных лиц благодаря более совершенной координации функций и более экономному расходованию энергетических ресурсов работа осуществляется более производительно.
§ 5. Показатели тренированности при выполнении предельно напряженной работы
Основные факторы, обеспечивающие высокие результаты в различных видах спорта. Достижение высоких спортивных результатов обусловлено многими факторами. Важнейшими из них являются: 1) совершенная техника движений; 2) быстрое и адекватное возникающим при работе требованиям повышение физиологических функций и одновременная их экономизация; 3) адаптация организма к продолжению работы при резко измененной внутренней среде. Каждый из этих факторов имеет относительно большее или меньшее значение в зависимости от вида спорта и индивидуальных особенностей человека.
В видах спорта, в которых оценка результата производится в баллах (спортивная гимнастика, акробатика, прыжки в воду и др.), высокая техника выполнения движений является решающим фактором. В циклических видах спорта она играет относительно меньшую роль. Например, по данным Р. Маргариа, улучшение техники бега на длинные дистанции повышает спортивный результат не более чем на 7%. Несмотря на это, при занятиях всеми видами спорта необходимо совершенствование техники движений, экономизирующее деятельность спортсмена.
В циклических видах физических упражнений (бег, плавание, гребля, лыжные гонки и др.) для достижения высоких результатов ведущее значение имеют разные факторы. Это обусловлено разной мощностью работы.
При работе субмаксимальной и максимальной мощности наибольшее значение имеют анаэробные процессы, освобождающие энергию. Очень велика здесь роль адаптации организма к деятельности при измененном составе внутренней среды.
411
При работе большой и умеренной мощности главным фактором, обеспечивающим высокую работоспособность организма, а следовательно, и высокий спортивный результат, является своевременная и по возможности удовлетворяющая потребность доставка кислорода.
При работе переменной мощности (спортивные игры, все виды единоборства и др.) наиболее важна способность организма стремительно повышать свои функции при увеличении требований к ним и снижать их в интервалах отдыха и при уменьшении мощности работы.
Потребление кислорода. Увеличение кислородной потребности происходит при любой спортивной деятельности, особенно при циклической работе субмаксималыюй и большой мощности, при которой у тренированных спортсменов потребление кислорода достигает 5—6 л/мин. Аэробные возможности организма при такой работе должны быть очень высоки. Об этих возможностях принято судить по величине МПК, которая определяется либо косвенным путем — по степени учащения сердцебиений при стандартных нагрузках умеренной мощности (см. выше), либо прямым, более точным, методом. В последнем случае испытуемые выполняют на велоэргометре 3—5-минутные повторные нагрузки постепенно повышающейся мощности. Определяемое при этом потребление кислорода повышается у них по мере увеличения мощности работы, достигает максимальной для данного спортсмена величины и далее остается постоянным, несмотря на увеличение мощности работы. Это и есть величина МПК данного спортсмена.
Определение МПК можно производить и в естественных условиях спортивной тренировки. Однако этот метод весьма сложен и не всегда точен.
Чем больше МПК, тем больше удовлетворяются потребности в кислороде. Это особенно важно при длительной циклической работе. Но потребление кислорода при спортивной деятельности редко достигает максимальных величин, так как при МПК можно работать весьма ограниченное время. Обычно при циклической работе потребление кислорода составляет около 8О°/0 от МПК данного спортсмена, временно повышаясь лишь при увеличении мощности работы (спурты, подъемы в гору и т. п.).
У спортсменов-стайеров высокой квалификации МПК составляет 5—6 л/мин (при расчете на 1 кг веса тела — 83—85 мл/мин). Максимальные величины этого показателя у спортсменов достигают почти 7 л/мин (или 90 мл/мин/кг). У тренирующихся к кратковременной циклической работе, а также к ациклическим и ситуационным видам мышечной деятельности МПК меньше. У лиц, не занимающихся спортом, эта величина не превышает 3—3,5 л/мин (менее 40 мл/м.ин/кг).
В лыжных гонках, беге на длинные дистанции, конькобежном и велосипедном спорте достижение высоких спортивных результатов в значительной мере зависит от уровня аэробных возможностей. Например, у спортсменов, достигающих в беге на 5 и 10 км резуль
412
татов, близких к международным рекордам, МПК составляет около 6 л/мин. Увеличение МПК на 1 мл/мин/кг способствует повышению результата в беге на 5 км в среднем на 3,5 сек. Лыжники-гонщики, имеющие МПК равное 80 мл/мин/кг, при передвижении по равнине выигрывают у спортсменов, МПК которых не превышает 65 мл/мин/кг, около 1 м за каждую секунду работы. Это означает, что на 10 км дистанции при прочих равных условиях первые выигрывают у вторых около 8 мин.
Высокие аэробные возможности организма наиболее важны в циклических видах спорта. Однако некоторую роль они играют и в ациклических, и в ситуационных видах. Увеличение МПК у представителей этих видов спорта достигается за счет большого объема тренировочной работы и использования средств общей физической подготовки. Данные, полученные при исследовании теннисистов и баскетболистов, свидетельствуют о достоверной связи между уровнем их мастерства и величиной МПК- Так, у квалифицированных баскетболистов этот показатель равен в среднем 5,1 л/мин, у неквалифицированных— лишь 3,66 л/мин (В. А. Данилов). При относительно одинаковой технической и тактической подготовленности лучших результатов в спортивных играх добиваются спортсмены, имеющие более высокие аэробные возможности, что обеспечивает лучшую выносливость к выполняемым нагрузкам.
Спортивная тренировка, особенно в циклических видах спорта, и некоторые упражнения общей физической подготовки способствуют повышению МПК. Величина его изменяется в процессе круглогодичной тренировки. Она повышается в подготовительном периоде, достигает максимума в соревновательном и несколько снижается в переходном. Например, наблюдения за тренирующимися велосипедистами показали, что в начале подготовительного периода МПК У них составляло в среднем 47 мл/мин/кг, в конце подготовительного — 58,9, в соревновательном — 64,3. У наиболее тренированных и успешно выступавших на соревнованиях оно достигло в это время 80—82 мл/мин/кг, у тренировавшихся нерегулярно не превышало 56.
Изменения МПК в годичном цикле тренировки более выражены у менее квалифицированных спортсменов, резко снижающих нагрузки в переходном периоде.
Для управления тренировочным процессом необходимо знать величины потребления кислорода при выполнении спортсменом нагрузок и процент этой величины по отношению к МПК. Однако исследовать газообмен при спортивной деятельности очень сложно. Поэтому о мощности (интенсивности) нагрузок и потреблении кислорода обычно судят косвенно по частоте сердцебиений. Известно, что в определенной зоне мощности работы имеется прямая зависимость между потреблением кислорода и сердечным ритмом. По степени учащения сердцебиений при работе у спортсмена можно определять величину потребления кислорода в процентном отношении к его МПК. Поэтому в спортивной практике получил широкое распространение метод радиотелеметрии, по-
413
зболяющий определять сердечный ритм рри любой деятельности спортсмена. /
Большинство исследователей считают, что при частоте сердцебиений 180—190 ударов в 1 мин. потребление кислорода составляет около 90—100% МП К. При менее частом сердечном ритме соответственно меньше и потребление кислорода. Работа при частоте сердцебиений 180—190 ударов в 1 мин. очень тяжела. Длительно ее могут выполнять лишь хорошо тренированные спортсмены. В связи с этим для оценки уровня выносливости спортсмена предложен тест, который заключается в определении длительности работы при сердечном ритме 180 ударов в 1 мин., выполняемой без снижения мощности. Чем длительнее может продолжаться такая работа, тем больше возможность спортсмена поддерживать потребление кислорода на уровне, близком к его МПК. Во многих видах спорта эта возможность важнее, чем сама величина МПК. Например, в беге на длинные дистанции лучший результат покажет бегун с относительно меньшим МПК, но способный более длительно поддерживать потребление кислорода на уровне, близком к этой величине.
Дыхательная система. В доставке кислорода тканям участвуют многие системы организма. При повышении кислородной потребности должна быть увеличена прежде всего вентиляция легких. Особенно больших величин она достигает при циклической работе субмаксимальной и большой мощности. По данным В. Б. Салтина и П. О. Астранда, легочная вентиляция при работе у тренированных спортсменов-мужчин может достигать 150— 200 л/мин, у женщин — 90—130 л/мин. Увеличение легочной вентиляции не должно сопровождаться значительным снижением коэффициента использования кислорода (КИО2). У нетренированных легочная вентиляция при работе не может достигать таких больших величин, как у тренированных. Но даже при относительно меньшем увеличении вентиляции легких КИО2 у нетренированных снижается. Эффективность легочной вентиляции при этом падает.
Сердечно-сосудистая система. К органам кровообращения при предельно напряженной работе также предъявляются очень большие требования. Особенно высоки они при циклической работе, когда потребление кислорода становится близким к МПК,- Деятельность сердца в таких условиях характеризуется значительным учащением ритма сокращений, укорочением почти всех фаз сердечного цикла, увеличением систолического и минутного объемов крови. Систолический объем крови при этом может нарастать до 150—200 мл, минутный — до 30—35 и более литров. Такой уровень деятельности сердца доступен лишь для квалифицированных спортсменов, тренирующихся к циклической работе большой и субмаксимальной мощности. В этом заключается одно из главных отличий спортсменов этой специализации от спортсменов других специализаций и лиц, не занимающихся спортом.
Для обеспечения кислородной потребности организма при предельно напряженной работе большую роль наряду с увеличением сердечного выброса играет происходящее при этом перераспределе-414
ние крови. Кровоснабжение органов брюшной полости при этом резко снижается (см. табл. 10 в гл. VII). То же происходит и в неактивных скелетных мышцах. О последнем можно судить по относительно большему увеличению жесткости артериальных стенок в неактивных областях тела. У более тренированных спортсменов сосудистые реакции, способствующие перераспределению крови при предельно напряженной работе, более эффективны, чем у нетренированных.
Система крови. При тяжелой работе у тренированных содержание эритроцитов и гемоглобина в крови несколько нарастает. Это способствует увеличению кислородной емкости крови (до 20— 22 мл). Однако если работа очень тяжела, то количество эритроцитов и гемоглобина может уменьшаться. Это происходит в результате интенсивного разрушения эритроцитов под влиянием некоторых продуктов обмена веществ. Кроветворная функция мозга при этом повышена. У нетренированных тяжела^ работа сопровождается более значительным уменьшением количества эритроцитов и гемоглобина в крови, что обусловлено разрушением эритроцитов и угнетением кроветворной функции.
Несмотря на усиление всех процессов, обеспечивающих доставку кислорода при предельно напряженной работе, особенно при работе субмаксимальной и большой мощности, потребность в кислороде не удовлетворяется и мышечная деятельность выполняется за счет анаэробных источников энергии. Это ведет к образованию значительного кислородного долга и изменению в кислую сторону pH крови в результате накопления в ней кислот, главным образом молочной. По величине максимального кислородного долга и по концентрации молочной кислоты в крови можно судить об уровне анаэробных возможностей организма. У нетренированных максимальный кислородный долг обычно не превышает 5—7 л. У тренированных же может достигать 20 и более литров. Соответственно повышается и концентрация молочной кислоты в крови (до 250—300 и более мг%)- В этом проявляется адаптация организма к работе в условиях резко измененной внутренней среды, что особенно важно при циклической работе субмаксимальной мощности.
При предельно напряженной мышечной деятельности происходят значительные изменения и в других системах организма. Обычно эти изменения более выражены у менее тренированных. Например, количество лейкоцитов в крови у них может достигать 30— 50 тыс. в 1 мм3. Значительно изменена при этом и лейкоцитарная формула крови (интоксикационная фаза). У нетренированных может резко уменьшаться содержание глюкозы в крови, что снижает их работоспособность.
Выделительные функции. Значительные изменения при напряженной работе происходят в деятельности почек. Перераспределение крови в организме и потери воды с потом ведут к уменьшению диуреза (вплоть до временного его прекращения). Уменьшение кровоснабжения почки вызывает ее кислородное голодание. В результате этого изменяется функциональное состояние почеч-
415
него эпителия и в моче резко повышается /одержание белка. В некоторых случаях при очень длительной ^напряженной работе в моче могут появляться эритроциты. Эти/изменения свидетельствуют о несоответствии функциональной подготовленности спортсменов выполняемой работе. /
Глава XVIII
ФИЗИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА ФИЗИЧЕСКИХ УПРАЖНЕНИИ С ЦИКЛИЧЕСКОЙ СТРУКТУРОЙ ДВИЖЕНИИ
§ 1. Общая характеристика циклических движений
При ходьбе, легкоатлетическом беге, скоростном беге на коньках, лыжных гонках, велогонках, гребле и плавании движения имеют циклический характер.
Особенности физиологических сдвигов при тренировке в каждом из этих видов спорта зависят от длины дистанции. Чем она больше, тем относительно меньше мощность работы. Циклическая работа по мощности и продолжительности делится на 4 зоны (см. гл. XIV, § 3). Однако это деление является условным. В действительности циклическая работа во всех видах спорта выполняется с некоторыми колебаниями мощности. На одной и той же дистанции она может меняться в зависимости от тактических задач, степени тренированности спортсмена и других факторов. Особенно резко изменяется мощность работы при кроссах.
Циклическая работа разной мощности предъявляет различные требования к организму и сопровождается разными морфофункциональными изменениями в его системах. Например, тренировка к работе умеренной мощности способствует развитию общей выносливости и повышению аэробной производительности. Тренировка же к спринту, развивая главным образом быстроту и силу, увеличивает анаэробные возможности организма.
Одни и те же циклические виды физических упражнений оказывают разное влияние на организм человека в зависимости от его возраста и уровня физической подготовленности. Кроме того, воздействие на организм тех или иных видов спорта зависит от условий, в которых проводятся тренировки и соревнования. Например, лыжные гонки, скоростной бег на коньках и особенно плавание дают больший закаливающий эффект, чем другие циклические упражнения.
§ 2. Спортивная ходьба
Соревнования по спортивной ходьбе проводят на дистанциях от 10 до 50 км. Таким образом, скороходы преимущественно выполняют циклическую работу умеренной мощности. Спортивная ходьба, так же как и обыкновенная, характеризуется чередованием одиночной и двойной опор. Отличия спортивной ходьбы от простой заклю
416
чаются в более сложной технике движений и в значительно большей скорости. Простая ходьба при тех скоростях, которые характерны для спортивной, неизбежно переходит в бег. Переход в бег является также наиболее частой ошибкой скороходов, не владеющих техникой спортивной ходьбы.
Двигательный аппарат. Мышцы спортсмена адаптированы к работе в аэробных условиях. Относительно длительная фаза опоры может влиять на морфофункциональное состояние мышц нижних конечностей. Например, скороходы, особенно высококвалифицированные, могут, как и бегуны, развивать очень большое произвольное мышечное напряжение. Способность же к произвольному расслаблению мышц у них несколько меньше, чем у бегунов.
Дыхание и расход энергии. Легочная вентиляция при спортивной ходьбе составляет в среднем 70—80 л/мин, потребление кислорода — около 4 л/мин. Однако эти величины при повышении мощности работы (спурты на дистанции, финиширование) могут быть значительно больше, в связи с чем при подготовке скорохода следует развивать его аэробные возможности.
Энерготраты при спортивной ходьбе очень велики. Например, при ходьбе на 50 км расход энергии может достигать 2000— 2500 ккал.
Кровообращение. Частота сердечных сокращений у скороходов на финише разных дистанций обычно составляет 150— 180 ударов в 1 мин. При увеличении мощности работы она может достигать 200 и даже 220 ударов в 1 мин.
Кровь. Количество эритроцитов и гемоглобина в крови при спортивной ходьбе, как правило, возрастает, но у недостаточно подготовленных спортсменов эти показатели могут снижаться. Миогенный лейкоцитоз обычно выражен значительно. Чаще наблюдается 2-я нейтрофильная фаза. Концентрация молочной кислоты в крови повышается нерезко. Щелочные резервы несколько снижаются.
Выделительные функции. Деятельность потовых желез при спортивной ходьбе очень интенсивна. Большие потери воды ведут к уменьшению мочеобразования. После напряженных соревнований в моче нередко обнаруживается белок.
§ 3. Легкоатлетический бег
Бег — естественная локомоция, в которой фаза одиночной опоры чередуется с фазой полета.
По технике движений наиболее сложен бег на короткие дистанции. Особенно трудным является разучивание старта и стартового разгона. При совершенной технике спринтерского бега энерготраты спортсмена значительно меньше, чем при нерациональной технике (по данным Г. И. Поповой, в среднем на 27%). Особенно сложна координация движений при барьерном беге.
Короткие, средние, длинные и сверхдлинпые дистанции легкоатлетического бега — типичные примеры циклической работы максимальной, субмаксимальной, большой и умеренной мощности. Осо-
417
большой мощности. Если же
Рис. 117. Функциональные свойства (в условных единицах) четырехглавой мышцы бедра у спринтеров разной квалификации (по В. В. Высо-чину).
Заштрихованные столбики — у мастеров спорта, белые столбики — у перворазрядников. А — скорость сокращения мышцы, Б — максимальная сила мышцы, В — скорость расслабления мышцы
бенности работы максимальной мощности наиболее ярко проявляются на дистанции 100 м, субмаксималр'ной— на дистанциях 800— 1500 м, большой — на дистанции 5000 м и умеренной — на марафонской дистанции. Остальные дистанции являются как бы промежуточными. В зависимости от скорости бега они могут быть отнесены либо к одной, либо к другой зоне мощности. Например, бег на 10000 м при результате равном 28—29 мин. близок к работе преодоление этой дистанции затрачивается более 30 мин., то по физиологическим сдвигам такой бег следует отнести к работе умеренной мощности. В действительности бег на всех дистанциях выполняется с переменной скоростью, а следовательно, и с изменяющейся мощностью, в некоторых случаях с переходом из одной зоны в другую.
Центральная нервная система. В процессе тренировки у бегуна формируются и закрепляются относительно однообразные динамические стереотипы нервных процессов, которые лежат в основе техники бега. При беге по гладкой дорожке структура движений изменяется лишь при ускорениях, беге по виражу и финишном броске.
Анализаторы. Сравнительно однообразная двигатель-
ная деятельность бегуна не предъявляет каких-либо особых требований к функциям анализаторов. Однако в условиях соревнований роль их повышается. В этих условиях бегуну необходимо быстро и точно воспринимать действия соперников и всю обстановку спортивной борьбы и тонко регулировать мышечные усилия. Значение зрительной и проприоцептивной рецепции увеличивается также при беге по пересеченной местности и особенно при беге с барьерами.
Двигательный аппарат. При беге на разные дистанции предъявляются различные требования к мышцам. Эффективность скоростного бега зависит главным образом от морфофунциональ-ного состояния опорно-двигательного аппарата. Мышцы спринтера должны обладать значительной силой, обеспечивающей мощность отталкивания от грунта, а также способностью очень быстро сокращаться (что определяет «взрывные» качества мышцы) и быстро расслабляться (что позволяет более эффективно использовать скоростно-силовые качества и достигать наибольшей скорости бега).
У спортсменов высокого класса сила мышц, их «взрывные» качества и особенно способность к быстрому расслаблению мышц бои
418
лее выражены, чем у менее квалифицированных (рис. 117). Спортсмены, отличающиеся хорошими «взрывными» качествами и относительно низкой скоростью расслабления мышц, как правило, опережают своих соперников на первой половине дистанции, но затем теряют достигнутое преимущество. У них быстрее развивается утомление и менее интенсивно происходит восстановление. Скорость сокращения и расслабления мышц определяет темп движений спринтера.
Оптимальная частота и длина шагов различны у разных бегунов. Частота шагов на дистанции — один из важнейших факторов, от которых зависит скорость. Наибольшей частоты движений, как правило, достигают более квалифицированные бегуны.
Мышцы спринтера должны быть адаптированы главным образом к работе в анаэробных условиях. При этом интенсивность восстановления АТФ играет решающую роль для поддержания скорости на протяжении всей дистанции.
При беге на средние дистанции требования к мышцам несколько иные, чем у спринтеров. Однако и на этих дистанциях и на более длинных умение быстро расслаблять мышцы является одним из важных качеств, обеспечивающих высокую работоспособность. При беге на средние дистанции мышцы работают в таком режиме, когда анаэробные процессы в организме сочетаются с аэробными. При этом чем длиннее дистанции, тем большую роль приобретают аэробные процессы. У бегунов на длинные и сверхдлинные дистанции эти процессы являются главными.
Ды хание и расход энергии. При беге на 100 м дыхание неглубокое и учащенное. Бегун производит 14—19 дыхательных циклов при средней глубине вдоха 420 мл (Е. М. Попова). Легочная вентиляция у квалифицированных бегунов достигает при этом в среднем 8 л. Кислородный запрос при беге на 100 м составляет в зависимости от скорости бега от 6 до 13 л. Кислородный долг при этом превышает 90% запроса. Такое соотношение величин кислородного запроса и кислородного долга указывает на то, что спринтеру необходимо развивать главным образом анаэробные возможности. Однако в последнее время экспериментально доказано большое значение и аэробных процессов при скоростном беге. Без наличия высоких аэробных возможностей удлиняется время восстановления и снижается способность к образованию кислородного долга (Н. И. Волков и др.). Кроме того, специфика тренировки спринтера (многократно повторяемая скоростная работа) требует высоких аэробных возможностей организма.
При беге на средние дистанции частота и глубина дыхания резко увеличиваются, в связи с чем легочная вентиляция может достигать 150 л/мин и более. Потребление кислорода при этом повышается до 4—5 л/мин. В конце бега на 1500 м оно может достигать максимальной для данного спортсмена величины. Суммарный кислородный запрос при беге на средние дистанции достигает 30 л и более. Кислородный долг, выраженный в % к запросу, тем больше, чем относительно короче дистанция. Кислородный же долг, выра
419
женный в литрах, наоборот, больше на 6oiiee длинных дистанциях. Например, при беге на 800 и 1500 м он достигает максимально возможных величин, т. е. 15—20 и даже--более литров.
У бегунов на средние дистанции должны быть хорошо развиты как анаэробные, так и аэробные возможности.
По данным Б. Салтина и П. О. Астранда, МПК у квалифицированных бегунов на средние дистанции составляет в среднем 76 мл/мин/кг, по данным В. Л. Карпмана— 69.
При беге на длинные дистанции частота дыхания и легочная вентиляция доходят почти до таких же величин, как при беге на средние дистанции. Потребление кислорода почти достигает предельного для данного спортсмена уровня и должно удерживаться на нем относительно длительное время. Несмотря на это, кислородный запрос полностью не удовлетворяется, и возникающее при беге на длинные дистанции устойчивое состояние является кажущимся (см. гл. XIV, § 4). В результате при такой работе образуется значительный кислородный долг. Его величина зависит от тактики бега. Если спортсмен бежит с ускорениями и резко финиширует, то кислородный долг достигает 12 и более литров. Суммарный кислородный запрос при беге на 5000 м составляет около 80—90 л, при беге 10 000 м — около 100—130 л. Восстановление АТФ при этой работе происходит главным образом аэробным путем. Поэтому для бегунов на длинные дистанции характерна большая величина МПК.
При беге на сверхдлинные дистанции дыхательные функции также значительно повышаются. Однако потребление кислорода не достигает столь высоких величин, как при работе большой мощности. Кислородный запрос почти полностью удовлетворяется, в связи с чем характерное для этой работы устойчивое состояние является истинным (см. гл. XIV, § 4). Кислородный долг образуется лишь при врабатывании и ускорениях. Обычно он составляет 4— 5 л. В целом же работа обеспечивается аэробными реакциями.
По величине МПК бегуны на сверхдлинные дистанции занимают одно из первых мест по сравнению с другими спортсменами.
Чем длиннее дистанция, которую пробегает спортсмен, тем относительно больше он расходует энергии. При беге на 100 м суммарный расход энергии составляет в среднем около 40—50 ккал, при беге на 800 м —около 150 ккал, при беге на 5000 м — около 450 ккал, при марафонском беге — около 2500 ккал.
Кровообращение. В состоянии покоя у бегунов часто наблюдается брадикардия. При этом чем длиннее дистанция, к которой готовится спортсмен, тем реже у него в покое сердечный ритм. Например, у бегунов-стайеров частота сердцебиений в покое равна в среднем 48 ударам в 1 мин., у бегунов на средние дистанции — 56, у спринтеров — 60. Сердечный ритм реже 50 ударов в 1 мин. среди стайеров наблюдался в 30% случаев, среди бегунов на средние дистанции— в 18%, среди спринтеров — лишь в 10% (В. В. Васильева). Брадикардия у бегунов часто сочетается с синусовой аритмией.
Непосредственно при беге сердечный ритм учащается в среднем до 170—190 ударов в 1 мин. (см. рис. 72). Лишь при ускорениях на
420
дистанции и при финишировании он может достигать 200—220 ударов в 1 мин. Восстановление сердечного ритма после окончания бега зависит от его длительности и интенсивности, а также от степени тренированности спортсмена. Обычно после бега на короткие дистанции оно происходит через 20—30 мин., после бега на средние и длинные дистанции — через несколько часов.
Размеры сердца, особенно у бегунов на длинные и сверхдлинные дистанции, как правило, увеличены.
Систолический и минутный объемы крови увеличиваются больше всего при беге на средние и длинные дистанции, достигая иногда 180—200 мл и 35—40 л/мин. Артериальное систолическое давление повышается до 180—220 мл рт. ст. Диастолическое давление при беге на длинные и сверхдлинные дистанции нередко понижается.
Кровь. Количество эритроцитов и гемоглобина в крови после бега оказывается увеличенным. Значительно возрастает и количество лейкоцитов, особенно после бега на сверхдлинные дистанции. Лейкоцитарная формула при этом изменяется. После длинных и сверхдлинных дистанций увеличивается число нейтрофилов, особенно палочкоядерных.
При беге на средние и длинные дистанции в крови резко повышается концентрация молочной кислоты (до 200—250 мг% и более). Это ведет к значительному снижению pH.
При беге на короткие и сверхдлинные дистанции содержание молочной кислоты в крови почти не изменяется. При сверхдлинных дистанциях может снижаться концентрация глюкозы в крови, что способствует развитию утомления.
Выделительные функции. После бега на длинные и сверхдлинные дистанции диурез в связи с усиленным потоотделением уменьшается. Удельный вес мочи при этом оказывается увеличенным. Концентрация молочной кислоты в моче после бега на средние дистанции может быть увеличена до 450 мг%, после длинных дистанций она меньше — 40—50 мг%.
После бега на средние, длинные и сверхдлинные дистанции может появляться белок в моче и даже эритроциты, особенно у нетренированных спортсменов.
Вес тела. После бега он уменьшается. Наибольшие потери его происходят при беге на сверхдлинные дистанции (до 4—5 кг).
Температура тела. Бег, особенно длительный, сопровождается усиленным теплообразованием. В жаркую погоду и при высокой влажности воздуха теплоотдача не обеспечивает полного освобождения организма от излишков тепла. В этих случаях температура тела может повышаться до 39—40°, в результате чего наступает перегревание организма и нарушение многих его функций.
§ 4. Плавание
Особенности двигательной деятельности плов-ц а. При нахождении тела пловца в роде земное притяжение является «топящей» силой. Однако этой силе противостоит «подъемная»
421
сила, определяемая потерей веса тела при погружении в воду. В пресной воде «топящая» сила превышает «подъемную» примерно на 0,8—1 кг. Таким и становится вес тела пловца в воде. В морской воде «подъемная» сила больше «топящей». Тело пловца в этих условиях становится еще легче. Снижение веса тела облегчает локомоции пловца. Однако передвижение в воде имеет и специфические трудности, которые обусловлены большей плотностью воды по сравнению с воздухом. При погружении в воду тело пловца испытывает гидравлическое давление, которое нарастает при увеличении глубины погружения.
Сопротивление воды движениям пловца очень значительно. Оно определяет величину мышечных усилий при плавании. Сопротивление воды рассчитывается по формуле: 7? = kV2, где R — сопротивление воды в кг, к — коэффициент сопротивления, зависящий от плотности воды, от коэффициента обтекаемости и площади наибольшего поперечного сечения тела, V — скорость движения в м/сек.
Сопротивление воды увеличивается пропорционально квадрату скорости движения. Например, при буксировке пловца со скоростью 1,3 м/сек оно составляет около 5 кг, при скорости 1,7 м/сек — 9,5 кг.
При плавании на различные дистанции совершается работа разной мощности: на отрезках 25—50 м — работа максимальной мощ ности, на дистанциях 100, 200 и 400 м — работа субмаксимальной, на 1500 м — большой мощности, на более длинных дистанциях — работа умеренной мощности.
Обычно при спортивной деятельности часть мышц выполняет статические напряжения, что необходимо для поддержания определенной позы тела. При плавании этого не требуется и все основные мышечные группы пловца выполняют динамическую работу.
Анализаторы. В процессе тренировки у пловцов формируется особое комплексное восприятие различных раздражителей, называемое «.чувством воды». Оно обусловлено ощущениями, возникающими при раздражении тактильного, температурного, проприоцептивного и вестибулярного рецепторов. При наличии «чувства воды» пловцы хорошо анализируют малейшие изменения в величине сопротивления воды, ее давление и температуру. Эти ощущения способствуют улучшению движений пловца.
Тренировка в плавании, особенно в плавании способом кроль, повышает функциональную устойчивость вестибулярного аппарата.Это обусловлено его многократными раздражениями при поворотах головы во время вдоха и выдоха и, кроме того, воздействием на вестибулярный аппарат холодовых раздражителей. При недостаточной вестибулярной устойчивости длительное воздействие ускорений и холодной воды может сопровождаться вегетативными реакциями, головокружением и потерей равновесия. Попадание холодной воды в наружный слуховой проход, особенно при нарушении целости барабанной перепонки, настолько сильно раздражает вестибулярный аппарат, что нередко является причиной несчастных случаев.
Двигательный аппарат. Под воздействием тренировки у пловцов развивается сила мышц. При плавании в работу вовлека
422
ются все основные мышечные группы. Способы плавания кроль и делофин предъявляют наибольшие требования к мышцам рук и плечевого пояса, вызывая преимущественное развитие этих мышц.
У пловцов, обладающих высокой техникой движений, отмечается строгая последовательность работы отдельных мышечных групп в каждом двигательном цикле. Она сохраняется и при повышении скорости плавания. При этом лишь нарастают мышечные усилия и удлиняется путь их приложения в фазе гребка. В фазе переноса длительность усилий, наоборот, уменьшается (Ю. В. Мельков).
Мышцы пловца должны быть адаптированы к работе как в аэробных, так и в анаэробных условиях. При этом чем длиннее дистанция, к которой готовится спортсмен, тем большее значение приобретают аэробные процессы.
Дыхание и расход энергии. При плавании дыхание осуществляется в необычных условиях. При вдохе и особенно при выдохе пловцу приходится преодолевать сопротивление воды. В связи с этим плавание является одним из эффективных средств развития дыхательной мускулатуры. По величине жизненной емкости легких пловцы занимают одно из первых мест среди спортсменов, специализирующихся в разных видах спорта. У пловцов — мастеров спорта (мужчин) она составляет в среднем 6055±410 мл, что превышает должную величину более чем на 30% (Г. И. Куренков).
Частота дыханий связана с частотой гребковых движений. При большей скорости плавания она может достигать 50—60 дыханий в 1 мин. По мере повышения скорости и, сл едовательно, учащения дыханий длительность дыхательного цикла уменьшается. Например, при увеличении скорости с 0,9 до 1,7 м дыхательный цикл укорачивается в среднем с 2,15 до 1,08 сек. Это происходит главным образом за счет укорочения вдоха. При медленном плавании длительность вдоха и выдоха почти одинаковы. При повышении скорости плавания время вдоха составляет не более */з дыхательного цикла. При максимальной скорости вдох осуществляется в течение 0,26— 0,31 сек. (Г. И. Куренков). Укорочение дыхательного цикла сочетается с повышением объемной скорости вдоха с 3,4 до 7,8 л/сек, объемной скорости выдоха — с 2,1 до 4,1 л/сек.
Легочная вентиляция при плавании может возрастать до 120— 150 л/мин. Однако даже такая величина ее недостаточна для удовлетворения потребности организма в кислороде. Коэффициент использования кислорода значительно увеличивается (до 5—6%). Однако при максимальных скоростях этот коэффициент снижается (в среднем до 4), что ведет к уменьшению потребления кислорода и преобладанию анаэробных процессов, обеспечивающих восстановление АТФ.
Потребление кислорода при плавании у квалифицированных спортсменов-мужчин составляет в среднем около 5—6 л/мин, что близко к величинам их МПК.
Значение аэробных возможностей организма при плавании очень велико. Это обусловлено прямой зависимостью между скоростью проплывания дистанции и уровнем аэробного обмена. Наибольшая
423
аэробная производительность отмечается у пловцов, тренирующихся в плавании на 400 и 1500 м. Наибольшие анаэробные возможности характеризуют пловцов-спринтеров. Так, у тренирующихся в плавании на 100 и 200 м МПК равно 65,2 мл/мин/кг, максимальный кислородный долг — 158 мл/кг, а у тренирующихся в плавании на 400 и 1500 м соответственно 72,6 и 138 (Н. И. Волков с соавт.).
Аэробные возможности несколько больше у пловцов, специализирующихся в плавании кролем. По данным Н. И. Волкова, у мастеров спорта — кролистов МПК достигает 6,26 л/мин (или 77 мл/мин/кг). У брассистов эти величины меньше—5,61 л/мин (или 69 мл/мин/кг). Максимальный кислородный долг, наоборот, больше у брассистов (14,3 л), чем у кролистов (11,5л).
Расход энергии при плавании несколько больше, чем при циклической работе в других видах спорта. Это обусловлено большими потерями энергии в виде тепла, что зависит от большей теплопроводности воды по сравнению с воздухом. Даже при неподвижном пребывании в воде 12° в течение 4 мин. расходуется 100 ккал тепла, т. е. столько же, сколько в воздушной среде за 1 час. Спокойное стояние в воде 24—25° в течение 3—4 мин. вызывает повышение энергетических трат более чем на 50%.
Расход энергии при плавании на различные дистанции зависит от их длины и мощности работы. На дистанциях 100—1500 м он составляет в среднем от 100 до 450 ккал и более.
Кровообращение. Горизонтальное положение тела при плавании облегчает работу сердца, так как в этих условиях отсутствует препятствие для продвижения крови — сила тяжести.
У человека в положении лежа систолический объем крови несколько больше, чем в положениях сидя и стоя. Поэтому при плавании увеличение систолического объема по отношению к исходному показателю несколько меньше, чем при других физических упражнениях. Однако абсолютные величины систолического объема крови при плавании могут быть очень большими. Увеличение систолического объема крови и учащение сердцебиений ведут к значительному повышению минутного объема крови.
Благоприятным фактором для деятельности сердца при плавании является отсутствие статических напряжений скелетных мышц. Их ритмичные сокращения, сочетающиеся с глубоким дыханием, усиливают венозный приток к сердцу.
Особенности работы сердца при плавании способствуют его развитию.
Кровь. Изменения в картине крови при плавании типичны для циклической работы субмаксимальной и большой мощности (увеличивается содержание эритроцитов, лейкоцитов, гемоглобина, повышается концентрация молочной кислоты, снижается резервная щелочность) .
Выделительные функции и терморегул яци я . При плавании почти отсутствует потоотделение. Продукты обмена веществ, которые при других физических упражнениях выводятся с потом, у пловцов могут выводиться только через почки. Это предъ
424
являет дополнительные требования к их функциям. Уменьшенное кровоснабжение почек при работе и необходимость выведения кислых продуктов обмена изменяют проницаемость почечного эпителия. В связи с этим у пловцов нередко после заплывов появляется в моче белок. Изменение деятельности почек является одной из специфических реакций организма на плавание. Эти реакции более выражены при плавании в холодной воде.
Длительное пребывание пловцов в прохладной воде может вести к значительным потерям тепла и переохлаждению тела. Однако у тренированных пловцов процессы, обеспечивающие сохранение постоянной температуры тела, более совершенны, чем у людей, не адаптированных к охлаждению. Таким образом, плавание является одним из эффективных средств закаливания организма.
§ 5. Гребля
Выполняемая гребцом работа относится преимущественно к работе субмаксимальной мощности и большой. Лишь при гребле на байдарках и каноэ (на дистанции 10 000 м) совершается работа умеренной мощности.
Анализаторы. Гребля предъявляет большие требования к анализаторам: двигательному, зрительному, вестибулярному, звуковому, осязательному. Импульсы от рецепторов двигательного аппарата обеспечивают информацию о степени напряжения мышц, амплитуде и скорости движения. Особенно велика роль этих импульсов во время подъезда и во время захвата воды веслами.
Зрительный контроль необходим при движении весел в воде (особенно при гребле в лодках с парными веслами). Это ведет к увеличению поля зрения. Расширению его способствует также необходимость следить во время гонок за лодками соперников.
Импульсы от вестибулярного аппарата обеспечивают сохранение равновесия в неустойчивых спортивных лодках. При гребле происходит постоянное раздражение вестибулярного аппарата, что совершенствует его функции.
В двойках и более крупных судах большую роль играет слаженность движений гребцов (их синхронность). Для достижения этого необходимо восприятие звуков, возникающих при захвате воды лопастью весла. Это предъявляет повышенные требования к органу слуха.
Двигательный аппарат. Работа мышц гребца, связанная со значительными силовыми напряжениями при быстрой проводке весла, является скоростно-силовой. Гребки совершаются очень часто. Например, квалифицированные гребцы на академических лодках производят до 46—48 рабочих циклов в 1 мин.
При академической гребле в продвижении лопасти весла в воде участвуют почти все мышцы тела. Однако основная нагрузка падает на мышцы ног и спины. Эти мышечные группы у гребцов на академических судах обычно хорошо развиты. При гребле на бай
425
дарке работают только мышцы рук и плечевого пояса и лишь частично мышцы спины. Мышцы ног участия в работе не принимают.
Мышцы гребцов должны быть адаптированы к работе в анаэробных и аэробных условиях. При тренировке на более длинные дистанции увеличивается значение аэробных процессов.
Телеметрические исследования показали, что биоэлектрическая активность мышц при гребле наиболее выражена при выполнении стартовых гребков и при ускорениях (С. П. Сарычев).
Дыхание. Частота дыхания при гребле обычно совпадает с ритмом движений и составляет в среднем 30—40 в 1 мин. и более. Исследования дыхательных движений при гребле, в частности путем радиотелсметрической регистрации, показали изменчивость их на протяжении дистанции. Например, при ускорениях и финишировании, т. е. при увеличении кислородной потребности, на фоне основных дыхательных движений у гребцов возникают еще и дополнительные. В связи с этим при интенсивной гребле на каждый двига-тельный'цикл может приходиться два дыхательных.
Своеобразная скоростно-силовая работа при гребле обусловливает особый характер дыхательных движений. В зависимости от мощности работы и квалификации спортсмена тип дыхания при гребле может быть разным. Проводка весел в воде происходит, как правило, при задержке дыхания и натуживании, что способствует повышению силы гребка. Вдох осуществляется при этом в начале проводки весел, выдох, усиленный к концу, производится при подъезде на банке и заносе весла.
Глубокое, соответствующее ритму движений дыхание имеет очень важное для работоспособности гребца значение. Поэтому параллельно с обучением гребцов технике движений необходимо обучать их наиболее рациональному дыханию.
Легочная вентиляция при гребле, как и при другой циклической работе субмаксимальной мощности, может достигать 100—150 л/мин. Это происходит главным образом за счет увеличения глубины дыхания, что способствует развитию дыхательных мышц. По величине жизненной емкости легких гребцы занимают одно из первых мест среди спортсменов различных специализаций.
Потребление кислорода на дистанции у квалифицированных спортсменов при академической гребле достигает 5 л/мин и более. Однако это не обеспечивает кислородной потребности и ведет к образованию значительного кислородного долга. Суммарный кислородный запрос на дистанциях 1500—2000 м составляет в среднем около 30—40 л. По данным Г. П. Пауперовой с соавт., суммарный кислородный запрос при гребле на байдарке на 1000 м равен в среднем 18 л, на каноэ—15 л. Величины кислородного долга — соответственно 5 и 4,3 л.
Аэробные и анаэробные возможности у квалифицированных гребцов очень высоки. Например, по данным Б. Салтина и П. О. Ас-транда, у гребцов на каноэ и академических судах средняя величина МПК превышает 5 л/мин. При расчете на 1 кг веса оно несколько больше у каноистов — соответственно 70 и 62 мл/мин/кг.
426
I
Расход энергии. При академической гребле на 1500—2000 м он равен в среднем 150—200 ккал., при гребле на 1000 м на байдарке — 86 ккал., на каноэ — 75 ккал.
Кровообращение. Частота сердцебиений у тренированных гребцов-мужчин в состоянии покоя равна в среднем 40—50 ударам в 1 мин. При тренировочной и соревновательной гребле сердечные сокращения резко учащаются. В лодке непосредственно перед стартом частота сердцебиений у гребцов нередко превышает 100 ударов в 1 мин., на финише может достигать 180—200 ударов в 1 мин.
Для многих гребцов характерна гипертрофия миокарда обоих желудочков.
При проводке весел в воде у гребцов возникает кратковременное натуживание. Оно затрудняет венозный приток к сердцу и этим несколько затрудняет его деятельность.
Выделительные функции. При тренировочных и соревновательных заездах у гребцов увеличивается концентрация молочной кислоты в моче, в связи с чем повышается ее кислотность и нередко появляется белок в моче.
Вес тела. После соревновательных заездов на 1500—2000 м вес тела гребца снижается в среднем на 200—300 г, после заездов на 25 км — до 2 кг и более.
§ 6. Скоростной бег на коньках
Циклическая работа конькобежцев на дистанциях 500—3000 м относится к зоне субмаксимальной мощности, на дистанциях же 5000—10 000 м — к зоне большой мощности.
Двигательный аппарат. Большая скорость, наличие скольжения, малая опорная поверхность конька и наклонное положение туловища усложняют двигательную деятельность спортсмена.
Для уменьшения сопротивления воздуха и увеличения разгибания в тазобедренных суставах (это позволяет производить отталкивание под меньшим углом) туловище конькобежца должно находиться почти в горизонтальном положении, что требует подавления врожденных выпрямительных рефлексов.
Хотя работа основных мышц конькобежца является динамической, обширные группы других мышц (спины, разгибатели бедра до начала отталкивания и др.) находятся при скоростном беге на коньках в статистическом напряжении.
Для овладения рациональной техникой и умением сохранять ее при большой скорости бега у конькобежца должна быть хорошо развита сила мышц. Углы сгибания в суставах при измерении силы должны быть такими же, как при беге на коньках во время отталкивания. Изометрическая сила мышц обычно больше их динамической силы. Например, у лучших шведских конькобежцев сила мышц ног, развиваемая в изометрических условиях, составляет 225—340 кг; а динамическая сила —200—230 кг. Однако эта динамическая сила, определяемая в лабораторных условиях, почти в 2
427
раза больше того усилия, которое производит конькобежец во время отталкивания (Н. И. Волков, Б. А. Стенин).
У конькобежцев, специализирующихся на длинных дистанциях, и у многоборцев показатели силы меньше, чем у спринтеров. При беге на длинные дистанции мышечные усилия относительно невелики, но они должны выполняться длительное время, что требует значительной выносливости.
Мышцы конькобежцев должны быть адаптированы к деятельности как в аэробных, так и в анаэробных условиях.
Анализаторы. Скоростной бег на коньках предъявляет высокие требования к двигательному, вестибулярному и зрительному анализаторам. Импульсы от них способствуют необходимому распределению мышечного тонуса и обеспечивают сохранение равновесия. Последнее особенно важно при прохождении виражей дорожки. На этих участках дистанции повышается роль вестибулярного аппарата и шейно-тонических рефлексов.
Дыхание. При скоростном беге на коньках у высококвалифицированных спортсменов легочная вентиляция может достигать 180 л/мин, потребление кислорода — 5 л/мин и более, кислородный долг— 8—9 л. Особенности газообмена, как и в других циклических видах спорта, зависят от длины дистанции и скорости бега. По мере увеличения дистанции повышается значение аэробных процессов и снижается роль анаэробных.
Максимальное потребление кислорода, по данным Б. Салтинаи П. О. Астранда, достигает у конькобежцев 5,8 л/мин (или 78 мл/мин/кг). Этот показатель повышается главным образом в подготовительном периоде тренировки. В это же время возрастают и анаэробные возможности организма, определяемые по максимальной величине кислородного долга. Так, на 1-м этапе подготовительного периода (май-июнь) средняя величина МПК у квалифицированных конькобежцев равна 66,4 мл/мин/кг, на 2-м этапе (сентябрь-октябрь) — 75,1, на 3-м этапе (ноябрь—декабрь)—76,1, средняя величина кислородного долга соответственно 111,5 мл/кг, 132,8 и 137,7 (Н. И. Волков, Б. А. Стенин).
Расход энергии. При скоростном беге на коньках суммарный расход энергии зависит от длины дистанции, мощности работы, степени тренированности спортсмена и других факторов. По приблизительным расчетам на дистанции 500 м он составляет в среднем около 45 ккал., 1500 м — около 80 ккал., 5000 м — более 200 ккал., 10 000 м — более 400 ккал.
Кровообращение. При скоростном беге на коньках, как и при другой циклической работе, предъявляются большие требования к сердечно-сосудистой системе.
Систематическая тренировка ведет к значительному увеличению размеров сердца. Например, по данным С. В. Шестакова, у конькобежцев с небольшим стажем тренировки поперечник сердца равен в среднем 12,9 см, длинник—13,8 см, при многолетней тренировке эти показатели соответственно увеличиваются до 13,7 и 14,8 см. По данным С. П. Летунова, гипертрофия левого желудочка наблюдает
428
ся у конькобежцев в 46% случаев, гипертрофия обоих желудоч* ков — в 19%.
Частота сердцебиений при беге на коньках зависит от мощности выполняемой работы. При беге с соревновательной скоростью сердечный ритм достигает 180—200 ударов в 1 мин. и более.
Кровь. При скоростном беге на коньках повышается содержание молочной кислоты в крови (до 200 мг% и более). Миогенный лейкоцитоз, как правило, характеризуется нейтрофильной фазой.
Выделительные функции. Бег на коньках оказывает значительное воздействие на функции почек. Кислотность мочи после бега оказывается резко увеличенной, нередко появляется белок в моче,
§ 7. Лыжные гонки
Хотя движения лыжника-гонщика имеют циклический характер, структура двигательных актов лыжника и мощность его работы, как и при других передвижениях по пересеченной местности, постоянно изменяются в зависимости от рельефа трассы.
Структура движений изменяется не только при преодолении подъемов и спусков, но и при переходе с одного способа передвижения на другой на равнинных участках дистанции. Как известно, лыжные ходы делятся в зависимости от характера работы рук на одновременные, попеременные и смешанные. Применение того или иного хода и смена их обусловлены рельефом местности и условиями скольжения. При использовании одного способа движения лыжника цикличны. В момент перехода на другой способ передвижения цикличность их нарушается. Затем они вновь становятся цикличными, но уже при иной структуре.
Мощность работы лыжника резко повышается на подъемах в гору и снижается при спусках.
Исходя из продолжительности работы гонки на лыжах относят к зоне большой (5 и 10 км) и умеренной (15, 30, 50 и более км) мощности. Однако в связи с резкой пересеченностью современных трасс правильнее характеризовать гонки на лыжах как работу переменной мощности.
Мощность работы лыжника, а также скорость его передвижения на дистанции зависят в значительной мере от метеорологических условий. Встречный ветер, большая влажность и относительно высокая или, наоборот, очень низкая температура воздуха, глубокий, свежий снег и т. п. сильно затрудняют движения спортсмена и снижают скорость. Уменьшение скорости при этом сочетается с увеличением энергетического расхода на каждый метр пути.
Двигательный аппарат. Лыжные гонки требуют развития общей и скоростной выносливости, а также силы. Эффективность работы мышц нижних и верхних конечностей обусловлена развитием их силовых качеств.
Движения лыжника на дистанции разнообразны и сложны по координации. Спортсмен должен не только владеть двигательными
429
навыками, обеспечивающими передвижение различными способами по равнине и позволяющими преодолевать подъемы и спуски, но и уметь использовать эти двигательные навыки в сложных условиях, иногда при мгновенном изменении рельефа и направлении трассы.
Техническая подготовка лыжника играет большую роль в достижении высоких результатов. При рациональной технике движений работа гонщика осуществляется более экономично. Энерготраты у квалифицированных гонщиков при одинаковой скорости меньше на 15—20%, чем у неквалифицированных.
При передвижениях на лыжах в работе участвуют почти все мышцы тела. Поэтому лыжники, как привило, отличаются равномерным развитием мускулатуры.
Мышцы лыжника должны быть адаптированы к работе как в аэробных, так и в анаэробных условиях.
Биохимические и морфологические изменения в скелетных мышцах, наступающие в процессе тренировки лыжника, сопровождаются сдвигами и в функциональном состоянии. Мышцы лыжника характеризуются повышенной возбудимостью и лабильностью.
Анализаторы. Сложность двигательных навыков и необходимость быстро изменять направление и скорость движений при изменениях рельефа трассы предъявляют большие требования к двигательному, зрительному и вестибулярному анализатору. Для ’ лыжников характерна высокая проприоцептивная чувствительность почти во всех звеньях тела. Зрительные функции, особенно периферическое зрение, очень важны для ориентировки лыжника при изменениях рельефа трассы. По данным А. Н. Крестовникова, квалифицированные лыжники имеют увеличенное поле зрения.
Удержание равновесия при спусках с гор, особенно при больших скоростях, требует высокой функциональной устойчивости вестибулярного анализатора.
Дыхание. Движения лыжника сочетаются с дыханием. При I попеременных ходах темп движений относится к частоте движений, как 1:1, 1:2, 2:1, 2:3. При одновременных ходах выдох всегда производится во время толчка палками и наклона туловища. Правильное сочетание дыхания и движений способствует повышению работоспособности лыжника.
Дыхательные мышцы выполняют при лыжных гонках интенсивную и длительную работу, что требует значительного их развития. В связи с этим лыжники имеют большую жизненную емкость легких. Развивая силу дыхательных мышц, тренировка способствует и повышению их выносливости. Поэтому у квалифицированных лыжников даже после тяжелых соревнований жизненная емкость не только не снижается, а может быть даже увеличенной.
Легочная вентиляция изменяется при беге на лыжах в зависимости от мощности работы. При ускорениях на дистанции и на подъемах легочная вентиляция может увеличиваться до 120—150 л/мин и более. Потребление кислорода на дистанции у квалифицированных гонщиков-мужчин составляет в среднем около 4—5 л/мин.
Для достижения высоких результатов в лыжных гонках спорт
430
смены должны иметь высокую аэробную производительность. У квалифицированных лыжников-мужчин, по данным Б. Салтина и П. О. Астранда, величина МПК равна около 75—86 мл/мин/кг, у женщин — около 60—65 мл/мин/кг. По данным Т. И. Раменской с соавт., у лыжников мастеров спорта МПК составляет в среднем 81,6 мл/мин/кг, у лыжников I разряда — 74,1 мл/мин/кг, II разряда— 64,1 мл/мин/кг. Многочисленные исследования показали, что для достижения высоких результатов на мировой арене лыжник должен иметь МПК не менее 80 мл/мин!кг, лыжница — не менее 70 мл /мин/кг.
Потребление кислорода при лыжной гонке не остается постоянным. На равнине оно поддерживается на уровне 80—82% от МПК, при подъемах повышается до 90—95%, на спусках, особенно длинных и пологих, становится ниже 80%. У квалифицированных лыжников, в совершенстве владеющих техникой, на подъемах потребление кислорода повышается в среднем до 87% от МПК, на спусках снижается до 62% (Т. И. Раменская).
Чем больше величина МПК у лыжника, тем относительно выше его соревновательная скорость (табл. 17). Однако несмотря на большое значение МПК, высокий результат в лыжных гонках не может быть достигнут без высоких анаэробных возможностей организма и совершенной техники движений.
Кислородный запрос при лыжных гонках больше, чем, например, при легкоатлетическом беге той же длительности и мощности. На дистанциях 30 и 50 км суммарный кислородный запрос достигает многих сотен литров. Это объясняется вовлечением в работу при беге на лыжах обширных мышечных групп, что не только увеличивает кислородный запрос, но и несколько облегчает потребление кислорода. Участие в работе больших мышечных групп обеспечивает при беге на лыжах лучшее использование кислорода артериальной крови, что ведет к повышению артерио-венозной разницы. Это облегчает поступление кислорода из альвеолярного воздуха в кровь. Образованию оксигемоглобина в легких способствует поступление в них холодного воздуха. Повышенная же температура в работающих мышцах способствует его диссоциации. Все эти факторы, облегчая потребление кислорода, обеспечивают возможность лыжнику работать при относительно устойчивом состоянии.
Таблица 17
Средняя соревновательная скорость (м/мин) на разных участках дистанции лыжной трассы у спортсменов, имеющих разную величину МПК
(по В. Ф, Громыко)
Гельеф местности Максимальное потребление кислорода (мл/мин/кг)
60 65 70 75 80
Подъем в гору 11—12° .... 108 114 126 132 145
Подъем в гору 3—4° 180 198 210 228 246
Равнина 240 254 270 306 320
431
Кислородный долг образуется у лыжников главным образом при подъемах в гору и увеличении скорости бега на равнине. Величина его зависит от разных условий (рельеф местности, скорость, уровень тренированности лыжника и др.).Он может достигать 8—12 л.
Об интенсивности анаэробных процессов при лыжных гонках можно судить по концентрации молочной кислоты в крови. Она повышается в основном при ускорениях и подъемах в гору; при малой скорости на пологом подъеме концентрация молочной кислоты в крови почти не увеличивается, но при повышенной скорости на этом же подъеме возрастает в среднем до 150 мг%.
Расход энергии. Он составляет при лыжных гонках в среднем около 20 ккал, в 1 мин. На отдельных участках дистанции повышается до 25 ккал, и более. Суммарный расход энергии в зависимости от длины дистанции достигает 350—4000 ккал, и более.
После длительной работы повышенный расход энергии может наблюдаться в течение нескольких суток. По данным В. В. Михайлова, на следующий день после участия в гонках основной обмен у лыжников остается повышенным на 25—26% по сравнению с должными величинами в состоянии покоя.
Большой расход энергии при лыжных гонках требует особого внимания к пищевому режиму спортсмена. В его суточном рационе должны быть полноценные белки, животные и растительные жиры, много углеводов и витаминов. В подготовительном и соревновательном периодах тренировки количество углеводов должно составлять в среднем 600—700 г в сутки. За несколько дней до гонок оно может быть увеличено до 800—900 г.
Кровообращение. Для тренированных лыжников, как правило, характерна в состоянии покоя резко выраженная брадикардия. У лыжников-мужчин частота сердцебиений снижается до 32—45 ударов в 1 мин., у женщин — до 44—48.
Брадикардия обычно сочетается у лыжников с синусовой аритмией, что является показателем увеличения воздействий на сердце блуждающего нерва. Об этом же свидетельствует изменение фазовой структуры сердечного цикла (увеличение фазы изометрического сокращения и др.).
Объем сердца у лыжников-гонщиков увеличен. По данным С. В. Хрущева, у квалифицированных лыжников-мужчин он равен в среднем 1010 + 34,7 см3, у женщин — 730+14,3 см3.
Частота сердцебиений на дистанции зависит от рельефа трассы и скорости передвижения лыжника. При наличии относительно коротких подъемов и спусков и большой скорости на равнине частота сердцебиений у лыжников может поддерживаться на стабильном уровне— 170—180 ударов в 1 мин., при резко выраженной пересеченности трассы она может значительно варьировать — от 140 до 200 раз в 1 мин.
Кровь. Количество эритроцитов и гемоглобина при лыжных гонках увеличивается. Возрастает и количество лейкоцитов (нейтрофильная фаза миогенного лейкоцитоза). В плазме крови повышается концентрация молочной кислоты. При недостаточном и нераци
432
ональном питании может резко снижаться содержание глюкозы в крови (до 50—40 мг°/о), что способствует развитию утомления.
Выделительные функции. Лыжные гонки вызывают в связи с увеличенным потоотделением временное уменьшение диуреза. Удельный вес мочи и ее кислотность при этом повышаются. В ней увеличивается содержание аммиака и креатина, что является следствием повышения интенсивности белкового обмена. У малотренированных спортсменов после бега содержание белка в моче иногда достигает 4—1О%о-
Температура тела. Низкая температура окружающей среды при лыжных гонках вызывает усиленную теплоотдачу. Повышенная теплопродукция при этом обусловлена включением в работу больших мышечных групп. Однако при малой скорости передвижения и нерациональной одежде гонщика теплоотдача может превышать теплопродукцию, что ведет к понижению температуры тела и снижению работоспособности.
Вес тела. После лыжных гонок вес тела уменьшается на 0,5— 5 кг. Это происходит в результате потери воды путем потообразования и испарения. Потери в весе зависят от длины дистанции, скорости передвижения, метеорологических условий и особенностей одежды лыжника.
§ 8. Велосипедный спорт
Передвижение на велосипеде обеспечивается мышечными усилиями через систему рычажных передач. Нижние конечности при этом совершают круговые движения, несвойственные естественным локомоциям. Пространственная структура движений велосипедиста относительно проста. Однако их внутренняя координационная структура, особенно при высоких темпах педалирования, отличается большой сложностью.
При езде на велосипеде мышцы ног выполняют динамическую работу, мышцы рук — статическую. Степень статического напряжения мышц рук зависит от квалификации гонщика (у менее квалифицированных она больше), от особенностей педалирования и посадки.
Рабочая поза велосипедиста характеризуется наклонным положением туловища, что целесообразно для уменьшения сопротивления воздуха, нарастающего при повышении скорости. Длительное сохранение наклонного положения тела очень утомительно. Поэтому на длинных дистанциях посадка гонщика более высокая.
При велогонках на треке может совершаться работа максимальной мощности (гиты 200 и 800 м), субмаксимальной (гит 1000 м и гонки преследования), большой и умеренной (гонки с лидером и др.). Велогонки на шоссе относятся к работе большой и умеренной мощности.
Шоссе, на которых проводятся гонки, обычно имеют разный рельеф, в связи с чем мощность работы велосипедиста может изменяться. Она несколько повышается при подъемах в гору и снижа
15—113
433
ется при спусках. Это приближает гонки на шоссе, особенно групповые, к видам спорта, в которых движения имеют ситуационный характер. Однако постоянство структуры движений гонщика, длительность работы и ее непрерывность позволяют отнести шоссейные гонки к циклическим видам спорта большой и умеренной мощности.
Анализаторы. Необходимость точно дозировать мышечные усилия, сохранять равновесие и ориентироваться в пространстве при очень высокой скорости предъявляет повышенные требования к функциям двигательного, вестибулярного и зрительного анализаторов велосипедиста. Поступающая от них информация способствует управлению движениями. Во время соревнований особое значение приобретает периферическое зрение, обеспечивающее ориентировку в пространстве и наблюдение за соперниками.
Двигательный аппарат. Как отмечалось выше, передача мышечных усилий через систему рычагов велосипеда не является сложным движением. Однако управление велосипедом усложняется при преодолении подъемов и спусков, при выполнении поворотов на дистанции, а также в связи с необходимостью сохранять равновесие при разной скорости передвижения.
Соревновательные нагрузки вызывают значительные изменения в функциональном состоянии мышц у велосипедистов (повышается реобаза и хронаксия, изменяются оптимум и пессимум частоты раздражения мышц). Эти сдвиги становятся меньше по мере развития специальной тренированности гонщиков (Д. А. Полищук).
Дыхание. Наклонное положение туловища, характеризующее рабочую позу велосипедиста, несколько затрудняет дыхание. Это особенно заметно при низкой посадке, когда гонщик держится за нижнюю часть руля. При высокой посадке велосипедиста, что характерно для работы на длинных дистанциях, условия для дыхания более благоприятны.
Соотношение частоты дыханий и движений гонщика зависит от скорости величины передачи. Оно может быть равным 3:1, 3,5:1, 4:1, 4,5:1 (И. М. Серопегин).
Особенностью дыхания квалифицированных гонщиков является большая продолжительность выдоха по сравнению со вдохом. При ускорениях и рывках у них могут возникать кратковременные задержки дыхания, что неблагоприятно отражается на работоспособности.
Несмотря на некоторые затруднения дыхания при езде на велосипеде, легочная вентиляция у спортсменов может достигать 120 л/мин и более. Потребление кислорода при этом составляет около 4—5 л/мин.
Степень насыщения крови кислородом у велосипедиста находится в прямой зависимости от частоты и глубины дыхания. При задержке дыхания, которая может иметь место во время стартового разгона и при спуртах, оксигенация крови уменьшается. Если дыхание ритмично и достаточно глубоко, то уменьшение незначительно (не более чем на 1—2%).
434
У велосипедистов, специализирующихся в гонках на шоссе, преобладают аэробные реакции. Однако при спуртах резко повышается и роль анаэробных процессов.
При скоростной работе на треке ведущее значение имеют анаэробные реакции.
Аэробные возможности велосипедистов, по данным разных авторов, различны. МПК варьирует у них, по средним данным, от 4,4 до 5,2 л (или от 63,0 до 74 мл/мин,/кг). Систематические наблюдения за величиной МПК у тренирующихся велосипедистов показали, что аэробные возможности у них повышаются в подготовительном пе-
риоде тренировки и остаются на постоянном уровне в соревновательном. Так, по данным В. В. Васильевой с сотр., в начале подготовительного периода МПК у велосипедистов в среднем равно 53,4 мл/мин/кг, в конце этого периода и в соревновательном периоде составляет 63 мл/мин/кг.
Кровообращение. Рабочая поза велосипедиста и статическое напряжение мышц верхних конечностей несколько затрудняют деятельность сердца и предъявляют особые требования к перераспределению крови. При работе гонщика должно быть усилено кровоснабжение нижних конечностей. У более квалифицированных велосипедистов сосудистые реакции, обеспечивающие
Рис. 118. Изменение скорости распространения пульсовой волны (в % к исходной величине) по артериям верхних (черные столбики) и нижних (заштрихованные столбики) конечностей у квалифицированных (Л) и менее квалифицированных (Б) велосипедистов под воздействием напряженной тренировки на треке (по Г. М. Тарасовой)
перераспределение крови,
совершеннее. Например некоторые наблюдения показывают, что после интенсивной тренировочной нагрузки на треке жесткость артериальных стенок на ногах у квалифицированных велосипедистов становилась ниже исходного уровня на 12%, У менее квалифицированных — на 8%; жесткость артериальных стенок
на верхних конечностях у квалифицированных повышалась, у менее квалифицированных оставалась почти неизменной (рис. 118).
У квалифицированных велосипедистов-шоссейников мужчин объем сердца составляет в среднем 1040±19 см3, у женщин 793± 14 см3 (С. В. Хрущев).
Частота сердцебиений при велогонках зависит от скорости передвижения, рельефа местности, метеорологических условий и других факторов. При скорости 30—35 км/час сердечный ритм не превы
15*
435
шает в среднем 120—130 ударов в 1 мин., при скорости 45— 50 км/час он достигает 170—190 ударов в 1 мин. При многодневных гонках такая скорость, а следовательно, и частота сердечных сокращений поддерживаются гонщиками на участках, составляющих 60—80% от дистанции. При повышении скорости до 50—55 км/час частота сердцебиений достигает 190—210 ударов в 1 мин. Такая скорость поддерживается (даже наиболее тренированными гонщиками) не более 1—3 мин. (Г. В. Мелленберг, А. В. Седов и др.).
У квалифицированных гонщиков-шоссейников частота сердечных сокращений в состоянии покоя равна 33—54 ударам в 1 мин.; на тренировочных занятиях — 152 ударам в 1 мин., на подъемах и при ускорениях повышается до 182—192 ударов в 1 мин.; на спусках снижается до 132—136 ударов (М. А. Артыков).
При участии в соревнованиях частота сердцебиений у велосипедистов несколько больше, чем на тренировках. По мере увеличения длины дистанции мощность работы, а следовательно, и сердечный ритм становятся меньше. В групповых гонках учащение сердцебиений несколько меньше, чем в командных и индивидуальных.
Кровь. Большая потребность в кислороде во время тренировочных и соревновательных нагрузок ведет к увеличению в крови количества эритроцитов и гемоглобина. По данным Л. Я. Евгеньевой, велосипедисты-шоссейники занимают по этим показателям первое место среди спортсменов-стайеров. Количество эритроцитов у них равно в среднем 5,3 млн. в 1 мм3 крови, в отдельных случаях — 6 млн., а количество гемоглобина — 15%.
Выделительные функции. Содержание хлоридов в моче после гонок уменьшается, что обусловлено большими их потерями с потом. В моче гонщиков находят повышенное количество недоокис-ленных продуктов и иногда белок.
Вес тела. В зависимости от длины дистанции и других условий потери веса тела у велосипедистов колеблются в пределах от 300 до 1800 г.
Глава XIX
ФИЗИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА ВИДОВ СПОРТА СО СТЕРЕОТИПНОЙ НЕЦИКЛИЧЕСКОЙ СТРУКТУРОЙ
ДВИЖЕНИЙ
Движениям стереотипной нециклической структуры свойственна строгая стандартность их формы, характера двигательных координаций конечностей и туловища. В этих движениях, в отличие от стереотипных циклических, нет монотонной повторяемости одного и того же ряда (цикла) фаз (элементов) движений. Элементы упражнения нециклического типа следуют друг за другом всегда в строго определенном порядке, но многократное повторение данного ряда элементов по его завершении отсутствует.
436
В спорте стереотипные движения нециклического характера делятся на две большие группы: 1) движения, результат которых выражается количественными мерами силы и пространства; 2) движения, оцениваемые по качеству их выполнения. Первая группа представляет собой ациклические однократные движения, в которых проявляется максимум силы, скорости или точности движений спортсмена: метания, прыжки, поднимание тяжестей и стрельба. Во вторую — движения смешанной структуры, в которых проявляются различные двигательные способности спортсмена: движения, характерные для спортивной и художественной гимнастики, акробатики, прыжков на батуте и в воду, фигурного катания на коньках.
§ 1. Общая характеристика однократных ациклических движений
В упражнениях со штангой, прыжках и метаниях мышцы должны развивать максимальную силу и скорость сокращения. Между силой (точнее, величиной преодолеваемого сопротивления) и скоростью сокращения существует зависимость, установленная А. Хиллом:
V = Ь(Р0—Р)(Р + а), где V — скорость сокращения; а и Ь — коэффициенты; Ра — максимальное изометрическое напряжение, при котором скорость сокращения равна нулю; Р — фактическое напряжение, зависящее от перемещаемой массы.
Согласно этой формуле, скорость сокращения мышц тем больше, чем меньше масса перемещаемого тела. Графически эта зависимость выражается кривой, показанной на рис. 119.
По второму закону Ньютона, сила F равна произведению массы m на сообщаемое ей ускорение а, т. е. F=m-a. Но в ациклических видах спорта максимум силы Fmax достигается либо за счет стремления к максимуму перемещаемой массы ттажпри относительно неизменном ускорении, либо за счет максимального повышения ускорения а-max при постоянстве величины перемещаемой массы.
Первый случай (Fmax= гптах-и) относится к упражнениям со штангой. Сила штангиста проявляется в увеличении веса поднимаемой штанги, со-
Рис. 119. Кривая зависимости скорости от силы мышечного сокращения (по А. В. Хиллу).
Ось абсцисс — отношение величины силы к максимальному (изометрическому) напряжению, ось ординат — скорость сокращения
437
общаемое же ей ускорение — величина относительно постоянная, от которой зависит лишь подъем штанги на определенную высоту. В связи с этим упражнения со штангой принято обозначать как собственно-силовые.
Во втором случае (77тах=Щ-атах) сила затрачивается главным образом на сообщение постоянной массе максимального ускорения. Упражнения такого рода называются скоростно-силовыми. К ним относятся прыжки и метания.
Рассмотрим участие различных физиологических систем в выполнении собственно-силовых и скоростно-силовых упражнений.
В центральной нервной системе процессы управления названными выше движениями характеризуются образованием динамического стереотипа. Поэтому в каждом упражнении видна своя форма и строгая последовательность движений. Но вместе с тем не все характеристики таких движений абсолютно стереотипны. Скорость и сила мышечных сокращений не должны быть постоянными, они должны возрастать до максимальной степени. В связи с этим управление силовыми и скоростно-силовыми упражнениями осложняется тем, что необходимо, сохраняя стереотипность их формы, добиваться увеличения силы и скорости мышечных сокращений.
Другой особенностью управления названной группой упражнений является сравнительно малое участие готовых, т. е. прирожденных, двигательных автоматов — безусловных двигательных рефлексов. Ациклические упражнения представляют собой специально вырабатываемый двигательный навык, который доводится до автоматизма лишь в результате многих повторений.
Группа упражнений ациклического типа — это движения однократные, кратковременные. Продолжительность силового или скоростно-силового акта измеряется секундами или даже долями секунды. Естественно, за такой короткий срок дыхание и кровообращение не могут значительно усилиться. Более того, дыхательные движения в момент силового напряжения обычно задерживаются, что в тяжелой атлетике сопровождается натуживанием и повышением внутригрудного давления. В связи с этим кровообращение в момент силового напряжения не только не усиливается, но даже уменьшается, в особенности в малом круге. Все это красноречиво свидетельствует о том, что силовые и скоростно-силовые упражнения относятся к анаэробной работе. Аэробные процессы совершаются уже после работы для погашения образовавшейся кислородной задолженности. Однако, несмотря на то, что мощность работы максимальна и сопровождается максимальным анаэробным распадом, общая величина кислородного долга сравнительно мала в связи с малой продолжительностью упражнения.
§ 2. Прыжки
По своей двигательной структуре прыжки представляют собой ациклическую локомоцию. По координации проще всего прыжки с места. Более сложные процессы управления характеризуют прыжки
438
с разбега. В них происходит резкое переключение циклической локомоции разбега на ациклическую локомоцию — прыжок. В прыжке в длину требуется, кроме того, точное попадание на брусок для отталкивания в условиях максимальной скорости разбега.
При прыжке в высоту должно вовремя, в конце разбега, произойти полное переключение горизонтальной составляющей усилий на вертикальную составляющую. Очень сложным является распределение движений при переходе через планку.
Еще более разнообразна координация при прыжке с шестом. Разбег усложнен здесь удерживанием в руках шеста, конец которого в момент отталкивания должен точно попасть в ямку, после чего следуют в условиях опоры на руки сложные акробатические движения при переходе через планку.
Вследствие кратковременности прыжков усиление вегетативных функций во время упражнения невелико. Вместе с тем многочисленные повторения прыжков на тренировках и на соревнованиях требуют выносливости и в целом вызывают значительное повышение функции органов кровообращения и дыхания.
§ 3. Метания
Управление движениями в метаниях представляет большую сложность, чем в простых прыжках. Это объясняется тем, что в основе прыжков лежат локомоции, т. е. филогенетически наиболее древний вид движения, управляемый частично автоматически действующими подкорковыми нервными центрами. Метания же — движения, не основывающиеся на автоматических локомоциях, совершаемые руками, т. е. присущие именно человеку.
Масса метаемого снаряда определяет степень развиваемого мышцами напряжения. При малом весе снаряда (в спортивном метании — копье) относительно незначительно напряжение сокращающихся мышц, но велика скорость сокращения. При большом весе (например, ядро) требуется для метания большая величина напряжения мышц, развивающих относительно меньшую скорость сокращения.
В координационном отношении сложным является не столько сам акт метания, сколько его сочетание с подготовительными движениями: разбегом в метании копья, вращениями в метании диска и молота, прыжковым движением в толкании ядра. Скорость конечного движения метающей снаряд конечности должна суммироваться с конечной скоростью движения туловища в подготовительном действии. Это требует от метателя очень точной координации движений.
Управление движениями в метаниях основывается почти исключительно на информации, поступающей от рецепторов двигательного аппарата. В метании копья, где требуется пространственная ориентировка в разбеге и в направлении полета копья, существенна роль зрения. Она несколько меньше в толкании ядра (известно, что
439
этот вид метаний доступен слепым легкоатлетам). В метании диска и молота зрительная ориентировка затруднена вследствие очень быстрых вращений и имеет значение главным образом в заключительном акте метания. Вращательные движения при метании диска и молота оказывают резкое воздействие на вестибулярный аппарат. Обычно метатели этих снарядов отличаются большой устойчивостью вестибулярного аппарата и после вращений сохраняют пространственную ориентировку.
§ 4. Поднимание тяжестей
Упражнения со штангой относятся, как было сказано выше, к движениям собственно-силового типа. Чем больше вес поднимаемой штанги, тем значительнее развиваемое мышцами напряжение. Последнее зависит, по закону Вебера, в первую очередь от поперечника мышц, что связано с массой тела штангиста. Это подтверждается почти линейной зависимостью веса поднимаемой штанги от веса штангиста и обусловливает наличие весовых категорий среди тяжелоатлетов.
То обстоятельство, что упражнения штангиста входят в рубрику собственно-силовых, а не скоростно-силовых движений, не означает исключение роли скорости сокращения мышц. В рывке и толчке требуется значительная скорость сокращения мышц для подбрасывания штанги, чтобы обеспечить «подсед» под нее.
Хотя штанга удерживается руками спортсмена, поднимание ее осуществляется не только мышцами рук. В рывке и в толчке главное усилие и скорость сокращения развивают мышцы туловища и ног.
В энергетическом отношении поднимание тяжестей занимает одно из первых мест по мощности выполняемой работы. Максимальную мощность работы штангиста не трудно подсчитать. Если исходить из рекордного веса штанги, поднятой в голчке, 240 кг и времени ее подъема 2 сек. на высоту вытянутых рук 2 м, мощность такой работы выразится цифрой 240 кгм/сек или свыше трех лошадиных сил. Отсюда можно рассчитать и общий расход энергии. Приняв коэффициент полезного действия при подъеме штанги за 20%, получим общую сумму работы за 2 сек. в 2400 кгм. При пересчете на калории эта величина составит 2400:427=5,6 ккал. Работа такой исключительно большой мощности совершается за счет анаэробных алактатных энергетических процессов. Поскольку при таких процессах потребление 1 л кислорода связано с освобождением 2,9 кал., общий кислородный запрос при рекордном подъеме штанги будет равен 5,6:2,9=около 2 л. Это — громадная величина, если учесть, что максимальное потребление кислорода у штангистов не превышает 4 л в 1 мин., а за 2 сек. — 0,13 л. Во время подъема штанги усиленного потребления кислорода практически не происходит и весь кислородный запрос выражается величинами кислородного долга.
440
§ 5. Стрельба
Стрельба, которая, как отмечалось выше, входит в группу однократных ациклических движений, не относится к скоростно-силовым действиям. Основное движение при стрельбе из огнестрельного оружия — нажатие пальцем на спусковой крючок — должно быть нерезким, плавным. Мышечная сила требуется лишь для удержания винтовки в стрельбе стоя, и в особености при стрельбе из лука. Натяжение тетивы, необходимое для пуска стрелы, может составлять у новичков 80% от максимального усилия, прикладываемого к тетиве, у мастеров спорта — 50% •
Главная задача в стрельбе из всех видов оружия — пространственная точность. Она определяется в основном двумя физиологическими факторами: устойчивостью позы тела и зрительным восприятием в прицеливании. Устойчивость необходима, чтобы при прицеливании исключались значительные колебания тела. Это зависит главным образом от состояния вестибулярного аппарата, степени мышечно-суставных ощущений и выраженности тремора (непроизвольного дрожания дистальных звеньев конечности).
Разные виды оружия, способы и условия стрельбы предъявляют специфические требования к физиологическим механизмам стрельбы. При многих видах стрельб необходимо очень точное соблюдение интервалов и тонкое распределение во времени движений конечности, удерживающей оружие. Соревнования в некоторых видах стрельбы могут длиться несколько часов. Это вызывает значительное утомление, как физическое (связанное со статическими усилиями при удержании необходимых поз), так и психическое.
§ 6. Другие прицельные движения
Однократные ациклические движения стереотипного характера, в которых на первый план выступает пространственная точность, встречаются и в нестереотипных, ситуационных видах спорта. Таковы подачи в волейболе и теннисе, штрафные броски в баскетболе, пенальти в футболе. Из них наиболее характерны по своей стереотипности штрафные броски в баскетболе. Здесь строго стандартно все: расстояние до кольца, его диаметр, высота. Движения баскетболиста по направленности, быстроте и силе должны быть также стандартными, чтобы обеспечить нужную траекторию полета мяча. Такие движения наиболее точно подходят под определение динамического стереотипа, как системы раздражителей, вызывающей определенную систему нервных процессов. Это, однако, не означает, что обучение штрафному броску должно сводиться только к бесконечному повторению таких бросков. Эксперименты показали, что варьирование условий (броски с разных точек площадки, с разных расстояний от кольца, в кольца, расположенные на разной высоте и разного диаметра) дает лучший эффект. Это подтверждает положение о динамичности стереотипа, т. е. изменчивости условий его проявления и образования.
441
§ 7. Общая характеристика упражнений, оцениваемых по качеству их выполнения
Многие упражнения, оценка которым дается по качеству выполнения, требуют большой силы, или быстроты, или пространственной точности, однако они оцениваются на соревнованиях не количественными мерами силы, времени и пространства, а в баллах. На первый план выступает в них способность спортсмена управлять своими движениями, тонко дозировать силу и скорость мышечных сокращений, обеспечивать координацию движений частей тела, в особенности способность управлять перемещениями тела в окружающем трехмерном пространстве, а также взаимными перемещениями звеньев тела. Во многих случаях необходимо умение управлять своими движениями в безопорном положении (во время фазы полета). Почти все упражнения этой группы требуют выразительности движений. Эстетическая, художественная сторона их выполнения играет существенную роль в оценке качества выступления спортсменов на соревновании.
Выполняемые упражнения представляют собой, как правило, сложные комбинации из отдельных двигательных актов (элементов), сначала разучиваемых по отдельности, а затем образующих связку в виде неразрывного целого.
Вся рассматриваемая здесь группа видов спорта, казалось бы весьма разнородных, имеет ряд общих черт. Все они представляют собой выраженный динамический стереотип. Стереотипна не только форма движений, их координационная структура, но и сила и скорость мышечных сокращений. Этим данная группа упражнений отличается от группы упражнений силового и скоростно-силового типа, в которых стереотипна лишь форма движений, характер двигательных координаций, в то время как сила и скорость мышечных сокращений не стандартны, а максимальны.
Для рассматриваемой группы упражнений характерно также многообразное участие различных афферентных систем. Необходимость сохранения равновесия в сложных условиях, совершение двигательных действий при различных положениях тела в пространстве, быстрые смены одних двигательных координаций другими — все это требует точного и быстрого анализа информации, поступающей от рецепторов двигательного аппарата, кожи, вестибулярного аппарата, от органов зрения.
Продолжительность упражнений этой группы различна — от нескольких секунд (акробатика, прыжки в воду) до многих десятков секунд (спортивная гимнастика) и нескольких минут (фигурное катание на коньках). Это позволяет отнести данные упражнения к зонам максимальной и субмаксимальной мощности, чем определяются и изменения в дыхании и кровообращении во время упражнений и после их завершения.
Характерной чертой всех упомянутых упражнений является их высокая эмоциональность, что связано с напряженностью функции эндокринной системы. При выступлениях в спортивной и художест-442
венной гимнастике, фигурном катании, где эмоциональная сторона движений выражена наиболее резко, может наблюдаться значительное усиление экскреции кортикостероидов.
Наряду с изложенными выше особенностями общего характера каждому упражнению, оцениваемому по качеству выполнения, присущи специфические физиологические особенности, которые освещаются ниже.
§ 8. Спортивная гимнастика
Спортивную гимнастику можно назвать наиболее универсальным видом спорта. При занятиях ею гармонично развиваются все мышцы тела. Так, в упражнениях на снарядах развивается сила мышц, в особенности рук и туловища; в опорных прыжках и вольных упражнениях — скоростно-силовые свойства мышц нижних конечностей, при этом увеличивается растяжимость мышц, определяющая гибкость в суставах; в упражнениях на бревне, в вольных упражнениях совершенствуется равновесие и т. д. Разнообразные упражнения требуют точного управления движениями при различных перемещениях тела в пространстве, когда голова и туловище занимают не только обычное (ортостатическое) положение в пространстве, но обращены книзу или находятся в наклонном положении к горизонту. Различные вращательные движения развивают устойчивость вестибулярного аппарата к действию адекватных раздражителей. Ряд акробатических элементов и соскоки со снарядов связаны с безопорным положением, создающим особые трудности для управления движением во времени и пространстве.
В энергетическом отношении гимнастика уступает циклическим движениям. Это связано с тем, что в ней встречаются паузы, статические усилия, задержка дыхания. Поэтому уровень максимального потребления кислорода у гимнастов высших разрядов ниже (около 4 л/мин), чем у высококвалифицированных представителей циклических видов спорта (5—6 л/мин). Продолжительность комбинаций упражнений на снарядах обычно не превышает 30—40 сек. и только в вольных упражнениях достигает 50—60 сек. Поэтому такие упражнения относятся к работе субмаксимальной мощности и связаны со значительным кислородным долгом гликолитического происхождения. Потребление кислорода во время комбинации сравнительно невелико и лишь при наиболее продолжительных вольных упражнениях достигает 2 л/мин.
Дыхание во время спортивной гимнастики неровное, связано с задержками и натуживанием, что ограничивает величину легочной вентиляции. Но его можно рационализировать, обучив гимнаста совершать выдох во время движений, требующих наибольших мышечных усилий, а вдох — при наименее трудных движениях.
§ 9. Художественная гимнастика
Управление движениями в художественной гимнастике определяется в первую очередь требованиями сложной координации движений. Необходима высокая пространственная точность. Она осо
443
бенно велика в упражнениях со снарядами. Точный глазомер имеет здесь большое значение, но совершенство, достигаемое во владении навыком, подчас таково, что некоторые точностные движения (с обручем, мячом) совершаются даже без зрительного контроля.
При выполнении больших прыжков характерна скоростно-силовая работа мышц нижних конечностей.
Не только при выпрыгивании, но и при приземлении необходимо точное управление движениями. В первом случае от него зависит дальность и высота полета тела гимнастки, во втором — мягкость и почти бесшумность приземления, обеспечиваемые амортизирующим напряжением мышц в короткий момент касания опоры.
Высокие требования предъявляются к гибкости тела. От нее зависит амплитуда движений в плечевых и тазобедренных суставах и позвоночника. Гибкость в суставах гимнастки проявляется не только при опорных ее положениях, но и в полете. Это значит, что растягивания мышц происходят не пассивно — под действием силы тяжести (как это часто бывает в вольных упражнениях спортивной гимнастики), а активно — под влиянием резких сокращений антагонистических мышц, которые для этого должны обладать большой силой.
Упражнения художественной гимнастики в исполнении мастеров этого вида связаны с относительно большими энерготратами. Во время выступления на соревновании частота сердечных сокращений достигает 180—210 ударов в 1 мин. Двухминутная длительность выполнения комбинации художественной гимнастики позволяет отнести ее к зоне субмаксимальной мощности. Это значит, что такая работа связана со значительным кислородным долгом. Действительно, по данным экспериментальных исследований, кислородный долг при выполнении комбинации художественной гимнастики превышает 4,1 л. В связи с этим можно считать, что по степени физиологической нагрузки художественная гимнастика мало чем уступает спортивной.
§10. Акробатика
Особенности управления движениями в акробатике сводятся в основном к равновесию и точному распределению движений в без-опорном положении. Эти движения совершаются -при изменении положения тела в пространстве, вращениях тела в сагиттальной и фронтальной плоскостях, что предъявляет высокие требования к устойчивости вестибулярного аппарата. Почти все движения акробата требуют большой гибкости как позвоночника, так и суставов конечностей. В парной акробатике спортсмен, находящийся внизу, должен иметь незаурядную мышечную силу и значительную выносливость в статических усилиях. Акробаты должны обладать совершенным равновесием, осложненным необходимостью удерживать сложные стойки на кистях или даже на одной кисти. Для акробатических прыжков характерны скоростно-силовые действия. По развитию прыгучести акробаты мало уступают прыгунам-легкоатлетам.
444
В связи с тем что продолжительность акробатической комбинации на соревновании не превышает обычно 10—15 сек., мощность выполняемой работы соответствует максимальной зоне и представляет собой анаэробную работу алактатного характера. Парные, групповые акробатические комбинации длятся около 1,5 мин., что характеризует зону субмаксимальной мощности. Однако здесь часты статические силовые позы, выполняемые на задержке дыхания. Но поскольку элементы бывают отделены друг от друга паузами, в это время происходит усиление дыхания и погашение части кислородного долга.
§ 11. Прыжки на батуте
Управление движениями в прыжках на батуте связано с двумя их фазами: опорной и полетной. Обе они отличаются от обычных акробатических прыжков своей продолжительностью. Благодаря упругим свойствам батута приземление оказывается нерезким. Прогибание батута в этот момент требует особой формы амортизирующих напряжений мышц нижних конечностей.
Спортсмен должен обладать способностью точно ощущать нарастание упругого сопротивления батута, чтобы синхронизировать максимальное толчковое усилие мышц с наибольшим выталкивающим усилием батута. В этом — главное отличие опорной фазы при прыжках на батуте от обычных прыжков на твердой опоре.
Полетная фаза при прыжках на батуте намного продолжительнее, чем при обычных прыжках. Она зависит, естественно, от высоты прыжка и достигает 2 сек. при высоте 3,5 м. Сложные изменения положения тела в полетной фазе и необходимость приземления в определенном участке батута требуют отличной ориентировки спортсмена в пространстве и во времени. Линейные и вращательные ускорения, возникающие при прыжках на батуте, оказывают сильное воздействие на отолитовую систему и полукружные каналы вестибулярного аппарата, который, с одной стороны, должен обладать устойчивостью к этим воздействиям, с другой — подавать точную информацию о всех изменениях положения тела в пространстве. Роль последнего особенно велика в связи с тем, что при быстрых вращениях тела зрительная ориентировка в окружающем пространстве сильно затруднена. Так же как в акробатических прыжках, соскоках в снарядовой гимнастике, прыжках с трамплина и прыжках в фигурном катании, первостепенное значение имеет «чувство времени». Специальные исследования показали, что квалифицированный прыгун на батуте может очень точно отмерять отрезки времени в полетной фазе.
Ощущение времени полета и его высоты приобретает еще большее значение в групповых прыжках, потому что от него зависит синхронизация прыжков партнеров. У квалифицированных прыгунов синхронность прыжков достигает такой степени, что расхождение во времени приземления, отталкивания и выполнения отдельных элементов в полете не превышает подчас десятой доли секунды.
445
На соревнованиях выполняется обычно 10 прыжков, что занимает 15—18 сек. Следовательно, совершаемая прыгуном работа находится в зоне максимальной мощности, а энергетически она характеризуется анаэробными реакциями алактатного характера.
§12. Прыжки в воду
Управление движениями при прыжках в воду связано в основном с тремя фазами: отталкиванием, полетом, приводнением.
Полетная фаза начинается с прыжка тела вверх. Ускорения, ко-
——.——-— ——___________ торыми сопровождается прыжок
Рис. 120. Циклограмма прыжка в воду (по К. Ю. Данилову).
Во время прыжка (соскок прогнувшись из передней стойки с места) испытуемый должен был включить лампочку в момент прохождения половины пути полета. Фактически включение произошло раньше — в момент половины времени полета
в воду, вызывают раздражение вестибулярного аппарата. С момента выпрыгивания скорость полета вверх быстро снижается и при достижении высшей точки полета равна нулю. Поэтому воздействие линейных ускорений на отолитовую систему вестибулярного аппарата резко падает. Свободное падение тела после достижения высшей точки полета (если только оно не связано с дополнительными вращательными движениями) не сопровождается раздражением вестибулярного аппарата, так как оно подобно состоянию невесомости. В то же время вращательные движения в полете являются специфическими раздражителями полукружных каналов, причем одновременное вращение в сложных прыжках вокруг продольной и поперечной осей тела комплексно раздражает все три пары полукружных каналов, расположенных в трех взаимно перпендикулярных плоскостях.
Сложные и разнообразные движения, преимущественно вращательные, должны завершаться выпрямлением туловища и погружением в воду вниз головой по возможности вертикально, чтобы всплеск воды был минимален.
Исследованиями показано, что прыгун в воду ориентируется главным образом на время полета и распределяет свои движения во
446
времени, а не в пространстве. Это характеризует механизмы управления движениями в полетной фазе не только при прыжках в воду, но и при всех других прыжках, в особенности тех, которые связаны с быстрыми вращениями тела, когда зрительное восприятие пространственных ориентиров затруднено (рис. 120).
Энерготраты во время прыжка в воду незначительны. Во всех движениях, за исключением выпрыгивания — первоначального скоростно-силового действия, отсутствуют антигравитационные моменты, требующие существенных энергетических затрат. Во время самого полета, не связанного с противодействием силы тяжести, расход энергии небольшой — идет только на сокращения мышц, осуществляющих сгибание или разгибание практически невесомых в этот момент частей тела. Значительно большего расхода энергии требуют плавательные движения после погружения в воду, выход из воды, подъем на вышку.
§ 13. Фигурное катание на коньках
В фигурном катании на коньках сочетаются элементы скоростного бега на коньках, художественной гимнастики, акробатики, хореографии. Богатство форм передвижений на коньках здесь очень велико. Ни в одном виде спорта управление движениями нижних конечностей не характеризуется такой сложностью, тонкостью и разнообразием, как в фигурном катании.
Непрерывность циклической локомоции сменяется ациклической в скоростно-силовых актах — прыжках. Сопровождающиеся многократными вращениями прыжки и мягкое приземление на одну ногу в скольжении требуют точного распределения движений во времени. В движениях ног, а также в движениях рук при поддержках проявляется большая мышечная сила. С нею связана подчас и гибкость, зависящая при определенных упражнениях не только от растяжимости мышц, но и от силы их антагонистов. Для поддержания трудных антигравитационных поз нужна статическая выносливость. При быстрых длительных вращениях на месте («винт», «волчок») необходима высокая устойчивость вестибулярного аппарата, в особенности при изменении положения головы («заклон»). Ни в одном виде спорта не встречаются такие быстрые и длительные вращения, как в фигурном катании.
Развитие способности к сложной координации движений конечностей— важный фактор в фигурном катании на коньках. Вместе с тем в связи с трудностью сохранения равновесия у начинающих фигуристов руки обычно находятся в статическом напряжении: согнуты в локтевых суставах и несколько разогнуты в плечевых и разведены в стороны. У более квалифицированных фигуристов наблюдается хорошее расслабление рук, что обусловливает возможность выполнения сложных выразительных движений.
В фигурном катании важнейшее значение имеет ориентировка в окружающем пространстве. Это ярко проявляется, в частности, в исполнении фигур обязательной программы. Вычерчивание лезвием конька определенных геометрических фигур, повторное «покрытие»
447
рисунка — все это основано на точных пространственных представлениях, на взаимодействии зрительного и двигательного анализаторов Это требует, кроме того, тонкой дозировки толчкового усилия, необходимого для выполнения на одном коньке той или иной части фигуры. Тонкое мышечно-суставное чувство, связанное главным образом с рецепторами голеностопного сустава, обеспечивает скольжение на ребре конька.
Расход энергии при исполнении фигур обязательной программы незначителен, так как оно требует точности, но не быстроты и поэтому несколько статично. Соответственно незначительны изменения в работе сердечно-сосудистой и дыхательной систем. Исполнение произвольной программы, которое чрезвычайно динамично, характеризуется большой быстротой, многочисленными прыжками и поэтому вызывает большие энерготраты. В связи с продолжительностью исполнения произвольной программы (4—5 мин.) оно относится к зоне работы субмаксимальной мощности. Усиление дыхания и кровообращения достигает близких к предельным значений. Несмотря на то что потребление кислорода приближается к максимальному, после окончания выступления остается значительный кислородный долг. Это говорит о том, что в этих видах фигурного катания работа совершается не только за счет аэробных реакций, но и при значительном участии анаэробных процессов.
Глава XX
ФИЗИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА НЕСТАНДАРТНЫХ ФИЗИЧЕСКИХ УПРАЖНЕНИЙ
К нестандартным, или ситуационным, физическим упражнениям относятся спортивные игры и все виды единоборств. Эти упражнения характеризуются тем, что структура, направление и сила при выполнении двигательных актов не программируются заранее, а зависят от ситуаций, возникающих в разные моменты спортивной деятельности. Своевременное и правильное решение двигательных задач при этом обусловлено способностью нервной системы к экстраполяции, четкой информацией, поступающей в центральную нервную систему от органов чувств, высокой подвижностью нервных процессов, соответствующей подготовленностью двигательного аппарата и обеспечивающих его деятельность вегетативных систем организма.
Разные виды нестандартных физических упражнений предъявляют различные требования к организму. Эти различия обусловлены вариативностью структуры и мощности движений на протяжении работы.
§ 1. Общая характеристика спортивных игр
Влияние различных спортивных игр на морфофункциональное состояние отдельных систем организма зависит от продолжительности игры, ее интенсивности, размеров площадки, количества
448
игроков в команде и некоторых других факторов. При этом чем больше беговых нагрузок в процессе игры, тем резче выражены вегетативные сдвиги.
Все спортивные игры характеризуются скоростно-силовыми движениями смешанного типа. Преобладающими являются ациклические движения. Большое значение имеет точное выполнение прицельных движений.
Работа мышц при спортивных играх в основном динамическая. Однако при силовых приемах мышцы развивают значительные, но кратковременные статические напряжения. Это предъявляет значительные требования к силовой подготовке спортсменов, занимающихся спортивными играми.
Мощность циклических движений при спортивных играх — переменная. В отдельные моменты игры она может быть большой, субмаксимальной и максимальной. Нередко в процессе игры происходит кратковременное прекращение деятельности спортсмена (нарушение правил, потеря мяча, одноминутный перерыв и пр.). При снижении мощности работы и при кратковременных остановках должны усиливаться восстановительные процессы. Они протекают более интенсивно у квалифицированных спортсменов, отличающихся высокой подвижностью нервных процессов. Спортивные игры предъявляют значительные требования к анаэробным возможностям организма. Если игра протекает в очень высоком темпе (например, хоккей с шайбой), то многие системы организма, функционируя в анаэробных условиях, не выдерживают длительного напряжения. В связи с этим правилами разрешается смена игроков. Для достижения высоких результатов в этих видах спорта наряду с анаэробными возможностями необходимо развитие и аэробных.
Спортивные игры способствуют развитию быстроты, силы, ловкости и специальной выносливости к выполнению работы переменной мощности. Большой объем тренировочных нагрузок требует также развития общей выносливости.
Двигательные навыки у занимающихся спортивными играми разнообразны. Сложность их обусловлена тем, что спортсмен должен вести мяч, выполнять передачи и другие игровые приемы непосредственно в момент быстрого передвижения по площадке. В спортивных играх, как и в других видах спорта, необходима автоматизация двигательных навыков. Чем она выше, тем эффективнее деятельность спортсмена. Автоматизированно должны осуществляться отдельные игровые приемы и даже компоненты сложных действий. Однако при игре нередко возникают такие ситуации, когда заученные привычные движения становятся неэффективными. Тогда стереотипная деятельность спортсменов должна изменяться в соответствии с создавшимися условиями. Это достигается путем экстраполяции, которая осуществляется как сознательно, так и автоматизированно. При этом у спортсменов образуются новые условнорефлекторные связи, обеспечивающие новые формы движений. Таким образом, при спортивных играх стереотипные движения должны быть легко изменяемыми. Это обеспечивается высокой подвиж-
449
ностыо нервных процессов, управляющих двигательном деятельностью.
Занятия спортивными играми совершенствуют подвижность нервных процессов. Один из косвенных показателей этого — укорочение латентного времени условнорефлекторных двигательных реакций. Более значительно при этом укорачивается латентное время сложных реакций, требующих выбора правильного ответа. Укорочение общего латентного времени реакций (на 5—20%, по данным С. М. Оплавина) происходит главным образом за счет уменьшения времени, затрачиваемого на выбор ответа.
Подвижность нервных процессов необходима не только для быстрой смены структуры и темпа движений, но и для соответствующего изменения деятельности органов дыхания и кровообращения. Эти органы должны быстро повышать свои функции при увеличении мощности работы и быстро восстанавливаться при ее снижении. Игровая деятельность требует быстрых, координированных и точных движений. Для этого необходима своевременная и четкая информация как о расположении игроков и мяча на площадке (обеспечиваемая органами зрения и слуха), так и о положении собственного тела и о состоянии его мышц, поступающая от вестибулярного и двигательного анализаторов.
Спортивные игры предъявляют специфические требования к двигательному аппарату. Подготовка игрока должна обеспечивать повышение «взрывной» силы мышц, «прыжковой» выносливости, быстроты стартовых ускорений, скоростной выносливости. Специальная силовая подготовка игроков сопровождается гипертрофией скелетных мышц. Но эта гипертрофия не должна снижать их скоростные качества.
§ 2. Баскетбол
Центральная нервная система. Для повышения квалификации баскетболиста необходимо совершенствовать управление движениями, в результате чего улучшается координация двигательной деятельности. Это можно наблюдать по изменению характера электрической активности мышц при выполнении некоторых игровых приемов. Например, у баскетболистов — мастеров спорта при передачах мяча и бросках его в кольцо относительно небольшие по частоте и амплитуде электрические потенциалы мышц рук и плечевого пояса возникают лишь непосредственно при выполнении движений, т. е. они сконцентрированы во времени. Только в более сложных условиях (например, при наличии третьего игрока, выполняющего роль защитника) электрическая активность мышц появляется у мастеров спорта еще до получения мяча. У баскетболистов низкой квалификации электрические потенциалы мышц не сконцентрированы во времени. Они возникают задолго до получения мяча и даже при обманных движениях партнера. Последнее объясняется отсутствием необходимых дифференцировок.
450
Анализаторы. Баскетбол предъявляет повышенные требования к многим анализаторам.
У квалифицированных баскетболистов поле зрения в обычных условиях имеет нормальные размеры. При выходе на игровую площадку и особенно после разминки оно увеличивается на 15—20% по сравнению с исходной величиной (рис. 121). Это происходит в
Рис. 121. Изменение поля зрения на бесцветный (черные стобикп) и зеленый (заштрихованные столбики) объекты у баскетболистов под влиянием разминки (в % к исходной величине).
Внизу — даты наблюдений и инициалы спортсменов
связи с увеличением возбудимости периферических элементов сетчатой оболочки глаза и зрительных центров.
Функции центра и периферии сетчатки при игре в баскетбол имеют одинаково большое значение. Периферические элементы сетчатки — палочки обеспечивают ориентировку на поле. Деятельность же ее центральных элементов — колбочек важна для выполнения точных бросков мяча в кольцо и т. п.
Большую роль во время игровой деятельности играет пространственное (глубинное) зрение. Пространственные восприятия у баскетболистов, как правило, лучше по сравнению со спортсменами других специализаций.
Систематическая тренировка баскетболистов совершенствует глазодвигательный аппарат. Если у лиц, не занимающихся спортивными играми, равномерное развитие мышц глаза (ортофория) от
451
мечается лишь в 40% случаев, то у баскетболистов — в 80% и более. Наличие ортофории снижает утомляемость глаза при зрительных нагрузках, которая очень велика при игре в баскетбол.
У квалифицированных баскетболистов хорошо развит двигательный анализатор, обеспечивающий информацию о силе, амплитуде, направлении выполняемых движений. Функциональное состояние этого анализатора оценивается по точности заданных движений, выполняемых сначала при зрительном контроле, а затем при закрытых глазах. Например, спортсмен получает задание выполнить поворот мяча, закрепленного на специальном градуированном штативе, на определенное число градусов. Сначала движение мяча контролируется зрением, затем выполняется при закрытых глазах. Чем меньше ошибка в амплитуде движения, допускаемая при этом, тем выше чувствительность двигательного анализатора в области лучезапястного сустава. У квалифицированных баскетболистов она намного лучше, чем у менее квалифицированных.
Д ы х а н и е. При игре в баскетбол частота дыхания достигает 50—58 и более дыхательных циклов в 1 мин. Дыхание при изменении темпа движений учащается быстрее, чем частота сердцебиений. Это свидетельствует о более высокой подвижности механизмов, регулирующих частоту дыхания.
Легочная вентиляция и потребление кислорода изменяются при игре в баскетбол по-разному, в зависимости от темпа игры и других условий.
Систематическая тренировка баскетболистов ведет к повышению анаэробных и в несколько меньшей степени аэробных возможностей организма. Наблюдения за баскетболистами показали, что 3-месячная тренировка сопровождалась увеличением максимального потребления кислорода в среднем с 4,3 до 4,72 л/мин, что составляет около 10%. Максимальный кислородный долг при этом возрос в среднем с 7,25 до 8,83 л, т. е. более чем на 20% (В. А. Данилов). Относительно небольшие величины максимального кислородного долга у баскетболистов обусловлены переменной мощностью работы, что обеспечивает возможность частичной его ликвидации непосредственно в процессе игры.
Кровообращение. В зависимости от темпа игры, активности игрока и других условий частота сердцебиений может достигать 200 и более ударов в 1 мин. В среднем этот показатель удерживается в процессе игры на уровне 170—190 ударов в 1 мин. Кратковременное снижение темпа движений и даже их прекращение на 5— 10 сек. не сопровождаются урежением сердечных сокращений. Лишь перерывы длительностью 20—60 сек. и более вызывают снижение сердечного ритма до 100—140 ударов в 1 мин. (Н. Б. Кичай-кина). При повышении мощности работы учащение сердцебиений наступает несколько быстрее, чем ее урежение при понижении мощности работы или в интервалах отдыха. Запаздывание изменений сердечного ритма по сравнению с изменением мощности работы свидетельствует о некоторой инертности механизмов, регулирующих этот показатель деятельности сердца.
452
По данным В. Л. Карпмана с соавторами объем сердца у квалифицированных баскетболистов-мужчин составляет в среднем 1201 см3. При работе значительной мощности в лабораторных условиях минутный объем крови у баскетболистов достигает 24 л/мин, систолический объем — 167 мл.
Выделительные процессы. В связи с увеличением деятельности потовых желез во время игры происходит уменьшение диуреза и увеличение удельного веса мочи. После игры повышается концентрация в моче молочной, мочевой и других кислот и нередко появляется белок.
Повышение концентрации глюкозы в крови у игроков в предстартовом периоде ведет к появлению ее в моче.
§ 3. Волейбол
При игре в волейбол передвижения на площадке менее интенсивны, чем при игре в баскетбол. Этим обусловлены менее выраженные вегетативные сдвиги.
Сложные по координации движения волейболистов требуют развития скелетных мышц, особенно их силовых и скоростных качеств.
Большую роль для достижения мастерства в этом виде спорта играют функции анализаторов. Несколько увеличенное у волейболистов поле зрения способствует лучшей ориентировке на площадке. Необходимость непрерывно следить за движениями мяча совершенствует глазодвигательный аппарат. Среди квалифицированных волейболистов ортофория наблюдается даже несколько чаще, чем у занимающихся другими видами спортивных игр.
Изменение дыхания и кровообращения при игре в волейбол зависит от темпа игры и активности игрока. По данным Н. Б. Кичай-киной, частота сердцебиений в процессе игры удерживается в среднем на уровне 170—190 ударов в 1 мин., частота дыханий составляет 42—48 дыхательных циклов в минуту.
§ 4. Футбол
Для футболиста необходима специальная подготовка двигательного аппарата. Производимая им работа является скоростно-силовой. Но большие размеры игрового поля и обусловленная этим значительная длительность пробежек, а также и продолжительность игры в целом требуют высокой адаптации мышц к работе в анаэробных и аэробных условиях.
Для повышения эффективности двигательной деятельности футболистов очень важна информация, поступающая от многих рецепторов тела. При уменьшенном поле зрения и сниженной функциональной устойчивости вестибулярного аппарата они, как правило, не могут достичь высоких спортивных результатов (А. Н. Крестовников и К. И. Лемешев).
Большое значение для результативности игры в футбол имеет быстрый бег. Ou осуществляется при наличии относительно корот
453
ких интервалов отдыха, что способствует развитию анаэробных возможностей организма. На протяжении игры футболист пробегает в общей сложности в среднем около 5—8 км. Это предъявляет высокие требования к аэробным возможностям организма. Однако показатель их относительно невысок (даже у квалифицированных тренированных футболистов). По данным В. Л. Карпмана, максимальное потребление кислорода у них составляет в среднем 4,4 л/мин (или 62,5 мл/мин/кг). Индивидуальные величины находятся в пределах от 3,2 до 5,3 л/мин. У футболистов в состоянии покоя отмечается умеренная брадикардия (48—54 удара в 1 мин.). По данным С. П. Летунова, размеры сердца у них в большинстве случаев увеличены. В 70% из них отмечается гипертрофия левого желудочка, в 21% —гипертрофия обоих желудочков. Более поздние исследования В. Л. Карпмана показали, что объем сердца у футболистов в среднем относительно невелик — 965 см3 (при индивидуальных колебаниях от 820 до 1140 см3).
Вегетативные сдвиги при игре в футбол зависят от темпа игры и других ее особенностей.
Частота сердцебиений у футболистов уже на первых минутах игры достигает 160—170 ударов в I мин. (Ю. А. Морозов). В дальнейшем этот показатель удерживается на уровне 160—180 ударов в 1 мин., лишь в отдельные моменты игры повышаясь до 200 и более ударов в 1 мин.
После игры, особенно если она проводится в холодную и влажную погоду, в моче у футболистов обнаруживается белок.
§ 5. Хоккей с шайбой
Хоккей отличается от других спортивных игр очень высоким темпом движений. Мышцы хоккеиста выполняют работу главным образом в анаэробных условиях. Динамическая, скоростно-силовая работа сочетается со значительными статическими напряжениями, что сопровождается гипертрофией мышц и развитием их силовых качеств.
Как и все спортивные игры, хоккей предъявляет большие требования к функциям анализаторов.
Вегетативные сдвиги в процессе игры очень значительны. По данным А. Н. Гуминского, за 6 месяцев тренировки максимальное потребление кислорода у хоккеистов увеличилось в среднем с 55,4 до 61,3 мл/мин/кг. У менее квалифицированных хоккеистов максимальное потребление кислорода в среднем составляет 4,2 л/мин (или 59 мл/мин/кг) при индивидуальных колебаниях от 3,2 до 5,1 л/мин (В. Л. Карпман). Таким образом, аэробная производительность хоккеистов не очень велика. Высокий темп игры и сравнительно кратковременное пребывание игрока на площадке способствуют развитию главным образом анаэробных возможностей.
Объем сердца у хоккеистов равен в среднем 927 см3 (при индивидуальных колебаниях от 685 до 1080 см3).
454
§ 6. Общая характеристика единоборств
Вариативность мощности работы и энергетических трат. Все виды единоборств характеризуются постоянной сменой ситуаций, каждая из которых требует от спортсмена новой программы нападающих или оборонительных действий. Вследствие этого работа совершается при постоянных изменениях как ее мощности, так и энергетических трат. Они могут быть весьма незначительными (сохранение некоторых поз при выжидании) и очень большими (броски в борьбе и т. п.).
Искусственное снижение веса тела. Соревнования по боксу и борьбе, как и по поднятию штанги, проводятся с учетом весовых категорий. Поэтому спортсмены, специализирующиеся в этих видах спорта, должны следить за весом своего тела и в случае необходимости принимать меры для искусственного его снижения. Спортсмену необходимо знать свой оптимальный вес, который характерен для состояния его наивысшей тренированности. Данные об оптимальном весе тела получают путем систематического взвешивания и наблюдений за его динамикой в разные периоды тренировки.
Определение так называемого «нормального» веса тела по формуле Брока (вес тела в кг равен росту в см минус 100), Бушара (вес в кг, умноженный на рост в см) и другим дает лишь приблизительные сведения. В каждом отдельном случае могут быть весьма существенные отклонения оптимального веса от «нормального», вычисляемого на основании роста, окружности груди и других антропометрических показателей.
В спортивной практике для снижения веса тела ограничивают прием пищи и питья, применяют специальные диеты, используют паровую баню, светотепловые ванны и другие средства. Применение диет с ограниченным содержанием углеводов и солей обеспечивает уменьшение веса на 2—3,5 кг в течение двух суток.
Пользование баней способствует быстрому и значительному снижению веса тела. Однако длительное (до 25—30 мин.) пребывание в парной (при температуре 50—60°), сочетаемое с ограничением приема жидкости, часто приводит к ухудшению самочувствия, общей слабости, беспокойному сну, повышению возбудимости нервной системы, учащению пульса, повышению кровяного давления. Все это ведет к снижению работоспособности спортсмена. Кратковременное пользование баней (10—15 мин.) хотя и менее эффективно для сгонки веса, но не вызывает неблагоприятных явлений.
Светотепловые ванны, почти совсем не снижающие работоспособность организма, при повторном применении с небольшими интервалами отдыха могут уменьшить вес тела на 2 кг и более. Для искусственного снижения веса тела можно также пользоваться воздушной баней, вызывающей обильное потоотделение без каких-либо неприятных ощущений, и горячим душем.
Физиологически наиболее целесообразным средством снижения веса тела является продолжительная физическая работа (напри
455
мер, бег) в теплой одежде, что вызывает обильное потоотделение. Однако снижение веса таким путем непосредственно перед участием в соревнованиях не рекомендуется.
Значительное снижение веса в краткие сроки (вне зависимости от применяемых способов), как правило, ведет к ухудшению самочувствия спортсмена, а в ряде случаев может снижать его работоспособность. Поэтому спортсмен, выступающий в определенной весовой категории, должен постоянно следить за своим весом, что исключит необходимость применения срочных мер для его снижения непосредственно перед участием в соревнованиях.
§ 7. Бокс
Двигательная деятельность боксера представляет собой скоростно-силовую динамическую работу переменной мощности. Движения боксера как при нанесении ударов, так и при защите против ударов противника ацикличны. Характер и структура их зависят от действий противника.
При некоторых раундах мощность работы очень велика. В кратковременные перерывы между ними не происходит полного восстановления, и боксер вновь выходит на ринг при наличии кислородного долга и других функциональных изменений, обусловленных предшествующим раундом.
Нервная система и двигательный а п п а р а т. Тренировка боксера способствует развитию силы, быстроты и специальной выносливости. Двигательные навыки боксера разнообразны. Во время боя движения выполняются в разной последовательности, с разной амплитудой и силой. Все это требует высокой подвижности нервных процессов и экстраполяции при программировании адекватных движений.
Для успешного ведения боя исключительно важна информация, поступающая от проприорецепторов двигательного аппарата и рецепторов сетчатки глаза. Для сохранения равновесия и точности движений необходима также высокая функциональная устойчивость вестибулярного аппарата.
Болевая и осязательная чувствительность участков кожи, которые подвергаются ударам, у квалифицированных боксеров несколько понижена. После прекращения занятий боксом она постепенно восстанавливается.
Систематическая тренировка боксера ведет к биохимическим, морфологическим и функциональным изменениям скелетных мышц, способствующим развитию их силы и быстроты сокращений.
Дыхание и расход энергии. Боксеры-новички обычно задерживают дыхание при ведении боя на соревнованиях и на тренировочных занятиях (например, при упражнениях с мешком). Квалифицированные боксеры сохраняют ритмичное глубокое дыхание на протяжении всего боя. Только в момент нанесения удара они производят кратковременную задержку дыхания на выдохе.
Повышенные требования к дыханию обусловливают развитие
456
дыхательных мышц. Жизненная емкость легких у боксеров ранил п среднем 4500 мм. После участия в соревновании она может временно уменьшаться на 100—300 мл. Легочная вентиляция у боксеров во время боя достигает 80—100 л/мин, потребление кислорода обычно не превышает 4 л/мин. Кислородный запрос при этом полностью не удовлетворяется, и при работе у боксера образуется значительный кислородный долг.
Максимальное потребление кислорода у боксеров равно в среднем 4,1 л/мин (или 60 мл/мин/кг). У отдельных наиболее тренированных и квалифицированных боксеров оно может достигать 6,5 л/мин. Расход энергии при ведении боя составляет 15—20 ккал, в 1 мин. Суммарные энерготраты в течение трех раундов в среднем равны около 200—250 ккал.
Сердечно-сосудистая система и кровь. В состоянии покоя у боксеров отмечается нерезко выраженная брадикардия. Объем сердца у них равен в среднем 948 см3 (при индивидуальных колебаниях от 615 до 1440 см3). Частота сердцебиений при ведении боя нарастает у боксера до 180—200 ударов в 1 мин. На тренировочных занятиях сердечный ритм учащается меньше.
Эмоциональное возбуждение, возникающее у боксера на соревнованиях перед боем, вызывает значительные сдвиги в функциональном состоянии всех систем организма. В частности, это проявляется в изменении состава крови. Уже в предстартовом периоде увеличивается содержание глюкозы в крови до 0,16—0,19% и более. У многих боксеров за 6—12 часов до соревнований снижается в крови число эозинофилов. Участие в боях ведет к дальнейшему их уменьшению, что свидетельствует о значительном напряжении всех функций организма. Одновременно с уменьшением эозинофилов под влиянием работы увеличивается содержание других форм лейкоцитов в крови. Возникает нейтрофильная фаза миогенного лейкоцитоза. После боя у боксеров увеличена концентрация молочной кислоты в крови и уменьшены щелочные резервы.
После участия в соревнованиях отмечается уменьшение диуреза, увеличение удельного веса мочи, повышение ее кислотности. В ней увеличивается содержание недоокисленных продуктов, а содержание хлоридов в связи с обильным потоотделением уменьшается. Гипергликемия на старте часто ведет к появлению глюкозы в моче. После соревнований в моче нередко находят белок (до 0,6—1,2%о).
Восстановительные процессы после соревновательных боев в связи с сильным эмоциональным возбуждением протекают несколько медленнее, чем после тренировочных нагрузок. О преобладании возбудительных процессов свидетельствует в это время нарушение ночного сна после соревнований.
Нокаут. При получении ударов в нижнюю челюсть, переносицу, область висков и сонных артерий, правое и левое подреберья и область солнечного сплетения у боксеров могут возникать нокдаун и нокаут, сопровождающиеся временным нарушением сознания, потерей равновесия тела и возникновением такого состояния, при котором спортсмен не может продолжать бой.
457
Нокдаун продолжается несколько секунд. Нокаут длится более продолжительное время. Если в течение 10 сек. после получения удара боксер не в состоянии возобновить бой, то он считается нокаутированным и ему засчитывают поражение.
Наиболее тяжелый вид нокаута возникает при ударе в область солнечного сплетения. Он сопровождается резкой болью, временной остановкой дыхания, замедлением деятельности сердца, падением артериального давления и потерей сознания. Возникновение нокаута при ударе в эту область имеет рефлекторный механизм. Происходящее при этом ударе раздражение окончаний чревного нерва изменяет состояние сосудодвигательного центра и центра сердечных нервов, что вызывает явление шока, характеризующееся, в частности, торможением деятельности сердца.
Нокауты при ударах в голову обусловлены сотрясением мозга, при ударах в нижнюю челюсть — сотрясением отолитового аппарата. Последние несколько менее болезненны, чем удары по животу, но вследствие резкого нарушения координации движений и равновесия действие их на боксера может быть также весьма неблагоприятным. Влияние нокаута на организм зависит от многих обстоятельств (область тела, куда нанесен удар, уровень тренированности спортсмена, особенности его индивидуальных реакций на нокаут, степень утомления, количество ударов, полученных при ведении боя, и др.). Повторные нокауты могут вызывать тяжелые расстройства нервной деятельности, в связи с чем боксеры, получившие нокаут, не допускаются в течение определенного времени к участию в соревнованиях.
§ 8. Борьба
Движения спортсменов во всех видах борьбы имеют разную структуру, но везде выполняются с переменной мощностью. Во время схваток динамическая скоростно-силовая, а в некоторых случаях и собственно-силовая работа чередуется со статическими напряжениями обширных мышечных групп. Соотношение между динамической работой мышц и статической в разных видах борьбы различно.
Нервная система и двигательный аппарат. Тренировка борцов ведет к развитию силы, быстроты, специальной выносливости и ловкости.
Для успешной деятельности борца необходимо развитие проприоцептивной чувствительности. У квалифицированных борцов она высоко развита во всех звеньях тела. Почти все мышцы борцов гипертрофированы. В процессе тренировки мышцы адаптируются главным образом к работе в анаэробных условиях.
Дыхание и расход энергии. Во время борьбы частота дыхания увеличивается до 40—50 раз в 1 мин. Ритм дыхания на протяжении схватки непостоянен. В моменты статических напряжений происходит задержка дыхания. После их окончания оно учащается.
458
Тренированные борцы могут хорошо регулировать дыхание. Задержки дыхания у них кратковрсмеины и феномен Лингардта про является в меньшей степени, чем у нетренированных борцов. Потребление кислорода при борьбе может быть различным. Оно i.i висит от мощности производимой работы. Кислородный запрос ом ре деляется мощностью работы и длительностью схваток. Потребность в кислороде во время борьбы полностью не удовлетворяется, в свя
зи с чем образуется кислородный долг. Он увеличивается в связи с задержкой дыхания во время статических напряжений.
У борцов наряду с развитием анаэробных возможностей существенное значение для поддержания высокого уровня специальной работоспособности имеет и повышение максимального потребления кислорода. Б. Салтин и П. Астранд показали, что у квалифицированных борцов максимальное потребление кислорода составляет в среднем 4,6 л/мин (или 57 мл/ мин/кг). По данным В. Л. Кар-пмана с соавторами, абсолютная велиична этого показателя
Рис. 122. Частота сердечных сокращений у борцов во время (I) и после (II) специальных упражнений (по Н. Г. Куликову и Е. А. Мирошникову).
Сплошная линия — максимальное число бросков партнера в течение 2,5 мии., прерывистая линия — максимальное число бросков чучела за то же время
у борцов I разряда в среднем несколько меньше — 4,1 л/мин, а при расчете на 1 кг веса тела — больше (59 мл). У отдельных борцов величина максимального потребления кислорода колеблется от 3,2 до 5,8 л/мин.
Кровообращение и кровь. В состоянии покоя частота сердцебиений у борцов равна 60—65 ударам в 1 мин.
Объем сердца у квалифицированных борцов, по результатам исследований Ю. А. Борисовой и Г. С. Туманяна, составляет в среднем 953 см3. По данным В. Л. Карпмана с соавторами, у борцов I разряда этот показатель в среднем равен 935 см3 (при индивидуальных колебаниях от 719 до 1248 см3). Объем сердца у борцов коррелирует с весом и ростом тела. Степень истинного увеличения объема сердца может быть выявлена лишь при расчете его объема на 1 кг веса и 1 см роста тела. Эта величина, обозначаемая как относительный объем сердца, составляет у борцов в среднем 69 см3/кг. см, у не занимающихся спортом — 50 см3/кг. см.
Во время проведения схваток частота сердцебиений у борцов достигает в зависимости от мощности усилий 170—200 ударов в 1 мин. Телеметрическая регистрация частоты сердцебиений у борцов на тренировочных занятиях показала, что при бросках чучела сердечный ритм может увеличиваться до 180 ударов в 1 мин., при бросках партнера — до 190 (рис. 122). Артериальное (систолическое) дав
459
ление повышается до 160—180 мм рт. ст., диастолическое также оказывается увеличенным.
При борьбе обычно возникают явления относительно продолжительного натуживания, что повышает требования к деятельности сердца и способствует его гипертрофии.
На тренировках и соревнованиях у борцов увеличивается количество эритроцитов и гемоглобина в крови. Как показали специальные исследования, в восстановительном периоде после схваток у них повыш на интенсивность деоксигенации эритроцитов, что благоприятно отражается на дыхательной функции крови. У борцов вольного и классического стиля скорость деоксигенации эритроцитов после схваток оказывается повышенной в среднем на 25% (В. И. Шубин с соавт.). После схваток наблюдается миогенный лейкоцитоз (нейтрофильная фаза). Концентрация глюкозы часто повышена. Увеличено и содержание молочной кислоты (до 130 мг% и более).
Во время борьбы резко усиливается потоотделение. При длительных схватках это ведет к большим потерям воды и снижению веса тела. В моче после напряженных схваток повышается концентрация недоокисленных веществ и иногда появляется белок.
§ 9. Фехтование
Фехтование по физиологической характеристике резко отличается от описанных выше нестандартных физических упражнений.
Нервная система и двигательный аппарат. Фехтование характеризуется сложной координацией движений. Для успешных выступлений в этом виде спорта необходима быстрота боевых действий, сочетаемая с правильностью выбора (боевая ориентировка) и точностью нанесения уколов рапирой или ударов саблей. Для этого необходима своевременная и предельно четкая зрительная информация и скорейшая ее переработка. Эффективное управление двигательной деятельностью фехтовальщика требует высокой подвижности нервных процессов, о которой можно судить по длительности простых и сложных (с выбором ответа) зрительно-двигательных реакций. По данным Ю. В. Варганова, скрытое время простых реакций с оружием у фехтовальщиков составляет в среднем 335 мсек. Сложные реакции при наличии выбора из двух раздражителей удлиняются в среднем до 375 мсек., из четырех — до 443 мсек., из восьми — до 552 мсек. У более квалифицированных фехтовальщиков скрытое время реакций уменьшено. Например, по данным И. Н. Сальченко, среднее время реакций в виде нанесения укола с выпадом у мастеров спорта составляет в среднем 407 мсек., а у спортсменов менее высоких разрядов — 422 мсек.
Мышцы спортсмена при ведении боя выполняют преимущественно динамическую скоростную работу. Однако необходимость поддержания специфической при фехтовании позы требует относительно длительных статических напряжений мышц туловища, а также верхних и нижних конечностей. Работа мышц производится преимущественно в анаэробных условиях.
460
Уже при подготовке защитных реакций у фехтовальщиков hoi-никает электрическая активность соответствующих мыши. Это явление наиболее выражено у спортсменов средней квалификации. Новички еще не умеют заранее готовиться к защите. У мастеров
спорта электрическая активность мышц сконцентрирована во времени, что обеспечивает более совершенную координацию движений.
Вегетативные сдвиги.
возникающие у фехтовальщика при ведении боя, типичны для скоростной, относительно кратковременной работы. Несмотря на переменную мощность работы и ацикличность движений у фехтовальщиков в процессе тренировочного урока, можно наблюдать период врабатывания, длящийся около 3—7 мин., и период наиболее высокой работоспособности, который обычно продолжается около 30—40 мин., после чего возникают первые признаки утомления. В период наиболее высокой работоспособности средняя длительность латентного периода двигательных реакций составляет около 354 мсек., моторный компонент реак-
М Продолжительность боя
Ф Уколы,полученные фехтовальщиком В.П< • Уколы,нанесенные фехтовальщиком В.Тй
Рис. 123. Частота сердечных сокращений у заслуженных мастеров спорта Г. С. (сплошная линия) и В. П. (прерывистая линия) на протяжении боя на турнире сильнейших фехтовальщиков СССР (по В. С. Келлеру с соавт.)
ций — около 405 мсек., точность уколов — 58%. С наступлением утомления длительность двигательных реакций нарастает, а точность уколов снижается (К. Ю. Ажицкий).
Дыхание. Необходимость пользоваться защитной маской несколько затрудняет дыхание, которое при ведении боя становится поверхностным и частым (И. Д. Карцев с соавт.).
Специфика двигательной деятельности фехтовальщика требует развития преимущественно анаэробных возможностей организма. Аэробные же возможности фехтовальщиков относительно невысоки. По данным В. Салтина и П. Астранда, максимальное потребление кислорода у них составляет в среднем 4,2 л/мин (или 59 мл/мин/кг).
Сердечно-сосудистая система и кровь. Радиоте-
леметрические исследования показали, что на тренировочных уроках учащение сердечного ритма не превышает 150—170 ударов в 1 мин. При выполнении отдельных движений частота сердцебиений зависит от количества работающих при этом мышц. Например, при выполнении укола одной рукой сокращения сердца учащаются до 96 ударов в 1 мин., при нанесении укола с шагом —до 120, при уколе с выпадом —до 120, при уколе с шагом вперед и выпадом —до
461
138 ударов в 1 мин. (Ю. В. Варганов). Таким образом, боевые действия фехтовальщиков, характеризующиеся активной работой мышц ног, вызывают большее учащение сердечного ритма, чем действия, выполняемые рукой.
При ведении фехтовательных боев ритм сердечных сокращений несколько выше, чем на тренировочных занятиях (рис. 123). В отдельных случаях он достигает 190—195 ударов в 1 мин. Наибольшая частота сердцебиений наблюдается у фехтовальщиков в боях, проводимых по системе прямого выбывания. Это свидетельствует о том, что сердечный ритм у фехтовальщиков во время боев обусловлен не только мощностью работы, но и значимостью соревнований.
У фехтовальщиков на тренировочных уроках часто наблюдаются микропаузы длительностью около 10—12 сек. После этих пауз, являющихся своеобразным отдыхом, частота сердцебиений оказывается сниженной. Уменьшается и длительность латентного периода двигательных реакций. Однако эти, казалось бы, благоприятные сдвиги сочетаются с повышением вариативности времени двигательных реакций и уменьшением точности нанесения уколов. Последнее снижает эффективность последующей деятельности спортсмена.
Быстрота и высокий темп движений фехтовальщика при ведении боя, а также его относительная продолжительность несколько нарушают гомеостаз. После выполнения стандартных нагрузок pH крови у него снижается в среднем на 0,16, буферные основания уменьшаются на 11,6 мэкв/л, парциальное давление углекислоты нарастает на 11,5 мм рт. ст. (И. В. Сивков с соавт.). По мере развития тренированности стандартные нагрузки вызывают относительно меньшие сдвиги в составе внутренней среды организма.
В связи с необходимостью пользоваться защитной одеждой и маской при ведении боев у фехтовальщика несколько затруднен тепловой обмен.
Глава XXI
ВЛИЯНИЕ ЭКСТРЕМАЛЬНЫХ УСЛОВИИ И АДАПТАЦИЯ
К НИМ СПОРТСМЕНОВ
Как известно, спортивная деятельность может происходить в самых различных условиях. В некоторых случаях спортсмены подвергаются воздействию экстремальных (extremum — крайний) факторов, т. е. сильно или необычно воздействующих на организм. Эти факторы, с которыми встречается в своей деятельности спортсмен, разнообразны. Ниже рассматривается влияние некоторых из них.
§ 1. Влияние на организм пониженного атмосферного давления
Общая характеристика влияния горного климата на организм человека. Земной шар окружен слоем воздуха — атмосферой. Под действием гравитационного поля вся масса воздуха оказывает дав
462
ление на поверхность земного шара. Если па уровне моря атмосферное давление соответствует 760 мм рт. ст., то с подъемом в юры оно пропорционально высоте будет снижаться при сохранении постоянства процентного состава сазов воздуха.
Принято считать высоты до 750—1000 м низкогорьем, от 1000 до 3000 м — среднегорьем, от 2500 м и выше — высокогорьем. Если низкогорье существенно не влияет на физическую работоспособность человека, то в условиях среднегорья, а тем более высокогорья человек испытывает дополнительные трудности при выполнении физических упражнений.
Действие горного климата на организм человека обусловлено многими природными факторами: усиленной солнечной радиацией, высокой ионизацией воздуха, резкими изменениями влажности и температуры, пониженным атмосферным давлением и связанным с ним низким парциальным давлением кислорода. Основным фактором, влияющим на функциональные вомзожности организма в условиях высоты, является гипоксия, т. е. снижение напряжения кислорода, что вызывает появление гипоксемии — снижения насыщения крови кислородом (табл. 18).
Понижение напряжения парциального давления кислорода в крови уменьшает градиент давления кислорода между капиллярной кровью и тканями, в результате чего ухудшается переход кислорода в ткани, снижается скорость окислительных процессов. В соответствии с кривой диссоциации оксигемоглобина (см. главу VII) на средних высотах в покое снижение парциального давления кислорода в альвеолярном воздухе не уменьшает процент оксигенации крови столь существенно, как на больших высотах.
Т аблица 18
Общее барометрическое давление, напряжение О2 в альвеолярном воздухе и артериальной крови и процент НВ02 в артериальной крови
Показатели Высота, м
100 2000 4300 7000 9000
Барометрическое давление 750 600 450 300 225
Альвеолярное рО2 103 76 54 30 26
Артериальное рО2 95 70 50 28 24
НВО2 в артериальной крови (%) 96 94 87 60 50
Влияние горного климата на организм человека необходимо учитывать при решении ряда спортивных задач. Такая необходимость, в частности, связана с проведением соревнований, в том числе международных, и с альпинистскими восхождениями в условиях высокогорья. Кроме того, как показали многочисленные исследования, спортсмены, тренирующиеся в среднегорье в условиях пониженного атмосферного давления, могут после этого более успешно выступать в местности на уровне моря.
463
Влияние среднегорья на физическую работоспособность. В состоянии покоя организм спортсмена сравнительно легко адаптируется к условиям среднегорья. При выполнении же напряженной мышечной работы вегетативные функции организма оказываются недостаточно приспособленными для обеспечения его необходимым количеством кислорода.
В первые дни пребывания в среднегорье происходит снижение величины максимального потребления кислорода и аэробных возможностей организма. Доставка одного и того же количества кислорода требует повышенного уровня деятельности органов дыхания и кровообращения, возрастания напряженности кислородного режима. Перестройка под влиянием новой среды координационных взаимоотношений тех функций, которые ответственны за поддержание гомеостаза организма, уменьшает эффективность и экономичность кислородного режима. Функциональные сдвиги в центральной нервной системе часто вызывают нарушение различных элементов координационной структуры движений. Все это увеличивает кислородный запрос при стандартной работе, требует большого расхода энергии.
Увеличение кислородного запроса и снижение рабочего уровня потребления кислорода при физической работе, неадекватность моторно-висцеральных координаций новым условиям обусловливают более выраженную в среднегорье двигательную гипоксию. Увеличение кислородного долга — одна из главных причин снижения показателей спортивной работоспособности и выносливости человека в горах, особенно при выполнении физических упражнений субмаксимальной и большой мощности. Однако в последующем отрицательное действие разреженной атмосферы на спортивную работоспособность уменьшается.
Скорость восстановления работоспособности зависит от состояния тренированности спортсменов, их горного стажа, характера двигательной деятельности в период пребывания в среднегорье, особенностей вида спорта. Сравнительно быстро повышается работоспособность в скоростно-силовых упражнениях, выполняемых в течение нескольких секунд. Например, на З-й неделе тренировок в среднегорье функциональные возможности гимнастов позволяют выполнять нагрузку, равную по объему и интенсивности тренировке в условиях на уровне моря. У пловцов же, бегунов на длинные и сверхдлинные дистанции при более длительных, чем 3 недели, тренировках в среднегорье не восстанавливается полностью исходный уровень работоспособности, который отмечался до переезда в горы.
Одним из важных механизмов адаптации и мышечной деятельности в начальный период пребывания в горной местности является увеличение альвеолярной вентиляции сверх исходных показателен. Это увеличение компенсирует недостаточное содержание кислорода в воздухе, но вместе с тем приводит к более быстрому выведению углекислого газа, нарушению кислотно-щелочного равновесия в сторону гипокапнии и даже к алкалозу, смещению кривой диссоциации
464
оксигемоглобина влево, т. е. уменьшению возможности отдавать кислород тканям организма. Кратковременно возникающий алкалоз может компенсироваться организмом за счет усиленного образования кислых продуктов, в частности за счет усиления анаэробного гликолиза.
При длительном воздействии специфических факторов горного климата в организме возникают иные пути приспособления, обусловленные изменением тканевой адаптации. При этом увеличивается артерио-венозная разность и меняется в лучшую сторону соотношение свободного и фосфорилирующего окисления — большая часть кислорода используется непосредственно на мышечную деятельность. Соответственно этому перестраиваются и регуляторные механизмы систем, обеспечивающие ткани кислородом.
Благодаря этим приспособительным реакциям повышаются функциональные возможности организма, эффективность и экономичность кислородного режима и биоэнергетика мышечной деятельности в целом, улучшается способность организма переносить кислородный долг и работать в неблагоприятных условиях.
Процесс адаптации к мышечной деятельности в среднегорье проходит ряд этапов, характеризующихся вначале понижением, а затем постепенным возрастанием спортивной работоспособности (в ряде случаев до исходного состояния и даже выше).
Высокий уровень работоспособности сохраняется в течение некоторого времени и после возвращения спортсменов в условия равнины. Как показали специальные наблюдения, во многих случаях спортсмены после тренировок в среднегорье устанавливали в условиях равнины личные, национальные и мировые рекорды.
Влияние климата высокогорья на организм человека. Акклиматизация. В высокогорье создаются весьма сложные условия для жизнедеятельности организма. При восхождении на большие высоты парциальное давление кислорода уменьшается до величин, при которых резко снижается насыщение крови кислородом. Чтобы сохранить в этих условиях достаточное снабжение тканей кислородом, организм мобилизует на «борьбу за кислород» физиологические реакции.
Основная реакция организма на влияние высоты заключается в усилении дыхания, увеличении минутного объема дыхания и в особенности альвеолярной вентиляции. Минутный объем дыхания возрастает пропорционально высоте. Под влиянием хеморецепторов синокаротидной зоны и аортальной, обладающих чувствительностью к снижению содержания кислорода в артериальной крови, возбуждается дыхательный центр; одновременно происходит усиление функций кровообращения.
В результате гипервентиляции напряжение кислорода в альвеолах падает в меньшей степени, чем это происходило бы при отсутствии усиленного дыхания. Но вследствие этого из легких в избыточном количестве удаляется углекислота, что затрудняет поддержание кислотно-щелочного равновесия и уменьшает степень утилизации кислорода из крови тканями.
16—113
465
Повышение функций сердечно-сосудистой системы характеризуется увеличением частоты сердечных сокращений, ускорением кровотока, снижением венозного давления, повышением артериального давления, способствующими улучшению кровоснабжения тканей.
Одной из особенностей кровообращения при пребывании на больших высотах является наличие легочной гипертензии (повышение кровяного давления в малом круге кровообращения), что затрудняет работу правого отдела сердца. При резком подъеме на значительную высоту может возникнуть отек легких. По мнению ряда исследователей, это происходит вследствие увеличения давления в малом круге кровообращения и повышенной проницаемости капилляров из-за недостаточного снабжения кислородом.
В результате гипоксии изменяется и деятельность различных анализаторов-, снижается острота зрения и слуха, уменьшается поле зрения, ослабляются восприятие цветности и аккомодация, нарушается баланс мышц глаза, понижается тактильная чувствительность, повышается болевая чувствительность.
Нервная система человека весьма чувствительна к недостатку кислорода, в особенности кора больших полушарий. Снижение насыщения крови кислородом приводит к возникновению фазовых изменений электрической активности коры головного мозга-, прежде всего обнаруживается активизация высокочастотных колебаний — происходит начальное возбуждение корковых нейронов, затем доминируют медленные волны высокой амплитуды — развивается процесс торможения и, наконец, происходит резкое угнетение биоэлектрической активности. Таким образом, на больших высотах нарушается баланс между возбуждением и торможением, снижается подвижность нервных процессов, страдает внутреннее торможение, особенно тонкие дифференцировки.
Гипоксия значительно влияет и на вегетативную нервную систему, нарушая нормальное равновесие между влияниями симпатического и парасимпатического отделов.
В результате столь значительных изменений функций организма, наблюдаемых в первые дни пребывания в высокогорье, существенно снижается умственная и физическая работоспособность человека.
Первые признаки недостаточной адаптации к высотной гипоксии проявляются в виде повышенной возбудимости нервной системы. Так, возникает первоначальная эйфория, выражающаяся, в частности, в ложном ощущении повышенных возможностей организма. Могут быть эмоциональные расстройства. Через некоторое время состояние первоначального возбуждения переходит в депрессию, которая сопровождается опасными приступами раздражительности, сонливостью. Ранними признаками являются и нарушения координации произвольных движений, внимания.
При более выраженной высотной гипоксии наблюдается ряд расстройств физиологических функций, известных под названием «горной болезни». Ее проявления разнообразны: одышка, приступы
466
удушья, цианоз и бледность кожных и слизистых покровов, ощущение сердцебиения, пульсации сосудов, носовое кровотечение, голо водружение, тошнота, рвота, нарушение сна. Выраженность этих симптомов зависит от степени тренированности, быстроты перемещений с одной высоты на другую, индивидуальных особенностей устойчивости к гипоксемии.
Проявление горной болезни при длительном пребывании на высоте уменьшается в результате акклиматизации. Под акклиматизацией к высокогорному климату понимают прежде всего приспособление, повышение стойкости организма к недостатку кислорода. Это происходит благодаря: 1) повышению доставки кислорода к тканям и 2) приспособлению тканей к существованию при пониженном содержании кислорода.
Адаптация к высокогорью характеризуется увеличением количества эритроцитов до 7—8 млн. в 1 мм3 крови, кислородной емкости крови — с 19—20 до 23—25 об. %. Благодаря этому кровь может транспортировать больше кислорода. В мышцах становится больше миоглобина. Наблюдается также повышение васкуляризации тканей, активности окислительно-восстановительных ферментов, резистентности тканей к различным повреждающим воздействиям.
В акклиматизации большую роль играют центральная нервная система, симпато-адреналовая система и адаптивные гормоны системы гипофиз — кора надпочечников.
Процессу акклиматизации способствует предсезонная усиленная физическая подготовка, ступенчатый подъем на большие высоты с соответствующими мышечными нагрузками и рациональное питание.
По мере акклиматизации работоспособность возрастает, но продолжает оставаться ниже исходного уровня, наблюдаемого на уровне моря. Подъемы свыше 7000 м сопровождаются резкими функциональными изменениями, особенно со стороны сердечной и дыхательной систем, и требуют от спортсмена максимального волевого напряжения. В этих условиях физическую работу значительно облегчает пользование кислородным прибором.
При длительной адаптации к высокогорью, наблюдаемой у местных жителей, отмечается снижение уровня вентиляции легких по отношению к первоначальным величинам, дыхание становится более глубоким, возрастает остаточный объем воздуха, что обусловливает менее выраженное снижение напряжения кислорода в альвеолах. Увеличивается минутный объем крови, умеренно снижается общее периферическое сопротивление артериальных сосудов, повышается уровень среднего артериального давления, венозного давления, уменьшается скорость движения крови. Количество эритроцитов в крови может возрастать до 6—7 млн. в 1 мм3. Со стороны кардиодинамики обнаруживается синдром регулируемой гиподинамии миокарда как показателя повышенного коэффициента полезного действия миокарда. Электрокардиографические данные указывают на развитие у коренных жителей высокогорья гипертрофии правого желудочка сердца.
467
16*
Часы суток
Условные обозначения:
@ Время обеда [о] Время тренировки
Рис. 124. Уровень двигательных функций при изменении времени тренировок и обеда в режиме дня (по С. Г. Харабурга)
§ 2. Ритмические изменения функциональной активности организма
Функциональная активность организма человека ритмически изменяется через определенные периоды. Ритмы функционирования организма, получившие название биологических ритмов, имеют различные периоды: сердечный ритм — секунды и доли секунды; дыхательный— несколько секунд; ритмика желудочной и кишечной секреции — часы; смена состояний бодрствования и сна —сутки; созревание яйцеклетки у женщин — десятки суток и т. д. Наиболее изучены суточные, или циркадные (от лат. cirke — приблизительно, около, dies — день), ритмы, период которых равен в среднем 24 часам.
Общие закономерности циркадных ритмов. Период циркадных ритмов приблизительно равен периоду вращения Земли вокруг своей оси. Циркадные ритмы присущи всем живым организмам, начиная от одноклеточных, и являются наследуемым свойством этих организ* мов. В многоклеточных организмах они поддерживаются на разных уровнях (клетка, орган, целый организм). Фазы ритмов могут быть сдвинуты при изменении внешних факторов, играющих роль синхронизаторов. Ритмы клеток, отдельных органов и систем синхронизируются с ритмами ведущих центров, локализующихся в головном мозгу. Согласно гипотезе Н. Клейтмана, циркадные ритмы различных органов привязываются к ритму сон — бодрствование, который должен рассматриваться как своеобразный ритм-ключ.
Факторы, лежащие в основе формирования биологических ритмов. Для человека первенствующее значение имеет режим труда — отдыха. Этот режим может изменяться в связи с особенностями производственной деятельности (ночные смены, дежурства и пр.) или при сдвиге по фазе светлого и темного времени суток (перемещение на несколько часовых поясов по географической долготе, например при перелете из Европы в Америку или от западной границы СССР к берегам Тихого океана). Изменение режима труда — отдыха может приводить к временному снижению работоспособнос
468
ти, а иногда к появлению ощущения недомогания. Это объясняется нарушением координации в работе отдельных органов и систем органов.
Для синхронизации циркадных ритмов функций разных систем органов в новых условиях (например, после быстрого перемещения с запада на восток) требуется определенный период. Он неодинаков для разных функций. Так, для ритма сон — бодрствование составляет от 2 до 9 дней; для ритма экскреции воды, калия и натрия — 6 дней и т. д. Люди, тренированные к частым сменам режима труда — отдыха (летчики, машинисты локомотивов и др.), адаптируются к новым условиям жизни быстрее.
Работоспособность в разные часы суток. В течение суток двигательные возможности людей возрастают и снижаются в полном соответствии с циркадным ритмом. В ночные часы суток у спортсменов отмечается значительное ухудшение результатов.
Периодические изменения работоспособности спортсменов на протяжении дневного времени в значительной мере определяются режимом тренировок. Наиболее высокий уровень различных показателей работоспособности (рис. 124) бывает в часы тренировочных занятий, наиболее низкий — в обеденное время (С. Г. Харабуга).
Изменение режима тренировок приводит вначале к выравниванию показателей, наблюдаемых в течение дня, с последующей полной перестройкой в соответствии с новым режимом. Для такой полной перестройки необходим срок примерно до 3 недель. Уменьшить его до 2 недель можно, увеличив нагрузку и интенсивность мышечной работы.
Работоспособность спортсменов изменяется также и от месяца к месяцу, и от сезона к сезону, т. е. имеются биоритмы с длительными периодами. Однако изучены они недостаточно, поэтому нет еще достаточных научно обоснованных предпосылок для использования этих биоритмов в тренерской практике.
В основе механизма действия главного синхронизатора суточных ритмов организма и ритмов изменения работоспособности в дневные часы лежат условные рефлексы на время, образующиеся в соответствии с режимом труда и отдыха человека.
Глава XXII
ФИЗИОЛОГИЧЕСКОЕ ОБОСНОВАНИЕ МАССОВЫХ ФОРМ оздоровительной физической культуры и производственной гимнастики
§ 1. Значение физических упражнений в условиях современной жизни
Развитие двигательных и вегетативных функций организма у детей и совершенствование и поддержание их на высоком уровне у взрослых и пожилых лиц обязательно требуют наличия двигатель
469
ной активности. Как известно, при современном уровне развития науки и техники производственный труд (управление и наблюдение за механизмами, научная, конторская и административно-хозяйственная работа и др.), ведение домашнего хозяйства, передвижения (механизированный транспорт, лифт и т. п.) не связаны со значительными мышечными наряжениями. Количество лиц, живущих в условиях недостаточной двигательной активности, с каждым годом увеличивается.
§ 2. Влияние на функции организма недостаточной двигательной активности
Недостаточность мышечных напряжений на производстве, в быту и при передвижении отрицательно влияет на физиологические функции. Об этом говорят как опыты на животных, так и наблюдения над людьми. Некоторые животные, поставленные в условия полного прекращения движений (акинезии — от греч. kinema — движение, а —частица отрицания) или резкого ограничения их (гипокинезии — от греч. hypo — приставка, указывающая на понижение). через несколько дней или недель погибают, у других же наблюдаются выраженные отрицательные изменения морфологической структуры тканей и функциональных свойств организма. Человек может жить и при отсутствии движений, но это приводит к значительному ухудшению его двигательных и вегетативных функций.
Функции двигательного аппарата. Гипокинезия всегда сопровождается атрофией и дегенерацией (от лат. degenera-ге—перерождение) скелетных мышц. Мышечные волокна становятся тоньше, вес мышц уменьшается. Об этом говорят, в частности, клинические наблюдения над больными при полиомиелите, накладывании на длительное время гипсовых повязок, ограничениях подвижности в суставах и т. д.
При полном выключении сократительной деятельности мышечных волокон в опытах на животных выявлено резкое ухудшение функциональных свойств мышц. Так, после 30 дней полного прекращения деятельности мышечная сила снизилась до */з исходной величины, длительность же цикла одиночного сокращения увеличилась в 1,5—2 раза (3. И. Барбашева, Е. К. Жуков).
Наряду с уменьшением силы и скорости при ограниченной подвижности у человека снижается и выносливость.
Нервная система и двигательные функции. Существенные изменения при гипокинезии происходят в деятельности нервной системы и сенсорных систем, проявляющиеся в возникновении при неврологическом исследовании ряда симптомов, не наблюдавшихся до гипокинезии. Это — расстройства двигательных функций (например, увеличение амплитуды колебаний центра тяжести и нарушение координации движений при ходьбе). Видимо, они объясняются не только изменением состояния самих мышц, но и нарушением координации их деятельности нервной системой.
Вегетативные функции. В результате длительной гипо-
470
кинезни происходят выраженные изменения в системе кровообращения: уменьшаются размеры сердца, снижается ударный и минутный объем крови, учащается пульс, уменьшается масса циркулирующей крови, удлиняется время ее кругооборота.
У спортсменов, для которых характерна интенсивная мышечная деятельность, при временном снижении двигательной активности до гипокинезии изменения в сердечно-сосудистой системе выражены । меньше, чем у спортсменов с меньшей двигательной активностью и в особенности у нетренированных лиц.
При гипокинезии в покое внешнее дыхание характеризуется уменьшением объема легочной вентиляции, а основной обмен — снижением на 5—20%- Во время мышечной работы ухудшается экономичность вегетативных функций, вследствие чего при тех же самых мышечных нагрузках увеличиваются как кислородный запрос, так и кислородный долг. Наблюдается также снижение функций эндокринных желез, в частности надпочечников.
Длительная неподвижность (например, при строгом постельном режиме) постепенно приводит в результате гипокинезии к резкому сглаживанию суточного ритма пульса, температуры и некоторых других функций организма. Но если для этих лиц вводятся ежедневные получасовые-часовые занятия физическими упражнениями, то проявления гипокинезии или не развиваются, или оказываются менее выраженными. Таким образом, физические упражнения являются эффективным средством, предохраняющим организм от неблагоприятных последствий недостаточной производственной и бытовой двигательной активности.
§ 3. Влияние на функции организма повышенной двигательной активности
Оздоровительное значение физической культуры общеизвестно. Имеется огромное количество исследований, показывающих поло-^ жительное влияние физических упражнений на опорно-двигательный аппарат, центральную нервную систему, функции кровообращения, дыхания, выделения, обмен веществ, теплорегуляцию, деятельность органов внутренней секреции. В результате физической тренировки значительно улучшается координация нервной системой двигательных и вегетативных функций; повышаются функциональные возможности многих органов и систем органов, в некоторых случаях в несколько раз. Например, при мышечных напряжениях у выдающихся спортсменов минутный объем крови может достигать 30—40 л, минутный объем легочной вентиляции — 150—180 л, максимальное поглощение кислорода — 5—G л. Повысившиеся функциональные возможности отдельных органов и систем органов человека позволяют значительно легче справляться с повышенными требованиями, предъявляемыми к сердечно-сосудистой, дыхательной и другим системам организма.
Велико значение физических упражнений как средства лечения, особенно при заболеваниях опорно-двигательного аппарата.
471
§ 4. Роль физических упражнений в повышении устойчивости организма к действию неблагоприятных факторов
ет к сеое в последние двадцать лет
Рис. 125. Максимальная длительность стандартной работы (ходьба по лесенке в заданном темпе — от 90 до ПО шагов в 1 мин.—«до отказа») в 3 сериях опытов (1, 2, 3) у нетренированных лиц и спортсменов при высокой температуре среды и 100%-ной влажности воздуха (по О. Г. Крюкову) .
Столбики — средние величины; белые прямоугольники внутри столбиков — диапазон индивидуальных колебаний. Цифры над столбиками — количество шагов в I мин., под столбиками — температура воздуха
Изменения в режиме мышечной деятельности могут отражаться как на отдельных двигательных и вегетативных функциях организма, так и на общей его устойчивости (резистентности) при действии неблагоприятных факторов внешней среды. Этот вопрос привлека-особое внимание.
Под влиянием сильных (чрезвычайных) раздражите лей в организме возникает напряжение, обозначаемое английским термином стресс. При этом развивается комплекс изменений (см. гл XIV), названный Г. Селье общим адаптационным синдромом (от греч. sindrome — совокупность, сочетание симптомов). Из трех стадий стресса патологические изменения в организме наблюдаются при первой (реакция тревоги) и третьей (истощение) стадиях. Вторая же стадия, повышающая устойчивость организма как к данному стрессору, так и к ряду других, представляет собой физиологическое явление.
Важнейшей особенностью влияния мышечных напряжений является то, что при постепенном увеличении нагрузок реакция тревоги проявляется слабо или отсутствует. В организме после нескольких тренировочных занятий начинает развиваться состояние повышенной резистентности как к специфическим, т. е. к тому же фактору, например мышечным нагрузкам, так и к неспецифическим, т. е. к целому ряду других неблагоприятных, воздействиям на организм. Третья стадия стресса — истощение— возникает только при чрезмерных для данного организма нагрузках. Таким образом, мышечная работа при весьма большом диапазоне нагрузок оказывает на организм только положительный эффект.
В частности, как было выявлено в опытах на животных и при наблюдениях над людьми, в результате мышечной деятельности неспецифически повышается устойчивость организма ко многим неблагоприятным воздействиям, которым подвергается человек в ус
472
ловиях современной жизни, например к действию гипоксии, некоторых ядов, радиоактивных веществ, инфекций, перегреванию (рис. 125), охлаждению и др. Установлено значительное снижение заболеваемости у лиц, занимающихся физической культурой и спортом (табл. 19).
Таблица 19
Данные о заболеваемости на одном из заводов Свердловска у занимающихся (3) и не занимающихся (НЗ) спортом (по В. В. Розенблату)
Показател ь Число заболеваний (%) Колич. дней нетрудоспособности Средняя длительность заболевания (колич. дней)
3 НЗ 3 НЗ 3 НЗ
Общая заболеваемость 53,7 79,1 318 731 5,9 9,2
Простудные заболевания органов дыхания 35,0 44,7 164 248 4,6 5,6
Травмы 6,0 10,9 82 166 13,7 15,2
Таким образом, рациональные занятия физическими упражнениями являются важным средством предохранения организма человека от ухудшения работоспособности при действии неблагоприятных факторов.
§ 5. Физиологическая характеристика утренней гигиенической гимнастики
Одной из наиболее регулярных и массовых форм оздоровительных занятий физическими упражнениями является утренняя гигиеническая гимнастика, представляющая собой важный элемент личной гигиены каждого человека на всех этапах возрастного развития.
Такая утренняя зарядка имеет разностороннее физиологическое значение. Она способствует, во-первых, более быстрому повышению работоспособности человека после сна, во-вторых, укреплению здоровья и закаливанию организма, в-третьих, совершенствованию путем регулярной физической тренировки мышечного аппарата, сердечно-сосудистой, дыхательной и других систем организма и развитию физических качеств, координации движений. В условиях все возрастающей в современном обществе гипокинезии утренняя гимнастика приобретает значение важного фактора укрепления здоровья и повышения работоспособности человека.
Ускорение полного восстановления работоспособности после сна. Во время сна интенсивность физиологических процессов в организме резко снижается, обмен веществ становится минимальным, температура тела несколько понижается,
473
ритм сердечных сокращений и дыхательных движений урежается, минутный объем сердца и объем легочной вентиляции уменьшаются, мышечные движения прекращаются совершенно или сводятся к минимуму. В тканях и органах человека, находящегося в состоянии сна, ослабляется циркуляция межтканевой жидкости, вследствие чего иногда обнаруживаются застойные явления, проявляющиеся, например, в виде отечности.
Нервная система во время сна, как известно, находится в состоянии торможения, в связи с чем многие нервные центры становятся недостаточно возбудимыми при обычной интенсивности действия световых, звуковых, тактильных, температурных, обонятельных и других раздражителей.
После пробуждения тормозное состояние в центральной нервной системе проходит не сразу и может проявляться спустя десятки минут и даже в первые часы после пробуждения. В связи с этим сразу после пробуждения пороги чувствительности анализаторов всегда повышены, производительность же умственной и физической работы несколько понижена.
Повышение возбудимости нервной системы и улучшение работоспособности после сна происходит тем быстрее, чем значительнее импульсации, поступающая из рецептивных зон в нервную систему. Вследствие этого утреняя гигиеническая гимнастика, сопровождающаяся поступлением в центральную нервную систему мощного потока импульсов от рецепторов многих сенсорных систем в особенности от проприоцепторов, способствуют быстрому повышению возбудимости центральной нервной системы и восстановлению нормальной работоспособности.
Если утренняя зарядка сочетается с применением воздействия на кожные рецепторы температурных раздражителей (например, в виде холода и водных процедур), то возбудимость нервной системы восстанавливается еще быстрее. Действие некоторых раздражителей внешней среды (температурный фактор, водные процедуры, действие свежего воздуха и солнца) наряду с повышением возбудимости нервной системы оказывает закаливающий эффект.
Утренняя гимнастика благодаря вовлечению в действие мышечных групп всех частей тела содействует усилению лимфообращения и тем самым быстрому устранению (у лиц с нормальным состоянием сердечно- сосудистой и выделительной систем) отечности тканей, в частности век, наблюдаемой иногда сразу после пробуждения.
Регулярность проведения утренней гимнастики является одной из предпосылок не только для сохранения достигнутого уровня физического совершенствования, но и для дальнейшего его повышения.
В связи с тем что утренняя гимнастика совершается до начала рабочего дня, интенсивность ее не должна быть чрезмерной, иначе она может ухудшить последующую работоспособность. Большой объем и высокая интенсивность утренней зарядки могут вызвать значительное утомление центральной нервной системы и снижение умственной и физической работоспособности после выполнения физических упражнений. Однако это в полной мере не относится к
474
тренированным лицам, так как благодаря систематической тренировке функциональные возможности человека в значительной степени совершенствуются. Выполнение ими утром даже таких упражнений, как бег, ходьба на лыжах, езда на велосипеде, верховая езда и плавание, не только не снижает, но и улучшает последующую работоспособность в течение дня.
§ 6. Физиологическая характеристика производственной гимнастики
Существенное влияние на организм и его работоспособность оказывают физические упражнения, проводимые в различных формах, непосредственно на производстве, в учреждениях, учебных заведениях. Эти упражнения, называемые производственной гимнастикой, проводятся как перед началом работы (вводная гимнастика), так и во время обеденного перерыва или специально организуемых перерывов (физкультпаузы). Систематические такие занятия способствуют производительности труда.
По своему физиологическому значению физкультпаузы являются видом активного отдыха, проводимого в форме гимнастических упражнений, спортивных игр и др. Как показано еще И. М. Сеченовым, физиологический механизм активного отдыха заключается в переключении с деятельности одних нервных центров, утомляющихся при работе, на деятельность других центров, связанных с регуляцией движений при физических упражнениях. Происходящее при этом углубление торможения в центрах работавших мышц способствует снятию утомления и повышению производительности труда.
Важное значение физические упражнения имеют для устранения застойных явлений, наблюдаемых при длительном пребывании в часы трудового дня в неподвижном или малоподвижном состоянии. Изменяются при регулярном применении физкультпауз показатели общего физического состояния организма: увеличивается мышечная сила, несколько повышается количество гемоглобина в крови и т. д.
Сочетание физических упражнений на производстве с закаливанием (в частности, с выполнением их на свежем воздухе) повышает сопротивляемость организма ко многим неблагоприятным факторам.
Производственная гимнастика особенно необходима в тех видах промышленности( химическая и др.), в которых организм человека подвергается различного рода неблагоприятным воздействиям.
Влияние физических упражнений на последующую работоспособность. Занятия физическими упражнениями во время работы, как и утренняя гигиеническая гимнастика, оказывают влияние на последующую работоспособность. Физиологическая сущность этого явления заключается в том, что под влиянием увеличения импульсации, поступающей из двигательного, висцерального, вестибулярного и других анализаторов, и изменения интенсивности секреции некоторых гормонов повышается возбудимость нервной системы до уровня, при котором работоспособность ее становится выше. С другой стороны, по физиологическому механизму активного отдыха бы-
475
стрее происходит восстановление нарушенных в результате развития утомления функций организма, связанных (^производственным трудом. /
Влияние физических упражнений, проводимых во время работы, на производительность труда зависит от целбго ряда факторов. Интенсивность и длительность физических упражнений перед работой или в интервалах трудового дня должны
соответствовать физической подготовленности
Рис. 126. Влияние бега перед работой на последующую работоспособность радиотелеграфистов: на передачу смыслового текста, многократную передачу стандартной комбинации из четырех букв (АПРО) и скорость постукивания — теппинг-тест (По Ю. К. Демьяненко)
занимающихся. Показано, что у хорошо подготовленных в этом отношении людей работоспособность улучшается даже после таких упражнений, как бег 30 мин. на 6 км и 60 мин. на 9 км.
Физические упражнения повышают последующую работоспособность (рис. 126) в сложной умственной деятельности (например, при передаче радиотелеграфистами.
смыслового текста) больше, чем при менее сложной работе (много-
кратная передача расположенных в стандартном порядке четырех букв или скорость постукивания). Результаты многочисленных исследований по вопросу о положительном влиянии специальных форм проведения физических упражнений на умственную работу свидетельствуют о большой целесообразности широкого использования лицами умственного труда вводной гимнастики и физкульт-
пауз.
Существенное значение имеет эмоциональность занятий. При прочих равных условиях такие эмоциональные упражнения, как спортигры, повышают работоспособность быстрее, чем гимнастические упражнения или бег. Однако если такие эмоционально насыщенные упражнения длятся долго, то влияние их на последующую производственную работу падает и они могут даже привести к понижению работоспособности.
Вводные производственные физические упражнения и физкульт-паузы полезны всегда. Но наибольшую эффективность в отношении повышения производительности труда они имеют в ранние утренние, поздние вечерние и ночные часы, когда тонус нервной системы несколько понижен.
В ряде экспериментов показано, что эффект от применения физических упражнений перед занятиями или в перерывах между ними продолжителен — не менее 2 часов, а в некоторых случаях — до 5 часов. . ......
476
Глава XXIII
ХАРАКТЕРИСТИКА ВОЗРАСТНЫХ ОСОБЕННОСТЕЙ
ФУНКЦИИ ОРГАНИЗМА ДЕТЕЙ ШКОЛЬНОГО ВОЗРАСТА
В СВЯЗИ С ФИЗИЧЕСКИМ ВОСПИТАНИЕМ
§ 1. Общие положения
В процессе развития организма человека происходит неравномерное развитие структур и функций различных его органов и тканей. Знание закономерностей этого развития имеет весьма существенное значение для рационального построения физического воспитания людей различного возраста. Теоретическое и прикладное значение эволюционного подхода в физиологии было сформулировано Л. А. Орбели. Он подчеркивал роль не только социальной, но и природной среды. Без должного внимания к этим условиям невозможно рациональное физическое воспитание. Педагогу необходимо учитывать возрастную ступень эволюции функций, на которой находятся его ученики, поскольку каждая фаза возрастного развития неповторима в жизни человека и требует соответствующего подхода к методам и средствам обучения.
В раннем периоде жизни человека преобладают процессы формирования тканей и образования новых координационных отношений в системах организма.
' С самого начала обучения детей в школе (с 7 лет) происходит сложный процесс сочетания особенностей возрастного развития с Характером обучения. За время обучения в школе организм ребенка претерпевает весьма значительные изменения, постепенно переходя из периода детского возраста в подростковый (мальчики — 13— 16 лет и девочки — 12—15 лет) и затем в юношеский возраст, завершающийся уже после окончания школы (юноши — 17—21 год, девушки — 16—20 лет). Для решения различных прикладных вопросов целесообразно пользоваться распределением детей на возрастные группы в соответствии с особенностями обучения в школе: 1) младший школьный возраст — с 7 до 11 лет, 2) средний школьный возраст— с 12 до 15 лет, 3) старший школьный возраст — с 16 до 19 лет.
Младший школьный возраст — это период детства, в котором происходят сложные процессы подготовки к половому созреванию и формируются начальные двигательные навыки в процессе систематического обучения. Средний школьный возраст совпадает с подростковым периодом — период начальной фазы полового созревания, когда завершается преобразование морфологических и функциональных особенностей организма детей с некоторым приближением к нормам взрослых. В старшем школьном возрасте у юношей отмечается еще большее приближение к функциям организма взрослых людей.
477
Возрастные периоды для сдачи норм комплекса/ГТО соответствуют этой классификации. Например, нормы многоборья комплекса ступени «Спортивная смена» сдаются детьми среднего школьного возраста, а ступени «Сила и мужество» — детьми старшего школьного возраста. '
При определении границ между возрастйыми периодами необходимо учитывать схематичность приведенных особенностей возрастных функций у детей. Эта схематичность связана с тем, что у них имеются значительные индивидуальные различия, при которых «физиологический» возраст и «паспортный» могут не совпадать. Чаще это обусловлено ускорением (акселерацией) физического развития по сравнению с паспортным возрастом. Наряду с ускорением физического развития происходит более раннее ускорение развития психической сферы. Акселерация проявилась особенно отчетливо в последнем столетии под влиянием резко изменившихся условий жизни людей. Такая «вековая» динамика (секулярный тренд, по Дж. Таннеру), несомненно, продолжается и в настоящее время, чему особенно способствует дальнейшее улучшение материального благосостояния людей.
§ 2. Развитие движений
На фоне безусловнорефлекторных движений ребенок начинает приобретать условнорефлекторные. Уже в первые дни после рождения формируются новые двигательные акты. Так, к 9-му дню возникают реакции на комплексные (тактильно-проприоцептивно-лабиринтные) раздражители, связанные с различными положениями тела при кормлении (М. М. Кольцова). В дальнейшем на протяжении первых 3 месяцев образуются и другие новые двигательные условные рефлексы, связанные с раздражением тактильного, двигательного, вестибулярного и зрительного анализаторов.
С 3 месяцев начинается целенаправленное развитие движений рук (захватывание предметов). К 5—6 месяцам появляется способность брать предмет одной рукой, а не двумя и удерживать одновременно каждой рукой по предмету. Со 2-го месяца ребенок поднимает голову лежа на животе, и до 6—7 месяцев осваивает подготовительные к ползанию движения — может вставать на четвереньки и к 8 месяцам обычно уже свободно ползает. В процессе ползания развивается мускулатура, удерживающая туловище при сидении, а затем и мускулатура, обеспечивающая сохранение равновесия тела при стоянии. После 8 месяцев начинается ранний период подготовки к стоянию и ходьбе. Однако в связи с недостаточным развитием силы мыши нижних конечностей и стопы нецелесообразно форсированное обучение сидению и стоянию. Это может привести к патологическим деформациям в различных звеньях тела — к нарушениям осанки и неправильному формированию свода стопы.
Ходьба. На протяжении примерно второго полугодия жизни у детей формируются предлокомоторные навыки. Важное значение
478
Время бега, сек.
Рис. 127. Изменение скорости бега у лиц разного возраста (по А. И. Васютиной, В. М. Король, А. Б. Тамбие-вой, В. С. Фарфелю и Л. Н. Жданову):
/ — 7—8 лет (50—70 м); II — 9—10 лет (70 и 100 м). III —11—12 лет (100 м). IV —13—14 лет (100 м). V — 15—16 лет (100 м), V/-17—18 лет (100 м), VII — 19—29 лет (100 м, мастера спорта)
имеют упражнения в ходьбе с поддержкой, которая часто начинается уже с б-месячнрго возраста. Самостоятельная ходьба в этом возрасте (без посторонней помощи, с наклоненным вперед туловищем, с согнутыми в коленях ногами) значительно отличается от ходьбы детей школьного возраста.
Длина шага у ребенка 4 лет равна 39,0—40,5 см, к 7 годам она достигает 51,7—53 см, а к 14 годам — 64,3—73,9 см. С возрастом скорость обычной ходьбы увеличивается. До 8 лет это в первую очередь связано с удлинением шага, а после 8 лет — с повышением темпа движений при постепенной стабилизации длины шага.
При обучении ходьбе с самого начала важно обращать внимание на правильную осанку, учить свободному, непринужденному шагу с параллельной постановкой стоп, с четким «перекатом» с пятки на носок и с конечным отталкиванием передней частью стопы.
Бег. Элементы полета, отличающие бег от ходьбы, наблюдаются уже у ребенка в возрасте около 2 лет. Полетное время в период с 2 до 10 лет увеличивается более чем в 2,5 раза. Это увеличение особенно выражено в возрасте от 2 до 5 лет. Максимальная скорость бега отчетливо повышается с возрастом. Так, у детей 7 лет она равна в среднем
4,55 м/сек, 10-—11 лет — 5,37 м/сек, 12—13 лет — 5,78 м/сек, 14— 15 лет — 6,07 м/сек, 15—16 лет — 7,59 м/сек, у 17—18-летних — 8 м/сек (у мастеров спорта в возрасте 19—29 лет — 9,77 м/сек). Скорость бега свыше 8 м/сек достигается только путем специальной тренировки в спринте, путем развития координационных механизмов центральной регуляции нервно-мышечных функций. Независимо от возраста максимум скорости на протяжении дистанций достигается к 5—6-й сек. (рис. 127).
Метания. Уже годовалый ребенок может не только захватывать предметы одной и двумя руками, но и бросать их. В 3 года дети бросают одной рукой маленький мяч вдаль на расстояние, превышающее 1 м. К школьному возрасту с развитием координации движений и силы мышц, особенно плечевого пояса и рук, способность к метаниям быстро прогрессирует, особенно у мальчиков. Так, по данным В. В. Леви-Гориневской, мальчики 5 лет метают (правой рукой), мешочки с песком весом 150—200 г на расстояние 7,73 м, а в 7 лет — на 10,80 м, девочки соответственно 4,57 и 5,70 м.
479
I
В школьном возрасте продолжает увеличиваться дальность и точность метаний в цель и становится еще заметнее,/различие между мальчиками и девочками, /
§ 3. Развитие физических качеств
Сила. Интенсивное развитие скелета у детей тесно связано с формированием мышц, сухожилий и связочно-суставного аппарата. Если вес мышц ребенка 8 лет отчетливо меньше */з веса тела, то к 15 годам он достигает почти ’/з, а к 17—18 годам — немногим менее половины общего веса тела. Наряду с увеличением веса и изменением физико-химических свойств мышц совершенствуется и их
Рис. 128. Изменение скорости движения, осуществляемого различными группами мышц, у нетренированных лиц разного возраста (по А. В. Коробкову).
Ось ординат — скорость движения (град/сек), ось абсцисс — возраст (в годах); •/ — палец, 2 — кисть, 3 — предплечье, 4 — плечо, 5 — туловище, 6 — шея, 7 — бедро, 8 — голеиь, 9 — стопа
функциональные свойства, обогащаются иннервационные отношения.
Увеличение веса различных мышц происходит неодинаково. Например, в первые 9 лет жизни вес прямой мышцы живота увеличивается в 88,9 раза, а вес наружной косой мышцы только в 66,6 раза. Неодинаково изменяется и вес мышц руки и ноги.
С возрастом увеличивается сила мышц. Так, у мальчиков 7 лет сила правой руки равна в среднем 4 кг, в 11 лет — 10,7 кг, а в 16 лет — 32,3 кг, а к старшему школьному возрасту, как правило, возрастает в 8 раз. Становая сила у мальчиков 8 лет равна в среднем 51 кг, в 11 лет — 64 кг, а в 16 лет — 110,2 кг. Увеличение силы различных мышц происходит избирательно: у детей различной спортивной специализации больше прирост силы наиболее тренируемых мышечных групп, которые играют особенно важную роль для спортивного результата.
480
Рис. 129. Возрастные изменения произвольного максимального темна движения (по Д. П. Букреевой).
Ось ординат — число движений в 1 сек., ось абсцисс — возраст (лет); I — мальчики, 11 — девочки
Увеличение мышечной массы отчасти объясняет прирост силы. Развитие силы зависит не только от морфологических сдвигов, но и от характера поступающего от нервных центров потока импульсов и увеличения числа двигательных единиц, сокращающихся в мышцах, осуществляющих те или иные движения.
Быстрота. Физиологическая природа различных проявлений быстроты — скорости перемещения звеньев тела, количества повторных движений в единицу времени (темп), времени латентного периода реагирования на различные сигналы и скорости перемещения в пространстве (например, при беге, гребле и др.) — весьма сложная.
Большое значение имеют подвижность нервных процессов, сократительные свойства и сила мышц, координация их деятельности. Длительность одиночного движения указательного пальца, кисти руки, предплечья, плеча, шеи, туловища, бедра, голени и стопы значительно возрастает в период от 4—5 до 13—14 лет, приближаясь к величинам, наблюдаемым у взрослых
людей (рис. 128). Затем к 16—17 годам она несколько снижается и к 20—30 годам достигает наибольших значений. Под влиянием тренировки скорость одиночных движений у детей в 9—11 лет изменяется мало, у подростков же в 13—14 лет, как и у лиц 20—30 лет, наблюдается отчетливое ее повышение.
Скорость однократного одиночного движения зависит от ряда причин. Определенное значение имеют биомеханические условия движений в суставах дистальных и проксимальных отделов. Только в раннем возрасте отсутствуют различия в скорости движений в разных суставах, в дальнейшем же движения в суставах дистальных частей тела совершаются быстрее, чем в проксимальных. Увеличение скорости движений в суставах с возрастом объясняется увеличением быстроты протекания физиологических процессов в нервно-мышечном аппарате и сонастраиванием ритмов возбуждения в мышечных группах, осуществляющих двигательный акт.
Тесно связан с быстротой однократного движения темп движений (частота их в единицу времени). Предельный темп при движениях в дистальных суставах выше по сравнению с проксимальными, что аналогично показателю темпа у взрослого человека. Максимальная частота движений с возрастом повышается неравномерно (рис. 129).
481
Наибольшее увеличение темпа движений происходит от 7 до 9 лет — средний годовой прирост равен 0,3—0,6 .Движения в 1 сек. У 10—11-летних он несколько ниже — 0,1—0,2 движения в 1 сек., а в 12—13 лет снова оказывается высоким — 0,3—0,4 движения в 1 сек. С 14 лет ежегодный прирост предельного темпа замедляется и к 16 годам почти совсем прекращается.
У детей отмечаются выраженные индивидуальные различия максимального темпа движений. В пределах одного и того же возрастного периода у одних могут наблюдаться весьма высокие показатели, у других — низкие. Однако имеется тесная связь между максимальным темпом движений разных мышц у одного и того же ребенка, что указывает на зависимость данного показателя от функций центральной нервной системы. Очевидно, это обусловлено способностью нервных центров к быстрой смене возбуждения и торможения, т. е. высокой подвижностью нервных процессов.
Латентные периоды при движениях различных мышечных групп до 13—14 лет укорачиваются, а затем стабилизируются.
Физические упражнения оказывают незначительное влияние на элементарные характеристики быстроты у детей. Очевидно, генетический фектор играет в развитии быстроты большую роль. Вместе с тем быстрота сложных движений при ходьбе, беге, гребле и т. д. зависит от развития двигательных навыков, структуры движений и должной мобилизации вегетативных функций.
Ловкость. Это — комплексное качество, компонентами которого являются точность движений в пространстве и во времени и способность изменять координацию движений в зависимости от внезапной смены внешних условий. Соответственно выделяются три стороны ловкости: первая характеризуется пространственной точностью движений, вторая — временной точностью и третья — точностью и быстротой двигательных решений в ответ на внезапные сигналы (В. С. Фарфель). При этом наиболее важной является способность адекватно приспосабливать свою деятельность к внезапной смене ситуаций. Пространственная и временная точность движений составляет необходимую предпосылку к развитию ловкости.
Точность смещений руки, определяемая по воспроизведению заранее заданного угла, наиболее заметно увеличивается в возрастной период от 7 до 10 лет и менее значительно — от 10 до 12 лет. В дальнейшем существенного изменения точности при простых движениях не обнаруживается. В последнее время отмечена высокая способность детей среднего школьного возраста (не уступающая взрослым) к синхронной точности выполнения работы симметричными частями тела.
При специальной тренировке точность движений при метаниях в цель, точность перемещения своего тела в пространстве (например, при прыжках с места) повышается и после 15—16 лет.
Развитие ловкости продолжается и за пределами школьного возраста, в котором только закладываются основные функциональные характеристики этого важнейшего качества. В раннем возрасте развитию ловкости особенно способствуют занятия играми
482
(П. Ф. Лесгафт) В игровой деятельности дети способны достигать хороших результатов уже в среднем школьном возрасте. В дальнейшем происходит совершенствование мастерства, что может продолжаться до пожилого возраста.
Диапазон проявления ловкости, возможность своевременно пред-: восхищать разнообразные внезапные ситуации зависят от накоплен-, ного двигательного опыта. При достаточном опыте уже начиная со среднего школьного возраста облегчается программирование необходимых ответных действий.
Выносливость к статическим усилиям. У детей школьного возраста выносливость различных мышц к статическим усилиям намного меньше, чем у взрослых. Значительное повышение этого показателя происходит к 13—14 годам. Важное значение имеет статическая выносливость мышц туловища, удерживающих позвоночный столб в правильном положении (так называемый «мышечный корсет»). От состояния этих мышц во многом зависит формирование хорошей осанки у детей, в частности кривизны позвоночника. Развитие статической выносливости мышц стопы и голени играет существенную роль в формировании свода стопы. У некоторых детей статическая выносливость различных мышечных групп тесно взаимосвязана, в чем проявляется центрально-нервная функция, позволяющая осуществить сменную работу функциональных нервно-мышечных единиц, увеличивая тем самым время поддержания той или иной позы, того или иного статического положения конечностей.
Физические качества и двигательные навыки. Возможность выполнения движений всегда тесно связана с развитием физических качеств. Так, например, в ходьбе, беге, прыжках и метаниях всегда в той или иной мере необходимы достаточно развитые сила, быстрота, ловкость и статическая выносливость. В единстве двигательных навыков и физических качеств проявляется не только возрастные особенности двигательной деятельности, но и половые различия. С возрастом все более выраженным становится некоторое ускорение темпа развития навыков и физических качеств у мальчиков по сравнению с девочками. Однако в ряде случаев девочки на некоторых этапах развития опережают мальчиков. Например, в возрасте от 11 до 13 лет у девочек максимальная частота движений, выполняемых по звуковым сигналам, больше, чем у мальчиков. В среднем школьном возрасте четко выявляются особенности движений девочек, характеризующиеся экономичностью, плавностью и мягкостью.
§ 4. Кровь
Кроветворение возникает еще внутриутробно. Функции печени, селезенки, лимфатических узлов и костного мозга достигают значительной активности уже в первые месяцы жизни. Кроветворная функция печени завершается на первом году жизни. На протяжении всего школьного возрастного периода продолжается развитие кроветворной функции селезенки.
483.
К моменту рождения полости всех костей заполнены красным костным мозгом, который в дальнейшем заменяется наполовину желтым костным мозгом. Красный мозг, заполняющий губчатое вещество плоских костей и эпифизы трубчать(х костей, осуществляет кроветворную функцию. Желтый (жировой )мозг, заполняющий к 7-летнему возрасту диафизы длинных трубчатых костей, в обычных условиях не несет кроветворной функции. В течение жизни в зависимости от особенностей питания и нейроэндокринных влияний количественные соотношения между красным и желтым мозгом могут значительно изменяться.
В системе кроветворных органов имеют существенное значение лимфатические узлы, в которых образуются лимфоциты, осуществляющие барьерную функцию. Лимфатические узлы окончательно оформляются как кроветворные органы уже к 12 годам. Таким образом, в начале периода среднего школьного возраста полноценно функционирует уже вся система органов, в которых образуются форменные элементы крови, выполняющие барьерную функцию путем поглощения (фагоцитоз) микробов и остатков погибших клеток.
У детей среднего школьного возраста в покое состав периферической крови такой же, как и у взрослых. Различия в отношении некоторых свойств крови— миогенного лейкоцитоза и др. — обнаруживаются только при мышечной работе. У подростков и юношей миогенный лейкоцитоз характеризуется большей степенью распада лейкоцитов (лойкоцитолизом) при длительных и тяжелых нагрузках. Аналогичная картина наблюдается в сдвиге числа эритроцитов. У юных спортсменов отмечается более выраженное их уменьшение (эритролиз), чем у взрослых. Весьма отчетливы различия в тромбоцитозе, возникающем при тяжелой работе (например, велогонки на 50 км). У юношей 16—18 лет наблюдается вторая фаза миогенного лейкоцитоза, тогда как у взрослых при таких же нагрузках возникает только начальная, т. е. первая лимфоцитарная, фаза. Кратковременные нагрузки у детей школьного возраста вызывают, как правило, такие же изменения числа форменных элементов в крови, как у взрослых. При продолжительных и тяжелых нагрузках миогенный лейкоцитоз и тромбоцитоз, а также изменения в числе эритроцитов (эритроцитоз) у детей наступают раньше и более выражены.
§ 5. Кровообращение
Развитие сердечно-сосудистой системы. Оно тесно связано с функциональными нагрузками и продолжается на протяжении всего школьного периода жизни. Темп роста сердца — более быстрый на первом году жизни. В дальнейшем его ускорение отмечается в возрасте 13—17 лет. Размеры сердца у девочек 12— 15 лет в среднем больше, чем у мальчиков того же возраста.
Объем сердца увеличивается пропорционально его размерам и весу и соответствует динамике физического развития детей. Отме-484
чается определенная связь объема сердца с весом тела, изменяющаяся с возрастом. Так, у новорожденных при весе 3,2 кг объем сердца составляет 22 мл, у детей в конце 1-го года жизни при весе 9,45 кг—42 мл, к концу 7-го года при весе тела 19 кг — 90 мл, у 13—14-летиих при весе 38,7 кг— 130 мл, тогда как у взрослых при весе тела 63,6 кг — 280 мл. (Е. А. Аркин). Уже к концу младшего школьного возраста завершается тканевая перестройка и размеры сердца у детей среднего школьного возраста близки к величине сердца взрослого. У 14-летних нервный аппарат сердца уже не отличается от нервного аппарата сердца взрослого человека (В. И. Пузик).
П риспособление сердечнососудистой системы к физическим нагрузкам происходит у детей иначе, чем у взрослых. Так, при нормальных условиях спортивной тренировки гипертрофия сердечной мышцы у них не достигает той степени, которая часто наблюдается у взрослых спортсменов. Адаптация к циклической длительности и тяжелой работе на выносливость происходит с большими трудностями, зависящими в основном от малой способности сердечной мышцы детей к обеспечению подобной нагрузки. Наблюдавшаяся же в некоторых случаях гипертрофия миокарда (например, у юных лыжников и велосипедистов) возникала в результате форсированной тренировки и отражала влияние чрезмерно длительных нагрузок, которые были не адекватны возможностям аппарата кровообращения у молодых спортсменов.
Частота сердцебиений. Этот показатель в условиях покоя отчетливо снижается у детей школьного возраста.
В начале периода младшего школьного возраста частота сердцебиений значительно больше (до 90 в 1 мин.) по сравнению с началом среднего школьного возраста (до 82 в 1 мин.). В начале же старшего школьного возраста ее показатель в покое приближается к величине взрослых людей (70—76 в 1 мин.). Это урежение частоты пульса обусловлено тем, что с возрастом снижается тонус симпатического нерва и усиливается тонус блуждающего нерва.
Значительное увеличение частоты сердцебиений у детей происходит при интенсивной мышечной работе —нередко до 200 в 1 мин., а иногда и более. Особенно высокая частота сердцебиений бывает при эмоциональном напряжении, соревновательных нагрузках.
Ударный (систолический) и минутный объемы сердца. С возрастом ударный объем сердца в покое увеличивается в большей степени, чем уменьшается частота сердцебиений. Этим обусловлено увеличение минутного объема сердца с возрастом. Так, в возрастной период 6—9 лет ударный объем в покое равен примерно 32 мл, в 10—12 лет — 44 мл, а в 13—16 лет — 59 мл, минутный объем соответственно 2,6, 3,2 и 3,8 л (М. А. Шалков). При предельно тяжелой работе ударный объем сердца также увеличивается с возрастом. У 12-летних он может достигать 104 мл, у 13-летних — 112 мл, у 14-летних— 116 мл. Эти цифры намного меньше показателей предельного увеличения минутного объема сердца у взрослых спортсменов (примерно до 190 мл). Соответственно возрастает и пре-
485
дельный минутный объем сердца при работе. У 12-летних он доходит до 19 л, у 13-летних — до 21 л и у 14-летних — до 22 л (у взрослых спортсменов — до 38 л).
Кровяное давление. У детей в покое артериальное давление обычно ниже, чем у взрослых. К 7—8 годам оно повышается до 99/67 мм рт. ст., к 9—12 годам — до 105/70, к 13—15 годам — до 117/73 и к 16—18 годам — до 120/70 (А. Б. Воловик). После спортивных упражнений отмечается значительный подъем максимального артериального давления— до 150—220 мм рт. ст. (А. И. Вер-хутина-Васютина). Минимальное артериальное давление при нагрузках изменяется мало, и поэтому увеличивается разница между максимальным и минимальным артериальным давлением: от 40— 45 мм рт. ст. в покое до 100—170 мм рт. ст. и более после работы. Эта величина при неизменном сопротивлении в сосудистом русле отражает величину ударного объема сердца.
§ 6. Дыхание
1 На протяжении возрастного периода от 7 до 19 лет происходят структурные изменения в легких. Так, размеры альвеол, почти одинаковые у детей от рождения до 7 лет, становятся больше в 2 раза у 12-летних и в 3 раза у взрослых по сравнению с размерами у новорожденных. Этим достигается значительное увеличение общей ловерхности альвеолярной мембраны.
Общая емкость легких. Измерение жизненной емкости легких (ЖЕЛ) можно проводить примерно с 4 лет. Остаточный объем начиная с 5 лет до 17-летнего возраста составляет примерно 20% от общей емкости легких, т. е. в относительных величинах к общей емкости легких аналогичен показателям его у взрослых Жизненная емкость легких увеличивается с возрастом. Например, у 4-летних она равна в среднем 1100 мл, у 6-летних — 1200 мл, у 10-летних— 1700 мл, у 14-летних — 2500 мл (Н. А. Шалков). Вариативность величины ЖЕЛ по отношению к должной у детей весьма велика. Существует зависимость величины ЖЕЛ от роста ребенка. Дети высокого роста, особенно занимающиеся упражнениями, требующими выносливости (плаванием, бегом, лыжными гонками, греблей и др.), нередко имеют ЖЕЛ более 3—4 л, т. е. до 130—150% от должной величины.
Вентиляция легких. С возрастом становится нсколько меньшей частота дыхательных движений в покое, а дыхательный объем увеличивается со 130—220 мл в 6—8 лет до 340'—450 в 16 лет. В старшем школьном возрасте дыхательный объем и частота дыхания близки к величинам их у взрослых. До 7 лет легочная вентиляция (произведение частоты дыханий на дыхательный объем) в покое у мальчиков и девочек почти одинакова, с 9-летнего возраста она становится отчетливо больше у мальчиков по сравнению с девочками (что соответствует их физическому развитию).
Как показали проведенные в последние годы исследования механики дыхания, у детей растяжимость ткани легкого меньше, чем 486
у взрослого. Высокая упругость легочной ткани плода уменьшается после рождения и с возрастом становится все меньше. Растяжимость легочной ткани, выраженную количеством миллилитров, па которое изменяется объем легких при изменении плеврального давления на 1 см водн. ст., относят обычно к определенной величине легочной вентиляции. Это связано с тем, что с увеличением вентиляции легких уменьшается и растяжимость легочной ткани. С возрастом она значительно увеличивается, однако и в 14 лет ее показатели намного меньше, чем у взрослых. С возрастом снижается величина работы дыхательных мышц — с 0,38 ±0,044 кгм/мин в 9 лет до 0,17±0,32 кгм/мин у взрослых. При этом уменьшается и относительное значение работы, необходимой для преодоления эластического сопротивления легочной ткани. Из всей работы дыхательных мышц на преодоление эластического сопротивления дыханию приходится у детей 8 — 9 лет 74%, у 14-летних — 68%, а у взрослых — 65%. При физических нагрузках с увеличением минутного объема дыхания работа дыхательных мышц у детей по сравнению со взрослыми значительно увеличивается, вследствие повышения сопротивления воздушному потоку в воздухоносных путях. Бронхиальное сопротивление в покое равно в 4—5 лет 9,6±2 см водн. ст. на 1 л вентиляции, в 6—7 лет — 8,7±2,3. В возрасте от 8 до 15 лет оно снижается до 5,8—3,3 см водн. ст. на 1 л вентиляции, однако не достигает величин, регистрируемых у взрослых (2—4 см водн. ст. на 1 л вентиляции). Эти возрастные особенности дыхания приобретают особенно большое значение при длительных циклических физических упражнениях со .значительной мобилизацией дыхательных функций. Например, при минутном объеме дыхания 60—100 л величина работы дыхательных мышц настолько возрастает, что это приводит к значительному снижению коэффициента полезного действия.
Использование кислорода из дыхательного воздуха у детей меньше, чем у взрослых. Дети школьного возраста значительно менее устойчивы к недостатку кислорода и избытку углекислого газа, чем взрослые. Это проявляется в уменьшении длительности времени задержки дыхания у детей (на вдохе — от 40 до 70 сек.) по сравнению со взрослыми (от 60 до 120 — 130 сек.). В среднем снижение насыщения крови кислородом при задержке дыхания или при дыхании в замкнутое пространство у детей наступает значительно быстрее и достигает критических величин в более короткий отрезок времени по сравнению со взрослыми. Это отчетливо отражает меньшую выносливость детей к недостатку кислорода и избытку углекислого газа, а также меньшие возможности к регуляции дыхания при тяжелой и длительной работе.
§ 7. Обмен веществ и энергии
Белковый обмен. Для роста и развития всех систем органов необходимо превышение поступления в организм белка над его выведением (положительный азотистый баланс). У детей 7—8 лет
487
суточная доза белка 2,2—2,5 г на 1 кг веса может лишь поддержать азотистое равновесие. Для удовлетворения же потребностей растущего детского организма эта доза должна быть не менее 2,8— 3 г на 1 кг веса. При этом принимаемые с пищей белки должны иметь соответствующий аминокислотный состав — с содержанием всех незаменимых аминокислот. Но если вводится с пищей избыточное количество белка, то организм усваивает из него не более 5— 6 г на 1 кг веса (А. Ф. Тур).
Углеводный обмен. Обеспечивая энергетический обмен» углеводы в известной мере участвуют и в пластических процессах при образовании соединительной ткани, клеточных оболочек и др. По сравнению со взрослыми людьми дети имеют меньше углеводных запасов в печени и мышцах, вследствие чего при работе у них быстрее снижается содержание сахара в крови (Н. Н. Яковлев).
Жировой обмен. Жир растворяет витамины A, D, Е и К» имеющие важное значение для роста и развития детского организма, и является источником энергии при мышечной работе.
Водно-солевой обмен. Вода, поступающая в организм через пищеварительный тракт, а также образующаяся при окислении жиров, углеводов и белков, выводится из организма у детей не только с мочой, но и экстраренально (внепочечно) — с потом и выдыхаемым воздухом (всегда полностью насыщенным водяными парами). Колебания веса тела у детей на протяжении суток обусловлены главным образом колебаниями баланса между поступающей в организм и выводимой из него водой.
Обмен воды тесно связан с солевым обменом. У детей вследствие меньшего совершенства нейрогуморальной регуляции обмен солей зависит в большей степени от поступления их с пищей, чем у взрослых.
Обмен энергии. Основной обмен, являющийся важным показателем интенсивности тканевых окислительных реакций при расчете на 1 кг веса тела у детей больше, чем у взрослых. Это объясняется прежде всего большей величиной показателя соотношения поверхности кожи к весу тела, а следовательно, и повышением теплоотдачи на 1 кг веса. Известное значение имеют также более высокая возбудимость детей и их типологические особенности.
С возрастом повышаются суммарные энерготраты при мышечной деятельности в связи с увеличением веса мышечной ткани. Однако при одинаковой мышечной работе при пересчете на 1 кг веса тела энерготраты у подростков больше, чем у взрослых. Суммарная величина максимального потребления кислорода (МПК), отражающая предельные энерготраты, у детей меньше по сравнению со взрослыми. Только в период завершения школьного возраста (к 18 годам) МПК приближается к показателям взрослых. У некоторых юношей-спортсменов величина МПК 5,5—6 л/мин, а при пересчете на 1 кг веса тела — 80—90 мл/мин, т. е. весьма близка к максимальным величинам у взрослых спортсменов.
Данные, показывающие динамику МПК у детей, представлены в табл. 20.
488
Таблица 20
Максимальное потребление кислорода (по данным велоэргометрической пробы) легочная вентиляция и частота пульса (по О. С. Елизаровой)
Показатели Возраст (в годах)
13 14 15 16 17 18 19 Взрослые
Вентиляции легких (л) . 65 95 109 127 142 153 161 156
Максимальное потребление кислорода (л) ... 2,5 3,5 3,9 4,7 5,2 5,5 5,6 6,0
То же на 1 кг веса тела (мл) 68 73 76 77 84 83 90 90
Частота пульса в 1 мин. 205 201 199 191 188 189 186 185
Как видно из данных табл. 20, в 13 лет, т. е. к началу периода полового созревания, МПК составляет менее половины по отношению к МПК в 18—19 лет. Соответственно меньше половины и величина минутного объема дыхания. При пересчете на 1 кг веса тела у 19-летних отмечается такая же величина МПК, как у взрослых мастеров спорта. Очевидно, это зависит от энергозатрат на работу дыхательных мышц при весьма большой у детей вентиляции легких.
Теплорегуляция. У детей температура кожи несколько выше, чем у взрослых. Этим объясняется большая теплоотдача у детей в условиях холода. Важно развивать у них путем постепенного закаливания адаптацию организма к условиям низкой температуры внешней среды.
§ 8. Железы внутренней секреции
На протяжении всего школьного периода развитие эндокринных функций у детей имеет важное значение для совершенствования физического развития и половой дифференциации. Период полового Созревания (пубертатный период) продолжается у девочек с 13— 14 до 18 лет, у мальчиков — с 15—16 до 19—20 лет.
Примерно с 10-летнего возраста начинается атрофия вилочковой железы (тимуса). Гормоны этой железы имеют большое значение для коррекции ростовых взаимоотношений и действуют как антагонисты по отношению к гормонам половых и других желез внутренней секреции. С 16 лет размеры тимуса уменьшаются, а к 20 годам вес этой железы становится меньше веса ее у новорожденных. С инволюцией (обратным развитием) вилочковой железы человека происходит развитие тех особенностей мужского и женского организма, которые свойственны взрослому человеку. С созреванием функций половых желез ускоряются темпы роста организма, повышается работоспособность, развиваются вторичные половые признаки. При этом повышаются функциональные возможности вегетативных систем организма. В периоде полового созревания не
489
сколько изменяется характер реакций организма на разнообразные раздражители. Так, в начале этого периода восстановительные процессы после мышечной деятельности протекают длительнее, чем у детей меньшего или большего возраста (К. М. Смирнов). В этом периоде наблюдается также снижение эмоциональной устойчивости, что следует учитывать при дозировке соревновательных спортивных нагрузок.
§ 9. Центральная нервная система
Основные структуры центральной нервной системы формируются в детские годы. Так, в 7 лет у детей уже намечены все борозды и извилины в коре больших полушарий (В. В. Бунак). В дальнейшем борозды углубляются, контуры их усложняются, поверхность коры значительно увеличивается, достигая к старшему школьному возрасту величин, близких величинам их у взрослых людей. Структурные элементы анализаторов закладываются уже к моменту рождения, причем их наиболее интенсивное развитие происходит на протяжении первых лет жизни. Особенно значительно в младшем и среднем школьном возрасте повышаются функциональные возможности коры больших полушарий, в том числе функциональные возможности центральных отделов анализаторов.
Важное значение для адаптации организма детей к разнообразным условиям внешней среды имеют реакции на словесные раздражители. Уже на 2-м году жизни формируются временные связи второй (словесной) сигнальной системы. Они тесно связаны с новыми двигательными навыками.
В младшем школьном возрасте относительно равномерно протекает совершенствование высшей нервной деятельности. При этом важную роль играет опыт, в том числе двигательный, приобретенный в течение первых 3 лет жизни. Условные двигательные рефлексы, образовавшиеся в раннем возрасте, являются весьма прочными. Поэтому в первые 3 года физическое воспитание играет важнейшую роль. Однако и после 3 лет имеются весьма широкие возможности для образования новых временных связей и закрепления уже возникших условных рефлексов. Легкость образования условных рефлексов и их прочность являются качественными особенностями, отличающими детей от взрослых.
Вследствие этого двигательные навыки — плавание, езда на велосипеде и другие, в том числе речедвигательные (фонетика иностранных языков), — закрепляются прочно. Такие навыки относительно легко восстанавливаются даже после длительных перерывов. Эти особенности детей при рациональном обучении делают возможным достижение высоких результатов в упражнениях, требующих высокой координации движений, в гимнастике, фигурном катании на коньках, плавании и др.
Совершенствование внутреннего торможения позволяет в возрасте 11—13 лет осуществлять дифференцирование весьма незначительных различий между раздражителями. Это способствует тонко
490
му различению временных и пространственных характеристик движения (В. С. Фарфсль).
В детском возрасте интенсивно развиваются анализаторы. Уже в 10—13 лет функции двигательного, зрительного, вестибулярного, тактильного и других анализаторов мало отличаются от функций их у взрослых.
Таким образом, уже в младшем школьном возрасте имеется все необходимые предпосылки к развитию спортивного мастерства во многих видах спорта, требующих высокой координаци движений. Вместе с тем следует учитывать сложность совершенствования двигательных навыков у детей, особенно в так называемом переходном возрасте, в котором обычно выделяют 2 фазы: первая фаза продолжается у мальчиков с 13 до 15 лет, у девочек — с 11 до 13 лет, вторая— у мальчиков с 15 до 17, у девочек с 13 до 15 лет. В первой фазе переходного периода нередко наблюдается временное ухудшение условнорефлекторной деятельности. Оно проявляется в генерализации двигательных рефлексов, в снижении точности дифференцировок, в замедлении реакций на второсигнальные (речевые) раздражители по сравнению с реакциями на первосигнальные. Возможны резкие вегетативные отклонения: учащение сердцебиений, сосудистые расстройства и др. Особено выражены эти явления у девочек.
Первая фаза переходного периода характеризуется высокой утомляемостью подростков; вторая фаза переходного периода (с 15—17 лет мальчики, с 13 до 15 лет девочки) —наибольшей неуравновешенностью поведения. Функции нервной системы могут осуществляться во второй фазе с преобладанием то возбуждения (экзальтации), то торможения (депрессии). Проявление психической неуравновешенности, чрезвычайная обидчивость и другие отклонения от обычных норм поведения детей требуют особого подхода к ним со стороны педагогов. В этом периоде при надлежащем воспитании имеются все возможности для двигательного совершенствования.
§10. Физиологическое обоснование спортивной тренировки детей школьного возраста
Как показывают физиологические исследования, физическое воспитание детей нужно начинать с самого раннего возраста. Двигательная активность в сочетании с закаливающими средствами укрепляет устойчивость детей к действию неблагоприятных воздействий внешней среды, совершенствуют координацию движений и улучшают функции всех систем организма. Занятия физическими упражнениями в дошкольных учреждениях и в школе, сдача школьниками норм ГТО подготавливают к систематическим занятиям спортом и облегчают рациональный выбор спортивной специализации. Возраст, в котором начинается спортивная специализация, зависит от вида спорта. Слишком ранняя спортивная специализация может нанести ущерб здоровью и физическому развитию детей.
Младший школьный возраст (7—II лет). В этом возрасте важ
491
ное значение имеет совершенствование координации движений, развитие быстроты и ловкости и стимулирование вегетативных функций. В связи с этим наряду с систематической подготовкой к освоению некоторых норм комплекса ГТО разрешается начинать занятия фигурным катанием на коньках (с 7 лет), теннисом и настольным теннисом (с 8 лет), акробатикой, художественной гимнастикой, лыжными гонками на 1—3 км (с 9 лет), слаломом и баскетболом (с 10 лет), спортивной гимнастикой, волейболом, легкой атлетикой (бегом на скорость), хоккеем и фехтованием (с 11 лет). Особую ценность для детей имеет плавание, которым целесообразно начинать заниматься еще в дошкольном возрасте (с 2—5 лет). Спортивное плавание разрешается позже — с 9 лет. Двигательная деятельность в воде совершается в условиях, близких к невесомости, что дает возможность весьма эффективно влиять на правильное формирование осанки и сводов стопы. Занятия плаванием способствуют совершенствованию анализаторных функций, развитию у детей тонкого «чувства воды». Движения в воде стимулируют процессы обмена вещества, благоприятно сказываются на адаптации организма к колебаниям внешней температуры.
Средний школьный возраст (12—15 лет). В этом возрасте, называемом переходным, происходит весьма сложный процесс перестройки структур и функций организма, связанный с половым созреванием. Для переходного возраста характерны ускорение роста тела в длину, наличие в ряде случаев диспропорций тела, высокая эмоциональность при неустойчивости функций вегетативных и двигательных систем, отчетливое проявление половых различий. Мальчики начинают превосходить девочек в силе, быстроте и выносливости. Девочки отличаются способностью к плавным, выразительным движениям.
Рациональные физические упражнения имеют важное значение для нормализации у подростков фазы полового созревания.
С 12-летнего возраста разрешается заниматься легкой атлетикой (в том числе бегом на средние дистанции), прыжками на лыжах, слаломом-гигантом (с ограничениями), ручным мячом, футболом, бегом на коньках, хоккеем и парусным спортом; с 13 лет — борьбой самбо, греблей на байдарке и каноэ и академической греблей, с 14 лет — велосипедным спортом, пулевой стрельбой и боксом, с 15 лет — тяжелой атлетикой. Боксом и некоторыми другими видами спорта можно заниматься только мальчикам. В ряде видов спорта имеются четкие различия в дозировке и характере физических нагрузок для мальчиков и девочек.
Старший школьный возраст (16—19 лет). Начиная с этого воз-роста, в котором завершается переходный период, для детей доступны занятия большинством видов спорта. Учет положительных особенностей этого возраста при рациональной системе обучения, эффективном использовании новых средств тренировки позволяет в этом возрасте достигнуть весьма высоких спортивных результатов.
492
ОГЛА ВЛ LIIИI
Введение (профессор // В. Зимкин)............................... 3
§ 1. Фи 1НОЛО1 и» и се содержание (стр. 3). § 2. Краткие сведения о развитии физиологии (стр. 5)
Часть первая. Общая физиология................................. 10
Глава I. Общие физиологические свойства и закономерности основных жизненных процессов (профессор А. А. Логинов)..................... 10
§ 1. Обмен веществ. Организм и внешняя среда. Гомеостаз (стр. 10).
§ 2. Раздражители и возбудимость (стр. 12). § 3. Функциональная активность (стр. 13). § 4. Биологические структуры и процессы обмена веществ (стр. 14). § 5. Мембранный потенциал (стр. 15) § 6. Распространение (проведение) возбуждения (стр. 20). § 7. Раздражение и основные свойства и параметры возбудимости (стр. 22). § 8. Изменение возбудимости при протекании одиночного цикла возбуждения (стр. 26). § 9. Функциональная подвижность (лабильность) (стр. 28). § 10. Оптимум и пессимум частоты (стр. 28). § 11. Парабиоз (стр. 29).
§ 12. Механизмы регулирования функций организма (стр. 31). § 13. Саморегулирование (стр. 33).
Глава II. Сенсорные (афферентные) системы (профессор Н. В. Зимкин) 35 § 1. Общая характеристика деятельности сенсорных систем (стр. 35). § 2. Зрительная сенсорная система (стр. 44). § 3. Слуховая сенсорная система (стр. 53). § 4. Вестибулярная сенсорная система (стр. 56). § 5. Двигательная сенсорная система (стр. 61). § 6. Висцеральная, болевая, температурная, тактильная, обонятельная и вкусовая сенсорные системы (стр. 63). § 7. Роль различных сенсорных систем при занятиях физическими упражнениями (стр. 66).
Глава III. Физиология нервно-мышечного аппарата (профессор Я- М. 1\оц) 67
§ 1. Строение нервно-мышечного аппарата (стр. 67). § 2. Нервно-мышечная передача (стр. 71). § 3. Механизм и энергетика мышечного сокращения (стр. 73). § 4. Формы мышечного сокращения (стр. 75). § 5. Режимы сокращения (стр. 78). § 6. Регуляция напряжения мышцы (стр. 82). § 7. Характеристические кривые мышцы (стр. 85). § 8. Работа мышц н ее механическая эффективность (стр. 89). § 9. Мышечная сила (стр. 93). § 10. Скоростные свойства мышц (стр. 101). § 11. Мышечная выносливость (стр. 102). § 12. Гладкие мышцы (стр. 107).
Глава IV. Физиология центральной нервной системы (доктор биологических наук Е. Б. Сологуб) ..................... 109
§ 1. Физиология нервной клетки (стр. ПО). § 2. Физиологические показатели функционального состояния нейрона (стр. 118). § 3. Системы нервных клеток (стр. 122). § 4. Свойства нервных центров (стр. 126). § 5. Координация деятельности центральной нервной системы (стр. 133). § 6. Специфические и неспецифическне системы (стр. 142). § 7. Основные рефлекторные центры нервной системы (стр. 145).
§ 8. Кора больших полушарий головного мозга (стр. 153). § 9. Нисходящие влияния головного мозга на двигательную деятельность (стр. 158). § 10. Нервная регуляция эмоций и вегетативных функций (стр. 164). § 11. Общие принципы регуляции двигательной деятельности (стр. 169). § 12. Позно-тонические реакции (стр. 172). § 13. Двигательная деятельность (стр. 178).
Глава V. Высшая нервная деятельность (условные рефлексы) (профессор
Е. К. Аганянц. профессор Н. В. Зимкин)........................ 185
§ 1. Роль высшей нервной деятельности в адаптации организма к изменяющимся условиям среды (стр. 185). § 2. Безусловные и условные рефлексы (стр. 187). § 3. Механизм и условия образования условных рефлексов (стр. 188). § 4. Сенсорные и оперантные условные рефлексы (стр. 193). §5. Виды условных рефлексов (стр. 195). §6. Первая и вторая сигнальные системы (стр. 199). § 7. Значение торможения в условно-рефлекторной деятельности и его виды (стр. 201). § 8. Динамический
493
стереотип (стр. 206). § 9. Экстраполяция в высшей нервной деятельности (стр. 208). § 10. Типы высшей нервной деятельности (стр. 209).
§ 11. Фазовые состояния (стр. 211). § 12. Особенности высшей нервной деятельности человека (стр. 213).
Глава VI. Кровь (профессор А. Б. Гандельсман, доктор биологических наук Л. Н. Евгеньева)..........................................
§ 1. Кровь как внутренняя среда организма (стр. 214). § 2. Количество крови в организме (стр. 215). § 3. Состав крови (стр. 215). § 4. Регуляция физико-химических процессов во внутренней среде организма (стр. 217). § 5. Защитные реакции крови (стр. 219). § 6. Изменения в крови при двигательной деятельности (стр. 220).
Глава VII. Кровообращение (профессор В. В. Васильева, профессор Г. Б. Сафронова) ......................
Сердце ......................................................
§ 1. Сердце и его физиологические свойства (В. В. Васильева) (стр. 223).
§ 2. Электрические процессы в сердце (Г. Б. Сафронова) (стр. 227). § 3. Частота сердечных сокращений в покое и при мышечной деятельности (В. В. Васильева) (стр. 229). § 4. Фазы сердечного цикла (В. В. Васильева) (стр. 231). § 5. Работа сердца как нагнетательного насоса (В. В. Васильева) (стр. 233). § 6. Механические и звуковые явления при сокращении сердца (В. В. Васильева) (стр. 234). § 7. Систолический и минутный объемы крови (В. В. Васильева) (стр. 235).
Кровеносные сосуды.........................................
§ 8. Основные сведения о гемодинамике (В. В. Васильева) (стр. 236).
§ 9. Общее периферическое сопротивление (В. В. Васильева) (стр. 238).
§ 10. Кровяное давление (В. В. Васильева) (стр. 239). § И. Артериальный пульс (В. В. Васильева) (стр. 243). § 12. Скорость распространения пульсовой волны по артериям (В. В. Васильева) (стр. 244). § 13. Кровообращение в артериолах и капиллярах (В. В. Васильева) (стр. 245). § 14. Кровообращение в венах (Г. Б. Сафронова) (стр. 246). § 15. Кровообращение в малом кругу (Г. Б. Сафронова) (стр. 247).
Регуляция кровообращения.....................................
§ 16. Регуляция деятельности сердца (В. В. Васильева) (стр. 248). § 17. Регуляция функционального состояния сосудов (В. В. Васильева) (стр. 251). § 18. Рабочая гиперемия (В. В. Васильева) (стр. 254). § 19. Кровообращение при физической работе (В. В. Васильева) (стр. 255) Обмен веществ между кровью и тканями тела....................
§ 20. Тканевая жидкость и обмен веществ через капиллярную стенку (В. В. Васильева) (стр. 256). § 21. Лимфа н ее течение (В. В. Васильева) (стр. 258).
Глава VIII. Дыхание (профессор А. Б. Гандельсман).............
§ 1. Внешнее дыхание (стр. 260). § 2. Вентиляция легких (стр. 264).
§ 3. Обмен газов между альвеолярным воздухом н кровью капилляров легких (стр. 266). § 4. Перенос газов кровью (стр. 267). § 5. Обмен газов между кровью н тканями (стр. 270). § 6. Регуляция дыхания (стр. 271). § 7. Дыхание при физической работе (стр. 275).
Глава IX. Пищеварение (профессор В. В. Васильева) ........ § 1. Общая характеристика пищеварительных процессов (стр. 281). § 2. Пищеварение в полости рта (стр. 282). § 3. Пищеварение в желудке (стр. 284). § 4. Пищеварение в кишечнике (стр. 288). § 5. Всасывание продуктов переваривания пищи (стр. 293). § 6. Печень и ее функции (стр. 293). § 7. Влияние мышечной работы на деятельность пищеварительных органов (стр. 294).
Глава X. Обмен веществ и энергии (профессор В. В. Васильева) . . . . § 1. Сущность обмена вещества и энергии (стр. 295). § 2. Обмен белков и его регуляция (стр. 296). § 3. Обмен углеводов и его регуляция (стр. 298). § 4. Обмен липидов и его регуляция (стр. 300). § 5. Водносолевой обмен (стр. 301). § 6. Витамины (стр. 302). § 7. Обмен энергии (стр. 303).
Глава XI. Выделительные функции (профессор А. А. Логинов)......
494
214
223
223
236
248
256
259
281
295
309
§ I. Общая характеристика выделительных процессии (сгр 301>) ( 2 Основные функции ночек и структура мочсобри.юиик явных единиц (стр 3(19). § 3 Механизм мочеобразования в почке (сгр 311) * 4 Регуляции мочеобразования (стр. 314). § 5. Мочевыведение (стп II >) § 6. Выделительная фуекцня потовых желез (сгр. 315) § 7. Влияние мышечной работы на функции выделения (стр. 316).
Глава XII. Теплообмен (доктор медицинских наук 3. Л. Бернинейн) , . 117
§ 1. Понкнлотермия и гомойотермия (стр. 317). § 2 Темпера)уриыг
«оболочка» и «ядро» тела (стр. 318). § 3. Температурные зоны внеш ней среды (стр. 319). § 4. Механизмы теплопродукции (стр. 320) § 5. Механизмы теплоотдачи (стр. 321). § 6. Центральная регуляция теплообмена (стр. 322). § 7. Особенности теплообмена при мышечной работе (стр. 323).
Глава XIII. Внутренняя секреция (профессор А. А. Виру)................324
§ 1. Общая характеристика желез внутренней секреции (стр. 324).
§ 2. Надпочечники (стр. 326). § 3. Половые железы (стр. 330). § 4. Поджелудочная железа (стр. 332). § 5. Щитовидная железа (стр. 332).
§ 6. Околощитовидные железы. Вилочная железа. Эпифиз (стр. 333).
§ 7. Гипофиз (стр. 334).
Часть вторая. Физиологические основы физического воспитания и спорта 336 Глава XIV. Физиологическая классификация поз и разных видов мышечной деятельности (профессор В. С. Фарфель)............................336
§ 1. Статическая и динамическая работа (стр. 336). § 2. Позы тела и мышечная деятельность (стр. 336). § 3. Классификация физических упражнений (стр. 338). § 4. Циклические движения (стр. 340). § 5. Ациклические движения (стр. 348). § 6. Прицельные движения (стр. 348). § 7. Упражнения, оцениваемые по качеству выполнения (стр. 348). § 8. Нестандартные движения (стр. 349).
Глава XV. Физиологическая характеристика состояний организма при спортивной деятельности (профессор В. В. Васильева, профессор Н. В. Зим-кин ).................................................................350
§ 1. Предстартовое н стартовое состояния (В. В. Васильева) (стр. 350).
§ 2. Изменение функционального состояния организма при разминке (В. В. Васильева) (стр. 353). § 3. Врабатывание (В. В. Васильева) (стр. 355). § 4. Устойчивое состояние (В. В. Васильева) (стр. 357)
§ 5. «Мертвая точка» и «второе дыхание» (В. В. Васильева) (стр. 359)
§ 6. Утомление (Н. В. Зимкин) (стр. 361). § 7. Восстановительные процессы (В. В. Васильева) (стр. 368).
Глава XVI. Физиологические механизмы двигательных навыков (профессор Н. В. Зимкин).........................................................376
§ 1. Временные связи как физиологическая основа формирования двн гательных навыков (стр. 376). § 2. Сенсорные и эффекторные компоненты двигательного навыка (стр. 377). § 3. Значение для формирования двигательных навыков ранее выработанных координаций (стр. 378).
§ 4. Характеристика формирования двигательного навыка по данным электромиографии (стр. 379). § 5. Вегетативные компоненты двигательного навыка (стр. 381). § 6. Афферентный синтез в двигательных навыках (стр. 381). § 7. Программирование в двигательном навыке и учет состояния исполнительных приборов (стр. 383). § 8. Стереотипность и изменчивость двигательного навыка (стр. 385). § 9. Экстраполяция в двигательных навыках (стр. 386). § 10. Автоматизация движений (стр. 387). §11. Стадии (фазы) формирования двигательного акта как навыка (стр. 389). § 12. Устойчивость навыков при разных состояниях организма и длительность сохранения их после прекращения трениров ки (стр. 389)
Глава XVII. Спортивная тренировка и развитие тренированности (профессор В. В. Васильева)..................................................390
§ 1. Спортивная тренировка. Тренированность. Перетренированность (стр. 390). § 2. Физиологические показатели тренированности (стр. 394).
§ 3. Реакция тренированного и нетренированного организма на стан дартные (тестирующие) нагрузки (стр. 406). § 4. Реакции отдельных
49Ь
систем организма на стандартные (тестирующие) нагрузки (стр. 409).
§ 5. Показатели тренированности при выполнении предельно напряженной работы (стр. 411).
Глава XVIII. Физиологическая характеристика физических упражнений с циклической структурой движений (профессор В. В. Васильева) ....
§ 1. Общая характеристика циклических движений (стр. 416). § 2. Спортивная ходьба (стр. 416). § 3. Легкоатлетический бег (стр. 417).
§ 4. Плавание (стр. 421). § 5. Гребля (стр. 425). § 6. Скоростной бег на коньках (стр. 427). § 7. Лыжные гонки (стр. 429). § 8. Велосипедный спорт (стр. 433).
Глава XIX. Физиологическая характеристика видов спорта со стереотипной нециклической структурой движений (профессор в. С. Фарфель) ....
§ 1. Общая характеристика однократных ациклических движений (стр. 437). § 2. Прыжки (стр. 438). § 3. Метания (стр. 439). § 4. Поднимание тяжестей (стр. 440). § 5. Стрельба (стр. 441). § 6. Другие прицельные движения (стр. 441). § 7. Общая характеристика упражнений, оцениваемых по качеству их выполнения (стр. 442). § 8. Спортивная гимнастика (стр. 443). § 9. Художественная гимнастика (стр. 443). § 10. Акробатика (стр. 444). § 11. Прыжки на батуте (стр. 445). § 12. Прыжки в воду (стр. 446). § 13. Фигурное катание на коньках (стр. 447) Глава XX. Физиологическая характеристика нестандартных физических упражнений (профессор б. В. Васильева) ............
§ 1. Общая характеристика спортивных игр (стр. 448). § 2. Баскетбол (стр. 450). § 3. Волейбол (стр. 453). § 4. Футбол (стр. 453). § 5. Хоккей с шайбой (стр. 454). § 6. Общая характеристика единоборств (стр. 455).
§ 7. Бокс (стр. 456). § 8. Борьба (стр. 458). § 9. Фехтование (стр. 460) Глава XXI. Влияние экстремальных условий и адаптация к ним спортсменов (профессор Д. А. Алипов, доцент В. П. Пономарев).........
§ 1. Влияние на организм пониженного атмосферного давления (Д. А. Алипов) (стр. 462). § 2. Ритмические изменения функциональной активности организма (В. П. Пономарев) (стр. 468).
Глава XXII. Физиологическое обоснование массовых форм оздоровительной физической культуры и производственной гимнастики (профессор Н. В. Зим-кин) ........................................................
§ Г. Значение физических упражнений в условиях современной жизни (стр. 469). § 2. Влияние на функции организма недостаточной двигательной активности (стр. 470). § 3. Влияние на функции организма повышенной двигательной активности (стр. 471). § 4. Роль физических упражнений в повышении устойчивости организма к действию неблагоприятных факторов (стр. 472). § 5. Физиологическая характеристика утренней гигиенической гимнастики (стр. 473). § 6. Физиологическая характеристика производственной гимнастики (стр. 475).
Глава XXIII. Характеристика возрастных особенностей функций организма детей школьного возраста в связи с физическим воспитанием (профессор А. Б. Гандельсман) .....................
§ 1. Общие положения (стр. 477). § 2. Развитие движений (стр. 478).
§ 3. Развитие физических качеств (стр. 480). § 4. Кровь (стр. 483).
§ 5. Кровообращение (стр. 484). § 6. Дыхание (стр. 486). § 7. Обмен веществ и энергии (стр. 487). § 8. Железы внутренней секреции (стр. 489). § 9. Центральная нервная система (стр. 490). § 10. Физиологическое обоснование спортивной тренировки детей школьного возраста стр. 491),
СОВЕТСКАЯ
УЧЕБНАЯ ЛИТЕРАТУРА ДЛЯ ВУЗОВ
SHEBA. SPB.liU/VUZ