


























































































































































































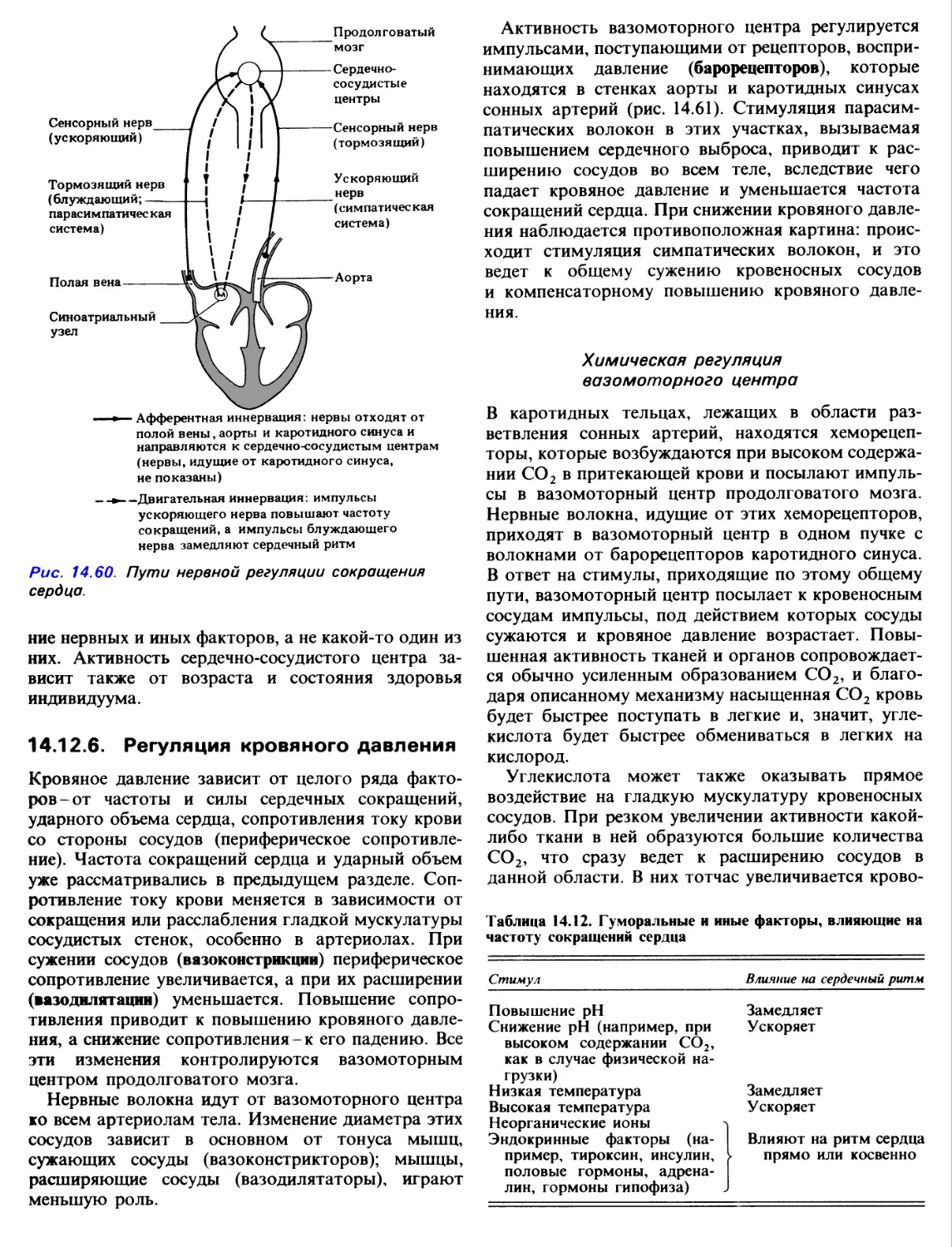


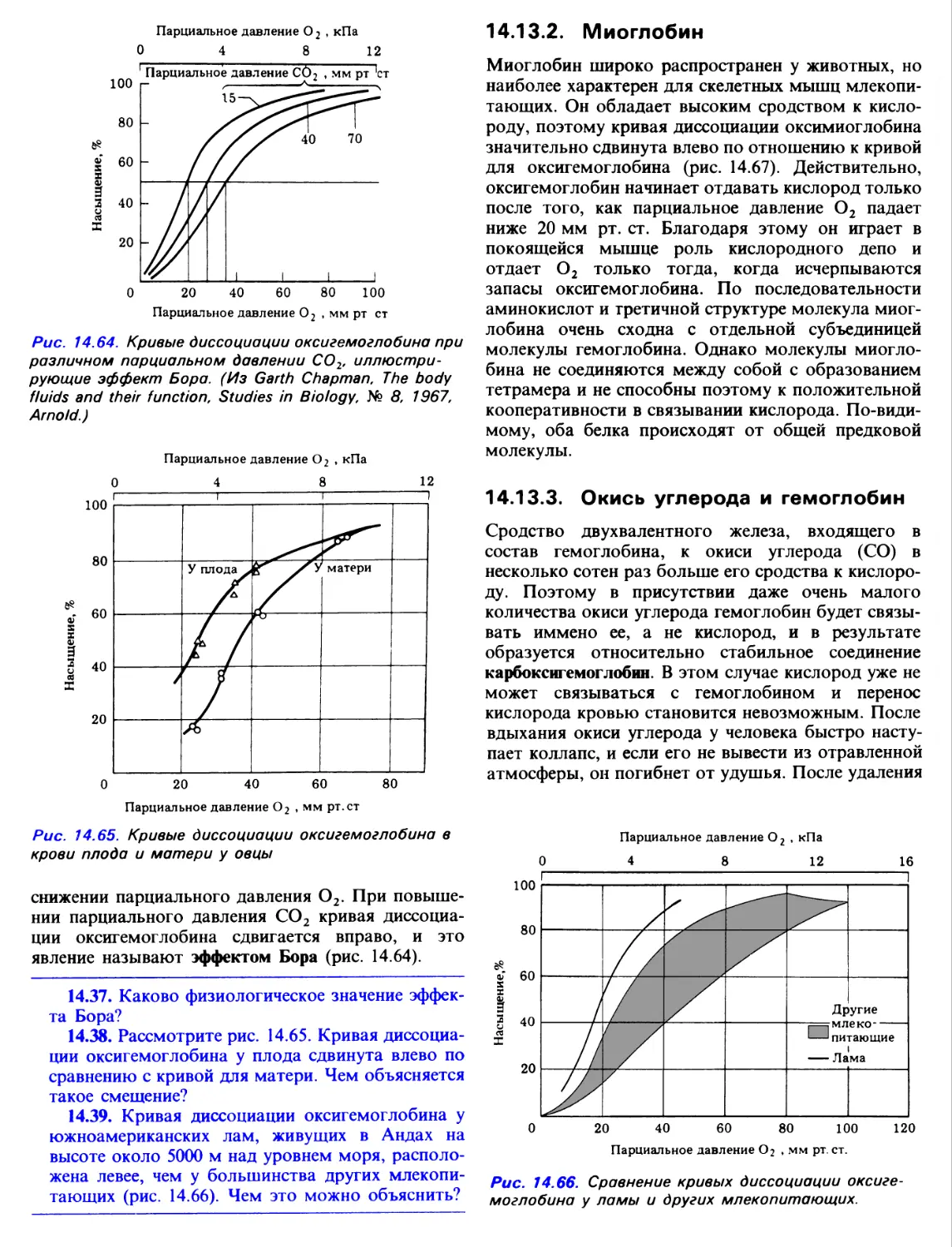








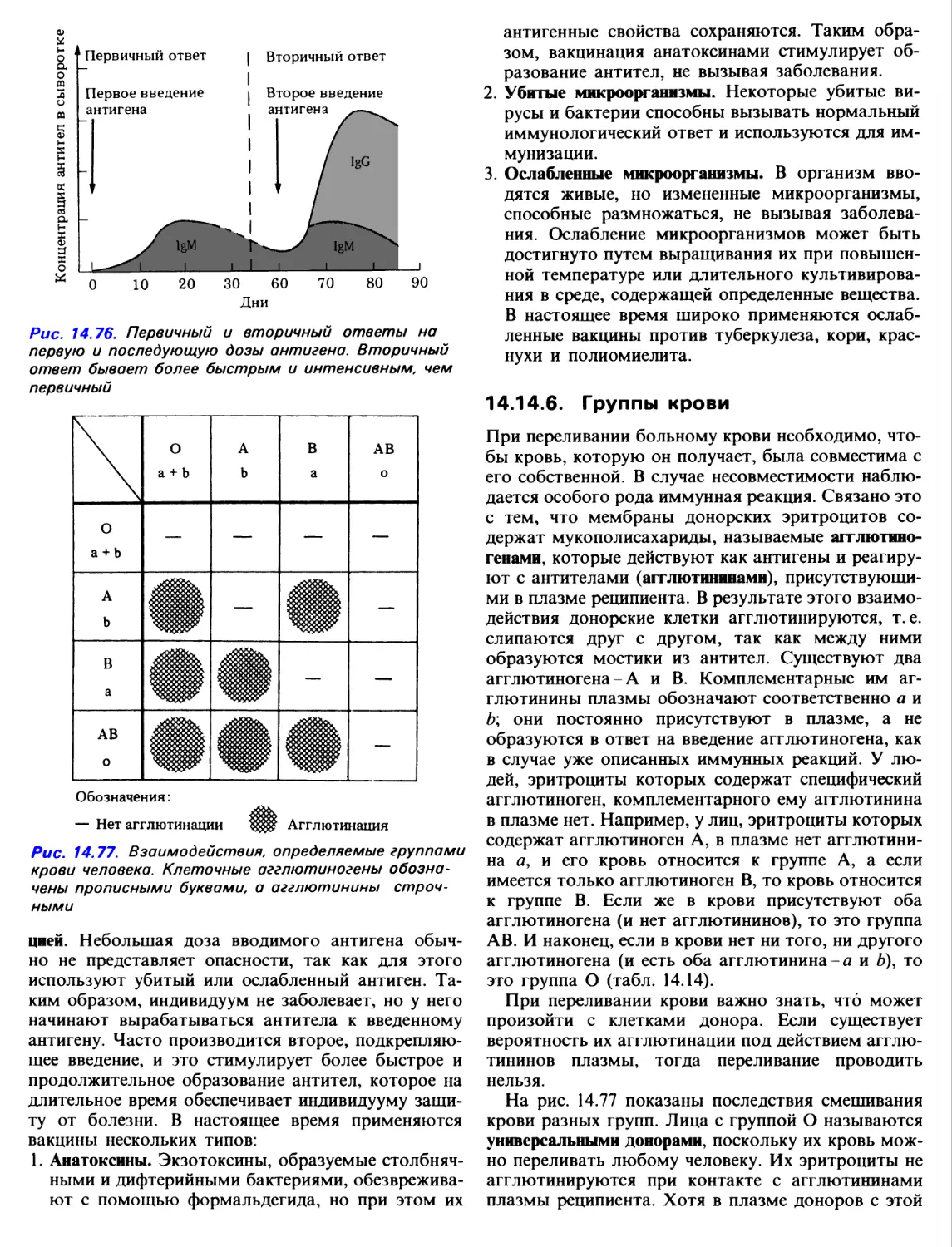









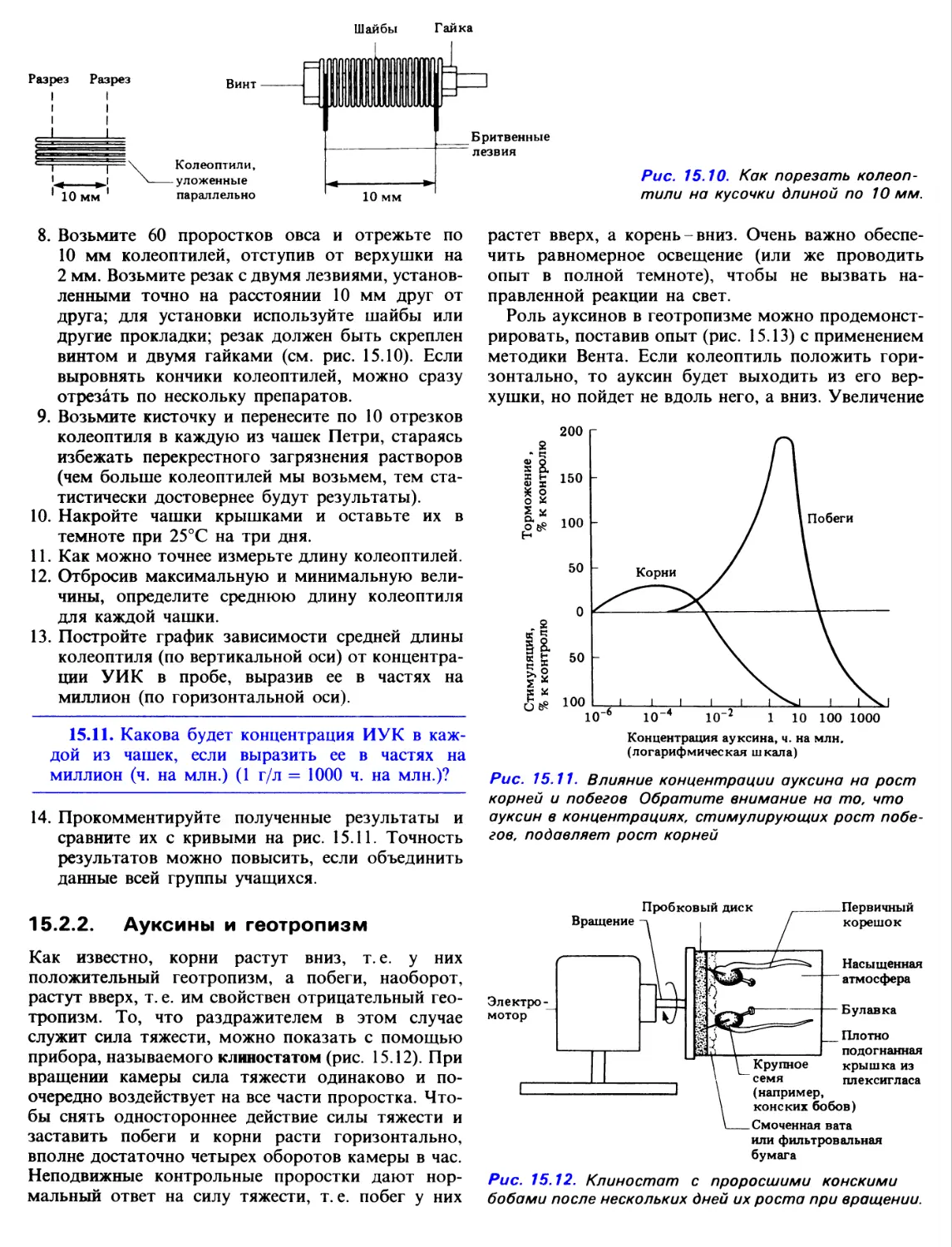






















































































Author: Грин Н. Стаут У. Тейлор Д.
Tags: общая экология биоценология гидробиология биогеография общая биология биология экология издательство мир учебник общей биологии координация регуляция жизненных процессов
ISBN: 5-03-001578-7
Year: 1990




Н.ГРИН, У СТАУТ, Д.ТЕЙЛОР
Биология
2
ИЗДАТЕЛЬСТВО «МИР»
BIOLOGICAL
SCIENCE
N.P.O. GREEN B.Sc, M.I.Biol
Headmaster
Sutton Manor High School for Boys,
Sutton, Surrey
G.W. STOUT B.Sc., M.Ed., M.I.Biol
Secretary for Biological Sciences
University of Cambridge Local Examinations Syndicate
D.J. TAYLOR B.Sc, Ph.D., M.I.Biol
Head of Biology
Strode's Sixth Form College, Egham
Editor
R. SOPER B.Sc, F.LBiol
Formerly Vice-principal and Head of Science
Collyers Sixth Form College, Horsham
CAMBRmGE UNIVERSITY PRESS
Cambridge
London New York New Rochelle
Melbourne Sydney
H. ГРИН
У СТАУТ
Д. ТЕЙЛОР
Биология
2 том
В 3-х томах
Под ред. P. Сопера
Перевод с английского
M. Г· Дуниной
канд. биол. наук
В. И. Mельгунова
канд. биол. наук
M. С. Морозовой
T. В. Никитиной
под редакцией
д-ра биол. наук
Б. M. Медникова
и д-ра биол. наук
А. А. Нейфаха
Москва «Мир» 1990
ББК 28.0
Г85
УДК 574.577
Грин H., Стаут У., Тейлор Д.
Г85 Биология: В 3-х т. T. 2.: Пер. с англ./Под ред. P. Сопера.- M.
Мир, 1990.-325 с, ил.
ISBN 5-03-001578-7
Современное руководство общей биологии, написанный английскими авторами. Весь
материал изложен на весьма высоком уровне, но простота изложения и удачное расположение
тем делают его доступным для неподготовленного читателя.
В т. 2 рассматриваются типы питания организмов, вопросы экологии, механизмы внутреннего
транспорта и способы координации и регуляции жизненных процессов.
Для студентов-биологов, преподавателей биологии в школе, биологов всех специальностей и
учащихся старших классов.
1901000000 005
Г 00 90, ч. 1
041(01) 90
ББК 28.0
Редакция литературы по биологии
ISBN 5-03-001578-7 (русск.)
ISBN 5-03-001576-0
ISBN 0-521-28407-4 (англ.)
ISBN 0-521-26951-2 (англ.)
© Cambridge University
Press 1984, 1985
This book was originally published in the English language
by Cambridge University Press of Cambridge England
First published 1984, 1985
Ninth printing 1989
© перевод на русский язык, коллектив переводчиков, 1990
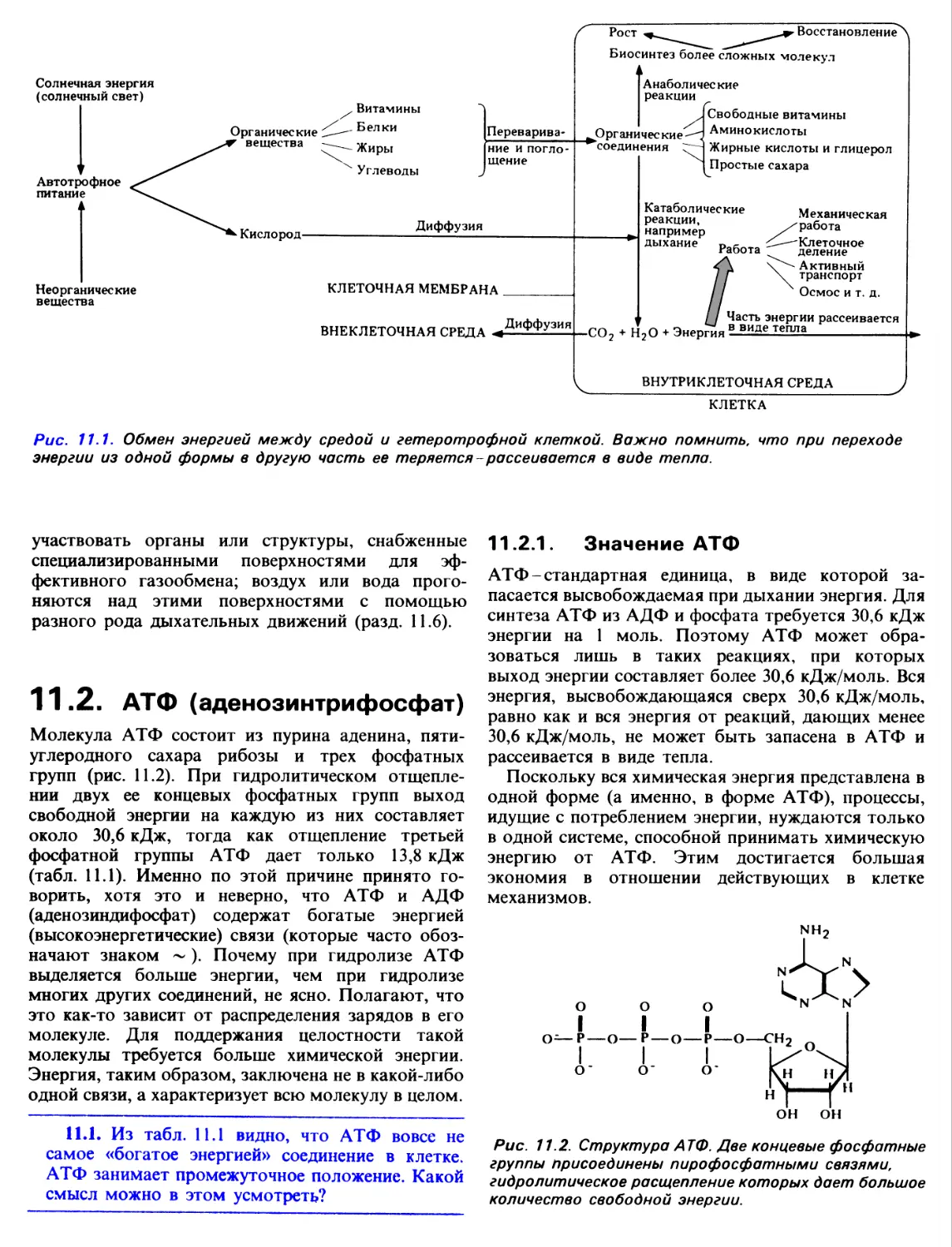
Глава 10
Гетеротрофное
питание
Гетеротрофами называются организмы,
использующие для питания готовые сложные органические
соединения. Эти соединения дают гетеротрофам
энергию, необходимую для их жизнедеятельности, а
также служат источником специфических атомов и
молекул, идущих на поддержание и возобновление
клеточной структуры и новообразование
протоплазмы в процессе роста. Вместе с пищей гетеротрофы
получают также коферменты и витамины, которые
не синтезируются в их организме, а между тем
абсолютно необходимы для целого ряда клеточных
процессов.
Выживание гетеротрофов прямо или косвенно
зависит от синтетической активности автотрофов. К
гетеротрофным организмам относятся все
животные, грибы и большинство бактерий. Некоторые
бактерии, например несерные пурпурные бактерии,
содержат бактериохлорофилл и способны к
фотосинтезу, но получаемую при этом энергию могут
использовать для построения собственных
органических соединений не из CO2, а из органического
«сырья». Такие бактерии называются фотогетеро-
трофами.
Способы добывания и поглощения пищи у
гетеротрофных организмов весьма разнообразны, но путь
превращения питательных веществ в усвояемую
форму у большинства из них очень сходен. По
существу это превращение состоит из двух
процессов, первый из которых - расщепление больших и
сложных молекул на более простые и растворимые
(переваривание), а второй - всасывание растворимых
молекул и транспорт их к собственным тканям
организма.
Известны четыре типа гетеротрофного питания:
гол озонный, сапрофитный, симбиотический и
паразитический.
10.1. Типы гетеротрофного
питания
10.1.1. Голозойный тип питания
Все организмы, питающиеся этим способом,
захватывают пищу внутрь тела, где она подвергается
перевариванию, превращаясь в небольшие
растворимые молекулы, которые могут всасываться и
усваиваться организмом. Свободноживущие голо-
зойные организмы обладают специальным
пищеварительным трактом, в котором и протекают эти
процессы. К голозойным организмам относится
большинство животных и насекомоядные растения.
Голозойный способ питания состоит из
следующих процессов:
Поглощение пищи. Потребление сложных
органических соединений.
Переваривание. Расщепление больших, сложных,
нерастворимых молекул органических соединений и
превращение их в небольшие, растворимые
молекулы, способные к диффузии. Переваривание
осуществляется путем механического измельчения и
ферментативного гидролиза пищи и может быть как
внеклеточным, так и внутриклеточным.
Всасывание. Перенос растворимых молекул через
мембрану из мест переваривания и доставка их к
тканям организма. Всосавшиеся питательные
вещества либо сразу поступают в клетки, либо
вначале попадают в кровеносное русло и с кровью
транспортируются к соответствующим участкам тела.
Ассимиляция. Использование организмом
всосавшихся молекул для получения энергии или для
пластических нужд.
Экскреция. Удаление из организма непереваренных
остатков пищи.
Процессы, составляющие голозойный способ
питания, представлены на рис. 10.1.
Животные, которые питаются растительной
пищей, называются травоядными, животные, которые
поедают других животных,-плотоядными, а
животные, которые едят смешанную
животно-растительную пищу,-всеядными. Если они поглощают пищу в
виде мелких частиц, то их называют микрофагами;
примером могут служить дождевые черви.
Животные, потребляющие пищу крупными кусками,
называются макрофагами. Существуют животные,
потребляющие только жидкую пищу, например тли и
комары. В табл. 10.1 перечислены все способы
голозойного типа питания.
10.1.2. Сапрофитный тип питания
Сапрофитами (от греч. sapros - гнилой и phyton-
растение) называются организмы, питающиеся
мертвым или разлагающимся органическим
материалом. Все сапрофитные организмы выделяют
ферменты непосредственно на потенциальный
продукт питания, который под воздействием этих фер-
Большие сложные нерастворимые
молекулы органических веществ пищи
Зубы
!(механическое
переваривание)
Гидролитичес/-
кие ферменты
(химическое
переварива- ^
ние)
Всосавшиеся
низ ко моле-^
кулярные И
простые рас}
творимые у
молекулы
ИСПОЛЬЗУЮТСЯ В
ОБМЕНЕ
Всасывание .
воды
Зубы
-(механическое '
переваривание)
-^Гидролитичеа
кие ферменты
"(химическое
переваривание)
ПОГЛОЩЕНИЕ
ПИЩИ
Всосавшиеся
низкомоле.|
кулярные
!ростые рас
творимые
молекулы
ИСПОЛЬЗУЮТСЯ В
ОБМЕНЕ
„ Всасьюание
воды
V ПЕРЕВАРИВАНИЕ
^ВСАСЫВАНИЕ
АССИМИЛЯЦИЯ
сапрофитам относятся многие бактерии и грибы,
типичным представителем которых является гриб
Mucor hiemalis.
Сапрофитный способ питания
гриба Mucor hiemalis
Питающие гифы гриба проникают в субстрат, на
котором он растет, и из кончиков гиф выделяются
ферменты, осуществляющие внеклеточное
переваривание (рис. 10.2). Карбогидразы и протеазы
катализируют расщепление крахмала и белков
соответственно до глюкозы и аминокислот. Состоящий из
тончайших нитей, сильно разветвленный мицелий
гриба обеспечивает большую поверхность
всасывания. Глюкоза используется грибом в процессе
дыхания, дающего энергию для метаболических
процессов, а аминокислоты используются для роста и
восстановления тканей. Избыток глюкозы
превращается в гликоген и жир, а избыток аминокислот - в
белковые гранулы, и эти запасные продукты
хранятся в цитоплазме гиф.
10.1. Кратко опишите возможные пути
использования мукора человеком
ДЕФЕКАЦИЯ
Рис. 10.1. Стадии голозойного питания у
млекопитающих.
Мицелий гриба
Субстрат
Гифа гриба
АМИНОКИСЛОТЫ
Π ротеаза-
Растворимые
/Ч продукты
'внеклеточного переваривания
транспортируются в другие участки
грибного мицелия
ЗЕЛОК
Ферменты,
секретируемые
кончиком гифы
ГЛЮКОЗА
Карбо-
гид раза
КРАХМАЛ
Рис. 10.2. Внеклеточное переваривание и всасывание
у мукора (Mucor hiemalis).
ментов подвергается перевариванию. Растворимые
конечные продукты такого переваривания
всасываются и ассимилируются сапрофитом. Питаясь
органическими остатками мертвых растений и
животных, сапрофиты участвуют в их уничтожении путем
разложения. Значительная часть образующихся при
этом низкомолекулярных веществ самими сапрофи-
тами не используется, но их поглощают растения.
Таким образом, деятельность сапрофитов является
важным звеном в круговороте веществ, обеспечивая
возвращение необходимых для жизни химических
элементов от мертвых организмов к живым. К
10.1.3. Симбиоз
Симбиоз (от греч. simbiosis - совместная
жизнь)-одна из форм совместного существования двух
различных организмов (см. разд. 2.2.5). Существуют две
разновидности симбиоза - мутуализм и
комменсализм.
Мутуализм
Мутуализмом называются взаимовыгодные
отношения между двумя организмами. Иногда этот
термин используют вместо термина «симбиоз». My-
Таблица 10.1. Разновидности и способы голозойного
питания
Микрофагия
1. С помощью псевдоподий
2 С помощью ресничек
3. Путем фильтрации
Макрофагия
1. С помощью щупалец
2. Путем соскабливания
3. Путем захвата добычи
4 Питание детритом
Питание жилкой пищей
1. Сосущий способ
2. Колюще-сосущий способ
Рис. 10.3. Актиния Calliactis parasitica, прикрепившаяся к раковине моллюска, в которой обитает
рак-отшельник Eupagurus bernhardus.
туализм может существовать между двумя
животными, двумя растениями или между растением и
животным. Так, например, актиния Calliactis
parasitica прикрепляется к раковине, в которой живет
рак-отшельник (рис. 10.3). Актиния питается
остатками пищи краба и перемещается вместе с ним с
одного места на другое. В свою очередь она
обеспечивает крабу маскировку, а ее стрекательные клетки
служат ему защитой. По всей видимости, актиния не
может существовать, не прикрепившись к раковине
краба, но и краб, лишившись актинии, будет искать
другую актинию и сам перенесет ее на раковину, в
которой обитает.
Растительноядные жвачные дают приют
обширной фауне ресничных, например Entodinium,
способных переваривать целлюлозу. Последние могут
существовать только в анаэробных условиях,
подобных тем, которые имеются в пищеварительном
тракте животных. Ресничные, обитающие в
пищеварительном тракте жвачных, питаются
содержащейся в пище хозяина целлюлозой, превращая ее в
такие соединения, которые уже сами жвачные
способны переваривать до продуктов, подвергающихся
всасыванию и ассимиляции.
Комменсализм
Комменсализм (лат. com-с, вместе и mensa-стол,
трапеза) - такая форма симбиоза, при которой один
из партнеров извлекает пользу из совместного
существования с другим партнером (иногда
называемым хозяином). При такой форме симбиоза хозяин
не получает от совместного существования ни
пользы, ни вреда. Так, например, колониальный полип
Hydractinia echinata прикрепляется к раковинам
брюхоногих моллюсков, в которых обитают раки-
отшельники. Полип получает от краба питание,
поглощая остатки его пищи, а для краба такое
совместное существование является совершенно
безразличным.
10.1.4. Паразитизм
Паразит (от греч. рага-около и sitos пища) обитает
внутри или на поверхности тела другого организма,
называемого хозяином, и получает от него пищу и,
как правило, местообитание. В данном случае
совместное существование выгодно только
паразиту, тогда как хозяину его присутствие может
приносить вред. Процветающий паразит может
сосуществовать с хозяином, не причиняя ему
серьезного вреда и обеспечивая таким образом собственное
будущее.
Паразиты, обитающие на поверхности тела
хозяина, называются эктопаразитами (например, клещи,
блохи и пиявки); такие организмы не всегда ведут
исключительно паразитический образ жизни.
Паразиты, обитающие внутри организма хозяина,
называются эндопаразитами (например, малярийный
плазмодий и цепень (Taenia). Паразитизм бывает
обязательным (облигатным) и необязательным
(факультативным). Если организм вынужден постоянно
вести паразитический образ жизни, его называют
облигатным паразитом. Примером факультативных
(необязательных) паразитов являются грибы,
которые вначале ведут паразитический образ жизни, а
после гибели хозяина переходят на сапрофитное
питание на мертвом теле. Некоторые зеленые
растения являются полупаразитами: обладая
способностью к фотосинтезу, микроэлементы тем не менее
они получают от хозяина. Примером таких
растений может служить омела белая, гаустории которой
проникают в древесину растения-хозяина и
высасывают из нее минеральные соли и воду.
Сама природа занимаемой паразитами ниши
свидетельствует о том, что они являются
высокоспециализированными организмами, обладающими
разнообразными приспособлениями, многие из
которых обусловлены видовой принадлежностью
хозяина и образом его жизни. В табл. 10.2 приведены
некоторые особенности строения, физиологии и
размножения паразитов. Эти особенности помогают
им приспосабливаться к суровым условиям
существования.
Таблица 10.2. Некоторые морфологические, физиологические и репродуктивные приспособления паразитов
M орфо
логические
Физиологические
Репродуктивные
Тип модификации
Отсутствие или редукция органов питания и
движения. Характерно для кишечных паразитов
Высокоспециализированный ротовой аппарат, как у
животных, питающихся жидкой пищей
Развитие гаусторий у некоторых паразитических
зеленых растений
Приспособления для пробуравливания покровов,
помогающие проникнуть в тело хозяина
Органы прикрепления типа крючьев или присосок
Резистентные наружные покровы
Атрофия органов чувств, связанная с постоянством
среды обитания паразита
Образование экзоферментов, осуществляющих
переваривание окружающих тканей хозяина
Образование антикоагулянта у видов, питающихся
кровью
Хемочувствительность, позволяющая находить
оптимальное местонахождение в организме
хозяина
Образование цитолитических веществ,
способствующих проникновению в организм хозяина
Образование антиферментов
Способность к дыханию в анаэробных условиях
Гермафродитизм, обеспечивающий
самооплодотворение
Огромное количество половых продуктов - яиц,
спор и цист
Резистентность половых клеток вне организма
хозяина
Наличие в жизненном цикле специальных
репродуктивных стадий
Использование промежуточного хозяина
Примеры
Fasciola, Taenia
Человеческая блоха (Pulex)
тля (Aphis)
Повилика (Cuscuta)
Нематоды, грибы
Цепень (Taenia), фасциола (Fasciola\ пиявка (Hirudo)
Цепень, фасциола
Цепень
Грибы, Plasmodium (простейшее из класса
споровиков, инфицирующее млекопитающих и птиц
и вызывающее малярию у человека)
Pulex, Hirudo
Plasmodium, Monocystis (простейшее из класса
споровиков, паразитирующее в семенных пузырьках
дождевого червя)
Cuscuta (цветковое растение, принадлежащее к сем.
Convolvulaceae, не имеющее хлорофилла и
паразитирующее на различных высших растениях)
Кишечные паразиты
Кишечные паразиты
Taenia, Fasciola
Taenia, Fasciola
Monocystis, Phytophthora
Fasciola
Taenia, Fasciola Plasmodium
10.2. Опишите особенности строения,
физиологии и размножения фасциолы (печеночной
двуустки), обеспечивающие ее процветание
10.2. Различные способы
питания животных
10.2.1. Животные, питающиеся
мелкими частицами
Питание с помощью псевдоподий
Амеба питается коловратками, десмидиевыми и
диатомовыми водорослями, бактериями,
жгутиковыми, ресничными и мельчайшими частичками
органических остатков, захватывая их путем
фагоцитоза. Псевдоподии амебы обтекают пищевую частицу
и включают ее в цитоплазму вместе с небольшим
количеством воды; образующийся при этом пузырек
называется пищевой вакуолью (рис. 10.4). Эта
вакуоль окружается множеством мельчайших лизо-
сом, которые в конечном счете сливаются с ее
мембраной и выделяют в вакуоль свое содержимое
(т. е. переваривание у амебы является
внутриклеточным). На этой стадии вакуоль называется
пищеварительной. Еще до слияния с лизосомами вакуоль
теряет некоторое количество воды и уменьшается в
размерах, а ее содержимое имеет кислую реакцию
(рН 5,6). Затем содержимое вакуоли становится
слабощелочным (рН 7.3).
Среди влившихся в вакуоль ферментов
содержатся карбогидразы, амино-, экзо- и эндопептидазы,
эстераза, коллагеназа и нуклеаза. Эти ферменты
поступают в вакуоль в разное время, поэтому их
переваривающее действие разделено во времени, а
не в пространстве, как у высших организмов. После
завершения переваривания мембрана пищевари-
1. Образуется пищевая вакуоль, содержащая
воду и пищевые частицы
2. Вакуоль слегка уменьшается в размерах
в результате потери воды.
Кислотность повышена (рН 5,6)
3. B пищевую вакуоль из окружающих ее
лизосом изливаются ферменты; теперь она
превращается в пищеварительную вакуоль,
в которой происходит переваривание
(рН становится 7,3)
4. От пищеварительной вакуоли во все стороны
отходят канальцы» по которым растворимые
продукты переваривания переходят
в окружающую протоплазму
тельной вакуоли расщепляется с образованием
множества тончайших канальцев. Растворимые
продукты переваривания поступают в эти канальцы, а из
них путем микропиноцитоза - в окружающую
цитоплазму. Непереваренные остатки пищи выводятся
из организма амебы путем экзоцитоза в каком-либо
участке ее поверхности.
Питание с помощью ресничек
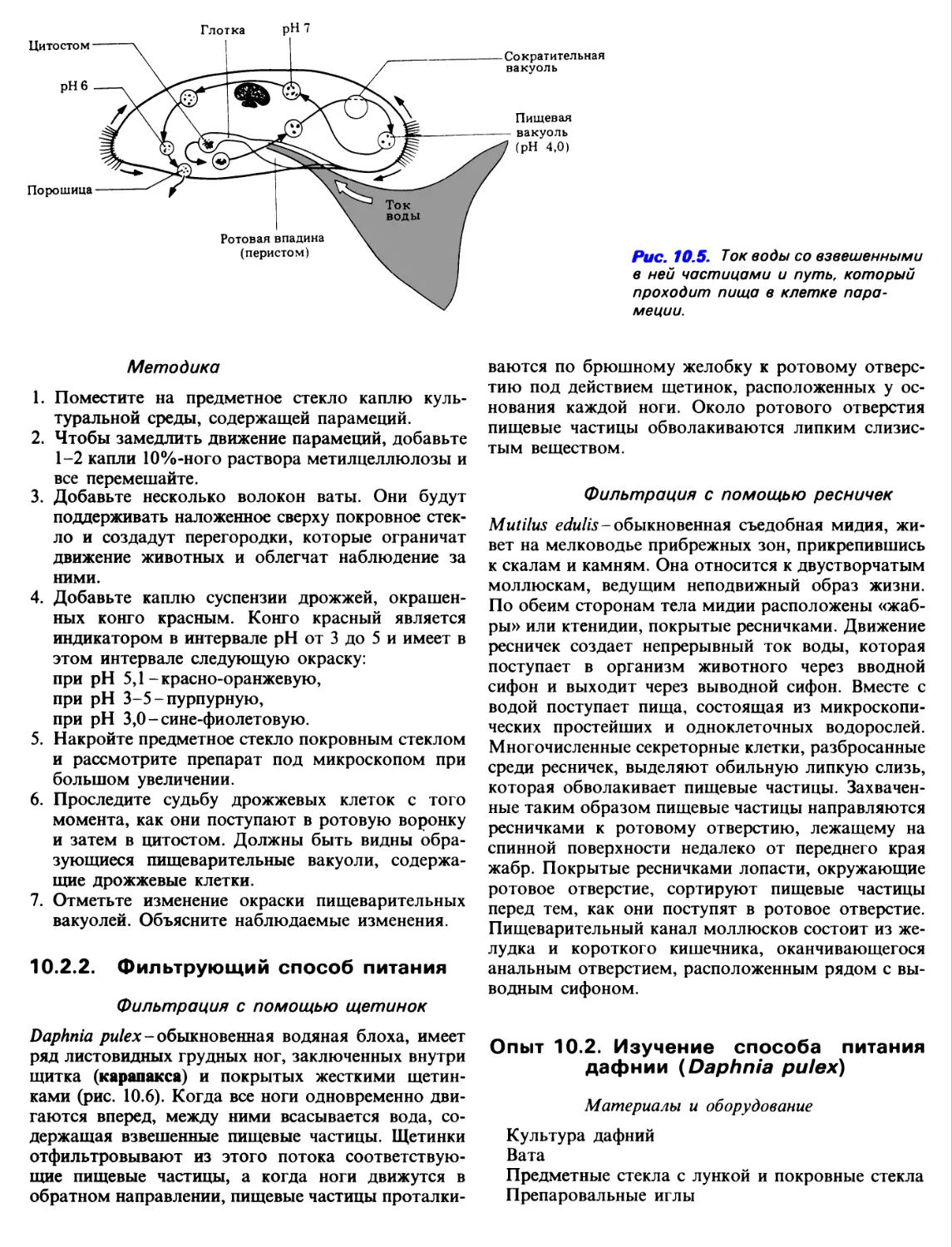
Основную пищу парамеции составляют бактерии.
Расположенные вдоль ротовой воронки
специализированные ряды ресничек гонят воду вместе со
взвешенными в ней микроорганизмами по направлению
к глотке (рис. 10.5). Вместе с током воды бактерии
поступают в глотку, а затем под действием ресничек
ундулирующей мембраны загоняются в рот (ци-
тостом)1. Вокруг рта находится ряд
специализированных, перекрестно расположенных ресничек,
которые играют роль фильтра, препятствуя
поступлению крупных частиц.
Мелкие частицы вместе с небольшим количеством
воды поступают в эндоплазму инфузории, где
вокруг них образуется пищеварительная вакуоль,
которая с током эндоплазмы проделывает
определенный путь вокруг всего тела инфузории. На
протяжении этого пути содержимое вакуоли приобретает
вначале кислую (рН 2-4), а затем слабощелочную
(рН 7-8) реакцию. В кислой среде происходит
умерщвление жертвы, а переваривание ее
осуществляется в основном с уменьшением кислотности
среды. В частности, протеолитические ферменты
обладают наибольшей активностью при рН 5,7-5,8.
Растворимые конечные продукты этого
внутриклеточного переваривания всасываются в
цитоплазму инфузории, а непереваренные частицы
выбрасываются путем экзоцитоза через порошицу (цито-
прокт).
Опыт 10.1. Переваривание дрожжевых
клеток и образование
пищеварительных вакуолей у
инфузории (Paramecium)
Материалы и оборудование
Культура парамеций
Покровные стекла и предметные стекла с лункой
Препаровальные иглы
10%-ный раствор метилцеллюлозы
Вата
Культура дрожжевых клеток, окрашенных конго
красным
Монокулярные микроскопы
5. Экзоцитоз нерастворимого
или неперевариваемого материала
Рис. 10.4. Поглощение, переваривание и всасывание
пищи у амебы.
1 По другим источникам (Жизнь животных./ Под ред.
Зенкевича, т. 1, стр. 136; Биологический
энциклопедический словарь, 1986, стр. 449) пища попадает сначала в рот,
а потом в глотку - Прим. перев.
Цитостом
рН6
Глотка
рН7
Сократительная
вакуоль
Пищевая
вакуоль
(рН 4,0)
Порошица
Ток
воды
Ротовая впадина
(перистом)
Рис. 10.5. Ток воды со взвешенными
в ней частицами и путь, который
проходит пища в клетке
парамеции.
Методика
1. Поместите на предметное стекло каплю куль-
туральной среды, содержащей парамеций.
2. Чтобы замедлить движение парамеций, добавьте
1-2 капли 10%-ного раствора метилцеллюлозы и
все перемешайте.
3. Добавьте несколько волокон ваты. Они будут
поддерживать наложенное сверху покровное
стекло и создадут перегородки, которые ограничат
движение животных и облегчат наблюдение за
ними.
4. Добавьте каплю суспензии дрожжей,
окрашенных конго красным. Конго красный является
индикатором в интервале рН от 3 до 5 и имеет в
этом интервале следующую окраску:
при рН 5,1-красно-оранжевую,
при рН 3-5-пурпурную,
при рН 3,0-сине-фиолетовую.
5. Накройте предметное стекло покровным стеклом
и рассмотрите препарат под микроскопом при
большом увеличении.
6. Проследите судьбу дрожжевых клеток с того
момента, как они поступают в ротовую воронку
и затем в цитостом. Должны быть видны
образующиеся пищеварительные вакуоли,
содержащие дрожжевые клетки.
7. Отметьте изменение окраски пищеварительных
вакуолей. Объясните наблюдаемые изменения.
10.2.2. Фильтрующий способ питания
Фильтрация с помощью щетинок
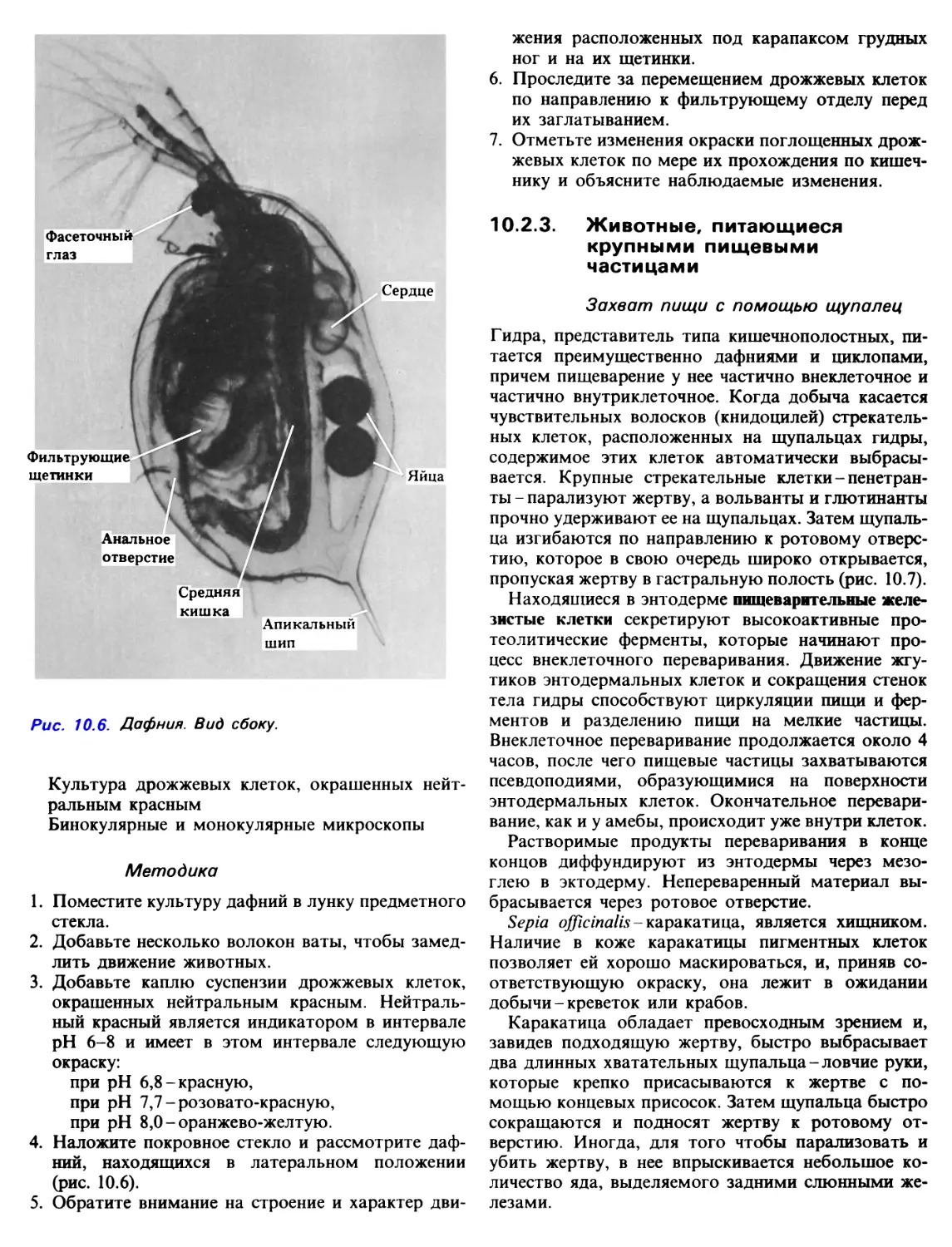
Daphnia /w/ex - обыкновенная водяная блоха, имеет
ряд листовидных грудных ног, заключенных внутри
щитка (карапакса) и покрытых жесткими
щетинками (рис. 10.6). Когда все ноги одновременно
двигаются вперед, между ними всасывается вода,
содержащая взвешенные пищевые частицы. Щетинки
отфильтровывают из этого потока
соответствующие пищевые частицы, а когда ноги движутся в
обратном направлении, пищевые частицы
проталкиваются по брюшному желобку к ротовому
отверстию под действием щетинок, расположенных у
основания каждой ноги. Около ротового отверстия
пищевые частицы обволакиваются липким
слизистым веществом.
Фильтрация с помощью ресничек
Mutilus edulis- обыкновенная съедобная мидия,
живет на мелководье прибрежных зон, прикрепившись
к скалам и камням. Она относится к двустворчатым
моллюскам, ведущим неподвижный образ жизни.
По обеим сторонам тела мидии расположены
«жабры» или ктенидии, покрытые ресничками. Движение
ресничек создает непрерывный ток воды, которая
поступает в организм животного через вводной
сифон и выходит через выводной сифон. Вместе с
водой поступает пища, состоящая из
микроскопических простейших и одноклеточных водорослей.
Многочисленные секреторные клетки, разбросанные
среди ресничек, выделяют обильную липкую слизь,
которая обволакивает пищевые частицы.
Захваченные таким образом пищевые частицы направляются
ресничками к ротовому отверстию, лежащему на
спинной поверхности недалеко от переднего края
жабр. Покрытые ресничками лопасти, окружающие
ротовое отверстие, сортируют пищевые частицы
перед тем, как они поступят в ротовое отверстие.
Пищеварительный канал моллюсков состоит из
желудка и короткого кишечника, оканчивающегося
анальным отверстием, расположенным рядом с
выводным сифоном.
Опыт 10.2. Изучение способа питания
дафнии (Daphnia p и lex)
Материалы и оборудование
Культура дафний
Вата
Предметные стекла с лункой и покровные стекла
Препаровальные иглы
Фасеточный
глаз
Сердце
Фильтрующие
щетинки
Яйца
Анальное
отверстие
Средняя
кишка
Апикальный
шип
Рис. 10.6. Дафния. Вид сбоку.
Культура дрожжевых клеток, окрашенных
нейтральным красным
Бинокулярные и монокулярные микроскопы
Методика
1. Поместите культуру дафний в лунку предметного
стекла.
2. Добавьте несколько волокон ваты, чтобы
замедлить движение животных.
3. Добавьте каплю суспензии дрожжевых клеток,
окрашенных нейтральным красным.
Нейтральный красный является индикатором в интервале
рН 6-8 и имеет в этом интервале следующую
окраску:
при рН 6,8-красную,
при рН 7,7-розовато-красную,
при рН 8,0-оранжево-желтую.
4. Наложите покровное стекло и рассмотрите
дафний, находящихся в латеральном положении
(рис. 10.6).
5. Обратите внимание на строение и характер
движения расположенных под карапаксом грудных
ног и на их щетинки.
6. Проследите за перемещением дрожжевых клеток
по направлению к фильтрующему отделу перед
их заглатыванием.
7. Отметьте изменения окраски поглощенных
дрожжевых клеток по мере их прохождения по
кишечнику и объясните наблюдаемые изменения.
10.2.3. Животные, питающиеся
крупными пищевыми
частицами
Захват пищи с помощью щупалец
Гидра, представитель типа кишечнополостных,
питается преимущественно дафниями и циклопами,
причем пищеварение у нее частично внеклеточное и
частично внутриклеточное. Когда добыча касается
чувствительных волосков (книдоцилей)
стрекательных клеток, расположенных на щупальцах гидры,
содержимое этих клеток автоматически
выбрасывается. Крупные стрекательные клетки - пенетран-
ты - парализуют жертву, а вольванты и глютинанты
прочно удерживают ее на щупальцах. Затем
щупальца изгибаются по направлению к ротовому
отверстию, которое в свою очередь широко открывается,
пропуская жертву в гастральную полость (рис. 10.7).
Находящиеся в энтодерме пищеварительные
железистые клетки секретируют высокоактивные про-
теолитические ферменты, которые начинают
процесс внеклеточного переваривания. Движение
жгутиков энтодермальных клеток и сокращения стенок
тела гидры способствуют циркуляции пищи и
ферментов и разделению пищи на мелкие частицы.
Внеклеточное переваривание продолжается около 4
часов, после чего пищевые частицы захватываются
псевдоподиями, образующимися на поверхности
энтодермальных клеток. Окончательное
переваривание, как и у амебы, происходит уже внутри клеток.
Растворимые продукты переваривания в конце
концов диффундируют из энтодермы через
мезоглею в эктодерму. Непереваренный материал
выбрасывается через ротовое отверстие.
Sepia officinalis -каракатица, является хищником.
Наличие в коже каракатицы пигментных клеток
позволяет ей хорошо маскироваться, и, приняв
соответствующую окраску, она лежит в ожидании
добычи - креветок или крабов.
Каракатица обладает превосходным зрением и,
завидев подходящую жертву, быстро выбрасывает
два длинных хватательных щупальца-ловчие руки,
которые крепко присасываются к жертве с
помощью концевых присосок. Затем щупальца быстро
сокращаются и подносят жертву к ротовому
отверстию. Иногда, для того чтобы парализовать и
убить жертву, в нее впрыскивается небольшое
количество яда, выделяемого задними слюнными
железами.
|Дафния I
ITrichodina sp.[
|Щупальце|
Рис. 10.7. А. Гидра, заглатывающая дафнию.
Б. Микрофотография, полученная с помощью
электронного микроскопа, на которой видна Trichodina,
застрявшая в щупальцах гидры.
Глаз
Мантия
|Щупальце|
[Боковой плавник!
Рис. 10.8. Каракатица (Sepia officinalis), у которой видны щупальца, окружающие ротовое отверстие.
С помощью восьми других, коротких щупалец
каракатица удерживает добычу около рта, где пара
клювообразных роговых челюстей разрывает
жертву на куски (рис. 10.8). Во рту у каракатицы
находится терка, перетирающая пищу в мелкую кашицу,
которая затем заглатывается. К механическому
дроблению пищи присоединяется действие протеаз,
секретируемых слюнными железами.
Питание путем соскабливания и
перетирания
Helix aspersa - обычная садовая улитка, питается с
помощью специального скребущего
органа-радулы, или терки, и роговой челюсти (рис. 10.9). Радула
представляет собой пластинку, на которой
расположено до 150 поперечных рядов зубчиков,
направленных вершинами назад. В каждом ряду
содержится более 100 зубчиков.
Когда улитка захватывает губами листья, радула
движется взад и вперед по листьям, прижимая их к
челюстной пластинке, и ее зубчики перетирают
пищу. В конечном счете образуются мельчайшие фраг-
Радула
Голова
Челюсть
Рот
Хрящ
Нога
Мешочек
радулы
Втягивающий
мускул глотки
Пищеварительная
трубка
!Центральный!
рубец
Рис. 10.9. А. Расположение радулы у садовой улитки
(Helix aspersa). Б. Микрофотография, полученная с
помощью сканирующего электронного микроскопа, на
которой виден участок радулы гигантской наземной
улитки.
Антенна'
Фасеточный глаз
Мандибула
(верхняя челюсть
или жвала)
Максилла
(нижняя
челюсть)
Нижняя губа
Простой глазок
Верхняя губа
Мандибула
Мандибула
Гипофа ринке
(по дг лоточник)
Рис. 10.10. Ротовые части обыкновенной кобылки
(Chonhippus)
менты растения, которые постепенно
проталкиваются в глотку. Зубчики на переднем конце терки от
употребления снашиваются, расшатываются и в
конце концов выпадают и проглатываются вместе с
пищей. Однако они быстро и непрерывно
замещаются вновь образующимися зубчиками. Благодаря
скребущему действию терки плотные целлюлозные
стенки растительных клеток разрушаются и их
содержимое становится доступным действию
гидролитических ферментов, и прежде всего протеаз, в
пищеварительном тракте.
Грызущие и пережевывающие
ротовые части
У кобылки (Chorthippus), представителя семейства
саранчовых, отростки четвертого, пятого и шестого
сегментов экзоскелета образуют ротовой аппарат,
который окружает рот, расположенный на брюшной
поверхности тела. Спереди рот ограничен верхней
губой (лабрумом), имеющей вид пластинки (рис. 10.10).
Под верхней губой лежит пара крепких и мощных
верхних челюстей - мандибул, или жвал. Вершины
жвал имеют зазубренные края, а их основания -
бугорчатую перетирающую поверхность. Работая
сообща, жвалы отрывают твердые частицы пищи,
разгрызают и перетирают их. За жвалами
располагается пара нижних челюстей, макенлл, каждая из
которых несет обонятельный щупик. Позади нижних
челюстей свешивается непарная складка покровов,
носящая название лабиума или нижней губы.
Нижняя губа способствует перемещению пищи, а также
выполняет сенсорную функцию. Кобылка -
травоядное насекомое и питается преимущественно
листьями растений. Она крепко зажимает лист между
губами и с помощью мандибул отрывает от него
кусочки. Движением нижних челюстей и нижней
губы пища направляется к ротовому отверстию и
заглатывается. В гипофаринксе пища смачивается
слюной, выделяемой слюнными железами. Слюна
содержит амилазу и сахаразу, поэтому
переваривание углеводов начинается сразу же.
Хватающие и заглатывающие
ротовые части
Scyliorhinus caniculus- обыкновенная кошачья акула,
относится к хищным плотоядным животным.
Питается ракообразными, кольчатыми червями,
мелкой рыбой и фрагментами тел мертвых или
умирающих животных. Широкий рот, расположенный
на брюшной стороне, позволяет акуле заглатывать
некоторых животных целиком. Обширная ротовая
полость имеет форму поперечной щели и снабжена
крупными направленными вершинами назад
кожными зубами, которые действуют как настоящие
зубы и удерживают во рту попавшую туда добычу.
Ротовая полость ведет в обширную глотку, в
которой находится плотный мышечный вырост-
язык. Язык помогает заглатыванию пищи,
передвигая ее вверх и назад по направлению к пищеводу.
Выстилка пищевода собрана в складки. При
попадании пищи в пищевод складки распрямляются и это
препятствует поступлению в кишечник слишком
большого количества воды. Желудок имеет
асимметричную форму и состоит из двух
отделов-расширенной кардиальной части, в которой протекает
кислая фаза пищеварения, и более узкой и короткой
пилорической части. За желудком следует
относительно короткая двенадцатиперстная кишка, в
которой происходит щелочная фаза пищеварения. В
двенадцатиперстную кишку открываются по
отдельности протоки поджелудочной железы и
желчного пузыря. Двенадцатиперстная кишка переходит
в подвздошную кишку, снабженную спиральным
клапаном. Слизистая подвздошной кишки образует
множество складок, которые замедляют
продвижение пищи и увеличивают всасывающую
поверхность. Вся всосавшаяся пища поступает
непосредственно в печень, имеющую очень большие размеры
(рис. 10.11).
По всей длине пищеварительного тракта
расположены многочисленные слизистые железы, секрет
которых облегчает продвижение пищи.
Непереваренные остатки пищи собираются в прямой кишке и
выбрасываются через анальное отверстие.
Животные, питающиеся детритом
Lumbricus terrestris - обыкновенный дождевой червь,
поедает частички свежего или разлагающегося
органического материала, главным образом растений,
находящихся на поверхности почвы или затянутых
червем в норку. Захваченные кусочки пищи
смачиваются в полости рта щелочным секретом,
выделяющимся в глотке и поступающим в ротовую
полость под действием накачивающих сокращений
мускулистой глотки. Затем с помощью
перистальтических движений пищеварительной трубки они
проталкиваются в глотку (рис. 10.12). Дождевой
червь питается органическими веществами почвы,
которую он заглатывает при прорывании ходов.
Пищеварительная трубка представляет собой
кишку, которая напрямую соединяет рот с
анальным отверстием, но при этом делится на участки,
каждый из которых играет определенную роль в
переваривании и всасывании поглощаемой червем
пищи. В табл. 10.3 указаны отделы
пищеварительной трубки червя и сегменты, в которых они
находятся, а также их строение и специфические
функции. Весь непереваренный материал проталкивается
под действием перистальтических сокращений в
заднюю кишку и выбрасывается через анальное
отверстие в виде характерных кучек - выбросов, или
копролитов.
Животные, питающиеся жидкой
пищей
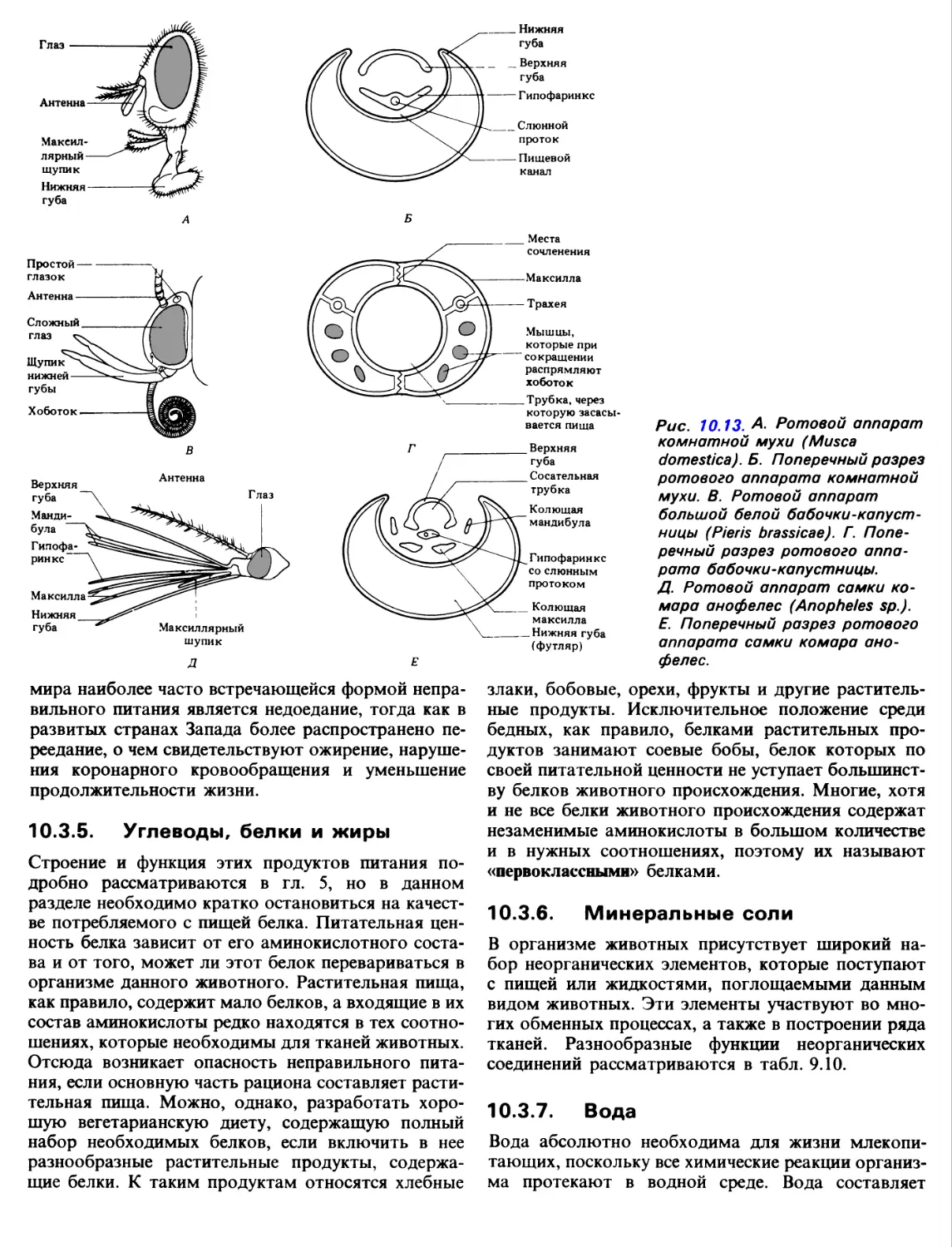
сосущий ротовой аппарат. Комнатная муха, Musca
domestica, имеет хоботок, образованный сильно
видоизмененной нижней губой. Жвалы у нее
отсутствуют, а нижняя челюсть редуцирована и
превращена в пару щупиков. На проксимальном конце
хоботка находится расположенный по центру рот, а
на дистальном конце-две сосательные лопасти,
носящие название лабелл. Каждая лабелла пронизана
многочисленными мельчайшими канальцами -
псевдотрахеями, которые в конечном счете соединяются
в центральный канал хоботка.
Обычно хоботок прижат к нижней стороне тела
насекомого, но, когда оно поглощает пищу, хоботок
под давлением крови выпрямляется и лабелла
присасывается к пищевой частице. Если пища твердая,
то из слюнных желез через отверстия,
расположенные над ротовым отверстием, выделяется слюна.
Слюна содержит ряд ферментов, под воздействием
которых пища становится растворимой. Ставшая
растворимой или бывшая изначально жидкой пища
проходит под действием капиллярных сил в
псевдотрахеи, а из них под действием перистальтических
сокращений засасывается в глотку (рис. 10.13,2T).
Бабочки, такие, как капустная белянка (Pieris bras-
sicae), питаются с помощью хоботка, который в
отличие от хоботка мух образуется из двух нижних
челюстей. Части каждой максиллы, именуемые га-
леа, соединяясь вместе, образуют хоботок, внутри
которого проходит желобок; по этому желобку и
засасывается пища. Жвалы у капустницы
отсутствуют, а максиллярные щупики или отсутствуют, или
недоразвиты.
В состоянии покоя хоботок скручен в спираль,
лежащую под головой. Во время сосания спираль
расправляется под действием рефлекторного
сокращения косых мышцы галеа. Хоботок проникает в
венчик цветка, и его конец непосредственно
погружается в пищевую массу, каковой является нектар-
разведенный раствор сахаристых веществ. Обычно
длина хоботка того или иного вида бабочек
соответствует глубине расположения нектара в тех
цветах, которые эти бабочки посещают. Благодаря
сокращениям мышц глотки нектар засасывается в
рот насекомого (рис. 10.13, Д).
Поджелудочная
железа
|Печень|
Кишечник,
в котором
имеется
спиральный
клапан
Кардиальный
отдел желудка
Пилорический
отдел желудка
Селезенка
Рис. 10.11. Вскрытая кошачья акула, видно строение пищеварительной трубки.
Глотка
Пищевод
Надглоточный
ганглий
Сегмен -
тарный
ганглий
Зоб
Мускульный
желудок
колюще-сосущие ротовые части. Самка комара
Anopheles питается кровью млекопитающих. Чтобы
добыть себе пищу, она должна проколоть кожу
животного, и проделывается это с помощью сильно
видоизмененных жвал или максилл,
превратившихся в четыре острых стилета. Стилеты располагаются
в желобке, образованном сильно вытянутой нижней
губой. Сверху желобок нижней губы прикрывается
желобком вытянутой верхней губы.
В состав образующегося таким образом хоботка
входит также гипофаринкс. Когда гипофаринкс
прижимается к верхней губе, образуется пищевой канал,
по которому и засасывается жидкая пища. По гипо-
фаринксу проходит слюнной проток. По нему в
кровь жертвы во время сосания выделяется слюна,
содержащая антикоагулянт, который препятствует
свертыванию крови, благодаря чему она может
засасываться в глотку через узкий пищевой канал
(рис. 10.13, E).
10.3. Питание млекопитающих
Нервная
цепочка
!Кишечник
Спинной
кровеносный
сосуд
10.3.1. Пищевой рацион
Каждое млекопитающее животное должно
ежедневно получать с пищей продукты, снабжающие его
энергией (углеводы и жиры) и строительным
материалом (белки), а также достаточное количество
минеральных солей, клетчатки и витаминов. Чтобы
пищевой рацион был адекватным и
сбалансированным, указанные компоненты должны находиться в
нем в правильных соотношениях. Такой рацион не
гарантирует от болезней, но в определенной степени
снижает вероятность заболевания, вызванного
неправильным питанием. Оптимальный пищевой
рацион должен заметно варьировать у разных
индивидуумов в зависимости от пола, возраста,
активности, размеров тела и температуры окружающей
среды (в жарком климате пищи потребляется меньше).
10.3. Почему мыши на единицу веса тела
требуется больше джоулей, чем человеку?
Рис. 10.12. Вскрытый дождевой червь; видно
строение пищеварительной трубки
10.3.2. Измерение энергетической
ценности пищевых продуктов
Адекватный пищевой рацион должен содержать
достаточное количество энергии, необходимой для
обеспечения ежедневных метаболических
потребностей организма. Энергетическая ценность пищевых
продуктов измеряется в виде тепловой энергии и
выражается в джоулях. Она может быть определена
путем сжигания известной массы продукта в
кислороде в калориметрической бомбе. Образующееся при
этом сжигании тепло передается определенной массе
воды, повышение температуры которой измеряется.
Исходя из того что для повышения температуры 1 г
воды на 1°С требуется 4,18 джоуля, можно рассчи-
тать количество джоулей, выделяющихся при
сгорании данного продукта питания (рис. 10.14).
10.4. Сколько выделится килокалорий, если при
сжигании в кислороде 1 г сахара температура
500 г воды повышается на 7,5°С?
В табл. 10.4 приведены рекомендуемые в Англии
суточные нормы потребления различных
питательных веществ для мужчин и женщин с учетом
возраста и физической активности. В табл. 10.5
приведен состав целого ряда продуктов питания,
рассчитанный на 100 г съедаемого продукта; величины
взяты из Manual Nutrition, HMSO, 1976.
10.3.3. Определение энергетических
затрат человека
Для определения энер1 етических затрат человека
применяют метод «непрямой калориметрии». Для
расчета энергетических затрат проводится точное
измерение поглощения кислорода и выделения CO2,
а иногда и выделения азота с мочой. В основе
метода лежит теоретическое предположение о том,
что при сгорании 1 г пищевого продукта в
организме поглощается такое же количество кислорода и
выделяется такое же количество CO2, теплоты и
воды, как при сгорании этого продукта на воздухе.
Однако определяемая таким путем величина
является приблизительной, поскольку полного окисления
пищевых продуктов в организме не происходит.
10.5. Рассчитано, что при взаимодействии 1 г
глюкозы с 774 мл кислорода выделяется 15,8 кДж
тепла, а при взаимодействии 1 г жирной кислоты
с длинной цепью с 2012 мл кислорода выделяется
39,4 кДж. Почему 1 г жирной кислоты дает в два с
лишним раза больше энергии, чем 1 г глюкозы?
10.6. Почему при сгорании белков и жиров в
калориметрической бомбе выделяется больше
тепла, чем при сгорании точно такого же их
количества в организме?
10.3.4. Неправильное питание
Неправильное питание имеет место в том случае,
когда в течение длительного времени организм
получает недостаточное количество пищи (недоедание)
или получает в избыточном количестве один или
несколько пищевых продуктов, богатых энергией
(переедание). Во многих слаборазвитых странах
Таблица 10.3. Строение и функции различных отделов пищеварительной трубки дождевого червя
Отдел
пищеварительного
тракта
Сегменты
Строение
Функция
Рот
С помощью рта червь захватывает пищу, затаскивает ее в нору и
отрывает от нее кусочки
Ротовая полость
1-3
Широкая тонкостенная
Пища смачивается и размягчается под действием секрета
глоточных желез. Секрет содержит слизь и протеолитические
ферменты. Периодически с помощью перистальтических движений
пища заглатывается
Глотка
4-5
Растягивающаяся
мышечная тонкостенная
Содержит скопления железистой ткани, выделения которой
способствуют размягчению пищи
Пищевод
6-13
Имеет вид узкой
тонкостенной трубки
В 10-м сегменте в пищевод открываются эзофагальные карманы.
В 11-ми 12-м сегментах имеются отверстия двух пар кальци-
фицирующих желез. Эти железы секретируют жидкость,
содержащую частички карбоната кальция, и выполняют скорее
выделительную, чем пищеварительную функцию, способствуя
удалению из организма избытка кальция
Зоб
14-16
Обширный
тонкостенный
Служит для хранения пищи и некоторого предварительного ее
переваривания
Мускульный
желудок
17-19
Сферический
тонкостенный твердый и
мускулистый
Содержит острые песчинки и служит для «пережевывания» пищи.
Кусочки пищи измельчаются путем перетирания между
песчинками и покрытой кутикулой выстилкой желудка
Кишечник
20
Стенка кишки
образована слоями продольных
и концевых мышечных
волокон. Площадь
поверхности увеличена за
счет тифлозоля
Имеет большую площадь поверхности для секреции ферментов и
всасывания переваренной пищи. Пищеварение внеклеточное.
Переваренная пища всасывается в сеть капилляров,
покрывающих кишечник. В кишечнике имеются три типа клеток:
1. Железистые клетки, секретирующие протеолитические,
амилолитические и липолитические ферменты; (обратите
внимание, что секретируется целлюлаза)
2. Ресничные клетки, способствующие перемешиванию пищи
с ферментами
3. Слизистые клетки, обеспечивающие смазывание пищи и
защиту кишечника от переваривания ферментами
Анальное
отверстие
Эвакуация каловых масс в виде копролитов
Глаз —
Антенна
Макеил-
лярный
щупик
Нижняя
губа
Нижняя
губа
Верхняя
губа
Гипофаринкс
Слюнной
проток
Пищевой
канал
Простой
глазок
Антенна
Сложный
глаз
Щупик
нижней
губы
Хоботок
Места
сочленения
Максилла
Трахея
Мышцы,
которые при
сокращении
распрямляют
хоботок
Трубка, через
которую
засасывается пища
Верхняя
губа
_ Сосательная
трубка
Колющая
мандибула
Гипофаринкс
со слюнным
протоком
_ Колющая
максилла
_ Нижняя губа
(футляр)
Верхняя
губа
Мандибула ^
Гилофа- '
ринке
Максилла:
Нижняя
губа
Антенна
Глаз
Максиллярный
шупик
Рис. 10.13. А. Ротовой аппарат
комнатной мухи (Musca
domestica). Б. Поперечный разрез
ротового аппарата комнатной
мухи. В. Ротовой аппарат
большой белой
бабочки-капустницы (Pieris brassicae). Г.
Поперечный разрез ротового
аппарата бабочки-капустницы.
Д. Ротовой аппарат самки
комара анофелес (Anopheles sp.).
E. Поперечный разрез ротового
аппарата самки комара
анофелес.
мира наиболее часто встречающейся формой
неправильного питания является недоедание, тогда как в
развитых странах Запада более распространено
переедание, о чем свидетельствуют ожирение,
нарушения коронарного кровообращения и уменьшение
продолжительности жизни.
10.3.5. Углеводы, белки и жиры
Строение и функция этих продуктов питания
подробно рассматриваются в гл. 5, но в данном
разделе необходимо кратко остановиться на
качестве потребляемого с пищей белка. Питательная
ценность белка зависит от его аминокислотного
состава и от того, может ли этот белок перевариваться в
организме данного животного. Растительная пища,
как правило, содержит мало белков, а входящие в их
состав аминокислоты редко находятся в тех
соотношениях, которые необходимы для тканей животных.
Отсюда возникает опасность неправильного
питания, если основную часть рациона составляет
растительная пища. Можно, однако, разработать
хорошую вегетарианскую диету, содержащую полный
набор необходимых белков, если включить в нее
разнообразные растительные продукты,
содержащие белки. К таким продуктам относятся хлебные
злаки, бобовые, орехи, фрукты и другие
растительные продукты. Исключительное положение среди
бедных, как правило, белками растительных
продуктов занимают соевые бобы, белок которых по
своей питательной ценности не уступает
большинству белков животного происхождения. Многие, хотя
и не все белки животного происхождения содержат
незаменимые аминокислоты в большом количестве
и в нужных соотношениях, поэтому их называют
«первоклассными» белками.
10.3.6. Минеральные соли
В организме животных присутствует широкий
набор неорганических элементов, которые поступают
с пищей или жидкостями, поглощаемыми данным
видом животных. Эти элементы участвуют во
многих обменных процессах, а также в построении ряда
тканей. Разнообразные функции неорганических
соединений рассматриваются в табл. 9.10.
10.3.7. Вода
Вода абсолютно необходима для жизни
млекопитающих, поскольку все химические реакции
организма протекают в водной среде. Вода составляет
К фильтру
насоса
Кислород —р£
Термометр
Мешалка
Тепл опере дающая
спираль
Вода
Асбестовая
подставка
Пища, сжигаемая ι
никелевом тигле
Рис. 10.14. А. Внешний вид калориметра,
предназначенного для определения энергетической
ценности пищевых продуктов. Для сжигания пищи
используется маленькая электрическая нагреватель
ная спираль. Б Схема строения калориметра
65-70% общей массы тела, а поскольку масса тела
изо дня в день остается относительно постоянной,
то те 2-3 л воды, которые организм ежедневно
теряет, должны возмещаться за счет жидкости или
пищи, которые животное ежедневно поглощает.
Насколько необходима вода для поддержания жизни,
видно из того, что без пищи человек может прожить
более 60 дней, а без воды-всего лишь несколько
дней. Полный перечень функций, выполняемых
водой в организме животных и растений, приводится в
гл. 5 и 14.
10.3.8. Грубоволокнистые пищевые
продукты
Грубая пища человека состоит из неперевариваемой
целлюлозы, входящей в состав клеточных стенок
растительных клеток. Целлюлоза, или клетчатка,
обладает способностью удерживать воду и
составляет основную массу содержимого кишечника,
особенно толстого. Растягивая стенки толстого
кишечника, клетчатка стимулирует его перистальтику,
способствуя тем самым продвижению содержимого
толстого кишечника к анальному отверстию и акту
дефекации. Отсутствие в рационе грубых волокон
может приводить к запорам и другим нарушениям
работы толстого кишечника.
10.3.9. Молоко
Молоко является единственной пищей
млекопитающих в первые недели их жизни. В этот период
развития они получают с молоком почти все
необходимые питательные вещества - углеводы, белки,
жиры, минеральные элементы (и прежде всего Ca,
Mg, P и К) и различные витамины. Из необходимых
элементов в молоке отсутствует только железо,
входящее в состав гемоглобина крови. Однако плод
преодолевает эту трудность, накапливая железо,
которое поступает от матери и которое к моменту
рождения сохраняется в его теле в достаточном
количестве. Эти запасы железа поддерживают
развитие плода и новорожденного до той поры, пока он
не начнет принимать твердую пищу.
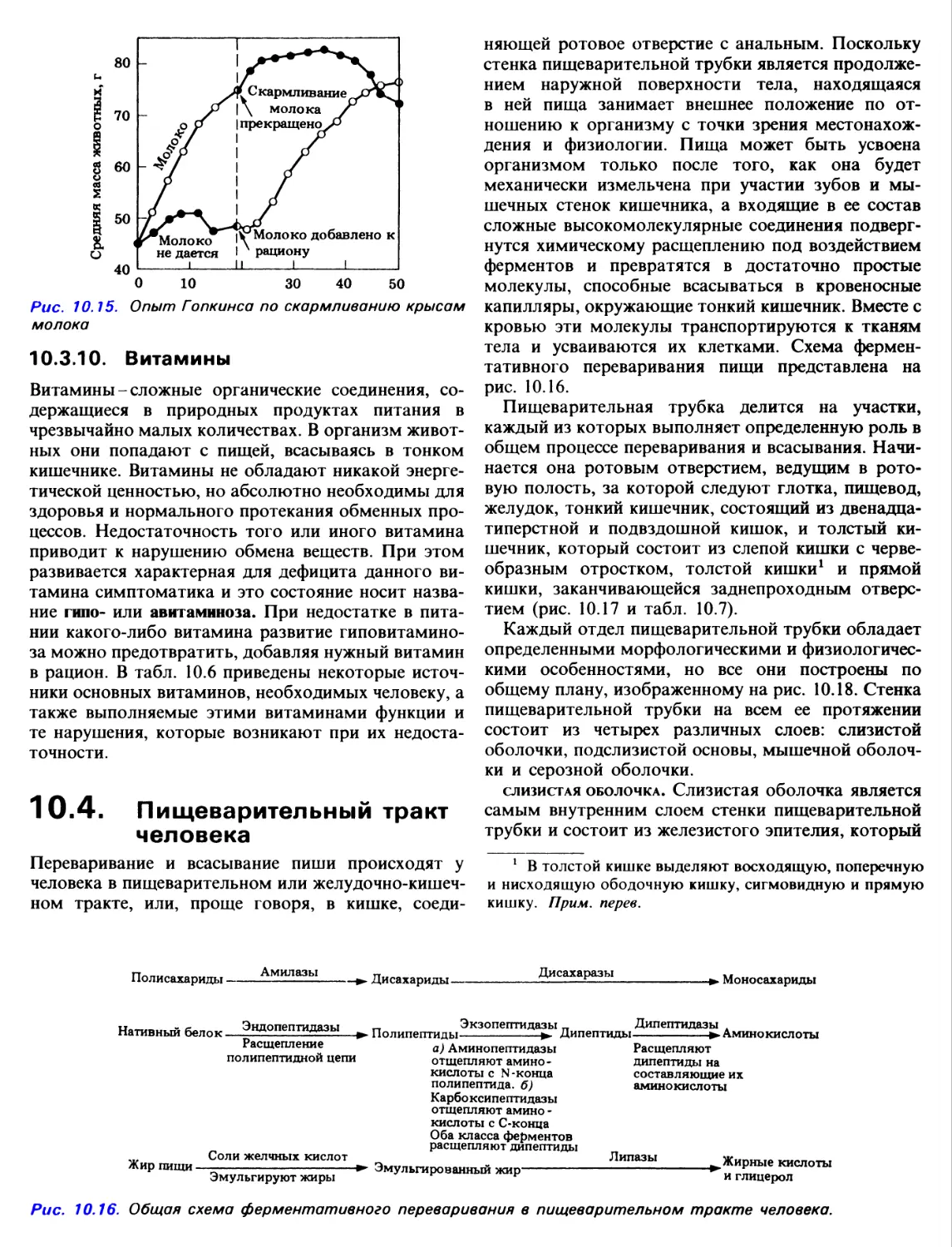
10.7. В начале нашего века Фредерик Гауленд
Гопкинс из Кембриджа провел известный опыт, в
котором две группы крысят (по 8 крысят в
каждой группе) кормили пищей, составленной
из чистого казеина, крахмала, сахарозы, сала,
неорганических солей и воды. Одна группа крысят
получала дополнительно 3 мл молока в день в
течение первых 18 сут. На 18-е сутки молоко было
исключено из рациона первой группы, но его
начали получать крысята второй группы.
Результаты приведены на рис. 10.15.
а) Какую гипотезу могли бы вы предложить на
основании рисунка?
б) Подкрепите ваш ответ пояснением.
в) Почему взрослые не могут питаться одним
молоком?
Таблица 10.4. Суточные нормы потребления энергии и питательных веществ для мужчин и женщин в зависимости от возраста
и активности, рекомендуемые министерством здравоохранения и социальною обеспечения Англии (1969 год)
Возраст и
образ жизни
Масса
тела, кг
Энергия
ккал
МДж
БелокХ),
г
Тиамин,
мг
Рибофлавин, мг
Никотиновая
кислота,
мг-экви-
валенты
Аскорбиновая
кислота,
мг
Витамины
А, мкг-
эквива-
ленты
ретинола^
мкг-экви-
валенты
кальциферола
Кальций,
мг
Железо,
мг
МАЛЬЧИКИ И
ДЕВОЧКИ
0-1 год
2-4 года
4-7 лет
7,3
13,5
20,5
800
1400
1800
3,3
5,9
7,5
20
35
45
0,3
0,6
0,7
0,4
0,7
0,9
5
8
10
15
20
20
450
300
300
10
10
2,5
600
500
500
6
7
8
МАЛЬЧИКИ
9-12 лет
12-15 лет
15-18 лет
31,9
45,5
61,0
2500
2800
3000
10,5
11,7
12,6
63
70
75
1,0
U
1,1
1,2
1,4
1,7
14
16
19
25
25
30
575
725
750
2,5
2,5
2,5
700
700
600
13
14
15
ДЕВОЧКИ
9-12 лет
12-15 лет
15-18 лет
33,0
48,6
56,1
2300
2300
2300
9,6
9,6
9,6
58
58
58
0,9
0,9
0,9
1,2
1,4
1,4
13
16
16
25
25
30
575
725
750
2,5
2,5
2,5
700
700
600
13
14
15
МУЖЧИНЫ
18 35 лет
Сидячий
Умеренно
активный
Очень
активный
65
2700
3000
3600
п,з
12,6
15,1
68
75
90
1,1
1,2
1,4
1,7
1,7
1,7
18
18
18
30
30
30
750
750
750
2,5
2,5
2,5
500
500
500
10
10
10
35-65 лет
Сидячий 65
Умеренно
активный
Очень
активный
2600
2900
3600
10,9
12,1
15,1
65
73
90
1,0
1,2
1,4
1,7
1,7
1,7
18
18
18
30
30
30
750
750
750
2,5
2,5
2,5
500
500
500
10
10
10
65-75 лет
75 и
более лет
Пред-
-
полагается
сидячий
образ
жизни
63
63
2350
2100
9,8
8,8
59
53
0,9
0,8
1,7
1,7
18
18
30
30
750
750
2,5
2,5
500
500
10
10
ЖЕНЩИНЫ
18-55 лет
Большинство
занятий
Очень
активный
55
2200
2500
9,2
10,5
55
63
0,9
1,0
1,3
1,3
15
15
30
30
750
750
2,5
2,5
500
500
12
12
55-75 лет
75 и
более лет
полагается
сидячий
образ
жизни
53
53
2050
1900
8,6
8,0
51
48
0,8
0,7
1,3
1,3
15
15
30
30
750
750
2,5
2,5
500
500
10
10
Беременность
2-й и 3-й
триместры
Лактация
2400
2700
10,0
п,з
60
68
1,0
1,1
1,6
1,8
18
21
60
60
750
1200
10,0
10,0
1200
1200
15
15
'Рекомендуется из расчета, что это количество белка дает 10% всей энергии
2)1 эквивалент ретинола = 1 мкг ретинола или 12 мкг других биологически активных каротиноидов.
Таблица 10.5. Состав некоторых продуктов питания в расчете на 100 г съедобной части. Величины взяты из Справочника
по питанию за 1976 год
Миндаль
Яблоки
Абрикосы
консервированные
Бекон,
ветчина
(вареные)
Бананы
Бобы
консервированные
в томатном
соусе
Фасоль
огненно-красная
Говядина
обычная
Говяжья
солонина
Пиво горькое
из бочки
Свекла
вареная
Печенье сухое
полусладкое
Хлеб черный
Хлеб белый
Хлеб из
непросеянной
муки
Капуста
брюссельская,
вареная
Масло
Капуста
кочанная,
вареная
Морковь
Капуста
цветная
Сыр чеддер
Цыпленок
жареный
Шоколад
молочный
Кокосовый
орех,
сушеный
Треска,
запеченная в
тесте
Треска,
пикша, белая
рыба
Кофе
растворимый
Энергия,
ккал
580
46
106
447
76
63
23
226
216
30
44
431
230
251
241
17
731
15
23
13
412
148
578
608
199
76
155
кДж
2397
197
452
1852
326
266
100
940
905
127
189
1819
981
1068
1025
75
3006
66
98
56
1708
621
2411
2509
834
321
662
Белок, г
20,5
0,3
0,5
24,5
1,1
5,1
2,2
18,1
26,9
0
1,8
7,4
9,2
8,0
9,6
2,8
0,5
1,7
0,7
1,9
25,4
24,8
8,7
6,6
19,6
17,4
4,0
Жир, г
53,3
0
0
38,8
0
0,4
0
17,1
12,1
0
0
13,2
1,4
1,7
3,1
0
81
0
0
0
34,5
5,4
37,6
62,0
10,3
0,7
0,7
Углеводы, г
4,3
12,0
27,7
0
19,2
10,3
3,9
0
0
2,3
9,9
75,3
48,3
54,3
46,7
1,7
0
2,3
5,4
1,5
0
0
54,5
6,4
7,5
0
35,5
Минеральные
элементы
Ca, мг
247
4
12
12
7
45
27
7
14
11
30
126
88
100
28
25
15
38
48
21
810
9
246
22
80
16
140
Fe, мг
4,2
0,3
0,7
1,4
0,4
1,4
0,8
1,9
2,9
0
0,7
1,8
2,5
1,7
3,0
0,5
0,2
0,4
0,6
0,5
0,6
0,8
1,7
3,6
0,5
0,3
4,0
Витамины
А, мкг
0
5
166
0
33
50
50
0
0
0
0
0
0
0
0
67
995
50
2000
5
420
0
6,6
0
0
0
0
D, мкг
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
1,25
0
0
0
0,35
0
0
0
0
0
0
Bj, мг
0,32
0,04
0,02
0,40
0,04
0,07
0,05
0,06
0,01
0
0,02
0,17
0,28
0,18
0,24
0,06
0
0,03
0,06
0,10
0,04
0,08
0,03
0,06
0,04
0,08
0
B2, мг
0,25
0,02
0,01
0,19
0,07
0,05
0,10
0,19
0,23
0,05
0,04
0,06
0,07
0,03
0,09
0,10
0
0,03
0,05
0,10
0,05
0,19
0,35
0,04
0,10
0,07
0,10
Никотиновая
кислота,
мг-эквиваленты
4,9
0,1
0,3
9,2
0,8
1,4
1,4
8,1
9,0
0,7
0,4
2,0
2,7
2,6
1,9
1,0
0,1
0,5
0,7
1,0
5,2
12,8
2,5
1,8
6,7
4,8
45,7
С, мг
0
5
5
0
10
3
20
0
0
0
5
0
0
0
0
41
0
23
6
64
0
0
0
0
0
0
0
Продолжение табл. 10.5
Кукурузные
хлопья
Сливки
двойные
Сливки
одинарные
Яйца
Рыбные
палочки
Мука
крупчатка
Фруктовый
кекс
Окорок
вареный
Мед
Мороженое
ванильное
Джем
Рыба копченая
Баранина
жареная
Салат
Печень
жареная
Мясо для
завтрака
Маргарин
Мармелад
Молоко
жидкое, цельное
Столовый жир
и
растительное масло
Лук
Апельсины
Пастернак
посевной
Персики
сервированные
Арахис
жареный
Груши свежие
Горох (свежий
или
замороженный)
вареный
Ананасы
сервированные
Энергия,
ккал
354
449
189
147
178
348
368
269
288
192
262
184
291
8
244
313
734
261
65
899
23
35
49
88
586
41
49
76
кДж
1507
1848
781
612
749
1483
1546
1119
1229
805
1116
770
1209
36
1020
1298
3019
1114
274
3696
98
150
210
373
2428
175
208
325
Белок,
IA
1,8
2,8
12,3
12,6
10,0
4,6
24,7
0,4
4,1
0,5
19,8
23,0
1,0
24,9
12,6
0,2
0,1
3,3
0
0,9
0,8
1,7
0,4
28,1
0,3
5,0
0,3
г Жир, г
0,4
48,0
18,0
10,9
7,5
0,9
15,9
18,9
0
п,з
0
11,7
22,1
0
13,7
26,9
81,5
0
3,8
99,9
0
0
0
0
49,0
0
0
0
Углеводы, г
85,4
2,6
4,2
0
16,1
80,0
55,0
0
76,4
19,8
69,2
0
0
1,2
5,6
5,5
0
69,5
4,8
0
5,2
8,5
11,3
22,9
8,6
10,6
7,7
20,0
Минеральные
элементы
Ca, мг
5
65
100
54
43
138
71
9
5
137
18
60
9
23
14
15
4
35
120
0
31
41
55
4
61
8
13
13
Fe, мг
0,3
0
0,1
2,1
0,7
2,1
1,6
1,3
0,4
0,3
1,2
1,2
2,1
0,9
8,8
1,0
0,3
0,6
0,1
0
0,3
0,3
0,6
1,9
2,0
0,2
1,2
1,7
Витамины
А, мкг
0
420
155
140
0
0
57
0
0
1
2
45
0
167
6000
0
900в)
8
44г)
37д)
0
0
8
0
41
0
2
50
7
D, мкг
0
0,28
0,10
1,50
0
0
0,80
0
0
0
0
22,20
0
0
0,75
0
8,0
0
0,05г)
0,01д)
0
0
0
0
0
0
0
0
0
B1, мг
1,13а)
0,04б)
0,02
0,03
0,09
0,09
0,30
0,07
0,44
0
0,05
0
0,02
0,10
0,07
0,27
0,07
0
0
0,04
0
0,03
0,10
0,10
0,01
0,23
0,03
0,25
0,05
B2, мг
1,41а)
0,10б)
0,08
0,13
0,47
0,06
0,03
0,07
0,15
0,05
0,20
0
0,30
0,25
0,08
4,30
0,12
0
0
0,15
0
0,05
0,03
0,09
0,02
0,10
0,03
0,11
0,02
Никотиновая
кислота,
мг-экви-
валенты
10,6а)
0,8б)
0,4
0,8
3,7
3,1
2,7
1,2
0,8
0,2
1,1
0
6,9
9,2
0,4
20,7
4,5
0,1
0
0,9
0
0,4
0,3
1,3
0,6
20,8
0,3
2,3
0,3
С, мг
0
0
0
0
0
0
0
0
0
1
10
0
0
15
20
0
0
10
1
0
10
50
15
4
0
3
15
8
Продолжение табл. 10.5
Кекс без
фруктов
Изюм
Свинина
обычная
Картофель
вареный
Картофель
фри
Картофель
жареный
Чернослив
Малина
Ревень
Рис
Рисовый
пудинг
Колбаса
свиная
Томатный суп
консервированный
Спагетти
Шпинат
Спирт 70%-
ный
Бифштекс и
пирог с
почками
Клубника,
земляника
Сахар-рафинад
Сахарная
кукуруза
сервированная
Сироп
Помидоры
свежие
Кресс-салат
Вино красное
Йогурт
фруктовый
Йогурт
натуральный
Энергия,
ккал
426
32
330
80
236
111
161
25
6
359
142
367
55
364
21
221
304
26
394
79
298
12
14
67
96
53
кДж
1785
137
1364
339
1028
474
686
105
26
1531
594
1520
230
1549
91
914
1266
109
1680
336
1269
52
60
277
410
224
Белок,
6,0
0,6
15,8
1,4
3,8
2,8
2,4
0,9
0,6
6,2
3,6
10,6
0,8
9,9
2,7
0
13,3
0,6
0
2,9
0,3
0,8
2,9
0
4,8
5,0
г Жир, г
24,0
0
29,6
0
9,0
1,0
0
0
0
1,0
7,6
32,1
3,3
1,0
0
0
21,1
0
0
0,8
0
0
0
0
1,0
1,0
Углеводы, г
49,7
7,9
0
19,7
37,3
27,3
40,3
5,6
1,0
86,8
15,7
9,5
5,9
84,0
2,8
0
14,6
6,2
105,0
16,1
79,0
2,4
0,7
0,3
18,2
6,4
Минеральные
элементы
Ca, мг
67
12
8
4
14
10
38
41
103
4
116
41
17
23
70
0
37
22
1
3
26
13
222
6
160
180
Fe, мг
1,4
0,3
0,8
0,5
1,4
1,0
2,9
1,2
0,4
0,4
0,1
1,1
0,4
1,2
3,2
0
5,1
0,7
0
0,1
1,4
0,4
1,6
0,8
0,2
0,1
Витамины
А, мкг
82
37
0
0
0
0
160
13
10
0
96
0
35
0
1000
0
126
5
0
35
0
117
500
0
10
10
D, мкг
1,20
0
0
0
0
0
0
0
0
0
0,08
0
0
0
0
0
0,55
0
0
0
0
0
0
0
0,02
0,02
B1, мг
0,08
0,05
0,58
0,08
0,10
0,10
0,10
0,02
0,01
0,08
0,05
0,04
0,03
0,09
0,12
0
0,11
0,02
0
0,05
0
0,06
0,10
0,01
0,05
0,05
B2, мг
0,11
0,03
0,16
0,03
0,04
0,04
0,20
0,03
0,07
0,03
0,14
0,12
0,02
0,06
0,20
0
0,47
0,03
0
0,08
0
0,04
0,16
0,02
0,23
0,26
Никотиновая
кислота,
мг-экви-
валенты
1,7
0,6
6,9
1,2
2,2
2,0
1,7
0,5
0,3
1,5
1,0
5,7
0,2
1,8
1,3
0
6,0
0,5
0
0,3
0
0,7
2,0
0,2
1,2
1,3
С, мг
0
3
0
4-15е)
6-20е)
6-23е)
0
25
10
0
1
0
6
0
60
0
0
60
0
4
0
21
60
0
1
0
Примечания
Углеводы даются как моносахариды
Витамин А дается в мкг-эквивалентах ретинола, т. е. величины соответствуют количеству ретинола, образующегося из его каротиновых
предшественников. 1 мкг ретинола эквивалентен 3,33 ME (международных единиц)
Витамин D дается в мкг; 1 мкг витамина D эквивалентен 40 ME.
Витамин B1 дается как тиамин, а витамин B2-как рибофлавин.
Величины никотиновой кислоты соответствуют ее содержанию как таковой или тому количеству никотиновой кислоты, которое
образуется из триптофана (60 мг триптофана дает 1 мг никотиновой кислоты).
а) добавлен;6) не добавлен,в' некоторые виды маргарина содержат каротин;г) летние вепичины; д) зимние величины;е) в свежем картофеле
содержание высокое, а при хранении оно снижается
Средняя масса животных, г
Скармливание
молока
прекращено
Молоко
не дается
Молоко добавлено к
рациону
Рис. 10.15. Опыт Гопкинса по скармливанию крысам
молока
10.3.10. Витамины
Витамины - сложные органические соединения,
содержащиеся в природных продуктах питания в
чрезвычайно малых количествах. В организм
животных они попадают с пищей, всасываясь в тонком
кишечнике. Витамины не обладают никакой
энергетической ценностью, но абсолютно необходимы для
здоровья и нормального протекания обменных
процессов. Недостаточность того или иного витамина
приводит к нарушению обмена веществ. При этом
развивается характерная для дефицита данного
витамина симптоматика и это состояние носит
название гипо- или авитаминоза. При недостатке в
питании какого-либо витамина развитие
гиповитаминоза можно предотвратить, добавляя нужный витамин
в рацион. В табл. 10.6 приведены некоторые
источники основных витаминов, необходимых человеку, а
также выполняемые этими витаминами функции и
те нарушения, которые возникают при их
недостаточности.
10.4. Пищеварительный тракт
человека
Переваривание и всасывание пиши происходят у
человека в пищеварительном или
желудочно-кишечном тракте, или, проще говоря, в кишке,
соединяющей ротовое отверстие с анальным. Поскольку
стенка пищеварительной трубки является
продолжением наружной поверхности тела, находящаяся
в ней пища занимает внешнее положение по
отношению к организму с точки зрения
местонахождения и физиологии. Пища может быть усвоена
организмом только после того, как она будет
механически измельчена при участии зубов и
мышечных стенок кишечника, а входящие в ее состав
сложные высокомолекулярные соединения
подвергнутся химическому расщеплению под воздействием
ферментов и превратятся в достаточно простые
молекулы, способные всасываться в кровеносные
капилляры, окружающие тонкий кишечник. Вместе с
кровью эти молекулы транспортируются к тканям
тела и усваиваются их клетками. Схема
ферментативного переваривания пищи представлена на
рис. 10.16.
Пищеварительная трубка делится на участки,
каждый из которых выполняет определенную роль в
общем процессе переваривания и всасывания.
Начинается она ротовым отверстием, ведущим в
ротовую полость, за которой следуют глотка, пищевод,
желудок, тонкий кишечник, состоящий из
двенадцатиперстной и подвздошной кишок, и толстый
кишечник, который состоит из слепой кишки с
червеобразным отростком, толстой кишки1 и прямой
кишки, заканчивающейся заднепроходным
отверстием (рис. 10.17 и табл. 10.7).
Каждый отдел пищеварительной трубки обладает
определенными морфологическими и
физиологическими особенностями, но все они построены по
общему плану, изображенному на рис. 10.18. Стенка
пищеварительной трубки на всем ее протяжении
состоит из четырех различных слоев: слизистой
оболочки, подслизистой основы, мышечной
оболочки и серозной оболочки.
слизистая оболочка. Слизистая оболочка является
самым внутренним слоем стенки пищеварительной
трубки и состоит из железистого эпителия, который
1 В толстой кишке выделяют восходящую, поперечную
и нисходящую ободочную кишку, сигмовидную и прямую
кишку. Прим. перев.
Полисахариды -
Амилазы
► Дисахариды-
Дисахаразы
»Моносахариды
Нативный белок энДопептидазы ^
Расщепление
полипептидной цепи
Полипептиды
Экзопептидазы
Дипептиды
Дипептидазы
^ Аминокислоты
Жир пищи ·
Соли желчных кислот
Эмульгируют жиры
а) Аминопептидазы
отщепляют
аминокислоты с N-конца
полипептида, б)
Карбо ксипепти дазы
отщепляют амино -
кислоты с С-конца
Оба класса ферментов
расщепляют дипептиды
Эмульгированный жир-
Расщепляют
дипептиды на
составляющие их
амино кислоты
Липазы
Жирные кислоты
и глицерол
Рис. 10.16. Общая схема ферментативного переваривания в пищеварительном тракте человека.
Таблица 10.6. Источники и функции основных витаминов, необходимых человеку
Название
витамина и обозначение
Основные источники
Жирорастворимые витамины
А
Ретинол
D
Кальциферол
E
Токоферол
К
Филлохинон
Жир из печени трески и
палтуса, печень
крупного рогатого скота,
молоко и молочные
продукты, морковь,
шпинат, кресс-салат
Жир из печени трески и
палтуса, яичный
желток, маргарин, молоко;
образуется в коже при
воздействии солнечного
света на липиды
Зародыши пшеницы,
ржаная мука, печень,
зеленые овощи
Шпинат, кочанная
капуста, брюссельская
капуста; синтезируется
микрофлорой
кишечника
Водорастворимые витамины
Тиамин
B6
Пиридоксин
B5
Пантотеновая
кислота
B3 (PP)
Никотиновая
кислота
M или Вс
Фолиевая
кислота
B12
Цианкобаламин
H
Биотин
С
Аскорбиновая
кислота
Зародыши пшеницы
или риса, экстракт
дрожжей, непросеянная
мука, печень, почки,
сердце
Яйца, печень, почки,
мука грубого помола,
свежие овощи
Широко распространен
во всех пищевых
продуктах
Мясо, хлеб грубого
помола, дрожжевой
экстракт, печень
Печень, белая рыба,
зеленые овощи
Мясо, молоко, яйца,
рыба, сыр
Дрожжи, печень, почки,
яичный белок;
синтезируется микрофлорой
кишечника
Цитрусовые, зеленые
овощи, картофель,
томаты
Функция
Необходим для нормального роста
и формирования эпителиальных
тканей. Альдегидная форма витамина А
необходима для образования
зрительного пигмента родопсина,
участвующего в темновой адаптации
Регулирует всасывание кальция в
пищеварительном тракте и
связанные с кальцием обменные процессы.
Необходим для образования костей
и зубов. Способствует всасыванию
фосфора
У крыс участвует в функциональной
активности мышц и половой
системы, препятствует гемолизу
эритроцитов. Функция у человека
неизвестна
Участвует на конечной стадии
синтеза протромбина в печени, являясь
незаменимым фактором
свертывания крови
Участвует в качестве кофермента
декарбоксилаз в химических
реакциях тканевого дыхания, прежде
всего в цикле Кребса
В фосфорилированной форме
участвует как кофермент в обмене
аминокислот и жирных кислот
Входит в состав кофермента А,
молекула которого активизирует кар-
боновые кислоты в клеточном
метаболизме
Незаменимый компонент кофермен-
тов НАД и НАДФ, играющих
роль акцепторов водорода в
составе ряда дегидрогеназ. Входит в
состав кофермента А
Участвует в синтезе нуклеопротеинов
и в образовании эритроцитов
Участвует в синтезе РНК.
Предупреждает развитие пернициозной
анемии
Играет роль кофермента в ряде
реакций карбоксилирования.
Участвует в синтезе белка и в трансамини-
ровании
Участвует в метаболизме
соединительной ткани и в образовании
здоровой кожи. Необходим для синтеза
коллагеновых волокон
Признаки недостаточности
Кожа становится сухой, развивается
сухость роговицы (ксерофтальмия) и
дегенерация слизистых оболочек.
Ухудшается, а при большом
дефиците полностью нарушается адаптация
к темноте. Если не добавлять
витамин А в пищу, то в результате ксеро-
фтальмии может развиться
непроходящая слепота
Рахит - нарушение кальцификации
растущих костей. У маленьких детей
характерным признаком рахита
являются кривые ноги, а у детей
постарше вывернутые внутрь
колени. У девочек-подростков
наблюдается деформация тазовых костей,
грозящая осложнениями при
беременности. Остеомаляция -
заболевание взрослых, выражающееся в
костных болях и спонтанных переломах
У крыс недостаточность может
вызвать бесплодие. Наблюдается
атрофия мышц и анемия, связанная
с гемолизом эритроцитов
При небольшой недостаточности
замедляется свертывание крови. При
большом дефиците кровь совсем не
свертывается
Бери-бери - поражение нервной
системы. Мышцы становятся слабыми и
болезненными. Возможны параличи
Сердечная недостаточность. Отеки.
Замедление роста у детей
В крови накапливаются кетокислоты, в
частности пировиноградная кислота
Депрессия и раздражительность.
Анемия. Диарея. Дерматиты
Нарушения нервно-мышечной
координации. Утомляемость. Мышечные
судороги
Пеллагра. Фото дерматиты. Сыпь
Диарея
Анемия, особенно выраженная у
женщин во время беременности
Пернициозная анемия
Дерматиты. Мышечные боли
Цинга. Десны становятся слабыми и
кровоточат. Не заживают раны. Не
образуются волокна соединительной
ткани.
Анемия. Сердечная недостаточность
Рот
Ротовая
полость
Пищевод
Печень
Двенадцати -
перегнал
кишка
Толстая
кишка
Слепая
кишка
Червеобразный
отросток
Анальное,
отверстие
Слюнные
железы
Глотка
Желудок
Селезенка
Поджелудочная
железа
Подвздошная
кишка
Прямая
кишка
Рис. 10.17. Общий план строения пищеварительного
тракта человека.
Железа
(например, печень или
поджелудочная железа)
Брыжейка
Брюшина
Серозная оболочка
Продольный мышечный слой
Ауэрбахово сплетение
Циркулярный мышечный слой
Мейснерово сплетение
Подслизистая основа
Мышечный слой слизистой
Собственная пластинка слизистой
Эпителий
Просвет кишки
Рис. 10.18. Общий план гистологического строения
стенок пищеварительной трубки. Серозная оболочка
образована рыхлой соединительной тканью;
ауэрбахово сплетение регулирует перистальтику;
мейснерово сплетение осуществляет регуляцию секреции
пищеварительных желез; подслизистая основа
образована главным образом коллагеновыми
волокнами; собственная пластинка слизистой образована
соединительной тканью, богата кровеносными и
лимфатическими сосудами.
секретирует большое количество слизи, а также
может секретировать ряд пищеварительных
ферментов. Слизь обволакивает пищу и облегчает
прохождение пищевого комка по пищеварительному
тракту, а также препятствует перевариванию стенок
кишечника его собственными ферментами.
Эпителиальные клетки лежат на базальной мембране, под
которой находится собственная пластинка
слизистой. Последняя состоит из соединительной ткани,
содержащей кровеносные и лимфатические сосуды.
Снаружи от нее лежит тонкий слой гладких мышц-
мышечная пластинка слизистой.
подслизистая основа. Состоит из соединительной
ткани, содержащей коллагеновые и эластические
волокна, а также кровеносные и лимфатические
сосуды и нервные сплетения. В этом слое могут
также располагаться слизистые железы,
выделяющие слизь через проток на поверхность слизистой
оболочки, как, например, бруннеровские железы
двенадцатиперстной кишки.
наружная мышечная оболочка. Состоит, как
правило, из двух слоев - внутреннего кольцевого и
наружного продольного. Координированные
сокращения этих слоев создают волнообразные
перистальтические движения стенок пищеварительного
тракта, которые способствуют продвижению пищевого
комка. В ряде участков пищеварительной трубки
слой кольцевых мышц утолщается, образуя
структуры, носящие названия сфинктеров. Расслабления и
сокращения сфинктеров контролируют
перемещение пищевого комка из одного отдела
пищеварительного тракта в другой. Сфинктеры находятся в
месте перехода пищевода в желудок (кардиальный
сфинктер), желудка в двенадцатиперстную кишку
(пилорический сфинктер), подвздошной кишки в
слепую (илеоцекальная заслонка) и вокруг
анального отверстия.
Между слоями кольцевых и продольных мышц
лежит ауэрбахово нервное сплетение, которое
содержит скопление нервных клеток вегетативной
нервной системы и контролирует перистальтику.
Импульсы, поступающие по симпатическим нервам,
вызывают расслабление мышц пищеварительного
тракта и сокращение сфинктеров, тогда как
импульсы, поступающие по парасимпатическим
нервам, вызывают сокращение мышечных стенок и
раскрытие сфинктеров (разд. 16.2). Между
циркулярным мышечным слоем и подслизистой основой
находится еще одно нервное сплетение (мейснерово
сплетение), которое регулирует секрецию желез, на-
Таблица 10.7. Функции различных отделов
пищеварительного тракта человека
Отдел
Ротовая полость
Глотка
Пищевод
Желудок
Двенадцатиперстная
кишка
Печень (желчь)
Поджелудочная железа
(панкреатический сок)
Подвздошная кишка
Толстый кишечник
Прямая кишка
Анальное отверстие
Функция
Прием пищи и ее
пережевывание
Проглатывание
Соединяет глотку с желудком
Хранение пищи и переваривание
белков
Переваривание и всасывание
Эмульгирование жиров
Переваривание крахмала,
белков и жиров
Завершение пищеварения и
всасывание образующихся
продуктов
Всасывание воды
Формирование и хранение
каловых масс
Дефекация
ходящихся в стенке пищеварительной трубки.
серозная оболочка. Образует самый наружный
слой кишечной стенки. Состоит из рыхлой,
волокнистой соединительной ткани.
Снаружи поверхность пищеварительной трубки
на всем ее протяжении (кроме пищевода) покрыта
брюшиной. Брюшина выстилает также брюшную
полость, в которой расположена большая часть
пищеварительного тракта, и образует брыжейку,
которая поддерживает и подвешивает к задней
стенке тела желудок и кишечник. Брыжейка образована
двумя слоями брюшины; в ней располагаются
нервы, кровеносные и лимфатические сосуды, идущие к
кишечнику и отходящие от него. Клетки брюшины
имеют влажную поверхность, благодаря чему
уменьшается трение различных отделов
пищеварительного тракта друг о друга и о другие органы.
10.4.1. Зубной аппарат человека
У человека имеется две челюсти-неподвижная
верхняя и подвижная нижняя. Обе челюсти снабжены
зубами, которые используются для измельчения
пищи (пережевывания). Жевание составляет
механическую стадию пищеварения, благодаря которой
увеличивается поверхность пищи, доступная
действию ферментов. Зубы представляют собой очень
твердые образования, идеально приспособленные
для выполнения своей функции. Человек имеет две
смены зубов (дифиодонтный тип). Вначале у него
появляются молочные зубы, которые постепенно
заменяются постоянными зубами. Зубы человека
имеют различную форму и размеры и обладают
разной жевательной поверхностью, поэтому
человека относят к гетеродонтным организмам в отличие
от рыб и рептилий, которые являются гомодонтны-
ми организмами, поскольку все зубы у них
одинаковые и, как правило, конусовидные. У человека
имеется 32 постоянных зуба, из них 8 резцов, 4
клыка, 8 ложнокоренных (премоляры) и 12
коренных (моляры). Эти зубы замещают 8 резцов, 4
клыка и 8 премоляров, составляющих молочный
ряд зубов. Состав зубов принято выражать зубной
формулой. У человека зубная формула имеет
следующий вид:
Г 2 1 2 31
2 i-c-pm-m- ,
L 2 Г 2 3_Г
где буквы являются начальными буквами латинских
наименований зубов (i-incisivi-резцы; c-canini-
клыки; pm-premolares-предкоренные и m-mola-
res - коренные), над чертой пишется число зубов
каждой группы в верхней, а под чертой-в нижней
челюсти с каждой стороны (рис. 10.19).
Число, размеры и форма зубов у человека и
других млекопитающих имеют характерные
видовые различия и соответствуют роду пищи и тем
функциям, которые они выполняют. Ниже
рассматриваются строение и функция зубов каждого типа.
Резцы, расположенные в ротовой полости спереди,
имеют ровные острые края, которые используются
для срезания и откусывания пищи (рис. 10.20).
Клыки имеют конусовидную форму (рис. 10.20).
У человека они развиты слабо, но очень хорошо
развиты у хищников, которым они служат для
захватывания и умерщвления жертвы и разрывания
ее на куски. Малые коренные зубы (премоляры,
предкоренные) имеют один или два корня и два
бугорка на жевательной поверхности (рис. 10.20).
Они приспособлены для раздавливания и
перетирания пищи, хотя у человека могут служить также для
разрывания пищи. Коренные зубы (моляры) имеют
несколько корней (верхние имеют три, а нижние-
два корня), а на вершине четыре или пять бугорков
(рис. 10.20). Эти зубы служат для дробления и
перемалывания пищи. В молочном ряду человека
коренные зубы отсутствуют.
10.4.2. Общий план строения зуба
Видимая часть зуба, называемая коронкой, покрыта
эмалью-самым твердым веществом организма,
относительно устойчивым к повреждению (рис. 10.21).
Шейка зуба окружена десной, а корень его погружен
в челюстную кость. Под эмалью находится дентин,
составляющий основную массу зуба. Будучи
довольно плотным, дентин, однако, менее тверд и
устойчив к повреждению, чем эмаль. Он пронизан
многочисленными дентинными канальцами,
содержащими цитоплазматические отростки одонтоблас-
тов-клеток, образующих дентин. Пульпарная
полость зуба содержит одонтобласты, окончания
чувствительных нервов и кровеносные сосуды, по
которым доставляются питательные вещества к живым
тканям зуба и удаляются конечные продукты их
обмена.
Корень зуба покрыт цементом, веществом,
похожим на кость. Многочисленные периодонтальные
волокна, прикрепленные одним концом к цементу, а
другим - к челюстной кости, прочно удерживают зуб
в альвеоле. Однако зуб обладает некоторой
подвижностью, благодаря чему меньше вероятность
того, что зуб сломается при жевании.
10.4.3. Развитие зубов у человека
У плода зубы начинают развиваться
приблизительно после 6-й недели внутриутробной жизни.
Эпителиальные клетки ротовой полости делятся, образуя
зубные зачатки. Зачатки увеличиваются в размерах
и внедряются в мезенхиму, образуя эмалевые
органы. Постепенно эмалевый орган становится
вогнутым и приобретает характерные очертания зуба.
Затем клетки эмалевого органа дифференцируются
на наружные и внутренние эпителиальные клетки.
Клетки внутреннего слоя окружают часть
мезенхимы-зубной сосочек, из которого развиваются
клетки, образующие дентин и зубную мякоть (пульпу).
Одновременно с началом образования дентина
|Резец|
Малый коренной
|зуб|
IKлык!
Коренной
зуб
■Корень зуба!
Рис. 10.19. А Рентгенограмма головы человека в профиль, на которой виден ряд постоянных зубов одной
стороны. Б. Рентгенограмма головы человека спереди, на которой виден полный ряд постоянных зубов.
Резец
Клык
Малый коренной
(премоляр)
Коренной
(моляр)
Рис. 10.20. Четыре типа зубов у человека
Эмаль
Дентин
Десна
Пулъпарная
полость
Цемент
Кость ■
Периодонтальные
волокна
Нервы-
Венозное
кровоснабжение
Артериальное
кровоснабжение
КОРОНКА
ШЕЙКА
КОРЕНЬ
Рис. 10.21. Вертикальный шлиф малого коренного
зуба.
внутренние клетки эмалевого органа
дифференцируются и превращаются в амелобласты - клетки,
образующие эмаль. Наконец, в челюстной кости
образуется углубление-альвеола зуба, в которой и
помещается растущий зуб. Стадии развития зуба
показаны на рис. 10.22. Первый зуб прорезывается у
младенца через десну приблизительно через 6
месяцев после рождения.
К трем годам обычно прорезываются уже все
молочные зубы, но челюсти продолжают расти, и
вскоре эти зубы становятся слишком мелкими. К 6
годам молочные зубы начинают шататься и
замещаться первыми постоянными зубами. К 13 годам
должны появиться уже 28 постоянных зубов.
Четыре остальные коренные зуба, или зубы мудрости,
обычно появляются после 17 лет.
У человека и плотоядных животных после того,
как зуб достигает определенного размера, отверстие
в основании зубной полости зарастает, в результате
чего дальнейший рост зуба прекращается. Однако у
травоядных животных это зарастание происходит
не в такой степени, как у плотоядных, и к зубу
продолжает поступать питание, благодаря чему он
постоянно растет.
10.4.4. Заболевания зубов
Существуют два основных вида поражения зубов-
пародонтоз и кариес. Причина обоих заболеваний-
возникновение зубного налета из бактерий и
веществ, содержащихся в слюне. Если позволить
бактериям скапливаться, они вызовут воспаление десен,
пародонтоз. Вследствие взаимодействия с
определенными химическими компонентами слюны
зубной налет становится твердым и кальцифицирован-
ным, в результате чего образуются отложения
зубных камней, которые уже нельзя удалить с
помощью зубной щетки. Некоторые бактерии,
присутствующие в зубном налете, способны
превращать сахар в кислоту, которая вызывает кариес
(рис. 10.23).
Пародонтоз
Пародонтоз - заболевание десен, вызываемое
микроорганизмами, которые присутствуют во рту в
составе зубного налета, особенно в области между
десной и зубом. Несоблюдение правил гигиены
полости рта создает благоприятные условия для
распространения этого заболевания. Сначала
пародонтоз вызывает воспаление десен. Обычно это
воспаление не сопровождается болью, но, если его
не прервать, оно может перейти на корень зуба и
Эмалевый орган
Эпителий
Зубная
пластинка
Клетки,
образующие
эмаль
1. Эпителий челюсти,
впячиваясь, образует
зубную пластинку
Клетки
мезенхимы
Зубной
сосочек -
2 На зубной пластинке
формируются зубные
сосочки
Клетки,
образующие
дентин
3. Дифференцируются
клетки, образующие
эмаль и дентин
Молочный зуб
Дентин
Клетки,
образующие эмаль
Эмаль
Периодонтальные
волокна
Пулъпарная
полость
Развивающийся
постоянный
зуб
Костная лунка
5 Зуб прорезывается через десну
4. Зубная пластинка разрушается
Образуется лунка зуба
Рис. 10.22. Стадии развития зуба.
привести к разрушению периодонтальных волокон,
которые удерживают зуб на месте. В конце концов
зуб расшатывается и может выпасть.
Кариес зубов
Микробы, входящие в состав зубного налета,
превращают присутствующий в ротовой полости сахар
в кислоту. Вначале эмаль разрушается под
действием кислоты медленно и безболезненно. Однако,
когда затрагиваются дентин и пульпа, появляется
сильная боль («зубная боль»), и это состояние
грозит потерей зуба.
В распространении кариеса зубов участвует ряд
факторов, в том числе продолжительное
употребление пищевых продуктов, содержащих сахар,
изменение состава слюны, несоблюдение правил гигиены
полости рта и низкое содержание фтора в питьевой
воде. Предупреждению кариеса способствует
добавление фтора к питьевой воде и некоторым
продуктам питания, в частности к молоку, употребление
детьми содержащих фтор таблеток, использование
для чистки зубов фторированных зубных паст,
тщательное соблюдение правил гигиены полости рта и
регулярные посещения зубного врача для лечения и
профилактического осмотра, а также внимательное
отношение к составу диеты.
Рис. 10.23. Развитие зубного налета. А. Первыми на
зубе поселяются кокки и, размножаясь, образуют
пленку. Б. Микроорганизмы погружаются в пленку,
состоящую из полимеров, образуемых бактериями
и содержащихся в слюне. В. Сообщество
микроорганизмов становится более сложным и
образуются палочковидные и нитевидные колонии. Г. В
полностью сформировавшемся сообществе можно
видеть множество необычных ассоциаций, в том числе
образования, напоминающие початки кукурузы.
Сосочек
Желоб, содержащий
вкусовые луковицы
Рис. 10.24.А. Микрофотография окруженного валом
сосочка трехнедельного щенка, полученная с помощью
сканирующего электронного микроскопа. В желобках,
окружающих сосочек, находятся вкусовые луковицы.
10.4.5. Ротовая полость
Ротовая полость образована челюстями, в ней
находится язык, и она выстлана многослойным плоским
эпителием. Язык является мышечным органом. Во
время жевания он переворачивает пищу во рту,
благодаря чему она перемешивается со слюной и
смачивается. Поверхность языка покрыта
вкусовыми луковицами (рис. 10.24), чувствительными к
сладким, соленым, кислым и горьким веществам, что
помогает определять характер пищи. У человека
язык выполняет также функцию органа речи.
Ежедневно у человека выделяется около 1,5 л
слюны, которая секретируется тремя парами
слюнных желез (рис. 10.25) и множеством щечных желез,
расположенных в слизистой полости рта. Слюна
представляет собой водянистый секрет, содержащий
ферменты амилазу и лизоцим, а также хлористый
натрий, бикарбонаты, фосфаты, карбонаты, Ca2 + ,
K + , Mg2 + , сульфоцианид и слизь. Слизь смачивает
и обволакивает пищу, облегчая тем самым ее
проглатывание. Под воздействием амилазы слюны
начинается расщепление крахмала, который
превращается сначала в декстрины - полисахариды с более
короткой цепью, а затем в дисахарид мальтозу. У
хищных млекопитающих, быстро заглатывающих
большие куски пищи, слюна не содержит никаких
пищеварительных ферментов. Лизоцим
способствует очищению ротовой полости от патогенных мик-
Сосочек
Вкусовые
луковицы
Глубокая
щель
Рис. 10.24. Б. Вертикальный разрез языка; видны вкусовые луковицы.
роорганизмов, катализируя расщепление их
клеточных стенок. В конечном счете полужидкая, частично
переваренная пища склеивается муцином, и при
участии языка из нее формируется пищевой комок,
который проталкивается языком по направлению к
глотке и в результате рефлекторного акта
заглатывается, попадая через глотку в пищевод.
10.4.6. Глотание
Глотание начинается как произвольный акт, но раз
начавшись, продолжается как непроизвольный
процесс вплоть до его завершения. На рис. 10.26
показаны стадии процесса глотания у человека.
10.4.8. Желудок
У человека желудок располагается под диафрагмой
в левой стороне брюшной полости. Он представляет
собой растяжимый мешок с мышечными стенками,
функция которого заключается в хранении и
частичном переваривании пищи. В отличие от других
отделов пищеварительного тракта мышечная
оболочка желудка состоит из трех слоев гладких мышц
(наружного продольного, промежуточного цирку-
10.4.7. Пищевод
Пищевод представляет собой узкую трубку,
имеющую мышечные стенки и выстланную
многослойным плоским эпителием, содержащим слизистые
железы (рис. 10.27). У человека длина пищевода
составляет около 25 см. Благодаря
перистальтическим движениям пищевода пища и жидкость быстро
транспортируются по нему из глотки в желудок. В
верхней части пищевода мышечный слой состоит из
поперечнополосатой и гладкой мускулатуры, а в
нижней части-только из гладкой. У хищников,
заглатывающих большие куски пищи, пищевод
содержит поперечнополосатые мышцы по всей своей
длине.
Слюнной
проток
Кубовидная
клетка с
большим ядром
Рис 10.25. Ткань слюнной железы.
Околоушная железа
Носовая полость
Пищевой комок
Язык
Подъязычная железа
Подчелюстная железа
Трахея
Мягкое нёбо
Глотка
Надгортанник
Позвоночник
Пищевод
1. Пища находится в полости рта. Надгортанник
поднят. Язык прижимает пищевой комок к
мягкому нёбу, благодаря чему вход в носовую
полость перекрывается
Рис. 10.26. Акты процесса глотания у человека.
Язык поднят
Пищевой комок
Гортань
Надгортанник
прижат
2. Гортань (открывающаяся в глотку)
закрывается надгортанником, благодаря
чему пищевой комок поступает не в трахею,
а в глотку
лярного и внутреннего косого), которые
обеспечивают механическое дробление пищи и
перемешивание ее с желудочным соком. Когда желудок нахо-
Собственная пластинка
слизистой
Циркулярный
мышечный слой
Многослойный
плоский
эпителий
Слизистая
железа
Продольный
мышечный слой
Подслизистая
основа
Просвет
пищевода
Лимфоидный
фолликул
дится в нерастянутом состоянии, его выстилка
(слизистая оболочка и подслизистая основа) собрана в
складки, а полностью растянутый желудок может
вмещать до 5 л пищевой массы. Толстая слизистая
оболочка желудка обильно снабжена
эпителиальными клетками, секретирующими слизь, и покрыта
многочисленными желудочными ямками (рис. 10.28
и 10.29). В этих ямках находятся главные и обкла-
дочные клетки, а также содержатся ферменты и
соляная кислота. Суммарный секрет желудка
называется желудочным соком. Слизь создает барьер
между слизистой желудка и желудочным соком и
таким образом препятствует самоперевариванию
желудка. Кардиальный сфинктер, расположенный
при входе пищевода в верхний, кардиальный, отдел
желудка, и пилорический сфинктер, расположенный
при выходе желудка в двенадцатиперстную кишку,
препятствуют бесконтрольной эвакуации пищи из
желудка. Оба сфинктера действуют как клапаны и
задерживают пищу в желудке приблизительно на
4 ч. При периодическом расслаблении пилорическо-
го сфинктера небольшие порции пищи переходят в
двенадцатиперстную кишку. Слизистая кардиально-
го отдела желудка содержит только слизистые
железы, тогда как в основном отделе желудка, в его дне,
содержится множество длинных трубчатых желез,
называемых знмогеннымн или главными (рис. 10.29).
Эти железы состоят из главных (зимогенных), об-
кладочных (париетальных) и аргирофильных (арген-
таффинных) клеток.
зимогенные, или главные клетки. Секретируют
неактивные ферменты пепсиноген и прореннин.
10.8. Почему необходимо, чтобы пепсин секре-
тировался в неактивной форме?
Рис. 10.27. Поперечный разрез пищевода человека.
ОБКЛАДОЧНЫЕ, ИЛИ ПАРИЕТАЛЬНЫЕ КЛЕТКИ. ЭТИ КЛеТКИ
секретируют 0,04-0,05 %-ный раствор соляной
кислоты, благодаря которой желудочный сок имеет рН
1-2,5 - оптимальный для ферментов желудочного
Желудочный сок
Соединительная ткань
собственной пластинки
слизистой
Ямка
Рис. 10.28. Поперечный разрез желудочных ямок
млекопитающих.
сока. Кислота оказывает бактерицидное действие,
разрыхляет ткани, размягчает волокна и
способствует превращению пепсиногена в активный
фермент-пепсин. Пепсин расщепляет белки на более
короткие полипептиды, а также превращает новые
молекулы пепсиногена в пепсин - процесс, носящий
название аутокатализа. Кроме того, соляная
кислота превращает прореннин в реннин, который в
присутствии Ca2+ вызывает коагуляцию растворимого
белка молока казеиногена и превращение его в
нерастворимую кальциевую соль, которая затем
переваривается пепсином. Наконец, под действием
соляной кислоты соли кальция и железа переходят в
такую форму, в которой они могут всасываться в
кишечнике, а также начинается расщепление
сахарозы на глюкозу и фруктозу и нуклеопротеинов на
нуклеиновые кислоты и белок.
АРГИРОФИЛЬНЫЕ, ИЛИ АРГЕНТАФФИННЫЕ, КЛЕТКИ. ЭТИ
клетки образуют внутренний фактор желудка,
необходимый для всасывания молекул витамина B12.
Благодаря сокращениям мышц желудочных
стенок пища хорошо перемешивается с желудочным
соком и в конце концов превращается в полужидкую
кашицу, носящую название химуса. Время от
времени пилорический сфинктер открывается, и
небольшое количество химуса выталкивается в
двенадцатиперстную кишку.
ЖЕЛУДОЧНАЯ^
ЖЕЛЕЗА А
Желудочная ямка
Слизистые клетки в шейке железы
Обкладочные клетки,
секретирующие соляную кислоту
Главные (зимогенные) клетки,
секретирующие пепсиноген
Мышечный слой слизистой
Подслизистая основа
Рис. 10.29. Продольный разрез стенки желудка, на
котором видна желудочная железа.
Мышечный
слой слизистой
Крипта
Продольный слой
гладких мышц
Ворсин
ки
Циркулярный слой
гладких мышц
10.4.9. Тонкий кишечник
Тонкий кишечник человека состоит из двух
отделов - двенадцатиперстной кишки длиной 20 см и
подвздошной кишки, имеющей в длину около 5 м. В
двенадцатиперстную кишку открываются протоки
поджелудочной железы и желчного пузыря (рис.
10.30, А, Б). Подслизистая основа тонкого кишечни-
Серозная
оболочка
Мышечный
слой
слизистой
Подслизистая
основа
Рис. 10.30. Вертикальные разрезы:
А-двенадцатиперстной кишки; Б - подвздошной кишки.
- Ворсинка
Клетки,
секрет ирующие слизь
Эпителии
Сеть кровеносных
капилляров
Лимфатический сосуд
Либеркюновы крипты
Клетки Панета
Артериола
Мышечный
слой слизистой
Венула
Подслизистая
Циркулярные мышцы
— Продольные мышцы
•^Ч— Серозная оболочка
Рис. 10.31. А. Поперечный разрез стенки тонкого
кишечника человека, на котором видна ворсинка.
Б. Микрофотография ворсинки тонкого кишечника,
полученная с помощью сканирующего электронного
микроскопа. В. Электронная микрофотография
слизистой клетки, на которой видны микроворсинки
Микроворсинки
Клеточная мембрана
Митохондрия
ка собрана в многочисленные складки. Слизистая
образует множество пальцевидных выростов,
называемых ворсинками, стенки которых обильно
снабжены капиллярами и лимфатическими сосудами и
содержат волокна гладких мышц. Ворсинки
способны непрерывно сокращаться и вытягиваться,
благодаря чему они находятся в тесном контакте с
присутствующей в тонком кишечнике пищей
(рис. 10.31). Расположенные на поверхности
ворсинок клетки несут на своих апикальных концах
крошечные микроворсинки.
10.9. а) Опишите особенности строения тонкого
кишечника, которые увеличивают площадь
поверхности.
б) Какие преимущества дает это животному?
По всей длине тонкого кишечника присутствуют
слизистые клетки, секретирующие слизь.
Расположенные в начальном отделе двенадцатиперстной
кишки бруннеровы железы тоже секретируют слизь,
а также щелочную жидкость, и этот секрет
защищает слизистую кишечника от поступающей из
желудка кислоты и создает рН 7-8, при котором
активно работают ферменты кишечника.
10.10. Что произошло бы с активностью
ферментов кишечника, если бы в двенадцатиперстной
кишке оставался рН 2,0?
Слизистая тонкого кишечника секретирует ряд
ферментов, составляющих кишечный сок. В состав
кишечного сока входят амилаза, превращающая
амилозу крахмала в мальтозу, мальтаза,
превращающая мальтозу в глюкозу, лактаза,
превращающая лактозу в глюкозу и галактозу, сахараза,
превращающая сахарозу в глюкозу и фруктозу, нуклео-
тидазы, превращающие нуклеотиды в нуклеозиды,
эрепсин - смесь аминопептидаз и дипептидаз,
превращающих пептиды и дипептиды в аминокислоты,
и энтерокиназа, которая не является
пищеварительным ферментом, а превращает неактивный трипси-
ноген панкреатического сока в активный трипсин. В
дополнение к собственной секреции тонкий кишеч-
Островок Лангерганса,
Протоки поджелудочной
железы разных размеров
Рис. 10.32. Поперечный разрез через ткань
поджелудочной железы, на котором видны протоки
и один островок Лангерганса.
ник получает щелочной сок из поджелудочной
железы и желчь из печени. Желчь, образуемая клетками
печени (разд. 18.4) и хранящаяся в желчном пузыре,
содержит смесь солей желчных кислот, главным
образом гликохолат и таурохолат натрия.
Последние уменьшают поверхностное натяжение жировых
глобул и способствуют их эмульгированию,
повышая таким образом общую площадь их
поверхности. В таком виде жиры активнее гидролизуются
липазами. Более подробное описание состава желчи
дается в разд. 18.4.
Поджелудочная железа представляет собой
крупную железу, экзокринная ткань которой напоминает
ткань слюнных желез (рис. 10.32). Она состоит из
групп клеток, называемых ацинусами. Ацинусы сек-
ретируют целый ряд ферментов, которые
изливаются в двенадцатиперстную кишку через общий проток
поджелудочной железы. В состав сока
поджелудочной железы входят следующие, перечисленные ниже
ферменты: амилаза - превращает амилозу в
мальтозу; липаза - расщепляет жиры на жирные кислоты
и глицерин; трипсиноген - под действием энтерокина-
зы превращается в трипсин, который катализирует
расщепление белков на более короткие
полипептиды, а также превращение новых молекул трипсино-
гена в трипсин; химотрипсиноген - превращаясь в
химотрипсин, расщепляет белки до аминокислот;
карбоксипептидазы - расщепляют пептиды на
аминокислоты; нуклеазы - превращают нуклеиновые
кислоты в нуклеотиды.
В табл. 10.8 перечислены все ферменты
пищеварительного тракта человека и те функции, которые
они выполняют. В табл. 10.9 указаны различия в
строении основных отделов пищеварительного
тракта человека.
Таблица 10.8. Пищеварительные ферменты и их действие
Секрет
Слюна (из
слюнных желез)
Желудочный сок
(из слизистой
желудка)
Кишечный сок
(из слизистой
кишечника)
Панкреатический
сок (из
поджелудочной
железы)
Желчь (из печени)
Фермент
Амилаза слюны
(Про)реннин
(у молодых)
Пепсин(оген)
Соляная кислота
(не фермент)
Амилаза
Мальтаза
Лактаза
Сахараза
Нуклеотидазы
Эрепсин
Энтерокиназа
Амилаза
Трипсин(оген)
Химотрипсин(оген)
Карбоксипептидаза
Липаза
Нуклеазы
Соли желчных
кислот (не ферменты)
Место действия
Ротовая полость
Желудок
Желудок
Желудок
Тонкий кишечник
» »
» »
» »
» »
» »
» »
» »
» »
» »
» »
» »
» »
» »
Оптимум рН
6,5-7,5
2,0
2,0
8,5
»
»
»
»
»
»
7,0
»
»
»
»
»
»
Субстрат
Амилоза
крахмала
Казеиноген
молока
Белки
Пепсиноген
Нуклеопротеины
Амилоза
Мальтоза
Лактоза
Сахароза
Нуклеотиды
Пептиды и дипеп-
тиды
Трипсиноген
Амилоза
Белки
Химотрипсиноген
Белки
Пептиды
Жиры
Нуклеиновые
кислоты
Жиры
Продукты
Мальтоза
Казеин
Пептиды
Пепсин
Нуклеиновая
кислота и белок
Мальтоза
Глюкоза
Глюкоза +
галактоза
Глюкоза -I-
фруктоза
Нуклеозиды
Аминокислоты
Трипсин
Мальтоза
Пептиды
Химотрипсин
Аминокислоты
Аминокислоты
Жирные
кислоты + глицерол
Нуклеотиды
Жировые капли
Таблица 10.9. Сравнение структуры основных отделов пищеварительного тракта человека
Слой
Слизистая
оболочка
а) Эпителий,
выстилающий
просвет
б) Собственная
пластинка
в) Мышечная
пластинка слизистой
Подслизистая
основа
Внутренний
циркулярный и
наружный
продольный мышечные
слои
Серозная
оболочка
Пищевод
Многослойный, плоский
Специализация:
немногочисленные
слизистые железы,
локализованные в собственной
пластинке и подслизис-
той
Некоторое количество
слизистых желез
Имеется
Некоторое количество
глубоких слизистых
желез
У человека в верхнем
отделе
поперечнополосатые мышцы, а в
нижнем - гладкие
Имеется
Желудок
Простой,
цилиндрический
Специализация:
желудочные железы,
локализованные в
ι собственной пластинке
и содержащие 4 типа
клеток:
1) слизистые
2) обкладочные
3) главные
4) эндокринные
Многочисленные
желудочные железы
Имеется
Имеется
С дополнительным
внутренним слоем
косых мышц. Мышцы
циркулярного слоя
образуют кардиальный
и пилорический
сфинктеры
Имеется
Тонкий кишечник
Однослойный,
цилиндрический, содержит
всасывающие и
слизистые клетки
Специализация:
1) кишечные железы в
либеркюновых
криптах
2) клетки Панета
3) эндокринные клетки
4) слизистые железы в
двенадцатиперстной
кишке
Кишечные железы и
выраженные
лимфатические сосуды
Имеется
Дуоденальные железы
Имеются
Имеется
Толстый кишечник
Однослойный,
цилиндрический, содержит
всасывающие и
слизистые клетки
Специализация:
кишечные железы,
расположенные в
собственной
пластинке
Трубчатые железы
Имеется
Кишечные железы
Имеются
Неполная
10.4.10. Всасывание пищи в тонком
кишечнике
Всасывание продуктов пищеварения в кишечнике
происходит через микроворсинки эпителиальных
клеток, выстилающих ворсинки подвздошной
кишки. Как видно на рис. 10.31, строение ворсинок
кишечника идеально приспособлено для
выполнения этой функции. Моносахариды, дипептиды и
аминокислоты всасываются в эпителий ворсинок, а
затем путем диффузии или активного транспорта
попадают в кровеносные капилляры (см. разд.
7.2.2).
10.11. Почему так важно участие активного
транспорта во всасывании моносахаридов, дипеп-
тидов и аминокислот пищи?
Кровеносные капилляры, выходящие из ворсинок,
соединяясь, образуют воротную вену печени, по
которой всосавшиеся продукты переваривания
поступают в печень. С жирными кислотами и
глицерином дело обстоит несколько иначе. Поступив в
цилиндрический эпителий ворсинок, они вновь
превращаются здесь в жиры, которые переходят затем
в лимфатические сосуды. Присутствующие в этих
лимфатических сосудах белки1 обволакивают
молекулы жира, образуя липопротеиновые шарики,
называемые хил омикронами. Через грудной
лимфатический проток хиломикроны поступают в
кровеносное русло. Впоследствии липопротеины гидро-
лизуются ферментами, присутствующими в плазме
крови, и образующиеся при этом жирные кислоты и
глицерин поступают в клетки, где они могут
использоваться в процессе дыхания или откладываются в
запас в виде жира в печени, мышцах, брыжейке и
подкожной жировой ткани.
В тонком кишечнике происходит также
всасывание неорганических солей, витаминов и воды.
10.4.11. Моторика пищеварительного
тракта человека
Находящаяся в пищеварительном тракте пища
подвергается воздействию целого ряда
перистальтических движений. В результате чередующихся ритми-
1 Имеются данные о том, что образование хиломик-
ронов происходит в самих энтероцитах, а не в
лимфатических сосудах. Физиология человека./ Под ред. Уилмера,
1986, т. 4, стр. 140.- Прим. перев.
ческих сокращений и расслаблений стенок тонкого
кишечника возникает его ритмическая сегментация,
при которой последовательно сокращаются
небольшие участки стенок, благодаря чему химус приходит
в тесное соприкосновение со слизистой кишечника.
Кроме того, кишечник совершает маятникообразные
движения, когда петли кишечника внезапно резко
укорачиваются, проталкивая пищу из одного конца
в другой, в результате чего она хорошо
перемешивается. Наконец, существует пропульсивная
перистальтика, продвигающая химус по
пищеварительному тракту. Илеоцекальная заслонка периодически
открывается и закрывается. При открывании
заслонки химус небольшими порциями поступает из
подвздошной кишки в толстый кишечник. Когда
заслонка закрыта, доступ химуса в толстый
кишечник прекращается.
10.4.12. Толстый кишечник
В толстом кишечнике человека происходит
всасывание основной массы воды и электролитов,
тогда как некоторые метаболические шлаки и
избыток электролитов, и прежде всего кальция и
железа, выделяются в виде солей. Слизистые клетки
эпителия секретируют слизь, которая смазывает
становящиеся все более твердыми остатки пищи,
называемые каловыми массами. В толстом
кишечнике обитает множество симбиотических бактерий,
синтезирующих аминокислоты и некоторые
витамины, в том числе витамин К, которые всасываются
в кровяное русло.
У человека червеобразный отросток представляет
собой слепо заканчивающийся отросток, отходящий
от слепой кишки и не выполняющий никакой
функции, тогда как у травоядных животных он имеет
большое значение (разд. 10.8.3). Каловые массы
состоят из мертвых бактерий, целлюлозы и других
растительных волокон, отмерших клеток слизистой,
слизи, холестерола, производных желчных
пигментов и воды. Они могут оставаться в толстой кишке в
течение 36 ч, прежде чем достигнут прямой кишки,
где они непродолжительное время хранятся, а затем
выделяются через анальное отверстие. Вокруг
анального отверстия имеются два сфинктера -
внутренний, образованный гладкими мышцами и
находящийся под контролем вегетативной нервной
системы, и наружный, образованный
поперечнополосатой мышечной тканью и находящийся под
контролем центральной нервной системы.
У маленьких детей рефлекс дефекации возникает
благодаря тому, что растяжение прямой кишки
вызывает расслабление внутреннего сфинктера. Со
временем ребенок приучается подчинять акт
дефекации контролю со стороны центральной нервной
системы, и дефекация происходит у него только
в том случае, когда расслабляется наружный
сфинктер.
Опыт 10.3.А. Изучение перистальтики
тонкого кишечника крысы
Материалы и оборудование
Только что забитая крыса (с помощью хлороформа)
Раствор Рингера для млекопитающих
Большая водяная баня на 37°С
Набор анатомических инструментов
Бинокулярный микроскоп
Методика
1. Забейте крысу и поместите ее в раствор Рингера
для млекопитающих при температуре 37°С.
2. Вскройте брюшную полость крысы и
рассмотрите кишечник.
3. Внимательно наблюдайте за тем, происходят ли
какие-либо движения стенки кишечника.
4. Зарегистрируйте и по возможности определите
все виды движений кишечника. Эти движения
могут быть трех типов:
а) пропульсивная перистальтика,
б) ритмическая сегментация,
в) маятникообразные движения.
(Более подробные сведения вы найдете в
тексте.)
Опыт 10.3.Б. Изучение анатомии
желудка, тонкого и толстого
кишечника и анализ аминокислот,
содержащихся в каждом отделе
Материалы и оборудование
Раствор Рингера для млекопитающих
Пробирки для образцов
Бинокулярный микроскоп
Набор анатомических инструментов
Дистиллированная вода
Пластинка для тонкослойной хроматографии
Проволочная петля
Хроматографический бачок с крышкой
н-Бутанол
Ледяная уксусная кислота
Вытяжной шкаф
Пульверизатор с нингидрином
Сушильный шкаф на 1000C
Методика
1. Используя крысу, препарированную в опыте
10.3,А, перевяжите и удалите по отдельности
желудок, тонкий кишечник и толстый кишечник.
Разрежьте их и смойте содержимое каждого
участка в отдельные, помеченные пробирки, в
которые предварительно налейте раствор
Рингера для млекопитающих.
Приготовление тонкослойной
хроматограммы
Прохождение
хрома гограм мы
Хроматограмма после
опрыскивания нингидрином
Готовая к использованию
тонкослойная пластинка
Проведите
карандашом
линию на
расстоянии
2 см от края
С помощью маленькой
проволочной петли
нанесите небольшую
каплю экстракта
Рис. 10.33. Приготовление тонкослойной хроматограммы.
Крышка
- Герметично
закрытый
бачок
Тонкослойная
пластинка
Стартовая линия
Растворитель
(толщина
слоя 1 см)
— Фронт растворителя
_ Пятна после
опрыскивания
нингидрином
Стартовая линия
2. Сделайте тонкие срезы со стенок каждого из
отделов и исследуйте их под бинокулярным
микроскопом. Рассмотрите и опишите вид их
внутренней поверхности.
3. Добавьте к содержимому пробирок небольшое
количество дистиллированной воды, встряхните
и оставьте на 15 мин отстаиваться.
4. Отберите из каждого образца каплю прозрачной
надосадочной жидкости, нанесите на отдельную
тонкослойную пластинку и подсушите.
Повторите процедуру, чтобы нанести на хромато-
грамму достаточное количество образца. После
того как пятна высохнут, поместите пластинки в
хроматографический бачок, на дно которого
налита слоем толщиной в 1 см смесь
растворителей, состоящая из н-бутанола, ледяной
уксусной кислоты и воды в соотношении 40:10:15, и
накройте бачок крышкой (рис. 10.33).
5. Оставьте пластинки в бачке на 90 мин, а затем
выньте их, высушите в вытяжном шкафу и
опрыскайте нингидрином.
6. Прогрейте пластинки в сушильном шкафу при
1000C.
7. Отметьте на пластинках пятна фиолетового и
других цветов. Эти пятна указывают на
присутствие в исследуемых образцах аминокислот.
(Данное практическое занятие взято из Nuffield
Α-level Biology Maintenance of the organism.)
Опыт 10.4. Изучение переваривания
и всасывания на модели кишки
Материалы и оборудование
Только что забитая крыса
Часовые стекла
1%-ный раствор крахмала
Три диализные трубки (Visking tubes) no 15 см
длиной
Вата
Пробирки для кипячения
Бумажные хомутики
Водяная баня на 300C
Таймер
Раствор Люголя
Раствор Бенедикта
Асбестовое полотенце
Пробирки и штативы
Пипетка
Методика
1. Забейте крысу, вскройте ее, перевяжите и
извлеките по отдельности желудок, тонкий и
толстый кишечник.
2. Поместите их порознь на часовые стекла и
залейте сверху 1%-ным раствором крахмала.
Размельчите ткань и перемешайте ее с раствором
крахмала.
3. Возьмите три диализные трубки диаметром
1 см и длиной 15 см и завяжите их с одного
конца узлом.
4. Слейте размельченную ткань каждого отдела
пищеварительного тракта порознь в три
диализные трубки и долейте сверху раствор
крахмала до общего объема 10 мл. Тщательно
все перемешайте.
5. Поместите диализные трубки по отдельности в
пробирки для кипячения, содержащие по 10 мл
дистиллированной воды, а чтобы их открытые
концы не упали в воду, прикрепите их к стенке
пробирки с помощью бумажных хомутиков.
6. Поставьте контрольный опыт с диализной
трубкой, в которую налито только 10 мл
1%-ного раствора крахмала.
7. Поместите все пробирки в водяную баню с
температурой 300C на 30 мин.
8. Через 30 мин проведите следующие пробы с
каждой пробиркой и ее содержимым:
а) Отберите каплю раствора из каждой
диализной трубки и с помощью раствора Люголя
проверьте его на содержание крахмала.
б) Отберите еще одну каплю содержимого
каждой трубки и определите в нем
содержание редуцирующих Сахаров с помощью
раствора Бенедикта. Полученные результаты
запишите.
в) Проведите в каждом случае пробу на крахмал
и редуцирующие сахара в воде, окружающей
диализную трубку.
9. Сведите полученные результаты в таблицу.
10. Имеются ли доказательства того, что ткань
пищеварительного тракта превращает крахмал
в редуцирующие сахара, и если такие
доказательства имеются, то в каком участке
пищеварительного тракта образуется наибольшее
количество Сахаров?
11. Что говорят ваши наблюдения о природе
диализных трубок?
10.5. Регуляция секреции
пищеварительных желез
Секреция пищеварительных ферментов требует
затраты энергии, и, если бы она происходила
непрерывно, тем более в отсутствие пищи, это было бы
слишком большим расточительством для
организма. В действительности же основная масса
пищеварительных соков образуется только тогда, когда
есть, что переваривать. Таким образом
осуществляются общая координация и регуляция процесса
пищеварения как упорядоченной
последовательности определенных стадий.
Секреция слюны в полость рта регулируется
двумя рефлексами-условным и безусловным.
Безусловный рефлекс, осуществляемый через черепно-
мозговые нервы, возникает под действием
присутствующей во рту пищи. Контакт пищи с вкусовыми
сосочками языка вызывает импульсы, идущие в
мозг, а из мозга в ответ поступают импульсы,
вызывающие секрецию. Условный рефлекс
возникает на вид, запах или вкус пищи.
Секреция желудочного сока протекает в три фазы.
Первая фаза-нервная, или вагусная. Присутствие
пищи в полости рта и ее проглатывание вызывают
импульсы, которые через блуждающий нерв
передаются в желудок и стимулируют секрецию
желудочного сока его слизистой оболочкой. Эта фаза
наступает еще до того, как пища достигнет желудка,
и, таким образом, подготавливает желудок к
приему пищи. Продолжительность вагусной фазы
составляет около 1 ч. Вторая фаза-фаза
растяжения, во время которой секреция желудочного сока
стимулируется растяжением желудка пищей. Третья
фаза - гастральная, или гуморальная, обусловлена
тем, что присутствующая в желудке пища
стимулирует образование в слизистой пилорического
отдела желудка гормона гастрина. Последний через
кровяное русло достигает слизистой других отделов
желудка и вызывает продолжающуюся около 4 ч
секрецию желудочного сока с высоким содержанием
соляной кислоты. Фазы желудочной секреции
изображены на рис. 10.34.
В ответ на присутствие в пище жирных кислот
слизистая желудка секретирует еще один гормон
энтерогастрон. Энтерогастрон, как правило, угнетает
Цефалическая
(вагусная) фаза
Общая фаза
Желудочная секреция
Желудочная
(гуморальная фаза)
Растяжение
Время, ч
Рис. 10.34. Фазы желудочной секреции. (Из
J. H. Green An introduction to human physiology, Oxford,
1968.)
секрецию соляной кислоты, замедляет
перистальтику желудка и задерживает его опорожнение.
Слизистая желудка вырабатывает также внутренний
фактор, способствующий всасыванию витамина
B12.
Когда кислый химус поступает в
двенадцатиперстную кишку и соприкасается с ее стенками,
слизистая двенадцатиперстной кишки начинает сек-
ретировать кишечный сок и два гормона - холецис-
токинин-панкреозимин и секретин. Панкреозимино-
вый компонент первого гормона доставляется с
кровью в поджелудочную железу, так же как и
секретин, который вместе с холецистокинином
попадает в печень. Панкреозимин вызывает
образование панкреатического сока с высоким
содержанием ферментов. Секретин стимулирует выделение
панкреатического сока с высоким содержанием
бикарбоната и синтез желчных кислот в печени. Холе-
цистокинин вызывает сокращения желчного пузыря
с последующим высвобождением желчи в
двенадцатиперстную кишку.
В табл. 10.10 суммированы гормоны, секретируе-
мые различными отделами пищеварительного
тракта, и те органы, на которые они действуют.
10.6. Судьба всосавшихся
питательных веществ
Углеводы и аминокислоты всасываются в
кровеносные сосуды, окружающие тонкий кишечник, и
поступают через воротную вену в печень. Большая
часть глюкозы откладывается в печени в виде
гликогена и жиров, но часть глюкозы выходит из нее с
кровью через печеночную вену и разносится по
тканям организма, где подвергается окислению в
процессе тканевого дыхания. Если между приемами
пищи организму потребуется повышенное
количество энергии, гликоген может превратиться в
глюкозу, и последняя транспортируется с кровью к тем
тканям, которые в ней нуждаются.
Аминокислоты используются для синтеза новых
Таблица 10.10. Сводная таблица гормонов пищеварительного тракта человека, секретирующих их органов и органов-
мишеней
Гормон
Гастрин
Энтерогастрон
Панкреозимин
Холецистоки-
нин
Секретин
Место образования
Слизистая желудка
Слизистая желудка
и тонкого кишечника
Слизистая
двенадцатиперстной кишки
Слизистая
двенадцатиперстной кишки
Слизистая
двенадцатиперстной кишки
Стимулы для секреции
Растяжение желудка
пищей
Жирные кислоты пищи
Присутствие пищи в
двенадцатиперстной
кишке
Присутствие жирной
пищи в
двенадцатиперстной кишке
Кислота и пища в
двенадцатиперстной
кишке
Орган-мишень
Желудок
Желудок
Поджелудочная железа
Желчный пузырь
Поджелудочная железа
Печень
Характер ответа
Повышение секреции HCl
Угнетение секреции HCl,
уменьшение
перистальтики и замедление
эвакуации
Повышенная секреция
ферментов с
панкреатическим соком
Сокращение желчного
пузыря
Повышенная секреция
HCOз с
панкреатическим соком
Образование желчи
белков цитоплазмы, восстановления поврежденных
участков тела и образования ферментов и гормонов.
Лишние аминокислоты не могут откладываться в
запас и подвергаются в печени дезаминированию. В
процессе дезаминирования от них отщепляется
аминогруппа (NH2), которая превращается в
мочевину. Мочевина доставляется с кровью в почки и
выделяется с мочой. Остатки молекул аминокислот
превращаются в гликоген и в таком виде
депонируются.
Всосавшиеся жиры минуют печень и поступают в
венозную кровь через грудной лимфатический
проток. Жиры являются основным энергетическим
депо организма. В нормальных условиях организм
получает достаточное количество глюкозы, и для
образования энергии жиры не требуются. В этой
ситуации они откладываются в подкожной жировой
клетчатке, в жировой ткани, окружающей сердце и
почки, и в брыжейке. Часть жиров встраивается в
плазматические и ядерные мембраны клеток.
10.7. Регуляция аппетита у
человека
У людей аппетит находится под контролем двух
центров, расположенных в гипоталамусе,-центра
голода и центра насыщения. Раздражение центра
голода заставляет индивидуума искать и
принимать пищу, а раздражение центра насыщения
подавляет аппетит. Наиболее важным фактором,
влияющим на оба центра, является концентрация
глюкозы в крови. Последняя находится под
контролем гипоталамуса и является хорошим
показателем степени насыщенности организма
питательными веществами.
Сразу после приема пищи концентрация глюкозы
в крови повышается, что приводит к возбуждению
центра насыщения и уменьшению аппетита. Через
определенное время после приема пищи
концентрация глюкозы в крови снижается, и это возбуждает
центр голода (рис. 10.35).
Это простое на первый взгляд объяснение ни в
коей мере не дает полной картины. На аппетит
влияет также целый ряд дополнительных факторов,
в том числе рефлексы растяжения
пищеварительного тракта, физиологическое и психическое
состояние индивидуума, нарушение психики или
повреждение головного мозга. Любой из этих факторов
может повлиять на активность гипоталамических
центров и привести к извращению аппетита.
Низкая концентрация
сахара в крови
Переваривания
I не происходит
Высокая концентрация
сахара в крови
!Дентр^
насыщения/
Центра
голода
Переваривание
'ГИПОТАЛАМУС^
Прекращение
приема пищи
Начало
приема пищи
Рис. 10.35. Гипоталамическая
регуляция аппетита.
10.8. Разновидности
пищеварительного тракта
млекопитающих
10.8.1. Плотоядные: кошка
Кошка-плотоядное животное, и ее зубы
приспособлены для того, чтобы схватывать добычу и
разрывать ее на куски. Зубная формула кошки
имеет следующий вид:
ГЗ 1 3 Л
2 i-c-pm-m- .
|_3 Г 2 IJ
Мелкие, плотно расположенные резцы имеют
долотообразную форму и служат для отрывания
мяса от костей. Клыки длинные, конусообразные,
изогнутые. Служат для захвата и умерщвления
жертвы и разрывания мяса. По два задних зуба с
каждой стороны, сильно увеличены и имеют
острые гребни, идущие параллельно линии челюсти.
Это так называемые хищные зубы (третий верхний
предкоренной и первый нижний коренной). Они
напоминают лезвия ножниц, поскольку при
разрывании жертвы на куски внутренняя поверхность
зубов верхней челюсти тесно примыкает к наружной
поверхности зубов нижней челюсти. Остальные
коренные зубы имеют уплощенные вершины и
острые края и используются для разрывания мяса и
разгрызания костей.
Челюстной сустав работает как хорошо
пригнанный шарнир и позволяет двигаться челюстям
только в вертикальном направлении. Задние зубы,
от которых требуется большее усилие, расположены
ближе других к челюстному суставу.
Нижняя челюсть поднимается благодаря
сокращению височной мышцы. Последняя
прикреплена к костному отростку нижней челюсти,
идущему вверх по направлению к ушам. Такое
строение создает эффективную систему рычагов для
перемещения пищи, когда кошка разгрызает мясо
или защелкивает зубы, умерщвляя жертву. Другая,
жевательная мышца поднимает вверх основание
нижней челюсти, уменьшая таким образом нагрузку
на челюстной сустав (рис. 10.36).
10.8.2. Травоядные: овца
Овца питается травой, и ее зубы прекрасно
приспособлены для ее образа жизни и способа питания.
Зубная формула овцы имеет следующий вид:
Г 0 0 3 31
2 i-c-pm-m- .
L 3 1F 2 3 J
Верхние резцы и клыки у овцы отсутствуют, а
вместо них имеется роговая пластинка, о которую
трутся нижние долотовидные резцы и клыки, когда
овца скусывает траву. Между передними и боко-
З-й малый коренной-,
(хищный) зуб \
Резец
Клык—
Малые коренные
зубы
Резец
Клык
Малые коренные
зубы
Височная мышца
Жевательная мышца
Коренной зуб
1-й коренной
(хищный)
зуб
Рис. 10.36. Челюсти, зубной ряд и мускулатура
челюстей у кошки.
Коренные
Малые коренные
Роговая.
пластинка
Диастема
Резец
Клык
Малые коренные
Височная
мышца
Жевательная
мышца
Коренные
Рис. 10.37. Челюсти, зубной ряд и мускулатура
челюстей у овцы.
выми зубами имеется большая щель диастема,
позволяющая языку манипулировать срезанной травой
таким образом, что та трава, которая
пережевывается, находится отдельно от новой порции
срезанной травы.
Боковые зубы обладают широкой перетирающей
поверхностью, площадь которой еще более
увеличивается за счет W-образных складок на
жевательной поверхности верхних зубов и М-образных
складок на поверхности нижних зубов. Гребни на
поверхности этих зубов состоят из прочной эмали, а
впадины между ними-из дентина. Челюсти
сочленены очень подвижно и могут двигаться вперед и
назад, вверх и вниз и из одной стороны в другую. Во
время жевания нижняя челюсть двигается из
стороны в сторону, и W-образные гребни верхних
коренных зубов плотно входят в желобки между
М-образными гребнями нижних зубов, благодаря
чему пища хорошо перетирается. Жевательная
мышца у овцы мощная, а височная - небольшая
по размеру (рис. 10.37), т.е. соотношение их
прямо противоположно тому, что наблюдается у
кошки.
10.8.3. Переваривание целлюлозы у
жвачных
Пищевод
К тонкому
кишечнику
Пищеварительный тракт жвачных имеет сложное
строение. Истинному желудку у них предшествует
несколько отделов, первым из которых является
рубец. Рубец выполняет роль ферментера, в
котором смешанная со слюной пища подвергается
ферментативному воздействию со стороны сим-
биотических микроорганизмов. Многие из этих
микроорганизмов синтезируют фермент целлюлазу,
которая расщепляет целлюлозу. Присутствие этих
микроорганизмов является для жвачного животного
жизненно необходимым, поскольку само животное
неспособно синтезировать целлюлазу. Конечными
продуктами ферментативного переваривания
целлюлозы является уксусная, пропионовая и
масляная кислоты, а также CO2 и метан. Кислоты
всасываются в кровь животного и, подвергаясь
окислительным превращениям, являются для
него основным источником энергии. В свою очередь
микроорганизмы используют энергию,
выделяющуюся в ходе химических реакций процесса
ферментации, и к тому же имеют оптимальную для их
жизнедеятельности температуру окружающей
среды.
Жвачное животное способно отрыгивать из рубца
частично переваренную пищу (это явление носит
название пережевывания или «жевания жвачки»), а
Сетка
Книжка
Сычуг
Рубец
Рис. 10.38. Сложное строение отделов,
предшествующих тонкому кишечнику, у жвачных.
затем пища вновь проглатывается и подвергается
дальнейшему ферментативному перевариванию.
Частично переваренная пища проходит через
начальные отделы пищеварительного тракта и в конце
концов попадает в сычуг, соответствующий желудку
человека. Начиная с этого отдела, дальнейшее
переваривание пищи происходит под действием общих
для всех млекопитающих пищеварительных
ферментов (рис. 10.38).
Глава 11
Использование
энергии
Каждая живая клетка-это сложная, высокоупо-
рядоченная система. Как показали эксперименты,
содержимое клетки находится в состоянии
непрерывной активности; различные вещества все время
входят в клетку и выходят из нее наружу. Все
реакции, протекающие в клетке, можно
подразделить на две группы. Анаболические реакции-это
реакции синтеза крупных молекул из более мелких и
простых; для этих процессов необходима затрата
энергии (т.е. это эндергонические процессы):
A+ B-* AB [+AG]
(где AG-изменение свободной энергии для данной
реакции).
Катаболические реакции-это реакции распада
крупных молекул на более мелкие и простые,
обычно с выделением энергии (т.е. это экзергони-
ческие процессы):
AB-A+ В [-AG].
Иногда образовавшиеся более простые молекулы
могут затем вновь использоваться для биосинтеза.
[Примечание. Важно отметить, что не во всякой
катаболической реакции высвобождается энергия.
Некоторые реакции распада, назначение которых
состоит в том, чтобы избавить клетку от
нежелательных веществ, являются эндергоническими.]
Совокупность катаболических и анаболических
реакций, протекающих в клетке в любой данный
момент, составляет ее метаболизм:
Катаболизм + Анаболизм = Метаболизм.
Поступающие в клетку органические вещества
служат для нее источником, во-первых, небольших
«строительных блоков», используемых для
биосинтеза новых клеточных компонентов или замены
компонентов, отслуживших свой срок, и, во-вторых,
источником химической энергии. Когда в клетке
происходит расщепление питательных веществ,
обычно высвобождается энергия. Значительную ее
часть клетка использует на поддержание своих
жизненных процессов. Энергия поступает в
различные участки клетки и переходит из одной формы в
другую. Каждая форма энергии служит затем для
выполнения в клетке определенной работы. Это
может быть биосинтез, механическая работа,
клеточное деление, активный транспорт, осмос, а в
некоторых специализированных клетках - мышечное
сокращение, биолюминесценция или электрические
разряды (рис. 11.1). Наиболее пригодна для
использования в живой клетке химическая энергия, так как
она может быстро передаваться из одной части
клетки в другую (а также из клетки в клетку) и
расходоваться экономно - строго отмеренными
порциями, там и тогда, где и когда это необходимо.
Первоисточником всей энергии служит Солнце. В
живых организмах энергия легко переходит из
одной формы в другую, но включиться в пищевые
цепи солнечная энергия может лишь после того, как
она будет поглощена зелеными растениями (авто-
трофами) и преобразована их содержащими
хлорофилл клетками в процессе фотосинтеза в химическую
энергию, заключенную либо в глюкозе (простой
сахар), либо в крахмале (полисахарид). Часть этой
энергии высвобождается и расходуется растениями
для их собственных нужд. Животные вынуждены
пользоваться готовым источником энергии (т.е.
пищей). Для некоторых (травоядных) животных
таким источником энергии служат растения,
которыми они питаются; другие (плотоядные) питаются
тканями травоядных животных (см. гл. 12).
11.1. Роль дыхания
Дыханием можно назвать практически любой
процесс, при котором окисление органических
веществ ведет к выделению химической энергии.
Когда этот процесс протекает в клетках, его
называют внутренним, тканевым или клеточным
дыханием. Если для него требуется кислород, то дыхание
называют аэробным; если же реакции идут в
отсутствие кислорода, то говорят об анаэробном
дыхании.
Органические молекулы (по большей части
углеводы или жиры) расщепляются последовательно,
связь за связью, в ряде ферментативных реакций. В
каждой из этих реакций высвобождается небольшое
количество энергии, и значительная часть этой
энергии запасается в молекулах нуклеотида, который
носит название аденозинтрифосфата (АТФ).
Тканевое дыхание не следует путать с процессами
поглощения кислорода из окружающей среды и
выделения CO2 в среду. В совокупности эти два
процесса называются внешним дыханием или
(лучше) газообменом. Во внешнем дыхании могут
Солнечная энергия
(солнечный свет)
Автотрофное
питание
Неорганичес кие
вещества
Органические:
~ вещества
, Витамины
. Белки
Жиры
4 Углеводы
Переварива-
ние и
поглощение
Кислород-
Диффузия
КЛЕТОЧНАЯ МЕМБРАНА .
ВНЕКЛЕТОЧНАЯ СРЕДА
Диффузия
Восстановление^
/ Рост
Биосинтез более сложных молекул
i
Анаболические
реакции
Свободные витамины
Аминокислоты
Жирные кислоты и глицерол
Простые сахара
Органические
соединения
Катаболические
реакции,
например
дыхание
Работа
Механическая
работа
Клеточное
деление
Активный
транспорт
Осмос и т. д.
Часть энергии рассеивается
в виде тепла
CO2 + H2O + Энергия
ВНУТРИКЛЕТОЧНАЯ СРЕДА
КЛЕТКА
Рис. 171. Обмен энергией между средой и гетеротрофной клеткой. Важно помнить, что при переходе
энергии из одной формы в другую часть ее теряется-рассеивается в виде тепла.
участвовать органы или структуры, снабженные
специализированными поверхностями для
эффективного газообмена; воздух или вода
прогоняются над этими поверхностями с помощью
разного рода дыхательных движений (разд. 11.6).
11.2. АТФ (аденозинтрифосфат)
Молекула АТФ состоит из пурина аденина, пяти-
углеродного сахара рибозы и трех фосфатных
групп (рис. 11.2). При гидролитическом
отщеплении двух ее концевых фосфатных групп выход
свободной энергии на каждую из них составляет
около 30,6 кДж, тогда как отщепление третьей
фосфатной группы АТФ дает только 13,8 кДж
(табл. 11.1). Именно по этой причине принято
говорить, хотя это и неверно, что АТФ и АДФ
(аденозиндифосфат) содержат богатые энергией
(высокоэнергетические) связи (которые часто
обозначают знаком ~ ). Почему при гидролизе АТФ
выделяется больше энергии, чем при гидролизе
многих других соединений, не ясно. Полагают, что
это как-то зависит от распределения зарядов в его
молекуле. Для поддержания целостности такой
молекулы требуется больше химической энергии.
Энергия, таким образом, заключена не в какой-либо
одной связи, а характеризует всю молекулу в целом.
11.1. Из табл. 11.1 видно, что АТФ вовсе не
самое «богатое энергией» соединение в клетке.
АТФ занимает промежуточное положение. Какой
смысл можно в этом усмотреть?
11.2.1. Значение АТФ
АТФ-стандартная единица, в виде которой
запасается высвобождаемая при дыхании энергия. Для
синтеза АТФ из АДФ и фосфата требуется 30,6 кДж
энергии на I моль. Поэтому АТФ может
образоваться лишь в таких реакциях, при которых
выход энергии составляет более 30,6 кДж/моль. Вся
энергия, высвобождающаяся сверх 30,6 кДж/моль,
равно как и вся энергия от реакций, дающих менее
30,6 кДж/моль, не может быть запасена в АТФ и
рассеивается в виде тепла.
Поскольку вся химическая энергия представлена в
одной форме (а именно, в форме АТФ), процессы,
идущие с потреблением энергии, нуждаются только
в одной системе, способной принимать химическую
энергию от АТФ. Этим достигается большая
экономия в отношении действующих в клетке
механизмов.
Рис. 11.2. Структура АТФ. Две концевые фосфатные
группы присоединены пирофосфатными связями,
гидролитическое расщепление которых дает большое
количество свободной энергии.
АТФ-постоянный источник энергии для клетки.
Он мобилен и может доставлять химическую
энергию в любую часть клетки. Когда клетка
нуждается в энергии, единственное, что требуется
для ее получения-это гидролиз АТФ. Поскольку
АТФ содержится во всех живых клетках, его часто
называют универсальным носителем энергии.
АДФ может быть рефосфорилирован в АТФ в
результате дыхательной активности (рис. 11.3) или
за счет другого высокоэнергетического соединения,
например креатинфосфата, присутствующего в
мышечных клетках. Если весь АДФ мышечной клетки
превращается в АТФ, то фосфат от АТФ
переносится на креатин с образованием креатинфосфата.
При этом вновь появляется некоторое количество
АДФ, который может, присоединив фосфат,
образовать АТФ. При понижении уровня АТФ
происходит обратный процесс: фосфат переносится от
креатинфосфата на АДФ, и запасы АТФ таким образом
восстанавливаются (рис. 11.4).
Третий путь рефосфорилирования АДФ-это
фосфорилирование, протекающее в хлорофилл-
содержащих клетках зеленых растений (разд. 9.4).
АТФ играет важную метаболическую роль
благодаря своему центральному положению в
клеточной активности. Он действует как связующее звено
между дыханием и процессами, требующими
затраты энергии. При этом его
высокоэнергетические фосфатные группы непрерывно
отщепляются и замещаются новыми.
11.2. Роль АТФ можно сравнить с ролью
аккумулятора. Объясните, в чем заключается это
сходство.
11.3. Биологическое окисление
В клетке происходят окислительные реакции
трех типов:
1. Прямое окисление молекулярным кислородом:
А + O2 -> AO2.
Таблица 11.1. Свободная энергия гидролиза некоторых
фосфатов
Соединение
Фосфоенолпировиноградная
кислота
1,З-Бисфосфоглицериновая кислота
Креатинфосфат
АТФ (до АДФ и фосфата)
АДФ (до АМФ и фосфата)
АМФ (до аденозина и фосфата)
Глюкозо-6-фосфат
AG (изменение
свободной энергии), кДж/молъ
-62,1
-49,5
-43,3
-30,6
-30,6
- 13,8
- 13,8
"Высокоэнергетическая" связь
Аденозин
+ H2O
Аденозин
Гидролиз
30,6 кДж/моль
1
Работа
Дыхательная активность
Аденозин
'30,6 кДж/моль
или более
(энергия,
поступающая
за счет
дыхания)
Аденозин
(АТФ)
Рис. 11.3. А. Гидролиз АТФ. Б. Рефосфорилирование
АДФ в результате дыхательной активности.
АДФ
АТФ
Креатин
■►Креатин
АДФ
АТФ
Рис. 11.4. Перенос высокоэнергетической
фосфатной группы между АТФ и креатином.
2. Реакции, в которых А окисляется за счет В:
AH2 + В - А + BH2.
11.3. Как называется этот тип окисления?
11.4. Как называются ферменты,
катализирующие такие реакции?
3. Реакции, в которых происходит перенос
электронов, например окисление одной ионной формы
железа (Fe2+) в другую (Fe3+):
Fe2+ - Fe3+ + е" .
Все эти три типа окисления встречаются в
последовательности реакций, составляющих вместе
процесс, который носит название аэробного дыхания.
11.3.1. Общая характеристика
клеточного дыхания
Клеточное дыхание-это окисление субстрата,
приводящее к получению химической энергии (АТФ).
Субстратами для дыхания служат органические
соединения-углеводы, жиры и белки.
углеводы. Большинство клеток использует в
первую очередь именно углеводы. !Слетки
головного мозга млекопитающих вообще не способны
использовать для дыхания ничего, кроме глюкозы.
Полисахариды вовлекаются в процесс дыхания
лишь после того, как они будут гидролизованы до
моносахаридов:
Крахмал (у растений) ^ξξτ—^
^^!П^=" Глюкоза
Гликоген (у животных)-——"""" 5^"
жиры. Жиры составляют «первый резерв» и
пускаются в дело главным образом тогда, когда
запас углеводов исчерпан. Впрочем, в клетках
скелетных мышц при наличии глюкозы и жирных
кислот предпочтение отдается жирным кислотам.
белки. Поскольку белки выполняют ряд других
важных функций, они используются лишь после
того, как будут израсходованы все запасы углеводов
и жиров, например при длительном голодании.
Окисление глюкозы-в тех случаях, когда
субстратом служит глюкоза,- подразделяется на три
четко различимые фазы: гликолиз (путь Эмбдена-
Мейергофа), окислительное декарбоксилирование
(цикл Кребса, иначе называемый циклом лимонной
кислоты или циклом трикарбоновых кислот) и
окислительное фосфорилирование (дыхательная
цепь, где происходит перенос водорода и
электронов). Гликолиз - фаза, общая для анаэробного и
аэробного дыхания, но две другие фазы можно
наблюдать только в аэробных условиях. Подробно
все эти процессы мы рассмотрим немного позднее, а
здесь скажем о них лишь несколько слов.
11.3.2. Гликолиз и цикл Кребса
При аэробном дыхании окисление глюкозы
происходит путем последовательных реакций
дегидрирования. При каждом дегидрировании
отщепляемый водород используется для восстановления
кофермента:
AH2
новленный
тельный
субстрат
+ в
Кофер-
мент
(акцептор
водорода)
Дегидрогеназа
>
А
ленный
тельный
субстрат
+ BH2
Вос-
станов-
лен-
ный
кофер-
мент
Большая часть этих реакций происходит в
митохондриях, где акцептором водорода служит обычно
кофермент НАД (никотинамидадениндинуклеотид):
НАД+ 2H-НАД H2,
или, в более точной записи,
НАД+ +2H-НАД H+ H + .
НАД H2 поступает затем в дыхательную цепь и
здесь снова подвергается окислению.
11.3.3. Дыхательная цепь и
окислительное
фосфорилирование
В дыхательной цепи НАД · H2 вновь окисляется до
НАД, а отщепившийся от него водород передается
Глюкоза
2АТФ
гликолизу
Пировиноградная
кислота
38АТФ
Ацетилкофермент
2АТФ
34АТФ
Пул
водорода
ДЫХАТЕЛЬНАЯ
ЦЕПЬ
H2O
/2 O2
ЦИКЛ КРЕБСА
CO2
Рис. 11.5. Схема аэробного дыхания.
6С-сахар
АТФ
Фосфорилированный бС-сахар
-АТФ
Расщепление
Фосфорилированный ЗС-сахар^== Фосфорилированный ЗС-сахар
2хНАД-
2хНАД.Н2
2х2АТФ
2 Пировиноградная кислота (3C)
Рис. 11.6. Схема гликолиза.
не менее чем через пять переносчиков к концу цепи,
где соединяется с молекулярным кислородом,
образуя воду. Переход водорода по этой дыхательной
цепи состоит из ряда
окислительно-восстановительных реакций. В некоторых из этих реакций
выделяется достаточно энергии для образования АТФ, и
такой процесс носит название окислительного фос-
форилирования. Чистый выход на одну молекулу
глюкозы при полном ее окислении до воды и CO2
составляет 38 молекул АТФ, синтезированного из
АДФ и неорганического фосфата. При этом две
молекулы АТФ дает гликолиз, две-цикл Кребса и
34 - дыхательная цепь (рис. 11.5).
11.3.4. Отдельные этапы гликолиза
Гликолизом называют последовательность
реакций, в результате которых одна молекула
глюкозы расщепляется на две молекулы пировино-
градной кислоты (рис. 11.6). Эти реакции протекают
не в митохондриях, а в цитоплазме, и для них не
требуется присутствия кислорода. Процесс можно
подразделить на два этапа: на первом из них
происходит превращение глюкозы в фруктозо-
1,6,-бисфосфат, а на втором - расщепление фрук-
тозо-1,6-бисфосфата на два трехуглеродных сахара,
которые позже превращаются в пировиноградную
кислоту. При этом на первом этапе две молекулы
АТФ потребляются в реакциях фосфорилирования,
а на втором - четыре молекулы АТФ образуются.
Поэтому чистый выход АТФ при гликолизе равен
двум молекулам. Кроме того, при гликолизе
освобождаются четыре атома водорода. Их судьбу
мы рассмотрим позднее. Суммарную реакцию
гликолиза можно записать так:
C6H12O6 - 2C3H4O3 + 4Н + 2АТФ .
Глюкоза
Пировиноградная
кислота
(Чистый
выход)
Потребление и выход различных веществ в
процессе гликолиза указаны в табл. 11.2.
Конечная судьба пировиноградной кислоты
зависит от присутствия кислорода в клетке. Если
кислород имеется, то пировиноградная кислота
переходит в митохондрии для полного окисления до
CO2 и воды (аэробное дыхание). Если же кислорода
нет, то она превращается либо в этанол, либо в
молочную кислоту (анаэробное дыхание).
11.5. Внимательно изучите рис. 11.7 и дайте
ответы на следующие вопросы:
а. Как называется процесс, происходящий на
этапах Б и Г?
б. К какому классу принадлежит фермент,
катализирующий реакцию В?
в. Как называется реакция, обозначенная
буквой Д?
г. Какой витамин входит в состав молекулы
НАД?
Таблица 11.2. Потребление и выход веществ в процессе
гликолиза
Общее потребление
1 молекула глюкозы
(6C)
2 АТФ
4 АДФ
2 χ НАД
2хФн
Общий выход
2 молекулы пировиноградной
кислоты (2 χ 3C)
4 АТФ
2 АДФ
2 χ НАД H2
2 χ H2O
Крахмал
(полисахарид
растений)
Гликоген
(полисахарид
животных)
Глюкоза
АТФ
АДФ
(-1 АТФ)
Глюкозо-6-фосфат
Фру ктозо-6-фосфат
АТФ
АДФ
(-1 АТФ)
Фру ктозо-1,6-бисфосфат
.Расщепление
Дигидрокси- ^
ацетонфосфат
± Глицеральдегид-
3-фосфат
(изомеры ЗС-сахара,
способные к взаимопревращению)
2 х НАД
-2 х НАД-H2-
2 х1,3-бисфосфоглицериновая кислота
2АДФ
Фн (включение неорганического
фосфата в этом пункте
важно, так как позже
он участвует в синтезе АТФ)
2АТФ-*-
(+2АТФ)
2 χ 3-фосфоглицериновая кислота
2АДФ
2АТФ
(несколько
этапов
пропущено)
(+2 АТФ)
H2O
2 χ Пировиноградная кислота (3C) ЧИСТЫЙ ВЫХОД=2АТФ
Рис. 11.7. Отдельные этапы гликолиза и общая
биохимия этого процесса. Важно отметить, что,
поскольку при расщеплении фруктозо-1,6-бисфос-
фата образуются два трехуглеродных соединения,
в последующих реакциях синтезируются четыре
молекулы А ТФ, по две на каждую молекулу треху г-
леродного соединения, превращающегося в
пировиноградную кислоту.
11.3.5. Аэробное дыхание
Аэробное дыхание распадается на две фазы. В
первой из них при достаточном количестве
кислорода каждая молекула пировиноградной кислоты
поступает в митохондрию, где она полностью
окисляется аэробным путем. Сначала происходит
окислительное декарбоксилирование
пировиноградной кислоты, т.е. отщепление CO2 с
одновременным окислением путем дегидрирования.
Во время этих реакций пировиноградная кислота
соединяется с веществом, которое называют ко-
ферментом А (сокращенно его часто обозначают
KoA или KoAS—H), в результате чего образуется
ацетилкофермент А. Количество выделяющейся при
этом энергии достаточно для образования в
молекуле ацетилкофермента А
высокоэнергетической связи. В действительности процесс гораздо
сложнее, чем следует из этого описания; в нем
участвуют пять разных коферментов и три фер-
Пировиноградная кислота (3C)
CO2
2Н
•НАД
.НАД H
• К дыхательной цепи
Ацетил-КоА (2C)
(CH3CO-S-KoA)
KoA (KoAS-H)
H2O
Щавелевоуксусная
кислота (4C)
Лимонная кислота (6C)
2Н+НАД
Яблочная
кислота (4C)
НАД -H2 НАД -H2
Все атомы водоро-
да направляются
в дыхательную
цепь
ФАД-Н2 НАД·H2
2Н+НАД
CO2
Фумаровая
кислота (4C)
2H+
ФАД
2H+N
НАД
а- Кетоглутаровая
кислота (5C)
НАД —накотинамид -
адениндинуклеотид
ФАД — флавин -
аденин дину кл еоти д
Янтарная
кислота (4C)
АТФ
CO2
Рис. 11.8. Упрощенная схема цикла
Кребса.
мента. Суммарная реакция имеет вид
CH3COCOOH + KoAS-H + НАД -
-> CH3CO - S-KoA + CO2 +
+ НАД H2 + Ацетил-КоА
НАД H2, являющийся продуктом этой реакции,
направляется в дыхательную цепь митохондрии.
Вторую фазу аэробного дыхания составляет цикл
Кребса (названный так в честь открывшего его
исследователя-сэра Ганса Кребса). Ацетильная
группа ацетил-КоА, содержащая два атома
углерода, включается в цикл Кребса при гидролизе
ацетил-КоА. Она присоединяется к щавелевоуксус-
ной кислоте - четырехуглеродному соединению, в
результате чего образуется шестиуглеродная
лимонная кислота. Для этой реакции требуется энергия; ее
поставляет высокоэнергетическая связь ацетил-КоА.
Далее следует цикл реакций, в которых ацетильные
группы, поступающие в цикл при гидролизе аце-
тил-КоА, дегидрируются с высвобождением четырех
пар атомов водорода и декарбоксилируются с
образованием двух молекул CO2. При декарбоксилиро-
вании для окисления двух атомов углерода до CO2
используется кислород, отщепляемый от двух
молекул воды. Этот процесс носит название
окислительного декарбоксилирования. В конце цикла
щавелевоуксусная кислота регенерируется. Теперь
она способна вступить в реакцию с новой
молекулой ацетил-КоА, и цикл повторяется. На каждую
окисленную молекулу ацетил-КоА образуются:
одна молекула АТФ, четыре пары атомов водорода
и две молекулы CO2. Водородные атомы
присоединяются к НАД или ФАД (разд. 6.2.3) и в конце
концов попадают в дыхательную цепь. Поскольку
из одной окисленной молекулы глюкозы
образуются две молекулы ацетил-КоА, для окисления
каждой молекулы глюкозы в процессе дыхания
требуются два оборота цикла. Поэтому в
конечном итоге при окислении одной молекулы
глюкозы синтезируются две молекулы АТФ,
выделяются четыре молекулы CO2 и
высвобождаются восемь пар атомов водорода, поступающие
затем в дыхательную цепь (рис. 11.8).
Суммарную реакцию для образования ацетил-
KoA и цикла Кребса можно записать так:
QH12O6 + 6H2O - 6CO2 + 4АТФ + 12AcH2,
где Ac-акцептор водорода.
11.3.6. Окислительное
фосфорилирование и
дыхательная цепь
Пары водородных атомов, отщепляемые от
промежуточных продуктов в реакциях
дегидрирования при гликолизе и в цикле Кребса, в конце
концов окисляются молекулярным кислородом до
H2O с одновременным фосфорилированием АДФ в
АТФ. Происходит это, когда водород,
отделившийся от НАД H2 или ФАД H2, передается по
цепи, включающей по меньшей мере пять пере-
носчиков-флавопротеин, кофермент Q и несколько
разных цитохромов. В конце цепи водород
соединяется с молекулярным кислородом и образует
воду. Промежуточные переносчики водорода
претерпевают при этом ряд окислительно-
восстановительных реакций. Переносчики
сгруппированы таким образом, что в трех пунктах цепи при
переходе водородных атомов от одного переносчика
к другому небольшое количество энергии
высвобождается и включается в молекулу АТФ. На
рис. 11.9 представлена дыхательная цепь, или цепь
переноса электронов. В действительности на
первом участке цепи происходит главным образом
перенос водорода, а на последнем - исключительно
перенос электронов. Переносчики X, Y и Z-цито-
хромы. XhY представляют собой белковые
пигменты с железосодержащей простетической
группой, которая называется гемом (такой же гем
содержится в гемоглобине). При каждой
окислительно-восстановительной реакции железо
оказывается попеременно в окисленной (Fe3 + ) и в
восстановленной (Fe2+) форме. На конечном этапе
медьсодержащий переносчик Z, называемый обычно
цитохромоксидазой (или цитохромом а + а3),
катализирует восстановление молекулярного
кислорода до воды. Цианид калия и окись углерода
блокируют клеточное дыхание на этом этапе.
11.3.7. Переносчики водорода и
электронов
НАД и НАДФ
(никотинамидадениндинуклеотидфосфат)
НАД и НАДФ-два очень близких по своей
структуре кофермента. Оба они представляют собой
производные никотиновой кислоты (одного из
витаминов группы В). Молекулы того и другого
кофермента электроположительны (у них
отсутствует один электрон) и могут играть роль
переносчика как электронов, так и атомов водорода.
Когда акцептируется пара атомов водорода, один
из атомов диссоциирует на протон и электрон:
H
Атом
водорода
H +
Протон,
или иоы
водорода
а второй присоединяется к НАД или НАДФ
целиком.
Суммарную реакцию можно записать так:
НАД(Ф)+ + H
Целый
атом
[H+ + <Г]
циированный
атом
НАД(Ф) H + H +
новленный
кофермент
Свободный
протон в
среде
Электрон
или проще
НАД(Ф) + Н2->НАД(Ф)-Н2.
Свободный протон позднее, при отщеплении
водорода, используется для обратного окисления
кофермента.
Флавопротеины
Флавопротеины - это коферменты, в состав которых
входит витамин B2. ФАД (флавинадениндинуклео-
тид) играет роль простетической группы, белковая
же часть молекулы флавопротеина действует как
фермент. В дыхательной цепи эта белковая часть
выступает в качестве НАД-дегидрогеназы,
катализируя окисление восстановленного НАД. Водород
переносится флавопротеином в виде целых
атомов.
В цикле Кребса белковая часть флавопротеина,
простетической группой которого служит ФАД,
действует как сукцинатдегндрогеназа. Она
катализирует окисление янтарной кислоты в фумаровую.
Восстановленный ФАД располагается в
дыхательной цепи после первого пункта окислительного
фосфорилирования (т. е. синтеза АТФ). Поэтому за
счет его обратного окисления синтезируются только
две молекулы АТФ (рис. 11.10).
Кофермент Q
Молекула этого кофермента содержит цикл из
шести атомов углерода. Кофермент Q принимает
водород от флавопротеина и передает его цито-
хрому Ъ.
над.н
НАД
Рис. 11.9. Схема дыхательной цепи, где происходит перенос водорода и электронов. Пункты, в которых
высвобождается достаточно энергии для синтеза А ТФ, не указаны. Кроме того, показаны не все переносчики,
входящие в дыхательную цепь.
2H
Янтарная кислота
Фумаровая кислота
2Н
НАД-H2
ФАД
ФАД-Н2
НАД
ФП
H2O
ΦΠ·Η2
2H +
KoQH2
2е-
KoQ
2Fe3 +
Цитохром b
2с"
2Fc2*
2Fe2>
2е
Цитохром с
2Fe3-
2Cu2+
Цитохром а + а 2
(цитохром-
оксидаза)
2Cu*
АДФ
АТФ
АДФ
АТФ
АДФ
АТФ
Рис. 11.10. Развернутая схема дыхательной цепи. Каждый цитохром способен передавать только один
электрон. Предполагается, что на каждом дыхательном пути действуют два ряда цитохромов. Здесь показан
только один, но цифры удвоены, чтобы количества образующихся конечных продуктов соответствовали
действительности. В энергетическом смысле электроны перемещаются «вниз».
Цитохромы
Все цитохромы - это белки с относительно
небольшой молекулярной массой. Они содержат гем в
качестве прочно связанной простетической группы и
переносят не водородные атомы, а электроны.
Роль переносящего электроны компонента в ци-
тохромах играет железо гема. Обычно оно
находится в окисленной форме (Fe3 + ), но после
присоединения электрона, переходит в
восстановленную форму (Fe2 + ). Каждый водородный
атом, поступающий от кофермента Q, распадается
на ион водорода и электрон:
H -> H+ + е~ .
Этот электрон присоединяется к иону железа:
Fe3 +
Окисленное
железо
+ е~^
Fe2 +
Восстановленное
железо
Ионы водорода поступают на время в окружающую
среду: они понадобятся вновь позднее, в конце
дыхательной цепи.
Электрон от цитохрома b переходит к цитохрому
с и далее к цитохрому a + аъ - прочному комплексу
двух цитохромов, называемому обычно цитохро-
моксидазой. Этот комплекс, помимо железа,
содержит еще и медь и вступает в окислительно-
восстановительную реакцию, когда цитохром аъ в
конце концов передает электроны кислороду
(рис. 11.10). Любой цитохром может переносить
только по одному электрону; поэтому полагают,
что во всякой дыхательной цепи имеются два ряда
цитохромов, которые и осуществляют перенос
электронных пар:
H2
2H+ + 2е-
е~ + Fc3* ^Fe2+
е~ + Fe3+ ^Fe2+
Теперь, когда читатель уже кое-что знает о
переносчиках водорода и электронов, ему будет
легче разобраться и в более подробной схеме
дыхательной цепи, приведенной на рис. 11.10.
Суммарные реакции аэробного
дыхания:
(I)C6H12O6 + 6H2O
Гликолиз
*-
Цикл Кребса
Гликолиз
>.
Цикл Кребса
6CO2 + 12H2 + 4АТФ
(2) 12H2 + 6O2
Дыхательная
>
цепь
Дыхательная
Ь
цепь
12H2O + 34АТФ
(») + (2):
C6H12O6 + 6O2 -> 6CO2 + 6H2O + 38АТФ
11.3.8. Анаэробное дыхание
Многие микроорганизмы (анаэробы) получают
большую часть своего АТФ за счет анаэробного
дыхания. Для некоторых бактерий сколько-нибудь
значительные количества кислорода вообще
губительны, так что они вынуждены жить там, где
кислород отсутствует. Такие организмы называют
облнгатнымн анаэробами (примеры - бактерии
Clostridium botulinum и С. tetani).
Известны и другие организмы, например дрожжи
и паразиты кишечного тракта (ленточные черви и
др.), которые могут существовать как без
кислорода, так и в его присутствии. Их называют
факультативными анаэробами. Некоторые клетки,
временами испытывающие недостаток кислорода (в
частности, мышечные клетки), тоже обладают
способностью к анаэробному дыханию.
В условиях, когда кислорода нет и, значит,
водородные атомы, освободившиеся в процессе
гликолиза, не могут быть ему переданы, вместо НАД
должен быть использован другой акцептор
водорода. Таким акцептором становится пировиноград-
ная кислота. При этом в зависимости от
метаболических путей, имеющихся у данного организма или у
самих клеток, конечные продукты анаэробного
дыхания оказываются различными. У дрожжей,
например, образуются этанол и CO2:
CH3COCOOH
Пировиноградная
кислота
CH3CHO + CO2
CH3CHO + HM-H2
CH3CH2OH
Этанол
+ НАД
(этот процесс называется спиртовым брожением), а
в животных клетках, испытывающих временный
недостаток кислорода, и у некоторых бактерий
Глюкоза
C6H12O6
к
2АДФ+2
2ΑΤΦ (чистый выход)
4Н через НАД
2 χ Пировиноградная кислота
происходит молочнокислое брожение, приводящее к
образованию молочной кислоты:
CH3COCOOH
Пировиноградная
кислота
+ НАД H2
Ch3CHOHCOOH
Молочная кислота
+ НАД
На рис. 11.11 представлены эти различные пути
анаэробного дыхания.
Ни тот, ни другой процесс не дает
дополнительного количества АТФ, так что выход энергии на
одну молекулу глюкозы, расщепленную путем
анаэробного дыхания, соответствует двум
молекулам АТФ. Значительная часть энергии,
заключенной в молекуле глюкозы, при этом так и не
извлекается (остается в конечных продуктах брожения-
этаноле и молочной кислоте). Поэтому анаэробное
дыхание (рис. 11.11) в сравнении с аэробным (см.
разд. 11.3.9) следует считать процессом
малоэффективным. Молочная кислота должна из мышечных
клеток удаляться кровью, чтобы не наступило
утомление. Позднее, в аэробных условиях, она вновь
превращается в глюкозу в печени, где глюкоза
запасается в форме гликогена. Подробно этот
процесс описан в разд. 16.4.
11.3.9. Эффективность превращения
энергии при аэробном и
анаэробном дыхании
Аэробное дыхание
C6H12O6 + 6O2 - 6CO2 + 6H2O + 38АТФ;
AG= - 2880 кДж/моль.
Эффективность =
38 (-30,6)
-2880
= 40,37%
СНзСОСООН g присутствии кислорода
В отсутствие
кислорода
Анаэробное дыхание
МИТОХОНДРИЯ
Животные
Растения
Молочная кислота
2*CH3CHOHCOOh
J- ^2CO2
Ацетальдегид
2*CH3CHO
1 Спиртовое
брожение
Этанол
2XCH3CH2OH
Рис. 11.11. Различные пути
анаэробного дыхания.
НАД. H2
X
НАД-Н2или
ΦΠ·Η2
Дыхательная
цепь
НАД
XH2
НАД или
ФП
Рис. 11.12. Модель челночной системы (подробное
объяснение см. в тексте). НАД - никотинамидаденин-
динуклеотид; ФП флавопротеин
( — 30,6 кДж-это величина свободной энергии,
образующейся при гидролизе АТФ до АДФ).
Анаэробное дыхание
1) Дрожжевое (спиртовое) брожение:
QHi2O6 -* 2C2H5OH + 2CO2 + 2АТФ;
AG = — 210 кДж/моль.
Эффективность =
2 (-30,6)
_-~2НГ~
= 29,14%.
2) Гликолиз в мышцах (молочнокислое брожение):
C6H12O6 - 2CH3CHOHCOOh + 2АТФ;
AG = — 150 кДж/моль.
Эффективность =
2 (-30,6)
-150
= 40,80%.
Приведенные цифры показывают, что
эффективность превращения энергии в каждой из этих систем
довольно высока по сравнению с бензиновым
(25-30%) или паровым (8-12%) двигателем.
Количество же энергии, запасаемой в виде АТФ при
аэробном дыхании, в 19 раз больше, чем при
анаэробном. Объясняется это тем, что значительная
часть энергии остается «запертой» в этаноле и
молочной кислоте. Энергия, заключенная в этаноле,
остается для дрожжей навсегда недоступной, и,
значит, спиртовое брожение в смысле получения
энергии - малоэффективный процесс. Из молочной
же кислоты довольно большое количество энергии
может быть извлечено позднее, если появится
кислород. В присутствии кислорода молочная
кислота превращается в печени в пировиноградную
кислоту. Последняя поступает затем в цикл Кребса
и полностью окисляется до CO2 и H2O, в
результате чего дополнительно образуется большое
число молекул АТФ (разд. 17.4.8).
11.4. Челночные системы
Хотя из описанной выше картины событий,
составляющих аэробное дыхание, видно, что на каждую
окисленную молекулу глюкозы образуется обычно
38 молекул АТФ, в разных тканях это число может
быть различным. В цитоплазме в процессе
гликолиза образуются два комплекса НАД H2. Ци-
топлазматический НАД не способен пройти сквозь
митохондриальную мембрану, поэтому
поставляемые гликолизом электроны должны поступать в
митохондрии не прямым путем, а при посредстве
особых челночных механизмов. В зависимости от
того, какой из этих механизмов действует,
окисление цитоплазматического НАД H2 дает либо
четыре, либо шесть молекул АТФ и, значит, общий
выход АТФ на одну окисленную молекулу глюкозы
составляет 36 или 38 молекул. Схема на рис. 11.12
показывает, как работает такой челночный
механизм. Вещество X играет роль переносчика-
транспортирует водород из цитоплазмы в
митохондрию; в отличие от НАД H2 оно способно
проходить через митохондриальную мембрану.
После реакции НАД-►НАД H2 в клетках
сердечной мышцы и печени обратное окисление
дает три молекулы АТФ.
После реакции ФП -> ФПН2 в мышечных и
нервных клетках обратное окисление дает две
молекулы АТФ. Следовательно, в сердечной мышце
и печени образуется в общей сложности 38
молекул АТФ, а в мышечных и нервных
клетках-36 молекул.
Челночные механизмы непрерывно переносят
электроны из цитоплазмы в митохондрии,
одновременно вновь окисляя цитоплазматический
НАД H2. Это надежно предотвращает накопление
водородных атомов в цитоплазме и позволяет
понять, почему при аэробном дыхании не
накапливается молочная кислота.
Опыт 11.1. Изучение окисления одного
из промежуточных продуктов
цикла Кребса
Наиболее эффективный способ извлечения энергии
из субстрата с запасанием ее для последующего
использования состоит в расчленении этого
процесса на ряд более простых обратимых реакций,
катализируемых ферментами. Одна из таких
промежуточных реакций-это окисление янтарной
кислоты в фумаровую путем отщепления водорода.
Известны вещества, способные присоединять
отщепленные водородные атомы и при этом
изменять свой цвет. Одно из таких веществ-2,6-
дихлорфенолиндофенол (ДХФИФ); его окисленная
форма окрашена в синий цвет, а восстановленная
бесцветна.
Если окисленная форма ДХФИФ при смешивании
с тканевым экстрактом обесцвечивается, то можно
предположить, что причина этого - присоединение
атомов водорода от янтарной кислоты. Добавим к
смеси янтарную кислоту. Если теперь скорость
обесцвечивания возрастает, то это подкрепит нашу
гипотезу о том, что ДХФИФ играет роль акцептора
атомов водорода, отщепляющихся от янтарной
кислоты.
Поскольку большинство реакций в живых
организмах катализируется ферментами, ясно, что
окисление будет происходить лишь в том случае,
если в среде имеется надлежащий фермент. Реакцию
окисления янтарной кислоты катализирует фермент
сукцинатдегидрогеназа, и дополнительные
исследования позволяют выявить присутствие этого
фермента. В нашем опыте источником фермента
будет служить суспензия митохондрий из
прорастающих семян маша (Phaseolus aureus). Важно
выделять митохондрии* как можно быстрее. После
разрушения клеток метаболизм почти сразу же
прекращается.
Опыт делится на две части: 1) выделение нужного
фермента и 2) использование его для окисления
янтарной кислоты. ДХФИФ служит индикатором,
который позволяет определить, идет ли реакция или
нет.
Все, что требуется для первой части опыта (т.е.
для приготовления ферментного экстракта),
желательно до начала работы не менее часа выдержать в
холодильнике.
Материалы и оборудование
4 центрифужные пробирки (по 15 мл)
2 стеклянные палочки
2 градуированные пипетки на 10 мл
2 лабораторных стакана емкостью 1 л
(желательно полиэтиленовые)
Лед.
Соль
Семена маша
Пробирки со штативом
1 градуированная пипетка на 1 мл
Таймер
Растворы (указания относительно приготовления
растворов даны после описания опыта):
Забуференный раствор сахарозы
Забуференный раствор сахарозы + раствор
янтарной кислоты
0,1%-ный ДХФИФ (приготовляется на забуфе-
ренном растворе сахарозы)
Дистиллированная вода
Методика
1. Прорастите семена маша (24 шт.), поместив их
на влажную вату и оставив в темноте на 3-4 дня.
2. Приготовьте ледяную баню: положите лед в
полиэтиленовый стакан емкостью 1 л и добавьте
к нему соли, чтобы понизить температуру.
3. Поместите в стакан со льдом забуференный
раствор сахарозы и две центрифужные
пробирки.
4. Удалите у 12 проросших семян оболочки и
корешки.
5. Поместите в каждую центрифужную пробирку
по шесть семян.
6. Добавьте в каждую пробирку по 1 мл забуферен-
ного раствора сахарозы, не содержащего
янтарной кислоты.
7. Тщательно размельчите семена с помощью
охлажденной стеклянной палочки; пробирки
должны при этом все время оставаться в стакане
со льдом.
8. Добавьте в каждую центрифужную пробирку
еще по 10 мл забуференного раствора сахарозы.
9. Поместите центрифужные пробирки в
центрифугу (друг против друга) и прокрутите их при
максимальной скорости в течение 3 мин.
10. Верните центрифужные пробирки в стакан со
льдом.
11. Внесите пипеткой в одну из лабораторных
пробирок 15 мл дистиллированной воды и отметьте
положение мениска.
12. Вылейте дистиллированную воду и заполните
пробирку до отметки надосадочной жидкостью
из центрифужных пробирок.
13. Следующее необходимо проделать очень
быстро: добавьте в ту же лабораторную пробирку
0,5 мл раствора ДХФИФ и перемешайте
содержимое, заткнув пробирку большим пальцем
и перевернув ее.
14. Во время перемешивания растворов включите
отсчет времени.
15. Отметьте цвет раствора через 20 мин.
16. Повторите весь эксперимент, использовав на
этот раз забуференный раствор сахарозы,
содержащий янтарную кислоту.
Проследить за ходом эксперимента можно и с
помощью колориметра.
1. Поставьте красный фильтр, включите
колориметр и дайте ему прогреться в течение 5 мин.
2. Добавьте 0,5 мл раствора ДХФИФ к 15 мл
надосадочной жидкости, как описано выше.
3. Смешайте растворы и включите отсчет времени.
4. Поместите пробирку в колориметр и установите
прибор на 0% пропускания.
5. Снимите показания колориметра через 1, 2, 5, 10 и
20 мин.
6. Повторите тот же опыт, но с забуференным
раствором сахарозы, содержащим янтарную
кислоту.
7. Постройте график зависимости пропускания (ось
ординат) от времени.
8. Сформулируйте выводы, которые можно сделать
на основании полученных результатов.
Приготовление растворов
Забуференный раствор сахарозы (100 мл)
Гидрофосфат натрия (Na2HPO4) 0,76 г
Дигидрофосфат калия (KH2PO4) 0,18 г
Сахароза 13,60 г
Сульфат магния 0,10 г
Забуференный раствор сахарозы + янтарная
кислота (100 мл)
Забуференный раствор сахарозы,
приготовленный, как описано выше, плюс:
Янтарная кислота 1,36 г
Гидрокарбонат натрия 1,68 г
Лучше всего сразу приготовить забуференный
раствор сахарозы в количестве, достаточном для обеих
частей опыта (раствор готовится на
дистиллированной воде). После этого следует разделить раствор на
две равные части и добавить к одной из них
янтарную кислоту и гидрокарбонат натрия (в
соответствующей концентрации).
Когда к чабуференному раствору сахарозы
добавляют янтарную кислоту и гидрокарбонат
натрия, жидкость вскипает. Раствор следует хорошо
встряхнуть, чтобы вся углекислота выделилась, так
как ее присутствие может сказаться на результатах
эксперимента.
Раствор ДХФИФ
Растворите 0,1 г дихлорфенолиндофенола в 10 мл
забуференного раствора сахарозы (без янтарной
кислоты для обеих частей эксперимента).
Растворяется Это вещество не слишком хорошо, поэтому
после тщательного перемешивания суспензию
следует профильтровать.
11.5. Митохондрии
Митохондрии имеются во всех эукариотических
клетках. Эти органеллы- главное место аэробной
дыхательной активности клетки. Впервые наблюдал
митохондрии в виде гранул в мышечных клетках
Кёлликер в 1850 г. Позднее, в 1898 г., Михаэлис
показал, что они играют важную роль в дыхании:
в его опытах митохондрии вызывали
изменение цвета окислительно-восстановительных
индикаторов.
Число митохондрий в клетке очень непостоянно;
оно зависит от вида организма и от природы
клетки. В клетках, в которых потребность в энергии
велика, содержится много митохондрий (в одной
печеночной клетке, например, их может быть около
1000). В менее активных клетках митохондрий
гораздо меньше. Чрезвычайно сильно варьируют
также размеры и форма митохондрий. Митохондрии
могут быть спиральными, округлыми, вытянутыми,
чашевидными и даже разветвленными; в более
активных клетках они обычно крупнее. Длина
митохондрий колеблется в пределах 1,5-10 мкм, а
ширина-в пределах 0,25-1,00 мкм.
11.6. Почему при столь больших различиях в
длине митохондрий их ширина сравнительно
постоянна?
Митохондрии способны изменять свою форму, а
некоторые могут также перемещаться в особо
активные участки клетки. Такое перемещение
(которому способствует ток цитоплазмы) позволяет
клетке сосредоточивать большее число
митохондрий в тех местах, где выше потребность в АТФ.
В других случаях положение митохондрий более
постоянно (как, например, в летательных мышцах
насекомых; рис. 11.13).
Полиморфные
митохондрии
| Миофибрилла]
| M итохондрияI
|Миофибрилла|
Рис. 11.13. Электронные микрофотографии лета-
тельной мышцы комнатной мухи (Musca), полученные
с помощью трансмиссионного (А) и сканирующего (Б)
электронного микроскопа. Видно, что каждая
миофибрилла окружена полиморфными
митохондриями.
11.5.1. Строение митохондрий
Митохондрии можно выделить из клеток в виде
чистой фракции с помощью гомогенизатора и
ультрацентрифуги. После этого их можно исследовать в
электронном микроскопе, используя для этой цели
различные методики, такие, как изготовление срезов
Криста
Метрике
Наружная
мембрана
Внутренняя
мембрана
Матрикс
Элементарная
частица
Основание
Компоненту
дыхательной цепи
Головка
элементарной
частицы
(АТФаза)
Ножка
Другие факторы
сопряжения
Гранула
в матриксе
Криста
Элементарная
частица
Наружная
мембрана
Матрикс
Внутренняя
мембрана
Митохондрия
Рис. 11.14. Строение митохондрий. А. Схематическое изображение митохондрии. Б Схема строения кристы
с ((элементарными частицами» внутренней мембраны В Строение элементарной частицы внутренней
митохондриальной мембраны. Г Электронная микрофотография митохондрии, полученная при малом
увеличении. Д. Электронная микрофотография, на которой видны элементарные частицы внутренней
митохондриальной мембраны (F1-F0 АТФаза) из разрушенных осмотическим шоком митохондрий комнатной
мухи.
и негативный контраст. Каждая митохондрия
окружена двумя мембранами; наружную мембрану
отделяет от внутренней расстояние в 6-10 нм.
Внутренняя мембрана заключает в себе полужесткий
матрикс митохондрии; эта мембрана образует
многочисленные гребневидные складки, так называемые
кристы (рис. 11.14). Обрабатывая митохондрии
ультразвуком и детергентами, можно отделить
наружную мембрану от внутренней, что позволяет изучать
структуру и функции каждой из них в отдельности.
Однако даже и с помощью такой методики нам пока
еще мало что удалось узнать о наружной
митохондриальной мембране. Полагают, что она
проницаема для веществ с молекулярной массой
меньше 21000 и что такие вещества через нее
диффундируют. Кристы внутренней мембраны
существенно увеличивают ее поверхность, обеспечивая
место для размещения мультиферментных систем и
облегчая доступ к ферментам, находящимся в ми-
тохондриальном матриксе. Внутренняя мембрана
отличается избирательной проницаемостью, т.е.
пропускает лишь определенные вещества. Известно,
что активный транспорт АДФ и АТФ через
внутреннюю митохондриальную мембрану осуществляют
особые ферменты, называемые транслоказами.
Метод негативного контрастирования, при котором
окрашенными оказываются не сами структуры, а
пространство вокруг них, позволил выявить
присутствие особых «элементарных частиц» на той
стороне внутренней митохондриальной мембраны,
которая обращена к матриксу (рис. 11.14, Б и Д).
Каждая такая частица состоит из головки, ножки и
основания. Хотя микрофотографии
свидетельствуют, казалось бы, о том, что элементарные частицы
выступают из мембраны в матрикс, считается, что
это артефакт, обусловленный самой процедурой
приготовления препарата, и что в действительности
они полностью погружены в мембрану. Головки
ЦИТОПЛАЗМА
МИТОХОНДРИЯ
ЯДРО
Рибосомная
РНК
Цитоплазматические
рибосомы
Структурный
белок
Матричнад
РНК
Ферменты цикла Кребса
Переносчики электронов
Элементы наружной
мембраны
Матричт
наяРНК
Митохондриальная
ДНК
Ядерная
ДНК
Рибосомная
РНК
Митохондриальные
рибосомы
Рис. 11.15. Процесс сборки митохондрии из ее компонентов. (По Tribe, Whittaker, Chloroplasts and mitochondria.
Series in Biology, № 31, Arnold.)
частиц ответственны за синтез АТФ. Это АТФаза
(ранее обозначавшаяся F1), обеспечивающая
сопряжение фосфорилирования АДФ с реакциями в
дыхательной цепи. В основании частиц, заполняя
собой всю толщу мембраны, располагаются
компоненты самой дыхательной цепи. Они размещены по
отношению друг к другу строго упорядоченным
образом. В митохондриальном матриксе
содержится большая часть ферментов, участвующих в цикле
Кребса и в окислении жирных кислот. Здесь же
находятся митохондриальные ДНК, РНК и
рибосомы, а также ряд различных не очень крупных
белков (разд. 7.2.12).
11.7. Какими химическими веществами
обмениваются цитоплазма и митохондрии? Укажите,
какие из них поступают в митохондрии и какие
переходят из митохондрий в цитоплазму.
11.5.2. Сборка митохондрий
Митохондриальная ДНК несет информацию для
синтеза приблизительно 30 белков. Этого, однако,
недостаточно, так как для построения новой
митохондрии требуется большее число белков. В
какой-то мере, следовательно, образование новых
митохондрий должно зависеть от ядерной ДНК, от
цитоплазматических ферментов и от некоторых
других молекул, поставляемых клеткой. На рис. 11.15
суммированы современные представления о
взаимодействии между митохондрией и другими частями
клетки в процессе сборки митохондрии.
11.5.3. Эволюция митохондрий:
эндосимбиотическая гипотеза
Существует гипотеза, согласно которой
митохондрии были некогда свободноживущими прокариоти-
ческими организмами, наподобие бактерий. Эти
прокариоты, случайно проникнув в клетку, вступили
затем с этой клеткой-хозяином во взаимовыгодный
симбиоз. По-видимому, условия внутри клетки
оказались благоприятными для прокариот, в обмен же
прокариоты своим присутствием резко повысили
«производительность» клетки в смысле синтеза АТФ
и придали ей способность к аэробному дыханию. В
пользу этой гипотезы свидетельствует ряд данных.
Во-первых, митохондриальная ДНК представлена
обычно кольцевой молекулой (рис. 11.16), а именно
такую ДНК мы находим у современных бактерий.
Во-вторых, митохондриальные рибосомы меньше
цитоплазматических и сходны по своим размерам с
бактериальными. В-третьих, движения
митохондрий напоминают движения некоторых бактерий.
И наконец, отмечено, что механизмы белкового
синтеза у митохондрий и бактерий, с одной
стороны, и в цитоплазме-с другой, чувствительны к
разным антибиотикам. Например, хлорамфеникол и
стрептомицин подавляют синтез белка в
митохондриях и у бактерий, а циклогексимид блокирует
синтез белка в цитоплазме (см. разд. 9.3.1 об эндо-
симбиотической гипотезе применительно к хлоро-
пластам).
11.5.4. Синтез АТФ
Изучением этого вопроса активно занимаются
больше 30 лет, однако четкого представления о
механизмах синтеза АТФ у нас пока еще нет. В последнее
время усиленно обсуждаются главным образом две
гипотезы: гипотеза химического сопряжения и хеми-
осмотическая гипотеза.
Гипотеза химического сопряжения
Согласно этой гипотезе, синтез АТФ сопряжен с
переносом электронов при посредстве одного или
нескольких «высокоэнергетических» промежуточных
продуктов. Энергия, высвобождаемая при переносе
электронов в окислительно-восстановительных
реакциях дыхательной цепи, используется в
нескольких ее звеньях для образования
высокоэнергетической связи в одном из таких продуктов. Затем при
фосфорилировании АДФ эта энергия переходит к
Рис. 11.16. Электронная микрофотография митохондриальной ДНК из пивных дрожжей Saccharomyces
carlsbergensis. Молекула представляет собой суперспирализованное кольцо двухцепочечной ДНК с длиной
покружности» 26 мкм. Она построена примерно из 75000 нуклеотидов.
ФАД
НАД· H2
ФАД H2
Дыхательная цепь
X
НАД-Х
НАД
-О-Ф
Х-О-Ф
X
АДФ
АДФ-Ф
Рис. 11.17. Гипотеза химического сопряжения.
X-пока еще не идентифицированный
промежуточный продукт. Здесь показан только один такой
продукт, но их могло быть и несколько.
высокоэнергетической связи АТФ (рис. 11.17). До
сих пор, однако, обнаружить подобные
промежуточные продукты не удалось, и до тех пор пока их
существование не подтвердится, эту гипотезу нельзя
считать убедительной.
Хемиосмотическая гипотеза
Большее признание завоевала гипотеза, выдвинутая
Митчеллом в 1961 г. Он полагал, что синтез АТФ
находится в тесной зависимости от того, каким
образом электроны и протоны передаются по
дыхательной цепи. Ниже перечислены условия,
соблюдения которых требует эта гипотеза.
1. Внутренняя митохондриальная мембрана должна
быть интактна и непроницаема для протонов
(ионов водорода), направляющихся снаружи внутрь.
2. В результате активности дыхательной (электрон-
транспортной) цепи ионы водорода поступают в
нее изнутри, из матрикса, а освобождаются на
наружной стороне мембраны.
3. Движение ионов водорода, направленное изнутри
наружу, приводит к их накоплению, вследствие
чего между двумя сторонами митохондриальной
мембраны возникает градиент рН. Это может
быть связано с тем, что ферменты, принимающие
и отдающие ионы водорода, расположены в
мембране определенным образом и поэтому могут
принимать ионы водорода только изнутри и
отдавать их только наружу.
4. Сам по себе градиент рН не мог бы
поддерживаться, так как ионы водорода
диффундировали бы обратно в митохондрию. Поддержание
такого градиента требует затраты энергии.
Предполагается, что энергию поставляет перенос
электронов по электронтранспортной (дыхательной)
цепи.
5. Эта энергия используется затем для синтеза АТФ.
Синтез АТФ, таким образом, поддерживается
наличием градиента рН.
6. АТФ образуется в результате фосфорилирования
АДФ:
АДФ + Фн *± АТФ + H2O.
По закону действующих масс удаление воды должно
ускорять реакцию, идущую слева направо, т. е.
благоприятствовать образованию АТФ. Согласно
теории Митчелла, фермент, ответственный за
образование воды при синтезе АТФ, ориентирован в
мембране таким образом, что ионы водорода
освобождаются с внутренней стороны мембраны, где
значение рН выше (т.е. концентрация H+ меньше), а
гидроксильные ионы (OH-)-с наружной стороны,
где рН ниже (т.е. концентрация H+ больше). Таким
образом, вода, образующаяся при синтезе АТФ,
быстро удаляется, и это стимулирует синтез.
Из гипотезы Митчелла ясно, почему мембрана
должна быть интактной (в ней расположены
ферменты, ответственные за прохождение ионов
водорода и за образование воды, так что любое
изменение структуры мембраны неминуемо
сказалось бы также на расположении ферментов и на их
структуре). Объяснимо и требование
непроницаемости мембраны для ионов водорода (в
направлении снаружи внутрь): в основе гипотезы лежит
представление о трансмембранном градиенте рН, а
такой градиент не мог бы поддерживаться, если бы
мембрана была полностью проницаемой. Следует
отметить также, что гипотеза обходится без каких
бы то ни было промежуточных продуктов в
процессе синтеза АТФ.
11.5.5. Другие пути дыхания
Пентозофосфатный
(гексозомонофосфатный) шунт
Этот метаболический путь требует наличия
кислорода и служит главным источником пятиуглерод-
ных Сахаров, входящих в состав важных нуклеоти-
дов (АТФ, НАД, ФАД) и нуклеиновых кислот. Шунт
может функционировать наряду с обычным гли-
колитическим путем, поставляя в разных клетках от
10 до 90% энергии, получаемой за счет расщепления
углеводов в процессе дыхания.
Суть превращений сводится к тому, что шесть
молекул глюкозо-6-фосфата сначала
дегидрируются, а затем декарбоксилируются. Акцептором
водорода служит НАДФ. В конечном итоге образуется
шесть молекул рибулозо-5-фосфаза (фосфорилиро-
ванного пятиуглеродного сахара) и в качестве
побочного продукта-шесть молекул CO2. Молекулы
рибулозо-5-фосфата претерпевают затем сложную
последовательность реакций, в результате которой
ресинтезируются пять молекул глюкозо-6-фосфата.
Кроме того, образуется глицеральдегид-3-фосфат,
который может быть направлен на путь гликолиза и
превращен в пировиноградную кислоту,
поступающую в конце концов в цикл Кребса.
Пентозофосфатный шунт дает в общем итоге 36 молекул АТФ.
Гликолиз вместе с циклом Кребса дает 38 молекул,
но шунт имеет то преимущество, что в нем меньше
реакций, а значит, меньше и число необходимых
ферментов. Суммарную реакцию можно записать
так:
6 χ глюкозо-6-Ф + 12 НАДФ+ + 6H2O
5хГлюкозо-6-Ф + 12 (НАДФ· H + H+) + 6CO2 + Φ
6O2- ¥
12 НАДФ + 12 H2O
Шунт поставляет также НАДФ H2, который
служит донором водорода и электронов
(восстановителем) при синтезе ряда веществ. В жировой ткани,
например, именно этот шунт дает большие
количества НАДФ H2, который используется для
восстановления ацетил-КоА до жирных кислот в
процессе синтеза липидов.
Глиоксилатный цикл
Глиоксилатный цикл, функционирующий в богатых
маслами семенах, обеспечивает превращение
запасных жиров в углеводы. Ферменты, участвующие в
этом цикле, сосредоточены главным образом в ор-
ганеллах, называемых глиоксисомами (они содержат
каталазу и, следовательно, представляют собой
разновидность пероксисом), и активных в период
прорастания семян; впрочем, этот цикл протекает и в
других органеллах. Он, однако, перестает
функционировать, после того как будет израсходован весь
запас жиров.
CH2OH
CHOH
I
CH2OH
Глицерол
АДФ
АТФ
Фосфорилирование
CH2OH
CHOH -
I
CH2O-@
Глицерол-3-
фосфат
НАД
СН20-(ф)
со
I
CH2OH
CHO
I
CHOH
CH2O <Ф)
Дигидро ксиацетон -
фосфат
Глицеральдегид-3-
фосфат
НАД-H2
Дыхательная
цепь
ЗАТФ
Рис. П.18. Превращение глицерола в глицеральдегид-3-фосфат.
Во время прорастания семян жиры гидролизу-
ются до жирных кислот и глицерола. Затем жирные
кислоты расщепляются путем β-окисления (разд.
11.5.6), что дает в итоге значительные количества
ацетил-КоА. Ацетил-КоА, как обычно, вступает в
цикл Кребса, но здесь к реакциям этого цикла
добавляется еще и глиоксилатный цикл. В каждом
обороте глиоксилатного цикла происходит
следующее суммарное превращение:
2Ацетил-КоА -> Янтарная кислота + 2Н + 2KoA.
2 χ 2С 4С
Пара отделившихся атомов водорода передается на
кислород через дыхательную цепь, в результате чего
синтезируется АТФ. Янтарная кислота может
использоваться как источник углеродных скелетов для
синтеза ряда различных соединений. Таким
образом, субстратом для глиоксилатного цикла служит
двухуглеродное соединение ацетил-КоА, а
поставляет этот цикл энергию и промежуточные четырех-
углеродные продукты для биологических синтезов.
11.5.6. Жиры как дыхательный субстрат
В некоторых животных тканях, например в печени, а
также в семенах растений, содержащих большие
количества жиров, эти жиры могут использоваться в
качестве дыхательного субстрата без
предварительного превращения их в углеводы. Сначала жиры при
участии ферментов, называемых липазами, гидро-
лизуются до жирных кислот и глицерола.
Глицерол
Глицерол (глицерин) сначала фосфорилируется под
действием АТФ, превращаясь в глицеролфосфат, а
затем дегидрируется под действием НАД с
образованием дигидроксиацетонфосфата. Этот последний
превращается в свой изомер-глицеральдегид-3-фос-
фат (рис. 11.18). В этих процессах, как видно из
рис. 11.18, одна молекула АТФ потребляется и три
образуются (при переносе водорода на кислород в
дыхательной цепи). Далее глицеральдегид-3-фосфат
вступает на путь гликолиза и в цикл Кребса, что
дает еще 17 молекул АТФ. Таким образом,
превращения одной молекулы глицерола в процессе
аэробного дыхания дают в общем итоге 20-1 = 19 АТФ.
Жирные кислоты
Каждая молекула жирных кислот окисляется в
результате процесса, называемого β-окислением. При
этом от молекулы жирной кислоты
последовательно отщепляются двухуглеродные фрагменты, так
что на каждом этапе эта длинная молекула
укорачивается на два атома углерода. Образовавшийся
ацетил-КоА может, как обычно, поступить в цикл
Кребса, чтобы окислиться до CO2 и воды (рис.
11.21). Этот процесс протекает в матриксе
митохондрий. Из каждой молекулы жирной кислоты
извлекается большое количество энергии: при
окислении стеариновой кислоты выход АТФ составляет,
например, 147 молекул. Неудивительно поэтому,
что жирные кислоты - важный источник энергии.
Около половины обычных энергетических затрат
сердечной мышцы, печени, почек и скелетных мышц
(в покое) покрывается именно за счет окисления
жирных кислот.
11.5.7. Белки как дыхательный субстрат
Белки используются как дыхательный субстрат
крайне редко-только после того, как будут
исчерпаны все имеющиеся запасы углеводов и жиров.
Сначала белки гидролизуются до аминокислот, из
которых состоят их молекулы, а затем
аминокислоты дезаминируются (от них отщепляются
аминогруппы). С отщеплением аминогрупп протекают
реакции двух типов: окислительное дезаминирова-
ние и трансаминирование.
Окислительное дезаминирование
Окислительное дезаминирование происходит в
клетках печени позвоночных. В результате
дегидрирования и гидролиза от аминокислоты отщепляется
молекула аммиака. Позже азот выводится из
организма в виде аммиака, мочевой кислоты или мо-
COOH
CH2
CH2+ НАД+ H2O
Глутаминовая
кислота
Дегидрогеназа
CHNH2
COOH
Глутаминовая
кислота
COOH
CH2
CH2 + NH3+ НАД-H2
со
COOH
α-Оксоглутаровая кислота
(направляется в цикл Кребса)
Рис. 11.19. Окислительное дезаминирование
глутаминовой кислоты.
чевины (у разных животных выводимый продукт
может быть различным; см. разд. 18.5).
Аминокислота при дезаминировании превращается в а-оксо-
кислоту. В зависимости от природы боковых групп
аминокислот их углеродные скелеты могут в
процессе дыхания расщепляться по пути, характерному
для углеводов или же для жирных кислот (рис.
11.19).
Трансаминирование
Эту реакцию, свойственную всем клеткам,
катализируют ферменты, называемые трансаминазами.
Трансаминирование-это перенос аминогруппы от
аминокислоты на оксокислоту. Таким образом, вместо
одной аминокислоты может получиться другая.
Одновременно образуются соответствующие а-оксо-
кислоты, которые могут затем подвергаться
обычному расщеплению в процессе дыхания. Несколько
примеров трансаминирования приведено на рис.
11.20. На рис. 11.21 представлены главные
метаболические пути, участвующие в дыхании.
11.6. Газообмен
Каким бы ни был процесс дыхания - аэробным или
анаэробным,-между организмом и средой
непрерывно должен происходить обмен газами. Аэробам
для окисления пищевых веществ и получения
энергии нужен поступающий из внешней среды
кислород, а в среду аэробы и большинство анаэробов
выделяют углекислоту (диоксид углерода
CO2)-конечный продукт («отходы») дыхания. Обмен CO2 и
O2 между средой и организмом называется
газообменом, а поверхность, на которой этот
обмен фактически происходит,-дыхательной
поверхностью. Осуществляется газообмен у всех
организмов путем диффузии. Для того чтобы диффузия
могла происходить, дыхательная поверхность
должна удовлетворять нескольким условиям:
1) она должна быть проницаемой, чтобы газы
могли сквозь нее проходить;
2) образующий ее слой должен быть тонким,
потому что диффузия эффективна на расстояниях
не более 1 см;
3) она должна быть влажной, так как оба газа-
кислород и CO2-диффундируют в растворе;
4) дыхательная поверхность должна быть
большой, чтобы через нее могли обмениваться
достаточные количества газов в соответствии с
потребностями организма.
Организмы получают необходимый им кислород
либо непосредственно из атмосферы, либо из воды,
в которой он растворен. Содержание кислорода в
воде и в воздухе далеко не одинаково. В воздухе в
единице объема содержится во много раз больше
кислорода, чем в таком же объеме воды. Отсюда
следует, что объем воды, который вынуждены
пропускать над дыхательной поверхностью для удов-
Аланин
a. -O ксо кисло га
Пировиноградная
кислота
Аминокислота
Глутаминовая кислота
α -О ксо кислота
α - О ксогл утаров ая
кислота
Аминокислота
Аспарагиновая
кислота
а-Оксокислота
Щавелевоуксусная
кислота
Аминокислота
Рис. 11.20. Примеры реакций трансаминирования.
Глюкоза
Гликоген
(углевод)
Белок
Глюкоза
Молочная
кислота
Глицеральдегид
Аминокислоты
Дезаминиро -
вание
α-О ксо кислоты
Глицерол
+
Жирная
кислота
Жир
Пировино-
градная
кислота
Этанол
► Алании-5'
^•Окисление
Ацетил-КоА
Трансаминирование
» Аспарагиновая
кислота·
Щавелевоуксусная
кислота
Лимонная
кислота
а-Оксо-
глутаровая
кислота
. Глутаминовая
кислота
летворения своих метаболических нужд водные
организмы, например рыбы, значительно больше
объема воздуха, достаточного для наземных
позвоночных животных.
11.6.1. Простейшие
У Amoeba proteus тело меньше 1 мм, поэтому
отношение поверхности к объему у нее очень велико.
Диффузия газов происходит у нее через всю
наружную клеточную мембрану, и этого вполне
хватает для ее метаболических нужд.
11.6.2. Кишечнополостные
У двуслойных многоклеточных Hydra и Obelia все
клетки находятся в контакте с водной средой, и у
каждой из этих клеток газообмен через клеточную
мембрану, соприкасающуюся со средой, достаточен
для удовлетворения ее потребностей.
11.6.3. Плоские черви
У таких свободноживущих плоских червей, как PIa-
naria, кислород поступает в организм путем
диффузии через всю поверхность тела. Этому
благоприятствует сильно уплощенная форма тела
(толщина тела не превышает, как правило, 0,06 см),
увеличивающая отношение поверхности к объему.
Способствует диффузии и то, что планарии обитают
обычно в хорошо аэрируемых водоемах.
Многие плоские черви, например
Taenia,-внутри с. 11.21. Главные
метаболические пути, участвующие в
процессе дыхания.
ренние паразиты, живущие в условиях, где
кислорода очень мало. В таком случае они ведут себя как
анаэробы. Ни размеры, ни форма их тела не
определяются при этом потребностью в диффузии
кислорода, хотя отношение поверхности к объему все еще
остается у них высоким.
11.8. Какое другое преимущество дает плоским
червям большое отношение поверхности тела к
объему?
11.6.4. Потребность
в специализированных
дыхательных структурах
и пигментах
С уэеличением размеров тела отношение
поверхности тела к объему уменьшается. Простая диффузия
теперь уже не может обеспечить достаточный приток
кислорода к тем клеткам, которые не находятся в
прямом контакте с внешней средой. К тому же
нередко и сама потребность в кислороде у таких
более крупных животных ввиду их высокой
метаболической активности оказывается выше.
Повышенная потребность в кислороде привела в
процессе эволюции к тому, что некоторые участки
тела превратились в специализированные
дыхательные поверхности. У разных животных типы
дыхательных поверхностей различны. В каждом случае
они приспособлены для работы в тех или иных
конкретных условиях. На рис. 11.22 показано, как их
можно в соответствии с этим классифицировать.
Клеточная мембрана
CO2
Кислород
Водная
среда
Наружная жабра
Полуводная
среда
Воздушная
среда
Вода
Жабры
(выросты
поверхности
тела)
Поверхность тела
Вода
Внутренняя жабра
Поверхность
тела
Вода
Эпителий
Кровеносный сосуд
Поверхность тела
Вода
Воздух
Трахеи
(впячивания
поверхности
тела)
Трахея
Ткань
Поверхность тела
Воздух
Легкие
(впячивания
поверхности
тела)
Кровь
Поверхность тела
Рис. 11.22. Типы дыхательных поверхностей.
Обычно площадь дыхательных поверхностей сильно
увеличена, и часто они связаны с какой-нибудь
транспортной системой, например кровеносной.
Транспортная система обеспечивает дыхательной
поверхности связь со всеми прочими тканями тела и
делает возможным непрерывный обмен кислородом
и углекислотой между дыхательной поверхностью и
клетками. Присутствие дыхательного пигмента в
крови еще больше повышает способность крови
переносить кислород. Кроме того, особые
вентиляционные движения ускоряют газообмен между
организмом и средой, поддерживая крутые
диффузионные градиенты.
Дыхательные пигменты
Кровь, содержащая какой-либо дыхательный
пигмент, служит гораздо более эффективным
переносчиком кислорода, нежели кровь без такого
пигмента. Пигмент позволяет поглощать и
транспортировать гораздо большие количества O2; он может
находиться в плазме крови или в особых клетках.
Любопытно отметить, что молекулы тех пигментов,
которые заключены в форменных элементах крови,
относительно невелики по сравнению с пигментами
плазмы. Пигменты плазмы представляют собой
агрегаты из многих небольших молекул, ведущие
себя как одна крупная молекула. Такая организация
дает возможность увеличивать количество пигмента
в крови без одновременного увеличения числа
частиц в растворе.
11.9. Почему это важно?
11.10. В чем преимущество системы, в которой
дыхательный пигмент сосредоточен в особых
клетках?
Все известные дыхательные пигменты
присоединены к белковым молекулам. Пигменты обладают
способностью обратимо связывать кислород. При
высоких концентрациях кислорода пигмент легко
его присоединяет, а при низких-быстро отдает.
Перенос кислорода гемоглобином более
подробно описан в гл. 14. В табл. 11.3 приведены
свойства распространенных дыхательных пигментов
животных.
11.6.5. Кольчатые черви
У кольчатых червей нет никаких систем,
предназначенных специально для газообмена, и газообмен
происходит путем диффузии через всю поверхность
тела. Специализированные системы кольчатым
червям в сущности и не нужны, так как благодаря
цилиндрической форме тела отношение поверхности
к объему у них велико, а при своей относительно
малой активности они расходуют мало кислорода
на единицу массы тела.
Кровеносная система у кольчатых червей имеется,
и в их крови растворен дыхательный пигмент
гемоглобин. Сокращения кровеносных сосудов
прогоняют кровь вместе с растворенными в ней газами по
всему телу, действуя как насос; это же способствует
и поддержанию крутых диффузионных градиентов.
У наземных олигохет (например, у Lumbricus)
кутикула постоянно увлажняется секретом
находящихся в эпителии желез и жидкостью, выделяемой
дорсальными порами. В эпителии непосредственно
под кутикулой расположены капилляры. Расстояние
между кровеносными сосудами и поверхностью
тела невелико, и это обеспечивает быструю диффузию
кислорода в кровь. Дождевые черви практически
ничем не защищены от высыхания и поэтому могут
существовать только во влажной среде.
У водных полихет (например, у Nereis sp.) на
каждом сегменте тела имеется по две параподии
(см. рис. 4.26). Это подвижные выросты стенки тела,
пронизанные многочисленными сосудами;
благодаря им площадь дыхательной поверхности
животного увеличивается. Диффузия газов ускоряется
благодаря тому, что кровь в этих сосудах проходит
совсем близко от поверхности тела.
11.6.6. Членистоногие
С системой, обеспечивающей газообмен у
членистоногих, можно ознакомиться на примере насекомых.
Газообмен осуществляется здесь через систему
трубочек-так называемых трахей. Благодаря этому
кислород из воздуха поступает прямо к тканям, и
необходимость в его транспортировке кровью
отпадает. Это гораздо более быстрый способ, нежели
диффузия растворенного кислорода сквозь ткани;
такой газообмен создает условия для высокой
интенсивности метаболизма.
Дыхальца-парные отверстия, имеющиеся на
втором и третьем грудном и на первых восьми
брюшных сегментах тела насекомого, ведут в воздушные
полости. От этих полостей отходят разветвленные
трубочки, называемые трахеями (рис. 11.23).
Каждая трахея выстлана плоским эпителием, секрети-
рующим тонкий слой хитинового материала.
Обычно этот жесткий слой дополнительно укреплен спи-
Таблица 11.3. Обычные дыхательные пигменты
Пигмент
Гемоцианин
Гемоэритрин
Хлорокруорин
Гемоглобин
Металл
Cu
Fe
Fe
Fe
Цвет
+ 02?± -O2
Синий ?± бесцветный
Красный *± бесцветный
Красный τ± зеленый
Оранжево-
красный
*± пурпурно-
красный
Группы животных
Некоторые улитки
Ракообразные
Головоногие моллюски
Некоторые кольчатые
черви
Некоторые кольчатые
черви
Некоторые моллюски
Некоторые кольчатые
черви
Рыбы
Амфибии
Рептилии
Птицы
Млекопитающие
Локализация в крови
В плазме
-»—
-»-
Всегда в клетках
В плазме
-»-
В плазме или в клетках
В клетках
-»-
-»—
-»-
-»—
Количество O1
(мл),
транспортируемого
в 100 млн.
крови1[)
2
3
8
2
9
2
7
9
11
10
18
25
!) При рассмотрении приведенных величин сопоставьте их с тем фактом, что в отсутствие связывающих кислород пигментов
растворимость кислорода составляет 0,2 мл O2 на 100 мл крови.
Грудной
воздушный мешок
Брюшные
воздушные мешки
Дорсальный
трахейный ствол
Вентральный
трахейный ствол
Дыхальца
Латеральный
трахейный ствол
Крупная
продольная трахея
Трахея
Трахеола
Мышечные
волокна
Рис. 11.23. А Вертикальный продольный разрез
тела прямокрылого Б Строение трохеи насекомого
ральными и кольцевыми утолщениями, благодаря
которым воздухоносные пути остаются открытыми,
даже если в просвете трахей давление оказывается
отрицательным (сравните с хрящевыми кольцами в
трахее и бронхах человека). В каждом сегменте тела
трахеи разветвляются на многочисленные более
мелкие трубочки, называемые трахеолами; трахеолы
тоже ветвятся, пронизывая ткани насекомого, и в
наиболее активных тканях, например в летательных
мышцах, оканчиваются слепо внутри отдельных
клеток. В трахеолах хитиновая выстилка
отсутствует; кроме того, степень их ветвления может
меняться, приспосабливаясь к метаболическим
нуждам ткани.
В состоянии покоя трахеолы наполнены
водянистой жидкостью (рис. 11.24); в это время
кислород диффундирует по ним к тканям (a CO2-B
обратном направлении) со скоростью, вполне
достаточной для удовлетворения потребностей
насекомого. В активном состоянии усиление
метаболической активности мышц ведет к накоплению
определенных метаболитов, в частности молочной
кислоты, и в тканях соответственно повышается
осмотическое давление. Когда это происходит, жидкость
из трахеол под действием осмотических сил
частично всасывается в ткани и в трахеолы поступает
больше воздуха, а значит, и больше кислорода,
причем этот кислород подается непосредственно к
тканям как раз тогда, когда они в нем нуждаются.
Общий поток воздуха, проходящий через тело
насекомого, регулируется механизмом,
закрывающим дыхальца. Отверстие каждого дыхальца
снабжено системой клапанов, управляемых очень
мелкими мышцами. Края этого отверстия покрыты
волосками, которые предотвращают попадание в
дыхальце чужеродных частиц и излишнюю потерю
влаги. Величина отверстия регулируется в
зависимости от количества CO2 в теле насекомого.
Усиленная активность ведет к усиленному
образованию CO2. Хеморецепторы улавливают это, и
дыхальца открываются. Тот же стимул может
вызывать и вентиляционные движения тела, особенно
у крупных насекомых. Дорсовентральные мышцы,
сокращаясь, делают тело насекомого более
плоским, вследствие чего объем трахейной системы
уменьшается и воздух выталкивается из нее наружу
(«выдох»). Всасывание воздуха («вдох») происходит
пассивно, когда сегменты тела благодаря своей
эластичности принимают исходную форму.
Судя по некоторым данным, грудные и брюшные
дыхальца открываются и закрываются
попеременно, и это в сочетании с вентиляционными
движениями тела создает однонаправленный поток
воздуха, который входит в тело насекомого через
грудной его отдел и выходит через брюшной.
Трахейная система, безусловно, весьма
эффективна в смысле газообмена, однако следует учитывать,
что газообмен определяется здесь исключительно
диффузией газообразного кислорода через ткани
насекомого. Диффузия же, как известно, эффективна
только на малых расстояниях, и это накладывает
жесткие ограничения на размеры, которых могут
достигать насекомые. Эти малые расстояния, на
которых диффузия достаточно эффективна, не
превышают 1 см; поэтому хотя и встречаются
насекомые длиной до 16 см, их тело не должно при
этом иметь в толщину более 2 см!
11.6.7. Рыбы
Хрящевые рыбы (например, акулы)
На каждой стороне глотки акулы расположено по
пять пар жаберных мешков, в каждом из которых
находится жабра. Обычно каждая жабра
поддерживается вертикальным хрящом-так называемой
жаберной дугой. От перегородки, лежащей над
жаберной дугой, отходит ряд горизонтальных складок-
жаберных лепестков. Каждый лепесток на верхней и
нижней поверхности в свою очередь образует
вертикальные складки, называемые вторичными
лепестками (рис. 11.25). Свободный край каждой
жаберной перегородки довольно сильно вытянут и
действует как надежный откидной (створчатый) клапан.
При дыхательных движениях он периодически
закрывает находящуюся непосредственно за ним
жаберную щель. Пространства между этими
клапанами и жаберными лепестками называются пара-
бранхиальными полостями.
Бедная кислородом кровь из брюшной аорты
поступает в каждую жабру по приносящей жаберной
артерии. В области жаберной пластинки эта артерия
многократно разветвляется на множество мелких
капилляров. Именно здесь происходит газообмен.
Капилляры затем вновь объединяются в выносящие
Жидкость
Дыхальце
Трахея
Воздух
Активность
Покой
Трахеола
Трахеолы окружены
гипотонической жидкостью.
Жидкость диффундирует
в трахеолы
Окружающая трахеолы жидкость
становится гипертонической вследствие
накопления молочной кислоты.
Жидкость из трахеол всасывается в ткани;
вместо нее в них поступает воздух
Рис. 11.24. Условия,
создающиеся в тканях
насекомого в покое и в
активном состоянии
(работа трахеол).
жаберные артерии, которые выходят из жабры у ее
основания.
Вентиляционные движения, действующие как
нагнетающий насос перед жабрами и отсасывающий
позади них, создают почти непрерывный ток воды
сквозь жабры. Вода входит через рот и брызгальца,
когда дно ротовой полости и глотки опускается.
Происходит это потому, что при увеличении объема
глоточной области давление в ней уменьшается и в
нее устремляется вода (рис. 11.26, Л). Одновременно
из-за снижения давления в глотке имеющиеся на
жабрах клапаны плотно прикрывают жаберные
щели, препятствуя поступлению воды с этой стороны.
При этом работает и отсасывающий насос: боковые
движения клапанов вызывают расширение пара-
бранхиальных полостей, так что гидростатическое
давление в них оказывается ниже, чем в глотке; из-за
этого перепада давлений вода не только поступает в
глотку, но одновременно проходит и между
жаберными лепестками, т.е. течет практически
непрерывно.
После того как полость глотки заполнится водой,
рот и брызгальца закрываются, клапаны
открываются, а дно ротовой полости и глотки поднимается.
Это проталкивает воду через жаберные мешки над
дыхательным эпителием жаберных лепестков, а
затем наружу через жаберные щели (рис. 11.26, Б).
Когда вода проходит через глоточную область,
сфинктер пищевода сокращается и закрывает
пищевод, препятствуя попаданию воды в
пищеварительный тракт. Растворенный в воде кислород
поглощается гемоглобином, находящимся в
эритроцитах, однако акулы извлекают из воды менее 50%
кислорода, тогда как костные рыбы-80%.
Объясняется это сравнительно небольшой поверхностью
жабер у акул (в сравнении с костными рыбами) и тем,
что значительная часть воды, проходящей через
жабры, движется в направлении, параллельном
направлению кровотока.
11.11. Почему газообмен происходит менее
эффективно, если ток крови параллелен току воды?
Костные рыбы
У костных рыб на каждой стороне глотки имеются
четыре жаберные дуги, разделяющие пять жаберных
щелей. На этих дугах находятся жабры. Каждая
жабра состоит из двух рядов тонких жаберных
Выносящий сосуд
Приносящий сосуд
Жаберная дуга
Ток воды
Вторичные
жаберные лепестки
Жаберные лепестки
Направление тока воды
Направление тока крови
Рис. 11.25. Ток воды около жаберных лепестков акулы.
Вода, входящая
через брызгальце
Низкое
давление
Вода, входящая через рот
Дно ротовой полости и глотки
опускается
Объем ротовой полости и глотки
увеличивается
Парабранхиальные
полости
Высокое
давление
Дно ротовой полости и глотки
поднимается
Объем ротовой полости и глотки уменьшается
Б
Рис. 11.26. Схема, поясняющая, как
происходит дыхание у акулы.
Изменения давления указаны
относительно наружного давления.
А. Наполнение ротовой полости и
глотки водой. Б. Изгнание воды
через жаберные щели.
лепестков, расположенных в виде буквы V (рис.
11.27). Лепестки, несущие жаберные пластинки и
имеющие примерно такое же строение, как у акулы,
пронизаны многочисленными капиллярами.
Подвижная жаберная крышка, укрепленная тонкими
костными слоями, служит наружной стенкой
жаберной полости и защищает жабры; она составляет
также часть вентиляционного механизма.
При вдохе ротовая полость расширяется и
давление в ней падает, вследствие чего внутрь
устремляется вода. Одновременно под напором воды извне
закрывается клапан на заднем конце жаберной
крышки, препятствуя поступлению воды с этой
стороны. В то же время, однако, сокращаются
мышцы жаберной крышки, вызывая расширение
жаберной полости. Давление в жаберной полости
ниже, чем в ротовой, и потому вода поступает из
ротовой полости в жаберную, проходя при этом над
жабрами. Газообмен, таким образом, может
продолжаться и тогда, когда рыба набирает в рот
новые порции воды.
При выдохе ротовая полость и входное отверстие
пищевода закрываются, а дно ротовой полости
поднимается. Вода вследствие этого проходит через
жаберные щели и выходит затем наружу около
заднего края открытой теперь жаберной крышки.
Благодаря согласованной активности ротовой
полости и мышц жаберной крышки жабры почти все
время омываются потоком воды.
Лепестки соседних жабер перекрываются своими
концами. Вместе с жаберными пластинками они
образуют своего рода сито; оно создает
сопротивление току воды, замедляя прохождение воды над
жаберными пластинками и тем самым увеличивая
время, на протяжении которого может происходить
газообмен. Кровь в жаберных лепестках течет в
направлении, противоположном току воды. В такой
противоточной системе кровь на своем пути все
время встречается с водой, в которой концентрация
растворенного кислорода относительно высока, и
градиент концентрации между кровью и водой
поддерживается по всей длине жаберного лепестка.
Благодаря этому костные рыбы могут извлекать до
80% растворенного в воде кислорода.
11.6.8. Амфибии
У лягушки газообмен может происходить тремя
разными путями: через кожу (кожное дыхание),
через эпителий, выстилающий ротовую полость
(ротовое дыхание), и через легкие (легочное дыхание).
кожное дыхание. Кожа лягушки обильно снабжена
капиллярами и все время остается влажной-ее
покрывает слизь, выделяемая слизистыми железами.
Кислород воздуха растворяется в этой слизи и затем
диффундирует в кровь. Это называется кожным
дыханием. Поглощение кислорода кожей остается
на протяжении всего года практически неизменным,
Костная
жаберная
дуга
Выносящий
сосуд
Приносящий
сосуд
Направление тока воды
через жаберную щель
Жаберный
лепесток
Вторичный лепесток
(жаберная пластинка)
Ток воды
Направление
тока крови
Кровеносные
сосуды
Рис. 11.27. Жаберный лепесток костной рыбы.
поскольку концентрация кислорода в воздухе
постоянна, а значит, остается постоянным и
диффузионный градиент. В зимнее время почти весь
необходимый животному кислород поступает в
организм именно таким путем, весной же, когда
лягушки наиболее активны, за счет кожного дыхания
удовлетворяется иногда лишь четверть всей
потребности в кислороде. Остальной кислород поступает
через ротовую полость и легкие.
ротовое дыхание. Заметные глазу движения горла у
лягушки поддерживают постоянный газообмен
между ротовой полостью и атмосферой. Это называется
ротовым дыханием. Для вдоха рот и голосовая
щель должны быть закрыты, ноздри открыты, а дно
ротовой полости должно опускаться, что
достигается сокращением грудино-подъязычных мышц,
прикрепленных к подъязычному хрящу (рис. 11.28, А).
Ротовая полость выстлана влажным, богатым
кровеносными сосудами эпителием; именно здесь и
происходит газообмен. Выдох наступает, когда
сокращаются каменисто-подъязычные мышцы, также
прикрепленные к подъязычному хрящу; при их
сокращении дно ротовой полости поднимается.
легочное дыхание. Легкие лягушки представляют
собой пару полых мешков. Стенки их образуют
многочисленные складки, но даже при этом по-
Рото-глоточная полость
Голосовая
щель
Гортань
Пищевод
Ноздря
Рот
m petrohyoideus
Подъязычный
хрящ
m sternohyoideus
Легкое
Воздух
Рото-глоточная
полость
Легкое
Ноздря
Дно ротовой
полости
Голосовая
щель
Воздух
Рис. 11.28. А. Вертикальный разрез через голову
лягушки, на котором виден подъязычный хрящ и
связанные с ним мышцы. Б. Вентиляция легких у
лягушки.
верхность их относительно невелика по сравнению с
поверхностью легких млекопитающего.
Выстилающий легкие эпителий увлажнен слизью и обильно
снабжается кровью. От каждого легкого отходит
короткая трубка, называемая бронхом. Два бронха
соединяются, образуя трахею. Трахея переходит в
небольшую полость-гортань, которая через
голосовую щель сообщается с ротовой полостью (рис.
11.2SV4).
У лягушки постоянно повторяются хотя и редкие,
но сильные глотательные движения,
обеспечивающие вентиляцию легких. Последовательность
событий при этом такова (рис. 11.28,UT, I—IV):
I. Рот закрыт, ноздри открыты, голосовая щель
закрыта, дно ротовой полости опущено.
Воздух входит в рото-глоточную полость.
II. Ноздри закрыты, голосовая щель открыта.
Воздух из легких в результате сокращения
мышц и эластической тяги легких
проталкивается в рото-глоточную полость, смешиваясь
со свежим воздухом, поступившим при вдохе.
III. Дно ротовой полости поднимается и
одновременно происходят усиленные глотательные
движения, в результате чего смешанный
воздух проталкивается в легкие. Когда легкие
заполнятся воздухом, голосовая щель
закрывается, и воздух на некоторое время
оказывается запертым в легких. В это время здесь и
происходит газообмен между капиллярами
легочного эпителия и воздухом,
заполняющим легкие.
IV. Через короткое время происходит выдох.
Ноздри закрываются, голосовая щель
открывается и дно ротовой полости снова
опускается. Воздух из легких засасывается в ротовую
полость. Затем ноздри открываются,
голосовая щель закрывается и дно ротовой полости
поднимается. Воздух выходит наружу через
ноздри.
11.6.9. Рептилии
Тело у рептилий одето роговым покровом, обычно
непроницаемым для газов, поэтому кожное дыхание
у них отсутствует. Газообмен происходит только в
легких. Их легкие тоже представляют собой, по сути
дела, мешки, но со значительно более сложной
складчатостью, чем у амфибий. У рептилий
имеются ребра, однако у них нет настоящей диафрагмы,
которая отделяла бы грудную полость от брюшной.
Вентиляция легких происходит за счет
движения ребер, вызываемого сокращением межреберных
мышц. Механизм дыхания очень сходен с таковым
у млекопитающих.
11.6.10. Птицы
Птицы - гомойотермные животные, и для
поддержания температуры тела им требуется весьма интен-
Межключичный
мешок
Передняя группа
воздушных мешков
Шейный мешок
Легкое
сивный метаболизм. Поэтому у них выработался в
процессе эволюции и высокоэффективный механизм
дыхания.
Легкие у птиц невелики; это компактные органы,
состоящие из многочисленных ветвящихся
воздухоносных трубочек, называемых бронхами.
Мельчайшие из них, парабронхи, пронизаны
многочисленными кровеносными сосудами; здесь и происходит
газообмен. С легкими сообщаются большие
тонкостенные воздушные мешки. Кровеносных сосудов
в них мало, и они не участвуют в газообмене. По
своему функциональному назначению воздушные
мешки делятся на две группы: передние и задние,
через которые воздух соответственно входит в
дыхательную систему и выходит из нее.
Вентиляционные движения у птиц сложны, но в
общем виде их можно свести к следующим этапам:
первый вдох-воздух поступает непосредственно в
задние мешки (рис. 11.29);
первый выдох - воздух из задних мешков поступает
в легкие;
второй вдох - воздух из легких переходит в
передние мешки;
второй выдох - воздух из передних мешков
выталкивается наружу.
Такой способ вентиляции обеспечивает
однонаправленный поток воздуха - из задних мешков через
легкие в передние мешки и затем наружу.
В состоянии покоя дыхательные движения
происходят у птиц так же, как и у
млекопитающих: межреберные мышцы, сокращаясь, производят
вдох, а брюшные-выдох. В полете вентиляция
легких усиливается в результате мощных сокращений
грудных мышц, управляющих движениями сильно
развитой грудины. В этих условиях легкие и
воздушные мешки птицы пропускают большой объем
воздуха, что необходимо для удовлетворения
возросших метаболических потребностей.
Брюшной мешок
Задняя группа
воздушных мешков
Рис. 11.29. Легкие и система
воздушных мешков птицы.
11.7. Газообмен
у млекопитающих
Дыхательная система млекопитающего состоит из
парных легких, расположенных в грудной полости,
и ряда воздухоносных трубок, связывающих их с
атмосферным воздухом. Путь воздуха включает
следующие разделы: носовые ходы, глотку, гортань,
трахею, бронхи, бронхиолы, легочные альвеолы.
Двенадцать пар костных ребер окружают
находящиеся в грудной полости сердце и легкие и служат
им защитой. Сзади каждое ребро сочленяется
с одним из грудных позвонков таким образом, что
оно может подниматься и опускаться. Спереди
десять пар ребер присоединены к костной пластине,
которая называется грудиной. Остальные две пары
называются свободными ребрами. Важными
частями этой системы служат также межреберные мышцы,
прикрепляющиеся к ребрам, и обширная диафрагма,
отделяющая грудную полость от брюшной.
Воздух поступает в организм через две наружные
ноздри, у каждой из которых имеется каемка
волосков, задерживающих посторонние частицы.
Носовые ходы выстланы ресничным эпителием, в
котором имеются бокаловидные клетки, секретирую-
щие слизь. Эта слизь выполняет две функции.
Во-первых, она улавливает любые частички,
которым удалось проскользнуть через каемку волосков,
окружающую ноздри. Биение ресничек направляет
затем эти частички к задней части ротовой полости,
и здесь они проглатываются, так что попасть в
воздухоносные пути уже никак не могут. Во-вторых,
слизь увлажняет вдыхаемый воздух; здесь же, в
носовых ходах, он и нагревается благодаря
неглубоко залегающим кровеносным сосудам. В крыше
задней части носовой полости находится
обонятельный эпителий, состоящий из нейросенсорных и
поддерживающих клеток, обильно снабжаемых кровью.
Гортань
Хрящ
Трахея .
Легкое
Легочная
артерия
Бронхиола
Легочная
вена
Висцеральная,
плевра
Плевральная
полость
Париетальная
плевра
Альвеолярный
мешочек
Ветвь
легочной
вены
Ветвь легочной
артерии
Бронхиола
Терминальная
бронхиола
Дыхательная
бронхиола
Альвеолярный ход
Альвеолы
Альвеолярный
мешочек, в котором
находятся альвеолы
Капиллярная сеть
в стенках альвеол
■Альвеолы
Рис. 11.30. Трахея и легкие
человека. (G. M. Hughes,
1973, The vertebrate lung,
Oxford Biology Readers,
№ 59.)
Именно этот участок ответствен за распознавание
запахов. Пройдя через носовые ходы, воздух
попадает в глотку через два внутренних отверстия. К
этому моменту он уже обычно освобожден от
имевшихся в нем частиц, согрет, увлажнен, и животное
распознало принесенные им запахи.
Далее воздух, прежде чем попасть в гортань,
должен пройти через глотку. Через глотку проходят
и воздух, и пища, поэтому щелевидное отверстие,
ведущее в гортань (оно называется голосовой
щелью), должно быть защищено от попадания
пищи, которая могла бы закупорить дыхательные
пути. Такой защитой служит имеющийся здесь
треугольный клапан из хрящевой ткани-так
называемый надгортанник.
Гортань-это полость перед входом в трахею,
образованная девятью хрящами. Прикрепленные к
ним мышцы позволяют этим хрящам двигаться
относительно друг друга. В гортани горизонтально
располагаются два ряда эластичных связок,
называемых голосовыми связками. Когда воздух с
силой проталкивается через голосовую щель,
возникают звуковые волны. С изменением натяжения
голосовых связок изменяется высота звука.
Из гортани воздух попадает в трахею-трубку,
которая лежит непосредственно перед пищеводом и
заканчивается в грудной полости. Стенки трахеи
укреплены С-образными хрящами; благодаря этим
хрящам она всегда остается открытой. Своей
незамкнутой стороной С-образные хрящи обращены к
пищеводу; они не позволяют трахее спадаться при
вдохе (рис. 11.30). Изнутри трахея выстлана
псевдомногослойным ресничным цилиндрическим
эпителием. В этом эпителии находятся секретирующие
слизь бокаловидные клетки. В слизи застревают
попавшие в трахею пылинки и микробы, а
ритмичные биения ресничек, направленные в сторону
ротовой полости (ее задней части), удаляют их из трахеи.
На нижнем конце трахея разделяется на два
бронха. Правый бронх в свою очередь разделяется на
три меньших бронха, каждый из которых
направляется в одну из трех долей правого легкого.
Аналогичным образом левый бронх разделяется на два
бронха, заканчивающиеся в двух долях левого
легкого (рис. 11.31). В обоих легких каждый бронх
многократно делится на еще более тонкие трубки,
называемые бронхиолами. Имеющиеся в бронхах
С-образные хрящи в более мелких трубках
замещены хрящевыми пластинками неправильной
формы, а в бронхиолах, внутренний диаметр
которых меньше 1 мм, хряща нет вовсе. Стенка
состоит здесь только из гладкой мускулатуры,
соединительной ткани с эластическими волокнами,
обеспечивающими возможность растяжения и
упругого сужения бронхиол, и выстилающего
бронхиолы ресничного эпителия с секретирующими слизь
клетками. Самые мелкие трубочки, называемые
дыхательными бронхиолами, имеют в диаметре около
0,5 мм. Они в свою очередь делятся на
многочисленные альвеолярные ходы, выстланные
кубическим эпителием и оканчивающиеся альвеолярными
мешочками, которые называются альвеолами (рис.
11.30, 11.32 и 11.33). Все вместе альвеолы и создают
ту поверхность, на которой у млекопитающего
происходит газообмен.
В легких млекопитающего могут быть сотни
миллионов альвеол с общей площадью поверхности в
десятки квадратных метров. Толщина альвеолярной
стенки составляет всего лишь около 0,0001 мм.
Наружная сторона альвеолярной стенки покрыта
Рис. 11.31. Легкое человека, в которое введен
специальный краситель, чтобы можно было видеть
воздухоносные пути и кровеносные сосуды.
густой сетью кровеносных капилляров; все они
берут начало от легочной артерии и в конце концов
объединяются, образуя легочную вену. Каждая
альвеола выстлана влажным плоским эпителием.
В альвеолярной стенке присутствуют также
коллаген и эластические волокна, придающие ей
гибкость и позволяющие альвеолам изменять свой
объем при вдохе и выдохе.
Кислород, растворившийся в слое влаги на
поверхности эпителия альвеол, диффундирует через
тонкий барьер, состоящий из этого эпителия и из
эндотелия капилляров, и поступает сначала в
плазму крови. Затем он соединяется в эритроцитах с
гемоглобином, который в результате этого
превращается в оксигемоглобин. Углекислый газ (диоксид
углерода) диффундирует в обратном направлении-
из крови в полость альвеол.
Диаметр альвеолярных капилляров меньше
диаметра эритроцитов, и эритроциты протискиваются
через эти капилляры под напором крови. При этом
большая доля их поверхности приходит в контакт с
поверхностью альвеол, на которой осуществляется
газообмен, и в эритроциты, таким образом,
поступает больше кислорода. Кроме того, эритроциты
движутся по капилляру относительно медленно, так
что газообмен может происходить дольше. Когда
кровь покидает альвеолы, парциальные давления
кислорода и диоксида углерода в ней те же, что и в
воздухе альвеолы.
Ядро эндотелиальной
клетки капилляра
Фагоцит
Кровеносный
капилляр
Рис. 11.32. Гистологическое строение легочных
альвеол.
1мкм
Пластинчатые
тельца
Интерстициальное
пространство
Рис. 11.33. Электронная микрофотография, на
которой виден альвеолярный капилляр легкого собаки
в продольном разрезе. (G. M. Hughes, 1973, The
vertebrate lung, Oxford Biology Readers, № 59.)
11.7.1. Строение грудной клетки
Легкие, находящиеся в грудной клетке, отделены от
ее стенок плевральной полостью - щелевидным
пространством, выстланным эластичной прозрачной
оболочкой (плеврой). Внутренний, висцеральный
листок плевры покрывает легкие, а наружный,
париетальный (пристеночный) листок выстилает стенки
грудной клетки и диафрагму. Плевральная полость
содержит жидкость, выделяемую плеврой. Эта
жидкость увлажняет плевру и тем самым уменьшает
трение между ее двумя листками при дыхательных
движениях. Плевральная полость непроницаема для
воздуха и давление в ней на 3-4 мм рт. ст. ниже, чем
в легких. Это существенно, так как благодаря этому
легкие заполняют практически всю грудную клетку.
Отрицательное давление в плевральной полости
поддерживается на протяжении всего входа, что
позволяет альвеолам расширяться и заполнять
любое дополнительное пространство, возникающее
при расширении грудной клетки.
11.7.2. Механизм вентиляции
Воздух поступает в легкие и выходит из них
благодаря работе межреберных мышц и диафрагмы; в
результате их попеременного сокращения и
расслабления объем грудной полости изменяется. Есть
две группы межреберных мышц: наружные
направлены под углом вниз и вперед, а внутренние - вниз и
назад (рис. 11.34). Диафрагма состоит из кольцевых
и радиальных мышечных волокон, расположенных
вокруг центрального сухожильного участка.
Вдох - это активный процесс. Наружные
межреберные мышцы сокращаются, а внутренние
расслабляются. Вследствие этого ребра отходят вперед,
удаляясь от позвоночника. Одновременно сокращается,
становясь более плоской, диафрагма. Оба этих
действия приводят к увеличению объема грудной
клетки. В результате давление в грудной клетке, а
поэтому и в легких становится ниже атмосферного,
так что воздух поступает внутрь и заполняет
альвеолы до тех пор, пока давление в легких не сравняется
с атмосферным (рис. 11.35).
Выдох - процесс в обычных условиях в основном
пассивный, происходящий под действием
эластического сокращения растянутой легочной ткани и
расслабления части дыхательных мышц. Наружные
межреберные мышцы и диафрагма расслабляются,
возвращаясь в прежнее положение и к прежним
своим размерам, а внутренние межреберные мышцы
сокращаются. Вследствие этого объем грудной
клетки уменьшается и давление в ней становится выше
атмосферного. Воздух поэтому выталкивается из
легких, и выдох таким образом заканчивается. При
физической нагрузке имеет место форсированное
дыхание. В действие вводятся дополнительные
мышцы, и выдох становится гораздо более активным
процессом, требующим расхода энергии.
Внутренние межреберные мышцы сокращаются более энер-
Внутренние
межреберные мышцы
Позвоночник
Грудина
Наружные
межреберные мышцы
Ребро
Рис. 11.34. Схема расположения межреберных мышц.
Позвоночник-
Положение
после вдоха
Положение
после выдоха
Грудина
Ребро
Диафрагма
Рис. 11.35. Схематическое изображение грудной
клетки, поясняющее, какие движения совершаются
при дыхании (вид сбоку; показано только одно ребро).
гично и резко отводят ребра вниз. Сильнее
сокращаются и брюшные мышцы, вызывая более активное
движение диафрагмы вверх.
11.7.3. Регуляция дыхания
Непроизвольную регуляцию дыхания осуществляет
дыхательный центр, находящийся в продолговатом
мозге (рис. 11.36). Вентральная часть дыхательного
центра ответственна за стимуляцию вдоха; ее
называют центром вдоха (инспираторным центром).
Дорсальная часть и обе латеральные тормозят вдох
и стимулируют выдох; они носят собирательное
название центра выдоха (экспираторного центра).
Дыхательный центр связан с диафрагмой диафраг-
мальными и грудными нервами. Бронхи и альвеолы
иннервируются ветвями одного из черепных
нервов - блуждающего.
Главным фактором, регулирующим частоту
дыхания, служит концентрация CO2 в крови. Когда
уровень CO2 повышается, хеморецепторы каротид-
ных и аортальных телец посылают нервные
импульсы в инспираторный центр. От него через диа-
Тормозящие
импульсы более
не поступают
Снова начинается
этап 1
Передний
мозг
Произвольная
регуляция
Подавление
вдоха
Тормозящие
импульсы от рецеп-
[торов растяжения]
в легких
Выдох
Легкие
спадаются!
Дыхательный центр
!Легкие рао-
ι ширяются H-
[Диафрагма и меж]
|реберные мышцы'
расслабляются
Импульсы
не поступают
Стимуляция

Накопление CO2
Импульсы
поступают
Межреберные
мышцы и
диафрагма сокращается
Вдох
Рис. 11.36. Регуляция дыхания.
фрагмальные и грудные нервы поступают импульсы
в диафрагму и межреберные мышцы, что ведет к их
сокращению. Таким образом автоматически
стимулируется вдох. При вдохе альвеолы расширяются и
находящиеся в них и в бронхиальном дереве
рецепторы растяжения посылают импульсы в
экспираторный центр, который автоматически подавляет
вдох. Дыхательные мышцы расслабляются, и
начинается выдох. После выдоха альвеолы уже не
растянуты, и рецепторы растяжения не
подвергаются больше стимуляции. Поэтому экспираторный
центр отключается и вдох может начаться снова.
Весь этот цикл непрерывно и ритмично повторяется
на протяжении всей жизни организма.
В известных пределах частота и глубина дыхания
могут регулироваться произвольно. При такой
регуляции импульсы, возникающие в головном мозге,
передаются в дыхательный центр, который и
выполняет соответствующие действия. Концентрация
кислорода тоже влияет на дыхание (разд. 11.8).
Однако в обычных условиях кислорода всегда бывает
достаточно, и потому его влияние относительно
невелико.
11.7.4. Объем легочного воздуха
Легкие человека вмещают в среднем около 5 л
воздуха (рис. 11.37). В состоянии покоя человек
вдыхает и выдыхает примерно 450 мл воздуха. Этот
объем воздуха называется дыхательным объемом.
Сверх этих 450 мл человек может вдохнуть еще
около 1500 мл (дополнительный воздух), а после
спокойного выдоха может выдохнуть
дополнительно около 1500 мл (резервный воздух). Если сделать
максимальный вдох, а затем максимальный выдох,
то общее количество выдыхаемого воздуха
(дыхательный + дополнительный + резервный) даст
величину, называемую жизненной емкостью легких.
После максимально глубокого выдоха в легких
остается еще 1500 мл воздуха. Выдохнуть его не
удается; он называется остаточным воздухом.
При вдохе из 450 мл вдыхаемого атмосферного
воздуха в легкие попадает лишь около 300 мл, а
приблизительно 150 мл остается в воздухоносных
путях и в газообмене не участвует. При выдохе,
который следует за вдохом, этот воздух выводится
наружу неизмененным, не отличающимся по своему
составу от атмосферного воздуха; его называют
поэтому воздухом мертвого, или вредного,
пространства. Воздух, достигающий легких,
смешивается здесь с 3000 мл воздуха, уже находящегося в
альвеолах. Вновь поступившая порция невелика по
сравнению с объемом, к которому она добавляется;
поэтому полное обновление всего находящегося в
легких воздуха-по необходимости медленный
процесс. Этот медленный и прерывистый обмен между
атмосферным и альвеолярным воздухом
сказывается на альвеолярном воздухе столь мало, что его
состав остается практически постоянным (13,8%
кислорода, 5,5% CO2 и 80,7% азота). Интересно
сравнить состав альвеолярного воздуха с составом
воздуха вдыхаемого и выдыхаемого (табл. 11.4). Из
таблицы видно, что одну пятую часть
поступающего кислорода организм удерживает для своих нужд,
тогда как выдыхаемое количество CO2 в 100 раз
больше того количества, которое поступает в
организм при вдохе. В тесный контакт с кровью вступает
альвеолярный воздух. По сравнению с вдыхаемым
воздухом он содержит меньше кислорода, но
больше CO2.
Таблица 11.4. Сравнение состава вдыхаемого,
альвеолярного и выдыхаемого воздуха (в объемных процентах)
Газ
Кислород
Диоксид
углерода
Азот
Во вдыхаемом
воздухе
20,95
0,04
79,01
В альвеолярном
воздухе
13,8
5,5
80,7
В выдыхаемом
воздухе
16,4
4,0
79,6
11.7.5. Измерение дыхания
Обычный прибор, которым в школах, лабораториях
и больницах измеряют объем водуха, поступающего
в легкие и выходящего из них,-это спирометр. Он
состоит из сосуда с водой и помещенного в него
вверх дном другого сосуда емкостью не менее 6 л, в
котором находится воздух. Ко дну этого второго
сосуда подведена система трубок. Через эти трубки
испытуемый дышит, так что воздух в его легких и в
сосуде составляет единую замкнутую систему.
Внутренний сосуд уравновешен, и, когда воздух
входит в него (если испытуемый делает выдох) или
выходит из него (при вдохе), он соответственно
поднимается или опускается. Писчик кимографа
записывает на медленно вращающемся барабане все
эти движения.
К спирометру прилагается подробная инструкция
по работе с прибором, и мы ее здесь обсуждать не
будем. Важно, однако, уметь анализировать записи
прибора и отдавать себе отчет в том, какую
информацию можно из них извлечь.
Спирометр дает возможность определять
интенсивность метаболизма, дыхательный коэффициент,
дыхательный объем, частоту дыхания и
потребление кислорода.
Под частотой дыхания понимают число циклов
дыхательных движений в минуту. Легочную
вентиляцию (JIB) определяют, умножая это число на
дыхательный объем (ДВ):
JlB = Частота дыхания χ Дыхательный объем.
Если, например, частота дыхания составляет 15/мин,
а дыхательный объем 400 мл, то JIB = 15 χ 400 мл =
= 6000 мл/мин (т. е. за 1 мин между организмом
испытуемого и средой обменивается 6000 мл
воздуха).
Под альвеолярной вентиляцией (AB) понимают
объем воздуха, фактически достигающего легких.
Альвеолярная вентиляция меньше легочной.
AB = Частота дыхания χ (Дыхательный объем —
— Объем мертвого пространства).
Если, например, ДО = 400 мл, объем мертвого
пространства = 150 мл и частота дыхания = 15/мин, то
AB = 15 χ (400 - 150) мл/мин =
= 15 χ 250 мл/мин =
= 3750 мл (т. е. за 1 мин между легкими
испытуемого и средой обменивается 3750 мл
воздуха).
11.12. Почему альвеолярная вентиляция
меньше легочной?
Измерение интенсивности
метаболизма
Поскольку почти любой вид метаболической
активности организма непосредственно связан с
дыханием, измеряя дыхание, мы можем судить об
интенсивности обмена веществ в организме. Ее можно
вычислить, определив потребление кислорода.
11.13. Ознакомьтесь с рис. 11.38. Нетрудно
заметить, что чем меньше животное, тем выше у
него интенсивность обмена. В чем причина этого?
11.14. Как можно сравнить интенсивность
обмена у млекопитающих, отличающихся по своим
размерам?
Уровень Г\максимального вдоха4
Дополнительный
воздух
Объем воздуха в легких, л
Вентиляция
в покое
Жизненная
емкость
легких
Дыхательный объем
Резервный
воздух
Уровень максимального выдоха
Функциональный
■остаточный
воздух
Остаточный
воздух
Время
Рис. 11.37. Соотношение объемов воздуха, находящегося в легких.
Интенсивность метаболизма,
мм3 02/(г-ч)
кМышь
Крыса
Кролик
fcСобака
k Человек
^Лошадь
кСлон
100 1000 10 000 1 000 000
100 000
Масса тела, г
Рис. 71.38. Интенсивность метаболизма в расчете
на 1 г массы тела (логарифмическая шкала).
11.7.6. Основной обмен
Основным обменом (основным метаболизмом)
называют минимальный уровень обмена веществ и
энергетических затрат, необходимый только для
поддержания жизни в состоянии полного покоя или
сна. Для определения основного обмена
необходимо, чтобы человек перед измерением в течение
12-18 ч находился в состоянии физического и
психического покоя и не принимал никакой пищи. Это
гарантирует, что к моменту измерения желудочно-
кишечный тракт испытуемого будет пуст. Величина
основного обмена зависит от возраста, пола, массы
тела и состояния здоровья индивидуума, а также
явным образом коррелирует с отношением
поверхности тела к его объему.
11.7.7. Дыхательный коэффициент (ДК)
Рассмотрим следующее уравнение:
C6H12O6 + 6O2 -► 6CO2 -I- 6H2O + Энергия.
Из этого уравнения видно, что объем двуокиси
углерода, образовавшейся за данное время при
окислении углевода в процессе дыхания, равен
объему потребленного за то же время кислорода
(напомним, что моль любого газа при данных
условиях температуры и давления занимает один и тот
же объем). Отношение CO2:02 называется
дыхательным коэффициентом. Для углеводов
дыхательный коэффициент равен 1,
т.е. ДК =
Объем выделенного CO2
Объем поглощенного O2
(из прямого наблюдения).
Или:
Число молей или молекул выделенной CO2
Число молей или молекул поглощенного O2
(из уравнений).
11.15. Окисление жира трипальмитина в
процессе дыхания описывается следующим
уравнением:
2C51H98O6 + 145O2 -► 102CO2 + 98H2O.
Чему равен ДК для трипальмитина?
11.16. Чему равен ДК, когда глюкоза
окисляется в анаэробных условиях до этанола и CO2?
Изучение дыхательных коэффициентов дает ценные
сведения о природе дыхательных субстратов и о
типе метаболизма (табл. 11.5).
11.17. Почему ДК у человека колеблется
обычно в пределах от 0,7 до 1,0?
11.18. Из спирограммы, приведенной на рис.
11.39, определите:
а) частоту дыхания;
б) дыхательный объем;
в) легочную вентиляцию;
г) потребление кислорода.
Опыт 11.2. Измерение потребления
кислорода у мелких
наземных беспозвоночных,
например мокриц
Поглощение кислорода беспозвоночными
измеряется в этом опыте при помощи манометра.
Используемый прибор изображен на рис. 11.40.
При дыхании кислород поглощается, a CO2
выделяется, поэтому для поглощения CO2 в прибор
помещают натровую известь.
Таким путем изучают влияние температуры на
поглощение кислорода. Температура воздуха, в
котором находятся исследуемые организмы,
поддерживается в течение опыта на заданном уровне при
помощи водяной бани.
Таблица 11.5. Дыхательные коэффициенты для различных
субстратов
Таким образом, из приведенного выше уравнения
ДК =
СО,
О,
-Г1"
ДК
> 1,0
1,0
0,9
0,7
0,5
0,3
Субстрат
Углевод, если происходит также и анаэробное
дыхание
Углеводы
Белки
Жир, например трипальмитин
Жир при наличии
синтеза углеводов
Углевод при наличии
синтеза
органичеДиоксид углерода,
образующийся при
дыхании, исполь-
► зуется для других
нужд и потому из
организма не
выделяется
ских кислот
Объем, мл
Время, мин
Рис. 11.39. Спирограмма человека,
находящегося в состоянии покоя.
Материалы и оборудование
Манометр
Манометрическая жидкость
Шприц на 1 мл
2 пробирки
2 куска цинковой сетки (размеры определяются
диаметром пробирок)
Стеклянные бусы (или любой другой не
поглощающий материал, занимающий тот же объем, что и
исследуемые организмы)
Зажимы и штативы
Водяная баня
Термометр
Таймер
Миллиметровая бумага
Винтовой зажим
Исследуемые
организмы
Цинковая сетка
Натровая известь
Мелкие беспозвоночные, например мокрицы или
личинки мясной мухи
Натровая известь
Методика
1. Заполните манометр до половины
манометрической жидкостью и к одному из его колен через
трехходовой кран присоедините шприц на 1 мл.
2. Поместите в обе пробирки равные по объему
количества натровой извести, а поверх
нее-цинковую сетку.
3. В одну из пробирок («Опыт») поместите
несколько мокриц или личинок мясной мухи, а в
другую («Контроль») - равное по объему
количество стеклянных бус. Животные не должны
Шприц на 1 мл
Трехходовой кран
Стеклянные бусы
Натровая известь
ОПЫТ
Капиллярная
U-образная трубка,
содержащая
манометрическую
жидкость
КОНТРОЛЬ
Рис. 11.40. Прибор, с помощью
которого можно измерять
потребление кислорода мелкими
наземными беспозвоночными.
соприкасаться с натровой известью; цинковая
сетка должна отделять их от натровой извести
вполне надежно.
4. Присоедините манометр к двум пробиркам, как
показано на рис. 11.40, и поставьте трехходовой
кран и винтовой зажим в такое положение,
чтобы манометр сообщался с атмосферным
воздухом.
5. Укрепите прибор так, чтобы пробирки
находились в водяной бане с температурой 200C, и
оставьте его при этой температуре с открытыми
кранами не менее чем на 15 мин.
6. Закройте трехходовой кран и винтовой зажим,
отметьте уровень манометрической жидкости и
включите отсчет времени.
7. Через одинаковые промежутки времени
отмечайте по шкале показания манометра.
8. По окончании опыта снова откройте
трехходовой кран и винтовой зажим.
9. Постройте график изменения уровня
манометрической жидкости в зависимости от времени.
10. Вычислите скорость поглощения кислорода.
11. Повторите тот же опыт несколько раз при
различных температурах, например при 20, 25, 30,
35 и 400C.
12. Постройте график зависимости потребления
кислорода от температуры.
Примечания
1. В качестве манометрической жидкости можно
использовать подкрашенную воду, масло или
ртуть. Чем меньше плотность жидкости, тем
больше смещение уровня ее мениска.
2. Смещение уровня мениска определяют по шкале.
Для этого V-образную трубку манометра
укрепляют на доске, на которую наклеена полоска
бумаги со шкалой. Можно наклеить шкалу и на
одно из колен манометра.
3. Прежде чем вести отсчет, следует проверить
герметичность прибора. Для этого в прибор с
помощью шприца нагнетают воздух, чтобы вызвать
смещение уровня манометрической жидкости.
После этого закрывают трехходовой кран,
отключая таким образом манометр от
атмосферного воздуха. Если прибор герметичен, то разность
уровней манометрической жидкости в двух
коленах не должна уменьшаться.
11.8. Дыхание в необычных
условиях
11.8.1. Влияние высоты над уровнем
моря и акклиматизация
При восхождении на высокие горы люди страдают
от недостаточной насыщенности крови кислородом.
Такое состояние называют аноксией или гипоксией.
Возникает оно вследствие того, что с возрастанием
высоты над уровнем моря парциальное давление
кислорода, так же как и других газов, содержащихся
в атмосферном воздухе, падает; на высоте 5450 м
атмосферное давление равно, например, уже только
0,5 бар, т. е. здесь оно вдвое меньше, чем на уровне
моря. И хотя воздух содержит здесь столько же
процентов кислорода, концентрация O2 на единицу
объема вдвое меньше.
Дыхательная активность стимулируется хеморе-
цепторами. На больших высотах усиление легочной
вентиляции, вызванное потребностью в большем
количестве кислорода, приводит к тому, что из
крови в легкие переходит больше CO2 и кислотность
крови соответственно снижается. Возрастание
щелочности (повышение рН) порождает состояние,
известное как алкалоз. При более высоком рН
активность хеморецепторов подавляется; легочная
вентиляция становится недостаточной, и это
вызывает недомогание и чувство сильной усталости.
Со временем дыхательная и кровеносная системы
могут в известной мере приспособиться к низкому
парциальному давлению кислорода,
существующему на больших высотах. По истечении нескольких
дней из организма начинает выводиться щелочная
моча, в результате чего алкалоз уменьшается.
Теперь, когда подавление хеморецепторов снято,
легочная вентиляция снова усиливается и
концентрация CO2 опять становится главным химическим
фактором, регулирующим интенсивность дыхания.
Одновременно стимулируется кроветворная
активность костного мозга-он начинает вырабатывать
больше эритроцитов. Вследствие этого возрастает
способность крови переносить кислород, и это
частично компенсирует неполное ее насыщение
кислородом в условиях пониженного парциального
давления O2. После того как все эти
приспособительные изменения произойдут, можно считать, что
организм акклиматизировался в новых условиях.
11.8.2. Млекопитающие, способные
долгое время оставаться
под водой
Тюлени могут оставаться под водой до 15 мин.
Способность их крови к переносу кислорода гораздо
выше, чем у человека: в 100 мл крови у них
переносится от 30 до 40 мл кислорода.
При нырянии в дыхательной и кровеносной
системах животного происходят значительные
изменения, обеспечивающие перераспределение кислорода
в теле животного и эффективное его использование.
Происходит это обычно следующим образом. В
начале ныряния рефлекторно снижается частота
сердечных сокращений и замедляется ток крови.
Кровяное давление в артериях остается при этом на
прежнем уровне, так как сосуды сужаются.
Некоторые сосуды сжимаются полностью, т. е.
отключаются вообще, и характер кровоснабжения меняется;
теперь кровью снабжаются только самые важные
для жизни органы: сердце, головной мозг и
некоторые другие части нервной системы. Эти измене-
ния означают, что кислород в крови используется
медленно, но при этом остается всегда доступным
для тех органов, которые наиболее чувствительны к
аноксии. В почти лишенных кислорода мышцах
тюленя идет процесс анаэробного дыхания, поэтому
в них накапливается молочная кислота. Однако,
поскольку из мышц в общий кровоток поступает
мало крови, эта молочная кислота не расходится по
телу животного в больших количествах и не
причиняет вреда. Когда животное, вынырнув на
поверхность, делает первый вдох, этот вдох служит
сигналом для повышения частоты сокращений
сердца и для возобновления нормального тока крови во
всех органах. Молочная кислота поступает теперь в
кровь и подвергается в печени соответствующим
превращениям. Воздух в легких быстро замещается
свежим, так как количество воздуха, обмениваемого
при каждом вдохе, составляет около 80% от его
общего количества в легких.
11.9. Цветковые растения
Растения расходуют на единицу массы меньше
энергии, чем животные, поскольку интенсивность
метаболизма у них ниже. У некоторых небольших
растений газообмен осуществляется путем диффузии
газов через всю поверхность, но у крупных цветковых
растений для этой цели служат устьица на листьях и
на зеленых стеблях (у травянистых форм), а также
чечевички и трещины в коре на одревесневших
стеблях (разд. 21.6.6).
Внутри растения распространение кислорода
определяется диффузионными градиентами в
воздухоносных межклетниках. По этим путям кислород
достигает клеток и растворяется во влаге,
покрывающей клеточные стенки. Отсюда он диффундирует
уже внутрь клеток. CO2 движется по растению тем
же путем, но в обратном направлении.
Сложнее обстоит дело с клетками, содержащими
хлорофилл, когда в них одновременно протекают и
дыхание, и фотосинтез. Здесь выделяемый хлоро-
пластами кислород может сразу же потребляться
митохондриями той же клетки, а продукт
дыхательного метаболизма митохондрий-CO2-может
использоваться для фотосинтеза хлоропластами.
Более подробные сведения о газообмене у
цветковых растений содержатся в гл. 9.
11.19. а. Перечислите (в виде таблицы)
основные различия между фотосинтезом и аэробным
дыханием, б. Укажите общие черты фотосинтеза и
аэробного дыхания (в том числе и
биохимические).
Глава 12
Организмы и окружающая среда
Экология-это наука о взаимоотношениях живых
организмов друг с другом и с окружающей средой.
Термин «экология» был впервые введен немецким
биологом Эрнстом Геккелем в 1869 г.; он образован
из двух греческих слов: oikos, что значит дом или
жилище, и logos - изучение или наука. Таким
образом, буквально «экология» означает нечто вроде
науки о земном «хозяйстве». Экология как наука
основывается на холистическом подходе, при
котором воссоздание общей картины важнее проработки
частных деталей. И хотя различные частности
нередко приходится анализировать порознь (см.
гл. 13), в рамках такого подхода осуществляется
синтез всей доступной информации для получения
всеобъемлющей картины живых систем и их
физического окружения.
Своими корнями экология уходит в
«естественную историю»; как самостоятельная биологическая
дисциплина она выделилась только с начала XX в.
наряду с физиологией, генетикой и др. С середины
пятидесятых годов ее значение и сферы приложения
стали значительно расширяться, и современную
экологию можно охарактеризовать как
междисциплинарную область, развивающуюся на стыке физики,
биологии и общественных наук. Взаимоотношения
экологии с другими биологическими науками
представлены на рис. 12.1, где показано, что живые
организмы можно изучать на разных уровнях
организации. Область экологии соответствует правой
части приведенной схемы и охватывает организмы,
популяции и сообщества. Экология рассматривает
их как живой (биотический) компонент системы,
называемой экосистемой; последняя включает также
неживой (абиотический) компонент - физическую
среду с ее веществом и энергией. Термины
«популяция», «сообщество» и «экосистема» имеют в
экологии точные определения, которые приводятся на
рис. 12.1. Различные экосистемы вместе образуют
биосферу, или экосферу, включающую все живые
организмы и всю физическую среду, с которой они
взаимодействуют. Таким образом, океан,
поверхность суши и нижние слои атмосферы - все это
входит в экосферу.
В экологических исследованиях по традиции
выделяют два направления - аутэкологию и синэколо-
гию. Аутэкология концентрирует свое внимание на
взаимоотношениях между организмом или
популяцией и окружающей средой, тогда как синэкология
занимается сообществами и средой. Например,
изучение отдельного экземпляра дуба или вида дуб
черешчатый (Quercus robur) или рода дуб (Quercus)
будет аутэкологическим исследованием, а
изучение сообщества дубового леса-синэкологическим
(разд. 13.4 и 13.5).
Как отмечает Саутвуд (1981) в обзоре,
резюмирующем происходящие в экологии изменения,
первая экологическая статья, опубликованная в
журнале New Scientist (1956), была посвящена реинтродук-
ции благородного оленя в Шотландию и
представляла собой в основном аутэкологическое
исследование. Однако позже акценты в экологической работе
переместились на изучение экосистем (т.е. сооб-
ГЕНЕТИКА
МОЛЕКУЛЯРНАЯ
БИОЛОГИЯ
цитология
гистология
АНАТОМИЯ
БИОХИМИЯ И ФИЗИОЛОГИЯ
Гены
Клетки
Ткани
> Органы
ОБЛАСТЬ, ПРЕДСТАВЛЯЮЩАЯ
ОСНОВНОЙ ИНТЕРЕС ДЛЯ
[ЭКОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ
Организмы -
►Популяции-
* Сообщества
Абиотические компоненты (Материя ♦ Энергия)
Экосистемы
Рис. 12.1. Уровни организации от генов до экосистем. Определения: Популяция-это группа организмов
одного вида, занимающая определенную территорию и обычно в той или иной степени изолированная от
других сходных групп. Сообщество - это любая группа организмов различных видов, сосуществующих в одном
и том же местообитании или на одной площади и взаимодействующих посредством трофических
и пространственных взаимоотношений Экосистема-это сообщество организмов с окружающей их
физической средой, взаимодействующих между собой и образующих экологическую единицу.
ществ и окружающей среды) и даже экосферы
(целой планеты). При этом существенный вклад в
понимание закономерностей, действующих на этих
уровнях организации, вносят и небиологические
науки, особенно химия, физика, почвоведение и
гидрология, а также различные социальные дисциплины.
Возрастающий авторитет экологии особенно в
проблемах, связанных с воздействием человека на
природу, виден из многочисленных примеров. Так, в
США перед реализацией любого проекта по закону
необходимо представить официальный доклад об
ожидаемом воздействии на окружающую среду.
В Великобритании многие местные планирующие
органы нанимают эколога или консультируются с
объединением натуралистов своего графства. Кроме
того, возрастает участие экологов в
правительственных комитетах, связанных с окружающей средой,
таких, как постоянная Королевская комиссия по
загрязнению среды (RCEP).
12.1. Подходы и методы
экологии
Для правильного понимания экологической
обстановки требуется одновременный учет всех
взаимодействующих в данном месте факторов, и уже сама
сложность такой задачи делает ее нелегкой. На
практике большинство экологов, предпринимая
новое исследование, применяют один из нескольких
основных подходов-экосистемный подход,
изучение сообществ (синэкология), популяционный
подход (аутэкология), анализ местообитаний,
эволюционный или исторический подходы.
Эти пять подходов в экологии частично
перекрываются и взаимодействуют друг с другом, и
одинаково подробное рассмотрение каждого из них
в нашей книге не представляется возможным.
Вместо этого сосредоточим внимание на экосистемном и
популяционном подходах и на изучении сообществ,
как на составляющих основную суть предмета
экологии. Анализ местообитаний и эволюционный
подход хотя и открывают ряд важных новых
перспектив, но нужны лишь для большего удобства и
по существу входят в три первых метода. Ниже
дается краткая характеристика всех пяти подходов и
вклада каждого из них в экологию.
Экосистемный подход
Впервые определение экосистемы как совокупности
живых организмов с их местообитанием было дано
Тэнсли в 1935 г. При экосистемном подходе в центре
внимания эколога оказываются поток энергии и
круговорот веществ между биотическим и
абиотическим компонентами экосферы. Его больше
интересуют здесь функциональные связи (такие, как
цепи питания) живых организмов между собой и с
окружающей средой, чем видовой состав сообществ
и определение редких видов или колебания
численности. Экосистемный подход выдвигает на первый
план общность организации всех сообществ,
независимо от местообитания и систематического
положения входящих в них организмов. Даже простое
сравнение водной и наземной экосистем,
представленных на рис. 12.2, подтверждает это. Обратите
внимание на сходство структуры и функциональных
единиц этих двух экосистем даже при резком
различии в среде обитания и в образующих систему
видах.
Вместе с тем в экосистемном подходе находит
приложение концепция гомеостаза (саморегуляции),
из которой становится понятным, что нарушение
регуляторных механизмов, например в результате
загрязнения среды, может привести к
биологическому дисбалансу. Экосистемный подход важен также
при разработке в будущем научно обоснованной
практики ведения сельского хозяйства.
Изучение сообществ
Экология сообществ уделяет особое внимание
биотическим компонентам экосистем. Синоним
экологии сообществ-синэкология (разд. 12.5 и 13.4). При
изучении сообществ исследуют растения, животных
и микроорганизмы, обитающие в различимых
биотических единицах, таких, как лес, луг или
вересковая пустошь. Могут быть выявлены и
лимитирующие факторы, но функциональные аспекты влияния
элементов физической среды, например климата,
обычно подробно не рассматриваются. Вместо
этого упор делается на определение и описание видов и
изучение факторов, ограничивающих их
распространение, в частности на конкуренцию и расселение.
Одним из аспектов подобных исследований
является представление о сукцессиях и климаксовых
сообществах, очень важное для решения вопросов
рационального использования природных ресурсов.
Популяционный подход
Популяционная экология занимается аутэкологичес-
кими проблемами (см. разд. 12.6 и 13.5). В
современных популяционных исследованиях
используются математические модели роста, самоподдержания
и уменьшения численности тех или иных видов.
Построение этих моделей связано с рядом важных
понятий, таких, как рождаемость, выживаемость и
смертность. Популяционная экология обеспечивает
теоретическую базу для понимания вспышек
численности вредителей и паразитов, имеющих значение
для сельского хозяйства и медицины, и открывает
возможности борьбы с ними при помощи
биологических методов (например, использование хищников
и паразитов вредителя), а также позволяет оценить
критическую численность вида, необходимую для
его выживания. Последнее особенно важно при
организации заповедников, ведении охотничьего
хозяйства, а в теоретическом плане-при изучении
вопросов эволюционной и исторической экологии.
Недавнее приложение экологических идей к па-
деонтологии позволило лучше понять
взаимоотношения видов в ископаемых сообществах. Популяци-
онная биология обеспечила теоретические основы
для анализа расселения и вымирания видов начиная
с самых ранних этапов эволюции жизни на нашей
планете.
Изучение местообитаний
Местообитание-это участок среды определенного
типа, где живет данный организм, например живая
изгородь, пресноводное озеро, дубовая роща или
каменистый берег. Организм приспособлен к
определенным физическим условиям местообитания. Но
в пределах последнего могут быть места с особыми
условиями (например, под корой гниющего ствола в
дубовой роще), иногда называемые
микроместообитаниями. Каждый вид занимает в своем
местообитании определенную экологическую нишу. Понятие
экологической ниши подразумевает не только
физическое пространство, где может быть обнаружен
данный вид, но также, что еще важнее,
определенную его роль в сообществе, в частности его питание
и взаимоотношения с другими видами. Когда два
вида занимают одну и ту же нишу, они обычно
конкурируют друг с другом, пока один из них не
будет вытеснен. Сходные местообитания включают
сходный набор экологических ниш, и в различных
частях земного шара можно встретить
морфологически близкие, хотя и различные по
таксономическому положению виды животных и растений.
Например, открытые луга, степи и заросли низкого
кустарника служат экологическими нишами для
быстро бегающих травоядных, но это могут быть
лошади, антилопы, бизоны, кенгуру и т.п.
Анализ местообитания особо выделяют в связи с
удобством проведения исследований, но он дает
мало дополнительной информации по сравнению с
тремя подходами, описанными ранее. Тем не менее
он широко распространен в полевых исследованиях,
поскольку местообитания легко поддаются
классификации. Приложения этого подхода в экологии
подробно описаны в гл. 13. Некоторые сообщества,
например сообщества песчаных дюн или засоленных
болот, так тесно связаны с конкретным
местообитанием, что их практически нельзя изучать в ином
контексте. Однако компетентное исследование
экологии песчаных дюн будет включать и остальные
четыре подхода.
Анализ местообитаний очень удобен также при
изучении физических факторов среды, таких,
как почва, влажность, освещенность, с которыми
тесно связана жизнь животных и растений. Здесь
связи с экосистемным подходом и изучением
сообществ особенно сильны. Развитие смежных на-
Солнце
(энергия)
Воздух
Почва
Материнская порода
Вода
Осадки
Материнская порода
Рис. 12.2. Простое сравнение макроструктуры водной (пресноводной или морской) и наземной экосистем.
Основные функциональные единицы: I-абиотические компоненты (основные неорганические и органические
соединения); Il - продуценты (растительность на суше, фитопланктон в воде); III - животные: HIA прямые или
«пастбищные» травоядные (настоящие саранчовые, калифорнийские полевки и т. д. на суше, зоопланктон
и т. д. в воде); HIB-непрямые или детритоядные консументы (почвенные беспозвоночные на суше, придонные
беспозвоночные в воде); IHC-конечные хищники (хищные птицы и крупная рыба), IV-редуценты (бактерии
и грибы, разлагающие органические вещества). Ill и IV относятся к консументам (Из Ю. Одум (1975) Ecology,
2nd ed., Holt, Rinehart and Wilson.)
ук-гидрологии, почвоведения, метеорологии,
климатологии, океанографии и др.-открыло новые
важные междисциплинарные области исследования.
К сожалению, это привело к тому, что выполнение
всестороннего исследования становится слишком
трудоемким для одного человека и требует создания
рабочей группы, где каждый отдельный эколог
обычно изучает лишь один аспект взаимодействия
животных или растений с окружающей средой,
например гидрологию леса, климатологию поля или
восстановление заброшенных земель. Опять-таки
можно ожидать, что и здесь будут использоваться
функциональные подходы (экосистемный, популя-
ционный, изучение сообществ).
Эволюционный и исторический
подходы
Изучая, как экосистемы, сообщества, популяции и
местообитания менялись во времени, мы получаем
важный материал для суждения о характере
вероятных будущих изменений. Эволюционная экология
рассматривает изменения, связанные с развитием
жизни на нашей планете, и позволяет понять
основные закономерности, действовавшие в экосфере до
того момента, когда важным экологическим
фактором, влияющим на большинство организмов и на
физическую среду, стала деятельность человека.
Эволюционная экология пытается
реконструировать экосистемы прошлого, используя как
палеонтологические данные (ископаемые остатки, анализ
пыльцы и др.), так и сведения о современных
экосистемах.
Историческая экология занимается изменениями,
связанными с развитием человеческой цивилизации
и технологии, с их возрастающим влиянием на
природу, и рассматривает период от неолита до
наших дней.
Используя эти подходы, можно выявлять
долговременные экологические тенденции, установить
которые только путем изучения современных
экосистем невозможно; таковы, например, изменения
климата, конвергентная эволюция (разд. 24.7.6),
расселение видов животных и растений (разд. 24.7.2).
Этот подход привносит больше новых
теоретических идей, чем анализ местообитаний.
Изучением древних сообществ и популяций
традиционно занималась палеонтология, применение
же экологических (экосистемных) идей к прошлому
началось сравнительно недавно. Эволюционная
экология - вес расширяющаяся и плодотворная область
исследования.
12.2. Экосистемы
12.2.1. Общая структура экосистем
Экосистемы состоят из живого и неживого
компонентов, называемых соответственно биотическим
и абиотическим. Совокупность живых организмов
биотического компонента называется сообществом.
На рис. 12.2 представлена упрощенная схема общей
структуры наземной и водной экосистем.
Исследование экосистем включает, в частности, выяснение и
описание тесных взаимосвязей, существующих
между сообществом и абиотическим компонентом. Здесь
эти два компонента будут рассмотрены кратко, а
более подробно речь пойдет о них в разделах
12.3 и 12.4.
Биотический компонент полезно подразделить на
автотрофные и гетеротрофные организмы. Таким
образом, все живые организмы попадут в одну из
двух групп, как было показано в табл. 9.1. Авто-
трофы синтезируют необходимые им органические
вещества из простых неорганических и делают это,
за исключением хемотрофных бактерий, с помощью
фотосинтеза, используя свет как источник энергии.
Гетеротрофы нуждаются в источнике органического
вещества и (за исключением некоторых бактерий)
используют химическую энергию, содержащуюся в
потребляемой пище. Далее будет показано, что
гетеротрофы в своем существовании зависят от
автотрофов и что понимание этой зависимости
необходимо для понимания экосистем.
Неживой, или абиотический, компонент
экосистемы в основном включает 1) почву или воду и 2)
климат. Почва и вода содержат смесь неорганических и
органических веществ. Свойства почвы зависят от
материнской породы, на которой она лежит и из
которой частично образуется. В понятие климата
входят такие параметры, как освещенность,
температура и влажность, в большой степени
определяющие видовой состав организмов, успешно
развивающихся в данной экосистеме. Для водных
экосистем очень существенна также степень солености.
При экосистемном подходе нам в конечном счете
нужен синтез знаний для воссоздания всего
сложного целого, и это следует иметь в виду, изучая
различные части системы по отдельности.
12.3. Биотический компонент
экосистемы:
взаимоотношения,
связанные с энергией
и питательными
веществами
12.3.1. Поток энергии и круговорот
питательных веществ
Организмы в экосистеме связаны общностью
энергии и питательных веществ, и необходимо четко
разграничить эти два понятия. В гл. 9-11 живые
организмы уже рассматривались как потребители
вещества и энергии, и мы очень рекомендуем
прочесть начальные разделы гл. 9.
Энергия
света
БИОТИЧЕСКИЙ
КОМПОНЕНТ
Тепловая
энергия
Солнце
Биогенные
элементы
АБИОТИЧЕСКИЙ КОМПОНЕНТ
Поток энергии
Круговорот биогенных элементов
Рис. 12.3. Поток энергии и круговороты биогенных
элементов в экосистеме.
В разд. 9.1 энергия определена как способность
совершать работу, и живые организмы нуждаются в
энергии для поддержания жизни точно так же, как
машины для того, чтобы работать. Всю экосистему
можно уподобить единому механизму,
потребляющему энергию и питательные вещества для
совершения работы. Питательные вещества первоначально
происходят из абиотического компонента системы,
в который в конце концов и возвращаются либо в
качестве отходов жизнедеятельности, либо после
гибели и разрушения организмов. Таким образом, в
экосистеме происходит постоянный круговорот
питательных веществ, в котором участвуют и живой и
неживой компоненты. Такие круговороты
называются биогеохимическими циклами, и они будут
описаны в разд. 12.4.6.
Движущей силой этих круговоротов служит в
конечном счете энергия Солнца. Фотосинтезирую-
щие организмы непосредственно используют
энергию солнечного света и затем передают ее другим
представителям биотического компонента. В итоге
создается поток энергии и питательных веществ
через экосистему, как показано на рис. 12.3.
Необходимо еще отметить, что климатические
факторы абиотического компонента, такие, как
температура, движение атмосферы, испарение и осадки,
тоже регулируются поступлением солнечной энергии.
Для того чтобы понять, почему имеет место
линейный поток энергии через экосистему, а не ее
круговорот и повторное использование (как в случае
питательных веществ), необходимо коротко
рассмотреть термодинамические соображения.
Энергия может существовать в виде различных
взаимопревращаемых форм, таких, как
механическая, химическая, тепловая или электрическая
энергия. Переход одной формы в другую, называемый
преобразованием энергии, подчиняется законам
термодинамики. Первый закон термодинамики, закон
сохранения энергии, гласит, что энергия может
превращаться из одной формы в другую, но не может
быть создана или уничтожена. Второй закон
утверждает, что при совершении работы энергия не
может быть использована на все 100% и часть ее
неизбежно превращается в тепло. Тепло есть
результат случайного движения молекул, тогда как работа
всегда означает неслучайное (т. е. упорядоченное)
использование энергии1. Понятие «работы» при-
ложимо к любому процессу, протекающему в живой
системе с потреблением энергии, начиная от
процессов на клеточном уровне, таких, как поддержание
электрических градиентов на мембране и синтез
белков, и кончая процессами на уровне целого
организма (рост, развитие, репарация,
размножение).
Таким образом, живые организмы - это
преобразователи энергии, и каждый раз, когда происходит
превращение энергии, часть ее теряется в виде тепла.
В конце концов вся энергия, поступающая в
биотический компонент экосистемы, рассеивается в виде
тепла. Можно было бы подумать, что, поскольку
и тепло способно совершать работу (например,
в паровозе), то нет причин, которые мешали бы
круговороту тепла. Однако процесс, производящий
тепло, требует больше энергии, чем может быть
возвращено путем вторичного использования этого
тепла; поэтому в целом происходит все же потеря
полезной энергии в системе. Фактически живые
организмы не используют тепло как источник энергии
для совершения работы - они используют свет и
химическую энергию.
Изучение потока энергии через экосистемы
называется энергетикой экосистем, и так как это
доминирующая тема в их изучении, нам важно
познакомиться с единицами, используемыми для измерения
энергии.
12.3.2. Единицы измерения энергии
Хотя в системе СИ энергию измеряют в джоулях,
все еще широко применяется и традиционная
единица-калория. Определения для обеих единиц даны
в табл. 12.1, в которую включены также данные
о содержании энергии в отдельных питательных
веществах и в живых организмах, а также дневная
потребность в энергии для некоторых из них.
12.1. Почему содержание энергии в табл. 12.1
дано в пересчете на сухую, а не сырую массу?
12.2. Чем можно объяснить большое различие
суточной потребности в энергии (на единицу
массы тела) у человека и у мелких птиц или
млекопитающих?
12.3.3. Солнце как источник энергии
Первоисточником энергии для экосистем служит
Солнце. Солнце-это звезда, излучающая в космос
огромное количество энергии. Энергия
распространяется в космическом пространстве в виде
электромагнитных волн, и небольшая часть ее, составляю-
1 Увеличение неупорядоченности, которым
сопровождаются преобразования энергии, измеряется как прирост
энтропии. Во Вселенной в целом суммарная энтропия
постоянно увеличивается.
щая примерно 10,5· 106 кДж/м2 в год, захватывается
Землей. Около 40% этого количества сразу
отражается от облаков, атмосферной пыли и поверхности
Земли без какого бы то ни было теплового эффекта.
Еще 15% поглощаются атмосферой (в частности,
озоновым слоем в ее верхних частях) и
превращаются в тепловую энергию или расходуются на
испарение воды. Оставшиеся 45% поглощаются
растениями или земной поверхностью. В среднем это
составляет около 5 106 кДж/м2 в год, хотя реальное
Таблица 12.1. Единицы энергии и содержание энергии в
некоторых живых организмах и питательных веществах
Единицы энергии
Калория (кал),
или
грамм-калория
Килокалория
(ккал или Кал)
Джоуль (Дж)
Килоджоуль (кДж)
Количество тепла (или энергии),
необходимое для повышения
температуры 1 г воды на ГС (от 14,5 до
15,5°С)
= 1000 кал
= 107 эрг; 1 эрг равен работе,
которую производит сила в 1 ньютон
на пути в 1 метр (1
ньютон-единица силы);
соответственно 981 эрг-работа,
совершаемая при поднятии веса 1 г
на высоту 1 см против силы
земного тяготения
= 1000 Дж
1 Дж = 0,239 кал; 1 кал = 4,186 Дж
Содержание энергии (средние или приблизительные вели-
чины) Дж/г сухой
массы (энергетическая
ценность)
Углеводы
Белки
Жиры
Наземные растения
Водоросли
Беспозвоночные (исключая
насекомых)
Насекомые
Позвоночные
16,7
20,9
38,5
18,8
20,5
12,6
22,6
23,4
(различия между этими группами организмов
обусловлены отчасти различным содержанием в них минеральных
веществ)
Ежедневная потребность в пище1*
Человек
Мелкие птицы или млекопитающие
Насекомые
кДж/кг живой
массы
167
(около 12 500 кДж
в день на взрослого
весом 70 кг)
4186
2093
Х) По данным из табл. 3.1 в работе E P Odum, Fundamentals
of Ecology, 3rd ed. (1971), Saunders.
количество энергии для данной местности зависит
от географической широты. Большая часть энергии
повторно излучается земной поверхностью и
нагревает атмосферу; приблизительно две трети
энергии поступает в атмосферу этим путем. И только
небольшая часть пришедшей от Солнца энергии
усваивается биотическим компонентом экосистемы.
12.3.4. Пищевые цепи
и трофические уровни
Внутри экосистемы содержащие энергию
органические вещества создаются автотрофными
организмами и служат пищей (источником вещества и
энергии) для гетеротрофов. Типичный пример: животное
поедает растения. Это животное в свою очередь
может быть съедено другим животным, и таким
путем может происходить перенос энергии через ряд
организмов - каждый последующий питается
предыдущим, поставляющим ему сырье и энергию. Такая
последовательность называется пищевой цепью, а
каждое ее звено-трофическим уровнем (греч. trophos-
питание). Первый трофический уровень занимают
автотрофы, или так называемые первичные
продуценты. Организмы второго трофического уровня
называются первичными консументами, третьего -
вторичными консументами и т. д. Обычно бывает четыре или
пять трофических уровней и редко больше шести-
по причинам, описанным в разд. 12.3.7 и очевидным
из рис. 12.12. Ниже приводится характеристика
каждого звена пищевой цепи, а их последовательность
показана на рис. 12.4.
Первичные продуценты
Первичными продуцентами являются автотрофные
организмы, в основном зеленые растения.
Некоторые прокариоты, а именно сине-зеленые водоросли
и немногочисленные виды бактерий, тоже фото-
синтезируют, но их вклад относительно невелик.
Фотосинтетики превращают солнечную энергию
(энергию света) в химическую энергию,
заключенную в органических молекулах, из которых
построены их ткани. Небольшой вклад в продукцию
органического вещества вносят и хемосинтезирующие
бактерии, извлекающие энергию из неорганических
соединений.
В водных экосистемах главными продуцентами
являются водоросли - часто мелкие одноклеточные
организмы, составляющие фитопланктон
поверхностных слоев океанов и озер. На суше большую
часть первичной продукции поставляют более
высокоорганизованные формы, относящиеся к
голосеменным и покрытосеменным. Они формируют леса
и луга.
Первичные консументы
Первичные консументы питаются первичными
продуцентами, т. е. это травоядные животные. На суше
Солнечная энергия
Тепловая энергия, теряющаяся при дыхании
Продуценты
Травоядные
Продукты
выделения и
неиспользованные
вещества
Хищники
:>[
Хищники
Детритофаги и редуценты
Продуценты = Первичные продуценты
Травоядные = Первичные консументы
Хищники = Вторичные консументы,
третичные консументы и τ д
Тепловая энергия,
теряющаяся при дыхании
Круговорот
веществ
Поток энергии
Рис. 12.4. Поток энергии и круговорот веществ в типичной пищевой цепи. Обратите внимание на.
существование двух путей обмена между хищниками, с одной стороны, и детритоядными и редуцентами-с
другой. Последние питаются мертвыми хищниками, хищники могут питаться живыми детритоядными
и редуцентами.
типичными травоядными являются многие
насекомые, рептилии, птицы и млекопитающие. Наиболее
важные группы травоядных млекопитающих - это
грызуны и копытные. К последним относятся
пастбищные животные, такие, как лошади, овцы,
крупный рогатый скот, приспособленные к бегу на
кончиках пальцев.
В водных экосистемах (пресноводных и морских)
травоядные формы представлены обычно
моллюсками и мелкими ракообразными. Большинство этих
организмов - ветвистоусые и веслоногие раки,
личинки крабов, усоногие раки и двустворчатые
моллюски (например, мидии и устрицы) - питаются,
отфильтровывая мельчайших первичных продуцентов
из воды, как описано в разд. 10.2.2. Вместе с
простейшими многие из них составляют основную
часть зоопланктона, питающегося фитопланктоном.
Жизнь в океанах и озерах практически полностью
зависит от планктона, так как с него начинаются
почти все пищевые цепи.
К первичным консументам относятся также
паразиты растений (грибы, растения и животные).
Консументы второго и третьего
порядка
Вторичные консументы питаются травоядными;
таким образом, это уже плотоядные животные, так же
как и третичные консументы, поедающие консумен-
тов второго порядка. Консументы второго и
третьего порядка могут быть хищниками и охотиться,
схватывать и убивать свою жертву, могут питаться
падалью или быть паразитами. В последнем случае
они по величине меньше своих хозяев. Пищевые
цепи паразитов необычны по ряду параметров (они
включены в пирамиды численности в разд. 12.3.6).
В типичных пищевых цепях хищников плотоядные
животные оказываются крупнее на каждом
следующем трофическом уровне:
Растительный материал (например, нектар) -►
муха -►паук -> землеройка -► сова
Сок розового куста -> тля -► божья коровка -* паук ->
насекомоядная птица -> хищная птица
В типичных пищевых цепях, включающих
паразитов, последние становятся меньше по размерам на
каждом следующем уровне (разд. 12.3.6). Приведем
еще несколько примеров пищевых цепей:
Морская:
Цепь
морского
побережья:
Пресноводная:
Лесная:
Диатомовые
(одноклеточные
водоросли,
составляющие
существенную часть
фитопланктона, например
Chaetoceros)
Морские
водоросли,
например Fu-
CUS
Диатомея,
например
Navicula
Растения,
например Ru-
bus sp.
(ежевика)
-> веслоногие
(мелкие
ракообразные,
составляющие
существенную
часть
зоопланктона,
например
Calanus)
-* береговая
улитка
Littorina
littoralis
-► личинка
поденки
Baetis
rhodani
-*· рыжая
полевка
Clethriono-
mys glareolus
-+ сельдь
Clupea harengus
-*■ кулик-сорока
Haematopus
ostralegus
-» личинка
ручейника
Rhyacophila
sp.
обыкновенная
неясыть
Strix aluco
Редуценты и детритофаги (детритные
пищевые цепи)
Существуют два главных типа пищевых цепей-
пастбищные и детритные. Выше мы привели
примеры пастбищных цепей, в которых первый
трофический уровень занимают зеленые растения, второй-
пастбищные животные (термин «пастбищные»
используется в широком смысле и включает все
организмы, питающиеся растениями) и третий -
хищники. Тела погибших растений и животных еще
содержат энергию и «строительный материал», так же как
и прижизненные выделения, например моча и
фекалии. Эти органические материалы разлагаются
микроорганизмами, а именно грибами и бактериями,
живущими как сапрофиты на органических
остатках. Такие организмы называются редуцентами. Они
выделяют пищеварительные ферменты на мертвые
тела или отходы жизнедеятельности и поглощают
продукты их переваривания. Скорость разложения
может быть различной. Органические вещества
мочи, фекалий и трупов животных потребляются за
несколько недель, тогда как упавшие деревья и
ветви могут разлагаться многие годы. Очень
существенную роль в разложении древесины (и других
растительных остатков) играют грибы, которые
выделяют фермент целлюлазу, размягчающий
древесину, и это дает возможность мелким животным
проникать внутрь и поглощать размягченный
материал.
Кусочки частично разложившегося материала
называют детритом, и многие мелкие животные
(детритофаги) питаются им, ускоряя процесс
разложения. Поскольку в этом процессе участвуют как
истинные редуценты (грибы и бактерии), так и
детритофаги (животные), и тех и других иногда
называют редуцентами, хотя в действительности
этот термин относится только к сапрофитным
организмам.
Детритофагами могут в свою очередь питаться
более крупные организмы, и тогда создается
пищевая цепь другого типа-цепь, начинающаяся с
детрита:
Детрит -► детритофаг -► хищник
Некоторые детритофаги лесных и прибрежных
сообществ изображены на рис. 12.5.
Приведем две типичные детритные пищевые цепи
наших лесов:
Листовая подстилка -> Дождевой червь -►
Lumbricus sp.
-► Черный дрозд -* Ястреб-перепелятник
Tur dm merula Л ccipiter nisus
Мертвое животное -► Личинки падальных мух -►
Calliphora vomitoria и др.
-» Травяная лягушка -► Обыкновенный уж
Rana temporaria Natrix natrix
Некоторые типичные наземные детритофаги - это
дождевые черви, мокрицы, двупарноногие и более
Дождевой червь
Лес
Мокрица
Личинка падальной мухи
Полихета
Прибрежная зона
Багрянка
Голотурия
Рис 12.5. Некоторые детритоядные, встречающиеся
в лесу и в прибрежной зоне.
мелкие (<0,5 мм) животные, такие, как клещи,
ногохвостки, нематоды и черви-энхитреиды.
Методы сбора и определения этих видов описаны в
разд. 13.2.
12.3.5. Пищевые сети
В схемах пищевых цепей каждый организм бывает
представлен как питающийся другими организмами
какого-то одного типа. Однако реальные пищевые
связи в экосистеме намного сложнее, так как
животное может питаться организмами разных типов из
одной и той же пищевой цепи или даже из разных
пищевых цепей. Это в особенности относится к
хищникам верхних трофических уровней.
Некоторые животные питаются как другими животными,
так и растениями; их называют всеядными (таков, в
частности, и человек). В действительности пищевые
цепи переплетаются таким образом, что образуется
пищевая (трофическая) сеть. На рис. 12.6
представлены примеры пищевых сетей в лесном и
пресноводном сообществах. На такой схеме могут быть
показаны только некоторые из многих возможных
связей, и она обычно включает лишь одного или
двух хищников каждого из верхних трофических
уровней. Такие схемы иллюстрируют пищевые связи
между организмами в экосистеме и служат основой
для количественного изучения экологических
пирамид и продуктивности экосистем (см. два
следующих раздела).
12.3.6. Экологические пирамиды
Пирамиды численности
Для изучения взаимоотношений между
организмами в экосистеме и для графического представления
этих взаимоотношений удобнее использовать не
Хищная птица,
например
сокол или сова
Хищное
млекопитающее,
например
лисица или горностай
Черный дрозд
Грызун,
например белка
Насекомоядная птица,
например синица
Обы кновенный
уж
Насекомоядное
млекопитающее, например
еж или землеройка
Лягушка
Паук
Насекомые,
питающиеся
нектаром и пыльцой
Дождевой червь
Жук
Слизень
Мокрица
Ж у ки-сверл илыцики
Гусеница
Плоды и семена
Цветковые
растения
Листья
Древесина
Кора
Насекомоядные
птицы
Взрослые
стрекозы
Взрослые
ручейни ки
Взрослые мелкие
двукрылые насекомые
Взрослые поденки
Колюшки
Хищные
водные жуки
Нимфы
стрекоз
Личинки
хищных водных
насекомых
Коловратки
Травоядные
водные жуки
Личинки
ручейников
Дафнии и
веслоногие
ракообразные
Пиявки
Мелкие
олигохеты
Нимфы
поденок
Водные улитки
Дв уство рчатые
моллюски
Нитчатые водоросли
Гумус и детрит
Мелкие зеленые жгутиковые,
одноклеточные водоросли,
диатомовые и десмидиевые
водоросли
Водные
покрытосеменные
растения
Бабочки
рода Nymphyla,
питающиеся на водных
растениях
Солнечная энергия
Л— личинки мелких двукрылых насекомых
Рис. 12.6. А Взаимоотношения между различными организмами в лесу, формирующие пищевую сеть.
Б Пищевая сеть в пресноводном местообитании (Рорпат (1955) Some aspects of life in fresh water, Heinemann )
Четвертый трофический уровень
Третий трофический уровень
Второй трофический уровень
Первый трофический
уровень
Третичные консументы
Вторичные консументы
Первичные консументы
Первичные продуценты
Плотоядные
Травоядные
Растения
Рис. 12.7. дво типа обычных пирамид численности (А и Б) и перевернутая пирамида (В) Объяснения см.
в тексте.
схемы пищевых сетей, а экологические пирамиды.
При этом сначала подсчитывают число различных
организмов на данной территории, сгруппировав их
по трофическим уровням. После таких подсчетов
становится очевидным, что численность животных
прогрессивно уменьшается при переходе от второго
трофического уровня к последующим. Численность
растений первого трофического уровня тоже
нередко превосходит численность животных,
составляющих второй уровень. Это можно отобразить в
виде пирамиды численности.
Для удобства количество организмов на данном
трофическом уровне может быть представлено в
виде прямоугольника, длина (или площадь)
которого пропорциональна числу организмов, обитающих
на данной площади (или в данном объеме, если это
водная экосистема). На рис. 12.7 показаны три типа
пирамид численности, отображающих реальные
ситуации в природе. Хищники, расположенные на
высшем трофическом уровне, называются
конечными хищниками.
12.3. В пирамиде А (рис. 12.7) первичные
продуценты (растения) - организмы малых размеров,
а численность их выше численности травоядных
животных. Опишите и объясните различия между
пирамидами А и Б.
12.4. Жгутиковые простейшие Leptomonas
паразитируют на мелких насекомых, тысячи их могут
быть найдены в одной блохе. Постройте
пирамиду численности на основе следующей пищевой
цепи:
Трава -* Травоядное млекопитающее -► Блоха -*
-> Leptomonas.
12.5. Дайте возможное объяснение различия
между пирамидами А и В на рис. 12.7.
Хотя данные, необходимые для построения
пирамид численности, можно сравнительно легко
получить путем прямого отбора образцов, существует
ряд неудобств, связанных с использованием этих
пирамид. Из этих неудобств наиболее важны
следующие:
1. Продуценты сильно различаются по размерам, а
между тем один экземпляр злака или водоросли,
например, имеет тот же статус, что и одно дерево.
Поэтому истинно пирамидальной формы часто
не получается. Цепи питания паразитов тоже
могут давать перевернутые пирамиды.
2. Диапазон численности разных видов настолько
широк, что часто трудно соблюсти масштаб при
изображении пирамиды (впрочем, в таких случаях
можно использовать логарифмическую шкалу).
Пирамиды биомассы
Неудобств, связанных с использованием пирамид
численности, можно избежать путем построения
пирамид биомассы, в которых учитывается суммар-
A . Заброшенное поле (Джорджия, США)
Зоопланктон
Фитопланктон
Б. Ла-Манш
Рис. 12.8. Пирамиды биомассы Тип А наиболее
распространен Тип Б относится к перевернутым
пирамидам (см. текст) Цифры означают продукцию,
выраженную в граммах сухой массы, приходящуюся на
7 м2. (E. Odum (1971). Fundamentals of Ecology, 3rd ed.,
W В. Saunders.)
ная масса организмов (биомасса) каждого
трофического уровня. Определение биомассы включает не
только учет численности, но и взвешивание
отдельных особей, так что это более трудоемкий процесс,
требующий больше времени и специального
оборудования. В идеале нужно было бы сравнивать сухую
массу, которая может быть или вычислена из общей
массы, или прямо определена после удаления воды
(см. опыт 13.1). Таким образом, прямоугольники в
пирамидах биомассы отображают массу
организмов каждого трофического уровня, отнесенную к
единице площади или объема. На рис. 12.8, Λ
показана типичная пирамида биомассы с характерным
уменьшением биомассы на каждом следующем
трофическом уровне.
При отборе образцов - иными словами, в данный
момент времени - всегда определяется так
называемая биомасса на корню, или урожай на корню.
Важно понимать, что эта величина не содержит
никакой информации о скорости образования
биомассы (продуктивности) или ее потребления; иначе
могут возникнуть ошибки по двум причинам:
1. Если скорость потребления биомассы (потеря
вследствие поедания) примерно соответствует
скорости ее образования, то урожай на корню не
обязательно свидетельствует о продуктивности,
т.е. о количестве энергии и вещества,
переходящих с одного трофического уровня на другой за
данный период времени, например за год.
Например, на плодородном, интенсивно
используемом пастбище урожай трав на корню может быть
ниже, а продуктивность выше, чем на менее
плодородном, но мало используемом для выпаса.
2. Продуцентам небольших размеров, таким, как
водоросли, свойственна высокая скорость
возобновления, т.е. высокая скорость роста и
размножения, уравновешенная интенсивным
потреблением их в пищу другими организмами и
естественной гибелью. Таким образом, хотя биомасса
_ Вторичные консументы .
_ Первичные консументы _
- Продуценты
Зима
Весна
Рис. 12.9. Сезонные изменения в пирамидах биомассы в одном из озер в Италии. Цифры означают продукцию,
выраженную в граммах сухой массы, приходящуюся на 1 м3. (Из E. Odит (1971). Fundamentals of Ecology, 3rd ed.,
W В. Saunders )
на корню может быть малой по сравнению с
крупными продуцентами (например, деревьями),
продуктивность при этом может быть не
меньшей, так как деревья накапливают биомассу в
течение длительного времени. Иными словами,
фитопланктон с такой же продуктивностью, как у
дерева, будет иметь намного меньшую биомассу,
хотя он мог бы поддерживать жизнь такой же
массы животных. Вообще популяции крупных и
долговечных растений и животных обладают
меньшей скоростью обновления по сравнению с
мелкими и короткоживущими и аккумулируют
вещество и энергию в течение более длительного
времени. Одно из возможных следствий этого
показано на рис. 12.8, 2>, где перевернутая
пирамида биомассы описывает сообщество Ла-Манша.
Зоопланктон обладает большей биомассой, чем
фитопланктон, которым он питается. Это
характерно для планктонных сообществ озер и морей в
определенное время года; биомасса
фитопланктона превышает биомассу зоопланктона во время
весеннего «цветения», но в другие периоды
возможно обратное соотношение. Подобных
кажущихся аномалий можно избежать, применяя
пирамиды энергии, как описано ниже.
12.6. Рассмотрите две пирамиды биомассы на
рис. 12.9. На них отображена биомасса планктона
в одном из озер Италии весной и зимой.
Объясните, почему в течение года пирамида
переворачивается.
12.7. На рис. 12.10 представлены данные о
биомассе на корню для продуцентов и первичных
консументов в озере на протяжении года и об
изменении некоторых параметров окружающей
среды.
а. В какие месяцы была бы получена перевернутая
пирамида биомассы?
б. Чем можно объяснить 1) весеннее увеличение
биомассы фитопланктона, 2) быстрое падение ее в
летние месяцы, 3) увеличение осенью и 4)
уменьшение зимой?
Пирамиды энергии
Наиболее фундаментальным и в определенном
смысле идеальным способом отображения связей
между организмами на разных трофических уровнях
служит пирамида энергии, обладающая рядом
преимуществ.
1. Она отражает скорость образования биомассы
в отличие от пирамид численности и биомассы,
описывающих только текущее состояние
организмов в отдельный момент времени. Каждая
ступенька пирамиды энергии отражает количество энергии
(на единицу площади или объема), прошедшей через
определенный трофический уровень за
определенный период. На рис. 12.11 представлена пирамида
энергии водной экосистемы. Обратите внимание,
что цифры здесь характеризуют поток энергии.
2. Как следует из табл. 12.1, при одинаковой
биомассе два вида совсем не обязательно содержат
одинаковое количество энергии. Поэтому
сравнение, основанное на биомассе, может ввести в
заблуждение.
3. Пирамиды энергии позволяют сравнивать не
только различные экосистемы, но и относительную
значимость популяций внутри одной экосистемы, не
получая при этом перевернутых пирамид. Это
иллюстрирует табл. 12.2, в которой сравнивается
поток энергии, проходящий через первичных
консументов разной биомассы. Обратите внимание,
например, на огромную важность почвенных бактерий
с точки зрения потока энергии, несмотря на их
ничтожную биомассу.
4. К основанию пирамиды энергии можно
добавить еще один прямоугольник, отображающий
поступление солнечной энергии.
Хотя пирамиды энергии - наиболее полезный из
трех типов экологических пирамид, получать
данные для их построения труднее всего, так как при
этом требуется даже больше измерений, чем при
построении пирамид биомассы. В частности,
необходима дополнительная информация об
энергетической стоимости данных масс организмов, а это
требует сжигания репрезентативных выборок. На
практике же пирамиды энергии иногда могут быть с
достаточной достоверностью получены из пирамид
биомассы путем пересчета, основанного на
проведенных ранее экспериментах.
12.8. Объясните связь между пирамидой
энергии и вторым законом термодинамики.
Продуценты
Биогенные элементы
Увеличение количества
Свет
Свет
Температура
Рис. 12.10. Изменение биомассы урожая на корню
продуцентов и первичных консументов и колебания
некоторых параметров окружающей среды в озере
в течение одного года (M A Tribe, M R Erant,
RK Snook (1974) Ecological principles, Basic Ecology
Course 4, Cambridge University Press )
От хищников к конечным хищникам
От травоядных к хищникам
От продуцентов к травоядным
Валовая продукция
Рис. 12.11. Пирамида энергии в системе
Силвер-Спрингс (Флорида) Цифрами обозначено
количество энергии на каждом трофическом уровне
в кДж м~ 2год~ г (E Odum (1971) Fundamentals of
Ecology, 3nd ed., W B Saunders )
12.3.7. Продуктивность
Изучая продуктивность экосистем, мы имеем дело с
потоком энергии, проходящим через ту или иную
экосистему.
Энергия поступает в биотический компонент
экосистемы первичных продуцентов. Скорость
накопления энергии первичными продуцентами в форме
органического вещества, которое может быть
использовано в пищу, называется первичной
продукцией. Это важный параметр, так как им определяется
общий поток энергии через биотический компонент
экосистемы, а значит, и количество (биомасса)
живых организмов, которые могут существовать в
экосистеме.
12.9. Какие группы организмов, помимо
растений, принимают участие в создании первичной
продукции экосистемы?
Как уже упоминалось в разд. 12.3.3, количество
солнечного света, падающего на растения,
различно. Оно зависит от географической широты и от
степени развития растительного покрова и на
Британских островах составляет (в энергетическом
выражении) 1 · 106 кДж/м2 в год. Примерно 95-99%
этого количества сразу отражается, поглощается с
переходом в тепло или расходуется на испарение
воды, и лишь 1-5% поглощается хлорофиллом и
используется для синтеза органических молекул.
Скорость, с которой растения накапливают
химическую энергию, называется валовой первичной
продуктивностью (ВПП). Примерно 20% этой энергии
расходуется растениями на дыхание и фото дыхание.
Скорость накопления органического вещества за
вычетом этого расхода называется чистой первичной
продуктивностью (ЧПП). Это энергия, которую
могут использовать организмы следующих
трофических уровней.
В примере, приведенном на рис. 12.12, ЧПП
составляет 8000 кДж/м2 в год, т. е. 0,8% от полученной
солнечной энергии (1% поглощенного излучения и
80% ВПП). В схеме указаны среднегодовые
показатели, так как продуктивность летом выше, чем
зимой.
При поедании одних организмов другими пища
(вещество и энергия) переходит с одного
трофического уровня на следующий. Непереваренная
часть пищи выбрасывается. Животные, обладающие
пищеварительным каналом, выделяют фекалии
(экскременты); в них содержится некоторое
количество энергии, так же как и в органических отходах
метаболизма (экскретах), например мочевине.
Животные, как и растения, теряют часть энергии
при дыхании. Энергия, оставшаяся после потерь,
связанных с процессами дыхания, пищеварения и
экскреции, идет на рост, поддержание
жизнедеятельности и размножение.
Количество органического вещества,
накопленного гетеротрофными организмами, называется
вторичной продукцией (на любом трофическом уров-
Таблица 12.2. Плотность, биомасса и поток энергии,
характеризующие пять популяций первичных консументов1'
Приблизительная
плотность
популяций, число
особей на 1 м2
Почвенные
бактерии 1012
Морские
веслоногие раки (Асаг-
tid) 105
Литоральные
брюхоногие
моллюски (Littorina) 200
Прямокрылые
соленых болот
(Orchelimum) 10
Полевки (Micro-
tus) 10~2
Олени (Odocoile-
us) 1O-5
1) Из E. P. Odum (1971).
Биомасса,
г/м2
0,001
2,0
10,0
1,0
0,6
1,1
Поток
энергии,
KjJ1TKJM2 в
сутки
4,2
10,5
4,2
1,7
2,9
2,1
2000
R
1 * 10 б Солнечная энергия
АВТОТРОФЫ
ТРАВОЯДНЫЕ
ПЕРВИЧНЫЕ
ХИЩНИКИ
-160
Вторичная
продукция
ВТОРИЧНЫЕ
ХИЩНИКИ
► 16
Вторичная
продукция
0,5-106
Фотосинтез
10 000
Поглощенная
Валовая первичная
продукция
8000
Чистая
первичная продукция
► 800
Вторичная
продукция
0,5· 10 б
Не поглощенная
(отраженная)
0,49· 10 6
Гибель
Гибель
R = Энергия, теряющаяся при дыхании
E = Энергия, заключенная в выделениях
(например, в моче) и экскрементах
(фекалиях) и переходящая
из пастбищной пищевой цепи к детритоядным
организмам и редуцентам
С = Потребление организмами
высшего трофического уровня
(Гибель
!Гибель
ДЕТРИТОЯДНЫЕ И РЕДУЦЕНТЫ
Все величины энергии выражены
в килоджоулях (кДж)
Рис. 12.12. Поток энергии через пастбищную пищевую цепь. Размерность цифр-кДжм 2год
не). Приведенное ниже словесное уравнение
суммирует расход потребленной животным энергии:
Потребленная пища = Рост + Дыхание +
+ Фекалии 4- Экскреты.
Некоторые члены этого уравнения нетрудно
определить на домашних или диких животных в
лабораторных условиях. Показателем роста за какое-то
время служит увеличение биомассы или же энергии,
заключенной в теле. Фекалии и экскреты можно
собрать, взвесить и вычесть их массу из общей
массы потребленной пищи, чтобы найти массу
усвоенной пищи, использованной для роста и других
процессов.
На рис. 12.12 ясно показано, что в каждом звене
пищевой цепи часть энергии теряется. Отсюда
очевидно, что длина пищевой цепи лимитируется
размерами этих потерь. Отношение количества
световой энергии, поглощаемой растениями, к чистой
первичной продукции очень велико. Последующие
переносы энергии по меньшей мере в десять раз
эффективнее, чем первоначальный перенос. Средняя
эффективность переноса энергии от растений к
травоядным животным составляет около 10%, а от
животного к животному - около 20%. В целом
травоядные менее эффективно усваивают пищу, чем
хищники. Это связано с тем, что растения содержат
большое количество целлюлозы, а часто и
древесины (содержащей целлюлозу и лигнин), которые
плохо перевариваются и поэтому не могут служить
источником энергии для большинства травоядных.
Энергия, теряющаяся при дыхании, не передается
другим организмам. Энергия же, заключенная в
экскрементах и отходах метаболизма, передается
детритофагам и редуцентам и, таким образом, не
теряется для экосистемы. Детритные цепи
начинаются с мертвых организмов и отмерших частей
растений (опавших листьев, ветвей). Доля чистой
первичной продукции, переходящей прямо в детрит
и в пищевые цепи редуцентов, в разных системах
неодинакова. В лесной экосистеме большая часть
первичной продукции поступает в детритные цепи, а
не в пастбищные. Поэтому лесная подстилка
оказывается местом значительной активности консумен-
тов, хотя эти организмы здесь обычно незаметны.
Однако в морских экосистемах и на интенсивно
используемых пастбищах больше половины
первичной продукции может поступать в пастбищную
пищевую цепь.
12.10. На рис. 12.13 показаны потоки энергии,
проходящие через небольшую часть луговой
экосистемы (цифры в килоджоулях на 1 м2 в год).
а) Какова валовая первичная продукция злаков и
разнотравья?
б) Какова эффективность фотосинтеза (т.е.
преобразования поступающей солнечной энергии
в общую продукцию?
в) Чему равна чистая продукция птиц,
питающихся семенами, паукообразных и кузнечиков
(по отдельности)?
г) Сколько энергии теряется при дыхании и
выделении фекалий у полевых мышей?
д) Какие организмы являются продуцентами,
первичными консументами и вторичными кон-
сументами?
ё) Какие организмы относятся к гетеротрофам?
ж) Какие еще могут быть пути для потоков
энергии? (Назовите три.)
[Из Tribe M. A. et al. (1975), Ecological Principles,
CUP.]
36· ΙΟ5
4
1856.ΙΟ6
4
Солнечный свет
1880· ΙΟ6
Дыхание
Травянистые
растения
Чистая
продукция
20,4· 10 6
Дыхание и фекалии 59,2 · 103
' 60-Ю3
Заглатывается
Птицы, питающиеся
семенами
Дыхание и фекалии
-374,3·103
500.
Дыхание и фекалии
^444-1O3
Заглатывается
Обычные зеленые
саранчовые
700
Пауки
116·103
Заглатывается
Apodemus sylvaticus
(полевая мышь)
19,8-Ю6
Гибель и другие
пути обмена
Чистая
продукция
-► 2000
Дыхание и фекалии
69-10J
Гибель и другие
пути обмена
Рис. 12.13. Поток энергии через небольшую часть луговой экосистемы. Размерность цифр кДж· м 2год 1.
(Из M. R. Tribe et all (1974). Ecological principles, Basic Ecology Course 4. Cambridge University Press.)
Детритные цепи сложнее и менее изучены, чем
пастбищные. Но роль их с точки зрения потока
энергии столь же велика, а часто и более
значительна, чем роль пастбищных цепей.
Приведенные в этом разделе
цифры-среднегодовые величины. Если экосистема стабильна и в ней не
происходит увеличение общей биомассы, то
биомасса в конце года будет такой же, как и вначале. Вся
энергия, заключенная в первичной продукции,
пройдет за это время через различные трофические
уровни, а чистая продукция будет равна нулю. Обычно,
однако, система изменяется. Например, молодой
лес к концу вегетационного периода накапливает
часть поступившей энергии в форме прироста
биомассы. Продуктивность удобно выражать в
среднегодовых показателях, так как при этом
учитываются сезонные изменения продуктивности, если
они есть. Например, первичная продукция больше в
то время года, когда начинается массовая вегетация
растений, а увеличение вторичной продукции
происходит позднее.
Одна из причин, которые побуждают изучать
поток энергии, проходящий через экосистему,-это
возможность применения полученных знаний для
удовлетворения потребности человека в пище и
энергии. Научный анализ позволяет оценивать
традиционные методы агротехники и вносить
усовершенствования с целью повышения их
эффективности. Поскольку на каждом трофическом уровне
происходит потеря энергии, ясно, что для всеядных
организмов (и в том числе для человека) наиболее
эффективный способ извлечения энергии из
экосистемы - потребление растительной пищи (табл.
12.3). Однако необходимо учитывать и другие
факторы. Например, животный белок содержит больше
незаменимых аминокислот, хотя некоторые
бобовые культуры (например, соя) лишь незначительно
уступают ему по своей ценности. Кроме того,
растительный белок переваривается труднее, чем
животный, так как, прежде чем извлечь его, необходимо
разрушить жесткие клеточные стенки. Наконец,
существует немало экосистем, где животные
добывают пищу на большой территории и где было бы
трудно выращивать культурные растения или
собирать их урожай. Таковы, например, неплодородные
земли, на которых пасутся овцы в Британии,
северные олени в Шотландии и Скандинавии, канны в
Восточной Африке.
Рациональное использование
экосистем
«Получение урожая» означает изъятие из
экосистемы тех организмов или их частей, которые
используются в пищу (или для иных целей). При этом
желательно, чтобы экосистема производила
пригодную для пищи продукцию наиболее эффективно.
Это может быть достигнуто путем повышения
урожайности культуры, уменьшения заболеваемости и
помех со стороны других организмов или
использования культуры, более приспособленной к условиям
данной экосистемы.
Для повышения урожаев культурных растений
могут применяться различные средства в
зависимости от конкретных условий. Можно, например,
вносить в почву удобрения, осуществлять
искусственное орошение или, наоборот, удалять избыток
воды через дренажную систему. Однако длительное
применение искусственных удобрений может
привести к ухудшению структуры почвы (разд. 12.4.1), а
это ведет к уменьшению ее плодородия. Внесение
удобрений и воды часто требует больших затрат
энергии.
Уменьшение вреда от насекомых и птиц, а также
от болезней достигается выведением генетически
устойчивых сортов и применением химических
средств, уничтожающих возбудителей болезней и
вредителей. Однако такие средства нужно
использовать осторожно. Применение стойких препаратов,
неспособных быстро разлагаться в экосистеме,
может иметь катастрофические последствия на других
трофических уровнях (разд. 12.3.8). Частое
применение эффективного пестицида может вызывать
вспышки численности устойчивых к нему форм
вредителей, с которыми труднее бороться. Такая
ситуация возникла в Африке при попытках
контролировать численность малярийного комара с
помощью ДДТ.
Примером использования культур, более
приспособленных к окружающей среде, служит разведение
в условиях относительно высокой интенсивности
света и температуры С4-растений, таких, как
сахарный тростник и кукуруза, а не С3-растений. В этих
условиях эффективность фотосинтеза и
соответственно продуктивность С4-растений выше, чем у
С3-растений (разд. 9.8.2). Следующий пример
объясняет, почему в Восточной Африке может быть
выгоднее использование диких копытных, чем разведение
домашнего скота. Дикие копытные, например канна,
имеют в течение года большую биомассу (82,2-
117,5· 105 кг/га по сравнению с 13,2-37,6· 105 кг/га
для домашних животных), так как они эффективнее
переваривают пищу и быстрее усваивают
питательные вещества, питаясь бедными по составу травами,
что особенно важно в течение сухого сезона в
саванне. Дикие копытные, вероятно, также более
устойчивы к заболеваниям, встречающимся в
районе их обитания.
Большую роль при разведении животных,
достигающих половой зрелости через несколько лет,
играет концепция максимального постоянного уровня
добычи. Это наибольшая скорость изъятия особей
из популяции без ущерба для ее дальнейшей
продуктивности. Для определения этой величины
необходимо иметь ясное представление о структуре и
динамике популяции (разд. 12.6), особенно о
рождаемости: ведь чем выше рождаемость, тем больше
особей можно изымать из популяции. Знание
структуры популяции позволяет установить, какие
возрастные группы играют наибольшую роль в
воспроизводстве стада, и соответственно отбирать для
убоя или отлова животных других возрастных
групп. В Англии, например, официально установлен
минимальный размер ячеек для сетей, которыми
можно пользоваться в промысловом рыболовстве.
Таким образом, вылавливается более крупная и
старая рыба, которая уже дала потомство, а не
достигшие половой зрелости особи в сеть, как
правило, не попадают. Вылов молоди привел бы к
уменьшению воспроизводства рыбного стада в
последующие годы и, таким образом, уменьшил бы
размеры популяций в будущем.
12.3.8. Эффекты концентрации
веществ в пищевых цепях
После второй мировой войны стало быстро
возрастать число различных синтетических веществ,
выбрасываемых в окружающую среду. Это прежде
всего гербициды и пестициды, предназначенные для
уничтожения организмов (особенно сорняков и
насекомых), наносящих вред урожаям, домашнему
скоту и самому человеку. Среди первых применявшихся
с успехом пестицидов была группа хлорированных
углеводородов, в том числе ДДТ (дихлордифенил-
трихлроэтан), диэлдрин и алдрин. Эти вещества
ядовиты для многих животных и человека, но
особенно вредоносны для птиц, рыб и беспозвоночных.
Таблица 12.3. Продукция сельскохозяйственных пищевых цепей в Соединенном Королевстве 1
Пищевая цепь
(а) Культурное растение -► человек
(б) Культурное растение -*■ домашний скот -►
->человек
(в) Интенсивно используемое пастбище -*
-► домашний скот -► человек
Мясо
Молоко
(г) Луг и культурные растения -► домашний
скот -* человек
Молоко
Примеры
Монокультуры пшеницы и
ячменя
Откармливаемый ячменем
мясной скот и поросята
Интенсивно откармливаемое
мясное стадо на тщательно
регулируемом пастбище
Энергия урожая,
потребляемого
человеком,
кДж-Ю3/га
7800-11000
745-1423
339
3813
1356
Содержание белка в
урожае,
потребляемом человеком,
кг/га в год
42
10-15
4
46
17
,} Данные из Duckham, A. N. & Mansfield, G. В. (1970) Farming Systems of The World, Chatto and Windus.
Птица (скопа)
Крупная рыба (сарган)
Мелкая рыба (атериновые)
Водные растения
(Cladophora)
Хищник 2
Хищник 1
Травоядное животное
Продуцент
Рис. 12.14. Количество ДДТ, заключенное в биомассе
организмов, находящихся на разных трофических
уровнях пищевой иепи Цифрами выражено
количество весовых единиц ДДТ, приходящееся на 1 млн
весовых единиц биомассы
В середине 60-х годов неожиданным для многих
ученых явилось сообщение о том, что ДДТ
обнаружен в печени пингвинов в Антарктиде-месте весьма
удаленном от районов возможного примене-
ния ДДТ.
12.11. На рис. 12.14 показано, какие количества
ДДТ содержатся на различных трофических
уровнях в пищевой цепи. Эти данные были
собраны в США.
а) Если концентрация ДДТ в воде, окружающей
водные растения, составляет 0,02 ч. на млн.,
то во сколько раз она возрастает при переходе
ДДТ 1) в первичных продуцентов, 2) в мелких
рыб, 3) в крупных рыб и 4) в тела хищных птиц,
завершающих пищевую цепь?
б) Какие заключения вы можете вывести из
вашего ответа на вопрос а?
в) 1) Ha каком трофическом уровне ДДТ скорее
всего оказывает наибольшее влияние? 2) На
каком уровне его легче всего обнаружить?
3) На каком уровне находятся насекомые-
вредители урожаев (обычная «мишень» для
ДДТ)?
г) Какими путями ДДТ мог попасть в печень
пингвинов?
д) Большое Чистое озеро в Калифорнии служило
местом отдыха, в частности рыбной ловли. В
1940-е годы нарушение естественной
экосистемы из-за эвтрофизации (обогащение
питательными веществами, см. разд. 3.2.8 и 12.4.6)
привело к увеличению популяций мелких
двукрылых насекомых. В 1949, 1954 и 1957 гг. эти
популяции были обработаны распыленным
ДДД (вещество, сходное с ДДТ). В результате
первой и второй обработки было уничтожено
около 99% этих насекомых, но они быстро
восстановили свою численность, а третья
обработка ДДД почти не оказала действия.
Анализ небольших рыб, выловленных в озере,
показал, что содержание ДДД в мышцах рыб,
употребляемых человеком в пищу, составляет
1-200 ч. на млн., а в жировой ткани 40-2500 ч. на
млн. Популяция западных поганок,
насчитывавшая около 1000 особей и кормившаяся на озере,
вымерла, а содержание ДДД в их жировых тканях
составляло 1600 ч. на млн.
1. Объясните, почему с помощью ДДД не
удалось уничтожить мелких двукрылых
насекомых и почему они так быстро восстановили
свою численность после повторного
применения ядохимиката.
2. Было замечено, что многие животные
погибают от отравления ДДТ в те периоды, когда
им не хватает пищи. Объясните это явление,
основываясь на ранее приведенных данных.
е) В Великобритании зимы 1946-1947 и 1962—
1963 гг. были особенно суровыми. Смертность
среди птиц была велика в обе зимы, но во
вторую из них была особенно высокой. Чем
это можно объяснить, учитывая приведенные
ранее данные о применении ДДТ?
От отравления пестицидами очень сильно
пострадали некоторые хищники верхних трофических
уровней, больше всего птицы. Например, из-за
отравления ДДТ на востоке США полностью исчез
сапсан. Птицы наиболее уязвимы потому, что ДДТ
вызывает гормональные изменения, влияющие на
обмен кальция. В результате скорлупа
откладываемых яиц становится тоньше, и они значительно
чаще разбиваются. Содержание ДДТ в жировых
тканях человеческого тела достигает в США 12-
16 ч. на млн., тогда как официально установленный
предел допустимого содержания его в пищевых
продуктах составляет 7 ч. на млн.
Позже был создан ряд сильнодействующих, но
быстро разрушающихся пестицидов, таких, как ор-
ганофосфаты (например, малатион), и применение
ДДТ резко сократилось. Однако производство ДДТ
обходится сравнительно дешево, и он по-прежнему
остается наиболее пригодным для определенных
целей, например для борьбы с малярией. Решая
вопрос о применении пестицидов, мы часто
вынуждены из двух зол выбирать меньшее. Благодаря
ДДТ во многих районах земного шара удалось
полностью искоренить малярию. На острове
Маврикий, например, хотя с 1900 г. рождаемость
значительно не изменилась, произошел взрыв
численности населения, так как гораздо меньше детей
стало умирать от этой болезни. За 10 лет в
результате распыления ДДТ смертность от малярии упала
со 150 до 50 случаев на 1 тыс. детей.
12.12. Какими свойствами в идеале должны
обладать, по вашему мнению, вновь создаваемые
пестициды?
12.4. Абиотический компонент
экосистемы
Абиотический, т. е. неживой, компонент экосистемы
подразделяется на эдафические (почвенные),
климатические, топографические и другие физические
факторы, в том числе воздействие волн, морских
течений и огня.
Международное общество по изучению почв
12.4.1. Эдафические факторы
Наука о почвах называется почвоведением. Уже в
ранних работах подчеркивалось значение почвы как
источника питательных веществ для растений.
Почву изучали также с геологической точки зрения, и
при этом внимание было сосредоточено на
физических и химических свойствах почвы,-ее описывали
как инертный неорганический материал. Русский
ученый Докучаев впервые стал рассматривать почву
как динамическую, а не инертную среду. Он сделал
это в своей классической работе о почвах России,
опубликованной в 1870 г. Докучаев доказал, что
почва постоянно изменяется и развивается, а в ее
активной зоне идут физические, химические и
биологические процессы. Он выявил пять главных почво-
образующих факторов, к которым относятся
климат, геологическая основа (материнская порода),
топография (рельеф), живые организмы и время.
Хотя мы включили почву в раздел об
абиотических факторах, правильнее считать ее важнейшим
связующим звеном между биотическими и
абиотическими компонентами наземных экосистем.
Почвой называют слой вещества, лежащий поверх
горных пород земной коры. Рост растений зависит
от содержания нужных питательных веществ в почве
и от ее структуры. Хотя современная технология
позволяет довольно успешно преодолевать
неблагоприятные почвенные условия, нужен верный
научный анализ проблемы, чтобы не допустить
побочных вредных воздействий на окружающую среду.
Примером может служить истощение запасов
питательных веществ в почве, зачастую связанное с
вырубкой лесов (особенно в тропиках), и избыточное
обогащение почвы удобрениями в районах
интенсивного земледелия. Оно ведет к вымыванию
питательных веществ и связанной с этим эвтрофизации
водоемов, как, например, в некоторых восточных
районах Англии.
В состав почвы входят четыре важных
структурных компонента: минеральная основа (обычно
50-60% общего состава почвы), органическое
вещество (до 10%), воздух (15-25%) и вода (25-35%).
Методы анализа этих компонентов описаны в разд.
13.1.1. Кроме того, имеется биотический компонент,
о котором говорилось в предыдущем разделе и
который будет рассмотрен подробнее в гл. 13.
Минеральный скелет (неорганический
материал)
Минеральный скелет почвы-это неорганический
компонент, который образовался из материнской
породы в результате ее выветривания.
структура почвы. Минеральные фрагменты,
образующие вещество почвенного скелета, различны - от
валунов и камней до песчаных крупинок и
мельчайших частиц глины. Скелетный материал обычно
произвольно разделяют на мелкий грунт (частицы
< 2 мм) и более крупные фрагменты. Частицы
Глина
Ил
Мелкий песок
Крупный песок
Гравий
0,002
0,02
0,2
2,0
Британская стандартная система (Soil Survey of England and Wales)
Глина
Ил
Мелкий
песок
Средний
песок
Крупный
песок
Камни
0,002
0,06 0,2
0,6
2,0
Диаметр почвенных частиц выражен в миллиметрах
(логарифмическая шкала)
Рис. 12.15. Классы почв по размеру частиц.
меньше 1 мкм в диаметре называют коллоидными.
Механические и химические свойства почвы в
основном определяются теми веществами, которые
относятся к мелкому грунту. Соотношение частиц
разных размеров в мелком грунте исследуют путем
механического анализа в лаборатории (разд. 13.1.1)
или при наличии опыта на ощупь в поле. На
рис. 12.15 представлены две наиболее известные
классификации для различения песка, ила и глины. В
обоих случаях глиной считают материал с
частицами меньше 0,002 мм (2 мкм) в диаметре.
Структура почвы определяется относительным
содержанием в ней песка, ила и глины. На рис. 12.16
изображен стандартный «треугольник почвенной
структуры» и указаны границы одиннадцати
структурных классов.
Структура почвы имеет большое значение для
сельского хозяйства. Средне- и мелкоструктурные
почвы, такие, как глины, суглинки и алевриты,
обычно более пригодны для роста растений, так как
содержат достаточно питательных веществ и
способны удерживать воду. Песчанистые почвы
быстрее дренируются и теряют питательные вещества
из-за выщелачивания, но их выгодно использовать
для получения ранних урожаев: их поверхность
высыхает ранней весной быстрее, чем у глинистых
почв, что приводит к лучшему прогреванию.
Степень каменистости почвы (частицы > 2 мм) тоже
имеет значение, так как влияет на быстроту износа
сельскохозяйственных орудий, а также на дренаж.
Обычно с увеличением содержания камней в почве
уменьшается ее способность удерживать воду.
Значение почвенной структуры можно отчасти
уяснить, сравнивая свойства чистого песка и глины
(табл. 12.4). Идеальная почва должна содержать
приблизительно равные количества глины и песка в
сочетании с частицами промежуточных размеров. В
этом случае образуется пористая, крупитчатая
структура, и почвы называются суглинками. Они
обычно обладают достоинствами двух крайних
типов почв и не имеют их недостатков. Таким
образом, легко выполнимый механический анализ почвы
дает полезное представление о ее вероятных
свойствах.
С сельскохозяйственной точки зрения почвы
иногда относят к тяжелым (глины) или легким (пески).
Таблица 12.4. Сравнение свойств песка и глины
Свойство
Структура
Сложение
Порозность
Водоудерживающая
способность
Температура
Удержание питательных
веществ
Песок
Грубая, размер частиц >0,06 мм
Бесструктурное
Поры относительно велики, хорошая
аэрация, быстрый дренаж
Слабо удерживает воду, мало удерживает
капиллярной воды (поверхностное
натяжение) или адсорбционной воды, не
заболачивается
Теплая, что обусловлено низким
содержанием влаги
Низкое, быстро выщелачивается
(песчаные почвы часто становятся кислыми,
так как основания выщелачиваются, а
гуминовые кислоты накапливаются)
Глина
Тонкая, размер частиц < 0,002 мм
Во влажном состоянии образует клейкую
массу (комья), при высыхании становится
твердой и растрескивается
Поры относительно малы, плохая аэрация,
медленный дренаж
Хорошо удерживает воду, сравнительно
много воды удерживается капиллярными
силами (поверхностное натяжение) и
адсорбцией, легко заболачивается
Холодная, что обусловлено высоким
содержанием влагиυ
Высокое, не выщелачиваются, частицы
глины притягивают катионы и некоторые
анионы
!) Вода отличается высокой теплоемкостью и большой скрытой теплотой парообразования.
Это разделение отражает те усилия, которые
необходимы для обработки почвы
сельскохозяйственными орудиями. Тяжелые почвы плохо
дренируются и обычно бывают сырыми и клейкими. Легкие
же почвы хорошо дренируются и прекрасно
поддаются обработке, так как их частицы легко
отделяются друг от друга.
химизм минерального скелета. Химизм почв час-
тично определяется минеральным скелетом,
частично органическим веществом, которое будет
описано ниже. Большая часть минеральных
компонентов представлена в почве кристаллическими
структурами. Это устойчивые продукты выветривания
материнской породы. Песок и алеврит состоят в
основном из очень инертного минерала - кварца
(SiO2), именуемого также кремнеземом. Кремнезем
служит источником силикат-ионов (SiO^"), которые
обычно соединяются с катионами, особенно с
катионами алюминия (Al3 + ) и железа (Fe3 + , Fe2 + ), и
образуют электронейтральные кристаллы.
Силикаты - преобладающие почвенные минералы.
Большую роль в удержании воды и питательных
веществ играет особенно многочисленная и важная
группа глинистых минералов. Большинство их
встречается в виде мельчайших плоских кристаллов,
часто шестиугольной формы, которые образуют в
воде коллоидную суспензию. Каждый кристалл
содержит слои силиката, объединенные со слоями
гидроокиси алюминия. Общая поверхность слоев,
граничащая с промежутками между ними, очень
велика по сравнению с объемом кристаллов
(5 - 800 м2 в 1 г глины). Важная особенность этих
минералов - наличие у них постоянного
отрицательного заряда, который нейтрализуется катионами,
адсорбированными из почвенного раствора.
Благодаря этому катионы не выщелачиваются из почвы и
могут обмениваться на другие катионы из
почвенного раствора и растительных тканей. Эта катионо-
обменная способность служит одним из важных
индикаторов плодородия почвы. Свободные
пространства втягивают также воду, что ведет к гидратации
и набуханию глины.
Глина
Алев ри
тистая
глина
Песчанистая
глина
Пылеватый
тяжелый
суглинок
Суглинок
Песчанистый
тяжелый суглинок
Песчанистый
суглинок .
Песчанистый
алеврит
Пылеватый
суглинок
Супесь
Песок
% песка 60-2000 мкм
Рис. 12.16. Треугольная диаграмма
классов почв по структуре,
использованная в Soil Survey of
England and Wales, 1974.
Органическое вещество
Органическое вещество почвы образуется при
разложении мертвых организмов, их частей (например,
опавших листьев), экскретов и фекалий. Мертвый
органический материал используется в пищу
совместно детритофагами, которые его поедают и
таким образом способствуют его разрушению, и
редуцентами (грибами и бактериями), завершающими
процесс разложения. Не полностью разложившиеся
органические остатки называются подстилкой, а
конечный продукт разложения - аморфное вещество, в
котором уже невозможно распознать
первоначальный материал,-получило название гумуса. Цвет
гумуса варьирует от темно-бурого до черного. В
химическом плане это очень сложная смесь
изменчивого состава, образованная органическими
молекулами различных типов; в основном гумус состоит
из фенольных соединений, карбоновых кислот и
сложных эфиров жирных кислот. Гумус, подобно
глине, находится в коллоидном состоянии;
отдельные частицы его прочно прилипают к глине и
образуют глино-гумусовый комплекс. Так же как и
глина, гумус обладает большой поверхностью частиц
и высокой катионообменной способностью. Эта
способность особенно важна для почв с низким
содержанием глины. Анионы в гумусе-это
карбоксильные и фенольные группы. Благодаря своим
химическим и физическим свойствам гумус
улучшает структуру почвы и ее аэрацию, а также
повышает способность удерживать воду и питательные
вещества.
Одновременно с процессом гумификации
(образования гумуса) жизненно важные элементы переходят
из органических соединений в неорганические,
например азот в ионы аммония (NH^"), фосфор в
ортофосфат-ионы (H2PO4^), сера в сульфат-ионы
(SO2^"). Этот процесс называется минерализацией.
Углерод высвобождается в виде CO2 в процессах
дыхания (см. рис. 9.2, где показан круговорот
углерода).
Для того чтобы формировался гумус того или
иного типа, необходим достаточный дренаж почвы:
в условиях переувлажнения разложение идет очень
медленно, так как нехватка кислорода ограничивает
рост аэробных редуцентов. В таких условиях
растительные и животные остатки сохраняют свою
структуру и, постепенно спрессовываясь, образуют торф,
который может накапливаться вплоть до больших
глубин. Метод определения содержания гумуса в
образцах почвы будет описан в следующей главе
(опыт 13.2).
Содержание воздуха
Почвенный воздух, так же как и почвенная вода,
находится в порах между частицами почвы. Пороз-
ность (объем пор) возрастает в ряду от глин к
суглинкам и пескам. Между почвой и атмосферой
происходит свободный газообмен, и в результате
этого воздух обеих сред имеет сходный состав.
Обычно в воздухе почвы из-за дыхания населяющих
ее организмов несколько меньше кислорода и
больше двуокиси углерода, чем в атмосферном воздухе.
Кислород необходим для корней растений,
почвенных животных и организмов-редуцентов.
Некоторые почвенные газы переходят в раствор, что
способствует газообмену. Если идет процесс
заболачивания, то почвенный воздух вытесняется водой и
условия становятся анаэробными. При этом такие
минеральные элементы, как железо, сера и азот,
проявляют тенденцию переходить в
восстановленное состояние (Fe2 + , сульфид, сульфит, нитрит), в
котором, окисляясь, они могут связывать еще
оставшийся кислород. Почва становится кислой, так
как анаэробные организмы продолжают
вырабатывать CO2. Обновление гумуса замедляется, и в
результате накапливаются гуминовые кислоты.
Почва, если она не богата основаниями, может
стать чрезвычайно кислой, а это наряду с
истощением запасов кислорода неблагоприятно
воздействует на почвенные микроорганизмы. Изменения в
степени окисленности железа влияют на цвет почвы.
Окисленная форма железа (Fe3 + ) придает почве
желтый, красный и коричневый цвета, тогда как
восстановленная форма (Fe2 + ) обусловливает серый
цвет, характерный для заболачиваемых почв.
Длительные анаэробные условия ведут к отмиранию
растений. Метод определения количества воздуха в
образцах почвы описан в следующей главе (опыт
13.3).
Содержание воды
Почвенные частицы удерживают вокруг себя
некоторое количество воды. Часть ее, называемая
гравитационной водой, может свободно
просачиваться вниз сквозь почву. Это ведет к
выщелачиванию, т. е. к вымыванию из почвы различных
минеральных веществ, в том числе азота.
Гравитационная вода проходит до уровня грунтовых вод,
глубина залегания которых колеблется в зависимости от
количества выпадающих осадков.
Вода может также удерживаться вокруг
отдельных коллоидных частиц в виде тонкой прочной
связанной пленки. Эту воду иногда называют
гигроскопической (рис. 12.17). Она адсорбируется за счет
водородных связей на поверхностях кварца и глины
или на катионах, связанных с глинистыми
минералами и гумусом. Эта вода наименее доступна для
корней растений, и именно она последней
удерживается в очень сухих почвах. Количество
гигроскопической воды зависит от содержания в почве
коллоидных частиц, и поэтому в глинистых почвах
ее намного больше (примерно 15% веса почвы), чем
в песчанистых (примерно 0,5%).
По мере того как накапливаются слои воды
вокруг почвенных частиц, она начинает заполнять
сначала узкие поры между этими частицами, а затем
распространяется во все более широкие поры.
Гигроскопическая вода постепенно переходит в
капиллярную, которая удерживается вокруг почвенных
частиц силами поверхностного натяжения (рис.
12.17). Капиллярная вода может подниматься по
узким порам и канальцам от уровня грунтовых вод.
Причиной этого служит высокое поверхностное
натяжение воды. Растения легко поглощают
капиллярную воду, которая играет наибольшую роль в
регулярном снабжении их водой. В отличие от
гигроскопической эта вода легко испаряется.
Тонкоструктурные почвы, например глины, удерживают
больше капиллярной воды, чем грубоструктурные,
такие, как пески.
Общее количество воды, которое может быть
удержано почвой (его определяют, добавляя
избыток воды и затем ожидая, пока она не перестанет
выходить каплями), называется полевой влаго-
емкостью (разд. 13.1.1).
Вода необходима всем почвенным организмам.
Она поступает в живые клетки путем осмоса (разд.
14.1). Вода также важна как растворитель для
питательных веществ и дыхательных газов,
поглощаемых из водного раствора корнями растений. Она
принимает участие в процессах разрушения
материнской породы, подстилающей почву (см. ниже).
12.4.2. Образование почвы
Почва образуется в результате взаимодействия
многих факторов, из которых пять наиболее
важных-это климат, материнская порода
(геологические условия), топография (рельеф или морфология
поверхности), живые организмы и время.
Рассмотрим каждый из этих факторов в отдельности.
Климат и выветривание
Физическое и химическое разрушение материнской
породы под влиянием климата и в меньшей степени
живых организмов называется выветриванием.
Наиболее важные факторы, вызывающие
выветривание,-это вода и температурные колебания, поэтому
роль климата удобнее рассматривать на примере
именно этих факторов. Влияние климата на
почвообразование в целом будет рассмотрено позже.
Физическое выветривание может происходить в
результате изменений температуры: горные породы
попеременно расширяются и сжимаются при ее
суточных колебаниях. Этот процесс ведет к
постепенному разрушению горных пород. Если в
трещинах имеется вода и если она периодически замерзает
и оттаивает, то при образовании льда может
создаваться огромное давление, также разрушающее
породу. Другими факторами физического
выветривания могут быть частицы разных размеров
(например, песчинки), которые перемещаются ветром,
водой и ледниками и вызывают эрозию
незащищенных поверхностей.
Химическое выветривание подвержено также
Частица почвы
Гигроскопическая вода
Капиллярная вода
Воздух или
гравитационная вода
Рис. 12.17. Три типа почвенной воды.
влиянию биологических и климатических факторов.
Оно обычно ускоряется с повышением температуры.
Вода действует как растворитель и как реагент в
процессе гидролиза некоторых минералов. Породы,
в некоторой степени растворимые в воде, мел и
известняк, состоящие в основном из карбоната
кальция, особенно подвержены эрозии. Эффективность
воды как растворителя может возрастать, когда в
ней содержится двуокись углерода (из атмосферы
или образующаяся в результате дыхания почвенных
организмов), так что получается разбавленный
раствор угольной кислоты. Лишайники относятся к
тем немногим организмам, которые могут расти на
обнаженной скальной поверхности, они тоже
способствуют химической эрозии, извлекая из
выветриваемых скал определенные питательные
вещества.
Температурный режим и осадки (дожди и снег)
являются двумя главными климатическими
факторами, вызывающими выветривание. Особенно
большую роль играет количество выпадающих осадков.
Если оно превышает испарение, в почве может идти
процесс выщелачивания, а в случае плохого дренажа
почвы заболачиваются. Если испаряется больше
влаги, чем выпадает осадков, происходит
капиллярное поднятие воды от уровня грунтовых вод; в
этом случае будет преобладать движение
растворенных веществ вверх, что тоже повлияет на свойства
развивающейся почвы. Растительный покров в
сухом климате скуден, и поэтому здесь образуется
меньше гумуса; в результате почвы становятся
менее кислыми. Недостаток воды замедляет
химическое выветривание и выщелачивание.
Влияние климата в его взаимодействии с другими
факторами отражено на приведенной схеме (рис.
12.18).
Материнская порода
Первичное происхождение всех горных пород
связано с застыванием расплавленной магмы,
внедрявшейся в верхние зоны земной коры из глубоких
недр. Породы, образовавшиеся непосредственно из
застывшей магмы, называются вулканическими.
Они могут быть кислыми (как гранит) или
основными (как базальт) в зависимости от их
минерального состава. При выветривании кислых пород боль-
КЛИМАТ
Выветривание
Влажность,
температура,
свет
ТОПОГРАФИЯ
Осадки, температура и τ д.
Вымывание, гидролиз,
скорость формирования
и т. д.
Микроклимат,
обусловленный
растительным покровом
Доступные
минеральные
питательные
вещества
и вода
ПОЧВА
Мощность почвы,
дренаж и смыв
Устойчивость
к выветриванию!
Выветривание
Химический состав
й размер частиц
Количество гумуса
и его природа,
степень перемешивания и
аэрации почвы
ОРГАНИЗМЫ
МАТЕРИНСКАЯ ПОРОДА
Рис. 12.18. Взаимодействие четырех факторов, непосредственно влияющих на процесс почвообразования.
Обратите внимание на большую роль климата. Время является шестым, косвенно влияющим фактором (в
схему не включено).
шая часть кремния остается в виде кварца (SiO2),
который представлен зернами песка. При
выветривании основных пород значительная часть кремния
связывается в различные силикаты, которые в
дальнейшем формируют алевриты и глины.
Осадочные породы образуются в результате
отложения продуктов выветривания других горных
пород или накопления остатков живых организмов.
Этот материал накапливается и уплотняется иногда
в течение миллионов лет. К типичным осадочным
породам относятся песчаники, мел и известняк; их
влияние на развитие почвы будет рассмотрено в
разд. 12.4.3.
Метаморфические горные породы образуются из
других пород в результате их нагревания и
перекристаллизации. К ним относятся, например,
сланцы и мрамор. Эти породы обычно более устойчивы
к выветриванию, чем другие.
Некоторые почвы образуются на перемещенных
материалах, таких, как эоловые пески,
аллювиальные отложения и ледниковые морены. В этом случае
для почвообразования существенно то, от какой
первоначальной породы происходят эти материалы.
Топография
Влияние топографии связано с тем, что на разной
высоте, при различной крутизне и экспозиции
склона условия различны. Рельеф местности влияет на
местный климат и дренаж. Дренаж обычно лучше на
склонах, а по направлению к низине или дну долины
он постепенно ухудшается, и там в случае
постоянного переувлажнения может образовываться торф.
Со склонов часть воды стекает, и при этом
смываются продукты выветривания и почва; поэтому
слой почвы на склонах обычно тоньше. Кроме того,
почва медленно сползает вниз под действием силы
тяжести. Этот процесс называется солифлюкцией и
ведет к скоплению почвы у основания склонов; его
может сдерживать растительный покров.
Рельеф местности, влияя на такие климатические
факторы, как выпадение осадков и температура, тем
самым косвенно сказывается и на процессе
почвообразования. Экспозиция и высота склона в
значительной мере определяют климат. На солнечных
склонах температура выше, поэтому почвы
развиваются быстрее, здесь они, как правило, более сухие,
что нередко влияет на растительность. Наветренные
склоны обычно более влажные, чем подветренные.
Организмы
При непосредственном участии живых организмов
образуются органические компоненты почвы
(подстилка и гумус), о физических и химических
свойствах которых уже говорилось. Растительность, так
же как климат и материнская порода, влияет на
развитие почв. Можно полагать, например, что
вероятность образования подзолов (разд. 12.4.3)
под степью и сообществами хвойных лесов выше,
чем под сообществами лиственных лесов. Это,
по-видимому, связано со свойствами веществ,
вымываемых из растительной подстилки, и их инги-
бирующим влиянием на процессы разложения
органического материала почвенными
микроорганизмами. Кислая подстилка степей и сосновых лесов в
отличие от подстилки лиственных лесов
неблагоприятна для дождевых червей.
Деятельность почвенных детритофагов, особенно
дождевых червей в зонах умеренного климата,
важна во многих отношениях и в свою очередь зависит
от растительности. Детритофаги ускоряют процесс
разложения, размельчая частицы подстилки и таким
образом увеличивая их поверхность. За счет фека-
лий и отходов метаболизма детритофагов
увеличивается масса перерабатываемого органического и
минерального вещества. Почва, проходя через
пищеварительный тракт дождевого червя,
измельчается, ее минеральные и органические компоненты
перемешиваются, и это улучшает структуру почвы.
Ходы дождевых червей улучшают аэрацию и
облегчают рост корней. Точно так же способствуют
перемешиванию и аэрации почвы ходы, прорытые
позвоночными животными - кротами, кроликами и
т.п.
влияние человека. Хрупкое динамическое
равновесие зрелой почвы может быть нарушено при
использовании ее человеком для сельскохозяйственных
целей. При этом часто происходит разрушение
почвенной структуры, истощение запасов питательных
веществ и эрозия почв.
Структура почвы особенно легко разрушается,
когда ее обрабатывают тяжелыми
сельскохозяйственными машинами. Уплотнение почвы приводит
к ряду последствий, в частности уменьшаются
размеры почвенных пор и в результате ухудшаются
аэрация и дренаж. Почва может уплотняться также
при неумеренном выпасе домашнего скота.
Повторное использование почвы, особенно под одни и те
же культуры, может понизить содержание в ней
важных питательных веществ. Распашка почвы
нередко усиливает ветровую и водную эрозию,
особенно на склонах. Точно так же уничтожение
защитного слоя растительности при чрезмерном выпасе
скота или при вырубке леса способствует эрозии.
Вырубка тропических дождевых лесов может иметь
очень серьезные последствия для обычно
маломощных почв таких районов. На обнаженной почве
усиливается поверхностный сток, что ускоряет не
только ее эрозию, но и вымывание питательных
веществ. Все это приводит к уменьшению
плодородия почвы, а иногда и к резкому падению
продуктивности экосистемы. Например, в некоторых
районах Малайи и бассейна Амазонки началась
массивная эрозия почв, сопровождающаяся заилением
рек. Таким образом, рациональное использование
почвы является одной из ключевых проблем как для
почвоведов, так и для земледельцев.
Время
Время имеет важное, хотя и косвенное значение для
образования почвы. Скорость этого процесса в
умеренном климате очень различна - может
потребоваться от нескольких десятилетий (при
формировании почвы на вулканическом пепле) до нескольких
тысяч лет (на обнаженной поверхности скальных
пород). В масштабах геологического времени
происходят климатические и топографические изменения,
которые, естественно, влияют на развитие почвы.
Таким образом, необходимо вновь подчеркнуть,
что рассмотренные почвообразующие факторы
действуют неизолированно. Каждый из них (исключая
время) непосредственно влияет на образование
почвы, но может оказывать также косвенное
воздействие, влияя на другие факторы. Схема на рис. 12.18
иллюстрирует взаимоотношения этих факторов.
12.4.3. Типы почв
Как мы видели, почвы представляют собой продукт
взаимодействия между биотическими и
абиотическими компонентами экосистемы. Это необычайно
сложные и разнообразные системы, поэтому их
очень трудно классифицировать. Из множества
описанных типов почв мы выбрали для примера три
весьма различных типа, которые встречаются во
влажных областях умеренной зоны, в том числе на
Британских островах. Это подзолы, буроземы и
рендзины. Почвы изучаются путем проходки
шурфов и использования буров, как описано в
разд. 13.1.1. При описании вертикального разреза
для характеристики почвенных слоев, или
горизонтов, применяются стандартные обозначения. Такие
обозначения для хорошо дренируемых почв, в том
числе трех выбранных для примера типов,
представлены на рис. 12.19 (там же даны объяснения).
Подзолы
Это название происходит от слова «зола».
Типичный вертикальный профиль подзола показан на
рис. 12.20.
Подзолы обычно образуются на песчаниках,
отложениях гравия (ледникового происхождения) и
аллювиальных отложениях и характерны в
основном для пустошей и хвойных лесов в областях с
прохладным влажным климатом. Подстилка
относительно мощная, так как ее материал (хвоя и т. п.)
отличается высоким содержанием фенолов,
устойчивых к микробному разложению. Кальция в таких
почвах мало, поэтому они слишком кислые для
того, чтобы в них могло обитать много дождевых
червей и других животных, заглатывающих почву. В
связи с этим главную роль в разложении подстилки
играют грибы, и разложение идет относительно
медленно; в результате образуется толстый слой
кислого гумуса (мора), который резко отделен от
нижележащих слоев.
Горизонт Еа по цвету напоминает серую золу и
ясно выделяется среди соседних слоев. Он состоит в
основном из крупинок песка. Это сильно
выщелоченный горизонт, содержание питательных
веществ в нем невелико из-за очень большого
количества осадков или высокой водопроницаемости
подстилающей материнской породы (или по обеим
этим причинам). Органические кислоты гумуса
способствуют выщелачиванию путем переноса
катионов вниз.
Под выщелоченным горизонтом накапливаются
железо, гумус и другие вещества, которые
вымываются из верхних горизонтов. Гумус и окись
железа могут объединяться в «твердый подпочвенный
пласт», или «ортштейн». Этот слой может быть
Горизонт А
(элювиальный горизонт,
т. е. горизонт,
из которого вымываются
минеральные вещества)
Горизонт В
(иллювиальный горизонт,
т. е. горизонт, в котором
накапливаются
питательные вещества)
Горизонт С
(материнская
порода, подвергающаяся
выветриванию)
Горизонт D
(материнская
порода, или коренная
подстилающая порода)
Свежий опад, образующий
органическую подстилку
Слой ферментации — частично
разложившаяся подстилка
Гумус (от темно-коричневого
до черного)
Поверхностный перегнойный
горизонт, состоящий из гумуса и
минеральных компонентов
Элювиальный (вымываемый)
кислый горизонт
(Еь~вымываемый основной горизонт)
ν Вариабельный слой
Слой,обогащенный железом,
с различным составом
Горизонт, обогащенный
окислами железа и алюминия
Рис. 12.19. Обобщенный профиль типичной легко дренируемой почвы с указанием почвенных горизонтов.
настолько твердым и плотным, что препятствует
проникновению корней в нижележащие слои. Между
ортштейном и материнской породой расположен
богатый питательными веществами слой, в котором
накапливается выщелачиваемый материал.
Выщелоченный горизонт, в котором расположена
большая часть корней, кислый (рН 3-4), содержит
мало нитратов и других питательных веществ,
необходимых растениям. Подзолы, таким образом,
относительно неплодородны. Большая часть
питательных веществ в хвойных лесах сохраняется в
биомассе деревьев и подстилки, а не в почве;
круговорот этих веществ и их разложение происходят
медленно. Такие почвы часто бывают главным
фактором, определяющим характер растительности
данного района, так как они благоприятны для
кислотолюбивых растений (кальцефобов).
Существует сравнительно немного кальцефобных видов,
к ним относятся многие типичные растения
пустошей, такие как верески (Calluna vulgaris, Erica tetrali,
E. cinerea), подмаренник скальный {Galium saxatile),
щавель малый (Rumex acetosella) и Rhododendron.
Буроземы (коричневые лесные почвы)
Буроземы имеют сравнительно несложный
профиль, образованный однородными коричневыми
или темно-коричневыми горизонтами А и В,
незаметно переходящими друг в друга (рис. 12.21).
Они характерны для лиственных лесов умеренной
зоны, расположенных, как правило, в более теплых
районах южнее области хвойных лесов с их
подзолистыми почвами. Скорость разложения здесь
выше, чем в подзолах, из-за более высоких
температур и меньшей устойчивости листьев к
разложению у лиственных пород; поэтому подстилка
обновляется быстрее. Она обычно маломощная, так
как почва достаточно богата кальцием (рН 4,5-8),
чтобы обеспечить обилие организмов (например,
дождевых червей), перемешивающих верхние слои
почвы. Образующийся гумус относится к типу,
известному под названием мулль. Содержание
питательных веществ возрастает по направлению к
горизонту В (как и в подзолах). В почве отсутствуют
ортштейны, поэтому корни растений могут сво-
ГОРИЗОНТЫ
Глубина залегания горизонта С различна,
обычно 80-120 см
ВНЕШНИЙ ВИД
Подстилка (остатки растений и животных,
сравнительно мощная (1—5 см),
рыхлая и пористая)
Темно-коричневый, активные редуценты
Черный гумусовый слой, кислый mod
Слой выщелачивания
Темный серо-коричн.
слой в пятнах
выщелачиваемого гумуса
Пепельно-серый
Слой с большим содержанием гумуса (рыхлый)
Железный ортштейн (твердый),
темного оранжево-коричневого цвета
Слой с большим содержанием железа,
- оранжевый (от красного до коричневого),
плотный песок, с высоким содержанием
биогенных элементов ,особенно катионов
Na+, K+, Ca2+
Выветриваемая материнская порода,
обычно это песок или гравий
Рис. 12.20. Профиль подзола.
ГОРИЗОНТЫ
ВНЕШНИЙ ВИД
Подстил ка
(сравнительно маломощная)
Темно-коричневый,
содержит гумус
Светло-коричневый,
частично выщелачиваемый
Коричневая почва
с тонкой текстурой
Выветриваемая
материнская порода
Рис. 12.21. Профиль бурозема.
бодно проникать в глубокие, более богатые
питательными веществами горизонты. Буроземы
обычно весьма плодородны. Созревая, они
приобретают суглинистую структуру.
Содержание элементов минерального питания в
биомассе лиственных лесов и в их почве выше, чем в
хвойных лесах. Круговорот этих веществ идет
быстрее.
Рендзины (дерново-карбонатные
почвы)
Название «рендзины» произошло от польского
rzedzic - дрожать или опасаться, так как почвы
маломощные и при их обработке
сельскохозяйственные орудия задевают подстилающую породу.
Рендзины-это очень маломощные почвы,
образующие тонкий темно-коричневый или черный слой,
богатый кальцием, над известковой материнской
породой (рис. 12.22). Формируются они, как
правило, на известняках и меловых отложениях.
Климат мало влияет на развитие этих почв,
поэтому они встречаются в разных климатических
условиях. Мел и известняк обычно образуют холмы
(такие, как Дауне и Котсуолдс в Англии). Поскольку
эти породы очень проницаемы, вода быстро
проходит сквозь них, не образуя потоков и не вызывая
эрозию.
Рендзины формируются обычно на крутых
склонах, где почва постоянно остается слаборазвитой,
что обусловлено главным образом ее сползанием
вниз. Из-за высокой водопроницаемости почвы
легко растворимый известковый материал со временем
вымывается, и в верхнем слое почвы остаются лишь
кремнистые вещества. Рендзины не содержат
горизонта В, слои почвы трудно различимы. С точки
зрения биологии важны две особенности этих почв:
высокое содержание карбоната кальция (до 80%),
придающее почве щелочной характер (рН > 8), и
быстрый дренаж, приводящий к высыханию.
Поэтому растения, произрастающие на рендзинах,
обычно имеют приспособления для уменьшения
потерь воды и для максимального ее использования.
Они часто обладают длинными боковыми корнями
или же глубинными корнями, нередко
проникающими в материнскую породу. Некоторые растения,
их называют кальцефильными, особенно пышно
развиваются в условиях высоких рН. Таковы,
например, Clematis (ломонос), истод (Polygala calcarea),
душица обыкновенная (Origanum vulgare) и душевка
чабрецевидная (Acinos arvensis). Флора рендзин
относительно богата. Как и подзолы, рендзины
служат примером почв, оказывающих особенно
сильное влияние на биотический компонент экосистемы.
12.4.4. Климатические факторы
Главными климатическими переменными в
экосистеме являются свет, температура, вода и ветер. Их
влияние на образование почвы было описано в
разд. 12.4.2. Ниже будет рассмотрено их прямое
влияние на биотический компонент экосистемы.
Свет
Свет необходим для жизни, так как это источник
энергии для фотосинтеза, однако есть и другие
аспекты его воздействия на живые организмы.
Рассматривая эти аспекты, полезно помнить, что
интенсивность света, его качество (длина волны, или
цвет) и продолжительность освещения (фотопериод)
могут оказывать различное влияние.
На интенсивность света влияет угол падения
солнечных лучей на земную поверхность; она
изменяется в зависимости от широты, сезона, времени
дня и экспозиции склона.
Длина дня (фотопериод) на экваторе более или
менее постоянна (около 12 ч), но в более высоких
широтах она изменяется в зависимости от времени
года. Для растений и животных таких широт
характерна реакция на фотопериод, которая
синхронизирует их активность с временами года.
Примерами могут служить цветение и прорастание
семян у растений (разд. 15.4), миграция, зимняя
спячка и размножение животных (разд. 16.8.5).
Необходимость света для растений существенно
ГОРИЗОНТЫ
ВНЕШНИЙ ВИД
Подстилка
- Слой от темно-коричневого
до черного цвета,
содержит органический и
известковистый матеоиал
Выветриваемый мел
или известняк
влияет на структуру сообществ. Распространение
водных растений ограничено поверхностными
слоями воды. В наземных экосистемах в процессе
конкуренции за свет у растений выработались
определенные стратегии, например быстрый рост в
высоту, использование других растений в качестве
опоры (у лиан), увеличение поверхности листьев. В
лесах это приводит к ярусной структуре сообщества
(разд. 12.5.1).
Некоторые важные процессы с участием света
(рассмотренные в других главах) перечислены в
табл. 12.5.
Температура
Главным источником тепла является солнечное
излучение; им могут также быть геотермальные
источники, но они играют важную роль только в
немногих местообитаниях, например в горячих
ключах, где развиваются бактерии и сине-зеленые
водоросли.
Данный организм может выживать только в
определенных температурных пределах, к которым
приспособлены его метаболизм и структура. Если
температура живой клетки падает ниже точки
замерзания, клетка обычно физически повреждается и
гибнет в результате образования кристаллов льда. Если
же температура слишком высока, происходит
денатурация ферментов. Между этими крайними
точками скорость реакций, контролируемых
ферментами, а значит, и интенсивность метаболизма
удваиваются с повышением температуры на каждые 10°.
Большинство организмов способно в той или иной
степени контролировать температуру своего тела с
помощью различных ответных реакций и
адаптации, которые могут смягчать воздействие
экстремальных условий и внезапных изменений среды
(разд. 18.3 и 18.4). В водной среде из-за высокой
теплоемкости воды не происходит резких изменений
температуры, так что в этом отношении условия
здесь более стабильны, чем на суше.
Температура, так же как интенсивность света, в
Рис. 12.22. Профиль рендзины
Таблица 12.5. Важнейшие процессы, протекающие у растений
и животных с участием света
Фотосинтез (гл. 9 и разд. 12.3.4)
В среднем 1-5% падающего на растения света
используется для фотосинтеза
Фотосинтез - источник энергии для всей остальной
пищевой цепи
Свет необходим также для синтеза хлорофилла
Транспирация (см. разд. 14.3)
Примерно 75% падающей на растения солнечной
радиации расходуется на испарение воды и таким образом
усиливает транспирацию; это важно в связи с проблемой
сохранения воды
Фотопериодизм (см. разд. 15.4 и 16.8.5)
Важен для синхронизации жизнедеятельности и
поведения растений и животных (особенно размножения) с
временами года
Движение (см. разд. 15.1 и гл. 17)
Фототропизм и фотонастии у растений; важны для того,
чтобы обеспечить растению достаточную освещенность
Фототаксис у животных и одноклеточных растений;
необходим для нахождения подходящего местообитания
Зрение у животных (см. разд. 16.5)
Одна из главных сенсорных функций
Прочие процессы
Синтез витамина D у человека
Длительное воздействие ультрафиолетовых лучей может
вызывать повреждение тканей, особенно у животных; в
связи с этим выработались защитные приспособления-
пигментация, поведенческие реакции избегания и т.п.
большой мере зависит от географической широты,
сезона, времени суток и экспозиции склона. Однако
часто встречаются и узколокальные различия в
температуре; это в особенности касается
микроместообитаний, обладающих собственным
микроклиматом. Растительность тоже оказывает некоторое
влияние на температуру. Например, иная
температура бывает под пологом леса (разд. 12.5.3) или в
меньшей степени внутри отдельных групп растений,
а также под листьями отдельного растения.
Влажность и соленость
Вода необходима для жизни и может быть важным
лимитирующим фактором в наземных экосистемах.
Вода поступает из атмосферы в виде осадков:
дождя, снега, дождя со снегом, града или росы. В
природе происходит непрерывный круговорот
воды-гидрологический цикл (разд. 12.4.6), от которого
зависит распределение ее на поверхности суши.
Наземные растения поглощают воду главным образом
из почвы. Быстрый дренаж, небольшое количество
осадков, сильное испарение или сочетание всех этих
факторов могут приводить к иссушению почв, а при
изобилии воды, напротив, возможно их постоянное
переувлажнение. Таким образом, количество воды в
почве зависит от водоудерживающей способности
самой почвы и от баланса между количеством
выпадающих осадков и совместным результатом
испарения и транспирации (эвапотранспирация).
Испарение происходит как с поверхности влажной
растительности, так и с поверхности почвы.
По способности переносить недостаток воды
растения можно разделить на ксерофиты (растения с
высокой выносливостью), мезофиты (со средней
выносливостью) и гидрофиты (с низкой
выносливостью, приспособленные к избытку воды).
Некоторые приспособления ксерофитов будут описаны в
разд. 14.3.7, где рассматривается транспирация, и в
разд. 19.3.2, а также перечислены в табл. 12.6.
Наземные животные тоже обладают
приспособлениями для добывания и сохранения воды, особенно в
засушливых местообитаниях (см. разд. 19.3.4 и
табл. 12.6).
Проблемы, связанные с водой, есть и у водных
организмов. Эти организмы можно подразделить
на пресноводные и морские-по степени солености
воды, в которой они обитают. Сравнительно
немногие растения и животные могут выдерживать
большие колебания солености. Такие виды обычно
обитают в эстуариях рек или в соленых маршах. К
ним относится, например, улитка Hydrobia ulvae,
способная выживать при изменениях концентрации
хлористого натрия от 50 до 1600 ммоль/мл. Co-
Таблица 12.6. Адаптации к засушливым условиям у растений
и животных
Примеры
Уменьшение потери воды
Листья превращены в
иглы или колючки
Погруженные устьица
Листья свернуты в
цилиндр
Толстая восковая
кутикула
Толстый стебель с
большим отношением
объема к поверхности
Опушенные листья
Сбрасывание листьев при
засухе
Устьица открыты ночью
и закрыты днем
Cactaceae, Euphorbiaceae
(молочаи), хвойные
деревья
Pinus, Ammophila
Ammophila
Листья большинства
ксерофитов; насекомые
Cactaceae и Euphorbiaceae
(«суккуленты»)
Многие альпийские
растения
Fouquieria splendens
Crassulaceae
(толстянковые)
Продолжение табл. 12.6
Эффективная фиксация
CO2 ночью при
неполностью открытых
устьицах
Выделение азота в виде
мочевой кислоты
Удлиненная петля Генле
в почках
Ткани выносливы к
высоким температурам из-
за уменьшения
потоотделения или
транспирации
Животные прячутся в
норах
Дыхательные отверстия
прикрыты клапанами
Увеличение поглощения
воды
Обширная поверхностная
корневая система и
глубоко проникающие
корни
Длинные корни
Прорытие ходов к воде
Запасание воды
В слизистых клетках и в
клеточных стенках
В специализированном
мочевом пузыре
В виде жира
(вода-продукт окисления)
Физиологическая
устойчивость к потере воды
При видимом
обезвоживании сохраняется
жизнеспособность
Потеря значительной
части массы тела и
быстрое ее
восстановление при наличии
доступной воды
«Уклонение» от проблемы
Переживают
неблагоприятный период в виде
семян
Переживают
неблагоприятный период в виде
луковиц или клубней
Распространения семян в
расчете на то, что
некоторые из них
попадут в благоприятные
условия
Поведенческие реакции
избегания
Летняя спячка в
слизистом коконе
Примеры
С4-растения, например Zea
mays
Насекомые, птицы и
некоторые рептилии
Пустынные
млекопитающие, например верблюд,
пустынная крыса
Многие пустынные
растения, верблюд
Многие мелкие пустынные
млекопитающие,
например пустынная крыса
Многие насекомые
Некоторые Cactaceae,
например Opuntia, и
Euphorbiaceae
Многие альпийские
растения, например Leontopo-
dium alpinum (эдельвейс)
Термиты
Cactaceae и Euphorbiaceae
Пустынная лягушка
Пустынная крыса
Некоторые эпифитные
папоротники и плауны,
многие мохообразные и
лишайники, осока Сагех
physoides
Lumbricus terrestris (теряет
до 70% массы), верблюд
(теряет до 30%)
Эшшольция
калифорнийская
Некоторые лилии
Различные растения
Почвенные организмы,
например клещи, дождевые
черви
Дождевые черви,
двоякодышащие рыбы
леность может иметь значение и в наземных
местообитаниях. Если испаряется больше воды, чем
поступает с осадками, почвы могут засоляться, и это
создает проблему в некоторых районах, где
применяется орошение.
Атмосфера
Атмосфера является важной частью экосферы, с
которой она связана биогеохимическими циклами,
включающими газообразные компоненты; это
главным образом круговороты углерода, азота,
кислорода и воды. Большое значение имеют также
физические свойства атмосферы: воздух оказывает лишь
незначительное сопротивление движению и не
может служить опорой для наземных организмов, и
это непосредственно сказалось на их строении. В то
же время некоторые группы животных стали
использовать полет как способ передвижения. В
атмосфере, так же как в океане, постоянно происходит
циркуляция, энергию для которой поставляет
Солнце.
Крупномасштабным результатом циркуляции
воздушных масс является перераспределение
водяных паров, так как атмосфера захватывает их в
одном месте (где вода испаряется), переносит и
отдает в другом месте (где выпадают осадки). Если
где-то в атмосферу поступают газы, в том числе
загрязняющие (например, двуокись серы в
промышленных районах), то система атмосферной
циркуляции перераспределит их и они выпадут в
других местах, растворенные в дождевой воде.
Ветер, взаимодействуя с другими факторами
окружающей среды, может влиять на развитие
растительности, особенно на деревья, растущие на
открытых местах. Это часто приводит к задержке их
роста и искривлению с наветренной стороны. Кроме
того, ветер увеличивает эвапотранспирацию в
условиях низкой влажности.
Ветер играет большую роль в распространении
спор, семян и т.п., расширяет возможности
распространения неподвижных организмов, например
растений, грибов и некоторых бактерий; он может
также влиять на миграцию летающих животных.
Еще одна атмосферная переменная - это
атмосферное давление, которое уменьшается с высотой. С
увеличением высоты снижается парциальное
давление кислорода и животные подвергаются действию
факторов, описанных в разделе 11.8.1; у растений
возрастает транспирация, поэтому у них
выработались адаптации для сохранения воды, как,
например, у многих альпийских растений.
Микроклимат
Климатические условия в каком-либо
местообитании или микроместообитании могут отличаться от
условий в окружающей области, тогда их и
называют микроклиматом. Понять всю сложность
экосистем невозможно без учета микроместообитаний
и их микроклиматических особенностей. Это можно
проиллюстрировать одним примером. В группе
ракообразных, называемых равноногими (мокрицы и
родственные животные), есть такие формы, как,
например, Porcellio, которые живут только во
влажном воздухе лесной подстилки, но есть и такие, как
Armadillidium, способные выдерживать более сухие
условия и поэтому перемещаться свободнее.
12.4.5. Топография
Влияние топографии тесно связано с другими
абиотическими факторами, так как она может сильно
сказываться на местном климате и развитии почвы,
о чем уже говорилось выше.
Главным топографическим фактором является
высота. С высотой снижаются средние
температуры, увеличивается суточный перепад температур,
возрастают количество осадков, скорость ветра и
интенсивность радиации, понижаются атмосферное
давление и концентрации газов. Все эти факторы
влияют на растения и животных. В результате
обычным явлением стала вертикальная зональность
(рис. 12.23).
Горные цепи могут служить климатическими
Высота, м
Пик Сан-Франциско
Север
Пик О'Лири
Альпийская зона
Зона верхней границы а
древесной растительности"
Зона еловых лесов φ
I Зона лесов из дугласии
Зона лесов из сосны
желтой
Зона лесов из сосны
однохвойной
Пустыня
Рис. 12.23. Зональное
распределение растительности на
юго-восточном склоне пика Сан-
Франииско. Аризона. (По Merriam
(1980). W. D. Billings (1972). Plants,
man and the ecosystem, 2nd ed.,
Macmillan: London and Basingstoke.)
(Эвапотранспирация)
Осадки,
выпадающие над сушей
Осадки,
выпадающие
в океан
Испарение
с поверхности
океана
Испарение
с суши,
озер и
рек
Транспирация
Океан
Поверхностный сток в океан
Почва
Грунтовые воды
Коренная порода
Цифры в диаграмме
рассчитаны, исходя из
допущения, что ежегодно
на Земле выпадает
100 единиц осадков
Вся вода
океаны 97%
пресная вода 3%
Пресная вода
пласты льда и
ледники 75%
грунтовая вода 25%
озера 0,3%
почвенная влага 0,06%
атмосфера 0,035%
реки 0,03%
Округлено до целого %
Рис. 12.24. Гидрологический цикл и накопление воды (R. J. Chorley, P. Haggett (eds.) (1967). Physical and
information models in geography, Methuen.)
барьерами. Воздух, поднимаясь над горами,
охлаждается, и при этом часто выпадают осадки. Уже
пролившиеся дождем массы воздуха поступают на
подветренную сторону гор, где воздух суше и
выпадает меньше осадков (дождевая тень). Это влияет
на экосистемы. Горы служат также барьерами для
распространения и миграции организмов и могут
играть важную роль изолирующего фактора в
процессах видообразования (разд. 25.7).
Еще один важный топографический фактор-
экспозиция склона. В Северном полушарии склоны,
обращенные на юг, получают больше солнечного
света, и поэтому интенсивность света и температура
здесь выше, чем на дне долин и на склонах северной
экспозиции (в Южном полушарии имеет место
обратная ситуация). Это оказывает поразительное
влияние как на естественную растительность, так и
на угодья, используемые человеком.
И наконец, важным топографическим фактором
является крутизна склона. Для крутых склонов
характерны быстрый дренаж и смывание почв,
поэтому почвы здесь маломощные и более сухие, с
ксероморфной растительностью. Если уклон
превышает 35°, почва и растительность обычно не
образуются, а создаются осыпи из рыхлого
материала.
12.4.6. Биогеохимические циклы
Биогеохимические циклы-это замкнутые пути, по
которым различные химические элементы
циркулируют из внешней среды в организмы и обратно во
внешнюю среду. Как и почвенные факторы, эти
циклы лучше всего рассматривать как связующее
звено между биотическим и абиотическим
компонентами экосистем. Здесь будут рассмотрены шесть
элементов, имеющих большое значение для жизни:
углерод, водород, кислород, азот, фосфор и сера.
Круговороты этих элементов, за исключением
водорода, были описаны в разд. 9.11.
Источником водорода служит вода (H2O), в
которой водород химически связан с кислородом.
Вода важна не только как источник водорода (при
фотосинтезе), но и сама по себе-как составная
часть живых клеток, как климатический фактор и
как среда для водных организмов. Круговорот воды
называется гидрологическим циклом. В этом цикле,
как показано на рис. 12.24, вода может находиться в
газообразном, жидком и твердом состоянии.
Общая картина круговоротов углерода, азота,
кислорода и воды представлена на рис. 12.25.
Из шести упомянутых элементов только углерод,
водород и кислород легко доступны в экосистемах.
Установлено, что азот, фосфор и калий могут
оказывать наибольшее положительное влияние на
урожаи культурных растений, и потому эти три
элемента в наибольших количествах вносят в почву с
удобрениями, применяемыми в сельском хозяйстве.
Поэтому азот и фосфор оказались главной
причиной ускоренной эвтрофизации озер в странах с
интенсивным земледелием. Эвтрофизация - это
процесс обогащения водоемов питательными
веществами. Она представляет собой естественное явление в
озерах, так как реки приносят питательные вещества
с окружающих дренажных площадей. Однако этот
процесс обычно идет очень медленно, в течение
тысяч лет. Неестественная эвтрофизация, ведущая к
стремительному увеличению продуктивности озер,
Лед
Солнечная энергия
Дыхание
Испарение
Осадки
CO2
Вулкан
Транспирация
Фотосинтез
Минеральные £
соединения
Органический
азот.
Известняк
Минеральные^
соединения
Сжигание
ископаемого
топлива
Азотфиксирующие
бактерии
H2O
Мочевина
Сине-зеленые
водоросли
Фотосинтез
Ca(HCO3) 2
H2O
Аммиак
Аммиак
H2O
Нитраты
Нитрит
CO2
(CH2O) „
Аммиак
CaCO з в
известковых
раковинах
Минеральные
соединения
Ископаемое топливо
Минеральные
частицы
Дождь детрита
Рис. 12.25. Круговороты основных элементов и воды в биосфере. (G. Evelyn Hutchinson (1970). The biosphere.
Scientific America, 50-1, Freeman.)
происходит в результате стока с
сельскохозяйственных угодий, которые могут быть обогащены
питательными веществами удобрений. Существуют
также два других важных источника фосфора-
сточные воды и моющие средства (детергенты).
Сточные воды, как в своем первоначальном виде,
так и обработанные, обогащены фосфатами (если
они не подверглись специальной обработке).
Бытовые детергенты содержат от 15% до 60%
биологически разрушаемого фосфата. Последствия
быстрой эвтрофизации обсуждались в разд. 3.2.8.
Кратко можно резюмировать, что эвтрофизация в
конце концов приводит к истощению ресурсов
кислорода и к гибели большинства живых
организмов в озерах, а в крайних ситуациях-и в
реках.
12.5. Экология сообществ и
экологические сукцессии
Сообщество - это совокупность взаимодействующих
популяций, занимающих определенную
территорию, живой компонент экосистемы. Сообщество
функционирует как динамическая единица с
различными трофическими уровнями, через него
проходит поток энергии и совершается круговорот
питательных веществ (разд. 12.3).
Некоторые типы взаимодействий уже
упоминались в разд. 12.3, например взаимоотношения
хищник - жертва (включая поедание растений) и
паразитизм. Существуют и другие типы
взаимоотношений, такие, как симбиоз (разд. 2.3.3) и
конкуренция (разд. 12.6.6).
Структура сообщества создается постепенно в
течение определенного времени. Примером,
который можно использовать как модель развития
сообщества, служит заселение организмами
обнаженной горной породы на недавно образовавшемся
вулканическом острове. Деревья и кустарники не
могут расти на голой скальной породе, так как здесь
нет необходимой для них почвы. Однако водоросли
и лишайники разными способами попадают на
такие территории и заселяют их, образуя пионерные
сообщества. Постепенное накопление отмерших и
разлагающихся организмов и эрозия горной породы
в результате выветривания приводят к
формированию слоя почвы, достаточного для того, чтобы
здесь смогли поселиться более крупные растения,
такие, как мхи и папоротники. В конце концов за
этими растениями последуют еще более крупные и
требовательные к питательным веществам формы -
семенные растения, включая травы, кустарники и
деревья. На рис. 12.26 представлена схема типичной
последовательности такого рода сообществ.
Такая смена одних видов другими за некоторый
период времени называется экологической
сукцессией. Завершающее сообщество-устойчивое,
самовозобновляющееся и находящееся в равновесии со
средой-называется климаксным сообществом. В
животном мире этих сообществ тоже происходит
смена одних видов другими, в значительной мере
обусловленная сменой растительности, но этот
процесс зависит еще и от того, какие животные могут
мигрировать из соседних сообществ.
Описанный выше тип сукцессии, начинающейся с
заселения обнаженной горной породы или другой
поверхности, лишенной почвы (например, песчаных
дюн или бывшего ложа ледника), называется
первичной сукцессией. В отличие от этого вторичной
называют сукцессию, начинающуюся там, где
поверхность полностью или в значительной степени
лишена растительности, но прежде находилась под
влиянием живых организмов и обладает
органическим компонентом. Таковы, например, лесные
вырубки, выгоревшие участки или заброшенные
сельскохозяйственные угодья. Здесь в почве могут
сохраняться семена, споры и органы вегетативного
размножения, например корневища, которые будут
оказывать влияние на сукцессию. Как при
первичной, так и при вторичной сукцессиях флора и фауна
окружающих территорий являются главным
фактором, определяющим типы растений и животных,
включающихся в сукцессию в результате случайного
расселения и миграций.
12.13. Какие факторы могут влиять на число и
разнообразие видов, появляющихся на
определенной территории?
Полную сукцессию иногда называют серией.
Серию образует ряд сериальных сообществ
(сериальных стадий). В сходных условиях среды серии
нередко бывают представлены сходными сукцессиями,
поэтому их можно классифицировать в
соответствии с этими условиями; например, гидросерия
развивается в водной среде в результате заселения
открытой водной поверхности, галосерия — на
соленых маршах.
Климаксные сообщества часто описывают как
имеющие один доминантный или несколько кодо-
минантных видов. Выбор доминантов часто
субъективен, но обычно к ним относят виды, обладающие
наибольшей совокупной биомассой или
продуктивностью, хотя при этом учитываются и физические
размеры отдельных организмов. Концепция
доминирования, однако, не имеет большого
практического значения (см. разд. 12.5.2).
Впервые теория сукцессии была детально
разработана в 1916 г. Клементсом. Он изучал сообщества
в Северной Америке и пришел к выводу, что
основным фактором, определяющим состав климаксного
сообщества, является климат. По представлениям
Клементса, в данных климатических условиях
может существовать только одно климаксное
сообщество, которое называется климаксом (концепция
моноклимакса). Любой перерыв в движении по
направлению к климаксу, вызванный местными
условиями рельефа, микроклимата, землепользованием
и т. д., тоже может привести к формированию
устойчивого сообщества, но его нельзя считать
климаксным, так как в конце концов действие
климатических факторов все-таки приведет к «теоретическо-
Крупное
нарушение в
окружающей среде
Голая земля
Сообщества серии (сериальные стадии)
Лишайники
+
Водоросли
Пионерное
сообщество
Мхи +
Папоротники
Травы-
Кустарники
Деревья
Луговая
растительность
Кустарниковая
заросль
Лес
Климаксное
сообщество
Рис. 12.26. Типичная наземная
сукцессия.
му» климаксу. Например, горы и холмы постепенно
будут разрушаться, что вызовет изменения в
структуре сообществ. Более современной является
концепция поликлимакса, согласно которой климакс
формируется под влиянием всех физических
факторов, причем один или несколько из них могут
доминировать (например, дренаж, почва,
топография, пожары). Сообщество считается настоящим
климаксом, если оно «устойчиво» длительный
период времени: любые изменения в нем происходят
относительно медленно по сравнению с временем,
необходимым для прохождения сукцессии до стадии
климакса.
Зональное распределение
В любое данное время виды внутри сообщества
могут быть пространственно распределены в
соответствии с различиями окружающей физической
среды. Это называется зональным распределением.
Хорошим примером служит распределение морских
водорослей и животных, встречающихся на
скалистых берегах между уровнями отлива и прилива и в
зоне попадания брызг. Физические условия этих зон
различны; особенно варьирует время их обнажения
между последовательными приливами. В каждой
зоне обитают виды, приспособленные к ее особым
условиям. Зональное распределение на морском
побережье описано и проиллюстрировано в разд. 13.4.
Еще один хороший пример такого рода -
вертикальная зональность с увеличением высоты в горах (см.
рис. 12.23). Внешне зональное распределение может
иметь сходство с сукцессиями, но необходимо
понимать основное различие: при зональном
распределении виды сменяются в пространстве, тогда как в
сукцессиях они сменяются во времени.
12.5.1. Примеры естественных и
искусственно созданных
климаксных сообществ
на Британских островах
Применение пыльцевого анализа позволило
воссоздать картину изменений растительности в
Британии с конца последнего ледникового периода, т. е.
примерно за последние 10 тыс. лет. В начале
неолита, примерно 4 тыс. лет назад, большая часть
низменностей была здесь покрыта лесными кли-
максными сообществами с преобладанием дуба.
Тогда впервые человек начал заниматься
земледелием и на больших площадях вырубал леса.
Обрабатывались в основном более сухие возвышенные
территории, такие, как меловые холмы Южного
Дауна и известняки Котсуолдса. Площади стали
использоваться как пастбища для домашнего скота
и иногда распахивались для посевов. В железном
веке здесь уже были широко распространены
возделывание растений и выпас скота. Типичным кли-
максным сообществом, например в античные
времена (когда в Британии заканчивался железный век),
был низинный лес, в котором доминировал дуб.
Там обитали травоядные (например, олени),
всеядные (например, медведи) и плотоядные животные
(волки и рыси). Такие леса покрывали, в частности,
область между северными и южными меловыми
холмами (Уилд) и Мидленд. Сосновые и березовые
леса росли на более бедных песчаных почвах на
большей высоте. На холмах из мела и известняка
повсеместно встречались ясень и бук.
Постепенно люди стали возделывать не только
холмы, но и более плодородные и легче
обрабатываемые низменности. Началось прогрессирующее
сведение лесов и все большее вмешательство
человека в естественные сообщества. Это
продолжается и до наших дней, когда сохранились лишь
небольшие участки лесов, которые можно считать
естественными. Некоторые сообщества с виду
естественной растительности, сохранившиеся
доныне, на самом деле уже не естественные, а созданные
человеком климаксные или «субклимаксные»
сообщества (их иногда называют плагиклимаксом).
Примером этого служат луга на мелах и верещатники
низменностей.
Для луговой растительности меловых холмов,
таких, как Южный и Северный Даун, характерно
богатое разнообразие цветов, многие из которых
(например, орхидные) сравнительно редки. Эту
растительность можно отнести к биотическому
климаксу, так как главную роль в поддержании ее
стабильности играет биологическое влияние
травоядных. К доминирующим травоядным относятся
овцы и крупный рогатый скот, а также кролики-с тех
пор как их завезли норманны. На отдельных
участках меловых холмов Дауна кролики стали
серьезными вредителями тех трав, которые не
использовались другими травоядными, так как были
слишком низкими для них. Но в 1953 г. в Англию было
«случайно» завезено вирусное заболевание миксо-
матоз. Болезнь так стремительно распространилась
в популяции кроликов, что к концу 1955 г. ее
размеры уменьшились в 10 раз. Вспышки заболевания
все еще временами повторяются, и популяция
кроликов сейчас намного меньше, чем прежде. В
результате уменьшения численности кроликов, а
также выпаса домашних травоядных стали выживать
молодые побеги кустарников и деревьев. Так или
иначе, восстановление леса сейчас затруднено,
особенно в некоторых областях, где любители природы
стараются сохранить меловые луга. Типичная
восстановительная последовательность (фактически
вторичная сукцессия) проходит через формацию, где
доминируют кустарники (такие, как ежевика нес-
ская, боярышник, кизил, можжевельник, терновник),
к климаксному сообществу, образованному
буковым лесом. Бук может иметь преимущество перед
дубом на более сухих меловых почвах и особенно
распространен на крутых склонах. Бук-сильный
конкурент отчасти и потому, что он затеняет другие
виды. На отлогих склонах с более глубокими и
Ясеневый лес
ПОЧВА
PH
ТИП
Буковый лес
(подлесник или
пролеска)
Кустарниковая
заросль (мож*
|жевельник (или)
боярышник с
различными
кальцефильны -
ми кустарника»·
ми)
Луг с
кальцефильными
травами
Тиссовый лес
Маломощная, образовалась
непосредственно
на меловых породах
7,5-8
Рендзина
Буковый лес
(мертвопокрож
ный или с μοχοί
вым покровом)
Дубовый лес
(Quercus robur
И(ИЛИ)
Q.petraea)
Березовый лес
\(Betula pendula
и(или)
B.pubescens)
Верещатник
(Calluna)
Сосновый лес
(развивается
редко)
Крупный песок, гравий
или оподзоленный
суглинок
3,5-4,5
Подзол
Рис. 12.27. Сукцессии β юго-восточной Англии. А. От мелового луга к лесу. Б. От верещатника к лесу.
(A. G. Tansley (1968). Britain's green mantle, 2nd ed., Allen & Unwin.)
более влажными почвами буку может
предшествовать ясень. На более тяжелых и влажных глинистых
почвах дуб получает преимущество перед буком и
становится доминантом. Большую роль в сукцессии
здесь играет почвенный фактор. На рис. 12.27, Л
представлена сукцессия от мелового луга к лесу в
юго-восточной Англии.
На рис. 12.27, Б показана сукцессия от
верещатника к лесу. Верещатники в низинах приурочены
главным образом к почвам определенного типа-к
кислым песчаным почвам. Как и меловые луга, они
относятся к климаксным сообществам, созданным
при участии человека. В прошлом они
использовались как пастбища для выпаса лошадей, крупного
рогатого скота, свиней и кроликов, и
доминирующими растениями здесь были верески, особенно
вереск обыкновенный (Calluna vulgaris). Рост вере-
щатников регулировался пожарами, во время
которых сгорали более старые особи, и это позволяло
прорастать молодым сочным побегам из подземных
частей, не пострадавших от огня. Огонь, таким
образом, является важным абиотическим фактором
в развитии этого сообщества. При отсутствии
пожаров верещатники постепенно сменяются лесом,
проходя через кустарниковую стадию, где могут
доминировать утесник обыкновенный, ракитник,
боярышник и другие виды. Первым укореняющимся
деревом часто бывает береза, вслед за ней
появляется дуб. Вся сукцессия занимает около 200 лет. С
дубом могут также конкурировать сосна и реже бук.
Для сохранения верещатников в низинах
необходимо удалять кустарники и деревья. Сохранение
верещатников желательно, так как поддерживает
разнообразие местообитаний на Британских
островах и способствует сохранению определенных
редких видов, которые здесь встречаются (например,
вонючей жабы и провансальской славки в
Дорсетшире).
Пример климаксного лесного
сообщества
К типичным наземным климаксным сообществам
относятся листопадные леса. Они отличаются
большим видовым богатством. Например, в лесу возле
Оксфорда, где проводились длительные подробные
исследования, было найдено около 4 тыс. видов
животных. Одна из причин такого богатства -
сложная структура лесного сообщества с большим
числом ниш и микроместообитаний.
На рис. 12.6, Λ была представлена трофическая
сеть в дубовом лесу. На рис. 12.28 показана
структура типичного лиственного леса. Важной
особенностью лесного сообщества является разделение его
на ярусы (ярусность). Большая часть первичной
продукции производится в древесном пологе,
наиболее интенсивное разложение идет на уровне
земли. Животные занимают ниши на всех уровнях.
Древесный ярус
(5-20 м +)
Кустарниковый
ярус (2 -5 м)
Травянистый ярус
(0-2м)
Приземный ярус
(на 3 см от почвы)
Доминантный вид растения, например дуб.
Несколько кодоминантных видов, например
клен белый. Птицы , например неясыть
обыкновенная, чеглок обыкновенный,
воробей, вяхирь, сорока, галка,
ворона, певчий дрозд, зяблик, большая
синица, лазоревка, сойка, дятел, поползень
обыкновенный, пищуха обыкновенная.
Личинки (гусеницы) пяденицы зимней
Серая белка.Большое количество насекомых
(свыше 1600 видов, в своей жизне -
деятельности тесно связанных с дубом)
Например, боярышник, терновник, кизил,
I бузина, калина обыкновенная,
| шиповник, растения рода Evonymus
ι крушина. Птицы, например зарянка,
* черный дрозд, мухолов ка-пеструшка,
I горихвостка, дятел, поползень
обыкновенный, пищуха обыкновенная
Много насекомых, таких же как в
травянистом ярусе Серая белка
Подстилка
Пахотный слой
(низший органический ярус)
Подпочва
(выветриваемая коренная порода)
Коренная порода
Травы, невысокие лесные растения,
подрост деревьев, папоротники Птицы,
например славки, крапивник, синица
московка, лесная завирушка Лань,
косуля. Мелкие млекопитающие,
например соня Бабочки, пчелы, осы,
журчалки, кровососущие насекомые,
мухи, комары, жуки, пауки
Мелкие прыгающие травяные беспозвоночные,
мухи, жуки, пауки, сенокосцы, муравьи,
полевки, землеройки, лесные мыши.
Лишайники, мхи, печеночники,
низкие травы
Мертвые и разлагающиеся организмы.
Редуценты (бактерии и грибы)
Дождевые черви, гусеницы, личинки мух,
жуки навозники и могильщики, губоногие
двупарноногие, мокрицы, ногохвостки,
клещи, нематоды, (см рис. 12.5) Кроты
Норы кроликов, барсуков, лисиц
Рис. 12.28. Вертикальная структура типичного сообщества широколиственного леса. Обратите внимание на
то, что некоторые животные могут перемещаться из одного яруса в другой. Например, серая белка кормится
на земле, а спит и выводит потомство на деревьях; птицы могут отдыхать на одном ярусе, а питаться на
других; например, неясыть обыкновенная охотится на млекопитающих в травянистом и приземном ярусах,
а гнездится в древесном пологе.
12.5.2. Основные особенности
сукцессии
Предпринималось несколько попыток найти и
понять основной принцип, предопределяющий ход
сукцессии по направлению к климаксу.
Экологическое доминирование
Наблюдения, касающиеся экологического
доминирования, наводят на мысль, что сукцессия всегда
идет в направлении к климаксу, где доминирует
один или несколько видов-кодоминантов, например
к дубовому лесу или к дубово-ясеневому лесу. В
таких лесах в связи с затенением и другими
причинами ограничен рост видов, которые могли бы
существовать рядом с доминантами. Однако в
некоторых случаях трудно применить концепцию
доминирования, например когда речь идет о
сообществах планктона, о животных или о тропических
дождевых лесах, где в приблизительно равных
количествах можно найти несколько сотен видов
деревьев.
Продуктивность и биомасса
Линдеман в 1942 г. высказал предположение, что
сукцессии сопровождаются повышением
продуктивности вплоть до климаксного сообщества, в
котором превращение энергии происходит наиболее
эффективно. Это логичная и привлекательная
гипотеза, так как поток энергии, проходящий через
экосистему, является одним из важных факторов,
лимитирующих численность и биомассу
организмов, жизнь которых она способна поддерживать.
Данные исследований показывают, что на поздних
стадиях сукцессии продуктивность действительно
возрастает, однако при переходе к климаксному
сообществу обычно происходит снижение общей
продуктивности.
Таким образом, продуктивность в старых лесах
ниже, чем в молодых, которые в свою очередь могут
Таблица 12.7. Основные изменения в экосистеме во время
типичной вторичной сукцессии
Энергия, выраженная
через общую продуктивность
Особенности
Отношение общей
продукции к дыханию
(P/R)
Чистая продуктивность
сообщества
Пищевые цепи
Общее количество
органического материала
(или биомасса)
Видовое разнообразие
Структура сообщества
Специализация по
нишам
Размеры организмов
Приспособительные
стратегии видов
(разд. 12.6.4)
Стадия развития экосистемы
незрелая
(ранняя)
Высокое (>1)
Высокая
Линейные, в
основном
пастбищные
Малое
Небольшое
Простая
Широкая
Малые
«г-стратегия»
зрелая (поздняя)
Приближается
к 1
Низкая
Трофическая
сеть, в
основном дет-
ритная
Большое
Большое0
Сложная
(ярусность,
много
микроместо-
обитаний)
Узкая
Крупные
«^-стратегия»
!) В большинстве растительных и в некоторых животных
сукцессиях максимум видового разнообразия достигается до
наступления климакса.
Достижение
климакса
Время
Рис. 12.29. Изменения общей продуктивности,
дыхания и биомассы в ходе типичной сукцессии. (Tribe
et a/. (1974). Ecological principles, Basic Biology Course 4.
Cambridge University Press.)
иметь меньшую продуктивность, чем
предшествовавшие им более богатые видами ярусы
травянистых растений. Сходное падение продуктивности
наблюдается в некоторых водных системах. Как
сейчас полагают, во многих сукцессиях максимум
продуктивности достигается очень быстро, хотя данных
об этом пока еще мало. Причины снижения
продуктивности остаются предметом гипотез, хотя,
если рассматривать в качестве примера лес, можно
ожидать, что продуктивность старых деревьев
действительно будет меньше, чем молодых. Для этого
есть несколько причин. Одна из них то, что
накопление питательных веществ в растущей биомассе
леса на корню может вести к уменьшению их
круговорота. Однако снижение общей
продуктивности могло быть просто результатом уменьшения
жизненности особей по мере увеличения их среднего
возраста в сообществе.
Хотя нет, по-видимому, оснований утверждать,
что сукцессии ведут к максимальной эффективности
превращения энергии (к максимуму общей
продуктивности), уже мож:но сказать, что они ведут к
максимальному накоплению биомассы. Это
наиболее очевидно в лесных сообществах, где в ходе
сукцессии растения становятся все крупнее и
крупнее. Накопленная биомасса в других климаксных
сообществах тоже обычно больше, чем в период
сукцессии. Изменения в общей продуктивности,
дыхании и биомассе в ходе типичной сукцессии
показаны на рис. 12.29. Можно видеть, что в кли-
максном сообществе эти параметры становятся
более или менее постоянными; видно также, что
верхний предел накопления биомассы достигается тогда,
когда общие потери на дыхание (R) становятся
почти равными общей первичной продуктивности
(P), т.е. отношение P/R приближается к единице.
Эти и другие особенности, вероятно, свойственные
сукцессиям, приведены в табл. 12.7.
По мере прохождения сукцессии все большая доля
доступных питательных веществ накапливается в
биомассе сообщества и соответственно уменьшается
их содержание в абиотическом компоненте
экосистемы (в почве или воде). Возрастает также количество
образующегося детрита, и главными первичными
консументами становятся не травоядные, а детри-
тоядные организмы. Соответствующие изменения
происходят и в трофических сетях, и детрит
становится основным источником питательных
веществ.
12.5.3. Взаимодействия между
организмами и абиотической
средой
На ранних стадиях сукцессии наиболее важны
взаимодействия между живыми организмами и
абиотической средой. В результате деятельности
организмов эта среда с течением времени изменяется.
Хорошим примером может служить образование
песчаных дюн вследствие накопления эолового
песка вокруг побегов песколюба (Ammophila, или
Psamma) (рис. 12.30). Время стабилизации дюн
составляет несколько лет. Хорошо изучена сукцессия
песчаных дюн вокруг озера Мичиган в Северной
Америке, где стабилизация дюн привела к
появлению более крупных и не требовательных к
питательным веществам растений (тополь, сосны). За
длительный период времени образуется почва, в
которой накапливается органический углерод,
поступающий из подстилки. Увеличение численности
азотфиксирующих организмов приводит к росту
количества азота в почве. Местами здесь появился
дуб, который стал доминирующим видом. После
того как дуб окончательно закрепился, почвенные
условия, по-видимому, остаются постоянными в
течение неопределенного времени: потребность в
питательных веществах удовлетворяется за счет
поступления их из лесной подстилки. В этом примере
почвенные факторы играют определяющую роль в
сукцессии и сами изменяются в процессе сукцес-
сионных смен. Может, однако, потребоваться
тысяча лет, для того чтобы химический состав почвы
стабилизировался и сукцессию можно было считать
законченной.
Этот пример иллюстрирует важный общий
принцип: ранние стадии сукцессии проходят значительно
быстрее, чем поздние. Дальнейшее развитие кли-
максного сообщества происходит только при
изменении условий окружающей среды. Хорошим
примером изменений климата, приведших к
коренной смене сообществ на больших площадях, служит
последовательность ледниковых периодов в
последние 1,8 млн. лет.
Биотическое сообщество может влиять на
физическую среду, а тем самым и на сукцессию
различными способами. Например, сообщества могут
локально изменять климат и формировать
микроклимат.
12.14. Каким образом могут изменяться под
кроной группы деревьев следующие условия:
а) свет, б) температура, в) ветер и г) влажность?
Воздушные побеги
заносятся песком
Рост Ammophila
продолжается
1 Укоренившаяся часть
корневища
Вокруг разросшейся
Ammophila
образуются дюны
Рис. 12.30. Образование дюны при постепенном
отложении переносимого ветром песка вокруг
воздушных побегов Ammophila.
Аналогичным образом создаются
микроместообитания, такие, как отмершая древесина (в которой
может обитать около 200 видов животных), навоз
(более 300 видов животных, в основном жуки и
мухи) и падаль (также заселяется жуками и мухами).
12.5.4. Взаимодействия между
организмами внутри
сообщества
На более поздних стадиях сукцессии повышается
роль биотических взаимодействий в формировании
детальной структуры сообщества. Возрастает
разнообразие живых организмов, и их взаимодействия
неизбежно становятся более сложными. Одно из
древнейших климаксных сообществ - тропический
дождевой лес - известно своим видовым богатством
и необычайной сложностью биотических
взаимоотношений. К экологии сообществ относят изучение
динамических взаимодействий между различными
видами, а не между особями одного и того же вида.
Встречаются различные типы взаимодействий.
Взаимоотношения типа «хищник - жертва» (в том
числе «травоядное животное - растение») уже были
упомянуты в связи с пищевыми цепями и всей сетью
трофических связей в разд. 12.3, где говорилось о
зависимости одних организмов от других. Большое
значение имеют также паразитические, симбиоти-
ческие и другие мутуалистические отношения. Как
показывают некоторые из примеров, приведенных в
разд. 12.6.6, исследование таких взаимоотношений
пересекается с изучением экологии популяций.
Все доступные экологические ниши постепенно
заполняются и может возникнуть конкуренция за
какую-либо нишу (разд. 12.1). Шансы на то, что
какой-то вид животных закрепится в данной
области, зависят от многих причин-от вероятности
миграции его в данный район, от наличия
пригодной для него пищи, от его способности отыскать
подходящую нишу и в случае необходимости за-
щитить ее от конкурентов. Чем больше
специализирован вид для использования данной ниши, тем
меньше вероятность прямой конкуренции.
С представлением об экологических нишах тесно
связано понятие разделения ресурсов, т.е.
распределения их между различными видами сообщества.
Специализация видов в отношении ресурсов
уменьшает конкуренцию и увеличивает стабильность
структуры сообщества. Существуют различные
типы разделения ресурсов:
1. Специализация морфологии и поведения в
соответствии с родом пищи, например клюв у птиц
может быть приспособлен для ловли насекомых,
долбления отверстий, раскалывания орехов,
разрывания мяса и т. п.
2. Вертикальное разделение, как, например, между
обитателями полога и лесной подстилки.
3. Горизонтальное разделение, например между
обитателями различных микроместообитаний.
Каждый из этих типов или их комбинация
приводит к разделению организмов на группы, которые
меньше конкурируют между собой, так как каждая
из них занимает свою нишу. Существует, например,
разделение птиц на экологические группы,
основанные на месте их питания (воздух, листва, ствол,
земля). Дальнейшее подразделение этих групп в
зависимости от основного типа пищи показано на
рис. 12.31. У растений для каждого вида существует
набор оптимальных, наиболее благоприятных
условий (влажность, высота над уровнем моря, рН
почвы и т.д.), что опять-таки приводит к
уменьшению прямой конкуренции.
Несмотря на то что каждый вид стремится
обзавестись собственной нишей, некоторая прямая
конкуренция между видами за доступные ресурсы
неизбежна. Подробнее это будет рассмотрено в
разд. 12.6.6.
12.6. Популяционная экология
Во введении к этой главе мы определили
популяцию как совокупность организмов одного вида,
обитающих в данном месте в данное время.
Исследование популяций имеет важные практические
приложения, например при изучении вредителей,
решении природоохранных задач и др. Изучая
популяции, интересуются не только их численностью в
данное время, но прежде всего их ростом,
поддержанием и уменьшением в зависимости от
факторов среды. Этот раздел популяционной экологии
называется динамикой популяций. Данные о том,
каким образом численность различных видов
изменяется во времени, используются также при
изучении потоков энергии и биомассы и помогают
построению теории экосистем.
Если не считать ряда длительных исследований по
динамике фитопланктона, большая часть работ по
динамике популяций проводилась на животных и
микроорганизмах. Имея дело с растениями, особен-
Парящие
Подстерегающие
Воздух
Питающиеся
нектаром
Листва
Насекомоядные
Питающиеся
семенами
Ствол
Ищущие
Долбящие
кору
Почва
Хищники
Насекомоядные
Питающиеся
семенами
Рис. 12.31. Экологическая классификация,
использованная при анализе трех сообществ птиц в
Калифорнии. (В. D. Collier et a/. (1974). Dinamic ecology.
Prentice Hall.)
но культурными, целесообразнее, а иногда и проще
учитывать биомассу, а не число особей. Динамика
популяций растений сильно влияет на динамику
популяций животных. Поэтому, например, при
изучении популяций человека (демография) обычно
учитывают и продовольственные ресурсы, основу
которых составляют растения. Некоторые сведения о
том, как собирают данные о численности животных
и растительных популяций и о распределении
особей, приводятся в разд. 13.2.
12.6.1. Рождаемость и смертность
Размеры популяции могут возрастать в результате
иммиграции из соседних популяций или за счет
размножения особей. Одним из показателей
размножения является плодовитость, которая
измеряется числом потомков одной женской особи.
Плодовитость можно выражать разными способами,
зависящими от целей изучения и от особенностей
изучаемого организма. Ее можно определить,
например, как среднее число оплодотворенных яиц,
производимых за один цикл размножения или в
течение всей жизни. Говоря о млекопитающих,
пользуются термином рождаемость, которую
определяют как число потомков, производимых одной
самкой за год. У человека рождаемость обычно
выражают числом рождений на 1000 человек за год
(табл. 12.8). В развивающихся странах с
недостаточным контролем рождаемости ее уровень
примерно вдвое выше, чем в развитых странах.
Социологи называют культурные изменения в обществе
при переходе с высшего уровня рождаемости на
низший демографическим переходом.
12.15. Если численность населения в данном
году составляла 500 000 человек и за год родилось
10 000, то какова была рождаемость в этом году
при пересчете на 1000 человек?
Размеры популяции могут уменьшаться в
результате эмиграции или смертности. В популяционной
биологии смертность означает вероятность смерти
и выражается либо в процентах от общего числа
особей, либо средним числом смертей на 1000
особей в год. Табл. 12.9 показывает, что в развитых
странах человеческая смертность в среднем ниже,
чем в развивающихся, что обусловлено лучшим
медицинским обслуживанием и питанием. Там, где были
введены программы медицинской помощи и
продовольственные программы (как, например, в
Индии после второй мировой войны), статистика
показывает снижение уровня смертности. Очень низкая
смертность в таких странах, как Япония и Коста-
Рика, обусловлена относительно большой долей
молодежи в населении.
Таблица 12.8. Статистика рождаемости в отдельных
странах и регионах. Цифры указывают число живых детей,
рождающихся за год, на тысячу человекп
Страны
Кения
Конго
Иран
Египет
Индия
Южная Африка
Мексика
Малайзия
Ямайка
Израиль
Чили
Китай
США
Куба
Япония
ГДР
Соединенное
Королевство
Италия
Швеция
ФРГ
53
45
44
41
36
36
33
31
27
25
22
18
16
15
14
14
13
12
12
10
υ Данные за 1981 г.
Регионы
Африка
Латинская
Америка
Азия
Северная
Америка
Австралия
Европа
46
32
29
16
16
14
Таблица 12.9. Статистика смертности в отдельных странах и
регионах. Цифры указывают число смертей за год,
приходящееся на тысячу человекп
Страны
Эфиопия
Конго
Индия
ГДР
Иран
Кения
Южная Африка
Соединенное
Королевство
ФРГ
Египет
Швеция
Италия
США
Мексика
Малайзия
Чили
Израиль
Китай
Куба
Ямайка
Япония
Коста-Рика
25
19
15
14
14
14
12
12
12
11
11
9
9
8
8
7
7
6
6
6
6
4
Регионы
Африка
Азия
Европа
Латинская
Америка
Северная Америка
Австралия
17
11
10
9
9
7
1} Данные за 1981 г.
12.6.2. Кривые выживания
Один из основных факторов, влияющих на размеры
популяции,-это процент особей, погибающих до
достижения половой зрелости (смертность в до-
репродуктивном периоде). В пределах данного вида
эта величина гораздо более изменчива, чем
плодовитость. Для того чтобы численность популяции
оставалась постоянной, в среднем только два
потомка каждой пары должны доживать до
репродуктивного возраста.
12.16. Ниже приведены данные о среднем числе
оплодотворенных яиц, производимых в течение
всей жизни самками различных животных (т.е. о
плодовитости)
Устрица
Треска
Камбала
Лосось
Трехиглая
колюшка
Зимняя
пяденица
100·
9-
35·
10·
5·
200
106
106
104
104
102
Мышь
Акула
Пингвин
Слон
Для сравнения:
английская женщина
эпохи королевы
Виктории
50
20
8
5
10
а. Сколько оплодотворенных яиц от одной
самки в среднем должно выжить, чтобы численность
популяции каждого из перечисленных видов
оставалась постоянной?
Выживание, %
Возраст
Рис. 12.32. Три типа кривых выживания. Объяснения
см. в тексте.
б. Напишите для каждого вида число потомков,
которые должны погибать до наступления
половой зрелости, чтобы численность популяции не
изменялась. Затем выразите это число в виде
процента от общего числа оплодотворенных яиц
(это даст величину смертности в дорепродуктив-
ном периоде).
в. Попробуйте объяснить, почему плодовитость
трехиглой колюшки и акулы намного меньше,
чем у других рыб, данные для которых приведены
в таблице.
Кривую выживания можно получить, если начать
с некоторой популяции новорожденных особей и
затем отмечать число выживших в зависимости от
времени. По вертикальной оси обычно
откладывают или абсолютное число выживших особей или
их процент от исходной популяции:
Число выживших
χ 100%
Численность исходной популяции
Каждому виду свойственна характерная кривая
выживания, форма которой отчасти зависит от
смертности неполовозрелых особей. Типичные
примеры приведены на рис. 12.32.
Большинство животных и растений подвержено
старению, которое проявляется в снижении
жизненности с возрастом после периода зрелости. Как
только начинается старение, вероятность
наступления смерти в определенный промежуток времени
возрастает. Непосредственные причины смерти
могут быть разными, но в основе их лежит уменьшение
сопротивляемости организма к действию
неблагоприятных факторов, например болезням. Кривая А
на рис. 12.32 очень близка к идеальной кривой
выживания для популяции, в которой старение
служит главным фактором, влияющим на смертность.
Примером может быть популяция человека в
современной развитой стране с высоким уровнем
медицинского обслуживания и рациональным
питанием. Большинство людей доживает до старости, но
среднюю ожидаемую продолжительность жизни
почти невозможно увеличить более чем до 75 лет.
Отклонение кривой от идеальной обусловлено
детской смертностью, отображаемой уклоном
начального участка. Хотя детская смертность в развитых
странах намного ниже, однако и здесь вероятность
смерти в раннем детстве выше средней. Кроме того,
на кривую выживания помимо старения влияет
фактор случайной гибели, причины которой с
возрастом могут изменяться. В Англии, например,
наибольшая смертность в результате
автомобильных катастроф падает на возраст 20-25 лет. Кривая,
сходная с кривой А, свойственна также однолетним
культурным растениям, например пшенице, когда
все растения на данном поле стареют
одновременно.
Кривая типа Б характерна для популяций
организмов с высокой смертностью в ранний период
жизни, например для горных овец или для
популяции человека в стране, где широко
распространены голод и болезни. Плавная кривая типа В может
быть получена, если смертность постоянна в течение
всей жизни организмов (50% за определенную
единицу времени). Это может быть тогда, когда
главным фактором, определяющим смертность,
становится случай, причем особи гибнут до начала
заметного старения. Кривая наподобие этой была
когда-то получена для «популяции» стеклянных
стаканов в кафетерии. Очень сходная кривая
характерна для популяций некоторых животных
(например, гидры), не подвергающихся особой опасности в
раннем возрасте. Для большинства беспозвоночных
и растений тоже характерна кривая такого типа, но
высокая смертность среди молодых особей
приводит к тому, что начальная часть кривой
спускается еще более круто.
12.17. В популяции какого типа -А или Б- нужна
большая скорость размножения для поддержания
стабильной численности? Объясните ваш выбор.
Существуют небольшие внутривидовые различия
в кривых выживания. Они могут быть обусловлены
разными причинами и нередко связаны с полом. У
людей, например, женщины живут несколько
дольше, чем мужчины, хотя точные причины этого
неизвестны.
Вычерчивая кривые выживания для различных
видов, можно определять смертность для особей
разного возраста и таким образом выяснять, в
каком возрасте данный вид наиболее уязвим.
Установив причины смерти в этом возрасте, можно
понять, как регулируется величина популяции.
12.18. Приводимые ниже средние цифры
относятся к нерке-рыбе из семейства лососевых,
обитающей в реках Канады. Осенью каждая
самка откладывает 3200 икринок на гравий в мелких
местах. Следующей весной 640 мальков, вывед-
шихся из отложенной икры, выходят в озеро
вблизи отмели; уцелевшие 64 серебрянки (мальки
постарше) живут в озере один год, а затем
мигрируют в море. Две взрослые рыбы (уцелевшие из
числа серебрянок) возвращаются к местам нереста
спустя 2,5 года; они нерестятся и умирают.
Подсчитайте процент смертности для нерки в каждом
из следующих периодов:
а) от откладки икры до переселения мальков в
озеро спустя шесть месяцев;
б) за 12 месяцев жизни в озере;
в) за 30 месяцев от выхода из озера до
возвращения к местам нереста.
Нарисуйте кривую выживания нерки в этой
водной системе (зависимость процента
выживших особей от возраста). Какова величина до-
репродуктивной смертности среди этих
лососевых?
12.6.3. Рост популяции и кривые роста
Если рождаемость в популяции превышает
смертность, то популяция будет расти, если, конечно,
изменения в результате иммиграции и эмиграции
незначительны. Чтобы понять закономерности
роста популяций, полезно вначале рассмотреть модель
(см. гл. 2), описывающую рост популяции бактерий
после посева их на свежую культу рал ьную среду. В
этой новой и благоприятной среде условия для
роста популяции оптимальны и наблюдается
экспоненциальный рост (см. рис. 2.7; см. также
разд. 21.1.2). Кривая такого роста-это
экспоненциальная, или логарифмическая кривая. Но, как
сказано в разд. 2.2.4 и 21.1.2, в конце концов
достигается такая точка, когда по нескольким причинам,
в том числе из-за уменьшения пищевых ресурсов и
накопления токсичных отходов метаболизма
экспоненциальный рост становится невозможным. Он
начинает замедляться так, что кривая роста
приобретает сигмоидную (S-образную) форму, как
показано на рис. 2.7 для бактерий и на рис. 21.1 для
дрожжей. Такой тип роста называют зависимым от
плотности, так как скорость роста зависит от
плотности популяции, которая влияет на истощение
пищевых ресурсов и накопление токсичных
продуктов, а потому на рост. С увеличением плотности
скорость роста популяции постепенно снижается до
нуля, а кривая выходит на плато. При нулевом
росте популяция стабильна, т.е. размеры ее не
меняются (напомним, что отдельные организмы
при этом могут расти и размножаться; нулевая
скорость роста популяции означает лишь то, что
скорость размножения, если оно происходит,
уравновешена смертностью). Такая сигмоидная кривая
роста получена для ряда одноклеточных и
многоклеточных организмов, например для клеток
водорослей в культуральной среде, для
фитопланктона озер и океанов весной, для насекомых, таких, как
мучные хрущаки или клещи, интродуцированные в
новое местообитание с обильными запасами пищи,
где нет хищников.
Кривая другого типа получается, когда
экспоненциальный рост продолжается вплоть до
внезапного падения плотности популяции в результате
исчерпания ресурсов среды. Эту кривую называют
«J-образной» или кривой типа «бум и крах». Такой
рост не зависит от плотности, так как его регуляция
не связана с плотностью популяции до самого
момента катастрофы. Крах может происходить по
тем же причинам, например из-за истощения
пищевых ресурсов, которое в случае сигмоидной
кривой роста заблаговременно оказывало
регулирующее влияние на рост. Миграция или расселение, так
же как и внезапное снижение скорости размножения,
может способствовать уменьшению численности
популяции. Расселение может быть связано с
определенной стадией жизненного цикла, например с
образованием семян. На рис. 12.33 показаны
примеры того и другого типа. Для обоих типов
характерна экспоненциальная фаза в начале роста.
Рассматривая вопрос об оптимальных размерах
популяции в данной среде, важно учитывать
поддерживающую емкость, или «кормовую
продуктивность», этой среды. Чем выше поддерживающая
емкость, тем больше максимальный размер
популяции, который может существовать в данном
местообитании неопределенно долгое время.
Дальнейшему росту популяции будет препятствовать
один или несколько лимитирующих факторов. Это
зависит от доступности ресурсов для данного вида.
Как показано на рис. 12.33, в случае J-образной
кривой роста популяция внезапно выходит за
пределы поддерживающей емкости среды. Эту
величину обозначают символом К, который можно
использовать также для обозначения максимальных
размеров стабильной популяции в данных условиях.
Рост, соответствующий сигмоидной и J-образной
кривым, можно описать алгебраически с помощью
простых дифференциальных уравнений (оба
уравнения относятся к популяциям, в которых
поколения полностью перекрываются, так что
популяция изменяется непрерывно; это и позволяет
использовать дифференциальные уравнения). Такие
уравнения приведены в табл. 12.10, там же дается
краткое объяснение.
Сигмоидная и J-образная кривые - это две модели
роста популяции. При этом предполагается, что все
организмы очень сходны между собой, имеют
равную способность к размножению и равную
вероятность погибнуть, так что скорость роста популяции
в экспоненциальной фазе зависит только от ее
численности и не ограничена условиями среды,
которые остаются постоянными. Но в отношении
природных популяций эти предположения часто
неверны. Например, скорость роста популяции в
естественном местообитании будет зависеть от
климатических изменений, от снабжения пищей и от
того, ограничено ли размножение определенным
Поддерживающая емкость среды
Число организмов
Дрожжи
Время, ч
Число организмов на 1 см 3
Поддерживающая
емкость среды
Резкое снижение
численности
Вспышка
численности
Б Возраст, дни
Рис. 12.33. Два типа кривых роста популяции.
(А) Сигмоидная (S-образная) кривая роста дрожжей
в культуре. Поддерживающая емкость среды
определяется изначальным количеством доступных
питательных веществ, содержащихся в среде.
(Б) J-образная кривая роста дафний в культуре
(A. S. Boughey (1971). Fundamental Ecology, International
Textbook Co.) См. также рис. 12.40-пример роста
популяции оленей на плато Кайбаб.
временем года. И все же модели роста популяций
способствуют лучшему пониманию естественных
популяций, и в случае надобности их можно
усовершенствовать.
12.6.4. Стратегии популяций
В предыдущем разделе в уравнениях, описывающих
рост популяции, были использованы символы г и К.
Виды, которые быстро размножаются, имеют
высокое значение г и называются r-видами. Это
обычно «оппортунистические» виды - типичные
пионерные виды нарушенных местообитаний. Такие
местообитания называют r-отбирающими, так как они
благоприятствуют росту численности г-видов.
Виды с относительно низким значением г
называют К-видами. Скорость их размножения
чувствительна к плотности популяции и остается близкой к
уровню равновесия, определяемому величиной К.
Они более характерны для поздних стадий
сукцессии. Об этих двух типах видов говорят, что они
используют соответственно r-стратегию и Х-стра-
тегию. Хотя существует и целый спектр
промежуточных стратегий, концепция двух стратегий
находит важное применение в популяционной
экологии и в экологии сообществ. Некоторые
характерные особенности крайних г- и К-стратегий
суммированы в табл. 12.11.
Дафния
По существу, эти две стратегии представляют два
различных решения одной задачи - задачи
длительного выживания вида. Виды с r-стратегией быстрее
заселяют нарушенные местообитания (характерные
для ранних стадий сукцессии, такие, как обнаженная
горная порода, лесные вырубки, выгоревшие
участки), чем виды с К-стратегией, так как они легче
распространяются и быстрее размножаются. Виды с
К-стратегией более конкурентоспособны, и в конце
концов они вытесняют г-виды, которые тем
временем перемещаются в другие нарушенные
местообитания. Высокий репродуктивный потенциал
г-видов означает, что, оставшись в каком-либо
местообитании, они быстро использовали бы доступные
ресурсы и превысили поддерживающую емкость
среды, а затем популяция погибла бы. Другими
словами, для них характерна J-образная кривая
роста с быстрым падением численности популяции в
конце (рис. 12.33,/>). Виды с r-стратегией занимают
данное местообитание лишь в течение жизни одного
Таблица 12.10. Уравнения для сигмойдной и J-образной
кривой роста
Если N - число особей в популяции и t - время, то скорость
изменения численности во времени, представленная
отношением dN/d/, пропорциональна N, т.е. άΝ/άί = rN, где г
константа - врожденная скорость роста численности
популяции, связанная с максимальной скоростью размножения
особи данного вида. Чем выше скорость размножения, тем
больше значение г
J-образная кривая (рост, не
зависящий от плотности)
Сигмоидная кривая (рост,
зависящий от плотности)
άΝ
~dt~
= rN
άΝ
~d7
= rN
(K-N)
K
Если г положительно,
численность популяции
увеличивается
экспоненциально
Если г отрицательно,
численность популяции
уменьшается
экспоненциально
Отсюда быстрые
увеличения и падения
численности популяции
Скорость роста каждого
организма не зависит от
плотности популяции.
Этот тип роста
популяции иногда
рассматривают как рост по сложным
процентам
Размеры популяции не
стабилизируются
где К - максимальное число
организмов, которое
может поддерживаться в
данных условиях среды.
Введение в уравнение К
означает, что влияние
среды на снижение роста
численности до какого-то
стационарного уровня
отражено в расчете. К
называют также
поддерживающей емкостью
среды
Если N > К, скорость
роста отрицательна
Если К> N, скорость
роста положительна, то
величина популяции
стремится к K = N, т. е.
приводится в соответствие с
поддерживающей
емкостью среды
Когда K = N скорость
роста популяции равна
нулю
Размеры популяции
остаются постоянными
Доля общей биомассы,%
Solidago speciosa в лесу
Solidago rugosa в лесу
Листья
Листья
Цветки
Цветки
Стебли
Стебли
Доля общей биомассы %
Solidago speciosa
в сухом местообитании
Solidago nemoralis
в сухом местообитании
Листья
Цветки
Цветки
Стебли
Стебли
Июнь Июль Авг. Сент Οκτ.
Июнь Июль Авг_ Сент. Окт,
Рис. 12.34. Процент биомассы,
приходящийся на листья, стебли
и цветки в четырех популяциях
золотарника (R. M. May (ed.)
(1976). Teoretical Ecology, B lack we//.)
Таблица 12.11. Характерные особенности г- и А-видов
r-Виды
(виды-«оппортунисты»)
Размножаются быстро
(высокая плодовитость,
время генерации короткое),
поэтому значение г
(врожденная скорость роста
популяции) высокое
Скорость размножения не
зависит от плотности
популяции
Энергия и вещество
распределяются между
многими потомками
Размеры популяции
некоторое время могут
превышать К
(поддерживающую емкость среды)
Вид не всегда устойчив на
данной территории
Расселяются широко и в
больших количествах; у
животных может
мигрировать каждое поколение
Размножение идет с
относительно большими
затратами энергии и
вещества
Малые размеры особей
Малая продолжительность
жизни особи
Могут поселяться на
открытом грунте
К-виды (с тенденцией к
равновесию)
Размножаются медленно
(низкая плодовитость,
продолжительное время
генерации), поэтому
значение г низкое
Скорость размножения
зависит от плотности
популяции, быстро
увеличивается, если плотность
падает
Энергия и вещество
концентрируются в
немногих потомках; родители
заботятся о потомстве
Размеры популяции близки
к равновесному уровню,
определяемому К
Вид устойчив на данной
территории
Расселяются медленно
Размножение идет с
относительно малыми
затратами энергии и вещества;
большая часть энергии и
вещества расходуется на
нерепродуктивный
(вегетативный) рост
Крупные размеры особей; у
растений деревянистые
стебли и большие корни
Большая
продолжительность жизни особи
Плохо приспособлены к
росту на открытых
местах
Продолжение табл 12.11
г-Виды
(виды-«оппортунисты»)
Местообитания
сохраняются недолго (например,
зрелые фрукты для
личинок Drosophild)
Слабые конкуренты
(способность к конкуренции
не требуется)
Защитные приспособления
развиты сравнительно
слабо
Не становятся
доминантами
Лучше приспособлены к
изменениям окружающей
среды (менее
специализированные)
Примеры
Бактерии
Парамеция
Тли
Мучные хрущаки
Однолетние растения
К-виды (с тенденцией к
равновесию)
Местообитания устойчивые
и сохраняются долго
(например, лес для обезьян)
Сильные конкуренты
Хорошие защитные
механизмы
Могут становиться
доминантами
Менее устойчивы к
изменениям условий среды
(высокая специализация для
жизни в устойчивых
местообитаниях)
Примеры
Крупные тропические
бабочки
Кондор (крупная хищная
птица)
Альбатрос
Человек
Деревья
или, самое большее, нескольких поколений. Затем
они переселяются на новое место. Отдельные
популяции могут регулярно вымирать, но вид при
этом перемещается и выживает. В целом эту
стратегию можно охарактеризовать как стратегию
«борьбы или бегства».
Как отмечает Саутвуд1, хотя для самых мелких
1 T. R. E. Southwood, Bionomic Strategies and Population
Parameters, in: Theoretical Ecology. Principles and
Applications, ed. R. M. May (1976), Blackwell.
Гусеницы пяденицы зимней
Число гусениц на 1 см 2
Годы
Рис. 12.35. Изменения численности популяции
гусениц пяденицы зимней (Operophthera brumata) в
Уитхемском лесу возле Оксфорда. Обратите
внимание, что численность откладывалась по
логарифмической шкале. (From Open University Science
Foundation Course (SWO) Unit 20 (1971). Open University Press.)
организмов, таких, как бактерии и насекомые,
характерна r-стратегия, а для самых крупных, таких,
как многие позвоночные и деревья,- К-стратегия,
большинству организмов свойственны стратегии
промежуточных типов. Даже в пределах таких
групп, как насекомые и позвоночные, можно
обнаружить различные стратегии. Среди птиц,
например, лазоревка (оппортунистический обитатель
лиственных лесов) проявляет резко выраженную
r-стратегию. Будучи маленькой, она тем не менее
откладывает сравнительно много яиц и способна
более чем вдвое увеличить свою численность за
один сезон. У одного из самых мелких попугаев-
волнистого попугайчика, кочующего по
Центральной Австралии,- очень небольшое время генерации,
поэтому этот вид тоже можно отнести к видам с
r-стратегией. Но существуют и птицы с резко
выраженной Х-стратегией, например кондор и
альбатрос. Размах крыльев у них превышает три метра,
но они откладывают только по одному яйцу каждые
два года. Альбатрос позднее всех других птиц
достигает половой зрелости (к 9-11 годам).
Стабильные местообитания способствуют отбору
на Х-стратегию. Хорошим примером служат
мясные (падальные) мухи, личинки которых питаются
падалью. У одного вида (Blaseoxiphia fletcheri)
личинки живут в жидкости кувшинчатых листьев
насекомоядного растения саррацении-более
долговечном местообитании, чем падаль, которую
используют другие виды. Самка этой мухи производит
всего лишь И личинок (тогда как самки
родственных видов-от 50 до 170); каждая такая личинка
имеет сравнительно крупные размеры.
12.19. Изучите рис. 12.34, где представлена доля
цветков в общей биомассе на корню у трех видов
золотарника (Solidago) на востоке США. На
рисунке указано, где растет каждый из этих видов - в
лесу или на открытых сухих местах (в
нарушенных местообитаниях на ранних стадиях
сукцессии):
а. Какие виды расходуют на размножение больше
биомассы?
б. Какие виды в большей степени подвержены
г-отбору?
в. Какие виды в большей степени подвержены
К-отбору?
г. Какой общий вывод можно сделать, сравнив
интенсивность цветения S. speciosa в лесу и на
открытых сухих участках?
12.6.5. Колебания и регуляция
численности популяций
В разд. 12.6.3 мы рассмотрели рост популяции.
После того как начальная фаза этого роста
закончилась, размеры популяции продолжают
колебаться от поколения к поколению. На рис. 12.35
показаны колебания численности гусениц зимней
пяденицы {Operophthera brumata) в дубовом лесу
возле Оксфорда.
Существенное влияние могут оказывать как
изменения климатических условий (например,
температуры), так и пищевые ресурсы, враги и т.п.
Иногда колебания происходят регулярно, и их
можно назвать циклами. Изучать их трудно, для
этого требуется длительное время, так как данные о
диких организмах часто приходится собирать в
течение нескольких лет. В ряде случаев для
получения данных использовались лабораторные
организмы с короткими жизненными циклами,
например дрозофилы, крысы или клещи. Их
популяции изучаются как модели, так как условия в
лаборатории можно контролировать более строго,
чем в поле.
Размеры популяции могут быть непостоянными в
результате изменений плодовитости или
смертности, а иногда того и другого. При изучении
размеров популяций и их изменений обычно стараются
отыскать ключевой фактор, т. е. фактор,
ответственный за наибольшую часть наблюдаемых сдвигов,
происходящих при смене поколений. В большинстве
исследованных случаев это фактор, влияющий на
смертность.
Хотя можно было бы ожидать, что колебания
размеров популяций будут носить чисто случайный
характер, в действительности есть ряд факторов,
удерживающих величину популяции в некоторых
пределах. Это факторы, которые сокращают
численность, способствуя повышению смертности или
снижению плодовитости. Они действуют более
эффективно при увеличении плотности популяции, т. е. это
факторы, зависимые от плотности. Иногда такими
факторами могут быть нехватка пищи или
возрастание численности врагов. Их непосредственное
влияние на смертность очевидно. К двум хорошо
изученным регулирующим механизмам, влияющим
на плодовитость, относятся территориальное
поведение и физические последствия перенаселения.
Территориальное поведение
Территориальное поведение, или территориальность,
встречается у широкого круга животных, в том
числе у некоторых рыб, рептилий, птиц,
млекопитающих и общественных насекомых. Это явление
особенно хорошо изучено у птиц. Один самец или
же самец и самка, образующие пару, могут
закрепить за собой гнездовую территорию, которую
они будут защищать от вторжения особей того же
вида. С помощью пения, а иногда визуальной
демонстрации (для которой служит, например,
красная грудь зарянки) птица утверждает свои права на
территорию, и незваные гости обычно удаляются,
иногда после короткой «ритуальной драки», в
которой конкуренты не причиняют друг другу
серьезного вреда (разд. 16.8.6); это имеет очевидные
преимущества над «настоящими» боями. Соседние
территории особей одного и того же вида совсем
или почти не перекрываются и обычно
обеспечивают достаточное количество пищи для родителей и
их птенцов. С ростом популяции отдельные
территории становятся меньше и на них может обитать
все меньшее число новых птиц. В крайних случаях
некоторым птицам не удается закрепить за собой
территорию, и они поэтому не размножаются.
Таким образом, регуляция здесь обусловлена
пространственными ограничениями.
Перенаселенность
Еще одна форма регуляции численности, в которой
большую роль играет пространство,-это действие
перенаселенности. Как показали лабораторные
эксперименты с крысами, когда плотность
популяции достигает определенной величины,
плодовитость животных сильно снижается, даже если нет
недостатка в пище. Происходят различные
гормональные сдвиги, которые влияют на половое
поведение; все чаще встречается неспособность к
спариванию, бесплодие, выкидыши, поедание
детенышей родителями. Родительская забота о потомстве
ослабевает, и детеныши покидают гнездо в очень
раннем возрасте, что снижает вероятность их
выживания. Усиливается агрессивность животных.
Подобные изменения характерны для ряда
млекопитающих и могут происходить также в природных
условиях вне лаборатории, например в природных
популяциях полевок.
Существует много примеров зависимости между
успешностью размножения и плотностью
популяции. Циклические изменения размеров популяции и
зависящего от ее плотности размножения были
выявлены у большой синицы в лесу Марли вблизи
Оксфорда (рис. 12.36).
Лабораторными исследованиями доказано, что
количество яиц, откладываемых плодовыми
мушками (Drosophila melanogaster) за день, уменьшается
с ростом плотности популяции. У растений число
семян, образующихся на каждой особи, тоже может
Числа от 47 до 63 означают
годы от 1947 до 1963
Число оперившихся птенцов
на размножающуюся пару
Число размножающихся взрослых особей
Рис. 12.36. Зависимость успеха размножения у
большой синицы в лесу Марли возле Оксфорда от
числа размножающихся взрослых пар. Данные
представлены за период с 1947 по 1963 год. (Из
M. E. Solomon (1976). Population dynamics. 2nd ed..
Studies in Biology № 18. Arnold.)
уменьшаться при возрастании плотности
популяции. В таких случаях не всегда ясно, какие именно
факторы играют регулирующую роль, но такими
факторами, несомненно, могут быть пищевые
ресурсы, конкуренция за территорию, физическое
беспокойство, которое доставляют друг другу
животные и которое может приводить к
гормональным сдвигам.
Регулирующие факторы помогают сглаживать и
компенсировать влияние других факторов
(например, климатических), которые оказывают
случайное, нерегулирующее действие на размеры
популяции. Например, очень низкий уровень размножения
больших синиц в лесу в 1947 г. (рис. 12.36),
возможно, был вызван высокой смертностью во время
холодной зимы 1946-1947 гг.; но падение
численности компенсировалось сравнительно
интенсивным размножением в 1947 и 1948 гг. Однако это
успешное размножение обеспечивалось,
по-видимому, целым комплексом причин.
Как уже отмечалось, ключевым фактором,
обусловливающим размеры популяции, часто бывает
фактор, влияющий на смертность. Это хорошо
видно на примере зимней пяденицы, жизненный цикл
которой показан на рис. 12.37. В дубовом лесу
около Оксфорда многие годы велись наблюдения за
ее численностью. В табл. 12.12 приведены средние
значения для числа особей, погибших под действием
каждого из шести факторов, влияющих на
смертность.
Как видно из таблицы, в среднем каждая пара
взрослых особей оставляет после себя двух
выживающих потомков, но, как показывает рис. 12.38,
в действительности размер популяции из года в год
варьирует. Если предположить, что плодовитость
постоянна, то по меньшей мере один из шести
факторов, влияющих на смертность, должен
изменяться. Он должен зависеть от плотности, так как
средняя величина популяции остается постоянной.
На рис. 12.38 показано, как факторы, влияющие на
смертность, изменялись в период с 1950 до 1961 г.
Ключевым фактором, влияющим на общую смерт-
Самки бескрылые и
влезают на ветви дуба ночью
Самки откладывают
яйца на ветвях дуба и
погибают
Самцы имеют
нормальные
крылья
Имаго
Яйцо
Активны в ноябре
и декабре (поэтому
"зимняя пяденица")
Взрослые особи
появляются в
ноябре и живут,
всего лишь
несколько
дней
Дек.
Янв.
Ноябрь
Февр.
Весной (в марте или
!апреле) из яиц появ·
[ляются зеленые гусе*·
^ницы; они питаются
^листьями дуба, затем
в мае спускаются
на тонкой нити на
землю, прячутся в
норках и окукливаются
Октябрь
Март
Гусеница
Сентябрь
Апрель
Август
Май
Июль
Июнь
Куколка
Рис. 12.37. Годичный жизненный
цикл пяденицы зимней. (Open
University Science Foundation Course
Unit 20 (1971). Open University Press.)
ность (в дорепродуктивном периоде), оказывается
зимняя гибель, так как именно зимой гибнет
большая часть особей, и это теснее всего коррелирует с
общей смертностью. Высокая смертность на этой
стадии развития объясняется тем, что вылупление
гусениц по времени не всегда точно совпадает с
распусканием почек и ростом молодых листьев
дуба. Если гусеницы выходят из яиц несколько
раньше, то многие гибнут от голода, а если
слишком поздно, то многие не успевают закончить свое
развитие до того, как листья станут непригодными в
пищу, т.е. жесткими с большим количеством
танина. Если же гусеницы выходят вовремя, то их
Таблица 12.12. Среднее число особей зимней пяденицы,
погибавших под действием шести факторов смертности11
Количество яиц, откладываемых самкой
Фактор смертности
Зимняя гибель (погибает часть яиц и
большое количество недавно вылупившихся
гусениц)
Мухи, паразитирующие на гусеницах
Другие паразиты гусениц
Болезни гусениц
Хищники, уничтожающие куколок в почве
(жуки, землеройки)
Осы, паразитирующие на куколках
Всего:
Число выживших взрослых особей,
способных размножаться
200
Число
погибших
184
1
1,5
2,5
8,5
0,5
198
2
11 По Open University SlOO Unit 20 См рис 12 37
популяция может достичь угрожающих размеров и
полностью уничтожить листья на деревьях.
Хотя зимняя гибель является ключевым
фактором, влияющим на смертность, размеры популяции
регулируются и какими-то другими факторами, так
как в большинстве лет колебания смертности были
менее разительны, чем колебания численности,
вызываемые зимней гибелью (исключение составляют
1960 и 1961 гг.). Таким образом, регулирующий
фактор способствует повышению смертности в те
годы, когда в результате уменьшения зимней гибели
популяция увеличивается. Иными словами,
регулирующий фактор зависит от плотности популяции.
Количество паразитов и хищников, уничтожающих
куколок, коррелирует с общей смертностью, и это
наводит на мысль, что именно оно служит
регулирующим фактором. Поедание куколок
увеличивается в годы, когда их численность высока (зимняя
гибель незначительна). Связь с паразитизмом более
сложная, поскольку увеличение плотности
популяции паразитов обычно не совпадает с ростом
плотности популяции хозяина. На куколках зимней
пяденицы паразитируют осы, личинки которых
питаются гусеницами. Паразит и хозяин связаны
сложными взаимоотношениями. Коротко можно
отметить, что увеличение плотности популяции
хозяина в данном году приводит к успешному
размножению паразитов и повышению плотности их
популяции в следующем году, и тогда соответственно
возрастает поражение куколок паразитами.
Существуют очень точные модели, позволяющие
прогнозировать численность зимней пяденицы.
Такого рода модели находят практическое
применение, когда изучаемый вид является вредителем, как,
например, зимняя пяденица в восточной Канаде.
Модель подсказывает способы борьбы с
вредителем, а в случае с зимней пяденицей она дала воз-
Общая смертность в
дорепродуктивном
периоде
Средний
уровень смертности ,
необходимый для того,
чтобы размер популяции
оставался неизменным
о
I
4)
Σ
О
Зимнее
исчезновение
Паразитизм
мух на гусеницах
Другие паразиты гусениц
Заболевания гусениц
Враги куколок хищники
паразиты
Годы
Рис. 12.38. Общая смертность в дорепродуктивном
периоде, влияние шести различных факторов на
общую смертность пяденицы зимней в Уитхемском
лесу в период с 1950 по 1961 год. (Open University
Science Foundation Course Unit 20 (1971). Open
University Press.)
Численность Eotetranychus
Жертва
Хищник
O
ί
1
I
Время, дни
Eotetranychus (жертва)
Typhlodromus (хищник)
Рис. 12.39. Колебания численности популяций
хищного клеща Typhlodromus и его жертвы -
растительноядного клеща Eotetranychus-в лабораторных
условиях. (M. К. Sands (1978) Problems in Ecology, Mills, Boon
Ltd.)
можность успешно прогнозировать результаты
интродукции паразитов как средства биологического
контроля.
Другой, зависимый от плотности фактор,
который может влиять на величину популяции,-это
миграция (или расселение). Например, у тлей при
высокой плотности популяции не только
замедляется размножение, но и у большей доли особей
развиваются крылья, что позволяет им покидать
растение, на котором они кормились.
Некоторые факторы, регулирующие величину
популяции, такие, как климат, обычно считались
независимыми от плотности, однако и они неизбежно
взаимодействуют с другими, зависимыми от
плотности факторами. Поэтому лучше не
разграничивать слишком резко эти две группы факторов, хотя
различать их иногда полезно. Следует
рассматривать отдельно каждый конкретный случай, учитывая
возможность сложных взаимодействий между
факторами того и другого типа. Изучение колебаний
численности и их регуляции - трудная область
экологии, и приведенные примеры служат только для
того, чтобы проиллюстрировать некоторые из более
очевидных путей воздействия этих факторов.
12.6.6. Взаимодействия между
популяциями
Редко изучают популяционную динамику только
одного вида. Как мы уже видели, для понимания
колебаний численности зимней пяденицы нужно
также кое-что знать о ее паразитах. Существует ряд
хорошо изученных типов взаимодействий между
популяциями различных видов (межвидовые
взаимодействия). На данном трофическом уровне это
может быть межвидовая конкуренция, т.е.
конкуренция между особями различных видов за
доступные ресурсы, например пищу и пространство.
Именно поэтому для экологии сообществ важно
изучение ниш. Популяции, находящиеся на разных
трофических уровнях, тоже могут
взаимодействовать - возможны, например, отношения типа
«хищник-жертва» и «хозяин-паразит». Существуют и
другие типы взаимоотношений; иногда они
трудноуловимы и сложны, как, скажем, некоторые сим-
биотические взаимоотношения, из которых оба
партнера извлекают пользу.
Взаимодействуют между собой и особи одного
вида (внутривидовые взаимодействия). Сюда
относятся территориальные отношения и другие формы
конкурентного поведения.
Взаимоотношения «хищник - жертва»
Для изучения межвидовых взаимодействий можно
создавать в лаборатории простые и легко
контролируемые ситуации, рассчитывая на то, что они
окажутся подходящими моделями реальных
ситуаций.
В качестве примера приведем простую и часто
используемую модель взаимоотношений «хищник-
жертва», реализуемую в экспериментах с двумя
клещами, один из которых хищный {Typhlodromus),
а другой травоядный {Eotetranychus). Ha рис. 12.39
показаны циклические колебания их численности.
Циклы этих двух видов несколько не совпадают по
фазе друг с другом.
Такие циклы можно объяснить следующим
образом: возрастание численности жертвы ведет к
увеличению популяции хищника, что в свою очередь
Численность популяции оленей, тыс.
Годы
Рис. 12.40. Изменения численности популяций оленей
на плато Кайбаб. вызванные хищничеством,
!-поддерживающая емкость среды в 1906 г; Il
поддерживающая емкость среды в 1940 г.
вызывает резкое снижение численности жертвы, а за
ним неизбежно следует уменьшение популяции
хищника; цикл замыкается, когда уменьшение числа
хищников позволяет снова вырасти числу жертв.
Каждый цикл включает несколько поколений.
Выводы, основанные на изучении таких лабораторных
моделей, можно лишь с большой осторожностью
переносить на отношения «хищник - жертва» в
природе, где могут действовать и другие факторы.
Однако регулярные циклические колебания
численности действительно характерны для популяций
некоторых травоядных млекопитающих. Например,
у леммингов в Северной Америке и Скандинавии
наблюдается четырехгодичный цикл. Поэтому
также циклически колеблется и численность их врагов -
песца и белой совы. Однако численность леммингов
периодически сокращается, скорее всего из-за
истощения кормовых ресурсов, а не из-за хищников.
Время от времени, когда популяции скандинавских
леммингов достигают исключительно высокой
плотности, они массами мигрируют, и многие при этом
гибнут в фиордах или тонут в реках. На циклические
изменения популяций других травоядных тоже
может в большей степени влиять наличие корма,
нежели хищники. Фактором, вызывающим колебания
численности, могут быть также болезни. Этот
фактор зависит от плотности популяций, так как
вероятность распространения эпизоотии возрастает с ее
увеличением. Судя по некоторым данным,
эпизоотии могут играть роль и в циклических колебаниях
популяции американского зайца-беляка в северной
Канаде. Здесь численность главного врага зайцев-
канадской рыси - колеблется сходным образом, но
со сдвигом по фазе. Зайцы составляют 80-90%
пищевого рациона рыси.
Хищники, несомненно, играют важную роль в
регулировании численности животных, хотя это,
возможно, и не единственный фактор. На примере
популяции оленей, обитающих на плато Кайбаб
в Аризоне, можно судить о важности отношений
«хищник - жертва» и о тех долговременных
преимуществах, которые эти отношения доставляют
популяции жертвы. В 1906 г. район плоскогорья был
объявлен заповедником, и, для того чтобы
защитить оленей от хищников (пумы, росомахи, койота)
последних систематически истребляли в течение
30 лет. До 1906 г. популяция оставалась стабильной
и состояла примерно из 4 тыс. особей, но
впоследствии произошел «демографический взрыв» (рис. 12.40),
и поддерживающая емкость среды, в которой по
оценкам могла существовать 30-тысячная
популяция оленей, оказалась недостаточной. В 1924 г.
численность оленей достигла 100 тыс. особей, т.е. рост
ее шел по J-образной кривой. Однако в результате
истощения кормовых ресурсов вскоре начался голод,
который вместе с болезнями привел к резкому
падению численности популяции. Растительность
была серьезно повреждена и уже не восстановилась
до уровня 1906 г.; в результате поддерживающая
емкость среды снизилась, и кормом теперь могло
быть обеспечено только 10000 оленей.
Взаимоотношения «хозяин-паразит»
Иногда наблюдаются циклические колебания
численности паразита и хозяина. Фазы циклов, как и в
примерах, описанных выше, не совпадают. Это
особенно характерно для случаев паразитизма одних
насекомых на других.
Межвидовая конкуренция
В экосистеме между популяциями возможна
конкуренция за любой из доступных ресурсов, например
за пищу, пространство, свет или убежища. Если два
вида находятся на одном трофическом уровне,
между ними весьма вероятна конкуренция за пищу. Со
временем адаптивная эволюция одного или обоих
видов может привести к тому, что каждый вид
в пределах одного трофического уровня будет
занимать отдельную нишу и конкуренция таким
образом будет сведена к минимуму. В других случаях,
когда конкурирующие виды занимают одинаковые
или сильно перекрывающиеся ниши, может быть
достигнуто равновесие, при котором ни один из
видов не будет развиваться столь же успешно, как
в отсутствие конкурента, или же численность одного
из видов начнет снижаться и он будет полностью
вытеснен. Последнее явление называют
конкурентным исключением. Изучать это явление в популяциях
диких организмов очень трудно, но был проведен
ряд классических экспериментов с лабораторными
популяциями. Первая такая работа принадлежит
советскому биологу Гаузе (1934), который изучал
конкурентные отношения между несколькими
видами Paramecium. Некоторые результаты этой работы
показаны на рис. 12.41.
Плотность популяции
Время, дни
Paramecium aurelia
Paramecium caudatum
Рис. 12.41. Рост численности двух
видов Paramecium. А. При
выращивании видов отдельно. Б. При
выращивании в одной культуре.
12.20. Рассмотрите рис. 12.41.
а) Какой тип кривой роста популяции характерен
для каждого из двух видов, когда они растут по
отдельности?
б) За какие ресурсы идет конкуренция между
двумя видами при их росте в одной культуре?
в) Какие факторы обеспечивают преимущество
P.aurelia в конкуренции с P.caudatum?
При выращивании двух видов инфузорий в одной
культуре вид P. aurelia оказывается более
конкурентоспособным в захвате пищи, чем P. caudatum. Через
пять дней численность P. caudatum начинает
уменьшаться, спустя примерно 20 дней этот вид
полностью исчезает, т.е. происходит его конкурентное
исключение. P. aurelia достигает стационарной фазы
роста позднее, чем при выращивании в отдельной
культуре. Хотя этот вид и более
конкурентоспособен, на него тоже отрицательно влияет конкуренция.
Это помогает объяснить давление отбора,
направленное на адаптацию конкурирующих видов к
отдельным нишам. В природных условиях менее
конкурентоспособный вид редко исчезает полностью-
просто его численность уменьшается, а иногда
может даже снова возрасти, прежде чем установится
равновесное состояние.
Принцип конкурентного исключения (или принцип
Гаузе) в дальнейшем был подтвержден другими
экспериментами на животных. Конкурентное
исключение наблюдается в популяциях растений,
например в смешанных культурах видов ряски (Lemna);
L. gibba способна вытеснять L. polyrrhiza.
Природные популяции изучать труднее, так как
здесь одновременно взаимодействует большее число
популяций, а параметры окружающей среды, такие,
как температура, влажность и пищевые ресурсы, не
поддаются контролю.
В популяциях растений одной из форм
конкурентных взаимодействий, вызывающей большой
интерес, является аллелопатия. Это частный случай
более общего явления - выработки растениями и
микроорганизмами разнообразных сложных
органических молекул, влияющих на рост других живых
организмов. Сюда входят антибиотики и
ингибиторы роста, такие, как пенициллин, который
вырабатывается грибом Penicillium и действует как
антибиотик на грамположительные бактерии (см. разд.
2.2.2). Химическая конкуренция между
микроорганизмами весьма сильна и взаимоотношения между
ними очень сложны. Когда выделяемые вещества
влияют на другие конкурирующие организмы того
же трофического уровня, такое взаимодействие
называют аллелопатией. Иногда при этом
используются летучие ароматические вещества - вторичные
метаболиты, выделяемые некоторыми растениями.
Например, Мюллер в 1966 г. показал, что летучие
терпены, выделяемые пахучими растениями чапа-
раля (тип кустарниковых зарослей) в Калифорнии,
адсорбируются на частицах почвы и задерживают
прорастание или подавляют рост соседних
растений. Фенольные соединения, которые
выщелачиваются в почву из подстилки некоторых растений,
оказывают такое же действие. Многие растения
меловых лугов являются ароматическими и
поэтому плохо поедаются травоядными. Это служит
дополнительным фактором в сохранении богатого
разнообразия травянистых растений. Недавнее
изучение аллелопатических взаимодействий позволяет
предположить, что они широко распространены как
в пределах одного трофического уровня, так и
между разными уровнями. К важным веществам,
выделяемым растениями, относятся фенол ьные
соединения, терпеноиды и алкалоиды. Интересно, что они
часто небезразличны и для животных. Примером
животного, извлекающего пользу из таких веществ,
может служить бабочка-данаида Danaus plexippus.
Ее гусеницы кормятся на ваточнике - растении,
которое содержит вещества, действующие на
позвоночных как сильный сердечный яд. Полагают, что
Рис. 12.42. Бобочка-данаида (Danaus plexippus).
ядовитость или неприятный вкус этого растения
предохраняет его от поедания травоядными
животными. Однако гусеницы данаид устойчивы к яду,
они могут накапливать его в себе и сохранять после
превращения во взрослую форму, и это в свою
очередь защищает бабочек от поедания птицами.
Яркая и характерная расцветка крыльев у данаиды
служит предостережением для потенциальных
врагов, т.е. действует как защитное приспособление
(рис. 12.42). Побочным следствием оказалось то,
что у некоторых других, уже неядовитых бабочек
выработалась подражательная окраска, которая им
тоже служит защитой от хищников. Этот случай
показывает, какими сложными могут быть
взаимодействия между видами. Растения нередко
содержат токсичные химические вещества, к которым
нечувствительны лишь немногие виды травоядных
животных.
Два других важных процесса, в которых
происходит взаимодействие между цветковыми растениями
и животными, - это распространение семян и
опыление (разд. 20.2). Последнее служит хорошим
примером распространенного явления, называемого
коэволюцией, когда виды приспосабливаются друг к
другу и со временем вырабатывают различные
адаптации, обусловливающие взаимную зависимость и
выгоду. В случае с ядовитыми растениями
наиболее полезным и надежным приспособлением для
устойчивых к яду животных может стать
«обслуживание» этих растений, например опыление их,
теми бабочками, гусеницы которых питаются
данным растением.
Глава 13
Количественная экология
Как указывалось в предыдущей главе, в основе
законов экологии лежат качественные и
количественные данные, полученные при изучении животных,
растений, микроорганизмов и абиотической среды.
В этой главе представлены как качественный, так и
количественный аспекты экологических
исследований и даны общие сведения относительно
некоторых методов и способов сбора, представления и
анализа результатов изучения абиотического и
биотического компонентов окружающей среды.
Прежде чем приступить к любому
экологическому исследованию, необходимо четко определить его
цели и задачи, а также требуемую степень точности.
Это в свою очередь позволит выбрать необходимые
методы исследования и обеспечит сбор данных,
достаточных для получения обоснованных
выводов. Часто это позволяет упростить используемые
методы, сократить время, деньги, ресурсы и усилия,
требуемые для проведения исследования. Однако
следует подчеркнуть, что нередко ход исследований
может изменяться в зависимости от того, какие
проблемы возникают по мере изучения объекта.
13.1. Методы измерения
факторов окружающей
среды
Для того чтобы дополнить анализ биотического
компонента, необходимо изучить основные
факторы окружающей среды - эдафические,
топографические и климатические (такие, как вода, влажность,
температура, свет и ветер). Ниже при описании
экспериментов приводятся различные методы
измерения факторов окружающей среды. Другие
методы количественного анализа описаны лишь в
общих чертах.
13.1.1. Эдафические факторы
Почвы значительно различаются по своей структуре
и химическому составу (см. разд. 12.4). Для
получения общего представления о структуре или
профиле почвы почвенный разрез делают таким
образом, чтобы он был строго вертикальным и были
четко видны отдельные слои. При этом можно
непосредственно измерить толщину ясно
различающихся по цвету и структуре горизонтов и отобрать
из них образцы для проведения различных
анализов, описанных ниже.
При работе с почвенным буром, который
представляет собой удлиненный инструмент с
пробковым винтом, бур ввинчивается в грунт на нужную
глубину и затем вынимается. Почву, попадающую в
резьбу винта с разных глубин, раскладывают по
отдельным полиэтиленовым пакетам для
проведения последующих анализов. При отборе почвенных
образцов данным методом необходимо записывать,
на какой глубине находился каждый взятый
образец. Эта запись должна быть сделана на
соответствующих пакетах.
Опыт 13.1. Определение содержания
воды в почвенном образце
Материалы и оборудование
Примерно 80 г почвы
Противень из алюминиевой фольги
Весы с точностью до 0,1 г
Сушильный шкаф с регулируемой температурой
Термометр с показаниями до 1500C
Эксикатор
Щипцы
Метод
1. Взвесьте пустой противень из алюминиевой
фольги. Запишите его массу (а).
2. Насыпьте в противень размельченный почвенный
образец и взвесьте. Запишите его массу (Ь).
3. Поместите противень с почвой на 24 ч в
сушильный шкаф при температуре 1100C.
4. Выньте образец из сушильного шкафа и охладите
его в эксикаторе.
5. Взвесьте остывший образец, запишите его массу.
6. Снова поместите образец на 24 ч в сушильный
шкаф при температуре 1100C.
7. Повторяйте операции 4 и 5 до тех пор, пока
результаты измерения не окажутся одинаковыми
(до постоянной массы). Запишите эту массу (с).
8. Рассчитайте процентное содержание воды по
следующей формуле:
Ъ-с
Ъ-а
χ 100.
9. До проведения опыта 13.2 храните почвенный
образец в эксикаторе.
Примечание
Полученное значение выражает процентное
содержание всей воды, присутствующей в образце. Эта
величина будет зависеть от количества осадков,
выпавших за последнее время. Альтернативными
оценками содержания воды являются полевая вла-
гоемкость и содержание доступной воды. Полевая
влагоемкость - это количество воды, сохраняющееся
в почве после того, как ее избыток дренируется под
влиянием гравитационных сил. Для получения этого
значения необходимо, чтобы почва в полевых
условиях была целиком затоплена в течение нескольких
минут. Затем через 48 ч можно отобрать образец
для проведения исследования. Доступная вода-это
та вода, которая может поглощаться растениями.
Ее содержание можно определить высушиванием
взвешенного образца при комнатной температуре
до постоянной массы. Разность масс влажной и
сухой почвы равна содержанию доступной воды.
Опыт 13.2. Определение содержания
органического вещества
(гумуса) в почвенном образце
Материалы и оборудование
Высушенный почвенный образец (из эксперимента
13.1) в эксикаторе
Тигель с крышкой
Штатив, бунзеновская горелка, асбестовая сетка,
огнеупорный треугольник
Эксикатор
Щипцы
Метод
1. Прокалите тигель и крышку на пламени горелки
для удаления остатков влаги. Для охлаждения
поместите в эксикатор. Взвесьте и запишите
массу (а).
2. Насыпьте из эксикатора в тигель высушенный
почвенный образец (возьмите из предыдущего
эксперимента) и взвесьте. Запишите массу (Ь).
3. Накалите тигель с почвенным образцом,
накрытым крышкой, до красного цвета и прокаливайте
в течение часа до полного сгорания всего
органического вещества. Остудите на воздухе 10 мин
и поместите в эксикатор.
4. Взвесьте остывший тигель с образцом.
5. Повторяйте операции 3 и 4 до получения
постоянной массы.
6. Подсчитайте процентное содержание
органических веществ по формуле:
Ъ-с
Ъ-а
χ 100.
7. Проведите этот эксперимент с почвенными
образцами, отобранными в разных районах, чтобы
показать различия в содержании органических
веществ.
Примечание
Полученная в этом эксперименте величина
соответствует процентному содержанию органических
веществ в сухой почве. Получить величину,
соответствующую содержанию органических веществ во
влажной почве, можно, используя данные опыта
13.1.
13.1. При анализе 60 г влажного почвенного
образца были получены следующие данные. После
неоднократного нагревания до 1100C и
охлаждения в эксикаторе почвенный образец приобрел
постоянную сухую массу, равную 45 г. Затем
сухой почвенный образец несколько раз
прокаливали в тигле до красного цвета, охлаждали в
эксикаторе и взвешивали. После чего его масса
стала равна 30 г. Подсчитайте содержание воды и
органических веществ во влажном почвенном
образце.
Опыт 13.3. Определение содержания
воздуха в почвенном образце
Материалы и оборудование
Жестяная банка емкостью примерно 200 см3
Стакан емкостью 500 см3
Вода
Стеклограф
Металлический снаряд
Методика
1. Поместите пустую банку открытым концом
вверх в стакан емкостью 500 см3 и наполните
стакан водой выше края банки. Отметьте уровень
воды в стакане.
2. Осторожно выньте банку, наполненную водой, и
измерьте объем воды мерным цилиндром.
Запишите объем (а). Уровень воды в стакане упадет
на величину, равную объему воды в банке.
3. Просверлите примерно восемь маленьких
отверстий на дне банки.
4. Банку открытым концом проталкивайте в почву,
с поверхности которой удалена растительность,
до тех пор пока почва не начнет выходить из
отверстий. Осторожно выкопайте банку,
переверните ее и удалите лишнюю почву,
находящуюся выше края банки.
5. Снова осторожно поместите банку с почвой
открытым концом вверх в стакан с водой и
разрыхлите почву снарядом, чтобы вытеснить из нее
воздух.
6. Уровень воды в стакане станет ниже, чем раньше,
потому что часть воды израсходуется на заме-
щение воздуха, который содержался в почве.
7. Из полного мерного цилиндра емкостью 100 см3
добавьте воду в стакан до прежнего уровня.
Запишите объем добавленной воды (Ь).
8. Процентное содержание воздуха в почвенном
образце можно определить по формуле:
Ъ
а
χ 100.
9. Повторите опыт с почвенными образцами из
разных районов.
Опыт 13.4. Определение
приблизительного
соотношения твердых частиц
в почвенном образце
(текстура почвы)
Материалы и оборудование
Мерный цилиндр емкостью 500 см3
100 см3 почвенного образца
300 см3 воды
Методика
1. Поместите почвенный образец в мерный цилиндр
и полностью залейте водой.
2. Энергично встряхните содержимое.
3. Для того чтобы частицы осели в соответствии с
их плотностью и площадью поверхности, дайте
смеси отстояться в течение 48 ч.
4. Измерьте объем различных фракций почвенного
образца.
Результаты
Наблюдается послойное распределение
составляющих почву частиц. Частицы органического
вещества плавают на поверхности воды, частицы
глины частично остались во взвешенном состоянии, а
более крупные осели в виде слоя поверх песка и
камней, распределившихся в соответствии со
своими размерами.
Опыт 13.5. Определение рН почвенного
образца
Материалы и оборудование
Длинная пробирка (145 мм) с пробкой
Штатив для пробирок
Сульфат бария
Раствор универсального индикатора и цветная
таблица
Почвенный образец
Шпатель
Дистиллированная вода
Пипетка на 10 см3
Методика
1. Поместите в пробирку примерно 1 см3 почвы
и 1 см3 сульфата бария, в результате чего
коллоидная глина выпадет в виде хлопьев.
2. Добавьте 10 см3 дистиллированной воды и 5 см3
раствора универсального индикатора. Закройте
пробирку пробкой, энергично встряхните и дайте
содержимому отстояться в течение 5 мин.
3. По цветной таблице сравните цвет жидкости в
пробирке с цветами индикатора и определите
соответствующее рН.
4. Повторите эксперимент с почвенными образцами
из разных районов.
Примечание
При исследовании почвы рН является одной из
наиболее важных характеристик. Несмотря на
простоту определения, значение рН зависит от
множества взаимодействующих факторов и служит
хорошим показателем содержания питательных
веществ в почве; кроме того, величина рН указывает
на то, какие виды растений (и соответственно
животных) могут успешно развиваться на данных
почвах. Кислые почвы, как правило, менее богаты
питательными веществами, поскольку в меньшей
степени способны удерживать катионы.
13.1.2. Климатические факторы
Вода, воздух и свет относятся к наиболее важным
климатическим факторам. В этом разделе кратко
описаны некоторые основные методы их измерения.
Опыт 13.6. Определение рН воды
Материалы и оборудование
Универсальная индикаторная бумага или рН-
метр
Проба воды
Методика
1. Опустите в исследуемую воду кусочек
универсальной индикаторной бумаги и сравните ее цвет
с цветной шкалой. Определите значение рН.
ИЛИ
2. Ополосните зонд рН-метра дистиллированной
водой, опустите его в пробу исследуемой воды и
снимите показания рН (этот метод более точен,
но до начала эксперимента необходимо
тщательно проверить рН-метр, используя готовые
растворы с известным значением рН). Вновь
ополосните зонд дистиллированной водой перед тем,
как поместить его на хранение в буферный
раствор.
3. Повторите опыт с пробами воды из различных
источников.
Опыт 13.7. Определение содержания
хлоридов в пробе
воды (приблизительная
оценка солености)
Материалы и оборудование
Проба воды
Пипетка объемом 10 см3
Бюретка
Дистиллированная вода в бутыли
3 конические колбы
Белая кафельная плитка
Калий-хроматный индикатор
50 см3 раствора нитрата серебра (2,73 г на 100
см3)
Методика
1. Налейте 10 см3 исследуемой воды в коническую
колбу и добавьте 2 капли калий-хроматного
индикатора.
2. Из бюретки оттитруйте раствором нитрата
серебра, постоянно встряхивая коническую колбу.
3. В конечной точке титрования осадок хлорида
серебра окрашивается в красный цвет.
4. Дважды повторите титрование с 10 см3
исследуемой воды. Подсчитайте среднее количество
израсходованного нитрата серебра.
5. Объем израсходованного раствора нитрата
серебра приблизительно равен содержанию
хлоридов в пробе воды (в г на 1л).
Опыт 13.8. Определение содержания
кислорода в пробе воды
Метод Винклера, который описан в этом разделе,
позволяет точно измерить содержание кислорода,
но для того чтобы пользоваться этим методом,
необходимы многочисленные реактивы.
Материалы и оборудование
10 см3 щелочного раствора иодида (3,3 г NaOH;
2,0 г KCl в 10 см3 дистиллированной воды)
(Обращаться с осторожностью!)
10 см3 раствора хлорида марганца (4,0 г MnCl2 в
10 см3 дистиллированной воды)
5 см3 концентрированной хлористоводородной
кислоты (Обращаться с осторожностью!)
Раствор крахмала (как индикатор)
Дистиллированная вода в бутыли
0,01 M раствор тиосульфата натрия
3 χ 5 см3 пипетки с делениями
Бюретка
Белая кафельная плитка
3 конические колбы
250 см3 исследуемой воды в стеклянном сосуде с
притертой пробкой
Методика
1. Осторожно, не расплескивая, наберите воду в
бутылку и оставьте ее под водой, чтобы в нее не
попадали пузырьки воздуха.
2. В пробу воды пипеткой добавьте 2 см3 хлорида
марганца и 2 см3 щелочного раствора иодида;
конец пипетки должен касаться дна бутылки.
Более тяжелый раствор солей вытеснит из
бутылки равное количество воды, находящейся в
верхнем слое.
3. Добавьте 2 см3 концентрированной
хлористоводородной кислоты и закройте бутылку так,
чтобы в ней не было пузырьков воздуха. Хорошо
потрясите бутылку для того, чтобы растворился
осадок. В результате образуется раствор в
избытке иодида калия. Теперь растворенный
кислород зафиксирован и бутылку можно вынуть из
воды.
4. Для исследования отлейте в коническую колбу
50 см3 воды. Из бюретки оттитруйте ее 0,01 M
раствором тиосульфата натрия следующим
образом:
а) постоянно встряхивая коническую колбу,
доливайте в нее раствор тиосульфата до тех пор,
пока желтый цвет не побледнеет;
б) добавьте 3 капли раствора крахмала и
продолжайте титровать, постоянно встряхивая колбу,
до тех пор, пока не исчезнет темно-синяя окраска
крахмала.
Запишите объем израсходованного тиосульфата
натрия.
5. Дважды повторите операцию 4 с 50 см3
исследуемой воды и вычислите средний объем
расходуемого тиосульфата (х).
6. При использовании этих растворов 1 см3 0,01 M
тиосульфата соответствует 0,056 см3 кислорода в
условиях НТД (нормальная температура и
давление).
7. Подсчитайте содержание кислорода в литре
воды, используя следующую формулу:
Содержание кислорода, см3/л =
0,056 ххх 1000
50
где χ -объем тиосульфата, расходуемый на
титрование 50 см3 воды.
Течение
Самым простым методом определения скорости
течения воды служит измерение времени,
необходимого какому-нибудь плавающему предмету для
того, чтобы пройти определенное расстояние.
Чтобы исключить влияние ветра, лучше использовать
предмет, который большей своей частью погружен в
воду. Или же поместить в поток воды L-образную
трубку высотой 50 см, длиной 10 см и диаметром
2 см таким образом, чтобы ее короткий конец был
обращен навстречу течению. Измерив высоту, на
Рис. 13.1. Аспирационный психрометр
которую вода поднялась в длинном конце трубки,
можно определить скорость течения, используя
следующую формулу:
ν = Jlhg
где ι;-скорость течения воды, см/с;
#-ускорение силы тяжести (981 см/с2);
Л-высота столба воды, см.
Влажность
Относительная влажность воздуха-это отношение
абсолютной влажности, выраженной в г/м3 к
максимальному содержанию водяных паров в воздухе;
относительная влажность выражается в процентах.
Относительная влажность изменяется с
температурой, так как воздух при нагревании расширяется и
может удерживать больше водяных паров. Их
содержание измеряется психрометром, состоящим из
влажного и сухого термометров, которые
закреплены в деревянной раме (рис. 13.1). По показаниям
термометров вычисляют влажность воздуха,
пользуясь психрометрическими таблицами.
Температура
Температуру воздуха, воды и почвы можно
измерить ртутным термометром. Однако определение
температуры в данной точке и в данное время имеет
небольшое значение для экологических
исследований. Более информативным является изменение
температуры за определенный период времени.
Поэтому обычно температуру измеряют в разное
время (по сложной временной схеме) или же
используют максимальный и минимальный термометры,
регистрирующие максимальную и минимальную
температуры соответственно. В
микроместообитаниях и труднодоступных местообитаниях, например
в сердцевине древесного ствола, температуру
измеряют термистором (рис. 13.2). Это электрический
Рис. 13.2. Термистор во
время работы.
прибор, который может иметь столь малые размеры,
что его можно вмонтировать в кончик шариковой
ручки. Электрическое сопротивление этого прибора
меняется с изменением температуры. Измеряя
сопротивление термистора и сравнивая его с ранее
составленной таблицей, где каждому
сопротивлению соответствует определенная температура,
можно установить температуру среды, в которую
помещен термистор. При проведении экологических
исследований полезно также определять
максимальные и минимальные температуры
микроместообитаний (микроклимат), так как часто именно
этим объясняется исчезновение из определенных
районов отдельных видов, например растений,
чувствительных к морозам.
Свет
Свет различается по своей интенсивности,
продолжительности и качественному составу (длине
волны). Чтобы получить полную информацию об этом
физическом факторе при проведении экологических
исследований, необходимо измерять все три
перечисленных параметра света. Для их измерения
используются специальные методы. В практических
целях для сравнения интенсивности падающего
света в различных районах обычно требуется некий
отнесенный к данной площади показатель
интенсивности света. Для этого достаточно обычного
фотоэкспонометра. Суммарная интенсивность света
за определенный период времени определяется с
помощью бумаги Ozalid, которая обладает
кумулятивной чувствительностью к свету.
Направление и скорость ветра
С экологической точки зрения важна не столько
скорость ветра в местообитании в данный момент
времени, сколько степень открытости
местообитания для ветров. Следовательно, важна частота
ветров, их интенсивность и направление. Однако для
большинства практических целей при сравнении
особенностей ветра в различных местообитаниях
достаточно простого прибора, показывающего
направление ветра и анемометра (рис. 13.3),
регистрирующего скорость ветра.
13.2. Биотический анализ
При исследовании организмов, населяющих данное
местообитание (биотический компонент
экосистемы), необходимо охарактеризовать структуру со-
общества, т.е. определить встречающиеся в
местообитании виды и численность популяции каждого
вида. Совершенно очевидно, что выявить и
подсчитать все организмы данного вида невозможно,
поэтому применяется такой метод отбора образцов,
который позволяет определить присутствующие
виды и их численность. Как правило, для получения
более точных результатов используются методы,
требующие больших затрат времени, поэтому
необходимо ясно представить цель предстоящих
исследований. И, если это возможно, применять
методы, не нарушающие естественных сообществ.
Всегда необходимо использовать достоверные
методы отбора и регистрации образцов и, как
говорится, «ни один камень не должен остаться
нетронутым» (при условии, что его вернут на место),
потому что организмы занимают почти все
доступные микроместообитания. Например, на
первый взгляд может показаться, что квадратный метр
травостоя, почвы, песка, скалистого берега или дна
реки заселен небольшим числом видов, но при более
внимательном рассмотрении, если при этом
перебрать руками почву, растения, перевернуть камни и
исследовать корни, стебли, цветы и плоды растений
и прикрепляющуюся часть таллома морских
водорослей, то можно обнаружить намного больше
видов.
При записи данных все виды растений и
животных, поддающиеся определению, необходимо
определить прямо в поле, пользуясь, если нужно,
определителями. Собирать надо только те виды, о
которых известно, что они широко распространены
в данной местности и не относятся к редким видам.
Массовый сбор видов может иметь серьезные
последствия для местных сообществ. При отборе
животных нужно постараться сохранить их живыми и
выпустить в местообитания, сходные с теми, из
которых они были изъяты. Организмы необходимо
определять с максимально возможной точностью,
т. е. до видов. Часто это сделать невозможно, тогда
надо постараться установить их принадлежность к
определенному классу, порядку или семейству.
Определение образцов зависит от степени знакомства с
определительными таблицами. Принципы
классификации, построение определительных таблиц и
подробное объяснение того, как ими пользоваться,
приведены в Приложении 3 (т. 1).
Список всех видов, населяющих данное
местообитание, дает некоторое представление о
многообразии структуры сообщества, о его видовом
разнообразии или богатстве.
(Существуют различные способы выражения
видового богатства с помощью математических
формул. Числовое значение называется индексом
видового разнообразия; подробно этот вопрос здесь
рассматриваться не будет.)
С помощью этих данных можно построить
возможные пищевые цепи и пищевые сети, но для
количественной характеристики сообщества этого
недостаточно. Степень разнообразия может
считаться полностью выявленной только тогда, когда
установлена численность организмов каждого вида
(размер популяции). Эти данные дают возможность
построить более детальную картину сообщества,
например пирамиду численности (разд. 12.3.6).
Полученные качественные и количественные дан-
Вид сбоку
Пластмассовые
фильтровальные
воронки
Поддерживающий
шест
Штифт, на котором
вращается
измерительный прибор
Деревянное крыло
с прикрепленной
воронкой
Вид сверху
Рис. 13.3. Простейший
анемометр, с помощью которого можно
измерять скорость ветра, измеряя
частоту оборотов деревянного
крыла, окрашенного в черный цвет.
ные о местообитании зависят от особенностей
методов отбора образцов и оценки численности
организмов. Метод выбирают в соответствии с образом
жизни, поведением и размерами организма.
13.2.1. Методы сбора организмов
Существует несколько моментов, которые
необходимо учитывать при сборе организмов:
1. Следует изучить свод законов данной области,
вышедших за последнее время.
2. Прежде чем начать экологические исследования,
необходимо получить разрешение от владельца
данной территории.
3. Не лишне получить консультацию в местном
краеведческом обществе, университете или в
обществе охраны природы о том, где и что вам
собирать.
4. Никогда не выносите организмы за пределы
местообитания и не уничтожайте их без особой
надобности.
5. Оставляйте местообитание по возможности
ненарушенным, верните, например, на прежнее
место камни, дерн, бревна и т. п.
6. При необходимости отобрать организмы для
определения берите как можно меньшее число
особей и, если это возможно, верните их на
прежнее место.
7. Животных во время доставки в лабораторию для
определения держите отдельно друг от друга для
того, чтобы предотвратить их порчу или
поедание хищниками. Например, не помещайте в один
сосуд многощетинковых червей и крабов. Поль-
Таблица 13.1. Бланк полевого дневника для записи эда-
фических, физиологических и климатических особенностей
Район Координаты .... Дата . . .
1. Подстилающая порода
2. Субстрат (почва)
а) особенности поверхности
б) мощность горизонта А
в) мощность горизонта В
г) мощность горизонта С
д) рН
е) температура
3. Топография
а) экспозиция склона угол
б) высота над уровнем моря
в) рельеф
г) дренаж
д) использование земель
е) высокий или низкий уровень воды, время
высота
4. Климат
а) температура воздуха, диапазон температур ....
б) осадки
в) облачность (освещенность)
г) относительная влажность
д) направление ветра
е) скорость ветра
ж) интенсивность света (по горизонтам), N ,
S ,E. . . . , W
з) время дня
зуйтесь приспособлениями для сбора образцов, в
том числе стеклянными банками,
полиэтиленовыми бутылками, пробирками и
полиэтиленовыми пакетами.
8. Во время сбора образцов всегда записывайте как
можно больше сведений о топографии
местообитания и климатических условиях, так как эта
информация может иметь отношение к
собранному материалу. Следует регистрировать такие
особенности местообитания:
а) характер горной породы или субстрата
(травостой, ил, почва и т.д.);
б) характер местности (например, плоская
поверхность, обращена на юг, угол наклона и т. д.);
в) дренаж;
г) профиль почвы, ила или песка;
д) температура субстрата, воды и воздуха;
е) рН субстрата или воды;
ж) облачность и осадки;
з) относительная влажность воздуха;
и) интенсивность освещенности (например, тень
или открытое пространство, показания можно
снять измерительным прибором);
к) скорость и направление ветра (тихий, легкий
бриз, буря, юго-западное направление);
л) время дня и дата.
Пример того, как следует записывать некоторые
из этих особенностей, приведен в табл. 13.1.
Существует множество методов сбора образцов.
Эти методы и их применение суммированы в табл.
13.2 и на рис. 13.4-13.9.
Попавшие в ловушку организмы необходимо
регулярно вынимать, определять, подсчитывать и,
если возможно, выпускать на свободу. Если
ловушка в виде ямы, то при попадании в нее организмов,
один из которых-хищник, а другой - жертва,
вполне вероятно, что к моменту сбора материала
жертвы в ловушке не окажется. Учитывая это, на дно
ловушки нужно налить 70% спирта, в котором будут
погибать попадающие в него организмы. При сборе
образцов требуются воображение и
изобретательность.
Место для сбора материала обычно выбирают не
случайно. В дальнейшем результаты, полученные
при обработке собранной коллекции, можно
соответствующим образом интерпретировать, что
позволит выбирать места образцов. Хотя место сбора
может не влиять на состав коллекции видов и
структура сообщества будет отображена правильно,
но при этом могут быть получены несколько
искаженные данные, касающиеся численности видов.
Например, использование наживок и приманок,
привлекающих организмы в клейкие ловушки,
применение ловушек в виде ям и ловушек для
млекопитающих повлияют на результаты, а
основанные на количественных данных выводы будут
необъективны. Поэтому при обсуждении
результатов необходимо четко показать, что выбор был не
случаен.
Таблица 13.2. Сводная таблица различных методов сбора организмов
Метод сбора
Стряхивание насекомых
на полотно
Ловля сачком в воздухе
Кошение скачком
Планктонная сеть
Клейкая ловушка
Ловчий цилиндр
Световая ловушка
Живоловка
Ловля водным сачком
Эксгаустер (всасыва-
тель)
Ручной разбор
Экстрагирование
Последовательность действий и назначение
Матерчатую простыню определенного размера прикрепляют к
складному каркасу и держат под веткой, по которой бьют
палкой или которую встряхивают. Животные падают на
простыню, их собирают с помощью всасывателя (см. примечание
далее)
Сетку из муслина прикрепляют к рукоятке и машут ею в воздухе.
Животные попадают в сетку. Методику отлова нужно
унифицировать, чтобы обеспечить единообразие собранных
образцов. Например, выбирать из сетки животных после восьми
взмахов, причем каждый взмах должен описывать восьмерку
Сетку из нейлона прикрепляют к жесткой рукоятке и резко
проводят по траве, кустарникам, в водной толще пруда или
реки
Просеивающую шелковую сетку прикрепляют к металлическому
обручу, привязывают к веревке и протягивают по воде. К
задней стороне прикрепляют небольшую банку для сбора
образцов
Темную патоку и сахар кипятят вместе, затем намазывают на
лист толстого полиэтилена, кнопками прикрепляют его к
отрезку доски из прессованных опилок. Этот лист можно
подвешивать в различном положении на различной высоте. К
клейкой массе в качестве привлекающих веществ можно
добавить джем и пиво
Стеклянную или консервную банку зарывают так, чтобы ее край
находился на уровне земли. Лучше закрыть банку так, чтобы
вокруг края образовался небольшой скат, это предотвратит
попадание в нее воды. Кусок шиферной плитки, установленной
на трех камнях, служит крышей, защищающей от дождя. В
ловушку необходимо положить приманку, например джем или
гниющее мясо. Ловушку необходимо регулярно чистить
(рис. 13.4)
Свет ртутной лампы привлекает летающих животных, которые
бьются о перегородки, падают вниз и попадают в картонную
коробку или бумажный пакет. Прежде чем осматривать
содержимое корооки, поместите в нее шерстяную нить,
пропитанную хлороформом, чтобы анестезировать или убить
животных (рис. 13.5)
Ловушку для млекопитающих (живоловку) (рис. 13.6) оставляют
на тропе, ведущей к водопою, предварительно заполнив ее
сухой травой. Приманку, например крупу или сухие фрукты,
можно оставить и снаружи и внутри ловушки. Первое время,
пока животные не привыкли к ловушке, ее можно не заряжать,
а затем зарядить. Животных ловят живыми, поэтому ловушку
необходимо регулярно осматривать. Одни животные очень
осторожны и никогда не заходят в ловушку, тогда как другие
попадают в нее регулярно. Различное поведение организмов
создает определенные трудности при использовании данного
метода для оценки численности популяции
Этот метод применяют для отбора образцов в пресных
проточных водах. Открытый сачок или планктонную сетку ставят
ниже по течению того места, которое намереваются
обследовать; на обследуемом участке переворачивают камни и
соскабливают с них организмы, которые заносятся течением в
сачок. Или же вода в месте сбора образцов взбалтывается
ударами ноги, так что организмы поднимаются со дна и
течением загоняются в сачок
Этот метод применяется для сбора мелких насекомых с поддонов
или прямо с растений для проведения более тщательного
исследования (или подсчета) (рис. 13.7).
Образцы почвы или растений, например злаки, лесную подстилку
или сачок с морскими водорослями, кладут на край подноса и
небольшие порции материала методично перебирают
пальцами. Найденные организмы помещают в банку для образцов,
а перебранный материал отодвигают на другой конец подноса.
Каждый образец исследуют и помещают на прежнее место
5 см3 4%-ного формальдегида добавляют к 50 см3 воды и этим
раствором поливают квадратный метр газона или луга. После
того как земляные черви выползут, их собирают и сразу же
обмывают водой, чтобы смыть формальдегид
Собираемые организмы
Нелетающие
насекомые, личинки, пауки
Летающие насекомые
Насекомые,
ракообразные
Планктон
Летающие насекомые
Ползающие насекомые,
многоножки,
ракообразные
Ночные насекомые,
особенно бабочки и
ручейники
Землеройки, полевки,
мыши
Водные насекомые и
ракообразные
Тли, мелкие насекомые
и пауки
Клещи, энхитреиды,
черви, мелкие насекомые
и их личинки
Дождевые черви
Продолжение табл. 13.2.
Метод сбора
Флотация
Эклектер
Тульгрена (сухая
экстракция)
Воронка Бсрмана
(мокрая экстракция)
Последовательность действий и назначение
Взвешенный комок почвы помещают в лабораторный стакан с
насыщенным раствором соли, несколько минут энергично
взбалтывают, затем дают почве осесть В концентрированном
соленом растворе организмы всплывают на поверхность.
Поверхностный слой жидкости сливают в чашку Петри и
исследуют под бинокулярным микроскопом. Затем все организмы
помещают в чашку Петри с 70%-ным спиртом, для того чтобы
убить и зафиксировать их. Каждый образец отдельно кладут на
предметное стекло и каплю глицерина, накрывают покровным
стеклом и определяют под бинокулярным микроскопом или
при малом увеличении обычного светового микроскопа
Многие организмы, населяющие почву и подстилку, избегают
источников тепла и перемещаются на влажные участки.
Образец почвы или подстилки помещают в сито примерно на
расстоянии 25 см под металлическим рефлектором с лампой
мощностью 100 Вт (рис. 13.8). Каждые два часа лампу
продвигают к образцу на 5 см, пока расстояние между лампой и
образцом не станет 5 см. В таком положении рефлектор
оставляют на 24 ч. Все мелкие членистоногие перемещаются
вниз и сквозь металлическую сетку падают в сосуд со спиртом
Образец почвы насыпают в муслиновый мешок, опускают в
воронку с водой и подвешивают на расстоянии 25 см под
металлическим рефлектором с лампой мощностью 100 Вт (рис. 13.9).
Оставляют на 24 ч. Вода и слабый нагрев способствуют тому,
что организмы перемещаются из образца в воду и оседают на
дне воронки. Через определенные интервалы зажим на дне
воронки открывают, и организмы падают в сосуд со спиртом.
Собираемые организмы
Клещи, насекомые, яйца
личинки и куколки
Мелкие членистоногие,
например двупарно-
ногие, губоногие,
клещи, ногохвостки и
коллемболы
Мелкие членистоногие,
энхитреиды и
нематоды
Шиферная плитка
Уровень земли
Поддерживающие
плитку камни
Стеклянная банка
Почва
Приманка,
привлекающая
насекомых и т. п.
Рис. 13.4. Простая ловушка типа «волчьей ямы».
Сделана из зарытой в землю стеклянной банки.
Рис. 13.5. Ртутная лампа для привлечения
насекомых .
13.2.2. Методы сбора образцов
на исследуемой территории
Для того чтобы унифицировать участки, на которых
проводятся исследования абиотического и
биотического компонентов экосистемы, обычно
закладывают трансекты и(или) квадраты и сбор образцов
ограничивают их площадью.
линейная трансекта. Она может быть использована
для сбора образцов на однородной площади, но на
практике линейную трансекту закладывают тогда,
когда полагают, что в пределах исследуемой
площади происходит переход одних местообитаний и
популяций в другие (рис. 13.10). Натянутая над
землей между двумя столбами тесьма или веревка
показывает положение трансекты. Собирают только
те виды, которые действительно соприкасаются с
линией трансекты.
ленточная трансекта. Ленточная трансекта - это
проложенная через изучаемое местообитание полоса
заданной ширины, образованная двумя линейными
трансектами, протянутыми на расстоянии
0,5 м или 1,0 м друг от друга, между которыми
производится учет видов. Для получения как
качественных, так и количественных данных проще
закладывать ленточную трансекту, а использовать
линейную трансекту и квадратную раму.
Изменения высоты, зарегистрированные вдоль
линейной или ленточной трансекты, образуют
профиль трансекты, который вычерчивают при
оформлении данных (рис. 13.19).
Рис. 13.6. Ловушка для млекопитающих.
Выбор типа трансекты зависит от качественного и
количественного характера исследования, от
требуемой степени точности, особенностей населяющих
территорию организмов, размеров площади,
которую необходимо исследовать и наличия времени.
На небольших расстояниях можно закладывать
линейную трансекту и записывать каждое растение,
находящееся непосредственно под ней. При
исследовании участков большей протяженности можно
записывать виды через каждый метр или другой
подходящий интервал.
квадрат. Этот инструмент представляет собой
металлическую или деревянную квадратную раму
определенной площади, например 0,25 м2 или 1 м2
(рис. 13.11), желательно, для облегчения переноса,
разборную. Раму кладут по одну сторону трансекты
и исследуют площадь, заключенную внутри рамы.
Потом раму переносят вдоль линейной трансекты в
другие точки. В зависимости от характера
исследования можно либо регистрировать находящиеся
внутри рамы виды, либо оценивать их численность
или обилие (разд. 13.2.6). В любом случае метод
учета видов должен быть последовательным,
например учитываются все виды, частично или полностью
находящиеся внутри квадрата. В соответствии с
требованиями исследования квадратную раму
можно видоизменить. Например, бечевкой или
проволокой ее можно разделить на определенные секции,
помогающие при подсчете численности или оценке
обилия видов (рис. 13.11). Это особенно удобно,
когда исследуемое местообитание заселено
несколькими видами растений.
Квадрат можно использовать и отдельно от
трансекты, если исследуемое местообитание имеет
явно однородный характер. В этом случае квадрат
помещают случайным образом. Один из методов
совершенно случайного отбора образцов состоит в
том, что квадрат бросают через плечо и записывают
виды, оказавшиеся внутри квадрата в месте его
падения. Эту процедуру повторяют несколько раз,
пока не будет охвачена достаточно большая
площадь участка. Определить место сбора образцов
можно, также выбрав на воображаемой сетке,
лежащей на участке, случайные координаты; для
этого используют либо таблицы случайных чисел,
либо получают эти числа с помощью некоторых
типов калькуляторов. Стороны сетки можно
отметить рулеткой. Результаты исследований
показывают, что если местообитание однородно, то
наступает момент, когда закладывать новые квадраты
уже не имеет смысла, так как при этом не
происходит увеличения числа зарегистрированных видов.
Эта зависимость представлена на рис. 13.12. Как
правило, если в пяти последних квадратах не
окажется ни одного нового вида, можно допустить,
что больше видов обнаружено не будет. Но, если
такое предположение сделано, его необходимо
записать, так как это может повлиять на
достоверность результатов.
рама со спицей (точечный метод). Это рама с
несколькими отверстиями, через которые можно
пропустить «стержень», например спицу для
вязания (рис. 13.13). Рама особенно удобна при
исследовании вдоль трансекты местообитаний с сильно
Рис. 13.7. Всасывающее устройство, применяемое для сбора мелких беспозвоночных.
разросшейся растительностью, где различные виды
растений частично перекрывают друг друга. Спицу
пропускают через каждое отверстие, при этом
записываются все виды, соприкасающиеся со спицей
по мере того, как она опускается к земле.
постоянный квадрат. При длительных
экологических исследованиях, включающих изучение смен
сообществ (сукцессии) или сезонных изменений,
используют постоянный квадрат или трансекту.
Натянутой на колышки нейлоновой веревкой
отмечают участок, на котором периодически исследуют
абиотические и биотические факторы. Результаты
представляют таким образом, чтобы
продемонстрировать не только направления изменений, но и сами
изменения, а также возможные факторы,
объясняющие или связанные с этими изменениями.
Металлический
рефлектор
Лампа (100 Вт)
Образец почвы
Металлическая сетка
Гладкая сторона
воронки
50%-ный спирт
Металлический
рефлектор
Лампа (100 Вт)
Стеклянный стержень
Бечевка
Муслиновый мешок
с почвенным образцом
Вода
Зажим
Соединяющая
резиновая трубка
50%-ный спирт
Рис. 13.8. Эклектор Тульгрена.
Рис. 13.9. Воронка Бермана.
Рис. 13.10. Положение линейной трансекты на каменистом берегу.
Гайка
Проволока
Металлическая или
деревянная рама
Рис. 13.11. Квадратная рама (1 м2), разделенная
проволокой на небольшие квадраты (каждый по
400 см2), образует размеченный квадрат
к
а
ill
υ
s
U
&
Число квадратов (или
исследуемая площадь)
Рис. 13.12. График зависимости между числом
видов, обнаруженных на участке, и числом описанных
квадратов (В количественных исследованиях нет
смысла описывать свыше определенного число
квадратов, это будет напрасной тратой времени.)
13.2.3. Методы оценки размеров
популяции
При проведении любых количественных
экологических исследований важно с большой степенью
точности дать оценку численности организмов,
населяющих определенную площадь на суше или
объем воды или воздуха. Как правило, это
равнозначно оценке величины популяции. Выбор метода
зависит от размеров и образа жизни организма и
площади исследуемой территории. На небольшом
участке можно непосредственно подсчитать число
или оценить проективное покрытие или обилие
растений и прикрепленных или медленно
передвигающихся животных. На больших же открытых
пространствах для оценки численности быстро
передвигающихся животных необходимы косвенные
методы учета. В местообитаниях, в которых
наблюдение за организмами затруднено вследствие
особенностей их поведения и образа жизни, приходится
оценивать численность организмов, используя либо
метод изъятия, либо метод мечения и повторного
отлова. Существуют объективные и субъективные
методы оценки численности популяции.
Объективные методы
Квадраты, прямое наблюдение и фотографирование
относятся к методам прямого учета, тогда как
метод изъятия и повторного отлова организмов
составляют косвенные методы учета.
квадрат. Если установлено число организмов в
пределах некоторого числа квадратов, занимающих
известную часть общей площади, то простым
умножением можно подсчитать численность
организмов на всей территории. Используя этот метод,
можно определить следующие три параметра
распределения видов.
1) Плотность вида. Плотность вида-это число
организмов определенного данного вида на данной
площади, например на Юм2. При определении
плотности подсчитывают число организмов в
брошенных случайным образом квадратах. Метод
имеет ряд преимуществ, а именно: он точен,
позволяет сравнивать различные участки и виды и
установить точное значение обилия. Недостатки
метода состоят в том, что он требует значительных
затрат времени и при пользовании им необходимо
четко определить категорию «особь». Например,
считать за одно растение всю дернину или каждый
побег?
2) Частота вида. Это вероятность обнаружения
определенного вида в пределах любого случайным
образом брошенного квадрата в данном районе.
Например, если вид встречается в одном из каждых
десяти квадратов, то его встречаемость равна 10%.
Чтобы получить эту величину, отмелают наличие
или отсутствие вида в каждом квадрате, брошенном
случайным образом. (Число присутствующих
особей не имеет значения.) При использовании этого
метода необходимо оговорить размер квадрата, так
как это будет влиять на результаты, и, кроме того,
условиться о том, какую частоту учитывать: побего-
вую или корневую? (В случае «побеговой» частоты
вид считается присутствующим, если его листья
попадают на площадь квадрата, хотя сама особь
находится вне его. При «корневой» встречаемости
вид считаетря присутствующим, если его особи
укореняются на площади, захваченной квадратом.)
Преимущества этого метода в том, что он прост и
не требует больших затрат времени. Его применяют
в определенных крупномасштабных экосистемах,
например в лесных массивах. Недостатки же
состоят в том, что на полученное значение частоты
влияют размеры квадрата, размеры растения и его
пространственное размещение (например,
случайное, равномерное или групповое; см. разд. П.2.8).
13.2. Какова частота вида, если он был
обнаружен в 86 квадратах из 200?
3) Проективное покрытие вида показывает, какая
часть почвы занята особями данного вида, и дает
оценку площади, покрытой этим видом в процентах
Стержень (спица
для вязания)
Другие отверстия
Гайка
Деревянная рама
Уровень
земли
Три вида растений,
которых коснется
спица
Рис. 13.13. Рама со стержнем.
Число животных в образце
Образец
1
2
3
4
Число
животных
120
93
60
35
Совокупный
размер
образца
0
120
213
273
Совокупный размер образца
Рис. 13.14. График зависимости числа животных
в каждом улове от общего числа пойманных ранее
животных. Продолжив линию к точке, в которой
число вновь пойманных животных равно 0, получим
оценку численности популяции.
от общей площади. Его определяют в нескольких
случайно выбранных точках путем регистрации
покрывающего почву вида, каждый раз субъективно
оценивая площадь квадрата, покрытую этим видом,
или используя раму со стержнем (рис. 13.13). Этот
метод удобен при оценке проективного покрытия
растений, особенно травянистых, когда посчитать
число особей трудно и не столь важно, как
определить проективное покрытие. Однако это достаточно
трудоемкий и утомительный метод.
13.3. Чему равно проективное покрытие
растений X, если рама с десятью спицами была
использована 10 раз и 36 раз было
зарегистрировано это растение?
прямое наблюдение. Прямой подсчет особей
применим не только к сидячим или медленно
передвигающимся животным, но и ко многим крупным
подвижным организмам. К таким, например, как
олени, дикие пони и львы, лесные голуби и летучие
мыши, в то время когда они покидают место
ночлега.
фотографирование. Прямым подсчетом особей на
фотоснимках можно установить размеры популяций
крупных млекопитающих и морских птиц,
собирающихся на открытых пространствах.
метод изъятия. Этот метод очень удобен для
оценки численности мелких организмов, особенно
насекомых, на определенном участке луга или в
определенном объеме воды. Взмахами специальной
сетки животных отлавливают, записывают число
пойманных и не выпускают до конца исследования.
Затем еще трижды повторяют отлов, при этом с
каждым разом число пойманных животных
уменьшается. При построении графика отмечают число
пойманных при каждом отлове животных против
общего числа пойманных ранее животных.
Продолжив линию графика к точке, в которой вновь
пойманных животных не окажется (т.е. их число в
последнем отлове равно нулю), можно оценить
общий размер популяции, как это показано на
рис. 13.14.
метод мечения и повторного отлова Этот метод
включает отлов животного, его мечение таким
образом, чтобы не причинить ему вреда и выпуск на
волю там, где его поймали, с тем чтобы оно могло
продолжить нормальную жизнедеятельность в
популяции. Например, на жаберные крышки
пойманной сетями рыбы прикрепляют алюминиевые
пластинки или на ноги пойманных сетями птиц
надевают кольца. Мелких млекопитающих можно
метить краской, надрезать ухо или остригать
пальцы, членистоногих также метят краской. В
любом из случаев можно применить такую форму
кодирования, которая позволяет различить
индивидуальные организмы. Пойманных животных
подсчитывают, метят репрезентативную выборку из
них, затем всех животных выпускают в то же самое
место. Через некоторое время животных снова
отлавливают и подсчитывают в выборке число
животных с меткой. Размер популяции оценивают,
используя следующее словесное уравнение:
Общий размер популяции =
Число животных χ Число животных во
в первом улове втором улове
Число животных с меткой во втором улове
Эта оценка размера популяции называется индексом
Линкольна. Индекс выводится исходя из ряда
приведенных ниже допущений.
1. Внутри популяции организмы размещены
случайным образом. (Это не всегда так,
поскольку некоторые организмы живут колониями,
стадами или стаями.)
2. Между первым и вторым отловом должно
пройти достаточное количество времени, чтобы
животные успели распределиться случайным
образом. Чем менее подвижны особи вида, тем
больше времени должно пройти между
отловами.
3. Индекс применим только к популяциям,
перемещение которых ограничено по географическим
причинам.
4. Организмы равномерно размещены в пределах
географического ареала популяции.
5. Изменения размеров популяции, вызываемые
иммиграцией, переселением, рождением и смертью
особей, незначительны.
6. Метки не должны мешать передвижению
животных и делать их заметными для хищников.
При изучении растений и мелких животных,
например усоногих раков, прямой подсчет
численности-очень трудоемкая процедура, поэтому ее в
зависимости от необходимой степени точности
можно заменить оценкой проективного покрытия
или обилия на площади квадратной рамы. В начале
работы, для того чтобы увеличить точность оценки,
рекомендуется пользоваться рамой, поделенной на
мелкие квадратики (рис. 13.11). Для определения
проективного покрытия или обилия можно освоить
различные методики, одни из которых целиком
субъективны, другие частично или полностью
объективны.
13.4. Чтобы оценить численность форели в
маленьком озере, 625 форелей были пойманы,
помечены и снова выпущены. Через неделю
поймали 873 форели, из них у 129 были
обнаружены метки. Определите примерные размеры
популяции.
Субъективные методы
Эти методы включают в себя определенный способ
оценки частоты, шкалу частоты или оценку обилия
через покрытие. Например, в условной шкале,
составленной Криспом и Саутвардом, для оценки
обилия живущих на скалистых берегах блюдечек
{Patella) используются следующие символы,
частоты и проценты.
А-обильный > 50%
С-обычный 10-50%
F-часто встречающийся 1-10%
О-редкий < 1%
R-единичный-за 30 минут поисков обнаружено
лишь несколько особей
Соотношение процентов и словесных обозначений
частоты условное, поэтому оно может меняться.
Например, в отдельных случаях обильным может
считаться вид с покрытием > 90%. Значения
приведенных выше пяти категорий можно использовать
при представлении данных, например при
построении кайт-диаграмм (разд. П.2.7.3). Главный
недостаток этого метода состоит в том, что он
необъективен и при использовании его наблюдается
тенденция к уменьшению величины покрытия для мелких
невзрачных видов по сравнению с цветущими,
бросающимися в глаза и растущими в куртинах
видами.
13.3. Экологические научные
программы и
исследования
Экологические программы предусматривают
изучение как населяющих определенную территорию
организмов (синэкология), так и изучение отдельных
видов (аутэкология). В обоих случаях требуется
время для прочтения и обсуждения вопросов,
связанных с программой, с тем чтобы уяснить ее
цели, характер и объем. Цель каждого
исследования-решение тех или иных насущных задач или
проверка какой-либо гипотезы.
Цели программы должны формулироваться
четко, и в ней должны содержаться как общие, так и
конкретные задачи. Например:
1) развить и закрепить любознательность и
пытливость;
2) развить способность планировать исследование,
выдвигать гипотезы и проектировать
эксперимент;
3) развить способность формулировать вопросы и
собирать соответствующие качественные и
количественные данные для получения ответа на
поставленные вопросы;
4) совершенствовать наблюдательность и
практические навыки в использовании аппаратуры и
определительных таблиц для идентификации
видов;
5) развить способность правильно записывать
данные;
6) развить способность применять имеющиеся
знания для объяснения полученных данных;
7) развить критическое отношение к данным,
способность оценивать обоснованность данных и
сделанных из них выводов;
8) развить способность переносить биологическую
информацию в таблицы и графики, делать
устные и письменные сообщения;
9) совершенствовать умение определять и хранить
организмы;
10) научиться оценивать связи между различными
организмами, между организмами и
окружающей их средой, а также понять динамический
характер экологии.
Этот перечень можно продолжить конкретными
задачами, представленными в разд. 13.5.
13.3.1. Подробное описание
программы или исследования
Независимо от качества исследования и полученных
данных программа или же само исследование будут
практически недоступны другим ученым до тех пор,
пока данные не будут представлены в виде
сообщения, которое необходимо оформить
соответствующим образом.
1. Введение: включает основную идею, проблемы,
гипотезы и цели (т.е. что вы хотели сделать и
почему).
2. Метод: стратегия программы (т.е. что вы
делали/сделали ранее), где и как это было
сделано, включая все существенные особенности
работы приборов и методов, применявшихся в
поле и в лаборатории).
3. Результаты и наблюдения: данные, сведенные в
таблицы, графики, гистограммы, профили,
диаграммы присутствия - отсутствия, кайт-диаграм-
мы, а также данные, представленные любыми
другими наглядными и информативными
способами.
4. Обсуждение результатов: включает анализ
результатов (желательно количественных) по
возможности пробные выводы, сделанные на основе
представленных данных и ссылок на уже
опубликованные материалы.
5. Обсуждение значения выводов: критическая
оценка применяемых методов, разбор источников
ошибок и предложения для дальнейших
исследований.
6. Список использованной литературы.
13.4. Синэкологическое
исследование
Синэкологическое исследование включает изучение
абиотического и биотического компонентов,
связанных с естественным сообществом (биотический
компонент экосистемы), обнаруженным в
определенном географическом районе (или экосистеме),
например в дубовом лесу или на скалистом берегу,
где может обитать несколько видов растений и
животных и, возможно, содержится несколько типов
местообитаний. При проведении такого
исследования необходимо сделать следующее:
1. Начертить план района и местообитания(й) и
при необходимости профиль.
2. Определить виды и оценить численность каждого
из них.
3. Измерить (или собрать и сделать анализы)
абиотические факторы в пределах
местообитания^).
Конечной целью такого исследования является
определение в пределах изучаемого района
качественных и количественных связей между
популяциями растений и животных и возможных
взаимоотношений между ними, а также изучение эдафичес-
ких, топографических и климатических факторов.
Имея эту информацию, можно объяснить природу и
значение факторов, регулирующих численность и
распределение организмов в пищевой сети, а в
случае детального исследования построить
пирамиды численности, биомассы и энергии.
13.4.1. Составление карты местности
Вид сверху
Описанный ниже простой метод предназначен
прежде всего для картирования небольших площадей,
например участка луга размером 10 χ Юм или
небольшого водоема, но его можно использовать
также на больших площадях, например при
картировании скалистого берега залива.
1. Выберите приблизительно участок для изучения
и натяните рулетку вдоль одной из его сторон.
Рулеткой отмечается базовая линия AY (рис. 13.15).
2. В направлении, перпендикулярном базовой
линии, отмерьте расстояния до определенных
естественных ориентиров, находящихся в
пределах изучаемого участка или до ограничивающих
участок колышков. Запишите эти значения.
3. На листе миллиметровой бумаги в
соответствующем масштабе начертите линию AY и перенесите
все отмеренные в перпендикулярном
направлении расстояния.
4. Приняв за основу уже начерченную базовую
линию AY и отложенные в перпендикулярном
направлении расстояния до естественных ориен-
.» тиров, закончите карту от руки.
5. Если исследуемая площадь сравнительно
невелика, то поделите базовую линию AY на равные
отрезки и проложите перпендикулярно к ней
линейные трансекты. Повторите все операции,
используя крайнюю левую трансекту AF как
новую базовую линию, для того чтобы натянуть
веревочную решетку, как это показано на
рис. 13.16. Нанесите эту решетку на карту и
отметьте ее буквами (А,В,С и т. д.) вдоль одного
края и цифрами вдоль другого.
6. Отметьте положение участков, ясно
различающихся по структуре и растительности.
7. В зависимости от характера изучаемой
территории с помощью рамы, рамы со стержнем или
сетки для отлова животных планомерно
исследуйте участок, скажем, слева направо и запишите
все встречающиеся виды, их численность или
обилие.
Рис. 13.15. Метод вычерчивания карты исследуемой
площади (например, небольшого водоема
неправильной формы) по полученным основным точкам.
Контрольный
столб
Рейка
Точка
А
Точка
В
Точка
С
Уровень отлива
Рис. 13.16. Карта исследуемой площади, разбитой
веревочной сеткой на квадраты, например А1 Е5
Сетка дает возможность точно обозначить
исследуемые участки при последующем изучении
Вид спереди,
показывающий
пересечение
нитей\
Местоположе
ние
перекрещивающихся
нитей
Контрольный столб
Картонная зрительная
труба
Деревянный брусок
Вынимающаяся
кнопка
Нить
Груз отвеса
Линия отвеса
Рис. 13.17. Простой нивелир, который можно
изготовить самому, прикрепленный к контрольному
столбу Положение столба выверяют до тех пор,
пока деревянный брусок не окажется в
горизонтальном положении, которое обозначается нитью,
показывающей 90° в точке отвеса на столбе
Удерживая столб в неизменном положении, наблюдатель
смотрит в зрительную трубу и называет человеку,
держащему рейку, отметку высоты на рейке,
соответствующую положению точки пересечения нитей
Эта высота записывается
8. Если участок очень велик и на нем необходимо
провести качественные и количественные
исследования, то проложите на участке через
определенные интервалы ленточные трансекты. Теперь
можно установить положение любой точки, в
которой проводятся стационарные исследования,
например методом квадрата, измерив правый
угол и расстояние до ленточной трансекты.
Необходимо как можно чаще проводить прямые
измерения свойств абиотической среды или же
отбирать образцы для последующего анализа.
Графическое изображение профиля
высот
В некоторых местах, например на скалистом
берегу, на размещение организмов могут влиять
факторы, связанные с высотой над уровнем моря.
Здесь продолжительность времени, которое каждый
Рис. 13.18. Методика измерения высот и расстояний
по прямой между точками, расположенными над
уровнем отлива.
участок берега свободен от воды, зависит от его
высоты. В этом случае необходимо начертить
профиль, показывающий, как изменяется высота
вдоль линии трансекты, например от высшей до
низшей отметок воды на скалистом берегу (см.
ниже). В каждой точке вдоль трансекты, где
проводятся исследования сообществ, необходимо
тщательно измерить высоту с помощью оптического
теодолита и контрольных точек. На небольших
расстояниях с помощью простого, заранее
сделанного нивелира, прикрепленного к контрольному
столбу, и рейки можно получить относительно
точные данные (см. ниже) (рис. 13.17).
1. Прикрепите нивелир на удобной высоте (Zz1),
например на высоте 1,5 м, к контрольному
столбу.
2. Проложите линейную трансекту от высшей
отметки воды до кромки воды.
3. Установите контрольный столб в определенной
точке, например на высшей отметке воды на
трансекте, а далее вниз по склону на
определенном расстоянии (х) установите рейку. Отметьте
их положение на трансекте и обозначьте как А и
В. Сохраняйте одну из сторон трансекты до тех
пор, пока идут измерения, чтобы предотвратить
затаптывание изучаемых организмов.
Таблица 13.3. Горизонтальные и вертикальные расстояния
между точками А - К на скалистом берегу. Нортумберленд,
1968 год
Точка
А
В
S
D
E
F
G
H
I
J
К
Горизонтальное расстояние,
м (х, Xx и
т.д.)
0
20
40
60
80
100
120
140
160
180
200
Вертикаль-
, ное расстояние
между
точками (h2 — A1 и
т.д.)
1,5
1,7
1,8
0,8
0,6
0,7
0,9
0,8
0,4
0,4
Высота над
уровнем отлива,
м (h2 — A1 и
т.д.)
9,6
8,1
6,4
4,6
3,8
3,2
2,5
1,6
0,8
0,4
0
Σ
л
η
S
о
Σ
α>
Da
а
χ
се
H
O
α
Ξ
03
Расстояние по горизонтали от уровня отлива, м
Рис. 13.19. На основе данных из табл 13 3 вычерчен профиль каменистого берега и отмечены положения
точек от А до J, в которых проводились исследования Обратите внимание, что ось χ имеет масштаб
10 м = / ед, а ось у 1 м = 1 ед Это дает искусственное искажение формы профиля, но при этом позволяет
показать расположение крутых и пологих участков берега
4. После того как деревянный зрительный брусок
установлен горизонтально, посмотрите через
зрительную трубу с визиром на рейку.
Совместите визир с рейкой и запишите высоту (A2).
Разница высот между точками А и В равна
A2-A1.
5. Перенесите контрольный столб в точку В, а рейку
на определенное расстояние (Jc1) в точку С.
Повторите операции 3 и 4 и запишите новую
высоту (A3) (рис. 13.18).
6. Далее на расстояниях jc3, X4. и т. д. отметьте точки
D, E и т. д. и измерьте соответствующие высоты
A4, A5 и т.д. Запишите все показания в виде
таблицы (табл. 13.3) и посчитайте разницу
высот между точками и расстояние между ними
по прямой.
7. Перенесите эти данные в масштабе, принятом
при изображении профиля берега на бумаге и
отметьте положение точек (рис. 13.19).
13.4.2. Определение и оценка
численности организмов по
видам
Как уже говорилось в разд. 13.2.2, для
систематического отбора образцов на исследуемой
территории используют линейные и ленточные трансекты,
квадратные рамы и рамы со стержнем. Организмы
идентифицируют с помощью определительных
таблиц, а их численность либо устанавливают путем
прямого подсчета, либо оценивают приблизительно
(см. разд. 13.2.3).
13.4.3. Регистрация и представление
данных
Данные необходимо заносить в полевой дневник
сразу после их получения. В случае проведения
синэкологических исследований морских
местообитаний данные удобно заносить в листы полевого
дневника, изображенные в табл. 13.4 и 13.5. Эти
листы лучше всего прикреплять на доску с
зажимами и вместе с карандашом держать в большом
полиэтиленовом пакете, чтобы предохранить от
дождя. Когда все данные собраны, их необходимо
представить в виде соответствующих диаграмм, в
которых будут ясно прослеживаться связи между
организмами и(или) между организмами и
характером окружающей среды. Способы представления
данных показаны в разд. П.2.7. Они включают
диаграммы присутствия - отсутствия, кайт-диаграм-
мы распределения, гистограммы и пищевые
(трофические) пирамиды, которые описаны в разд. 12.3.6.
Некоторые примеры использования всех
четырех методов представления данных показаны на
рис. 13.20-13.22.
13.4.4. Сбор и анализ абиотических
факторов
Количество времени, необходимое для проведения
этой части синэкологического исследования, зависит
от характера изучаемой территории. Такое исследо-
Таблица 13.4. Образец записи в полевом дневнике
Полевой дневник - Морская экология
1. Название местности и координаты
2. Характер профиля (скалистый, песчаный, илистый,
дюна)
3. Схематичная карта местности, включающая
исследуемую площадь (показывающая положение трансект)
4. Другие особенности (местоположение, экспозиция и
т.д.)
5. Дата
6. Погодные условия
7. Время прилива и отлива:
прогнозный прилив
прогнозный отлив
наблюдаемый прилив
наблюдаемый отлив
прогнозная амплитуда прилива и отлива
8. Примечание и условные обозначения при записи данных
(шкала обилия - % покрытия, указание высоты уровня,
например Yi19 и т.д.)
Высота над уровнем отлива
Зона
брызг
Верхняя часть
берега
Средняя
часть
берега
Ниж
няя часть
берега
Сублиторальная
зона
Приливно-
отливная зона
L neritoides
L sax at ilis
L littorea
L littoralis
5
0
5
0
5
0
5
0
5
Рис. 13.20. Койт-диогроммы, показывающие частоту и характер размещения четырех обычных видов
улиток Littorina на каменистом берегу полуострова Дейл, Пембрукшир, апрель 1976 г. Диаграммы построены
по данным табл. 13.6.
вание целесообразнее проводить в районах, где
доминируют эдафические факторы, например в
лесу, на лугу или солончаке, чем на каменистых
берегах водоемов. Абиотические факторы, которые
необходимо исследовать, и методы их изучения
описаны в разд. 13.1.
13.5. Аутэкологическое
исследование
Аутэкологическое исследование включает изучение
всех экологических факторов, влияющих на
отдельный вид растения или животного на протяжении его
жизненного цикла. Целью исследования является
возможно более точное описание экологической
ниши вида. Выбранный для изучения вид должен
быть распространенным и доступным. На первой
стадии исследования необходимо как можно шире
ознакомиться с литературой о выбранном виде.
Изучая литературу, необходимо обращать внимание
на все аспекты биологии вида, а также на черты,
удобные для его практического изучения.
Исследование может включать в себя уже выполненную
другими работу или решать новые задачи.
Удобнее всего составить исчерпывающий список
вопросов, для того чтобы ответив на них, выявить
все, что известно об изучаемых видах. Литературу
необходимо изучать очень внимательно, а
поставленные вопросы принять за программу
исследования. Программа частично должна состоять из
новых, ранее не проводимых исследований,
включающих наблюдения, измерения и эксперименты.
Она не должна представлять собой простой
перечень сведений, почерпнутых из прочитанных книг
и журналов. Сведения об изучаемом виде
необходимо собирать в течение всего года.
Ниже приведен образец вопросов, которые
обычно ставят при изучении животного.
1. Систематическое положение. Как называется
вид? Какие другие группы организмов наиболее
сходны с ним? Каково сходство и каковы
различия между близкородственными видами?
Каково его полное таксономическое описание?
2. Местообитание. Где оно находится? Каковы
особенности абиотической среды обитания? Как
изменяются абиотические факторы в течение
года?
3. Морфология. Каково строение взрослой особи?
Каковы характерные внешние признаки?
Каковы размеры и масса организма?
4. Передвижение. Как перемещается животное?
Какие его части принимают участие в движении
и каковы функции этих частей?
5. Питание. Чем питается организм? В какое
время? Сколько пищи поедает? Каким образом
ловит и заглатывает пищу? Какие черты строе-
Таблица 13.5. Данные, которые необходимо внести в полевой дневник при проведении синэколо!ических исследований
морских местообитаний
Название
места
Горизонтальное
расстояние от
берега, м
Обозначение
уровня (A2
и т.д.), м
Разница
высот
и т.д.), м
Высота над
уровнем
отлива, м
Время
нахождения
над водой
Время
нахождения
под водой
ЖИВОТНЫЕ
(виды и
обилие)
РАСТЕНИЯ
(виды и
обилие)
Для
заметок
Высота над уровнем отлива, м
Расстояние от берега по горизонтали, м
Место
обитание А
В Зона
Зона
брызг
Верхняя
часть берега
Средняя часть берега
Нижняя
часть
берега
Сублиторальная
зона
Г Виды растений
Caloplaca sp.
Verrucaria maura
Pelvetia canaliculata
Chaetomorpha sp.
Enteromorpha intestinalis
Fucus spiralis
Porphyra umbilical is
Cladophora sp.
Chondrus crispus
Ceramium rubrum
Fucus vesiculosus
Ectocarpus sp.
Fucus ceranoides
Ascophyllum nodosum
Polysiphonia sp.
UIva lactuca
Fucus serratus
Corallina officinalis
L i thothamnion sp.
Rhodvmenia palmata
Gigartina stellata
Laurencia sp.
Furcellaria Jastigiata
Lomentaria articulata
Laminaria saccharina
Lam ι nana digita
Местообитания
Общее
число
видов
Рис. 13.21. Зональное распределение растительности на скалистом берегу, изображенное в виде ряда
диаграмм. Графически представлены следующие данные: А-профиль берега; Б-положение местообитаний от
А до Y с интервалами в 10 м; В-береговые зоны, возникающие в результате деятельности
прилива; Г-диаграмма присутствия-отсутствия видов растений; Д-диаграмма с вертикальными столбцами,
показывающая общее количество видов в каждом местообитании. Эти данные были получены группой
студентов шестого курса, использовавших ленточную трансекту и квадратную раму площадью 1 м2
на каменистом берегу полуострова Гоуэр, Южный Уэльс, июль, 1981 г.
Pelvetia canaliculata
Масштаб
Fucus spiralis
Масштаб
Ascophillum nodosum
Масштаб
Fucus serratus
Fucus vesiculosus
Масштаб
Масштаб
Рис. 73.22. Трофические пирамиды биомассы «урожая на корню» для пяти сообществ каменистого берега.
Первичными продуцентами каждого сообщества являются водоросли разных видов. Цифры обозначают
величину биомассы «урожая на корню» в гм~ 2. Площадь, окрашенная в серый цвет, соответствует биомассе
первичных продуцентов; заштрихованная площадь-биомассе первичных консументов (травоядные и дет-
ритофаги); вертикальные линии-биомасса вторичных консу ментов (плотоядных). Не следует, однако,
считать, что каждый трофический уровень поддерживается только нижележащим уровнем. Побережье
Нортумберленд, март 1969 г. (D. A S. Smith (1970) School Science Review, ASE.)
ния способствуют заглатыванию? Обладает ли
вид какими-либо особенностями переваривания
и всасывания пищи?
6. Дыхание. Где находятся поверхности для
газообмена? Как происходит газообмен? Сколько
кислорода требуется организму?
Выделение. Каковы конечные продукты обмена?
Каким образом они выводятся из организма?
Какими специальными органами выделения
обладает организм?
8. Размножение. Являются ли особи
раздельнополыми? Какие видимые внешние различия
существуют между полами? Встречаются ли
какие-нибудь формы в поведении ухаживания?
Защищает ли животное свою территорию? Как
происходит спаривание? Когда происходит
спаривание? Как часто животные спариваются?
Сколько вырабатывается половых клеток? Где
происходит оплодотворение?
9. Жизненный цикл. Сколько времени идет
развитие организма? В какой мере родители
заботятся о потомстве? Имеется ли личиночная стадия?
Когда взрослые особи достигают
половозрелого состояния? Чему равна средняя продолжи-
Таблица 13.6. Распределение четырех обычных видов
улиток Littorina на каменистом берегу полуострова Дейл,
Пембрукшир. Апрель 1976 года. Эти данные представлены
графически в виде кайт-диаграммы на рис. 13.20
Высота
над
уровнем
отлива, M
9-10
8-9
7-8
6-7
5-6
4-5
3-4
2-3
1-2
0-1
L. neritoides
число
63
54
7
—
—
—
—
—
—
баллы
по
шкале
обилия
10
10
4
—
—
—
—
—
—
L. saxatalis
число
3
8
17
6
1
—
—
баллы
по
шкале
обилия
2
4
8
4
1
—
—
L. littorea
число
—
1
3
13
6
2
1
баллы
по
шкале
обилия
—
1
2
6
4
2
1
L. Hit oralis
число
—
—
2
9
16
5
1
баллы
по
шкале
обилия
—
—
2
4
8
4
1
Шкала обилия: ^20=10; 19-15 = 8; 14-10 = 6;
9-5 = 4; 4-2 = 2; 1 = 1
тельность жизни индивидуума данного вида?
10. Поведение. Каким образом животное
воспринимает раздражители? На какие раздражители
главным образом реагирует животное? Как
основные органы чувств приспособлены к
образу жизни животного? В какой мере животное
поддается обучению? Как реагирует животное
на неблагоприятные погодные условия? Как
общаются животные между собой?
11. Экология. Какова численность популяции?
Какие еще животные обитают в данном
местообитании? Каким образом различные виды
размещены в пределах местообитания? Как
изучаемый вид связан с другими видами,
населяющими то же местообитание, в пищевых
цепях и пищевых сетях? Является ли животное
хозяином, паразитом или симбионтом? Какова
экологическая ниша вида?
Ниже приведен образец аналогичных вопросов,
которые должен поставить перед собой
исследователь при изучении цветкового растения.
1. Систематическое положение. Как называется вид?
Какие существуют подвиды, разновидности и
экотипы данного вида? Каковы сходства и
различия между близкородственными видами? Каково
полное таксономическое описание вида?
2. Местообитание
а) Эдафические факторы. К какому типу
относится материнская порода? К какому типу
относится почва? Какова толщина различных
горизонтов? Каково процентное содержание воды в
почве (полевая влагоемкость)? Каково
процентное содержание органического вещества в почве?
Каков минеральный состав почвы? Чему равен
рН почвы? Как связан уровень грунтовых вод и
его сезонные изменения с жизненным циклом и
распределением особей вида?
б) Климатические факторы. Чему равны
экстремальные и средние температуры в
местообитании? Чему равна средняя относительная
влажность воздуха в местообитании? Сколько осадков
выпадает ежегодно? Каково направление
господствующих ветров? Сколько света получает
растение?
в) Топографические факторы. На склонах какой
экспозиции обычно встречается вид?
Предпочитает ли вид открытые или закрытые места?
Предпочитает ли вид наклонные или равнинные
местообитания? Влияет ли высота над уровнем
моря на распределение вида?
3. Морфология. Насколько развита корневая
система? Каков тип корневой системы? Каков тип
ветвления? Сколько листьев на каждой ветви?
Какой формы листья? В каких пределах
изменяются длина и ширина листьев? Чему равна
высота растения?
4. Физиология. Какие пигменты содержатся в
листьях и лепестках? На какой поверхности листа
транспирация идет интенсивнее? Как темнота
влияет на скорость транспирации? Происходит
ли изменение содержания воды в листьях в
течение суток?
5. Размножение:
а) Цветки. Сколько цветков в среднем образуется
на растении? Сколько чашелистиков, лепестков,
тычинок, плодолистиков, или пестиков, в цветке?
Какой они формы и размера? В каких пределах
изменяется цвет лепестков? Какие пигменты
содержатся в листьях? Когда начинается цветение?
Как долго оно продолжается? Как происходит
опыление? Какие существуют приспособления
для опыления насекомыми или ветром?
б) Плоды и семена. Как образуются плоды?
Каково строение плодов? Сколько семян
образуется в среднем на каждый цветок? Как
распространяются плоды и семена? На какое
расстояние распространяются плоды и семена?
в) Вегетация. Как происходит вегетативное
размножение? Какие органы растения сохраняют
жизнеспособность в неблагоприятный период? С
какой скоростью вид расселяется по территории?
6. Жизненный цикл. Какого типа семена образуются
у растения? Какие условия необходимы для
прорастания семян? Когда прорастают семена?
Какой процент семян прорастает? Какие формы
прорастания встречаются? С какой скоростью
развиваются побеги? Сколько времени идет
развитие растения и каких размеров оно
достигает? (Почему часть проростков не становятся
взрослыми?)
7. Экология. Растут ли особи вида как отдельные
растения или куртинами? Каковы размеры
куртин? Какие еще виды растут в данном
местообитании? Какова степень интенсивности
конкуренции между изучаемым видом и другими
видами растений? Является ли вид паразитом,
хозяином или симбионтом? Как вид связан с
животными с точки зрения его положения в
трофической сети? Служат ли растения вида
защитой или убежищем для животных? Если да,
то какие животные и как используют растение в
качестве убежища? Какова экологическая ниша
вида?
Объектами аутэкологического исследования
могут являться также грибы, водоросли, мхи,
печеночники или голосеменные растения. В этом случае
приведенные выше вопросы можно видоизменить в
соответствии с природой изучаемого вида.
Глава 14
Транспорт
Индивидуальные клетки обмениваются веществами
с окружающей средой в результате физического
процесса диффузии (сюда относится и осмос) и
активных процессов-активного транспорта, эндо-
цитоза и экзоцитоза. Внутри клеток вещества
обычно перемещаются за счет диффузии, однако
происходят и активные процессы, например токи
цитоплазмы. При небольших расстояниях эти средства
транспорта достаточно быстро и хорошо
функционируют у одноклеточных и у тех многоклеточных
организмов, у которых достаточно велико
отношение поверхности тела к его объему; в таких случаях
нет нужды в специальных транспортных системах.
Например, у дождевых червей и других мелких
животных дыхательный газообмен с окружающей
средой происходит путем диффузии газов через всю
поверхность тела.
У более крупных и более сложно устроенных
организмов клетки могут быть настолько удалены и
друг от друга, и от окружающей среды, что одной
диффузии становится недостаточно. Возникает
необходимость в специализированных системах
транспорта на дальнее расстояние, гарантирующих
быстрое перемещение нужных веществ. При этом
вещества обычно передвигаются за счет системы
массового потока. Массовый, или общий, поток-это
массовый перенос веществ из одного места в другое
за счет разности давления в этих двух местах. Для
массового потока характерно то, что все вещества,
находятся ли они в растворе или суспензии,
переносятся с одинаковой скоростью, как в реке, и это
отличает массовый поток от диффузии, при которой
молекулы передвигаются независимо друг от друга
в соответствии с их диффузионными градиентами.
Некоторые системы массового потока у растений и
животных перечислены в табл. 14.1.
Транспорт у растений
В табл. 14.2 перечислены важнейшие группы
веществ, которые транспортируются у растений; там
же даны некоторые сведения об основных путях и
механизмах поглощения, транспорта и удаления
веществ.
Передвижение веществ по проводящим тканям
растений называют транслокацией. У сосудистых
растений (папоротникообразных и семенных)
проводящие ткани крайне специализированы и
подразделяются на ксилему и флоэму. По ксилеме
осуществляется главным образом транслокация воды,
минеральных солей, некоторых органических
соединений азота и гормонов, и транспорт при этом
направлен от корней к надземным частям растения.
По флоэме переносятся самые разные органические
и неорганические вещества, направляемые из
сформировавшихся листьев в другие части растения.
Изучение транслокации имеет большое
практическое значение. Например, очень важно знать, как
попадают в растение гербициды, фунгициды,
регуляторы роста и питательные элементы и по каким
путям они перемещаются внутри растения. Это
нужно и для того, чтобы решить, как лучше всего
применять такие вещества, и для того, чтобы уметь
предвидеть их возможные эффекты. Кроме того,
транслокация иногда приводит к распространению
таких патогенных для растений агентов, как грибы,
бактерии и вирусы, и от этого может зависеть выбор
лечебных или предупредительных мер. Одним из
сравнительно недавних новшеств явилось
применение так называемых системных фунгицидов,
которые разносятся по всему растению и обеспечивают
более длительную и более надежную защиту от
таких опасных болезней, как милдью (ложная
мучнистая роса).
14.1. Водный режим растений
14.1.1. Осмос
Чтобы понять, как поддерживается водный режим
растения, нужно прежде всего разобраться в таких
физических процессах, как осмос и диффузия,
которые объясняются в Приложении 1.5 (т. I). Там особо
подчеркнуто, что осмос можно рассматривать как
своеобразную диффузию, при которой
диффундируют только молекулы воды, а молекулы всех
остальных веществ и взвешенные в воде частицы не
проходят через полупроницаемую мембрану.
Молекулы воды передвигаются из того места, где их
много (т. е. из разбавленного раствора), туда, где их
концентрация ниже (т.е. в более
концентрированный раствор), и при этом мембрана избирательно
пропускает только молекулы воды. Известно, что
этот процесс идет быстрее, чем следовало бы из
Таблица 14.1. Некоторые системы массового потока у животных и растений
Система массового потока
Растения
Проводящие ткани:
ксилема (гл. 14)
флоэма (гл. 14)
Животные
Пищеварительная
система (гл. 10)
Дыхательная система
(гл. 11)
Кровеносная система
(гл. 14)
Лимфатическая система
Переносимые вещества
Главным образом вода и
минеральные соли
Главным образом
органические питательные
вещества, например сахароза
Пища и вода
Воздух или вода
Кровь
Лимфа
Движущая сила
Дыхание и корневое
давление; механизм не
совсем понятен
Мускулатура
пищеварительного тракта
Дыхательная
мускулатура
Сердце или
сокращающиеся кровеносные
сосуды
Общая активность всех
мышц тела
Группы организмов
Сосудистые растения: Pte-
ridophyta и Spermatophy-
ta
Annelida, Mollusca, Arthro-
poda, Chordata
Mollusca, Chordata
Annelida, Mollusca, Arthro-
poda, Chordata
Mammalia
Таблица 14.2. Передвижение веществ у растений
Вода
Путь
Механизм
Растворенные вещества
Путь
Механизм
Газы1*
Путь
Механизм
Поглощение
Корневые волоски
Осмос
Корневые волоски
Диффузия или
активный транспорт
Устьица, чечевички,
корневой эпидермис
Диффузия
Транспорт
Ксилема
Массовый поток
(корневое давление)
Ксилема (главным
образом неорганические
вещества)
Массовый поток
Межклеточные
пространства и клетки
Диффузия
Удаление
Устьица (кутикула и чечевички)
Диффузия водяных паров
Сбрасывание листьев, коры, плодов
и семян; в противном случае
вещества сохраняются до отмирания
всего растения или переходят к
следующему поколению вместе с
зародышем семени
Контролируется ростовыми
веществами
Устьица, чечевички, корневой
эпидермис
Диффузия
1) Передвижение газов более подробно описано в гл. 11.
расчетов для обычной диффузии, и что в нем
участвует массовый поток. Тем не менее такое
передвижение воды вполне объяснимо с помощью законов
диффузии.
14.1.2. Осмотическое давление (ОД) и
осмотический потенциал (π)
Осмотическое давление-это давление1, которое
создается в растворе, помещенном в осмометр,
1 Прежде давление измеряли в атмосферах (атм), а
сейчас-в паскалях (Па) (см. Приложение 4.3).
1 Па = 1 Hm"2 (H-ньютон)
1 бар = 0,987 атм = 105 Па = 100 кПа
1 атм = 1,0132 бар = 1,0132· 105 Па = 101,32 кПа.
после того как будет достигнуто равновесие с
чистой водой. Чем концентрированнее раствор, тем
выше ОД.
В наше время чаще используют термин
осмотический потенциал, так как при обычных
обстоятельствах речь идет лишь о потенциальном
(теоретическом) давлении, которое становится реальным
только в особых условиях-при измерении
осмометром (см. Приложение 1.5.2). Осмотический
потенциал условно считают отрицательным, а
осмотическое давление-положительным. Для обозначения
осмотического давления в этой главе мы
используем сокращение ОД, имея в виду положительную
величину, так как отрицательные значения могут
затруднить понимание.
14.1.3. Водный потенциал
(символ ψ, греч, буква «пси»)
В наши дни физиологи растений пользуются
термином «водный потенциал»1, ψ, когда хотят описать
тенденцию молекул воды к перемещению из одного
места в другое. В литературе могут встретиться и
другие, более традиционные термины, о которых
будет сказано в разд. 14.1.10.
Вода перемещается из участка с более высоким
водным потенциалом к участку с более низким
водным потенциалом.
Условно принято считать, что при обычном
атмосферном давлении водный потенциал чистой
воды равен нулю. В этих же условиях все растворы
будут иметь более низкий водный потенциал, а
следовательно, величина ψ будет отрицательной.
Теперь мы можем определить понятие «осмос»
по-иному: это перемещение молекул воды через
полупроницаемую мембрану, направленное из
области с более высоким водным потенциалом в
область с более низким водным потенциалом.
Если приложить давление выше атмосферного,
водный потенциал чистой воды или раствора
возрастет. Так можно поднять водный потенциал
системы и до положительной величины, как,
например, в жидкости, находящейся внутри почечных
клубочков. При нормальном атмосферном
давлении водный потенциал раствора обычно
определяют по осмотическому давлению раствора и
выражают его в единицах давления:
Ψраствора ^^раствора ·
Преимущество, связанные с
использованием водного потенциала
Водный потенциал можно рассматривать как
стремление воды покинуть некую систему. Чем выше (т. е.
чем менее отрицателен) будет водный потенциал,
тем больше будет это стремление. Если две системы
находятся в контакте друг с другом, вода будет
переходить из системы с более высоким водным
Полупроницаемая
мембрана
1 Следует сказать несколько слов о происхождении
термина «водный потенциал». Этот термин взят из
термодинамики, где означает свободную энергию воды в
данной системе, например в почве или в клетке. Молекулы
воды, как и все другие молекулы, всегда передвигаются по
градиенту свободной энергии - от более высокого значения
к более низкому в соответствии с законами энтропии. Этот
процесс и называется диффузией. Водный потенциал
системы-это разность между значениями свободной энергии
в данной системе и в чистой воде при обычном
атмосферном давлении. Биологи обычно выражают эту величину в
единицах давления, хотя ее можно выражать и в единицах
энергии.
Дальнейшие подробности можно найти в статьях Brad-
beer, Thomason, Bradbeer, 1980 (School Science Review, 62,
272-273) н Hutchinson CS., SutclilTe J.F. 1983 (J. Biol, Ed.,
17, 123-130), написанных в расчете на преподавателей
биологии и успешно занимающихся студентов.
Раствор А
I Раствор Б
О Молекулы или ионы
растворенных веществ,
например сахара или
соли
о Молекулы растворителя
(воды)
Растворенное вещество
+ растворитель=раствор
Рис. 14.1. Два раствора, разделенные
полупроницаемой мембраной
потенциалом в систему с более низким
потенциалом.
Таким образом, термин «водный потенциал»
позволяет оценить стремление воды к перемещению
между любыми двумя системами-не только из
одной клетки растения в другую, но, например, и из
почвы в корень, из листа в воздух или из почвы в
воздух. Можно сказать, что внутри растения вода
передвигается по непрерывному градиенту водного
потенциала от почвы до воздуха.
Чем круче будет градиент потенциала, тем
быстрее пойдет поток воды по градиенту.
14.1.4. Передвижение воды между
растворами за счет осмоса
Все упомянутые выше термины можно уверенно
использовать только в том случае, если они
правильно поняты. Чтобы проверить это,
попробуйте ответить на вопросы 14.1 и 14.5-14.8.
14.1. Внимательно рассмотрите рис. 14.1, где
изображены два раствора, разделенные
полупроницаемой мембраной. Ответьте на вопросы:
а. В каком из растворов выше концентрация
молекул воды?
б. Какой из растворов более
концентрированный, т.е. в каком из них выше концентрация
растворенного вещества?
в. В каком направлении пойдет осмос?
г. У какого раствора более высокое
осмотическое давление?
д. Какая из двух приведенных ниже величин ψ
выше:
1) - 2000 кПа; 2) - 1000 кПа;
е. В каком из растворов более высокий водный
потенциал (ψ)?
ж. Какова связь между ОД и ψ раствора при
нормальном атмосферном давлении?
14.1.5. Осмос и растительная клетка
На рис. 14.2 показаны полупроницаемые мембраны,
имеющие особое значение для водного режима
Протопласт
— живая
часть клетки
Клеточная стенка — полностью проницаемая
для небольших молекул
( Плазматическая
мембрана
Цитоплазма
Тонопласт
Вакуоль
— содержит вакуолярныи
сок, т. е. раствор Сахаров,
солей и других веществ
Ядро
Плазматическая
мембрана
— окружает цитоплазму
Тонопласт— мембрана,
окружающая вакуоль -
Полупроницаемые
мембраны
Рис. 14.2. Полупроницаемые мембраны типичной вакуолизированной растительной клетки. Обратите
внимание, что в нормальных условиях плазматическая мембрана плотно прижата к клеточной стенке.
растительной клетки. Клеточная стенка обычно
полностью проницаема для веществ, находящихся в
растворе, так что ее нельзя назвать осмотическим
барьером. В клетке находится большая центральная
вакуоль, содержимое которой (клеточный сок)
способствует поддержанию осмотического давления в
клетке. Две очень важные мембраны-это
плазматическая мембрана и тонопласт. В своем влиянии на
водный режим растений плазматическая мембрана,
цитоплазма и тонопласт выступают как единое
целое-действуют как одна полупроницаемая
мембрана.
Опыт 14.1. Изучение осмоса в живых
растительных клетках
Материалы и оборудование
Эпидермис (кожица) лука или молодого ревеня
Микроскоп
2 предметных и 2 покровных стекла
Скальпель и пинцет
Дистиллированная вода
1 M раствор сахарозы
2 пипетки с грушей
Фильтровальная бумага
Методика
Оторвите полоску эпидермиса с нижней
поверхности одной из мясистых чешуи луковицы или черешка
молодого листа ревеня. Ревень удобнее потому, что
клеточный сок у него окрашен, а у лука зато легче
отделяется эпидермис. Эпидермис можно содрать,
если сначала сделать небольшой надрез, а затем
взяться за край надреза рукой или пинцетом и
оторвать один слой клеток. Быстро перенесите
оторванную полоску эпидермиса на предметное
стекло и капните на него две или три капли
дистиллированной воды. Осторожно накройте
покровным стеклом и посмотрите, как выглядят клетки
под микроскопом. Зарисуйте несколько клеток.
Отделите другую полоску эпидермиса и повторите
всю процедуру, взяв вместо воды 1 M раствор
сахарозы. Понаблюдайте за полоской в течение
15 мин при большом увеличении и зарисуйте
изменения, которые вы заметите в одной или нескольких
типичных клетках. Можно посмотреть, нельзя ли
повернуть наблюдаемый процесс вспять. Для этого
под покровное стекло капают дистиллированную
воду и отмывают препарат от раствора сахарозы;
избыток жидкости удаляют фильтровальной
бумагой.
Результаты
На рис. 14.3 показано, как выглядят клетки
эпидермиса лука после того, как их подержали разное
время в 1 M растворе сахарозы.
Протопласт, отстающий
от клеточной стенки
1,5 мин
1,75 мин
2 мин
2,25 мин
Сжавшийся
протопласт
Плазмолизированные
клетки через 10 мин
Клетка на разных
стадиях плазмолиза
(сначала плазмолиз
идет быстро, затем
замедляется)
Плазматическая
мембрана
Клеточная
стенка
Рис. 14.3. Внешний вид эпидермальных клеток лука
во время плазмолиза. Полоски эпидермиса оставляли
в 1 M растворе сахарозы на разное время.
14.1.6. Плазмолиз и тургорное
давление
Если клетка находится в контакте с
гипертоническим раствором, т.е. с раствором, имеющим более
низкий водный потенциал, чем собственное
содержимое клетки, вода начинает выходить из нее путем
осмоса через плазматическую мембрану. Сначала
теряется вода цитоплазмы, а затем через тонопласт
выходит вода и из вакуоли. Протопласт, т. е. живое
содержимое клетки, окруженное клеточной стенкой,
сморщивается и в конце концов отстает от
клеточной стенки. Этот процесс называют плазмолизом, а
про такую клетку говорят, что она плазмолизиро-
вана. При начинающемся плазмолизе протопласт
только-только перестает оказывать какое-либо
давление на клеточную стенку, и клетка становится
вялой. Вода выходит из протопласта до тех пор,
пока его содержимое не приобретает такой же
водный потенциал, что и окружающий раствор.
После этого клетка перестает сморщиваться
дальше.
14.2. Чем занято пространство между
клеточной стенкой и сморщенным протопластом в
плазмолизированных клетках?
Процесс плазмолиза обычно обратим, клетка при
этом не получает никаких стойких повреждений.
Если плазмолизированную клетку поместить в
чистую воду или в гипотонический раствор, т.е. в
раствор с более высоким, чем у содержимого
клетки, водным потенциалом, вода начинает поступать в
клетку путем осмоса. По мере того как
увеличивается объем протопласта, он начинает давить на
клеточную стенку и растягивав! ее. Клеточная
стенка сравнительно жесткая, поэтому давление внутри
клетки растет очень быстро. Давление, оказываемое
протопластом на клеточную стенку, называется тур-
горным давлением (ТД). Иногда вместо этого
говорят о противодавлении клеточной стенки (ПД), имея
в виду давление клеточной стенки на протопласт.
Оба давления равны по величине, но
противоположны по направлению. При постепенном
увеличении тургорного давления из-за того, что вода
поступает в клетку за счет осмоса, клетка
становится тургесцентной. Полное набухание клетки, т.е.
максимальное ТД, наблюдается только тогда, когда
клетку помещают в чистую воду. Когда стремление
воды войти в клетку и тургорное давление в
точности уравновешивают друг друга, из клетки
выходит ровно столько воды, сколько в нее входит, и
клетка теперь находится в равновесии с
окружающим ее раствором. Осмотическое давление в клетке,
вероятно, все еще выше, чем в окружающем
растворе, так как для того, чтобы поднять тургорное
давление до точки равновесия, нужно очень малое
количество воды; его явно недостаточно, чтобы
существенно разбавить содержимое клетки. Тот
факт, что в состоянии равновесия осмотическое
давление в растительной клетке может быть
несколько выше, чем у окружающего раствора,
объясняется именно наличием тургорного давления.
Тургорное давление - это уже не потенциальное, а
реальное давление, оно создается только при
наличии клеточной стенки. У животных клеток нет
клеточной стенки, а плазматические мембраны
слишком нежны, чтобы защитить клетку от набухания и
разрыва в гипертоническом растворе. Поэтому в
животных клетках должна быть система защиты,
основанная на осморегуляции (см. гл. 19).
14.3. Каково тургорное давление в вялой
клетке?
14.4. У каких еще организмов, кроме растений,
есть клеточные стенки?
14.1.7. Осмотическое передвижение
воды из клетки в раствори
обратно
Если хотят определить, куда будет переходить
вода-из растительной клетки в окружающий раствор
или наоборот,-нужно сравнить водный потенциал
клетки и окружающего раствора, так как вода
всегда пойдет в направлении от более высокого водного
потенциала к более низкому. В состоянии
равновесия водный потенциал клетки должен быть равен
водному потенциалу окружающего раствора.
Как мы уже говорили, при нормальном
атмосферном давлении водный потенциал данного
раствора определяется его осмотическим давлением:
Ураствора = ~ ОД
раствора *
Водный потенциал клетки определяют два
фактора-ее осмотическое давление и тургорное
давление. Как и для любого раствора, водный
потенциал клетки будет тем ниже, чем выше
осмотическое давление раствора внутри клетки. Однако
всякое повышение тургорного давления усиливает
стремление воды выходить из клетки (или
увеличивает сопротивление поступлению воды извне), т.е.
при повышении тургорного давления повышается и
водный потенциал клетки. Все это можно описать
следующим уравнением:
Уклетки = ТДклетки ~ °Д клетки
В состоянии равновесия уклетки = Ураствора, или
Увнутр = Унаружн» гДе «внутр» означает внутренний
раствор, а «наружи» - наружный раствор.
Ниже мы даем несколько вопросов для проверки
понимания тех терминов, которые мы только что
использовали. При этом не следует забывать, что
1) вода всегда движется от более высокого водного
потенциала к более низкому;
2) для того чтобы две системы находились в рав-
1 Уклеей лучше всего выразить как \\fw = ψρ 4- ψ5 (см.
разд. 14.1.10).
новесии, у них должен быть одинаковый водный
потенциал;
3) в большинстве случаев количества воды,
поступающей в растительную клетку или выходящей
из нее, недостаточно для того, чтобы заметно
изменить осмотическое давление в клетке
(поэтому при дальнейших расчетах исходят из того,
что осмотическое давление остается
постоянным), хотя тургорное давление сильно
изменяется.
14.5. Опишите, что произойдет, если клетку в
состоянии начинающегося плазмолиза с ОД
2000 кПа поместить в раствор с ОД 1200 кПа.
Каковы будут величины ψ и ТД клетки в
состоянии равновесия?
14.6. Если ОД клеточного сока в клетке,
уравновешенной с чистой водой при нормальном
атмосферном давлении, составляет 110 кПа, каково
будет ТД?
14.7. Если клетку сначала уравновесить с
чистой водой, а затем перенести в раствор
сахарозы, водный потенциал которого составляет
-800 кПа, то
а) Какова будет разность величин ψ между
содержимым клетки и наружным раствором в
момент переноса?
б) Куда пойдет вода-внутрь клетки или из
клетки?
в) Возрастет или понизится ТД?
14.1.8. Осмотическое передвижение
воды из клетки в клетку
Рассмотрите ситуацию, изображенную на рис. 14.4,
когда две клетки с вакуолями, имеющие разный
водный потенциал, контактируют друг с другом.
14.8. а) В какой клетке водный потенциал
выше?
б) В каком направлении пойдет вода в результате
осмоса?
в) Каков будет водный потенциал в этих клетках
в состоянии равновесия?
г) Какими будут величины осмотического и тур-
горного давления в этих клетках при достижении
равновесия?
Опыт 14.2. Определение среднего
осмотического давления
клеточного сока в препарате
растительных клеток методом
начинающегося плазмолиза
Осмотическое давление в растительных клетках
можно измерять различными методами, но самый
удобный из них-это метод начинающегося плаз-
Полностью проницаемые
клеточные стенки
КЛЕТКА А
T
^ = -1200кПа|
и
| ТД = 800 кПа
1
ОД=2000кПа
КЛЕТКА Б
У=-800кПа
(ТД = 600кПа|
ОД= 1400кПа
Полупроницаемые
'
плазматические
мембраны
Рис. 14.4. Две соседние вакуолизированные клетки.
молиза. Он основан на следующих соотношениях:
0) Ψ™™ = ТД - ОД; i|/pacTBOpa = -ОД;
(2) Уклетки = ¥раствора » *ОГДа ЭТИ ДВв СИСТвМЫ
находятся в равновесии.
Препараты исследуемой ткани уравновешивают в
растворах разной концентрации (с разным водным
потенциалом); цель этого - подобрать раствор,
который вызывает начинающийся плазмолиз, т.е.
заставляет протопласты сморщиваться настолько,
что едва-едва начинается отделение протопласта от
клеточной стенки. В этот момент тургорное
давление равно нулю, поскольку протопласт уже не давит
на клеточную стенку, так что \|/клеТ1СИ = -ОДклетки =
ΨΡ
= -одп
[из уравнений (1) и (2),
приведенных выше]. Другими словами,
осмотическое давление раствора, который вызывает
начинающийся плазмолиз, будет таким же, как и у
клеточного сока.
На самом деле осмотическое давление в клетках
одной и той же ткани сильно варьирует, так что
одни клетки плазмолизируются в более
разбавленных растворах, чем другие. Считают, что плазмолиз
начинается тогда, когда плазмолизируется 50%
клеток. В этот момент остальные 50% клеток еще не
плазмолизированы, и можно сказать, что «средняя»
клетка находится в состоянии начинающегося
плазмолиза. Полученная величина отражает среднее
значение осмотического давления в ткани.
Материалы и оборудование
Черешок ревеня или луковица
6 чашек Петри
6 пробирок
Штатив для пробирок
Этикетки или восковой карандаш
2 градуированные пипетки на 10 или 25 мл
2 стакана на 100 мл
Тонкая кисточка для рисования
Дистиллированная вода
1 M раствор сахарозы
Тонкий пинцет
Пастеровские пипетки
Предметные и покровные стекла
Микроскоп
Миллиметровка
Бритвенное лезвие или острый скальпель
Методика
(После этого первого метода мы опишем другую
методику для свеклы.)
1. Подпишите шесть чашек Петри и шесть
пробирок для следующих растворов сахарозы: 0,3; 0,35;
0,4; 0,45; 0,5 и 0,6 M.
2. Возьмите градуированную пипетку на 10 или
25 мл, стакан дистиллированной воды и стакан
1 M раствора сахарозы и приготовьте в
пробирках по 20 мл раствора различной концентрации.
Объемы, которые нужно взять для этого,
указаны в табл. 14.3.
Таблица 14.3. Приготовление разбавленных растворов
сахарозы для опыта 14.2
Концентрация
раствора, M
0,30
0,35
0,40
0,45
0,50
0,60
Объем
дистиллированной воды, мл
14
13
12
11
10
8
Объем
1 M раствора
сахарозы, мл
6
7
8
9
10
12
Таблица 14.4. Осмотическое давление растворов сахарозы
при 20 С
Концентрация
раствора
сахарозы, M
0,05
0,10
0,15
0,20
0,25
0,30
0,35
0,40
0,45
0,50
0,55
0,60
0,65
0,70
0,75
0,80
0,85
0,90
0,95
1,00
1,50
2,00
Осмотическое
давление, к Π а
130
260
410
540
680
820
970
1120
1280
1450
1620
1800
1980
2180
2370
2580
2790
ЗОЮ
3250
3510
6670
11810
Осмотическое
давление, атм
1,3
2,6
4,0
5,3
6,7
8,1
9,6
11,1
12,6
14,3
16,0
17,8
19,5
21,5
23,3
25,5
27,5
29,7
32,1
34,6
65,8
116,6
3. Хорошо перемешайте каждый раствор путем
встряхивания пробирки. Это очень важно.
Вылейте полученные растворы в соответствующие
чашки Петри.
4. Лук. Оторвите одну из мясистых чешуи
луковицы. Пока внутренний эпидермис еще находится
на месте, возьмите лезвие или скальпель и
вырежьте в нем 6 квадратиков размером
приблизительно 5x5 мм. Тонким пинцетом отделите
квадратики и сразу же положите их по одному в
каждую чашку Петри. Осторожно покачайте
чашки, чтобы ткань была полностью погружена
и хорошо промылась раствором сахарозы.
Оставьте чашки примерно на 20 мин.
Ревень. Прорежьте в наружном эпидермисе 6
квадратиков размером около 5x5 мм, отделите
их и т.д., как описано для лука.
5. С помощью кисточки выньте ткань из 0,60 M
раствора и перенесите ее на предметное стекло в
раствор сахарозы той же концентрации.
Накройте покровным стеклом и рассмотрите под
микроскопом.
6. При малом увеличении выберите подходящую
зону клеток. Затем используйте объектив для
среднего или большого увеличения и
просмотрите выбранную зону, передвигая стекло.
Запишите, в каком состоянии (плазмолизированном
или нет) находятся первые 100 просмотренных
клеток. Клетку считают плазмолизированной,
если протопласт в ней хотя бы чуть-чуть отошел
от клеточной стенки.
7. Повторите всю процедуру с остальными
квадратиками ткани в соответствующих растворах.
8. Подсчитайте процент плазмолизированных
клеток в каждом растворе. Постройте график,
отложив по вертикальной оси процент
плазмолизированных клеток, а по горизонтальной - мо-
лярность раствора сахарозы.
9. По графику определите молярность раствора
сахарозы, при котором плазмолизируется 50%
клеток.
10. Постройте график зависимости осмотического
давления (по вертикальной оси) от молярности
раствора сахарозы (по горизонтальной оси) по
данным, приведенным в табл. 14.4.
11. Из полученного графика определите
осмотическое давление того раствора, который вызывает
50%-ный плазмолиз. Эта величина будет
соответствовать усредненному осмотическому
давлению клеточного сока.
Результаты
На рис. 14.5 показана типичная кривая для
эпидермиса лука. Сходные результаты получаются и с
эпидермисом ревеня.
14.9. Какова величина осмотического давления
в клетках эпидермиса лука, если 50% клеток
плазмолизируется в 0,38 M растворе сахарозы?
Доля плазмолизированных
клеток, %
Молярность раствора сахарозы
Рис. 14.5. Доля плазмолизированных клеток в
эпидермисе лука при разных концентрациях сахарозы
в растворе.
Использование тканей корня свеклы
Корень свеклы - гораздо менее удобный материал;
однако, если этот опыт объединить с опытом
14.3, можно будет определить тургорное давление в
клетках свеклы, хотя следует подчеркнуть, что у
разных корней водный потенциал и осмотическое
давление могут быть разными. В обычных условиях
осмотическое давление у свеклы выше, чем у лука
или ревеня, так как в вакуолях свеклы больше
сахара и неорганических солей.
Методику нужно слегка модифицировать: этапы
1-3 те же, что и в опыте с луком и ревенем.
Единственное отличие-это концентрации
растворов: 0,4; 0,45; 0,5; 0,55; 0,6 и 0,7 M. Затем (4) вырежьте
из корня прямоугольный «столбик» квадратного
сечения размером примерно 5x5 мм. Возьмите
лезвие и сделайте тонкие квадратные срезы (не более
0,5 мм толщиной). Чем тоньше будут срезы, тем
легче будет подсчитать плазмолизированные
клетки. Окрашенный сок облегчает подсчет. Срезы
следует сделать перед самым занятием и подержать их
в дистиллированной воде. Положите по нескольку
срезов в каждый раствор и оставьте их в сахарозе
примерно на 30 мин. Тем временем рассмотрите
под микроскопом срезы, залитые дистиллированной
водой, чтобы познакомиться, как выглядят неплаз-
молизированные клетки. Скорее всего, срезы будут
тоньше с краю, и там их легче рассматривать.
Некоторые поврежденные клетки могут оказаться
обесцвеченными, а около проводящей ткани иногда
будут видны более мелкие клетки; их не надо брать
для подсчета. Этапы 5-11 те же, что и в предыдущем
опыте; начните с ткани, помещенной в раствор
0,7 M.
Результаты
Серия данных, полученных для свеклы, приведена в
табл. 14.5.
14.10. Каково усредненное осмотическое
давление в клетках свеклы, использованных в этом
эксперименте? (Чтобы ответить на этот вопрос,
вам нужно будет построить график.)
Таблица 14.5. Изменение доли плазмолизированных клеток
в ткани из корня свеклы при различной концентрации
сахарозы в растворе
Концентрация раствора
сахарозы, M
0,30
0,40
0,45
0,50
0,55
0,60
Доля плазмолизированных
клеток, %п
2,5
3,5
13,5
74,0
100,0
100,0
!) Размер выборки -200 клеток
Опыт 14.3. Определение водного
потенциала растительной
ткани
Водный потенциал - это мера стремления молекул
воды перемещаться из одного места в другое.
Принцип опыта состоит в подборе раствора с известным
водным потенциалом, в котором исследуемая ткань
не будет ни поглощать, ни терять воду. Препараты
ткани помещают для уравновешивания в растворы
различной концентрации. Ткань будет иметь тот же
водный потенциал, что и раствор, в котором не
произойдет никаких изменений (увеличения или
уменьшения) массы или объема ткани. Описанный
ниже метод основан на регистрации изменений
объема, а не массы.
Материалы и оборудование
Свежий клубень картофеля или корень свеклы
6 чашек Петри
5 пробирок
Штатив для пробирок
Этикетки или восковой карандаш
2 градуированные пипетки на 10 или 25 мл
Кафельная плитка
Дистиллированная вода
1 M раствор сахарозы
Скальпель или нож
2 стакана на 100 мл
Миллиметровка
Методика
1. Подпишите шесть чашек Петри для
дистиллированной воды и для 0,1; 0,25; 0,75 и 1,0 M
растворов сахарозы. Подпишите пять пробирок,
по одной для каждого раствора сахарозы.
2. Возьмите градуированную пипетку, стакан с
дистиллированной водой и стакан с 1 M
раствором сахарозы и приготовьте в отдельных
пробирках по 20 мл раствора сахарозы
различной концентрации. Для разведения удобно
пользоваться таблицей 14.3.
3. Встряхивая пробирки, хорошо перемешайте
каждый раствор.
4. Вылейте приготовленные растворы в
соответствующие чашки Петри. В 6-ю чашку налейте
20 мл дистиллированной воды.
5. Проверьте, не мокрые ли чашки (если надо,
протрите их снизу), и поставьте на
миллиметровку.
6. Возьмите нож или скальпель, сделайте срез
толщиной 2 мм из середины большого корня
свеклы или большой картофелины и вырежьте
из него 12 прямоугольных полосок толщиной
2 мм, шириной 5 мм и максимальной длины
(около 5 см). Работать надо быстро, чтобы
избежать потери воды в результате испарения,
так как это понизит водный потенциал ткани.
7. В каждую чашку Петри положите по две
полоски, полностью погрузите их в раствор и
тотчас же измерьте длину полосок по
миллиметровке, на которой стоят чашки. Покачайте
каждую чашку, чтобы промыть полоски.
8. Закройте чашки крышками и оставьте их хотя
бы на час, а лучше на сутки.
9. Снова измерьте длину полосок, рассчитайте
среднее изменение длины в процентах.
Постройте график зависимости среднего изменения
длины полоски (по вертикальной оси) от
молярной концентрации раствора сахарозы (по
горизонтальной оси). Изменение длины полосок
означает и изменение объема.
10. По полученной кривой определите ту
концентрацию раствора сахарозы, при которой
длина полосок совершенно не изменяется.
11. Постройте график зависимости осмотического
давления (по вертикальной оси) от молярной
концентрации раствора сахарозы (по
горизонтальной оси), воспользовавшись данными,
приведенными в табл. 14.4.
12. По этому графику определите, при каком
осмотическом давлении раствора длина полосок не
меняется. Водный потенциал ткани вычисляют с
помощью уравнения
Ψ клетки Ψ наружного раствора ^r-^наружного раствора *
13. Если использовался корень свеклы и если опыт
14.2 уже проведен (т.е. уже определено
осмотическое давление), рассчитайте тургорное
давление по формуле ψ = ТД — ОД.
Результаты
Более точные цифры обычно получаются, если
свести воедино результаты всей группы учащихся.
Возможные результаты представлены в табл. 14.6.
14.11. Каково будет усредненное значение
водного потенциала в клетках свеклы, если
воспользоваться данными табл. 14.6? [Вам нужно
будет рассчитать среднее изменение длины (в
процентах) и построить график.]
14.12. Зачем в каждую чашку Петри надо
класть по меньшей мере две полоски ткани?
14.13. Зачем накрывают чашки Петри, когда их
оставляют стоять?
14.14. Если осмотическое давление в клетках
свеклы 1400 кПа, а водный потенциал равен
— 950 кПа, то каково будет усредненное
тургорное давление?
14.15. Рассмотрите эксперимент,
представленный на рис. 14.6. Полый стебель одуванчика
{Taraxacum officinale) разрезают вдоль на 4
полоски по 3 см длиной, а затем эти полоски
погружают в дистиллированную воду или в
растворы сахарозы разной концентрации.
а) Почему при продольном разрезе стебля
отрезанные полоски тотчас же отгибаются
наружу?
б) Почему полоска Б в дистиллированной воде
еще больше закручивается наружу?
в) Почему полоска В в концентрированном
растворе сахарозы закручивается внутрь?
г) Почему полоска Л сохраняет свою кривизну
в разбавленном растворе сахарозы?
д) Что можно определить в таком опыте:
осмотическое давление, водный потенциал или
тургорное давление клеток стебля?
Спланируйте постановку эксперимента для
определения данной величины, полностью опишите все
детали процедуры.
14.1.9. Влияние нагревания и спиртов
на мембраны
Некоторые воздействия, например обработка
этанолом или нагревание, нарушают
избирательную проницаемость клеточных мембран. Хотя мем-
Таблица 14.6. Длина полосок, вырезанных из корня свеклы,
после 24 ч выдерживания их в дистиллированной воде или в
растворах сахарозы различной концентрации
Концентрация
сахарозы,
M
0,00
тиллированная
вода)
0,10
0,20
0,25
0,30
0,40
0,50
0,60
0,75
0,90
1,00
1,50
Длина полоски в начале
опыта, см
1
4,8
5,1
5,1
5,2
4,9
4,9
5,0
4,8
4,9
4,9
4,8
4,9
2
5,0
4,8
4,9
4,8
4,9
5,0
4,8
5,0
4,9
5,0
4,9
4,9
3
5,3
4,9
4,9
5,0
5,0
4,8
5,1
5,0
5,0
4,9
4,9
4,9
Длина полоски через
сутки, см
1
5,0
5,3
5,2
5,2
4,9
4,9
4,8
4,6
4,6
4,5
4,7
4,5
2
5,3
4,9
4,9
4,9
5,0
5,0
4,7
4,9
4,7
4,7
4,6
4,1
3
5,6
5,1
5,0
5,0
5,1
4,8
5,0
4,9
4,8
4,7
4,4
4,5
Полоски разрезанного
стебелька тотчас
закручиваются назад
Толстостенные
клетки эпидермиса
Центральная
полость
Тонкостенные
клетки коры
Через 15 с после препарирования
Плоскость разреза
Поперечный разрез стебелька
Эпидермис
Эпидермис
Во время препарирования
Л, Б и В — полоски,
выдержанные до состояния
равновесия (около 30 мин) в
растворах сахарозы разной
концентрации или в
дистиллированной воде
В разбавленном растворе
сахарозы
В дистиллированной
воде
В концентрированном
растворе сахарозы (1 M)
или после подсушивания
на воздухе
Рис. 14.6. Опыт со стебельками
одуванчика. Исследование влияния
дистиллированной воды и
растворов сахарозы на скручивание
полосок, вырезанных из стебелька.
браны и сохраняются, но они ведут себя так, будто в
них образовались отверстия, и при этом исчезает
барьер проницаемости для таких крупных молекул,
как сахароза. Высокие температуры и спирты
денатурируют белки мембран и увеличивают текучесть
мембранных липидов; кроме того, при высокой
концентрации спирты растворяют липиды.
14.16. Вакуоли клеток свеклы окрашены в
красный цвет. Исходя из этого, спланируйте
эксперимент для исследования влияния
нагревания и этанола на избирательную проницаемость
клеточных мембран свеклы.
14.1.10. Дополнение
(1) Термин «водный потенциал» был предложен для
замены терминов «дефицит диффузионного
давления» и «сосущая сила». Смысл последних
объясняется ниже.
Дефицит диффузионного давления (ДДД) и
сосущая сила (CC). Как мы уже говорили, осмос можно
рассматривать как особый род диффузии, когда
переносятся только молекулы воды. Раньше
стремление молекул воды к перемещению измеряли
величиной диффузионного давления (ДД). Чем выше
концентрация молекул воды в растворе, тем выше
ДД данного раствора. Поэтому при нормальном
атмосферном давлении чистая вода будет иметь
максимальное ДД, а растворенные в ней вещества
будут понижать ДД раствора. В стандартных
условиях ДД раствора всегда будет ниже, чем у
чистой воды; чем концентрированнее будет раствор,
тем меньше в нем будет концентрация воды и тем
ниже будет ДД раствора.
Поэтому раньше говорили, что по отношению к
чистой воде раствор имеет дефицит диффузионного
давления (ДДД), при этом ДДД чистой воды был
произвольно принят за нуль. Чем
концентрированнее раствор, тем выше ДДД для этого раствора.
При нормальном атмосферном давлении ДДД для
любого раствора будет равен его осмотическому
давлению. Термин ДДД явился заменой термина
сосущая сила (CC). Происхождение последнего
термина ведет свое начало от представления о том,
что при осмосе гипертонический раствор
присасывает воду из гипотонического раствора. Поэтому
говорили, что гипертонический раствор имеет
большее всасывающее давление. Теперь стало
понятно, что движущей силой для передвижения
воды служит более высокий водный потенциал
(или иначе диффузионное давление)
гипотонического раствора.
Связь между рассмотренными терминами можно
выразить следующим образом:
-ψ = ДДД = CC;
а для раствора в стандартных условиях
-ψ = ДДД = ОД = CC.
(2) Вместо уравнения
Уклетки = ТД
клетки
-од
клетки
физиологи растений предпочитают использовать
уравнение
где \|/w- водный потенциал клетки, \|/5-доля,
вносимая растворенным веществом в общий водный
потенциал (ψ5 имеет отрицательную величину,
равную величине осмотического давления), а ψρ-
доля тургорного давления в общем водном
потенциале (она либо равна нулю, либо имеет
положительную величину).
14.2. Передвижение воды у
цветковых растений
Вода, находящаяся в растении, непосредственно
контактирует с водой почвы и с парами воды в
окружающем растение воздухе. Мы уже говорили о
том, что вода движется от более высокого водного
потенциала к более низкому. Поэтому, как
полагают физиологи растений, вода передвигается по
растению из области с высоким водным
потенциалом (т.е. из почвы) в область с более низким
водным потенциалом (т.е. в атмосферу) в
соответствии с градиентом водного потенциала. Водный
потенциал в умеренно влажном воздухе на
несколько десятков тысяч килопаскалей ниже, чем в
растении; отсюда и большее стремление воды
выходить из растения.
Основная масса воды поступает в растение через
корневые волоски. Вода проходит через кору корня
в ксилему, поднимается по ксилеме к листьям и
испаряется с поверхности клеток мезофилла, а затем
диффундирует наружу через устьица. Последний
процесс называется транспирацией, а поток воды из
корня к транспирирующей поверхности - транспира-
ционным током. Установлено, что растение в
среднем использует менее 1% поглощаемой им воды.
Куда расходуется вода, мы уже рассматривали в
разд. 5.1.2.
14.3. Транспирация и
передвижение воды
в листьях
В обычных условиях вода выходит из растения в
виде пара. Для перехода воды из жидкого состояния
в парообразное нужна дополнительная энергия,
которую называют скрытой теплотой
парообразования. Ее дает солнце (солнечная энергия), и
согласно наиболее принятой теории транспирации,
теории сцепления - натяжения, именно эта энергия
обеспечивает общий поток воды через растение.
Транспирация - это испарение воды с поверхности
растения; ниже перечислены три основные
структуры, участвующие в этом испарении (в скобках
указана относительная потеря воды через них при
открытых устьицах).
1. Устьица - поры, через которые диффундирует
вода, испарившаяся с поверхности клеток; такие
поры есть в эпидермисе листьев и зеленых стеблей
(около 90% воды).
2. Кутикула - восковой слой, покрывающий
эпидермис листьев и стеблей; через него проходит
вода, испаряющаяся с наружных стенок клеток
эпидермиса (около 10%; больше или меньше в
зависимости от толщины кутикулы).
3. Чечевички: обычно их роль в испарении воды
очень мала, но у листопадных деревьев после
сбрасывания листвы через чечевички теряется
основная масса воды.
Потери воды в результате транспирации могут
быть очень велики. Травянистые растения,
например хлопчатник или подсолнечник, ежедневно
теряют до 1-2 л воды, а старый дуб-более 600 л.
14.17. Почему транспирация идет главным
образом через листья?
Вода поступает в листья по ксилеме. В листьях
есть своя собственная, сильно разветвленная сеть
проводящих пучков, охватывающая весь лист.
Ее ветви оканчиваются одним или несколькими
сосудами или трахеидами, которые почти не лиг-
нифицированы. Поэтому вода легко проходит через
их целлюлозные стенки и поступает к снабжаемым
ими клеткам мезофилла. На рис. 14.7 показаны три
основных пути, по которым вода идет дальше:
апопластный (через клеточные стенки), симпласт-
ный (через цитоплазму и плазмодесмы) и вакуо-
лярный (от вакуоли к вакуоли).
14.3.1. Путь через апопласт
Аоопласт-это система, образованная из
примыкающих друг к другу клеточных стенок, создающих
непрерывное целое (за исключением поясков
Каспари в корнях, разд. 14.5.2). «Свободное
пространство», которое может быть заполнено
водой, занимает до 50% объема клеточных стенок.
Испарение воды из апопласта мезофилла в
межклеточное воздушное пространство создает
натяжение в непрерывном потоке воды через апопласт, и
вода массовым потоком проходит через стенки
в результате сцепления (когезии) ее молекул
(разд. 14.4). В апопласт вода поступает из ксилемы.
14.3.2. Путь через симпласт
Симпласт-это система связанных между собой
протопластов растения. Цитоплазма соседних
клеток соединена плазмодесмами - цитоплазма-
тическими тяжами, проходящими через поры в
примыкающих друг к другу клеточных стенках
(рис. 14.7, Б). Детали строения плазмодесм до сих
пор окончательно не выяснены, поэтому не
известно, в какой мере они служат каналами для
передвижения веществ. Кажется, однако,
вероятным, что если уж вода и растворенные в ней
вещества проникнут в цитоплазму одной из клеток,
то дальше они могут переходить из клетки в клетку,
минуя мембраны. Как и при вакуолярном
переносе (разд. 14.3.3), вода идет по градиенту водного
потенциала. Возможно, что этому способствуют и
токи цитоплазмы. Передвижение воды по симпласту
имеет большее значение, чем путь через вакуоли.
14.3.3. Вакуолярный путь
В этом случае вода переходит из вакуоли в
вакуоль через другие компоненты смежных клеток;
Лигнин
Целлюлоза
Сосуд ксилемы
или τ pax ей да
СИМПЛАСТНЫЙ
ПУТЬ
Плазматическая
мембрана \
Тонопласт J
Клеточная стенка—
полностью проницаемая
Цитоплазма
Клеточный сок внутри
вакуоли
АПОПЛАСТНЫЙ
ПУТЬ
Полупроницаемые
ВАКУОЛЯРНЫЙ
ПУТЬ
Клетка
мезофилла
Испарение воды с
поверхности клетки и
последующая диффузия
водяных паров через
воздушные пространства
Подустьичное
воздушное
пространство
Транспирация через
устьица (диффузия
+ массовый поток
молекул, вызванный
движением воздуха)
Транспирация через
кутикулу (испарение)
Эпидермальная
клетка
Замыкающая
клетка
Устьице
-Кутикула
Клеточная
стенка
Плазмодесма
Цитоплазма
Тонопласт Вакуоль
Плазматическая
мембрана
Апопластный путь — через клеточные стенки
Симпластный путь — через цитоплазму и плазмодесмы
Вакуолярный путь - через плазматические мембраны, цитоплазму и
тонопласты вакуолей (по этому пути переносится сравнительно
мало воды)
Рис. 14.7. А. Схема путей
передвижения воды в листе.
Существуют три возможных пути:
симпластный и вакуолярный пути
показаны слева, а апопластный
путь - справа. Клетки 1, 2, 3
упоминаются в тексте. Толщина
клеточных стенок намеренно
преувеличена. Б. Схематическое
изображение группы клеток, на
котором суммированы все
возможные пути передвижения
воды (и растворенных в ней
веществ). Одновременно может
использоваться несколько
путей. Такие пути могут
функционировать и в листе, и в коре
корня. Перенос ионов по ва-
куолярному пути обязательно
включает активный транспорт.
Наибольшее значение имеет
апопластный путь, а
наименьшее-вакуолярный (им
можно пренебречь).
при этом она пересекает симпласт и апопласт и
проходит через плазмалемму и тонопласт путем
осмоса (рис. 14.7, £). Она движется по градиенту
водного потенциала, который создается как описано
ниже.
Вода испаряется с увлажненной поверхности
клеточных стенок мезофилла и попадает в
межклетники, в частности в околоустьичные воздушные
пространства. Рассмотрим, например, клетку 1 на
рис. 14.7, А. Потеря ею воды приводит к тому, что в
ней падает тургорное давление и понижается
водный потенциал. В результате этого водный
потенциал клетки 2 становится выше, чем у клетки 1 (хотя
в состоянии равновесия он был одинаков). Поэтому
вода идет из клетки 2 в клетку 1, и водный
потенциал клетки 2 понижается по сравнению с
потенциалом клетки 3. Так в листе создается
градиент водного потенциала на всем пути от сосудов
ксилемы с ее более высоким водным потенциалом
до клеток мезофилла, потенциал которых гораздо
ниже. За счет осмоса вода поступает из ксилемы в
клетки мезофилла. Хотя нам удобнее рассматривать
передвижение воды шаг за шагом, следует
подчеркнуть, что градиент водного потенциала,
который создается в листе, на самом деле непрерывен, и
вода равномерно течет по градиенту, подобно тому
как жидкость движется по фитилю.
Иногда создается неверное представление, что
вода передвигается в листе под действием
градиента осмотического давления. Однако нет
никаких данных, которые бы позволили
предположить, что осмотическое давление в
соответствующих клетках сильно различается, хотя градиент
водного потенциала, несомненно, существует.
Разность водного потенциала возникает в основном
из-за разности тургорного давления (вспомните, что
потеря даже небольшого количества воды в первую
очередь влияет на тургорное давление, а не на
осмотическое). То же самое можно сказать и о корне
(разд. 14.5), в котором есть градиенты тургорного
давления и водного потенциала, но не всегда
существуют градиенты осмотического давления.
14.3.4. Выход воды через устьица
Все три пути переноса воды завершаются
испарением ее в воздушные пространства листа. Отсюда
пары воды диффундируют через устьица по пути
наименьшего сопротивления, т. е. от более высокого
водного потенциала внутри листа к более низкому
за его пределами. У двудольных растений устьица
обычно располагаются в основном на нижней
поверхности листа, на верхней же их меньше или нет
вовсе. Регуляцию открывания и закрывания устьиц
мы рассмотрим в разд. 14.3.9. Все листья окружены
неподвижным слоем воздуха, толщина которого
определяется размерами листа, особенностями его
строения, например опущенностью, и скоростью
Нижний эпидермис
Устьице
Диффузионная
оболочка в
неподвижном
воздухе
Подвижный воздух
Стрелками показано отклонение молекул
воды во время диффузии
Рис. 14.8. Диффузия молекул воды из устьица.
Обратите внимание на то, что у краев отверстия
диффузионный градиент круче.
Линиями показаны области одинаковой концентрации
молекул воды (одинакового водного потенциала), чем круче
градиент водного потенциала, тем ближе друг к другу
расположены эти линии и тем быстрее происходит
диффузия Поэтому наибольшей будет скорость диффузии у
краев устьичной щели Этот «краевой эффект» означает,
что потеря влаги и газообмен будут более быстрыми при
большом числе малых отверстий, чем при малом числе
больших отверстий той же суммарной площади
ветра. Пары воды сначала диффундируют через
этот слой и лишь затем уносятся воздушными
потоками. Чем тоньше неподвижный слой, тем
быстрее идет транспирация. Существует
диффузионный градиент, направленный от
неподвижного слоя назад к клеткам мезофилла. Теоретически
каждое устьице должно иметь свой собственный
диффузионный градиент - свою «диффузионную
оболочку» (рис. 14.8). На самом же деле, если
воздух неподвижен, диффузионные оболочки
соседних устьиц перекрываются и образуют одну
общую оболочку.
14.3.5. Измерение скорости
транспирации
Транспирацию легче всего продемонстрировать,
если взять растение в горшке и накрыть его
стеклянным колпаком, а горшок завязать в
полиэтиленовый пакет, чтобы избежать прямого испарения
почвенной влаги. Если растение транспирирует, на
внутренней поверхности колпака будут собираться
капельки жидкости. Проверив эту жидкость
индикатором, можно убедиться, что это вода.
Индикатором могут служить сухая бумага, пропитанная
хлоридом кобальта (H), которая в воде из синей
становится розовой, или белые кристаллы
безводного сульфата меди (II), в воде синеющие.
Измерить абсолютную скорость транспирации
довольно трудно. Ниже описаны два простых
эксперимента, дающие приличные результаты (по
крайней мере для целей сравнения).
Опыт 14.4. Изучение и измерение
действия различных
факторов на транспирацию
с помощью потометра
Потометр-это прибор, предназначенный для
измерения скорости поглощения воды срезанными
побегами или молодыми сеянцами. Саму транспирацию
он не измеряет, но поскольку основная масса
поглощенной воды теряется за счет транспирации, эти два
процесса тесно взаимосвязаны. Можно купить
готовый потометр, а можно сделать его самому; один из
простейших вариантов изображен на рис. 14.9.
Материалы и оборудование
Потометр (рис. 14.9: колба Бунзена, короткая
резиновая трубка, резиновая пробка с одним
сквозным отверстием, шприц с иглой для
подкожных инъекций, градуированные
капиллярные трубки)
Большой черный полиэтиленовый мешок
Большой прозрачный полиэтиленовый мешок
Небольшой электрический вентилятор
Штатив для реторт и зажим
Запорный кран
Термометр
Вазелин
Стебель с листьями, например ветка сирени
Ведро
Методика
1. Подберите подходящее растение с листьями,
срежьте стебель и тотчас же погрузите срез в
ведро с водой, чтобы свести к минимуму
возможность попадания воздуха в ксилему. После этого
сразу обрежьте стебель еще раз, но под водой,
сделав косой срез на несколько сантиметров выше
первоначального среза. Толщина стебля должна
быть такой, чтобы он туго входил в отверстие в
резиновой пробке потометра.
2. Погрузите колбу Бунзена в раковину,
заполненную водой, и наберите в колбу воду. Перенесите
стебель с листьями из ведра в раковину и тотчас
же сделайте новый косой срез на несколько
сантиметров выше предыдущего. Под водой вставьте
стебель в отверстие пробки, а затем плотно
заткните этой пробкой колбу.
3. Опустите в раковину градуированный капилляр
вместе с прикрепленной к нему резиновой
трубкой, заполните его водой и подсоедините к
боковому отростку колбы.
4. Выньте всю систему из раковины и проткните
резиновую трубку иглой шприца, как показано на
рис. 14.9. Шприц укрепите в вертикальном
положении. То место, где стебель входит в
отверстие пробки, следует замазать вазелином, чтобы
не проходил воздух.
5. По мере всасывания воды стеблем мениск
жидкости в капилляре будет смещаться. Его можно
вернуть на старое место, добавив немного воды
из шприца. Дайте ветке постоять 5 мин и в это
время регулярно подкачивайте воду в систему.
6. Засеките время, за которое столб воды в
капилляре сместится на определенное расстояние, и
оцените скорость поглощения воды, выразив ее в
подходящих единицах, например в см/мин.
Следует сделать несколько замеров, чтобы убедиться,
что скорость достаточно постоянна, а затем
рассчитать среднее значение. Не забудьте записать
температуру воздуха вокруг растения.
7. Каждый раз, как только пузырек воздуха дойдет
до конца градуированного отрезка капилляра,
возвращайте его в исходное положение, подкачав
воду шприцем.
8. Можно выяснить, как влияют на скорость
поглощения воды такие факторы, как
а) ветер (для этого используйте небольшой
электрический вентилятор; струя воздуха не
должна сильно бить по листьям, иначе
закроются устьица);
Облиственный
стебель
Резиновая
пробка
Вода
Колба Бунзена
Штатив для
перегонки
Шприц с водой для
оттеснения воздуха
к правому концу
капилляра
Градуированная
капиллярная трубка
Воздух
Резиновая трубка
Рис. 14.9. Простейший потометр.
б) влажность (поместите ветку в мешок из
прозрачного пластика);
в) темнота (закройте ветку черным
полиэтиленовым мешком);
г) удаление половины листьев (уменьшится ли
в два раза скорость транспирации?);
д) смазывание нижнего или верхнего эпидермиса
листьев вазелином с целью уменьшить
потерю воды.
В каждом случае надо подождать, чтобы скорость
транспирации снова стала постоянной. Не всегда
можно изменить только одно из условий;
например, закрыв растение в прозрачном мешке, мы в
то же время уменьшим, хотя и незначительно,
интенсивность освещения.
Абсолютная скорость поглощения
воды
Полученные результаты можно выразить как
истинный объем воды, поглощенной за единицу времени,
например в мл/ч; для этого нужно знать объем,
соответствующий одному делению на шкале
капилляра.
Основная масса поглощенной воды теряется через
листья. Скорость потери воды на единицу
поверхности листа можно определить, если сначала
измерить потерю воды, как описано выше, а затем
оборвать все листья и определить площадь их
поверхности. Эту площадь можно измерить, если
положить листья на миллиметровку и очертить их
контуры, а затем подсчитать, сколько квадратиков
это займет. Если такие данные есть, полученные
результаты можно выразить в мл/ч на 1м2
поверхности листа.
Результаты
Влияние ветра, температуры, влажности и темноты
будут рассмотрены в разд. 14.3.6.
14.3.6. Влияние факторов
окружающей среды на
транспирацию
У растений имеется ряд морфологических и
анатомических особенностей, уменьшающих потерю
воды в засушливых условиях. Такие признаки
называются ксероморфными, и сейчас мы их
рассмотрим. Растения, которые растут в засушливых местах
и потому приспособлены к нехватке влаги,
называются ксерофитами. У них очень много
специфических ксероморфных черт, которые мы подробнее
обсудим в разд. 19.3.2. Растения, приспособленные к
условиям достаточной влажности, называются
мезофитами, однако у них могут быть и некоторые
ксероморфные признаки.
Свет
Свет влияет на транспирацию потому, что устьица
обычно открываются на свету и закрываются в
темноте. Поэтому потери воды ночью невелики и
связаны в основном с транспирацией через
кутикулу, а может быть, и через чечевички. Как только
утром открываются устьица, скорость
транспирации возрастает.
Растения сем. толстянковых (Crassulaceae)-o,£tHa
из групп суккулентов - уменьшают потери воды,
закрывая устьица днем и открывая их на ночь.
Двуокись углерода, поступая ночью через устьица,
связывается в виде органических кислот, а затем
снова высвобождается, но уже внутри листа; так
обеспечивается дневной фотосинтез.
Температура
На свету самое сильное влияние на транспирацию
оказывает температура. Чем выше температура, тем
быстрее испаряется вода из клеток мезофилла и тем
больше насыщен воздух внутри листа водяными
парами. В то же время повышение температуры
приводит к уменьшению относительной влажности
воздуха, окружающего растение. Эти два явления
повышают крутизну градиента молекул воды между
воздухом в листе и окружающей атмосферой. Чем
круче этот градиент, тем быстрее идет диффузия.
Можно сказать и по-другому: водный потенциал
внутри листа возрастает, а вне его-убывает.
Солнечное излучение повышает температуру
листа. Бледнее окрашенные листья отражают
больше солнечной радиации и поэтому не так быстро
обогреваются. Более бледная окраска листьев
обычно связана с тем, что они покрыты густым
слоем волосков, чешуек или воска, и является ксеро-
морфным признаком.
Влажность и давление водяного пара
Пониженная влажность воздуха (более низкое
давление водяного пара) около листа обычно
способствует транспирации, так как градиент водяного
пара (или водного потенциала) между увлажненным
воздухом внутри листа и окружающей атмосферой
становится более крутым. А с повышением
концентрации водяного пара в воздухе, т.е. с
увеличением влажности, диффузионный градиент,
наоборот, становится более пологим. Давление
водяных паров в атмосфере уменьшается с увеличением
высоты над уровнем моря; это связано с
понижением атмосферного давления. Поэтому у
высокогорных растений часто наблюдаются различные
ксероморфные приспособления, предназначенные для
уменьшения транспирации.
Один из ксероморфных признаков, свойственных
некоторым растениям,-это погруженные устьица,
т.е. устьица, которые утоплены в углублениях или
складках эпидермиса, способствующих локальному
повышению влажности и уменьшению потерь воды.
В некоторых случаях весь лист скручивается так, что
в образовавшейся полости создается влажная
атмосфера, например у Ammophila (песчаного тростника)
(рис. 14.10). Шуба из эпидермальных волосков или
Флоэма
Сосуд ксилемы
Толстая кутикула
нижнего эпидермиса.
В эпидермисе нет
устьиц
Фотосинтезиру-
ющие клетки
мезофилла
"Шарнирные"
клетки
Влажная
атмосфера
Одноклеточные волоски^
захватывающие
влажный воздух
Устьице-^
Верхний эпидермис
с тонкой кутикулой
и устьицами
-"Шарнирные"
клетки
Волокна мезофилла
Рис. 14.10. Поперечный разрез ксероморфного листа песчаного тростника (Ammophila). Показано
расположение тканей. Лист изображен в скрученном состоянии.
чешуек задерживает влагу, создавая неподвижный
слой увлажненного воздуха у самой поверхности
листа, что тоже уменьшает транспирацию.
Ветер
Когда нет ветра и воздух спокоен, у поверхности
листа создается воздушная оболочка, насыщенная
парами воды. Это уменьшает крутизну
диффузионного градиента между внутренней атмосферой
листа и наружным воздухом. При малейшем
движении воздуха эта оболочка сдувается. Поэтому
скорость транспирации при ветреной погоде
возрастает; при этом ускорение транспирации выражено в
наибольшей степени, когда ветер слабый. При
сильном ветре устьица закрываются и транспирация
через них прекращается.
Как мы уже говорили, всевозможные волоски и
чешуйки создают зону спокойного воздуха и таким
образом несколько замедляют транспирацию.
Доступность воды в почве
При высыхании почвы вода, как правило, прочнее
связывается с частицами почвы. К тому же (хотя это
имеет меньшее значение) почвенный раствор
становится более концентрированным, т.е. уменьшается
водный потенциал. Эти факторы снижают
осмотическое поглощение воды корнями. Вскоре
замедление поглощения воды приводит и к уменьшению
Рис. 14.11. Суккулентные растения. Слева - кактус
Opuntia echinocarpa; справа - Kalanchoe verticillata.
транспирации, так как возникает сопротивление
передвижению воды в растении, а градиент водного
потенциала в направлении почва -»растение -►
воздух становится более пологим.
14.3.7. Влияние особенностей самого
растения (внутренних
факторов) на скорость
транспирации
Мы уже видели, как влияют некоторые ксеро-
морфные приспособления на скорость
транспирации. Сейчас мы еще на нескольких примерах
покажем, как действуют такие «внутренние» факторы и
чем они отличаются от «внешних», т.е. факторов
окружающей среды.
Поверхность листа и соотношение
объем/поверхность
Транспирация возрастает с увеличением общей
поверхности листьев и уменьшением соотношения
объем/поверхность. Листовая поверхность может
быть уменьшена, если листья превратятся в иглы,
как у сосны и других хвойных, или в колючки, как
это произошло у кактусов. Еще один способ -
просто уменьшение всей поверхности листвы при
недостатке влаги. У листопадных растений это
достигается путем сбрасывания листьев в период
засухи или похолодания. Листопад-это тоже
ксероморфное приспособление. Зимой вода
замерзает и становится недоступной; кроме того, при
низких температурах замедляется и передвижение
воды в растении.
Соотношение поверхность/объем может
уменьшиться, если главным фотосинтетическим органом
будет весь стебель, как, например, у кактусов. На
рис. 14.11 показаны примеры характерного
уменьшения листовой поверхности у кактусов и других
суккулентов.
Кутикула
Транспирация через кутикулу обычно идет тем
быстрее, чем тоньше кутикула, хотя важен и ее
химический состав. Если кутикула тонкая (как,
например, у папоротников), через нее может
теряться до 30-45% всей транспирируемой воды.
Это может иметь особое значение, когда
транспирация через устьица сведена к минимуму, а воды
мало. У двудольных растений кутикула на верхней
поверхности листьев обычно толще, чем на нижней;
это связано с тем, что верхняя поверхность, во-
первых, открыта лучам солнца и, во-вторых, хуже
защищена от воздушных потоков. Усиленное
отложение воска на листьях может практически
полностью устранить транспирацию через
кутикулу. К тому же листья, покрытые воском,
обычно имеют блестящую поверхность и поэтому
отражают больше солнечных лучей.
Устьица
Чем больше устьиц приходится на единицу
поверхности листа, тем быстрее идет транспирация
через устьица, хотя важно также и их расположение.
Например, у двудольных растений на нижней
поверхности листа устьиц обычно больше, чем на
верхней (табл. 14.7), а у однодольных, листья у
которых обычно торчат вертикально, устьица
распределяются одинаково и на верхней, и на
нижней поверхности (см. данные о кукурузе и овсе в
табл. 14.7). У растений, адаптированных к
засушливым условиям, устьиц в среднем намного
меньше; поэтому число устьиц варьирует даже у
растений одного и того же вида.
Таблица 14.7. Число устьиц на единицу поверхности листа у
некоторых обычных растений. (По Weier T. E., Stocking C. R.,
Barbour M. G., 1970, Botany, an Introduction to Plant Biology,
4th ed., John Wiley a. Sons, p. 192.)
Растение
Однодольные
Кукуруза (Zea mays)
Овес (A vena sativa)
Двудольные
Яблоня (Malus spp.)
Фасоль (Phaseolus vulgaris)
Капуста (Brassica spp.)
Люцерна {Medicago sativa)
Nasturtium
Число устьиц на 1 см2
верхний
эпидермис
5200
2500
0
4000
14100
16900
0
Дуб (Quercus spp.) 0
Картофель (Solarium tuberusum) 5100
Томат (Lycopersicon esculentum) 1200
нижний
эпидермис
6800
2300
29400
28100
22600
13800
13000
45000
16100
13000
Опыт 14.5. Изучение распределения
устьиц
Материалы и оборудование
Прозрачный лак для ногтей
Предметные и покровные стекла
Тонкий пинцет
Свежие полностью развитые листья
Микроскоп
Методика
Для того чтобы изучить распределение устьиц,
удобно будет воспользоваться отпечатками
(репликами), сделанными с поверхности листа. Для
этого нанесите на лист тонкий слой лака с помощью
кисточки, вделанной в крышку пузырька. Дайте
лаку подсохнуть, а затем возьмите тонкий пинцет и
осторожно стяните тонкую пленку с отпечатком
поверхности. Положите реплику на предметное
стекло и накройте покровным стеклом. Рассмотрите
препарат в микроскоп. Сосчитайте, сколько устьиц
видно в поле зрения. Повторите эту процедуру
несколько раз с различными участками препарата.
Рассчитайте среднюю величину. Определите
площадь круга, видимую в поле микроскопа,
измерив диаметр с помощью калиброванного стеклышка
или прозрачной линейки. (Площадь круга равна кг2,
где г-радиус, а π = 3,142.) Затем можно подсчитать,
сколько устьиц приходится на 1 квадратный
сантиметр поверхности.
Сравните плотность распределения устьиц в
верхнем и нижнем эпидермисе одного и того же листа
или у разных видов. Есть ли какая-нибудь
корреляция между количеством устьиц и местообитанием
растения?
14.18. Опишите, какая зависимость существует
между тремя переменными, представленными
на рис. 14.12, и объясните ее.
Относительная величина,
% от максимума
Температура
воздуха на
уровне посева
Скорость
транспирации
Интенсивность
освещения
Время, часы суток
Рис. 14.12. Связь между интенсивностью света,
температурой воздуха и скоростью транспирации в
листьях люцерны. (По данным L. J. Briggs, H. L. Shantz
[1916], J. Agr. Res., 5, 583-649, цит. по А. С. Leopold
[1964], Plant growth and development, p. 396,
McGraw-Hill.)
14.3.8. Физиологическая роль
транспирации
Транспирацию называют «неизбежным злом», так
как это неминуемое, но потенциально вредное
следствие того, что клеточные стенки должны быть
влажными и что с них испаряется вода. Пары воды
выходят из растения в окружающую среду по тем
же самым путям газообмена, которые необходимы
для фотосинтеза и дыхания. Как мы уже говорили,
основной газообмен идет через устьица. Если бы не
было кутикулы, устьица были бы совсем не нужны,
а газообмен происходил бы даже более эффективно.
Но тогда нельзя было бы контролировать потерю
воды. Кутикула уменьшает эту потерю, а
дальнейшую регуляцию осуществляют устьица, которые
у большинства растений очень чувствительны к
недостатку воды и закрываются, например, при
засухе. Обычно они закрыты и ночью, когда
прекращается фотосинтез. В засушливых условиях
потеря воды может вести к завяданию, опасному
обезвоживанию, а затем и к гибели растения. Есть
достаточно данных о том, что даже незначительный
водный стресс замедляет рост, а это приводит к
экономическим потерям из-за снижения
урожайности сельскохозяйственных культур.
Несмотря на явную неизбежность транспирации,
интересно было бы знать, не дает ли она растению
какую-то выгоду? Существуют две возможности:
1. Высказано предположение, что транспирацион-
ный ток нужен для доставки минеральных солей
всем частям растения, т.е. что эти вещества
переносятся вместе с водой. Может быть, это так, но,
по-видимому, для передвижения минеральных солей
вполне хватило бы очень малой скорости
транспирации. Например, поступление минеральных солей в
листья ничуть не снижается ночью, когда транспи-
рация невелика; это связано с тем, что растительный
сок, находящийся в ксилеме, ночью более
концентрирован, чем днем. Всасывание минеральных
солей не зависит существенным образом от тран-
спирационного тока, хотя возможно, что быстрое
поглощение воды позволяет корням извлекать воду
и растворенные в ней вещества из более отдаленных
участков почвы.
2. Транспирации сопутствует испарение воды из
клеток мезофилла. Этот процесс идет с затратой
энергии, поэтому испарение приводит к охлаждению
листьев точно так же, как пот охлаждает кожу у
млекопитающих. Иногда это очень важно,
особенно на открытом солнце, когда листья
поглощают много лучистой энергии и могли бы
перегреваться, а сильный перегрев затормаживает процессы
фотосинтеза. Однако вряд ли такой охлаждающий
эффект имеет какое-либо значение в нормальных
условиях. В странах с жарким климатом у растений
существуют другие способы защиты от теплового
стресса.
Ι^^^ΜΰΚ'ΐηίΗΗ
!Устьице!
!Замыкающие
клетки
Рис. 14.13. Микрофотография устьица на нижней
поверхности листа, полученная с помощью
сканирующего электронного микроскопа.
14.3.9. Устьица: строение и механизм
открывания и закрывания
Устьица-это поры в эпидермисе, через которые
происходит газообмен. Они имеются главным
образом в листьях, но есть и на стебле. Каждое устьице с
двух сторон окружено замыкающими клетками,
которые в отличие от других эпидермальных клеток
содержат хлоропласта. Замыкающие клетки
контролируют величину отверстия устьица за счет
изменения своей тургесцентности. На рис. 14.13
показан вид устьиц и замыкающих клеток на
микрофотографии, полученной с помощью
сканирующего электронного микроскопа.
В разд. 8.1 мы уже говорили, как выглядят
клетки эпидермиса, замыкающие клетки и устьица,
если рассматривать их сверху в световом
микроскопе. На рис. 14.14 приведено схематическое
изображение устьица в разрезе. Здесь видно, что
клеточные стенки замыкающих клеток неоднородны по
толщине: та стенка, которая ближе к отверстию
устьица, явно толще, чем противоположная стенка.
К тому же целлюлозные микрофибриллы, из
которых состоит клеточная стенка, расположены
таким образом, что стенка, обращенная к
отверстию, менее эластична, а некоторые волокна
образуют своего рода обручи вокруг замыкающих
клеток, похожих на сардельки (см. рис. 14.14, ^). По
мере того как клетка всасывает воду и становится
тургесцентной, эти обручи не дают ей расширяться
дальше, позволяя лишь растягиваться в длину.
Поскольку замыкающие клетки соединены своими
концами, а более тонкие стенки вдали от устьичной
щели растягиваются легче, клетки приобретают
полукруглую форму (рис. 14.14). Поэтому между
замыкающими клетками появляется отверстие.
(Такой же эффект мы получим, если будем
надувать колбасовидный воздушный шарик с липкой
лентой, приклеенной к нему вдоль одной из его
сторон.)
И наоборот, когда вода выходит из замыкающих
клеток, пора закрывается. Каким образом
происходит изменение тургесцентности клеток, пока не
ясно.
В одной из традиционных гипотез - «сахаро-
крахмальной» гипотезе - предполагается, что днем
в замыкающих клетках возрастает концентрация
сахара, а в результате повышается осмотическое
давление в клетках и поступление в них воды.
Однако никому еще не удалось показать, что в
замыкающих клетках накапливается достаточное
количество сахара, чтобы вызвать наблюдаемые
изменения осмотического давления. Недавно было
установлено, что днем на свету в замыкающих
клетках накапливаются ионы калия и
сопутствующие им анионы; такого накопления ионов вполне
достаточно, чтобы вызвать наблюдаемые
изменения. В темноте ионы калия (K + ) выходят из
замыкающих клеток в прилегающие к ним клетки
эпидермиса. До сих пор неясно, каким анионом
уравновешивается положительный заряд иона калия. У
некоторых (но не у всех) изученных растений
отмечалось накопление большого количества
анионов органических кислот типа малата.
Одновременно уменьшаются в размере крахмальные
зерна, которые появляются в темноте в хлоро-
пластах замыкающих клеток. Это позволяет
предполагать, что крахмал на свету превращается в
малат. Один из возможных путей такого
превращения:
ФЕП
(фосфоенол пируват)
+ CO2
ФЕП-карбоксилаза
Оксалоацетат
Мала тдег идрогеназа
Малат
НАДФ-Н2
НАДФ
Серия реакций
Крахмал
(Сравните с процессом фотосинтеза у С4-расте-
ний, разд. 9.8.2.)
У некоторых растений, например у Allium сера
(лук), в замыкающих клетках нет крахмала.
Поэтому при открытых устьицах малат не
накапливается, а катионы, по-видимому, поглощаются
вместе с неорганическими анионами типа хлорида
(СГ).
Некоторые вопросы остаются нерешенными.
Например, почему для открывания устьиц нужен
свет? Какую роль играют хлоропласты, кроме
запасания крахмала? Превращается ли малат в
темноте обратно в крахмал? В 1979 г. было показано,
что в хлоропластах замыкающих клеток Vicia faba
(конские бобы) отсутствуют ферменты цикла
Кальвина и система тилакоидов развита плохо, хотя
хлорофилл и имеется. В результате не работает
Устьице
Хлоропласт
Ядро
Небольшая
вакуоль
Цитоплазма
клетки (вид сверху)
Толстая вентральная стенка
замыкающей клетки
Тонкая дорсальная стенка
замыкающей клетки
Микрофибриллы
целлюлозы
Расположение микрофибрил
целлюлозы вокруг
замыкающих клеток
Эпидермальная клетка
Рис. 14.14. А. Вертикальный
разрез устьица (видна часть нижней
поверхности листа). Б. Структура
микрофибрилл целлюлозы в
стенках замыкающих клеток.
обычный С3-путь фотосинтеза и не образуется
крахмал. Это могло бы помочь объяснить, почему
крахмал образуется не днем, как в обычных фото-
синтезирующих клетках, а ночью. Другой
интересный факт - отсутствие плазмодесм в
замыкающих клетках, т.е. сравнительная
изолированность этих клеток от остальных клеток эпидермиса.
14.4. Подъем воды по ксилеме
Ксилема цветковых растений состоит из двух
типов структур, переносящих воду,- трахеид и
сосудов. В разд. 8.2.1 мы уже говорили о том, как
выглядят соответствующие клетки в световом
микроскопе, а также на микрофотографиях,
полученных с помощью сканирующего электронного
микроскопа (рис. 8.11). Строение вторичной
ксилемы (древесины) мы рассмотрим в разд. 21.6.6.
Ксилема вместе с флоэмой образует проводящую
ткань высших растений. Эта ткань состоит из так
называемых проводящих пучков, которые состоят из
особых трубчатых структур. На рис. 14.15 показано,
как устроены проводящие пучки и как они
располагаются в первичном стебле у двудольных и
однодольных растений.
14.19. Резюмируйте в виде таблицы различия в
строении первичного стебля у двудольных и
однодольных растений.
14.20. Какова трехмерная форма следующих
тканевых компонентов: а) эпидермиса; б)
ксилемы; в) перицикла двудольных и г) сердцевины?
То, что вода может подниматься по ксилеме,
очень легко продемонстрировать, погрузив нижний
конец срезанного стебля в разбавленный раствор
красителя, например эозина. Краситель
поднимается по ксилеме и распространяется по всей сети
листовых жилок. Если сделать тонкие срезы и
просмотреть их в световом микроскопе, краситель
будет обнаружен в ксилеме.
То, что ксилема проводит воду, лучше всего
показывают опыты с «кольцеванием». Такие опыты
проводили задолго до того, как стали применяться
радиоактивные изотопы, позволяющие очень легко
проследить путь вещества в живом организме. В
одном из вариантов опыта вырезают кольцо коры с
флоэмой. Если опыт не очень продолжителен, такое
«кольцевание» не влияет на подъем воды по стеблю.
Однако, если отслоить лоскут коры и вырезать
ксилему, не повреждая лоскута коры, растение
быстро завянет.
Любая теория, объясняющая передвижение
воды по ксилеме, не может не учитывать
следующие наблюдения:
1. Сосуды ксилемы - мертвые трубки с узким
просветом, диаметр которого варьирует в пределах
от 0,01 мм в «летней» древесине примерно до
0,2 мм в «весенней» древесине.
2. Большие количества воды переносятся
относительно быстро: у высоких деревьев
зарегистрирована скорость подъема воды до 8 м/ч, а у
других растений она часто составляет около
1 м/ч.
3. Чтобы поднять воду по таким трубкам к вершине
высокого дерева, необходимо давление порядка
4000 кПа. Самые высокие деревья -
калифорнийские гигантские секвойи (хвойные, у которых нет
сосудов и есть только трахеиды) и австралийские
эвкалипты - бывают выше 100 м. Вода
поднимается по тонким капиллярным трубкам
благодаря высокому поверхностному натяжению под
действием капиллярных сил; однако только за
счет этих сил даже по самым тончайшим сосудам
ксилемы вода не поднимется выше 3 м.
Все эти наблюдения удовлетворительно объясняет
теория сцепления (когезии), или теория натяжения.
Согласно этой теории, подъем воды от корней
обусловлен испарением воды из клеток листа.
Как мы уже говорили в разд. 14.3, испарение
приводит к снижению водного потенциала клеток,
примыкающих к ксилеме. Поэтому вода входит
в эти клетки из ксилемного сока, у которого более
высокий водный потенциал; при этом она
проходит через влажные целлюлозные клеточные
стенки сосудов ксилемы на концах жилок, как показано
на рис. 14.7.
Сосуды ксилемы заполнены водой, и по мере того
как вода выходит из сосудов, в столбе воды
создается натяжение. Оно передается вниз по стеблю на
всем пути от листа к корню благодаря сцеплению
(когезии) молекул воды. Эти молекулы стремятся
Внутрипучковый камбий
Межпучковый камбий
Проводящий пучок
Эпидермальный волосок
(у Helianrhus многоклеточный)
Кутикула
(образована из кутина)
Поперечный разрез
Эпидермис
(однослойный)
Кора (в
наружных клетках мо-'
гут быть хлоро -
пласты)
Колленхима-
Паренхима
Эндодерма-
толщиной в одну клетку;
клетки могут содержать
крахмал)
Сердцевина
(паренхима)
Перицикл
(волокна склеренхимы)
Флоэма (ситовидные трубки,
клетки-спутницы, паренхима, волокна)
Внутрипучковый камбий
Мета ксилема
(зрелые сосуды ксилемы, волокна
и паренхима)
Поперечный разрез
проводящего пучка
Πротоксилема эндархная, т. е. находится ближе к
центру, чем метаксилема; сосуды с кольчатыми и
спиральными утолщениями
Продольный
разрез
Кутикула
(как у двудольных)
Эпидермис
(как у двудольных)
Гиподерма
(колленхима, в наружных
клетках могут быть
хлоро пласты)
Основная ткань
(паренхима; нет деления
на кору и сердцевину)
Сосудистые пучки
(многочисленные,
разбросанные, к центру крупнее)
Перицикл, или обкладка-
проводящего пучка
(склеренхима, окружающая пучок)/
Флоэма
(только ситовидные трубки
и клетки-спутницы)
Метаксилема
(два больших сосуда)
Прото ксилема
Поперечный разрез
проводящего пучка
Лизигенный канал
(образовался в результате
разрушения прото ксилемы)
Эпидермис
Эпидермис -
Колленхима-
Проводящий
пучок
(см. рис. 14.15, Л)
Паренхима коры
Колленхима
Пробка
(начинается
вторичный рост)
Паренхима
сердцевины
Эпидермальный
волосок
Паренхима
(основная
ткань)
Проводящие
пучки (см. рис
14.15,E)
Рис. 14.15. А. Главные особенности анатомического
строения стебля у типичного двудольного растения
Helia nth us annuus (подсолнечника). Б. Анатомическое
строение стебля у типичного однодольного растения
lea mays (кукурузы). В. Часть поперечного среза
стебля подсолнечника при малом увеличении. Г.
Часть поперечного среза стебля Zea при малом
увеличении. Д. Поперечный срез проводящего пучка из
стебля подсолнечника при большом увеличении. E.
Поперечный срез проводящего пучка из стебля
кукурузы при большом увеличении. Ж. Микрофотография
продольного среза стебля подсолнечника. 3.
Микрофотография продольного среза стебля кукурузы.
Перицикл из
волокон
склеренхимы
Флоэма
Камбий
Сосуды
метаксилемы
Прото ксилема
Перицикл
из волокон
склеренхимы
Клетка-спутница
Ситовидная трубка
Флоэма
Сосуды
метаксилемы
Прото ксилема
Лизигенная
полость,
возникшая
в результате
распада
прото ксилемы
Волокна
склеренхимы
Спиральные сосуды
Кольчатый сосуд
Сетчатый сосуд
«прилипнуть» друг к другу, потому что они
полярные и притягиваются друг к другу электрическими
силами, а затем удерживаются вместе за счет
водородных связей (разд. 5.1.2). Кроме того, они
стремятся прилипнуть к стенкам сосудов под действием
сил адгезии. Высокая когезия молекул воды
означает, что для того, чтобы разорвать столб воды,
требуется сравнительно большое растягивающее
усилие; иными словами, столб воды обладает
высокой прочностью на разрыв. Натяжение в сосудах
ксилемы достигает такой силы, что может тянуть
весь столб воды вверх, создавая массовый поток;
при этом вода поступает в основание такого столба
в корнях из соседних клеток корня. Необходимо,
чтобы стенки сосудов ксилемы тоже обладали
высокой прочностью и не вдавливались внутрь.
Такую прочность обеспечивают лигнин и
целлюлоза. Данные о том, что содержимое сосудов
ксилемы находится под воздействием большой
растягивающей силы, были получены при
измерении суточных изменений диаметра ствола у
деревьев с помощью прибора, называемого
дендрометром. Минимальные значения
регистрировались в дневные часы, когда скорость тран-
спирации максимальна. Крохотное сжатие
отдельных сосудов ксилемы суммировалось и давало
вполне измеримое уменьшение диаметра всего
ствола.
Оценки прочности на разрыв для столба кси-
лемного сока варьируют в пределах примерно от
3000 до 30 000 кПа, при этом более низкие значения
получены позднее. В листьях зарегистрирован
водный потенциал порядка —4000 кПа, и
прочность столба ксилемного сока, вероятно,
достаточна, чтобы выдержать создающееся натяжение.
Не исключено, конечно, что столб воды может
иногда разрываться, особенно в сосудах большого
диаметра.
Критики этой теории указывают на то, что любое
нарушение непрерывности столба сока должно
немедленно останавливать весь поток, так как сосуд
должен заполняться воздухом и парами воды
(явление кавитации). Кавитацию может вызвать
сильное сотрясение, сгибание ствола или недостаток
воды. Хорошо известно, что на протяжении лета
содержание воды в стволе дерева постепенно
уменьшается, а древесина заполняется воздухом.
Это используется в лесной промышленности,
потому что такое дерево обладает лучшей
плавучестью. Однако разрыв водного столба в части
сосудов не очень сильно влияет на скорость
переноса воды. Это можно объяснить тем, что вода
переходит из одного сосуда в другой или же
обходит воздушную пробку, передвигаясь по
соседним клеткам паренхимы и их стенкам. Кроме
того, согласно расчетам, для поддержания
наблюдаемой скорости потока вполне достаточно, чтобы в
каждый момент времени функционировала хотя бы
небольшая часть сосудов. У некоторых деревьев и
кустарников вода передвигается только по самому
молодому наружному слою древесины, который
называют заболонью. У дуба и ясеня, например,
вода движется главным образом по сосудам
текущего года, а остальная часть заболони выполняет
функцию водного резерва. В течение вегетационного
сезона все время прибавляются новые и новые
сосуды, но больше всего их образуется в начале
сезона, когда скорость потока гораздо выше.
Вторая сила, которая участвует в передвижении
воды по ксилеме,- это корневое давление. Его можно
обнаружить и измерить в тот момент, когда
отрезают крону, а штамб с корнями продолжает
выделять сок из сосудов ксилемы. Этот процесс эксу-
дации подавляется цианидом и другими
ингибиторами дыхания и прекращается при недостатке
кислорода или понижении температуры. Для
работы такого механизма, по-видимому, нужна
активная секреция в ксилемный сок солей и других
водорастворимых веществ, снижающих водный
потенциал. Затем в ксилему поступает вода за счет осмоса
из соседних клеток корня.
Одного положительного гидростатического
давления около 100-200 кПа (в исключительных
случаях до 800 кПа), создаваемого за счет корневого
давления, обычно недостаточно, чтобы обеспечить
передвижение воды вверх по ксилеме, но его вклад у
многих растений несомненен. У медленно транспи-
рирующих травянистых форм этого давления,
однако, вполне хватает для того, чтобы вызвать
гуттацию. Гуттация-это выведение воды в виде
капель жидкости на поверхности растения (тогда
как при транспирации вода выходит в виде пара).
Все условия, уменьшающие транспирацию, т.е.
слабая освещенность, высокая влажность и т.п.,
способствуют гуттации. Она весьма обычна у
многих растений влажных тропических лесов и часто
наблюдается на кончиках листьев молодых
проростков.
14.21. Перечислите те свойства ксилемы,
благодаря которым она обеспечивает транспорт
воды и растворенных в ней веществ на большие
расстояния.
14.5. Поглощение воды
корнями
На рис. 14.16 показано первичное строение корня у
типичных двудольных и однодольных растений.
14.22. Составьте таблицу главных различий
в первичном строении корня у двудольных и
однодольных растений.
Основная масса воды поглощается более
молодыми частями корня в зоне корневых волосков.
Когда корень растет в почве, на нем непрерывно
Слой клеток с корневыми
волосками
(видоизмененный эпидермис
толщиной в одну клетку, без кути
кул ы, у двухдольных быстро исчезает;
часто отсутствует в готовых препаратах)
Экзодерма
(защитный слой толщиной в одну или
несколько клеток, род гиподермы
иногда по своей структуре и функции
напоминает эндодерму корня)* У
Кора (паренхима,
может запасать крахмал)
Поперечный разрез
(1/2-1/4 корня)
Корневой волосок
(вырост одиночной клетки)
Камбий (имеется
у некоторых
двудольных)
Эндодерма
(толщиной в одну клетку, у клеток
есть пояски Каспари из суберина
(рис. 14.17,5) клетки могут образ<
вывать крахмалоносное влагалище)]
Перицикл (паренхима
толщиной в одну или несколько
клеток; у двудольных сохраняет
меристематическую активность,
участвует в образовании боковых
корней и процессах вторичного
роста)
Прото ксилема (экзархная,
сосуды с кольчатыми и спиральными
утолщениями; у двудольных до 2—8
групп, т. е. мест дифференциации,
в соответствии с чем корни называют
диархными, триархными и т.д.;
в данном случае корень тетрархный)
Продольный разрез
Флоэма
(чередуется
с ксилемой)
M era ксилема I
(у двудольных имеет
несколько лучей,
в данном случае четыре)
Корневой волосок
(как у двудольных)
Слой клеток с корневыми волосками
(как у двудольных, но сохраняется
постоянно)
Экзодерма (как у двудольных)
— Кора
(как у двудольных)
Эндодерма
(как у двудольных)
Перицикл (как у
двудольных, но не сохраняет мери-
стематической активности)
Флоэма (чередуется с
ксилемой)
Прото ксилема (экзархная,
свыше восьми групп, т. е.
корень полиархный)
Метаксилема
Сердцевина (паренхима,
может быть, а может и
отсутствовать)
Экзодерма
(эпидермис утрачен)
Паренхима коры
Зерна крахмала
Флоэма
Корневой
волосок
Слой клеток с
корневыми
волосками
Волокна
экзодермы
Паренхима коры
Эндодерма
с U-образными
утолщениями
Перицикл
Метаксилема
Прото ксилема
Пропускные
клетки эндодермы
Эндодерма
Перицикл
Рис. 14.16. А. Главные особенности анатомического строения корня у типичного двудольного растения
Ranunculus (лютика). Б. Главные особенности анатомического строения корня у типичного однодольного
растения lea mays (кукурузы). В. Поперечный срез корня лютика при малом увеличении. Г Поперечный срез
корня кукурузы при малом увеличении.
Метаксилема
Прото ксилема
Паренхима сердцевины
Флоэма
образуются новые корневые волоски, а старые
отмирают. Новые корневые волоски возникают на
некотором расстоянии позади зоны растяжения.
Корневые волоски-это трубчатые выросты эпидер-
мальных клеток (рис. 14.16), намного
увеличивающие поверхность, которая может поглощать воду и
минеральные соли. Они вступают в тесный контакт
с частицами почвы.
На рис. 14.17, Λ, схематически показано, как вода
проходит по корню. В корне существует градиент
водного потенциала - от более высокого уровня в
клетках, образующих корневые волоски, к более
низкому в клетках, примыкающих к ксилеме.
Градиент водного потенциала поддерживается
двумя способами:
1) за счет передвижения воды вверх по ксилеме,
при котором, как мы уже говорили, в ксилеме
создается натяжение и таким образом понижается
водный потенциал ксилемного сока;
2) за счет того, что осмотическое давление в
ксилемном соке намного выше, чем в более
разбавленном почвенном растворе.
Вода передвигается по корню по тем же путям,
что и в листьях, а именно по апопласту, симпласту и
через вакуоли.
14.5.1. Симпластный и вакуолярный
пути
По мере того как вода поднимается вверх по
корневой ксилеме, ее замещает вода из соседних
клеток паренхимы, скажем таких, как клетка 1 на
Клеточная
стенка
Цитоплазма
Слой клеток,
образующих
корневые волоски
Кора-
Вакуоль
Эндодерма
Перицикл
Корневой
волосок
Воздух + вода
Сосуды ксилемы
Поясок
Каспари
Частичка
почвы
Тонкая пленка
воды, прочно связанная
с частичкой глинозема
Б
Поясок Каспари,
видимый на поперечном
разрезе, в световом
микроскопе выглядит
как "пятнышко "
Поперечный
разрез
Старые
клетки.
Поперечный
разрез
Пропускная
клетка
Наружная
тангенциальная
стенка
Молодые
клетки
АПОПЛАСТНЫЙ ПУТЬ
(прерывается в эндодерме)
СИМПЛАСТНЫЙПУТЬ
ВАКУОЛЯРНЫЙ ПУТЬ
Поясок \
Каспари
охватывает"
всю клетку
Добавочное
вторичное
утолщение ~
из суберина
Добавочный
суберин
Радиальная
стенка
Пропускная
клетка без
добавочных
утолщений
\Внутренняя
тангенциальная
стенка
Поперечная
стенка
Тангенциальная
стенка
Трехмерное изображение
клетки эндодермы
Молодая клетка
Старые клетки
Передвижение воды и
растворенных веществ,
например минеральных солей
Трехмерное изображение
трех соседних клеток эндодермы
В трехмерном
изображении
Рис. 14.17. А. Схема передвижения воды и ионов в корне (корень изображен в поперечном разрезе). Толщина
клеточных стенок для ясности преувеличена. Клетки 1, 2, 3 упоминаются в тексте. Наибольшее значение для
передвижения воды и растворенных в ней веществ имеет апопластный путь. Менее важен симпластный путь,
если не считать переноса солей в области эндодермы. Передвижение по вакуолярному пути ничтожно мало.
Б. Структура и функция эндодермы корня. Показаны пояски Каспари в молодых эндодермальных клетках и
отложение добавочного суберина в более старых клетках, за исключением «пропускных» клеток. В. Названия
стенок: поперечные и радиальные стенки расположены антиклинально (т. е. под прямым углом к
поверхности корня), α тангенциальная стенка - периклинально (т.е. параллельно поверхности корня).
рис. 14.17, А. В результате потери воды водный
потенциал клетки 1 уменьшается, и в нее начинает
поступать вода из клетки 2. Она переходит туда
путем осмоса или по симпласту точно так же, как
это происходило в таких же клетках 1 и 2 в листе
(см. разд. 14.3.2). Тогда в свою очередь снижается и
водный потенциал клетки 2, в нее начинает
поступать вода из клетки 3 и так далее через весь
корень до самого эпидермиса, образующего
волоски.
Почвенный раствор обычно бывает достаточно
разбавленным и потому водный потенциал его
выше, чем в клетках, образующих корневые
волоски, в том числе и в самих волосках. В
результате вода входит в корень из почвы за счет осмоса.
14.23. Расположите по порядку в соответствии
с величиной ψ почвенный раствор, ксилемный сок,
клетку 1, клетку 2, клетку 3, клетку корневого
волоска. (Используйте символ >.)
14.5.2. Апопластный путь
Апопластный путь в корне функционирует в
принципе так же, как в листе (разд. 14.3.1), но с одним
существенным отличием. Когда вода, передвигаю-
щаяся в клеточных стенках, достигает эндодермы, ее
дальнейшему продвижению препятствует
водонепроницаемый материал суберин, который
откладывается в клеточных стенках в виде полосок,
называемых поясками Каспари. Эти пояски
препятствуют передвижению воды по апопласту
(рис. 14.17, £), и поэтому вода и растворенные в ней
соли должны проходить через плазматическую
мембрану под контролем цитоплазмы эндодермальной
клетки. Как полагают, благодаря этому живые
клетки могут регулировать передвижение воды и
минеральных солей из почвы в ксилему. Возможно,
что этот механизм служит также для защиты от
проникновения токсичных веществ, патогенных
грибов и т. п. Интересно отметить, что даже при
плазмолизе цитоплазма эндодермальной клетки не
отстает от клеточной стенки в области пояска
Каспари, хотя она и сжимается в остальной части
клетки. С возрастом опробковение клеток
эндодермы в корне часто усиливается (рис. 14.17, Б). Это
препятствует нормальному выходу воды и
минеральных солей через внутренние тангенциальные
клеточные стенки. Однако в таких стенках могут
сохраняться поры в виде плазмодесм и, кроме того,
остаются так называемые «пропускные» клетки, у
которых не происходит добавочного утолщения
стенки и через которые свободно проходят вода и
растворенные вещества.
Относительная роль апопластного, симпластного
и вакуолярного путей не выяснена.
накоплению ионов против концентрационного
градиента (разд. 7.2.2).
3. Существует непрерывная система клеточных
стенок - апопласт, который распространяется
внутрь от слоя клеток, образующих корневые
волоски. Вода и любые растворенные в ней
вещества поступают в эту систему из почвы путем
массового потока и в меньшей степени путем
диффузии.
4. Передвижение воды по апопласту составляет
часть общего транспирационного тока.
На рис. 14.18 представлено поглощение ионов
калия молодыми корнями злаков, тщательно
отмытыми в чистой воде. Через 90 мин в раствор
добавляли ингибитор дыхания - цианид калия.
14.24. а) Опишите, как происходит поглощение
ионов калия при 00C и при 25°С (рис. 14.18).
б) Объясните выявленные различия и влияние
цианистого калия (KCN).
Результаты, сходные с представленными на
рис. 14.18, можно получить и с изолированными
тканями; обычно используют ткани запасающих
органов, например корней моркови. Кривые на
рис. 14.19 подтверждают, что цианистый калий
ингибирует дыхание.
14.25. На рис. 14.19 показано, что скорость
дыхания в вырезанных из моркови дисках
возрастает, когда их переносят из чистой воды
14.6. Поглощение
минеральных солей
и их транспорт в корнях
Для питания растений необходимы не только
углеводы, образующиеся при фотосинтезе, но и
определенные минеральные элементы. Для чего
нужны эти элементы, указано в табл. 9.10. У высших
растений в поглощении минеральных веществ из
почвы или окружающей воды участвуют корни.
Максимальное поглощение происходит в зоне
корневых волосков.
Минеральные элементы входят в состав
различных солей; в растворе эти соли диссоциируют и
образующиеся ионы свободно передвигаются. В
любой попытке объяснить поглощение и
передвижение минеральных ионов должны учитываться
следующие факты:
1. Мембраны клеток, включая плазматическую
мембрану и тонопласт, строго говоря, не
полупроницаемы, а обладают дифференциальной
проницаемостью - в разной степени пропускают ионы и
другие вещества, растворенные в воде.
2. Возможен активный транспорт веществ
через клеточные мембраны. Для этого нужна
энергия в виде АТФ, образующегося в процессе
дыхания. Активный транспорт может приводить к
Поглощение калия, мкг
на пробу ткани корня
Добавлен цианид калия
Время, мин
Рис. 14.18. Поглощение ионов калия корнями
молодых растений злаков, находящимися в аэрируемом
растворе.
Скорость дыхания
или поглощения KCl
Перенос
из чистой воды
в раствор
KCl
Дыхание
Добавлен
цианид
,калия
Содержание KCl
Время, мин
Рис 14.19. Дыхание и поглощение хлорида калия
кружочками из корня моркови. (По данным Robertson,
Turner, 1945.)
в раствор хлористого калия. Объясните причину
этого, пользуясь представленными данными.
14.26. Почему содержание хлористого калия
перестает возрастать после добавления KCN?
14.27. Если поставить опыт, аналогичный
опыту на рис. 14.18, но с поглощением фосфата,
то окажется, что 16% фосфата, поглощенного
корнями ячменя при кратковременной
инкубации, легко отмывается после переноса корней
снова в чистую воду. Объясните, почему.
14.28. Могут ли ионы дойти до ксилемы
только путем передвижения по апопласту?
Подводя итоги, можно сказать, что поглощение
ионов корнями - это сочетание пассивного
поглощения, когда ионы движутся за счет
массового потока и диффузии через апопласт, с
активным поглощением, или активным транспортом,
при котором ионы переносятся в клетки против
градиента концентрации за счет энергии
дыхания.
Активный транспорт избирателен и зависит от
дыхания, а диффузия неизбирательна и не зависит
от дыхания. В результате пассивного поглощения
каждая клетка в коре корня омывается раствором
такого же состава, как и почвенный раствор.
Так создается большая поверхность для поглощения
ионов.
Ионы, передвигающиеся по апопласту, доходят
только до эндодермы, где, как уже говорилось в
разд. 14.5.2, пояски Каспари препятствуют
дальнейшему передвижению ионов. Чтобы пересечь
эндодерму, ионы должны пройти путем диффузии
или активного транспорта через плазматические
мембраны эндодермальных клеток и попасть в их
цитоплазму, а возможно, и в вакуоли. Так растение
«следит» за тем, какие ионы в конце концов
попадают в ксилему, и контролирует этот
процесс.
14.29. Как бы вы показали с помощью
радиоактивного иона и радиоавтографии, что
эндодерма является барьером для передвижения
ионов через клеточные стенки?
Ионы могут передвигаться также по симпласт-
ному пути. Как только они попадут в цитоплазму
какой-нибудь клетки, они начинают передвигаться
дальше по симпласту, не проходя больше через
плазматические мембраны. Непрерывный сим-
пластный путь идет от корневых волосков до
самой ксилемы. На рис. 14.17, Λ показаны все
возможные пути транспорта ионов в корне.
Конечный этап передвижения минеральных солей
по корню - высвобождение ионов в ксилему. Чтобы
попасть туда, ионы должны на каком-то этапе
выйти из живых клеток, т.е. снова пересечь
плазматическую мембрану, но в обратном
направлении. Это происходит либо путем диффузии,
либо за счет активного транспорта.
14.7. Транспорт минеральных
солей внутри растения
Мы только что рассмотрели путь минеральных
солей в корне от его поверхности до ксилемы, но это
лишь самый первый этап их передвижения. Попав в
ксилему, соли разносятся дальше по всему
растению с транспирационным током,
представляющим собой массовый поток жидкости. Как мы
уже говорили, передвижение минеральных
элементов по ксилеме можно продемонстрировать,
поставив опыты с кольцеванием растений. При этом
удаление тканей, расположенных ближе к
поверхности ствола, т.е. флоэмы и т.д., не влияет на
передвижение ионов вверх по растению. Анализы
ксилемного сока говорят о том, что значительная
доля азота переносится в виде аминокислот и
других близких к ним органических соединений, хотя
часть азота мигрирует в виде неорганических ионов
нитрата или аммония. Следовательно, эти ионы
частично превращаются в корнях в
аминокислоты. Точно так же и небольшие количества
фосфора и серы транспортируются в виде
органических соединений.
Поэтому, хотя традиционно ксилему и флоэму
рассматривают как две ткани, проводящие
соответственно минеральные и органические
вещества, это различие не вполне четкое. К тому же
обычно происходит некоторый обмен веществами
между ксилемой и флоэмой и, кроме того, как мы
увидим дальше, по флоэме переносятся
значительные количества минеральных элементов из
других органов растения.
Главными потребителями минеральных
элементов, т.е. местами их использования, являются
растущие части растения, т. е. верхушечные и
пазушные меристемы, молодые листья, развивающиеся
плоды, цветки и запасающие органы. Растворенные
вещества «выгружаются» из ксилемы на концах
тончайших жилок, при этом они поступают в клетки
путем диффузии и активного поглощения. Иногда в
этом процессе участвуют особые передаточные
клетки (см. разд. 14.8.6).
14.7.1. Рециркуляция и
ремобилизация
Очень часто транслокация минеральных элементов
по ксилеме и флоэме не завершается переносом
веществ из корня к органу-потребителю. В общем
ксилема только доставляет тот или иной элемент к
определенному органу, и если этот элемент здесь не
нужен, то флоэма уносит его дальше вверх или вниз
по растению. Такая рециркуляция была изучена с
помощью радиоактивных изотопов и, как
выяснилось, это довольно обычное явление. В одном из
экспериментов корни, идущие от одного стебля
кукурузы, размещали в двух стаканах с
питательным раствором, при этом в одном из стаканов был
радиоактивный фосфор. За 6 ч этот фосфор про-
делал путь по ксилеме вверх по стеблю, затем
спустился вниз по флоэме и оказался во втором
стакане. Это только часть данных, говорящих о
том, что у растений фосфор быстро и непрерывно
мигрирует по ксилеме и флоэме. Другие элементы
менее подвижны. Сера, например, в основном
удаляется из проводящих тканей за время первой
циркуляции, а кальций известен своей неподвижностью
и практически весь остается в том органе, в который
был доставлен, так как он плохо передвигается по
флоэме.
Очень часто элемент остается внутри органа
только на определенное время, а затем вновь
мобилизуется, т. е., выполнив какую-то полезную функцию
в данном органе, выходит из него и направляется в
другую часть растения. Это происходит при
неодновременном старении листьев, когда более старые,
отмирающие листья экспортируют значительную
часть своих минеральных веществ в более молодые
листья. Точно так же у листопадных деревьев и
кустарников полезные вещества листьев перед
сбрасыванием листвы снова мобилизуются и
сохраняются, откладываясь где-нибудь в другом месте.
Ремобилизация происходит также при развитии
цветков, плодов, семян и запасающих органов.
Направленное передвижение питательных веществ
контролируется ростовыми веществами, особенно
цитокининами (разд. 15.2.7). Из всех элементов
легче всего ремобилизуются фосфор, сера, азот и
калий.
14.8. Транслокация
органических веществ по
флоэме
У тех многоклеточных растений, у которых корни и
другие органы удалены от мест фотосинтеза, для
распределения продуктов фотосинтеза нужна
специальная транспортная система. У сосудистых
растений эти органические продукты переносятся из
главных органов фотосинтеза - листьев - ко всем
остальным частям растения по флоэме. Флоэма
состоит из ситовидных элементов,
клеток-спутников, паренхимы, волокон и склереид. Строение
флоэмы по данным световой микроскопии мы уже
рассматривали в разд. 8.2.2. Ситовидные элементы
располагаются торец в торец и образуют
ситовидные трубки, в которых каждый элемент отделен от
соседнего ситовидной пластинкой. Связи между ав-
тотрофными клетками, образующими органические
питательные вещества, и клетками, получающими
эти вещества, показаны на рис. 14.20. Обратите
внимание на то, что, как видно из этого рисунка,
органические вещества у растений
транспортируются и вверх, и вниз, т. е. в двух направлениях. Это
отличает флоэму от ксилемы, по которой вещества
передвигаются только вверх. Запомните также, что
запасающие органы в разное время выступают то
Фотосинтезирующая ткань
(главным образом листья)
Место запасания
(стебель, корни,
луковицы, клубни и т. п.)
Центры роста
(меристемы, молодые
листья, цветки, плоды)
Рис. 14.20. Передвижение органических веществ в
зеленом растении.
как экспортеры питательных веществ, то как их
импортеры.
Как правило, около 90% растворенных веществ,
переносимых по флоэме, составляет дисахарид
сахароза. Это сравнительно инертный и хорошо
растворимый углевод, который не играет почти никакой
роли непосредственно в самом метаболизме и
поэтому служит идеальной транспортной формой
сахара, так как маловероятно, чтобы он расходовался
в процессе переноса. Основное предназначение
сахарозы-вновь превратиться в более активные
моносахариды. Концентрация сахарозы во флоэме
может быть очень высокой, и у сахарного тростника
она достигает 25% (вес/объем).
Следует отметить, что по флоэме в различной
форме переносятся и некоторые минеральные
элементы, например азот и сера-в форме
аминокислот, фосфор-в виде фосфорилированных Сахаров и
ионов неорганического фосфата, калий-в виде
ионов. Кроме того, в небольшом количестве в ней
могут присутствовать витамины, ростовые
вещества (ауксины, гиббереллины и др.), искусственно
привнесенные химикалии, вирусы и иные
компоненты. Значение флоэмы для рециркуляции и ремо-
билизации минеральных веществ уже обсуждалось в
разд. 14.7.1.
Данные о циркуляции углерода в растении можно
получить, если ввести в листья CO2 с
радиоактивным изотопом 14C. Радиоактивная углекислота
фиксируется в процессе фотосинтеза, и 14C
переходит в растворенные органические вещества,
например сахарозу. Передвижение метки в растении
можно проследить самыми разными методами,
предназначенными для выявления радиоактивных
изотопов, например с помощью радиоавтографии,
сканирования поверхности счетчиком Гейгера или
экстракции изотопа из разных частей растения. В
конечном счете и флоэма, и ксилема будут
непосредственно участвовать в циркуляции углерода.
Так, например, углерод в форме сахарозы может
попадать в корни и там использоваться для
превращения нитратов в аминокислоты; а затем
синтезированные аминокислоты, содержащие этот
углерод, могут транспортироваться в ксилемном
соке вверх по стеблю.
Опыт 14.6. Изучение распределения б
продуктов фотосинтеза
у растений гороха
В основе описанного ниже опыта лежат данные,
использованные в учебной телевизионной
программе «Исследование фотосинтеза и транспорта асси-
милятов». В этом опыте фотосинтезирующим
растениям гороха (Pisum sativum) давали углекислоту,
содержавшую радиоактивный изотоп углерода 14C.
Это весьма удобный изотоп, так как у него очень
большой период полураспада (5570 лет), поэтому
его радиоактивность практически не меняется на
протяжении всего эксперимента (сравните с 11C, у
которого период полураспада составляет всего
лишь 20,5 мин). К тому же с ним относительно
безопасно работать, так как его излучение -
слабопроникающее (β-частицы с низкой энергией).
Растение использует 14CO2 для фотосинтеза
точно так же, как и обычную углекислоту (12CO2), и
включает ее в органические продукты (ассимиляты).
Можно экспериментально проследить перемещение
таких ассимилятов, потому что в них содержится
14C; о таких веществах говорят, что они помечены
14C.
Рис. 14.21. Этапы эксперимента по изучению
распределения ассимилятов, меченных 14C, у гороха.
Фото А.
Растение гороха, один из листьев которого помещен
в прозрачный, хорошо освещенный мешок с 14CO2
Общее время экспозиции 15—30 мин.
Лист вынимают из мешка, а растение
оставляют на 24 ч, чтобы меченные
14C ассимиляты (главным образом
сахароза) могли транспортироваться
из листьев в другие части растения.
Радиоавтография целого растения
Радиоавтография растений,
меченных 14C
Количественное определение содержания
14Cb разных частях растения
Растение разрезают на части
Фото Б. I
Растение после высушивания Апикальная
и расправления на бумаге; зона стебля
стрелкой показан тот лист,
который инкубировался с 14CO2
Лист, Другие Стебель Боб
обрабо- листья (если
тайный есть)
14С
Корни
Фото В.
Радиоавтограф, полученный
с помощью рентгеновской пленки,
которую кладут сверху на высушенное
растение и оставляют затем в темноте
на четыре дня; β -Излучение 14C засвечивает
пленку; чем чернее участок, тем больше в нем С
Рис. 14.21. (продолжение).
Каждую часть сжигают в печи с О 2
вещества, содержащие 14C, дают
при этом меченую углекислоту
I4CO2, которая поглощается
сцинтиляционной жидкостью,
применяемой в счетчиках
Радиоактивная проба + сцинтил-
ляционная жидкость. При
распаде 14C энергия излучаемых
β -частиц заставляет сцинтилля-
ционную жидкость
флуоресцировать; пробу помещают в прибор,
называемый сцинтилляционным
счетчиком, где слабые вспышки
света преобразуются в усиленный
электрический сигнал; частота
таких сигналов пропорциональна
количеству 14C в пробе
Методика
На рис. 14.21 представлены основные этапы
эксперимента, в котором 14CO2 поступает извне в
нижний или в верхний лист отдельных растений гороха,
имеющих по одному бобу. После этого в течение
суток меченные 14C ассимиляты транспортируются
из листьев в другие части растения. По прошествии
этих суток исследуют распределение
радиоактивности методом радиоавтографии или же путем
измерения количества 14C в каждой из частей
растения. Можно сначала получить радиоавтографы, а
затем разрезать растение на части и определить в
них содержание 14C после сжигания.
Результаты
Результаты радиоавтографии представлены на
рис. 14.21. Результаты подсчета
радиоактивности 14C в разных частях растения приведены в
табл. 14.8.
14.30. а) Взяв данные из табл. 14.8, рассчитайте
долю радиоактивности (в %) в различных частях
обоих растений и прямо сравните распределение
меченых ассимилятов в этих двух случаях.
Каковы основные различия и в чем сходство картины
экспорта 14C из верхних и из нижних листьев?
б) Оцените физиологическое значение
наблюдаемых различий или сходств в связи с ростом всего
растения и отдельных его частей. (Попробуйте
нарисовать простейшую схему растения гороха,
изобразив на ней около восьми листьев и
поместив боб в районе третьего от вершины листа, а
затем указать направление и интенсивность
движения ассимилятов от верхнего и от нижнего
листа к бобу и к корневой системе.)
14.8.1. Характерные особенности
транслокации по флоэме
Если для транспорта веществ по ксилеме имеется
вполне удовлетворительная гипотеза, то в
отношении флоэмы вопрос остается спорным. Прежде чем
рассмотреть возможные механизмы транслокации
по флоэме, полезно будет перечислить некоторые
факты, которые сильно затрудняют решение
проблемы, но тем не менее должны учитываться в любой
гипотезе.
1. Количество переносимого материала может быть
очень большим. Подсчитано, например, что за
вегетационный период вниз по стволу большого
дерева может пройти около 250 кг сахара.
2. Скорость движения велика, обычно она составляет
20-100 см/ч. Зарегистрированы максимальные
скорости более 600 см/ч.
14.31. Если сахароза перемещается по
ситовидным трубкам со скоростью 100 см/ч, а длина
ситовидного элемента 200 мкм, за какое время
молекула сахарозы пройдет один ситовидный
элемент?
В стеблях двудольных растений скорость
передвижения нередко составляет около 10—25 г
сухого вещества на 1 см2 поперечного сечения
ситовидных трубок в час.
3. Возможен перенос на очень большие расстояния.
Самые высокие деревья, например эвкалипты,
могут быть высотой до 100 м и выше. Листья
эвкалипта находятся в основном у верхушки
ствола, поэтому ассимиляты должны
продвигаться по всей длине ствола, а часто еще и на
большое расстояние по корням.
4. Относительная масса флоэмы невелика. Толщина
слоя функционально активной флоэмы,
расположенного по окружности древесного ствола,
близка к толщине почтовой открытки. Из
флоэмы состоит самый внутренний слой коры
одревесневших стеблей и корней; при этом более
Таблица 14.8. Радиоактивность (ими. мин) различных частей
растения гороха через 24 ч после включения 14CO21'
Часть растения После включения в После включения в
верхний лист нижний лист
Верхушечная зона 1 123 759
стебля
Лист, обработан- 11 325 11 372
ный 14C
Другие листья 234 168
Стебель 819 1 160
Боб 9055 4937
Корни 842 2 700
!) За вычетом естественного фона, обусловленного
космическим излучением.
старые слои флоэмы растягиваются и отмирают
по мере роста органа и увеличения его
окружности.
5. Ситовидные трубки очень тонки, их диаметр не
превышает 30 мкм. Последняя величина сравнима
с толщиной очень тонкого человеческого волоса.
Ситовидные трубки через определенные
промежутки перегорожены ситовидными пластинками
с порами еще меньшего диаметра. Чем уже
трубки и мельче поры, тем выше их
сопротивление протеканию жидкости и тем большая сила
нужна для того, чтобы преодолеть это
сопротивление.
14.32. Через сколько ситовидных пластинок
пройдет молекула сахарозы, если она
переместится на 1 м по ситовидным трубкам, состоящим
из ситовидных элементов длиной 400 мкм
каждый?
6. Помимо ситовидных пластинок ситовидные
трубки имеют и ряд других особенностей, о
которых тоже не следует забывать.
14.8.2. Ультраструктура ситовидных
трубок
В отличие от сосудов ксилемы - мертвых пустых
трубок практически без каких-либо внутренних
препятствий - ситовидные трубки флоэмы являются
живыми образованиями и определенно содержат
рят препятствий, мешающих току раствора, а
именно ситовидные пластинки и (в меньшей степени)
цитоплазму. Поскольку механизм передвижения до
сих пор не выяснен, в поисках новых сведений очень
важно изучить ультрамикроскопическую структуру
ситовидных трубок. На рис. 14.22 представлена
электронная микрофотография зрелого ситовидного
элемента, а на рис. 14.23-схема, на которой
указаны все основные детали ситовидных элементов и
примыкающих к ним клеток-спутников.
В процессе развития ситовидного элемента из
меристематической клетки ядро дегенерирует, и
перед нами оказывается необычный пример
живой клетки, не имеющей ядра; в этом отношении
она сходна с эритроцитом млекопитающего. В то
же самое время происходят и другие глубокие
изменения, результаты которых можно видеть на
рис. 14.23. Торцовые клеточные стенки
преобразуются в ситовидные пластинки; при этом плазмо-
десмы, проходящие через эти стенки, сильно
увеличиваются в размерах и превращаются β
ситовидные поры. На рис. 8.12 показано, как выглядит
ситовидная пластинка при взгляде сверху. В
результате всех этих изменений остается трубкооб-
разная структура, состоящая из большой
внутренней полости и очень тонкого, почти незаметного
периферического слоя живой цитоплазмы,
ограниченного плазматической мембраной.
Каждый ситовидный элемент тесно связан с од-
Клетка-спутница
ФБ
Клеточная
стенка
Срединная
пластинка
Каллоза
КС
Срединная пластинка
ситовидной пластинки
Ситовидная трубка
КС — клетка-спутница
Π — ситовидная пора
Пл — пластида
ФБ — флоэмный белок
Рис. 14.22. Электронная микрофотография зрелого
ситовидного элемента.
ной или несколькими клетками-спутниками, т.е.
паренхимными клетками, которые возникают из
той же самой родительской клетки, что и соседний
ситовидный элемент. Клетки-спутники имеют очень
плотную цитоплазму, содержащую небольшие
вакуоли и обычные клеточные органеллы. Эти клетки
обладают очень высокой метаболической
активностью, на что указывает присутствие
многочисленных митохондрий и рибосом (рис. 14.23); в
структурном и физиологическом отношении они очень
тесно связаны с ситовидными элементами и
совершенно необходимы для их функционирования: в
случае гибели клеток-спутников погибают и
ситовидные элементы.
У двудольных и некоторых однодольных
растений в ситовидных элементах образуются большие
количества фибриллярного белка, который
называют флоэмным белком (Ф-белком). Этого белка
иногда откладывается так много, что его можно
видеть даже в световой микроскоп. Когда-то такие
отложения называли «слизистыми тельцами» или
«слизистыми пробками», но они не состоят из
углеводов и поэтому их нельзя считать настоящей
слизью. Много спорят о том, присутствует ли этот
белок в ситовидных порах в нормальных условиях.
Иногда, хотя и не во всех случаях, волокна Ф-белка
выявляются в ситовидных порах при электронной
микроскопии. Одна из больших трудностей при
подготовке препарата флоэмы для электронной
микроскопии связана с тем, что содержимое
ситовидных трубок, как полагают, находится под
высоким гидростатическим давлением - возможно, до
3000 кПа. При подготовке среза для фиксации такое
давление могло бы внезапно сбрасываться, и в
результате флоэмный белок вместе с остальным
содержимым ситовидного элемента мог бы
продавливаться через поры и закупоривать их. В этом
случае появление флоэмного белка в порах
ситовидных пластинок было бы артефактом. Чтобы
решить этот вопрос, пробовали, например, совсем
устранить или уменьшить гидростатическое
давление, подсушивая растение перед приготовлением
срезов. Электронная микроскопия увядших
растений показала, что ситовидные поры у них иногда
закупорены полностью, иногда лишь частично, а
иногда не закупорены вовсе. Таким образом, этот
прием, так же как и другие методы, не позволил
решить проблему.
14.8.3. Данные о передвижении
веществ по флоэме
Очень важно, особенно в связи с вопросом о
механизме транспорта, иметь полную уверенность в том,
что растворенные органические вещества
действительно передвигаются по ситовидным трубкам
флоэмы. Самые первые данные о передвижении
Сахаров и других соединений по флоэме были
получены в опытах с кольцеванием, когда вырезали
кольцо ткани, содержащей флоэму, а ксилему не
трогали. Еще в 1675 г. Мальпиги показал, что вода
идет вверх по древесине, а питательные
вещества-вниз по «коре». Он делал кольцевые вырезки
коры у деревьев (в коре находится флоэма) и
обнаружил, что листья при этом не вянут, а рост ниже
кольца сильно замедляется.
Очень много экспериментов по кольцеванию
продел ами Мэзон и Маскелл, работавшие в 20-е и 30-е
годы на Тринидаде с растениями хлопчатника. Один
из этих опытов представлен на рис. 14.24; его
результаты привели исследователей к выводу, что,
когда флоэма перерезана, но остается в контакте с
ксилемой, между обеими тканями может
происходить некоторый обмен сахарами (рис. 14.24,/!), но
передвижение веществ вниз по стволу происходит
по флоэме (рис. 14,24,5" и В).
Чтобы продемонстрировать перенос сахарозы по
флоэме, ставились простые эксперименты двух
типов. В 1945 г. в растение вводили 13C в виде 13CO2 и
затем выявляли этот нерадиоактивный изотоп
углерода при помощи масс-спектрометрии. Кольцо
флоэмы ошпаривали тонкой струей перегретого
пара, чтобы убить клетки, и после этого
транслокация меченной 13C сахарозы через ошпаренный
участок прекращалась. Однако такая обработка не
влияла на передвижение минеральных элементов по
ксилеме. Эксперименты второго типа были
основаны на выявлении радиоактивности во флоэме
ЭЛЕМЕНТ СИТОВИДНОЙ ТРУБКИ, или
СИТОВИДНЫЙ ЭЛЕМЕНТ, в зрелом состоянии
не имеет ядра, рибосом и аппарата Гольджи;
митохондрии мелкие, цитоплазмы очень мало;
диаметр 10 — 15 мкм, длина 150—1000 мкм
Митохондрия
Гладкий эндоплазматический
ретикулум
Цитоплазма образует тонкий
периферический слой под клеточной
стенкой и плазматической мембраной
Клеточная стенка
Пластида-
Крахмальное зерно
Эндоплазматический //
ретикулум —
обычно стопки
цистерн
Флоэмный белок
(в живых ситовидных
клетках может присутствовать
в ситовидных порах, но
может и отсутствовать)
Ситовидная пора
Каллоза
(полимер глюкозы)
Плазматическая
мембрана
Целлюлозная
клеточная стенка
Срединная
пластинка
Клеточная
стенка
Небольшая
вакуоль
Свободные
рибосомы
(во всей
цитоплазме)
Аппарат
Гольджи
Рибосомы
Шероховатый
эндоплазматический
ретикулум
Плазмодесма
(крупная)
Ядро
Небольшая
вакуоль
Митохондрия
КЛЕТКА-СПУТНИЦА
с плотной цитоплазмой,
многочисленными
рибосомами и
митохондриями, очень активная
Флоэмный белок
Рис. 14.23. Схематическое изображение элементов ситовидной трубки и клетки-спутника, как они выглядят
на продольном срезе в электронном микроскопе. Если ситовидная трубка повреждена (например, травоядным
животным), в ситовидной пластинке быстро откладывается каллоза, что препятствует потере ценных
веществ, содержащихся в ситовидной трубке.
после инкубации растений с 14CO2 и последующей
микрорадиоавтографии срезов стебля. Введение в
практику радиоактивных изотопов сильно
стимулировало работы по транслокации в 30-е и 40-е
годы.
То, что передвижение идет по ситовидным
трубкам, было потверждено в изящных экспериментах, в
которых была использована способность тлей
питаться переносимыми по флоэме сахарами. Тля
прокалывает ткани растения с помощью своего
специализированного ротового аппарата; в этот
аппарат входят чрезвычайно тонкие трубчатые
«стилеты», которые медленно проталкиваются
через ткани растения до тех пор, пока не достигнут
флоэмы. Как видно из рис. 14.25, такие стилеты
могут проникать в индивидуальную ситовидную
трубку.
Если обездвижить тлю двуокисью углерода и
затем удалить тело, оставив стилеты внутри
растения, содержимое ситовидной трубки будет
продолжать проталкиваться через колюще-сосущий
ротовой аппарат гидростатическим давлением в
ситовидной трубке; выходящую наружу жидкость
можно собрать с помощью микрокапилляра. Этот
прием нашел ряд полезных приложений, например
для измерения скорости тока жидкости в
ситовидных трубках (по скорости ее выхода из капилляра) и
для анализа их содержимого.
Наконец, повышение чувствительности пленки,
используемой для микрорадиоавтографии,
позволило точно локализовать соединения, меченные
слабо излучающим изотопом водорода тритием
(3H), именно в ситовидных трубках, а не в
каких-либо иных клетках флоэмы. Изотоп вводили в
составе той или другой аминокислоты или же
сахарозы.
Все эти опыты позволили также установить, что
самые разные вещества переносятся по флоэме
одновременно и вверх и вниз, хотя такое двустороннее
движение происходит, вероятно, не в одной и той же
ситовидной трубке, а в разных трубках,
расположенных по соседству друг с другом.
МЕТОДЫ
Один из четырех
лоскутов, лоскуты
привязывают обратно
на свое место, как
показано на схеме Б,
но без вазелина
Разрезы
длиной
16,5 см
Кольцо
шириной)
1,3 см
Лоскуты, вырезанные
как на схеме А \ перед
их привязыванием на
место ксилему
замазывают вазелином,
поэтому ксилема и
флоэма не могут
обмениваться
веществами
Как на
схеме Л,
но ствол
кольцуют
с двух
сторон
разреза
РЕЗУЛЬТАТЫ
Углеводы обнаружи
ваются и в ксилеме,
и во флоэме
Углеводы обнаруживаются
главным образом
во флоэме
И в ксилеме, и во флоэме
очень мало углеводов
Рис. 74.24. Опыты по кольцеванию растений хлопчатника, проведенные Мэйсоном и Маскеллом.
Ситовидная
трубка
,флоэмы
Волокна
Хоботок
тли
Рис. 14.25. А. Тля, хоботок которой проник через эпидермис листа. Б. Хоботок тли, вошедший в отдельную
ситовидную трубку.
14.8.4. Механизм транслокации
веществ по флоэме
В разд. 14.8.1 были перечислены факты, которые
должны быть учтены в любой гипотезе. В
последующих разделах мы нарисовали картину активной
флоэмы, по которой через очень тонкие ситовидные
трубки с относительно большой скоростью
переносятся большие количества материала. Внутри
трубок имеются добавочные препятствия - ситовидные
пластинки; кроме того, есть и другие структурные
особенности (в частности, наличие флоэмного
белка), о роли которых ничего не известно. Если мы
учтем еще один факт, а именно то, что вся система
очень хрупка и легко повреждается при всяком
постороннем вмешательстве, станет понятно, что
исследователям было очень трудно выяснить,
механизм транслокации по ситовидным трубкам.
Многие сейчас считают, что по ситовидным
трубкам идет массовый поток раствора, т.е.
перемещение всей массы воды и растворенных в ней веществ в
одном направлении, в отличие от диффузии, при
которой различные молекулы и ионы перемещаются
независимо друг от друга. Сама по себе диффузия-
слишком медленный процесс, чтобы им можно
было объяснить наблюдаемую скорость передвижения
веществ. Сейчас мы перечислим данные, которые
указывают на то, что по ситовидным трубкам идет
массовый поток вещества.
1. Когда флоэму перерезают, начинает вытекать
сок; по-видимому, он движется путем массового
потока. Иногда это используют в практических
целях для получения сахара. Так, например,
сахарная пальма (аренга) «плачет», выделяя
ежедневно по 10 л сахаристого сока.
2. Длительное вытекание раствора сахарозы через
хоботок тли (см. разд. 14.8.8) говорит о наличии
в ситовидных трубках гидростатического
давления.
3. Некоторые вирусы переносятся по флоэме, что
Массовый поток раствора
Перемещение воды
Полупроницаемая
мембрана
Раствор с высоким ОД,
например
концентрированный раствор сахарозы
указывает на массовый поток, а не диффузию,
так как вирус не находится в растворе и
переносится пассивно.
Основные споры касаются механизма самого
массового протока, и сейчас мы рассмотрим важнейшие
гипотезы.
Гипотеза Мюнхо о перемещении под
давлением
В 1930 г. Мюнх предложил чисто физическую
гипотезу, объясняющую, как происходит транслокация в
ситовидных трубках. Эта гипотеза
проиллюстрирована с помощью модели на рис. 14.26.
В этой модели вода исходно стремится
переходить путем осмоса из D в А и в С, но это стремление
для А больше, так как раствор в А более
концентрирован, чем в С. По мере того как вода входит в А,
в замкнутой системе А—В—С создается тургорное
(гидростатическое) давление, заставляющее воду
выходить из С. Массовый поток раствора проходит
через В в соответствии с установившимся
градиентом гидростатического давления. Кроме того,
существует и градиент осмотического давления,
направленный от А к С. В конце концов вся система
приходит в равновесие, когда содержимое А
разбавляется водой, а в С накапливаются растворенные
вещества. Эту модель можно применить и к живым
растениям. Отсек А соответствует тканям листа,
образующим сахар в процессе фотосинтеза, в
результате чего повышается осмотическое давление в
клетках листа. Вода, поступающая к листьям по
ксилеме (D), входит в клетки листа за счет осмоса и
повышает тургорное давление в них. Одновременно
сахара расходуются потребителями ассимилятов,
например корнями (С), для самых разных целей, в
том числе для дыхания и синтеза целлюлозы. Это
понижает осмотическое давление в клетках таких
тканей. Между листьями и корнями (или, в более
общей форме, между источниками ассимилятов и их
Манометр, прикрепленный к трубке В,
позволяет регистрировать гидростатическое
давление
Полупроницаемая
мембрана
Раствор с низким ОД,
например
разбавленный раствор сахарозы
Вода
Рис. 14.26. Физическая модель, иллюстрирующая гипотезу Мюнха о том, что транслокация по флоэме
происходит за счет массового потока. Моделируемые части живого растения: А - источник ассимилятов,
например лист; В флоэма, С ткани, потребляющие ассимиляты, например корни, меристемы, плоды,
D-ксилема, клеточные стенки и межклеточные пространства.
потребителями) создается градиент
гидростатического давления, что приводит к массовому потоку
жидкости. Равновесие никогда не достигается, так
как растворенные вещества все время расходуются
тканями-потребителями (С) и непрерывно
образуются в ассимилирующих тканях (А).
Гипотеза Мюнха дает чисто физическое
объяснение току под давлением, но она не объясняет,
почему ситовидные трубки должны быть живыми и
метаболически активными. Кроме того, она не
позволяет объяснить тот факт, что клетки листа
способны загружать ситовидные трубки ассимилятами
против градиента концентрации, т.е. факт более
высокого осмотического давления в ситовидных
трубках по сравнению с клетками листа. Поэтому
гипотеза была видоизменена - в нее включили
активный механизм переноса растворенных веществ в
ситовидные трубки. В этом случае градиент
осмотического и гидростатического давления начинается
не в фотосинтезирующих клетках, а в ситовидных
трубках. Кроме того, полагают, что разгрузка
флоэмы на уровне потребителей-тоже активный
процесс.
Донные в пользу гипотезы Мюнха
1. Согласно этой гипотезе, по ситовидным трубкам
идет массовый поток вещества, и это скорее всего
верно (соответствующие данные приводились
выше).
2. Гипотеза исходит из существования
осмотического градиента и высокого тургорного
(гидростатического) давления во флоэме. Оба этих
фактора продемонстрированы у ряда растений.
Данные против гипотезы Мюнха
1. Для того чтобы растворенные вещества
передвигались с наблюдаемой скоростью, нужны
относительно высокие градиенты гидростатического
давления. Если ситовидные поры полностью
открыты, достаточно будет градиента около
100 кПа/м, однако существование столь крутых
градиентов до сих пор остается сомнительным.
Если же поры ситовидных пластинок заполнены
цитоплазмой или флоэмным белком, т.е.
имеются значительные препятствия для потока,
расчетные величины нужного давления
становятся совершенно нереальными.
2. Гипотеза Мюнха не объясняет, для чего нужны
ситовидные пластинки и флоэмный белок,
которые, по-видимому, создают дополнительные
препятствия для массового потока.
Электроосмос
Механизм, основанный на электроосмосе,
предложили независимо друг от друга Фенсом в 1957 г. и
Спэннер в 1958 г. Явление электроосмоса
иллюстрирует рис. 14.27.
Электроосмос обусловлен наличием поверхности
Катод
Анод
Исходный уровень
раствора
Массовый поток
раствора
Ионный раствор
Пористая мембрана, тонкие трубочки или
макромолекулы; они закреплены на месте
и имеют заряженную поверхность
Рис. 14.27. Электроосмос. В случае, показанном на
рисунке, закрепленная поверхность заряжена
отрицательно, а раствор несет соответствующий
суммарный положительный заряд.
с фиксированным зарядом, находящейся в ионном
растворе. Эта поверхность приобретает заряд, когда
молекулы, из которых она состоит, начинают
отдавать ионы. Например, если нерастворимые
молекулы (скажем, молекулы фибриллярного белка),
помещенные в воду, начинают освобождать ионы
водорода, то эти положительно заряженные ионы
могут свободно перемещаться в растворе, а в том
месте, где они были раньше, создается
отрицательно заряженная поверхность, состоящая из
неподвижных молекул. В результате раствор несет
подвижный положительный заряд, который
сбалансирован с неподвижным отрицательным зарядом.
Если к заряженной поверхности приложить
разность потенциалов, положительные ионы будут
двигаться к отрицательно заряженному электроду
(катоду). Каждый ион окружен оболочкой из
молекул воды (рис. 5.5) и движется вместе с нею, а также
с другими ионами и незаряженными молекулами
типа сахарозы в массовом потоке всего
раствора, проходящего мимо фиксированного заряда
(рис. 14.27).
В ситовидных трубках такими фиксированными
заряженными поверхностями могли бы служить
боковые клеточные стенки, ситовидные пластинки и
флоэмный белок; все они были бы заряжены
отрицательно при тех щелочных условиях, которые обычно
создаются внутри флоэмы. Гипотеза Спэннера
видоизменялась несколько раз, но «окончательная
новая форма», опубликованная в 1979 г.1, может быть
вкратце изложена так:
1. Массовый поток возникает в результате загрузки
ситовидных трубок ассимилятами и
осуществляется так, как это описано Мюнхом.
Spanner D С. (1979). Plant, Cell and Environment, 2.
2. Ф-белок рыхло упакован внутри ситовидных
пор-видимо, под действием массового потока.
3. Концентрация ионов калия (K+) в растворе,
находящемся внутри ситовидной трубки,
достаточно высока. Эти и другие катионы проходят через
ситовидную пластинку, а анионы отталкиваются
отрицательным зарядом флоэмного белка (и
клеточных стенок).
4. На ситовидной пластинке возникает разность
потенциалов, так как выше ее начинают
накапливаться анионы. Выше пластинки создается
суммарный отрицательный заряд, а
ниже-положительный.
5. Как только достигается критическая разность
потенциалов, происходит местное истечение
ионов H+ (протонов) со стенки ситовидной
трубки; протоны проходят через плазматическую
мембрану в просвет трубки и, увеличивая
концентрацию H + , понижают в ней рН.
6. Протонов с избытком хватает для нейтрализации
отрицательного заряда, в результате чего
разность потенциалов на пластинке меняет знак и
местное окружение над пластинкой на время
приобретает положительный заряд.
7. Появление такого положительного заряда
приводит к электроосмосу ионов K+ через
ситовидную пластинку мимо фиксированного
отрицательного заряда Ф-белка. В результате
передвижения калия и других катионов индуцируется
массовый поток раствора.
8. Протонный насос мембраны ситовидной трубки
быстро восстанавливает высокое значение рН
внутри трубки, активно откачивая из нее
протоны. Энергию для этого доставляют в форме АТФ
клетки-спутники и ситовидные трубки.
9. Снова начинает создаваться исходная разность
потенциалов.
Конечный результат состоит в том, что
транслокация по ситовидным трубкам происходит за счет
массового потока, предположенного Мюнхом, но
она ускоряется электроосмосом на ситовидных
пластинках. Ускорение происходит за счет
кратковременных толчков-импульсов. Совсем не
обязательно, чтобы оно происходило одновременно на
всех ситовидных пластинках; может существовать
определенная последовательность «включения»
разных пластинок, но ее может и не быть. Поэтому
поток в ситовидных трубках может носить не
равномерный, а пульсирующий характер.
Данные в пользу
электроосмотической гипотезы
1. Установлена роль клеток-спутников в снабжении
АТФ.
2. В ситовидных трубках обнаружены высокие
концентрации ионов калия.
3. Давление, необходимое для транслокации, было
бы намного ниже, чем по гипотезе Мюнха.
4. Намечена роль для ситовидных пластинок: они
создают заряженную пористую структуру,
позволяющую растворенным веществам передвигаться
за счет электроосмоса. К тому же выясняется и
возможное предназначение флоэмного белка,
если он находится в ситовидных порах.
Данные против
электроосмотической гипотезы
Эта гипотеза основана по существу на косвенных
соображениях - прямых ее подтверждений пока нет.
Так, например, не удалось измерить
предполагаемые в ней изменения разности потенциалов. Не
показано также и то, что стенки ситовидных трубок
служат резервуаром ионов водорода.
Эстафетная гипотеза
Существенный недостаток гипотезы Спэннера
состоит в том, что для поддержания наблюдаемой
скорости потока понадобились бы очень большие
затраты метаболической энергии, так как
«закачивание» раствора происходит только на сравнительно
небольшой поверхности ситовидных пластинок. В
1979 г. Александр Лэнг высказал мысль, что
закачивание могло бы происходить на гораздо большей
поверхности, если бы в нем участвовали также
мембраны смежных перекрывающихся ситовидных
трубок в области перекрывания. Процесс
закачивания раствора мог бы протекать так же, как при
загрузке или разгрузке ситовидных трубок у
источника или потребителя ассимилятов. Растворенные
вещества по-прежнему передвигались бы по
градиенту давления между источником и потребителем,
как предполагал Мюнх, но в этом случае требовался
бы гораздо меньший градиент давления. Такой
«эстафетный» механизм по своей сути почти ничем
не отличается от течения под давлением, но он
позволяет примирить некоторые противоречивые
данные, приводимые в пользу и против гипотезы
Мюнха. В поддержку новой гипотезы можно
привести данные о том, что растворенные вещества
легко входят и выходят из ситовидных трубок по
всей их длине, например из флоэмы в ксилему и
обратно.
Другие гипотезы
Есть и другие гипотезы о флоэмном транспорте, в
частности учитывающие движение цитоплазмы, а
также наличие цитоплазматических «трансцеллю-
лярных» тяжей, прикрепленных к ситовидным
пластинкам и тянущихся на большие расстояния вдоль
ситовидных трубок; последнее наводит на мысль,
что растворы могут каким-то образом
перекачиваться и вдоль всего тяжа. Так же как и теория
электроосмоса, эти идеи интересны, но пока не
подкреплены почти никакими экспериментальными
данными.
14.8.5. Механизмы «первой помощи»:
возможная роль
ситовидных пластинок,
флоэмного белка и пластид
Одна из опасностей, с которой сталкиваются все
растения,-это повреждение травоядными
животными. Если при этом будет нарушена целость
ситовидных трубок, то утечка таких
высокоэнергетических веществ, как сахароза, дорого обойдется
растению. Поврежденные ситовидные трубки
обычно закупориваются уже через несколько минут в
результате отложения каллозы на ситовидных
пластинках и закрытия пор. Возможно, что это одна из
функций ситовидных пластинок. Было высказано
предположение, что у двудольных растений роль
флоэмного белка как еще одного средства «первой
помощи» связана с быстрым блокированием
ситовидных пор сразу же после повреждения трубки; при
этом может использоваться возникающий перепад
давления, о котором упоминалось в разд. 14.8.2. У
однодольных, у которых флоэмного белка обычно
очень мало, ту же роль могли бы играть пластиды,
находящиеся в ситовидных трубках.
14.8.6. Загрузка ситовидных трубок
ассимилятами
Было показано, что концентрация сахарозы в
ситовидных трубках листа обычно находится в пределах
от 10 до 30%, тогда как в фотосинтезирующих
клетках, где этот сахар образуется, концентрация
его в растворе составляет всего лишь около 0,5%.
Значит, загрузка ситовидных трубок происходит
против концентрационного градиента. На коротком
пути от хлоропласта до ситовидной трубки,
составляющем не более 3 мм, вещества могут двигаться и
по симпласту, и по апопласту.
В 1968 г. Ганнинг и его сотрудники описали
новый тип видоизмененных клеток-спутников. В
результате дополнительного утолщения клеточные
стенки у них образуют многочисленные внутренние
выступы, и это примерно в десять раз увеличивает
площадь плазматической мембраны, выстилающей
такую стенку. Как полагают, такое видоизменение
клеток связано с необходимостью активно
поглощать вещества из соседних фотосинтезирующих
клеток и активно же передавать их ближайшим
ситовидным элементам через сложно устроенные,
сильно развитые плазмодесмы. Энергию для всех
этих активных процессов доставляют
многочисленные митохондрии, лежащие в цитоплазме таких
клеток. Эти видоизмененные клетки называются
передаточными клетками. Позднее сходного типа
клетки с выростами клеточной стенки были найдены
и в других местах, где происходит транспорт
веществ на короткое расстояние, например в
паренхиме ксилемы. Такие клетки очень легко рассмотреть
на сделанных вручную срезах узла традесканции
после окраски толуидиновым синим. Эти клетки
есть не у всех растений, но они довольно обычны у
бобовых и некоторых других семейств. На рис. 14.28
показано, как они выглядят в электронном
микроскопе.
Ситовидные
трубки
Выросты
клеточной стенки
в передаточной
клетке
Сосуды
ксилемы
Хлоропласты
Плазмодесма
Стрелками
показан
возможный
перенос
растворенных
веществ
из ксилемы
во флоэму
Рис. 14.28. Поперечный разрез одной из тонких
жилок листа Senecio vulgaris. B зоне флоэмы (вверху)
видны шесть клеток паренхимы, окружающих два
ситовидных элемента. Сверху и снизу от ситовидных
элементов две пары клеток-спутников,
видоизмененных в передаточные клетки типа А. Сбоку от них
находится по одной передаточной клетке типа В.
Клетки этих двух типов резко различаются по
относительной плотности иитоплазмы, по
полярности зон, в которых находятся внутренние выросты
клеточных стенок, и по способу соединения с
ситовидными элементами; клетки типа А, как правило,
соединены плазмодесмами. тогда как между
клетками типа В и ситовидными элементами
плазмодесмы втречаются очень редко Внизу видны два
элемента ксилемы, а по обеим сторонам от них
находятся две большие клетки обкладки проводящего
пучка, они видны лишь частично Стрелками показаны
некоторые из возможных путей возврата
растворенных веществ внутрь жилки после их доставки к
апопласту по ксилеме (см в тексте)
Увеличение χ 6560.
Транспорт у животных
Почти у всех животных имеется специальная
система для транспорта и распределения веществ в
организме. Исключение составляют простейшие,
кишечнополостные и плоские черви. У представителей
этих типов очень велико отношение поверхности
тела к его объему, поэтому для их нужд вполне
достаточно простой диффузии газов через
поверхность тела. Да и внутри организма веществам
приходится проходить такие малые расстояния, что они
вполне могут перемещаться путем диффузии или с
током цитоплазмы (разд. 7.2.4).
С увеличением размеров и сложности строения
организмов возрастают и количества веществ,
поступающих внутрь и выходящих наружу.
Расстояния, которые приходится проходить этим
веществам в организме, тоже увеличиваются, и для их
распределения уже недостаточно простой диффузии.
Поэтому здесь нужен какой-то иной способ
транспорта веществ из одной части тела в другую, и
таким способом становится перенос их с током
жидкости.
14.9. Общие особенности
циркуляторной системы
Функция циркуляторной системы состоит в
быстром переносе веществ из одной части тела в другую
на такие расстояния, при которых диффузия была
бы слишком медленным способом
транспортировки. Достигнув места назначения, вещества должны
выходить через стенки циркуляторной системы в
органы или ткани, а другие вещества, которые в
этих тканях образуются, должны поступать в цирку-
ляторную систему.
В состав любой циркуляторной системы должны
входить следующие три компонента:
1) циркулирующая жидкость, которой обычно
служит кровь;
2) сократимый орган, который действует как насос
и обеспечивает движение жидкости по всему телу;
таким органом может быть сердце или
видоизмененный кровеносный сосуд;
3) трубки, называемые кровеносными сосудами, в
которых циркулирует жидкость.
У высших беспозвоночных и у позвоночных
известны циркуляторные системы двух типов -
незамкнутые и замкнутые.
незамкнутая система (у большинства
членистоногих, некоторых головоногих моллюсков,
оболочников). Кровь накачивается сердцем в аорту,
которая разветвляется на ряд артерий. Последние
открываются в систему полостей, образующих гемо-
цель. Находясь под низким давлением, кровь
медленно движется между тканями и постепенно вновь
собирается в сердце через открытые концы венозных
сосудов. Распределение крови между тканями почти
не регулируется.
замкнутая система (у иглокожих, головоногих
моллюсков, кольчатых червей, позвоночных).
Нагнетаемая сердцем кровь быстро обходит тело,
постоянно находясь под высоким давлением, и
возвращается обратно в сердце. На всем пути кровь
заключена в специальные сосуды и не вступает в
прямой контакт с тканями тела. Распределение
крови между тканями может регулироваться. Вход в
эту замкнутую систему и выход из нее
осуществляются только через стенки кровеносных сосудов.
Кровеносные сосуды носят разные названия в
зависимости от их строения и функции. Сосуды,
несущие кровь от сердца, называются артериями.
Артерии разветвляются на более мелкие сосуды,
называемые артериоламн. Артериолы многократно
ветвятся, образуя микроскопические капилляры,
проходящие между клетками практически всех
тканей тела. Именно в капиллярах происходит обмен
веществами между кровью и тканями.
Внутри органа или ткани капилляры сливаются,
образуя венулы, по которым начинается возврат
крови к сердцу. Венулы, соединяясь, образуют все
более крупные вены. Строение кровеносных сосудов
каждого из этих типов будет детально рассмотрено
в разд. 14.12.
14.10. Эволюция транспортных
систем у животных
У простейших вещества перемещаются в основном с
током протоплазмы. У кишечнополостных
циркуляция веществ осуществляется благодаря движению
стенок тела, создающему в гастроваскулярной
полости токи воды, с которыми и переносятся пища,
вода и растворенные в ней газы. Этому
перемещению способствуют мышечно-эпителиальные и
жгутиковые клетки энтодермы. Плоские черви имеют
очень тонкое уплощенное тело, благодаря чему
обмен веществами внутри тела и с окружающей
средой может осуществляться путем диффузии.
14.10.1. Кольчатые черви
Кольчатые черви относятся к вторичнополостным
животным. Благодаря наличию вторичной полости
тела (целома) стенки тела отделены у них от
внутренних органов, что обеспечивает независимость
движения таких образований, как кишка. Однако с
появлением вторичной полости тела возникла
необходимость в приспособлении, которое связывало
бы разделенные ею области и обеспечивала обмен
питательными веществами, газами и отходами
метаболизма между кишкой и окружающей средой.
Поэтому именно на уровне вторичнополостных
впервые появляется кровеносная система,
осуществляющая связь между пищеварительной трубкой и
стенками тела.
СБОР
РАСПРЕДЕЛЕНИЕ
Выносящий
сосуд кишки
Кольцевой
сосуд
Выносящий
сосуд нефридия
Выносящий
сосуд стенки тела
Субневральный
сосуд
Спинной cocyz
Приносящий
сосуд стенки
тела
Приносящий
сосуд кишки
Приносящий
сосуд
нефридия
. Брюшной
сосуд
Боковой
■ невральный
сосуд
Рис. 14.29. Распределение крови в
отдельном сегменте дождевого
червя в участках, расположенных
позади пояска. (Из A. E. Vines,
N. Rees, Plant and animal biology, v. 1,
4th ed, 1972, Pitman.)
Замкнутая кровеносная система
дождевого червя
У дождевого червя имеется хорошо развитая
кровеносная система, и кровь циркулирует по телу в
замкнутой системе сосудов. Самым большим
кровеносным сосудом является продольный спинной
сосуд, имеющий мышечные стенки и расположенный
над кишкой. Перистальтические сокращения,
возникающие на заднем конце спинного сосуда, гонят
кровь к переднему концу тела животного, а
движению крови в обратном направлении препятствует
серия клапанов, образованных складками
эндотелия. В спинной сосуд собирается кровь от стенки
тела, кишки, нервного ствола и нефридиев.
Названия основных сегментарных сосудов приведены на
рис. 14.29 и 14.30.
Спинной сосуд соединяется с меньшим по
размеру, но обладающим большей сократительной
активностью брюшным сосудом через пять пар
пульсирующих «ложных сердец», расположенных в
сегментах 7-11. Каждое «сердце» имеет по четыре клапана,
пропускающие кровь только в сторону брюшного
сосуда.
По брюшному сосуду кровь течет к заднему
концу тела и через серию сегментарных сосудов
направляется к нефридиям, нервной цепочке, кишке
и стенкам тела. Нервная цепочка дополнительно
снабжается кровью через расположенный под нею
продольный субневральный сосуд.
Внутри всех органов червя имеется сеть
капилляров, в которых происходит обмен веществами
между кровью и тканями. В конце концов кровь опять
собирается в спинной сосуд и снова начинает свой
путь по замкнутой кровеносной системе.
Кровь дождевого червя имеет красный цвет, так
как содержит гемоглобин, растворенный в плазме
(разд. 14.13.1). Она разносит по телу кислород,
углекислоту, растворимые отходы метаболизма и
питательные вещества. В крови циркулируют также
бесцветные амебоидные клетки, выполняющие
защитную функцию.
14.10.2. Членистоногие
У членистоногих объем целома резко уменьшен и
его место занимает гемоцель. Последний
представляет собой сеть заполненных кровью незамкнутых
полостей, называемых синусами, в которых лежат
внутренние органы. Газообмен у большинства
членистоногих происходит через систему трахей (разд.
14.10), поэтому кровеносная система не
используется для транспорта дыхательных газов. Кровь
(гемолимфа) у них бесцветная; она не содержит
гемоглобина и служит для транспорта питательных
веществ и отходов, подлежащих экскреции. В крови
циркулируют бесцветные амебоидные лейкоциты.
Незамкнутая кровеносная система
таракана
У таракана имеется только один кровеносный
сосуд-спинной; его задний участок превратился в
хорошо выраженное «сердце», а передний носит
название аорты. Сердце расположено в большом
синусе, представляющем собой видоизмененный
участок гемоцеля, перикардиальном синусе (рис.
14.31). Сердце состоит из тринадцати расширений
или камер, три из которых находятся в грудных
сегментах, а десять-в брюшных. Каждая камера, за
Спинной сосуд
Пять пар "сердец"
Кольцевой сосуд
Брюшной сосуд
Субневральный сосуд
Рис. 14.30. Главные кровеносные сосуды дождевого
червя (Lumbricus terrestris). (Из E. G. Springthorpe, An
introduction to functional systems in animals, 1973,
Longman.)
Сердечная связка
Остия
Перикардиальная
мембрана
Перивисцеральная
полость
Перикардиальная
полость
Крыловидная
мышца
Кишка
Направление
кровотока
Рис. 14.31. Схематический поперечный разрез тела,
показывающий строение незамкнутой кровеносной
системы у таракана. (По Ramsey, Physiological
approach to lower animals, 1968, Cambridge Univ. Press.)
исключением последней, имеет пару боковых
отверстий, называемых остиями. Остии закрываются
клапанами, позволяющими крови входить в камеру,
но не выходить из нее. Между соседними камерами
тоже имеются клапаны, препятствующие обратному
току крови. К брюшной стенке сердца прикреплены
сегментарные крыловидные мышцы. При
сокращении этих мышц объем сердца увеличивается, в нем
создается отрицательное давление и кровь
насасывается в него из перикардиального синуса через
остии. При расслаблении крыловидных мышц
сердце сокращается и кровь выталкивается в аорту.
Аорта делится на несколько артерий с открытыми
концами, и кровь изливается из них в кровеносные
синусы, где непосредственно омывает внутренние
органы насекомого. В конце концов кровь опять
собирается в перикарде и отсюда поступает в
сердце.
14.10.3. Циркуляторные системы
позвоночных
У всех позвоночных имеется хорошо развитое
мышечное сердце, занимающее вентральное положение
в области плечевого пояса. Благодаря сердцу кровь
быстро разносится по всему телу животного. От
сердца кровь течет по артериям, а возвращается к
нему по венам. В дополнение к кровеносной системе
у позвоночных есть еще одна циркуляторная
система - лимфатическая.
Данные сравнительной эмбриологии показывают,
что у эмбрионов всех позвоночных от брюшной
аорты отходят шесть пар артериальных дуг,
которые, соединяясь, образуют две боковые спинные
аорты, сливающиеся в конце концов в одну
срединную аорту (рис. 14.32). Во взрослом состоянии
строение кровеносной системы, сходное с
эмбриональным, сохраняется только у рыб. У всех
остальных позвоночных эмбриональная кровеносная
система претерпевает большие изменения.
Акула
Сердце акулы, лежащее в полости перикарда, имеет
S-образную форму и состоит из двух главных
камер - предсердия и желудочка. Кровь от всего тела
собирается в предшествующую сердцу
тонкостенную камеру-венозный синус, откуда благодаря
отрицательному давлению в перикарде поступает в
предсердие с относительно слабой мышечной
стенкой. Из предсердия кровь проходит в желудочек,
имеющий более толстые мышечные стенки. При
сокращении желудочка кровь выталкивается через
артериальный конус в брюшную аорту. Клапаны,
расположенные между предсердием и желудочком и
между желудочком и артериальным конусом,
препятствуют обратному току крови. Артериальный
конус весьма эластичен и легко растягивается под
давлением крови, поступающей из желудочка. В
свою очередь его упругие стенки давят на кровь и
проталкивают ее вперед с более равномерной
скоростью, обеспечивая таким образом непрерывный
ток крови в брюшной аорте.
От брюшной аорты у акулы отходят пять пар
приносящих жаберных артерий, несущих дезоксиге-
нированную кровь к жабрам (рис. 14.33, Л). В
жабрах эти артерии многократно ветвятся, образуя
множество тончайших капилляров. Капилляры
соединяются, образуя в конечном счете выносящие
жаберные артерии. Последние несут оксигенирован-
ную кровь и образуют с каждой стороны тела по
четыре петли, окружающие с внутренней стороны
по периметру каждую из первых четырех жаберных
щелей. Одиночная выносящая жаберная артерия,
отходящая от передней части последней полужабры,
впадает в четвертую петлю. С каждой стороны тела
от этих петель отходит по четыре наджаберных
артерии, которые несут кровь назад и, сливаясь по
средней линии тела, образуют спинную аорту. От
спинной аорты отходят вперед внутренние сонные
Атриовентрику-
лярный клапан
Артериальный
конус
Брюшная
аорта
-Брюшная аорта
_ Боковая аорта
Спинная аорта
Предсердие
Синусно-
-предсердный
клапан
Кювьеров
проток
Желудочек
Венозный
синус
Рис. 14.32. А. Строение эмбриональной кровеносной
системы позвоночных. От брюшной аорты отходят с
каждой стороны шесть артериальных дуг, которые
затем сливаются, образуя боковую аорту. Б.
Строение сердца у эмбриона.
артерии, несущие оксигенированную кровь к голове.
В задней части тела от спинной аорты отходят
многочисленные артерии, снабжающие оксигениро-
ванной кровью остальные органы.
Поскольку кровь у акулы за один обход тела
проходит через сердце лишь один раз, принято
говорить, что у нее имеется один круг
кровообращения (рис. 14.33, В). При прохождении крови через
капилляры жабр ее давление падает, и в спинной
аорте она течет уже под низким давлением. Когда
кровь проходит через органы, то из-за
сопротивления в капиллярных сетях ее давление падает еще
сильнее, поэтому возврат крови к сердцу
происходит очень медленно. Этот видимый недостаток
преодолевается благодаря большим полостям,
заполненным кровью,-кардинальным синусам, в
которых сопротивление току крови очень мало. По
задним кардинальным синусам кровь возвращается
к сердцу и поступает в венозный синус через парные
кювьеровы протоки. Обратному току крови в венах
препятствуют клапаны, движению крови вперед
способствуют сокращения мускулатуры тела,
сжимающей эти сосуды.
В заднем отделе тела у рыб имеются кровеносные
сосуды, носящие название портальных (воротных)
вен. Эти сосуды образуют капиллярные сети на
обоих концах. Кровь, оттекающая от тонкого
кишечника, поступает в печень через воротную вену
печени, а кровь, оттекающая от хвостового отдела,-
в почки через воротные вены почек. Таким образом,
у акулы кровь, уже прошедшая через жаберные
капилляры, может проходить еще через две
капиллярные сети одной из портальных систем, т.е. в
сумме-через три капиллярные сети.
Кровь акулы содержит взвешенные в плазме
эритроциты овальной формы, имеющие ядро, и
амебоидные лейкоциты. Лимфатическая система
состоит из лимфатических сосудов, впадающих в два
продольных спинных лимфатических протока,
которые в конце концов изливают свое содержимое (т. е.
лимфу) в кровь через кардинальные синусы.
Земноводные: лягушка
У взрослой лягушки в связи с ее полуназемным
образом жизни сердце и аортальное
кровообращение претерпели сильные изменения. В качестве
органов дыхания у нее развились легкие, заменившие
Брызгальце
Наджаберная артерия
Выносящая жаберная артерия
Приносящая жаберная артерия
Брюшная аорта
Голова
Жабры
Сердце
Печень
Кишка
Туловище
Почки
Хвост
Спинная аорта
А Артериальная система
Предсердие
Синусно-предсердный
клапан
Венозный синус
Предсердно-желудочковый
(атриоветрикулярный)
клапан
Желудочек
Перикард
Артериальный
конус
В Общая схема
циркуляции
Один круг кровообращения
Б Сердце с двумя камерами — одним предсердием и
одним желудочком.
Рис. 74.33. Артериальная система акулы. Из шести артериальных дуг осталось только пять; первая
жаберная щель превратилась в брызгальце. Прямые жаберные сосуды разделились на выносящую, приносящую
и наджаберную артерии, что обеспечивает большую поверхность кровеносных сосудов в жабрах, а значит, и
большую поверхность для газообмена.
ГОЛОВАСТИК
ЛЯГУШКА
Каротидная
дуга
Системная
дуга
Брюшная
аорта
Легочная
дуга
Спинная
аорта
Оксигениро-
ванная кровь
Дезоксиге-
нированная
кровь
Голова и
передние
конечности
Сердце
Легкие
Кожа
Печень
Кишечник
Туловище
Почки
Хвостовой
отдел
Первая и вторая артериальные
дуги исчезли; дуги 3—6 несут
кровь к жабрам
Правое предсердие
Левое предсердие
Три функционирующие дуги
— 3, 4 и 6 жаберные капилляры
шунтированы; 6-я дуга несет кровь
Брюшная аорта
Каротидная дуга"
Системная дуга-
Легочная дуга"
Легочные вены
Передние полые
вены
Задняя полая
вена
Венозный синус
Артериальный
конус
Желудочек
Рис. 14.34. Кровеносная система лягушки.
жабры, и между сердцем и легкими сформировался
новый круг кровообращения-легочный. В связи с
этим отпала необходимость в сохранении всех дуг
аорты, снабжающих жабры, и некоторые из них
получили новое применение, а другие были просто
утрачены. Третья дуга аорты превратилась в
сонную артерию, по которой кровь течет к голове
(рис. 14.34, А). Четвертая дуга стала системной
дугой, несущей оксигенированную кровь, которая по
спинной аорте доставляется к остальным частям
тела. По шестой дуге дезоксигенированная кровь
поступает из сердца в легкие через легочную
артерию. Первая, вторая и пятая дуги исчезли.
Сердце лягушки состоит из трех основных
камер-двух предсердий и одного желудочка (рис.
14.34, Б). Кровь, насыщенная в легких кислородом,
поступает по легочной вене в левое предсердие, а
дезоксигенированная кровь собирается со всего тела
в переднюю и заднюю полые вены, по которым она
попадает в венозный синус и затем в правое
предсердие. В предсердиях венозная и артериальная
кровь никогда не смешивается, но оба предсердия
одновременно выталкивают кровь в единственный
желудочек. Смешиванию крови в желудочке
частично препятствуют мышечные гребни, образуемые
внутренней поверхностью дорсальной и
вентральной стенок желудочка; а когда кровь поступает из
желудочка в артериальный конус, то здесь ее
смешивание затрудняет спиральный клапан, который не
полностью подразделяет артериальный конус на
два протока. Дорсальный проток, называемый кож-
но-легочным стволом, переходит в легочные
артерии, от которых ответвляются кожные артерии. А
из вентрального протока кровь поступает в общую
сонную артерию и в системную дугу аорты.
Таким образом, очевидно, что у лягушки имеются
два раздельных круга кровообращения, один из
которых идет к легким, а другой-к остальным
органам тела. Однако эти два круга у лягушки еще
не вполне обособлены. Кровь поступает из сердца в
легкие и возвращается из них в сердце, не проходя
через сосуды остального тела (рис. 14.34, B)9 но в
ней еще нет полного разделения артериальной и
венозной крови.
Кровеносная система лягушки имеет очень
специфическое строение, приспособленное к
земноводному образу жизни. Когда лягушка находится в
относительном покое, кислород диффундирует в
капилляры кожи и переносится с кровью через
подключичную вену в правое предсердие. Легкие при этом
не используются - они начинают функционировать
только при резком повышении активности
животного. Таким образом, обычно левое предсердие
получает кровь, бедную кислородом.
В венозной системе лягушки синусы заменены
венами. Кровь возвращается в венозный синус по
передней и задней полым венам, собирающим кровь
из вен, отходящих от важнейших органов. У
лягушки, так же как у рыб, имеются воротные вены почек
и печени.
Лимфа (разд. 14.12.1) из межклеточных ходов
собирается в лимфатические сосуды и в конце концов
изливается в венозную систему в двух участках -
переднем и заднем. В каждом из этих участков лимфу
перекачивает в вены пара маленьких лимфатических
сердец.
Кровь лягушки состоит из плазмы и взвешенных в
ней клеток-эритроцитов, имеющих ядро и
содержащих гемоглобин, и лейкоцитов; последние делятся
на макрофаги, лимфоциты, гранулоциты и
моноциты (разд. 14.11.2).
Пресмыкающиеся: крокодил
Сердце у крокодила четырехкамерное и состоит из
двух предсердий и двух желудочков, причем правая
и левая половины сердца практически полностью
разделены (рис. 14.35). В связи с таким разделением
у крокодила имеются два круга кровообращения, и
артериальная кровь, находящаяся в левой половине
сердца, почти полностью отделена от венозной,
находящейся в правой половине. Вместо
артериального конуса у крокодила имеются три сосуда,
самостоятельно отходящих от желудочков: правая дуга
аорты с отходящим от нее общим стволом сонных
артерий, левая дуга аорты и легочная артерия.
Птицы и млекопитающие. Общий
план строения
У птиц и млекопитающих сердце четырехкамерное.
Правые предсердие и желудочек полностью
отделены от левых предсердия и желудочка (рис. 14.36).
Это означает, что оксигенированная и дезоксигени-
рованная кровь тоже полностью разделена. Чтобы
из правой половины сердца попасть в левую, кровь
Каротидная дуга
Брюшная аорта
Правая системная дуга
аорты
Левая системная
дуга, несущая
дезоксигениро-
ванную кровь
Легочная дуга
Артериальная система
Рис. 14.35. Артериальная система
пресмыкающегося (крокодила).
должна пройти через легкие, а из левой половины
сердца в правую кровь может попасть, только
совершив полный обход всего тела. Благодаря
такому строению кровеносной системы
оксигенированная кровь, идущая к тканям, находится под высоким
давлением, а не под низким, как у рыб. К тому же
перед поступлением в различные ткани кровь
проходит через легкие, поэтому к активно дышащим
органам поступает кровь, богатая кислородом.
14.33. Почему следует считать преимуществом
высокое давление артериальной крови,
поступающей к тканям, у птиц и млекопитающих?
У птиц и млекопитающих артериальная система
подверглась дальнейшей редукции. У птиц
сохранилась только правая половина системной дуги, а у
млекопитающих - только левая (рис. 14.36). Третья
дуга превратилась в ствол сонных артерий, а
шестая - в легочную дугу. В венозной системе воротную
систему почек заменила задняя полая вена, и кровь
из задних отделов тела возвращается прямо в
сердце, благодаря чему увеличилась скорость возврата
крови.
14.11. Состав крови
млекопитающих
Кровь у млекопитающих состоит из клеток,
взвешенных в жидкой среде, называемой плазмой.
Клетки составляют около 45%, а плазма-55% общего
объема крови.
14.11.1. Плазма
Плазма-это бледно-желтая жидкость, 90% которой
составляет вода, а 10%-растворенные и
взвешенные в ней вещества. В норме концентрации одних
растворенных веществ остаются постоянными, а
содержание других может колебаться в узких
пределах в зависимости от скорости их удаления из крови
или поступления в нее из тех или иных органов.
Основные компоненты плазмы и их функции
приведены в табл. 14.9.
14.11.2. Форменные элементы крови
Эритроциты
Эритроциты - красные кровяные клетки. У человека
это мелкие клетки, лишенные ядра и имеющие
форму двояковогнутых дисков. Средний диаметр
эритроцитов составляет 7-8 мкм и приблизительно
равен диаметру кровеносных капилляров.
Специфическая форма эритроцита обусловливает более
высокое отношение поверхности к объему, что
увеличивает возможности газообмена. Толщина
отдельного эритроцита очень мала, и это облегчает
диффузию газов с поверхности внутрь клетки. Благодаря
Артериальное
кровообращение
ПТИЦЫ
МЛЕКОПИТАЮЩИЕ
Каротидная
дуга — к
голове и передним
конечностям
Левая системная
дуга исчезает
егочная дуга —
к легким
Правая системная
дуга— к органам тела
Оксигенирован-
ная кровь
Дезоксигени-
рованная
кровь
Голова и
передние
конечности
Сердце
Пегкие
Печень
Пищеварительный
тракт
Туловище
Почки
Хвостовой
отдел
Легочная артерия
Правое предсердие
Правый желудочек
Сердце четырехкамерное,
правая и левая половины
полностью разделены
Левая системная
дуга — к органам тела
Легочная дуга —
к легким
Правая системная
дуга исчезает
Аорта
Левое предсердие
Левый желудочек
Общая схема циркуляции
с двумя кругами
кровообращения
Рис. 14.36. Артериальные системы птиц (А) и млекопитающих (Б).
эластичности своей мембраны эритроцит может
проходить через капилляры, просвет которых
меньше его диаметра.
Процесс образования эритроцитов носит название
гемопоэза (эритропоэза) или кроветворения, а ткань,
в которой он происходит, называют кроветворной
или гемопоэтической. У младенцев кроветворная
ткань содержится во всех костях, а у взрослых
людей главными местами образования эритроцитов
являются кости черепа, рёбра, грудина, позвонки,
ключицы и лопатки. В 1 мл крови содержится около
5 млн. эритроцитов, но эта величина варьирует в
зависимости от возраста, пола и состояния
здоровья.
Важнейшая особенность эритроцитов -
присутствие в них гемоглобина, который обратимо
связывает кислород (превращаясь в оксигемоглобин) в
участках с высокой концентрацией O2 и отдает его в
участках с низкой концентрацией O2. В эритроцитах
содержится также фермент карбоангидраза,
участвующий в транспорте углекислоты.
Продолжительность жизни эритроцитов у
взрослых людей составляет около трех месяцев, после
чего они разрушаются в печени или селезенке.
Белковые компоненты эритроцита расщепляются на
составляющие их аминокислоты, а железо,
входящее в состав гема, удерживается печенью и хранится
в ней в составе ферритина (белка, связывающего
железо). Железо может в дальнейшем
использоваться повторно при образовании новых эритроцитов и
при синтезе цитохромов. Остальная часть молекулы
гема расщепляется с образованием желчных
пигментов билирубина и биливердина. Оба пигмента в
конце концов выводятся с желчью в кишечник.
Каждую секунду в организме человека
разрушается от 2 до 10 млн. эритроцитов. Скорость распада
эритроцитов и замещения их новыми зависит от
содержания в атмосфере кислорода, доступного для
переноса кровью. Низкое содержание кислорода в
крови стимулирует костный мозг, и в нем
образуется больше эритроцитов, чем разрушается в печени.
Этот механизм служит одним из путей
акклиматизации млекопитающих к пониженному содержанию
кислорода в атмосфере на больших высотах. При
высоком содержании кислорода наблюдается
противоположная картина.
Белые кровяные клетки -
лейкоциты
Лейкоциты крупнее эритроцитов, но содержатся в
крови в гораздо меньшем количестве (около 7000 в
1 мл крови). Они играют важную роль в защите
организма против болезней. Все лейкоциты имеют
ядро. Несмотря на наличие ядра,
продолжительность жизни лейкоцитов в кровяном русле
составляет в норме всего лишь несколько дней.
Существуют две основные группы лейкоцитов - гранулоциты
(зернистые лейкоциты) и агранулоциты
(незернистые лейкоциты).
гранулоциты (зернистые, или полиморфноядер-
ные, лейкоциты) образуются в костном мозге, но не
из тех клеток, из которых образуются эритроциты.
Все гранулоциты содержат разделенное на лопасти
ядро и зернистую цитоплазму (табл. 14.10) и
обладают способностью к амебоидному движению.
Гранулоциты можно далее подразделить на нейтрофи-
лы, эозинофилы и базофилы.
Нейтрофилы (фагоциты) составляют 70% всех
лейкоцитов. Они обладают способностью
проходить между клетками, образующими стенки
капилляров, и проникать в межклеточные пространства
тканей (процесс, называемый диапедезом) и
направляться к инфицированным участкам тела.
Нейтрофилы - активные фагоциты, они поглощают и
переваривают болезнетворные бактерии (разд. 14.23.5).
Эозинофилы содержат цитоплазматические
гранулы, окрашиваемые эозином в красный цвет. Обычно
они составляют всего лишь 1,5% общего числа
лейкоцитов, но при аллергических состояниях
(например, астме или сенной лихорадке) их количество
возрастает. Как полагают, эозинофилы обладают
Таблица 14.9. Компоненты плазмы крови и их функции
антигистаминным действием. Содержание эозино-
филов в крови контролируется гормонами коры
надпочечников.
Базофилы составляют 0,5% популяции
лейкоцитов. Они вырабатывают гепарин и гистамин;
содержат гранулы, которые окрашиваются в синий цвет
основными красителями типа метиленового синего.
агранулоциты (одноядерные лейкоциты) содержат
ядро овальной формы и незернистую цитоплазму.
Существуют два основных типа агранулоцитов:
моноциты и лимфоциты.
Моноциты (4%) образуются в костном мозге и
содержат ядро бобовидной формы. Активно
фагоцитируют (поглощают) бактерий и другие крупные
частицы. Способны мигрировать сквозь стенки
капилляров в очаги воспаления, где действуют так же,
как нейтрофилы.
Лимфоциты (24%) образуются в тимусе (вилоч-
ковой железе) и лимфоидной ткани из
клеток-предшественниц, поступающих из костного мозга.
Имеют округлую форму и содержат очень мало
цитоплазмы. Способность к амебоидным движениям
ограниченна. Главная функция - индукция
иммунных реакций или участие в них (образование анти-
Компонент
Функция
Компоненты, присутствующие в постоянной концентрации
Вода
Белки плазмы
Сывороточный альбумин
Сывороточные глобулины
α-Глобулин
β-Глобулин
γ-Глобулин
Протромбин
Фибриноген
Ферменты
Минеральные ионы
Na + , K + , Ca2 + , Mg2 + ,
H2PO4", PO^, СГ, HCO3-, SO2/
Основной компонент лимфы. Служит источником воды для клеток.
Разносит по телу множество растворенных в ней веществ. Способствует
поддержанию кровяного давления и объема крови
Содержится в очень большом количестве. Связывает присутствующий в
плазме кальций
Связывает тироксин и билирубин
Связывает железо, холестерол и витамины A, D и К.
Связывает антигены и играет важную роль в иммунологических реакциях
организма (γ-глобулины обычно называют антителами). Связывает
также гистамин
Каталитический фактор, участвующий в свертывании крови
Участвует в свертывании крови
Участвуют в метаболических процессах
Совместно участвуют в регуляции осмотического давления и рН крови.
Оказывают ряд других воздействий на клетки организма; например, Ca2 +
может участвовать в свертывании крови, а также в регуляции мышечного
сокращения и чувствительности нервных клеток, влияет на коллоидное
состояние клеточного содержимого
Компоненты, концентрации которых изменяются
Растворимые продукты пищеварения
Растворимые продукты, подлежащие
экскреции
Витамины
Гормоны
Постоянно транспортируются в клетки и выделяются из них
Таблица 14.10. Форменные элементы крови
Клетки
Эритроциты
Лейкоциты
а) Гранулоциты (72% общего
количества лейкоцитов)
Нейтрофилы (70%)
Эозинофилы (1,5%)
Базофилы (0,5%)
б) Агранулоциты (28%)
Моноциты (4%)
Лимфоциты (24%)
Тромбоциты (кровяные пластинк!
Место образования
Костный мозг
Костный мозг
Костный мозг
>
КОСТНЫЙ M03I
Костный мозг и
лимфоидные ткани,
селезенка
i) Костный мозг
Число в 1 мм3
5000000
4900
105
35
280
1680
250000
Функция
Переносят кислород и частично CO2
Захватывают бактерий
Антигистаминное действие
Образуют гнетамин и гепарин
Захватывают бактерий
Вырабатывают антитела
Инициируют свертывание крови
Строение
тел, отторжение трансплантата, уничтожение
опухолевых клеток). Продолжительность жизни
варьирует от нескольких дней у грызунов до десяти и более
лет у человека.
14.11.3. Тромбоциты (кровяные
пластинки)
Тромбоциты представляют собой фрагменты
клеток, имеющие неправильную форму, окруженные
мембраной и обычно лишенные ядра. Они
образуются из крупных клеток костного мозга,
называемых мегакариоцитами. Играют важную роль в
инициации свертывания крови. В 1 мл крови
содержится около 250000 тромбоцитов.
Опыт 14.7. Приготовление мазка крови
Материалы и оборудование
Полотенца одноразового пользования
Ватные тампоны
70%-ный спирт
Стерильные ланцеты одноразового
пользования
Два предметных стекла
Герметичные контейнеры
Краситель Лейшмана
Дистиллированная вода
Глицерол (глицерин)
Покровное стекло
Свежеприготовленный раствор хлорида
натрия
Методика
1. Тщательно вымойте руки водой с мылом. Те, у
кого будут брать кровь, должны особенно
хорошо промыть место взятия крови. Руки следует
вытирать только полотенцем одноразового
пользования.
2. Несколько раз встряхните кистью опущенной
руки, чтобы кровь прилила к пальцам.
3. Простерилизуйте участок кожи мизинца на
расстоянии 5-10 мм от нижнего края ногтя с
помощью ватного тампона, смоченного 70%-ным
спиртом, и дайте ему просохнуть (рис. 14.37, А).
4. Непосредственно перед использованием
извлеките из пакетика новый стерильный ланцет
одноразового пользования. Ни к чему не
прикасайтесь острым концом ланцета.
Ноготь
Направление движения
Предметное
стекло
Капля крови
Рис. 14.37. А. Участок пальца, из которого берут
кровь. Удобнее делать прокол, если палец согнут в
последнем суставе. Б. Приготовление мазка крови.
5. С помощью ланцета проколите кожу в
намеченном участке пальца, а затем уберите ланцет в
контейнер. Ланцеты должны использоваться не
более одного раза.
6. Перенесите выступившую каплю крови на конец
стерильного предметного стекла. Избегайте при
этом всякого контакта между проколотым
участком кожи и стеклом.
7. Протрите место укола чистым ватным
тампоном, смоченным в 70%-ном спирте; если нужно,
слегка прижмите тампон, чтобы остановить
кровотечение. Все использованные тампоны
кладите в тот же контейнер, куда складываются
ланцеты.
8. Возьмите второе предметное стекло и, держа
его под углом 45°, прикоснитесь им к капле
крови, а затем проведите по первому стеклу в
направлении от капли, чтобы получился мазок
(рис. 14.37,5).
9. В течение нескольких минут подсушите мазок,
чтобы клетки приклеились к стеклу. Подсушить
мазок можно, подержав его над абажуром
лампы или помахав им в воздухе.
10. Добавьте 6 капель красителя Лейшмана и
оставьте на 60 с.
11. Добавьте 6 капель дистиллированной воды и,
покачивая стекло, перемешайте ее с красителем.
12. Оставьте мазок на 15 мин, а затем сполосните
водой.
13. Добавьте к препарату каплю глицерола,
накройте его покровным стеклом и рассмотрите под
микроскопом.
14. Эритроциты будут окрашены в розовый цвет, а
тромбоциты и ядра лейкоцитов-в синий.
15. Кровь, попавшую во время опыта на стол или
еще куда-нибудь, необходимо немедленно смыть
свежеприготовленным водным раствором
хлората натрия.
16. Необходимо принимать все меры
предосторожности, чтобы кровь от одного человека не
попала к другому; если же это произойдет,
загрязненный участок следует тщательно протереть
хлоратом натрия, разведенным в 10 раз водой, а
затем хорошо промыть его водой с мылом.
17. В конце практического занятия следует вымыть
руки с мылом и вытереть их полотенцем
одноразового пользования.
18. В связи с опасностью загрязнения через
поврежденную кожу в этом занятии не должны
участвовать лица с какими-либо открытыми
ранками.
14.1 2. Кровеносная система
млекопитающих
Кровь, непрерывно циркулируя в организме,
проходит через артерии, капилляры и вены. Стенки
артерий и вен построены из трех слоев: внутреннего
(интима, tunica intima), состоящего из плоского
эндотелия, среднего (tunica media), состоящего из
гладкой мускулатуры и эластических волокон, и
наружного (tunica externa), состоящего из
фиброзной соединительной ткани, содержащей коллагено-
вые волокна (табл. 14.11 и рис. 14.39, А).
Крупным артериям, расположенным вблизи
сердца (аорта, подключичные и сонные артерии),
приходится выдерживать большое давление крови,
выталкиваемой левым желудочком сердца. Эти сосуды
имеют толстые стенки, средний слой которых
состоит главным образом из эластических волокон.
Благодаря этому во время систолы они могут
растягиваться, не разрываясь (разд. 14.12.3). После
окончания систолы стенки артерий сокращаются,
что обеспечивает непрерывный ток крови на всем их
протяжении (рис. 14.40).
Артерии, расположенные дальше от сердца,
имеют сходное строение, но содержат больше гладких
мышечных волокон в среднем слое. Они иннерви-
руются волокнами симпатической нервной системы,
и поступающие по этим волокнам импульсы
регулируют их диаметр, что имеет важное значение для
регуляции кровотока в различных участках тела.
Из артерий кровь поступает в более мелкие
сосуды, называемые артериолами. Во всех артериолах,
за исключением легочных, средний слой стенок
целиком состоит из гладкой мускулатуры, иннервиро-
ванной симпатическими волокнами (разд. 16.2). Во
многих артериолах в местах перехода их в
капилляры находятся прекапиллярные «сфинктеры» (рис.
14.41 и 14.42), при сокращении которых поступление
крови в капиллярную сеть прекращается. В
определенных участках тела имеются также артериовеноз-
ные анастомозы, которые действуют как шунты
между артериолами и венулами и служат для
регуляции количества крови, протекающей через
капиллярные сети, в соответствии с нуждами организма.
Из артериол кровь поступает в капилляры - самые
мелкие из всех кровеносных сосудов тела.
Капилляры образуют обширную сеть сосудов, пронизываю-
Агранулоцит
(моноцит)
Эритроцит
Эритроциты,
имеющие вид
круглых
двояковогнутых
дисков
Гранулоцит
Агранулоцит
(лимфоцит)
Рис. 14.38. А Мазок крови, на котором видны эритроциты и три типа лейкоцитов Б. Эритроциты
млекопитающего. Микрофотография, полученная с помощью сканирующего электронного микроскопа.
щих все части тела. Плотность их расположения так
велика, что расстояние от клеток до ближайшего
капилляра не превышает 0,5 мм. Диаметр
капилляров составляет 7-10 мкм, а их стенки, состоящие из
одного лишь эндотелия, проницаемы для воды и
растворенных в ней веществ. Именно в капиллярах
происходит обмен веществами между кровью и
клетками тела.
Из капилляров кровь собирается в венулы, стенки
которых состоят из тонкого слоя коллагеновых
волокон. Из венул кровь поступает в вены, по
которым в конце концов возвращается к сердцу.
Вена
Кровяные
клетки
Клапан
Толстая стенка
артерии
Коллагеновые
волокна
Кровяные
клетки
Эластические
волокна
и гладкая
мускулатура
Венозная
стенка
Рис. 14.39. А Поперечный срез через артерию и вену. Б. Продольный срез через вену, в которой виден клапан.
Желудочек
сокращается
Клапан открыт
Клапан закрыт
Желудочек
расслабляется
Происходит
упругое сжатие
растянутого
сосуда
Рис. 14.40. Эта схема показывает, каким образом
артерии, расположенные вблизи сердца,
способствуют непрерывному току крови, несмотря на
перерывы в поступлении крови из желудочков сердца (Из
Clegg, Clegg, Biology of the mammal, 2nd ed, Heinemann
Medical Books )
В венах средний слой стенки содержит меньше
мышечных и эластических волокон, чем в артериях,
а диаметр просвета больше, чем в артериях. В венах
имеются полулунные клапаны (рис. 14.43),
образованные складками внутренней оболочки, которые
пронизаны эластическими волокнами. Клапаны
препятствуют обратному току крови и таким образом
обеспечивают ее движение только в одном
направлении. Некоторые вены расположены между
крупными мышцами (например, в руках и ногах). При
своем сокращении эти мышцы давят на вены и
сжимают их (рис. 14.44), способствуя таким
образом возврату венозной крови к сердцу. Общая схема
двух кругов кровообращения у млекопитающих
представлена на рис. 14.45.
14.12.1. Образование межклеточной
(тканевой) жидкости
Интерстициальная, или тканевая, жидкость
образуется при прохождении крови по капиллярам. Стенки
капилляров проницаемы для всех компонентов
крови, за исключением эритроцитов и белков плазмы.
Артериовенозный
анастомоз
Венула
Артериола
Ток крови
(о ксигенированной)
Прекапиллярный
сфинктер
Капиллярная сеть
Дезо ксигенирован-
ная кровь
Венула
Рис. 14.41. Возможные пути
перехода крови из артериолы в венулу.
Аретриолы|
Капилляры|
А Клапан открыт
Б Клапан закрыт
Рис. 14.42. Сеть капилляров в ткани
Рис. 14.43. Механизм действия полулунного клапана
в вене. А Давление крови, направленное вверх,
заставляет клапан открыться, и кровь течет по
направлению к сердцу Б Обратное движение крови
приводит к закрытию клапанов, поэтому кровь не
может течь по направлению от сердца
Таблица 14.11. Сравнение структуры и функций артерий, капилляров и вен
Просвет
Эндотелий
Средний слой
из гладкой
мускулатуры
и эластических
волокон
Эндотелиальные
клетки
Артерии
Несут кровь от сердца
Средний слой стенки толстый,
состоит из эластических и мышечных
волокон
Полулунные клапаны отсутствуют
Давление крови высокое и
пульсирующее
Кровь течет быстро
Кровь оксигенированная, за
исключением легочных артерий
Наружный слой из
коллагеновых волокон
Капилляры
Соединяют артерии с венами.
Служат местом обмена веществами
между кровью и тканями
Средний слой отсутствует. Стенки
состоят только из эндотелия и не
содержат эластических волокон
Полулунные клапаны отсутствуют
Давление крови понижающееся,
непульсирующее
Течение крови замедляется
Смешанная оксигенированная и де-
зоксигенированная кровь
Вены
Несут кровь к сердцу
Средний слой относительно тонкий и
содержит мало мышечных элементов.
Эластические волокна
немногочисленны
По всей длине имеются полулунные
клапаны, препятствующие обратному току
крови
Давление крови низкое, непульсирующее
Кровь течет медленно
Кровь дезоксигенированная, за
исключением легочных вен
Наружный слой из
коллагеновых волокон
Просвет
Эндотелий
Средний слой
из гладкой
мускулатуры
и эластических
волокон
Верхний
клапан
Нижний
клапан
Мышца
Движение крови
к сердцу
Вена
Клапан
Мышцы расслаблены,
клапаны закрыты
Мышцы сокращаются, верхний
клапан открывается и кровь
выталкивается верх; нижний
клапан остается закрытым
Мышцы расслабляются, верхний
клапан закрывается, а нижний
открывается вследствие
сокращения мышц в нижележащих
участках, и кровь проходит
через него
Рис. 74.44. Эта схема показывает, каким образом сокращение окружающих вену мышц способствует
однонаправленному движению крови к сердцу.
Осмотическое давление, создаваемое белками
плазмы, составляет около 25 мм рт. ст.* и намного
превышает осмотическое давление тканевой
жидкости. В этих условиях можно было бы ожидать, что
тканевая жидкость будет переходить в плазму
крови, однако этого не происходит. Кровяное давление
В медицине за единицу давления до сих пор
принимают 1 мм рт.ст. Сравнение с единицами системы СИ см.
в Приложении 4.3.3.
в артериальном конце капилляров составляет около
32 мм рт.ст., поэтому жидкость выходит из
капилляров в мельчайшие промежутки между клетками и
образует тканевую жидкость. Именно через эту
жидкость происходит обмен веществами между
кровью и тканями. Кровь не может постоянно
терять так много жидкости, и значительная часть
этой жидкости возвращается обратно в кровяное
русло. Это возвращение происходит двумя путями.
Часть тканевой жидкости поступает обратно в кро-
Яремная вена
Голова и шея
η Сонная артерия
Подключичная
вена
Передние
конечности
Подключичная
артерия
Передняя
полая вена
Легочная
артерия
ПП
-+
ПЖ
лп
+
лж
Спинная аорта
Легкое
Легкое
Легочная
вена
Печеночная вена
,Печеночная артерия
Печень
Воротная вена
печени
Задняя
полая вена
Сегментарные
поясничные вены
варительный
тракт
Желудочная артерия
Передняя «
брыжеечная артерия
Задняя брыжеечная
Туловище
артерия ^—
Сегментарные
■•поясничные артерии]
Почечная вена
Почки
Почечная артерия
Половая вена
Гонады
Половая артерия
Подвзодшая
вена
Задний отдел
туловища и
задние
конечности
Подвздошная
артерия
Рис. 14.45. Общая схема кровеносной системы
млекопитающих, состоящей из двух кругов Показаны
главные кровеносные сосуды без капиллярных сетей
ПП-правое предсердие, ЛП-левое предсердие,
ПЖ правый желудочек; ЛЖ - левый желудочек.
веносные капилляры за счет того, что в венозном
конце капилляров давление крови падает до
12 мм рт ст., т.е. становится ниже осмотического
давления, создаваемого белками плазмы (рис. 14.46).
Остальная часть тканевой жидкости
диффундирует в замкнутые с одного конца лимфатические
капилляры и с этого момента называется лимфой.
Лимфатические капилляры, соединяясь между
собой, образуют более крупные лимфатические
сосуды. Лимфа движется по этим сосудам благодаря
сокращению окружающих мышц, а обратному ее
току препятствуют клапаны, которые имеются в
крупных сосудах и действуют подобно клапанам вен
(рис. 14.48). Лимфатические сосуды, идущие от ног,
объединяясь с лимфатическими сосудами
пищеварительного тракта, образуют грудной проток.
Последний открывается в кровяное русло в области шеи
через левую подключичную вену. Правый
лимфатический проток изливает лимфу в кровь через правую
подключичную вену (рис. 14.47).
По ходу лимфатических сосудов в некоторых
местах расположены лимфатические железы, или
узлы (рис. 14.49). В лимфатических узлах
задерживаются и скапливаются лимфоциты,
циркулирующие в крови и лимфе. Они образуют антитела и
составляют важную часть иммунной системы
организма. Кроме того, лимфатические узлы
отфильтровывают присутствующие в лимфе бактерии и
чужеродные частицы, которые в конце концов
поглощаются фагоцитами.
14.12.2. Сердце млекопитающих
У млекопитающих сердце расположено в грудной
клетке между легкими, за грудиной. Оно окружено
конусовидным мешком - околосердечной сумкой,
или перикардом, наружный слой которого состоит
из нерастяжимой белой фиброзной ткани, а
внутренний-из двух листков, висцерального и
париетального. Висцеральный листок сращен с сердцем, а
париетальный-с фиброзной тканью. В щель между
этими листками выделяется перикардиальная
жидкость, которая уменьшает трение между стенками
сердца и окружающими тканями. Неэластичный в
целом характер перикарда препятствует излишнему
растяжению сердца или переполнению его кровью.
Сердце состоит из четырех камер: двух верхних-
тонкостенных предсердий и двух нижних -
толстостенных желудочков (рис. 14.50). Правая половина
сердца полностью отделена от левой. Функция
предсердий состоит в том, чтобы собирать и на короткое
время задерживать кровь, пока она не перейдет в
желудочки. Расстояние от предсердий до
желудочков очень мало, поэтому от предсердий не требуется
большой силы сокращения. В правое предсердие
поступает дезоксигенированная кровь из системного
круга, а в левое - насыщенная кислородом кровь из
легких. Мышечные стенки левого желудочка по
меньшей мере втрое толще стенок правого
желудочка. Эта разница связана с тем, что правый
желудочек снабжает кровью только легочный (малый) круг
кровообращения, тогда как левый гонит кровь по
системному (большому) кругу, снабжающему
кровью все тело. Соответственно кровь,
поступающая в аорту из левого желудочка, находится под
значительно большим давлением (приблизительно
105 мм рт.ст.), чем кровь, поступающая в легочную
артерию (16 мм рт.ст.).
14.34. Какие другие преимущества дает более
низкое давление крови в легочном круге по
сравнению с большим кругом?
Гидростатическое
давление,
мм рт. ст
Артериальный
конец
капилляра
Ток крови
Артериальный
конец капилляра
Венозный конец
капилляра
Онкотическое
давление плазмы
,Онкотическое
давление
плазмы
25 мм
DT. CT
Гидростатическое
—давление
12 мм рт. ст.
ι идростатическое
давление
32 мм рт. ст.
H2Oi
Венозный
конец
капилляра
Лимфатический капилляр
"Лимфа-
Тканевая
-жидкость
.Тканевая
клетка
При сокращении предсердий кровь выталкивается
в желудочки, и при этом кольцевые мышцы,
расположенные при впадении полых и легочных вен в
предсердия, сокращаются и перекрывают устья вен,
благодаря чему кровь не может оттекать обратно в
вены. Левое предсердие отделено от левого
желудочка двустворчатым клапаном, а правое
предсердие от правого желудочка - трехстворчатым
клапаном. К створкам клапанов со стороны желудочков
прикреплены прочные сухожильные нити, которые
другим концом прикреплены к конусовидным сосоч-
ковым (папиллярным) мышцам, представляющим
собой выросты внутренней стенки желудочков. При
сокращении предсердий клапаны открываются, а
при сокращении желудочков створки клапанов
плотно смыкаются, препятствуя возврату крови в
предсердия. Одновременно сокращаются сосочко-
вые мышцы, натягивая сухожильные нити и
препятствуя выворачиванию клапанов в сторону
предсердий. У основания легочной артерии и аорты
находятся соединительнотканные карманы - полулунные
Лимфатический
сосуд
Рис. 14.46. Образование тканевой жидкости и
лимфы. Часть тканевой жидкости всасывается обратно
в кровь на венозном конце капилляров, а остальная
часть собирается в лимфатические капилляры. (По
К. Schmidt-Nielsen, Animal physiology, 2nd ed., 1980,
Cambridge.)
Клапан
Шейные
лимфатические
узлы
Яремная
вена
Подключичная вена
Подмышечные
лимфатические
узлы
Грудной проток
Брыжеечные
лимфатические
узлы
Кишечник
Паховые
лимфатические
узлы
Рис. 14.47. Лимфатическая система человека. (Из
E. G. Springthorpe, An introduction to functional systems in
animals, 1973, Longman.)
Рис. 14.48. Продольный срез через лимфатический
сосуд, в котором виден внутренний клапан
Капсула
Лимфатический
узел
клапаны, пропускающие кровь в эти сосуды и
препятствующие ее возвращению в сердце.
Стенки сердца состоят из сердечных мышечных
волокон, соединительной ткани и мельчайших
кровеносных сосудов. Каждое мышечное волокно
содержит одно или два ядра, миофиламенты и
множество крупных митохондрий. Мышечные волокна
разветвляются и соединяются между собой
концами, образуя сложную сеть. Это обеспечивает
быстрое распространение волн сокращения по волокнам,
так что каждая камера сокращается как одно целое.
В стенках сердца не содержится никаких нейронов
(рис. 14.51 и 14.52).
14.12.3. Сердечный цикл
Сердечным циклом называют последовательность
событий, происходящих во время одного
сокращения сердца. Цикл состоит из трех фаз:
1. В правое предсердие поступает под низким
давлением дезоксигенированная кровь, а в левое - окси-
генированная кровь. Постепенно предсердия
растягиваются. Вначале двустворчатый и
трехстворчатый клапаны остаются закрытыми, но по мере
заполнения предсердий кровью давление в них
растет; когда оно становится выше, чем в
желудочках, клапаны открываются. При этом
некоторое количество крови переходит в расслабленные
желудочки. Этот период покоя всех камер сердца
называется диастолой (рис. 14.53, А).
2. Когда диастола заканчивается, оба предсердия
одновременно сокращаются. Эта фаза носит
название систолы предсердий и приводит к тому, что
в желудочки выталкивается дополнительное
количество крови (рис. 14.53,5). Почти тотчас же
после систолы предсердий сокращаются
желудочки (рис. 14.53 В), и это сокращение носит название
систолы желудочков. Во время систолы
желудочков двустворчатый и трехстворчатый клапаны
закрываются. Давление в желудочках возрастает
Рис. 74.49. Срез лимфатической железы.
Легочные вены
(от легких)
Полая вена
Правое
предсердие-
Трехстворчатый
клапан
Сосочковая
мышца
Правый
желудочек
Межжелудочковая
перегородка
Аорта
Легочная
артерия
Легочные вены
(от легких)
Левое
предсердие
Полулунный
клапан
Двустворчатый
(митральный)
клапан
Сухожильные
нити
Левый
желудочек
Рис. 14.50. Сердце
млекопитающего (в разрезе)
Поперечная
исчерченность
миофибрилл
Ядра в
саркоплазме
Ветвящееся
мышечное волокно
Вставочный
диск
Рис. 14.51. Строение сердечной мышцы.
Миофибриллы
с поперечной
исчерченностью
Вставочный
диск
Анастомозы
между
волокнами
Промежутки между
волокнами,
заполненные
соединительной тканью и
капиллярами (последние
не видны)
Поперечные
полосы
Ядро
Ядрышко
Вставочный
диск
Рис. 14.52. Микрофотография среза сердечной
мышцы.
и вскоре оказывается выше, чем в аорте и
легочной артерии, в результате чего открываются
полулунные клапаны и кровь выталкивается в эти
эластичные сосуды. Во время систолы
желудочков кровь ударяет в закрытые атриовентрикуляр-
ные клапаны и в результате этого удара
возникает первый тон сердца («лаб»).
3. Систола желудочков заканчивается, и за ней
следует диастола желудочков (рис. 14.53,7"). Под
действием высокого давления, создавшегося в
аорте и легочной артерии, часть крови
устремляется обратно в сторону желудочков, кровь
заполняет полулунные клапаны и они закрываются,
препятствуя возвращению крови в желудочки.
При ударе этого обратного тока крови о
полулунные клапаны возникает второй тон сердца
(«дап»):
систола желудочков - «лаб»
диастола желудочков - «дап»
Во время систолы желудочков стенки эластичных
артериальных сосудов растягиваются, а во время
диастолы возвращаются в исходное состояние и
выталкивают кровь, благодаря чему поступление
крови в большой и малый круги кровообращения
носит пульсирующий характер. По мере удаления
крови от сердца пульсация становится менее
выраженной, пока совсем не затухает в капиллярах и
венах, где кровь течет равномерно (рис. 14.55 и
14.56).
Один полный цикл состоит из одной систолы и
одной диастолы и продолжается около 0,8 с
(рис. 14.54).
Полые вены.
Правое
предсердие
Правый
желудочек
Легочная
вена
Левое
предсердие
Левый
желудочек
А. Предсердия в фазе диастолы,
заполняются кровью
Трехстворчатый
клапан
Сократившаяся
стенка
предсердия
Двустворчатый
клапан
Б. Систола предсердий. Кровь выталкивается в
желудочки. Двустворчатый и трехстворчатый
клапан открыты. Сфинктеры полой и легочной
вен закрыты
Аорта
Легочная
артерия^
В. Предсердия расслабляются, желудочки
сокращаются. Кровь выталкивается в
аорту и легочную артерию.
Г. Клапаны аорты и легочной артерии закрыты.
Предсердия начинают вновь наполняться кровью
Желудочки в фазе диастолы
Рис. 14.53. Последовательность фаз одного
сердечного цикла.
Предсердия
Желудочек
Митральный
клапан
Аортальный
клапан
Изометрическая
фаза
Левая
половина
сердца
Давление,
мм рт. ст.
Систола
Диастола-
Открыт
Открыт
Левый
желудочек
Аорта
Правая
половина
сердца
Давление,
мм рт ст
Правый
желудочек
Наполнение
100%
70%
0%
Ударный
объем
Объем
желудочка
ЭКГ
Тоны
сердца
Время, с
Рис. 14.54. Последовательность событий на
протяжении сердечного цикла начиная с систолы
предсердий Показана также часть следующего цикла
ПА кровяное давление в легочной артерии, ПП в
правом предсердии, ПП в левом предсерции (Из
J H Green, An introduction to human physiology, 1968,
Oxford Univ Press )
14.12.4. Механизм возбуждения и
сокращения сердца
Когда сердце извлекают из тела животного и
помещают в хорошо оксигенированный раствор Рин-
гера, оно в течение длительного времени
продолжает сокращаться, несмотря на отсутствие каких-либо
нервных или эндокринных стимулов. Этот факт
свидетельствует о миогенной природе сердечного
ритма, т.е. о том, что оно имеет собственный,
«встроенный» в него механизм, возбуждающий
сокращения мышечных волокон.
Миогенный контроль сердечного
ритма
Импульсы, вызывающие ритмические сокращения
сердца, возникают в особом участке правого
предсердия, называемом сшюатриальным (синусно-пред-
сердным) узлом и расположенном у места впадения
верхней полой вены. Этот узел является
рудиментарным остатком венозного синуса низших
позвоночных животных. Он состоит из небольшого числа
беспорядочно расположенных сердечных мышечных
волокон, бедных миофибриллами и иннервирован-
ных окончаниями вегетативных нейронов.
В клетках синоатриального узла за счет разности
концентраций ионов поддерживается мембранный
потенциал порядка —90 мВ. Мембране этих клеток
всегда свойственна высокая проницаемость для
натрия, поэтому ионы натрия непрерывно
диффундируют внутрь клетки. Поступление Na+ ведет к
деполяризации мембраны, в результате чего в
клетках, соседствующих с узлом, возникают
распространяющиеся потенциалы действия. Волна
возбуждения проходит по мышечным волокнам сердца и
заставляет их сокращаться. Синоатриальный узел
называют водителем сердечного ритма (пейсмейке-
ром), так как именно в нем зарождается каждая
волна возбуждения, которая в свою очередь служит
стимулом для возникновения следующей волны.
Раз начавшись, сокращение распространяется по
стенкам предсердия через сеть сердечных мышечных
волокон со скоростью 1 м/с. Оба предсердия
сокращаются более или менее одновременно. Мышечные
волокна предсердий и желудочков полностью
разделены соединительнотканной предсердно-желудочко-
вой перегородкой, и связь между ними
осуществляется только в одном участке правого предсердия -
атриовентрикулярном (предсердно-желудочковом) узле.
Ткань этого узла сходна с тканью
синоатриального узла. От атриовентрикулярного узла отходит
пучок специализированных волокон (антриовентри-
кулярный пучок)-единственный путь, по которому
волна возбуждения передается от предсердий к
желудочкам. Передача импульсов от синоатриального
узла к антриовентрикулярному происходит с
задержкой, составляющей около 0,15 с, благодаря
чему систола предсердий успевает закончиться
раньше, чем начинается систола желудочков. Атрио-
вентрикулярный пучок переходит в пучок Гиса,
который состоит из видоизмененных сердечных
мышечных волокон и от которого отходят более
тонкие веточки - волокна Пуркинье. Импульсы
проходят по пучку со скоростью 5 м/с и
распространяются в конце концов по всему миокарду желудочков.
Оба желудочка сокращаются одновременно, причем
волна их сокращения начинается в верхушке сердца
и распространяется вверх, выталкивая кровь из
желудочков в артерии, которые отходят от сердца
вертикально вверх (рис. 14.57 и 14.58).
Сердечная мышца обладает рядом особенностей,
Аорта
Артерии
Артериолы
Капилляры
Венулы
Вены
Полая вена
Объем, мл
Давление, мм рт. ст.
Скорость, см/с
Рис. 14.55. Распределение объема крови, давления и скорости кровотока в кровеносной системе человека. (Из
К. Schmidt-Nielsen, Animal physiology, 2nd ed., Cambridge Univ. Press, 1980.)
Артерии
Артериолы
_ ι Капилляры
Вены
Кровяное давление, мм рт. ст.
Пульсирующие
Непульсирующие
Рис. 14.56. Кровяное давление в различных участках
кровеносной системы человека. (Из J. H. Green, An
introduction to human physiology, 1968, Oxford Univ.
Press.)
позволяющих ей выполнять роль насоса, который
гонит кровь по всему телу в течение всей жизни
животного. Начав сокращаться, сердечная мышца
уже не может отвечать ни на какие другие стимулы
до тех пор, пока она не начнет расслабляться. Эта
стадия носит название рефрактерного периода, а
отрезок времени, в течение которого мышца не
отвечает ни на какие стимулы, называется периодом
абсолютной рефрактерное™ (рис. 14.59). У
сердечной мышцы этот период более продолжителен, чем
у мышц других типов, и это позволяет ей энергично
и быстро сокращаться, не испытывая утомления.
Благодаря этой особенности сердечная мышца не
способна к длительному сокращению, называемому
тетанусом, и в ней не создается кислородной
задолженности.
14.12.5. Регуляция сердечного ритма
Как уже отмечалось, собственный ритм сокращений
сердца задается синоатриальным узлом. Даже после
удаления из тела и помещения в искусственную
среду сердце продолжает ритмично сокращаться,
хотя и более медленно. Однако в организме к
сердечно-сосудистой системе предъявляются
постоянно меняющиеся требования, а соответственно
должна меняться и частота сердечных сокращений.
Эти изменения достигаются благодаря динамичной
и согласованной работе двух регуляторных
механизмов - нервного и гуморального,
осуществляющих тот гомеостатический контроль, который
поддерживает достаточное кровоснабжение тканей при
непрерывно меняющихся условиях.
Количество крови, протекающей через сердце за
1 мин, называется минутным объемом; оно зависит
от объема крови, выбрасываемой сердцем за одно
сокращение, и от частоты сокращений. Эти три
переменные связаны между собой следующим
уравнением:
Минутный объем = Ударный объем · Частота
сокращений.
Минутный объем, или сердечный выброс,-очень
важная переменная величина, и одним из способов
ее регуляции служит изменение частоты сокращений
сердца.
Полая вена
Синоатриальный
узел
Атриовентрикуляр-
ный узел
Правое предсердие
Правый желудочек
Аорта
Левое предсердие
Пучок Гиса,
содержащий
волокна Пуркинье
Левый желудочек
Межжелудочковая
перегородка
Рис 14.57. Расположение синоатриального и атрио-
вентрикулярного узлов и пучка Гиса.
Время, с
Рис. 14.58. Электрокардиограмма (ЭКГ)-запись
электрической активности сердца в течение одного
сердечного цикла. P-деполяризация мышцы
предсердия и распространение возбуждения от синоатри-
ального узла во время систолы предсердий; Q, R и
S-систола желудочков; T-начало диастолы
желудочков.
Сокращение
Время
Возбудимость
Нормальная
Период
абсолютной
рефрактерное™
Период
относительной рефрак
терности
Повышенная
Нормальная
Рис. 14.59. Период рефрактерности сердечной
мышцы. Верхняя кривая отражает сокращение мышцы, а
нижняя-изменения ее чувствительности к
стимуляции. (Из Clegg, Clegg, Biology of the mammal, 2nd ed.,
1963, Heinemann Medical Books.)
Нервная регуляция сердечного ритма
В продолговатом мозге-одном из отделов заднего
мозга-имеется ряд участков, регулирующих
деятельность сердечно-сосудистой системы, в том
числе частоту сокращений сердца. От
расположенного здесь кардиоингибиторного центра отходит
пара блуждающих нервов, содержащих
парасимпатические волокна и направляющихся по обеим
сторонам трахеи к сердцу. В сердце
парасимпатические волокна подходят к синоатриальному узлу,
атриовентрикулярному узлу и пучку Гиса, и
поступающие по этим волокнам импульсы уменьшают
частоту сердечных сокращений. В продолговатом
мозге находится также прессорный участок сосудо-
двигательного (вазомоторного) центра, от которого
берут начало нервы симпатической системы.
Пройдя вдоль позвоночника, эти нервы подходят в
грудном отделе к сердцу и посылают импульсы
синоатриальному узлу. Импульсы, поступающие по
симпатическим волокнам, ускоряют работу сердца.
Совместная деятельность тормозящих и
стимулирующих центров продолговатого мозга
контролирует частоту сердечных сокращений.
К сердечно-сосудистым центрам продолговатого
мозга подходят чувствительные нервные волокна от
рецепторов растяжения (барорецепторов),
расположенных в стенках дуги аорты, каротидного синуса и
полых вен. Импульсы, поступающие от аорты и
каротидного синуса, замедляют работу сердца,
тогда как импульсы, приходящие от полых вен,
ускоряют ее. При увеличении количества крови в
этих сосудах их стенки растягиваются, в результате
чего возрастает число импульсов, посылаемых от
них в сердечно-сосудистые центры продолговатого
мозга.
Например, при интенсивной физической работе
мышцы сильно сокращаются, что ускоряет
возвращение крови к сердцу по венам. Поступление
большого количества крови в полые вены вызывает
их растяжение, а это приводит к ускорению работы
сердца. Одновременно повышенный приток крови к
сердцу вызывает растяжение сердечной мышцы, и в
ответ на это сердце сильнее сокращается и
выбрасывает больше крови во время систолы (увеличение
ударного объема). Эта зависимость между объемом
поступающей в сердце крови и ударным объемом
сердца получила название закона Старлинга по
имени открывшего его английского физиолога.
Увеличение ударного объема приводит к
растяжению стенок аорты и сонных артерий и к
возникновению импульсов, которые поступают в кардиоин-
гибиторный центр и вызывают замедление работы
сердца. Таким образом, существует автоматический
механизм саморегуляции, который препятствует
слишком частым сокращениям сердца и позволяет
так изменять его активность, чтобы в любой
момент оно могло эффективно справляться с
объемом притекающей крови (рис. 14.60).
Другие факторы, регулирующие
сердечный ритм
Существует целый ряд гуморальных и иных
факторов, действующих непосредственно на сердечную
мышцу или на синоатриальный узел. Эти факторы
перечислены в табл. 14.12.
На сердечно-сосудистый центр, так или иначе,
влияют многие факторы, в том числе эмоции,
связанные, например, с гневом или воздействием
слуховых и зрительных стимулов и выражающиеся
в покраснении или побледнении кожи. В этих
случаях сенсорные импульсы передаются в мозг и
через внутримозговые связи в сердечно-сосудистый
центр, который на них соответствующим образом
реагирует (табл. 14.12).
Следует заметить, что на этот центр в каждый
данный момент всегда влияет определенное сочета-
Продолговатый
мозг
Сердечнососудистые
центры
Сенсорный нерв
(тормозящий)
Ускоряющий
нерв
(симпатическая
система)
Аорта
Сенсорный нерв
(ускоряющий)
Тормозящий нерв
(блуждающий;
парасимпатическая
система)
Полая вена
Синоатриальный
узел
Афферентная иннервация: нервы отходят от
полой вены, аорты и каротидного синуса и
направляются к сердечно-сосудистым центрам
(нервы, идущие от каротидного синуса,
не показаны)
Двигательная иннервация: импульсы
ускоряющего нерва повышают частоту
сокращений, а импульсы блуждающего
нерва замедляют сердечный ритм
Рис. 14.60. Пути нервной регуляции сокращения
сердца.
ние нервных и иных факторов, а не какой-то один из
них. Активность сердечно-сосудистого центра
зависит также от возраста и состояния здоровья
индивидуума.
14.12.6. Регуляция кровяного давления
Кровяное давление зависит от целого ряда
факторов-от частоты и силы сердечных сокращений,
ударного объема сердца, сопротивления току крови
со стороны сосудов (периферическое
сопротивление). Частота сокращений сердца и ударный объем
уже рассматривались в предыдущем разделе.
Сопротивление току крови меняется в зависимости от
сокращения или расслабления гладкой мускулатуры
сосудистых стенок, особенно в артериолах. При
сужении сосудов (вазоконстрикции) периферическое
сопротивление увеличивается, а при их расширении
(вазодилятации) уменьшается. Повышение
сопротивления приводит к повышению кровяного
давления, а снижение сопротивления - к его падению. Все
эти изменения контролируются вазомоторным
центром продолговатого мозга.
Нервные волокна идут от вазомоторного центра
ко всем артериолам тела. Изменение диаметра этих
сосудов зависит в основном от тонуса мышц,
сужающих сосуды (вазоконстрикторов); мышцы,
расширяющие сосуды (вазодилятаторы), играют
меньшую роль.
Активность вазомоторного центра регулируется
импульсами, поступающими от рецепторов,
воспринимающих давление (барорецепторов), которые
находятся в стенках аорты и каротидных синусах
сонных артерий (рис. 14.61). Стимуляция
парасимпатических волокон в этих участках, вызываемая
повышением сердечного выброса, приводит к
расширению сосудов во всем теле, вследствие чего
падает кровяное давление и уменьшается частота
сокращений сердца. При снижении кровяного
давления наблюдается противоположная картина:
происходит стимуляция симпатических волокон, и это
ведет к общему сужению кровеносных сосудов
и компенсаторному повышению кровяного
давления.
Химическая регуляция
вазомоторного центра
В каротидных тельцах, лежащих в области
разветвления сонных артерий, находятся хеморецеп-
торы, которые возбуждаются при высоком
содержании CO2 в притекающей крови и посылают
импульсы в вазомоторный центр продолговатого мозга.
Нервные волокна, идущие от этих хеморецепторов,
приходят в вазомоторный центр в одном пучке с
волокнами от барорецепторов каротидного синуса.
В ответ на стимулы, приходящие по этому общему
пути, вазомоторный центр посылает к кровеносным
сосудам импульсы, под действием которых сосуды
сужаются и кровяное давление возрастает.
Повышенная активность тканей и органов
сопровождается обычно усиленным образованием CO2, и
благодаря описанному механизму насыщенная CO2 кровь
будет быстрее поступать в легкие и, значит,
углекислота будет быстрее обмениваться в легких на
кислород.
Углекислота может также оказывать прямое
воздействие на гладкую мускулатуру кровеносных
сосудов. При резком увеличении активности какой-
либо ткани в ней образуются большие количества
CO2, что сразу ведет к расширению сосудов в
данной области. В них тотчас увеличивается крово-
Таблица 14.12. Гуморальные и иные факторы, влияющие на
частоту сокращений сердца
Стимул
Повышение рН
Снижение рН (например, при
высоком содержании CO2,
как в случае физической
нагрузки)
Низкая температура
Высокая температура
Неорганические ионы
Эндокринные факторы
(например, тироксин, инсулин,
половые гормоны,
адреналин, гормоны гипофиза)
Влияние на сердечный ритм
Замедляет
Ускоряет
Замедляет
Ускоряет
Влияют на ритм сердца
\ прямо или косвенно
Продолговатый мозг
Вазомоторный
центр
Наружная
сонная
артерия
Каротидное>
тельце —
регистрирует изменение в
соотношении
CO2/О 2 ирН
Общая сонная
артерия
Внутрен^у
няя сонная
артерия
Ток крови
Много- ]
численные
живающие
импульсы
Каротидный синус —
регистрирует
изменения давления
Много импульсов
Мало импульсов
Малочисленные
сосудорас-
ширяющие
импульсы
Малочисленные
сосудосуживающие!
импульсы
Сужение сосудов
Многочисленные
сосудорас -
ширяющие
импульсы
Расширение сосудов
Рис. 14.61. Взаимоотношения между каротидным тельцем, каротидным синусом, вазомоторным центром и
общей системой кровообращения.
ток, и к активным клеткам поступает больше
кислорода и глюкозы. Следует, однако, помнить,
что углекислота, поступившая из этой области в
общий кровоток, будет влиять на активность
вазомоторного центра, способствуя сужению сосудов в
других частях тела. Эта ситуация ясно
показывает, каким динамичным и гибким может быть
регулирование кровяного давления, а значит, и
всего процесса циркуляции и распределения крови.
Другие факторы, в частности различные виды
эмоционального стресса (возбуждение, боль и т. п.),
повышают активность симпатической нервной
системы, и в результате кровяное давление
повышается. Выброс адреналина при стимуляции
мозгового вещества надпочечников импульсами,
поступающими из высших отделов центральной нервной
системы, приводит к ускорению ритма сердца и
способствует общему сужению сосудов и
повышению кровяного давления. Значение регуляции
сердечного ритма и кровяного давления подробнее
обсуждается в разд. 18.1.5.
14.35. При ранении животного общее кровяное
давление повышается, но вокруг раны образуется
отек в результате местного расширения сосудов.
Почему это происходит?
14.36. Опишите основные изменения сердечного
ритма и кровеносной системы перед, во время и
после пробега на 100 м.
14.13. Функции крови у
млекопитающих
Кровь млекопитающих выполняет множество
важных функций. В приведенном ниже списке первые
пять функций принадлежат только плазме.
1. Перенос растворимых органических веществ от
тонкого кишечника к различным органам и
тканям, где эти вещества откладываются в запас
или участвуют в метаболизме, а также доставка
питательных веществ из мест их хранения к
местам использования.
2. Транспорт подлежащих экскреции растворимых
отходов из тканей, где они образуются, к
органам выделения.
3. Перенос побочных продуктов метаболизма из
мест их образования к другим участкам тела.
4. Транспорт гормонов из желез, где они
образуются, ко всем органам и тканям или к
определенным органам-мишеням для передачи
информации внутри организма.
5. Перенос тепла от глубоко расположенных
органов, предупреждающий перегрев этих органов и
поддерживающих равномерное распределение
тепла в организме.
6. Доставка кислорода из легких ко всем тканям
организма и перенос в обратном направлении
углекислоты, образующейся в тканях.
7. Защита от болезней, в которой участвуют три
механизма:
а) свертывание крови, предотвращающее
излишнюю потерю крови и проникновение в
организм болезнетворных агентов;
б) фагоцитоз, осуществляемый гранулоцитами,
которые захватывают и переваривают
попавших в кровеносное русло бактерий;
в) иммунная защита, осуществляемая
антителами и/или лимфоцитами.
8. Поддержание постоянного осмотического
давления и рН с помощью белков плазмы. Белки
плазмы и гемоглобин содержат как кислые, так и
основные аминокислоты, поэтому они могут
связывать и освобождать ионы водорода и таким
образом снижать до минимума изменения рН.
Рис. 14.62. Строение молекулы гема.
S
§
1
Напряжение
при насыщении
Напряжение
при полу-
насыщении
Напряжение O2 , мм рт. ст.
Рис. 14.63. Этот график поясняет термины,
принятые для описания кривых диссоциации пигментов,
переносящих кислород. Напряжение насыщения-это
то напряжение кислорода, при котором насыщено
кислородом 95% пигмента, а напряжение полуна -
сыщения-то, при котором насыщено кислородом
50% пигмента. (Из Florey, An introduction to general and
comparative physiology, 1966, W. B. Saunders and Co.)
14.13.1. Транспорт кислорода
Кислород переносят молекулы гемоглобина,
содержащиеся в эритроцитах. Гемоглобин представляет
собой тетрамерный белок с мол. массой 68000. В
его состав в качестве простетических групп входят
четыре молекулы гема, которые присоединены к
четырем полипептидным цепям глобина и придают
крови характерный красный цвет. Каждый гем
содержит один атом двухвалентного железа,
непрочно связывающего одну молекулу кислорода
(рис. 14.62):
Hb + 4O2 τ± HbO8.
Присоединение кислорода к гемоглобину с
образованием оксигемоглобина происходит при
высоком парциальном давлении кислорода-таком, как в
капиллярах легочных альвеол. При низком
парциальном давлении кислорода (как в капиллярах
метаболически активных тканей) связь между
кислородом и гемоглобином становится непрочной,
кислород освобождается и диффундирует в
окружающие клетки.
Количество кислорода, которое может связаться с
гемоглобином, зависит от напряжения кислорода в
среде. Последнее выражают как парциальное
давление кислорода в атмосфере, находящейся в
равновесии с данной средой. Парциальное давление до
сих пор выражают в миллиметрах ртутного столба.
Например, атмосферное давление на уровне моря
составляет 760 мм рт. ст. Кислород составляет
около 1/5 атмосферного воздуха; следовательно, его
парциальное давление на уровне моря равно
примерно 1/5-760 = 152 мм рт. ст.
Определяя процент насыщения крови кислородом
при различном парциальном давлении O2,
получают S-образную кривую, называемую кривой
диссоциации оксигемоглобина (рис. 14.63).
Анализ этой кривой показывает, что в
физиологическом плане гемоглобин можно считать полностью
насыщенным кислородом при таком напряжении
O2, при котором фактически насыщается 95%
гемоглобина. В примере, приведенном на рис. 14.63,
такое напряжение составляет около 73 мм рт. ст.
При более высоком парциальном давлении O2
может происходить дальнейшее его поглощение, но
100%-ное насыщение гемоглобина достигается
редко. При парциальном давлении кислорода около
30 мм рт. ст. только 50% гемоглобина находится в
форме оксигемоглобина, а при нулевом
парциальном давлении O2 молекулы гемоглобина вообще не
связывают кислород. В области крутого наклона
кривой уже при небольшом снижении парциального
давления O2 процент насыщения гемоглобина
значительно уменьшается. Кислород,
освобождаемый пигментом, становится доступным для тканей.
Почему же кривая насыщения имеет S-образную
форму? Такая форма кривой объясняется аллостери-
ческим характером взаимодействия гемоглобина с
кислородом (разд. 6.6). Когда молекула O2
связывается с атомом двухвалентного железа одного из
гемов, конфигурация этого гема слегка изменяется.
Это изменение отражается на конфигурации всей
молекулы гемоглобина, которая определенным
образом меняется. По мере присоединения
кислорода к второй и третьей группам гема происходит
дальнейшее изменение конфигурации гемоглобина,
и в результате каждая последующая молекула O2
присоединяется легче, чем предыдущая (явление
кооперативности), так что последний гем связывает
кислород в несколько сотен раз легче, чем первый.
Обратный процесс происходит тогда, когда
оксигемоглобин оказывается в области низкого
парциального давления O2, как это бывает в
активно дышащих тканях. Первую молекулу кислорода
оксигемоглобин отдает тканям очень легко, но
вторая, третья и четвертая молекулы отделяются
значительно труднее и только при очень сильном
Парциальное давление О 2 , кПа
Парциальное давление CO2 , мм рт 'ст
Насыщение, %
Парциальное давление O2 , мм рт ст
Рис. 14.64. Кривые диссоциации оксигемоглобина при
различном парциальном давлении CO2,
иллюстрирующие эффект Бора. (Из Garth Chapman, The body
fluids and their function. Studies in Biology, № 8, 1967,
Arnold.)
Парциальное давление O2 , кПа
4 8
Насыщение, %
У плода
У матери
Парциальное давление O2 , мм рт.ст
Рис. 14.65. Кривые диссоциации оксигемоглобина в
крови плода и матери у овцы
снижении парциального давления O2. При
повышении парциального давления CO2 кривая
диссоциации оксигемоглобина сдвигается вправо, и это
явление называют эффектом Бора (рис. 14.64).
14.37. Каково физиологическое значение
эффекта Бора?
14.38. Рассмотрите рис. 14.65. Кривая
диссоциации оксигемоглобина у плода сдвинута влево по
сравнению с кривой для матери. Чем объясняется
такое смещение?
14.39. Кривая диссоциации оксигемоглобина у
южноамериканских лам, живущих в Андах на
высоте около 5000 м над уровнем моря,
расположена левее, чем у большинства других
млекопитающих (рис. 14.66). Чем это можно объяснить?
14.13.2. Миоглобин
Миоглобин широко распространен у животных, но
наиболее характерен для скелетных мышц
млекопитающих. Он обладает высоким сродством к
кислороду, поэтому кривая диссоциации оксимиоглобина
значительно сдвинута влево по отношению к кривой
для оксигемоглобина (рис. 14.67). Действительно,
оксигемоглобин начинает отдавать кислород только
после того, как парциальное давление O2 падает
ниже 20 мм рт. ст. Благодаря этому он играет в
покоящейся мышце роль кислородного депо и
отдает O2 только тогда, когда исчерпываются
запасы оксигемоглобина. По последовательности
аминокислот и третичной структуре молекула миог-
лобина очень сходна с отдельной субъединицей
молекулы гемоглобина. Однако молекулы миогло-
бина не соединяются между собой с образованием
тетрамера и не способны поэтому к положительной
кооперативности в связывании кислорода.
По-видимому, оба белка происходят от общей предковой
молекулы.
14.13.3. Окись углерода и гемоглобин
Сродство двухвалентного железа, входящего в
состав гемоглобина, к окиси углерода (СО) в
несколько сотен раз больше его сродства к
кислороду. Поэтому в присутствии даже очень малого
количества окиси углерода гемоглобин будет
связывать иммено ее, а не кислород, и в результате
образуется относительно стабильное соединение
карбоксигемоглобин. В этом случае кислород уже не
может связываться с гемоглобином и перенос
кислорода кровью становится невозможным. После
вдыхания окиси углерода у человека быстро
наступает коллапс, и если его не вывести из отравленной
атмосферы, он погибнет от удушья. После удаления
Парциальное давление O2 , кПа
Насыщение,%
Другие
млекопитающие
ι
Лама
Парциальное давление О2 , мм рт. ст.
Рис. 14.66. Сравнение кривых диссоциации
оксигемоглобина у ламы и других млекопитающих.
Миоглобин
Гемоглобин
Насыщение, %
Парциальное давление О 2 , мм рт. ст.
Рис. 14.67. Сравнение кривых диссоциации оксиге-
моглобина и оксимиоглобина. Миоглобин остается
насыщенным O2 на 80% до тех пор, пока парциальное
давление кислорода не падает ниже 20 мм рт. ст.
Это означает, что миоглобин удерживает кислород
в покоящемся мышечном волокне, но отдает его,
когда в результате интенсивной мышечной работы
используется весь кислород, доставленный
гемоглобином.
пострадавшего из атмосферы, содержащей окись
углерода, необходимо провести искусственное
дыхание смесью чистого кислорода с CO2, так как
напряжение кислорода в воздухе недостаточно для
быстрого вытеснения окиси углерода, связанной с
гемоглобином.
14.13.4. Транспорт углекислоты
Существуют три способа переноса углекислоты
(двуокиси углерода, CO2) кровью.
перенос в растворенном виде (5%) Большая часть
переносимой этим способом углекислоты
транспортируется в физически растворенном виде, и лишь
очень небольшая часть находится в виде угольной
кислоты (H2CO3).
ПЕРЕНОС B СОЕДИНЕНИИ С БЕЛКОМ (10 20%) CO2 ПрИСОе-
диняется к аминогруппам гемоглобина с
образованием карбаминогемоглобина. Количество CO2,
способное связаться с гемоглобином, зависит от
количества уже связанного гемоглобином кислорода.
Чем меньше кислорода связано с гемоглобином,
тем больше CO2 может быть перенесено этим
способом:
HHbH2
Гемоглобин
+ CO2-
Карбаминогемоглобин
Перенос в виде карбоната (85%). Образующаяся в
тканях углекислота проникает путем пассивной
диффузии в кровеносное русло и поступает в
эритроциты, где, соединясь с водой, образует
угольную кислоту. Этот процесс катализируется
содержащимся в эритроцитах ферментом карбоангидразой
(угольной ангидразой) и протекает менее чем за
одну секунду. Угольная кислота диссоциирует затем
на ионы водорода и гидрокарбоната (бикарбоната):
CO2 + H2O *± H2CO3 τ± H+ + HCO3".
Когда эритроциты покидают с кровью легкие,
содержащийся в них оксигемоглобин (HbO2) имеет
слабокислую реакцию и связан с ионами калия
(обозначается как KHbO2). В участках с высокой
концентрацией CO2 (в тканях) оксигемоглобин
легко отдает килород, приобретая при этом
сильнощелочную реакцию. В этом состоянии он теряет
ионы калия и легко принимает от угольной кислоты
ионы водорода, превращаясь в гемоглобиновую
кислоту (H · Hb). В свою очередь ионы калия
связываются с ионами гидрокарбоната, образуя
гидрокарбонат калия:
KHbO2 т± KHb + O2;
H + HCO3 + KHb «± H Hb + KHCO3.
(Гемогло-
биновая
кислота)
Присоединяя ион водорода, гемоглобин
действует как буферная система; поэтому большое
количество угольной кислоты может переноситься к
легким без значительного изменения рН крови.
Плазматическая мембрана эритроцитов
относительно непроницаема для ионов натрия и калия, но
имеется натриевый насос, который откачивает из
эритроцитов в плазму большое количество ионов
натрия. Основная масса ионов гидрокарбоната,
образующихся в эритроцитах, выходит в плазму путем
диффузии по градиенту концентрации и, соединяясь
здесь с ионами натрия, образует гидрокарбонат
натрия. Выход ионов гидрокарбоната из
эритроцитов компенсируется поступлением туда хлорид-
ионов, диффундирующих из плазмы. Таким
образом поддерживается электронейтральность. Это
явление носит название хлорндного сдвига. Тот
гидрокарбонат калия, который образуется в
эритроцитах, тоже способен диссоциировать, и часть
хлорид-ионов, проникающих в эритроциты,
соединяется с освобождающимися ионами калия, образуя
KCl. Когда ионы гидрокарбоната покидают
эритроцит, оставшиеся в нем в избытке ионы водорода
снижают рН, вызывая дальнейшую диссоциацию
калиевой соли оксигемоглобина KHbO2 на
кислород и KHb (рис. 14.68).
Когда эритроциты попадают в легкие, в них
происходит обратный процесс.
14.40. Опишите кратко, каким образом
содержащаяся в крови углекислота удаляется в легких в
газообразной форме.
14.13.5. Защитные функции крови
Каждое млекопитающее обладает сложной
системой защитных приспособлений, позволяющей ему
отражать атаки болезнетворных агентов, удалять из
Плазма
Тканевая
[жидкость.
CO2 + H2O
H2CO1
H++ HCO3,
CO2+ HHbNHT
(Гемоглобин)
HHbNHCOOH
(Карбамино-
гемоглобин)
HHb + K+
KHCO3
K+ + НСОГ
NaHCO3
Плазма
Na+
+ i
СГ
NaCl
Хлоридный
сдвиг
JCCl
Эритроцит
CO2-
O2
Тканевая
жидкость'
O7+ KHb
KHbO2
"(Оксигемоглобин)'
Митохондрия
Рис. 14.68. Перенос углекислоты плазмой и эритроцитами; показана роль CO2 в высвобождении кислорода в
тканях.
организма чужеродные материалы и т. п. В этом
разделе будут рассмотрены три защитных
механизма:
1) свертывание крови;
2) фагоцитоз;
3) иммунный ответ на инфекцию.
Два первых механизма участвуют в заживлении ран.
Свертывание крови
При повреждении ткани из нее вытекает кровь,
которая вскоре свертывается, образуя кровяной
сгусток. Последний препятствует дальнейшей
потере крови и проникновению в организм
болезнетворных микроорганизмов. Очевидно, что этот
процесс имеет большое значение для выживания
животного. Не менее важно и то, что в неповрежденных
сосудах кровь не свертывается. Чрезвычайная
сложность многостадийного процесса свертывания
служит одновременно для того, чтобы этот процесс не
происходил тогда, когда в нем нет необходимости.
Для его осуществления нужны по меньшей мере 12
различных факторов, действующих согласованным
образом; мы рассмотрим только важнейшие из этих
факторов.
Кровь, вытекающая на поверхность раны,
соприкасается с воздухом и смешивается с веществами,
выделяющимися из поврежденных клеток и
разрушенных тромбоцитов. Высвобождающийся из
поврежденных тканей липопротеин тромбопластин
вместе с факторами свертывания VII и X (ферменты
плазмы) и ионами кальция катализирует
превращение неактивного белка плазмы протромбина в
тромбин. Тромбин - протеолитический фермент; он
расщепляет крупную молекулу растворимого
глобулярного белка плазмы фибриногена на более мелкие
единицы - мономеры, которые затем полимеризу-
ются и образуют сеть, состоящую из длинных
перепутанных нитей фибрина - нерастворимого
фибриллярного белка (рис. 14.69). После удаления
фибриногена из плазмы остается жидкость, которая
называется сывороткой. В сети, образованной
волокнами фибрина, задерживаются форменные
элементы крови (рис 14.70) и в результате образуется
кровяной сгусток. Позднее он сжимается, подсыхает
и образует струп, который препятствует дальнейшей
потере крови и создает механический барьер для
проникновения патогенных агентов.
При такой сложности всего процесса отсутствие
или низкая концентрация любого из незаменимых
факторов свертывания крови могли бы вызвать
чрезмерно сильное кровотечение. Такое состояние
известно под названием гемофилии. Например, если
нет или слишком мало какого-либо из факторов,
необходимых для действия тромбопластина, то
малейший порез приведет к длительному
кровотечению. Гемофилия связана с мутацией
определенного гена и передается с Х-хромосомой. Скрытыми
носителями дефектного гена бывают женщины, но
страдают гемофилией только мужчины. Дело в том,
что дефектный аллель рецессивен по отношению к
нормальному аллелю (см. разд. 23.6.1). Поэтому он
не проявляется у женщин, имеющих две Х-хромо-
сомы, в том числе одну с нормальным аллелем, но
вызывает болезнь у мужчин, у которых второй
Х-хромосомы нет.
Ранение
Разрушение
' тромбоцитов
Соприкосновение"
с воздухом
.Повреждение ткани
Тромбопластин
Протромбин
плазмы
(образуется
в печени)
Тромбин
Ферменты
плазмы
Ca2 +
Фибриноген
плазмы
(образуется
в печени)
Фибрин
(сгусток)
Рис. 14.69. Основные этапы процесса свертывания
крови
|Эритроцит|
|Фибриновая сеть|
Рис. 14.70. Эритроцит, захваченный нитями фибрина при образовании кровяного сгустка
В неповрежденных кровеносных сосудах кровь не
свертывается, поскольку выстилка сосудов обладает
очень гладкой поверхностью и не вызывает
разрушения тромбоцитов или клеток крови. К тому же
в крови имеются активные вещества,
препятствующие свертыванию. Одно из таких веществ - гепарин,
присутствующий в небольшом количестве в плазме
и выделяемый тучными клетками, находящимися в
соединительной ткани и печени. Гепарин
препятствует превращению фибриногена в фибрин и
поэтому широко используется в клинике как
антикоагулянт.
Если сгусток образуется внутри кровеносного
сосуда, он называется тромбом, и его образование
ведет к острому нарушению кровообращения-
тромбозу. Тромбоз может произойти в результате
повреждения эндотелия кровеносного сосуда, так
как шероховатость поврежденного участка
способствует разрушению тромбоцитов и таким образом
запускает процесс свертывания крови. Особенно
опасен и может привести к внезапной смерти
коронарный тромбоз, т. е. тромбоз венечной артерии
сердца.
Фагоцитоз
Фагоцитоз осуществляют главным образом нейтро-
филы. Это амебоидные клетки, мигрирующие в
места повреждения клеток и тканей. Стимулами для
их миграции служат, по-видимому, какие-то
вещества, освобождаемые разрушенными клетками
крови и тканями. Нейтрофилы обладают способностью
распознавать любые бактерии, проникшие в
организм. Эту способность усиливают плазменные
белки, называемые опсонинами, которые
прикрепляются к поверхности бактерий и каким-то образом
делают их легче узнаваемыми. Обнаружив
бактерию, нейтрофил захватывает ее путем фагоцитоза, и
в нем формируется фагосома (рис. 14.71). С фаго-
сомой сливаются мелкие лизосомы, образуя фаго-
лизосому. В фаголизосому из лизосом изливаются
лизоцим и другие гидролитические ферменты, под
действием которых бактерия переваривается. В
конце концов растворимые продукты переваривания
бактерий поглощаются окружающей цитоплазмой
нейтрофил а.
Нейтрофилы способны проходить через стенки
кровеносных капилляров (этот процесс называется
диапедезом) и передвигаться в межклеточных
пространствах. В таких органах, как печень, селезенка и
лимфатические узлы, имеются крупные
неподвижные фагоциты, называемые обычно макрофагами.
Роль макрофагов состоит в поглощении токсичных
чужеродных частиц и микроорганизмов и
длительном, а часто и постоянном их удерживании внутри
себя. Таким путем нередко приостанавливается
распространение инфекции. Вместе с нейтрофилами
макрофаги образуют ретикулоэндотелиальную
систему организма.
Воспаление
При ранении какого-либо участка тела возникает
местная реакция окружающих тканей, которая
проявляется в опухании и болезненности. Это состояние
называется воспалением и связано с выделением из
поврежденных тканей определенных веществ, в
частности гистамина и серотонина. Эти вещества
вызывают местное расширение капилляров, в
результате чего усиливается приток крови к
поврежденному участку и повышается его
температура. Возрастает также проницаемость капилляров,
и в результате плазма выходит в окружающие ткани
и вызывает их набухание - отёк. Плазма содержит
бактерицидные факторы, антитела и нейтрофилы,
которые все вместе противодействуют
распространению инфекции. Кроме того, в плазме
присутствует фибриноген, способствующий в случае
необходимости свертыванию крови, а избыток
тканевой жидкости снижает концентрацию и
токсичность любых потенциально опасных раздражающих
агентов.
Заживление ран
К концу воспалительного процесса появляются
клетки, называемые фибробластами, которые секре-
тируют коллаген. Это фибриллярный белок,
который, соединяясь с полисахаридами, образует сеть
из беспорядочно переплетающихся волокон рубцо-
вой ткани. Для образования коллагена необходим
витамин С, так как без него молекулы коллагена не
гидроксилируются и остаются недостроенными.
Примерно через две недели после ранения
беспорядочная масса волокон реорганизуется и собирается
в пучки, расположенные вдоль линий натяжения в
области раны. Здесь начинают также прорастать
многочисленные мелкие кровеносные сосуды,
которые снабжают кислородом и питательными
веществами клетки, участвующие в заживлении раны.
Одновременно к процессу заживления подключается
и окружающий эпидермис. Отдельные клетки
эпидермиса мигрируют в рану и поглощают остатки
тканей и фибрин образовавшегося кровяного
сгустка. Встречаясь друг с другом, эпидермальные клетки
соединяются, образуя сплошной слой под струпом.
Когда формирование этого слоя заканчивается,
струп отпадает и открывается эпидермис.
Перечень стадий процесса заживления
1. В поврежденном участке возникает
кровотечение.
2. Происходит свертывание крови.
3. Начинается воспалительный процесс.
4. В рану мигрируют лейкоциты, которые
поглощают чужеродный материал, бактерий и
остатки клеток.
Бактерия
Нейтрофил
Миграция нейтрофила
по направлению
к бактерии
Прилипание бактерии
к нейтрофилу
Образование
фагосомы
Лизосома
Фагоцитоз
Фаголизосома
Лизосома
Переваривание
Всасывание
переваренных
частей бактерии
Рис. 14.71. Фагоцитоз: захват и переваривание
бактерии нейтрофилом.
5. В ране собираются фибробласты,
синтезирующие коллаген, из которого формируется рубцо-
вая ткань.
6. Эпидермальные клетки поглощают последние
остатки разрушенных клеток и начинают
разрушать рубец.
7. Эпидермис образует в области раны новую
кожу.
8. Струп слущивается.
Если рана небольшая, то для борьбы с инфекцией
бывает достаточно одного лишь фагоцитоза, но при
значительных размерах повреждения в действие
вступает иммунная система организма.
14.14. Иммунная система
Макфарлейн Бёрнет-один из создателей
современной теории иммунитета - определяет его как
«способность распознавать вторжение в организм
чужеродного материала и мобилизовать клетки и
образуемые ими вещества на более быстрое и
эффективное удаление этого материала».
Основные понятия
Антитело - молекула, синтезируемая организмом
животного в ответ на присутствие чужеродного
вещества, к которому эта молекула обладает
высоким сродством. Все антитела - белки, называемые
иммуноглобулинами. Молекула иммуноглобулина
состоит из двух тяжелых цепей (H-цепей) с мол.
массой 50000-60000 и двух легких цепей (L-цепей) с
мол. массой 23000. В функциональном отношении
молекула антитела подразделяется на константные
и вариабельные участки, причем последние
действуют подобно ключу, который подходит к
определенному замку (рис. 14.72). Каждый организм способен
производить тысячи видов антител различной
специфичности, которые могут распознавать
всевозможные виды чужеродных веществ.
Чужеродное вещество, вызывающее образование
антител, называют антигеном или иммуногеном.
Обычно антиген представляет собой белковую или
полисахаридную молекулу, находящуюся на
поверхности микроорганизма или в свободном виде. У
млекопитающих сформировались две системы
иммунитета - клеточный и гуморальный иммунитет.
Такое разделение функций иммунной системы
связано с существованием двух типов
лимфоцитов - Т-клеток и В-клеток. Клетки обоих типов
образуются в костном мозге из
клеток-предшественниц. В формировании иммунологической
компетентности Т-клеток решающую роль играет тимус
(вилочковая железа). Что касается В-клеток, то
полагают, что аналогичное влияние на их развитие
оказывают плацента или костный мозг и печень
плода. Клетки каждого из этих двух типов
обладают колоссальной способностью «узнавать»
какой-либо из миллионов существующих антигенов.
Реакция антиген - антитело направлена на то, чтобы
связать антиген, инактивировать его и
предотвратить таким образом его вредное воздействие на
организм.
Клеточный иммунитет. При взаимодействии с
антигеном Т-лимфоциты, несущие на мембране
рецепторы, способные распознавать этот антиген,
начинают размножаться и образуют клон таких же
Т-клеток. Клетки этого клона вступают в борьбу с
несущими антиген микроорганизмами или
вызывают отторжение чужеродной ткани.
Гуморальный иммунитет. В-лимфоциты
распознают антиген таким же образом, как и Т-клетки, но
реагируют по-иному. Размножаясь при стимуляции,
они образуют клон плазматических клеток, которые
синтезируют антитела и выделяют их в кровь или
тканевую жидкость. Здесь антитела связываются с
антигенами на поверхности бактерий и ускоряют их
захват фагоцитами или присоединяются к
бактериальным токсинам и нейтрализуют их.
14.14.1. Тимус и развитие Т-клеток
Тимус (вилочковая железа) расположен в грудной
клетке под грудиной, около вентральной (у
человека-передней) стороны сердца. Он начинает
функционировать в период внутриутробного
развития и проявляет наибольшую активность в
момент рождения и в последующее время. После
окончания вскармливания материнским молоком
тимус уменьшается в размерах и вскоре перестает
функционировать.
Константные участки,
где последовательность-
аминокислот одинакова
во всех молекулах
данного типа
Антиген-
связывающий
участок
Вариабельные участки,
где последовательность
аминокислот различна
в разных молекулах
Легкая цепь
Тяжелая цепь
Шарнирная область
Дисульфидная
связь
Рис. 14.72. Молекула
иммуноглобулина. Антиген связывается
между вариабельными участками
легкой и тяжелой цепей. (Из M. Cooper,
A Lav/ton, The development of the
immune system. Scientific American,
7972)
Костный мозг
Стволовые клетки
Тимус
Лимфоциты
Тимоциты
Тимозин
Лимфокины
Иммунокомпе-
тентные
Т-лимфоциты
Стимулируют
образование
антител в
В-лимфоцитах
Лимфоидная
ткань
Клоны
Селезенка
Клоны
Общая циркуляция
Рис. 14.73. Дифференциация и активность
T-лимфоцитов.
На важную роль тимуса в развитии иммунного
ответа указывают следующие экспериментальные
данные:
1. Удаление тимуса у новорожденных мышат
приводит к гибели от хронической недостаточности
лимфоцитов в крови и тканевой жидкости.
2. Новорожденные мышата с удаленным тимусом
не распознают ткань, пересаженную от других
мышей, и не дают на нее иммунного ответа.
3. Удаление тимуса у мышей в значительно более
позднем возрасте не вызывает каких-либо
неблагоприятных последствий.
Будущие Т-лимфоциты, образовавшиеся из
стволовых клеток костного мозга, приобретают
способность функционировать лить после того, как
пройдут через ткань тимуса. Механизм созревания
Т-лимфоцитов еще не вполне ясен. Известно, что
тимус выделяет гормон, называемый тимозином,
который, возможно, способствует созреванию
Т-клеток, однако роль тимуса как эндокринного
органа до сих пор изучена плохо. Небольшое число
лимфоцитов имеется при рождении в селезенке и
других лимфатических железах.
Корковое вещество тимуса набито лимфоцитами,
которые называют тимоцитами, чтобы отличить их
от Т-лимфоцитов, циркулирующих в крови и
тканевой жидкости (рис. 14.73). Большинство тимоци-
тов-незрелые клетки, но некоторые из них
способны взаимодействовать с антигеном. В результате
такого взаимодействия Т-лимфоциты начинают
размножаться и синтезировать сложные молекулы-
лимфокины, которые помогают атаковать и
уничтожать чужеродные частицы. Зрелые, иммунокомпе-
тентные Т-лимфоциты способствуют также
созреванию В-клеток и выработке ими антител.
Т-клетки все время выходят из тимуса и
поступают в лимфатические узлы и селезенку, где в
случае встречи со специфическим антигеном узнают
его и начинают делиться; так образуются клоны
идентичных клеток, способных узнавать этот
антиген и реагировать с ним (рис. 14.74). Т-лимфоциты
могут атаковать чужеродные клетки благодаря
специфическому рецептору для антигена,
встроенному в плазматическую мембрану. Когда этот
«сенсибилизированный» лимфоцит узнаёт
комплементарный к его рецептору антиген, он присоединяется
к нему (подобно тому как ключ входит в замок) и
разрушает его. Т-клетки регулярно переходят из
лимфоидных тканей в циркулирующую кровь и
тканевую жидкость, благодаря чему увеличивается
вероятность их встречи с антигеном и его
уничтожения.
14.14.2. Образование В-клеток
Стволовые клетки, которым предстоит стать
В-лимфоцитами, должны подвергнуться
дальнейшей дифференцировке за пределами костного мозга.
Эта дифференцировка может происходить в печени,
селезенке или лимфатических узлах.
Когда поверхностные рецепторы
(иммуноглобулины) В-лимфоцитов узнают комплементарные им
антигены, В-лимфоциты начинают делиться и
дифференцироваться, образуя клоны плазматических
клеток и «клетки памяти» (рис. 14.74).
Плазматические клетки, генетически идентичные друг
другу, синтезируют большие количества антител
одного вида.
Тимус
Костный мозг
Лимфоидная
ткань
Периферические
Т-лимфоциты
^ ^ Периферические
В-лимфоциты
•Антиген
Активированные
Т-лимфоциты
(иммуноциты)
Реакции,
осуществляемые
клетками
КЛЕТОЧНЫЙ
ИММУНИТЕТ
- Клетки
памяти
Плазматические
клетки
(В-иммуноциты)
Антитела
ГУМОРАЛЬНЫЙ
ИММУНИТЕТ
Рис. 14.74. Сравнение клеточного и гуморального
иммунитета.
Клетки, образующие антитела, живут всего лишь
несколько дней, но в это время они могут
синтезировать и выделять около 2000 идентичных молекул
антител в секунду.
Клетки памяти играют важную роль в активации
иммунного ответа организма при повторном
внедрении того же антигена. О клетках памяти
известно очень немногое - лишь то, что они
существуют и каким-то образом позволяют организму,
уже встречавшемуся с данным антигеном, при
повторной встрече реагировать на него быстрее и
энергичнее. Эта реакция носит название вторичного
иммунного ответа и приводит к массовому выбросу
антител, которые быстро нейтрализуют вредное
действие антигена (см. рис. 14.76). Так формируется
иммунитет. Следует, однако, помнить, что
иммунитет к одному антигену не защищает организм от
других антигенов. Каждый раз при попадании в
организм нового патогенного агента заболевание
может быть предотвращено только в том случае,
если образуются соответствующие антитела нового
типа.
14.14.3. Классы иммуноглобулинов
и их биологическая активность
Известно пять классов иммуноглобулинов,
принадлежность к которым определяется типом тяжелых
Таблица 14.13. Классы иммуноглобулинов и их
биологическая активность
Иммуноглобулин
IgM
IgG
IgA
IgD
IgE
Тяжелая цепь
μ (мю)
γ (гамма) 1
» 2
» 3
» 4
α (альфа) 1
» 2
δ (дельта)
ε (эпсилон)
Активность
Первый класс антител,
появляющийся в сыворотке после
введения антигена; начинает первичный
иммунный ответ
Основной класс антител в
сыворотке; начинает вторичный
иммунный ответ
Основной класс антител,
выделяемых с такими секретами, как слюна,
слезная жидкость, бронхиальная и
кишечная слизь; составляет первую
линию обороны организма против
бактериальных и вирусных
антигенов
Почти не секретируются; связаны с
мембранами. Функция неизвестна
Возможно, участвуют в
аллергических реакциях. Другие функции
неизвестны
цепей. Все классы различаются по биологической
активности. Легкие цепи иммуноглобулинов
относятся к типу κ (каппа) или λ (лямбда). Здесь
нет необходимости более подробно рассматривать
строение иммуноглобулинов. Основные функции
различных классов иммуноглобулинов указаны в
табл. 14.13.
14.14.4. Клонально-селекционная теория
образования антител
Клонально-селекционная теория образования
антител была разработана в 1950-е годы Ерне, Бёрнетом,
Толмеджем и Ледербергом и в настоящее время
общепринята как рабочая модель образования
антител (рис. 14.75). В основе этой теории лежат
следующие положения:
1. У каждого индивидуума имеется чрезвычайно
широкий набор лимфоцитов, каждый из которых
способен распознавать только один
специфический антиген.
2. Специфичность антитела (которое представляет
собой белок) зависит от его аминокислотной
последовательности, которая в свою очередь
определяется кодирующей ее последовательностью
ДНК. Таким образом, способность клетки к
синтезу данного специфического антитела
предопределена еще до ее встречи с антигеном.
3. Каждая клетка во время созревания образует
небольшие количества антител, которые
встраиваются в ее плазматическую мембрану и играют
роль рецепторов для комплементарного
антигена.
4. Предполагается, что если в период
внутриутробной жизни незрелый лимфоцит встречает
соответствующий ему антиген, то он погибает.
Благодаря этому у животного не синтезируются
антитела к собственным макромолекулам, и оно
становится толерантным к собственным антигенам.
5. Взаимодействие антигена с рецептором зрелого
лимфоцита каким-то образом стимулирует
клетку, заставляя ее вырабатывать антитела. Контакт
с антигеном необходим также для того, чтобы
лимфоцит начал дифференцироваться и делиться
с образованием клона плазматических клеток,
синтезирующих антитела, и клеток памяти.
6. Поскольку все плазматические клетки в пределах
данного клона генетически идентичны, все они
производят одни и те же антитела.
7. Клетки памяти продолжают жить и после
исчезновения антигена, сохраняя каким-то образом
способность стимулироваться антигеном, когда
он появляется вновь. Это явление называется
иммунологической памятью (рис. 14.76), и именно
оно ответственно за вторичный ответ организма,
который и обеспечивает иммунитет
животного против того или иного специфического
антигена.
14.14.5. Типы иммунитета
Естественный пассивный иммунитет
При иммунитете этого типа предобразованные
антитела передаются от одного индивидуума к
другому того же вида. Такой иммунитет обеспечивает
лишь кратковременную защиту от инфекции, так
как по мере выполнения антителами их функции, а
также вследствие естественного разрушения в
организме их количество и защитное действие
постепенно уменьшаются. Примером иммунитета этого типа
служит иммунитет новорожденного. Антитела
матери могут проходить через плаценту и попадать в
организм плода, обеспечивая ребенку защиту до тех
пор, пока не сформируется полностью его
собственная иммунная система. Пассивный иммунитет
может также обеспечиваться антителами, которые
содержатся в молозиве (первичном секрете молочных
желез) и всасываются в кишечнике новорожденного.
Приобретенный пассивный иммунитет
Этот тип иммунитета создается искусственно путем
введения готовых антител. Для этого выделяют
антитела, образовавшиеся в организме одного
индивидуума, и вводят их в кровь другому индивидууму
того же или иного вида. Например, специфические
антитела против столбняка или дифтерии получают
от лошадей и затем вводят их людям. Эти антитела
действуют профилактически, предупреждая
заболевание столбняком или дифтерией соответственно.
Иммунитет этого типа тоже непродолжителен.
Естественный активный иммунитет
При инфицировании каким-либо агентом у
животного вырабатываются собственные антитела.
Поскольку клетки иммунологической памяти,
образующиеся при первой встрече с антигеном, способны
стимулировать выработку больших количеств
антител при повторном воздействии того же антигена,
иммунитет этого типа наиболее эффективен и
сохраняется, как правило, в течение длительного времени,
а иногда и всю жизнь (рис. 14.76).
Приобретенный активный
иммунитет
Иммунитет этого типа создают, вводя в организм
небольшие количества антигена в виде вакцины.
Этот процесс называется вакцинацией или иммуниза-
Будущие
лимфоциты
Во время эмбрионального
развития собственные
незрелые лимфоциты
плода реагируют
с антигеном
Ar
Каждый будущий лимфоцит содержит гены, кодирующие
синтез только одного специфического антитела
Превращаются в
лимфоциты,
неспособные синте
зировать антитела]
против
собственных антигенов,и
гибнут
После
рождения
Клон В-иммуноцитов
Ar.
Превращаются в
лимфоциты,
неспособные
синтезировать антитела
против
собственных антигенов,и
гибнут
Митоз
Митоз
Митоз
Образование антител
Клетка
памяти
Рис. 14.75. Клонольно-селекционная
теория образования антител
Первичный ответ
Вторичный ответ
Концентрация антител в сыворотке
Первое введение
антигена
Второе введение
антигена
IgG
IgM
IgM
Дни
Рис. 14.76. Первичный и вторичный ответы на
первую и последующую дозы антигена. Вторичный
ответ бывает более быстрым и интенсивным, чем
первичный
О
а + Ь
А
b
В
а
AB
о
О
а + Ь
А
b
В
а
AB
о
Обозначения:
— Нет агглютинации
Агглютинация
Рис. 14.77. Взаимодействия, определяемые группами
крови человека. Клеточные агглютиногены
обозначены прописными буквами, а агглютинины
строчными
цией. Небольшая доза вводимого антигена
обычно не представляет опасности, так как для этого
используют убитый или ослабленный антиген.
Таким образом, индивидуум не заболевает, но у него
начинают вырабатываться антитела к введенному
антигену. Часто производится второе,
подкрепляющее введение, и это стимулирует более быстрое и
продолжительное образование антител, которое на
длительное время обеспечивает индивидууму
защиту от болезни. В настоящее время применяются
вакцины нескольких типов:
1. Анатоксины. Экзотоксины, образуемые
столбнячными и дифтерийными бактериями,
обезвреживают с помощью формальдегида, но при этом их
антигенные свойства сохраняются. Таким
образом, вакцинация анатоксинами стимулирует
образование антител, не вызывая заболевания.
2. Убитые микроорганизмы. Некоторые убитые
вирусы и бактерии способны вызывать нормальный
иммунологический ответ и используются для
иммунизации.
3. Ослабленные микроорганизмы. В организм
вводятся живые, но измененные микроорганизмы,
способные размножаться, не вызывая
заболевания. Ослабление микроорганизмов может быть
достигнуто путем выращивания их при
повышенной температуре или длительного
культивирования в среде, содержащей определенные вещества.
В настоящее время широко применяются
ослабленные вакцины против туберкулеза, кори,
краснухи и полиомиелита.
14.14.6. Группы крови
При переливании больному крови необходимо,
чтобы кровь, которую он получает, была совместима с
его собственной. В случае несовместимости
наблюдается особого рода иммунная реакция. Связано это
с тем, что мембраны донорских эритроцитов
содержат мукополисахариды, называемые агглютино-
генами, которые действуют как антигены и
реагируют с антителами (агглютининами),
присутствующими в плазме реципиента. В результате этого
взаимодействия донорские клетки агглютинируются, т.е.
слипаются друг с другом, так как между ними
образуются мостики из антител. Существуют два
агглютиногена - А и В. Комплементарные им
агглютинины плазмы обозначают соответственно а и
Ь\ они постоянно присутствуют в плазме, а не
образуются в ответ на введение агглютиногена, как
в случае уже описанных иммунных реакций. У
людей, эритроциты которых содержат специфический
агглютиноген, комплементарного ему агглютинина
в плазме нет. Например, у лиц, эритроциты которых
содержат агглютиноген А, в плазме нет
агглютинина а, и его кровь относится к группе А, а если
имеется только агглютиноген В, то кровь относится
к группе В. Если же в крови присутствуют оба
агглютиногена (и нет агглютининов), то это группа
AB. И наконец, если в крови нет ни того, ни другого
агглютиногена (и есть оба агглютинина - а и Ь), то
это группа О (табл. 14.14).
При переливании крови важно знать, что может
произойти с клетками донора. Если существует
вероятность их агглютинации под действием
агглютининов плазмы, тогда переливание проводить
нельзя.
На рис. 14.77 показаны последствия смешивания
крови разных групп. Лица с группой О называются
универсальными донорами, поскольку их кровь
можно переливать любому человеку. Их эритроциты не
агглютинируются при контакте с агглютининами
плазмы реципиента. Хотя в плазме доноров с этой
Таблица 14.14. Группы крови
Группа крови
Частота в
популяции
Агглютиногены
Агглютинины
О
46%
-
a + b
Л
42%
А
Ь
В
9%
В
а
AB
3%
А + В
-
группой содержатся агглютинины а и Ь, они будут
вызывать лишь очень слабую агглютинацию
эритроцитов реципиента, так как донорская плазма
будет настолько сильно разбавлена кровью
реципиента, что практически не будет обладать
агглютинирующей активностью. Лица с группой AB могут
получать кровь любой группы и поэтому их
называют универсальными реципиентами; но кровь
этой группы можно переливать только лицам с
группой AB.
14.14.7. Резус-фактор
У 85% людей эритроциты содержат особый агглю-
тиноген, называемый резус-фактором (Rh), и этих
лиц называют резус-положительными (Rh+). У
остальных людей резус-фактор отсутствует, и их называют
резус-отрицательными (Rh-). Плазма
резус-отрицательной крови обычно не содержит
резус-агглютининов. Однако если резус-положительная кровь
попадает в организм человека с резус-отрицательной
кровью, то у последнего образуются
резус-агглютинины.
Практическое значение этого факта становится
очевидным в тех случаях, когда
резус-отрицательная мать беременна резус-положительным плодом.
На поздних стадиях беременности фрагменты
резус-положительных эритроцитов плода могут
проникать в кровеносную систему матери и вызывать у
нее образование резус-агглютининов, а эти антитела
могут проходить через плаценту и разрушать
эритроциты плода. При первой беременности титр
резус-агглютининов обычно бывает недостаточно
высок, и у плода не возникает серьезных
осложнений. Однако у последующих резус-положительных
младенцев может наблюдаться хронический распад
эритроцитов. Обычно такие дети рождаются
недоношенными и страдают анемией и желтухой.
В этих случаях может потребоваться полная замена
крови младенца нормальной донорской кровью.
В настоящее время есть возможность производить
такую процедуру еще до рождения ребенка.
Возможности предотвращения
резус-конфликта
1. Рассмотрим случай, когда резус-отрицательная
мать с кровью группы О беременна
резус-положительным плодом, имеющим любую из групп
крови, кроме группы О. При попадании
эритроцитов плода в кровь матери а- и ^-агглютинины
вызовут их разрушение раньше, чем у матери
успеют образоваться резус-агглютинины.
2. Если резус-отрицательной матери в течение 72 ч
после родов ввести внутривенно антитела к
резус-агглютининам, называемые анти-D (анти-D-
глобулин)1, то этим будет предотвращена
сенсибилизация резус-отрицательной матери
резус-положительными эритроцитами плода. По всей
видимости, анти-D связываются с эритроцитами
плода, попавшими в кровь матери, и так
изменяют их, что материнские клетки, образующие
антитела, не могут их распознать, благодаря чему
антитела к ним не образуются (рис. 14.78). Это
может предотвратить резус-реакцию при
следующей беременности.
14.14.8. Трансплантация тканей
Трансплантацией, или пересадкой, называют замену
пораженных тканей или органов здоровыми. В
настоящее время трансплантация все шире
применяется в хирургии. Однако при пересадке чужеродной
ткани она отторгается организмом реципиента,
поскольку действует как антиген и вызывает
иммунный ответ. Ниже приводятся названия различных
видов трансплантатов.
Аутотрансплантат-ткань, пересаженная из одного
участка тела в другой тому же индивидууму.
Изотрансплантат-ткань, пересаженная от
генетически идентичного организма, например от
однояйцового близнеца.
Аллотрансплантат - ткань, пересаженная от одной
особи другой того же вида, но иного генотипа.
Ксенотрансплантат - ткань, пересаженная от
организма другого вида, например от свиньи человеку.
В настоящем разделе мы рассмотрим только алло-
трансплантацию.
Поскольку кровь является тканью, любое
переливание крови можно рассматривать как аллотранс-
плантацию. Как уже отмечалось, отторжение в этом
случае происходит путем агглютинации донорских
эритроцитов. При отторжении ткани, например
кожи, наблюдаются следующие стадии:
1. Вначале, в течение первых двух-трех дней, в
кожный трансплантат прорастают кровеносные
сосуды и он выглядит вполне здоровым.
2. В течение последующих шести дней васкуляриза-
ция трансплантата уменьшается и вокруг него
собирается множество T-лимфоцитов и
моноцитов (разд. 14.14.1 и 14.11.2).
3. Еще через два дня клетки трансплантата
начинают отмирать, и в конце концов трансплантат
отпадает.
1 D-глобулин - один из нескольких резус-агглютино-
генов, обладающий наиболее выраженными антигенными
свойствами. - Прим. перев.
Кровь плода
Кровь матери
Резус-конфликт
иммуноцит
Эритроцит
Антирезусные
агглютинины (анти-D)
aD
Разрушение эритроцитов
плода и анемия
Плацента
Предотвращение
резус-конфликта
Введение анти-D
в течение 72 ч
после родов
Rh-
иммуноцит
не узнает эритроциты плода,
и антитела не образуются
Рис. 14.78. Механизм развития
резус-конфликта и способ его
предотвращения.
Предупреждение отторжения
трансплантата
В настоящее время существует несколько способов,
позволяющих предотвратить отторжение
трансплантата:
1) подбор наиболее совместимого донора -
очевидная и необходимая мера предосторожности,
которую следует соблюдать при любой пересадке;
2) облучение рентгеновскими лучами костного
мозга и лимфатических тканей, подавляющее
образование лейкоцитов и таким образом замедляющее
процесс отторжения;
3) использование иммунодепрессантов, т.е.
агентов, угнетающих активность всей иммунной
системы. При этом отторжение трансплантата
задерживается, но возникает другая серьезная
проблема, состоящая в том, что реципиент
становится восприимчивым ко всем видам инфекции.
Показано также, что подавление иммунной системы
может повысить вероятность развития рака.
Для преодоления недостатков, присущих методу
неспецифической иммунодепрессии, необходимо
найти способ избирательного подавления только тех
Т-клеток, которые реагируют на антигены данного
трансплантата. В этом случае остальная часть
иммунной системы будет продолжать нормально
функционировать. Наиболее перспективный
подход-введение больному в кровь (или
непосредственно в костный мозг) антител, которые узнают и
разрушают только те Т-лимфоциты, которые
ответственны за отторжение трансплантата.
14.14.9. Система интерферона
Интерферон - групповое название, под которым в
настоящее время объединяют целый ряд белков со
сходными свойствами. Интерфероны человека
подразделяют на три группы-α, β и γ-в зависимости
от типа клеток, в которых они образуются. а-Ин-
терфероны включают нескольких видов белков с
мол. массой около 20000.
Наиболее изученным свойством интерферона
является его способность препятствовать
размножению вирусов. Он образуется в клетках
млекопитающих и птиц в ответ на вирусную инфекцию;
по-видимому, это активный противовирусный агент,
характерный для большинства типов клеток и
действующий в большей или меньшей степени против всех
вирусов.
При заражении клетки вирусом события
развиваются следующим образом. Вирус начинает
размножаться, и одновременно клетка-хозяин начинает
продуцировать интерферон (рис. 14.79). Интерферон
выходит из клетки, вступает в контакт с соседними
клетками и делает их невосприимчивыми к вирусу.
Он действует, запуская цепь событий, приводящих к
подавлению синтеза вирусных белков и (в
некоторых случаях) сборки и выхода вирусных частиц.
Таким образом, интерферон не обладает прямым
противовирусным действием, но вызывает такие
изменения в клетке, которые препятствуют
размножению вируса. Образование интерферона могут
стимулировать не только интактные вирусы, но и
различные другие агенты, например некоторые
Клетка стала
резистентной
к вирусу
Высвобо*
ждение
интерферона'
Образование
интерферона
Ядро
Стимуляция"
синтеза
интерферона
Размножение
вируса
Высвобождение
интерферона
Клетка стала
резистентной^
к вирусу
Вирусная атака
Рис. 14.79. Образование и
действие интерферона.
инактивированные вирусы, двухцепочечные РНК,
синтетические двухцепочечные олигонуклеотиды и
бактериальные эндотоксины. Создается
впечатление, что клетка образует интерферон как бы в ответ
на нанесенную ей «обиду».
Биологическая активность интерферона
чрезвычайно высока: у мышиного интерферона она
составляет 2· 109 ед./мг, а одна единица снижает
образование вирусов примерно на 50%. Это означает,
что достаточно одной молекулы интерферона,
чтобы сделать клетку резистентной к вирусной
инфекции.
Интерферон вызывает и целый ряд других
биологических эффектов, в том числе подавляет
размножение клеток. Недавние исследования показали,
что в определенных условиях он может
препятствовать развитию рака. Установлено также, что
интерферон действует на иммунную систему и
вызывает изменение клеточных мембран. Таким
образом, интерфероновая система, вероятно, может
играть важную роль в защите организма от
вирусов.
Глава 15
Координация и регуляция
у растений
Для того чтобы растения росли и развивались
упорядоченным образом и адекватно реагировали
на условия среды, им так же, как и животным,
нужны какие-то формы внутренней координации.
Но у растений нет нервной системы, и потому они
могут использовать лишь химические
координирующие факторы. В связи с этим растения отвечают
на раздражители гораздо медленнее, и ответ часто
выражается только в изменении роста; в свою
очередь рост может приводить к перемещению того
или иного органа. В этой главе мы рассмотрим
сначала движения растений, а затем - различные
способы координации функций.
15.1. Движения растений
Почти для всех растений, кроме некоторых
одноклеточных форм, характерно то, что они не способны
к локомоции (т. е. к передвижению всего организма).
Однако у растений возможны движения отдельных
органов, и они определяются тем, насколько
чувствительно данное растение к внешним стимулам.
Движения, вызываемые внешними воздействиями,
можно разделить на три категории: тропизмы,
настии и таксисы.
15.1. Какова основная причина того, что
животные могут передвигаться, а растения-нет?
15.1.1. Тропизмы
Тропизм-это перемещение части растения,
вызываемое и направляемое внешним стимулом. Такое
перемещение - это почти всегда ростовое движение.
Тропизмы называют положительными или
отрицательными в зависимости от того, куда направлено
ответное движение - по направлению к фактору,
стимулирующему рост, или от него. В табл. 15.1
приведены некоторые примеры тропизмов.
15.2. Дополните табл. 15.1 еще одной,
четвертой, колонкой и впишите в нее, какую выгоду
приносит растению ответная реакция на тот или
иной раздражитель.
15.3. Как должен видоизмениться рост усика,
чтобы усик обвился вокруг твердой опоры?
Фототропизм и геотропизм будут более подробно
рассмотрены позднее (разд. 15.2.1 и 15.2.2).
Таблица 15.1. Примеры тропизмов
Раздражитель
Свет
Сила
тяжести
Химические
факторы
Вода
Твердая
поверхность;
прикосновение
Воздух
(кислород)
Тип тропизма
Фототропизм
Геотропизм
Хемотропизм
Гидротропизм
(хемотропизм
особого рода)
Гаптотро-
пизм (тигмо-
тропизм)
Аэротропизм
(хемотропизм
особого рода)
Примеры
Побеги и колеоптили
обладают положительным
фототропизмом, а
некоторые корни -
отрицательным фототропизмом
(например, придаточные
корни вьющихся растений
вроде плюща)
Побеги и колеоптили
проявляют отрицательный
геотропизм, а
корни-положительный геотропизм;
корневищам, отпрыскам,
усам, листьям
двудольных свойствен диагеотро-
пизм1*, а боковым корням
и разветвлениям стебля-
пл агиогеотропизмl}
Положительный
хемотропизм характерен для
гиф некоторых грибов,
например Мисог.
Пыльцевые трубки проявляют
положительный
хемотропизм в ответ на вещества,
образуемые в микропиле
семязачатка
Положительным
гидротропизмом обладают
корни и пыльцевые
трубки
Положительный гапто-
тропизм свойствен
усикам вьющихся растений,
например гороха.
Центральные чувствительные
волоски росянки (одного
из насекомоядных
растений) проявляют
положительный гаптотропизм
(разд. 9.12.2)
Отрицательный
аэротропизм проявляют
пыльцевые трубки
υ Диагеотропизм - рост под углом в 90° к направлению силы
тяжести, т.е. горизонтальный рост. Плагиогеотропизм - рост под
определенным углом к направлению силы тяжести, но не
горизонтальный и не в сторону раздражителя или от него.
15.1.2. Таксисы
Таксис-это перемещение всей клетки или всего
организма (т.е. локомоторная реакция),
вызываемое и направляемое определенным внешним
стимулом. Подобно тропизмам, таксисы тоже делятся
на положительные и отрицательные; кроме того, их
можно классифицировать в соответствии с
природой раздражителя (табл. 15.2).
15.4. Продумайте, как поставить опыт, чтобы
определить по фототаксису, какую интенсивность
света предпочитает эвглена или хламидомонада.
15.5. На рис. 15.1 показано, как распределяются
подвижные бактерии через 10 мин после того, как
их поместят под покровное стекло вместе с нитью
зеленой водоросли.
а) Назовите, какого рода водоросли подходят для
этого опыта, б) Предложите гипотезу,
объясняющую картину окончательного распределения
бактерий, в) Как можно проверить вашу гипотезу?
15.1.3. Настии
Настия-это ненаправленное движение части
растения в ответ на внешний раздражитель. Направление
перемещения определяется структурой
соответствующего органа. Движение происходит в результате
роста или же изменения тургора; при этом
ничтожное смещение отдельных клеток обычно приводит
к значительному движению органа благодаря
специфическому положению этих клеток.
«Сонные движения» (никтинастия) некоторых
цветков и листьев, когда они раскрываются или
закрываются в ответ на изменение освещенности (фо-
тонастия) или температуры (термонастия) относятся
к настическим потому, что внешние стимулы только
запускают их, а направление зависит от внутренних
факторов. Некоторые цветки, например у крокуса
или тюльпана, закрываются ночью потому, что
лепестки снизу растут быстрее (гнпонастия), а
открываются в результате того, что начинает быстрее
расти верхняя часть лепестков (эпинастия). У многих
растений, особенно у бобовых (таких, как клевер
или мимоза), в листьях и листочках имеются особые
структуры, называемые листовыми подушечками.
Листовая подушечка - это особое вздутие у
основания черешка или листочка, в котором находятся
крупные паренхимные клетки. Быстрое изменение
тургорного давления в таких клетках приводит к
тому, что листовая подушечка начинает работать
как шарнир, с помощью которого и происходит
движение.
Гаптонастические движения, происходящие в ответ
на прикосновение, принадлежат к числу самых
удивительных движений, свойственных растениям, так
как реакция в этом случае бывает очень быстрой и
весьма специфичной. Всем известное
«чувствительное» растение Mimosa pudica реагирует на прикосно-
Бактерии
Предметное
стекло
Покровное
стекло
Нить
зеленой
водоросли
Рис. 15.1. Распределение подвижных бактерий на
предметном стекле.
вение и на целый ряд других раздражителей. У этого
вида есть и обычные сонные движения, но он очень
резко реагирует на «шоковые» воздействия (сейсмо-
настия), например на резкий порыв ветра,
повреждение, внезапное изменение температуры или
освещенности. Если раздражать кончики листочков, они
складываются через несколько секунд. При большой
силе или длительности раздражения начинают скла-
Таблица 15.2. Примеры таксисов
Раздражитель
Свет
Химические
факторы
Воздух
(кислород)
Сила
тяжести
Магнитное
поле
Сопротивление среды
Таксис
Фототаксис
Хемотаксис
Аэротаксис
(хемотаксис
особого
рода)
Геотаксис
Магнитотак-
сис
Реотаксис
Примеры
Положительный фототаксис:
эвглена плывет к свету, хлоро-
пласты перемещаются к свету.
Отрицательный фототаксис:
дождевые черви, личинки
мясных мух, мокрицы и тараканы
убегают от света
Положительный хемотаксис:
сперматозоиды печеночников,
мхов и папоротников плывут в
сторону веществ, выделяемых
яйцеклеткой; подвижные
бактерии движутся в сторону
различных пищевых субстратов.
Отрицательный хемотаксис: комары
избегают репеллентов
Положительный аэротаксис:
подвижные аэробные бактерии
движутся в сторону кислорода
Положительный геотаксис:
личинки-планулы некоторых
кишечнополостных опускаются на
дно моря. Отрицательный
геотаксис: личинки-эфиры
некоторых кишечнополостных
всплывают к поверхности моря
Некоторые подвижные бактерии
Положительный реотаксис:
планарии движутся против тока
воды, ночные и дневные бабочки
летят против ветра
Черешок
Листочки в
нормальном
положении
Листовая
подушечка
Поникший
черешок
Листочки,
сложенные
вверх
Листовые
подушечки
у основания
листочков
Рис. 15.2. Реакция «чувствительного» растения мимозы (Mimosa pudica) на шоковое воздействие. Слева-до
воздействия, справа-после воздействия.
дываться и соседние листочки; в конце концов
действие стимула распространяется на весь лист, и
черешок поникает (рис. 15.2). Если воздействовать
на стебель, реакция будет распространяться в
обратном направлении. На рис. 15.2 указаны те три
участка, в которых находятся листовые подушечки. Как
полагают, раздражение передает гормон,
переносимый по ксилеме; передача сигнала связана и с
электрическими изменениями, хотя нервных клеток
у растений нет.
Самые сложные движения наблюдаются у
насекомоядных представителей растительного царства,
часть таких движений -гаптонастические.
15.1.4. Кинезы
Еще один тип двигательных реакций, о котором мы
не упоминали,-это кинезы. Поскольку этот тип
движений фактически свойствен только животным,
мы рассмотрим его в главе 16, посвященной
поведению животных.
15.2. Ростовые вещества
растений
Химическая координация у животных
осуществляется с помощью гормонов, т. е. органических веществ,
которые синтезируются в одном месте, а действуют,
причем в очень малых концентрациях, в других
местах. У растений физиологические процессы
координируются веществами, которые вовсе не
обязательно переносятся куда-то из того места, где
синтезируются, и поэтому их не всегда можно назвать
гормонами. Учитывая это обстоятельство и то, что
они обычно в той или иной мере влияют на рост, их
обычно называют ростовыми веществами. Кроме
того, важно подчеркнуть, что конкретный механизм
действия тех ростовых веществ, которые уже
открыты, далеко не ясен и что аналогия с гораздо
лучше изученными гормонами животных может
ввести в заблуждение. Очевидно, что ростовые
вещества необходимы для развития растений, но
остается неясным, в какой мере их действие состоит в
«запуске» ростовых изменений или же в
«интеграции» процессов, запускаемых другими,
неизвестными пока событиями. Не следует забывать и то, что
процесс роста складывается из трех этапов - деления
клеток, их растяжения и их дифференцировки
(специализации) и что эти этапы по-разному протекают
в разных частях растения (разд. 21.6). Поэтому
можно ожидать, что все это будет отражаться и на
действии, и на распределении различных ростовых
веществ в растении. Выделяют пять основных
классов ростовых веществ: ауксины, гиббереллины, ци-
токинины, абсцизовую кислоту и этилен (этен).
В общем можно сказать, что цитокинины связаны с
делением клеток, ауксины и гиббереллины-с
увеличением размеров клеток и их дифференцировкой,
абсцизовая кислота-со стадиями покоя (например,
в боковых почках), а этилен-со старением.
В этой главе мы сначала рассмотрим по
отдельности все классы ростовых веществ, а затем
обсудим ключевые моменты жизненного цикла
растений, особо выделив тот факт, что ростовые
вещества часто вызывают тот или иной эффект, действуя
совместно.
Поверхность
почвы
Первый лист,
появляющийся-
на конце
колеоптиля
Колеоптиль
Колеоптиль
Зерно
Колеоптиль — полый
не окрашенный
защитный чехол
Первый лист —
скрученный и
зеленый
Верхушка стебля
Узел
Мезокотиль
Рис. 15.3. Прорастание типичного
проростка злака. А, Б и
В-отдельные стадии прорастания; Г-разрез
колеоптиля на стадии Б.
15.2.1. Ауксины и фототропизм
Открытие ауксинов
Ауксины были открыты в результате изучения
фототропизма, которое было начато еще в опытах
Чарлза Дарвина и его сына Френсиса. Взяв ко-
леоптили овса, они показали на этом очень удобном
объекте (рис. 15.3), что рост проростков в сторону
света обусловлен тем, что от верхушки стебля
передается какое-то «влияние» на лежащую позади зону
роста. Часть этих экспериментов представлена на
рис. 15.4, где каждая схема отражает результаты не
одного, а многих опытов.
15.6. а) Подробно запишите выводы, которые
вы могли бы сделать на основании опытов А -Г
на рис. 15.4, учитывая, что изгибы стебля
обусловлены изменениями роста в зоне, расположенной
ниже верхушки стебля.
б) Почему нужно было поставить опыт В, узнав
результаты опыта Б1
Если ответную реакцию представить схемой:
раздражитель -> рецептор -* передача сигнала -►
эффектор -> реакция, то оказывается, что меньше всего
мы знаем о природе передачи сигнала. В 1913 г.
датский физиолог растений Бойсен-Йенсен впервые
исследовал этот вопрос. Некоторые из его опытов
отображены на рис. 15.5.
15.7. Какую дополнительную информацию
дали опыты Бойсен-Йенсена?
15.8. Какие результаты будут получены, если
повторить эти опыты при равномерном
освещении со всех сторон? Нарисуйте схемы, объясните
ответ.
В 1928 г. датский физиолог растений Вент
окончательно доказал, что существует специфический
химический передатчик. Вент поставил перед собой
цель перехватить и собрать это вещество в тот
момент, когда оно распространяется из верхушки
назад, а затем показать его эффективность в
различного рода тестах. Он рассуждал, что небольшая
диффундирующая молекула должна
беспрепятственно проникать внутрь небольшого блока из
агарового геля, в котором между молекулами агара
остаются довольно большие свободные пространства.
Некоторые опыты Вента представлены на рис. 15.6.
15.9. Какие выводы позволяют сделать
результаты, показанные на рис. 15.6?
15.10. Какой результат будет получен, если
обработанный блок поместить на правую сторону
декапитированного колеоптиля в варианте Б1
На рис. 15.7 представлен еще один эксперимент
Вента, который заслуживает особого внимания.
В контрольных опытах кончик колеоптиля, поме-
Нео бработанный
контроль
Кончик удален
Cb етонепроницаемый
колпачок
Мелкий черный песок
Свет
Рис. 15.4. Опыты Дарвина по
фототропизму колеоптилей овса. А,
Б, В и Г-разные опыты; слева в
каждом случае показано
воздействие, справа-его результат.
Необработанны й
контроль
Слюдяная пластинка —
тонкий непроницаемый барьер
Свет
Рис. 15.5. Опыты Бойсен-Йенсена по фототропизму
колеоптилей овса. А, Б и В-разные опыты; слева в
каждом случае показано воздействие, справа его
результат.
щенный на два агаровых блока А и Б, инкубировали
при равномерном освещении или в темноте, а затем
переносили агаровые блоки на колеоптиль с
удаленным кончиком; величина изгиба, индуцируемого
блоками А и Б, в этом случае была одинакова.
Одностороннее же освещение верхушки колеоптиля
приводило к неравномерному распределению
активного вещества между блоками А и Б (рис. 15.7). Это
не только подтверждает выводы Бойсен-Йенсена
о влиянии света на распределение активного
вещества, но и показывает, что можно определять
количество этого вещества биологическим методом (с
помощью «биотеста»). Вент установил, что
величина изгиба колеоптилей овса прямо
пропорциональна концентрации активного вещества (в диапазоне
ее нормальных физиологических величин).
Впоследствии это вещество было названо
«ауксином» (от греч. auxein - увеличивать). В 1934 г. оно
было идентифицировано как индолилуксусная
кислота (ИУК). Вскоре выяснилось, что ИУК широко
распространена у растений и что с нею тесно
связано увеличение размеров клеток. На рис. 15.8
показано, как, по современным представлениям,
передвигается ИУК при одностороннем освещении
колеоптилей. Следует, однако, отметить, что
колеоптиль-это самая простая из изученных до сих
пор систем и что другие системы, по-видимому,
устроены гораздо сложнее. Кроме того, нет почти
никаких данных о том, что градиент ауксина
создается еще во время критического периода, до
проявления ответной реакции.
Строение ИУК
Строение ИУК показано на рис. 15.9.
Вскоре были выделены и другие вещества,
сходные с ИУК по структуре и активности; удалось
также синтезировать несколько соединений
подобного типа, так что появился целый класс ростовых
веществ, называемых ауксинами. Некоторые из них
будут рассмотрены в разд. 15.2.5.
Синтез и распределение ауксинов
Ауксины непрерывно образуются в точке роста
стебля и в молодых листьях. Перемещение ауксинов
от верхушки можно назвать базипетальным (от
точки роста к основанию органа) и полярным (т.е.
идущим в одном направлении). По-видимому, они
движутся от клетки к клетке путем диффузии и в
конце концов инактивируются и разрушаются
ферментами. Транспорт на дальние расстояния
происходит по проводящей системе (в основном по
флоэме) и направлен от побегов к корням.
Вероятно, в корнях ауксины почти не образуются. Влияние
различных концентраций ауксина на рост побега
можно оценить, поставив опыт вроде описанного
ниже.
Агаровый блок
Берут
нормальный
калеоптиль
Отрезают
кончик
Переносят кончик на
агаровый блок и оставляют
на несколько часов
ίЗатем переносят блок на срезы
колеоптилей с только что
удаленной верхушкой
Необработанный
агаровый блок
Стимулятор роста
Необработанный
агаровый блок
Рост
Роста
почти нет
Роста
почти
нет
Контроль для А
Контоль для Б
Рис. 15.6. Опыты Вента. А и Б разные опыты; слева в каждом случае показано воздействие, справа его
результат. Рядом показаны контрольные эксперименты. Все процедуры проводились в темноте или при
постоянном освещении.
Верхушка колеоптиля
Степень изгиба пропорциональна
количеству препарата
Свет
Тонкая
металлическая I
пластинка
А, Б — два
агаровых блока
Изгиб меньше,
чем в опыте
с блоком А
Рис. 15.7. Опыт Вента,
демонстрирующий влияние
одностороннего освещения на распределение
химического фактора (ауксина).
Опыт 15.1. Изучение влияния
индолилуксусной кислоты
(ИУК) на рост колеоптилей
овса
Цель эксперимента - исследовать влияние разных
концентраций ИУК на рост колеоптилей овса.
Белый свет влияет на рост растений, поэтому резать и
переносить колеоптили во время опыта нужно при
красном свете или при возможно более слабом
освещении. В опыте используют раствор сахарозы,
так как для роста нужна энергия, а сахароза служит
ее источником. Верхушку (3 мм) колеоптиля
удаляют, чтобы природные ауксины, которые в нем
образуются, не влияли на рост.
Материалы и оборудование
Проросшие семена овса с колеоптилиями не менее
1,5 см длиной [замочите 100 зерен овса в воде и
оставьте их на ночь; замоченные семена
положите на мокрую фильтровальную бумагу в
чашке Петри, накройте чашку алюминиевой
фольгой и поместите в темное место (семена
проращивают пять дней в термостате при
200C); для того чтобы получить 60 колеоптилей,
нужных для каждого опыта, следует замачивать
по меньшей мере 100 зерен, так как часть их
может не прорасти]
6 пробирок вместе со штативом
6 чашек Петри с крышками
5 градуированных пипеток на 5 мл
Мерный цилиндр на 25 мл или градуированная
пипетка на 10 мл
Резак для колеоптилей (рис. 15.10)
Кисточка
Исходный раствор ИУК (1 г/л). (ИУК с трудом
растворяется в воде, поэтому ее сначала
растворяют в этаноле: растворите 1 г ИУК в 2 мл
этанола и разбавьте раствор дистиллированной
водой до 900 мл; нагрейте раствор до 800C и
подержите его 5 мин при этой температуре;
доведите раствор дистиллированной водой до
1 л. Количество раствора готовится в
соответствии с тем, сколько его нужно для опыта)
2%-ный раствор сахарозы
Дистиллированная вода
Методика
1. Возьмите 6 пробирок и 6 чашек Петри и
подпишите их буквами от А до E.
2. В каждую пробирку внесите 18 мл 2%-ного
раствора сахарозы.
3. С помощью чистой пипетки на 5 мл добавьте
2 мл исходного раствора ИУК в пробирку А и
тщательно перемешайте оба раствора.
4. Взяв новую пипетку, перенесите 2 мл раствора из
пробирки А в пробирку Б и тщательно
перемешайте содержимое пробирки Б.
5. Используя каждый раз новую пипетку,
перенесите 2 мл из пробирки Б в пробирку В,
перемешайте; затем-2 мл из пробирки В в
пробирку Г, перемешайте; затем - 2 мл из пробирки
Г в пробирку Д.
6. В пробирку E добавьте 2 мл дистиллированной
воды.
7. Перенесите растворы из пробирок A-E в чашки
Петри A-E.
Зона
растяжения
Расщепление
ауксина
ферментами
Свет
Перемещение
ауксина
Ауксин движется в сторону от света.
Он не инактивируется светом.
Повышенная концентрация ауксина
на теневой стороне стимулирует
растяжение клеток, и растение
изгибается.
Рис. 15.8. Гипотеза, объясняющая влияние
одностороннего освещения на распределение ауксина в
колеоптиле.
CH2COOH
Индол
Уксусная кислота
Рис. 15.9. Строение индолилуксусной кислоты.
Шайбы
Гайка
Разрез
Разрез
10 мм
Винт -
Колеоптили,
уложенные
параллельно
10 мм
Бритвенные
~ лезвия
Рис. 15.10. Как порезать
колеоптили на кусочки длиной по 10 мм.
8. Возьмите 60 проростков овса и отрежьте по
10 мм колеоптилей, отступив от верхушки на
2 мм. Возьмите резак с двумя лезвиями,
установленными точно на расстоянии 10 мм друг от
друга; для установки используйте шайбы или
другие прокладки; резак должен быть скреплен
винтом и двумя гайками (см. рис. 15.10). Если
выровнять кончики колеоптилей, можно сразу
отрезать по нескольку препаратов.
9. Возьмите кисточку и перенесите по 10 отрезков
колеоптиля в каждую из чашек Петри, стараясь
избежать перекрестного загрязнения растворов
(чем больше колеоптилей мы возьмем, тем
статистически достовернее будут результаты).
10. Накройте чашки крышками и оставьте их в
темноте при 25°С на три дня.
11. Как можно точнее измерьте длину колеоптилей.
12. Отбросив максимальную и минимальную
величины, определите среднюю длину колеоптиля
для каждой чашки.
13. Постройте график зависимости средней длины
колеоптиля (по вертикальной оси) от
концентрации УИК в пробе, выразив ее в частях на
миллион (по горизонтальной оси).
15.11. Какова будет концентрация ИУК в
каждой из чашек, если выразить ее в частях на
миллион (ч. на млн.) (1 г/л = 1000 ч. на млн.)?
14. Прокомментируйте полученные результаты и
сравните их с кривыми на рис. 15.11. Точность
результатов можно повысить, если объединить
данные всей группы учащихся.
15.2.2. Ауксины и геотропизм
Как известно, корни растут вниз, т.е. у них
положительный геотропизм, а побеги, наоборот,
растут вверх, т. е. им свойствен отрицательный
геотропизм. То, что раздражителем в этом случае
служит сила тяжести, можно показать с помощью
прибора, называемого клиностатом (рис. 15.12). При
вращении камеры сила тяжести одинаково и
поочередно воздействует на все части проростка.
Чтобы снять одностороннее действие силы тяжести и
заставить побеги и корни расти горизонтально,
вполне достаточно четырех оборотов камеры в час.
Неподвижные контрольные проростки дают
нормальный ответ на силу тяжести, т.е. побег у них
растет вверх, а корень-вниз. Очень важно
обеспечить равномерное освещение (или же проводить
опыт в полной темноте), чтобы не вызвать
направленной реакции на свет.
Роль ауксинов в геотропизме можно
продемонстрировать, поставив опыт (рис. 15.13) с применением
методики Вента. Если колеоптиль положить
горизонтально, то ауксин будет выходить из его
верхушки, но пойдет не вдоль него, а вниз. Увеличение
Торможение,
% к контролю
Побеги
Корни
Стимуляция,
% к контролю
Концентрация ауксина, ч. на млн.
(логарифмическая шкала)
Рис. 15.11. Влияние концентрации ауксина на рост
корней и побегов Обратите внимание на то, что
ауксин в концентрациях, стимулирующих рост
побегов, подавляет рост корней
Вращение
Пробковый диск
Электро
мотор
.Первичный
корешок
Насыщенная
атмосфера
Булавка
Плотно
подогнанная
крышка из
плексигласа
Крупное
семя
(например,
конских бобов)
Смоченная вата
или фильтровальная
бумага
Рис. 15.12. Клиностат с проросшими конскими
бобами после нескольких дней их роста при вращении.
концентрации ауксина на нижней поверхности ин-
тактного колеоптиля приведет к усиленной
стимуляции растяжения клеток в этом месте, и в результате
рост будет направлен вверх.
Декапитация корня тоже делает его
нечувствительным к земному тяготению, но передвижение
ауксинов в корне продемонстрировать не так легко,
потому что их здесь очень мало, и результаты
описанного выше биотеста будут не особенно
убедительными. Тем не менее можно получить
интересные результаты, проделав опыт, показанный на
рис. 15.14.
15.12. Какие выводы можно сделать из опыта,
показанного на рис. 15.14?
Наблюдения подобного рода привели к гипотезе,
согласно которой диаметрально противоположная
реакция корней и стеблей обусловлена разной
чувствительностью к ауксину; эта гипотеза представлена
на рис. 15.15. Несколько позже мы обсудим
различные модификации этой гипотезы, учитывающие
данные последних лет.
Отдельные случаи отрицательного фототропизма
корней тоже можно объяснить различной
чувствительностью к ауксину (рис. 15.11); повышенное
накопление ауксина на затененной стороне должно
подавлять рост, и в результате этого клетки на
освещенной стороне будут растягиваться быстрее,
заставляя корень удаляться от света.
Таким образом, выявляется один из важных
аспектов регуляции роста растений: важна не только
природа действующего ростового вещества
(качественный контроль), но и его концентрация
(количественный контроль).
Механизм восприятия силы тяжести
Возникает вопрос: как же воспринимается сила
тяжести? Еще Дарвин показал, что геотропическая
реакция исчезает после удаления корневого чехлика,
т.е. группы крупных паренхимных клеток, которые
защищают кончик корня, растущий в почве. На
срезе корневого чехлика видны большие
крахмальные зерна, лежащие в амилопластах внутри клеток
(рис. 15.16).
Еще в 1900 г. было высказано предположение о
том, что эти клетки ведут себя как статоциты, т.е.
как рецепторы, воспринимающие силу тяжести, а
сами крахмальные зерна играют роль статолитов -
структур, перемещающихся под действием этой
силы. Согласно «гипотезе крахмальных статолитов»,
крахмальные зерна осаждаются на нижней стороне
клетки (рис. 15.16). Это каким-то образом влияет на
распределение ростовых веществ, которые, как мы
знаем, иногда образуются в точке роста корня,
иногда в корневом чехлике, а иногда и там и тут.
Существует много данных в пользу этой гипотезы.
Во всех органах, воспринимающих силу тяжести,
есть и статоциты. Так, например, они находятся в
Верхушка Агаровый блок
колеоптиля,
лежащая
горизонтально
(Распределение
ауксина изучали
с помощью
пробы Вента)
Рис. 15.13. Влияние силы тяжести на распределение
ауксина в лежащей горизонтально верхушке
колеоптиля.
B левой части блока
концентрация ауксина
выше
Декапити-
рованный
КОЛЕОПТИЛЬ
ИЛИ ПОБЕГ
Агаровый
блок
Верхушка
колеоптиля,
положенная
горизонтально
Ауксин
КОРЕНЬ
Де капитированный
ВОЗДЕЙСТВИЕ
РЕЗУЛЬТАТ
Рис. 15.14. Влияние неравномерного распределения
ауксина на рост декапитированных колеоптилей и
корней.
ПОБЕГ
КОРЕНЬ
Высокая концентрация
ауксина на нижней
стороне стимулирует
рост, поэтому побег
растет вверх
Высокая концентрация
ауксина на нижней
стороне тормозит рост,
поэтому корень
растет вниз
Рис. 15.15. Гипотеза, объясняющая ростовые
эффекты перераспределения ауксина в положенном
горизонтально сеянце.
обкладке проводящих пучков стебля. Растения, у
которых тем или иным способом удалены зерна
крахмала, теряют и чувствительность к силе
тяжести, но она восстанавливается, если дать им
возможность накопить крахмал.
15.13. В чем сходны механизмы восприятия
силы тяжести у растений и животных?
Современные представления
о геотропизме
Как мы только что говорили, реакция на силу
тяжести у колеоптилей, по-видимому, опосредована
ауксинами, однако у большинства растений
геотропическая реакция стебля наблюдается даже после
удаления верхушки, и поэтому до сих пор не ясно,
связана ли она с передвижением ауксинов. В корнях
перераспределение ауксинов действительно
происходит, но оно, по всей видимости, не столь уж
велико, чтобы полностью объяснить наблюдаемые
изменения скорости роста. Было показано, что
какой-то ингибитор роста и в самом деле передается
из корневого чехлика в зону растяжения, но это,
быть может, и не ауксин. Несколько групп
исследователей пытались найти ауксин в корневых чехликах
проростков кукурузы - одного из самых обычных
экспериментальных объектов. Однако вместо
ауксина была найдена абсцизовая кислота, тоже
хорошо известный ингибитор роста. Не исключено,
что здесь действует еще один ингибитор - этилен.
И наконец, было обнаружено, что концентрация
гиббереллинов (стимуляторов роста) на быстро
растущей стороне стеблей или корней при
геотропической стимуляции намного превышает норму.
15.14. Какие выводы можно сделать из опытов,
показанных на рис. 15.17? В контроле, где агар не
обработан, изгиба нет. Если вместо абсцизовой
кислоты берут ИУК, изгиб тоже незначителен.
15.2.3. Механизм действия ауксинов
Мы более или менее понимаем, каким образом
ауксины способствуют росту клеток. Для
растяжения клетки нужно, чтобы жесткая целлюлозная
основа клеточной стенки была ослаблена. Увеличение
размеров клетки связано с набуханием ее за счет
осмотического поступления воды и одновременным
отложением нового материала клеточной стенки.
Направление растяжения клетки, вероятно, зависит
от ориентации уже существующих микрофибрилл
целлюлозы. «Разрыхление» клеточной стенки
индуцируется повышенной кислотностью и действием
ауксинов.
В 1973 г. четыре группы исследователей
одновременно показали, что ауксины стимулируют
секрецию ионов водорода (протонов). Это приводит к
понижению внеклеточного рН, а тем самым и к
разрыхлению клеточной стенки; возможно, в этом
процессе участвует фермент с низким оптимумом
рН. Кроме того, нужно, чтобы внутри клетки
поддерживался высокий осмотический потенциал и
было достаточно воды, которая входила бы в клетку и
создавала тургорное давление.
Аппарат Гольджи
Клеточная
стенка
Эндопл азматичес кий
ретикулум
Митохондрии
Крахмальные зерна
Точка
роста корня
Амилопласты
IO мкм
Ядро
Рис. 15.16. Электронная микрофотография среза корневого чехлика, видны амилопласты с крахмальными
зернами, лежащие в нижних участках клеток
15.2.4. Другие эффекты ауксинов
Помимо стимуляции растяжения клеток, а значит, и
роста побегов, ауксины выполняют и ряд других
очень важных для растения функций; эти функции
перечислены в табл. 15.4. С ролью ауксинов в диф-
ференцировке, доминировании верхушки, опадении
листьев и созревании плодов мы познакомимся в
соответствующих разделах.
15.2.5. Практическое применение
ауксинов
Открытие ИУК привело к тому, что химики
синтезировали большой набор активных соединений с
очень сходной структурой. Синтетические ауксины
оказались в некоторых отношениях лучше
природных. Их дешевле получать, чем ИУК, и они часто
оказываются более активными, так как у растений
обычно нет ферментов, способных их расщеплять.
В табл. 15.3 приведено несколько примеров таких
веществ с указанием их структуры и способов
применения. Часто активность ауксинов возрастает при
введении в молекулу хлора. На рис. 15.18 показано,
как влияет ауксин на завязывание плодов у
обработанных им томатов.
15.2.6. Гиббереллины
Открытие гиббереллинов
В 20-х годах группа японских ученых, работавших в
Токийском университете, исследовала широко
распространенное во всем мире и весьма вредоносное
Таблица 15.3. Практическое применение ауксинов
Тип ауксина
Индолы и нафтилы
Феноксиуксусные
кислоты *
Бензойные кислоты
Примеры
НУК (нафтилуксусная кислота (сравните с
ИУК на рис. 15.19)
Индолилпропионовая кислота
2,4-Д (2,4-дихлорфеноксиуксусная
кислота)
2,4,5-Т (2,4,5-трихлорфенокси-
уксусная кислота) - строение почти такое
же, но есть лишний атом хлора в
положении 5 кольца; МХФА (2-метил-4-хлор-
феноксиуксусная кислота) строение как
у 2,4-Д, но вместо хлора в положении 2
кольца находится метильная группа (CH3)
2,3,6-трихлорбензойная кислота
2,4,6-трихлорбензойная кислота - строение
как у предыдущей кислоты, но атом хлора
находится не в положении 3 кольца, а в
положении 4
Применение
Способствуют естественному завязыванию
плодов, а иногда и завязыванию их без опыления
(партенокарпия); содействует укоренению
черенков
Избирательные гербициды вызывают гибель
широколистных растений (двудольных).
Применяются в посевах зерновых и на газонах, а также
в питомниках хвойных деревьев для очистки от
кустарников (сами хвойные нечувствительны).
[Смесь 2,4-Д с 2,4,5-Т применялась США во
время войны во Вьетнаме (дефолиант «Эйджент
Орандж»).]
Подавляют прорастание глазков картофеля
при его хранении; предотвращают
преждевременное опадение незрелых плодов (замедляют их
отделение)
Мощные гербициды
п Вызывают некоординированный рост и чрезмерно интенсивное дыхание растений 2,4,5-Т вышла из употребления из-за того, что
содержит примесь диоксина одного из самых токсичных веществ, известных человеку Диоксин вызывает, например, рак, аномалии
внутриутробного развития плода и тяжелые кожные заболевания
Дека питированный
корень кукурузы
Агаровый блок с абсцизовой
кислотой (такие же блоки
используются в опытах Б—Г)
Барьер
Сильный
изгиб
вниз
Барьер
Слабый
изгиб
вверх
Сильный
изгиб
вверх
Рис. 15.17. Влияние абсцизовой кислоты на
геотропическую реакцию декапитированных корней (По
данным PiIeI 1975 )
заболевание проростков риса, которое вызывал
гриб Gibberella (сейчас его относят к роду Fusarium).
Зараженные сеянцы вытягивались в длину,
обесцвечивались и в конце концов погибали или давали
очень плохой урожай. В 1926 г. был получен
экстракт гриба, вызывавшего все эти симптомы у
растений риса. В 1935 г. было получено в
кристаллическом виде первое активное вещество, а к
1938 г.-еще два. Эти вещества были названы гнб-
береллннамн по названию гриба. Языковые барьеры,
а затем и вторая мировая война задержали развитие
работ на Западе, но сразу же после окончания войны
началось соревнование между английскими и
американскими учеными, старавшимися выделить эти
вещества. В 1954 г. английские исследователи
выделили одно из активных веществ, которое они
назвали гибберелловой кислотой. Оно оказалось
третьим и наиболее активным гиббереллином (ГК3)
из числа тех, что были выделены в Японии. Вскоре
были выделены гиббереллины из ряда высших
растений, однако химическая структура ГК3
была окончательно установлена только в 1959 г.
(рис. 15.19). В настоящее время известно свыше 50
природных гиббереллинов, и все они лишь
незначительно оличаются от ГК3.
Строение гиббереллинов
Все гиббереллины относятся к терпенам, очень
сложной группе растительных веществ, близкой к
липидам; все они-слабые кислоты и все содержат
«гиббановый» скелет (рис. 15.19).
Синтез гиббереллинов
и распределение их в растении
Больше всего гиббереллинов в молодых,
растущих органах, и синтезируются они главным
образом в молодых апикальных листьях (возможно, в
хлоропластах), почках, семенах и кончиках корней.
Отсюда они мигрируют вверх и вниз по растению-
передвижение из листьев носит неполярный
характер. Перемещаются они по флоэме и ксилеме.
Действие гиббереллинов
Подобно ауксинам, гиббереллины вызывают
главным образом удлинение стебля, в основном путем
Рис. 15.18. После опрыскивания томата бета-
нафтоксиуксусной кислотой были получены три
большие кисти плодов Небольшая гроздь в нижнем
левом углу снимка не была опрыскана, и на ней
сформировался только один плод нормальной
величины
Гиббановый скелет
Гибберелловая кислота (ГК 3 )
Рис 15.19. Структура гиббанового скелета и
гибберелловой кислоты (77C3)
Рис. 15.20. Влияние гибберелловой кислоты (ГК) на рост карликовой фасоли сорта Метеор. Крайнее
растение слева не обрабатывалось ГК и имеет типичный карликовый габитус. Все остальные растения
обрабатывали ГК, для каждого растения указана доза (в микрограммах) Увеличение дозы вплоть до 5 мкг
приводит к усилению роста стебля На этом основан биотест на гиббереллины с использованием карликовой
фасоли.
растяжения клеток. С их помощью можно
восстановить нормальный рост у карликовых сортов гороха
и кукурузы или превратить карликовую фасоль во
вьющуюся лиану (рис. 15.20), а также
стимулировать рост побегов у обычных растений.
Взаимодействие гиббереллинов с ауксинами будет
рассмотрено в разд. 15.3.
Один из классических эффектов гиббереллинов,
который очень интенсивно исследовали, пытаясь
понять его механизм,-это выход из покоя семян
некоторых растений, в особенности злаков.
Прорастание семян вызывают, замачивая их в воде.
После того как зародыш впитывает влагу, он
начинает выделять гиббереллин, который
диффундирует в алейроновый слой и стимулирует
образование ряда ферментов, в том числе α-амилазы (рис.
15.21). Ферменты начинают расщеплять пищевые
запасы эндосперма, и продукты их переваривания
диффундируют в зародыш, где используются для
его роста.
15.15. а) Что служит субстратом а-амилазы?
б) Что является продуктом реакции, которую она
катализирует? в) Какой другой фермент нужен
для полного расщепления субстрата а-амилазы?
г) Почему α-амилаза столь важна для
прорастания семян хлебных злаков?
15.16. Объясните роль белков алейронового
слоя, рассмотрев рис. 15.21.
Опыт 15.2. Проверка двух гипотез:
а) что гиббереллин
стимулирует расщепление
крахмала в прорастающих
зернах ячменя и
б) что гиббереллин
образуется в зародыше
Амилазу в семенах ячменя можно обнаружить,
поместив разрезанное семя на поверхность агара,
содержащего крахмал. Если смочить поверхность
агара, амилаза диффундирует из семени и начинает
катализировать расщепление крахмала. Если
добавить иод, оставшийся крахмал окрасится в
сине-черный цвет, а вокруг семени останется прозрачная
зона, где крахмал переварен. По размеру этой
круглой зоны можно приближенно оценить количество
амилазы в семени. При этом лучше вести работу в
стерильных условиях, так как амилазу могут
синтезировать также различные бактерии и грибы.
При надлежащей постановке эксперимента можно
проверить и другие гипотезы (см., например, Сорра-
ge J., Hill T. А. (1973), J. Biol. Ed., 7, 11-18).
Материалы и оборудование
(в расчете на одного учащегося)
Белая кафельная плитка
Скальпель
Пинцет
Стакан на 50 мл
Наклейки или карандаш по стеклу
Раствор иода с йодистым калием
Стерильная дистиллированная вода в стерильных
колбах (χ 3)
5%-ный раствор гипохлорита натрия или
продажный препарат для стерилизации, например
раствор Милтона или 70%-ный спирт
две чашки с агаром и крахмалом (стерильные):
1%-ный агар с 0,5% крахмала заливают в
стерильные чашки Петри слоем толщиной около
0,25 см
ДВЕ ЧАШКИ С АГАРОМ, СОДЕРЖАЩИМ КРАХМАЛ И ГИББЕРЕЛ-
лин: в агар перед автоклавированием добавляют
гиббереллин (ГК3)-1 мл 0,1%-ного раствора
ГК3 на 100 мл агара (конечная концентрация
гиббереллина 10 ч. на млн.) (ГК3 плохо
растворима в воде и лучше всего растворяется в
этаноле; часть ГК3 разрушается при автоклави-
ровании, но семена чувствительны к гибберел-
лину даже в концентрации Ю-5 ч. на млн.)
вылущенные семена ячменя: для удаления шелухи
семена замачивают в 50%-ном (по объему)
водном растворе серной кислоты на 3-4 часа,
затем тщательно промывают (до десяти раз) в
дистиллированной воде. Большая часть шелухи
отделяется при энергичном встряхивании зерен
в колбе Эрленмейера; семена тотчас же
используют в опыте, так как после набухания они
начинают прорастать. Если это невозможно,
семена следует разделить на «зародышевую» и
«незародышевую» половинки и положить в
холодильник; в сухих условиях семена можно
хранить не более 2-3 дней. У пшеницы зерна
голые, и удалять шелуху не нужно; можно взять
их вместо ячменя.
Методика
1. Возьмите две чашки с крахмальным агаром и две
чашки с агаром, содержащим крахмал и
гиббереллин, и пометьте их соответствено + Г и
— Г. Чашки должны быть простерилизованы.
Одну чашку из каждой пары подпишите
«зародышевая», а другую-«незародышевая».
2. Разрежьте поперек хотя бы два сухих зерна
ячменя (рис. 15.22) на кафеле. Так получают
«зародышевую» и «незародышевую» половинки.
3. Простерилизуйте половинки в 5%-ном растворе
гипохлорита натрия в течение 5 мин. Затем
промойте их в трех сменах стерильной
дистиллированной воды, разлитой по трем стерильным
колбам.
4. Простерилизуйте пинцет в 70%-ном этаноле, а
затем пинцетом перенесите половинки зерен в
соответствующие чашки и положите их срезом
вниз. Крышки чашек при этом приподнимают как
можно меньше. Получаются такие варианты:
-Г
зародышевая
половинка
+г
зародышевая
половинка
-Г
незародышевая
половинка
+ г
незародышевая
половинка
5. Проинкубируйте препараты в течение 24-48 ч при
20-300C.
6. Проверьте каждую чашку на наличие крахмала:
залейте агар раствором I2 /KI. Зарисуйте, как
будут выглядеть чашки после пробы на крахмал.
Обсудите полученные результаты.
Рис. 15.21. Роль гиббереллина в
мобилизации резервных
питательных веществ в зерне ячменя при
выходе из состояния покоя
Диффузия
Биохимическая
реакция
Шелуха или оболочка
плода (околоплодник),
слившаяся с семенной
оболочкой
Алейроновый слой
(у ячменя состоит из
трех слоев клеток,
содержит белки)
Крахмалистый
эндосперм
Скутеллум
(всасывающий орган)
Колеоптиль
+ побег
Корень
Зародыш
Вода
Крахмал -
Мальтоза
► Мальтоза
Глюкоза
Синтез
гиббе-
релли7
1на
Разрез
Зародыш
"Незародышевая
половина"
" Зародышевая
половина"
Рис. 15.22. Как разрезать зерно ячменя для опыта
15.2.
15.17. При работе с крахмальным агаром
активность амилазы часто выявляется и в
отпечатках пальцев. Подумайте, с чем это может быть
связано.
15.18. Какой эксперимент можно поставить,
используя те же материалы, чтобы доказать, что
гиббереллины вызывают синтез нового фермента,
а не активируют предсуществующую амилазу?
15.19. Как вы докажете, что синтез амилазы
происходит в алейроновом слое?
15.20. Как можно использовать действие гиб-
береллина на семена ячменя для разработки
биотеста на активность гиббереллина?
Другие эффекты гиббереллинов
О том, как влияют гиббереллины на цветение, рост
плодов и состояние покоя у растений и как с ними
связаны фотопериодизм и яровизация, мы
поговорим позднее в соответствующих разделах. Все эти
эффекты перечислены в табл. 15.4.
Механизм действия гиббереллинов
Механизм действия гиббереллинов до сих пор не
выяснен. Было установлено, что в семенах хлебных
злаков ГК3 стимулирует синтез нового белка,
особенно α-амилазы; при этом ГК3 эффективна в столь
малой концентрации (около 10"5 мкг/мл), что
действует, вероятно, на таком глубоком уровне
клеточного метаболизма, как «включение» или
«выключение» генов в процессе дифференцировки клеток
(разд. 22.7). До сих пор, однако, убедительных
подтверждений этого не получено, и к тому же для
других эффектов требуются гораздо большие
концентрации гиббереллинов. Влияние гиббереллинов
на растяжение клеток зависит от присутствия
ауксинов.
Практическое применение
гиббереллинов
Для коммерческих целей гиббереллины получают из
культуры грибов. Гиббереллины способствуют
завязыванию плодов, и их используют для
выращивания бескосточкового винограда (партенокарпия).
ГК3 применяют в пивоварении для стимуляции
образования α-амилазы у ячмены, т.е. для более
эффективного осолаживания. Некоторые
синтетические ретарданты роста действуют как «антигиббе-
реллины», т.е. подавляют эффект гиббереллинов.
Применение таких веществ очень часто приводит к
получению коротеньких (карликовых), но плотных
растений с темно-зелеными листьями, а иногда и с
повышенной устойчивостью к вредителям и
болезням. Они занимают меньше места и, возможно,
будут давать более высокий урожай в расчете на
единицу площади; к тому же они меньше полегают
при сильном ветре.
15.2.7. Цитокинины
Открытие цитокининов
В 40-е и 50-е годы предпринимались попытки
усовершенствовать методы выращивания культур
растительной ткани. Создание улучшенных методов
позволило бы изучать процессы развития в условиях,
когда не влияют другие части растения, и дало бы
возможность оценивать действие различных
химических препаратов. Сохранять клетки живыми
удавалось, но стимулировать их рост было очень
трудно. В 1954-1956 гг. Скуг, работавший в США,
обнаружил, что в молоке кокосового ореха есть
какой-то ингредиент, который способствует
делению зрелых сердцевинных клеток табака в культуре.
«Кокосовое молоко»-это жидкий эндосперм (запас
питательных веществ), и первые данные о том, что в
нем содержатся ростовые вещества, были получены
уже в начале 40-х годов, когда его исследовали
как возможный источник веществ, стимулирующих
рост зародыша. Испытывая другие активные
вещества, Скуг и его группа обнаружили, что сходной
активностью обладают старые препараты ДНК,
тогда как в свежих пробах ДНК такой активности не
было. Потом, однако, выяснилось, что и свежий
препарат ДНК активен, если он был подвергнут
автоклавированию. Было установлено, что
активный компонент по химическому строению сходен с
аденином - одним из оснований, входящих в состав
ДНК (разд. 5.6). Этот компонент назвали кинетн-
ном. Термин кинин был предложен Скугом для
обозначения веществ, участвующих в регуляции
клеточного деления. Позднее был принят термин цито-
кинин; отчасти это было связано с понятием
цитокинеза, означающим деление клеток, отчасти с тем,
что термин «кинин» уже был использован в
физиологии животных и имел там совершенно иное
значение (кинин-это один из полипептидов крови).
Первый природный цитокинин, который был химически
идентифицирован, выделили из молодых зерен
кукурузы (Zea mays) в 1963 г.; поэтому его назвали
зеатином. Обратите внимание на то, что он тоже
сходен по строению с аденином (рис. 15.23).
Кинетин (6-фурфуриладенин) —
синтетический цитокинин
Зеатин — природный цитокинин
Аденин (близок к цитокининам)
Рис. 15.23. Структура кинетина, зеатина и аденина.
Синтез и распределение цитокининов
Больше всего цитокининов там, где происходит
быстрое деление клеток, особенно в незрелых
плодах и развивающихся семенах,-цитокинины
связаны здесь с развитием зародыша. Судя по
имеющимся данным, в зрелых растениях они часто
образуются в корнях и переносятся оттуда к растущим
побегам вместе с транспирационным током (по
ксилеме). Затем цитокинины могут вновь
экспортироваться из листьев по сосудам флоэмы.
Эффекты, вызываемые цитокининами
Цитокинины, если судить по их названию, должны
способствовать делению клеток. Однако так они
действуют только в присутствии ауксинов. Какую-
то роль могут играть и гиббереллины, как,
например, в камбии. Взаимодействие цитокининов с
другими ростовыми веществами мы рассмотрим в
разд. 15.3.2.
Одно из самых интригующих свойств
цитокининов-это их способность замедлять нормальный
Таблица 15.4. Роль ростовых веществ в процессах роста и развития растений. (В таблице представлены самые общие
сведения. Ростовые вещества не обязательно всегда оказывают то влияние, которое здесь указано; довольно обычны
различия в реакции у разных растений. Наибольшее внимание лучше обращать на положительные эффекты; они выделены
полужирным шрифтом и наиболее важны для студентов.)
Затрагиваемый процесс
Рост стебля
Рост корня
Инициация роста корней
Инициация образования по
чек (побегов)
Рост листьев1)
Рост плодов
Доминирование верхушки
Покой почекυ
Покой семян u
Цветение0
Старение листьев
Созревание плодов
Опадение
Влияние на устьица
Ауксины
Содействуют увеличению клеток в области
позади точки роста. Способствуют
делению клеток камбия
В очень низких концентрациях стимулируют.
В более высоких - ингибируют (пример:
геотропизм?)
Способствуют образованию корней у
черенков и каллусов
- У некоторых растений способствуют
образованию побегов, но иногда действуют
как антагонисты цитокининов и
ингибируют этот процесс. Иногда стимулируют
рост побегов у интактного растения, если
снято доминирование верхушки (см.
ниже)
Неактивны
Стимулируют. Иногда вызывают партено-
карпию
Усиливают (т.е. подавляют рост боковых
почек)
Неактивны
Неактивны
Обычно неактивны (стимулируют у
ананаса)
У некоторых растений замедляют
-
Ингибируют. Иногда стимулируют,
особенно если опадение уже началось или если
их наносить со стороны отделительного
слоя, обращенной к стеблю
Неактивны
Гиббереллины
Содействуют увеличению клеток в
присутствии ауксина. Кроме того, способствуют
делению клеток верхушечной меристемы и
камбия. У некоторых розеточных растений
способствуют «выходу в стрелку»
Обычно неактивны
Ингибируют
Способствуют росту побегов у каллуса
хризантемы. Иногда стимулируют рост
побегов у интактного растения, если снято
доминирование верхушки.
Стимулируют
Стимулируют. Иногда вызывают партено-
карпию
Усиливают действие ауксинов
Нарушают
Нарушают (например, у хлебных злаков,
ясеня)
Иногда могут заменить красный свет; поэтому
стимулируют у растений длинного дня,
ингибируют у растений короткого дня
У некоторых растений замедляют
-
Неактивны
Неактивны
Продолжение табл. 15.4
Затрагиваемый
процесс
Рост стебля
Рост корня
Инициация роста корня
Инициация образования
почек (побегов)
Рост листьев2)
Рост плодов
Доминирование верхушки
Покой почекυ
Покой семянυ
Цветение
Старение листьев
Созревание плодов
Опадение
Устьица
Цитокинины
Стимулируют деление клеток в верхушечной
меристеме и камбии. Иногда подавляют
растяжение клеток
Неактивны или ингибируют рост
первичных корней
Неактивны или способствуют росту
боковых корней
Стимулируют (пример-протонема мхов)
Стимулируют
Стимулируют. Изредка могут вызывать
партенокарпию.
Антагонисты ауксинов-стимулируют рост
боковых почек
Нарушают
Нарушают
Обычно неактивны
Замедляют
-
Неактивны
Способствуют открыванию
Абсцизовая кислота
Ингибирует,
особенно при
физиологическом стрессе,
например при засухе
или подтоплении
Ингибирует
(например, при
геотропизме?)
-
—
-
—
Стимулирует
(примеры-платан,
береза)
Стимулирует
Иногда стимулирует
у растений
короткого дня и
подавляет у растений
длинного дня
(антагонист гиббе-
реллинов)
Иногда стимулирует
-
Стимулирует
Стимулирует
закрывание в условиях
водного стресса
(завядания)
Этилен
Ингибирует, особенно
при физиологическом
стрессе
Ингибирует
(например, при
геотропизме?)
—
—
-
—
—
Нарушает
-
Стимулирует у
ананаса
-
Стимулирует
-
Неактивен?
1} Кроме того, участвуют свет и температура-см. разделы о фотопериодизме и яровизации.
процесс старения листьев. Если лист отделить от
растения, он обычно очень быстро стареет, что
проявляется в его пожелтении и в утрате им белка,
РНК и ДНК. Однако, если нанести на такой лист
каплю кинетина, в нем останется зеленый островок
активной ткани, окруженный со всех сторон
пожелтевшей тканью, и затем наблюдается
передвижение питательных веществ из окружающих клеток в
этот зеленый островок (рис. 15.24).
15.21. Рассмотрите рис. 15.24 и затем ответьте
на следующие вопросы:
а) Чем различается судьба нанесенной
аминокислоты в старом и в молодом листе?
б) Почему должна быть такая разница?
в) В чем выражается влияние кинетина на
распределение радиоактивной аминокислоты в старых
листьях?
Сходный, хотя и более слабый эффект наблюдается
даже в том случае, когда кинетин наносят на
отмирающие листья прямо на растении. Было
установлено, что в стареющих листьях резко снижается
содержание природных цитокининов. Поэтому
возможно, что программа естественного старения
включает перенос цитокининов из более старых листьев в
более молодые (по флоэме).
Вероятно, цитокинины участвуют и во многих
других стадиях роста и развития растений (см.
табл. 15.4 и разд. 15.3).
Механизм действия цитокининов
Структурное сходство цитокининов с аденином-
компонентом РНК и ДНК-наводит на мысль, что
цитокинины могут играть очень важную роль в
метаболизме нуклеиновых кислот. Было показано,
что некоторые необычные основания, выделенные
из транспортных РНК, обладают цитокининовой
активностью, и поэтому возникает вопрос о
возможном участии цитокининов в синтезе
транспортных РНК. Однако это предположение, даже если
оно подтвердится, не исключает и иных механизмов
действия этих ростовых веществ; сейчас ведутся
дополнительные исследования.
Кинетин
Кинетин
Молодой
Старый
Рис. 15.24. Влияние ки нети но на транслокацию аминокислот в листьях табака. Вносили радиоактивную
аминокислоту, а затем через определенное время, необходимое для транслокации, листья переносили на
фотопленку. На полученных радиоавтографах видны черные зоны, содержащие меченую аминокислоту.
Практическое применение
цитокининов
Цитокинины продлевают жизнь свежей капусты,
салата и некоторых других зеленых овощей (так как
задерживают старение листьев) и срезанных цветов.
Кроме того, их применяют для прерывания покоя
у некоторых семян.
15.2.8. Абсцизовая кислота
Открытие абсцизовой кислоты
Позднее физиологи растений получили данные о
том, что для нормальной регуляции роста важны не
только ауксины, гиббереллины и цитокинины,
стимулирующие рост, но и ингибиторы роста. Уже
давно подозревали, что состояние покоя
обусловлено действием специфических ингибиторов. Именно
из этого исходила группа ученых, работавших под
руководством Уэринга в Аберистуитском
университете в Уэльсе, когда они начали в конце 50-х годов
поиски в этом направлении. В 1963 г. было
показано, что экстракт из листьев березы индуцирует
состояние покоя у березовых почек. Чтобы
имитировать наступление зимы, листья выдерживали на
коротком световом дне. В 1964 г. из листьев платана
были выделены чистые кристаллы активного
вещества. Полученное вещество назвали дормином.
Оно оказалось идентично соединению, выделенному
другой группой ученых в 1963 г. из молодых
коробочек хлопчатника. Дормин ускорял опадение (по-ла-
тыни-abscisio) коробочек и поэтому был назван
абсцизином II (абсцизин I действует сходным
образом, но отличается по химическому строению и
менее активен). В 1967 г. было решено называть это
вещество абсцизовой кислотой (АБК). Абсцизовая
кислота была найдена у всех высокоорганизованных
растений, начиная с мхов, а у водорослей и
печеночников было найдено другое вещество - лунуларовая
кислота, которая играет сходную роль.
Строение абсцизовой кислоты
Подобно гиббереллинам, АБК относится к терпе-
ноидам и имеет сложное строение (рис. 15.25). Это
единственное ростовое вещество данного класса.
Синтез и распространение
абсцизовой кислоты
АБК образуется в листьях, стеблях, плодах и
семенах. Тот факт, что ее могут синтезировать
изолированные хлоропласты, еще раз указывает на
возможную связь АБК с каротиноидными пигментами,
которые тоже образуются в хлоропластах. Как и все
остальные ростовые вещества, АБК
транспортируется по проводящей системе, в основном по флоэме.
Кроме того, она может распространяться из
корневого чехлика путем диффузии (см. раздел о
геотропизме).
Эффекты, вызываемые АБК
В табл. 15.4 указано, как влияет АБК на рост и
развитие растений. Это важный ингибитор роста, и
он действует как антагонист веществ всех остальных
трех классов, способствующих росту. Можно
упомянуть такие классические эффекты АБК, как ее
влияние на покой семян, зимний покой почек
(включая и доминирование верхушки) и опадение листьев
(см. разд. 15.3.4). Кроме того, АБК каким-то
образом участвует в увядании, цветении, старении
листьев и, возможно, геотропизме. Она связана и со
стрессовыми воздействиями, в особенности с
потерей воды при засухе. Так, например, концентрация
АБК в завядших листьях томата в 50 раз выше, чем
в нормальных листьях, и полагают, что АБК
участвует в закрывании устьиц. Высокие концентрации
АБК полностью останавливают рост растения.
Механизм действия АБК неизвестен.
Практическое применение
абсцизовой кислоты
АБК используют для опрыскивания плодовых
деревьев в конце сезона, чтобы вызвать
одновременное опадение плодов. Это позволяет намного
сократить время сбора урожая.
15.2.9. Этилен (этен)
Открытие еще одного ростового
вещества - этилена
Еще в начале 30-х годов стало известно, что газ
этилен ускоряет созревание плодов цитрусовых и
оказывает различное влияние на рост растений.
Позднее было установлено, что некоторые спелые
плоды, например бананы, выделяют газ,
обладающий сходным действием. В 1934 г. выяснилось, что
желтеющие яблоки испускают этилен, а еще позже
было показано, что этот газ выделяют самые
разные спелые плоды и другие органы растений,
особенно поврежденные. Следовые количества этилена
выделяют практически все органы.
Строение этилена показано на рис. 15.26.
Синтез и распространение этилена
Как мы уже говорили, очень многие, если не все,
органы растений образуют этилен. Хотя этилен-
газ, он обычно плохо распространяется по системе
Рис. 15.25. Строение абсцизовой кислоты.
Рис. 15.26. Строение этилена
внутренних воздушных пространств, так как
значительная часть его прямо выходит наружу через
поверхность растения. Однако было показано, что
водорастворимый предшественник этилена
переходит по ксилеме из затопленных корней в стебли.
Действие этилена
Из эффектов, вызываемых этиленом, известны
главным образом его влияние на созревание плодов и
связанное с ним ускорение дыхания, наблюдаемое у
некоторых растений (разд. 15.3.5). Подобно АБК,
этилен действует как ингибитор роста, и в
некоторых случаях он способствует опадению плодов и
листьев. Эффекты этилена перечислены в табл. 15.4.
Практическое применение этилена
Этилен вызывает зацветание ананасов и
стимулирует созревание томатов и цитрусовых. Чтобы
замедлить созревание плодов, их очень часто хранят в
бескислородных условиях, а созревание
контролируют, вводя этилен с кислородом.
Промышленность выпускает препарат «этефон», который во
время обработки им растений распадается с
выделением этилена; это соединение применяют для
стимуляции вытекания латекса у каучуконосов.
15.3. Синергизм и антагонизм
После того как были изучены отдельные ростовые
вещества, стало ясно, что обычно они действуют
совместно и что нет таких аспектов роста, которые
бы специфически регулировались лишь каким-то
одним веществом. Существует, видимо, два способа
регуляции. В одних случаях два или несколько
веществ дополняют друг друга в своем действии, и
часто оказывается, что суммарный эффект намного
выше, чем был бы результат простого сложения их
индивидуальных эффектов. Это явление называют
синергизмом, а о веществах, действующих подобным
образом, говорят, что они синергисты. В других
случаях два регулирующих вещества оказывают
противоположное влияние на один и тот же
процесс-одно стимулирует его, а другое ингибирует.
Это явление называют антагонизмом, а такие
вещества - антагонистами. В этом случае ответная
реакция определяется балансом между данными
веществами.
Теперь мы можем обратиться к некоторым
наиболее изученным стадиям роста и развития растений и
показать, какое значение имеют явления синергизма
и антагонизма.
15.3.1. Рост побега
Действие гиббереллинов на удлинение стеблей,
черешков, листьев и гипокотилей зависит от
присутствия ауксинов.
Высокое отношение
кинетин: ауксин
!Рост побегов
Промежуточное отношение
кинетин: ауксин
Рост самого каллуса
Низкое отношение
кинетин: ауксин
|Рост корней
Рис. 15.27. Культуры каллусов табака В каждом случае в культуральной среде присутствовала ИУК (2 мг/л).
Культура, содержавшая 0,2 мг кинетина на 1 л (в середине), продолжала расти в виде каллуса; в меньшей
концентрации (0,02 мг/л) кинетин вызывал образование корней, а в более высокой концентрации (0,5 мг/л)
образование стеблей.
15.22. Как это можно было бы показать
экспериментальным путем?
15.3.2 Деление и дифференцировка
клеток
Цитокинины способствуют делению клеток только
в присутствии ауксинов. Иногда какую-то роль
играют и гиббереллины, как, например, в камбии, в
который ауксины и гиббереллины поступают из
находящихся поблизости почек и листьев.
Взаимодействие цитокининов с другими ростовыми
веществами было продемонстрировано в 50-е годы в
классических экспериментах Скуга, о которых мы уже
говорили. Его группа показала, как влияют
различные концентрации кинетина и ауксина на рост
каллуса из сердцевинных клеток табака. При высоком
соотношении ауксин/кинетин наблюдалось
образование корней, а при высоком соотношении кине-
тин/ауксин просыпались боковые почки, из которых
вырастали побеги с листьями. В случае
сбалансированного воздействия ростовых веществ шло
образование недифференцированной ткани (рис. 15.27).
15.3.3. Апикальное доминирование
Апикальное доминирование (доминирование
верхушки) состоит в том, что присутствие растущей
верхушечной почки подавляет рост боковых почек.
Точно так же рост главного корня ингибирует
образование боковых корней. Удаление верхушки
побега приводит к тому, что начинает развиваться
боковая почка, т. е. происходит ветвление стебля.
Это используют при обрезке, когда хотят, чтобы
растение росло кустом, а не тянулось вверх.
15.23. а) Какое ростовое вещество образуется в
точке роста стебля? 6) Продумайте, как поставить
опыт, чтобы доказать, что именно оно
ответственно за доминирование верхушки.
Интересно отметить, что содержание ауксина в
боковых почках часто бывает не настолько
высоким, чтобы подавить их рост. Как действуют при
этом ауксины, неизвестно; возможно, что они
каким-то образом «привлекают» питательные
вещества к точке роста. У дурнишника резкое падение
уровня ауксина в стебле после удаления верхушки,
по-видимому, как-то позволяет боковым почкам
инактивировать АБК, которая в них содержится в
большом количестве. Ответную ракцию на ИУК
часто усиливают гиббереллины. С другой стороны,
обработка боковых почек кинетином нередко
выводит их из состояния покоя, хотя бы на некоторое
время. Кинетин плюс ИУК полностью снимают
состояние покоя. Обычно цитокинины образуются в
корнях и затем переносятся по ксилеме к побегам.
Поэтому в нормальных условиях они, вероятно,
направляются туда, где образуются ауксины, и там
эффекты тех и других совместно стимулируют рост
почек.
Доминирование верхушки-это классический
пример того, как одна часть растения контролирует
другую при помощи того или иного ростового
вещества. Эти явления называют ростовой
корреляцией (рис. 15.28).
Градиент ауксинов и гиббереллинов,
направленный от верхушечной почки,
"привлекает" цитокинины и питательные
вещества
-Лист
Узел <
-Питательные
'вещества из листа
Цитокинины и
минеральные соли
(из корней к точке
роста)
Спящая боковая почка
с низким содержанием
цитокининов,ауксинов
и гиббереллинов;
может также содержать
ингибитор роста,
например абсцизовую
кислоту
-Верхушечная почка
удалена
Боковая почка
активируется и
образует собственные
цитокинины, ауксины
и гиббереллины; если
здесь есть ингибитор
роста, то он инактиви-
руется
-Питательные
вещества
из листа
"Цитокинины и
минеральные
соли
Рис. 15.28. Возможное участие ростовых веществ в
доминировании верхушки: А-в присутствии
верхушечной почки, Б-после ее удаления.
15.3.4. Опадение
Опадение-это организованное сбрасывание
определенных органов у растений; обычно опадают
листья, неопыленные цветки и плоды. У основания
всех этих органов, в так называемой зоне отделения,
находится слой живых клеток, которые разделяются
в результате частичного растворения срединной
пластинки, а иногда и клеточных стенок. Такие
клетки образуют отделительный слой (рис. 15.29).
Окончательное сбрасывание всего органа
происходит тогда, когда механически порвутся проводящие
пучки, например при порыве ветра. Для защиты
рубца от инфекции и от потери воды под
отделительным слоем образуется защитный слой, который
участвует в Закупорке проводящего пучка. У
древесных растений этот слой пробковый и
формируется из той ткани, которую образует пробковый
камбий, а именно из перидермы (разд. 21.6).
Опадение листьев у листопадных деревьев и
кустарников обычно связано с наступлением зимы,
но в тропиках оно часто бывает связано с приходом
засушливого сезона. И в том и в другом случае оно
дает возможность уберечься от возможной нехватки
воды, поскольку главным органом, который теряет
воду в результате транспирации, являются листья.
Зимой, например, вода, находящаяся в почве, может
замерзать и становиться недоступной для растений.
У вечнозеленых растений листья постепенно
опадают и заменяются новыми в течение всего года, и при
этом они обычно видоизменяются таким образом,
чтобы уменьшить потерю влаги.
Как было установлено, по мере старения листа
содержание ауксина в нем снижается. Влияние
ауксина на опадение листьев показано на рис. 15.30.
Следует отметить, что позже, когда опадение
листьев уже началось, ауксины, по-видимому, ускоряют
его.
Абсцизовая кислота (АБК) действует как
антагонист ауксина и иногда способствует опадению
плодов. В незрелых семенах образуются ауксины,
но по мере созревания ауксинов синтезируется все
меньше и меньше, и начинается образование АБК.
В развивающихся коробочках хлопчатника,
например, наблюдаются два пика накопления АБК.
Первый из них соответствует «июньскому
сбрасыванию», когда происходит самоистощение растений;
на этой стадии АБК накапливается только в
абортивных недозрелых мелких плодах. Второй пик
совпадает с созреванием коробочек.
Сомнительно, чтобы АБК вызывала и опадение
листьев. Этот эффект достигается при высоких
концентрациях АБК, но он может быть связан с
действием этилена, образование которого стимули-
. Эпидермис
Перидерма
Стебель
Пазушная
почка
Черешок
Ксилема
Флоэма
Проводящий
пучок
Отделительный
слой
Защитный слой
Отделительная
зона
Перидерма
(наружный слой —
пробковый)
Отделительный слой показан в тот момент,
когда только что началось отделение
Пробковый наружный слой
перидермы образует рубец
Слегка выступающее место
соответствует прежнему
проводящему пучку
Рис. 15.29. Отделительная зона листа в процессе
его отделения (А) и после отделения (Б).
Опадение
черешка
Удаление листовой
пластинки
ИУК в ланолиновой
пасте
Рис. 15.30. Влияние ауксина (ИУК) на опадение
листового черешка. Удаление листовой пластинки
ведет к опадению черешка. ИУК заменяет собой
листовую пластинку.
руется при такой обработке. Этилен образуется в
стареющих листьях и созревающих плодах, и если
им обработать зрелые части растения, он всегда
способствует их опадению. У некоторых
листопадных кустарников и деревьев АБК образуется в
листьях как раз перед самой зимой (разд. 15.5.1), но
это может быть нужно просто для того, чтобы
перевести почки в состояние покоя.
Сбрасывание органов имеет огромное значение
для садоводства, так как с ним связано опадение
плодов. Это находит свое отражение в том, как
применяются ауксины и АБК на практике (об этом
мы уже говорили в начале главы).
Неопыленные цветки обычно сбрасываются (см.
ниже).
15.3.5. Рост пыльцевой трубки,
завязывание плода
и развитие завязи
Прорастающие пыльцевые зерна не только сами
служат богатым источником ауксинов, но и
стимулируют образование ауксинов в тканях столбика
и завязи. Этот ауксин необходим для «завязывания
плода», т.е. для сохранения завязи, из которой
после оплодотворения развивается плод. Если этого
не происходит, цветок, как правило, опадает. После
оплодотворения завязь и зреющие в ней семена
продолжают вырабатывать ауксины, которые
стимулируют дальнейшее развитие плода.
Известно несколько примеров, когда плод
развивается без оплодотворения и потому не образуется
семян, как, скажем, у банана, ананаса и некоторых
бессемянных сортов апельсинов и винограда. Такое
развитие называется партенокарпическим. При этом
в завязи накапливается необычайно большие
количества ауксинов. Иногда партенокарпию можно с
помощью ауксинов вызывать искусственно,
например у томатов, тыквы или перца. Так же легко
можно получить бессемянные бобы у гороха! У ряда
растений, на которые не действуют ауксины,
например у вишни, абрикоса и персика, точно такой
же эффект вызывают гиббереллины. Развивающиеся
семена - богатейший источник не только ауксинов и
гиббереллинов, но и цитокининов (разд. 15.2.7). Эти
ростовые вещества связаны главным образом с
развитием зародыша и накоплением пищевых
запасов в семени, а иногда и в околоплоднике (стенке
плода), куда питательные вещества поступают из
других частей растения.
Созревание плодов-это фактически один из
процессов старения, и оно часто сопровождается
вспышкой дыхательной активности, которую называют
климактерическим подъемом дыхания. Эта вспышка
связана с образованием этилена. О том, какую роль
играют этилен и АБК в опадении плодов, мы уже
говорили в разд. 15.3.4.
15.4. Влияние света
на развитие растений
и роль фотохрома
О значении факторов окружающей среды для роста
и ориентации растения и отдельных его частей мы
уже говорили, когда обсуждали движения
растений. Важнейшим внешним стимулом,
оказывающим многообразное влияние на рост растений,
является свет. Он не только дает энергию для
фотосинтеза и вызывает двигательные реакции, но и
непосредственно влияет на процессы развития.
Индуцируемые светом изменения в развитии называют
фотоморфогенезом.
15.4.1. Этиоляция
Вероятно, самый лучший способ
продемонстрировать, как важен свет для растения,-это вырастить
растение в темноте! Такое растение не накапливает
хлорофилл и поэтому остается белым или бледно-
желтым (как говорят, хлоротичным), но не
становится зеленым; междоузлия стебля становятся
тонкими и длинными. Все это называют этиоляцией,
а само растение -этиолированным. У двудольных
растений при прорастании с выносом семядолей
удлиняется подсемядольное колено, а при
прорастании без выноса семядолей - надсемядольное колено
(разд. 21.6.2); при этом верхушка почечки
искривляется. Листья остаются маленькими и неразвитыми.
У однодольных растений при прорастании
удлиняется мезокотиль, а листья остаются в закрученном
состоянии. Во всех случаях хлоропласты в листьях
не образуют нормальных мембранных систем и
называются в таком виде этиопластами. У растений
образуется мало опорной ткани, и они становятся
ломкими и легко полегают. В конце концов
расходуются все запасы питательных веществ, и если
растение не выйдет на свет и не приступит к фотосинтезу,
то оно гибнет. Но если такое растение поставить на
свет, оно будет расти нормально. Значение
естественной этиоляции в том, что она позволяет
максимально расти в длину с минимальной затратой
углеродных запасов, которые без света не
восстанавливаются за счет фотосинтеза.
15.24. Каким образом морфология
этиолированного растения способствует его прорастанию
через слой почвы?
15.4.2. Открытие фитохрома
На первой стадии любого процесса,
чувствительного к свету, свет поглощается тем или иным
пигментом-так называемым фоторецептором. Пигмент
поглощает характерный для него набор световых
волн, образующих спектр поглощения данного
пигмента (разд. 9.3.2); остальные световые волны
отражаются и придают пигменту присущий ему цвет
(хлорофилл, например, поглощает красный и синий
свет и отражает зеленый).
Семена многих растений прорастают только на
свету. В 1937 г. было установлено, что красный свет
стимулирует, а дальний красный (т.е. свет с
большей длиной волны), наоборот, подавляет
прорастание семян латука. Бортвик и Хендрик, работавшие в
50-е годы в Департаменте сельского хозяйства
США, определили спектр действия для процесса
прорастания (спектр действия-это кривая
относительной эффективности света как стимулятора
данного процесса в зависимости от длины волны). Как
видно из рис. 15.31, лучше всего семена прорастают
при длине волны около 660 нм (красный свет), а
ингибирующее действие света максимально при
длине волны около 730 нм (дальний красный свет).
Кроме того, эти исследователи показали, что
достаточно кратковременного освещения и что
эффект красного света снимается дальним красным
светом и наоборот. Поэтому при попеременном
освещении растения красным и дальним красным
светом эффект будет определяться длиной волны
при последней экспозиции. В 1960 г. американским
ученым удалось в конце концов выделить пигмент,
ответственный за эту реакцию, и он был назван
фитохромом. Как они и предсказывали, фитохром
оказался сине-зеленым пигментом, который
существует в двух взаимопревращающихся формах. Одна
форма Фдк (Ф730) поглощает дальний красный свет, а
вторая Фк (Фббо)-красный. Поглотив свет,
фитохром быстро переходит из одной формы в другую
(за несколько секунд или минут, это зависит от
интенсивности освещения):
Красный свет
(660 нм)
Биохимический
процесс
Дальний красный
свет (730 нм)
Эффективность
660 нм 730 нм
Содействует прорастанию
семян латука, увеличению/
листьев, выпрямлению
согнутой верхушки
гипокотиля у
этиолированных проростков
фасоли
Ингибирует
прорастание
семян латука,
подавляет рост
листьев, вызывает
искривление
верхушки у
этиолированных
проростков
фасоли
Длина волны, нм
Рис. 15.31. Типичные спектры действия для реакции,
контролируемой фитохромом.
В обычном солнечном свете преобладают красные
лучи, их больше, чем дальних красных, поэтому
днем преобладает форма Фдк. Это физиологически
активная форма, но ночью она медленно переходит
в более устойчивую, но неактивную форму Фк. Было
показано, что фитохром состоит из белка и
присоединенной к нему пигментной части. Он
содержится во всех органах растения в ничтожных
количествах (поэтому его не видно, хотя он и окрашен),
но в растущих верхушках его концентрация
повышена. Спектр поглощения фитохрома показан на
рис. 15.32.
15.25. В чем различие между спектром
поглощения и спектром действия?
Целый ряд процессов развития инициируется
слабым красным светом, и этот эффект можно снять
темнотой или дальним красным светом; все это
говорит об участии фитохрома (рис. 15.31 и табл.
15.5). Самый важный из таких процессов - цветение,
о котором речь пойдет в следующем разделе. Об
участии фитохрома в том или ином процессе судят
по тому, насколько спектр действия для данной
реакции на свет совпадает со спектром поглощения
фитохрома.
15.4.3. Фотопериодизм и цветение
Один из важных путей воздействия света на живой
организм связан с восприятием изменений длины
светового дня (фотопериода). На экваторе длина дня
почти постоянна и составляет примерно 12 часов, но
чем дальше от экватора, тем больше будут сезонные
колебания длины дня. Поэтому продолжительность
дня является очень важным внешним фактором в
умеренных широтах, где она изменяется в течение
года в пределах примерно от 9 до 15 часов.
(Действие фотопериода на животных мы рассмотрим в
Поглощение
Длина волны, нм
Рис. 75.32. Спектры поглощения двух форм фито-
хрома.
разд. 16.8.5.) У растений такие явления, как
цветение, образование плодов и семян, переход почек и
семян в состояние покоя, листопад и прорастание
семян тесным образом связаны с сезонными
изменениями длины дня и температуры.
Наиболее глубокие изменения происходят во
время цветения, когда меристема побегов
переключается с образования листьев и боковых почек на
формирование цветков. Значение фотопериода было
открыто еще в 1910 г., но впервые было четко описано
только в 1920 г. Гарнером и Аллардом. Эти авторы
показали, что растения табака зацветают лишь
после выдерживания на коротком фотопериоде в
течение нескольких дней. В естественных условиях это
происходит осенью, но короткий день
(продолжительностью 7 ч) можно создать искусственно и
летом, особенно в теплице. Когда был исследован и
ряд других растений, выяснилось, что некоторым
растениям для цветения нужен длинный день
(растения длинного дня), а некоторые зацветают, как
только созреют, независимо от длины дня
(растения, нейтральные в отношении фотопериода).
Позже обнаружились и другие сложности. Так,
например, некоторые растения при одной
температуре ведут себя как нейтральные, а при другой - нет;
некоторым нужно, чтобы одна длина дня сменялась
другой; а есть и такие, у которых определенная
длина дня только ускоряет наступление цветения, но
не является обязательным условием.
Важным шагом вперед было выяснение того, что
на самом-то деле решающее значение имеет не
длина дня, а продолжительность темного периода.
Поэтому фактически растения короткого дня
оказываются растениями длинной ночи. Если их
выращивать в условиях короткого дня и долгой ночи, но
ночь прерывать коротким периодом освещения,
растения не зацветут. А растения длинного дня
зацветают и в условиях короткого дня, если
прерывать ночной период темноты. Эффект длинного
дня не снимается, однако, при кратковременном
затемнении растений днем. Три основные группы
растений перечислены в табл. 15.6.
15.4.4. Качество и количество света
Теперь остается только установить, сколько и
какого (по спектральному составу) света нужно
растениям. Эксперименты с дурнишником-растением
короткого дня, которое не зацветает в условиях
длинной ночи, прерываемой светом,-позволили
выяснить, что красный свет эффективно препятствует
цветению, а дальний красный свет снимает эффект
красного. Следовательно, фоторецептором служит
фитохром. Эти опыты фактически и были
выполнены в ходе программы исследований, приведших к
открытию фитохрома. Как и следовало ожидать,
длиннодневные растения в условиях короткого дня
зацветают, если длинную ночь прерывать
непродолжительным воздействием красного света. И эту
стимуляцию цветения опять-таки можно снять
дальним красным светом. Ответная реакция
растений во всех случаях определяется тем, какое
воздействие было последним.
Иногда, хотя и не всегда, достаточно
кратковременного (всего несколько минут) воздействия
слабого света, что тоже типично для реакции,
контролируемой фитохромом. Чем ярче свет, тем короче
нужное время экспозиции на таком свету.
15.26. Дурнишник (Xanthium)-растение
короткого дня. Поэтому в обычных условиях цветение у
него индуцируется длинными темными
периодами. В опытах было исследовано, как влияет на
Таблица 15.5. Некоторые реакции растений, контролируемые
фитохромом
Затрагиваемая
область
физиологических процессов
Прорастание u
Фотоморфогенез
(контролируемое
светом развитие
формы и
структуры)
Фотопериодизм
Конкретный процесс, которому
способствует красный свет
Прорастание некоторых семян,
например семян определенных сортов
латука; прорастание спор папоротников
Расширение листовой пластинки у
двудольных. Раскручивание листьев у
злаков (однодольных). Развитие хлоро-
пластов (этиопласты превращаются в
хлоропласты: см. разд. 15.4.1).
Позеленение (протохлорофилл
превращается в хлорофилл). Подавление роста
междоузлий, включая надсемядольное
колено, подсемядольное колено и ме-
зокотиль (т. е. красный свет
предотвращает этиоляцию). Выпрямление
ростовой почечки у двудольных
Цветение у растений длинного дня
(однако цветение у растений короткого
дня подавляется)
п Постановка опытов для иследования действия света на
прорастание семян описана в статье: Hannay J. W., J. Biol. Ed. (1967),
1, 65-73. Автор предлагает использовать сорт латука «Grand
Rapids», но его теперь уже не достать; правда, был подобран ряд
подходящих современных сортов: «Dandie», «Kloek» и «Kweik».
Вероятно, наиболее надежен первый из них, но его семена стоят
довольно дорого, так как это сорт для зимней выгонки в тепличных
условиях Его можно приобрести у Suttons (в мелкой расфасовке)
или у E. W. King a. Co, Coggeshall (Эссекс) в упаковках по 10 г
(этого количества достаточно для засева 250 чашек Петри из
расчета по 50 семян на каждую). Интересно взять также семена
фацелии, это полная противоположность латуку. Примечание во
всех этих опытах в качестве светофильтра, поглощающего дальний
красный свет, можно использовать зеленые листья.
цветение прерывание длинной (12 ч) ночи красным
светом или же сначала красным, а затем дальним
красным светом в течение трех следующих друг за
другом ночей. Интенсивность цветения оценивали
особым индексом (от 1 до 8); полученные
результаты представлены в табл. 15.7.
Постройте график по этим данным, отложив по
вертикальной оси индекс цветения, а по
горизонтальной - продолжительность освещения (на
одном рисунке можно построить оба графика).
Прокомментируйте полученные результаты.
15.4.5. Восприятие и передача сигнала
В середине 30-х годов было показано, что световой
сигнал воспринимается листьями, а не апексом
(верхушкой), где образуются цветки.
Таблица 15.6. Подразделение растений по фотопериоду,
необходимому для цветения
Растения короткого дня
Примеры: дурнишник
(Xanthium pensylvanicum),
хризантемы, соя, табак 1\
земляника
Цветение индуцируется,
когда темный период
превысит критическую
величину, например для
дурнишника 8,5 ч, для табака
ΙΟΙ 1 ч. (В естественных
условиях это эквивалентно
длине дня меньше
критической, например для
дурнишника 15,5 ч, для
табака 13-14 ч)
Растения длинного дня
Примеры: белена
(Hyoscyamus niger)l\
львиный зев, капуста, яровая
пшеница, яровой ячмень
Цветение индуцируется,
когда темный период станет
короче критического,
например для белены 13 ч. (В
естественных условиях это
эквивалентно длине дня
превышающей критическую,
например для белены 11 ч)
Растения, нейтральные в отношении фотопериода
Примеры: огурцы, томаты, садовый горошек, кукуруза,
хлопчатник. Цветение не зависит от фотопериода
п Табак и белена зацветают при длине дня около 12-13 ч
Таблица 15.7. Влияние прерывания длинной ночи красным и
дальним красным светом на зацветание дурнишника. (По
Downs R. J. [1956], Plant Physiol., 31, 279 284.)
Красный свет
Индекс
цветения
6,0
5,0
4,0
2,6
0,0
Время
воздействия
красного света, с
0
5
10
20
30
Две минуты на красном свету и
затем освещение дальним
красным светом
Индекс
цветения
0,0
4,0
4,5
5,5
6,0
Время
воздействия дальнего
красного света,
с
0
12
15
25
50
15.27. Как это можно доказать для растения
короткого дня?
В дополнение к этому оказалось, что дурнишник
зацветает, даже если всего лишь один лист был
помещен в условия, индуцирующие цветение, а все
остальное растение находилось в неиндуцирующих
условиях. Это означает, что какой-то агент (гормон)
вызывает цветение, переходя из такого листа в
точку роста. В пользу этого говорят данные о том,
что фактор, вызывающий цветение, можно передать
от индуцированного растения неиндуцированному
путем прививки и что такой фактор, по-видимому,
один и тот же для короткодневных, длиннодневных
и нейтральных растений, так как он успешно
передается при самых разных вариантах прививки. Это
вещество - гипотетический гормон цветения -
назвали флоригеном1, но его еще не удалось выделить.
Некоторые физиологи растений даже сомневаются в
существовании такого гормона.
15.4.6. Механизм действия фитохрома
Каким же образом осуществляется регулирующее
действие фитохрома? В конце светлого периода он
существует в активной форме Фдк. Во время
короткого темнового периода он медленно превращается
в неактивную форму Фк, но такое превращение,
происходящее только в темноте, может не дойти до
конца. Поэтому можно предположить, что Фдк
способствует цветению длиннодневных растений и
подавляет цветение короткодневных. В последнем
случае для цветения нужны достаточно длинные ночи,
чтобы успела исчезнуть форма Фдк. К сожалению,
кратковременное воздействие дальнего красного
света, которое должно было бы давать тот же
эффект, лишь отчасти воспроизводит действие
длинной ночи; значит, последнее требует более сложного
объяснения. К тому же в какой-то степени важен и
сам фактор времени.
Мы, конечно, знаем, что гиббереллины в
некоторых случаях достаточно хорошо имитируют
действие красного света. Гибберелловая кислота (ГК3)
вызывает цветение у ряда растений длинного дня, в
основном у розеточных форм вроде белены,
которые перед цветением выходят в стрелку («выход в
стрелку»-это быстрое увеличение длины стебля).
Кроме того, ГК3 подавляет цветение и у некоторых
короткодневных растений. Антигиббереллины
(ретарданты роста) снимают эти эффекты.
Итак, стимулирует ли Фдк образование гибберел-
линов и не служат ли последние гормонами
цветения? К сожалению, исключений из общего правила
слишком много, чтобы можно было сделать такой
вывод. Абсцизовая кислота ингибирует цветение у
Lolium и некоторых других длиннодневных расте-
1 Термин предложен советским исследователем
M. X. Чайлахяном, который разработал
соответствующую теорию в 1930-х годах.- Прим. перев.
Рис. 15.33. Растения моркови (сорт Early french
forcing). Слева-контроль, в середине - растение,
которое росло при 17°С, но в течение 4 недель
ежедневно получало по 10 мг гиббереллино, справа
растение, подвергшееся яровизации (6 недель на
холоду).
ний, но при этом индуцирует зацветание земляники
и некоторых других растений короткого дня.
Короче говоря, наши знания о регуляции цветения все
еще неполны.
15.5. Яровизация и цветение
Некоторые растения, особенно двулетние и
многолетние, зацветают после выдерживания при
низкой температуре. Это явление называется
яровизацией. В этом случае действующий фактор
воспринимается не листьями, как при фотопериодизме, а
точкой роста зрелого стебля или же зародышем
семени. Точно так же как и надлежащий
фотопериод, яровизация может быть абсолютно
необходима (например, у белены) или же просто ускоряет
переход к цветению (как у озимых хлебных злаков).
Яровизация нужна и растениям длинного дня
(например, капусте), и растениям короткого дня
(вроде хризантемы), и нейтральным в отношении
фотопериода (скажем, крестовнику). Необходимая
длительность охлаждения варьирует в пределах от
четырех дней до трех месяцев, при этом наиболее
эффективны обычно температуры около 4°С. Xo-
лодовой стимул, так же как и фотопериодический,
можно передать от растения к растению путем
прививки. В этом случае гипотетический гормон
был назван верналином. Позднее было установлено,
что в ходе яровизации возрастает уровень гибберел-
лина и что холодовую обработку можно заменить
обработкой неяровизированных растений гибберел-
линами (рис. 15.33). Сейчас полагают, что «верна-
лин»-это какой-то гиббереллин. Мы теперь знаем,
что фотопериодизм и яровизация служат для
синхронизации репродуктивного поведения растений с
изменениями в окружающей среде и что так
обеспечивается размножение в самое благоприятное время
года. Кроме того, фотопериодизм и яровизация
помогают одновременному зацветанию растений
данного вида и таким образом способствуют
перекрестному опылению и перекрестному
оплодотворению со всеми вытекающими отсюда
преимуществами, связанными с генетическим разнообразием.
15.5.1. Фотопериодизм и регуляция
покоя
Образование зимних почек у деревьев и кустарников
умеренного пояса обычно связано с
фотопериодической реакцией на осеннее уменьшение длины дня;
так обстоит дело, например, у березы, бука и
платана. Внешний стимул воспринимается листьями, и,
как мы уже говорили, начинает возрастать
количество абсцизовой кислоты; она переносится к
меристемам и подавляет рост. Кроме того, наступление
коротких дней индуцирует опадение листвы у
листопадных деревьев. Зимний покой почек можно
бывает снять только воздействием холода
(«раскрывание почек»). Точно так же холодовая обработка
иногда нужна для прорастания набухших в воде
семян (такое воздействие на семена называют
стратификацией): это предохраняет семена от
преждевременного прорастания сразу же после созревания (об
этом речь пойдет в разд. 21.10). Холодовой стимул
можно заменить гиббереллинами, а при
естественном раскрывании почек наблюдается повышение
уровня гиббереллинов и во многих случаях
снижение уровня абсцизовой кислоты. Выход почек из
состояния зимнего покоя у березы и тополя, как
выяснилось, совпадает по времени с повышением
уровня цитокининов. Помимо почек и семян
состояние покоя характерно и для запасающих органов, и
здесь опять-таки большое значение имеет
фотопериод. Например, короткий день индуцирует
образование клубней у картофеля, а длинный день
способствует образованию луковиц у лука.
15.28. Некоторые почки даже летом остаются в
состоянии покоя. Чем здесь обусловлено такое
состояние?
Глава 16
Координация и регуляция
у животных
Характерная черта всего живого - раздражимость,
или чувствительность. Всем организмам нужна
определенная степень внутренней координации и
регуляции; надлежащая взаимосвязь между
стимулом и реакцией необходима для поддержания
стационарного состояния (см. гл. 18) и выживания.
Животные в отличие от растений имеют две
различные, но взаимосвязанные системы
координации - нервную и эндокринную. Нервная система
действует очень быстро, ее эффекты четко
локализованы, а в основе ее деятельности лежит
электрическая и химическая передача. Эндокринная система
действует более медленно, ее эффекты носят
диффузный характер, а в основе ее действия лежит
химическая передача сигнала через систему
кровообращения. Как полагают, у большинства
многоклеточных животных обе системы развивались
параллельно.
16.1. Нервная система
Нервная система состоит из высокодифференциро-
ванных клеток, функция которых заключается в
том, чтобы воспринимать сенсорную информацию,
кодировать ее в форме электрических импульсов и
передавать (нередко на значительные расстояния)
другим дифференцированным клеткам, способным
отвечать надлежащим образом.
У многоклеточных животных вся сенсорная
информация воспринимается видоизмененными
нервными клетками, носящими название рецепторов,
структура и функция которых описана в разд. 16.5.
Воспринимаемая рецепторами информация
передается в конце концов эффекторным клеткам и
вызывает в них реакцию, определенным образом
связанную со стимулом. Краткое описание строения и
функций эффекторов дается в разд. 16.7 и в гл. 17.
Связь между рецепторами и эффекторами
осуществляют проводящие клетки нервной системы-
нейроны. Они являются основными структурными и
функциональными элементами нервной системы, и
их разветвленные отростки пронизывают весь
организм, образуя сложную систему связей. Типы
нейронов и их структура были описаны в разд. 8.6.
Сложность строения отдельных нейронов и всей нервной
системы зависит от филогенетического положения
животного и сильно варьирует-от примитивной
нервной сети кишечнополостных (разд. 16.3) до
сложнейшей центральной нервной системы
млекопитающих (разд. 16.2). Если в первом случае
нервная система выполняет чисто связующую роль, то
во втором обеспечивает также хранение
информации, ее извлечение в нужный момент, переработку и
интеграцию.
16.1.1. Природа нервного импульса
О том, что нервные сигналы передаются по
нейронам в виде электрических импульсов, влияющих на
сокращение мышц и секреторную активность желез,
было известно уже более 200 лет назад. Однако
механизм возникновения и распространения этих
импульсов был выяснен только в последние 40 лет,
после того как у кальмара были открыты гигантские
аксоны толщиной около миллиметра. Они иннерви-
руют мускулатуру мантии и вызывают ее быстрое
сокращение, когда животному нужно спасаться от
врага. Большая толщина этих аксонов позволила
провести на них некоторые из самых ранних
электрофизиологических исследований.
На рис. 16.1 изображен прибор, применяемый в
настоящее время для изучения электрической
активности нейронов. Его важнейшую часть составляет
микроэлектрод - стеклянная трубочка, вытянутая на
конце в капилляр диаметром 0,5 мкм и заполненная
раствором, проводящим ток, например 3 M KCl.
Этот микроэлектрод вводят в аксон, а второй
электрод, имеющий вид маленькой металлической
пластинки, помещают в солевой раствор,
омывающий исследуемый нейрон. Электроды подсоединены
к усилителю, замыкающему цепь. Сигнал,
усиленный примерно в 1000 раз, передается на двухлучевой
осциллограф. Все перемещения микроэлектрода
осуществляются с помощью
микроманипулятора-специального устройства, позволяющего с большой
точностью регулировать положение
микроэлектрода. Когда кончик микроэлектрода проходит сквозь
плазматическую мембрану аксона, лучи
осциллографа раздвигаются. Расстояние между лучами
показывает разность потенциалов между двумя
электродами. Эта разность называется потенциалом покоя
аксона и составляет у всех исследованных видов
приблизительно —65 мВ. Таким образом, мембрана
Стимулятор
Выключено
Стимули-^
рующие
электроды
Включено
3 M раствор
KCl
Микро электрод
Усилитель
Катодный двухлучевой
осциллограф
Внешний,
референтный электрод
Омывающий
солевой
раствор
Аксон
Мембрана
аксона
Рис. 16.1. Схема аппаратуры для регистрации электрической активности аксона отдельного нейрона.
Стимулятор подает ток, возбуждающий потенциал действия, который отмечается и записывается с
помощью внутриклеточных микроэлектродов и двулучевого катодного осциллографа.
аксона поляризована, а минус перед потенциалом
покоя означает, что с внутренней стороны она
заряжена отрицательно по отношению к наружной
поверхности. В сенсорных клетках, нейронах и
мышечных волокнах эта величина во время активности
изменяется, поэтому такие клетки называются
возбудимыми. На мембранах всех остальных живых
клеток тоже существует подобная разность
потенциалов, известная как мембранный потенциал, но в
этих клетках она остается постоянной, поэтому их
называют невозбудимыми клетками.
Потенциал покоя
У большинства нейронов млекопитающих
потенциал покоя остается постоянным до тех пор, пока
клетка находится в неактивном состоянии из-за
отсутствия стимула. Кертис и Коул в США и
Ходжкин и Хаксли в Англии в конце 30-х годов
Таблица 16.1. Концентрации ионов во внеклеточной среде и
внутриклеточной жидкости аксона кальмара (приближенные
величины; выражены в миллимолях на 1 кг H2O; из
Hodgkin, 1958)
Ион
K +
Na +
Cl
A- (органические
анионы)
Внеклеточная
концентрация
20
460
560
0
Внутриклеточная
концентрация
400
50
100
370
установили, что потенциал покоя имеет физико-
химическую природу и обусловлен разностью
ионных концентраций по обе стороны мембраны аксона
и избирательной проницаемостью мембраны для
ионов. Анализ жидкости, находящейся внутри
аксона, и омывающей аксон морской воды показал, что
между обеими жидкостями существуют
электрохимические ионные градиенты (табл. 16.1).
В аксоплазме, находящейся внутри аксона,
больше ионов К+ и меньше Na+, тогда как в жидкости,
омывающей аксон, наоборот, больше ионов Na+H
меньше K+ (распределение ионов Cl- в
последующем описании не учитывается, так как оно не играет
существенной роли в интересующих нас явлениях).
Указанные градиенты поддерживаются за счет
активного переноса ионов против их
электрохимических градиентов, который осуществляют
определенные участки мембраны, называемые катионными
или натриевыми насосами. Эти непрерывно
действующие транспортные механизмы работают за счет
энергии, освобождающейся при гидролизе АТФ; при
этом происходит выведение из аксона Na + ,
сопряженное с поглощением K+ (рис. 16.2, А).
Активному транспорту этих ионов противостоит
их пассивная диффузия, поскольку они постоянно
перемещаются «вниз» по электрохимическим
градиентам, как показано на рис. 16.2, Б. Скорость
диффузии определяется проницаемостью мембраны
аксона для данного иона. Ионы K+ более
подвижны, и проницаемость мембраны для них в 20 раз
больше, чем для Na +, поэтому К + легче выходит из
аксона, чем Na+ входит в него, и в результате в
аксоне становится меньше катионов и создается
отрицательный заряд. Величина потенциала покоя
определяется главным образом электрохимическим
градиентом K + .
Изменение проницаемости мембраны
возбудимых клеток для ионов K+ и Na+ приводит к
изменению разности потенциалов на мембране, к
возникновению потенциалов действия и
распространению нервных импульсов по аксону.
Потенциал действия
При стимуляции аксона электрическим током (рис.
16.3) потенциал на внутренней поверхности
мембраны меняется с — 70 мВ до + 40 мВ. Это изменение
полярности носит название потенциала действия
(спайка) и регистрируется на двухлучевом
осциллографе в виде кривой, представленной на рис. 16.3.
Потенциал действия возникает в результате
внезапного кратковременного повышения
проницаемости мембраны аксона для ионов Na+H входа
последних в аксон. Вследствие увеличения
проводимости (электрический эквивалент проницаемости)
для натрия число положительно заряженных ионов
внутри аксона возрастает, и мембранный потенциал
снижается по сравнению с величиной покоя,
составляющей около —70 мВ. Такое изменение мем-
СНАРУЖИ
Мало K+,много Na +
Мембрана аксона
(7,5 нм)
Транспорт за счет
энергии ΑΤΦ
Много K+, мало Na+
ВНУТРИ
Рис. J 6.2. Активное (А) и пассивное (Б) перемещение
ионов, связанное с созданием отрицательного
потенциала внутри аксона. Натриево-калиевый насос
осуществляет активный перенос ионов (А), которые в
то же время проходят через мембрану путем
пассивной диффузии в направлении их
электрохимических градиентов (Б)
Разность потенциалов на
мембране аксона, мВ
Время, мс
Рис. 16.3. Типичный потенциал действия в аксоне
кальмара.
Мембранный потенциал, мВ
Деполя -
ризация
Реполя-
ризация
Гиперполяризация
Направление
распространения
импульса
Изменение
ионной проводимости
Разность
потенциалов
на мембране
аксона +T+
В
Аксоплазма
Мембрана аксона
Время, мс
Рис. 16.4. Схема взаимоотношений между
мембранным потенциалом (А), ионной проводимостью (Б) и
распределением потенциалов на обеих сторонах
мембраны аксона (В) во время потенциала действия.
бранного потенциала называется деполяризацией.
Повышение натриевой проводимости и
деполяризация влияют друг на друга по принципу
положительной обратной связи, т.е. взаимно усиливают
друг друга, и в результате возникает фаза крутого
подъема потенциала действия. Как показали
расчеты, при деполяризации порядка 110 мВ,
происходящей при развитии потенциала действия, в аксон
входит сравнительно немного ионов Na+ (около
10_6% от общего количества Na + ,
присутствующего в клетке, в зависимости от диаметра аксона).
На пике потенциала действия проницаемость для
натрия начинает падать (инактивация натриевых
каналов), а примерно через 0,5 мс после начала
деполяризации повышается проницаемость для К +,
и калий выходит из аксона. По мере выхода ионов
K+ положительный заряд с внутренней стороны
мембраны меняется на отрицательный. Эта фаза
реполяризации мембраны, представленная
нисходящей областью пика потенциала действия, приводит
к восстановлению исходного мембранного
потенциала.
Из приведенного объяснения видно, что если
потенциал покоя определяется главным образом
ионами K + , то потенциал действия зависит в основном
от ионов Na+ (рис. 16.4).
Мембранный потенциал, мВ
Время, мс
Рис. 16.5. Запись мембранных потенциалов в аксоне
кальмара, омываемом обычной и разведенной морской
водой.
16.1. Приведите две причины резкого
увеличения входа Na+ в аксон при повышении
проницаемости мембраны аксона для Na + .
16.2. Как изменился бы потенциал действия,
если бы одновременно повысилась проницаемость
мембраны для инов Na+ и K+?
16.3. В 1949 г. Ходжкин и Катц исследовали
влияние ионов Na+ на потенциалы действия в
аксонах кальмара. С помощью внутриклеточных
микроэлектродов их регистрировали в аксонах,
омываемых изотоничной морской водой и той же
водой в разведениях 1:2 и 1:3. Результаты
представлены на рис. 16.5. Какой среде соответствует
каждая из приведенных трех кривых? Объясните
влияние различных растворов на потенциалы
действия.
Характерные особенности
потенциалов действия
возникновение потенцилла действия. Стимуляция сен-
сорных клеток приводит к деполяризации их
мембран, и если эта деполяризация достигнет
определенной пороговой величины, она вызовет потенциал
действия. Амплитуда потенциала действия (рис. 16.3)
постоянна для каждого нейрона, и увеличение силы
или частоты стимуляции на нее не влияет; поэтому
говорят, что потенциал действия подчиняется
закону «всё или ничего».
передача нервных импульсов. В нервной системе
информация передается как серия нервных
импульсов - распространяющихся потенциалов действия.
Нервный импульс пробегает по аксону в виде волны
деполяризации. Такое распространение потенциала
действия происходит благодаря натриевому току:
когда ионы Na+ входят в аксон, в данном участке
аксона создается область положительного заряда и
в местной цепи возникает ток между этим активным
участком и отрицательно заряженной областью,
находящейся непосредственно перед ним.
Возникающий в местной цепи ток снижает мембранный
потенциал в этой области, и в результате
деполяризации здесь повышается проницаемость
мембраны для Na+ и в свою очередь возникает потенциал
действия, развивающийся по принципу «всё или
ничего». Последовательная деполяризация все
новых и новых участков мембраны ведет к тому, что
потенциал действия распространяется по аксону все
дальше без изменения амплитуды. Таким образом
потенциалы действия могут передаваться на любые
расстояния, т. е. носят незатухающий характер (без-
декрементное проведение). Причина этого
заключается в том, что изменение в каждой точке аксона
происходит за счет локального запаса энергии в
виде ионного градиента, хотя и запускается
изменением в соседней точке. До тех пор пока существует
необходимая разность ионных концентраций внутри
и вне аксона, потенциал действия, возникающий в
одном участке, будет генерировать потенциал
действия в соседнем участке.
период рефрактерности. Нервные импульсы
проходят по аксону только в одном направлении. Это
связано с тем, что участок, бывший ранее активным,
переходит в восстановительную фазу. Во время этой
фазы мембрана аксона не может отвечать на
деполяризацию изменением натриевой проводимости
даже в том случае, если интенсивность стимула
увеличилась. Эта фаза носит название периода
абсолютной рефрактерности и продолжается около
1 мс. За нею следует период относительной
рефрактерности, продолжающийся 5-10 мс, в течение
которого сильные стимулы могут вызывать
деполяризацию. Период рефрактерности ограничивает
возможную частоту нервных импульсов.
16.4. Опишите изменения, происходящие в
аксоне во время периода рефрактерности.
скорость проведения. В немиелинизированных
аксонах (разд. 16.2), характерных для беспозвоночных,
скорость распространения потенциалов действия
зависит от сопротивления аксоплазмы вдоль аксона.
Это сопротивление в свою очередь зависит от
диаметра аксона-чем меньше диаметр, тем больше
сопротивление. В тонких аксонах (<0,1 мм)
большое сопротивление аксоплазмы влияет на
проведение тока и уменьшает длину местной цепи, так что в
нее включается только тот участок, который
расположен непосредственно впереди потенциала
действия. Эти аксоны проводят импульсы со скоростью
около 0,5 м/с. Гигантские аксоны, типичные для
многих червей, членистоногих и моллюсков и
имеющие диаметр около 1 мм, проводят импульсы со
скоростью до 100 м/с, оптимальной для передачи
жизненно важной информации.
16.5. Объясните, опираясь на сведения о
сопротивлении аксоплазмы и местных токах, почему
гигантские аксоны проводят импульсы с большей
скоростью, чем тонкие нервные волокна.
У позвоночных большинство нервных волокон,
особенно в спинномозговых и черепномозговых нер-
вах, имеют миелиновую оболочку, образованную
мембранами шванновских клеток (разд. 8.6.3).
Миелин-вещество липидной природы, оно обладает
большим электрическим сопротивлением и
действует как изолятор подобно резиновому или
пластиковому покрытию электрического провода.
Суммарное сопротивление мембраны аксона и миелиновой
оболочки очень велико, но там, где в миелиновой
оболочке имеются разрывы, называемые
перехватами Ранвье, сопротивление току между аксоплаз-
мой и внеклеточной жидкостью меньше. Только в
этих участках и замыкаются местные цепи, и именно
здесь через мембрану аксона проходит ток,
генерирующий следующий потенциал действия. В
результате импульс перескакивает от одного перехвата к
другому и пробегает по миелинизированному
аксону быстрее, чем серия меньших по величине местных
токов в безмиелиновом нервном волокне. Такой
способ распространения потенциала действия,
называемый сальтаторным (от лат. saltare-прыгать),
может обеспечивать проведение импульса со
скоростью до 120 м/с (рис. 16.6).
На скорость проведения нервных импульсов
влияет температура, и по мере ее повышения до 400C эта
скорость увеличивается.
16.6. Почему миелинизированные аксоны
лягушки, имеющие диаметр 3,5 мкм, проводят
импульсы со скоростью 30 м/с, а аксоны кошки
такого же диаметра-со скоростью 90 м/с?
кодирование нервной информации. Нервные
импульсы распространяются в нервной системе в виде
потенциалов действия, подчиняющихся закону «всё
или ничего» и имеющих постоянную для данного
вида нейронов амплитуду; в гигантском аксоне каль-
Направление
распространения
импульса
Перехват
Ранвье
Местная цепь
Миелиновая
оболочка
Мембрана
аксона
Рис. 16.6. Схема, показывающая разную длину
местных цепей, возникающих в миелинизированном (А) и
немиелинизированном (В) аксоне Проведение тока в
первом случае называют сальтаторным, так как
потенциал действия быстро «перескакивает» от
одного перехвата к другому
Мембранный потенциал, мВ
Направление
распространения
импульса
Диффузия ионов.
Катионный насос
(постоянно активный)
Аксон
Местные цепи
Аксон!
Состояние
аксона
Тип
потенциала
Мембрана
Движение
ионов
Внутренний
потенциал
Наружный
потенциал
Ток
Покой
Потенциал,
покоя ι
Поляризо-,
вана ι
Сохраняет-'
ся стабиль-|
ная раз- ,
ность ион-1
ных кон- ι
центраций '
Отрица- ι
тельный |
Положи- [
тельный ;
Активность
Потенциал
действия
Деполяризована
Повышенная
проницаемость
для Na+ и K +
ι
Положитель- .
ный I
I
Отрицатель- '
ный '
Возникают местные цепи между
неактивными и активными
участками
Восстановительный
период
Потенциал
покоя
Реполяризу-
ется
Восстанавливается
исходная
разность
потенциалов
Отрицатель -
ный
Положитель -
ный
Отсутствует
Рис. 16.7. Общая схема событий, сопровождающих
возникновение местных цепей в аксоне и
распространение потенциала действия в немиелинизированном
аксоне А Потенциал действия и направление его
распространения Б Перемещение ионов через
мембрану аксона В. Токи в местных цепях
мара, например, она составляет 110 мВ. В связи с
этим информация не может кодироваться
амплитудой, а используется только частота импульсов.
Впервые этот факт был установлен в 1926 г. Эдриа-
ном и Зоттерманом, которые показали, что частота
нервных импульсов находится в прямой
зависимости от силы вызывающего их стимула.
16.1.2. Синапс
Синапс представляет собой место функционального,
а не физического контакта между нейронами; в нем
происходит передача информации от одной клетки к
другой. Обычно встречаются синапсы между
концевыми веточками аксона одного нейрона и дендрита-
ми (аксодендритные синапсы) или телом (аксосома-
Синапсы
Ядро
Тело клетки
Аксон
Рис. 16.8. Синапсы на двигательном нейроне
(электронная микрофотография)
тические синапсы) другого нейрона. Число синапсов,
как правило, очень велико, что обеспечивает
большую площадь для передачи информации.
Например, на дендритах и телах отдельных мотонейронов
спинного мозга находится свыше 1000 синапсов.
Некоторые клетки головного мозга могут иметь до
10000 синапсов (рис. 16.8).
Существуют два типа синапсов - электрические и
химические - в зависимости от природы проходящих
через них сигналов. Между окончаниями
двигательного нейрона и поверхностью мышечного волокна
существует нервно-мышечное соединение,
отличающееся по строению от межнейронных синапсов, но
сходное с ними в функциональном отношении.
Структурные и физиологические различия между
обычным синапсом и нервно-мышечным
соединением будут описаны несколько позже.
Строение химического синапса
Химические синапсы - наиболее распространенный
тип синапса у позвоночных. Это луковицеобразные
утолщения нервных окончаний, называемые синап-
тическими бляшками и расположенные в
непосредственной близости от окончания дендрита.
Цитоплазма синаптической бляшки содержит
митохондрии, гладкий эндоплазматический ретикулум, мик-
рофиламенты и многочисленные синаптические
пузырьки. Каждый пузырек имеет в диаметре около
50 нм и содержит медиатор-вещество, с помощью
которого нервный сигнал передается через синапс.
Мембрана синаптической бляшки в области самого
синапса утолщена в результате уплотнения
цитоплазмы и образует пресинаптическую мембрану.
Мембрана дендрита в области синапса также
утолщена и образует постсинаптическую мембрану. Эти
мембраны разделены промежутком - синаптической
щелью шириной около 20 нм. Пресинаптическая
мембрана устроена таким образом, что к ней могут
прикрепляться синаптические пузырьки и
выделяться в синаптическую щель медиаторы. Постсинапти-
ческая мембрана содержит крупные белковые
молекулы, действующие как рецепторы медиаторов, и
многочисленные каналы и поры (обычно закрытые),
через которые в постсинаптический нейрон могут
поступать ионы (см. рис. 16.10,/!).
Синаптические пузырьки содержат медиатор,
который образуется либо в теле нейрона (и попадает в
синаптическую бляшку, пройдя через весь аксон),
либо непосредственно в синаптической бляшке.
В обоих случаях для синтеза медиатора нужны
ферменты, образующиеся в теле клетки на
рибосомах. В синаптической бляшке молекулы медиатора
«упаковываются» в пузырьки, в которых они
хранятся до момента высвобождения. Основные
медиаторы нервной системы позвоночных - ацетилхо-
лин и норадреналин, но существуют и другие
медиаторы, которые будут рассмотрены позже.
Ацетилхолин - аммонийное производное,
формула которого приведена на рис. 16.9. Это первый из
известных медиаторов; в 1920 г. Отто Леви
выделил его из окончаний парасимпатических нейронов
блуждающего нерва в сердце лягушки (разд. 16.2).
Структура норадреналина подробно
рассматривается в разд. 16.6.6. Нейроны, высвобождающие
ацетилхолин, называются холинэргическими, а
высвобождающие норадреналин - адренэргическими.
Механизмы синаптической передачи
Как полагают, прибытие нервного импульса в
синаптическую бляшку вызывает деполяризацию преси-
наптической мембраны и повышение ее
проницаемости для ионов Ca2 + . Входящие в синаптическую
бляшку ионы Ca2+ вызывают слияние синаптиче-
ских пузырьков с пресинаптической мембраной и
выход их содержимого из клетки (экзоцитоз), в
результате чего оно попадает в синаптическую
щель. Весь этот процесс называют
электросекреторным сопряжением. После высвобождения медиатора
материал пузырьков используется для образования
новых пузырьков, заполняемых молекулами
медиатора. Каждый пузырек содержит около 3000
молекул ацетил хо л ина.
Молекулы медиатора диффундируют через
синаптическую щель (этот процесс занимает около 0,5 мс)
и связываются с находящимися на постсинаптичес-
кой мембране рецепторами, способными узнавать
молекулярную структуру ацетилхолина. При связы-
Puс. 16.9. Структурная формула ацетилхолино.
Аксон пресинапти-
ческого нейрона
Митохондрия -
Синаптический
пузырек
Мембрана постсинапти-
ческого нейрона
Пресинаптическая
мембрана
Область
рецепторов
Синаптическая
бляшка
Синаптическая
щель
Постсинаптическая
мембрана
Синаптические пузырьки,
сливаются с пресинапти -
ческой мембраной
и разрываются
Молекулы медиатора-
Проницаемость
пресинаптической
мембраны для Ca2 +
повышается
(1) Диффузия молекул
медиатора через
щель
(2) Молекулы медиатора
связываются
с рецепторными
молекулами
Распространяющийся
потенциал действия
(3) Открываются каналы,
и по ним поступают
ионы из синаптической
щели (повышается
проницаемость постинап -
тической мембраны)
Местная
деполяризация
Гидролитические ферменты,
расщепляющие медиаторы,
например ацетилхолинэстераза
Рис. 16.10. Механизм химической передачи импульсов в нервном синапсе; от А до Д-последовательные этапы
процесса.
вании молекулы рецептора с медиатором ее
конфигурация меняется, что приводит к открытию ионных
каналов и поступлению в постсинаптическую клетку
ионов, вызывающих деполяризацию или
гиперполяризацию (рис. 16.4,^4) ее мембраны в зависимости от
природы высвобождаемого медиатора и строения
молекулы рецептора. Молекулы медиатора,
вызвавшие изменение проницаемости постсинаптической
мембраны, сразу же удаляются из синаптической
щели либо путем их реабсорбции пресинаптической
мембраной, либо путем диффузии из щели или
ферментативного гидролиза. В случае холинэрги-
ческих синапсов находящийся в синаптической щели
ацетилхолин гидролизуется ферментом ацетилхо-
линэстеразой, локализованным на
постсинаптической мембране. В результате гидролиза образуется
холин, он всасывается обратно в синаптическую
бляшку и вновь превращается там в ацетилхолин,
который хранится в пузырьках (рис. 16.10).
В возбуждающих синапсах под действием ацетил-
холина открываются специфические натриевые и
калиевые каналы, и ионы Na+ входят в клетку, а
ионы K+ выходят из нее в соответствии с их
концентрационными градиентами. В результате
происходит деполяризация постсинаптической
мембраны. Эту деполяризацию называют возбудительным
постсинаптическим потенциалом (ВПСП).
Амплитуда ВПСП обычно невелика, но продолжительность
его больше, чем у потенциала действия. Амплитуда
ВПСП меняется ступенчатым образом, и это
позволяет предполагать, что медиатор освобождается
порциями, или «квантами», а не в виде отдельных
молекул. По-видимому, каждый квант
соответствует освобождению медиатора из одного синапти-
ческого пузырька. Одиночный ВПСП не способен,
как правило, вызвать деполяризацию пороговой
величины, необходимой для возникновения
потенциала действия. Но деполяризующие эффекты
нескольких ВПСП складываются, и это явление носит
название суммации. Два или больше ВПСП,
возникших одновременно в разных синапсах одного и
того же нейрона, могут сообща вызвать
деполяризацию, достаточную для возбуждения потенциала
действия в постсинаптическом нейроне. Это
называют пространственной суммацией. Быстро
повторяющееся высвобождение медиатора из пузырьков
одной и той же синаптической бляшки под
действием интенсивного стимула вызывает отдельные
ВПСП, которые следуют так часто один за другим
во времени, что их эффекты тоже суммируются и
вызывают в постсинаптическом нейроне потенциал
действия. Это называется временной суммацией.
Таким образом, импульсы могут возникать в
одиночном постсинаптическом нейроне либо как результат
Аксон моторного нейрона, покрытый
шванновской клеткой
Двигательная концевая
пластинка
Вторичная
синаптическая щель
Шванновская клетка
Митохондрия
Синаптические
пузырьки
"Поверхность мышечного волокна
Пресинаптическая мембрана
моторного нейрона
Синаптическая щель
Соединительная складка сарколеммы
(постсинаптической мембраны)
Митохондрия
Трубочка
Т-сисгемы
Миофибрилла
Диск
Саркомер
Рис. 16.11. Схема строения двигательной концевой пластинки и нервно-мышечного соединения. В верхней
части рисунка (над линией X X) изображено окончание аксона моторного нейрона на поверхности
мышцы - двигательная концевая пластинка. В нижней части (под линией X X) показана ультраструктура
нервно-мышечного соединения
слабой стимуляции нескольких связанных с ним
пресинаптических нейронов, либо как результат
повторной стимуляции одного из его
пресинаптических нейронов. В тормозных синапсах
высвобождение медиатора повышает проницаемость
постсинаптической мембраны за счет открытия
специфических каналов для ионов K+ и Cl". Перемещаясь
по концентрационным градиентам, эти ионы
вызывают гиперполяризацию мембраны, называемую
тормозным постсинаптическим потенциалом (ТПСП).
Медиаторы сами по себе не обладают
возбуждающими или тормозящими свойствами.
Например, ацетилхолин оказывает возбуждающее
действие в большинстве нервно-мышечных соединений и
других синапсов, но вызывает торможение в
нервно-мышечных соединениях сердца и висцеральной
мускулатуры. Эти противоположные эффекты
обусловлены теми событиями, которые
развертываются на постсинаптической мембране. От
молекулярных свойств рецептора зависит, какие ионы
будут входить в постсинаптический нейрон, а эти ионы
в свою очередь определяют характер изменения
постсинаптических потенциалов, как описано выше.
Электрические синапсы
У многих животных, в том числе у
кишечнополостных и позвоночных, передача импульсов через
некоторые синапсы осуществляется путем
прохождения электрического тока между пре- и постсинап-
тическими нейронами. Ширина щели между этими
нейронами составляет всего лишь 2 нм, и
суммарное сопротивление току со стороны мембран и
жидкости, заполняющей щель, очень мало.
Импульсы проходят через синапсы без задержки, и на их
передачу не действуют лекарственные вещества или
другие химические препараты.
Нервно-мышечное соединение
Нервно-мышечное соединение представляет собой
специализированный вид синапса между
окончаниями двигательного нейрона (мотонейрона) и эндо-
мизием мышечных волокон (разд. 17.4.2). Каждое
мышечное волокно имеет специализированный
участок - двигательную концевую пластинку, где
аксон моторного нейрона (мотонейрона)
разветвляется, образуя немиелинизированные веточки
толщиной около 100 нм, проходящие в неглубоких
желобках по поверхности мышечной мембраны.
Мембрана мышечной клетки - сарколемма - образует
множество глубоких складок, называемых постсина-
птическими складками (рис. 16.11). Цитоплазма
окончаний мотонейрона сходна с содержимым си-
наптической бляшки и во время стимуляции
освобождает ацетилхолин с помощью того же
механизма, о котором говорилось выше. Изменения
конфигурации молекул-рецепторов, находящихся на
поверхности сарколеммы, ведут к изменению ее
проницаемости для Na+ и K + , и в результате
происходит местная деполяризация, называемая
потенциалом концевой пластинки (ПКП). Эта деполяризация
по величине вполне достаточна для возникновения
потенциала действия, который распространяется по
сарколемме в глубь волокна по системе поперечных
трубочек (Т-системе) (разд. 17.4.7) и вызывает
сокращение мышцы.
Функции синапсов и
нервно-мышечных соединений
Основная функция межнейронных синапсов и
нервно-мышечных соединений состоит в передаче
сигнала от рецепторов к эффекторам. Кроме того,
строение и организация этих участков химической
секреции обусловливают ряд важных особенностей
проведения нервного импульса, которые можно
резюмировать следующим образом:
1. Однонаправленность передачи. Высвобождение
медиатора из пресинаптической мембраны и
локализация рецепторов на постсинаптической
мембране допускают передачу нервных сигналов по
данному пути только в одном направлении, что
обеспечивает надежность работы нервной системы.
2. Усиление. Каждый нервный импульс вызывает
освобождение в нервно-мышечном синапсе
достаточного количества ацетилхолина, чтобы вызвать
распространяюпгайся ответ в мышечном волокне.
Благодаря этому нервные импульсы, приходящие к
нервно-мышечному соединению, как бы они ни
были слабы, могут вызвать реакцию эффектора, и это
повышает чувствительность системы.
3. Адаптация, или аккомодация. При непрерывной
стимуляции количество освобождающегося в
синапсе медиатора постепенно уменьшается до тех пор,
пока запасы медиатора не будут истощены; тогда
говорят, что синапс утомлен, и дальнейшая передача
им сигналов тормозится. Адаптивное значение
утомления состоит в том, что оно предотвращает
повреждение эффектора вследствие
перевозбуждения. Адаптация имеет место также на уровне
рецепторов. (См. описание в разд. 16.4.2.)
4. Интеграция. Постсинаптический нейрон может
получать сигналы от большого числа
возбуждающих и тормозных пресинаптических нейронов
(синаптическая конвергенция); при этом
постсинаптический нейрон способен суммировать сигналы от
всех пресинаптических нейронов. Благодаря
пространственной суммации нейрон интегрирует
сигналы, поступающие из многих источников, и выдает
координированный ответ. В некоторых синапсах
имеет место облегчение, состоящее в том, что после
каждого стимула синапс становится более
чувствительным к следующему стимулу. Поэтому
следующие друг за дутом слабые стимулы могут вызывать
ответ, и это явление исцользуется для повышения
чувствительности определенных синапсов.
Облегчение нельзя рассматривать как временную сумма-
цию: здесь происходит химическое изменение
постсинаптической мембраны, а не электрическая сум-
мация постсинаптических мембранных
потенциалов.
5. Дискриминация. Временная суммация в синапсе
позволяет отфильтровывать слабые фоновые
импульсы, прежде чем они достигнут мозга.
Например, экстероцепторы кожи, глаз и ушей постоянно
получают из окружающей среды сигналы, не
имеющие особого значения для нервной системы: для нее
важны лишь изменения интенсивности стимулов,
приводящие к увеличению частоты импульсов,
которое обеспечивает их передачу через синапс и
надлежащую реакцию.
6. Торможение. Передача сигналов через синапсы
и нервно-мышечные соединения может
затормаживаться определенными блокирующими агентами,
воздействующими на постсинаптическую мембрану
(см. ниже). Возможно и пресинаптическое
торможение, если на окончании аксона чуть выше данного
синапса оканчивается другой аксон, образующий
здесь тормозный синапс. При стимуляции такого
тормозного синапса уменьшается число синапти-
ческих пузырьков, разряжающихся в первом,
возбуждающем синапсе. Такое устройство позволяет
изменять воздействие данного пресинаптического
нейрона с помощью сигналов, приходящих от
другого нейрона.
Химические воздействия на синапс
и нервно-мышечное соединение
Химические вещества выполняют в нервной системе
множество различных функций. Воздействия одних
веществ широко распространены и хорошо изучены
(как, например, возбуждающее действие
ацетилхолина и адреналина), тогда как эффекты других носят
локальный характер и пока еще недостаточно ясны.
Некоторые вещества и их функции приведены в
табл. 16.2.
Полагают, что некоторые лекарственные
препараты, используемые при таких психических
нарушениях, как тревожность и депрессия, воздействуют
на химическую передачу в синапсах. Многие
транквилизаторы и седативные средства (трициклический
антидепрессант имипрамин, резерпин, ингибиторы
моноаминоксидазы и др.) оказывают свой лечебный
эффект, взаимодействуя с медиаторами, их
рецепторами или отдельными ферментами. Так,
например, ингибиторы моноаминоксидазы подавляют
фермент, участвующий в расщеплении адреналина и
норадреналина, и скорее всего оказывают свой
лечебный эффект при депрессии, увеличивая
продолжительность действия этих медиаторов.
Галлюциногены типа диэтиламида лизерговой кислоты и
мескалина, воспроизводят действие каких-то
природных медиаторов мозга или же подавляют
действие других медиаторов.
Проводившееся недавно изучение действия
некоторых болеутоляющих веществ - опиатов героина и
морфина-показало, что в мозгу млекопитающих
присутствуют природные (эндогенные) вещества,
вызывающие сходный эффект. Все эти вещества,
взаимодействующие с опиатными рецепторами, по-
лучили общее название эндорфинов. К настоящему
времени открыто много таких соединений; из них
лучше всего изучена группа относительно
небольших пептидов, называемых энкефалинами (мет-энке-
фалин, β-эндорфин и др.). Считается, что они
подавляют болевые ощущения, влияют на эмоции и
имеют отношение к некоторым психическим
заболеваниям.
Все это открыло новые пути для изучения
функций мозга и биохимических механизмов, лежащих в
основе воздействия на боль и лечения с помощью
таких различных методов, как внушение, гипноз и
акупунктура. Предстоит выделить еще много других
веществ типа эндорфинов, установить их строение и
функции. С их помощью можно будет получить
более полное представление о работе мозга, и это
лишь вопрос времени, так как методы
выделения и анализа веществ, присутствующих в столь
малых количествах, непрерывно
совершенствуются.
Таблица 16.2. Вещества, влияющие на синапсы и нервно-
мышечные соединения млекопитающих
Вещество
Ацетилхолин
Гамма-аминомас-
ляная кислота
Дофамин, серо-
тонин, норад-
реналин
Диэтиламид ли-
зерговой
кислоты, меска-
лин
Столбнячный
токсин
Ботулинический
токсин
Никотин
Эзерин, стрихнин,
фосфоорганиче-
ские гербициды
и инсектициды
Кураре
Атропин
Мускарин
Место действия
Нервная система
позвоночных
Мозг
млекопитающих
Мозг
млекопитающих
Мозг
млекопитающих
Π ресинаптическая
мембрана
Пресинаптическая
мембрана
Постсинаптиче-
ская мембрана
Постсинаптиче-
ская мембрана
Постсинаптиче-
ская мембрана
нервно-мышечного
соединения
Постганглионар-
ные
парасимпатические
окончания
Постганглионар-
ные
парасимпатические
окончания
Функция
Возбуждение или
торможение
Торможение
Возбуждение
Вызывают
галлюцинации,
действуя подобно
медиаторам или
как их
антагонисты
Препятствует
освобождению
тормозных
медиаторов
Препятствует
освобождению аце-
тилхолина
Воспроизводит
действие ацетил-
холина
Препятствуют
расщеплению аце-
тилхолина, инак-
тивируя ацетил-
холинэстеразу
Блокирует
действие ацетилхо-
лина
Блокирует
действие ацетилхо-
лина
Воспроизводит
действие ацетил -
холина
16.2. Нервная система
позвоночных
Нервная система позвоночных отличается большим
структурным и функциональным разнообразием
нейронов и сложностью их взаимодействий.
Существует несколько способов классификации
компонентов нервной системы, и каждый из них
имеет свои достоинства и недостатки. Согласно
схеме, приведенной на рис. 16.12, периферическая
нервная система подразделяется в соответствии с
зонами иннервации. В эту классификацию не
включены вставочные нейроны (интернейроны),
видоизмененные нервные клетки, нейроны мозга и органов
чувств.
16.2.1. Периферическая нервная
система
Спинномозговые (спинальные) нервы берут начало в
спинном мозге и выходят из него между соседними
позвонками почти по всей длине позвоночника. В их
состав входят и сенсорные, и моторные нейроны,
поэтому их называют смешанными нервами. Ниже
будет дано более подробное описание спинальных
нервов и спинного мозга. Черепномозговые нервы
отходят от вентральной поверхности мозга и, за
одним исключением, иннервируют рецепторы и
эффекторы головы. У низших позвоночных имеется 10
пар черепномозговых нервов, а у
млекопитающих-12 пар, и их обозначают римскими цифрами
I-XII. Не все черепномозговые нервы являются
смешанными (табл. 16.3).
Десятый черепномозговой нерв - блуждающий -
включает очень важный моторный нерв
вегетативной нервной системы, иннервирующий сердце,
бронхи и пищеварительный тракт.
16.2.2. Рефлексы и рефлекторные
дуги
Простейшей реакцией нервной системы является
рефлекс. Он представляет собой быструю,
автоматическую, стереотипную реакцию на раздражение,
его называют непроизвольным актом, так как он не
находится под контролем сознания. Нейроны,
образующие путь нервных импульсов при рефлекторном
акте, составляют рефлекторную дугу. Простейшая
рефлекторная дуга у животных включает один
нейрон и имеет следующий вид:
Нейрон
Стимул -► Рецептор Эффектор -► Реакция
Такой уровень организации характерен для
нервной системы кишечнополостных. Рефлекторные
дуги всех групп животных с более высоким уровнем
структурной и функциональной организации
состоят по меньшей мере из двух нейронов - афферент-
Нервная система позвоночных
Центральная нервная система
Периферическая нервная система
Головной
мозг
Спинной
мозг
Висцеральная
нервная система
(вегетативная)
Соматическая
нервная
система
Симпатическая
нервная система
Парасимпатическая
нервная система
Только
спинномозговые
нервы
Спинномозговые
нервы
Черепно -
мозговые
нервы
Спинно-
мозговые
нервы
Черепно-
мозговые
нервы
Рис 16.12. Важнейшие составные части нервной системы позвоночных. (Все спинномозговые и некоторые
черепномозговые нервы содержат и сенсорные, и двигательные волокна )
ного, или сенсорного (чувствительного),
проводящего импульсы от рецептора, и эфферентного, или
двигательного (моторного), передающего импульсы
к эффектору. Между этими двумя нейронами могут
быть еще вставочные нейроны, находящиеся в
скоплении нервных клеток - ганглии, нервной цепочке
или центральной нервной системе (рис. 16.13).
Существует огромное множество рефлексов различной
структурной и функциональной сложности, но все
они могут быть разделены на следующие четыре
группы:
1. Моносинаптические рефлексы. Это рефлексы с
простейшей дугой, встречающейся у позвоночных.
Сенсорный нейрон непосредственно контактирует с
телом моторного нейрона. В такой дуге участвует
только один синапс, находящийся в центральной
нервной системе. Такие рефлексы весьма обычны у
всех позвоночных, они участвуют в регуляции
мышечного тонуса и позы (таков, например, коленный
рефлекс - разгибание ноги в коленном суставе). В
этих рефлекторных дугах нейроны не доходят до
головного мозга, и рефлекторные акты
осуществляются без его участия, так как они стереотипны и
не требуют обдумывания или сознательного
решения. Они экономны в отношении числа
участвующих центральных нейронов и обходятся без
вмешательства головного мозга, который может
«сосредоточиться» на более важных делах.
2. Полисинаптические спинномозговые рефлексы.
В таких рефлексах участвуют по меньшей мере два
Таблица 16.3. Черепномозговые нервы млекопитающих, их функции и иннервируемые органы
Черепномоз
нерв
I
II
III
IV
V
VI
VII
VIII
IX
X
XI
XII
teoeou Название
Обонятельный
Зрительный
Глазодвигательный
Блоковый
Тройничный
Отводящий
Лицевой
Слуховой
Языкоглоточный
Блуждающий
Добавочный
Подъязычный
Тип
Сенсорный
Сенсорный
Моторный
Моторный
Смешанный
Моторный
Смешанный
Сенсорный
Смешанный
Смешанный
Моторный
Моторный
Иннервируемый орган
Орган обоняния
Сетчатка
Четыре глазные мышцы
Одна глазная мышца
Мышцы челюстей, зубы, кожа
лица
Одна глазная мышца
Щеки, мышцы лица, язык
Улитка, полукружные каналы
Язык, мускулатура глотки
Гортань, глотка, сердце,
кишечник
Голова и шея
Язык
Функция
Обоняние
Зрение
Движения глаз
Движения глаз
Движения челюстей, рецепторы
прикосновения и боли
Движения глаз
Слюноотделение, мимика,
восприятие сладкого, кислого и
соленого
Слух, равновесие
Восприятие горького вкуса,
глотание
Речь, глотание, замедление
ритма сердца, стимуляция
перистальтики
Движения головы
Движения языка
спинной
мозг
(1) Стимул
Белое
вещество
Серое
вещество
Стимул
U) Рецептор
(3) Сенсор
ный
нейрон
Вставочный нейрон
(5) Эффектор
(4) Моторный
нейрон
(6)Реакция
Реакция
Рис. 16.13. Схема двух видов
простых рефлекторных дуг. А.
Моносинаптическая рефлекторная
дуга, включающая основные
элементы всякой рефлекторной дуги
(1 6). Б. Простая полисинапти-
ческая рефлекторная дуга
(P-рецептор, Э-эффектор).
синапса, находящиеся в ЦНС, так как в дугу
включен третий нейрон-вставочный, или промежуточный
(интернейрон). Синапсы имеются здесь между
сенсорным и вставочным нейронами и между
вставочным и двигательным нейронами (рис. 16.13,2^).
Этот вид рефлекторного акта служит примером
простого рефлекса, замыкающегося в спинном
мозгу. На рис. 16.14 представлен в сильно упрощенном
виде рефлекс, возникающий при уколе пальца
булавкой.
Простые рефлекторные дуги типа 1 и 2 позволяют
организму осуществлять автоматические
непроизвольные реакции, необходимые для приспособления
к изменениям внешней среды (например, зрачковый
рефлекс или сохранение равновесия при
передвижении) и к изменениям в самом организме (регуляция
частоты дыхания, кровяного давления и т.п.), а
также предотвращать повреждение тела, например
ранение или ожог.
3. Полисинаптические рефлексы с участием как
спинного, так и головного мозга. В рефлекторных
дугах этого типа сенсорный нейрон образует в
спинном мозгу синапс со вторым нейроном,
который посылает импульсы в головной мозг. Таким
образом, эти вторые сенсорные нейроны образуют
восходящие нервные пути (рис. 16.15,/I). Головной
мозг истолковывает эту сенсорную информацию и
сохраняет ее для дальнейшего использования.
Наряду с этим он в любой данный момент может
инициировать двигательную активность, и тогда
импульсы будут передаваться двигательными
нейронами по нисходящему нервному пути прямо на
спинальные мотонейроны через синапсы,
расположенные в той же области, что и выходные синапсы
вставочных нейронов (рис. 16.15).
4. Условные рефлексы. Условные рефлексы
представляют собой тип рефлекторной активности, при
которой характер ответа зависит от прошлого
опыта. Эти рефлексы координируются головным
мозгом. Основу всех условных рефлексов (таких, как
привычка к совершению туалета, слюноотделение
при виде и запахе пищи, осознание опасности)
составляет научение (разд. 16.9).
Существует множество ситуаций, когда возникает
одна из двух возможных рефлекторных реакций с
участием определенной группы мышц, которые
могут либо сокращаться, либо расслабляться, что
приводило бы к противоположным результатам. В
(3) Сенсорный
нейрон
Тело клетки
Ганглий заднего корешка
Синапс
Направление
импульса
-Белое
вещество
Вставочный
нейрон
(2)Окончания \
чувствительного
нерва в коже
(1) Булавка
(4) Моторный
нейрон
Серое вещество
(тела клеток
и передаточные
нейроны)
Передний
корешок
(6) Палец отдергивается
от булавки
Синапс(передача
от нейрона к нейрону)
Спинной мозг
в поперечном
разрезе
(5) Двуглавая мышца сокращается
(5) Трехглавая мышца расслабляется
Рис. 16.14. Упрощенная схема рефлекторного акта и рефлекторной дуги (цифрами в скобках указаны
основные компоненты рефлекторной дуги, обозначенные теми же цифрами на рис 16.13.)
Кора мозга
Базальный
ганглий
(а) Восходящий
сенсорный нейрон
Задний рог
Боковой рог
Таламус
(б) Нисходящий
моторный нейрон
Белое вещество
Серое вещество
Задний корешок
Передний рог
Центральный
канал
Ганглий
заднего
корешка
Передний
корешок
Спинномозговой
нерв
Рис. 16.15. Упрощенная схема поперечных разрезов
головного и спинного мозга, на которой показаны: (а)
путь сенсорных импульсов от рецептора через
спинной мозг в кору головного мозга и (б) путь
двигательных импульсов, возникающих в коре и
направляющихся через спинной мозг к эффектору
этой ситуации обычный спинномозговой рефлекс
осуществлялся бы рефлекторной дугой,
изображенной на рис. 16.14, однако «условия», в которых
действует стимул, могут изменять ответ. В таких
случаях действует более сложная рефлекторная
дуга, включающая и возбуждающие, и тормозные
нейроны. Например, если мы схватим рукой пустую
металлическую сковороду, которая окажется
слишком горячей и будет обжигать пальцы, мы ее,
вероятно, тотчас же выпустим из рук, но столь же
горячую пищу на обжигающем пальцы дорогом
блюде осторожно и быстро поставим на место.
Разница в реакции указывает на то, что мы имеем
дело с условным рефлексом, в котором участвуют
память и сознательное решение, принятое мозгом. В
этой ситуации ответ осуществляется по более
сложному рефлекторному пути, показанному на рис. 16.16.
В обоих случаях стимул вызывает импульсы,
идущие к сенсорному отделу головного мозга по
восходящему нервному пути. Когда эти импульсы
поступают в мозг, он анализирует их, учитывая
информацию, поступающую от других органов чувств,
например от глаз, и устанавливает причину стимула.
Входящая в мозг информация сопоставляется с той,
которая в нем уже хранится,- с информацией о том,
что скорее всего произойдет, если спинномозговой
рефлекс осуществится автоматически. В случае с
металлической сковородой мозг вычислит, что
если она будет брошена, то это не причинит
никакого вреда телу или сковороде, и пошлет импульсы по
возбуждающему пути. Этот путь идет вниз по
спинному мозгу до уровня, где в спинной мозг поступил
стимул, и образует связи с телами моторных
нейронов, осуществляющих данный рефлекс. Скорость
проведения импульсов по этому пути такова, что
импульсы от возбуждающего мотонейрона
головного мозга достигают специального мотонейрона
одновременно с импульсами от вставочного
нейрона простой рефлекторной дуги. Эффекты тех и
других импульсов суммируются, и к мышечному
эффектору по аксону спинального мотонейрона
поступают возбуждающие импульсы, заставляющие
бросить сковороду.
А вот в случае с горячим блюдом головной мозг
быстро вычислит, что если бросить его, то можно
обварить ноги, а к тому же будет испорчена пища и
разбито дорогое блюдо. Если же блюдо удержать и
осторожно поставить на место, это не вызовет
сильного ожога пальцев. После принятия мозгом
такого решения в нем возникнут импульсы, которые
тоже будут переданы к спинальным мотонейронам,
но на этот раз уже по тормозному пути. Они
прибудут одновременно с возбуждающими
импульсами от вставочного нейрона и погасят их действие.
В результате по мотонейронам к соответствующим
мышцам не будет поступать никаких импульсов и
блюдо будет удержано в руках. Одновременно мозг
может дать мышцам иную программу действий, и
блюдо будет быстро и осторожно поставлено на
место.
Сделанное выше описание рефлекторных дуг,
естественно, сильно упрощено. Ведь процесс
координации, интеграции и регуляции функций в организме
намного более сложен. Так, например,
определенные нейроны связывают между собой разные
уровни спинного мозга, контролирующие, скажем, руки
и ноги, так что активность одного уровня
координируется с активностью другого, а еще какая-то
Тормозный моторный нейрон
Возбуждающий моторный нейрон
Восходящий сенсорный нейрон
Вставочный
нейрон
(возбуждающий)
Эфферентный
моторный
нейрон
Афферентный
сенсорный нейрон
Рис 16.16. Взаимоотношения между нейронами
спинного мозга и модифицирующее действие
возбуждающего и тормозного нейронов головного мозга
(упрощенная схема)
Висцеральные
сенсорные нейроны
ЦНС
Эффектор
IAX
Парасимпатический ганглий
(расположен на периферии)
Симпатический ганглий
(расположен в ЦНС)
Преганглионарные
нейроны
Эффектор
Ад I
Преганглионарный нейрон
Постганглионарный нейрон
Ацетилхолин
Норадреналин
Рис. 16.17. Основные особенности парасимпатической (А) и симпатической (Б) нервной системы
(висцеральные сенсорные нейроны не входят в состав вегетативной нервной системы).
группа нейронов осуществляет общий контроль со
стороны мозга.
В то время как совместная деятельность мозга и
эндокринной системы играет важную роль в
согласовании многих видов нервной деятельности,
описываемых далее в настоящей главе, регуляцию
вегетативных функций осуществляет другая
рефлекторная система, в основе которой лежит исключительно
нервная деятельность. Эту систему называют
вегетативной или автономной нервной системой.
16.2.3. Вегетативная нервная система
Автономная (autos-сам; nomos-управляющий),
или вегетативная, нервная система является частью
периферической нервной системы и регулирует
непроизвольную активность внутренних органов, в
том числе частоту сердечных сокращений,
перистальтику и потоотделение. Она состоит из
двигательных нейронов, иннервирующих гладкую
мускулатуру. Активность вегетативной системы
регулируется в основном с помощью местных
висцеральных рефлексов, замыкающихся в спинном или
головном мозгу, без сознательного контроля со
стороны высших отделов мозга. Однако некоторые
функции, например активность мышечных
сфинктеров анального отверстия и мочевого пузыря,
находятся под контролем сознания, и этим функциям
приходится обучаться. Существует мнение, что
усилие воли позволяет управлять и многими другими
видами вегетативной активности; управление
вегетативными функциями составляет физиологическую
основу многих форм медитации и расслабления, и
некоторые люди уже достигли значительных
успехов в регуляции сердечного ритма и в снижении
кровяного давления с помощью сознания или «силы
воли». Однако общий контроль активности
вегетативной нервной системы осуществляется центрами,
расположенными в продолговатом мозгу и в
гипоталамусе (см. разд. 16.2.4). Эти центры получают и
интегрируют сенсорную информацию и,
координируя ее с информацией от других отделов нервной
системы, формируют надлежащую ответную
реакцию.
Вегетативная нервная система состоит из
нейронов двух типов - преганглионарных и постганглио-
нарных. Тела преганглионарных нейронов лежат в
головном или спинном мозгу, а их немиелинизиро-
ванные аксоны покидают центральную нервную
систему в составе вентральных (у человека - передних)
корешков сегментарного нерва и образуют синапсы
с дендритами постганглионарных нейронов. Тела
постганглионарных нейронов находятся в ганглии, а
немиелинизированные аксоны направляются к
органу-эффектору.
Вегетативная нервная система подразделяется на
две системы - симпатическую и парасимпатическую;
они различаются прежде всего своей структурной
организацией, и эти различия показаны на рис. 16.17.
В симпатической нервной системе синапсы и тела
постганглионарных нейронов расположены в
ганглиях, находящихся рядом со спинным мозгом. Как
показано на рис. 16.18, каждый симпатический
ганглий связан со спинным мозгом белой соединительной
ветвью, а со спинальным нервом-серой
соединительной ветвью. Соседние сегментарные
симпатические ганглии соединены между собой
симпатическим нервным стволом и образуют две цепи,
идущие по обе стороны спинного мозга вдоль всей
его длины. В парасимпатической нервной системе
ганглии расположены вблизи эффекторного органа
или внутри него.
Остальные различия между двумя системами
касаются природы химических медиаторов,
высвобождающихся в постганглионарных синапсах с
эффекторами, общего воздействия всей системы на
организм и тех условий, в которых она
активируется. Эти различия суммированы в табл. 16.4.
Таблица 16.4. Различия между симпатической и парасимпатической системами
Особенность или свойство
Происхождение нервных
волокон
Расположение ганглиев
Длина волокон
Число волокон
Распределение волокон
Зона влияния
Медиатор
Общие эффекты
Суммарный эффект
В каких условиях
активируется
Симпатическая система
Выходят из черепного, грудного и
поясничного отделов ЦНС
Рядом со спинным мозгом
Короткие преганглионарные и длинные
постганглионарные волокна
Многочисленные постганглионарные
волокна
Преганглионарные волокна иннервируют
обширные области
Действие генерализованное
В эффекторе освобождается норадреналин
Повышает интенсивность обмена
Усиливает ритмические формы активности
Снижает пороги чувствительности
Возбуждающий
• Доминирует во время опасности, стресса и
активности; контролирует реакции на
стресс
Парасимпатическая система
Выходят из черепного и крестцового отделов
ЦНС
Поблизости к эффектору
Длинные преганглионарные и короткие
постганглионарные волокна
Немногочисленные постганглионарные
волокна
Преганглионарные волокна иннервируют
ограниченные участки
Действие местное
В эффекторе освобождается ацетил холин
Снижает интенсивность обмена или не влияет
не нее
Снижает ритмические формы активности
Восстанавливает пороги чувствительности до
нормального уровня
Тормозящий
Доминирует в покое; контролирует обычные,
«повседневные» физиологические функции
Как правило, симпатическая и парасимпатическая
системы оказывают противоположное воздействие
на иннервируемые органы, что позволяет организму
быстро и точно регулировать деятельность
внутренних органов, поддерживая таким образом
устойчивое состояние. Например, учащение сердечного
ритма, вызванное высвобождением норадреналина из
симпатических нейронов, компенсируется
высвобождением ацетилхолина из парасимпатических
нейронов. Этот эффект препятствует слишком
частым сокращениям сердечной мышцы, и в конечном
счете, когда секреция обоих медиаторов будет
уравновешена, восстанавливается нормальный
сердечный ритм. Антагонистические эффекты двух
отделов вегетативной нервной системы
резюмированы в табл. 16.5.
Серое
вещество
Белое
вещество
Постганглионарный
симпатический
нейрон
Преганглио-
нарный
симпатический
нейрон
Передний корешок
Серая
соединительная ветвь
Симпатический
ганглий
Белая
соединительная
ветвь
Рис. 16.18. Упрощенная схема расположения
симпатического ганглия и его взаимосвязей со спинным
мозгом и спинномозговым нервом.
16.2.4. Центральная нервная система
Центральная нервная система (ЦНС) развивается из
впячивающейся внутрь складки эктодермы,
лежащей непосредственно над эмбриональной хордой.
При смыкании краев этой складки образуется полая
спинная нервная трубка, проходящая по всей длине
тела. В процессе дальнейшего развития нервная
трубка дифференцируется, формируя расширенный
передний конец-головной мозг-и длинный
цилиндрический спинной мозг.
На всем своем протяжении центральная нервная
система покрыта тремя мозговыми оболочками и
заключена в защитную костную капсулу,
состоящую из черепа и позвоночника. Снаружи мозг
покрыт прочной твердой мозговой оболочкой (dura
mater), которая сращена с надкостницей черепа и
позвоночника. Непосредственно к ткани мозга
прилегает мягкая мозговая оболочка (pia mater). Между
твердой и мягкой оболочками находится паутинная
оболочка (arachnoidea), образующая сеть из
перекладин соединительной ткани, благодаря которым
между мягкой и паутинной оболочками образуется
подпаутинное пространство, заполненное
спинномозговой (цереброспинальной) жидкостью. Большая
часть спинномозговой жидкости содержится в
центральном канале спинного мозга, а в головном мозге
она заполняет четыре расширенных
участка-мозговых желудочка. Спинномозговая жидкость
омывает мозг снаружи и изнутри, и с ней соприкасаются
кровеносные сосуды, обеспечивающие снабжение
нервных тканей питательными веществами и
кислородом и удаление продуктов обмена (рис. 16.19). В
крыше мозга находятся переднее и заднее
сосудистые сплетения, клетки которых выделяют
спинномозговую жидкость и осуществляют связь между
жидкостью, находящейся внутри мозга и снаружи.
Твердая оболочка
Паутинная оболочка
Подпаутинное
пространство
Кровеносный сосуд
Арахноидальная
трабекула
Мягкая оболочка
Периваскулярное
пространство
Нервная ткань
Рис. 16.19. Схема строения мозговых оболочек и их
взаимоотношений с кровеносными сосудами.
Обратите внимание на то, что каналы, по которым
проходят сосуды в нервную ткань, выстланы мягкой
мозговой оболочкой
Объем спинномозговой жидкости составляет около
100 мл; помимо питательной и выделительной
функций она выполняет также опорную функцию и
защищает нервные клетки от механических ударов о
твердую костную поверхность. Ресничные клетки,
выстилающие полость желудочков и центрального
канала, поддерживают непрерывную циркуляцию
спинномозговой жидкости.
В функции центральной нервной системы входят
координация, интеграция и регуляция почти всех
видов нервной активности; при этом ЦНС работает
в тесном контакте с периферической нервной
системой. Высшие формы нервной деятельности,
свойственные высокоорганизованным животным,-
память и интеллектуальные функции-связаны,
возможно, с увеличенными размерами определенных
участков головного мозга.
Спинной мозг
Спинной мозг представляет собой уплощенный в
дорсовентральном направлении цилиндр из нервной
ткани, который идет от основания головного мозга
до крестцового отдела и на всем протяжении
защищен позвоночником. Он состоит из ткани двух
типов: во внутренней массе серого вещества,
имеющей в поперечном разрезе Η-образную форму,
находятся тела нервных клеток, дендриты и синапсы,
а лежащее снаружи белое вещество образуют пучки
нервных волокон, цвет которых связан с наличием
миелиновой оболочки. От спинного мозга отходит
31 пара сегментарных спинномозговых нервов,
каждый из которых сразу по выходе из спинного мозга
разделяется на вентральные и дорсальные (у
человека - передние и задние) корешки. В составе
дорсальных корешков в спинной мозг вступают аксоны
сенсорных нейронов, тела которых находятся в
ганглиях дорсальных (задних) корешков, расположенных
рядом со спинным мозгом и образующих вздутия. В
спинном мозге эти аксоны направляются в
дорсальные (задние) рога серого вещества, где они
образуют синапсы со вставочными нейронами
(интернейронами). Последние в свою очередь образуют
синапсы с мотонейронами, лежащими в
вентральных (передних) рогах спинного мозга, аксоны
которых покидают спинной мозг в составе вентральных
корешков (см. рис. 16.15). Так как вставочных
нейронов намного больше, чем двигательных, в сером
веществе спинного мозга должна осуществляться
какая-то интеграция. Некоторые сенсорные нейроны
образуют синапсы непосредственно с
мотонейронами, лежащими в вентральных рогах, как в случае
уже знакомого нам коленного рефлекса (рис. 16.13,^4).
Таблица 16.5. Эффекты симпатической и парасимпатической систем в организме
Орган
Голова
Сердце
Легкие
Кишечник
Кровеносная
система
Кожа
Почки
Мочевой
пузырь
Половой член
Железы
Симпатическая система
Расширяет зрачки
Угнетает слюноотделение
Повышает амплитуду и частоту сокращений
Расширяет бронхи и бронхиолы
Усиливает вентиляцию легких
Угнетает перистальтику
Угнетает секрецию пищеварительных соков
Усиливает сокращение анального сфинктера
Сужает артериолы кишечника и гладких мышц;
расширяет артериолы мозга и скелетных мышц
Повышает кровяное давление
Увеличивает объем крови за счет сокращения
селезенки
Вызывает сокращение мышц, приподнимающих
волосы
Сужает артериолы в коже конечностей
Усиливает потоотделение
Уменьшает диурез
Усиливает сокращение сфинктера мочевого
пузыря
Вызывает эякуляцию
Вызывает выброс адреналина из мозгового слоя
надпочечников
Парасимпатическая система
Сужает зрачки
Стимулирует слезотечение
Стимулирует слюноотделение
Уменьшает амплитуду и частоту сокращений
Сужает бронхи и бронхиолы
Уменьшает вентиляцию
Усиливает перистальтику
Стимулирует секрецию пищеварительных соков
Угнетает сокращение анального сфинктера
Поддерживает постоянный тонус артериол
кишечника, гладких мышц, мозга и скелетных мышц
Снижает кровяное давление
Расширяет артериолы в коже лица
Расслабляет сфинктер мочевого пузыря
Стимулирует эрекцию
Таблица 16.6. Отделы головного мозга у позвоночных
Отделы мозга
у эмбриона
Отделы мозга у взрослого
животного
Структурно-функциональные участки
у рыб, амфибий, рептилий и птиц
у млекопитающих
Передний мозг
Средний мозг
Задний мозг
Конечный мозг
Промежуточный мозг
Средний мозг
Задний мозг
Продолговатый мозг
Обонятельные доли (у рыб) |
Неопаллиум (у рептилий) J
Полосатые тела (у птиц)
Таламус
Гипоталамус
Зрительные доли (у амфибий)
Крыша среднего мозга
Мозжечок
Мост
Продолговатый мозг
Большие полушария
Базальные ганглии
Таламус
Гипоталамус
Четверохолмие
Мозжечок
Мост
Продолговатый мозг
В грудном, верхнепоясничном и крестцовом отделах
спинного мозга серое вещество образует боковые
рога (рис. 16.15), содержащие тела преганглионар-
ных нейронов вегетативной нервной системы. Белое
вещество состоит из пучков нервных волокон,
образующих проводящие пути (тракты), которые
идут от серого вещества спинного мозга к
головному мозгу и осуществляют связь между спинальными
нервами и мозгом. Восходящие пути несут
головному мозгу сенсорную информацию, а по нисходящим
путям от головного мозга спинному передаются
двигательные сигналы.
Функция спинного мозга заключается в том, что
он служит координирующим центром простых спи-
нальных рефлексов (вроде коленного рефлекса) и
автономных рефлексов (например, сокращения
мочевого пузыря), а также осуществляет связь между
спинальными нервами и головным мозгом.
Головной мозг
Головной мозг-это расширенный передний конец
нервной трубки позвоночных, роль которого
состоит в координации и регуляции деятельности всей
нервной системы. В целом головной мозг состоит из
скоплений тел нервных клеток, нервных трактов и
кровеносных сосудов. Нервные тракты образуют
белое вещество мозга и состоят из пучков нервных
волокон, проводящих импульсы к различным
участкам серого вещества - «ядрам» или «центрам» - или
от них. Проводящие пути связывают между собой
различные ядра, а также головной мозг со спинным.
В процессе филогенетического развития
позвоночных число проводящих путей и сложность их
взаимосвязей возрастали. «Ядра» весьма различны по
своим размерам-от небольших групп, состоящих
из нескольких сотен нейронов, до таких обширных
участков, как кора большого мозга и кора
мозжечка, включающих у человека миллиарды клеток.
строение головного мозга позвоночных. В процессе
эмбрионального развития головной мозг у
позвоночных сначала делится на три пузыря - передний,
средний и задний мозг. Первоначально эти отделы
были связаны соответственно с функциями
обоняния, зрения и равновесия. Последующее развитие
мозга идет в различных классах позвоночных и даже
внутри одного и того же класса по-разному, но во
всех случаях исходная трехчастная структура
утрачивается, так как передний, средний и задний мозг
подвергаются дальнейшему подразделению. Мозг
взрослого позвоночного состоит из пяти отделов,
значение которых у представителей каждого класса
зависит от образа жизни и уровня структурной и
функциональной сложности, достигнутой этим
классом. Некоторые участки мозга увеличиваются в
размерах, что указывает на их особую важность,
тогда как другие уменьшаются. Общая схема
строения мозга у позвоночных представлена на рис. 16.20,
а в табл. 16.6 суммировано развитие отделов мозга
у млекопитающих и остальных позвоночных.
Рыбы в поисках пищи сильно зависят от
обоняния, и у них большие обонятельные доли,
перерабатывающие сенсорные сигналы. Крупный
мозжечок осуществляет у них координацию движений, а
весьма развитые зрительные доли мозга хотя и
участвуют в зрительных реакциях, но в основном
служат главным координирующим центром мозга.
Зрительные доли составляют главную часть мозга и
у амфибий, у которых они выполняют ту же роль,
что и у рыб. У рептилий размеры среднего мозга
уменьшаются, а переднего-увеличиваются. Эта
тенденция продолжается и у птиц, у которых
увеличиваются размеры таламуса и полосатых тел,
координирующих сложные инстинктивные формы
поведения. И наконец, для мозга млекопитающих
характерно наличие больших полушарий,
прикрывающих хорошо развитый таламус. Эти две
структуры, содержащие ядра и проводящие пути,
отражают потребность млекопитающих в хранении
сенсорной информации и в интегрировании всех
произвольных действий.
Относительные размеры продолговатого мозга в
ходе эволюции позвоночных менялись мало, и это
говорит о существенной роли, которую играет про-
Передний мозг
Средний
мозг
Задний мозг
Конечный мозг
Большой мозг
Промежуточный мозг
Переднее cocy-j
дистое сплетение
Средний
мозг
Задний
мозг
Продолговатый
мозг
Таламус
Эпифиз!
Зрительная
доля ι
(тектум) ]
Мозжечок
Заднее сосудистое
сплетение
Боковые
желудочки
Спинной
мозг
Обонятельная
доля
Гипоталамус I
Мост
Продолговатый
мозг
Рис. 16.20. Схема строения мозга позвоночных (продольный разрез). Цифры указывают положение III и IV
мозговых желудочков.
долговатый мозг в рефлекторной регуляции всех
жизненно важных функций, таких, например, как
работа сердца.
Методы исследования функций
головного мозга
Простое изучение анатомии и гистологии
различных участков головного мозга мало что дает для
понимания их функций. Доставляя ценные сведения
о структурных взаимоотношениях, такие
исследования ничего не говорят о физиологической
активности мозга. Активность мозга изучают с
помощью электрофизиологических методов, в том
числе электроэнцефалографии. Для получения
электроэнцефалограммы на коже головы укрепляют
электроды (хороший электрический контакт
обеспечивается с помощью электродного клея), которые
регистрируют электрические импульсы,
отражающие активность многочисленных клеток мозга. С
электродов импульсы передаются на усилитель и
записываются с помощью писчика.
Обнаружены три основных вида электрических волн-
α-, β- и δ-волны. α-Волны регистрируются, когда
человек расслаблен и глаза его закрыты. Обычно
α-волны маскируют более частый ритм β-волн,
которые всегда присутствуют, но выявляются лучше
всего, когда человек находится под наркозом.
Отсутствие β-волн или полное отсутствие
электрической активности мозга является признаком
«мозговой смерти» и наряду с остановкой сердца и
дыхания считается клиническим показателем смерти.
δ-Волны имеют наименьшую частоту и наибольшую
амплитуду и регистрируются во время сна. Хотя
электроэнцефалограммы мало что дают для
непосредственного понимания функций мозга, они
имеют большое диагностическое значение, так как
позволяют определять локализацию очаговых
нарушений, например различных форм эпилепсии.
Эпилептический припадок - результат
неупорядоченной повышенной активности мозга. При одной
форме эпилепсии (grand mal) возникают сильные
судорожные припадки, длящиеся от нескольких
секунд до нескольких минут, а при другой форме (petit
mal) судороги бывают очень слабыми и
кратковременными.
Деятельность мозга изучают также при
операциях, проводимых под местной анестезией. В
исследованиях одного типа больные описывают
ощущения, которые они испытывают при раздражении
различных участков мозга электрическим током. В
других исследованиях выясняются двигательные
функции тех или иных участков коры мозга
(рис. 16.23). Аналогичным образом при стимуляции
сенсорных клеток и органов чувств запись
электрической активности с помощью микроэлектродов,
введенных в тела или аксоны нейронов
определенных участков мозга, позволяет картировать участки,
выполняющие сенсорные функции.
Мозолистое тело
Боковые
желудочки
Таламус
Гипоталамус
Гипофиз
Красное ядро
Мост
Полушарие
большого мозга
Кора мозга
Подпаутинное
пространство
Переднее
сосудистое сплетение
Эпифиз
Четверохолмие
Мозжечок
Заднее сосу -
дистое сплетение
Спинной мозг
Центральный
канал
Продолговатый мозг
Рис. 16.21. Упрощенная схема строения мозга
человека (вертикальный разрез). Цифрами указаны
мозговые желудочки
Строение и функции головного мозга
В настоящем разделе дается лишь краткое описание
мозга, суммирующее современные представления о
его строении и функциях. В нем не приводятся
детали и соответствующие экспериментальные
данные, за исключением тех случаев, когда это
потребуется ради ясности. Мы последовательно
рассмотрим строение и функциональную активность
каждого из отделов, указанных в табл. 16.6. Такое
описание неизбежно будет сильно упрощенным, так
как во многих видах деятельности участвует
несколько отделов мозга. Во всяком случае следует
иметь в виду, что в функциональном отношении
мозг можно подразделить на три главных
взаимосвязанных отдела: большой мозг (большие
полушария), мозжечок и ствол мозга, который состоит
из продолговатого мозга, моста, среднего мозга и
таламуса. Ствол является продолжением спинного
мозга и содержит сложные нервные структуры,
контролирующие деятельность сердечно-сосудистой
системы и желудочно-кишечного тракта, дыхание,
движения глаз, равновесие и большую часть
стереотипно выполняемых функций организма.
Конечный мозг
Конечный мозг-это самый передний участок
головного мозга; он состоит из большого мозга и
базальных ганглиев. Большой мозг составляет
крышу и стенки конечного мозга и достигает крупных
размеров, образуя левое и правое полушария,
покрывающие сверху большую часть головного мозга.
Базальные ганглии расположены в основании
конечного мозга.
большой мозг. Полушария большого мозга состоят
из коры и лежащей под ней центральной массы
белого вещества. Кора представляет собой тонкий
(3 мм) слой серого вещества, образованного плотно
расположенными нервными клетками, число
которых может составлять более 109. Белое вещество
состоит из проводящих путей. Левое и правое
полушария соединены широким нервным трактом,
носящим название мозолистого тела. Поверхность
коры сильно увеличена за счет многочисленных
складок, называемых извилинами. Каждое
полушарие для удобства делят на четыре доли, показанные
на рис. 16.23. С помощью электрофизиологических
методов установлено, что в коре можно различить
области трех типов в соответствии с функциями,
которые выполняют находящиеся в них клетки: 1)
сенсорные зоны, получающие импульсы от
рецепторов (входные сигналы); 2) ассоциативные зоны,
которые интерпретируют и хранят получаемую
информацию и вырабатывают ответ с учетом
сходного прошлого опыта; 3) двигательные зоны,
посылающие импульсы к эффекторам (выходные
сигналы).
Взаимосвязи между этими зонами позволяют
коре большого мозга контролировать и координи-
Кора мозга
Мозолистое тело
Боковой желудочек
Подпаутинное
пространство
Базальный ганглий
Третий
желудочек
Гипоталамус
Таламус
Рис. 16.22. Упрощенная схема строения мозга
человека (поперечный разрез по линии X-X на рис. 16.21)
Двигательная
соматическая зона
Премоторная зона
Сенсорная
соматическая
зона
Ступня
Нога
Туловище
Рука
Лицо
/Грудовыё\
навыки
Ассоциативная
зона речи
Зрительная
сенсорная зона
Моторный
центр речи
Память
Височная доля
Слуховая
ассоциативная зона
Зрительная
ассоциативная
зона
Слуховая
сенсорная зона
Рис. 16.23. Схема расположения четырех главных
долей коры мозга-лобной, теменной, затылочной
и височной Показана локализация сенсорных,
ассоциативных и двигательных зон и центров, а также
отмечены некоторые зоны, связанные со
специфическими функциями и участками тела.
ровать все произвольные и некоторые
непроизвольные формы деятельности, включая такие высшие
функции, как память, научение, сознание и свойства
личности. Полное разрушение коры не привело бы к
смерти, но такой больной не проявлял бы никакой
спонтанной активности. Он мог бы реагировать на
определенные раздражители, но был бы неспособен
к обучению или логическому мышлению; у него
исчезли бы все видимые признаки интеллекта и
сознания - остались бы только те проявления,
которые контролируются в основном продолговатым
мозгом, например желание есть и спать.
Сенсорные зоны-это входные участки коры,
которые через восходящие нервные пути получают
сенсорную информацию от большинства
рецепторов тела. Они занимают отдельные участки коры,
связанные с определенными видами ощущений, как
показано на рис. 16.23. Размеры этих зон
коррелируют с числом рецепторов в соответствующей
сенсорной системе.
Ассоциативные зоны называются так по ряду
причин. Во-первых, они связывают вновь поступающую
сенсорную информацию с полученной ранее и
хранящейся в блоках памяти, благодаря чему новые
стимулы «узнаются». Во-вторых, информация от
Мышление!
одних рецепторов сопоставляется здесь с сенсорной
информацией от других рецепторов. В-третьих,
здесь же сенсорные сигналы интерпретируются,
«осмысливаются» и при надобности используются
для «вычисления» наиболее подходящей ответной
реакции, которая выбирается в ассоциативной зоне
и передается в связанную с ней двигательную зону.
Таким образом, ассоциативные зоны участвуют в
процессах запоминания, научения и мышления, и
результаты их деятельности составляют то, что не
очень точно называют «интеллектом».
Отдельные крупные ассоциативные области
расположены в коре рядом с соответствующими
сенсорными зонами. Например, зрительная
ассоциативная зона расположена в затылочной доле
непосредственно впереди сенсорной зрительной зоны и
осуществляет описанные выше ассоциативные функции,
связанные со зрительными ощущениями.
Некоторые ассоциативные зоны выполняют лишь
ограниченную специализированную функцию и связаны
с другими ассоциативными центрами, способными
подвергать информацию дальнейшей обработке.
Например, слуховая ассоциативная зона анализирует
звуки, разделяя их на обширные категории, а затем
передает сигналы в более специализированные
зоны, такие как речевая ассоциативная зона, где
воспринимается смысл услышанных слов. Собственная
речь формируется в двигательном центре речи,
который относится уже к третьему типу
функциональных зон, имеющихся в коре.
Двигательные зоны-это выходные области коры.
В них возникают двигательные импульсы, идущие к
произвольным мышцам по нисходящим путям,
которые начинаются в белом веществе больших
полушарий.
Многие двигательные импульсы идут прямо в
спинной мозг через два больших пирамидных (кор-
тикоспинальных) тракта, проходящих в стволе
мозга. Остальные двигательные импульсы передаются
по экстрапирамидным путям; здесь идут
двигательные импульсы от таких частей головного мозга, как
базальные ганглии и мозжечок. В продолговатом
мозгу все пути перекрещиваются, так что импульсы,
идущие от коры левого полушария, иннервируют
правую половину тела и наоборот.
Тела нейронов, участвующих в образовании
пирамидных трактов, лежат в моторных зонах коры, а
их аксоны образуют синапсы непосредственно с
мотонейронами спинного мозга в том его сегменте,
где эти нейроны выходят на периферию. В головном
мозгу нет никаких промежуточных синапсов,
поэтому импульсы и последующие ответы на них по
пути не задерживаются и не видоизменяются.
Функциональные участки двигательных зон коры
картированы, и примеры таких участков приведены
на рис. 16.23. Размеры каждого участка зависят от
сложности контролируемых им движений.
Главным экстрапирамидным трактом является
ретикулоспиналъный тракт, переключающий
импульсы от ретикулярной формации, которая лежит в
стволе мозга между таламусом и продолговатым
мозгом. Из различных отделов головного мозга,
контролирующих двигательную активность,
импульсы поступают в определенные участки
ретикулярной формации, где они модифицируются под
воздействием импульсов, идущих из коры, и
становятся либо возбуждающими, либо тормозными.
Например, импульсы от мозжечка и премоторной
зоны коры, управляющей координированными
движениями, поступают в ту область ретикулярной
формации, которая находится в продолговатом
мозгу и посылает импульсы, стимулирующие
тормозные мотонейроны. Последние подавляют
активацию определенных мышц, что дает возможность
осуществлять сложные координированные
движения тела при такой деятельности, как, например,
ужение рыбы, игра в теннис или крокет, игра на
скрипке и т.п. Другие комбинации двигательных
импульсов, напротив, стимулируют возбуждающие
нейроны, и общее воздействие ретикулярной
формации на двигательную активность оказывается
возбуждающим.
Большинство волокон сенсорных нейронов на
своем пути через таламус к коре отдает коллатерали
(боковые ветви) в ретикулярную формацию,
участвуя в образовании ретикулярной активирующей
системы, которая тонизирует кору и участвует в
пробуждении организма от сна. Недостаточная
активность этой системы или ее разрушение приводит
соответственно к глубокому сну или коме. Как
полагают, многие вещества, вызывающие общий
наркоз, оказывают свое действие, временно
блокируя синаптическую передачу в этой системе.
Предполагается также, что ретикулярная
активирующая система ответственна за возникновение и
поддержание побуждений к действию и к
концентрации внимания, т.е. за взаимоотношения между
утомлением и активностью.
И наконец, функции некоторых участков коры, в
частности обширных передних областей-префрон-
тальных зон,- остаются еще неясными. Эти области,
а также ряд других участков мозга, называют
немыми, так как при раздражении их электрическим
током не возникает никаких ощущений или реакций.
Предполагают, что эти зоны ответственны за наши
индивидуальные особенности, или личность.
Удаление этих зон или перерезку проводящих путей,
идущих от них к остальному мозгу (префронтальную
лоботомию), применяли для снятия у больных
острого возбуждения, но от этого пришлось
отказаться из-за таких побочных эффектов, как
снижение уровня сознания и интеллекта, способности к
логическому мышлению и творчеству. Эти
побочные эффекты косвенно указывают на функции,
выполняемые префронтальными зонами.
базальные ганглии. Эти участки переднего мозга
содержат тела клеток, на которых оканчиваются
идущие из коры аксоны двигательных нейронов, и
здесь же происходит переключение импульсов на
ретикулярную формацию. Функции базальных
ганглиев многообразны; например, один из базальных
ганглиев посылает тормозные импульсы для ре-
ципрокной регуляции мышечного тонуса при
медленных движениях. Повреждение этого ганглия
вызывает дрожание рук при паркинсонизме - одной из
форм мышечного паралича.
Промежуточный мозг
Промежуточный мозг-это задний отдел переднего
мозга; его дорсальный и боковые участки образуют
таламус, а вентральная часть-гипоталамус. Здесь
же находится эпифиз, функции которого будут
описаны в разд. 16.6.3.
таламус. В таламусе оканчиваются аксоны
большинства сенсорных нейронов, несущих импульсы в
кору мозга. Здесь анализируются происхождение и
характер этих импульсов, и они передаются в
соответствующие сенсорные области коры по
волокнам, берущим начало в таламусе. Таким образом,
таламус играет роль перерабатывающего,
интегрирующего и переключающего центра для всей
сенсорной информации, и в этом отношении он сходен
с телефонным коммутатором. Кроме того, в
таламусе модифицируется информация, поступающая из
определенных зон коры, и полагают, что он
участвует в ощущении боли и удовольствия. В таламусе
начинается та область ретикулярной формации,
которая, как уже говорилось, имеет отношение к
регуляции двигательной активности. Дорсальный
участок, лежащий непосредственно перед таламу-
сом,-переднее сосудистое сплетение - ответствен за
транспорт веществ между спинномозговой
жидкостью, находящейся в третьем желудочке, и
жидкостью, заполняющей подпаутинное пространство
(см. рис. 16.21).
гипоталамус Это главный координирующий и
регулирующий центр вегетативной нервной системы.
К нему подходят волокна сенсорных нейронов от
всех висцеральных, вкусовых и обонятельных
рецепторов. Отсюда через продолговатый и спинной
мозг информация передается на эффекторы и
используется для регуляции сердечного ритма,
кровяного давления, дыхания и перистальтики. В
других участках гипоталамуса лежат специальные
центры, от которых зависят голод, жажда и сон, а также
поведенческие реакции, связанные с агрессивностью
и размножением.
Обладая богатой сетью кровеносных сосудов,
гипоталамус «следит» за концентрацией метаболитов
и гормонов в крови, а также температурой крови.
На основании получаемой информации он вместе с
расположенным под ним гипофизом регулирует
секрецию большинства гормонов и поддерживает
постоянство состава крови и тканей. Более
подробно эндокринные функции гипоталамуса будут
рассмотрены в разд. 16.6.2.
Средний мозг
Средний мозг связывает два передних отдела мозга
с двумя задними, поэтому все нервные пути
головного мозга проходят через эту область,
составляющую часть ствола. Крышу среднего мозга
образует четверохолмие, где находятся центры
зрительных и слуховых рефлексов. Верхняя пара бугорков
четверохолмия получает сенсорные импульсы от
глаз и мышц головы и контролирует зрительные
рефлексы, например движения головы и глаз,
позволяющие фиксировать взором тот или иной объект.
Нижняя пара бугорков получает импульсы от ушей
и мышц головы и контролирует слуховые рефлексы,
такие как движения головы при определении
направления и источника звука.
В вентральной части среднего мозга расположены
многочисленные центры или ядра, управляющие
разнообразными бессознательными стереотипными
движениями, такими как наклоны или повороты
головы и туловища. Например, стимуляция
определенного участка одного из этих ядер - красного
ядра - заставляет голову и верхнюю часть туловища
откидываться назад.
Задний мозг
Дорсальная область заднего мозга образует
мозжечок, а вентральная - варолиев мост.
мозжечок. Мозжечок состоит из двух полушарий
и, подобно большому мозгу, имеет кору из серого
вещества. Серое вещество содержит характерные
крупные грушевидные клетки Пуркинье, от которых
отходит множество дендритов. Эти клетки
получают импульсы, связанные с мышечной активностью,
из множества разнообразных источников, в том
числе от рецепторов вестибулярного аппарата, от
проприоцепторов суставов, сухожилий и мышц и от
моторных центров коры. Как полагают, мозжечок
интегрирует всю эту информацию и обеспечивает
координированную работу всех мышц,
участвующих в том или ином движении или в поддержании
определенной позы. При повреждении мозжечка
движения становятся резкими и плохо
управляемыми. Мозжечок абсолютно необходим для
координации быстрых мышечных движений, как,
например, при беге, печатании на машинке и даже
разговоре. Все функции мозжечка осуществляются
без участия сознания, но на ранних этапах
тренировки могут включать элемент научения. На этих
этапах мозжечком управляет кора, и нужны
определенные волевые усилия - например, когда мы
учимся ходить, плавать или ездить на велосипеде.
После выработки навыка мозжечок берет на себя
функцию рефлекторного контроля.
мост. Мост (или варолиев мост) составляет часть
мозгового ствола. Он лежит на вентральной
стороне заднего мозга, и в нем проходят восходящие и
нисходящие нервные пути. Кроме того, здесь
имеются ядра, переключающие импульсы на мозжечок.
Продолговатый мозг
Продолговатый мозг-самый задний участок
головного мозга, непосредственное продолжение
спинного мозга. На его дорсальной стороне
расположено заднее сосудистое сплетение. В продолговатом
мозгу восходящие и нисходящие пути переходят с
левой стороны на правую и наоборот. Здесь берут
начало черепномозговые нервы VIII-XII. В этом же
отделе мозга находятся важные центры
рефлекторной регуляции вегетативных функций, в том числе
ритма сердца (гл. 14 и 18), кровяного давления
(гл. 14 и 18), дыхания (гл. 11 и 18), глотания,
слюноотделения, чихания, рвоты и кашля.
16.3. Филогенетическое
развитие нервной
системы
Изучение филогенеза животного мира
свидетельствует о постепенном усложнении структур и
функций в ряду от простейших до млекопитающих. Как
показывает изучение раздражимости у простейших,
кишечнополостных, червей, членистоногих и
млекопитающих, в процессе эволюции наметилось
несколько направлений и типов организации систем
органов, и это нашло свое четкое отражение в
развитии нервной системы.
У простейших способность отвечать на стимулы
присуща одной-единственной клетке. У них нет
пространственного разобщения между стимулом и
реакцией, и клетка функционирует одновременно как
рецептор и как эффектор. Изучение возбудимости
амебы позволило предположить, что в ответе на
раздражение, например на укол тупой иглой,
участвуют плазматическая мембрана и зернистая
эндоплазма. Вероятно, механизм этого процесса
включает систему АТФ-АТФаза, которая доставляет
энергию для амебоидного движения (разд. 17.6.1),
позволяющего амебе уклоняться от раздражителя.
Появление многоклеточной организации у
кишечнополостных привело к пространственному
разделению стимула и реакции, рецептора и эффектора.
К счастью, возникновение многоклеточности
сопровождалось дифференциацией тканей и образованием
нервных клеток, связывающих рецептор с
эффектором, что позволило преодолеть трудности,
связанные с пространственной разобщенностью. Нервная
система примитивных кишечнополостных,
например гидры, представлена нервной сетью или
сплетением, состоящим из одного слоя нейронов.
Многочисленные короткие отростки нейронов
соединяются друг с другом, образуя сеть, пронизывающую
все тело животного. Импульсы распространяются
от места раздражения во всех направлениях, и в
каждом синапсе часть импульсов затухает; эти
затухающие импульсы используются для
«подзарядки» синапсов, чтобы через них могли проходить
следующие импульсы (процесс, называемый
облегчением). Такой механизм называется декрементным
(т. е. затухающим) проведением. Передача нервных
сигналов у таких организмов осуществляется
медленно, так как на ее пути много синапсов, и
ограничена в пространстве, поскольку импульсы
затухают по мере удаления от исходного пункта. Такая
система годится для локальных реакций, например
для реакции одного щупальца, но от нее мало
пользы всему организму в целом, если стимуляция
не будет достаточно сильной или продолжительной.
У большинства высших кишечнополостных, таких
как медузы или актинии, помимо нервной сети
имеется система удлиненных биполярных нейронов,
образующих цепочки - «транзитные пути»,
способные быстро передавать импульсы на значительные
расстояния без заметного их угасания. Такая
система позволяет организму довольно быстро
осуществлять общую, нелокальную реакцию на
угрожающие стимулы (например, отдергивать все
щупальца), и является прообразом объединения нейронов в
нервы, как у высших организмов.
У кольчатых червей объединение нейронов
привело к образованию нервной системы, состоящей из
одного продольного тяжа, идущего по всей длине
тела,-брюшной нервной цепочки. Эта цепочка
состоит из парных сегментарных узлов, соединенных
пучками нервных волокон - коннективами - и
посылающих сегментарные нервные волокна к органам и
тканям каждого данного сегмента (см. рис. 17.37).
Благодаря однонаправленному способу
передвижения червей у них обособилась голова. Эта часть
тела помогает червю находить пищу. Будучи тем
участком тела, который первым сталкивается с
новыми элементами окружающей среды, голова
содержит все чувствительные органы (рецепторы),
необходимые для восприятия внешних стимулов.
Повышенный приток информации от этих рецепторов
в нервную систему привел к утолщению переднего
конца нервной цепочки. Это концентрирование
органов чувств и нервной ткани в одном участке около
ротового аппарата называется цефализацией.
Следует подчеркнуть, что этот термин относится к
развитию всех особенностей, отличающих голову от
остального тела, а не только к нервным структурам.
Степень цефализации зависит от степени
структурной сложности организма и от его образа жизни.
Нервные системы всех других беспозвоночных по
общему плану более или менее сходны с нервной
системой кольчатых червей. Расширенный передний
конец нервной цепочки образует пару церебральных
(мозговых) ганглиев, расположенных над глоткой и
связанных с брюшной цепочкой окологлоточными
коннективами. У членистоногих нервная система
имеет почти такое же строение, как у кольчатых
червей, за исключением того, что церебральные
ганглии лежат у них над пищеводом. Эти ганглии
аналогичны головному мозгу позвоночных, но не
играют такой доминирующей роли, как у
последних. Так, например, удаление головы у беспозво-
ночного мало влияет на его движения, тогда как у
позвоночных все движения инициируются и
контролируются головным мозгом. По-видимому, у
беспозвоночных церебральные ганглии действуют
просто как передаточные центры между
рецепторами и эффекторами, и их роль в интеграции и
координации сводится к нескольким нейроэндо-
кринным воздействиям, влияющим, например, на
время размножения у кольчатых червей или на
линьку у членистоногих (разд. 21.7.3).
16.4. Сенсорные рецепторы
Координированная деятельность организма
основана на непрерывном притоке информации из
внешней и внутренней среды. Если какой-то сигнал
вызывает изменение в активности или поведении
животного, он называется стимулом (раздражителем).
Специальные органы или клетки, воспринимающие
стимулы, называются рецепторами.
Рецептор простейшего, наиболее примитивного
типа состоит из одной неспециализированной
нервной клетки - первичного сенсорного нейрона,
периферическое окончание которого способно
воспринимать раздражения и порождать нервный импульс,
направляющийся в мозг; примером могут служить
кожные механорецепторы типа телец Пачини
(разд. 16.5.1). Более сложные рецепторы
называются вторичными сенсорными клетками и представляют
собой видоизмененные эпителиальные клетки,
способные воспринимать стимулы. Эти клетки
образуют синапсы с соответствующими сенсорными
нейронами, передающими импульсы в мозг; таковы,
например, клетки вкусовых луковиц млекопитающих
(рис. 16.31). Наиболее сложными рецепторами
являются органы чувств, состоящие из большого
числа чувствительных клеток и связанных с ними
вспомогательных приспособлений. Примером
сложности, которой достигают органы чувств, может
служить устройство глаза или уха. В глазу имеются
вторичные воспринимающие клетки двух типов
(палочки и колбочки), множество вставочных нейронов
и целый ряд вспомогательных структур, таких как
хрусталик и радужная оболочка. Вспомогательные
структуры часто выполняют двойную функцию-
препятствуют действию посторонних стимулов и
усиливают действие нужного стимула.
В зависимости от расположения в теле и
характера воспринимаемых стимулов все рецепторы
делятся на три типа:
1) экстероцепторы реагируют на стимулы,
поступающие из внешней среды (как, например, ухо,
воспринимающее звуки);
2) интероцепторы воспринимают стимулы,
поступающие из внутренней среды организма (таковы,
например, рецепторы сонных артерий,
реагирующие на изменения кровяного давления и
содержания CO2 в крови);
3) проприоцепторы отвечают на стимулы, связанные
с положением и движением частей тела и
сокращением мышц.
В табл. 16.7 приведена другая и, пожалуй, более
существенная схема классификации рецепторов,
основанная на природе воспринимаемых ими
стимулов.
Животные воспринимают стимулы в виде одного
из видов энергии, перечисленных в табл. 16.7.
Структуры, преобразующие энергию раздражителя
в электрические сигналы, возникающие в аксонах,
называются преобразователями, и в этом смысле
рецепторы действуют как биологические
преобразователи.
Все рецепторы преобразуют энергию стимула в
локальный нераспространяющийся электрический
сигнал, который возбуждает нервные импульсы в
сенсорном нейроне. Таким образом, рецепторы
кодируют многообразные стимулы в виде нервных
импульсов, которые передаются в мозг, где они
расшифровываются и используются для выработки
нужной ответной реакции, как это показано на
рис. 16.13,ZT. Характер ответа, его величина и
продолжительность находятся в прямой зависимости
от природы стимула.
16.4.1. Механизм преобразования
Все рецепторные клетки являются возбудимыми
клетками и, подобно нейронам и мышечным
волокнам, могут отвечать на соответствующие
стимулы быстрым изменением электрических
характеристик. В отсутствие раздражителя они способны
поддерживать потенциал покоя, описанный в
разд. 16.1, но при стимуляции их мембранный
потенциал изменяется. В 1950 г. Бернард Катц,
используя специализированный рецептор растяжения,
называемый мышечным веретеном, смог выявить
наличие деполяризации в участке, непосредственно
примыкающем к окончанию сенсорного нерва. Эта
местная деполяризация обнаруживается только в
рецепторной клетке, и ее называют генераторным
или рецепторным потенциалом. В последующих
работах путем отведения электрических потенциалов
(с помощью внутриклеточных электродов) от ре-
Таблица 16.7. Типы рецепторов и воспринимаемые ими
стимулы
Тип рецепторов
Фоторецепторы
Электрорецепторы
Механорецепторы
Терморецепторы
Хеморецеп-
торы
Энергетическая
природа стимула
Электромагнитная
Электромагнитная
Механическая
Тепловая
Химическая
Тип стимула
Свет
Электричество
Звук, прикосновение,
давление, гравитация
Изменение температуры
Влажность, запах, вкус
Давление
(стимул)
Усилитель
Микро-
электрод
Катодный
осциллограф
Тонкая
стеклянная
палочка
Микро-,
электрод II
Тельце
Пачини
Аксон
Миелиновая
оболочка
Слабое
давление
Разность
потенциалов
на
мембране,
мВ
Запись с микро-
электрода I
Среднее
давление
Сильное
давление
Немиелинизированное окончание
- Концентрические слои
соединительной ткани
Запись с микро -
электрода II
цепторных клеток мышечных веретен и кожных
механорецепторов - телец Пачини - были получены
следующие данные о механизме преобразования:
1) рецепторный потенциал возникает в результате
того, что под действием стимула возрастает
проницаемость мембраны сенсорной клетки для
ионов Na+ и K + , которые перемещаются в
направлении их электрохимических градиентов;
2) величина рецепторного потенциала варьирует в
зависимости от силы раздражителя;
3) когда рецепторный потенциал достигает
определенной пороговой величины, он возбуждает
распространяющийся потенциал действия в
сенсорном нервном волокне, отходящем от
рецептора (рис. 16.24);
4) частота нервных импульсов в этом волокне
находится в прямой зависимости от силы
раздражения.
Последняя закономерность была установлена еще в
1926 г. Эдрианом (Кембридж), который показал,
что сенсорная информация передается
потенциалами действия типа «всё или ничего» в форме
частотного кода. Теперь мы знаем, что рецепторный
потенциал кодирует силу стимула своей амплитудой,
но после достижения определенной пороговой ам-
Рис. 16.24. Запись
электрической активности с помощью двух
микроэлектродов (I и II), один из
которых введен в окончание
аксона сенсорного нейрона,
находящееся внутри тельца
Пачини, а второй-в тот же аксон
уже вне тельца Пачини.
Надавливание тонкой стеклянной
палочкой, действующее как
стимул, вызывает в окончании
аксона локальный, нераспростра-
няющийся рецепторный
потенциал, который по мере
увеличения давления возрастает и,
достигнув определенной
пороговой величины, возбуждает в
сенсорном нейроне
распространяющийся потенциал действия.
плитуды в нейроне возникает распространяющийся
потенциал действия (импульс), который
подчиняется закону «всё или ничего».
16.7. Рассмотрите графики на рис. 16.25,
полученные при исследовании мышечного веретена
лягушки, и опишите взаимоотношения между
стимулом, рецепторным потенциалом и частотой
нервных импульсов.
16.4.2. Свойства рецепторов
Поступление в ЦНС сигналов от рецепторов
обеспечивает организм всей информацией, необходимой
для выживания. Как правило, одна рецепторная
клетка не может воспринимать все множество
стимулов данного типа; организму, однако, нужна
информация относительно силы (интенсивности)
каждого стимула, чтобы правильно на него
реагировать. Рецепторы обладают двумя очень
важными свойствами, которые повышают их
эффективность и надежность. Эти свойства -
чувствительность и способность к различению, и их
обеспечивают особые структурные и функциональные
приспособления, описанные ниже.
Вес, г (стимул)
Частота импульсов, с 1
Частота импульсов, с"'
Рецепторный потенциал, мВ
Рис. 16.25. Графики, полученные при исследовании мышечного веретена лягушки. (А-по Adrian. Zotterman, 1926,
5-по Katz, 1950)
Параллельные сенсорные клетки
с различными порогами возбуждения
Некоторые рецепторные органы, например
рецепторы растяжения в мышцах, состоят из множества
чувствительных клеток, имеющих разные пороги
возбуждения. Клетки с низким порогом
возбуждаются под действием слабых стимулов, а по мере
возрастания силы раздражителя в отходящем от
клетки нервном волокне частота импульсов
увеличивается. В определенной точке наступает
насыщение, и дальнейшее усиление стимула уже не
повышает частоту импульсов; однако при этом
возбуждаются сенсорные клетки с более высоким
порогом чувствительности, и теперь эти клетки
тоже посылают импульсы, частота которых
пропорциональна силе действующего стимула. Таким
образом диапазон эффективного восприятия
расширяется (рис. 16.26).
Адаптация
При длительном воздействии сильного
раздражителя большинство рецепторов вначале возбуждает в
сенсорном нейроне импульсы с большой частотой,
но постепенно частота их снижается, и это
ослабление ответа во времени называют адаптацией.
Например, войдя в комнату, вы можете сразу обратить
внимание на тиканье часов, но затем перестанете его
замечать. Скорость наступления и степень
адаптации рецепторной клетки зависят от ее функции. В
этом отношении существуют два типа рецепторов -
медленно и быстро адаптирующиеся.
Быстро адаптирующиеся (фазные) рецепторы в
момент «включения» или «выключения» стимула
отвечают на изменения его интенсивности
высокочастотным разрядом импульсов. Именно так
действуют, например, тельца Пачини, реагирующие на
прикосновение, и другие рецепторы, отвечающие на
внезапные изменения стимула: эти рецепторы
доставляют сведения о его динамике.
Медленно адаптирующиеся (тонические) рецепторы
отвечают на постоянный стимул постепенно
уменьшающейся частотой импульсов. Например,
рецепторы растяжения речного рака регистрируют
статические аспекты стимулов, связанных с более или
менее стабильными условиями.
Полагают, что адаптация связана с уменьшением
проницаемости мембраны рецептора для ионов
вследствие непрерывной стимуляции. В результате
амплитуда и продолжительность рецепторного
потенциала постепенно убывают, и когда его величина
Интенсивность стимула
(в произвольных единицах)
Стимул включается
Время,с-10"3
Электрическая активность
(потенциалы действия)
в сенсорный нейронах
Рис. 16.26. Частота потенциалов действия,
возникающих в нейронах, отходящих от трех сенсорных
клеток Д Б и В с разными порогами возбуждения
У клеток Б и В точка активации совпадает с точкой
насыщения для сенсорной клетки с более низким
порогом
становится ниже пороговой, импульсация в
чувствительном нейроне прекращается.
Значение адаптации сенсорных клеток состоит в
том, что она позволяет животному получать
точную информацию об изменениях в окружающей
среде. Когда этих изменений нет, клетки находятся в
покое, что предотвращает перегрузку центральной
нервной системы ненужной информацией.
Благодаря этому повышается общая эффективность и
экономность работы центральных механизмов - они
могут игнорировать статическую фоновую
информацию и сосредоточиться на восприятии внешних
событий, имеющих жизненно важное значение.
Конвергенция и суммация
Во многих сенсорных органах высокая
чувствительность достигается благодаря особому
анатомическому расположению рецепторов и нейронов-так
называемой конвергенции. В таких случаях
выходные пути от нескольких рецепторных клеток
сходятся (или, как говорят, конвергируют) к одному
сенсорному нейрону. Как правило, эти клетки
невелики по размерам, присутствуют в большом числе
и чрезвычайно чувствительны к стимуляции. В то
время как воздействие стимула на одну из этих
клеток не могло бы вызвать ответ в сенсорном
нейроне, одновременная стимуляция нескольких
клеток дает достаточный суммарный эффект. Их
совместное воздействие на сенсорный нейрон
называется суммацией и сходно в функциональном
отношении с суммацией в синапсах (разд. 16.1.2) и в
эффекторах (разд. 16.6 и 17.4.5). Яркий пример
конвергенции и суммации - функция палочек в сетчатке
млекопитающего. Некоторые из этих клеток
способны реагировать даже на один квант света, но
создающийся в них рецепторный потенциал
недостаточен для возбуждения распространяющегося
потенциала действия в волокне зрительного нерва.
Однако несколько палочек (от 2-3 до нескольких
сотен) связаны через биполярные нейроны с одним
волокном зрительного нерва. Чтобы вызвать в нем
импульс, требуется стимуляция по меньшей мере
шести палочек. Повышенная чувствительность к
свету, обусловленная таким совместным действием
палочек, служит прекрасным приспособлением для
сумеречного зрения, хорошо развитого у животных,
ведущих ночной образ жизни, например у сов,
барсуков и лисиц. Однако эта высокая
чувствительность связана с уменьшением остроты зрения, в чем
мы можем легко убедиться, если попробуем читать
при слабом освещении. В глазу человека и
животных, ведущих дневной образ жизни, этот дефект
палочкового зрения преодолевается благодаря
присутствию колбочек, которым (за немногими
исключениями) не свойственна конвергенция, приводящая
к суммации. Жертвуя чувствительностью, система
колбочек обеспечивает большую остроту зрения
(разд. 16.5.4).
Спонтанная активность
В некоторых органах чувств нервные импульсы
возникают в сенсорных нейронах и в отсутствие
стимула. Эта система не так бессмысленна, как
может показаться: она дает два важных
преимущества. Во-первых, она повышает чувствительность
рецептора, обеспечивая немедленный ответ на
стимуляцию, которая в противном случае была бы
слишком мала, чтобы вызвать реакцию сенсорного
нейрона; любое, даже очень малое, изменение
интенсивности стимула будет вызывать изменение
частоты импульсов в этом нейроне. Во-вторых, эта
система позволяет регистрировать направление
изменения стимула в виде увеличения или уменьшения
частоты разрядов сенсорного нейрона. Например, в
рецепторах инфракрасного света, расположенных в
ямках на голове гремучей змеи и действующих как
локаторы при обнаружении жертвы или врага,
существует спонтанная активность, благодаря
которой они способны улавливать повышение или
понижение температуры всего на О, ГС.
Обратная связь в регуляции
рецепторов
В некоторых органах чувств порог
чувствительности может понижаться или повышаться под
действием эфферентных импульсов, поступающих из
центральной нервной системы, благодаря чему
рецептор может проявлять одинаковую
чувствительность к стимулам разной интенсивности. Во многих
случаях эта регуляция осуществляется по принципу
обратной связи с рецептором и вызывает изменения
во вспомогательных структурах, благодаря чему
рецепторная клетка функционирует в ином
диапазоне величин стимула. Такие изменения происходят,
например, в мышечном веретене и в радужной
оболочке глаза.
Латеральное торможение
Важную роль в повышении чувствительности
рецептора и его разрешающей способности играет
латеральное торможение. Оно состоит в том, что
соседние сенсорные клетки, возбуждаясь, оказывают
друг на друга тормозящее воздействие.
Исследования, проведенные на отдельных глазках (оммати-
диях) фасеточного глаза мечехвоста (Limulus)
показали, что смежные омматидии, если они
подвергаются стимуляции одновременно, дают более
слабый ответ, чем в том случае, если они
стимулируются по отдельности. Латеральное
торможение усиливает контраст между двумя соседними
участками, различающимися по интенсивности
стимула. На рис. 16.27 представлена модель
зрительного восприятия, показывающая, каким образом
разница в освещении двух соседних рецепторов
«преувеличивается» и край светлого участка кажется
более светлым, чем он есть на самом деле. Этот
Светочувствительные
клетки
Потенциалы
действия
СВЕТ
Сенсорные нейроны
А Без латерального торможения
Световой
стимул
СВЕТ
Тормозные
нейроны
Б При латеральном торможении
Световой
стимул
Рис. 16.27. Схема возникновения потенциалов
действия в четырех соседних омматидиях 7, 2, 3 и 4
Омматидии 2 и 3 непосредственно стимулируются
узким лучом света, а 7 и 4 слабо стимулируются
рассеянным светом от луча, падающего на 2 и 3. Если
латерального торможения нет (А), омматидии
7 и 4 возбуждаются и в их сенсорных нейронах
возникают потенциалы действия. При наличии
латерального торможения прямой свет стимулирует
не только 2 и 3, но и тормозные нейроны,
соединяющие их с 7 и 4 соответственно, в результате
потенциалы действия в последних не возникают.
Этот механизм усиливает контраст,
воспринимаемый церебральными ганглиями при разном
освещении соседних фоторецепторов. Этот же принцип
применим к колбочкам и палочкам глаза
млекопитающих (По Lamb, Ingram, Johnson, Pitman, 1980)
эффект лежит в основе зрительной иллюзии,
показанной на рис. 16.28.
В глазу человека латеральное торможение
увеличивает разрешающую способность, или остроту
зрения. Разрешающая способность системы-это
способность воспринимать два или более стимулов
одинаковой интенсивности как раздельные. Ее
можно измерять наименьшим угловым расстоянием,
при котором две темные линии одинаковой
плотности еще могут восприниматься не как одна, а как
две раздельные линии. Острота зрения
человеческого глаза очень велика, и это связано отчасти с
латеральным торможением, но в основном-с
расположением колбочек в сетчатке. Свет, падающий
из двух источников на одну и ту же колбочку или на
две соседние колбочки, не воспринимается как
поступающий из двух точек; но если он падает на
колбочки, разделенные всего лишь одной
колбочкой, он может уже восприниматься как идущий из
разных точек. Глаз человека содержит около 7 млн.
колбочек. В центральной ямке сетчатки-участке
диаметром 1 мм-95% фоторецепторов составляют
колбочки, плотность которых здесь достигает
150 тыс. на 1 мм2. Каждая из колбочек связана с
биполярной клеткой, соединяющей колбочку с ее
«собственным» сенсорным нейроном зрительного
нерва. Именно отсутствие конвергенции и тесное
расположение колбочек в центральной ямке
обеспечивают большую остроту зрения.
16.5. Строение и функции
рецепторов
В животном мире существует огромное
разнообразие в строении и функции рецепторов. В настоящем
разделе будут кратко описаны некоторые рецепторы
млекопитающих и более подробно - глаз и ухо
млекопитающих и глаз членистоногих.
16.5.1. Механорецепторы
Механорецепторы считаются наиболее
примитивным типом рецепторов; они воспринимают
разнообразные механические стимулы, такие как
давление, сила тяжести, перемещение и вибрация.
Прикосновение и давление
Различие между прикосновением и давлением чисто
количественное, и восприятие этих стимулов
зависит от расположения рецепторов в коже. Рецеп-
Рис. 16.28. Пример зрительной иллюзии, связанной
с латеральным торможением Кажущиеся серые
пятна на пересечении белых полос позникают,
по-видимому, в связи с тем, что рецепторы,
воспринимающие белые полосы, затормаживают соседние
рецепторы, препятствуя их стимуляции
Отсутствие стимуляции со стороны темных участков
и стимуляция со стороны белых совместно создают
ощущение серых пятен на пересечении белых полос
Ороговевшие
эпителиальные
клетки
Плоский
многослойный
эпителий
Мальпигиев
слой
Тельце
Мейснера
Рис. 16.29. Схема строения рецептора
прикосновения-тельца Мейснера, показывающая его
взаимоотношения с кожей
Ороговевший
слой
Свободные
нервные
окончания
дермис
Дерма
Рис. 16.30. Свободные нервные окончания в
эпидермисе и дерме, реагирующие на температуру
торы прикосновения, чувствительные к слабому
давлению, расположены близко к поверхности кожи
и представляют собой многочисленные тончайшие
свободные окончания сенсорных нейронов
(первичных сенсорных клеток). Они могут лежать в
эпидермисе или прикрепляться к волоскам в волосяных
фолликулах и реагируют на деформацию кожи или
волоска. Рецепторы прикосновения сосредоточены в
определенных участках тела и обусловливают
повышенную чувствительность этих участков. Например,
кончик языка может различать два стимула,
разделенные расстоянием всего лишь 1 мм, тогда как в
середине спины это возможно только в том случае,
если расстояние между раздражаемыми точками
составляет не менее 60 мм.
На прикосновение отвечают также
специализированные рецепторы, называемые тельцами
Мейснера, которые лежат под самым эпидермисом
(рис. 16.29). Они состоят из одного извитого
нервного окончания, заключенного в капсулу,
заполненную жидкостью. В коже, суставах, сухожилиях,
мышцах и брыжейке имеются тельца Пачини,
состоящие из окончания одного нейрона, окруженного
соединительнотканными пластинками. Эти
рецепторы реагируют на давление (см. рис. 16.24).
Полагают, что во всех рецепторах,
воспринимающих прикосновение и давление, рецепторный
потенциал возникает в результате деформации
клеточной мембраны, приводящей к повышению ее
проницаемости для ионов.
Мышечное веретено
В мышцах млекопитающих, амфибий,
ракообразных и насекомых имеются специализированные про-
приоцепторы, называемые мышечными веретенами.
Они действуют как рецепторы растяжения во всех
процессах, связанных с регуляцией мышечного
сокращения. Мышечные веретена выполняют три
главные функции, одна из которых является
статической, а две другие-динамическими:
1) посылают в ЦНС информацию о состоянии и
положении мышц и связанных с ними структур
(статическая функция);
2) рефлекторно усиливают сокращение мышцы и
тем устраняют ее удлинение при увеличении
растягивающей нагрузки (динамическая
функция);
3) изменяют напряжение мышцы и «настраивают»
ее на новую длину (динамическая функция).
Структура и функции мышечного веретена
подробно описаны в разд. 17.5.4.
16.5.2. Терморецепторы
В дерме имеются два типа рецепторов, которые
обычно считают ответственными за температурную
чувствительность. Полагают, что на тепло
реагируют тельца Руффини, а на холод - колбочки Краузе.
Однако главными температурными рецепторами
кожи скорее всего являются многочисленные
свободные нервные окончания (рис. 16.30).
16.5.3. Хеморецепторы
Ощущения вкуса и запаха связаны с действием
химических веществ, возбуждающих специфические
хеморецепторы и играющих важную роль в
питании, избегании опасности и других поведенческих
реакциях, включая ухаживание, спаривание и
социальную активность.
Вкус
Многие ощущения, воспринимаемые как вкусовые,
на самом деле обусловлены раздражением как
вкусовых, так и обонятельных рецепторов. Если
человек с завязанными глазами и зажатым носом
(чтобы химические стимулы не действовали на
орган обоняния) будет жевать лук или яблоко, вряд ли
он сможет их различить. Это свидетельствует о
малой чувствительности вкусовых рецепторов.
Вкусовые раздражители взаимодействуют со
специфическими молекулами, находящимися в
микроворсинках сенсорных клеток, погруженных в
бокаловидные образования - вкусовые почки-на верхней и
боковых поверхностях языка (рис. 16.31).
Пора, заполненная
жидкостью
Лицевой нерв
Опорная
клетка
Эпителиальная
клетка
Микро-
ворсинки
Сенсорная
клетка
Языкогло-
точный
нерв
Базальная
клетка
Сладкое
Соленое
Кислое
Горькое
Рис. 16.31. А Схема строения вкусовой почки. Обладающие вкусом вещества растворяются в жидкости,
омывающей микроворсинки, и диффундируют в рецепторные клетки. (Воспроизведено с разрешения из Schmidt,
1978.) Б. Симметричное распределение вкусовых рецепторов и их иннервация в языке человека.
Существуют четыре типа вкусовых ощущений, и
неравномерное распределение воспринимающих их
рецепторов можно продемонстрировать, нанося на
разные участки языка различные вещества: сладкое
(сахарин), кислое (слабую кислоту), соленое
(морскую соль) и горькое (хинин). Распределение
соответствующих рецепторных зон показано на
рис. 16.31, £. Однако до сих пор не удалось
обнаружить какую-либо корреляцию между химическим
строением веществ и их вкусом. Предполагают, что
рецепторный потенциал возникает в рецепторной
клетке под действием веществ, проникающих через
ее мембрану или взаимодействующих с рецептор-
ными молекулами на поверхности этой мембраны.
Для стимуляции вкусовых рецепторов требуются
более высокие концентрации веществ, чем для
стимуляции обонятельных рецепторов.
Обоняние
Обонятельные ощущения обладают большей
остротой, чем вкусовые, и вызываются пахучими
веществами, находящимися в воздухе. Эти вещества
растворяются в слизи и стимулируют обонятельный
эпителий, расположенный в верхней части носовой
полости. В каждой половине носа имеется
обонятельная область площадью около 2,5 см2 с
клетками трех типов, показанных на рис. 16.32.
В 1963 г. Аморе (Amoore) выдвинул стереохими-
ческую теорию обоняния, согласно которой
существует соответствие между свойствами
рецепторных молекул, находящихся в ресничках
обонятельных клеток, и конфигурацией и размерами молекул
пахучих веществ. Он предположил, что существует
семь основных запахов, таких как гнилостный,
острый, едкий и т.д., каждому из которых
соответствуют свои специфические рецепторы. Всего
можно различать около 10 000 запахов, что, по
мнению Аморе, связано с разной степенью
стимуляции рецепторов тех или иных типов. Наиболее
сильные обонятельные ощущения вызывают
вещества, растворимые в воде и жирах.
Вкус и обоняние у насекомых
Органы вкуса у насекомых обладают
исключительно высокой чувствительностью и довольно
обстоятельно изучены. Эти органы - вкусовые волоски
расположены на усиках, на максиллярных и губных
щупиках и на лапках. Они представляют собой
полые образования, содержащие окончания
сенсорных нейронов, и чувствительны как к химическим
веществам, так и к механическим смещениям.
У падальной мухи Calliphora существует вкусовой
рефлекс, который состоит в вытягивании хоботка,
когда лапки прикасаются к съедобному веществу.
Пороговая концентрация сахарозы для
возникновения этого рефлекса составляет у мухи всего лишь
0,8 10"4M, тогда как для человека она равна
0,2-101 M.
Как ни поразителен столь низкий порог реакции
вкусовых рецепторов, чувствительность насекомых
к запахам еще больше. Обычно обонятельные
рецепторы расположены на усиках (антеннах); их
интенсивно изучали у бабочек. Самки тутового
шелкопряда (Bombyx mori) способны привлекать самцов
того же вида, находящихся на расстоянии
нескольких километров, с помощью выделяемого ими
специфического вещества. Такие вещества, которые,
подобно этому половому аттрактанту, действуют
как средство внутривидовой связи, называются
феромонами. В одном из экспериментов единственная
самка тутового шелкопряда, находившаяся в
муслиновом садке, привлекла 40% большой популяции
самцов, выпущенных на расстоянии 4 км от нее.
Насекомые, использующие феромоны для столь
дальней связи, имеют сложно разветвленные
антенны, несущие множество (например, у самцов
Слой слизи
Обонятельная
булава
Обонятельная
ресничка
Микротрубочка
Митохондрия
Опорная
клетка
Сенсорная
клетка
Ядро
сенсорной
клетки
Базальная
клетка
Базальная
мембрана
Аксон
сенсорного
нейрона
Рис. 16.32. Упрощенная схема взаимоотношений
между сенсорными, опорными и базальными
клетками в обонятельном эпителии
Молекулы-рецепторы для пахучих веществ находятся на
поверхностных мембранах ресничек обонятельного эпителия.
бабочки Те lea polyphemus 15· 104) сенсорных клеток.
Молекулы полового аттрактанта улавливаются
антеннами из обдувающего их воздуха.
Вопреки распространенному мнению, самцы
бабочек не летят в поисках самки прямо по
концентрационному градиенту феромона. Они просто
устремляются против ветра, несущего феромон, и
начинают пересекать воздушный след зигзагами, всякий
раз меняя направление при исчезновении запаха;
только оказавшись вблизи самки, они определяют
ее местонахождение по градиенту концентрации
феромона.
16.5.4. Глаз млекопитающих
Глаз млекопитающих - орган чувств, состоящий из
большого числа рецепторных клеток (палочек и
колбочек сетчатки), сенсорных нейронов,
образующих зрительный нерв, и сложной системы
вспомогательных приспособлений. Такое устройство
позволяет глазу воспринимать свет с различной
длиной волны, отражаемый объектами, находящимися
в поле зрения на разных расстояниях, и
преобразовывать его в электрические импульсы, которые
направляются в головной мозг и порождают
удивительно точное восприятие.
Свет распространяется в виде волн
электромагнитного излучения, и волны, воспринимаемые
человеческим глазом, составляют узкую, так
называемую видимую часть спектра (длины волн 380-
760 нм; см. Приложение 1.7). Свет-один из видов
энергии, он испускается и поглощается дискретными
порциями - квантами, или фотонами. Каждый квант
в видимой части спектра несет энергию,
достаточную для того, чтобы вызвать фотохимическую
реакцию в чувствительных клетках глаза. Работа
глаза основана на тех же перечисленных ниже
принципах, что и фотокамера, а именно он 1)
контролирует количество света, проходящее внутрь; 2)
фокусирует изображения предметов внешнего мира
с помощью системы линз; 3) регистрирует
изображение на чувствительной поверхности; 4)
перерабатывает невидимое изображение во внутренний образ
видимой картины мира.
Строение и функция человеческого
глаза
Глаза расположены во впадинах черепа,
называемых глазницами; глаз укреплен здесь при помощи
четырех прямых и двух косых мышц, управляющих
его движениями. Глазное яблоко человека имеет
диаметр около 24 мм и весит 6-8 г. Большую часть
глаза составляют вспомогательные структуры,
назначение которых в том, чтобы проецировать поле
зрения на сетчатку - слой фоторецепторных клеток,
выстилающий глазное яблоко изнутри.
Стенка глаза состоит из трех концентрических
слоев: 1) склеры (белковой оболочки) и роговицы;
2) сосудистой оболочки, ресничного тела,
хрусталика и радужки; 3) сетчатки. Форма глаза
поддерживается за счет гидростатического давления
(25 мм рт. ст.) водянистой влаги и стекловидного
тела. Схема строения человеческого глаза приведена
на рис. 16.33. Ниже дается краткое перечисление
различных его частей и выполняемых ими функций.
Склера-самая наружная оболочка глаза. Это очень
плотная капсула, содержащая коллагеновые
волокна; защищает глаз от повреждения и помогает
глазному яблоку сохранять свою форму.
Роговица - прозрачная передняя сторона склеры.
Благодаря искривленной поверхности действует как
главная светопреломляющая структура.
Конъюнктива - тонкий прозрачный слой клеток,
защищающий роговицу и переходящий в эпителий
век. Конъюнктива не заходит на участок роговицы,
прикрывающий радужку.
Веко - защищает роговицу от механического и
химического повреждения, а сетчатку-от слишком
яркого света.
Сосудистая оболочка - средняя оболочка; пронизана
сосудами, снабжающими кровью сетчатку, и
покрыта пигментными клетками, препятствующими
отражению света от внутренних поверхностей глаза.
Ресничное (цилиарное) тело-место соединения
склеры и роговицы. Состоит из эпителиальных клеток,
Верхнее веко
Конъюнктива
Роговица
Водянистая
влага
Зрачок
Хрусталик
Радужная
оболочка
Циннова
связка
Цилиарное
Склера
Сосудистая
оболочка
^ Сетчатка
Стекловидное
тело
Центральная
ямка
Слепое пятно
Зрительный
нерв
Цилиарная
мышца
Рис. 16.33. Строение глаза
млекопитающего
кровеносных сосудов и цилиарной мышцы.
Цилиарная мышца-кольцо, состоящее из гладких
мышечных волокон, кольцевых и радиальных,
которые изменяют форму хрусталика при
аккомодации.
Цилиарная (циннова) связка - прикрепляет хрусталик
к цилиарному телу.
Хрусталик - прозрачное эластичное двояковыпуклое
образование. Обеспечивает тонкую фокусировку
лучей света на сетчатке и разделяет камеры,
заполненные водянистой влагой и стекловидным телом.
Водянистая влага - прозрачная жидкость,
представляющая раствор солей. Секретируется цилиарным
телом и переходит из глаза в кровь через шлеммов
канал.
Радужка - кольцевая мышечная диафрагма,
содержит пигмент, определяющий цвет глаз. Разделяет
пространство, заполненное водянистой влагой, на
переднюю и заднюю камеры и регулирует
количество света, проникающего в глаз.
Зрачок - отверстие в радужке, через которое свет
проходит внутрь глаза.
Стекловидное тело - прозрачное полужидкое
вещество, поддерживающее форму глаза.
Сетчатка - внутренняя оболочка, содержащая фото-
рецепторные клетки (палочки и колбочки), а также
тела и аксоны нейронов, образующих зрительный
нерв.
Центральная ямка - наиболее чувствительный
участок сетчатки, содержащий только колбочки. В этом
участке наиболее точно фокусируются лучи света.
Зрительный нерв-пучок нервных волокон,
проводящих импульсы от сетчатки в мозг.
Слепое пятно-место на сетчатке, где из глаза
выходит зрительный нерв; оно не содержит ни палочек,
ни колбочек и потому не обладает
светочувствительностью.
16.8. Перечислите по порядку структуры, через
которые проходит свет по пути к сетчатке.
Аккомодация
Аккомодация - это рефлекторный механизм, с
помощью которого лучи света, исходящие от объекта,
фокусируются на сетчатке. Он включает два
процесса, каждый из которых будет рассмотрен
отдельно.
РЕФЛЕКТОРНОЕ ИЗМЕНЕНИЕ ДИАМЕТРА ЗРАЧКА. При Яр-
ком свете кольцевая мускулатура радужки
сокращается, а радиальная расслабляется; в результате
происходит сужение зрачка и количество света,
попадающего на сетчатку, уменьшается, что
предотвращает ее повреждение (рис. 16.34). При слабом
свете, наоборот, радиальная мускулатура
сокращается, а кольцевая расслабляется. Дополнительное
преимущество, доставляемое сужением зрачка,
состоит в том, что увеличивается глубина резкости, и
поэтому различия в расстоянии от объекта до глаза
меньше сказываются на изображении.
преломление (рефракция) света. От объекта,
удаленного на расстояние больше 6 м, в глаз поступают
практически параллельные лучи света, тогда как
лучи, идущие от более близких предметов, заметно
расходятся. В обоих случаях для того, чтобы свет
сфокусировался на сетчатке, он должен быть
преломлен (т. е. его путь изогнут), и для близких
предметов преломление должно быть более сильным.
Нормальный глаз способен точно фокусировать
Яркий свет
Слабый свет
Кольцевая мышца
сокращается
Зрачок
суживается
Радиальная
мышца
расслабляется
Радиальная
мышца
сокращается
Зрачок
расширяется
Кольцевая
мышца
расслабляется
Рис. 16.34. Реакция радужной
оболочки (и тем самым зрачка) на
изменение освещенности.
Свет от дальнего объекта
Свет об ближнего объекта
1. На глаз падают параллельные лучи
2. Роговица преломляет лучи света
3. Кольцевая мышца расслабляется
4. Циннова связка натягивается
5. Хрусталик уплощается
6. Свет фокусируется на сетчатке
1. В глаз попадают расходящиеся лучи
2 Роговица преломляет лучи света
3. Кольцевая цилиарная мышца сокращена
4 Циннова связка расслаблена
5. Эластичный хрусталик становится
более выпуклым
6. Свет фокусируется на сетчатке
Рис. 16.35. Аккомодация при
восприятии лучей света от
предметов, находящихся на разных
расстояниях А. Вид глаза сбоку. Б. Вид
глаза спереди
свет от объектов, находящихся на расстоянии от
25 см до бесконечности. Преломление света
происходит при переходе его из одной среды в другую,
имеющую иной коэффициент преломления, в
частности на границе воздух - роговица и у поверхностей
хрусталика. Форма роговицы не может изменяться,
поэтому рефракция здесь зависит только от угла
падения света на роговицу, который в свою очередь
зависит от удаленности предмета. В роговице
происходит наиболее сильное преломление света, а
функция хрусталика состоит в окончательной
«наводке на фокус». Форма хрусталика регулируется
цилиарной мышцей: от степени ее сокращения
зависит натяжение связки, поддерживающей
хрусталик. Последняя воздействует на эластичный
хрусталик и изменяет его форму (кривизну поверхности), а
тем самым и степень преломления света. При
увеличении кривизны хрусталик становится более
выпуклым и сильнее преломляет свет. Полная картина
этих взаимоотношений представлена в табл. 16.8.
На рис. 16.35 показаны изменения, происходящие в
глазу при аккомодации для восприятия отдаленных
и близких предметов.
На сетчатке изображение получается
перевернутым, но это не мешает правильному восприятию,
так как все дело не в пространственном положении
изображения на сетчатке, а в интерпретации его
мозгом.
Строение сетчатки
Сетчатка развивается как вырост переднего мозга,
называемый глазным пузырьком. В процессе
эмбрионального развития глаза фоторецепторный участок
пузырька впячивается внутрь до соприкосновения с
сосудистым слоем. При этом рецепторные клетки
оказываются лежащими под слоем тел и аксонов
нервных клеток, связывающих их с мозгом
(рис. 16.36).
Сетчатка состоит из трех слоев, каждый из
которых содержит клетки определенного типа. Самый
наружный (наиболее удаленный от центра глазного
яблока) светочувствительный слой содержит
фоторецепторы-палочки и колбочки, частично
погруженные в пигментный слой сосудистой оболочки. Затем
идет промежуточный слой, содержащий биполярные
нейроны, которые связывают фоторецепторы с
клетками третьего слоя. В этом же промежуточном
слое находятся горизонтальные и амакриновые
клетки, обеспечивающие латеральное торможение.
Третий слой - внутренний поверхностный
слой-содержит ганглиозные клетки, дендриты которых
соединены синапсами с биполярными клетками, а
аксоны образуют зрительный нерв.
Строение и функция полочек и
колбочек
Палочки и колбочки очень сходны по своему
строению: в тех и других светочувствительные пигменты
находятся на наружной поверхности
внутриклеточных мембран наружного сегмента; и те и другие
состоят из четырех участков, строение и функции
которых кратко описаны ниже.
наружный сегмент. Это тот светочувствительный
участок, где световая энергия преобразуется в ре-
цепторный потенциал. Весь наружный сегмент
заполнен мембранными дисками, образованными
плазматической мембраной и отделившимися от
нее. В палочках число этих дисков составляет
Таблица 16.8. Взаимоотношения между структурами,
участвующими в изменении формы хрусталика, и степенью
преломления света
Цилиарная
мышца
Сокращена
Расслаблена
Циннова
связка
Не
натянута
Натянута
Кривизна хрусталика
Увеличена
(хрусталик более
выпуклый)
Уменьшена
Преломление
света
Усилено
Ослаблено
600-1000, они представляют собой уплощенные
мембранные мешочки и уложены наподобие стопки
монет. В колбочках мембранных дисков меньше, и
они представляют собой складки плазматической
мембраны.
перетяжка. Здесь наружный сегмент почти
полностью отделен от внутреннего впячиванием
наружной мембраны. Связь между двумя сегментами
осуществляется через цитоплазму и пару ресничек,
переходящих из одного сегмента в другой. Реснички
содержат только 9 периферических дублетов
микротрубочек: пара центральных микротрубочек,
характерных для ресничек, отсутствует.
внутренний сегмент. Это область активного
метаболизма; она заполнена митохондриями,
доставляющими энергию для процессов зрения, и
полирибосомами, на которых синтезируются белки,
участвующие в образовании мембранных дисков и
зрительного пигмента. В этом же участке
расположено ядро.
синаптическая область. В этом участке клетка
οδό клера
Эпителий сосу -
диетой оболочки
с пигментным
слоем
Палочка
Мембранные
диски
Колбо
Складки
плазматической мембра- у
ны V\/,
Параресничек(
Mитохонд
рии
Наруж-j
ный
сегмент
Сужение
Слой
фоторецепторов
Внутренний
сегмент
Ядро
Концевое
утолщение
► Синап-t:
тический
участок
Горизонтальная
клетка
Синаптические
окончания
Биполярная
клетка
межуточный
слой
Внутренний
поверхностный
слой
Нейроны
зрительного
нерва
Амакриновая
клетка
Направление
лучей света
Рис. 16.36. Схематический разрез сетчатки глаза с
деталями ультраструктуры палочек и колбочек.
Показаны связи между сенсорными клетками и
нейронами зрительного нерва. Лучи света должны
пройти через слои ганглиозных клеток и других
нейронов, прежде чем достигнут палочек и колбочек.
разует синапсы с биполярными клетками.
Диффузные биполярные клетки могут образовывать
синапсы с несколькими палочками. Это явление,
называемое синаптической конвергенцией, уменьшает
остроту зрения, но повышает
светочувствительность глаза. Моносинаптические биполярные клетки
связывают одну колбочку с одной ганглиозной
клеткой, что обеспечивает большую по сравнению с
палочками остроту зрения. Горизонтальные и ама-
криновые клетки связывают вместе некоторое число
палочек или колбочек. Благодаря этим клеткам
зрительная информация еще до выхода из сетчатки
подвергается определенной переработке; эти клетки,
в частности, участвуют в латеральном торможении.
Различия между палочками и
колбочками
Палочек в сетчатке содержится больше, чем
колбочек (120 · 10е и 6-7* 106 соответственно).
Распределение палочек и колбочек тоже неодинаково.
Тонкие, вытянутые палочки (размеры 50 χ 3 мкм)
равномерно распределены по всей сетчатке, кроме
центральной ямки, где преобладают удлиненные
конические колбочки (60 χ 1,5 мкм). Так как в
центральной ямке колбочки очень плотно
упакованы (15· 104 на 1 мм2), этот участок отличается
высокой остротой зрения (разд. 16.4.2). В то же
время палочки обладают большей
чувствительностью к свету и реагируют на более слабое освещение.
Палочки содержат только один зрительный
пигмент, не способны различать цвета и
используются преимущественно в ночном зрении.
Колбочки содержат три зрительных пигмента, и это
позволяет им воспринимать цвет; они используются
главным образом при дневном свете. Палочковое
зрение отличается меньшей остротой, так как
палочки расположены менее плотно и сигналы от них
подвергаются конвергенции, но именно это
обеспечивает высокую чувствительность, необходимую
для ночного зрения.
16.9. Объясните, почему конвергенция
должна повышать чувствительность глаза к слабому
свету.
16.10. Объясните, почему ночью предметы
видны лучше, если не смотреть прямо на них.
Механизм фоторецепции
Палочки содержат светочувствительный пигмент
родопсин, находящийся на наружной поверхности
мембранных дисков. Родопсин, или зрительный
пурпур, представляет собой сложную молекулу,
образующуюся в результате обратимого связывания
липопротеина скотопсина с небольшой молекулой
поглощающего свет каротиноида-ретиналя.
Последний представляет собой альдегидную форму
витамина А и может существовать (в зависимости
от освещения) в виде двух изомеров (рис. 16.37).
Установлено, что при воздействии света на
родопсин один фотон способен вызывать
изомеризацию, показанную на рис. 16.37. Ретиналь играет
роль простетической группы, и полагают, что он
занимает определенный участок на поверхности
молекулы скотопсина и блокирует реактивные группы,
участвующие в генерации электрической активности
в палочках. Точный механизм фоторецепции пока
неизвестен, но предполагается, что он включает два
процесса. Первый из них-это превращение \\-цис-
ретиналя в полностью-т/?яноретиналь под
действием света, а второй - расщепление родопсина через
ряд промежуточных продуктов на ретиналь и ско-
топсин (процесс, называемый выцветанием):
Выцветание
Родопсин > Ретиналь + Скотопсин.
После прекращения воздействия света родопсин
тотчас же ресинтезируется. Вначале полностью-
т/?днс-ретиналь при участии фермента ретиналь-
изомеразы превращается в ΙΙ-ί/моретиналь, а затем
последний соединяется со скотопсином. Этот
процесс лежит в основе темповой адаптации. В полной
темноте требуется около 30 мин, чтобы все палочки
адаптировались и глаза приобрели максимальную
чувствительность. Однако во время этого процесса
проницаемость мембраны наружного сегмента для
Na+ уменьшается, в то время как внутренний
сегмент продолжает откачивать ионы Na+ наружу, и в
результате внутри палочки возрастает
отрицательный потенциал, т.е. происходит гиперполяризация
(рис. 16.38). Это прямо противоположно тому, что
обычно наблюдается в других рецепторных клетках,
где раздражение вызывает деполяризацию, а не
гиперполяризацию. Гиперполяризация замедляет
высвобождение из палочек возбуждающего
медиатора, который в темноте выделяется в наибольшем
количестве. Биполярные клетки, связанные через
синапсы с палочками, тоже отвечают
гиперполяризацией, но в ганглиозных клетках, аксоны
которых образуют зрительный нерв, в ответ на сигнал
от биполярной клетки возникает
распространяющийся потенциал действия.
Цветовое зрение
В видимой части спектра человеческий глаз
поглощает свет всех длин волны, воспринимая их в
Таблица 16.9. Цвета видимого спектра и приблизительно
соответствующие им длины волн
Цвет
Красный
Оранжевый
Желтый
Зеленый
Синий
Фиолетовый
Длина волны, нм
Более 620
590-620
570-590
500-570
440-500
Менее 440
Свет
11- цис -ретиналь
Полностью- транс -ретиналь
Рис. 16.37. Переход 11-цис-ретиналя в полностью-
транс-ретиналь под действием света.
Свет
Нет света
Наружный
сегмент
Внутренний
сегмент
Активный
транспорт
Рис. 16.38. Схема строения палочки,
иллюстрирующая предполагаемые изменения проницаемости
наружного сегмента для Na+ под действием света.
Отрицательные заряды на правой стороне палочки
соответствуют потенциалу покоя, а на левой
стороне - гиперполяризации
виде шести цветов, каждый из которых
соответствует определенному участку спектра (табл. 16.9).
Существуют три типа колбочек - «красные», «зеленые»
и «синие», которые содержат разные пигменты и, по
данным электрофизиологических исследований,
поглощают свет с различной длиной волны.
Цветовое зрение объясняют с позиций трехком-
понентной теории, согласно которой ощущения
различных цветов и оттенков определяются степенью
раздражения каждого типа колбочек светом,
отражаемым от объекта. Так, например, одинаковая
стимуляция всех колбочек вызывает ощущение
белого цвета. Первичное различение цветов
осуществляется в сетчатке, но окончательный цвет, который
будет воспринят, определяется интегративными
функциями мозга. Эффект смешения цветов лежит в
основе цветного телевидения, цветной фотографии и
живописи.
цветовая слепота. Полное отсутствие или
недостаток колбочек какого-либо типа может приводить к
различным формам цветовой слепоты или
аномалиям цветоощущения. Например, люди, у которых
нет «красных» или «зеленых» колбочек, не
различают красный и зеленый цвета, а те, у кого имеется
недостаточное количество колбочек одного из этих
двух типов, плохо различают некоторые оттенки
красного и зеленого цвета. Для выявления дефектов
цветового зрения применяют тестовые таблицы
типа таблиц Исахари, на которых нанесены пятнышки
разных цветов. На некоторых таблицах из этих
пятнышек составлены цифры. Человек с
нормальным цветовым зрением легко различает эти цифры,
а лица с нарушенным цветоощущением видят
другое число или вообще не видят никакой цифры.
Цветовая слепота передается по наследству как
рецессивный признак, сцепленный с Х-хромосомой.
Среди мужчин около 2% не различают красный
цвет и 6%-зеленый, тогда как среди женщин
аномалиями цветового зрения страдают только 0,4%.
16.11. Испытуемый помещает перед одним
глазом зеленый фильтр, а перед другим - красный и
смотрит на предмет. Используя данные,
приведенные в табл. 16.9, опишите его цветовые
ощущения.
Бинокулярное зрение и
стереоскопическое зрение
Бинокулярное зрение имеет место в том случае,
когда зрительные поля обоих глаз перекрываются
таким образом, что их центральные ямки
фиксируются на одном и том же объекте. Бинокулярное
зрение имеет ряд преимуществ по сравнению с
использованием одного глаза, в том числе
расширяет поле зрения и дает возможность
компенсировать повреждения одного глаза за счет другого.
Кроме того, бинокулярное зрение снимает эффект
слепого пятна и, наконец, лежит в основе
стереоскопического зрения. Стереоскопическое зрение
обусловлено тем, что на сетчатках двух глаз
одновременно возникают слегка различающиеся
изображения, которые мозг воспринимает как один образ.
Чем больше глаза направлены вперед, тем больше
стереоскопическое поле зрения. У человека,
например, общее поле зрения охватывает 180°, а
стереоскопическое-140°. У лошади глаза расположены по
бокам головы, поэтому их фронтальное
стереоскопическое поле зрения ограниченно и используется
лишь для рассматривания удаленных предметов.
Чтобы лучше рассмотреть близкий предмет,
лошадь поворачивает голову и пользуется
монокулярным зрением. Для хорошего стереоскопического
зрения необходимы глаза, направленные вперед, с
центральными ямками, лежащими посередине их
полей, что обеспечивает большую остроту зрения. В
этом случае стереоскопическое зрение позволяет
получать более точное представление о размерах и
форме предмета, а также о расстоянии, на котором
он находится. В основном стереоскопическое зрение
характерно для хищных животных, которым оно
абсолютно необходимо, если они ловят добычу,
внезапно набрасываясь на нее или пикируя с
высоты, как это делают представители семейства
кошачьих, ястребы или орлы. У животных, которым
приходится спасаться от хищников, глаза, напротив,
расположены по бокам головы, благодаря чему они
имеют более широкий обзор, но ограниченное
стереоскопическое зрение. Например, у кролика общее
поле зрения охватывает 360°, а фронтальное
стереоскопическое поле-всего 20°. Анализ изображений,
получаемых на сетчатке при стереоскопическом
зрении, осуществляется в двух симметричных участках,
составляющих зрительную кору.
Зрительные пути и зрительная кора
Нервные импульсы, возникающие в сетчатке,
поступают по миллиону или около того волокон
зрительного нерва в зрительную кору, расположенную в
задней части затылочных долей. В этой зоне
спроецированы все мельчайшие участки сетчатки,
включающие, возможно, всего лишь по нескольку
палочек и колбочек, и именно здесь зрительные сигналы
интерпретируются и мы «видим». Однако то, что
мы видим, приобретает смысл только после обмена
сигналами с другими участками коры и прежде
всего с височными долями, где хранится
предшествующая зрительная информация и где она
используется для анализа и идентификации текущих
зрительных сигналов (разд. 16.2.4). В мозгу человека
аксоны от левых половин сетчатки обоих глаз
направляются к левой половине зрительной коры, а
аксоны от правых половин сетчатки обоих глаз-к
правой стороне зрительной коры. Аксоны, идущие
от носовых половин обеих сетчаток, пересекаются;
место их пересечения называется зрительным
перекрестом или хиазмой (схема зрительных путей пред-
Правый
глаз
Левый
глаз
Нос
Зрительный
нерв
Зрительный
перекрест
Боковое
коленчатое
тело
Зрительная
кора
Рис. 16.39. Схема зрительных путей человека. Вид
с нижней стороны мозга.
ставлена на рис. 16.39). Около 20% волокон
зрительного нерва не доходит до зрительной коры, а
вступает в средний мозг и участвует в рефлекторной
регуляции диаметра зрачка и движений глаз.
16.5.5. Глаза членистоногих
У членистоногих бывают глаза двух типов - простые
глаза, свойственные личинкам некоторых
насекомых, и более типичные сложные, или фасеточные,
глаза. Простой глаз состоит из единственной линзы,
прикрывающей небольшое число
светочувствительных клеток. Он может различать свет и темноту, но
не способен создавать изображение. Нередко такие
глаза присутствуют вместе с большими по размеру
и функционально более важными фасеточными
глазами.
Каждый фасеточный глаз состоит из тысяч
отдельных зрительных единиц, называемых оммати-
диями. У стрекозы, например, число омматидиев
составляет около 30000. Каждый омматидий имеет
две светопреломляющие структуры -
двояковыпуклую роговичную линзу и кристаллический конус,
являющийся производным особых клеток. Эти
структуры фокусируют свет на скоплении
светочувствительных клеток ретинулы. Внутренние
мембраны этих клеток несут микроворсинки, которые,
сливаясь, образуют рабдом. В микроворсинках рабдома
содержится зрительный пигмент, и здесь же
возникают нервные импульсы, которые по зрительному
нерву передаются в мозг. Каждый омматидий
окружен и отделен от соседних омматидиев
удлиненными пигментными клетками (рис. 16.40).
Поскольку поле зрения каждого отдельного ом-
матидия очень мало, общее изображение,
возникающее в целом глазу, состоит из множества отдельных
перекрывающихся изображений и называется
мозаичным. Этот тип глаза не обладает такой большой
разрешающей способностью, как глаз
млекопитающего, но наложение изображений придает ему
большую чувствительность и позволяет хорошо
улавливать движение объектов.
16.5.6. Ухо млекопитающих
Ухо млекопитающих представляет собой орган
чувств, содержащий механорецепторы,
чувствительные к силе тяжести, к перемещению в пространстве
и к звукам. Движения и положение головы
относительно направления силы тяжести фиксируются
вестибулярным аппаратом, состоящим из
полукружных каналов и двух небольших мешочков - круглого
(sacculus) и овального (utriculus). Все остальные
структуры предназначены для восприятия, усиления
и преобразования звуковой энергии в электрические
импульсы, которые, поступая в слуховые зоны
мозга, порождают слуховое ощущение.
Строение и функции уха
Ухо состоит из трех отделов - наружного, среднего
и внутреннего уха (рис. 16.41). Наружное ухо состоит
из ушной раковины, представляющей собой
покрытый кожей хрящевой вырост, который фокусирует и
направляет звуковые волны в наружный слуховой
проход. Звуковые волны вызывают колебания
барабанной перепонки, отделяющей наружное ухо от
среднего. В среднем ухе находятся три слуховые
косточки - молоточек, наковальня и стремя, которые
передают колебания барабанной перепонки на
перепонку овального окна, отделяющего среднее ухо от
Роговичная
линза
- Кристаллический
конус
- Кристаллическая
клетка
-Проксимальная
пигментная клетка
-Дистальная
пигментная клетка
-Клетка ретинулы
Пигментная клетка
Клетка ретинулы
-Рабдом
Рабдом
Нейроны
зрительного
нерва
Рис. 16.40. Схема строения отдельного омматидия.
А - продольный разрез, Б- поперечный разрез.
Стремя
Наковальня
Молоточек
Барабанная
перепонка
Овальное,
окно
Полукружный
канал
Ампула
Овальный
мешочек
Круглый
мешочек
Лестница
улитки
Направление
звуковых волн
Среднее ухо
Круглое
окно
Лестница \
преддверия
Барабанная
\ лестница
Наружный
слуховой проход
Евстахиева труба
(Все структуры
расположены
в костной капсуле)
Перилимфа
| (peri —вокруг)
Эндолимфа
(endo—внутри)
Улитка
(в
действительности
закручена
в спираль)
Рис. 16.41. Схематическое изображение основных
структур уха млекопитающего, образующих органы
равновесия и слуха.
внутреннего. При этой передаче колебания
усиливаются в 22 раза благодаря системе рычагов,
образуемой слуховыми косточками, и относительным
площадям контакта между молоточком и
барабанной перепонкой (60 мм2) и между стременем и
овальным окном (3,2 мм2). Заполненное воздухом
среднее ухо соединяется через евстахиеву трубу с
глоткой, и это предохраняет барабанную перепонку
от повреждения при перепадах атмосферного
давления. И наконец, внутреннее ухо находится в сложной
системе каналов и полостей, образующих костный
лабиринт (внутри височной кости) и заполненных
жидкостью, называемой перилимфой; внутри
костного лабиринта находится перепончатый лабиринт,
заполненный эндолимфой и содержащий
чувствительные рецепторы. Слуховые рецепторы находятся
в улитке, а рецепторы органа равновесия-в
овальном и круглом мешочках и в ампулах полукружных
каналов. Овальное и круглое окна ограничивают
перемещение перилимфы.
Природа звука
Звук-это колебания частиц, из которых состоит
среда. Он распространяется в виде
волн-чередующихся участков высокого и низкого давления-и
способен проходить через жидкости, твердые тела и
газы. Расстояние между двумя идентичными
точками соседних волн называется длиной волны, и эта
величина обратно пропорциональна частоте
колебаний, от которой зависит слышимая высота звука.
Человеческое ухо воспринимает звуки с частотой от
40 гц до 20000 Гц (циклов в секунду). У собак
предел слышимости достигает 40 000 Гц, а у летучих
мышей-100000 Гц. Частота звуков человеческой
речи варьирует от 500 до 3000 Гц. С возрастом
чувствительность к высоким звукам уменьшается.
Характер музыкального тона определяется
набором входящих в него частот. Например, скрипка и
труба, на которых берется одна и та же нота,
скажем среднее «до», дают одну и ту же основную
Перилимфа
Эндолимфа
Перилимфа
Лестница
преддверия
Рейснерова
мембрана
Лестница
улитки
Текториаль -'
нал мембрана
Базилярная
мембрана
Волосковая
клетка
Слуховой
нейрон
Барабанная
лестница
Кортиев
орган
Рис. 16.42. Схема поперечного разреза улитки, на
котором виден кортиев орган
частоту (256 Гц), но звуки получаются разные; это
зависит от издаваемых инструментом обертонов
(гармоник), которые определяют качество звука,
или его тембр. Тот же принцип применим к
человеческому голосу и определяет его индивидуальное
звучание.
Интенсивность (громкость) звука зависит от
амплитуды звуковых волн и является мерой
содержащейся в них энергии.
Улитка и слух
Улитка представляет собой спиральный канал
длиной 35 мм, разделенный продольными перепонками
на три части (рис. 16.42).
Два канала - лестница преддверия и барабанная
лестница - содержат перилимфу и соединяются
между собой в верхушке улитки через маленькое
отверстие - геликотрему. Средний канал-канал улитки-
содержит эндолимфу. Он отделен от барабанной
лестницы базилярной мембраной, на которой
расположены чувствительные волосковые клетки,
которые могут приходить в соприкосновение с
расположенной над ними текториальной мембраной. Все это
образование, состоящее из базилярной мембраны,
чувствительных клеток и текториальной мембраны,
носит название кортиева органа и является тем
местом, где происходит преобразование звуковых
волн в электрические импульсы.
Звуковые волны, приходящие по слуховому
проходу, передаются на перепонку овального окна и
вызывают колебания перилимфы в лестнице
преддверия, а те в свою очередь передаются через
рейснерову мембрану эндолимфе, заполняющей
средний канал, а затем базилярной мембране и
перилимфе барабанной лестницы и в конце концов
гасятся в воздушной среде среднего уха,
передаваемые ей колебаниями перепонки круглого окна.
Механизм преобразования волн давления в
нервные импульсы в точности неизвестен, но
предполагают, что он основан на взаимном перемещении
базилярной и текториальной мембран. В результате
колебаний базилярной мембраны, вызванных
волнами давления, две мембраны скользят
относительно друг друга, и при этом чувствительные волоски
трутся о текториальную мембрану. Вызванная этим
деформация чувствительных волосков приводит к
деполяризации сенсорных клеток и возникновению в
них рецепторного потенциала, порождающего
потенциалы действия в аксонах слухового нерва.
Различение высоты и силы звука
Способность различать высоту звуков зависит от
того, что звуки данной частоты вызывают
колебание базилярной мембраны и возбуждение
сенсорных клеток в строго определенном участке
кортиева органа. Возбужденные клетки посылают
импульсы в соответствующие участки слуховой зоны
Ютоконии
Пучки волосков
чувствительных
клеток
Рис. 16.43. Структуры внутреннего уха Микрофотография, полученная с помощью сканирующего
электронного микроскопа. Л Отоконии. Б. Слой отокониев, часть которого удалена, чтобы показать лежащие под ним
чувствительные клетки макулы
коры головного мозга, и в результате возникает
слуховое ощущение. По мере удаления от основания
улитки и приближения к ее вершине базилярная
мембрана становится все более широкой и
эластичной, и чувствительность ее меняется таким
образом, что до верхушки доходят только низкие звуки.
Поэтому звуки высокой частоты стимулируют
чувствительные клетки только в основании улитки, а
звуки низкой частоты - только в ее верхушке.
Чистый тон, представляющий собой звук одной
частоты, будет стимулировать только один небольшой
участок базилярной мембраны; однако
большинство звуков состоит из разных частот и стимулирует
одновременно много участков. Слуховая зона коры
мозга интегрирует стимулы, поступающие из
разных участков базилярной мембраны, и в результате
возникает ощущение одного смешанного звука.
Различение интенсивности, или громкости, звука
зависит от того, что каждый участок базилярной
мембраны содержит набор сенсорных клеток с
разными порогами чувствительности к вибрации.
Например, тихий звук данной высоты может
стимулировать всего лишь несколько сенсорных клеток, а
более громкий звук той же частоты будет
возбуждать и некоторые другие клетки-с более высоким
порогом для вибрации, и в итоге будет происходить
пространственная суммация.
Равновесие
Поддержание равновесия тела в покое и при
движении связано с тем, что мозг все время получает
информацию о положении различных частей тела.
Импульсы от проприоцепторов, находящихся в
суставах и мышцах, сообщают о положении и
состоянии конечностей, но жизненно важная информация о
положении и движениях головы поступает из
вестибулярного аппарата внутреннего уха, который
состоит из овального и круглого мешочков и
полукружных каналов.
В этих структурах находятся рецепторные клетки,
снабженные волосками,- волосковые клетки,
которые расположены на плотных структурах,
омываемых эндолимфой. Движение головы приводит к
отклонению волосков и возникновению в волос-
ковой клетке рецепторного потенциала.
Участки поверхности овального и круглого
мешочков, называемые макулами, содержат
рецепторные клетки, волоски которых покрыты сверху
студенистой массой, содержащей кристаллы карбоната
кальция-так называемые отоконии (рис. 16.43).
Отоконии, отклоняясь под действием силы тяжести,
деформируют волоски, и благодаря этому
сенсорные клетки могут сообщать мозгу о движениях
головы относительно вертикали.
Овальный мешочек реагирует на движения
головы в вертикальном направлении, и отоконии
вызывают максимальную стимуляцию, когда
оттягивают рецепторные волоски вниз, как, например, при
положении тела вниз головой (рис. 16.44).
Круглый мешочек реагирует на наклоны головы в
стороны. Когда голова находится в вертикальном
положении, волосковые клетки круглого мешочка
расположены горизонтально. При наклоне головы
влево в левом и правом мешочках возникает разный
ответ. Левый мешочек испытывает повышенную
стимуляцию, так как отоконии оттягивают волоски
вниз, а на правой стороне стимуляция уменьшается.
Это ведет к изменению импульсов, направляющихся
в мозжечок, и позволяет чувствовать положение
головы.
Три полукружных канала расположены в трех
плоскостях под прямыми углами друг к другу. Они
воспринимают направление и скорость изменения в
положении головы. В основании каждого из каналов
имеется расширение - ампула, содержащая
студенистую массу-купулу. Купула покрывает волоски ре-
цепторных клеток и почти вплотную примыкает к
противоположной стенке ампулы (рис. 16.45). При
начале вращения головы и полукружных каналов
движению купулы противодействует инерция эндо-
лимфы, стремящейся остаться неподвижной. Это
приводит к относительному смещению купулы,
которая отгибается в сторону, противоположную
повороту головы. В сенсорных клетках в ответ на
раздражение возникает рецепторный потенциал,
возбуждающий потенциалы действия в
вестибулярных нейронах. Линейное ускорение воспринимается
и макулами, и купулами.
16.6. Эффекторы: эндокринная
система
Эффектор - это дифференцированная структура
(клетка, ткань, орган или система органов),
осуществляющая специфическую реакцию в ответ на
стимулы, поступающие из нервной системы.
Наиболее распространенными и важными являются те
эффекторы, которые участвуют в движении и
секреции.
Организмы, отвечающие на внешние стимулы
двигательной активностью, могут в зависимости от
их строения и образа жизни использовать
амебоидное движение или локомоцию с помощью ресничек,
жгутиков или мышц. Эти механизмы будут
подробно описаны в главе 17. У многоклеточных
организмов в реакции на внешние стимулы могут
участвовать все виды движения. Это может быть,
например, амебоидное движение лейкоцитов,
проходящих через стенки капилляров к месту инфекции в
тканях; усиленное движение ресничек трахеи при
инфекции в легких; активация жгутиков у
сперматозоидов под влиянием выделений влагалища;
рефлекторное сокращение пищевода при поступлении в
него пищевого комка.
Ответом на другие стимулы, как внешние, так и
внутренние, является секреция экзокринных или
эндокринных желез (разд. 8.3.3). В качестве примера
реакции стороны экзокринных желез может
служить слезотечение при попадании в глаз частичек
пыли, выделение поджелудочного сока при
попадании кислой пищи в двенадцатиперстную кишку или
усиленное образование синовиальной жидкости при
повреждении сустава. Эндокринные железы тоже
отвечают на разнообразные стимулы. Примеры:
секреция инсулина при повышении уровня глюкозы
в крови; выделение хорионического гонадотропина
стенками матки в начале беременности после
имплантации бластоцисты; повышение секреции
тироксина при длительном воздействии холода.
16.6.1. Эндокринная система
Эндокринная и нервная системы действуют
координирование, поддерживая постоянство внутренней
среды организма. Нервная система передает
сигналы в виде нервных импульсов, а эндокринная
использует для этого вещества, переносимые кровью.
При очевидном различии в механизме передачи
информации общим для обеих систем является
высвобождение химических веществ в качестве средств
коммуникации между клетками. Как полагают, обе
системы возникли и развивались параллельно, по
мере того как межклеточные связи усложнялись
вместе с увеличением размеров и сложности орга-
Частота импульсов,
приходящих от
сенсорных клеток
Положение головы
(как показано внизу)
Овальный
мешочек
Сенсорная клетка
Волоски
Отолит
Постепенное изменение
положения головы
(слева начальное положение,
справа конечное)
Левый круглый
мешочек
Рис. 16.44. Влияние положения головы на
активность рецепторных клеток овального и круглого
мешочков
Ампула
Купула
Эндолимфа
Чувствительный
волосок
Сенсорная
клетка
Вестибулярный
нейрон
Рис. 16.45. Схема строения ампулы полукружного
канала (поперечный разрез)
Таблица 16.10. Сводная таблица основных эндокринных желез человека, их функций и путей регуляции их активности
Железа
Гипоталамус
Задняя доля
гипофиза
Передняя доля
гипофиза
Паращитовид-
ная железа
Щитовидная
железа
Кора надпочеч
НИКОВ
Мозговой слой
надпочечников
Островки
Лангерганса
Желудок
Двенадцатиперстная
кишка
Гормоны
Либерины и статины; из них семь
идентифицированы; общее
число неизвестно (см. табл. 16.2.)
Здесь же образуются гормоны
задней доли гипофиза
Здесь не образуются никакие
гормоны, а хранятся и секретиру-
ются следующие:
Окситоцин
Антидиуретический гормон (ва-
зопрессин)
Фолликулостимулирующий
гормон (ФСГ)
Лютеинизирующий гормон (ЛГ)
Пролактин
Тиреотропный гормон (ТТГ)
Адренокортикотропный гормон
(АКТГ)
Гормон роста (соматотропный
гормон, СТГ)
Паратгормон
Трииодтиронин (T3) и тироксин
(T4)
Тиреокальцитонин
- Глюкокортикоиды (кортизол)
Минералокортикоиды (альдосте-
рон)
[ Адреналин (эпинефрин)
Норадреналин (норэпинефрин)
Инсулин (бета-клетки)
Глюкагон (альфа-клетки)
Гастрин
Секретин
Функции (или эффекты)
Регуляция секреции
специфических гипофизарных гормонов
Стимуляция активного выведения
молока молочной железой и
сокращений матки при родах
Уменьшение диуреза
У мужчин-стимуляция
сперматогенеза
У женщин - стимуляция роста
яйцевого фолликула
У мужчин-стимуляция секреции
тестостерона
У женщин-стимуляция секреции
эстрогенов и прогестерона, а
также овуляции; поддержание
существования желтого тела
Стимуляция образования и
секреции молока
Стимуляция синтеза и секреции
тиреоидных гормонов и роста
щитовидной железы
Стимуляция синтеза и секреции
гормонов коры надпочечников,
а также роста этой железы
Стимуляция белкового синтеза и
роста, особенно костей
конечностей
Повышение уровня Ca2+ и
снижение уровня PO45- в плазме
Регуляция основного обмена,
роста и развития
Снижение уровня Ca2+ в крови
Стимуляция расщепления белков,
синтеза глюкозы и гликогена.
Адаптация к стрессу.
Противовоспалительное и
антиаллергическое действие
Задержка Na+B почках,
повышение отношения Na+/K+ во
внеклеточной и внутриклеточной
жидкостях. Повышение
кровяного давления
Повышение частоты и силы
сердечных сокращений, сужение
капилляров в коже и
внутренних органах Расширение арте-
риол в сердце и скелетных
мышцах Повышение уровня
глюкозы в крови
Общее сужение мелких артерий;
повышение кровяного давления
Снижение уровня глюкозы в
крови, повышение поглощения и
использования клетками
глюкозы и аминокислот
Повышение уровня глюкозы в
крови, усиленное расщепление
гликогена до глюкозы в печени
Секреция желудочного сока
Секреция панкреатического сока;
угнетение секреции
желудочного сока
Факторы, регулирующие активность
Секреция регулируется уровнем
метаболитов и гормонов по
принципу обратной связи
Механизм отрицательной
обратной связи с участием
гормонов и нервной системы
Осмотическое давление крови
Уровни эстрогенов и
тестостерона в плазме; действуют через
гипоталамус
Уровень тестостерона в плазме;
действует через гипоталамус
Уровень эстрогенов в плазме,
действует через гипоталамус
Гормоны гипоталамуса
Уровень тиреоидных гормонов
(T3 и T4) в плазме; действует
через гипоталамус
Уровень АКТГ и кортикостеро-
идов в плазме; действует через
гипоталамус
Гормоны гипоталамуса
Уровень Ca2+ и PO4- в плазме
ТТГ
Уровень Ca2+ в крови
АКТГ
Уровни Na+ и K+ в плазме,
низкое кровяное давление
Симпатическая нервная система
Нервная система
Уровень глюкозы и
аминокислот в крови
Уровень глюкозы в крови
Присутствие пищи в желудке
Присутствие кислой пищи в
двенадцатиперстной кишке
Продолжение табл. 16.10.
Железа
Почки
Яйцевой
фолликул
Желтое тело
Плацента
Семенники
Гормоны
Холецистокинин (панкреозимин)
Ренин
Эстрогены (17р-эстрадиол)
Прогестерон
Прогестерон и эстрогены
Хорионический гонадотропин
Плацентарный лактоген
Тестостерон
Функции (или эффекты)
Сокращение желчного пузыря и
выведение панкреатического
сока в двенадцатиперстную
кишку
Превращение ангиотензиногена в
ангиотензин
Развитие вторичных женских
половых признаков, регуляция
менструального цикла
Поддержание беременности,
подавление овуляции
Стимуляция роста и развития
матки, продолжение развития
плода
Поддержание желтого тела
Стимуляция роста молочных
желез
Развитие вторичных мужских
половых признаков
Факторы, регулирующие активность
Присутствие жирных кислот и
аминокислот в
двенадцатиперстной кишке
Уровень Na+B плазме, низкое
кровяное давление
ФСГ и ЛГ
ЛГ
ЛГ
Развивающийся плод
Развивающийся плод
Развивающийся плод
ЛГ и ФСГ
низма. Главная роль обеих систем состоит в
регулировании, интеграции и координации важнейших
форм жизнедеятельности.
Железа-это структура, выделяющая
специфические вещества. В организме имеются железы двух
типов - экзокринные и эндокринные. Эндокринные
железы секретируют гормоны (от греч. hormao-
двигаю, побуждаю)-специфические химические
соединения, которые образуются в каком-то одном
участке тела, поступают в кровеносное русло и
доставляются с кровью к удаленным органам,
тканям или группам клеток, где проявляют свое
регулирующее действие. Важнейшие эндокринные
железы и выделяемые ими гормоны приведены в табл.
16.10.
Методы изучения желез и гормонов
Большая часть первоначальных сведений о
функциях эндокринных желез и их секретах была
получена при изучении тех изменений, которые
возникали в организме при заболеваниях, связанных с
недостаточной или избыточной активностью той
или иной железы. В некоторых случаях для
установления функции железы производили ее
экспериментальное удаление. Разработка методов выделения,
очистки, анализа и синтеза гормонов дала биологам
возможность снимать симптомы, вызванные
удалением железы, путем введения экстрактов этой
железы или синтетических препаратов ее гормона. Эти
методы позволили выяснить роль многих желез и их
секреции, но не могли быть применены ко всем
железам, например к печени или почкам, поскольку
их удаление вызвало бы нежелательные
последствия, не связанные с их эндокринной функцией.
Для измерения в крови, моче и других средах
концентраций гормонов, лекарственных препаратов,
ферментов, вирусов, бактериальных и опухолевых
антигенов и других органических материалов,
представляющих интерес для биологии, широко
применяется радиоиммунологический метод. Принцип
этого метода довольно прост. Нужно иметь
специфическую антисыворотку к исследуемому веществу
(антигену, в частности гормону), само это вещество
и, наконец, то же вещество, меченное
радиоактивным изотопом. Немеченый антиген конкурирует с
меченым за ограниченное число связывающих
участков антитела. Чем больше в данном материале
немеченого антигена, тем больше в его присутствии
уменьшается связывание меченого антигена
антителом. Для определения степени этого уменьшения
антиген, связанный с антителами, отделяют
(обычно путем осаждения) от свободного антигена.
Сравнивая полученные результаты с результатами
опытов, в которых использовался ряд пробирок с
известными концентрациями антигена,
рассчитывают его содержание в исследуемом образце.
Схематически это можно представить следующим
образом:
Ат 4- Ar + Ar* ±+ АтАг + Ат-Аг*
Специфичес- Немеченый Меченый Немеченый Меченый
кое антитело антиген антиген комплекс
антиген - антитело
В настоящее время в этой области наблюдается
тенденция заменять радиоактивную метку фермент-
но-флуоресцентной или хемолюминесцентной, что
избавляет от работы с радиоактивным материалом.
Механизмы действия гормонов
По химической природе все гормоны
позвоночных можно разделить на четыре группы: 1)
производные аминов (например, тирозина), 2) пептиды и
белки, 3) стероиды и 4) жирные кислоты (табл.
16.11). В регулировании секреции гормонов могут
участвовать следующие механизмы:
А. Присутствие специфического метаболита в
крови. Например, избыток в ней глюкозы вызывает
секрецию поджелудочной железой инсулина, ко-
Гипоталамус
Количество
вещества
Ко рти кол и берин
0,1 мкг
Передняя
доля гипофиза
Адренокор -
тикотропный
гормон
1,0 мкг
Кора
надпочечников
Кортизол
40 мкг
ПЕЧЕНЬ
Глюкоза -
-► Гликоген 5600 мкг
Рис. 16.46. Пример «каскадного» эффекта в
регуляции превращения глюкозы в гликоген в результате
освобождения кортиколиберина. Исходный эффект
усилен в 56000 раз (По данным Bradley, 1976.)
торыи снижает уровень глюкозы в крови.
Б. Присутствие в крови другого гормона.
Например, многие гормоны, выделяемые передней
долей гипофиза, стимулируют секрецию гормонов
другими железами организма.
В. Стимуляция со стороны вегетативной нервной
системы. Так, например, при беспокойстве,
стрессе или опасности клетки мозгового слоя
надпочечников начинают выделять адреналин и
норадреналин.
В первых двух случаях время секреции гормона и
его количество регулируются по принципу обратной
связи. В регуляции активности железы участвует и
положительная, и отрицательная обратная связь, но
положительная, повышающая нестабильность
системы, действует лишь как часть общего регулятор-
ного механизма. Например, высвобождение люте-
инизирующего гормона (ЛГ) под влиянием
эстрогенов осуществляется по принципу положительной
обратной связи, но его чрезмерную секрецию
предотвращает в надлежащий момент прогестерон,
выделяющийся под действием Л Г.
Секреция гормонов, стимулируемая каким-то
другим гормоном, обычно находится под
контролем гипоталамуса и гипофиза, и конечный
метаболический или трофический эффект может быть
результатом секреции трех различных гормонов, как
это показано на рис. 16.46. Такого рода каскадный
механизм имеет большое значение, так как
благодаря ему в эндокринной цепи во много раз
усиливается действие малых количеств исходного
гормона.
Гормоны обладают специфичностью и
воздействуют только на те клетки-мишени, которые
обладают специальными рецепторами белковой или ли-
попротеиновой природы, реагирующими с данным
гормоном. У клеток, не являющихся мишенями для
данного гормона, таких рецепторов нет, и поэтому
гормон не оказывает на них влияния.
Многие гормоны имеют в своей молекуле
специфические участки, ответственные за присоединение к
рецептору. Будучи связан рецептором, гормон
может воздействовать 1) на плазматическую
мембрану, 2) на фермент, находящийся в этой мембране,
3) на клеточные органеллы или 4) на гены. Ниже
приводятся примеры каждого из перечисленных
механизмов действия гормонов.
1. Один из эффектов инсулина - усиленное
поглощение глюкозы клеткой - обусловлен тем, что
гормон, связавшись с рецептором, изменяет
проницаемость клеточной мембраны для глюкозы.
2. Адреналин и многие пептидные гормоны при-
Таблица 16.11. Химическая природа важнейших гормонов
Химическая группа
Амины
<
Катехо-
ламины
Производные
тирозина
Белки и пептиды
Стероиды
Жирные кислоты
Гормон
Адреналин
Норадреналин
Тироксин
Трииодтиронин
Либерины и ста-
тины
Фолликуло-
стимулирующий
гормон
Лютеинизирую-
щий гормон
Пролактин
Тиреотропный
гормон
АКТГ
Гормон роста
Окситоцин
Вазопрессин
Паратгормон
Основной источник
Симпатическая
нервная система,
мозговой слой
надпочечников
Щитовидная
железа
Гипоталамус
Передняя доля
гипофиза
Задняя доля
гипофиза
Паращитовидные
железы
Тиреокальцитонин Щитовидная
железа
Инсулин
Глюкагон
Гастрин
Секретин
Тестостерон
Эстрогены
Прогестерон
Кортикостероиды
Простагландины
Островки Лангер-
ганса
(поджелудочная
железа)
Слизистая желудка
Слизистая
двенадцатиперстной
кишки
Семенники
Яичники и
плацента
Кора
надпочечников
Многие ткани
соединяются к поверхностным рецепторам
клеточной мембраны и вызывают освобождение «второго
посредника», который начинает цепь
ферментативных реакций, приводящих к специфическому
конечному эффекту. Во многих случаях этим вторым
посредником является нуклеотид циклический аде-
нозинмонофосфат (цАМФ), который образуется из
АТФ под действием фермента аденилатциклазы
после ее активации гормон-рецепторным
комплексом. В упрощенной форме этот механизм показан на
рис. 16.47.
3. Один из эффектов тироксина осуществляется
на уровне митохондрий, где гормон влияет на
ферменты, участвующие в транспорте электронов,
сопряженном с синтезом АТФ (разд. 11.5.4). Под
влиянием тироксина значительная часть энергии,
выделяющейся в процессе транспорта электронов,
теряется в виде тепла.
4. Стероидные гормоны, в том числе гормон
линьки насекомых (ждизон), проходят через
плазматическую мембрану и связываются с рецепторами в
цитоплазме. Образовавшийся комплекс
транспортируется в клеточное ядро, где воздействует
непосредственно на хромосомы путем активации генов и
стимуляции транскрипции (т. е. образования
информационной РНК).
По-видимому, и во многих других случаях
гормоны оказывают свое действие, влияя на ферменты,
связанные с мембранами, или на геном.
16.6.2. Гипоталамус и гипофиз
Как уже говорилось, связь между клетками
осуществляют нервная и эндокринная системы,
действующие независимо или совместно. Главными
центрами координации и интеграции функций этих двух
регуляторных систем служат гипоталамус и
гипофиз. Гипоталамус играет ведущую роль в сборе
информации от других участков головного мозга и
от собственных кровеносных сосудов. Эта
информация передается в гипофиз, который путем секреции
специфических гормонов прямо или косвенно
регулирует активность всех других эндокринных желез.
Гипоталамус
Гипоталамус расположен в основании переднего
мозга непосредственно под таламусом и над
гипофизом. Он состоит из нескольких дискретных
участков, называемых ядрами и представляющих собой
скопления тел нейронов, аксоны которых
оканчиваются на кровеносных капиллярах в срединном
возвышении и в задней доле гипофиза (рис. 16.48).
Регуляция многих физиологических функций (голод,
жажда, сон, выработка и отдача тепла)
осуществляется нервным путем с помощью импульсов,
поступающих из гипоталамуса по нервам
вегетативной системы. В то же время контроль над
эндокринной секрецией со стороны гипоталамуса
основан на его способности регистрировать содержание
метаболитов и гормонов в крови. Информация,
поступающая при этом в гипоталамус, вместе с
информацией от множества других отделов мозга
передается в гипофиз-либо путем выделения в
кровеносное русло специальных гормонов, либо через
нейроны. В последнем случае передатчиками служат
специализированные нейроны, называемые нейро-
секреторными клетками.
Все нервные клетки выделяют в синаптических
окончаниях химические вещества - медиаторы, но у
Адреналин
КРОВЬ
Глюкоза
Клеточная
мембрана
Рецептор
Адени-
\ латци -
. клаза .
АТФ-
^ цАМФ
(несколько
этапов)
Неактивная —
фосфорилаза Ъ
Активная
фосфорилаза а
Гликоген ■
Глюкоза
Рис. 16.47. Механизм действия адреналина,
вызывающего освобождения глюкозы из клеток печени
(упрощенная схема). Активаиия аденилатциклазы,
связанной с мембраной, приводит к образованию
циклического АМФ (цАМФ); цАМФ активирует
ферментные системы, расщепляющие гликоген до
глюкозы, которая диффундирует из клеток в кровеносное
русло.
Первичная сеть
(в нее секретируются
либерины и статины)
Нейросекреторные
клетки
Срединное
возвышение
Гипоталамо-
гипофизарная
портальная
система
Вторичная сеть
(в нее высво
бождаются
гормоны)
Передняя доля
гипофиза
Гипоталамо-
гипофизарный
нервный тракт
Нейрогемальный
участок
Задняя доля
гипофиза
Ветви
гипофизарной
вены
Рис. 16.48. Схема взаимоотношений между нейро-
секреторными клетками и кровеносными сосудами в
гипоталамусе и гипофизе.
нейросекреторных клеток эта способность достигла
особенно высокого развития. Вещества,
образующиеся в телах этих клеток, упаковываются в
гранулы или пузырьки и с током аксоплазмы
транспортируются вниз по аксону. Нервные окончания
этих клеток образуют синапсы на капиллярах, в
которые они и высвобождают свой секрет под
действием нервных импульсов, приходящих по аксону.
Гипофиз
Гипофиз - небольшая железа красновато-бурого
цвета, которая у человека весит около 0,5 г и
связана с головным мозгом ножкой. Гипофиз имеет
двойное происхождение, и эта особенность
проявляется в его функциональной активности. Он
состоит из двух долей - передней и задней.
Передняя доля гипофиза, или аденогипофиз. Этот
отдел образуется из направленного вверх выроста
крыши первичной ротовой полости. Он связан с
гипоталамусом кровеносными сосудами гипотала-
мо-гипофизарной воротной системы, которая состоит
из первичной капиллярной сети, расположенной в
срединном возвышении, и вторичной капиллярной
сети, находящейся в аденогипофизе.
Нервные окончания специализированных
нейросекреторных клеток гипоталамуса выделяют в
первичную капиллярную сеть две группы веществ,
получивших название либеринов (рилизинг-факторов)
и статинов. Из первичной капиллярной сети эти
факторы поступают во вторичную капиллярную
сеть и, воздействуя на переднюю долю гипофиза,
стимулируют или тормозят высвобождение одного
или нескольких из шести «тропных» гормонов,
которые здесь образуются и хранятся. Тропные
гормоны выводятся клетками аденогипофиза в
кровоток, разносятся с кровью по всему организму и
действуют на специфические органы-мишени (табл.
16.12).
Секреция первых двух из указанных в таблице
гормонов аденогипофиза находится под двойным
контролем гипоталамуса - стимулирующим и инги-
бирующим. Выделение четырех остальных
гормонов регулируется по принципу отрицательной
обратной связи гормонами желез-мишеней,
действующими через посредство рецепторов гипоталамуса и
гипофиза. В общем виде это происходит следующим
образом: гипофизарный гормон стимулирует
соответствующую периферическую железу, и уровень ее
гормонов в крови повышается; это приводит к
подавлению секреции гипоталамического фактора
(либерина) и гипофизарного гормона; в результате
активность периферической железы снижается, и
когда концентрация ее гормонов в крови падает
ниже определенного уровня, их угнетающее
действие на гипоталамус и гипофиз уменьшается и
последние снова усиливают секрецию гормонов.
Этот регуляторный механизм будет подробнее
описан в разд. 18.4.4 и 19.6.
Задняя доля гипофиза, или нейрогипофиз. Этот
отдел развивается как нижний вырост
гипоталамуса. Он не синтезирует никаких гормонов, а лишь
хранит и высвобождает два гормона -
антидиуретический гормон (вазопрессин) и окситоцин.
Антидиуретический гормон выделяется в кровь при
уменьшении содержания воды в плазме; он усиливает
обратное всасывание воды в дистальных канальцах
и собирательных трубках почек, и она удерживается
в плазме. При этом уменьшается количество мочи и
возрастает ее осмотическая концентрация (разд. 19.6).
Окситоцин вызывает сокращения матки при родах и
активное выведение молока из сосков (разд. 20.3.8).
Антидиуретический гормон и окситоцин
образуются в телах нейросекреторных клеток, лежащих в
ядрах гипоталамуса, и транспортируются по их
аксонам, находясь в комплексе с
белком-носителем - нейрофизином. Эти нейросекреторные клетки
являются значительно более специализированными,
чем те, которые выделяют либерины и статины. В
задней доле гипофиза расширенные окончания их
аксонов образуют структуры, называемые нейроге-
мальными органами: здесь эти окончания,
окруженные соединительной тканью, прилегают к
кровеносным капиллярам (рис. 16.49).
Афферентные нервные сигналы, приходящие к
телам нейросекреторных клеток из других участков
мозга, передаются по их аксонам к нейрогемальным
органам. Под действием этих сигналов гормон,
хранящийся в нейрогемальном органе в пузырьках,
высвобождается в кровеносное русло и
транспортируется кровью к органам-мишеням.
Физиологическая реакция, к которой в конечном счете приводит
такое преобразование сенсорного стимула в
секрецию гормона, носит название нейроэндокринного
ответа. Многие виды нейроэндокринных ответов
проявляются в поведенческих реакциях - нейроэндо-
Таблица 16.12. Главные гипоталамические факторы,
регулируемые ими гормоны гипофиза и органы-мишени этих
гормонов
Гипоталамиче-
ский фактор
Соматоли-
берин
Соматоста-
тин
Пролактоли-
берин
Пролакто-
статин
Люлиберин
(фоллибе-
рин)
Тиреолибе-
рин
Кортиколи-
берин
Действие на аденогипофиз
Стимулирует секрецию
гормона роста
Тормозит секрецию
гормона роста
Стимулирует секрецию
пролактина
Угнетает секрецию
пролактина
Стимулирует секрецию
фолликулостимулирующе-
го и лютеинизирующего
гормонов
Стимулирует секрецию
тиреотропина
Стимулирует секрецию
АКТГ
Мишени
соответствующего
гипофизарного
гормона
Большинство
тканей
Яичники и
молочные
железы
Яичники и
семенники
Щитовидная
железа
Кора
надпочечников
Афферентные
нервные
импульсы
Тело нейро-
секреторной
клетки
Нервные
импульсы,
идущие
по аксону
Митохондрия
Нейро -
гемальный
пузырек
Базальная
мембрана
Нейроге-
мальный
участок
Гормоны, высвобождаемые
из нейросекреторных
гранул в кровь
Ядро эндо-
телиальной
клетки
Стенка капилляра
Рис. 16.49. Схема строения нейросекреторной
клетки и нейрогемального органа (единый масштаб не
выдержан).
кринных рефлексах, которые часто бывают связаны
с размножением (ухаживание, лактация и др.).
16.6.3. Шишковидная железа, или
эпифиз
Эпифиз-это крошечная железа, образующаяся из
крыши промежуточного мозга и прикрытая сверху
мозолистым телом и большими полушариями. Вес
эпифиза составляет около 150 мг. Древние римляне
считали эпифиз «вместилищем души», однако
теперь известно, что это эндокринная железа. Эпифиз
не имеет никакой прямой связи с центральной
нервной системой, но богато снабжен кровеносными
сосудами. Полагают, что он секретирует несколько
гормонов, в том числе мелатонин. Эпифиз иннерви-
руют симпатические нейроны, которые находятся
под влиянием ядер головного мозга, получающих и
передающих сигналы от фоторецепторов сетчатки.
В темноте синтез мелатонина повышается, и этот
гормон, действуя через мозг, изменяет активность
щитовидной железы, надпочечников и гонад.
Имеются данные, позволяющие предполагать, что
мелатонин воздействует на мозг и влияет на время
наступления ряда физиологических процессов, таких
как половое созревание, овуляция и сон. Опухоли
эпифиза вызывают у мальчиков преждевременное
половое созревание. Кроме того, эпифиз,
по-видимому, играет роль «биологических часов» и
действует как орган, преобразующий периодическую
нервную активность, вызываемую светом, в
эндокринную секрецию. Для выяснения многочисленных
функций этой железы нужны дальнейшие
исследования.
16.6.4. Паращитовидные железы
У человека имеются четыре небольшие
паращитовидные железы, погруженные в ткань щитовидной
железы. Эти железы секретируют только один
гормон - паратгормон, который представляет собой
полипептид, состоящий из 84 аминокислот.
Паратгормон и гормон щитовидной железы кальцитонин
действуют как антагонисты, регулирующие
концентрацию кальция и фосфата в плазме. Секреция
паратгормона поддерживает концентрацию кальция
в плазме на нормальном уровне около 2,5 ммоль/л
и снижает концентрацию фосфора. Функция пара-
щитовидных желез регулируется простым
механизмом отрицательной обратной связи, показанным на
рис. 16.50.
Пониженная активность паращитовидных желез -
гипопаратиреоз - приводит к снижению уровня
кальция в плазме и тканях в результате выведения его с
мочой; вследствие этого может развиться тетания-
патологическая склонность к длительному
сокращению мышц. Экскреция фосфата при этом
уменьшается, а уровень его в плазме возрастает.
16.6.5. Щитовидная железа
У человека щитовидная железа имеет форму
галстука-бабочки. Она находится в области шеи и состоит
из двух долей, расположенных по обеим сторонам
трахеи и соединенных тонким перешейком. Вся
железа весит около 25 г. Она секретирует три
активных гормона - трииодтиронин (T3), тироксин (T4) и
тиреокальцитонин. T3 и T4 участвуют в регуляции
основного обмена, роста и развития, а
тиреокальцитонин регулирует концентрацию кальция в плазме.
Строение щитовидной железы
Щитовидная железа состоит из множества
фолликулов диаметром 0,1 мм, которые заполнены
прозрачным «коллоидом», содержащим гликопротеин ти-
реоглобулин. Стенка каждого фолликула состоит из
одного слоя кубических клеток. При активации
щитовидной железы тиреотропным гормоном адено-
гипофиза клетки фолликулов становятся цилиндри-
Много Ca2^4
Много PO-^
4
Кальцитонин
Паратгормон
Мало PO3.
4
''Мало Ca2+"
Рис. 16.50. Роль паратгормона и кальцитонина в
регуляции уровня кальция в плазме.
3,5,3-трииодтиронин (T )
3,5,3,5-тетраиодтиронин (T )
Тироксин
Рис. 16.51. Структурные формулы главных тиреоид-
ных гормонов.
ческими и на их внутренней поверхности
появляются микроворсинки.
Образование и секреция
тиреоидных гормонов
Иод поступает в щитовидную железу в форме ионов
I-, активно поглощаемых клетками фолликулов из
крови многочисленных капилляров, окружающих
фолликулы. Затем эти ионы окисляются до
молекулярного иода пероксидазами, и молекулярный иод
реагирует с аминокислотой тирозином, входящей в
состав тиреоглобулина - белка, выделяемого
фолликулярными клетками в просвет фолликула.
Дальнейшее иодирование молекул тирозина и
последующая их конденсация приводит к образованию двух
главных тиреоидных гормонов, структура которых
показана на рис. 16.51.
Функция T3 и T4
T3 и T4 оказывают большое влияние на многие
метаболические процессы, включая обмен
углеводов, белков, жиров и витаминов. Их главное
физиологическое действие состоит в повышении
интенсивности основного обмена - в так называемом калори-
генном эффекте. Интенсивность основного обмена,
составляющая у человека 160 кДж на 1 м2
поверхности тела в 1 ч, поддерживается на стабильном
уровне тироксином, который способствует
расщеплению глюкозы и жиров с образованием легко
сгорающих продуктов (разд. 18.4.2). Калоригенный
эффект связан с повышением поглощения кислорода
и скорости ферментативных реакций, участвующих
в транспорте электронов в митохондриях. В
конечном результате это приводит к повышенному
образованию АТФ и тепла в тканях. Конкретный
механизм действия тироксина неизвестен.
Вместе с гормоном роста T3 и T4 стимулируют
синтез белка, что приводит к ускорению роста. Этот
эффект отчетливо проявляется при метаморфозе у
лягушки (разд. 21.7.4) и подтверждается задержкой
роста у детей, страдающих недостаточностью
тиреоидных гормонов.
Во многих метаболических процессах,
подверженных влиянию тироксина, его роль, по-видимому,
состоит в усилении действия других гормонов,
таких как инсулин, адреналин и глюкокортикоиды
(разд. 18.5.2).
Регуляция секреции T3 и T4
Тиреоидные гормоны оказывают более длительное
действие, чем большинство других гормонов,
поэтому поддержание постоянного уровня T3 и T4
имеет жизненно важное значение для организма.
Это одна из причин, почему T3 и T4 запасаются в
железе и в любой момент готовы для выведения в
кровь.
Высвобождение T3 и T4 из щитовидной железы
регулируется их концентрацией в крови. Эта
регуляция осуществляется на уровне гипофиза и
гипоталамуса по принципу отрицательной обратной
связи. Когда концентрация T3 и T4 в крови становится
выше той, которая необходима для поддержания
постоянного уровня основного обмена, они
подавляют секрецию тиреолиберина гипоталамусом и ти-
реотропного гормона гипофизом. На этот механизм
накладывается влияние внешних факторов
(например, температуры), которые через посредство
вышележащих центров головного мозга стимулируют
выделение тиреолиберина; в результате этого
изменяется порог чувствительности гипофиза к
сигналам отрицательной обратной связи. Эти механизмы
регуляции представлены на рис. 16.52.
Повышенная активность
щитовидной железы (гипертиреоз)
Как чрезмерная, так и недостаточная активность
щитовидной железы может сопровождаться
увеличением ее размеров - образованием зоба. При
повышенной секреции тироксина увеличенной
щитовидной железой развивается гипертиреоз. Его
симптомы: ускорение ритма сердца (тахикардия, см. разд.
18.1.5), учащение дыхания и повышение
температуры тела. Интенсивность основного обмена может
быть повышена на 50%; параллельно повышаются
потребление кислорода и теплопродукция. Больные
становятся очень нервными и раздражительными, и
при вытягивании рук у них дрожат кисти. Крайняя
степень гипертиреоза, называемая тиреотоксикозом,
сопровождается повышенной возбудимостью
сердечной мышцы, что может привести к сердечной
недостаточности. Лечение состоит в удалении
большей части железы хирургическим путем или в
разрушении ее путем введения радиоактивного иода.
Пониженная активность
щитовидной железы (гипотиреоз)
Гипотиреоз может быть связан с недостаточной
секрецией тиреотропного гормона гипофизом, с де-
фицитом иода в пищевом рационе или с
недостаточностью ферментных систем, участвующих в
образовании тироксина. Недостаток тироксина с момента
рождения приводит к кретинизму, который
выражается в задержке роста и умственного развития. Если
выявить заболевание на ранней стадии, можно
добиться восстановления нормального роста и
развития путем введения тироксина. Недостаточность
тироксина во взрослом состоянии приводит к
состоянию, называемому микседемой, симптомы
которого состоят в снижении основного обмена,
потребления кислорода, легочной вентиляции,
частоты сокращений сердца и температуры тела.
Снижается также умственная активность, становятся
замедленными движения. Вес тела увеличивается за
счет накопления под кожей полужидкого эксудата. В
результате лицо и веки отекают, язык набухает,
кожа становится грубой, выпадают волосы и
редеют брови. Заболевание лечат таблетками с
тироксином, при этом все перечисленные симптомы
исчезают.
Рецепторы
Тормозящее
воздействие
Стимуляция
Кора
Холод
Тепло
Гипоталамус
Регулирует
"заданный
уровень"
тироксина
в крови
Передняя доля
гипофиза
Щитовидная
железа
Кровь/ткани
Рис. 16.52. Общая схема взаимодействий,
регулирующих секрецию тироксина и участвующих в го-
меостатической регуляции интенсивности
метаболизма
Кальцитонин
В щитовидной железе наряду с фолликулярными
клетками имеются еще так называемые С-клетки.
Они вырабатывают полипептидный гормон
кальцитонин, который действует как антагонист парат-
гормона - снижает уровень кальция в крови.
16.6.6. Надпочечники
Надпочечники-парный орган. Эти железы
прилегают к верхушкам почек, и каждая весит около 5 г.
Надпочечник состоит из двух слоев, которые имеют
разное происхождение и функционируют
независимо друг от друга. Наружный слой - кора -
образуется из клеток нервной пластинки и составляет 80%
всей массы железы. Кора имеет плотную
консистенцию и покрыта фиброзной капсулой; ее функция
необходима для поддержания жизни. Внутренний
слой надпочечника - мозговой - образуется из
клеток нервного гребешка и сохраняет сходство с
нервной тканью. Жизненно важного значения он не
имеет.
Кора надпочечников
По гистологическому строению кора надпочечников
делится на три зоны, которые состоят из клеток,
группирующихся вокруг капилляров. Она выделяет
стероидные гормоны двух типов. Данные о
гистологическом строении зон коры, секретируемых ими
гормонах и их функциях приводятся в табл. 16.13.
Все стероиды образуются из общего
предшественника - холестерола (холестерина), который может
синтезироваться в самой коре и поглощаться из
крови, когда он поступает в организм с пищей. В
разных зонах коры холестерол превращается в
специфические для них стероидные гормоны.
Будучи жирорастворимыми соединениями,
стероидные гормоны диффундируют через клеточные
мембраны и связываются с белками-рецепторами в
цитоплазме. Образующиеся комплексы
транспортируются в ядро, где они присоединяются к
определенным участкам хромосом и дерепрессируют
(активируют) гены, вызывая образование
информационных РНК.
Размеры надпочечников в большой степени
связаны с секрецией АКТГ и со способностью организма
выдерживать «стресс». Во время продолжительного
стресса надпочечники увеличиваются. Изучение
поведения животных показало, что с увеличением
плотности популяции выброс кортикостероидов при
стрессе повышается. У тех животных, у которых
существует социальная иерархия, отмечается
прямая связь между положением особи на
иерархической лестнице и размерами надпочечников.
Регуляция активности коры
Как видно из табл. 16.13, секреция минералокор-
тикоидов стимулируется ренином и ангиотензином, а
глюкокортикоидов - адренокортикотропным
гормоном (АКТГ). Соответствующие механизмы
показаны на рис. 16.53.
АКТГ - полипептид, состоящий из 39
аминокислот. Он присоединяется к рецепторам на
поверхности кортикальных клеток и активирует аденилат-
циклазу, которая превращает АТФ в цАМФ. цАМФ
участвует в активации ферментов протеинкиназ,
которые способствуют превращению холестерола в
прегненолон. Кроме того, АКТГ стимулирует
поглощение клетками холестерола и ответствен за
поддержание размеров коры надпочечников и
активности ферментов, участвующих в биосинтезе
стероидов.
Мозговой слой надпочечников
Мозговой слой надпочечника образует его
сердцевину. Это мягкая ткань, состоящая из клеточных
тяжей, окруженных капиллярами и обильно иннер-
вированных. Клетки этого слоя представляют собой
видоизмененные постганглионарные симпатические
нейроны и при стимуляции со стороны преганглио-
нарных нейронов выделяют норадреналин и
адреналин (разд. 16.6.1). Мозговой слой надпочечников не
является необходимым для жизни, так как его роль
сводится лишь к усилению действия вегетативной
нервной системы.
Норадреналин и адреналин образуются из
аминокислоты тирозина и принадлежат к группе
биологически активных соединений, называемых катехол-
аминами (рис. 16.54). Клетки мозгового слоя
надпочечников секретируют оба гормона, тогда как в
постганглионарных синапсах вегетативной нервной
системы выделяется только норадреналин. В
основном действие обоих гормонов одинаково (табл.
16.14), но на кровеносные сосуды они влияют
по-разному. Норадреналин вызывает сужение всех
сосудов, тогда как адреналин, сужая сосуды кожи и
кишечника, расширяет сосуды мозга и мышц. В
органах-мишенях оба гормона взаимодействуют с
рецепторами двух типов-а- и β-адренэргическими
рецепторами. Присоединяясь к ним, катехоламины
активируют аденилатциклазу и таким образом
вызывают специфическую реакцию ткани подобно
тому, как это показано на рис. 16.53 для АКТГ. В
большинстве органов имеются и а-, и β-рецепторы,
причем α-рецепторы обладают, по-видимому,
большим сродством к норадреналину, а β-рецепторы - к
адреналину.
Действие катехоламинов распространяется на
весь организм. Они подготавливают животное к
реакции типа «борьбы или бегства» и облегчают
адаптацию к неожиданному стрессу, вызванному,
например, болевым ощущением, холодом,
гипогликемией, понижением кровяного давления,
раздражением или гневом.
Таблица 16.13. Гистологическое строение и функции трех зон коры надпочечников (сводная таблица)
Зона
Клубочко-
вая зона
Пучковая
зона
Сетчатая
зона
Гистологические
особенности
Состоит из
округлых
скоплений
мелких
клеток,
содержащих
удлиненные
митохондрии
Два-три ряда
параллельных
колонок
клеток,
расположенных
под прямым
углом к
поверхности
железы и
содержащих
округлые
митохондрии
Состоит из
нерегулярно
расположенных колонок
клеток,
содержащих
удлиненные
митохондрии
Гормоны
Минерало-
кортикоиды
(альдосте-
рон и др.)
Глюко-
кортикои-
ды (кор-
тизол и
др.)
Функции гормонов
Контролируют водно-солевой обмен
путем активации катионных насосов
в мембранах, в результате чего ионы
Na+ и Cl- удерживаются, a K+
выводится. Предотвращают
избыточную потерю Na+ с потом, слюной и
мочой и поддерживают
осмотическую концентрацию жидкостей тела
на постоянном уровне
A. Влияние на углеводный обмен: 1)
усиливают глюконеогенез; 2)
способствуют образованию гликогена
в печени; 3) повышают уровень
глюкозы в крови
Б. Белковый обмен: 1) стимулируют
расщепление белков плазмы; 2)
увеличивают запас свободных
аминокислот, доступных для синтеза
белка в печени
B. Другие функции: 1) подавляют
воспалительные и аллергические
реакции; 2) уменьшают образование
антител
Примечания
Ренин, выделяемый юкстагломеруляр-
ным аппаратом почек, катализирует
образование ангиотензина, а
последний стимулирует секрецию альдосте-
рона, который повышает реабсорб-
цию Na+ в почках и вызывает
освобождение антидиуретического
гормона, повышающего реабсорбцию
воды в почечных канальцах. АКТГ
не влияет на секрецию альдостерона
Повышенная активность приводит к
развитию синдрома Кушинга
(отложение жира в области живота,
атрофия мышц, повышенное кровяное
давление, диабет, чрезмерное об-
волосение). Сходный синдром,
связанный с повышенной секрецией
АКТГ гипофизом, называют
болезнью Кушинга. Пониженная
активность приводит к развитию ад-
дисоновой болезни, выражающейся
в мышечной слабости, низком
кровяном давлении, слабой
сопротивляемости инфекции, потемнении
кожи
Низкая
концентрация
Na+ в плазме
Лихорадка
Боль
"Биологические
часы"
Болезни
Тревога
Сон
Стресс
Почки
(юкстагломе-
рулярный
аппарат)
Гипоталамус
Кортиколиберин
Ренин
Передняя доля
гипофиза,цАМФ
Ангиотен -
зиноген
плазмы
АКТГ-секрети -
рующие клетки
Ангиотензин
Адренокорти -
котропный гормон
(АКТГ)
Отрица-
тельная
обратная
связь
Клубочковая зона
Пучковая
и сетчатая зоны
Альдостерон
(минералокортикоид)
Кортизол
(глюкокортикоид)
Рис. 16.53. Общая схема взаимодействий,
регулирующих секрецию главных гормонов коры
надпочечников.
Норадреналин
Адреналин
Рис. 16.54. Структурные формулы норадренолина и
адреналина.
16.6.7. Поджелудочная железа
Поджелудочная железа выполняет и экзокринную, и
эндокринную функцию и связана с
пищеварительным трактом (разд. 10.4). Основную массу железы
составляет экзокринная ткань, состоящая из ациноз-
ных клеток; эти клетки образуют округлые
структуры - ацинусы, в полость которых выводятся
пищеварительные ферменты зимогеновых гранул (разд.
10.4.9). Между ацинусами разбросаны островки Лан-
герганса, содержащие небольшое число крупных
альфа-клеток, многочисленные мелкие бета-клетки
и кровеносные капилляры. Альфа-клетки секретиру-
ют глюкагон, а бета-клетки - инсулин, и эти два
гормона оказывают противоположное действие на
уровень глюкозы в крови.
Инсулин
Инсулин - белок, состоящий из 51 аминокислоты.
Он выделяется в ответ на повышение уровня
глюкозы в крови (более 100 мг на 100 мл) и на увеличение
концентрации глюкагона. В плазме крови инсулин
связан с бета-глобулином. Он усиливает процессы
анаболизма во всех органах и тканях тела. Гормон
присоединяется к рецепторам на поверхности
клеточной мембраны, и это приводит к изменению ее
проницаемости и активности ряда ферментных
систем, что вызывает в клетке следующие эффекты:
1) увеличение скорости превращения глюкозы в
гликоген (гликогенеза);
2) ускорение переноса глюкозы через клеточные
мембраны в мышцах и жировой ткани;
3) усиление синтеза белка и липидов;
4) повышение скорости синтеза АТФ, ДНК и РНК.
Инсулин необходим для жизни, так как это
единственный гормон, снижающий концентрацию
глюкозы в крови. Недостаточная секреция инсулина
приводит к нарушению метаболизма, известному
под названием сахарного диабета. При этой болезни
содержание глюкозы в крови превышает тот
максимальный уровень, при котором глюкоза еще может
реабсорбироваться в почках (так называемый
почечный порог), и она начинает выводиться с мочой.
Связь сахарного диабета с функцией поджелудочной
железы была установлена почти столетие назад, но
только в 1921 г. канадские исследователи Бантинг и
Бест добились успеха в лечении диабета с помощью
инсулина, выделенного из поджелудочных желез
животных. В 1956 г. Сэнгер в Кембридже
расшифровал первичную структуру инсулина, открыв таким
образом возможность получать его синтетическим
путем. Эффекты, вызываемые недостатком или
избытком инсулина, суммированы в табл. 16.15.
Глюкагон
Глюкагон - полипептид, состоящий из 29
аминокислот. Он выделяется вместе с несколькими другими
гормонами в ответ на снижение уровня глюкозы в
крови, вызванное усиленным ее использованием.
Глюкагон стимулирует образование углеводов за
Таблица 16.14. Физиологические эффекты адреналина и
норадреналина
Расширяют зрачки
Заставляют волосы вставать дыбом
Расширяют бронхиолы, улучшая таким образом легочную
вентиляцию
Угнетают перистальтику и пищеварение
Препятствуют сокращению мочевого пузыря
Увеличивают силу и частоту сокращений сердца
Вызывают генерализованное сужение кровеносных сосудов
Повышают кровяное давление
Стимулируют превращение гликогена в глюкозу в печени
Снижают пороги реакции органов чувств
Повышают настороженность
Таблица 16.15. Некоторые симптомы недостатка и избытка
инсулина
При недостатке
Высокий уровень глюкозы
в крови (гипергликемия)
Распад мышечной ткани
Потеря веса
Утомляемость
При избытке
Низкий уровень глюкозы в
крови (гипогликемия)
Повышенный аппетит
Потливость
Раздражительность
Двоение в глазах
счет расщепления белков и жиров (глюконеогенез;
разд. 18.5.2). Он связывается в печени с
рецепторами, находящимися на поверхности клеточных
мембран, и активирует аденилатциклазу,
катализирующую синтез цАМФ. Действие глюкагона сходно с
действием адреналина: оба гормона активируют
фосфорилазу, расщепляющую гликоген с
образованием глюкозо-6-фосфата (см. рис. 16.47). На
гликоген мышц глюкагон не влияет.
Общие схемы действия инсулина и глюкагона
приведены на рис. 18.7 и 18.23.
16.6.8. Различия между нервной и
эндокринной регуляцией
Нервная и эндокринная системы могут действовать
порознь, но могут и совместно регулировать многие
виды жизнедеятельности организма. В этой главе
были описаны обе ситуации, и теперь полезно будет
резюмировать различия между этими двумя
системами, так как это позволит яснее увидеть
преимущества каждой системы в связи с функцией,
которую она выполняет. Рассмотрите следующую
таблицу:
Таблица 16.16. Различия между нервной и эндокринной
регуляцией
Нервная регуляция
Информация передается по
аксонам в виде
электрических импульсов
(химическая передача в
синапсах)
Передача быстрая
Ответ наступает тотчас
Ответ кратковременный
Ответ четко локализован
Эндокринная регуляция
Информация передается
химическими веществами
через кровеносное русло
Передача медленная
Ответ обычно развивается
медленно (как, например,
рост)
Ответ продолжительный
Ответ обычно
генерализованный
16.7. Изучение поведения
(этология)
Поведение можно определить как направленные
вовне действия организма в ответ на стимулы,
возникающие в той или иной ситуации. Эти
действия каким-то образом изменяют
взаимоотношения организма с окружающей средой и имеют
адаптивное значение - способствуют сохранению вида.
Все живые существа обладают различными
формами поведения, которые зависят от способности
отвечать на стимулы.
Реакции могут быть самыми различными - от
таких относительно простых, как рост стебля растения
по направлению к источнику света, до таких
сложных форм поведения, как зищита территории,
ухаживание и спаривание у птиц и млекопитающих.
Поведение растений ограничено движениями,
вызываемыми ростом или изменением тургора, и
бывает стереотипным и предсказуемым. Две основные
формы поведения растений - тропизмы и настии-
подробно описаны в разд. 15.1.
Поведение животных намного сложнее и
разнообразнее, поэтому его очень трудно изучать и
объяснять с какой-либо степенью научной достоверности.
Существуют три главных подхода к изучению
поведения животных: виталистический, механистический
и это логический.
виталистический подход Сторонники такого
подхода стремятся объяснить поведение животных
лишь на основе тех действий, которые можно
наблюдать, и пытаются связать эти действия с
изменениями в окружающей среде. Они полностью
отрицают возможность изучения животных вне их
природной среды обитания. Этот подход коренится
в традициях «естественной истории», и с его
помощью получено очень много ценных сведений,
однако их нельзя считать вполне научными, так как
это лишь описания прошлых событий, которые
нельзя проверить экспериментальным путем.
механистический подход Этот подход является
экспериментальным и основан на изучении той или
иной формы поведения в лаборатории в
контролируемых условиях. Ему можно поставить в упрек
искусственность условий эксперимента, характера
поведенческих реакций и интерпретации
результатов. Однако этот метод, основоположником
которого был И. П. Павлов, широко применяется в
зоопсихологии.
этологический подход Это наиболее современный
подход к изучению поведения животных,
включающий оба описанных выше метода. Сторонники
этого направления пытаются объяснять поведенческие
реакции, наблюдаемые в естественных условиях,
связывая их с определенными стимулами.
Основоположники этологии - Лоренц, фон Фриш и Тин-
берген.
Изучая поведение животных, необходимо
соблюдать величайшую осторожность при интерпретации
результатов наблюдений во избежание
субъективности. Следует, например, остерегаться ставить себя
на место животного (антропоцентризм),
интерпретировать наблюдения с точки зрения человеческого
опыта (антропоморфизм) или объяснять причину
наблюдаемого явления, исходя из заданности его
результата (телеология).
Изучению поведенческих реакций животных во
многом способствовали последние достижения в
технике видео- и звукозаписи. Фотография в
инфракрасных лучах сделала возможной киносъемку
животных ночью. Замедленная съемка позволяет
регистрировать медленные процессы (например,
линьку насекомых), а ускоренная - очень быстрые
(например, полет птицы) и затем рассматривать их при
скорости, более подходящей для анализа.
Использование миниатюрных кассетных магнитофонов для
записи звуков и последующий анализ записей с
помощью звукоспектрографов и компьютеров
помогли в изучении звуковой сигнализации у
животных. Миграцию животных изучают в настоящее
время с помощью имплантированных
миниатюрных датчиков, подающих сигналы, которые можно
улавливать специальными приборами
(определяющими также направление, откуда приходит сигнал)
или же с помощью радиолокаторов. Оба метода
успешно используются для того, чтобы следить за
миграцией млекопитающих, птиц и саранчи.
Какой бы подход или метод ни применялся при
изучении поведения, объяснение поведенческих
реакций должно охватывать всю цепь событий от
стимула до реакции, все уровни внутренней организации,
связывающие причину и следствие.
В самом широком плане поведение можно
подразделить на два типа - врожденное и приобретенное,
но между ними нет четкой границы, и большинство
поведенческих реакций высших организмов,
несомненно, содержит элементы того и другого типа.
Однако для большей простоты в нашем вводном
очерке эти два типа поведения будут рассмотрены
по отдельности.
16.8. Врожденное поведение
Врожденные формы поведения составляют не одну
какую-то четко отграниченную категорию, а скорее
разнородную совокупность реакций, наследуемых
вместе со специфическими нервными или цитоплаз-
матическими связями у многоклеточных и
бесклеточных организмов соответственно. Благодаря
существованию этих «встроенных» связей данный
стимул всегда будет вызывать одну и ту же реакцию.
Эти формы поведения развивались и
совершенствовались на протяжении многих поколений путем
отбора, и главное их приспособительное значение
состоит в том, что они способствуют выживанию
вида. Другая важная особенность врожденного
поведения-то, что оно обеспечивает экономное
использование нервных путей у многоклеточных
организмов, так как не предъявляет слишком
больших требований к высшим отделам нервной
системы.
Существует градация сложности врожденных
поведенческих реакций, отражающая сложность
нервных путей, участвующих в их осуществлении. К
врожденным формам поведения относятся
ориентация, таксисы, кинезы, простые рефлексы и
инстинкты. Последние могут быть чрезвычайно сложными
и включают биологические ритмы,
территориальное поведение, ухаживание, спаривание, агрессию,
альтруизм, социальную иерархию и социальную
организацию. У растений все формы поведения
являются врожденными.
Таксисы
Таксис-это направленное движение всего
организма в целом, вызванное внешним стимулом. Такое
движение может быть направлено в сторону
стимула (положительный таксис), в противоположную
сторону (отрицательный таксис) или под
определенным углом по отношению к стимулу. Таксисы
классифицируют в соответствии с природой
стимула (фототаксис, хемотаксис и т. д.). Некоторые виды
таксисов перечислены в табл. 15.2. Известно,
например, что определенные виды муравьев находят
путь к своему муравейнику, ориентируясь по
солнцу. Другие организмы ориентируются в
пространстве таким образом, чтобы какая-то сторона тела
(обычно спинная) всегда находилась сверху;
подобная реакция наблюдается, скажем, у таких рыб, как
камбала, которая всегда держится определенной
стороной вверх.
Многие организмы определяют направление
действия стимула, двигая из стороны в сторону
головой, на которой находятся главные органы чувств.
Это явление, называемое клинотаксисом, позволяет
воспринимать раздражитель с помощью
рецепторов, расположенных на голове симметрично,
например фоторецепторов. Если одинаково
стимулируются оба рецептора, организм будет двигаться
вперед приблизительно по прямой линии. Этот тип
поведения можно видеть у планарии, движущейся
по направлению к источнику пищи, и у личинок
мясной мухи, движущихся в обратном направлении
от источника света. Полагают, что во всех случаях
клинотаксиса необходимо достаточно частое
возбуждение рецепторов на обеих сторонах тела, чтобы
«мозг» все время получал информацию, так как
долговременной памяти у этих организмов нет.
Кинезы
Кинез-это ненаправленная двигательная реакция,
при которой скорость движения зависит от
интенсивности стимула, а не от направления его действия.
Например, щупальца гидры в поисках пищи
медленно движутся в разных направлениях, но если в
непосредственной близости от гидры поместить
слюну, глутатион или водяных блох, щупальца
начинают двигаться быстрее.
Кинез и таксис можно наблюдать у мокриц в
опыте, описанном ниже.
Лейкопластырь
Натянутая ткань
Мокрицы
Основания двух
чашек Петри
Отверстие для внесения мокриц
Мокрая вата
Полоска пластилина
Гранулы безводного
хлористого кальция
Рис. 16.55. Прибор для изучения реакции на
влажность у мокриц.
Опыт 16.1 Изучение ориентации у
мокриц с применением
простой камеры
Материалы и оборудование
Одна пара колготок
Две пластиковые чашки Петри
Липкая лента
Горячая металлическая палочка
Вата
Безводный хлористый кальций
Лейкопластырь
Десять мокриц
Пластилин
Методика
1. Вырежьте из старых колготок кружок диаметром
10 см, натяните его на чашку Петри диаметром
8,5 см и закрепите с помощью кольца из липкой
ленты.
2. В дне этой чашки с помощью нагретой
металлической палочки проплавьте отверстие диаметром
1 см.
3. Дно второй чашки разделите на две половинки с
помощью полоски пластилина длиной 8,5 см,
шириной 0,5 см и высотой 1,4 см.
4. В одну половину этой чашки поместите
пропитанную водой вату, а в другую - гранулы
безводного хлористого кальция.
5. Укрепите первую чашку над второй с помощью
лейкопластыря, как показано на рис. 16.55.
6. Через отверстие в верхней чашке Петри внесите в
полученный прибор десять мокриц и с
интервалом в 1 мин записывайте общее число животных
и число активных (передвигающихся) животных в
каждой половине камеры в таблицу типа табл.
16.17.
7. Через 20 мин постройте график числа мокриц в
каждой половине камеры в зависимости от
времени.
8. Рассчитайте процент активных мокриц в сухой
половине и составьте график его зависимости от
времени.
9. Объясните полученные результаты, опираясь на
представление о кинезах и таксисах.
В простых опытах подобного типа можно
исследовать реакцию животных на условия среды,
соответствующие крайним значениям изучаемой
переменной. Проявлением таксиса в таком случае будет
предпочтение животными определенной среды.
Например, мокрицы в камере, где они могут выбирать
высокую или низкую влажность, скапливаются в
большем числе во влажной половине, проявляя
таким образом положительный гидротаксис.
Используя сочетание разных переменных, можно
усложнить эксперимент, чтобы выяснить, какая из них
сильнее влияет на конечную реакцию.
Регистрируя активность мокриц в каждой
половине камеры, скажем, с 30-секундными интервалами,
мы могли изучать кинез. Результаты такого
исследования показывают, что сразу после помещения в
камеру на границу двух участков с разными
условиями одни мокрицы беспорядочно ползают, а
другие остаются неподвижными. Однако вскоре все
мокрицы начинают двигаться, причем скорость их
движения всегда больше на сухой половине. Эту
повышенную подвижность на сухой половине,
носящую, видимо, беспорядочный характер, можно
понять как попытку найти оптимальные условия, и
когда мокрицы находят эти условия, их
подвижность уменьшается. Такие реакции служат
примером ортокинеза. Во влажной половине камеры
мокрицы движутся медленнее и в результате, как
правило, скапливаются именно здесь. Предпочтение
Таблица 16.17. Примерный вид таблицы с результатами
Время, мин
0
1
2
20
Влажная половина
Общее число
животных
4
5
Число активных
особей
1
2
и т.д.
Сухая половина
Общее число
животных
6
5
Число активных
особей
4
5
и т.д.
— Доля активных на сухой
половине
40%
50%
влажной камеры указывает на положительный
таксис по отношению к влажности.
Не все подобные реакции жестко
детерминированы, некоторые из них могут варьировать в
зависимости от других факторов, например от степени
голода, жажды, освещенности, темноты, тепла,
холода или влажности.
16.8.1. Простые рефлексы
позвоночных
Простой рефлекс - это непроизвольная стереотипная
реакция какой-то части организма на тот или иной
стимул. Она определяется наследственно
детерминированными связями нейронов, образующих спи-
нальные и черепномозговые рефлекторные дуги,
строение и функции которых были описаны в разд.
16.2.
В поведенческом плане простые спинальные
рефлексы представляют собой либо сгибательные
движения, приводящие к отдергиванию конечности от
причиняющего боль раздражителя, либо реакции на
растяжение, поддерживающие равновесие и позу. Те
и другие в своей основе непроизвольны и чаще всего
не требуют какой-либо координации или
интеграции помимо той, которая осуществляется на уровне
спинного мозга. Однако реакции обоих типов могут
видоизменяться в зависимости от условий и
прошлого опыта под влиянием головного мозга. Когда
это происходит, врожденные и приобретенные
поведенческие реакции накладываются друг на друга, и в
этом случае рефлекторный акт будет уже
называться условным (см. табл. 16.18). Многие простые
рефлексы, замыкающиеся в головном мозгу, тоже
могут стать условными, как, например, моргание в
ответ на внезапное резкое движение.
16.8.2. Инстинкты
Инстинкты - это сложные врожденные стереотипные
формы поведения, возникающие в ответ на
определенные изменения окружающей среды и имеющие
большое значение для выживания организма.
Инстинкты специфичны для каждого вида и
отличаются от простых рефлексов степенью сложности.
Лауреат Нобелевской премии этолог Конрад Лоренц
определял инстинкты как «врожденные видоспеци-
фические комплексы двигательных реакций».
Инстинктивное поведение играет
преобладающую роль у беспозвоночных животных, в частности
у насекомых, у которых короткий срок жизни не
позволяет изменять поведение путем «проб и
ошибок». Обеспечивая организм набором готовых
поведенческих реакций, инстинкты у насекомых и
позвоночных позволяют экономно использовать
нервные клетки. Эти реакции передаются из поколения в
поколение, выдерживая жесткий естественный
отбор, и, несомненно, имеют важное значение для
выживания.
Однако, прежде чем делать вывод, что
инстинктивные формы поведения жестко детерминированы
благодаря своей наследственной природе, следует
сразу сказать, что это не так. Все аспекты развития
организма - анатомические, биохимические,
физиологические, экологические или поведенческие -
являются результатом влияния постоянно меняющихся
факторов среды на наследственную основу.
Поэтому ни одну поведенческую реацию нельзя
рассматривать как только инстинктивную (т. е.
наследственную) или как только приобретенную (т.е.
выработанную условиями жизни), и любая из описанных
ниже форм поведения, на первый взгляд как будто
бы чисто инстинктивная или чисто приобретенная,
зависит от обоих факторов. Некоторые
исследователи предпочитают говорить о видоспецифическом
поведении вместо «инстинктивного» и об
индивидуально-специфическом поведении вместо
«приобретенного». Но какова бы ни была терминология, в
обоих случаях сохраняет свою силу принцип
взаимодействия наследственности и условий среды. Это
ясно продемонстрировал профессор Торп,
изучавший пение зяблика. Он установил, что зяблики,
выращенные в изоляции или лишенные слуха сразу
после вылупления, издают звуки, которые ухо
человека распознает как типичное пение зяблика. Однако
спектрофонограммы показали, что их пение носит
лишь «зачаточный» характер. Дело в том, что
зяблики, выращенные родителями, прислушиваются к
их пению и пению других зябликов той же
популяции, и у них вырабатывается такой же характер
звучания, как у старших птиц, т. е. характерный для
местной популяции. Оказалось, что пение птиц
одного вида имеет «местные диалекты». Песни глухих
птиц или выращенных в неволе оставались
рудиментарными, и из этого было видно, что условия жизни
могут значительно изменять инстинктивную форму
поведения.
16.8.3. Мотивация
На степень проявления и характер любой
поведенческой реакции влияет множество факторов, и
совокупность таких влияний называют мотивацией. У
одного и того же организма один и тот лее стимул
не всегда вызывает одинаковый ответ. Разница
всегда бывает обусловлена внешними или
внутренними факторами. Например, у голодного
животного реакция на пищу будет совсем не такой, как у
сытого. Между этими двумя крайностями
возможны ответы разной интенсивности в зависимости от
того, насколько животное голодно. А если бы
потребление пищи грозило голодному животному
опасностью быть схваченным хищником, то оно
сдержало бы свое желание поесть до того момента,
когда опасность минует. Многие поведенческие
реакции, связанные с размножением, зависят от
мотивации. Например, самки многих
млекопитающих восприимчивы к притязаниям самцов только в
Рис. 76.56. Самка шимпанзе, демонстрирующая самцу свою половую рецептивность.
определенное время года. Эти периоды совпадают у
них с периодом течки, и такое совпадение имеет
приспособительное значение, так как благодаря ему
спаривание происходит в оптимальный для
оплодотворения момент и потомство появляется в
наиболее благоприятный сезон. Такие периодические
изменения поведенческих реакций называются
биологическими ритмами; они описаны в разд. 16.8.5. У
многих животных степень мотивации, или
«влечения», у самцов и самок совпадает, но у других видов
для выражения степени мотивации нужна
определенная система сигналов между особями разного
пола. У многих приматов наступление течки у самок
сопровождается набуханием и изменением цвета
кожи около половых органов, и самка
демонстрирует эти изменения самцу (рис. 16.56). Такое поведение
уменьшает вероятность того, что самец будет
делать попытки к спариванию в то время, когда самка
к этому не готова. Сигналы, используемые для
воздействия на поведение, называются сигнальными
раздражителями, и в зависимости от их
функциональной роли могут подразделяться на мотивацион-
ные, пусковые или терминирующие.
мотивационные стимулы. Это могут быть как
внешние факторы (например, продолжительность
светового дня, обусловливающая у птиц
территориальное поведение и ухаживание), так и внутренние
(например, истощение запасов питательных веществ
в организме во время зимней спячки, приводящее к
пробуждению животного и поискам пищи).
Мотивационные стимулы создают «потребность» или
«цель», подготавливающую организм к активности,
которая может быть вызвана сигнальным
раздражителем второго типа-пусковым.
пусковые стимулы (релизеры). Релизер - это либо
простой стимул, либо ряд последовательных
стимулов, которые исходят от одной особи и вызывают
поведенческую реакцию у другой особи того же
вида. Термин «релизер» был предложен Лоренцом,
а роль таких стимулов в поведении широко
изучалась Тинбергеном.
Действие пускового стимула было
продемонстрировано Тинбергеном в исследованиях по кормлению
птенцов у серебристых чаек. Обычно эти птенцы
тычутся клювиком в красное пятно, расположенное
на конце желтого подклювья родителя, требуя,
чтобы тот отрыгнул им рыбу, которую они и
проглатывают. В серии модельных опытов с использованием
муляжей головы взрослой чайки Тинберген и Пер-
дек установили, что пусковым стимулом для
реакции птенцов служит наличие контрастной окраски
на клюве родителя. Как видно из рис. 16.57, сила
действия этого стимула такова, что заостренная
палочка с чередующимися цветными полосками
вызывала у птенцов реакцию, причем даже более
сильную, чем родительский клюв.
терминирующие стимулы. Как показывает их
название, терминирующие стимулы вызывают окончание
поведенческой реакции. Они могут быть как
внешними, так и внутренними. Например, вид успешно
построенного гнезда (внешний стимул) заставляет
птиц прекратить работу; внутреннее удовлетворение
или «пресыщение», сопровождающее эякуляцию у
самца, приводит к прекращению спаривания; при
полном желудке животное перестает есть.
Другие примеры сигнальных раздражителей и их
роль в поведенческих реакциях обсуждаются в разд.
16.8.4-16.8.9.
16.8.4. Врожденные пусковые
механизмы
Лоренц высказал предположение, что должны
существовать способы фильтрации, позволяющие
отделять несущественные стимулы от тех, которые
имеют значение для правильной поведенческой
реакции. Проведенные исследования указывают на
то, что такая фильтрация может быть
периферической, осуществляемой на уровне рецепторов, или
центральной - на уровне центральной нервной
системы. Так, например, Шнейдер установил, что хемо-
рецепторы, находящиеся на антеннах у самцов
бабочек, чувствительны к половым аттрактантам
(феромонам), выделяемым самками только того же, а не
какого-либо другого вида. Новые варианты опытов,
проведенных Тинбергеном и Пердеком на
серебристых чайках, позволяют думать, что реакции на
сигнальные раздражители, как и предполагал
Лоренц, контролируются нейросекреторными
механизмами центральной нервной системы.
16.8.5. Биологические ритмы
Многие формы поведения повторяются с
регулярными интервалами и служат одним из проявлений
биологических ритмов, или биоритмов. Хорошо
известны такие примеры, как периоды ухаживания и
гнездования у птиц весной и перелеты определенных
видов осенью. Интервалы между периодами
активности могут варьировать в пределах от нескольких
минут до нескольких лет в зависимости от
характера активности и вида животного. Например, много-
щетинковый червь пескожил (Arenicola marina),
живущий в норе в илистом или песчаном дне, каждые
6-7 минут совершает движения, связанные с
питанием. Этот цикл питания не имеет никаких внешних
или внутренних физиологических мотивационных
стимулов. По-видимому, он регулируется
«биологическими часами»-механизмом, который в данном
случае зависит от водителя ритма (пейсмейкера),
находящегося в глотке, откуда стимулы
распространяются вдоль тела червя по брюшной нервной
цепочке.
Ритмы, задаваемые внутренними «часами» или
водителями ритма, называются эндогенными - в
отличие от экзогенных, которые регулируются
внешними факторами. Если не считать таких случаев, как
цикл питания пескожила, большинство
биологических ритмов являются смешанными, т.е. частично
эндогенными и частично экзогенными.
Во многих случаях главным внешним фактором,
регулирующим ритмическую активность, служит
фотопериод, т. е. продолжительность светового дня.
Это единственный фактор, который может быть
надежным показателем времени года, и он
используется для установки «часов». Конкретная природа
«часов» неизвестна, но нет сомнений, что здесь
действует какой-то физиологический механизм,
который может включать как нервные, так и
эндокринные компоненты. Влияние фотопериода
широко исследовалось в связи с поведением
млекопитающих, птиц и насекомых. Хотя он, несомненно,
играет важную роль в контроле таких видов активности,
как подготовка к зимней спячке у млекопитающих,
перелеты у птиц и диапауза у насекомых, это не
единственный внешний фактор, регулирующий
биологические ритмы. У некоторых животных
существуют лунные ритмы, как, например, у много-
щетинкового червя палоло. Во всей южной части
Тихого океана этот червь выделяет яйца и сперму
Рис. 16.57. А. Горизонтальные столбики показывают
число клевательных ответов птенцов серебристой
чайки на картонные муляжи головы взрослой чайки,
имеющие серые клювы и пятна разных оттенков
(Hinde R. А.. 1966, по Tinbergen, 1961.) S.
Искусственный клюв, окрашенный в красный цвет с тремя
белыми полосками, вызывал на 20% больше
клевательных ответов, чем точная трехмерная модель
головы взрослой чайки, имеющая желтый клюв
с красным пятном. (Tinbergen, Per deck. Behavior, 1950,
3, 1)
ежегодно один день, приуроченный к определенной
фазе луны (в среднем около 2 ноября).
Хорошо известна связь приливов с лунными
циклами, и было показано, что эти два внешних
фактора определяют периодическую активность
комара Clunio maritimus. Личинка этого насекомого
питается красными водорослями, растущими на
нижней границе отлива, которая не бывает покрыта
водой только два раза в каждый лунный месяц. В
естественных условиях вылупление личинок,
спаривание взрослых особей и откладка яиц происходят в
течение всего лишь двух часов (это весь период
жизни насекомого), пока водоросли вновь не
накроет приливная волна. В лабораторных условиях
при постоянном фотопериоде (12 ч света и 12 ч
темноты) личинки продолжали вылупляться с
примерно 15-дневными интервалами; это указывало на
существование эндогенных часов,
запрограммированных на приблизительно половинный лунный
цикл, совпадающий с 14,8-дневным циклом
приливов и отливов.
Поведение многих насекомых, ведущих
полностью наземный образ жизни, контролируется,
по-видимому, эндогенными ритмами, связанными с
чередованием света и темноты. Например, плодовая
мушка дрозофила выводится из куколки на
рассвете, а тараканы становятся наиболее активными с
наступлением темноты и перед рассветом. Эти
регулярные биологические ритмы с периодом около 24 ч
называются циркадианными (лат. circa-около, dies -
день) или околосуточными ритмами. Исследование
активности таракана Periplaneta при двух различных
световых режимах (в течение 10 дней 12 ч свет и 12 ч
темнота, а в последующие 10 дней-полная темнота
на протяжении суток) показало, что при втором
режиме активность таракана ограничена временем,
приблизительно совпадающим с периодом
активности после наступления темноты при первом
режиме. Результаты этого исследования, приведенные на
рис. 16.58, означают, что суточный ритм
сохраняется и без каких-либо внешних временных сигналов,
хотя начало активности изо дня в день слегка
варьирует. Эти результаты подтверждают
представление о том, что суточные ритмы контролируются
каким-то эндогенным механизмом -
«биологическими часами», ход которых настраивается внешними
факторами.
Предполагают, что циркадианные ритмы имеют
многообразное адаптивное значение, специфичное
для каждого вида и, в частности, связанное с
ориентацией. Такие животные, как рыбы, черепахи, птицы
и некоторые насекомые, мигрирующие на большие
расстояния, используют в качестве компаса солнце и
звезды. Другие животные (пчелы, муравьи и рач-
ки-бокоплавы) ориентируются по солнцу при
поисках пищи и при возвращении домой. Ориентация по
солнцу или луне надежна только в том случае, если
животное способно каким-то образом определять
время, чтобы учитывать суточные перемещения
солнца и луны. Примером ситуации, когда
присущий человеку физиологический циркадианный ритм
отклоняется от естественного чередования дня и
ночи, может служить «сдвиг фаз», с которым все
чаще приходится сталкиваться пассажирам дальних
авиалиний.
16.8.6. Территориальность
Территория - это участок, который занимает какая-
то особь или популяция и который она защищает от
других представителей этого же вида или иных
видов. Территориальность широко распространена
у всех позвоночных животных, кроме земноводных,
но у беспозвоночных встречается редко. Характер и
биологическую роль территориальности интенсивно
изучали у птиц и приматов. У приматов
территориальность составляет важную часть
общественного поведения.
Для каждого вида животных территориальность
имеет, вероятно, свое специфическое значение, но во
всех случаях она обеспечивает условия, при которых
каждая пара особей и ее потомство имеют
достаточно места для пропитания и размножения. Таким
путем виду удается оптимально использовать
природную среду. Размер территории, занимаемой
каким-либо видом, варьирует от сезона к сезону в
зависимости от обилия пищевых ресурсов. Хищным
птицам и крупным плотоядным млекопитающим
для того, чтобы прокормиться, нужна территория в
несколько квадратных миль. В то же время
серебристые чайки и пингвины (рис. 16.59) занимают
территории всего в несколько квадратных метров,
так как для добывания пищи выходят за пределы
Световой
режим
Дни 1-10
Дни 11-20
-24 ч-
Рис. 16.58. Результаты регистрации активности
тараканов в течение 20 дней. С 1-го по 10-й день
световой режим состоял из 12 ч света и 12 ч
темноты, а с 11-го по 20-й день тараканы находились
в постоянной темноте. Черными участками
представлены периоды активности в течение каждого
дня
своей территории, используя ее только для
размножения.
Обычно поисками территории занимается перед
размножением самец. Наиболее активная защита
территории ведется в период размножения, и
наибольшей остроты достигают столкновения между
самцами одного и того же вида. Существует
множество поведенческих реакций, связанных с защитой
индивидуальных участков, в том числе
демонстрации угрозы владельцам соседних участков. Эти
демонстрации включают определенные стимулы,
которые вызывают соответствующую реакцию.
Например, Лэк показал, что взрослый самец зарянки
(Erithacus rubecula) атакует чучело взрослого самца
того же вида, обладающего красной грудкой, или
просто пучок его красных перьев, но не трогает
чучело молодого самца, не имеющего красного
оперения на груди.
Чем ближе к центру участка, тем агрессивнее
ведет себя его владелец. Агрессивность самцов
определяется отчасти содержанием тестостерона в
крови, которое может влиять и на размеры территории,
охраняемой самцом. Например, введя тестостерон
самцу шотландской куропатки, можно добиться
увеличения занимаемого им участка. На рис. 16.60
показано, как меняются размеры участков,
занимаемых тремя самцами, при введении им тестостерона.
Животные, занимающие соседние участки, на грани-
Рис. 16.59. Территории, занимаемые пингвинами
(снимок с воздуха)
Рис. 16.60. Сплошные линии-границы участков,
занимаемых самцами шотландской куропатки
Прерывистыми линиями показаны изменения границ,
происшедшие после введения тестостерона птицам
A, X и Y Птицы X и Y до введения тестостерона не
имели собственных участков (По A Watson,
J Reprod Fert, 1970, Suppl, 71, 3)
це этих участков проявляют одинаково сильную
защитную реакцию. Несмотря на видимость
конфликта и агрессивность, связанные с охраной
территории, настоящие драки, которые могли бы быть
пагубными для вида, случаются редко и заменяются
обычно угрожающими звуками, телодвижениями и
позами. Представители многих видов, захватив
территорию, метят ее границы, оставляя пахучий след.
С этой целью они мочатся на различные предметы,
расположенные вдоль границы территории
(«пограничные столбы») или трутся о них тем местом, где
находятся пахучие железы. Хотя территориальность
предполагает разделение мест обитания и ресурсов
между членами популяции, всегда каким-то особям
не удается занять и защищать индивидуальный
участок. У многих птиц, например у куропаток,
более слабые особи оттесняются на периферию
местообитания, где они не находят партнера для
спаривания. Очевидно, это одна из сторон
адаптивного значения территориальности, поскольку лишь
наиболее «приспособленные» особи могут
захватывать территорию, приносить потомство и
передавать свои гены следующему поколению. Таким
образом, еще одна функция территориальности
связана с внутривидовой конкуренцией и может
действовать как механизм, регулирующий размеры
популяции.
16.8.7. Ухаживание и спаривание
Существует множество сложных ритуальных видо-
специфических форм поведения, связанных с
ухаживанием и спариванием. У птиц, млекопитающих и
некоторых рыб эти два процесса часто происходят
сразу же после закрепления территории самцом.
Ухаживание-это комплекс поведенческих реакций,
предназначенных для стимуляции половой
активности партнера. У тех видов, у которых в
выращивании потомства участвуют оба родителя
(например, у дроздов), оно связано с образованием
брачных пар. У стадных животных самцы и самки
находятся в одном стаде, как у павианов. Для
большинства таких видов характерна периодическая
половая активность такого типа, как описанная в
разд. 16.8.5.
Ухаживание контролируется в основном моти-
вационными и пусковыми стимулами и приводит к
спариванию (копуляции), которым и завершается.
При спаривании поведенческие реакции
инициируются пусковыми стимулами, а заканчиваются под
действием терминирующих стимулов, связанных с
выделением спермы самцом.
У большинства видов ухаживание регулируется
внешними мотивационными стимулами (например,
фотопериодом), вызывающими повышение уровня
половых гормонов и созревание гонад. У
большинства видов это приводит к резким изменениям
вторичных половых признаков и поведенческих
реакций, в том числе таким, как изменение окраски
(например, появление красной окраски брюшка у
самцов трехиглой колюшки), увеличение размеров
определенных частей тела (оперения у райских птиц
и т. п.), брачное пение (у соловья и др.),
демонстрационные позы (как у чомги, рис. 16.61), а также
использование химических половых аттрактантов
(как у бабочек).
Среди множества сигналов, используемых в
процессе ухаживания для привлечения особи
противоположного пола, наиболее важную роль играют
зрительные, звуковые и обонятельные сигналы.
Например, самец манящего краба (Uса) использует
зрительный сигнал и привлекает самку, размахивая
огромной клешней и совершая при этом ритмичные
движения, сходные с движениями скрипача. По мере
приближения самки движения клешни становятся
все более энергичными.
Многие насекомые, амфибии, птицы и
млекопитающие используют при ухаживании акустические
сигналы. Самки некоторых видов комаров
привлекают самцов звуками, возникающими в результате
вибрации крыльев, тогда как кузнечики, сверчки и
цикады во время ухаживания стрекочут.
Стрекочущий звук создается либо при трении задних
конечностей друг о друга или о надкрылья (твердые
передние крылья), либо при трении между
надкрыльями. Стрекотание по своему звуковому
«рисунку» видоспецифично и вызывает ответную
реакцию только у представителей того же вида.
Некоторые пауки используют для привлечения
партнера механические приспособления. Самец
приближается в самке, сидящей в центре паутины, и
перебирает нити паутины с частотой, характерной
для данного вида. Эти колебания паутины
«умиротворяют» самку и уменьшают ее природную
агрессивность, что позволяет самцу приблизиться к ней и
оплодотворить ее. Если же самец по ошибке
приблизится к самке не того вида или «возьмет
неверную ноту», он будет атакован и убит!
Рис. 16.61. Эпизод ухаживания у чомги. Самец и
самка демонстрируют друг другу гнездовой материал
(По Huxley J S., Proc Zoo/ Lond., 1914, 2. 491.)
Поведение самки
Появляется
Поведение самца
Преследует
в воздухе
Летает
Нагоняет
самку
и прикасается
к ней кисточками
из волосков
Садится
на растения"
Складывает
крылья
Парит над
самкой,
задевая ее
кисточками
Садится
сбоку
Прини -
мает
ухаживания
Копули-
рует
Послебрачный
полет
Поведение
самки
Поведение
самца
Рис. 16.62. Ухаживание и копуляция у
бабочек-данаид. (Стрелки идут от стимулов к вызываемым ими
реакциям). (В rower L. Р., В rower J. V. Z., Cranston F. P.,
Zoologica, 1965, 50, 18.)
Для ухаживания и спаривания (а также, как мы
увидим, для регуляции поведения в сообществах)
используется выделение особями малых количеств
химических веществ, вызывающих специфическую
физиологическую или поведенческую реакцию у
других особей того же вида. Эти вещества,
называемые феромонами, обычно представляют собой
весьма летучие соединения с небольшой
молекулярной массой. Многие из них действуют как
природные половые аттрактанты. Первыми среди них были
идентифицированы цибетон, выделяемый
африканской циветтой, и мускон, выделяемый кабаргой; оба
эти вещества являются секретами анальных желез
самок и используются в парфюмерной
промышленности. Кобылы, коровы и суки выделяют феромоны
во время течки. Эти вещества не воспринимаются
обонятельным эпителием человека, но
улавливаются самцами тех же видов. Бомбикол - феромон,
выделяемый выворачивающейся железой на кончике
брюшка неоплодотворенной взрослой самки
тутового шелкопряда,-может привлекать самцов того
же вида, находящихся на значительных расстояниях.
Обонятельные рецепторы на антеннах
бабочек-самцов улавливают феромоны, присутствующие в
чрезвычайно низких концентрациях; при этом самцы
проявляют реотаксис, летая против ветра до тех
пор, пока не достигнут самки. В настоящее время
феромоны находят все более широкое применение
как биологические средства борьбы с вредными
насекомыми. Например, для борьбы с непарным
шелкопрядом искусственно распространяют в
воздухе феромон джиплур, который привлекает самцов
к источнику феромона, где они могут быть пойманы
и уничтожены. При этом не только сразу же за счет
самцов уменьшаются размеры популяции, но и
снижается численность следующего поколения, так
как пойманные самцы уже не могут спариваться.
Феромоны используются животными и при самом
спаривании, как, например, у бабочек-данаид Danaus
gilippus. У этих бабочек самец выделяет феромон и
наносит его на антенны самки с помощью пары
кисточек, выпускаемых из кончика брюшка. Вся
последовательность фаз ухаживания и спаривания
этих бабочек показана на рис. 16.62.
У некоторых животных ухаживание
сопровождается конфликтным поведением со стороны одного
или обоих партнеров. У тех видов, особи которых
обычно ведут одиночный образ жизни, конфликт
ухаживания может быть связан с изменением
отношения к другим особям того же вида в результате
повышения секреции гормонов. Другой смысл
такого поведения может состоять в том, что оно
обеспечивает более прочную связь между
партнерами и синхронизацию развития гонад, с тем чтобы их
половые клетки созревали одновременно. У
некоторых видов пауков, например у тарантула, конфликт
между самцом и самкой утихает только к моменту
копуляции, которая, однако, заканчивается тем, что
самка убивает самца.
16.8.8. Агрессия (агонистическое
поведение)
Агрессия-это комплекс поведенческих реакций,
включающий угрожающие позы, ритуальные атаки,
а иногда и физическое нападение на других особей,
которое, однако, не имеет отношения к
хищничеству. Обычно все эти действия бывают
направлены против представителей того же вида и
выполняют различные функции, такие как изгнание других
особей с какого-то участка (обычно с территории
или места питания), защита партнера или потомства
и установление ранга в социальной иерархии.
Слово «агрессия» вызывает определенные эмоции
и наводит на мысль об ожесточенных схватках
внутри групп животных. Пожалуй, более
правильным было бы слово «агонизм». Агонистическое
поведение играет адаптивную роль, так как
способствует ослаблению внутривидовых конфликтов и
предотвращает настоящие драки, которые совсем не
в интересах вида. У большинства видов
«агрессивность» находит выход в ритуальных поединках,
демонстрациях силы и в угрожающих позах,
хорошо знакомых всем особям данного вида. Например,
рогатые животные (олени, американские лоси,
горные альпийские козлы и серны) устраивают
«турниры», которые разыгрываются по определенным
«правилам». Партнеры лишь сталкиваются рогами
и никогда не используют их для удара в
незащищенный и легко ранимый бок соперника. Сиамская
бойцовая рыбка Betta splendens принимает
угрожающие позы, при которых ее кажущиеся размеры
увеличиваются (рис. 16.63).
Угрозы, выражаемые двумя особями в агонисти-
ческой конфликтной ситуации, всегда
заканчиваются тем, что одна из особей (как правило, более
слабая) уступает и выходит из поединка, принимая
позу подчинения или умиротворения. У собак и
волков умиротворяющая поза выражается в том,
что животное ложится на спину или подставляет
победителю свое горло.
Во время драк животные обычно избегают
применять против представителей того же вида свое
наиболее эффективное оружие. Например, жирафы,
выясняя отношения, бодаются рожками, тогда как в
случае защиты от других животных они используют
свои мощные копыта.
Чтобы агонистические поведенческие реакции
были наиболее эффективны, они должны быть
стереотипными для каждого вида. Тинберген исследовал
некоторые из таких реакций у трехиглой колюшки.
В одной серии опытов он показал, что угрожающая
поза самца эффективна только в том случае, если он
находится в горизонтальном положении с
оттопыренными грудными плавниками и приподнятыми
колючками. Когда же самца помещают в пробирку,
где он находится в вертикальном положении, он уже
не имеет для других самцов такого устрашающего
вида, как в том случае, когда он плавает свободно
или находится в горизонтальной пробирке (рис.
16.64).
В другой серии опытов Тинберген показал, что
воинственное поведение самца колюшки,
защищающего свою территорию, может быть
спровоцировано объектами почти что любой формы, если их
нижняя сторона окрашена в красный цвет. Эти
фигуры действуют как модели самца колюшки, у
которого в брачный сезон краснеет брюшко и
которого владелец участка считает потенциальным
противником (рис. 16.65).
В условиях стресса, например в конфликтной
ситуации или во время ухаживания и спаривания,
животные могут производить какие-то
малозначащие действия, не имеющие отношения к данной
ситуации. Это явление носит название смещенной
активности и возникает в тех случаях, когда при
наличии сильной мотивации два разных стимула
действуют в противоположных направлениях.
Например, одна из двух птиц, участвующих в
территориальном конфликте, оказавшись перед выбором
«драться или бежать», вдруг начинает собирать
травинки, т.е. заниматься деятельностью,
связанной с устройством гнезда. Такая реакция как бы
дает выход сдерживаемому возбуждению. У
человека многие действия в определенных условиях тоже
можно рассматривать как смещенную активность
(например, сжимание кулаков, удары кулаком по
столу, кусание ногтей, расправление одежды,
барабанящие движения пальцев и т. п.). Сходная форма
поведения называется «вакуумной активностью»
или «переадресовкой реакции» и наблюдается в тех
случаях, когда при наличии сильной мотивации
отсутствует пусковой стимул. Тогда возникает
нормальный ответ, но он не может быть направлен на
соответствующий объект или ситуацию и служит,
таким образом, лишь способом уменьшить
внутреннее напряжение (например, раздражение против
кого-то, кто в действительности не является
виновником раздражения, но оказывается как бы его
заместителем).
16.8.9. Социальная иерархия
У многих насекомых и у большинства позвоночных
животных существуют разнообразные
поведенческие реакции, связанные с существованием
постоянных или временных сообществ. Совокупность этих
реакций носит название общественного поведения, а
достигаемая при этом сплоченность и кооперация
имеют приспособительное значение, так как
увеличивают жизнеспособность вида и дают ему
преимущество перед другими видами. В сообществах
такого рода очень важна система коммуникаций; при
этом эффективность возрастает и за счет того, что
каждая особь играет в сообществе определенную
роль. Отсюда вытекает один из главных аспектов
Рис. 16.63. Демонстрация угрозы у сиамской
бойцовой рыбки А и 6- отсутствие демонстрации, β и
Г-угрожающие позы, жаберные крышки (о) и
плавники оттопырены, что создает впечатление больших
размеров тела. (Hinde R А., 1970, Simpson, 1968)
Рис. 16.64. А. Угрожающая поза самца колюшки. Б
Находясь в вертикальном сосуде, самец имеет менее
угрожающий вид для других самцов (По N. Tinbergen,
1951).
Рис. 16.65. Модели, использованные в качестве
стимулов для агрессивного поведения самца колюшки,
занявшего территорию. А. Точная модель самца, не
имеющая красного брюшка, не вызывает агрессивной
реакции. БД. Модели самцов, не имеющие точного
сходства с колюшкой по форме, но окрашенные снизу
в красный цвет, вызывают у самца агрессивную
реакцию. (По Tinbergen, 1951.)
общественного поведения - существование
социальной иерархии, или «порядка клевания», как ее в
некоторых случаях называют.
«Порядок клевания» представляет собой
доминантную иерархию, иерархию по степени
доминирования-иными словами, животные располагаются в
сообществе по рангам. Например, в курятнике
существует линейный порядок, согласно которому
курица А будет клевать любую другую курицу,
курица Б будет клевать всех кур, кроме курицы А и
т. д. Положение в иерархии обычно устанавливается
с помощью каких-то агонистических форм
поведения, помимо драк. Сходная иерархия наблюдается
также у других птиц, у мышей, крыс, коров и
павианов. В принципе и все существующие
человеческие сообщества основаны на той или иной
иерархии.
«Порядок клевания» возможен только в том
случае, если животные способны различать друг друга
индивидуально и обладают некоторой способностью
к научению. Положение животного на
иерархической лестнице обычно зависит от его размеров, силы,
выносливости и агрессивности и у птиц остается
более или менее постоянным на протяжении всей
жизни особи. Самцы более низкого ранга могут
перейти в более высокий ранг, если им вводить
тестостерон, который повышает их агрессивность.
Если мышей низкого ранга отделить от группы и
кормить без ограничения, их вес увеличится, они
станут активнее и после возвращения в прежнюю
группу могут занять в ней более высокое положение.
Точно так же, если поместить мышей невысокого
ранга в другие группы, где они будут занимать
доминирующее положение, это как бы вселит в них
определенную уверенность (говоря
антропоморфным языком), которую они сохранят и после
возвращения в первоначальную группу, где займут
теперь более высокое положение.
Одно из преимуществ иерархии состоит в том, что
она уменьшает агрессивность особей, связанную с
питанием, выбором полового партнера и места для
выведения потомства. Кроме того, она избавляет
животных от травм, которые они могли бы
получить, если бы завоевание доминантного положения
не обходилось без драк. Еще одно преимущество
иерархической системы то, что благодаря ей
ресурсы распределяются таким образом, что выживают
самые жизнеспособные. Например, если группа из
100 кур получает корм, достаточный для
пропитания только 50 кур, то с точки зрения интересов вида
лучше, если 50 кур будут получать достаточное
питание, а 50 более слабых кур погибнут, чем если
будут жить все куры, получая половинный рацион,
так как это может помешать успешному
размножению. Короче говоря, социальная иерархия
повышает генетическую жизнеспособность сообщества
благодаря тому, что наиболее сильные и
генетически приспособленные животные имеют
преимущество, когда приходит время размножаться.
Общественная организация
Когда животные образуют прочные сообщества,
отдельные особи часто выполняют в них
специализированные функции, что повышает
жизнеспособность сообщества в целом (рис. 16.66). Такими
функциями могут быть добывание пищи,
размножение, забота о потомстве или защита от врагов.
Кооперация между членами сообщества при
подобном разделении функций основана на стереотипных
формах поведения и эффективных средствах
коммуникации. Эти формы поведения и средства
коммуникации варьируют у разных видов и очень сильно
различаются в сообществах приматов и насекомых.
У приматов сообщества довольно гибки в том
смысле, что роли между членами группы могут
перераспределяться, тогда как роли в сообществах
насекомых определяются строением тела и
способностью к размножению (это явление называют
полиморфизмом).
Рис. 16.66. Взаимная чистка шерсти у взрослых
шимпанзе способствует сплочению особей в группе.
Муравьи, термиты и пчелы являются
общественными насекомыми и живут колониями (семьями),
организованными по принципу кастовой системы. В
семье пчел имеется одна плодовитая самка-матка,
несколько тысяч рабочих особей (стерильных самок)
и несколько сотен плодовитых самцов. Пчелы
каждого типа выполняют свойственный им круг
обязанностей, определяемых в основном тем, из каких
яиц - оплодотворенных или неоплодотворенных -
эти пчелы вывелись. Оплодотворенные яйца
диплоидны, и из них развиваются самки. Неопло-
дотворенные яйца гаплоидны и дают самцов. Кто
выведется из оплодотворенного яйца-матка или
рабочая особь - зависит от характера пищи,
которую будет получать личинка. Особая пища для
личинки, из которой выводят матку, называется
маточным кормом; это один из примеров того, какое
важное значение имеют химические вещества в
организации сообществ. Внутри колонии информация
передается либо с помощью пахучих веществ и
феромонов при разнообразных формах
облизывания и «чистки» (такая передача веществ называется
трофаллаксисом), либо с помощью особых
демонстраций, называемых танцами.
Немецкий зоолог, лауреат Нобелевской премии
Карл фон Фриш изучал природу этих танцев,
используя меченых рабочих пчел и специально
сконструированные ульи со стеклянными стенками,
позволявшие вести наблюдения. Рабочие пчелы
разыскивают источники нектара и сообщают о
направлении и расстоянии, на котором они находятся,
другим рабочим пчелам с помощью «танца»,
обычно на вертикальных сотах. Если расстояние
составляет (приблизительно) менее 90 м, пчела исполняет
круговой танец (рис. 16.67), уведомляющий о том,
что источник нектара находится недалеко, но не
дающий никаких указаний относительно
направления. Если же расстояние больше 90 м, пчела
исполняет виляющий танец, который указывает
направление к источнику нектара по отношению к улью и к
солнцу. При этом пчела описывает путь,
напоминающий восьмерку, и во время пробега по средней
линии этой восьмерки виляет брюшком. Согласно
фон Фришу, скорость движения пчелы обратно
пропорциональна расстоянию от улья до источника
нектара, а угол между средней линией восьмерки и
вертикалью равен углу между нужным
направлением полета и направлением на солнце.
Интенсивность «виляния» отражает количество корма в
найденном месте (рис. 14.67,5). Предполагают, что
поправка на перемещение солнца вносится у пчел с
помощью «биологических часов» и что при
неполной облачности пчелы могут определять положение
невидимого солнца по поляризации света от
отдельных участков голубого неба.
Данные, полученные в последнее время,
позволяют предположить, что для сообщения другим
рабочим особям о местонахождении нектара пчелы
могут использовать высокочастотные звуки. Пока
неизвестно, является ли этот способ основным
средством связи, но его открытие не обесценивает
наблюдений и выводов Фриша, поскольку это может
быть добавочной сигнализацией, подкрепляющей
зрительные сигналы-танцы. Известно, однако, что
возвращающаяся с добычей пчела может сообщить
другим пчелам о типе цветков, которые она
посетила, скармливая им некоторое количество
собранного нектара.
А Круговой танец
Источник
пищи внутри
этого круга
Улей
Б Виляющий танец
Солнце
Источник
пищи
К пище
Виляющий
танец
К пище
Улей
Улей
Рис. 76.67. Танцы пчел. А. Круговой танец, который
исполняется, когда корм находится на расстоянии
менее 90 м от улья. Б. Виляющий танец, указывающий
на взаимное расположение улья, солнца и источника
корма.
Среднее число атак
Пробы
16.9. Поведение, связанное с
научением
16.9.1. Память
Память-это способность хранить и извлекать
информацию о прошлом опыте, и научение без
памяти невозможно. Прошлый опыт, включающий
стимулы и реакции, фиксируется в виде «следов
памяти», или энграмм, а так как способность к
научению у млекопитающих пропорциональна
величине больших полушарий, очевидно, что именно
они служат местом образования и хранения энграмм.
Природа энграммы неизвестна, о ней существуют
лишь гипотезы, основанные на противоречивых
данных. Эти гипотезы можно разделить на две главные
группы: согласно одной из них, в основе памяти
лежат изменения в структуре нейронов и их
организации в центральной нервной системе, а согласно
другой-стойкие изменения в биохимии мозга.
Гистологическое исследование ткани головного
мозга указывает на существование замкнутых
нейронных цепей, что позволило выдвинуть
представление о реверберирующих цепях как элементах
энграммы. Согласно этому представлению, ре-
верберирующие цепи позволяют возбуждению
циркулировать по кругу, сохраняя таким образом
информацию. Кажется, однако, сомнительным,
чтобы такая активность могла продолжаться
сколько-нибудь долго, и эксперименты
показывают, что память обладает большей емкостью и
прочностью, чем можно было бы достичь с
помощью одного только этого механизма. Так,
например, при охлаждении мозга крысы до 00C вся
электрическая активность в нем прекращается, но
после возвращения температуры к норме никакого
разрушения следов памяти не обнаруживается.
Полагают, однако, что такие цепи могут играть роль в
кратковременной памяти, т.е. в хранении
информации в пределах нескольких минут (как, например,
при запоминании человеком шестизначного номера
телефона с тем, чтобы тут же его набрать) и в
облегчении передачи сигналов по определенным
нервным путям. После сотрясения мозга память на
предшествующие ему события может ослабевать
или исчезать; кратковременная память постепенно
ухудшается также в процессе старения.
Долговременная память более прочна, и поэтому
полагают, что в ее основе лежат какие-то стойкие
изменения в ткани головного мозга.
Можно думать, что образование следов памяти
связано с действием биохимического механизма,
включающего синтез в мозгу определенных
веществ. Экстракты из центральной нервной ткани
обученных плоских червей или крыс при введении
необученным червям или крысам соответственно
сокращали время, необходимое для усвоения тех же
задач. Активным фактором во всех этих опытах
была, по-видимому, РНК.
Рис. 16.68. Типичная кривая научения. На графике
показано число нападений каракатицы на креветку,
находящуюся в стеклянном сосуде. Креветку
показывали каракатице 8 раз, каждый раз по 3 мин. Так как
попытки напасть на креветку не приводили к успеху,
число атак уменьшалось, по мере того как
каракатица «усваивала» то, что она не может схватить
креветку. Приведенная кривая основана на усреднении
результатов, полученных на 40 каракатицах.
(Messenger J. В., Symp. Zoo. Soc. Lond., 1977, 38, 347.)
Есть также данные в пользу того, что состав РНК
нейронов в процессе научения изменяется и что это
может приводить к синтезу специфических «белков
памяти». Было показано, что введение ингибитора
белкового синтеза - пуромицина - вызывает
нарушения памяти. Например, у мышей, только что
обученных выбирать направление в лабиринте,
после введения в мозг пуромицина эта способность
утрачивалась, тогда как у контрольных животных, в
мозг которых вводили физиологический раствор,
она сохранялась.
В заключение следует сказать, что мы далеки еще
от понимания механизмов памяти, но, по всей
видимости, в образовании следов памяти участвуют
изменения электрических свойств нейронов,
проницаемости синаптических мембран,
эффективности синаптической передачи и синтеза
ферментов, имеющих отношение к синапсам. Вряд ли
можно сомневаться в том, что память тесно
связана с процессами, происходящими в синапсах.
16.9.2. Научение
Научение-это адаптивное изменение
индивидуального поведения в результате предшествующего
опыта (рис. 16.68). Устойчивость вновь
приобретенных форм поведения зависит от памяти,
хранящей полученную в прошлом информацию. У
человека запоминание «фактов», например для
сдачи экзамена, может быть кратковременным,
тогда как навыки координированной двигательной
активности (такой, как совершение туалета, езда на
велосипеде или плавание) сохраняются в течение
всей жизни. Способность к научению обычно
ассоциируется у нас с поведением позвоночных, и
прежде всего млекопитающих, однако она
обнаружена у всех животных, за исключением
простейших, кишечнополостных и иглокожих, у которых
нервная система отсутствует или организация ее
Таблица 16.18. Основные типы научения, основанные на классификации, предложенной Торпом (1963)
Тип научения
Привыкание
Ассоциативное научение
Классическое
обусловливание
(выработка условного
рефлекса)
Инструментальное
обусловливание
(научение путем
проб и ошибок)
Латентное научение
Инсайт
(«постижение»)
Импринтинг (за-
печатление)
Характеристика поведения
При продолжительном повторении стимулов, не подкрепляемых поощрением или наказанием,
реакция на них постепенно угасает; например, птицы перестают бояться пугала (т.е.
научаются не обращать на него внимания). Это имеет важное значение для формирования
поведения у молодых животных-помогает им распознавать нейтральные элементы
окружающей среды (движения предметов, вызванные ветром, тень от облаков, движение волн и
т.п.). Привыкание-результат изменения в нервной системе, а не форма сенсорной
адаптации, так как новое поведение здесь устойчиво и после привыкания стимул никогда уже не
вызывает реакции
Выявлено в исследованиях Павлова, проводившихся на собаках. Метод состоял в выработке
условного рефлекса слюноотделения: животное начинает реагировать не только на
безусловный раздражитель (вид пищи), но и на новый условный раздражитель (удары метронома),
который предъявлялся собаке в сочетании с безусловным раздражителем. Животные
научаются связывать безусловный стимул с условным и давать ответ на любой из них.
Например, птицы не едят черных и оранжевых гусениц одного вида бабочек из-за их
неприятного вкуса и избегают есть других гусениц того же цвета, даже если они вполне
съедобны
Выявлено в исследованиях Скиннера, проведенных на голубях. Всякий раз, когда птица
производила определенное действие, ее за это вознаграждали (положительное подкрепление)
или подвергали «наказанию» (отрицательное подкрепление). Сочетание данного действия с
наградой или наказанием соответственно повышает или уменьшает вероятность повторения
этого действия в дальнейшем. Эффективность такого научения может возрастать при
проведении ряда повторных опытов (пример-эксперименты с каракатицей, рис. 16.68)
Не все формы поведения бывают явно направлены на удовлетворение непосредственных нужд
или получение награды. Иногда животные исследуют новую обстановку и получают
информацию, которая может пригодиться или даже приобрести жизненно важное значение
лишь в дальнейшем. Например, знание непосредственного окружения норки может помочь
мыши убежать от хищника, тогда как в момент приобретения этого знания оно, казалось, не
имело никакой ценности. Поэтому такое научение называют латентным. По-видимому, к
этой категории можно отнести и усвоение видоспецифической песни зябликом (см. разд.
16.8.2)
Вероятно, инсайт-это высшая форма научения. Он не является результатом прямого научения
методом проб и ошибок, а основан на информации, полученной ранее при других в чем-то
сходных обстоятельствах. Инсайт возможен лишь при достаточном развитии
интеллектуальных функций. Примером могут служить наблюдения Кёлера над шимпанзе. Когда
подвешивали слишком высоко бананы и помещали отдельно деревянные ящики, обезьяна
ставила ящики друг на друга и, взобравшись на них, доставала бананы. Наблюдения
показали, что этим действиям предшествовал некоторый период «размышления».
Правильное решение в данном случае могло быть в значительной степени облегчено
предшествующими играми с ящиками (т.е. латентным научением)
Простая специализированная форма научения, которая проявляется во время критического
(чувствительного) периода развития животного. Приобретаемое поведение становится
относительно стойким и с трудом поддается изменению. Импринтинг состоит в том, что в мозгу
детеныша (или птенца) запечетлевается образ другого индивидуума, обычно родителя, или
какого-то крупного объекта и создается особая «привязанность» к нему. Лоренц наблюдал,
как лишенные родителей гусята и утята следовали за ним по пятам, принимая его за мать.
Так же ведут себя прирученные и выкормленные из бутылки ягнята, что может оказывать
глубокое и не всегда желательное влияние на всю последующую жизнь животного,
затрудняя установление нормальных взаимоотношений с другими особями своего вида. В
естественных условиях импринтинг, несомненно, имеет приспособительное значение,
помогая детенышам быстро перенимать необходимые навыки от родителей (например,
учиться летать) и запоминать характерные особенности окружающей среды (например, для
лососей это может быть «запах» реки, в которой они вывелись и куда они будут
возвращаться для нереста).
весьма примитивна. Психологи пытались
установить общие «законы научения», но до сих пор все
эти попытки были неудачными. По-видимому,
научение представляет собой индивидуальный процесс
и осуществляется у разных видов и в разных
обстоятельствах по-разному. Приведенная в настоящей
главе классификация форм и особенностей
приобретенного поведения является искусственной, и об
этом не следует забывать. Однако она охватывает
весь спектр существующих представлений о типах
научения и подкреплена экспериментальными
данными. Основные типы приобретенного поведения
суммированы в табл. 16.18, где, однако, даются
лишь вводные сведения.
Ответы и обсуждение
Глава 10
10.1. 1) Разлагает органическое вещество и,
следовательно, способствует возврату химических элементов
мертвых организмов к живым.
2) Перемешивает органические остатки.
3) Вызывает порчу пищевых продуктов (например,
образует плесень на хлебе).
4) На Дальнем Востоке Мисог использовали для
выработки спирта. В смесь Мисог и дрожжей добавляли
рис. Мисог превращал крахмал риса в сахара, которые
затем дрожжи превращали в спирт.
10.2. См. разд. 4.5.3 (т. 1).
10.3. Из-за постоянной потери тепла с относительно
большой поверхности тела мыши.
10.4. 4,18 Дж повышают температуру 1 г воды на ГС;
7,5*4,18 Дж повышают температуру Ir воды на
7,5°С; 7,5-4,18-500 Дж повышают температуру
500 г воды на 7,5°С; следовательно, при сгорании
1 г сахара в кислороде вырабатывается 15,675 кДж.
10.5. Жиры намного богаче водородом, чем углеводы, а
поскольку большая часть высвобождающейся в
организме энергии образуется при окислении водорода в
H2O, жиры дают больше теплоты, чем углеводы.
10.6. В организме высвобождается меньше энергии, так
как оба эти вещества окисляются не полностью;
например, азот белка выделяется в виде мочевины в моче, не
успевая окислиться до двуокиси азота, следовательно, с
мочевиной организм теряет часть энергии.
10.7. а) В пище в небольших количествах должны
содержаться определенные «факторы» (сейчас их
называют витаминами). Они необходимы для нормального
роста и развития.
б) «Факторы роста» должны были содержаться в той
порции молока (3 мл), которую получали крысы.
Этим подтверждается то, что «факторы роста» нужны
лишь в очень малых количествах. Когда молоко
перестали давать, рост быстро прекратился. Крысы,
не получавшие молока, вначале росли; значит, у них был
в организме небольшой запас витаминов.
в) Взрослые будут испытывать недостаток железа,
витамина В и грубой пищи.
10.8. Активный фермент переваривал бы
вырабатывающие его клетки, так как в зимогенных железах нет
защитного барьера, создаваемого слизью.
10.9. а) Слизистая тонкого кишечника образует складки,
покрытые ворсинками и микроворсинками.
6) Это во много раз увеличивает секретирующую и
всасывающую поверхность тонкого кишечника и
повышает эффективность этих процессов.
10.10. Активность ферментов снижалась в результате
денатурации под действием кислой среды.
10.11. Этим обеспечивается переход в кровь
растворимых веществ пищи даже в том случае, если их
концентрация ниже, чем в крови.
Глава 11
11.1. Те вещества, при гидролизе которых выделяется
больше энергии, чем при гидролизе АТФ, способны
переносить свои фосфатные группы на АДФ с
образованием АТФ; а те, при гидролизе которых
выделяется меньше энергии, чем при гидролизе
АТФ, будут присоединять фосфатные группы от АТФ.
11.2. АТФ можно сравнить с аккумулятором в том
смысле, что его производство требует энергии и он
служит удобным кратковременным накопителем
энергии. АТФ вырабатывается в процессе дыхания и может
перемещаться в любую часть клетки, где необходима
энергия, «разряжаться» (превращаясь в АДФ), а затем
при дыхании вновь «заряжаться» (когда АДФ снова
превращается в АТФ).
11.3. Дегидрогенирование.
11.4. Дегидрогеназы (см. табл. 6.3).
11.5. а) Фосфорилирование.
б) Изомеразы.
в) (1) Дегидрогенирование (или окисление).
(2) Фосфорилирование.
г) Один из витаминов группы В.
11.6. Для быстрой диффузии промежуточных продуктов
из цитоплазмы в митохондрии и обратно.
11.7. Поступающие:
Пировиноградная кислота
Кислород
Восстановленный
переносчик водорода
АДФ
Фосфат
Выходящие:
Двуокись углерода
Окисленный
переносчик водорода
АТФ
Вода
11.8. Обеспечивает увеличение поверхности, всасывающей
продукты переваривания пищи.
11.9. С повышением концентрации растворенных молекул
будет повышаться осмотическое давление плазмы, а это
может повлиять на многие физиологические процессы.
11.10. 1) В клетке пигмент отделен от более изменчивой
химической среды плазмы крови.
2) Заключение пигмента внутрь клеток уменьшает
вязкость крови и тем самым облегчает работу сердца.
11.11. Вначале при соприкосновении крови с водой
градиент концентрации кислорода между ними будет
велик. Однако по мере дальнейшего параллельного
течения обеих жидкостей градиент будет
уменьшаться до тех пор, пока относительное насыщение
кислородом крови и воды не станет одинаковым. В
результате насыщение крови будет значительно
ниже максимальной точки и поэтому будет
недостаточным (рис. 11.11 отв.).
11.12. Потому что часть объема легочной вентиляции
составляет мертвое пространство.
11.13. У мелких животных велико отношение
поверхности тела к его объему, что увеличивает потерю
тепла; поэтому им требуется больше кислорода для
поддержания постоянной температуры тела.
11.14. Для этого нужно сравнить потребление ими
кислорода (в единицу времени), приходящееся на
1 г массы тела.
11.15. ПДК =
CO2
102
145
= 0,70.
11.16. ПДК =
CO2
2
О
= оо .
11.17. Потому что на дыхание у человека обычно
расходуются углеводы и жиры.
11.18. а) Частота дыхания равна около 17 в 1 мин.
б) Объем воздуха, обмениваемого за один вздох,
равен 450 мл (в среднем).
в) Легочная вентиляция составляет 17-450 мл = 7,65 л в
1 мин.
г) Потребление кислорода выражено наклоном линии
AB. Таким образом, потребление кислорода равно
1500 мл за 4 мин, или 375 мл за 1 мин.
11.19.(а)
Аэробное дыхание
Это процесс диссимиляции,
в результате которого
молекулы углеводов
расщепляются до простых
неорганических
соединений
Энергия запасается в виде
АТФ
Кислород расходуется
Двуокись углерода и вода
выделяются
Происходит уменьшение
сухой массы
У эукариот процесс
протекает в митохондриях
Происходит непрерывно в
течение жизни во всех
клетках независимо от
наличия хлорофилла и
света
Фотосинтез
Анаболический процесс, в
результате которого из
простых неорганических
соединений синтезируются
молекулы углеводов
Энергия накапливается и
запасается в углеводах.
Образуется также
некоторое количество АТФ.
Кислород выделяется
Двуокись углерода и вода
потребляются
Происходит увеличение
сухой массы
У эукариот процесс
протекает в хлоропластах
Происходит только в
клетках, содержащих
хлорофилл, и только на свету
б) Перечень черт сходства между фотосинтезом и
аэробным дыханием
Оба процесса приводят к преобразованию энергии.
Для обоих необходим механизм, обеспечивающий
обмен двуокиси углерода и кислорода.
Для протекания обоих процессов у эукариот необходимы
специальные органеллы, а именно митохондрии-для
дыхания и хлоропласты-для фотосинтеза; митохондрии и
Насыщение, %
Насыщение, %
Расстояние вдоль
вторичного лепестка
Расстояние вдоль
вторичного лепестка
Рис. 11.11 (отв) А. Противоток воды и крови. 6.
Параллельный ток воды и крови.
хлоропласты сходны с прокариотическими организмами
тем, что обладают кольцевой ДНК и прокариотическим
типом белоксинтезирующей системы. Световые реакции
фотосинтеза сходны с процессами клеточного дыхания в
следующих отношениях:
1) в обоих случаях происходит фосфорилирование (т.е.
синтез АТФ из АДФ и Ф„);
2) те и другие процессы связаны с потоком электронов
вдоль цепи переносчиков электронов;
3) для сопряжения переноса электронов с фосфорилирова-
нием нужна определенная организация системы
переносчиков в мембранах; в митохондриях такими мембранами
служат кристы, в хлоропластах-тилакоиды.
Оба процесса связаны с циклическими цепями реакций,
которые протекают в окружающем мембрану растворе (с
циклом Кребса при дыхании, с циклом Кальвина при
фотосинтезе). Часть ферментов гликолиза является общей
для обоих процессов.
Глава 12
12.1. Сухую массу используют потому, что содержание
воды в различных пищевых продуктах или организмах
может варьировать, но оно не влияет на количество
энергии.
12.2. У мелких птиц и млекопитающих отношение
поверхности тела к его объему больше, чем у человека;
поэтому они быстрее теряют тепло. Так как мелкие
млекопитающие и птицы, так же как и человек, являются
гомойотермными («теплокровными») организмами, они
должны потреблять больше энергии для поддержания
определенной температуры тела. (У птиц интенсивность
метаболизма и температура тела выше, чем у
млекопитающих.)
12.3. В пирамиде Б первичные продуценты представлены
крупными организмами, такими, как деревья, поэтому
численность их меньше численности питающихся ими
травоядных. Так же обстоит дело, когда, например, тли
кормятся на розовом кусте или гусеницы-на дереве.
12.4.
Leptomonas I
Блоха
Травоядное
Трава
Перевернутая
пирамида
12.5. В основе пирамиды В лежит паразитическая цепь
питания, в которой одни паразиты паразитируют на
других. Пирамида численности имеет необычный
перевернутый вид потому, что по ходу пищевой цепи
размеры организмов постепенно уменьшаются, а
численность их возрастает. На первом трофическом уровне
здесь может находиться дерево или кустарник. Иногда
паразитическую цепь представляют как начинающуюся
с хозяина, и тогда это может быть как растение, так и
животное.
12.6. Весной, с наступлением благоприятных условий,
начинается быстрый рост и размножение водорослей,
образующих фитопланктон (цветение воды) Биомасса
фитопланктона в это время превышает биомассу
поедающих его организмов. Затем, по-видимому,
происходит увеличение численности и биомассы
первичных консументов, а вслед за этим и вторичных
консументов. При этом вещество и энергия
переносятся с одного трофического уровня на другой.
Биомасса фитопланктона уменьшается с увеличением
биомассы его консументов и с наступлением
неблагоприятных зимних условий. В некоторый момент
биомасса более долгоживущих консументов
становится больше, чем биомасса продуцентов Такие
сезонные изменения пирамид биомассы характерны для
озерных и морских экосистем, в которых первичным
продуцентом служит фитопланктон.
12.7. а) В мае, июне и июле.
б) (1) Увеличением интенсивности света и длины дня,
повышением температуры в сочетании с наличием
питательных веществ. Все это благоприятствует
фотосинтезу и росту.
(2) Поеданием первичными консументами, например
зоопланктоном, и уменьшением продуктивности из-за
истощения запасов питательных веществ (последнее
обусловлено тем, что остатки отмерших продуцентов
опускаются в холодные слои воды, не участвующие
в циркуляции).
(3) Падением численности зоопланктона. Увеличением
пищевых ресурсов (циркуляция питательных веществ
улучшается осенью, когда поверхностные слои
воды охлаждаются и лучше смешиваются с более
глубокими, холодными слоями). Температурные и
световые условия еще благоприятны.
(4) Свет и температура неблагоприятны для
фотосинтеза и роста.
12.8. Перенос энергии с одного уровня на другой не может
происходить со 100%-ной эффективностью, поэтому до
последующих уровней доходит меньше энергии и на них
может располагаться в среднем меньшее число
животных.
12.9. К фотосинтезирующим организмам относятся также
сине-зеленые водоросли и некоторые бактерии (это уже
прокариоты, а не растения). Хемосинтезирующие
бактерии тоже являются автотрофами (табл. 9.1) и участвуют
в создании первичной продукции. Общий вклад всех этих
организмов по сравнению с растениями невелик.
12.10. а) 24106кДжм-2год-1
б) 1,3%.
в) 800, 200 и 69 700 кДжм~2год-1 соответственно.
г) 114-Ю3 кДж-м"2тод"1.
д) Злаки и другие травы (продуценты), зерноядные
птицы, прямокрылые и лесные мыши (первичные
консументы), пауки (вторичные консументы).
е) Все, за исключением злаков и других трав.
ж) Другие первичные или вторичные консументы,
редуценты, детритофаги (возможна также эмиграция).
12.11. а) 1) χ 2; 2) χ 500; 3) χ 2500; 4) χ 3750.
б) Концентрация ДДТ увеличивается в процессе
передачи его по пищевой цепи. Из этого можно заключить,
что ДДТ - устойчивое вещество, мало подверженное
разрушению. Оно накапливается в живых организмах
быстрее, чем расщепляется в процессах метаболизма.
(Фактически ДДТ в почве сохраняет активность в
течение 10-15 лет.)
в) 1) и 2) На четвертом трофическом уровне
(хищники высшего порядка); 3) на втором трофическом
уровне (травоядные).
г) ДДТ распространился по всему миру по двум
причинам. Во-первых, он, хотя и в очень
незначительных количествах, переносится водой. Если ДДТ
смывается с сельскохозяйственных угодий, то по
рекам часть его попадает в моря и концентрируется в
морских пищевых цепях. Пингвины питаются рыбой и
входят в пищевые цепи. Во-вторых, ДДТ может
распространяться в атмосфере, так как относится к летучим
веществам и его распыляют в виде мелкого порошка,
который может переноситься воздушными потоками на
большие расстояния.
д) (1) Исходно небольшая часть мелких двукрылых
насекомых была устойчива к ДДД и не погибла после
его распыления. В промежутках между распылениями их
численность увеличилась. После дальнейших
распылений они продолжали размножаться и со временем стали
составлять большую часть популяции. Другими
словами, популяция подверглась интенсивному давлению
отбора (см. гл. 25).
(2) Приведенные данные указывают на то, что ДДД
(и соответственно ДДТ) накапливается
преимущественно в жировых тканях. (Это происходит потому,
что ДДД и ДДТ в жирах растворяются лучше, чем в
воде.) Во время нехватки пищи жиры мобилизуются и
используются организмом; при этом ДДД или ДДТ,
накопившийся за длительный период времени,
переходит в кровь в сравнительно высоких концентрациях.
е) Можно предположить, что высокая смертность
среди птиц зимой 1962/63 г. по сравнению с 1946/47 г.
была вызвана дополнительным воздействием ДДТ,
ранее запасенного в жировых тканях. В 1946-47 г. г.
ДДТ использовался в ограниченных масштабах,
а в конце 1950-х и в начале 1960-х годов он уже
получил широкое применение.
12.12. Они должны 1) обладать максимально возможной
специфичностью, т.е. оказывать минимальное
воздействие на виды, не являющиеся вредителями, 2) быть
недорогими и 3) легко разрушаться.
12.13. Географические барьеры, например океаны;
экологические барьеры, например неподходящие для
данного вида зоны, разделяющие области с
благоприятными условиями; расстояние, на которое происходит
расселение; воздушные и водные течения; размеры и
характер заселяемой территории.
12.14. Могут измениться многие факторы окружающей
среды, а) Свет. Интенсивность света под пологом
леса может составлять всего лишь 1-6% от
интенсивности света, падающего на кроны деревьев.
Может изменяться также качество света: свет,
прошедший через листья, обогащается лучами дальней
красной области спектра (более коротковолновая
часть красного света задерживается). Это имеет
физиологическое значение для системы фитохромов
(см. гл. 15).
б) Температура. Суточные и сезонные колебания
температуры в лесу меньше, чем за его пределами.
В лесу максимальные температуры обычно ниже,
а минимальные-выше. Средняя температура летом
ниже, а зимой выше по сравнению с температурой
воздуха вне полога.
в) Ветер. Растения под кронами деревьев защищены
от ветра, скорость которого здесь составляет в
среднем 40-80% от скорости ветра над кронами.
г) Влажность. Перехватывание осадков и последующее
испарение с крон деревьев приводит к уменьшению
количества влаги, достигающей растительности
нижних ярусов. Относительная влажность в лесу обычно
выше, чем за его пределами, отчасти из-за более низких
температур в течение дня.
12.15. Рождаемость =
10000
500000
• 1000 = 20 особей на тысячу
особей в год
12.16. а) От каждой самки в среднем должно выжить два
яйца.
б)
Устрица
Треска
Камбала
Лосось
Трехиглая колюшка
Зимняя пяденица
Мышь
Акула
Пингвин
Слон
Английская женщина
эпохи королевы
Виктории
Число
оплодотворенных яиц,
которые должны
погибать, чтобы
размер популяции не
изменялся
(100106)-2
(9 106)-2
(35 104)-2
(10104)-2
498
198
48
18
6
3
8
Дорепродуктив-
ная смертность,
%
>99,9
>99,9
>99,9
>99,9
498/500 = 99,6
99,0
96,0
90,0
75,0
60,0
80,0
в) Трехиглая колюшка и акула рождают живых мальков,
т. е. это живородящие организмы. Таким образом, в тех
случаях, когда родители принимают большое участие в
созревании потомства, требуется меньшее число яиц.
Кроме того, самки фактически не могут тогда
производить потомство в большем числе.
12.17. В популяции ZT, так как в ней большая доля
особей погибает, не достигнув репродуктивного
возраста. В популяции А высокая выживаемость сочетается
с низкой рождаемостью, так что размеры популяции
остаются неизменными.
12.18. а) Из 3200 яиц 640 выживают, а 2560 погибают,
т.е. смертность равна 80%.
б) Из 640 мальков 64 выживают, а 576 погибают-
смертность равна 90%.
в) Из 64 серебрянок 2 выживают, а 62 погибают-
смертность равна примерно 97%.
Общая дорепродуктивная смертность у лосося
составляет 3198 особей из 3200, т.е. 99.97% (см.
рис. 12.18 (отв.).
12.19. a) S. nemoralis;
б) S. nemoralis;
в) S. rugosa.
г) Результаты опытов со S. speciosa показали, что
этот вид обладает пластичной реакцией на
условия среды. При заселении стабильного лесного
местообитания, где действует /f-отбор, жизненная
стратегия вида больше напоминает /f-стратегию:
вид расходует меньше биомассы на размножение,
чем при жизни в нарушенном местообитании и на
более ранней стадии сукцессии.
Можно было бы сделать общий вывод, что некоторые
виды обладают переменным уровнем «г», зависящим
от условий окружающей среды. Это, по-видимому,
действительно верно для популяций некоторых
растений и животных. Если та или иная стратегия
отбирается в течение долгого периода времени,
может происходить видообразование, как это было в
случае S. rugosa и S. nemoralis.
12.20. а) Сигмоидная (S-образная) кривая роста.
б) За пищу и пространство. В данном случае скорее
всего за пищу.
в) Большая скорость размножения. Более обильное
питание. Более высокая устойчивость к токсичным
продуктам жизнедеятельности Paramecium или
бактерий, растущих в той же культуре (как было
показано, P. aurelia более устойчива, чем P. caudatum).
Выделение отравляющего вещества или ингибитора
роста (аллелопатия). Хищничество.
Глава 13
13.1. Сырая масса почвы 60 г
Сухая масса почвы 45 г
Следовательно, масса воды 60 — 45 = 15 г
Следовательно, содержание воды
в сырой почве
15
60
• 100 = 25%
Сухая масса почвы 45 г
Сухая масса почвы после
сжигания 30 г
Следовательно, вес органического
вещества 15 г
Следовательно, содержание
органического вещества в сырой почве
15
60
• 100 = 25%
13.2. 43%.
13.3. 36%.
13.4. 4230.
Глава 14
14.1. а) В растворе Б. 6) Раствор А. в) Из Б в А.
г) В растворе А. д) (2) —1000 кПа. Если это вас
смущает, вспомните, что —1000 ближе к нулю, чем —2000.
Нуль-это максимальное, наивысшее значение ψ.
Число выживших, %
Время, годы
Рис. 12.18. (отв.). График, показывающий дорепро-
дуктивную смертность среди нерки.
ё) В растворе Б. ж) Чем выше ОД раствора, тем
ниже ψ (практически —ОД = ψ для раствора при
нормальном атмосферном давлении).
14.2· Гипертоническим раствором. Вспомните, что
клеточная стенка легко проницаема для растворов.
14.3. Нулевое. Протопласт не оказывает давления на
клеточную стенку.
14.4. У прокариот (бактерий и сине-зеленых
водорослей) и грибов. Хотя у прокариот нет вакуолей,
их клетки обладают такими же осмотическими
свойствами, что и клетки растений и грибов.
14.5. В начале ¥клетки = ТД - ОД, a VpacTBOpa = - ОД =
= —1200 кПа; ТД = 0, так как клетка находится в
состоянии начинающегося плазмолиза; ОД = 2000 кПа;
таким образом, ψ^™ = ° кПа - 2000 кПа =
= —2000 кПа. Вначале ψ раствора выше, чем ψ клетки.
Поэтому вода за счет осмоса пойдет из раствора в
клетку. Единственный член уравнения, который
существенно изменяется при переходе воды в клетку -
это ТД. По мере поступления воды ТД, а
следовательно и ψ, возрастает до тех пор, пока оно не станет
препятствовать дальнейшему входу воды в клетку.
В этот момент ψ клетки становится равным ψ
раствора, и клетка становится тургесцентной.
Поэтому в состоянии равновесия ψ^ηχη — — ^200 к^а.
ψ = ТД - ОД, отсюда ТД = ψ + ОД = - 1200 кПа +
+ 2000 кПа = 800 кПа.
14.6. ОДклетки = 1100 кПа; ψ чистой воды = 0; В
состоянии равновесия \|/клетки = 0. ТД = ψ + ОД = 0 кПа +
+ 1100 кПа= 1100 кПа.
14.7. α) ψ,^βχκΗ во время переноса = ψ чистой воды = 0,
а ψ раствора сахарозы = — 800 кПа; отсюда разность
ψ между содержимым клетки и наружным
раствором равна 800 кПа.
б) Вода будет выходить из клетки (от более высокого
ψ к более низкому).
в) ТД должно уменьшиться.
14.8. а) Клетка Б. б) Из клетки Б в клетку А.
в) В состоянии равновесия клетки должны иметь
одинаковый ψ, он будет равен среднему от величин
двух исходных ψ, т.е. —1000 кПа.
г) Клетка А в состоянии равновесия:
ТД = ψ + ОД
= - 1000 кПа + 2000 кПа
= 1000 кПа
Клетка Б в состоянии равновесия:
ТД = ψ + ОД
= - 1000 кПа + 1400 кПа
= 400 кПа
14.9. 1060 кПа. Для величин, занимающих
промежуточное положение между значениями, приведенными в
табл. 14.4, постройте график зависимости
осмотического давления от молярной концентрации
раствора сахарозы.
14.10. Усредненное ОД в клетках свеклы будет
составлять около 1400 кПа.
14.11. ψ в клетках свеклы составит примерно —940 кПа.
14.12. Можно получить более точный результат, взяв
среднюю величину из двух или более повторностей.
Некоторое представление о размерах вариаций,
которых можно ожидать в этом случае, дают цифры,
приведенные в табл. 14.6.
14.13. Чтобы предотвратить испарение воды и связанное
с ним повышение концентрации растворов сахарозы,
а также возможное высыхание полосок ткани.
14.14. ТД = ψ + ОД
= - 950 кПа + 1400 кПа
= 450 кПа
Обратите внимание на то, что у разных корнеплодов
свеклы могут быть разные значения ОД и ψ.
14.15. а) Клетки интактного стебелька тургесцентны,
поэтому их стенки стремятся растянуться под
действием тургорного давления. Толстые стенки
эпидермальных клеток менее растяжимы, чем
тонкие стенки клеток коры, и таким образом
препятствуют расширению этих клеток. Клетки коры
сжаты. Разрез эпидермиса снимает такое
ограничение, все клетки коры слегка расширяются, общий
объем коры увеличивается, и это приводит к тому,
что полоски закручиваются наружу.
б) Дистиллированная вода имеет более высокий
водный потенциал, чем клетки стебелька. Поэтому
вода поступает в ткань за счет осмоса, еще больше
растягивая клетки коры и увеличивая изгиб.
в) Концентрированный раствор сахарозы имеет более
низкий водный потенциал, чем клетки стебелька.
Поэтому вода в результате осмоса выходит из ткани,
клетки коры сжимаются сильнее, чем клетки
эпидермиса, и ткань изгибается внутрь.
г) Разбавленный раствор сахарозы должен иметь точно
такой же водный потенциал, что и клетки стебелька;
поэтому-то и не происходит чистого поступления воды
или чистого выхода ее из раствора или из ткани.
д) Водный потенциал. План такого эксперимента
может быть следующим.
Приготовьте ряд разбавленных растворов сахарозы,
начиная с 1 M раствора и кончая дистиллированной
водой (например: дистиллированная вода-0,2 M-
0,4 M-0,6 M-0,8 M-1,0 M). Типичный изгиб
свежесрезанных стебельков одуванчика следует
зарегистрировать, зарисовав их; затем в каждый из растворов,
разлитый по отдельным подписанным чашкам Петри,
помещают по две полоски стебелька (лучше взять по две
полоски, чтобы определить средний результат). После
того как будет достигнуто равновесие (примерно через
30 минут), тщательно зарегистрируйте изгиб стебельков
(например, зарисовав их). Тот раствор, в котором не
произойдет никаких изменений, будет иметь тот же
самый ψ, что и усредненная клетка стебелька в момент
его среза.
14.16. План двух подходящих экспериментов может
быть следующим.
Влияние температуры. Вырежьте кубики из свежего
корнеплода свеклы, промойте их, чтобы удалить
красный пигмент из поврежденных клеток, и
поместите в стаканчики с водой разной температуры,
скажем, в пределах от 20 до 1000C. Появление
красного пигмента в воде будет указывать на нарушение
избирательной проницаемости тонопласта (вакуоляр-
ной мембраны) и плазматической мембраны,
приводящее к диффузии пигмента из клеточного сока в
воду. Время, необходимое для появления
определенного количества пигмента, позволит судить о
скорости разрушения структуры мембран.
Интенсивность окрашивания можно измерять на колориметре
или же оценивать визуально.
Влияние этанола. Методика та же самая, только
вместо изменения температуры используют ряд
концентраций этанола.
14.17. а) Листья имеют множество устьиц, служащих для
газообмена, и нет почти никаких препятствий для
передвижения водяных паров через эти поры.
б) У листьев большая поверхность (для того чтобы
улавливать солнечный свет и для газообмена). Чем
больше поверхность, тем значительнее потеря воды в
результате транспирации.
14.18. Чем выше поднимается солнце, тем больше
интенсивность света; она достигает максимума в полдень,
когда солнце находится в верхней точке своего пути.
Точно так же возрастает и температура воздуха, но с
некоторым запаздыванием-примерно на два часа
позже (главным образом потому, что сначала
нагревается почва, а затем она излучает тепло в
воздух). Первоначальное ускорение транспирации
между 3 и 6 часами утра, еще до того, как повысится
температура воздуха, обусловлено открытием устьиц
на свету. С 6 часов утра нарастающая скорость
транспирации тесно коррелирует с температурой
(причины этого объясняются в тексте). Она слабо
коррелирует с интенсивностью света, так как устьица
в это время открыты полностью, и всякое дальнейшее
повышение освещенности не оказывает никакого
действия.
После полудня интенсивность света убывает, так
как солнце начинает снижаться. Понижается и
температура, но с такой же задержкой примерно в два
часа. Транспирация замедляется и в результате
понижения температуры, и в результате уменьшения
освещенности, но она гораздо больше коррелирует
со светом, ослабление которого приводит к
постепенному закрыванию устьиц. Примерно к 19.30
наступает темнота, и устьица, вероятно, уже закрыты.
Вся остальная транспирация, по-видимому, происходит
через кутикулу, и на нее по-прежнему влияет
температура.
14.19. См. табл. 14.19 (отв.).
14.20. а) Тонкостенный полый цилиндр.
б) Массивный стержень (цилиндр), обеспечивающий
опору.
Таблица 14.19 (отв.)
Двудольные
Сосудистые пучки
образуют кольцо
Эндодерма имеется
Раздельные сердцевина и
кора
Перицикл образует чехлик
над сосудистым пучком
Метаксилема состоит из
множества сосудов
Протоксилема имеется
Камбий имеется, и позднее
может происходить
вторичное утолщение (у
некоторых травянистых
растений вроде Helianthus
формируются вторичные
сосудистые пучки; у
многолетних образуются
древесина, лучи, годичные
кольца, пробка)
Однодольные
Сосудистые пучки
разбросаны
Эндодерма отсутствует
Основная ткань не разделена
на сердцевину и кору
Перицикл полностью
окружает сосудистый пучок
Метаксилема состоит из
небольшого числа сосудов
(часто из двух или трех)
Большая часть протоксиле-
мы разрушается, образуя
лизигенную полость
Камбия нет, поэтому нет и
вторичного утолщения (за
редкими исключениями,
когда стебли однодольных
образуют древовидную
форму)
Таблица 14.22 (отв.)
Двудольные
Зона корневых волосков
быстро исчезает
Мало групп протоксилемы
(мало лучей метаксиле-
мы)-2-8, обычно четыре
(тетрархная
протоксилема) или пять (пентархная).
Сердцевины обычно нет.
Ксилема образует
сплошной цилиндр в центре
корня
Часто имеется камбий, что
делает возможным
вторичное утолщение
Однодольные
Зона корневых волосков
сохраняется
Много групп протоксилемы
(протоксилема полиарх-
ная)
Центральная сердцевина.
Ксилема образует
столбики вокруг сердцевины
Камбий присутствует редко;
поэтому нет вторичного
утолщения
в) Массивный стержень (цилиндр), обеспечивающий
опору.
г) Массивный цилиндр.
14.21. 1) Ксилема состоит из длинных трубок,
образованных в результате слияния соседних клеток и
разрушения поперечных стенок между ними.
2) В трубках нет живого содержимого, отсюда
меньшее сопротивление потоку.
3) Трубки обладают высокой прочностью, поэтому они
не спадаются.
4) Тонкие трубки необходимы для того, чтобы водяной
столб в них не разрывался.
14.22. См. табл. 14.22 (отв.).
14.23. Почвенный раствор > клетка корневого волоска >
> клетка 3 > клетка 2 > клетка 1 > ксилемный сок.
14.24. а) При обеих температурах происходит быстрое
начальное поглощение K+ (в первые 10-20 мин).
Через 20 мин при 25°С продолжается непрерывное
постепенное поглощение K + , а при 00C никакого
дальнейшего поглощения не происходит.
Поглощение при 250C ингибируется KCN.
б) Есть две ясно различимые фазы поглощения. Во
время начальной быстрой фазы K+ диффундирует в
свободные пространства корня. Он проходит через
клеточные стенки зоны корневых волосков, которые
обращены к раствору, и диффундирует через апопласт
(возможно, проникая в межклеточные пространства там,
где в них находится раствор). Эксперименты
показывают, что эта фаза мало зависит от температуры и,
следовательно, не связана с обменом веществ (реакции,
контролируемые ферментами, протекали бы при 25°С
быстрее, чем при 00C). Значит, эта первая фаза-
пассивный процесс.
Вторая фаза зависит от температуры и не осу-
ществлятся при 00C, когда интенсивность
метаболизма очень низка. Это позволяет предполагать,
что поглощение требует затраты энергии, а его
ингибирование KCN указывает на то, что оно зависит
от дыхания. Таким образом, вторая фаза представляет
собой активный транспорт через клеточную мембрану
внутрь клетки.
14.25. Увеличение интенсивности дыхания
сопровождается усилением поглощения KCl. Если KCl достаточно,
он, по-видимому, поглощается путем активного
транспорта за счет энергии, которую доставляет
дыхание.
14.26. KCN, ингибируя дыхание, тем самым подавляет
и активный транспорт KCl в высечки из моркови.
14.27. Значительная часть фосфата внутри корня
находилась в свободном пространстве и могла поэтому
диффундировать обратно в окружающую воду,
обращая вспять пассивное поглощение
14.28. Нет. Эндодерма - это барьер для передвижения
воды и растворенных в ней веществ по апопластному
пути (см. разд. 14.5.2).
14.29. Радиоавтография выявляет локализацию иона в
тонких срезах. Ингибируйте у одного из растений
активный транспорт (например, с помощью KCN
или низкой температуры), а другое растение возьмите
в качестве контроля. Пусть теперь оба растения
поглощают радиоактивный ион. У растения с
подавленным активным транспортом ионы будут
передвигаться только пассивно через клеточные
стенки. Радиоавтография должна показать, что
радиоактивный ион почти не проникнет здесь дальше
эндодермы, тогда как в контроле должно быть видно
гораздо большее проникновение ионов в ткань за
эндодермой.
14.30. а) См. табл. 14.30 (отв.).
Черты сходства в экспорте 14C из верхнего и из нижнего
листа. Оба листа экспортируют одинаковую долю асси-
милятов (сравните результаты для «листа,
обработанного 14С»), и одинаковая доля переходит в другие
листья. Главным местом назначения ассимилятов в
обоих случаях является боб.
Различия в экспорте 14C из верхнего и из нижнего
листа. На экспорт ассимилятов из того или иного
листа влияет расположение этого листа на стебле.
Верхний лист экспортирует большую долю своих
ассимилятов в боб и апикальную зону стебля, чем
нижний лист, и меньшую долю-в корни.
б) Когда боб уже образовался, он становится
существенным потребителем ассимилятов, особенно из тех
листьев, которые находятся в непосредственной
близости к нему. У взрослого растения есть еще два
важных места потребления ассимилятов, а именно
апикальная зона стебля и корни. Нижние листья
экспортируют ассимиляты главным образом в корни,
а верхние-в основном к верхушке стебля.
14.31. 0,72 с. Этот ответ получается таким образом:
100 см = 1000 мм = 1 000000 мкм = 106 мкм;
следовательно, сахароза движется со скоростью 106 мкм/ч
= 1 мкм за 1/106 ч
= 200 мкм за 200/106 ч = 2/104 ч
= (2-3600)/104с = 2-0,36 с
= 0,72 с.
Таблица 14.30 (отв.)
Часть растения Доля радиоактивности в этой части, %
Апикальная зона
стебля
Лист,
обработанный 14C
Другие листья
Стебель
Боб
Корни
после обработки
верхнего листа
4,8
48,4
1,0
3,5
38,7
3,6
после обработки
нижнего листа
3,6
53,9
0,8
5,5
23,4
12,8
14.32. Через 2500 ситовидных пластинок на каждый
метр:
1 м = 106 мкм,
400 мкм = 4-102 мкм,
106/(4 102)= 104/4 = 2500.
14.33. Благодаря высокому давлению оксигенированная
кровь может поступать в ткани достаточно быстро,
чтобы удовлетворить метаболические потребности
организма. Кроме того, на высоком гидростатическом
давлении крови основана нормальная работа таких
органов, как почки и кровеносные капилляры.
14.34. В большом круге кровообращения кровь
поступает в капилляры под значительно более высоким
давлением, а относительно низкое давление крови
в легочной артерии предотвращает разрыв нежных
легочных капилляров.
14.35. Благодаря расширению кровеносных сосудов в
поврежденном участке улучшается его снабжение
оксигенированной кровью и питательными веществами
и быстрее идет процесс заживления. Повышение
общего кровяного давления подготавливает организм
животного к более быстрому и эффективному ответу на
любой последующий стресс.
14.36. Перед стартом. Ожидание старта вызывает
выброс адреналина. Под действием адреналина
сужаются все кровеносные сосуды, кроме тех, которые
снабжают кровью жизненно важные органы, и как
следствие - повышается кровяное давление.
Повышается частота сокращений сердца. Из селезенки
в общую циркуляцию выбрасывается дополнительное
количество крови. Во время бега. В это время
повышается метаболическая активность, особенно в
скелетных мышцах. Углекислота, образующаяся
в мышцах в повышенных количествах, вызывает
местное расширение кровеносных сосудов. В то же
время повышение концентрации CO2 в системной крови
регистрируется хеморецепторами аорты и каротидных
телец, и сигналы от них стимулируют сосудодви-
гательный центр. Стимуляция этого центра приводит
к сужению сосудов, повышению кровяного давления
и ускорению кровотока. Увеличивается также
частота и сила сердечных сокращений, и желудочки
сердца полнее освобождаются от крови. К концу
пробега в мышцах преобладает анаэробное дыхание и
накапливается молочная кислота (разд. 11.3.8). Сильные
сокращения мышц приводят к ритмическому
сдавливанию вен, что способствует ускорению венозного
возврата крови в сердце. Восстановительный период.
Полностью ликвидируется кислородная
задолженность, и молочная кислота удаляется из крови,
уменьшается активность тканей и снижается содержание
CO2. В результате этого частота сердечных
сокращений и кровяное давление возвращаются к норме.
14.37. В активно дышащих тканях существует высокое
парциальное давление CO2, которое приводит к
снижению рН и смещению кривой диссоциации окси-
гемоглобина вправо. Анализ рис. 14.34 показывает,
что в этих условиях облегчается переход кислорода
из крови в ткани, где он может быть использован для
выработки энергии в процессе дыхания. При низком
парциальном давлении O2 гемоглобин связывает его
слабо, но, будучи насыщен кислородом, легко его
отдает.
Высокая метаболическая активность приводит также к
повышению температуры в данном участке тела, а это
ведет к уменьшению сродства гемоглобина к O2 и
усиленной диссоциации оксигемоглобина. В результате
кривая диссоциации тоже сдвигается вправо, и это имеет
физиологический смысл, так как в активные участки
поступает больше кислорода из крови. Кривые
диссоциации оксигемоглобина у разных видов не совсем
одинаковы; например, у мелких млекопитающих они
сдвинуты вправо по сравнению с человеком. У мелких
млекопитающих метаболическая активность намного
выше, чем у человека, поэтому кислород должен у них
освобождаться намного быстрее.
14.38. Сдвиг кривой диссоциации у плода по отношению к
ее положению у матери означает, что кровь плода
обладает большим сродством к O2, чем кровь матери, и
это вполне естественно, поскольку плод должен
получать весь кислород через плаценту из материнской
крови. Поэтому при любом парциальном давлении
кислорода кровь плода будет поглощать его из
материнской крови и всегда будет сильнее насыщена
кислородом, чем материнская кровь
14.39. У южноамериканских лам кровь отличается
высоким сродством к кислороду и способна связывать
его при низком парциальном давлении, существующем
на больших высотах. Это еще один яркий пример
физиологической адаптации.
14.40. 1. Карбоксигемоглобин попадает в легкие,
насыщается кислородом и превращается в оксигемоглобин.
2. Оксигемоглобин, будучи более слабым
основанием, чем карбоксигемоглобин, освобождает ионы
H + .
3. Ионы H+ присоединяются в эритроцитах к ионам
гидрокарбоната с образованием угольной кислоты.
4. Угольная кислота диссоциирует на CO2 и воду.
5. В результате потери ионов гидрокарбоната
эритроцитами в эритроциты из плазмы диффундируют
новые ионы гидрокарбоната.
6. Из гидрокарбоната в эритроцитах образуются новые
молекулы угольной кислоты, которые опять
диссоциируют на CO2 и воду
7. CO2 диффундирует из эритроцитов и в конечном
счете выводится из организма через легкие.
Глава 15
15.1. Передвижение прежде всего обусловлено
необходимостью искать пищу (и тесно связано с развитием
нервной системы). Зеленые растения -автотрофные
организмы, они способны сами синтезировать нужные
им органические вещества, так что им нет надобности
искать органическую пищу.
15.2. См. табл. 15.2 (отв.).
15.3. Рост мог бы ингибироваться на стороне контакта или
же стимулироваться на противоположной стороне (или
могло бы происходить то и другое) Фактически рост
замедляется (т.е клетки меньше удлиняются) в зоне
контакта, а рост на противоположной стороне
ускоряется в 40-200 раз.
15.4. Возможны самые разные методики. Простейший
эксперимент показан на рис. 15.4 (отв ).
15.5. a) Spirogyra (или любая другая нитчатая зеленая
водоросль).
б) Бактерии- аэробы и проявляют положительный
аэротаксис. Поэтому они движутся в сторону кислорода по
градиенту от низкой концентрации O2 к более высокой.
Наивысшая концентрация кислорода по краям
покровного стекла, где кислород диффундирует в воду из
воздуха, и в непосредственной близости от нити
водоросли, где кислород выделяется как побочный
продукт фотосинтеза.
в) Можно оставить препарат в темноте примерно на
30 мин и снова рассмотреть его. Все бактерии должны
теперь скопиться у краев покровного стекла, так как
водоросль в темноте не фотосинтезирует.
15.6. а) Световой стимул воспринимается кончиком ко-
леоптиля Затем какой-то сигнал передается от кончика
(рецептора) в область ниже кончика (эффектор).
б) Опыт В был нужен для проверки результатов опыта Б,
которые могли быть следствием повреждения коле-
оптиля.
15.7. Были получены дальнейшие данные о
существовании какого-то сигнала - по-видимому, химического
вещества (гормона). Это вещество не может пройти
через непроницаемый барьер. Оно передвигается
главным образом вниз по затененной стороне колеоптиля. В
опыте Б слюда мешает такому передвижению. Поэтому
свет либо ингибирует образование гормона, либо
вызывает его инактивацию (стимулирует его распад), либо
заставляет его перераспределяться в латеральном
направлении.
Таблица 15.2 (отв.)
Примеры
Побеги и колеоптили
обладают положительным
фототропизмом, а корни-
отрицательным
Побеги и колекоптили
проявляют отрицательный
геотропизм, а
корни-положительный
Корневищам, усам и т.п.
свойствен диагеотропизм;
листьям двудольных - тоже
Боковые корни и
разветвления стебля проявляют
плагиогеотропизм
Гифы грибов и пыльцевые
трубки обнаруживают
положительный
хемотропизм
Корням и пыльцевым
трубкам свойствен
положительный гидротропизм
Усики лиан проявляют
положительный гаптотро-
пизм; чувствительные
волоски росянки-тоже
Пыльцевым трубкам
свойствен отрицательный
аэротропизм
Преимущества
Листья оказываются на
свету, который служит
источником энергии для
фотосинтеза. Обнаженные корни растут
в сторону почвы или иного
подходящего субстрата
Побеги и проростки растут
вверх, т.е. к свету, а корни
врастают глубже в почву
Это помогает растениям
заселять новые участки почвы;
листья растут горизонтально
и получают максимум света
Корни используют больший
объем почвы, и их
расположение усиливает их опорную
функцию; листья на ветвях
занимают больше
пространства для использования света
Гифы растут по
направлению к пище, а пыльцевые
трубки-к семязачатку (где
происходит оплодотворение)
Вода нужна для всех
жизненных процессов
Это позволяет усикам
выполнять опорную функцию, а
волоскам - улавливать
насекомых
Трубка сразу же начинает
расти в сторону ткани
пестика (дальше от воздушной
среды)
Полупрозрачная крышка постепенно
изменяющейся толщины; спустя
примерно 30 мин чашку открывают
для исследования под бинокуляром
СВЕТ
Зачерненная
чашка Петри
Организмы скапливаются
в области оптимальной
интенсивности света
Рис. 15.4 (отв.) Опыт, позволяющий выяснить,
какую интенсивность света предпочитает эвглена или
хламидомонада.
Колеоптиль
Слюда
КОНТРОЛЬ -
нормальный
рост
Гормон стимулирует рост только
на той стороне, где путь ему
не преграждает пластинка слюды
Рис. 15.8 (отв.). Повторение опытов Бойсен-
Йенсена при равномерном освещении. Показаны три
эксперимента; во всех случаях слева представлены
условия опыта, справа- результат
15.8. См. рис. 15.8 (отв.).
15.9. Кончик колеоптиля образует какое-то химическое
вещество, которое диффундирует в агар. Оно может
стимулировать рост в зоне, расположенной ниже
кончика, и восстанавливает нормальный рост (опыт Л). В
условиях равномерного освещения или в темноте
практически не происходит никакой латеральной передачи
этого вещества (опыт Б).
15.10. Колеоптиль должен будет расти в левую сторону
15.11. А - 100 ч. на млн.; Б - 10 ч. на млн ; В - 1 ч. на млн.;
Г-0,1 ч. на млн.; Д-0,01 ч. на млн.; E-нуль.
15.12. Относительно высокая концентрация ауксина
стимулирует рост колеоптилей (или побегов), но ингиби-
рует рост корней. Это подтверждает выводы, сделанные
из опыта 15.1.
15.13. См. разд. 16.5.6.
15.14. а) Абсцизовая кислота может транспортироваться
из кончиков корней вверх, претерпевать латеральный
транспорт в тканях корня в ответ на воздействие силы
тяжести и подавлять рост.
б) ИУК, вероятно, не участвует в геотропической
реакции корня кукурузы, поскольку она, по-видимому,
не транспортируется из кончика корня вверх.
15.15. а) крахмал;
б) мальтоза;
в) мальтаза.
г) Крахмал, запасенный в эндосперме, служит главным
питательным веществом семян хлебных злаков.
15.16. Запасные белки перевариваются (гидролизуются) с
освобождением аминокислот - основных структурных
единиц белка. Из этих аминокислот синтезируются
ферменты (все ферменты - белки), например α-амилаза; эти
ферменты используются затем для переваривания
питательных веществ эндосперма.
15.17. Амилазная активность может быть связана с
присутствием микроорганизмов на коже рук или же с
попаданием слюны на пальцы изо рта. Поэтому в
подобного рода опытах очень важно не брать в руки
семена, после того как их поверхность простерили-
зована.
15.18. Можно инкубировать семена с радиоактивными
(мечеными 14C) аминокислотами. Это приведет к
образованию меченой амилазы. Можно также инкубировать
семена с ингибиторами белкового синтеза (например, с
циклогексимидом); это помешает синтезу амилазы, и
тогда не будет обнаружено никакой амилазной
активности.
15.19. Разделив семена на алейроновую и неалейроновую
части, можно показать, что меченая амилаза появляется
сначала в алейроновом слое. Можно также раздельно
инкубировать эндосперм с алейроновым слоем и
эндосперм без этого слоя на агаре с крахмалом и гибберел-
лином; тогда амилаза должна синтезироваться только в
первом случае (практически, однако, достичь такого
результата нелегко).
15.20. Одна из лучших биологических проб на гиббе-
реллин (быстрая, надежная и чувствительная) основана
на инкубации зародышевых половинок семян ячменя с
исследуемым материалом. Через два дня содержание
редуцирующих Сахаров в зародышах будет
пропорционально количеству гиббереллина в этом материале.
15.21. а) В молодом листе аминокислота задерживается и
не уходит далеко от места ее нанесения. В старом листе
часть ее экспортируется по жилкам и черешку
б) Молодой лист использует аминокислоту в процессе
роста для построения белков. Старый же лист уже не
растет и поэтому экспортирует питательные вещества в
другие части растения, например в корни и молодые
листья.
в) Ткань, обработанная кинетином, удерживает
аминокислоты или даже «притягивает» их. (Причины этого
неизвестны; возможно, это связано с тем, что кинетин
либо поддерживает, либо стимулирует нормальную
активность клетки.)
15.22. Можно, например, взять такое растение, о котором
известно, что добавленный гиббереллин влияет на рост
его стебля, и удалить собственный источник ауксинов,
отрезав верхушку стебля. Тогда гиббереллин должен
оказаться неэффективным. Очень важно показать, что
ответную реакцию растения можно восстановить,
добавив ауксин (например, ИУК в ланолиновой пасте), так
как отсутствие реакции на гиббереллины могло бы быть
обусловлено повреждением растения или же влиянием
какого-то другого фактора. Такие опыты действительно
демонстрируют полную зависимость реакции от
ауксина.
15.23. а) Ауксин (ИУК).
б) См. рис. 15.23 (отв.).
15.24. Мелкие листья легче проходят сквозь почву
(листья злаков остаются внутри колеоптиля).
Изогнутые крючком почечки зародыша у двудольных
растений предохраняют нежную верхушечную
меристему от повреждения частицами почвы. Удлиненные
междоузлия дают листьям максимальные шансы выйти
на поверхность, к свету.
Ланолиновая
паста
Ланолиновая
паста с ИУК
Верхушечная
почка
Боковая почка
Нормальный
рост
После удаления
верхушечной
почки
апикальное
доминирование
прекращается
ИУК
в осстанав лив ает
апикальное
доминирование
Рис. 15.23 (отв.). Эксперимент, показывающий роль
ИУК в доминировании верхушки.
Дальний красный свет
S
H
О)
§
9
H
U
Красный свет
Длительность освещения, с
Рис. 15.26 (отв). Влияние прерывания «длинной»
ночи красным и дальним красным светом на цветение
дурнишника.
15.25. См. гл. 9.
15.26. График представлен на рис. 15.26 (отв.). Он
демонстрирует противоположное действие красного и
дальнего красного света. 30-секундная экспозиция на
красном свету (при интенсивности освещения,
использованной в данном эксперименте) полностью аннулирует
индуцирующее действие длинной ночи. Эффективность
красного света возрастает с увеличением времени
экспозиции до 30 с. Эффект красного света можно полностью
снять дальним красным светом, хотя для этого нужна
более длительная экспозиция (50 с). Эти результаты
позволяют предполагать, что фоторецептором служит
фитохром.
15.27. Можно использовать несколько методов. Один из
простейших опытов показан на рис. 15.27 (отв.).
Прямоугольниками обведены части растения, укрываемые от
света для создания эффекта короткого дня.
15.28. Подавление роста пазушных почек, или
доминирование верхушки, обусловлено в основном
действием ауксинов. (О доминировании верхушки см. в
разд. 15.3.3).
Глава 16
16.1. а) Между наружной и внутренней поверхностями
мембраны аксона существует крутой градиент
концентрации Na + , и ионы Na+ быстро диффундируют по этому
градиенту.
б) Поступление в аксон положительно заряженных
ионов Na+ поддерживается также относительно
высоким отрицательным потенциалом внутри аксона.
16.2. Если бы выход ионов Na+ уравновешивался
поступлением в него ионов К+ , ничего не изменилось бы или,
возможно, слегка уменьшился бы потенциал покоя. Но
этого было бы недостаточно для достижения пороговой
величины, необходимой для возникновения потенциала
действия.
16.3. (а) Нормальная морская вода;
(б) разведенная до 1/2;
(в) разведенная до 1/3.
Амплитуда потенциала действия в (а) и степень
деполяризации мембраны в (б) и (в) определяются числом
ионов Na+, поступающих в аксон из внеклеточной
жидкости. В растворах, соответствующих графикам на
рис. 16.5, ионы Na+ содержались в постепенно
уменьшающихся концентрациях.
16.4. Ионы Na+ откачиваются из аксона, а ионы K+
накачиваются в него. Перемещение этих ионов
взаимосвязано (сопряжено) и осуществляется катионным
насосом.
16.5. Чем больше диаметр аксона, тем меньше
сопротивление его аксоплазмы продольному току. С
уменьшением этого сопротивления увеличивается длина того
участка мембраны, на который влияет местная цепь, и
это ведет к увеличению расстояния между соседними
деполяризующимися участками и к более быстрому
проведению импульсов.
16.6. Лягушка - холоднокровное (пойкилотермное)
животное, активное при температурах 4-25°С, а
кошка-теплокровное (гомойотермное) животное и имеет постоянную
температуру тела около 35°С. При таком повышении
температуры скорость проведения импульса
увеличивается в 3 раза.
16.7. График А показывает, что частота импульсов в
сенсорном нейроне находится в прямой зависимости от
силы стимуляции мышечного веретена. Известно, что
Короткие дни
Цветки
Длинные
дни
Цветки
Верхушеч-1
ная почка
Листья
.Короткие
дни
Длинные
дни
Цветков
нет
Короткие
дни
Цветков
нет
Длинные
дни
Рис. 15.27 (отв.). Эксперимент, позволяющий
установить, какие части растения-листья или цветковый
апекс-чувствительны к тому фотопериоду, который
стимулирует цветение.
стимул вызывает деполяризацию мембраны,
называемую генераторным или рецепторным потенциалом, и
величина этого потенциала, как видно из графика />,
прямо пропорциональна частоте сенсорных импульсов.
Эти данные позволяют заключить, что усиление
стимуляции ведет к увеличению рецепторных потенциалов,
а оно в свою очередь - к повышению частоты импульсов
в сенсорном нейроне.
16.8. Попадая в глаз, луч света проходит следующий
путь: конъюнктива -* роговица -* водянистая влага -►
-> хрусталик -► стекловидное тело -► сетчатка.
16.9. Свет, падающий от объекта на несколько палочек,
связанных с мозгом через разные нейроны, может не
обладать достаточной энергией, чтобы вызвать
распространяющиеся потенциалы действия в каком-либо из
этих нейронов, и тогда он не будет воспринят. Если же
этот свет падает на три палочки, связанные с мозгом
через один общий нейрон, то отдельные рецепторные
потенциалы, индуцируемые палочками, будут
суммироваться и смогут возбудить распространяющийся
потенциал действия, который будет восприниматься
мозгом как свет
16.10. Когда мы смотрим на предмет прямо, отраженный
от него свет проходит вдоль оптической оси глаза и
падает на сетчатку в центральной ямке, содержащей
только колбочки. Днем благодаря большой
интенсивности света, падающего на колбочки, в мозгу будет
возникать детальное изображение предмета. Однако
ночью сила света слишком мала для активации
колбочек. Если же направить взгляд немного в сторону, свет
от предмета будет падать не на центральную ямку, а на
какой-то другой участок сетчатки, где имеются палочки,
обладающие большей чувствительностью; палочки
будут активироваться даже слабым светом, и
изображение будет воспринято мозгом.
16.11. Казалось бы, предмет будет казаться желтым.
Сетчатка каждого глаза будет воспринимать только один
цвет. В одном глазу свет с длиной волны 530 нм будет
стимулировать зеленые колбочки, а в другом глазу
свет с длиной волны 620 нм-красные колбочки. В
мозгу сигналы от обоих глаз будут действовать
совместно, и предмет будет казаться окрашенным в цвет,
соответствующий средней длине волн (
530 + 620
2
= 575 нм I, т.е. в желтый цвет. В действительности,
однако, смешение контрастных цветов, предъявляемых
правому и левому глазу, затруднено из-за так называемой
«борьбы полей зрения»; результатом может быть
чередующееся восприятие красного и зеленого цвета.
Научное издание
ИБ № 7295
Найджел Грин, Уилер Стаут, Деннис Тейлор
БИОЛОГИЯ
Под ред. P. Сопера
В трех томах
Том 2
Сдано в набор 06.02.89. Подписано к печати 27.07.90. Формат
84 χ 108Viе- Бумага офсетная № 2. Печать офсетная. Гарнитура
тайме. Объем 10,25 бум. л. Усл.печ.л. 34,44. Усл.кр.-отт. 69,73.
Уч.-изд.л. 38,57. Изд. № 4/5707. Тираж 100000 экз. Зак. 7. Цена
4 р. 80 к.
Заведующий редакцией чл.-корр. АН СССР T. M. Турпаев
Зам. зав. редакцией M. Д. Гроздова
Ст. научный редактор Ю. И. Лашкевич
Научный редактор M. Б. Николаева
Мл. редактор И. А. Деменцова
Художник В. В. Перельман
Художественные редакторы А. Я. Мусин, Л. M. Аленичева
Технический редактор 3. И. Резник
Корректор E. В. Морозова
Издательство «МИР» В/О «Совэкспорткнига» Государственного
комитета СССР по печати, 129820, ГСП, Москва, И-110, 1-й
Рижский пер., 2.
Можайский полиграфкомбинат В/О «Совэкспорткнига»
Государственного комитета СССР по печати.
г. Можайск, ул. Мира, 93.
Уважаемый читатель!
Ваши замечания о содержании книги, ее оформлении,
качестве перевода и другие просим присылать по
адресу:
129820, Москва, ГСП. И-110,
1-й Рижский пер , д. 2,
издательство «Мир»