
































































































































Author: Добровольский Г.В.
Tags: сельское хозяйство в целом физика химия почвы почвоведение академия наук ссср издательство наука журнал почвоведение
ISBN: 0032-180Х
Year: 1996




Text
Номер 6
Июнь 1S96
ISSN 0032-180Х
РОССИЙСКАЯ АКАДЕМИЯ НАУК
ПОЧВОВЕДЕНИЕ
Главный редактор
Г.В. Добровольские
Журнал основан в январе 1899 г. На его страницах публикуются оригинальные обзоры,
отражаются различные аспекты теоретических и экспериментальных исследований
генезиса, географии, физики, химии, биологии, плодородия почв; освещаются результа¬
ты теоретических и экологических исследований в глобальном и региональном аспектах.
МАИК “НАУКА”
“НАУК А”
Российская академия наук
ПОЧВОВЕДЕНИЕ
№ 6 1996 Июнь
Основан в январе 1899 г.
Выходит 12 раз в год
ISSN: 0032-180Х
Главный редактор
Г.В. Добровольский
Заместители главного редактора
А.Д. Воронин, C.B. Зонн
Ответственный секретарь
А.Н. Геннадиев
Редакционная коллегия:
Б.Ф. Апарин, P.B. Арнольд (США), B.E.X. Блюм (Австрия),
И.М. Гаджиев, Е.А. Дмитриев, B.H. Ефимов, B.T. Емцев,
Ф.Р. Зайдельман, А.Н. Каштанов, B.H. Кудеяров,
В.В. Медведев (Украина), Д.С. Орлов, И. Сабольч (Венгрия),
Н.И. Смеян (Белоруссия), И.А. Соколов,
А.Д. Фокин, Ф.Х. Хазиев, JI.JI. Шишов, А.П. Щербаков
Зав. редакцией Е.В. Достовалова
Адрес редакции: 109017, Москва, Ж-17, Пыжевский пер., 7, тел. 230-80-66
Москва
Издательство “Наука”
Международная академическая
издательская компания “Наука”
© Российская академия наук
Отделение общей биологии РАН
Общество почвоведов при РАН, 1996
СОДЕРЖАНИЕ
Номер 6,1996
ПОЧВА И БИОРАЗНООБРАЗИЕ
От редколлегии
693
Значение почв в сохранении биоразнообразия
Г В. Добровольский
694
Почвы и микробное разнообразие
Т. Г. Добровольская, Л. В. Лысак, Д. Г. Звягинцев
699
Разнообразие грибов и актиномицетов и их экологические функции
Д. Г. Звягинцев, И. П. Бабъева, Г. М. Зенова,Л. М. Полянская
705
Адаптивные стратегии освоения животными почвенного яруса
Б. Р. Стриганова
714
Структура почвенного покрова и разнообразие лесных фитоценозов
Л. О. Карпаневский
722
Влияние дренажа и агромелиорации на почвенную биоту
Ф. Р. Зайделъман
728
Значение биоразнообразия для диагностики почв
Ю. Г. Гелъцер, А. С. Яковлев
735
ГЕНЕЗИС И ГЕОГРАФИЯ ПОЧВ
Роль окисления сульфидов в генезисе дерновых шунгитовых железистых почв Карелии
/7. В. Красильников, А. М. Володин
743
Эволюция и развитие почв в Приаралье при антропогенном опустынивании
Г. С. Куст
754
ХИМИЯ почв
Фазовый состав соединений железа в почвах и гидротермальных отложениях Абхазии
в связи с проблемами латеритообразования
JI. С. Ильина, Т. С. Гендлер, В. В. Лежава
Содержание серы и микроэлементов в пахотных вулканических почвах Камчатки
В. И. Голову С. М. Бахова
763
775
ФИЗИКА ПОЧВ
Формирование агрегатного состава почв и оценка его изменения
С. Ю. Булыгин, Ф. Н.Лисецкий
783
Определение коэффициента эффективной диффузии водяного пара в соломенной мульче
Е. М. Гусев, О. Е. Бусарова
789
МИНЕРАЛОГИЯ И МИКРОМОРФОЛОГИЯ почв
Микроморфология почв Восточно-Европейского сектора Субарктики
Г. В Русанова
793
ЭРОЗИЯ почв
гтипичного чернозема
Вынос питательных веществ поверхностным стоком из тиии
в почвозащитном севообороте
В. П. Веретельников, В. Л. Рядовой -—
ХРОНИКА
Юбилей профессора Ильяса Мамедовича Гаджиева
Фангат Хаматович Хазиев (к 60-летию со дня рождения и 35-летию научной деятельности)
Благодарность
Международный симпозиум “Засоление лагунных экосистем”
Е. И. Панкова, И. Н. Любимова
803
810
811
812
РЕЦЕНЗИИ
Е.С. Мигунова. Леса и лесные земли (количественная оценка взаимосвязей). М.: Экология, 1993.
Е.С. Мигунова. Лесоводство и почвоведение (исторические очерки). М.: Экология, 1994
Д. Г. Тихоненко
Contents
Number 6,1996
Simultaneous English language translation of the journal is available fromMAHK Hayica/Interperiodica Publishing (Russia).
Eurasian Soil Science ISSN 1064-2293.
SOIL AND BIODIVERSITY
Editorial Note
693
The Role of Soils in Biodiversity Conservation
G. V. Dobrovol’skiy
694
Soils and Microbial Diversity
T. G. Dobrovol'skaya, L. V. Lysak, D. G. Zvyagintsev
699
Diversity of Fungi and Actinomyces in Soils and their Ecological Functions
D. G. Zvyagintsev, I. P. Bab’ieva, G. M. Zenova, L. M. Polyanskaya
705
Adaptive Strategies of Animal Populations to Soil Layer
B. R. Striganova
714
The Soil Cover Structure and Diversity of Forest Phytocoenoses
L. 0. Karpachevskiy
722
Drainage and Agromelioration Effects on Soil Biota
F. R. Zaidelman
728
The Importance of Biodiversity for Soil Diagnostics
Yu. G. Gel’tser, A. S. Yakovlev
735
SOIL GENESIS AND GEOGRAPHY
The Role of Sulfide Oxidation in Genesis of Ferrous Sod-Shunghite Soils in Karelia
P. V. Krasilnikov, A. Af. Volodin
743
Soil Evolution under Conditions of Anthropogenic Desertification
in the Near-the Aral Sea Areas
G. S. Kust
754
SOIL CHEMISTRY
Phase Composition of Iron Compounds in Soils and Hydrothehnal Deposits of Abkhazia
as Related to Problems of Laterization \
L. S. Ilyina, T. S. Gendler, V. V. Lezhava
763
Content of Sulfur and Microelements in Arable Volcanic Soils of Kamchatka
V. 1. Golov, S. M. Bakhova
775
SOIL PHYSICS
The Formation of the Soil Aggregate Composition and its Assessment
S. Yu. Bulygin, F. N. Lisetskiy
783
Calculation of Effective Diffusion Coefficient of Water Stream in Straw Mulch
Ye. M. Gusev, 0. Ye. Busarova
789
SOIL MINERALOGY AND MICROMORPHOLOGY
Micromorphology of East European Subarctic Soils
G. V. Rusanova
793
SOIL EROSION
Nutrients Removal from Typical Chernozem in Soil-protecting Crop Rotation
V. P. Veretelnikov, V. A. Ryadovoi
803
CHRONICLE
Jubilee Celebration of Prof. Dr. Gadzhiev Ilias Mamedovich
808
Fangat Khamatovich Khaziev (to his 60th Birthday and 35 Years of Creative Activities)
810
International Symposium “Soil Salinity of Lagoon Ecosystems”
Ye. /. Pankova, /. N. Lyubimova
812
Ye.S. Migunova. Forests and Forest Lands (Quantitative Assessment of Interrelations). М.: Ekologiya, 1993.
Ye.S. Migunova. Forestry and Soil Science (Historical Essay). М.: Ekologiya, 1994.
D.T.Tikhonenko 814
ПОЧВОВЕДЕНИЕ, 1996, № 6, с 693
ПОЧВА
И БИОРАЗНООБРАЗИЕ
УДК 631 46
ОТ РЕДКОЛЛЕГИИ
В связи с обострением угрозы глобального
экологического кризиса все большее внимание
ученых и широкой общественности привлекает
проблема сохранения жизни. На состоявшейся в
1992 г. в Рио-де-Жанейро Конференции ООН
по окружающей среде и развитию была принята
специальная “Конвенция о биологическом раз¬
нообразии”.
В ней подчеркивается, что сохранение биоло¬
гического разнообразия является общей задачей
всего человечества.
С точки зрения почвоведения очень важным
представляется формулировка в Конвенции во¬
проса о том, что “основным условием сохранения
биологического разнообразия является сохране¬
ние in situ экосистем и естественных мест обита¬
ния, поддержание и восстановление жизнеспо¬
собных популяций видов в их естественных усло¬
виях”.
Не менее важно и определение понятия экоси¬
стемы. В Конвенции оно дано в следующем виде:
“Экосистема означает динамичный комплекс со¬
обществ растений, животных и микроорганиз¬
мов, а также их неживой окружающей среды, вза¬
имодействующих как единое функциональное
тело”.
Приведенные выдержки явно свидетельству¬
ют о том, что почва как среда обитания всего раз¬
нообразия жизни на суше рассматривается в Кон¬
венции как неотъемлемый компонент наземных
экосистем.
Учитывая важность проблемы сохранения
биоразнообразия, Министерство науки и техничес¬
кой политики Российской Федерации приняло Го¬
сударственную научно-техническую программу
“Биологическое разнообразие” и сформировало
Научно-технический совет по этой программе в со¬
ставе 35 человек во главе с академиком В.Е. Соко¬
ловым. В программе предусмотрен ряд проектов
по изучению роли почв в сохранении биоразнооб¬
разия.
Следует, однако, заметить, что вопросы о роли
и значении почв в сохранении биоразнообразия
изучены еще совершенно недостаточно ни почво¬
ведами, ни биологами. И это несмотря на то, что
именно почва является основной средой обитания
подавляющего числа видов и подвидов животно¬
го мира, особенно беспозвоночных и микроорга¬
низмов. Недостаточно полно была представлена
эта проблема и на XV Международном конгрессе
почвоведов в Мексике.
А между тем у нас есть все основания для ак¬
тивной совместной разработки почвоведами и би¬
ологами вопроса о роли почв.в биологическом
разнообразии. Основы учения по этой проблеме
были заложены еще Ч. Дарвиным, В.В. Докучае¬
вым, Г.Н. Высоцким, Н.А. Димо, а позже развиты
М.С. Гиляровым, Н.А. Красильниковым, Е.Н. Ми-
шустиным, их учениками и последователями.
Уместно подчеркнуть, что именно М.С. Гиля¬
рову принадлежит приоритет в изучении и оценке
почвы как уникальной среды обитания и ее роли
в эволюции беспозвоночных.
Нельзя не вспомнить в связи со сказанным и
очень многозначительное утверждение В.И. Вер¬
надского о том, что “Значение почв в истории
планеты гораздо больше, чем это обычно кажет¬
ся” (Избр. соч. Т. V. М.: Изд-во АН СССР, 1960.
С. 97).
Думается, что исследования по программе
“Биоразнообразие” могут послужить основанием
для разработки такой фундаментальной научной
проблемы, как “Роль почвы в эволюции жизни на
Земле”. Такая постановка вопроса в полной мере
соответствовала бы научным заветам Докучаева
и Вернадского.
Надо надеяться, что организованная факуль¬
тетом почвоведения МГУ научная конференция
на тему “Почвы и биологическое разнообразие”
привлечет внимание почвоведов и биологов к ак¬
тивной ее разработке.
693
ПОЧВОВЕДЕНИЕ, 1996, № 6. с 694-698
ПОЧВА
И БИОРАЗНООБРАЗИЕ
УДК 631 46
ЗНАЧЕНИЕ ПОЧВ В СОХРАНЕНИИ БИОРАЗНООБРАЗИЯ*
© 1996 г. Г. В. Добровольский
Почвенный факультет МГУ им. М.В. Ломоносова
Поступила в редакцию 05.02.96с.
Рассмотрены примеры тесных связей различных почв с характерными для них видами растений и
почвообитающих животных. Утверждается, что невозможно сохранить биологическое разнообра¬
зие, не сохраняя разнообразие почв.
Среди документов, принятых Конференцией
ООН по окружающей среде и развитию, состояв¬
шейся в Бразилии в 1992 г., очень важное значение
имеет Конвенция по биоразнообразию. Впервые
человечество в полной мере осознало свою ответ¬
ственность за ежегодное исчезновение 10-15 ты¬
сяч разновидностей биологических организмов.
Если этот разрушительный процесс не прекра¬
тить, то в течение 50 будущих лет с лица планеты
Земля исчезнет от четверти до половины всего
биологического разнообразия, созданного эво¬
люцией жизни за многие сотни миллионов лет.
Именно поэтому принятая Конвенция гласит, что
“... сохранение биологического разнообразия яв¬
ляется общей задачей всего человечества”.
Основным условием сохранения биологичес¬
кого разнообразия является сохранение in situ
экосистем и естественных мест обитания, поддер¬
жание и восстановление жизнеспособных популя¬
ций видов в их естественных условиях. При этом
понятие экосистемы определяется в Конвенции
как “... динамичный комплекс сообществ расте¬
ний, животных и микроорганизмов, а также их
неживой окружающей среды, взаимодействую¬
щих как единое функциональное тело”.
Если не забывать, что почва является основ¬
ной средой обитания наземных организмов и вхо¬
дит в экосистемы как неотъемлемый их компо¬
нент, то совершенно очевидной становится невоз¬
можность сохранения биоразнообразия без
одновременного сохранения почв.
К сожалению, эта достаточно простая истина
еще не нашла должного признания и внимания сре¬
ди экологов и специалистов по охране природы.
На состоявшемся в 1994 г. XV Международном
конгрессе почвоведов в Мексике был проведен
специальный симпозиум “Почвы и биоразнооб¬
разие”, но докладов было представлено немного.
Непосредственно проблеме связи почв с биораз¬
нообразием были посвящены доклады Д.Г. Звя¬
гинцева [24], Г.В. Добровольского [22]. Руководи¬
тель симпозиума д-р Sombroek [23] выразил на¬
дежду, что в ближайшее время будет выработана
’Работа выполнена в рамках программы “Биоразнообразие”.
программа исследований почв как генератора и
хранителя биологического разнообразия.
Почвоведами и биологами накоплены обшир¬
нейшие материалы, доказывающие тесноту свя¬
зей между разнообразием почв и разнообразием
характерных для них биоценозов, отдельных ви¬
дов растений и почвообитающих животных.
Установление этих связей послужило основой
развития специальных направлений ботаничес¬
ких и зоологических исследований. Речь идет об
индикационной геоботанике [3] и зоологическом
методе диагностики почв [9].
В индикационной геоботанике индикаторами
называются такие виды растений или целые их
сообщества, которые достаточно четко приуро¬
чены в своем распространении к определенным
экологическим условиям и прежде всего к опре¬
деленным типам и видам почв со свойственными
им гидротермическими и физико-химическими
режимами. Особенно четко эти экологические
связи проявляются в аридных ландшафтах, где по
наличию растений-индикаторов можно судить не
только о гранулометрическом составе почв, вы¬
соте и степени минерализации почвенно-грунто¬
вых вод, но даже о химическом их составе. Изве¬
стно, например, что такие виды растений, как со¬
лерос (Solicomia herbacea), сарсазан (Holocnemum
strobilaceum), кермек Гмелина (Limonium Gmeli-
ni), тамарикс солончаковый (Tamarix hispida), со¬
лянки (Salsola) указывают на хлоридно'е и суль-
фатно-хлоридное засоление почв, а биюргун
(Anabasis salsa), боялыч (Salsola arbuscula), шведка
(Suaeda), поташник (Kalidium caspicum) - на суль¬
фатное или хлоридно-сульфатное [18].
На близость незаселенных (пресных) грунто¬
вых вод указывает присутствие таких растений-ин-
дикаторов, как рогоз или куга (Tupha angustifolia),
чий (Stipa splendens), солодка (Glicyrrhiza glabra),
ива южная (Salix australior), лох или джида (Elea-
gnus angustifolia).
В надежности индикаторов нам постоянно
приходилось убеждаться во время работы в полу¬
пустынных степях и пустынях Прикаспийской
низменности и Казахстана [13].
694
Виды Lumbricidae
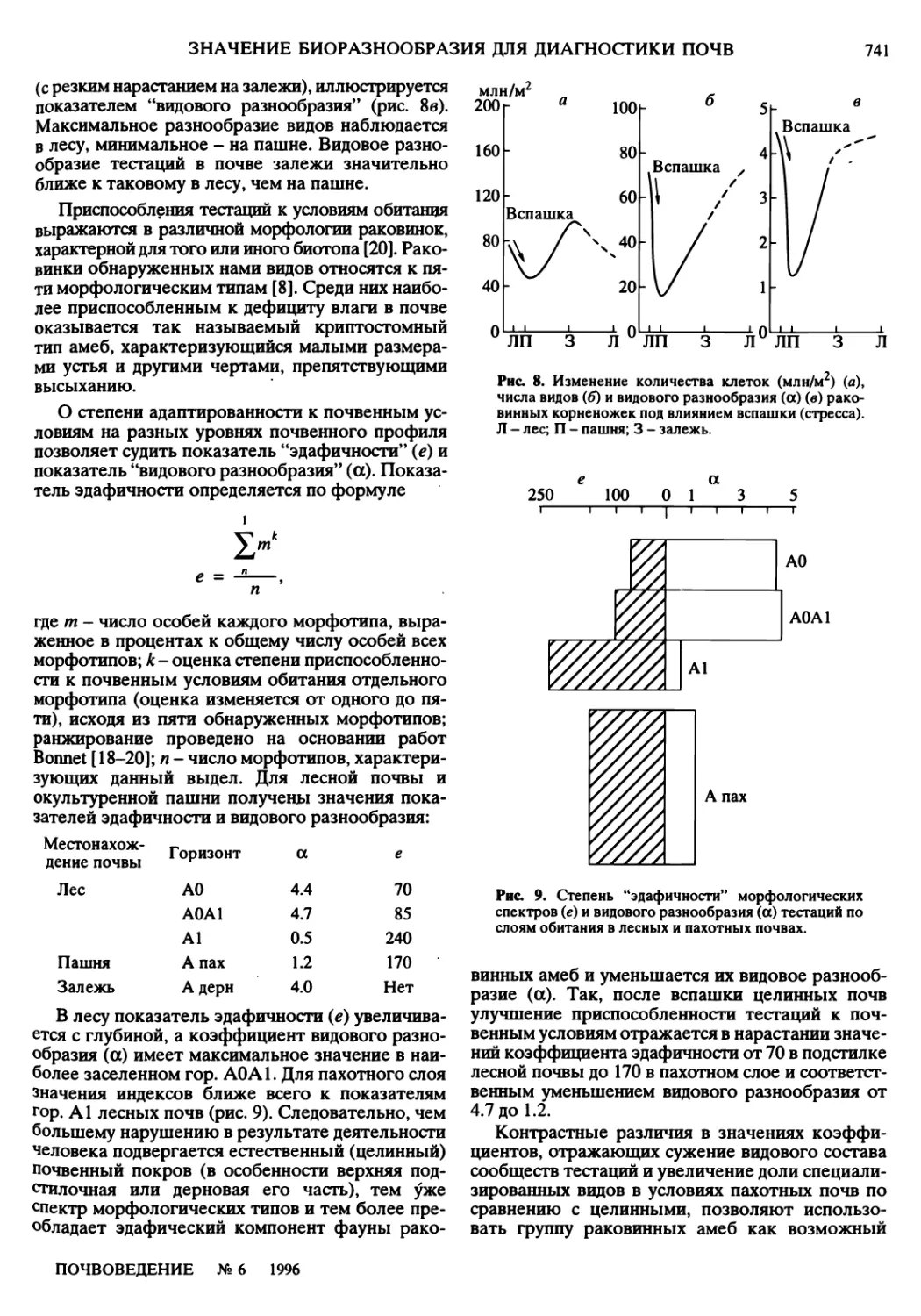
а — б в г
д е tvvvvvvl ж I I з
Распределение видов Lumbricidae (% общего числа) в почвах Княгинкинской поймы р. Клязьмы. 7-7,9 - опытные пло¬
щадки. Почвы: 1,4,9- дерново-луговые и луговые центральной поймы; 2,3- дерновые прирусловья; 5, 6 - лугово-
болотные и болотные притеррасья; 7 - дерново-подзолистые коренного берега. Виды Lumbricidae: a- Allolobo-
phora caliginosa (Sav); б - Eisenia nordenskialdi Mich; в - Lumbricus rubellus Hoffm; г - Bimastus tenuis (Eisen); d - Den-
drobaena oktaedra (Sav); e - Octolasium lacteum (Oerley); ж - Eisenia rosea (Sav); з - прочие виды.
ж
Индикационная геоботаника широко исполь¬
зуется при почвенно-картографических, почвен-
но-мелиоративных и почвенно-геохимических
исследованиях, в том числе и с применением дис¬
танционных аэрокосмических методов.
Нельзя не упомянуть, что основы учения о тес¬
ноте связей разнообразия почв и растений, теоре¬
тические основы индикационной геоботаники
были заложены трудами выдающихся почвове¬
дов, ботаников и геохимиков - В.В. Докучаева,
A.П. Виноградова, В.Н. Сукачева, Б.А. Келлера,
Н.А. Димо, И.В. Ларина, Л.Г. Раменского,
B.А. Ковды, а в наше время С.В. Викторова,
Б.В. Виноградова, Е.А. Востоковой.
Связи животного мира с почвой сложнее, ди¬
намичнее, они в большей мере опосредованы че¬
рез многоступенчатые трофические связи. И все
же они настолько глубоки и показательны, что
легли в основу зоологического метода диагностики
почв, блестяще разработанного Гиляровым [9].
Плодотворность этого метода для решения слож¬
нейших вопросов генезиса и диагностики почв по¬
казана М.С. Гиляровым при изучении подзолис¬
тых, болотных и темноцветных почв южно-таеж¬
ной зоны европейской России, лесостепных и
степных черноземов центральных областей Рос¬
сии, буроземов Молдавских Кодр и орехово-пло-
довых лесов южной Киргизии, terra rossa, бурозе¬
мов, коричневых и горно-луговых почв Крыма
и Кавказа, горных своеобразных темно-серых
почв байрачных лесов в степях Украины, в полеза¬
щитных лесных полосах юга России и др. [7, 8,9].
При изучении пойменных почв мы также
убедились в том, насколько закономерно состав
почвенной фауны соответствует разным типам
пойменных почв, отчетливо различающихся по
экологическим условиям, прежде всего по водно¬
воздушному режиму, гранулометрическому со¬
ставу, количеству и распределению по профилю
органического вещества (гумуса) [12]. Соответст¬
венно этим различиям в почвах изменяется и со¬
став мезофауны в них. Особенно показательна
приуроченность к разным типам почв разных ви¬
дов дождевых червей (Lumbricidae) (рисунок).
В дерновых и дерново-луговых супесчаных, суг¬
линистых почвах прирусловой поймы почти все
дождевые черви относятся к виду Allolobophora
caliginosa, всегда предпочитающих рыхлые, хоро¬
шо аэрируемые почвы. Полуболотные и особен¬
но болотные почвы притеррасной поймы харак¬
теризуются господством мелких червей, принад¬
лежащих виду Dendrobaena octaedra, причем почти
все они сосредоточены в самом верхнем горизон¬
те почвы и заторфованной дернине. В луговых
и лугово-лесных почвах центральной поймы, об¬
ладающих наиболее благоприятным водно-воз-
душным режимом и богатством органических ве¬
ществ, видовой состав дождевых червей очень
разнообразен. Здесь в значительных количествах
встречаются Eisenia nordenskioldi, Octolasium lac¬
teum, Lumbricus rubellus, Alloloborhora caliginoza,
Dendrobaena octaedra и др.
Экологическая приуроченность определен¬
ных видов люмбрицид к дерновому, луговому и
болотному типам пойменных почв выражена на¬
столько четко, что может служить целям генети¬
ческой и агрономической диагностики почв. В то
же время потеря разных типов пойменных почв
при нарушении пойменного режима (например,
вследствие заболачивания и пересушки при гид¬
ротехническом строительстве) будет сопровож¬
даться утратой биоразнообразия почвообитаю¬
щих животных в долинах рек. А с ними связано
в свою очередь разнообразие видового состава
растительности высокопродуктивных поймен¬
ных лугов и лесов.
С точки зрения сохранения биологического
разнообразия особый интерес представляет изу¬
чение биоценозов, характерных для древних и ре¬
ликтовых почв. Приведем некоторые примеры.
ПОЧВОВЕДЕНИЕ № 6 1996
696
ДОБРОВОЛЬСКИЙ
Соотношение средиземноморских (по зоогеографичес-
кой характеристике) и широкораспросграненных видов
жуков и муравьев в разных почвах Южного берега Кры-'
ма (по [8])
Почвы, местонахождение,
растительность
Соотношение
видов, %
среди¬
земно-
морские
широко*
распрост¬
раненные
Terra rossa, Гурзуф, “шибляко-
вый” кустарник
96
4
Terra rossa, мыс Мартьян,
можжевеловый лес
83
17
Красноцветная почва на диорите,
Аю-Даг, “шибляковый” кустарник
80
20
Бурозем, над Гурзуфом,
буковый лес
25
75
Аллювиальная темноцветная,
Магарач
22
78
Степной маломощный чернозем
на массивно-кристаллических
породах, с. Коккозы
0
100
Среди почв Южного берега Крыма особое ме¬
сто занимают древние красноцветные почвы (terra
rossa), образовавшиеся под древесно-кустарнико-
вой растительностью с участием многих реликто¬
вых вечнозеленых видов древовидного можже¬
вельника (Yuniperus excelsa), земляничного дерева
(Arbutus andrachne), кустарников - иглицы (Ruscus
ponticus), ладанника (Cistus tauricus) и др. [19].
Южнобережным почвам свойственна не только
растительность средиземноморского типа, но и
средиземноморская почвенная фауна. По наблю¬
дениям Гилярова [8, 9], фауна южнобережных
красноцветных почв (как на известняках, так и на
' массивно-кристаллических породах) представле¬
на главным образом средиземноморскими вида¬
ми (таблица).
Красноцветные почвы на массивно-кристал¬
лических породах (диорите) отличаются от terra
rossa на известняках отсутствием или меньшей
численностью калькофильных форм беспозво¬
ночных (улитки, мокрицы, многоножки-диплопо-
ды)[11].
Из таблицы видно, что в буроземах под сосно¬
выми и буковыми лесами, расположенными зна¬
чительно выше по южному склону Крымских
гор, средиземноморская фауна резко обеднена
и представлена более “северными” и влаголюби¬
выми видами, широко распространенными в ши¬
роколиственных лесах Европы.
Большая часть красноцветных почв южного
Крыма подвергается сильной эрозии, антропо¬
генному загрязнению и распространены они срав¬
нительно небольшими участками. Если они не бу¬
дут сохранены, то вместе с ними исчезнут и среди¬
земноморские виды растений и животных в самой
северной части их ареала.
Не менее интересны почвенно-биологические
исследования, выполненные Гиляровым в горах
Тянь-Шаня [7]. Здесь под реликтовыми ореховы¬
ми (Yuglans fallax) лесами на уникальных черно¬
коричневых многогумусных почвах обнаружена
очень богатая и разнообразная мезофауна, мно¬
гие виды которой характерны для почв средизем¬
номорских стран.
Генезис и систематическое положение этих
почв явились предметом длительной дискуссии.
По новейшим исследованиям [4,17] они отнесены
к типу коричневых субтропических почв, харак¬
терных для Средиземноморья.
Эти почвы содержат от 8 до 23% гумуса, мощ¬
ность гумусовых горизонтов колеблется от 60 до
150 см, карбонаты обнаруживаются на глубине
80-110 см, реакция нейтральная или очень слабо¬
щелочная. Более 50% структурных агрегатов
в этих почвах представлены водопрочными коп-
ролитами дождевых червей и других беспозво¬
ночных животных. На 1 м2 почвы насчитывается
в среднем свыше 100 крупных (преимущественно
дождевые черви) и до 40 тысяч мелких беспозво¬
ночных животных (клещи, ногохвостки и др.).
Очень интенсивны в этих почвах микробиологи¬
ческие процессы. Эти ценнейшие реликтовые
орехово-плодовые леса и уникально биогенные и
богатые почвы очень страдают от эрозии, бесси¬
стемной вырубки леса, нерегулируемого выпаса
скота. Если в ближайшее же время не принять не¬
обходимых противоэрозионных и вообще приро¬
доохранных мер, то погибнут и ценнейшие релик¬
товые леса и уникальные почвы с их богатейшей
средиземноморской фауной на самом восточном
ареале их распространения.
Очень интересны и важны для полноты пред¬
ставлений о связях биологического разнообразия
с разнообразием почв новые материалы о специ¬
фике видов и жизненных форм организмов в поч¬
вах с экстремальными условиями жизни - край¬
ней аридностью, низкой или избыточно высокой
температурой и т.п. Так, Гельцер описал осо¬
бый способ выживания у инфузории Colpoda
maupasii в крайне аридных почвах Заалтайской
Гоби (Монголия) [16]. При полном отсутствии
влаги в почвах этого края в течение длительного
времени инфузории этого вида переходят во вре¬
менное нежизнеспособное инцистированное со¬
стояние и превращаются в неподвижные оваль¬
но-угловатые кристаллоподобные тела с тонкой
контрастной оболочкой и грубозернистым содер¬
жимым. Такой способ перехода в неактивное со¬
стояние Ю.Г. Гельцер назвал “псевдокрисгаллиза-
цией”, поскольку скопление неподвижных обезво¬
женных колпод напоминает друзу кристаллов. При
ПОЧВОВЕДЕНИЕ № 6 1996
ЗНАЧЕНИЕ ПОЧВ В СОХРАНЕНИИ БИОРАЗНООБРАЗИЯ
697
появлении влаги через 10-15 мин происходит
“оживление” и гидратация цитоплазмы клеток.
“Псевдокристаллизацию” автор рассматривает
как типичный случай ксероанабиоза.
Не менее интересны результаты исследований
Д.Г. Звягинцева, Д.А. Гиличинского и других о со¬
хранении в течение более миллиона лет жизнеспо¬
собности клеток микроорганизмов в постоянно
мерзлых грунтах и погребенных в них почв [14].
По-видимому, по морфологическим и функцио¬
нальным особенностям этих микроорганизмов воз¬
можны палеоэкологические реконструкции [20].
Приведенные примеры свидетельствуют о ши¬
роте и разнообразии адаптаций жизни к экстре¬
мальным почвенно-экологическим условиям.
Для изучения роли и значения почвы в биоло¬
гическом разнообразии очень важное значение
имеют детальные стационарные почвенно-био-
логические исследования. Об этом можно судить,
например, по результатам исследований, проведен¬
ных С.Я. Трофимовым, А.А. Бобровым, А.В. Го-
ловченко и О.В. Стариковой в Центральном лес¬
ном биосферном заповеднике в Тверской обл. [16].
Здесь изучалось таксономическое и функцио¬
нальное разнообразие микроорганизмов, про¬
стейших и беспозвоночных животных в двух типах
леса - в ельнике сфагново-черничном на подзолис¬
той почве и в ельнике сложном на буроземной поч¬
ве. Установлено, что в вертикально-ярусных сис¬
темах обоих биогеоценозов, т.е. в наземной части
растительности, подстилках и почвенных гори¬
зонтах, наибольшее разнообразие бактерий в ве¬
сенний и осенний период обнаружено в лесных
подстилках, особенно в ферментативных их сло¬
ях. Летом показатели наибольшего разнообразия
бактерий смещаются в нижележащие горизонты:
в ельнике черничнике - в гумусированный слой
подстилки, в ельнике сложном - в гумусово-акку-
мулятивный горизонт почвы.
Буроземная почва под ельником сложным ха¬
рактеризуется по сравнению с подзолистой поч¬
вой под ельником-черничником более благопри¬
ятным для большинства почвообитающих орга¬
низмов водно-воздушным режимом, меньшей
выщелоченносгью биофильных элементов. Соот¬
ветственно этим различиям буроземная почва ха¬
рактеризуется ббльшим количеством бактерий и
большим разнообразием видов микроартропод
(преобладают орибатиды сапрофаги).
В подзолистой почве ельника-черничника вы¬
ше, чем в буроземной почве ельника сложного,
оказалась плотность населения раковинных амеб
(тестаций). Это вполне соответствует большей
потребности тестаций во влаге, особенно среди
видов бриофильной группы.
Нет сомнения в том, что и по другим видам ми-
кро- и мезофауны различия между названными
биогеоценозами и свойственными им почвами
окажутся еще более яркими.
Принятая международная Конвенция по био¬
логическому разнообразию означает серьезный
поворот в осознании людьми своей ответственно¬
сти за все живое на Земле. Конвенция привлекла
внимание всех ученых и специалистов, работаю¬
щих в области охраны природы и природопользо¬
вания. Она содействует также дальнейшему раз¬
витию теории и методов исследования богатства
органического мира [1, 5, 21].
По мере накопления результатов сопряжен¬
ных почвенно-биологических исследований мы
все в большей мере убеждаемся в справедливости
определения В.И. Вернадским почвенного покро¬
ва Земли как особой биосферной пленки сгуще¬
ния жизни. В такой же мере обоснованными
представляются взгляды биологов М.С. Гиляро¬
ва, Д.А. Криволуцкого на почвенный покров как
хранилище генетического разнообразия жизни
на Земле [10, 15].
Из этих обобщающих положений неизбежно
следует мысль о том, что нельзя сохранить био¬
логическое разнообразие, не сохраняя разнообра¬
зие почв.
СПИСОК ЛИТЕРАТУРЫ
1. Биологическая диагностика почв: Тез. докл. Все-
союз. совещ. “Проблемы и методы биологической
диагностики и индикации почв”. М.: Наука, 1976.
360 с.
2. Биоразнообразие. Степень таксономической изу¬
ченности. М.: Наука, 1994.
3. Викторов С.В., Ремезова ГЛ. Индикационная гео¬
ботаника. М.: Изд-во МГУ, 1988. 168 с.
4. Владыченский А.С. Особенности горного почво¬
образования и формирования почвенного покрова
гор: Автореф. дис.... д-ра наук. М., 1994.
5. Гельцер Ю.Г. Биологическая диагностика почв.
М.: Изд-во МГУ, 1986. 82 с.
6. Гельцер Ю.Г. Псевдокристаллизация - особый спо¬
соб перенесения неблагоприятных условий суще¬
ствования у инфузорий Colpoda maupasii из крайне
аридных почв Заалтайской Гоби (Монголия) //
Цитология. 1988. № 30.11. С. 1386-1388.
7. Гиляров М.С. Почвенная фауна орехово-плодовых
лесов Ферганского хребта // Вестн. МГУ. 1947. № 1.
С. 97-102.
8. Гиляров М.С. Диагностика и география почв в све¬
те почвенно-зоологических исследований // Успе¬
хи современной биологии. 1949. № 28. Вып. 3(6).
С. 339-353.
9. Гиляров М.С. Зоологический метод диагностики
почв. М.: Наука, 1965. 268 с.
10. Гиляров М.С., Криволуцкий Д. А. Жизнь в почве.
М.: Молодая гвардия, 1985. 190 с.
11. Добровольский Г.В. О красноцветных почвах юж¬
ного берега Крыма // Вестн. МГУ. 1950. № 9.
С. 123-139.
12. Добровольский Г.В., Гельцер Ю.Г. Почвенно-фау-
нистические наблюдения в пойме р. Клязьмы //
Вестн. МГУ. Сер. Почвоведение. 1958. №4.
ПОЧВОВЕДЕНИЕ № 6 1996
698
ДОБРОВОЛЬСКИЙ
13. Добровольский Г.В., Федоров К.Н., Стасюк Н.В.
Геохимия, мелиорация и генезис почв дельты Те¬
река. М.: Изд-во МГУ, 1975. 247 с.
14. Звягинцев Д.Г., Гилининский Д.А., Благодат¬
ский С.А. и др. Длительность сохранения микро¬
организмов в постоянно мерзлых осадочных поро¬
дах и погребенных почвах // Микробиология. 1985.
Т. 54. № 1.С. 155-161.
15. Криволуцкий ДА. Почвенная фауна в экологичес¬
ком контроле. М.: Наука, 1994. 270 с.
16. Отчет по теме “Структурно-функциональная ор¬
ганизация почвенной биоты в ненарушенных био¬
ценозах южной тайги” / ИЭМЭЖ им. А.Н. Север-
цова, проект № 93-04-6138 РФФИ. 1995.
17. Розанов А.Б. Почвы ореховых и плодовых лесов
Тянь-Шаня: Автореф. дис. ... канд. наук МГУ. М.,
1993. 24 с.
18. Справочник-определитель литологического со¬
става поверхностных отложений и глубины зале¬
гания подземных вод / Под. ред. Н.Г. Верейского,
Е.А. Востоковой. М., 1963.
19. Станков С.С. Южный берег Крыма. Н. Новгород,
1926. 148 с.
20. Хлебникова Г.М., Гилининский ДА., Федоров-Да-
выдовД.Г. и др. Количественная оценка микроор¬
ганизмов в многолетнемерзлых отложениях и по¬
гребенных почвах // Микробиология. 1990. Т. 59.
№ 1. С. 148-155.
21. Чернов ЮМ. Биологическое разнообразие: сущ¬
ность и проблемы // Успехи современной биоло¬
гии. 1991. Т. III. Вып. 4. С. 499-507.
22. Dobrovolsky G. The Importance of soil sustaining Biodi¬
versity //Transaction 15-th World Congress of Soil Sci.
Acapulco, Mexico. 1994. V. 4b. P. 75-76.
23. Sombroek W.G. Soils and biodiversity // Transaction 15-th
World Congress of Soil Sci. Acapulco, Mexico. 1994.
V. 4a. P. 184-185.
24. Zvyagintsev D. Biodiversity of microorganism in differ¬
ent soil types // Transaction 16-th World Congress of
Soil Sci. Acapulco, Mexico. 1994. V. 4a. P. 187-189.
The Role of Soils in Biodiversity Conservation
G. V. DobrovoPskiy
Under consideration are some examples permitting to show a close correlation between different soils and prop¬
er species of plants and soil inhabitants. One cannot but admit that the conservation of soil diversity is required
for biodiversity conservation.
ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВОВЕДЕНИЕ, 1996, № 6, с. 699-704
ПОЧВА
■ И БИОРАЗНООБРАЗИЕ
УДК 631.46
ПОЧВЫ И МИКРОБНОЕ РАЗНООБРАЗИЕ*
© 1996 г. Т. Г. Добровольская, JI. В. Лысак, Д. Г. Звягинцев
Факультет почвоведения МГУ им. М.В. Ломоносова
Поступила в редакцию 13.02.96 г.
Рассматриваются в историческом аспекте различные подходы к оценке микробного разнообразия
почв. Продемонстрировано влияние разных экологических факторов на структуру и функциониро¬
вание бактериальных сообществ почв. Показано, что в почвах разных природно-климатических зон
содержатся разные типы бактериальных сообществ.
Палеомикробиология располагает прямыми
свидетельствами развития бактерий в очень отда¬
ленном геологическом прошлом. К ним относятся
строматолиты и микрофоссилии. Строматолиты -
слоистые осадочные породы, представляющие
собой продукты жизнедеятельности сообществ си¬
незеленых водорослей. Анализ современных циа¬
нобактериальных матов позволил заключить [6],
что они аналогичны таким же цианобактериаль¬
ным сообществам прошлого. Микрофоссилии - ми¬
нерализованные остатки микроорганизмов - также
свидетельствуют о консервативности прокариот
в плане их эволюции. Установлено, что в течение
докембрия сложилось большинство известных
ныне морфологических типов бактерий. Трофи¬
ческим окружением центрального ядра из циано¬
бактерий были представители класса Proteobacteri-
ае, включающего разнообразные формы грамот-
рицательных бактерий. Цианобактериальное
сообщество безраздельно господствовало на Земле
около 2.7 млрд. лет, т.е. 2/3 времени эволюции
биосферы, и изменилось очень мало до настоя¬
щего времени [7, 12].
В современных амфибиальных ландшафтах
протеобактерии остаются доминирующими груп¬
пами прокариотных сообществ [3]. В наземных
экосистемах, где основной средой обитания явля¬
ется почва, а не вода, эволюционно сложились
другие, более разнообразные типы бактериоце-
нозо'в.
Почва благодаря своей трехфазности способ¬
на обеспечивать условия для существования в ней
разнообразных микроорганизмов, одни из кото¬
рых находятся на почвенных частицах, другие -
в водных пленках, капиллярах или почвенном
растворе. Для адсорбции на почвенных частицах
у бактерий существуют специальные приспособле¬
ния: капсулы, простеки, фимбрии и др. Большая
часть всех почвенных микроорганизмов в естест¬
венных условиях адгезирована на твердофазном
Работа выполнена в рамках программы “Биоразнообразие”.
субстрате [8]. Микрозональность распределения
микроорганизмов в почве с адгезией клеток на
твердых поверхностях имеет принципиальное
значение, так как в адсорбированном состоянии у
микроорганизмов меняются морфология клеток,
циклы развития, продолжительность лаг-фазы,
скорость размножения. В отдельных микрозонах
создаются совершенно различные условия и по¬
является возможность развития в почве - в чрез¬
вычайно структурированном биотопе - самых
различных по своим свойствам групп микроорга¬
низмов.
Определение микробного разнообразия в поч¬
вах невозможно без решения многих вопросов,
связанных с методами выделения и идентифика¬
ции микроорганизмов, составом питательных
сред, способом отбора и предварительной обра¬
ботки образцов, что также служило предметом
постоянных дискуссий среди почвенных микро¬
биологов. Наиболее полный обзор методов и
приемов, используемых для выявления и учета
разных форм почвенных микроорганизмов, в ис¬
торическом аспекте дан Аристовской [1] и Ники¬
тиным с соавт. [15]. Наиболее идеальной, но до
сих пор неосуществленной, представляется мето¬
дология, предложенная Аристовской, утверждав¬
шей, что “от микробного пейзажа следует идти к
отдельным микроорганизмам и микробным про¬
цессам, а затем от них снова к пейзажу” [1]. Пред¬
ставляется, что для оценки микробного разнооб¬
разия следует использовать весь накопленный
в практике микробиологии опыт и следовать
принципу дополнительности Бора, а не взаимоис¬
ключения. Метод чистых культур с использова¬
нием элективных питательных сред для каждой
группы микроорганизмов не исключает метода
наблюдения за почвенными микроорганизмами
in situ с его многочисленными модификациями
(стекла обрастания, педоскопы, мембранные
фильтры и др.). Широкие возможности открылись
для изучения микроорганизмов при использова¬
нии люминесцентной и электронной сканирую¬
щей микроскопии. Разработаны сукцессионные
699
700
ДОБРОВОЛЬСКАЯ и др.
подходы, позволяющие следить в модельных
опытах за изменением состава микробных сооб¬
ществ во времени [19]. Такой подход был удачно
использован для изучения экологии простекобак-
терий [17] и основывался на том, что олиготроф-
ные простекобактерии являются партнерами
аэробных целлюлозоразрушающих бактерий и
могут быть обнаружены в их колониях на заклю¬
чительных этапах деструкции растительных ос¬
татков в почве.
На примере анализа распространения ана¬
эробных бактерий в почвах было показано, что
группировки доминирующих видов анаэробных
бактерий не только различны в почвах разных
климатических зон [17], но и обладают разными
экологическими функциями и разной активнос¬
тью метаболических процессов. Отмечена также
температурная адаптация этих бактерий к усло¬
виям климата и почвы.
Выход за пределы почвенного яруса при изу¬
чении микробных сообществ, т.е. биоценологи-
ческий подход, оказался весьма перспективным,
так как он позволил выявить разнообразные ис¬
точники формирования почвенных микробоце-
нозов, проследить пути миграции микроорганиз¬
мов, отделить ярусы и локусы активного размно¬
жения микробов от мест их переживания и
сохранения, связать местоположение определен¬
ных микроорганизмов с их функциями и типом
стратегии [3,5,9,10]. В результате были установ¬
лены закономерности вертикального распрост¬
ранения микроорганизмов, относящихся как к эу-
кариотным, так и прокариотным, в почвах и со¬
пряженных субстратах разных природных зон
СНГ. В этих публикациях был использован но¬
вый подход для количественного исследования
таксономической структуры микробных сооб¬
ществ, разработанный на основе общепринятых
синэкологических методов. В настоящей статье
в соответствии с ее тематикой, отраженной в за¬
главии, сознательно делается акцент на характе¬
ристике микробного разнообразия в пределах
почвенного яруса разных типов почв, включая,
однако, подстилки, без которых невозможно
формирование почвенного профиля.
Сама постановка вопроса о приуроченности
определенных групп микроорганизмов к опреде¬
ленным типам почв осложняется тем, что боль¬
шинство микроорганизмов обнаруживаются во
всех типах почв, т.е. являются космополитами.
Это положение утверждается первой частью пра¬
вила Бейеринка: “Все есть всюду...”. Однако вто¬
рая часть девиза: “но среда отбирает...” - обнаде¬
живает и предполагает, что исследователь может
и должен найти те экологические факторы, кото¬
рые определяют формирование микробных сооб¬
ществ, адаптированных к данному типу почв.
Ведущими экологическими факторами, опреде¬
ляющими структуру бактериальных сообществ,
являются тип субстрата и влажность. Влияние
типа субстрата на бактериальное разнообразие на¬
иболее четко прослеживается при анализе верти¬
кального распределения бактерий в биогеоцено¬
зах (БГЦ) в ряду: филлоплана растений-подстил-
ка-почвенные горизонты. Такие вертикальные
ряды являются сукцессионно-хорологически-
ми, так как отражают смену сообществ во вре¬
мени в процессе разложения растительных ос¬
татков, реализованную в пространстве в виде
стратифицированных слоев подстилки и почвен¬
ных горизонтов. В таксономическом отношении
смена бактериальных ярусных доминантов соот¬
ветствует схеме: псевдомонады-бациллы-бакте-
рии актиномицетной линии-олиготрофы. В це¬
лом такая тенденция в смене бактериальных
доминантов прослеживается во всех типах зо¬
нальных почв, однако, в каждой природно-клима¬
тической зоне вырисовываются свои детали
в структуре бактериальных комплексов разных
ярусов.
Влияние типа субстрата на состав бактериаль¬
ных сообществ можно также проследить, анализи¬
руя соотношение между содержанием органичес¬
кого вещества в почве и встречаемостью некото¬
рых бактериальных таксонов. В примитивных
почвах, на поверхности которых поселяются пер¬
воначально лишь лишайники, пионерами среди
бактерий являются представители актиномицет¬
ной линии - Корине- и нокардиоподобные бакте¬
рии. О доминировании этой группы бактерий,
объединяемых ранее под названием “микобакте¬
рии”, в таких примитивных БГЦ сообщалось в ра¬
ботах Красильникова [13], Сушкиной [18], Арис¬
товской [2]. Эти авторы показали роль микобак¬
терий в процессах выветривания и первичного
почвообразования. Если перейти на современный
уровень таксономического лексикона, то следует
вычленить из этой сборной группы артробактер
(род Arthrobacter) как организм, доминирующий в
олиготрофных местообитаниях. Это получило
подтверждение и при изучении бактериальных
комплексов в почвах тундры Западного Таймыра.
Бактерии рода Arthrobacter преобладали на по¬
верхности голых пятен, в глеевых горизонтах, по¬
родах, т.е. в олиготрофных эконишах'. В диамет¬
рально противоположных по содержанию орга¬
нического вещества биотопах - очёсе мха,
торфянистых почвах трещин - доминировали
спорообразующие бактерии рода Bacillus. Встре¬
чаемость артробактера и бацилл в зависимости
от типа субстрата была прослежена нами при ис¬
следовании бактериальных сообществ в ряде
почв сухих субтропиков. Было показано, что арт¬
робактер - типичный обитатель примитивных
каменистых почв и сероземов, его доля составля¬
ла 55-80% общего числа бактерий сапротрофного
блока. При переходе к более гумусированным се¬
ро-коричневым и коричневым почвам содержание
ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВЫ И МИКРОБНОЕ РАЗНООБРАЗИЕ
701
Биомы
Почвы
Arthro¬
bacter
Bacillus
Azoto-
Aquaspi-
Entero-
Rhodo¬
Geoder-
bacter
rillum
bacter,
coccus,
matophi-
'
Plesio-
Micro-
lus
monas
coccus
I
Halobac-
terium,
Halo-
coccus
Тундра
Тайга
Болота
Степь
Полупу¬
стыни
Пустыни
Глеевые тунд¬
ровые
Подзолистые
Дерново-подзо¬
листые
Торфяники
Черноземы
Каштановые
Сероземы
Серо-бурые
Солончаки
# #
# #
# #
# #
# #
# #
* * *
♦
♦
♦
♦
♦
♦
@
@
@
@
♦
*
♦
@
@
# #
# #
# #
# #
# #
X
* • * - ширина соответствует доле таксона в различных почвах.
# #
Бактерии в почвах различных биомов.
артробактера уменьшалось, а доля бацилл значи¬
тельно увеличивалась. Из других форм коринепо-
добных бактерий в почвах сухих субтропиков бы¬
ли обнаружены бактерии родов Rhodococcus и
Micrococcus. Однако их содержание составляло
не более 2-10%. В почвах тундр артробактер был
единственным представителем группы коринепо-
добных бактерий.
Другая пара родов контрастных по местооби¬
танию и физиологии - Arthrobacter и Pseudomonas.
Если обратиться опять к тундровым почвам, то
для псевдомонад в качестве благоприятных мес¬
тообитаний называются влажные почвы со сла-
боразложившимся опадом - осоково-моховые и
ерниково-моховые тундры, а для артробактера -
щебнистые, лишайниковые и куртинные тундры
с почвами, лишенными моховой дернины [17].
Доминирование артробактера в почвах или гори¬
зонтах, характеризующихся низким содержанием
органического вещества, объясняется его способ¬
ностью не только выживать в условиях голода¬
ния, но и функционировать за счет экономного
энергетического обмена. Анализ стратегии всей
группы коринеподобных бактерий, проведенный
на основании изучения кинетических параметров
и особенностей метаболизма этих бактерий, дан
в обзоре “Экология коринеподобных бактерий”
[16]. В результате показана корреляция между ме¬
стообитанием коринеподобных бактерий (КЛБ) и
их жизненной стратегией.
Не менее важным фактором, определяющим
качественное разнообразие бактериальных ком¬
плексов почв, является влажность. Наиболее на¬
глядно это демонстрируется при сравнении бак¬
териальных сообществ в почвах, расположенных
в контрастных по влажности биомах: болотах и
пустынях. В почвах с постоянным переувлажнени¬
ем - торфяниках - и сопряженных с ними почвах
окраин болот доминируют грамотрицательные
формы бактерий, выделенные недавно в самосто¬
ятельный класс - Proteobacteriae. Это подвижные
палочковидные либо спиральные и вибриоидные
формы бактерий, апигментные, с простым жиз¬
ненным циклом. Большинство протеобактерий,
обитающих в торфяниках, представлены спирил¬
лами либо факультативно-анаэробными форма¬
ми, относящимися к пяти родам из семейств Entero-
bacteriaceae и Vibrionaceae (рисунок), т.е. типичны¬
ми гидробионтами. В почвах пустынь (серо-бурые,
примитивные каменистые, литосоли и ксеросоли,
серые пустынные почвы) и опаде пустынных рас¬
тений доминируют бактерии актиномицетной ли¬
нии - континентальная ветвь прокариот (рису¬
нок). Эти бактерии со сложным жизненным цик¬
лом характеризуются мицелиальной стадией хотя
бы на одной из стадий развития, либо тенденцией
к мицелиальному росту как приспособлению к оби¬
танию в твердофазном субстрате. Все они устой¬
чивы к высушиванию и повышенной инсоляции,
чему способствует образование ими конидий, ка-
ротиноидных и меланоидных пигментов. У боль¬
шинства представителей родов актинобактерий
колонии окрашены в желтые, оранжевые, крас¬
ные или черные цвета. Все бактерии актиноми¬
цетной линии представляют собой довольно мо¬
нолитную группу бактерий с высоким содержанием
ГЦ (гуанин + цитозин) в ДНК - 60-75%. Считается,
что генофоры, содержащие ДНК ГЦ-типа, более
устойчивы к ультрафиолетовому облучению [11].
ПОЧВОВЕДЕНИЕ № 6 1996
702
ДОБРОВОЛЬСКАЯ и др.
Итак, все перечисленные признаки бактерий
можно рассматривать в качестве адаптивных,
сложившихся в результате их приспособления к
условиям окружающей среды. Недаром в спект¬
рах потенциальных доминантов болотных и пус¬
тынных экосистем нет ни одного общего доми¬
нанта:
Пустыни
Болота
Протеобактерии
Aquaspirillum
Klebsiella
Proteus
Enterobacter
Aeromonas
Vibrio
Plesiomonas
Актинобактерии
Cellulomonas
Micrococcus
Clavibacter
Geodermatophilus
Rhodococcus
Arthrobacter
Streptomyces
Представляется, что бактериальные комплек¬
сы этих контрастных экосистем можно рассматри¬
вать как сообщества, отражающие ступени эво¬
люции микробного мира при переходе от водного
к наземному образу жизни. Следует напомнить,
что самые древние из амфибиальных ландшаф¬
тов - это прибрежные цианобактериальные ма¬
ты докембрия, в которых в качестве постоянных
симбионтов были обнаружены протеобактерии,
т.е. те группы, которые и сейчас доминируют
в современных амфибиальных экосистемах. Пус¬
тыни являются примером ландшафтов, где адап¬
тация организмов к условиям суши выражена на¬
иболее резко. Актиномицеты и родственные им
бактерии служат примером приспособления к на¬
земному существованию на прокариотном уровне.
Влажность является тем фактором, который
определяет и сезонные изменения в таксономиче¬
ском составе бактериальных комплексов почв,
расположенных в пределах одной природно-кли¬
матической зоны. Нами исследовалась динамика
структуры бактериальных сообществ в почвах
Центрального государственного лесного био¬
сферного заповедника (бело-подзолистая почва и
бурозем) и в почвах Окского биосферного запо¬
ведника (дерново-подзолистая, торфянисто-под¬
золистая, дерново-луговая почвы). Было показано,
что в периоды длительного переувлажнения почв,
наблюдающегося в результате затяжных дож¬
дей, в бактериальных сообществах доминируют
типичные представители водной фазы почв, т.е.
гидробионты. Они представлены большей час¬
тью спириллами, миксобактериями и другими
формами подвижных грамотрицательных бакте¬
рий. И, наоборот, в те сезоны, когда происходило
иссушение почв, доминантами становились ба¬
циллы и бактерии актиномицетной линии. Таким
образом, в одном и том же типе почв в зависимо¬
сти от степени насыщенности их влагой таксоно¬
мический состав бактериальных комплексов бу¬
дет разным. Поэтому сравнивать разные почвы
по составу бактериальных сообществ и делать
выводы о различиях в их бактериальном разнооб¬
разии следует с большой осторожностью и толь¬
ко на основании сезонных наблюдений с обяза¬
тельным отбором образцов во влажные и сухие
периоды года.
Кислотность почв является одним из селектив¬
ных факторов, которые либо ограничивают, ли¬
бо способствуют распространению бактерий оп¬
ределенных таксономических групп. Так, извест¬
но, что азотобактер обнаруживается только
в нейтральных либо слабощелочных почвах, его
нет ни в почвах тундр, ни в болотных, ни в подзо¬
листых. Максимальная концентрация бактерий
этого рода зарегистрирована в богатых, с нейт¬
ральной реакцией среды почвах - черноземах и
каштановых. Артробактер также чувствителен
к pH среды и не обнаруживается в подзолистых
почвах и торфяниках (кроме низинного высоко¬
зольного торфяника с нейтральной реакцией сре¬
ды). В то же время спириллы и факультативно¬
анаэробные бактерии с успехом развиваются и
часто доминируют в кислых почвах таежной зо¬
ны (рисунок).
Многие формы бактерий чувствительны к по¬
вышенной концентрации солей в почве. В солон¬
чаках - почвах, где концентрация солей достигает
наивысших значений, бактериальные комплексы
характеризуются низким разнообразием. Галото-
лерантными являются лишь бациллы, микрокок¬
ки и некоторые стрептомицеты. Из этих биото¬
пов выделяются и настоящие галофилы - бакте¬
рии, требующие для роста 20-25% соли в среде.
Это представители родов Halobacterium и Halococ-
cus (рисунок).
Таким образом, бактериальное разнообразие
разных типов почв определяется многими эколо¬
гическими факторами: содержанием органичес¬
кого вещества, влажностью, кислотностью сре¬
ды, концентрацией солей. От сочетания этих
факторов, определяющих тип почвообразования,
зависит соотношение таксонов в бактериальных
комплексах почв. Мы не упомянули температуру
в числе экологических факторов, так как она ма¬
ло влияет на качественный состав бактериоцено-
зов. Она определяет лишь интенсивность и ско¬
рость биологических процессов. Известно также,
что в южных почвах по сравнению с северными
выше максимальные температуры роста многих
групп бактерий.
Характеристика бактериального разнообра¬
зия почв на видовом уровне осложняется прежде
всего трудностями, существующими в определе¬
нии бактерий до вида. Дифференциация видов на
основании физиолого-биохимических признаков
по таблицам, приводимым в определителях Берги,
представляется весьма условной, так как многие
ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВЫ И МИКРОБНОЕ РАЗНООБРАЗИЕ
703
виды по этим признакам практически невоз¬
можно разделить. Сложные генетические методы,
которые представляются наиболее надежными
при установлении видового диагноза бактерий,
невозможно использовать в экологических ис¬
следованиях в связи с их трудоемкостью, необхо¬
димостью привлечения к работе специалистов-
генетиков, отсутствием реактивов и приборов и
т.д. Видовая дифференциация несколько упроща¬
ется лишь в тех группах бактерий, где первона¬
чальное деление на группы видов возможно на
основании сочетаний таких простых признаков,
как цвет колоний и некоторые микроморфологи-
ческие особенности цикла развития. Коринепо-
добные бактерии являются именно такой груп¬
пой. На примере распределения родов и видов ко¬
ринеподобных бактерий по профилю чернозема
типичного охарактеризовано на видовом уровне
это сообщество (таблица). Максимальное разно¬
образие бактерий этой группы обнаруживается
в подстилке, где происходит интенсивное накоп¬
ление и разложение растительного материала.
К этому ярусу БГЦ приурочены представители
четырех родов КПБ, окрашенные в желто-оран-
жево-красные тона. При этом представители ро¬
да Mycobacterium представлены пятью видами,
Rhodococcus - четырьмя видами. Обращает на се¬
бя внимание значительная доля среди КПБ пред¬
ставителей рода Mycobacterium. Следует отме¬
тить, что о широком распространении микобак¬
терий, представленных видами М. fortuitum,
М. smegmatis, М. phlei, в степных почвах Украины
сообщали и другие авторы [14]. В нижележащем
дерновом горизонте микобактерии практически
не обнаруживались, здесь наиболее часто встре¬
чались артробактерии, представленные тремя ви¬
дами, родококки и представители родов Cellu-
lomonas и Promicromonospora. В почвенных гори¬
зонтах доминировали бактерии рода Arthrobacter.
Следует отметить, что в подстилке и дерновом го¬
ризонте преобладали виды с яркой окраской коло¬
ний, в то время как в почвенных горизонтах - апиг-
ментные виды артробактера. Таким образом, раз¬
ные виды приурочены к разным ярусам БГЦ.
По таксономическому составу бактериаль¬
ных сообществ можно составить представление
об их экологических функциях. Так, например,
доминирование протеобактерий в торфяниках и
почвах, испытывающих временное избыточное
увлажнение, свидетельствует о замедленной ско¬
рости минерализации растительных остатков,
так как среди протеобактерий практически от¬
сутствуют гидролитики (кроме скользящих бак¬
терий). Преобладание в пустынных почвах бакте¬
рий актиномицетной линии, большинство кото¬
рых обладает гидролитической активностью,
коррелирует с быстрыми темпами деструкции
растительного опада в пустынных экосистемах.
В качестве примера корреляции между таксоно-
Распределение некоторых родов и видов коринеподоб¬
ных бактерий по профилю чернозема
Ярус
Горизонт
почвы
Роды и виды*
Наземный (подстилка)
АО
Mycobacterium phlei
fortuitum
parafortui-
tum
gordonae
Rhodococcus luteus
erythropolis
longus
terrae
Promicromonospora citrea
Cellulomonas sp.
Почвенный
Ad
Arthrobacter aurescens
globiformis
pascens
Rhodococcus erythropolis
luteus
longus
Promicromonospora citrea
Cellulomonas sp.
А
Arthrobacter globiformis
pascens
Rhodococcus erythropolis
АВ
Arthrobacter globiformis
В
Arthrobacter pascens
* Виды в таблице расположены в порядке убывания их обилия.
мическим составом бактерий, их местообитанием
и функциями могут быть приведены следующие:
доминирование представителей родов Cytophaga,
Cellulomonas и порядка Myxobacterales в подстилке
свидетельствует об участии бактерий этих таксо¬
нов в разрушении целлюлозы; обилие родококков
в нижних слоях подстилок связано с их способнос¬
тью использовать всевозможные ароматические
соединения, образующиеся в качестве промежу¬
точных продуктов в процессе деструкции лигнина;
обнаружение бактерий рода Hyphomicrobium
в торфяниках позволяет предположить участие
этих бактерий в использовании метанола - продук¬
та окисления метана, образующегося в болотах
в результате деятельности метаногенных бакте¬
рий; доминирование бацилл в нижних слоях лес¬
ных подстилок и верхних почвенных горизонтах
связано с их способностью разрушать крахмал,
пектин, лизировать клеточные стенки грибов и
дрожжей, т.е. реутилизировать микробную био¬
массу.
ПОЧВОВЕДЕНИЕ № 6 1996
704
ДОБРОВОЛЬСКАЯ и др.
Таким образом, изучение бактериального разно¬
образия в почвах разных природно-климатических
зон позволит составить представление о функциях
бактериальных сообществ, связанных с их учас¬
тием в процессах разложения растительного опа-
да и круговорота различных элементов в БГЦ.
Однако разнообразие бактерий столь велико,
а функции их столь многогранны, что собрать всю
информацию о составе и функциях бактериаль¬
ного мира почв практически невозможно, так как
для этого потребовались бы усилия всех бактери¬
ологов - специалистов по разным таксономичес¬
ким группам бактерий. В настоящей статье были
изложены материалы, касающиеся экологии бак¬
терий сапротрофного блока, включающего аэ¬
робные и факультативно-анаэробные формы
(около 50 родов), являющиеся характерными оби¬
тателями растений, подстилок и почв. Все они
растут на среде, предложенной нами для выделе¬
ния эпифитных и сапротрофных бактерий из
почв и растительных субстратов [4]. Использова¬
ние синэкологических методов анализа для оцен¬
ки таксономической структуры этого деструк¬
тивного блока бактерий позволило выявить
четкие различия в бактериальном населении
почв и сопряженных субстратов в разных типах
экосистем.
СПИСОК ЛИТЕРАТУРЫ
1. Лристовская Т.В. Микробиология подзолистых
почв. М.; Л.: Наука, 1965. 186 с.
2. Лристовская Т.В. Микробиология процессов поч¬
вообразования. Л.: Наука, Ленингр. отд-ние, 1980.
187 с.
3. Головченко А.В., Полянская JI.M., Доброволь¬
ская Т.Г. и др. Особенности пространственного
распределения и структуры микробных комплек¬
сов болотно-лесных экосистем // Почвоведение.
1993. № 10. С. 78-89.
4. Добровольская Т.Г., Скворцова И.Н., Лысак J1.B.
Методы выделения и идентификации почвенных
бактерий. М.: Изд-во МГУ, 1989. 72 с.
5. Добровольская Т.Г., Чернов И.Ю., Лысак Л.В. и др.
Бактериальные сообщества пустыни Кызыл-Ку-
мы: пространственная дисперсия и таксономичес¬
кий состав // Микробиология. 1994. Т. 63. Вып. 2.
С. 334-343.
6. Заварзин Г.А. Микробное сообщество в прошлом
и настоящем // Микробиол. журн. 1989. Т. 51. № 6.
С. 3-14.
7. Заварзин Г А. Протеобактерии: экологический
принцип в систематике прокариот // Природа.
1990. №5. С. 8-17.
8. Звягинцев Д.Г. Почва и микроорганизмы. М.:
Изд-во МГУ, 1987. 256 с.
9. Звягинцев Д.Г., Добровольская Т.Г., Лысак Л.В.
Вертикальный континуум бактериальных сооб¬
ществ в наземных биогеоценозах // Журн. общей
биологии. 1991. Т. 52. С. 162-171.
10. Звягинцев Д.Г., Бабьева И.П., Добровольская Т.Г.
и др. Вертикально-ярусная организация микроб¬
ных сообществ лесных биогеоценозов // Микроби¬
ология. 1993. Т. 62. С. 5-36.
11. Калакуцкий Л.В., Агре Н.С. Развитие актиномице-
тов. М.: Наука, 1977.286 с.
12. КацЛ.Н. Палеомикробиология: проблемы и пер¬
спективы // Успехи микробиологии. 1990. Т. 24.
С. 194.
13. Красильников НА. Микроорганизмы почвы и выс¬
шие растения. М.: Изд-во АН СССР, 1958.463 с.
14. Нестеренко О.А., Квасников ЕЛ., Ногина Т.М.
Нокардиоподобные и коринеподобные бактерии.
Киев: Наукова думка, 1985. 334 с.
15. Никитин Д.И., Васильева Л.В., Лохмачева Р.А.
Новые и редкие формы почвенных микроорганиз¬
мов. М.: Наука, 1966. 71 с.
16. Паников Н.С., Добровольская Т.Г., Лысак Л.В.
Экология коринеподобных бактерий // Успехи ми¬
кробиологии. 1989. № 23. С. 51.
17. Паринкина О.М. Микрофлора тундровых почв. Л.:
Наука, Ленингр. отд-ние, 1989. 160 с.
18. Сушкина Н.Н., Цюрупа И.Г. Микрофлора и пер¬
вичное почвообразование. М.: Изд-во МГУ, 1973.
156 с.
19. Polyanskaya L.M., Zvyagintsev D.G. Microbial succes¬
sion in soil // Sov. Sci. Rev. F. Physiol. Gen. Biol. 1995.
V. 9. Part. 1. P. 1-67.
Soils and Microbial Diversity
T. G. Dobrovol’skaya, L.V. Lysak, D. G. Zvyagintsev
Several approaches to estimate the soil microbial diversity are considered under historical aspect. The influence
of different environmental factors on the structure and functioning of bacteria communities in soils is con¬
vincingly demonstrated. Diverse types of bacteria communities are shown to be inherent to soils of various
natural-climatic zones.
ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВОВЕДЕНИЕ, 1996, № 6, с. 705-713
ПОЧВА
И БИОРАЗНООБРАЗИЕ
УДК 631.46
РАЗНООБРАЗИЕ ГРИБОВ И АКТИНОМИЦЕТОВ
И ИХ ЭКОЛОГИЧЕСКИЕ ФУНКЦИИ*
© 1996 г. Д. Г. Звягинцев, И. П. Бабьева, Г. М. Зенова, Jl. М. Полянская
Факультет почвоведения МГУ им. М.В. Ломоносова
Поступила в редакцию 13.02.96 г.
Приводятся данные по географическому расселению микроорганизмов разных групп в основных
типах почв СНГ, полученные на основании многолетних исследований. Показана четкая корреля¬
ция между таксономическим составом микробных комплексов, экологическими функциями микро¬
организмов и их приуроченностью к определенным типам почв. Продемонстрировано влияние раз¬
ных экологических факторов на структуру микробных комплексов, отражающую адаптацию мик¬
роорганизмов к природно-климатическим условиям почвенных зон. Показано, что полнота учета
микробного разнообразия в почвах, определяемая числом видов, увеличивается во много раз в ре¬
зультате использования сукцессионного подхода.
Теория коэволюционного освоения суши [9]
предполагает существенную роль симбиотической
ассоциации грибов с низшими и высшими растени¬
ями в виде микориз и лишайников в становлении
наземной флоры. Прокариотные мицелиальные
организмы также способны образовывать актино-
ризу на корнях многих кустарников и трав. Акти-
номицетный мицелий был обнаружен на древних
водорослях - вендотениях [8, 10]. Такое партнер¬
ство растений и мицелиальных организмов позво¬
лило решить проблемы питания и переживания
в условиях суши. Все основные экологические
группы растений (кроме цветковых) сложились
уже в девоне, т.е. наблюдался быстрый скачок от
водорослей ордовика до развитой флоры девона
благодаря симбиотическим ассоциациям грибов и
актиномицетов с растениями. Это партнерство
сохранилось и до сих пор - 9/10 видов всех расте¬
ний микотрофны, т.е. имеют на корнях микоризу.
Более 160 видов растений имеют на корнях акти-
норизу. В основе симбиоза лежит снабжение рас¬
тения актиномицетами рода Frankia связанным
азотом в обмен на получаемые продукты фото¬
синтеза [8]. Древние симбиозы растений с микроор¬
ганизмами не только способствовали выходу расте¬
ний на сушу, но и определили направление почвооб¬
разовательного процесса. Свою теорию дернового
почвообразовательного процесса В.Р. Вильямс по¬
строил на основе симбиотрофного питания луго¬
вых злаков [9]. Образование эктомикориз на кор¬
нях древесных пород также напрямую связано со
стадией почвообразовательного процесса. Так,
гигантские девонские леса были свободны от ми¬
коризы, поскольку почвы были еще плохо сфор¬
мированы, они стали более развитыми лишь
в карбоне. Современные эктомикоризы харак-
Работа выполнена в рамках программы “Биоразнообразие”.
терны для мулей и модеров. Грибы распространя¬
лись на сушу не только под покровом сосудистых
растений в виде различных микориз. Наиболее
ранние наземные сапротрофные грибы известны
с позднего силура и раннего девона [9]. Поступле¬
ние растительных остатков, содержащих лигно-
целлюлозу, способствовало выходу на первый
план микроорганизмов - активных деструкторов
этих сложных полимеров, т.е. грибов. Таким об¬
разом, прежняя деструкционная бактериальная
цепь, сложившаяся во времена господства циано¬
бактерий, в составе которых не было лигноцел-
люлозы, была заменена на комплекс сапротроф-
ных грибов - разрушителей лигноцеллюлозы [3].
Недаром именно грибы составляют, по последним
данным, 90% и более от общих запасов микробной
биомассы во всех типах почв [15]. Благодаря их де¬
ятельности в комплексе с другими группами мик¬
роорганизмов осуществляется конвейерная пере¬
работка растительного опада, наглядным отраже¬
нием которой являются стратифицированные
слои подстилок и почвенных горизонтов.
С другой стороны, сама почва как сложная ге-
терофазная система явилась той благоприятной
средой обитания, в которой происходила эволю¬
ция микроорганизмов, и в особенности мицели¬
альных. Считается, что эволюция мицелиальной
организации у грибов и актиномицетов тесней¬
шим образом связана с адаптацией к условиям су¬
ществования в почве. При развитии в почве мице¬
лиальные организмы располагают следующими
преимуществами: они способны проникать через
поверхности раздела фаз и колонизировать но¬
вые пространства, осуществлять транспорт пита¬
тельных веществ на расстояние [6]. Резкие колеба¬
ния влажности, температуры, радиации, временные
перерывы в снабжении организмов водой и пита¬
тельными веществами, т.е. режимы, характерные
2 ПОЧВОВЕДЕНИЕ № 6 1996
706
ЗВЯГИНЦЕВ и др.
для почв, послужили главными факторами эво¬
люции грибов и актиномицетов. В результате у
них сформировались специализированные пере¬
живающие стадии в цикле развития - споры
и плодовые тела с их функциями репродукции
и распространения спор в условиях суши [6, 9].
АКТИНОМИЦЕТЫ В ПОЧВАХ
Актиномицеты (порядок Actinomycetales) со¬
ставляют весьма обширную группу грамположи-
тельных бактерий, в пределах которой можно об¬
наружить разнообразные случаи перехода от до¬
статочно простой клеточной дифференциации к
более сложной. Понятие “актиномицеты” соби¬
рательное, оно объединяет мицелиальные и неми-
целиальные организмы. Следует отметить, что
если генетический (и, возможно, филогенетичес¬
кий) “вес” такого признака, как образование ми¬
целия, подвергается в настоящее время пересмот¬
ру, то внимание к экологической его значимости
лишь усиливается [8]. Мицелиальная организация
актиномицетов обусловливает схожесть экологи¬
ческих стратегий этих прокариотных организмов
с эукариотными микроорганизмами - грибами.
Сказанное побуждает нас выделить эту группу
мицелиальных прокариот среди прочих бактерий
и описать пространственно-функциональные за¬
кономерности распределения ее в почвах.
Широкое распространение актиномицетов в
природе определяется их устойчивостью к высу¬
шиванию, временному отсутствию питательных
веществ в среде обитания и эффективностью рас¬
селения их спор. Многие актиномицеты в лабора¬
торных культурах способны медленно расти на
водном агаре и других средах с низким содержа¬
нием питательных веществ. В почве активно рас¬
тущий вегетативный мицелий обнаруживается
преимущественно в микрозонах с повышенным
содержанием органического вещества. Корреля¬
цию между содержанием актиномицетов в почве
и количеством органического вещества в ней от¬
мечал Красильников [11]. Обладая небольшой
скоростью роста, актиномицеты обычно не ока¬
зываются в первом эшелоне г-стратегов при по¬
падании в почву легкодоступных многим микро¬
организмам веществ, и начинают доминировать
лишь тогда, когда создаются условия для исполь¬
зования сравнительно труднодоступных субстра¬
тов. Большинство актиномицетов нейтрофилы
(оптимальные для роста значения pH среды 6-8),
однако, существуют сведения о присутствии
в почве ацидофильных и алкалофильных форм [7].
В настоящее время представители почти всех
известных родов актиномицетов выделены из
почвы или обнаружены в почве. Однако законо¬
мерности распределения мицелиальных прокари¬
от в почвах до недавнего времени были исследова¬
ны только в отношении представителей родов
Streptomyces и Streptoverticillium [4]. Введение поня¬
тия комплекса почвенных актиномицетов, струк¬
тура которого отражает пространственно-вре-
меннбе соотношение отдельных таксонов (родов,
видов) мицелиальных прокариотных микроорга¬
низмов, способствовало выявлению общих зако¬
номерностей распределения этих организмов
в природных субстратах и, в первую очередь, в
почве [5]. Структура комплекса почвенных акти¬
номицетов определяется составом и численнос¬
тью типичных родов и видов и величиной видово¬
го спектра. Выделить представителей всех изве¬
стных в настоящее время родов актиномицетов
не представляется возможным, поскольку для вы¬
явления многих представителей порядка Actino¬
mycetales требуются специальные методы, включа¬
ющие ряд селективных приемов, связанных со
свойствами как спор, так и мицелия этих организ¬
мов, имеющих сложные жизненные циклы.
В своих исследованиях мы использовали тра¬
диционный метод поверхностного посева из раз-
ведений на плотные питательные среды: казеин-
глицериновый агар и среду с пропионатом натрия.
В отдельных случаях для выделения некоторых
малоизученных родов актиномицетов применяли
предпосевную обработку почвенных образцов
(нагреванием при температуре 100°С в течение 1 ч
или при 45°С - 16 ч).
Структура комплексов почвенных актиноми¬
цетов специфична для каждого биогеоценоза
(БГЦ). Лесной БГЦ характеризуется наличием
одного доминирующего рода Streptomyces с ти¬
пичными доминирующими видами только одной
секции и серии. Виды почвенных актиномицетов
лесного БГЦ функционируют в условиях, где ос¬
новными средообразующими факторами являют¬
ся подстилка, невысокое содержание гумуса, низ¬
кое значение pH среды. Эти условия определяют
заполнение основной части пространства эколо¬
гических возможностей (65%) в лесной почве
только стрептомицетами секции Cinereus Achro-
mogenes. Показано, что экологические ниши изу¬
ченных актиномицетных популяций дерново-
подзолистой почвы не разобщены полностью, а в
той или иной мере перекрываются. Виды секций
и серий Cinereus Achromogenes и Albus Albus име¬
ют сходные экологические позиции, степень пе¬
рекрывания их экологических ниш более 70%.
То же можно сказать и о видах секций и серий Ro-
seus Lavendulde-roseus и Cinereus Chromogenes.
Если степень перекрывания ниш рассматривать
как коэффициент конкуренции видов, то конку¬
рентные взаимоотношения в почве наиболее ве¬
роятны между представителями секций и серий
Cinereus Achromogenes и Albus Albus; Roseus Laven-
dulae-roseus и Cinereus Chromogenes. Следует напом¬
нить, что концепция экологической ниши как мно¬
гомерного пространства, в пределах которого воз¬
можно существование вида, широко используется
ПОЧВОВЕДЕНИЕ № 6 1996
РАЗНООБРАЗИЕ ГРИБОВ И АКТИНОМИЦЕТОВ
707
Биомы
Почвы
Strep-
Strepto-
Micro¬
Saccha¬
Saccha-
Actino-
Strepto¬
tomyces
verti-
mono¬
romono¬
ropoly-
madura
sporan¬
cillium
spora
spora
spora
gium
Nocardia
Тайга
Болото
Степь
Пустыня
Подзолистые
Дерново-подзо-
л истые
Торфяники
Черноземы
Каштановые
Серо-бурые
Такыры
# #
# #
□I
# #
# #
# #
* - ширина соответствует доле таксона в различных почвах.
Рис. 1. Актиномицеты в почвах различных биомов.
в общей экологии для характеристики экологи¬
ческих различий между популяциями.
Почва является тем природным субстратом, из
которого актиномицеты выделяются в наиболь¬
шем количестве и разнообразии, составляя чет¬
вертую часть по численности от всех бактерий,
выделяемых из почв на питательные среды. Од¬
нако представители некоторых родов актиноми¬
цетов (Streptoverticillium, Micromonospora) обнару¬
живаются в подстилке в значительно (на поря¬
док) большем количестве, чем в почве. Что
касается представителей рода Micromonospora, то
в лесных подзолистых, в том числе и дерново-
подзолистых почвах эти актиномицеты являются
минорными компонентами актиномицетного
комплекса. Они становятся доминантами в мохо¬
вой подстилке ельников, состоящей из листосте¬
бельных зеленых мхов, где их доля в актиноми-
цетном комплексе достигает 95%, а в нижних сло¬
ях лесной подстилки - 80%. Известно, что мхи
обладают высокой водоудерживающей способнос¬
тью и для них характерна теплая забуференная сре¬
да. Для микромоноспор - обладателей гидрофиль¬
ных спор - в мохостое создаются благоприятные
условия существования. В нижних слоях подстил¬
ки - этого специфического биоценотического
горизонта, для которого характерно ежегодное
пополнение органическим веществом, - требо¬
вательные к содержанию питательных элемен¬
тов в субстрате микромоноспоры также находят
оптимальные условия для жизни.
Низкие значения pH лесных подзолистых почв
способствуют развитию в них представителей ро¬
да Streptosporangium, тяготеющих к слабокислым
условиям среды. Стрептоспорангии, обычно
представляющие минорный компонент актино-
мицетных комплексов наземных экосистем, в
лесных подзолистых почвах составляют до 70%
всех актиномицетов (рис. 1).
В торфяниках актиномицеты сосредоточены
в дернине на залуженных участках территорий,
в нижних слоях лесной подстилки, где идут актив¬
ные процессы деструкции растительных остат¬
ков, в напочвенных разрастаниях водорослей,
мхов, лишайников. Специфическая особенность
микробных сообществ торфяных почв определя¬
ется во многом обилием микромоноспоровых ак¬
тиномицетов, численность которых во все сроки
определения, как правило, на 1-2 порядка превы¬
шает численность стрептомицетов. В большинст¬
ве же зональных типов почв микромоноспоры,
как и представители других “редких” родов акти¬
номицетов, по численности уступают стрептоми-
цетам. Типичными доминирующими представи¬
телями комплекса во всех рассматриваемых тор¬
фяных почвах явились актиномицеты родов
Streptomyces и Micromonospora. Стрептомицеты
представлены значительным видовым разнооб¬
разием, включающим виды секций и серий Ci-
nereus Achromogenes, С. Chromogenes, С. Chryso-
malus, С. Aures, Helvolo-flavus Helvolus. В актино-
мицетном комплексе целинной торфяной почвы
представители почти всех выделяемых на среде
с пропионатом натрия родов - Streptomyces,
Micromonospora, Saccharomonospora, Streptospo¬
rangium, Saccharopolyspora) являются доминиру¬
ющими или часто встречающимися, исключение
составляют актиномицеты рода Streptoverticilli¬
um, случайные в торфяных почвах. По широте
родового спектра комплекс актиномицетов в це¬
линной торфяной почве сходен с комплексом
чернозема и резко отличается от комплекса дер¬
ново-подзолистой почвы. Другой особенностью
ПОЧВОВЕДЕНИЕ № 6 1996
2*
708
ЗВЯГИНЦЕВ и др.
таксономического состава актиномицетного ком¬
плекса торфяников является доминирование са-
харомоноспоровых и сахарополиспоровых акти¬
номицетов, редко встречающихся в почвах зо¬
нального типа.
Почвы степных биогеоценозов - черноземные
и каштановые - отличаются от подзолистых лес¬
ных почв тем, что максимальное количество ак¬
тиномицетов в них приурочено к почвенному яру¬
су, причем доминантами, кроме представителей
рода Streptomyces, являются также представители
родов Micromonospora, Actinomadura, Nocardia
(в черноземе), меньшую долю составляют предста¬
вители родов Saccharomonospora, Saccharopolyspora,
Streptosporangium. Среди стрептомицетов в черно¬
земных почвах встречаются виды, принадлежащие
к секции Cinereus сериям Achromogenes, Chromoge¬
nes, Violaceus и Aureus. На основании изучения ди¬
намики численности почвенных актиномицетов
в ходе микробных сукцессий в черноземе опреде¬
лены показатели перекрывания экониш и уста¬
новлены наиболее благоприятные условия для
выявления популяций определенного таксономи¬
ческого спектра. Наиболее близки оказались
экониши у видов секции Cinereus серий Violaceus
и Aureus, наиболее далекими - у Cinereus Achro¬
mogenes и Cinereus Chromogenes. Установлено,
что для большинства видов актиномицетов наи¬
более благоприятные условия складываются на
последних этапах сукцессии.
Серо-бурые почвы пустынных ландшафтов
обладают специфическими особенностями в от¬
ношении распределения актиномицетов. Наши
исследования [5] показали, что существует чет¬
кая вертикальная стратификация сообществ ак¬
тиномицетов по ярусам пустынных экосистем с
максимальной концентрацией актиномицетов в
наземном ярусе - в поверхностной корочке песча¬
ной пустынной почвы, покрытой водорослями и
мхами, в ризосфере пустынных растений и в зоо-
генных субстратах. Для пустынных почв характер¬
но постоянное присутствие в них представителей
родов Streptomyces, Streptoverticillium, Nocardia; вы¬
сокая доля микромоноспоровых актиномицетов;
обилие темноокрашенных форм актиномицетов,
что можно рассматривать как адаптацию мицели¬
альных прокариот к пустынным условиям. Серо¬
бурые пустынные почвы отличаются актиноми-
цетным комплексом с большим разнообразием
стрептомицетных форм, принадлежащих почти
ко всем известным секциям и сериям.
Одним из характерных местообитаний акти¬
номицетов являются выходы карбонатных пород.
В наших исследованиях показано, что в формиру¬
ющихся на поверхности этих пород альгобакте-
риальных ценозах актиномицеты находятся в ак¬
тивной жизненной форме (в виде мицелия) и вместе
с водорослями выполняют роль ценозообразовате-
лей. В разных географических регионах (на вы¬
ходах скальных карбонатных пород в Прибалти¬
ке, Приднестровье, на Тянь-Шане, на известняко¬
вых карьерах Подмосковья) альгобактериальные
ценозы однотипны по таксономическому составу,
характеризуются доминированием альгокомпо-
нентов в биомассе ценозов (биомасса водорослей
составляет 96-98% общей массы ценоза) и акти¬
номицетного мицелия в биомассе сапротрофного
компонента ценоза (мицелий составляет более
50% биомассы бактерий). Актиномицеты в аль-
гобактериальных ценозах на выходах карбонат¬
ных пород представлены родами Streptomyces и Mi¬
cromonospora; среди стрептомицетов доминируют
представители секций Helvolo-flavus и Roseus. Вме¬
сте с другими группами микроорганизмов, входя¬
щими в микробные сообщества, складывающиеся
на поверхности пород, актиномицеты участвуют в
биохимическом выветривании и первичном поч¬
вообразовательном процессе.
Актиномицеты представляют собой единое
звено в трофической цепи любой экосистемы,
осуществляя в ней функции микробов-редуцен¬
тов. Основная роль мицелиальных прокариот со¬
стоит в разложении таких сложных полимеров,
как лигнин, хитин, целлюлоза, гумусовые соеди¬
нения. Актиномицеты участвуют также в накоп¬
лении в почве биологически активных веществ и
формировании азотного баланса почв. Посколь¬
ку большинство актиномицетов - гидролитики,
с их ферментативной деятельностью связана ло¬
кализация этих организмов в нижних слоях под¬
стилки, в кишечном тракте почвенных беспозво¬
ночных животных. В почве, где сосредоточена
основная масса стрептомицетов, актиномицеты
разлагают не только растительные субстраты и
гумусовые соединения; благодаря их способности
к образованию литических ферментов, они реу-
тилизируют микробную биомассу, прежде всего
грибную; растут на копролитах беспозвоночных
животных; участвуют в деструкции почвенных
минералов. Во всех этих случаях актиномицеты
проявляют себя как L, либо К-стратеги.
В почве и подстилке актиномицеты вступают
во взаимодействие с животными и растениями,
в том числе с водорослями, участвуя в формиро¬
вании трофических цепей биогеоценозов и сим¬
биотических ассоциаций.
МИКОБИОТА ПОЧВ
Грибы обнаруживаются во всех почвах, вклю¬
чая примитивные, на поздних этапах деградации
или в вечномерзлом состоянии. Экологическая
значимость грибов в разных условиях почвенной
среды различается в зависимости от факторов,
определяющих их численность, динамику разви¬
тия, таксономический состав.
ПОЧВОВЕДЕНИЕ № 6 1996
РАЗНООБРАЗИЕ ГРИБОВ И АКТИНОМИЦЕТОВ
709
О почвенных грибах можно сказать, что они -
главные редуценты в экосистемах суши. Эта их
основная функция осуществляется в почве. Одна¬
ко среди почвенных грибов есть специфические
группы, занимающие иное положение в трофиче¬
ских цепях: например, грибы-хищники, питающи¬
еся почвенными микроскопическими беспозвоноч¬
ными, грибы-паразиты, вызывающие корневые
гнили и другие болезни растений, факультативные
патогены, микоризообразователи и др. [13].
Рассматривая проблему биоразнообразия поч¬
венного микроскопического мира, мы будем ка¬
саться только наиболее многочисленной и широко¬
распространенной во всех типах почв группы мик-
ромицетов-сапротрофов, представители которой
осуществляют громадную по масштабам и одну
из важнейших экологических функций - разло¬
жение органических веществ, начиная от самых
простых углеводов и кончая такими сложными
биополимерами, как целлюлоза, хитин, лигнин, а
также создаваемых человеком полимерных соеди¬
нений, в огромном количестве поступающих в поч¬
ву, загрязняющих и отравляющих среду жизни.
Почвенные сапротрофные микромицеты как
в таксономическом отношении, так и по связи
с разными субстратами и по приспособленности
к факторам физической среды, чрезвычайно раз¬
нообразны. Их образ жизни определяется типом
роста (одноклеточный или мицелиальный), ха¬
рактером обмена веществ, наличием разного
рода приспособительных реакций, связанных с
внешними условиями, устойчивостью к стрессо¬
вым воздействиям [14]. Основной экологический
фактор, определяющий разнообразие грибов, -
тип субстрата как среды обитания. Он разделяет
три основных комплекса дрожжевых грибов в
биогеоценозе - фитобионтов, сапробионтов и пе-
добионтов, которые населяют, соответственно,
филлосферу, подстилку и почвенные горизонты
[1, 17]. Среди фитобионтов преобладают одно¬
клеточные дрожжевые грибы. Для них характер¬
ны наличие защитных от радиации каротиноид-
ных пигментов или меланинов, окислительный
тип обмена как более экономный для жизни за
счет экссудатов на поверхности листьев, образо¬
вание баллистоспор, которые активно рассеива¬
ются в воздушной среде. Наиболее характерные
черты видов-сапробионтов - образование гидро¬
литических экзоферментов и мицелиальный тип
роста, направленный на поиск нового субстрата.
Истинные педобионты обладают экономичным
обменом, накоплением резервных источников уг¬
лерода, образованием полисахаридных капсул,
которые обеспечивают особую межклеточную
сРеДУ> где концентрируются элементы питания и
сорбируется вода при дефиците их в окружающей
почве [2].
Наиболее типичными представителями ука¬
занных трех комплексов дрожжей могут быть ви¬
ды родов Sporobolomyces, Bullera и Rhodotorula -
для фитобионтов; род Trichosporon - для обитате¬
лей лесной подстилки (сапробионтов) и роды
Lipomyces и Cryptococcus - для педобионтов. Видо¬
вая структура перечисленных комплексов дрож¬
жей подчиняется определенным географическим
закономерностям.
Таксономический состав дрожжевых сооб¬
ществ меняется не только при переходе от расти¬
тельных к минеральным субстратам по вертика¬
ли БГЦ. В почвах одного типа, но под разными
растительными ассоциациями, комплексы доми¬
нантных видов могут быть различными. Так,
в почвах под хвойными и смешанными лесами зо¬
ны умеренного климата доминируют в подстил¬
ках представители родов Trichosporon и Cystofilo-
basidium, в минеральных горизонтах - Lipomyces
starkeyi и некоторые виды рода Cryptococcus.
В этой же зоне под травянистыми ассоциациями
наиболее характерны для почв, наряду с крипто¬
кокками, некоторые виды аскоспоровых дрож¬
жей родов Debaryomyces u Williopsis. В работе
Мирчинк [12] было показано, что на территории
Малинского стационара Московской обл. в дер-
ново-подзолистой почве под ельником доминирую¬
щими видами были Aureobasidium pullulans, Chrisos-
porium sulfureum, Mortierella ramanniana и Verticilli-
um terrestre, а в почве под березняком - Aposphaera
pulviscula, Cladosporium herbarum, Penicillium dale-
ae и Trichoderma koningi.
Из других факторов на грибное население
почвы значительно влияют температурные коле¬
бания в ее верхних слоях. Большинство грибов -
мезофилы, облигатные психрофилы - описаны
только среди дрожжевых грибов и в роде Acremo-
nium. И, наоборот, среди почвенных дрожжей нет
термофильных и термотолерантных видов, но
они широко известны среди видов p. Aspergillus.
Отношение к температуре определяет в значи¬
тельной степени местообитание многих видов
микромицетов или долю их популяций в сообще¬
ствах в разные периоды года. Так, например, об-
лигатно-психрофильные дрожжи p. Mrakia (Leu-
cosporidium) в дерново-подзолистых почвах Подмо¬
сковья могли быть обнаружены только в холодные
периоды года, когда численность их популяций
превысила критические значения, а конкуренция
со стороны других видов была низкой [1].
Детальное сравнение структуры грибного на¬
селения разных почв удобно сделать на биомах
резко различных географических зон - тундро¬
вых и пустынных (рис. 2). И те, и другие характе¬
ризуются отсутствием древесной растительности,
но резко различаются тепловым и световым ре¬
жимами, а также количеством и режимом влаги.
В зоне тундр надземная фитомасса значительно
превышает подземную ее часть, а отсюда следует
и высокая экологическая значимость микробной
биоты эпифитного комплекса фитобионтов, в
ПОЧВОВЕДЕНИЕ № 6 1996
710
ЗВЯГИНЦЕВ и др.
Биомы
Почвы
Lipomyces
starkeyi
lipofer
tetra-
albidus
aerius
diff-
gilves¬
sporus
luens
cens
Cryptococcus
podzo-
licus
Willi-
opsis
satur-
nus
Schw.
occi-
den-
talis
Тундра
Тайга
Болото
Степь
Полупу¬
стыня
Пустыня
Глеевые
тундровые
Подзолистые
Дерново-
подзолистые
Торфяники
Луговые
черноземы
Черноземы
Каштановые
Сероземы
Серо-бурые
# #
# #
.
# #
# #
# #
* * * - ширина соответствует доле таксона в различных почвах.
# #
Рис. 2. Дрожжи в почвах различных биомов.
частности, дрожжевых грибов. В тундровых поч¬
вах их очень мало и по численности, и по видово¬
му разнообразию. В основном, это виды, обычно
развивающиеся на зеленых частях растений, либо
в подстилках. Все виды - базидиомицетового
аффинитета: домгнируют криптококки (Cr. gilve-
scens, Cr. laurentii, Cr. albidus) и облигатно-психро-
фильные анаморфы рода Mrakia. В почвах тунд¬
ры полностью отсутствуют не только типичные
для многих более южных почв дрожжи p. Lipomy¬
ces, но и все другие аскоспоровые дрожжи.
Во всех почвах тундры, принадлежащих к разным
типам: тундровых глеевых, арктотундровых, арк¬
тических, примитивных, торфянистых, дерновых
почвах тундровых лугов формируется один и тот
же тип дрожжевого населения с Cr. gilvescens
в качестве доминанта и Mrakia gelida, Cr. albidus
в качестве сопутствующих видов [1]. Среди мице¬
лиальных микромицетов доминантами в почвах
тундр Таймыра выступают такие виды, как Peni-
cillium kapuscinskii, Geomyces pannorum и Sporor-
miella minimoides, достигающие в некоторых поч¬
вах 100% встречаемости, а также S. intermedia,
Podospora tetraspora и Mortierella sp. с встречаемо¬
стью 60-80%.
Своеобразие микобиоты тундровых почв состо¬
ит в том, что представители многих таксонов, ши¬
роко распространенные и доминирующие в других
районах, здесь отсутствуют. С другой стороны,
виды-доминанты в тундре обычны и для почв
умеренных широт. Большой процент стерильных
форм в Арктике может быть обусловлен непод¬
ходящими условиями для споруляции, и различ¬
ные виды микромицетов могут попадать в группу
Mycelia sterilia. Собственно педобионтный ком¬
плекс видов микромицетов в криоморфных поч¬
вах беден и характеризуется высокой долей пси-
хротолерантных видов [13]. Таким образом, под¬
водя итоги исследованиям грибного населения
тундровые почв, можно отметить следующие ха¬
рактерные для него черты: редукция таксономи¬
ческого состава, выпадение крупных групп
(например, аскоспоровых дрожжей), малая пред¬
ставленность собственно почвенных видов, высо¬
кая доля психрофильных и психротолерантных
форм. Экологические функции грибов в экосис¬
темах тундры сводятся, в основном, к иммобили¬
зации и трансформации легкодоступных соедине¬
ний в короткий период вегетации растений. Эти
процессы протекают в фитоярусе и на поверхно¬
сти почв, что приводит к накоплению неразло-
жившихся растительных остатков и торфообра-
зованию.
В почвах полупустынь, пустынь и сухих суб¬
тропиков Средней Азии и Южного Казахстана ус¬
тановлены другие закономерности распростране¬
ния и иные экологические функции грибов-мик-
ромицетов. Число обнаруженных в почвах
мицелиальных грибов значительно выше, чем в
тундре, а доля дрожжевых грибов среди них чрез¬
вычайно низка. В составе типичных форм - ас-
пергиллы, пенициллы и темноокрашенные виды
(рис. 3). Частота встречаемости типичных форм
не превышает 60%. Среди аспергиллов в серозе¬
мах доминируют виды A. flavus, A. ustus, A. terrens,
A. flavus, A. fumigatus. Пенициллы представлены
видами секции Asymmetrica: P. simplicissicum,
P. fellutanum, P. verrucosum, P. nigricans. Для зоны
сухих субтропиков показательно разнообразие
темноокрашенных грибов семейства Dematiaceae.
ПОЧВОВЕДЕНИЕ № 6 1996
РАЗНООБРАЗИЕ ГРИБОВ И АКТИНОМИЦЕТОВ
711
Биомы
Почвы
Aspergillus
terreus ustus wentii
Penicillium (seria)
chrisogenum
nigncans
funicu-
losum
Fusa-
rium
spp.
Tricho-
derma
spp.
Chryso-
sporium
spp.
Mortie-
rella
raman-
niana
Тундра
Тайга
Степь
Полупу
стыня
Глеевые тунд¬
ровые
Дерново-подзо¬
листые
Серые лесные
Черноземы
Каштановые
Сероземы
* * * * * *
# #
# #
# #
# #
# #
□г
# #
# #
I
¥
и
# #
* * *
* * *
# #
* * *
# #
# #
# #
♦ *
* *
И
# #
ширина соответствует доле таксона в различных почвах.
Рис. 3. Грибы в почвах различных биомов.
Мукоровые грибы малочисленны, род Trichoder-
ша представлен единичными видами, что харак¬
терно для засушливых почв.
Из дрожжевых грибов абсолютными доминан¬
тами выступают криптококки, представленные в
этих регионах почти исключительно видами Сг.
diffluens, Cr. albidus и Cr. laurentii. Психрофильные
дрожжи ни в почвах, ни на растениях не обнару¬
жены. Типичные педобионты - липомицеты -
в пустынных почвах не найдены.
Сравнение дрожжевых комплексов тундровых
и пустынных экосистем выявляет общие и специ¬
фические для каждого из биомов особенности.
Сходство дрожжевого населения, обусловленное
экстремальностью климата на крайнем севере
и юге, заключается в явном преобладании видов
эпифитного комплекса, в отсутствии собственно
почвенных дрожжей рода Lipomyces и в абсолют¬
ном доминировании криптококков. Различия
в таксономической структуре дрожжевых ком¬
плексов определяются разностью в основных ли¬
митирующих факторах в этих регионах: недо¬
статком тепла в тундре, недостатком влаги в пус¬
тыне. Поэтому в сходных группировках
происходит замена одних видов на другие - таксо¬
номически близкие, но экологически разные.
По сравнению с такими полноразвитыми экосис¬
темами, как лесные и степные БГЦ, экологичес¬
кие функции дрожжевых грибов в экстремаль¬
ных условиях резко сужаются, и дрожжи высту¬
пают в роли типичных эпифитов, в то время как
комплекс видов-гидролитиков и почвенных оби¬
тателей - липомицетов - отсутствует.
РАЗНООБРАЗИЕ МИКРООРГАНИЗМОВ
В ПОЧВАХ В ХОДЕ СУКЦЕССИИ
Состав доминирующих форм микроорганиз¬
мов динамичен и изменяется во времени (микроб¬
ные сукцессии) [16, 18]. При инициировании сук¬
цессий в почве оттаиванием, увлажнением или
внесением различных субстратов первыми повы¬
шают свою численность грибы, затем развивают¬
ся бактерии и актиномицеты. Каждая группа поч¬
венных микроорганизмов претерпевает в ходе
общей сукцессии микробиоты свою собственную
сукцессию. В составе комплекса почвенных гри¬
бов можно выделить два крупных блока, разли¬
чающихся по ряду экологических характеристик:
это быстрорастущие и медленнорастущие попу¬
ляции. Медленнорастущие популяции преоблада¬
ют практически на всех стадиях сукцессии. Эти
грибы отличаются наибольшей гидролитической
активностью и играют ведущую роль в разложе¬
нии полимеров растительного опада. Быстрорас¬
тущие грибы также способны осуществлять гид¬
ролиз, но в силу особенностей своей кинетики мо¬
гут эффективно перехватывать легкодоступные
субстраты, поступающие из зон активного гидро¬
лиза, осуществляемого медленнорастущими гри¬
бами. Быстрорастущие грибы на протяжении
всей сукцессии существенно уступают в числен¬
ности медленнорастущим грибам. Видовой ана¬
лиз комплекса почвенных грибов в ходе сукцес¬
сии показывает, что как среди медленнорастущих
грибов, так и среди быстрорастущих происходит
постоянная смена доминирующих популяций.
При многократном анализе инкубируемого поч¬
венного образца удается выделить в 4-5 раз
больше видов грибов, чем при обычном одно¬
кратном анализе [16].
При внесении в почву различных субстратов
ход сукцессии изменяется: целлюлоза резко по¬
вышает численность целлюлозоразрушающих
грибов, внесение сахаров - грибов-сахаролитиков
и т.д. В варианте с простым увлажнением чернозе¬
ма доминирующим был вид Penicillium ochrochlo-
ron, в варианте с внесением хитина на 6-13 сутки -
Aspergillus fumigatus, Mortierella alpina, начиная с
ПОЧВОВЕДЕНИЕ № 6 1996
712
ЗВЯГИНЦЕВ и др.
гга з
Рис. 4. Динамика численности некоторых доминиру¬
ющих видов (%) при инициации сукцессии увлажнени¬
ем (а), увлажнением и внесением хитина (б) и целлю¬
лозы (в).
/ - P. ochrochloron; 2 - Humicola grisea; 3 - Mycogone
nigra.
"I 1 1—^
13 30 51 120 140
. Сутки
Рис. 5. Количество выявленных видов (ось ординат)
грибов в ходе сукцессии.
30 суток абсолютным доминантом становится
Mycogone nigra. В сукцессии, инициированной ув¬
лажнением с внесением целлюлозы, на ранних
стадиях доминировали микромицеты из рода
Acremonium, а с 30 суток опыта в структуре ком¬
плекса почвенных грибов доминирующее поло¬
жение занимали грибы Mycogone nigra и Humicola
grisea (рис. 4а-4в). Четко прослеживается приуро¬
ченность развития грибов разных родов и видов
к определенным этапам сукцессии. При этом в
доминанты могут выходить популяции тех видов,
которые вообще не удается выделить из данной
почвы в обычных условиях, что позволяет значи¬
тельно расширить информацию о таксономичес¬
ком спектре микромицетов, обитающих в иссле¬
дуемой почве. На рис. 5 наглядно продемонстри¬
ровано нарастание количества видов грибов,
выделяемых на протяжении 140 суток сукцессии,
запущенной в черноземе путем увлажнения и вне¬
сения целлюлозы и хитина. Число видов, кото¬
рые удалось выделить в этом опыте, достигло
к концу сукцессии 90.
Изучение видового состава микромицетов в про¬
цессе сукцессии в дерново-подзолистой почве по¬
казало, что комплекс микромицетов представ¬
лен, в первую очередь, видами рода Penicillium,
встречаются представители родов Fusarium, Mu-
cor, Trichodeima. При этом на 0-6 сутки в структу¬
ре комплекса микромицетов доминировали виды
P. nigricans, P. simplicissium и Fusarium solani. В хо¬
де сукцессии возрастала численность быстрорас¬
тущих видов Trichodeima koningii и Т. harsianum.
Общее количество видов, обнаруженных в про¬
цессе сукцессии, достигло 42, в то время как при
однократном анализе этой почвы удалось вы¬
явить лишь 17 видов. Таким образом, сукцессион-
ный подход позволил выявить в 3-5 раз больше
видов микромицетов как в черноземе, так и дер¬
ново-подзолистой почве, и получить информа¬
цию об их месте в сукцессии, а следовательно,
и об их экологических функциях.
Сукцессия прокариотных микроорганизмов
в различных почвах характеризуется, как прави¬
ло, преобладанием вначале грамотрицательных
бактерий и увеличением доли актиномицетов на
более поздних стадиях. Увеличение численности
актиномицетов наблюдается в тот период, когда
биомасса грибов начинает уменьшаться. Можно
полагать, что актиномицеты специализируются
на использовании отмершего мицелия грибов.
Широко представленную у актиномицетов хити-
назную активность логично связать с высоким со¬
держанием хитина в клеточной стенке грибного
мицелия. Изучение сукцессии комплекса почвен¬
ных актиномицетов показало, что на начальных
стадиях доминируют медленнорастущие актино¬
мицеты. В ходе сукцессии доля быстрорастущих
актиномицетов увеличивается, и в ряде почв быс¬
трорастущие популяции доминируют на поздних
стадиях. Общая закономерность изменений, на¬
блюдаемая в ходе сукцессии в разных почвах и
с различными воздействиями (внесение мономе¬
ра или полимера), состоит в том, что на ранних
этапах сукцессии разнообразие актиномицетов
уменьшается, а в дальнейшем - возрастает. Попу¬
ляции актиномицетов, выделяемые на разных
ПОЧВОВЕДЕНИЕ № 6 1996
РАЗНООБРАЗИЕ ГРИБОВ И АКТИНОМИЦЕТОВ
713
этапах сукцессии, различаются не только по ра¬
диальной скорости роста и трофическим потреб¬
ностям, но и по проявлению антибиотической ак¬
тивности. Это позволяет судить о характере меж-
микробных взаимодействий, происходящих на
разных этапах сукцессии. Становятся понятными
специфические законы функционирования ком¬
плекса почвенных микроорганизмов, такие, на¬
пример, как избыточное многообразие микроб¬
ного пула и при этом ограниченный набор доми¬
нирующих видов, ведущих процесс в каждый
заданный момент [6].
ЗАКЛЮЧЕНИЕ
Для разных групп почвенных микроорганиз¬
мов разные экологические факторы являются ве¬
дущими: для актиномицетов - это режим влажно¬
сти и значение pH; для микромицетов - тип субст¬
рата и гидротермический режим почв. Роль и
экологическое значение отдельных групп микро¬
биоты почв меняется в зональной проекции, что
можно было проследить на примере распростра¬
нения как группировок мицелиальных и дрожже¬
вых грибов, так и комплексов актиномицетов.
Таксономический состав микробных сообществ
меняется во времени. Модельные опыты показа¬
ли, что полнота учета микробного разнообразия
в почвах, определяемая числом видов, увеличива¬
ется во много раз в результате использования
сукцессионного подхода. Растянутость во време¬
ни сукцессионных изменений, определяемая се¬
зонностью и сменой климатических факторов,
затрудняет их учет.
Для сохранения и изучения микробного разно¬
образия, его структуры и функций необходимо
сохранить все многообразие почв, поскольку в
разных почвах структура микробных комплексов
различна. Необходимо создание коллекций раз¬
личных почвенных образцов, которые хранились
бы при температуре жидкого азота, когда в них со¬
храняются естественные микробные сообщества.
СПИСОК ЛИТЕРАТУРЫ
1. Бабьева И.П. Дрожжи в биогеоценозах разных
природных зон // Почвенные организмы как компо¬
ненты биогеоценоза. М.: Наука, 1984. С. 131-140.
2. Бабьева И.П., Горин С.Е. Почвенные дрожжи. М.:
Изд-во МГУ, 1987. 78 с.
3. Заварзин Г. А. Протеобактерии: экологический
принцип в систематике прокариот // Природа.
1990. № 5. С. 8-17.
4. Зенова Г.М. Актиномицеты в биогеоценозах //
Почвенные организмы как компоненты биогеоце¬
ноза. М.: Наука, 1984. С. 162-170.
5. Зенова Г.М., Звягинцев Д.Г Актиномицеты в на¬
земных экосистемах // Журн. общей биологии.
1994. Т. 55. № 2. С. 198.
6. Калакуцкий Л.В., Агре Н.С. Развитие актиномице¬
тов. М.: Наука, 1977. 286 с.
7. Калакуцкий Л.В., Зенова Г.М. Экология актино¬
мицетов // Успехи микробиологии. 1984. № 19.
С. 203-221.
8. Калакуцкий Л.В., Шарая Л.С. Актиномицеты и
высшие растения // Успехи микробиологии. 1990.
№ 24. С. 26-65.
9. Каратыгин И.В. Коэволюция грибов и растений.
СПб.: Гидрометеоиздат, 1993.
10. Кац Л.Н. Палеомикробиология: проблемы и пер¬
спективы // Успехи микробиологии. 1990. Т. 24.
С. 194.
11. Красильников Н.А. Микроорганизмы почвы и выс¬
шие растения. М.: Изд-во АН СССР, 1958. 463 с.
12. Мирчинк Т.Г. Почвенные грибы как компонент
биогеоценоза // Почвенные организмы как компо¬
ненты биогеоценоза. М.: Наука, 1984. С. 114-130.
13. Мирчинк Т.Г. Почвенная микология. М.: Изд-во
МГУ, 1988. 224 с.
14. Мирчинк Т.Г., Бабьева И.П. Грибы, образующие
мицелий, и дрожжи в природных экосистемах //
Журн. общ. биологии. 1981. Т. 42. № 3. С. 390-398.
15. Полянская Л.М., Гейдебрехт В.В., Степанов AJI.,
Звягинцев Д.Г. Распределение численности и био¬
массы микроорганизмов по профилю зональных
типов почв // Почвоведение. 1995. № 3. С. 322-328.
16. Полянская Л.М., Мирчинк Т.Г., Кожевин П.А.,
Звягинцев Д.Г. Изменение структуры комплекса
почвенных микромицетов в ходе микробных сук¬
цессий//Микробиология. 1990. Вып. 2. С. 349-354.
17. Bab’eva I.P., Chernov I.Yu. Geographical aspects of
yeast ecology // Sov. Sci. Rev. F. Physiol. Gen. Biol.
1995. V. 9. Part. 3.
18. Polyanskaya L.M., Zvyagintsev D.G. Microbial succes¬
sion in soil // Sov. Sci. Rev. F. Physiol. Gen. Biol. 1995.
V. 9. Part. 1. P. 1-67.
Diversity of Fungi and Actinomyces in Soils and their Ecological Functions
D. G. Zvyagintsev, I. P. Bab’ieva, G. M. Zenova, L. M. Polyanskaya
For many years the authors investigated numerous microorganisms with the view of their geographical popu¬
lation in the main soil types of the country. A clearly expressed correlation is shown between taxonomic com¬
position of microbial complexes, ecological functions of microorganisms and their attribution to definite soil
types. Several environmental factors prove to be responsible for structure of microbial complexes, which re¬
flects adaptability of microorganisms to natural and climatic conditions of soil zones. Further discussed is the
succession pattern, capable to make more complete accounting the microbial diversity of soils.
ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВОВЕДЕНИЕ, 1996, № 6. с. 714-721
УДК 631.46
ПОЧВА
И РАЗНООБРАЗИЕ
АДАПТИВНЫЕ СТРАТЕГИИ
ОСВОЕНИЯ ЖИВОТНЫМИ ПОЧВЕННОГО ЯРУСА
© 1996 г. Б. Р. Стриганова
Институт проблем экологии и эволюции им. А.Н. Северцова РАН, Москва
Поступила в редакцию 31.08.95 г.
Гетерогенность почвенной среды рассматривается как фактор формирования таксономического и
экологического разнообразия животного населения почвы в процессе эволюции. Анализируются
различия пространственных ниш разных размерных группировок беспозвоночных в связи с особен¬
ностями их дыхания. Обсуждаются пути реализации трех механизмов газообмена на примере поч¬
венных животных. Рассматриваются этолого-экологические адаптации крупных почвенных обита¬
телей к формированию и поддержанию аэрированного пространства в минеральной массе почвы.
Наличие макропор аэрации является основным условием освоения аэробионтными животными
глубоких горизонтов почвенного профиля. Обосновывается гипотеза об освоении почвенного яру¬
са наземными формами с поверхности почвы.
В 40-х гг. была выдвинута гипотеза о почве как
особой специфической среде обитания, через кото¬
рую осуществлялся выход животных на сушу [5].
Подавляющее число современных групп назем¬
ных беспозвоночных связано с почвой в течение
всего жизненного цикла, или в почве проходит
развитие их ювенильных стадий (яйца, личинки,
нимфы, куколки). Практически лишь представи¬
тели двух классов членистоногих - пауки и кры¬
латые насекомые - оказались способными пол¬
ностью приспособиться к жизни в открытой ат¬
мосфере.
М.С. Гиляров показал, что у почвенных и под¬
стилочных обитателей нет эффективных меха¬
низмов защиты от испарения воды через покровы
и они способны к нормальной жизнедеятельности
лишь в условиях высокой влажности окружающе¬
го воздуха. Почва представляет для таких форм
благоприятную среду: влажность почвенного воз¬
духа постоянно близка к насыщению, а колебания
температуры сглажены по сравнению с наземной
средой. Насыщенность воздуха водяными пара¬
ми, присутствие капельной и пленочной влаги,
выровненность температурных условий, варьи¬
рование содержания кислорода и углекислоты -
признаки, сближающие почву с водной средой [5].
Эти особенности почвы дали повод некоторым ав¬
торам сравнивать ее обитателей с планктоном [28].
В то же время наличие воздушного пространства
в почве и действие сил гравитации - признаки на¬
земной среды.
Выход животных из воды на сушу сопровож¬
дался кардинальными перестройками функцио¬
нальной морфологии различных систем органов,
связанными прежде всего с условиями газо- и во¬
дообмена, а также с передвижением в воздушной
среде [6]. Была высказана гипотеза о возможнос¬
ти выхода из почвы в наземный ярус через расти¬
тельные остатки (мертвая древесина) или живые
растительные ткани [5]. Проблема перехода бес¬
позвоночных животных к обитанию в почве и в
надземном ярусе рассматривалась, в основном, с
позиций их приспособлений к поддержанию вод¬
ного баланса в воздушной среде с колеблющимся
содержанием влаги.
Однако не менее важной проблемой при пере¬
ходе животных к сухопутному образу жизни явля¬
ется приспособление к дыханию атмосферным
воздухом. В настоящем сообщении рассмотрены
адаптивные стратегии животных, определившие
начальные стадии эволюционного процесса вы¬
хода на сушу, а именно, приспособления к дыха¬
нию в почвенной среде.
ДИФФЕРЕНЦИАЦИЯ
ПРОСТРАНСТВЕННЫХ НИШ
ПОЧВЕННЫХ БЕСПОЗВОНОЧНЫХ
Наличие в почве микро- и макрополостей и аг¬
регатов разного размера обеспечивает высокое
разнообразие пространственных ниш, заселяе¬
мых разными группами организмов. Структурная
сложность почвенного яруса формируется и за
счет органического материала, поступающего в
виде наземного опада и массы отмерших корней.
Поэтому почва представляет среду, где могут
обитать организмы, кардинально различающие¬
ся по эко-физиологическим требованиям.
Размеры почвообитающих беспозвоночных
варьируют от 10 мкм до 20-30 см. Традиционно
животное население почвы разделяется на четы¬
ре размерных класса [7]. Эта классификация от¬
ражает не только формальные различия разме¬
ров, но и вытекающие из этого эколого-физиоло-
гические различия отдельных группировок.
714
АДАПТИВНЫЕ СТРАТЕГИИ ОСВОЕНИЯ ЖИВОТНЫМИ ПОЧВЕННОГО ЯРУСА
715
Группы, относящиеся к эумикрофауне пред¬
ставлены по существу физиологически водными
животными, дышащими кислородом, растворен¬
ным в воде. Они обитают в капиллярных водных
полостях и при недостатке капельной влаги адге-
зируются к поверхности твердых частиц почвы,
окруженных водной пленкой. Тончайший слой
в одну молекулу воды достаточен для погруже¬
ния этих организмов, и представители эумикро-
фауны могут переживать длительные неблаго¬
приятные условия в неактивном состоянии. При
насыщении почвы влагой они способны свободно
мигрировать по системе водных микроводоемов.
Для эумикрофауны почва представляет разветв¬
ленную систему сообщающихся водных каналов.
Простейшие и нематоды концентрируются в оча¬
гах разложения органического материала, где об¬
разуются обводненные продукты распада. Там они
дают вспышки размножения, которые заканчива¬
ются при истощении пищевых резервов (микроб¬
ная масса), либо при пересыхании этих очагов.
Более крупные размерные группировки бес¬
позвоночных являются настоящими сухопутны¬
ми формами. Мелкие черви и микроартроподы
живут в почвенных полостях, заполненных возду¬
хом, и свободно передвигаются по системе внут-
рипочвенных ходов. Для этой группы почва пред¬
ставляет некую систему полостей, поверхность
которых они используют для передвижения и
сбора пищи. Суммарная поверхность стенок по¬
лостей определяет величину пространства для ак¬
тивной жизнедеятельности животных.
Лишь наиболее крупные беспозвоночные, от¬
носящиеся к мезофауне, встречаются с фактором
плотности почвенной среды и должны решать
ряд проблем, связанных с передвижением в плот¬
ном субстрате, дыханием, нахождением пищи, по¬
лового партнера, реализацией размножения.
Таким образом, для мелких водных форм пе¬
реход из водоемов в почвенную влажную среду не
представлял никаких проблем, и они сохранили все
физиологические черты водных обитателей. Объ¬
ем водных микрополостей в почве лимитирует раз¬
меры таких физиологически-водных животных, их
величина измеряется десятками микрон.
Фаунистический состав простейших в водо¬
емах и в почве сходен на уровне надвидовых так¬
сонов. Но население простейших в почве как прави¬
ло представлено более мелкими видами, размеры
которых в 5-10 раз меньше, чем у систематически
близких представителей водных форм [14]. Брод¬
ский [1] отмечал заселение почвенными простей¬
шими аридных почв по всей глубине профиля.
Помимо резких периодических колебаний об¬
щего запаса капельной влаги, водные формы в
почве встречают ряд других факторов, отсутству¬
ющих в водоемах: особенности солевого и газового
состава почвенной воды, сила поверхностного
натяжения водных пленок, более резкие темпера¬
турные колебания. В процессе эволюции почвен¬
ные формы приспособились успешно переживать
экстремальные воздействия этих факторов в не¬
активном состоянии. Для некоторых форм отме¬
чено сокращение длительности жизненного цик¬
ла в сравнении с близкими водными видами [34].
Более крупные размерные группировки аэро-
бионтных почвенных беспозвоночных включают
таксономические группы, имеющие водных пред¬
ков и близкородственные водные формы в совре¬
менной фауне (олигохеты, мокрицы, амфиподы,
моллюски), а также группы, сформировавшиеся в
наземной среде (представители паукообразных,
онихофоры, многоножки, энтогнаты, насекомые).
Мелкие беспозвоночные (микрофауна), оби¬
татели воздушных полостей, заселяют в основ¬
ном подстилку и гумусовый слой почвы. Энхитреи-
ды концентрируются в верхних 10 см почвы и лишь
в полевых почвах более равномерно распределя¬
ются по всему пахотному горизонту. Наличие
доступного органического материала и влаж¬
ность - основные факторы, определяющие харак¬
тер пространственного распределения энхитреид
[23]. Но на фоне этого имеет значение и аэриро-
ванность почвы, т.е. количество воздушных по¬
лостей. Как правило, в песчаных почвах числен¬
ность энхитреид выше, чем почвах тяжелого ме¬
ханического состава [30]. При уплотнении почвы
вследствие пастбищной или рекреационной на¬
грузки численность энхитреид резко снижается
[33], что связано с уменьшением объема воздуш¬
ных полостей в почве.
Микроартроподы (клещи и коллемболы) так¬
же заселяют в основном самый верхний горизонт
почвы, где концентрируется до 90% их биомассы.
Среди клещей и коллембол выделен ряд жизнен¬
ных форм, различающихся по ярусному преферен-
думу в почвенном профиле и по ряду морфологиче¬
ских особенностей, определяющих их адаптации, с
одной стороны, к обитанию в почве, а с другой -
к выходу в подстилочный и напочвенный ярусы
[20, 37]. В характере вертикального распределе¬
ния этих беспозвоночных нет четко выраженной
зависимости от суммарного объема воздушных
пор [23]. Более важную роль для их пространствен¬
ного распределения играет концентрация доступ¬
ного органического материала, обогащенного ми¬
крофлорой, служащей основным пищевым ресур¬
сом микроартропод, а также фактор влажности.
Формы, обитающие в глубоких слоях почвы,
отличаются мелкими размерами: глубокопочвен¬
ные виды Oppiidae (Oribatei), Mesaphorura, Folsomia,
Anurida (Collembola) имеют размеры менее 1 мм.
У внутрипочвенных жизненных форм коллембол
имеются признаки редукции прыгательной вилки,
особенно хорошо развитой у поверхностно живу¬
щих групп, а также частичная или полная утрата
органов зрения [20]. В то же время у них отмече¬
но усложнение антеннального и постантенналь-
ного органов, свидетельствующее об усилении
развития тактильных и хеморецепторов. Таким
ПОЧВОВЕДЕНИЕ № 6 1996
716 СТРИГАНОВА
Таблица 1. Содержание углекислоты в почвенном воздухе на разных глубинах (Центрально-черноземный запо¬
ведник) (по [15])
Глубина, см
Содержание углекислоты, %
Глубина, см
Содержание углекислоты, %
Степь
Лес
Степь
Лес
25
0.36
0.70
125
0.62
0.74
50
0.48
0.74
150
0.58
0.82
75
0.54
0.78
200
0.57
0.94
100
0.54
0.82
300
0.55
1.08
образом, морфологические изменения у внутрипоч-
венных форм микроартропод носят явно вторич¬
ный характер. Освоение этими мелкими обитателя¬
ми воздушных полостей минеральных горизонтов
почвы, очевидно, связано с более постоянным ре¬
жимом влажности почвенного воздуха.
Крупные роющие беспозвоночные встреча¬
ются до глубины 1.5-2 м. Было отмечено совпа¬
дение границ гумусового горизонта, распростра¬
нения корней и зоотической активности в почвен¬
ном профиле разных типов почв [4]. Как и для
микрофауны, основным фактором, определяю¬
щим вертикальную локализацию крупных беспо¬
звоночных в почве, является обеспеченность пи¬
щевыми ресурсами и влажность.
Для животного населения почвы характерно
четкое разделение пространственных ниш в поч¬
венном профиле. Среди них выделяются обитате¬
ли подстилки и поверхностного горизонта, обита¬
тели минерального слоя почвы и формы, совер¬
шающие регулярные вертикальные миграции.
Животные локализуются в водных или воздуш¬
ных порах, что обеспечивает их пространствен¬
ную сегрегацию в цределах одного горизонта.
Среди обитателей воздушных пор выделяются
группы, использующие уже имеющиеся ходы, и
роющие формы, способные создавать и поддер¬
живать длительно действующую систему ходов,
активно регулируя тем самым степень аэрирован-
ности нижних горизонтов почвы. Роющая деятель¬
ность и вертикальные миграции животных играют
существенную роль и в обогащении глубоких гори¬
зонтов органическими остатками, которые зано¬
сятся в кишечнике животных или погребаются в
ходах. Это способствует заселению глубоких слоев
микрофлорой, а зоогенная аэрация почвы стиму¬
лирует развита» аэробных микробиальных процес¬
сов. Кроме того, прокладывая систему ходов, рою¬
щие формы создают специфические ниши, годные
для обитания более мелких нероющих форм. Они
усиливают пространственную дифференциацию
почвенной среды и способствуют повышению раз¬
нообразия и обилия животного населения.
ОСОБЕННОСТИ ДЫХАНИЯ
ПОЧВЕННЫХ БЕСПОЗВОНОЧНЫХ
Приспособление, животных к обитанию в поч¬
ве было сопряжено с резким повышением уровня
энергообмена для преодоления сил гравитации и
возможности активного передвижения внутри
плотного субстрата. Условия дыхания животных
в почве определяются особенностями газового
состава почвенного воздуха. В почве с глубиной
напряжение кислорода падает и возрастает кон¬
центрация углекислоты. В табл. 1 показана дина¬
мика нарастания содержания углекислоты с глу¬
биной в черноземной почве. Напряжение угле¬
кислоты значительно выше, чем в атмосферном
воздухе, а в лесу почти вдвое выше, чем в степной
почве, даже в поверхностном слое. Причиной
этого является высокая концентрация разлагаю¬
щейся подстилки. Постепенное нарастание содер¬
жания углекислоты несколько модифицируется
активностью дыхания корней, и на глубине наи¬
большего скопления массы сосущих корней ее со¬
держание выше, чем в нижележащих слоях поч¬
вы. Таким образом, в почве на большой глубине
животные испытывают недостаток кислорода.
Для нормальной жизнедеятельности, в том числе
и для рытья ходов, они нуждаются в формирова¬
нии эффективных механизмов газообмена.
Существуют три типа реализации газообмена:
первый - простая диффузия газов через поверх¬
ность тела. Этот тип дыхания характерен для мел¬
ких и малоактивных животных. Второй тип - это
формирование трахейной системы, по которой
кислород, поступающий через дыхальца, достав¬
ляется непосредственно ко всем клеткам тела.
Развитая трахейная система обеспечивает кисло¬
родом все ткани тела независимо от их удаленно¬
сти от дыхалец. Третий тип - обеспечение кисло¬
родом с помощью дыхательных пигментов крови
или гемолимфы. Кислород из органов аэрации
(жабры или легкие) или от поверхности тела
связывается пигментами и от них поступает в
клетки тела. Это - наиболее прогрессивный ме¬
ханизм газообмена, характерный для всех позво¬
ночных животных.
У почвенных и напочвенных беспозвоночных
встречаются все три типа газообмена. Простая
диффузия наблюдается у простейших, коловра¬
ток и нематод, дышащих кислородом, растворен¬
ным в воде. При относительно высоком обмене
обеспечение тканей кислородом возможно лишь на
глубину 0.1 см. При расстоянии между источником
кислорода и тканями более 0.5 см необходим допол¬
нительный механизм переноса кислорода [31].
Таким образом, малые размеры представителей
ПОЧВОВЕДЕНИЕ № 6 1996
АДАПТИВНЫЕ СТРАТЕГИИ ОСВОЕНИЯ ЖИВОТНЫМИ ПОЧВЕННОГО ЯРУСА 717
Таблица 2. Распространение дыхательных пигментов у беспозвоночных
Группа организмов
Гемоглобин
Гемоцианин
Группа организмов
Гемоглобин
Гемоцианин
Infusoria
+
-
Arachnoidea
+
+
Plathelminthes
+
-
Crustacea
+
+
Nemertini
+
-
Entognatha
+
-
Nematoda
+
-
Insecta
+
-
Oligochaeta
+
-
Mollusca
+
-
эумикрофауны лимитируются и особенностями их
газообмена.
Трахейная система, встречающаяся только
у членистоногих, обеспечивает сравнительно вы¬
сокий уровень газообмена. Активный обмен
у многоножек-диплопод может в 300 раз превы¬
шать уровень стандартного обмена в состоянии
покоя. Возникновение трахейной системы в про¬
цессе эволюции рассматривалось как ароморфоз,
обеспечивший в дальнейшем членистоногим ос¬
воение надземного яруса [5].
В процессе эволюции у животных возникло че¬
тыре дыхательных пигмента; у полихет отмечено
три пигмента, при этом хлорокруорин и гемире-
трин встречаются только в этой группе. У мол¬
люсков, ракообразных, мечехвостов и некоторых
паукообразных распространен гемоцианин, кото¬
рый также, очевидно, возник в группе червей, дав¬
ших начало моллюскам и ракообразным. Гемогло¬
бин распространен наиболее широко в животном
мире. У позвоночных имеется только гемоглобин.
Он встречается, кроме того, и у некоторых сосуди¬
стых растений и грибов. Коржуев [11] предполага¬
ет неоднократное и независимое возникновение
гемоглобина в разных группах животных. Струк¬
тура гема-железо-порфиринового кольца одина¬
кова у всех, но структура белков, с которыми свя¬
зан гем, различается в разных группах.
Гемоглобин может насыщаться кислородом
при очень низком парциальном давлении послед¬
него, чем объясняется его высокая эффектив¬
ность и прогресс в животном мире. Но сродство
гемоглобинов к кислороду у разных групп живот¬
ных широко варьирует и, кроме того, зависит от
ряда внешних факторов - температуры, pH сре¬
ды, наличия СО, который инактивирует гемогло¬
бин. Хлорокруорин переносит кислород только
при высоком парциальном давлении, а гемирит-
рин выполняет функции получения кислорода из
морской воды и передачи его гемиритрину гемо¬
лимфы. Гемоцианин - единственный пигмент,
в состав которого входит не железо, а медь. У чле¬
нистоногих он не насыщается кислородом полно¬
стью. У ракообразных содержание меди (и гемо-
Цианина) в гемолимфе относительно низкое [18].
Его носители нуждаются в атмосферном воздухе
с высоким содержанием кислорода.
У почвообитающих беспозвоночных встреча¬
ется два дыхательных фермента - гемоглобин и
гемоцианин (табл. 2). Последний характерен
лишь для мокриц. У беспозвоночных гемоглобин
служит переносчиком кислорода при нормаль¬
ном его парциальном давлении. При низком дав¬
лении пигмент функционирует как депо кисло¬
рода при временной гипоксии или прекращении
дыхания. Сродство гемоглобина к кислороду выра¬
жается через парциальное давление, при котором
половина молекул насыщена кислородом (Р^).
Величина Р50 сильно колеблется у разных видов и
групп в зависимости от характера их местообита¬
ний. Наиболее высокое сродство к кислороду отме¬
чено у паразитических форм (Р^ аскариды - 0.05,
у личинок лошадиного овода - 0.02 мм рт. ст.). Наи¬
более высокие показатели Р^ установлены у лито¬
ральных обитателей Eupolymnia - 36 мм рт. ст. [18].
У дождевых червей Р50 составляет 1-6 мм рт. ст.
Эта величина сравнима с Р50 у водных полихет, за¬
рывающихся в грунт (у пескожила Arenicola Р50
составляет 1.6-4.0 мм рт. ст.).
Гемоглобин играет особенно большую роль
в дыхании дождевых червей. Дождевые черви ды¬
шат поверхностью тела, их покровы, пронизанные
многочисленными сосудами, смачиваются слизис¬
тыми выделениями. Адаптация червей к дефициту
кислорода направлена на усиление синтеза гемо¬
глобина не только в гемолимфе, но и в мышечной,
и нервной тканях.
У мокриц имеются разные модификации ды¬
хательной системы. У водных форм развиты лис¬
товидные жабры, образовавшиеся из эндоподитов
(внутренних лопастей) брюшных конечностей.
Жаберные листочки прикрыты снизу лопастью
из сросшихся экзоподитов (наружных ветвей
брюшных конечностей). На суше многие мокри¬
цы остались водно-дышащими формами. Для по¬
лучения кислорода им достаточно капли воды.
Если капля воды попадает на их покровы, то по
сиртеме скульптурированных образований на
панцире капля проводится к жабрам. Такая же
капиллярная система имеется и на уроподах.
У некоторых форм увлажнение жабер произво¬
дится жидкостью, выбрасываемой из анального
отверстия. Поэтому большинство мокриц ведет
приводный образ жизни, и в отсутствие капель¬
ной влаги они погибают. В этой группе имеются
и настоящие наземные формы с воздушным ды¬
ханием. У них в толще экзоподитов образуется
воздушная полость со слепыми псевдотрахеями.
Трахейное поле хорошо развито у Oniscus, Philos-
cia; при этом у них функционируют и жабры.
ПОЧВОВЕДЕНИЕ № 6 1996
718
СГРИГАНОВА
У Porcellio, Armadillidium жабры редуцированы,
и развиты только воздушные мешки на экзоподи-
тах двух передних плеопод [29].
Среди паукообразных скорпионы и жгутоно¬
гие имеют легочные мешки, сформировавшиеся
на основе жаберных пластинок их водных пред¬
ков. У ложноскорпионов, сенокосцев и части кле¬
щей имеются настоящие трахеи. Трахейная сис¬
тема характерна для протрахеат, многоножек,
насекомых, некоторых пауков.
У наземных беспозвоночных четко дифферен¬
цируются два основных типа приспособлений
к дыханию на суше - формирование трахейной
системы, представленной в наиболее развитой
форме у насекомых, и сосредоточение дыхатель¬
ных функций в сосудистой системе при наличии
дыхательных пигментов в крови или гемолимфе,
характерное для дождевых червей. Оба типа при¬
способления к дыханию атмосферным воздухом
обеспечивают относительно высокий уровень
энергообмена, позволяющий реализовать полет
в воздухе (крылатые насекомые) и передвижение
в плотном субстрате (почва, растительные тка¬
ни). В то же время почвенные беспозвоночные
нуждаются в воздухе, в котором парциальное дав¬
ление кислорода близко к атмосферному. Лишь
немногие формы способны дышать воздухом
с повышенным содержанием углекислоты.
ФОРМИРОВАНИЕ АЭРИРОВАННОГО
ПРОСТРАНСТВА В ПОЧВЕ
Среди глубокопочвенных обитателей наибо¬
лее многочисленными и активными группами яв¬
ляются дождевые черви, пустынные мокрицы,
муравьи и термиты, личинки жесткокрылых.
Потребности в кислороде у обитателей мине¬
рального горизонта почвы относительно высоки.
Пустынные мокрицы роют глубокие верти¬
кальные норки глубиной до 1 м. Их распределе¬
ние четко связано с гипсоносными сероземами
тяжелого гранулометрического состава [8, 12].
В глинистых почвах ходы мокриц долго сохраня¬
ются, насмотря на то, что их вертикальные стен¬
ки не укрепляются выделениями животных. Экс¬
кременты мокриц очень сухие. Необходимость
поддержания режима аэрации на дне ходов вы¬
звала формирование сложного комплекса пове¬
денческих адаптаций: ежедневная чистка, вынос
на поверхность обвалившейся земли, экскремен¬
тов, остатков пищи, проветривание хода и закры¬
вание его собственным телом [12, 13]. Сходные
формы поведения описаны у австралийских пус¬
тынных мокриц, относящихся к другому роду [26].
Пустынные мокрицы питаются на поверхности
почвы наземным опадом, однако, в жаркий сезон
они уносят корм в норы [22]. У мокриц питание и
экскреция связаны с поверхностью почвы, т.е. ос¬
новные функции жизнедеятельности реализуют¬
ся в наземном ярусе. Это позволяет говорить об
относительно недавнем переходе к обитанию в
минеральном горизонте, связанном с освоением
аридных ландшафтов. Некоторые показатели
роющей активности мокриц Hemilepistus cristatus
Ulj., исследованных нами в Бадхызском заповед¬
нике (Туркменистан), приведены ниже:
Частота норок на 1 м2 67.5 ±1.8
Глубина норок, см 80-90
Диаметр норок, см 0.45
Общий объем норок в монолите почвы 858.4
объемом 100 х 100 х 80, см3
Масса почвы, выброшенной на 79.2 ± 2.4
поверхность вокруг норки, г
Общий объем почвы, поднятой на поверх- 5.3
ность из глубокого горизонта, кг/м2/год
При невысокой для агрегации этих мокриц плот¬
ности они выбрасывают наверх до 5 кг почвы. При
этом объем их нор составляет около 800 мл, что су¬
щественно влияет на режим аэрации этих тяже¬
лых почв. Вместе с тем у этих мокриц имеются
адаптации к сохранению влаги в норах и поддер¬
жанию влажности воздуха около 85-90% (закры¬
вание входов в нору).
В почвах аридных районов большое значение
имеет роющая деятельность муравьев и терми¬
тов, строящих подземные гнезда. Ниже приводят¬
ся несколько параметров их гнездостроительной
активности в почвах Туркмении [9,10] (табл. 3).
Глубина гнезд муравьев варьирует у разных
вчдов и семей от 30 до 170 см. Муравьи роют
очень глубокие ходы, достигающие водоносного
слоя. По ним в гнездо проходит холодный влаж¬
ный воздух, где он блокируется благодаря плот¬
ной структуре почвенного купола. Уровень влаж¬
ности в гнезде регулируется открыванием вход¬
ных отверстий. Строя и поддерживая гнездо,
муравьи существенно изменяют структуру и
гранулометрический состав почвы,, сдвигая его в
сторону увеличения содержания физической глины
(табл. 4). Муравьи выносят глинистые частицы из
нижних горизонтов и укрепляют ими стенки хо¬
дов, обмазывая своими выделениями. Устраивая
гнезда в слабозакрепленных песках, они способ¬
ствуют их закреплению. Повышение содержания
физической глины имеет место и в гнездах терми¬
тов (табл. 5). Таким образом, используя глинистую
фракцию почвы, муравьи и термиты строят в оби¬
таемом горизонте систему макропор, улучшая аэ¬
рацию минерального слоя. Концентрируя пище¬
вые остатки в определенных камерах, они регу¬
лируют поток углекислоты в гнезде. В табл. 6
показаны результаты оценки воздействия рою¬
щей деятельности беспозвоночных на пористость
почвы, найденной по отношению плотности поч¬
вы к плотности твердой фазы.
Дождевые черви, создавая почвенные ходы, так¬
же разрушают почвенные капилляры и формиру¬
ют систему воздушных пор. Здесь приводятся лишь
некоторые данные, характеризующие частоту
ПОЧВОВЕДЕНИЕ № 6 1996
АДАПТИВНЫЕ СТРАТЕГИИ ОСВОЕНИЯ ЖИВОТНЫМИ ПОЧВЕННОГО ЯРУСА
719
почвенных ходов червей на разных глубинах
(табл. 7). Количество ходов учитывали на зачи¬
щенной горизонтальной поверхности разных гори¬
зонтов при снятии верхнего пласта почвы [32].
У дождевых червей, многоножек и личинок
жесткокрылых, обитающих в засушливых райо¬
нах, описаны случаи строительства специальных
камер из частиц почвы, экскрементов и слюнных
выделений, в которых происходит диапауза или
развитие ювенильных стадий. Камеры жестко¬
крылых находили на разной глубине в минераль¬
ном горизонте пустынных сероземов [3]. В неко¬
торых местообитаниях масса камер составляла до
40% массы почвы. У некоторых пластинчатоусых
жуков и филлоцерид камеры имеют двойную
стенку. Непроницаемость стенок камер для вод¬
ных паров позволяла рассматривать такие обра¬
зования как защиту от дефицита влаги в почве.
Однако возможна и иная трактовка назначения
таких зоогенных структур, в частности, сохране¬
ние большой воздушной полости, позволяющей
длительно существовать в ней относительно
крупным беспозвоночным. Пользуясь промерами
величины камер [36], мы приводим расчеты воз¬
душного пространства внутри камер некоторых
жуков: Phylloceridae до 140 см3, Lethnis - 4-8 см3,
Copris hispanus - 25 см3, Synaptus tmolus - 100 см3.
В местообитаниях с высокой численностью таких
форм строительство камер оказывает существен¬
ное влияние на структуру почвы и режим аэрации
ее минеральных горизонтов. При этом в камерах
с двойной стенкой влажность воздуха поддержи¬
вается на уровне около 100% в течение длитель¬
ного времени.
Таким образом, переходя к жизни во влагообес¬
печенных горизонтах почвы, членистоногие живот¬
ные активно формируют в них условия воздушного
Дыхания, приближающиеся к наземным. В про¬
цессе эволюции у них сложились этолого-физио-
логические и морфологические адаптации, обес¬
печивающие высокий уровень газообмена в ми¬
неральной толще почвы. Переход в подземный
ярус для активно передвигающихся представите¬
лей мезофауны был неизбежно связан с много¬
кратным увеличением уровня энергообмена, не¬
обходимого для рыхления и рытья. Это вызвало
также существенные морфологические перестрой¬
ки мускулатуры и хитиновых структур с образова¬
нием специальных органов рытья (модифицирован¬
ные конечности, верхние челюсти, передняя часть
головной капсулы) и опорных органов на заднем
конце тела [5, 21]. Эти структуры рассматрива¬
ются как апоморфные, сформированные уже
в наземных условиях.
В то же время дождевые черви способны к нор¬
мальному газообмену в условиях частичной гипо¬
ксии, создающейся в минеральном горизонте.
Сродство гемоглобина к кислороду существен-
1,0 различается у представителей разных эколо¬
гических группировок. У почвенно-подстилочных
Таблица 3. Глубина и объем ходов муравьев в пустын¬
ных экосистемах Туркмении [10]
Вид
Глубина
гнезда, см
Суммарный
объем ходов, см3
Monomorium barbatulum
65-115
11-20
Tetramorium schneideri
30-37
83-279
Cataglyphis aenescens
78
787
Messor aralocaspius
170-330
7764-14075
Camponotus xerxes
240-390
7279-40560
Таблица 4. Содержание физической глины (%) в почве
гнезда и в окружающей почве [10]
Глубина,
см
Cataglyphis setipes
Messor variabilis
Гнездо
Контроль
Гнездо
Контроль
0-10
57
54
28
10
10-20
48
36
40
10
20-30
60
50
29
9
30-40
55
52
20
8
40-50
61
50
18
22
50-60
55
51
20
20
Таблица 5. Влияние роющей деятельности термитов
на гранулометрический состав почвы (по [3,8,19])
Вид термитов
Содержание физической глины, %
Гнездо
Окружающая почва
Acanthoteimes
turkestanicus
20.8
9.4
A. ahngerianus
37.1
15.3
Тоже
36.8
32.3
Таблица б. Влияние роющих беспозвоночных на порис¬
тость почвы (%) (по [32])
Группа животных
Плотность/плотность
твердой фазы
Гнезда, скоп¬
ления ходов
Контроль
Термиты:
Acanthotermes ahngerianus
56.0
52.0
A. turcestanicus
57.8
54.3
Дождевые черви:
пастбище (Австралия)
44.9
21.5
пастбище (Франция)
48.0
-
ПОЧВОВЕДЕНИЕ № 6 1996
720
СТРИГАНОВА
Таблица 7. Частота ходов дождевых червей на разной
глубине в почвах агроценозов[32]
Местообитание
Глубина, см
Число ходов червей
Пастбище
Поверхность
800
Фруктовый сад
10
3-25
50
25-33
Пашня
7
200-600
2
27-117
60
348-363
Lumbricus Р50 составляет около 8 см рт. ст., а у поч¬
венных Aporrectodea - всего 0.7-6 см рт. ст. Это го¬
ворит о наличии физиологических адаптаций к ды¬
ханию при пониженном давлении кислорода.
Пашенный червь A. caliginosa способен поддер¬
живать нормальный уровень стандартного обме¬
на при снижении парциального давления кисло¬
рода, характерного для минерального горизонта
почвы [2].
Способность к дыханию в условиях частичной
гипоксии характерна для водных полихет, зары¬
вающихся в грунт (Arenicola) и олигохет, обитаю¬
щих в донном иле (Tubificidae). Таким образом,
при освоении минерального горизонта почвы,
дождевые черви использовали стратегию водных
близкородственных групп, зарывающихся в ми¬
неральный грунт и делающих в нем ходы. Воз¬
можность постоянного пребывания в выровнен¬
ных гидротермических условиях глубоких слоев
почвы определяется наличием гемоглобина, поз¬
воляющего дышать воздухом с повышенным пар¬
циальным давлением углекислоты.
ЗАКЛЮЧЕНИЕ
Освоение почвы животными как яруса обита¬
ния в процессе эволюции шло очевидно разными
путями: (1) непосредственным переходом из ин¬
терстициальных местообитаний прибрежной зо¬
ны в микрополостй, заполненные водой (эуми-
крофауна); (2) заселением глубоких горизонтов
с поверхности путем создания системы аэриро¬
ванных макрополостей (микро- и мезофауна).
Наиболее древние ходы животных обнаружены
в палеопочвах верхнего ордовика еще до появле¬
ния на суше сосудистых растений [17]. Высказано
предположение о возможности обитания первых
наземных животных в водорослевых корках на
поверхности почвы. Колонизация суши низшими
растениями началась в середине ордовика. В ран¬
несилурийских экскрементах беспозвоночных
найдены гифы грибов [16, 17]. Это говорит о воз¬
можности использования первыми наземными
животными низших растений в качестве ресурс¬
ной базы, а также о том, что их уход в почву не
был прямо связан с появлением там корней. При
первичном почвообразовании на рыхлых продук¬
тах выветривания в настоящее время колониза¬
ция почвы беспозвоночными начинается также
с поверхности. Пионерные формы как правило
связаны с почвенными водорослями и лишайни¬
ками [35].
Появление воздушного дыхания стимулирова¬
лось недостатком кислорода в мелких заболочен¬
ных водоемах в условиях высокой температуры в
позволяло резко повысить уровень энергообмена
и компенсировать затраты на преодоление сил
гравитации в наземной среде. Высокая влажность
приземного слоя воздуха и поверхности грунта
способствовали развитию сухопутной низшей
растительности и сапротрофной микрофлоры,
утилизирующей растительные остатки. Водорос¬
ли сохраняют большой запас влаги и защищают
поверхность почвы от высыхания. Определен¬
ную роль играли очевидно и береговые выбросы
водной растительности. Возможно предполо¬
жить, что возможность существования беспозво¬
ночных на поверхности грунта обусловливалась
достаточной влажностью климата, аккумуляцией
первичной продукции, растительных остатков и
сапротрофной микрофлоры на поверхности поч¬
вы, а также отсутствием хищников.
Развитие трахейного дыхания, позволившее на¬
земным членистоногим резко повысить уровень
энергообмена, сопровождалось широкой адаптив¬
ной радиацией и освоением разнообразных сухо¬
путных местообитаний, в том числе и почвенного
яруса. Уход в минеральный горизонт почвы, оче¬
видно, стимулировался, с одной стороны, незащи¬
щенностью животных от пересыхания в условиях
резких перепадов влажности, а с другой - появлени¬
ем хищников (стегоцефалы, рептилии). Шмальгау-
зен [24,25] рассматривает биотические отношения,
и особенно пресс хищничества, как важные фак¬
торы, определяющие темпы и направление эво¬
люции отдельных групп. Появление насекомояд¬
ных подвижных позвоночных поставило беспо¬
звоночных в положение жертв. Это привело к
последующему измельчанию размеров и перехо¬
ду многих групп к скрытой жизни в почве и расти¬
тельных тканях. Освоение новой среды обеспечи¬
ло сухопутным беспозвоночным возможность
прогрессивной эволюции, сопровождавшейся ак¬
тивной дивергенцией форм и развитием разнооб¬
разных алломорфных приспособлений к пита¬
нию, локомоции и ориентации в почве.
Таким образом, адаптации аэробионтных жи¬
вотных к дыханию в почве позволяют предполо¬
жить, что этот ярус заселялся не из воды, а с по¬
верхности суши. Освоение минерального гори¬
зонта почвы наземными формами шло путем (1)
приспособления к дыханию в условиях частичной
гипоксии и (2) создания системы аэрации мине*
рального горизонта. В то же время при уходе
в почву животным не требовались специальные
адаптации для удержания влаги в теле. Наоборот»
при выходе в надземный ярус основные адаптаций
животных были сосредоточены на сохранений
ПОЧВОВЕДЕНИЕ № 6 1996
АДАПТИВНЫЕ СТРАТЕГИИ ОСВОЕНИЯ ЖИВОТНЫМИ ПОЧВЕННОГО ЯРУСА
721
влаги и формировании непроницаемых покровов.
Это обусловило и разные направления адаптиоге-
неза почвенных и напочвенных форм.
СПИСОК ЛИТЕРАТУРЫ
1. Бродский А.Я. Protozoa почвы и их роль в почвен¬
ных процессах // Бюл. Среднеаз. гос. ун-та. 1935.
Вып. 20. С. 3-19.
2. Бызова Ю.Б. Дыхание почвенных беспозвоночных //
Экология почвенных беспозвоночных М.: Наука,
1973. С. 3-39.
3. Валиахмедов Б.В. Роль почвенных беспозвоноч¬
ных в генезисе и плодородии основных типов почв
Таджикистана // Почвенная фауна и почвенное
плодородие. М.: Наука, 1987. С. 510-511.
4. Гиляров М.С. Почвенная фауна буроземов орехо-
во-плодовых лесов Ферганского хребта и ее значе¬
ние для диагностики почв // Вестн. МГУ. 1947. № 1.
С. 97-102.
5. Гиляров М.С. Особенности почвы как среды оби¬
тания и ее роль в эволюции насекомых. М.: Изд-во
АН СССР, 1949.
6. Гиляров М.С. Закономерности приспособления
членистоногих к жизни на суше. М.: Наука, 1970.
7. Гиляров М.С. Условия обитания беспозвоночных
разных размерных групп в почве // Методы поч¬
венно-зоологических исследований. М.: Наука,
1975. С. 7-11.
8. Димо НА. Мокрицы и их роль в почвообразовании
пустынь // Почвоведение. 1945. № 2. С. 115-121.
9. Длусский Г.М. Муравьи пустынь. М.: Наука, 1981.
10. Длусский Г.М., Союнов О.С., Забелин С.И. Мура¬
вьи Туркменистана. Ашхабад: Ылым, 1989.
11. Коржуев П.А. Гемоглобин. М.: Наука, 1964.
12. Лащак ТА. Материалы к биологии пустынной мо¬
крицы Hemilepistus cristatus B.-L. // Изв. АН
ТуркмССР. 1952. Т. 5. С. 52-57.
13. Лащак ТА. Мокрицы подрода Hemilepistus как
почвообразователи // Уч. зап. Туркм. ун-та. 1954.
Т. 1.С. 108-118.
14. ЛепинисА.К., Гельцер Ю.И., Чибисова О.И., Гепт-
нер В.А. Определитель Protozoa почв Европейской
части СССР. Вильнюс: Минтис, 1973. 171 с.
15. Мацкевин В.Б. Цит. по: А.А. Роде. Почвоведение.
М.; Л.: Гослесбумиздат, 1955.
16. Мейен С.В. Первые “сухопутные” растения // При¬
рода. 1989. №5. С. 17-20.
17. Пономаренко А.Г. Наземные беспозвоночные //
Природа. 1989. № 5. С. 20-23.
18. Просер ЛБраун Ф. Сравнительная физиология
животных. М.: Мир, 1967.
19. Союнов О.С. Комплексы насекомых северных Ка¬
ракумов Ашхабад: Ылым, 1991.
20. Стебаева С.К. Жизненные формы ногохвосток
(Collembola) // Зоол. журн. 1970. Т. 49. Вып. 10.
С.1437-1454.
21. Стриганова Б.Р. Закономерности строения орга¬
нов питания личинок жесткокрылых. М.: Наука,
1966.
22. Стриганова Б.Р., Валиахмедов Б.В. Роль пустын¬
ных мокриц в разложении растительных остатков //
Зоол. журн. 1975. Т. 54. Вып. 10. С. 372-378.
23. Тишлер В. Сельскохозяйственная экология. М.:
Колос, 1971.
24. Шмальгаузен И.И. Происхождение наземных по¬
звоночных. М.: Наука, 1964.
25. Шмальгаузен И.И. Факторы эволюции. М.: Наука,
1968.
26. Edney Е.В. Woodlice and the land habitat // Soil review.
1954. №29. P. 185-219.
27. Edwards C.A., Lofty J.R. Biology of earthworms. Lon¬
don: Chapman & Hall, 1977.
28. France R.H. Das Edaphon. Munchen: Verl. Deutsch.
Mikrobiol. Ges., 1937.
29. Herold W. Beitrage zur Anatomie und Physiologie der
Landisopoden // Zool. Jahrb. 1913. Bd. 35. Abt. Anat.
S. 457-526.
30. Jegen С. Die Bedeutung der Enchytraiden fur die Hu-
musbildung // Landw. Jahrb. Schweiz. 1920. Bd. 34.
S. 55-71.
31. Krogh A. The comparative physiology of respiratory
mechanisms. Philadelphia: Univ. Pennsylvania Press,
1941.
32. Lee K.E. Earthworms. Sydney: Acad. Press, 1985.
33. StockliA. Uber das Vorkommen der Enchytraeiden in Wie-
sen, Weiden, Ackerland und Waldboden der Schweiz //
Landw. Jahrb. Schweiz. 1957. Bd. 71 (6). S. 171-188.
34. Stout J.D., Heal O.W. Protozoa // Soil Biology (A. Burg¬
es & F. Raw Eds.). London-New York: Acad. Press,
1967. P. 149-194.
35. Striganova B. Effect of temperature on the feeding activ¬
ity of Sarmatiulus kessleri (Diplopoda) // Oikos. 1972.
V. 23. №2. P. 197-199.
36. Valiachmedov B. Spuren der Lebenstatigkeit von Boden-
virbellosen als diagnostische Merkmale fur Subtypen
von Grauen Wustenboden in Tadshikistan // Pedobiolo-
gia. 1977. Bd. 17. H. 2. S. 60-69.
37. Wallwork JA. Acari // Soil Biology (A. Burges &
F. Raw Eds.). London-New York: Acad. Press, 1967.
P. 363-396.
Adaptive Strategies of Animal Populations to Soil Layer
B. R. Striganova
Heterogeneity of the soil stratum is considered as a determining factor of taxonomic and ecological diversity of an¬
imal populations in the course of soil evolution. A comprehensive analysis reveals distinctions between spatial nich¬
es for different invertibrates in terms of their peculiar respiration. Under discussion are three mechanisms of gas-
exchange on the example of soil animals. Both etological and ecological adaptations of large-sized inhabitants to
the formation and maintenance of available air space in the soil mineral mass is also discussed. Macroporosity ap¬
pears to be an indispensable condition for aerobiont animals in deeper horizons of the soil profile. The development
of soil stratum by terrestrial forms from the soil surface is substantiated as a hypothesis proposed.
3 почвоведение №6 1996
ПОЧВОВЕДЕНИЕ, 1996, № 6, с. 722-727
ПОЧВА
И БИОРАЗНООБРАЗИЕ
УДК 631.46
СТРУКТУРА ПОЧВЕННОГО ПОКРОВА
И РАЗНООБРАЗИЕ ЛЕСНЫХ ФИТОЦЕНОЗОВ
© 1996 г. JI. О. Карпачевский
Почвенный факультет МГУ им. М.В. Ломоносова
Поступила в редакцию 15.02.96 г.
Формирование современных почв определяется не столько почвообразующими породами, или ли¬
тогенетическими факторами, сколько почвами предыдущих БГЦ, или экосистем. Можно выделить
два основных ряда в формировании почв: элювиальный, или нормальный по Докучаеву, и наносной,
анормальный. В первом случае разнообразие почвенного покрова заметно ниже, чем во втором,
что, возможно, влияет на биологическое разнообразие как растений, так и педофауны.
Почва как природное тело обладает опреде¬
ленной степенью разнообразия, что учитывается
многочисленными ее классификациями. Связь
типов почв и растительных группировок на высо¬
ком таксономическом уровне - известный факт.
И хотя некоторые исследователи сомневаются
в зональности почв, справедливо отмечая, что
в пределах одной формации и даже одного биогео¬
ценоза (БГЦ) можно встретить несколько типов
почв, но и они признают зональность процессов
почвообразования (в понимании П.С. Коссовича
и К.Д. Глинки) и корреляции их с растительными
зонами.
Варьирование почв в нижних таксонах органи¬
зации биосферы связано с пестротой почвообра¬
зующих пород, микроклиматом и воздействием
живых организмов. В первую очередь следует
выделить роль неоднородного состава почвооб¬
разующих пород. Кодебания в валовом химичес¬
ком составе, отражающем минералогический и
гранулометрический состав, даже близко залегаю¬
щих почв, составляют 20-100% и больше (табл. 1).
При этом отмечается, что почти для всех окси¬
дов, кроме А1203, варьирование максимально
в гор. А1 и А1А2. В то же время валовой состав
ила варьирует заметно меньше не только в одной
почве, но и в группе почв (содержание Si02 в иле
разных тундровых, подзолистых, коричневых
почв, черноземов, вертисолей колеблется в пре¬
делах 50-57% от массы ила). Содержание Si02
в суглинистых почвах в целом варьирует в боль¬
шей степени - в пределах 50-80%. Значительное
варьирование валового состава в верхних слоях
почвы в пределах одного типа леса может быть
связано с воздействием живых организмов, в пер¬
вую очередь растений.
Существует много работ по влиянию растений
на почву. Доказана связь анизотропности почв
в тессере с парцеллярной структурой БГЦ. Эти
данные основаны в большинстве случаев на ана¬
лизе образцов почв, взятых под разными ассоци¬
ациями растений. Меньше работ с анализом вли¬
яния конкретных растений. При этом основная
часть этих исследований посвящена воздействию
деревьев на почвы и лишь отдельные работы -
травяным растениям [3]. Исследователи устано¬
вили, что для травяных растений в лесу при мощ¬
ном влиянии деревьев-эдификаторов обнаружи¬
вается варьирование свойств верхних почвенных
горизонтов, связанное с локальным воздействием
отдельных растений (табл. 2-4).
Не менее интенсивно локальное влияние на
свойства почвы животных и микроорганизмов.
Формирующаяся структура БГЦ определяется
биогеоценотическими полями конкретных эди-
фикаторов (в лесу - деревьями, кустами, в травя¬
ных ценозах - куртинами трав одного вида или
относительно постоянными микроассоциациями,
парцеллами). На эту, в общем, закономерную
структуру БГЦ и парцеллы накладывается влия¬
ние случайных факторов, приводящих к общей
стохастичности экосистем. Состав парцелл по¬
стоянен, инвариантен для данного типа БГЦ, но
реальное расположение парцелл, а внутри них
распределение отдельных растений - вероятност¬
но. Оно определяется историей БГЦ, микрорель¬
ефом, погодными условиями з период прораста¬
ния семян растений, случайностью прохода таких
животных, как кабан, лось, олень, медведь и пр.
Ветровал с созданной им западиной и возвышени¬
ем может обновить отдельные популяции лесных
видов, почвенный профиль и послужить основой
для формирования самых разных, но свойствен¬
ных данному БГЦ, парцелл.
Влияние почвенных условий на биоценоз, его
разнообразие в первую очередь определяют эко¬
логические функции почв [1]. Они влияют на хи¬
мические и биохимические процессы в экосисте¬
мах, физические ее свойства, в том числе формы
корневых систем, их мощность, скорость диффу¬
зии питательных веществ и пр.
722
СТРУКТУРА ПОЧВЕННОГО ПОКРОВА 723
Таблица 1. Содержание различных компонентов в дерново-подзолистых почвах двух типов ельников, % на про¬
каленную навеску
Горизонт
М
т
а
V, %
Горизонт
М
т
а
V,%
Si02
к2о
А1
75.28
3.37
5.73
7.6
А1
2.81
0.23
0.41
14.7
А1А2
79.94
0.45
1.01
1.3
А1А2
2.78
0.05
0.12
2.8
А2
81.19
1.01
2.03
2.5
А2
2.90
0.12
0.24
8.3
А2В
79.01
1.74
3.99
5.2
А2В
2.86
0.08
0.19
6.52
В1
74.44
0.59
1.03
2.7
В1
2.97
0.03
0.05
1.6
В2
77.62
1.64
3.28
4.2
В2
2.74
0.11
0.22
8.1
Fe203
СаО
А1
3.08
0.68
1.18
35.2
А1
0.74
0.11
0.19
25.7
А1А2
2.45
0.45
1.01
41.6
А1А2
0.50
0.06
0.13
26.0
А2
3.53
0.32
0.65
18.4
А2
0.55
0.03
0.05
9.1
А2В
3.93
0.25
0.56
14.2
А2В
0.59
0.03
0.07
11.9
В1
4.49
0.07
0.14
3.1
В1
0.56
0.03
0.05
8.9
В2
5.25
0.15
0.31
6.9
В2
0.62
‘ 0.025
0.05
8.1
А1203
MgO
А1
9.83
0.25
0.45
4.6
А1
0.45
0.18
0.81
68.9
А1А2
9.22
0.17
0.38
4.1
А1А2
0.35
0.012
0.024
6.9
А2
8.43
1.22
2.45
29.1
А2
0.32
0.08
0.14
43.8
А2В
11.63
0.74
1.65
14.2
А2В
0.48
0.04
0.09
18.8
В1
12.62
0.23
0.40
3.2
В1
0.53
0.05
0.09
17.0
В2
9.89
0.68
1.26
12.7
В2
0.52
0.019
0.039
7.71
Таблица 2. Плотность и поры разного диаметра в гор. А1 дерново-подзолистой почвы под разными растениями,
% общего объема пор
Растение
Плотность, г/см3
Порис¬
тость, %
Диаметр пор, мм
почвы
твердой
фазы
>0.3
0.3-0.15
0.15-0.1
0.1-0.06
0.06-0.04
0.04-0.03
Хвощ лесной
0.67
2.44
72.4
11.3
4.1
2.8
2.9
3.3
2.7
Живучка
0.63
2.47
74.5
6.5
4.7
3.3
2.8
2.6
2.0
Сныть
0.55
2.43
77.4
9.2
3.8
1.8
2.4
1.9
-
Кислица
0.68
2.39
71.7
7.9
3.2
2.7
3.9
3.8
3.3
Копытень
0.71
2.52
71.8
18.1
2.6
2.1
2.3
2.4
2.9
Папоротник мужской
0.55
2.36
76.6
12.4
2.1
1.5
2.3
1.9
3.2
Костяника
0.67
2.41
72.3
10.0
4.5
4.3
2.6
2.9
2.9
Осока волосистая
0.87
2.40
63.9
5.4
3.9
3.1
3.6
3.6
2.9
Щитовник Линнея
0.74
2.40
69.0
13.5
4.1
4.1
3.2
3.2
3.3
Установленная связь почв и растений (или
приуроченность растений к почвам с определен¬
ными свойствами), даже если эта связь вероятно¬
стная, все равно требует ответа на сакраменталь¬
ный вопрос: растение поселилось на почве с опре¬
деленными свойствами или почва изменилась в
необходимом для растения направлении под воз¬
действием самого растения? В общей форме веро¬
ятен следующий ответ, который вытекает из ана¬
лиза связи между почвой и растением (табл. 2).
Достаточно небольшого сектора почвы с подхо¬
дящими для данного растения свойствами, чтобы
оно смогло успешно развиваться и изменять ос¬
ПОЧВОВЕДЕНИЕ № 6 1996
тальную часть занятой им площади в Соответст¬
вующем направлении. Если же растение поселя¬
ется на почве, полностью соответствующей ему
по свойствам, то оно поддерживает эти свойства
на необходимом уровне (pH, гумус, плотность, об¬
менные основания).
Литогенная неоднородность может проявиться
в пределах одного БГЦ, но в лесных экосистемах
влияние геологической породы может затушевы¬
ваться тем, что дерево лишь частично использует
другую породу или тем, что глубина залегания
литогенно отличающегося слоя (например, кар¬
бонатной морены) может ограничивать влияние
3*
724
КАРПАЧЕВСКИЙ
Схема основных функций почвы как природного тела
Почва
Функции: регуляция химического
состава других тел
Объекты: гидросфера, другие
твердые биокосные тела,
атмосфера
трансформация других тел
литосфера,
гидросфера
атмосфера
живые организмы
регуляция циклов
веществ в биосфере
биологический
круговорот,
геологический круговорот
Форма воздействия: химические (трансформация природных веществ, синтез новых веществ, включая
комплексы и смеси, каталитическая активность);
биохимические (деструкция органических веществ, воспроизводство гумуса);
биологические (накопление биологических и органических стимуляторов и ингибиторов, среда обита¬
ния живых организмов, источник питательных веществ, стимулятор и ингибитор живых организмов,
выделение газов в атмосферу);
физико-химические (сорбция и десорбция веществ, диффузия ионов, окислительно-восстановительный
режим);
физические (влияние на остаточные электромагнитные поля, формирование структуры экосистемы).
его на дерево. Если слой лежит слишком глубоко
или слишком близко к поверхности, то он пере¬
стает влиять на пестроту почвенного покрова
в заметной степени. Визуальные описания лесов
обычно не выявляют влияние таких вариантов на
почвы и растения. Так, в суглинистых дерново-
подзолистых почвах на покровных суглинках
(Малинки, Московская обл.) на глубине 40 см
встречаются в виде “жил” треугольные в сечении
включения красно-бурой опесчаненной суглинис¬
той морены. С поверхности почвы эти включения
себя ничем не обнаруживают, за исключением
одной детали: к местам с залеганием этих жил с
большей вероятностью приурочены вывалы де¬
ревьев. Варьирование свойств верхних горизон¬
тов над покровным суглинком и красно-бурой мо¬
реной не различается.
Таблица 3. Специфические свойства гор. А1 дерново-подзолистой почвы под разными растениями
Растение
Плотность почвы, г/см3
Водопро¬
ницаемость,
мм/мин
pH почвы
Обменные катионы,
мг-экв/100 г
пределы
средняя
Са2+
Mg2+
Костяника
0.5-1.1
0.80
0-75
4.6-4.9
3-5
1-2
Кислица
0.5-1.1
0.71
25-200
4.3-4.6
1-3
1-2
Хвощ лесной
0.7-1.1
0.85
25-150
3.4-5.5
3-5
6-7
Копытень
0.7-0.9
0.88
0-100
4.6-5.5
7-13
1
Осока волосистая
0.4-1.1
0.65
75-150
4.0-4.6
5-7
0-2
Щитовник Линнея
0.3-0.8
0.61
0-150
4.6-5.2
1-3
-
Папоротник мужской
0.5-0.9
0.70
0-150
4.0-:.2
3-5
4-5
Ландыш
0.3-0.5
0.4
-
-
11-15
3^
Живучка
0.6-0.7
0.65
25-50
4.9-5.2
Не опр.
Сныть
0.4-0.5
0.45
25
4.0^.3
»
Мертвопокровная парцелла
0.3-0.5
0.40
125-150
4.0-4.3
»
Таблица 4. Динамика значений pH почвы в гор. А1 под разными растениями
Растение
13.06
20.06
27.06
12.07
21.07
31.07
Осока
волосистая
4.4
5.2
4.9
4.3
4.3
4.1
Папоротник мужской
4.9
4.5 •
4.6
4.3
4.1
4.1
Щитовник
Линнея
4.8
4.9
5.2
5.0
5.1
4.9
ПОЧВОВЕДЕНИЕ № 6 1996
СТРУКТУРА ПОЧВЕННОГО ПОКРОВА
725
Гранулометрический состав почв, который в
основном является литогенетическим признаком
почв, варьирует очень сильно (табл. 5). Меньше
варьируют содержание гумуса и плотность.
Влажность почв и ее колебания в летний период
определяются растениями, потреблением ими
воды из почвы. Это подтверждается данными по
изменению потенциала почвенной влаги: в верх¬
нем слое 0-10 см на 21 июля он соответствовал
-1 ... -12 атм [3].
О роли растений и сопутствующих им микро¬
организмов, животных, грибов говорит выделе¬
ние С02 из почвы (чисто ценогенетическое свой¬
ство почвы). В этом случае именно современная
парцелла определяет активность биоценоза, ко¬
торую можно оценить по выделению углекисло¬
го газа.
Разнообразие свойств почв должно проявиться
также в формировании новых почв, когда неод¬
нородность палингенетических свойств, унасле¬
дованная от предыдущего БГЦ, проявится в поч¬
ве и структуре нового БГЦ. В связи с этим стоит
вспомнить теорию Личкова [5], что все современ¬
ные почвы, включая красноземы и ферраллит-
ные, сформировались не из почвообразующих
пород, а из предыдущей почвы. Если не происхо¬
дит катастрофического нарушения почвенного
покрова (эрозия, погребение), то почвы нового
БГЦ формируются из почв предыдущего БГЦ,
“старые” почвы служат почвообразующими поро¬
дами для новых почв. При этом в новых почвах со¬
храняется множество палингенетических свойств,
унаследованных от предыдущих почв. Таким обра¬
зом, современные почвы несут на себе неодно¬
родность, унаследованную от почвообразующей
породы (литогенетическая), от предыдущей поч¬
вы (палингенетическая) и от современного БГЦ
(ценогенетическая).
Итак, сегодня возможны два случая формирова¬
ния почв: непосредственно из почвообразующей
породы и из почвы, сформировавшейся раньше.
В первом случае формируются вначале примитив¬
ные почвы и упрощенные по составу растений и
животных биоценоза. Почвы обычно слабо диф¬
ференцированы с одним четким горизонтом -
гумусовым. Они в принципе интразональны и
различаются в основном валовым составом, унас¬
ледованным от породы, величиной и pH. В амери¬
канской классификации примитивные почвы всех
Таблица 5. Содержание (%) гранулометрических фракций, мм, и коэффициент вариации (V, %) в некоторых почвах
Почва
Гори¬
зонт
1-0.05
0.05-0.005
0.05-0.01
<0.005
<0.01
Повторность об¬
разцов для оценки
содержания ила
%
V
%
V
%
V
%
V
%
V
Дерново-подзолистая
А1
15
21
63
8
51
11
22
16
34
4
18
суглинистая
А2
8
7
74
5
60
7
19
9
33
2
9
А2В
9
23
60
14
49
14
34
15
45
5
11
Чернозем типичный
А
11
20
58
8
44
10
30
10
45
6
6
тяжелосуглинистый, лес
А1
11
37
55
5
39
5
33
8
49
3
6
В
10
8
53
2
38
3
36
3
50
2
4
ВС
10
5
52
2
36
4
36
3
52
2
4
Чернозем типичный,
А1
6
18
57
12
45
12
37
7
49
2
Нет
поле
В
6
84
60
9
47
11
37
9
48
7
»
ВС
5
38
57
3
48
2
38
8
47
3
• »
Темно-каштановая
А
16
14
71
8
54
5
13
19
30
10
13
В
12
7
51
12
35
9
37
13
54
5
11
с
11
16
45
9
30
7
44
7
58
3
5
Серозем
А
12
14
39
9
33
7
39
9
9
13
16
ААВ
17
18
39
10
27
16
33
16
11
8
9
АВВ
17
13
38
7
25
6
32
10
12
11
14
Солонец
А
17
19
66
5
51
6
17
22
32
11
21
В1
15
14
54
12
41
14
31
23
45
15
23
ВС
13
15
61
21
47
23
27
46
41
27
64
Светло-каштановая
А1
19
9
66
6
50
9
15
20
30
14
18
АВ
15
16
58
12
42
13
27
27
43
11
30
ВС
15
18
58
12
43
25
27
22
42
15
21
Темноцветная западины
А1
19
8
67
3
52
7
15
15
29
12
11
(лугово-каштановая)
АВ
17
9
60
10
46
12
22
25
38
18
26
ВС
16
14
55
7
42
11
29
19
42
14
16
ПОЧВОВЕДЕНИЕ № 6 1996
726
КАРПАЧЕВСКИЙ
т 4
□ 5
Два пути эволюции почв: / - при поступлении матери¬
ала сверху; II-в условиях сноса. / - гумусовый гори¬
зонт; 2 - погребенный гумусовый гор. АВ; 3 - гор. В;
4 - гор. ВС; 5 - гор. С.
зон объединены в азональный порядок Entisols.
Часть песчаных почв также относится к этой
группе. В них почвообразование привело лишь
к формированию гумусового и дернового горизон¬
тов. Смагин [6] приводит пример эволюции почв
сосняков на свежих аллювиальных отложениях.
На них образуются дерновые маломощные поч¬
вы с очень слабо выраженным гор. А2 и форми¬
руется простой сосняк. Второе поколение сосня¬
ка вырастает уже на сформированных почвах,
превращая их в дерново-подзолистые полнопро¬
фильные. При этом сосняк становится сложным,
в его составе встречаются дубы, лещина, разно¬
травье с более богатым набором видов. Анало¬
гичную картину формирования почв приводит
Зоин для Камчатки [2]. На вулканическом аллю¬
вии р. Пахчи сначала появляются кусты кедрово¬
го стланика и отдельные деревья лиственницы.
Под ними в основном образуется лесная подстил¬
ка. Второе поколение БГЦ на этих почвах пред¬
ставляет собой уже лиственничник брусничный с
грубогумусными почвами. Третье поколение ли¬
ственничника представляет собой уже багульни¬
ковый тип с брусничными парцеллами и с ксеро-
морфными грубогумусными почвами с более
мощным гор. А1 и хорошо выраженным гор. В.
В почвах возрастают емкость поглощения, содер¬
жание гумуса, дифференциация профиля.
Предложенная схема не охватила еще один ас¬
пект, выдвинутый в свое время В.В. Докучаевым:
формирование нормальных и анормальных почв.
К первым Докучаев отнес почвы, развивающиеся
на элювии пород, ко вторым - наносные. Сам
принцип формирования анормальных почв хоро¬
шо освещен Неусгруевым [5], но, к сожалению,
не был разработан более детально. Две стороны
этого процесса приводят к формированию двух
рядов почв. Эрозия и дефляция сносят материал с
водоразделов и склонов, выравнивая рельеф и об¬
разуя эродированные почвы, а иногда просто обна¬
жая почвообразующие или даже подстилающее
породы. В этом случае почвообразование идет щ
“обезглавленной” почве (рисунок). В.П. Лидов счи¬
тал, что выделенные в Смоленской обл. бурые
лесные почвы на полях с сельскохозяйственной
обработкой представляют собой эродированные
дерново-подзолистые, у которых снесены гор. А1
и А2 (устное сообщение). Обработка этих почв,
внесение удобрений, перегнивание жнивья пра¬
вело к образованию специфического верхнего
слоя - А пахотного, аналога гор. А1 природных
почв. Вторая сторона процесса - привнос эроди¬
рованного материала в другие БГЦ. В этом слу¬
чае, если привнос достаточно равномерный и уме¬
ренный, то он не нарушает существующего БГЦ.
Приносимый мелкозем включается в подстилку и
постепенно - в гор. А1 существующей почвы.
Гумусовый слой наращивается кверху, и в резуль¬
тате формируются глубокогумусные почвы.
Именно поэтому во всех почвах с глубиной воз¬
раст гумуса увеличивается. В этом плане чернозе¬
мы, бурые лесные, пойменные луговые и пере-
гнойно-дерновые почвы, как и все другие глубо¬
когумусные почвы, формируются одинаково:
в результате привноса материала. Разница в том,
в каких климатических условиях это происходит,
какими БГЦ он задерживается и каков способ пе¬
реноса - ветром или водой. Именно эти факторы
влияют на скорость минерализации гумуса в по¬
гребенных горизонтах, его состав и содержание
в профиле. В соответствии с условиями почвооб¬
разования меняются значения pH, состав обмен¬
ных катионов и пр. Выделенные два процесса
эволюции почв - эрозия и аккумуляция - влияют
на биологическое разнообразие почв и БГЦ.
В частности, эродированные почвы обедняются
видовым составом растений, общей численнос¬
тью и видовым составом педофауны. Наносные
почвы, напротив, поддерживают высокое биоло¬
гическое разнообразие не только в результате
приноса семян растений, мелких представителей
педофауны, но и в результате постоянного обога¬
щения почв БГЦ, что создает условия для форми¬
рования более богатых типов БГЦ. Процесс об¬
разования глубокогумусных почв может быть
прерван катастрофическим приносом материала
в результате резкого усиления эрозии или извер¬
жений вулканов. В этом случае в почве отмечает¬
ся перерыв в почвообразовании, который марки¬
руется соответствующим слоем. Возвращение к
нормальной скорости процессов приноса матери¬
ала приводит к образованию на маркирующем
слое новой глубокогумусной почвы. О преобла¬
дании эрозионных или аккумулятивных процес¬
сов в истории почвы можно судить по их биологи¬
ческому разнообразию. Возможно, что разница
ПОЧВОВЕДЕНИЕ № 6 1996
СТРУКТУРА ПОЧВЕННОГО ПОКРОВА
727
в количестве педофауны черноземов и серых лес¬
ных почв Северо-Украинской и Окско-Донской
провинций связано не с различиями в бассейнах
Днепра и Дона, как это считают зоологи, а имен¬
но с разным участием процессов эрозии и аккуму¬
ляции в формировании почв этих провинций в те¬
чение голоцена: эрозия, даже очень постепенная,
приводит к уменьшению биологического разно¬
образия, а аккумуляция - к увеличению. Очевид¬
но, что предложенная схема (рисунок) представ¬
ляет собой крайние - “чистые” варианты. В при¬
роде эрозия и аккумуляция могут чередоваться на
одной и той же поверхности, поэтому история ре¬
альных почв может быть сложнее. Но остается
главный вывод: варьирование свойств почвы го¬
раздо шире, чем тех же свойств почвообразую¬
щих пород. Формирование неоднородного поч¬
венного покрова приводит к увеличению биоло¬
гического разнообразия. Эрозия уменьшает,
а аккумуляция мелкозема в почвах увеличивает
биологическое разнообразие БГЦ.
СПИСОК ЛИТЕРАТУРЫ
1. Добровольский Г.В., Никитин Е.Д. Экологичес¬
кие функции почвы. М.: Изд-во МГУ, 1986. 136 с.
2. Зонн С.В., Карпачевский JI.O., Стефин В.В. Лес¬
ные почвы Камчатки. М.: Изд-во АН СССР, 1963.
254 с.
3. Карпачевский JI.O. Пестрота почвенного покрова
в лесном биогеоценозе. М.: Изд-во МГУ, 1977.312 с.
4. Неуструев С.С. Почвы и циклы эрозии // Генезис
и география почв. М.: Наука, 1977. С. 129-141.
5. Переписка В.И. Вернадского с Б.Л. Дичковым
1918-1939. М.: Наука, 1979. 270 с.
6. Смагин А.В. Почва как результат самоорганиза¬
ции биогеоценоза // Докл. АН СССР. 1989. Т. 308.
№3. С. 729-731.
7. Структура и функционирование почвенного насе¬
ления дубрав Среднерусской лесостепи. М.: Наука,
1995. 152 с.
The Soil Cover Structure and Diversity of Forest Phytocoenoses
h. O. Karpachevskiy
Recent soil formation is determined not so much by soilfoiming rocks or lithogenetic factors as by soils of pre¬
ceding biogeocoenoses or ecosystems. Two basic ways of soil formation are proposed to distinguish: eluvial or
normal according to Dokuchaev’s statement and illuvial or abnormal. In the first case the soil cover is diversi¬
fied to a lesser extent as compared to the second one. The latter may provide better conditions for biodiversity
of plants and soil fauna.
ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВОВЕДЕНИЕ, 1996, № 6, с. 728-734
УДК 631.46:631
ПОЧВА
И БИОРАЗНООБРАЗИЕ
ВЛИЯНИЕ ДРЕНАЖА И АГРОМЕЛИОРАЦИИ
НА ПОЧВЕННУЮ БИОТУ
© 1996 г. Ф. Р. Зайдельман
Факультет почвоведения МГУ им. М.В. Ломоносова
Поступила в редакцию 0S.02.96 г.
Дренаж и агромелиоративные мероприятия оказывают различное влияние на почвенную биоту.
В минеральных почвах супераквальных ландшафтов дренаж и глубокое рыхление способствуют
увеличению численности водорослей, простейших, микроорганизмов и их проникновению в глубо¬
кие горизонты профиля. Осушение и глубокое мелиоративное рыхление на фоне дренажа резко
снижают численность и биомассу червей и насекомых. Глубокое рыхление особенно неблагоприят¬
но отражается на численности молоди дождевых червей.
В субаквальных ландшафтах осушение после устранения постоянного затопления оказывается
определяющим фактором заселения торфяных почв мезофауной и, в частности; дождевыми чер¬
вями. Показана необходимость исследования почвенной биоты с учетом степени заболоченности
почв и неоднородности (первичной и вторичной) структуры.
Мелиорация - неотъемлемый элемент земле¬
делия - оказывает глубокое влияние на гидротер¬
мический режим почв и их свойства. Следствием
их трансформации в результате орошения, дрена¬
жа и других мероприятий является резкое изме¬
нение условий местообитания растений, живот¬
ных, микроорганизмов. Сведения об эволюции
растительных ценозов (лесных и травянистых)
изучены весьма полно. Особое внимание в этом
случае привлекали данные, отражающие новые
условия для роста и развития различных сельско¬
хозяйственных культур. Менее полно раскрыта
изменчивость почвенной микро-, мезофито- и зоо¬
биоты почв, вовлеченных в современное сельско¬
хозяйственное использование.
На мелиорированных территориях почвенная
зообиота и ее изменения в результате антропоген¬
ного воздействия изучалась весьма неравномерно.
В аридных регионах влияние ирригации на почвен¬
ную фауну изучали Димо [4], Валиахмедов [2] и
другие. Они исследовали разные группы беспозво¬
ночных в орошаемых и неорошаемых сероземах
и выявили роль термитов и других беспозвоноч¬
ных в современном почвообразовании аридной
зоны. В гумидной зоне наблюдения за изменения¬
ми микрофлоры и фауны под действием мелиора¬
ции были начаты сравнительно недавно. Их ре¬
зультаты до настоящего времени остаются весь¬
ма ограниченными.
Вместе с тем очевидно, что влияние мелиора¬
ции на биоту весьма существенно и не однознач¬
но. Оно может по-разному проявляться в разных
природных условиях. Ниже предпринята попытка
оценить влияние закрытого дренажа и агромели¬
оративных мероприятий на различные группы
почвенной биоты в Нечерноземной зоне России.
Нами систематизированы материалы, получен¬
ные при изучении дренированных почв этого ре¬
гиона, выполненных под руководством автора
в 1977-1987 гг. [5, 6] в южнотаежной подзоне ев¬
ропейской территории Российской Федерации.
Цель работы заключалась в изучении влияния за¬
крытого керамического дренажа и глубокого ме¬
лиоративного рыхления на гидротермический ре¬
жим, физические и химические свойства тяжелых
болотно-подзолистых и дерново-глеевых почв.
Исследования были приурочены к почвам, обра¬
зованным на озерно-ледниковых тонкослоистых
кислых тяжелых глинах, легких лёссовидных кис¬
лых глинах и красноцветных карбонатных тяже¬
лых суглинках и глинах. Во всех случаях причи¬
ной гидроморфизма почв были поверхностные
неминерализованные намывные склоновые во¬
ды. Одновременно с работами мелиоративного и
гидрофизического цикла на этих же массивах ис¬
следовалась почвенная альгофлора, простейшие,
черви, личинки и имаго насекомых, другие орга¬
низмы почвенной биоты. Наряду с изложением
оригинальных данных, здесь рассмотрены также
литературные материалы, представляющие важ¬
ное значение при трактовке полученных данных.
В целом дренаж повышает общую биологиче¬
скую активность осушенных почв. Об этом свиде¬
тельствует рост интенсивности дыхания осушен¬
ных почв по сравнению с недренированным кон¬
тролем, резкое увеличение массы корней и
урожая. По данным Козлова [10], увеличение сте¬
пени канализации территории существенно повы¬
шало интенсивность дыхания (табл. 1) и биологи¬
ческой активности почв. Из этого наблюдения
вероятен вывод о том, что дренаж создает благо¬
приятные условия для развития почвенной биоты.
728
ВЛИЯНИЕ ДРЕНАЖА И АГРОМЕЛИОРАЦИИ НА ПОЧВЕННУЮ БИОТУ
729
Таблица 1. Влияние дренажа на интенсивность дыха¬
ния и рост корневой и наземной массы многолетних
трав. Олонецкая равнина, Карелия [10]
Средняя за
июнь-сен¬
тябрь ин¬
тенсивность
выделения
С02, кг/га ч
Масса
корней
Урожай¬
ность (сено)
Вариант
ц/га
Без осушения
Закрытый дренаж:
2.7
70
14
расстояния меж¬
ду дренами 36 м
3.5
143
44
расстояния меж¬
ду дренами 10 м
4.2
146
60
Однако такой казалось бы справедливый вывод
имеет слишком общий характер. Реальная карти¬
на происходящих изменений оказывается значи¬
тельно более сложной. Рассмотрим это на приме¬
ре суглинистых и глинистых оглеенных почв,
осушенных закрытым гончарным дренажом с
применения глубокого (до 80 см) мелиоративного
рыхления и без него. Глубокое рыхление, как по¬
казали многолетние исследования, вызывает де¬
формацию в зоне обработки всех горизонтов
почвенного профиля [6]. При этом изменяются
морфология, сложение и структура почв, улуч¬
шается их аэрация; значительный объем мелко¬
зема пахотного горизонта, обогащенного гуму¬
сом, поступает в глубокие слои профиля. В них
проникает значительная масса корней и увеличи¬
вается общая масса перегноя. Все это стимулиру¬
ет развитие альгофлоры и микроорганизмов.
В нашей программе водоросли и микрофлора бы¬
ли изучены (табл. 2) профессором Штиной совме¬
стно с Марковой и Кондаковой [11, 12]. Такие на¬
блюдения были выполнены на дерново-глеевых
почвах, образованных на пермских карбонатных
красноцветных глинах, в Котельническом районе
Кировской обл. В результате было установлено
следующее. В осушенных и рыхленых почвах аб¬
солютное количество водорослей оказалось вы¬
ше и разнообразнее по видовому составу по срав¬
нению с почвами дренированных контрольных
вариантов. На всех участках наблюдалось “цвете¬
ние” почвы, вызванное, преимущественно, сине-
зелеными водорослями. Среди доминантов - ви¬
ды рода Cylindrospermum. Максимальная чис¬
ленность и биомасса водорослей отмечена в
дренированных и рыхленых глееватых почвах
(466.5 тыс. клеток на 1 г абсолютно сухой почвы
и 0.45 мг/г биомассы водорослей). В дренирован¬
ной, но нерыхленой глееватой почве, численность
и биомасса водорослей была существенно ниже
(136 тыс. клеток и 0.205 мг/г). В недренированных
и нерыхленых почвах (природные условия, кон¬
троль) численность составила 58 тыс. клеток, био¬
масса - 0.011 мг/г. Таким образом, дренаж и рых¬
ление обусловили значительное увеличение чис¬
ленности водорослей и их биомассы по всему
профилю - до нижней границы обработки почв
рыхлителями. Установлена существенная разни¬
ца в видовом составе водорослей по профилю
рыхленых и нерыхленых почв. В рыхленых поч¬
вах дренированного участка водоросли, как и
корни высших растений, более глубоко проника¬
ют по профилю, особенно по призмам рыхления.
На дренированном участке с нерыхлеными поч¬
вами разнообразие водорослей с глубиной резко
уменьшается (в пахотном горизонте - всего 4 ви¬
да, на глубине 35-40 и 60-65 см водоросли не об¬
наружены). Одновременно на дренированном уча¬
стке с рыхлеными почвами на глубине 0-24 см
идентифицировано 24 вида водорослей (по приз¬
мам рыхления), а на глубине 35-40 и 60-65 см обна¬
ружено 14 и 14 видов соответственно. При этом
один вид водорослей проник на глубину 85-90 см
рыхленой дренированной почвы. Аналогичные
закономерности распределения альгофлоры и
влияние мелиоративных мероприятий ка абсо¬
лютное число клеток водорослей по горизонтам
было установлено на этих почвах в других райо¬
нах Кировской обл. Все это позволяет признать
обнаруженные закономерности достоверными.
Важно и то, что была установлена четкая реак¬
ция альгофлоры на гидрологический режим почв.
В тех случаях, когда глубокое рыхление дерновых
глееватых почв осуществлялось без дренажа, на¬
блюдался длительный застой влаги и вторичное
Таблица 2. Изменение общей численности, массы и видового состава водорослей в дерново-глееватых почвах на
пермских глинах под влиянием дренажа и глубокого мелиоративного рыхления
Осушение
Контроль
(без осушения)
Гончарный
дренаж
Гончарный
дренаж + рыхление
Численность, тыс. клеток/г абсолютно сухой
58.0
136.0
465.0
почвы, гор. А1
Масса, мг/г абсолютно сухой почвы, гор. А1
0.0110
0.0205
0.0447
Число видов на глубине, см:
0-24
4
Не опр.
24
35-40
0
»
14
60-65
0
»
14
ПОЧВОВЕДЕНИЕ № 6 1996
730
ЗАЙДЕЛЬМАН
Таблица 3. Изменение видового состава и численности
(тыс. клеток/г абс. сухой почвы) простейших в гор. В.1
(50-80 см) дерново-подзолистых суглинистых глеева-
тых почв под влиянием дренажа и глубокого мелиора¬
тивного рыхления
Вид простейших
Контроль
(без осу¬
шения)
Дренаж
Дренаж +
+ рыхление
Жгутиконосцы:
Oicomonas Sp.
Нет
0
193
Bodo Edax
»
57
0
Bodo Uncinatus
»
0
193
Инфузории:
Colpoda in Flata
Нет
57
58
Количество клеток
Нет
114
444
переувлажнение почв [6]. В этом варианте было
обнаружено в налетах на поверхности почвы и в
воде понижений преобладание Witschia palea и дру¬
гих видов, указывающих на повышенную влаж¬
ность. Существенно, что эти виды не были иденти¬
фицированы в дерново-глееватых дренирован¬
ных, а также дренированных и рыхленых почвах.
Следует признать, что глубокое мелиоратив¬
ное рыхление заболоченных почв на фоне дрена¬
жа значительно увеличивает ареал вертикального
распространения альгофлоры и является основной
причиной увеличения видового разнообразия во¬
дорослей. Это общее заключение, по-видимому,
справедливо для большинства или для всех мик¬
роорганизмов почвенной биоты.
Так, микробиологический анализ почвенных
образцов на наших двух стационарах (“Ивакин-
ские пашни” и “Горевский массив”) в Котельни¬
ческом и Свечинском районе Кировской обл. по¬
казал, что в осушенных рыхленых почвах наблю¬
дается более высокая численность всех основных
групп микроорганизмов по профилю [11]. Как и
для водорослей, наибольшая разница между вари¬
антами отмечалась в подпахотном горизонте, где
общая численность микроорганизмов на контро¬
ле была на порядок ниже, чем на дренированной
рыхленой почве. Наиболее высокая насыщен¬
ность микроорганизмами наблюдается по призме
рыхления.
Близкие результаты были получены и при
изучении некоторых групп простейших. Эти ис¬
следования были выполнены в западной части
Московской обл. на дерново-подзолистых глее-
ватых почвах, образованных легкими кислыми
лёссовидными глинами под многолетними трава¬
ми. Наблюдения осуществлялись на третий год
последействия дренажа (Е 20 м) и на второй после
активного глубокого рыхления. Изучение реак¬
ции жгутиконосцев и инфузорий на дренаж и об¬
работку почв глубокими активными мелиоратив¬
ными рыхлителями [3] показало, что в окульту¬
ренных почвах простейшие концентрируются
в пахотном горизонте. Иллювиальные оглеенные
горизонты дерново-подзолистых почв в условиях
естественного режима неблагоприятны для их
развития: представители жгутиконосцев и инфу¬
зорий в них отсутствуют (табл. 3). В слое 50-80 см
глееватых почв они появились только после осуше¬
ния. В осушенных оглеенных почвах увеличивается
глубина распространения корней, по ходам кото¬
рых простейшие обычно проникают из пахотно¬
го в иллювиальные горизонты. Рыхление осушен¬
ных почв способствует увеличению (в 4 раза) чис¬
ленности простейших в рассматриваемом слое
(в исходном состоянии - 0; после осушения - 144;
после глубокого рыхления осушенных почв -
444 тыс. клеток/г абсолютно сухой почвы). Реак¬
ция разных видов и родов этих животных на глу¬
бокое рыхление оказалась весьма неоднознач¬
ной. После рыхления исчез вид Bodo edax, но
появился вид Bodo uncinatus рода Oicomonas. Чис¬
ленность инфузорий не изменилась.
В наших исследованиях общую микробиологи¬
ческую активность почв на качественном уровне
оценивали по накоплению нингидринположитель-
ных веществ на полотнах. В слое 30-80 см рыхле¬
ных почв зон накопления таких веществ оказа¬
лось значительно больше, чем в нерыхленых.
Эти зоны приурочены к крупным порам, затекам
или включениям (после рыхления) гуму сиров эн¬
ного мелкозема, в которых концентрировались
корни растений. В них поселялись простейшие.
Таким образом, изучение видового состава, био¬
массы и распределения водорослей, простейших,
микробов позволяет утверждать, что дренаж и осо¬
бенно сочетание дренажа и глубокого мелиоратив¬
ного рыхления резко активизируют жизнедеятель¬
ность этих представителей почвенной биоты.
Влияние этих распространенных мелиоратив¬
ных мероприятий на мезофауну оказывается су¬
щественно иным. По наблюдениям Гельцера [3],
они вызвали принципиально иную трансформа¬
цию численности и видового состава дождевых
червей, членистоногих и крупных насекомых.
Дождевые черви на контроле (неосушенная за¬
лежь с осоково-злаковой растительностью, ис¬
пользуемая под сенокос) принадлежали к пяти ви¬
дам. Существенных различий в видовом составе
червей из почв на целинном (1, контроль), осу¬
шенном (2), осушенном и рыхленом (3) участках
не обнаружено. На всех вариантах доминируют
Nicodrilus caliginosus и Lumbrix teeiestris. В мень¬
шем количестве на залежи и в осушенной нерыхле-
ной почве встречен Eisenia nordenskiodi. После
рыхления этот вид не идентифицирован. В почве,
обработанной глубокими рыхлителями, выше чис¬
ленность Lumbricus rubellus по сравнению с двумя
другими вариантами (контроль и дренаж). Вид
Nicodrius roseus на всех вариантах эксперимента
обнаружен единично.
ПОЧВОВЕДЕНИЕ № 6 1996
ВЛИЯНИЕ ДРЕНАЖА И АГРОМЕЛИОРАЦИИ НА ПОЧВЕННУЮ БИОТУ 731
Таблица 4. Изменение численности и динамика люмбрицид (экз/м3) в дерново-подзолистых глееватых тяжело¬
суглинистых почвах под влиянием дренажа и глубокого мелиоративного рыхления. Московская область
Структура сообщества
1979 г.
1980 г.
1981 г.
09
05
06
07
09
05
Залежь
Половозрелые
Не опр.
52
30
50
114
80
Неполовозрелые
»
213
180
126
240
320
Общее количество
»
265
210
176
354
400
Дренаж (без рыхления)
Половозрелые
54
66
64
58
Не опр.
30
Неполовозрелые
114
138
70
100
»
156
Общее количество
168
204
134
158
»
186
#
Дренаж + глубокое рыхление
Половозрелые
5
10
16
22
40
20
Неполовозрелые
19
16
10
72
72
40
Общее количество
24
26
26
94
112
60
Дождевые черви - наиболее многочисленные
представители почвенной мезофауны. На залежи
без дренажа и глубокого рыхления их общая чис¬
ленность составила 250-400 экз./м2. Зоомасса
червей была равна 5.5-9.5 ц/га сырого органичес¬
кого вещества. В почвах, вовлекаемых в актив¬
ное земледелие, изменение численности люмбри¬
цид происходило в два этапа. После осушения и
распашки глееватых почв общее количество
дождевых червей уменьшилось на 30-50% и бо¬
лее, а их общая биомасса - до 3.5-4.0 ц/га. Сниже¬
ние плотности населения червей на осушенном
участке произошло, в основном, за счет уменьше¬
ния количества молоди. В осушенных не рыхле-
ных почвах количество взрослых особей в мень¬
шей степени подвержено изменениям.
Глубокое рыхление осушенной почвы резко
снизило численность люмбрицид (табл. 4). В сен¬
тябре 1979 и в мае 1980 гг. общее количество поло¬
возрелых особей в рыхленой почве по сравнению с
осушенной нерыхленой в первые годы последейст¬
вия рыхления осушенных почв оказалось ниже
в 6-8 раз, а с залежью (неосушенной и нерыхле¬
ной почвой) - в 12-18 раз. При этом, однако, мож¬
но проследить общую тенденцию восстановления
популяции люмбрицид к концу второго-третьего
года последействия. Это неметившееся увеличе¬
ние, однако, не ликвидирует резкое различие в
численности дождевых червей на трех вариантах
эксперимента даже в конце третьего года. Зоо¬
масса дождевых червей в рыхленых и осушенных
почвах оказалась минимальной и не превышала
1.2 ц/га сырого органического вещества.
К столь резкому уменьшению численности
люмбрицид в нерыхленых почвах приводит дей¬
ствие нескольких факторов: агротехнические ме¬
роприятия по подготовке поверхности почвы к
ПОЧВОВЕДЕНИЕ № 6 1996
глубокому рыхлению, движение тракторов по
вспаханному полю, воздействие на поверхност¬
ные горизонты почвы мощностью до 80 см ножа
и лемеха рыхлителя. Люмбрициды попадают в
новые экологические условия, основная особен¬
ность которых заключается в появлении разрых¬
ленного слоя почвы до глубины 80 см, в котором
нарушаются природные высокая плотность, низ¬
кая пористость и сеть ходов почвенных живот¬
ных. Вертикальное передвижение дождевых чер¬
вей в таком слое затруднено. Норники из глуби¬
ны почвы не могут подняться на поверхность за
пищей, среднеярусные черви лишены возможно¬
сти мигрировать в глубь почвы при возникнове¬
нии неблагоприятных условий. В этих условиях
вероятна гибель люмбрицид.
В дальнейшем при уплотнении разрыхленной
толщи до 1.2-1.3 г/см3 и выше численность дож¬
девых червей возрастала. Тем не менее в сентяб¬
ре 1980 и в мае 1981 гг., т.е. на третий-четвертый
год после рыхления при плотности слоя 20-80 см
1.46 г/смэ численность люмбрицид в рыхленой
почве все еще оставалась в 3-7 раз ниже, чем в
осушенной нерыхленой почве и на залежи. В этой
связи важно подчеркнуть, что после проведения
агротехнических мероприятий численность дож¬
девых червей под многолетними травами и на
культурном пастбище восстанавливается через
5-6 лет [1,13 и др.]. По-видимому, такой же более
продолжительный срок необходим для восста¬
новления численности люмбрицид, нарушенной
глубоким рыхлением. В почвах под многолетни¬
ми травами создаются лучшие условия для обра¬
зования более богатого биоценоза почвенных
беспозвоночных, чем под пашней при интенсив¬
ном севообороте [15,17]. Поэтому целесообразно
использовать мелиорированные почвы в системе
732
ЗАЙДЕЛЬМАН
Таблица 5. Численность насекомых (экз/м2) в дерново-подзолистых глееватых суглинистых почвах и ее изменение
под влиянием дренажа и глубокого мелиоративного рыхления. Московская область
Состав
Контроль (без осушения)
Дренаж
Дренаж + рыхление
05
06
07
09
05
06
07
09
05
06
07
09
Личинки:
проволочники
22
36
Нет
12
4
8
Нет
2
Нет
2
10
16
жужелицы
6
Нет
12
Нет
10
12
4
Нет
»
Нет
6
Нет
Имаго:
жужелицы
Нет
12
8
»
12
6
10
»
»
2
8
2
стафилины
»
8
Нет
»
2
4
Нет
»
»
Нет
Всего:
»
20
8
»
14
10
10
»
»
2
8
2
Личинки + имаго
28
56
20
12
28
30
14
2
»
4
24
18
севооборотов с высокой насыщенностью полями
многолетних трав.
На территориях, обработанных глубокими
рыхлителями (особенно после сплошного
рыхления), могут создаться условия, когда для
восстановления почвенной биоты и особенно чис¬
ленности дождевых червей, необходимо кратко¬
срочное залужение полей с использованием их в
качестве зеленых угодий. В этом случае можно
предполагать высокую экологическую и эконо¬
мическую эффективность инокуляции люмбри-
цид в рыхленые почвы. Последнее оправдано и
тем, что эффект последействия этого приема со¬
храняется 12-18 лет и более [6, 18]. Вторым при¬
емом восстановления численности мезофауны
может оказаться орошение, особенно зеленых
угодий [16].
Реакция насекомых на дренаж и глубокое ме¬
лиоративное рыхление оказалась близкой или
тождественной реакции дождевых червей. Как
показали наблюдения [3] (табл. 5), наибольшее
количество личинок и имаго обитало на залежи,
меньшее - в осушенной нерыхленой почве и еще
меньшее - в дренированных и рыхленых почвах.
Многоножки и пауки обитали на залежи (кон¬
троль, неосушенная почва) и в осушенной нерых¬
леной почве. В почвах с дренажом и глубоким
рыхлением они отсутствовали.
Необходимо отметить близкий состав членис¬
тоногих на залежи, на осушенном нерыхленом и
осушенном рыхленом участках. Проволочники от¬
носятся к двум родам - Agriotes и Athous. Личинки
жужелиц представлены тремя родами - Clivina,
Pteroctichum и Harpalus, личинки двухкрылых -
тремя семействами - Empididae, Dolichorus, Rha-
gionidae. Родовой состав имаго жужелиц более
многочисленный, чем личинок - Clivina, Amara,
Bembidion, Harpalus, Pteroctichus. Среди многоно¬
жек обнаружены многосвязы Polydesmus denticu-
latus, кивсяки Brachyinlus jawlowskai и геофилы
Geophilus proximus.
В заключение необходимо остановиться на не¬
скольких общих вопросах изучения почвенной
биоты мелиорируемых территорий. В этой связи,
во-первых, следует отметить, что при оценке ро¬
ли мелиорации в изменении видового состава и
численности почвенной биоты особое значение
имеет степень переувлажнения почв. Дренаж и
агромелиоративные мероприятия могут оказы¬
вать принципиально различное влияние на биоту
почв суб- и супераквальных ландшафтов. Так,
в сильнозаболоченных почвах (торфяных и глее-
вых) на фоне естественного гидротермического
режима отсутствуют или весьма ограничены не¬
обходимые условия для формирования мезозоо-
биоты. По данным Кипенварлиц [9], люмбрици-
ды заселяют торфяные почвы только после осу¬
шения. С другой стороны, по наблюдениям
Смирновой [14], максимальное развитие жуже¬
лиц отмечено в дубраве черничной, тогда как
в условиях более сухого местообитания (дубрава
орляковая) их видовой состав и численность су¬
щественно уменьшаются. Во-вторых, в настоя¬
щее время широко распространено представле¬
ние о том, что дренаж и агромелиорация “вырав¬
нивают” гидротермический режим осушаемых
почв и их разнообразие как среды обитания.
Поэтому анализ биоты в любой точке поля ре¬
презентативен его общему состоянию. Реальная
ситуация значительно сложнее. Как показали на¬
ши исследования [7, 8], мелиорированное поле
оказывается весьма неоднородным. Существуют
длительно и постоянно действующие факторы,
которые определяют его экологическую и агро-
экологическую пестроту.
Разнообразие экологических условий осушен¬
ного поля определяется прежде всего первич¬
ными факторами гидрологической, педогенной и
литологической пестроты. Гидрологический фак¬
тор обусловлен разнообразием исходной степени
заболоченности почв. Гидрологические различия
могут быть устранены за относительно короткое
ПОЧВОВЕДЕНИЕ № 6 1996
ВЛИЯНИЕ ДРЕНАЖА И АГРОМЕЛИОРАЦИИ НА ПОЧВЕННУЮ БИОТУ
733
время при правильном проектировании, строи¬
тельстве и эксплуатации дренажа.
Педогенный фактор связан с исходной генети¬
ческой неоднородностью почвы. Он особенно от¬
четливо проявляется при комплексном сочетании
минеральных и органических почв и обычно
оказывает свое действие на протяжении весьма
длительного времени. Литологическая первичная
пестрота обусловлена разнообразием грануломе¬
трического состава почв и является постоянно
действующим фактором их разнообразия, кото¬
рый практически не может быть нивелирован с
помощью мелиоративных мероприятий.
Наряду с этими естественными факторами
экологическая неоднородность осушаемой тер¬
ритории определяется и вторичными гидрологи¬
ческими и литологическими факторами, оказы¬
вающими прямое влияние на почвенную биоту
и урожай. Возникновение вторичной гидрологи¬
ческой пестроты осушенного поля обусловлено
формированием депрессионных кривых грунто¬
вых вод или верховодки между дренажными ли¬
ниями. Поэтому в пределах междренного рассто¬
яния постоянно сохраняется разное увлажнение
(минимальное - у дрен и максимальное - в середине
междренного пространства). Такое дискретное
увлажнение носит вторичный характер, возника¬
ет только после осушения и всегда свойственно
осушаемой территории. Его невозможно устра¬
нить. Вторичная экологическая пестрота, связан¬
ная с действием литологического фактора, обус¬
ловлена особенностями строительства дренажных
систем. В этом случае на осушенных массивах воз¬
никают вертикальные литологические структу¬
ры траншейных или щелевых засыпок, которые
закономерно и постоянно функционируют на
всей площади массива. Они повторяются со стан¬
дартным шагом, соответствующим принятым в
проекте междренным расстоянием. Актуальны в
этом случае и новые образования, связанные с пе¬
ремещением значительных земляных масс на осу¬
шаемой территории, - разравниванием кавалье¬
ров вдоль каналов, планировкой поверхности и
др. Все эти особенности мелиорированного поля
(первичные и вторичные) необходимо тщательно
учитывать при оценке почвенной биоты мелио¬
рируемых массивов.
Эти методологические аспекты изучения поч¬
венной биоты имеют актуальное значение для
правильной организации и выполнения экологи¬
ческих наблюдений и мониторинга. Результаты
наших исследований, выполненных на тяжелых
дерново-подзолистых и дерново-глеевых глеева¬
тых почвах, а также анализ опубликованных дан¬
ных позволяет признать, что реакция почвенных
организмов на осушение и агромелиоративные ме¬
роприятия существенно различна. После дренажа
и глубокого рыхления уменьшается количество
люмбрицид, насекомых, многоножек, пауков.
Одновременно возрастает численность простей¬
ших, микроорганизмов, повышается микробиоло¬
гическая активность почв. В этой связи, однако,
остается нерешенным вопрос об экологических и
агроэкологических критериях происходящих из¬
менений различных представителей биоты, их
связи с экологическим состоянием и плодородием
почв. По-видимому, на основе полученных дан¬
ных может быть дан общий ответ, основанный на
интегральной оценке. В тех случаях, когда целе¬
сообразность осушения и рыхления обоснованы
эколого-гидрологическим состоянием почв [5, 8],
эти мероприятия повышают урожай и таким об¬
разом благоприятно влияют на их плодородие.
Однако рассмотренные результаты определенно
показывают, что в этой весьма разноплановой
проблеме существуют и нерешенные вопросы.
К ним в первую очередь следует отнести пробле¬
му регулирования численности дождевых червей,
проволочников и других представителей мезо-
(и микро-) фауны.
СПИСОК ЛИТЕРАТУРЫ
1. Атлавините О.П. Экология дождевых червей и их
влияние на плодородие почвы в Литовской ССР.
Вильнюс: Москлас, 1974.200 с.
2. Валиахмедов Б.В. Влияние орошения и террасиро¬
вания склонов на почвенную фауну беспозвоноч¬
ных // Тр. НИИ почвоведения Госагропрома
ТаджССР. 1987. Т. 26. С. 75-81.
3. Ггльцер В.Ю. Влияние дренажа и глубокого рых¬
ления на физические свойства, водный режим и би¬
оту дерново-подзолистых почв на лёссовидном
суглинке (на примере южнотаежной подзоны): Ав-
тореф. дис. ...канд. биол. наук/МГУ. М., 1983.20 с.
4. Димо Н.А. Наблюдения и исследования по фауне
почв. Кишинев: Госиздат Молдавии. 1955.137 с.
5. Зайдельман Ф.Р. Гидрологический режим почв
Нечерноземной зоны. Л.: Гидрометеоиздат, 1985.
328 с.
6. Зайдельман Ф.Р. Эколого-гидрологические осно¬
вы глубокого мелиоративного рыхления. М.: Изд-
во МГУ, 1986. 193 с.
7. Зайдельман Ф.Р. Структура почвенного покрова
осушенных территорий и урожай // Почвоведение.
1988. № 10. С. 5-15.
8. Зайдельман Ф.Р. Эколого-мелиоративное почво¬
ведение гумидных ландшафтов. М.: Агропромиз-
дат, 1991. 320 с.
9. Кипенварлиц А.Ф. Изменения почвенной фауны
низинных болот под влиянием мелиорации и сель¬
скохозяйственного освоения. Минск: Сельхозгиз,
1961.200 с.
10. Козлов Г.В. Некоторые особенности воздушного
режима дерново-подзолистых глеевых почв под
сеяными лугами // Современные почвенные про¬
цессы и плодородие почв Карелии. Петрозаводск,
1979.
11. Кондакова Л.В. Влияние глубокого мелиоративно¬
го рыхления на биологическую активность почв //
ПОЧВОВЕДЕНИЕ № 6 1996
734
ЗАЙДЕЛЬМАН
Мелиорация, использование и охрана почв Нечер¬
ноземной зоны. М.: Изд-во МГУ, 1980.
12. Маркова Г.И. Влияние осушения переувлажнен¬
ных минеральных почв на группировки почвенных
водорослей // Мелиорация, использование и охра¬
на почв Нечерноземной зоны. М.: Изд-во МГУ,
1980.
13. Перель Т.С. Распространение и закономерности
распределения дождевых червей фауны СССР. М.:
Наука, 1979. 270 с.
14. Смирнова Т.П. Изменения населения жужелиц не¬
которых типов дубрав Белорусского Полесья в
связи с осушением // Биологическая диагностика.
М.: Изд-во МГУ, 1976. С. 258-259.
15. Страздане В.М. Личинки насекомых // Влияние
агротехники на почвенных беспозвоночных жи¬
вотных. Вильнюс: Изд-во АН ЛитССР, 1974.
С. 141-170.
16. Шляпетене ЮЛ. Влияние мелиорации и агротех¬
ники на биоценоз почвенных беспозвоночных в
дерново-глеевых почвах Литвы. Нематоды (Nema-
toda) // Влияние агротехники на почвенных беспо¬
звоночных животных. Вильнюс: Изд-во АН
ЛитССР, 1974. С. 19-40.
17. Эйтминавичюте И.С., Страздене И.К., Сукацке-
не И.К. и др. Соотношение между отдельными
группами почвенных беспозвоночных животных //
Влияние агротехники на почвенных беспозвоноч¬
ных животных. Вильнюс: Изд-во АН ЛитССР,
1974. С. 171-183.
18. Schroder D., Schulte-Karring Н. Nachweis 20-jahriger
Wirksamkeit von Tieflockerungsmassnahmen in loss-
beeinflussten Graulehm-Pseudogleyen // Z.f. Pflanzen-
emahr., Diing. und Bodenkunde. 1984. Bd. 147. № 5.
Drainage and Agromelioration Effects on Soil Biota
F. R. Zaidelman
Drainage and agromeliorative management practices exeit different influence on soil biota. In superaqueous
landscapes drainage and deep loosening of mineral soils promote the increased number of algae. Protozoa and
microorganisms, being penetrated into deeper horizons of the soil profile. The amount and biomass of worms
and insects become drastically declined due to these reclamation facilities. Deep loosening is especially un¬
favourable for population of young earthworms. In subaqueous landscapes drainage after the long-duration
floodin'; in particular appears to be a determining factor of mesofauna population in peat soils. Detailed studies
of soils biota necessitate taking into complete account the soil wetting degree and heterogeneity of primary and
secondary structure.
ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВОВЕДЕНИЕ. 1996, № 6, с. 735-742
ПОЧВА
И БИОРАЗНООБРАЗИЕ
УДК 631.46
ЗНАЧЕНИЕ БИОРАЗНООБРАЗИЯ ДЛЯ ДИАГНОСТИКИ ПОЧВ*
© 1996 г. Ю. Г. Гельцер, А. С. Яковлев
Факультет почвоведения МГУ им. М.В. Ломоносова
Поступила в редакцию 09.02.96 г.
В диагностике почв и почвенных процессов широко используют биохимические, микробиологичес¬
кие, ботанические и зоологические показатели состояния среды. Биологический метод диагностики и
индикации почв базируется на разнообразии видов, жизненных форм и физиологических функций
почвенной биоты, а также на гомеостазе организмов и популяций. Множественность задач биотесги-
рования почв, грунтов и искусственных субстратов предполагает использование как разнообразных
биологических объектов, как и методических приемов исследования. Свободноживущие почвен¬
ные простейшие - обязательный компонент лесных биогеоценозов и агроценозов. Разнообразие
видового состава, эколого-морфологических типов и физиологических функций клеток - четкие
показатели состояния среды обитания организмов. Исследуется новое направление - цитологичес¬
кая диагностика почв.
Биологический метод диагностики и индика¬
ции почв базируется на разнообразии видов, жиз¬
ненных форм и физиологических функций поч¬
венной биоты, а также на гомеостазе организмов
и популяций. Гомеостаз есть совокупность слож¬
ных приспособительных реакций организма, на¬
правленных на устранение или максимальное ог¬
раничение действия различных факторов внеш¬
ней или внутренней среды [14]. Биотестирование
позволяет уловить присутствие стрессирующего
воздействия раньше, чем многие обычно исполь¬
зуемые анализы физического состояния и хими¬
ческих свойств почвы.
Основные принципы зоологического метода ди¬
агностики почв изложены академиком М.С. Гиля¬
ровым в монографии [10], где он пишет: “Участие
в комплексных экспедиционных исследованиях и
совместная работа с почвоведами и геоботаника¬
ми на ряде стационаров... привели меня к выводу,
что данные изучения комплексов почвообитаю¬
щих животных, их экологические и зоогеографи-
ческие характеристики могут быть с успехом
привлечены для решения спорных вопросов диа¬
гностики и географии почв, для выявления изме¬
нений почвенных условий и направления почвооб¬
разовательного процесса под влиянием хозяйст¬
венной деятельности человека и т.п.” [ 10, с. 3-4].
Разнообразие растительных сообществ, от¬
дельные виды растений и их внутривидовые фор¬
мы широко используются в индикационной гео¬
ботанике для определения условий среды. Расти¬
тельные сообщества хорошо заметны на аэро- и
космофотоснимках, что позволяет использовать
их связь со средой для дешифрирования почв,
Работа выполнена при содействии Государственной науч¬
но-технической программы России “Биологическое разно¬
образие”.
горных пород и подземных вод, а также распозна¬
вать проявления естественных экзогенных и эн¬
догенных процессов и различные формы дея¬
тельности человека [1].
Академик Г.В. Добровольский развивает уче¬
ние о функциональном многообразии почв. Осо¬
бое внимание он уделяет функциям почвы как
среды обитания разнообразных организмов и
фактора биологической эволюции почвообита-
телей. Почва - связующее звено биологического
и геологического круговоротов, и в этом ее важ¬
нейшая общебиологическая функция. Сохране¬
ние видового разнообразия в экосистемах - важ¬
нейшая функция почвы, привлекающая в настоя¬
щее время пристальное внимание почвоведов,
биологов, экологов. Наиболее полно биологиче¬
ский метод представлен в монографии Г.В. Доб¬
ровольского “Почвы речных пойм центра Рус¬
ской равнины” [13].
Оценивая устойчивость почвы к техногенным
воздействиям, ведущий почвовед-геохимик про¬
фессор Глазовская [11] придает большое значе¬
ние ответным реакциям почвы как биокосного
тела и сложноорганизованной биокосной системы,
проявляющимся в изменении э колого-геохимичес-
кого состояния среды обитания педобиоценозов.
При этом интегральным показателем эколого-гео-
химической устойчивости является нормальное
функционирование биопедоценозов, населяющих
внутрипочвенное и напочвенное экологическое
пространство.
Биологическое разнообразие (% от исходного),
а также численность и биомасса (% от нормы)
почвенных беспозвоночных - зоологические ин¬
дикаторы зон экологической нормы, риска, кризи¬
са и бедствия экосистем [2]. Примером разнообра¬
зия в биологической диагностике почв, грунтов и
735
736
ГЕЛЬЦЕР, ЯКОВЛЕВ
Рис. 1. Amoeba proteus L.: а - трофозоит (контроль);
б, в - изменение клеток под действием диурона; г -
лизис клетки.
субстратов могут служить одноклеточные почво¬
обитающие животные - простейшие (Protozoa).
Среди разнообразных населяющих почву орга¬
низмов простейшие занимают особое положение:
по размерам клеток, численности, характеру мета¬
болизма, интенсивности размножения, повсемест¬
ности расселения, некоторым культуральным при¬
знакам и многим другим показателям - это типич¬
ные микроорганизмы; по способам питания и
ряду физиологических функций - представители
животного царства. Простейшие - наиболее мно¬
гочисленная и повсеместно распространенная
группа почвообитающих животных. В настоящее
время из почв выделено порядка 450 видов и ва-
риететов простейших, относящихся к трем боль¬
шим широко распространенным группам - сарко-
довым, жгутиконосцам и инфузориям [5, 7, 15].
Четкая функциональная дифференциация клет¬
ки, наличие разнообразных хорошо различимых
в обычный световой микроскоп органоидов и
сложная структура почвообитающих популяций
позволяют успешно использовать протисгов для
биотестирования экологического состояния среды.
В лабораторных условиях в опытах in vitro изу¬
чали влияние различных доз гербицида диурона
на вегетативные клетки свободноживущих про¬
стейших: Hartmannelia rhysodes Singh, Amoeba pro¬
teus L., Paramaecium caudatum Ehren., Euglena viri-
dis Ehren., Colpoda cucullus O.F.M., Oxytricha sp.
При концентрации диурона 0.6 мг/л (так называе¬
мая полевая или “производственная” доза) Н. rhy¬
sodes Singh начали инцистироваться через 96 ч, а на
контроле (физиологический раствор) - через двое
суток. При 6 мг/л цисты мелких амеб появились
через 72 ч. Через сутки появились цисты на среде
с 60, 120, 300 мг/л диурона; развитие амеб шло
медленно, наблюдались лизирующиеся клетки.
При дозе 600 мг/л у некоторых амеб лизис насту¬
пал через 6 ч; есть двойные клетки, которые как
бы остановились в процессе деления, а часть кле¬
ток находилась в стадии покоя. Через 9 ч почти
все клетки инцистируются, остаются единичные
покоящиеся особи.
При концентрации диурона 120, 300 и 600 мг/л
у Amoeba proteus L. наблюдали лизис клеток че¬
рез 3-4 ч, а при 1200 мг/л гибель наступала мгно¬
венно (рис. 1). У многих амеб произошло сокраще¬
ние псевдоподий, они стали широкими, короткими
и неподвижными; у некоторых амеб имелась масса
очень мелких псевдоподий. Отдельные клетки
представляли собой шарообразные образования
без видимых выростов.
У активных инфузорий и жгутиконосцев при¬
нимается во внимание такой показатель, как ос¬
тановка движения 50% особей в поле зрения мик¬
роскопа (BOjo). Диурон токсичен для парамеций и
эвглен в кислой среде, в щелочной среде время
остановки у этих организмов больше, чем в нейт¬
ральной. Протистоцидное действие диурона в дозе
120 мг/л проявляется на парамеций через 84 ч и че¬
рез 20 ч на эвглен при значениях pH 5.5. Большие
дозы (1200 мг/л) действуют на клетки простей¬
ших почти моментально. При величине pH 7.8
(600 мг/л) лизис клеток наступал быстрее, время
остановки 50% особей было равно 4-8 мин. При
действии диурона на P. caudatum Ehrbg. и Е. viridis
Ehrbg. в концентрации 300 мл/л через 22 и 12 ч
(pH 6.8) и 24 и 8 ч (pH 7.8) наблюдали образование
округлых клеток, выпячивание пелликулы и за¬
тем полную гибель животных (рис. 2, 3).
Концентрация 600 мг/л вызывала скорую ги¬
бель всех простейших. Имели место нарушение
физиологических функций, деформация клеток,
отслаивание пелликулы, коагуляция цитоплаз¬
мы, разрушение ядра, полный лизис клеток.
Внесение гербицидов в почву вызывает изме¬
нение количества и видового состава простей¬
ших. Реакция различных видов Protozoa зависит
от вносимого гербицида, времени его воздействия
на почвенную биоту, реакции и ряда экологичес¬
ких параметров среды. Представляется перспек¬
тивным подобрать виды-маркеры простейших на
определенные гербициды. По активности и мор¬
фологическим изменениям клеток можно судить
о концентрации ядохимикатов в почве.
Простейшие реагируют на биологически ак¬
тивные метаболиты почвенной микрофлоры.
Однако три вида инфузорий неодинаково реаги¬
ровали на экзотоксины исследованных грибов
(таблица). Культуральная жидкость Penicillium
cyclopium проявляла наибольшую токсичность.
Наименее устойчивой оказались Stylonychia myti-
lus, гораздо выше резистентность Colpoda fastiga-
ta, Paramaecium caudatum занимала промежуточ¬
ное положение. При высоких концентрациях ток¬
сина парамеции одновременно останавливались,
прекращали работу все реснички, тело сжима¬
лось и принимало лимоновидную или грушевид¬
ную форму (рис. 4); происходило выпячивание
пелликулы, либо последняя отставала от цитоплаз¬
мы, вздуваясь пузырями. Затем наступал цитолиз:
пелликула разрывалась, и цитоплазма изливалась
наружу. При более низких концентрациях токсиче¬
ского вещества (разведение 1: 32,1:64) остановка
ПОЧВОВЕДЕНИЕ № 6 1996
ЗНАЧЕНИЕ БИОРАЗНООБРАЗИЯ ДЛЯ ДИАГНОСТИКИ ПОЧВ
737
а б в г д
Рис. 2. Paramaecium caudatum Ehrenberg: а - трофозоит (контроль); 6-г - выпячивание пелликулы под действием диу¬
рона; г) - лизис клетки.
Рис. 3. Euglena viridis Ehrenberg; а - трофозоит (контроль); б-д - изменение формы клетки под действием диурона;
е- лизис клетки.
инфузорий происходила не одновременно, неко¬
торое время еще можно было наблюдать беспо¬
рядочное движение ресничек. Дольше других
функционировали реснички перистомальной зо¬
ны. Наблюдалось увеличение числа и объема со¬
кратительных вакуолей; на поверхности инфузо¬
рий появлялись светлые гомогенные пузырьки
Цитоплазмы.
Стилонихии в неразведенной культуральной
жидкости мгновенно останавливались и полностью
распадались. При меньших концентрациях внешне
никаких морфологических изменений заметить
не удавалось, но через некоторое время клетки
распадались.
Колподы в разведениях культуральной жидко¬
сти 1 : 32 - 1 : 128 быстро округлялись и постепен¬
но замедляли движение до полной остановки, ко¬
торая наступала у разных экземпляров в разное
время. Затем часть особей распадалась, а некото¬
рые успевали инцистироваться.
Почва - гетерогенная и полидисперсная систе¬
ма. В нормально увлажненной почве микроорга¬
низмы развиваются в гуматных пленках, облека¬
ющих отдельные частицы мелкозема, либо в за¬
полненных почвенным раствором капиллярах и
микролакунах. В настоящее время разрабатыва¬
ются методы биотестирования непосредственно в
почвенной среде, либо в имитирующих почву ус¬
ловиях, так называемых микрокосмах [3-6]. По¬
следние более соответствуют почвенной среде,
чем водные суспензии. Метод проращивания поч¬
венного мелкозема на тонко агаризованных и по¬
мещенных во влажную камеру покровных стек¬
лах позволяет исследовать трофозоиты простей¬
ших непосредственно в их природной среде
обитания, часто внедренными в микроколонии
бактерий, дрожжей и водорослей, которыми они
питаются.
При больших увеличениях светового микро¬
скопа протозойная клетка может дать богатую
информацию о состоянии среды обитания. Так,
на окрашенном молибденовым гематоксилином
препарате почвенного мелкозема можно наблю¬
дать у амебы Vahlkampfia sp. крупное интерфаз¬
ное ядро с кариосомой и периферическими хро-
матиновыми зернами. Множество вакуолей в
клетке указывает на паранекротическое состоя¬
ние цитоплазмы, сопутствующее обезвоживанию
организма (рис. 5).
4 ПОЧВОВЕДЕНИЕ № 6 1996
738 ГЕЛЬЦЕР, ЯКОВЛЕВ
Реакция различных видов свободноживущих инфузорий на нативные жидкости почвенных грибов (время оста¬
новки 50% инфузорий ВО50, мин)
№ и вид штамма грибов
Вид инфузорий
Протистоцидное действие на инфузорий (ВО50)
разведений культуральной жидкости грибов
0
1:2
1:4
1:8
1 : 16
1 :32
1 :64
1 : 128
37. Penicillium cycloppium
Paramaecium caudatum
0.4
2
5.5
7
8
23
37
81
Stylonychia mytilus
0.5
1
2
3
5
10
24
Нет данных
Colpoda fastigata
1
6
12
24
49
79
130
»
202а. Penicillium cycloppium
Paramaecium caudatum
2
7
105
180
>720
Нет данных
Stylonychia mytilus
2
6
87
180
>720
>:
►
Colpoda fastigata
1.5
61
180
>720
-
>:
►
2088. Trichoderma lignorum
Paramaecium caudatum
4
8
112
>120
Нет данных
Stylonychia mytilus
5
8
66
>120
»
Colpoda fastigata
3
33
>120
>:
►
224. Стерильный мицелий
Paramaecium caudatum
5
62
134
>180
|
Нет данных
Stylonychia mytilus
8
99
>180
>:
►
Colpoda fastigata
3.5
97
>180
»
По аналогии с зоологическим методом диагно¬
стики почв, разработанным М.С. Гиляровым,
оценку экологических условий среды по состоя¬
нию клеток микроорганизмов мы назвали цито¬
логическим методом диагностики, или сокращен¬
но - цитодиагностикой.
Разнообразие форм переживания неблагопри¬
ятных условий у простейших - хороший тест для
оценки степени опустынивания ландшафтов.
Обычная почвенная инфузория Colpoda maupasii
Enriquez - убиквист, т.е. повсеместно распростра¬
ненный организм. Однако способы образования
стадий переживания неблагоприятных условий у
нее совершенно различны в гумидных и аридных
условиях. Инцистирование С. maupasii, выделен¬
ных в культуру из дерново-подзолистой почвы,
протекает по известной схеме (рис. 65, в): трофо-
зоит прекращает движение, клетка округляется,
сократительная вакуоль интенсивно выводит во¬
ду из заметно уплотняющейся цитоплазмы; орга-
неллы клетки дедифференцируются, неперева¬
ренные пищевые частицы выбрасываются нару¬
жу. Округлившаяся клетка заметно уменьшается
в объеме и образует внешний бесструктурный
слизистый слой, со временем уплотняющийся в
относительно тонкую и слегка рефрактирующую
оболочку. Оболочка дифференцируется на два
слоя - эндоцисту и эктоцисту; эктоциста морщи-
мкм
Рис. 4. а — инфузория Paramaecium caudatum Ehrenberg, экспонированная в нативной жидкости гриба Penicillium cyclo-
pium West, и окрашенная ядерным красителем по Роскину (объектив х60, окуляр х 15); б-вегетативная клетка амебы
Hartmannelia rhysodes Singh, окрашенная нейтральным красным (водный раствор 1 : 100000) после суточной экспози¬
ции в культуральной жидкости гриба Penicillium cyclopium West.; в - нормальная вегетативная клетка амебы Hartman¬
nelia rhysodes, окрашенная водным раствором нейтрального красного (1 : 100000).
ПОЧВОВЕДЕНИЕ № 6 1996
ЗНАЧЕНИЕ БИОРАЗНООБРАЗИЯ ДЛЯ ДИАГНОСТИКИ ПОЧВ
739
не Заалтайской Гоби (Монголия). Здесь форми¬
руются так называемые гаммады - щебнистые
пустынные территории, практически лишенные
растительности; температура поверхности в лет¬
ние месяцы достигает 75°С, сумма годовых осад¬
ков в районе Эхийнгольской сельскохозяйствен¬
ной станции, где проводились исследования, не
превышает 20 мм. Но и в этих жестких условиях в
почве обитают жгутиконосцы, амебы и инфузо¬
рии, проявляя определенные черты приспособле¬
ния к экстремальным условиям существования.
Переход от активного состояния (рис. 6а) к не¬
активному у Colpoda maupasii, выделенных из пу¬
стынных почв МНР, проходил иначе, чем у инфу¬
зорий из почв российского Нечерноземья. При
подсыхании капли с культурой колпод трофозои-
ты не прекращают движения, и дегитротация ци¬
топлазмы начинается у еще подвижных форм
(рис. 6г); стадии покоя с ротирующей цитоплаз¬
мой отсутствуют. Инфузории сжимаются, прини¬
мают угловатую форму (рис. 6д, 6е); дедиффе-
ренцирование содержимого клетки происходит
очень быстро, однако, слабое движение ресничек
сохраняется. Через 10-15 мин клетка превраща¬
ется в неподвижное прямоугольное или овальное
угловатое кристаллоподобное тело с тонкой кон¬
трастной оболочкой и грубозернистым содержи¬
мым (рис. 6е, 6ж). Такой способ перехода в неак¬
тивное состояние мы назвали “псевдокристалли¬
зацией”, поскольку скопление неподвижных
обезвоженных колпод напоминает друзу крис¬
таллов (рис. 6ж). Псевдокристаллизация у С. mau¬
pasii не сопровождается процессом деления, хотя,
по мнению Laminger [23], при обычном инцисти-
ровании он обязателен.
10 мкм
I
Рис. 6. Colpoda maupasii из пустынной крайнеаридной почвы Заалтайской Гоби (МНР).
а - трофозоит, 40 мкм; 6 - стадии покоя; в - циста, 20 мкм; г-е - этапы псевдокристаллизации; ж - скопление (“друза”)
псевдокристаллов колпод. Об. х60, ок. х 10.
Рис. 5. Вегетативная клетка Vahlkampfia sp. среди
почвенных частиц. Рисовальный аппарат. хбОО.
Я - ядро с крупной кариосомой и периферическими
зернами хроматина. Препарат мелкозема на агаризо-
ванном покровном стекле, окрашенном гематоксили¬
ном; 1 - плазмалемма; 2 - почвенная частица.
нистая, складчатая, от чего циста Colpoda имеет
часто звездчатую форму. Во время образования
оболочки цитоплазма оживленно ротирует.
Эксцистирование Colpoda происходит путем
разрыва оболочки и длится достаточно долго, ино¬
гда до 1 ч. У инфузорий, обитающих в хорошо ув¬
лажненных почвах умеренных широт, эксцистиро-
ванию предшествует длительный период покоя.
Пример экстремальных условий существова¬
ния простейших - крайнеаридные почвы в пусгы-
ПОЧВОВЕДЕНИЕ № 6 1996
4*
740
ГЕЛЬЦЕР, ЯКОВЛЕВ
" О. f?"® О
'©<о ©(О
я б
Л
"Оо
А б
Рис. 7. Схема экологических морфотипов раковин те¬
стаций (по [20]). / - апланатный; П — трахелостомный;
/// - акростомный (а - без латерального сжатия, б-с ла¬
теральным сжатием); IV - центростомный с уплощен¬
ной вентральной поверхностью; V: А - плагиостомный
простой; Б - с козырьком; VI: А - криптостомный
простой; Б- с козырьком; а - план, 6 - профиль.
Псевдокрисгаллизация - особый способ перехо¬
да в состояние временной нежизнеспособности, ти¬
пичный случай ксероанабиоза (ангидроанабиоза)
у свободноживущих простейших в экстремаль¬
ных условиях [12]. Вышеописанный способ инци-
стирования у пустынных колпод связан с резкими
колебаниями температуры и кратковременными
увлажнениями почвы редкими дождями и диагно¬
стирует аридные процессы почвообразования.
Раковинные амебы - тестации (Testacida) пред¬
ставляют перспективный объект биоиндикаци-
онных исследований, поскольку раковины имеют
комплекс константных таксономических призна¬
ков, значительное время сохраняются в почве,
что позволяет определять состав населения и его
изменения даже после гибели оргайизмов. Харак¬
терная особенность тестаций - наличие у многих
видов экологических преферендумов по отноше¬
нию к среде обитания, в результате которых фор¬
мируются четко различающиеся комплексы этих
простейших.
Все разнообразие форм раковин тестаций
можно свести к нескольким эколого-морфологи-
ческим типам раковины - экоморфотипов (рис. 7).
Для российских почв пока названы; апланатный
(дисковидный), трахелостомный, акростомный,
центростомный, плагиостомный и криптостом¬
ный экоморфотипы. Каждый из них соответству¬
ет определенному местообитанию; от водной сре¬
ды болот до минеральных горизонтов почвы.
Преобладание в популяции тех или иных экомор¬
фотипов позволяет судить о сформированности
почвенного горизонта. Таким образом, по набору
морфотипов раковины и экологических групп
в образце можно в общих чертах дать характери¬
стику субстрата и происходящих в нем процессов:
в развитых автоморфных почвах без подстилки
преобладают крупные плагиостомные и криптос-
томные виды; в лесной почве значительна доля ак-
ростомных мелких населяющих подстилку видов;
в комплексах гидроморфных почв - гидробион-
тов.
Примером использования принципа биоразно¬
образия в диагностике почв могут служить иссле¬
дования по влиянию окультуренности дерново-
подзолистых почв на фауну раковинных корнено¬
жек [9]. Так, структурированность сообществ,
обитающих в разных по окультуренности биото¬
пах (их внутренняя организация, соотношение ви¬
дов внутри сообщества), может быть хорошо ил¬
люстрирована показателями рассеяния (доминиро¬
вания) видов с, а также отношением о/а [18, 21],
где а - показатель видового разнообразия [22],
а а определяется по формуле
/Цт,-т)2
Ч S-1 ’
где 5 - число родов на пробной площадке; mt -
число видов каждого рода, % от общего количе¬
ства; in - число видов в среднем на один род, %.
Получены следующие величины показателей:
Лес Залежь Пашня
а 10 11 24
ст/а 2 3 25
Максимальную степень доминирования огра¬
ниченной группировки видов наблюдали на паш¬
не, минимальную - в лесу. Залежь по степени до¬
минирования значительно ближе к лесу, чем к
пашне. Объясняется это более жесткими услови¬
ями обитания (в частности, отсутствием подстил¬
ки), в результате чего наблюдается преобладание
узкой группы специализированных чисто почвен¬
ных видов Centropyxis, Plagiopyxis [17].
Виды-педобионты, которые доминируют в ус¬
ловиях пашни, наиболее активно развиваются и
на залежи, где не наступило еще равновесие био¬
логических факторов, наблюдаемое в лесу. За
счет быстрого роста указанной группировки ви¬
дов общее количество организмов, зафиксиро¬
ванных на залежи, выше, чем в лесу и на пашне
(рис. 8а), а общее количество видов меньше, чем
в лесу, но больше, чем на пашне (рис. 86).
Общая картина изменения населения раковин¬
ных амеб, выражающаяся в уменьшении количе¬
ства видов и снижении обилия раковинных амеб
сразу после “стресса” [16], вызванного вспашкой
ПОЧВОВЕДЕНИЕ № 6 1996
ЗНАЧЕНИЕ БИОРАЗНООБРАЗИЯ ДЛЯ ДИАГНОСТИКИ ПОЧВ
741
(с резким нарастанием на залежи), иллюстрируется
показателем “видового разнообразия” (рис. 8в).
Максимальное разнообразие видов наблюдается
в лесу, минимальное - на пашне. Видовое разно¬
образие тестаций в почве залежи значительно
ближе к таковому в лесу, чем на пашне.
Приспособления тестаций к условиям обитания
выражаются в различной морфологии раковинок,
характерной для того или иного биотопа [20]. Рако¬
винки обнаруженных нами видов относятся к пя¬
ти морфологическим типам [8]. Среди них наибо¬
лее приспособленным к дефициту влаги в почве
оказывается так называемый криптостомный
тип амеб, характеризующийся малыми размера¬
ми устья и другими чертами, препятствующими
высыханию.
О степени адаптированности к почвенным ус¬
ловиям на разных уровнях почвенного профиля
позволяет судить показатель “эдафичности” (е) и
показатель “видового разнообразия” (а). Показа¬
тель эдафичности определяется по формуле
млн/м2
200 г а
160
120
80
40
0
Вспашка
ЛП
100
80
60
'ч 40
ч
20
0
Вспашка
\\
Л ЛП
-I pi L
Ли ЛП
л
Рис. 8. Изменение количества клеток (млн/м2) (а),
числа видов (б) и видового разнообразия (а) (в) рако¬
винных корненожек под влиянием вспашки (стресса).
Л - лес; П - пашня; 3 - залежь.
е а
250 100 0 1 3 5
1 1—I—I—|—I—I—I—I—г
т
е =
где т - число особей каждого морфотипа, выра¬
женное в процентах к общему числу особей всех
морфотипов; к- оценка степени приспособленно¬
сти к почвенным условиям обитания отдельного
морфотипа (оценка изменяется от одного до пя¬
ти), исходя из пяти обнаруженных морфотипов;
ранжирование проведено на основании работ
Bonnet [18-20]; п - число морфотипов, характери¬
зующих данный выдел. Для лесной почвы и
окультуренной пашни получены значения пока¬
зателей эдафичности и видового разнообразия:
Местонахож¬
дение почвы
Лес
Горизонт
а
е
АО
4.4
70
А0А1
4.7
85
А1
0.5
240
А пах
1.2
170
А дерн
4.0
Нет
Пашня
Залежь
В лесу показатель эдафичности (е) увеличива¬
ется с глубиной, а коэффициент видового разно¬
образия (а) имеет максимальное значение в наи¬
более заселенном гор. А0А1. Для пахотного слоя
значения индексов ближе всего к показателям
гор. А1 лесных почв (рис. 9). Следовательно, чем
большему нарушению в результате деятельности
человека подвергается естественный (целинный)
почвенный покров (в особенности верхняя под¬
стилочная или дерновая его часть), тем уже
спектр морфологических типов и тем более пре¬
обладает эдафический компонент фауны рако-
Рнс. 9. Степень “эдафичности” морфологических
спектров (е) и видового разнообразия (а) тестаций по
слоям обитания в лесных и пахотных почвах.
винных амеб и уменьшается их видовое разнооб¬
разие (а). Так, после вспашки целинных почв
улучшение приспособленности тестаций к поч¬
венным условиям отражается в нарастании значе¬
ний коэффициента эдафичности от 70 в подстилке
лесной почвы до 170 в пахотном слое и соответст¬
венным уменьшением видового разнообразия от
4.7 до 1.2.
Контрастные различия в значениях коэффи¬
циентов, отражающих сужение видового состава
сообществ тестаций и увеличение доли специали¬
зированных видов в условиях пахотных почв по
сравнению с целинными, позволяют использо¬
вать группу раковинных амеб как возможный
ПОЧВОВЕДЕНИЕ № 6 1996
742
ГЕЛЬЦЕР, ЯКОВЛЕВ
объект для оценки почв разной степени окульту-
ренности.
Значительное биологическое разнообразие на¬
селяющих почву организмов, объединенных по¬
нятием почвенная биота, предоставляет исследо¬
вателям - почвоведам, агрохимикам, экологам -
широкие возможности для применения метода
биотестирования при диагностике почв и почвен¬
ных процессов.
СПИСОК ЛИТЕРАТУРЫ
1. Викторов С.В., Ремезова ГЛ. Индикационная гео¬
ботаника. М.: Изд-во МГУ, 1988. 168 с.
2. Виноградов Б.В., Орлов В.П., Снакин В.В. Биоти¬
ческие критерии выделения зон экологического
бедствия России // Изв. РАН. Сер. Географ. 1993.
№ 5. С. 77-89.
3. Гельцер Ю.Г. О новой методике изучения почвен¬
ных простейших // Вестн. МГУ. Сер. Биология.
1960. № 6. С. 67-75.
4. Гельцер Ю.Г. Прибор для наблюдения за ростом
корневых систем и почвенными микробами // Био¬
логия в школе. 1961. № 5. С. 28-33.
5. Гельцер Ю.Г'. Protozoa почвы и методы их обнару¬
жения //Успехи современной биологии. 1962. Т. 53.
Вып. 2. С. 237-245.
6. Гельцер Ю.Г. О методах изучения почвенных амеб //
Микроорганизмы в сельском хозяйстве. М.: Изд-во
МГУ, 1963. С. 441-458.
7. Гельцер Ю.Г. Простейшие (Protozoa) как компо¬
нент почвенной биоты (систематика, экология).
М.: Изд-во МГУ, 1993. 175 с.
8. Гельцер Ю.Г., Корганова Г.А., Яковлев А.С., Алек-
сеев ДА. Раковинные корненожки (Testacida) почв //
Почвенные простейшие. Сер. Протозоология.
1980. Вып. 5. С. 108-142.
9. Гельцер Ю.Г, Яковлев А.С. Влияние окультурен-
ности почв на фауну раковинных корненожек (Pro¬
tozoa, Testacida) // Фауна и экология почвенных
беспозвоночных Московской обл. М.: Наука, 1983.
С. 156-162.
10. Гиляров М.С. Зоологический метод диагностики
почв. М.: Наука, 1965. 278 с.
11. Глазовская М.А. О принципах оценки устойчивос¬
ти почв к техногенным воздействиям (устное сооб¬
щение). МГУ, 1995.
12. Голдовский А.М. Анабиоз. Л.: Наука, 1981. 136 с.
13. Добровольский Г.В. Почвы речных пойм центра
Русской равнины. М.: Изд-во МГУ, 1968. 296 с.
14. Захаров В М., Кларк Д.М. Биотест. Интегральная
оценка здоровья экосистем и отдельных видов. М.,
1993. 68 с.
15. Николюк В.Ф., Гельцер Ю.Г. Почвенные простей¬
шие СССР. Ташкент: Фан, 1972. 311 с.
16. Одум Ю. Основы экологии. М.: Мир, 1975. 740 с.
17. Яковлев А.С. Биологическая активность дерново-
подзолистых и болотно-подзолистых почв и во¬
просы диагностики их свойств: Автореф. дис. ...
канд. биол. наук / МГУ. М., 1981.22 с.
18. Bonnet L. Le peuplement th6camoebien des sols // Rev.
Ecol. Biol. Sols. 1964. V. 1. № 2. P. 123-408.
19. Bonnet L. Le peuplement thecamoebien de quelques sols
du Spitzbergem // Bull. Soc. Hist. Natue. Touluse. 1965.
V. 100. №3/4. P. 281-293.
20. Bonnet L. Types morphologiques, ecologie et evolution
de la th6que cher les Th6camoebiens // Protistologica.
1975. V. ll.№3. P. 363-378.
21. Couteaux M.-M. Dynamisme de L’6quilibre des ТЬё-
camoebiens dans quelques sols climaciques // Мёт.
Mus. Nat. Hist. Natur. Paris, 1976. Nuov. serie, ser. A,
Zool. V. 96. P. 1-183.
22. Fischer R.A., Corbet AS., Williams C.B. The relation be¬
tween number of individuals and the number of species
in a randoms of animal population // J. Animal Ecol.
1943. V. 12. P. 42-58.
23. Laminger H. Bodenprotozoologie // Mikrobios. 1980.
Bd. 1. S. 1-142.
The Importance of Biodiversity for Soil Diagnostics
Yu. G. Gel’tser, A. S. Yakovlev
In characterizing the soils and soil processes biochemical, microbiological, botanical and zoological indices of
the soil medium are widely used. Biological method of soil diagnostics and indication is based on a diversity
of species, life forms and physiological functions of soil biota as well as on homeostasis of organisms and pop¬
ulations. The major tasks as related to biotesting the soils, grounds and artificial media point to the need for a
better understanding of diverse biological objects and methods to study them. Soil Protozoa appear to be a main
component of forest biogeocoenoses and agrocoenoses. Diversity of species composition, ecologo-morpholog-
ical types and physiological functions of cells proves to be very specific for living organisms habitat. Cytolog-
ical diagnostics of soils as a new trend has being elaborated.
ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВОВЕДЕНИЕ, 1996, № 6, с. 743-753
ГЕНЕЗИС
И ГЕОГРАФИЯ ПОЧВ
УДК 631.48
РОЛЬ ОКИСЛЕНИЯ СУЛЬФИДОВ В ГЕНЕЗИСЕ ДЕРНОВЫХ
ШУНГИТОВЫХ ЖЕЛЕЗИСТЫХ ПОЧВ КАРЕЛИИ
© 1996 г. П. В. Красильников, А. М. Володин
Институт биологии Карельского НЦ РАН, Петрозаводский государственный университет
Поступила в редакцию 30.08.95 г.
На сульфидсодержащих углеродистых (шунгитовых) сланцах Карелии формируются охристые и
красноцветные почвенные профили. Окисление сульфидов приводит к выщелачиванию углерода
шунгитов, обеднению поглощающего комплекса обменными основаниями и увеличению кислотно¬
сти. Окраска почв определяется наличием гетита, гематита и ярозита. В илистой фракции присутст¬
вуют биотит, вермикулит и смешанослойный слюда-вермикулит с тенденцией к упорядоченности.
Дерновые шунгитовые почвы Карелии издав¬
на привлекали внимание исследователей своим
своеобразием. По данным Марченко [10], первые
упоминания о темноцветных почвах Олонецкой
губернии относятся к началу XIX в., а первые их
систематические исследования были проведены
в конце того же века академиком Левинсон-Лес-
сингом и местным агрономом Быстровым. Подроб¬
ное исследование дерновых шунгитовых почв
проводилось в 50-60-х гг. сотрудниками кафедры
почвоведения Петрозаводского государственно¬
го университета Тойкка и Володиным [4, 14, 15].
Эти авторы выяснили распространение дерновых
шунгитовых почв на территории Карельской
АССР, что было отражено на карте-схеме мас¬
штаба 1 : 500000. Ими были выделены типичные
дерновые шунгитовые почвы, неполноразвитые,
оподзоленные и железистые, что внесло коррек¬
тивы во взгляды на дерновые шунгитовые почвы
как на слабодифференцированные по окраске.
Тогда же эти авторы предположили, что ожелез-
нение профилей некоторых дерновых шунгито¬
вых почв вызывается окислением сульфидов же¬
леза, присутствие которых в шунгитовых сланцах
известно [20]. Из-за отсутствия данных специаль¬
ных исследований это предположение не было
отражено в публикациях. Впоследствии Ивано¬
вой [8] в классификации почв Карелии были отра¬
жены только дерновые литогенные шунгитовые и
Дерновые литогенные шунгитовые оподзоленные
почвы. Дерновые литогенные шунгитовые желе¬
зистые почвы были в дальнейшем включены
в классификацию в качестве подрода оподзолен-
ных шунгитовых почв [11]. В последнее время по¬
явились работы, которые подробно характеризу¬
ют свойства почв на породах, обогащенных шунги-
товым материалом [18,19], однако, ожелезненные
профили не описывались и не анализировались.
Морфологическое описание ожелезненного поч¬
венного профиля на элювии шунгитовых сланцев
принадлежит Пудовкину [12], но приводимые
аналитические данные касаются только прочно¬
стных свойств шунгитовых сланцев. Таким обра¬
зом, до настоящего времени генезис и свойства
дерновых шунгитовых железистых почв не были
изучены. Актуальность подобного исследования
диктуется как достаточно широким распростра¬
нением этих почв (рис. 1), так и их своеобразием.
ОБЪЕКТЫ И МЕТОДЫ
Исследования проводились в Медвежьегор¬
ском районе Республики Карелия к югу от пос.
Толвуя (рис. 1). Территория относится к Приозер¬
ному подрайону Южного агроклиматического
района республики [2]. Количество осадков за год
около 650 мм, сумма температур выше +5°С за ве¬
гетационный период 1850-1900°С, средняя темпе¬
ратура воздуха в январе -10°С, в июле около
+ 16°С. Почвообразующими породами для иссле¬
дованных почв служат шунгитовые сланцы, отно¬
сящиеся преимущественно к группе IV (т.е. содер¬
жащие собственно шунгитового материала от 10
до 20%), типу Б (т.е. содержащие 67-71% Si02)
[20]. По составу силикатной основы шунгиты IV-Б
отличаются преобладанием биотита и хлорита.
Кроме того, для них характерна обогащенносгь
сульфидами, иногда образующими сплошные
прослои руды.
Почвенные разрезы закладывались возле карь¬
еров “Зажогино” и “Максово”. Карьер “Зажогино”
достигает глубины 10 м. Порода по всей стенке
карьера трещиноватая, в верхней части с гнезда¬
ми щебня. По граням отдельностей и по трещи¬
нам до глубины 2.5 м порода покрыта тонкими
пленками красного (10 R 4/8) цвета [24]. Ниже
красные пленки сменяются интенсивно-бурыми
(7.5 YR 5/8) и бледно-желтыми (5 Y 8/4). Переход к
непрокрашенной породе постепенный, с глубины
4 м интенсивность прокраски убывает, на глубине
743
744
КРАСИЛЬНИКОВ, ВОЛОДИН
Рис. 1. Фрагмент карты-схемы распространения дер¬
новых шунгитовых почв в Заонежье [15]. Дополнена
по материалам исследований 1994 г. П о ч в ы: а -тем¬
ноцветные дерновые шунгитовые; б - железистые;
1 - участок “Зажогино”; 2 - участок “Максово”.
Рис. 2. Обломки шунгитовой породы: а - из карьера
“Зажогино”; б- из разреза 6ш/95; / - прослои пирита;
2 - неизмененная черная (N 2/) порода; 3 - выщело¬
ченная светло-серая (N 7/) порода; 4 - глинистая жел¬
товато-красная (5 YR 5/6) пленка.
6 м остаются единичные бурые и желтые пленки
по трещинам. Неизмененный шунгитовый сланец
представляет собой черную массивную породу,
пронизанную тонкими прожилками халцедона
и сульфидов (рис. 2а). На сколе сульфиды имеют
золотистую окраску. Порода пронизана сульфи¬
дами неоднородно: местами они образуют густую
сеть прожилок, а местами отсутствуют.
Карьер “Максово” достигает глубины 7 м. По¬
роды отличаются от пород в карьере “Зажогино”
большей твердостью, отсутствием макроскопи¬
ческих прожилок сульфидов и обилием халцедо¬
новых прожилок и стяжений. В верхней части по¬
рода покрыта бурыми, желтыми и красноватыми
пленками по трещинам. Отчетливой зональнос¬
ти, как в карьере “Зажогино”, не наблюдается.
Было заложено 6 почвенных профилей. Раз¬
резы 1ш/94, 2ш/94 и Зш/94 морфологически не
различаются, поэтому приводится описание толь¬
ко одного из них. При определении цветов в поле
мы пользовались атласом Манселла. Цвета при¬
водятся для почвы во влажном состоянии.
Разрез 1ш/94. Карьер “Зажогино”. Разрез
заложен в 30 м от верхнего среза карьера. Склон
около 5° северной экспозиции. Территория ранее
распахивалась. Растительность луговая: клевер
(Trifolium medium), ромашка (Matricaria matricario-
ides), овсяница (Festuca pratensis), ежа (Dactylis glom-
erata), тимофеевка (Phleum pratense), одуванчик
(Taraxacum officinale), чертополох (Carduus crispus).
Адерн 0-5 см. Очень темно-серый (N 3/); свежий;
дернина; уплотненный; минеральная часть по грануло¬
метрическому составу соответствует среднему суглин¬
ку; структура бусовидная по корням, непрочная; встре¬
чаются обломки шунгитовых пород размером от 1 до
200 мм; некоторые фрагменты выщелочены до полно¬
го исчезновения шунгитового вещества так, что оста¬
ется сероватая силикатная основа; граница ровная, пе¬
реход ясный по содержанию корней.
А1 5-25 см. Старопахотный горизонт; очень темно¬
серый (N 3/); влажноватый; уплотненный; средний суг¬
линок; структура крайне непрочная комковатая; мно¬
го корней; обломки породы разнообразных размеров
как выветрелые до полной выщелоченности, так и
слабоизмененные; на некоторых обломках имеются
красные и бурые пленки; граница ровная по плужной
подошве; переход заметный по плотности и количест¬
ву корней.
А1В 25-55 см. Очень темно-серый (N 3/); влажнова¬
тый; плотный; тяжелый суглинок; структура выраже¬
на слабо; единичные корни; много обломков шунгито¬
вой породы как выщелоченных, так и неизмененных,
частично покрытых красными и бурыми пленками;
подстилается трещиноватой шунгитовой породой.
Разрез 4ш/94. Карьер “Максово”. Разрез за¬
ложен непосредственно над стенкой карьера. Рас¬
тительность естественная: ель (Picea abies), ольха
(Alnus in с ап а), кустарничковый ярус представлен
единичной черникой (Vaccinium myrtillus), на по¬
верхности почвы зеленые мхи и лишайники
Chladonia. У ели отмечены признаки хлороза
(светло-зеленая к желтой хвоя).
L 1-0 см. Лиственный и хвойный опад; единичные
корни кустарничков; угли; отмершие и живые мхи и
лишайники; граница ровная; переход ясный.
ПОЧВОВЕДЕНИЕ № 6 1996
РОЛЬ ОКИСЛЕНИЯ СУЛЬФИДОВ
745
А1 0-11 см. Черный (N 2/); влажноватый; средний
суглинок; структура мелкокомковатая; слабоуплот¬
ненный; много корней; угли; много обломков шунги-
товой породы; граница волнистая; переход ясный.
В 11-30 см. Темно-бурый (10 YR 3/3); влажнова¬
тый; огравленный тяжелый суглинок; структура не¬
прочная ореховатая; уплотненный; мало корней; мно¬
го обломков шунгитовых пород, некоторые из облом¬
ков имеют бурые пленки; граница волнистая; переход
заметный.
ВС 30-35 см. Очень темно-серовато-бурый (2.5 Y 3/2);
влажноватый; огравленный средний суглинок; струк¬
тура глыбистая непрочная; слабоуплотненный; еди¬
ничные корни; очень много обломков шунгита; под¬
стилается трещиноватой шунгитовой породой.
Разрез 5ш/94. Заложен в 150 метрах к югу
от карьера “Максово” на выровненном участке
у подножья небольшого (2-3 м в высоту) полого¬
го склона. Растительность переходная от луговой
к лесной: подрост березы (Betula pendula), сосны
(Pinus sylvestris), клевер (Trifolium medium), мыши¬
ный горошек (Picea сгасса), костер (Bromus inermis),
ежа (Dactyllus glomerata), нивянник (Leucanthemum
vulgaris), звездчатка (Stellaria sp.), многочислен¬
ные злаки без генеративных органов. Почва ста¬
ропахотная.
Адерн 0-4 см. Бурый (10 YR 4/3); свежий; мелкозем
среднесуглинистого состава; структура бусовидная по
корням; уплотненный; дернина; граница ровная; пере¬
ход ясный по количеству корней.
А1 4—15 см. Бурый (10 YR 4/3); свежий; опесчанен-
ный средний суглинок; структура непрочная комкова¬
тая; рыхлый; много корней; много обломков шунгито¬
вых пород различного размера, покрытых бурыми и
ржавыми железистыми пленками; граница волнистая;
переход ясный.
Bfl 15-50 см. Темно-желтовато-бурый (10 YR 4/6);
влажноватый; рыхлый; тяжелый суглинок; структура
мелкоореховатая непрочная; уплотненный; мало кор¬
ней: много обломков шунгита длиной от 5 до 300 мм,
местами дезинтегрированных так, что режутся ножом
и лопатой; выделяются пятна желтовато-бурого цвета
(10 YR 5/8), диагностируемые как полностью выветре-
лые обломки шунгитовых сланцев; граница волнистая;
переход постепенный.
Bj (ярозитовый) 30-50 см. Неправильной формы
морфон на правой боковой стенке разреза; бледно-
желтый (5 Y 6/4) с крупными, до 5 см диаметром, жел¬
тыми (5 Y 8/6) пятнами; влажноватый; легкий сугли¬
нок; структура призмовидная непрочная; рыхлый; еди¬
ничные корни; много обломков шунгитовых сланцев,
в том числе значительно дезинтегрированных; пере¬
ход к окружающим горизонтам заметный.
Bf2 50-95 см. Интенсивно-бурый (7.5 YR4/6); влаж¬
новатый; огравленный тяжелый суглинок; структура
мелкоглыбистая непрочная; уплотненный; единичные
корни; очень много обломков шунгитовых сланцев
размером от дресвы до крупных камней, покрытых
бурыми и ржавыми железистыми пленками; граница
волнистая; переход заметный.
ВС 95-115 см. Темно-желтовато-бурый (10YR 3/4);
влажноватый; огравленный средний суглинок; струк¬
тура мелкоглыбистая непрочная; очень много облом¬
ков шунгитовых сланцев, переходящих в нижней части
горизонта в трещиноватую породу.
Разрез 6ш/95. Карьер “Зажогино”. Разрез за¬
ложен в 300 м к северу от карьера возле распахан¬
ного поля. Растительность луговая: тимофеевка
(Phleum pratense), осот (Sonchus arvensis), мятлик
(Роа sp.), овсяница (Festuca pratensis), одуванчик
(Taraxacum officinale), клевер (Trifolium medium).
Адерн 0-3 см. Очень темно-серый (N 3/); свежий;
дернина; уплотненный; минеральная часть по грануло¬
метрическому составу соответствует среднему суглин¬
ку; структура бусовидная по корням, непрочная; встре¬
чаются обломки шунгитовых пород размером от 1 до
200 мм; некоторые фрагменты выщелочены до полно¬
го исчезновения шунгитового вещества так, что оста¬
ется сероватая силикатная основа; граница ровная, пе¬
реход ясный по содержанию корней.
А1 5-35 см. Старопахотный горизонт; очень темно¬
серый (N 3/); влажноватый; уплотненный; средний суг¬
линок; структура крайне непрочная комковатая; мно¬
го корней; обломки породы разнообразных размеров
как выветрелые до полной вьпцелоченности, так и
слабоизмененные; на многих обломках имеются
красные пленки; местами примазки белого и красно¬
го цвета; граница ровная по плужной подошве; пере¬
ход ясный.
Bf 35-52 см. Желтовато-красный (5 YR 5/6); влажно¬
ватый; тяжелый суглинок; структура комковато-глы-
бистая; уплотненный; единичные корни; в профиле
множество обломков шунгитовых сланцев, большей
частью выщелоченных до светло-серой (N 7/) окраски
и часто дезинтегрированных до такой степени, что
легко режутся ножом и лопатой; на поверхности об¬
ломков желтовато-красные глинистые кутаны толщи¬
ной до 5 мм (рис. 26); граница языковатая; переход за¬
метный.
ВС 52-85 см. Очень темно-серовато-бурый
(2.5 Y 3/2); влажноватый; состоит преимущественно
из щебня и дресвы шунгитовых сланцев с красными,
бурыми и желтыми пленками; мелкозем занимает не
более 20% по объему; средний суглинок; плотный;
единичные корни; подстилается трещиноватой шунги¬
товой породой.
Из почв были отобраны образцы по генетиче¬
ским горизонтам для химических и минералоги¬
ческих анализов.
В лаборатории определяли значения pH вод¬
ной и солевой суспензии, гидролитическую кислот¬
ность (по Каппену), обменные водород и алюминий
(по Соколову), обменные основания (по Шоллен-
бергеру), органический углерод (по Тюрину) [1].
Также определяли железо в дитионит-цитратной
вытяжке по Мера-Джексону, железо, алюминий
и кремний в оксалатной вытяжке по Тамму и же¬
лезо, алюминий и органический углерод в пиро-
фосфатной вытяжке по Баскомбу [5]. Анализы
выполнялись сотрудниками лаборатории эколо¬
гии и географии почв Института биологии КНЦ
РАН А.А. Матулайтине и Г.И. Черкашиной.
Сульфиды и мелкозем из нескольких почвен¬
ных горизонтов профиля 5ш/94 исследовали мето¬
дом мессбауэровской спектроскопии (ядерного
ПОЧВОВЕДЕНИЕ № 6 1996
746 КРАСИЛЬНИКОВ, ВОЛОДИН
Таблица 1. Некоторые химические и физико-химические свойства дерновых шунгитовых почв
Горизонт,
глубина, см
рНн2о
РНкС!
ГК,
смоль (+)/кг
Обменная кислотность
по Соколову, смоль (+)/кг
Обменные основания по
Шолленбергеру, смоль (+)/кг
Н+
А13+
Са2+
Mg2+
К+
Na+.
Разрез 1ш/94
А1
5-25
'6.4
5.9
2.6
0.05
0.15
33.0
7.0
0.5
0.5
В
25-55
6.8
6.2
2.2
Сл.
0.05
20.8
7.2
0.5
0.8
Разрез Зш/94
А1
5-20
6.4
6.2
2.6
Сл.
0.05
41.0
7.0
0.3
0.5
В
20-50
6.9
6.4
1.3
»
0.05
21.5
1.0
0.5
0.4
Разрез 4ш/94
А1
0-11
4.6
3.9
10.9
0.30
3.20
2.5
2.5
0.2
0.3
В
11-30
4.4
3.4
24.9
7.10
5.70
1.5
1.5
0.1
0.3
ВС
30-35
4.3
3.2
17.5
3.00
5.80
2.0
2.0
0.1
0.3
Разрез 5ш/94
А1
4-15
. 6.0
4.7
8.7
0.10
0.20
.24.0
13.0
0.7
0.4
Bfl
15-50
5.0
3.8
10.5
0.10
6.90
7.0
6.5
0.4
0.3
Bj
30-50
4.5
2.9
24.1
15.70
5.80
1.5
11.0
0.4
0.2
Bf2
50-95
5.0
3.8
10.5
0.10
5.20
6.0
8.5
0.4
0.2
ВС
95-115
5.1
4.0
8.7
0.05
3.70
9.0
5.3
0.2
0.2
гамма-резонанса - ЯГР) для идентификации со¬
единений железа. Спектры были получены и ин¬
терпретированы сотрудником лаборатории ра¬
диоспектроскопии и магнитных методов ВИМС
В.В. Коровушкиным. Мессбауэровские измере¬
ния проводили при комнатной температуре на
спектрометре ЯГРС-4М с источником Со2+ в мат¬
рице хрома. Использовали порошковые пробы
крупностью 0.05-0.07 мм. Калибровку шкалы
скоростей проводили цо ЯГР-спектру a-Fe203.
Изомерный сдвиг рассчитывали относительно
нержавеющей стали. Спектры были разложены
на составляющие с использованием лоренцов-
ской формы линий поглощения. Вероятности ре¬
зонансного эффекта железа присутствующих
фаз и валентных форм принимались равными.
Илистые фракции из почв выделяли отмучи-
ванием по Горбунову, насыщали магнием и ис¬
следовали в ориентированных образцах в воз¬
душносухом состоянии, после прокаливания до
температуры 550°С и после насыщения глицери¬
ном. Рентген-дифрактометрические исследования
проведены на универсальном рентген-дифрактоме¬
тре ДРОН-3, излучение медное, Ка, 2. Применялся
графитовый монохроматор. Режим работы аппара¬
та: напряжение на трубке 30 кВ, анодный ток
25 мА, скорость вращения гониометра град/мин
(26), скорость протяжки диаграммы 600 мм/ч,
предел измерения 400 имп./с, постоянная време¬
ни 2 с. Съемку производил научный сотрудник
лаборатории физических методов исследований
Института геологии КНЦ РАН А.М. Софронов.
РЕЗУЛЬТАТЫ
Химические и физико-химические свойства.
Реакция среды темноцветных почв из разрезов
1ш/94 и Зш/94 слабокислая, типичная для дерно¬
вых шунгитовых почв (табл. 1). Значения pH мор¬
фологически ожелезненных почв из разрезов
4ш/94 и 5ш/94 значительно ниже. Минимальные
значения pH наблюдаются в гор. Bj разреза 5щ/94.
Гидролитическая кислотность также значительно
выше в железистых почвах, чем в темноцветных.
Существенно выделяются железистые почвы по
содержанию обменного алюминия: во всех их го¬
ризонтах его количество на два порядка выше,
чем в темноцветных почвах. Содержание обмен¬
ного водорода в большей части горизонтов же¬
лезистых почв соотносимо с его содержанием в
темноцветных шунгитовых почвах, и только в
гор. В и ВС разреза 4ш/94 и гор. Bj разреза 5ш/94
оно выше на два порядка. Содержание обменно¬
го кальция высоко во всех почвах, но в темно-
цветных оно выше в несколько раз. В железис¬
тых почвах высоким содержанием обменного
кальция выделяется старопахотный гор. А1 разреза
ПОЧВОВЕДЕНИЕ № 6 1996
РОЛЬ ОКИСЛЕНИЯ СУЛЬФИДОВ 747
Таблица 2. Содержание оксидов в селективных вытяжках из дерновых шунгитовых почв, %
Горизонт,
Fe203 по
Оксиды по Тамму
Оксиды по Баскомбу
глубина, см
Мера-Джексону
Fe203
ai2o3
Si02
Fe203
А1203
С
Разрез 1ш/94
А1
5-25
1.34
0.47
0.59
0.10
0.26
0.03
0.60
В
25-55
1.22
0.34
0.27
0.13
0.19
0.03
0.28
°азрез Зш/94
А1
5-20
1.54
0.50
0.63
0.33
0.16
0.04
0.76
В
20-50
2.02
0.41
0.30
0.18
0.18
0.03
0.35
Разрез 4ш/94
А1
0-11
5.44
1.45
0.74
0.11
0.42
0.04
0.47
В
11-30
12.80
2.92
0.93
0.11
1.34
0.08
0.80
ВС
30-35
7.04
2.00
0.50
0.11
0.94
0.04
0.35
Разрез 5ш/94
А1
4-15
Не опр.
1.15
0.41
0.13
0.42
0.03
0.12
Bfl
15-50
»
0.55
0.43
0.09
0.53
0.03
Сл.
Bj
30-50
»
0.88
0.43
0.31
0.24
Сл.
»
Bf2
50-95
0.88
0.32
0.09
0.26
»
»
ВС
95-115
»
2.00
0.40
0.14
0.41
»
»
5ш/94. Аккумулятивное распределение обменно¬
го кальция в темноцветных почвах подтверждает
преимущественно биогенное его накопление. Об¬
менного магния содержится меньше, чем каль¬
ция, во всех почвах. В железистых и темноцвет¬
ных почвах магний содержится в поглощающем
комплексе в соотносимых количествах. Содер¬
жание обменных калия и натрия незначительно,
от ожелезнения почв не зависит. Содержание ор¬
ганического углерода велико во всех профилях, в
темноцветных оно несколько выше. Однако ин¬
терпретировать цифры, полученные при опреде¬
лении органического углерода в данных почвах,
следует с большой осторожностью, поскольку в
шунгитовых почвах при окислении углерода ор¬
ганического вещества частично окисляется и уг¬
лерод шунгитов [18]. Кроме того, известно, что
при определении органического углерода методом
мокрого озоления в сульфидсодержащих почвах
на результаты влияет окисление сульфидов [29].
Содержание железа, извлекаемого дитионит-
цитратной вытяжкой, в темноцветных шунгито¬
вых почвах, составляет от 1.2 до 2% (табл. 2).
В железистом профиле оно достигает 12% в гор. В.
Содержание оксалат-извлекаемого железа (отно¬
симого к тонкодисперсным оксидам и гидрокси¬
дам) в железистых почвах в 2-5 раз выше, чем в
темноцветных. Существенная часть оксидов и ги¬
дроксидов находится в наиболее тонкодисперс¬
ном и слабоокрисгаллизованном виде и извлека¬
ется пирофосфатной вытяжкой [21]. Алюминия
извлекается оксалатной вытяжкой соотносимое
количество из темноцветных и железистых почв.
Органического углерода пирофосфатной вытяж¬
кой извлекается существенное количество из
темноцветных почв. В железистых почвах (разр.
5ш/94), напротив, органический углерод в пиро¬
фосфатной вытяжке обнаруживается в следовых
количествах.
Мессбауэровская спектроскопия. ЯГР-спектр
сульфида из прожилок в шунгитовой породе
представляет собой дублет с изомерным сдвигом
0.45 мм/с и квадрупольным расщеплением 0.65 мм/с
(рис. За). Сульфид интерпретируется как пирит
[7]. Железосодержащих примесей не идентифи¬
цируется.
Поскольку в образцах мелкозема предполага¬
лось наличие ярозита (KFe3(S04)2(0H)6), а в лите¬
ратуре ЯГР-спектров ярозита нам не удалось об¬
наружить, был отснят образец ярозита гипер-
генного происхождения с поверхности скалы,
прилегающей к жиле гидротермальной сульфид¬
ной минерализации на мысе Картеш (Канда¬
лакшский залив Белого моря) [9]. Спектр пред¬
ставляет собой дублет с изомерным сдвигом
0.58 мм/с и квадрупольным расщеплением 1.18 мм/с
(рис. 36).
ЯГР-спектры образцов мелкозема из гор. Bf2
и ВС разреза 5ш/94 близки (рис. 4). Преимущест¬
венным в них является дублет с изомерным сдви¬
гом 0.42-0.45 мм/с и квадрупольным расщеплением
0.6 мм/с. В обоих спектрах отмечается появление
ПОЧВОВЕДЕНИЕ № 6 1996
748
Л/x 1(H
320 -
310
300
290
-2
КРАСИЛЬНИКОВ, ВОЛОДИН
a N x 10_3
320
310
4
v, мм/с
300
290
-2
4
V, мм/с
Рис. 3. Мессбауэровские (ЯГР) спектры: а - пирита из прожилки в шунгитовой породе (карьер “Зажогино”); б - яро-
зита (гипергенный, Северная Карелия [9]).
N х 10'3
Рис. 4. Мессбауэровские (ЯГР) спектры мелкозема из профиля дерновой шунгитовой железистой почвы (5ш/94):
а - гор. Bf2; б - гор. ВС.
магнитной сверхтонкой структуры (СТС), не¬
сколько большей в гор. ВС. По параметрам дуб¬
летов их можно связать с тонкодисперсными гид¬
роксидами железа (гетитом, гидрогетитом),
в которых часть частиц имеет размер выше кри¬
тического, соответствующего переходу из супер-
парамагнитного состояния в магнитоупорядочен¬
ное, чем и обусловлено появление СТС. В гор. ВС,
соответственно, таких частиц несколько больше.
Кроме того, в образце из гор. ВС линии дублета
уширены, что может быть вызвано присутствием
других железистых фаз (гидрослюд, ярозита,
пирита).
ЯГР-спектр мелкозема из гор. Bj разреза
5ш/94 представляет собой асимметричный дублет
с изомерным сдвигом 0.SS мм/с и квадрупольным
расщеплением 1.15 мм/с (рис. 5). При ошибке
0.02 мм/с он может быть идентифицирован как
принадлежащий ярозиту (рис. 36). Кроме дублета
ярозита, в ЯГР-спектре фиксируется дублет Fe2+
силикатной фазы с изомерным сдвигом 1.4 мм/с и
квадрупольным расщеплением 2.45 мм/с, кото¬
рый можно приписать гидрослюдам.
Рентгеновские исследования. Профиль 1ш/94
слабо дифференцирован по минералогическому
составу илистой фракции (рис. 6, 7,2). Доминиру¬
ющей минеральной фазой является кварц, что,
очевидно, связано с его преобладанием в силикат¬
ной основе шунгитовых сланцев района карьера
“Зажогино” над слоистыми силикатами. Присут¬
ствуют также слюда и вермикулит. По соотноше¬
нию первого и второго базального рефлексов
слюда диагностируется как триоктаэдрическая
с небольшим содержанием железа в октаэдрах
ПОЧВОВЕДЕНИЕ № 6 1996
РОЛЬ ОКИСЛЕНИЯ СУЛЬФИДОВ
749
N х КГ1
Рис. 5. Мессбауэровские (ЯГР) спектры мелкозема из
горизонта Bj дерновой шунгитовой железистой поч¬
вы (5ш/94).
(биотит, близкий к флогопиту). В гор. А1 интен¬
сивность рефлексов биотита и вермикулита ни¬
же, чем в нижележащем горизонте. Это может
быть связано как с потерей структуры указанны¬
ми минералами в верхнем горизонте почвы, так и
с тем, что частицы минералов покрыты пленками
органики. Относительное содержание биотита
в гор. А1 также ниже, чем в нижележащем гори¬
зонте. В гор. А1 отмечается присутствие незначи¬
тельного количества смешанослойного слюда-
вермикулита, диагностируемого по “плечу” в об¬
ласть больших углов на рефлексе 14.2 А.
Илистая фракция почвы гор. А1 ожелезненного
профиля 4ш/94 (рис. 6,3) сходна по минералогиче¬
скому составу с таковой из профиля 1ш/94. Здесь
также представлены кварц, биотит и вермику¬
лит. Относительное содержание кварца в образ¬
цах из разреза 4ш/94 также меньше, что связано
с меньшим содержанием кварца в шунгитовых
сланцах района карьера “Максово”. В илистой
фракции гор. В разреза 4ш/94 биотит отсутст¬
вует (рис. 6, 4). Преобладающей минеральной
фазой, помимо кварца, является вермикулит.
По слабому рефлексу 11 А в прокаленном образ¬
це диагностируется присутствие смешанослойно¬
го хлорит-вермикулита с незначительным содер¬
жанием хлоритовых пакетов. В илистой фракции
гор. ВС того же профиля присутствуют кварц,
вермикулит, биотит, а также смешанослойный
слюда-вермикулит, диагностируемый по рефлек¬
су 12.4 А. Высокое содержание слюды в гор. А1 дан¬
ного профиля было нами приписано эоловому при¬
внесу и необратимой дегидратации вермикулита
Рис. 6. Рентгеновские дифрактограммы илистых (< 0.001 мм) фракций дерновой шунгитовой темноцветной (1ш/94)
и дерновой шунгитовой железистой почв: I - разр. 1ш/94, гор. А1 (10-20 см); 2 - разр. 1ш/94, гор. А1В (30-40 см);
3 - разр. 4ш/94, гор. А1 (0-10 см); 4 - разр. 4ш/94, гор. В (20-30 см); J - разр. 4ш/94, гор. ВС (30-35 см); а - воздушносухой
препарат; 6 - прокаленный при температуре 550°С; в - насыщенный глицерином.
ПОЧВОВЕДЕНИЕ № 6 1996
750
КРАСИЛЬНИКОВ, ВОЛОДИН
Рис. 7. Рентгеновские дифрактограммы илистых (<0.001 мм) фракций дерновой шунгитовой железистой почвы
(5ш/94): 1 - гор. А1 (4-15 см); 2 - гор. Bfl (15-50 см); 3 - гор. Bj (30-50 см); 4 - гор. Bf2 (50-95 см); 5 - гор. ВС (95-115 см);
а, б, в-то же, что и на рис. 6.
в результате пожара (диагностируемого по оби¬
лию свежих углей в горизонте).
В иле из гор. А1 разреза 5ш/94 (рис. 1,1) обна¬
руживается вермикулит и существенно упорядо¬
ченный смешанослойный слюда-вермикулит
с содержанием вермикулитовых пакетов 60-70%.
В меньших количествах содержатся биотит и
кварц. Наличие лепидокрокита (рефлекс 6.27 А)
и гетита (рефлекс 4.18 А> предполагается. Однако
те же рефлексы могут быть кажущимися базаль¬
ными отражениями смешанослойного минерала.
В тонкой пыли из того же горизонта (рис. 8,1) при¬
сутствуют биотит, вермикулит, кварц и гетит.
Смешанослойного минерала не обнаруживается.
То, что рефлекс вермикулита в илистой фракции
сжимается при прокаливании до 10 А, а в тонко¬
пылеватой - до 9.7 А, связано с неполной
дегидратацией вермикулита в илистых части¬
цах [16].
Илистая фракция из гор. Bfl того же разреза
(рис. 7,2) содержит вермикулит, гетит и незначи¬
тельные количества биотита и кварца.
Ил из гор. Bj (рис. 7, 3) содержит вермикулит,
незначительные количества биотита и кварца и
существенное количество слюда-вермикулита с
тенденцией к упорядоченности. Содержание вер¬
микулитовых пакетов определяется в 60-70%.
В тонкопылеватой фракции из того же горизонта
(рис. 8,2) упорядоченность смешанослойного ми¬
нерала возрастает, на что указывает наличие сум¬
марного рефлекса 2S.2 А. Интенсивный рефлекс
3.08 А был отнесен нами к ярозиту, однако, не ис¬
ключено, что это кажущееся отражение смеша¬
нослойного минерала.
Ил из гор. Bf2 (рис. 7,4) содержит вермикулит,
биотит, кварц, гетит и незначительные количества
хлорита и смешанослойного слюда-вермикулита.
Ил из гор. ВС (рис. 7, 5) состоит преимущест¬
венно из вермикулита. Биотит и кварц присутст¬
вуют в малых количествах. В тонкопылеватой
фракции того же горизонта (рис. 8, 3) обнаружи¬
вается смешанослойный слюда-вермикулит.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Окисление пирита в почвах приводит к обра¬
зованию своеобразных ожелезненных профилей.
При окислении пирита выделяется серная кисло¬
та. Реально процесс окисления сульфидов более
сложен [26], однако, выделение серной кислоты
на определенном этапе происходит практически
всегда. Действие серной кислоты на почву двоя¬
ко. С одной стороны, внесение сильной минераль¬
ной кислоты приводит к подкислению почвенно¬
го раствора и, следовательно, к усилению гидроли¬
за силикатов и увеличению активности ряда
элементов, таких как алюминий и железо. С другой
ПОЧВОВЕДЕНИЕ № 6 1996
РОЛЬ ОКИСЛЕНИЯ СУЛЬФИДОВ
751
Рис. 8. Рентген-дифрактограммы тонкопылеватых (0.001-0.005 мм) фракций дерновой шунгитовой железистой
почвы: / - гор. А1 (4-15 см); 2 - гор. Bj (30-50 см); 3 - гор. ВС (95-115 см); а, б, в- то же, что и на рис. 6.
стороны, серная кислота как сильныи окислитель
вызывает интенсивную минерализацию органи¬
ческого вещества и окисление структурного же¬
леза в слоистых силикатах. Окисление органиче¬
ского углерода шунгитов серной кислотой оче¬
видно из рис. 1. Окисление пиритных прожилок
приводит к “выгоранию” углерода шунгитов на
контакте с пиритом. Иногда углерод из обломков
шунгита полностью выщелочен. Аналитически
окисление углерода диагностируется по его низ¬
кому содержанию в пирофосфатной вытяжке из
всех, кроме старопахотного, горизонтов разреза
5ш/94. Окисление углерода приводит к ускоре¬
нию образования мелкозема, т.к. углерод являет¬
ся одним из цементирующих агентов в шунгито¬
вых сланцах.
Действие серной кислоты на слоистые силика¬
ты также определяется ее кислотными и окисли¬
тельными свойствами. При незначительных кон¬
центрациях ее действие сводится к ускорению де-
градационной трансформации слюд и хлоритов.
Для молодых приморских почв Швеции, содержа¬
щих седиментогенный пирит, сообщалось о пре¬
образовании хлорита и слюд в вермикулит [27].
В канадских почвах на сульфидсодержащей ло¬
кальной морене обнаружено преобразование слюд
в разбухающий минерал [22]. Однако при высо¬
ких концентрациях серной кислоты происходит
быстрое окисление октаэдрического железа в
триоктаэдрических слюдах, что приводит к поте¬
ре структуры кристаллической решеткой [28].
В нашем случае органический углерод шунгитов
служит своеобразным буфером, на который расхо¬
дуется окислительный потенциал серной кислоты.
Морфологический облик дерновых шунгито¬
вых железистых почв определяется присутствием
гетита, гематита и ярозита. Последние два мине¬
рала являются типичными именно для почв, фор¬
мирующихся при окислении сульфидов. Ярозит
является типичным метастабильным продуктом
окисления сульфидов железа. Нго формирование
связано с тем, что он стабилен при низких значени¬
ях pH [25] и высокой активности серы. Гематит,
редко формирующийся в гипергенных условиях
в холодных гумидных областях, отмечался как ха¬
рактерный для современного сульфидсодержаще¬
го элювия даже в очень холодных условиях [17].
Это связано с тем, что при формировании зоны
окисления сульфидов химические реакции, при¬
нимающие участие на стадиях формирования
сульфатов, карбонатов и оксидов железа, являют¬
ся экзотермическими [3]. По морфологии эти поч¬
вы и подстилающий элювий могут рассматривать¬
ся как особая кора выветривания - зона окисления
сульфидных месторождений [13], поскольку на¬
блюдается характерная последовательность суль¬
фидной, сульфатной (ярозитовой) и гетит-гемати-
товой зон. Однако силикатная часть элювия име¬
ет характерный для этих широт гидрослюдистый
состав [6], почти не зависящий от преобразования
сульфидов. Очевидно, в данном случае следует
говорить о наложении двух кор выветривания;
обломочно-гидрослюдистой и зоне окисления
сульфидов.
Слоистые силикаты в темноцветных и желези¬
стых профилях трансформированы практически
в равной степени. Отмечается возрастание доли
ПОЧВОВЕДЕНИЕ № 6 1996
752
КРАСИЛЬНИКОВ, ВОЛОДИН
вермикулита за счет биотита в образцах из желе¬
зистых профилей. Некоторая асимметрия ре¬
флекса 10 А в область малых углов в образцах из
ряда ожелезненных горизонтов (рис. 6, 46, 7, 26)
указывает на некоторую хлоритизацию вермику¬
лита. Высокая активность алюминия в профилях,
подверженных действию продуктов окисления
сульфидов, теоретически может приводить к воз¬
никновению фрагментарных гиббситовых просло¬
ев в вермикулите, что и фиксируется на дифрак-
тограмме как хлоритизация. Также отмечается
наличие смешанослойного слюда-вермикулита
в разрезе 5ш/94, причем максимальное его содер¬
жание наблюдается в ярозитовом горизонте, где
процесс окисления сульфидов еще не завершен.
Это позволяет предположить, что смешанослой-
ный минерал является промежуточным продук¬
том деградационной трансформации биотита.
С другой стороны, в профиле 4ш/94 подобного
минерала не обнаружено. Возможно, образова¬
ние слюда-вермикулитов с тенденцией к упорядо¬
ченности является характерным для железистых
почв, однако без массовых определений минера¬
логического состава говорить об этом было бы
преждевременным.
Высокое содержание железа в дерновых шун¬
гитовых железистых почвах связано как с его вы¬
делением при окислении пирита, так и с тем, что
при деградационной трансформации биотита
происходит диоктаэдризация последнего с вы¬
бросом избытка железа из октаэдров.
Высокая кислотность и сравнительно невысо¬
кое содержание обменных оснований связаны
с происходившим (и отчасти происходящим сей¬
час) выделением серной кислоты. Поглощающий
комплекс насытился водородом и алюминием.
Кроме того, углерод шунгитов, определяющий
поглотительную способность дерновых шунгито¬
вых почв, в основном окислился. Большую часть
коллоидной фракции по,чвы составляют гидро¬
ксиды железа, имеющие низкую поглотительную
способность.
Несмотря на то, что морфология и свойства
изученных нами почв определяются процессом
окисления сульфидов, выделять их как сульфат¬
нокислые не следует. Из формальных признаков
сульфатнокислых почв [23] присутствует ярози-
товый горизонт, но он встречается не во всех
профилях и pH водной вытяжки из образца
почвы этого горизонта выше 3.5. Поэтому пред¬
лагается включать эти почвы, как и предлага¬
лось ранее [8, 11], в подтип дерновых литогенных
ненасыщенных почв, но выделять в отдельный
род дерновых шунгитовых железистых.
СПИСОК ЛИТЕРАТУРЫ
1. Аринушкина Е.В. Руководство по химическому
анализу почв. М.: Изд-во МГУ, 1970. С. 103, 311,
327.
2. Атлас Карельской АССР. М., 1989. С. 14.
3. Бугелъский Ю.Ю. Гипергенная миграция рудных
компонентов в различных климатических районах //
Кора выветривания. Вып. 4. М.: Изд-во АН СССР.
С. 261-287.
4. Володин А .М., Тойкка МА. К вопросу о географи¬
ческой закономерности распространения дерно¬
вых шунгитовых почв Заонежского района Ка¬
рельской АССР // Уч. зап. Петрозаводского гос-
университета. Биол. науки. 1956. Вып. 7. N° 3.
С. 138-157.
5. Воробьева ЛА., Гриндель Н.М. Подготовка почв к
валовому и фазовому анализу. М.: Изд-во МГУ,
1989. С. 66.
6. Градусов Б.П., Черняховский А.Г Генетико-гео¬
графические закономерности состава почвообра¬
зующих пород на карте Мира //Глобальная геогра¬
фия почв и факторы почвообразования. М., 1991.
С. 196-301.
7. Дериватограммы, инфракрасные и мессбауэров-
ские спектры стандартных образцов фазового со¬
става: Дополнение к каталогу. СПб., 1992. 128 с.
8. Иванова Е.Н. Классификация и диагностика почв
Карелии // Классификация и диагностика почв
СССР. М.: Наука, 1976. С. 104-144.
9. Красильников П.В., Сафонова В.М. Сульфатно¬
кислый процесс в почвах Карелии // Контроль со¬
стояния и регуляция функций биосистем на раз¬
ных уровнях организации. Петрозаводск, 1993.
С. 126-134.
10. Марченко А.И. Почвы Карелии. М.; Л.: Изд-во АН
СССР, 1962. С. 12.
11. Почвы Карелии. Петрозаводск, 1981. С. 39.
12. Пудовкин В.Г., Гульмалиева Т.Е. Влияние процес¬
сов выветривания на строительные свойства шун¬
гитов Максово-Зажогинского месторождения //
Комплексное и рациональное использование ми¬
нерального сырья Карелии. Петрозаводск, 1986.
С. 133-139.
13. Смирнов С.С. Зона окисления сульфидных место¬
рождений. М.; Л.: Изд-во АН СССР, 1955. С. 57.
14. Тойкка МА. Физические и химические свойства
темноцветных (шунгитовых) почв Карело-Фин-
ской ССР // Уч. зап. Карело-Финского госунивер-
ситета. Биол. науки. 1954. Т. 6. Вып. 3. С. 149-182.
15. Тойкка М.А.У Володин А.М. Дерновые слабопод¬
золистые шунгитовые почвы Кондопожского рай¬
она// Вопр. почвов. Карельской АССР. 1961. Вып. 2.
С. 3-23.
16. Уокер Г.Ф. Вермикулитовые минералы // Рентге¬
новские методы изучения и структура глинистых
минералов. М.: Мир, 1965. С. 284-344.
17. Черняховский А.Г. Элювий и продукты его пере-
отложения. М.: Наука, 1966. 85 с.
18. Шекк Г.В., Градусов Б.П., Урусевская И.С., Сквор¬
цова И.Н. К структурно-генетической неоднородно¬
сти гумуса шунгитовых почв // Вестн. МГУ. Сер. 17.
Почвоведение. 1993. № 4. С. 9-24.
19. Шекк Г.В., Градусов Б.П., Урусевская И.С. Осо¬
бенности химического состава тонкодисперсного
ПОЧВОВЕДЕНИЕ № 6 1996
РОЛЬ ОКИСЛЕНИЯ СУЛЬФИДОВ
753
материала почв, развитых на шунгитах и шунгитсо¬
держащих моренных отложениях Карелии // Вестн.
МГУ. Сер. 17. Почвоведение. 1993. № 3. С. 32-45.
20. Шунгиты Карелии и пути их комплексного исполь¬
зования. Петрозаводск: Карелия, 1975. С. 34.
21. Borggaard O.K. Phase identification by selective disso¬
lution techniques // Iron in soils and clay minerals, ed.
J.W. Stucki, B.A. Goodman & U. Schwertmann. NATO
ASI. 1987. Ser. С. V. 217. P. 83-98.
22. De Kimpe C., Miles N. Formation of swelling clay min¬
eral by sulfide oxidation in some metamorphic rocke and
related soil of Ontario, Canada // Can. J. Soil Sci. 1992.
V. 72. № 3. P. 263-270.
23. Keys to Soil Taxonomy. USDA. Soil Concervation Ser¬
vice. 1994. P. 13.
24. Muncell Soil Color Charts. Baltimore: Md., 1975.
25. Nickel E.H. An unusual pyrite-sulphur-jarosite assem¬
blage from Arkaroola, South Australia // Miner. Mag.
1984. V. 48. №2. P. 139-142.
26. Nordstrom D.K. Aqueous pyrite oxidation and the con¬
sequent formation of secondary iron minerals // Acid
sulfate weathering. SSSA Special Publ. 1982. V. 10.
P. 37-56.
27. Obom /. Some effects of chemical weathering in three
cultivated acid sulfate soils in Sweden // Dev. Plant Soil
Sci. 1991. V. 45. P. 55-63.
28. Ross G.J., Eilers R.G., lvarson K.C. Influence of acid
sulfate weathering on chemical and mineralogical prop¬
erties of an acid sulfate soil of Manitoba // Can. J. Soil
Sci. 1988. V. 68. № 3. P. 629-644.
29. Wilett l.R. Determination of organic carbon in pyritic
and acid sulfate soils // Commun. Soil Sci. Plant Anal.
1987. V. 18. №7. P. 715-724.
The Role of Sulfide Oxidation in Genesis
of Ferrous Sod-Shunghite Soils in Karelia
P. V. Krasilnikov, A. M. Volodin
Ochric and red-coloured soil profiles are mainly formed on sulfide-beaming carbonic (shunghite) shists in
Karelia. Sulfide oxidation leads to shunghite carbon leaching, exchange bases loss and the acidity increase. Soil
colouring is explained by presence of goethite, hematite and jarosite. Biotite, vermiculite and partially ordered
interstratified mica-vermiculites occur in clay fractions of soils under study.
5 ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВОВЕДЕНИЕ, 1996, № 6, с. 754-762
УДК 631.4(213.5)
ГЕНЕЗИС
И ГЕОГРАФИЯ ПОЧВ
ЭВОЛЮЦИЯ И РАЗВИТИЕ ПОЧВ В ПРИАРАЛЬЕ
ПРИ АНТРОПОГЕННОМ ОПУСТЫНИВАНИИ*
©1996 г. Г. С. Куст
Факультет почвоведения МГУ им. М.В. Ломоносова
Поступила в редакцию 17.09.93 г.
Методологическая база и понятийный аппарат теории эволюции почв применены к анализу меха¬
низмов устойчивости и движущих сил изменения почв и почвенного покрова в Приаралье, террито¬
рия которого испытывает в настоящее время интенсивное воздействие антропогенного опустыни¬
вания. Проанализированы отличия эволюционных состояний почв и почвенного покрова. Для оцен¬
ки устойчивого состояния почвенного покрова предложено использовать понятие гомеостаза
почвенного покрова. Предложен принцип определения границ гомеостатических территорий.
Изучение устойчивости почв и почвенного по¬
крова к антропогенным воздействиям относится
к числу важнейших теоретических и прикладных
проблем современного естествознания. В почвове¬
дении накоплен обширный материал, позволяю¬
щий судить в разных конкретных случаях о степени
изменения почв, скорости происходящих измене¬
ний, обусловленных теми или иными воздействи¬
ями. Однако, к сожалению, вероятно, в связи с
чрезвычайным разнообразием антропогенных
воздействий, действующих на фоне исходно мно¬
гообразных почв, до настоящего времени отсут¬
ствуют методологическая база и понятийный ап¬
парат для оценки устойчивости почв и почвенного
покрова. Поэтому, хотя понятия устойчивости, ста¬
бильности, равновесия, толерантности, буфернос-
ти и т.п. стали достаточно общеупотребительны¬
ми в науке о почвах, до сих пор смысловые грани¬
цы этих понятий остаются недостаточно четко
очерченными. Критерий и индикаторы, которы¬
ми пользуются при диагностике степени устойчи¬
вости, оказываются, как правило, чрезвычайно
индивидуальными и применимыми только в кон¬
кретных целях какого-либо одного исследования,
для какой-то одной территории.
Вместе с тем, как нам представляется, в неко¬
торых областях почвоведения на сегодняшний
день существует ряд фундаментальных разрабо¬
ток, напрямую не связанных с теорией устойчи¬
вости почв, однако, разработанный понятийный
аппарат которых позволяет приблизиться к пони¬
манию механизмов устойчивости почв.
В данной работе нами сделана попытка приме¬
нить методологическую базу и понятийный аппа¬
рат теории эволюции почв, разрабатываемой в
* Работа выполнена при финансовой поддержке Российско¬
го фонда фундаментальных исследований, ЮНЕСКО и
BMFT Германии, ГНТП России.
работах [8,9,10], к анализу механизмов устойчиво¬
сти и движущих сил изменения почв и почвенного
покрова на примере такой чрезвычайно нестабиль¬
ной территории, как Приаралье, испытывающей
в настоящее время интенсивное воздействие антро¬
погенного опустынивания. Проанализированы от¬
личия эволюционных состояний почв и почвенно¬
го покрова. Для оценки устойчивого состояния
почвенного покрова предложено использовать
понятие гомеостаза почвенного покрова.
ДВИЖУЩИЕ СИЛЫ
ЭВОЛЮЦИОННЫХ ИЗМЕНЕНИЙ
В более ранних работах [4, S] мы уже отмечали,
что разнообразие почвенного покрова в Приара¬
лье тесно связано не только с особенностями ли¬
тологических, геоморфологических и гидрологи¬
ческих условий их формирования, но и с их эво¬
люцией.
Эволюционные изменения, в силу относитель¬
ной молодости почв дельтовых равнин, охватыва¬
ют практически все почвенные разности, практи¬
чески все территории. Так, на территории обсох¬
шего дна Аральского моря в настоящее время
происходит становление молодых почв на постак-
вальных поверхностях. На территории современ¬
ных неорошаемых дельт Амударьи и Сырдарьи
наблюдается сложный комплекс эволюционных
изменений почв в направлении их аридизации, ме¬
стами и временами осложненных нерегулярным
обводнением старых русел и межрусловых пони¬
жений. На территории древнедельтовых опусты¬
ненных равнин в зависимости от времени, про¬
шедшего с момента прекращения их обводнения
или орошения, прослеживаются завершающие
этапы эволюции почв в направлении их отакыри-
вания или опесчанивания. Здесь же получают
754
ЭВОЛЮЦИЯ И РАЗВИТИЕ ПОЧВ В ПРИАРАЛЬЕ
755
начальное развитие эволюционные и деградацион-
ные изменения, вызванные такими комплексными
антропогенными воздействиями, как отгонное жи¬
вотноводство, техногенное воздействие, сведение
древесной и кустарниковой растительности. Даже
на территориях современных песчаных пустынь
Кызылкумов и Каракумов, столовых и останцо-
вых возвышенностей, предгорных наклонных
равнин эволюция почв не прекращается. Здесь
почвы также никогда не находятся в равновесии,
будучи подвержены циклическим изменениям де¬
градации-восстановления в связи с периодически¬
ми или спорадическими антропогенными воздей¬
ствиями, перечисленными выше. Эволюционные
изменения почв в ежегодном цикле отмечаются
и на орошаемых территориях. В связи с близким
залеганием минерализованных грунтовых вод на
орошаемых массивах циклы засоления-рассоле¬
ния обусловливают ежегодные эволюционные
изменения почв в направлении образования солон¬
чаков. После ежегодных осенних и/или весенних
промывок циклы возобновляются. На орошае¬
мых землях нового освоения эволюция почв до¬
полняется формированием агроирригационных
горизонтов.
Особенности движущих сил эволюционных
изменений почв Приаралья можно проследить на
примере отношения этих изменений к состоянию
равновесия почв, почвенного покрова и природ¬
ных систем в целом.
Все разнообразие почв Приаралья, в том числе
и промежуточные эволюционирующие вариан¬
ты, было сведено нами к нескольким генерализо¬
ванным эволюционным эколого-генетическим
рядам. Анализ этих рядов показывает, что любые
трансформации природных систем в целом и поч¬
венного покрова, в частности, в аридных зонах
обусловлены изменением динамического равно¬
весия между условиями почвообразования. При
смещении равновесия в сторону какого-либо из
них возникает некий эволюционный градиент,
обусловливающий напряженность и скорость
эволюционных процессов. Чем более противоре¬
чивы факторы, в комплексе обусловливающие
формирование природных систем и почвенного
покрова той или иной территории, тем более кар¬
динальные изменения охватывают экосистемы в
Целом и почвенный покров и почвы, в частности,
в ходе их эволюции.
В Приаралье основные противоречия, опреде¬
ляющие эволюцию почв и почвенного покрова,
как правило, состоят в различии, с одной сторо¬
ны, аутоэволюционных онтогенетических изме¬
нений почв и почвенного покрова в сторону до¬
стижения равновесия с аридным макроклиматом,
Который выступает здесь в роли постоянного и
повсеместно действующего фактора зонального
почвообразования; а с другой - аллоэволюцион-
Гидрогенный
кваэиклимакс
(а)
(б)
кваэиклимакс
-~7 — S - -9 тю □ и
■12
Рис. 1. Основные направления эволюции почв и поч¬
венного покрова в современных “живых” и опустыни¬
вающихся дельтах (а) и современных пустынях (б):
а: 1 - гидрогенные скачки; 2 - направление аутоэво¬
люции; 3 - направление гидрогенной аллоэволюции;
4 - циклоидные процессы; 5 - промежуточные состо¬
яния почв и почвенного покрова, в том числе - поли-
генетические квазиклимаксы; 6 - направленность
опустынивания; б: 7 - пасторальные и техногенные
скачки; 8 - направление аутоэволюции; 9 - направ¬
ления пасторальной и техногенной аллоэволюции;
10 - циклоидные процессы; // - промежуточные со¬
стояния почв и почвенного покрова, в том числе - по-
лигенетические квазиклимаксы; 12 - направленность
опустынивания.
ных изменении, протекающих под воздействием
антропогенных или естественных причин локаль¬
ного действия, смещающих равновесие почв и
почвенного покрова с аридным макроклиматом
или осложняющих ход указанных аутоэволюци¬
онных изменений.
С этой точки зрения движущие силы практи¬
чески всех видов эволюционных изменений почв
и почвенного покрова в Приаралье являются од¬
нотипными (рис. 1).
На рисунке показано, что на фоне единой для
всех неравновесных или квазиравновесных поч¬
венных систем тенденции к достижению устойчи¬
вого равновесия с наиболее мощным в регионе и
повсеместно действующим фактором - аридным
макроклиматом - постоянно возникают дополни¬
тельные локальные воздействия, как бы "отбра¬
сывающие'’ почвы в их аутоэволюционном раз¬
витии “назад”. При этом, если эти дополнитель¬
ные действия носят циклоидный характер, то
возникают условия для поступательной аллоэво¬
люции под влиянием дополнительного локально¬
го фактора или факторов, а если эти действия
ПОЧВОВЕДЕНИЕ № 6 1996
5*
756 КУСТ
[виды состояний;
I
I
т
•аугоэволюция1
'АЛЛОЭВОЛЮЦИЯ1
1 СКАЧОК 1
1 1
1 КЛИМАКС 1
1 1
' КВАЗИКЛИМАКС1
1 1
1 1
| Изменения ' I
1 1
I Изменения I
1 1
I Изменения i
1 1
| Изменения i
1 1
I Изменения I
1 циклоидные 1
1 циклоидные 1
1 резкие 1
1 медленные 1
1 циклические 1
1 1
1 1
, моментальные,
1 циклические ,
А
1 1
I 1 I I I * t
1 Моногенетическая 1
1 Полигенетическая 1
1
1 Моногенетический 1
'Моногенетический \
1 Полигенетический |
1 Гидрогенная
1
I Гидрогалогенная
1
I Гидрогенный
1
I Гидрогенный
1
1
1 Гидрогалогенный
1
1 Галогенная
1
1 Техно галогенная
1 Пасторальный
1 Галогенный
1
1 Техногалогенный
. Пасторальная
[ Другие
, Техногенный
1 Пасторальный
! Другие
1 Техногенная
1 Техногенный
Рис. 2. Виды эволюционных состояний почв и почвенного покрова Приаралья.
носят или приобретают циклический характер, то
возникает состояние квазиравновесия (или квази¬
климакса), которое будет продолжаться неопреде¬
ленно долгое время вплоть до изменения эволю¬
ционного градиента в сторону какого-либо фак¬
тора.
Таким образом, для понимания механизмов из¬
менения почв и почвенного покрова Приаралья
при опустынивании следует различать следую¬
щие понятия (рис. 2):
Аутоэволюция - основная тенденция самораз¬
вития автономных почв в направлении достиже¬
ния равновесия с аридным макроклиматом. Как
правило, аутоэволюция сопровождается рядом
циклоидных процессов, характеризующихся на¬
правленной тенденцией изменения почвенных
свойств.
Климакс - окончательное состояние равнове¬
сия почв и почвенного покрова при завершении
мыслимой аутоэволюции. Это понятие, как пра¬
вило, не имеет природного воплощения, но ока¬
зывается удобным при раскрытии движущих сил
эволюции почв и почвенного покрова, а также
для понимания явления опустынивания.
Полигенетическая (или моногенетическая) ал¬
лоэволюция - локальные или (реже) региональ¬
ные тенденции изменения почв и почвенного по¬
крова в направлении достижения квазиравнове¬
сия (квазиклимакса), отвечающего совокупному
действию как зонального макроклимата, так и
дополнительных локальных антропогенных или
природных воздействий. В зависимости от типа
воздействия можно различать антропогенную и
естественную аллоэволюцию, а в зависимости от
направленности эволюционных изменений - гид¬
рогенную, галогенную, пасторальную, техноген¬
ную. Гидрогенная аллоэволюция обусловлена воз¬
действием периодического или спорадического
обводнения; галогенная - возникновением засо¬
ления почв; пасторальная - воздействием выпаса;
техногенная - транспортными и другими техно¬
генными сбоями. Полигенетичность аллоэволю-
ционных процессов находит отражение и в терми¬
нологий. Например, орошение часто приводит
к гидрогалогенной аллоэволюции. Очевидно, что
любой вид аллоэволюции сопровождается цикло¬
идными процессами.
Полигенетический (или моногенетический)
квазиклимакс (квазиравновесие) - состояние ди¬
намического равновесия почв и почвенного по¬
крова, отвечающее законченному совокупному
действию фактора зонального макроклимата и
противодействующих ему локальных антропо¬
генных или природных воздействий. Основной
атрибут любого квазиклимакса - активные цик¬
лические процессы. В соответствии с видами
аллоэволюции в Приаралье нами выделяются
следующие виды моногенетических квазиклимак¬
сов - гидрогенный, галогенный, пасторальный,
техногенный. А также соответствующие виды
полигенетических квазиклимаксов - гидрогалоген¬
ный, техногалогенный и т.п.
Скачок - тип резких моментальных (револю¬
ционных) изменений почв и почвенного покрова,
отличающийся от аллоэволюции отсутствием
циклоидных направленных процессов, а также
отсутствием ряда промежуточных стадий. Как
правило, скачок не приводит к достижению дли¬
тельного состояния квазиклимакса, а лишь вызы¬
вает новую цепь аллоэволюционных трансфор¬
маций или “отбрасывает” почвенные системы
ПОЧВОВЕДЕНИЕ № 6 1996
ЭВОЛЮЦИЯ И РАЗВИТИЕ ПОЧВ В ПРИАРАЛЬЕ
Таблица 1. Эволюционные изменения почв и почвенного покрова в Приаралье
757
Тип территории
Объект
Совре¬
менная
опусты¬
ниваю¬
щаяся
дельта
Древняя
опустыненная
дельта
Обсох¬
шее дно
моря
Песчаные пустыни
Орошаемые территории
Живая
дельта (в
настоящее
время прак¬
тически от¬
сутствует)
без регу¬
лируемого
выпаса
с регу¬
лируемым
выпасом
новое
орошение
древнее
и дли¬
тельное
орошение
Почвы
Состоя¬
Состояние ауто¬
Состоя¬
Состояние
Состояние
Состояние ау¬
Полиге-
Состояние
ние ауто-
эволюции, ос¬
ние ауто¬
аутоэволю¬
аутоэволю¬
тоэволюции,
нетичес-
аутоэволю¬
эволю¬
ложненное пас¬
эволю¬
ции, ослож¬
ции, ослож¬
осложненное
кий ква¬
ции, ослож¬
ции
торальной алло¬
ции
ненное пас¬
ненное пас¬
гидрогенной и
зикли¬
ненное гид¬
эволюцией
торальной
торальной
галогенной ал¬
макс
рогенной
аллоэволю¬
аллоэволю¬
лоэволюцией
аллоэволю¬
цией
цией. Места¬
и гидрогенны¬
цией и гид¬
ми - полиге-
ми скачками
рогенными
нетический
скачками
квазикли¬
макс
Почвен¬
Состоя¬
Состояние ауто¬
Состоя¬
Состояние
Полигенети-
Состояние ау¬
Полиге-
Полигене-
ный по¬
ние ауто-
эволюции, ос¬
ние ауто¬
аутоэволю¬
ческий ква¬
тоэволюции,
нетичес-
тический
кров
эволю¬
ложненное пас¬
эволю¬
ции, ослож¬
зиклимакс
осложненное
кий ква¬
квазикли¬
ции
торальной алло¬
ции
ненное пас¬
гидрогенной и
зикли¬
макс
эволюцией.
торальной
галогенной ал¬
макс
Местами - по-
аллоэволю¬
лоэволюцией
лигенетический
цией
и гидрогенны¬
квазиклимакс
ми скачками
Почвен¬
Актив¬
Заторможен¬
Актив¬
Циклоид¬
Цикличес¬
Активные
Актив¬
Активные
ные про¬
ные цик¬
ные циклоид¬
ные цик¬
ные
кие
циклоидные
ные цик¬
цикличес¬
цессы
лоидные
ные
лоидные
лические
кие
к начальным этапам аутоэволюционных измене¬
ний. На территории Приаралья нами выделяется,
в отличие от видов аллоэволюции, три вида скач¬
кообразных изменений: 1) гидрогенный - обуслов¬
ленный внезапным обводнением ранее обсохших
территорий, в отличие от гидрогенной аллоэволю¬
ции, поддерживаемой за счет периодического
и/или поступательного подъема фунтовых вод;
2) техногенный - обусловленный внезапным тех¬
ногенным воздействием; 3) пасторальный - обус¬
ловленный резким усилением пастбищной диг¬
рессии. Как правило, в Приаралье эти скачкооб¬
разные трансформации отмечаются чаще, чем
соответствующие виды аллоэволюционных из¬
менений.
При обсуждении эволюционных изменений
почв и почвенного покрова Приаралья с точки
зрения их отношения к состоянию равновесия
особое значение занимают вопросы, связанные с
отличием в этом ключе собственно почв от поч¬
венного покрова (табл. 1).
ЭВОЛЮЦИОННЫЕ СОСТОЯНИЯ ПОЧВ
Как мы указывали выше, в Приаралье практи¬
чески нет таких почв, которые находились бы в
состоянии равновесия. Постоянное присутствие
дополнительных антропогенных или естествен¬
ных воздействий локального характера, создаю¬
щих эволюционный градиент на фоне квазикли-
максных почв или изменяющих направление
аутоэволюции, обусловливает перманентные из¬
менения почвенных свойств в результате цикло¬
идных или активных контрастных циклических
процессов. Иначе говоря, почвы Приаралья ни¬
когда не достигают состояния полного (мыслимо¬
го) климакса. Перечень квазиравновесных состо¬
яний отдельных почвенных индивидуумов также
крайне ограничен. Это связано с неравномернос¬
тью локальных антропогенных и природных воз¬
действий на почвенный покров, и, в первую оче¬
редь, со своего рода “переложностью” земельных
участков по отношению к любому из этих воздей¬
ствий, вызывающих скачкообразные или аллоэво-
люционные изменения. Иначе говоря, в Приаралье
редко можно описать какой-либо из почвенных ин¬
дивидуумов, не находящихся в состоянии эволю¬
ции и испытывающий только циклические изме¬
нения.
На участках, функционирующих в режиме
“живой” дельты, это связано с неравномерным и
непостоянным функционированием тех или иных
рукавов и протоков. На землях, находящихся
ПОЧВОВЕДЕНИЕ № 6 1996
758
КУСТ
в пастбищном использовании, степень пастбищ¬
ной нагрузки крайне неравномерна как по годам -
в зависимости от условий увлажненности, так и
в связи с нерегулируемой нагрузкой, которая вре¬
мя от времени приводит к дигрессии и вынуждает
забрасывать пастбища до их полного или частич¬
ного восстановления. Неравномерность освоения
характерна также для орошаемых земель, кото¬
рые издавна в связи с высоким развитием засоле¬
ния находились в Приаралье в режиме “перелож¬
ного” земледелия.
Редкие состояния квазиравновесия отдельных
почвенных индивидуумов удается описать только
в тех случаях, когда циклы саморазвития почв
совпадают по времени с циклами локальных ант¬
ропогенных и природных воздействий. При этом
общая длительность периода, в течение которого
отмечается это совпадение, не должно быть ко¬
роче того временного промежутка, в течение ко¬
торого завершается период аллоэволюционного
развития (характерное время данной полигенети-
ческой или моногенетической аллоэволюции).
К числу подобных состояний можно отнести
следующие:
- длительноорошаемые луговые почвы с мощ¬
ным агроирригационным горизонтом в условиях
хорошего искусственного или естественного от¬
тока грунтовых вод;
- развитые песчаные пустынные почвы под
сложными сформированными эфемерово-бело-
полынно-белосаксауловыми сообществами в ус¬
ловиях регулируемого выпаса;
- солончаки по периферии массивов длитель¬
ного орошения в условиях неизменности границ
орошаемых территорий и условий дренажа.
Все другие варианты почвенных индивидуу¬
мов в Приаралье подвержены циклоидным изме¬
нениям и эволюционируют, различаясь только
скоростью и степенью завершенности эволюци¬
онных процессов, а также контрастностью состо¬
яний в пределах циклоидных изменений.
ЭВОЛЮЦИОННЫЕ СОСТОЯНИЯ
ПОЧВЕННОГО ПОКРОВА
Поскольку почвы Приаралья постоянно под¬
вержены аллоэволюционным или скачкообраз¬
ным изменениям, то и почвенный покров в целом
никогда не достигает состояния климакса.
Вместе с тем для многих территорий Приара¬
лья, как в современный период, так и, в особенно¬
сти, до начала современного интенсивного опус¬
тынивания, характерно такое состояние, когда
почвы могут постоянно эволюционировать в том
или ином направлении, а такие параметры поч¬
венного покрова, как его состав (разнообразие
составляющих его почв) и соотношение площа¬
дей, занятых разными почвами, остаются посто¬
янными. Структура почвенного покрова и кон¬
кретные почвенные индивидуумы при этом могут
находиться в состоянии эволюции.
Для удобства дальнейшего изложения для ха¬
рактеристики такого состояния мы использовала
термин гомеостаз почвенного покрова в отличие
от состояния стабильности почвенного покрова,
при котором неизменной остается также и струк¬
тура почвенного покрова. Состояние гомеостаза
почвенного покрова уже по определению следует
характеризовать не как климаксное, а как квази-
климаксное (квазиравновесное). Наиболее инте¬
ресным при анализе состояния гомеостаза поч¬
венного покрова является вопрос об определении
границ территорий, состояние почвенного покро¬
ва которых можно характеризовать как гомео¬
статическое. Наиболее ярко это можно проиллю¬
стрировать на примере почвенного покрова тер¬
риторий, функционирующих в режиме “живой”
дельты. Нами были проанализированы материа¬
лы повторных инвентаризаций почвенных ресур¬
сов дельт Амударьи, Сырдарьи, а также, для под¬
тверждения полученных результатов, дельт ряда
других рек.
С учетом различий классификационных под¬
ходов при инвентаризации почв, проводимых раз¬
ными исследователями и в разные годы, и степе¬
ни хозяйственного освоения территорий, нами
была отмечена любопытная закономерность: не¬
смотря на довольно существенные изменения почв
в пределах различных участков дельт, связанные с
миграцией рукавов и протоков и сопровождающи¬
еся определенными изменениями в структуре поч¬
венного покрова, состав его и соотношение пло¬
щадей образующих его почв остается практичес¬
ки постоянным при условии сохранения среднего
многолетнего стока рек в створе вершины дель¬
ты. Это характерно для почвенного покрова
дельт Амударьи и Сырдарьи до 1964 г., а также
для дельт таких крупных рек, как Или, Чу, Терек
и другие (рис. 3, 4). Почвенный покров таких
дельт и был охарактеризован нами как гомеоста¬
тический.
Дальнейший анализ показал, что постоянство
состава почвенного покрова и соотношения площа¬
дей слагающих его почвенных разностей в такого
рода динамически равновесных гомеостатических
естественных дельтовых экосистемах поддержива¬
ется в результате сбалансированного противодей¬
ствия двух основных факторов: 1) аридного зо¬
нального макроклимата; 2) дополнительного ко¬
личества воды извне, приносимого питающими
дельту реками.
Совокупное действие этих факторов приводит
к тому, что естественные процессы развития
дельтовых экосистем и природных комплексов в
засушливых условиях носят ярко выраженный
динамический характер при условии сохранения
ПОЧВОВЕДЕНИЕ № б 1996
ЭВОЛЮЦИЯ И РАЗВИТИЕ ПОЧВ В ПРИАРАЛЬЕ
759
объемов среднемноголетнего речного стока.
Цикличность развития экосистем связана с перио¬
дическим новообразованием, развитием и последу¬
ющим отмиранием дельтовых протоков и малых
русел. Развитие и отмирание малых русел сопро¬
вождается обсыханием и/или засолением террито¬
рий, приуроченных (подкомандных) к этим прото¬
кам. Это проявляется в снижении уровня и увели¬
чении минерализации грунтовых вод, изменении
почвенного и растительного покровов в сторону
уменьшения степени гидроморфизма и ксерофи-
тизации соответственно. Вместе с тем периоди¬
ческое или спорадическое обводнение дельт па¬
водковыми водами, миграция русел и протоков
приводили к периодическому возобновлению
циклов эволюции дельтовых экосистем и обеспе¬
чивали общее динамическое равновесие в. дель¬
тах, поддерживаемое за счет поступления воды
извне.
Таким образом, вода, поступающая в дельто¬
вые экосистемы, выступает, с одной стороны, как
воздействие, обеспечивающее постоянное возоб¬
новление циклов гидрогенной аллоэволюции на
уровне подсистем современной дельты, что обус¬
ловливает их динамичность и неустойчивость, а
с другой - как фактор, обеспечивающий гомеостаз
почвенного покрова дельт в целом. Иначе говоря,
равновесие почвенного покрова дельтовых эко¬
систем крайне неустойчиво и поддерживается не
за счет сил и средств саморегуляции, а лишь за
счет внешних дополнительных источников массы
и энергии. Прекращение действия этих источни¬
ков ведет к смещению баланса условий почвооб¬
разования в сторону преобладания воздействия
фактора аридного макроклимата. Отсюда следу¬
ет, что площадь территорий, характеризующихся
состоянием гомеостаза почвенного покрова, ог¬
раничивается тем уровнем “командования”, на
котором обеспечивается длительное постоянство
(сопоставимое с собственными характерными
временами развития подкомандных систем) при¬
чин, вызывающих аллоэволюцию “подкоманд¬
ных” (подвешенных) систем. Для почвенного по¬
крова дельт в целом этим уровнем командования
является створ вершины дельты при условии со¬
хранения среднемноголетнего стока в этом створе.
Логично предположить, что принцип опреде¬
ления размера гомеостатических территорий но¬
сит универсальный характер. Он подтверждается
не только для дельтовых систем в целом, но и для
систем более мелкого и более крупного уровней.
Так нами, а также в литературе [2, 3,7], описаны
более мелкие внутридельтовые природные комп¬
лексы, функционирующие в режиме естественно¬
го или искусственного паводкового затопления,
в пределах которых такие параметры, как состав
почвенного покрова и соотношение площадей,
занятых разными почвами, аналогичны или весь¬
ма близки к этим же параметрам, характерным
для “живой” дельты в целом. Такого рода терри-
%
Рис 3. Почвенный покров Приаралья: а - дельта
Амударьи, 1953 г.; б-то же, 1980 г.; в - ключ Бозатау,
1990 г. П о ч в ы: / - луговые и болотно-луговые пой-
менно-аллювиальные; 2 - луговые аллювиальные;
3 - луговые орошаемые; 4 — болотные остаточные;
5 - луговые остаточные; 6 - лугово-остаточные ту¬
гайные; 7 - луговые такыровидные; 8 - луговые та¬
кыровидные орошаемые; 9 - такыры; 10 - такыро¬
видные; 11 - песчаные пустынные, развеваемые пес¬
ки; 12 - солончаки типичные, луговые, болотные;
13 - солончаки остаточно-болотные; 14 - солонча¬
ки остаточно-луговые; 15 - солончаки остаточные;
16 - серо-бурые; 17 - водная поверхность.
О 1 О) 2
Рис. 4. Почвенный покров Гуляевской дельты р. Чу
(по [1]) до (/) и после (2) зарегулирования. 1 -1963 г.;
2 - 1983 г. П о ч в ы: Б - болотные; Бзас - болотные
засоленные; БЛ - болотно-луговые; БЛзас - болот¬
но-луговые засоленные; БЛост - болотно-луговые
остаточные; ЛБ - лугово-болотные; ЛБзас - лугово¬
болотные засоленные; А - аллювиальные; Азас - ал¬
лювиальные засоленные; С - солончаки; ТВсол - та¬
кыровидные солончаковые.
тории подкомандны какому-либо из рукавов или
протоков, в верхнем створе которого обеспечи¬
вается постоянство среднемноголетнего стока
(рис. Зв - ключ Бозатау).
Действенность указанного принципа на более
высоком уровне фактически подтверждается со¬
временным состоянием интенсивного опустыни¬
вания в Приаралье. Попытка человека изменить
структуру почвенного покрова в бассейне Араль¬
ского моря за счет увеличения площадей гидро-
морфных почв в верхнем и среднем течении рек
неизбежно вызывала уменьшение площадей этих
почв в дельтах. В данном случае в качестве команд¬
ного уровня рассматривается среднемноголетний
ПОЧВОВЕДЕНИЕ № 6 1996
760
КУСТ
тыс. км2
Е21 1 Ш 2 IZ2 3
Рис. 5. Изменение соотношения обводняемых площа¬
дей в бассейне Аральского моря. Площадь:/ - мор¬
ской поверхности; 2 - болот и водоемов в дельтах;
3 - орошения в бассейне Аральского моря.
сток рек бассейна Аральского моря, подкоманд-
ным по отношению к которому является почвен¬
ный покров всего бассейна. Иллюстрацией этого
является рис. 5, составленный по данным, полу¬
ченным из нескольких независимых источников.
Аналогичным образом действует принцип оп¬
ределения гомеостатических территорий и в слу¬
чае сбалансированного противодействия между
аридным макроклиматом и антропогенными воз¬
действиями, как то: отгонное животноводство,
техногенное воздействие, сведение древесной и
кустарниковой растительности. При этом коли¬
чество вариантов такого рода гомеостатических
состояний природных систем в целом и почвенно¬
го покрова, в частности, может быть весьма зна¬
чительным. Все зависит от состояния баланса,
обусловленного интенсивностью постоянно дей¬
ствующего того или иного воздействия, вызыва¬
ющего аллоэволюцию.
Так, в условиях постоянно интенсивной паст¬
бищной нагрузки в песчаных пустынях формиру¬
ются квазиравновесные природные комплексы,
в которых доминирующее положение занимают
разбитые или развеваемые пески, а также слабо¬
развитые и деградированные песчаные пустын¬
ные почвы. В условиях умеренной пастбищной
нагрузки формируется почвенный Покров, состо¬
ящий почти нацело из развитых пустынных пес¬
чаных почв под полночленными сообществами
белого саксаула. В условиях недовыпаса возника¬
ют природные комплексы с песчаными пустынны¬
ми почвами, в которых в зависимости от степени
недовыпаса участвуют или доминируют деградиру¬
ющие песчаные пустынные почвы с моховым по¬
крытием (Tortula desertorum). Размер таких гомео¬
статических территорий зависит от степени паст¬
бищной нагрузки, не изменяющейся длительное
(сопоставимое с периодом, требующимся для до¬
стижения квазиравновесного состояния) время.
В частности, для комплексов из разбитых песков,
характерных для экосистем, формирующихся
вокруг длительно используемых водопоев,
“командным” уровнем может быть количество
голов скота, ежегодно приходящееся на данный
водный источник. Если на пастбищах, на которых
длительное время выпасается определенное ко¬
личество скота, используется в качестве водопо¬
ев несколько водных источников, функциониру¬
ющих в переменном режиме, то гомеостатической
следует считать всю территорию данного пастби¬
ща, а участки приколодезных сбоев в этом случае
рассматриваются только как один из составных
элементов этих гомеостатических территорий.
Для такого рода территорий, равно как и для
природных комплексов с доминированием замо-
ховелых или развитых пустынных песчаных почв
в качестве командного уровня следует рассматри¬
вать ежегодное количество голов скота, приходя¬
щееся на единицу площади, которое остается не¬
изменным в течение длительного промежутка
времени, сопоставимого с периодами эволюцион¬
ных циклов. Например, гомеостатическими мо¬
гут быть как большие по площади территории,
многие пастбищные участки которых периодиче¬
ски находятся в перелоге для восстановления по¬
сле дигрессии, так и незначительные площади, на
которых ведется умеренный выпас.
Из принципа определения границ гомеостати¬
ческих территорий вытекает важное следствие:
только та территория может считаться го¬
меостатической в отношении почвенного по¬
крова, для которой можно определить границы
и командный уровень причин, вызывающих цик¬
лические трансформации внутри систем.
Если границы системы невозможно опреде¬
лить, значит она не относится к гомеостатичес¬
кой, а является подчиненной по отношению к го¬
меостатической системе более высокого уровня.
Так, если нельзя определить командный уровень
для системы почвенного покрова, приуроченной
к обсыхающему или обсохшему рукаву или про¬
току дельты, значит почвенный покров этой сис¬
темы целиком находится в состоянии эволюции и
необходимо говорить об эволюции, а не о гомео¬
стазе почвенного покрова в целом.
К негомеостатическим системам почвенного
покрова в Приаралье относятся такие крупные
территории, как древние опустыненные дельты,
современные опустынивающиеся дельты, обсох¬
шее дно моря, многие орошаемые массивы. Для
этих территорий характерно нарушение одного или
обоих условий гомеостаза почвенного покрова:
1) наличие устоявшегося сбалансированного про¬
тиводействия аридного макроклимата и воздейст¬
вия, вызывающего циклические трансформации
внутри систем (вызывающего различные виды
аллоэволюционных изменений и инициирующего
новые изменения в направлении аутоэволюции);
2) достаточная длительность устоявшегося сба¬
лансированного действия упомянутых выше
фактора климата и дополнительного внешнего
ПОЧВОВЕДЕНИЕ № 6 1996
ЭВОЛЮЦИЯ И РАЗВИТИЕ ПОЧВ В ПРИАРАЛЬЕ
Таблица 2. Гомеостатичность различных типов территорий Приаралья
761
Тип территории
Условия
гомеостаза
Современ¬
Древняя
Обсох¬
шее дно
моря
Песчаные пустыни
Орошаемые территории
Живая дель¬
та (в настоя¬
щее время
практически
отсутствует)
почвенного
покрова
ная опусты¬
нивающая¬
ся дельта
опусты¬
ненная
дельта
без регу¬
лируемо¬
го выпаса
с регу¬
лируемым
выпасом
новое
орошение
древнее и
длительное
орошение
Устоявшийся
баланс аридного
климата и допол¬
-
+
—
—
+
+
+
+
нительного внеш¬
него воздействия
Завершение цик¬
лоидных транс¬
+
+
формаций поч¬
венного покрова
Гомеостатич¬
ность территории
Т
т
Примечание. + - наличие условия; — отсутствие условия.
воздействия, не меньшая, чем время, необходи¬
мое для завершения трансформаций почвенного
покрова, носящих циклоидный характер (табл. 2).
Еще одно следствие из принципа определения
границ территории, гомеостатической в отношении
почвенного покрова, заключается в том, что если
существует гомеостатическая система почвен¬
ного покрова, то всегда можно определить го¬
меостатическую систему почвенного покрова,
подкомандную более высокому уровню. Иначе
говоря, любая гомеостатическая система всегда
является подсистемой другой гомеостатической
системы. Это следствие объясняет, почему в При¬
аралье гомеостаз почвенного покрова в большей
мере присущ более крупным по площади терри¬
ториям, и реже идентифицируется для менее
крупных. В целом же высказанное соображение
аналогично известному положению биоэкологии
о том, что вид более устойчив чем особь, а экоси¬
стема более устойчива чем вид [6].
ВЫВОДЫ
1. Показано, что основные противоречия, обус¬
ловливающие эволюцию почв и почвенного по¬
крова в Приаралье, как правило, состоят в разли¬
чии, с одной стороны, аутоэволюционных изме¬
нений почв и почвенного покрова в направлении
Достижения равновесия с аридным макроклима¬
том, который выступает здесь в роли постоянного
и повсеместно действующего фактора почвообра¬
зования; а с другой - аллоэволюционных измене¬
ний, протекающих под воздействием антропоген¬
ных или природных причин локального действия,
смещающих равновесие почв и почвенного по¬
крова с аридным макроклиматом или осложня¬
ющих ход указанных аутоэволюционных изме¬
нений.
. 2. Предложено для понимания механизмов из¬
менения почв и почвенного покрова Приаралья
при опустынивании использовать понятия: ауто¬
эволюция; климакс; полигенетическая (или моно-
генетическая) аллоэволюция (в зависимости от
типа воздействий различается антропогенная и
естественная аллоэволюция, а в зависимости от
направленности эволюционных изменений раз¬
личается гидрогенная, галогенная, пасторальная,
техногенная аллоэволюция); полигенетический
или моногенетический квазиклимакс (квазирав¬
новесие) (различаются гидрогенный, галоген¬
ный, пасторальный, техногенный моногенетиче-
ские квазиклимаксы, а также соответствующие
виды полигенетических квазиклимаксов - гидро¬
галогенный, техногалогенный и т.п.); скачок
(различаются гидрогенный, техногенный, пасто¬
ральный скачки).
3. Для характеристики такого состояния поч¬
венного покрова, при котором отдельные поч¬
венные индивидуумы и структура почвенного по¬
крова в целом могут постоянно меняться в том
или ином направлении, а такие параметры поч¬
венного покрова, как его состав (разнообразие
составляющих его почв) и соотношение площа¬
дей, занятых разными почвами, остаются посто¬
янными, предложено использовать термин го¬
меостаз почвенного покрова.
4. Предложен принцип определения границ го¬
меостатических территорий.
ПОЧВОВЕДЕНИЕ № 6 1996
762
КУСТ
СПИСОК ЛИТЕРАТУРЫ
1. Аханов Ж.У. Почвообразование в дельтовых рав¬
нинах Южного Казахстана. Алма-Ата: Наука,
1987. 240 с.
2. Жоллыбеков Б. Изменение почвенного покрова
приморской дельты Амударьи при аридизации.
Нукус: Билим, 1991. 132 с.
3. Кабулов С.К. Изменение фитоценозов пустынь
при аридизации. Ташкент: Фан, 1990. 240 с.
4. Куст Г.С. О путях эволюции почвенного покрова
дельтовых территорий при опустынивании // Вестн.
МГУ. Сер. Почвоведение. 1992. № 2. С. 35-43.
5. Куст Г. С. О постгидроморфной эволюции почвен¬
ного покрова водно-аккумулятивных равнин засу¬
шливых территорий // Почвоведение. 1994. № 2.
С. 16-29.
6. Одум Ю. Экология. Т. 1. М.: Мир, 1986. 328 с.
7. Попов В.А. Проблема Арала и ландшафты дельты
Амударьи. Ташкент: Фан, 1990. 112 с.
8. Самойлова ЕМТолчельников Ю.С. Эволюция
почв. М.: Изд-во МГУ, 1991. 90 с.
9. Соколов И А. Почвообразование и время: поли-
климаксность и полигенетичность почв // Почво¬
ведение. 1984. № 2. С. 102-112.
10. Соколов И.А., Караваева Н.А., Александров-
скийА.Л., Иванов И.В. Эволюция почв: понятия и
термины (опыт разработки) // Эволюция и возраст
почв СССР. Пущино, 1986. С. 5-18.
Soil Evolution under Conditions of Anthropogenic Desertification
in the Near-the Aral Sea Areas
G. S. Kust
Methodologies and concepts of the soil evolution theory are applied to study mechanisms responsible for
changes in soils and soil cover within the Near-the Aral Sea areas, which are highly desertified due to man-
made effects now. Evolutional state of soils and soil cover is shown as changed to a certain extent. To estimate
the sustainable state of the soil cover the concept of homeostasis is proposed to use as well as a principle to
determine boundaries of homeostatic territories.
ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВОВЕДЕНИЕ, 1996, № 6, с. 763-774
ХИМИЯ
~ почв
УДК 631.4
ФАЗОВЫЙ СОСТАВ СОЕДИНЕНИЙ ЖЕЛЕЗА В ПОЧВАХ
И ГИДРОТЕРМАЛЬНЫХ ОТЛОЖЕНИЯХ АБХАЗИИ
В СВЯЗИ С ПРОБЛЕМАМИ ЛАТЕРИТООБРАЗОВАНИЯ
© 1996 г. JI. С. Ильина*, Т. С. Гендлер**, В. В. Лежава***
*Факулыпет почвоведения МГУ им. М.В. Ломоносова,
**Институт физики Земли РАН,
***Тбилисский государственный университет
Поступила в редакцию 23.01.91 г.
Исследован фазовый состав соединений железа методами ядерной у-резонансной спектроскопии и
термомагнитного анализа в конкреционных горизонтах “желтоземно-подзолистых” почв Абхазии
и железистых отложений современных гидротермальных источников этого региона. Установлено
присутствие тонкодисперсного гетита, пирита, пирротинов - промежуточных продуктов окисления
пирита, магнетита. Близость фазового состава обеих групп образцов и описываемы^ в литературе
некоторых сульфидсодержащих Fe-Mn-конкреций донных морских отложений гидротермального
генезиса свидетельствует об относительной древности геологических процессов, приведших в обра¬
зованию конкреционного ожелезненного слоя. Предполагается два возможных пути его образова¬
ния - палеогидротермальное воздействие на отложения, связанное с аккумуляцией железа, и палео-
гидрогенная аккумуляция железа из гидротермальных растворов. Погребение ожелезненного лате-
ритного горизонта под слоем суглинистых отложений произошло после его формирования.
Конкреционный горизонт препятствует нор¬
мальному развитию корневой системы чайного ку¬
ста и представляет собой конгломерат, в основном,
крупных (5-20 см) темноокрашенных Fe-Mn-кон-
креций [9,10,13-15] с примесью гальки. Конкреци¬
онный мощный слой может залегать над разными
породами - галечниками, суглинками, песгроцвет-
ными и зебровидными глинами. Характер его зале¬
гания определяется литолого-геоморфологически-
ми факторами, а мощность, степень выраженности
и сцементированности заметно варьируют.
Решение вопроса о происхождении этого оже¬
лезненного слоя тесно связано с мелиорацией
почв и дальнейшим их сельскохозяйственным ис¬
пользованием. Наличие латеритного горизонта
является фактором, лимитирующим возможнос¬
ти сельскохозяйственного освоения этих почв.
Зонн [6] связывает формирование мощных
конкреционных слоев с древними процессами ги¬
дрогенной аккумуляции железа в период форми¬
рования галечников, которые часто составляют
основу этих слоев. Вопросы генезиса латеритов и
почв, содержащих конкреционные горизонты,
остаются остродискуссионными до настоящего
времени. Вместе с тем данные о фазовом составе,
размерах частиц оксидов железа могут в опреде¬
ленной степени прояснить картину их генезиса:
являются ли они литогенными образованиями
“допочвенной” стадии ожелезнения или образова¬
лись в результате мощной педогенной трансфор¬
мации исходного материала, т.е. сформировались
в результате почвообразовательного процесса.
Объектами исследования послужили образцы
почв с латеритными горизонтами, мощность ко¬
торых составляет ~60 см и более. Залегают гори¬
зонты на глубине от 30 до 80 см. Почвы с такими
образованиями, как правило, развиты на дву¬
членных отложениях, часто несут признаки огле-
ения с поверхности до подстилающих пород, рас¬
пространены в северо-западной части Колхиды
и приурочены к волнистым и выровненным пред¬
горным равнинам, холмам и террасам.
Современное формирование ортштейнов в ре¬
зультате сегрегации в условиях избыточного по¬
верхностного увлажнения не исключено, оно так¬
же возможно при частичном разрушении латери¬
та в современных условиях.
Очевидно, гипотеза палеообразования латери¬
та в результате гидрогенной или гидротермаль¬
ной аккумуляции Fe и его существование на фоне
современных процессов сегрегации и ортштейно-
образования требуют объективных доказа¬
тельств. Обобщение литературных материалов и
наши исследования [2, 7, 8,10,11, 18,20,22] дают
основание предположить, что происхождение
мощного конкреционного железистого горизон¬
та связано с гидротермальными процессами. При
этом фазовый состав соединений Fe и их дисперс¬
ность могут быть важными диагностическими
признаками этого процесса.
Объекты исследования. Образцы для изуче¬
ния отобраны в Очамчирском районе Абхазии.
Они представляют собой конкреции, взятые из
нескольких разрезов с латеритным горизонтом
763
764
ИЛЬИНА и др.
(10, 11 , 12*, 13*, 14, 15). Образец 14 (диамет¬
ром 12 см) взят из латеритного горизонта одной
из древних морских террас и представляет собой
конгломерат конкреций темно-коричневого цве¬
та, на поверхности которого наблюдаются охрис¬
тые железистые отложения современного гидро¬
термального источника. Для изучения использова¬
лись темная (14) и осветленная (14’) центральные
части, охристый поверхностный слой (14м) кон¬
гломерата.
Для более надежного определения источника
аккумуляции железа были также отобраны об¬
разцы из современных гидротермальных источ¬
ников того же района (образцы 2,5,6,7), а также
образец 1, представляющий собой железистые
корочки с поверхности гальки из почвы.
Для диагностики минералов, содержащихся
в описанных выше объектах, дополнительно бы¬
ло изучено несколько образцов природных пири-
тов и синтезированного лепидокрокита. Образцы
пиритов отбирали из пород верхне- и среднеэоце-
нового возраста (Учамбо, Вараза - Абхазия) и из
прожилок ивкрапленийместорождения Неждан-
ка (Восточная Якутия) .
Приборы и методы исследования. Для выяв¬
ления степени дисперсности и типа соединений
железа были применены ядерный гамма-резо¬
нанс (ЯГР) на изотопе 57Fe и магнитные измере¬
ния. Использование этих методов для изучения
железосодержащих соединений почв является на¬
иболее оптимальным. ЯГР на изотопе 57Fe выяв¬
ляет все железосодержащие соединения как маг¬
нитные, так и немагнитные при данной температу¬
ре измерения. Однако чувствительность метода
составляет 2-3%, в связи с чем на фоне парамаг¬
нитных соединений с большим содержанием же¬
леза (обычно в почвах - это силикаты или тонко¬
дисперсные гидроксиды железа) не проявляются
спектры ферромагнитных фаз, концентрация ко¬
торых существенно меньше. Магнитные измере¬
ния, напротив, избирательны к ферримагнитным
компонентам, содержащимся в образцах. Чувст¬
вительность магнитных методов существенно
выше - десятые-сотые доли процента в зависи¬
мости от удельных величин намагниченностей.
ЯГР-спектры получены при комнатной темпе¬
ратуре и температуре кипения жидкого азота на
спектрометре электродинамического типа с ана¬
лизатором LP-4000. Источником у-квантов слу¬
жил 57Со(Сг). Величины изомерного сдвига при¬
ведены относительно нитропруссида Na
комплекс магнитных измерений при изучении
фазового состава почв сводится к получению
ряда характеристик:
*Эти образцы описаны в работе [13] под № 1,2, 3.
Образцы из коллекции С.Ю. Бродской.
Измерения проводились в лаборатории ядерных методов
физического факультета МГУ.
1. JSo, JrSo - удельные величины намагничен¬
ности насыщения и остаточной намагниченности
насыщения, соответственно.
2. J /J , JrSj/JrSo - отношение тех же величин
после первого и второго последовательных на¬
гревов в диапазоне температур 20-700°С.
3. JS(T), JTS(Т)- температурные зависимости
J, и Jrs в процессе непрерывных последователь¬
ных лабораторных нагревов.
4. Тс,Тб- температуры Кюри и блокирующие
температуры ферримагнитных фаз.
5. Т„р - температуры превращений (фазовых
переходов), происходящих при нагреве натураль¬
ных образцов.
Все магнитные измерения проводились в лабо¬
ратории геомагнетизма Института физики Земли
РАН. Величины Js и температурные зависимости
JS(T) измеряли на вибромагнетометре с чувстви¬
тельностью 10~5 А/м в магнитном поле Я = 0.25 Тл
в интервале температур 20-700°С. Аналогичные
измерения остаточной намагниченности насыще¬
ния проводили на спин-магнетометре с чувстви¬
тельностью 10-7 А/м в том же диапазоне темпера¬
тур. Образцы предварительно намагничивались
в магнитном поле Я = 2.0 Тл.
Основную информацию о железистых магнит¬
ных минералах, содержащихся в натуральных об¬
разцах, дает анализ температурных зависимостей
JS(T) и Jrs(T)- По Тс и Тб на этих кривых диагности¬
руются ферримагнитные минералы. По харак¬
терным температурам необратимых фазовых из¬
менений определяются неустойчивые к нагревам
пара- и ферримагнитные соединения. Для несколь¬
ких образцов выборочно был проведен валовой хи¬
мический анализ и получены рентгеновские ди-
фрактограммы.
ЭКСПЕРИМЕНТАЛЬНЫЕ РЕЗУЛЬТАТЫ
Данные валового химического анализа трех
образцов из современных гидротермальных ис¬
точников (2, 5, 6), конкреций (10) и железистых
корочек на гальке (1) приведены в табл. 1. Образ¬
цы гидротермального происхождения обеднены
Si02; А1203; К20, МпО и обогащены СаО и S03 по
сравнению с конкрециями (10) и образцом 1. Же¬
лезо в гидротермальных осадках и конкрециях, в ос¬
новном, представлено в виде несиликатных форм.
Конкреции из почв. На дифрактограммах двух
конкреций (11, 14) наблюдаются только линии
кварца, следовательно, железосодержащие фазы
представлены либо аморфными формами, либо
тонкодисперсными частицами. ЯГР-спектры об¬
разцов 11-13, как было показано ранее [13], по¬
добны и типичны для многих красноцветных
ПОЧВОВЕДЕНИЕ № 6 1996
ФАЗОВЫЙ СОСТАВ СОЕДИНЕНИЙ ЖЕЛЕЗА 765
Таблица 1. Данные валового химического анализа, % на прокаленную навеску
Объект исследования
Na20
MgO
ai2o3
Si02
p2o5
so3
Cl
к2о
Ti02
MnO
Fe203
CaO
Образцы из современных
гидротермальных источников:
2
0.00
0.85
1.34
23.41
0.11
1.65
0.00
0.28
0.07
0.06
33.90
38.24
5
0.85
2.17
2.93
15.76
0.34
2.95
0.03
0.60
0.11
0.04
11.14
63.00
6
0.00
1.07
0.88
18.44
0.19
2.70
0.00
0.13
0.02
0.03
17.96
58.38
Корочки на гальке из почвы (1)
0.13
1.46
9.48
35.32
0.18
0.39
0.00
0.85
1.08
0.36
47.37
3.16
Конкреции из почвы (10)
0.10
0.50
13.30
58.80
0.05
0.16
0.03
1.03
1.29
5.86
18.58
0.23
почв и кор выветривания, океанических осадков
и их конкреций [2, 8,25]. При комнатной темпера¬
туре спектры являются парамагнитными дубле¬
тами различной интенсивности с параметрами
ЬЕ = 0.61 и ДЕ = 0.57 мм/с. При 80 К они трансфор¬
мируются в сумму дублетов и секстета с уширен¬
ными компонентами (рис. 1), что подтверждает
предположение о тонкодисперсности железосодер¬
жащих фаз, входящих в конкреции. Менее интен¬
сивный дублет на рис. 1а с параметрами 5Е = 1.95,
АЕ = 3.31 мм/с относится к ионам Fe2+ и составля¬
ет около 5%. Эти ионы стабильны и не окисляют¬
ся при хранении. Их спектры характеризуются
низкой температурной зависимостью /(Г), рас¬
считанной по величинам площадей (5) при 300 и
80 К (5 2,/S г» =1). Величины 8Е и АЕ близки к
FeW) Few,
таковым, например, для ионов двухвалентного
железа в октаэдрическом слое смектитов [5].
Появление при 80 К сверхтонкого магнитного
расщепления с Яэфф ~ 455 кэ при уменьшении ин¬
тенсивности центрального дублета, позволяет от¬
нести большую часть железистых фаз к гетиту
с размером частиц 7-12 нм [1]. Доля этих частиц,
по парциальным площадям спектров ЯГР, для
конкреций 11-13 составляет 66-81% [13].
Сохранившийся при 80 К центральный дублет
может отвечать ультрадисперсным частицам гид¬
роксида железа (£7 нм), пириту и ионам трехва¬
лентного железа в составе силикатов [1, 9, 24].
Наибольшая относительная площадь централь¬
ного дублета при 80 К наблюдается для образца
12 (-30%).
Предварительные лабораторные
эксперименты. Выше было отмечено, что
ЯГР-спектры, приведенные на рис. 1, типичны
для различных типов почв, континентальных и
морских конкреций и металлоносных осадков.
В связи с неоднозначностью интерпретации пара¬
метров центрального дублета для определения
железосодержащих соединений и принципиаль¬
ными трудностями применения минералогичес¬
кого и рентгеновского анализов для тонкодис¬
персных почвенных частиц, рассмотрим возмож¬
ности магнитоминералогической диагностики
предполагаемых соединений. Для этой цели были
проведены лабораторные определения темпера¬
турной зависимости намагниченности насыщения
JS(T) нескольких образцов природного пирита и ле-
пидокрокита. Гамма-резонансные спектры этих
образцов при 300 и 80 К имеют близкие параме¬
тры [2, 23].
Известно, что оба этих минерала испытывают
превращения после 300°С [3,21]. Окисление пири¬
та представляет собой сложную реакцию, которая
в зависимости от целого ряда свойств исходных
зерен может быть как одно-, так и многостадий¬
ной [23-26]. Окисление пирита приводит к обра¬
зованию сильномагнитного, достаточно устойчи¬
вого магнетита и может сопровождаться образо¬
ванием промежуточных продуктов окисления:
моноклинного и/или гексагонального пирротина
и других. В зависимости от сложности и стадийно¬
сти реакции кривые JS(T) имеют различные тем¬
пературы Кюри промежуточных фаз и темпера¬
туры фазовых превращений [17,21,25,26].
Наиболее интенсивно в лабораторных услови¬
ях пирит окисляется при температурах 400-500°С.
Однако этот интервал, а также стадии окисления
могут меняться в зависимости от размера зерна,
происхождения и рассеянности пирита в породе.
На рис. 2 приведены нормализованные кривые
JJX) трех последовательных нагревов для различ¬
ных природных образцов пирита. Кривые перво¬
го нагрева (рис. 2а, 2Ь) близки в первом прибли¬
жении для всех образцов и демонстрируют резкое
возрастание намагниченности в форме пика в ин¬
тервале температур 300-600°С, а также возраста¬
ние величины намагниченности насыщения по¬
сле охлаждения до комнатной температуры. Это
связано с образованием при нагреве сильномаг¬
нитных соединений.
Кривые J,(T) второго и третьего нагревов
(рис. 2c-2h) различны и демонстрируют различие
в типах протекания процесса окисления. Эти кри¬
вые сложные, многофазные с пиками и “площад¬
ками”. Четко проявляются две или три ферримаг-
нитные фазы с ГС| ~ 310-320, TCi ~ 360-450°С,
ТС} - 600-620°С. Первая точка Кюри (ГС|) отно¬
сится к моноклинному пирротину, вторая -
к неидентифицированной фазе переменного со¬
става, очевидно, окисленному пирротину проме¬
жуточного типа, третья - к частично окисленному
магнетиту (Fe304+y).
ПОЧВОВЕДЕНИЕ № 6 1996
766
ИЛЬИНА и др.
** ч~ '♦♦V'4* ♦ ♦«✓>*4*»* _ _v5v>..*.' V*/*.v*'*v Vv
* .♦♦ V ~
■v
100.00
99.93
99.86
Каналы
Рис. 1. Спектр ЯГР образца 12: а - 300 К; б-80 К.
Пики и “площадки” в области температур
250-320°С отвечают фазовым превращениям гек¬
сагонального и моноклинного пирротинов в про¬
цессе лабораторных нагревов [21]. Сглаженный
пик при 400-600°С отвечает незаконченному
окислению пирита. В некоторых случаях значи¬
тельно менее заметно, но также устойчиво, про¬
являются Тс ~ 110-200°С (рис. 2g), отвечающие
слабомагнитному гексагональному пирротину с
частично упорядоченными вакансиями [25, 26].
Таким образом, процесс окисления пирита в
лабораторных условиях, отраженный на кривых
температурной зависимости величины намагни¬
ченности насыщения (рис. 2a-2h), можно запи¬
сать следующим образом:
mFeS2—► aFeS2 + £FeS, + cFeS, + d¥eS. +
L L x\ *2 *3
+ Fe304+r
где величины xit x2, x3 варьируют и относятся к
пирротинам различного состава и структуры:
гексагональному, моноклинному и промежуточ¬
ному окисленному, соответственно.
Количественное соотношение фаз может ме¬
няться в значительных пределах, вплоть до пол¬
ного отсутствия некоторых промежуточных про¬
дуктов реакции. Например, окисление пирита
с образованием только одной фазы - магнетита -
приведено на рис. 2h. Следовательно, лаборатор¬
ные нагревы пирита можно рассматривать как
моделирование природных процессов окисления.
Существование сглаженных пиков и площадок на
кривых намагниченности насыщения исходных
образцов является надежным признаком естест¬
венного (природного) окисления пирита.
Обобщенные результаты литературных дан¬
ных и лабораторных экспериментов можно пред¬
ставить в виде схемы температур Кюри соедине¬
ний, возникающих при окислении пирита, и обла¬
стей температурных превращений в этих фазах
(рис. За). Температурные интервалы, указанные
на схемах, существенно варьируют в зависимости
от исходного состояния пирита и состава пирро¬
тинов.
Поскольку при лабораторном нагреве (или
окислении) пирита образуются сильномагнитные
соединения, то характерным признаком такого
превращения является значительное возрастание
величины У. ( Js /JSfj > 1). Для ряда последова¬
тельных нагревов, как правило, наблюдается воз¬
растание величины намагниченности насыщения
У, и выполняется условие У,о < JS) < Jh ,...,< JSj.
Поэтому для диагностики самого пирита и других
сульфидов железа разной степени окисленности
по кривым JS(T) существенны характеристики
У,/У* и У /У (табл. 2), а также появление про¬
межуточных фаз с температурами Тс - 100-200,
ПОЧВОВЕДЕНИЕ № 6 1996
ФАЗОВЫЙ СОСТАВ СОЕДИНЕНИЙ ЖЕЛЕЗА 767
Т,°С
•1 *2 »3
Рис. 2. Термомагнитные кривые JS(T) образцов пиритов из месторождений Учамбо (а, е, #); Вараза (Ъ, Л,/); Неждан-
ка (с, d) и синтезированного лепидокрокита (к% /). Кривые Js (7) первого (/), второго (2) и третьего (5) нагревов. Средний
размер зерна образцов (мм): Вараза: Ъ ~ 0.2;/~ 0.2; Учамбо: а ~ 0.1; g ~ 0.1; е ~ 0.2; Нежданка с ~ 0.1; d ~ 0.1 (в сростках).
ПОЧВОВЕДЕНИЕ № 6 1996
768
ИЛЬИНА и др.
Гексагональный
пирротин
г
2-
Моноклинный Окисленный Магнетит Окисленный Гематит
пирротин пирротин | магнетит
Моноклинный
X-Пирротин пирротин
В
Пирит
я
100
200
300
400
500
600
700
Г, °С
Гексагональный
пирротин
Моноклинный Окисленный Магнетит и окисленный
пирротин пирротит магнетит
,_и.
Пирит
L
100
200
Гексагональный
пирротин
. ?7Л .
300
Моноклинный
пирротин
400
500
600
700
т, °с
Окисленный
пирротин
_|_ЕЁЭ 1
Окисленный
магнетит
JL
Моноклинный
пирротин
Пирит
100
200
300
400
500
600
700
г,° с
Рис. 3. Схемы областей точек Кюри (1) для различных Fe-фаз, возникающих при окислении пирита и температурных
превращений в системе Fe-S (2): а - литературные данные [7, 8,16,17,21,24-27] и модельные эксперименты настоя¬
щей работы; b - образцы конкреций из почвы (латеритный горизонт); с - железистые аккумуляции современных ги¬
дротерм, Абхазия.
310-320, 360-450°С. Эти особенности окисления
сульфидов железа резко отличают их от лепидо-
крокита.
При термомагнитном анализе лепидокрокита
кривая J4 (Т) первого нагрева (рис. 2к, 21) анало¬
гична таковой пирита и имеет широкий пик в ин¬
тервале температур 350-450°С, отражающий пе¬
реход y-FeOOH —► y-Fe^. Однако кривые вто¬
рого нагрева резко отличаются - они всегда
однофазны с Тс = 570°С. Для лепидокрокита вели¬
чина удельной намагниченности насыщения по¬
сле первого прогрева возрастает (Js /К > 1),
а после второго, в отличие от пирита, - снижает¬
ся (Js/Js, ^ О- Это отражает последователь¬
ность фазового перехода лепидокрокита через
маггемит в гематит:
y-FeOOH -^2 y-Fe203 a-Fe203 (рис. 2к,
21). В случае тонкодисперсного лепидокрокита
переход у —► a-Fe203 может наблюдаться уже
при первом нагреве.
Мерой концентрации парамагнетика и показа¬
телем его трансформации в исходном соединении
могут служить величины Js и Jrs после серии нагре¬
вов и высота максимума (Л) на кривой JS(T) (табл. 2).
Магнитные свойства образцов конкреций из
почвы. По результатам у-резонансной спектро¬
скопии в образцах конкреций был установлен
тонкодисперсный гетит (</ — 7—12 нм) и предполо¬
жено присутствие пирита или лепидокрокита.
Рассмотрим теперь магнитные характеристики
образцов конкреций (табл. 2, рис. 4).
Кроме пиков в соответствующих интерва¬
лах температур, кривые JS(T) характеризуются
тремя температурами Кюри Тс ~ 170-200°С,
ТСг - 300-310°С, Тч - 575-600°С (для образца I
TCj ~ 675°С).
По величинам J4, У„о и стабильности к нагре¬
ву исходные образцы делятся на две группы 10—14
и 1, 15. Для образцов первой группы величины
удельной намагниченности насыщения (У1о) ко¬
леблются от 4.4 до 11 А/м, для второй группы со¬
ставляют, соответственно, 40 и 30 А/м. После
прогрева образцов первой группы до 700°С ве¬
личины 7Jo и У„о возрастают в 4-12 раз, во вто¬
рой группе меняются слабо (JSf / JSo = 1.3 и 0.85,
соответственно), последующие прогревы не ме¬
няют величины намагниченности насыщения
(Л3 /л2 = 1).
Кривые температурной зависимости остаточ¬
ной намагниченности насыщения Jrs(T) для образ¬
цов конкреций существенно отличаются от кри¬
вых JS(T) и подобны для всех образцов. На них
более или менее четко проявляются фазы с бло¬
кирующими температурами Т6 ~ 150-180,300-340
и 580-600°С.
ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВОВЕДЕНИЕ № 6 1996
Таблица 2. Магнитные характеристики образцов пиритов, лепидокрокита, конкреций латеритного горизонта и из современных гидротермальных
источников
Образец.
Месторождение
10-2,
А/м
Js Us
Sl s0
Js Us
s2 s,
T пика,
°C
sr0
io-3,
A/m
JfsxUrs0
tCo a6o)°c
О
n
Пирит. Нежданка:
21 (d~ 0.1 мм)
2.0
95.0
2.1
550
He onp.
225
480
625
300
400; 625
22 (d ~ 0.1 мм)
2.3
300.0
3.2
525
»
650
340
450; 650
23 (d~ 0.1 мм)
0.6
210.0
1.3
525
»
650
425
650
Вараза:
24 (d ~ 0.2 мм)
0.6
508.0
1.6
450
»
125
600
425
600
Учамбо:
25 (d - 0.1 мм)
21.0
100.0
1.0
460
»
125
375
525
580
300
550; 600
26 (d ~ 0.2 мм)
9.2
161.3
1.3
475
»
550
320
550
Лепидокрокит
16.0
25.6
0.13
300
»
550
550
Конкреции из почвы:
ю
11.0
5.7
1.0
375
3.7
12.7
125
300
420
600
600; 700
11
4.7
12.3
1.0
365
7.7
4.0
100
480
580
560
12
5.8
1.1
1.0
365
580
550
13
5.9
4.6
1.0
365
575
550
14
4.4
10.5
1.0
360
5.3
3.0
175
300
480
600
550
600
15
30.0
1.3
1.0
He onp.
600
600
Образцы из современных
гидротермальных источников:
2
1.38
32
1
570
4.1
0.72
150
325
460
600
125;325
600; 700
5
1.57
7.5
1
585
0.7
12.4
180
280
340
500; 600
150; 280
350; 625
6
16.1
0.88
He onp.
550
1.02
1.62
150
300
500
600
650; 700
14'
2.7
36.4
»
550
1.04
1.73
150
325
625
650
14"
4.3
1.5
1.2
550
700
700
7
4.0
1.48
He onp.
550
nc uiip.
600
He onp.
Корочки на гальке из почвы:
1
40
0.85
He onp.
350
17.8
1.1
170
375
550
600
600
Примечание. Js^ - величина намагниченности насыщения исходных образцов при Г = 20°С; - величины намагниченности насыщения образцов при 20°С после
первого и второго нагревов до 700°С соответственно; Гпика - максимальная температура пика на кривой JS(T); Jrs^; - остаточная намагниченность насыщения при
Т = 20°С в исходных образцах и после первого нагрева; TCq ; Т6q ; ГС(; - температуры Кюри и блокирующие температуры для исходных образцов и после одного
нагрева до 700°С. ^
ФАЗОВЫЙ СОСТАВ СОЕДИНЕНИЙ ЖЕЛЕЗА
770
ИЛЬИНА и др.
Г,°С
Рис. 4. Кривые термомагнитного анализа намагниченности насыщения JS(T) (а, с) и остаточной намагниченности на¬
сыщения Jrs(T) (b, d) образцов конкреций из почвы: а,Ь- первый нагрев; с, d - второй нагрев. Цифры на кривых -
номера образцов.
Кривые повторного нагрева J. (Г) и Jrs (Г)
одинаковы для всех конкреций (рис. 4с, 4d), одно¬
фазны, Тб = 580°С и подобны кривой JS(T) для об¬
разца 15. Следовательно, в процессе уже первого
лабораторного нагрева до 700°С в них происхо¬
дит полное превращение всех промежуточных
фаз и образование магнетита.
Таким образом, сравнивая кривые Js (Т) на
рис. 4 и рис. 2, можно с уверенностью заключить,
что в образцах конкреций присутствует не липи-
докрокит, а пирит в различной степени окислен¬
ный: гексагональный, моноклинный и промежу¬
точный пирротины.
Образцы четко различаются по содержанию
пирита. Наибольшее его количество содержится
в образцах 11 и 14, тогда как в образце 15 он пол¬
ностью исходно окислен до магнетита. В образце 1
гипергенное окисление пирита зафиксировано на
промежуточной стадии пирротинов различного
состава и гематита.
Магнитные свойства отложений современ¬
ных гидротерм. Типичные кривые JS(T) и 7„(Л
ПОЧВОВЕДЕНИЕ № 6 1996
ФАЗОВЫЙ СОСТАВ СОЕДИНЕНИЙ ЖЕЛЕЗА
771
Г,°С
Рис. 5. Кривые термомагнитного анализа намагниченности насыщения JS(T) (а, с) и остаточной намагниченности на¬
сыщения Jrs(T) (b, d) образцов современных гидротермальных отложений: а,Ь- первый нагрев; с, d - второй нагрев.
Цифры на кривых - номера образцов.
образцов из современных гидротерм (2,5,6,7,14',
14") приведены на рис. 5а-5с, а основные их маг¬
нитные характеристики - в табл. 2. Как правило,
на этих кривых отмечаются два пика - небольшой
при 300-350°С и основной при 550-580°С. Эти пи¬
ки имеют существенно меньшую интенсивность,
чем в случае конкреций, часто вырождены в пло¬
щадки. После прогрева до 700°С величина Js воз¬
растает также в различной степени.
Как и в конкрециях, интенсивность пиков h на
кривых J4(T) и отношения J /У5(| и JrS[ /У„о об¬
ратно пропорциональны намагниченности исход¬
ных образцов. Это отражает степень окисленности
до магнетита. Так, наибольшей начальной
намагниченностью и стабильностью к последую¬
щим нагревам обладают образцы 6,7,14" (табл. 2),
они же исходно наиболее окислены. Во всех об¬
разцах отложений современных гидротерм изу¬
чаемого региона в том или ином количестве со¬
держится пирит. На кривых JS(T) он диагностиру¬
ется при наличии устойчивой величины JS[ по
пикам или “площадкам” при Т = 500-600°С.
ПОЧВОВЕДЕНИЕ № 6 1996
б*
772
ИЛЬИНА и др.
Кроме двух пиков (или площадок), для ряда об¬
разцов (2,5,6,14") на кривых JS(T) отмечается еще
несколько характерных точек Тх = 150-180°С;
Т2 =* 280-325 и Г3 = 420-480°С. Эти особенности
либо исчезают при повторном нагреве, либо со¬
храняются, указывая на точки Кюри определен¬
ных устойчивых магнитных фаз.
Кривые Jrs(T) образцов гидротерм и латерит-
ных горизонтов подобны, наиболее явно это про¬
является в наличии блокирующих температур
в соответствующих температурных интервалах.
Как и ранее, блокирующие температуры могут
быть отнесены к трем типам пирротинов:
1) Т6 = 150-180°С - гексагональному, слабо¬
магнитному, частично упорядоченному; 2) Т6 =
= 300-350°С - моноклинному ферримагнитному;
3) Тб - 450-480°С - окисленному промежуточному.
Кривые JS(T) и Jrs(T) повторного нагрева при¬
ведены на рис. 5с и 5d. Они демонстрируют посте¬
пенное сглаживание пиков и “площадок” в темпе¬
ратурном интервале 550-580°С и трансформацию
кривых в соответствии с устойчивостью проме¬
жуточных продуктов окисления пиритов. Однако
однородные магнетитовые кривые JS(T) и Jrs(T),
подобные кривым, полученным для конкреций
(рис. 4с и Ad), для образцов из гидротерм при по¬
вторных лабораторных нагревах не наблюдают¬
ся. В то же время присутствие незначительного
количества частично окисленного магнетита
фиксируется на кривых для всех образцов из гид¬
ротерм по Тс ~ 620°С (рис. 5а); по-видимому, он
является тонкодисперсным, так как на кривых
Jrs(T) его блокирующие температуры существен¬
но ниже, чем Тс, и не превышают 550-570°С.
Магнетит, образующийся в результате лабора¬
торных нагревов до 700°С, вызывает дополни¬
тельное увеличение намагниченности образцов
после прогрева.
ОБСУЖДЕНИЕ
Проведенные исследования показали, что об¬
щий состав железосодержащих фаз в конкрециях
латеритного горизонта и образцах современных
гидротермальных ожелезненных образований
достаточно близок и может быть записан следую¬
щим образом:
a-FeOOH
гетит
aFeS?
bFeSr
пирит и окислен- гексагональный
ный пирит слабомагнитный
пирротин
cFeSr
dFeS
моноклинным окисленным частично
ферримагнит- пирротин окисленный
ный пирротин магнетит
При очень большом сходстве по фазовому со¬
ставу образцов конкреций и современных гидро¬
термальных образований наблюдаются и некото¬
рые различия по свойствам отдельных фаз. Для
конкреций температура начала окисления пирита
(возможно, и пирротина) в лабораторных услови¬
ях при прогреве ниже, чем для гидротермальных
образований. По-видимому, за счет этого, уже
в течение первого лабораторного нагрева пирит
конкреций полностью окисляется до магнетита.
Обращает на себя внимание фаза с перемен¬
ной точкой Кюри Тс от 420 до 480°С, проявляю¬
щаяся в той или иной мере во всех образцах либо
в исходном состоянии, либо при лабораторных
нагревах. Мы предположительно отнесли ее к
окисленному промежуточному пирротину, одна¬
ко, структурные характеристики и состав этой
фазы не известны и нуждаются в дополнитель¬
ных исследованиях.
Схемы фазового состава океанических суль¬
фидсодержащих конкреций по литературным
данным и результатам модельных экспериментов
настоящей работы, а также состав почвенных
латеритных конкреционных горизонтов и желе¬
зистых отложений современных гидротерм пред¬
ставлены на рис. 3. Фазовый состав конкреций
также близок к составу “черных курильщиков” -
уникальных сульфидных многометровых построек
гидротермального происхождения, обнаруженных
в зонах спрединга в океанах.
Присутствие в конкреционном латеритном го¬
ризонте почв сульфид-магнетитовой ассоциации
минералов, продуктов окисления пирита и пирро¬
тина в комплексе с гетитом (d ~ 7-12 нм) свиде¬
тельствует об относительной древности геологи¬
ческих процессов, приведших к образованию
ожелезненного слоя. Суть этих процессов - гидро¬
термальное воздействие на отложения, связанное
с аккумуляцией железа; не исключено воздействие
низкотемпературных гидротерм и палеогидроген-
ная аккумуляция железа. Гидротермальные источ¬
ники известны в данном районе и сейчас. Они вы¬
носят на поверхность значительное количество
Fe и Si02. Можно предположить, что подобные
процессы были широко распространены в геоло¬
гическом прошлом территории, когда поствулка-
ническая активность проявлялась ярче. Об этом
свидетельствуют горизонтальные слои ожелез-
ненного сцементированного галечника в обнаже¬
ниях морских террас на глубине 3, 6, Юм, кото¬
рые чередуются с прослоями обычного галечни¬
ка с песчаным или супесчаным материалом.
Как известно, общим фактором, приводящим
к нарушению равновесия и выпадению сульфидов
металлов из раствора, является снижение их ще¬
лочности и повышение окислительно-восстанови¬
тельного потенциала системы. В рассматривае¬
мом случае такие процессы могли происходить
при смешении глубинных щелочных растворов.
ПОЧВОВЕДЕНИЕ № 6 1996
ФАЗОВЫЙ СОСТАВ СОЕДИНЕНИЙ ЖЕЛЕЗА
773
обогащенных сероводородом и поверхностных
вод, имеющих более высокий окислительно-вос-
становительный потенциал. Такая обстановка
могла существовать у зеркала грунтовых вод, и
непосредственно на поверхности почвы вокруг
источников. Фактически вся зона аэрации и самая
верхняя часть горизонта грунтовых вод представ¬
ляет собой геохимический барьер для глубинных
растворов. Вместе с тем исследования в кальдере
Узона (Камчатская обл.) показали, что сульфиды
остаются на месте образования лишь в том слу¬
чае, если взаимодействие происходило в свобод¬
ной обстановке, например, в приповерхностном
горизонте на дне озера, в бессточной котловине.
Лишь в зоне растекания раствора (при смешива¬
нии глубинных и поверхностных окисленных вод
наблюдается образование аурипигмента As2S3,
скородита (FeAs04 • 2Н20) или тонкодисперсных
пирита и марказита [12].
В связи с этим накопление сульфидов (как и
других рудных веществ) наиболее вероятно в ус¬
ловиях, когда сравнительно рыхлые породы на¬
ходятся на уровне грунтовых вод или крупных
плоскодонных “полей” с многочисленными выхо¬
дами ювенильных вод, в том числе низкотемпера¬
турных гидротерм. В этом случае происходит вза¬
имодействие ювенильных и поверхностных вод.
Возможен был и поток растворов по галечнику,
их взаимодействие с осажденными вокруг гальки
слоями Fe-соединений. Но в этом случае поток
должен был быть очень медленным, и формиро¬
вание латеритного горизонта шло в относительно
спокойной геологической обстановке на по¬
верхности глин, гальки или суглинков. Иногда
в конкрециях латерита встречаются остатки рас¬
тительных тканей [15], что является дополни¬
тельным свидетельством их образования в припо¬
верхностном корнеобитаемом слое палеопочвы
или донных отложений. В пользу того, что некогда
конкреционный латеритный горизонт был припо¬
верхностным свидетельствует вертикальная неод¬
нородность почвенного профиля. Под латеритом
на границе с глинистыми отложениями и в верхней
толще современной почвы встречаются погребен¬
ные горизонты, выделяющиеся по содержанию гу¬
муса, более высокому значению pH (5.5 по сравне¬
нию с 4.5 в других слоях), а также по данным мик¬
роморфологии и рентгеноструктурного анализа
крупных и илистых фракций [15]. Такая неодно¬
родность над конкреционным слоем и под ним сви¬
детельствует о захоронении конкреционного гори¬
зонта под слоем глинисто-суглинистых наносов.
Процессы, идущие в современной поверхност-
но-глеевой почве, гумусообразование, сегрегация
железа и формирование мелких Fe-гумусовых но¬
вообразований в верхней оглеенной толще поч¬
вы, оглеение и миграция в пределах горизонта
тонкодисперсного материала не могут способст¬
вовать накоплению сульфидов железа в самом
латеритном слое.
Поскольку содержание магнетита до и после
прогрева конкреций латерита прямо коррелиру¬
ет с содержанием сульфидной фазы Fe (чем боль¬
ше сульфида железа в исходном образце, тем
больше величина пика и содержание магнетита
после прогрева / JSo > 1, табл. 2), можно с уве¬
ренностью утверждать, что из сульфидов Fe вну¬
три конкреций при их постепенном окислении об¬
разуется магнетит. Этим можно объяснить, в ча¬
стности, полученные ранее данные о том, что
более мелкие литогенные конкреции, как прави¬
ло, содержат больше магнетита, чем более круп¬
ные [15].
Присутствие более мелких “современных”
Fe-новообразований в латеритном слое и в верх¬
ней толще почвы свидетельствует о том, что в на¬
стоящее время происходит постепенное разруше¬
ние древнего ожелезненного слоя под воздейст¬
вием почвообразования, в том числе, оглеения,
накопление аморфной гидроокиси Fe и вторичная
сегрегация более мелких частиц по остаткам
ожелезненного слоя при высушивании.
ВЫВОДЫ
1. В латеритном конкреционном горизонте почв
установлено присутствие гетита в качестве ос¬
новной фазы, а также пирита разной степени
окисленности и концентрации, промежуточных
продуктов его окисления - пирротинов и окис¬
ленного магнетита.
2. Фазовый состав конкреций латеритного го¬
ризонта близок к составу металлоносных донных
океанических отложений зон спрединга и гидро¬
термальной деятельности, а также ожелезненных
осадков современных гидротермальных источни¬
ков и рудных месторождений исследованного реги¬
она. Сходство состава указывает на определенную
генетическую связь всех этих образований.
Поэтому латеритный конкреционный горизонт
не является продуктом почвообразования в зоне
гипергенеза.
3. Фазовый состав латеритного горизонта - на¬
личие пирита и серии метастабильных фаз его
окисления до магнетита и гематита - свидетель¬
ствует о древности геологических процессов, от¬
вечающих за его образование. Возможны два пу¬
ти образования этого комплекса: а) в результате
палеогидротермального воздействия на отложе¬
ния, связанного с аккумуляцией железа на суше;
б) палеогидротермальная аккумуляция железа из
гидротерм при формировании донных отложений
(морских).
4. Древний ожелезненный слой был впоследст¬
вии погребен под слоем суглинистых наносов -
последующих отложений различного генезиса.
В настоящее время происходит постепенное раз¬
рушение латеритного горизонта под воздействи¬
ем почвообразования.
ПОЧВОВЕДЕНИЕ № 6 1996
774
ИЛЬИНА и др.
СПИСОК ЛИТЕРАТУРЫ
1. Багин В.И., Гендлер Т.С., Авилова Т.Е. Магнетизм
а-окислов и гидроокислов железа. М.: Наука, 1988.
180 с.
2. Багин В.И., Гендлер Т.С., Деков В.М., Полищук О.Б.
Мессбауэровские исследования металлоносных
осадков восточно-тихоокеанского поднятия
(20о30'-22°00' ю.ш.) // Океанология. 1993. Т. 33.
№ 5. С. 763-770.
3. Буров Б.В., Ясонов П.Г. Введение в дифференци¬
альный анализ горных пород. Казань: Изд-во Ка¬
зан. гос. ун-та, 1979. 280 с.
4. Водяницкий Ю.Н., Манавариани Л.Г. Лепидокро-
кит в подзолисго-желтоземных почвах // Докл.
ВАСХНИЛ. 1990. № 8. С. 16-18.
5. Гендлер Т.С., Ершова JI.C., Карпачевский Л.О.,
Кузьмин Р.Н. Исследование поведения железа в
структуре монтмориллонита и на его поверхности
методом ядерного гамма-резонанса // Почвоведе¬
ние. 1980. № 12. С. 86-95.
6. Зонн С.В. Железо в почвах. М.: Наука, 1982. 206 с.
7. Жабин А.Г., Самсонов Н.С., Абесадзе Г.Н. Золото¬
носные диагенетические пиритовые конкреции //
Литология и полезные ископаемые. 1993. № 4.
С. 117-120.
8. Железо-марганцовые конкреции Центральной кот¬
ловины Индийского океана / Под ред. И.О. Мурд-
маа, Ю.Ю. Житковский, Н.С. Скорнякова. М.: На¬
ука, 1989. 223 с.
9. Ильина Л.СЛежава В.В. Применение ЯГРС и
термомагнитного анализа при изучении Fe-ново¬
образований в почвах // Мат. Всесоюз. совещания.
Тбилиси-Кобулети, 1988. С. 19-20.
10. Ильина Л.С., Лежава В.В., Гендлер Т.С. Особенно¬
сти фазового состава Fe-соединений в конкрецион¬
ных горизонтах почв Западной Грузии // Мат. Все¬
союз. конференции. Тбилиси, 1990. С. 7-8.
11. Капщук М.П., Градусов Б.П., Чижикова Н.П. Или¬
сто-минералогические признаки почв мандарино¬
вых плантаций разной продуктивности // Науч. тр.
ВАСХНИЛ, 1988. С. 5-17.
12. Карпов Г.А. В кальдере вулкана. М.: Наука, 1980.
94 с.
13. Лежава В.В., ИлъинаЛ.С., Гендлер Т.С. Состав кон¬
креций в субтропических подзолистых почвах Абха¬
зии//Сообщение АН ГрузССР. 1984. Вып. 114. № 1.
С. 141-144.
14. Лежава В.В. О почвенных конкрециях Западной
Грузии // Тр. Геогр. об-ва Грузии. 1989. Т. XVIII.
С. 50-56.
15. Манавариани Л.Г. Микроморфология и микрохи¬
мия подзолисго-желтоземных почв Грузии: Дис....
канд. с.-х. наук / Тбилисский гос. ун-т. Тбилиси,
1989. 250 с.
16. Нагата Т. Магнетизм горных пород. М.: Мир,
1965. 347 с.
17. Нгуен Тхи Ким Тхоа. Кристаллизационная и хими¬
ческая остаточная намагниченность магнетита:
Дис. ... д-ра физ.-мат. наук / ИФЗ АН СССР. М.,
1985.
18. Разумова В.Н. Древние коры выветривания и гид¬
ротермальный процесс. М.: Наука, 1977.156 с.
19. Соколов И А., Лежава В.В. О проблемах генезиса
и эволюции почв влажных субтропиков на приме¬
ре Западной Грузии // Мат. Всесоюз. конференции.
Пущино, 1984. С. 164-166.
20. Соколов И.А., Лежава В.В. О происхождении ла¬
терита в Западной Грузии // Мат. Всесоюз. конфе¬
ренции. Тбилиси, 1990. С. 14—15.
21. Соппа И.В. Химическая намагниченность про¬
дуктов превращения пирита и титаномагнетита:
Дис. ... канд. физ.-мат. наук / ИФЗ АН СССР. М.,
1989.
22. Шатский Г.В., Зайцевская Ю.В. Проблемы изото¬
пии и сульфатредукции осадочно-вулканических
комплексов Кукасозерской зоны // Литология и
полезные ископаемые. 1993. № 4. С. 91-104.
23. Garg V.K., Lin Y.S., Puri S.P. Mossbauer electric field
gradient study in FeS2 (pyrite) // J. Appl. Phys. 1974. V. 45.
№ 1. P. 70-72.
24. KruczykJ. Phase transitions in iron oxides and sulphides
and their relations to mangetic properties of rocks // Pub¬
lications of the institute of geophysics. Polish academy
of sci. Warszawa-Lodz, 1983. P. 34-56.
25. Schwarz EJ. Magnetic property of pyrrhotite and their
use in applied geology and geophysics // Geol. Survey
Canada. 1975. P. 59-74.
26. Schwarz EJ.t Vaughan D.J. Magnetic Phase Relations
of Pyrrhotite. J. Geomag. Geoelectr. 1972. V. 24.
P. 441-458.
27. Krs М., Navak F„ Krsova М., Prunor P., Kouklikova L.,
Jans a S. Magnetic properties and metastability of greig-
ite - smythite mineralization in brown-coal basins of the
Krusnehory. Piedmont, Bohemia. Phys. Earth Planet. In¬
ter, 1992. V. 70. P. 273-287.
Phase Composition of Iron Compounds in Soils and Hydrothermal Deposits
of Abkhazia as Related to Problems of Laterization
L. S. Ilyina, T. S. Gendler, V. V. Lezhava
The phase composition of iron compounds is studied by Messbauer’ spectroscopy and thermo-magnetic tech¬
nique of lateritic horizons in yellow podzolic soils as well as in Fe-containing deposits of recent hydrothermal
sources of Abkhazia. The data so obtained testify the presence of fine-dispersed goethite, pyrite, pyrrhotine
products of pyrite and magnetite oxidation. The phase composition of investigated samples is found to be sim¬
ilar to that of some sulfide-containing Fe-Mn concretions in sea bottom sediments of hydrothermal origin,
being indicative of ancient geological processes resulted in the formation of ferruginous layer. Two possible
ways of its formation are assumed to consider: paleohydrothermal effects on deposits associated with Fe accu¬
mulation and paleohydrogenic accumulation of Fe from hydrothermal solutions. Ferruginous laterite horizon
proves to be burned by loamy deposits after its formation.
ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВОВЕДЕНИЕ. 1996, № 6. с. 775-782
ХИМИЯ
ПОЧВ
УДК 631.416.3+631.416.9(571.6)
СОДЕРЖАНИЕ СЕРЫ И МИКРОЭЛЕМЕНТОВ
В ПАХОТНЫХ ВУЛКАНИЧЕСКИХ ПОЧВАХ КАМЧАТКИ
© 1996 г. В. И. Голов, С. М. Бажова
Биолого-почвенный институт ДВО РАН. Владивосток
Камчатская проектно-изыскательская станция химизации сельского хозяйства
Поступила в редакцию 08.06.94 г.
Обследованы пахотные почвы Камчатки (58 тыс. га) на содержание валовых и подвижных форм се¬
ры и основных микроэлементов (Mn, Zn, Си, В, Мо и Со). Содержание валовых форм изучаемых
элементов за исключением серы находится в пределах кларка. Большинство почв относится к низ¬
кообеспеченным подвижными формами соединений S (73%), В (51%), Zn (46%), Mn (43%), Со и
Мо (38%), около пятой их части слабо обеспечена Си (20%).
Почвы, используемые в сельскохозяйствен¬
ном производстве Камчатки, практически все
подвержены влиянию вулканической деятельнос¬
ти, так как приурочены, в основном, к восточно¬
му и южному побережью полуострова, а также
центральной Камчатской депрессии, незначи¬
тельно удаленных от действующих вулканов.
Почвообразующие породы пахотных почв со¬
стоят главным образом из пирокластических от¬
ложений различной мощности, которая по мере
удаления от действующих вулканов колеблется
от 20 до 150 см. Вулканическая деятельность яв¬
ляется причиной однообразия почвообразующих
пород, представленных, в основном, базальтами,
андезито-базальтами и андезитами. Такой состав
магмы характерен для всего Тихоокеанского вул¬
канического пояса. В минералогическом составе
вулканических почв обычно преобладают сред¬
неосновное вулканическое стекло и полевые
шпаты. В меньшем количестве присутствуют слю¬
ды, пироксены, кварц, роговая обманка. Среди
вторичных минералов, образующихся при вывет¬
ривании названных пород, преобладают аморф¬
ные минералы - аллофаны, гидроксиды железа и
алюминия. С этим связана высокая сорбционная
способность рассматриваемых почв по отноше¬
нию к фосфатам [15, 18].
В пределах речных пойм встречаются аллюви¬
альные отложения, на которых формируются лу¬
гово-дерновые аллювиальные, а на слабодрен-
нированных участках (что связано с наличием
вечной мерзлоты) - торфяно-болотные почвы.
В составе аллювиальных, аллювиально-пролю-
виальных и реже делювиальных отложений, на¬
ряду с продуктами выветривания коренных поч¬
вообразующих пород, существенную роль играет
переотложенный вулканогенный материал, в со¬
ставе которого присутствует больше окристалли-
зованных минералов, а не аморфных, что харак¬
терно для аэральных отложений. В составе тонко¬
дисперсной фракции аллювиальных отложений
больше содержится вторичных минералов, что
предопределяет их ббльшую гидроморфносгь.
Химический состав аллювия отличается от аэраль¬
ных вулканических отложений более низким со¬
держанием щелочных и щелочноземельных эле¬
ментов, особенно Са и Na [7,11,15, 18].
Физические свойства вулканических почв до¬
вольно устойчивы и однообразны, слабо зависят
от мощности пирокластического материала. Как
правило, все почвы и почвообразующие породы
легкого гранулометрического состава. Для них
характерны низкая плотность, высокая порис¬
тость, хорошая аэрация и фильтрационная спо¬
собность. Несмотря на хорошую фильтрацию,
вулканическим почвам свойственна высокая гид-
рофильность, которая объясняется значительным
содержанием рыхлосвязанной воды, присутствую¬
щей в аморфном вулканическом пепле, обладаю¬
щем большой адсорбционной поверхностью, спо¬
собной удерживать кристаллическую влагу.
Поэтому в засушливый период растения крайне
редко теряют влагу до пределов, близких к коэф¬
фициенту завядания [11, 15]. г
Пахотные почвы Камчатки по валовым запа¬
сам основных питательных элементов и cep^i
сравнимы с наиболее плодородными почвами за¬
падных областей России (серыми лесными и чер¬
ноземами). Высокое содержание гумуса и азота
в почвах, залегающих под лесной и луговой рас¬
тительностью, объясняется обильным поступле¬
нием органического вещества за счет древесного
и лугового опада (до 10 т/га). Минерализация ор¬
ганического вещества за счет активной микроби¬
ологической деятельности, которая отмечается
в теплый период (июль-август), идет быстро, не¬
смотря на суровый и влажный климат [7, 18].
Быстрой минерализации органического вещества
почв способствует наличие вулканического пес¬
ка, отличающегося хорошей проницаемостью и
775
776 ГОЛОВ, БАХОВА
Таблица 1. Шкала обеспеченности почв подвижными формами микроэлементов, использованная в настоящей работе
Микро¬
элемент
Экстрагент
Дифференциация почв по содержанию микроэлементов, мг/кг
низкое
среднее
высокое
Мп
0.1 н. H2S04 ацетатно-аммонийный
30
31-70
71
Zn
Буферный раствор pH 4.8
2.0
2.1-5
5.1
Си
1 н. НС1
1.5
1.6-3.3
3.4
Со
1 н. HN03
1.0
1.1-2.2
2.3
В
Н20, горячая
0.33
0.34-0.70
0.71
Мо
Оксалатный буфер pH 3.3
0.1
0.11-0.22
0.23
аэрацией. Особенно это заметно при осушении и
сельскохозяйственном использовании торфяни¬
ков, широко распространенных на восточном по¬
бережье Камчатки. До 50-х гг. XX в. торфяники
Камчатки считались непригодными для сельско¬
хозяйственного освоения из-за слабой минерали¬
зации растительного опада и наличия мерзлоты.
Однако разработанные В.В. Бурлака агротехни¬
ческие приемы осушения и обработки болотной
целины позволили освоить и использовать в даль¬
нейшем обширные торфяные массивы Восточно¬
го побережья, а также бассейнов рек Авача,
Паратунка и др. [4, 15]. Большинство почв Кам¬
чатки, включая торфяно-болотные, характеризу¬
ются большими запасами слабогумифицирован-
ного органического вещества и питательных эле¬
ментов как органогенных (С, N, S), так и зольных
(Р, К, Mg и др.). Однако содержание подвижных
форм основных питательных элементов, вклю¬
чая микроэлементы, очень низкое. Отсутствие
подвижных соединений азота объясняется корот¬
ким вегетационным периодом для автоморфных
почв и слабой микробиологической деятельностью
для торфяников. Низкое содержание фосфора
объясняется энергичным связыванием его аморф¬
ными гидроксидами Fe и А1, а калия и других эле¬
ментов, включая серу и микроэлементы, - высокой
промываемостью вулканических почв [11,15].
Легкий гранулометрический состав большин¬
ства пахотных почв Камчатки, а также относи¬
тельная бедность почвообразующих вулканичес¬
ких пород основными питательными элементами
способствуют формированию неустойчивого пи¬
тательного режима, дефицитного практически по
всем элементам питания. В связи с этим здесь на¬
блюдается высокая эффективность минеральных
удобрений [15, 17]. С другой стороны, высокие
дозы питательных элементов, вносимых с удоб¬
рениями, могут оказывать негативное воздейст¬
вие на окружающую среду и качество получае¬
мой продукции в силу невысокой адсорбционной
способности камчатских почв. Поэтому в данном
регионе наблюдается весьма специфичная реак¬
ция возделываемых растений на вносимые удоб¬
рения, требующая более глубокого и тщательно¬
го изучения.
В основу настоящей статьи положены данные,
полученные лабораторией почвоведения и эко¬
логии почв Биолого-почвенного института по со¬
держанию валовых форм изучаемых элементов,
а также материалы 5-летних исследований (с 1987
по 1992 г.) Камчатской проектно-изыскательской
станцией химизации сельского хозяйства по опре¬
делению подвижных форм соединений серы и ос¬
новных микроэлементов в пахотных почвах Кам¬
чатской области.
Побуждающим мотивом этих исследований по¬
служили, с одной стороны, практически полное от¬
сутствие систематических данных по содержанию
соединений серы и основных микроэлементов в
почвах, используемых в сельскохозяйственном про¬
изводстве, с другой стороны, противоречивые
факты (высокой и незначительной) эффективно¬
сти внесения борных, молибденовых и других ми¬
кроудобрений в почвы Камчатки [19, 20]. Кроме
того, в последние годы наблюдались случаи высо¬
кого накопления нитратов в сельскохозяйствен¬
ной продукции, в основном в овощных и кормо¬
вых культурах. Это зачастую происходит из-за
недостаточного питания элементами, участвую¬
щими в азотном метаболизме растений (серы, мо¬
либдена, меди и др.).
Определение содержания подвижных форм
соединений микроэлементов проводили по мето¬
ду Пейве-Ринькиса, модифицированному и реко¬
мендованному ЦИНАО. Валовые формы микро¬
элементов определяли спектральным методом
в образцах почв из разрезов, заложенных на ос¬
новных типах почв. Всего было проанализирова¬
но 28 разрезов и 2500 средних образцов почв со
всей пахотной площади Камчатской области
(58 тыс. га). Отбор почвенных образцов для опре¬
деления доступных форм соединений элементов
проводили по методике, разработанной ЦИНАО
для агрохимических лабораторий России [12].
Оценка обеспеченности пахотных почв прово¬
дилась по шкале, разработанной на основе вытя¬
жек, предложенных Ринькисом (1963) в модифи¬
кации ЦИНАО (табл. 1).
Сульфатную серу определяли в солевой вы¬
тяжке по методике Айдиняна и др. [1, 8], согласно
ПОЧВОВЕДЕНИЕ № 6 1996
СОДЕРЖАНИЕ СЕРЫ И МИКРОЭЛЕМЕНТОВ
777
которой шкала обеспеченности почв серой вы¬
глядит следующим образом: <6 мг/кг - слабообе¬
спеченная почва, от 6.1 до 12 мг/кг - средне- и
> 12.1 мг/кг - хорошо обеспеченная.
Поскольку приватизация пахотных земель и
передача их в аренду на Камчатке идут значи¬
тельно медленнее, чем в других регионах России,
в статье сохранены старые названия хозяйств.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Сера. Содержание серы в почвах в значитель¬
ной мере зависит от содержания органического
вещества [1, 6, 10]. Плодородие основных почв
Камчатки (охристых вулканических), используе¬
мых в сельскохозяйственном производстве, очень
низкое, поэтому устойчивые и хорошие урожаи
можно получать только при внесении достаточно¬
го количества удобрений [15,17]. Кислая реакция
среды, недостаточное количество тепла, особен¬
но в начале вегетации, являются причиной низ¬
кой микробиологической активности почв в на¬
чале лета, и, как следствие, незначительного на¬
копления в них нитратов и других форм азота из
гумусовых веществ. О низкой микробиологичес¬
кой активности почв свидетельствует отсутствие
в них азотобактера и небольшое количество цел¬
люлозоразлагающих микроорганизмов [11]. Ана¬
логичным образом обстоят дела с серой, поэтому
в почвах Камчатки ее не хватает, особенно в на¬
чальные фазы развития растений.
Содержание валовой серы колебалось от 0.33
до 1.3% (в расчете на S03), что значительно выше
кларка для почв средней полосы России. Это
можно объяснить высоким содержанием серы в
вулканических пеплах (0.14-0.53%) и органичес¬
ком веществе [11]. Причем с пеплопадами сера,
как правило, поступает в почву преимущественно
в виде газообразных соединений, адсорбированных
пепловыми частицами, поэтому в первый год по¬
сле извержения вулканов бывает доступной для
растений. Однако содержание подвижных форм
соединений серы (в солевой вытяжке) небольшое и
колеблется от следов до 16 мг/кг (в расчете на S).
Большинство пахотных почв Камчатки (73%)
следует отнести к категории низкообеспеченных
(менее 6 мг/кг) и только 27% к средне- и хорошо
обеспеченным. Это обстоятельство позволяет
сделать предположение о положительной реак¬
ции возделываемых на Камчатке культур на вне¬
сение серных удобрений. Большая разница в со¬
держании различных форм соединений серы (вы¬
сокое - валовой и низкое - подвижной) можно
объяснить тем, что в составе вулканических пеп-
лов сера находится в сульфидной форме, которая
весьма устойчива к факторам выветривания в зо¬
не гипергенеза и недоступна в качестве источни¬
ка питания для растений [6, 10, 11]. Кроме того,
сульфатная форма соединений серы, образую¬
щихся в конечном счете в результате минерали¬
зации органического вещества и медленного вы¬
ветривания сульфидных минералов, легко вымы¬
вается за пределы почвенного профиля, т.е.
анионная поглотительная способность данных
почв ничтожна.
Как видно из данных, приведенных в табл. 2,
около 3/4 исследованных почв характеризуются
низким содержанием сульфатной серы. Наиболее
низким содержанием серы отличаются почвы сов¬
хозов: “Заречный” (67%), “Камчатский” (85%),
“Карагинский” (100%), “Ключевской” (92%),
“Корякский” (76%) и “Елизовский” (100%). Более
благополучная ситуация по содержанию серы от¬
мечена в почвах совхозов “Лазовский” (36%),
“Моховский” (10%) и “Рассвет” (22%). Высокое
содержание подвижных форм соединений серы
в почвах (точнее, грунтах) тепличного совхоза
“Термальный”, где все почвы (40 га) попадают в
категорию хорошо обеспеченных, следует вос¬
принимать с некоторой поправкой, так как для
закрытого грунта существуют другие градации
с более высоким содержанием как серы, так и
других элементов. Для выяснения связи содержа¬
ния серы и микроэлементов с агрохимическими
свойствами почв мы приводим статистически об¬
работанные агрохимические показатели и содер¬
жание определяемых элементов в основных поч¬
вах территории землепользования совхоза “Кам¬
чатский”, который является наиболее типичным
в почвенно-климатическом отношении (табл. 3).
Лучше других обеспечены серой аллювиаль¬
ные пойменные (лугово-дерновые, по Соколову)
почвы, отличающиеся повышенным содержанием
гумуса и более тяжелым гранулометрическим со¬
ставом по сравнению с более распространенными
охристыми вулканическими почвами (табл. 3 и 4).
Следует подчеркнуть, что наименее обеспечены
серой почвы северной части Камчатки, особенно
горных территорий - торфянистые иллювиаль¬
но-гумусовые вулканические, охристо-подзоли¬
стые и примитивно-литоморфные (совхозы:
“Тигильский”, “Карагинский”, “Корфский”, “Та-
ловский”, “Паланский”, обладающие небольши¬
ми площадями пахотных угодий). Более благопо¬
лучная ситуация по содержанию доступной серы
складывается в почвах равнин и предгорных ува¬
лов южной и центральной частей области (совхо¬
зы: “Моховский”, “Рассвет”, “Петропавловский”
и др.) в зоне распространения лугово-дерновых,
перегнойно-глеевых, пойменных и торфянистых
почв [15,18].
Марганец. Содержание валового марганца
в обследованных почвах колебалось от 0.01 до
0.3% (в расчете на элемент), причем в основном оно
было ниже кларкового (кларк 0.085%). По содер¬
жанию подвижного марганца почти половина
ПОЧВОВЕДЕНИЕ № 6 1996
778 ГОЛОВ, БАХОВА
Таблица 2. Агрохимические показатели основных почв совхоза “Камчатский”
Агрохимический показатель
Статис¬
тический
показатель
Почвы
лесные вулканические
охристые дерновые
аллювиальные
старопойменные
аллювиальные
пойменные
Гумус, %
М
6.5
7.3
7.3
±т
0.14
0.52
0.55
V
12.3
25.5
18.5
pH солевой суспензии
М
5.3
5.3
5.5
±т
0.08
0.10
0.19
V
8.3
6.5
8.2
Гидролитическая кислотность,
М
4.5
5.1
4.1
мг-экв/100 г
±т
0.35
0.56
0.72
V
45.7
39.8
43.1
Сумма поглощенных
М
7.6
8.3
9.0
оснований, мг-экв/100 г
±т
0.42
0.56
1.22
V
32.2
24.2
33.2
Обменный кальций, мг-экв/100 г
М
6.9
7.6
8.2
±т
0.41
0.55
1.19
V
34.6
26.0
35.6
Степень насыщенности
М
62.0
62.2
67.6
основаниями, мг-экв/100 г
±т
2.88
3.73
6.53
V
27.0
21.5
23.6
Примечание. Здесь и далее: М - среднеарифметическая, т - ошибка среднего, V, % - коэффициент вариации.
Таблица 3. Содержание микроэлементов в основных почвах совхоза “Камчатский”
Статистический
показатель
Почвы
Элемент
лесные вулканические,
охристые дерновые
аллювиальные
старопойменные
аллювиальные
пойменные
Сера
М
4.6
4.1 .
7.8
±т
0.64
0.54
2.88
V
51.3
47.2
90.4
Марганец
М
36.3
40.9
36.3
±т
0.95
1.26
2.12
V
15.3
11.1
14.3
Цинк
М
1.25
1.46
1.42
±т
0.116
0.242
0.36
V
53.2
59.5
62.6
Медь
м
2.8
2.3
2.9
±т
0.27
0.17
0.44
V
55.5
26.7
37.3
Бор
М
0.52
0.62
0.65
±т
0.042
0.058
0.107
V
46.2
33.4
40.3
Кобальт
М
0.48
0.98
0.61
±т
0.143
0.328
0.376
V
175.2
145.5
151.5
Молибден
М
0.15
0.16
0.14
±т
0.008
0.013
0.017
V
33.7
30.0
29.9
ПОЧВОВЕДЕНИЕ № 6 1996
СОДЕРЖАНИЕ СЕРЫ И МИКРОЭЛЕМЕНТОВ
779
Таблица 4. Обеспеченность пахотных почв Камчатки подвижными формами соединений серы, марганца и цин¬
ка, % от обследованной площади
Сельскохозяйст¬
Обследо-
D О UllO Q ПТТЛ-
Количество про¬
анализирован¬
ных образцов
Сера
Марганец
Цинк
венное предприятие
ОйППйЛ 11 л и
щадь, га
I
II
III
I
II
III
I
II
III
“Заречный”
5559
121
67
7
26
65
35
0
100
0
0
“Камчатский”
2818
40
85
5
10
20
80
0
78
22
0
“Ключевской”
2161
87
92
8
0
1
86
13
1
84
15
“Корякский”
2843
97
76
10
14
0
20
80
2
98
0
“Лазовский”
4009
127
36
20
44
49
38
13
95
5
0
ОПХ “Октябрьское”
3132
67
80
19
1
65
35
0
63
37
0
“Петропавловский”
4155
94
48
20
32
4
81
15
24
74
2
“Рассвет”
2088
103
21
22
57
89
11
0
70
29
1
“Большерецкий”
3814
76
71
18
11
79
21
0
92
8
0
“Крутобереговский”
2870
57
82
3
15
50
50
0
96
4
0
“Пограничный”
4539
93
59
19
22
81
15
4
100
0
0
“Комсомольский”
6188
106
75
14
11
19
81
0
99
1
0
Военный J4® 146
1850
50
71
29
0
32
68
0
0
0
100
“Мильковский”
6322
130
90
6
4
47
52
1
99
1
0
"Начикинский”
2851
120
97
3
0
60
40
0
75
25
0
“Тигильский”
1129
246
96
3
1
96
4
0
0
48
52
Военный № 7
976
71
59
4
37
0
37
69
4
89
7
“Соболевский”
907
205
94
0
6
35
65
0
38
62
0
Итого (в среднем по области)
58483
2474
73
10
17
43
38
19
46
35
19
Примечание. Здесь и в табл. 5: 1. Обеспеченность: I - низкая; II - средняя; III - высокая (см. табл. 1). 2. В итоговую цифру
вошла вся обследованная площадь пахотных почв, а в таблице приведены только те хозяйства, на которые приходится 1000 га
пахоты и более.
(43%) почв попадает в группу слабообеспечен¬
ных, 38% - среднеобеспеченных и только 19% - в
категорию хорошо обеспеченных. Наименьшее
содержание марганца обнаружено в гумидных
почвах западного побережья и севера Камчатки в
зоне распространения охристо-подзолистых, тор-
фянисто-глеевых, торфянистых и торфянисто-
перегнойных почв. Например, на западном побе¬
режье к группе слабообеспеченных марганцем
почв относится 96% их площади в совхозе “Ти-
гильский”, 79% в совхозе “Анавгайский”, 75%
в совхозе “Быстринский”, а на севере Камчатки
в нее входят 100% почв совхоза “Карагинский”.
Хорошо обеспечены подвижным марганцем поч¬
вы (100%) под многолетними насаждениями (де¬
коративные и ягодные питомники). В основном
же обследованные почвы содержат незначитель¬
ное количество подвижного марганца, что пред¬
определяет положительное воздействие марган¬
цевых удобрений на урожай многолетних трав и
некоторых овощных культур [11, 20, 22].
Цинк. Содержание валового цинка колебалось
от 30 до 120 мг/кг и составило в среднем для верх¬
них горизонтов 75 мг/кг, что в 1.5 раза больше
кларка (50 мг/кг). Содержание подвижного цин¬
ка, так же как и марганца, невысокое (табл. 4).
Половина пахотных почв (46%) характеризуется
низким содержанием цинка (<2 мг/кг), 35% - сред¬
ним и только 19% - высоким (>5 мг/кг). Менее
всего обеспечены подвижным цинком почвы сов¬
хозов: “Заречный”, где 100% пашни попадает
в группу с низким содержанием этого элемента,
“Пограничный” (100%), “Мильковский” (99%),
“Комсомольский” (99%), “Крутобереговский”
(96%), “Камчатский” (78%). Все эти хозяйства
расположены вдоль восточного побережья Кам¬
чатки по долине одноименной реки, а также бо¬
лее мелких рек: Сторож, Озерная, Ука и других,
впадающих в Беренгово море. На территории
землепользования этих хозяйств, в основном, рас¬
пространены охристые вулканические, лугово¬
дерновые и аллювиальные почвы. Лучше других
обеспечены цинком почвы совхозов “Быстрин¬
ский”, “Тигильский”, “Паланский”, расположен¬
ных в долинах рек, впадающих в Охотское море,
где более половины обследованных почв попада¬
ют в группу хорошо обеспеченных. Хорошо обес¬
печены цинком почвы под многолетними насаж¬
дениями, где регулярно вносятся большие дозы
органических удобрений. Надежных результатов
ПОЧВОВЕДЕНИЕ № 6 1996
780
ГОЛОВ, БАХОВА
Таблица 5. Обеспеченность пахотных почв Камчатки подвижными формами соединений меди, бора, кобальта
и молибдена, % от обследованной площади4*
Сельскохозяйст¬
венное предприятие
Медь
Бор
Кобальт
Молибден
I
II
III
I
II
1П
I
II
Ш
I
II
III
“Заречный”
6
58
36
74
22
4
77
21
2
64
36
0
“Камчатский”
8
77
15
11
63
26
85
8
7
17
83
0
“Ключевской”
54
44
2
100
0
0
100
0
0
90
10
0
“Корякский”
42
54
4
100
0
0
97
3
0
36
20
44
“Лазовский”
31
9
60
16
7
77
50
28
22
36
49
15
ОПХ “Октябрьское”
1
40
59
23
50
27
14
76
10
55
45
0
“Петропавловский”
3
33
64
100
0
0
100
0
0
55
42
3
“Рассвет"
74
0
26
1
1
98
60
39
1
73
26
1
“Большерецкий”
5
85
10
21
46
33
14
80
6
18
77 ’
5
“Крутобереговский”
0
56
44
0
13
87
4
54
42
64
36
0
“Пограничный”
0
19
81
5
49
46
17
70
13
1
75
24
“Комсомольский”
1
56
43
7
16
77
4
50
46
71
29
0
Военный № 146
0
34
66
20
35
45
28
56
16
100
0
0
“Мильковский”
26
68
6
49
34
17
10
82
8
40
59
1
“Начикинский”
19
79
2
1
31
68
33
40
27
40
58
2
“Тигильский”
1
88
11
14
36
50
0
19
81
37
60
3
Военный № 7
46
46
8
100
0
0
97
3
0
19
69
12
“Соболевский”
11
89
0
14
33
53
10
76
14
35
65
0
Итого (в среднем по области)
20
41
39
51
18
32
42
41
16
38
51
11
* Обследованная площадь и количество проанализированных образцов см. в табл. 4.
по эффективности цинковых удобрений на поч¬
вах Камчатки пока не получено.
Бор. Количество валового бора в почвах Кам¬
чатки колебалось от 10 до 51 мг/кг и составляло в
среднем для верхних горизонтов пахотных почв
17 мг/кг. Это несколько выше значения кларка
для почв западных областей России (10 мг/кг).
Что касается содержания подвижных соединений
бора, то создается впечатление, что из всех эле¬
ментов питания для большинства почв он являет¬
ся лимитирующим фактором урожайности возде¬
лываемых культур. Пожалуй, нет в стране да и в
мире таких сельскохозяйственных регионов,
в почвах которых невозможно было бы обнару¬
жить хотя бы следы этого элемента (во всяком,
случае авторы таких не встречали), так как метод
анализа и индикатор (хинализарин) довольно чув¬
ствительны. И тем не менее в почвах 10 хозяйств
(11.5 тыс. га) из 35, числившихся до 1992 г. на
Камчатке, бора не было обнаружено совсем. Тер¬
ритория землепользования этих хозяйств приуро¬
чена к склонам действующих или потухших вул¬
канов. Всего 51% пахотных почв попадает в груп¬
пу слабообеспеченных (29.3 тыс. га), на долю
среднеобеспеченных приходится 18% и хорошо
обеспеченных - 31%. Наиболее благополучная
ситуация по содержанию подвижного бора в поч¬
вах складывается в совхозах “Карагинский”,
“Таловский”, расположенных в долинах рек Та-
ловка и Карага (север Камчатки), где все пахот¬
ные почвы (всего 332 га) относятся к группе хоро¬
шо обеспеченных. В совхозах “Рассвет” и “Круто-
береговский” также большинство почв относится
к этой группе (98 и 87%). Вообще же строгих свя¬
зей между содержанием подвижных соединений
бора и местоположением, генезисом почв и их ли¬
тологическим составом не обнаружено, за исклю¬
чением отмеченных выше. В одном и том же хо¬
зяйстве могут быть почвы и с низким, и с высо¬
ким его содержанием (табл. 5).
Первые опыты по выявлению эффективности
борных удобрений были проведены на Камчатке
в начале 60-х гг. с дикорастущими и сеяными тра¬
вами [19]. Увеличение урожая зеленой массы раз¬
нотравного луга от двукратного опрыскивания
растворами солей бора и молибдена составило
38%. Позднее положительные результаты от
применения одних борных удобрений, а также
борных удобрений по фону извести были получе¬
ны в опытах со свеклой, морковью и однолетни¬
ми травами [5,20, 22, 24].
ПОЧВОВЕДЕНИЕ № 6 1996
СОДЕРЖАНИЕ СЕРЫ И МИКРОЭЛЕМЕНТОВ
781
Молибден. Почвы с низким содержанием по¬
движного молибдена занимают 22 тыс. га или 38%
всего пахотного фонда, со средним - 29.3 тыс. га
(51%) и с достаточным - 6.3 тыс. га (11%). Хуже
всего обеспечены подвижным молибденом поч¬
вы хозяйств, расположенных в северной части
полуострова и имеющих небольшие пахотные
фонды (общая, площадь 572 га), которые все без
исключения (100%) попадают в группу малообес¬
печенных. Много таких почв в совхозах “Ключев¬
ской” (90%) и “Рассвет” (73%). Достаточно хоро¬
шо обеспечены почвы птицефабрики (231 га) и
совхоза “Тепличный” (40 га), где все они попада¬
ют в третью группу, по-видимому, в результате
внесения повышенных доз органических и мине¬
ральных удобрений.
Содержание валового молибдена колебалось
от 1 до 3.5 мг/кг и составило в среднем для верх¬
них горизонтов исследованных почв 1.4 мг/кг, т.е.
несколько ниже кларка (2.0 мг/кг). Следователь¬
но, в целом пахотные почвы Камчатки характе¬
ризуются низкими как потенциальными, так и по¬
движными запасами молибдена.
Молибденовые удобрения были эффективны
при внесении под однолетние травы, особенно
бобовые [19, 20]. Следует подчеркнуть, что при¬
менение молибденовых удобрений при возделы¬
вании сельскохозяйственных культур в условиях
Камчатки может иметь положительное значение
в борьбе с избыточным накоплением нитратов
в кормовых и овощных культурах, так как молиб¬
ден входит в состав ферментов нитратредуктазы
и нитрагеназы, которые участвуют в превраще¬
нии нитратных форм азота в аммиачные и далее
в амидные. То же самое можно сказать и о других
элементах, участвующих в метаболизме растений
(сере, в отсутствие которой замедляется синтез
белка, меди и др.) [3, 6,9, 14].
Медь. Исследованные почвы по содержанию
подвижной меди в основном можно отнести к
средне- (42%) и хорошо обеспеченным (38%) и
только пятая часть пахотного фонда (20%) отно¬
сится к категории слабообеспеченных. Более
других обеднены медью почвы птицефабрики
“Пионерская”, 94% которых (из 231 га) относится
к первой группе, и совхоза “Рассвет”, 74%
(из 2080 га) почв которого слабо обеспечены
медью. Особенно бедны медью торфянистые и
торфяно-болотные почвы, составляющие значи¬
тельную долю пахотного фонда в совхозах “Рас¬
свет” и “Лазовский”. Низким содержанием меди
характеризуются лесные вулканические слоисто-
пепловые почвы, которых много в совхозе “Клю¬
чевской” (1071 из 2161 га), поэтому здесь 54% па¬
хотных почв попадают в первую группу (табл. 5).
Повышенное содержание подвижных соединений
меди отмечено в почвах совхозов “Карагинский”,
где 100% пашни попадают в третью категорию, а
также “Корфский” (92%) и “Таловский” (82%).
Все эта почвы находятся на севере Камчатки и
характеризуются также низким содержанием по¬
движных соединений молибдена.
Есть факты, свидетельствующие о том, что из¬
быток меди в почвах усугубляет недостаток мо¬
либдена в растениях; особенно это сказывается
при скармливании их животным, в организме ко¬
торых соотношение меди и молибдена имеет жиз¬
ненно важное значение [2, 9, 13]. Данное обстоя¬
тельство позволяет рекомендовать внесение мо¬
либденовых удобрений в эти почвы под кормовые
культуры для улучшения их питательной ценности
независимо от влияния на их урожайность. То же
самое можно сказать и о применении удобрений,
содержащих серу и микроэлементы, практикуе¬
мом в других странах [23], что, безусловно, требу¬
ет дополнительных исследований.
Содержание валовой меди колебалось от 13 до
100 мг/кг и составило в среднем 33 мг/кг, что поч¬
ти в 1.5 раза превышало кларковое (20 мг/кг). На¬
именьшее количество меди как валовой, так и по¬
движной наблюдалось в почвах торфянистых и
аллювиальных на галечниковых отложениях,
больше - в лугово-дерновых и охристых вулкани¬
ческих с признаками оподзоливания.
Проведенные опыты (весьма немногочислен¬
ные) по выявлению эффективности медных удо¬
брений дали противоречивые результаты [15,20],
что, по-видимому, объясняется отсутствием или
недостатком информации о содержании меди в
изучаемых почвах.
Кобальт. Содержание валового кобальта в
проанализированных образцах колебалось от 4 до
22 мг/кг. Среднее его значение превысило незна¬
чительно (13 мг/кг) величину кларка (10 мг/кг).
Большинство почв по содержанию подвижного
кобальта следует отнести к бедным (43%) и сред¬
необеспеченным (41%). Повышенное его содер¬
жание отмечено только для 16% почв. Практиче¬
ски все хозяйства имеют в составе почвенного по¬
крова почвы с низким содержанием подвижного
кобальта, за исключением территорий земле¬
пользования совхозов “Быстринский”, “Анавгай-
ский” и “Тигильский”, где почвы в основном ха¬
рактеризуются высоким содержанием этого эле¬
мента. Все эти хозяйства находятся на западном
побережье, в зоне распространения охристо-под-
золистых, перегнойно-глеевых и торфянистых
вулканических почв. Опытов по применению
удобрений, содержащих кобальт, на Камчатке не
проводилось, поэтому трудно прогнозировать
или тем более давать конкретные рекомендации
по применению этого элемента в растениеводст¬
ве и животноводстве.
ВЫВОДЫ
1. Большинство пахотных почв Камчатки (73%)
содержат незначительное количество подвижных
соединений серы, которое зависит, в основном, от
ПОЧВОВЕДЕНИЕ № 6 1996
782
ГОЛОВ, БАХОВА
содержания органического вещества и интенсив¬
ности его минерализации, а также выветривания
сульфидных минералов, которые протекают очень
медленно. Применение серных удобрений перспек¬
тивно в первую очередь на требовательных к ней
культурах (крестоцветные, бобовые и др.).
2. Содержание валовых и подвижных форм со¬
единений марганца, цинка, кобальта и молибдена
и особенно бора недостаточное, поэтому необхо¬
димо изучение эффективности этих видов удоб¬
рений с учетом полученных данных по обеспе¬
ченности пахотных почв этими элементами.
3. Содержание меди в почвах, в основном, до¬
статочное, за исключением торфянистых почв.
Внесение медных удобрений перспективно на
торфянистых и аллювиальных вулканических
почвах.
СПИСОК ЛИТЕРАТУРЫ
1. Лйдинян Р.Х., Иванова М.С., Соловьева Т.С. Ме¬
тоды извлечения и определения различных форм
серы в почвах и растениях / Почв, ин-т им. В.В. До¬
кучаева ВАСХНИЛ. М., 1975. 21 с.
2. Буркин И.А. О молибдене как микроудобрении и
его токсичности в кормах для животных // Агрохи¬
мия. 1964. № 1. С. 123-126.
3. Буркин И.А. Физиологическая роль и сельскохо¬
зяйственное значение молибдена. М.: Наука, 1968.
294 с.
4. Бурлака В.В. Растениеводство Дальнего Востока.
Хабаровск, 1965.494 с.
5. Голов В.И. Влияние известкования на подвиж¬
ность микроэлементов в почвах Дальнего Востока
и эффективность микроудобрений // Пути повы¬
шения продуктивности растениеводства, кормо¬
производства и садоводства на Дальнем Востоке.
Владивосток, 1987. С. 28-37.
6. Голое В.И., Прокопова Т.К. Об эффективности
серных удобрений на почвах Дальнего Востока //
Пути повышения продуктивности растениеводства
на Дальнем Востоке. Владивосток, 1981. С. 44-56.
7. Зонн С.В., КарпачевскийЛ.О., Стефин В.В. // Лес¬
ные почвы Камчатки. М.: Изд-во АН СССР, 1963.
254 с.
8. Ильковская З.Г. Определение серы и железа в
почвах // Агрохимические методы исследования
почв. М.: Наука, 1965. С. 28-36.
9. Каталымов М.В. Микроэлементы и микроудобре¬
ния. М.: Химия, 1965. 330 с.
10. Коллингс Г.Х. Удобрения, содержащие второсте¬
пенные элементы питания - серу, кальций, магний //
Промышленные удобрения, их производство и
применение / Пер. с англ. Под ред. Э.И. Шконде.
М.: Сельхозгиз, 1960. С. 328-363.
11. Ливеровский Ю.А. Почвы равнин Камчатского по¬
луострова. М.: Изд-во АН СССР, 1959. 130 с.
12. Методические указания колориметрического оп¬
ределения подвижных форм микроэлементов в
почвах, кормах и растениях методом атомно-аб-
сорбционной спектрофотометрии / ЦИНАО,
МСХ СССР. М., 1985. 95 с.
13. Муравин Э.М., Журавлева С.В. Взаимодействие
фосфора, молибдена и меди в процессе питания
растений // Изв. ТСХА. 1970. № 4. С. 92-100.
14. Пейве Я.В. Микроэлементы и ферменты. Рига:
Изд-во АН ЛатвССР, 1960. 136 с.
15. Прохорова З.А., Соколов И.А. Агрохимическая
характеристика почв Камчатской области // Агро¬
химическая характеристика почв СССР. М.: На¬
ука, 1971. С. 170-239.
16. Ринькис Г.Я. Определение микроэлементов в поч¬
вах и растениях // Микроэлементы в СССР. 1963.
№ 5. С. 26-44.
17. Система земледелия в Камчатской области / Под
ред. А.М. Ярушина. Новосибирск, 1982. 151 с.
18. Соколов И.А. Вулканизм и почвообразование
(на примере Камчатки). М.: Наука, 1973. 224 с.
19. Степанова К Д. Первые опыты применения мик¬
роэлементов в культуре многолетних трав на Кам¬
чатке // Микроэлементы в Восточной Сибири и на
Дальнем Востоке. Улан-Удэ: Изд-во АН СССР.
Сиб. отд-ние, 1962. С. 64-65.
20. Стружкина Г.М., Тужилина И.Б., Скрипченко А.Ф.
и др. Микроэлементы в почвах Камчатки и эффек¬
тивность их как удобрений // Микроэлементы в
почвах Дальнего Востока: Уч. зап. ДВГУ. 1969.
Т. XXVII. С. 66-71.
21. Чичканов В.П. Дальний Восток, стратегия эконо¬
мического развития. М.: Экономика, 1988. 246 с.
22. ЯрушинА.МЧудина К.А. К вопросу о семеновод¬
стве тимофеевки луговой на Камчатке // Вопр. ин¬
тенсификации с.-х. производства Камчатской об¬
ласти: Сб. науч. тр. 1979. № 6. С. 46-50.
23. During С. The amelioration of volcanic ash soils in New
Zeland // Meeting on the Classification and Corralation
of Soils from Volcanic Ash. Tokyo, L964.
24. Golov V.I., Bakhova S.M. Content of sulphur and micro¬
elements in plowable volcanic soils in Kamchatka //
Bridges of the Sci. between North America and the Rus¬
sian Far East. 45th Arctic Sci. Conf.: Abs. Vladivostok:
Dalnauka, 1994. Book 50. P. 143-144.
Content of Sulphur and Microelements
in Arable Volcanic Soils in Kamchatka
V. I. Golov, S. M. Bakhova
Plowable soils of Kamchatka (58 ha) were studied on content of the total and movable forms of sulphur and
basic microelements (Mn, Zn, Си, B, Mo and Co). The content of total forms of the investigated elements, with
the exception of sulphur is within of Klark. Majority of plowable soils belongs to poorly supplied movable forms
of sulphur (73%) and boron (51%). The soils are poorly supplied with movable form of Zn (46%), Mn (43%),
Co (42%), Mo (38%) and Cu (20%).
ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВОВЕДЕНИЕ, 1996, № 6. с. 783-788
ФИЗИКА
ПОЧВ
УДК 631.4
ФОРМИРОВАНИЕ АГРЕГАТНОГО СОСТАВА ПОЧВ
И ОЦЕНКА ЕГО ИЗМЕНЕНИЯ
© 1996 г. С. Ю. Булыгин, Ф. Н. Лисецкий
Институт почвоведения и агрохимии им. А.Н. Соколовского, Харьков
Одесский государственный университет им. И.И. Мечникова
Поступила в редакцию 22.09.93 г.
Обоснован выбор показателей, отражающих иерархические уровни агрегатного состава почвы.
Выявлена зависимость формирования высокой агрегированности пахотных почв от особенностей
их биологии и гумусного состояния. Предложены двухуровневая система управления агрегатным
составом и принципы его оперативного мониторинга.
Почвенный агрегат можно рассматривать как
объект, аккумулирующий результат многих поч¬
вообразовательных процессов. Совокупность аг¬
регатов различных размеров и форм образует аг¬
регатный состав. Оценка, контроль и поддержа¬
ние оптимального агрегатного состава почвы,
безусловно, должны стать непременными усло¬
виями любой земледельческой деятельности че¬
ловека.
Агрегатный состав почвы обычно разделяют
на два иерархических уровня: макроагрегатный и
микроагрегатный. Разделение на макроагрегаты
диаметром >0.25 мм и микроагрегаты <0.25 мм
подкрепляется определенным физическим смыс¬
лом, в частности, их различной устойчивостью
к разрушению. Силы сцепления между элемен¬
тарными почвенными частицами (ЭПЧ) в микро¬
агрегатах на порядок, а чаще на 2-3 порядка вы¬
ше, чем в макроагрегатах, в которых соединены
между собой главным образом микроагрегаты.
Очевидны различия и в организации макро- и ми¬
кроагрегатов. Придерживаясь концепции иерар¬
хии структурных уровней [6] на фоне взаимосвя¬
занности всех уровней в систему, необходимо в то
же время признать их относительную самостоя¬
тельность. Поэтому важное теоретическое и при¬
кладное значение имеет выбор показателей, ко¬
торые наиболее полно и адекватно отражают
суть и диагностируют тенденции развития каждо¬
го из отмеченных иерархических уровней.
Воронин [6] констатирует, что неагрегирован-
ные ЭПЧ содержатся во всех фракциях микроаг-
регатного состава, а их соотношение с микроаг¬
регатами в той или иной фракции зависит от осо¬
бенностей почвообразующей породы и типа
почвообразования. Следовательно, таким соот¬
ношением в обобщенном виде можно охаракте¬
ризовать микроагрегатный уровень агрегатного
состава почвы. Активные разработки в направле¬
нии как поиска формы выражения этого концеп¬
туального положения, так а в апробации показа¬
телей на разнообразных объектах позволили
остановить наш выбор на коэффициенте агреги¬
рованности [1, 2]. Впервые он был предложен
еще в 1932 г. Бэйвером и Роадесом и вычисляется
по формуле (в нашей модификации)
КА = 100^-=Д
где а - содержание частиц и микроагрегатов диа¬
метром от 0.05 до 0.25 мм; б - содержание ЭПЧ
тех же размеров.
Для определения содержания неагрегирован-
ных ЭПЧ и свободных (не связанных в макроаг¬
регаты) микроагрегатов в отдельных фракциях
использовали метод прямого подсчета в отражен¬
ном свете на микроскопе МБС-9. Почвенные об¬
разцы предварительно рассеивали в колонке сит
с диаметром ячеек не только 0.25 мм и более, но
и 0.20,0.16,0.10,0.065,0.05 мм на приборе “Уста¬
новка лабораторная для определения фракцион¬
ного состава. Модель 029” в течение 30 с. После
рассева пропорционально доли каждой фракции
макроагрегатов (L > 0.25 мм) составляли образец
общей массой 25 г для мокрого просеивания на
приборе Бакшеева [3].
По результатам этого анализа рассчитывали
средний взвешенный диаметр водопрочных агре¬
гатов. Объектом исследований служили зональ¬
ные почвы регионов Украины, характеризующих¬
ся довольно разнообразными почвенно-климати¬
ческими условиями: Левобережной Лесостепи,
Донецкой Степи, сухой степи Побужья. Для осве¬
щения почвенно-эволюционных аспектов про¬
блемы использованы археологические датировки
памятников почв сельскохозяйственной округи
Ольвии - античного государства, имеющего деся¬
тивековую историю.
783
784
БУЛЫГИН, ЛИСЕЦКИЙ
20 30
КА/Г
Рис. 1. Зависимость среднего взвешенного диаметра
водоустойчивых агрегатов d от агрегирующей эф¬
фективности гумуса Кд/Г в слое 0-20 см черноземов
южных и темно-каштановых почв Побужья: I - це¬
линные полнопрофильные; 2 - молодые разновозра¬
стные; 3 - залежные; 4 - старопахотные в античное
время и ныне; 5 - пахотные в последние 130-150 лет.
При земледельческой оценке почвы важней¬
шая характеристика ее макроагрегатного уров¬
ня - устойчивость макроагрегатов к разрушению.
Причем истинный агрегат должен быть прежде
всего водопрочным, что прямо зависит от содер¬
жания гумусовых соединений [4]. Одним из наибо¬
лее интегральных и информативных показателей
водопрочности агрегатов считается средний взве¬
шенный диаметр водоустойчивых агрегатов [7],
поэтому он может быть выбран в качестве базо¬
вого показателя макроагрегатного уровня агре¬
гатного состава.
С учетом изложенного предлагается двухуров¬
невая система управления агрегатным составом
почвы. Для каждого уровня управления, отража¬
ющего конкретную иерархическую степень орга¬
низации агрегатного состава почвы, используется
контролирующий параметр или их система. На няш
взгляд, достаточно информативные и сравни¬
тельно несложные в определении показатели мак-
ро- и микроагрегатного уровней: d - средний взве¬
шенный диаметр водоустойчивых агрегатов (мм)
и КА - коэффициент агрегированности (рис. 1).
Почвы, находящиеся в разных условиях сель¬
скохозяйственного использования, имеющие су¬
щественные различия в гумусовом состоянии,
в итоге характеризуются относительной незави¬
симостью макро- и микроагрегатных уровней.
Для пахотных почв эффективность участия гуму¬
са в микроагрегированности не сопряжена с изме¬
нением диаметра водопрочных агрегатов, а для
почв, формирующихся под естественной расти¬
тельностью, повышение водопрочности структу¬
ры не обеспечивается усилением микроагрегиро¬
ванности и увеличением эффективности гумуса
в этом процессе. Очевидно, для почв, находящих¬
ся в таких контрастных условиях почвообразова¬
ния, уже недостаточно сопоставить лишь физико¬
химические показатели.
Принципиальные различия имеются и в био¬
логии этих почв. О значении корневой системы
зональных ассоциаций в фрагментации и структу-
рообразовании почв (на примере степной зоны)
дает представление отношение массы живых кор¬
ней к массе почвы (табл. 1). По сравнению с па¬
хотным горизонтом верхний 20-сантиметровый
слой почвы под естественной растительностью
насыщен корнями в 3-4 раза больше. Целинные и
залежные почвы по сравнению с пашней обладают
значительно большей общей пористостью (ходы
дождевых червей, отмерших корней), а также
внутриагригатной пористостью [6]. В природных
условиях почвообразования все это создает
Таблица 1. Фитомасса подземных органов в целинных ассоциациях и агроценозах теплой зоны (чернозем южный)
Фитоценоз, агроценоз
Слой
почвы, см
Корневи¬
ща (Afi)
Корни
деятельные (К2)
Корни
недеятельные
К1+К2
W
г/м2
Разнотравная типчаковая ассоциация
0-10
48.5
834.5
283.0
7.3
0-20
65.0
1074.5
413.0
4.8
Типчаково-ковыльная ассоциация
0-10
72.5
717.5
1159.5
6.6
0-20
87.0
870.5
1465.5
4.0
Люцерна 4 года пользования
0-10
275.8
202.8
61.6
4.0
0-20
392.0
376.0
97.6
3.2
Пшеница озимая
0-10
Нет
204.0
Нет
2.0
0-20
»
пол
»
1.4
Кукуруза
0-10
»
160.4
»
1.6
0-20
»
243.3
»
1.2
Примечание. IV - масса почвы в соответствующем слое, кг/м .
ПОЧВОВЕДЕНИЕ № 6 1996
ФОРМИРОВАНИЕ АГРЕГАТНОГО СОСТАВА ПОЧВ
785
благоприятные предпосылки для развития гриб¬
ных гифов, участие которых в повышении ста¬
бильности почвенных агрегатов установлено экс¬
периментами [12] и подтверждено хорошо извест¬
ной динамикой водоустойчивости структуры [4].
Дождевые черви также ежегодно могут создать
на поверхности почвы слой копролитов мощнос¬
тью до 5 мм, усиливая в ходе почвообразования
агрегированность верхнего горизонта [5].
В широком зональном ряду пахотных почв
(от дерново-подзолистых до темно-каштановых)
выявлена тесная зависимость КА слоя почвы
0-20 см от содержания в нем гумуса (г - 0.89 ± 0.19).
При рассмотрении аналогичной зависимости во
внутризональном разрезе (пахотные почвы: чер¬
ноземы южные и темно-каштановые Нижнего
Побужья), очевидно, вследствие резкого сужения
диапазона варьирования содержания гумуса (от 1
до 2.8%), его связи с величиной КА не отмечается
(рис. 2). При этом пахотные почвы имеют высокую
степень микроагрегированности (КА = 31-48%),
что позволяет предположить более сложный ме¬
ханизм агрегирования, в котором участвует не
только гумус и/или гумус иного качества.
Следует отметить, что пока речь о водоустой¬
чивости агрегатов не ведется. По схеме Хана [11]
агрономически благоприятная агрегированность
почвенной массы считается результатом образо¬
ваний и действия органо-минеральных соедине¬
ний при непременном наличии трех компонентов:
новообразованного гумуса, высокодисперсных
глинистых минералов и обменных оснований.
В интенсивно используемых пахотных почвах при
небольшом поступлении растительных остатков
участие новообразованного гумуса в агрегиро¬
ванное™ крайне невелико, и потому отмеченный
уровень гумусированности (1.1-2.8%) можно отне¬
сти на счет прочных органо-минеральных коллои¬
дов, консервирующих гумус, по словам JI.H. Алек¬
сандровой, на “сотни и тысячи лет”. Таким образом,
микроагрегированность пахотных и старопахот¬
ных почв может быть связана главным образом
с действием полимерных коллоидов, которые,
как было установлено Кульманом [8], имеют аг¬
регирующую эффективность, аналогичную гуму¬
совым веществам. Важным представляется так-'
же поиск особенностей группового и фракцион-*
ного состава гумуса пахотных почв.
Для почв, формирующихся под естественной
растительностью (целина, залежь) в условиях по¬
стоянного воспроизводства новообразованного
гумуса, различия в гумусированности находят
свое отражение в различной микроагрегирован¬
ности (рис. 2). Сопоставление коэффициентов ре¬
грессии уравнений связи КА с содержанием гумуса
на зональном и внутризональном уровнях позво¬
ляет сделать вывод о разнокачесгвенности гуму¬
са пахотных почв, содержащих значительное ко¬
личество неспецифических гумусовых веществ,
КА
Г,%
Рис. 2. Зависимость коэффициента агрегированнос-
ти КА от содержания гумуса Г в слое 0-20 см пахот¬
ных и новообрабатываемых почв. Кд = 6.2 Г - 10.43;
г = 0.63 ± 0.34. Обозначения те же, что на рис. 1.
что определяет необходимость учета агрегирую¬
щей эффективности гумуса. Поэтому мы предла¬
гаем оценивать его по отношению КА/Г, показы¬
вающему, какую агрегированность (%) обеспечи¬
вает 1% гумуса данной почвы.
В пахотных почвах отмечается уменьшение
содержания гумуса по сравнению с целинными
полнопрофильными, а также молодыми и залеж¬
ными почвами. Однако различия в агрегирующей
эффективности гумуса не соответствуют дости¬
гаемой степени дегумификации (табл. 2), поэтому
принципиальным моментом является увеличение
прежде всего микроагрегированности пахотных
почв. Анализ группового и фракционного соста¬
ва гумуса показывает, что выявленную законо¬
мерность усиливает фракция свободных и связан¬
ных с подвижными полуторными оксидами гуми-
новых кислот, содержание которой в пахотных
почвах возрастает в 2-3 раза по сравнению с це¬
линной. На наш взгляд, совместно с водораство¬
римым гумусом эта фракция может играть суще¬
ственную роль в насыщении гумусовых веществ
функциональными группами, создающими пери¬
ферические элементы для образования гетеропо-
лярных органо-минеральных соединений, образу¬
ющих, в свою очередь, “подвижные” ассоциаты
и микроагрегаты [9, 10].
Но особенно примечательна, по нашему мне¬
нию, четко выявляемая зависимость агрегирую¬
щей эффективности гумуса от содержания фрак¬
ции “агрессивных” фульвокислот, т.е. свобод¬
ных и связанных с подвижными полуторными
оксидами. Хотя это лишь частичный ответ на во¬
прос “как?” Механизм этого влияния еще не ясен.
Несмотря на то, что в сумме кислоторастворимых
7 ПОЧВОВЕДЕНИЕ № 6 1996
786 БУЛЫГИН, ЛИСЕЦКИЙ
Таблица 2. Агрегирующая эффективность гумуса почв при различной интенсивности использования
Почвы
Количество определений
КА
Г,%
КА/Г
■*min ■*max/*cp
Целинные
6
22-43/31
2.78-4.21/3.47
8-11/9.5
Молодые
9
21-39/31
2.16-5.35/3.55
5-13/9
Залежные
10
18-38/29
2.28-3.87/3.16
7-13/9.5
Пахотные
17
34-48/42
1.51-2.76/1.95
17-25/22
Старопахотные
7
27-48/42
1.11-2.19/1.89
11-43/24
органических веществ в целинных и залежных поч¬
вах большое значение могут иметь неспецифичес-
кие органические соединения, в наших исследова¬
ниях все пахотные почвы отличались наиболее
высоким содержанием “агрессивных” фульвокис-
лот (15-26%). Очевидно, эта особенность тесно
связана с ранее установленным фактом [10], объ¬
ясняющим формирование агрономически опти¬
мальных структур в пахотных черноземах преиму¬
щественно фульвокислотной фракцией лабильной
части гумуса, что подтверждается результатами
стационарного опыта. Так, замена отвальной
вспашки ежегодной поверхностной обработкой
под все культуры севооборота уже через 5 лет на
фоне общего увеличения содержания гумуса в чер¬
ноземе обыкновенном достоверно повышала до¬
лю фульвокислот. Это позволяет утверждать,
что существует принципиальная возможность уп¬
равления процессом агрегирования ЭПЧ. Следо¬
вательно, в пахотных почвах не только интенсив¬
нее (по сравнению с целиной) воспроизводятся
лабильные фракции гумусовых веществ, но и
полнее реализуется их агрегирующий потенциал.
В этой связи вполне объяснима, на первый взгляд
парадоксальная, закономерность уменьшения
d, мм
г, годы
Рис. 3. Зависимость среднего взвешенного диаметра
водоустойчивых агрегатов d от длительности обра¬
ботки почвы. Обозначения те же, что на рис. 1.
суммы неагрегированных ЭПЧ в зависимости от
длительности обработки почвы (от 18-34% на це¬
лине до 6-20% на пашне разной длительности
земледельческого пользования).
На основании археологических и историчес¬
ких датировок можно представить зависимость
величины d от длительности обработки почвы
(рис. 3). Уже через 100-150 лет обработки сред¬
ний взвешенный диаметр водоустойчивых агре¬
гатов уменьшится в 2-3 раза и в дальнейшем
(1000 лет) стабилизируется на уровне 0.50-0.75 мм.
Это позволяет предположить наличие у пахот¬
ных почв устойчивого зонального показателя
противоэрозионной стойкости, но зависящего не
от сравнительно изменчивых показателей агро¬
физического состояния пахотного горизонта, а от
почвенно-генетических характеристик, к кото¬
рым может быть отнесен коэффициент агрегиро¬
ванное™. Естественно, при другой интенсивнос¬
ти использования почвенного покрова и разной
длительности возможен иной диапазон значений.
Диапазон изменений параметра d в условиях
определенной культуры земледелия зависит от
многих причин, в том числе от технологии обра¬
ботки почвы. Так, по нашим исследованиям, ра¬
зовое применение различных способов основной
обработки почвы обусловливает статистически
доказуемое различие величин d уже осенью.
К весне эти различия усиливаются, но к уборке
возделываемой культуры влияние основной об¬
работки на параметр не фиксируется. Значитель¬
но труднее изменить значения экстремумов уста¬
новившегося диапазона d. Особенно важно под-
нять значение нижнего экстремума, которое
может служить одним из количественных крите¬
риев системы земледелия, обязанной быть эконо¬
мически эффективной, экологически безопасной
и сохранять плодородие почвенного покрова при
проявлении почвенно-климатических факторов
не ниже 10%-ной обеспеченности. Поэтому зна¬
чение нижнего дециля (dot) может в достоверной
степени характеризовать нижний экстремум ус¬
тановившегося диапазона колебаний.
ПОЧВОВЕДЕНИЕ № 6 1996
ФОРМИРОВАНИЕ АГРЕГАТНОГО СОСТАВА ПОЧВ
787
В полевом стационарном опыте на черноземе
обыкновенном (Донецкая Степь) испытывали
качественно различные технологии обработки поч¬
вы под культуры 10-польного полевого севооборо¬
та: отвальную, плоскорезную, щелевую (чизель-
ную) и поверхностную. Через 5 лет к началу весны
значение 5o.i слоя почвы 0-20 см составило по ва¬
риантам соответственно: 0.34; 0.43; 0.36; 0.45 мм; на
участке целины косимой - 2.00 мм; летом в пери¬
од наибольшей опасности ливневой эрозии - 0.40;
0.40; 0.50; 0.63 мм; на целине изменение do.i не
было установлено. Таким образом, улучшение гу-
мусного состояния почвы в опыте обусловило уве¬
личение нижнего экстремума диапазона колебаний
водоустойчивости макроагрегатов. Это свидетель¬
ствует о возможности в процессе земледельческой
деятельности не только управлять сезонной дина¬
микой водоустойчивости макроагрегатов, но и
целенаправленно изменять ее пределы, т.е. улуч¬
шать стабильную, генетически обусловленную
водоустойчивость почвенных агрегатов. Поэтому
разработка рациональных почвоохранных техно¬
логий возделывания культур требует глубокого на¬
учного обеспечения. Необходимо иметь количест¬
венные представления о влиянии на агрегатный
состав почвы всех отдельных операций э+их тех¬
нологий, т.е. следует установить оперативный
мониторинг агрегатного состава почвы при ее
сельскохозяйственном использовании.
Безусловно, мониторинг можно было бы про¬
водить по значениям КА и d. Но сенсорносгь этих
показателей недостаточна, чтобы по ним оцени¬
вать отдельные технологические операции. Для
более оперативного мониторинга агрегатного со¬
става почв нужен дополнительный показатель.
Установлено, что содержание неагрегированных
ЭПЧ моментально изменяется, причем в опреде¬
ленном диапазоне при любом агротехническом
воздействии [1, 2]. Постоянно поддерживая со¬
держание неагрегированных ЭПЧ возле верхнего
или нижнего экстремума, мы тем самым создаем
направленность в изменении агрегатного состава
почвы. Для почв Донецкой Степи с интервалом
значений параметров d 0.50-0.75 мм была уста¬
новлена соответствующая амплитуда содержания
всех неагрегированных ЭПЧ от 15 до 5%, т.е.
можно говорить об обратной связи между вели¬
чиной d и содержанием неагрегированных ЭПЧ,
которое обозначим символом С.
В условиях стационарного полевого опыта оп¬
ределена хорошо выраженная цикличность изме¬
нения значений С в течение сельскохозяйствен¬
ного года, но технология обработки оказывала
решающее воздействие на амплитуду и фазы этой
цикличности. Практически любая технологичес¬
кая операция ведет к увеличению величины С,
которая после проведения операции начинает
уменьшаться. Видимо, как и при рассмотрении
ряда d, значение нижнего дециля С01 будет наи¬
более полно и точно характеризовать воздейст¬
вие технологии обработки почвы на содержание
неагрегированных ЭПЧ. Этот показатель говорит
о степени восстановления агрегированности после
очередной технологической операции. И чем его
величина ниже, тем лучше, так как он характери¬
зует степень разрушения агрегатов до ЭПЧ.
Хорошей иллюстрацией сказанному будут значе¬
ния С0слоя почвы 0-20 см по вариантам: 8.4; 6.2;
4.1 и 4.0%. При отвальной технологии отмечается
наибольшее воздействие на агрегаты, которые не
успевают восстановиться до исходного состоя¬
ния; при этом их водоустойчивость снижается.
Применение технологий обработки почвы, под¬
держивающих значение С0л близким к 4%, обес¬
печило рост водоустойчивости макроагрегатов
по сравнению с отвальной технологией, что, разу¬
меется, произошло из-за общего увеличения со¬
держания гумусовых соединений в почве. Следо¬
вательно, поддержание чрезмерно высокого
уровня неагрегированных ЭПЧ при отвальной
вспашке ведет к увеличению скорости минерали¬
зации гумуса, а следовательно, к ухудшению агре¬
гатного состава почв. Отметим, что это всего
лишь гипотеза. Необходимы непосредственные
измерения.
Для обеспечения научно обоснованного мони¬
торинга агрегатного состава по содержанию не¬
агрегированных ЭПЧ, который позволял бы оце¬
нивать рациональность отдельных операций и
технологий возделывания в целом, нужны норма¬
тивы предельно допустимого их содержания. Нам
представляется, что для этого есть довольно на¬
дежный и простой способ, а именно: наличие не¬
агрегированных ЭПЧ почвы под пластом много¬
летних трав 3-4 года жизни может служить нор¬
мативом для близких по генезису почвенных
разновидностей. К этому времени в почве проис¬
ходит “стирание” тех особенностей, которые
обусловлены обработкой до посева трав, и уста¬
навливается минимальное содержание ЭПЧ, ха¬
рактеризующее почвы, находящиеся в интенсив¬
ном использовании.
Таким образом, мониторинг агрегатного со¬
става почвы предлагается проводить по показате¬
лю, отражающему состояние предшествующего
по степени организации иерархического уровня, -
уровню неагрегированных ЭПЧ. Вероятно, уста¬
новленный принцип (отражение результата про¬
явления деградационных процессов прежде всего
в подчиненном иерархическом уровне структур¬
ной организации) носит более универсальный ха¬
рактер и может быть рассмотрен как концепту¬
альная база мониторинга почвенного покрова.
ПОЧВОВЕДЕНИЕ № 6 1996
7*
788
БУЛЫГИН, ЛИСЕЦКИЙ
СПИСОК ЛИТЕРАТУРЫ
1. Булыгин С.Ю., Лисецкий Ф.Н. Оценка агрегиро-
ванности почв степной зоны УССР // Б юл. Почв,
ин-та им. В.В. Докучаева. 1989. Вып. 51. С. 74-75.
2. Булыгин С.Ю., Комарова Т.Д. К оценке влияния
механической обработки на почву // Почвоведе¬
ние. 1990. №6. С. 135-138.
3. Вадюнина А.Ф., Корчагина З.А. Методы исследо¬
вания физических свойств почв. М.: Агропромиз-
дат, 1986. 416 с.
4. Вершинин П.В. и др. Основы агрофизики почв /
Под ред. А.Ф. Иоффе, И.Б. Ревута. М.: Физматгиз,
1959. 904 с.
5. Возраст и эволюция черноземов / Н.Я. Марголина,
А.Л. Александровский, Б.А. Ильичев и др. М.: На¬
ука, 1988. 144 с.
6. Воронин АД. Основы физики почв. М.: Изд-во
МГУ, 1986. 244 с.
7. Кузнецов М.С. Протиаоэрозионная стойкость
почв. М.: Изд-во МГУ, 1981.136 с.
8. Кульман А. Искусственные структурообразовате-
ли почвы. М.: Колос, 1982. 158 с.
9. Орлов Д.С. Химия почв. М.: Изд-во МГУ, 1985.
376 с.
10. Сорочкин В.М., Орлова Л.П., Кучеряева Е.В.
К механизму образования структуры обрабатывае¬
мых лесостепных почв // Почвоведение. 1990. № 6.
С. 51-58.
И. Хан Д.В. Органоминеральные соединения и струк¬
тура почв. М.: Наука, 1989. 140 с.
12. Molope М.В., Grive /.С., Page ЕЯ. Contributions by
fungi and bacteria to aggregate stability of cultivated
soils //J. Soil Sci. 1987. V. 38. № 1. P. 71-77.
The Formation in the Soil Aggregate Composition and its Assessment
S. Yu. Bulygin, F. N. Lisetskiy
The choice of indices is substantiated to characterize hierarchical levels of the soil aggregate composition.
A higher aggregation of arable soils appears to be dependent on peculiar features of their biology and humus
state. Two-level system of governing the soil aggregate composition is proposed as well as principles of its op¬
erative monitoring.
ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВОВЕДЕНИЕ, 1996. № 6. с. 789-792
УДК 631.436
ФИЗИКА
ПОЧВ
ОПРЕДЕЛЕНИЕ КОЭФФИЦИЕНТА ЭФФЕКТИВНОЙ ДИФФУЗИИ
ВОДЯНОГО ПАРА В СОЛОМЕННОЙ МУЛЬЧЕ
© 1996 г. Б. М. Гусев, О. Е. Бусарова
Институт водных проблем РАН, Москва
Поступила в редакцию 15.09.92 г.
Получено уравнение для оценки коэффициента эффективной диффузии водяного пара в мульче.
Оно может быть рекомендовано для использования при расчетах испарения воды почвой, покрытой
соломенной мульчей, в частности, при определении эффективности мульчирования с точки зрения
повышения влагообеспеченности сельскохозяйственных посевов.
Использование мульчирующего покрытия
почвы в качестве депрессора непродуктивного
испарения является одной из ресурсосберегаю¬
щих технологий в сельском хозяйстве. Мульча
значительно уменьшает непродуктивное испаре¬
ние воды почвой, создавая благоприятные усло¬
вия водообеспеченности даже в таких подвержен¬
ных засухам районах, какими являются степная и
лесостепная зоны России [2]. Кроме того, исполь¬
зование в качестве мульчи растительных остат¬
ков, в частности, соломы, приближает сельскохо¬
зяйственную экосистему к естественной, для ко¬
торой свойственно наличие на ее поверхности
подстилки и войлока из отмершей растительнос¬
ти. С точки зрения взаимодействия человека с ок¬
ружающей средой этот факт имеет важное значе¬
ние, поскольку наиболее экологически благо¬
приятны те технологии, которые максимально
близки к эволюционным тенденциям ненарушен¬
ной биосферы [6].
Поскольку основная область фазового пере¬
хода вода в почве-пар отделена от прилегающего
слоя атмосферы слоем мульчирующего покры¬
тия, затрудняющего перенос пара от поверхности
почвы, то эффективность мульчирования с точки
зрения уменьшения непродуктивного испарения
воды почвой определяется как толщиной слоя
мульчи 6, так и коэффициентом эффективной
диффузии водяного пара в ней dk. Определение
этого параметра для соломенной мульчи и яви¬
лось основной целью настоящей работы. Здесь
необходимо отметить следующее обстоятельст¬
во. Использование мульчирующего слоя обычно
предполагается в течение длительного периода,
в результате чего может происходить накопле¬
ние влаги в глубоких слоях почвы от года к году.
Поэтому достаточно очевидно, что эффект по¬
вышения влагообеспеченности посевов при муль¬
чировании может усиливаться на второй, третий
годы ее применения. Возникает необходимость
исследования изменения величины dk во времени,
обусловленного изменением структуры соломен¬
ной мульчи, связанным с уплотнением и перегни-
ванием слоя соломы. Этот вопрос также был за¬
тронут в настоящем исследовании.
В основу методики определения dk была поло¬
жена следующая связь параметров Ьи dk с вели¬
чиной испарения воды почвой Е [4]
Е =
Ф'
С„ + L<p'
В
D_
(1)
где
En =
C„ + L(p'
c„ + Li#
- так называемая испаряемость, характеризую¬
щая испарение при достаточном увлажнении под¬
стилающей поверхности.
Здесь R - радиационный баланс; В - поток теп¬
ла в почву от ее поверхности; D - коэффициент
турбулентного обмена между подстилающей по¬
верхностью и высотой наблюдения гидрометео¬
рологических параметров z6 (обычно равной 2 м),
Du = dk/6 - коэффициент обмена для переноса па¬
ра в мульче, Ср, рв, d(j — соответственно теплоем¬
кость, плотность и дефицит влажности воздуха на
уровне г#", L- теплота фазового перехода вода-
пар; ф' - производная по температуре зависимос¬
ти насыщающей влажности воздуха от его темпе¬
ратуры.
Уравнение (1) может служить основой экспе¬
риментального определения dk в случае независи¬
мого измерения всех остальных входящих в него
789
790
ГУСЕВ, БУСАРОВА
Рис. 1. Опытный комплекс по определению коэффи¬
циента эффективной диффузии водяного пара в
мульчирующем слое:
1 - лизиметр с увлажненной почвой; 2 - подпитываю¬
щее устройство; 3 - термопары; 4 - электроизмери¬
тельное устройство; 5 - балансомер; 6 - анемометр;
7 - психрометр; 8 - мульчирующий слой.
с помощью поправочного коэффициента. Ука¬
занный коэффициент находился из результатов
сопоставления значений Е0 и Епо, рассчитанных
по уравнению (1) при значениях радиационных
балансов, измеренных одновременно над соло¬
менной мульчей и оголенной увлажненной поч¬
вой (рис. 1). Результатом сопоставления явилась
эмпирическая связь
Е0 = 0.78£по. (3)
Другая часть испарителей, содержащих почву,
покрытую соломенной мульчей, на основе изме¬
рений расхода воды на подпитывание этих лизи¬
метров давала возможность за те же периоды из¬
мерений определить величину Е.
Величину Ь находили по данным наблюдений
за динамикой температуры почвы с помощью
термопар, расположенных на нескольких глуби¬
нах в почвенных испарителях с соломенной муль¬
чей (рис. 1). Поток тепла В оценивали при этом
следующим образом:
величин. Выражение для dk примет при этом сле¬
дующий вид:
В
dJb = D= D
I- ■
(2)
Приведенное выражение было использовано в
настоящей работе для оценки dk соломенной
мульчи. Оценка была проведена на основе серии
экспериментов по определению испарения с ого¬
ленной смоченной почвы Е„0 и с почвы, покрытой
слоем мульчи, - Е. При этом, естественно, одно¬
временно измерялись величины D, В, 5.
Методика измерений указанных характерис¬
тик, проводимых на Курской биосферной стан¬
ции ИГ АН СССР, заключалась в следующем.
Специально изготовленные лизиметры [2] содер¬
жали оголенную почву (типичный чернозем) и поч¬
ву, покрытую слоем соломенной мульчи (рис. 1).
Все лизиметры обеспечивались с помощью сосу¬
да Мариотта постоянным подпитыванием водой,
находясь в условиях достаточного увлажнения.
По расходу подпитывающей воды в первой
группе испарителей - лизиметрах без мульчи оп¬
ределялись ежесуточные, а затем и средние за пе¬
риод измерения (порядка нескольких суток) зна¬
чения испаряемости для оголенной почвы Еп0, от¬
личающиеся от Е0, поскольку радиационный
баланс для поверхности соломенной мульчи, вхо¬
дящий в выражение (1), отличается от радиацион¬
ного баланса для оголенной почвы. Поэтому зна¬
чения Епо, полученные на основе водного балан¬
са, пересчитывались для получения величин £0
= f C,ATdz - A.jGradT’l
Д tJ '■
(4).
где Сэ, X, - соответственно теплоемкость и коэф¬
фициент теплопроводности почвы, АТ - измене¬
ние температуры почвы за период измерения Д/,
z„ - высота испарителя, Grad Г| - средние за пе¬
риод измерения градиенты температуры у нижне¬
го конца испарителя, определяемые по разнице
температур двух соседних термопар.
Для задания параметров X, и С, использова¬
лись сведения, содержащиеся в работе [5], и дан¬
ные о плотности и влажности почвы, получаемые
непосредственно при проведении эксперимен¬
тальных исследований.
За период измерения определялась также и
средняя скорость ветра на уровне z6 (рис. 1), необ¬
ходимая для оценки D в соответствии Со следую¬
щим эмпирическим выражением [2]:
D =
0.76и«
1 + 0.82 7^’
(5)
где иб - скорость ветра на уровне г* = 2 м. Здесь D
выражен в см/с, а и6- м/с.
Эксперименты были проведены в течение двух
летних сезонов (1988,1989 гг.). Всего было постав¬
лено около 35 серий опытов при различной вы¬
соте слоя соломенной мульчи (от 2 до 8 см) и при
разных значениях средней за период наблюдения
скорости ветра (от 0.5 до 2.5 м/с).
Кроме того, были использованы варианты
соломенной мульчи различной плотности
(0.019-0.066 г/см3). Как было отмечено выше,
ПОЧВОВЕДЕНИЕ № 6 1996
ОПРЕДЕЛЕНИЕ КОЭФФИЦИЕНТА ЭФФЕКТИВНОЙ ДИФФУЗИИ
791
исследование возможного влияния вариации плот¬
ности мульчи рм на величину коэффициента эф¬
фективной диффузии водяного пара в ней обус¬
ловлено тем, что в реальных условиях плотность
соломенного покрытия значительно меняется во
времени. Связано это с уплотнением мульчи из-за
ее оседания под действием собственной тяжести,
ветра, осадков, а также из-за перегнивания соло¬
мы. Теоретический анализ показывает, что при
том диапазоне плотности соломы, который имеет
место в реальных условиях, ожидать заметного
влияния рм на интенсивность переноса пара не
приходится. Действительно, коэффициент эф¬
фективной диффузии пара в соломе можно пред¬
ставить в следующем виде:
die = <1ф1, (6)
где d/a - коэффициент эффективной диффузии
водяного пара в воздухе; £ - пористость мульчи;
I - коэффициент извилистости пор в мульче. При
этом
(7)
где рс - плотность твердой фазы (непосредствен¬
но соломы), которая в первом приближении мо¬
жет быть принята равной плотности древесины
(~0.5 г/см3); в - объемное влагосодержание мульчи.
Последняя характеристика может варьиро¬
вать примерно от 0 до 0.04 [3]. Таким образом,
при указанных выше значениях плотности муль¬
чи значения £ будут в общем случае лежать в ди¬
апазоне от 0.83 до 0.96, а для сухой мульчи - от
0.87 до 0.96, т.е. варьировать очень незначитель¬
но (отклонения величины £ от ее среднего значе¬
ния лежат в пределах 7% в общем случае и в преде¬
лах 5% для сухой мульчи). Также следует отметить,
что при таких больших значениях пористости, ко¬
торые указаны выше, значение коэффициента
извилистости / во всех случаях близко к 1. Таким
образом, в соответствии с уравнением (6) вариа¬
ция плотности мульчи должна очень слабо сказы¬
ваться на значениях коэффициента эффективной
диффузии пара в ней.
Экспериментальная вариация плотности муль¬
чи достигалась как ее искусственным уплотнени¬
ем, так и использованием в качестве мульчирую¬
щего покрытия соломы различной степени све¬
жести. В частности, было выделено две группы
соломенного покрытия. Первая группа опытов
была проведена с сухой, достаточно свежей соло¬
мой, другая - с соломой, подвергшейся значи¬
тельному перегниванию.
Проведенные измерения показали отсутствие
заметной корреляции величин D/DM со скоро¬
стью ветра, и, как и ожидалось, с плотностью со¬
ломы в тех диапазонах изменений этих характе¬
ристик, какие были в опытах. Было установлено
6, см
Рис. 2. Зависимость отношения интегрального коэф¬
фициента обмена водяного пара между поверхностью
мульчи и высотой измерения метеорологических эле¬
ментов к интегральному коэффициенту обмена водя¬
ного пара в мульче от высоты мульчирующего слоя
соломы: 1 - солома свежая; 2 - солома гнилая; 3 - ли¬
нейная регрессия D/DM от 8.
также, что величина D/Du примерно пропорцио¬
нальна 5 (рис. 2), хотя и с не очень высоким коэф¬
фициентом корреляции (=0.6). Таким образом,
учитывая выражение (2), можно сделать вывод,
что величина dk в первом приближении зависит от
скорости ветра примерно так же, как и D, т.е.
в соответствии с равенствами (2), (5) и рис. 1
kD = к
0.76иб
1 + 0.82 7«6
(8)
а
где к » (1.0 ± 0.1) см (здесь dk выражен в см2/с,
иб - в м/с).
Выражение (6) в первом приближении спра¬
ведливо как для свежей, так и для полусгнившей
соломы. Отсюда следует, что эффективность
уменьшения соломенной мульчей непродуктив¬
ного испарения в значительной мере сохраняется
даже при перегнивании соломы. Полученное для
оценки коэффициента эффективной диффузии
пара в мульче выражение (6) может быть реко¬
мендовано для использования при проведении
расчетов испарения воды почвой, покрытой соло¬
менной мульчей, в частности, при определении
эффективности мульчирования с точки зрения
повышения влагообеспеченности сельскохозяй¬
ственных посевов.
СПИСОК ЛИТЕРАТУРЫ
1. Будаговский А.И. Методы оценки параметров мо¬
делей испарения почвенных вод // Водные ресур¬
сы. 1986. № 6. С. 3-15.
ПОЧВОВЕДЕНИЕ № 6 1996
792
ГУСЕВ, БУСАРОВА
2. Будаговский А.И.У Григорьева Н.И. Пути повыше¬
ния эффективности использования ресурсов поч¬
венных вод//Водные ресурсы. 1991. № 1.С. 131-142.
3. Гусев Е.М., Ясинский С.В. Влияние мульчи из рас¬
тительных остатков на формирование термичес¬
кого режима почвы в зимний период // Водные ре¬
сурсы. 1990. № 12. С. 46-54.
4. Гусев ЕМ.У Бусарова О.Е.у Шурхно АЛ., Ясин¬
ский С.В. Влияние соломенной мульчи на терми¬
ческий режим почвы после схода снежного покро¬
ва // Почвоведение. 1992. № 5. С. 49-59.
5. Калюжный ИЛ., Павлова К.К. Формирование по¬
терь талого стока. Л.: Гидрометеоиздат, 1981. 160 с.
6. Каммонер Б. Замыкающийся круг. Л.: Гидрометео¬
издат, 1974. 274 с.
Calculation of Effective Diffusion Coefficient
of Water Stream in Straw Mulch
Ye. M. Gusev, O. Ye. Busarova
An equation is offered to calculate effective diffusion coefficient of water stream in straw mulch. This equation
can be used for calculation of the soil evaporation in fields, covered by straw mulch and particularly for assess¬
ment of efficient mulching with the view of increasing the water supply of agricultural crops.
ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВОВЕДЕНИЕ, 1996, № 6, с. 793-802
МИНЕРАЛОГИЯ
И МИКРОМОРФОЛОГИЯ почв
УДК 631.43(1-924.81)
МИКРОМОРФОЛОГИЯ почв
ВОСТОЧНО-ЕВРОПЕЙСКОГО СЕКТОРА СУБАРКТИКИ*
© 1996 г. Г. В. Русанова
Институт биологии Кольского научного центра Уральского отделения РАН, г. Сыктывкар
Поступила в редакцию 12.04.94 г.
Дана характеристика микростроения и морфогенетических свойств основных типов почв (подбуры,
тундровые глеевые и тундровые болотные) северной части Большеземельской тундры.
Рассматриваются эволюционные стадии от свежего пятна до зрелой почвы.
Несмотря на длительную историю исследова¬
ний почв тундровой зоны, к настоящему времени
лучше изучен Азиатский сектор. Подзоне суб¬
арктической или типичной тундры Восточно-Ев-
ропейского сектора посвящена обобщающая ра¬
бота Игнатенко [7], а горные тундры Кольского
п-ва и Полярного Урала освещены в работе Таргу-
льяна [12]. В почвенно-микроморфологическом от¬
ношении изученность этих почв крайне слабая [3],
а материалы по микроморфологии почв на песча¬
но-супесчаных и каменисто-мелкоземистых от¬
ложениях отсутствуют. Также в недостаточной
степени разработана классификация [2,6,7,9,12].
Район исследований автора расположен в Ар¬
хангельской обл., в типичной тундре северной ча¬
сти Большеземельской тундры, представляющей
собой слабопокатую к северу оконечность Рус¬
ской равнины. Территория характеризуется по¬
логоувалистым рельефом с высотами 25-60 м
ближе к побережью Баренцева моря и холмис¬
тым с высотами 150-200 м к югу от побережья.
Небольшие реки (Варкнивъяха, Лабанъяха, На-
улъяха), в бассейнах которых проводились иссле¬
дования, впадают в оз. Б. Торавей и Хайпудыр-
скую губу Баренцева моря. Район сплошного рас¬
пространения многолетней мерзлоты, мощность
которой колеблется в пределах 150-200 м. Глубина
оттаивания болотных торфяных почв составляет
20-40 см, суглинистых - 40-60 см, а супесчано-пес-
чанных - 60-80 см. Зональная растительность -
ивняково-мелкоерниковые зеленомошные ассоци¬
ации. Площадь болот достигает 40%, господству¬
ют гипново-осоковые и осоково-сфагновые ком¬
плексы.
Согласно ландшафтному районированию [1],
на территории выделяются холмистые равнины на
элюво-делювии плотных, часто карбонатных
пород с каменистыми тундрами горного типа,
возвышенные холмистые, а также волнистые и
'Работа выполнена при финансовой поддержке Междуна¬
родного научного фонда.
пологохолмистые равнины с плоскобугристыми
и полигональными тундрами, травяно-осоковы¬
ми болотами. Ближе к побережью заболоченные
территории преобладают. Почвы формируются
на моренных суглинках и супесях, верхнечетвер¬
тичных морских, ледниково-морских и озерно¬
аллювиальных отложениях, элюво-делювии ко¬
ренных пород. В работе использована классифи¬
кация почв восточно-европейской тундры, разра¬
ботанная Игнатенко [7].
ПОЧВЫ СО СВОБОДНЫМ
ВНУТРЕННИМ ДРЕНАЖОМ
Тип подбуров, формирующихся на песках и су¬
песях, включает подтипы типичных и надмерз-
лотно-глееватых почв. В условиях Таймырской
тундры на древних надпойменных террасах на пе¬
счаных отложениях выделены тундровые дерно¬
вые почвы [2], отличающиеся от подбуров.
Подбуры типичные равнинных тундр форми¬
руются в условиях холмистого рельефа на морен¬
ных супесях и песчаных гривах высотой 1.5-2 м
вдоль озер, как это наблюдалось и на Таймыре [2],
под кустарничково-мохово-лйпайниковым покро¬
вом (карликовая ива, брусника, водяника, мхи, ли¬
шайники, злаки).
Характерной особенностью морфологического
строения подбуров (рис. 1) является четкая диф¬
ференциация профиля: маломощная подстилка
(АО 0.5 см), темно-серый или бурый, рыхлый био¬
генно-аккумулятивный горизонт (А0А1), буро¬
коричневый бесструктурный иллювиально-гуму¬
совый горизонт (Bh), светло-бурый уплотненный
иллювиальный горизонт (Bhf), плотный ортзанд
со следами турбаций (S), слоистая почвообразую¬
щая порода (С). Криогенные процессы в подбурах
песчаных реализуются в криогенных деформаци¬
ях ортзандов.
В микросгроении гор. А0А1 отмечаются следу¬
ющие черты: округлые агрегаты из растительных
остатков с пылеватыми зернами по периферии,
793
794
РУСАНОВА
50-
т°гт
47V
ги
(а) А0А1
АО
А1
Bh В1
В1
• &
«д
S
ВС
ВС
о • «
а » о
Разр. 32(a)
-1.5 м-
20 м
Bh
о о#
(б)
(в)
Bg
в
I 118 12 I я> I ? i 6 14 вин5 I а 16 I»»I 7 ДЕ 81 ж i 9
Рис. 1. Морфология и микростроение субарктического подбура (а), почвы голого пятна (б) и покрытого водорослевой
пленкой (в): I - пленки; 2 - органогенные агрегаты; 3 - сгустки плазмы; 4 - скопления зерен скелета; 5 - агрегаты
в порах и вокруг зерен; 6 - растительные остатки; 7 - зерна скелета; 8 - агрегаты; 9 - коагуляционные агрегаты.
гроздьевидные сгустки гумусовой плазмы между
зернами и на их поверхности; обрывки расти¬
тельных остатков, местами мелко фрагментиро¬
ванных, скелетные зерна с желто-бурыми плен¬
ками и без них; глинистые включения (литоре-
ликты) бурые, овальной формы, с чешуйчатой
плазмой и околопоровыми глинистыми пленка¬
ми. Характерным признаком иллювиально-гуму-
Таблица 1.
почвах
Характер пленок и натеков в тундровых
Подзона
Почва
Гори¬
зонт
Пленки, натеки
Типичная
Подбур
А0А1
Гроздьевидные сгустки гу¬
тундра
типич¬
ный
Bh
S
мусовой плазмы, пылева¬
тые частицы вокруг органо¬
генных агрегатов
Бурые (Fe-гумусовые)
пленки, коричнево-бурые
гроздьевидные сгустки
вокруг скелетных частиц
Бурые (Fe-гумусовые)
пленки на зернах
Тундро¬
Bgtx
Пылевато-глинистые и гли¬
вая
нистые натеки, папулы, вее¬
глеевая
BCg
рообразные прожилки глин
в основе
Гипокутаны, скелетаны в
трещинах, пылевато-гли-
нистые натеки и пленки на
стенках пор
Южная
Поверх-
GB
Обломки красноватых
тундра
ностно-
глеевая
B2
Fe-глинистых натеков со
слабым двупреломлением
Глинистые и Fe-глинистые,
бурые и красно-бурые, ред¬
кие, мелкие, скорлуповатые
натеки
сового гор. Bh является формирование как Fe-гу¬
мусовых пленок с пылеватыми частицами, так
и округлых гроздьев, слипшихся сгустков корич-
нево-бурого органического вещества вокруг зе¬
рен скелета и между ними, на стенках пор (табл. 1).
Мелкие коагуляционные агрегаты образуют бо¬
лее крупные округлые и угловатые агрегаты.
Микрозональность основы диагностирует слабое
оглеение над ортзандом, заметное только в шли¬
фах и не являющееся, по-видимому, достаточным
основанием для перевода почвы в подтип над-
мерзлотно-глееватых. Отличия в микростроенш
ортзанда - темно-серая изотропная плазма, ком¬
пактное сложение. Таким образом, микросгроение
показывает, что в подбурах тонкодисперсный ма¬
териал сконцентрирован, в основном, в виде пле¬
нок на зернах, в криогенно-коагуляционных агре¬
гатах, в промежутках между зернами и на их по¬
верхности. Последние'отражают роль криогенных
процессов в организации почвенной массы. В ре¬
зультате коагуляции коллоидов при проморажива¬
нии и их склеивания образуются сложные округлые
aiperaibi. Характерен аккумулятивно-иллювиаль¬
ный тип профильного распределения тонкодис-
персной массы в подбурах. Признаки элювииро-
вания (удаление пленок с части зерен) в гор. А0А1
не вызывают осветления массы горизонта.
Подбуры имеют среднекислую реакцию среды,
кислотность уменьшается с глубиной. Аккумуля¬
ция гумуса происходит как в иллювиально-гуму¬
совом горизонте, так и в ортзанде (табл. 2).
Основными процессами почвообразования в
подбурах являются: криокоагуляционная агрега¬
ция тонкодисперсной массы, Al-Fe-гумусовая ми¬
грация, криотурбация. Механизм формирования
ортзанда - наложение иллювиирования на лито-
генный слой. Согласно Таргульяну [12], в подбо¬
рах происходит ожелезнение и алюминирование
ПОЧВОВЕДЕНИЕ № 6 1996
МИКРОМОРФОЛОГИЯ почв
795
Таблица 2. Физико-химические свойства подбуров
Разрез. Почва
Гори¬
зонт
Глуби¬
на, см
pH
Гумус
по Тю¬
Обменные
основания, мг-экв/100 г
водный
солевой
рину, %
Са2+
Mg2+
32. Подбур типичный
А0А1
0-4
4.9
3.9
-
1.6
1.0
Bh
4-10
5.2
4.1
0.7
0.8
0.6
Bhf
10-29
5.8
4.3
0.4
1.5
0.9
S
29-34
5.7
4.3
1.1
2.7
0.7
С
34-62
6.1
4.7
0.6
2.3
0.6
4. Подбур надмерзлотно-глееватый
(водорослевая пленка)
0-1
5.0
4.3
0.4
0.5
0.3
2. То же, заросшее пятно
А0А1
0-1
4.9
4.2
2.5
0.4
0.3
5. Подбур, горная тундра
А0А1
0-10
6.0
5.6
4.2
13.4
3.4
Bh
10-14
5.9
4.9
0.4
1.8
1.5
8. Подбур, почва зарастающего пятна
A0A1
0-5
5.9
5.4
7.6
19.4
9.7
В
5-10
5.1
4.2
2.5
4.0
2.6
профиля, обеднение Si02 вследствие различной
геохимической подвижности этих соединений
в кислой среде.
Подбуры надмерзлотно-глееватые формиру¬
ют трехчленный комплекс: почвы бугорков, пятен
и ровных участков на супесчано-песчаных породах.
Занимают незаболоченные вершины и склоны ува¬
лов, приурочены к пятнисто-бугорковатым лишай¬
никовым тундрам с ерником, ивой, брусникой, зла¬
ковыми и политриховыми мхами в покрове.
При близком залегании мерзлоты (30 см) в ус¬
ловиях пятнисто-бугорковатой тундры формиру¬
ются надмерзлотно-глееватые подбуры с недиф¬
ференцированным (пятно) и слабодифференци¬
рованным (бугорок, ровный участок) профилем.
Наблюдается эоловое разрушение бугорков и за¬
растание образующихся пятен. Наиболее молодая
стадия почвообразования - полностью разрушен¬
ный бугор, на месте которого образовалось пятно
(дефляционное обнажение). На его поверхности -
темная водорослевая пленка толщиной 1 мм.
Подобные почвы выделяются в самостоятель¬
ный тип “почв пятен” [2] или почв-пленок [9].
В профиле присутствуют признаки оглеения в ви¬
де ржавых, охристых, белесых пятен и охристой
каймы на глубине 3-5 см. Профиль, заложенный
через бугорок (минеральный), слабо заросший
ягелем, политриховыми мхами, обнаруживает сла¬
бые признаки дифференциации (однородный свет-
ло-бурый песок с пятнами внизу). Мощность
поверхностного темно-бурого слоя (А0А1 + Bh)
здесь составляет 5 мм, а на слабо заросших
мхом, с водорослевыми корками пятнах между
бугорками, на ровных участках - 5 см (глубже -
строение сходное). Последние отличаются луч¬
шей дифференциацией, более кислой средой и
высоким содержанием гумуса. В поверхностном
слое мощностью 1 см этой почвы ровного участ¬
ка накапливается до 2-5% гумуса (анализы вы¬
полнены для поверхностных горизонтов). Над¬
мерзлотно-глееватые подбуры характеризуются
усилением криодинамических процессов и оглее¬
ния при близком залегании мерзлоты.
В центральной части Большеземельской тунд¬
ры расположен Большеземельский хребет в на¬
правлении от Хайпудырской губы до устья р. Циль-
мы с высотами 150-200 м над ур. м. [13].
Подбуры горных тундр, названные горными
подбурами [2], горно-тундровыми слаборазвиты¬
ми [5], ортоподбурами [12], примитивно-щебнис-
тыми [9], распространены ограниченно, наблю¬
даются на вершинах, сложенных элюво-делюви-
ем коренных пород, с каменистыми пятнистыми
тундрами горного типа, под кустарничково-мохо-
во-лишайниковой растительностью с примесью
дриад, злаков, разнотравья. Формируется ком¬
плекс, состоящий из зрелого с развитым профи¬
лем подбура (ровньщ участок) и почв пятен (све¬
жего и зарастающего).
Горные подбуры отличаются маломощным
профилем (14 см). В зрелых почвах под хорошо
разложившимся органо-аккумулятивным горизон¬
том (А0А1) залегает черно-коричневый горизонт,
опесчаненный суглинок, щебнистый, пропитанный
гумусом (Bh), подстилаемый крупнообломочным
материалом пород с мелкоземом в промежутках.
Почвы незаросших пятен не имеют признаков
дифференциации. В зарастающих пятнах мощ¬
ность гор. А0А1 меньше, чем в зрелых подбурах.
Залегающий ниже гор. B(g) - сизовато-бурого
цвета, опесчаненный суглинок, содержит обло¬
мочный материал и подстилается неоглеенным
коричневато-бурым легкосуглинистым гор. ВС.
ПОЧВОВЕДЕНИЕ № 6 1996
796
РУСАНОВА
Мощность профиля 17 см. В профиле почвы све¬
жего пятна обнаруживаются тиксотропия и сле¬
ды оглеения, связанные с пылевато-суглинистым
составом (массообмен при пятнообразовании).
Обломочный материал вследствие криогенных
процессов также переносится к поверхности.
Слабые следы оглеения еще сохраняются на ста¬
дии зарастания пятна. На стадии зрелой почвы
происходят пропитка подвижными гумусово-Fe
соединениями, формирование структуры, что обес¬
печивает хороший дренаж и отсутствие следов
оглеения.
Микростроение почвы свежего пятна характе¬
ризуется неагре гированн остью, наличием окислен¬
ных и восстановленных микрозон и пылевато-гли¬
нистыми пленками на скелетных зернах. Зарастаю¬
щее пятно с водорослевой пленкой на поверхности
отличается большим количеством мелкораздроб¬
ленных растительных остатков (0.2-0.05 мм),
гумусовыми (коричневыми) коагуляционными
агрегатами между зернами, слипающимися и об¬
разующими агрегаты П порядка, гумусово-глини¬
стыми пленками на зернах. Гумусовые агрегаты
образуют пленку на поверхности зерен скелета,
обломков породы.
Микростроение зрелой почвы ровного участ¬
ка характеризуется наличием крупных фрагмен¬
тов остатков растений в верхней части профиля.
Гумусовые коагуляционные агрегаты в проме¬
жутках между скелетными зернами и бурые гуму¬
сово-Fe пленки и натеки придают почве темно¬
бурый цвет. Основные диагностические признаки
этих почв следующие:
1) Морфологически недифференцированный
бурый монотонный профиль, отсутствие границы
между органо-аккумулятивным и иллювиально¬
гумусовым горизонтами. Коагуляционные бурые
агрегаты в промежутках между скелетными зер¬
нами. 2) Бурые гумусово-Fe пленки и натеки
в средней и нижней частях профиля. 3) Включе¬
ния обломков пород по всему профилю.
К особенностям этих почв относятся высокое со¬
держание гумуса и слабокислая среда, значитель¬
ное количество обменных оснований (табл. 2).
Аккумулятивный характер профильного распре¬
деления гумуса обнаружен и другими исследова¬
телями подобных почв [2,12]. Гор. Bh диагности¬
руется по микростроению.
Богатство пород (кристаллических сланцев) ос¬
нованиями и легковыветривающимися минерала¬
ми придает почвообразованию аккумулятивную
направленность.
Ведущими процессами в почве свежего пятна
являются криогенные. На следующей стадии сук-
цессионного развития (зарастающее пятно с во¬
дорослевой пленкой) начинают проявляться эле¬
ментарные почвенные процессы метаморфизма
органического вещества, переорганизации массы
(агрегация) с участием продуктов метаболизма ми¬
кроорганизмов, Al-Fe-гумусовой миграции. В мик¬
ростроении зрелой почвы обнаруживаются при¬
знаки иллювиально-гумусового процесса.
ПОЧВЫ С ЗАТРУДНЕННЫМ
ВНУТРЕННИМ ДРЕНАЖОМ
Среди минеральных почв тундровой зоны
наиболее широко распространены суглинисто-гли¬
нистые почвы с затрудненным внутренним дрена¬
жом. На исследованной территории типичной
тундры описаны почвы с недифференцирован¬
ным или слабодифференцированным профилем,
относящиеся к типу тундровых глеевых.
1. Автоморфные почвы. Тундровые глеевые
почвы формируются в комплексе с остатонно-
глеевыми пятен и торфянисто-глеевыми бугор¬
ков на плоских водоразделах и увалах, сложен¬
ных суглинисто-глинистыми породами (рис. 2).
Приурочены к ландшафтам пятнисто-бугоркова-
тых тундр с ерниково-ивняковым кустарничково-
моховым покровом. Разрез тундровой глеевой
почвы сделан в 30 км от побережья Баренцева
моря на возвышенности, сложенной пылевато¬
суглинистыми отложениями, подстилаемыми ко¬
ренными породами. Растительность представле¬
на нанополигональной дриадово-мохово-лишайни-
ково-кустарничковой тундрой. Пятнисто-бугорко-
ватый комплекс состоит из пятен различных стадий
зарастания и моховых бугорков высотой 15 см.
Организация полигонов (10 х 10 м) следующая: бу¬
горки высотой 15 см и размером 20 х 30 и 30 х 50 см,
заросшие лишайниками, мхами, осоками, травя¬
нистыми растениями, занимают полигон. По пе¬
риферии полигона формируются голые и слабо-
зарастающие пятна, окруженные валиками из
мелких бугорков, сходных со срединными.
Зарастающее водорослями, хвощом, мхами
пятно с обломками пород на поверхности. В тем¬
но-коричневой рыхлой торфянистой подстилке
(АО) обнаружен дождевой червь длиной 5 см.
Гор. А0А1 (0-6 см) темно-бурый, криокоагуляци-
онного генезиса. Гор. Gtx (6—14 см) грязно-сизый
суглинок с обломками породы; гор. Bgtx (14-35 см)
сизовато-бурый тиксотропный суглинок; гор.
BCg (35-57 см) темно-сизо-бурый суглинок, под¬
стилается обломками породы. Мерзлота на глу¬
бине 60 см (рис. 2).
Анализ микросгроения показывает, что для
гор. А0А1 характерна агрегированность биоген¬
ного характера (копролиты, состоящие из мелких
растительных остатков с пылеватыми частица¬
ми), наряду с криогенной в виде коагуляционных
мелких гумусовых сгустков, образующих слож¬
ные округло-угловатые агрегаты. Фрагментар¬
ность растительных тканей, что отмечается
и другими исследователями в континентальных
тундрах [3], и гумусово-глинйстые пленки на зер¬
нах скелета объясняются также криогенными
ПОЧВОВЕДЕНИЕ № 6 1996
МИКРОМОРФОЛОГИЯ почв
797
процессами. Гор. Gtx грязно-сизого цвета, ком¬
пактный, с окисленными и восстановленными мик¬
розонами с гумусово-глинистыми пленками на зер¬
нах скелета. Отличием нижележащего гор. В gtx
являются редкие глинистые пленки на стенках
пор и на зернах, чешуйчато-волокнистая плазма,
свидетельствующие о возможности очень слабой
миграции тонкодисперсной массы. Отчетливы
микрозоны с криотурбированным гумусом, папу¬
лами, ооидами. Надмерзлотный горизонт (BCg)
обнаруживает реликтовые признаки (темный гу¬
мус) и пылевато-глинистые пленки в порах (рис. 3),
свидетельствующие о прошлых этапах почвооб¬
разования, глинистые, бурые, тусклые натеки
и псевдоморфозы по корням (оглиненные корни
с выбросами клещей).
О реликтовом характере гумуса свидетельст¬
вуют большое количество мелкораздробленных
черных растительных остатков и темноокрашен-
ный сгустковый гумус вокруг скелетных частиц.
Пылевато-глинистые пленки и натеки на той же
глубине в тундровых глеевых почвах Субарктики,
вероятнее всего, также не связаны с современным
режимом почвообразования и более свойствен¬
ны поверхностно-глеевым (элювиально-глеевым)
почвам южной тундры. Как и фрагменты релик¬
тового гумуса, натеки и пылевато-глинистые
пленки могли сохраниться от прошлых фаз диф¬
ференциации профиля. Этот вывод, сделанный
на основе изучения микростроения, подтвержда¬
ет предположение Игнатенко [7] о дифференциа¬
ции (хотя и слабой) этих почв по илу. Возмож¬
ность современного очень слабого лессиважа
предполагается в связи с образованием сети мел¬
ких трещин при промерзании почвогрунтов и гра¬
витационным выносом по ним суспензий после
вытаивания линз льда. Слабая дифференциация
профилей по распределению ила объясняется ре¬
ликтовым элювиально-глеевым почвообразова¬
нием в период среднеголоценового климатичес¬
кого оптимума [7]. Ливеровский [9] считает, что
тундровое почвообразование наложилось на лес¬
ное. Гумусовые педореликты, гетерогенные на¬
теки в нижней части профиля могут свидетельст¬
вовать о ранних периодах почвообразования. По
радиоуглеродным датировкам торфяника вблизи
г. Воркуты [8] слои торфа примерно на той же
глубине имеют среднеголоценовый возраст.
Тундровые глеевые почвы отличаются силь¬
ным оглеением минеральных горизонтов, ярко
выраженной тиксотропностью деятельного слоя,
слабым проявлением гумусово-аккумулятивных
процессов и дифференциации профиля, лучше за¬
метной в микростроении. Ведущими процессами
являются глеевый и криогенные.
Сильнокислая реакция наблюдается в почвах,
формирующихся на моренных суглинках. На мор¬
ских и элювиально-делювиальных (карбонатных)
отложениях реакция по всему профилю нейтраль-
*
Б
АО
G
Bg
Вс
&
77^
г©
£=> Q
1 ^ 121 <==? Ы • * | 4\ О I 5
4№/\б I * 171 Г I 8\ I яЕПП] 10
Рис. 2. Морфология и микростроение тундровой глее-
вой (А) и поверхностно-глеевой (Б) почв: а - слой 0-4 см
голого пятна: б - гор. А0А1 зарастающего пятна;
1 - пленки; 2 - конкреции; 3 - растительные остатки;
4 - коагуляционные агрегаты; 5 - биогенные агрега¬
ты; 6 - поры; 7 - прожилки глин; 8 - гумусовые педо¬
реликты; 9 - натеки в порах; 10 - плазма.
ная. Незначительное содержание гумуса коррели¬
рует с маломощной подстилкой, сумма обменных
оснований (Са, Mg) также невысокая. В крипто¬
гамных корках пятен мощностью 0.5-1 см количе¬
ство гумуса выше - 0.8% (табл. 3). Эти черные во¬
дорослевые корки аккумулируют органическое
вещество, обогащают почву азотом, благодаря
фиксации из атмосферы, и увеличивают содер¬
жание питательных веществ [15].
ПОЧВОВЕДЕНИЕ № 6 1996
798
РУСАНОВА
(а) (б)
Рис 3. Микрофотографии генетических горизонтов подбура (а) и тундровой глеевой почвы (б).
Горизонты, а: / - А0А1; 2 - Bh; 3 - S; б: 4- А0А1; 5-Glx; 6- BCg.
ПОЧВОВЕДЕНИЕ № 6 1996
МИКРОМОРФОЛОГИЯ почв
Таблица 3. Физико-химические свойства тундровых глеевых почв
799
Разрез. Почва
Гори¬
зонт
Глуби¬
на, см
рн
Гумус
по Тю¬
рину, %
Обменные основа¬
ния, мг-экв/100 г
водный
солевой
Са2+
Mg2+
41. Тундровая глеевая
АО
0-6
7.1
7.0
-
35.8
15.1
Gtx
6-14
7.0
6.9
0.6
8.6
2.2
Bgtx
14-35
7.1
6.9
0.4
7.3
0.7
Пятно
0-1
6.7
5.7
0.8
—
—
21. Тундровая перегнойно-глеевая
А0А1
0-7
5.7
5.2
-
20.1
10.0
Bgtx
7-14
6.0
4.5
1.4
7.7
2.5
49. Тундровая поверхностно-глеевая
АО
0-3
6.1
6.0
-
43.9
16.6
G
3-14
5.9
4.6
1.7
8.8
4.5
Bg
14-34
5.7
4.6
3.2
8.4
4.4
На дренированных бровках склонов преиму¬
щественно южной экспозиции встречены поверх-
ностно-глеевые почвы, отнесенные Игнатенко [7]
к типу элювиально-глеевых, и характерные для
подзоны южной кустарниковой тундры. В подзоне
типичной тундры эти почвы приурочены к ланд¬
шафтам мелкобугорковатых тундр с мохово-осо-
ково-разнотравным напочвенным покровом, при
преобладании разнотравья, злаков. Характеризу¬
ются отсутствием оглеения нижней части профи¬
ля (34-80 см), слабокислой реакцией органоген¬
ной и кислой - минеральной толщи. Наблюдается
более высокое содержание гумуса по сравнению
с тундровыми глеевыми почвами и увеличение
его количества с глубиной (табл. 3). В данном
конкретном случае это может быть связано с бо¬
ковым привносом как мелкофрагментированных
растительных остатков, так и дисперсных форм
гумуса.
Микростроение поверхностно-глеевой почвы
четко отражает разницу с тундровой глеевой.
Прежде всего усиливаются элювиально-иллюви-
альные процессы. Об этом свидетельствует от-
мытость от глинистых пленок скелетных зерен
верхней части профиля (гор. G) и присутствие их и
натеков (в большем количестве по сравнению с тун¬
дровой глеевой почвой) в нижней (гор. В, 34-80 см).
В гор. Gtx тундровой глеевой почвы глинистые
пленки сохраняются. Также отчетливо видно
формирование маломощного гумусового гори¬
зонта криокоагуляционного генезиса в контакт¬
ной с минеральной толщей зоне. Этот горизонт
обладает округлой и сгустковой формами гумусо¬
вой плазмы, как и в тундровой глеевой почве.
В гор. G отмечены округлые гумусово-Fe сгустки
и дисперсный гумус, а в гор. Bg (14-34 см) - про-
гумусированные бурые микрозоны, что согласу¬
ется с данными содержания гумуса в этой почве
(табл. 3).
На современный характер лессиважа (очень
слабого) указывают “свежие” глинистые натеки
и пленки на стенках пор и чешуйчато-волокнис¬
тая плазма в гор. В (34-80 см) тундровых поверх-
ностно-глеевых почв.
Анализ микростроения автоморфных тундро¬
вых глеевых почв показал, что ведущими процес¬
сами в органогенных горизонтах (А0А1) являются
1) криогенная переработка материала и 2) акку¬
муляция грубого гумуса на границе с минераль¬
ной толщей. В верхней части профиля (гор. Gtx)
преобладают криогенные и глеевые процессы.
В иллювиальных горизонтах (гор. Bg) отчетливы
признаки 1) иллювиирования (глинистые пленки
на зернах, на стенках пор и натеки), 2) оглеения
(микрозональность, конкреции), 3) криогенных
процессов (ооиды, папулы, турбированный гумус);
в гор. BCg - унаследованные признаки (гумусо¬
вые педореликты, гетерогенные натеки, псевдо¬
морфозы). Сохранность палеопочвенных призна¬
ков имеет место благодаря слабой переработке
почвообразованием материала нижней части
профиля (на глубине 50-60 см над мерзлотой).
В тип тундровых глеевых почв включены на
уровне подтипа перегнойно-глеевые [7], широко
распространенные в арктической тундре и встре¬
чающиеся локально в типичной. Перегнойно-
глеевая почва формируется на средней и нижней
частях склона увала южной экспозиции, менее
подверженных влиянию ветра, со злаково-бобо-
выми компонентами в напочвенном покрове, со¬
стоящем из ивы карликовой, мхов, осок, мятлика,
овсяницы, дриады, копеечника. Характеризуется
наличием интенсивно черного грубогумусового
перегнойного горизонта (7-20 см) с более высо¬
ким, чем в тундровых глеевых почвах, содержа¬
нием гумуса. Мощность его увеличивается вниз
по катене. Подстилается сизо-бурым тиксотроп-
ным гор. Bgtx с охристой каймой в нижней части.
Увеличение мощности перегнойного горизон¬
та в нижней части склона связано и с поверхност¬
ным стоком, приносящим мелкие растительные
остатки и растворимые соединения, ил. Горизонт
темно-бурый с коричневыми зонами, форма
которых напоминает гумусовые затеки. Фраг¬
ПОЧВОВЕДЕНИЕ № 6 1996
800
РУСАНОВА
Таблица 4. Физико-химические свойства тундровых болотных почв
Разрез. Почва
Горизонт
Глуби¬
на, см
pH
Гумус
по Тю¬
рину, %
Обменные основания,
мг-экв/100 г почвы
водный
солевой
Са2+
Mg2+
15. Торфянисто-глеевая
Ат
0-9
4.4
3.7
Не опр.
9.4
-
Ржавая кайма
9-11
5.1
4.0
2.9
4.4
-
G
11-25
5.2
3.8
0.2
2.9
3.1
36. Торфяно-глеевая
Ат
0-10
3.9
3.0
Не опр.
0.6
-
15-29
4.3
3.6
»
0.3
0.5
G
29-31
4.8
3.6
5.0
0.8
0.8
38. Болотная низинная
Ат
0-39
5.9
5.3
Не опр.
56.0
13.5
40. Болотная верховая
Ат
0-39
4.1
3.2
»
14.4
10.5
39. Перегнойно-торфяная бугров
Ат
1-35
4.1
3.1
»
13.6
3.8
менты растительных остатков с глубиной меня¬
ют окраску (от желтой до коричневой). Сложе¬
ние горизонта становится более плотным. 6 ни¬
жней его части появляются округлые агрегаты из
бурых растительных остатков, окаймленные ста¬
билизирующими их гумусово-глинистыми плен¬
ками, и черно-коричневые аморфные гумусовые
сгустки, хлопья. Много склероций, выбросов эн-
хитреид внутри растительных тканей. Коричне¬
во-желтый дисперсный гумус проникает в мине¬
ральную толщу, маскируя ориентацию тонкодис¬
персной массы.
В почвах накапливаются слаборазложившиеся
грубые растительные остатки, углистые частицы.
С глубиной степень разложения выше, на 60-70%
растительные остатки состоят из мелкодисперс¬
ных фрагментов, не различимых на макроуровне
исследований. Гумус плазмы тонкодисперсный,
перемешанный с минеральной составляющей.
2. Полугидроморфные и гидроморфные почвы.
Тундровые болотные почвы. Комплекс почв по¬
лигональной ивняково-пушицево-моховой тунд¬
ры широко распространен на изученной террито¬
рии. Он приурочен к пологим склонам и выров¬
ненным водоразделам, надпойменным террасам,
депрессиям рельефа.
Характерный микронанорельеф образован
полигонами, валиками и трещинами. Полигоны
имеют 5-6-угольную форму с бугорками в сере¬
дине высотой 10-15 см, разделены трещинами
шириной 0.3-0.4 м. Вдоль трещин формируются
валики высотой 15-20 см и шириной 0.5 м. Иногда
полигоны окружены по периферии голыми и за¬
растающими пятнами с валиками из мелких бу¬
горков, сходных со срединными. Компоненты
комплекса полигональной тундры различаются
по мощности торфяной толщи и степени ее раз-
ложенности. В трещинах полигонов развиваются
торфяно-глеевые почвы, а на валиках и полиго¬
нах в дренированных условиях - тундровые тор-
фянисто-глеевые. В торфяно-глеевых почвах
слой торфа мощностью от 30 см и выше отлича¬
ется высокой степенью минерализации. Глубина
мерзлоты в торфяно-глеевых почвах 30 см, в тор-
фянисто-глеевых - 50 см. В профиле последней
под торфом наблюдается ржавая оторочка, в ко¬
торой аккумулированы соединения Fe с гумусом.
Ниже залегает глеевый тиксотропный горизонт.
Почвы кислые, с невысоким содержанием гумуса,
за исключением ржавой каймы (9-11 см). Обмен¬
ных оснований небольшое количество (табл. 4).
Минеральные горизонты по микростроению
близки гор. Gtx тундровых глеевых почв, особен¬
но торфянисто-глеевых, в которых также отчет¬
лива микрозональность под органо-аккумулятив-
ным горизонтом.
Комплекс остаточно-болотных мерзлотных
перегнойно-торфяных почв бугров приурочен
к плоскобугристым лишайниковым болотам, за¬
нимающим северные склоны невысоких плоских
водоразделов. Представляет собой комплекс буг¬
ров и мочажин. Высота торфяных бугров достига¬
ет 40 см на севере, 70 см на юге. Поверхность плос¬
кая, нередко подвержена водной эрозии, имеет
трещины. Растительный покров состоит из кус-
тарничково-мохово-лишайниковых сообществ,
в мочажинах - травяно-сфагновых. Торф на буг¬
рах и в мочажинах черно-коричневый, кислый.
Мерзлота на глубине 20 см, под бугорками - 30 см.
Анализ микростроения показывает слабую
биогенную переработку торфа, которая наблю¬
дается лишь в слое 0-3 см благодаря лучшей
прогреваемости. Отчетливо заметно морозное аг¬
регирование растительного материала, окантовка
агрегатов органо-Fe плазмой. Большие скопления
ледяных кристалов в торфяных горизонтах спо¬
собствуют расчленению отдельных зон с орга¬
ническими остатками, сглаживанию их краев
и формированию округлых агрегатов (0.5-0.07 мм).
Последние окантованы (ориентация при промо¬
раживании) органо-железистой плазмой, способ¬
ствующей повышению устойчивости агрегата,
его механической прочности. Доминирует меха¬
ническое дробление растительных остатков в ре¬
зультате криогенных процессов.
ПОЧВОВЕДЕНИЕ № 6 1996
МИКРОМОРФОЛОГИЯ почв
Рис. 4. Микрофотографии подбура (а) и тундровой глеевой почвы (б).
Обводненные понижения между буграми пред¬
ставляют собой торфяники, на которых произра¬
стают осока, пушица. Реликтовые бугристые тор¬
фяники южной подзоны тундры нередко содержат
остатки древесных растений, что подтверждается
анализом ботанического состава торфа. В опти¬
мальные периоды голоцена дренированные плос¬
кие бугры заселялись древесной растительнос¬
тью [11]. Химическими анализами [10] обнаруже¬
ны сильнокислая реакция и бедность обменными
основаниями. Вновь образующихся низинных
торфяников в тундре не встречается.
ЭВОЛЮЦИОННЫЕ РЯДЫ ПОЧВ
Изучение микростроения эволюционных ря¬
дов тундровых почв (подбуры, тундровые глее-
вые) от свежего пятна до зрелой почвы показало,
что структурная организация поверхностных сло¬
ев возникает на самых начальных этапах разви¬
тия под действием не только мерзлотного факто¬
ра [4], но и биогенного. Поселение на поверхности
пятна с момента его появления микроорганизмов
(бактерий и водорослей), продуцирование ими ге¬
леобразного материала, слизи и органических со¬
единений бактерий [14] способствуют связыва¬
нию почвенных частиц в гетерогенную массу, обра¬
зованию первичных почвенных агрегатов (рис. 4).
Следующий этап в структурной организации осу¬
ществляют почвенные водоросли (табл. 5), синте¬
зирующие от 25 до 37% углерода и фиксирующие
атмосферный азот (3.6%). В черных криптогам¬
ных корках происходит аккумуляция Са и Mg орга¬
нического вещества [15]. Таким образом, механизм
формирования микроагрегатов в поверхностном
слое - биогенный и коагуляционный (округлые
и округло-угловатые агрегаты), обусловленный
гумусированностью плазмы. Последнее обязано
активной деятельности бактерий и водорослей.
В более глубоких слоях профиля преимуществен¬
ную роль в агрегации, очевидно, играет мерзлот¬
ный фактор.
Итак, пространственное разнообразие, ком¬
плексность почвенного покрова тундры являют¬
ся отражением не только сложной ледниково¬
морской истории формирования четвертичных
отложений и современного рельефа, развития
растительного покрова, особенностей ландшафт¬
но-экологической обстановки, но и динамичнос¬
ти природной среды, проявления криогенных
процессов.
Микроморфологические исследования пока¬
зали, что ведущими процессами почвообразова¬
ния, ответственными за формирование подбуров,
следует считать криогенную агрегацию массы,
Al-Fe-гумусовую миграцию, криотурбацию, мета¬
морфизм органического вещества.
Подбуры равнинных тундр на песчаных поро¬
дах отличаются от Al-Fe-гумусовых почв отсутст¬
вием гор. А2, аккумулятивно-иллювиальным типом
распределения тонкодисперсной массы, представ¬
ленной не только в виде пленок, но и коагуляцион¬
ных агрегатов. В отличие от дерновых [2,9], в этих
почвах формируются поверхностный грубогуму¬
совый гор. А0А1 и темно-бурый гор. Bh.
Таблица 5. Содержание С и N (%) в почвах пятен (бас¬
сейн р. Кары)
Тундра
Образец
С
N
C/N
Кустарничково-моховая
с дриадами:
а) водорослевая корка
Корка
37.18
3.60
10
б) водорослево-мохово¬
Корка
25.46
1.23
20
лишайниковая корка
Почва
2.90
0.21
14
в) заросшее пятно
Корка
10.95
0.51
21
Почва
2.57
0.15
17
8 ПОЧВОВЕДЕНИЕ № 6 1996
802
РУСАНОВА
Горно-тундровые подбуры на щебнисто-мел-
коземистых отложениях не имеют аналогов среди
равнинных почв. Согласно одной классификацион¬
ной схеме [9], они разделяются на примитивно-щеб-
нистые и криогенно-структурные комплексы, а
в других классификациях [2,12] эти почвы названы
подбурами. В отличие от равнинных, они недиф-
ференцированы, криогенно-оструктурены, щеб-
нисгы, с различным содержанием мелкозема в за¬
висимости от степени выветрелости. Наиболее
удачным для этих почв является предложенное
Таргульяном [12] название “ортоподбуры”.
Основными процессами в почвах, формирую¬
щихся в условиях затрудненного внутреннего дре¬
нажа, являются: аккумуляция грубого гумуса на
границе с минеральной толщей, криогенная пере¬
работка материала, оглеение.
6 качестве разграничительных диагностичес¬
ких микроморфологических признаков тундровых
глеевых почв с поверхносгно-глеевыми можно
назвать следующие: 1) гумусово-глинистые пленки
на зернах скелета, отсутствие агрегатов и конкре¬
ций в гор. G и Bg, свидетельствующие об усиле¬
нии криогенеза и оглеения; 2) уменьшение коли¬
чества натечных форм глинистого вещества в ни¬
жней части профиля, ослабление элювиально¬
иллювиальных процессов. Гумусовые педорелик-
ты встречаются повсеместно на северо-востоке
Русской равнины. Чтобы судить, являются ли они
диагностическими для типа тундровых глеевых
почв, достаточных литературных данных пока
нет. Подтипы тундровых глеевых почв выделяют¬
ся по особенностям формирования и состава орга-
ногенно-аккумулятивных горизонтов [2,6,7], сте¬
пени оглеения. Отличительными признаками пе-
регнойно-глеевой почвы являются органогенная
агрегация грубогумусового перегнойного горизон¬
та и ослабление оглеения минеральной толщи.
К почвам автономного ряда относятся также ос-
таточно-болотные мерзлотные перегнойно-тор¬
фяные реликтовых бугристых торфяников [7].
Тундровые болотные почвы (торфяно- и торфя-
нисто-глеевые) не отражают специфики тундро¬
вых процессов, кроме образования в них органо¬
генных агрегатов, диагностируемых на микро¬
уровне.
Самостоятельный тип почв пятен выделяет¬
ся [2] на основании изучения их морфологии, хи¬
мизма и состава почвенных растворов. Поскольку
формирование пятен связано с потерей биоген¬
но-активного слоя почвы, а образующаяся почва-
пленка резко отличается от исходной, выделение
самостоятельного типа представляется целесооб¬
разным. Преобладающая роль в восстановлении
почв пятен, образовании поверхностной почвен¬
ной корки, агрегации и обогащении гумусом при¬
надлежит биологическому фактору (пионерным
формам бактерий и водорослей). Первые стадии
преобразования грунта и становления почвы под
криптогамной коркой особенно отчетливо наблю¬
даются при изучении микростроения этих обра¬
зований.
СПИСОК ЛИТЕРАТУРЫ
1. Атлас Архангельской области. М.: ГУ ГК, 1976.
2. Васильевская В Д. Почвообразование в тундрах
Средней Сибири. М.: Наука, 1980.234 с.
3. Герасимова М.И., Губин С.В., Шоба С.А. Микро¬
морфология почв природных зон СССР. Пущино,
1992. 214 с.
4. Губин С.В. Динамика формирования структуры
тундровых мерзлотных неглеевых почв (тундровых
криоземов) // Почвоведение. 1993. № 10. С. 62-70.
5. Забоева И.В. Почвы и земельные ресурсы Коми
АССР. Сыктывкар: Коми кн. изд-во, 1975. 343 с.
6. Иванова Е.Н., Караваева Н.А., Забоева И.В., Тар-
гульян В.О. Основные подтипы тундровых глее¬
вых почв СССР // Биол. основы использования
природы Севера. Сыктывкар: Коми кн. изд-во,
1970. С. 94-99.
7. Игнатенко И.В. Почвы восточно-европейской тун¬
дры и лесотундры. М.: Наука, 1979.278 с.
8. Красовская Т.М. Пространственно-временные зако¬
номерности накопления тяжелых металлов в экоси¬
стемах Воркутинского промышленного района //
Биоиндикация природной среды Воркутинской тунд¬
ры. Сыктывкар. 1995.
9. Ливеровский Ю.А. Проблемы генезиса и геогра¬
фии почв. М.: Наука, 1987.247 с.
10. Полынцева О А. Почвы тундры и лесотундры вдоль
Печорской ж.д. (от ст. Абезь до ст. Воркута) //
Тр. Коми филиала АН СССР. Сер. геогр. 1952.
Вып. 1. С. 33-72.
11. Пьявченко Н.И. Бугристые торфяники. М.: Изд-во
АН СССР, 1955.278 с.
12. Таргульян В.О. Почвообразование и выветрива¬
ние в холодных гумидных областях. М.: Наука,
1971.267 с.
13. Толмачев А .И. Большеземел ьская тундра // Хозяй¬
ство Севера. 1931. № 5-6. С. 64-74.
14. Bolter М. Organic matter and its availability to microor¬
ganisms in Antarctic soils // Proc. Ith Intern. Conf. on
cryopedology. Pushchino, 1992. P. 189-198.
15. Sletten R.S., Ugolini F.C. Pedogenic evolution in polar
desert soils - the biotic factor // Abstr. Ith Intern. Conf.
on cryopedology. Pushchino, 1992. P. 55-56.
Micromorphology of East European Subarctic Soils
G. V. Rusanova
Major soil types of Bol’shezemel’skaya tundra northern part are described in terms of their genesis and micro-
morphology. Soil formation in followed at different stages of soil evolution - from a bare spot to a mature soil.
ПОЧВОВЕДЕНИЕ № 6 *1996
ПОЧВОВЕДЕНИЕ. 1996. № 6. с. 803-807
ЭРОЗИЯ
почв
УДК 631.431.1
ВЫНОС ПИТАТЕЛЬНЫХ ВЕЩЕСТВ ПОВЕРХНОСТНЫМ СТОКОМ
ИЗ ТИПИЧНОГО ЧЕРНОЗЕМА В ПОЧВОЗАЩИТНОМ СЕВООБОРОТЕ
© 1996 г. В. П. Веретельников, В. А. Рядовой
Сумское научно-производственное объединение "Элита”
Поступила в редакцию 31.10.95 г.
Рассматриваются объемы стока и количество теряемых почвой элементов питания растений в зави¬
симости от погодных условий, системы обработки, экспозиции склона, дозы вносимых удобрений.
Исследования показали, что в среднем ежегод¬
но с 1 га севооборотной площади терялось с по¬
верхностным стоком на зяби 174 м3 воды, S.4 т/га
почвы, 0.806 кг азота, 0.100 кг фосфора и 0.429 кг
калия, а на посевах озимой ржи - 118 м3, 0.596,
0.099 и 0.314 кг соответственно.
Водная эрозия способствует потере питатель¬
ных веществ не только самой почвы, но и вноси¬
мых удобрений [1,2].
В северо-восточной части Украины, в ближай¬
ших районах Курской и Белгородской обл. на¬
блюдается интенсивное проявление эрозии. В ле¬
состепной зоне Сумской обл. широко распростра¬
нены эродированные почвы, доля которых в
хозяйствах составляет 17.6%, а в отдельных райо¬
нах-до 30-40% [1]. В Центрально-Черноземном
экономическом районе, где около 57% пахотных
почв расположено на склонах различной крутиз¬
ны, проблема охраны их от водной эрозии стоит
особенно остро [4]. Поэтому, разрабатываемая
для этих регионов технология обработки почвы
должна прежде всего обеспечивать их высокую
противоэрозионную устойчивость. Этой цели от¬
вечает плоскорезная (безотвальная) обработка
почвы [2,3,6]. Однако данных о количественных
потерях азота, фосфора и калия вследствие вод¬
ной эрозии в условиях почвозащитного земледе¬
лия практически не имеется. Именно поэтому при
расчетах баланса питательных веществ в сево¬
оборотах эта статья расхода чаще всего не учиты¬
вается.
Начиная с 1976 г. на опытном поле Кировского
отделения Сумской с.-х. опытной станции НПО
“Элита” нами ведется длительный полевой опыт
по изучению различных уровней насыщенности
6-польных почвозащитных севооборотов мине¬
ральными и органическими удобрениями на агро¬
фонах, создаваемых обработками (отвальной
вспышкой, плоскорезным рыхлением и минималь¬
ной обработкой). Чтобы определить размер потерь
питательных веществ с поверхностным стоком во¬
ды на опытных делянках и стоковых площадках
вдоль склона на протяжении 150 м осенью напа¬
хивали валики с основанием 30-35 см.
Опытный участок расположен на склоне кру¬
тизной 3°-9° юго-восточной экспозиции. Почва -
чернозем типичный малогумусный среднесугли¬
нистый слабой и средней степени смытости.
Агрохимическая характеристика пахотного слоя
на стоковых площадках следующая: содержание
гумуса 3.7 и 3.3%, подвижного фосфора 9.2 и 8.7,
калия 6.6 и 6.0 мг/100 г почвы, рН^ 6.3 и 6.9.
Обработку почвы, посев и уход за растениями
проводили поперек склона. С 1978 по 1980 г. в од¬
ном севообороте на делянках опыта и стоковых
площадках высевали кукурузу на силос (гибрид Бу-
ковинский 3), затем в 1981 г. ячмень (Нутанс 244)
с подсевом травосмеси (клевер, люцерна, эспар¬
цет, кострец, овсяница). В последующие годы
следовала травосмесь первого-третьего года
пользования. Во втором севообороте вместо ку¬
курузы высевали озимую рожь. Под основную
обработку почвы при возделывании кукурузы
на силос вносили N120P90K60 и N180P120K90,
ячменя - N40P40K40 и N60P60K60, в подкормку
травосмеси первого-второго года пользования -
N30P30K30 и N45P45K45, третьего года -
N60P60K60 и N90P90K90. Аналогичные дозы
удобрений вносили и под культуры второго сево¬
оборота.
Смыв почвы определяли по Соболеву, плот¬
ность сложения-буром Качинского. Весной, перед
снеготаянием определяли запасы воды в снеге.
Пробы отбирали в начале стока и по его оконча¬
нии и анализировали на содержание водораствори¬
мых форм питательных веществ N-N03, N-NH4,
р2о5,к2о.
Размеры потерь питательных веществ из почвы
и внесенных удобрений определяются в основном
поверхностным стоком воды и смывом мелкозе¬
ма, объемы которых зависят в свою очередь от
запасов воды в снеге, характеристики склона,
промерзания и увлажнения почвы, применяе¬
мых противоэрозионных мероприятий, агрофона,
803
8*
804 ВЕРЕТЕЛЬНИКОВ, РЯДОВОЙ
Таблица 1. Поверхностный сток талых вод и смыв почвы
Ва¬
риант
Агрофон
Осадки за с.-х. год, мм
Средняя t° воздуха
Запасы
воды в
снеге
Объем
стока
Коэф¬
фициент
стока
Смыв
почвы,
т/га
м3/га
1978 г., перед посевом кукурузы на силос (*) и на посевах озимой ржи (**)
1
Зяблевая вспашка на 20-22 см
636
5.7
530*
530**
108*
75**
0.20*
0.14**
20.4*
Нет
2
Плоскорезное рыхление на 20-22 см
636
5.7
550*
550**
150*
161**
0.27*
0.29**
11.6*
Нет
3
Минимальная обработка
636
5.7
1979 г., перед посевом я*
550*
550**
шеня
178*
117**
0.32*
0.21**
9.0*
Нет
1
Зяблевая вспашка на 20-22 см
653
6.5
360
70
0.19
Не опр.
2
Плоскорезное рыхление на 20-22 см
653
6.5
380
169
0.44
0.5
3
Минимальная обработка
1980,
653
6.5
г., перед посевом кукуруз
450
1ы на силос
282
0.63
3.3
1
Зяблевая вспашка на 20-22 см
753
5.4
601
220
0.36
14.2
2
Плоскорезное рыхление на 20-22 см
753
5.4
627
254
0.40
16.9
3
Минимальная обработка
753
5.4
1981 г. перед посевом ян
627
меня
294
0.47
1.2
1
Зяблевая вспашка на 20-22 см
743
7.2
370
70
0.19
1.3
2
Плоскорезное рыхление на 20-22 см
743
7.22
380
162
0.42
1.4
3
Минимальная обработка
743
7.2
390
136
0.35
0.4
Среднее за 1978-И
)81 гг.
496
163
0.33
5.4
1
Зяблевая вспашка на 20-22 см
478
109
0.22
7.2
2
Плоскорезное рыхление на 20-22 см
497
179
0.36
6.1
3
Минимальная обработка
513
201
0.40
2.8
метеорологических условий снеготаяния и других
факторов водной эрозии [4].
Гидрометеорологические условия формирова¬
ния стока талых вод по годам были неодинаковы¬
ми. Наиболее влажным и прохладным был 1980 г.
(табл. 1). Количество осадков составило 753 мм за
год (среднее многолетнее 508 мм) при средней тем¬
пературе воздуха 5.4°С (средняя многолетняя 6°С).
Почва под зиму ушла увлажненной и непромерз¬
шей. На месяц позже наступило похолодание и
замерзание почвы (14.12) и спустя два дня устано¬
вился устойчивый снежный покров (средняя мно¬
голетняя дата 01.12), который всю зиму лежал ус¬
тойчиво (средняя максимальная высота до 40 см).
К концу зимы почва промерзла на глубину 47 см
(максимальная многолетняя глубина 160 см).
Устойчивый переход температуры воздуха через
0° произошел 28 марта, т.е. в обычные сроки.
ПОЧВОВЕДЕНИЕ № 6 1996
ВЫНОС ПИТАТЕЛЬНЫХ ВЕЩЕСТВ ПОВЕРХНОСТНЫМ СТОКОМ
805
Перед весенним снеготаянием запасы воды в снеге
на стоковых площадках достигали 601-627 м3/га.
Поскольку потепление наступало очень медлен¬
но, период стока талых вод был растянутым.
Снег, пролежавший на делянках почти 4 месяца
сошел 07.04. В сложившихся условиях объем сто¬
ка на вспашке, плоскорезном рыхлении и мини¬
мальной обработке составил 220, 254 и 294 м3/га,
а смыв почвы соответственно 14.2, 16.9 и 1.2 т/га.
В следующем 1980/81 с.-х. году по сравнению
с 1979/80 с.-х. годом осадков выпало на 10 мм
меньше, а средняя температура воздуха повыси¬
лась на 2.2°. Устойчивый переход среднесуточных
температур от положительных к отрицательным
и одновременное замерзание почвы произошли
на 10 дней раньше обычного срока (01.11). В этот
период образовался первый снежный покров.
Зима была теплой. Среднезимняя температура
воздуха была -3.3°С, что на 3.7° выше среднемно¬
голетней. Из 90 календарных дней зимы 45 бы¬
ло с оттепелями, максимальная температура возду¬
ха в отдельные дни повышалась до +4°С. В услови¬
ях положительных температур и выпадения жидких
осадков снег на полях таял и снежный покров во¬
зобновлялся 4 раза (01.11, 05.01, 19.01, 09.02). Для
этого периода было характерно обилие осадков
(в основном, за счет жидких): 180.5 мм или 205%:
нормы. Запасы воды в снеге перед каждым снего¬
таянием на делянках с обработками достигали
370-390 м3/га. По вспашке, плоскорезному рых¬
лению и минимальной обработке объем стока со-*
ставил соответственно 70, 162 и 136 м3/га, а смыв
почвы - 0.4—1.4 т/га, т.е. в пределах допустимого.
В 1977/78 с.-х. году почва замерзла на 15 дней
позже обычного срока (29.11). Тогда же образовал¬
ся устойчивый снежный покров высотой до 32 см.
Осадки выпадали часто, но в небольшом количе¬
стве. Потепление, начавшееся в конце февраля,
в марте постепенно усиливалось, среднесуточная
температура воздуха в отдельные дни второй де¬
кады марта достигала +5 ... +8°С. Снежный по¬
кров сошел 28.03 и началось интенсивное оттаи¬
вание почвы, и к 08.04 она оттаяла на всю глубину
промерзания (85 см). Запасы воды в снеге перед
снеготаянием достигали 530-550 м3/га, объем
стока по вспашке, плоскорезной и минимальной
обработкам составил 107, 150 и 178 м3/га, смыв
почвы соответственно 20.4, 11.6 и 9.0 т/га.
В 1978/79 с.-х. году замерзание почвы про¬
изошло на 20 дней позже обычного, тогда же
(03-04.12) образовался и устойчивый снежный
покров. Вплоть до середины третьей декады ян¬
варя удерживалась холодная погода. С 26.01 тем¬
пература воздуха резко повысилась, достигая
днем +4 ... +5°С. Снег почти полностью сошел к
концу третьей декады января. С возвратом холодов
в первой декаде февраля на почве образовалась
ледяная корка толщиной 4-5 см. В целом зима бы¬
ли снежной, осадков выпало 206% нормы. К концу
зимы почва промерзла на глубину 70-76 см. Пол¬
ностью снег сошел 24.03. Запасы воды в снеге
были небольшими (360-450 м3/га), объем стока
по обработкам колебался в значительных преде¬
лах (70-282 м3/га). Поверхностный сток талых
вод по отвальной вспашке составил 70 м3/га, по
плоскорезной и минимальной обработкам повы¬
сился соответственно на 98 и 212 м3/га (табл. 1).
Результаты 4-летних (1978-1981 гг.) наблюде¬
ний на стоковых площадках показали, что даже
в условиях почвозащитного севооборота и проти-
воэрозионной агротехники с 1 га склоновых зе¬
мель в среднем по обработкам терялось 163 м3/га
или 33% от общего запасы воды в снеге и 5.4 т/га
почвы. В условиях сильного проявления эрозии
(1980 г.) поверхностный сток талых вод по мини¬
мальной обработке достигал 294 м3/га или 47% от
запасов воды в снеге, а смыв почвы - 1.2 т/га.
Потеря влаги в размере 163-294 м3/га приводит
к недобору 1.5-3.0 ц/га зерна. В зависимости от
объема стока и смыва почвы колебались по го¬
дам и размеры потерь азота в нитратной и амми¬
ачной формах, подвижных форм фосфора и ка¬
лия (табл. 2). Максимальные потери питательных
веществ (кг/га) с водой отмечены в 1980 г. по мини¬
мальной обработке: нитратной и аммиачной форм
азота соответственно 0.970 и 0.441 кг/га, фосфора
0.323 и калия 0.794 кг/га. В среднем за 1978, 1979,
1981 гг. вследствие меньшего объема стока поте¬
ри питательных веществ на зяби были примерно
в 1.2-1.3 раза ниже, чем в 1980 г., и составили
(по агрофонам): азота 0.654, фосфора 0.097 и ка¬
лия 0.334 кг/га. В 1978 г. по плоскорезной и мини¬
мальной обработкам по сравнению со вспашкой
на посевах озимой ржи отмечены аналогичные
концентрации нитратного и аммиачного азота,
подвижных форм фосфора и калия в поверхност¬
ном стоке талых вод. В среднем с продуктами
водной эрозии с 1 га севооборотной площади те¬
рялось (кг) перед посевом кукурузы на силос нит¬
ратного и аммиачного азота 0.663 и 0.322, фосфо¬
ра 0.100 и калия 0.462 соответственно, а на посе¬
вах озимой ржи - 0.413 и 0.183, 0.099 и 0.314.
Несмотря на незначительные различия в объ¬
емах стока, на удобренных стоковых площадках в
1978 г. содержание питательных веществ было
практически одинаковым, а в 1979 г. оно было
больше там, где вносили полуторную дозу мине¬
ральных удобрений (табл. 2). Большая концент¬
рация N03, NHj,, Р205, К20 в поверхностном стоке
талых вод и несколько ббльшие величины их по¬
тери в течение периода стока из почвы, в кото¬
рую вносили полуторную дозу минеральных удо¬
брений, служат еще одним доказательством того,
что в результате водной эрозии теряются пита¬
тельные вещества не только почвы, но и вноси¬
мых удобрений. Наши наблюдения показали, что
за период снеготаяния содержание N03, NH4,
ПОЧВОВЕДЕНИЕ № 6 1996
806 ВЕРЕТЕЛЬНИКОВ, РЯДОВОЙ
Таблица 2. Концентрация питательных веществ в поверхностном стоке вод, мг/л
Вариант
N03
nh4
Р205
k2o
На зяби перед посевом кукурузы на силос, среднее за 1978,1980 гг.
Отвальная вспашка:
N120P90K60
1.3
6.2
2.5
3.1
m |cn
old
1.5
1.3
N180P120K90
1.4
9.9
1.0
1.4
0.4
0.3
1.4
1.6
Плоскорезное рыхление:
N120P90K60
1.0
1.9
1.9
2.1
PIP
u>l
1.9
1.9
N180P120K90
1.0
4.9
1.6
0.7
0.8
0.3
2.8
3.0
Минимальная обработка:
N120P90K60
1.0
4.9
1.0
1.5
1.4
0.5
3.1
2.3
N180P120K90
1.0
5.6
1.0
0.7
0.8
0.5
3.1
3.4
НСР0$ для удобрений
1.1
0.3
0.2
0.2
НСР05 для обработки почвы
1.3
0.4
0.2
0.3
На посевах озимой ржи за 1978 г.
Отвальная вспашка:
N120P90K60
3.1
4.2
1.1
1.1
0.4
1.0
1.3
2.8
N180P120K90
2.7
5.6
0.7
1.7
0.6
0.2
1.4
1.5
Плоскорезное рыхление:
N120P90K60
2.0
5.1
1.0
1.0
0.9
0.3
1.9
2.2
N180P120K90
2.1
6.3
1.1
2.3
1.1
0.6
2.7
2.8
Минимальная обработка:
1.0
4.6
1.5
2.2
1.1
0.5
OIO
cn lc«*i
N180P120K90
0.5
4.8
1.6
3.0
2.6
0.4
4.3
4.2
HCPqj для удобрений
0.4
0.3
0.4
0.4
HCPqs для обработки почвы
0.6
0.3
0.6
0.6
Примечание. Над чертой - начало стока, под чертой - его окончание.
Р205, К20 в стоке непостоянно. Особенно четко
эти различия по обработкам выражены по азоту
на зяби и на посевах озимой ржи. Так, при весен¬
нем снеготаянии на разных агрофонах (зябь и по¬
севы озимой ржи) потери азота на зяби были ни¬
же на 0.085 кг/га по плоскорезной обработке и
выше на 0.152 кг/га по минимальной, а на посевах
озимой ржи они были выше на 0.470 и 0.179 кг/га
по плоскорезной и минимальной обработкам в
сравнении с потерями по отвальной вспашке
(0.784 кг/га на зяби и 0.383 кг/га на посевах ози¬
мой ржи). Менее четко эти зависимости просле¬
живаются с фосфором и калием. Наибольшие по¬
тери подвижных форм фосфора и калия наблю¬
дали за весь период поверхностного стока как на
зяби (0.178 и 0.669 кг/га), так и на посевах озимой
ПОЧВОВЕДЕНИЕ № 6 1996
ВЫНОС ПИТАТЕЛЬНЫХ ВЕЩЕСТВ ПОВЕРХНОСТНЫМ СТОКОМ
807
ржи (0.140 и 0.421 кг/га) по минимальной обработ¬
ке по сравнению с другими обработками, что объ¬
ясняется заделкой вносимых удобрений в верх¬
нюю часть (0-10 см) пахотного слоя и последую¬
щим их вымыванием с поверхностным стоком.
Как показали наши данные, в среднем за
1978-1981 гг. потери почвы на зяби с поверхност¬
ным стоком талых вод составили 5.4 т/га, мине¬
рального азота - 0.806, фосфора 0.100 и калия
0.429 кг/га, а на посевах озимой ржи они умень¬
шались по азоту и калию соответственно на 0.210
и 0.115 кг/га. Потери фосфора как на зяби, так и
на посевах озимой ржи были практически одина¬
ковыми (0.099-0.100 кг/га).
ВЫВОДЫ
1. В среднем по обработкам с 1 га чернозема
типичного на склоне юго-восточной экспозиции
в среднем ежегодно терялось с поверхностным
стоком на зяби 174 м3 воды (36% общего запаса
воды в снеге) и 5.4 т почвы, а на посевах озимой
ржи -118 м3 (22% от общего запаса воды в снеге).
2. На зяби наибольшие потери азота отмечены
по отвальной вспашке (0.784 кг/га) и минималь¬
ной обработке (0.936 кг/га), меньше - по плоско¬
резной обработке (0.699 кг/га). На посевах ози¬
мой ржи больше всего азота терялось по плоско¬
резной обработке (0.853 кг/га), меньше - по
минимальной (0.562 кг/га) и наименьшее количе¬
ство - по отвальной вспашке (0.383 кг/га).
3. Наибольшие потери Р205 (0.140-0.178 кг/га)
и К20 (0.314-0.446 кг/га) отмечались на обоих аг¬
рофонах по минимальной обработке.
4. Потери питательных веществ на посевах
озимой ржи уменьшались на 0.326 кг/га по срав¬
нению с их потерями на зяби (1.335 кг/га).
СПИСОК ЛИТЕРАТУРЫ
1. Веретельников В.П. и др. Улучшение гумусового
состояния эродированного чернозема // Химиза¬
ция сельского хозяйства. 1991. № 8. С. 77-80.
2. Каштанов А.Н. Основные направления дальней¬
шего развития почвозащитного земледелия в
СССР//Земледелие. 1989. № 2. С. 2-7.
3. Миронов Г.И. На склонах лесостепи Украины //
Земледелие. 1984. № 3. С. 36-37.
4. Рындыч Л.П., Явтушенко В.Е. Смыв питатель¬
ных веществ из выщелоченного чернозема в поч¬
возащитном севообороте // Агрохимия. 1987. № 4.
С. 117-123.
5. Рядовой В.А. и др. Влияние плотности сложения
чернозема на сток и смыв питательных веществ
при его различных обработках // Почвоведение.
1990. № з. с. 147-152.
6. Шикула Н.К. Земледелие без плуга // Земледелие.
1983. № 11. С. 5-9.
7. Юркин С.Н., Макаров Н.Б., Пименов Е.А. Потери
азота, фосфора и калия из почвы и удобрений с по¬
верхностным стоком // Агрохимия. 1978. № 11.
С. 133-141.
Nutrients Removal from Typical Chernozem
in Soil-protecting Crop Rotation
V. P. Veretelnikov, V. A. Ryadovoi
Under examination are runoff volume and soil loss of nutrients depending on the weather conditions, soil man¬
agement options, slope exposition and the rates of fertilizers applied.
ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВОВЕДЕНИЕ, 1996, № 6, с. 808-809
ХРОНИКА
УДК 631.4
ЮБИЛЕЙ ПРОФЕССОРА ИЛЬЯСА МАМЕДОВИЧА ГАДЖИЕВА
14 ноября 199S г. исполнилось 60 лет видному
почвоведу директору Института почвоведения и аг¬
рохимии СО РАН Ильясу Мамедовичу Гаджиеву.
Выпускник Горского сельскохозяйственного
института (Владикавказ), Alma Mater таких изве¬
стных почвоведов, как С.В. Зонн, Р.В. Ковалев,
Е.В. Рубилин, Ильяс Мамедович связал свою
судьбу с проф. Р.В. Ковалевым и с Сибирью.
В конце 50-х гг. Роман Викторович Ковалев был
заведующим отделом географии и генезиса почв
Института почвоведения и агрохимии АН Азер¬
байджана. Ему предложили организовать акаде¬
мический почвенный институт СО РАН. Р.В. Ко¬
валев взял на себя эту трудную миссию - объеди¬
нить почвоведов Сибири. И благодаря своему
молодому коллективу, “кавказскому десанту”, пе¬
реехавшему вместе с ним в Новосибирск, он достиг
своей цели. Среди первопроходцев был и Ильяс
Мамедович. Он был заместителем Ковалева,
т.е. тем человеком, на котором лежала вся органи¬
зационно-административная ответственность за
выполнение созидательных планов. И.М. Гаджи-
ев с честью оправдал все возлагавшиеся на него
надежды. Начиная с Западно-Сибирского сове¬
щания с экскурсией (1967 г.) и кончая последним
Съездом Всесоюзного общества почвоведов
(1989 г.) - все совещания в Сибири проходили под
знаменем Института почвоведения и агрохимии и
были прекрасно организованы в первую очередь
благодаря И.М. Гаджиеву. Из почвенного отдела
Биологического института вырос мощный инсти¬
тут, где сплав мудрости и молодости дал самый
прекрасный урожай - урожай мысли. Институт
расширил географию исследований: кроме тради¬
ционных Кулунды и Барабы исследованиями были
охвачены Васюганье, Салаир, Горный Алтай, Куз¬
басс. Именно в ИПА начались глубокие работы
по рекультивации нарушенных ландшафтов.
Институт взялся за составление разномас¬
штабных карт Сибири. Были организованы ста¬
ционарные исследования. Расширилась издатель¬
ская деятельность института. Ильяс Мамедович
нес основную нагрузку как заместитель директо¬
ра при проведении в жизнь всех планов институ¬
та. Рос институт, защищались молодые доктора,
многим их них становились тесными стены род¬
ного института и они переезжали в другие науч¬
ные организации Сибири и смежных регионов.
ИПА по праву стал кузницей кадров для Сибири
и Дальнего Востока. И сегодня Ученый совет
ИПА, возглавляемый И.М. Гаджиевым, - один из
самых авторитетных в России. Результаты рабо¬
ты института впечатляют. Когда-то окрашенная
в 5-6 цветов почвенная карта Сибири заиграла
множеством цветов и оттенков. Были выделены
совершенно новые типы почв. Вскрылись особые
закономерности формирования почвенного покро¬
ва Сибири. И.М. Гаджиев был пионером этого но¬
вого почвоведения. Он поставил почвоведение на
строгую историческую, эволюционную основу.
Он показал, что современный почвенный покров
образовался из покрова предыдущей стадии эво¬
люции биосферы. Современные почвы формиру¬
ются не из материнских (почвообразующих) по¬
род. Почвообразующие породы становятся уже
косвенным фактором почвообразования, они бук¬
вально отходят в прошлое, а современные почвы
формируются из ранее развивавшихся в данном ре¬
гионе почв предыдущей ландшафтной эпохи.
Идея глобальной и региональной эволюций поч¬
венного покрова оказалась именно той теорети¬
ческой базой, которая позволила по-новому
взглянуть на почвы и оценить их свойства и фор¬
мирование, особенно в таком сложном регионе,
как Сибирь.
Географо-генетический подход к эволюции
почв И.М. Гаджиев сопровождал глубокими ис¬
следованиями самого механизма эволюции почв.
Были заложены специальные опыты с переносом
разных типов почв в несвойственные им условия
почвообразования. Эти опыты длятся уже около
30 лет и дают очень многое для понимания поч¬
венных процессов.
Вместе со своей сотрудницей, д. б. н. М.И. Дер-
гачевой И.М. Гаджиев выдвинул одну из самых
оригинальных новых гипотез - о матричном вос¬
производстве гумуса в почвах. В этой гипотезе
скрыт ключ для понимания эволюции почв, пре¬
вращения гумуса, формирования почвенных
агрегатов. В настоящее время институт Почвове¬
дения и агрохимии занят обобщением всех матери¬
алов по почвам Сибири. Это дело требует большо¬
го опыта, мудрости, оригинальности взглядов
и отсутствия предвзятости к полученным данным.
И мы уверены, что институт обладает громадным
научным потенциалом, который позволит ему до¬
вести начатое дело до успешного окончания.
С 1986 г., когда И.М. Гаджиев сменил на директор¬
ском посту Р.В. Ковалева, он еще более упрочил
авторитет ИПА. Именно в это время был проведен
Съезд Всесоюзного общества почвоведов, а Ильяс
Мамедович был избран его вице-президентом.
808
ЮБИЛЕЙ ПРОФЕССОРА ИЛЬЯСА МАМЕДОВИЧА ГАДЖИЕВА
809
Он член редколлегии журнала “Почвоведение”.
Он умело направляет работу своих сотрудников
на решение актуальных задач почвоведения. Одна
из самых прекрасных черт И.М. Гаджиева - умение
привлекать к себе прекрасных специалистов, про¬
фессионалов. Он периодически приглашает в
ИПА почвоведов самого разного профиля для
чтения лекций. На заседаниях Ученого совета ин¬
ститута заслушивают самые злободневные тео¬
ретические работы. Институт связан в своих ра¬
ботах с учеными Омска, Томска, Барнаула, и во
многом благодаря ему традиционные научные
школы в этих городах продолжают существовать
и радовать нас своими научными достижениями.
Расширились международные связи ИПА: рос-
сийско-японские, российско-германские, россий¬
ско-монгольские и др. И в большинстве этих ра¬
бот И.М. Гаджиев принимает непосредственное
участие.
Отмечая 60-летие Ильяса Мамедовича, позд¬
равляем юбиляра и желаем ему творческих и ор¬
ганизаторских успехов, новых монографий, ус¬
пешного завершения многолетних трудов по поч¬
вам Сибири.
Институт почвоведения и агрохимии СО РАН
Общество почвоведов при РАН
и его Новосибирское отделение
Редколлегия журнала “Почвоведение”
ПОЧВОВЕДЕНИЕ № б 1996
ПОЧВОВЕДЕНИЕ, 1996, № 6, с. 810-811
ХРОНИКА
УДК 631.4
ФАНГАТ ХАМАТОВИЧ ХАЗИЕВ
(К 60-ЛЕТИЮ СО ДНЯ РОЖДЕНИЯ
И 35-ЛЕТИЮ НАУЧНОЙ ДЕЯТЕЛЬНОСТИ)
Фангат Хаматович Хазиев известен широкому
кругу почвоведов своими фундаментальными
трудами в области почвенной энзимологии и эко¬
логии почв.
Он родился 26 апреля 1936 г. в деревне
“Буляк”, расположенной на границе лесов и сте¬
пей Башкирии, на берегу чистой и быстрой реч¬
ки. Красота родной природы и определила его на¬
учные интересы, связанные с экологией почв.
По окончании биолого-почвенного факульте¬
та Казанского государственного университета
в 1960 г. он работал в почвенной экспедиции Уп¬
равления сельского хозяйства Пермской обл., где
получил большие навыки почвоведа-полевика.
Позже, на кафедре почвоведения Башкирского
сельскохозяйственного института и в лаборато¬
рии почвоведения Института биологии Башкир¬
ского филиала АН СССР, он увлекся почвенной
энзимологией. Его учитель - профессор М.Н. Бу-
рангулова помогла выявить в молодом ученом
особенности хорошего методиста-аналитика и
Ф.Х. Хазиевым были разработаны принципиаль¬
ные положения и методы изучения почвенных
ферментов и впервые в стране изданы научно-ме-
тодические руководства (Ферментативная актив¬
ность почв. М.: Наука, 1976; Методы почвенной
энзимологии. М.: Наука, 1990).
Целеустремленные исследования, начатые во
время обучения в аспирантуре (1963-1966 гг.),
кандидатская работа “Активность фосфогидро-
лаз и динамика фосфорорганических соединений
в почвах Башкирии” (1966 г.), многолетняя работа
в качестве старшего научного сотрудника (с 1970 г.)
и заведующего лабораторией (с 1977 г.) получили
свое развитие и логическое завершение в доктор¬
ской диссертации “Системно-экологический ана¬
лиз ферментативной активности почв” (1983 г.).
Ф.Х. Хазиев разработал эколого-генетический
подход в изучении ферментативной активности
почв; он выдвинул и обосновал системно-эколо¬
гическую концепцию формирования и функцио¬
нирования ферментативного потенциала почв,
которая признана в качестве научной основы но¬
вого направления в биохимии почвы - экологиче-
• ской почвенной энзимологии.
Ф.Х. Хазиев внес большой вклад в изучение
почв Башкирии, им проведены комплексные ис¬
следования изменений почв при интенсификации
земледелия, в результате эрозии, мелиорации, за¬
грязнения нефтью, по характеристике биологи¬
ческой активности и гумусного состояния почв,
экологической оптимизации агроэкосистем и т.д.
По его инициативе разработана и реализуется
комплексная программа повышения плодородия
почв Башкирии.
Ф.Х. Хазиев - автор и соавтор ряда фундамен¬
тальных работ по почвам Башкортостана: “Чер¬
ноземы Башкирии” (1969), “Почвы Башкирии”
(1975), “Морфогенетическая и агропроизводст-
венная характеристика почв Башкирской АССР”
(1985), “Органическое вещество почв Башкирии”
(1991) и др., которые являются научной основой
рационального использования почв республики,
воспроизводства их плодородия. Им опубликова¬
но более 300 научных работ, в том числе 17 моно¬
графий, 8 работ в зарубежных изданиях, под его
редакцией издано 17 монографий и сборников, он
соавтор 9 изобретений по ингибиторам нитрифи¬
кации азота в почве и биологической рекультива¬
ции нефтезагрязненных почв.
Организаторский талант и глубокие знания
почв республики позволили Ф.Х. Хазиеву объе¬
динить ученых-почвоведов различных учрежде¬
ний в авторский коллектив для написания фунда¬
ментальной двухтомной монографии “Почвы
Башкортостана”, первый том которой “Эколого¬
генетическая и агропроизводственная характери¬
стика почв” вышел в 1996 г.
Ф.Х. Хазиев - опытный организатор науки, за¬
ведующий лабораторией почвоведения, работал
заместителем директора Института биологии,
в составе оргкомитетов принимал участие в про¬
ведении ряда всесоюзных и региональных конфе¬
ренций, им создана научная школа по изучению
почвенных ферментов, под его научным руковод¬
ством подготрвлены один доктор наук, 12 кандида¬
тов наук. Он принимает участие в педагогической
работе в Башкирском государственном универси¬
тете. Ф.Х. Хазиев активно участвует в подготов¬
ке кадров, выступая в качестве официального оп¬
понента кандидатских и докторских диссертаций
(МГУ, ТСХА, Институт почвоведения и агрохи¬
мии СО РАН, Воронежский университет и др.).
В его лаборатории проходят стажировку по освое¬
нию методов почвенной энзимологии аспиранты и
810
ФАНГАТ ХАМАТОВИЧ ХАЗИЕВ
811
научные работника из различных научно-иссле¬
довательских институтов и вузов России.
Ф.Х. Хазиев ведет большую научно-общест-
венную работу: он - член Научного совета РАН
по проблемам почвоведения, член Центрального
совета и председатель Башкирского отделения
Общества почвоведов, член Международного об¬
щества почвоведов, и.о. академика-секретаря
Отделения биологических наук и председатель
Научного совета АН Республики Башкортостан,
член редколлегии журнала “Почвоведение”.
За заслуги в развитии науки Ф.Х. Хазиеву при¬
своено почетное звание “Заслуженный деятель
науки Башкирии”. Он награжден почетными гра¬
мотами Президиума Верховного Совета Башки¬
рии, Президиумов АН СССР и БНЦ РАН, медаля¬
ми быв. ВДНХ. Он лауреат премии им. акад.
В.Р. Вильямса за работы в области почвенной эн-
зимологии.
Поздравляя Фангата Хаматовича с 60-летием,
коллеги, друзья и ученики юбиляра желают ему
дальнейших творческих свершений на благо
нашей науки.
Отделение биологических наук
АН Башкортостана
Институт биологии УНЦ РАН
Башкирское отделение
Общества почвоведов при РАН
Редколлегия журнала “Почвоведение”
Благодарность
Не имея возможности ответить лично всем, поздравившим меня с 70-летием научной деятельности
и 90-летием со дня рождения, я через журнал приношу свою искреннюю благодарность за внимание,
уважение и память.
Искренне Ваш С. Зонн
ПОЧВОВЕДЕНИЕ № б 1996
ПОЧВОВЕДЕНИЕ, 1996, № 6, с. 812-813
УДК 631.4
ХРОНИКА
МЕЖДУНАРОДНЫЙ симпозиум
“ЗАСОЛЕНИЕ ЛАГУННЫХ ЭКОСИСТЕМ”
© 1996 г. Е. И. Панкова, И. Н. Любимова
Почвенный институт им. В.В. Докучаева, РАСХН
Поступила в редакцию 25.12.95 г.
Международный симпозиум проходил с 18 по
25 сентября 1995 г. в г. Валенсия (Испания). Орга¬
низаторами симпозиума были Университет г. Ва¬
ленсии, Estudi General и подкомиссия засоленных
почв Международного общества почвоведов.
В работе симпозиума приняли участие 129 деле¬
гатов из 30 стран: Австралии, Австрии, Аргенти¬
ны, Болгарии, Бразилии, Венгрии, Германии,
Египта, Индонезии, Ирака, Ирана, Испании, Ита¬
лии, Кении, Китая, Колумбии, Коста-Рики, Мекси¬
ки, Нидерландов, Пакистана, Португалии, Рос¬
сии, Румынии, США, Таиланда, Узбекистана, Фи¬
липпин, Франции, Швеции, Югославии, а также
ФАО, ЮНЭПа и генеральный секретарь ISSS.
Торжественное открытие симпозиума состоялось
18 сентября в зале заседаний Университета Ва¬
ленсии - старейшего в Испании. На открытии
присутствовали представители Правительства
страны, города Валенсии, международных орга¬
низаций (FAO, UNEP, ISSS), а также руководство
университета.
На пленарном заседании было заслушано 6 до¬
кладов. Первый доклад председателя подкомис¬
сии М. Редли (Redly, Венгрия) был посвящен ана¬
лизу 30-летней работы подкомиссии засоленных
почв. Она рассмотрела истоки создания подко¬
миссии, показала научную и практическую значи¬
мость работ по изучению засоленных почв, на¬
звала основные направления и результаты иссле¬
дований, дала информацию о международных
симпозиумах, которые проводились в разных
странах по темам, представляющим большой ин¬
терес для страны, организующей симпозиум, и
для широкой научной аудитории. Всего с 1964 по
1992 г. было проведено 12 заседаний. Докладчик
подчеркнула огромную роль проф. И.Сабольча в
работе подкомиссии. Два пленарных выступления
были посвящены теоретическим вопросам почво¬
ведения. Это доклад проф., докт. В.Блюма (Blum,
Австрия) на тему “Soil Resilience” и проф., докт.
В.О. Таргульяна (Россия) на тему “Soil as Recording
System: Recent and Inherithed Soil Memory”. Три сооб¬
щения были связаны с основной проблемой симпо¬
зиума - засолением почв лагун. Проф., докт. И. Со-
больч (Венгрия) обратил внимание на разнообра¬
зие засоления почв лагунных экосистем мира;
докт. С. Арунин (Arunin, Таиланд) говорила о за¬
солении почв побережья Юго-Восточной Азии; ру¬
ководитель симпозиума проф., докт. Дж. Батлле-
Салеса (Batlle-Sales, Испания) осветил особенности
засоления почв лагунных экосистем Испании.
На симпозиуме было заслушано 53 устных
и 45 стендовых докладов. Тезисы их были опуб¬
ликованы до начала работы симпозиума и вруче¬
ны всем участникам. Полный текст выступлений
планируется издать после завершения работы
симпозиума. Доклады были посвящены трем ос¬
новным проблемам: 1) изучению засоленных
почв лагун, их генезису, экологии, использованию;
2) засоленным почвам других экосистем, включая
вопросы генезиса, классификации, картирования и
мелиорации; 3) проблеме использования минера¬
лизованных вод для орошения.
Засолению почв лагун было уделено значи¬
тельное внимание в работе симпозиума. Большой
интерес представляли сообщения, характеризую¬
щие особенности и генезис кислых солончаков
морских побережий, которые долгое время не
включали в разряд засоленных почв, но которые,
как показали исследования последних лет, имеют
широкое распространение в разных странах и
в разных природных зонах.
Многие выступающие рассматривали почвы
лагун и прибрежных экосистем с хлоридным,
сульфатным и содовым засолением. Как показа¬
ли заслушанные доклады, анализирующие эколо¬
гические условия формирования этих почв, причи¬
ны их разнообразия и прогноз эволюции лагунных
экосистем, эти проблемы требуют дальнейшего
изучения, особенно в связи с освоением и исполь¬
зованием этих почв.
Большой интерес представили доклады, по¬
священные солеустойчивости культур и рацио¬
нальному использованию засоленных земель. Дис¬
куссию вызвали выступления, касающиеся исполь¬
зования морских вод для орошения рендзин.
На заключительном заседании были подведе¬
ны научные итоги симпозиума, дан анализ стендо¬
вых докладов и намечены планы дальнейших ра¬
бот подкомиссии. Следующий симпозиум планиру¬
ется провести в сентября 1997 г. в Каире (Египет).
Была также дана информация о проведении
812
МЕЖДУНАРОДНЫЙ СИМПОЗИУМ “ЗАСОЛЕНИЕ ЛАГУННЫХ ЭКОСИСТЕМ”
813
международных рабочих заседаний, которые бу¬
дут проходить в 1995-1996 гг. по проектам, фи¬
нансируемым UNEP.
В ходе работы симпозиума и после его заверше¬
ния были организованы полевые экскурсии, кото¬
рые позволили его участникам увидеть засоленные
почвы лагун, районы распространения гипсонос¬
ных почв и пород и районы освоенных засоленных
почв. Первая экскурсия (20 сентября) включала
знакомство с ландшафтами и почвами прибрежной
зоны национального парка La Albufera, а также по¬
сещение рисовых полей. Трехдневная послесим-
позиумная экскурсия проходила с 23 по 25 сентя¬
бря. Экскурсанты ознакомились с торфяными за¬
соленными почвами побережий; освоенными
почвами лагунных экосистем, используемыми
в настоящее время под сады и виноградники, и
с зоной распространения древних гипсоносных
пород. Обсуждались проблемы проявления вто¬
ричного засоления на орошаемых землях под са¬
довыми культурами.
Участники симпозиума выразили глубокую
благодарность его организаторам и в первую оче¬
редь проф., докт. Дж. Батлле-Салесу, который
создал доброжелательную и рабочую обстановку
для научного общения людей, имеющих общие
научные интересы.
ПОЧВОВЕДЕНИЕ № 6 1996
ПОЧВОВЕДЕНИЕ, 1996, № 6, с. 814-816
РЕЦЕНЗИИ
УДК 634.0.12:634.0.114
Е.С. МИГУНОВА. ЛЕСА И ЛЕСНЫЕ ЗЕМЛИ
(КОЛИЧЕСТВЕННАЯ ОЦЕНКА ВЗАИМОСВЯЗЕЙ).
М.: ЭКОЛОГИЯ, 1993.
Е.С. МИГУНОВА. ЛЕСОВОДСТВО И ПОЧВОВЕДЕНИЕ
(ИСТОРИЧЕСКИЕ ОЧЕРКИ).
М.: ЭКОЛОГИЯ, 1994
© 1996 г. Д. Г. Тихоненко
Харьковский аграрный университет им. В.В. Докучаева
Поступила в редакцию 17.05.95 г.
В издательстве “Экология” вышли в свет две
книги доктора сельскохозяйственных наук Е.С. Ми-
гуновой. Первая посвящена исследованиям взаимо¬
связей леса и почв, вторая - анализу взаимодейст¬
вия наук, изучающих их, - лесоводства и почвове¬
дения. Автор многие годы изучал взаимосвязи леса
и почв и шире - леса и лесных земель (с учетом
почвообразующих, подстилающих пород и грун¬
товых вод), в том числе историю этого вопроса
и его развитие почвоведами и лесоводами.
Исследования, результаты которых обобще¬
ны в книгах, проводились не традиционными ме¬
тодами, а на основе разработок школы лесной ти¬
пологии, созданной ближайшими соратниками
В.В. Докучаева - Г.Ф. Морозовым и Г.Н. Высоц¬
ким. Ее основными принципами являются призна¬
ние полной обусловленности характера лесных на¬
саждений в пределах однородного по климату реги¬
она особенностями почвенно-грунтовых условий
и использование сопряженной классификации почв
и насаждений - эдафической сетки, - построен¬
ной в координатах основных факторов плодоро¬
дия почв: уровня обеспеченности элементами пи¬
тания (трофности) и уровня водообеспеченности.
Однако разделение почв на типы в данной класси¬
фикации производится не по этим “внутренним”
их свойствам, а по изменению состава, структуры
и продуктивности произрастающих на них насаж¬
дений, по наличию в последних растений разных
экологических групп - олиго-, мезо- или мегатро-
фов; ксеро-, мезо- или гигрофитов, т.е. по типу
леса. Почвы таким образом разделяются по их спо¬
собности формировать разные типы леса, вместе
с которыми они образуют единства - экосистемы,
биогеоценозы (боры, субори, дубравы и др.).
Первая книга объемом 23.5 печ. л. состоит из
семи глав. Пять из них посвящены плодородию
лесных почв, характеризуемому по их обеспечен¬
ности влагой и элементами питания, с учетом та¬
ких негативных факторов, как ограничение кор¬
необитаемой зоны и наличие токсичных соедине¬
ний. Такое направление в изучении плодородия
почв берет начало от П.А. Костычева. Многие
годы его развивал А.Н. Соколовский, а в даль¬
нейшем успешно продолжили его ученики и по¬
следователи. Исходным материалов для этих глав
явились данные более 1.5 тыс. пробных площадей
с таксационными описаниями и полными физико¬
химическими анализами почв, грунтов и грунто¬
вых вод, заложенных в основном в южной части
лесной зоны и в лесостепи Русской равнины, но
также почти во всех других физико-географичес-
ких зонах бывшего Союза ССР - от предтундро-
вых лесов до пустынных тугаев. '
Автором очень убедительно показана строгая
зависимость лесных насаждений от уровня водо¬
обеспеченности почв, обусловливаемой не только
количеством атмосферных осадков, но и их пере¬
распределением рельефом и водно-физическими
свойствами грунтов. Охарактеризовано значение
объема корнеобитаемого слоя почвогрунта для
роста леса и резко негативное влияние его огра¬
ничения при неглубоком залегании плотных, ток¬
сичных, вечномерзлых пород и грунтовых вод:
Значительное внимание уделено оценке богатства
почв элементами питания. Предложены методы
определения доступных для древесной раститель¬
ности количеств фосфора и калия, чаще всего ли¬
митирующих их рост, и уровня обеспеченности
ими пород, обладающих разной требовательнос¬
тью к почвенному плодородию. Таким методом
оказалась вытяжка Гинзбург, в которую перехо¬
дят валовые фосфор и калий, кроме калия, за¬
ключенного в кристаллических решетках поле¬
вых шпатов. Олиготроф сосна способна расти на
почвах, содержащих в минеральных горизонтах
менее 0.01% Р205 и 0.03% К20. Мегатрофы ясень,
клены, липа удовлетворительно растут лишь при
содержании в почвах более 0.06% Р205 и 0.80%
К20, переходящих в вытяжку Гинзбург. На боль¬
шом материале охарактеризовано влияние токсич¬
ных легкорастворимых солей на древесные поро¬
ды разной степени солевыносливости. Выделено
пять категорий лесопригодности засоленных почв.
814
ЛЕСА И ЛЕСНЫЕ ЗЕМЛИ
815
Е.С. Мигунова показала, что в эдафической
сетке уровень водообеспеченности почв хорошо
отражает их генетический тип, а обеспеченность
элементами питания - гранулометрический со¬
став, обусловленный минералогическим соста¬
вом почвообразующих пород. Автор утверждает,
что в целом эдафическая сетка характеризует из¬
менения состава и строения поверхностных отло¬
жений, а потому она может называться геологиче¬
ской. При размещении таких геологических сеток
отдельных регионов в глобальной климатичес¬
кой сетке создается единая классификационная
модель всех основных природных факторов -
классификация экосистем, точнее геоэкосистем.
Координатами ее являются ведущие абиотические
факторы - климат и земля, зависимыми перемен¬
ными - биологические и биокосные - раститель¬
ность, животные, почвы. Основу этой классифика¬
ции составляет признание единства организмов
и среды. Организмы в ней - прежде всего это выс¬
шая растительность - характеризуются составом
и продуктивностью, среда - плодородием клима¬
та и грунтов. Последние оцениваются по их лими¬
тирующим жизнь параметрам - уровню обеспе¬
ченности теплом, влагой и элементами питания.
Установлены количественные величины этих па¬
раметров, что дает возможность прогнозирования
растительности (ее состава и продуктивности)
и почв (их типа и производительности) по задан¬
ным параметрам климата и грунтов, а также мас¬
штабов их изменений при тех или других наруше¬
ниях природной среды. Это положение представ¬
ляет особый интерес, поскольку позволяет подойти
к количественному решению уравнения В.В. Доку¬
чаева связи почв с факторами почвообразования.
Специальная глава посвящена почвообразова¬
нию в лесах, в том числе изменению степных почв
под влиянием древесной растительности. Однако
главной темой монографии является анализ вли¬
яния почв на состав и продуктивность лесной рас¬
тительности, оценка лесорастительного потенци¬
ала почв, их плодородия. Это направление дове¬
дено до установления оптимальных составов и
разработки уравнений, позволяющих рассчиты¬
вать возможные запасы насаждений в возрасте
спелости на разных видах лесных почв, а также
предельные высоты посадок на почвах степной
зоны. На представленных в книге моделях раз¬
ных зон почвы по их плодородию, оцениваемому
запасом или высотой древостоев, размещаются
очень закономерно. Выделено 10 классов произ¬
водительности почв - от наиболее высокопроиз¬
водительных до практически бесплодных.
Выявлена исключительная роль почвообразу¬
ющих и подстилающих пород, их гранулометри¬
ческого состава (отражающего минералогию,
химизм) и строения, предопределяющих обеспе¬
ченность почв элементами питания и влагой.
В связи с этим автор замечает, что лесоводы мо-
розовской школы практически не пользовались
термином “почва”, а только “почвогрунт”, что по
отношению к древесной растительности, безус¬
ловно, справедливо.
Вторая монография “Лесоводство и почвове¬
дение” объемом 15.5 печ. л. состоит из 18 очер¬
ков, посвященных истории изучения взаимосвя¬
зей леса и почв и некоторым другим вопросам
развития лесоводства и почвоведения. Книга име¬
ет три раздела. В первом помещены четыре очер¬
ка, содержащих краткий анализ материалов одного
из старейших специальных научных изданий Рос¬
сии - “Лесного журнала” (1834-1918 гг.). В этом
журнале нашла отражение не только история
развития лесоводственной науки на протяжении
XIX-начала XX вв., но и многих сопредельных ес¬
тественно-научных дисциплин. В очерках основ¬
ное внимание уделено публикациям, развиваю¬
щим представления о связи леса с окружающей
средой, в том числе с почвами, которые занимали
на страницах журнала с самых первых его номе¬
ров очень большое место.
Во втором разделе “Почвоведение и лесовод¬
ство” два очерка посвящены характеристике науки
о почве - почвопознания - первой половины XIX в.,
также в основном по материалам “Лесного жур¬
нала”. Следует заметить, что этот додокучаев-
ский период развития почвоведения нигде в России
не получил, по-видимому, такого полного освеще¬
ния, как на страницах этого журнала. Отмечается,
что все методы изучения почв в то время были на¬
правлены на оценку их плодородия, и в этом от¬
ношении имелись заметные достижения. Охарак¬
теризован вклад лесоводов морозовской школы
в изучение почв.
Один из очерков посвящен Особой экспедиции
Лесного департамента, руководимой В.В. Доку¬
чаевым. В работах этой экспедиции, представляв¬
шей первый опыт комплексного изучения приро¬
ды России, единение лесоводов и почвоведов про¬
явилось особенно ярко. Автор, однако, отмечает,
что В.В. Докучаев, настойчиво доказывавший не¬
обходимость познания природных взаимосвязей,
сам не предложил метода их изучения. В одной из
своих последних работ он писал, что формирова¬
ние науки о взаимосвязях органической и нежи¬
вой природы еще только начинается. Действи¬
тельно, в начале XX в. такие науки - экология
и ландшафтоведение - появились. Тем не менее
представленные в третьем разделе книги очерки
об основоположниках лесной типологии как на¬
уке о единстве леса и почв дают автору основание
считать ее наиболее близкой той науке, появле¬
ние которой предсказал В.В. Докучаев. Особый
интерес представляет очерк о А.А. Крюденере.
Вклад этого крупного лесовода, соратника Г.Ф. Мо¬
розова и Г.Н. Высоцкого, в оформление лесной ти¬
пологии огромен. Однако его эмиграция в 1918 г.
ПОЧВОВЕДЕНИЕ № 6 1996
816
ТИХОНЕНКО
в Германию обусловила тот факт, что этот вклад
был предан забвению. Е.С. Мигуновой удалось
собрать биографические данные и научные тру¬
ды этого ученого, созданные в России и Герма¬
нии. Оказалось, что именно ему принадлежит за¬
слуга создания, по-видимому, первой в истории
мировой науки сопряженной классификации раз¬
ных природных факторов - лесов и почвогрун-
тов. Особо подчеркивается, что идея этой класси¬
фикации была воспринята Крюденером в народ¬
ной среде, он многие годы собирал народные
природоведческие знания.
В связи с этим стоит вспомнить, что В.В. Доку¬
чаев настойчиво доказывал необходимость обоб¬
щения народных знаний о почвах. Этот завет на
рубеже XIX-XX вв. в значительной мере осуще¬
ствили русские лесоустроители, вынужденные
в силу специфики своей работы тесно общаться
с сельским населением. Главным среди народных
знаний о почвах была оценка их плодородия по
гранулометрическому составу грунтов (“тощие”
пески - “жирные” глины) и синтетический взгляд
на природу, отраженный в следующем постулате:
“каков грунт земли, таков и лес”. На этом прин¬
ципе и построена классификация А.А. Крюдене-
ра: в ней выделены типы почвогрунтов, обеспе¬
чивающие формирование на них разных типов
леса. Отмечается определенное сходство клас¬
сификации А.А. Крюденера с классификацией
Н.М. Сибирцева (1895), построенной в координа¬
тах климата (генетические типы) и грануломет¬
рического состава грунтов. При этом ему удалось
объединить классификации В.В. Докучаева и
П.А. Косгычева. Типологический принцип отли¬
чается тем, что лесоводы создали две классифи¬
кации, представляющие модели плодородия кли¬
мата и почвогрунтов.
В заключении книги Е.С. Мигунова пишет
о большой роли знания истории науки для ее посту¬
пательного развития. Это облегчает выбор наи¬
более перспективных направлений исследований
и кратчайших путей для их развития. Оно позво¬
ляет избегать “тупиковых” путей, не столь уж
редких в истории науки.
Представленный краткий анализ работ Е.С. Ми¬
гуновой свидетельствует, что развиваемое ею на¬
учное направление, основы которого заложили
русские лесоводы, представляет безусловный инте¬
рес для почвоведов. Оно дает возможность по-ново-
му осмыслить многие устоявшиеся положения,
найти решение ряда сложных и спорных вопро¬
сов, таких, в частности, как оценка пригодности
почв для разных древесных пород и сельскохо¬
зяйственных культур, количественная оценка их
плодородия, их лесорастительного потенциала.
Совершенствование классификации почв на ос¬
нове более полного учета почвообразующих по¬
род позволит совместить генетические классифи¬
кации с их группировками по плодородию. Рецен¬
зент, хорошо знакомый с принципами и методами
лесной типологии, разделяет сожаление автора
книг по поводу того, что это перспективное на¬
правление до сих пор не получило широкой изве¬
стности и признания.
Сдано в набор 01.03.96 г. Подписано к печати 29.04.96 г. Формат бумаги 60 х 88'/g
Офсетная печать Уел. печ. л. 16.0 Уел. кр.-отт. 13.7 тыс. Уч.-изд. л. 16.0 Бум. л. 8.0
Тираж 833 экз. Зак. 4260
Отпечатано в Московской типографии № 2 РАН, 121099, Москва, Г-99, Шубинский пер., 6