

























































































































































































































































































































































































Author: Шунтов В.П.
Tags: aves птицы в целом орнитология биология зоология морские животные
Year: 1972




Text
ТИХООКЕАНСКИЙ
НАУЧНО-ИССЛЕДОВАТЕЛЬСКИЙ
ИНСТИТУТ РЫБНОГО ХОЗЯЙСТВА
И ОКЕАНОГРАФИИ (ТИНРО)
В. П. ШУНТОВ
МОРСКИЕ ПТИЦЫ
И БИОЛОГИЧЕСКАЯ
СТРУКТУРА ОКЕАНА
ДАЛЬНЕВОСТОЧНОЕ
КНИЖНОЕ ИЗДАТЕЛЬСТВО
ВЛАДИВОСТОК, 1972
УДК 598.2/9+ 577.472
В. П. ШУНТОВ
МОРСКИЕ ПТИЦЫ И БИОЛОГИЧЕСКАЯ
СТРУКТУРА ОКЕАНА
Работа является итогом многолетних исследований ав-
тора тю морским птицам Тихого и Индийского океанов.
Рассматривается видовой состав, сезонное распределение,
миграции, плотность населения, численность и географиче-
ское распространение морских птиц на фоне существующей
схемы биологической структуры океана, на фоне выявлен-
ных закономерностей в распределении океанологических
характеристик и количественного распределения других
морских животных.
Большое внимание обращается на анализ особенностей в
составе, сезонных явлениях, количественном распределении,
географическом распространении, характерных для различ-
ных географических и ландшафтных эон. Делается попытка
оценить численность морских птиц в Мировом океане и их
роль как потребителей других животных.
Рис. 73, табл. 37, бнблиогр. 713 наимен.
Редакционная коллегия: И. В. КИЗЕВЕТТЕР (ответст-
венный редактор), Б. Н. АЮШИН, И. П. ЛЕВАНИДОВ, В. Г. ОСИ-
ПОВ, ю. В. НОВИКОВ, И. А. ПИСКУНОВ, Н. С. ФАДЕЕВ, Е. В. КУ-
ЧЕРЯВЕНКО.
3
... Великолепная и загадочная красота жизни
Мирового океана надежно сокрыта от глаз
восхищенного наблюдателя внешней яркой обо-
лочкой его...
Зато птицы, безраздельно владеющие Вели-
ким Воздушным Океаном, в полной мере воз-
дают пытливому уму натуралиста и его эсте-
тическим притязаниям.
И. Л. Гончаров. Из путевих записок в круго-
светном путешествии на фрегате .Паллада*.
ВВЕДЕНИЕ
Последние десятилетия характеризуются возрастающими
темпами в комплексном изучении Мирового океана. Важное ме-
сто при этом отводится биологическим исследованиям, наиболее
характерной и важной особенностью которых в данный период
является получение и накопление количественных показателей
процессов, определяющих биомассу и продукцию планктона,
донной фауны и нектона. Синтезом работ этого направления
явилось учение о биологической структуре океана — системе
взглядов о количественном распределении жизни и особенно-
стях биологических явлений в Мировом океане в плане геогра-
фической и вертикальной зональности (Зенкевич. 1948; Богоров,
1959; Богоров и Зенкевич, 1966). На основании полученных
представлений о количественных связях между живыми сущест-
вами различных трофических уровней даны общие глобальные
оценки продукционных свойств океана (Богоров. 1967, 1967а;
Моисеев, 1969; и др.).
В настоящее время все большее внимание биологов привле-
кает изучение биогеоценозов океана. Продолжение исследова-
ний в этом направлении в итоге приведет к моделированию
структур биогеоценозов отдельных районов, систем течений и,
наконец, целостной биогеоценотической системы океана. Акаде-
мик Л. А. Зенкевич (1970) назвал следующие десять основных
групп растительного и животного населения, биогеоценотиче-
ское значение которых должно обязательно учитываться. Это
бактерии, фитопланктон, фитобентос, зоопланктон, плейстон,
головоногие моллюски, рыбы, зообентос, морские млекопитаю-
щие и морские птицы. Для оценки слагаемых общего биологиче-
ского баланса океана по каждой из перечисленных групп долж-
ны быть получены данные по биомассам, продуктивности, а так-
же количественные показатели биогеоценотических связей и их
энергетических характеристик.
В связи с изложенным видна важность изучения морских
птиц, которым посвящена настоящая работа. Значение морских
птиц, конечно, не ограничивается их ролью в биологическом ба-
4
ВВЕДЕНИЕ
лансе моря. Они являются индикаторами водных масс, различ-
ных биологических явлений в море, продуктивных зон и районов
скоплений различных морских промысловых животных. Некото-
рые из них сами являются промысловыми животными. Само-
стоятельное значение имеет изучение закономерностей их рас-
пространения, распределения и миграций. Будучи одной из
групп животных, изучением которой занимается специальная
дисциплина — орнитология, морские птицы дают информацию
по вопросам общего порядка, разработкой которых занимается
данная наука. Известно также, что класс птиц является хорошей
«модельной» группой, на основе изучения которой многие зооло-
ги (Ф. Склэтер, А. Уоллес, М. А. Мензбир, П. П. Сушкин, Г. П.
Дементьев. Э. Майр, Д. Лэк, Б. К. Штегман и другие) решали
вопросы общебиологического и биогеографического плана. Из-за
доступности для наблюдений морские птицы, очевидно, могут
быть удобным материалом для аналогичных разработок в изу-
чении океана.
Состав группы морских птиц в понимании различных авто-
ров несколько различен. На море встречаются многие виды
птиц, но не все из них морские. Настоящие морские птицы сушу
используют только как субстрат для гнездования. Все осталь-
ные их связи, и в первую очередь трофические, с морем (Каф-
тановский, 1951). Всего таких видов около 250. Исключительно
морские — семейства альбатросов — Diomedeidae, буревестни-
ков — Procellariidae, качурок — Hydrobatidae. ныряющих буре-
вестников — Petecanoididae, пингвинов — Sphenisci'dae, чисти-
ковых — Alcidae, фрегатов — Fregatidae, олуш — Sulidae, фа-
этонов — Phaethontidae. Настоящие морские виды есть также
в семействах поморников — Stercorariidae, крачек — Sternidae,
чаек — Laridae, бакланов — Phalacrocoracidae, пеликанов —
Pelecanidae. В пяти последних семействах, а также среди кули-
ков, уток и гагар, есть много видов, которые значительную часть
времени проводят на море, но вместе с этим сохранили тесную
связь с сушей и пресными водоемами. Многочисленные на море
с осени до весны нырковые утки, кулики, гагары, некоторые
чайки, крачки и другие птицы одновременно мигрируют по ре-
кам и озерам. Значительную часть летнего периода они прово-
дят на внутренних водоемах, где размножаются. Характерно,
что на море фактически все они ведут прибрежный образ жиз-
ни. Здесь они нередко доминируют в видовом и количественном
отношении над настоящими морскими птицами. Общее количе-
ство таких полуморских видов не меньше, чем настоящих мор-
ских птиц. Конечно, они должны учитываться при исследовании
морской орнитофауны, за исключением вопросов, связанных с
орнитогеографическнм районированием.
ВВЕДЕНИЕ
В настоящее время морские (птицы далеко не самая изучен-
ная группа морских животных. Правильнее сказать — это очень
неравномерно изученная группа, хотя список литературы, по-
священной морским птицам, насчитывает тысячи наименований.
Большая часть имеющихся работ касается гнездового, т. е. бе-
регового периода жизни птиц. Неплохо известен видовой состав
птиц, размножающихся в различных районах. У большинства из
них известны границы гнездовой части ареала. Можно с уверен-
ностью сказать, что сейчас описаны почти все виды морских
птиц. Имеется большое количество работ, описывающих биоло-
гию размножения многих видов. Определена численность птиц,
состав и величина их рационов в ряде районов колониального
гнездования.
Успехи в изучении морских >птиц нашли отражение в ряде
серьезных сводок. Можно назвать книги Мэрфи (Murphy,
1936) об океанических птицах Южной Америки, Фишера (Fisher,
1952) и Така (Tuck, 1960) о глупыше и кайре, Фишера и Локли
(Fisher and Lockley, 1954) о морских птицах Атлантического
океана. Л. О. Белопольского (1957) о птицах Баренцева моря,
С. М. Успенского (1969) о птицах советской Арктики и некото-
рые другие.
За исключением времени размножения большую часть жизни
морские птицы проводят в кочевках, используя для этих целей
всю свободную от льдов площадь Мирового океана- Этот пери-
од жизни птиц наименее исследован. Дело в том, что в много-
численных морских экспедициях, проводившихся различными
странами, нечасто принимали участие орнитологи. Еще два де-
сятилетия назад публикации с описанием поведения, распреде-
ления и миграций птиц в открытом море появлялись в значи-
тельной степени эпизодически. В связи с этим были мало изве-
стны сама картина послегнездового распределения и миграций
лтиц. общие вопросы количественного размещения и факторы
среды, определяющие их закономерности. В последние двадцать
чет положение заметно изменилось, появилось значительное ко-
личество работ, при этом по районам, наименее изученным, в
частности по Антарктике и тропическим водам Тихого и Индий-
ского океанов. Сильно расширились масштабы кольцевания
морских птиц. Особого упоминания заслуживают работы по
кассовому кольцеванию птиц учеными различных стран на ме-
стах гнездования в Южном океане и Антарктиде, районах,
•ставившихся до этого слабо изученными с орнитологической
точки зрения.
Большой объем накопленной информации позволяет делать
•'^которые общие выводы и заключения. Тем не менее изучение
''орского периода (и не'только морского) должно продолжать-
6
ВВЕДЕНИЕ
ся. Мы еще далеки от того, чтобы иметь полную и целостную
картину жизни морских птиц в это время. Для примера можно
сослаться на степень изученности питания птиц. Фактически все
имеющиеся обширные материалы по данному вопросу относятся
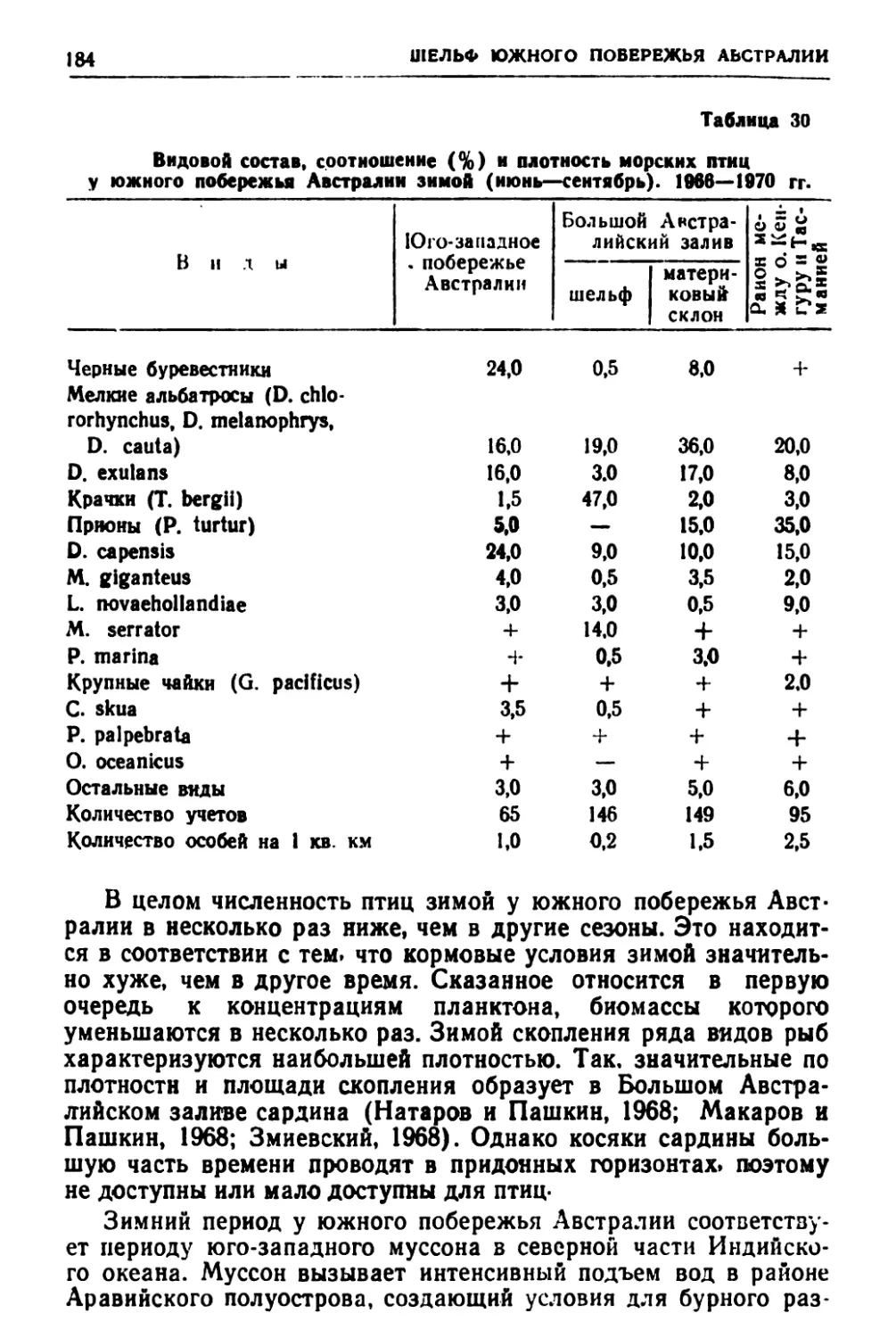
к гнездовому периоду. Состав питания, а также рационы во вре-
мя кочевок по большинству видов не исследовались. Ясно, что
для конкретных суждений о роли птиц в биологическом балан-
се того или иного района эти сведения нужны. По-прежнему
слабо изученными остаются многие районы и особенности ми-
граций большой группы видов. Недостаточно данных о числен-
ности птиц- Важными являются вопросы межгодовой изменчи-
вости, по которым вообще сведений очень мало. Здесь нужны
наблюдения, повторяющиеся ежегодно по стандартным мето-
дикам, как это делается в гидрологии, ихтиологии и гидробиоло-
гии.
Два последних десятилетия характеризуются бурным рас-
ширением океанского рыболовства на Тихом океане. Разработ-
кой рекомендаций по использованию сырьевой базы Для разви-
вающейся рыбной промышленности Дальнего Востока занимал-
ся и занимается ТИНРО, многочисленный исследовательский
флот которого проводит научно-поисковые работы во всех райо-
нах Тихого и восточной части Индийского океанов.
Занимаясь поиском и изучением рыбных ресурсов в период
с 1959 по 1968 г., я имел возможность принимать участие и ру-
ководить работой 15 научно-поисковых экспедиций ТИНРО, про-
водивших работы в Беринговом, Охотском, Японском, Восточ-
но-Китайском, Южно-Китайском, Арафурском, Тиморском, Тас-
мановом морях, в северо-западной части Тихого, в юго-восточ-
ной части Индийского океанов, на Новозеландском плато.
В перечисленных районах, а также на переходах между ними
в течение всех экспедиций проводились учеты морских птиц.
В других районах Тихого океана по моей просьбе проводили
учеты В. М. Шпак. П. Л. Кантемиров, Г. Г. Солодовников,
А. И. Чигиринский, В. Г. Царев, О. Е. Комраков, Ю. К. Деми-
денко и другие. В обшей сложности я располагаю данными
около 17 500 часовых и получасовых визуальных учетов, охва-
тывающих всю акваторию Тихого и восточную часть Индий-
ского океанов. Так как наблюдения проводились в определенные
промежутки времени и охватывали определенную полосу учета
(на ходу судна), было возможно полученные данные пересчи-
тывать на площадь — 1 кв. км.
Визуальные учеты морских птиц, как и сухопутных, конечно,
дают весьма приближенные цифры по плотности населения.
Многие виды не отличимы или почти не отличимы на расстоя-
нии. При этом определенную ошибку в расчеты вносят птицы.
ВВЕДЕНИЕ
7
которые привлекаются судами и подолгу их сопровождают. Но
ценность результатов учетов увеличивается благодаря сравни-
мости полученных данных в плане сравнения районов, зон, сезо-
нов и т. д. Материалы этих исследований дают возможность
описать не только сезонные аспекты в орнитофауне и изменение
плотности населения птиц в отдельных районах, но и дать ори-
ентировочную оценку их численности в масштабе Мирового
океана.
Наблюдения над морскими птицами проводились параллель-
но с изучением биологии, запасов и условий обитания рыб. Вы-
явленные закономерности в распределении рыб в связи с гидро-
логическими условиями и характером кормовой базы способст-
вовали формированию представлений, характеризующих зако-
номерности видового состава, поведения и распределения мор-
ских птиц, а также сезонных изменений их численности в тех
пли иных районах. Таким образом, от констатации фактов во
многих случаях удалось перейти к объяснению причинности на-
блюдаемых явлений. Весьма способствовало этому и наличие
обширной литературы по гидрологии, планктону и рыбам Тихо-
го и Индийского океанов.
Слабой стороной полученных материалов является ограни-
ченное количество данных из непосредственно прибрежной
5—10-мильной зоны, а также полное отсутствие собственных
береговых наблюдений. Последнее обстоятельство в какой-то
степени удалось компенсировать литературными сведениями.
Все затрагиваемые ниже вопросы — распределение и мигра-
ции отдельных видов, сезонные аспекты в орнитофауне, общие
закономерности распределения и миграций в масштабе всего
океана, орнитогеографическое деление океана и другие—я пытал-
ся рассматривать на фоне выявленных закономерностей в харак-
тере и распределении океанологических характеристик, а также в
связи с кормовой базой (планктон, нектон). Если говорить крат-
ко, то основная цель настоящей работы — попытка «вписать»
морских птиц в существующую схему биологической структуры
океана. Рассматриваемые, однако, вопросы не охватывают все
етороны темы, связанные с биологической структурой океана.
Упор в работе делается на морской период жизни птиц.
При обработке материалов учетов, а также подготовке ру-
кописи к печати мне большую помощь оказывала моя постоян-
ная помощница В. А. Костикова. Предварительно рукопись бы-
ла просмотрена докторами биологических наук Л. О. Белополь-
ским и К. А. Юдиным, а также сотрудниками ТИНРО В. В. Фе-
доровым и В. А. Снытко, сделавшими некоторые замечания и
Давшими полезные советы. Всем перечисленным я признателен
за их труд.
8
ГЛАВА I
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ
ОТДЕЛЬНЫХ ВИДОВ МОРСКИХ ПТИЦ
В этой и следующей главах излагается большое количество
фактических материалов, необходимых для дальнейших обобще-
ний. В настоящей главе я не имею в виду описать распростране-
ние и миграции всех видов морских птиц. По значительной части
видов подробной ннформации, характеризующей распределение
во все сезоны, сейчас пока нет. Здесь подобраны отдельные ви-
ды, по которым я располагаю обширными собственными материа-
лами, а также есть литературные сведения. В основном это хоро-
шо различимые в полевых условиях птицы, некоторые из них при-
влекаются судном.
1. Северотихоокеанские альбатросы.
В северной части Тихого океана обитает три вида альбатро-
сов — темноспинный — Diomedea immutabilis Rothsch., черноно-
гий — D. nigripes Audub. и белоспинный — D. albatrus Pall. Не-
сколько десятков лет назад последний вид имел высокую числен-
ность и гнездился на девяти островах, расположенных южнее
Японии и в Восточно-Китайском море. К настоящему времени он
почти истреблен,.сохранилось несколько десятков особей, раз-
множающихся лишь на острове Торисима (Austin, 1949; Austin
and Kuroda, 1953; Rice and Kenyon, 1962; Kuroda, 1964). Ha
шести островах уничтожен также черноногий альбатрос и на пя-
ти—темноспинный. Но на Гавайских островах пока оба вида мно-
гочисленны.
Около 10—12 лет назад численность темноспинного аль-
батроса оценивалась в 1 500000, черноногого в 300 000 осо-
бей (Rice and Kenyon, 1962). Однако сообщения последних лет
(Fisher, 1963, 1966; и др.) показывают, что истребление альба-
тросов продолжается и сейчас.
Темноспинный и черноногий альбатросы в настоящее время
размножаются в основном на Гавайских островах. Небольшое
количество черноногого альбатроса гнездится также на островах
Торисима и Марианских (Jouanin, 1959; Rice and Kenyon, 1962).
Основываясь на большом количестве собственных наблюде-
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
9
ний, а также данных других авторов по отдельным районам
(Miller. 1940; Arnold, 1948; Kenyon, 1950; Thompson, 1951; Kuro-
da, 1955, 1960, 1963; McHugh, 1955; Motova and Fujii, 1956; Ha-
milton, 1958; Nakamura, 1963; Holmes, 1964; Слепцов, 1959; Ку-
рочкин, 1963; и др.), я уже делал попытку построить схемы се-
зонного размещения темноспинного и черноногого альбатросов
для всей северной части Тихого океана (Шунтов, 1968). В по-
следние годы я получил новые данные, которые позволяют не-
сколько уточнить и конкретизировать особенности распределе-
ния обоих видов.
Все имеющиеся в моем распоряжении материалы разбиты
на три. периода: первый — с ноября по март, второй — с апреля
по июнь, третий — с июля по октябрь. Первый период охваты-
вает время с ноября (начало гнездования) (Munro, 1960; Rice
and Kenyon, 1962a; Hsher, 1969) до начала весны. Второй пе-
риод охватывает весну и начало лета до окончания размноже-
ния. Разлет с колоний происходит в середине июня у черноного-
го альбатроса и в середине июля у темноспинного (Munro, 1960;
Rice and Kenyon, 1962a). Третий период — это время' кочевок
всех возрастных групп альбатросов. Приведенные ниже схемы
распространения альбатросов построены с учетом выделенных
периодов.
Наиболее характерной особенностью в распределении рас-
сматриваемых видов альбатросов в океане является заметная
разобщенность районов их обитания. На рис. 1—3 показано со-
отношение темноспннного и черноногого альбатросов в различ-
ных районах океана. Обращает на себя внимание факт приуро-
ченности кочевок темноспинного альбатроса к северо-западной
части Тихого океана. Наоборот, доля черноногого альбатроса
закономерно увеличивается в юго-восточном и восточном на-
правлениях.
В зимний период темноспинный альбатрос преобладает в
учетах между Алеутскими островами и 45° с. ш., в заливе Аляс-
ка, а также на обширной акватории севернее 30° с. ш. между
Японией и 175е з. д. (см. рис. 1). Весной и особенно летом об-
ласть преобладания в учетах темноспинного альбатроса сокра-
щается в северо-западном направлении (см. рис. 2 и 3).
Известно, что северо-западная часть Тихого океана, занятая
субарктическими водами с холодным промежуточным слоем
(Uda, 1955; Радзиховская, 1965; Радзиховская и Леонтьева,
1968). значительно холоднее северо-восточной. Поверхностные
изотермы в водах к востоку от Японии, Курильских островов и
Камчатки под влиянием холодных течений имеют скорее мери-
лиональное направление, чем широтное (Панфилова, 1968).
включая летний период, то же самое можно сказать и о темпе-
ратуре воздуха.
20
J
vy
000000600 ЭЭ
7'[0оооооооооооэе«э«»г
7 Foooooq®®®®®®®®®®»® I
y'.bGOGGGG ••••
GG®O OGG
••••®®®®®<э
•• ®9®G
18O
ISO
Pik*. 1. Соотношение в учетах темносппнного— D. iinmutabilis и черноногого D. riigripes альбатросов в
Тихим оксане зимой (ноябрь—март). Томноспинный— светлые секторы и кружки, черноногий — черные
Рис. 2. Соотношение в учетах темноспинного и черноногого альбатросов в Тихом океане весной (ап-
рель—июнь). Обозначения, как на рис. I
Риг. 3. Соотношение в учетах темноспииного и черноногого альбатросов в Тихом океане в летне-осенний
период (июль—октябрь). Обозначения, как на рис. I
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
13
По осредненным за все сезоны данным, темноспинный аль-
батрос встречается на кочевках в районах, имеющих температу-
ру воздуха от минус 2 до 26° (чаще 6—12) и температуру воды
от 2 до 28* (чаще 7—14°), а черноногий — при температуре воз-
духа от 6 до 26° (чаще 12—20’) и воды от 6 до 30° (чаще
10—20*). Таким образом, темноспинный альбатрос в период
кочевок придерживается более холодных, а чернояогий — болсг
теплых районов северной части Тихого океана. .
Тяготение черноногого альбатроса во все сезоны к более теп-
лым районам океана логично связывать с теплолюбивостью это-
го вида. Аналогичные взгляды высказывали также другие ав-
торы (Kuroda, 1960; Sanger, 1970). Темноспинный альбатрос,
как и черноногий, размножается на субтропических и тропиче-
ских островах, где очень тепло, поэтому приуроченность этого
вида к северо-западной части океана не приходится объяснять
его холодолюбивостью. По-видимому, наиболее правдоподобно
объяснять раздельность в обнтанин альбатросов межвидовыми
отношениями двух близких видов. Характерно, что в периоды,
когда область распространения темноспинного альбатро-
са наибольшая, у черноногого она наименьшая, и наоборот
(см. рис. 1—3). Некоторым подтверждением такого объяснения
могут служить особенности распределения альбатросов в райо-
нах, где обычны или многочисленны оба вида, например у Га-
вайских островов в гнездовой период. В таких случаях, как
правило, в отдельных учетах доминирует один из видов и редко
в равных соотношениях отмечаются оба вида.
В зимний период1 темноспинный альбатрос встречается на
обширной акватории: от Японии до Калифорнии и от южной ча-
сти Берингова моря до Гавайских островов (рис. 4). В среднем
южная граница его распространения проходит между 28 и
30° с. ш. на западе океана и около 35* с. ш. на востоке. Правда,
отдельные особи у тихоокеанского побережья Америки могут
проникать до п-ова Калйфорниж. В районе Гавайских островов
граница распространения Проходит значительно южнее. В не-
больших количествах он проникает здесь в тропики — до
15° с. ш. (King, 1967). В это время темноспинный альбатрос
отсутствует на большей Части Берингова и Охотского морей’, 1 2
1 В это время темноспипный и чернонэгий альбатросы размножаются.
Часть особей обоих видов впервые приступает к размножению в возрасте
' лет; ежегодно гнездится только 73% взрослых. Особи в возрасте 1—2 лет
совсем не появляются в районах размножения. Начиная же с 3-летнего возра-
ста, неполовозрелые особи, а также неразмножаюшиеся взрослые, посещают
гнездовЬЯ (Rice and Kenyon, 1962, 1962а).
2 Раньше не было известно о проникновении темноспинного альбатроса зи-
мой в суровое Охотское море. 25 декабря 1967 г. один экземпляр его был ветре-
нон, у юго-западного побережья Камчатки в координатах 51°29' с. ш.
в. д.
Рис. 4. Распределение темноспинного альбатроса Тихом океане зимой. !— поверхностные изотермы. 2 ме-
ста гнездования. 3 — редок, 4 - обычен. 5 — многочислен
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
15
что, несомненно, связано не со льдами, которые закрывают глу-
боководные районы этих морей только частично (Булгаков,
1965), а с сильным охлаждением воздуха и воды. Средняя ме-
сячная температура воздуха в январе & северных районах Бе-
рингова моря понижается до минус 23,3е, а в различных районах
Охотского моря — от минус 6 до минус 25° (Леонов, 1960).
Основные концентрации темноспинного альбатроса в рас-
сматриваемый период располагаются между Японией, Гавайски-
ми и Алеутскими островами. Севернее 40° с. ш. местами обычен
он и в восточной части Тихого океана. В советских водах зимой
темноспинный альбатрос постоянно встречается только у север-
ных Курильских островов и у юго-восточного побережья Кам-
чатки (см. рис. 4).
Черноногий альбатрос зимой откочевывает в южном и юго-
восточном направлениях. Северная граница его распространения
проходит в это время от середины острова Хонсю на северо-
восток к южной части зал. Аляска. В ноябре он еще встречается
у побережья северной части острова Хонсю, но уже в декабре
здесь остается только темноспинный альбатрос (Kuroda, 1963;
Шунтов, 1966). Наиболее часто встречается черноногий альба-
трос в водах между Гавайским районом и восточным побе-
режьем США (рис. 5). По мере движения на запад приблизи-
тельно от 170е в. д. количество его закономерно уменьшается,
при этом область распространения здесь простирается в широт-
ном направлении всего на 1000—1200 миль, в то время как на
востоке океана — на 2000 миль. Немногочислен черноногий аль-
батрос и в южных районах ареала. Южная граница его распро-
странения проходит в районе 25—30° с. ш. на западе океана,
опускается до 10—15е с. ш. в центральной части и вновь подни-
мается до 23—25° с. ш. на востоке океана.
Занятые выведением потомства взрослые птицы, видимо, мо-
гут улетать за кормом за сотни миль. Однако вдали от мест
гнездования зимой кочуют главным образом неполовозрелые и не-
размножающиеся в данном году особи. Так как разлет с мест
гнездовий происходит только летом, весной в открытом океане
численность альбатросов не увеличивается. Основное отличие
весеннего (апрель-июнь) распределения от зимнего состоит в
том, что по мере потепления черноногий альбатрос расширяет
область кочевок на север и северо-запад, а темноспинный со-
кращает в тех же направлениях. Особенно заметно сокращает-
ся количество темноспинного альбатроса в восточной части Ти-
хого океана. Правда, отдельные особи .могут быть встречены
здесь в течение всего весеннего периода. Так, 26 мая 1967 г.
один темноспинный альбатрос был встречен у полуострова Ка-
лифорния на широте 25’11' с. ш. Как и зимой, темноспинный ал»-
Рис. 5. Распределение яерноногого альбатроса в Тихом океане зимой. Обозначения, как «на рис. 4
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЕНЬЕ ВИДОВ
17
батрос весной наиболее часто встречается в открытых водах
между Японией, Алеутскими и Гавайскими островами.
Черионогий альбатрос в июне на западе океана достигает
широты островов Хоккайдо и Курильских, но держится на значи-
тельном удалении от них в зоне более теплых вод (Kuroda,
1960; Шунтов, 1968). В восточной части Тихого океана он в это
время уже обычен в зал. Аляска и у Алеутских островов (Arnold.
1948; Kenyon, 1950). По-прежнему наиболее многочисленным
весной черноногий альбатрос остается в восточной части Тихого
океана, у Гавайских островов, а также к северо-западу от них
до 150е в. д. и к северу до 40—45° с. ш.
Летом и в первой половине осени кочуют все возрастные
группы альбатросов, поэтому численность их в открытых водах
в это время наибольшая. Темноспинный альбатрос в летне-осен-
ний период концентрируется главным образом севернее 40° с. ш.
между Камчаткой, Курилами и Хоккайдо на западе и 175* з. д.
на востоке, проникает при этом в северные районы Охотского и
Берингова морей (рис. 6). В советских водах летом темноспин-
ный альбатрос наиболее многочислен у Камчатки, Командорских
и Курильских островов.' В восточной части Тихого океана он
малочислен и встречается в основном севернее 45* с. ш. в водах,
прилегающих, к Алеутским островам и зал. Аляска (см. рис. 6).
В период разлета с гнездовий летом и во время возвращения к
местам размножения осенью темноспинный альбатрос встре-
чается между Гавайскими островами и областью кочевок, пока-
занной на рис. 6. Очевидно, через эти районы он пролетает не
задерживаясь. Поэтому летом южнее 40* с. ш. на западе океана
и 45° с. ш. на востоке он почти никем не отмечался (Motoda and
Fujii, 1956; Hamilton, 1958; Kuroda, 1960; Wilhoft, 1961; Nakamu-
ra, 1963; Шунтов, 1968).
Черноногий альбатрос летом занимает очень обширную об-
ласть распространения. Он проникает в Охотское и Берингово
моря, а по наблюдениям М. М. Слепцова (1959), даже в Япон-
ское море в районе пролива Лаперуза. В западной часта Тихого
океана этот вид весьма обычен в водах восточнее Японии и Ку-
рильских островов. В советских водах в летне-осенний период
он наиболее многочислен у северных Курильских и у Коман-
дорских островов. Основные его концентрации наблюдаются ле-
том на акватории, ограниченной линиями, соединяющими се-
верную часть Хонсю с восточной частью Алеутских островов и
Калифорнией'(рис. 7).
Во второй половине осени в связи с приближением сезона
размножения (ноябрь) и похолоданием начинается откочевка
части альбатросов в более южные районы.
Все приведенные и описанные выше схемы распределения
Рис. 6. Распределение темноопинного альбатроса в Тихом океане в летне-осенний период.
Обозначения, как на рис. 4
Рис. 7, Распределение черноногого альбатроса в Тихом океане в летне-осеипнй период. Обо-
значения, как на рис. 4
20
СЕВЕРОТИХООКЕАНСКИЕ АЛЬБАТРОСЫ
альбатросов построены на основании осреднения данных за ряд
лет. При этом каждый из рассмотренных периодов охватывает
несколько месяцев. Естественно, что в отдельные годы и в каж-'
дый данный момент могут наблюдаться некоторые, иногда за-
метные, отклонения от средней картины*. Так, я не отмечал чер-
ноногого альбатроса в западной части Берингова моря. Но его
встречали здесь М. М. Слепцов (1959) и Е. Н. Курочкин (1963).
В водах залива Аляска черноногий альбатрос появляется обыч-
но в конце весны, но есть одно наблюдение о встрече его у ост-
рова Кадьяк 19 апреля (Gabrielson and Lincoln, 1959). В разные
годы изменяется положение южной границы распространения
альбатросов. В частности, на востоке океана темноспинный аль-
батрос достигает полуострова Калифорния только в отдельные
годы (Blake, 1953; мои данные). Наблюдаются некоторые раз-
личия в разные годы и в размещении районов концентраций
альбатросов. Оба вида нередко питаются отходами рыболовно-
го и китобойного промысла, поэтому часто собираются в боль-
ших количествах в районах концентрации промыслового флота.
В 1967—1970 гг. впервые проводился промысел рыбы в цент-
ральных районах северной части Тихого океана на банках Им-
ператорского и Гавайского подводных хребтов. Раньше здесь
также отмечалась повышенная плотность альбатросов, в этот
же период их здесь было особенно много.
Раньше уже была показана связь распределения скоплений
альбатросов с продуктивными водами (Miller, 1940; Thompson,
1951; McHugh, 1955; Шунтов, 1968; и др.) . Районы концентраций
альбатросов у Японских, Курильских, Алеутских островов, в
зал. Аляска, около Камчатки, тихоокеанского побережья Кана-
ды и США, как будет показано в следующих разделах, харак-
теризуются наиболее высокой продуктивностью в северной час-
ти Тихого океана. Но повышенная плотность альбатросов наб-
людается и в удаленных от берегов водах (см. рис’ 4—7), хотя
открытые океанские районы характеризуются в среднем более
низкой биологической продуктивностью, чем прибрежные. Здесь
меньше планктона и рыб. К сожалению, по этим районам ниче-
го неизвестно о кальмарах, одних из основных кормовых объ-
ектов альбатросов. Вполне возможно, что в некоторых районах
они многочисленна Впрочем, альбатросы могут быть обычными
и в районах, не отличающихся высокой биологической продук-
тивностью. Для них характерен длительный поисковый полет-’,
1 Уилхофт (Wilhoft, 1961) сообщает о встрече значительного количества
черноногого альбатроса в августе 1960 г. v Филиппинских островов, в районе,
не входящем в ареал этого вида. По-видимому, здесь была ошибка в опре-
делении. С 1961 по 1968 г. в этом районе я был 10 раз, но ни одного аль-
батроса наблюдать не пришлось.
2 Все альбатросы значительную часть жизни проводят в воздухе.
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
2!
во время которого они обследуют огромные акватории. Это дает
им возможность обитать в районах с невысокой концентрацией
кормовых объектов. Как показывают исследования ТИНРО в
последние годы, в открытом океане на фоне общей сравнитель-
ной бедности выделяются локальные районы с высокой продук-
цией вод. Одним из таких районов является зона Император-
ского и Гавайского подводных хребтов, где наблюдается увели-
чение биомасс планктона, количества рыбы и, как я уже отме-
чал, альбатросов.
Из изложенного выше видно, что ряд особенностей в распре-
делении альбатросов в северной части Тихого океана находится
в зависимости от продуктивности вод, сезонных изменений тем-
пературы воды и воздуха, а также межвидовых отношений.
Однако, если рассматривать характер распространения толь-
ко в связи с этими факторами, остается неясным, почему альба-
тросы малочисленны летом в Охотском море, на большей час-
ти Берингова моря, а в Японском отсутствуют вообще. Темпера-
тура воды и воздуха в летние месяцы здесь оптимальна для
альбатросов, а по богатству жизнью дальневосточные моря за-,
нимают одно из первых мест среди других районов Тихого океа-
на. Очевидно, это объясняется тем, что черноногий и темноспин-
ный альбатросы тяготеют в своем распространении к океаниче-
ским водным массам. Так, в Беринговом море, соединенным с
океаном широкими проливами, через которые наблюдается ин-
тенсивное поступление океанических вод, альбатросы сравни-
тельно многочисленны (в основном на юге моря близ проли-
вов). Охотское море более изолировано от океана, и в нем зна-
чительно меньше альбатросов; они здесь часто встречаются
только в районе проливов северной части Курильской гряды, че-
рез которые в Охотское море поступают океанические воды.
Японское море почти полностью изолировано от океана, соеди-
няется с ним узкими проливами; альбатросы его почти не посе-
щают. Показательно и то, что наиболее редки альбатросы на-
шел ьфах.
Зависимость распространения в связи с определенными вод-
ными массами была показана мною также на примере распро-
странения черноногого альбатроса в районе полярного фронта в
северо-западной части Тихого океана (Шунтов, 1968). В районе
Японии наблюдается четко выраженный гидрологический по-
лярный фронт, где встречаются и взаимодействуют субтропиче-
ские водные массы, связанные происхождением с течением Ку-
росио, и субарктические, связанные с Курильским (Ойясио) те-
чением (Бурков и Арсеньев, 1958; Радзиховская и Леонтьева.
1968). Зона стыка различных водных масс является важной
биогеографической границей. Весной и в начале лета черноног
22
АЛЬБАТРОСЫ ЮЖНОГО ОКЕАНА
гий альбатрос встречается здесь только к югу от фронта Ойясио
(в том числе в зоне смещения вод), не выходя за поверхност-
ную изотерму 11'. хотя в других районах он наблюдается и при
температуре 6*. Характерно, что северная граница его распро-
странения проходит в это время, как и поверхностные изотер-
мы, не в широтном направлении, а с юго-запада на северо-вос-
ток. По мере прогрева вод и смещения фронта на север чер-
ноногий альбатрос также расширяет область распространения.
Непосредственно у южных Курильских островов он становится
обычным только во второй половине лета, когда сюда прибли-
жается фронт Ойясио.
Сравнивая в этом плане схемы ареалов альбатросов со схе-
мами течений, легко заметить, что темноспинный альбатрос кочу-
ет в основном в зоне холодных Курильского, Камчатского и
Алеутского течений, с которыми связано формирование суб-
арктических вод с холодным промежуточным слоем (Радзихов-
ская, 1965;* Радзиховская и Леонтьева, 1968). В зимнее время
его численность увеличивается в водах Куросио, Северотихооке-
анского (продолжение Куросио) и Калифорнийского течений1.
Черноногий альбатрос зимой придерживается вод теплых Ку-
росио, Северотихоокеанского, и Калифорнийского течений. Ле-
том увеличивается его численность в зоне Аляскинского и Але-
утского течений.
2. Альбатросы Южного океана.
В Южном океане обитает 9 видов альбатросов. В различных
районах распределение и миграции их изучейя неодинаково.
Так как я располагаю многочисленными собственными данны-
ми только по Австрало-Новозеландскому району и сопредельно-
му сектору Антарктики, где встречаются все 9 видов альбатро-
сов, ниже в очерках по миграциям птиц южного полушария
рассматриваются преимущественно эти районы.
Странствующий альбатрос — Diomedea exulans L. Стран-
ствующий альбатрос размножается на островах Южная
Георгия, Тристан-да-Кунья, Гоф, Марион, Принца Эдуарда.
Крозе, Кергелен, Окленд, Антиподов. Макуори, Кэмпбелл
(Murphy, 1936; Alexander, 1954; Oliver, 1955; и др.), Фолькленд-
ских (Cawkell and Hamilton, 1961) и Амстердам (Tickell, 1968).
Все перечисленные острова, за исключением Южной Георгии и
Амстердама, располагаются между субтропической и антарк-
тической конвергенциями, т. е. находятся в пределах южной
умеренной зоны. Область кочевок странствующего альбатроса
1 Название течений по В. А. Буркову (1966).
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
23
более обширна п включает не только воды Южного океана, но
также субтропические и частично тропические.
Гнездовой цикл странствующего альбатроса, как и королев-
ского, продолжается около года, поэтому значительная часть
его особей размножается только раз в два года (Richdale, 1952;
Tickell, 1968; и др.)'. К размножению он, по-видимому, впервые
приступает в возрасте около 10 лет, хотя часть молодых начина-
ет появляться на гнездовых колониях с 4 лет (Tickell, 1968;
Tickell and Gibson, 1968). Таким образом, в любое время года в
открытом океане находится не только несколько генераций не-
половозрелых птиц, а также около половины половозрелых
особей.
Северная и южная границы распространения странствующе-
го альбатроса претерпевают заметные изменения в сезонном
аспекте. Летом с прогревом воды и воздуха, как и многие дру-
гие животные, он повсеместно смещается в сторону высоких
широт. На юг отдельные особи мигрируют до паковых льдов,
но, как правило, граница распространения проходит несколь-
ко севернее льдов (Murphy, 1936; Jameson, 1961; и др.). Поло-
жение южной границы распространения зависит от интенсивно-
сти прогрева вод и положения кромки льдов. В западно-тихо-
океанском секторе Южного океана от течения западных вет-
ров происходит ответвление в море Росса, вызывающее прогиб
в южном направлении изотерм и кромки льда (Нейман, 1961;
Трешников, 1964; Григорьев, 1969; и др.). Здесь странствующий
альбатрос проникает в антарктические воды наиболее далеко
(рис. 8 и 9). Так, летом 1967/68 г. он был ’ обычным здесь до
65е ю. ш. (крайняя встреча 65’51' ю. ш.), а в 1968/69 г. до
67° ю. ш. (крайняя встреча на 69’15' ю. ш.)1 2. Восточнее рас-
сматриваемого сектора южная граница проходит заметно се-
вернее, и за 65° ю. ш. проникают только редкие одиночки (край-
няя встреча в 1968/69 г. на 67’18' ю. ш.).
В индоокеанском секторе Антарктики, как известно, берега
Антарктиды и соответственно льды имеют более северное поло-
жение. Здесь количество странствующего альбатроса в связи с
этим резко уменьшается уже в районе 63° ю. ш. (крайняя встре-
ча на 63с44' ю. ш.). В целом южная граница распространения
подвижна и в течение лета по мере потепления постепенно сме-
щается на юг. Наблюдаются некоторые различия в ее положе-
нии и в разные годы (см. рис. 9).
1 Есть сообщения о том, что на островах Кроле каждая пара размно-
жается в три года (Voisin, 1969). Особи, которые в текущем году теря-
ют кладку, размножаются только на следующий год (Tickell, 1968).
2 Как южный предст распространения другие авторы отмечали широты
•68—68°40' ю. iu.t(Murpny, 1936; Jameson. 1961; Tickell. 1968; и др.).
Рис. 8. Распределение странствующего альбатроса — D. exulans в районе
Австралии, Новой Зеландии и сопредельных водах Антарктики. 1 — грани-
цы распространения зимой, 2 — границы распространения летом, 3 — по-
верхностные изотермы зимой, 4 — поверхностные изотермы летом, 5 —
районы основных концентраций зимой, 6 — районы основных концентра-
ций летом, 7 — места гнездования
Рис. 9. Распределение странствующего альбатроса в новозеландском сек-
торе Южного океана летом 1967—1969 гг. 1—южная граница распростра-
нения в 1967/68 г.. £ —южная граница распространения в 1968/69 г.,
5—районы концентрации, 4 — места гнездования, 5 — поверхностные изо
термы в 1967/68 г.
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
25
Северная граница распространения странствующего альба-
троса еще более непостоянна, чем южная. Изменяет она свое
положение и в течение летних месяцев, постепенно опускаясь на
юг. Кроме того, она изменяется в течение коротких промежут-
ков времени при колебаниях температуры воздуха, связанных
с ветрами различных направлений. Так было отмечено, что у
восточного побережья Австралии распространение альбатросов
характеризуется наиболее северным положением в дни сильных
ветров южных напра1влений, особенно при прохождении поляр-
ных циклонов (Amiet, 1958). Летом на мелководье Большого
Австралийского залива странствующий альбатрос также появ-
ляется в основном в дни ветров южных направлений, когда
температура воздуха понижается до 17—2Г (Шунтов, 1968а)!.
Следовательно, границы распространения даже для одного се-
зона являются довольно широкими зонами. Анализ наи-
более северных встреч странствующего альбатроса, по
собственным наблюдениям, а также данным других
авторов (Fleming, 1950; Gibson, 1951; Mac Donald and Lawford,
1954; Laird, 1956; Amiet, 1958; Harrison, 1962; и др.), показыва-
ет, что северная граница его распространения летом (декабрь-
март) проходит в восточной части Индийского океана и в Ти-
хом океане между 29°30z и 35* ю. ш.
Таким образом, область летнего распространения странству-
ющего альбатроса очень обширна и охватывает океан поясом
шириной около 2100—2200 миль. Среднее положение границ
этой зоны в общем соответствует поверхностным изотермам 20
и 0° (см. рис. 8)1 2.
По мере приближения к северному и южному пределам рас-
пространения численность странствующего альбатроса умень-
шается. Основные его концентрации летом в Тихом океане, на
востоке Индийского океана и сопредельных водах Антарктики
наблюдаются между 35—37 и 55—60° ю. ш. (табл. 1, рис. 8),
т. е. занимают зону, охватывающую океан поясом шириной око-
ло 1000—1200 миль. При этом в течение лета происходит посте-
пенное смещение концентраций на юг. Странствующий альба-
трос весьма обычен в пределах очерченных широт от Австралии
До Южной Америки, но наиболее часто встречается в новозе-
ландском секторе Южного океана, где нередко судно сопровож-
дает по 5—20 особей (Gibson, 1967; Ozawa et al., 1968; Шунтов,
1967, 1967a). Только в этом районе рассматриваемый вид бы-
1 Странствующий альбатрос обычно встречается при температуре возду-
ха не выше 18—20°; предельной же температурой для этого вид.а является
21—23° (Laird, 1956; Harrison, 1962; Rand, 1962; мои наблюдения).
2 Положение -средних поверхностных изотерм 20 и 0° довольно близко
положению изотерм воздуха 20 и 0*.
26
АЛЬБАТРОСЫ ЮЖНОГО ОКЕАНА
вает многочислен до 65—67° ю. ш. К западу и востоку он имеет
более северное распространение и встречается гораздо реже.
В осенний период (апрель-май) численность странствую-
щего альбатроса заметно увеличивается в более северных ши-
ротах. В это время северная граница его распространения про-
ходит приблизительно между 27 и 34’ ю. ш., а основные кон-
центрации располагаются между 30—35 и 50’ ю. ш. (см. табл. 1).
Следовательно, как и летом, альбатрос многочислен осенью
в поясе шириной около 1000—1200 миль. Разница состоит лишь
в том, что зона обилия сдвигается к северу в среднем на 300
миль.
Таблица 1
Среднее количество особей странствующего альбатроса на часовой учет
на разных широтах в районе -Австралии, Новой Зеландии я сопредельных
водах Антарктики. 1964—1969 гг.
Районы Месяцы Ш и р о т а
-25 - 30 - 35- 40 -45-50- 55-60 -65 70-75
Декабрь— 0 март 0 ,0.3- 3,0 4,1 2,8 3,2 1,0 0,5 0,4 0
Глубоко- водные Апрель- май 0 + 2.4 3,5 6,6 5.0 0,9 Нет данных
Июнь- сентябрь 0,7 3,5 3,8 5,6 7,0 1,8 1.0 Нет данных
Декабрь- март 0 0,1 0,1 0.4 1.2 3,0 Нет шельфа
Шельфовые Апрель—' май 0 + 0,3 22 0,7 2,1 Нет шельфа
Июнь- сентябрь -г 0.1 0,3 2.5 1,9 1.0 Нет шельфа
Начавшееся с осенним похолоданием смещение странстую-
щего альбатроса на север продолжается зимой. Однако проник-
новение на север в различных районах происходит с различной
интенсивностью. В Индийском океане у южного побережья Ав-
стралии количество странствующего альбатроса осенью увели-
чивается (Шунтов, 19686), но вдоль западного побережья на
север он заходит лишь немного дальше (29—30“ ю. ш.), чем
летом. Лишь в конце зимы граница его распространения при-
ближается к северо-западному углу Австралии'.
1 Австралийские орнитологи (Serventy and Whittell, 1951; Cayley, 1965)
северным пределом называют здесь только широту порта Фримантл, распо-
ложенного в районе 33° ю. ш.
ОЧГРКИ ПО FAC ПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
27
В западной части Тихого океана (моря Тасманово, Корал-
ловое и Фиджи) странствующий альбатрос мигрирует наибо-
лее далеко на север, достигая Большого Барьерного рифа и
Новой Каледонии. По многочисленным регистрациям наиболее
дальних залетов (Laird, 1956; Amiet, 1958; Cayley, 1965; мои
данные) северная граница распространения колеблется здесь
в различные зимние месяцы и годы между 19°30' и 25° ю. ш.
В центральной и восточной частях Тихого океана, по дан-
ным ряда авторов (Murphy, 1936; Fleming, 1950; Harrison, 1962),
странствующий альбатрос доходит лишь до 28’50'—32’ ю. ш-
Однако, по некоторым непроверенным сообщениям, в июне-
июле 1968 г. у побережья Южной Америки он отмечался до
26° ю. ш. По-видимому в зоне холодного Перуанского течения
он проникает еще севернее.
Сведений о наиболее южных встречах альбатроса зимой ма-
ло. Связано это с тем, что в данное время в штормовом Южном
океане плавает мало кораблей. В тихоокеанском секторе Юж-
ного океана странствующий альбатрос встречался в райо-
не 58°57' ю. ш. 159°5Г о. Д. и 57°19' ю. ш. 125’22' з. д. (Szijj,
1967). Таким образом, и в зимний период, как и летом, область
распространения странствующего альбатроса с севера на юг
простирается не менее чем на 2100—2200 миль.
В сравнении с осенним и особенно летним периодом числен-
ность альбатроса зимой в северных широтах значительно уве-
личивается. В этот сезон основные его концентрации находятся
между 30 и 45’ ю. ш., но в Тасмановом море он довольно обы-
чен также и между 25 и 30° ю. ш. Таким образом, в юго-запад-
ной части Тихого, восточной части Индийского океанов и со-
предельных водах Антарктики странствующий альбатрос мно-
гочислен в течение всего года. Летом здесь в глубоководных
районах в среднем на учет приходится около 4, а осенью и зи-
мой около 5 особей. •
Распределение странствующего альбатроса по широтам вес-
ной в целом аналогично распределению в осенний период. Раз-
ница в основном заключается в том, что весной отдельные осо-
би проникают на север почти также далеко, как и зимой. Так,
2 ноября 1968 г. одиночки странствующего альбатроса наблюда-
лись у Новой Каледонии на 19°37' ю. ш.
В описанную выше схему сезонных перемещений странствую-
щего альбатроса, составленную на основании большого количе-
ства учетов, укладываются наблюдения многих других орнито-
логов, выполненные в отдельных районах, например, восточнее
Новой Зеландии (Fleming, 1950; Harrison, 1962; и др.,), в Тасма-
новом море (Gibson and Sefton, 1955; Hindwood, 1955; Amiet
1958; Gibson, 1967) и южнее Новой Зеландии в водах Антаркти-
28
АЛЬБАТРОСЫ ЮЖНОГО ОКЕАНА
ки (Ozawa et al., 1968; Darby, 1970). Вместе с этим некоторые
данные отдельных авторов не совсем точны. В научной и попу-
лярной литературе стало привычным называть область распро-
странения странствующего альбатроса акваторию океана между
30 и 60° ю. ш. Как показано выше, район кочевок альбатроса
обширнее в долготном направлении по крайней мере на 900
миль. Как видно из табл. 1, требуют детализации и данные
Диксона (Dixon, 1933), который писал, что летом и осенью
странствующий альбатрос концентрируется в основном между
30 и 603 ю. ш., а зимой между 30 и 50* ю. ш. Этот автор отме-
чал, что зимой странствующего альбатроса мало в восточной
части Тихого океана. По моим данным, он здесь обычен, хотя
частота встречаемости заметно ниже, чем в западной половине
океана. Саммерхейс (Summerhayes, 1969) писал, что в восточ-
ной части Тасманова моря северная граница распространения
рассматриваемого вида зимой проходит значительно южнее,
чем у пооережья Австралии. Но как было показано выше, гра-
ница проходит от Большого Барьерного рифа к Новой Каледо-
нии строго в широтном направлении, лишь дальше в океане она
опускается несколько к югу. Норрис (Norris, 1965) предпола-
гал, что странствующий альбатрос, откочевывая на юг, в нояб-
ре покидает воды восточного и северо-восточного побережий
острова Северный (Н. Зеландия). Численность этого вида ле-
том у острова Северный, расположенного между 35 и 4 Г ю. ш.,
действительно уменьшается, но он регулярно встречается
здесь все лето (Шунтов, 1967). Несколько искусственной пред-
ставляется схема Гибсона (Gibson, 1967) о перемещениях аль-
батроса в течение зимы в Тасмановом море. Этот автор гово-
рит о смещении скоплений странствующего альбатроса в сере-
дине зимы от побережья Австралии в центральные районы Тас-
манова моря, где в начале зимы он встречается реже. По мо-
им наблюдениям, такой правильной картины в кочевках альба-
троса нет. Он может быть многочисленным в центральных рай-
онах Тасманова моря осенью и в начале зимы, а у побережья
Австралии — во второй половине зимы.
Все изложенное выше о кочевках странствующего альбатро-
са из более высоких широт в низкие и обратно говорит о том,
что перемещения этого типа регулярны и носят массовый харак-
тер. Наряду с описанными перемещениями странствующий аль-
батрос совершает миграции и в широтном направлении, при
этом часть особей с попутными западными ветрами огибает да-
же земной шар (Dixon, 1933; Jameson, 1961).
Много нового в познании миграций странствующего альба-
троса дает кольцевание, весьма интенсивно проводимое в по-
следние годы (Gibson and Sefton, 1959, 1960; Hitchcock, 1961;
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
29
Murray, 1962; Tickell, 1962, 1968; Gibson, 1963, 1967; Sladen et
al., 1968; Tickell and Gibson, 1968).
Особи, занятые выведением потомства, в период размноже-
ния могут улетать на большие расстояния, но обычно держатся
по соседству с островами, на которых располагаются гнезда. Об
этом свидетельствуют наблюдения за окрашенными- краской
альбатросами на Южной Георгии (Tickell, 1968а). Неполовозре-
лые и неразмножающиеся в данном году особи широко разле-
таются по Южному океану, при этом наиболее отчетливо про-
слеживается движение в восточном направлении. Воды Австра-
лии, например, посещают альбатросы, размножающиеся на остро-
вах Южная Георгия, Марион, Крозе, Кергелен. Часть особей
с мест гнездования мигрирует в северном и северо-западном на-
правлениях. Об этом свидетельствуют возвраты из района Ав-
стралии птиц, окольцованных на острове Окленд, а из района
Южной Америки и Африки—окольцованных на Южной Георгии.
Очевидно, для отдельных популяций странствующего альба-
троса существуют традиционные места зимовок и кочевок. В Тас-
мановом море, например, во время работ по кольцеванию среди
отлавливаемых особей большой процент приходился на долю
особей, окольцованных здесь в прошлом году (Gibson, 1967;
Tickell and Gibson, 1968; Tickell, 1968). Здесь же было установ-
лено, что часть особей может держаться в ограниченном райо.
4е по 2—3 недели (Gibson and Setton, 1959).
ib; Как видно из табл. 1, во все сезоны плотность концентраций
Занствующего альбатроса на шельфе в среднем в 5 раз ниже,
Г в глубоководных районах океана. У берегов в значитель-
t количествах он наблюдается только в тех местах, где
шельф узкий и большие глубины подходят близко к берегу.
В таких районах странствующий альбатрос залетает даже в бух-
ты. В частности, постоянно отмечается он в бухте порта Веллинг-
тон (Oliver, 1955; Harrison, 1962; и др.).
Особенно мало странствующего альбатроса на обширных
мелководьях. Сказанное можно наглядно проследить на разрезе
от западного побережья Австралии до Новой Зеландии. В боль-
ших количествах здесь он встречается по кромке шельфа у юго-
западного, юго-восточного побережий Австралии и вокруг Тас-
мании, где мелководья узкие. Наоборот, на обширных шельфах
Большого Австралийского залива и Бассова пролива странствую-
щий альбатрос редок. По выходе из Бассова пролива в Тасмано-
во море он становится опять многочисленным. Количество его
резко снижается на обширном новозеландском мелководье, рас-
положенном западнее пролива Кука; далее к востоку от пролиза
Кука численность снова резко увеличивается'.
Известно, что оперение странствующего альбатроса с возра-
стом светлеет: молодые птицы почти полностью бурого цвета,
30
АЛЬБАТРОСЫ ЮЖНОГО ОКЕАНА
а старые — в основном белые с черными концами крыльев. Осо-
би, начинающие размножение, особенно самки, имеют еще до-
вольно много пестрин на корпусе и голове (Matthews, • 192?);
Murphy, 1936; Rankin, 1951; Oliver, 1955; Holdgate, 1958; Юдин,
1968; и др.). Странствующий альбатрос по окраске подразде-
ляется на 5—6 групп (Fleming, 1950; Murray, 1962; Gibson.
1967; Tickell, 1968). Столь дробная градация несколько громозд-
ка для наблюдений в море, поэтому при учетах я выделял толь-
ко три группы: 1 — в буром оперении, 2 — в промежуточном
оперении, так называемая «леопардовая стадия» (с многочис-
ленными пестринами на корпусе, особенно спине), 3 — в белом
•оперении (спина и остальная часть тела белые, все крылья или
концы их черные).
Как видно из рис. 10, в течение летнего и осеннего сезоноз
в районе Австралии к Новой Зеландии процент молодых птиц в
буром оперении изменяется в сравнительно небольших пределах.
Летом на долю бурых птиц в различных районах приходится
10—24, осенью 10—20%, при этом нельзя выделить места, к ко-
торым явно тяготеют молодые особи. Количество пестрых особей
закономерно увеличивается с запада на восток. В водах южного
и восточного побережий Австралии на их долю приходится ле-
том н осенью 20—25%, в Тасмановом море у Новой Зеландии ле-
том — 20—40, осенью 30—35%. К востоку от Новой Зеландии
.они особенно многочисленны (около 60%). Как писал Соренсен
«(Bailey a. Sorensen, 1962), расположенные в этом районе остро-
ва Антиподов являются основным местом размножения пестрых
•особей. Наоборот, количество белых особей увеличивается* с во-
•стока на запад. Восточнее Новой Зеландии на их долю прихо-
дится в рассматриваемые периоды лишь 20—25%, у южного по-
бережья Австралии — 55—65%, в Тасмановом море увеличивает-
•ся с запада на восток с 40—45 до 60—65%.
Зимой наблюдается, несколько другая картина (рис. 11). Об-
ращает на себя внимание тяготение молодых к более северным
районам1. Их процент составляет между южной частью Большо-
го Барьерного рифа и Новой Каледонией — 63, между Корал-
ловым и Тасмановым морями — 25—48, в Тасмановом море —
10—20, у южного побережья Австралии — 10—22, к востоку от
Новой Зеландии — 1—5. Восточнее Новой Зеландии значитель-
но меньше становится зимой и пестрых особей (у острова Юж-
ный 3%, у острова Северный 45%), но сильно увеличивается про-
цент (50—96%) белых. Процент белых птиц в Тасмановом мо-
ре и у южного побережья Австралии зимой изменяется меньше*
1 Очевидно, это характерно и для других районов. В частности, по наблю-
дениям Ранда (Rand, 1963), в западной части Индийского океана зимой мо-
лодые отмечались в основном только к северу от 39е ю. ш.
Рис. 10. Соотношение различных возрастных групп странствующего альбатроса в районе Ав-
стралии и Новой Зеландии летом и осенью. Вверху — лето, внизу — осень, 1—молодые в буром
оперетт, 2— особи в промежуточном пестром оперении, 3—взрослые в белом оперении
Рис. 11. Соотношение различных возрастных групп странствующего альбатроса в районе
Австралии и Новой Зеландии зимой. Обозначения, как па рис. 10
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
33
соответственно 55—60 и 60—75%. То же наблюдается и в отно-
шении пестрых особей (в Тасмановом море 25—30, у южного по-
бережья Австралии 10—30%).
Дымчатый альбатрос — Phoebetria palpebrata (Forst). В ти-
хоокеанском секторе Южного океана дымчатый альбатрос гнез-
дится на островах Антиподов, Окленд, Кэмпбелл и Макуори (Ale-
xander, 1954; Oliver, 1955; и др.). В летний период у Новой Зе-
ландии он не поднимается севернее южной оконечности острова
Южный. К югу от Большого Австралийского залива в марте
1968 г. он отмечался до 42’40' ю. ш. В период наибольшего про-
грева вод этот вид проникает на юг дальше других альбатросов
В «юре Росса он отмечался до 77’50' ю. ш. (Siple and Lindsey,
1937); обычно же дымчатый альбатрос держится несколько се-
вернее и южная граница его распространения повторяет поло-
жение кромки сплошных льдов. Это значит, что дымчатый аль-
батрос летом достигает побережья Антарктиды.
В летний период дымчатый альбатрос обычен или многочис-
лен у мест размножения, к югу от Тасмании, а также в других
районах, расположенных южнее (рис. 12). В водах Антарктики
Рис. 12 Распределение дымчатого альбатроса — Р. palpebrata s -районе
Австралии, Новой Зеландии и сопредельных водах Антарктики. 1 — граня-
«ы распространения зимой, 2 — границы распространения летом, 3 — по-
верхностные изотермы зимой, 4 — поверхностные изотермы летом, 5 —
районы основных концентраций летом, 6 — места гнездования
34
АЛЬьАТРОСЫ ЮЖНОГО ОКЕАНА
он распространен непрерывно, однако в центральных и восточ-
ных районах тихоокеанского сектора Южного океана имеет
очень неравномерное распределение. В открытом море наиболее
многочислен в антарктических водах между 60 и 70* ю. ш.
(табл. 2).
Таблица 2
Среднее количество особей дымчатого альбатроса на часовой учет
иа различных широтах в австрало-новозеландском секторе Южного океана
в декабре—марте 1964—1968 гг.
Широта 30 — 35 — 40 — 45 — 50 — 55 — 60 - 65 — 70 — 75
Количество
особей _ — _ 0,02 0,3 0,5 0,4 0,9 1.0 0,6
В море дымчатый альбатрос держится рассредоточенно, обыч-
но не более чем по 1—2 экземпляра. Но близ ледовой кромки, у
островов Балени и у мест гнездования за учет можно встретить
по 5—20 особей.
- Осенью дымчатый альбатрос постепенно откочевывает к се-
веру. В апреле отдельные особи достигают Большого Австралий-
ского залива, а в районе Новой Зеландии дымчатый альбатрос
становится обычным до широты пролива Кука (Шунтов, 1967а,
1968а).
В зимний период в северных районах области распростране-
ния дымчатый альбатрос немногочислен, даже редок. У Австра-
лии он не проникает дальше вод южного побережья. У Новой
Зеландии, как и осенью, встречается, по моим данным, в основ-
ном к югу от пролива Кука, но отдельные особи достигают
30° ю. ш. (Sorensen, 1964; Summerhayes, 1969).
Основные концентрации дымчатого альбатроса зимой, как и
летом, остаются южнее 45* ю. ш. В июле на разрезе от пролива
Кука на юго-восток он был обычным между 46*4 Г и 59*18' ю, ш.
(Szijj, 1967). В июне-августе 1968 г. за три месяца наблюдений
в районе острова Южный (Тасманово море, залив Кантербери,
Чатамскнй хребет) я встретил всего 2 экземпляра дымчатого
альбатроса, при этом оба в Тасмановом море.
Таким образом, дымчатый альбатрос в сравнении с другими
альбатросами во все сезоны характеризуется наиболее южным
распространением. Поэтому его миграции изучены хуже, чем у
других альбатросов.
Чернобровый альбатрос — D. melanophrys Tetnm. Самый
многочисленный из альбатросов в Мировом океане. В районе
Австралии и Новой Зеландии гнездится на островах Окленд, Ан-
типодов, Кэмпбелл и Макуори (Alexander, 1954; Oliver, 1955;
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИ! РАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
35
и др.), расположенных в пределах южной умеренной зоны. Об-
ласть кочевок так же обширна, как и у странствующего аль-
батроса.
Распределение чернобрового альбатроса в океане в различ-
ные сезоны претерпевает значительные изменения. Размножаю-
щиеся в данном году особи сравнительно рано заканчивают ко-
чевки и мигрируют* * на юг к районам гнездования. На местах
размножения они появляются в конце августа—сентябре (Oliver,
1955; Bailey and Sorensen, 1962; и др.). Весной по мере прогрева
вод и воздуха постепенно откочевывают в более южные широты
также неполовозрелые и неразмножающиеся в данном году
особи.
Смещение на юг продолжается в течение всего весеннего и
летнего периодов (сентябрь—март). В среднем северная граница
распространения чернобрового альбатроса летом проходит в
районе юго-западного угла Австралии (35° ю. ш.), Большого
Австралийского залива (33’20' ю. ш.), между 33 и 35° ю. ш. в Тас-
мановом море, опускаясь приблизительно до 38—40° ю. ш. во-
сточнее Новой Зеландии (рис. 13) *. На юг он, как и странствую-
щий альбатрос, проникает далеко, достигая паковых льдов. Юж-
нее Новой Зеландии и Австралии чернобровый альбатрос отме-
чался в 1968/69 г. до 65—69’1 Г ю. ш. (рис. 14).
Так как в летние месяцы продолжается гнездовой период и
большое количество особей концентрируется у островов, числен-
ность чернобрового альбатроса в открытых водах заметно ниже,
чем в другие сезоны. Обычно за учет наблюдается не более 2—3
его особей, лншь местами, чаще всего в районе островов, где
есть гнездовья, у судна собирается по 10—20 птиц. Наиболее
многочислен чернобровый альбатрос летом на обширной аква-
тории к югу от Тасманова моря и Новой Зеландии (см. рис.
13-—14). В центральных и восточных районах тихоокеанского
сектора Южного океана численность и плотность населения на-
много ниже.
В апреле и мае количество чернобрового альбатррса в райо-
не Новой Зеландии и южного побережья Австралии увеличи-
вается. Сначала это связано с появлением особей, кочевавших в
южных широтах, а затем — с начавшимся в апреле разлетом от
мест гнездовий. Осенью (апрель-май) чернобровый альбатрос
концентрируется главным образом между 35—45° ю. ш. Чаще все-
го (обычно 3—15 за учет, изредка несколько десятков) он встре-
1 При установлении границ распространения чернобрового и других альба-
тросов, кроме собственных материалов, использованы данные но крайним
встречам альбатросов, приводимые в сообщениях других авторов (Fleming.
*950; Serventy and Whittell, 1951; Gibson, 1951; MacDonald and Lawford,
*954; Hindwood, 1955; Laird, 1956; Amiet, 1958; Harrison, 1962; Cayley, 1965;
" ДР.).
»$0 190 <50
Рис. 13. Распределение чернобрового альбатроса — D. melanophrys в рай-
оне Австралии, Новой Зеландгж и сопредельных водах Антар.стики. 1 —
границы распространения зимой, 2 — границы распространения летом,
3 — поверхностные изотермы зимой, 4 — поверхностные изотермы летом.
5 — районы основных концентраций зимой, 6 — районы основных кон-
центраций летом. 7 — места гнездования
Рис. 14. Распределение чернобрового альбатроса в новозеландском секто-
ре Южного океана летом 1967-1969 гг. 1 южная граница распростране-
ния в 1967/68 г., 2 — южная граница распространения в 1968/69 г., 3 —
районы концентраций, 4 — места гнездования, 5 — поверхностные изо-
термы в 1967/68 г.
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ II МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
37
чается в это время в Тасмановом море, вокруг Новой Зеландии и
у южного побережья Австралии между Тасманией и Большим
Австралийским заливом. В апреле и мае северная граница рас-
пространения в Тасмановом море поднимается до 28*30'—
29*00' ю. ш.. а восточнее Новой Зеландии — до 33*30' ю. ш.
В течение зимних месяцев продолжается распространение
чернобрового альбатроса на север. В Тасмановом море он до-
стигает 24с07'—24*30' ю. ш. Восточнее Новой Зеландии грани-
ца проходит несколько южнее, правда, отдельные особи слу-
чайно достигают острова Питкэн — 25°04' ю. ш. (Williams,
1960). В сравнении с осенним периодом зимой в Тихом океане
заметно увеличивается его численность севернее 40° ю. ш. Зна-
чительно чаще он встречается в это время и у южного побе-
режья Австралии, однако вдоль западного побережья матери-
ка проникает только до 33* ю. ш. Основные концентрации черно-
брового альбатроса зимой, как и осенью, тяготеют к водам юж-
ного побережья Австралия, Тасманова моря и Новой Зелан-
дии (см. рис. 13). Здесь за учет нередко можно встретить 5—20
его особей.
Таким образом, как и странствующий, чернобровый альба-
трос совершает регулярные сезонные перемещения в долготном
в юго-западной
распространения
500—600 миль
открытых водах,
направлении. Наиболее четко они выражены
части Тихого океана, где северная граница
смещается в течение года примерно на
(см. рис. 13).
Рассматриваемый вид многочислен как в
так и у побережий. Однако на обширных мелководьях — Бас-
сов пролив. Большой Австралийский залив, шельф между ост-
ровами Северный и Южный — численность его значительно ни-
же, чем в глубоководных районах.
Желтоносый альбатрос — D. chlororhynchus Gmel. Гнез-
довья этого вида находятся на островах Тристан-да-
Кунья и Гоф в Атлантическом океане, а также на острове Сен-
Прльв Индийском океане (Murphy, 1936; Alexander, 1954; и др.,
Воды Австралии и Новой Зеландии он посещает во время коче-
вок.
В течение всего лета желтоносый альбатрос весьма обычен,
а местами многочислен в Большом Австралийском заливе
(Шунтов, 1968а) и прилегающих к нему районах южного побе-
режья Австралии. Наиболее далеко на север — до 32е ю. ш. он
проникает в это время в Большом Австралийском заливе. У за-
падного побережья Австралии летом желтоносый альбатрос до-
ходит только до юго-западного угла материка (35* ю. ш.). При-
близительно до этой же широты (между 34 и 35° ю. ш.) встре-
чается он летом и у восточного побережья Австралии. Южиая
38
АЛЬБАТРОСЫ ЮЖНОГО ОКЕАНА
граница распространения проходит, по-видимому, где-то в райо*
не 50° ю. ш. В Тихом океане желтоносый альбатрос обычен
только у юго-восточного побережья Австралии (Amiet, 1958).
В открытых водах Тасманова моря численность его резко
уменьшается. Восточнее Новой Зеландии этот вид пока не от-
мечался.
В осенние месяцы желтоносый альбатрос встречается в ос-
новном к северу от 36° ю. ш., при этом по-прежнему наиболее,
многочислен в районе Австралии. Вдоль ее западного побе-
режья в мае он поднимается уже до залива Шарк —
25°40'—26’00' ю. ш., а у восточного побережья материка — до
26’16'—28°00' ю. ш. Основные концентрации его в осенние меся-
цы находятся в Большом Австралийском заливе, прилегающих
к нему водах, а также вдоль западного побережья Австралии
(нередко 3—15. временами по нескольку десятков за учет).
Распространяясь на север, зимой желтоносый альбатрос до-
стигает тропиков. У западного побережья Австралии он отме-
чается до северо-западного угла материка (Serventy and Whit-
tell, 1951; Cayley. 1965; Pocklington, 1967) ’, а у восточного по-
бережья — до 24*24' ю. ш. (Amiet, 1958). Положение южной
границы распространения в зимние месяцы не установлено.
В западной части Индийского океана, по наблюдениям Ранда
(Rand, 1962, 1963), он держится зимой севернее 40° ю. ш. Кон-
центрации в зимний период наблюдаются в тех же районах,
что и осенью; примерно на одном уровне остается и их плот-
ность.
В районе Австралии этот вид более часто, чем чернобровый
альбатрос, встречается в прибрежной зоне. Однако в Большом
Австралийском заливе, где ширина шельфа достигает нескольких
десятков миль, плотность населения его в прибрежных водах
(глубины менее 80 м) в 10—20 раз меньше, чем во внешней
части залива.
Пугливый альбатрос — D. cauta Gould. Эндемик Австрало-
Новозеландского района. Размножается на островах Чатам,
Сиэре, Баунти, Окленд, Альбатрос, Педро Бланка (Alexander,
1954; Oliver, 1955; Cayley, 1965; Hill, 1968).
Летом пугливый альбатрос обычен или многочислен только
в районах гнездования: южнее и восточнее острова Южный, а
также у юго-восточного побережья Австралии (штат Виктория)
и вокруг Тасмании. Северная граница распространения у Но-
вой Зеландии проходит в летние месяцы приблизительно по 40°
ю. ш., а у Австралии — в районе ее юго-восточного побережья
1 В июле 1941 г. мертвый желтоносый альбатрос был обнаружен на ост-
рове Кокос-Кил.тннг — 12° ю. ш. 97° в. д. (Serventy and Whittell, 1951). Не
сомненно, это был залетный экземпляр.
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
39
(крайняя точка встречи 35°52' ю. ш. 138°40' в. д. — Condon.
1946).
Осенью (апрель), закончив размножение, значительная
часть особей, гнездящихся на островах, расположенных южнее
Новой Зеландии, откочевывает к главным островам Новой Зе-
ландии. В апреле и мае у восточного побережья острова Юж-
ный, а также на мелководье, расположенном к западу от про-
лива Кука, он весьма многочислен, и за кормой рыболовного
судна нередко можно наблюдать по нескольку десятков особей
(Шунтов, 1967а). В открытом море пугливый альбатрос регу-
лярно отмечается осенью только южнее и восточнее острова
Южный (не южнее острова Окленд). В водах Австралии он
также обычен преимущественно в пределах шельфа н материко-
вого склона. С мест гнездования, расположенных у Тасмании,
ок распространяется, по-видимому, в основном вдоль южного
побережья.
Распределение пугливого альбатроса в течение зимнего пь
риода имеет много общего с осенним. По-прежнему он немного-
числен в открытых водах. На значительном удалении от бере-
гов вокруг Австралии, в Тасмановом море и восточнее Новой
Зеландии встречаются лишь отдельные экземпляры. Довольно
обычен он в районе шельфа и материкового склона у южного
побережья Австралии. Здесь в учетах встречается регулярно, но
значительных скоплений, по-видимому, не образует.
Как и осенью, основные концентрации пугливого альбатро-
са зимой располагаются у Новой Зеландии. Распределение его
здесь характеризуется следующими особенностями. Вдоль во-
сточного побережья острова Южный, особенно в заливе Каи-
тербери, в течение всей зимы пугливый альбатрос многочислен.
У рыболовных судов здесь нередко собирается по 20—100 осо-
бей. Резко уменьшается его количество у восточного побережья
острова Северный и за пределами свала глубин у острова Юж-
ный. Зимой много пугливого альбатроса наблюдается также в
пределах шельфа и свала глубин Новой Зеландии со стороны
Тасманова моря. У ведущих здесь лов рыбы судов обычно мож-
но видеть 2—15 его особей. Концентрации пугливого альбат-
роса в этом районе держатся всю зиму. В течение осенне-зим-
него периода наибольшая их плотность наблюдается на мелко-
водье .между островами Северный и Южный. Во второй полови-
не зимы здесь количество его несколько уменьшается, но замет-
но увеличивается вдоль всего западного побережья острова Се-
верный. Северная граница распространения пугливого альбат-
роса зимой проходит в районе 33° ю. ш. у юго-западного побе-
режья Австралии и между 29’15' и 33’ ю. ш. в Тасмановом море.
Альбатросы сероголовый — D. chrysostoma Forst, и Буллера
40
АЛЬБАТРОСЫ ЮЖНОГО ОКЕАНА
— D. bulleri Rothsch. Распределение этих видов значительно
отличается друг от друга, однако рассматриваются они вместе,
так как из-за большого внешнего сходства в учетах фиксирова-
лись без разделения. Альбатрос Буллера — эндемик Новой Зе-
ландии; он размножается на островах Чатам и Сиэре. Имею-
щий широкое распространение в Южном океане, сероголовый
альбатрос в Австрало-Новозеландском районе гнездится на ост-
ровах Кэмпбелл и Макуори (Alexander, 1954; Oliver, 1955; и др.).
В летние месяцы эти виды обычно не встречаются севернее
40° ю. ш. У Австралии отдельные особи сероголового альбатро-
са встречаются в это время, очевидно, лишь в районе Тасмании.
Не встречаются оба вида летом и на большей части акватории
Тасманова моря.
Обычен, а местами многочислен альбатрос Буллера у побе-
режья острова Южный и в прилегающих к нему водах, особен-
но у островов Чатам. Южнее количество его значительно умень-
шается; численность же сероголового альбатроса, наоборот,
увеличивается. Последний вид при наблюдениях в 1967—1969 гг.
оказался весьма обычным в открытом море к югу от Но-
вой Зеландии до 65—67° ю. ш.
Осенью, как и другие альбатросы, рассматриваемые виды
расширяют область распространения на север. В апреле и мае
сероголовый альбатрос достигает южного побережья Австра-
лии, а в Тасмановом море отмечается в районе 32*30'—34*00'
ю. ш. Однако эти перемещения не носят такого массового ха-
рактера, как у чернобрового, желтоносого и странствующего
альбатросов. В открытом море в Австрало-Новозеландском
районе осенью они обычны только южнее Новой Зеландии. Ос-
новные концентрации альбатроса Буллера располагаются у во-
сточного побережья острова Южный. В заливе Кантерберм у
рыболовных судов в это время нередко собирается по 10—100
(по'видимому, вместе с сероголовыми) особей. Обычен альбат-
рос Буллера осенью и на мелководье между островами Север-
ный и Южный.
Распределение обоих видов в зимний период мало отличает-
ся от описанного для осени. Это относится к положению,райо-
нов концентраций и к северному пределу распространения.
Известны случаи проникновения сероголового альбатроса вдоль
западного побережья Австралии до залива Шарк (Serventy and
Whittell, 1951; Cayley, 1965). Но постоянно он встречается в во-
дах Австралии, очевидно, только у южного побережья матери-
ка. В течение всей зимы сохраняются концентрации альбатро-
сов у восточного побережья острова Южный (по 10—100 экземп-
ляров у рыболовных судов). Заметно увеличивается их количе-
ство и вдоль западного побережья острова Северный, где У су-
дов обычно держится по 1—5 их особей. Многочислен зимой аль-
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОН
41
батрос Буллера и вдоль западного побережья острова Южный
(Dawson, 1962). В открытом море, как и в другие сезоны, они
обычны только южнее острова Южный.
Некоторые общие закономерности в распределении и мигра-
циях альбатросов южного полушария. Из обзора распределения
отдельных видов видно, что для всех альбатросов характерно
смещение зон их обилия по сезонам в долготном направлении.
В летний период, когда все виды размножаются и много особей
концентрируется у берегов, в открытом море они, как правило,
не многочисленны1. Наиболее часто в это время в Южном океа-
не встречаются мелкие виды альбатросов между 40 и 60° ю. ш.
а также у южного побережья Австралии между 33 и 40” ю. ш.
(табл. 3).
Таблица 3
Среднее количество мелких альбатросов на часовой учет на различных
широтах в районе Австралии, Новой Зеландии и сопредельных водах
Антарктики в декабре—марте 1964—1968 гг.
Широта —30 —35 —40 —45 —50 —55 —60 —65 —70 Всего
Количество
особей + 2,0 1,5 2,1 3,8 2,8 2,4 1,1 0,9 1,9
Начиная с апреля, численность альбатросов в открытых водах
в Австрало-Новозеландском районе увеличивается приблизитель-
но в 4,5 раза (за счет откочевки с юга и разлета с мест гнездова-
ния). При этом зона их концентраций постепенно смещается на
север. В апреле и мае они наиболее многочисленны между 30 и
50* ю. ш. (табл. 4)
Таблица 4
Среднее количество мелких альбатросов на часовой учет
на разных широтах в районе Австралии и Новой Зеландии
в апреле—мае. 1965—1968 гг.
Виды Ш и р о т а
— 30 — 35 — 40 -45 -50 -55 Всего
D. chlororhynchus 1,0 3,2 0,1 + — — 1,2
D. melanophrys + 0,5 4,4 3,0 0,4 + 2,1
D. cauta —— + 0,1 3,6 0,4 0,3 М
D. chrysostoma, D. bulleri — + 0,1 3,3 2,2 0,2 1,1
Неопределенные виды — 1,7 5,0 3,7 0,5 0,2 3,0
Все виды 1,0 5,4 9,7 13,6 3,5 0,7 8Л
1 Все мелкие виды альбатросов размножаются ежегодно, очевидно, только
«ероголовый альбатрос размножается через сезон (Tickell and Pinder, 1967).
Возвраты окольцованных особей чернобрового и сероголового альбатро-
сов показывают, что молодые особи этих видов в небольшом количестве на-
чинают появляться на местах гнездования в возрасте 3 лет, но в основном
в 5 лет, правда, в этом возрасте они еще не размножаются (Tickell, 1967).
42
АЛЬБАТРОСЫ ЮЖНОГО ОКЕАНА
Распределение альбатросов зимой имеет много общего с’рас-
пределением их в осенний период. В зимние месяцы все виды
распространяются наиболее далеко на север, однако основные
концентрации они образуют по-прежнему между 30 и 50° ю. ш.
На одном уровне остается и общая численность альбатросов в
районе (табл. 5). В среднем зона обилия альбатросов смещает-
ся в течение года с севера на юг и обратно примерно на десять
градусов.
Таблица 5
Среднее количество мелких альбатросов на часовой учет
на разных широтах в районе Австралии и Новой Зеландии
в июне—сентябре. 1966—1968 гг.
Виды Широта
— 25 — 30 — 35 — 40 — 45 — 50 — 55 Всего
D. chlororhunchus 0,1 0,6 I»2 0,1 — — — од
D. melanophrys + 0,5 1.2 7<5 4,0 1.3 1.8 2,9
D. cauta — + + 0,9 5,3 + — 1Л
D. chrysostoma,
D. bulleri — + + 0,4 8,6 0,7 оз 23
Неопределенные виды — — 0,8 13 2,9 — — 1.3
Все виды 0,1 1.1 3,3 103 20,8 2,0 2,0 8,4
Наряду с кочевками в долготном направлении, которые вы-
ражены весьма четко, все рассматриваемые виды совершают миг-
рации в широтном направлении, при этом часть особей, несом-
ненно, огибает земной шар.
Существование миграций в широтном направлении обнару-
живается даже без привлечения данных по кольцеванию. Лишь
миграциями с запада на восток можно объяснить наличие кон-
центраций в водах Австралии гнездящегося на западе Индий-
ского океана и в Атлантике желтоносого альбатроса. О мигра-
циях такого рода свидетельствуют и кочующие в водах Южной
Америки особи альбатросов пугливого, королевского—Diomedea
epomophora Less, и Буллера, размножающихся только в Австра-
ло-Новозеландском районе. Почти все виды альбатросов встре-
чаются на обширных пространствах Тихого океана между Новой
Зеландией и Южной Америкой, хотя ни один вид здесь не раз-
множается из-за отсутствия островов.
Регулярность миграций в широтном направлении подтверж-
дена кольцеванием. О результатах кольцевания странствующего
альбатроса говорилось выше. Особи чернобрового и сероголово-
го альбатросов, окольцованные на Южной Георгии, добывались
у южного побережья Австралии и Новой Зеландии (Tickell, 1967;
Sladen et al., 1968); почти все возвраты королевского альбатроса.
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
43
окольцованного на острове Кэмпбелл, приходятся на западное и
восточное побережья Южной Америки (Kinsky, 1963; Robertson,
1964; Tickell, 1968; и др.).
Все имеющиеся данные свидетельствует о том, что при кочев-
ках в широтном направлении преобладающим движением яв-
ляется восточное с попутными западными ветрами. В этом пла-
не не является парадоксальным тот факт, что у Австралии, рас-
положенной сравнительно недалеко от Новой Зеландии, новозе-
ландские альбатросы королевский и Буллера отмечались только
по одному разу (Gibson and Sefton. 1962; Strong, 1964), а у на-
ходящейся за тысячи миль Южной Америки альбатрос Буллера
встречается регулярно, королевский же местами даже многочис-
лен. Конечно, изложенное не является отрицанием существова-
ния миграций с востока на запад. Примером таких перемещений
являются обратные миграции того же желтоносого альбатроса к
местам размножения из вод Австралии, королевского—от Амери-
ки к Новой Зеландии, странствующего — от Австралии на Юж-
ную Георгию.
Говоря о перемещениях мелких альбатросов с запада на во-
сток, нужно заметить, что по массовости и расстояниям они усту-
пают миграциям странствующего альбатроса и тем более цир-
кумполярным перелетам молодых особей гигантского буревест-
ника. Так, желтоносый альбатрос на восток в основном не идет
дальше Австралии, лишь отдельные особи достигают Новой Зе-
ландии. Кольцеванием доказано, что большая часть особей чер-
нобрового альбатроса с Фольклендских-островов мигрирует в во-
ды восточного побережья Америки, а с Южной Георгии —в во-
ды южного и юго-западного побережья Африки (Tickell, 1962,
1967; Sladen et al., 1968). Аналогичная картина, то есть преоб-
ладание кочевок в пределах определенного сектора Южного
океана над циркумполярными миграциями, несомненно, харак-
терна и для мелких видов альбатросов Австрало-Новозеланд-
ского района. Как было показано выше, наибольшие концентра-
ции чернобровый альбатрос образует здесь во внегнездовое вре-
мя на обширной акватории между южной Австралией и Новой
Зеландией. Здесь плотность его концентраций остается пример-
но на одном уровне с осени до весны; восточнее и западнее чис-
ленность чернобрового альбатроса уменьшается. Местом основ-
ных концентраций альбатроса Буллера в течение всего осенне-
го и зимнего сезонов являются воды острова Южный и частич-
но острова Северный. К востоку и западу от Новой Зеландии он
встречается хотя и регулярно, но редко.
Основные концентрации пугливого альбатроса, размножающе-
гося в Новозеландском районе, во внегнездовое время распола-
гаются вдоль восточного побережья острова Южный и в Тасма-
44
АЛЬБАТРОСЫ ЮЖНОГО ОКЕАНА
новом море у побережья Новой Зеландии. Значительная часть
особей пугливого альбатроса, размножающегося в районе Тасма-
нии и Бассовом проливе, по-видимому, не мигрирует дальше вот
юго-восточного и южного побережий Австралии. Пока только в
этих районах обнаруживались особи, окольцованные на острове
Альбатрос в Бассовом проливе (Macdonald and Green, 1963).
Выше рассматривались распределение и миграции альбатро-
сов в пределах их постоянных, так сказать, нормальных обла-
стей распространения. Вместе с этим известно много случаев
обнаружения отдельных особей альбатросов за пределами их
ареалов, в том числе в водах северной Атлантики и Пацифики.
В дополнение к известным ранее залетам (Murphy, 1936; Alexan-
der, 1954; и др.) в последние годы появляются заметки о новых
встречах альбатросов в северных водах (Slipp. 1952; Orlando.
1958; Bull, 1961; Richmond, 1967: Waterston, 1968; Paxton, 1963;
и др.). Обстоятельный обзор дальних залетов трубконосых,
включая альбатросов, недавно дал Буэн (Bourne, 1967).
По частоте залетов в северное полушарие на первом месте
стоит самый массовый из альбатросов — чернобровый, затем
желтоносый (Атлантический океан). Другие виды — странствую-
щий и пугливый, отмечаются редко. Как правило, такие залеты
приходятся на период с осени до весны южного полушария1, ког-
да распределение альбатросов характеризуется наиболее север-
ным положением.
При анализе залетов за пределы ареалов предлагаются раз-
личные объяснения. Во многих случаях попадание в несвойст-
венные районы объясняется заносом птиц штормовыми ветрами,
особенно при прохождении сильных циклонов. В других — свя-
зывается с залетами бродячих или молодых особей, для которых
вообще характерны странствия вдоль побережий материков и
постоянных течений. Через тропическую зону альбатросов могут
перевозить и моряки, которые сопровождающих судно особей ча-
сто ловят на крючки. Мне кажется, что все эти объяснения впол-
не приемлемы. Конечно, встреча южного альбатроса, скажем, у
берегов Европы — факт сам по себе интересный. Тем не менее
подобные залеты не могут рассматриваться как регулярные
трансэкваториальные миграции, свойственные для некоторых
других трубконосых.
Очевидно, все виды альбатросов дальше на север проникают
вдоль побережий. В открытом океане границы распространения
проходят несколько южнее. Воды, прилегающие к побережьям,
отличаются более высокой продуктивностью, и во все сезоны
численность альбатросов здесь выше. Естественно полагать, чь>
там, где концентрируется много особей, больше вероятность про-
никновения части из них в низкие широты.
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
45
Хорошо известно, что холодолюбивые виды птиц проникают
наиболее далеко в низкие широты с холодными течениями, а теп-
лолюбивые виды — в высокие широты с теплыми течениями
(Murphy, 1936; Alexander, 1954; и др.). О такой особенности в
миграциях странствующего альбатроса говорит Джеймесон
(Jameson, 1961). Действительно, во многих случаях наблюдения
подтверждают это правило. В пользу высказанного говорят наи-
более дальние проникновения альбатросов в Антарктику в сек-
торе моря Росса (см. рис. 8, 9, 12, 13, 14). О зависимости распре-
деления от гидрометеорологических условий свидетельствует и
пульсация границ распространения в сезонном аспекте по мере
прогрева или охлаждения вод, а также изменение северной гра-
ницы распространения при ветрах различных направлений. Од-
нако некоторые особенности в распространении альбатросов не
находят объяснения с этих позиций. Выше обращалось внимание
на то, что странствующий и чернобровый альбатросы наиболее
далеко заходят на север в западной части Тихого океана, где на-
правлено на юг теплое Восточно-Австралийское течение. У за-
падного же побережья Австралии в эоне идущего на север хо-
лодного Западно-Австралийского течения они встречаются за-
метно южнее. В этом районе только желтоносый альбатрос в
зимние месяцы достигает тропиков.
Некоторая часть особей альбатросов отличается значитель-
ной эвритермностью. Об этом свидетельствует проникновение их
в тропики, а также преодоление экваториальной полосы бродя-
чими и унесенными штормами птицами. Имея в виду сказанное,
нужно подчеркнуть, что это не отрицает избирательности к темпе-
ратурам и наличия у них определенного температурного диа-
пазона обитания. Так. область распространения странствующего
и чернобрового альбатросов удовлетворительно совпадает с поя-
сом, ограниченном поверхностными изотермами 0 и 20° (см.
рис. 8 и 13). Температуре воды и- воздуха как фактору, опреде-
ляющему границы распространения южных альбатросов, нужно
отвести решающую роль.
В связи с этим нужно заметить, что нередко, особенно в на-
учно-популярной литературе, приуроченность альбатросов к
Южному океану связывается с тяготением их к зоне постоянных
и устойчивых западных ветров. Такое объяснение выглядит не
совсем убедительным. Достаточно сказать, что в обширной пас-
сатной зоне ветры как раз и отличаются наибольшим постоян-
ством и устойчивостью. Альбатросы же в небольшом количест-
ве проникают в южную часть этой зоны лишь зимой, когда тем-
пература воздуха значительно понижается.
Если границы распространения альбатросов определяются
в первую очередь температурой воды и воздуха, то на количе-
46
ГИГАНТСКИЙ буревестник
ственное размещение накладывает отпечаток распределение
продуктивных зон. Правда, как и северотихоокеанские альбат-
росы, благодаря способности к длительному поисковому полету
(и днем, и ночью), эти птицы могут быть довольно многочислен-
ны в районах с невысокими концентрациями корма (странст-
вующий, чернобровый, дымчатый). В этом отношении они отли-
чаются от планктоноядных трубконосых, скопления которых-
приурочены только к высокопродуктивным районам. Тем не ме-
нее тяготение к богатым жизнью зонам (а не локальным уча-
сткам) заметно у всех видов и этой группы.
В летний период, в сезон размножения, альбатросы наибо-
лее многочисленны близ мест гнездования. В другие сезоны са-
мые большие концентрации они образуют в водах шельфа и
свала глубин у восточного (особенно залив Кантербери) и за-
падного побережий Новой Зеландии. Высокая плотность аль-
батросов и в водах, омывающих южное побережье Австралии
и Тасманию. Хотя здесь нередко можно встретить несколько де-
сятков альбатросов за учет, плотность их концентраций и общая
численность ниже, чем у Новой Зеландии. Тяготение основных
концентраций альбатросов к водам шельфа и свала глубин
легко объясняется высокой биологической продуктивностью
этих районов, на чем более подробно я остановлюсь в следую-
щей главе.
3. Гигантский буревестник —
Macronectes giganteus (Gmel).
Широко распространенный в Южном океане вид; имеет циркум-
полярный ареал. В тихоокеанском секторе гнездится на распо-
ложенных южнее Новой Зеландии островах Чатам, Стюарт,
Окленд, Кэмпбелл, Антиподов, Макуори, а также местами в Ан-
тарктике — на Южных Шетландских островах, мысе Адэр
(Alexander, 1954; Oliver. 1955; Reid, 1962; и др.).
Гигантский буревестник размножается ежегодно. Гнездовой
цикл продолжается не менее полгода, при этом сроки яйцеклад-
ки и разлета молодых несколько отличается в различных райо-
нах. На островах Окленд и Кэмпбелл откладка яиц начинается
в августе, на Макуори отдельные особи откладывают яйца в
конце августа, но в основном в начале октября, на Южной Ге-
оргии — в середине октября; на острове Хед, на земле Адели,
в Антарктиде — в конце октября. Разлет молодых наблюдается
в основном в марте и апреле, раньше в тех районах, где рань-
ше откладываются яйца (Mathews. 1929; Oliver, 1955; Downes
et al; 1959; Bailey and Sorensen, 1962; Warham, 1962; Prevost,
1964; и др.). Половозрелым гигантский буревестник становится,
очевидно, не раньше 6—8-летнего возраста (Sladen. 1965; и др.).
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ К МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
47
хотя часть молодых начинает появлятся на островах на 3-ем и
4-ом годах жизни (Ingham, 1959; и др.). Следователь»*?, у ги-
гантского буревестника, как и у альбатросов, в гнездовое вре-
мя в море кочует значительное количество молодых особей.
Распределение в море характеризуется заметной изменчи-
востью по сезонам, очень непостоянной является и северная
граница распределения. В течение лета она постепенно смеща-
ется к югу, в среднем ее положение для лета соответствует ши-
роте южного побережья Австралии и северо-восточной оконеч-
Ррс. 15. Распределение гигантского буревестника — Macronectes giganteus
в районе Австралии, Новой Зеландии и сопредельных водах Антарктики.
1 — границы распространения зимой. 2 — границы распространения ле-
том, 3 — поверхностные изотермы зимой, 4 — поверхностные изотермы
летом, 5 — районы основных концентраций зимой, 6 — районы основных
концентраций летом. 7 — места гнездования, 8 — граница залета отдель-
ных особей зимой
ности Новой Зеландии; к востоку от Новой Зеландии она опу-
скается до 38—40° ю. ш. (рис. 15). Севернее случайно встреча-
ются, по-видимому, только одиночки. Так, в январе у западного
побережья Австралии один окольцованный экземпляр был об-
наружен на широте ЗГ57' ю. ш., а в декабре — у восточного
побережья Австралии, в районе 34“ ю. ш. (Tickell and Scotland,
1961). На юге гигантский буревестник достигает берегов Ан-
тарктиды, где он гнездится. В море Росса отдельные особи про-
никают до 77°45' ю. ш. (Darby, 1970).
48
гигантский буревестник
Основные концентрации гигаитского буревестника распола*
гаются летом между 40 и 70° ю. ш. (табл. 6). Повышенная плот-
ность рассматриваемого вида наблюдается в это время в районе
Новой Зеландии южнее пролива Кука, а также местами в во-
дах Антарктики (см. рис. 15). Наиболее многочислен (до 10—
30 и более за учет) он у островов, на которых расположены
гнездовья. В открытом море, в районах концентраций, отме-
ченных на рис. 15, гигантский буревестник обычен, но, как пра-
вило, за учет наблюдается не более 1—5 особей. К востоку и
западу от Новой Зеландии плотность населения его сильно
уменьшается. Вновь количество гигантского буревестника за-
метно увеличивается на востоке Тихого океана у берегов Юж-
ной Америки, а также в море Беллинсгаузена у Земли Грейа-
ма и Южных Шетландских островов.
Таблица 6
Среднее количество особей гигантского буревестника на часовой учет
на разных широтах в районе Австралия, Новой Зеландии
и сопредельных водах Антарктики. 1964—1968 гг.
Широта
—30 — 35 — 40 — 45 — 50 — 55 — 60 — 65 — 70 — 75 Всего
Декабрь —
март — — 0,2 1,3 1,2 1,6 0,4 0,3 0,5 0,2 0,7
Июнь —
сентябрь 0.1 0,2 1,2 5,5 2,4 1,8 Нет данных 1,5
Таким образом, гигантский буревестник, имея непрерывное
распространение в Южном океане, образует концентрации в
основном в водах, прилегающих к берегам. О тяготении этого
вида к побережьям писали многие авторы, в частности Холгер-
сен (Holgersen, 1957). С одной стороны, эта особенность в рас-
пространении связана с более высокой кормностью прибрежных
вод, а с другой, — с тем, что гигантский буревестник часто пи-
тается падалью (в частности, трупами и плацентами тюленей)
и отбросами разного рода береговых баз и предприятий. На-
глядным примером зависимости распределения буревестника от
деятельности береговых баз может служить изменение его чис-
ленности в бухте порта Веллингтон. Максимальное количество
его обычно наблюдается здесь весной и летом. В это время
происходит массовый забой овец, и в бухту поступает много от-
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
49
бросав. Зимой гигантский буревестник становится здесь немно-'
гочисленным; он концентрируется на другой стороне пролива
Кука в районе Новозеландской китобойной базы, функциони-
рующей в этот сезон. В годы слабого китобойного промысла ги-
гантский буревестник многочислен в бухте Веллингтона и зимой
(Mcllwaine, 1964).
Осенью, в апреле и особенно в мае, количество гигантского
буревестника в связи с разлетом с мест гнездовий, а также
с откочевкой части особей с юга в более северных широтах уве-
личивается. Это очень заметно у Новой Зеландии. Например, в
районе залива Тасман в течение лета, а также в апреле он
встречается регулярно, но за учет обычно отмечается не более
1—2 особей. В мае количество его здесь увеличивается не ме-
нее чем в 3—5 раз.
В зимний период основные концентрации гигантского буре-
вестника в Австрало-Новозеландском районе располагаются
между 35 и 55° ю.ш. Наибольшая плотность (обычно 3—15 за
учет) наблюдается в течение всей зимы на шельфе Новой Зе-
ландии и прилегающих к нему открытых водах. Сравнительно
многочислен он и южнее острова Южный (см. рис. 15). Весь-
ма обычным становится гигантский буревестник зимой и в во-
дах Австралии, но количество его и плотность в целом несрав-
нимо ниже, чем у Новой Зеландии. Восточнее Новой Зеландии,
примерно от 160—170° з. д., а также в открытых водах Тасма-
нова мбря количество его сильно уменьшается, хотя, как и ле-
том, он встречается по всему Южному океану. У берегов Юж-
ной Америки платность населения вновь увеличивается.
Северная граница распространения поднимается зимой до
25—29* ю. ш., но отдельные особи проникают значительно даль-
ше (см.- рис. 15). В восточной части Индийского океана он встре-
чается до северо-западного угла Австралии (Serventy and Whit-
tell, 1951; Cayley, 1965; и др.). Есть сообщение о гигантском бу-
ревестнике с островов Фиджи и Туамоту (Hitchcock and Carrick.
1960; Morgan and Morgan, 1965). У побережья Южной Амери-
ки он достигает Перу (Murphy, 1936).
О распределении гигантского буревестника в водах Антарк-
тнкн зимой известно мало. Южная граница распространения
проходит, очевидно, между 60 и 65° ю. ш. К юго-востоку от Но-
вой Зеландии он отмечался у льдов на широте 62’25' ю. ш.
(Szijj, 1967).
Выше отмечалось, что гнездовой период у гигантского буре-
вестника начинается рано — в августе. Следовательно, откочев-
ка на юг половозрелых особей должна быть еще зимой. Что же
касается молодых особей, то они мигрируют на юг позднее.
Например, у северо-восточного побережья острова Северный их
50
ГИГАНТСКИМ БУРЕВЕСТНИК
количество резко уменьшается только в начале ноября (Norris,
1965).
В последние годы гигантский буревестник весьма интенсив-
но кольцевался на местах гнездования в южной части Атланти-
ческого, Индийского и Тихого океанов (Howard, 1956; Carrick
et al., 1957; Hitchcock and Carrick, 1958, 1960; Sladen and Tickell
1958; Stonehouse, 1958; Ingham, 1959; Tickell and Scotland, 1961;
Tickel, 1962; Hudson, 1963; Sladen, 1965; Sladen et al., 1968; Sib-
son, 1969; и др.). Было показано, что после разлета с островов
молодые особи в течение двух лет совершают протяженные цир-
кумполярные миграции, о чем предполагал раньше Мэрфи
(Murphy, 1936). Большая часть особей при этом смещается в
восточном и северо-восточном направлениях. За 1—2 месяца
некоторые птицы из южной Атлантики достигают Австралии, а
с острова Хед и от Новой Зеландии — Южной Америки. Обыч-
но же перемещение происходит более медленно- Большая часть
особей с островов Южной Атлантики достигает вод Австралии
в середине (июнь-июль), а Новой Зеландии — во второй поло-
вине зимы. Многие особи летят на север или северо-восток, а
некоторые, очевидно, и на северо-запад. Об этом говорят мно-
гочисленные случаи обнаружения в водах Новой Зеландии и
Австралии особей, окольцованных на острове Макуори, а в во-
дах Африки — окольцованных на островах в южной части Ат-
лантического океана.
Таким образом, кольцеванием было подтверждено широкое
рассеивание гигантского буревестника после размножения по
всему Южному океану. Эти данные свидетельствуют, в частно-
сти, о том, что ежегодно в зимние месяцы через Австрало-Но-
возеландский район проходит волна мигрантов из Индийского
и Атлантического океанов. Такие дальние миграции совершают
преимущественно молодые в первые два года жизни. Для моло-
дых же характерно проникновение в более низкие широты. Раз-
множающиеся ежегодно половозрелые особи не совершают
столь протяженных миграций, а остаются в течение зимы в пре-
делах секторов океана, где имеются острова, на которых они
гнездятся. Небольшое количество буревестника держится в те-
чение всей зимы даже у берегов далеко расположенных на юге
островов Макуори (Oliver, 1955; Warham, 1962), Хед (Downes
et al., 1959), Южная Георгия (Mathews, 1929).
Преимущественный разлет молодых буревестников в восточ-
ном направлении справедливо связывается с устойчивыми за-
падными ветрами. В зависимости от устойчивости и преоблада-
ния ветров определенных направлений в отдельные периоды
преобладает передвижение или в восточном, или в северо-вос-
точном направлениях. Например, в годы сильных устойчивых
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОЗ
51
западных ветров с островов южной Атлантики мало буревест-
ников достигает вод южного побережья Африки, а с острова Ма-
куори — вод Новой Зеландии и Австралии, то есть в такие го-
ды в миграциях преобладает восточное направление (Ingham,
1959; Tickell and Scotland, 1961). Имея в виду сказанное, все
же вряд ли можно согласиться с существующими мнениями
(Dorst, 1962) о том, что миграции такого великолепного летуна,
каким является гигантский буревестник, носят пассивный ха-
рактер. Правильнее говорить о том, что птицы используют для
своих миграций ветер. В этой связи нужно заметить, что дан-
ные кольцевания говорят не только о циркумполярных мигра-
циях гигантского буревестника. Они свидетельствуют также о
том, что многие особи, найдя кормный район, могут оставаться
в одном месте длительное время и «не сноситься» ветрами на
восток. Равным образом половозрелые особи, обитая в эоне тех
же западных ветров, не следуют с ними вокруг земного шара.
Известно, что гигантский буревестник диморфен. В северной
половине умеренной зоны светлая морфа редка, по мере про-
движения на юг процент светлых птиц увеличивается. На ост-
рове Хед на долю белых приходится 0,5%, на островах Макуорн
н Южных Оркнейских — 8—10%, в Антарктиде — 12—18%
(Oliver, 1955; Downes et al, 1959; Warham, 1962; Hudson, 1968;
и др.)1. В водах Австралии белые особи не встречаются совсем,
а у Новой Зеландии редкие одиночные птицы достигают остро-
рова Северный (Oliver, 1955). По моим наблюдениям, летом на
широте островов Южный и Северный белая морфа не встреча-
ется совсем, а осенью и зимой на ее долю приходится здесь
0,2—0,3%. при этом белые особи не проникают севернее южной
части острова Северный. Таким образом, особи белой морфы в
зимние месяцы, хотя и мигрируют на север, основной массой
по-прежнему кочуют в более высоких широтах. Аналогичные*
выводы получены при помощи колцевания на Южных Оркней-
ских островах (Hudson, 1968).
4. Капский голубь —
Daption capense (L).
Капский голубь — один из самых массовых и.известных бу-
ревестников в Южном океане. Он постоянно сопровождает ко-
рабли, особенно в больших количествах собирается у промысло-
вых судов и баз.
1 Шогнеоси (Shaughnessy, 1971) приводит несколько другие цифры. По
его Данным в некоторых районах Антарктидой белых птиц около 0,3%, в за-
падных колониях на о. Maxvopn 1—6,8%, в восточной колонии этого остро--
м — 15,1%.
52
КХПСКИИ ГОЛУБЬ
В австрало-новозеландском секторе Южного океана гнездо-
вья располагаются на островах Баунти, Антиподов, Сиэре и на
земле Адели в Антарктиде (Alexander, 1954; Oliver, 1955; Pre-
vost, 1964, 1964а; и др.). Возможно гнездование на острове
Кэмпбелл (Bailey and Sorensen, 1962), Балени и других остро-
вах.
В открытых водах он многочислен во многих районах, в
том числе в сезон размножения, который продолжается с октяб-
ря-ноября до марта—апреля (Downes et al., 1959; Prevost.
1964, 1964 а; Сыроечковский, 1966). Летом в большинстве райо-
нов капский голубь явно тяготеет к водам Антарктики. В част-
Таблица 7
Среднее количество особей капского голубя на часовой учет па различных
широтах в районе Австралии, Новой Зеландии я сопредельных водах
Антарктики. 1964—1969 гг.
UJ и р о т а
' " 25 - 30 — <5 — 4<* — 45 5() 55 - 60 -65-70 - 75 Всего
— 1 _ — --------------------------
.Декабрь-март — — — + 25 8,0 3.0 2,0 7,0 12,0 3,0 6,0
Апрель-май — — — 0,5 5,0 7,0 9,0 нет данных 5,4
Июнь-сентябрь 0,1 1,3 1,5 6,5 16,5 20,0 12,0 нет данных 8,1
ности, в !шдийском секторе Южного океана основные концент-
рации его располагаются в это время вдоль ледовой кромки
(Routh, 1949). Это характерно и для тихоокеанского сектора.
"Однако в районе Новой Зеландии капский голубь в значитель-
ных количествах встречается и в более северных водах — в райо-
не островов Южный, Окленд, Кэмпбелл, Антиподов и Баунти,
где гнездится подвид D. с. australis (табл. 7, рис. 16)-
В декабре отдельные особи капского голубя могут быть еще
встречены в южной половине Тасманова моря, но в разгар лета
граница его распространения опускается здесь до шпроты юж-
ной оконечности острова Южный. Вдоль восточного побережья
Новой Зеландии отдельные особи встречаются до острова Се-
верный (в феврале 1965 г. наблюдались в заливе Хок). Вос-
точнее и западнее Новой Зеландии северная граница распрост-
ранения капского голубя опускается к югу и во второй полови-
не лета севернее 50—60° ю. ш., очевидно, встречаются только
53;
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
.___________________________________________*______
Рис. 16. Распределение капского голубя — Daption capensis в районе Ав-
стралии, Новой Зеландии и сопредельных водах Антарктики. 1 — грани*
цы распространения зимой, 2 — границы распространения летом, 3 —
поверхностные изотермы зимой, 4 — поверхностные изотермы летом,
5 — районы основных концентраций зимой, 6 — районы основных кон-
центраций летом, 7 — известные места гнездования
случайные одиночки. Это объясняет, почему капский голубь не
фигурировал в учетах Диксона (Dixon, 1933), проводившего
наблюдения в Тихом и Атлантическом океанах севернее
60° ю. ш. между меридианами 180 и 0°.
Концентрации капского голубя в умеренных водах Новой
Зеландии и в Антарктике (60—70° ю. ш.) устойчиво держатся
все лето. Главное отличие в распределении в различные месяцы
лета заключается в том, что по мере прогрева вод концентрации
голубя, как и северная граница его распространения, постепен-
но смещаются на юг. Особенно это четко выражено в водах Ан-
тарктики у ледовой кромки, отступающей с потеплением в те-
чение всего лета к югу. В антарктических водах в поясе от 60—
63 до 68—70° ю. ш. капский голубь наиболее многочислен меж-
ду землей Адели и приблизительно 170° з. д. Концентрации здесь
образует южный подвид D. с. capensis (Ozawa et al., 1968)- Он
весьма обычен, а местами многочислен в этих широтах и далее
на восток по всему тихоокеанскому сектору Южного океана. В
Умеренных водах в летний период капский голубь, как уже от-
54
КАПСКИП ГОЛУБЬ
мечалось, в значительных количествах встречается только в
районе Новой Зеландии. Как видно из рис. 16, между районами
скоплений D. с. australis и D. с. capensis существует зона, где го-
лубь редок. Это отмечают и другие авторы (Ozawa et al., 1968;
Darby, 1970). Правда, Озава с соавторами не совсем точен, го-
воря о полном отсутствии голубя в районе, разделяющем кон-
центрации двух подвидов.
С осенним похолоданием первым проникает наиболее далеко
на север D. с. australis. В небольших количествах в апреле и срав-
нительно регулярно в мае он начинает встречаться в открытых
водах южной части Тасманова моря. В это же время вдоль все-
го восточного побережья острова Северный капский голубь уже
обычен, и, как правило, за судном можно видеть по 5—10 его
особей. Через пролив Кука еще в апреле он проникает на мел-
ководье между островами Северный и Южный, при этом в тече-
ние всей осени количество его здесь постепенно увеличивается.
В водах Тасмании отдельные особи капского голубя появляются
в конце апреля, в мае его уже можно встретить у юго-восточно-
го побережья Австралии. Однако обычным и многочисленным у
южного побережья Австралии он становится только к июлю.
В течение первой половины зимы количество капского голубя
в умеренных и субтропических водах продолжает увеличивать-
ся. У западного побережья Австралии северная граница его
распространения поднимается, по данным австралийских орни-
тологов (Serventy and Whittell, 1951; Cayley, 1965; Hill, 1968),
до Джералдтона — 29° ю. ш., а у восточного побережья — до
южной части Большого Барьерного рифа. По моим данным, во
второй половине зимы капский голубь достигает северо-запад-
ного угла Австралии. Здесь же (2Г00' ю. ш.) в июле отмечал
его Поклингтон (Pocklington, 1967).
Восточнее Австралии граница несколько опускается к югу;
в районе Новой Каледонии капский голубь отмечался мною до
2346' ю. ш. В центральной части Тихого океана граница прохо-
дит между 24 и 26* ю. ш. На востоке океана она поднимается
до побережья Эквадора (Harris, 1968)-
Капский голубь отмечался у Маркизских островов (Alexan-
der, 1954; Oliver, 1955), расположенных севернее очерченного
выше предела распространения. Очевидно, столь далеко на се-
вер случайно проникают лишь отдельные бродячие (или зане-
сенные штормами) особи. Это вполне можно допустить, если
учесть, что изредка капский голубь отмечается в северном по-
лушарии (Bourne, 1967; и др).
Южная граница распространения капского голубя зимой
повторяет положение ледовой кромки. Основные концентрации
.его зимой в Австрало-Новозеландском районе располагаются
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
55
на обширной акватории, включающей воды южного побережья
Австралии значительную часть Тасманова моря и воды к югу
и востоку от Новой Зеландии (см. рис. 16). По плотности его
населения первое место ореди этих районов занимают шельф
Новой Зеландии и прилегающие к нему с юга воды океана. Во-
сточнее Новой Зеландии капский голубь многочислен, по-види-
мому, только до 150° з. д.Как показали наблюдения на разре-
зе от Южной Америки к Новой Зеландии в июле 1969 г. (меж-
ду 40 и 45е ю. ш.), между 150 и 80е з- д. капский голубь был
немногочислен; непосредственно у побережья Южной Америки
количество его вновь увеличивалось.
В отмеченных на рис. 16 местах концентраций капского го-
лубя за судном обычно можно наблюдать по 5—20 его особей,
нередко — по 30—100 и более особей.
В связи с наступлением сезона размножения откочевка на
юг части особей происходит рано. На земле Адели этот вид
появляется в середине октября (Prevost, 1964, 1964а). В более
северных районах, в частности на острове Хед, увеличение коли-
чества и активности наблюдается уже в конце августа (Dow-
nes et al., 1959). Однако многие особи капского голубя, очевид-
но, в первую очередь неполовозрелые, остаются в северных во-
дах значительную часть весны. Для примера отметим, что в во-
дах Австралии он довольно многочислен почти до конца октяб-
ря. По наблюдениям Норриса (Norris, 1965), в водах острова
Северный количество капского голубя сильно уменьшается толь-
ко в первые две недели ноября.
Миграции капского голубя в сравнении с миграциями альба-
тросов и гигантского буревестника изучены меньше. Сравни-
тельно мало этот вид и кольцевался. Однако даже небольшое
количество возвратов окольцованных особей позволило полу-
чить интересные выводы- С одной стороны, кольцевание показы-
вает существование протяженных миграций в широтном направ-
лении. Летом на Южных Оркнейских островах было обнаруже-
йо 12 особей, окольцованных в районе Новой Зеландии. Один
голубь, окольцованный в море Уэделла в 1938 г., через 20 лет
был пойман у Новой Зеландии (Hudson, 1963). Несомненно,
для этого вида свойственны циркумполярные миграции, о чем
можно предполагать также исходя из непрерывного распрост-
ранения голубя в Южном океане. С другой стороны, кольцева-
ние показывает тяготение значительной части особей к опреде-
ленным локальным районам. Так, молодые и взрослые особи
капского голубя с Южных Оркнейских островов зимуют глав-
1 В|7,оль ледовой кромки капский голубь, по-видимому, как и летом, зимой
°бычен или многочислен пл всему тихоокеанскому сектору Антарктики. Это
подтверждено наблюдениями в западно-тихоокеанском секторе (Szijj, 1967).
56
ТОНКОКЛЮВЫЙ И СЕРЫЙ 6УРЕВГ.СТНИКИ
ным образом у Чилийского побережья (Beck, 1969). Об этом
же говорят многочисленные случаи обнаружения особей капско-
го голубя у восточного побережья Новой Зеландии, окольцован-
ных здесь в предыдущие годы (Robertson, 1964).
5. Тонкоклювый и серый буревестники — Puffinus
tenuirostris (Temm.), Р. gn'seus (Gmel.).
Среди видов, совершающих трансэкваториальные миграции,
особое место занимают тонкоклювый и серый буревестники.
Ежегодно, когда в северном полушарии наступает весна, мил-
лионы особей этих видов, закончив размножение, мигрируют из
южного полушария в воды северной части Тихого океана.
В начале пятидесятых годов А- М. Судиловокая (1950, 1951)
и Сервенти (Serventy, 1953) независимо друг от друга дали
схемы миграций тонкоклювого буревестника, согласно которым
этот вид в Тихом океане совершает перелеты по часовой стрел-
ке: весной на север — по западной части океана, а осенью на
юг по восточной. В связи с очень ограниченным количеством
наблюдений, выполненных к тому времени в морских условиях,
схемы данных авторов оказались, во многом неточными. Не-
сколько лет назад я предложил новую схему миграций тонко-
клювого буревестника, при этом использовал результаты мор-
ских наблюдений, проведенных в северной части Тихого океана
(Шунтов, 1964). Однако накопленные мною в последние годы
дополнительные материалы (наблюдения от Чукотки и Аляски
до Антарктиды), а также новые литературные данные показы-
вают, что и эта схема нуждается в значительном уточнении.
Филлипс (Phillips, 1963) и Ричдейл (Richdale, 1963) сделали
обзоры пелагического распространения и миграций серого буре-
вестника, из которых следует, что миграции этого вида изучены
значительно меньше в сравнении с миграциями тонкоклювого
буревестника. Филлипс при этом полагает, что часть особей но-
возеландских популяций серого буревестника весной летит по
западной части Тихого океана, а возвращается по восточной..
В целом нужно признать, что морской период жизни обоих
видов известен сравнительно меньше, чем гнездовой, которому
посвящена серия работ (Marshall and Serventy, 1956, 1959; Ser-
venty, 1957, 1958, 1961, 1967; Earner and Serventy, 1959; Richdale,
1963; и др.).
Тонкоклювый буревестник размножается только в районе
Австралии — на юго-восточном побережье материка, в Тасма-
нии и на прилегающих островах (Serventy and Whittell, 1951;
Alexander, 1954; Lane, 1962; Cayley, 1965; и др.). Основные гне-
здовья серого буревестника расположены на побережье и при-
легающих к Новой Зеландии островах (в том числе на Чатам,
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
57
Снэрс, Стюарт, Окленд, Кэмпбелл, Антиподов, Макуоры), а так-
же в районе южной оконечности Южной Америки и на Фоль-
клендских островах. В небольших количествах серый буревест-
ник размножается в районе Тасмании и юго-восточного побе-
режья Австралии (Murphy, 1936; Alexander, 1954; Oliver, 1955;
Richdale, 1963; Cayley, 1965; и др.).
Оба рассматриваемых вида размножаются в весенне-летний
период южного полушария. Половозрелые особи начинают воз-
вращаться на места гнездования в последнюю неделю сентяб-
ря. Откадка яиц происходит в конце ноября — начале декаб-
ря, вылупление птенцов — в январе, отлет с колоний взрос-
лых — в течение апреля, молодых данного года — в конце ап-
реля — начале мая. Тонкоклювый и серый буревестники стано-
вятся половозрелыми в возрасте 5—8 лет. Однако на гнездовых
колониях в больших, количествах они начинают появляться уже
в возрасте 3—4 лет (изредка в возрасте 2 лет). при этом прибыва-
ют они обычно позднее (чем старше, тем раньше) взрослых, а уле-
тают раньше — в феврале-марте (Marshall and Serventy. 1956;
Serventy, 1956; Serventy, 1957. 1958, 1961. 1967; Richdale, 1963,
1964 и др.).
Из изложенного следует, что в период размножения тонко-
клювый и серый буревестники должны быть многочисленными
на море в том оекторе океана, где располагаются их колонии.
Непосредственные наблюдения подтверждают это. В течение
всех весенних и летних месяцев тонкоклювый буревестник мно-
гочислен у юго-восточного побережья Австралии, особенно в
Бассовом проливе и у Тасмании. Концентрации здесь образуют
я взрослые, проводящие дневное время на море, и неполовозре-
лые особи, часть из которых в некоторые периоды выходит на бе-
рег. В этом районе нередко встречаются стаи, насчитывающие
сотни и тысячи экземпляров, а плотность достигает нескольких
десятков и даже сотен птиц на квадратный километр. Харрисон
(Harrison, 1962) ссылается на капитана Флиндерса, который
встреченную в Бассовом проливе стаю оценивал в 12 миллио-
нов особей. По-видимому, такие данные нужно признать чрезмер-
но завышенными, однако они подчеркивают мощность концент-
раций тонкоклювого буревестника в районе гнездования. Столь
же значительные концентрации образует и серый буревестник.
Основная масса его особей, связанных с гнездовыми колония-
ми, концентрируется вдоль восточного побережья острова Юж-
ный, а также в районе островов Стюарт, Снэрс, Окленд, Кэмп-
белл (Bailey, and Sorensen, 1962; Richdale, 1963; Шунтов, 1967;
и др.). Севернее широты пролива Кука численность его резко
сокращается, хотя гнездовья есть и у северной оконечности
острова Северный.
58
ТОНКОКЛЮВЫЙ И СЕРЫЙ БУРЕВЕСТНИКИ
В открытых водах Тасманова моря в летние месяцы числен-
ность обоих видов невелика. К востоку от Новой Зеландии се-
рый буревестник встречается в это время на расстоянии не-
скольких сот миль (Fleming, 1950; Harrison, 1962), однако срав-
нительно многочислен он здесь только до островов Чатам.
Значительная часть особей обоих видов концентрируется
летом в Южном океане в водах, расположенных южнее Австра-
лии и Новой Зеландии. О наличии скоплений серого буревест-
ника в водах Антарктики близ ледовой кромки сообщал ряд
авторов (Falla, 1937; Oordt van and Kruijt, 1953; Phillips, 1963;
Richdale, 1963; Darby, 1970). Есть такие сообщения и по тонко-
клювому буревестнику (Routh, 1949; Ozawa et al., 1968)*.
Я также располагаю данными многих учетов (три летних сезо-
на — с 1967 по 1970 г.), говорящими о том, что в австралий-
ском и новозеландском секторах Антарктики в летние месяцы
скапливаются огромные количества тонкоклювого и серого бу-
ревестников. В восточном направлении буревестники сравни-
тельно обычны в Южном океане до 160—170° з. д. (в основном
серый). Но многочисленны они лишь к западу от 160—170° в. д.
Концентрации их известны на запад до 70’ в. д. (Oordt van and
Kruijt, 1953; Phillips, 1963). Здесь они отмечаются почти в каж-
дом учете, часто можно наблюдать стаи по нескольку десятков
и сот особей. На юг они проникают в значительных количест-
вах до 65’ ю- ш., а отдельные — до 70’ ю. ш. (в секторе моря
Росса). В воды между 55 и 60° ю- ш. отдельные стаи буревест-
ников проникают рано, в середине ноября1 2. В течение лета их
количество увеличивается. В районе ледовой кромки основные
концентрации они образуют в январе—марте.
Скопления буревестников в высоких широтах Антарктики в
гнездовой период образуют, конечно, в первую очередь нетолово-
зрелые особи. В море в период размножения у обоих видов на-
ходится несколько генераций неполовозрелых особей, часть из
которых совсем не появляется у гнездовых колоний или появ-
ляется на непродолжительное время. Как увидим ниже, время
появления стай буревестников в водах Антарктики согласуется
со сроками отлета их из северного полушария. Нужно, однако,
заметить, что в воды Антарктики проникает, по-видимому, и
часть половозрелых особей. Особенно это относится к популя-
ции серого буревестника с острова Макуори, расположенного
на расстоянии всего около 600 миль от ледовой кромки. Оба
вида, являясь великолепными летунами, в период гнездования
1 Филлипс (Phillips, 1963) полагает, что наблюдения Раута (Routh, 1949)
относятся не к тонкоклювому, а к серому буревестнису.
2 В обзорах Филлипса (Phillips, 1963) и Рнчдейла (Richdale, 1963) отме-
чается, что в вады Антарктики серый буревестник мигрирует в период с ян-
варя-февраля по март.
ОЧЕРКИ IiO РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
59
способны улетать на значительные расстояния от островов, на ко-
торых они гнездятся (Phillips, 1963; Richdale, 1963; Serventy, 1967).
Некоторые авторы (Falla, 1937; Oordt van and Kruijt, 1953),
исходя из фактов встречи серого буревестника в индийском сек-
торе Антарктики, предполагали его миграции через Южный
океан в Атлантику. С этим трудно согласиться. Как будет пока-
зано дальше, с наступлением южной осени серый буревестник
мигрирует в северную часть Тихого океана. В южной же Атлан-
тике между 0 и 25° з. д. в летний период он не отмечается
(Bierman and Voous, 1950).
С наступлением в южном полушарии осени тонкоклювый и
серый буревестники начинают миграцию в воды северного по-
лушария. Отлет основной массы птиц на север продолжается
около полутора месяцев. Первыми мигрируют неполовозрелые
особи. Речдейл (Richdale, 1963) наблюдал летящие на север
стаи серого буревестника вдоль восточного побережья Новой
Зеландии в конце марта, когда половозрелые особи еще вы-
кармливали птенцов. В это же время начинаются миграции и у
тонкоклювого буревестника. Родители обоих видов оставляют
птенцов и улетают на север в среднем в середине апреля, хотя
этот процесс растянут приблизительно на месяц (Serventy,
1953, Marshall and Serventy, 1956; Richdale, 1963). Таким обра-
зом, с начала апреля к мигрирующему потоку неполовозрелых
птиц присоединяются закончившие размножение половозрелые
особи. Молодые данного года оставляют острова и улетают
вслед за родителями в конце апреля — начале мая. Из изло-
женного следует, что в течение всего осеннего периода перерыва в
миграциях не наблюдается. Однако преобладание в потоке от-
летающих буревестников сначала неполовозрелых, затем поло-
возрелых и, наконец, молодых данного года позволяет говорить
о трех миграционных волнах.
Численность тонкоклювого и серого буревестников в Авст-
рало-Новозеландском районе заметно уменьшается уже к кон-
цу первой половины апреля, когда отлетает часть половозрелых
особей. Особенно это заметно в районе Бассова пролива и
Тасмании- У восточного побережья Новой Зеландии (остров
Южный) серый буревестник сравнительно многочислен до сере-
дины мая. К началу второй половины мая подавляющая часть
особей обоих видов оставляет воды Австралии и Новой Зелан-
дии. Не встречаются они с этого времени и восточнее Новой
Зеландии (Harrison, 1962). Правда, отдельные особи обоих ви-
дов остаются зимовать в южном полушарии. В отношении вод
Австралии на это указывал Д. Сервенти (Serventy, 1953) в пер-
вом обзоре миграций тонкоклювого буревестника. У Новой Зе-
ландии я также встречал в небольших количествах серого бу-
60
ТОНКОКЛЮВЫП И СЕРЫЙ Г>УРЕПЕСГНИКИ
ревестника в течение всех зимних месяцев. На протяжении
всей зимы мертвые экземпляры серого буревестника обнаружи-
вают и на пляжах Новой Зеландии (Bull and Boeson, 1961).
В период миграций оба вида могут быть встречены в несвой-
ственных для них районах. Есть сообщение об обнаружении
двух особей тонкоклювого буревестника на Цейлоне и в Запад-
ном Пакистане (Phillips, 1951; Jouanin, 1957). Проникает в не-
больших количествах в Индийский океан и серый буревестник
{Gill, 1967).
Пролет буревестников происходит широким фронтом, и
лишь в отдельных случаях представляется говорить о пролет-
Рис. 17. Миграции тонкоклювого — Puffinus tenuirostris и серого —
Р. griseus буревестников из южного полушария в северное. Сплошные
стрелки — тонкоклювый, прерывистые - серый Крупные стрелки — ос-
новные направления, мелкие — второстепенные направления. Заштрихо-
ваны основные районы концентрации
0uri KII ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
61
ных путях (рис. 17). Настоящий поток мигрирующего серого,
буревестника наблюдается, например, в апреле вдоль побе-
режья острова Южный. Однако в это же время серый буревест-
ник встречается на несколько сот миль к востоку от Новой Зе-
ландии, а также в Тасмановом море. Миновав широты Новой
Зеландии и Австралии, серый буревестник широким фронтом пе-
ресекает тропическую зону Тихого океана. Во время миграций
он может быть встречен по всей тропической части Тихого оке-
ана, но большая часть пролетает между Маршалловыми и Мар-
кизскими островами (King, 1967)-
Тонкоклювый буревестник из района размножения, а также
из Антарктики, мигрирует сначала через Тасманово море на севе-
ро-восток. Вдоль восточного побережья Австралии в период
миграций он также встречается, но в небольших количествах.
Основная его масса летит между Новой Зеландией и Новой Ка-
ледонией, затем поворачивает на север ( см. рис. 17). В отли-
чие от серого буревестника при пролете через тропическую зо-
ну на север тонкоклювый буревестник тяготеет к восточному
полушарию (King, 1967).
Тропическую зону оба вида преодолевают быстро, при этом,
по наблюдениям Кинга (King, 1967), они почти не питаются.
В умеренных водах северной части Тихого океана тонкоклювый
буревестник появляется в середине апреля. 22 апреля он наблю-
дался на юге Охотского моря (Austin and Kuroda, 1953), а в се-
редине месяца — несколько южнее Алеутских островов. В за-
ливе Аляска, по данным Габрмельсона и Линкольна (Gabrielson
and Lincoln, 1959), а также Мури (Murie, 1959), тонкоклювый
буревестник прилетает во второй декаде мая. По-видимому, это
слишком поздние сроки; по моим наблюдениям, он еще в кон-
це апреля проникает в Берингово море (Шунтов, 1961). В нача-
ле второй декады апреля отмечался в водах Японии и серый
буревестник (Austin and Kuroda, 1953). В районе Калифорнии
этот вид встречается в течение всего года (Loomis, 1918; Dawson,
1940; и др.). У расположенного севернее острова Ванкувер он
появляется в конце марта (Martin, 1942).
Основная масса обоих видов прибывает в северную часть
Тихого океана в период с конца апреля до конца мая или на-
чала июня. Пролет буревестников проходит здесь также широ-
ким фронтом. В умеренные воды большая их часть проникает
между Японией и Гаванскими островами (см. рис. 17). Особен-
но плотные концентрации на пролете наблюдаются между Хон-
сю и 160° в. д., при этом к берегам Японии буревестники при-
ближаются с юго-востока. В Восточно-Китайское море, по-види-
мому, попадают только отдельные особи обоих видов. Не на-
блюдается заметного пролета буревестников и через Корейский
02 ТОНКОКЛЮВЫЙ И СЕРЫЙ БУРЕВЕСТНИКИ
пролив*. В Японское море значительное количество их проника-
ет через пролив Сангарский и особенно пролив Лаперуза.
Через проливы Курильской гряды много особей серого и
тонкоклювого буревестников проникает в Охотское море. Боль-
шие количества серого буревестника в течение всего лета дер-
жатся на широте Японии; большая же часть стай тонкоклювого
буревестника продвигается на север и северо-восток в Беринго-
во море, в приалеутские воды и залив Аляска- Характерно, что
в апреле н начале мая более выражено движение тонкоклюво-
го буревестника в северо-восточном направлении, в результате
чего в это время он становится очень многочисленным в восточ-
ной части Берингова моря, но в несравнимо меньших количест-
вах встречается в западной части моря и в водах Камчатки.
В последних районах его количество значительно увеличивает-
ся во второй половине мая.
О весенних миграциях тонкоклювого буревестника в северо-
восточной части Тихого океана данных меньше. Раннее появле-
ние его на востоке Берингова моря, на основании чего я делал
вывод о пролете вдоль тихоокеанского побережья Северной
Америки (Шунтов, 1964), можно объяснить миграцией через
западную часть Тихого океана. Все же некоторое количество
тонкоклювого буревестника, по-видимому, приникает на север
между Гавайскими островами и побережьем Северной Амери-
ки. По наблюдениям Гамильтона (Hamilton, 1958), пересекав-
шего в первой половине июня Тихий океан от Японии к Ванку-
веру, оба вида встречались на всем пути; на восточном отрезке
пути преобладал серый буревестник.
О встречаемости в апреле тонкоклювого буревестника в во-
дах Калифорнии упоминал Даусон (Dawson, 1940); правда, он
уточнял, что в последующие месяцы до осени этот вид в рас-
сматриваемом районе не наблюдается. Что касается серого бу-
ревестника, то этот вид, преодолев широким фронтом тропиче-
скую зону, мигрирует весной и через восточную половину север-
ной части Тихого океана. Восточнее Гавайских островов в зна-
чительных количествах отмечал его Кинг (King, 1970). У тихо-
океанского побережья Северной Америки птицы новозеландских
популяций смешиваются с южноамериканскими, которые мигри-
руют по восточной части океана (см. рис. 17)1 2.
Итак, к началу северного лета многомиллионные популяции
1 20 июня 1963 г. в районе Корейского залива были добыты два околь-
цованных тонкоклювых буревестника (Лим Чун, 1963). В Японское маре
они, 1*о-шидртмам1у, проникли через Корейский пролив.
2 Серый буревестник, гнездящийся у южной оконечности Южной Амери-
ки, совершает миграции вдоль побережий в северные части Тихого и Атлан-
тического океанов. В настоящем обзоре он не рассматривается.
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
63
тонкоклювого и серого буревестников, миновав тропическую зо-
ну, сосредоточиваются в северной части Тихого океана и даль-
невосточных морях. В общих чертах распределение их здесь
выглядит следующим образом. В Японском море они встреча-
ются в основном в его северо-восточной части и Татарском про-
ливе- Наибольшие концентрации наблюдаются у входа и в
проливе Лаперуза. В Охотском море в больших количествах
встречаются оба вида. Крупные стаи их особенно часто наблю-
даются в районе пролива Лаперуза и Курильских островов.
На юге Охотского моря более многочислен серый, а в северной
половине моря чаще встречается тонкоклювый буревестник.
Однако численность его здесь не очень высокая, хотя встреча
стай по неокольцу десятков или сот особей не может считаться
необычной. Большие скопления буревестников летом держатся
у восточного побережья Японии, а также у Курильских остро-
вов. Многочисленные наблюдения показывают, что вся обшир-
ная акватория северо-западной части Тихого океана между
Японией и западными островами Алеутской гряды характеризу-
ется повышенной плотностью населения буревестников. Нужно
заметить, что на широте Японии и южных Курильских островов
серый буревестник преобладает над тонкоклювым, ио севернее
в водах Камчатки, Алеутских и северных Курильских островов
соотношение в пользу тонкоклювого. О скоплениях серого буре-
вестника у южных Курильских островов, а тонкоклювого меж-
ду Алеутскими островами и Камчаткой ранее сообщали также
Курода (Kuroda, 1955, 1960) и М. М. Слепцов (1959).
Восточнее 170° в. д. количество буревестников в открытых
водах северной части Тихого океана сильно уменьшается; вновь
плотность их населения увеличивается вдоль побережья Север-
ной Америки. В течение всех летних месяцев у Калифорнии,
Орегона, Ванкувера и восточной части залива Аляска скопле-
ния, которые достигают здесь большой мощности, состоят преиму-
щественно из серого буревестника (Loomis, 1918; Dawson, 1940;
Blake, 1953; Gabrielson and Lincoln, 1959)- В западной части за-
лива Аляска и особенно у Алеутских островов численность се-
рого буревестника уменьшается. Большие скопления в этих рай-
онах образует тонкоклювый буревестник (Arnold, 1948; Murie,
1959; Gabrielson and Lincoln, 1959; мои данные). Особенно мно-
гочислен тонкоклювый буревестник в Беринговом море (Слеп-
цов, 1959; Курочкин, 1963; Шунтов, 1965; Murie, 1959; Gabriel-
son and Lincoln, 1959; и др.). Здесь во многих местах он обра-
зует крупные скопления и резкопреобладаетнад серым буревест-
ником. Последний вид встречается только в южной части моря.
Весной и в начале лета наибольшие концентрации тонкоклю-
вого буревестника наблюдаются на востоке Берингова моря,
где располагаются его линные скопления (Шунтов, 1961). В за-
64
ТОНКОКЛЮВЫЙ и серый буревестники
ладной части моря он становится сравнительно многочислен-
ным только в конце весны. Во второй половине лета тонкоклю-
вый буревестник по-прежнему очень многочислен в восточной
части Берингова моря, правда, основные концентрации смеща-
ются с мелководья в глубоководную часть между островами
Прибылова и Унимаком. Сопоставимые по плотности скопле-
ния в это время наблюдаются и на северо-западе моря в Олкр
торско-Наваринском районе (Шунтов, 1965). При этом значи-
тельное количество его через западную часть моря и Берингов
пролив проникает в Чукотское море, где он известен до остро-
вов Врангеля, Геральда и мыса Барроу (Портенко, 1947; Gab-
ridson and Lincoln. 1959; Swartz. 1967; и др.).
В то время, когда стаи тонкоклювого буревестника еще ле-
тят в Чукотское море, половозрелые особи обоих видов начина-
ют миграции на юг. Отлет половозрелых буревестников, по-ви-
димому, начинается еще в конце августа, но основная масса
отлетает из северного полушария в начале сентября- Период
отлета неполовозрелых птиц сильно растянут: начинается в
сентябре, массовый характер носит в октябре, после чего коли-
чество буревестников в северном полушарии сильно уменьшает-
ся. В Чукотском море у мыса Барроу тонкоклювый буревест-
ник наблюдался даже 19 ноября, а в заливе Аляска в неболь-
ших количествах — в декабре (Gabridson and Lincoln, 1959).
В северо-западной части Тихого океана тонкоклювый буревест-
ник в больших количествах пролетает в сентябре и октябре
(Слепцов, 1959; мои данные). У побережья Калифорнии серый
буревестник отмечается в течение всего года, а тонкоклювый —
с сентября по апрель. Основная масса особей серого проле-
тает здесь в сентябре и октябре, тонкоклювого — в декабре
(Loomis, 1918; Dawson, 1940; и др.). Отдельные особи тонкоклю-
вого буревестника наблюдаются зимой в северо-западной части
Тихого океана (Шунтов, 1966) и у’Гавайских островов (King,
1970).
Серый буревестник осенью мигрирует и по восточной, и по-
западной половинам северной части Тихого океана. Филлипс
(Phillips, 1963) полагает, что часть особей серого буревестника,
как и тонкоклювого, летевшая весной по западной части Тихо-
го океана, осенью возвращается назад вдоль побережья Северной
Америки. По-видимому, это соответствует действительности, од-
нако конкретных данных, подтверждающих сказанное, пока
нет. Тонкоклювый буревестник в сентябре и октябре летит в
основном по западной половине океана, при этом стаи его на-
блюдаются на всей обширной акватории между Курильскими и
Гавайскими островами. В конце осени и начале зимы пролет
происходит в основном по восточной части северной Пацифики.
От побережья Калифорнии стаи тонкоклювого буревестника по-
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
65
ворачивают на юго-запад. В ноябре и декабре восточнее Гавай-
ских островов его наблюдал Кинг (King, 1970)-
Поток серого буревестника у Калифорнии разделяется на
два — часть птиц следует вдоль побережья на юг к гнездовьям
у южной оконечности Америки, а другие поворачивают на юго-
запад в сторону Новой Зеландии (рис. 18). Тропическую часть
Тихого океана серый буревестник пересекает широким фронтом,
но, как и весной, основная масса его пролетает в сентябре и
октябре между Маршалловыми и Маркизскими островами. Через
центральную часть Тихого океана проходит в октябре и ноябре
также главный поток тонкоклювого буревестника (King, 1967).
Так как отлет буревестников из северного полушария про-
исходит в течение нескольких месяцев, прилет их в районы раз-
Рис. 18. Миграции тонкоклювого — Puffinus tenuirostris и серого —
Р. griseus буревестников из северного полушария в южное.
Обозначения, как ва рис. 17
66
ТОНКОКЛЮВЫЙ И СЕРЫЙ БУРЕВЕСТНИКИ
множения сильно растянут. Две стайки тонкоклювого буревест-
ника, летевшие в юго-западном направлении, я наблюдал 16
сентября 1968 г. между Новой Каледонией и Новой Зеландией.
Стаи серого и тонкоклювого буревестников в районах размно-
жения появляются в конце сентября, но пролег продолжается
до декабря (Serventy, 1953, Marshall and Serventy, 1956; Oliver,
1955; Richdale, 1963; и др.)1. Оба вида приближаются к местам
размножения широким фронтом (см. рис. 18).
Мощный пролет тонкоклювого буревестника ежегодно наб-
людается вдоль восточного побережья Австралии (Serventy,
1953; и др.), а серого — вдоль восточного побережья Новой Зе-
ландии (Richdale, 1963; и др). Оба вида на осеннем пролете
встречаются на обширном пространстве между Австралией и
Новой Зеландией, а также на несколько сот миль к востоку от
Новой Зеландии. Кроме собственных наблюдений, об этом сви-
детельствуют данные, приводимые в работах других авторов
(Fleming, 1950; Serventy, 1941, 1953; Harrison, 1962; Phillips,
1963; Oliver, 1955; Norris, 1965; и др.). Часть неполовозрелых
особей, достигнув вод Австралии м Новой Зеландии, по-видимо-
му, сразу следует южнее — в Антарктику. Выше уже отмеча-
лось, что в водах Антарктики они появляются еще в ноябре.
При рассмотрении трансэкваториальных миграций буревест-
ников, -в особенности при объяснении положения путей пролета,
большое значение отводится господствующим ветрам, течениям
(Судиловская, 1950; Kuroda, 1957; Serventy, 1953; и др.) и разме-
щению продуктивных эон (Шунтов, 1964). Несомненно, все эти
факторы имеют большое значение, однако влияние нх иногда
Трактуется упрощенно. При пересечении тропической зоны бу-
ревестники почти не питаются (King, 1967) и преодолевают ее
быстро, без задержки. Поэтому на данном этапе миграций
вряд ли есть смысл искать связи 'путей миграций с расположе-
нием продуктивных зон.
Другое дело — распределение буревестников в районах кон-
центрации в северной и южной частях Тихого океана, а также в
Южном океане. В этом случае наблюдается четкая зависимость
мощности и устойчивости концентраций рт кормноста вод. Так-
же не оправданно на основании совпадении направления переле-
та на отдельных отрезках пути с направлением ветров и тече-
ний делать выводы о том, что миграции буревестников происхо-
дят с попутными ветрами и течениями. Как видно из рис. 17 и
18,^ построенных на основании большого количества наблюде-
ний и литературных данных, преобладающими направлениями
* 14 августа 1968 г. я наблюдал две стайки (по 10 штук) серого буреве-
стника и заливе Кентербери, летящие в юго-западном напра1влании. Впрочем,
это могли быть особи, зимовавшие в южном полушарии
0ЧИРКИ ПО РАСПРЕДЕЛЕНИЮ 1< МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
67
движения между умеренными зонами обоих полушарий являют-
ся направления с юга на север и с севера на юг. Напомним, что
течения и ветры между 40е с. ш. и 40е ю. ш. в центральных рай-
онах океана, где мигрируют буревестники, имеют общее направ-
ление или с востока на запад, или с запада на восток. Следо-
вательно, на этом участке, когда миграции буревестников пра-
вильнее считать перелетом, а не кочевками, пути их проходят
поперек основных течений -и воздушных потоков.
Трансэкваториальные миграции буревестников являются
приспособлением для освоения кормовых ресурсов наиболее
продуктивных вод умеренных зон обоих полушарий, максималь-
ная кормность которых приходится на противоположные
сезоны года (Шутов, 1964). В следующих разделах работы бу-
дут подробно описаны условия обитания в различных районах
Тихого океана. Здесь только следует подчеркнуть, что сроки при-
бытия и пребывания буревестников в обоих полушариях совпа-
дают с максимальным развитием жизни в их водах. В апреле и
мае, когда буревестники покидают воды Австралии и Новой Зе-
ландии, поверхностные слои моря еще богаты планктоном и ры-
бой (Marchall and Serventy, 1956). Но в нюне картина меняется;
уменьшаются биомассы планктона, мелкие пелагические рыбы
реже появляются у поверхности. Зато в умеренных водах север-
ного полушария в мае и нюне, когда сюда прибывает основная
маоса буревестников, наблюдается бурное развитие жизни. Осе-
нью из северного полушария, когда здесь кормовые условия зна-
чительно ухудшаются, улетают на юг не только половозрелые осо-
би, которым предстоит выводить потомство, но и неполовозре-
лые. Они спешат в южное полушарие, в воды умеренной и ан-
тарктической эон, где в это время наступает благоприятный
для размножения и откорма весенне-летний сезон.
Таким образом, в объяснении происхождения и целого ряда
особенностей трансэкваториальных миграций серого и тонко-
клювого буревестников, очевидно, нужно исходить из решающе-
го влияния кормового фактора. Кроме приведенных выше дан-
ных, значение обеспеченности пищей может быть показано и на
Других примерах. Д. Сервенти (Serventy, 1953, 1967) отмечает,
что в годы, когда у восточных берегов Австралии мало планк-
тона, в октябре-январе наблюдается очень высокая смертность
возвращающихся с севера молодых буревестников. Как приспо-
собление к уменьшению пищевой конкуренции, по-видимому,
нужно рассматривать разобщенность мест основных концентра-
ций серого и тонкоклювого буревестников в течение всего года.
Влияние других факторов на распределение рассматриваемых
видов менее существенно. Так, температура воды и воздуха, оп-
ределяющая особенности распределения многих видов, в данном
С8
ГЛУПЫШ
случае заметной роли не играет. Оба вида за короткий проме-
жуток времени способны преодолевать все климатические поя-
са от Антарктики до Арктики.
6. Глупыш — Fulmarus gladalis (L).
Амфибореальный вид, многочисленный и широкораспрост-
раненный в Тихом и Атлантическом океанах. Глупышу посвяще-
на обстоятельная монография Фишера (Fisher, 1952). Но тихо-
океанскому подвиду в этой книге отводится лишь небольшой
раздел.
Известные в Тихом океане гнездовья глупыша не выходят за
пределы умеренной зоны. Большая часть гнездовых колоний
располагается на Курильских, Командорских, Алеутских остро-
вах, в западной части залива Аляска, в Беринговом море на
островах Прибылова и Святого Матвея (Судиловская, 1951;
Fisher, 1952; Gabrielson and Lincoln, 1959; и др.). Ю. В. Аверин
(1948) указывает его гнездование на Камчатке, Н. Е. Кабанов
(I960) — на острове Святого Ионы в Охотском море. Периоди-
чески глупыш размножается на Сахалине на мысе Терпения
(Гизенко, 1955). По всей вероятности, в настоящее время изве-
стны еще не все места гнездования глупыша*. Ниже будет пока-
зано, что есть все основания предполагать его гнездование в се-
веро-восточной части Охотского моря.
Глупыш — полиморфный вид- Многочисленные цветовые ва-
риации его обычно подразделяются на две морфы — светлую и*
темную. Как в Атлантическом, так и в Тихом океане в распро-
странении светлого и темного глупышей наблюдается опреде-
ленная географическая локализация. На гнездовых колониях,
по Фишеру (Fisher, 1952), обобщившему наблюдения ряда ав-
торов, в различных популяциях процент темных птиц составля-
ет: на Курильских островах — 100, на Командорских — 99, в
западной части залива Аляска — 95—99, в восточной части
Алеутской гряды — не менее 60, на островах Прибылова —.
0—5, Святого Матвея — 0.
Таким образом, в более северных районах размножаются в
основном белые особи, в южных — темные. Меньше известно
распространение светлой и темной морф в море. О том, что в
северной части Берингова моря доминирует светлая морфа,
а в южной и прилегающих к ней 'районах Тихого океана — тем-
ная, отмечали многие авторы (Судиловская, 1951; Слепцов,
1 Александер (Alexander, 1954) к местам размножения относит также ост-
рова Врангеля, Геральда, Святого Лаврентия, а А. М. Судиловская (1951) —
острова в Беринговом проливе. 'Возможность гаезд.ования глупыша на севе-
ре Берингова и в южной части Чукотского моря допускает и Фишер (1952).
Эго, однако, ие подтверждается другими авторами.
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ II МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
69
1959; Курочкин, 1963; Шунтов, 1965, 1966; Fisher, 1952; Kuroda,
1955; Gabrilson and Lincoln, 1959; и др.). Однако распределе-
ние морф в море в пределах всего ареала, тем более в сезонном
аспекте, в этих работах детально не рассматривается. Накоп-
ленный мною материал позволяет в настоящее время этот про-
бел заполнить. Осреднение имеющихся данных дает следующую
картину.
В летний период в Чукотском море, а также на шельфе се-
верной и восточной частей Берингова моря, встречается исклю-
чительно белая морфа (рис. 19). Граница распространения тем-
Рпс. 19. Соотношение в учетах светлых п темных особей глупыша —
Fulmarus glacialis летом
ной морфы проходит в это время вдоль материкового склона и
внешней кромки мелководья от Наваринского района к полу-
острову Аляска (Шунтов, 1965). В первой половине лета (июнь-
июль) в районе материкового склона на долю темной морфы
приходится 35—70% (увеличиваясь на юго-восток), а в авгус-
те— начале сентября уменьшается до 5—15%. По мере движе-
ния на юг процент темной морфы увеличивается. В юго-запад-
ной глубоководной половине Берингова моря на ее долю прихо-
дится уже от 90 до 100% (за исключением Олюторского райо-
на).
В северной части Тихого океана повсеместно доминирует
темная морфа. Между Камчаткой, Командорами и централь-
ной частью Алеутской гряды доля белых особей колеблется от
70
ГЛУПЫШ
0 до 5%. В районе восточной части Алеутских островов процент
их увеличивается до 10—30 (соответствует увеличению количе-
ства светлых особей, гнездящихся на островах Прибылова и
Чагулак). По заливу Аляска я располагаю ограниченным коли-
чеством данных. По Фишеру (Fisher. 1952), здесь на долю тем-
ных особей приходится не менее 80—95%, у побережья Канады
процент белых уменьшается до 0,5—2. Южные районы летней
области распространения занимает исключительно темная мор-
фа. Отдельные светлые особи отмечаются приблизительно до
47—48’ с. ш.
В Охотском море распределение морф аналогично распреде-
лению в Беринговом море. На юге летом встречается только
темная морфа, с широты южной оконечности Камчатки появ-
ляется белая (1—3%), доля которой быстро увеличивается в се-
верном направлении, достигая на широте середины полуостро-
ва 50%- Еще севернее процент светлых птиц увеличивается до
95—98 (см. рис. 19). Абсолютное доминирование на севере
Охотского моря белой морфы, а на известных гнездовых колони-
ях (преимущественно Курильские острова) темной, говорит
о том, что в северной части моря должны быть неизвестные
гнездовья этого вида1.
Осенью наблюдается заметное смещение на юг и светлой, и
темной морф (рис. 20). В это время на большей части Беринго-
ва моря доминирует белая1 морфа: на шельфе северо-восточной
половины моря, а также в Олюторско-Наваринском районе —
100%, в районе материкового склона — 70—95%, в централь-
ной части моря и у Командорских островов — 65—85%, в при-
алеутских водах (севернее гряды) — 15%. Значительно увели-
чивается количество светлых птиц у восточного побережья Кам-
чатки (14% у южной оконечности полуострова и северных Ку-
рильских островов, 45—80% между Авачинским заливом и
Командорскими островами). Меньше заметно изменение соотно-
шения светлых и темных птиц в тихоокеанских водах у Алеут-
ских островов: в районе западной и центральной частей гряды
светлых 0—15%, в районе восточной части — 10—30%, в запад-
ной части залива Аляска — 7—12%. В восточной части залива
Аляска белых, по Фишеру (Fisher, 1952), около 25%. В Охот-
ском море светлый глупыш по-прежнему доминирует в север-
ной части моря. На юг он распространяется до широты средней
части Курильской гряды (2—8%).
Зимой светлый глупыш вдоль побережья Америки достигает
Калифорнии (Fisher. 1952), а вдоль Курильских островов — до
। Очевидно, часть белых особен гнездится на острове Святого Ионы, нэ &
этом районе/весьма высок процент и темных птиц (45—90%).
Рис. 20. Соотношение в учетах светлых н темных особей глупыша Fulmarus facialis осенью
72
ГЛУПЫШ
южной части гряды и острова Хоккайдо. В Беринговом море в
это время в районе материкового склона (у льдов) встречают-
ся только светлые особи, их процент уменьшается на юге моря
до 70. Лишь в районе западной и центральной частей гряды зна-
чительна доля темных — 50—85%. Между Камчаткой и Алеут-
скими островами процент светлой морфы колеблется от 35 до
55. У северных Курильских островов на долю светлой приходит-
ся 35%, в районе средней части гряды — 20%, южной —
10—15%. Южнее и восточнее Курильских островов встречаются
только темные особи. В западной части залива Аляска белых
зимой около 30% (рис. 21).
Рис. 21. Соотношение в учетах светлых и темных особей глупыша —
— Fulmarus glacialis зимой
Весной светлый глупыш быстро откочевывает в Берингово и
Охотское моря. В апреле вдоль Курильских островов его остает-
ся не более 1—2%. Южнее Командорских островов и вдоль Але-
утских соотношение морф весной близко к тому, что наблюдает-
ся летом; белых здесь от 2 до 8%. Заметно увеличивается доля
белых (до 30%) в водах восточного побережья Камчатки.
В Беринговом море весной, как и зимой, почти повсеместно пре-
обладают светлые особи (рис. 22). Темная морфа доминирует
только в районе западной и центральной частей Алеутской гря-
ды (85%).
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
73
Рис. 22. Соотношение в учетах светлых и темных особей глупыша —
Fulmarus glacialis весной
Таким образом, во все сезоны в распределении светлой и тем-
ной морф глупыша наблюдается заметная разобщенность. В те-
чение всего года наиболее северные районы ареала занимает
светлая морфа, а южные — темная. При этом в северной поло-
вине Берингова и Охотскогб морей всегда доминирует светлая
морфа, в большинстве районов северной части Тихого океана —
темная. В южной глубоководной половине Берингова моря, а
также между Камчаткой и Командорскими островами, летом до-
минирует темная морфа, осенью и зимой — светлая. В целом
можно заключить, что при откочевке одной морфы на север или
на юг, другая распространяется в этом же направлении.
Географические вариации окраски птиц и млекопитающих со-
ответствуют определенным климатам, отражают различную ин-
тенсивность обмена веществ в различных ландшафтных зонах и
определяются степенью окисления пропигмента меланина.
В этом плане считается; что светлая окраска в условиях сурово-
го климата севера обусловлена сверхокислением, а черная —
высокой степенью окисления пропигмента. Умеренные климаты
дают темную окраску, отражающую средние ступени окисления
пропигмента (Ларионов, 1939; Дементьев и Ларионов, 1944; Де-
ментьев, 1957). В пределах Палеарктики посветление окраски
обычно увеличивается в северном и восточном направлениях
74
ГЛУПЫШ
(Успенский, 1969; и др.). Аналогичная тенденция наблюдается и
в распределении светлого и темного глупышей в Тихом океане.
Однако в Атлантическом океане наблюдается совершенно иная
картина: темный глупыш обитает в северных высокоарктических,
а светлый — в южных, бореальных районах (Fisher 1952; и др.)
В настоящее время это не находит объяснения.
Полиморфизм окраски связан с другими, более глубокими,
различиями и свойствами организмов. В этом смысле цветовые
вариации являются скорее внешним проявлением (индикатора-
ми) полиморфизма (Huxley, 1955; Дементьев, 1957; Майр, 1968).
Нередко распространение полиморфизма имеет параллельный
с географической изменчивостью характер и контролируется од-
ними и теми же факторами1. В таких случаях морфам свойствен-
ны, кроме морфологических, также экологические отличия и гео-
графическая локализация (Дементьев и Ларионов, 1944; Де-
ментьев, 1957). Глупыш может служить хорошим примером,
иллюстрирующим это правило. Как показал Саломонсен (Salo-
monsen, 1965), белый и темный глупыши Атлантики относятся
даже к различным подвидам: темный — F. g. glacial is, светлый—
F. g. auduboni. Распространение первого подвида ограничено
преимущественно высокоарктической зоной, а второго — низко-
арктической и бореальной. Тихоокеанский глупыш в этом отно-
шении не должен представлять исключения. Правда, в таком
плане он пока не исследовался.
Описанное выше размещение в море светлого и темного глу-
пышей в различные периоды года не дает полного представления
о его сезонном распределении. Кратко остановимся на сезонных
перемещениях без разделения на светлых и темных особей.
Зимой, когда мелководные районы дальневосточных морей
покрываются льдами, северная граница распространения глупы-
ша повторяет очертания* ледовой кромки, постепенно смещаясь
в течение зимы на юг. Южная граница в это время проходит
между 30 и 25° с. ш. На западе океана зимой он встречается
вдоль островов Нампо, а на востоке — у полуострова Калифор-
ния. Отдельные особи могут проникать даже в тропики (Fisher,
1952; Austin and Kuroda, 1953; Blake, 1953; King, 1967). Зани-
мая столь обширную область распространения, глупыш распре-
деляется в ее пределах очень неравномерно. Обычен или много-
числен он только к северу от 35—40° с. ш., при этом зона его
основных концентраций огибает северную часть Тихого океана
от Хонсю до тихоокеанского побережья США, включая южные
части Охотского и, особенно, Берингова моря (рис. 23)2. Таким
1 В данном случае климатические и океанографические факторы.
г В связи с изложенным видно, что замечание Б. А. Подкоеыркина (1955)
о том, что зимой глупыша нет у северной части Курильской гряды, неточно.
Рис. 23. Распределение глупыша — Fulrnarus glacialis з Тихом океане зимой. 1— льды, 2—поверхностные изо-
термы, 3—редок, 4—обычен, 5 — многочислен, 6— места гнездования
76
ГЛУПЫШ
образом, несмотря на то, что зимой глупыш рассредоточивается
на очень обширной акватории, значительная часть особей ос-
тается в пределах нескольких сот миль от мест гнездования.
В этом же убеждают рассмотренные выше особенности распре-
деления светлой и темной морф.
Распределение глупыша весной в общих чертах близко к то-
му, что наблюдается зимой. В сравнении с зимним периодом об-
ращает на себя внимание увеличение его количества в Охот-
ском, Беринговом морях и прилегающих к ним водах Тихого
океана у Курильских, Командорских и Алеутских островов. Со
льдами в течение весны смещается и северная граница распро-
странения рассматриваемого вида. Однако в связи с поздним
отступлением льдов в крайние, северные районы глупыш прони-
кает поздно.
В мае, по-видимому, все размножающиеся в этом году особи
концентрируются у мест гнездования. Кладка происходит в
в конце мая — начале июня (Fisher, 1952; Гизенко, 1955; Маре-
ков, 1966; и др.). Как и для других трубконосых, для глупыша
характерно позднее половое созревание. В связи с этим в гнез-
довой период в открытом море остается большое количество
особей. Несомненно, неполовозрелые и холостые особи проника-
ют летом в высокие широты Арктики. В Атлантическом секторе
Арктики по разводьям глупыш почти достигает Северного полю-
са (Портенко, 1946; Успенский, 1969; и др.). Тихоокеанский под-
вид проникает к северу от Берингова пролива не далее 500 миль
(Fisher, 1952).
Летняя южная граница распространения глупыша в видеду-
гн огибает северную часть Тихого океана от северного побе-
режья Хонсю к Калифорнии (рис. 24). Таким образом, в сравне-
нии с зимним периодом область распространения глупыша в
океанских водах сокращается примерно вдвое. Основные кон-
центрации его здесь, как и зимой, тяготеют к курильским, кам-
чатским, командорским и алеутским водам. Очевидно, не мень-
ше половины общего поголовья глупыша кочует летом в Охот-
ском и Беринговом морях. В Охотском море он многочислен по
всей глубоководной части моря, а также в районе Пенжинского
залива, Тауйской губы и на шельфе Камчатки. В-Беринговом
море глупыш чаще всего встречается в районе материкового
склона, прилегающих к нему с севера и юга водах, между остро-
вами Прибылова, Святого Матвея и Чукотским полуостровом, а
также на юге моря вдоль островной гряды (см. рис. 24). Наибо-
лее редок в Беринговом море он в районе заливов Нортон-Саунд
и Бристольском. Это вызывает некоторое недоумение: ведь ря-
дом, на островах Прибылова и Святого Матвея, располагаются
его большие колонии. В Японском море летом глупыш встречает-
ся только в Татарском проливе и прилегающих к нему водах.
Рис. 24. Распределение глупыша — Fuhnarus glacialis в Тихом океане летом. 1—«говоряностные изотермы, 2 — мс-
< ia гнездования, 3 — редок, 4 — обычен, 5 — многочислен
78
ГЛУПЫШ
Сезон размножения глупыша продолжается до сентября —
начала октября, когда с колоний происходит разлет молодых
особей (Fisher, 1952; Гизенко, 1955; Мараков, 1966). В это же
время начинается расширение области распространения на юг.
Первые признаки этого могут быть замечены еще раньше. На-
пример, в центральной части Берингова моря заметно смещение
на юг и светлого, и темного глупышей уже в августе (Шунтов,
1965).
Расширение области распространения глупыша на юг проис-
ходит довольно быстро. В конце октября и ноябре отдельные
особи уже достигают широты южного побережья Японии и полу-
острова Калифорния. Но основная масса в течение осени, как и
летом, продолжает концентрироваться в Охотском и Беринговом
морях, в заливе Аляска, а также в океанских водах, прилегаю-
щих к Курильским, Командорским, Алеутским островам и Кам-
чатке. Выше отмечалось, что в этих районах глупыш многочис-
лен и в другие сезоны, в том числе зимой. Из изложенного вид-
но, что утверждение А. И. Гизенко (1955) о том, что со второй
половины сентября глупыш уже не встречается в районе север-
ной части Курильской гряды, не соответствует действитель-
ности.
Глупыш более чем другие трубконосые северного полушария
использует отходы рыболовного и китобойного промыслов. В этом
отношении он сравним с чайками. Большие его стаи сопровож-
дают суда, ведущие лов рыбы в различных районах северной
части Тихогр океана. Аналогичная картина наблюдается в Ат-
лантике. Привязанность к судам увеличивается в зимний период,
когда кормовые условия ухудшаются. Эта особенность в поведе-
нии глупыша имеет большое значение для поддержания высо-
кий численности и ее роста. Как известно, в Атлантическом
океане в связи с развитием китобойного и рыболовного промыс-
ла глупыш, используя дополнительный источник пищи, за по-
следние 200 лет сильно увеличил численность и расширил гнездо-
вую часть ареала. В пределах умеренной зоны появилось огром-
ное количество новых гнездовых колоний (Fisher, 1952; Larsson,
1960; Holm, 1962; и др.). В Тихом океане точного учета колоний
и численности глупыша не проводилось, тем более в течение дли-
тельного периода времени. Как отмечалось выше, в настоящее
время недостаточно полно выявлено даже размещение гнездо-
вых колоний. Поэтому говорить конкретно о том, как отразилось
на численности глупыша расширение здесь промысла, достигше-
го в последние десятилетия огромных масштабов, не приходится.
Скорее всего в Тихом океане, как и в Атлантическом, произош-
ло увеличение численности глупыша. По мнению М. М. Слепцо
ва (1959), например, на Курильских и Командорских островах
с 1948 по 1954 г. численность глупыша увеличилась в 3—4 раза.
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
7»
7. Обыкновенная моевка — Rissa tridatyla (L).
Арктобореальный вид. Гнездовая часть ареала почти совпа-
дает с таковой толстоклювой кайры. В Тихом океане она гнез-
дится по побережьям и островам от Чукотского моря до верши-
ны залива Аляска н южной части Курильской гряды (Дементьев,
1951; Gabrielson and Lincoln, 1959; и др.). Наиболее крупные ко-
лонии располагаются в Беринговом море.
В летний период морская область распространения моевки
примыкает к районам размножения. Южная граница распрост-
ранения в это время почти не выходит за широты южных гнездо-
вий: она проходит от южной части Курильской гряды вдоль
Алеутских островов в залив Аляска (рис. 25).
Рис. 25. Распределение обыкновенной моевки — Rissa tridactyla в Тихом
океане летом. 1 — поверхностные изотермы, 2 — места гнездования, 3 —
редка, 4 — обычна или многочисленна
В Японском море летом моевки нет, за исключением север-
ной части Татарского пролива, где встречаются редкие одиноч-
ки. Редка моевка и на большей части Охотского моря, особен-
но в его глубоководных районах. Количество ее несколько уве-
личивается в водах шельфа, но многочисленной она становится
здесь только в прибрежной зоне. То же самое можно сказать и о
северной части Тихого океана. Здесь, за исключением прибреж-
ных вод у Курильских, Алеутских островов и залива Аляска, мо-
евка сравнительно обычна только между Камчаткой и западной
частью Алеутской гряды. В последнем районе в июне ее регу-
лярно отмечал также Курода (Kuroda, 1955, 1960). Наиболее
80
ОБЫКНОВЕННАЯ МОЕВКА
часто за пределами прибрежной зоны моевка встречается в Бе-
ринговом море. Здесь она обычна, а местами многочисленна и
образует скопления от Олюторского района до Бристольского
залива и Берингова пролива (Шунтов, 1963). По наблюдениям
Шварца (Swartz, 1967), летом обычна моевка и в свободной от
льдов южной части Чукотского моря. Часть ее особей залетает
далеко на север от ледовой кромки. Проникновение летом в
центральную Арктику вплоть до приполюсных районов для это-
го вида считается довольно обычным явлением (Портенко,
1946; Успенский, 1969; и др.).
Д. Коулсон и Е. Уайт (Coulson and White, 1956, 1959) при
изучении моевки в Англии установили, что этот вид начинает
гнездиться на 3—4 году, при этом в гнездовых колониях в сезон
размножения* держится большое количество неразмножающихся
особей. Наблюдения в дальневосточных морях показывают, что
неполовозрелые и неразмножающиеся в данном году особи мо-
евок концентрируются у берегов не только в тех местах, где есть
колонии.. В больших количествах они собираются у низменных
побережий, в частности в устьях рек. Летом моевка имеет наи-
меньшую в сравнении с другими сезонами область распростра-
нения. Но в связи с тем, что половозрелые и значительная часть
неполовозрелых особей держится у побережий, плотность насе-
ления в открытом море в это время наиболее низкая.
Завершение периода размножения обыкновенной моевки при-
ходится на конец августа в южной и на первую половину сен-
тября в северной части ареала (Дементьев, 1951; Гизенко, 1955;
Kenyon and Phillips, 1965; и др.). На сентябрь приходится и на-
чало осенних кочевок моевки. Наиболее характерным в период
послегнездовых кочевок является быстрое рассредоточение в
сторону открытого моря, а также весьма раннее смещение в об-
щем южном направлении. В сентябре и октябре моевка стано-
вится многочисленной *на всей акватории Берингова моря, в том
числе и в глубоководных районах моря. Большая часть ее осо-
бей мигрирует у азиатского побережья, а также вдоль шельфа
северо-восточной половины моря, где плотность заметно выше,
чем в центральных районах моря. В Охотском море в сентябре
и октябре моевка также становится обычной в глубоководной
части моря, но основные потоки мигрантов направлены вдоль
шельфов по западной и восточной частям моря к проливу Лапе-
руза, Курильским островам и Хоккайдо. В Японское море моев-
ка проникает преимущественно через пролив Лаперуза (часгъ
через проливы Татарский и Сангарский). С середины октября
она становится многочисленной вдоль Приморья, Хоккайдо, но
пролет наблюдается и через центральную глубоководную часть
моря. В конце октября моевка достигает Корейского пролива, а
затем распространяется в Восточно-Китайское и Желтое моря.
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
81
Большое количество особей моевки, откочевавших из Берин-
гова и Охотского морей, в октябре рассеивается по северной
части Тихого океана. Уже в начале октября ее стаи встречаются
в нескольких сотнях миль южнее Алеутской гряпы. В течение
осенних месяцев моевка обычна или многочисленна в открытом
океане на всей огромной акватории между Японией, Камчаткой
и восточной частью Алеутских островов. В северо-восточной ча-
сти Тихого океана (приблизительно восточнее долготы острова
Унимак) наблюдается другая картина. Здесь в открытом океане
моевка немногочисленна. Основные пути ее миграций в этом
районе проходят вдоль шельфа залива Аляска, Британской Ко-
лумбии и далее к Калифорнии. Раньше к таким же выводам
пришел Кеньон (Kenyon, 1949).
К началу зимнего периода моевка в основном покидает райо-
ны, где расположены ее гнездовья. Как видно из рис. 26, не
только гнездовая часть ареала, но и область летнего распростра-
нения моевки в море почти полностью разобщены с районом
зимнего распространения Северная граница распространения
зимой проходит от южной части полуострова Корея к острову
Хоккайдо, затем на северо-восток к западной части Алеутской
гряды и далее к островам Прибылова. Южная граница распро-
странения в виде дуги пересекает Тихий океан от южной части
Восточно-Китайского моря к северной части полуострова Кали-
форния. Отдельные птицы иногда проникают южнее очерченно-
го предела. В частности, залетные особи моевки неоднократно
отмечались на. Гавайских островах (King, 1967; Sibley and
McFarlane, 1968). В пределах очерченных границ распределе-
ние моевки на зимовках неоднородно. Основные концентрации
располагаются в океане южнее Алеутских островов и восточнее
Японии, в юго-восточной части Японского моря, а также у тихо-
океанского побережья Америки (см. рис. 26).
Тот факт, что моевка на зимний период покидает наиболее
суровую по климату и гидрологическому режиму северо-запад-
ную часть ареала, говорит о значении температуры в распреде-
лении данного вида в это время. Этот вывод подтверждается и
конфигурацией северной границы распространения, положение
которой соответствует направлению изотерм (см. рис. 26).
Начало весенних миграций моевки соответствует началу ве-
сеннего прогрева вод. В южной части района зимовок движение
на север начинается со второй половины марта. С самых первых
чисел апреля заметно увеличивается количество моевкн в Берин-
говом море, особенно в его восточной части. Однако в дальней-
шем распространение на север замедляется из-за льдов. Так, на
остров Святого Лаврентия первые птицы прилетают только в
конце первой декады мая (Fay and Cade, 1959), а на остров
Диомида в Беринговом проливе — в середине мая (Kenyon and
1»о ' ito ito
Рис. 26. Распределение обыкновенной моевки — Rissa tridactyla в Тихом океане зимой. I—льды, 2 — поверхно-
стные изотермы, 3—редка, 4 — обычна или многочисленна, 5 - места гнездовании
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
83
Brooks, 1960). Как и осенью, весенний пролет моевки происходит
широким фронтом. Однако в некоторых районах, особенно у по-
бережий и в пределах шельфов, он выражен наиболее интенсив-
но. В Японском море при движении на север птицы тяготеют к
восточной части моря. Очень интенсивные миграции моевки на-
блюдаются в восточной части Берингова моря (в том числе
вдоль ледовой кромки), куда пролетные стайки этого вида про-
никают через проливы восточной части Алеутской гряды. В юж-
ной части ареала завершение отлет'а на север приходится на
май. В Беринговом и Охотском морях и прилегающих к ним во-
дах океана весенние миграции заканчиваются в июне, но и после
этого, как было показано выше, значительная часть особей ос-
тается в открытом море.
По дальности проникновения на юг моевка не выделяется
среди других чаек. Так, вилохвостая чайка — Хета sabinei (Sa-
bine) мигрирует даже в южное полушарие, а многие крупные
чайки (например, полярная — Larus hyperboreus Gunn и сере-
бристая — Larus argentatus Pont.) достигают в небольших коли-
чествах тропиков. Однако по массовости миграций за пределы
гнездовой части ареала и по степени тяготения в период кочевок
к открытым водам моевка резко выделяется среди других чаек.
В этом смысле она классифицируется как пелагический вид.
Как и у других птиц, у моевки наиболее широкие перемеще-
ния совершают молодые особи. Окольцованные в Англии особи
в возрасте 2—3 лет добывались на расстоянии 1360 км от гнез-
довых колоний летом и 2580 км зимой; половозрелые же в воз-
расте 4 лет и старше — не далее 93 км летом и 760 км зимой
(Coulson, 1966). Непосредственные наблюдения показывают, что
у тихоокеанской моевки имеет место аналогичная тенденция. Мо-
лодые, как правило, мигрируют дальше, а возвращаются на се-
вер весной позднее половозрелых.
8. Толстоклювая и тонкоклювая кайры (Uria lomvia L.
U. aalge Pontop.).
Кайры* — одни из наиболее многочисленных морских птиц
в северном полушарии. По ориентировочным подсчетам Така
(Tuck, 1960), общая их численность, по-видимому, не менее
50 млн. особей. Кайры больше других морских птиц используют-
ся человеком в промысловых целях. Это всегда определяло к
ним интерес, в итоге к настоящему времени онм изучены лучше,
чем многие другие виды. Вопросы биологии кайр рассматри-
ваются во многих работах (Кафтановский, 1951; Дементьев,
1 В полевых условиях эти близкие виды не отличимы, поэтому их распреде-
ление и миграции рассматриваются вместе.
84 1ОЛСТОКЛЮВАЯ И ТОНКОКЛЮВАЯ КЛИПЫ
1961а; Гизенко, 1955; Успенский, 1956; Белопольский, 1957; Коз-
лова, 1957; Tuck, 1960; и др.). Нужно, однако, заметить, что све-
дения по тихоокеанским кайрам накоплено несравнимо меньше,
чем по атлантическим. В настоящее время здесь неполностью
выяснено даже распределение их гнездовых колоний. Слабо ис-
следованным, естественно, остается и размещение кайр в море.
Все это нашло отражение в неточности схем в ряде работ, в част-
ности, в наиболее полной, сводке Така (Tuck, 1960). Только в по-
следние годы появилось несколько работ, в которых приводятся
сведения по распределению и миграциям кайр в различных райо-
нах северо-западной части Тихого океана и в дальневосточных
морях (Kuroda, 1955, 1960; Шунтов, 1961, 1965, 1965а, 1966; Гуд-
ков, 1962; Курочкин, 1963).
Граница гнездовой части ареала рода Uria в Тихом океане
почти совпадает с границами тихоокеанской умеренной подобла-
сти. В западной части Тихого океана гнездовья тонкоклювой
кайры известны до Корейского залива и Сангарского пролива,
а толстоклювой — до острова Монерон и южной части Куриль-
ской гряды. В восточной части океана тонкоклювая кайра на
гнездовьях доходит до нижней Калифорнии (острова Санта
Барбара), а толстоклювая — до острова Кодьяк. Толстоклювая
кайра, кроме умеренной зоны, гнездится местами в Арктике, а
тонкоклювая заходит только в южную часть Чукотского моря.
Таким образом, на значительном протяжении гнездовые области
рассматриваемых видов перекрываются, при этом в колониях
'более южных районов преобладает тонкоклювая кайра, а более
северных — толстоклювая (Дементьев, 1951а; Козлова. 1957;
Dawson, 1940; Austin and Kuroda, 1953; Gabrielson and Lincoln,
1969; Tuck, 1960; Udvardy, 1963; и др.).
Южный предел распространения кайр в море в летний пе-
риод почти совпадает и широтой наиболее южных гнездовий.
Ют Корейского залива и Сангарского пролива граница распро-
странения проходит на северо-восток к мористой части залива
Аляска, поворачивая затем вдоль побережья Америки на юг
(рис. 27). За пределами очерченных границ, по-видимому, встре-
чаются только случайные одиночки. Так, Гамильтон (Hamilton,
1958) наблюдал в июне двух кайр в открытом океане в 750 ми-
лях от Алеутских островов.
Летом, то есть в период размножения, за пределами узкой
прибрежной зоны кайры многочисленны только в районах рас.
положения больших гнездовых колоний, при этом распределе-
ние их связано с размерами прилегающих шельфов. Крупные
колонии кайр располагаются на Курильских, Командорских. Але-
утских островах, в заливе Аляска и местами у тихоокеанского
побережья Америки. Во всех этих районах шельф очень узкий.
Соответственно концентрации кайр размещаются узкой полосой
150 180 '^С
Рис. 27. Распределение кайр(13па lomvia и U. aalge) в Тихом океане летом. 1—повераностные изотермы, 2 — ме-
ста гнездования, 3—южная граница гнездования толстоклювой кайры, 4 — редки, 5—обычны, 6 — многочисленны.
86
ТОЛСТОКЛЮВАЯ И ТОНКОКЛЮВАЯ КАЙРЫ
в основном в пределах мелководья и свала глубин; мористее
плотность их населения резко падает. Другая картина наблю-
дается на обширных мелководьях Охотского и особенно Берин-
гова морей. Здесь повышенная плотность наблюдается не толь-
ко у берегов, но и на всей площади шельфов (см. рис. 27).
Многочисленны кайры на несколько десятков миль от берегов и
в Чукотском море, где шельф также большой (Swartz, 1967).
Очень редки кайры на большей части Японского моря. Очень
мало их также в глубоководных районах Охотского моря и поч-
ти везде в открытых водах Тихого океана. Над большими глу-
бинами они сравнительно обычны в это время только в водах
между Камчаткой, Алеутскими и Командорскими островами
(Kuroda, 1955, I960) и в Беринговом море. В этих двух районах
возможна даже встреча небольших стаек кайр.
Описанная схема летнего размещения кайр в море является,
конечно, осредненной. При сравнении распределения в различ-
ные месяцы летнего периода обращает на себя внимание более
частая встречаемость их в открытом море в начале лета, в июне.
В этой связи июнь нужно рассматривать как месяц завершения
весенних миграций к берегам и на север.
Гнездовья кайр располагаются от арктической до южно-
бореальной зоны. Ясно, что в ритмах их годичных циклов в раз-
личных зонах в связи с различием условий обитания должны
быть существенные отличия. В первую очередь нужно отметить
различия в сроках гнездования. В районе Японии, а также в во-
сточной части Тихого океана с их мягкими климатом и гидроло-
гическим режимом кладка яиц начинается еще в первой полови-
не мая. В Охотском и в северной половине Берингова моря в
это время еще льды. В этих районах кайры откладывают яйца
в основном в июне (Дементьев, 1951а; Гизенко, 1955; Козлова,
1957; Kenyon and Brooks, 1960; Tuck, 1960; и др.). В связи с
этим весенние миграции в южной части ареала заканчиваются
раньше, чем на севере. Соответственно на юге раньше начинают-
ся послегнездовые кочевки, к которым кайры приступают после
спуска молодых на воду. В 1963 г. я наблюдал кайр с птенцами
в проливе Лаперуза еще шестого июля. В более северных райо-
нах, где располагаются основные колонии кайр, молодые поки-
дают берег во второй половине августа и в начале сентября.
Первое время молодые и сопровождающие их взрослые держат-
ся вблизи побережий, затем постепенно рассредоточиваются и
открытое море1. В это время распределение кайр близко к тому,
что наблюдается летом. Заметные различия отмечаются со вто-
1 Отдельные особи откочевывают в открытое пюре сразу. Так, 19 августа я
встречал кайр с птенцами в 15 милях от Чукотского полуострова, а 12 сен-
тября 1960 г. — в ста милях от островов Святого Матвея и Прибылова.
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
87
рой половины сентября, когда кайры более быстрыми темпами
начинают распространяться в сторону открытого моря. Харак-
терно, что миграции кайр в начале осени не обязательно ориен-
тированы на юг. В,некоторых районах наблюдается даже дви-
жение на север (Tuck, 1960; Holgersen, 1961).
Осенние миграции, кайр проходят постепенно, часто вплавь и
в основном в пределах шельфов. В Охотском море наблюдается
движение кайр вдоль побережья Камчатки и Сахалина. В част-
ности, кайры, мигрирующие вдоль восточного побережья Саха-
лина ( часть из них гнездится на острове Тюлений), уже во вто-
рой половине октября достигают залива Анива и пролива Лапе-
руза. В Беринговом море осенью, как и летом, кайры наиболее
многочисленны на обширном шельфе северо-восточной половины
моря от залива Анадырского до Бристольского залива и от Бе-
рингова пролива до материкового склона. Здесь наиболее отчег-
ливо прослеживается в конце осени общее смещение кайр в юго-
восточном направлении. Значительно меньше мигрирует их
вдоль побережья Олюторско-Наваринского района и восточного
побережья Камчатки.
Так как откочевка кайр происходит постепенно и медленно,
в северной части ареала они задерживаются до образования
льдов. Например, в Чукотском море у мыса Барроу толстоклю-
вая кайра наблюдалась девятого декабря (Gabrielson and Lin-
coln, 1959). Интенсивное льдообразование в прибрежной зоне
начинается обычно в ноябре. С этого времени распределение
кайр в северных районах дальневосточных морей начинает во
многом зависеть от степени ледовитости и положения ледовой
кромки. В декабре значительная часть мелководий Берингова и
Охотского морей еще свободна от льда. Но в январе уже во мно-
гих районах кромка льда достигает материкового склона (Бул-
гаков, 1965). Не откочевавшие раньше кайры со льдами посте-
пенно отступают на юг. В зимний период их распределение вы-
глядит следующим образом.
В Японском море большое количество кайр зимует в районе
Хоккайдо и пролива Лаперуза, куда они откочевывают из за-
падной части Охотского моря. Здесь их количество увеличивает-
ся во второй половине зимы, когда пролив Лаперуза и приле-
гающие районы Охотского моря замерзают полностью и кайры
вынуждены откочевывать в соседние районы. По восточной ча-
сти Японского моря кайры опускаются зимой только до широты
середины острова Хонсю (Austin and Kuroda, 1953). В западной
части моря отдельные особи (очевидно, тонкоклювая кайра)
проникают почти до Корейского пролива. Сильно увеличивается
их количество зимой в сравнении с другими, сезонами вдоль все-
го Приморья, где сплошным льдом покрывается только при-
88
ГОЛСГОКЛЮВАЯ И ТОНКОКЛЮВАЯ КАЙРЫ
брежная эона и заливы. Характерно, что кайры обычны здесь
не только на шельфе, но и в пределах нескольких десятков миль
от материкового склона. Столь значительное увеличение числен-
ности кайр зимой в западной части Японского моря может быть
объяснено только при условии допущения миграций некоторого
количества их особей из Охотского моря.
В Охотском море в начале зимы (декабрь) кайры многочис-
ленны у кромки льда вдоль Сахалина, Камчатки и, особенно, в
районе пролива Лаперуза. Однако в середине и конце зимы, ког-
да льды закрывают большую часть моря, кайры остаются мно-
гочисленными только в районе Курильских островов и Кам-
чатки.
Много кайр зимует вдоль восточного побережья Хоккайдо,
Курильских островов, восточного побережья Камчатки и в при-
легающих к ним водах Тихого океана. Здесь зимуют кайры,
гнездящиеся в этих районах, а также из Охотского, западной
части Берингова морей и, по-видимому, часть особей, размно-
жающихся на Командорских островах.
В Беринговом море зимой льды покрывают почти все мелко-
водье. Свободные от льда участки на шельфе остаются только
в восточной части Берингова моря. Здесь на чистой воде, а так-
же в разводьях располагаются самые мощные в Тихом океане
зимние концентрации кайр. Высокая плотность населения кайр
(часты стаи, насчитывающие сотни птиц) наблюдается здесь
на обширной акватории от района островов Прибылова до Бри-
стольского залива и пролива Унимак (рис. 28). Положение ле-
довой кромки в течение зимы меняется постоянно. Во многом
оно зависит от направления и силы ветра. При устойчивых се-
верных ветрах льды закрывают почти все мелководье, а при юж-
ных — уходят севернее широты острова Прибылова. В первом
случае это вызывает увеличение концентраций кайр, во втором—
их рассредоточение. В восточной части Берингова моря зимуют
кайры, гнездящиеся на островах северной половины моря, а так-
же часть кайр из Чукотского моря. В свободных от льда глубо-
ководных районах Берингова моря кайры встречаются повсеме-
стно, но в незначительных количествах. Численность их увеличи-
вается у Алеутских островов, особенно в восточной части гря-
ды. Небольшое количество кайр зимует в разводьях на севере
моря.
Многочисленны зимой кайры в заливе Аляска, куда откоче-
вывает часть особей из Берингова моря. В восточной части Ти-
хого океана зимовки кайр располагаются в основном в пределах
шельфа, который здесь имеет небольшую ширину.
В зимний период южная граница распространения кайр за-
нимает крайнее южное положение. В водах Японии она доходит
Pile. 28. Раилгределенме кайр (LJria lorn via и V. aalge) в Тихом океане зимой. 1—поверхностные изотермы,
2 — льды, 3 — места гнездования, 4 — редки, 5— обычны, 6 — многочисленны
90
ГОЛСТОК.1ЮВАЯ И ТОНКОКЛЮВАЯ КАЙРЫ
до шпроты середины острова Хонсю (Austin and Kuroda, 1953),
в виде дуги огибает северную часть Тихого океана и на востоке
опускается до нижней Калифорнии. В открытом океане кайры,
как правило, редки. Весьма обычны они только в полосе не-
скольких десятков миль вдоль Алеутской гряды, а также на об-
ширной акватории между северной частью острова Хонсю и за-
падной частью Алеутских островов (см. рис. 28). Таким обра-
зом, в зимний период, в связи с образованием льдов на значи-
тельной площади шельфа, кайры более часто, чем летом, встре-
чаются в глубоководных районах. Однако их основные концен-
трации, как и в другие сезоны, располагаются в мелководных и
прибрежных районах (см. рис. 28).
Начало массовых весенних миграций кайр приходится на
апрель. Если осенние миграции кайр являются, как правило, по-
степенной откочевкой, совершаемой часто вплавь, то весенние в
ряде районов напоминают массовый пролет перелетных птиц.
Интенсивный весенний пролет кайр в восточной части Берингова
моря был описан мною раньше (Шунтов, 1961). В этом районе
пролет кайр, часто большими стаями, происходит вдоль ледовой
кромки в апреле. Весьма оживленный пролет наблюдается в
апреле и мае также вдоль обоих побережий Камчатки, по за-
падной части Берингова моря, у островов восточной части Але-
утской гряды (особенно, пролив Унимак) и в других районах.
Весной мигрирующие стайки кайр могут быть встречены и в глу-
боководных районах. Так как на весенних миграциях кайры ча-
ще, чем осенью, пользуются крыльями, скорость их перемеще-
ний в это время, конечно, выше. Правда, тон в быстрых весен-
них передвижениях задают особи, которые в данном сезоне бу-
дут размножаться. Неполовозрелые особи, по крайней мере
часть их, мигрируют медленнее. Выше отмечалось, что еще в
июне, когда даже на севере Берингова моря гнездовой период в
разгаре, весенние миграции только заканчиваются. Несомненно,
что в открытом море в это время концентрируются преимущест-
венно неполовозрелые и неразмножающиеся в данном году осо-
би.
Как было видно из изложенного, даже в период зимовок юж-
нее широт гнездования кайры проникают в незначительных ко-
личествах. Кочевки их происходят в основном в пределах гнез-
довой части ареала. Наименьшую протяженность при этом име-
ют миграции у южных популяций, наибольшую — у северных.
Самые дальние перемещения совершают кайры, гнездящиеся в
Чукотском море, на севере Берингова и Охотского морей, кото-
рые покрываются сплошным льдом.
Анализ имеющихся материалов позволяет выявить некото-
рые общие особенности в распределении кайр в море. В опреде-
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
9Г
лении крайних границ распространения обоих видов определен-
ное значение имеет температура, при этом тонкоклювая кайра
является более теплолюбивой. Так (Tuck, 1960) показал, что
южная зимняя граница тонкоклювой кайры соответствует фев-
ральской поверхностной изотерме 15е, а толстоклювой 5е. Как
видно из рис. 28, в районах побережий кайры действительно не
проникают южнее изотермы 15°. Правда, толстоклювая кайра в
небольших количествах проникает в воды с температурой вы-
ше 5е. Зимой она встречается в водах Хонсю и Британской
Колумбии (Austin and Kuroda, 1953; Gabrielson and Lincoln,
1959).
Кайры питаются в основном массовыми видами рыб (сайка,
мойва, песчанка, молодь сельди, трески и другие) и в меньшей
степени — планктоном (Дементьев, 1951а; Успенский, 1956; Бе-
лопольский, 1957, 1971; Tuck, 1960; и др.). Пища добывается
исключительно при нырянии, при этом кайры способны нырять
на глубину в несколько десятков метров. При таком способе пи-
тания, когда птицы лишены возможности в поисках пищи обсле-
довать большие акватории, важное значение приобретает оби-
лие пищи. Хорошо известно, что именно шельфовые воды, а
также воды материкового склона имеют наиболее богатую в
качественном и количественном отношении фауну рыб. За пре-
делами материкового склона плотность концентраций рыб во
много раз уменьшается. Отсюда становится понятным тяготение
основных скоплений кайр к мелководьям во все сезоны года.
Обитая в мелководных районах, кайры могут добывать пищу и в
придонных слоях, когда ее мало в толще воды. В связи с тяготе-
нием к шельфовой зоне южная граница распространения кайр,
в открытом океане не повторяет положение изотерм воды
(см. рис. 27 и 28).
Во многих работах (Storer, 1952; Gabrielson and Lincoln,
1959; Tuck, 1960; Козлова, 1957; и др.) делаются выводы о том,
что многие южные популяции кайр оседлы, а миграции наблю-
даются главным образом в северных районах, где море зимой
замерзает, и что главная причина миграций — образование
льдов. Основанием для таких суждений являются наблюдения
кайр среди льдов в полыньях, а также круглогодичная встре-
чаемость их во многих незамерзающих районах. Действительно,
ледовитость в северных районах играет большую роль в разме-
щении гнездовых колоний, сроках гнездования, характере мигра-
ций кайр (Успенский, 1956, 1969; Tuck, 1960; и др.).
Выше уже отмечалось, что в Тихом океане наиболее протя-
женные миграции совершают кайры, гнездящиеся в Чукотском
море и на севере Берингова и Охотского морей. Здесь именно
ледовая обстановка определяет сроки появления кайр весной,
92
ТОПОРОК
а зимой делает эти районы совершенно непригодными для их
обитания. Однако считать образование льдов единственной
причиной миграции кайр, значит подходить к этому вопросу од-
носторонне. Мне кажется, правильнее рассматривать миграции
кайр, как и других птиц, в связи с сезонной изменчивостью, на-
блюдаемой в кормовой базе, а также в климато-гидрологическом
режиме, где деловитость является только одним из существен-
ных факторов. В пользу такой трактовки говорят многие дан-
ные. В северных районах кайры начинают кочевки еще осенью,
когда море свободно от льда. Выше отмечалось, например, что
в заливе Анива кайры начинают миграции с шельфа восточного
побережья Сахалина еще в октябре. Море у Командорских ост-
ровов всегда свободно от льда. Однако основная масса кайр
этот район зимой покидает. Толстоклювая кайра гнездится не
южнее южной части Курильской гряды, зимой же встречается
в водах Хонсю. В восточной части Тихого океана этот вид гнез-
дится только до острова Кодъяк, а зимой мигрирует до Британ-
ской Колумбии, хотя залив Аляска никогда не замерзает.
Совершают миграции кайры и у тихоокеанского побережья
•США, где зимой очень тепло. Так (Tuck, (1960) приводит дан-
ные о результатах кольцевания тонкоклювой кайры в районе
Орегона, где температура воды зимой не бывает ниже 10°. Из
54 возвращенных колец 25 пришлось на район Орегона, 12 —
Вашингтона, 14 — Британской Колумбии и 3 — Калифорнии.
Аналогичные результаты были получены путем кольцевания
кайр и в Атлантике (Tuck, 1960; Holgersen, 1961; и др.). Таким
образом, если в каком-либо южном районе кайры держатся зи-
мой, то из этого не следует вывод, что это одни и те же осед-
лые особи. В определенные периоды это могут быть особи из
других районов, затем откочевывающие отсюда. Выше, исходя
из непосредственных наблюдений в море, при рассмотрении рас-
пределения кайр на зимовках предположительно назывались ме.
ста зимовок кайр из определенных районов. Однако конкретно
о расположении на зимовках и путях миграции отдельных попу-
ляций представится возможным говорить только после массово-
го кольцевания, которое на Дальнем Востоке пока почти не про-
водилось.
9. Топорок — Lunda drrhata (Pall.).
Характерный тихоокеанский бореальный вид. Гнездовая
часть ареала топорка огромной дугой огибает северную часть
Тихого океана с прилежащими морями от залива Петра Вели-
кого и северной части Хоккайдо до островов Санта Барбара у
Нижней Калифорнии. На севере его гнездовья встречаются до
мыса Лисберн и острова Колючин в южной части Чукотского
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
9S'
моря (Дементьев, 1951а; Козлова, 1957; Gabrielsen and Lincoln,.
1959; Udvardy, 1963; и др.).
Среди чистиковых птиц топорок имеет наиболее широкое рас-
пространение в открытых водах. Даже в летний период, когда’
его основные концентрации смещаются в Охотское и Берингово-
моря, он встречается на всей обширной акватории- океана к се-
веру примерно от 40—45° с. ш. (рис. 29). При этом в некоторых
удаленных от побережий районов он встречается весьма часто.
Курода (Kuroda, 1955, 1960) нашел его обычным в июне между
Камчаткой и Алеутскими островами, а Гамильтон (Hamilton,
1958) — в центральных районах северной части Тихого океана.
Наблюдения этих авторов подтверждаются моими данными
(см. рис. 29).
В летний период, как и в другие сезоны, топорок редок в
Японском море. Здесь он встречается преимущественно в север-
ных водах у побережий. В Охотском море его очень мало толь-
ко в юго-западной глубоководной половине моря. В северной
мелководной половине моря и у Курильских островов он обы-
чен, а местами многочислен. Аналогичная картина наблюдается
и в Беринговом море. В открытых водах океана и морей встре-
чаются главным образом молодые неполовозрелые особи, хоро-
шо отличаемые по коротким косицам и менее массивному и яр-
кому клюву.
Гнездовой период топорка (за исключением южной части
ареала) заканчивается в конце августа — начале сентября (Ги-
зенко, 1955; Мараков, 1966; Gabrielson and Lincoln, 1959; Fay
and Cade, 1959 и др.). В это время после спуска на воду моло-
дых начинаются осенние кочевки. В сентябре топорок еще обы-
чен на большей части Охотского моря, правда, уже с середины
месяца в крайних северных районах моря численность его
уменьшается значительно. В середине сентября начинается от.
кочевка топорка и из Берингова моря. Характерно, что чис.
ленность его в первую очередь сокращается не только на севере
моря, но и в его северо-западной части (Олюторско-Наварин-
ский район), охлаждающейся быстрее центральных и восточных
районов моря. С конца сентября и особенно в октябре топорок
становится немногочисленным и на юге моря.
Из Охотского и Берингова морей подавляющая часть особей
топорка откочевывает в Тихий океан, где они рассеиваются на
огромной акватории. Размещение вида на зимовках выглядит
следующим образом. Северная граница распространения прохо-
дит от южной оконечности Сахалина по Курильским островам,
затем от южного побережья Камчатки к Командорским остро-
вам и далее через южную часть Берингова моря к островам
Прибылова (рис. 30). Положение границы определено в основ-
Рис. 29. Распределение топорка — Lunda cirrhata ib Тихом океане летом. 1 — поверхностные нзо1срмы. 2 .мел а
гнездования, 3 — редок, 4 — обычен, 5 — многочислен
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ
95
ном по литературным источникам (Аверин, 1948; Дементьев,
1951а; Гизенко, 1955; Козлова, 1957; Мараков, 1963; и др. Нужно
заметить, что у северного предела распространения топорок зи-
мой очень редок. В годы исследований я не встречал его зимой,
например, у Курильских островов и Камчатки (Шунтов, 1966).
Как видно из рис. 30, зимовки топорка располагаются преиму-
щественно в водах северной части Тихого океана, севернее при-
близительно 40° с. ш. О зимовках рассматриваемого вида в
открытом океане предполагалось и раньше (Bent, 1919; Hami-
lton, 1958; Шунтов, 1966). Эти предположения подтвердились, но
с чем связано тяготение к открытым водам вида, который, как
и кайры, питается рыбой, добывая ее при нырянии, не по-
нятно.
У побережья Японии южная граница распространения про-
ходит в районе северной половины острова Хонсю (немного юж-
нее 40° с. ш.), а на востоке Тихого оекана опускается до Ниж-
ней Калифорнии (Dawson, 1940; Austin and Kuroda, 1953). От-
дельные особи проникают на юг дальше. Так, зимой 1968 г.
один топорок был встречен к северо-западу от Гавайских остро-
вов на широте 32'30' с. ш. Как видно из сопоставления рис. 29 и
30, положение южной границы распространения топорка в про-
тивоположные сезоны отличается мало; его кочевки ограничи-
ваются широтами размещения их гнездовий. Но необходимо от-
метить два обстоятельства, подчеркивающие значительную про-
тяженность кочевок. Для топорка, как указывалось выше, боль-
ше других чистиковых характерен пелагический образ жизни;
зимой он обитает преимущественно в океанических водах. Ха-
рактерно также, что наибольшие сезонные перемещения свой-
ственны для популяций, населяющих Охотское, Берингово моря
и прилегающие районы северо-западной части Тихого оекана.
Эту часть ареала, где летом вид наиболее многочислен, зимой
он покидает. В результате оказывается, что основная часть райо-
на зимовок только примыкает к дальневосточным морям.
Ранняя откочевка топорка из дальневосточных морей, отсут-
ствие его зимой в наиболее охлажденной части ареала, а также
тяготение на зимовках к наиболее теплым районам бореальной
области говорят об относительной теплолюбивости и влиянии на
его распределение температурного фактора. В связи с этим на-
ходится и позднее возвращение топорка весной в северные воды.
Если у Калифорнии рассматриваемый вид начинает размножать-
ся в апреле (Dawson, 1940), то на местах гнездования на Кам-
чатке, Курильских, Командорских островах и островах Прибы-
лова он появляется только в конце апреля — начале мая (Аве-
рин, 1948; Гизенко, 1955, Мараков, 1963; Gabrielson and Lincoln,
1959), а на севере Еерингова моря — лишь в конце мая и начз-
Рис. 30. Распределение топорка — Lunda cirrhata в Тихом океане зимой. 1 — поверхностные изотермы, 2 — льды,
3 — места гнездования, 4 — область кочевок
ОЧЕРКИ ПО РАСПРЕДЕЛЕНИЮ И МИГРАЦИЯМ ОТДЕЛЬНЫХ ВИДОВ 97
пе нюня (Козлова, 1957; Fay and Cade, 1959; Kenyon and Brooks,
‘i960).
Как и зимой, в период весенних кочевок в открытых водах
топорок держится рассредоточенно, мигрируя широким фрон-
том. Значительные скопления отмечаются только в мелковод-
ных районах, например, в водах полуострова Аляска, восточной
части Алеутской гряды, северных Курильских островов и южно-
го побережья Камчатки. Как отмечалось выше, в районы гне-
здования мигрируют далеко не все особи топорка. После за-
вершения весенних перемещений много их (преимущественно
неполовозрелых) держится все лето в открытом океане.
98
ГЛАВА II
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ РАЗЛИЧНЫХ
РАЙОНОВ ТИХОГО И И НДИЙСКОГО ОКЕАНОВ
1. Берингово море
Берингово море с его очень богатой фауной в течение по-
следних двадцати лет было местом интенсивных рыбохозяйст-
венных иоследований. Многочисленные экспедиции, большая
часть которых проводилась ТИНРО, внесли большой вклад в
дело изучения гидрологического режима, планктона, бентоса и
ихтиофауны моря. Вслед за научными экспедициями сюда дви-
нулись флотилии промысловых судов; в итоге Берингово море к
началу шестидесятых годов стало одним из важнейших промыс-
ловых районов на Дальнем Востоке.
В ряде экспедиций, работавших в Беринговом море, прово-
дились наблюдения над морскими птицами, что нашло отраже-
ние в серии посвященных им статей (Слепцов, 1959; Шунтов,
1961, 1963, 1965, 1966; Гудков, 1962; Курочкин, 1963). Обобще-
ние опубликованных данных, а также результатов дополнитель-
ных наблюдений, дает возможность осветить особенности соста-
ва, распределения и миграций птиц в связи с океанографиче-
скими факторами в сезонном аспекте, а также приблизительно
оценить их численность в этом водоеме.
В Беринговом море, как и во многих других районах, зима
— наименее благоприятный период для обитания морских
птиц. Почти над всей акваторией моря господствуют отрица-
тельные температуры воздуха, в связи с чем выхолаживаются
поверхностные воды, почти вся шельфовая зона покрывается
льдами. Изменения в гидрологическом режиме вызывают ухуд-
шения кормовых условий для птиц- Вследствие уменьшения чис-
ленности мезапланктона, а также ухода его под влиянием ох-
лаждения вод ниже границы зимней конвекции, биомассы в до-
ступном для птиц поверхностном слое резко уменьшаются и,
как правило, не превышают 50 мг/м3 (Виноградов, 1956, 1968;
Гейнрих, 1959). Зимой в поверхностных слоях не встречается в
заметных количествах и макропланктон, в частности эвфаузии-
ды (Пономарева, 1963). Низкие температуры воды являются
причиной миграций в более глубокие районы и большей части
видов рыб, в том числе пелагических. В связи с ухудшением
ctJ3OliriUE АСПЕКТЫ В ОРНИТОФАУНЕ
99
кормовой базы (в первую очередь доступность кормовых орга-
низмов), сильным охлаждением воздуха и воды, а также льдооб-
разованием в мелководных районах большая часть морских птиц
из Берингова моря откочевывает в северную часть Тихого океана.
Основным местом зимовок птиц в Беринговом море является
его юго-восточная часть, включающая шельф с прилегающими
водами, к востоку от долготы острова Святого Матвея (рис. 31).
Рис. 31. Основные районы зимовок морских птиц в Беринговом море. 1 — льды,
2 — основные районы концентраций птиц, 3 — теплые течения, 4 — холодные
течения, 5 — изобата 200 м, 6 — поверхностные изотермы
Над другими видами здесь резко доминируют кайры (преиму-
щественно толстоклювая) и серокрылая чайка — Larus glau-
cescens Naum. Обычны в районе материкового склона глупыш
и обыкновенная моевка, в разводьях среди ледяных полей —
белая чайка — Pagophila ebumea Phipps, по всему району —
полярная чайка. Увеличение плотности населения птиц наблю-
дается также у Алеутских и Командорских островов. Здесь
кроме чаек (серокрылая и моевка) и кайр многочисленны мел-
кие чистиковые, преимущественно конюги. Обычен также глу-
пыш и временами темноспинный альбатрос.
Как видно из табл. 8, суммарная плотность на юго-восточ-
ном мелководье и в приалеутских водах, по данным визуаль-
100
БЕРИНГОВО МОРЕ
инн учетов, довольно высокая — 10 птиц на 1 кв. к.м. При ис-
пользовании этой плотности для оценки общей численности
пчнц в рассматриваемом районе получено около 3.5 миллиона
особей. Следует, однако, заметить, что в учетах, проводившихся
в основном за пределами непосредственно прибрежной зоны,
мало представленными оказались утки, мелкие чистиковые и
бакланы, многочисленные на зимовках у берегов Алеутских и
Командорских островов (Gabrielson and Lincoln, 1959; Murie,
1959; Мараков, 1966; и др.). С учетом их, особенно уток, зи-
мующих также местами на севере моря в разводьях (Fay and
Cade, 1959), численность птиц в рассматриваемых районах бу-
дет. конечно, выше.
Таблица 8
Видом* света*, ммщмммм (%> м плотность морских птиц
* Бериагомм маре эимоВ (декабрь-март). 1963—1965 гг.
Виды Район шельфа и материкового склона Глубоководная часть моря
Кайры (U. lomvia) 68,0 10,0
Другие чиепшовые 4,0 7,0
L. glaucescens 17,0 19,0
R. tridactyla 2,0 3,0
Другие и неопределенные чайки 3.0 9.0
F. glacialis 2,0 48,0
Другие трубконосые + 4,0
Утки 3,0
Бакланы (Р. pelagicus) 1,0 —
Количество особей на 1 кв. км. 10,0 0,8
Количество учетов 170 64
Примечание. В этой и следующих таблицах в скобках — наиболее массо-
вые виды в каждой группе.
Восточный шельф и прибрежные воды островной гряды край-
него юга моря фактически единственные районы, благоприятные
для зимовок птиц в Беринговом море. Только здесь шельф не
закрывается или закрывается льдами частично (Атлас гидроме-
теорологических и промысловых данных. Восточная часть Берин-
гова моря, 1969). Данный район, находящийся под отепляющим
влиянием вод Тихого океана, выгодно отличается от западной
части моря, находящейся под влиянием сильно охлаждающегося
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ Ю|
зимой материка Азия, более мягким климатом и гидрологичес-
ким режимом (Леонов, I960). В северной и. западной частях
.моря часты северные штормовые ветры, нагоняющие холодный
арктический воздух. В юго-восточной части моря и у Алеутских
островов преобладают ветры неустойчивых направлений, более
редки штормы. На севере моря средняя температура воздуха ко-
леблется от минус 14 до минус 22°, в юго-западной части моря—
от минус 6 до минус 14е, в юго-восточной — от минус 2 до ми-
нус 10е (Карпова. 1963). Средняя температура воды поверхно-
стного двухсотметрового слоя в юго-восточной части моря на
3° выше, чем в юго-западной (Васюкова, 1964). Значительно вы-
ше в поверхностных слоях биомассы зоопланктона (Атлас ги-
дрометеорологических и промысловых данных, 1969). Очень су-
щественно и то, что на юго-восточном мелководье в районах,
полностью или частично свободных от льда, образуют мощные
концентрации пелагические и придонные рыбы и их молодь —
минтай, сельдь (Дудник и Усольцев, 1964; Шабанеев. 1965; Се-
робаба. 1968, 1970; и др.), песчанка, корюшка и мойва. Косяки
рыб, находящиеся у грунта, на нижней части шельфа, конечно,
недоступны для птиц. Однако, как показывают наблюдения,
значительная часть рыбы держится в толще воды, при суточных
вертикальных миграциях поднимаясь почти до поверхности. Не-
сомненно, эти рыбы доступны для кайр, которые способны ны-
рять на глубину в несколько десятков метров (Tuck, 1960). Из
табл. 8 видно, что именно кайры имеют наибольшую числен-
ность зимой в Беринговом море.
На мелководье и свале глубин в восточной части моря, а
также у Алеутских островов в течение всего зимнего периода
работает многочисленный траловый флот, добывающий сельдь,
минтая, треску, камбалу, окуня, палтусов и угольную рыбу. От-
ходы промысла составляют важный источник питания крупных
чаек, большая часть которых концентрируется в районах сосре-
доточения добывающего флота.
В глубоководных (центральная и западная части) районах
моря, бедных кормом (мало планктона, нет массовых виде*
рыб) и имеющих более суровый климат и гидрологический ре-
жим. плотность населения и численность птиц значительно ни-
же, чем на шельфе (0,8 на 1 км2; 0,68 млн. особей). За немно-
гим исключением (белая чайка, утки, бакланы) видовой состаа
здесь такой же. как и в шельфовых водах, только основу насе-
ления составляют трубконосые, главным образом глупыш
(см. табл. 8).
Из изложенного видно, что зимой обычны и многочисленны
в Беринговом море только хорошо ныряющие виды или виды,
потребляющие отходы рыбного промысла. К первым относятся
кайры, нырковые утки, бакланы и чистик — Cepphus grylle (L),
102
БЕРИНГОВО МОРЕ
ко вторым — крупные чайки: серокрылая, полярная и тихооке-
анская — Larus schistisagus Stejn. (последний вид только у по-
бережья Камчатки). Места зимовок видов первой группы приу-
рочены к шельфу, ледовой кромке или прибрежным водам, где
легче добывать пищу ныряющим птицам, где меньше влияние
сильных ветров и крупной штормовой волны, столь частых в Бе.
ринговом море зимой. В течение всей зимы довольно много ны-
ряющих птиц остается на севере моря в разводьях. В частности,
у берегов Святого Лаврентия в полыньях зимует большое ко-
личество морянки — Clangula hyemalis (L) и гаг. Обычны здесь
толстоклювая кайра и чистик (Fay and Cade. 1959).
Для размещения и численности крупных чаек особенно боль-
шое значение имеет деятельность промыслового флота, который,
впрочем, также концентрируется главным образом в пределах
мелководья и материкового склона. Характерно, что среди зи-
мующих в Беринговом море всех видов чаек преобладают взрос-
лые особи. На долю светлых особей самой массовой серокрылой
чайки приходится 80% в восточной части моря и 85% — в запад-
ной.
Для видов, добывающих пищу в верхних слоях моря, зимой
кормовые условия в Беринговом море особенно неблагоприят-
ны. Поэтому-то мало в это время здесь трубконосых и мелких
чистикоцых, которые очень многочисленны здесь с весны до
осени. Особенно резко эти различия заметны на примере серой
качурки — Oceanodroma furcata (Gmel.) и темного глупыша.
Зимой в небольших количествах они встречаются только в юж-
ной части моря, а летом нередко доминируют над другими ви-
дами по всей юго-западной половине моря.
Для некоторых видов, например, обыкновенной моевки и то-
порка необходимо допустить, что основной причиной отлета из
Берингова моря являются низкие температуры воды и воздуха.
В пользу этого говорит их быстрый отлет, особенно из наиболее
рано охлаждающихся районов, и характер размещения на зи-
мовках. Если обыкновенная моевка довольно обычна зимой на
юге моря, тр топорок редок даже у Алеутских островов. Основ-
ная масса обоих видов, как было показано выше, широко рас-
сеивается по северной части Тихого океана. Покидает Бе-
рингово море и значительная, а может, и большая часть особей
красноногой моевки — Rissa brevirostris (Bruch).
По климатическим и гидрологическим условиям март в Бе-
ринговом море почти не отличается от февраля, однако уже в
конце его наблюдаются первые признаки наступающей весны.
В глубоководной части моря, а также в районе материкового
склона более многочисленным становится глупыш (в основном
светлый), а на востоке моря — оба вида моевок. Начало мас-
совых миграций птиц приходится на апрель — месяц больших
СЕЗОННЫЕ аспекты В ОРНИТОФАУНЕ ЮЗ
изменений в гидрометеорологическом режиме и биологии моря.
В апреле средняя температура воздуха на севере моря заметно
повышается и колеблется от минус 4 до минус 10°, а на юге мо-
ря — от 2 до минус 2° (Карпова, 1963). В связи с прогревом
вод и увеличением повторяемости южных ветров постепенно
отступают в северном направлении льды (Булгаков, 1964; Атлас
гидрометеорологических и промысловых данных, 1969). Хотя в
апреле льды по-прежнему покрывают большую часть шельфа,
они становятся менее плотными, сокращается площадь полей,
увеличиваются разводья. В апреле, по мере прогрева поверхно-
стных вод, обогащенных в период зимней конвекции биогенны-
ми элементами, начинается бурное развитие фитопланктона.
В это время для питания и размножения поднимается из
нижних горизонтов в верхние мезо- и макропланктон, в резуль-
тате чего биомассы его в верхних слоях значительно увеличи-
ваются (Виноградов, 1956, 1968; Гейнрих, 1969; Пономарева,
1963; Мещерякова, 1970; и др.). В апреле и мае совершают не.
рестово-кормовые или кормовые миграции на меньшие глу-
бины основные промысловые рыбы Берингова моря (Фадеев,
1963, 1970; Новиков, 1964; Дудник и Усольцев, 1964; Качина и
Прохоров, 1966; Серобаба, 1968; Румянцев и Дарда, 1970; и др.).
Таким обазом, прогрев вод и воздуха, уменьшение ледови-
тости и особенно увеличение в верхних слоях моря количества
планктона,, рыбы и ее молоди резко улучшают условия для оби-
тания морских птиц. В сравнении с зимним периодом наиболее
заметно повышается обеспеченность кормом планктоноядных
птиц. Это находит отражение в составе весенней орнитофауны
моря (табл. 9). В сравнении с зимним периодом особенно силь-
но увеличивается численность трубконосых и мелких чистико-
вых, в питании которых планктон играет основную роль. Одна-
ко период, наиболее благоприятный в кормовом отношении для
йланктоноядных птиц, наступает только в мае. В апреле наблю-
дается массовое цветение, но биомассы зоопланктона в верхних
слоях еще не достигают наибольших величин. По-видимому,
в связи с этим трубконосые становятся доминирующей группой
только с мая. В апреле же самые массовые в Беринговом море,
как и зимой, чистиковые и чайки.
Первыми начинают весенние миграции виды, зимовки кото-
рых располагаются в Беринговом море и прилегающих водах
Тихого океана. С первых чисел апреля (некоторые виды с конца
марта) начинают перемещение зимовавшие в Беринговом море
кайры, глупыш, утки, моевки., чистик, крупные чайки и другие.
В это же время стайки уток, кайр, моевок и бакланов, а также
глупыш, начинают проникать в Берингово море из Тихого океа-
на. Прогрев вод наступает раньше в относительно теплой юго-
восточной части моря. В связи с этим здесь несколько раньше
км
БЕРИНГОВО МОРЕ
Таблица 9
Видовой состав, соотношение (%)н плотность морских птиц
в Беринговом море весной (апрель-май). 1М1—1965 гг.
Виды Районы шельфа и материкового склона Глубоководная часть моря
Буревестники (Р. tenuirostris) 36,0 10,0
F. glacialis 9,0 32,0
Другие трубконосые 1,0 4,0
Кайры (U. lomvia) 23,0 5,0
Мелкие чистиковые 8,0 32,0
Другие чистиковые 1,0 2,0
Моевки (R. tridactyla) 6,0 8.0
L. glaucescens 4,0 3,0
Другие и неопределенные чайки 1,0 1.0
Утки 5,0 1,0
Бакланы (Р. pelagicus) 1.0 +
Поморники + 2.0
Плавунчики (Р. fulicarius) 4,0 - •
Остальные виды 1,0 —
Количество особей на 1 кв. км 20,0 4,0
Количество учетов 170 40
начинаются и миграции птиц. Пролет перечисленных выше
птиц продолжается весь апрель и захватывает часть мая. При-
близительно с середины апреля заметно увеличивается количе-
ство качурок, мелких чистиковых, топорка и ипатки — Fratercu-
la corniculata (Naum.). В первой половине мая численность
птиц в Беринговом море вновь значительно увеличивается.
В это время начинается массовое прибытие буревестников из
южного полушария (преимущественно тонкоклювый, меньше—
серый). В основном в мае происходит пролет вдоль побережий
гагар. В мае же появляется в приалеутских водах черноногий
альбатрос. Во второй половине мая и даже в начале июня, ког-
да миграции основной массы птиц заканчиваются, происходит
пролет видов, зимовки которых простираются до тропиков или
южного полушария. Это — поморники, плавунчики, полярная
крачка — Sterna paradisea Pontop. и вилохвостая чайка.
С конца апреля и в течение всего мая регулярно в открытом
море встречаются сухопутные птицы и кулики. Чаще они наблю-
даются при прохождении проливов или пересечении заливов.
Следует заметить, что пролет сухопутных птиц в Беринговом
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
105
море по массовости уступает тому, что наблюдается в более юж-
ных морях, и в частности Охотском и Японском. Несомненно, это
связано с меньшей численностью сухопутных птиц в высоких
широтах.
Берингово море в широтном направлении протянулось на
900 миль, весна в его северных районах начинается приблизи-
тельно на месяц позднее, чем в южных. Соответственно здесь
смещены сроки миграций птиц и их гнездования. Если на распо.
ложенные в южной части моря острова Прибылова. Командор-
ские и Алеутские прилет основной массы птиц приходится на
апрель — начало мая, то на острова северной части моря —
только во второй половине мая — начале июня (Fay and Cade,
1959; Gabrielson and Lincoln, 1959; Kenyon and Brooks, 1960;
Мараков, 1963, 1966; и др.).
При весенних миграциях многие прибрежные виды встре-
чаются (часто в заметных количествах) над глубоководной ча-
стью моря (см. табл. 9). Но, как и зимой, имеются различия в
соотношении групп и, главное, в доминирующих видах в шель-
фовых и глубоководных районах, В шельфовых водах концен-
трируется основная масса буревестников, кайр, мелких чистико-
вых (в основном конюги), чаек, уток, бакланов, светлого глупы-
ша, плавунчиков и других. В глубоководной части наиболее
многочислен темный глупыш; местами (в основном на западе
моря) многочисленны также мелкие чистиковые, обычны чайки,
кайры, качурки и буревестники.
Различия в составе птиц имеются между западной и восточ-
ной частями моря. К восточной части моря тяготеют буревест-
ники, кайры, серокрылая чайка, плосконосый плавунчик —
Phalaropus fulicarius (L). Основное направление миграций в
этом районе отчетливо прослеживается от восточной части
Алеутской гряды на север и северо-запад через мелководье к
островам Прибылова, Святого Матвея, Святого Лаврентия и Бе-
рингову проливу. На западе моря основной поток мигрантов
проходит вдоль азиатского побережья (шельф и материковый
склон). Здесь больше, чем на востоке моря, мигрируют конюги;
только здесь встречается тихоокеанская чайка. В западной ча-
сти моря больше круглоносого плавунчика — Lobipes lobatus
(L), но меньше кайр и буревестников. Миграции обыкновенной
моевки, уток, гагар, топорка и поморников проходят, по-видимо-
му, с одинаковой интенсивностью по обеим половинам моря.
Соотношение видов птиц весной в Беринговом море может
характеризовать табл. 9. Здесь нужно, однако, учитывать, что
относительно верно она отражает только состав орнитофауны
в открытых водах, исключая непосредственно прибрежную зо-
ну. В связи с этим, если говорить о всем море, то численность
и доля некоторых групп (в первую очередь буревестников)
106
БЕРИНГОВО МОРЕ
оказывается завышенной, а доля уток, мелких чистиковых и
бакланов — заниженной.
Общая численность птиц весной в Беринговом море намно-
го выше, чем зимой. Соответственно увеличивается и плотность
населения. Хотя в мелководных районах моря плотность возра-
стает только в два раза, а в глубоководных — в пять раз, воды
над мелководьем и материковым склоном по-прежнему оказы-
ваются более населенными. Здесь концентрируется более'80%
птиц, а средняя плотность в пять раз выше, чем в глубоковод-
ных районах. Для объяснения этого обратимся к данным, харак.
теризуюшим кормность различных районов моря.
Б глубоководных районах Берингова моря, как отмечалось
выше, весной биомассы планктона увеличиваются в несколько
раз. В связи с этим не является случайным пятикратное “увели-
чение здесь и плотности птиц, в первую очередь планктонояд-
ных. Тем не менее, шельфовые воды в рассматриваемый пе-
риод, как и в другие сезоны, в целом более благоприятны в кор-
мовом отношении. Достаточно отметить, что первичная продук-
ция неритической группировки фитопланктона, занимающей
шельфовук) зону, в 3—5 раз больше океанической, занимающей
глубоководные районы моря (Иваненков, 1964). В шельфовых
водах более обилен макропланктон, в частности эвфаузииды
(Пономарева, 1963). На мелководье концентрируется основная
масса рыб, среди них имеющие высокую численность сельдь,
сайка, песчанка, корюшка, минтай и мойва. Значительная часть
шельфа Берингова моря характеризуется весьма высокими био-
массами бентоса (Нейман, 1963; и др.).
Полученные для весеннего периода цифры плотности птиц,
несомненно, довольно внушительные. Такое количество птиц
может концентрироваться только в очень продуктивном водое-
ме. Здесь, однако, нет противоречия. Как показывают опреде-
ления содержания в водах необходимых для фотосинтеза био-
генных элементов, а также расчеты продукции, Берингово море
может быть поставлено в ряд с наиболее продуктивным райо-
нами Мирового океана (Давидович, 1963; Федосов и Давидович,
.1963; Азова, 1964; Иваненков, 1964). В связи с этим, конечно, не
является случайным, что значительная часть многомиллионного
населения буревестников, совершающих миграции через экватор,
собирается на откорм и линьку именно в Беринговом море.
В начале июня весенние миграции птиц заканчиваются. Оп-
ределившиеся весной тенденции в изменении состава орнито-
фауны и особенностях распределения птиц сохраняются в тече-
ние всего лета. Из табл. 10 видно, что в сравнении с другими се.
зонами доминирование трубконосых в это время особенно за
метно. В течение лета значительная часть особей размножаю-
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
107
Таблица 10
Видовой состав, соотношение (%) и плотность морских птиц
в Беринговом море летом (нюнь—август). 1961—1965 гт.
Виды Районы шельфа и материкового склона Глубоководная часть моря
Буревестники (Р. tenuirostris) 39,0 20,0
Качурки (О. furcata) 15,0 50,0
F. glacialis 12,0 17,0
Другие трубюоносые + 1.0
Кайры (U. lomvia) 15.0 2,0
Мелкие чистиковые 10,0 1.0
L. cirrhata 4- . 1,0
Моевки (R. tridactyla) 3,0 3,0
Крупные чайки (L. glaucescens) 1,0 2,0
Плавунчики (Р. fulicarius) 1.0 ’ +
Остальные виды 4,0 3,0
Количество особей на 1 кв. км 18,0 2,7
Количество учетов 280 79
щихся здесь видов концентрируется у берегов. В открытом море
уже в конце мая мало крупных чаек. С июня до третьей декады
августа встречи их за пределами прибрежной зоны очень редки.
Богатство литоральной зоны обуславливает концентрацию в
прибрежной зоне и значительной части особей пелагической чай*
ки — обыкновенной моевки. За пределами узкой прибрежной
зоны не встречаются утки й гагары, очень редки бакланы.
Несмотря на тяготение многих видов к береговой полосе,
а части их к пресным водам (утки, гагары, поморники), плот-
ность населения птиц в Беринговом море остается высокой и ма-
ло отличается от весенней. Определяется это следующими об-
стоятельствами. Убыль в море ряда видов компенсируется уве-
личением количества трубконосых (буревестники, качурки и
альбатросы). Существенно и то, что многие виды в гнездовой
период удаляются за кормом от мест размножения на десятки
миль (трубконосые, кайры, моевки) и таким образом учиты.
ваются при наблюдениях. Наконец, в открытом море остается
значительное количество неполовозрелых и неразмножающих-
ся в данном году особей. В связи с этим получена довольно
внушительная цифра общей летней численности птиц в Берин-
говом море — 23,8 млн. особей (20,7 млн. в районе шельфа и
материкового склона, 3,1 млн. в глубоководной части моря)'.
1 При оценке общей численности птиц площадь акватории прибрежной
эоны также учитывается, при этом на нее переносилась плотность, получен-
ная для шельфовых районов. В связи с этим цифра общей численности птиц
является, по>внднмому, заниженной.
108
БЕРИНГОВО МОРЕ
При анализе табл. 10 нужно иметь в виду замечания, сделанные
к табл. 9: процент трубконосых для моря в целом завышен,
а чистиковых и чаек—занижен.
В начале лета особенно много птиц на мелководье в юго-
восточной части Берингова моря. Здесь над всеми видами абсо-
лютно доминирует собирающийся на линьку тонкоклювый бу-
ревестник. Обычны в этом районе кайры, обыкновенная моевка,
местами топорок и конюги. Севернее островов Прибылова и Ну-
нивак птиц меньше. Основу населения в этих районах составля-
ют кайры, конюги, светлый глупыш и обыкновенная моевка. Вы.
сокая плотность птиц наблюдается в районе материкового
склона центральной части моря. Фон населения здесь определя-
ют тонкоклювый буревестник, глупыш (светлый и темный), се-
рая качурка, обыкновенная и красноногая моевки. В централь-
ных глубоководных районах моря птиц меньше. Хотя здесь
временами и можно встретить скопления, однако они не быва-
ют здесь такими мощными- и стабильными, как на шельфе. Как
и в другие сезоны, летом в глубоководных районах моря встре-
чаются' в основном пелагические виды — буревестники (тонко-
клювый и серый), темный глупыш, качурки, моевки, топорок,
альбатросы и тайфунник Пила — Pterodroma inexpectata
(Forst). Сведения о встречаемости последнего вида в Беринго-
вом море появились только в последние годы (Kenyon ana Phil-
lips, 1965). По моим наблюдениям, этот новозеландский вид
встречается летом по всей глубоководной части Берингова мо.
ря к югу от линии мыс Наварин — острова Прибылова.
Повышенная плотность населения птиц летом отмечается
вдоль Алеутской гряды и у азиатского побережья Берингова
моря. Видовой состав птиц здесь очень пестрый: обычны или
многочисленны виды, перечисленные для глубоководной часта
моря, а также гнездящиеся в этих районах различные чистико-
вые, чайки и другие. Нужно только отметить, что вместо серо-
крылой чайки, которая доминирует среди крупных чаек к во
стоку от Командорских стровов, у западного побережья Берии
гова .моря преобладает тихоокеанская чайка.
Хотя весенние миграции птиц в начале июня заканчиваются,
это не значит, что в составе орнитофауны во всех районах уста-
навливается стабильность. Уже приблизительно с середины
июля наблюдаются заметные изменения в распределении ряда
видов. Закончив линьку на юго-восточном мелководье, устрем-
ляется на юг тонкоклювый буревестник. Ранний отлет полово,
зрелых особей связан с тем, что у этого вида уже в сентябре
начйнается период размножения. С этого времени на юго-во-
сточном мелководье буревестник в заметных количествах нс
встречается.
СЕЗОННЫЕ аспекты в орнитофауне
109
В июле начинается отлет на юг части особей плосконосого
плавунчика и других куликов. В море небольшие группы плос-
коносого плавунчика встречаются в течение всего июня, но в
середине июля его численность заметно увеличивается; в это
время нередки стайки по нескольку десятков и даже сотен птиц.
Наиболее оживленный пролет проходит по восточной части Бе-
рингова моря. Пролет плосконосого плавунчика быстрый: позд-
нее последней декады августа в открытом море он почти не
встречается. Судя по срокам, летящие в это время птицы не мо-
гут быть молодыми данного года. Так как в более поздние сро-
ки этот вид в открытом море не встречается, можно предполо-
жить, что его пролет осенью проходит у побережий. В конце ле-
'та начинает постепенно откочевывать на юг из центральной ча-
!сти Берингова моря темный глупыш.
J Таким образом, уже в начале второй половины лета часть
'особей некоторых видов имеет тенденцию к отлету на юг. Не
'менее характерными, однако, для этого времени являются чег-
; ко выраженные миграции, ряда массовых планктоноядных птиц
'в более северные районы. Из них в первую очередь нужно на-
звать серую качурку. В конце июля — начале августа ее стаи,
нередко насчитывающие сотни и тысячи особей, перемещаются
с юга в Олюторско-Наваринский район, а с юго-востока — в
район материкового склона центральной части Берингова моря,
где они держатся до конца сентября — начала октября. В за-
падной половине Берингова моря в более северные районы сме-
щаются и концентрации буревестников. Миграции тонкоклюво-
го буревестника во второй половине лета вдоль западного побе-
режья моря по массовости не уступают серой качурке и отли-
чаются более дальним проникновением на север (до Чукотско-
го моря). Он обычен в это время в районе материкового склона
центральной части моря, но особенно много концентрируется в
Олюторско-Наваринском районе. Миграции на север планктоно-
’ ядных трубконосых птиц во второй половине лета не случайны
и связаны с сезонными изменениями в планктоне. Во второй по-
ловине лета биомассы зоопланктона в поверхностном слое в
южной части Берингова моря уменьшаются под влиянием вые-
дания и опускания в более низкие горизонты. На севере же, в
связи, с более поздним прогревом вод, в это время наблюдается
годовой максимум численности планктона (Виноградов, 1956;
Мещерякова, 1970а).
В августе в открытых водах вновь появляются одиночные осо-
би серокрылой, а на севере моря—полярной и серебристой чаек,
которые почти не наблюдаются вдали от берегов в июне и июле.
Летом, как и в другие сезоны, более высокая плотность на-
селения птиц наблюдается у побережий, в зоне шельфа и мате-
рикового склона (рис. 32). В среднем она здесь приблизительно
но
БЕРИНГОВО МОРЕ
в семь раз выше, чем в глубоководной части моря. Связано это
с тем, что воды шельфа более продуктивны.
Скопления птиц приурочены к районам, где благодаря тем
или иным особенностям гидрологии моря создаются особенно
благоприятные условия для развития высокой биологической
продуктивности. В этом смысле общеизвестно богатство жиз-
нью в летний период вод (планктон, бентос, рыба) непосредст-
венно прибрежной зоны. В открытых водах Берингова моря,
как и в других районах, повышенной продуктивностью отли-
чаются зоны усиленного вертикального движения вод. Одним
Рис. 32. Распределение концентраций морских птиц в Беринговом море летом.
1 — низкие, 2 — средние, 3 — высокие, 4 — течения
из таких районов является материковый склон, где в результа-
те встречи идущих на север тихоокеанских вод с круто подни-
мающимся дном возникает интенсивная макротурбулентность.
Вертикальная циркуляция обогащает биогенными элементами
поверхностные слои не только над материковым склоном, но и
прилегающие участки шельфа; в итоге создаются необходимые
условия для высокой биологической продуктивности этих уча-
стков моря, в том числе для образования высоких концентра-
ций планктона и рыб (Натаров. 1963; Мещерякова, 1964; Мои-
сеев, 1964; Новиков. 1970; Шунтов. 1966а, 1971; и др.). В связи
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
111
с этим в течение летнего периода район материкового склона
является местом устойчивых концентраций ряда видов птиц.
Аналогичная картина наблюдается и при прохождении течений
через Алеутскую гряду. Условия для больших скоплений птиц
создаются на шельфе Берингова моря по краям холодных пятен
вод, занимающих летом среднюю часть мелководья и прости-
рающихся от Анадырского залива до Бристольского. Хотя воды
холодных пятен имеют высокое содержание биогенных элемен-
тов, массового развития планктона из-за низкой температуры
в них не происходит. Однако по мере прогрева и развития пере-
мешивания вод в течение лета эти пятна постепенно отдают ак-
кумулированные в них вещества, необходимые для развития
жизни, создавая по краям зону плодородия (Натаров, 1963; Me.
щеря ков а, 1964). Холодные пятна накладывают существенный
отпечаток на распространение рыб. пути их миграций и разме-
щение скоплений (Фадеев, 1963, 1970; Шунтов, 1963а, 1970;
Новиков, 1964; и др.). Продуктивная зона, формирующаяся пр
к?жной границе холодных пятен, смыкается с аналогичной зо-
.Ной, возникающей в районе материкового склона. В итоге вся.
, обширная акватория, включающая воды, материкового склона '
i и нижней половины шельфа от Олюторско-Наваринского райо-
; на до Бристольского залива и полуострова Аляска, рассматри-
. вается как весьма продуктивная. Летом здесь обилен планктон;
: мощные концентрации образуют различные виды рыб и, как по-
। казано выше, морские птицы (см. рис. 32). Имеют место скоп-
ления птиц местами и по северной границе холодных пятен вод.
В частности, на восточном мелководье вдоль северной границы
пятен располагается в июне часть линных скоплений тонкоклю-
вого буревестника (Шунтов, 1961).
В видовом составе птиц западной и восточной половин Бе-
рингова моря резких различий нет. Различия касаются в основ-
ном соотношения видов, о чем говорилось выше. Другая кар-
тина наблюдается при сопоставлении состава орнитофауны вон
шельфа и глубоководной центральной части моря. Преимущест->
вен но в шельфовых водах встречается основная масса видов
чистиковых, светлый глупыш, бакланы, а также прибрежные
виды — утки, гагары и некоторые чайки (сизая — Larus canus
L., озерная — Larus ridibundus L.). Значительная группа ви-
дов — качурки серая и северная — Oceanodroma leucorrhoa
(Vieillot), темноспинный и черноногий адьбатросы, тайфунник
Пила, темный глупыш, красноногая моевка тяготеют к глубоко-
водным районам моря*. Там, где шельф узкий (Алеутские и
1 Такие массовые виды как тонкоклювый буревестник, обыкновенная мо-
евка и топорок обычны или многочисленны и в мелководных, н в глубоковод-
ных районах моря.
112
БЕРИНГОВО МОРЕ
Командорские острова, западная часть моря), наблюдается бо-
лее пестрый видовой состав. Здесь встречаются представители
обеих основных группировок. В центральной части моря, где ма-
териковый склон разделяет море на мелководную и глубоковод-
ную половины, основные комплексы птиц разделены. Границей
между ними является зона кромки шельфа от мыса Наварин к
полуострову Аляска (рис. 33). Если многие шельфовые виды и
небольших количествах встречаются южнее этой границы, то
все виды, перечисленные для глубоководной части моря, захо-
дят севернее нижней части шельфа лишь случайно.
Рис. 33. Распределение неритических и океанической группировок морских
птиц в Беринговом море летом. 1 — неритические, 2 — океаническая
Описанное различие в составе орнитофауны находится в
связи со спецификой гидрологических условий в шельфовой и
глубоководной половинах моря. Глубоководная половина Бе-
рингова моря заполнена тихоокеанскими водами, поступающи-
ми сюда через проливы между островной грядой. По своим фи-
зико-химическим характеристикам эти воды мало отличаются от
вод северной части Тихого океана. В районе материкового
склона океанские воды поворачивают на северо-запад, но часть
их проникает на шельф, а затем через Берингов пролив в Чу-
котское море. На мелководье эти воды трансформируются; во
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
113
все сезоны они имеют более низкую температуру и соленость.
Отличаются они и по пидрооптическим характеристикам —
прозрачности и цвету. Характерно, что изолинии всех характе-
ристик водных масс параллельны изобатам материкового скло-
на (Натаров, 1963; Ароеньев, 1967; Арсеньев и Войтов, 1968; Ат-
лас гидрометеорологических и промысловых данных, 1969; и
др.). Различия в характеристиках водных масс накладывают
существенный отпечаток на распределение фитопланктона и вод-
ных животных и определяют существование у них неритических
и океанических группировок. По данным Г. И. Семиной (1955),
граница между неритической группировкой фитопланктона,
приуроченной к шельфовым водам, и океанической, приурочен-
ной к глубоководной части моря, проходит, как в случае с пти-
цами, в районе материкового склона. Океаническая группировка
зоопланктона проникает со струями течений на шельф несколь-
ко дальше, в результате чего на значительной части мелководья
наблюдается смешение океанических и неритических видов
(Виноградов, 1956).
При объяснении тяготения группировок морских птиц к рай-
онам с определенными водными массами, по-видимому, следует
иметь в виду комплексное влияние различных факторов и общий
фон океанографических условий. Вместе с этим нужно подчерк-
нуть значение кормового фактора, в частности, доступность пи-
щи в различных водных массах и районах моря. В следующей
главе эти вопросы будут рассматриваться.
Осенью, уже с сентября, почти над всей акваторией Беринго-
ва моря начинают преобладать ветры северных румбов- Посте-
пенно охлаждаются водные и воздушные массы. Средняя месяч-
ная температура понижается на севере моря до 3—7* в сентяб-
ре и до О минус 12° — в ноябре (Карпова, 1953). Осенью за-
канчивается вегетация фитопланктона и на большей части Бе-
рингова моря биомассы зоопланктона значительно уменьшаются
(Виноградов, 1956); исчезают из планктона личинки донных
животных (Гейнрих, 1959) и отходят на глубины наиболее мас-
совые рыбы и их молодь. Таким образом, осенью происходит за-
кономерное обеднение поверхностных и прибрежных вод кор-
мом для птиц, в связи с чем в сентябре начинается смещение
многих видов их на юг. В это время у большинства видов закан-
чивается период размножения, после чего они отходят от бере-
гов. Рассеивание птиц, начавших кочевать после гнездования,
происходит во всех направлениях, но преобладающим является
южное.
В сентябре на северо-восточном мелководье становится обыч-
ной, а у островов Прибылова многочисленной серокрылая чай-
ка. Осенние кочевки ее происходят в основном по восточной
114
БЕРИНГОВО МОРЕ
мелководной половшие моря. В центральных глубоководных
районах Берингова моря она немногочисленна, хотя и встречает-
ся регулярно. Разлет с мест гнездования происходит не только
в южном, но и северном направлении1. Первыми откочевывают
в открытые воды молодые серые особи. В сентябре на их долю
приходится около 75%. В октябре и ноябре до 40% увеличивает-
ся доля взрослых светлых птиц. В это же время в открытом мо-
ре появляются серебристая и полярная чайки- В отличие от пе-
речисленных видов тихоокеанская чайка по-прежнему остается
в западной части моря. Она почти не выходит за пределы шель-
фа и материкового склона; миграции этого вида происходят
вдоль побережий. В основном в сентябре проходит отлет вило-
хвостой чайки и крачек. Рано отлетают поморники. В первой
половине сентября они еще обычны, затем становятся редкими,
а после первой декады октября не встречаются. Уже в первой
половине Сентября уменьшается в Беринговом море количество
Топорка. В начале октября он становится редким. К началу ок-
тября резко сокращается в большинстве районов моря и числен-
ность качурок. Откочевка многих массовых видов происходит
в течение более значительного промежутка времени. До конца
октября много в Беринговом море тонкоклювого буревестника,
мелких чистиковых, уток, моевок.
Осенние миграции по сравнению с весенними происходят бо-
лее постепенно. Численность птиц в Беринговом море особенно
сильно уменьшается лишь к началу ноября. В сентябре же, не-
смотря на отлет значительной части особей ряда видов, плот-
ность населения мало отличается от летней. Нужно заметить,
что условия обитания птиц в начале осени еще нельзя назвать
неблагоприятными- В это время еще сравнительно тепло, а в се-
верных районах наблюдается даже годовой максимум планк-
тона.
Как и весной, многие шельфовые виды (чистиковые, бакла-
ны, утки) нередко встречаются осенью в глубоководных райо-
нах моря. Однако основные потоки мигрантов проходят над
мелководьем и вдоль побережий. Особенно интенсивный пролет
происходит вдоль азиатского побережья. В больших количест-
вах здесь встречаются тонкоклювый буревестник, конюги, обык-
новенная моевка, утки, кулики и другие Много мигрирует птиц
и по восточной мелководной части- Берингова моря. В отличие
от западного побережья здесь значительно больше кайр и се-
рокрылой чайки, но меньше конюг и буревестников. В цент-
1 Аналоганная картина наблюдается у этого видд в районе Британской
Колумбии. Здесь откочевка части особей в северном направлении подтверж-
дается кольцеванием (Pearse, 1963).
СЕЗОННЫЕ аспекты в ОРНИТОФАУНЕ
иг-
ральной глубоководной части Берингова моря осенью по-преж-
нему доминируют трубконосые (табл. 11).
Таблица 11
Видовой состав, соотношение (%) и плотность морских птиц
в Беринговом море осенью (сентябрь—ноябрь), 1959—1866 гг.
13 И л ы Районы шельфа и материкового склона Глубоководна» часть моря
Бурезестники (Р. teniiirostris) 20,0 20,0
Качурки (О. funcata) 15,0 8,0
F. glacialis 9,0 39,0
Другие трубканосые — 4.0
- Кайры (U. lomvia) 17,0 IX)
Мелкие чистиковые 10,0 4,0
Другие чистиковые 1.0 —
Моевки (R. tridactyla) 10,0 11,0
L. glaucescens 10,0 2,0
Другие и неопределенные чайки 3,0 6,0
Утки 2,0 3,0
Остальные виды 3,0 ЪО
Катичеспво особей на 1 кв. км 12,0 2.8
Количество учетов 130 70
Наиболее быстро охлаждается северо-западная часть моря.
Несколько раньше здесь заканчивается и откочевка птиц. При
осредненных данных за весь осенний период получена плотность
населения 12 особей на 1 кв. км для района шельфа и матери-
кового склона и 2,8 особей на 1 кв. км — для глубоководной
части моря-
2. Охотское море.
Орнитофауна Охотского моря в сравнении с Беринговым
изучена меньше. Материалы, имеющиеся в моем распоряжении,
позволяют сравнительно подробно описать только летний и осен-
ний периоды. По другим сезонам есть только отрывочные дан-
ные.
Охотское море глубоко вдается в сильно охлаждающийся
азиатский материк, поэтому зимой по климатическим условиям
оно мало отличается от полярных морей. В январе средняя тем-
пература воздуха на севере моря составляет минус 20—25*;
на юге — минус 6* (Леонов, I960)- Большая часть моря
зимой замерзает. Свободным от льда остается только не-
116
ОХОТСКОЕ МОРЕ
большой участок, прилегающий к Курильским островам
и западному побережью Камчатки (Булгаков. 1965). В свя-
зи с этим большая часть морских птиц на зимний пе-
риод покидает Охотское море. Совсем не встречаются
зимой здесь такие относительно теплолюбивые ви-
ды, как моевка, чернохвостая чайка — Larus crassirostris Vieill.,
очковый чистик — Cepphus carbo (Pall.), топорок, рогатый ту-
пик — Cerorhinca monocerata (Pall), качурки, черноногий альба-
трос- Особенно резко в сравнении с другими сезонами умень-
шается численность трубконосых. Обычным в свободных от
льда прикурильских водах остается из этой группы только глу-
пыш. Изредка сюда проникают из океана отдельные особи тем-
носпинного альбатроса.
В начале зимы (декабрь), когда льды закрывают только
часть шельфа, численность птиц в Охотском море еще большая.
Они концентрируются у Курильских островов, а также на мно-
гих участках мелководья вдоль кромки льдов и в разводьях. В
районе пролива Лаперуза, у восточного побережья Сахалина,
Курильских островов и юго-западного побережья Камчатки мно-
гочисленны в это время кайры и мелкие чистиковые. В разво-
дьях в северной части моря кайр меньше, но очень многочис-
ленны мелкие чистиковые (по-видимому, конюги). Абсолютное
доминирование чистиковых во всех отмеченных районах бес-
спорно. Значительные скопления они образуют и у Курильских
островов, но здесь многочисленны также утки и бакланы. За
исключением глубоководной части моря во всех перечисленных
выше районах обычна или многочисленна тихоокеанская чайка.
Зимой в Охотском море остаются главным образом половозре-
лые светлые особи, на долю которых на севере моря приходится
около 98%. а у юго-западного побережья Камчатки и Северных
Курил — не менее 90%. Другие чайки — полярная, серебрис-
тая и белая1 встречаются на зимовках в Охотском море, но чис-
ленность их невелика.
В январе—марте, когда льды закрывают всю северную и во-
сточную часть моря, значительная часть птиц, концентрировав-
шаяся в начале зимы в этих районах, откочевывает через про-
лив Лаперуза в Японское море, а также в прикурильские воды
Тихого океана. Таким образом, в Охотском море, как и в Берин-
говом, фон зимней орнитофауны составляют ныряющие птицы
( чистиковые, утки, бакланы) и крупные чайки (тихоокеанская)-
На размещение скоплений чаек существенное влияние оказыва-
ет большой промысловый флот, добывающий минтая на шель-
1 О встречаемости этой чайки в Охотском море сообщил мне Шереметь-
ев Ю. В.
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
117
фе юго-западного побережья Камчатки. Большие стаи чаек в
течение всей зимы концентрируются у судов, ведущих лов рыбы.
Плотность населения птиц в водах шельфа и прилегающих
к нему водах материкового склона соответствует тому, что наб-
людается в Беринговом море и составляет около 10 особей на
I кв. км.
Весной наблюдений в Охотском море почти не проводилось.
Известно только, что основные потоки мигрантов из гнездящих-
ся на севере моря видов проходят в апреле и мае вдоль запад-
ного побережья Камчатки и восточного побережья Сахалина.
Прилетающие в мае в больших количествах из южного полуша-
рия серый и тонкоклювый буревестники концентрируются в это
время в южной части моря, особенно в районе пролива Ла-
перуза.
Летний период характеризуется значительным преобладани-
ем трубкбносых в открытых водах как на шельфе, так и в глу-
боководной части Охотского моря (табл. 12). Состав доминиру-
ющих видов в различных участках моря несколько отличается.
На крайнем юге моря вдоль Хоккайдо, в проливе Лаперуза,
у Южных Курил резко преобладают буревестники (серый, тон-
коклювый, меньше бледноногий — Puifinus carneipes Gould) и
серая качурка. Все эти виды образуют здесь крупные скопле-
ния. Высокая плотность птиц наблюдается в прикурильских во-
Таблица 12
Видовой состав, соотношение (%) и плотность морских птиц
в Охотском шоре летом (июль—август). 1963— IMS гг.
Виды Шельф н прилегающие воды Глубоководная часть моря
F. glacialis 24.0 50.0
Качурки (О. funcata) 20,0 35,0
Буревестники (Р. griseus, Р. tenuirostris) 23.0 1,0
Кайры 6,0 +-
Мелкие чистиковые 10,0 2.0
L. cirrhata 4,0 6.0
R. tridactyla 6,0 1.0
L. schistisagus 1,0 4-
Утки 3,0 —
Остальные виды 3,0 5,0
Количество особей на 1 кв. км 20,0 2.7
Количество учетов 47 82
118
ОХОТСКОЕ МОРЕ
дах- В этом районе фон населения составляют качурки, глупыш
и топорок. Гнездящиеся на Курильских островах в больших ко-
личествах другие виды (кайры, бакланы, моевка) тяготеют здес:>
к узкой прибрежной зоне.
На шельфе западного побережья Камчатки птиц меньше.
Основу населения составляют большая конюга — Aethia
•cristatella Pall.), глупыш, серая качурка и топорок. На северном
мелководье, включая залив Шелихова, плотность населения
значительно возрастает. Здесь преобладают светлый глупыш,
кайра, топорок, конюги и обыкновенная моевка. В глубоковод-
ных районах основу населения составляют серая качурка, тем-
ный глупыш и в меньшей степени — топорок. Кроме них могут
быть встречены и другие виды — альбатросы, буревестники,
поморники, моевки и другие; все они здесь немногочисленны.
Таким образом, в Охотском море, как и в Беринговом, сос-
тав морских птиц шельфовых и глубоководных районов в значи-
тельной степени различается. Здесь представляется возможным
говорить о неритических и океанических птицах и соответствую-
щих их группировках. Правда, характер распределения выде-
ленных группировок несколько отличается. Шельфовая зона в
Охотском море уже, чем в Беринговом; в связи с этим нерити-
ческая группировка фито- и зоопланктона имеет ограниченное
распространение. В летний период область распространения не-
ритического комплекса планктона в виде узкой полосы вдоль по-
бережий окаймляет Охотское море. Наоборот, океанический
комплекс планктона распространен широко: он занимает всю
глубоководную часть моря, с течениями вдоль глубоководных
жолобов проникает далеко на север в направлении залива Ше-
.лихова и острова Святого Ионы (Смирнова, 1959; Лубны-Гер-
цык, 1959)' Нечто аналогичное наблюдается и у морских птиц
(рис. 34). Виды, тяготеющие к мелководным районам, имеют
здесь летом более ограниченное распространение. В северной
части моря, где шельф сравнительно широкий, область их рас-
пространения заметно шире. За пределами шельфа почти не
встречаются тихоокеанская и чернохвостая чайки, кайры, коню-
ги, ипатка. Летом за пределы мелководья редко выходят помор-
ники и обыкновенная моевка, которая в Беринговом море обыч-
на в глубоководной части. Такие птицы, как бакланы, очковый
и обыкновенный чистики, утки, пыжики, крачки, рогатый тупик,
вообше концентрируются в непосредственно прибрежной зоне.
1 Интенсивное поступление океанических вод по жолобам в северо-запад-
ном и северо-восточном направлениях накладывает большой отпечаток на
распространение и распределение скоплений беспозвоночных и рыб не толь-
ко в поверлностных слоях, но и в батиали (Ушаков, 1963; Шунтов, 19656,
.19676).
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
119
Рис. 34. Распределение неритических и океанической группировок морских
птиц в Охотском море летом. 1 —неритические, 2 — океаническая
Океаническая группировка насчитывает меньше видов, но
занимаемая ею область более обширна. Она включает всю глу-
боководную часть моря и районы нижней части шельфа, находя-
щиеся под воздействием океанических вод. Характерный и мно-
гочисленный вид этой группировки — серая качурка. В северо-
западной части моря она почти не заходит севернее изобаты
200 м, но вдоль жолоба. по которому происходит наиболее да-
лекое проникновение океанических вод на север, доходит до
залива Шелихова. Район нижней части мелководья, а также
прикурильские воды, где шельф почти не выражен, являются
местом смешанного обитания видов рассматриваемых комплек-
сов.
Таким образом, различия в составе птиц в северной и юж-
ной частях моря во многом определяются большим развитием
120
OXOICKOE МОРЕ.
на севере неритической, а на юге — океанической эон. Однако
имеют здесь значение и климатические различия этих районов
.моря
Различия эти существенны, так как Охотское море в мери-
диональном направлении простирается больше, чем на тысячу
миль (Леонов, 1960). Ограниченны в распространении более
теплой южной частью моря чернохвостая чайка, рогатый тупик,
бледноногий буревестник, тайфунннк Пила, уссурийский баклан
— Phalacrocorax filamentosus (Temm. et Schleg.) С юга на север
уменьшается численность темного глупыша, альбатросов, качу-
рок. На севере моря меньше тонкоклювого и серого буревестни-
ков, хотя отдельные стаи их могут достигать залива Шелихова.
Наоборот, с юга на север увеличивается плотность и числен-
ность светлого глупыша, кайр, топорка, ипатки, коиюг,
поморников, обыкновенной моевки тихоокеанской чайки и
других.
Факт обитания большей части рассматриваемых выше ви-
дов в Охотском море известен. Ранее не отмечались здесь крас-
ноногая моевка, бледноногий буревестник и тайфунннк Пила,
которые посещают Охотское море во время летнего и осеннего
сезонов. По моим наблюдениям, бледноногий буревестник мо-
жет быть отнесен к вполне обычным птицам южной части моря.
Красноногая моевка редка, однако залеты ее в Охотское море
не являются случайными. Она отмечалась у восточного побере-
жья Камчатки, северо-восточного побережья Сахалина и у Ку-
рильских островов. По-видимому, регулярны проникновения в
глубоководную часть моря и тайфунника Пила.
Охотское море — очень продуктивный водоем. Некоторые
его участки более продуктивны, чем в Беринговом море Но в
целом, судя по содержанию биогенных элементов, величинам
первичной продукции, биомассам планктона, бентоса и концент-
рациям рыб, эти два моря вполне сопоставимы (Ушаков, 1953;
Лубны-Герцык, 1959; Смирнова, 1959; Бруевич и др., 1960; Сэ-
вилов, 1961; Зенкевич, 1963; и др.). Это находит отражение в
плотности населения и численности морских птиц. Как видно из
табл. 12, плотность населения на 1 кв. км — 20 для мелковод-
ных и 2,7 особей для глубоководных районов — фактически
идентична плотности в Беринговом море. Как и в Берин-
говом море, плотность населения птиц в мелководных райо-
нах в несколько раз выше, чем в глубоководных, что находится
в связи с большей продуктивностью неритической зоны. При вы-
числении численности птиц в Охотском море полученная для
шельфа плотность была перенесена на прибрежную зону, в ко-
торой проводилось ограниченное количество учетов. Цифра об-
щей численности птиц оказалась довольно внушительной —
СЕЗОННЫЕ аспекты в орнитофауне
121
16,4 млн. в мелководной и 2,1 млн. особей в глубоководной ча-
сти моря.
На фоне значительной плотности, характерной для шельфо-
вых и прибрежных вод, выделяются три района, где скопления
пищ наиболее мощны и устойчивы: пролив Лаперуза, северо-
восточная часть моря и прикурильские воды (рис. 35)- Пролив
Лаперуза и прилегающие к нему воды являются очень оживлен-
ным районом. Эта часть моря характеризуется сложной дина-
микой вод; с севера сюда подходит холодное Восточно-Саха-
линское течение, а из Японского моря — теплое течение Соя
(Леонов, 1960; Морошкин, 1964). В течение всего летнего пе-
риода здесь очень обилен планктон (Пономарева. 1954, 1961), с
юга сюда мигрируют кальмары, анчоус, сайра. Уже в мае в этом
районе начинают собираться стаи серого и тонкоклювого буре-
вестников, затем серой качурки.
Рис. 35. Распределение концентраций морских птиц в Охотском море летом.
1 — низкие, 2 — средние, 3 — высокие, 4 — течения
122
ОХОТСКОЕ МОРЕ
Исключительно высокой биологической продуктивностью ха-
рактеризуется северо-восточная часть Охотского моря от Тауй-
ской губы до залива Шелихова. Этот район интересен тем, что
здесь наблюдается мощный выход на поверхность холодных
глубинных океанических вод, поступающих на север по впадине
ТИНРО. В связи с этим даже в разгар лета у входа в залив
Шелихова наблюдаются поверхностные температуры порядка
1,5—2,0°. Непрерывному процессу перемешивания вод способст-
вует летом фронтальная зона, формирующаяся южнее Тауйской
губы в результате встречи теплых поверхностных тихоокеанских
вод с холодными водами шельфа. В итоге по богатству подни-
маемых течениями в слои фотосинтеза биогенных элементов се-
веро-восточный угол Охотского моря относится к одной из са-
мых. продуктивных акваторий Мирового океана (Бруевич и др.,
I960; Винокурова, 1965; Морошкин, 1966; Чернявский, 1970,
1970а; и др.). Как показывают исследования разных лет, это
находит подтверждение в мощном развитии планктона и бенто-
са (Кун, 1951; Смирнова, 1959; -Лубны-Герцык, 1959; Микулич,
1960; Савилб®, 1961;.Котляр, 1965,1967, 1970; и др.)-
Рассматриваемый район является местом обитания и кон-
центраций двух весьма крупных стад сельди — охотского и ги-
жигииско-камчатского (Аюшин, 1956; Прайоторова, 1965; Хари-
тонова, 1965; и др.). Таким образом, большая численность
птиц (чистиковые, чайки, трубконосые) в северо-восточной ча-
сти Охотского моря находится в полном соответствии с высокой
кормностью этого района. В связи с поздним прогревом и по-
стоянным поступлением в верхние слои биогенов, развитие
планктона и его высокие биомассы наблюдаются и в осенние
месяцы. До глубокой осени нагуливается здесь сельдь. В тече-
ние всего осеннего периода сохраняются и концентрации птиц.
Общеизвестно богатство вод прикурильского района. Благо-
приятные условия для обитания морских птиц здесь также соз-
дают многочисленные острова, на которых располагаются гнез-
довые колонии.
Гнездовой период большей части размножающихся по бе-
регам Охотского моря птиц заканчивается во второй половице
августа — первой половине сентября. Но некоторые виды еще
раньше начинают осенние перемещения. Со второй половины
июля появляются стайки круглонооого плавунчика, а затем и
другие кулики. Массовый пролет куликов наблюдается с нача-
ла августа до начала сентября. В третьей декаде августа начи-
нается интенсивный пролет сухопутных птиц, который продол-
жается в сентябре и октябре. С конца августа в открытом море
появляются пролетные стайки уток, количество которых значи-
тельно увеличивается в сентябре. Основные пролетные пути
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
123
ут<ж и куликов проходят вдоль побережий. Однако в больших
количествах они встречаются и вдали от берега. Весьма интен-
сивный пролет уток, куликов, а также сухопутных птиц наблю-
дается от северного побережья на юг и юго-запад в направлении
Сахалина.
Тонкоклювый, серый и бледноногий буревестники встреча-
ются в Охотском море в течение всей осени. Но уже к сентябрю,
в связи с отлетом на юг размножающихся в данном году осо-
бей, численность этих видов заметно уменьшается. В результа-
те этого, а также увеличения в открытых водах других птиц,
трубконосые с начала осени перестают быть доминирующей
группой в мелководных районах. В глубоководной части моря
они по-прежнему преобладают, -хотя доля их уменьшается
(табл. 13).
Таблица 13
Видовой состав, соотношение (%) и плотность морских птиц
в Охотском море осенью (сентябрь—ноябрь). 1959—1965 гг.
Виды Шельф и приле- гающие воды Глубоководная часть моря
Кайры 40,0 3,0 ч
Мелкие чистиковые 15,0 8,0
L. cirrhata 1,0 6,0
F. glacialis 9,0 28,0
Буревестники (Р. griseus,
Р. tenuirostris) 9,0 20,0
Качурки (О. furcata) + 14.0
R. tridactyla 8,0 4,0
L. schistisagus 4,0 .7,0
Другие и неопределенные чайки 3,0 2,0
Поморники 1,0 1.0
Бакланы (P.pelagicus) 1,0 +
L. lobatus 4,0 +
Утки 1,0 4,0
• Остальные виды 4,0 3,0
Количество особей на 1 кв. км 14,0 3,0
Количество учетов 45 40
Закончив размножение, большая часть видов рассредоточи-
вается в сторону открытого моря, а некоторые сразу начинают
откочевку на юг. Естественно, что в связи с этим в сентябре за
пределами прибрежной эоны заметно увеличивается количество
чаек и чистиковых (см. табл. 13). У тихоокеанской чайки первы-
ми рассеиваются в море молодые особи. Так, летом в северной.
124
ЯПОНСКОЕ МОРЕ
половине моря на долю особей в сером оперении приходится не
более 5% из встречаемых в открытом море чаек, а в сентябре
их процент увеличивается до 50. В Беринговом море и у восточ-
ного побережья Камчатки эта чайка во все сезоны тяготеет к
шельфовой зоне. В Охотском море в период осенних кочевок
она широко рассеивается по всему морю.
В начале сентября начинают откочевку на юг поморники.
На севере моря их количество сразу уменьшается, а на юге, осо-
бенно у пролива Лаперуза, во второй половине сентября и в
первой декаде октября они становятся многочисленными.
Уменьшается в сентябре в Охотском море численность серой ка-
чурки, а на севере моря — топорка. В это же время по всему
морю встречаются мигрирующие на юг особи обыкновенной мо-
евки. Сравнительно обычными становятся серебристая и по-
лярная чайки.
Таким образом, к октябрю, в результате начавшегося отлега
на юг, кроме буревестников, уменьшается численность топорка,
поморников, моевки, качурок. Обычными или многочисленными
в октябре они становятся только на юге моря. Большая часть
видов чистиковых птиц, крупных чаек и глупыш в сентябре
также рассредоточиваются, выходя из прибрежной зоны. Но за-
метное смещение нх на юг начинается только в октябре. Мигра-
ции этих птиц происходят постепенно и, как было показано вы-
ше, много кайр, мелких чистиковых и крупных чаек встречается
в Охотском море и зимой. Осенью, как и летом, плотность птиц
в шельфовых районах в несколько раз выше, чем в глубоковод-
ных (см. табл. 13). Это связано с тем, что основные потоки ми-
грантов проходят вдоль побережий: на востоке моря вдоль
шельфа Камчатки к Курильским островам и на западе моря
вдоль Сахалина к проливу Лаперуза и Хоккайдо, а затем в
Японское море и Тихий океан.
3. Японское море.
В общих чертах сезонные аспекты в распределении птиц в
Японском море мною описаны несколько лет назад (Шунтов,
1965а). Накопленные после этого новые материалы позволяют
уточнить и дополнить некоторые из полученных выводов.
По климатическим и гидрологическим условиям Японское
море исключительно контрастно. В связи с этим в видовом со-
ставе и особенностях распределения морских животных, в том
числе морских птиц, в различных районах и в различные сезо-
ны наблюдаются большие различия. В северной части моря
(в основном в Татарском проливе) гнездятся преимущественно
северо-бореальные виды. У берегов Приморья. Кореи, Сахалина
н Хоккайдо вместе с широко распространенными бореальными
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
126
видами обитают также южнобореальные виды» а в юго-восточ-
ной части моря среди гнездящихся видов основу населения со-
ставляют субтропические птицы (Austin, 1948; Austin a. Kuroda,
1953; Воробьев, 1964; Гизенко, 1955; Успенский, (959; и др.).
Северная и северо-западная части Японского моря, находя-
щиеся под влиянием холодного дыхания азиатского материка,
зимой сильно охлаждаются. В январе средняя месячная изотер-
ма воздуха 0° проходит примерно от Корейского залива к Сан-
гарскому проливу. У берегов Приморья в этом месяце средняя
температура составляет около минус 7°, а на севере Татарского
пролива — минус 19°. В северных районах моря до отрицатель-
ных температур охлаждаются зимой поверхностные воды (Лео
нов, I960), почти весь Татарский пролив и часть шельфа При-
морья покрываются льдами (Крындин, 1961). Южная и юго-во-
сточная части моря, находящиеся под отепляющим влиянием
Цусимского течения и Тихого океана, значительно теплее. Сред-
няя температура января здесь 5°, а поверхностные воды вдоль
Хонсю и на юге моря имеют температуру 7—13’ (Леонов, 1960).
В связи со значительным охлаждением воздуха и воды теп-
лолюбивые виды откочевывают в южную часть Японского моря,
а также в южнее расположенные районы. Совсем покидают на
зимний период Японское море субтропические виды. Лишь в
начале зимы только в районе Корейского пролива в небольшом
количестве встречается пестролицый буревестник — Puffinus
leucomelas Temm. Основная масса особей этого многочисленно-
го вида улетает в Восточно-Китайское, Австрало-азиатские мо-
ря и прилегающие воды Тихого океана. Из холодной северном
части моря к берегам полуострова Корея, Японии и частично
в Восточно-Китайское море откочевывают южно-бореальные ви-
ды — чернохвостая чайка, очковый чистик, рогатый тупик, уссу.
рийский баклан (отдельные особи последнего, возможно, зиму-
ют). Не задерживаются здесь на зимовку и некоторые относи-
тельно теплолюбивые северные виды, которые обычны или мно-
гочисленны у берегов Приморья и Сахалина во время осенних
миграций: обыкновенная моевка, поморники, гагары и другие.
Несмотря на откочевку многих видов на юг, в течение всего
зимнего периода численность птиц в северной части Японского
моря остается высокой. Сюда мигрирует довольно большое ко-
личество особей из Охотского моря и других северных районов.
Часть нх особей распространяется по всему Японскому морю, а
также мигрирует южнее.
В северной части Японского моря у берегов Приморья, Са-
халина и Хоккайдо основу населения составляют чистиковые
птицы' (конюги, кайры и другие), утки (морянка, каменушка —
Histrionicus histrionicus (Ь.)и другие), тихоокеанская чайка н
бакланы. На их долю здесь приходится не менее 90% птиц
126
ЯПОНСКОЕ МОРЕ
(табл. 14). В южной и юго-восточной частях моря у берегов
Хонсю и юго-восточного побережья полуострова Корея все эти
группы также представлены, но в меньших количествах. Здесь
увеличивается численность относительно теплолюбивых видов,
в том числе представителей южно-бореального комплекса. Как
видно из табл. 14, в этих районах над всеми видами преобла-
дают чайки, из которых наиболее многочисленны обыкновенная
моевка и чернохвостая чайка. Преимущественно чайки состав-
ляют фон зимнего населения птиц и в открытых глубоковод-
ных районах Японского моря. В водах, прилегающих к мелко-
водью и материковому склону, еще могут быть встречены кай-
ры и другие чистиковые, но большая часть глубоководных райо-
нов моря особенно пустынна.
Таблица 14
Видовой состав, соотношение (%) и плотность морских птиц
в Яроиском море зимой (декабрь—март). 1959—1966 гг.
Виды Шельф и прилегаю- щие воды Глубоковод- ная часть моря
севернее 40° с. ш. южнее 40° с. ш.
L. schistisagus 8.0 4.0 60,0
L. crassirostris 4.0 20,0 10,0
R. tridactyla 2.0 40,0
Другие и неопределенные чайки 4.0 18,0 9,0
Мелкие чистиковые 37,0 13,0 3.0
Кайры (U. aalge) 15.0 + 9.0
Другие чистиковые 3.0 + +
Утки 21,0 + —
Бакланы (Р. pelagicus) 5,0 + —
Р. leucomelas — 4,0 —
Остальные виды 1.0 1,0 9,0
Количество особей на 1 кв. км 8,0 4,0 0,2
Количество учетов 69 28 28
Среди чаек в северной части Японского моря преобладают
взрослые светлые особи. Так, на их долю у тихоокеанской чайки
в водах Приморья и Сахалина приходится в среднем около 90%
При этом в течение зимы доля светлых птиц постепенно увели-
чивается. В южной части моря доля серых птиц в 2—3 раза вы-
ше. Аналогичная картина наблюдается и у чернохвостой чайки.
Таким образом, в течение большей части зимнего периода
орнитофауну Японского моря слагают только бореальные эле-
менты. На севере моря — это северо-бореальные и широко рас-
СЕЗОННЫЕ аспекты в ОРНИТОФАУНЕ
127
пространенные в бореальной зоне виды, а в южной половине
моря к ним добавляются виды южно-бореального комплекса.
Выше отмечалось, что решающими факторами, вызывающими
рткочевку птиц и особенности их распределения в Японском мо-
ре на зимовках, являются климатические и гидрологические ус-
ловия. Однако велика роль и кормового фактора. В сравнении
с другими сезонами кормовые условия для птиц зимой в Япон-
ском море менее благоприятны. С осени значительно умень-
шаются биомассы планктона (Пономарева, 1954а; Ивлева, 1960;
Мещерякова, 1960; и др.); осенью же в связи с охлаждением
вод. откочевывают на юг Японского или в Восточно-Китайское
моря многие субтропические пелагические рыбы (сайра, анчоус,
сардина, скумбрия), их молодь, а также массовый пелагическйй
тихоокеанский кальмар — Todarodes pacificus.
Условия для существования птиц зимой в Японском море
более благоприятны, чем во многих районах Берингова и Охот-
ского морей. Здесь мягче климат и гидрологический режим,
лучше кормовые условия. В северных морях зоопланктон в зим-
ний период в основном уходит ниже слоя зимней конвенции.
В Японском же море, наоборот, он концентрируется в верхних го-
ризонтах. И хотя в сравнении с летним сезоном его биомасса
уменьшается, все же остается более высокой, чем в Беринговом
И Охотском морях (Пономарева, 1954а; Мещерякова, 1960;
В др.). У ряда бореальных видов рыб в южно-бореальных райо-
нах, в том числе в Японском море, время размножения и мигра-
ций на меньшие глубины приходится на осенние и зимние ме-
сяцы (Моисеев, 1953). В частности, в это время происходит
мощный нерест самой массовой рыбы Японского моря — мин-
тая, и в ряде мелководных районов наблюдаются большие
скопления его икры в поверхностных слоях (Веденский, 1949;
Кагановская, 1950, 1954; и др.). Видимо, поэтому значительное
количество птиц, гнездящихся в более северных районах и. по-
кидающих их на зимний период, избирают Японское море для
зимовок. Это большая и крошечная — Aethia pusilia (Pall.)
конюги, пыжики, кайры, нырковые утки, тихоокеанская, сизая,
серебристая и полярная чайки, берингов баклан — Phalacroco-
rax pelagicus Pall., обыкновенная моевка и другие.
Общая численность птиц зимой в Японском море значитель-
на. Как видно из рис. 36, концентрации птиц располагаются в
прибрежной зоне с прилегающими водами и как бы окаймляют
море. Плотность населения здесь в 20—40 раз выше, чем в глу.
боководных районах моря (см. табл. 14). Пересчет на соответ-
ствующие площади дал 2,1 млн. особей для мелководных райо-
нов и 0,12 млн. для глубоководных. По-видимому, цифра для
мелководья занижена. Дело в том, что при расчетах на непо-
средственно прибрежную зону перенесена плотность, получен-
Рис. 36. Основные районы зимовок морских птиц в Японском море.
1 — льды, 2 — теплые течения, 3 — холодные течения, 4 — основные
районы концентраций птиц
СЕЗОННЫЕ аспекты В ОРНИТОФАУНЕ 129
ная для всего шельфа. В то Жевремя некоторые виды, в первую
очередь утки, тяготеют к узкой прибрежной полосе (чаще бух-
ты, заливы), где проводилось мало наблюдений. Плотность же
их в этих районах значительно выше, чем на остальной части
шельфа.
Различие в плотности н численности птиц в шельфовых и
глубоководных частях моря вполне закономерны и объяснимы.
Связано оно в основном с различной продуктивностью и соот-
ветственно кормностыо рассматриваемых .районов. В глубоко-
водных открытых районах зимой почти нет рыбы. Птицы могут
здесь питаться только планктоном, биомассы которого невели-
ки, и отбросами с пересекающих море судов. Другое дело —
мелководье с его обильным рыбным населением, бентосом и
планктоном. Существенно также то, что фактически вдоль все-
го побережья Японского моря работает многочисленный рыбо-
ловный флот, отбросами которого в течение зимы кормится
большое количество чаек.
Потепление поверхностных вод Японского моря начинается
со второй половины марта (Панфилова, 1961). Весной усили-
вается поступление через Корейский пролив тихоокеанских вод
с Цусимским течением (Сизова, 1961). Именно через Корейский
пролив раньше всего начинается весенний пролет птиц. Во вто-
рой половине марта в значительных количествах здесь мигри-
руют зимовавшие южнее особи чернохвостой чайки и обыкно-
венной моевки. В марте в Корейский пролив и южную часть
Японского моря проникает пестролицый буревестник. В начале
весны три перечисленных вида составляют основу населения
южной части моря. Встречающиеся здесь также другие чайки,
пойорники и чистиковые более редки.
Зимовавшие в северной части Японского моря конюги, кай-
ры, утки, тихоокеанская чайка и другие во второй половине
марта начинают откочевку на север. На смену им прилетают из
более южных районов другие виды, в том числе южно-бореаль-
ные. Особенно заметна смена доминирующих видов у чаек.
У берегов Приморья преобладающую зимой тихоокеанскую
чайку заменяет чернохвостая, а в непосредственно прибрежной
зоне — также озерная. Многочисленная в марте на юге моря
обыкновенная моевка в апреле здесь становится редкой, но на
широте Сангарского пролива остается обычной. Пролет этого
вида на север происходит преимущественно по восточной части
моря вдоль берегов Японии. Чернохвостая чайка наоборот бо-
лее многочисленна на пролете вдоль побережья полуострова
Кореи. В открытых водах юго-западной половины моря в апре-
ле она становится самой многочисленной птицей. Среди мигри-
рующих на север особей чернохвостой чайки преобладают свет-
лые половозрелые, на долю которых приходится в разных райо.
130
ЯПОНСКОЕ МОРЕ
нах от 95 до 100%. Лишь в Корейском проливе соотношение
светлых и серых особей обратное.
В апреле в Японском море увеличивается численность пе-
стролицего буревестника. Этот теплолюбивый вид сначала кон.
центрируется на крайнем юге моря в районе Корейского проли-
ва и островов Оки. Здесь он резко доминирует над другими
птицами, при этом тяготеет к восточной части моря к зоне дей-
ствия Цусимского течения (рис. 37). В основном через восточ-
ную часть Японского моря мигрируют на север и поморники,
весьма обычные в течение апреля.
В конце марта — начале апреля в Японское море прилетает
бледноногий буревестник. В течение всего апреля он держится
на крайнем юге моря в районе Корейского пролива, а в мае
смещается ненамного севернее. Самые массовые из трансэква-
ториальных мигрантов — тонкоклювый и серый буревестники-
через Корейский пролив, как отмечалось выше, проникают ма-
ло. Их пролетные пути проходят восточнее. В конце апрели,
когда стаи этих видов приближаются к Хонсю и Хоккайдо,
часть их проникает через Сангарский пролив в северо-восточ-
ную часть Японского моря. В мае тонкоклювый и серый буреве.
стники попадают в Японское море также через пролив Лаперу-
за из Охотского моря. У входа в проливы Сангарский и Лапе-
руза рассматриваемые виды могут встречаться в значительных
количествах. Однако весной и в дальнейшем их распростране-
ние ограничивается главным образом северо-восточной частью
моря.
В течение мая пролет чаек и поморников через южную по-
ловину Японского моря в основном заканчивается. В открытом
море сравнительно обычной остается только чернохвостая чай-
ка, которая регулярно встречается в полосе нескольких десят-
ков миль вдоль всего побережья полуострова Кореи и вокруг
острова Дажелет. Кроме чернохвостой чайки, в южной половине
моря в это время можно встретить бродячих особей серебри-
стой, полярной чаек, моевки и отдельных поморников. К началу
лета в открытом море остаются почти одни буревестники.
Весной через Японское море проходит интенсивный пролет
куликов и сухопутных птиц. Из куликов самым многочисленным
является круглоносый плавунчик, массовый пролет которого
происходит в первой половине мая. Основной его поток прохо-
дит вдоль побережья Хонсю. Во время миграций сухопутные
птицы встречаются по всему Японскому морю, но наиболее ожив-
ленный пролет происходит через южную часть моря и Корейский
пролив от полуострова Корея к Японии. Много сухопутных птиц
и куликов летит также через северо-западную часть моря от се-
верного побережья полуострова Корея к Южному Приморью.
Рис. 37. Распределение пестрошцепо буревестника — Puffinus leucomelas в Японском море. Слева — весна,
справа — лето, 1 — холодные течения, 2 — теплые течения, 3 — поверхностные изотермы, 4—«редок, 5 — обычен.
6 — многочислен
132
ЯПОНСКОЕ МОРЕ
Соотношение видов весной в открытых водах Японского мо-
ря может характеризовать очень осредненная таблица 15. Пр:;
сравнении с зимним периодом видно, что весной происходит
смена доминирующей группы: чистиковых и чаек заменяют
трубконосые. Убыль птиц, откочевавших на север, компенси-
руется прилетевшими с юга буревестниками. В итоге плотность
населения и общая численность птиц весной почти не отличает-
ся от того, что наблюдается зимой (2,3 млн. особей в мелковод-
ных районах и0,2 млн. в глубоководных).
По некоторым признакам западная половина Японского мо-
ря более продуктивна^ чем восточная. Концентрация биогенных
элементов в водах Приморского течения примерно в два раза
выше, чем в водах Цусимского течения (Мокиевская, 1961).
В связи с этим наиболее высокие биомассы планктона наблю-
даются именно в северных и западных районах моря (Брод-
ский, 1941; Мещерякова, 1960; и др.). Раньше, когда на север
моря совершали массовые миграции иваси и скумбрия, у бере-
гов полуострова Корея и Приморья эти виды были более мно-
гочисленными,, чем у Японии (Кагановский, Г9ЭД: 1951; Веден-
ский, 1954). В настоящее время здесь более обильны такие
планктоноядные рыбы, как минтай, сайра, а также кальмар. Од-
нако сравнение общей плотности населения птиц не дает осно-
вания для противопоставления западной и восточной половин
Японского моря. На весенних миграциях только чернохвостая
чайка, бледноногий буревестник и некоторые чистиковые более
многочисленны в западной части моря. Наоборот, пестролицый
буревестник, моевка, поморники и круглоносый плавунчик тяго-
теют на пролете к восточной части моря. Очевидно, обеспечен-
ность птиц пищей на востоке моря удовлетворительная. Действи-
тельно, до сих пор у побережья Хонсю в заметных количествах
обитает сардина, многочисленны здесь в весенне-летний период
ставрида, скумбрия, летучие, рыбы, анчоус, сайра, кальмар и
другие- Цусимское течение приносит сюда с юга большое коли-
чество молоди различных рыб.
Как видно из табл. 15, плотность птиц в глубоководных рай-
онах моря в сравнении с зимним периодом увеличивается в 2 ра-
за. Тем не менее она остается в 15 раз ниже, чем в водах шель-
фа и материкового склона. Все это находится в соответствии с
распределением кормовых организмов. Прибрежная зона и при-
легающие к ней воды более продуктивны. Здесь обильнее планк-
тон. К водам шельфа тяготеют концентрации всех массовых ви-
дов рыб*. В открытом море сравнительно обычны только сайра п
кальмар. Но я эти животные и их молодь также больше тяготе-
ют к краевым зонам моря (Соеда, 1956; Fukataki. 1959; Наша-
be a. Shimizu, 1966; Шунтов. 1964а, 1965b, 1967b; и др.).
СЕЗОННЫЕ АСПЕКТЫ D ОРНИТОФАУНЕ
133
Таблица 15
Видовой состав, соотношение (%) и плотность морских птиц
в Японском море весной (апрель—май). 1961—1965 гг.
В и .1 ы Шельф и прилегаю- щие воды Глубоководная часть моря
р, leucomelas 41,0 22,0
р. tenuirostris, Р. griseus 10,0 1.0
Р. carneipes 5,0 1.0
L. crassirostris 15,0 47,0
R. tridactyla 10,0 5,0
Крупные чайки 4,0 5,0
Чистиковые 3,0 4,0
L lobatus 10,0 +
Поморники 1.0 3,0
Остальные виды 1.0 12,0
Количество особен на 1 кв. м 6,0 0.4
Количество учетов 124 72
Примечание. Большая часть наблюдений проводилась в южной половине
моря. В связи с зги-м в таблице оказались недостаточна или совсем непред-
ставленными чистиковые, утки, бакланы.
Время миграций птиц, гнездовья которых располагаются се-
вернее Японского моря, определяется не столько условиями на
местах зимовок, сколько сложившимися сроками, размножения.
В самом деле, миграции северных видов заканчиваются еще до
наступления наиболее благоприятного в кормовом отношений
летнего периода. Условия на местах гнездования во многом оп-
ределяют и.срргки миграций птиц, размножающихся в Япон-
ском море. Другое дело — характер миграции бледноногого бу-
ревестника, прилегающего в Японское море на откорм. Переме-
щение на север этого вида хорошо согласуется с общим ходом
сезонных изменений биологии открытых вод Моря. Максималь-
ная продукция планктона в рассматриваемом районе наблюдает-
ся между изотермами 13 и 18°. Синхронно с сезонным изменени-
ем расположения этих изотерм смещаются и зоны повышенных
биомасс планктона (Matsudaria et al.. 1956). Весной наиболее
богата зоопланктоном южная часть моря; севернее в это время
обилен фитопланктон1 (Мещерякова, 1960). Только на юге моря
отмечаются весной концентрации сайры и тихоокеанского каль-
мара.
Как отмечалось выше, в апреле и мае бледноногнй буреве-
стник не выходит за пределы южной части моря- Максимальное
развитие зоопланктона в северо-западной части Японского мо-
ря приходится на июнь (Бродский, 1941; Мещерякова, 1960; и
ЯПОНСКОЕ МОРЕ
др.). В тоне в эти районы смещаются концентрации сайры,
кальмара и бледноногого буревестника.
Летом плотность наделения птиц в Японском море уменьша-
ется. Примерно « два -раза уменьшается и общая численность
птиц (0,8 млн. в мелководных районах и 0,2 млн. в глубоковод-
ных) . Происходит это за счет откочевавших на север особей ря-
да видов. Уменьшение плотности населения происходит также
в связи с концентрацией размножающихся птиц непосредствен-
но у побережий.
В летний период в Японском море доминируют в основном
те же виды, что и в конце весны. Это, главным образом, трубко-
носые (табл. 16). Увеличивается в это время численность глу-
пыша. В небольшом количестве глупыш встречается в северной
половине Японского моря весь год. Летом его становится боль-
ше за счет особей, проникающих через пролив Лаперуза из
Охотского моря. Вместе с глупышом через пролив Лаперуза
проникает также серая качурка. Лётом через Корейский пролив,
в Японское море могут залегать тропические птицы (в 1962 г.
мною, например, добывался фрегат). В течение всего лета, хотя
и в небольших квличесгаак, но регулярен) встречается южно-
пойярнмй еюмеримс — Caftharacta skua Briton. С прогревом вед
в течение первой половины лета расширяют на север области
распространения пестролицый и бледаоногий буревестники. В се-
редине лета они достигают вод Приморья. Затем бледноногий
буревестник поднимается до Татарского пролива, а пестроли-
цый. как и весной, основной массой тяготеет к юго-восточной
половине моря (см. рис. 37)
В -юмшай ломаине моря в течение всего лета преобладаю-
щим видам остается нвстролицый буревестник. В меньших коли-
чествах здесь встречается бледаоногий буревестник. За преде-
лами «рифрежных >вод могут быть изредка встречены чернохво-
стая чайка и вилоавостая качурка — Oceanodroma monodiis
(Swtoh.). в северо-западном углу моря основу населения сос-
тавляют бледненогий. «остролицый буревестники и чернохвостая
чайка, но здесь не может считаться случайной встреча тонко-
клювой кайры, старика — Synthliboramphus anliguus (Gmell.)
(на мелководье — других чистиковых и бакланов) или бродяче-
го номоринка. По данным В. А. Нечаева и А. Г. Юдакова (1967.
4968). на островах н заливе Петра Великого гнездится более 20
тысяч особей вилонвостой качурки1. Учитывая позднее созрева-
ние трубконосых, нужно полагать, что общая численность этой
качурки в северо-западном углу моря должна быть выше. В
* Б. Г. Масленников (1965) «авшаегг другую цифру — всего окаю 300
качурок.
СЕЗОННЫЕ аспекты н орнитофауне
135
Таблица 1$
Видовой состав, соотамкте (%) мотмсть морских птиц
в Яванском море летом (омов август). 1ВИ—1МВ гт.
Виды Шельф и прилегающие воды Глубоковод- ная часть
севернее 40° с. ш. южнее 40° с. ш.
Р. canneipes 28,0 15,0 41,0
Р. leucomelas 9,0 70,0 41,0
Р. griseus, Р. tenuirostris 11,0 — —
F. glacialis 9,0 — —
Качурки (О. lureata) О. nronorhis) 1,0 1,0 +
L. cras&irostris 19,0 4,0 9,0
Другие чайки ЗЛ> + — J
Чистиковые 9,0 + + )
Утки 3,0 — —
Осталшые виды 3,0 10,0 9,0
Количество особей на 1 кв. км 2,0 2Д 0,3
Количество учетов 33 39 58
этой связи я не могу объяснить, почему при наблюдениях весной
и летом в рассматриваемом районе отмечались только редкие
одиночки вилохвостой качурки. В гнездовой период качурка ве-
дет ночной образ жизни, поэтому при дневных наблюдениях она
может выпасть из поля зрения. Но такой образ жизни характе-
рен и для других качурок, тем не менее в других районах, где
они гнездятся в значительных количествах, в гнездовой период
нх всегда можно встретить на море (хотя бы неполовозрелых
особей).
В северной части Японского моря состав фоновых видов дру-
гой. Это тонкоклювый, серый буревестники, глупыш, местами
серая качурка, поморники, крупные чайки, а в прибрежной зоне
также гнездящиеся здесь чистиковые и бакланы.
При описании летнего распределения морских птиц в Берин-
говом и Охотском морях подчеркивалось различие в распрост-
ранении неритических и океанических видов- В Японском море
картина несколько иная. Прибрежные птицы (в основном чисти-
ковые и бакланы) летом в Японском море имеют ограниченное
распространение, что связано с узостью шельфа и соответствен-
но неритической зоны. Область их распространения узкой поло-
136
ЯПОНСКОЕ МОРЕ
сой окаймляет Японское море (рис. 38). В северной половине
моря — это кайры, очковый чистик, рогатый тупик, ипатка,
обыкновенный .старик, уссурийский иберингов бакланы, а в юж-
ной — старики обыкновенный и хохлатый — Synthliboramphus
wumizusume (Temm.), местами рогатый тупик. Столь же ограни-
ченное распространение в Японском море имеет и неритический
комплекс планктона (Кун и Мещерякова, 1954). Океаническая
группировка птиц в Японском море не выражена. Преобладаю-
щие в глубоководных районах моря буревестники и чайки бо-
лее часто встречаются в водах шельфа.
Вместе с этим заслуживает внимания вопрос о различии в
составе морских птиц в северо-западной и юго-восточной поло-
винах Японского моря. Выше уже подчеркивались различия в
климатическом и гидрологическом режимах данных частей мо-
ря. Эти различия накладывают решающий отпечаток на состав
и особенности распределения вСех групп морских животных. В по-
верхностной япономорской водной массе1, формирование которой
связано с холодным Приморским течением, обитает комплекс
холоднолюбивых и умеренных видов планктона. Южная граница
распространения этих вод проходит приблизительно от Корей-
ского залива к южной оконечности Сахалина- В поверхностной
тихоокеанской водной массе, формирование которой связано с
теплым Цусимским течением, обитают теплолюбивые субтропи-
ческие и тропические виды. В зоне смещения вод встречаются
виды обоих комплексов (Киселев, 1947; Кун и Мещерякова.
1954; Кун. 1969; и др.). В общих чертах то же самое можно ока?
зать и о составе фаун рыб рассматриваемых районов. Нечто по-
добное наблюдается и у птиц (рис. 38). В юго-восточной поло-
вине моря как в прибрежной зоне, так и в открытых водах, до-
минирует субтропический вид — пестролицый буревестник. Пе-
Стролицый буревестник, а также другой субтропический вид
вилохвостая качурка — встречаются летом до широты Примо-
рья. Область распространения в море бореальных видов летом
ограничивается водами северной части моря. Вдоль восточного
побережья полуострова Кореи, а также у Южного Приморья и
Хоккайдо орнитофауна носит смешанный характер. .Здесь вмес-
те с пестролицым буревестником встречаются .чернохвостая чай-
ка и несколько видов чистиковых (см. рис. 38).
Японское море менее продуктивно, чем Берингово и Охот-
ское. Его. воды менее насыщены биогенными элементами (Мо-
ки ев ска я, 1961)- В целом менее обилен здесь зоопланктон и ни-
же численность рыб. Это находит отражение в плотности насе-
ления и численности птиц. Во все сезоны в Японском море они
1 Номенклатуоа водных масс по М. А. Радзиховской (1961).
Рис. 38. Распределение различных группировок морских птиц в Японском
море летом. 1 — область распространения бореальных и юЖ|»о-бореаль-
ных видов, 2 — область распространения субтропических видов. 3 — нери-
тическая группировка
138 ЯПОНСКОЕ МОРЕ
значительно ниже. Летом довольно большие концентрации лтиц
наблюдаются только у восточного побережья Хонсю (пестроли-
цый буревестник) и в районе пролива Лаперуза (серый и тон-
коклювый буревестники).
Говоря о более низкой продуктивности Японского моря, нуж-
но. однако, отметить, что некоторым исключением в этом смысле
являются воды Татарского пролива. Этот район по продуктивно-
сти. по-видимому, сопоставим с богатыми районами Охотского и
Бершгова морей. Здесь очень обилен планктон, биомассы кото-
рого достигают нескольких тысяч миллиграммов на кубометр
воды. При этом местами большие концентрации образуют такие
ценные в кормовом отношении планктеры, как эвфаузииды
(Бродский, 1959; Ивлева, 1960; Мещерякова, 1960; Пономарева.
1963; и др.).
В северной части Японского моря нагуливалось имевшее
раньше очень высокую численность сахалино-хоккайдское стадо
сельди. В больших количествах на нагул сюда поднимались ко-
сяки сардины и скумбрии. В настоящее время высокую числен-
ность здесь имеют такие планктоноядные рыбы, как минтай, тер-
пуг, песчанка и другие. Летом мощные концентрации образует
тихоокеанский кальмар. Богато развита на литорали и сублито-
рали северной части Японского моря бентическая фауна (Мо-
киевскин, 1960; Зенкевич, 1963). Численность же гнездящихся
здесь птиц нельзя назвать высокой. В Японском море нет мощ-
ных птичьих базаров. На мой взгляд, главная причина этого —
ограниченность мест, пригодных для размножения колониально
гнездящихся птиц. По-видимому, особенно существенно то. что
на севере Японского моря мало скалистых островов. Например,
в заливе Петра Великого, где довольно много островов, еще есть
сравнительно большие базары.
Описанная для летнего периода картина распределения птиц
в общих чертах сохраняется и в сентябре. По климатическим
и гидрологическим условиям сентябрь, особенно его первая по-
ловина, в Японском море напоминает летний месяц. Тем не ме-
нее, в это время уже намечаются изменения в распределении
птиц, свойственные для осени. Первыми признаками приближа-
ющейся осени могут служить появляющиеся в море со второй по-
ловины августа пролетные кулики и сухопутные птицы. Вдоль
побережий Татарского пролива в сентябре начинают появлять-
ся летящие в южном направлении крачки, озерная чайка, стайки
уток- В сентябре в северной части Японского моря увеличивает-
ся численность крупных чаек (тихоокеанская и серебристая) и
поморников. Отчетливо прослеживается перемещение этих птиц
из Охотского моря через проливы Лаперуза и Татарский.
К началу сентября к местам гнездования в южное полуша-
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
139
рие отлетвет часть бледионогого буревестника. Но, очевидно,
неполовозрелые особи этого внда встречаются в Японском море
в течение веего октября. В эти же сроки отлетают к местам гнез-
дования тонкоклювый и серый буревестники- Большая часть
проникших в северо-восточную часть Японского моря их осо-
бей сначала возвращается в Охотское море, а затем летит на
юг. Через Корейский пролив, по-видимому, отлетают только от-
дельные птицы или группы. В октябре откочевывает на юг и
часть пестролицего буревестника, при этом сокращается область
его распространения в Японском море. В начале октября он
встречается только к югу от 4(Г с. ш.
В октябре за счет мигрантов из северных районов количест-
во птиц в Японском море увеличивается- Обычными или много-
численными в северной части моря становятся крупные чайки,
обыкновенная моевка, поморники, мелкие чистиковые> бакланы,
утки, гагары. Наиболее интенсивный пролет всех перечисленных
птиц начинается со второй половины октября и продолжается
в ноябре. Лишь основная масса поморников пролетает до нача-
ла ноября- В октябре происходит рассредоточение из прибреж-
ной эоны чернохвостой чайки; в открытом море преобладают в
это время ее молодые серые особи.
Ряд видов, в первую очередь чайки и поморники, широко рас-
сеиваются во время миграции по Японскому морю. В связи с
этим плотность в глубоководных районах увеличивается в срав-
нении с летом в два раза (табл. 17). Но основные потоки миг-
рантов проходят, как и в другие сезоны, у побережий. Это отно-
сятся особенно к уткам, бакланам, чистиковым, гагарам, опер-
ной и сизой чайкам. У берегов Приморья пролет перечисленных
птиц временами достигает большой интемснвиости.
Таблице 17
Видовой состав, соотношение (•%) и плотность морсмш птиц
в Японском море осенью (сентябрь—ноябрь). 1383—1386 гг.
Виды Шельф и ярмлегающме «о» Глубоковод- ная часть моря ।
севернее 40 с. ш. южнее 40° с. ш. ।
R. tridactyla 11,0 7,0 17,0 L crassirostris 8,0 7,0 14,0 Крупные чайки (L. schistisagus) 7,0 10,0 15,0 Мелкие чистиковые 27,0 + 23,0 Утки 34,0 4- 7,0 Бакланы 11,0 + + Р. leucomelas — 73,0 16,0 Поморники + + 2,0 Остальные виды 2,0 3JD 6,0 Количество особей на 1 кв. км 7,1 1,5 0,6 Количество учетов 44 16 54
140
СЕВЕРНАЯ ЧАСТЬ ТИХОГО ОКЕАНА
В теплую южную часть Японского моря в октябре, проника-
ет относительно небольшое количество представителей мощной
волны осенних мигрантов. Здесь, как и летом, преобладает Суб-
тропический вид — пестролицый буревестник (см. табл. 17).
Зимний облик орнитофауна южной части Японского моря при-
нимает только в ноябре—декабре. Время интенсивных мигра-
ций через Японское море совпадает с периодом' существенных
изменений в климатическом, гидрологическом и биологическом
режимах моря. В октябре наблюдается быстрое охлаждение вод-
ных и воздушных .масс, значительное обеднение поверхностных
вод планктоном, а субтропические пелагические рыбы и кальма-
ры быстро отходят в южную часть Японского моря-
В среднем плотность населения птиц осенью в Японском мо-
ре сопоставима с весенней. Соответственно была получена сход-
ная цифра и общей численности — 2,8 млн. особей (2,4 млн. в
прибрежных районах. 0,4 млн. в глубоководных).
В заключение характеристики сезонных аспектов в орнито-
фауне Японского моря нужно подчеркнуть две особенности, от-
личающие рассматриваемый район от Берингова и Охотского
морей. В Японском море более резко меняется по сезонам сос-
тав, общее количество видов и распределение -птиц. Достаточно
вспомнить, что летом в южной половине моря доминируют суб-
тропические виды, а зимой — бореальные. В северных морях
минимальная плотность и численность птиц приходится на зим-
ний период, а в Японском море — на летний. При объяснении
этих различий существенно то, что Японское море находится на
южном пределе бореальной и северном пределе субтропической
эон. Это определяет контрастность климатических и гидрологи-
ческих условий как в пространственном, так и сезонном аспекте.
Находя в Японском море зимой сравнительно благоприятные ме-
ста для зимовок, сюда откочевывает значительное количество
массовых видов с севера. Именно время нахождения северных
видов в Японском море является периодом наибольшего оби-
лия птиц. Заменяющие их летом представители южно-бореаль-
ного и субтропического комплексов по ряду причтен не имеют
такой большой численности -
4. Северная часть Тихого океана
(к северу от 30° с. ш.).
По северной части Тихого океана я располагаю данными бо-
лее двух тысяч учетов, выполненных большей частью к западу о:
175° з. д. Восточная часть океана охвачена наблюдениями недо-
статочно. Имеющиеся материалы позволяют охарактеризовать
состав, распределение и миграции птиц в общих чертах, при
этом ‘почти не затрагивая восточной части океана.
Воды северной части Тихого океана являются местом зимо-
СЕЗОННЫЕ аспекты в орнитофауне
141
вок большого количества особей многих видов, гнездящихся в
дальневосточных морях. Выше отмечалось, что зимой из Берин-
гова и Охотского морей откочевывают все или почти все особи
качурок, моевок, топорка, мелких чистиковых, темного глупыша,
поморников, гагар. Некоторые из них рассредоточиваются в при-
брежной зоне океана, другие широко рассеиваются как в при-
брежных. так и в открытых водах северной части Тихого океана.
Откочевывает сюда также значительное количество особей ви-
дов, многочисленных на зимовках в дальневосточных морях - -
крупных чаек. кайр, светлого глупыша, уток, бакланов.
В пределах прибрежной зоны располагаются зимовки основ-
ной массы уток, бакланов, гагар, обыкновенного и очкового чис-
тиков, рогатого тупика, старика, пыжиков, сизой и озерной ча-
ек. Все они почти не выходят за край шельфа, а утки, бакланы,
сизая и озерная чайки вообще тяготеют к узкой прибрежной по-
лосе. К водам шельфа приурочены скопления кайр и конки. Но
эти птицы весьма обычны местами и за пределами мелководья
на удалении от побережий в несколько десятков и даже сотен
миль. В частности, большие скопления конюг ежегодно наблю-
даются в полосе около 200 миль вдоль Курильских островов и
Хоккайдо (рис. 39).
мо _____________ <бо___________________ieo________________
Рис. 39. Распределение мелких чистиковых в дальневосточных морях и
северо-западной части Тихого океана. 1 — основлце районы концентраций
летом. 2 — основные районы концентраций зимой
142
СЕВЕРНАЯ ЧАСТЬ ТИХОГО ОКЕАНА
Тяготеет к шельфу и прилегающим водам основная масса
особей крупных чаек — серонрылой. тихоокеанской, западной —
Larus occidentalis Audub., полярной, серебристой и чернохвостой.
Часть особей этих видов широко рассеивается по умеренным
районам северной части Тихого океана и в меньшей степени —
в пределах субтропической эоны (рис. 40). Отдельные особи
крупных видов чаек как вдоль побережий, так и в открытом оке-
ане, проникают в северную часть тропиков (Austin and Kuroda,
1953; Blake, 1953; King, 1967; Sibley and McFarlane, 1968). Одна-
ко за пределами 100—200-мнльной зоны численность всех круп-
ных чаек невелика, и обычно встречаются лишь отдельные особи
При этом в открытом океане они особенно редкими становятся
южнее приблизительно 40° с. ш. Из перечисленных видов серо-
крылая и серебристая чайки имеют наиболее обширные области
зимовок. Они часто встречаются в открытом океане и далеко
проникают на юг. Вольте других видов к окраинным водам
океана тяготеют чернохвостая, тихоокеанская и западная чайки.
Характерно распределение молодых и взрослых чаек- Как при-
мер на рис. 41 Дано соотношение в разных районах молодых и
взрослых особей тихоокеанской чайки. Водно, что в южных, сле-
довательно, в более теплых районах зимует основная часть мо-
лодых бурых, а в северных — взрослых светлых птиц. Такая
особенность в распределении наблюдается и у других видов.
С этим вполне согласуется тот факт, что на островах северной
части тропической зоны добывались только неполовозрелые
особи (Sibley and McFarlane, 1968). Более значительное по
сравнению с взрослыми рассеивание молодых особей показано
при помощи кольцевания крупных чаек в районе северной части
Атлантики (Smith, 1959. 1962; Radford, 1960; Bruun, 1963;
и др.), а также серокрылой чайки в Британской Колумбии
(Pearse, 1963). К птицам, распространение которых не свя-
зано с водами шельфа, относятся черноногий и темно-
спинный альбатросы, глупыш, серая и северная качурки,
обыкновенная моевка, топорок и некоторые другие. Область
зимнего распространения этих птиц охватывает обширную ак-
ваторию северной части Тихого океана между Курильскими л
Японскими островами и побережьем Северной Америки. Места-
ми они обычны или многочисленны в прибрежных водах, ко боль-
шая часть особей рассеивается в зимний период в открытых во-
дах.
В связи с изложенным вполне естественно видеть отчетливые
различия в видовом составе и соотношении групп птиц в при-
брежной зоне, с одной стороны, и открытом океане, — с другой.
Из табл. 18 видно, что в водах, прилегающих к Японии, Куриль-
ским. Алеутским островам, Камчатке и Аляске (в полосе около
Рис. 40. Распределение крупный чаек и северной части Тихого океана. 1 — распространение в море летом,
2, 3, 4 — распространение % море зимой (интенсивность штрмхотаси соответствует обилию), 5 — южная гра-
ница единичных залетов
144
НЕВЕРНАЯ ЧАСТЬ ТИХОГО ОКЕАНА
150 миль); приблизительно 50% приходится на долю чистико-
вых птиц. Затем нужно назвать обыкновенную моевку и крупя-
ных чаек (тихоокеанская на западе, серокрылайна севере райо;
на), уток и бакланов. Из трубконосых здесь многочислен толь-
ко глупыш. Таким образом, в соотношении видов и доминирую-
щих .групп птиц в прибрежных районах северной части Тихого
океана наблюдается большое сходство- с дальневосточными
морями.
В открытом океане картина,, иная. Здесь основу населения
составляют пелагические виды — трубконосые (альбатросы, ка-
чурки, глупыш), на долю Которых приходн+ся больше 50%, и
обыкновенная моевка.
Различия в орнитофауне, наблюдаемые при сравнения за-
падной части с восточной, и северной части с южной во все
сезоны связаны, с одной стороны, с некоторыми различиями в
составе гнездящихся здесь видов, а с другой, — с тем, что ми-
грирующие с севера птицы для зимовок избирают определен-
ные районы. В частности, к западной части Тихого океана тяготе-
ют или встречаются исключительно в этом районе темноспинный
альбатрос, пестролицый буревестник, тихоокеанская и чернохво-
/ДООННЫВ АСПЕКТЫ В ОРНИТОФАУНЕ
145
• стая чайки, уссурийский баклан, очковый, чистик: а в восточной*—
червоногий альбатрос, серокрылая и восточная чайки, алеутский
. пыжик — Ptychoramphus aleuticus (Pall.), бакланы Брандта —
Phalacrocorax penicillatus (Brandt) и ушастый — Р. auritas
(Less.). К северо-западным и северным районам (от Японии до
залива Аляска) рассматриваемой акватории приурочены основ.
' ные зимовки конюг. В восточной части Тихого океана их меньше.
Кайры наиболее крупные скопления образуют в северо-восточной
части океана (залив Аляска, восточная часть Алеутской гряды).
При сопоставлении состава птиц в северных и южных райо-
нах северной части Тихого океана обращает на себя внимание
уменьшение доли чистиковых птиц в более юЖных районах. Осо-
бенно это заметно с широты северной части Хонсю. Количество
трубконосых, за исключением глупыша, наоборот, по мере дви-
жения на юг,увеличивается. Это касается, в первую очередь, алъ-
1 батросов и Качурок, а также буревестников — пестролицёго, се-
v- ’ • • '
1 Таблица ‘18>к
• I. : т
Видовой состав,.соотношение (%) и плотность морских птнд-
в северной части Тихого океада (севернее 30° с. ш<).» зимний период
(декабрь—март). I960—1965 у. ’ , ,, ,
♦ В н д ы . ^Р1}бре>|сные районы > фткрьу, , •тый i I океан, г 1 * * ’ S
11 f ’О ' Хоккайдо . Курилы , А:1*?™’., и Хонсю и Камчатка
•: у , . » . • ’ J V.! редкие чистиковое. • i (в основном конюги) 50,0 36,0 9,0 , , ДО Кайры 2,0 , 12,0 40,0 . 3,0 . L. cirrhata 1,0 4- ' д.‘ 2,0 - U schistisagus 7,0 12,0 " 2,0 ' .L. glaucescens + 2,0 t 18,0 4,0 Моевки (R, trldactiyla) 15,0 ,.+ 15.0 , 18,0 t.- crassirostris 3,0 — — —, * Неоорзделепные чайки 8,0 6j0 + 4,0 ЕЬморвдки 1,0 — — 1,0 F. glacialis 1,0 12,0 13,0 14,0 D. immutabilis + +,. + 25,0, D. nigripes — — — 14,0 Качурки + '-r - 5,0 ужи 5,0 12,0 - + Ьдкланы (P. pelagicus) 1,0 6,0 2,0 — Гагары 2,0 + + — Остальные виды *4,0 2,0 3,0 3,0 Количество особей на 1 кв. км 10,0 12,0s 11,0 0,4 Количество учетов 87 122 25 598
146
СЕВЕРНАЯ ЧАСТЬ ТИХОГО ОКЕАНА
рого, тонкоклювого и других. Только в южных районах северной
части Тихого океана встречаются зимой поморники. Значитель-
ная часть нх особей на зиму мигрирует, как известно, в южное
полушарие. В связи с уменьшением на юге численности массовой
группы чистиковых доля чаек в учетах здесь увеличивается, хотя
плотность их населения соответствует плотности на севере.
В целом почти на всей акватории к северу от 30° с. ш. зимой
доминируют представители умеренного комплекса птиц.
В сравнении с другими сезонами зимний период в этом отноше-
нии отличается низким процентом субтропических видов и ви-
дов из южного полушария. Как было показано в первой главе,
буревестники, совершающие трансэкваториальные миграции, н
это время концентрируются в основном в нотальных и антаркти-
ческих водах, в северном же полушарии остается небольшое чис-
ло особей. Субтропические виды — пестролицый буревестник,
качурки вилохвостая и мадейрская — Oceanodroma castro
(Нагс.) и другие птицы, многочисленные летом южнее пример-
но 40° с. ш., зимой откочевывают в основном в субтропическую
и тропическую зоны. Поэтому численность их в водах, располо-
женных к северу от 30° с. ш., сильно уменьшается. Показателен
в этом смысле рис. 42, где дано сезонное распределение пестро-
лицего буревестника. Из видов, гнездящихся в субтропической
и тропической зонах к северу от 30° с. ш., зимой многочислен-
ны, по-вндимому, только альбатросы.
Условия для зимовок птиц во многих районах северной части
Тихого океана более благоприятны, чем на боямней площади
Берингова и Охотского морей. В зимнее время здесь ограничен,
ное распространение имеют льды, которые образуются в основ-
ном только в узкой прибрежной полосе на западе океана (Бул-
гаков, 1968). Над большей частью акватории зимой наблюдают-
ся положительные температуры воздуха. Нулевая январская
изотерма проходит от северной части Хонсю к западной око-
нечности Алеутской гряды. На западе океана между 40 и
30° с. ш. и на востоке океана между 50 и 40° с. ш. средняя ян-
варская температура воздуха составляет 5—10* (Морской ai-
лас, 1953). Только в западной части океана вдоль Курильских
островов и Камчатки поверхностные воды охлаждаются до от-
рицательных температур. Но уже на широге острова Хоккайдо
на западе океана и в южной части залива Аляска на востоке,
поверхностная температура поднимается до 5, а в районе
40е с. ш. — до 10—12е (Панфилова, 1968). Таким образом, кли-
матические и гидрологические условия в северной части Тихого
океана менее суровы в сравнении с северными дальневосточны-
ми морями. Весьма существенным при этом является различие
в климатическом и гидрологическом режимах в западной и во-
сточной частях океана. В первой главе на примере обыкновен-
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
447
Рис. 42. Сезонное распределение пестролинего буревестника — Puffinus
leucomelas. I — места размножения. 2 — область распространення летом.
3 — основные районы концентраций летом. 4 — область распространения
зимой
ной моевки, черноногого альбатроса и топорка было показано,
что сравнительно теплолюбивые виды в наиболее суровой севе-
ро-западной части океана зимой не встречаются или встречают-
ся в небольших количествах. Границы распространения их здесь
ориентированы с юго-запада на северо-восток, т. е. параллельно
направлению изотерм воды и воздуха.
В кормовом отношении зимний период в северной части Ти-
хого океана, как и в дальневосточных морях, наименее благо-
приятный. В поверхностных слоях количество кормовых организ-
мов сильно сокращается, хотя в большинстве районов сезонные
изменения в гидрологическом режиме не так контрастны, как
в дальневосточных морях. Из-за недостатка освещенности и
неустойчивости верхних слоев (конвективное перемешивание)
148
СЕВЕРНАЯ ЧАСТЬ ТИХОГО ОКЕАНА
вегетация фитопланктона почти прекращается (Семина, 1967;
и др.). В связи с охлаждением поверхности моря наиболее мас-
совые виды зоопланктона зимой скапливаются в теплом проме-
жуточном слое ниже границы зимней конвекции. Поэтому; а
также в результате выедания, концентрация планктона в верх-
них слоях зимой очень сильно уменьшается (Виноградов, 1968;
и др.). Особенно заметны описанные изменения в умеренных
водах к северу от полярного фронта (приблизительно до
40° с. ш.). Для этих районов характерно уменьшение в верхних
слоях моря и на малых глубинах количества рыб, что является
следствием их миграций на глубину или на юг. В частности, по-
кидают умеренные воды на зимний период такая массовая пе-
лагическая рыба, как сайра, и многочисленные пелагические
кальмары.
Таким образом, в умеренных водах северной части Тихого
океана, как и в дальневосточных морях, условия для обитания
птиц, питающихся с поверхности моря, ухудшаются наиболее
заметно. С этим хорошо согласуется сильное уменьшение чис-
ленности в северных водах трубконосых. Здесь, как видно из
табл. 18, преобладают чистиковые, добывающие пищу при ны-
рянии, и всеядные чайки. Большое количество чаек в течение
всей зимы кормится в портах’ и у рыболовных судов, ведущих
промысел у побережий по всей дуге шельфа и материкового
склона от Японии до Калифорнии. Вместе с чайками в районах
лова рыбы питается отходами промысла глупыш, единственный
из трубконосых, встречающийся зймой в больших количествах
в северных водах.
Южнее полярного фронта сезонные колебания в количестве
планктона выражены-менее резко. В-зимнее время биомассы
планктона в поверхностных слоях здесь выше, чем-в умеренных
водах (Каредин. 1969; Кун, 1969а; Кун и др., 1969; и др.). Юж-
нее полярного фронта зимой концентрируется основная масса
субтропических пелагических рыб (сайра, анчоус, скумбрия),
их молодь и пелагические кальмары. У некоторых из них зимой
в субтропических водах начинается нерест, в результате к концу
зимнего периода в поверхностных слоях появляется значитель-
ное количество молоди.
Изложенное помогает понять, почему большая часть трубко-
носых, покидающих на зимний период северо-умеренные воды
Тихого океана, откочевывает в южную часть умеренной, субтро-
пическую и тропическую зоны, а также в южное полушарие.
О благоприятности кормовых условий для птиц в субтропиче-
ских водах зимой приходится судить в сравнении с охлажден-
ными водами умеренной зоны. Зимой биомассы планктона в
субтропических водах заметно ниже, чем весной и. летом, а пло-
щади с повышенной концентрацией планктона меньше. По-ви-
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
149
димому, в связи с этим птицы, питающиеся на поверхности мо-
ря, и в первую очередь трубконосые, держатся здесь рассредото-
чение и рассеиваются на больших акваториях. Исключение мо-
гут представлять только некоторые прибрежные районы (напри-
мер, у Японии и Калифорнии), где располагаются зимовки и не-
рестилища массовых пелагических рыб и где наблюдаются скоп-
ления птиц.
Из табл. 18 видно, что в прибрежных водах различных райо-
нов северной части Тихого океана (в полосе около 150 миль)
цифры средней плотности птиц вполне сопоставимы. Из-за ог-
раниченного количества учетов в таблице не приведены данные
по восточной части океана. В общем плотность здесь имеет так-
же сходные показатели. Приведенные цифры говорят о том, что
плотность концентраций птиц в прибрежных водах Тихого
океана, как и на шельфе дальневосточных морей, равна около
10—12 особей на 1 кв. км. В открытом океане она, примерно, в
25 раз ниже. Как будет показано далее, в другие сезоны это
различие сохраняется, хотя и. несколько сглаживается. На объ-
яснении этого явления остановимся несколько подробнее. Как и
в морях, высокая плотность птиц у окраин океана в сравнении
с открытыми водами связана с тем, что прибрежные районы ха-
рактеризуются более высокой продуктивностью, а следователь-
но и корм костью. Сказанное подтверждается расчетами первич-
ной продукции (Кобленц-Мишке, 1967; и др.), распределением
биомасс планктона (Богоров, 1967; Семина, 1967; и др.) и бен-
тоса (Зенкевич, 1963).
Распространение подавляющего количества массовых видов
рыб и особенно их концентраций также ограничивается приле-
гающими к побережью водами. В северо-западной части Тихого
океана это минтай, японская скумбрия, сайра, японский анчоус,
японская ставрида и большая группа других донных и придон-
ных рыб (Моисеев, 1953; Новиков Н., 1965; Полутов и др., 1966;
Новиков Ю., 1967; Kawasaki, 1966; Hayasi 1967; и др.), а в се-
веро-восточной — хек, северный анчоус, калифорнийская сар-
дина, симметричная ставрида и многие донные и придонные ви-
ды (Alverson et al., 1964, 1969; Ahlstrom, 1967; Baxter, 1967;
Blunt, 1969; Messersmith et al., 1969; Nelson and Larkins, 1970;
Ермаков, 1970; и мн. др.). В верхних слоях открытого океана
обычны, а местами многочисленны, по-видимому, только сайра,
лососи, светящиеся анчоусы (Беккер, 1967; Бирман, 1967; Виг-
дорович, 1967; Парин, 1967; и др.). Из прибрежной зоны сюда
могут проникать скумбрия, анчоус, ставрида и другие виды
(Бирман, 1958; Blunt, 1969; и др.), однако численность их здесь
невелика, а присутствие непостоянно. По всей вероятности, в
открытом океане местами многочисленны кальмары. Но основ-
150
СЕВЕРНАЯ ЧАСТЬ ТИХОГО ОКЕАНА
ные концентрации этих животных наблюдаются у окраин океана.
Говоря о более низкой продукции и кормности вод открытого
океана, следует иметь в виду, что в ряде районов здесь имеются
весьма благоприятные условия для формирования высокой про-
дуктивности. Одним из таких участков является район Импера-
торского подводного хребта, протянувшегося от Гавайских до
Командорских островов. В южной части хребта, где имеется
большое количество гайотов, благодаря высокой динамике вод
создаются благоприятные условия для формирования высоких
концентраций планктона и рыб. На ряде банок имеют место
исключительно мощные по плотности концентрации кабан-рыбы.
В этом районе отмечается и повышенная плотность населения
птиц. В некоторые периоды увеличение плотности птиц наблю-
дается в районе субполярной дивергенции, проходящий несколь-
ко южнее Алеутских островов.
В характере и сроках весенних миграций птиц в северной
части Тихого океана есть много общего с дальневосточными мо-
рями. Начало весенних миграций приходится на март, но ос-
новные потоки мигрантов проходят в апреле и мае. Первыми на-
чинают перемещения виды, многочисленные на зимовках и в
более северных водах. Это — чистиковые (в первую очередь
кайры) и крупные чайки. В апреле в постоянном движении на
север находятся все зимующие в северной половине океана ви.
ды. В это время начинают прибывать из южного полушария
трансэкваториальные мигранты — буревестники, поморники,
плавунчики, вилохвостая чайка, основной пролет которых про-
ходит в мае и частично в июне. Виды, которые на зимовках тя
готеют к окраинам океана (большинство чистиковых, крупные
чайки, бакланы, утки), мигрируют вдоль побережий, повторяя
очертания шельфа и материкового склона. Многочисленны в
прибрежной зоне и другие птицы — моевка, глупыш, альбатро-
сы, качурки и буревестники. Однако большое количество нх осо-
бей мигрирует и через открытую часть Тихого океана. В частно-
.сти, большие стаи буревестников встречаются в это время на
всей огромной акватории между Японскими и Гавайскими ост-
ровами н заливом Аляска. В связи с тяготением видов к опре-
деленным районам океана сохраняются свойственные для зимы
черты различия, наблюдаемые в составе птиц в прибрежных и
океанических водах. Здесь имеется в виду преобладание в при-
брежных водах чистиковых, чаек и уток, а в океанических —
трубконосых. Из табл. 19 видно, что весной доля трубконосых
увеличивается и в прибрежных районах. В океанических же во-
дах доминирование данной группы становится особенно замет-
ным.
Плотность населения птиц весной в прибрежной зоне почти
не отличается от зимней. Убыль отлетевших на север птиц ком-
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
151
пенснруется подлетающими : юга. В открытом же океане она
увеличивается в несколько раз (табл. 19). Это увеличение
связано главным образом с прибывающими из южного полуша-
рия буревестниками, которые преобладают здесь над другими
птицами.
Таблица 19
Видовой состав, соотиошеине (%) и плотность морских птиц
в северной части Тихого океана (севернее 30° с. ш.) в весеиний период
(апрель—май). 1N3-1M8 гг.
Виды Прибрежные районы Открытый океан
Хоккайдо и Хонсю Курилы и Камчатка
Р. tenuirostris, Р. griseus 30,0 9.0 43,0
Р. leu cornel as 40,0 — +
F. glacial is + 4.0 4,0
0. furcatu, 0. leucarhoa + 28,0
D. immutabilis + + 3,0
D. nigripes -г + 5,0
Мелкие чистиковые 8,0 24,0 4,0
Кайры 3,0 15,0 +
Моевки (R. tridactyla) 11,0 1,0 3,0
L. schistisagus 1,0 10,0 +
L. crassirostris 1,0 -4- —
Бакланы (P. pelagicus) 1,0 9,0 —
Утки 2,0 23,0 —
Плавунчики + + 4,0
Остальные виды 3,0 5,0 6,0
Количество учетов 20 30 331
Количество особей ив 1 кв. км 10,0 15,5 3,0
В результате откочевки в дальневосточные моря, Арктику, а
также к побережьям, уже со второй половины весны в открытом
океане почти не остается крупных чаек. Уменьшается в откры-
тых водах численность и многих других видов. Тем не менее, в
море вне связи с берегами в течение всего периода размно-
жения остается часть особей всех видов. Летняя область рас-
пространения в тихоокеанских водах кайр, чистиков, конюг, пы-
жиков, крупных чаек, моевок, бакланов, крачек и других пред-
ставляет собой прибрежную полосу, обычно не шире нескольких
десятков миль и простирающуюся от Японии вдоль Курильских
островов, Камчатки, Командорских и Алеутских островов, по-
луострова Аляска и тихоокеанского побережья Северной Аме-
152
СЕВЕРНАЯ ЧАСТЬ ТИХОГО ОКЕАНА
рики.-За.лределами этой полосы могут быть встречены очей»
редкие одиночки перечисленных видов. Шире распространены
северные поморники. Отдельные бродячие особи их встречаются
по всей северной части океана к северу от полярного фронта.
Впрочем, более часто они встречаются также у побережий.
.Как и в другие сезоны, летом широкое распространение
имеют альбатросы, качурки, глупыш, буревестники, тайфунни-
ки, топорок, местами ипатка. В большинстве районов северной
части Тихого океана шельф развит слдбо, нетрансформирован-
ные океанические воды располагаются близко от берега. Поэто-
тому представители океанической группировки, главным обра-
разом трубконосые, обитают не только в открытых водах, но и
близ побережий. А вся область их распространения намного об-
ширнее области, занимаемой прибрежными видами. Наиболее
ощутима эта разница именно в летний период, когда многие
прибрежные виды почти не выходят за пределы шельфа. В свя-
зи с этим в летний период не только в открытом океане, но и
в 150-мильной прибрежной полосе почти повсеместно наблю-
дается доминирование трубконосых (табл. 20) — особенно тон-
коклювого и серого буревестников. Хотя летом большое коли-
чество их особей откочевывает в Берингово и Охотское моря,
в северной части океана они остаются по-прежнему многочи-
сленными. В начале и в конце лета во многих районах численное
преобладание над другими видами определяют их пролетные
стаи; в июне — следующие на север особи-, так называемой
третьей волны буревестников, состоящей из молодых данного
года, в августе — начавшие движение на юг на места гнездо-
вания стаи половозрелых особей.
До сих пор речь шла о видах, составляющих летний фон на-
селения в водах океана к северу от полярного фронта. Поляр-
ный фронт является границей умеренных вод, он пересекает се-
верную часть Тихого океана в основном между 40 и 45е с. ш.
(Uda, 1963; Мороз, 1969; и др.). За немногим исключением, пе-
речисленные выше виды летом не встречаются южнее фронта, а
если и встречаются (альбатросы, северная качурка), то центр
обилия их находится в умеренных водах. Южнее полярного
фронта в субтропических водах видовой состав в значительной
мере другой. Здесь весьма характерны буревестники пестроли-
цый, Буллера — Puffinus bulleri Salvin и бледноногий, качур-
ки — О. castro, вилохвостая, бонинский тайфунннк — Ptero-
droma hypoleuca (Salvin) и другие виды. В основном южнее
фронта встречается южнополярный поморник и хохлатый ста-
рик. Летом в южную -часть рассматриваемой акватории прони-
кают и тропические птицы. У берегов Японии это — клинохво-
стый буревестник — Puffinus pacificus (Gmel.), фаэтоны, фре-
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
163
Таблица 20
Видовой состав, соотношение (%) и плотность морских птиц
в северной части Тихого океана (севернее 30° с. ш.) в летний период
(нюнь—август). I960—1968 гг.
Виды Прибрежные районы Откры- тый океан
Хоккайдо и Хонсю Курилы и Кам- чатка Алеуты и Ко- мандоры
Р. tenuirostris, Р. griseus 30,0 35,0 28,0 50,0
Р. leucomelas 30,0. 4- — 2,0
Качурки 27,0 12,0 5,0 22,0
F. glacialis 1,0. 18,0 18,0 7.0
D. immutabilis 2,6 2,0 1,0 5,0
D. nigripes . kQ 4- 1.0 6,0
L. cirrhata 4; 3,0 4,0 1,0
Кайры , 4- 2,0 17,0, 1,0
Мелкие чистиковые .4- 8,0 2.0 1,0
Моевед (R. trida^tyUy + 4,0 15,0 1,0
L. schistisagus 4- г. 3,0 — —
L. glaucescens , . , — + 6,0 —
Неопределенные, чайки 4-’ 9,0 — 4-
Бакланы ,(P. pe(agicus) 4- 1>0 . , т —
Остальные воды. 9,0 3.0 2,0 . 4,0
Количество учетов 25 59 10 421
Количество оробей на 1 кв. км 13,0 11,0 11,0 1,6
гаты, олуши, тропические крачки и другие (Austin and Ku-
roda, 1953; и др.), Аналогичная картина наблюдается) и. в во-
сточной части океана. Здесь летом многочисленны буревестники
розовоногий — Puffinus creatopits Coues и Puffinus opisthomela»
Coues (Dawson, 1940; и др.). (Здесь я.хочу еще раз напомнить,
что в настоящем разделе в связи, с ограниченным, количеством
учетов из восточной части океана рассматривается в>основном
западная половина северной части Тихого океана).
Умеренные воды северной части Тихого океана, а также зо-
на полярного фронта характеризуются в весенне-летннй'период
обильным развитием планктона (Богоров, 1967; Куи, 1969;
Кун и др., 1969; и мн. др.). На нагул в умеренные воды и в
район фронта мигрируют массовые пелагические рыбы (сайра,
скумбрия, анчоус) и кальмары. Кормовые условия1 здесь для
птиц в связи с этим становятся более благоприятными, чем к
югу от фронта. Этому способствует, несомненно, и тот факт,* что
в весенне-летний период концентрации планктона тяготеют к
154
СЕВЕРНАЯ ЧАСТЬ ТИХОГО ОКЕАНА
верхним слоям моря (Виноградов, 1967; 1968; Кареднн. 1969;
и мн. др.). Высокие биомассы зоопланктона в умеренных водах
наблюдаются не только в прибрежной зоне, но и в открытом
океане, в частности южнее Алеутских островов в районе суб-
полярной дивергенции (Миклухииа, 1969). Здесь же образует
концентрации и сайра (Вигдорович, 1967). Богатство умерен-
ных вод кормом является одной из основных причин смещения
сюда концентраций птиц с юга, особенно планктоноядных птиц.
Зимой, как отмечалось выше, их концентрации тяготеют к бо-
лее южным водам. Летом же плотность населения в открытых
водах к северу от 40° с. ш. приблизительно в два раза выше,
чем между 30 и 40“ с. ш.
Прибрежные северные и южные воды больших различий в
плотности населения, как правило, не имеют (табл. 20). В при-
брежных водах Японии и Калифорнии благодаря особенностям
гидрологического режима (высокая динамика вод) создаются
благоприятные условия для высокой биологической продуктив-
ности в летний период. Наличие концентраций птиц в водах
Японии в летний период отмечалось другими авторами (Kuroda
1955; I960; Hotta et al., 1961; и др.). Из табл. 20 видно, что по
данным имеющихся в моем распоряжении учетов, плотность
птиц у Курильских и Алеутских островов оказалась немного
ниже, чем у Японии. Здесь сказалось недостаточное количество
учетов непосредственно у островов, где, как известно, много
крупных гнездовых колоний птиц. В действительности плот-
ность населения в этих районах, по-видимому, равнозначна или
даже несколько выше, чем у Японии.
Летом, как и в другие сезоны, плотность населения в при-
брежной зоне в несколько раз выше, чем в открытом океане.
Здесь она мало отличается от того, что наблюдаехсв зимой и
весной. В открытых же водах она уменьшается в сравнении с
весной почти в два раза (см. табл. 20), что связано с откочевкой
части буревестников в дальневосточные моря и в прибрежную
зону (Курильские, Алеутские острова и залив Аляска).
Уже в конце августа начинается откочевка части особей бу-
ревестников в южное полушарие. Однако они остаются много-
численными в северной части Тихого океана в течение всего
сентября и начале октября, когда происходит отлет на юг основ-
ной массы их особей. В сентябре начинаются миграции и дру-
гих птиц. Основными тенденциями, ведущими к установлению
описанной выше зимней картины, кроме уменьшения численно-
сти трансэкваториальных мигрантов, является, с одной сторо-
ны, откочевка на юг субтропических и тропических птиц, а так-
же бореальных трубконосых, а с другой, — постепенное увели-
чение количества видов, гнездящихся в Охотском, Беринговом
морях и Арктике. В связи с этим, как видно из табл. 21, осенью
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
Мб
в соотношении групп птиц в водах северней части Тимого -океа-
на вполне отчетливо вырисовываются черты, свойственные зим-
нему периоду.
В конце сентября заметно увеличивается в водах северной
части Тихого океана количество обыкновенной моевки, топорка,
поморников и других. Что касается основной массы видов чи-
стиковых птиц н крупных чаек, то явное увеличение их числен-
ности н расширение областей распространения происходит в ос-
новном в октябре и ноябре.
Таблица 21
Видовой состав, еитиошенне (%) и ывмюсть мсрсках птиц
в северной части Тихого океана (севернее 30° с. .) а осенний период
(сентябрь—ноябрь). 198В— ТМ8 гг.
Виды Прибрежные районы Открытый океан
F. glacralis 11,0 15,0
Р. termirostris, Р. griseus 10,0 11,0
Качурки to 23,Ю
О. imfiratabilis 2,0 8,0
D. nigripes 1.0 12;0*
Моевки (F. tridactyla) 19,0 18,0
L. schiStisagus 11,0 +
L. glaucescens 5,0 2,0
Кайры 18,0 1,0
L ci rr hat a + 2,0
Мелкие чистиковые 16.0 2,0
Поморники + 1.0
Утки 5,0 +
Бакланы (Р. pelagicus) 3,0 -L
Плавунчики 2,0 1,0
Остальные виды 1;0 4,0
Количество учетов 63 323
Количестве особей на 1 кв. км 5,0 0,7
Из табл. 21 видно, что осенью наблюдается уменьшение
плотности птиц и в прибрежных, и в океанических районах. Ха-
рактерно. что для прибрежных вод была получена наиболее
низкая плотность по сравнению с другими сезонами. Уменьям,
ние связано с тем. что в это время отлетает на юг большая
часть буревестников. Убыль же их компенсируется только зи-
мой, когда заканчивается откочевка птиц из дальневосточных
морей.
156.
ВОСТОЧНО-КИТАЙСКОЕ МОРЕ
5. Восточно-Китайское море.
В Восточно-Китайском море гнездится сравнительно неболь-
шое количество, преимущественно субтропических видов, мор-
ских птиц. Однако видовой состав этого района сильно увеличи-
вается за счет птиц бореальной и тропической зон, встречаю-
щихся здесь в определенные сезоны. Как и в других районах,
особенности распределения птиц и их численность в Восточно-
Китайское море определяются спецификой гидрологического
режима — характером распределения температуры воды и воз-
духа, теплых и холодных течений, фронтов и продуктивных зон
(Шунтов, 1970а).
В конце осени и зимой большое количество птиц откочевы-
вает в Восточно-Китайское море с севера, главным образом че-
рез Корейский пролив. Фон зимней орнитофауны составляют пе-
стролицый буревестник, обыкновенная моевка, чернохвостая, ти-
хоокеайская, серебристая чайки, поморники (по-видимому, ча-
ще короткохвостый — Stercorarius parasiticus L.) и мелкие чи-
стиковые (по-видимому, в основном обыкновенный и хохлатый
старики). В начале зимы (декабрь) во время пролета на юг
здесь еще встречаются стайки круглоносого плавунчика и
бледноногого буревестника. В январе и феврале эти виды не
отмечаются совсем, но в марте с наступлением весны они вновь
появляются в Восточно-Китайском море. Встречаются в рас-
сматриваемом районе зимой также вилохвостая качурка, аль-
батросы (все три северотихоокеанских вида), полярная чайка,
тонкоклювая кайра, рогатый тупик . — Qerorhinca monocerata
(Pall.), очковый чистик, уссурийский баклан и некоторые дру-
гие. Однако все эти виды везде редки, Из перечисленных видов
только пестролицый буревестник, старики й- качурка гнезДйтся
в Восточно-Китайском море.. Гнездовья других, за исключением
альбатросов и бледноногого буревестника, располагаются в
пределах бореальной зоны.
Как видно из табл. 22, видовой состав, а главное, соотноше-
ние видов, в северной и южной половинах моря различны. В се-
верной половине моря наиболее многочисленны чайки, пестро-
лицый буревестник, мелкие чистиковые и поморники. Все чи-
стиковые не заходят далеко на юг и встречаются только в райо-
не Японских островов и полуострова Корея. В южной полови-
не моря над другими видами резко доминирует пестролицый бу-
ревестник. Весьма обычны здесь поморниик, но количество
всех видов чаек сильно сокращается.
Еще больше различий обнаруживается при сравнении видо-
вого состава птиц в северо-западной мелководной и юго-восточ-
ной глубоководной половинах моря. Эти различия находятся в
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТЦФАУНЕ
Т57
связи со спецификой гидрологических условий в рассматривае-
мых районах моря. Резюмируя данные ряда авторов об особен-
ностях гидрологического режима Восточно-Китайского моря,
(Tsujita, 1957, 1963; Kato, 1958, 1959; Kikuchi, 1959; Miyazaki
and Abe, 1960; Харченко, 1968; и др.), нужно отметить' следую-
щее. Юго-восточная глубоководная часть моря во все сезоны’
более теплая, что связано с прохождением здесь течения Куро-
сио. Куросио входит в Восточно-Китайское море с юга, затем
следует на северо-восток вдоль материкового склона. Южнее ост-
рова Кюсю Куросио дивергирует на две ветви: одна уходит в.
океан, другая—на север, давая начало Цусимскому течению. Из
Желтого моря в западную половину Восточно-Китайского выхо-
дят менее соленые холодные воды, которые распространяются
на большей части шельфа. Холодное течение следует также на
юг вдоль берегов Китая. По кромке мелководья между Куросио.
и желтоморскими холодными водами формируется четко выра-
женная фронтальная зона с градиентом температуры в зимний
период около 9° на 60 миль. Положение фронта, приблизитель-
но определяемое зимой изотермой 15°, варьирует по сезонам.
Наиболее далёко на юг холодные желтоморские воды проника-
ют в период развития .северо-западного муссона (зимой). Таким
образом, по условиям обитания мелководная и глубоководная
части Восточно-Китайского моря оказываются различными.
Особенно большие различия наблюдаются в температуре воды,
которая на севере моря опускается зимой до 6—8’, на юге — не
бывает ниже 20—24*, Весьма характерно, что изотермы имеют
не широтное направление, а ориентированы с юго-запада на
северо-восток, т. е. параллельно фронту и материковому склону.
/ Таблица 22
Видовой состав, соотношение (%) и плотность морских птиц
в Восточно-Китайском море зимой (декабрь—март). 1961—1968 гг.
В н д ы . . а 35 - 30° с. ш. 30—25° с. ш. Все море
Р. leucomelas 29,0 82.0 53.5
Крупные чайки 18.0 2,0 11.0
R. tri dactyls 17.0 0,5, 9,5
L. crassirostris 11,0 1,0 6.5
Поморники 7.0 10.0 8.5
L. lobatus 4.0 2.5 3,0
Мелкие чистиковые 9,0 — 4,5
P. carneipes 4.0 -г 2,0
0. monorhis — 0.7 0,3
Альбатросы + 0,3 0,2
Неопределенные виды 1,0 1,0 1,0
Количество особей на 1 кв. км 1,4 1.3 1,4
Количество учетов 75 100 175
«56
ВОСТОЧНО-КИТАЙСКОЕ МОРЕ
Различия в условиях обитания накладывают существенный
отпечаток на распределение и. видовой состав морских живот-
ных в отдельных районах моря. Анализируя данные по распре-
делению планктона, В. В. Крылов (1969) показал, что воды Ку-
росио являются биотопом океанического биоценоза, а желтомор.
<ские водные массы — неритического. Зона смешения вод яв-
ляется переходным биотопом, населенным представителями ос-
новных биоценозов. Размещение водных масс и положение
фронта определяют многие особенности в распределении рыб
(Tsujita, 1957, 1961, 1963; Tsujita and Kondo, 1957; Чигирин-
ский. 1964, 1969; и др.). Морские птицы в этом отношении не
представляют исключения. Большинство видов, мигрирующих
из умеренной зоны, южнее Корейского пролива концентрируют-
ся в пределах области, занятой холодными водами (шельф).
Рис. 43. Распространение моевки — Rissa tridactyla и пестролнцето —
буревестника — Puffinus leucomelas в Восточно-Китайском море в январе-
феврале. 1 — поверхностные изотермы, 2 — теплые течения, 3 — холод-
ные течения, 4 — граница распространения буревестника, 5 — граница
распространения моевки, 6 — район распространения моевки, 7 — район
распространения буревестника
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
15©
Рис. 44. Распространение чаек р. Larus и поморников р. Stercorarius в
Восточно-Китайском море в январе-феврале. 1 — поверхностные изотер-
мы, 2 —теплые течения, 3 —холодные течения, 4 —граница распрост-
ранения чаек, 6 — район распространения чаек. 7 — район распростра-
нения поморников
Такая особенность в распределении характерна для моевки,
чернохвостой, тихоокеанской, серебристой и полярной чаек. За
пределами шельфа чайки встречаются только случайно. По-
морники и особенно субтропические виды — пестролицый бу-
ревестник и вилохвостая качурка, а также альбатросы тяготеют
зимой к водам Куросио. В зоне смешения вод встречаются ви-
ды обеих группировок (рис. 43 и 44).
Одним из ведущих факторов, определяющих отмеченные
особенности в распределении птиц, является температура. Так,
пестролицый буревестник в начале зимы (декабрь), а также в
другие сезоны встречается в Корейском проливе. Лишь в пе-
риод наибольшего охлаждения (январь-февраль) северная
граница его распространения опускается до южной части остро-
ва Кюсю. В марте он снова расширяет область распростране-
ния па север. Однако только температурой все особенности в
160
восточно-китаиское море
распространении объяснить, очевидно, нельзя.. Тот же пестроли-
цый буревестник даже в летний период, когда повсеместно тем-
пература воды и воздуха повышается, продолжает тяготеть к
зОнё действия Куросио и его ветвей, в частности Цусимского те-
чения (включая Японское море и Тихий океан). Чайки в Во-
сточно-Китайском море почти не выходят за пределы шельфа.
В то же время, как отмечалось выше, зимой они регулярно за-
летают на океанические острова в тропической части Тихого
океана (Sibley and Farlane, 1968), где температура выше, чем
в восточной части Восточно-Китайского моря. При объяснении
тяготения группировок птиц к районам с определенными водны-
ми массами правильнее говорить о комплексном влиянии раз-
личных факторов, об Общем фоне океанографических условий
(климатические, гидрологические, кормовые), как это делается
в отношении водных животных.
Рис. 45. Распределение скоплений морских птиц в Восточно-Китайском
море в декабре—марте. 1 — скопления чаек, 2 — скопления буревестни-
ков, 3 — теплые течения, 4 — холодные течения, 5 — конвергенция, 6 —
* изобаты
СЕЗОННЫЕ АСПЕК1Ы В ОРНИТОФАУНЕ
191
В течение всего зимнего периода повышенная плотность на-
селения птиц наблюдается в двух основных районах: в водах,
прилегающих к Корейскому проливу, Кюсю и полуострову Ко-
рея, а также в центральной части моря в зоне конвергенции
(рис. 45). В каждом из этих участков средняя плотность населе-
ния в период исследований составила по 1,7 особей на 1 кв. км.
В районе Корейского пролива концентрации образуют преиму-
щественно чайки, а в центральной части моря — пестролицый
буревестник. На остальной акватории моря плотность населения
птиц примерно в два раза ниже: в водах Куросио — 0,7, в жел-
томорских водах — 0.6 особей на 1 кв. км.
Распределение скоплений птиц хорошо согласуется с разме-
щением продуктивных эон; последние же приурочены к райо-
нам усиленного вертикального перемешивания или подъема глу-
бинных вод. Наиболее стационарным и большим по протяжен-
ности является район подъема вод вдоль кромки мелководья и
материкового склона. Это связано с наличием здесь конверген-
ции и с так называемыми каскадными явлениями, возникающи-
ми на склоне глубин при прохождении потока. Другие зоны
подъема вод располагаются на севере моря в районах, приле-
гающих к Корейскому проливу и острову Кюсю. Их формирова-
ние связано с дивергенциями ветвей Куросио. а также образо-
ваниями завихрений и круговоротов.
Все отмеченные районы характеризуются повышенными био-
массами планктона и являются основными местами концентра-
ций пелагических рыб и кальмаров. В районе Корейского про-
лива располагаются концентрации сардины, ставриды, анчоуса,
сайры, кальмара и других, а в центральной части моря — став-
риды, скумбрии, и сайры (Tsujita, 1957, 1961, 1963; Mukai, 1953;
Uda and Ishino, 1958; Ямада. 1962; Харченко, 1968; Крылов, 1969;
и др.). Приведенные данные показывают, что не только распро-
странение птиц, но и размещение их концентраций зависит и на-
ходится в соответствии с особенностями гидрологического режи-
ма моря. В перечисленных районах в течение всего зимнего пе-
риода работает многочисленный японский рыболовный флот
(сотни судов), отходы промысла с которых существенно улуч-
шают кормовую базу морских птиц.
Заканчивая описание зимнего периода, нужно отметить, что
среди поморников в Восточно-Китайском море приходится на
долю светлой морфы около 90%, темной — 10%, среди чер-
нохвостой чайки 1/% светлых взрослых, 83% — серых молодых;
среди крупных чаек 67% — светлых, 33% — серых.
Весенний прогрев вод Куросио начинается в конце февраля
— начале марта, а желтоморских холодных вод — с середины
марта (Kikuchi, 1959). В марте, как отмечалось выше, в Восточ-
1бе
' ВОСТОЧНО-КИТАЯСКОВ МОРЕ
но-Китайоком море появляется бледноногий буревестник, зиму-
ющие же здесь виды начинают перемещение на север. В это вре-
мя пестролицый буревестник, мигрирующий в основном по вос-
точной половине моря, проникает в Корейский пролив и в юж-
ную часть Японского моря. Довольно многочисленны в марте в
Корейском пролнве '(на пролете на север) и чайки, мигрирую-
щие главным образом по западной половине Восточно-Китай-
ского моря (рис. 46). Признаком наступления весны служат так-
же появляющиеся с начала марта и сухопутные птицы, доволь-
но интенсивный пролет которых широким фронтом продолжает-
ся с середины марта до середины мая со стороны Китая в об-
щем направлении на Японские острова. Здесь летят различные
кулики, ласточки, трясогузки, коньки, сорокопуты, овсянки, цап-
ли, голуби, дневные и ночные хищники, кукушки и другие.
Как следствие весенних перемещений, численность всех видов
морских птиц в Восточно-Китайском море заметно изменяется.
К апрелю сильно уменьшается количество всех чаек, при этом
lap t26 йУ
Рис. 46. Основные пути и направления весенних, миграций в Восточно-Ки-
тайском море. 1—чайки, 2 — буревестники и поморники, 3 —изобаты.
* 4 — поверхностные изотермы
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
163
встречаются они преимущественно только в районе Корейского
пролива. Уменьшается численность и других гнездящихся в се-
верных районах видов, а также альбатросов (табл. 23). Чис-
ленность пестролицего буревестника весной в Восточно-Китай-
ском море увеличивается, хотя довольно много его стай в это
время уже проникает в Японское море. Увеличение происходит
за счет прилета особей, кочевавших зимой на юге за пределами
Восточно-Китайского моря. Как видно из табл. 23. весной во
всех районах наблюдается абсолютное доминирование этого ви-
да над всеми остальными. В апреле и мае продолжается пролет
в сторону Корейского пролива бледноного буревестника. На
юге моря в это время можно, наблюдать и тропические виды, из
которых наиболее обычным является буревестник Бульвера —
Bulweria bulweri (Jard. andSelb.).
Таблица 23
Видовой состав, соотношение (%) н плотность морских птиц
в Восточно-Китайском море весной (апрель—май). 1961—1968 гг.
Виды 35—30 с. in. . 0 -25' с. in. Все море
Р. leucomelas 89,0 93,0 91,0
Р. carneipes 1,0 1,2 1,2
Крупные чайки 1,1 — 0,5
L crassirostris 0,3 — 0,1
R. tridactyla 0.5 — 0,3
В. bulweri — 1,4 0,7
Поморники 0,1 0,9 0,5
Крачки 0,2 0,5 0,3
Мелкие чистиковые 1,1 — 0,5
С. monocerata 0,7 — 0,3
L. lobatus 3,3 0,1 1,7
Неопределенные виды 2,7 2,9 2,9
Количество особей на I кв. км 3,3 1,5 2,3
Количество учетов 54 72 126
Несмотря на то, что многие виды
весной откочевывают на
север, общая плотность населения птиц увеличивается за счет
пролетных стай пестролицего буревестника (см. табл. 23). В это
время от широты острова Тайвань до Корейского пролива на фо-
не в общем-то разрозненного распространения регулярно встре-
чаются его стаи, насчитывающие по нескольку десятков или со-
тен особей- Характерно, что пролегающие через Восточно-Китай-
ское море на север стаи пестролицего буревестника, как прави-
ло, придерживаются края мелководья и материкового склона.
164
ВОСТОЧНО-КИТАЙСКОЕ море
где зимой наблюдалось увеличение плотности населения птиц.
В связи с этим нужно заметить, что особенности циркуляции
вод весной в общем сходны с зимним периодом (Kikuchi, 1959;
Харченко, 1968). При этом конвергенция, как и зимой, проходит
с юго-запада на северо-восток вдоль края шельфа. Соответст-
венно сохраняет свое положение и продуктивная эона.
Гнездовой сезон пестролицего буревестника приходится на
весенне-летний период (Иосида, 1962). К июню пролет заканчи-
вается, и л открытых водах Восточно-Китайского моря его чис-
ленность сильно уменьшается. По-видимому, в Японское море
откочевывают и многие неполовозрелые и неразмножающиеся
в данном году особи. В несколько раз уменьшается летом по
сравнению с зимним и весенним периодами и общая плотность
населения птиц (табл. 24). Сравнительно обычен летом пестро-
лицый буревестник лишь в районе Корейского пролива и остро-
ва Кюсю. Здесь он, как и весной, доминирует над другими пти-
цами, ио приблизительно южнее ЗГ с. ш. в течение всего лета он
редок, как редки и другие виды.
Таблица 24
Видовой состав, соотношение (%) н плотность морских нтнц
в Восточно-Китайском море летом (июнь—сентябрь). 1961—1968 гг.
В н д ы • 5 — O'J с. ш. 30 - 25° с. ш. Вег море
Р. leucomelas 46,5 42,0 46,0
Р. carneipes. Р. pacificus 3.0 17,0 4.0
В. bulweri 1.0 21,0 2.5
L. lobatus 36,0 — 33.5
Крачки — 4.0 0.5
Олуши — 2,0 +
Неопределенные виды 13,5 14.0 13.5
Количество особей на 1 кв. км 0,6 0.2 0.4
Количество учетов 42 30 72
Примечание. L. lobatus отсутствует в июне—августе и появляется только
в сентябре.
Таким образом, летом Восточно-Китайское море наиболее
бедно птицами- Это связано, как было показало выше, с откочев-
кой на север и к берегам гнездящихся в северном полушарии,
видов. Буревестники южного полушария, миллионы особей кото-
рых заполняют просторы Берингова, Охотского морей и приле-
гающих к ним районов океана, мигрируя на север, основной
массой минуют Восточно-Китайское море (пролетают восточ-
нее). Они следуют на откорм в более продуктивные районы. В
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
165
Восточно-Китайском же море кормовые условия для птиц летом,
очевидно, мало благоприятны. Зоны повышенной биологической
продуктивности с прогревом вод смещаются на север. В частно-
сти, на север моря смещается конвергенция Куросио и желтомор-
ских вод, а следовательно, и наиболее продуктивный переход-
ный биоценоз (Kato, 1958; Kikuchi, 1959; Tsujita, 1963; Харченко,
1968; Крылов, 1969 и др.)- Как следствие этого, из Восточно-
Китайского в Японское и Желтое моря уходят основные массо-
вые виды рыб — скрумбрня, анчоус, сайра, сардина- ставрида,
а также кальмар.
Низкая численность птиц сохраняется в течение всех летних
месяцев и в первой половине сентября- который по гидрологи-
ческим условиям относится к летнему сезону. Вместе с этим с
начала сентября все же наблюдаются первые признаки прибли-
жающейся осени: появляются сухопутные птицы и стайки круг-
лоносого плавунчика.
Состав орнитофауны и распределение птиц осенью во многом
сходны с таковыми весной. Снова многочисленным становится
пестролицый буревестник, который постепенно откочевывает с
севера. С севера в октябре (отдельные экземпляры раньше, регу-
лярно — с конца месяца) начинают прибывать чайки, поморни-
ки и другие виды. В результате этого заметно увеличивается и
общая плотность населения птиц (табл. 25)-
Таблица 25
Видовой состав, соотношение (%) и влотиость морских птиц
в Восточно-Китайском море осенью (октябрь—ноябрь). fMI—1М8 гг.
Виды 35—30 с. ш 30—25° с. ш. Все море
Р. leucomelas 80,0 96.0 87,0
L. crassirostris 10,0 — 5,7
Поморники 2,7 0.2 1,6
Крупные чайки р. Larus 2,3 — 1,3
Олуши 2.2 1.3
Р. carneipes 0,2 0,1
D. nigripes — 0.8 0,3
О. monorhis — 0.8 0,3
R. tridactyla 0.3 — 0,2
Неопределенные виды 2,5 2,0 2,2
Количество особей на 1 кв. klM 2,3 1.1 1,7
Количество учетов 42 42 84
166 ТРОПИЧЕСКИЕ ВОДЫ .МЕЖДУ АВСТРАЛИЕК И ЗОНДСКИМИ ОСТРОВАМИ
Заканчивая изложение данных по распределению и мигра-
циям морских птиц Восточно-Китайского моря, нужно подчерк-
нуть следующее. В рассматриваемом районе, как и в других мо-
рях, в составе орнитофауны, численности и распределении птиц
наблюдается большая сезонная изменчивость. Только пестроли-
цый буревестник является массовым видом; во все сезоны года
он резко доминирует над другими видами. Птицы умеренных
широт (чайки, поморники, чистиковые) сравнительно многочис-
ленны в Восточно-Китайском море лишь в холодное время, а тро-
пические виды — в теплое. На распределение птиц существен-
ный отпечаток накладывают большие различия в гидрологиче-
ском режиме, наблюдаемые в мелководной и глубоководной ча-
стях моря. Большая часть бореальных видов концентрируется
в районе корейского пролива и в северо-западной мелководной
половине моря в зоне желтоморских холодных вод, а субтропи-
ческие виды и поморники — в юго-восточной глубоководной, в
зоне теплого течения Куросио. В связи с этим миграции первых
проходят по западной части моря, миграции вторых — по восточ-
ной части.
Основные скопления морских птиц располагаются в районе
Корейского пролива и в зоне смешения желтоморских холодных
вод с водами Куросио. то есть районах концентраций пелагиче-
ских рыб и рыболовного флота. Наиболее высокая плотность
населения птиц в Восточно-Китайском море наблюдается с осе-
ни до конца весны (октябрь—май), то есть в период миграций
и зимовок. Летом плотность уменьшается в несколько раз, что
связано с откочевкой большей части особей многих видов на се-
вер. Средняя плотность населения, по данным визуальных уче-
тов, составила летом 0,4, зимой 1,4, осенью 1,7, весной 2,3 особи
на 1 кв. км. Пересчет этих данных на всю площадь моря (752
тыс- кв. км.) дает следующие цифры общей численности птиц:
300 800, 1 052 800, 1 278 400 и 1 729600 особей. Сравнивая эти
данные с другими районами, нужно заметить, что плотность на-
селения птиц в Восточно-Китайском море значительно ниже, чем
в умеренных широтах, но несколько выше, чем во многих тро-
пических районах.
6. Тропические воды между
северным побережьем Австралии
и Зондскими островами
До недавнего времени распределение и миграции птиц в тро-
пических водах Индийского океана были очень слабо исследова-
ны. Появившиеся в последние годы публикации до некоторой
степени восполняют этот пробел (Phillips, 1954; Lavery, 1964;
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
107
Bailey, 1965, 1966, 1968; Ozawa at el., 1966; Ozawa and Naka-
mura, 1966; Pocklington, 1965, 1967; Gill, 1967; Шунтов, 19686; и
др.). Большая часть имеющихся работ касается западной поло-
вины Индийского океана.
В настоящем разделе описывается распределение птиц в ме-
нее изученной восточной части Индийского океана на акватории,
ограниченной на севере Зондскими островами, на юге — север-
ным и северо-западным побережьем Австралии. Рассматривае-
мый район целиком расположен в тропической зоне. Основу ор-
нитофауны составляют гнездящиеся здесь типичные тропические
виды — многие крачки, олуши, фрегаты и фаэтоны.
Несмотря на низкоширотное положение района, сезонность
климата и гидрологического режима здесь выражены вполне
отчетливо. Основным фактором, определяющим изменчивость в
условиях обитания, является смена преобладающих ветров: юго-
восточного пассата, господствующего с мая по сентябрь (зимний
период), и северо-западного муссона, господствующего с ноября
по март (летний период)-
У северного побережья Австралии и расположенных на этих
широтах островах в Индийском океане, в частности островах
Рождества и Кокосовые, птицы размножаютсй круглогодично,
но преобладающим является зимний тип гнездования (Serventy,
1952; Pearson, 1966), то есть большая часть особей гнездится в
период юго-восточного пассата. На островах у северного побе-
режья Австралии самые массовые из гнездящихся птиц — крач-
ки и, как видно из табл. 26, они преобладают здесь в учетах на
море. В Тиморском, в западной части Арафурского моря и у се-
веро-западного побережья Австралии среди крачек резко доми-
нирует темная крачка — Sterna fuscata L., в восточной части
Арафурского моря и в заливе Карпентария — несколько видов
крачек со светлой окраской, в том числе Sterna sumatrana Raff-
les, S. bengalensis Less., S. bergii Licht., S. albifrons Vroeg., S. dou-
galli Mont, и др. Весьма обычны, а местами многочисленны на
североавстралийском шельфе олуши (чаще бурая — Sula leuco-
gaster (Bodd). значительно реже S. sula L., S- dactylatra (Less.),
фрегаты и фаэтоны (чаще Phaethon rubricaudus (Bodd). У се-
верного побережья Австралии гнездятся и другие птицы, в част-
ности бакланы. Однако во все сезоны они тяготеют к узкой!
прибрежной зоне и выпали из моих наблюдений, проводившихся
не ближе 15—20 миль от берега. Это же можно сказать и о
красноклювой серебристой чайке—Larus novaehollandiae Steph.,
только отдельные особи которой выходят за пределы прибреж-
ной полосы.
Численность всех гнездящихся в районе североавстралий-
ского мелководья видов резко уменьшается за пределами мел-
168 ТРОПИЧЕСКИЕ ВОДЫ МЕЖДУ АВСТРАЛИЕЙ И ЗОНДСКИМИ ОСТРОВАМИ
ководья. Плотность населения некоторых из них вновь увеличи-
вается в зоне Южно-Пассатного течения, особенно к северу
от 12—13° ю. ш. Это — олуши (S. sula, S. leucogaster), фаэтоны
(Р. rubricaudus. Р. lepturus Daud), фрегаты (Fregata minor-
(Gmel.), F. andrewsi Math.), гнездящиеся на островах Рождест-
ва и Кокосовые и преобладающие над другими видами между
5 и 15’ ю. ш. (см. табл. 26).
Таблица 26
Видовой состав, соотношение (%) и плотность населения морских птиц
в восточной части Индийского океана в период юго-восточного пассата
(июнь—сентябрь). 1965—1970 гг.
Виды Арафурское море Тиморское море Шельф у северо- западного побе- режья Австралии Открытый океан
1П ш oSl—S 15—25° ю. ш.
Крачки 74,0 90,0 15,0 -L +
Черные буревестники (Р. pacificus) 6,0 + 3,0 14,0 61,0
Олуши (S. leucogaster) 13,0 7,0 10,0 63,0 +
Фрегаты 3,0 2,0 2,0 10,0 +
Фаэтоны (Р. lepturus, Р. rubricaudus) 3,0 + + 12,0 6,0
Медеие белобрюхие буревестники
(Р. assimiiis) — 1,0 64,0 1.0 20,0
Качурки (О. oceanicus) 1,0 + 1,0 4- 12,0
Альбатросы (D. chlororhynchus) — — 2,0 — 1,0
D. capensis — — 2,0 — —
L. novaehollandiae + + 1.0 — —
Количество учетов 221 98 115 60 51
Количество особей на 1 кв. км 0,2 1,6 0,7 1,0 0,1
Наряду с размножающимися тропическими видами для рас-
сматриваемого района Индийского океана в период юго-восточ-
ного пассата характерна также большая группа птиц, гнездя-
щихся южнее. В первую очередь нужно назвать клинохвостого
буревестника, многочисленного летом на гнездовье вдоль запад-
ного побережья Австралии (Serventy and Whittell, 1951; Cayley,
1965; Hill. 1968). В знмний период он кочует в открытых водах
Индийского океана, регулярно встречается между островом Ява
и северо-западным побережьем Австралии, но в сравнении с
летним периодом численность его на востоке Индийского океа-
на сильно уменьшается (см. табл. 26). Проникает в тро-
пические ‘воды гнездящийся у юго-западного побережья
Австралии сумеречный буревестник — Puffinus assimiiis
СЕЗОННЫЕ аспекты в орнитофауне
169
Gould и, по-видимому, другие мелкие белобрюхие буревестники.
У северо-западного побережья Австралии они доминируют в это
время над другими птицами, а в открытом океане встречаются
регулярно (см. табл. 26) *. В небольших количествах здесь мо-
жет быть встречен и другой субтропический вид, гнездящийся у
юго-западного побережья Австралии, — белолицая качурка —
Pelagodroma marina (Lath.). Большая часть особей этого вида
на зимовки откочевывает в северо-западную часть Индийского
океана (Morzer Bruyns and Voous, 1964; Bailey, 1966, 1968; Cill.
1967; и др.).
В районе между Индонезией и северным побережьем Авст-
ралии, по-видимому, регулярно встречаются качурки из Тихого
океана. Судя по наблюдениям ряда авторов (Pocklington, 1967;
Bailey, Pocklington, Willis, 1968), это в основном Oceanodroma
matsudairae.
В период юго-восточного пассата температура воды и воз-
духа у северного побережья Австралии заметно понижается,
В это время до северо-западного угла Австралии поднимается
поверхностная изотерма 20°, что дает возможность некоторым
нотальным и антарктическим видам с зональным распростране-
нием проникать в южную часть тропической зоны. До северо-
западного угла Австралии расширяют область своего распро-
странения желтоносый и странствующий альбатросы, капский
голубь, гигантский буревестник, прионы и некоторые другие.
Не зависят от температуры в период миграций качурка
Вильсона и южно-полярный поморник. Качурка Вильсона в те-
чение всего зимнего периода многочисленна в северной части
Индийского океана (Gibson-Hill, 1948; Phillips, 1950, 1954; Bai-
ley, 1966, 1968; и др.). Встречается она зимой и в водах, приле-
гающих к северному побережью Австралии (Serventy, 1952а;
Pocklington, 1967; и др.), но более многочисленна здесь, по-ви-
димому, в периоды миграций. Только на миграциях, то есть
осенью и весной, встречается в рассматриваемом районе южно-
полярный поморник. В тропической части Индийского океана
севернее экватора он встречается регулярно (Morzer Bruyns
and Voous, 1965; Bailey, 1966, 1968; и др.).
Таким образом, видовой состав птиц в тропиках восточной
части Индийского океана в период юго-восточного пассата до-
1 У меня нет полной уверенности в правильности определения видовой
принадлежности мелких белобрюхих буревестников у северо-западного побе-
режья Австралии. Раньше я предполагал (Шунтов, 19686), что это Puffinus
gavia (Forst.), но этот вид не называется для данного района австралийски-
ми орнитологами (Serventy and Whittell, 1951; Cayley, 1965; Hill, 1968).
Часть особей относится, видимо, к Р. mollis (Gould), напоминающему
Р. assimilis или Р. gavia в полевых условиях; этот вид отмечен для данного
района Поклингтоном (Pocklington, 1967).
170 ТРОПИЧЕСКИЕ РОДЫ МЕЖДУ АВСТРАЛИЕЙ И ЗОНДСКИМИ ОСТРОВАМИ
вольно пестрый. Здесь встречаются гнездящиеся на этих широ-
тах тропические виды, гнездящиеся у западного и юго-западно-
го побережья Австралии субтропические виды и виды из но-
тальной и антарктической зон. На большей части акватории
между Зондскими островами и побережьем Австралии преобла-
дают тропические виды (см. табл. 26). Примесь южных видов
существенна только у северо-западного побережья Австралии и
расположенных на этой широте открытых водах Индийского
океана (исключая воды Южно-Пассатного течения).
Как видно из рис. 47, основные скопления птиц наблюдают-
ся на шельфе Австралии В этом районе, за исключением Ара-
фурского моря, плотность в 7—16 раз выше, чем в прилегающих
водах океана. Повышение плотности обусловлено хорошей
кормностью отмеченных районов, а также наличием octdobob, на
которых птицы размножаются.
Зимой в период юго-восточного пассата под влиянием гос-
подствующих ветров на шельфе Австралии преобладает движе-
ние вод с востока на запад (Wyrtki, 1961). В результате сгона
поверхностных вод с мелководья па запад и северо-запад в при-
донных слоях наблюдается поступление глубинных вод на
шельф. Эти воды содержат значительное количество биогенов
(Rochford, 1962; Kirkwood, 1967). Подъем вод на шельф созда-
ет благоприятные условия для развития высоких биомасс планк-
тона здесь и в прилегающих к северо-западному побережью
районах (Богоров и Виноградов, 1961; Виноградов и Воронина,
1962; Суханова, 1962, 1964; Маркина, 1969; Tranter, 1962; и др.).
Во все сезоны на шельфе многочисленны мелкие пелагические
рыбы. На этом фоне не понятна бедность птицами большей
части Арафурского моря (см. табл. 26). В Арафурском море
подъем и выход глубинных вод на шельф выражен особенно
сильно, что находит отражение в очень обильном развитии
планктона (Маркина, 1969). В глубоководных районах здесь
очень многочисленны летучие рыбы (Шунтов, 1968в), которые
являются одним из основных кормовых объектов многих мор-
ских птиц в открытых водах тропиков. На малых глубинах в
больших количествах обитают различные мелкие пелагические
рыбы — сельди, ставриды, сребробрюшки и другие (Шунтов и
Демиденко, 1970; Шунтов, 1971а). Сопоставимы по плотности
населения птиц с Тиморским морем и шельфом у северо-запад-
ного побережья Австралии только прибрежные воды залива
Карпентария (см. рис. 47).
Открытые воды Индийского океана между субтропической
конвергенцией (около 40° ю. ш.) и 15—16’ ю. ш. отличаются
низкой продукцией планктона, что связано со слабым верти-
кальный перемешиванием вод (Богоров и Виноградов, 1961;
Tranter, 1962). В общем не многочисленны здесь и летучие ры-
Рис. 47. Распределение скоплений морских птица восточной части Индийского океана в период юго-восточного
пассата (май—сентябрь). 1—редки, 2 — обычны, 3 — многочисленна 4 — поверхностные течения
172 ТРОПИЧЕСКИЕ ВОДЫ МЕЖДУ АВСТРАЛИЕЙ И ЗОНДСКИМИ ОС1РОВАМИ
бы (Шунтов, 1968в). Как видно из табл. 26, южнее 15° ю. ш.
плотность населения птиц очень небольшая - всего 0,1 на
I кв. км. Севернее, то есть в зоне Южно-Пассатного течения,
плотность птиц в 10 раз выше. Воды к югу от острова Ява (на
широте острова Рождества) по плотности птиц сопоставимы с
шельфом Австралии. Этот район, как известно, является местом
формирования Южно-Пассатного течения, уносящего воды на
запад. Как следствие переноса вод, наблюдается мощный
подъем богатых биогенами вод к поверхности, (Rochford, 1962;
Wirtki, 1962), являющийся причиной высокой продуктивности
этого района (Кобленц-Мишке, Кабанова, 1964). Здесь же отме-
чается высокая для открытых тропических вод биомасса зо-
опланктона (Богоров и Виноградов, 1961; Виноградов и Ворони-
на, 1962; Богоров и Воронина, 1967; Маркина, 1970). Значитель-
ные концентрации образуют в водах экваториальных течений
летучие рыбы (Шунтов, 1968в), являющиеся основный кормом
олуш, фрегатов и фаэтонов, доминирующих здесь над другими
птицами.
В период северо-западного муссона в связи с изменением
океанографического окружения в распределении птиц и составе
их населения наблюдаются существенные изменения. Еще до
окончательной смены преобладающих ветров и установления
северо-западного муссона покидают тропическую часть Индий-
ского океана виды умеренных и антарктических широт. Соби-
раются в это время у южного и юго-западного побережий Ав-
стралии в районе мест гнездования белолицая качурка и блед-
ноногий буревестник. Бледноногий буревестник от Австралии
мигрирует на зимовки- в северо-западную часть Индийского и
на север Тихого океана. Пути миграций особей, зимующих в
Индийском океане, проходят западнее рассматриваемого райо-
на. Через восточную часть Индийского океана летят особи, зи-
мующие в Тихом океане. Весной они пролетают здесь в октябре
и ноябре. Основной поток мигрантов проходит через восточную
часть Зондского архипелага между Целебесом и Молуккскими
островами.
В летний период размножается клипохвостый буревестник,
который в больших количествах гнездится вдоль всего западно-
ного побережья Австралии (Serventy and Whittell, 1951; Cayley,
1965; и др.). В связи с этим его количество в восточной части
Индийского океана сильно увеличивается (табл. 27). Он стано-
вится многочисленным не только в тропических водах восточной
части Индийского океана, но и вдоль западного побережья Ав.
стралии (Ozawa and Nakamura, 1966; Шунтов, 19686).
Заслуживает упоминания пестролицый буревестник, не из-
вестный раньше для австралийских морей. Наблюдения пока-
зывают, что в период северной зимы этот вид совершает регу-
СЕЗОННЫЕ ЛСПЕК1Ы В ОРНИТОФАУНЕ
173
лярные миграции в воды шельфа северного побережья Австра-
лии, где становится весьма обычным в Тиморском и Арафур-
ском морях (см. табл. 27).
Что касается североавстралийских крачек и веслоногих, то
в летний период эти птицы, как и раньше, доминируют на шель-
фе над другими группами. У многих из них к этому времени! за-
канчивается размножение, и они чаще начинают встречаться в
открытом океане. В небольших количествах летом они распро-
страняются до южного побережья Австралии. Однако основные
концентрации их по-прежнему наблюдаются у североавстралий-
ского побережья. У северного побережья Австралии и прилегаю-
щих водах, по-видимому, проводит зиму часть особей темной
крачки, в больших количествах гнездящейся у западного побе-
режья. В тропиках этот вид, как и другие, гнездится в период
зимнего пассата. У западного побережья размножение протека-
ет в весенне-летний период (Serventy and Whittell, 1951; Serven-
ty, 1952). В это время крачка здесь многочисленна (Ozawa and
Nakamura, 1966; Шунтов, 19686).
Более широкое распространение имеют летом и птицы, гнез-
дящиеся на острове Рождества. В связи с этим плотность кон-
центраций южнее острова Ява несколько уменьшается, хотя и
остается, выше, чем в более южных районах открытого океана
(см. табл. 27).
Таблица 27
Видовой состав, соотношение (%) и плотность населения морских птиц
в восточной частя Индийского океана в период северо-западного муссона
(декабрь—март). 1965—1970 гг.
В и д ы Арафурское море Тиморское море Шельф у севере- западною побе- режья Австралии Открытый океан
“? а Is 15—2; ° ю. ш.
Крачки 77,0 77,0 10,0 5,0 52,0
Черные буревестники (Р. padficus) t 2,0 33,0 43,0 25,0
Олуши 13,0 17,0 48,0 28,0 17,0
Фрегаты 3,0 + 2,0 10,0 +
Фаэтоны 1,0 4- 11,0 5,0
Крупные белобрюхие бу рент пики (Р. leucomelas) 1,0 3,0 — —
Остальные виды 5,0 1.0 7,0 3,0 1,0
Количество учетов 149 38 84 120 63
Количество особей на 1 кв. км 2,3 0,3 1.1 0,6 0,3
174 ТРОПИЧЕСКИЕ ВОДЫ МЕЖДУ АВСТРАЛИЕЙ И ЗОНДСКИМИ ОСТРОВАМИ
Описанные выше общие черты количественного распределе-
ния птиц сохраняются в период северо-западного муссона, хотя
и наблюдается некоторое перераспределение видов (рис. 48).
Южная граница Южно-Пассатного течения летом опускается до
20* (Муромцев, 1959). Подъем вод южнее острова Ява в это
время сохраняется, поэтому сохраняются предпосылки, для фор-
мирования повышенной продуктивности района. Этому способ-
ствует, по-видимому, и зона дивергенции между Южно-Пассат-
ным течением и Экваториальным противотечением, которая на
восток прослеживается до района острова Рождества (Иванов-
Францкевич, 1961; Овчинников, 1961). Из рисунка 48 и таблицы
27 видно, что здесь, как и зимой, наблюдается повышенная
плотность населения птиц. В сравнении с периодом юго-восточ-
ного пассата продуктивность вод и биомассы планктона летом
ниже (Маркина, 1970а). Уменьшается в данном районе в это
время и численность летучих рыб. Они. рассредоточиваются ><
южном направлении (Шунтов, 1968в). Как видно из табл. 27.
плотность птиц в северной части Южно-Пассатного течения
(до 15° ю. ш.) несколько уменьшается, но южнее 15е увеличи-
вается. Последнее, очевидно, можно связывать со смещением на
юг Южно-Пассатного течения.
Для летнего типа циркуляции вод на северном шельфе Ав-
стралии характерно общее движение вод в восточном направ-
лении. Вдоль всего северо-западного побережья прослеживает-
ся Западно-Австралийское течение (Wyrtki, 1961, 1962). В это
время подъем глубинных вод на шельф прекращается; в связи с
этим заканчивается массовое развитие планктона. Особенно по-
нижаются биомассы планктона в Тиморском море. При этом
характерно смещение зон высоких биомасс планктона в при-
брежные воды. Несмотря на уменьшение количества планктона,
кормовые условия для птиц на мелководье остаются благопри-
ятными. Дело в том, что многочисленные мелкие пелагические
рыбы не покидают шельфа в течение всего года. Летом они об-
разуют концентрации в прибрежной зоне вдоль всего побе-
режья, особенно в Арафурском море. Кроме того, у большинства
видов рыб в весен не-летний период происходит нерест, и в при-
брежной зоне появляется большое количество молоди рыб.
Больше всего молоди наблюдается в Арафурском море, куда
она заносится и из Тиморского моря (Шунтов, 1971а). Именно
в Арафурском море особенно заметно увеличивается плотность
населения птиц (см. табл. 27, рис. 48). Происходит это, по-видн-
мому, за счет миграций птиц из районов, расположенных запад-
нее, и откочевки после размножения крачек и веслоногих из при-
брежной 15—20-мильной зоны, нс охваченной наблюдениями.
Высокая плотность птиц в Арафурском море (за пределами при-
брежной зоны) бывает только летом. Весной (октябрь) и
Рис. 4ft. Распределение скоплений морских птиц в восточной части Индийского океана <в период северо-западное
муссона (ноябрь—март). 1 — редки, 2 —обычны, 3—многочисленны, 4 — поверхностные течения
176
ШЕЛьФ ЮЖНОГО ПОБЕРЕЖЬЯ АВСТРАЛИИ
осенью (апрель—май) она составляет, как и зИмой, всего 0,2
особи на I кв. км. У северо-западного побережья Австралии
плотность населения птиц также несколько увеличивается. Про*
исходит это в основном за счет клинохвостого буревестника и
бурой олуши (см. табл. 27). Тиморское море — единственный
район североавстралийского шельфа, где плотность пт|1ц летом
уменьшается значительно — в пять раз. •
7. Шельф южного побережья Австралии i
и прилегающие воды
Южное побережье Автралии расположено в субтропической
зоне. Среди гнездящихся птиц здесь абсолютно доминируют суб-
тропические виды. Как видно из табл. 28, летом основу населе-
ния в водах юго-западного побережья и в Большом Австралий-
ском заливе составляют субтропические виды. Это бледноногий
буревестник, длиннокрылый тайфунник — Pterodroma' macrop-
tera (Smith.), хохлатая крачка, белолицая качурка и другие
Все они гнездятся у южного побережья, при этом большая
часть видов в теплое время года (Serventy and Whittell, 1951;
Cayley, 1965; Hill, 1968). К северу от юго-западного угла Ав-
стралии бледноногий буревестник заменяется тропическим кли-
нохвостым буревестником (Ozawa and Nakamura, 1966; Шун-
тов, 19686). В этом районе располагаются к его гнездовья
(Serventy and Whittell, 1951; Storr, 1964; и др.).
Летом у южного побережья Австралии тепло. В Большом
Австралийском заливе температура воды на поверхности подни-
мается до 20—22*. В это время здесь можно встретить настоя-
щих тропических птиц — темную крачку, фаэтонов, фрегатов.
Из птиц умеренных широт сопоставим по численности с субтро-
пическими видами только желтоносый альбатрос. В небольших
количествах, но регулярно встречаются также чернобровый и
странствующий альбатросы. Южнее в открытых водах восточ-
ной части Индийского океана они весьма обычны. У побережья
Австралии в течение лета их численность не остается постоян-
ной. При продолжительных ветрах южных направлений увели-
чивается, например, количество странствующего альбатроса,
правда, в прибрежные воды он не заходит и держится в районе
кромки шельфа и материкового склона. В мористых районах
Большого Австралийского залива эпизодически могут быть
встречены и другие виды, гнездящиеся в умеренных и антаркти-
ческих широтах, в том числе пугливый альбатрос, южно-поляр-
ный поморник, качурка Вильсона.
Иная картина наблюдается у юго-восточного побережья
Австралии и Тасмании. Здесь резко над другими видами доми-
нирует размножающийся в этом районе тонкоклювый буревест-
СЕЗОННЫЕ аспекты в орнитофауне
177
ник. Многочисленны также огненный прион — Pachyptila turtur
(Kuhl.) и альбатросы. Среди последних, кроме желтоносого,
обычны пугливый, чернобровый и странствующий.
Шельф южного побережья Австралии узкий, и свал глубин
проходит вблизи берегов; только в Большом Австралийском
заливе и Бассовом проливе мелководье простирается на десятки
миль. Детальные исследования в Большом Австралийском зали-
ве (Шунтов, 1968а) показали, что в видовом составе мелковод-
ной и глубоководной частей залива наблюдаются заметные раз-
личия. В мористых участках залива в районе материкового
склона (глубже 100 м) абсолютно преобладают трубконосые —
бледноногий буревестник, белолицая качурка, желтоносый аль-
батрос (см. табл. 28). Трубконосые в целом более многочислен-
ны и на шельфе, но здесь они обычны только на западе и восто-
ке залива, где наблюдается поступление океанических вод на
мелководье. Очень характерны для мелководья крачки, среди
которых преобладает хохлатая. На востоке залива у острова
Кенгуру обычен короткохвостый поморник, мигрирующий сюда
из северного полушария.
Таблица 28 отражает состав птиц за пределами 15-мильной
зоны. Тенденция изменения з соотношении видов еще больше
выражена в прибрежной зоне. Как показали учеты во время за-
хода в порт Аделаида, у берегов очень многочисленны серебри-
стая красноклювая чайка и бакланы. Вместе с крачками они
составляют основу населения птиц прибрежной полосы. К во-
стоку и западу от Большого Австралийского залива шельф бо-
лее узкий, и птицы, тяготеющие к океаническим водным массам,
встречаются близко у берегов вместе с прибрежными видами
(рис. 49). При этом за пределами 15-мильной зоны везде пре-
обладают океанические виды — буревестники и альбатросы.
Состав доминирующих видов в различных районах во мно-
гом определяется составом и характером кормовой базы. Так,
в мелководной части Большого Австралийского залива основу
биомассы планктона слагает мозопланктон; здесь имеют высо-
кую численность мелкие пелагические рыбы — сардина, анчоус,
шпрот, а также молодь многих рыб, обитающих в глубоковод-
но* части залива. В глубоководной части кроме мезопланктона
сравнительно обилен макропланктон, питаясь которым образу-
ют концентрации многие придонные и крупные пелагические
рыбы — тунцы, ставрида, скумбрия (Натаров и Пашкин, 1968;
Макаров к Пашкин, 1968; Змиевский, 1968; Шунтов, 1969; Мар-
кина, 1969; Шунтов и Демиденко, 1970). Концентрации рыб, в
том числе тунцов, вдоль кромки мелководья имеют место также
к западу и востоку от залива (Robins, 1962; Jones, 1966; Hynd
and Robins, 1967; и др.). Многочисленные или обычные в при-
178
ШЕЛЬФ ЮЖНОГО ПОБЕРЕЖЬЯ АВСТРАЛИИ
Таблица 28
Видовой состав, соотношение (%) и плотность морских птиц
у южного побережье Австралии летом (декабрь—март). 1666—1970 гт.
В и д ы Юго-западное побережье Ав- стралии Большой Австралийский залив
шельф материковый склон
Черные буревестники (Р. carneipes) 85,0 60,0 75,0
Мелкие альбатросы (D. chlororhynchus) 5,0 7,0 10,0
Р. marina 5,0 1Д ПЛ
S. bergii 0,5 21.0 2,0
S. parasiticus + 4,5 4-
D. exulans 1,5 — 0,5
L. novaehollandiae 0,5 +
M. serrator 4- 1,0 4-
Остальные виды 3,0 5,0 1,5
Количество учетов 10 72 241
Количество особей на 1 кв. км 3,0 0,8 4,0
брежных районах хохлатая крачка, австралийская олуша —
Moris senator (Gray) и серебристая чайка питаются мелкими
пелагическими рыбами. В глубоководных районах залива они
немногочисленны. Здесь, как отмечалось выше, доминируют пи-
тающиеся макропланктоном трубконосые — буревестники, ка-
чурки и питающиеся кальмарами и более крупной рыбой аль-
батросы.
Заслуживает специального рассмотрения вопрос о размеще-
нии скоплений птиц в летний период. Как показывают исследо-
вания В. Н. Пашкина в Большом Австралийском заливе (На-
таров и Пашкин, 1968; Макаров и Пашкин, 1968; Пашкин, 1970),
летняя циркуляция вод здесь формируется под влиянием цикло-
нического круговорота, образующегося южнее материкового
склона. Результатом этого является перенос вод вдоль присва-
ловой части мелководья и материкового склона. На этом фоне
в ряде районов материкового склона наблюдается сложная
картина активных динамических процессов (круговороты, за-
вихрения), обуславливающих вертикальное перемешивание вод,
благодаря чему создаются условия для развития повышенной
биологической продуктивности. Вдоль материкового склона фор-
мируется своего рода зона жизни. Это подтверждается, в част-
ности, и распределением пелагических (Шунтов, 1969) и при-
донных рыб (Kesteven and Stark, 1967; Шунтов и Демиденко.
1970: и др.).
Рис. 49. Распределение морских птиц у южного побережья Австралии в летний период. 1 — течения, 2 —районы по-
вышенной плотности птиц, 3—неритические виды, 4 — океанические «яды
480
ШЕЛЬФ ЮЖНОГО ПОБЕРЕЖЬЯ АВСТРАЛИИ
Около 40 лёт назад Д. Сервенти (Serventy, 1935) отмечал
тяготение бледнсногого буревестника к кромке мелководья. Эти
было подтверждено моими наблюдениями (Шунтов, 1968а); при
этом было показано, что основные скопления птиц (буревестни-
ки и альбатросы) тяготеют к западной и восточной частям .зали-
ва, где на мелководье наблюдается выход глубинных вод. Как
видно из табл. 28, плотность птиц в глубоководной части залива
в 4 раза выше, чем на мелководье. Хотя летом в мелководной
части залива концентрации рыб неплотные, в частности, рассре-
доточение держится самый массовый вид -- сардина, постоян-
ная низкая плотность птиц вызывает некоторое недоумение.
Повышенная плотность населения птиц вдоль кромки мелю-
водья и материкового склона наблюдается к востоку и западу
от Большого Австралийского залива (см. рис. 49). Преимуще-
ственно вдоль этой зоны распространяется почти до Тасмании
часть неполовозрелых особей бледноногого буревестника, воз-
вращающихся на лето к Австралии из северной части Индийско-
го и Тихого океанов. Южнее зоны материкового склона ' птиц
меньше; вновь плотность несколько увеличивается южнее при-
близительно 40° ю. ш., где проходит субтропическая конверген-
ция (см. рис. 49). -
В табл. 28 не представлен район Тасмании и Бассова проли-
ва. За летний период по этому району я располагаю ограничен-
ным количеством учетов. Однако нужно заметить, что пло+ность
птиц здесь выше, чем в Большом Австралийском заливе. Уже
неоднократно упоминалось об огромных концентрациях тонко-
клювого буревестника в этой районе. Высокая продуктивность
данного района, являющаяся предпосылкой значительной, плот-,
ности птиц, находит подтверждение и в больших концентрациях
различных рыб (Blackburn, 1950, 1950а; Blackburn and Gartner.
1954; Roughley, 1961; и др.).
Осенью, начиная с апреля, в составе населения птиц наб-
людаются большие изменения, связанные с отлетом некоторы..
видов в другие районы, а также с откочевкой к берегам Австра-
лии особей многих видов из южных широт. В апреле, закончив
размножение, улетает на север Тихого океана тонкоклювый бу-
ревестник. В районе Тасмании и Бассова пролива в апреле—
мае на его долю в учетах приходится еще почти половина осо-
бей (табл. 29). но в целом плотность и численность птиц
у юго-восточного побережья Австралии заметно уменьшается.
В апреле заканчивается размножение и у бледноногого буре-
вестника (Warham, 1958). В апреле данный вид еще многочис-
лен у южного побережья Австралии (см. табл. 29), но
в это время его стаи уже начинают появляться у Маль-
дивских и Лакадивских островов й в Аравийском море
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
181
(Bourne, 1960; Phillips, 1963; Bailey. 1965, 1966, 1968;
Gill, 1967). В мае, по-видимому, из района Австралии он исче-
зает полностью. Аналогичные изменения происходят и в распре-
делении белолицей качурки. Она откочевывает на север Индий-
ского океана, но некоторое количество особей остается в водах
Австралии. В апреле покидают воды Австралии северные по-
морники — короткохвостый и средний1.
Таблица 29
Видовой состав, соотношение (%) и плотность морских птиц
у южного побережья Австралии осенью (апрель-май). 1966—1970 гг.
В и л i.i Юго-западное побережье Австралии Большой Австра- лийский залив Район ме- жлу о. Кен гуру и Тас
шельф матери- ковый склон
Черные буревестники (Р. cameipes, Р. tenuirostris) Мелкие альбатросы 65,0 9.0 58,0 40,0
(D. chlororhynchus, D. mela- nophrys, D. cauta) 11,0 7,0 29,0 17,5
Крачки (S. bergii) Прионы (P. turtur) 1,0 + 76,0 4,0 + 2,0 20,0
D. exulans 10,0 1,0 2,5 5,5
L. novaehollandiae 5,0 2,0 4,0
M. serra tor -г 1,0 4- 0,5
€. skua 4- 0,5 4- 0,5
S. parasiticus + 4- -j- 0,5
0. oceanicus 4,0 — 1,0 0,5
P. marina т — 3,0 1,0
P. palpebrata 4- — 4- 4
D. ca pensis — — + 4-
M. giganteus •f- — 4- +
Остальные виды 4,0 3,5 2,0 8,0
Количество учетов 35 49 229 146
Количество особей на 1 кв. км 1,5 0,5 2,8 4,7
Очевидно, большая часть видов, тяготеющих к прибрежной
зоне, после сезона размножения остается в водах Австралии,
совершая кочевки вдоль побережий- При помощи кольцевания
установлено распространение вдоль побережий красноклювой
серебристой чайки (Carrick, et al., 1957а; Murray and Carrick.
1960; Wheeler and Watson, 1963), малого пингвина — Eudyptula
minor (Forst) (Phillips, 1961), хохлатой крачки (Carrick et
1 Оба вида наиболее многочисленны на зимовках у юго-восточного побе-
режья Австралии (восточнее острова Кенгуру). В Большом Австралийском
заливе они редки, у юго-западного побережья Австралии обычны. Более мно-
гочисленный короткохвостый поморник представлен в основном темной мор-
фой
182
ШЕЛЬФ ЮЖНОГО ПОБЕРЕЖЬЯ АВСТРАЛИИ
al., 1957; Hitchcock and Carrick, 1960; Hitchcock, 1961). Как вид-
но на примере Большого Австралийского залива (см. табл. 29),
различие в составе птиц в районе шельфа и . материкового
склона осенью сохраняется.
Факт посещения зимой вод Австралии птицами из мотальной
и антарктической зон достаточно хорошо известен (Serventv
and Whittell, 1951; Storr, 1964a; Cayley, 1965; Hill, 1968; Шунтов,
1968a, 19686; и др.). Табл. 29 показывает изменения их количе-
ства и соотношения в различные сезоны. Осенью в сравнении
с летом количество и численность южных видов значительно
увеличивается. Особенно заметно увеличение количества альба-
тросов, качурки Вильсона и южно-полярного поморника. Таким
образом, осенью орнитофауна южного побережья Австралии
имеет смешанный характер, но, как и летом, за исключением
района Тасмании, везде преобладают субтропические виды (см.
табл. 29).
Определившаяся осенью тенденция в изменении видового сос-
тава имеет развитие в течение всей зимы. Как видно из табл. 30,
во всех районах остается очень мало черных буревестников. У
юго-западного побережья и в Большом Австралийском заливе
.из них наиболее часто вствечается. по-видимому, длиннокрылый
тайфунннк. В отличие от бледноногого буревестника размноже-
ние этого вида приходится на зимний период (Warham, 1956).
А так как численность его ниже, то он не может компенсиро-
вать уменьшение в учетах доли буревестников с черной окраской,
вызванное откочевкой бледноногого буревестника.
За исключением прибрежных вод (в частности, прибрежной
зоны Большого Австралийского залива), зимний период харак-
теризуется преобладанием у южного побережья Австралии ви-
дов из нотальной и антарктической зон (рис. 50, табл. 30). Это
— альбатросы, прионы, капский голубь, гигантский буревестник
и другие. Численность одних из них (альбатросы) в течение
всей зимы изменяется мало, другие (капский голубь) становят-
ся более многочисленными во второй половине зимы, а качурка
Вильсона, откочевав на север Индийского океана, почти исче-
зает из района (см. табл. 30).
Зимой океанические виды чаще заходят в прибрежную зону.
На обширный шельф Большого Австралийского залива прони-
кает, в частности, странствующий альбатрос. С другой стороны,
прибрежные виды чаще встречаются за пределами мелководья.
Тем не менее различие в составе доминирующих видов на мел-
ководье и в глубоководных районах сохраняется. В прибреж-
ных водах Большого Австралийского залива, например, почти
половина особей в учетах приходится на долю многочисленной
в течение всего года хохлатой крачки, (см. табл. 30).
Рис. 50. Распространение зональных групп морских птиц в (районе Австралии. Слева — лето, справа — зима. 1 — тро
лические. 2 — субтропические, 3 — потальные и потальн о-антарктические
184
ШЕЛЬФ ЮЖНОГО ПОБЕРЕЖЬЯ АВСТРАЛИИ
Таблица 30
Видовой состав, соотношение (%) и плотность морских птиц
у южного побережья Австралии зимой (июнь—сентябрь). 1966—1970 гг.
Виды Юго-западное . побережье Австралии Большой Австра- лийский залив матери- шельф новый склон Район ме- жду о. Кен- гуру и Тас- манией
Черные буревестники 24,0 0,5 8,0 4- Мелкие альбатросы (D. сМо- rorhynchus, D. melanophrys, D. cauta) 16,0 19,0 36,0 20,0 D. exulans 16,0 3.0 17,0 8,0 Крачки (T. bergii) 1,5 47,0 2,0 3,0 Прионы (P. turtur) 5,0 — 15,0 35,0 D. ca pen sis 24,0 9,0 10,0 15,0 M. giganteus 4,0 0,5 3,5 2,0 L. novaehollandiae 3,0 3,0 0,5 9,0 M. senator 4- 14,0 + + P. marina 4- 0,5 3,0 + Крупные чайки (G. pacificus) 4- 4- 4- 2,0 C. skua 3,5 0,5 4* 4- P. palpebrata 4- 4- 4- 4- O. oceanic us 4- — 4- 4- Остальные виды 3,0 3,0 5,0 6,0 Количество учетов 65 146 149 95 Количество особей на 1 кв. км 1,0 0,2 1,5 2,5
В целом численность птиц зимой у южного побережья Авст-
ралии в несколько раз ниже, чем в другие сезоны. Это находит-
ся в соответствии с тем, что кормовые условия зимой значитель-
но хуже, чем в другое время. Сказанное относится в первую
очередь к концентрациям планктона, биомассы которого
уменьшаются в несколько раз. Зимой скопления ряда видов рыб
характеризуются наибольшей плотностью. Так. значительные по
плотности и площади скопления образует в Большом Австра-
лийском заливе сардина (Натаров и Пашкин, 1968; Макаров и
Пашкин, 1968; Змиевский, 1968). Однако косяки сардины боль-
шую часть времени проводят в придонных горизонтах, поэтому
не доступны или мало доступны для птиц-
Зимний период у южного побережья Австралии соответству-
ет периоду юго-западного муссона в северной части Индийско-
го океана. Муссон вызывает интенсивный подъем вод в районе
Аравийского полуострова, создающий условия для бурного раз-
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
185
вития планктона. На севере Индийского океана во время летне-
го муссона количество планктона приблизительно в два раза
выше, чем зимой (Богоров и Воронина. 1967). В этих продуктив-
ных районах проводят зиму многие мигрирующие с юга планк-
тоноядные птицы, в том числе бледноногий буревестник и бело-
лицая качурка (Bourne, 1960; Bailey, 1966, 1968; и др.).
Нотальные и антарктические виды доминируют в водах юж.
ного побережья Австралии до начала весны (до конца сентяб-
ря). В это время южные виды проникают наиболее далеко на
север (зональные виды вдоль западного побережья до северо-
западного угла Австралии).
В конце сентября в воды южного побережья начинают воз-
вращаться массовые мигранты из северного полушария: тонко-
клювый буревестник — в район юго-восточного побережья, блед-
ноногий — к юго-западному побережью и в Большой Австра-
лийский залив. В октябре происходит массовый пролет этих ви-
дов. В это время южные виды постепенно откочевывают в более
высокие широты.
8. Район Новой Зеландии и
сопредельные воды Антарктики
По обилию и видовому разнообразию морских птиц мало
районов могут сравниться с юго-западной частью Тихого океана
с сопредельными водами Антарктики. По этому району в по-
следние годы появилось значительное количество статей, отра-
жающих видовой состав и распределение птиц в отдельные се-
зоны и годы (Dell, 1960; Harrison, 1962; Norris, 1965; Gibson.
1967; Ozawa et al., 1968; Summerhayes, 1969; Darby, 1970; Шун-
тов, 1967, 1967a; и др.).
Миграции и распределение некоторых характерных и много-
численных видов (альбатросы, буревестники) в данном районе
подробно описаны выше. В настоящем разделе рассматривают-
ся общие аспекты сезонного распределения и изменения в сос-
таве и численности птиц в связи с изменчивостью в условиях
обитания.
Наибольшим обилием птиц рассматриваемый район харак-
теризуется в летний период — сезон размножения большинства
видов. Видовой состав птиц и плотность их населения в летние
месяцы на различных участках отражены в табл. 31—33. Даже
при беглом анализе таблиц видны существенные различия в на-
селении птиц на различных широтах, а также между шельфом
и открытыми водами. Изменение видового состава с изменением
широты связано с характером распределения водных масс теп-
лых и холодных течений. Новая Зеландия расположена на пути
двух мощных течений: теплого Тасманова (продолжение Вос-
18$
РАЙОН НОВОЙ ЗЕЛАНДИИ И СОПРЕДЕЛЬНЫЕ ВОДЫ АНТАРКТИКИ
точно-Австралийского) и холодного течения Западных ветров.
Граница между ними — субтропическая конвергенция — прохо-
дит по Чатамскому хребту, вдоль восточного побережья остро-
ва Южный, затем по южной части Тасманова моря. Таким об-
разом, остров Северный и северная часть острова Южный омы-
ваются субтропическими водами, плато Кэмпбелл — субантарк-
тическими, а значительная часть восточного побережья острова
Южный находится в зоне смешения водных масс (Burling, 1961;
Garner, 1961, 1962; Garner and Ridgway, 1965; и др.).
Из таблиц 31 и 32 видно, что в мелководных районах вокруг
острова Северный (к северу от 40° ю. ш.) преобладают субтро-
пические виды птиц. Наиболее многочислеными фоновыми вида-
ми являются буревестники Буллера и бледноногий, а также ав-
стралийская олуша. Весьма обычны белолицая качурка, длин-
нокрылый тайфунник, сумеречный и порхающий буревестники
и другие виды. Многие из перечисленных субтропических видов
характерны и для вод Австралии (в Тасмановом море), но
здесь над всеми видами резко доминирует тонкоклювый буре-
вестник, огромные колонии которого располагаются на островах
Бассова пролива и у Тасмании. На мелководье и в открытом
океане также встречаются весьма характерные для субтропиче-
ской зоны тайфунники Кука — Pterodroma cooki (Gray), бонин-
ский, Гоулда — Р. leucoptera. (Gould), Штейнегера — Р. longi-
rostris (Stejn.).
Таблица 31
Вцдовой состав, соотношение (%) и плотность морских птиц
в Тасмановом море летом (декабрь—март). 1964—1970 гг.
Вилы ( Шельф Австралии Шельф Н. Зеландии Открытые воды
35—40" 1 40—45° 35— 40" 40— 45э 30-40° 40- 45е
Прионы (Р. turtur) 5,0 9,0 6,5 66,0 6,5 26,0 Р. tennuirostris 77,0 86,0 — — 10,0 5,0 Р. carneipes 1,5 — 15,0 4,0 34,0 4,0 Р. bulleri 4- — 9,0 0,5 4- — М. giganteus 4- 4- 0,5 4- + 7,0 Dj capensis — — — — + 7,5 Ныряющие буревестники 4- 4-2,0 4- — — D. exulans, D. epomophora 4- 0,5 1,5 4- 33,0 32,5 D. melanophrys + 4- 4- 4- 2,5 5,5 D. cauta 2,0 0,5 — + — 3,0 L. domonicanus 4- 4- 20,0 4,5 — — L. novaehollandiae 5,0 4- 4- 4- — — S. parasiticus 4,0 + 4- 4- 4- + M. senator 1,0 + 42,0 0,5 1,0 — Остальные виды 4,5 4,0 3,5 5,5 13,0 9,5 Количество Учетов 21 25 31 44 36 33 Количество особей на 1 кв. км 4,5 25,0 5,5 22,0 1,0 1,0
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
1в7
Рассматривая, орнитофауну субтропической зоны, нельзя
не заметить, что в большинстве районов как на шельфе, так и в
открытых водах в значительных количествах встречаются и по-
тальные виды. На шельфе Новой Зеландии в Тасмановом море
наиболее характерны из них прионы (самый массовый — ог-
ненный). В южной части субтропической зоны (к югу от
40° ю. ш.) они преобладают над всеми видами. На шельфе вос-
точного побережья Новой Зеландии южнее пролива Кука, то
есть в зоне смешения вод, кроме прионов, многочисленны также
серый буревестник, капский голубь (нотальный подвид —
D. capensis australis) и альбатросы. Почти повсеместно много-
численна на шельфе Новой Зеландии доминиканская ча^са —
Larus dominicanus Licht. Заметное преобладание потальных ви-
дов в южной части субтропической зоны и в открытых водах
Тасманова моря» Здесь основу населения составляют, альбатро-
сы и прионы (см. табл. 31). В течение летнего периода соотно-
шение видов (в частности доля нотальных видов), не остается
неизмененным. В начале лета все эти виды имеют более север-
ное распространение и в субтропических водах встречаются ча-
ще. Во второй половине лета с прогревом вод многие особи от-
кочевывают в более южные широты.
Субтропический виды в отличие от нотальных имеют 6ojfte
ограниченное распространение и почти не заходят в смежную
зону. На рис. 51 показано распределение только двух субтропи-
ческих видов — буревестников Буллера и бледноНогого.Но ог-
раниченность в распространении в южном направлении зоной
субтропической конвергенции характерна и для других видов.
Нотальные виды активно проникают за пределы умеренной
зоны и в южном направлении. Из рис. 51 и табл. 33 видно, что
область их кочевок простирается далеко к югу от антарктиче-
ской конвергенции (в антарктическую эону). В открытых водах
Южного океана в зоне циркумполярного течения Зацддных вет-
ров, южнее Тасманова моря и Новой Зеландии преобладают та-
кие нотальные виды, как серый и тонкоклювый буревестники,
прионы, капский голубь (северный подвид), качурки <(Garrodia
nereis (Gould) и Fregetta tropica (Gonld), гигантский буревесг-
ник и альбатросы (наиболее массовые странствующий и черно-
бровый). Менее многочисленные, но весьма характерные для
умеренных вод буревестники белогорлый — Proceliana aeuqinoc-
tialis L., голубой — Halobaena caerulea (Gmel.), тайфунники бе-
логоловый — Pterodroma lessoni (Garn.) и Пила. Часть видов
многочисленна или обычна в пределах всей умеренной зоны,
другие тяготеют к определенному сектору Южного океана. Так,
среди черных буревестников южнее Тасмании преобладает тон-
коклювый буревестник, а у Новой Зеландии — серый. К Новозе-
ландскому району тяготеют также альбатросы королевский и
Рис. 51. Распространение морских птиц в районе Австралии, Новой Зеландии и сопредельных водах Ан-
тарктики летом. 1 — бледнонопий буревестник — Р. carneipes, 2 — серебристо-серый буревестник — г. gla
cialoides, 3 — основная область летних кочевок шоталь ных видов, 4 — снежный буревестник — Р. nivea.
5 Гпргвссгиик Буллера - Р. ntilleri, 6 -- антарктический буревестник — Т. antarctica, 7 — субтропическая
к.1 вер! цп. 8 - ан । арктическая конверт еиния
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ 188> Таблица 32 Видовой состав, соотношение (%) и плотность морских птиц у северного и восточного побережий Новой Зеландии летом (декабрь—март). 1965—1970 гг.
Виды о. Северный о. Южный
35—40е 40-45° 45—50
„ 1 9.0 60.0 niXTiP. turtur) 0Л 1.0 210 Ж’ ™ й = Р. carneipes 27.0 • 5,5 °? 15 05 М. giganteus + ,’5 Качурки *.0 т • D. exulans. D. epomophora 0.5 J.u J*» Альбатросы неопределенные + J’X £0 L. dominicanus 7.0 200 2Л М. senator у Ныряющие буревестники + + £ Остальные виды 2,0 /.• • Количество учетов *2 « Количество особен на 1 кв. км 12,0 10,0 Таблица 33 Видовой состав, соотношение (%) и плотность морских птиц в открытых водах у восточного побережья Новой Зеландии и сопредельных вод Антарктнкн летом (декабрь-март). 1967—1971 гг.
Виды Широта
30— 40° 40- 50- 50° 60° 60- 70- 70 80'-
Черные буревестники Hft _ (Р. griseus, Р. tenuirostris) 14.0 24Д 68.0 11,0 D. capensis 3,5 5,5 4,0 21.0 75 Прионы 20.0 17,0 9,5 26, Т. antarctica - ~ ~ 1. F. glacialoides - _ + g ((+ М. giganteus - 4.5 2,0 1,5 05 D. exulans, D. epomophora U5 11,0 3,0 05 D. melanophrys 20.0 3.5 4,0 1,0 - D. chrysostoma. D. bulleri 3,0 1,5 0,5 0, P. palpebrata - 0,5 1,0 25 15 1$ 19.0 9.0 Ы 10.0 качурки . go 16,0 c“sZaH“ - Й 2’5 H Остальные виды U.0 6.0 3.5 2,5 Количество учетов 40 200 393 747 Количество особей на 1 кв. км 0,5 2,о 4,5 4Д ,
190
РАЙОН НОВОЙ ЗЕЛАНДИИ И СОПРЕДЕЛЬНЫЕ ВОДЫ АНТАРКТИКИ
Буллера, капский голубь и другие размножающиеся здесь
виды.
Многие из массовых видов умеренной зоны многочисленны и
южнее 60° ю. ш. (табл. 33). Но для этих районов, особенно для
наиболее высоких широт, очень характерна целая группа антарк-
тических видов: антарктический — Thalassoica antarctiea
(Gmel.), серебристо-серый — Fulmarus glacialoides (Smith.)
и снежный — Pagodroma nivea (Gmel.) буревестники, южный
подвид капского голубя (D. с. capensis). Здесь заметно умень-
шается численность серого и тонкоклювого буревестников, а
также альбатросов, за исключением дымчатого. Также, как и
субтропические, антарктические виды ограничены в распростра-
нении в широтном направлении. Летом они не доходят даже до
антарктической конвергенции (см. рис. 51).
Положение границы распространения нотальных и антаркти-
ческих видов в водах Антарктики изменяется в течение летнего
периода и во многом определяется ходом прогрева вод и поло-
жением ледовой кромки. Естественно, что наиболее южным рас-
пространением характеризуются все виды во второй половине
лета — в феврале и марте, когда ледовая кромка приближается
к Антарктиде и среди льдов открывается проход в море Росса.
Течение Западных ветров, встретив на своем пути Новозеланд-
ское плато, отклоняется на юг, что вызывает прогиб в этом на-
правлении антарктической конвергенции. В этом районе от те-
чения Западных ветров происходит ответвление в море Росса,
вызывающее прогиб изотерм и кромки льдов (Нейман,. 1961:
Трешников, 1964; Григорьев, 1969; и др.). Здесь у входа в море
Росса, нотальные виды наиболее глубоко проникают на юг, а
.северная граница антарктических видов опускается к югу.
Распространение льдов и положение ледовой кромки имеют
большое значение в распределении многих видов птиц (Routh,
1949; Cline et al., 1969; Darby, 1970; и др.). За пределы льдов нс
выходит снежный буревестник. В отличие от антарктической
крачки — Sterna vittata Gmel., встречающейся к северу от ледо-
вой кромки, арктическая крачка Sterna paradisea Brunn, как и
.снежный буревестник, встречается только среди ледовых полей
(Salomonsen, 1967; и др.). К северу от ледовой кромки не встре-
чаются летом антарктические виды пингвинов и антарктиче
<ский подвид поморника С. skua maccormicki1. Серебристо-ct .
рый и антарктический буревестники, южный подвид капского
голубя, качурка Вильсона чаще всего встречаются среди льдов,
1 Некоторое количество особей антарктического подвида поморника зна-
чительную часть лета проводит в районах колоний других птиц, чаше пиш-
:вмнов, питаясь их яйцами и птенцами. Другие добывают корм (криль, рыба)
в разводьях и у кромки льдов (Eklund, 1961, 1964; Young, 1963, 1963а).
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
191
но они многочисленны в пределах нескольких десятков миль и к
северу от ледовой кромки. Широко распространен в открытых во-
дах Южного океана дымчатый альбатрос, он многочислен и сре-
ди льдов. Местами у ледовой кромки плотность его даже увеличи-
вается. Большая часть умеренных видов, в частности, альбатро-
сы странствующий, чернобровый, сероголовый, буревестники
серый, тонкоклювый, голубой, тайфунники Пила, белоголовый,
прионы и другие виды более многочисленны в. открытых водах.
В районе ледовой кромки их численность уменьшается, и в раз-
водья большая их часть далеко не заходит.
Шельфовые участки в юго-западной части Тихого океана и
прилегающих к ней водах Антарктики развиты слабо. Это нахо-
дит отражение в распределении океанических и прибрежных
видов морских птиц. Так как прибрежные воды окаймляют ост-
рова узкой полосой, а океанические водные массы подходят к
берегам, большая часть океанических видов обычна или много-
численна на значительной части шельфов1. Резко уменьшаете'!
численность ряда океанических видов (например, странствую-
щего и чернобрового альбатросов) только на таких обширных
шельфах, как Бассов пролив и район залива Тасман у Новой
Зеландии, а также непосредственно в прибрежной зоне. Ограни-
ченное распространение прибрежных видов соответствует сла-
бо развитому шельфу. Не выходят или почти не выходят за пре-
делы мелководья ныряющие буревестники, красноклювая сере-
бристая и доминиканская чайки, все бакланы и пингвины.'
Южный океан называют царством трубконосых и пингвинов.
В районе Новой Зеландии и на островах, расположенных к югу
от нее, располагается большое количество колоний различных
видов пингвинов. Однако за пределами 15-мильной зоны на не-
сколько тысяч учетов пришлось всего несколько встреч пингви-
нов. По-видимому, большая их часть в течение всего года кон-
центрируется близ районов размножения. Более подвижны ан-
тарктические виды пингвинов, которые могут встречаться на
значительном удалении от мест расположения колоний, но в по-
давляющем большинстве случаев они концентрируются в разво-
дьях и у кромки льдов. Это заметно по увеличению их доли юж-
нее 60° ю. ш. (см. табл. 33).
Летние концентрации птиц в рассматриваемом районе отли-
чаются большой плотностью и наблюдаются на различных ши-
ротах. Размещение их в общем соответствует распределению
продуктивных зон. Упоминания заслуживает в первую очередь
Новозеландское плато. В основе повышенной биологической
продуктивности этого района лежит высокая динамика вод, ко-
1 Здесь имеются в виду участки мелководья за пределами 15-мильаой
прибрежной эоны.
192
РАЙОН НОВОЙ ЗЕЛАНДИИ И СОПРЕДЕЛЬНЫЕ ВОДЫ АНТАРКТИКИ
торая определяется сложным рельефом дна и расположением
его на пути сильных течений. В районе свала глубин интенсив-
ное вертикальное перемешивание прослеживается, например,
на глубину в несколько сот метров. Существенным является и
то, что значительная часть плато расположена в зоне субтропи-
ческой конвергенции, где, как известно, процессы вертикального
перемешивания выражены особенно сильно.
В летние месяцы биомассы планктона здесь значительно
увеличиваются и на обширных акваториях составляют по не-
скольку сот миллиграммов на кубометр воды. Особенно богаты
некоторые мелководные районы, кормность которых увеличи-
вают скопления рыб, наблюдающиеся на многих участках шель-
фа (Graham, 1953; Шунтов, 19706). Большие устойчивые скоп-
ления птиц наблюдаются на мелководье между островами Се-
верный и Южный (к западу от пролива Кука). В течение всего
лета здесь держатся большие стаи прионов, гнездящихся на ост-
ровах у северной оконечности острова Южный. Этот район ин-
тересен тем, что здесь на шельф поступают воды с трех направ-
лений: с запада и севера из Тасманова моря, а с востока — че-
рез пролив Кука. Высокая динамика вод создает условия для
формирования кормовых полей, на которых откармливаются не
только планктоноядные птицы, но и многочисленные косяки пе-
лагических рыб, в частности ставрида (Шунтов, 19706). Вдоль
северного побережья острова Северный все лето наблюдаются
стаи размножающихся здесь буревестников Буллера, бледно-
ногого и австралийской олуши. Много птиц держится вдоль во-
сточного побережья острова Южный, который, как отмечалось,
находится в зоне субтропической конвергенции. Здесь, особенно
к югу от залива Кантербери, концентрируется большое количе-
ство стай серого буревестника. Многочисленны и другие птицы,
в частности прионы и порхающий буревестник.
Сопоставим по плотности птиц с новозеландским мелководь-
ем шельф Тасмании и Бассова пролива, где скопления образует
тонкоклювый буревестник. Но в открытых водах Тасманова мо-
ря, характеризующихся более низкой продуктивностью, плот-
ность птиц в 5—25 раз ниже, чем на шельфе (см. табл.31).
В открытых водах к югу Новой Зеландии и Тасманова моря
(южнее 40е ю. ш.) плотность птиц в среднем в 3—4 раза выше,
чем в Тасмановом море. Особого упоминания заслуживает
участок, включающий Чатамский хребет и плато Кэмпбелл.
Здесь существенны два обстоятельства: во-первых, наличие мно-
гих островов, на которых располагаются большие гнездовья ко-
лоний трубконосых и пингвинов, во-вторых, высокая кормность
ряда районов. Плотность жизни здесь местами соответствует
тому, что наблюдается на шельфе. Это подтверждается высоки-
ми биомассами планктона и значительными концентрациями рыб
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
1вв
(Шунтов, 19716). Скопления птиц в водах, омывающих Ново-
зеландское плато, обязаны не только значительным концентра-
циям планктона и рыб; судя по траловым уловам, здесь много-
численны кальмары, которые играют большую роль в питании
альбатросов.
Южнее плато Кэмбелл в открытых водах Южного океана,
птиц также много (рис. 52), а в некоторых районах плотность со-
поставима с шельфовыми районами. Высокая продуктивность и
кормность вод Южного океана общеизвестна. Гидробиологи, за-
нимающиеся изучением планктона (Виноградов и Наумов, 1961;
Воронина и Наумов, 1968; Воронина, 1966; Долженков, 1970;
и др.), выделяют две основные зоны высоких биомасс планкто-
на, как бы опоясывающих Антарктиду. Одна связана с антарк-
тической конвергенцией и располагается несколько южнее ее,
другая — с антарктической дивергенцией. Обеим зонам соответ-
ствует увеличение плотности птиц (см. рис. 52).
Наибольшим богатством кормовых ресурсов характеризует-
ся антарктическая дивергенция, на которой наблюдается интен-
сивный выход глубинных вод, создающий условия для развития
фитопланктона. Подъем вод усиливается в местах малоподвиж-
ных атмосферных циклонов (Иванов и Тареев, 1959). Весеннему
состоянию в планктоне здесь способствует и то, что дивергенция
расположена летом у края льдов или на небольшом удалении or
них. Обильное развитие фитопланктона в районе антарктиче-
ской дивергенции создает условия для формирования скоплений
криля — Euphausia superba — основного корма всех антаркти-
ческих животных. На полях криля откармливаются киты, тю-
лени и морские птицы. Наиболее мощные скопления криля при-
урочены к участкам зоны дивергенции, где отмечаются кругово-
роты вод, связывающие течения Западных и Восточных ветров.
Особое значение имеют круговороты вод, соответствующие цен-
трам циклонической деятельности в атмосфере, а также подъе-
мы вод вблизи островов, расположенных на пути течений (Бе-
клемишев, 1961; Шевцов и Макаров, 1969; Ярогов, 1969).
До последнего времени значительные скопления криля бы-
ли известны только для атлантического и индийского секторов
Южного океана, при этом предполагалось, что в тихоокеанском
секторе нет условий для развития высокой численности этого
рачка (Магг, 1962; Пономарева. 1966). Однако исследованиями
ТИНРО в последние годы было показано, что ряд районов ти-
хоокеанского сектора Антарктики располагает значительными
ресурсами криля. Большие поверхностные скопления его отме-
чаются к северо-западу и северо-востоку от моря Росса. На
этих участках на дивергенции постоянно имеют место кругово-
роты вод (Трешников, 1964; Григорьев, 1969; и др.), соответст-
вующие зоне усиленной циклонической деятельности (Ярогов,
Рис. 52. Распределение концентраций морских птиц летом в районе Австралии, Новой Зеландии и сопре-
дельных водах Антарктики. 1 — низкие, 2 — средние, 3—(высокие, 4 —течения, 5 — субтропическая конвер-
генция, 6 — антарктическая конвергенция
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
195
1969)- Анализ результатов трехлетних наблюдений показывает,
что наиболее плотные скопления птиц в зоне антарктической ди-
вергенции отмечаются именно к северо-западу и северо-востоку
от моря Росса. К северо-востоку от моря Росса основу скопле-
ний составляют прионы. В районе островов Балени скопления
образуют капский голубь, антарктический, серебристо-серый и
снежный буревестники; весьма обычны здесь прионы и альба-
тросы. К западу от островов Балени вдоль зоны дивергенции на-
чинают преобладать серый и тонкоклювый буревестники. Та-
ким образом, обильные кормовые ресурсы высоких широт Ан-
тарктики привлекают сюда не только стаи антарктических, но и.
нотальных видов. Из табл. 33 видно, что на долю нотальных ви-
дов в зоне антарктической дивергенции (приблизительно между
60 и 70° ю. ш ) приходится почти половина особей. В водах Ан-
тарктики проводит северную зиму антарктическая крачка. Про-
никают сюда и отдельные особи других северных мигрантов —
короткохвостый и средний поморники (Sladen, 1952, 1954). Но в
отличие от крачки большая часть их особей остается в пределах
субтропической и северной части нотальной зоны, в частности
у южного побережья Австралии (где они наиболее многочис-
ленны) и у Новой Зеландии. Выше отмечалось, что в районах
гнездования птиц в умеренной зоне (Тасмания, острова на Но-
возеландском плато) в течение всего лета держаться большие
количества буревестников, прионов и альбатросов. Многие из
них половозрелы и размножаются в это время. Что касается кон-
центраций серого и тонкоклювого буревестников, прионов, аль-
батросов и других видов умеренной зоны в Антарктике, то они,
несомненно, состоят в основном из неполовозрелых и нераз-
множающнхся в данном году особей.
Распространение нагульных концентраций на юг продолжа-
ется до конца марта, после чего начинается обратный процесс.
Некоторые виды птиц, размножающихся в субтропической зоне,
заканчивают гнездовой период во второй половине лета. Так,
молодые особи австралийской олуши из популяций, гнездящих-
ся у северного побережья Новой Зеландии, в конце января и
феврале начинают появляться в водах Австралии; разлет же из
более южных колоний происходит в марте—мае (Wodzicki
and Stein, 1958; Wodzicki, 1967). Завершение размножения и на-
чало послегнездовых кочевок у большинства видов нотальной и
антарктической зон приходится на апрель. В это время проис.
ходит весьма существенное перераспределение птиц и измене-
ния их численности в различных районах.
В апреле и начале мая отлетает в северное полушарие ос-
новная масса трансэкваториальных мигрантов — буревестни-
ков серого, тонкоклювого, бледноногого, Буллера, тайфунника
Пила- Это уменьшает общую плотность и численность птиц в водах
196
РЛПОИ НОВОЙ ЗГ.ЛЛНДИИ И СОПРЕДЕЛЬНЫЕ ВОДЫ АНТАРКТИКИ
Южного океана к югу от Новой Зеландии, на шельфе острова
Южный, Тасмании и в Бассовом проливе. Наоборот, пролетаю-
щие на север стаи, особенно серого и тонкоклювого буревестни-
ков, создают временное увеличение плотности птиц в открытых
водах Тасманова моря (табл. 34). На шельфе Новой Зеландии
в Тасмановом море численность птиц осенью остается высокой.
Как и летом, между островами Северный и Южный кон-
центрируется огромное количество прионов, которые доминиру-
ют над всеми видами. Некоторое увеличение здесь плотности
птиц обусловлено и распространяющимся на север капским
голубем, альбатросами, гигантским буревестником и другими
видами. Постепенное увеличение численности этих видов наблю-
дается в течение всей осени. Накопление птиц на шельфе запад-
ного побережья Новой Зеландии происходит в результате их
миграций через пролкв Кука (капский голубь, гигантский буре-
вестник, альбатросы пугливый и Буллера). И наоборот, здесь
уменьшается количество австралийской олуши (в связи с миг-
рациями к Австралии) и буревестников Буллера и бледноногого
(в связи с миграциями в северное полушарие).
Таблица 34
Видовой состав, соотношение (%) и плотность морских птиц
в Тасмановом море осенью (апрель—май). 1996—1970 гг.
В н л ы Шельф Открытые воды
35- 40J 40-455 30 -40 40 45е
Прионы (Р. turtur) 52,0 75,0 -L 4,0
Черные буревестники (Р. griseus, Р. tenuirostris) + + 60,0 84,0
D. capensis 4- 3,5 — +
D. cauta 4,0 4,5 + +
D. exulans, D. epomophora 0,5 0,5 8,0 3,0
D. melanophris 8,0 2,0 8,0 2,5
D. bulteri, D. ch г у sos tom a + 0,5 1,0 4- -
Альбатросы неопределенные 2,5 2,0 1.0 -г
P. carneipes 4,0 0,5 + 4-
P. bulleri 7,0 0,5 10,5 —
Качурки 4- 1,5 10,5 3,5
M. giganteus — 1,0 — 4-
L. doniinicanus 1,0 3,0 " —
M. serrator 18,0 0.5 + 4-
Остальные виды 3,0 4,5 1,0 3,0
Количество учетов 11 91 29 40
Количество особей на 1 кв. км 5.0 33.0 2,0 5.7
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
1вГ
На шельфе у восточного побережья Новой Зеландии увели-
чивается численность целого ряда видов — капского голубя,
всех альбатросов, гигантского буревестника. Чаще за предела-
ми прибрежных вод встречаются красноклювая* и доминикан-
ская чайки. Но в связи с отлетом самого массового вида — се-
рого буревестника общая плотность птиц, за исключением зали-
ва Кантербери, уменьшается (табл. 35).
Таблица 35
Видовой состав, соотношение (%) и плотность морских птиц
у восточного побережья Новой Зеландии осенью (апрель—май).
1966—1970 гг.
Виды Ill е л ь ф Открытые воды
35-40° | 40-4?° 1 45—50° 40-50° а? ° 1
Прионы (Р. turtur) 31,0 2,5 11,0 7,0
D. capensis 20,0 8,0 29,0 14,0 22,0
D. bulleri, D. chrysostoma 5,0 4,5 1.5 5,5
D. exulans, D. epomophora 4.0 2,5 7,5 7.5 5,5
D. melanophrys 12,0 U 3,0 2,0 +
D. cauta + 1.0 + + +
Альбатросы неопределенные — 6,0 12,0 6,5 15,0
M. giganteus 5,0 0.5 4.5 3,0 1.0
Черные буревестники (Р. griseus) 5.0 6,5 25,0 19,0 13,0
Р. bull er 1.0 3,0 — 1.5 —.
Качурки + 0,5 2,0 25,0 23,0
Р. palpebrata -U + 2.5 1.5 1,5
С. skua — + + 1,5 1,0
L. novaehollandiae + 30,0 2,5 — —
L. dominicanus 35,0 2.0 + — —
M. serra tor 15,0 •+• — —
Остальные виды 3,0 2,5 5,0 6,0 5,5
Количество учетов 7 77 34 78 53
Количество особей на 1 кв. км 1.0 11,5 2,5 1.0 1.6
В открытых водах к югу от Новой Зеландии в связи с завер-
шением сезона размножения на островах Окленд, Кэмпбелл,
Чатам, Макуори, Баунти, Антиподов и разлетом буревестников
и альбатросов численность птиц в целом уменьшается. Чаще,
чем летом, здесь встречается, по-видимому, только небольшая
часть видов, в частности, качурка Вильсона, дымчатый альба-
1 Значительная часть гнездовых колоний красноклювон чайки распо-
лагаются v восточного побережья Новой Зеландии, где во многих районах
благодаря сложному подводному рельефу создаются условия для формиро-
вания скоплений планктона, в частности Nictiphanes australis (Gurr and Kins-
ky, 1965; Mills, 1969). Осенью в заливе Кантербери у восточного побережья
острова Южный N. australis образует поверхностные концентрации, на кото-
рых красноклювая чайка откармливается вместе с трубконшгыми.
198
РАЙОН НОВОЙ ЗЕЛАНДИИ И СОПРЕДЕЛЬНЫЕ ВОДЫ АНТАРКТИКИ
трос, белоголовый тайфунннк. Я не располагаю осенними дан-
ными по составу, численности и распределению птиц в водах
Антарктики. Однако можно с уверенностью говорить о том, что
в это время в связи с откочевкой на север многих нотальных ви-
дов и такой массовой антарктической птицы, как качурка Виль-
сона, общая численность птиц значительно уменьшается. Кро-
ме того, с этого времени до следующего лета южнее антаркти-
ческой конвергенции должны абсолютно доминировать антарк-
тические виды птиц.
Описанные изменения в сезонном распределении птиц нахо-
дятся, как и в других районах, в соответствии и в связи с изме-
нениями, происходящими в гидрологическом режиме и кормо-
вой базе. Летнее смещение всех видов в южном направлении
хорошо согласуется с постепенным прогревом вод и воздуха, а
также со смещением на юг зон повышенной продукции планк-
тона.
Летом в районе припая планктон находится в зимнем состо-
янии, севернее, в зоне плавучих льдов — в весеннем (бурное
развитие фитопланктона), в умеренных водах — в летнем. По
мере прогрева вод и смещения ледовой кромки на юг смещает-
ся в этом направлении и зона массового цветения. Таким обра-
зом, положение летнего максимума планктона постепенно пере-
двигается в южном направлении. В Южном океане в летнее
время планктон концентрируется в верхних слоях (Виноградов,,
и Наумов, 1961; Воронина, 1966; Воронина и Наумов, 1968; и
другие). Летний максимум планктона в поверхностных слоях
наблюдается и в районе Новозеландского плато.
Количество планктона в верхних слоях Южного океана на-
чинает уменьшаться в апреле и происходит это в значительной
мере за счет его опускания в более глубокие горизонты (Виног-
радов и Наумов, 1961). В апреле и мае очень высокие концент-
рации планктона в поверхностных слоях наблюдаются только
в отдельных районах. К таким районам относятся, в частности,
два участка мелководья Новой Зеландии: залив Кантербери и
район залива Тасман, где в верхних слоях моря в течение всей
осени концентрируются стаи эвфаузиид (по-аидимому, N. austra-
lis). На скоплениях планктона в районе залива Тасман откарм-
ливаются в это время многочисленные косяки ставриды и скоп-
ления прионов, а в заливе Кантербери — косяки снека, шпрота,
скопления прионов, красноклювой чайки, альбатросов (Шунтов,
1967а, 19706). Однако и на этих широтах в большинстве райо-
нов концентрации планктона в поверхностных слоях осенью
уменьшаются, поэтому птицы держатся рассредоточение. Ни в
одном из богатых кормом районов не задерживаются буревест-
ники, которые улетают на север- Выше отмечалось, что их сроки
СЕЗОННЫЕ АСПЕКТЫ В ОРНИТОФАУНЕ
1Ю
отлета хорошо согласуются с сезонными изменениями в биоло-
гии моря в северном полушарии.
В течение зимнего периода по мере дальнейшего охлаждения
продолжается распределение птиц в северном направлении.
Почти полностью исчезают из юго-западной части Тихого океа-
на совершающие перелеты за экватор буревестники серый, тон-
коклювый, бледноногий, Буллера. Многие виды широко рассеи-
ваются в открытых водах южной части Тихого океана.
На шельфе Новой Зеландии очень сильно уменьшается чис-
ленность прионов. В то же время в открытом океане они стано-
вятся весьма многочисленными. Область кочевок нх зимой рас-
ширяется’до южной части тропической зоны. По данным Фле-
минга (Fleming, 1950) и Харрисона (Harrison, 1962), прионы
зимой многочисленны к востоку от Новой Зеландии на протяже-
нии нескольких суток пути.
Уменьшается к зиме в районе Новой Зеландии численность
австралийской олуши. Молодые и часть взрослых особей улета-
ют к берегам Австралии. Из молодых лишь отдельные особи
остаются, зимовать в районах размножения (Stein, 1961, 1962).
Миграций к Австралии совершают и другие виды, гнездящиеся
у Новой Зеландии, в частности белолобая крачка — Sterna
striata Gmel- (Clark and Dawson. 1957).
В течение всей зимы на мелководье Новой Зеландии в Тас-
мановом море многочисленны альбатросы, капский голубь, ги-
гантский буревестник, доминиканская чайка, ныряющие буреве-
стники. На шельфе Австралии кроме перечисленных видов мно-
гочисленны красноклювая чайка и длиннокрылый тайфуниик.
Многие из указанных выше видов обычны или многочисленны и
в открытых водах Тасманова моря. Здесь нет только чаек, олу-
ши и наряющих буревестников, редки многочисленные на шель-
фе альбатросы — пугливый и Буллера (табл. 36). Значительная
часть многочисленных в открытых водах Тасманова моря видов,
как и прионы, проникает зимой далеко на север, достигая юж-
ной части тропической зоны.
Преобладание нотальных и нотально-антарктических видов
характерно и для шельфа восточного побережья Новой Зелан-
дии. Капский голубь, альбатросы, гигантский буревестник и чай-
ки составляют в большинстве участков мелководья основу на-
селения птиц. Эти же виды, за исключением чаек, преобладают н
открытых водах к югу и востоку от Новой Зеландии (табл. 37).
Южнее плато Кэмпбелл наблюдения зимой почти не проводи-
лись. Мне известна только одна работа, в которой описывается
состав и распределение птиц в это время в Южном океане К
югу от Новозеландского плато (Szijj, 1967). По наблюдениям
этого автора, в умеренных водах к северу от антарктической
конвергенции преобладают капский голубь, странствующий, се-
200
РАЙОН НОВОЙ ЗЕЛАНДИИ И СОПРЕДЕЛЬНЫЕ ВОДЫ АНТАРКТИКИ
роголовый, дымчатый альбатросы, гигантский буревестник, бе-
логоловый тайфунник, буревестники голубой и бурый — Adama-
stor dnereus (Gmel.), белолицая качурка. Южнее антарктиче-
ской конвергенции из перечисленных видов обычны или много-
числены только капский голубь, гигантский и голубой буревест-
ники. Кроме них для вод Антарктики характерны антарктиче-
ский, серебристо-серый, снежный буревестники и пингвины
(только у льдов). В отличие от летного периода распростране-
ние антарктических видов характеризуется более северным по-
ложением. К северу от антарктической конвергенции проника-
ют антарктический и серебристо-серый буревестники. Залеты
последнего отмечаются в это время даже к берегам Новой Зе-
ландии (Oliver. 1955).
Таблица 36
Видовой состав, соотношение (%) н плотность морских птиц
в Тасмановом море зимой (июнь—сентябрь). 1966—1870 гг.
Виды Шельф Ав- стралии Шельф Н. Зе- ландии Открытые воды
30*40° 35-40° 40—45* 30—40е 40—50°
Прионы (Р. turtur) 1,0 29,0 30,0 34,5 25,0 D. capensis 2,0 19.0 19,0 11.0 23,0 D. melanophrys 10,0 7,0 1,0 14,0 13,0 D. exulans, D. epomophora 10,0 4,0 1,0 13,0 12,0 D. cauta 6,0 4.0 4- + D. bulleri, D. chrysostoma 4- 2,0 1,0 0,5 2,0 At giganteus 1,0 5,0 6,0 1,0 2,0 Черные буревестники (P. macroptera) 24,0 3,0 1,5 19,0 — Качурки 1.0 + 4- 2,0 11,0 L. novaehollandiae 31,0 0,5 4- — — L. dominicanus 4- 19,0 0,5 — — M. serrator 4- 1,0 0,5 — — Ныряющие буревестники 4- + 30,0 — — Остальные виды 20,0 4,5 5,5 5,0 12,0 Количество учетов 4 38 78 68 47 Количество особей на 1 кв. км 4,5 3,0 5,0 1,0 1,2
Таким образом, большинство видов птиц в зимний период
характеризуется большой подвижностью. Преимущественно это
океанические трубконосые, собирающие пищу с поверхности
или в верхних слоях моря. Зимой биомассы планктона в верх-
ний х слоях уменьшаются в несколько раз, и даже в продуктивном
Новозеландском районе биомассы обычно не превышают 100
миллиграммов на кубометр воды. В то же время в придонных
сезонные аспекты в орнитофауне
201
горизонтах на шельфе акустические приборы фиксируют много-
численные стаи эуфаузнид. Больше, чем летом, тяготеют здесь
к глубинным слоям и пелагические рыбы (Шунтов» 19706).
В водах восточного побережья Австралии зимой стаи самой
массовой пелагической рыбы сардины, наоборот» наиболее часто
появляются у поверхности (Blackburn, 1950, 1960). По-видимо-
му, это обстоятельство играло большое значение в формирова-
нии перелетов австралийской олуши от Новой Зеландии к Авст-
ралии. Питается этот вид исключительно пелагическими рыба-
ми (Wodzicki and Moreland, 1966; и др).
Таблица 37
Видовой состав, соотношение (%) и плотность морских птиц
у восточного побережья Новой Зеландии зимой (июнь—сентябрь).
1966-1970 гг.
В и д ы Шельф Открытые воды
35—40° 40-45° 45—50° 40—50° 50-55°
D. capensis 35,0 29,0 54,0 45,0 54,0
D. bulleri, D. chrysostoma 1,0 18,0 8,0 7,5 14,0
D. cauta + 5,5 + 0,5 —
D. melanophrys 3,0 7,0 3,0 7,0 2,0
D. exulans, D. epomophora 7,0 5,0 4,0 7,5 4,0
Альбатросы неопределенные + 6,0 6,0 4,0 3,0
Прионы (P. turtur) 15,0 3,5 + 11,0 4“
M. giganteus 6,0 3,5 6.5 8,5 9.0
P. griseus 1.5 4- 2,0 —
P. macroptera 20,0 0,5 — 0,5 —
M. serrator 2,0 + + —. —
L. novaehollandiae + 11,0 + — —
L. dominicanus 3,0 6,5 8,5 — —
Ныряющие буревестннкп 2,0 + 3,0 — —
Остальные виды 6,0 3,0 7,0 6,5 14,0
Количество учетов 7 141 39 88 50
Количество особей на 1 кв. км 1,1 7,1 1,6 0,8 0,6
Как и в другие сезоны, менее подвижны прибрежные виды,
добывающие пищу при нырянии, и чайки, питающиеся в при-
брежной полосе, а также у судов* Несомненно, ограничены пе-
редвижения пингвинов. За пределами 15-мильной прибрежной
зоны, где проводились наблюдения, они встречаются, как и ле-
том, очень редко. Мало выходят зимой за пределы шельфа и ны-
ряющие буревестники. Много их держится в заливах, в частно-
сти в заливе Тасман, где меньше влияние сильного зимнего вол-
нения. Преимущественно вдоль берегов на ограниченные рас-
стояния кочуют чайки. Доминиканская чайка, например» в райо-
2оа
РАЙОН НОВОЙ ЗЕЛАНДИИ И СОПРЕДЕЛЬНЫЕ ВОДЫ АНТАРКТИКИ
не Велингтона начинает концентрироваться у мест размножения
с середины зимы. В сезон размножения у колоний держатся и
половозрелые особи. Разлет после размножения обычно не пре-
вышает нескольких десятков миль (Fordham, 1964, 1964а, 1968).
В сравнении с летним периодом численность птиц на боль-
шинстве участков шельфа Новой Зеландии и Тасмании умень-
шается в 5—20 раз. В 3—7 раз уменьшается численность птиц
и в открытых водах океана к югу от Новой Зеландия. И только
в Тасмановом море и в водах океана к востоку от него плотность
птиц остается такой же (см. табл. 36). Убыль, вызванная отко-
чевкой части видов на север, компенсируется нотальными вида-
ми из более южных широт и с шельфа Новой Зеландии. В от-
личие от летного периода зимой менее резко выражена разница
в плотности населения птиц на шельфе и в открытом море.
Весной наблюдается картина, обратная описанной для осен-
него периода. Поэтому нет необходимости подробно останавли-
ваться здесь на этом сезоне. Следует отметить только, что за-
метные изменения в составе птиц начинаются со второй поло-
вины сентября. В это время начинают прибывать мигранты с
севера. Весьма характерно то, что на фоне начавшегося общего
движения на юг многие нотальные виды в начале весны имеют
наиболее северное распространение. По-видимому, это связано
с тем, что конец зимы и начало весны — время максимального
охлаждения поверхностных вод.
зов
ГЛАВА III
НЕКОТОРЫЕ ОБЩИЕ ЗАКОНОМЕРНОСТИ
В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ
В предыдущей главе были рассмотрены особенности распре-
деления птиц в различных районах океана, расположенных в
бореальной (дальневосточные моря и сопредельные воды океа-
на), субтропических (Восточно-Китайское море, мелководье у
южного побережья Австралии, район острова Северный у Новой
Зеландии), тропической (воды между Индонезией и северным
побережьем Австралии), южной умеренной и антарктической
(Новая Зеландия и сопредельные воды) зонах. Все районы,
естественно, имеют свои особенности в видовом составе, разме-
щении птиц, численности и ее изменчивости- Но все зоны объ-
единяет то, что в каждой из них наиболее существенными из-
менениями в орнитофуане являются сезонные.
В массе работ, посвященных миграциям птиц, показано, что
в основе сезонных перемещений лежат сезонные колебания ус-
ловий обитания. Миграции позволяют птицам использовать
районы, благоприятные для существования только в определен-
ные периоды года, и, таким образом, осваивать большие терри-
тории. Не должны представлять исключения здесь и морские
птицы, имеющие в этом отношении принципиальное сходство со
многими морскими рыбами и морскими млекопитающими. Од-
нако во многих работах по морским птицам причинная сторова
миграций трактуется несколько упрощенно. Так, основной при-
чиной откочевки на юг ряде видов из северных широт указы-
вают образование ледового покрова, южные же популяции этих
птиц, живущих в незамерзающих районах, относят к оседлым.
Как было показано выше на примере дальневосточных морей и
других районов, картина миграций имеет более сложный харак-
тер и определяется комплексом факторов, при этом миграции
не являются свойством только северных птиц-
По дальности и характеру сезонных перемещений птицы
обычно подразделяются на оседлых, кочующих н перелетных
(Дементьев, 1940; и др.). Это подразделение в известной мере
условно, так как масса видов совмещает в себе признаки осед-
лых и кочующих, кочующих и перелетных птиц. Поэтому А. В.
Михеев (1964) выделяет оседлых, полуоседлых. кочующих, пе-
релетно-кочующих. слабо-перелетных и настоящих перелетных.
204 ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ
Оседлые птицы не совершают регулярных сезонных перемеще-
ний. Считается, что перелетные птицы имеют определенные и на
следственно фиксированные области зимовок, кочующие же
птицы фиксированных мест зимовок не имеют, весь внегнездо-
вой период они находятся в непрерывном движении.
Мне кажется, что мнение об отсутствии у кочевых птиц на-
следственно закрепленных мест зимовок и кочевок нуждается в
уточнении. Ниже, при обсуждении путей миграций этот вопрос
еще будет затрагиваться. Впрочем, А. В. Михеев (1964) подчер-
кивает ненужность противопоставления кочевок и перелетов,
так как те и другие имеют единую биологическую основу и еди-
ное происхождение. Справедливость этого замечания подтверж-
дается особенностями миграций морских птиц. Вот несколько
примеров. Дальневосточные бакланы (берингов, краснолиций.
японский) относятся к кочующим видам, но нередко осенью и
весной можно наблюдать весьма интенсивный их пролет стая-
ми вдоль побережий- В таких случаях они напоминают нырко-
вых уток, в массе совершающих перелеты у побережий. Часть
этих бакланов зимует в пределах широт гнездовой части ареа-
ла, но значительное количество отлетает южнее в воды полу-
острова Корея. Китая и Японии. То же самое можно оказать о
некоторых чистиковых, в частности, конюгах. Довольно боль-
шая группа буревестников южного полушария совершает на-
стоящие перелеты в северные части Тихого и Атлантического
океанов. Но, зимуя здесь, они находятся в непрерывном движе-
нии, перекочевывая из одного района в другой.
По многим особенностям сезонных перемещений все альбат-
росы должны быть отнесены к кочующим птицам. Но в характе-
ре движения и размещения мест кочевок у некоторых из них
наблюдаются элементы, характерные для перелетных птиц. Все
они имеют область кочевок (и зимовок), выходящую за преде-
лы гнездовой части ареала. У ряда видов и популяций выявле-
ны вполне фиксированные места зимовок и кочевок. У королев-
ского альбатроса, гнездящегося на островах Новозеландского
плато, это — воды, омывающие Южную Америку, у желтоно-
сого — район Австралии, у темноспинного альбатроса с Гавай-
ских островов — северо-западная часть Тихого океана, у чер-
ноногого — северо-восточная. Чернобровый альбатрос широ-
ко разлетается в период кочевок по Южному океану, но вместе
с этим установлено вполне определенное тяготение особей с
Фольклевдских островов к водам восточного побережья Южной
Америки, а с Южной Георгии — к водам южного побережья
Африки. Часть особей австралийской олуши,..размножающейся
у Новой Зеландии, зимой кочует в пределах гнездовой части
ареала; большая же часть молодых совершает перелеты в воды
Австралии, где кочует вдоль побережий.
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ I; МИГРАЦИЯХ МОРСКИХ П(ИЦ 905
Настоящих оседлых видов среди морских птиц очень мало,
по-видимому, не более 5—6%- Если учесть виды, у которых
оседлы только некоторые популяции, то эта цифра несколько
увеличится. К полностью оседлым видам, очевидно, нужно от-
нести гнездящихся на островах Галапагос и совсем не встречаю-
щихся в других районах нелетающего баклана*— Nannopterum
harrisi (Rothsch.), чаек — Larus fuliginosus Gould, Greagrus
furcatus (Neboux), пингвина — Spheniscus mendiculus Sundev.
Другие примеры несомненно оседлых видов или популяций так-
же относятся к прибрежным птицам изолированных океаниче-
ских островов. Много таких островов разбросано в Южном океа-
не. где они разделены огромными пространствами воды, являю-
щимися преградой для распространения прибрежных видов.
Только на островах Чатам встречается баклан — Phalacrocorax
featherstoni Bull., а на островах Кергелен — баклан Р. verruco-
sus (Cab.). Аналогичная картина наблюдается у видов или по-
пуляций пингвинов, ныряющих буревестников, доминиканской и
красноклювой чаек, гнездящихся на изолированных, далеко от-
стоящих от побережий островах. В этом отношении очень пока-
зателен район Новозеландского плато, где мне удалось провести
детальные наблюдения в течение всех сезонов. Здесь на остро-
вах Чатам, Антиподов, Баунти. Окленд и Кэмпбелл гнездятся
несколько видов пингвинов, бакланов, ныряющие буревестники,
доминиканская и красноклювая чайки, некоторые из них в боль-
ших количествах (Oliver, 1965). Наблюдения показывают, что в
разделяющих острова открытых водах все эти птицы фактиче-
ски не встречаются, тем более не может быть речи о каких-либо
направленных и регулярных миграциях- В то же время популя-
ции данных видов, гнездящиеся непосредственно у побережий
Новой Зеландии и Австралии, совершают вдоль побережий ко-
чевки различной протяженности.
Оседлость, однако, не нужно понимать в абсолютном смыс-
ле. Например, отдельные особи островных популяций несколь-
ких видов пингвинов регулярно достигают вод Австралии и Но-
вой Зеландии. Совсем не имеют строго оседлых видов семейст-
ва Procellariidae- Diomedeidae. Hydrobatidae. Stercorarriidae,
Alcidae, Phaethontldae, Fregatidae, Sulidae, Pelecanidae. Stemidae.
По-видимому, только некоторые из них, в частности сем. Sterni-
dae, имеют отдельные полуоседлые популяции (тропическая зо-
на). За исключением чистиковых, птицы перечисленных се-
мейств относятся к хорошим летунам.
Немного (7—10%) среди морских птиц и настоящих перелет-
ных1. Регулярные перелеты совершают буревестники Буллера,
* При подсчете не учитывались чайки и крачки, гнездящиеся на внутренних
водоемах, но зимующие и совершающие перелеты вдоль морских побережий.
206
МИГРАЦИИ В РАЗЛИЧНЫХ КЛИМАТИЧЕСКИХ ЗОНАХ ТИХОГО ОКЕАНА
бледноногий, тонкоклювый, серый, большой — Puffinus gravis
(O‘Reilly), розовоногий, тайфунники Штейнегера и Пила, три ви-
да северных поморников, вилохвостая чайка, качурка Вильсона,
арктическая крачка и некоторые другие.
Подавляющая часть видов (порядка 85%) относится к коче-
вым. Однако многие из них, как отмечалось выше, имеют при-
знаки. характерные для перелетных птиц. При современной сла-
бой изученности миграций морских птиц трудно говорить о точ-
ном количестве таких видов. По ориентировочным подсчетам их
не менее 20%•
Большая подвижность морских птиц, в частности стремление
к кочевкам, находит объяснение в специфике водной среды, с
которой они тесно связаны. Я имею в виду большую подвиж-
ность водной среды. Подвижность не только в смысле перемеще-
ния поверхностных водных масс в виде течений. Изменчивостью
характеризуются температурные и другие характеристики вод-
ных масс, продукция и определяемая ею кормовая база птиц.
Например, формирование и размещение кормовых полей, где
обильно развивается планктон и концентрируется рыба, зави-
сит от временных и стационарных циркуляционных образова-
ний, преобладающих ветров и траекторий циклонов, образова-
ния эон подъема вод. пульсации течений и т. д. В умеренных и
высоких широтах большое значение имеет также перемещение
в течение всего весеннего и летнего времени зон максимального
обилия планктона, связанное с распространяющимся от низких
широт к высоким прогревом вод. В связи с этим даже перелет-
ные виды на зимовках, как правило, не остаются постоянно в.
локальных районах, а кочуют, перемещаясь от одной продук-
тивной зоны к другой.
1. Особенности сезонных миграций
морских птиц в различных климатических
зонах Тихого океана.
А. В. Михеевым (1964) на примере птиц Палеарктики убе-
дительно показано, что особенности миграций находятся в тес-
ной зависимости от характера и степени приспособленности
птиц к переживанию неблагоприятного периода года, а также
от степени изменчивости условий обитания в том или ином рай-
оне. Этот автор подчеркивает важность в определении мест зи-
мовок состава питания, способов и места добычи пищи. Резуль-
тативно рассмотрение в таком плане и миграций морских птиц.
Арктическая зона. Среди всех климатических зон наиболее
суровыми условиями отличается Арктика с ее деловитостью и
низкими температурами. В связи с этим большая часть птиц на
зимний период покидает районы, закрывающиеся плотными
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 207
льдами. В атлантическом секторе Арктики массовые зимовки
птиц располагаются, например, в Баренцевом море к югу от ле-
довой кромки (Белопольский. 1957; Успенский, 1969). К северу
от кромки льдов в разводьях и на полыньях остается на зимов-
ках часть особей чистика, люрика — Plautus alle (L.), толсто-
клювой кайры, белой, розовой — Rhodostethia rosea (Macgill.)*,
полярной чаек и морянки (Успенский. 1969)- Однако, учитывая
суровость условий, трудно допустить, что численность зимующих
во льдах птиц высокая. Этот вывод относится и к тихоокеанско-
му сектору Арктики, где ледовая кромка зимой уходит далеко
на юг в умеренную зону.
В тихоокеанском секторе Арктики наиболее богато морски-
ми птицами Чукотское море, где гнездится значительное коли-
чество бореальных и аркто-бореалвных видов. Осенью большая
часть особей всех видов откочевывает южнее. Некоторые виды,
в частности, поморники, моевка, полярная крачка, вилохвостая
чайка начинают движение на юг довольно рано, до образования
льдов. Процесс откочевки многих чистиковых и крупных чаек
растянут. Часть особей постепенно отступает на юг вместе с ле-
довой кромкой. Зимовки всех чистиковых, откочевывающих че-
рез Берингов пролив, по-видимому, располагаются в северной
части умеренной зоны, в частности в Беринговом море1 2. Не идет
южнее ледовой кромки белая чайка, весьма обычная зимой в
разводьях в Беринговом море- Другие чайки совершают более
протяженные миграции. В пределах умеренной и субтропической
зон зимуют обыкновенная моевка, полярная и серебристая чай-
ки- Плотность их населения в субтропических водах не ниже, а
местами выше, чем в умеренных. Отдельные особи этих видов
проникают в северную часть тропической зоны. Полностью в
тропиках у берегов Южной Америки находятся зимовки вило-
хвостой чайки. От северной субтропической до южной умерен-
ной зоны располагаются зимовки поморников. Арктическая
крачка, как известно, зимует в Антарктике.
Таким образом, морские птицы, гнездящиеся в Арктике, ис-
пользуют для зимовок все зоны океана, но основная масса осо-
бей и видов зимует в смежной умеренной зоне (рис. 53). Важное
значение, как место для зимовок, имеют и северные субтропики,
во их роль несопоставима с более северными водами. В умерен-
1 Я располагаю несколькими сообщениями о встречая зимой разовой чай-
ки в Беринговом и Охотском морях. Описаны такие случаи и в литературе
(Дементьев, 1951; Нечаев, 1969).
2 До последнего времени в Тихом океане не отмечался люрик. Судя по
наблюдениям Бедарда (Bedard, 1965), часть особей этого вида может про-
никать из Арктики в Берингово море. Бедара даже допускает возможность
гнездования вида в Беринговом море.
208
МИГРАЦИИ В РАЗЛИЧНЫХ КЛИМАТИЧЕСКИ* ЗОНАХ ТИХОГО ОКЕАНА
Рис. 53. Районы кочевок и зимовок морских птиц, гнездящихся в аркти-
ческой и умеренной зонах. 1 — птицы арктической зоны, 2—4 — птицы
умеренной зоны (интенсивность штриховки соответствует значению райо-
нов как мест кочевок)
ных и в меньшей степени в субтропических водах находятся зи-
мооки многих размножающихся в Арктике нырковых уток
Среди морских птиц (без учета уток), гнездящихся в аркти-
ческой зоне, около четверти приходится на долю перелетных, у
которых зимовки отстоят на большом расстоянии от гнездовой
части ареала. Остальные виды кочующие, но по крайней мере
около двух третей из них в характере сезонных перемещений
ймеют признаки перелетных птиц. Оседлых видов в Арктике
нет.
Бореальная зона. В бореальной зоне условия обитания зна-
чительно изменяются по сезонам, ио в зимний период они менее
экстремальны, чем в Арктике. В связи с этим большая часть
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 209
гнездящихся здесь видов и особей птиц, смещаясь в южном на-
правлении, тем не менее, остается зимовать в умеренной зоне.
За пределы умеренной зоны не выходят зимовки подавляю-
щей части особей чистиковых, хотя многие из видов этой груп-
пы (рогатый тупик, старик, пыжики, очковый чистик, топорок,
тонкоклювая кайра) нередки в северной части субтропической
зоны. В умеренной и субротической зонах располагаются зимов-
ки северных бакланов. Вдоль побережий они встречаются не-
прерывно от Нижней Калифорнии, Китая и южной части Япо-
нии до льдов Берингова и ОхотскоТго морей. На обширных про-
странствах умеренных и субтропических зон рассеиваются в пе-
риод осенне-зимних кочевок чайки. Часть из них (красноногая
моевка, серокрылая чайка) тяготеет к умеренным водам, дру-
гие (чернохвостая, западная) — к субтропическим, а третьи
(обыкновенная моевка, тихоокеанская, серебристая) часто
встречаются в обеих зонах. Некоторые из них (чернохвостая,
западная, серебристая, обыкновенная моевка) в небольших ко-
личествах заходят в северную часть тропиков. Полностью отле-
тает в тропики, как отмечалось выше, вилохвостая чайка, кото-
рая гнездится не только в Арктике, но и в северной части уме-
ренной зоны. В пределах умеренной и субтропической зон зи-
муют глупыш и серая качурка. Северная качурка зимой распро-
странена шире, она встречается и в северной половине тропиче-
ской зоны. В южное полушарие отлетают поморники (часть
остается в субтропиках и тропиках) и арктическая крачка, о ко-
торых упоминалось выше при описании районов зимовок птиц
Арктики. В южной части умеренной зоны гнездится несколько
субтропических видов (вилохвостая качурка, пестролицый буре-
вестник, малая крачка), все они на зимний период мигрируют в
субтропическую и тропическую зоны.
Таким образом, морские птицы, гнездящиеся в бореальной
зоне, для зимовок используют, как и птицы Арктики, все клима-
тические зоны океана (см. рис. 53). Однако размах разлета
здесь все же выражен меньше, чем в Арктике, и основные зи-
мовки располагаются в бореальной и северной субтропической
зонах*. По существу, за экватор мигрируют только поморники,
полярная крачка и вилохвостая чайка, гнездящиеся в Арктике
и в прилегающей к ней северной части умеренной зоны. В целом
для бореальной области процент настоящих перелетных птиц
невысокий, по-видимому, около 10—15%. Остальная часть —
кочевые виды. Из них не менее 50% в характере сезонных ми-
граций имеют признаки, свойственные перелетным птицам. Стро-
1 В северной Атлантике иногда отмечаются северогихоокеа некие чистико-
вые — большая конюга и белобрюшка — Cvdorrhvnchus p^ittacula (Pallas)
(•Makalsch, 1969). Несомненно, это залетные илч занесенные штормами особи.
210
МИГРАЦИИ В РАЗЛИЧНЫХ КЛИМАТИЧЕСКИХ ЗОНАХ ТИХОГО ОКЕАНА
го оседлых видов (и, наверное, популяций) в умеренной зоне
нет-
Аналогичная картина ‘наблюдается в северной Атлантике.
В этом районе изучение морских птиц проводится весьма интен-
сивно, и многие детали миграций и размещения районов коче-
вок птиц выяснены кольцеванием. Можно сослаться, например,
на результаты кольцевания тонкоклювой кайры в Норвегии
(Holgersen, 1961), большой морской чайки — Larus marinus L.
в Дании (Bruun, 1963) > малого буревестника — Puffinus puffinus
(Briinn), большого поморника и моевки в Британии (Thomson,
1965, 1966; Coulson, 1966). Дальние миграции за экватор здесь
совершают преимущественно те же виды, которые были перечис-
лены для Тихого океана. Мигрирует в южное полушарие также
малый буревестник, но в небольших количествах (Spencer, 1962,
1963; Thomson, 1965).
Основными факторами, вызывающими ухудшение условий
обитания в умеренной зоне зимой, являются изменения в кормо-
вой базе, понижение температуры воды и воздуха, а в северных
районах также образование льдов. Льды делают непригодными
для существования обширные пространства в Беринговом и
Охотском морях. В разводьях к северу от ледовой кромки оста-
ются зимовать немногие виды. Например, в районе острова Свя-
того Лаврентия, расположенного недалеко от Чукотского полу-
острова. зимуют морянка, гага-гребенушка — Somatena specta-
bilis (L.), обыкновенная гага — S. mollissima (L.). толстоклювая
кайра, чистик и полярная чайка (Fay and Cade, 1959)- Однако
все эти, а также другие виды морских птиц начинают осенние
кочевки, в том числе миграции на юг еще до образования льда.
Поэтому последний не следует рассматривать в качестве причи-
ны. вызывающей сезонные миграции.
Непосредственной причиной полной откочевки ряда видов из
некоторых северных районов умеренной области является зна-
чительное понижение температуры воды и воздуха. Так как на-
иболее сильно зимой охлаждаются районы, прилегающие к
Азии, то изотермы на севере Тихого океана в это время имеют'
направление с юго-запада на северо-восток. Аналогичную кон-
фигурацию имеют северные границы распространения ряда от-
носительно теплолюбивых видов, в частности, обыкновенной мо-
евки. чернохвостой чайки, топорка, ипатки, качурок. Однако
главным фактором, определяющим характер зимнего распреде-
ления птиц, является изменение в кормовой базе. Особое значе-
ние имеет сильное обеднение зимой верхних слоев моря планк-
тоном и рыбой. Уменьшение в верхних горизонтах биомасс
планктона в бореальной зоне связано с сокращением численно-
сти и опусканием его в теплый промежуточный слой (Гейнрнх,
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ГНИЦ 2U
1961. 1967; Виноградов. 1967, 1968). Большинство пелагических
рыб и их молодь переходят к обитанию .в промежуточные или
придонные слои вод. Многие из них образуют при этом малопо-
движные зимовальные скопления. На большие глубины смеща-
ются также донные и придонные рыбы.
При анализе сезонных аспектов в орнитофуане дальневосточ-
ных морей и соиоеделыных водах океана было показано, что ус-
ловия зимой в северных районах умеренной зоны значительно
более суровы, чем в южно-бореальных районах. В южной части
умеренной зоны не бывает или почУи не бывает льдов, здесь
значительно теплее, сезонные изменения в концентрациях планк-
тона л рыбы в поверхностных слоях выражены несколько мень-
ше Таким образом, каждая популяция морских птиц, откочевы-
вая из более северных районов в южном направлении, даже не
выходя за пределы бореальной области, попадает в более бла-
гоприятные условия для зимовки.
Естественно, что обеднение поверхностных слоев кормом ска-
зывается, в первую очередь, на птицах, добывающих пищу с по-
верхности моря или ныряющих на небольшую глубину. С этим
хорошо согласуется откочевка из северных районов умеренной
зоны большей части планктоноядных трубконосых. Гнездящиеся
в умеренных широтах глупыш, серая и северная качурки широ-
ко рассеиваются в пределах умеренной и субтропической зон, а
последний вид кроме того — в тропической. Мелкие планктоно-
ядные чистиковые — конюги кормятся и в толще воды, но ны-
ряют неглубоко. Протяженность миграций их также значитель-
на. Как и трубконосые, рассеивается на огромных пространствах
океана мелкая пелагическая чайка моевка. Меньший размах
имеют сезонные миграции бакланов и крупных чистиковых (оба
вида кайр, чистиюи). Эти птицы способны нырять на большую
глубину, поэтому влияние обеднения поверхностных слоев моря
кормом для них менее ощутимо. Многочисленны на зимовках в
северных умеренных районах крупные чайки — серокрылая,
тихоокеанская, полярная. Чайки питаются с поверхности моря,
но свойственная им эврибионтность (всеядность многообразие
приемов охоты, применение длительного поискового полета) по-
зволяет существовать в условиях низких концентраций корма
(Юдин. 1965). Пищей для чаек являются и отходы с многочис-
ленных судов, ведущих промысел рыбы на шельфе и материко-
вом склоне во всех районах умеренной зоны. Что касается не
идущей южнее льдов белой чайки, то она вообще в состоянии
длительное время обходиться без чистой воды (Успенский,
1969).
Перечисленные птицы вместе с большим количеством нырко-
вых уток в прибрежных водах составляют основу зимней орни-
тофуаны в северной части умеренной зоны. В южной половине
212
МИГРАЦИИ В РАЗЛИЧНЫХ КЛИМАТИЧЕСКИХ ЗОНАХ ТИХОГО ОКЕАНА
заметно увеличивается количество планктоноядных птиц —
трубконосых, мелких чистиковых.
В предыдущей главе достаточно подробно были описаны
особенности весенних миграций в отдельных районах северной
части Тихого океана. Здесь нужно подчеркнуть только некото-
рые моменты. Начало весенних миграций приходится на конец
марта, разгар — на апрель и май, завершение — на первую по-
ловину июня. Первыми мигрируют виды, наиболее приспособ-
ленные к обитанию в суровых условиях, последними — наиболее
специализированные планктоноядные и теплолюбивые птицы, а
также виды, зимовки которых расположены на больших рас-
стояниях от мест гнездования, в частности в южном полушарии.
Время весенних миграций соответствует периоду наиболее су-
щественных изменений, происходящих в гидрологии и биологии
моря (прогрев вод, вегетация фитопланктона, подъем в верх-
ние слои моря и размножение зоопланктона, миграции на мень-
шие глубины рыб и размножение многих из них).
Весенне-летний период, несомненно, наиболее благоприятный
для обитания сезон. Достаточно сказать, что плотность концент-
раций планктона увеличивается в верхних слоях моря в десятки
и сотни раз. Здесь же много пелагических рыб и их молоди. Ин-
тенсивного развития достигает жизнь в литоральной зоне. Это,
а также суровость зимнего сезона являются причиной размно-
жения всех птиц в умеренной эоне в весенне-летнее время. Осен-
не- и зимнеразмножающихся видов птиц, гнездящихся в \ мерен-
ной зоне, нет.
Сезонные перемещения в умеренной зоне не ограничиваются
миграциями на зимовки и обратными перемещениями весной.
Летом относительно строго придерживаются определенных рай-
онов только размножающиеся в данное время особи. В море же,
как неоднократно подчеркивалось выше, в сезон размножения
остается большое количество неполовозрелых особей. В умерен-
ных водах, кроме них, концентрируется масса трубконосых из
южного полушария. Все они образуют подвижные скопления,
перемещающиеся из одного продуктивного района в другой.
На фоне этого ненаправленного движения, обусловленного осо-
бенностями формирования и размещения кормовых полей, обра-
щает на себя внимание постепенное смещение многих видов в
северном направлении в течение всего лета и даже в начале
осени. В основе этих перемещений лежат изменения, происходя-
щие в биологии и гидрологии моря. Характерно, что после ве-
сенне-летнего максимума планктона во второй половине лета в
результате выедания и начавшегося опускания в более нижние
горизонты биомассы планктона в поверхностных слоях умень-
шаются. А так как весенние изменения более рано начинаются
ЗЛКОНС.’ШРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 213
в южЬой части умеренной зоны, то на протяжении летнего пери-
ода зона летнего максимума планктона постепенно смещается на
север. На севере Берингова моря, например, наибольшие био-
массы планктона наблюдаются только в конце лета. Поэтому в
августе и начале сентября, когда некоторые виды (плавунчики,
поморники, половозрелые буревестники) уже мигрируют на юг,
значительные количества особей серой качурки и тонкоклювого
буревестника (неполовозрелые) перемещаются в северном на-
правлении. В конце лета смещаютсй в северном направлении
не только планктоноядные трубконосые, но и часть особей, на-
чавших лослегнездовые кочевки кайр и чаек-
Воды умеренной зоны используют для кочевок многие виды,
гнездящиеся в более южных широтах. Из них заслуживают, в
первую очередь, упоминания несколько видов трубконосых, раз-
множающихся в северной субтропической и тропической (темно-
спинный и черноногий альбатросы, буревестники пестролицый
и Р.opisthomelas, вилохвостая качурка), а также в южной суб-
тропической и ноталыюй зонах (буревестники серый, тонкоклю-
вый, бледноногий, Буллера, роэовоногий. тайфунники Пила и
Шлегеля). Общее число видов-мигрантов в бореальной области
из южных широт точно не известно, но, очевидно, их не меньше
20. Характерно, что большинство южных видов в умеренной зо-
не кочует в период с весны до осени. Зимой из них многочислен-
ны только черноногий и темноспинный альбатросы. Поэтому в
зимний период на всем протяжении умеренной зоны по числу ви-
дов и особей абсолютно преобладают бореальные и аркто-боре-
альные виды. Летом картина меняется. Даже если рассматри-
вать всю летнюю орнитофауну умеренной зоны, то на долю юж-
ных мигрантов приходится около одной трети видов. Что каса-
ется южной части умеренной зоны, то здесь соотношение видов
нередко в пользу южных мигрантов. По числу особей в целом,
особенно в большинстве шельфовых районов, преобладают се-
верные виды. Но в мелководных районах в южной части умерен-
ной зоны, а также в открытых водах океана обычно на долю
южных мигрантов приходится больше половрны особей.
Северная субтропическая зона. Сезонные явления в субтро-
пических широтах выражены, но отличия между сезонами здесь,
менее контрастны, чем в умеренных водах. В северной субтро-
пической зоне температура воды почти нигде не опускается ни-
же 10°. Менее резко и четко, чем на севере, изменяется количе-.
ство планктона и его вертикальное распределение (Гейнрих,
1961а. 1967; Виноградов, 1968; и др.). Кормовые условия для
птиц зимой здесь довольно благоприятны. Как показано многи-
ми гидробиологами (Marumo et al, 1954; Marumo, 1957; Motoda
and Marumo, 1963; Карелин, 1969; Кун, 1969; Кун и др.. 1969; ц
214
МИГРАЦИИ В РАЗЛИЧНЫХ КЛИМАТИЧЕСКИХ ЗОНАХ ТИХОГО ОКЕАНА
ми. др.), в субтропических водах Японии биомассы планктона
в зимний период ниже, чем летом, но эти отличия не очень зна-
чительны. К тому же в конце зимы здесь начинается весеннее
развитие планктона. В зимний и зимне-весенний периоды проис-
ходит нерест некоторых пелагических рыб, в том числе такого
массового вида, как сайра, и в поверхностных слоях много их
молоди. Аналогичная картина наблюдается в восточной части
Тихого океана у берегов Калифорнии. Зимнее понижение био-
масс планктона здесь не очень значительно (Thrailkill. 1963;
Fleminger, 1964; Alvarino, 1965; Abbott and Albee, 1967; Brinton,
1967; Strickland, 1967; и мн. др.), в течение большей части зимы
в поверхностных слоях много молоди рыб, особенно анчоуса
(Kramer and Ahlstrom, 1968; и др.).
Мигрирующие в субтропические воды северные виды находят
здесь, несомненно, благоприятные кормовые условия. Мягкость
же климата и термического режима вод в этих широтах общеиз-
вестна. Однако условия, благоприятные для северных видов, не
обязательно должны быть благоприятными для субтропических.
Для последних, по-видимому, имеет значение зимнее понижение
температуры воды и воздуха. Неблагоприятная синоптическая
обстановка, определяемая частыми зимой штормами уменьшает
доступность пищи. Все это вместе с некоторым общим уменьше-
нием концентраций планктона, очевидно, и определяет заметные
сезонные изменения в распределении и биологии субтропических
птиц. В субтропической зоне, как и в умеренной, ясно выраже-
на сезонность в размножении. Правда, мягкий климатический
режим и нерезкие колебания в кормовой базе позволяют размно-
жаться в зимний период пяти видам (три альбатроса, бонинский
тайфунннк, качурка — О. tristrami) *• Но размножение большей
части видов (примерно около 90%) приходится на весенне-лет-
ний период. При этом у многих видов, особенно у видов север-
ного происхождения (чистиковые), оно начинается рано — в
марте. Более раннее, чем в умеренной зоне, размножение птиц
хорошо согласуется с более ранним началом весны в этих ши-
ротах. Обычно весенняя вспышка цветения приходится иа конец
зимы — начало весны. Вскоре после этого начинается нараста-
ние биомасс зоопланктона (Магштю et al., 1954; Abbott and Albee,
1967; и др.).
Зимовки большинства вадов птиц, размножающихся в се-
верной субтропической эоне, располагаются в северной части
тропической и в южной части субтропической зоны (рис. 54).
Только альбатросы частично остаются зимой в бореальной об-
ласти. В тропики откочевывают крачки (субтропические виды и
1 Приуроченность размножения к зямяему периоду у альбатросов и тай-
«фуниика, несомненно, связана с их южным происхождением.
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 215
популяции тропических видов, гнездящихся в субтропиках), олу-
ши. Качурки частично остаются в пределах южной части суб-
тропической зоны, но вместе с этим широко рассеиваются в во-
дах Северо-Пассатного течения, а два вида — вилохвостая и
О-matsudairae — проникают в Индийский океан. Аналогичное
распространение имеют буревестники. В частности пестролицый
буревестник достигает северного побережья Австралии и архи-
пелага Бисмарка. Наименьшую .протяженность миграций име-
ют чистиковые (калифорнийские пыжики, хохлатый старик, суб-
тропические популяции нескольких бореальных видов), бакланы
и чайки. Перемещения большей их части ограничиваются суб-
тропическими водами. В связи с отсутствием резкой контраст-
Рис. 54. Районы кочевок и зимовок морских птиц, гнездящихся в субтро-
пических и тропической зонах. I — птицы северной субтропической зоны,
2 — птицы южной субтропической эоны, 3—4 — птицы тропической эо-
ны (интенсивность штриховки соответствует значению районов как мест
кочевок)
216
МИГРАЦИИ В РАЗЛИЧНЫХ КЛИМАТИЧЕСКИХ ЗОНАХ ТИХОГО ОКЕАНА
ности в условиях обитания в субтропической зоне нет или поч-
ти нет перелетных птиц. Нет здесь и оседлых видов. Фактически
все виды в зоне кочующие, при этом, по крайней мере, у третьей
части видов в сезонных миграциях известны признаки, свойст-
венные перелетным птицам.
Таким образом, птицы северной субтропической эоны ис-
пользуют для внегнеэдового обитания в основном две зоны. Они
в ограниченных количествах проникают за экватор и почти
не бывают многочисленны в бореальной области. В умеренные
воды летом, когда температура воды и воздуха значительно по-
вышается, кроме альбатросов, проникают также субтропические
виды буревестников и качурок. Распространение последних ог-
раничивается здесь южно-бореальной зоной. В северных водах
их привлекает обилие планктона, развивающегося в это время.
Имеет значение и то, что на нагул в богатую планктоном юж-
ную часть умеренной зоны с прогревом вод поднимаются многие
массовые пелагические рыбы (сайра, анчоус, сардина, скумбрия
и другие) и кальмары-
Северная субтропическая зона занимает промежуточное по-
ложение между бореальной и тропической. Это накладывает
большой отпечаток на состав орнитофауны в различные сезоны.
В зимний период, когда большая группа бореальных и арктобо-
реальных видов расширяет область распространения на юг, во
многих районах субтропической зоны они доминируют по числу
видов и особей. В прибрежных водах в северной части зоны
вместе с ними многочисленны различные нырковые утки и гага-
ры. Летом северные виды здесь немногочисленны. Зато с кха
сюда проникают многие виды из тропической, южной субтропи-
ческой и нотальной зон. И вновь во многих районах субтропи-
ческой зоны местные виды как бы теряются среди мигрантов из
других широт. Из изложенного видно, что рассматриваемая зо-
на характеризуется наиболее смешанным, а следовательно, на-
именее оригинальным составом орнитофауны.
Тропическая зона. Сезонные явления в гидрологии и биоло-
гии моря в этой самой обширной климатической зоне выражены
менее резко, чем во всех других. Общеизвестны сравнительно
небольшие сезонные колебания температуры воды и воздуха;
обилие света и тепла является основной предпосылкой кругло-
годичной продукции планктона, в связи с чем сезоны в тропи-
ческом планктоне имеют менее резкие границы (Гейнрих, 1961а,
1967). Отсутствие резких сезонных колебаний в температурном
режиме поверхностных водных масс и мало меняющаяся в те-
чение года стратификация вод определяют отсутствие у тропи-
ческого планктона правильных сезонных вертикальных мигра-
ций, важнейшей причины изменения концентраций планктона в
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 217
поверхностных слоях (Виноградов, 1967, 1968)- В пределах рас-
сматриваемой зоны в течение всего года остается большая часть
тропических рыб. При этом для обитающих в поверхностных
слоях видов обычно нельзя выделить какой-либо сезон, когда
они переходят к обитанию в более глубоких слоях, т. е. они не
образуют такие подвижные придонные скопления, как рыбы
более высоких широт зимой.
Морские тропические птицы, постоянно обитая в условиях
сравнительно небольших сезонных, климатических изменений, в
основном являются стенобионтными видами. Это, а также отсут-
ствие большой контрастности в условиях обитания объясняют
не очень значительный размах сезонных миграций в долготном
направлении. Здесь, по-видимому, нет или почти нет перелетных
видов. Приблизительно половина видов зоны при кочевках не
выходит за пределы тропиков. К этой группе птиц относятся
бакланы, чайки, многие тропические крачки и трубконосые. В то
же время в летний период, когда в субтропиках температура
воды и воздуха достигает значений, характерных для тропиче-
ской зоны, многие виды расширяют область кочевок в смежные
субтропические зоны (в обоих полушариях до широты ЭО и да-
же 40°). Регулярно достигает вод Японии, Новой Зеландии, Ка-
лифорнии, южного побережья Австралии некоторое количество
фаэтонов, фрегатов, олуш, крачек. Более массовый характер но-
сят подобные перемещения значительной группы трубконосых,
особенно буревестников (клннохвостый, Одюбона — Puffinus
lherminieri, Бульвера, рождественский — Puffinus nativitatis
Streets и другие) и тайфунников (бонинский, гавайский —Pte-
rodroma phaeopygia (Salvin) кермадекский — Pterodroma
neglecta (Schleg) и другие). Миграции в умеренную зону север-
ного и южного полушарий для тропических птиц в общем не
характерны. Чаще всего это отдельные залетные особи. Исклю-
чение могут составить, очевидно, только темноспинный я черно-
ногий альбатросы, гнездящиеся на Гавайских островах и мигри-
рующие в умеренную зону океана. Впрочем, указанные альбат-
росы не являются настоящими тропическими видами.
Таким образом, морские тропические птицы для кочевок ис-
пользуют в основном одну биогеографическую область. Основ-
ное значение, как место для кочевок, имеет тропическая эона;
существенно также значение субтропических эон (см. рис. 54).
Обитая в условиях относительного постоянства условий, тро-
пические птицы тем не менее в подавляющем большинстве не
являются оседлыми. Оседлы всего несколько видов (по-видимо-
му, не более 4—5%). Такие виды есть на островах Галапагос, о
них говорилось выше. Бытовавшие ранее представления об осед-
лости тропических птиц были основаны на круглогодичной
218
МИГРАЦИИ В РАЗЛИЧНЫХ КЛИМАГИЧЕСКИХ ЗОНАХ ТИХОГО ОКЕАНА
встречаемости многих видов на местах гнездования- Морских
же наблюдений проводилось мало. В частности, считалось, что
оседлы фрегаты и держатся они только вблизи суши. Кольце-
вание же на островах центральной части Тихого океана показа-
ло, что после гнездования происходит разлет их на тысячи ки-
лометров (Sibley and Clapp, 1967). Обширность областей рас-
пространения в море многих видов тропической части Тихого
океана хорошо иллюстрируют схемы Кинга (King, 1967). Подав-
ляющая часть тропических видов относится к кочевым. Только
у незначительной части из них в характере сезонных перемеще-
ний есть элементы, свойственные перелетным птицам.
Отсутствие очень резких сезонных контрастов в условиях
обитания в тропической зоне, конечно, не означает неизмен-
ность условий существования вообще. В первую очередь это от-
носится к сезонным изменениям в кормовой базе, циркуляции
вод и преобладающим ветрам. Если говорить о шельфовых
районах и водах, омывающих океанические архипелаги, то, по-
жалуй, трудно найти район, где бы не наблюдались значитель-
ные сезонные различия в количестве планктона. Основным фак-
тором, ограничивающим продуцирование тропического планкто-
на, является содержание в поверхностных слоях биогенов, кото-
рое в свою очередь зависит от интенсивности подъема глубин-
ных вод. Условия для подъема вод, связанного с характером
циркуляции вод, определяют особенности синоптической обста-
новки (Гейнрих, 1961а, 1967; Гурьянова, 1970; и др.)- Обширные
области подъема вод чаще всего зависят от сгонно-нагонных
явлений, вызываемых муссонами и пассатами. Так как в дейст-
вии ветров определенных направлений наблюдается периодич-
ность, обильное развитие планктона характерно только для оп-
ределенных сезонов. Так, в районах формирования Южно-Пас-
сатного течения в Индийском океане на фоне круглогодичного,
довольно интенсивного подъема вод и соответствующего разви-
тия планктона, особенно выделяется зимний период, когда ин-
тенсивно дует юго-восточный пассат. Аналогичная ситуация наб-
людается, как отмечалось во второй главе, на североавстралий-
ском шельфе. В западной части Аравийского моря подъем вод
и обильное развитие планктона происходит летом во время юго-
западного муссона, а в восточной части моря — зимой, во вре-
мя северо-восточного муссона. Такая же картина наблюдается
в других азиатских тропических морях.
У северного и южного пределов тропической зоны опреде-
ленное значение для развития планктона и обитания питающих-
ся им рыб и птиц имеют заметные сезонные колебания темпера-
туры воды и воздуха. Для примера можно отметить, что север-
ная и южная границы распространения летучих рыб, являющих-
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 219
ся важнейшими пищевыми объектами многих тропических птиц,
смещаются на 700—800 миль. В летний период северного полу-
шария границы поднимаются в северном направлении. В это
время в северном полушарии они становятся многочисленными
не только в северной части тропической зоны, но и в субтропи-
ческих водах (до 35’ с. ш.)- В южном полушарии в это время зи-
ма, и в результате миграций в более низкие широты летучие ры-
бы многочисленны только до 15’ ю. ш. Летом в южном полуша-
рии летучие рыбы многочисленны до 30° ю. ш., а в северном по-
лушарии в это время они обычны или многочисленны только
южнее 20’с. ш. (Шунтов, 1969а).
Таким образом, говоря о тропической области, важно под-
черкнуть, что в любой сезон одни районы благоприятны в кор-
мовом отношении для птиц, в других концентрации кормовых
организмов уменьшаются. В этом еще одно существенное отли-
чие тропической эоны от более высоких широт. Там, как отме-
чалось выше, в неблагоприятный зимний сезон ухудшение усло-
вий обитания происходит в пределах всей зоны.
Рассматриваемые особенности сезонных изменений в гид-
рологии и биологии моря в низких широтах, очевидно, и яви-
лись основными предпосылками формирования миграций тропи-
ческих птиц в пределах тропической зоны и сопредельных суб-
тропических вод. Птицы здесь постепенно перекочевывают из
одного продуктивного района в другой- Важную роль для от-
корма имеют не только обширные стационарные районы подъ-
ема вод. например, у побережья Аравии, на востоке Тихого и
Индийского океанов, в зоне экваториальной дивергенции. Су-
щественно значение также локальных, часто временных, облас-
тей подъема вод, которые формируются в открытом океане, а
также в районах океанических архипелагов в результате тех
или иных особенностей в циркуляции.
В настоящее время многие детали миграций тропических
морских птиц не известны. Для неритических видов, очевидно,
наиболее характерными являются кочевки вдоль побережий. В
Тихом океане, где побережья ориентированы в общем с севера
на юг, миграции прибрежных птиц происходят в этом направ-
лении и обратно. Так, кольцеванием подтверждены миграции
вдоль побережья Южной Америки массовых веслоногих с пе-
руанских птичьих базаров (Jordan, 1958. 1960). Для океаниче-
ских видов характерны кочевки как в меридиальном, так и ши-
ротном направлениях. Миграции с востока на запад и обратно
в системе пассатных и экваториальных течений совершают
многие виды, гнездящиеся на океанических архипела-
гах. Такие миграции, как правило, ограничиваются пре-
делами одного океана, но для некоторых видов известно
проникновение из Тихого в Индийский океан. Выше упомина-
220
МИГРАЦИИ В РАЗЛИЧНЫХ КЛИМАТИЧЕСКИХ ЗОНАХ ТИХОГО ОКЕАНА
лись такие миграции двух качурок. Большинство океанических
видов совершает кочевки также с севера ла юг и обратно. Осо-
бенно характерным в таких перемещениях является проникнове-
ние в субтропические воды в летний период- В этом случае тро-
пические птицы, как и летучие рыбы, в периоды наибольшего
распространения в смежные широты в северном полушарии в
южном полушарии имеют наиболее ограниченное распростране-
ние.
Неодновременность сезонных изменений в условиях обита-
ния в различных участках обширной тропической зоны является
причиной того, что различные популяции (особенно, у видов с
большой гнездовой частью ареала) одного и того же вида име-
ют специфичные ритмы биологических циклов. С другой сторо-
ны, в связи с особенностями в экологических потребностях наб-
людаются большие отличия в годичных циклах и сроках мигра-
ций у видов, обитающих в одном и том же районе. Это особенно
заметно при анализе сезонов размножения тропических птиц.
В настоящее время по размножению морских тропических
птиц имеется обширная литература (Moreau, 1950; Richardson,
1957; Kuroda, 1960а; Brosset, 1962; Ashmole, 1965; Harris. 1969;
и мн. др.). Анализ данных показывает, что в каждом районе
тропической зоны имеются свои особенности. Общим является
то, что во всех районах размножение наблюдается в течение все-
го года. Несомненно, это связано с отсутствием в тропиках рез-
ких климатических изменений. Однако круглогодично размно-
жается только часть видов. У большинства сроки размножения
фиксированы, хотя и сильно растянуты'. Во всех или почти во
всех районах на фоне круглогодичного размножения представ-
ляется возможным говорить о периодах наиболее массового
гнездования. Эти периоды приурочены к сезонам, наиболее бла.
гоприятным по кормовым и климатическим условиям. У север-
ного побережья Австралии большая часть видов размножается
зимой в период юго-восточного пассата (Serventy, 1952) —
время интенсивного подъема глубинных вод и обильного разви-
тия планктона. Несомненно, имеет значение и небольшое коли-
чество осадков. Преимущественное размножение в период юго-
восточного пассата характерно для многих океанических остро-
вов Индийского океана, в частности. Рождества, Кокосовых
(Pearson, 1966) и Сейшельских (Ridley, 1957; Ridley and Percy.
1958). На Гавайских островах интенсивное размножение прихо-
дится на весенне-летний период, наиболее благоприятный в кор-
мовом отношении, а также по синоптическим условиям (Richard-
son, 1957). У восточного побережья Африки по интенсивности
размножения выделяется период юго-западного муссона (Мо-
1 Например, на островах Галапагос фрегаты только птенцов выкармливают
в течение шести месяцев (Nelson, 1966).
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 221
reau, 1950), вызывающий подъем глубинных вод в северо-запад-
ной части. Индийского океана. Лишь в отдельных районах по име-
ющимся материалам нельзя выделить сезон, когда размножает-
ся большая часть видов. По-видимому, к таким районам отно-
сятся острова Галапагос (Brosset, 1962; Harris, 1969)*.
Таким образом, в одно и то же время, когда часть видов или
особей кочует, другие размножаются и концентрируются у мест
гнездования. Здесь нет. следовательно, как в высоких широтах,
однонаправленного для всей эоны хода гнездовых и миграцион-
ных циклов у всех представителей орнитофауны.
Так как большая часть видов и особей тропических птиц в те-
чение всего года не покидает пределы рассматриваемой зоны,
они, как правило, составляют здесь основу населения во все се-
зоны. В то же время в определенные сезоны в ряде районов фон
определяют мигранты из других зон. В зимний период в южной
части тропической зоны, когда сюда мигрируют с юга субтропи-
ческие, нотальные и нотально-антарктические виды, а часть тро-
пических птиц откочевывает севернее, южные мигранты нередко
преобладают по количеству видов и особей. Во второй главе это
было показано в отношении района северо-западного побережья
Австралии, куда мигрирует много южных трубконосых. Анало-
гичная картина наблюдается к востоку от Австралии. Здесь до
Кораллового моря проникают с юга в значительных количествах
буревестники, качурки, странствующий альбатрос. Много особей
южных видов трубконосных поднимается в тропики вдоль восточ-
ного побережья Южной Америки. Выше уже упоминалось о мас-
совых миграциях субтропических, нотальных и нотально-антарк-
тических птиц в период южной зимы в Аравийское море. Некото-
рые из них имеют высокую численность (бледноногий буревест-
ник, качурки Вильсона и белолицая), поэтому не удивительно,
что местами они составляют здесь основу населения. В течение
летнего периода, когда южные виды откочевывают в свои эоны,
в тропиках южнее экватора повсеместно преобладают тропиче-
ские виды.
В северном полушарии в зимний период субтропические виды,
откочевывая в северную часть тропической зоны, в ряде районов
создают временное преобладание над тропическими видами. Так,
в Филиппинском море основу населения в это время нередко со-
1 В приведенном обзоре сезонов размножения отмечены только основные
факторы, определяющие сроки размножения тропических птиц. Вообще же
проблема эта значительно сложнее. В частности, в ряде случаев, кроме обилия
корма н особенностей климата, большое значение имеет происхождение вида,
конкурентные межвидовые взаимоотношения. Частота размножения некоторых
видод (или особей) определяется только временем необходимым для после-
гнездовой лишки оперения крыльев и хвоста (Richardson, 1967; Kuroda,
1960а; Harris, 1960; и др.).
222
МИГРАЦИИ В РАЗЛИЧНЫХ КЛИМАТИЧЕСКИХ ЗОНАХ ТИХОГО ОКЕАНА
ставляют пестролицый буревестник и качурки, гнездящиеся
в районе Японии. Аналогичная картина наблюдается в водах
мексиканского побережья, куда откочевывают птицы из района
полуострова Калифорния. Летом, когда субтропические виды от-
кочевывают, здесь повсеместно начинают преобладать тропиче-
ские птицы. Особого упоминания заслуживают массовые буре-
вестники и тайфунники, совершающие миграции за экватор из
южного полушария. В период миграций на север и обратно во
многих районах тропической зоны, в том числе в приэкватори-
альных, большие стаи этих птиц определяют временное их преоб-
ладание над другими видами. Птицы бореальной и арктической
зон нигде и никогда в тропиках не встречаются в таких больших
количествах, как птицы нотальной и антарктической зон. В райо-
не холодного Перуанского течения довольно многочисленной на
зимовках бывает вилохвостая чайка (Chapman, 1969), местами
обычны зимой и на пролете поморники*, по-видимому, крачки.
Однако говорить о их преобладании над тропическими птицами
не приходится.
Южная субтропическая зона. Сезонные явления в южной суб-
тропической зоне выражены более отчетливо, чем в тропической.
Как и в северной субтропической зоне, зимнее понижение темпе-
ратур воды и воздуха здесь не очень значительно. Но сезонные
изменения в биологии моря выражены почти так же, как в уме-
ренной зоне (по крайней мере сильнее, чем в северной субтропи-'
ческой зоне). Наиболее существенным является обеднение верх-
них слоев моря планктоном и рыбой. Во второй главе при описа-
нии сезонных аспектов в орнитофауне субтропических вод у юж-
ного побережья Австралии и у Новой Зеландии уже отмечалось,
что в зимние месяцы биомассы планктона в несколько раз ниже,
чем летом. Многие пелагические рыбы переходят к обитанию
в придонных горизонтах, а некоторые из них образуют малопод-
вижные придонные скопления. В весенне-летний период обеспе-
ченность птиц кормом улучшается, что определяется не только
обилием в поверхностных слоях планктона, рыб и местами каль-
маров. Условия добычи пищи улучшаются с прекращением зим-
них штормов. Существенно также то, что большая часть видов
рыб в это время нерестится, в результате чего в ряде районов
много икры и молоди рыб.
. В южной субтропической зоне в связи с изменчивостью в ус-
ловиях обитания довольно четко выражена сезонность в размно-
жении птиц. Большая часть их размножается в весенне-летний
период. Правда, мягкие климатические условия дают возмож-
1 Летом, когда пролет северных поморников заканчивается, в тропиках мо-
гут оставаться отдельные бродячие особи. В частности, средний поморник
в нюне наблюдался в Панамском заливе (Robins, 1958).
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 223
ность некоторым видам гнездиться в зимний период. Это харак-
терно, например, для длиннокрылого тайфунника, сумеречного
буревестника, некоторых популяций красноклювой чайки в
Австралии.
Ухудшение зимой кормовых условий для птиц, добывающих
пищу в поверхностном слое моря, явилось причиной формирова-
ния у ряда планктоноядных видов перелетов за экватор. Всего
таких видов в зоне не менее 12 (около 20—25%). До бореальной
зоны мигрируют тайфунник Штейнегера, буревестники розовоно-
гий, бледноногий, Буллера, а также субтропические популяции
серого и тонкоклювого буревестников (до Чукотского моря) и
тайфунника Пила. По-видимому, только до северной субтропиче-
ской зоны поднимаются тайфунники Кука, белошейный —
Pterodroma externa (Salvin), Соландера — Pterodroma solandri
Gmel.
Много субтропических птиц зимует в тропиках. От южного
побережья Австралии в тропическую часть Индийского океана
мигрирует значительная часть особей бледноногого буревестника
и белолицей качурки. В тропические воды Тихого океана расши-
ряют область зимнего распространения бонинский тайфунник,
буревестники порхающий и черный — Procellaria parkinsoni
Gray, белолицая качурка, многие крачки, а также субтропиче-
ские популяции некоторых нотальных видов, например, прионы.
Большое количество субтропических птиц кочует зимой в пре-
делах субтропической зоны. В основном это птицы, для которых
зимнее обеднение поверхностных вод планктоном менее ощути-
мо, чем для планктоноядных трубконосных. Это — или ныряю-
щие, или всеядные, или виды, питающиеся молодью рыб. В тече-
ние всей зимы многочисленны у южного побережья Австралии
крачки. Не совершают значительных перемещений в широтном
направлении размножающиеся в субтропической зоне пингвины
(Гумбольда — Spheniscus humboldti Меуеп, малый и др.), ны-
ряющие буревестники, чайки (Gabianus pacificus (Lath.), красно-
клювая), бакланы (белогрудый — Phalacrocorax fuscescens
(Vieill.), красноногий — Р. gaimardi (Less.), малый черный —
Р. sulcirostris Less, и др.). Большие по расстояниям миграции со-
вершают олуши. Австралийская олуша совершает перелеты к
Австралии. Здесь она концентрируется в мелководных районах
в пределах субтропических вод, лишь небольшая часть мигран-
тов проникает в тропики. Весьма протяженные миграции совер-
шает южноафриканская олуша {Moris capensis (Licht.), но и
здесь в тропики попадает только часть молодых (Broekhuysen et
al., 1961).
Во второй главе было показано, что субтропические виды
к югу от субтропической конвергенции проникают мало. Но в
рассматриваемой зоне гнездится часть популяций некоторых ви-
224
МИГРАЦИИ В РАЗЛИЧНЫХ КЛИМАТИЧЕСКИХ ЗОНАХ ТИХОГО ОКЕАНА
дов, например, серого и тонкоклювого буревестников, тайфунни-
ка Пила, прионов. Летом они проникают в высокие широты до
Антарктики.
Таким образом, морские птицы, гнездящиеся в южной субтро-
пической зоне, для миграций используют все зоны от Арктики до
Антарктики (см. рис. 54). Но основное значение имеют три зоны
— южная и северная субтропические и тропическая, где кочуют
настоящие субтропические виды. В целом преобладающим на-
правлением миграций являются перемещения с севера на юг и
обратно.
Южная субтропическая зона занимает промежуточное поло-
жение между тропической и нотальной. Это накладывает суще-
ственный отпечаток на состав орнитофауны в различные сезоны.
Зимой, когда часть субтропических видов откочевывает, а птицы
нотальной и антарктической зон расширяют область распростра-
нения на север, последние в большинстве районов преобладают
над субтропическими и составляют основу населения. Как было
показано выше на примере района Новой Зеландии, в южной
части субтропической зоны нотальные виды по числу особей пре-
обладают и летом. В северной части субтропической зоны в это
время существенна примесь тропических видов. В некоторых
районах на их долю приходится более половины особей. Напри-
мер, у западного побережья Австралии летом доминируют клино-
хвостый буревестник и темная крачка. Таким образом, южная
субтропическая зона, как и северная субтропическая, характери-
зуется во все сезоны наиболее смешанным составом.
Нотальная зона. Различия в условиях обитания птиц в зим-
ний и летний сезоны выражены в нотальной зоне более отчетли-
во, чем в субтропиках. Правда, климатический и гидрологический
режимы этой зоны в сравнении с бореальной менее суровы и
контрастны. Здесь не бывает отрицательных температур воды
и никогда не замерзает океан. Сравнительная мягкость зим дает
возможность в это время даже размножаться нескольким видам
морских птиц. На зимние месяцы приходится размножение бак.
лана — Phalacrocorax carunculatus (Gmel.), пингвина — Eudyptes
pachyrhynchus (Gray), бурого и сумеречного буревестников, длин-
нокрылого тайфунника, части особей белогорлого буревестника.
Зимой продолжают выкармливание птенцов странствующий и ко-
ролевский альбатросы.
Среди факторов, вызывающих ухудшение условий обитания
в умеренных широтах в зимний период, нужно подчеркнуть два:
уменьшение в поверхностных слоях планктона, рыб и кальмаров,
а также неблагоприятную синоптическую обстановку.
По характеру сезонных изменений в планктоне Южный океан
имеет большое сходство с бореальной зоной. Зимой прекращает-
ся вегетация фитопланктона (ослабление интенсивности освеще-
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 225
ния), уменьшаются биомассы зоопланктона, при этом концентри-
руется он глубже 200—250 м. Различия в количестве планктона
в противоположные сезоны в верхних слоях достигают десяти-
краткой величины (Гейнрих, 1967; Виноградов, 1968). Опреде-
ленное значение для части видов птиц, очевидно, имеет зимнее
понижение температуры воды и воздуха.
Как и в южной субтропической зоне, сильное обеднение по-
верхностных слоев планктоном явилось причиной формирования
у некоторых массовых планктоноядных трубконосых перелетов
за экватор, в Арктобореальную область, В Тихом океане это тон-
коклювый и серый буревестники, тайфунннк Пила, а в Атланти-
ке — серый и большой буревестники, качурка Вильсона. Список
таких мигрантов небольшой, но все они имеют высокую числен-
ность. Большая часть видов нотальной зоны относится к кочевым.
У некоторых из них в характере сезонных перемещений есть при-
знаки, свойственные для перелетных птиц. Осенью и зимой после
завершения размножения они широко рассеиваются в океане,
расширяют область распространения в субтропическую, а часть
— в тропическую области. Для многих из них характерны мигра-
ции через Южный океан в системе течения Западных ветров. Та-
кие миграции свойственны, в первую очередь, океаническим труб-
коносым — альбатросам, буревестникам, тайфунникам, прионам,
качуркам. Сезонные перемещения некоторых из них подробно
описаны в первой главе. Не покидают или почти не покидаюг
пределы нотальной зоны чайки, бакланы и пингвины. Две по-
следние группы в рассматриваемой зоне хорошо представлены
видами и имеют огромную суммарную численность. Именно сре-
дн этих групп есть оседлые и полуоседлые островные популяции
или виды. Для видов же, гнездящихся у побережья Новой Зелан-
дии и Южной Америки, характерны перемещения вдоль побере-
жий, не имеющие, однако, большой протяженности.
Летом в период размножения численность птиц в нотальной
зоне максимальная. Возвращаются сюда в это время также не-
половозрелые особи, численность которых значительна, если
учесть позднее созревание морских птиц вообще и трубконосых
в особенности. Для неразмножающихся особей характерно посте-
пенное продвижение на юг в течение всего летнего периода. Это
хорошо согласуется с закономерным смещением в высокие широ-
ты областей повышенных концентраций планктона. В умеренных
водах максимум планктона приходится на весну и начало лета
(ноябрь—декабрь), а в водах Антарктики на разгар и конец ле-
та (январь—март). У ледовой кромки, постепенно отступающей
на юг, наблюдается постоянное весеннее состояние планктона,
Гейнрих, 1967; Виноградов 1968). В середине и второй половине
лета большое количество неполовозрелых особей многих ноталь-
ных видов мигрирует в воды Антарктики, где они распростра-
226
МИГРАЦИИ В РАЗЛИЧНЫХ КЛИМАТИЧЕСКИХ ЗОНАХ ТИХОГО ОКЕАНА
няются до ледовой кромки. До осени многочисленны в антаркти-
ческой зоне альбатросы, прионы, несколько видов нотальных бу-
ревестников, особенно тонкоклювый и серый. Во многих районах
Антарктики они доминируют в это время над антарктическими
видами. Осенью и зимой почти все они мигрируют в более север-
ные воды.
Таким образом, птицы, гнездящиеся в нотальной зоне, для ко-
чевок используют все зоны океана от Арктики до Антарктики
(рис. 55). Основное значение имеет нотальная зона. Зимой очень
большое значение имеет также южная субтропическая зона, я
летом — антарктическая. Менее существенно, но также весомо
место бореальной зоны, в которой проводит зиму несколько
очень массовых видов.
Во всех рассмотренных выше зонах, даже таких, как тропиче-
Рис. 55. Районы кочевок и зимовок морских птиц, гнездящихся в южной
умеренной и антарктической зонах. 1 г- птицы антарктической зоны,
2—4 — птицы южной умеренной зоны (интенсивность штриховки соответ-
ствует значению районов как мест кочевок)
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 23Т
ская и бореальная, в отдельные сезоны наблюдается очень пест-
рый видовой состав. Мигрирующие из других зон птицы местами
могут доминировать над местными видами как по числу видов,
так и по числу особей. Нотальная зона в этом смысле яв-
ляется исключением. Во все сезоны основу населения
и видового состава составляют гнездящиеся здесь птицы. От-
меченная особенность объясняется, с одной стороны, тем, что в
нотальную зону в ограниченных количествах проникают птицы
из других широт (с севера летом и с юга зимой), а с другой, —
большим видовым разнообразием и высокой численностью но-
тальных видов, многие из которых кочуют в пределах зоны и зи-
мой. Здесь нет необходимости более подробно описывать измене-
ние состава птиц в рассматриваемой зоне, особенности миграций
ряда характерных представителей потальной зоны детально рас-
смотрены в первой главе. Во второй главе в разделе, посвящен-
ном Австрало-Новозеландскому району, описано распростране-
ние и изменение доли в общей численности основных видов из
различных комплексов.
Антарктическая зона. В Антарктической зоне сезонные коле-
бания в гидрологии и биологии моря выражены более резко, чем
в нотальной зоне, однако они менее контрастны, чем в Арктике.
В этом отношении антарктическая зона сопоставима с северно-
бореальными районами Тихого океана. Зимой она только частич-
но закрывается льдами; даже в период наибольшего охлаждения
на значительной части зоны сохраняются низкие положительные
температуры воды.
В зимний, наименее благоприятный для обитания период, пти-
цы Антарктики кочуют, часть из них проникает в другие зоны. В
северное полушарие совершает регулярные перелеты массовая
вилохвостая качурка Вильсона. В Атлантическом океане в зим-
ний период она концентрируется в пределах бореальной (Dorst.
1962а), в Индийском и Тихом — тропической зоны. До бореаль-
ной зоны в Тихом океане и в тропики в Индийском и Атлантиче-
ском мигрирует часть особей южно-полярного поморника. Но
этот вид в значительных количествах зимует и в Антарктике в
районе ледовой кромки (Eklund, 1961, 1964). До южной субтро-
пической зоны распространяются чернобрюхая качурка, капский
голубь, гигантский буревестник. Два последних вида в неболь-
ших количествах проникают и в южную часть тропической зоны.
В нотальную зону расширяют зимой область распространения
антарктический, серебристо-серый и снежный буревестники, прн-
он — Pachyptila desolata (Gmel). Правда, большая часть первых
трех видов, особенно снежного буревестника, остается в преде-
лах Антарктики. Наименее выражены перемещения за пределы
антарктической зоны у пингвинов. На север их миграции, как
правило, ограничиваются границей распространения льдов, хо-
228
МИГРАЦИИ Б РАЗЛИЧНЫХ КЛИМАТИЧЕСКИХ ЗОНАХ ТИХОГО ОКЕАНА
тя отдельные особи могут достигать не только островов в ноталь-
ной зоне, но даже вод Австралии. Из изложенного видно, что в
Антарктике, как и в других зонах, наибольшая протяженность
миграций свойственна видам, добывающим пищу в поверхност-
ном слое, наименьшая — ныряющим.
В летний период антарктические птицы за пределами рас-
сматриваемой зоны не встречаются. Основные их концентрации
в это время в районе ледовой кромки.
Таким образом, антарктические птицы для кочевок использу-
ют все зоны Мирового океана, за исключением Арктики (см.
рис. 55). Основное значение имеют две зоны — антарктическая
и нотальная. Кроме миграций с севера на юг и обратно для час-
ти видов характерны перемещения вдоль зоны. Миграции этого
вида выявлены при помощи кольцевания. Показан, в частности,
весьма значительный разлет вдоль побережья южно-полярного
поморника (Eklund, 1961, 1964). В первой главе цитировались
многие работы, в которых описываются аналогичные миграции
гигантского буревестника и капского голубя.
Бурное развитие планктона в Антарктике в летний период хо-
рошо известно. Огромные скопления планктона и особенно криля
делают этот сезон исключительно благоприятным в кормовом от-
ношении для птиц и других животных. Но в связи с коротким ле-
том этот благоприятный период имеет сравнительно небольшую
продолжительность. Поэтому в районы гнездования антарктиче-
ские птицы прилетают рано — в октябре и ноябре1. Вылупление
птенцов приходится на январь, а в конце февраля — в марте
птицы уже покидают гнездовья (Maher. 1962; Prevost, 1964.
1964а; Прайор, 1964; Сыроечковскнй, 1966; и др.). Таким обра-
зом, время выкармливания птенцов приходится на наиболее бла-
гоприятный в кормовом отношении период. Выше уже подчерки-
валось, что максимальные концентрации планктона в поверхност-
ных слоях в Антарктике наблюдаются в середине и второй поло-
вине лета. Опускание его в нижние горизонты начинается в апре-
ле (Виноградов и Наумов, 1961; Виноградов, 1968). В это время
птицы начинают откочевывать на север.
Огромные пищевые ресурсы антарктических вод привлекают
сюда большое количество птиц из соседней нотальной зоны. Ле.
том нотальные буревестники, альбатросы и прионы доминируют
над антарктическими видами во многих районах зоны. Среди
льдов местами обычна северная арктическая крачка. Зимой, ког-
да все они откочевывают на север, на протяжении всей зоны пре-
обладают антарктические птицы.
Краткий обзор особенностей сезонных перемещений птиц в
1 Хорошо известным исключением из этого правила -является император-
ский пингвин — Aptenodytes forsteri Gray, размножающийся в зимний период.
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 229
различных зонах океана позволяет сделать несколько общих за-
ключений. Размах сезонных миграций морских птиц увеличи-
вается с увеличением широты, в связи с усилением контрастно-
сти в сезонной изменчивости условий обитания (климат, гидро-
логия, кормовая база). Во всех зонах преобладают кочевые пти-
цы. Особенно много (не менее 90%) их в тропической зоне. По
мере движения от экватора на юг и север увеличивается процент
перелетных и перелетно-кочевых птиц. Оседлых видов среди мор-
ских птиц мало вообще, и встречаются они только в тропической,
южной субтропической и нотальной зонах. Во всех зонах, за ис-
ключением тропической, климатические и кормовые условия оби-
тания птиц наиболее благоприятны в весенне-летний период. В
это время размножается большая часть видов птиц. В крайних
северных и южных районах в связи с коротким летом сроки вы-
ведения потомства очень сжаты. В более низких широтах в свя-
зи с мягким климатом и менее резкими колебаниями в обеспечен-
ности кормом размножение более растянуто, часть видов гнездит-
ся и в зимний период. В тропической зоне размножение птиц про-
исходит круглогодично, при этом в одних районах преобладает
гнездование в летний, а в других — в зимний периоды.
Из всех изменений среды, определяющих характер и масшта-
бы миграций птиц, наиболее важны изменения в обеспеченности
пищей и ее доступности. Этот фактор играет решающую роль во
всех зонах, в том числе в тропиках. С увеличением широты, осо-
бенно в северном полушарии, на распределение и распростране-
ние птиц существенный отпечаток накладывает непосредственное
влияние климатических и гидрологических факторов.
Во всех зонах обеспеченность пищей зависит главным обра-
зом от сезонных изменений в численности морских беспозвоноч-
ных и рыб вообще, а в поверхностных слоях и на малых глуби-
нах -г- в особенности. Имеет значение также образование льдов
и синоптическая обстановка (частота и сила штормов), опреде-
ляющие доступность кормовых организмов. Наиболее чутко реа-
гируют на изменения в кормовой базе пелагические виды, добы-
вающие пищу в поверхностном слое моря. Виды, способные до-
бывать пищу на различных горизонтах, меньше пелагических за-
висят от сезонных изменений в кормовой базе. Миграции таких
видов (чистиковые, пингвины, бакланы) во всех зонах имеют
меньшую по сравнению с пелагическими птицами протяженность.
С другой стороны, перемещения пелагических трубконосых огра-
ничиваются площадью Мирового океана.
Имея в виду вынужденность сезонных перемещений птиц,
нельзя не видеть, что в них отсутствует хаос. Миграции носят
приспособительный характер. Благодаря им, птицы имеют воз-
можность переживать неблагоприятное для обитания в том или
яао
МИГРАЦИИ Б РАЗЛИЧНЫХ КЛИМАТИЧЕСКИХ ЗОНАХ ТИХОГО ОКЕАНА
ином районе время, а также максимально использовать кормо-
вые ресурсы океана. Яркой иллюстрацией последнего положения
может служить довольно большая группа южных трубконосых*
совершающая регулярные миграции в северное полушарие.
Время пребывания их в каждом полушарии соответствует пе-
риоду наибольшего обилия корма в поверхностных водах. Та-
кие миграции, несомненно, дают большую выгоду данным ви-
дам, о чем свидетельствует их высокая численность. Перелеты до-
вольно большой группы видов с высокой численностью приводят
к заметному перераспределению плотности населения птиц меж-
ду северным и южным полушариями в противоположные сезоны.
Это хорошо видно на рис. 56 и 57, отражающих распределение
плотности населения птиц в Тихом океане в различные сезоны.
Все рассмотренные выше основные факторы внешней среды
Рис. 56- Средняя плотность населения птиц в Тихом океане и сопредель-
ных водах, оо данным визуальных счетов в декабре—марте. I — менее 0,5;
2 — от 0.5 до I; 3 — от 1 до 10; 4 — более 10 особей на 1 кв. км
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 231
Рис. 57. Средняя плотность населения птиц в Тихом океане и сопредель-
ных водах, по данным визуальных учетов в июне—сентябре. Обозначения,
как на рис. 56
(кормовые, климатические и гидрологические), вернее, их из-
менчивость, являются первопричинами миграций птиц. Хорошо
известно и значение факторов другого рода, а пинаю—внутрен-
них и внешних факторов, вызывающих перелетное состояние
птиц или являющихся непосредственными стимулами миграций.
Я не имею возможности останавливаться здесь на этих сложных
и менее изученных вопросах. Работы в этом направлении по
сухопутным птицам в настоящее время весьма успешно разви-
ваются, в том числе в Советском Союзе. Особого упоминания
заслуживают исследования В. Р. Дольника (1965, 1967, 1968).
Установлено, что миграционное поведение, имеющее наследст-
венную сигнальную природу, определяется гармональными, ме-
таболическими, погодными, пищевыми и психическими воздей-
ствиями. Важнейшим элементом миграционного состояния яв-
232
МИГРАЦИИ В РАЗЛИЧНЫХ КЛИМАТИЧЕСКИХ ЗОНАХ ТИХОГО ОКЕАНА
ляется установление особых отношений между жировым и угле-
водным метаболизмом. Накопление в теле жира усиливает ме-
таболическую ситуацию, являющуюся основой миграционного
поведения.
Эти выводы, очевидно, могут быть перенесены и на морских
птиц, совершающих перелеты или в сезонных перемещениях ко-
торых есть элементы перелетности. Однако для большинства
кочевых птиц непрерывные миграции, исключая гнездовый пе-
риод, являются нормой. Их концентрации в районах гнездова-
ния связаны только с размножением. Послегнездовое рассеива-
ние является возвращением к нормальному образу жизни. Оче-
видно, для этого не нужно особого физиологического состояния
или сигнала из внешней среды. Несомненно, в условиях плотных
концентраций, какие бывают в районах колоний, прокормиться
и выжить труднее, чем осваивая ресурсы более обширных аква-
торий. Не нужно особого физиологического состояния и для то-
го. чтобы перекочевывать из одного кормного района в другой.
Непосредственным фактором таких перемещений является
ухудшение в обеспеченности пищей; птицы при обеднении кор-
мом одного района просто вынуждены искать другой. О конт-
роле сигнальными факторами, а также гормональными и мета-
болическими процессами поведения таких птиц, по-видимому,
представляется возможным говорить только относительно эта-
пов годичного цикла, связанных с размножением.
Все изложенное выше показывает исключительную роль со-
временных экологических условий, определяющих основные осо-
бенности сезонных перемещений морских птиц в различных
климатических зонах- В общих чертах это соответствует выво-
дам А. В. Михеева (1964, 1967, 1969), который больше других
подчеркивает, что современная картина перелетов обусловлена
ныне существующими условиями среды. Во многом соглашаясь
с этим автором, трудно, однако, разделить его точку зрения, ког-
да от отрицает или почти отрицает роль наследственных и исто-
рических факторов в формировании направлений и путей миг-
раций, мест зимовок, поведения птиц во время перелетов. Мас-
са доказательств в этом смысле приводится во многих работах,
написанных в различное время. (Мензбир, 1934; Тугаринов, 1937,
1950, 1952; Промптов, 1941; Штейнбахер. 1956; Гриффин, 1966;
Dorst, 1962а; и мн. др.). Правда, в работах более раннего перио-
да значение исторических причин слишком абсолютизирова-
лось, а значение современных условий недооценивалось. Ненуж-
ное противопоставление современных и исторических причин пе-
релетов, наследования миграционных направлений, их измен-
чивости и явилось причиной длительной дискуссии в орнитоло-
гии о перелетах птиц.
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 233
Невозможность объяснить современными экологическими
условиями все особенности миграций морских птиц видна во
многих случаях. Трудно допустить, например, что для полярной
крачки нет благоприятных условий для зимовок ни в одной из
зон, кроме льдов Антарктики, куда она совершает ежегодно
столь протяженные миграции. Не очень далеко от гнездовой
части ареала этого вида, а именно — в северной субтропиче-
ской зоне, она могла бы найти обильные кормовые условия в
виде массовой молоди сайры, анчоуса и других пелагических
рыб, размножающихся зимой- По-видимому, Уильямс (Williams,
1952) был прав, предполагая сравнительно недавнее распрост-
ранение ее из Антарктики, где обитает очень близкий вид —
антарктическая крачка.
Примеров, когда район зимовки вида является местом, отку-
да шло расселение вида, немного. В прошлом значение путей
расселения в формировании направлений миграций, конечно, пе-
реоценивалось. Путь миграций повторяет путь расселения,
очевидно, только первоначально, затем, в связи с влиянием дру-
гих факторов и изменением условий, пути миграций и места
зимовок претерпевают большие изменения. Это, однако, не дает
оснований для выводов, отрицающих значение наследственной
основы в выборе направлений и путей миграций, а также мест
зимовок. Ведь, как бы не изменялись в зависимости от измене-
ний в гидрологическом режиме сроки миграций и перераспре-
деление птиц на зимовках, общий характерный план распреде-
ления остается. Тонкоклювый буревестник, например, от Авст-
ралии летает ежегодно только в северную часть Тихого океана,
а бледноногий, кроме того, — в Аравийское море. Вилохвостая
чайка в Тихом океаме мигрирует и зимует только на востоке
Тихого океана, хотя аналогичные условия обитания есть и в за-
падной части океана. Даже альбатросы, которых считают веч-
ными странниками, не кочуют беспорядочно. Кольцеванием до-
казано, что характерные и относительно постоянные места ко-
чевок и зимовок свойственны не' только каждому виду альбат-
росов, но и отдельным популяциям (см. выше). Еще больше та-
ких примеров известно о сухопутных птицах. Но, признавая, что
птицы имеют свойственные видам или популяциям направления
перелетов и места кочевок или зимовок, просто невероятно до-
пускать, что такая сложная, но относительно постоянная карти-
на миграций иа огромных пространствах создается каждым по-
колением заново, как это полагает А. В. Михеев. Она могла
сформироваться только в течение длительного ряда поколе-
ний на основе преемственности между поколениями (пусть
даже случайно). Вывод о том, что миграции птиц про-
исходят на наследственной основе под влиянием современных
234
К ВОПРОСУ О ПРОЛЕТНЫХ ПУТЯХ МОРСКИХ птиц
условий, является признанием того, что современная картина
сезонных перемещений отражает и историю их формирования.
Конечно, наследуемое постоянство направлений и мест коче-
вок не нужно понимать абсолютно. Это постоянство может
быть наиболее относительно среди всех других наследуемых яв-
лений и признаков. Здесь можно сослаться, например, на пока
недостаточно изученную межгодовую изменчивость, наблюдае-
мую в распределении птиц. Известно, что в теплые годы тепло-
любивые виды наиболее далеко проникают в высокие широты,
а в холодные годы холодолюбивые — в низкие. Заметные пе-
рераспределения птиц вызывают изменения в интенсивности
течений, особенно в тех районах, где холодные и теплые тече-
ния направлены вдоль побережий на север или на юг (напри-
мер, Куросио, Перуанское). Несомненно, изменения в окружаю-
щей среде, имеющие однонаправленный характер, т. е. имеющие
тенденцию развиваться в одном направлении в течение очень
длительного периода, способны вызвать большие и кординаль-
ные изменения в распределении птиц. Что же касается измен-
чивости в распределении и миграциях, вызываемой межгодовы-
ми изменениями в климате и гидрологии моря в аспекте корот-
коцикловых колебаний или имеющих неправильный ход, то она
носит частный характер и совершается на фоне средней типич-
ной картины.
Нужно заметить, что консервативность многих черт и явле-
ний вообще свойственна птицам, особенно морским. Очень чет-
ко проявляется у морских птиц, например, гнездовой консерва-
тизм. Очевидно, в связи с ограниченностью пригодных для
гнездований мест и их большой разобщенности гнездовой кон-
серватизм у морских птиц выражен более резко, чем у многих
сухопутных1.
2. К вопросу о пролетных путях
морских птиц
Стремление к классификации миграционных путей, как из-
вестно, имело широкое распространение в прошлом. При этом
большое внимание уделялось прибрежным путям, по которым
мигрирует большое количество береговых и водных птиц. Океа-
1 Это показано в цитированных выше работах, освещающих результаты
кольцевания разных видов морских птиц. Можно, кроме того, сослаться на ре-
зультаты кольцевания пингвинов (Sladen and Tickell, 1958; Lacan et al. 1969;
и др.). Гнездовой консерватизм также не нужно понимать в полном смысле
слова. Абсолютный консерватизм был бы отрицанием расселения. В то же
время у некоторых видов отмечено изменение размеров гнездовой части ареа-
ла за сравнительно небольшие отрезки времени. Выше уже упоминалось о рас-
ширении ареала и увеличении численности глупыша в северной Атлантике.
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 235
нические птицы в то время почти не рассматривались, так как
их миграции Фактически не были известны- Такие попытки име-
ли место позднее. Сервенти (Serventy, 1953),-например,разделил
трубконосых, встречающихся в Тихом и Индийском океанах,
на оседлых, мигрирующих в долготном и в широтном направле-
ниях. По-видимому, из-за отсутствия соответствующих данных
в группу оседлых птиц попали такие дальние мигранты, как ко-
ролевский альбатрос, тайфунннк Штейнегера, белолицая качур-
ка и другие. Что касается миграций с севера на юг, с юга на
север, с запада на восток и с востока на запад, то они действи-
тельно имеют место- Однако такая классификация мало что да-
ет. Граница между миграциями в широтном и долготном нап-
равлениях весьма условна. Выше было показано, что даже та-
кие циркумполярные мигранты, как странствующий альбатрос
и гигантский буревестник, совершают регулярные перемещения
с севера на юг и обратно на тысячи километров.
10 миграционных путей у трубконосых выделил Курода
(Kuroda, 1957). Анализ этой классификации обнаруживает ее
большую искусственность и неточность. По данному автору,
северотихоокеанские альбатросы летают в пределах Северной
Пацифики по часовой стрелке по круговому пути. Но, как по-
казано в первой главе, в характере миграций этих птиц нет я
намека на такой путь. Трансэкваториалыные мигранты из райо-
на Австралии и Новой Зеландии, по Курода, совершают пере-
леты по западно-тихоокеанскому и центрально-тихоокеанскому
путям. На его схеме, а также у Сервенти, опять же иллюстри-
руется круговой путь по часовой стрелке. В первой главе в этом
плане подробно рассмотрены только тонкоклювый и серый бу-
ревестники. Из представленных схем видно, что буревестники
широким фронтом пересекают океан и в данных маршрутах не-
возможно уловить подобие предполагаемых путей. Этот вывод
можно перенести и на других, мигрирующих за экватор, труб-
коносых Тихого океана. Не подтвержденным остается и индо-
океанский круговой путь, предложенный Гибсон-Хиллом (Gib-
son-Hill, 1953). В частности, бледноногий буревестник, которо-
му приписывался этот путь, осенью от Австралии летит прямо
на северо-запад в Аравийское море (Bailey, 1966, 1968). Ана-
логичные замечания можно сделать и по другим путям схемы
Курода.
В некоторых районах, например в Англии, увеличила численность моевка. По-
казано, что новые колонии образуют молодые особи, значительный процент
которых ежегодно рассеивается в другие колонии (Coulson and White, 1958,
1960; Coulson, 1963). Аналогичная картина наблюдается с красноклювой чай-
кой в Австралии (Sharland, 1956). В последнем районе в настоящее время,
по-видимому, начал размножаться новозеландский буревестник Буллера
(D’ombrain and Gwynne, 1962}.
236
К ВОПРОСУ О ПРОЛЕТНЫХ ПУТЯХ МОРСКИХ птиц
Большая пестрота, наблюдаемая в миграциях птиц, и рас-
средоточение многих из них на обширных акваториях, по-види-
мому, будут делать бесперспективными попытки разработки
стройной классификации пролетных путей и в будущем. Этот
вывод относится и к пелагическим перелетным птицам, которые
в большинстве мигрируют широким фронтом. Делая такое зак-
лючение, я не хочу отрицать существование пролетных путей
вообще. У некоторых видов они. несомненно, существуют.
Вопрос о пролетных путях птиц много дискутировался в ли-
тературе- Много внимания ему уделяли отечественные орнито-
логи. В результате представления о формах перелета птиц пре-
терпели существенные изменения. После стройных схем и пред-
ставлений о пролетных путях (Мензбир, 1934; Тугаринов, 1937;
и др.) широкое признание получили мнения о миграциях широ-
ким фронтом (Кумари, 1957, 1959; Михеев, 1964; и др.). Харак-
терно, что в какой степени раньше абсолютизировались пролет-
ные пути, нередко столь же решительно они отрицаются в на-
стоящее время. Такой подход к рассматриваемой проблеме вряд
ли правилен. Я согласен с И. И. Пузановым (1960), который
на Первой Всесоюзной орнитологической конференции подчер-
кивал, что пролетные Пути предавать забвению рано. Сущест-
вование их у ряда видов отрицать невозможно. Правильна, на
мой взгляд, следующая трактовка данного явления, которую
высказывали в разное время некоторые авторы (например,
Промптов, 1941; Штейнбахер, 1956; и др.)- Большая часть птиц
совершает перелеты широким фронтом. Такая форма миграций
позволяет говорить о перелетных потоках, которые то сужива-
ются, то расширяются и разветвляются в зависимости от гео-
графических и экологических условий- В местах наложения по-
токов различных видов, особенно там, где фронт сужается, об-
разуются пути массового пролета птиц. Сужение фронта ггроле-
та имеет место у побережий морей, вдоль долин рек, в районе
узких проливов, гряд островов, горных хребтов и т. д. Переле-
ты только узким фронтом — явление более редкое. Однако они
имеют место и свойственны в первую очередь наиболее специа-
лизированным видам, в частности водным.
Подробнее остановимся на особенностях миграций в этом
плане морских птиц. Предварительно, однако, нужно сделать
некоторое отступление для характеристики экологических груп-
пировок морских птиц, миграции представителей которых за-
метно отличаются.
При описании сезонных аспектов в орнитофауне различных
районов уже говорилось о прибрежных, неритических и океа-
нических видах и группировках птиц. В работах иностранных
авторов морские птицы подразделяются на три группы: при-
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 237
брежные (inshore), дальне-береговые (offshore) и океанические
(Murphy, 1936; Baker. 1951; и др.). Птицам перечисленных
группировок свойственны некоторые биологические особенности.
Так, для океанических видов в сравнении с прибрежными ха-
рактерно более позднее созревание, более длинный гнездовой
период, меньшая плодовитость (Баск, 1967).
Классификация морских животных в таком плане довольно
распространена. Например, Гловер (Glover, 1961) в планктоне
Северного моря выделил неритическую, промежуточную и океа-
ническую группы. В масштабе всего Мирового океана разрабо-
тал эту проблему применительно к планктону К. В. Беклеми-
шев (1967. 1969). Этот автор выделяет неритические, дальне-
неритические и океанические виды и группировки планктона,
распространение которых приурочено к определенным водным
массам. Океанические виды обитают преимущественно в пер-
вичных водных массах, связанных с крупномасштабными зо-
нальными круговоротами. Стациями дальне-неритических видов,
по К. В. Беклемишеву, являются вторичные водные массы нейт-
ральных областей. Наконец, неритические виды обитают в при-
брежных модифицированных водных массах. Нечто аналогичное
наблюдается и у рыб. Например, Н. В. Парии (1968) говорит о
неритических, нерито-океанических и океанических рыбах. Ю. Ю.
Марти (1970) показал различие в протяженности миграций и
других особенностях, наблюдаемых у рыб различных группиро-
вок.
Для перечисленных выше группировок птиц, а также всех
других животных, по-видимому, целесообразно использовать
терминологию, применяемую К. В. Беклемишевым, а именно, го-
ворить о неритических, дальне-неригаческих и океанических
видах и группах. Правда, содержание этих терминов примени-
тельно к различным систематическим группам может быть не-
сколько различным. Для планктона к мелких рыб большое зна-
чение имеет, в частности, непосредственное влияние течений, оп-
ределяющих распространение видов и фор-му их ареалов.
Ю. Ю. Марти (1970). имея это в виду, называет районы основ-
ных крупномасштабных круговоротов областями вечного дрей-
фа. Решающее значение непосредственному влиянию водных
потоков на распространение пленктеров придает и К. В. Бекле-
мишев. Характер связей активно мигрирующих водных живот-
ных, в том числе большинства рыб. с водными массами, конеч-
но, иной, тем более не могут рассматриваться «рабами» тече-
ний морские птицы. Для всех истинно водных животных, неза-
висимо от того, пассивные или активные это мигранты, большое
значение имеют физико-химические характеристики водных
масс. Общеизвестно огромное значение в распространении жи-
238
К ВОПРОСУ О ПРОЛЕТНЫХ ПУТЯХ МОРСКИХ птиц
вотных температуры, солености и других факторов, определяю-
щих специфику водных маос.
Для морских птиц эти факторы также имеют значение. На-
пример, температура воды играет решающую роль в формиро-
вании широтной зональности распространения. Что касается
рассматриваемых группировок птиц, то их тяготение к опреде-
ленным водным массам, очевидно, нужно объяснять, как уже
отмечалось выше, влиянием комплекса факторов, формирую-
щих общий фон океанографического окружения. Наряду с
этим предоставляется возможным говорить, что в формировании
группировок решающую роль имели, с одной стороны, различия
в способах добычи, связанных с морфологическими особенностя-
ми видов, а с другой. — различия в условиях добычи пищи в
различных водных массах. В этом плане исключительное значе-
ние имеют гидрооптические свойства вод, а также плотность
концентраций в них кормовых организмов. Изменение данных
характеристик по мере движения из открытого океана к бере-
гу связано с трансформацией вод, обусловленной взаимодейст-
вием океанов и континентов. Результат этого взаимодействия
выражается в наличии неритических зон и дальне-неритических
областей (Беклемишев, 1969).
К прибрежной или неритической группе относится большая
часть видов бакланов, некоторые чайки, крачки, пингвины, по-
видимому, отдельные чистиковые (очевидно, обыкновенный и оч-
ковый чистики), а также многие полуморские птицы — нырко-
вые утки, гагары, водорезы. Характерным для этой группы яв-
ляется обитание в узкой, не более 4—5 миль, прибрежной зо-
не. За ее пределы, как правило, не выходят постоянно обитаю-
щие здесь неритические морские виды, а также зимующие пти-
цы с пресных водоемов. Большей частью все они мигрируют
вдоль побережий. Таким образам, рассматриваемая группа да-
ет значительное количество примеров миграций узким фронтом.
Неритические птицы имеют наибольшее распространение в се-
верном полушарии, особенно в Аркто-бореальной области
(рис. 58). Здесь они наиболее многочисленны. Миграции при-
брежных птиц вдоль азиатского и американского побережий в
Тихом океане носят очень массовый характер. После наблюде-
ний здесь многочисленных верениц мигрирующих прибрежных
птиц вдоль берегов материков и островных дуг не остает-
ся сомнений в реальности существования пролетных пу-
тей. Несомненно, когда Пальмен и М. А. Мензбир выделяли ма-
рино-литоральные и океаническо-береговые пролетные пути,
они имели для этого все основания.
К дальне-неритическим видам нужно отнести поморников,
большую часть чистиковых, почти всех морских чаек, многих
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 239
Рис. 58. Распространение неритической (1), дальне-неритической (2) и
океанической (3) группировок морских пищ в Тихом океане и сопредель-
ных водах
крачек, ныряющих буревестников, пингвинов, часть олуш, фре-
гатов и несколько видов трубкоиосых Дальне-неритические виды
имеют более обширную область распространения, охватываю-
щую всю площадь шельфа и прилегающие к нему глубоко-
водные районы. Таким образом, виды этой группы встречаются
на десятки и даже сотни миль от берегов. Так как большая
часть их обнаруживает четкую связь с водами шельфа, они
имеют широкое распространение в тех районах, где мелководья
довольно обширны. В этом смысле вполне объяснимо большое
видовое разнообразие и широкое распространение дальне-нери-
тических видов в северном полушарии и, в первую очередь, в
Аркто-бореальной области, (см. рис. 58). Наиболее ограниченное
распространение дальне-неритические виды имеют в летний
К ВОПРОСУ О ПРОЛЕТНЫХ ПУТЯХ МОРСКИХ птиц
период, когда за пределами материкового склона многие из них
не встречаются совсем. Зимой область распространения значи-
тельно шире. На севере Тихого океана в это время многие даль-
не-неритические виды обычны в глубоководных районах дальне-
восточных морей и в открытом океане на сотни миль от Япон-
ских, Курильских и Алеутских островов. В первой и второй гла-
вах было показано, какое широкое распространение имеют зи-
мой многие чистиковые и чайки.
В большинстве районов южного полушария дальне-неритиче-
ские виды имеют менее широкое распространение. К этой груп-
пе относятся многие виды птиц (веслоногие, крачки, чайки),
гнездящиеся у побережья Центральной и Южной Америки.
Здесь, как известно, шельф узкий, и эти виды тяготеют к приб-
режной эоне. Основными направлениями сезонных миграций в
этих районах являются перемещения вдоль побережий. В цент-
ральной части Тихого океана, где нет шельфовой зоны, поведе-
ние некоторых из них или близких видов другое. Как обратил
внимание Бекер (Baker, 1951), гнездящиеся здесь фрегаты,
олуши, некоторые крачки не отличаются по распространению
от океанических видов. На западе океана, где сконцентрирова-
но огромное количество островов, архипелаги которых прости-
раются на восток от Австралии и Индонезии, дальне-неритиче-
ские виды имеют более широкое распространение (см. рис. 58).
В этом птицы значительно отличаются от планктона. Дальне-
неритические виды планктеров в связи с более обширными нейт-
ральными областями на востоке океана имеют здесь и более
широкое распространение (Беклемишев, 1969).
Особую подгруппу составляют так называемые ледово-нери-
тические птицы. В северном полушарии характерным ледово-
неритическим видом является белая чайка, в южном — снеж-
ный буревестник, антарктические виды пингвинов. Распростра-
нение этих птиц, как правило, ограничено плавучими льдами.
Черты, свойственные ледово-неритическим видам, характеризуют
распространение и некоторых других антарктических птиц, в
частности антарктического и серебристо-серого буревестников.
При объяснении распространения ледово-неритических видов
планктона допускается, что лед в данном случае играет роль
берега (Беклемишев, 1969). О правдоподобности такой трактов-
ки говорит хотя бы такой пример. Обыкновенный чистик обыч-
но обитает в узкой прибрежной зоне. Зимой в период широко-
го распространения льдов он становится обычным у кромки
льдов в центральной части Берингова и Охотского морей. Ледо-
во-неритические виды птиц, по-видимому, должны рассматри-
ваться в группе дальне-неритических птиц.
В обеих неритических группировках по количеству видов и
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 241
особей преобладают ныряющие итицы- Многие из них являются
посредственными или плохими летунами, что, конечно, ограни-
чивает размеры обследуемой площади при поиске пищи. С этим
вполне согласуется обитание таких птиц в прибрежных, более
насыщенных жизнью водах. На малых глубинах для ныряющих
птиц оказываются доступными и бентические животные. Мень-
ше, но также хорошо представлены в прибрежных группах
птицы, добывающие пищу при помощи поискового полета. Наи-
более характерны из них чайки, которые являются в основном
всеядными, способными кормиться и в литоральной зоне, и в
открытом море.
Анализ особенностей миграций птиц дальне-неритической
группировки не дает данных о миграциях узким фронтом. В
большинстве случаев возможно выделять только сгущения миг-
рационных потоков. Такие сгущения обычно имеют место в райо-
нах пролетных путей неритических видов, а именно: вдоль бе-
регов, островных гряд и в проливах. В двух первых главах при
описании отдельных видов и районов это подчеркивалось неод-
нократно. В большинстве случаев, однако, трудно говорить о
пролетных путях в классическом понимании. Скажем, миграции
чаек и пестролицего буревестника через Корейский пролив на-
поминают оживленный пролетный путь. Но вое эти виды, осо-
бенно пестролицый буревестник, к югу и северу от пролива
мигрируют фронтом шириною в сотни миль- Миграционные по-
токи чистиковых и чаек вдоль побережий дальневосточных мо-
рей не уступают по массовости волнам прибрежных видов. Но
одновременно они встречаются на всей* акватории не только мо-
рей, но и на обширных пространствах сопредельных вод север-
ной части Тихого океана. Аналогичная картина наблюдается и
в южном полушарии, правда, здесь дальне-неритические виды
встречаются в более узкой прибрежной полосе. В категорию
птиц, летающих узкими пролетными путями, не могут быть
включены и такие трансэкваториальные мигранты, как помор-
ники.
Итак, миграции большой части дальне-неритических видов
происходят широким фронтом, ширина которого чаще всего со-
ставляет сотни, реже десятки миль.
Настоящие океанические виды есть лишь в некоторых се-
мействах. Только океаническими видами представлены семейст-
ва альбатросов, качурок, по-видимому, фаэтонов. В основном
океанические виды — в семействе буревестников. Есть океани-
ческие виды среди крачек, олуш, фрегатов. С сушей и прибреж-
ной зоной океанические птицы бывают связаны только в период
размножения- Область распространения этой группы наиболее
обширна. Она включает фактически весь Мировой океан, иск-
лючая только непосредственно прибрежную зону (см. рис. 58).
242
К ВОПРОСУ О ПРОЛЕТНЫХ ПУТЯХ МОРСКИХ птиц
Океанические виды встречаются, таким образом, не только
в первичных водных массах, по и в трансформированных водах
окраинных морей и нейтральных областей. В морях, сильно
изолированных от океана, а также в районах с обширными
шельфами приуроченность рассматриваемых видов к струям
океанских вод проявляется особенно четко. В местах, где
шельф узкий и нетраисформированные океанические воды под-
ходят непосредственно к берегу, океанические виды проникают
наиболее близко к побережьям. С этим хорошо согласуется
распространение океанических птиц среди многочисленных ост-
ровов в центральной части Тихого и Индийского океанов. Океа-
ническая группа морских птиц представлена только видами, ко-
торые добывают пишу в поверхностных слоях. Для них харак-
терен длительный поисковый полет — необходимое условие
для обитания в условиях низких концентраций кормовых орга-
низмов. Наиболее широкое распространение океаническая груп-
пировка имеет в Южном полушарии, где слабо развиты шель-
фовые зоны. Эти районы характеризуются и наибольшим видо-
вым разнообразием океанических птиц.
Имея столь обширное распространение, океанические пти-
цы, в том числе совершающие дальние миграции через Южный
океан или из южного полушария в северное и обратно, не дают
ни одного примера миграций узким фронтом. В периоды коче-
вок или дальних однонаправленных перемещений все они
встречаются на акваториях, простирающихся на тысячи миль.
В продуктивных районах и зонах они могут образовывать зна-
чительные скопления.
Таким образом, как и береговые птицы, большая часть мор-
ских птиц совершает миграции широким фронтом. Но процент
видов, мигрирующих узким фронтом, здесь выше, чем у берего-
вых птиц. Последнее связано с многочисленностью прибрежной
группировки птиц.
* • •
Подведем некоторые итоги обсуждения закономерностей в
распределении и миграциях морских птиц. Наиболее общие
черты в распространении и распределении связаны с климатиче-
ской зональностью. Каждой крупномасштабной зоне свойствен-
ны характерный видовой состав и особенности в биологии, рас-
пределении и миграциях морских птиц. Именно климатические
факторы определяют границы распространения зональных ви-
дов. В пределах каждой зоны на распределение птиц наклады-
вают отпечаток особенности океанографических условий зоны.
Здесь очень важным является размещение водных масс и тече-
ний, определяющих условия добычи пищи и распределение кор-
мовых организмов. Этим вопросам выше уделено достаточно
ЗАКОНОМЕРНОСТИ В РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 243
много внимания. Ниже в главе по орнитогеографическому райо-
нированию проблема широтной зональности будет освещена
дополнительно в несколько другом аспекте.
В связи с тем, что условия обитания претерпевают сущест-
венные изменения по сезонам, птицы находятся в непрерывном
движении. Перемещения их по сезонам, а также в период каж-
дого сезона нужно рассматривать не только как вынужденные,
вызываемые ухудшением климатических и кормовых условий.
Периодические миграции имеют приспособительный характер в
условиях жизни непостоянной среды и, что самое главное,
направлены на максимальное использование кормовых ресур-
сов океана. В этом смысле районы кочевок и зимовок птиц
должны быть благоприятными не только по климатическим и
гидрологическим, но и кормовым условиям. На почве использо-
вания кормовых ресурсов птицы неизбежно вступают в конку-
рентные отношения друг с другом, что должно оказывать влия-
ние на их численность и распределение. Это, а также зависи-
мость количества и плотности населения морских птиц от про-
дуктивных зон и концентраций кормовых организмов будут
предметом специального рассмотрения в следующих главах.
Таким образом, распределение птиц в Мировом океане конт-
ролируется комплексом факторов, влияние которых в большом
ряду поколений обусловило относительно постоянные районы
кочевок и зимовок видов. В этом плане общие направления миг-
раций определяются взаимным расположением мест гнездова-
ния и летнего обитания с местами' зимних кочевок. Однако неко-
торые факторы на том или ином участке пути определяют за-
метные отклонения от общего генерального направления, соеди-
няющего районы обитания в противоположные сезоны. Болыпое-
значение здесь имеют очертания береговой линии и материко-
вой отмели. Значительная часть неритических и дальне-нерити-
ческих видов при миграциях повторяет очертания или берегов,
или шельфов. Отклонение от основного направления может вы-
зываться расположением продуктивных зон, во многом опреде-
ляемом особенностями циркуляции вод. В ряде случаев опреде-
ленное значение может иметь ветер. Но влияние ветра на раз-
мещение птиц и пути их миграций нередко переоценивается.
Достаточно сказать, что вся схема миграционных путей Курода
(Kuroda, 1957) построена исключительно на основе схемы вет-
ровой циркуляции. Слов нет, ветры и особенно сильные цикло-
ны имеют большое значение в жизни птиц. Они могут ухудшать
условия для питания и миграций- Сильные ветры иногда раз-
носят птиц за пределы обычной области распространения и не-
редко вызывают гибель их. После штормов обнаруживается
большое количество мертвых птиц, например, на побережье
244
СУХОПУТНЫЕ ПТИЦЫ В МОРЕ
Австралии и Новой Зеландии (Goddard et al. 1951; Hindwood
and McGill. 1955; Mills, 1958; Kinsky, 1968; Imber and Boeson,
1969; и мн. др.). Погодные условия, конечно, отражаются на
перераспределении птиц в пределах района обитания, оказыва-
ют влияние на скорость перемещения птиц1. Ветры определяют
направление движения птиц в каждом данном моменте, но не
в смысле формирования постоянных миграционных путей и
районов кочевок. Представления о том, что именно ветры опре-
деляют пути миграций птиц, основаны на выборочных случаях,
когда основное направление перемещения соответствует направ-
лению господствующих ветров. Обычно для примеров исполь-
зуются миграции трубконосых в Южном океане с попутными
западными ветрами. Но, как было показано, не менее харак-
терным здесь являются миграции с севера на юг и обратно. За-
бывается при этом, что те же альбатросы и другие птицы, ко-
торые из Атлантики и Индийского океана мигрируют к Австра-
лии по ветру, назад вынуждены лететь против него. Кроме то-
го, непосредственные полевые наблюдения показывают, что мор-
ские птицы, особенно трубконосые, летают не по ветру, а по i
углом к нему (нередко поямым) вдоль волн и под углом к ним.
3. Сухопутные птицы в море
Ежегодно весной и осенью в ряде районов Тихого океана су-
хопутные птицы встречаются в больших количествах. Они при-
влекают к себе внимание, хотя и не входят в морские сообщества
В литературе хорошо освещены перелеты в Атлантическом океа-
не, по Тихому океану таких данных значительно меньше.
В период моих эскпеднционных работ все случаи встреч су-
хопутных птиц в море фиксировались. Наблюдения показали,
что наиболее оживленный пролет наблюдается осенью и весной
в дальневосточных морях, при этом количество наблюдаемых
птиц уменьшается в роду Восточно-Китайское—Японское—
Охотское—Берингово моря (Шунтов, 1961а; 1962; 1965а; 1970а).
Это обстоятельство связано с тем, что через Берингово море ле-
тят -в основном птицы, гнездящиеся в тундровой зоне, а в более
южных морях встречаются, кроме них, птицы с обширных про-
странств востока азиатского материка, а также крупных остро-
вов (Японские, Сахалин, Курильские). Зимовки большей ча-
стя птиц восточной Палеарктики располагаются не южнее Ки-
тая. В связи с этим >к югу от Восточно-Китайского моря коли-
чество сухопутных птиц в море сильно уменьшается. Через Ав-
страло-азиатские моря в больших количествах летят, главным
1 Важность в изучении воля пролета птиц синоптической обстановки на-
глядно показана, например, И. И. Пузановым и Л. Ф. Назаренко (1965).
ЗАКОНОМЕРНОСТИ Ь РАСПРЕДЕЛЕНИИ И МИГРАЦИЯХ МОРСКИХ ПТИЦ 245.
образом, кулики и ласточки. В тропической зоне миграции име-
ют, как известно, меньший размах- Однако тропические птицы
встречаются в море, хотя и в меньших количествах, чем север-
ные. Регулярные перелеты тропических птиц существуют через
моря, отделяющие Австралию от Новой Гвинеи и Зондских ост-
ровов. При работах в этих районах я имел возможность наблю-
дать такие миграции. Хорошо известны перелеты некоторых но-
возеландских птиц (Мауг. 1953; и др).
Сухопутные птицы встречаются в море, главным образом, в
весенний и осенний периоды, т- е. во время сезонных миграций.
В западной части Тихого океана пролет более оживленный, чем
на востоке океана. Причина этого заложена уже в самой конфи-
гурации азиатского побережья, где много краевых морей и
гряд островов, вытянутых в общем направлении с севера на юг.
Естественно, птицы более часто встречаются на пролете через,
проливы или заливы, которые они пересекают, сокращая путь.
Очень оживленный продет птиц проходит, в частности, через се-
верную часть Восточно-Китайского моря и Корейский пролив,
между северной частью полуострова Корея и Южным Приморь-
ем, между северным побережьем Охотского моря и Сахалином,
вдоль Курильских островов к Японии и обратно1- При сильных
ветрах (а осенью постоянны ветры от материка в море) часть
мигрирующих птиц может просто сноситься в открытый океан,
где они в большинстве погибают. Очевидно, такова же участь
многих особей, сбившихся с пути во время тумана.
Важность наличия для мигрирующих птиц островных гряд
показана на примере западной и центральной частей Тихого
океана (В<акег, 1953; Мауг, 1953; Kuroda, 1961). В центральные
районы Тихого океана мигрируют только кулики и утки1 2. Но на
западе океана на несколько сот миЛь от Японии, Филиппинских
и Зондских островов регулярно встречаются и настоящие берего-
вые птицы (в основном воробьиные). На востоке океана остро-:
вов почти нет, и береговая линия слабо изрезана. Отсюда в рай-
он Полинезии проникают только кулики и утки, но в меньших
количествах, чем со стороны Азин.
Осенью сухопутные птицы в открытом море наблюдаются
чаще, чем весной, и встречаются на более обширных акватори-
ях. Дело в том, что осенью мигрирует много молодых, более
слабых и хуже ориентирующихся особей. Большое значение в
этом случае имеют постоянные ветры с материка (северо-за-
падный муссон), уносящие птиц в море.
1 Я не рассматриваю адесь хорошо известный факт наиболее интенсив*
ных миграций вдоль непосредственно материкового побережья.
2 Многие кулики регулярно залетают и на острова тропической части Ин-
дийского океана (Bailey, МВ7).
246
ГЛАВА IV
РАСПРЕДЕЛЕНИЕ ПТИЦ В СВЯЗИ С МЕЖВИДОВОЙ
КОНКУРЕНЦИЕЙ ЗА ПИЩУ
Вывод Ч. Дарвина о решающей роли биотических связей ь
эволюции в настоящее время является вполне обоснованным л
доказанным. При этом особое значение придается определяющей
роли конкурентных отношений между организмами, которые
предопределяются ограниченностью многих ресурсов среды.
Как правило, ограниченными бывают пищевые ресурсы, а изме-
нение и развитие пищевых связей и конкуренция за пищу имеют
наибольшие эволюционные последствия (Лэк. 1957; Майр, 1968;
Давиташвили. 1969; Шмальгаузен, 1969; и мн. др.).
Конкуренция принимает более острые формы между живот-
ными с наиболее сходными экологическими потребностями. Этот
вывод является основной мыслью известного «закона Гаузе»
(Gause, 1934) и различных его интерпретаций (Hutchinson and
Deevey, 1949; Gilbert et al., 1952; Hardin, 1960; Макфедьен, 1965;
и др.).
Любая конкуренция, даже слабая, создает серьезное давле-
ние естественного отбора; последний вызывает дивергенцию
признаков, уменьшающую или устраняющую конкуренцию меж-
ду видами. Устранение или ослабление конкуренции достигает-
ся географическим и биотопическим расхождением в местах
обитания, а также всевозможными экологическими различиями,
касающимися использования ниш в одном и том же биотопе
(Лэк, 1949, 1957; Майр, 1968 и мн. др.). В результате этого при
изучении сложившихся биоценозов и фаунистических комплек-
сов нередко, даже чаще всего, бросается в глаза не проявление
острой конкуренции как таковой, а ее последствия в виде раз-
личных взаимоисключений. Данные выводы во многом обяза-
ны наблюдениям за птицами, дающими огромное количество
случаев взаимоисключений.
В последние два десятилетия начали развиваться исследова-
ния такого направления и по морским птицам. В серии обстоя-
тельных работ (Fisher and Lockley, 1954; Wynne-Edwards, 1962;
Ashmole. 1963; Bourne, 1963; Rown, 1965; Ashmole and Ashmole.
1967; Lack, 1967; Белопольский, 1957; и др.) показано, что пища
за немногими исключениями является основным фактором, вы-
РАСПРЕДЕЛЕНИЕ ПТИЦ В СВЯЗИ С КОНКУРЕНЦИЕЙ ЗА ПИЩУ 347
зывающим конкурентные взаимоотношения с вытекающими из
них последствиями, а также фактором, определяющим общий
уровень численности птиц. При этом, конечно, не отрицается
большое значение и других факторов: хищников, климатических
условий, наличия мест для гнездования и других*.
Одной из характерных биологических особенностей морских
птиц является колониальность пнеэдования. Среди морских
птиц на долю колониальных приходится 93%. в то время как
для орнитофауны земли в целом эта цифра составляет всего
13% (Lack, 1967). Концентрируясь в больших количествах на
ограниченных площадях, морские птицы в колониях вступают
в весьма сложные и разнообразные Отношения-
H. Ашмол и М. Ашмол (Ashmole and Ashmole, 1967), прово-
дившие исследования сообщества гнездящихся птиц на распо-
ложенном в тропической части Тихого океана острове Рождест-
ва, показали, что несмотря на то, что здесь многие виды близки
в систематическом отношении и по экологическим потребностям,
конкуренция за пищу уменьшена до уровня, когда возможно
совместное существование. Это достигается различиями в мето-
дах питания, что определяет использование видами различной
пищи или позволяет одним видам ловить добычу, не доступную
для других; различиями в морфологических характеристиках
близких видов (размеры тела, клюва и т. д.), определяющих
размеры используемой пищи; несовпадением гнездовых сезонов
у некоторых родственных видов, а также питанием в разны»1
часы суток и на различном удалении от побережья.
Аналогичные выводы получены и по птицам умеренных ши-
рот, гнездовые колонии которых располагаются в зоне более
продуктивных вод, размножающимся к тому же в сезон (весен-
не-летний) наибольшего обилия вод кормовыми организмами.
Вопрос межвидовых отношений колониальных птиц в северных
морях наиболее обстоятельно рассмотрен Л. О- Белопольским
(1955, 1957, 1957а) в серии работ, являющихся итогом весьма
интенсивных отечественных исследований в Баренцевом море в
предвоенный и послевоенный периоды. Л. О. Белопольский под-
черкивает первостепенное значение кормового фактора, опреде-
ляющего численность и размещение птиц. Его анализ питания
птиц, способов добычи корма, путей развития различных на-
правлений в освоении кормовых биотопов (специализация при
освоении ресурсов открытого моря, «универсализм» при исполь-
зовании ресурсов литорали) выявляет экологическую радиацию,
1 В колониях птиц, интенсивно эксплуатирующихся человеком, о большом
значении естественных факторов в изменении численности говорить не при-
ходится. В связи с этим вспоминается известная драматическая судьба бес-
крылой гагарки в Атлантике и белоспинного альбатроса в Тихом океане.
248
РАСПРЕДЕЛЕНИЕ ПТИЦ В СВЯЗИ С КОНКУРЕНЦИЕЙ ЗА ПИЩУ
направленную на наиболее полное использование кормовых ре-
сурсов. Пищевые ниши большинства морских птиц в Баренце-
вом море не совпадают с таковыми кайр, численно преобладаю-
щих здесь над другими видами. Расхождение достигается раз-
личиями в способах добычи (глупыш, моевка), в местах кормле-
ния (чистики, бакланы), морфологических особенностях, опре-
деляющих специфичность пищевых спектпов (гагарка, люрик) ’•
Л. О. Белопольским показано, что большое значение имеют и
другие формы взаимоотношений, в которые вступают птицы при
совместном гнездовании. Так, в результате конкуренции за ме-
ста в колониях изменяется видовой состав, численность птиц,
характер гнездования, а также происходит изменение окружаю-
щих природных условий1 2.
Детальных исследований по экологии птиц в других морях
бореальной зоны проводилось мало. Но, по-видимому, многие
из вскрытых в Баренцевом море биологических особенностей
морских птиц носят характер общих закономерностей. Изучение
колониальных сообществ морских птиц, естественно, нуждается
в продолжении. Не иначе как следствие недостаточной изучен-
ности надо рассматривать противоречивые мнения ряда авторов
(Ashmole, 1963; Rown, 1965) о значении конкуренции за пищу у
хорошо обеспеченных кормом птиц умеренных областей. При об-
суждении рассматриваемой проблемы, как правило, имеются
в виду конкурентные отношения между птицами в период размно-
жения, когда они концентрируются на ограниченных площадях.
Районы кочевок у всех видов морских птиц по площади значи-
тельно превосходят районы, где они добывают пищу в гнездо-
вой период. Конкуренция за пищу в это время, естественно, вы-
ражена меньше А некоторые авторы наличие ее в данном слу-
чае вообще отрицают (Ashmole, 1963; Rown. 1965; и др.).
Океан действительно располагает колоссальными кормовыми
ресурсами. Тем не менее большой ряд факторов свидетельству-
ет о том, что корм во многих случаях является лимитирующим
численность фактором и во внепнездовой период, а конкуренция
за пищу определяет важные особенности в распределении птиц.
Сравнительные данные о питании разных видов птиц во вне-
1 О значении различий мест кормления в уменьшении межвидовой кон-
куренции недавно писал Пирсон (Pearson, 1968), изучавший питание птиц,
гнездящихся на острове Фарне (Северная Атлантика).
2 Состав населения гнездовых колоний зависит от рельефа и геологиче-
ского строения берегов. По мере изменения нх геофизической природы изме-
няется видовой состав птиц в колониях. В изменении условий гнездования
заметная роль принадлежит и птицам (изменение химического состава поч-
вы, уничтожение или распространение растительности и т. д.). Таким образом,
сами птицы принимают участие в создании условий, благоприятных для
гнездования одних видов и (неблагоприятных для других (Гнэенко, 1955;
Бреслина и Карпович, I960).
РАСПРЕДЕЛЕНИЕ ПТИЦ В СВЯЗИ С КОНКУРЕНЦИЕЙ ЗА ПИЩУ
249
гнездовой период по большинству районов Мирового океана
очень ограничены. Поэтому о пищевой конкуренции, вернее, о ее
последствиях приходится судить по косвенным признакам. Важ-
ным доводом в пользу признания большого значения кормового
фактора является зависимость плотности концентраций птиц
от продуктивности вод, о чем неоднократно подчеркивалось в
предыдущих разделах. Об ограниченности в морях и океанах
доступной для птиц пищи, особенно для видов, питающихся с
поверхности моря, свидетельствует склонность многих видов
(чайки, альбатросы, некоторые буревестники) питаться отброса-
ми с судов и отходами промысла. Тенденция концентрироваться
у судов наиболее проявляется в малокормный зимний период-
Именно с улучшением кормовой базы в виде большого количе-
ства отходов рыболовного и китобойного промысла связывается
значительное увеличение численности ряда видов морских птиц,
например, глупыша и чаек (Fisher, 1952; Fisher and Lockley,
1954; Lockley, 1958; и др.). Наконец, в пользу большого значе-
ния конкуренции говорит огромное количество пространствен-
ных взаимоисключений между близкими по потребностям видов.
Описанию таких случаев и будет посвящена остальная часть
настоящей главы.
Элтон (Elton, 1946) для выяснения значения конкурентных
отношений сравнивал списки большого количества животных и
растительных сообществ различных биотопов. Было показано»
что в сообществах на 84—86% представлены роды с одним ви-
дом. Эти соотношения сильно отличаются от фаунистических
списков больших областей. В этом автор справедливо видит ре-
зультат влияния на формирование состава сообщества конку-
рентных отношений. Так (Tuck, I960), исследуя состав морской
орнитофауны на зимовках в водах Ньюфаундленда, пришел к
заключению, что конкуренция за корм между видами здесь в
значительной мере ослаблена- Это обеспечивается подбором ви-
дов. занимающих различные пищевые ниши. Более тщательный
анализ группировок морских птиц в двух районах тропической
зоны — в Аравийском море и водах Атлантики у западного по-
бережья Африки, провел Буэн (Bourne, 1963). В обоих рас-
смотренных районах концентрируется большое количество мор-
ских птиц, представленных как размножающимися здесь вида-
ми, так и видами, мигрирующими сюда на зимовки из более вы-
соких широт северного и южного полушарий. Проведенный ана-
лиз дал несколько важных выводов. Группировка птиц в каж-
дом районе включает набор видов, способных эксплуатировать
все уровни кормовой базы — от мелкого зоопланктона до круп-
ных рыб и кальмаров. В одном районе, как правило» держатся
виды, отличающиеся способом питания. Если совместно ком-
250 РАСПРЕДЕЛЕНИЕ ПТИЦ В СВЯЗИ С КОНКУРЕНЦИЕЙ ЗА ПИЩУ
центрируются родственные виды» то они отличаются размерами
тела, длиной и формой клюва, что находит отражение в видо-
вой специфике добываемой пищи. Виды со сходными потребно-
стями, т. е. занимающие близкие экологические ниши, обычно
разобщены пространственно или встречаются в одном районе в
различные сезоны. Значительное количество накопленных в по-
следние годы сведений по распределению и миграциям птиц да-
ет возможность рассмотреть в таком плане группировки птиц н
во многих районах Тихого океана. Сопоставление распределения
родственных видов птиц дает здесь много примеров простран-
ственных взаимоисключений. Рассмотрим несколько таких
примеров.
В дальневосточных морях и в северо-западной части Тихого
океана из планктоноядных птиц в значительных количествах
встречаются мелкие чистиковые (наиболее массивы конюги), ка-
чурки, буревестники, глупыш и плавунчики. На море плавунчи-
ки наблюдаются только на пролете, при этом у азиатского по-
бережья преобладает круглоносый, у американского — плоско-
носый. Трубконосые и мелкие чистиковые обитают в рассматри-
ваемом районе круглогодично. Первые питаются в поверхност-
ных слоях моря и тяготеют к открытым водам, вторые добыва-
ют пищу при нырянии и тяготеют к шельфовым водам- Мелкие
качурки и имеющие средние размеры глупыш и буревестники
в значительной степени используют животных различных тро-
фических уровней, следовательно, занимают несходные эколо-
гические ниши.
Области распространения в море родственных серой и север-
ной качурок (гнездовые части ареалов перекрываются) заметно
отличаются. На большей части Берингова. Охотокосо и севера
Японского морей доминирует серая качурка, а для океанских
вод более характерна северная. В южной половине Японского
моря и на этой широте у восточных берегов Японии данные ви-
ды замещаются вилохвостой качуркой. Зимой, когда и северная,
и серая качурки держатся в океанских водах, серая тяготеет к
более северным, а северная — к южным районам. В северной
части Тихого океана -кроме трех отмеченных видов качурок оби-
тает еще шесть. По-видимому, в связи с этим сюда не соверша-
ют массовых трансэкваториальных миграций качурки южного
полушария. Интересно, что в северную часть Атлантического
океана, где гнездится только четыре вида качурок, до умеренных
широт из южного полушария проникает качурка Вильсона, а в
северную часть Индийского океана, где качурки нс гнездятся
совсем, на зимний период мигрируют в больших количествах- из
южного полушария три вида, а из Тихого океана — два вида
качурок (Bourne. 1963, Bailey et al-, 1968).
РАСПРЕДЕЛЕНИЕ ПГИЦ В СВЯЗИ С КОНКУРЕНЦИЕЙ ЗА ПИЩУ
ttl
Другая картина наблюдается в миграциях черев экватор бу-
ревестников. В умеренных водах дальневосточных морей и се-
верной части Тихого океана, за исключением глупыша, нет гнез-
дящихся видов буревестников. Зато из южного полушария сюда
регулярно совершает перелеты целая группа видов и среди них
такие массовые, как тонкоклювый и серый. Районы основных
концентраций этих двух видов в основном не совпадают. Тонко-
клювый буревестник, как было показано выше, откармливается
в более северных водах, а серый — в более южных. Оба вида
почти не заходят в Восточно-Китайское и южную половину
Японского моря> где многочислен пестролицый буревестник.
В значительной мере разобщенно с данными видами обитает и
бледноногий буревестник. В Японское море бледноногий и пест-
ролицый буревестники мигрируют весной через Корейский про-
лив. Первый вид тяготеет к западной, второй — к восточной
части моря. Оба вида немногочисленны на севере моря, куда
через проливы Сангарский и Лаперуза проникают серый и тон-
коклювый буревестники.
Глупыш и буревестники занимают различные ниши, однако, с
их пищевых спектрах есть и большое сходство: все они в боль-
ших количествах потребляют макропланктон, особенно эвфау-
зиид. Но буревестники посещают северную часть Тихого океана
в период, когда глупыш размножается и многие его особи кен-
центрируюкя у островов. Буревестники образуют особенно
плотные скопления там, где численность глупыша сравнительно
невысокая. Буревестники очень многочисленны летом на юге
Охотского моря, в районе Японии, южной части Курильской гря-
ды, шельфе восточной части Берингова моря, в Олюторсвю-На-
варинском районе, в заливе Аляска, местами в открытом океане.
Глупыш же наиболее многочислен летом в Охотском море, аа
севере Берингова моря в районе островов Святого Лаврентия,
Святого Матвея. Прибылова, вдоль Алеутских, Командорских и
северной части Курильских островов.
Вне конкуренции с другими трубконосыми находятся в север-
ной части Тихого океана питающиеся кальмарами альбатросы.
Как отмечалось выше, .северотихоокеанские альбатросы — чеп,-
ноногий и теммоспинный — размножаются в одних и тех же
районах (в настоящее время в основном на Гавайских остро-
вах). Во время кочевок во все сезоны темноспинный альбатрос
тяготеет к северо-западной, а черноногий — к северо-восточной
части Тихого океана. В районах наложения областей их распро-
странения в учетах нередко отмечаются одновременно оба вида.
Как правило, один из них явно преобладает.
К сожалению, сейчас еще мало конкретных данных о степе-
ни сходства экологических ниш конюг. нередко в больших коли-
252
РАСПРЕДЕЛЕНИЕ ПТИЦ В СВЯЗИ С КОНКУРЕНЦИЕЙ ЗА ПИЩУ
•чествах встречающихся совместно, особенно на севере Беринго-
ва моря. Есть косвенные данные о некотором различии потреб-
ностей мелких чистиковых. Об этом, в первую очередь, говорят
заметные морфологические особенности, наблюдаемые у разных
видов. Эти различия касаются и строения клюва (Козлова,
1957), что особенно важно. Здесь можно также сослаться на
пример, описанный Бедардом (Bedard, 1969). На острове Свя-
того Лаврентия совместно гнездятся в больших количествах кро-
шечная и большая конюги. Крошечная конюга предпочитает
участки с камнями размерами 20—40 см, большая — 60—80 см.
Это обеспечивает видовую разобщенность при совместном засе-
лении одной и той же каменистой россыпи.
Специфичны экологические ниши чаек. По берегам северной
части Тихого океана гнездится несколько крупных чаек: черно-
хвостая, тихоокеанская, серокрылая, полярная, серебристая, во-
сточная. Гнездовые части ареалов этих видов почти не перекры-
ваются (рис. 59). В период кочевок все виды широко рассеива-
ются по северной части Тихого океана- Несмотря на то. что об-
ласти их кочевок в значительной степени перекрываются, основ-
ные районы концентраций остаются разобщенными. Чернохво-
стая чайка зимует в основном на юге Японского, в Восточно-
Китайском море и вдоль тихоокеанского побережий Хонсю. Ти-
хоокеанская чайка концентрируется в основном в северной ча-
сти Японского моря, в Охотском море, а также вдоль восточно-
го побережья Камчатки, Курильских островов и Хоккайдо. Лю-
бопытно* что в период кочевок этот вид не идет далеко в откры-
тые воды в районе восточного побережья Камчатки, где много-
численна серокрылая чайка. В Охотском же море она обычна
в это время по всей акватории моря. Серокрылая чайка преоб-
ладает на обширной акватории от Командорских островов до
Калифорнии (исключая крайнюю северную часть Берингова мо-
ря). Южнее, вдоль берегов южной Калифорнии и Мексики зи-
мует восточная чайка. Занимающие на гнездовье наиболее се-
верные области полярная и серебристая чайки сравнительно
равномерно рассредоточиваются в районах зимовок перечислен-
ных выше видов* 1.
Крупные чайки рода Larus являются птицами шельфовых
вод. Зимой основная их масса концентрируется в пределах не-
। Интересные выводы получил Смит (Smith, 1966), изучавший естествен*
ные изоляционные механизмы крупных чаек L. hyperboreus, L. argentatus,
L. glaucoides, L. thayeri в арктической Канаде. Было показано, что гнездовые
части ареалов аллопатричных видов почти не (налегают друг на друга. У сим-
патричных же водов репродуктивная изоляция достигается гнездованием в
различных биотопах, несовпадением пиков половой активности и определен-
ными морфологическими различиями, влияющими на формирование пар и ко-
пуляцию.
Ри-с. 59. Распространение крупных чаек в северной части Тихого океана. Гнездовая часть ареала: 1 —
L. crassirostris, 2— L. schistjsagus, 3 — L. hyperboreus, 4 — L. glaucescens, 6 — L. occidentals
254
РАСПРЕДЕЛЕНИЕ ПТИЦ В СВЯЗИ С КОНКУРЕНЦИЕЙ ЗА ПИЩУ
скольких десятков миль, но в это время нередко их можно встре-
тить и далеко в открытом океане (особенно серокрылую). По-
являющаяся же осенью на морском побережье сизая чайка зи-
му проводит только в узкой прибрежной эоне- Тяготение к ли-
торальной зоне характерно и для некоторых мелких чаек (озер-
ная, Бонапарта). Наоборот, как правило, избегает зоны литора-
ли обыкновенная моевка — самая массовая из чаек, имеющая
наиболее обширную зону кочевок, включающую всю северную
часть Тихого океана.
Вместе с обыкновенной моевкой в центральной части Берин-
гова моря и прилегающих водах океана встречается красноно-
гая моевка. Характер их отношений не совсем ясен. Но нужно
иметь в виду, что красноногая моевка, несомненно, угасающий
вид. Большая часть ее земного поголовья (несколько тысяч
пар) концентрируется на острове Святого Георгия (Kenyon and
Phillips, 1965). Посещают северную часть Тихого океана и дру-
гие чайки, например, вилохвостая и белая. Но вилохвостая
встречается только во время пролета на зимовку в южное полу-
шарие и обратно, а белая появляется только зимой и держится
среди льдов, когда здесь нет других мелких чаек.
Из массовых видов, питающихся в основном рыбой и добы-
вающих ее при нырянии, в северной части Тихого океана обита-
ют бакланы, два вида кайр и топорок. Специфика пищевых
спекторов этих птиц кроме морфологических различий определя-
ется известной разобщенностью в местах их обитания. Бакланы
держатся только в прибрежной зоне, кайры многочисленны по
всему шельфу за пределами непосредственно прибрежных вод,
а топорок обычен как на шельфе, так и в глубоководной части
моря-
Сложнее объяснить совместное обитание очень сходных по
потребностям близких видов. Выше в таком плане затрагива-
лись конюги- Показательны в этом отношении и противоречи-
вые мнения о межвидовых отношениях толстоклювой и тонко-
клювой кайр, одних из наиболее многочисленных рыбоядных
морских птиц в умеренных водах северного полушария. Эти два
вида представляют собой пример аллопатрического формообра-
зования. Толстоклювая кайра обособилась в самостоятельный
вид от исходной формы в Арктике, а тонкоклювая в южных рай-
онах умеренной зоны (Sergent, 1951; Storer, 1952). В настоящее
время ареалы их в значительной мере перекрываются; во мно-
гих колониях они гнездятся совместно. Это, а также известное
сходство в пищевых спектрах послужили основой для выводов
об экологической идентичности данных видов (Sergent, 1951;
Tuck, 1960; и др.). Имеется, однако, много данных, свидетель-
ствующих о неполном совпадении экологических ниш толсто-
РАСПРЕДЕЛЕНИЕ ПТИЦ В СВЯЗИ С КОНКУРЕНЦИЕЙ ЗА ПИЩУ
256
клювой и тонкоклювой кайр. Л. О Белопольский (1957) пока-
зал, что тонкоклювая кайра является более специализирован-
ным ихтиофагом. В питании же толстоклювой кайры в Барен-
цевом море значительную роль играют беспозвоночные. Недав-
но Шварц (Swartz, 1966, 1967) установил, что в Чукотском мо-
ре пищевые спектры этих видов также заметно отличаются: в
питании толстоклювой кайры сравнительно большое значение
имеют беспозвоночные и донные животные. Наблюдается неко-
торое различие и в местах добычи корма. В гнездовых колони-
ях на побережье Аляски на долю толстоклювой кайры прихо-
дится около 60%, а в открытых водах — более 90%. Это гово-
рит о более пелагическом образе жизни толстоклювой кайры.
Последний вывод для других дальневосточных морей и северной
части Тихого океана подтверждают и мои наблюдения. Таким
образом, говорить о полной идентичности ниш двух видов кайр
не приходится.
Этот случай, по-видимому, может рассматриваться как тре-
тий случай «закона Гаузе». предусматривающий ситуацию, при
которой существует зона экологичеоюбго неперекрывания, допу-
скающая сосуществование двух близких по требованиям видов-
При этом оказывается, что в некоторых биотопах преимущест-
во имеет один вид. а в других — другой. Действительно, как
показано на примере кайр Баренцева моря, в более северных
районах в колониях преобладает толстоклювая, а в более юж-
ных ее вытесняет тонкоклювая (Белопольский, 1955, 1957; Ус-
пенский, 1956 и др.). Такая же особенность в распределении
двух видов наблюдается и в Тихом океане. В южной половине
умеренной зоны преобладает тонкоклювая, а на севере — тол-
стоклювая кайра (Дементьев. 1951; Гизенко, 1955; Козлова,
1957; Gabrielson and Lincoln, 1959; и др.).
Есть все основания полагать, что и в других случаях со-
вместного обитания близких видов дальнейшие исследования об-
наружат неполную идентичность их потребностей, т. е. только
частичное перекрытие ниш. При этом важное значение могут
иметь тонкие экологические различия, часто незаметные при бег-
лом анализе.
В таком же плане, как было рассмотрено население птиц в
северной части Тихого океана, остановимся на орнитофауне юго-
западной части Тихого океана (район Новой Зеландии и Авст-
ралии). Основу населения в данном районе составляют различ-
ные виды планктоноядных трубконосых. Низшие трофические
уровни кормовой базы используются здесь мелкими видами: ка-
чурками, ныряющими буревестниками и прионами. Ныряющие
буревестники занимают экологическую нишу, аналогичную та-
ковой мелких чистиковых. Другие трубконосые добывают пищу
356
РАСПРЕДЕЛЕНИЕ ПТИЦ В СВЯЗИ С КОНКУРЕНЦИЕЙ ЗА ПИЩУ
в поверхностных слоях моря. Находясь в этом смысле вне кон-
куренции, ныряющие буревестники тем не менее имеют неболь-
шую численность. Эти виды, как известно, — прибрежные пти-
цы. Может быть, их низкая численность находится в связи с ог-
раниченностью в данном районе шельфовой эоны.
Качурки используют ресурсы с более обширных акваторий.
Но в Новозеландском районе, отличающемся высокой продук-
тивностью. где гнездится несколько видов качурок, численность
их в целом невелика. Есть основание связывать это с присутст-
вием здесь огромного количества прионов, с которыми по чис-
ленности сопоставим разве только серый буревестник- В водах
острова Северный и у южного побережья Австралии, где прио-
нов меньше, численность качурок (белолицая качурка) выше.
Род прионов насчитывает шесть видов. Все они встречаются
у Новой Зеландии, при этом четыре вида гнездятся (Falla, 1940;
Oliver, 1955). Экология прионов изучена мало, в полевых усло-
виях они не отличимы, поэтому в настоящее время трудно объяс-
нить совместное существование нескольких близких видов. По-
видимому, существенное значение имеют отличия в форме и ве-
личине клюва, выраженные у различных видов весьма заметно
(Falla, 1940; Oliver, 1955). Последнее обстоятельство, как под-
черкивалось многими авторами (Лэк, 1949; 1957; Schoener, 1965;
и др.), имеет большое значение в расхождении ниш.
Из-за недостаточной изученности не представляется возмож-
ным подробно рассматривать и буревестников, большое количе-
ство видов которых совместно обитает в районе Австралии и
Новой Зелавдии. В этой группе есть и примеры явных прост-
ранственных взаимоисключений. Так, тонкоклювый буревестник
гнездится только в районе Тасмании и юго-восточного побе-
режья Австралии, где он образует огромные колонии. Серый
буревестник имеет здесь низкую численность- В больших коли-
чествах он гнездится в районе Новой Зеландии и Южной Аме-
рики (Murphy, 1936; Serventy and Whitley, 1951; Oliver 1955;
Cayley. 1965; и др.). В значительной степени разобщенно с дан-
ными видами обитает и бледноногий буревестник, гнездовья ко-
торого располагаются у юго-западного побережья Австралии,
на острове Лорд Хау и вокруг острова Северный. В последнем
районе гнездится и серый буревестник, но он здесь немногочис-
лен; его огромные колонии располагаются южнее. В северную
часть Тихого океана совершает перелеты только часть особей
бледноногого буревестника. От юго-западного побережья Авст-
ралии многие его особи мигрируют в северо-западную часть
Индийского океана.
Лучше изучены особенности размещения альбатросов, хоро-
шо различимых в полевых условиях. В Австрало-Новозеланд-
РАСПРЕДЕЛЕНИЕ ПТИЦ В СВЯЗИ С КОНКУРЕНЦИЕЙ ЗА ПИЩУ
257
ском районе многочисленны восемь видов альбатросов. В их
размещении на море наблюдаются заметные различия, о чем
можно судить по данным, изложенным в первой главе. Особен-
ности их распределения в связи с обсуждаемыми вопросами
кратко сводятся к следующему. Концентрации альбатросов жел-
тоносого, пугливого и Буллера в период кочевок располагаются
преимущественно в водах, прилегающих к побережьям (рис. 60).
Желтоносый альбатрос в больших количествах встречается толь-
ко у южного и западного побережий Австралии, альбатрос Бул-
лера — у острова Южный и западного побережья острова Се-
верный, а пугливый — вокруг Новой Зеландии и в районе Тас-
мании. Из перечисленных видов большие совместные скопления
образуют только альбатросы Буллера и пугливый в районе Но-
вой Зеландии. Однако эти виды нельзя считать- экологически
идентичными. Они отличаются размерами, при этом пугливый
альбатрос имеет более крушшй клюв. К открытым водам тяго-
теют основные концентрами странствующего, чернобрового,
сероголового и дымчатого альбатросов. Из трех последних ви-
дов наиболее северным распрострайением характеризуется чер-
нобровый альбатрос, наиболее южным — дымчатый, а сероголо-
вый занимает промежуточное положение. Очень четкая разоб-
щенность в местах поиска корма свойственна для очень близ-
ких в систематическом отношении чернобрового и сероголового
альбатросов и в водах атлантического сектора Южного океана;
следствием этого являются значительные расхождения в пище-
вых спектрах (Tickell, 1964). Районы скоплений странствующе-
го альбатроса нередко совпадают с таковыми чернобрового.
Но нужно иметь в ввду, что первый вдвое крупнее второго.
Пока не представляется возможным оконтурить районы
скоплений королевского альбатроса, очень близкого странству-
ющему по размерам и в систематическом отношении. Но в све-
те рассматриваемых закономерностей, очевидно, не случайно,
что из района Новой Зеландии, где многочислен на кочевках
странствующий альбатрос (в том числе особи, прилетающие из
индийского и атлантического секторов), значительное количе-
ство особей королевского мигрирует в воды Южной Америки, о
чем можно судить по данным кольцевания (Robertson, 1964;
Tickell, 1968; и др.). Странствующий и королевский альбатросы
совместно гнездятся на островах Кэмпбелл и Окленд; на острове
Кэмпбелл, где многочислен королевский альбатрос, странствую-
щего насчитывается всего несколько пар. На острове Окленд
соотношение обратное (Oliver, 1955; Westerskov, 1960; Bailey
and Sorensen, 1962). Королевский альбатрос в отличие от стран-
ствующего больше тяготеет к прибрежным водам (Westerskov,
1960а).
Рис. 60. Основные районы концентраций мелких альбатросов рода Diomedea в районе Австралии и Новой Зеландии
зимой. 1 — D. melanophrys, 2 — D. chlororhynchus, 3 — D. caula, 4 —* D. bulleri
РАСПРЕДЕЛЕНИЕ ПТИЦ В СВЯЗИ С КОНКУРЕНЦИЕЙ ЗА ПИЩУ
259
Чайки представлены в районе Австралии и Новой Зеландии
несколькими видами. Очень широко в южном полушарии рас-
пространена доминиканская чайка. Она гнездится в районе по-
бережий Южной Америки, Африки, Новой Зеландии и на мно-
гих островах, разбросанных в Южном океане. В зоне занимае-
мых ею широт она до последнего времени отсутствовала толь-
ко у Австралии (Alexander, 1954; и др.). Здесь обитает близкая
по габитусу Gabianus pacificus. Лишь в последние годы доми-
никанская чайка освоилась на гнездовьях в районе юго-восточ-
ного побережья Австралии (Thomas, 1967; Hill, 1968). Совмест-
но с обоими видами обитает красноклювая серебристая чайка.
Однако экология данного вида специфична, к тому же он вдвое
мельче.
Выше рассмотрены только два района и только часть оби-
тающих в них видов- Перечень подобных примеров пространст-
венного и экологического взаимоисключения можно было бы
продолжить.
Майр (1968) подчеркивает важность'отличать случаи, когда
географическое замещение связано с конкуренцией и когда рас-
пространению вида препятствует экологическая непригодность
области. Но все перечисленные выше примеры пространственно-
го взаимоисключения касаются видов, области кочевок, которых
перекрываются, а разобщенными бывают лишь основные кон-
центрации.
Все изложенное показывает, что закон конкурентного взаи-
моисключения находит вполне определенное выражение в рас-
пространении и распределении морских птиц. Его действие
проявляется как в гнездовой, так и в негнездовой период жизни
птиц. Это убеждает в том, что птицы в целом ограничены дос-
тупной пищей не только во время гнездования, но и в период
кочевок, когда они рассеиваются на больших пространствах.
Не представляется возможным в этом смысле противопостав-
лять птиц тропической и умеренных зон.
Заключая анализ распределения и состава сообществ мор-
ских птиц в аспекте влияния лимитирующего численность кор-
мового фактора, можно суммировать некоторые общие выводы.
Состав кормовой базы в различных районах Мирового океана
значительно отличается. Это находит отражение в количествен-
ном соотношении птиц, потребляющих животных различных
трофических уровней. Так, северные умеренные воды Тихого оке-
ана, как известно, богаты планктоном, рыбой и кальмарами.
Хорошо выражены здесь насыщенная жизнью литораль и бога-
тый бентосом шельф. Соответственно здесь имеют высокую чис-
ленность планктонофаги (мелкие чистиковые, качурки, буревест-
ники, глупыш), ихтиофаги (крупные и среднего размера чисти-
260
РАСПРЕДЕЛЕНИЕ ПТИЦ В СВЯЗИ С КОНКУРЕНЦИЕЙ 3> ПИЩУ
«Юбые бакланы), питающиеся кальмарами альбатросы, потреб-
ляющие бентос и >ыбу нырковые утки и чайки.
В умеренных и атарктичмких водах южного полушария
основной источник кормовой базы планктон (в том числе криль}.
Здесь, как нигде, многочисленны яланктоыоядные трубконосые
В основном макроклммтмгам в Южном океане питаются и
пингвины, имеющие высокую численность. В пределах этих зон
у побережья НомА Зеландии ж Южной Америки, где в значи-
тельных количествах встречаются многие виды рыб, увеличи-
вается доля ихтиофагов (чайки, бакланы, олуши).
В продуктивном перуанском районе, где из кормовых объ-
ектов наиболее обилен анчоус, фон птичьего населения
определяют рыбоядные веслоногие — бакланы, олуши, пе-
ликаны.
Для субтропических и тропических шельфовых вод Австра-
лии характерно большое обилие мелких пелагических рыб и их
молоди. Основу населения птиц здесь слагают питающиеся ими
крачки. Подобное соответствие характера кормовой базы и чис-
ленности определенного вида потребителей имеет место во всех
рейпаах океана. О еомоветствии кормовой базы и потребителей
говорит и различие в численности птиц в зависимости от того,
какого трофического уровня животными питаются они. Извест-
но, что в пищевом ряду фитопланктон — мелкий планктон —-
крупный планктон — мелкие рыбы — крупные рыбы и т. д, при
переходе с одного трофического уровня на другой происходит
большое уменьшение численности и объема продукции. Объем
цродукдаи смежных уровней может разниться в десятки и да-
же сотни раз (Бегоров, 1967; Моисеев, 1969; и др.). В связи с
этим не вызывает, удивления то, что самые многочисленные из
морских птиц являются(планктонофагами. Это буревестники, ка-
чурки, мелкие чистиковые, некоторые пингвины. Численность
птиц, использующих животных более высоких трофических уров-
ней, значительно ниже. Среди ихтиофагов наиболее многочис-
ленны виды, питающиеся мелкими пелагическими видами рыб.
Это тропические крачки, веслоногие, некоторые чистиковые.
Вполне логичен в этом плане в среднем невысокий уровень чис-
ленности крупных морских птиц, питающихся крупными рыбами,
кальмарами и другими животными.
Орнитофауна каждого района состоит из набора видов, по-
зволяющего максимально использовать кормовые ресурсы на
всех уровнях — от мезопланктона до сравнительно крупных
рыб и кальмаров. Плаактонофаги, в значительной мере питаю-
щиеся мезоилаактоном, мовут быть представлены в том или
ином районе различными мелками трубконосыми или медшми
чистиковыми и плавунянками; питающиеся мамропланктоном —
РАСПРЕДЕЛЕНИЕ ПТИЦ В СВЯЗИ С КОНКУРЕНЦИЕЙ ЗА ПИЩУ
261
это среднего размера буревестники, тайфунники, пингвины; их-
тиофаги, питающиеся мелкой рыбой и кальмарами, — крачки,
мелкие чайки, чиспгковые, крупной рыбой и кальмарами, —
крупные чистиковые, веслоногие, альбатросы, некоторые пингви-
ны, крупные чайки. В питании части видов последней группы
(чайки, чистиковые), а также уток существенное место занима-
ют бентические животные.
В те районы, где нет гнездящихся видов из потребителей опре-
деленной группы, они мигрируют из других. Например, в пре-
делах тропической части Индийского океана нет своих мелких
планктоноядных птиц. Зато сюда мигрируют качурки и плавун-
чики из других районов. В бореальной зоне летом, когда гнез-
дящиеся здесь потребители планктона в значительной мере тя-
готеют к побережью, богатые ресурсы планктона открытых вод
ими недоиспользуются. Это дает возможность откармливаться
здесь миллионам особей трубконосых из южного полушария.
Подбор в каждом районе видов с различными потребностя-
ми, а также пространственная разобщенность экологически
близких видов направлены на уменьшение конкуренции за пи-
щу и во многом определяются ею. Воды всех зон Мирового
океана в этом смысле оказываются как бы поделенными между
различными видами птиц для наиболее рационального исполь-
зования его кормовых ресурсов- Степень насыщенности раз-
личных районов океана потребителями ресурсов разных трофи-
ческих уровней и наличие в них незанятых экологических ниш,
несомненно, имели большое значение при формировании сооб-
ществ на местах гнездования, а также миграций птиц, их путей,
определении мест кОчевок и зимовок.
Известно, что более разнообразные сообщества наиболее ус-
тойчивы к вторжениям извне (Элтон, 1960). Хорошим подтверж-
дением этого правила могут быть примеры о миграциях качу-
рок и буревестников из южного полушария в северное. Анало-
гичные примеры можно указать и в других группах. Так, по-
морники р. Stecorarius, гнездящиеся только на севере, во время
кочевок мигрируют за экватор в южные умеренные воды в Ат-
лантическом, Индийском и Тихом океанах. Южно4юлярный по-
морник, обитающий в Южном океане, в северное полушарие
проникает только в Индийском и Тихом океанах. В северной
Атлантике его заменяет близкий подвид (или вид), который
также не мигрирует за экватор (Thomson. 1966). Все эти и дру-
гие аналогичные примеры могут быть рассмотрены в более об-
щем плане, с позиций различия в распространении и разнообра-
зии неритических и океанической группировок птиц в южном н
северном полушариях.
Как было показано выше, океаническая группировка наибо-
202
РАСПРЕДЕЛЕНИЕ ПТИЦ В СВЯЗИ С КОНКУРЕНЦИЕЙ ЗА ПИЩУ
лее представлена в южном полушарии. С этим хорошо согла-
суются миграции многих южных пелагических видов в северное
полушарие. Массовых миграций гнездящихся в северном полу-
шарии птиц океанической группировки в плотно заселенные пе-
лагическими птицами воды южного полушария не наблюдается.
С другой стороны, неритические группы птиц хорошо представ-
лены в северном полушарии. Некоторые из них при миграциях
проникают на юг за экватор. Это, например, три северных по-
морника, вилохвостая чайка, пестролицый буревестник, аркти-
ческая крачка и. видимо, другие. Аналогичных примеров сре-
ди птиц этих группировок, гнездящихся в южном полушарии,
почти нет. Очевидно, можно назвать только перелеты южно-по-
лярного поморника в северную часть Тихого океана.
Всевозможные приспособления для уменьшения конкуренции
за пищу не могут ее невелировать полностью. Так, даже в та-
ком высокопродуктивном водоеме, каким является Баренцево
море, в годы, неблагоприятные в кормовом отношении, конку-
ренция за пищу может достигать остроты, вызывающей измене-
ние численности видов (Белопольский, 1957). Об этом свиде-
тельствуют и другие примеры. В частности, выше упоминалось,
что в связи с появлением дополнительного источника питания в
виде отбросов и отходов промысла некоторые птицы заметно
увеличили свою численность-
Таким образом, если конкуренция остается, хотя и в смягчен-
ной форме, сохраняются и развиваются предпосылки, определяй
ющие давление естественного отбора, направленного на расхож-
дение признаков и уменьшение противоречий. Следовательно,
сложившиеся соотношения в численности и распределении ви-
дов не являются застывшими. Они должны меняться в связи и
в соответствии с изменяющимися условиями среды. Должны на
этом фоне наблюдаться изменения и в пространственном разме-
щении птиц. Н. А. Гладков (1960) отмечал, что устойчивость
пространственных связей у птиц несомненна, но она не абсо-
лютна, а относительна. К сожалению, орнитологические иссле-
дования по морским птицам в данном направлении имеют пока
ограниченный ряд лет наблюдений. Морская ихтиология, к при-
меру, располагает огромными материалами, отражающими ана-
логичные изменения в сообществах рыб.
263
ГЛАВА V
ОРНИТОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ
МИРОВОГО ОКЕАНА
Морские птицы еще не совсем нашли свое место в схемах
биогеографического деления океана. Нередко наравне с сухо-
путными птицами их используют для выделения и характеристи-
ки различных регионов суши. Делается это на том основании,
что для размножения они используют берега — пограничную
полосу между двумя резко различными средами — сушей и мо-
рем. Правильнее, однако, распространение морских птиц, явля-
ющихся настоящими морскими животными и, следовательно,
имеющих отличные от сухопутных птиц условия существования,
расселения и пути развития, рассматривать в связи и в зави-
симости от океанографических факторов. Более 30 лет назад на
это обращали внимание Р. Мэрфи (Murhy, 1936) и Ю. М. Каф-
таноеский (1951).
Действительно, на всех этапах жизненного цикла связи мор-
ских птиц с морем более тесны и важны в сравнении со связя-
ми с сушей- Это справедливо и для периода размножения, ког-
да, будучи привязанными к берегам, они с морем сохраняют
трофические связи. Нужно также учитывать, что и климатиче-
ские условия непосредственно береговой полосы, где распола-
гаются гнездовья птиц, находятся под решающим влиянием
моря, особенно течений.
Морские птицы, как и другие морские животные, дают ха-
рактерные примеры амфибореального (F. glacialis, U. aalge,
О. leuoorhoa), амфипацифического (О. пкхюгшв, С. monocerata)
и биполярного (Р. marina, С. skua, Р. mollis, Р. hypoleuca, Р. as-
simiiis) типов распространения. Зависимость от условий моря
иллюстрируют освещенные в предыдущих главах закономерно-
сти в распределении и миграциях, определяемые особенностя-
ми сезонных явлений в гидрологии и биологии моря-
Хорошо известно, что многие морские птицы в период коче-
вок, имея большие возможности для передвижения, широко
рассеиваются по океану, и часто вся область распространения
вида охватывает несколько климатических и гидрологических
эон. В связи с этим при районировании приходится учитывать
только репродуктивную часть ареала птиц. Таким образом,
'ЭД ОРНИ ГОГЕОГРАФИЧЕСКИЕ РЕГИОНЫ ТИХОГО И ИНДИЙСКОГО ОКЕАНОВ
предлагаемая ниже система регионов основана на распростра-
нении птенцов'. Это представляется вполне логичным, так как
период размножения — наиболее уязвимый момент в жизнен-
ном цикле животных; в это время птицы, как никогда, находят-
ся в большой зависимости от гидрометеорологических условий.
Далее, при анализе населения птиц того или иного района ис-
пользовались только данные по настоящим морским птицам, а
именно: чайкам, крачкам, поморникам, трубконосым, веслоно-
гим, пингвинам и чистиковым. Не учитывались встречающиеся
на море в больших количествах во внегнездовое время утки, ку-
лики, гагары. Из списка исследуемых видов исключены также
те виды из семейств чаек, крачек, бакланов и пеликанов, кото-
рые размножаются на внутренних водоемах, следовательно, не
зависят или мало зависят в это время от морских условий. Пос-
ле перечисленных олраннчений список использованных для райо-
нирования видов сократился до 250.
Распределение морских птиц в период размножения характе-
ризуется четко выраженной крупномасштабной зональностью.
Анализ распространения птиц позволяет выделить в Мировом
океане семь крупных регионов, сменяющих друг друга: аркти-
ческий, бореальный, северный субтропический, тропический,
южный субтропический, мотальный и антарктический (рис. 61).
Границы, разделяющие регионы, выявлены путем анализа
соотношений видов, характерных для различных зон. Ниже в
этом плане подробно рассмотрены только Тихий и Индийский
океаны.
1. Границы орннтогеографическнх регионов
в Тихом и Индийском океанах.
Распространение морских птиц бореального комплекса огра -
ничено в основном умеренным поясом. Северная граница боре-
ального орннтогеографического региона проходит по южной ча-
сти Чукотского моря (см. рис. 61). На участке, ограниченном се-
вером Чукотского полуострова, островом Врангеля й
мысом Барроу на Аляске, гнездится 19 видов морских птиц:
9 арктобореальных, 7 бореальных и 3 арктических1 2. 7 видов —
Phalacrocorax pelagicus, Aethia pusilia, A. cristatella, Brachyra-
mphus brevirostris, Cyclorhynchus psitacula, Lunda cirrhata,
Eratercula corniculata — определяют соотношение в этом районе
в пользу бореальных видов. При этом нужно отметить, что в
1 й орнитологии такой подход считается общепрвэнаяным (Дементьев,
1968; Лортенко, 1965 и др..
2 Все три арктических вида относятся к чайкам — L. hyperboreus, X. sa-
binei, Р. eburnea. Однако Р. eburnea гнездится в этом районе только на остро-
ве Геральд.
Рис. 61. Орнитогеографическое районирование Мирового океана. Орнитогеографические регионы: 1 — аркти-
ческий и антарктический, 2 — бореальный и нотальный, 3 — северный н южный субтропические, 4 — тропи-
ческий
2в6 ОРНИТОГЕОГРЛФИЧЕСКИЕ РЕГИОНЫ ТИХОГО И ИНДИЙСКОГО ОКЕАНОВ
данном районе все они имеют северный предел распростране-
ния.
В южной половине умеренной зоны по мере движения на юг
постепенно исчезают многие характерные для бореального ре-
гиона виды. До полуострова Корея с севера доходит только
6 видов — Phalacrocorax filamentosus, Larus crassirostris, Cero-
rhinca monocerata, Synthliboramphus antiquus, Cepphus carbo,
Uria aalge. Однако они составляют здесь основу (75%) видово-
го состава морских птиц. Южных элементов в районе восточного
побережья полуострова Корея, очевидно, всего 3 — Oceanodro-
ma monorhis, Sterna albifrons и Puffinus leucomelas.
У Японских островов южная граница бореального региона
проходит приблизительно на пять градусов севернее. От южной
части Корейского полуострова она поднимается к северной ча-
сти острова Хонсю. Острова в Корейском проливе, а также Кю-
сю, Сикоку и Хонсю, за исключением крайней северной части
последнего, относятся к субтропическому региону. Здесь гнездит-
ся 5 (72%) субтропических и тропических видов (Puffinus leuco-
melas, Oceanodroma monorhis, О. castrd, Synthliboramphus
wumisuzume, Sterna albifrons) и только 2 — южно-бореаль-
ных — Cerorhinca monocerata и Larus crassirostris. У северной
части о. Хонсю соотношение видов уже в пользу бореального ре-
гиона. Здесь обитает только три южных вида (Puffinus leucome-
las, Oceanodroma monorhis, Oceanodroma castro); все они име-
ют в этом районе северный предел распространения. Бореаль-
ных видов здесь 5 (63%), и у 3 из них (Phalacrocorax pelagicus,
Cepphus carbo, Uria aalge) южная граница распространения
проходит в районе северной части Хонсю, а 2 — Larus crassiro-
stris, Cerorhinca monocerata проникают на юг лишь немного
дальше. На острове Хоккайдо субтропических птиц нет, здесь
гнездятся только бореальные и южно-бореальные виды. Таким
образом, в районе северной части Хонсю наблюдается довольно
отчетливый стык фаун двух зон.
Аналогичная картина имеет место и у Нижней Калифорнии
в районе островов Санта Барбара, где гнездится 16 видов мор-
ских птиц: 8 бореальных, 5 субтропических и тропических и
3 общих для бореальной и субтропической зон. 5 бореальных
видов — Lunda cirrhata, Cepphus grylle, Uria aalge, Brachyra-
mphus marmoratus, Pelecanus erythrorhynchus — имеют здесь
южный предел распространения. С другой стороны, не идут се-
вернее островов Санта Барбара все 5 встречающихся здесь суб-
тропических и тропических видов — Brachyramphus hypoleucus,
Oceanodroma homochroa, Puffinus opisthomelas, Pelecanus occi-
dentalis, Thalasseus maximus.
Район американского побережья между островами Санта
Барбара и основанием полуострова Калифорния относится к
ОРЛНТОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
267
субтропичесокму региону. Из 14 встречающихся здесь видов
8 субтропических и тропических (Halocyptena microsoma, Осе-
anodroma monorhis, О. meiania, О. homochroa, Puffinus opistho*
melas, Brachyramphus hypoleucus, Pelecanus occidentalis, Thalas-
seus maximus) и только три бореальных (Oceanodroma leucor-
hoa, Ptychoramphus aleuticus, Pnalacrocorax pelagicus).
Учитывая все изложенное, бореальный орнитогеографиче-
ский регион можно очертить с юга границей, проходящей от
южной части Корейского полуострова через северную часть
Хонсю к Нижней Калифорнии в район островов Санта Барбара
(см. рис. 61).
Северная граница северного субтропического региона
очерчена при анализе распространения орнитофауны уме-
ренной зоны. В западной части Тихого океана к субтропиче-
скому региону, кроме юго-восточной части Японского моря, от-
носятся Восточно-Китайское и Желтое моря, а также район
островов Идзу и Рю-Кю. Южная граница региона проходит
здесь от северной части острова Тайвань, при этом острова Рю-
Кю остаются к северу от нее. Правомочность проведения грани-
цы в данном районе видна из анализа, состава гнездящихся
птиц. До Тайваня, где гнездится, по-видимому, не менее 11 ви-
дов птиц (5 тропических, 4 субтропических, 2 общих для тропи-
ческой и субтропической зон), с севера доходят и имеют предел
распространения субтропические Thalasseus zimmermanni,
Puffinus leucomelas, Oceanodroma monorhis и Diomedea albat-
rus1, а с юга — тропические Sterna anaethetus, Puffinus pacifi-
cus, Bulweria bulweri. Два тропических вида — Sterna sumat-
rana и Anous stolidus проникают немного севернее. На островах
Рю-Кю гнездится 10 видов морских птиц: 2 тропических (Bulwe-
ria bulweri, Sterna sumatrana), 5 субтропических (Puffinus le-
ucomelas. Pterodroma hypoleuca, Oceanodroma monorhis, Synth-
liboramphus wumisuzume, Diomedea albatrus) и 3 общих для
тропической и субтропической зон (Sterna albifrons, S. dougal-
li, Thalasseus bergii).
На островах Идзу соотношение видов еще больше в пользу
субтропических: здесь гнездятся один тропический (Sula leuco-
gaster), 4 субтропических (Puffinus leucomelas, Oceanodroma
monorhis, Synthliboramphus wumisuzume, Diomedea albatrus)
и 3 общих для тропической и субтропической зон (Diomedea
immutabilis, Diomedea nigripes, Sterna albifrons). Острова Бо-
нин, расположенные южнее, имеют в основном тропический со-
став морских птиц: тропических — 6 (Puffinus pacificus, Bulwe-
1 D. albatrus к настоящему времени на островах Пескадорских и Рю-Кю
истреблен.
268 ОРНИТОГЕОГРАФИЧВСКИЕ РЕГИОНЫ ТИХОГО И ИНДИЙСКОГО ОКЕАНОВ
ria. bulweri, Anous stolidus, Sula leucogaster, Phaethon rubricau-
da, Puffinus lherminieri), субтропических — 2 (Diomedea albat-
rus, Pterodroma hypoleuca), общих для тропической и субтропи-
ческой зон — 2 (Diomedea nigripes, Thalasseus bergii).
Таким образом, граница, разделяющая тропический и суб-
тропический регионы, от («верной оконечности острова Тайвань
и южной части островов Рю-Кю должна быть продолжена на
северо-восток между островами Идзу и Бонин. В центральной
части северной Пацифики она поднимается почти до 30’ с. ш.,
оставляя в пределах тропического региона всю гряду Гавайских
островов, на которых из 22 гнездящихся видов 19 являются тро-
пическими.
В восточной части Тихого океана граница вновь опускается
южнее. К северному субтропическому региону на востоке океа-
на относится район побережья между островами Санта-Барба-
ра и полуостровом Калифорния, о чем говорилось выше, а также
весь полуостров Калифорния с расположенными на его широте
островами (см. рис. 61). В районе полуострова Калифорния гнез-
дится 23 вида морских птиц: 10 субтропических (Brachyramp-
hus hypoleucus, В. craveri, Oceanodroma macrodactyla, О. mela-
nia, O. monorhis, O. homochroa, Halocyptena microsoma, Puffi-
nus opisthomelas, Larus heermanni, Thalasseus elegans), 5 тро-
пических (Sula leucogaster, S. nebouxii, S. dactylatra, Phaethon
aethereus, Fregata magnificens), 3 общих для тропической и суб-
тропической зон (Gelochelidon nilotica, Sterna albifrons, Peleca-
nus occidentalis), 2 бореальных (Oceanodroma leucorhoa, Pty-
choramphus aleuticus) и 3 общих субтропической и бореальной
зонам (Phalacrocorax auritus, Р. penicillatus, Larus occidenta-
lis). Из приведенного перечня видно, что состав гнездящихся
здесь видов довольно пестрый, что говорит о переходности райо-
на. Тем не менее преобладание субтропической группы над дру-
гими хорошо заметно. При этом нужно учитывать, что все пять
настоящих тропических видов не идут севернее рассматривае-
мого района.
Южнее на участке между полуостровом Калифорния и
островами. Ревилья Хихедо картина уже иная; здесь основу на-
селения составляют тропические виды: Sula dactylatra, S. sula,
S. nebouxii, S. leucogaster, Fregata magnificens, Phaethon aethe-
reus, Sterna fuscata, Anous stolidus. Здесь гнездится только
3 субтропических вида — Oceanodroma monorhis, О. melania,
Larus heermanni.
Учитывая соотношение видов различных комплексов в раз-
личных участках Калифорнийско-Мексиканского района, грани-
цу, разделяющую субтропический и тропический регионы, оче-
видно, нужно провести по южной оконечности полуострова Ка-
лифорния.
ОРНИТОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
369
Тропический орнитогеографический регион, располагающийся
к югу от северной оконечности Тайваня и полуострова Калифор-
ния, очень обширный; он занимает всю тропическую эону обоих
полушарий.
В юго-западной части Тихого океана у восточного побережья
Австралии граница, разделяющая тропический и южный субтро-
пический регионы, проходит в районе Сиднея (около 34е ю. ш.).
Из 20 гнездящихся здесь видов основная часть относится к тро-
пическим (7) и общим для тропической и субтропической зон
(7). Все тропические виды -• Puffinus pacificus, Sterna fuscata,
S. anaetheta, Anous stolidus, Sula dactylatra, Phaethon rubricau-
da, Pterodroma leucoptera не идут на юг дальше этого района.
С юга до юго-восточного побережья Австралии поднимаются
субтропически^ и мотальные виды — Gabianus pacificus, Puffi-
nus griseus, P. bulleri и P. tenuirostris.
У западного побережья Австралии граница тропического ре-
гиона проходит приблизительно на 300 миль севернее — в райо-
не Джеральдтона (29° ю. ш.). На расположенных здесь скалах
Хаут.ман гнездится 18 видов морских птиц: 6 тропических
(Sterna fuscata, Anous tenuirostris, A. stolidus. Phaethon rubri-
cauda, Puffinus pacificus. Sterna anaetheta), из которых 4 не
встречаются южнее (в списке первые), 8 общих для тропиче-
ской и субтропической зон (Phalacrocorax sulcirostris, Sterna
dougalli, S. nereis, Phalacrocorax varius, Ha lie tor melanoleucus,
Pelecanus conspicillatus, Gelochelidon nilotica, Thalasseus bergii),
2 субтропических (Puffinus assimilis, Gabianus pacificus), один
общий для субтропической и нотальной зон (Pelagodroma ma-
rina) и один межзональный (Larus novaehollandiae).
От юго-восточного побережья Австралии южная граница
тропического региона проходит на восток, оставляя острова
Лорд-Хау и Керм а дек к северу от нее. На островах Лорд-Хау из
15 видов 7 тропических и лишь 2 субтропических, а на островах
Кермадек из 14 видов 9 тропических и 3 субтропических.
В восточной части Тихого океана у побережья Южной Аме-
рики граница региона приблизительно на 1750 миль поднимает-
ся к северу в район северного побережья Перу (5—6' ю. ш.).
Здесь па сравнительно ограниченном участке вполне отчетливо
выражен стык орнитофаун тропического и субтропического ком-
плексов. С севера до указанного района проникают и имеют
здесь границу распространения 5 тропических видов — Fregata
magnificens, Pelecanus occidentalis, Phaethon aethereus, Suia
nebouxi и, очевидно, Sula dactylatra. С юга сюда доходят и
также имеют предел распространения 9 субтропических видов —
Phalacrocorax bougainvillei. Sterna lorata, Larostema inca, Sula
variegata, Pelecanus thagus, Pelecanoides garnoti. Spheniscus
270 ОРНИТОГЕОГРАФИЧЕСКИЕ РЕГИОНЫ ТИХОГО И ИНДИЙСКОГО ОКЕАНОВ
humboldti, Larus belcheri, Oceanodroma tethys и один межзо-
нальный — Larus dominicanus.
К южному субтропическому региону относятся районы юго-за-
падного, юго-восточного и всего южного побережья Австралии.
Орнитофауна побережья Тасмании и Бассова пролива по соста-
ву довольно пестрая. Здесь гнездится целая группа нотальных
видов: Pachyptila turtur, Puffinus griseus, Pelecanoides urinatrix,
Diomedea cauta, Puffinus tenuirostris. Тем не менее и в этом
районе более многочисленной является группа видов, свойствен-
ных субтропической и тропической зонам — Moris serrator, Ga-
bianus pacificus. Sterna nereis, Thalasseus bergii, Gelochelidon
nilotica, Phalacrocorax fuscescens, P. sulcirostris, P. varius,
Halictor melanoleucus, Pelecanus conspiciliatus.
Орнитофауна островов Новой Зеландии (острова Северный
и Южный) еще более смешанная. На острове Северный и при-
легающих к нему островах гнездятся следующие виды: субтро-
пические — Puffinus carneipes, Р. bulleri, Р. gavia, Р. assimilis,
Pterodroma longirostris, Procellaria parkinsoni, Moris serrator
общие для тропической и субтропической зон — Halietor mela-
noleucus, Phalacrocorax varius, P. sulcirostris, Sterna nereis;
нотальные — Puffinus griseus, Pachyptila turtur, Pelecanoides
urinatrix; общие для нотальной и субтропической зон — Eudyp-
tula minor, Pterodroma cooki, Pelagodroma marina, Pterodroma
macroptera, Pterodroma inexpectata. Sterna striata, Phalacroco-
rax punctatus; межзональные — Phalacrocorax carbo, Larus
dominicanus, Larus novaehollandiae.
В проливе Кука и в районе северной части острова Южный
гнездится 18 видов: субтропические — Procellaria parkinsoni,
Puffinus gavia; общие для тропической и субтропической зон —
НаПёЬг melanoleucus, Phalacrocorax varius; межзональные —
Phalacrocorax carbo, Larus dominicanus, L. novaehollandiae;
нотальные — Pelecanoides urinatrix, Procellaria aeguinoctialis,
Puffinus griseus. Pachyptila turtur, Eudyptes pachyrhynchus;
общие для субтропической и нотальной зон — Pterodroma inex-
pectata, Р. cooki, Pelagodroma marina, Eudyptula minor, Sterna
striata, Phalacrocorax punctatus. При большом количестве об-
щих видов у острова Северный соотношение все же в пользу
субтропических видов, чего не наблюдается в районе пролива
Кука. Флеминг (Fleming, 1956) говорит даже об абсолютном
доминировании в проливе Кука нотальных видов'. Учитывая
изложенное, границу южного субтропического региона нужно
проводить по южной оконечности острова Северный. Южнее
границы, следовательно, остается весь остров Южный, а также
1 Флеминг (Fleming, 1956), как и Мэрфи (Marphy, 1936), натальные виды
называет субантарктическими.
ОРНИТОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
271
расположенные восточнее острова Чатам, где, как ранее отмечал
Флеминг (Fleming, 1939), почти нет субтропических видов.
На востоке Тихого океана у побережья Южной Америки гра-
ница, разделяющая южный субтропический и нотальный регио-
ны, довольно размыта. По-видимому, она проходит где-то з
районе 37—38° ю. ш.1. До этих широт доходят с севера и имеют
здесь границу распространения субтропические Pelecanoides
garnoti, Phalacrocorax bougainvillei, Pelecanus thagus. До
30—33’ ю. ш. распространены на юг также другие субтропиче-
ские птицы — Larus belcheri, Larosterna inca, Spheniscus hum-
boldti. С юга до 37—38’ ю. ш. доходят, но не проникают севернее
3 нотальных вида — Puffinus griseus, Catharacta skua, Sphenis-
cus magellanicus и один нотально-антарктический — Phalacroco-
rax atriceps. До 40—42’ ю. ш. доходит с юга еще 5 нотальных
видов — Larus maculipennis, Gabianus scoresbyi, Pelecanoides
magellani, P. urinatrix, Phalacrocorax magellanicus.
Все побережье Южной Америки к югу от очерченной грани-
цы относится к нотальному региону. На Огненной Земле и при-
легающих к ней островах из 17 гнездящихся видов 11 являются
потальными — Puffinus griseus, Spheniscus magellanicus, Apte-
nodytes patagonica, Eudyptes crestatus, Phalacrocorax magel-
lanicus, Phalacrocorax albiventer, Diomedea melanophrys,
D. chrysostoma, Larus maculipennis, Pelecanoides magellani,
Gabianus scoresbyi. На расположенном южнее пролива Дрейка
Антарктическом архипелаге почти нет нотальных видов. Из 16
гнездящихся видов 7 антарктических — Pagodroma nivea, Ful-
marus glaciloides, Pygoscelis adeliae, Pygoscelis antarctica, Oce-
anites oceanicus, Aptenodytes forsteri, Thalassoica antarctica и 7
общих для. нотальной и антарктической зон — Pygoscelis papua,
Eudyptes chrysolophus, Phalacrocorax atriceps, Daption capensis,
Fregetta tropica, Macrosnectes giganteus Catharacta skua.
Учитывая характерное соотношение видов в двух рассмотрен-
ных районах, границу между потальным и антарктическим ре-
гионами нужно проводить через пролив Дрейка.
На западе Тихого океана к нотальному региону отходят все
острова в районе Новой Зеландии к югу от пролива Кука до
острова Макуори включительно. На острове Макуори, как и на
Огненной Земле резко доминируют потальные виды — 15 из 21:
Phalacrocorax albiventer, Pterodroma lessoni, Puffinus griseus,
Adamastor cinereus, Procellaria aeguinoctialis, Pelecanoides uri-
natrix, P. georgicus, Sterna vittata, Aptenodytes patagonica,
Eudyptes schlegeli, Eudyptes crestatus, Diomedea chrysostoma,
D. exulans, D. melanophris, Phoebetria palpebrata. Располо-
женные приблизительно на этой широте в Индийском
1 Острова Сан-Амбросио я Хуан-Фернандес остаются в пределах южного
субтропического региона.
272 ОРНИТОГЕОГРАФИЧЕСКИЕ РЕГИОНЫ И БИОГЕОГРАФИЧЕСКИЕ ОБЛАСТИ
океане острова Хед и Кергелен, а в Атлантическом — Южная
Георгия я также отношу к нотальному региону. Здесь больше,
чем на Макуори, антарктических и- общих для антарктической
и умеренной зон видов, однако, основу составляют нотальные ви-
ды: на острове Хед — 10% антарктических, 55 — нотальных,
30% общих для антарктической и умеренной зон; на острове Кер-
гелен соответственно 7, 63, 23%; на Южной Георгии — 14, 43.
33%. К антарктическому региону, следовательно, отходят побере-
жье Антарктиды и прилегающие острова (см. рис. 61).
2. Место орнитогеографических регионов
в системе биогеографических областей
Естественная система областей и других биогеографических
подразделений должна строиться на данных по распространению
всего живого населения. На такой позиции стоят многие био-
логи. Можно сослаться, например, на высказывания Е. Ф. Гурь-
яновой (1945) и Д. Д. Дарлингтона (1966), подчеркивающих,
что система фаунистических областей должна быть средним
всех типов распространения различных групп животных, у каж-
дой из которых имеются свои особенности распределения.
Тем не менее большая часть существующих в настоящее вре-
мя биогеографических схем основана на распространении от-
дельных групп животных. При этом почти все авторы выделен-
ным таким путем регионам присваивают ранги областей, подоб-
ластей, провинций и. д. А так как в распределении различных
групп имеются некоторые различия, то существует большое ко-
личество вариантов областей и других биогеографических под-
разделений. Показательна в этом смысле, например, статья
Е. Н. Курочкина (1964) об орнитогеографическом делении се-
верной части Тихого океана. В последней работе К. В. Бекле-
мишева (1969) в отличие от многих других исследователей для
целей биогеографического районирования океана используются
данные по распространению очень большого числа видов (713),
но это опять же только планктон.
Еще со времени основоположника морской биогеографии Орт-
мана (Orthmann, 1896) районирование океана проводится от-
дельно для каждого крупного вертикального раздела моря —
шельфа, пелагиали и глубоководной зоны. Делается это на ос-
новании больших различий в условиях существования и рассе-
ления в каждой вертикальной зоне. Учитывается также различ-
ная способность к передвижению и преодолению преград у пе-
лагических и донных животных.
Так как в распределении очень многих групп морских жи-
вотных наблюдаются черты принципиального сходства, есть
предпосылка к тому, что в будущем, по-видимому, будет разра-
ОРНИТОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
273
ботана единая для океана система биогеографических областей,
обобщающая распространение всех групп животных и растений*.
Пока же приходится придерживаться раздельного районирова-
ния для донных и пелагических животных. Морские птицы,
имеющие большие возможности для передвижения, а также свя-
занные в основном с поверхностными слоями моря, должны рас-
сматриваться в системе областей пелагической зоны жизни.
Для пелагической зоны жизни Ортман выделил четыре об-
ласти: Арктическую, Индопацифическую, Атлантическую и Ан-
тарктическую. Арктическая область разделена им в свою оче-
редь на три подобласти: Циркумполярно-арктическую, Атланти-
ческую бореальную, Тихоокеанскую бореальную, а Антарктиче-
ская область — на Нотально-циркумполярную и Циркумполяр-
но-антарктическую подобласти. Представления Ортмана с опре-
деленными изменениями и дополнениями до сих пор лежат в ос-
нове воззрений биогеографов. Современные схемы отличаются в
основном положением границ областей, а также объединением
двух тропических областей в одну (Ekman, 1963; Наумов, 1969).
В новейших схемах районирования океана, построенных на
основании распределения фитопланктона (Семина, 1967), зоо-
планктона (Беклемишев, 1965, 1967, 1969) и рыб эпипелагиали
(Парии, 1967а, 1968), арктическая, а также умеренные зоны Ти-
хого и Атлантического океанов объединяются в Аркто-бореаль-
ную область, тропическая и субтропические зоны всех океанов
— в Тропическую, а нотальная и антарктическая — в Антаркти-
ческую (Нотально-антарктическую)1 2.
Арктический и бореальный орнитогеографические регионы.
В арктическом регионе гнездится 18 видов морских птиц, от-
носящихся к 12 родам. Из них только 3 рода (25%) (Pagophila.
Хета, Plautus) и 5 видов (28%) (Pagophila eburnea, Larus leu-
copterus, Хета sabinei, Plautus alle, Larus hyperboreus) являют-
ся арктическими. Больше, половины (55%) видов относится к
аркто-бореальным (Stercorarius pomarinus, S. longicaudus.
S. parasiticus, Fulmarus glaciais, Larus argentatus, Rissa
tridactyla, Sterna paradisea, Cepphus grylle, Uria lom-
via, Fratercula arctica). Такое соотношение говорит о большой
подчиненности орнитофауны Арктики фауне птиц умеренной зо-
ны. Орнитофауна арктического региона в общем представляет
единое целое. Из 15 арктических и аркто-бореальных видов 1)
1 В учебнике А. Г. Воронова (1963) приводится схема районирования
океана, в которой сделана попытка объединить распространение донных и
пелагических животных. Эта схема мне кажется не совсем удачной, так как
в ней литоральная и пелагическая эоны искусствен зо объединяются в ранге
подобластей.
2 Н. В. Ларин о возможности объединения потального и антарктического
нхтногеографических регионов в одну область говорит предположительно.
274 ОРНИТОГЕОГРАФИЧЕСКИЕ РЕГИОНЫ И БИОГЕОГРАФИЧЕСКИЕ ОБЛАСТИ
(73%) имеют циркумполярное распространение. Некоторый
специфический оттенок на восточный и западный секторы Аркти-
ки накладывают только проникающие сюда отдельные бореаль-
ные виды. Однако их влияние сказывается только в самых юж-
ных частях арктической зоны.
Большое сходство, основанное на значительном количестве
общих видов и родов, наблюдается между орнитофаунами уме-
ренных зон Тихого и Атлантического океанов. В тихоокеанском
умеренном регионе насчитывается 20 родов, из них нет в соот-
ветствующих районах Атлантического океана 9 — Aethia, Вга-
chyramphus, Lunda, Synthiliboramphus, Ptychoramphus, Cerorhi-
пса, Cyclorrhynchus, Pelecanus, Хета. В умеренной зоне Атлан-
тики насчитывается 18 родов, из них в бореальном регионе Ти-
хого океана отсутствуют 7 — Hydrobates, Plautus, Alca, Moris,
Thalasseus, Gelochelidon, Catharacta. Общих родов для обоих
регионов 11. Однако роды Moris, Thalasseus, Catharacta, Gelo-
chelidon, представители которых отсутствуют в умеренной зоне
Тихого океана, есть в других районах этого океана, а роды
Pelecanus и Хета, которые отсутствуют в умеренной Атланти-
ке, обитают в соседних регионах этого океана. Из 43 видов, гнез-
дящихся в пределах умеренной зоны Тихого океана, 15 (35%)
встречаются в Северной Атлантике, а 28 (65%) — только здесь.
В умеренной зоне Атлантического океана гнездятся 28 видов, из
них 15 обитают в Тихом океане, что составляет 54%.
Проведенный анализ свидетельствует о значительном родст-
ве орнитофаун арктической, умеренной тихоокеанской и уме-
ренной атлантической зон. Ни в одном из рассмотренных регио-
нов нет эндемичных семейств, процент эндемичных родов везде
менее 50. Ясно, что более высоким оказывается видовой энде-
мизм. Но все же процент видов, общих с фауной умеренных вод
Тихого океана в Арктике и в северной Атлантике, более 50: соот-
ветственно 58 и 54%.
Несмотря на то, что бореальная и субтропическая зоны
имеют некоторое количество общих видов, их обособленность
вполне четкая. Из 8 семейств, представители которых обитают в
умеренной зоне Тихого океана, в субтропических широтах отсут-
ствует семейство Stercorariidae, а из семейства Alcidae проника-
ют только отдельные виды. Из 11 семейств, встречающихся в суб-
тропиках, представители четырех (36%) — Diomedeidae, Suli-
dae, Phaethontidae, Fregatidae не гнездятся в бореальной зоне.
Аналогичная картина наблюдается и в северной Атлантике. Из
8 семейств бореальной зоны 2 — Alcidae и Stercorariidae от-
сутствуют в субтропической, а из 9 семейств субтропической зо-
ны 3 (33%) — Pelecanidae, Fregatidae, Phaethontidae отсутству-
ют в бореальной. Из 43 гнездящихся в пределах тихоокеанского
умеренного региона видов только 3 (7%) — Larus occidentalis
ОРНИТОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
275
Phalacrocorax penicillatus и Р. auritus общи для обоих регио-
нов1. 6 (14%) бореальных видов (Oceanodroma leucorhoa, Syn-
thliboramphus antiquus, Ptychoramphus aleuticus, Cerorhinca
monocerata, Larus crassirostris, Phalacrocorax pelagicus) прони-
кают в северные районы субтропической зоны, 4 (9%) субтро-
пических вида — Oceanodroma monorhis, О. homochroa, Sterna
albifrons и Puffinus leucomelas заходят в южные районы умерен-
ной зоны. 67,5% видов бореальной и 66,5% видов субтропиче-
ской зон совсем не переходят границу, разделяющую соответ-
ствующие регионы.
Выделенные выше арктический и бореальные орнитогеогра-
фические регионы при сравнении с соответствующими зональ-
ными подразделениями по планктону и пелагическим рыбам об-
наруживают очевидное сходство. Отличия состоят в основном в
некотором расхождении границ регионов. К. А. Бродский (1957»
1960) на основании анализа распределения зоопланктона юг
Чукотского моря приблизительно до широты острова Врангеля
относит к бореальной области1 2 3. Обычно же границу, разделяю-
щую арктическую и северо-тихоокеанскую фауны, проводят не-
сколько южнее — по Берингову проливу (Андрияшев, 1939;
Гурьянова, 1951; Ekman, 1953; Физико-географический атлас,
1964; и др.) или по северной части Берингова моря (Пария,
1967а, 1968; и др.). Сдвиг границы к Берингову проливу, rfo-види-
мому, оправдан в отношении донных животных. Если в летний
период поверхностные воды северной части Берингова моря и
юга Чукотского прогреваются сравнительно хорошо (что спо-
собствует проникновению сюда бореальных животных), то в
придонных слоях сохраняются низкие температуры, позволяю-
щие арктическим видам выходить через Берингов пролив.
Положение южной границы северотихоокеанской подобласти
(или области у некоторых авторов) в схемах различных авто-
ров близко к очерченной выше границе бореального орнитогео-
графического региона. Особенно много сходства обнаруживает-
ся при сравнении со схемами К. А. Бродского (1957, 1960; Физи-
ко-географический атлас, 1964). Этот автор границу северо-
тихоокеанской области проводит от южной части Корейского по-
луострова к северной оконечности острова Хонсю и далее в
район Нижней Калифорнии. От северной части Хонсю к Нижней
Калифорнии проводит границу бореального ихтиогеографиче-
ского региона Н. В. Ларин (1967, 1968).
Учитывая большое сходство в видовом составе птиц арктиче-
1 Общим для обоих регионов является и Phalacrocorax carbo, но это ши-
роко распространенный вид, понти космополит.
2 Арктическую и умеренные зоны К. А. Бродский рассматривает в качестве
самостоятельных областей.
276 ОРНИТОГЕОГРАФИЧЕСКИЕ РЕГИОНЫ И БИОГЕОГРАФИЧЕСКИЕ ОБЛАСТИ
ского и атлантического умеренного регионов с тихоокеанским
умеренным, а также общее совпадение границ орнитогеографи-
ческих регионов с границами подразделений океана по рыбам
и планктону, приходим к выводу о соответствии арктического и
бореальных орнитогеографических регионов Аркто-бореальнэй
биогеографической области. Следовательно, каждый из перечис-
ленных регионов приобретает ранг подобласти — Арктической,
Тихоокеанской бореальной и Атлантической бореальной.
Орнитофауну Аркто-бореальной области характеризуют
представители 9 семейств. Наиболее многочисленными и харак-
терными, главным образом, для этой области являются семей-
ства Alcidae, Laridae и Stercorariidae. По нескольку представи-
телей имеют в рассматриваемой области семейства Phalacroco-
racidae, Procellariidae, Hydrobatidae, Sternidae, по одному —
Sulidae и Pelecanidae. Область характеризуется значительным
количеством свойственных только ей родов, относящихся пре-
имущественно к семействам Alcidae и Laridae. Эндемичны для
Аркто-бореальной области 12 родов из 27 встречающихся в ней—
Хеша, Rissa, Pagophila, Stercorarius, Plautus, Cepphus. Uria,
Fratercula, Lunda, Alca, Aethia и Cerorhinca.
Наиболее новыми являются схемы биогеографического райо-
нирования Тихого океана, предлагаемые Г. И. Семиной (1967)
по фитопланктону и К. В. Беклемишевым (1965, 1967, 1969) по
зоопланктону. Сравнение схем этих авторов с другими затрудне-
но, так как обширные зоны океана они выделяют как переход-
ные без определенного биогеографического ранга. Так, в север-
ной части Тихого океана к переходной зоне они относят пояс
океана от Сангарского пролива и Ванкуверо-Орегонского райо-
на до южной части Хонсю и полуострова Калифорния. Ширина
этого переходного пояса достигает нескольких сот миль. Еще
более широкая переходная зона выделяется в южном полуша-
рии. Конечно, даже на очень резко выраженном стыке двух
фаун всегда существует определенная зона, где обитают одно-
временно животные разных областей, поэтому нельзя границу,
разделяющую биогеографические области, представлять линией.
Что же касается переходных зон шириною в сотни миль, то труд-
но допустить, чтобы на таких огромных пространствах были оди-
наково представлены виды разных зон. При учете всего видово-
го состава (в некоторых случаях также численности) неизбежно
определяется граница преобладания видов одной из фаун. Хуже
заметная на одних группах, граница обнаруживается более оп-
ределенно при анализе распространения других животных. При
использовании же распространения всех систематических групп,
что пока не применяется, проблема огромных переходных зон,
очевидно, отпадает. В частности, в районе восточного побережья
Японии, относимого К. В. Беклемишевым и Г. И. Семиной к пе-
ОРНИТСГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
277
реходной зоне, стык умеренного и тропического планктона вы-
ражен вполне четко (Богоров и Виноградов, 1955; Бродский,
1955). Нужно заметить, что введение в схемы районирования пе-
реходных эон, по площади не уступающих некоторым подобла-
стям, делает их неудобными для использования. Особенно это
становится очевидным при более дробном делении океана на
провинции и округа.
Несколько лет назад попытку орнитогеографического де-
ления северной части Тихого океана сделал Е. Н. Курочкин
(1964). Северную границу бореальной области он провел по югу
Чукотского моря по линии мыс Барроу — остров Врангеля, но-
южную сдвинул далеко на юг. К бореальному региону Е. Н. Ку-
рочкин отнес весь полуостров Калифорния, весь остров Хонсю
и, судя по схеме, приводимой в его работе, острова Идзу. Осно-
ванием для этого послужило присутствие здесь отдельных боре-
альных видов. При этом оказался игнорированным факт гнездо-
вания в этих районах большого количества тропических и суб-
тропических видов. Выше было показано, 'что в субтропическую
зону действительно заходят некоторые представители бореаль-
ной фауны, но уже на участке между полуостровом Калифор-
ния и островами Санта Барбара встречается только 3 (21%) бо-
реальных вида. Что же касается района полуострова Калифор-
ния, то из 23 гнездящихся здесь видов только 2 — бореальные,
но 18 (78%) субтропических и тропических. Здесь гнездятся та-
кие характерные для тропиков виды, как Sula leucogaster,
S. nebouxii, S. dactylatra, Phaethon aethereus, Fregata magni-
ficens. Такое соотношение северных и южных элементов харак-
терно и для района острова Хонсю. За исключением его север-
ной части, основу гнездящихся видов составляют, как уже под-
черкивалось, субтропические и тропические виды. Что же ка-
сается островов Идзу, то на них бореальные виды совсем не
гнездятся. Мне кажется, вряд ли правильно для характеристики
умеренной области называть Brachyramphus hypoleucus, В. сга-
veri и Synthliboramphus wumisuzume. Семейство Alcidae своим
происхождением и развитием действительно связано с умерен-
ной зоной. Однако все три перечисленных вида являются, не-
сомненно, субтропическими. Гнездовая часть ареала первых
двух ограничена районами полуострова Калифорния, а послед-
него — островами, расположенными в Корейском проливе и у
южной части Хонсю.
Субтропические и тропический регионы. Северный субтропи-
ческий регион, имея вполне четкую обособленность от бореаль-
ного, обнаруживает большое сходство с тропическим. Говоря об
отличиях на уровне семейств, здесь нужно только отметить, что-
в субтропики проникают отдельные виды бореального семейства
Alcidae, а в тропики — одни вид мотального семейства Sphenis-
ХП ЭРНИТОГЕОГРАФИЧЕСКИЕ РЕГИОНЫ И БИОГЕОГРАФИЧЕСКИЕ ОБЛАСТИ
cidae (Spheniscus mendiculus)1; все остальные семейства об-
щие.
В северо-тихоокеанском субтропическом регионе из 20 встре-
чающихся здесь родов эндемичен только один (5%) — Halocyp-
tena. Даже при учете четырех бореальных родов — Synthlibo-
ramphus, Brachyramphus, Cerorhinca, Ptychoramphus, представи-
тели которых обитают в субтропиках, количество общих с тро-
пиками родов достигает 15 (75%). В северо-атлантическом
субтропическом регионе из 17 родов только два — Hydrobates
и Pelagordoma не встречаются в тропической зоне. Большое
сходство наблюдается и при сравнении видового состава. Из
42 видов тихоокеанского субтропического региона 15 (36%)
субтропические. 55% видов встречается в субтропическом и
тропическом регионах (сюда включены субтропические, заходя-
щие в тропики, и тропические, заходящие в субтропики). Для
северной Атлантики эта цифра еще выше — 65%.
Большинство биологоп, анализируя распространение живог-
ных различных систематических групп, рассматривают фауну
северной субтропической зоны в составе тропической области.
Как можно судить из изложенного, орнитофауна северного суб-
тропического региона не представляет в этом смысле исключе-
ния. Нужно подчеркнуть также ее большую подчиненность бо-
лее разнообразной орнитофауне тропического региона. Н. В.
Парии (1967, 1968) южную границу субтропического ихтиогео-
графического региона проводит от южной части Южно-Китай-
ского моря на восток (Гавайские острова остаются в тропиче-
ском регионе) к оконечности полуострова Калифорния. Таким
образом, она почти полностью совпадает с южной границей суб-
тропического орнитогеографического региона (см. рис. 61).
В составе тропической области рассматривается также суб-
тропическая зона южного полушария (Бродский, 1960; Бекле-
мишев, 1965, 1967, 1969; Семина, 1967; Парии, 1967, 1968)а. Этот
вывод можно перенести и на морских птиц. Однако орнитофау-
на южного субтропического региона находится под большим
влиянием также фауны нотальной зоны. Здесь обитают пред-
ставители 10 семейств, из них 9 характерны для тропической
зоны, и 9 — для нотальной. Правда, семейство Sulidae, харак-
терное в основном для тропиков, но хорошо представленное и
в субтропической зоне, в нотальной имеет всего один вид •<-
Moris serrator, который в незначительном количестве проникает
только в самую северную часть умеренной зоны. Из 25 родов 19
встречаются в тропическом и 16 — в нотальной регионах. Эндеми- 1 2
1 Эти раличия касаются только Тихого океана. В Атлантике все семейст-
<ва общие.
2 К. В. Беклемишев часть субтропических вод относит к переходной зоне.
ОРНИТОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
279
чей для региона лишь один род — Larosterna. С севера в юж-
ный субтропический регион проникают представители таких теп-
лолюбивых родов, как Pelecanus, Sula, Thalasseus, Gelochelidon,
Halietor, Procelsterna, Anous, а с юга — типичных нотальных —
Pelecanoides, Eudyptula, Pachyptila, Procellaria.
В южном субтропическом регионе размножаются около ЬО
видов морских птиц, из них 28 субтропических; 23 встречаются
в тропической и субтропической зонах (виды общие для обеих
зон, субтропические, проникающие в тропическую, и тропиче-
ские, проникающие в субтропическую); 17 — в нотальной и суб-
тропической (общие для обеих зон, субтропические, проникаю-
щие в нотальную и нотальные, проникающие в субтропиче-
скую). В тихоокеанском секторе южного субтропического ре-
гиона гнездится 50 видов; из них 24 субтропических, 19 встре-
чаются в тропической и субтропической, 15 — в субтропической
и нотальной зонах.
Выше было показано, что границы арктического, бореально-
го и субтропического орнитогеографических регионов в северной
части Тихого океана удовлетворительно совпадают с границами
соответствующих подразделений по планктону, пелагическим
рыбам, а в некоторых случаях и по донным животным. Прибли-
зительное сходство в положении границ регионов различных
групп животных имеет место и в южном полушарии. Границу,
разделяющую тропический и южный субтропический ихтиогео-
графические регионы. Н. В. Парим (1967, 1968) проводит от се-
верного побережья Перу (полное совпадение с птицами) к
30° ю. ш. у восточного побережья Австралии (у птиц, 34е ю. ш.).
Граница распространения тропической мелководной донной
фауны проводится в Тихом океане по 6° ю. ш. у побережья Аме-
рики и в районе Сиднея (34° ю. ш.) у побережья Австралии
(Ekman, 1953; Физико-географический атлас, 1964). Здесь видно
практически полное совпадение границ распространения донной
фауны и птиц, хотя последние рассматриваются в системе обла-
стей для пелагических животных.
Южную границу субтропического ихтиогеографического ре-
гиона Н. В. Парии (1967, 1968) проводит по широте северного
побережья Тасмании, середины острова Северный у Новой Зе-
ландии и 30° ю. ш. у берегов Чили. Эта граница близка к гра-
нице южной умеренной зоны К. А. Бродского (1960, Физико-гео-
графический атлас, 1964) с той только разницей, что К. А. Брод-
ский к умеренной зоне относит весь район Бассова пролива, а
у Новой Зеландии границу проводит по проливу Кука. Граница
орнитогеографического региона (см. рис. 61) в некоторых райо-
нах проходит чуть южнее, чем у цитированных авторов: через
Тасманию и южную часть острова Северный к 37—38° ю. ш. у
побережья Южной Америки.
280 ОРНИТОГЬОГРАФИЧЕСКИЕ РЕГИОНЫ И БИОГЕОГРАФИЧЕСКИЕ ОБЛАСТИ
Выше отмечалось, что тропическая и субтропическая зоны
Тихого, Атлантического и Индийского океанов рассматриваются
в составе единой пелагической тропической области. Данные
по распределению морских птиц, как было показано, также не
противоречат объединению тропической н субтропических зон.
Кратно остановимся на сходстве и различии орнитофауны со-
ответствующих регионов различных океанов. В первую очередь
нужно отметить большое сходство на уровне семейств. Говоря
о различиях, можно лишь сказать, что в Тихом океане в южный
субтропический регион из нотальной зоны проникают представи-
тели семейства Pelecanoididae, а в северный субтропический ре-
гион — виды бореального семейства Alcidae. Только в Тихом
океане в субтропические и тропические широты заходят и виды
семейства Diomedeidae. Однако все три семейства не являются
эндемичными для Тихого океана в целом. Они представлены и в
других океанах, но только в умеренных широтах. В тропиках
Индийского океана не гнездятся виды семейства Hydrobatidae,
но это семейство представлено здесь в субтропиках (Pelagodro-
ma marina). Таким образом, ни в одном из океанов нет энде-
мичных семейств1.
Несколько иная картина наблюдается при сравнении спис-
ков родов в различных океанах. Для тепловодных районов Ти-
хого океана эндемичны роды Greagrus, Procelsterna, Larosterna,
Nesofregetta, Halocyptena, Nannopterum. В тропических и суб-
тропических районах Атлантического и Индийского океанов нег
целой группы родов, представители которых проникают в тропи-
ки и субтропики Тихого океана из умеренных широт Diomedea,
Pelecanoides, Pachyptila, Procellaria, Fregetta, Oceanites, Syn-
thliboramphus, Brachyramphus, Cerorhinca, Ptychoramphus.
В Индийском океане эндемичных родов нет совсем, а в Атланти-
ческом только один — Hydrobates. Из 20 родов, представлен-
ных в Индийском океане, все (100%) есть в Тихом и 18 (90%) —
в Атлантическом. В Атлантическом океане также 20 родов, из
них 18 (90%) общие с Индийским и 19 (95%) — с Тихим океа-
нами. Как увидим ниже, во всех океанах есть значительное ко-
личество и общих видов.
Таким образом, данные по распределению морских птиц
подтверждают правильность выделения единой тропической
биогеографической области для пелагических животных. Учиты-
вая значительное сходство фаун трех океанов, а также принци-
пиальное сходство границ описанных нами регионов с соответ-
ствующими подразделениями по планктону и рыбам, можно счи-
тать, что выделенные орнитогеографические регионы — тропи-
1 Здесь не учитывается семейство Rynchopidae, распространение которого
связано с реками я их устьями.
ОРИИТОГЬОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
281
ческий, южный и северный субтропические — являются состав-
ными частями единой тропической биогеографической обла-
сти.
Тропическую пелагическую область характеризуют предста-
вители 12 семейств. Исключительно или преимущественно в
этой области живут представители семейств Phaethontidae, Fre-
gatidae, Sulidae, Sternidae, Pelecanidae. Хорошо здесь представ-
лены семейства Procellariidae (много видов отдельных родов),
Hydrobatidae. Phaiacrocoracidae, меньше Laridae. Отдельных
представителей, главным образом, на периферии области, име-
ют семейства Diomedeidae, Spheniscidae и Alcidae. Для тропиче-
ской области характерно значительное количество эндемичных
родов, относящихся к семействам, имеющим в ней большое ко-
личество представителей. Эндемичны для области 12 родов из
37 — Fregata, Sula, Phaethon, Bulweria, Halocyptena, Anous,
Gygis, Procelsterna, Creagrus, Nesofregetta, Nannopterum и La-
rosterna.
Тропическая пелагическая область на основании данных по
распределению планктона и рыб подразделяется на две подоб-
ласти — Индо-пацифическую и Атлантическую. Рассмотрим, на-
сколько соответствует такому подразделению распространение
морских птиц.
Наибольшее количество родов (28) и видов (71) известно в
тропической области из западной части Тихого океана. Несом-
ненно, с этим районом должны быть объединены Микронезия,
Полинезия и Гавайские острова, имеющие общие с западной ча-
стью Тихого океана 93% родов и 69% видов. Особенно близ-
ки орнитофауны западной части Тихого океана и восточной ча-
сти Индийского океана, имеющие 100% общих родов и 90% ви-
дов. Этот вывод распространяется и* на западную часть Индий-
ского океана. Последний район имеет общие с Тихим океаном
93% родов и 55% видов. С восточной частью Атлантического
океана на западе Индийского океана количество общих родов
и видов меньше — соответственно 75% и 35%.
Донная тепловодная мелководная фауна тихоокеанского по-
бережья Америки более близка к фауне Атлантического, но не
к фауне западной части Тихого океана, что является следствием
влияния на распространение, с одной стороны, так называемого
восточнотихоокеанского глубоководного барьера, с другой, сво-
бодного обмена между океанами в связи с отсутствием в прош-
лом Панамского перешейка (Ekman, 1953; и др.). Для пелагиче-
ских животных большие глубины восточной части Тихого океана
не являются преградой для распространения, и поэтому фауны
восточной и западной частей Тихого океана не отличаются
большой обособленностью.
Морские птицы имеют большие возможности для распростра-
282 ОРНИТОГЕОГРАФИЧЕСКИЕ РЕГИОНЫ И БИОГЕОГРАФИЧЕСКИЕ ОБЛАСТИ
нения. Тем не менее орнитофауна восточной части Тихого океа-
на характеризуется некоторой обособленностью. Из 61 гнездя-
щихся здесь видов 31 (51%) не встречается в других районах.
По составу более широкораспространенных видов восточная
часть Тихого океана занимает промежуточное положение меж-
ду западными частями Тихого и Атлантического океанов. Из
25 родов 18 (72%) характерны для западной половины Тихого
океана, а 14 (56%) — для западной части Атлантического. Ко-
личество общих видов составляет соответственно 39 и 32%. Осо-
бенно большое сходство наблюдается при сравнении рассматри-
ваемых районов в пределах тропического региона: соответствен-
но 57 и 48%.
Орнитофауны субтропических регионов отличаются весьма
заметной обособленностью. Это видно как при сравнении фаун
разных океанов, так и фаун противоположных побережий одно-
го океана. Среди субтропических видов Калифорнийского райо-
на нет ни одного, встречающегося в Атлантическом океане и
только один (Oceanodroma monorhis) — на западе Тихого
океана. Встречаются в обоих океанах только широко распрост-
раненные виды: тропические, общие для тропической и субтро-
пической зон или общие для субтропической и бореальной зон.
Аналогичная картина наблюдается и при сравнении районов
южного субтропического региона. В субтропическом регио-
не у западного побережья Южной Америки из 28 видов 14 (50%)
встречаются в западной части Тихого океана, но только 2 из
них — субтропические, остальные являются широкораспростра-
ненными тропическими или общими для тропической и субтро-
пической зон. Общих субтропических видов для западного и во-
сточного побережья Южной Америки нет совсем. Общими яв-
ляются только 8 (28%) широкораспространенных, в основном
тропических, птиц.
Из приведенных данных видно, что больше половины родов
и тропических видов, известных в восточной части Тихого океа-
на, есть и в пределах Индо-Вест-Пацифики. На этом основании
было бы логично этот район рассматривать в составе Индо-па-
цифической подобласти.
Атлантическая часть тропической области отделена от Ин-
дийского и Тихого океанов материками. Тем не менее основу ви-
дового состава здесь составляют виды, характерные для Тихого
или Индийского океана (58% встречаются в Тихом и 51% — в
Индийском). Эндемичны для Атлантического океана только
30% (15 из 50) видов и один род (Hydrobates). Обращая внима-
ние на большую подчиненность орнитофауны Атлантического
океана орнитофауне Индо-тихоокеанской подобласти, я, тем не
менее, не имею в виду ставить под сомнение правомочность вы-
ОРНИТОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
283-
деления Атлантической подобласти, что сделано на основании
распространения планктона и рыб.
Потальный и антарктический регионы. Прежде, чем перейти'
к анализу орнитофауны умеренных широт и Антарктики, нужно*
заметить, что умеренный пояс южного полушария часто непра-
вильно называют субантарктическим. Этот термин распростра-
няется и на животных, в том числе морских птиц. Субантаркти-
ческими называли птиц умеренной (нотальной или антиборе-
альной) зоны, например, Мэрфи (Murphy, 1936)*, Флеминг
(Fleming, 1939, 1952, 1956) и Фолл (Falla, 1952, 1964). На са-
мом деле Субантарктика является частью Антарктики и зани-
мает зону между антарктической дивергенцией и антарктиче-
ской конвергенцией (Таубер, 1956; Марков, и др. 1968). Такое
подразделение имеется в виду ниже при анализе распростране-
ния морских птиц.
Донная фауна Антарктики сильно обособлена от фауны уме-
ренных вод (Regan, 1914; Norman, 1938; Ekman, 1953; De Witt,
1965; Андрияшев, 1964; и др.). Фауны Ясе пелагических живог-
ных антарктической и нотальной зон не имеют такой обособлен-
ности. о чем уже говорилось выше. Поэтому в настоящее время
нх объединяют в одну Антарктическую или Нотально-антаркти-
ческую область, с выделением Антарктической и Нотальной под-
областей, как это делал еще Ортман. Орнитофауна выделенно-
го выше потального орнитогеографического региона весьма
разнообразна, специфична и насчитывает 74 вида, относящихся
к 29 родам. Эндемичны для этой зоны 5 (17%) родов — Ada-
mastor, Phoebetria, Halobaena, Garrodia, Magadyptes и 39 видов
(52%). Всего нотальных видов 45 (60%), но часть из них захо-
дит или в субтропическую, или в антарктическую зону.
В пределах антарктического орнитогеографического региона
гнездится только 18 видов, относящихся к 15 родам. Эндемичны
здесь лишь 2 (13%) рода — Pagodroma и Thalassoica. Из 18 ви-
дов антарктическими являются 7 (но 4 из них заходят в южную
часть потального региона), а 9 следует рассматривать, как свой-*
ственных и нотальной и антарктической зонам. Таким образом,
больше половины видов орнитофауны Антарктики встречаются
и в пределах умеренных вод. Следовательно, кроме не очень
значительной обособленности, приходится говорить и о большой
зависимости антарктической фауны от южной умеренной. Таким
образом, данные по распространению морских птиц не противо-
речат объединению двух зон в единую Нотально-антарктиче-
скую область. Это, а также общее сходство границ орнитогео*-
графических регионов с границами распространения других жи-
1 Позднее Мэрфи (Murphy, 1964) птиц умеренной зоны .называет антибо*-
реальными.
ЗОН ОРНИТОГЕОГРАФИЧЕОИЕ РЕГИОНЫ И БКОГЕО! РАФИЧЕСКИЕ ОБЛАСТИ
вотных позволяют считать потальный и антарктические регио-
ны составными частями Нотально-антарктической области.
Нотально-антарктическую область характеризуют представи-
тели 10 семейств. Наиболее характерны из них семейства
Spheniscidae. Procellariidae, Pelecanoididae, Diomedeidae. По не-
скольку представителей имеют семейства Hydrobatidae, Phaia-
crocoracidae, Sternidae, Laridae, Stercorariidae. Семейство Su-
iidae имеет только одного представителя, проникающего в се-
верную часть области в районе Новой Зеландии. Из 31 рода,
известного для Нотально-антарктической области, эндемичны
12, относящиеся к трубконосым и пингвинам — Megadyptes,
Eudyptes, Aptenodytes, Pygoscelis, Phoebetria, Garrodia, Macro-
nectes, Halobaena, Adamastor, Daption, Thalassoica и Pagod-
roma.
Уже давно было замечено, что для многих пелагических жи-
вотных Южного океана характерно циркумполярное распростра-
нение. Относительно морских птиц обстоятельно этот вопрос
раньше "был рассмотрен Р. Мэрфи (Murphy, 1936). О слабой
обособленности фаун морских птиц отдельных районов Южного
океана можно судить по следующим сопоставлениям. Из 72 ви-
дов, характерных для умеренной и антарктической зон. 34
(47%) встречаются в тихоокеанском, индоокеанском и атланти-
ческом секторах Южного океана, при этом процент циркумпо-
лярных видов увеличивается в более высоких широтах. Относи-
тельной обособленностью характеризуется фауна умеренной зо-
ны западно-тихоокеанского сектора. Свойственными только для
этого района являются 12 (27%) видов; в то же время в индо-
океанском секторе таких видов всего 4 (10%), атлантическом —
2 (5%), восточно-тихоокеанском — 1 (4%). 82% видов, обитаю-
щих в индоокеанском секторе, встречаются также в атлантиче-
ском и 72% — в тихоокеанском. В атлантическом секторе общие
с индоокеанским — 76%, с тихоокеанским 83% видов. В восточ-
но-тихоокеанском секторе общие с атлантическим — 92%, с за-
падно-тихоокеанским 58% видов.
Такое большое сходство в составе видов, гнездящихся в раз-
личных районах и океанах, не дает материалов для выделения
подобластей, охватывающих какой-либо сектор Южного океана.
Различия, свойственные орнитофаунам южной умеренной и ан-
тарктической зон, как об этом можно судить по приведенным
сопоставлениям, гораздо выше; в них есть даже эндемичные
роды.
Таким образом, в этом смысле морские птицы также дают
данные, подтверждающие правильность выделения зональных
Нотальной и Антарктической пелагических подобластей. Этот
вывод не является принципиально новым. Некоторые авторы
(Murphy, 1936; Fleming, 1939, 1952, 1956; Falla, 1952. 1964;
ОРНИТС ГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
28В
и др.).. рассматривая! орнитофауну Южного океана, уже гово-
рили о комплексах антарктическом и субантарктическом и со-
ответственно антарктических и субантарктических видах птиц.
Как уже отмечалось, субантарктическими они называла виды
умеренной зоны.
Есть и другие, на мой взгляд, менее удачные попытки райо-
нирования Южного океаоа на основании распространения мор-
ских птиц. Это можно сказать об орнитогеографическом райо-
нировании Антарктики, приводимом Е. С. Короткеничем и Bu Н.
Петровым в «Атласе Антарктики» (1966). Эти авторы выдаля-
ют три зоны — антарктическую, субантарктическую и умерен-
ную, исходя из существования соответствующих физико-геог
графических поясов.
Я провел сравнение видового состава морских итиц каждояо-
из поясов, при этом принимались границы поясов, приводимые
в «Атласе Антарктики» (1966) и «Географии Антарктиды»
(Марков и др-. 1968). Оказалось, что субантарктический пояс
не имеет ни одного эндемичного вида/ Всего здесь гнездится
28 видов морских птиц: 15 — нотальных, 7 — нотально-антаркти-
ческих. 5— антарктических и 1 межзональный. Казалось бы,
соотношение видов.— в пользу нотальных. Однако 12 из 15 уме-
ренных видов в субантарктический пояс заходят только в райо-
не островов Южная Георгия и Хед. Если рассматривать эти ост-
рова в составе нотального региона, что я и делаю, то в остав-
шихся частях субантарктического пояса соотношение видов бу-
дет уже. несомненно, в пользу антарктических. Нужно также
заметить, что острова Принца Эдуарда, Крозе, Кергелен и Ма-
куори относятся к умеренному физико-географическому поясу
(Атлас Антарктики, 1966; Марков и др., 1968). Но на схеме рас-
пространения птиц Антарктики Е. С. Короткевича и В. Н. Пет-
рова северная граница орнитогеографической субантарктиче-
ской зоны отодвинута несколько к северу, в результате чего ост-
рова Макуори, Кергелен и Крозе оказались в пределах субан-
тарктической зоны. Вместе с этим 70% видов (14 из 20), встре-
чающихся на острове Макуори, гнездятся также на расположен-
ных севернее островах Окленд, Кэмпбелл, Антиподы и Боунти,
которые правильно относятся данными авторами к умеренной
зоне. Характерно также то, что 63% (19 из 30) морских птиц,
обитающих на островах Окленд, Кэмпбелл, Антиподы и Боунти,
встречаются на островах Крозе, Кергелен, Хед и Принца Эду-
арда.
3. Некоторые закономерности географического
распространения морских птиц
Много видов морских птиц в период кочевок встречается В;
нескольких климатических зонах. Тем не менее, как было покат
286
ЗАКОНОМЕРНОСТИ ГЕОГРАФИЧЕСКОГО РАСПРОСТРАНЕНИЯ
зано при описании закономерностей сезонного распределения
птиц, общие черты зонального распространения орнитофауны
.сохраняются во все периоды года. Что касается географического
распространения птиц в гнездовой период, то в данном случае
зональность вырисовывается весьма четко. В пределах несколь-
зких из очерченных на рис. 61 орнитогеографичеоких регионов
располагаются гнездовые части ареалов всего у 2% видов мор-
ских птиц, в пределах двух соседних регионов — у 19%• У 79%
видов основная гнездовая часть ареала не выходит за границы
одного региона.
Приведенные цифры станут еще более показательными, если
учесть, что птицы чаще других животных попадают в другие зо-
ны. Ураганами и штормами они регулярно и в больших коли-
чествах уносятся далеко за пределы областей их постоянного
обитания. Не представляют здесь исключения и виды, в совер-
шенстве освоившие воздушную среду. По-видимому, только в
очень редких случаях такие заносы могут привести к заселению
новой территории. По крайней мере, достоверных случаев по
морским птицам в настоящее время не известно. Таким обра-
зом, морские птицы не представляют исключения среди других
морских животных. Их распространение подчиняется закону ми-
ровой зональности Гумбольдта, Докучаева, Берга.
В основе географической зональности и на суше, и в океанах
лежит неравномерное распределение по поверхности планеты
солнечной радиации*. В океанах последовательная цепь клима-
тической, географической и биологической зональности выраже-
на не менее отчетливо, чем на суше (Зенкевич, 1948; Богоров,
1964; Богоров и Зенкевич, 1966; Богоров, 1970; и др.)- Каждую
климатическую зону океана занимает определенная первичная
водная масса, большая часть воды которой циркулирует в пре-
делах крупномасштабных круговоротов. Формирование физи-
ко-химических характеристик водных масс определяется кли-
матическими факторами данной зоны (Добровольский, 1961).
Поэтому наиболее общей и характерной чертой в размещении
структур вод в океане является широтная зональность (Радзи-
ховская и Леонтьева. 1968).
Водные массы являются биотопами пелагических биоцено-
зов. Отсюда следует, что на границах водных масс должна про-
исходить смена их населения, что и имеет место в действитель-
ности (Беклемишев, 1969). Наиболее резко градиенты физико-
химических свойств вод. особенно их температурных характери-
стик, выражены в местах схождения различных водных масс, то
1 В серии работ А. А. Григорьева и М. И. Будыко (Григорьев, 1966)
обосновано, что на суше большое значение имеют также распределение осад-
ков и соотношение тепла и влаги.
ОРНИТОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
287
есть в эонах конвергенций. В связи с этим в районе конверген-
ций находят предел распространения многие животные, а смена
населения происходит особенно резко. Поэтому по конвергенци-
ям нередко лроводятся границы крупных биогеографических
подразделений.
Неплохо совпадают с соответствующими географическими
эонами и выделенные выше орнитогеопрафические регионы-
Подчеркнем это на примере Тихого океана.
Бореальный орнитогеографический регион соответствует уме-
ренной или, как ее иногда называют, субарктической зоне. Эта
эона занята субарктической структурой вод. Границей между
поверхностными субарктической и субтропической водными
массами является субарктическая конвергенция, проходящая
между 40 и 45° с. ш. Северный субтропический орнитогеографи-
ческий регион соответствует северной субтропической зоне, за-
нятой субтропической структурой вод. Границей между поверх-
ностными субтропической и тропической водными массами явля-
ется субтропическая конвергенция, проходящая в районе 23—25°
с. ш. Тропический орнитогеографический регион соответствует
пассатным и межпассатной зонам, занятым тропическими и эква-
ториальными структурами вод. Границей между поверхностны-
ми южной тропической и южной субтропической водными мас-
сами является тропическая конвергенция, проходящая в райо-
не 30° ю. ш. Южный субтропический орнитогеографический ре-
гион соответствует южной субтропической зоне, занятой суб-
тропической структурой вод. Границей между поверхностными
южной субтропической и субантарктической водными массами
является субтропическая конвергенция, проходящая примерно
в районе 40° ю. ш. Нотальный орнитогеографический регион со-
ответствует южноумеренной зоне, занятой субантарктической
структурой вод. Границей между поверхностными субантаркти-
ческой и антарктической водными массами является антарктиче-
ская конвергенция, проходящая в западной части Тихого океана
между 55 и 56° ю. ш., в восточной — в районе 63° ю. ш. Антарк-
тический орнитогеографический регион соответствует антарктиче-
ской зоне, занятой антарктической структурой вод*. В Тихом
океане не представлен арктический орнитогеографический ре-
гион, соответствующий арктической зоне.
Не следует, однако, понимать, что границы регионов различ-
ных групп животных и единых биогеографических подразделе-
ний для всех животных должны абсолютно совпадать с конвер-
генциями. Разговор идет о совпадении зон и регионов в общем
среднем плане. Это замечание, в первую очередь, необходимо
1 Номенклатура и расположение водных масс приводится по сводке
tTnxnfl океан» (Радзиховская и Леонтьева, 1968).
288
ЗАКОНОМЕРНОСТИ ГЕОГРАФИЧЕСКОГО РАСПРОСТРАНЕНИЯ
отнести к гнездящимся на или у побережий континентов мор-
ским птицам, на распределение которых в период размножения
некоторое влияние оказывают условия суши. В связи с этим
нужно заметить, что при биогеографическом делении пел аги а ли
океана по планктону К. В. Беклемишевым (1969) установлено
два типа зональности: океанического и дальне-неретического.
Существование последнего обусловлено тем, что благодаря
взаимодействию океанов и континентов на широтную климати-
ческую зональность накладывается цнркумконтинеитальная зо-
нальность.
Специфичность и сложность условий в прибрежной зоне яв-
ляется одной из причин смешанного состава орнитофауны в не-
которых районах. Так, в восточной части Тихого океана в райо-
не полуострова Калифорния и побережья Перу в результате ох-
лаждающего влияния холодных Калифорнийского и Перуанско-
го течений воды имеют более низкую температуру, чем в откры-
том океане на этих широтах. В то же время температура возду-
ха здесь отличается от среднеширотной не так значительно, что
является результатом влияния прогретой суши. В итоге веду-
щие в основном воздушный образ жизни тропические фрегаты,
фаэтоны, олуши и крачки обитают у полуострова Калифорния
вместе с чистиковыми, а у перуанского побережья — с пингвина-
ми и ныряющими буревестниками, проводящими большую
часть времени в воде. На отмеченную особенность в .распреде-
лении птиц на востоке Тихого океана более сорока лет назад
обратил внимание Александер (Alexander, 1954).
Изложенное выше, конечно, не является исчерпывающим
объяснением и обоснованием смешанного состава фаун на гра-
ницах зон и областей. Известно, что границ зон и биогеографи-
ческих областей достигают многие виды, характеризующиеся
широким распространением. Такие виды отличаются большой
экологической пластичностью, и это позволяет им проникать в
смежные зоны. Для водных животных большое значение имеет
и сам факт смешения различных водных масс, что создает ка-
кой-то средний тип среды обитания.
Несмотря на то, что в течение миллионов лет положение
климатических зон в общих чертах соответствует современному,
при похолоданиях и потеплениях наблюдается смещение границ
эон с севера на юг и обратно. В этом смысле не случайно Удвар-
дн (Udvardy, 1969) свою книгу по зоогеографии назвал «Дина-
мическая зоогеография». Смещение зон является причиной под-
вижности границ фаунистических комплексов в общем и границ
ареалов видов, в частности. Такая пульсация границ зон и кон-
вергенций имеет различную периодичность. Заметна она и на
коротких отрезках времени. Напомню хорошо известные факты
ОРНИТОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА 289
проникновения в умеренные воды Атлантического и Тихого
океана многих тропических животных в периоды потепления. В
историческом плане при смещении границ какого-либо фауни-
стического (или флористического) комплекса большая часть
видов этого комплекса изменяет область своего распростране-
ния в соответствии с изменением конфигурации климатической
зоны, с которой связан этот комплекс*. Большая часть, но не
все. Небольшое количество видов или только некоторые попу-
ляции отдельных видов могут оставаться и, таким образом, ока-
зываться уже в пределах новой зоны. Это реликты. О таком
пути образования реликтов писали, например. Б. К. Штегман
(1936) при обсуждении распространения типов фаун в Палеарк-
тике и Флеминг (1956) при анализе биогеографической принад-
лежности морских птиц залива Кука (Новая Зеландия). При-
сутствие реликтов в районах границ биогеографических облас-
тей является еще одной причиной смешанного состава фаун у
пределов областей. <
Взятый мною для орнитогеаграфического районирования
принцип широтной зональности имеет в орнитологии свою исто-
рию. В таком плане птицы Южного океана в тридцатые годы
рассматривались Мэрфи (1936), а чистиковые — Ю. М. Кафта-
новским (1951). Позднее о связях в распространении комплек-
сов морских птиц с определенными водными массами и значе-
нии конвергенций в определении границ различных видов птиц
писали и другие авторы (Fleming, 1939, 1952, 1956; Bourne, 1963;
Salomonsen, 1965). Буэн (Bourne, 1963) в отличие от других
рассмотрел птиц не в отдельном районе, а в пределах всего Ми-
рового океана. В каждом полушарии он выделяет полярное»
субполярное, субтропическое и тропическое сообщества птиц, а
их границы определяет по конвергенциям. Буэн ограничился пе-
речислением зональных сообществ, их место в системе биогеог-
рафических областей он не рассматривал.
Из изложенного в данном разделе видно, что связь геогра-
фических комплексов птиц с определенными ландшафтными зо-
нами океана в общем и с водными массами, в частности, очевид-
на. Не всегда ясен в деталях характер этой связи. Выше при об-
суждении распределения неритической, дальне-неритической и
океанической группировок говорилось о значении условий добы-
чи пищи, а также общего океанографического окружения. Что
касается зонального распределения видов и комплексов птиц, то
из всех физико-химических характеристик водных масс наи-
> Напомню, что именно на признании соответствия природных зон (в том
числе зоогеографических и геоботанических) климатическим, а также син-
хронности пространственной соразмерности и однонаправленности первых от
вторых основано восстановление климатов прошлого (Синицын, 1965—1967).
'290
3 ХКОНОМЕРНОСТИ ГЕОГРАФИЧЕСКОГО РАСПРОСТРАНЕНИЯ
большее значение имеет температурный фактор. Это объясняет,
почему многие зональные виды в определенные периоды пере-
секают фронты. Так, зимой, когда температура воды и воздуха
понижается, арктические и антарктические птицы в больших ко-
личествах проникают в умеренные воды, виды умеренной зоны
— в субтропические, а виды субтропической эоны — в тропиче-
скую.
Большая зависимость от температурного фактора, по-види-
мому, и является тем основным общим моментом, который опре-
деляет черть» сходства в географическом распространении жи-
вотных таких различных групп, как планктон, нектон и мор-
ские птицы. Как отмечает Н. В. Парии (1968), совпадение райо-
нирования пелагиали по планктону и нектону является вполне
обоснованным, так как температурные границы, служащие
пределами распространения географических группировок нек-
тона, довольно точно соответствуют границам основных круго-
воротов вод, определяющих ареалы планктона. Это станет еще
очевиднее, если учесть, что ареалы планктона определяются не
только циркуляцией, но И физико-химическими характеристика-
ми водных масс (в том числе температурой). Решающая роль
температуры бросается в глаза при анализе конфигурации гра-
ниц зональных комплексов птиц. Там, где теплые течения нап-
равлены в сторону высоких широт (Куросио, Восточно-Австра-
лийское, Цусимское), в этом же направлении наблюдается сдвиг
границ регионов. Наоборот, в сторону экватора наблюдается
сдвиг границ в зонах холодных течений — Перуанского, Кали-
форнийского, Приморского, Западно-Австралийского.
Несмотря на значительные отличия в условиях обитания
между всеми географическими зонами, различия между такими
группами зон, как умеренная и арктическая, нотальная и ан-
тарктическая, субтропическая и тропическая, гораздо больше,
чем между зонами в каждой группе. На этом основании все зо-
ны можно сгруппировать в три: одну теплую (тропики и суб-
тропики) и две умеренно-холодные (аркто-бореальная и ноталь-
но-антарктическая). Выше было показано, что и орнитофауны
арктической и умеренной, субтропической и тропической, но-
тальной и антарктической зон имеют между собой много обще-
го- Поэтому, как и у других морских животных, они рассматри-
ваются в системе единых пелагических областей — Аркто-боре-
альной, Циркум-тропической и Нотально-Антарктической.
Ф. Дарлингтон (1966) в результате анализа распределения фа-
ун суши резюмирует, что все основные региональные фауны
отделяются друг от друга или границей между тропиками и
умеренной зоной, или солеными водами. И далее он подчерки-
вает, что зональный климат и соленые воды — самые важные
ОРНИТОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
291
факторы неживой природы, вызывающие дифференциацию ре-
гиональных фаун и способствующие ей. Конечно, о преградах,
создаваемых солеными водами, для морских животных говорить
не приходится. Вывод же о влиянии климатичееяой зональности,
как видно из изложенного выше, всецело можно перенести и на
морских животных, в том числе морских птиц.
4. О необходимости сочетания при биогеографическом
районировании экологогеографнческнх,
исторических и фаунистических данных
Зонально-географический подход к биогеографическому
районированию не является общепризнанным, при этом биогеог-
рафия, несмотря на свою сравнительно длительную историю,
еще далека от утверждения единого метода районирования. По
мнению одних, система областей должна основываться не на ти-
пе современного распространения, а на . истории фауны. Этого,
так сказать, фауно-генетического принципа придерживаются
многие биологи (Гептнер, 1936; Пузанов, 1938; Авдрияшев, 1939;
Гурьянова, 1945, 1951; и др.). Другие авторы в основу райони-
рования кладут ландшафтно-климатические зоны, то есть совре-
менные условия. Зональные биогеографические подразделения
фигурируют в работах и по морским (Murphy, 1936; Bourne, 1963;
Salomonsen, 1965; Бродский, 1957; Парни, 1968; и др.). и по су-
хопутным животным (Штегман, 1938; Кузнецов, 1950; Дементь-
ев, 1958; Воинственский. 1960; Портенко, 1965; Реймерс, 1966;
Успенский, 1969; и мн. др.). Любопытны при этом некоторые
категоричные и одновременно противоречивые высказывания.
Так, Ф. Дарлингтон (1966) повторяет мнение Уоллеса о том,
что объединение прошлого и настоящего при выделении облас-
тей, якобы, приводит к путанице, хотя причины, приведшие к
современному распространению, заложены в прошлом. И здесь
же он пишет, что как прошлое, так и настоящее одинаково важ-
ны для зоогеографии и одно нельзя понять без другого.
Недавно критический анализ двух основных принципов био-
географического районирования суши провел К). И. Чернов
(1968), который подчеркивает, что необходимо разграничивать
зональные подразделения и их границы, обусловленные градиен-
том современных экологических факторов, а также региональ-
ные подразделения с их границами, связанные с историческими
причинами. С другой стороны, этот автор говорит о неизбежно-
сти синтеза историко-регионального и зонального подходов
при дробном зоогеографическом районировании.
Наконец, К. В. Беклемишев (1969) считает, что при биогеог-
рафическом районировании надо класть в основу ареал, а не
экологические и исторические характеристики видов и, тем бо-
292
О ПРИНЦИПАХ БИОГЕОГРАФИЧЕСКОГО ; РАЙОНИРОВАНИЯ
лее, деление океана на водные массы и физико-географические
зоны. Тем не мерее, этот автор в своих работах демонстрирует
не что иное, как зонально-экологический подход к райониро-
ванию океана по планктону. Это и не удивительно, так как фор-
мы и размеры ареалов планктонных организмов во многом оп-
ределяются экологическими условиями, в том числе течениями,
— фактором, которому К. В. Беклемишев придает первостепен-
ную роль.
Мне кажется, что проблема противопоставления двух прин-
ципов в биогеографическом районировании весьма искусствен-
на. Интересно здесь то, что, как отмечает К. А. Бродский (1957),
два принципа, представляющихся многим авторам противопо-
ложными, дают нередко сходный результат в районировании,
особенно для крупных подразделений. Я совершенно согласен
с этим автором в том,'что правильным путем к разрешению
противоречий является рассмотрение районирования, как фор-
мы проявления закона мировой зональности Гумбольдта, До-
кучаева, Берга. При районировании исходным материалом дол-
жны быть закономерности в распределении животных, которые
отражают и настоящие, и прошлые условия. Аналогичные
взгляды высказывали также И. В. Кожанчиков (1946) и Г. П.
Дементьев (1958). В частности, Г. П. Дементьев подчеркивал на
примере птиц СССР, что развитие авиафауны протекало в
строгом соответствии со становлением, развитием и изме-
нением основных природных зон. Зональное распростра-
нение птиц, таким образом, соответствует определенным
комплексам физико-географических и фаунистических ус-
ловий, при этом отражает историю фауны. Чтобы пока-
зать это на примере морских птиц, необходимо сделать некото-
рое отступление и охарактеризовать распространение семейств.
Семейство пингвинов — Spheniscidae. Семейство объединяет
17 видов, относящихся к 6 родам. Обитают в основном в южно-
умеренной зоне, где встречается 12 видов (все 6 родов), из ко-
торых больше половины эндемичны для этой зоны. В антаркти-
ческой зоне гнездится шесть видов, относящихся к трем родам.
Из них только 3 являются антарктическими — A. forsteri,
Р. adeliae, Р. antarctica. В субтропическую зону проникают
лишь 3 вида, относящихся к двум родам, а в тропическую —
один (S- mendiculus).
Во время сезонных кочевок пингвины не выходят за пределы
области гнездования. Таким образом, распространение семейст-
ва пингвинов в целом характеризуется отчетливо выраженной
зональностью (рис. 62). Зональность распространения семейст-
ва нарушают только гнездящийся на островах Галапагос
S. mendiculus. а на побережье Перу — S. humboldti. Сдвиг гра-
Рис. 62. Распространение пингвинов (Sphentscidae) в Мировом океане. 1 — места гнездования
294
О ПРИНЦИПАХ БИОГЕОГРАФИЧЕСКОГО РАЙОНИРОВАНИЯ
ницы распространения семейства на север в восточной части Ти-
хого океана связан, как и у других животных, с холодным Пе-
руанским течением.
Наибольшим видовым разнообразием характеризуется рай-
он Южной Америки, где обитает 10 видов, относящихся к 4 ро-
дам (Spheniscus, Pygoscelis, Eudyptes, Aptenodytes). и западно-
тихоокеанский сектор. Южного океана, где гнездится 11 видов,
относящихся к 5 родам (Eudyptes, Eudyptula, Megadyptes, Ap-
tenodytes, Pygoscelis). Большая часть ископаемых видов извест-
на из Южной Америки. Этот район, очевидно, и является местом
происхождения пингвинов. Другой важный центр развития груп-
пы — район Новой Зеландии- Все известные ископаемые пинг-
вины (18 родов, 25 видов) обитали в пределах области .распро-
странения современных видов (Murphy, 1959).
Семейство альбатросов — Diomedeidae. Большинство авторов
склонны предполагать, что альбатросы по происхождению яв-
ляются южно-умеренной группой (Murphy, 1936; Williams, 1952;
Fisher and Peterson, 1964; и др.). Из 13 современных видов толь-
ко 3 гнездятся на севере Тихого океана, один — в тропической
части Тихого океана, 9 — в умеренной зоне Южного океана, при
этом в южном полушарии обитают представители двух родов —
Diomedea и Phoebetria, а в севермом — только одного — Diome-
dea. Время размножения альбатросов южного полушария при-
ходится на весенне-летний период. Этот цикл календарно сохра-
нился и у северо-тихоокеанских альбатросов, которые размно-
жаются зимой северного полушария. Вместе с этим, основы-
ваясь на фактах находок ископаемых остатков в Европе, а так-
же некоторых примитивных чертах в характере размножения.
Курода (Kuroda. 1954, 1960а) допускает возможность северного
происхождения альбатросов. В прошлом альбатросы имели
действительно более широкое распространение. Но не вы-
зывающий сомнения факт происхождения отряда трубконо-
сых в целом в южном полушарии, где в настоящее время оби-
тает большая часть их видов, заставляет склоняться к мнению
авторов первой группы.
Как видно из рис. 63, распределение альбатросов во время
гнездования имеет вполне выраженный зональный характер.
Исключение представляет только D. irrorata, размножающийся
на островах Галапагос. Как и в случае с пингвинами, наруше-
ние зональности в восточной части Тихого океана связано с хо-
лодным Перуанским течением, проникающим до экваториаль-
ной зоны. Во время кочевок альбатросы распространяются на
огромные акваюрин. но у южных альбатросов общие черты зо-
нального распределения сохраняются.
Семейство ныряющих буревестников — Pelecanoididae. Се
мейство включает один род с четырьмя видами, из которых тр:г
Рис. 63. Распространение альбатросов (Diomedefdae) в Мировом океане. 1 — места гнездования, 2 — область
кочевок
296
О ПРИНЦИПАХ БИОГЕОГРАФИЧЕСКОГО РАЙОНИРОВАНИЯ
нотальные (Р. urinatrix заходит в субтропическую зону), один—
субтропический (Р. garnoti). Местом происхождения и развития
группы предполагается район западного побережья Южной
Америки (Murphy, 1936; Williams, 1952). Только в этом районе
встречаются эндемичные виды — Р. garnoti и Р. magellani.
Ныряющие буревестники не совершают протяженных мигра-
ций. Во время сезонных кочевок они фактически не выходят за
пределы очерченной -на рис. 64 области распространения семей-
ства в гнездовой период. Таким образом, распространение се-
мейства ныряющих буревестников характеризуется широтной
зональностью, несколько нарушаемой только у западного побе-
режья Южной Америки.
Семейство буревестников — Procellariidae. Большое семей-
ство, объединяющее 64 вида, относящегося к 12 родам. Обла-
стью распространения семейства является весь Мировой океан,
от льдов Арктики до льдов Антарктики. В пределах тропической,
а также обеих умеренных и субтропических зон гнездятся ви-
ды рода Puffinus (19 видов), в тропической, нотальной и обоих
субтропических — виды рода Pterodroma (26 видов). Большая
часть видов этих родов концентрируется в южном полушарии в
южной части тропической, южной субтропической и нотальной
зонах. Что касается остальных 10 родов, то их распространение
характеризуется четко выраженной зональностью. В нотальной
н антарктической зонах гнездятся виды родов Pachyptila,
Pagodroma, Daption, Macronectes, в антарктической — Thalas-
soica, в нотальной — Adamastor, Procellaria, Halobaena. Род
Fulmarus имеет более широкую область, но его распространение
также зонально: арктическая и умеренная зоны северного полу-
шария и антарктическая и мотальная зоны южного полуша-
рия. Зонально распространение и рода Bulweria, ограниченно-
го тропической зоной (рис. 65).
Многие виды широкораспространенных родов Puffinus и
Pterodroma в период сезонных кочевок мигрируют за пределы
зон, где они размножаются. Некоторые из них совершают мас-
совые перелеты через экватор. Что же касается остальных родов,
то их миграции в долготном направлении ограничены. Так, буре-
вестники эндемичных южных родов подобно альбатросам совер-
шают дальние миграции в широтном направлении, на север же
их распространение зимой ограничивается у одних южной ча-
стью тропической, у других — южной субтропической или
нотальной зонами.
Современное распространение буревестников дает много
доводов о происхождении этой группы в южном полушарии, где
гнездится 81% известных видов. В южном полушарии обитают
представители всех 12 родов, из которых 8 родов эндемичны.
В северном полушарии только 4 рода, все они имеют представи-
Рис. М. Распространение ныряющих буревестников (Pelecanoididae) в Мировом океане. I — места гнездова-
ния
Рнс. 65. Распространение некоторых буревестников (Procellariidae) в Мировом океане. 1 — места гнездо-
вания буревестников родов Pachyptila, Pagodroma, Daption, Macronectes, Thalassoica, Adamastor, Procella-
ria, Halobaena; 2 — общая область их кочевок; 3 — места гнездования буревестников рода Bulweria
ОРНИТОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
299
телей в южном полушарии. Существенно и то, что эндемичные
роды есть только в нотальной (2 из 9) и антарктической зонах
(2 из 6). Обе зоны вместе располагают 6 эндемичными родами.
Здесь нет только представителей рода Bulweria. Наибольшее ви-
довое разнообразие свойственно нотальной (25). тропической
(25) и южной субтропической (20) зонам. Но в тропической зо-
не это представители всего трех, а в субтропической — четырех
родов. Эти данные позволяют сузить предполагаемый район
происхождения семейства умеренной зоной Южного океана с
сопредельными водами. Небольшое количество видов у побере-
жий Южной Америки (9) и большое разнообразие буревестников
в районе Новой Зеландии (27) дают возможность ограничить
предполагаемый район юго-западной частью Тихого океана с
прилегающими с юга и запада водами Южного океана, характе-
ризующимися наибольшим количеством видов вообще и энде-
мичных видов, В частности. Уильямс (Williams. 1952) предпола-
гает следующие пути проникновения представителей семейства
из Тихого океана в другие районы: на запад в Индийский океан
и на восток через пролив Дрейка в южную часть Атлантики;
в северную Атлантику из южной Атлантики, через район Панам-
ского перешейка (в период его отсутствия), море Тетис и Арк-
тику.
Как видно из современного распространения, большая часть
родов расселялась в широтном направлении, поэтому не слу-
чайно, что почти все роды и многие виды в Южном океане име-
ют циркумполярное распространение. Иное дело — роды Puffi-
nus и Pterodroma. В основном именно их представители засели-
ли другие зоны и районы.
Семейство качурок — Hydrobatidae. Широко распространен-
ное семейство, насчитывающее 21 вид, относящийся к 8 родам.
Представители семейства гнездятся от северной части бореаль-
ной зоны до Антарктики. Распространение семейства в целом
не зональное. Из 8 родов строго зональны только Garrodia (но-
тальная зона), Halocyptena (северная субтропическая зона),
Nesofregetta (тропическая эона).
Рассматриваемое семейство, несомненно, тихоокеанского про-
исхождения. Здесь гнездятся все виды за исключением Н. pela-
gicus, 13 из них эндемичны. В индийском и атлантическом сек-
торах Южного океана гнездится по четыре вида (О. oceanicus.
G. nereis, Р. marina, F. tropica), встречающихся и в тихоокеан-
ском секторе. В тропической части Индийского океана качурки
совсем не гнездятся, а в тропиках Атлантики только О. castro.
В Северной Атлантике гнездится 4 вида — Н. pelagicus. О. leu-
corhoa, О. castro, и Р. marina. Лишь Н. pelagicus эндемичен для
этого района. Остальные обитают в Тихом океане. Очевидно,
нужно согласиться с Уильямсом (Williams, 1952), который пред-
300
О ПРИНЦИПАХ БИОГЕОГРАФИЧЕСКОГО РЛИОНИРС-Вл'1ИЯ
полагает те же, что и у буревестников, пути расселения качурок
из Тихого океана в Атлантический.
Семейство фаэтонов — Phaethontidae. Тропическое семейст-
во, включающее три вида, объединяемые в один род Phaethon.
Большая часть гнездовий располагается в тропической эоне,
лишь местами размножаются в субтропиках- За исключением
редких залетных и занесенных штормами особей, фаэтоны не
проникают за пределы тропических и субтропических широт и
в период кочевок (рис. 66). Зональность в распределении се-
мейства сохранялась, очевидно* и в прошлом. Этому не противо-
речат ископаемые находки из эоценовых отложений в Англии.
В первой половине третичного периода в данном районе был
субтропический климат.
Местом происхождения и дифференциации отряда веслоно-
гих, в том числе фаэтонов, предполагается восточная часть мо-
ря Тетис, что соответствует современному району между Инди-
ей и Австралией (WilHams, 1952; Fisher and Lockley, 1954;
Fisher and Peterson, 1964).
Семейство фрегатов — Fregatidae. Тропическое семейство по
распространению сходное с фаэтонами. За исключением полу-
острова Калифорния все районы размножения находятся в тро-
пической зоне. Известны случаи залетов фрегатов в умеренные
широты, но область регулярных кочевок не выходит за пределы
субтропических зон (рис- 67). Семейство включает один род
Fregata, объединяющий пять видов. Расселение фрегатов в тро-
пики северной Атлантики происходило, по-видимому, через
район Панамского перешейка из Тихого океана (F. magnificens),
а в южную Атлантику (F. ariel, F. minor) — из Индийского
океана вокруг Африки (Williams, 1952).
Семейство олушей — Sulidae. Включает два рода — Moris с
тремя видами и Sula с шестью видами. Род Moris биполярен:
М. bassanus гнездится в северной Атлантике, М. capensis — в
районе южной Африки, М. serrator — в районе Новой Зеландии
и Тасмании. Род Sula имеет зональное распространение: гнездо-
вые части ареалов всех его видов не выходят за пределы суб-
тропических широт. В пределах тропической и субтропической
зон виды р. Sula совершают и сезонные кочевки (рис. 68). Наи-
большее видовое разнообразие олуш р. Sula свойственно Тихо-
му (5) и Индийскому (4) океанам. В Атлантическом океане
гнездится 3 вида. Все атлантические виды — S. sula, S. dacty-
latra, S. leucogaster есть в Индийском и Тихом океанах, откуда
они проникли, очевидно, через район Панамского перешейка
(Williams, 1952). Ископаемые остатки олуш р- Sula известны из
южной части Франции.
Семейство пеликанов — Pelecanidae. Тропическое по прайс-
Рис. 66. Распространение фаэтонов (Phaethontidae) в Мировом океане. 1 — места гнездования, 2 — область
кочевок
Рис. 67. Распространение фрегатов (Fregatidae) в Мировом океане. 1 - места гнездования, 2 — область
кочевок
Рис. 68. Распространение олуш (Sulidae) в Мировом океане. 1 — места гнездования видов рода Sula,
2 — область их кочевок, 3 — места гнездования видов рода Moris
304
О ПРИНЦИПАЛ БИОГЕОГРАФИЧЕСКОГО РАЙОНИРОВАНИЯ
хождению семейство. В настоящее время большая часть видов
(из восьми известных) обитает в пределах тропиков и субтро-
пиков на побережьях, а также на озерах и реках. Только
Р. erythrorhynchus и Р. crispus гнездятся в южной части умерен-
ной зоны. По берегам Индийского океана обитает четыре вида,
в восточной части Тихого океана— три, в западной Атлантике—
один, в восточной — два.
Семейство бакланов — Phalacrocoracidae. Семейство вклю-
чает 29 видов, объединяемых в три рода. Род Nannopterum
(1 вид) эндемичен для Галапагоских островов, распространение
видов рода Halietor (4 вида) ограничено на западе Средизем-
ным морем и Африкой, на востоке Новой Зеландией и Новыми
Гебридами, представители рода Phalacrocorax (24 вида) обита-
ют во всех зонах от Арктики до Антарктики. Таким образом,
распространение семейства бакланов почти космополитно. Наи-
большим разнообразием характеризуется южная часть Тихого
океана- В Австрало-Новозеландском районе обитает 9, в южной
Атлантике — 8, в тропической части Ицдийокого океана — 8,
в юго-восточной части Тихого океана — 7, на севере Тихого оке-
ана — 6, в северной Атлантике — 4, в Антарктике — 1 вид.
Уильямс (Williams, 1952) называет следующие направления и
пути расселения семейства из Австрало-Азиатского района,
предполагаемого места его происхождения: а) на север (пре-
имущественно виды черного цвета) в Европу и Азию, откуда —
в Северную Америку; б) на юго-восток (в основном белобрю-
хие ввды) через Австралию в Южный океан и затем вдоль Ан-
тарктиды к Южной Америке; в) вдоль Антарктиды от Южной
Америки к Новой Зеландии (виды серого цвета). Расселение
бакланов от Австралии вдоль Антарктиды к Южной Америке
предполагал и Мэрфи (Murphy, 1936). Расселение бакланов про-
исходило вдоль побережий, а ряда видов и по внутренним водое-
мам. По этой причине до сих пор остались не заселенными ими
океанические архипелаги в тропической зоне всех океанов.
Семейство чаек Laridae. Включает 7 родов, объединяющих
44 вида. В аркто-умеренной зоне обитают представители 5 ро-
дов — Larus, Rissa, Хета, Pagophila, Rhodostethia, в тропиче-
ской 2 — Larus и Creagrus (последний эндемик островов Галапа-
гос) в южной умеренной — также 2 — Larus и Gabianus. Пред-
ставители рода Larus, объединяющего 36 видов, встречаются от
Арктики до Антарктики, остальные роды по распространению
вполне зональны. Почти половина видов рода р. Larus, а также
R. rosea гнездятся на внутренних водоемах и посещают морские
побережья только во время кочевок.
Виды, связанные в период размножения с внутренними во-
доемами, наиболее многочисленны в умеренной и субтропической
ОРНИТОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
305
зонах северного полушария. Чайки, гнездящиеся на морских по-
бережьях, наиболее многочисленны в северной полушарии, точ-
нее в умеренной и арктической зонах, где, как отмечалось выше,
обитают представители 5 родов. Так, в северной Атлантике раз-
множается 10 видов, относящихся к четырем родам, в Арктике
соответственно 5 и 4, в северной части Тихого океана — 12 и 3.
В тропиках чайки представлены мало: в Атлантическом океане
1 вид, в Индийском — 3, в западной части Тихого — 1, в восточ-
ной части Тихого — 3. Небольшим разнообразием морских ви-
дов чаек отличается и умеренная зона южного полушария: Юж-
ная Америка — 3, Австрало-Новозеландский район — 3, остро-
ва в Южном океане — 2 вида.
Основываясь на современном распределении чаек, ряд авто-
ров (Fisher and Lockley, 1954; Fisher and Peterson, 1964)
полагает, что центром происхождения этой группы является
район северного полушария, включающий Арктику и северные
части Тихого и Атлантического океанов. Большое видовое раз-
нообразие и четыре эндемичных рода в этих зонах говорят в поль-
зу данного предположения. Однако эта трактовка затрудняет
объяснение распространения обитающих южнее большого коли-
чества чаек рода Larus на внутренних водоемах. В связи с этим
интересна гипотеза Уильямса (Williams, 1952), который связы-
вает происхождение чаек с морем Тетис. Он полагает, что бело-
головые виды чаек могли иметь местом своего развития север-
ное побережье Тетис, а черноголовые — южное. Половина со-
временных видов чаек является белоголовыми. Почти все они
обитают только в северном полушарии. Черноголовые виды мно-
гочисленны в тропиках, субтропиках и в прилегающих районах
умеренных зон, при этом значительная часть гнездится на внут-
ренных водоемах. Может быть, гипотезу Уильямса следует
принять только для объяснения происхождения и расселения
видов рода Larus. Вместе с этим нельзя отрицать наличие оча-
гов видообразования чаек на севере Тихого океана и в северной
Атлантике.
Морские виды чаек, за немногим исключением, — прибреж-
ные виды. Вдоль побережий шло их расселение и в прошлом. По
этой причине они не заселили большинство океанических архи-
пелагов в тропической зоне.
Семейство крачек — Sternidae. Преимущественно тропиче-
ское семейство, насчитывающее 42 вида, относящихся к 10 ро-
дам. Представители трех родов Hydroprogne, Chlidonias, Phae-
tusa, а также некоторые виды других родов (в частности Sterna)
— всего 12 видов — гнездятся в основном на внутренних водое-
мах.
Виды родов Anous Gygis, Procelsterna, Larosterna ограниче-
ны тропической и субтропической зонами, роды Thalasseus и Ge-
306
О ПРИНЦИПАХ БИОГЕОГРАФИЧЕСКОГО РАЙОНИРОВАНИЯ
lochelidon заходят в умеренные широты; только род Sterna рас-
пространен от Арктики до Антарктики. К тропическим и субтро-
пическим районам приурочено распространение и большей части
видов крачек, при этом на первом месте по разнообразию стоят
западная часть Тихого океана и Индийский океан. Без учета ви-
дов внутренних водоемов в тропиках Индийского и западной ча-
сти Тихого океанов обитает 17, Атлантике — 11, восточной части
Тихого океана — 11 видов. В умеренных районах северной ча-
сти Тихого океана гнездится 2, северной Атлантике — 5, южной
Атлантике — 3, южной части Тихого океана — 5 видов.
Наиболее вероятное место происхождения семейства —
район между Индией и Австралией (Williams, 1952). Таким об-
разом, происхождение крачек, как и других тропических се-
мейств, связано с районом восточной части моря Тетис. Радиа-
ция и расселение семейства происходили главным образом в ши-
ротном направлении. Они заселили не только прибрежные, но
и океанические районы. В Атлантический океан крачки расселя-
лись как с запада через район Панамского перешейка, так и с
востока через западную часть моря Тетис. С востока через Па-
намский перешеек и с запада через Океанию присходило за-
селение и восточной части Тихого океана. Расселение в северном
и южном направлениях имело ограниченные масштабы, при
этом в умеренные районы проникли представители, главным об-
разом, рода Sterna.
Семейство поморников — Stercorariidae. Биполярная группа.
В Аркто-бореальной зоне обитает 4 вида, относящихся к родам
Stercorarius и Catharacta, в нотально-антарктической — 1 вид
рода Catharacta (многие авторы антарктического поморника
рассматривают как самостоятельный вид — С. moccormicki). Та-
ким образом, распространение семейства в гнездовой период но-
сит зональный характер. В период сезонных миграций все виды
совершают трансэкваториальные миграции. Наиболее вероятно,
что по происхождению поморники — северная группа (Fisher
and Peterson, 1964). В пользу предположения о происхождении
рассматриваемого семейства в южном полушарии (Williams,
1952) данных очень мало.
Семейство чистиковых — Alcidae. В настоящее время семей-
ство включает 22 вида, объединяемых в 12 родов. Чуть менее
ста лет назад в Атлантике был истреблен еще один вид —
Pinguinus impennis. Гнездовая область трех видов — В. craveri,
В. hypoleucus и S. wumisuzume не выходит за пределы субтро-
пиков. Один вид арктический, все остальные являются бореаль-
ными и арктобореальными. Лишь три из них — Р. aleuticus,
S. antiquus, С. monocerata — проникают в северную часть суб-
тропической зоны. Почти не выходят чистиковые за пределы
умеренной зоны и во время сезонных кочевок (рис. 69).
Рис. 69. Распространение чистиковых (Alcidae) в Мировом океане. 1 — места гнездования, 2 — область
кочевок
зов
О ПРИНЦИПАХ БИОГЕОГРАФИЧЕСКОГО РАЙОНИРОВАНИЯ
Происхождение чистиковых связано с северной частью Ти-
хого океана. Данный вопрос не вызывает разногласий. Разли-
чия в мнениях авторов сводятся в этом смысле лишь к тому, сле-
дует ли рассматривать местом происхождения чистиковых
район Берингова моря или другой район северной части Тихого
океана. За тихоокеанское происхождение говорят раннетретич-
ные находки чистиковых на тихоокеанском побережье Америки
и современное видовое разнообразие этой группы в Тихом океа-
не. В Тихом океане обитает 18 видов, относящихся к 10 родам,
из которых 6 родов эндемичны, в Атлантическом — 6 видов, от-
носящихся к 5 родам.
Распространение семейства чистиковых в целом характери-
зуется зональностью. В основном в широтном направлении про-
исходило их распространение и в прошлом. В северную Атлан-
тику они проникли, по-видимому, в олигоцене или начале мио-
цена, где обособились такие роды, как Uria, Alca, Pinguinus,
Cepphus, Plautus. Позднее в плиоцене и плейстоцене вновь про-
исходил обмен между фаунами Тихого и Атлантического океа-
нов. Из Атлантического в Тихий океан проникли роды Uria. Cep-
phus, из Тихого в Атлантический — Fratercula. Во все периоды
расселение чистиковых шло через Арктику и Берингов пролив
(Козлова, 1957; Udvardy, 1963).
* * •
Современное распространение морских птиц в гнездовой пе-
риод известно довольно хорошо. Вместе с этим испытывается
большой недостаток в ископаемых данных, в связи с чем непол-
но разработаны вопросы происхождения и .расселения птиц.
Однако ряд конкретных выводов, представляющих интерес
< биогеографической точки зрения, все же можно сделать.
Эволюция морских птиц, как и других птиц, имеет продолжи-
тельность не менее 70 млн. лет. В конце мела — начале кайно-
зоя уже наметились современные отряды, а к середине кайнозоя
— современные семейства. Большинство современных родов и
многие виды появились в конце плиоцена. Вместе с этим неко-
торые роды морских птиц — Moris, Uria, Procellaria — извест-
ны с миоцена, a Puffinus, Pelecanus, Phalacrocorax, Sula — даже
с олигоцена (Lambrecht, 1933 Wetmore, 1940, 1951; Howard,
1950; Storer, 1960; Дементьев, 1964).
Значительная часть времени существования птиц характери-
зовалась более теплым, чем сейчас, климатом. Однако в настоя-
щее время может считаться доказанным, что уже на протяже-
нии мезозоя происходила дифференциация термического режи-
ма и постепенное усиление контрастности климата планеты в
целом. Развитие и усиление этой тенденции имело место в кай-
нозое, особенно с конца палеоцена, когда конфигурация Клима-
ОРНИТОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКСАНА
309'
тическн.х зон в общих чертах соответствовала современному
(Berry, 1937; Axelrod, 1952; Barghoorn, 1953; Страхов, 1962;.
Синицын. 1965—1967; Тахтаджян, 1970; и др.). В частности, бо-
реальная область в западной части Тихого океана в начале
неогена имела очертания, близкие к современным; она включала
север Японского, Охотского, Берингово моря и сопредельные-
воды океана (Синицын, 1965—1967). Таким образом, представ*
ляется возможным говорить о том, что эволюция семейств мор-
ских птиц происходила на фоне отчетливо выраженной широтной,
климатической зональности. Это, конечно, наложило свой отпе-
чаток на распространение птиц.
По-видимому, вполне определенно можно говорить о проис-
хождении фактически всех отрядов и семейств морских птиц в;
Тихом и Индийском океанах. Здесь происходила основная ра-
диация семейств, отсюда шло их расселение. Орнитофауна Ат-
лантического океана более молодая, менее разнообразна и яв
ляется производной от орнитофаун Индийского и Тихого океа-
нов. Формообразование здесь также имело место, но протекало
на уровне видовой дифференциации. В некоторой степени ис-
ключение могут представлять только чистиковые. Северная Ат-
лантика является местом формирования нескольких родов этого
семейства. Вывод о молодости и происхождении орнитофауны
Атлантического океана объясняет подчиненность всех его орни-
тогеографических регионов соответствующим подразделениям.
Тихого океана.
Из обзора семейств видно, что распространение 6 из 14-
(Spheniscidae, Alcidae, Stercorariidae, Phaethontidae, Fregatidae,
Pelecanoididae) имеют зональный характер. Кроме того, не рас-
сматривались семейства, связанные в основном с внутренними
водоемами, Anhingidae и Rynchopidae; размножение представи-
телей этих семейств происходит в тропической и субтропических
зонах. Распространение других восьми семейств морских птиц,
более широкое. Однако незональность распространения в этих
случаях определяется обычно широким распространением одно-
го-двух родов. В семействе Sternidae—это род Sterna, Laridae—
Larus, Procellariidae — Puffinus и Pterodroma, Sulidae — Moris
Распространение же большей части родов зонально. Незональ-
ным может быть названо распространение только 29% (всего 65<
родов) родов морских птиц, из них только три — Phalacrocorax,.
Larus и Sterna характерны для всех зон и являются космополи-
тами.
Зональное распространение значительного количества се-
мейств и большей части родов связано с наличием климатиче-
ской зональности, на фоне которой происходила радиация отря-
дов и семейств в течение третичного периода. Существование
зон, характеризующихся существенными различиями в уело-
310
О ПРИНЦИПАХ БИОГЕОГРАФИЧЕСКОГО РАЙОНИРОВАНИЯ
виях обитания, создавало большие препятствия для расселения
в северном и южном направлениях. Не случайно при сравнении
орнитофаун океанов, отделенных друг от друга массивами су-
ши, наблюдается меньше различий, чем при сравнении крупно-
масштабных зон в пределах одного океана. Действительно, пред-
ставители всех семейств морских птиц известны во всех океа-
нах. Но каждая биогеографическая область имеет свои харак-
терные семейства: Аркто-бореальная — Alcidae, Циркумтропи-
ческая — Phaethontidae, Fregatidae, Sulidae, Нотально-Антарк-
тическая — Spheniscidae, Pelecanoididae. Существование такой
ситуации определялось преимущественным расселением птиц в
широтном направлении (рис. 70). Наличие сообщений между
океанами во всех зонах облегчало эту задачу.
В широтном направлении через Полярный бассейн происхо-
дило расселение чистиковых, многих видов чайковых, отдельные
трубконосых. Время и пути обмена между фаунами птиц север-
ной части Тихого и Атлантического океанов находятся в соот-
ветствии с разработанными, главным образом, отечественными
зоологами (Андрияшев, 1939; Световидов, 1948, 1952; Шмидт,
1950; Гурьянова, 1951; Берг, 1953; Линдберг, 1955; и мн. др.), кон-
цепциями, освещающими закономерности происхождения и рас-
селения морских фаун арктической и бореальной зон. В преде-
лах тропических и субтропических широт в широтном направле-
нии, в том числе через море Тетис и район Панамского пере-
шейка, происходило расселение многих веслоногих, в частности,
фаэтонов, олушей и фрегатов, большинства крачек, некоторых
трубконосых. Возможно, что расселение в этих районах продол-
жалось после образования узких сухопутных перешейков (Па-
намский и Суэцкий). Через Южный океан, также в широтном
направлении происходило расселение большей части родов
пингвинов и многих родов трубконосых. Преимущественное
расселение в долготном направлении, т. е. вдоль побережий, бы-
ло характерно в основном для бакланов и, по-видимому, пелика-
нов. Расселившиеся в обособившиеся в различных зонах чай-
ки и трубконосые распространялись впоследствии также в пре-
делах определенных зон.
Таким образом, изложенные данные говорят о том, что эво-
люция почти каждого семейства морских птиц проходила в пре-
делах определенной крупномасштабной климатической зоны, где
это семейство возникло. За пределы этих зон распространились
представители только отдельных родов. Широтное направление
расселения птиц (и других животных) на стадии формирова-
ния и радиации семейств создало предпосылки для формирова-
ния крупных биогеографических областей, объединяющих одно-
типные зоны всех океанов. Разобщенность Атлантического океа-
на от Тихого и Индийского материками создало вместе с этим
Рис. 70. Предполагаемые центры происхождения и направления расселения морских птиц. 1 — основные
направления, 2 — второстепенные направления
312
ПРЕДПОСЫЛКИ РАЙОНИРОВАНИЯ ОКЕАНА на уровне провинция
предпосылки для развития черт различия, достаточных для вы-
деления подобластей: Атлантической бореальной и Тихоокеан-
ской бореальной, Атлантической и Индопацифической тропиче-
ских. В Арктике и Южном океане нет сухопутных барьеров; в
связи с этим здесь часто встречается циркумполярный тип рас-
пространения. Поэтому в основу выделения регионов в ранге
подобластей учитываются зональные отличия фаун: между арк-
тической и бореальной и между нотальной и антарктической.
Все это говорит в пользу подчеркнутого выше утверждения о
том, что современное зональное распространение животных, и
морских птиц в частности, соответствует настоящим экологогео-
графическим условиям и отражает историю фаун. И далее, под-
ход к биогеографическому районированию, как форме проявле-
ния всеобщей зональности, не противоречит ни одному из проти-
вопоставляемых ему принципов — историческому, фаунистиче-
скому, экологическому, наоборот, он является синтезом данных
принципов и естественно вытекает из разумного их сочетания.
5. Предпосылки к орнитогеографическому
районированию океана на уровне провинций
Абсолютных преград для распространения птиц на Земле,
очевидно, нет. Относительно морских птиц нужно отметить, что
именно их представители проникают в приполюсные районы
Арктики и Антарктики. Однако свои большие способности к рас-
пространению и расселению используют далеко не все птицы.
Здесь я вновь возвращаюсь к вопросу о консерватизме террито-
рии у птиц. У разных видов и групп птиц стремление к родине и
явление дисперсии выражены различно. По А. С. Мальчевскому
(1968, 1969), в случае заметного преобладания одной из этих
тенденций следует говорить о дисперсном или консервативном
типах эволюции. При консервативном типе эволюции территори-
альное постоянство преобладает над стремлением к дисперсии.
В этих случаях даже при широком распространении и высокой
численности вид не способен быстро заселять новые пространст-
ва. При дисперсном типе эволюции происходит постоянное рас-
селение части особей, чаще всего молоди. Птицы, развивающие-
ся по данному типу, способны сравнительно быстро заселять
новые пространства.
В связи с существованием данных типов эволюции стоит во-
прос различия в экологической пластичности видов. Несомненно,
был прав Дарлингтон (1966), когда подчеркивал, что расселяю-
щиеся группы — это доминирующие группы, характеризующие-
ся относительной многочисленностью по числу особей и видам,
разнообразием адаптаций и большей конкурентоспособностью.
В связи с этим не случайно, что низкая численность и ограни-
ченное распространение, определяющиеся во многом низкой эко-
ОРНИТОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
313
логической пластичностью, встречаются среди птиц, развиваю-
щихся по консервативному типу эволюций.
В различных группах морских птиц две рассматриваемые
противоположные тенденции выражены в различной степени.
Факт наличия во многих семействах родов, широко расселивших-
ся в Мировом океане, то есть развивающихся по дисперсному
типу эволюции, определяет большое сходство орнитофаун одно-
типных зон в различных океанах, на основании чего они объеди-
няются в единые биогеографические области. Благодаря таким
широко распространенным группам значительным оказывается
процент родов и семейств, общих для различных областей. Без
же, если говорить о морских птицах в целом, в первую очередь,
отмечаются признаки эволюции по консервативному типу. Это
проявляется в том, что распространение большинства родов, тем
более видов, ограничено определенными зонами, что в Атланти-
ческий океан проникли представители только относительно не-
большой части родов и видов. Наконец, что проявляется в том,
что лишь около 25—30% морских птиц имеют очень обширный
ареал. В Аркто-бореальной области это, например, обыкновен-
ная моевка, чистик, кайра, глупыш, поморники и другие; в Тро-
пической области. — фаэтоны, олуши, некоторые крачки, фре-
гаты, клинохвостый буревестник, буревестник Бульвера и другие,
в Нотально-антарктической области — странствующий, черно-
бровый и сероголовый альбатросы, гигантский буревестник,
капский голубь, доминиканская чайка, некоторые пингвины и
другие.
Больше двух третей (около 70%) видов птиц ограничены в
распространении только определенной частью одного из океа-
нов. При этом 20% видов, гнездовая часть ареала которых огра-
ничена одним или несколькими мелкими островами, являются
локальными эндемиками. Так, чайки — С. furcatus и L. fuligino-
sus, баклан — N. harrisi, альбатрос — D. irrorata, пингвин —
S. mendiculus гнездятся только на островах Галапагос, бакла-
ны — Р. verrucosus — на островах Кергелен, Р. featherstoni —
на островах Чатам, тайфунники — Р. cohow — на Бермудских
островах, Р. caribbaea — на острове Ямайка, буревестник —
Р. auricularis — на островах Ревилья Хихедо, пингвин —
Е. schlegeii — на остроНе Макуори.
Наличие видов с ограниченным распространением создает
предпосылки более дробного районирования океана, то есть вы-
деления регионов второго порядка, соответствующих рангу про-
винций. Здесь в основу районирования должны быть положены
не отличия в составе птиц между зонами, а различия в ком-
плексах птиц в различных районах в пределах каждой зоны.
Рассмотрим в таком плане птиц Тихого океана.
Тихоокеанская бореальная подобласть. Население морских
314
ПРЕДПОСЫЛКИ районирования океана на уровне провинция
птиц в различных районах умеренной зоны Тихого океана не-
однородно. Характерно, что по мере движения с севера на юг
увеличиваются различия в видовом составе птиц у американско-
го и азиатского побережий. Наряду с этим, у обоих побережий
наблюдаются некоторые различия в составе населения птиц в
южно-бореальных районах, с одной стороны, и в северо-боре-
альных — с другой. Аналогичные картины в составе фаун на-
блюдаются и у других животных (Андрияшев, 1939; Шмидт,
1950; Гурьянова, 1951).
В западной части Тихого океана к самостоятельному южно-
бореальному региону (азиатскому), по-видимому, должны быть
отнесены районы Корейского полуострова, Приморья, Хоккайдо
и северной части Хонсю. Однако, здесь нет ни одного характер-
ного только для этого региона вида. Все типичные южно-бореали-
ные виды — Larus crassirostris, Cerorhinca monocerata, Cepphus
carbo, Phalacrocorax fiiamentosus проникают на север Японско-
го моря и в Охотское море. Но если на юге Охотского моря н<.
долю перечисленных видов приходится лишь около 17%, то в
южной части Японского моря — 50%. Значительное количество
видов — Brachyramphus marmoratus, Fratercula corniculata,
Larus schistisagus, Rissa tridactyla, Uria lomvia не гнездится
южнее Татарского пролива. Не идут южнее района северо-во-
сточного побережья Хоккайдо и Южных Курильских островов
Larus schistisagus, Phalacrocorax urile, Lunda cirrhata, Brachy-
ramphus marmoratus, Synthliboramphus antiquus, Oceanodroma
leucorhoa, Uria lomvia, Cepphus grylle, Aethia cristatella, Fra-
tercula corniculata. В результате этого видовой состав южно-
бореального региона в очерченных выше границах оказывается
обедненным в сравнении с более северными районами.
В восточной части Тихого океана к южно-бореальному ре-
гиону (американскому) нужно отнести район тихоокеанского
побережья Америки, по-видимому, южнее островов Королевы
Шарлотты и Ванкувер. В районе островов Королевы Шарлотты
и Ванкувер проходит южная граница распространения Larus
argentatus. Larus glaucessens, Fratercula corniculata, Synthlibo-
ramphus antiquus, Sterna paradisea, Brachyramphus brevirostris.
С другой стороны, только до этих районов поднимаются с юга
Phalacrocorax penicillatus, Larus occidentalis, Pelecanus erythro-
raynchus, Cerorhinca monocerata.
Для американского южно-бореального региона очень харак-
терны Pelecanus erythrorhychus, Cerorhinca monocerata, Pty-
choramphys aieuticus, Larus occidentalis, Phalacrocorax auritus,
P. penicillatus, а также некоторые широко распространенные в
бореальной зоне виды.
В составе населения птиц азиатского и американского южно-
бореальных регионов общих видов немного. При этом большая
ОРНИТОГЕОГРАФНЧЕСКОБ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
SIS
часть их — это широко распространенные виды, обитающие о г
Приморья или северного побережья Хоккайдо до тихоокеан-
ского побережья США (Brachyramphus marmoratus, Uria aalge,
Lunda cirrhata, Oceanodroma leucorhoa, Phalacrocorax pelagi-
cus). Из южно-бореальных видов общим является только один—
Cerorhinca monocerata.
Что касается северо-бореальных районов, то здесь картина
иная. В северной части Японского моря гнездится 14 видов
птиц. Все они есть на юге Охотского моря. В южной части
Охотского моря обитает 23 вида, 19 из них есть и в его северной
части. На севере Охотского моря также 23 вида, 22 из них есть
и в Беринговом море. В Беринговом море 31 вид, 28 из них ха-
рактерны его западной и восточной половинам. В заливе Аляс-
ка также 23 вида птиц, все они гнездятся и в Беринговом море.
17 видов встречаются в заливе Аляска, Беринговом и Охотском
морях. На основании большого сходства в составе птиц в пре-
делах рассматриваемых районов северную часть Тихого океана,
включая Татарский пролив, все Охотское, Берингово моря и за-
лив Аляска до островов Королевы Шарлотты, по-видимому, нуж-
но объединить в один северо-бореальный регион. Проникающие
в южную часть Охотского моря и в залив Аляска южно-боре-
альные виды составляют здесь незначительный процент. Так,
на Южных Курилах на полю четырех южно-бореальных видов
приходится всего около 26%, а заливе Аляска — около 9%.
Объединение северной части Тихого океана от Охотского
моря до островов Королевы Шарлотты в один северо-бореаль-
ный регион не отрицает существования азиатского и американ-
ского центров развития л распространения фаун, что убедитель-
но показано и на других группах морских животных. Типичными
азиатскими видами являются, например, Cepphus carbo, Aethu
cristatella, Larus schistisagus, американскими — Brachyramphus
brevirostris, Larus glaucessens, Phalacrocorax auritus, Ptychora-
mphus aleuticus. Третьим центром формирования фауны северо-
бореальной зоны, очевидно, является район современного Берии-'
гова моря. Примерами берингийских видов могут быть Rissa
brevirostris и Aethia pusilia.
К настоящему времени значительная часть азиатских видов
распространилась на восток до американского побережья, аме-
риканских — на запад до азиатского побережья, следствием
чего и является большое сходство в составе морских птиц, гнез-
дящихся от Охотского мэря до залива Аляска.
Средн всех других районов северной части Тихого океана
(в пределах бореальной зоны) Берингово море отличается наи-
более разнообразной орнитофауной (31 гнездящийся вид). С од-
ной стороны (в первую очередь), это следствие смешения фаун
Двух побережий; именно через Берингово море происходило рас-
Об ПРЕДПОСЫЛКИ районирования океана на уровне ПРОВИНЦИЙ
ширение ареалов американских видов на запад, а азиатских на
восток. С другой стороны — следствие того, что район Беринго-
ва моря был (и, очевидно, есть) одним из центров формообра-
зования.
Тихоокеанская часть Тропической области. В видовом соста-
ве птиц, гнездящихся у азиатского и американского побережий
в пределах северной субтропической зоны, наблюдаются значи-
тельные различия. Здесь всего семь общих видов, шесть из них
относятся к широкораспространенным в тропической области.
Среди настоящих субтропических видов общим является только
Oceanodroma monorhis. Для азиатской части характерны —
Diorriedea albatrus, Puffinus leucomelas, Thalasseus zimmer-
manni, Synthliboramphus wumisuzume, Pterodroma hypoleuca
и другие, для американской — Puffinus opisthomelas^ Larus
.heermanni, Thalasseus elegans, Brachyramphus hypoleucus, Bra-
chyramphus craveri, Oceanodroma homochroa, O. macrodactyh,
O. melania, Halocyptena microsoma и другие. Таким образом,
в субтропиках еще больше, чем в южно-умеренных районах, вы-
ражено различие в составе птиц у противоположных побережий
океана. Исходя из изложенного, район Японии и Восточно-Ки-
тайское море, с одной стороны, и район полуострова Калифор-
ния, с другой, должны быть выделены в регионы второго поряд-
ка. Эти регионы разделены открытыми водами океана, где в
пределах северной субтропической зоны нет островов, на кото-
рых могли бы гнездиться птицы.
Аналогичная картина наблюдается и в южной субтропиче-
ской зоне. Среди- субтропических видов, пожалуй, только Ptero-
droma longirostris и Р. externa гнездятся и в восточной, и в за-
падной частях Тихого океана. Общими здесь являются, главным
образом, широкораспространенные в тропической зоне виды и
их довольно много — 12. Характерны для западной части Тихо-
го океана, но отсутствуют в восточной следующие субтропиче-
ские, а также общие для субтропической и нотальной зон виды—
Moris serrator, Puffinus assimilis, P. tenuirostris, P. carneipes,
P. bulleri, P. gavia, Procellaria parkinsoni, Pelagodroma marina,
Pelecanus conspicillatus, Sterna nereis, Thalasseus bergii, Halie-
tor melanoleucus, Phalacrocorax varius, P. sulcirostris, P. puncta-
tus, P. fuscescens, Pterodroma inexpectata, P. hypoleuca, P. ma-
croptera, P. cooki, Eudyptula minor, Larus novaehollandiae,
Gabianus pacificus. Для восточной части Тихого океана харак-
терны отсутствующие в западной — Larosterna inca, Phalacro-
corax bougainvillei, P. gaimardi, Puffinus creatopus, Pelecanoides
garnoti, Oceanites gracilis, Spheniscus humboldti, Larus belcheri,
Oceanodroma tethys, Sterna lorata, Pelecanus thagus, Sula varie-
gata, Sterna hirundinacea. Таким образом, в южном субтропиче-
ском регионе в Тихом океане также могут быть выделены два
ОРНИТСГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
317
региона второго порядка, разделенных открытыми без островов
водами океана. При этом в составе австрало-новозеландского'
региона нужно рассматривать субтропические районы западной
части Тихого океана (юго-восточное побережье Австралии, Но-
вая Зеландия с прилегающими субтропическими островами) и
юго-восточной части Индийского океана (южное и юго-западное
побережья Австралии с прилегающими островами). Все виды,
гнездящиеся в субтропической зоне юго-восточной части Ин-
дийского океана, есть и в'Тасмановом море.
При сравнении видового состава птиц в восточной и запад-
ной частях Тихого океана в границах тропического региона об-
наруживается много сходства. На этом основании, как отмеча-
лось выше, они рассматриваются в составе одной подобласти.
Наряду с этим, в каждом из данных районов, а также в цент-
ральной части океана (приблизительно в границах Океании)
имеется достаточное количество видов, не встречающихся в
других районах. Для восточной части Тихого океана специфич-
ны отсутствующие в других его районах — Spheniscus mendicu-
lus, Oceanodroma tethys, О. melania, Oceanites gracilis, Phala-
crocorax auritus, P. penicillatus, Nannopterum harrisi, Diomedea
irrorata, Sula nebouxii, Pelecanus occidentalis, Thalasseus maxi-
mus, Phaethon aethereus, Tregata magnificens, Larus fuliginosus,.
Creagrus furcatus, Pterodroma externa, Puffinus auricularis.
Для Океании характерны отсутствующие в западной и восточ-
ной частях Тихого океана Nesofregetta albigularis, N. moestissi-
ma, Fregetta lineata, Pterodroma rostrata, P. brevipes, P. ultima,
P. alba, Puffinus newelli, P. nativitatis, Bulweria macgillivrayi.
Для западной части Тихого океана специфичны Phalacrocorax
sulcirostris, Р. varius, Р. carbo, Ha^tor niger, Pelecanus conspi-
cillatus, Sterna nereis, S. dougalli, Thalasseus bengalensis, Fre-
gata andrewsi, Larus novaehollandiae, Pterodroma becki, Puffi-
nus heinrothi.
Таким образом, тропический регион Тихого океана можно
подразделить на три региона второго порядка — восточный
центральный и западный. К последнему должна быть отнесена
и восточная часть Индийского океана (Бенгальский залив,
Австрало-азиатские моря), имеющая 90% общих с западной
частью Тихого океана видов.
Тихоокеанская часть Нотально-Антарктической области.
Распространение птиц в антарктическом регионе не дает дан-
ных для более дробного районирования. Почти все антарктиче-
ские и потально-антарктические виды имеют циркумполярное
распространение. Многие нотальные виды распространены че-
рез весь Южный океан, но в различных секторах здесь есть и
значительное количество специфичных видов. В частности, за-
метны различия в видовом составе птиц в западной и восточной.
3ts ПРЕДПОСЫЛКИ РАЙОНИРОВАНИЯ океана НА УРОВНЕ ПРОВИНЦИЙ
частях Тихого океана. Характерны для западной части Тихого
океана (район Новой Зеландии) отсутствующие в восточной —
Eudyptes pachyrhynchus, Е. sclateri, Е. schlegeli, Megadyptes
antipodum, Endyptula albosignata, E. minor, Phalacrocorax
corunculatus, P. featherstoni, P. punctatus, Diomedea epomopho-
та, D. bulleri, D. cauta, Pachyptila turtur, Pterodroma inexpecta-
ta и другие. В западной части Тихого океана отсутствуют из
обитающих у побережья Южной Америки Eudyptes chrysolop-
hys, Spheniscus magellanicus, Sterna hirundinacea, Gabianus
scoresbyi, Larus maculipennis. Phalacrocorax atriceps, Phalacro-
corax magellanicus, P. gaimardi, P. olivaceus, Pelecanoides ma-
gellani. Эти различия позволяют западную и восточную части
Тихого океана относить к различным регионам второго порядка,
разделенным огромными пространствами открытых (нет остро-
вов) вод.
Новозеландская часть нотального региона имеет большое
количество (19) специфичных видов, не встречающихся в соот-
ветствующих широтах Индийского океана (острова Кергелен,
Крозе, Хед, Принца Эдуарда, Марион). С другой стороны, 13
видов, встречающихся на перечисленных островах в Индийском
океане, отсутствуют в районе Новой Зеландии. В то же время
большая часть видов, встречающихся в восточной части Индий-
ского океана, есть в Атлантическом. В некоторой степени обо-
собленно выглядят комплексы гнездящихся птиц побережья
Южной Америки. Острова южной части Атлантического океана
(Южная Георгия, Тристан-да-Кунья, Гоф) населяют главным
образом виды, общие для Индийского и западной части Тихого
океана. В итоге потальный орнитогеографический регион, по-
видимому, может быть разделен на три региона второго порядка.
Таким образом, шесть больших регионов Тихого океана, со-
ответствующих основным крупномасштабным эонам, могут быть
подразделены на 13 регионов второго порядка (рис. 71). Так
как выделенные крупные регионы соответствуют определенным
частям биогеографических пелагических областей и подобла-
стей, можно суммировать следующее. Тихоокеанская бореаль-
ная подобласть может быть подразделена на три региона — севе-
ро-бореальный, азиатский южно-бореальный и американский
южно-бореальный. Тихоокеанская часть Индо-тихоокеанской
тропической подобласти может быть подразделена на семь ре-
гионов второго порядка — субтропические: азиатский, северо-
американский, австрало-новозеландский, южно-американский:
тропические: австрало-азиатский, американский, центрально-ти-
хоокеанский. Тихоокеанская часть Нотальной подобласти мо-
жет быть подразделена на два региона: новозеландский и южно-
американский. Антарктическая подобласть, как отмечалось вы-
ше, является весьма однородной.
ОГНИТОГЕОГРАФИЧЕСКОЕ ДЕЛЕНИЕ МИРОВОГО ОКЕАНА
319
Данные по распределению птиц дают основания для еще
более дробного деления. Эти вопросы я не рассматриваю в на-
стоящей работе. Не проводится здесь также сопоставление ор-
нитогеографических регионов второго порядка с соответствую-
щими подразделениями по другим животным. Дело в
том, что в дробных делениях на уровне провинций, округов
и т. д. у различных авторов наблюдаются большие различия.
Регионы второго порядка в пределах одной и той же подоб-
Рис. 71. Орнитогеографическое районирование Тихого океана. Крупно-
масштабные регионы; 1 — арктический и антарктический, 2 — бореаль-
ный и нотальный, 3 — северный и южный субтропические, 4 — тропиче-
ский. Регионы второго порядка: 1 — азиатский южно-бореальный, 2 —
северо-бореальный, 3 — американский южно-бореальный, 4 — азиатский
субтропический, 5 — северо-американский субтропический, 6 — австрало-
азиатский тропический, 7 — центрально-тихоокеанский тропический, 8 —
американский тропический, 9 — австрало-новозеландский субтропический,
10 — южно-американский субтропический, 11 — новозеландский ноталь-
ный, 12 — южноамериканский нотальный
320 ПРЕДПОСЫЛКИ РАЙОНИРОВАНИЯ ОКЕАНА НА УРОВНЕ ПРОВИНЦИИ
ласти объединяет значительное количество широкораспростра-
ненных видов. Тем не менее население каждого из них характе-
ризуется заметным своеобразием. Каждый регион имеет значи-
тельное число видов, гнездящихся только в пределах данного
региона. Это связано с тем, что, по-видимому, почти во всех ре-
гионах имели место очаги формообразования различного масш-
таба.
321
ГЛАВА V!
КОЛИЧЕСТВЕННОЕ РАСПРЕДЕЛЕНИЕ МОРСКИХ ПТИЦ
И БИОЛОГИЧЕСКАЯ СТРУКТУРА ОКЕАНА
Обилие животных в том или ином районе океана, в первую
очередь, определяется кормностью вод. Это, конечно, не отрицает
определенного значения и других факторов среды, в том числе
условий для размножения и физико-химических характеристик
вод. Относительно морских птиц представление о зависимости
их количества от продуктивности вод утвердилось еще в три*
дцатые годы (Jesperson, 1930; Brooks, 1934; Белопольский.
1933; и др.). Удовлетворительная изученность основных океано-
логических и биологических процессов в океане позволяет в на-
стоящее время проводить анализ количественного распределе-
ния птиц в более широком плане на фоне биологической струк-
туры океана.
Особенности биологической структуры океана определяют за-
коны биологического продуцирования. Биологическая структура
океана отражает количественное распределение жизни и харак-
тер биологических явлений в Мировом океане в аспекте геогра-
фической и вертикальной зональности. В основе биологиеской
зональности и биполярности лежит широтная географическая
зональность и сходный характер изменений условий среды от
экваториальной к холодным зонам, проявляющийся в распре-
делении многих геохимических и геофизических характеристик.
Наблюдается хорошо выраженная зональность, а также симмет-
рия по отношению к экватору и в характере динамики вод —
важнейшем условии, определяющем величину биологической
продуктивности. А так как в количественном распределении
первичной продукции, планктона, бентоса, рыб и морских птиц
обнаруживается закономерное сходство, то можно говорить о
следующем общем правиле: обилие жизни в океане от умерен-
ных вод уменьшается в направлении тропиков и вновь несколько
увеличивается в приэкваториальных районах (Зенкевич, 1948;
Богоров, 1967а, 1970а; Богоров и Зенкевич, 1966; Кобленц-Миш*
ке, 1967; Кобленц-Мишке и др., 1970; Моисеев, 1969; Шунтов,
1970в; и др.).
Во второй главе я неоднократно возвращался к сопоставле-
ниям величин плотности населения птиц в зависимости от корм*
322 ЗОНАЛЬНОСТЬ И СИММЕТРИЯ в распределении плотности птип
ности вод в том или ином районе. Ниже эти вопросы рассматри-
ваются в плане крупномасштабной широтной зональности на
примере Тихого и Индийского океанов.
1. Широтная зональность и симметрия
в распределении плотности населения
морских птиц.
На рис. 56 и 57 показано распределение плотности насе-
ления морских птиц в летний и зимний периоды. В результате
осреднения их. а также аналогичных данных за весенний и
осенний периоды была построена общая схема количественного
распределения морских птиц в Тихом и восточной части Индий-
ского океана (рис. 72). Для этого использовались материалы
около 17500 визуальных учетов и некоторые литературные дан-
ные из ряда цитированных в предыдущих главах работ. Пред-
варительно океан был разбит на квадраты по пять градусов по
широте и долготе. Путем осреднения данных всех учетов для
каждого квадрата было определено среднее количество особей
птиц на 1 кв. км. Эти данные легли в основу для проведения
изолиний на рис. 56, 57 и 72. Приближенность полученных цифр
не отрицает их сравнимость при сопоставлении различных райо-
нов и зон.
Из представленных схем видно, что распределение плотности
населения морских птиц вполне подчиняется законам широтной
зональности-
Многочисленны птицы в умеренной зоне. Общеизвестны
большие гнездовые колонии различных видов на Курильских,
Командорских и Алеутских островах, в районе побережий в
Охотском и Беринговом морях, а также в заливе Аляска и у тихо-
океанского побережья Канады. Все эти районы расположены в
северной бореальной зоне. В южно-бореальных районах также
есть колонии птиц, но по мощности они не идут в сравнение с
массовыми северными гнездовьями.
Повышенная плотность населения птиц в течение всего года
наблюдается и в открытых водах умеренной зоны. В северной
части Тихого океана и расположенных в умеренной эоне дальне-
восточных морях средняя плотность в районе шельфа и матери-
кового склона, как правило, составляет около 10—20. а в глу-
боководных районах — 0,5—2,5 особей на 1 кв. км. Плотность
особенно больших концентраций достигает внушительных вели-
чин — сотен и даже тысяч птиц на 1 кв. км. Места значитель-
ных концентраций показаны на рис. 56, 57 и 72; более подробно
о них говорилось во второй главе при описании сезонных аспек-
тов в отдельных районах. Особенно большие концентрации наб-
людаются на мелководье к востоку от островов Прибылова,
КОЛИЧЕСТВЕННОЕ РАСПРЕДЕЛЕНИЕ МОРСКИХ ПТИЦ
323
Рис. 72. Средняя плотность населения птиц в Тихом океане, по данным
визуальных учетов (осредненные данные за весь год). 1 — не менее 0,5;
2 — от 0,5 до 1; 3 — от 1 до 10; 4 — более 10 особей на 1 кв. м
в прибрежных водах в Олюторско-Наваринском районе, у Чу-
котского полуострова, в западной части залива Аляска, в север-
ной части Охотского моря, в проливах Лаперуза и Сангарском.
Высокая численность и плотность птиц в рассматриваемых
широтах соответствуют повышенной биологической продуктив-
ности вод северной умеренной зоны. Как было показано выше,
здесь многочисленны не только гнездящиеся виды (чистиковые,
чайки, трубконосые). Богатые пищевые ресурсы зоны привлека-
ют сюда большое количество птиц океанической группировки
из южных районов. Это вызывает заметное увеличение плотнос-
ти птиц на обширных акваториях зоны (см. рис. 56, 57 и 73).
Сопоставление общих карт раопределения первичной продук-
ции, планктона, бентоса, рыб (БогороВ и Зенкевич, 1966; Бого-
324' ЗОНАЛЬНОСТЬ И СИММЕТРИЯ В РАСПРЕДЕЛЕНИИ ПЛОТНССТН ПТИН
ров, 1967а, 1970а; Кобленц-Мишке и др.. 1970; Моисеев, 1969)’
с распределением плотности морских птиц на севере Тихого
океана, как нигде, обнаруживает большое сходство- На схемах
распределения во всех группах населения выделяются зоны
обилия жизни, окаймляющие окраины океана.
Таким образом, высокая численность и плотность населения
морских птиц в северной части Тихого океана в первую очередь
связана с хорошими кормовыми условиями. Кроме того, здесь
благоприятные климатические и гидрологические условия и мно-
гочисленны скалистые острова, что очень важно для успешного
гнездования размножающихся в этих районах птиц.
Большое значение в жизни морских птиц умеренной зоны
отводится полярному фронту (Кафтановский, 1951; Белополь-
ский, 1957; Судиловская, 1962; Успенский, 1969; и др.). Выше
было показано, что зона полярного фронта (субарктическая
конвергенция), проходящая в Тихом океане между 40 и 45’ с. ш.,
является важной биогеографической границей. В этом районе
находится предел распространения ряда видов птиц. Подавляю-
щее большинство птиц умеренной зоны не гнездится к югу от
фронта- Но влияние фронта на количественное распределение
птиц сказывается только непосредственно в тех районах, где о i
проходит1 2. Фактически в течение всего года значительные кон-
центрации птиц наблюдаются у восточного побережья Японии.
В формировании повышенной биологической продуктивности
этого района особенно велика роль конвергенции между тече-
ниями Куросио и Ойясио. Что касается большей части умерен-
ной зоны, то формирование в ней продуктивных районов нахо-
дится вне связи с полярным фронтом. Понятно, что фронт, про-
ходящий на широте Японии, не может оказывать существенного
влияния <на птиц, находящихся в Охотском, Беринговом, Чукот-
ском морях или в заливе Аляска.
Как и в других районах, важнейшим условием высокой
продуктивности умеренных вод северной части Тихого океана
является повышенная динамика вод. Велико значение осенне-
зимнего конвективного перемешивания, благодаря которому в
период отсутствия вегетации фитопланктона на всей акватории
зоны происходит накопление биогенных элементов в поверхност-
ных слоях. На этом фоне благодаря тем или иным особенностям
1 Я не привожу для сравнения эти схемы в работе, так как в настоя-
щее время они вошли в ряд общеизвестных обобщающих сводок и обзор-
ных статей.
2 Благоприятные кормовые условия для птиц в районах конвергенций
создаются не только благодаря повышенной биологической продуктивности
этих зон. Увеличению концентраций кормовых организмов способствует ме-
ханический сиос планктона течениями. В отношении нейстона на это обра-
щает внимание Ю. П. Зайцев (1970).
КОЛИЧЕСТВЕННОЕ РАСПРЕДЕЛЕНИЕ МОРСКИХ ПТИЦ
325
циркуляции вод формируются участки интенсивного верти-
кального перемешивания и подъема вод» определяющие особен-
но высокий уворень биологической продукции. Такие участки
могут создаваться в районах стыка течений, например, Восточ-
но-Сахалинского и Соя в проливе Лаперуза, теплых океанских
вод с холодными шельфовыми водами на севере Охотского и
Берингова морей, при прохождении течений через район со
2
Рис. 73. Средняя плотность населения птиц на разных широтах, а — при-
брежные воды (заштриховано — июнь—сентябрь, черным — декабрь—
март), б — прибрежные воды (осреднено за год), в — океанические
районы (заштриховано — июнь— сентябрь, черным — декабрь—март),
г — океанические районы (осрелнено зз гол)
326 ЗОНАЛЬНОСТЬ И СИММЕТРИЯ В РАСПРЕДЕЛЕНИИ ПЛОТНОСТИ ПТИЦ
сложным рельефом дна, например, в проливах Курильских и
Алеутских островов или в районе материкового склона цент-
ральной части Берингова моря- Велико в этом смысле значение
стационарных циклонических круговоротов, в которых происхо-
дит подъем к поверхности глубинных вод.
По мере продвижения на юг плотность населения птиц за-
метно уменьшается (см. рис. 72, 73). В Восточно-Китайском мо-
ре средняя плотность в два-три раза ниже, чем в расположен-
ном севернее Японском море. В целом по прибрежным водам
вместе с краевыми морями в субтропической зоне (между 40
и 25° с. ш.) средняя плотность населения птиц в 3—4 раза ниже,
чем в бореальных районах (см. рис. 73). Значительные скопле-
ния птиц, сравнимые по плотности с таковыми умеренной зоны,
в субтропиках известны только в водах Японии, а также у бере-
гов Калифорнии и Мексики (см. рис. 72). Аналогично уменьша-
ется плотность населения птиц и в субтропических водах откры-
того океана. Таким образом, обращает на себя внимание до-
вольно высокая плотность птиц в прибрежной зоне и низкая в
открытом океане. Все это находит объяснение в анализе распре-
деления первичной продукции и обилия других морских живот-
ных.
В прибрежных водах на востоке и на западе Тихого океана
в пределах субтропической зоны благодаря повышенной дина-
мичности вод создаются благоприятные условия для биологиче-
ского продуцирования. Это подтверждают схемы распределения
первичной продукции, фито- зоопланктона и бентоса (Богоров
и Зенкевич, 1966; Богоров, 1967а, 1970а; Семина. 1967; Кобленц-
Мишке и др.. 1970). В Восточно-Китайском море, Калифорний-
ско-Мексиканском районе, прибрежных водах Японии многочис-
ленны такие пелагические рыбы, как сайра, анчоусы, скумбрии,
ставриды, сардины, а также пелагические кальмары.
Зимой, когда в результате охлаждения в поверхностных во-
дах в умеренной зоне уменьшается количество рыб и планктона,
прибрежные субтропические воды становятся местом зимовок
многих птиц, мигрирующих с севера. Летом здесь концентриру-
ется значительное количество птиц из более южных широт.
В тропической эоне наиболее плотно заселены птицами во-
ды у побережья Южной Америки. Несколько выделяются также
восточная часть Зондского архипелага, североавстралийский
шельф и восточная часть Индийского океана (см. рис. 56. 57,
72). В восточных частях Тихого и Индийского океанов, в зоне
формирования пассатных течений наблюдается очень интенсив-
ный выход богатых биогена1ми глубинных вод к поверхности,
что является основной предпосылкой высокой продуктивности
этих районов (Богоров и Виноградов. 1961; Богоров и Зенкевич,
1966; Богоров, 1967а, 1970а; Кобленц-Мишке и др., 1970). В
КОЛИЧЕСТВЕННОЕ РАСПРЕДЕЛЕНИЕ МОРСКИХ ПТИЦ 397
рассматриваемых районах обычны и местами образуют скопле-
ния различные пелагические рыбы — сельдевые, анчоусы, став-
риды. скумбриевые, летучие рыбы и другие- Однако очень рез-
кое (по сравнению с окружающими районами) увеличение плот-
ности морских птиц наблюдается только непосредственно у бе-
регов Южной Америки. За пределами 200-мильной рыболовной
эоны скопления птиц уже не отмечаются*. В прибрежных райо-
нах Индо-Вест-Пацифики плотность птиц приблизительно в 25
раз ниже, чем в северных умеренных, и в 5—7 раз ниже, чем в
субтропических водах (см. рис. 73).
В океанических районах тропической зоны птиц также мало,
как и в открытых водах субтропиков. Средняя плотность на
разных широтах, составляет, как правило, 0,1—0,4 особи на
I кв. км. Субтропические зоны и прилегающие к ним области
тропической зоны соответствуют обширным антициклональным
круговоротам вод. В этих районах слабо выражена вертикаль-
ная циркуляция вод (преобладает опускание вод), поэтому по-
верхностные слои бедны биогенными элементами (Гусарова,
и др., 1967; и др.). Это в свою очередь служит причиной бедно-
сти данных районов жизнью. Здесь мало планктона, а из пела-
гических рыб обычными можно назвать, пожалуй, только лету-
чих рыб, тунцов и некоторых батнпелагических рыб. В итоге
обширнейшие районы открытого океана, охватывающие субтро-
пические (в обоих полушариях) и прилегающие к ним районы
тропической эоны, могут быть охарактеризованы как области
двух океанических биологических минимумов. Центры этих двух
бедных районов совпадают с двумя обширными антициклони-
ческими круговоротами вод.
Области отмеченных минимумов разделяются более насыщен-
ной жизнью приэкваториальной полосой, обязанной своим про-
исхождением заметному перемешиванию вод, возникающему в
системе пассатных и межпассатных течений. Все это находит
наглядное подтверждение на схемах распределения первичной
продукции, планктона, бентоса, рыб, кальмаров (Богоров и Зен-
кевич, 1966; Уда, 1966; Богоров, 1967а, 1970а; Моисеев. 1969;
Беляев, 1970; Богданов, 1970; Кобленц-Мишке и др., 1970; и
др.) и морских птиц (ом. рис. 56, 57. 72, 73)- Большая роль в
обогащении межпассатных районов биогенными элементами от-
родится подъему вод на экваториальной дивергенции. При этом
показано, что в результате переноса меридиональными состав-
ляющими течений зоны обилия планктона и его потребителей
располагаются параллельно экваториальной дивергенции на
значительном удалении от нее (Виноградов и Воронина, 1965;
1 На рис. 73 данные по тропическим прибрежным водам относятся толь-
ко к Индо-Вест-Паиифнке.
328 ЗОНАЛЬНОСТЬ И СИММЕТРИЯ В РАСПРЕДЕЛЕНИИ ПЛОТНОСТИ ПГИЦ
и др.). По наблюдениям Кинга и Пила (King and Pyle, 1957),
в центральной части Тихого океана птицы наиболее часто встре-
чаются не в районе дивергенции, а к северу от нее> приблизи-
тельно, в районе 10° с. ш. Правда, такая характерная картина
наблюдается далеко не всегда. Дело в том, что на распределе-
ние плотности птиц большое влияние оказывают многочислен-
ные архипелаги островов, расположенные в центральных и за-
падных районах Тихого и Индийского океанов- К районам архи-
пелагов птицы тяготеют не только в период гнездования. У ост-
ровов формируются локальные продуктивные зоны, где птицы
концентрируются и во внегнездовое время. А так как океаниче-
ские острова разбросаны и в межпассатной, и пассатных зонах»
увеличение плотности птиц может наблюдаться в различных
участках тропиков.
В связи с влиянием островов на распределение птиц числен-
ность и плотность их в западной и восточной половинах океани-
ческой зоны тропиков Тихого океана различны. В западных и
центральных районах в целом птиц больше, чем на востоке оке-
ана, где мало островов (см. рис. 72). Это, конечно, не исключает
возможности встречи в последнем районе в отдельные периоды
скоплений птиц. Так, в 1967 г. повышение концентрации птиц
наблюдалось в открытых вадах океана к югу от мексиканского
побережья до 5° с. ш. (Eastropac Atlas, 1970, 1971).
В распределении первичной продукции и планктона наблю-
дается несколько иная картина. В связи с мощным подъемом
вод на востоке океана концентрации биогенных элементов, и
соответственно планктона, значительно выше. По мере продви-
жения на запад наблюдается закономерное их уменьшение.
В открытом океане в пределах южной субтропической зоны
птиц очень мало» при этом не наблюдается увеличения плотнос-
ти по сравнению с тропическими широтами. В прибрежных же
районах в южном полушарии, как и северном, картина совсем
другая. В западной части Тихого и в восточной части Индийско-
го океанов плотность населения птиц в 5—7 раз выше, чем в при-
брежных тропических районах Индо-Вест-Пацифики, т. е. она со-
вершенно сопоставима с плотностью в северной субтропической
зоне.
Южнее, в умеренных водах она увеличивается еще в не-
сколько раз, в итоге в районе Тасмании и Новой Зеландии плот*
ность достигает величин, характерных Охотскому и Берингову
морям. Здесь нередко можно встретить огромные скопления
птиц. Плотность в таких случаях достигает нескольких сотен и
тысяч особей на 1 кв. км. На многих островах в этих районах
располагаются большие гнездовые колонии. Плотно заселены
птицами прибрежные воды в пределах субтропической и уме-
КОЛИЧЕСТВЕННОЕ РАСПРЕДЕЛЕНИЕ МОРСКИХ ПТИЦ
329
ренной зон на востоке Тихого океана. В 4—5 раз в
сравнении с субтропической зоной выше плотность птиц и в
открытых умеренных водах, где в среднем около двух птиц
ча I кв. км. Такая же величина плотности характерна и для
Антарктики. >
Из изложенного видно, что в аналогичных широтах северно-
го и южного полушарий цифры плотности населения птиц впол-
не сопоставимы. Следовательно» в самых общих чертах можно
говорить о зеркальном отражении картины, наблюдаемой в се-
верном полушарии. Увеличение количества птиц повсеместно в
умеренной зоне и в прибрежных водах субтропиков южного по-
лушария вполне согласуется с увеличением продуктивности вод
этих районов. На формирование условий для повышенной био-
логической продуктивности в умеренных и антарктических ши-
ротах южного полушария влияют те же факторы, что и в соот-
ветствующих зонах северного полушария- На обобщающих схе-
мах распределения биогенных элементов, первичной продукции,
фито-зоопланктона и бентоса обращает на себя внимание за-
метное увеличение показателей всех характеристик при продви-
жении из субтропических районов в умеренные (Богоров и Зен-
кевич, 1966; Богоров, 1967а, 1970а; Гусарова и др., 1967; Семи-
на. 1967; Кобленц-Мишке и др., 1970). Однако наряду с прин-
ципиальным сходством обнаруживаются и некоторые несоответ-
ствия по отдельным районам. В частности, на общих схемах фи-
гурирует как малопродуктивный район юго-западная часть Ти-
хого океана. При описании сезонных аспектов в орнитофауне
Австрало-Новозеландского района отмечалось, что, по-видимо-
му, это связано с недостаточной изученностью данной аквато-
рии. Исследования ТИНРО в последние годы показывают, что
у южного побережья Австралии и на Новозеландском плато
весьма обилен планктон и имеют высокую численность многие
виды рыб.
Выше описаны основные особенности количественного рас-
пределения птиц относительно экваториальной оси симметрии.
Кроме экваториальной, может быть проведена также меридио-
нальная ось симметрии, делящая океан на западную и восточную
половины. Наиболее характерной особенностью в распределе-
нии жизни относительно меридиональной плоскости симметрии
является уменьшение количества животных при продвижении от
прибрежных зон в центральные части океана- В целом нерити-
ческие и шельфовые районы характеризуются значительно бо-
лее высокими показателями первичной продукции., биомасс
планктона, бентоса и рыб. Биомасса планктона в поверхност-
ном слое океана при продвижении от прибрежной эоны к цент-
ральной части уменьшается в 20—30 раз (Богоров и Зенкевич,
330 ЗОНАЛЬНОСТЬ И СИММЕТРИЯ В РАСПРЕДЕЛЕНИИ ПЛОТНОСТИ ПТИЦ
1966). На рисунке 73 плотность птиц в прибрежной эоне ниже,
чем в океанической, в среднем в 5—10 раз. Однако здесь нужно
учитывать два обстоятельства. Во-первых, я располагаю ограни-
ченным количеством учетов в 15-<мильной прибрежной зоне, где
птицы весьма многочисленны; во-вторых, при расчетах к при-
брежной эоне относились все краевые моря и довольно обшир-
ные акватории окраин океанов. Если же сравнить непосредст-
венно прибрежную эону с центральными океаническими райо-
нами, разница выразится цифрами того же порядка* что и в слу-
чае с планктоном.
На основании приведенных данных о распределении плотно-
сти птиц в Тихом и Индийском океанах по отношению к эквато-
риальной и меридиональной плоскостям симметрии можно со
всей определенностью говорить, что птицы не представляют ис-
ключения среди других морских животных; их количественное
распределение соответствует особенностям распределения жиз-
ни в океане в целом и укладывается в схему биологической
структуры океана.
Наряду с симметричным расположением в океане биологи-
ческих характеристик имеют место явные нарушения правиль-
ности широтной зональности и симметрии, обусловленные осо-
бенностями циркуляции вод, конфигурацией побережий, релье-
фом дна и так далее (Богоров и Зенкевич, 1966). В отношении
морских птиц можно также отметить несколько отклонений от
симметричного распределения и изменения плотности их насе-
ления.
1. Увеличение плотности птиц в прибрежных районах на-
блюдается и на востоке, и на западе Тихого океана. Однако в
пределах тропических широт в восточной части океана концен-
трации птиц значительно выше, чем в западной. Это общее пра-
вило для всех групп животных и увеличение плотности населе-
ния птиц следует здесь за увеличением количества других жи-
вотных. Как отмечалось выше, богатство района связано с мощ-
ным подъемом глубинных вод на восточной периферии Тихого
океана. Противопоставлять западную и восточную части Индий-
ского океана в этом смысле не представляется возможным. На
востоке Индийского океана значительный подъем вод происхо-
дит между Зондскими островами и Австралией. В западной
части Индийского океана аналогичные зоны подъема вод фор-
мируются у побережий вод влиянием муссонов. В обоих слу-
чаях наблюдаются повышенные концентрации птиц, сопостави-
мые по плотности.
2. Открытие воды значительной части тропической зоны, а
также обеих субтропических зон, как показано выше, отли-
чаются большой бедностью жизни, в том числе птицами. Эту
КОЛИЧЕСТВЕННОЕ РАСПРЕДЕЛЕНИЕ МОРСКИХ ПТИЦ
331
общую картину нарушают острова, которые в огромном количе-
стве разбросаны в Тихом и Индийском океанах. На островах
гнездится большое количсстро крачек, олуш, фрегатов, трубко-
носых. Многие гнездовые колонии (особенно крачек) достигают
внушительных размеров.
Рассматриваемые острова расположены, как правило, на
пути пассатных течений, поэтому естественно, что вблизи остро-
вов наблюдаются завихрения вод и создаются условия для пе-
ремешивания. Это основная причина повышенной продуктивно-
сти вод в районе островов (Sette, 1955; и др.). Большинство ост-
ровов в низких широтах — коралловые или окружены корал-
ловыми рифами, которые по продукции органического вещества
не уступают наиболее продуктивным районам умеренной зоны
и являются среди тропических морей «оазисами в пустыне»
(Yonge, 1963). Высокая продуктивность вод у островов, конеч-
но, — основная предпосылка больших концентраций здесь птиц.
Определенную роль в увеличении продуктивности прибрежных
вод в районе гнездовых колоний должны играть и сами птицы.
Такой вывод напрашивается в связи с данными, полученными
при изучении колониальных птиц Баренцева моря (Головкин и
Позднякова, 1964, 1965; Головкин и Зеликман, 1965; Головкин,
1970). Установлено, что в результате удобрения прилежащих к ко-
лониям вод экскрементами птиц увеличивается содержание био-
генных элементов, что создает условия для формирования ста-
бильных районов с высокой биологической продуктивностью. При
этом локальные круговороты и завихрения течений в районах ко-
лоний уменьшают скорость разноса биогенных элементов.
Следует заметить, что открытый океан на удалении от остро-
вов на несколько десятков миль на этих широтах чаще всего
поражает своей пустынностью. Приуроченность к островам
гнездящихся птиц показана в ряде работ по центральной части
Тихого океана (Dixon and Starrett, 1952; Murphy and Ike-
hara, 1955; Waldron, 1964; King, 1970). Характерно это и для
других районов. Так, в Коралловом море установлено наличие
гнездовых колоний на многих океанических островах (Hindwood
et al., 1963). В открытом же море птицы здесь также немного-
численны, как и в центральной части Тихого океана.
3. Временные нарушения в Описанное распределение плотно-
сти вносят и трансэкваториальные мигранты. Весной и осенью
большие количества мигрантов могут быть встречены в бед-
ных птицами водах океана. Однако эти районы они преодолева-
ют обычно в короткие промежутки времени и не образу-
ют здесь устойчивых скоплений. Большую часть года они прово-
дят е продуктивных районах северного и южного полушарий.
4. Хотя в целом в распределении плотности населения .мор-
ская птиц наблюдается крупномасштабная зональность, зоны
332 ЗОНАЛЬНОСТЬ И СИММЕТРИЯ В РАСПРЕДЕЛЕНИИ ПЛОТНОСТИ ПТИЦ
различного обилия птиц расположены не строго по определен-
ным широтам. Такого рода отклонения от симметричного распо-
ложения характерны и .тля других морских животных, и соз-
даются они существующими системами циркуляции вод (Бого-
ров и Зенкевич. 1966).
5. Плотность концентраций птиц в Антарктике соответст-
вует плотности в бореальной и нотальной зонах Тихого океана.
Меньше заселена птицами Арктика — аналог антарктической
зоны в северном полушарии. Причина этого — неблагоприятный
климатический и гидрологический режимы данной зоны. Исклю-
чение представляют только атлантический сектор Арктики, на-
ходящийся под влиянием Гольфстрима, и крайний юг Чукотско-
го моря, находящийся под влиянием тихоокеанских вод.
Все отмеченные нарушения в симметричном распределении
плотности населения птиц лишний раз иллюстрируют зависи-
мость численности птиц и плотности их концентраций от продук-
тивности района.
Исходя из положения, что биологическая структура океана
определяется законами биологического продуцирования, Л. А.
Зенкевич (1970) выделяет в океане восемь типовых зон с типо-
вым населением, обуславливающих особенности биогеоценозов
первого порядка. Эти зоны в общем плайе являются отражени-
ем деления океана на основные ландшафты, населенные специ-
фичными сообществами. При характеристике океанических
ландшафтов наиболее существенны следующие их элементы:
климат с соответствующей атмосферной циркуляцией и особен-
ностями синоптических процессов, циркуляция вод, физико-хи-
мические характеристики водных масс, видовой состав, особен-
ности распределения и продуктивность населения (Беклемишев,
1967, 1969; Богоров, 1970). Согласно цитированным авторам,
основные ландшафты океана: полярноморские (арктический и
вьгсокоаитарктический), субполярные (северный и южный уме-
ренные) , субтропические и экваториальный.
Изложенное относится к океанической зоне. Неритическая
зона в этом смысле специфична. Тем не менее сходный харак-
тер зависимости от крупномасштабной климатической зонально-
сти определяет и наличие общих черт в биологических процес-
сах в океанических районах, с одной стороны, и в прибрежных
— с другой.
В предыдущих главах было показано, что каждая зона от
арктической до антарктической характеризуется специфичной
орнитофауной. Степени различия и сходства легли в основу
предложенного оргнитогеографического районирования океана.
Для каждой зоны удалось установить особенности сезонного
распределения и миграций птиц. Было показано, что характер
КОЛИЧЕСТВЕННОЕ РАСПРЕДЕЛЕНИЕ МОРСКИХ ПТИЦ
333
сезонных изменений в орнитофауне, а также особенности годич-
ного цикла находятся в первую очередь в зависимости от сезон-
ных изменений в биологии и гидрологии моря. Анализ количе-
ственного распределения птиц в данной главе проводился так-
же с учетом зональных подразделений океана- Такой подход,
как мне кажется, является результативным.
По величине показателей биологического продуцирования
типовые зоны Л. А. Зенкевича могут быть объединены в три
группы. Высокие показатели продукции имеют сообщества ниж-
неарктической, антарктической, северной и южной умеренной
зон, а также прибрежных участков и коралловых рифов в тро-
пической области. Средним уровнем продуцирования характери-
зуются открытые воды умеренных широт, а также пояс эквато-
риальных течений. Низкие показатели продуцирования свойст-
венны открытым районам тропической области, включая воды
субтропических широт.
Изложенное выше показывает, что данные по количественно-
му распределению птиц вполне соответствуют рассмотренной
градации зон.
2. Численность морских птиц и их роль как
потребителей других морских животных
Абсолютная численность птиц на земном шаре неизвестна.
Ориентировочно она оценивается в 100 млрд, особей (Fisher
and Peterson, 1964). Используя данные учетов в море, я делал
попытку определить общую численность птиц в Мировом океа-
не (Шунтов, 1970в) *. Проведенные расчеты позволили допус-
тить, что общая численность морских птиц находится в преде-
лах от 1 до 3 млрд, особей. Цифры эти в общем внушительные,
но, по-видимому, не завышенные. Об этом свидетельствуют ре-
зультаты учета птиц на местах гнездования в ряде районов. Вот
некоторые примеры.
По подсчетам С. М. Успенского (1959, 1969), в Баренцевом
море гнездится около 3 млн. морских птиц, главным образом
кайр. Численность гнездящихся птиц на островах у побережья
Перу оценивается в различные годы от 15 до 25 млн. особей
(Dorsi. 1956; Ullmann, 1963; Jordan and Fuentes, 1964; Jordan,
1967). Эти цифры достаточно большие, но истинная численность
птиц в этих районах выше. При подсчете гнездящихся особей
не учитываются находящиеся в море неполовозрелые особи.
Численность последних также высокая, так как большая часть
морских птиц созревает поздно, в возрасте нескольких лет. Так,
। При издании этой статьи допущено много опечаток.
334 ЧИСЛЕННОСТЬ И РОЛЬ ПТИЦ КАК потребителей других животных
общая численность королевского альбатроса определена в
19 тыс. особей В то же время половозрелых птиц всего 9200
особей (Westerskov, 1960, 1963).
Общая численность кайр, по-видимому, достигает 50—
60 млн. (Tuck. 1960). а только взрослых тупиков — 15 млн. осо-
бей (Fisher and Lockley, 1954). Многомиллионно население
темной крачки, широко распространенной в тропиках. Только на
острове Рождества гнездится около 1 млн. ее особей (Ashmole
and Ashmole, 1967). Такой же цифрой оценивается численность
крошечной конюги на острове Малый Диомид (Kenyon and
Brooks, I960)- Столь же массовые гнездовья этого вида, а так-
же большой конюги есть и в других местах на севере Беринго-
ва моря, например, на острове Святого Лаврентия (Fay and Ca-
de, 1959).
Допускается, что на острове Гоф гнездится по 5—10 млн.
особей ширококлювогэ приона — Pachvptila vittata (Gmel.) и
тайфунника — Pterodroma mollis (Gould) (Holdgate. 1958), а на
острове Хед — 4 млн. особей пингвина — Eudyptes cristatus
Forst. (Swales, 1965). О таких же больших колониях и популя-
циях сообщают и многие другие авторы, в частности о птицах
островов Фольклецдских, Южных Сандвичевых, Тристан-да-Ку-
нья и Антарктиды (Voous and Wattell, 1963; Strange, 1965; Har-
per, 1966; Коновалов и Шулятин, 1964 и др.). Перечисленные
работы не ограничивают описок публикаций, где есть сведения
о численности птиц в отдельных районах Мирового океана. Пе-
речень аналогичных указаний можно было бы продолжить. Но
приведенные здесь данные, даже учитывая некоторую их завы-
шенность, являющуюся следствием эмоционального восприятия
при наблюдениях грандиозных скоплений птиц на гнездовьях,
говорят о больших порядках цифр, характеризующих общую
численность морских птиц.
Раньше для оценки средней плотности и численности птиц
я использовал данные около 8 тыс. учетов. Путем осреднения
всех учетов было получено, что средняя плотность птиц состав-
ляет около четырех особей на 1 кв. км (Шунтов, 1970в). Пере-
счет с привлечением большого количества дополнительных уче-
тов (всего около 17500 учетов) дал несколько меньшую цифру
— около 3,5 особи на 1 кв. км. Уменьшение вызвано поступле-
нием большого количества учетов из бедных птицами океаниче-
ских районов Тихого океана. Несомненно, эта цифра должна
быть признана несколько заниженной. Как уже отмечалось, по
большинству районов в учетах слабопредставленными оказа-
лись птицы непосредственно прибрежных вод, где нередко наб-
людаются их плотные концентрации. К примеру, в учетах почти
не фигурировали пингвины, десятки миллионов которых населя-
ют прибрежные воды южного полушария. Показательно в этом
КОЛИЧЕСТВЕННОЕ РАСПРЕДЕЛЕНИЕ МОРСКИХ ПТИЦ
335
смысле и следующее сопоставление. В летний период, по моим
данным, средняя плотность населения птиц иа шельфе Берин-
гова моря составляет около 18 особей на 1 кв. км. По наблю-
дениям Н. Н. Карташова (1961), в 1—5 км от берега острова
Медный (Командорские острова) только на 1 км маршрутного
учета с лодки приходится 175 птиц. Такая же закономерность
наблюдается и во многих других районах- Учитывая сказанное,
при расчетах я оставляю среднюю плотность 4 птицы на 1 кв. км,
переношу ее и на Атлантический океан1.
Площадь Мирового океана составляет около 361 млн. кв. км.
Около 30 млн. кв. км приходится на приполярные районы, за-
крытые льдом (Назаров, 1970). Расчеты на указанных исходных
материалах дают 1,3 млрд, морских птиц для всего океана. Учи-
тывая предполагаемую заниженность данных по плотности, я
думаю, что цифра 1,3—1,5 млрд, особей является только воз-
можным нижним пределом численности птиц в Мировом океа-
не. Однако мало вероятно, что занижение средней плотности по
данным визуальных учетов более чем двукратное. Таким обра-
зом, при дальнейших обобщениях я исхожу из того, что числен-
ность птиц в Мировом океане находится в пределах от 1,3—1,5
до 3 млрд, особей. Из этого следует, что цифру 100 млрд, птиц
(она, на мой взгляд, несколько завышена) слагают, главным об-
разом, сухопутные виды. Известные данные по населению птиц
различных ландшафтов суши дают материалы для подтвержде-
ния этой .мысли.
В последние десятилетия накоплено значительное количест-
во сведений о количественном распределении птиц в различных
биотопах. Широкий размах такие исследования получили в Со-
ветском Союзе- Количественные учеты свидетельствуют о том,
что плотность населения птиц на суше намного выше, чем в
океане. В лесных биотопах Палеарктики плотность, как прави-
ло, находится в пределах от 200 до 4000 (чаще 400—2000) осо-
бей на 1 кв. км (Дубинин и Торопа-нова, 1956; Новиков, 1960;
Реймерс, 1966; Пузаченко, 1967; и др.). Столь же высокие кон-
центрации птиц характерны и для культурных открытых ланд-
шафтов. В неосвоенных открытых ландшафтах (тундра, пусты-
ни, степи), птиц меньше, ио и в этом случае плотность населе-
ния колеблется от нескольких десятков до нескольких сот осо-
бей на 1 кв. км (Чельцов-Бебутов, 1959; Злотин, 1963; Дроздов,
1967; Второв, 1968; Успенский, 1969; и др). Близкие показате-
лям концентраций птиц в океане Ю. Г. Пузаченко (1962) при-
водит цифры плотности птиц для пустыни Кара-Кум.
< Этому не противоречат данные по продуктивным свойствам всех океа-
нов. По средним величинам первичной продукции Атлантический океан не-
сколько уступает Индийскому, но превосходит Тихнй (Кобленц-Мишке и
др., 1970).
336 ЧИСЛЕННОСТЬ И РОЛЬ ПТИЦ КАК ПОТРЕБИТЕЛЕЙ ДРУГИХ ЖИВОТНЫХ
Результаты сопоставлений материалов, цитированных выше
авторов, с количественным распределением птиц в океане (см.
рис. 73) первоначально несколько удивляют. Ведь привычно го-
ворить об огромных биологических ресурсах океана. Однако
описанные различия вполне объяснимы. Более высокая числен-
ность и плотность концентраций птиц на суше находится в связи
со значительно более высокой продуктивностью сухопутных
сообществ- Н. И. Базилевич, Л. Е. Родин и Н. Н. Розов (1971)1
показали, что живое вещество суши Земли по весу в 750 раз пре-
вышает живое вещество Мирового океана, биомасса консумен-
тов и редуцентов суши более чем в шесть раз выше биомассы
консументов и редуцентов океана. При этом надо иметь в виду,
что площадь суши более чем в два раза меньше площади океа-
на (соответственно 149 и 361 млн. кв. км). Первичная продук-
ция Мирового океана в три раза меньше продукции наземных
растительных сообществ. Согласно Одуму (Odum, 1959), продук-
тивность открытых океанических районов, занимающих боль-
шую часть океана, почти не отличается от пустынных и полу-
пустынных наземных ландшафтов. Одум отмечает, что продук-
тивность вод шельфов сопоставима с экосистемами смешанных
лесов и лугов, но значительно уступает таким сообществам, ка-
кими являются вечнозеленые леса и выращиваемые сельскохо-
зяйственные культуры.
Хотя по продуктивности сообщества вод шельфа сопостави-
мы с некоторыми сообществами суши, плотность птиц в них, как
показано выше, значительно ниже плотности птиц в открытых
ландшафтах суши. Редко в местах особенно больших скоплений
плотность концентраций морских птиц достигает нескольких сог
и тысяч особей на 1 кв- км. Здесь большое значение имеет раз-
личие в доступности пищи в условиях суши и моря. На суше
пищу добывать, конечно, проще. В этом отношении в худших
условиях по сравнению с сухопутными птицами находятся даже
ныряющие виды, не говоря уже о видах, добывающих корм на
поверхности моря.
Таким образом, более низкая продуктивность океана в срав-
нении с сушей, а также недоступность для птиц большей части
кормовых организмов, скрытых водной толщей, рассматривают-
ся мною как основные причины сравнительно низкой плотности
и численности птиц в Мировом океане. Лишь в районах очень
крупных колоний плотность населения птиц может превосходить
плотность наиболее заселенных птицами биотопов суши. Но
большие колонии располагаются только в продуктивных райо-
нах. Особый смысл приобретают здесь и выводы о большом
1 По океану указанные авторы использовали некоторые данные В. Г. Бо-
горова (1969).
КОЛИЧЕСТВЕННОЕ РАСПРЕДЕЛЕНИЕ МОРСКИХ ПТИЦ
337
I значении в увеличении продуктивности прилегающих к колони-
ям вод удобрения экскрементами птиц. Продолжение исследо-
ваний этого направления, начатых на Мурмане (Головкин и
Позднякова, 1964, 1965; Головкин и Зеликман, 1965; Головкин,
1970). в будущем, несомненно, даст важные результаты.
Для определения роли птиц, как потребителей других мор-
ских животных, наряду с численностью важно знание величины
и состава их рационов. В этом отношении не все виды изучены
достаточно полно. Вернее, исчерпывающие данные есть только
по отдельным видам и районам- Установлено, например, что
баклан Бугенвиля — Phalacrocorax bougainvillei (Less.) в сут-
ки съедает 430—495 г рыбы, главным образом анчоуса (Avila,
1954; Jordan, 1967). Весьма интенсивно изучаются численность,
питание и роль колониальных птиц как потребителей рыбы
у южного побережья Африки (Davies, 1955, 1956, 1958; Rand,
1959, 1960, 1960а, 1963а, 19636). Полученные здесь результаты
относительно изъятия рыбы капской олушей, капским бакланом
— Phalacrocorax capensis (Sparrm) и ослиным пингвином —
Spheniscus demersus (L.) довольно противоречивы. Дэвис назы-
вает суточную норму для баклана — около 560 г, а для олуши
и пингвина — цифры близкие к весу тела- Ранд суточную норму
для баклана определяет 280 г, пингвина — 320 г, олуши —
350 г. Естественно, у этих авторов разными оказались и общие
изъятия рыбы. Большое значение изучению питания рыбоядных
птиц уделялось и уделяется отечественными орнитологами. Ис-
следования показали, что суточная норма мелких птиц состав-
ляет обычно несколько десятков граммов. Так, различные крач-
ки потребляют в сутки 35—65 г корма (Бородулина, 1965; Мар-
кузе, 1965). С. М. Успенский (1959) полагает, что мелкие чисти-
ковые и трубконосые в сутки потребляют около 100 г корма.
Птицы средних размеров имеют суточную норму обычно между
100 и 200 г. Рацион моевки определен в 108—140 г, среднего
размера чистиковых — в 150 г. Крупные чайки и чистиковые в
сутки потребляют в среднем 200—300 г, крупные бакланы —
около 700 г (Успенский, 1959; Головкин, 1963; Гераси-
мова. 1965; Марков. 1965; и др.). Таким образом, средние ра-
ционы колеблются от нескольких десятков у мелких видов до
нескольких сот граммов у крупных видов морских птиц.
Больше килограмма корма в сутки потребляют, очевидно,
только такие крупные птицы, как пеликаны. Много пищи за
один раз могут поглощать также другие крупные птицы в тех
случаях, когда находят массовые скопления корма. Так, явно
повышенные количества пищи потребляют чайки, альбатросы,
глупыш и другие птицы в моменты их концентраций у ведущих
лов рыбы судов. В таких случаях нередко можно видеть, как
338 ЧИСЛЕННОСТЬ И ЮЛЬ ПТИЦ КАК ПОТРЕБИТЕЛЕЙ ДРУГИХ ЖИВОТНЫХ
птицы заглатывают рыбу таких размеров, что это мешает нор*
мальному полету. В обычных же природных условиях такие кар-
тины наблюдать не приходится. В морских условиях добывать
пищу довольно трудно. В процессе длительных полевых наблю-
дений у меня сложилось мнение, что значительная часть мор-
ских птиц (особенно виды океанической группировки) большую
часть жизни проводят в поисках корма.
Основу населения мороких птиц в Мировом океане состав-
ляют мелкого и среднего размера виды. Это прионы, качурки,
крачки, мелкие чистиковые, мелкие чайки, буревестники, тай-
фунники. Высокую численность имеют также относительно
крупные виды — крупные чистиковые, бакланы, пингвины, олу-
ши. Как отмечалось выше, суточный рацион большинства видов
этих групп не превышает 400—500 г.
Исходя из того, что численность морских птиц в Мировом
океане составляет 1,3—3 млрд, особей, а среднесуточный раци-
он одной птицы — 150 г, то ежегодное изъятие планктона, ры-
бы, кальмара и бентоса выразится величинами как минимум 0,7,
как максимум — 1,6 млрд. ц. Эти цифры не претендуют на абсо-
лютную точность, но они вполне могут характеризовать масш-
табность явления.
Так как точное соотношение планктоноядных и рыбоядных
птиц в океане не известно, трудно в конкретных цифрах гово-
рить о том, какая доля из потребляемого корма приходится на
рыб. Однако не будет преувеличением сказать, что как мини-
мум 50%, а вернее — больше половины потребляемой пищи
приходится на долю планктона. Преимущественно плактоном
питаются наиболее массовые виды морских птиц — мелкого и
среднего размера трубконосые, мелкие чистиковые. Планктон
является основным кормам также пингвинов в Южном
океане. Но даже если на долю нектона (рыбы и кальмары)
приходится несколько меньше половины пищи, потребляемой
птицами в океане, суммарная цифра остается внушительной.
Она не меньше количества рыбы и кальмаров, ежегодно добы-
ваемых в океане человеком*.
Морские птицы питаются, главным образом, наиболее мно-
гочисленными и доступными для добычи водными, животными2.
Среди планктонных организмов это эвфаузиевые и копеподы.
Из рыб наиболее часто встречаются массовые пелагические ви-
ды или молодь придонных видов, обитающая в верхних слоях.
1 В 1967 г. мировой улов рыбы и нерыбных объектов (без китов) достиг
0.6 млрд, ц (Моисеев, 1969).
г Характерно, что состав питания птиц отражает сезонные изменения в
численности кормовых объектов в море (Белопольский, 1967; Ashmole and
Ashmole, 1968; и др ).
КОЛИЧЕСТВЕННОЕ РАСПРЕДЕЛЕНИЕ МОРСКИХ ПТИЦ
339
Например, у берегов Южной Америки — это преимущественно
анчоус (Avila, 1954; Jordan, 1967), у южного побережья Африки
— сардина и ставрида (Davies. 1958; Rand, 1959, 1960, 1960а), в
северных умеренных морях и Арктике — сельдь, тресковые,
мойва, песчанка и другие (Белопольский, 1957; Успенский, 1969;
Swanz, 1966; и др.). Кроме планктона и рыб велико значение в
питании морских птиц, особенно трубкодосых, пелагических
кальмаров.
Изложенное противоречит выводам Ю. П. Зайцева (1970),
который в обстоятельной книге о морском нейстоне говорит о
том. что морские птицы существуют главным образом или ис-
ключительно за счет животного населения верхнего, менее чем
пятисантиметрового слоя пелагиали. Перечисленные выше, а
также другие массовые кормовые объекты морских птиц не яв-
ляются нейстонными организмами. Нейстон, как подчеркивает
Ю. П. Зайцев, специфичное сообщество пограничной поверх-
ности раздела вода—'воздух. Наряду с комплексом приспособ-
лений. обеспечивающих существование в области пленки по-
верхностного натяжения, для нейстонных организмов характер-
но количественное преобладание в слое 0—5 см по сравнению
с нижележащими горизонтами.
В связи с этим из списка птиц, в питании которых нейстон иг-
рает заметную роль, нужно исключить ныряющих на значитель-
ную глубину видов — пингвинов, чистиковых, большинство
веслоногих. Нейстонные организмы чаще должны встречаться в
питании птиц, добывающих пищу с поверхности моря, — чаек,
крачек, трубконосых. Это, однако, не значит, что они питаются
главным образом нейстонными животными. Основу питания их
составляют виды макропланктона, кальмаров и рыб с довольно-
широким распространением по вертикали, но поднимающихся
в поверхностные слои во время суточных вертикальных мигра-
ций. Как хорошо известно, суточные вертикальные миграции
всех пелагических и части донных животных носят всеоб-
щий и массовый характер- В этом смысле большое значение име-
ет способность многих морских птиц питаться в темное время
суток, когда поверхностные слои наиболее населены планкто-
ном, рыбой п кальмарами.
Названные выше цифры предполагаемого изъятия из океана
птицами других водных животных — 0,7—1,6 млрд, ц — сами
по себе мало о чем говорят. Они приобретают смысл при сопо-
ставлении их с величинами продукции потребляемых ими жи-
вотных.
В настоящее время наибольшим признанием пользуются ве-
личины биологической продуктивности, определенные В. Г. Бо-
горовым (1967, 1970а) и П. А. Моисеевым (1969). В. Г. Бог.)-
’340 ЧИСЛЕННОСТЬ И РОЛЬ ПТИЦ КАК ПОТРЕБИТЕЛЕЙ ДРУГИХ ЖИВОТНЫХ
ров полагает, что ежегодная продукция зоопланктона в Миро-
вом океане составляет 530 млрд, ц (биомасса 215 млрд, ц), зоо-
бентоса 30 млрд, ц (биомасса 100 млрд, ц), нектона 2 млрд, ц
(биомасса 10 млрд, ц)- Продукция всех консументов составляет
562 млрд. ц. П. А. Моисеев, во многом соглашаясь с В. Г- Бого-
ровым, называет несколько более низкие цифры.
Таким образом, от указанной продукции всех морских жи-
вотных доля потребляемого птицами корма составляет всего
около 0.2—0.3% • На этом основании можно сделать самый об-
щий вывод о том, что роль морских птиц в биологическом ба-
лансе Мирового океана в целом не должна быть очень замет-
ной.
Если же говорить о морских птицах как о потребителях нек-
тона, то здесь роль их, конечно, более существенна. К сожале-
нию, вопрос о продукции нектона в настоящее время изучеi
хуже, чем о продукции планктона. Недавно обстоятельный об-
зор существующих воззрений о рыбопродуктивности Мирового
океана сделал П. А. Моисеев (1969).
Обращает на себя внимание следующая тенденция в измене-
нии представлений на продуктивность нектона и прежде всего
рыб. Если в недалеком прошлом бытовали представления о не-
исчерпаемых биологических ресурсах океана, то сейчас на их
место выступают трезвые расчеты- Большинство специалистов
склоняются к мнению о том. что возможный вылов рыбы и про-
мысловых беспозвоночных не может превысить 1 млрд. ц. Прав-
да, здесь допускается другая крайность: некоторые ученые пре-
делом промыслового изъятия называют 0,55 млрд, ц, — вылов,
превзойденный еще в 1966 г. По В. Г. Боторову (1970а), еже-
годная продукция нектона составляет около 2 млрд- ц. П. А.
Моисеев (1969) отмечает, что суммарная продукция ценных
для человечества водных объектов составляет не более 3—3,2
млрд. ц. Оба автора на основании своих расчетов называют
возможное промысловое изъятие 0,8—0.9 млрд, ц в год. Эти
взгляды разделяют многие гидробиологи и ихтиологи. Однако
некоторые сопоставления склоняют к мысли, что эти оценки так-
же являются заниженными. По подсчетам И. И. Акимушкина
(1970), только кашалоты в год съедают около 0,55 млрд, ц голо-
воногих моллюсков, а А. Г. Томилин (1970) называет следую-
щие цифры суточного потребления корма морскими млекопита-
ющими: усатые киты — 3 млн. ц криля и рыбы, кашалоты —
4,5 млн. ц головоногих, дельфины — 17 млн. ц рыбы и голово-
ногих. В пересчете на год получается несколько миллиардов
центнеров. Конечно, ряд моментов в расчетах цитированных ав-
торов навряд ли можно принять. Нельзя, например, согласить-
ся с перенесением И. И. Акимушкиным данных по 30 видам на
КОЛИЧЕСТВЕННОЕ РАСПРЕДЕЛЕНИЕ МОРСКИХ ПТИЦ
341
всю фауну головоногих Мирового океана, в результате чего об-»
щая биомасса этих животных оказалась равной 22 млрд. ц.
Большие возражения вызывают также перенесение А. Г. Томи-
линым плотности дельфинов для Черного моря на весь Мировой
океан. Но даже имея в виду завышенность этих оценок, все же
масштабы описываемого явления будут велики.
Таким образом, если сложить ежегодный вылов человеком
рыбы и промысловых беспозвоночных — более 0,6 млрд, ц, приб-
лизительно такое же количество нектона, потребляемое морски-
ми птицами, и по крайней мере не менее 1 млрд, ц нектона, по-
требляемое морскими млекопитающими, станет видно, что в эти
масштабы не укладываются названные выше величины продук-
ции нектона в Мировом океане. В связи с этим нужно напом-
нить. что существуют более оптимистичные оценки продуктив-
ных свойств океана, пользующиеся, однако, меньшим признани-
ем. Называются величины продукции нектона и вылова, превос-
ходящие указанные выше в несколько раз (Chapman, 1964;
Schaefer, 1965).
Из изложенного видно» что представления различных авто-
ров о величине рыбопродуктивности океана весьма противоре-
чивы. По этой причине говорить о конкретной доле изъятия
птицами нектона от его годичной продукции пока не представ-
ляется возможным. Если принять продукцию нектона 2 млрд, ц
то на долю птиц придется 20—30%, что, конечно, немыслимо.
При принятии наиболее оптимистической оценки продукции
нектона, на долю птиц придется всего несколько процентов. Ес-
ли второй подход более близок к действительности, то пока
можно говорить только о заметной, но ие очень большой ролн
морских птиц, как потребителей нектона. Действительно, труд-
но допустить» что на площади одного квадратного километра
роль четырех особей морских птиц, как потребителей других
животных, будет очень значительной. Но роль птиц существен-
но увеличивается в тех районах, где они концентрируются в
больших количествах и плотность населения составляет не-
сколько десятков, сотен и тысяч на 1 кв. км. О таких районах
говорилось выше. Это, в первую очередь, прибрежные районы
во всех зонах океана и некоторые океанические районы в уме-
ренных водах обоих полушарий.
Так. по подсчетам А. Н. Головкина (1963), кайры и моевка
за лето в Баренцевом море выедают 760 тыс. ц рыбы. По С. М.
Успенскому (1959. 1969), все морские птицы в Баренцевом мо-
ре съедают за год 617 тыс. ц рыбы и 757 тыс. ц беспозвоноч-
ных. Столь же внушительные цифры называются по району юж-
ного побережья Африки (Davies, 1958; Rand, 1959, 1960, 1960а).
342 ЧИСЛЕННОСТЬ И РОЛЬ ПТИЦ КАК потребителей других животных
Нельзя здесь снова не вспомнить и перуанский район. Только
гнездящиеся особи трех видов (баклан, олуша и пеликан) по-
требляют до 28 млн. ц рыбы в год (Jordan, 1967).
В последние годы все большее развитие получают работы
по изучению биогеоценозов океана и сообществ определенных
групп животных. В сравнительно недалеком будущем по мере
накопления данных можно предвидеть построение моделей
структур биогеоценозов отдельных районов океана*. Учитывая
важную роль морских птиц, как потребителей других морских
животных, в этих моделях должны найти место количественные
показатели их биогеоценотических связей. Разумеется, здесь
имеются в виду районы, где птицы многочисленны. Что касается
обширных открытых пространств тропической и субтропической
зон, где птицы немногочисленны, то здесь их роль в общем не-
значительна.
При изучении питания птиц, как было показано выше, во
многих случаях получаются довольно большие цифры потреб-
ляемой ими рыбы. Отсюда нередко следуют выводы о так назы-
ваемом хозяйственном значении птиц. При этом имели и имеют
место призывы к уничтожению птиц, как конкурентов человека
(смотрите, например, А. И. Пахульский (1957). А. И. Гизенко
(1955). Подробное рассмотрение этих вопросов не входит в за-
дачу настоящей работы- Здесь я кратко остановлюсь только на
некоторых моментах.
Проблема динамики и состояния запасов рыб успешно раз-
рабатывается уже несколько десятков лет. Сейчас совершенно
ясно, что изменения в численности определяются в основном
гидрологическими и кормовыми условиями, а состояние перело-
ва и истощения запасов связано исключительно с хозяйствен-
ной деятельностью человека и, в первую очередь, сю слабо ре-
гулируемым промыслом. Характерно, что «проблема хозяйст-
венного значения птиц» ни в одном из районов не возникла, по-
ка люди не подрывали запасы рыб. Но она нередко всплывала
уже после подрыва запасов. Характерным примером этого могут
служить наши внутренние южные моря. Показателен здесь и
пример с перуанским анчоусом- У берегов Перу, за исключени-
ем отдельных лет. когда здесь распространяется теплое тече-
ние Эль-Нино, вызывающее резкое сокращение численности ан-
чоуса и птиц, всегда было много птиц и рыбы. Ни у кого не воз-
никал вопрос о необходимости уничтожения птиц. Как произво-
дителей гуано, их охраняли и пока охраняют сейчас. Но в по-
следнее время запасы анчоуса имеют тенденцию к сокращению
1 В ТИНРО, например, изучение сырьевых ресурсов в последние годы на-
чинает проводиться с учетом специфики сообществ рыб определенных райо-
нов или систем течений.
КОЛИЧЕСТВЕННОЕ РАСПРЕДЕЛЕНИЕ МОРСКИХ ПТИЦ
343
под влиянием чрезмерного промысла. Немедленно появилось и
предложение о существенном сокращении поголовья птиц (Мои-
сее®, 1969). Нужен ли этот свермпрактицизм? Уместность дан-
ного вопроса очевидна сейчас, когда на глазах только одного
поколения под влиянием человеческой деятельности во много
раз сокращается численность одного за другим видов живот-
ных, когда дело охраны природы становится острой необходи-
мостью в масштабе планеты. В связи с этим нельзя не сослать-
ся на «Решение заседания Бюро Ихтиологической Комиссии» и
выводы изданного по ее решению сборника «Рыбоядные птицы
и их значение в рыбном хозяйстве» (1965). Основную их идею
может отражать заключение Н. А. Гладкова (1965) о том, что
проблема роли рыбоядных птиц в рыбном хозяйстве — второ-
степенная проблема.
Здесь возникает, правда, одно кажущееся несоответствие. В
то время, когда мировой промысловый вылов еще не достигал
6,5 млрд. ц. местами уже наблюдались признаки сокращения
запасов рыбы и перелова. Отсюда следовали сверхзаниженные
оценки рыбопродуктивности океана. В то же время более зна-
чительное изъятие рыбы морскими птицами и млекопитающими
к такому результату не приводит. Объясняется это принципи-
альными различиями в характере воздействия на нектон про-
мысла, с одной стороны, и морских животных, — с другой.
Промысел, как известно, базируется на запасах отдельных
.видов и групп рыб, имеющих к тому же хорошие пищевые или
технологические качества- Большей частью это виды среднего,
нередко крупного размеров — сельдевые, тресковые, ставридо-
вые, скомброидные, скорпеновые, камбаловые, лососевые. Иск-
лючением могут быть только мелкие анчоусовые (Мартинсен,
1966; Моисеев, 1969). Именно среди этих групп есть виды, за-
пасы которых подорваны чрезмерным промыслом1.
Птицы в отличие от промысла потребляют всех доступных
им рыб. в том числе мелкие виды, многие из которых являются
сорными. Например, в бореальной и арктической зонах высо-
кую численность имеют мойва, песчанки, корюшки, сайка и
другие мелкие виды, недостаточно используемые промыслом.
В питании же птиц они играют очень большую роль. В большин-
стве районов океана слабо развит промысел головоногих мол-
1 После сокращения численности некоторых видов, вызванного переловом,
освободившиеся кормовые ресурсы эффективно используют другие рыбы, не
испытывающие такою давления промысла. В морях Дальнего Востока в свя-
зи с сокращением количества камбал резко увеличили численность непро-
мысловые бентосоядные рыбы — бычки, ликоды и другие, а также краб-
стригун. Параллельно с уменьшением количества сельди и окуней сильно
увеличил численность другой планктонофаг — минтай, ставший самой мас-
совой рыбой на севере Тихого океана.
344 ЧИСЛЕННОСТЬ И РОЛЬ ПТИЦ КАК ПОТРЕБИТЕЛЕЙ ДРУГИХ ЖИВОТНЫХ.
люсков, численность которых велика. В питании морских птиц
эти животные занимают большое место-
Промысел использует, главным образом, половозрелых осо-
бей, имеющих средние и крупные размеры. Птицы потребля-
ют преимущественно молодь этих рыб. Это обстоятельство очень
существенно, так как численность и естественная смертность
младших возрастных групп значительно выше. Учитывая это, а
также то, что в литании большого количества видов птиц основ-
ное место занимает планктон, можно сделать вывод о том, что
в сравнении с человеком птицы используют ресурсы в целом бо-
лее низких (на 1—2 порядка) трофических уровней. Как хорошо
известно, наиболее эффективное в количественном отношении
использование биологических ресурсов возможно при условии
использования продукции животных более низких трофических
уровней.
В настоящее время особенно интенсивный промысел развит
в умеренных и субтропических водах северного полушария, но
далеко не во всех продуктивных районах тропиков и южного по-
лушария1. Можно напомнить о слабо развитом промысле в Ин-
дийском океане и в большинстве районов южной умеренной зо-
ны. Птицы используют ресурсы всех зон Мирового океана-
Промысел интенсивен только в местах больших концентра-
ций рыб. Облов современными орудиями лова рассредоточенных
концентраций не рентабелен* 2. Птицы «собирают урожай» с бо-
лее обширных пространств. Их местом добычи пищи является
весь Мировой океан.
Наконец, не нужно забывать, что птицы потребляют также
многочисленные отходы промысла.
Все изложенное говорит против упрощенных суждений о ры-
боядных птицах как о конкурентах человека в использовании
биологических ресурсов морей и океанов. Вопрос о роли птиц
как потребителей других водных животных должен рассматри-
ваться в аспекте изучения сложившихся связей в сообществах,
ииаче говоря — это биогеоценологическая проблема-
Освоение промыслом многих южных районов рассматривается как важ-
нейший путь увеличения мирового вылова морских животных.
2 Общие запасы рыбы, обитающей рассредоточенно, в Мировом океане
огромны. Они являются важным резервом для увеличения вылова в буду-
щем, когда будут разработаны эффективные способы облова неплотных кон-
центраций. *
345
ЗАКЛЮЧЕНИЕ
Распространение, распределение, численность и многие осо-
бенности биологии морских птиц контролируются целым рядом
факторов, вернее, комплексом факторов, формирующих общий
фон океанографического окружения.
Наиболее общий и характерный отпечаток на пространствен-
ное размещение морских птиц и состав орнитофауны наклады-
вает климатическая зональность. В Мировом океане последова-
тельная цепь климатической, географической и биологической
зональности выражена столь же четко, как и на суше. Морские
птицы не представляют здесь исключения среди других морских
животных. Космополитов, в строгом смысле этого слова, среди
морских птиц нет. Очень широкое распространение характерно
для немногих видов. Репродуктивная часть ареала подавляю-
щего количества видов морских птиц не выходит за пределы од-
ной климатической зоны. Зональное распространение находит
объяснение не только в связи видов и комплексов с современны-
ми ландшафтными зонами океана. Формирование зональности
в распространении морских птиц происходило в процессе диффе-
ренциации семейств на фоне становления климатических и геог-
рафических зон в течение третичного периода. Если учесть, что
сказанное справедливо и для других групп, то можно утверж-
дать, что это создавало предпосылки для формирования круп-
ных пелагических биогеографических областей.
Велико значение в распространении и распределении морских
птиц структурной неоднородности водных масс Мирового океа-
на. Приуроченность к определенным водным массам хорошо за-
метна уже при анализе зонального распространения птиц. Это
we случайно, так как наиболее существенной чертой в размеще-
нии структур вод в океане является широтная зональность, про-
являющаяся в том, что каждую климатическую зону занимает
характерная первичная водная масса. Четкая приуроченность
ряда видов к зональным водным массам делает их индикатора-
ми этих масс. Важен и другой аспект влияния водных масс на
распределение морских птиц- Наличие океанических, а также
трансформированных вторичных и прибрежных водных масс —
346
ЗАКЛЮЧЕНИЕ
одна из основных предпосылок формирования океанических,
дальне-неритических и неритических группировок и видов мор-
ских птиц. Распространение этих масс определяет ряд важных
особенностей в распространении соответствующих видов птиц.
Факторы среды, формирующие климатическую и географи-
ческую зональность, а также специфику водных масс, определя-
ют области и границы распространения видов птиц. Числен-
ность и плотность населения птиц в пределах этих областей в
первую очередь определяются кормностью вод. В связи с этим
вполне закономерно видеть принципиальное сходство в количе-
ственном распределении первичной продукции, планктона, бен-
тоса, рыб и морских птиц. Обилие жизни в океане и морских
птиц от умеренных вод уменьшается в направлении тропиков
и вновь несколько увеличивается в приэкваториальных районах.
Следовательно, распределение плотности населения морских
птиц вполне подчиняется законам широтной зональности. Это и
понятно, так как в характере динамики вод — важнейшем ус-
ловии величины биологической продуктивности — также наблю-
дается хорошо выраженная зональность-
Морские птицы в целом ограничены доступной пищей, в том
числе в период кочевок. Состав кормовой базы в том или ином
районе обычно находит отражение в количественном соотноше-
нии птиц, потребляющих животных различных трофических
уровней. Наиболее многочисленны среди морских птиц виды,
питающиеся животными более низких трофических уровней.
Конкуренция за пищу является важнейшей предпосылкой подбо-
ра в каждом районе Мирового океана видов с различными по-
требностями, а также пространственной разобщенности эколо-
гически близких видов. Орнитофауна каждого района в этом
смысле состоит из набора видов, позволяющего максимально ис-
пользовать его кормовые ресурсы. Эта мысль может быть вы-
ражена более конкретно: океан «поделен» между различными
видами морских птиц для наиболее рационального использова-
ния его кормовых ресурсов. Степень насыщенности различных
районов океана потребителями ресурсов различных трофических
уровней и наличие в них незанятых экологических ниш имело
и имеет большое значение при формировании сообществ на мес-
тах гнездования, а также миграций птиц, их путей, мест коче-
вок и зимовок.
Из всех изменений в биологии моря наиболее существенны
сезонные. Климатические, океанографические и кормовые усло-
вия претерпевают большие изменения в сезонном аспекте. Эти
изменения лежат в основе сезонных миграций птиц, определя-
ют все основные особенности сезонных изменений в составе ор-
нитофауны, численности и биологических циклах морских птиц.
ЗАКЛЮЧЕНИЕ
347
Зональность в пространственном размещении птиц в период
сезонных миграций, когда птицы рассеиваются на огромных
пространствах, выражена менее заметно, чем в гнездовой пери-
од. Вместе с этим специфичность изменчивости условий обита-
ния в каждой физико-географической зоне накладывает сущест-
венный отпечаток на особенности миграций птиц, гнездящихся
в ней. Так, размах миграций увеличивается с увеличением ши-
роты в связи с усилением контрастности в сезонной изменчивос-
ти условий обитания. Под влиянием ухудшения условий обита-
ния птицы не просто перемещаются в смежные области. Мигра-
ции носят приспособительный характер, в них отсутствует хаос-
Благодаря им птицы имеют возможность не только переживать
неблагоприятное для обитания время, но и осваивать районы,
благоприятные для существования только в определенные пери-
оды года, следовательно, максимально использовать кормовые
ресурсы океана.
Основной вывод данной работы можно сформулировать сле-
дующим образом: особенности и характер изменчивости видово-
го состава, численности, географического распространения, рас-
пределения, сезонных миграций и годовых циклов морских
птиц, определяемые климатической, географической и биологи-
ческой зональностью, соответствуют представлениям о биологи-
ческой структуре океана и отражают ее закономерности.
В настоящей работе рассмотрены только некоторые аспекты
распространения, распределения, миграций, численности и био-
логии морских птиц на основе представлений о биологической
структуре океана. Вообще же эта проблема более многоплано-
вая. Здесь я не имею в виду останавливаться на всех пробле-
мах дальнейшего изучения морских птиц. Однако в плане темы
«Морские птицы — биологическая структура океана» нужно
подчеркнуть следующее. Большой интерес представляют иссле-
дования по изучению годовых циклов птиц, особенно размно-
жения, с упором на выявление закономерностей, характерных
для определенных зон. В этом плане должны быть исследованы
изменение плодовитости, особенности гнездования и гнездовых
циклов, характерные как для популяций одного вида, обитаю-
щих в разных зонах, так и для зональных комплексов птиц.
Нуждаются в значительном углублении и многие вопросы, за-
тронутые в данной работе. В первую очередь, это относится к
изучению питания птиц, но, конечно, не в узком смысле «рыбо-
хозяйственного значения птиц». Морские птицы имеют внуши-
тельную численность. В местах их постоянных больших концент-
раций они играют заметную роль в биоэнергетическом балансе
морских биогеоценозов, выявление этой роли немыслимо без
хорошего знания питания и численности птиц.
348
ЗАКЛЮЧЕНИЕ
Все соотношения в биогеоценозах не являются застывшими.
Они постоянно изменяются, отсюда следует необходимость изу*
чения биогеоценозов в плане межгодовой изменчивости. Все эти
исследования также должны вестись с учетом значительной
специфичности, свойственной ландшафтным зонам и зональным
комплексам животных-
349
ЛИТЕРАТУРА
Аверин Ю. В. 1948. Наземные позвоночные Восточной Камчатки.
Труды Кроноцкого гос. запое., вып. 1.
Азова Н. В. 1964. Первичная продукция Прибылово-Бристольского
района Берингова моря. Известия ТИНРО, т. 52.
Акимушкин И. И. 1970. Головоногие моллюски (Cephalopoda), нх
распространение и трофические связи с остальным нектоном. В сб. «Биогео-
ценозы морей и океанов». Изд-во «Наука».
Андрияшев А. П. 1939. Очерк зоогеографии и происхождения фау-
ны рыб Берингова моря и сопредельных вод. Изд. ЛГУ.
Андрияшев А. П. 1964. Обзор фауны рыб Антарктики. Исслед.
фауны морей, II (X). Результ. биолог, исслед. Советск, антарктич. экспед.
(1955—1958 гг.), 2.
Арсеньев В. С. 1967. Течения и водные массы Берингова моря. Изд-
во «Наука», М.
Арсеньев В. С. и Войтов В. И. 1968. Относительная прозрач-
ность и цвет вод Берингова моря. «Океанология», т. 8, вып. 1.
Атлас Антарктики, 1966, т. 1. Главн. упр. геодезии и картографии
МГ СС£Р, М.-Л.
Атлас гидрометеорологических и промысловых данных. Восточая часть Бе-
рингова моря, 1969.
Аюшин Б. Н. 1956. Разведка сельди в северной части Охотского мо-
ря. Примизцат, Владивосток.
Базилевич Н. И., Родия Л. Е., Розов Н. Н. 1971. Сколько
весит живое вещество планеты. «Природа», № 1.
Беккер В. Э. 1967. Светящиеся анчоусы (сем. Myctophidae). В книге
«Тихий океан. Биология Тихого океана. Рыбы». Изд-во «Наука», И.
Беклемишев К. В. 1961. Влияние антарктических цикланов на по-
ля питания китов в Антарктике. Труды ИО АН СССР, т. 51.
Беклемишев К. В. 1965. Биогеографическое деление верхних слоев
пелагиали Тихого океана и его зависимость от течений и распределения вод-
ных масс. Океанологии, исслед. по прогр. МГГ, № 13.
Беклемишев К. В. 1967. Биогеографическое деление пелагиали
Тихого океана. В книге «Тихий океан. Биология Тихого океана. Планктон».
Изд-во «Наука», М.
Беклемишев К. В. 1969. Экология и биогеография пелагиалж
Изд-во «Наука», М.
Белопольский Л. О. 1933. К вопросу о количественном распре-
делении Rissa rissa и Fulmarus glaciMis по Кольскому и 38чму меридианам
в Баренцевом море. Труды Арктяч. инет., 8.
Белопольский Л. О. 1956. Роль межвидовых взаимоотношений в
развитии колониальное™ птиц. «Зоологический журнал», т. 34, вып. 3.
Белопольский Л. О. 1957. Экология морских колониальных птиц
Баренцева моря. М.—-Л.
350
Белопольский Л. О. 1957а. Кормовые биотопы и состав пищи мор-
ских колониальных птиц Баренцева моря. Труты АНИИ, т. 205.
Белопольский Л. О. 1971. Состав кормов морских птиц Барен-
цева моря. Зоологические исследования северо-западных районов СССР. Ка-
лининградский гос. университет, Ученые зап., 6. Калининград.
Беляев Г. М. 1970. Клювы кальмаров в донных осадках Тихого океа-
на. Труды ИО АН СССР, т. 88.
Берг Л. С. 1953. Об амфибореальном распространении морской фау-
ны в северном полушарии. В об. «Очерки по общим вопр. ихтиологии». Изд.
АН СССР, М.-^1.
Бирман И. Б. 1958. О распространении некоторых пелагических рыб
в северной части Тихого океана. «Зологический журнал», т. 37, вып. 7.
Бирман И. Б. 1967. Лососи в морском периоде жизни. В книге «Ти-
хий океан. Биология Тихого океана. Рыбы». Изд-во «Наука», М.
Богданов Д. В. 1970. Океанографические условия тропической зо-
цы Атлантического и Индийского океанов. Отдел НТИ ВНИРО, М.
Богоров В. Г. 1959. Биологическая структура океана. Докл.
.АН СССР, т. 128, № 4.
Богоров В. Г. 1964. Единство природы океана. В сб. «Соврем, пробл.
географии». Изд-во «Наука», М.
Богоров В. Г. 1967. Биологическая трансформация и обмен энергии
и веществ в океане. «Океанология», т. 7, № 5.
Богоров В. Г. 1967а. Биомасса зоопланктона и продуктивные райо-
ны Тихого океана. В книге «Тихий океан. Биология Тихого океана. Планк-
тон». Изд-во «Наука», М.
Богоров В. Г. 1969. Жявнь океана. М.
Богоров В. Г. 1970. Биогеоценозы пелагиали океана. В сб. «Про-
грамма и методы изучения биогеоценозов водной среды». Изд-во «Наука», М.
Богоров В. Г. 1970а. Биологическая продуктнняость океана и осо-
бенности ее географического распределения. «Вопросы географии», № 84,
Изд-во «Мысль».
Богоров В. Г. и Виноградов М. Е. 1955. О зоопланктоне севе-
ро-западной части Тихого океана. Докл. АН СССР, т. 102, № 4.
Богоров В. Г., Виноградов М. Е. 1961. Некоторые черты рас-
пределения биомассы планктона в поверхностных водах Индийского океана
зимой 1959/60 г. «Оке ано л. исслед. по прогр. МГГ», № 4.
Богоров В. Г. и Зенкевич Л. А. 1966. Биологическая струк-
тура океана. В сб. «Экология водных организмов». Изд-но «Наука», М.
Богоров В. Г. и Воронина Н. М. 1967. Распределение биомас-
сы планктона в продуктивном слое Индийского океана. В сб. «Биология и
распределение планктона южных морей». Изд-во «Наука», М.
Бородулина Т. Л. 1965. Рыбохозяйственное значение чаек и кра-
чек Азово-Черноморского и Каспийского бассейнов. В сб. «Рыбоядные пти-
цы и их значение в рыбном хозяйстве». Изд-во «Наука».
Бреслина И. П. и Карпович В. Н. 1909. Развитие раститель-
ности под влиянием жизнедеятельности колониальных птиц. «Ботанический
журнал», 54, № 5.
Бродский К. А. 1941. Обзор количественного распределения и соста-
ва зоопланктона северо-западной части Японского моря. Труды Зоол. ин-та
АН СССР, т. 7, вып. 2.
Бродский К. А. 1955. Планктон северо-западной части Куросио и
прикурилыжих вод Тихого океана. Труды ИО АН СССР, т. 18.
Бродский К. А. 1957. Фауна веслоногих рачков (Calanofdae). Изд.
АН СССР, М.-Л.
Бродский К. А. 1959. Зоопланктон морских вод Южного Сахалина
и Южных Курильских островов. Иссл. Дальневост, морей. СССР, вып. 6.
Бродский К. А. 1960. Зоогеографические зоны в Южной части Ти-
351
хого океана и биполярное распространение некоторых Calonoidae. Труды
океаногр. Комисе. АН ССОР, т. 10, вып. 4.
Бруевич С. В., Богоявленский А. Н., Мокиевски! В. В.
Ю60.р Гидрохимическая характеристика Охотского моря. Труды ИО АН
Булгаков Н. П. 1964. О распространении льда в Беринговом море.
«Океанология», т. 9, вып. 5.
Булгаков Н. П. 1965. Предельная зимняя граница льдов в дальне-
восточных морях. «Океанология, исслед. по прогр. МГГ», Xs 13.
Булгаков Н. П. 1968. Льды. — В мните «Тихий океан. Гидрология
Тихого океана». Ивд-во «Наука», М
Бурков В. А. 1966. Структура течений Тихого океана и их номенкла-
тура. «Океанология», т. 6, вып. 1.
Бурков В. А. и Арсеньев В. С. 1958. Опыт выделения водных
масс в эоне соприкосновения Куросио и Курильского течения. Труды ИО АН
СССР, т. 27.
Васюкова Н. Г. 1964. Тепловой баланс поверхностных вод в неко-
торых рыбопромысловых районах Берингова моря. Известия ТИНРО, т. 51.
Веденский А. П. 1949. Опыт поисков скоплений минтая по плава-
ющей икре. Известия ТИНРО, т. 29.
Веденский А. П. 1964. Биология дальневосточной скумбрии в Япон-
ском море. Известия ТИНРО, т. 42.
Вигдоровяч О. П. 1967. О распределении и биологии сайры в при-
алеутских водах Тяхогю океана. Известия ТИНРО, т. 56.
Виноградов М. Е. 1956. Распределение зоопланктона в западных
районах Берингова моря. Труды Всес. гидробиол. об-ва, т. 7.
Виноградов М. Е. 1967. Вертикальное распределение зоопланктона
в Тихом океане. В книге «Тихий океан. Биология Тихого океана. Планктон».
Изд-во «Наука», М.
Виноградов М. Е. 1968. Вертикальное распределение океаническо-
го зоопланктона. Изд-во «Наука», М.
Виноградов М. Е., Наумов А. Г. 1961. Количественное рас-
пределение планктона в антарктических водах Индийского и Тихого океанов.
«Океанология, исслед. по прогр. МГГ», № 3.
Виноградов М. Е., Воронина Н. М 1962. Некоторые черты
распределения зоопланктона северной части Индийского океана. Труды
ИО АН СССР, т. 58.
Ви но градов М. Е., Воронина Н. М. 1965. Некоторые особен-
ности распределения планктона в водах экваториальных течений Тихого и
Индийского океанов. «Океанология. исслед. по прогр. МГГ», № 13.
Винокурова Т. Т. 1965. Изменчивость температурных условий вод
северной части Охотского моря. Известия ТИНРО, т. 59.
Воииственский М. А. 1960. Птицы степной полосы европейской
части СССР. Соврем, состояние орнитофауны и ее происхождение. Киев,
АН УССР.
Вор о б ье® К. А. .1954. Птицы Уссурийского края. Изд. АН СССР. М.
Воронина Н. М. 1966. Распределение биомассы зоопланктона Юж-
ного океана. «Океанология», т. 6, вып. 6.
Воронина Н. М., Наумов А. Г. 1968. Количественное распределе-
ние и состав мезоплаиктона Южного океана. «Океанология», т. 8, вып. 6.
Воронов А. Г. 1963. Биогеография с элементами биологии. Изд.
МГУ. М.
Второв П. П. 1968. Птицы культурных ландшафтов восточного При-
иссыкуль'и. «Орнитология», вып. 9.
Гейнрих А. К. 1959. Биологические сезоны в планктоне Берингову
моря и горизонтальное распределение биомассы зоопланктона. Труды ИОАН
СССР, т. 30.
352
Гейнрих А. К. 1961 Сезонные явления в планктоне Мирового океа-
на. 1. Сезонные явления в планктоне средних и высоких широт. Труды ИО
АН СССР, т. 51.
Гейнрих А. К. 1961а. Сезонные явления в планктоне Мирового океа-
на. II. Сезонные явления в планктоне низких широт. «Океанология», т. I,
вып. 3.
Гейнрих А. К. 1967. Сезонные явления в зоопланктоне Тихого
океана. В книге «Тихий океан. Планктон». Изд-во «Наука», М.
Гептнер В. Г. 1936. Общая зоогеография. Биомедгиз, М.—Л.
Герасимова Т. Д. 1965. Питание чаек Мурманского побережья. —
В сб. «Рыбоядные птицы и их значение в рыбном хозяйстве». Изд-во
«Наука», М
Гизенко А. И. 1955. Птицы Сахалинской области. Изд. АН СССР,М.
Гладков Н. А. 1960. Птицы и пространство. «Орнитология», вып. 3.
Гладков Н. А. 1965. Рыбоядные птицы и их возможное рыбохозяй-
ственное значение. В сб. «Рыбоядные птицы и их значение в рыбиом хозяй-
стве». Изд-во «Наука», М.
Головкин А. Н. 1963. О выедании рыбы кайрами и моевками в гнез-
довой период в Баренцевом море. «Зоологический журнал», т. 42, № 3.
Головкин А. Н. 1970. Гидрологические предпосылки увеличения про-
дуктивности сообществ близ птичьих базаров Баренцева моря. Тезисы док-
ладов II съезда ВГБО, Кишинев.
Головкин А. Н., Позднякова Л. Е. 1964. Влияние морских
колониальных птиц на режим биогенных элементов в прибрежных водах
Восточного Мурмана. Труды Мурм. морск. биол. ин-та, вып. 6 (10).
Головкин А. Н.» Позднякова Л. Е. 1965. Влияние морских
колониальных птиц на режим биогенных солей в прибрежных водах Мурма-
на. В сб. «Рыбоядные птицы и их значение в рыбном хозяйстве». Изд-во
«Наука», М.
Головкин А. Н., Зеликман Э. А. 1965. Развитие калянуса в
районе гнездовий морских колониальных птиц в прибрежье Мурмана. «Океа-
нология», т. 5, вып. 1.
Григорьев А. А. 1966. Закономерности строения и развития геогра-
фической среды. Изд-во «Мысль», М. '
Григорьев К). А. 1969. Горизонтальная циркуляция вод в Тихо-
океанском секторе Южного океана. Проблемы Арктики и Антарктики, Гид-
рометиздат, вып. 30.
Гриффнн Д. 1966. Перелеты птиц. Изд-во «Мир», М.
Гудков В. М. 1962. О связях в распределении зоопланктона, мор-
ских птиц и усатых китов. Труды ИО АН СССР, т. 58.
Гурьянова Е. Ф. 1945. Принципы биогеографического районирова-
ния Мирового океана. Науч. бюлл. ЛГУ, № 4.
Гурьянова Е. Ф. 1951. Бокоплавы морей СССР и сопредельных
вод. Изд. АН СССР, М.-^Л.
Гурьянова Е. Ф. 1970. О годовом цикле развития планктона в не-
ритических районах тропической зоны. В сб. «Вопр. эволюц. морф, и биоце-
иол.». Казань.
Гусарова А. Н., Иваненков В. Н., Коннов В. А., Сапож-
ников В. В., Чернякова А. М. 1967. Основные закономерности
распределения химических элементов в водах Тихого океана. В книге «Тихий
океан», т. 3, вып. 1. Изд-во «Наука», М.
Давидович Р. Л. 1963. Гидрохимические черты южной н юго-вос-
точной частей Берингова моря. Известия ТИНРО, т. 50.
Давиташвили Л. Ш. 1969. Причины вымирания организмов. Изд.
«Наука», М.
Дарлингтон Ф. Д. 1966. Зоогеография. Географическое распростра-
нение животных. Изд. «Прогресс», М.
353
Дементьев Г. П. 1940. Руководство по зоологии. Птицы. Изд. АН
СССР,
Дементьев Г. П. 1951. Отрад чайки. «Птицы Советского Союза»,
Изд-во «Наука», т. 3, М.
Дементьев Г. П. 1951а. Отряд чистики, Alcae или Alciformes. «Пти-
цы Советского Союза». Изд-во «Наука», т. 2, М.
Дементьев Г. П. 1957. Исследования по окраске позвоночных жи-
вотных IV. Морфизм окраски птиц фауны СССР, «зоологический журнал»,
т. 36, № 7.
Дементьев Г. П. 1958. К вопросу об истории фауны птиц Советско-
го Союза. Уч. зап. Моск, ун-та, 197. Орнитология.
Дементьев Г. П. 1964. Основы палеонтологии. Класс Aves. Птицы.
Изд-во «Наука», М.
Дементьев Г. П., Ларионов В. Ф. 1944. Исследования по ок-
раске позвоночных животных. I. О возникновении географических вариаций
окраски. «Зоологический журнал», т. 23, № 5.
Добровольский А. Д. 1961. Об определении водных масс. «Океа-
нология», т. 1, вып. 1.
Долженков В. Н. 1970. О распределении планктона в тихоокеан-
ском секторе Антарктики. Известия ТИНРО, т. 70.
Дольник В. Р. 1965. Физиологические основы миграций птиц. В сб.
«Биологическое значение и функциональная детерминация миграционного
поведения животных». Изд-во «Наука», М.—Л.
Дольник В. Р. 1967. О взаимосвязи, жировых резервов перелетных
птиц и миграции. Труды Зоол. ин-та АН СССР, т. 40.
Дольник В. Р. 1968. Таинственные перелеты. Изд-во «Наука», М.
Дроздов Н. Н. 1967. Фауна и население птиц культурных ландшаф-
тов. «Орнитология», вып. 8.
Дубинин Н. П. и Торопаиова Т. А. 1956. Птицы лесов доли-
ны р. Урал. Части 2 и 3. Труды И-та леса, т. 32.
Дудник Ю. И., Усольцев Э. А. 1964. О сельди восточной части
Берингова моря. Известия ТИНРО, т. 51.
Ермаков Ю. К. 1970. Некоторые данные о биологии тихоокеанского
хека. — В сб. вып. 2. «Исследования по биологии рыб и промысловой океа-
нографии», Владивосток.
Зайцев Ю. П. 1970. Морская нейстонология. Изд-во «Наукова Дум-
ка», Киев.
Земкевич Л. А. 1948. Биологическая структура океана. «Зоологиче-
ский журнал», т. 27, вып. 2.
Зенкевич Л. А. 1963. Биология морей СССР. Изд. АН ССОР, М.
Зенкевич Л. А 1970. Общая характеристика биогеоценозов океана
и сравнение их с биогеоценозами суши. — В кн. «Биогеоценозы морей и
океанов», Изд-во «Наука», М.
3логин Р. И. 1963. Зимнее население птиц Шнрванской степи.
«Орнитология», вып. 6.
3 м и е в с к и й В. А. 1968. О распределении сардины в Большом Австра-
лийском заливе. В сб. НТИ .ВНИРО, вып. 7.
Ивлева Н. А. 1960. Характер распределения зоопланктона у юго-за-
падного берега Сахалина. Известия ТИНРО, т. 46.
Иваненков В. Н. 1964. Гидрохимия Берингова моря. Изд-во «Нау-
ка», М.
Иванов Ю. А., Тар ее в В. А. 1959. О структуре зоны антарктиче-
ской дивергенции. Известия АН ООСР, серия географ., № 6.
Иване в-Ф ранцкевич Г. Н. 1961. О некоторых особенностях
гидрологической структуры в водных массах Индийского океана. «Океано-
логия. исслед. по прогр. МГГ», № 4.
Иосида Наотси 1962. Экология гнездования буревестника Calo-
354
nectris leucomelas в Камуридзиме. I. Майдзуру, префектура Киото, Tori,
Bull. Ornithol. Soc. Japan», 17, № 79—80 (Яп.).
Кабанов H. E. 1960. На уединенной скале. «Природа», № 5.
Кагаиовская С. М. I960. Материалы к познанию минтая. Изве-
стия ТИНРО, т. 32.
Кагаиовская С. М. 1954. О распределении икры и личинок неко-
торых рыб в зал. Петра Великого. Известия ТИНРО, т. 42.
Кагановский А. Г. 1939. Дальневосточная сардина. Дальгиз.
Кагановский А. Г. 1951. Миграции скумбрии в Японском море.
Известия ТИНРО. т. 35.
Каредин Е. П. 1969. Сезонные изменения в распределении макро-
планктона в зоне течения Куросио. Известия ТИНРО, т. 68.
Карпова Л. А. 1963. Основные черты климата Берингова мрря. Из-
вестия ТИНРО, т. 50.
Карташев Н. Н. 1961. Птицы Командорских островов и некоторые
предложения по рациональному их использованию. «Зоологический журнал»,
т. 40, вып. 9.
К афт а нов ск ий Ю. М. 1951. Чистиковые птицы Восточной Атлан-
тики. Материалы к познанию фауны и флоры СССР, нов. сер. отд. зоол.,
вып. 28 (XIII).
Качина Т. Ф., Прохоров В. Г. 1966. Корфо-карагинская сельдь.
Рыбное хозяйство, Xs 11—<12.
Киселев И. А. 1947. Фитопланктон дальневосточных морей как по-
казатель некоторых особенностей их гидрологического режима. — Труды
ГОИ, вып. 1 (13).
Кобленц-Мишке О. И. 1967. Первичная продукция. В кн. «Тихий
океан. Биология Тихого океана. Планктон». Изд-во «Наука», М.
К о б л е н ц-М и ш к е О. И., Кабанова Ю. Г. 1964. О первичной
продукции в северо-восточной части Индийского океана в период летнего
муссона. Труды ИО АН СССР, т. 65.
Коб л ей ц-М и шке О. И., Во л ко винский В. В., Кабано-
ва Ю. Г. 1970. Первичная продукция планктона Мирового океана. В сб.
«Биогеоценозы морей и океанов». Изд-во «Наука», М.
Ко ж а н ч и к о в И. В. 1946. Экологические предпосылки для зоогеогра-
фических делений Евразии. «Жури. общ. биологии», т. VII, № 1.
Козлова Е. В. 1957. Подотряд чистиковые. Фауна СССР, Птицы,
т. 2, вып. 3.
Коновалов Г. В., Шулятин О. Г. 1964. Уникальный птичий ба-
зар в Антарктиде. «Природа», Xs 10.
Котляр Л. К- 1965. Особенности распределения планктона в заливе
Шелихова в июне 1963 г. Известия ТИНРО, т. 59.
Котляр Л. К- 1967. Распределение зоопланктона в северной части
Охотского моря в августе 1964 г. Известия ТИНРО, т. 61.
Котляр Л. К. 1970. Закономерности развития и количественного рас-
пределения зоопланктона как кормовой базы сельди в северо-восточной час-
ти Охотского моря. Известия ТИНРО, т. 71.
Крылов В. В. 1969. Распределение планктона в Восточно-Китайском
море. Труды ВНИРО, т. 65.
Крындин А. Н. 1961. Льды. В сб. «Основные черты геологии и гид-
рологии Японского моря». Изд. АН СССР.
Кузнецов Б. А. 1950. Очерк зоогеографического районирования
СССР. Изд. Моск, об-ва испыт. природы, М.
К у м а р и Э. В. 1957. К теории пролетных путей и миграций широким
фронтом. Труды Второй прибалт, орнитол. конфер. Изд. АН СССР, М.
Кумари Э. В. 1959. Миграции птиц как зоогеографическая проблема.
Тр. Третьей Прибалт, ориитол. конфер. АН Литовск. ОСР, Вильнюс.
355
. Кун М. С. 1951. Распределение планктона й питание сельди в север-
ной части Охотского моря. Известия ТИНРО, т. 35.
Кун М. С. 1969. Географическая изменчивость некоторых видов рода
Calanus в Японском море. «Зоологический журнал», т. 48, вып. 7.
Кун М. С. 1969а. Сезонные изменения в составе и распределении ме-
зопланктона в зоне течения Куросно. Известия ТИНРО, т. 68.
Кун М. С. и Мещерякова И. М. 1954. Распределение типов
зоопланктона в Японском море. Известия ТИНРО, т. 39.
Кун М. С., Гладких Г. Н., К а р е д и н Е. П., Павлы-
чев В. П., Рачков В. И., Стародубцев Е. Г. 1969. Гидрологиче-
ские условия и биологическая характеристика вод Куросио. Известия
ТИНРО, т. 68.
Курода Н. 1963. Зимний учет морских птиц между Токио и Кусиро,
о-в Хоккайдо (Японск.). — Misc. Repts. Jamashinas Inst. Ornithol. and
Zool., 3, 4.
Курочкин E. H. 1963. Распределение некоторых видов морских
птиц в северной части Тихого океана. «Зоологический журнал», т. 42, вып. 8.
Курочкин Е. Н. 1964. К орнитогеографии северной части Тихого
океана. Труды 3-ей Всесоюзн. орнитол. коифер., Львов.
Ларионов В. Ф. 1939. Региональное™ окраски птиц и факторы, ее оп-
ределяющие. Докл. АН СССР, т. 23, № 2.
Леонов А. К. 1960. Региональная океанография. Гидрометеоиздат, Л.
Лим Ч у н 1963. О находке окольцованных буревестников Р. tenuirOB-
tris Temm. Сэнмунь, жури, биол., науки, 2, № 4, 47 (корейск. яз.).
Линдберг Г. У. 1955. Четвертичный период в свете биогеографиче-
ских данных. Изд. АН СССР, М.—Л.
Л у б н ы-Г е р ц ы к Е. А. 1959. Состав и распределение зоопланктона
Охотского моря. Труды ИО АН СССР, т. 30.
Лэк Д. 1949. Дарвиновы вьюрки. Изд-во «Иностранная литерату-
ра», М.
Лэк Д. 1957. Численность животных и ее регуляция в природе. Изд-во
«Иностранная литература», М.
Майр Э. 1968. Зоологический вид и эволюция. Изд-во «Мир», М.
Макаров В. Н. и Пашкин В. Н. 1968. Общие закономерности
распределения рыб Большого Австралийского залива «Рыбное хозяйство», № 3.
Макфедьен Э. 1965. Экология животных. Изд-во «Мир», М.
Мальчевский А. С. 1968. О консервативном и дисперсном типах
эволюции популяций у птиц. «Зоологический журнал», т. 47, вып. 6.
Мальчевский А. С. 1969. Дисперсия особей и эволюция видов и
популяций у птиц. В сб. «Орнитология в СССР», книга 1. Изд-во «Ылым»,
Ашхабад.
Мараков С. В. 1963. Птичьи базары острова Медного и возможно-
сти их практического использования. Сб. НТИ. Всесоюзн. н-и. инс-та жив.
сырья н пушнины, вып. 5 (8).
Мараков С. В. 1966. Край непуганых птиц. Изд-во «Наука», М.
Маркина Н. П. 1969. Некоторые особенности распределения планкто-
на у побережья Австралии в различные сезоны года. Тезисы докладов кон-
ференции по тропической зоне Мирового океана, 24—28 ноября 1969 г., М.
Маркина Н. П. 1970. Сведения о распределении планктона в севе-
ро-восточной части Индийского океана (за 1963—1965 гг.). Аннотации на-
учных работ, выполненных ТИНРО в 1967 г. Владивосток.
Маркина Н. П. 1970а. О распределении планктона в северо-восточ-
ной части Индийского океана. В сб. вып. 2. «Исследования по биологии
рыб и промысловой океанографии», Владивосток.
Марков В. И. 1965. Заметки о пеликанах и бакланах Аральского
моря. В сб. «Рыбоядные птицы и их значение в рыбном хозяйстве». Изд-вр
«Наука», М.
356
Марков К. К.» Бардин В. Н, Л е б е д е в В. Л., Орлов А. И.
я Суетова И. А. 1968. География Антарктиды. Изд-во «Мысль», М.
Маркузе В. К- 1965. Рыбоядные птицы в нерестово-выростных хо-
зяйствах дельты Волги и их значение. В сб. «Рыбоядные птицы и их зна-
чение в рыбном хозяйстве». Изд-во «Наука», М.
Маслеников Б. Г. 1965. Новые данные о малой вилохвостой качурке
(О. monorhis Swinh). Записки приморск. фил. Географ, общ-ва СССР № 1 (24).
Марти Ю. Ю. 1970. Рыбы и биогеоценозы океана. В сб. «Биогеоце-
нозы морей и океанов». Изд-во «Наука», М.
Мартинсеи Г. В. 1966. Современный мировой промысел рыбы и не-
рыбных объектов. М.
Мензбир М. А. 1934. Миграции птиц. Биомедгиз, М.—Л.
Мещерякова И. М. 1960. Сезонные изменения планктона в откры-
тых водах Японского моря. Известия ТИНРО, т. 46.
Мещерякова И. М. 1964. Количественное распределение планктона
в юго-восточной части Берингова моря летом 1968 и 1959 г. Известия
ТИНРО, т. 51.
Мещерякова И. М. 1970. Зимне-весенний планктон юго-восточной
части Берингова моря. Известия ТИНРО, т. 72.
Мещерякова И. М. 1970а. Планктон восточной части Берингова
моря в весенний и осенний гидрологические сезоны. Известия ТИНРО, т. 72.
Микл у хин а А. П. 1969. Распределение и видовой состав планктона
в зоне тихоокеанского дрейфа. Известия ТИНРО, т. 68.
Микулич Л. В. 1960. О распределении планктона в северной части
Охотского моря летом 1955 г. Известия ТИНРО, т. 46
Михеев А. В. 1964. Роль факторов среды в формировании сезонных
миграций птиц Восточной Палеарктики. Ученые записки Моск. гос. пед.
инст-та им. Ленина (227).
Михеев А. В. 1967. Некоторые вопросы происхождения перелетов
птиц Палеарктики. «Итоги орнитол. исслед. в Прибалтике». Изд-во «Валгус»,
Таллин.
Михеев А. В. 1969. Происхождение перелетов птиц Палеарктики.
«Жури, общей биологии», т. 30, Ns 1.
Моисеев П. А. 1953. Треска и камбалы дальневосточных морей.
Известия ТИНРО, т. 40.
Моисеев П. А. 1964. Некоторые итоги исследований Беринговоморской
научно-промысловой экспедиции. Известия ТИНРО, т. 52.
Моисеев П. А. 1969. Биологические ресурсы Мирового океана. Изд.
«Пищевая промышленность», М.
Мокиевсхая В. В. 1961. Химическая характеристика водных масс.
В сб. «Основные черты геологии и гидрологии Японского моря». Изд.
АН СССР, М.
Мокиевскнй О. Б. 1960. Фауна литорали северо-западного побере-
жья Японского моря. Труды ИО АН СССР, т. 34.
Мороз И. Ф. 1969. Общая характеристика субарктического фронта
Тихого океана. Известия ТИНРО, т. 68.
Морошкин К. В. 1964. Новая схема поверхностных течений Охот-
ского моря. «Океанология», т. 4, вып. 4.
Морошкин К. В. 1966. Водные массы Охотского моря. Изд-во
«Наука», М.
Морской Атлас, 1953. Изд. Главного штаба ВМС.
Муромцев А. М. 1959. Основные черты гидрологии Индийского
океана, Л.
Назаров В. С. 1970. Ледяной покров океанов и направленность кли-
матических процессов Земли. «Вопросы географии», Изд-во «Мысль», № 84.
< Натаров В. В. 1963. О водных массах и течениях Берингова моря.
Известия ТИНРО, т. 50.
357
Натаров В. В. и Пашкин, В. Н. 1968. Влияние океанографиче-
ских характеристик на формирование промысловых районов в водах, приле-
гающих к Австралии. Труды ВНИРО, т. 66.
Наумов Г. В. 1969. Краткая история биогеографии. Изд-во «Нау-
ка*, М.
Нечаев В. А. 1969. Птицы Южных Курильских островов. Изд-во
«Наука», Л.
Нечаев В. А., Юдаков А. Г. 1967. На островах залива Петра Ве-
ликого. «Природа», № 5.
Нечаев В. А., Юдаков А. Г. 1968. О гнездовании морских птиц
на островах залива Петра Великого (Южное Приморье). Известия Сиб. отд.
АН СССР, сер. бнол.-мед. наук, № 15, вып. 3.
Нейман В. Г. 1961. Динамическая карта Антарктики. «Океанологи-
ческие исследования», № 3,
Нейман А. А. 1963. Количественное распределение бентоса иа шель-
фе и верхних горизонтах склона восточной части Берингова моря. Известия
ТИНРО, т. 50.
Новиков Г» А. I960. Географическая изменчивость плотности насе-
ления лесных птиц в Европейской части СССР и сопредельных странах.
«Зоологический журнал», т. 39, вып. 3.
Новиков Н. П. 1964. Основные черты биологии тихоокеанского бе-
локорого палтуса (Hippoglosuss hippoglosus stenolepis Schmidt) в Беринго-
вом море. Известия ТИНРО, т. 61.
Новиков Н. П. 1965. Сырьевые ресурсы материкового склона север-
ной части Тихого океана. «Рыбное хозяйство», № 8.
Новиков Н. П. 1970. Биология малоглазого долгохвоста Chalinura
pectoralis в северной части Тихого океана. Известия ТИНРО, т. 72.
Новиков Ю. В. 1967. Основные черты биологии и состояние запа-
сов тихоокеанской сайры. Известия ТИНРО, т. 56.
Овчинников И. М. 1961. Циркуляция вод северной части Индий-
ского океана в период зимнего муссона. «Океанологии, исслед. по прогр.
МГГ», №4.
Панфилова С. Г. 1961. Температура воды. В сб. «Основные черты
геологии и гидрологии Японского моря». Изд. АН СССР, М.
Панфилова С. Г. 1968. Температура вод. В книге «Тихий океан.
Гидрология Тихого океана». Изд-во «Наука», М.
Парии Н. В. 1967. Сарганообразные рыбы открытого океана. В кни-
ге «Тихий океан. Биология Тихого океана. Рыбы». Изд-во «Наука», М.
Парии Н. В. 1967а. Основные особенности географического распро-
странения рыб эпипелагиали. В книге «Тихий океан. Биология Тихого океа-
на. Рыбы». Изд-во «Наука», М.
Парии Н. В. 1968. Ихтиофауна океанской эпипелагиали. Изд-во
«Наука», М.
Пахульский А. И. 1967. Роль птиц в рыбном хозяйстве. Пище-
промиздат.
Пашкин В. Н. 1970. Особенности формирования продуктивных зон
иа Австрало-Новозеландском шельфе. В сб. «Современное состояние биол.
ресурсов Мирового океана и перспективы их использования». Калининград.
Подковыркин В. А. 1955. Список птиц северной части Курильской
гряды. «Зоологический журнал», 34, № 6.
Полутон И. А., Лагунов И. И., Никулин П. Г., Вер-
сии В. Д., Дроздов В. Г. 1966. Промысловые рыбы Камчатки. Пет-
ропавловск-Камчатский.
Пономарева Л. А. 1954. О сезонных изменениях зоопланктона
пролива Лаперуза. Труды ИО АН СССР, т. Г1.
Пономарева Л. А. 1954а. Зимний зоопланктон северной части
Японского моря. Труды ИО АН СССР, т. 9.
358
Пономарева Л; А. . 1961. Зоопланктон залива Анива. Труды ИО
АН СССР, т. 51.
Пономарева Л. А. 1963. Эвфаузниды северной половины Тихого
океана, их распространение и экология массовых видов. Изд. АН СССР, М.
Пономарева Л. А. 1966. Эвфаузниды антарктических вод. Кали-
нинград.
Портенко Л. А. 1946. Птицы Северного Ледовитого океана. Труды
дрейф, экспед. Главсевморпути на ледок, пароходе «Г. Седов» 1937—
1940 гг. Л.
Портенко Л. А. 1947. Особенности перелета птиц в Арктике. «При-
рода», 4.
Портенко Л. А. 1965. Орнитогеографическое районирование террито-
рии СССР. Труды Зоол. ин-та АН СССР, т. 35.
Правоторова Е. П. 1965. Некоторые данные по биологии гижигин-
ско-камчатской сельди в связи с колебаниями ее численности и изменени-
ем ареала нагула. Известия ТИНРО, т. 59.
Прайор М. Э. 1964. Серебристо-серый и антарктический буревестни-
ки о. Хасуэлл. Бюлл. Сов. Антаркт. экспедиции, Хе 50.
Промптов А. Н. 1941. Сезонные миграции птиц. Изд. АН СССР,
М.-Л.
Пузанов И* И. 1938. Зоогеография. Учпедгиз, М.
Пузанов И. И. 1960. М. А. Мензбир как зоогеограф и проблемы раз-
вития советской зоогеографии. Труды проблем, и тематяч. совещаний ЗИН,
вып. 9.
Пузанов И. И. и Назаренко Л. Ф. 1965. Применение данных
синоптической метеорологии при изучении перелетов птиц. В сб. «Миграции
птиц и млекопитающих». Изд-во «Наука», М.
1 Пузаченко Ю. Г. 1962. Еще о зимней.орнитофауне Репетека. «Ор-
нитология», вып. 5.
Пузаченко Ю. Г. 1967. Географическая изменчивость обилия и струк-
туры населения птиц лесных биоценозов. «Орнитология», вып. 8.
Радзиховская М. А. 1961. Водные массы Японского моря. В сб.
^Основные черты геологии и гидрологии Японского моря». Изд. АН СССР.
Радзиховская М. А. 1965. Основные черты структуры вод север-
ной части Тихого океана. «Океанологии, исслед. по прогр. МГГ», № 13.
Радзиховская М. А. и Леонтьева В. В. >1968. Структура
вод и водные массы. В книге «Тихий, океан. Гидрология Тихого океана».
Изд-во «Наука», М.
Реймерс Н. Ф. 1966. Птицы и млекопитающие южной тайги средней
Сйбири. Изд-во «Наука», М.—Л.
Румянцев А. И. и Дар да М. А. 1970. Летняя сельдь восточней
части {Берингова моря. Известия ТИНРО, т. 72.
Савилов А. И. 1961. Экологическая характеристика донных сооб-
ществ беспозвоночных Охотского моря. Труды ИО АН СООР, 46.
Световидов А. Н. 1948. Трескообразные. Фауна СССР. Рыбы, т. 9,
вып. 4, М.—Л. m
• С в е т ов и д о в А. Н. 1962. Сельдевые. .Фауна СССР. Рыбы, т. 2,
вып. 1, М.—Л.
Семина Г. И. 1965. О двух зональных группировках фитопланктона
(на примере Берингова моря). Докл. АН СССР, т. 101» № 2,
Семина Г. И. 1967. Фитопланктон. В книге , «Тихий океан. Биоло-
гия Тихого океана. Планктон». Изд-во «Наука», М.
Серобаба И. И. 1968. Нерест минтая в северо-восточной части Бе-
рингова моря. «Вопросы ихтиологии», т. 8, вып. 6.
Серобаба И. И. 1970. Распределение минтая Theragra chalcogramma
(Pall.) в восточной части Берингова моря и возможности его промысла. Из-
вестия ТИНРО, т. 72.
359
Сизова Ю. В. 1961. Циркуляция вод Японского моря. В сб. «Основ-
ные черты геологии и гидролорш Яновского моря». Изд. АН СССР, М.
Синицын В. М. 1965—1967. Древние климаты Евразии. Изд. Ле-
нингр. университета.
Слепцов М. М. 1959. Трубконосые в районах китобойного промыс-
ла на северо-западе Тихого океана. «Орнитология», вып. 2.
Смирнова Л. И. <1959. Фитопланктон Охотского моря и прикуриль-
ского района. Труды ИО АН СССР, 30.
Сое да Д. 1956. Изучение экологии и размножения тихоокеанского
кальмара. Бюлл. Хюккайд. н-исолед. ла борат, рыбол. № 14.
Страхов Н. М. 1962. Основы теории литогенеза. 1, М.
Судиловская А. М. .1950. Новые данные о миграциях и кочевках
тонкоклювого буревестника. Бюлл. Моск, об-ва вспыт. природы, отд. биоло-
гии, т. 55 (5).
Судиловская А. М. 1951. Отряд трубконосые (Tubinares). В книге
«Птицы Советского Союза», т. 2. Изд-во «Наука», М.
Судиловская A. iM. 1962. Основные черты строения и распростра-
нения чистиков и пингвинов. Научи, докл. высш, школы. Биол. науки, № 1.
Суханова И. Н. 1962. О видовом составе и распределении фито-
планктона в северной части Индийского океана. — Труды ИО АН СССР, 58.
Суханова И. Н. <1964. Фитопланктон северо-восточной части Индий-
ского океана в период юго-западного муссона. Труды ИО АН СССР, 65.
Сыроечковский Е. £. 1966. Орнитологические наблюдения в Ан-
тарктиде и некоторые вопросы биогеографии суши Антарктики. В сб. «Ан-
тарктика». Докл. комиссии 4965.
Таубер Г. М. 1956. Антарктика. Основные черты климата и погоды.
Гидрометеоиздат, Л.
Т а х т а д ж я н А. Л. 1970. (Происхождение и расселение цветковых
растений. Изд-во «Наука», Л.
Т о м н л и н А. Г. 1970. Морские млекопитающие и их биогеоценетиче-
ские связи с другими группами морской фауны. В сб. «Биогеоценозы морей
и океанов». Изд-во «Наука», М.
Трешников А. Ф. 1964. Циркуляция поверхностных вод Южного
Ледовитого океана. Информационный бюллетень Советской антарктической
экспедиции, № 45. Изд-во «Транспорт», М.—Л.
Тугаринов А. Я. 1937. Миграции птиц на территории СССР в свете
четвертичной истории страны. Известия АН СССР, сер. биол., № 4.
Тугаринов А. Я. 1950. Происхождение миграций птиц Палеарктики.
В сб. «Памяти акад. П. П. Сушкина». Изд. АН ССОР, М.—Л.
Тугаринов А. Я. 1962. Пути перелетов птиц как свидетельство бы-
лых ландшафтов и территориальных связей. «Зоологический журнал», т. 31,
вып. 5.
Уда М. 1966. Подъем субполярных промежуточных вод в тропической
зоне Мирового океана и его связь с расположением областей высокой биоло-
гической продуктивности, в том числе районов, благоприятных для лова тун-
ца. Труды 2-го Международного океаногр. конгресса. Изд-во «Наука», М.
Успенский С. М. <1956. Птичьи базары Новой Земли. М.
Успенский С. М. 1959. Морские колониально гнездящиеся птицы
северных и дальневосточных морей СССР, их размещение, численность и
роль как потребителей планктона и бентоса. Бюлл. Моск, об-ва нспыт. при-
роды, отд. биологии, т. 64 (2).
Успенский С. М. 1969. Жизнь в высоких широтах. Изд-во «Мысль», М,
У ш а к о в П. В. 1953. Фауна Охотского моря и условии ее существо-
вания. Изд. АН СССР.
Фадеев Н. С. 1963. Желтоперая камбала восточной части Берингова
моря. Известия ТИНРО, т. 30.
360
Фадеев Н. С. 1970. Промысел и биологическая характеристика жел-
топерой камбалы восточной части Берингова моря. Известия ТИНРО, т. 72.
Федосов М. В., Давидович Р. Л. 1963. Некотооые особенности
гидрохимического режима Берингова моря. Известия ТИНРО, т. 50.
Физико-географический Атлас Мира. 1964. АН СССР и Главное упр. гео-
дезии и картографии МГ СССР, М.
Харитонова О. А. 1965. К вопросу о распределении нагульной сель-
ди в северной части Охотского моря. Известия ТИНРО, т. 59.
Харченко А. М. 1968. Течения и водные массы Восточно-Китайского
моря. «Океанология», т. 8, вып. 1.
Чельцов-Бебутов А. М. 1959. Опыт количественной оценки птичь-
его населения открытых ландшафтов. «Орнитология», вып. 2.
Чернов Ю. И. 1968. Географическая зональность и животный мир
суши. Сообщение 1 и 2. «Зоологический журнал», т. 47, вып. 7—8.
Чернявский В. И. 1970. О причинах, высокой биологической про-
дуктивности северной части Охотского моря. Известия ТИНРО, т. 71.
Чернявский В. И. 1970а. Гидрологический фронт северной части
Охотского моря. Известия ТИНРО, т. 71.
Чигиринский А. И. 1964. О нересте ставриды Trachurus japonicus
(Temm. et Schl.) в Восточно-Китайском море. Известия ТИНРО, т. 55.
Чигиринский А. И. 1969. О нерестовых популяциях и миграциях
ставриды в Восточно-Китайском море. «Рыбное хозяйство», № 2.
Шабонеев И. Е. 1965. О биологии и промысле сельди восточной ча-
сти Берингова моря. Известия ТИНРО, т. 53.
Шевцов В. В. и Макаров Р. Р. 1969. К биологии антарктиче-
ского криля. Труды ВНИРО, т. 66.
Шмальгаузен И. И. 1969. Проблемы дарвинизма. Изд-во «Нау-
ка», Л.
Шмидт П. Ю. 1950. Рыбы Охотского моря. Труды Тихоокеанской
комиссии АН СССР, 4.
Штегман Б. К- 1936. О принципах зоогеографического деления Па-
леарктики на основе изучения типов орнитофауны. Известия АН СССР,
№ 2—3 (сер. биол.).
Штегман Б. К. 1938. Основы орнитогеографического деления Пале-
арктики. Фауна СССР, Птицы, т. 1, вып. 2, М.—Л.
Штейнбахер И. 1956. Перелеты птиц и их изучение. Изд-во «Иност-
ранная литература», М.
Шунтов В. П. 1961. Миграции и распределение морских птиц в юго-
восточной части Берингова моря в весенне-летний период. «Зоологический
журнал», т. 40, вып. 7.
Шунтов В. П. 1961а. Сухопутные птицы в море. «Природа», № 12.
Шунтов В. П. 1962. Сухопутные птицы и кулики в Беринговом мо-
ре. «Орнитология», вып. 4.
Шунтов В. П. 1963. Летнее распределение моевок в Беринговом мо-
ре. «Орнитология», вып. 6.
Шунтов В. П. 1963а. Особенности распределения ихтиофауны в юго-
восточной части Берингова моря. «Зоологический журнал», т. 42, вып. 5.
Шуитов В. П. 1964. Трансэкваториальные миграции тонкоклювого
буревестника — Puffinus tenuirostris (Temm.). «Зоологический журнал»,
т. 43, вып. 4.
Шунтов В. П. 1964а. Распределение н миграции тихоокеанского
кальмара в Японском море. Известия ТИНРО, т. 55.
Шунтов В. П. 1965. О трубконосых и чистиковых птицах Беринго-
ва моря. «Орнитология», вып. 7.
Шунтов В. П. 1965а. Сезонные аспекты в распределении морских
цтиц в открытых водах Японского моря. «Зоологический журнал», т. 44,
вып. 3.
361
Шунтов В. П. 19656. Вертикальная зональность в распределении
рыб в верхней батиали Охотского моря. «Зоологический журнал», т. 44,
вып. 11.
Шунтов В. П. 1965в. • Сайра Японского моря. В сб. «Сайра», Влади-
восток.
Шунтов В. П. 1966. О зимовках птиц в дальневосточных морях н се-
верной части Тихого океана. «Зоологический журнал», т. 45, вып. 11.
Шунтов В. П. 1966а. Вертикальное распределение черного н стрело-
зубых палтусов в северной части Тихого океана. «Вопросы ихтиологии»,
т. 6, вып. 1 (38).
Шунтов В. П. 1967. Характеристика летнего населения морских птиц
в юго-западной части Тихого океана (район Новой Зеландии). «Орнитоло-
гия», вып. 8.
Шунтов В. П. 1967а. Осеннее население морских птиц в юго-запад-
ной части Тихого океана в районе Новой Зеландии. «Зоологический журнал»,
т. 46, вып. 9.
Шунтов В. П. 19676. Новые данные о распределении глубоковод-
ных рыб Охотского моря. «Вопросы ихтиологии», т. 7, вып. 1.
Шунтов В. П. 1967в. Сайра Японского моря. Известия ТИНРО, т. 56.
Шунтов В. П. 1968. Некоторые закономерности в распределении
альбатросов (Tubinares, Diomedeidae) в северной части Тихого океана. «Зоо-
логический журнал», т. 47, вып. 7.
Шунтов В. П. 1968а. Характеристика летне-осеннего населения мор-
ских птиц и их численность в Большом Австралийском заливе. «Орнитоло-
гия», вып. 9.
Шунтов В. П. 19686. Количественный учет морских птиц в восточ-
ной части Индийского океана. «Океанология», т. 8, вып. 3.
Шунтов В. П. 1968в. Количественный учет летучих рыб в восточной
части Индийского океана. «Вопросы ихтиологии», т. 8, вып. &
Шунтов В. П. 1969. Некоторые черты экологии пелагических рыб
Большого Австралийского залива. «Вопросы ихитиологии», т. 9, вып. 6 (59).
Шунтов В. П. 1969а. О закономерностях в распределении летучих
рыб в Тихом и Индийском океанах. Тезисы докл. коиф. по троп, зоне Миро-
вого океана, ноябрь 1969 г. М.
Шунтов В. П. 1970. Сезонное распределение черного и стрелозубых
палтусов в Беринговом море. Известия ТИНРО, т. 72.
Шу и то в В. П. 1970а. Морские птицы в отрытых водах Восточно-
Китайского моря. «Экология», № 4.
Шунтов В. П. 19706. Некоторые аспекты сезонного распределения
шельфовых рыб Новой Зеландии. «Вопросы ихтиологии», т. 10, вып. 3.
Шунтов В. П. 1970в. Морские птицы и биологическая структура океа-
на. В сб. «Программа и методика изучения биогеоценозов водной среды».
Изд-во «Наука».
Шуитов В. П. 1971. Некоторые закономерности распределения черно-
го и стрелозубых палтусов в северной части Тихого океана. Известия
ТИНРО, т. 75.
Шунтов В. П. 1971а. Некоторые данные о биологии рыб Арафурско-
го моря и условиях их обитания. «Вопросы ихтиологии», т. 11, вып. 1.
Шунтов В. П. 19716. * Рыбы верхней батиали Новозеландского пла-
то. «Вопросы ихтиологии», т. 11, вып. 3.
Шунтов В. П., Демиденко Ю. К. 1970. Рыбные ресурсы райо-
нов Австралии и Новой Зеландии. В сб. «Современное состояние биол. ре-
сурсов Мирового океана и перспективы их использования». Калининград.
Элтон Ч. 1960. Экология нашествий животных и растений. Изд-во
«Иностранная литература», М.
362
Юдин К. А. 1965. Филогения и классификация ржанкообразных. Фау-
на СССР, ПЙгам, т. 2, вып. 1, часть I. Изд-во «Наука», М.—Л.
Юдин К. А. 1968. Краткий определитель трубконосых птиц (РгосеЬ
lariiformes) Антарктики я Субантарктики. Исследования фауны морей,
VI (XIV). Результаты бнологич. иссл. Советской антарктической экспедиции
<1935—«1958 гт). Зкюлогяч. инет. АН СССР, 4.
Я м а д а Т. 1962. Промысел ставриды кошельковыми неводами в Восточ-
но-Китайском море. Жури. «Суй санкай», № 9 (перевод с я поиск.).
Ярогой Б. А. 1969. О физико-географических условиях ареала
Euphausia superba Dana. Труды В НИРО, т. 66.
Abbott D. and Albee R. 1967. Summary of thermal conditions and
phytoplankton volumes measured in Monterey Bay, California 1961—1966. Ca-
lif. Cooper. Oceanic. Fish. Investig. Rep. vol. 11.
AhlatT о m E. H. 1967. Cooccurrences of sardine and anchovy larvae in
the California current region off California and Baja California. Calif. Cooper.
Oceanic. Fish. Investig. Reports, vol. II.
Alexander W. B. 1954. Birds of the Ocean. 2 nd edition. New York.
A1 v a r i n о A. 1965. Distributional Atlas of Chaetognatha in the Califor-
nia Current Region. Calif. Cooper. Oceanic Fish.. Investig, Atlas n. 3.
Alverson D. L., Pruter A. T. and RonholtL. L. 1961.
A Study of Demersal Fishes and Fisheries of the Northeastern Pacifis Ocean.—
Instit. Fish. Univer. British Columbia, Vancouver.
Alverson D. and Larkins H., 1969, Status of knowledge of the
Pacific Hake resource. Calif. Cooper. Ocenjc. Fish. Investig. Reports, vol. 13.
Amiet L. U958. The distribution of Diomedea in Eastern Australian waters:
north of Sydney. Notornis, vol. 7, n. 8.
Arnold L. W. 4948. Observations on populations of North Pacific pela-
gic birds. The Auk, vol. 65, n. 4.
A s h m о 1 e N. P. 1963. The regulation of number of tropical oceanic
birds. The Ibis, vol 103, n. 3.
Ash mole N. 1965. Adaptive variation in the breeding regime of a
tropical sea bird. Proc. Nat. Acad. Sci. U. S. A. 53, n. 2.
Ash mole N. P. and Ash mole M. J. 1967. Comparative Feeding
Ecology -of Sea Birds of a Tropical Oceanic Island. Peabody Museum of Nat.
Mist. Yale Univ., bull. 24.
As h mо 1 e M. J. and Ash mo 1 e X. P. 1968. The use of food samp-
les from sea birds in the study of seasonal variation in the surface fauna of tro-
pical oceanic areas. Pacif. Sci., vol. 22, n. I.
Austin O. L. J. 1948. The Birds bf Korea. Bull, of the Museum of Compara-
tive Zoology at Harvard College. Vol. 101, n. I.
Austin O. L. J. 1949. The status of Steller’s Albatross. Pacific Sci., vol. 3.
Austin O. L. J. and Kuroda N. 1953. The birds of Japan. Their sta-
tes and distribution. Bull. Mus. Camp. Zool. Harv. Coll. voll. 109, n. 4.
Avila E. 1954. Potencia deyectiva del guanay (Phalacrocorax bouga-
invillii). Biol. Compania administr. Guano. 1, n. 2.
Axelrod D. 1. 1952. A theory of angiosperm evolution. Evolution., 4.
Bailey R. S. 1965. Cruise of R. R. S. Discovery in the Indian Ocean.
Sea swallow, 17.
Bailey R. S. 1966. The sea-birds of the Southeast coast of Arabia. The
Ibis, vol, 108, n. 2.
Bailey R. S. 1967. Migrant waders in the Indian Ocean. The Ibis, vol.
109, n. 3.
Bailey R. S. 1968. The pelagic distribution of sea-birds in the Western
Indian Ocean. The Ibis, vol, 110, n. 3.
363
Bailey R. S., Pocklington R. and Willis Pk R. 1968. Storm-
petrels Oceanodroma spp. in the Indian Ocean. The Ibis. vol. 110, n. 1.
Bailey A. M. and Sorensen J. H. 1962. Subantarctic Campbell
island. Denver Mus. of Natur. Proc. n. 10.
Baker R. H., 1951. The avifauna of Micronesia, its origin, evolution and
distribution. Univ. Kansas Publ. Mus. N^t. Hist., 3, n. 1.
Baker R. H. 1953. Migrations of shore birds in the Central Pacific. Pro-
ceed. Seventh Pacif. Scien. Congr.
Barghoorn E. S. 1953. Climatic changes in the light of the geological
past of the plant life. In: H. Shapley (ed. by). Climatic change. Cambridge.
Baxter J. L 1967. Summary of biological information on the northern
anchovy Engraulis mordax Girard. Calif. Cooper. Ocenic. Eish. Investig. Re-
ports, vol. 11.
Beck J. R. Food, moult and age of first breeding in the cape pigeon,
Daption capensis. Brit. Antarct. Surv. Bull., n. 21.
Bedard J. 1966. New records of aicids St. Lawrence Island, Alaska. The
Condor, vol. 68, n. 5.
Bedard J. 1969. The nesting of the crested least and parakeet auk*
Jets on St. Lawrence Island, Alaska. The Condor, vol. 71, n. 4.
Bent A. 1919. Life histories of North American diving birds. Smiths. Instit.
U. S. Nat. Museum. 107.
Berry E. W. 1937. Tertiary Погда of Eastern North America. Bot. Rev., 3.
Bierman W. H. and V о о u s К. H. 1950. Birds observed and colle-
cted during the whaling expeditions of the Willem Barendsz in the Antarctic,
1946—1947 and 1947—1948. — Ardea, 37.
Blackburn M. 1950. Studies on the Age, Growth, and Life History o;
the Pilchard, Sardinops neopilchardus (Steindachner), in Southern and Western
Australia. Austr. Journ. Mar. Freshw. Res. vol. 1, n. 2.
Blackburn M. 1950a. A biological study of the Anchovy, E. australis
(White) in Australian Waters. Aust. Journ. of Mar. and Freahw. Res. 1, n. 1.
Blackburn M. 1960. Synopsis of biological information on the Austra-
lian and New Zealand Sardine Sardinops neopilchardus (Steindachner). FAO
Fish. Biol. Synopsis, n. 12.
Blackburn M. and Gartner P. E. 1954. Populations of barraco-
uts, Thyrsites atun (Euphrasen), in australian waters. Austr. Journ of Mann,
and Freshw. Res. 5, n. 3.
Blake E. R. 1953. Birds of Mexico. Chicago.
Blunt С. E. 1969. The Jack mackerel (Trachurus symmetricus) resource
of the eastern North Pacific. Calif. Cooper. Oceanic. Fish. Investig. Reports,
vol. 13.
Bourne W. R. P. 1960. The petrels of the Indian Ocean. Sea Swallow, 13.
Bourne W. R. P. 1963. A Review of Oceanic Studies of the Biology of
Sea birds. Proc. 13 Intern. Ornithol. Congress, vol. 2.
Bourne W. R. P. 1967. Long-distance vagrancy in the petrels. The
Ibis, vol. 109, n. 2.
Brin ton E. 1967. Distributional atlas of Euphausiacea (Crustacea) in
the California Current region, part I. Calif. Cooper. Oceanic Fish. Investig.
Atlas n. 5.
Broekhuysen G. J., Livers idge R., Rand R. W. 1961. The
South African gannet Morus capensis. I. Distribution and movements. Ostrich,
32, n. 1.
Brooks S. C. 1934. Oceanic currents and the migration of pelagic birds.
The Condor, vol. 36. n. 5.
Brosset A. 1962. La reproduction des oiseaux de mer des iles Galapagos
en 1962. Alauda 31, n. 2.
Bruun Bertel 1963. Svartbagen (Larus marinas L): Danmark. En
364
undersogerse baseret pa ringsemaerkninger of skandinaviske Svartbage. Dansk
ornithol. foren. tidsskr. 57, n. 2.
Bull J. H. 1961. Yellow-nosed Albatross off the coast of Long Island.
New York. The Auk, vol. 78, n. 4.
Bull P. C. and Boe son B. W. 1961. Preliminary analysis of records
of “storm-killed” sea birds from New Zealand* 1939—59. Notornis 9.
Burling R. W. 1961. Hydrology of Circumpolar waters South of New
Zealand. N. Z. Dep. Sci. Industr. Res. Bull. 143.
Carrick R., Keith N. and Keith K. 1957. Third annual report of
the Australian bird-banding scheme, lune, 1957. C. S. I. R. 0. Wildlife Res. 2, n. 2.
Carrick R.. Wheeler W. R. and Murray M. D. 1957a. Se-
asonal dispersal and mortality in the silver gull, Larus novae-holandiae Ste-
phens, and, crested tern. Sterna bergii Lichtenstein in Australia. C. S. I. R. O.
Wildlife Res. 2. n. 2.
C a w к e 11 E. M. and Hamilton! E. 1961. The birds of the Folk-
land. The Ibis, vol. 103a, n. I.
Cayley N. W. 1965. What Bird is that? Sydney.
Chapman S. 1969. The Pacific winter quarters of Sabine's Gull. The
Ibis, vol. Ill, n. 4.
Chapman W. M. 1964. The management of ocean fisheries. Governor’s
Advisory Commission on Ocean Resources, California.
Clark W. C. and Dawson E. W. 1957. The trans-Tasman dispersal of
the white-fronted tern (Sterna striata Gm.). Notornis, 7, n. 3.
Cline D. R., Si niff D. B. and Erickson A. W. 1969. Summer
birds of the pack ice in the Weddell sea, Antarctica. The Auk, vol. 86, n. 4.
Condon H.T. 1946. South Australian Birds. The South Australian Na-
turalist, vol. 24.
Coulson J. C. 1963. The status of the Kittiwake in the British Isles.
Bird Study, vol. 10, n. 3.
Coulson J. C. 1966. The movements of the Kittiwake. Bird Study, vol.
13 n. 2.
Coulson J. C. and White E. 1956. A study of colonies of the Kitti-
wake Rissa tridactyla (L.). The Ibis, vol. 98, n. 1.
Coulson J. C. and White E. 1958. The effect of age on the bree-
ding biology of the Kittiwake Rissa tridactyla. The Ibis, vol .100, n. 1.
Coulson J. C. and White E. 1959. The postfledging mortality of
the Kittiwake. Bird Study, 6, n. 3.
Coulson J. C. and Whitte E. 1960. The effect of age and density
of breeding birds on the time of breeding of the Kittiwake Rissa tridactyla.
The Ibis, vol. 102, n. 1.
Darby Marie 1970. Summer Sea birds between New Zealand and
McMurdo Sound. Notornis, 17, n. 1.
Davies D. H 1955. The South African pilchard (Sardinops ocellata); Bird
predators, 1953—4. Commerce and Industry, Investig. rep. n. 18.
Davies D. H. 1956. The South African pilchard (Sardinops ocellata)
and maasbanker (Trachurus trachurus); Bird predators, 1954—55. Commerce
and Industry, Investig. rep. n. 23.
Davies D. H. 1958. The South African pilchard (Sardinops ocellata)
and maasbanker (Trachurus trachurus); the predation of Sea birds in the com-
mercial fishery. Commerce and Industry, Investig. rep. n. 31.
Dawson E. W. 1962. A west coast South Island, sea bird log in winter,
1961. Notornis, vol 10, n. 2.
Dawson W. L. 1940. The birds of California. New York.
Dell R. K. 1960. Sea-bird logs between New Zeeland and the Ross Sea.
Rec. Dominion Mus. 3 (IT).
363
DeWitt Hugh H. 1965. Antarctic ichthylogy. Bio Scienoe, 15, n. 4.
Dixon С. C. 1933. Some observations on the albatrosses and other birds
of the Southern oceans. Trans. Roy. Canad. Inst. vol. 19, pt. 1.
Dixon K. L. and Starrett W. C. 1952. Offshore observations of
Tropical sea birds in the western Pacific. The Auk. vol. 69, n. 3.
D‘ о mbr a i n A. and Gwynn e'A. 1962. Buller's Shearwater on Cab-
bage Tree Island, New South Wales. The Emu, vol. 61, n. 4.
D о г s t J. 1956. Les oiseaux a guano des cotes peruviennes. Sci. et nature,
n. 14.
D о г s t J. 1962. Les immenses migrations des oiseaux de mer naguere
insoupcon-nees. Nature Sci. progr. n. 3322.
D о г s t J. 1962a. The Migration of Birds. Heinemann, 1962.
Downes M. С.» Ealey E. H. M., Gwynn A. M. and Young
P. S. 1959. Birds of Heard Island. ANARE Reports, Melbourne, ser. B., vol. I.
Eastropac Atlas, 1970—1971. Vol. 2, 4. U. S. Depart. Commerce, Cir-
cular 330, Washington.
Eklund C. R. 1961. Distribution and life history studies of the south-
polar skua. Bird-Band, 32, n. 4.
Eklund C. R. 1964. The Antarctic skua.«Scient. Amer. 210, n. 2.
E к m a n S. 1953. Zoogeography of the Sea. London.
Elton C. 1946. Competition and the structure of ecological communites—J.
Anim. Ecol., 15.
Falla R. A. 1937. Birds. B. A. N. Z. A. R. E. Reps., Ser.B., 2.
Falla R. A. 1940. The genus Pachyptila liliger. The Emu, vol. 40, n. 3.
Falla R. A. 1952. Antarctic Birds. The Antarctic today.
Falla R. A. 1964. Distribution patterns of birds in th antarctic and
high-latitude Subantarctic. Biol, antarctique, Paris.
F a r n e r D. S. and Serventy D. L. 1959. Body temperature and
the ontogeny of thermoregulation in the Slender — billed Shearwater. The
Condor, vol. 61, n. 6.
F a у F. H. and C a d e T. J. 1959. An ecological analysis of the avi-
fauna of St. Lawrence island, Alaska. Univ. Calif. Pubis. Zool., 63, n. 2.
Fisher H. I. 1963. Airplane-albatross collisions on Midway Atoll. The
Condor, vol. 68, n. 3
Fisher H. I. 1966. Aerial census of Laysan Albatrosses breeding on Mid-
way Atoll in December. The Auk, vol. 83, n. 4.
Fisher H. I. 1969. Eggs and egg-laying in the Laysan Albatross, Dio-
medea immutabilis. The Condor, vol. 71, n. 2.
Fisher J. 1952. The Fulmar. London.
Fisher J. and Lockley R. M. 1954. Sea-Birds. London. Collins.
Fisher J. and Peterson R. T. 1964. The World of Birds. London.
Fleming C. A. 1939. Birds of the Chatham Islands. The Emu, 38.
Fleming C. A. 1950. Some South Pacific Sea-bird Logs. The Emu,
vol. 49. n. 3.
Fleming C. A. 1952. The Seas between. The Antarctic today.
Fleming C. A. 1956. Biogeography of Cook strait sea-birds. Proc. N. Z.
Ecol. Soc. n. 4.
Fleminger A. 1964. Distributional Atlas of Calanoid Copepods in the
California Current Region, Part I. Calif. Cooper. Oceanic Fish. Invest., Atlas n. 2.
Fordham R. A. 1964.Breeding,biology of the southern black-backed
gull. I. Pre-egg and egg stage. Notornis, 11, n. L
Fordham R. A. 1964a. Breeding biology of the southern black-backed
gull. II. Incubation and the chick stage. Notornis, 11, n. 2.
Fordham R. A. 1968. Dispersion and dispersal of the Dominican gull
in Wellington, New Zealand. Proc. N. Z. Ecol. Soc., n. 15.
366
F u к a t a к i H. 1959. On the Year Round Occurrence and Ecology of Eggs
and Larwae of the Principal Fishes in the Japan Sea. II. Occurence and Distri*
bution of Eggs and Larwae of the Saury in the Tsushima Current Region.
Bull. Jap.Sea Reg. Fish. Res. Lab. n. 6.
Gabrielson I. N. and Lincoln E. C. 1959. The birds of Alaska.
Washington.
Garner D. M. 1961. The Hydrology of New Zealand Coastal Waters.
1955. N. Z. Dep. Sci. Industr. Res. Bull., 138.
Garner D. M. 1962. Analysis of Hydrological observations in the New
Zealand Region, 1874—1955. N. Z. Dep. Sci. Industr. Res. Bull, 144.
Garner D. M. and Ridgway N. M. 1965. Hydrology of New Ze-
aland off shore Waters. N. Z. Dep. Sci. Industr. Res. Bull., 162.
Gause G. F. 1934. The struggle for existence. Williams and Wilkins,
Baltimore.
Gibson J. D. 1963. Third Report of the New South Wales Albatross
Study Group (1962). Summarizing activities to date. The Emu, vol. 63, n. 3.
Gibson J. D. 1967. The wandering albatross (Diomedea exulans): in
New South Wales coastal Waters and the Tasman sea. Notornis, vol. 14, n. 1.
Gibson J. D. and Sefton A. R. 1955. Notes on some albatrosses
of coastal New South Wales. The Emu, vol. 55, n. 1.
Gjbson J. D. and Sefton A. R. 1959. First Report of the New So-
uth Wales Albatross study group. The Emu, vol. 59, n. 2.
Gibson J. D. and Setton A. R. 1960. Second report of the New
South Wales Albatross study group. The Emu, vol. 60, n. 2.
Gibson J. and Sefton A. R. 1962. First Australian Record of
the Royal Albatross. The Emu, vol. 62, n. 3
Gibson-Hi 11 C. A. 1948. The Storm-petrels occurring in the northern
Indian Ocean and adjacent seas. J. Bombay Nat. Hist. Soc., 47.
G i b s о n - H i 11 C. A. 1953. Hotes on the sea birds of the ord. Procella-
riiformes and Pelicaniformes recorded as strays or visitors to the Ceylon coast.
Spolia Zeylanica, 27.
Gilbert O. et al., 1952. Gause's Hypothesis: an examination. J. Anim.
Ecol., 21.
Gill F. B. 1967. Observations on the Pelagic Distribution of Seabirds in
the Western Indian Ocean. Proc. Unit. Stat. Nat. Mus. Vol. 123, n. 3605.
Glover R. S. 1961. Biogeographical boundaries: the shapes of distri-
bution. In: Oceanography, Amer. Assoc. Adv. Sci., Publ. n. 67, Ed. M. Sears,
Washington.
Goddard M. T., Dorrigo N. S. W. and Hindwood K. A.
1951. Sea-birds and Cyclones: Some Interesting New South Wales Records. The
Emu, vol. 51, n. 2.
Graham D. M. 1953. A treasury of New Zealand Fishes. Wellington.
GurrL. and Kinsky F. C. 1965. The. distribution of breeding colo-
nies and status of the red-billed gull in New Zealand and its outlying island.
Notornis, 12, n. 4.
Hamabe M. and Shimizu T. 1966. Ecological Studies on the
Common Squid, Todarodes pacificus Steenstrup, mainly in the Southwestern
Waters of the Japan Sea. Bull. Jap. Sea Reg. Fish. Res. Lab. (16).
Hamilton W. J. 1958. Pelagic birds observed on a North Pacific cro-
ssing. The Condor, vol. 60, n. 3.
Hardin G. 1960. The competitive exclusion principle. Science, 131.
Harper P. C. A. 1966. New Zealand ornithologist on ”Eltani“. Antarctic,
4, n. 8.
Harris M. P. Black-browed albatross (Diomedea melanophris) and other
sea-birds near Guayaguil. Ecuador. Ardea, vol. 56, n. 3—4.
Harris M. P. 1969. Breeding seasons of seabirds in the Galapagos Is-
lands. J. Zool. vol. 1959, n. 2.
____________________________________________:__________________________367
Harrison Р. Р. О. 1962. Sea Birds of the South Pacific. Narberth.
H a у a s i S. 1967. A note on the biology and Fishery of the Japanese
«anchovy Engraulis japonica (Houttuyn). Calif. Cooper. Oceanic. Fish. Investig.
Reports., vol. 11.
Hill R. 1968. Australian Birds. Melbourn.
Hindwood K. A. 1955. Sea-birds and sewage. The Emu, vol. 58, n. 3.
Hindwood K. A. and McGill A. R. 1955. Sea-bird mortality in
coastal New South Wales during July, 1954. The Emu, vol. 55, n. 2.
Hi nd wood K., A. Keith K. and Serventy D. L. 1963. Birds
of the South-West Coral Sea. CSIRO, Divis. Wildl. Res. Techn. paper n. 3.
Hitchcock W. B. and Carrick R. 1958. First report of banded
birds migrating between Australia and other parts of the world. C. S. 1. R .0.
Wildlife Res. 3, n. 1.
Hitchcock W. B. and Carrick R. 1960. Fifth Annual Report of
the Australian Bird-Banding scheme, July 1958 to June 1959. C. S. I. R. O.
Wildlife Res. A n. 1.
Hitchcock W. B. 1961. Sixth Annual Report of the Australian Bird-
Banding scheme, July 1959 to June 1960. C. S. I. R. O. Wildlife Res. 6, n. 1.
H о 1 d g a t e M. 1958. Mountains in the sea. St. Martin’s Press Co., New
York.
Holgersen H. 1957. Ornithology of the Brategg Expedition. Scient.
Results of the Brategg Expedition 1947—48, n. 4.
Holgersen H. 1961. Norske lomviers vandzinger. Sterna, vol. 4, n. 6.
Holm J. A. 1962. Mollemukken. Vor vjden, n. 14.
Holmes R. T. 1964. Notes on the occurrence of the laysan albatross
near the California coast. The Condor, vol. 60. n. 4.
Hotta H., Fukushima S., Odate S. andAizawa Y. 1961.
Observations of Fish schools and sea-bird flocks in the Tohoku sea area of
Japan. Bull. Tohoku Reg. Fish, Rep. Lab., n. 19.
Howard H. 1950. Fossil evidence of avian evolution. The Ibis, vol.
92, n.. 1.
Howard P. F. 1956. Banding of Giant Petrels at Heard Island and
Macquarie lsland-II. The Emu, vol. 56, n. 4.
Hudson R. 1963. Bird ringing in British Antarctic territory 1957—1962.
Ring. Internal. Ornitol. Bull., n. Й.
Hudson R. 1968. The white-phase giant petrels of the South Orkney
Islands. Ardea, vol. 56, n. 1—2.
Hutchinson G. E., D e e v e у E. S. 1949. Ecological studies on po-
pulations. Surv. Biol. Progr., 1.
Huxley J. 1955. Morphism in birds, Acta XI Congressus Int. Ornithol.
Hynd J. S. and Robins J. P. 1967. Tasmanian Tuna Survey Re-
port of First Operations Period. Division of Fish and Oceanogr. Techn. Paper
n. 22 C. S. I. R. O.
Imber M. J. and Boeson B. W. 1969. Seabirds found dead in New
Zealand in 1964. Notornis, 16, n. 1.
Ingham S. E. 1959. Banding of Giant Petrels by the Australian Natio-
nal Antarctic Research Expeditions, 1955—1958. The Emu, vol. 59, n. 3.
Jameson W. 1961. The wandering albatross. New York.
Jesperson P. 1930. Ornithological observations in the North Atlantic
Ocean. The Danish "Dana" Exped., 1920—1922, Oceanogr. Repts., n. 7.
Jones S. 1966. A preliminary report on the commercial sea Fisheries of
the western sector the IPFC region Indo-Рас. Fish, council F. A. O., Proc.
12 th session 1966.
Jordan R. 1958. Breve nota sobre la anillacion de piqueros. Bol. Compa-
nia admin, guano, 34, n. 11.
Jordan R. 1960. Some results of the banding of guano producing birds
in Peru. Ring, 2, 23.
368
Jordan R. 1967. The predation of Guano birds on the Peruvian anchovy
(Engrauljs ringens Jenyns). Calif. Cooperat. Oceanic Fish. Investig. Repts, 11.
Jordan R. and Fuentes 1964. Resultados de los censos graficos de
aves guaneras efectuados durante el ciclo reproductive 1962—1963. Inst. In*
vest. Recurs. Mar., Informes (22).
J о u a n i n C. 1957: Les Procellariidfcs melaniques signales en mer d'Oman.
Oiseau 27.
J о u a n i n C. 1959. Une colonie тёсоппие d'Albatros a pieds noirs, Diome*
dea nigripes, dans les iles Mariannes. Bull. Musdum nat. histoire natur., 31, 6.
Kato T. 1958. Analysis of distribution and correlation of water masses on
the East Chine Sea in Summers, 1954—1956. Journ. of Meteorol. Res. vol. 10,
n. 8.
Kato T. 1959. Oceanographic Conditions of the East China Sea in winters.
Journ. of Meteorol. Res. vol. 11, n. 9.
Kawasaki T. 1966. Structure of the Pacific Population of the Mackerel.
Bull. Tokai Reg. Fish. Res. Lab. n. 47.
Kenyon K. W. 1949. Distribution of the Pacific Kittiwake in November
and December of 1948. The Condor, vol. 51, n. 4.
Kenyon K. W. 1950. Distribution of albatrosses in the North Pacific and
adjacent waters. The Condor, 52, n. 3.
Kenyon K. W. and Brooks J. W. 1960. Birds of little Diomede
island, Alaska. The Condor, vol. 62, n. 6.
Kenyon K. and Phillips R. 1965. Birds from the Pribilof Islands
and vicinity. The Auk, vol. 82, n. 1.
Kesteven G. L. and Stark A. E. 1967. Demersal Fish Stocks of
the Great Australian Bight as Estimated from the Results of Operations of
F. V. Southern Endeavour. Div. of Fish, and Oceanogr. Tech, paper, n. 24.
C. S. I. R. O.
Kikuchi S. 1959. Research of the Sea Surface temperature of the East
China Sea (1 st Report). Journ. of Meteorol. Res vol. 11, n. 3.
К i n g W. B. 1967. Seabirds of the tropical Pacific Ocean. U. S. National
Museum Smithsonian Institution, Washingon.
King W. B. 1970. The Trade Wind Zone Oceanography Pilot Study. Part
VII: Observations of Sea Birds March 1964 to June 1965. U. S. Fish. Wildl.
Serv. Spec. Sci. Rep. Fish. n. 586.
К i n g J. E. a n d P у 1 e R. L. 1957. Observations on sea birds in the
tropical Pacific. The Condor, vol. 59, n. 1.
Kinsky F. C. 1963. 13 th Annual Report of the New Zealand Bird Ba-
nding Scheme for the year ending 31 st March 1963. Dominion Museum. Wel-
lington.
Kinsky F. C. 1968. An unusual seabird mortality in the southern North
Island (New Zealand) april. Notornis, vol. 15, n. 3.
Kirkwood L. F. 1967. Inorganic Phosphate, Organic Phosphorus, and
Nitrate in Australian waters. Divis. of Fish and Ooeanogr. Tech, paper n. 25,
C. S. I. R. O.
Kramer D. and Ahl strom E. 1968. Distributional atlas of fish larvae
in the California Current region: Northern Anchovy, Engraulis mordax Girard,
1951 through 1965. Calif. Cooper. Oceanic. Fish. Investig., Atlas n. 9.
Kuroda N. 1954. On the classification and phylogeny of the order Tu-
binares (Osteology and habit differentiation), Tokyo.
Kuroda N. 1955. Observations on pelagic birds of the northwest Paci-
fic. The Condor, vol. 57, n. 5.
Kuroda N. 1957. A brief note on the pelagic migration of thej Tubinares.
Misc. Repts. Jamashina's Inst. Ornithol. and zool. n. 11.
Kuroda N. 1960. Analysis of sea birds distribution in the northwest Pa-
cific Ocean. Pacif. Sci., 14, n. 1.
3694
Kuroda N. 1960a. Notes on the breeding seasons in the Tubinares. Japa-
nese Journal of Zool. vol. 12, n. 4.
K u г о d a N. 1961. The over-sea crossings of land birds in the Western’
Pacific. Misc. Repts Yamashina's Inst. Ornithol. and Zool. 3, n. 1.
Kuroda N. 1964. Uber den Bestand seltener Stelzvogel und des Kurz-
schwans — Albatros in Japan 1963—64x Vogelwarte, 22, n. 3--4.
Lacan F., Prevost J. and Van Beveren M. 1969. Etude des
populations doiseaux de Urchipel de Pointe Gedlogie d£ 1965 a 1968. Oiseau
et Rev. franc, ornithol., 39, num. spec.
Lack D. 1967. Interrelationships in breeding adaptations as shown by
marine birds. Proc, of the 14 Intern. Ornithol. Congress.
Laird M. 1956. Biological notes on a sea vayage from Auckland to Si-
ngapore. Notornis, vol. 6, n. 8.
Lambrecht K. 1933. Handbuch der Palaornithologie. Berlin.
Lane S. G. 1962. Nesting of the short tailed shearwater on Brush Is-
land and other coustal islands of N. S. W. The Emu, vol. 62, n. 3.
Larsson E. 1960. Stormfagelfynd fran svenska vastkusten varen 1959’
Fauna och flora, n. 1—2.
Lavery H. J. 1964. Notes on Birds in the Gulf of Corpentaria. The
Emu, vol.63, n. 4.
L о с к 1 e у R. M. 1958. Sea birds and their protection. Bird Notes, 28, n. 4.
Loomis L. M. 1918. Review of albatrosses, petrels and diving petrels.
Proc. Calif. Acad. Sci. (4) 2, pt. 2.
MacDonald J. D. and Lawford P. A. 1954. Sight Pecords of
Birds in the Pacific: Compiled from the Bird Log Kept during the Recent Grui-
ses of H. M. S. Challenger. The Emu, vol. 54, n. I.
MacDonald D. and Green R. H. 1963. Albatross Island. The*
Emu, vol. 63, n. 1.
Maher W. J. 1962. Breeding biology of the Snow petrel near Cape Hal-
lett, Antarctica. The Condor, vol. 64. n. 6.
M a к a t s c h W. 1969. Wir bestimmen die Vogel Europas. Neumann Ver-
lag.
Marr G. W. S. 1962. The Natural History and Geography of the Antarc-
tic krill (Euphausia superba Dana). Discovery Reports, vol. 32.
Marshall A. J. and Serventy D. L. 1956. The breeding cycle of
the Short-tailed shearwater, Puffinus tenuirostris (Temm), in relation to trans-
equatorial migration and its environment. Proc. Zool. Soc. Lond. vol. 127, p. 4.
Marshall A. J and Serventy D. L. 1959. Experimental demon-
stration of an internal rhythm of reproduction in a trans-equatorial migrant
(the short-tailed shearwater Puffinus tenuirostris). Nature, vol. 184.
Martin P. W. 1942. Notes on some pelagic birds of the coast of British-
Columbia. The Condor, vol. 44, n. 1.
Marumo R. 1957. On the distribution of plankton settling volumes in-
the neighbouring seas of Japan. I—II. Bull. Japan Soc. Sci. Fish. vol. 23, n. 4.
Marumo R., К i t о u M. and A s а о к a O. 1954. The productivity
and its seasonal variations of main plankton groups in the open sea. J. Oaea-
nogr. Soc. Japan. 10, n. 4.
Matsudaria Ch., Iwasaki H., and Tsuda T. 1956. The
Quantitative Ecology of the Marine Populations in the Boundary Zone. Bull, ol
the Japan. Soc. of Scien. Fish. vol. 22, n. 3.
Matthews L. H. 1929. Birds of South Georgia. Discovery Reports, I.
Cambridge. •
Мауг E. 1953. On the origin of bird migration in the Pacific. Proceed,
seventh Pacif. Scien. Congr.
McHugh J. L. 1955. Distribution of black-footed albatross, D. nigripes
on the west coast of North America, 1949 and 1950. Pacif. Sci., 9, 4.
370
М с 11 w a i п е С. P. 1964. Fluctuations in the Number of Giant Petrels
at Ngauranga, Wellington Harbour, New Zealand. The Emu, vol. 64, n. 1.
Messershmith J. D., Baxter J. L. and Roedal P. H. 1969.
The anchovy Resources of the California Current Region off California and
Baja California. Calif. Cooper. Oceanic. Fish. Investig. Reports, vol. 13.
Miller L. 1940. Observations on the black.-footed albatross. The Condor,
vol. 42, n. 2.
Mills J. A. R. 1958. A die-off of shags in north Otago. — Notornis.
8, n.l
Mills J. A. R. 1969. The distribution of breeding red-billed gull colonies
in New Zealand in relation to areas of plankton enrichment. Notornis. 16, n. 3.
Miyasaki M. and Abe S. 1960. On the Water Masses in the Tsu-
shima Current Area. J. Oceanogr. Soc. Jap. 16, n. 2.
Moreau R. E. 1950. The breeding seasons oi African birds. 2. Sea
birds. The Ibis, vol. 92, n. 3.
Morgan B. and Morgan J. 1965. Some notes on birds ob the Fiji
Island. Notornis, vol. 12, n. 3.
Morzer Bruyns W. F. J. and Voous К. H. 1964. White-faced
Storm Petrels (Pelagodroma marina) in the Indian Ocean. Ardea, 52.
Morzer Bruyns W. F. J. and Voous К. H. 1965. Great Skuas
(Stercorarius skua) in Northern Indian Ocean. Ardea, 53.
Motoda S. and Fujii T. 1956. Report from the Oshoro Maru on
Oceanographic and biological investigations in the Bering Sea and Northern
North Pacific in the summer of 1955. Bull. Fac. Fish. Hokkaido Univ., 6, 4.
Motoda S. and Marumo P. 1963. Plankton of the Kuroshio water.
Proceeding of Simposium on the Kuroshio, 1963. October, 29, Tokyo.
Mukai M. 1958. On the Relation between Sardine and Sea Conditions in
the West Sea Area of Kyushi. J. Meteorol. Res., vol. 10, n. 8.
Munro G. C. 1960. Birds of Hawaii. Bridgewav Press.
Murie O. J. 1959. Fauna of the Aleutian Island and Alaska Peninsula.
Washington.
Murphy R. C. 1936. Oceanic birds of South America, vol. 1. 2. New
York.
Murphv R. C. 1959. History of the penguins, part II. Natur. History,
68, n. 3.
Murphy R. C. 1964. Systematics and distribution of Antarctic petrels.
Biol, antarctique, Paris.
Murphy G. and Ik eh ar a I. 1955. A Summary of Sightings of Fisn
Schools and Bird Flocks and Trolling in the Central Pacific. U. S. Fish and
Wildl. Serv. Spec. Scien. Rep.: Fisheries, n. 154.
Murray M. D. 1962. The. wandering albatross. Austral, Natur. Hist.
14, n. 3.
Murray M. D. and Carrick R. 1960. Seasonal movements and
habitats of the silver gull, Larus novaehollandiae Stephens, in southeastern
Australia. C. S. I. R. O. Wildlife Res., 9, n. 2.
Nakamura T. 1963. Distribution of the black-footed albatross in the
north Pacific Ocean. Misc. Repts. Yamashinas Inst. Ornithol. and Zool., 3, 4.
Nelson B. 1966. The man-o-war bird. Natur. History, vol. 75, n. 5.
Nelson M. O. and Larkins H. A. 1970. Distribution and Biology
of Pacific Hake. A Synopsis. Pacific Hake. U. S. Fish and Wilblife serv. Bu-
reau commercial Fish., Circular 332.
Norman J. R. 1938, Coast fishes. РаИ. III. The Antarctic Zone. Disc.
Rep., 18.
Norris A. Y. 1965. Obsevations of seabirds in the Tasman Sea and in
New Zealand waters in October and November, 1962. Notornis, vol. 12, n. 2.
Odum E. P. 1959. Fundamentals of ecology. Philadelphia-London, W. B.
Sounders Co.
37!
О 1 i v е г W. R В. 1955. New Zealand birds, 2 nd edition, Wellington.
Oordt G. J. Van and Kruijt J. P. 1953. On the pelagic distri-
bution of some Procellariiformes in the Atlantic and Southern Oceans. The
Ibis, vol. 95, n. 4.
Orlando C. 1958. Cattura di un Albatross Urlatore (Diomedea exulans
exulans) in Sicilia. Riv. Ital. Orn., 28, n. 2.
Orthmann A. 1896. Grundzuge der marine Tiergeographie. Jena.
Ozawa К.. Ko take J. and Matsuike K. 1966. Observations of
Birds in the Indian Ocean (I). J. Tokyo Univ. Fish., vol. 8, n. 1.
Ozawa K. and Nakamtfra Y. 1966. Observations of birds in
the Indian Ocean (II). J. Tokyo Univ. Fish., vol. 8, n. 2.
Ozawa K., Yamada T., Kira M. and Shimizu T. 1968».
Observations of sea-birds in the Southern Ocean (III). J. Tokyo Univ. Fish.,
vol. 9, n. 2.
Paxton R. O. 1968. Wandering Albatrc^s in California. The Auk, vol.
85, n. 3.
Pearse Theed 1963. Pesults from banding -Glaucous-winged gulls in
the northern gulf of Georgia, В. C. from 1922 to 1949. Bird-Band, vol. 34, n. 1.
Pearson A. J. 1966. The birds of Christmas Island (Indian Ocean).
Bull. Brit. Ornithol. Club., 86, n. 4.
Pearson T. H. 1968. The feeding biology of sea-bird species breeding
on the Fame Islands. Northumberland. J. Animal Ecol., vol. 37, n. 3.
Phillips A. 1961. Recovery in Victoria of a fairy penguin, Eudyptala
minor novaeftollandia (Forster), banded in Southern Tasmania. Austral. J. Sci.,
23, n. 7.
Phillips J. H. 1963. The pelagic distribution of the sooty shearwater Pro-
cellaria grisea. The Ibis, vol. 105, n. 3.
Phillips W. W. A. 1950. Wilsons Stormpetrels, Shearwaters and other sea
Birds in the Gulf of Aden and Indian Ocean. J. Bombay Nat. Hist. Soc., 49
Phillips W. W. A. 1951. On the occurrence of the slender-billed she-
arwater (Puffinus tenuirostris) in Ceylon. Spolia Zeylanica, vol. 26, p. 2.
Phillips W. W. A. 1954. Petrels, shearwaters and other oceanic birds tn
the north Indian Ocean. J. Bombay Nat. Hist. Soc. 52.
Phillips W. W. A. 1963. The Birds of the Maidive Islands, Indian
Ocean. Journ. of the Bombay Natur. History Soc. vol. 60, n.3.
Pocklington R. 1965. A summary of birds seen over the western In-
dian Ocean. — August — November 1963. The Ibis, vol. 107, n. 3.
Pocklington R. 1967. Birds seen at sea off N. W. Australia. July
23—30, 1965. The Emu, vol. 76, n. 1-2.
Prevost J. 1964. Quelques aspects de Tecologie des procellariens an-
tarctques. Sci. et nature, n. 62.
Prevost J. 1964a. Remorques fccologiques sur quelques procellariens
antarctiques. Oiseau et Rev. franc, ornitol., Й, Num. spec.
Radford M. C., Mrs. 1960. Common gull movements shown by
ringing returns. Bird Stuby, vol. 7, n. 2.
Rand R. W. 1959. The biology of guanoproducing sea-birds. Commerce
and Industri, Investig. rep. n. 39.
Rand R. W. 1960. The biology of guanoproducig sea-birds. Commerce
and Industry, Investig. rep., n. 41.
Rand R. W. 1960a. The biology of guanoproducing sea-birds. Commerce
and Industry, Investig. rep., n. 42. •
Rand R. W. 1962. Sea-birds south of Madagascar. Ostrich, vol. 33, n. 3.
Rand R. W. 1963. Seabirds in the southern Indian Ocean. Ostrich, vol.
34, n. 3.
Rand R. W. 1963a. The biology oi guanoproducing seabirds. Commerce
and Industry, Investig. rep. n. 43.
.'372
Rand R. W. 1963b. The biology of guanoproducing sea-birds. Commerce
>and Industry, Investig. rep. n. 46.
Rankin N. 1951. Antarctic islet. Collins, London.
Regan С. T. 1914. Fishes. British Antarct. (’’Terra Nova") Exp. 1910.
Nat. Hist. Rep., Zool. 1.
Reid B. 1962. An assessment of the size of the Cape Adare Adelie Penguin
rookery and skuary-with notes on petrels. Notornis, vol. 10, n. 3.
Rice D. W. and Kenyon K. W. 1962. Breeding distribution, histo-
ry and populations of North Pacific albatrosses. The Auk, vol. 79, n. 3.
Rice D. W. and Kenyon K. W. 1962a. Breeding cycles and be-
havior of laysan and blackfooted albatrosses. The Auk, vol. 79, n. 4.
Richardson F. 1957. The breeding cycles of Hawaiian sea birds. Bull.
В. P. Bishop. Museum, n. 218.
Rich d ale L. E. 1952. Post-egg period in albatrosses, Bjol. Monographs
n. 4, Otago Daily Times, etc., Dunedin.
R i c h d a 1 e L. E. 1963. Biology of the sooty shearwater Puffinus gri-
seus. Proceed, of the Zoological society of* London, vol. 141, p. 1.
Richdale L. E. 1964. Notes on the mottled petrel, Pterodroma inex-
pectata and other petrels. The Ibis, vol. 106, n. 1.
Richmond K. 1967. An albatross in Scotland. Animals, vol. 10, n. 6.
Ridley M. W. 1957. Terns of the Seychelles island. J. Bombay Natur.
History Soc., 54, n. 4.
Ridley M. W. and Percy L. R. 1958. The exploitation of Sea
Birds in Seychelles. Colonial Res. Stud. n. 25.
Robertson C. J. R. 1964. 14 th Annual. Report of the New Zealand
Bird Banding Scheme for the Year ending 31 st March 1964, Dominion Mu-
seum. Wellington.
Robins R. C. 1958. Observations on oceanic birds in the Gulf of Pa-
nama. The Condor, vol. 60, n. 5.
Robins J. P. 1962. Synopsis of Biological Date on Bluefin tuna Thun-
nus thynnus maccoyii (Castelnau).
Rochford D. J. 1962. Hydrology of the Indian Ocean II. The surface
water of the South-East Indian Ocean and Arafura Sea in Spring and Sum-
mer. Austr. J. Marine and Freshwater Res., vol. 13, n. 3.
Roughley T. C. 1961. Fish and Fisheries of Australia. Angus Robert-
son. Holstead Press, Sydney.
Routh M. 1949. Ornithological observations in the antarctic seas,
1946—47. The Ibis, vol. 91, n. 4.
Rown M. K. 1965. Regulation of Sea-Bird numbers. The Ibis, vol. 107, n. к
Salomonsen F. 1965. The geographical variation of the Fulmar (Ful-
marus glacialis) and the zones of marine environment in the North Atlantic.
The Auk. vol. 82, n. 3.
Salomonsen F. 1967. Migratory movements of the Arctic Tern (Ster-
na paradisea Pont.) in the Southern ocean. Det Kong. Danske Videns.
Biol. Medd., 24, n. 1.
Sanger G. A. 1970. The seasonal distribution of some seabirds off
Washington and Oregon, with notes on their ecology and behavior. The Condor,
vol. 72, n. 3.
Schaefer M. B. 1965. The potential harvest of the sea. Trans. Amer.
Fish. Soc., vol. 94, n. 2.
Schoener T. W. 1965. The evolution of bill size differences among
sympatric congeneric species of birds. Evolution, vol. 19, n. 1.
Serventy D. L. 1935. Birds in the Great Australian Bight. The Emu,
vol. 35, n. 1.
Serventy D. L. 1941. Mating in petrels. The Emu, vol. 41, n. 1.
373
Serventy D. L. 1952 The Bird Island of the Sahul Shelf. The Emu
vol. 52. n. 1.
Serventy D. L. 1952a. Movements of the Wilson Storm-Petrel on Aust-
ralian Seas. The Emu, vol. 52, n. 2. „ , t .. j
Serventy D. L. 1953. Movements of pelagic sea-birds in the Indo-
Pacific region. Proc. 7 th Pacific Science Congress, 4.
Serventy D. L. 1957. The banding programme on Puffinus tenuirist-
ris (Temm.). 1. First report C, S. I. R. O. Wildlife Research, vol. 2, n. 1.
Serventy D. L. 1958. Recent studies on the Tasmanian mutton-bird.
Austral. Museum Mag. 12, n. 10. '
Ser ventv D. L. 1961. The banding programme on Puffinus tenuirost-
ris (Temm). 2.’Second report. C. S4 1. R. O. Wildlife Research, vol. 6, n. 1.
S e r v e n t у D. L. 1967. Aspects of the population ecology of the Short-
tailed shearwater Puffinus tenuirostris. Proc of the 14 Internal. Ornithol.
Congress. >
Serventy D. L. and Whittell H. W. 1951, Birds of Western
Australia. Perth, W. A.
Sergent D. E. 1951. Ecological Relationships of the Guillemots Uria
aalge and Uria lomvia. Proc, of the 10 Intern. Ornithol. Congress.
S e 11 e О. E. 1955. Consideration of midocean fish production as related
to ocean circulatory systems. J. Marine Res., vol. 12, n. 4.
S h a r 1 a n d M. 1956. Population rise in two sea -birds. The Emu, vol.
56, n. 1.
Shaughnessy P. D. 1971. Frequency of the white phase of the sout-
hern giant petrel, Macronectes giganteus (Gmelin). “Austral. J. Zoo!.”, 19, n. I.
Sibley F. C., Clapp К. B. 1967. Distribution and dispersal of
Central Pacific Lesser Frigatebirds Fregata ariel. The Ibis, vol. 109, n. 3.
Sibley F. C. and McFarlane W. 1968. Gulls in the Central
Pacific. Pacific Science, vol. 22, n. 3.
Sibson R. B. 1951. A Tasman Bird Log. Notornis, vol. 4, n. 6.
S i b s о n R. B. 1969. Giant Petrels as migrants to northern New Zea-
land. Notornis, vol. 16, n. 1.
Siple and Lindsey. 1937. Ornithology of the Second Bird Antar-
ctic Expedition. The Auk, vol. 54, n. 2.
S 1 a d e n W. J. L. 1952. Arctic skua in the Antarctic. The Ibis, vol.
94, n. 3.
S 1 a d e n W. J. L. 1954. Pomarine skua in the Antarctic. The Ibis, vol.
vol. 96, n. 2.
S laden W. J. L. 1965. Ornithological reseach in Antarctica. Bio Sci-
ence, vol. 15, n. 4.
SI a den W. J. L. and Tickell W. L. N. 1958. Antarctic Bird-
banding by the Falkland Islands dependencies survey, 1945—1954. Bird-Ban-
ding, vol. 29, n. 1.
Sladen W. J. L, Wood R. C. and Monagham E. P. 1968.
Th USARP bird banding Program, 1958—1965. Antarstic Res. series, vol. 12.
S 1 i p p J. W. 1952. A record of the White-capped Albatross (Diomedea cau-
ta cauta) in American North Pacific Waters. The Auk, vol. 69, n. 4.
Smith W. J. 1959. Movements of Michigan herrning gulls. Bird-Band,
30, h. 2.
Smith W. J. 1962. Drifting gulls. Natur. History, 71, n. 1.
Smith N. G. 1966. Evolution of some arctic gulls (Larus) an experi-
mental study of isolating mechanisms. Ornithol. monograph, n. 4. Publ. Amer.
Ornithol. Union.
Sorensen J. H. 1364. Birds of the Kermadec Islands. Notornis, vol.
11, n. 2.
374
Spencer R. 1962. British-ringed Manx Shearwater recovered in Austra-
lia. Brit Birds, 55, n. 2.
Spencer R. 1963. The bird ringing scheme, 1958—1961. Bird Study,
10, n. 2.
Stein P. 1961. Winter occurrence of juvenile gannets in New Zealand
Waters. Notornis, vol. 9, n. 5.
Stein P. 1962. Winter sighting of gannets. Notornis, vol. 9, n. 8.
Stonehouse B. 1958. Notes on the ringing and the breeding distri-
bution of the Giant petrel Macronectes giganteus. The Ibis, vol. 100, n. 2.
S t о r r G. M. 1964. The Avifauna of Rott nest Island Western Australia.
I. Marine Birds. The Emu, vol. 64, n. 1.
S t о г г G. M. 1964a. Zonation and Seasonal Occurrence of Marine Birds
in the Seas off Fremantle, Western Australia. The Emu, vol. 63, n. 4.
Storer R. W. 1952. A comparison of variation, behaviour and evolu-
tion in the sea-bird genera Uria and Cepphus. Univ. Calif. Publ. Zool. 52.
Storer R. W. 1960. Evolution of the diving birds. Proc. 12 Intern. Orni-
thol.Congr. Helsinki.
Strange I. F. 1965. The Falkland Islands — a south atlantic bird ha-
ven. Animals, 6, n. 20.
Strickland J. D. 1967. The ecology of the plankton off La jolla, Ca-
lifornia, in the period april through September. Bull. Scrips. Inst. Ocean.,
vol. 17.
Strong P. 1964. Buller's Albatross: an Addition to the Australian List.
The Emu, vol. 64, n. 2.
Summerhayes С. P. 1969. Seabirds seen in the northern Tasman sea
in winter. N. Z. Journ. Mar. and Freshw. Res. 3, n. 4.
Swales M. K. 1965. The sea-birds of Gough Island. The Ibis,
vol. 107, n. 2.
Swartz L. G. 1966. Sea-cliff birds. Chap. 23, pp. 611—678. In: Environ-
ment of the Cape Thompson Region, Alaska, U. S. Atomic Energy Commission,
Oak Ridge.
Swartz L. G. 1967. Distribution and Movements of Birds in the Bering
and Chukchi Seas. Pacific Science, vol. 21, n. 3.
S z i j j L. J. 1967. Notes on the winter distribution of Birds in the wes-
tern Antarctic and adjacent Pacific waters. The Auk, vol. 84, n. 3.
Thomas D. G. 1967. The Dominican Gull in Tasmania. The Emu,
vol. 66, n. 3.
Thomson A. L. 1965. The transeguatorial migration of the manx she-
arwater (Puffin des anglais). Oiseau Rev. franc, ornithol. 35, num. spec.
Thomson A. L. 1966. An anlysis of recoveries of Great Skuas ringed
in Shetland. Brit. Birds., vol. 59, n. 1.
Thompson D. Q. 1951. Notes on distribution of the North Pacific al-
batrosses. The Auk, 68, n. 2.
Thrailkill J. R. 1963. Zooplankton volumes off the Pacific Coast,
1959. U. S. Fish, and Wildl. Serv. Spec. Scien. Rep. Fish. n. 414.
Tickell W. L. N. 1962. Report of the bird-banding expedition to Bird
island South Georgia 1960—61. Ring. Internal Ornithol. Bull. n. 30—31.
Tickell W. L. N. 1964. Feeding preferences of the albatrosses D. me-
lanophris and D. chrysostoma at South Georgia. Biol, antarctique. Paris.
Tickell W. L. N. 1967. Movement of Black-browed and Grey-headed
Albatrosses in the South Atlantic. The Emu, vol. 66, n. 4.
Tickell W. L. N. 1968. The biology of the Great Albatrosses, D. exu-
lans and D. epomophora. Antarctic Res. Series, vol. 12.
Tickell W. L. N. 1968a. Colorgyeing Albatrosses. Bird-Banding,
vol. 39, n. 1. •
375
. Tickell W. L. N. and Gibson J. D. 196& Movements of Wande-
ring Albatrosses Diomedea exulans. The Emu, vol. 68. n. 1.
Tickell W. L. N. and Pinder R. 1967. Breeding frequencies in
the albatrosses D. melanophris and D. chrysostoma. Nature, 213.
Tickell W. L. N. and Scotland C. D. 1961. Recoveries of rin-
ged giant petrels Macronectes. giganteus. The Ibis. vol. 103a, n. 2.
Tranter D. J. 1962. Zooplancton abundance Australasian water. Aust-
ral J. Marine and Freshwater Res., vol. 13, n. 2.
Tsujita T. 1957. Fisheries oceanography of the East China Sea and
Tsushima Strait. 1. Hydrographic Structure of the fishing grounds. Bull. Seikai
Peg. Fish. Res. Lab. n. 13.
Tsujita T. 1961. A Brief Report on Progress in the Studies on the
Exploeted Pelagic Fish Populations in the Waters off Western Japan during
jthe Years 1950—1959. Bull. Seikai Reg. Fish. Res. Lab., n. 21.
Tsujita T. 1963. The fisheries oceanography of the East China Sea
and the Tsushima Strait. Bull. Seikai Reg. Fish Res. Lab., n. 28.
Tsujita T. and Kondo M. 1957. Some contributions to the ecology
of the mackerel and the oceanography of the fishing grounds in the East China
Sea. Bull. Seikai Reg. Fish. Res. Lab., n. 14.
Tuck L. M. 1960. The murres, their distribution, populations and biology.
Ottawa.
Uda M. 1956. Research on the fluctuation of the North Pacific circula-
tion. Res. Oceanogr. Works Japan, vol. 2, n. 2.
Uda M. 1963. Oceanography of Subarctic Pacific Ocean. Journal Fish.
Res. Board of Canada, 20. *
Uda M. and Ishino 1958. Enrichment pattern resulting from eddy
systems in relation to fishing graunds. J. Токио Univ. Fish. 44 (1—2).
U d v a r d у M. D. F. 1963. Zoogeographical study of the Pacific Alci-
dae/ Pacific Basin Biogeogr. Honolulu.
U d v a r d у M. D. F. 1969. Dynamic Zoogeography. V. N. R. Company,
New York.
Ullmann H. S. 1963. The most prosperous bird colony. Audubon Mag.,
vol. 65, n. 3.
Voisin J. F. 1969. L‘albatros Hurleur Diomedea exulans a rile de la
Possession. Oiseau et Reiv. franc, ornithol., 39, num. sp4c.
Voous К. H. and Wattel J. 1963. Distribution and migration of
the greater shearwater. Ardea, vol. 51, n. 2—4.
Waldron K. D. 1964. Fish schools and Bird Flocks in the central Pa-
cific Ocean, 1950—1961. U. S. Fish and Wildl. Serv. Spec. Scien, Rep.: Fishe-
ries, n. 464.
W a r h a m J. 1956. The breeding of the great-winged petrel Pterodroma
macroptera. The Ibis, vol. 98. n. 2.
Warham J. 1958. The nesting of the shearwater Puffinus carneipes. The
Auk, vol. 75, n. 1.
W a r h a m J. 1962. The biology of the Giant petrel Macronectes gigan-
teus. The Auk, vol. 79, n. 2.
Waterston G. 1968. Black-browed albatross on the Bass Rock. Brit.
Birds, vol. 61, n. I.
Westerskov K. I960. Birds of Campbell Island. Dept. Internal Alfa-
irs. Wildlife Pabl. 61. Wellington. , t
Westerskov K. 1960a. Field indentification and sex determination of
the Royal albatross. Notornis, vol. 9, n. 1.
Westerskov K. 1963. Ecological Factors Affecting Distribution of a
Nesting Royal Albatross Population. Proc. XIII Intern. Ornithol. Congress,
vol. 2.
376
Wetmore A. 1940. A check-list of the fossil birds of North America.
Smith. Inst. Misc. Coll., n. 4.
Wetmore A. 1951. Recent additions to our knowledge of prehistoric
birds, 1933—1949. Proc. X Intern. Ornithol. Congr. Upsala, 1950.
Wheeler W. R. and Watson 1. 1963. The Silver Cull Larus no-
vaehollandiae Stephens. The Emu, vol. 63, n. 2.
Wilhoft D. C. 1961. Birds observed during two crossings of the Pa-
cific Ocean. The Condor, vol. 63, n. 3.
Williams G. G. 1952. The origins and dispersal of oceanic birds. Te-
xas J. Sci., 4.
Williams G. R. 1960. — The birds of the Pitcairn islands central
South Pacific ocean. The Ibis, vol. 102, n. 1.
Wodzicki K. 1967. The Gannets at Cape Kindnapper. 2. Dispersal and
Movements. Trans. Royal Soc. N. Z. Zoology, vol. 9, n. 2.
Wodzicki K. and Moreland J. 1966. A note on the food of
New Zealand Gannets. Notornis, vol. 13, n. 2.
Wodzicki K. and Stein P. 1958. Migration and Dispersal of New
Zealand Gannets. The Emu, vol. 58, n. 4.
Wynne-Edwards. 1962. Animal Dispersion in Relation to Social Be-
haviour. Edinburgh: Oliver a. Boyd.
Wyrtki K. 1961. Physical Oceanography of the Southeast Asian Waters.
Naga Report, vol. 2.
Wyrtki K. 1962. The upwelling in the region between Java and Aust-
ralia during the South-East Monsoon. Austr. J. Marine and Freshw. Res. vol.
13, n. 3.
Yonge С. M. 1963. The biology of coral reefs. Advances Marine Biol.,. 1.
Young E. C. 1963. Feeding hubits of the South Polar Skua Catharacta
maccormicki. The Ibis, vol. 105, n. 3.
Young E. C. 1963a. The breeding behaviour of the South Polar Skua
Catharacta maccormicki. The Ibis. vol. 105. n. 2.
ОГЛАВЛЕНИЕ
Введение ......................................................... 3
Глава I. Очерки по распределению и миграциям отдельных видов мор*
ских птиц...........................................................8
1. Северотихоокеанские альбатросы...............................8
2. Альбатросы Южного океана....................................22
3. . Гигантский буревестник.................................. 46
4. Капский голубь..............................................5!
5. Тонкоклювый и серый буревестники............................56
6. Глупыш......................................................68
7. Обыкновенная моевка.........................................79
8. Толстоклювая и тонкоклювая кайры........................... 83
. 9. Топорок.....................................................92
Глава П. Сезонные аспекты в орнитофауне различных районов Тихого
и Индийского океанов ...............................98
1. Берингово море..............................................98
2. Охотское море...............ч.................................Н5
3. Японское море..............................................124
4. Северная часть Тихого океана (к северу от 30° с. ш.).......140
Б. Восточно-Китайское море.....................................156
6. Тропические воды между северным побережьем Австралии
и Зондскими островами.........................................166
7. Шельф южного побережья Австралии и прилегающие воды . . <176
8. Район Новой-Зеландии и сопредельные воды Антарктики < < • 185
Глава III. Некоторые общие закономерности в распределении и мигра-
циях морских птиц.................................................293
1. Особенности сезонных миграций морских птиц в различных
климатических зонах Тихого океана.............................206
2. К вопросу о пролетных путях морских птиц.....................234
3. Сухопутные птицы в море.....................................
Глава IV. Распределение птиц в связи с межвидовой конкуренцией за
пищу.................................................................246
Глава V. Орнитогеографическое деление Мирового океана .............263
1. Границы орнитогеографическях регионов в Тихом и Индийском
океанах 264
2. Место орнитогеографическжх регионов в системе
биогеографических областей ....................................272
3. Некоторые закономерности географического распространения
морских птиц ...................................................285
4. О необходимости сочетания при биогеографическом районировании
экологогеографическех, исторических и фаунистических данных . 291
5. Предпосылки к орнитогеографическому районированию океана
на уровне провинций.............................................312
Глава VI. Количественное распределение морских птиц и биологическая
структура океана.....................................................321
1. Широтная зональность и симметрия в распределении плотности
населения морских птиц...........................................322
2. Численность морских птиц и нх роль как потребителей
других морских животных....................................... . 333
Заключение . 345
Литература . 349
ВЯЧЕСЛАВ ПЕТРОВИЧ ШУНТОВ
МОРСКИЕ ПТИЦЫ
И БИОЛОГИЧЕСКАЯ СТРУКТУРА ОКЕАНА
Редактор В. Чашкова. Техмич. редактор И. Маслов
Корректоры В. Ладинова, В. Анисимова
ВД 00201 от 26 сентября 1972 г. Сдано в набор 29.11.1972 г.
Подписано к печати 26.IX.1972 г. -Формат бОХЭО1/^ Объем 23,75 печ. л.
Уч.-изд. л. 33,5. Заказ 683. Тираж 600. Цена 1 руб. 66 коп.
Дальневосточное книжное рздательство Комитета по печати
при Совете Министров РСФСР. Владивосток, Ленинская, 43.
Типография издательства «Боевая вахта», Владивосток, Посьетская, 22.