





























































































































































































































Author: Лихарев И.М. Виктор А.Й.
Tags: mollusca моллюски биологические науки в целом зоология фауна ссср
Year: 1980




Text
ФАуНА
СССР
МОЛЛЮСКИ
III
выпуск
5
ЛКАЛЕМИЯ Н АУК ССС Р
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ
НОВАЯ СЕРИЯ, № 122
ФАУНА СССР
МОЛЛЮСКИ
Том III, ВЫП. 5
И. М. ЛИХАРЕВ и А. Й. ВИКТОР
СЛИЗНИ ФАУНЫ СССР И СОПРЕДЕЛЬНЫХ СТРАН
(GASTROPODA TERRESTRIA NUDA)
в
ЛЕНИНГРАД
«НАУК А»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
1980
УДК 594.382(47+57} (983.71)
Слизни фауны СССР и сопредельных стран (Gastropoda terrestria nuda). Лиха-
рев И. М., Виктор А. Й. (В серии: Фауна СССР. Моллюски. Т. Ш, вып. 5). Л.,
«Наука», 1980. 438 с.
Комплекс наземных легочных моллюсков, лишенных наружной раковины и объединяемых
понятием слизни — это сборная группа, многие виды которой являются опасными вре-
дителями сельского хозяйства как на открытом, так и на закрытом грунте. В книге пред-
ставлен систематический обзор всех видов, обитающих на- территории СССР и сопредель-
ных стран. Установлено 102 вида, принадлежащих к 29 родам и 8 семействам. На базе
морфо-функционального сравнения строения слизней и раковинных улиток выявлены
черты параллелизма в организации слизней; впервые сделана попытка обосновать фило-
генез родов и семейств слизней и их положения в пределах надотряда стебельчатоглазых
брюхоногих моллюсков; предлагается новая классификация слизней Голарктики. Да-
ется обзор основных особенностей физиологии и экологии слизней, особенно видов, яв-
ляющихся вредителями сельского хозяйства. Приводятся таблицы для определения
семейств, родов и видов. Лит. — 488 назв., табл. — 2, ил. — 576.
THE FAUNA OF SLUGS OF THE USSR AND ADJACENT COUNTRIES
(GASTROPODA TERRESTRIA NUDA)
I. M. LIKHAREV A. WIKTOR
(Zoological Institute (Museum of Natural History
of Academy of Sciences, of Wroclaw University)
Leningrad)
Главный редактор серии «Фауна СССР»
директор Зоологического института АН СССР'
О. А. Скарлато
Редакционная коллегия:
И. М. Лихарев (отв. ред. серии), О. Л. К рыжановский (зам., отв. редактора),.
3. И. Баранова, И. М. Громов, В. Ф. Зайцев, Л. А. Кутикова,
Г. С. Медведев, М. Е. Тер-Минасян, Н. А. Филиппова
Ответственный редактор
Я, И. Старобогатов
_ 21008-682
Ф' „ . ™ о 536-80 2005000000
055 (02)-80
© Издательетво «Наука», 1980 г-
ПРЕДИСЛОВИЕ
Среди брюхоногих моллюсков (Gastropoda), большинство которых об-
ладает наружной раковиной, нередко очень характерной для соответствую-
щего семейства, рода и вида, встречаются отдельные роды и семейства,
утратившие ее и в связи с этим обычно называемые слизнями, так как тело
их очень скользкое из-за обильной слизи, выделяемой их покровами.
Если наземные улитки благодаря разнообразным и нередко красивым
раковинам с давних пор привлекали взимание как зоологов, так и коллек-
ционеров-любителей, то сухопутные слизни с их довольно однообразной
и нередко отталкивающей внешностью не пользовались такой популярностью
и являлись «пасынками» у малакозоологов. Другая причина более слабой
по сравнению с улитками изученности слизней — это необходимость при
их исследовании и диагностике широко использовать анатомию, так как
многие важные таксономические признаки связаны с особенностями внут-
реннего строения.,
, Между тем изучение слизней имеет большое как теоретическое, так и
практическое значение. Будучи гетерогенной группой, объединяющей комп-
лекс жизненных форм разного происхождения, слизни представляют собой
прекрасный пример параллельной эволюции систематически далеких друг
от друга групп. Среди 77 семейств брюхоногих легочных моллюсков отряда
'Geophila известны 11 семейств, в основном или полностью состоящих из
слизней, и, кроме того, отдельные «слизневые» роды в семействах, в основном
•состоящих из улиток. Таким образом, слизни как морфологический тип
строения широко распространены в пределах указанного отряда и изучение
их с позиций эволюционного учения представляет несомненный интерес.
Кроме того, эти животные являются превосходными модельными объек-
тами при исследовании различных сторон физиологии наземных моллюсков,
в некоторых случаях более удобными, чем улитки.
Среди многих видов слизней отчетливо прослеживается тенденция
к синантропии. А так как это обычно растительноядные полифаги, то боль-
шинство их являются серьезными вредителями многих культурных расте-
ний, в частности столь важных для человека, как пшеница, рожь, многие
огородные и полевые культуры, цитрусы и некоторые технические культуры.
С другой стороны, некоторые слизни служат передатчиками (промежуточ-
ными хозяевами) целого ряда гельминтозов, опасных для многих домашних
животных. Поэтому сведения об их экологии, физиологии и фауне необхо-
димы для эффективного регулирования и прогноза численности популяций
этих вредных видов..
Первые серьезные попытки классифицировать отдельные группы слиз-
ней относятся к концу XIX и началу XX века. Особенно значительный
вклад в познание этих животных сделал Зимрот (Н. Simroth, 1851—1917).
Его перу принадлежит большой список работ, посвященных слизням мно-
гих стран и континентов нашей планеты, но особенно много им сделано в по-
знании фауны Евразии. Среди этих работ бесспорно самой значительной
является монография «Фауна слизней Российской империи», изданная на
немецком языке в Петербурге Российской академией наук в 1901 г. До вы-
4
ПРЕДИСЛОВИЕ
хода этой сводки о слизнях России было известно очень мало, главным
образом из работ Калениченко (Kaleniczenko, 1851а, Ь) и О. Бётгера (Boett-
ger О., 1881—1889). Как и некоторые другие, менее значительные исследова-
ния, где в числе наземных моллюсков европейской части России упоминались
некоторые виды слизней, эти сообщения основывались на определении слиз-
ней по внешнему виду, без попытки использовать данные по их анатомии.
В упомянутой монографии Зимрот охарактеризовал как внешний облик,
так и внутреннее строение исследованных видов. Особенное значение в плане
диагностики и классификации он придавал окраске тела и строению гени-
талий. Для написания этой книги Зимрот использовал коллекционный
материал, в основном полученный из Зоологического музея Российской
академии наук (ныне Зоологический институт Академии наук СССР —
ЗИН АН СССР), а также немногие сборы из Зенкенбергского музея
(Франкфурт-на-Майне) и Гельсингфорского музея (ныне Зоологический
музей Хельсинкского университета). После выхода упомянутой моногра-
фии Зимрот опубликовал три работы меньшего объема по фауне слизней
Кавказа и Средней Азии, тоже в основном по материалам Петербургского
зоологического музея (1910а, 1912а, Ь), которые послужили дополнением
к его монографии.
Эти исследования вместе с тем, что было известно о слизнях западной
и средней Европы (в значительной степени тоже благодаря Зимроту), впервые
дали сравнительно четкую характеристику фауны сухопутных слизней не-
тропических областей Евразии. Вместе с тем выяснился значительный
эндемизм этой фауны на Кавказе и в Средней Азии. Основные выводы Зим-
рота в области классификации палеарктических семейств слизней дали воз-
можность Гессе (Hesse Р., 1926) уточнить и разработать классификацию
этих семейств, которой большинство исследователей придерживается и в на-
стоящее время.
Несмотря на несомненную ценность, исследования Зимрота имеют и ряд
существенных недостатков, которые проистекают из скудости изученного
материала. В связи с этим автор не мог разобраться не только в экологи-
ческой изменчивости видов, но даже в возрастных изменениях, происходя-
щих с окраской и гениталиями. Многие виды были описаны по 1—2 экземп-
лярам, нередко молодым, неполовозрелым или плохой сохранности. Не слу-
чайно составленные им таблицы для определения слизней Кавказа и Средней
Азии (1912а) нечетки и непригодны для использования.
После смерти Зимрота (1917 г.) исследования слизней в масштабе Па-
леарктики не проводились. Однако появилось немало сводок и отдельных
статей по некоторым областям Европы: Британские о-ва (Quick, 1960),
Норвегия (Okland, 1923), средняя Европа (Hudec, 1960—1970; Wiktor,
1973), юго-западная Европа (Altena, 1950—1971), юго-восточная Европа
(Grossu, 1955—1970; Grossu, Lupu, 1957—1966; Lupu, 1968—1977; Urbanski,
Wiktor, 1968; Wagner, 1929—1935). Естественно, что эти региональные ис-
следования не ставили задачей ревизию хотя бы одного семейства.
Наряду с этим опубликовано немало статей, где описания новых для науки
видов выполнены на основании изучения гениталий единичных, нередко
неполовозрелых экземпляров слизней, без попытки проанализировать из-
менчивость этих животных и возрастные изменения гениталий. Это, в част-
ности, относится к описаниям новых видов Deroceras, Limax и Lehmannia
из Румынии (см. работы Grossu и Lupu).
Главная задача авторов настоящей книги заключалась в подготовке
сводки по фауне слизней СССР и сопредельных стран, т. е. Финляндии,
Польши, Чехословакии, Румынии, Венгрии, Турции, Ирану, Афганистану,
Монголии, соседних провинций Китая и КНДР. В связи с этим были иссле-
дованы наиболее значительные коллекции: по территории СССР —
ЗИН АН СССР (наиболее крупная коллекция!), Зоологического музея
ПРЕДИСЛОВИЕ
5
Московского университета, Львовского научно-природоведческого музея
АН УССР, Института зоологии АН АрмССР, Института зоологии
АН КазССР, Института зоологии и паразитологии АН ТаджССР; по Фин-
ляндии — некоторые материалы Зоологического музея Хельсинкского уни-
верситета; по Польше — коллекции Зоологического института ПАН (Вар-
шава), Естественно-исторического музея Вроцлавского университета;
по ЧССР, Венгрии и Румынии — материалы, в том числе и типы видов,
присланные коллегами; по Турции . и Ирану — материалы, собранные
в конце XIX и в начале XX века и хранящиеся в ЗИН АН СССР и Венском
естественно-историческом музее, а также новые сборы из Лейденского музея;
по Афганистану — сборы К. Линдберга (Лихарев, Старобогатов, 1967);
по Монголии и КНДР — сборы зоологов ЗИН АН СССР и ЗИН ПАН;
по КНР — сборы конца XIX и начала XX века, хранящиеся в ЗИН
АН СССР.
Вне исследования осталась фауна слизней западных и южных областей
Европы и фауна Северной Африки, откуда авторы имеют лишь случайные
сборы. Из литературы известно, что в этих областях обитают виды трех родов
подсемейства Arioninae (из них два рода возможно подроды Arion) и неко-
торое число видов, принадлежащих к родам, рассмотренным в этой книге,—
Limax, Lehmannia (Limacidae), Deroceras (Agriolimacidae), Parmacella (Par-
mac ellidae), Milax, Tandonia (Milacidae). Слизни Балканского п-ова хотя
не включены в данную сводку, но учтены при составлении характеристик
соответствующих родов, так как они являются предметом текущего иссле-
дования А. Виктора.
В процессе изучения слизней и разработки их классификации авторы
настоящей книги стремились не только максимально учитывать экологи-
ческую и возрастную изменчивость, но и расширить диапазон исследуемых
признаков внутреннего строения, а в их оценке придерживаться экологи-
ческой и функциональной интерпретации. В частности, с новых позиций
был рассмотрен мантийный комплекс органов, а изучение гениталий не огра-
ничивалось описанием их внешнего облика, но исследовалось внутреннее
строение копулятивных органов, что давало возможность судить о меха-
низме копуляции.
Поскольку в организации слизней имеются многочисленные конверген-
ции, связанные со сходным образом жизни, при оценке систематического
положения отдельных семейств авторы стремились выявить анцестральные
признаки, не затронутые параллельной эволюцией. Благодаря этому уда-
лось не только наметить положение отдельных семейств в общей системе
отряда Geophila (Шилейко, 1979), но и сделать выводы об основных эволю-
ционных тенденциях внутри семейства.
В итоге авторы пришли к заключению, что фауна слизней Палеарктики
слагается из 4 надсемейств и 8 семейств. На изученной территории она пред-
ставлена 29 родами и 102 видами. Наиболее разнообразной фауной, богатой
эндемиками видового и родового рангов, обладают Кавказ и сопредельные
горно-лесные области Турции (Понтийские горы) и Ирана (Эльбурс). Только
здесь обитают Trigonochlamydidae. Другим центром эндемизма, но меньшим
по масштабу (2 рода, 8 видов), являются горные области Средней Азии.
К началу настоящего исследования считалось,1 что на территории СССР
обитает 125 видов, принадлежащих к 23 родам и 4 семействам. В данной
книге с той же территории указаны 70 видов, из 27 родов и 8 семейств, в том
числе 12 видов, новых для фауны СССР, и 2 вида, новых для науки. Кроме
того, описаны 4 новых рода и подрода и 3 подсемейства. В то же время 68 ви-
дов и 3 рода, упомянутые у Лихарева и Раммельмейер, рассматриваются
1 Согласно книге Лихарева и Раммельмейер (1952), с учетом работы Левушкина
и Матекина (1965),
6
ПРЕДИСЛОВИЕ
здесь как младшие синонимы; 2 рода переведены в подроды, а 3 подрода —
в ранг родов.
Следует особо отметить, что при написании этой работы авторы распола-
гали большинством типов видов, описанных Зимротом с территории СССР
и сопредельных стран, а также другими материалами по слизням, определен-
ным тем же автором. Большинство этих слизней находятся в ЗИН АН СССР,
значительно меньшая часть в Венском естественно-историческом музее и
Зоологическом музее Хельсинкского университета. К сожалению, в Зен-
кенбергском музее (Франкфурт-на-Майне) не сохранились типы зимротов-
ских видов, как не сохранилось многих типов слизней, описанных О. Бёт-
гером. По-видимому, в настоящее время ЗИН АН СССР обладает наиболее
крупной коллекцией типов видов слизней, описанных Зимротом, и других
материалов, определенных тем же автором.
За годы, прошедшие после выхода в свет последних работ Зимрота по
фауне слизней России, коллекция ЗИН АН СССР сильно пополнилась сбо-
рами по этим животным. Обработка поступлений, равно как исследование
аналогичных коллекций ЗИН ПАН и Вроцлавского музея, а также поступ-
ления от многих коллег послужили основой, на которую опираются выводы
авторов этой книги.
Работа между авторами распределялась следующим образом. «Общая часть»,
а также характеристики семейств выполнялись совместно. В «Системати-
ческой части» совместно написаны разделы, посвященные родам Deroceras
и Eumilax. Роды Krynickillus, Mesolimax, Boettgerilla, Milax, Tandonia,
Meghimatium и Arion написацы А. Й. Виктором; остальные группы —
И. М. Лихаревым. Им же составлена таблица для определения семейств и
осуществлена стилистическая унификация всей работы.
Хотя обработанный материал, по мнению авторов, по объему значительно
превосходит тот, которым располагал Зимрот, список видов, живущих на
исследованной территории, не следует считать завершенным. Огромные
пространства Сибири и Средней Азии таят в себе еще некоторое число неопи-
санных видов. Однако родовой состав фауны вряд ли может сильно изме-
ниться. В связи с тем что некоторые виды и роды из сопредельных с СССР
стран исходно описаны недостаточно полно, а авторы не располагали необ-
ходимыми материалами, чтобы определить свою точку зрения, эти таксоны
приводятся в данной книге как «Дополнения» к соответствующим родам
и семействам.
Настоящая книга снабжена 576 штриховыми рисунками, одной цветной
таблицей и 18 фотографиями радул. Эти фотографии выполнены и любезно
предоставлены Ю. Юнгблутом (J. Jungbluth, Heidelberg), за что авторы
приносят ему благодарность. Большая часть рисунков оригинальные и вы-
выполнены с натуры; для заимствованных иллюстраций указан источник,
откуда они взяты. Все рисунки переведены в тушь художником Н. Н. Фу-
зеевой, ею же выполнен акварельный оригинал цветной таблицы.
В процессе работы авторы получили неоценимую помощь от своих
многочисленных коллег. В первую очередь она выражалась в сборе и при-
сылке необходимых коллекционных материалов. Такие сборы были осуществ-
лены: на Кавказе — Н. Н. Акрамовским (Ереван), 3. И. Калитиной (Орджо-
никидзе), А. А. Шилейко (Москва); в Средней Азии и Казахстане —
В. В. Гороховым (Москва), 3. И. Иззатуллаевым (Душанбе), П. В. Матё-
киным (Москва), А. Б. Мухитдиновым (Ленинабад), К. К. Увалиевой (Алма-
Ата), А. А. Шилейко; в Приморском и Хабаровском краях — И. М. Москви-
чевой (Хабаровск).
Огромную помощь в получении необходимого музейного материала ав-
торы получили от И. Валовирта (I. Valovirta, Helsinki), Е. Гиттенбергера
(Е. Gittenberger, Leiden), А. Гроссу (A. Grossu, Buguresti), Р. Килиаса
ПРЕДИСЛОВИЕ.
7
(R. Kilias, Berlin), О. Паджета (0. Paget, Wien), А. Риделя (A. Riedel,
Warszawa), P. Янсена (R. Janssen, Frankfurt a. M.), а также от покойного
Регтерен Альтена (С. О. van Regteren Altena, Leiden).
В процессе написания разделов книги, посвященных вопросам конвер-
генции у слизней, их филогении и классификации, авторы получили полез-
ные советы от Я. Рафальского (J. Rafalski, Рогпай), А. Риделя, Я. И. Старо-
богатова (ЗИН АН СССР) и А. А. Шилейко..Последний, кроме того, выпол-
нил ряд сложных натурных рисунков по анатомии некоторых слизней.
Всем упомянутым коллегам авторы приносят живейшую благодарность.
Очень признательны они также В. Е. Аренс, любезно согласившейся пере-
водить на русский язык разделы, написанные А. Й. Виктором на польском
языке, а также А. С. Довгялдо и Л. Л. Ярохнович за повседневную помощь
при работе с коллекциями и при подготовке рукописи данной книги к пе-
чати. ’
Со смешанным чувством грусти и благодарности авторы вспоминают по-
койного профессора А. А. Стрелкова (ЗИН АН СССР), который живо ин-
тересовался их работой и дал много ценных советов в период написания этой
книги.
В заключение авторы очень признательны Я. И. Старобогатову за науч-
ное редактирование их работы.
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
1. Надсемейство LIMACOIDEA Rafinesque, 1815
I. Семейство AGRIOLIMACIDAE Wagner, 1935
1. Род Deroceras Rafinesque, 1820
1. Подрод Deroceras s. str.
1. D. (D.) laeve (Muller, 1774)............................................ 129
2. D. (D.) sturanyi (Simroth, 1894)...............'......................... 132
3. D. (D.) pollonerae (Simroth, 1889)....................................... 135
4. D. (D.) osseticum (Simroth, 1901)........................................ 136
5. D. (D.) ilium (Simroth, 1901)............................................ 139
2. Подрод Agriolimax Morch, 1865
6. D. (A.) agreste (Linnaeus, 1758)......................................... 142
7. D. (A.) altaicum (Simroth, 1886) ....................................... 146
8. D. (A.) reticulatum (Muller, 1774) . .................................... 149
9. D. (A.) turcicum (Simroth, 1894)......................................... 152
3. Подрод Plathystimulus Wiktor, 1973
10. D. (Pl.) subagreste (Simroth, 1892).................................... 154
11. D. (Pl.) bakurianum (Simroth, 1912)..................................... 158
12. D. (Pl.) tauricum (Simroth, 1901)....................................... 163
13. D. (Pl.) praecox Wiktor, 1966 ....................................... 165
14. D. (Pl.) rodnae Grossu et Lupu, 1965 .................................. 167
4. Подрод Liolytopelte Simroth, 1901
15. D. (L.) caucasicum (Simroth, 1901)...................................... 169
16. D. (L.) bureschl (Wagner, 1934) 174
17. D. (L.) moldavicum (Grossu et Lupu, 1981)............................... 176
18. D. (L.) occidentals (Grossu et Lupu, 1966)............................. 178
Дополнение к роду Deroceras!
D. (?) absoloni (Simroth, 1916).............................................. 180
D. (Agriolimax) callatis Grossu et Lupu, 1963 ............................... 180
D. (?) crimense (Simroth, 1901).............................................. 180
D. (Agriolimax) dobrogicum Grossu, 1969 ..................................... 182
D. (A.) forcarti Grossu et Lupu, 1961........................................ 183
D. (?) jaeckeli Grossu, 1969 ................................................ 183
D. (Plathystimulus)! padisii Lupu, 1969 ..................................... 185
D. (Agriolimax?) pentheri (Babor, 1905)...................................... 185
D. (A.) ponori Lupu, 1972 ................................................... 187
D. (A.) schbeschi Grossu et Lupu, 1961....................................... 187
D. (?) steidachneri (Wagner, 1931)........................................... 187
D. (Agriolimax) vranceanum Grossu, 1969 ..................................... 189
D. (Plathystimulus?) zilchi Grossu, 1969 .................................... 189
2. Род Krynickillus Kaleniczenko, 1851
1. K. melanocephalus Kaleniczenko, 1851..................................... 191
2. K. urbanskii (Wiktor, 1971)............................................. 193
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ 9
3. Род Lytopelte О. Boettger, 1886
L. maculata (Koch et Heynemann, 1874) . .................................. 196
4. Род Megalopelte Lindholm, 1914
M. simrothi Lindholm, 1914................................................ 202
5. Род Mesolimax Pollonera, 1888
1. M. brauni Pollonera, 1888 ............................................. 205
2. M. escherichi Simroth, 1899 ......................................... 207
Дополнение к роду Mesolimax:
M. reibischi Simroth, 1892 . ............................................. 207
Дополнение к семейству Agriolimacidae:
Род Toxolimax Simroth, 1899
T. hoplites (Simroth, 1899).............................................. 208
II. Семейство BOETTGERILLIDAE Goethem, 1972
Род Boettgerilla Simroth, 1910
1. B. pallens (Simroth, 1912)............................................. 214
2. B. compressa Simroth, 1910........................................... 217
III. Семейство LIMACIDAE Rafinesque, 1815
1. Подсемейство Limacinae Rafinesque, 1815
1. Род Gigantomilax O. Boettger, 1883
1. Подрод Gigantomilax s. str.
1. G. (G.) lederi (O. Boettger, 1883) . . . ............................. 223
g|| 2. Подрод Vitrinoides Simroth, 1891
2a. G. (P.) monticola armeniacus (Simroth, 1886) . . ..................... 226
2h. G. (P.) monticola monticola (0. Boettger, 1881)....................... 230
3. G. (P.) koenigi (Simroth, 1912).......•............................... 231
4. G. (P.) daghestanus (Simroth, 1898)................................... 233
3. Подрод Monochroma Simroth, 1896
5. G. (M.) brunneus (Simroth, 1901) .................................... 235
6. G. (M.) lenkoranus Simroth, 1912...................................... 238
Щ 2. Род Caucasolimax Likharev et Wiktor, gen. nov.
C. caucasicus (Simroth, 1898)............................................. 241
3. Род Caspilimax P. Hesse, 1926
C. keyserlingi (Martens, 1880) ........................................... 244
4. Род Turcomilax Simroth, 1901
1. Подрод Turco-milax s. str.
1. T. (T.) nanus (Simroth, 1901)........................................ 249
2. T. (T.) ferganus (Simroth, 1910)............................... . . 251
10
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ
2. Подрод Michaelisia Likharev et Wiktor, subgen. nov.
3. T. (JIT.) natalianus (Michaelis, 1892) ....................... • . . 252
3. Подрод Taulimax Likharev et Wiktor, subgen. nov.
4. T. (T.) turkestanus (Simroth, 1898)................................ 255
5. T. (T.) tzvetkovi Likharev et Wiktor sp. nov. ....'.................. 258
5. Род Malacolimax Malm, 1868
M. tenellus (Muller, 1774) .......................................... ' 259
6. Род Lehmannia Heynemann, 1862
1. L. marginata (Muller, 1774).......................................... 263
2. L. macroflagellata Grossu et Lupu, 1962 ......”...................... 265
3. L. horezia Grossu et Lupu, 1962 ..................................... 267
4. L. medioflagellata Lupu, 1968 ....................................... 267
5. L. valenttana (Ferussac, 1823)....................................... 268
6. L. jaroslaviae Grossu, 1967 ......................................... 270
7. L. nyctelia (Bourguignat, 1861)..................................... 271
7. Род Limax Linnaeus, 1758
1. Подрод Limax s. str.
1. L. (L.) cinereoniger Wolf, 1803 .................................... 274
, 2. L. (L.) maxtmus Linnaeus, 1758 ................................... 278
3. L. (L.) dobrogicus Grossu et Lupu, 1960 ............................ 280
2. Подрод L i m a c u s Lehmann, 1864
4. L. (L.) flavus Linnaeus, 1758 ....................................... 281
5. L. (L.) maculatus (Kaleniczenko, 1851)............................... 284
2. Подсемейство Bielziinae Likharev et Wiktor, subfam. nov.
8. Род Bielzia Clessin, 1887
B. coerulans (M. Bielz, 1851) .......................................... 288
3. Подсемейство Eumilacinae Likharev et Wiktor, subfam. nov.
9. Род Eumilax 0. Boettger, 1881
1. E. brandtt (Martens, 1880) .......................................... 293
2. E. intermittens (O. Boettger, 1883) ................................ 297
10. Род Metalimax Simroth, 1896
1. M. varius (O. Boettger, 1884)........................................ 301
2. M. elegans Simroth, 1901 ........................................... 305
2. Надсемейство T RIGONOCBLAM YDOI DE A P. Hesse, 1882
I. Семейство TRIGO NOCIILAMYDIDAE P. Hesse, 1882J
1. Подсемейство Trigonochlamydinae P. Hesse, 1882
1. Род Trigonochiamys O. Boettgery 1881
Tr. imitatrix O. Boettger, 1881 . ... . ........... 312
2. Род Hyrcanolestes Simroth, 1901 ]
H. velitdrls (Martens, 1880) '. . ; . .................. . 314
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ 11
3. Род Parmacellilla Simroth, 1910
Р. filipowitschi Simroth, 1910 ........................................ 320
4. Род Troglolestes Ljovuschkin et Matekin, 1965
Tr. sokolovi Ljovuschkin et Matekin, 1965 .............................. 322
5. Род Drilolestes Lindholm, 1925
Dr. retowskii (0. Boettger, 1884) .................................... 324
2. Подсемейство Selenochlemydinae Likharev et Wiktor, subfam. nov.
6. Род Selenoehlamys 0. Boettger, 1883
S. pallida 0. Boettger, 1883 .......................................... 330
3. Надсемейство Z 0 N I T QI DE A Morch, 1864
I. Семейство PARMACELLTDAE Cutfier, 1804
1. Род Parmacella Grau, 1860
P. ibera Eichwald, 1841 .............................................. 335
2. Род Candaharia Godwin-Austen, 1888
1. Подрод Candaharia s. str.
1. С. (C.) rutellum (Hutton, 1849)...................................... 342
2. С. (C.) aethiops (Westerlund, 1896)................................... 345 •
2. Подрод Levanderia Likharev et Wiktor, subgen. nov.
3. C. (L.) levanderi (Simroth, 1901)................................... 348 .
4. C. (L.) izzatullaevi Likharev et Wiktor, sp. nov...................... 352 .
II. Семейство MILACIDAE Ellis, 1926
1. Род Milax Gray, 1855
Подрод Milax s. str.
1. M. (M.) caucasicus (Simroth, 1912)................................... 358
2. M. (M.) gagates (Draparnaud, 1801) ....'............................. 362
2. Род Tandonia Lessona et Pollonera, 1882
Подрод Tandonia s. str.
1. T. (T.) cristata (Kaleniczenko, 1851)................................ 365
2. T. (T.) kaleniczenkoi (Clessin, 1883) .’............................. 368
3. T. (T.) rustica (Millet, 1843)............... . . ................. 372
4. T. (T.) kusceri (Wagner, 1931)....................................... 374
5. T. (T.) budapestensis (Hazay, 1881).................................. 376
Дополнение к семейству Milacidaes
Milax samsunensis Forcart, 1942 . . . . ................................ 379
4. Надсемейство A R I 0 NOIDEA Gray, 1840
I. Семейство PHILOMYCIDAE Gray, 1847
Род Meghimatium Hasselt, 1823
M. btlineatum (Benson, 1842)............................................ 382
12
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ
П. Семейство ARIONIDAE Gray, 1840
1. Подсемейство Arioninae Gray, 1840
Род Arion Ferussac, 1819
1. Подрод Arion s. str.
1. A. (A.) ruf us (Linnaeus, 1758).......................................... 390
2. A. (A.) ater (Linnaeus, 1758) . .......................................... 394
3. A. (A.) lusitanicus Mabille, 1868 .................................... 395
2. Подрод M e s ar i on P. Hesse, 1926
4. A. (M.) subfuscus (Draparnaud, 1805)................ ........ 396
5. A. (M.) sibiricus Simroth, 1901 ......................................... 400
3. Подрод Carinarion P. Hesse, 1926
6. A. (C.) circumscriptus Johnston, 1828 .................................. 403
7. A. (C.) stlvaticus Lohmander, 1937 . . .................................. 405
8. A. (C.) fasciatus (Nilsson, 1823)......................................... 407
4. Подрод K o b e 11 i a -Seibert, 1873
9. A. (K.) hortensis Ferussac, 1819 .............. 409
5. Подрод Microarion P. Hesse, 1926
10. A. (M.) -intermedius (Normand, 1852) .................................... 411
ОБЩАЯ ЧАСТЬ
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
Как известно, подавляющее большинство видов брюхоногих моллюсков
обладает хорошо развитой наружной раковиной. Тем не менее от этого ос-
новного и исходного в пределах класса плана строения имеются многочис-
ленные отклонения, связанные с определенным образом жизни. Так, виды,
ползающие по дну водоемов и поверхности суши, чаще всего имеют спирально
закрученную наружную раковину, в которую они могут втянуть голову
и ногу и где всегда помещается так называемый внутренностный, или висце-
ральный, мешок. Однако у видов, живущих в других условиях, например •
у планктонных и паразитических форм, внешний облик кардинально иной.
В частности, раковина может испытывать полную или частичную редук-
цию.
Редукция раковины и перестройка типичного для Gastropoda плана строе-
ния иногда имеет место и у ползающих форм. При этом туловище (цефало-
подиум) в общем сохраняет обычный облик, но внутренностный мешок
погружается в толщу ноги, раковина подвергается редукции, и животное при-
обретает вытянутую цилиндрическую или веретеновидную форму, уплощен-
ную снизу, т. е. со стороны подошвы. Таких брюхоногих моллюсков с дав-
них пор называют слизнями (limaces — фр.; slugs — англ.; Nacktschnecken—
нем.).1 В то же время формы с типичным для класса строением, т. е. со спи-
рально закрученным внутренностным мешком, закрытым хорошо развитой
наружной раковиной, в которую могут втягиваться голова и нога, называют
улитками (escargots — фр.; snails — англ.; Gehauseschnecken — нем.).
Слизни встречаются как среди морских, так и среди наземных брюхо-
ногих моллюсков. В данной работе рассматриваются сухопутные слизни из
отряда Geophila (надотряд Stylommatophora, подкласс Pulmonata). Даже
в пределах этого отряда слизни независимо возникали в разных группах.
Следовательно, и здесь это понятие объединяет разные по происхождению
группы моллюсков, приобретших под влиянием сходного образа жизни
ряд общих черт. Поэтому в данной работе под понятием «слизень» подразуме-
вается определенный морфологический тип организации и комплекс сходных
жизненных форм, независимо повторяющихся в генетически далеких друг
от друга группах. Аналогичное содержание вкладывается и в понятие
«улитка» — другой, еше более распространенный среди Gastropoda морфо-
логический тип организации. Однако в отличие от слизней улитки являются
первичным типом организации и, следовательно, генетически связаны друг
с другом.
Ниже, излагая материал о строении и физиологии слизней, мы стремимся
выявить особенности, общие не только для всех групп, но и свойственные
только отдельным семействам и родам, а также показать принципиальные
1 В русской литературе их иногда называли «голыми слизнями». Однако это название
неправомочно, так как альтернативой понятию «слизень» является понятие «улитка».
Недаром в немецкой литературе слизней называют «голыми улитками».
14
ОБЩАЯ ЧАСТЬ
отличия слизней от улиток. Подобный подход преследует несколько за-
дач: а) установить сравнительно-морфологические и физиологические пути:
перехода от плана строения улитки к плану строения слизня; б) раскрыть,
коррелятивные связи между отдельными системами органов; в) выявить
черты сходства в строении и физиологии слизней, связанные с параллелиз-
мами в эволюции разных групп; г) обнаружить наиболее характерные осо-
бенности отдельных групп слизней, которые можно считать, близкими
к анцестральным, дающие возможность судить об их положении в системе-
отряда Geophila.
Естественно, что при анализе морфологии слизней совершенно необ-
ходим функциональный и экологический подход.
Внешний вид
Форма тела и размеры. Тело слизней сильно вытянуто»
в длину, так что при рассматривании сверху боковые контуры почти парал-
лельны. Впереди, к голове, они плавно и закругленно смыкаются, сзади
сходятся под более или менее острым углом (у большинства семейств) или
тоже плавно и закругленно смыкаются (Philomycidae, Arionidae). В общем
тело слизней, особенно во время ползания, имеет цилиндрическую или слабо-
веретеновидную форму, уплощенную снизу, т. е. со стороны' подошвы..
При сокращении, особенно после жесткой фиксации, тело слизней приобре-
тает более вздутую и массивную веретеновидную или булавовидную форму
за счет общего сжатия в длину и втягивания головы внутрь тела.
Тело слизней в отличие от улиток было бы внешне строго симметричным'
благодаря отсутствию наружного внутренностного мешка, если бы не поло-
жение пневмостома, ануса, выделительной поры и полового отверстия, ко-
торые лежат на правой стороне. Цефалоподиум слизней состоит из следую-
щих основных отделов (рис. 1): голова, шея, мантия, спина, нога с подошвой..
Среди слизней встречаются как «гиганты», длина которых при движении;
превышает 20 см (Eumilax brandti, Limax maximus, Arion ater), так и «кар-
лики» — не более 2 см (Arion intermedins, Deroceras laeve).
Голова (рис. 2). У слизней она построена по тому же плану, что и
у улиток Geophila. Впереди, в центре «лицевой» поверхности, располага-
ется рот с двумя ротовыми лопастями по сторонам и складчатыми верхней
и нижней губами. Снизу, между головой и передним концом подошвы, име-
ется широкая щель, в которую открывается наружу педальная железа.
Несколько отступя от ротовых лопастей, на голове располагается пара не-
больших губных щупалец; их иногда еще называют передними, или ниж-
ними, щупальцами. Выше и немного сзади губных щупалец располагается
пара более крупных глазных щупалец (омматофоры); иногда их еще называют
задними, или верхними, щупальцами. Участок головы между глазными щу-
пальцами называют затылком.
Шея. Сзади голова плавно переходит в шею. Под последней понима-
ется вся передняя часть тела между головой и местом прирастания мантии'
к верхней стороне цефалоподиума. На правой стороне шеи недалеко от ноги
находится половое отверстие. У большинства слизней оно лежит немного»
позади правых щупалец. В некоторых группах половое отверстие сильно»
смещено назад и лежит вблизи от пневмостома (Arion) или более или менее-
на полпути между щупальцами и пневмостомом (Milaeidae).
Мантия. В отличие от улиток, у которых мантия по форме повторяет
внутренностный мешок и раковину, у слизней она имеет вид плоского утол-
щения на верхней стороне тела. Чаще она удлиненно-овальная, реже более
или менее круглая. Нередко задний край мантии посередине вытянут в ост-
рый или закругленный угол; передний край мантии обычно широко закруг-
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
'___• V
лен. У Trigonochlamydidae мантия имеет округло-треугольную или асиммет-
ричную щитовидную форму.
Вблизи от правого края мантия пронизана дыхательным отверстием
или пневмостомом, который с краем мащгии связан глубокой щелью или вы-
резкой. Как пневмостом, так и мантийная щель гомологичны и сходны с
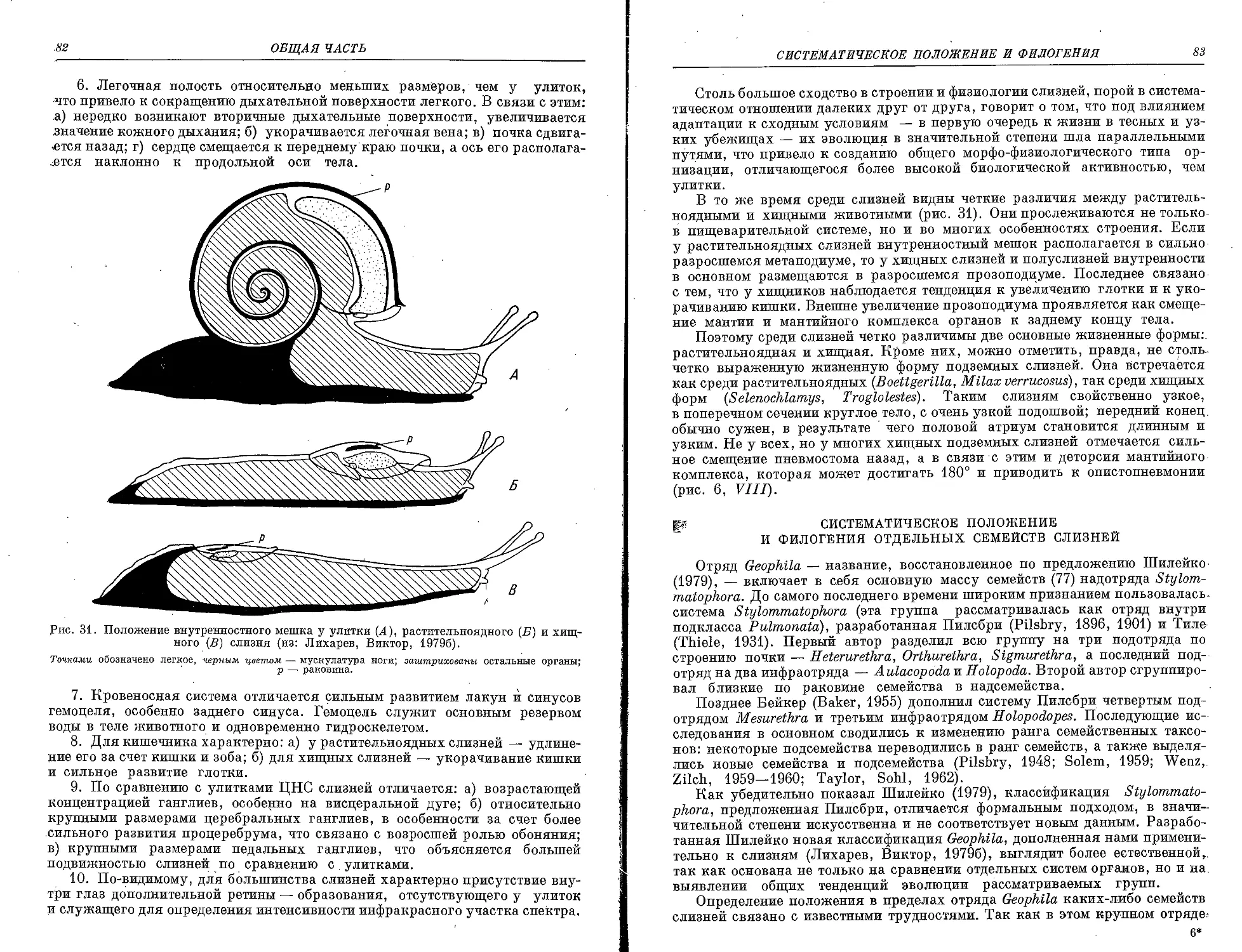
Рис. 1. Основные отделы тела улитки (/), полуслизня (II) и слизня (III).
йРаковина обозначена точками, а контуры ее, закрытые мантией, — прерывистой линией} мантия за-
штрихована в клетку; метаподиум — косой штриховкой; прозоподиум — не ^штрихован.
такими же образованиями у улиток, причем щель образована за счет сбли-
жения левой и правой лопастей мантийного края раковинных легочных мол-
люсков. В мантийной щели рядом с пневмостомом располагаются анус и
выделительная пора. Пневмостом.нередко окружен колцевым удолщением
или валиком, внутри которого залегает сфинктер.
Только у Philomycidae и некоторых Trigonochlamydidae мантия прира-
стает к телу всей нижней поверхностью, не образуя свободных лорастей.
У большинства остальных слизней она заканчивается. впереди более или
менее широкой свободной лопастью — капюшоном. Это образование играет
защитную роль, так как призвано закрывать шею, а у многих и голову при
сокращении животного. Кроме того, периодически втягивая шею под капю-
шон, слизень очищает с помощью слизи эту часть тела от пыли и гряаи.
16
ОБЩАЯ ЧАСТЬ
У Рarmacellidae капюшон достигает особенно крупных размеров — он
больше, чем задняя приросшая часть мантии. У Arionidae и Milacidae это
образование составляет меньшую часть мантии, но тем не менее хорошо вы-
ражено. В большинстве же семейств капющон составляет около половины
всей мантии.
Чаще всего мантия располагается ближе к голове и далеко от заднего
конца тела. Однако имеются исключения. Так, у Philomycidae мантия пол-
Рис. 2. Передний конец тела Limax.
1 — глазное щупальце; 2 — рот; 3 — губное щупальце; 4 — ротовая лопасть; 5 — выходное отверстие
педальной железы; в — половое отверстие; 7 — боковые доли подошвы; 8 — срединная доля подошвы;
9 — нога; 10 — перипедальная борозда; 11 —• оторочка; 12 — супрапедальная борозда; 13 — боковая
борозда; 14 — капюшон мантии; 15 — затылочные борозды.
ностью закрывает всю верхнюю сторону животного почти до ноги, и край
ее плотно прирастает к телу по всему периметру. У Megalopelte (Agrioli-
macidae) мантия тоже закрывает всю верхнюю сторону животного, но к телу
она прирастает только в центральной части и широкая свободная лопасть
закрывает не только шею, но и спину и бока слизня.
Для хищных слизней характерно уменьшение размеров мантии и смеще-
ние ее Kj заднему концу тела. Так, у Trigonochlamydidae можно видеть все
ступени.’обоих процессов. Если у Troglolestes крупная мантия закрывает
всю верхнюю сторону животного и лишь при ползании из-под крупного ка-
пюшона выступает часть шеи и голова, то у Drilolestes мантия значительно
меньших размеров, с очень узким капюшоном и располагается более или ме-
нее .на середине тела. У Trigonochlamys мантия относительно еще меньше,
без капюшона и лежит за серединой тела. Наконец, у S elenochlamys от ман-
тии остается едва заметный щиток, лежащий у самого заднего конца живот-
ного.
крупных размеров — он
irionidae и Milacidae это
тем не менее хорошо вы-
тавляет около половины
ове и далеко от заднего
diilomycidae мантия пол-
5
пасть; 5 — выходное отверстие
8 — срединная доля подошвы;
альная борозда; 13 — боковая
борозды.
почти до ноги, и край
7 Megalopelte (Agrioli-
1 животного, но к телу
сая свободная лопасть
«еров мантии п смеще-
idae можно видеть все
1ая мантия закрывает
и из-под крупного ка-
?s мантия значительно
гагается более или ме-
штельно еще меньше,
Selenoehlamys от ман-
задпего конца жпвот-
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
1Г
Характерно, что в этом же семействе одновременно с уменьшением раз-
меров мантии и смещением ее к заднему концу тела происходит сдвиг пнев-
мостома к заднему краю мантии.
Спина. Участок верхней стороны тела слизней, лежащий позади ман-
тии, называется спиной. Она соответствует той части верхней стороны тела
улиток, которая располагается за внутренностным мешком. К заднему
концу спина в различной степени снижается и сужается. У многих слизней
вдоль средней линии спины тянется киль. У одних он имеет вид четкой муску-
листой складки, простирающейся от мантии к заднему концу тела, а у других
выражен менее ясно — как легкая угловатость, порой ограниченная -только-
задней частью спины.
У большинства Arionidae всякие следы киля отсутствуют и спина на всем
протяжении плавно закруглена. У Arion на заднем конце тела имеется ха-
рактерный орган — хвостовая ямка или каудальная железа. Внешне она
имеет вид треугольной ямки, лежащей над ногой и прикрытой небольшой
лопастью (см. рис. 538).
Нога. У улиток под ногой подразумевают мускулистую часть цефа-
лоподиума, впереди располагающуюся под внутренними органами (глотка,
зоб, гениталии), а сзади лежащую за внутренностным мешком и заполнен-
ную мускулатурой (рис. 1, /). У слизней в связи со смещением органов внут-
ренностного мешка в толщу цефалоподиума нога как впереди, так и сзади
располагается под внутренностями, т. е. является мускулистым утолщением
нижней стенки тела (рис. 1, III). Внешне от остального тела она отграни-
чена перипедальной бороздой (рис. 2, 10). Нижнюю поверхность ноги назы-
вают подошвой.
Нога повторяет общие очертания тела слизня, т. е. впереди она широко-
закруглена, боковые края ее почти параллельны, а сзади они смыкаются,
плавно закругляясь или под более или менее острым углом. У большинства
слизней при рассмотрении сверху нога почти не выступает за боковые кон-
туры спины и шеи, но у некоторых, в особенности у Arionidae и Philomycidae,
нога заметно шире верхней стороны тела. G другой стороны, у видов, веду-
щих преимущественно подземный образ жизни (Trigonochlamydidae, Boett-
gerillidae и др.), нога обычно значительно уже остального тела.
Рельеф. Как верхняя сторона тела слизней, так и нижняя, т. е.
подошва, обладают определенным рельефом, слагающимся из различной
формы возвышений (морщин) и углублений (борозд). Все основные формы
рельефа кожи слизней встречаются и у улиток.
Рельеф верхней стороны (кроме мантии) образован сетью бороздок, между
которыми возвышаются ряды полигональных, морщин. Основу этой сети
составляют бороздки, довольно регулярно радиально расходящиеся от ман-
тии к ноге. Число их, по-видимому, более или менее постоянно для каждого
рода, а в некоторых случаях для определенного вида.1 2 На шее наиболее
характерны две пары бороздок. Одна пара — затылочные бороздки
(рис. 2, 15) — простирается вдоль средней линии шеи. Она берет начало от
передней точки прирастания мантии к телу (т. е. у видов с капюшоном —
от основания его) и тянется вперед к затылку, где распадается на сеть мел-
ких лицевых бороздок.
Другая пара — боковые бороздки — располагается наклонно (рис. 2,
13). Из них правая берет начало у пневмостома и сходит на нет околи
правого губного щупальца. Левая бороздка располагается соответственна
на другой стороне шеи.
1 При характеристике таксонов обычно указывают число рядов морщин, между сред-
ней линией спины (или килем) и мантийной щелью.
2 И. М. Лихарев, А. Й. Виктор
18 ОБЩАЯ ЧАСТЬ
Как уже упоминалось, над ногой по всему периметру тела (с перерывом
только у головы) залегает четкая перипедальная борозда. У многих слизней
немного выше ее лежит вторая, но не столь четкая борозда — супрапедаль-
ная. Между ними имеется невысокая складйа кожи — оторочка (рис. 2, 11).
Только у Philomycidae- и у большинства -Arionidae супрапедальная борозда
и оторочка отсутствуют.
Кроме радиальных и продольных бороздок, между ними находятся корот-
кие и менее глубокие бороздки, в итоге создающие общую сеть. В ячеях
этой сети располагаются возвышения — морщины — гексагональной или по-
лигональной формы. Указанный рельеф при жизни обычно менее четок,
чем после фиксации, особенно после жесткой фиксации (с. 116). Тогда в связи
с сокращением тела он. выражен особенно сильно. При этом морщины не
только вспухают, но нередко на них возникают серии вторичных бугорков.
Поверхность мантии имеет свой рельеф. У одних слизней — Agriolima-
cidae, Вoetigerillidae, многие L'imacidae (за исключением Eumilacinae) —
при жизни по ней пробегают волны концентрических сокращений или мор-
щин. После фиксации эти морщины исчезают и мантия становится гладкой.
У других групп — Parmacellidae, Milacidae, Trigonochlamydidae, Philomy-
cidae, Arionidae и у Eumilacinae (Limacidae) — при жизни по мантии про-
бегают многочисленные сосочки, которые после фиксации обычно исчезают
и мантия приобретает зернистый рельеф. Только у Troglolestes (Trigonochla-
mydidae) эти сосочки столь велики, что сохраняются и после смерти живот-
ного.
Кроме указанных форм рельефа мантии, у некоторых групп слизней на
ее поверхности имеются 1—2 бороздки. Так, для Parmacellidae и Milacidae
характерна одна, изогнутая дугой (выпуклостью вперед), подкововидная
бороздка (см. рис. 468). Она сохраняется и после фиксации животного. В двух
других группах — Boettgerilla (Boettgerillidae) и Krynickillus (Agriolimacidae)
на мантии имеются небольшие бороздки иного типа, отходящие от пневмо-
стома. У первой их две — одна впереди, а другая позади дыхательного от-
верстия (см. рис. 246); у второго только одна, лежащая позади пневмостома
(см. рис. 209).
Подошва в отличие от верхней стороны тела имеет более простой рельеф
(рис. 2). При жизни он слагается из двух глубоких продольных бороздок
(педальные бороздки), разделяющих подошву на три доли: срединную и две
боковых. Подобный рельеф имеют все рассматриваемые здесь группы слиз-
ней, за исключением Philomycidae и рода Arion (Arionidae). Однако и в этих
двух группах, несмотря на отсутствие педальных бороздок, срединная доля
подошвы микрорельефом заметно отличается от боковых долей.
Та или иная дифференциация подошвы на три продольных доли связана
с особой активностью срединной доли — при движении по ней спереди
назад пробегают волны сокращений. В связи с этим средний участок ноги
отличается от боковых расположением мускулатуры и иннервацией.
Кроме двух основных педальных бороздок, у некоторых слизней (Parma-
cellidae, Mesolimax) ттекпся еще несколько менее четких дополнительных
продольных бороздок, пролегающих на боковых долях подошвы.
После фиксации на подошве появляются многочисленные поперечные
бороздки. Они могут пересекать всю подошву или ограничиваться только
боковыми долями. У Milacidae поперечные бороздки не прямые, а V-образно
изогнутые назад (рис. 3), а у Agriolimacidae на боковых долях они прямые,
а на срединной V-образно изогнутые.
Рельеф как верхней стороны тела, так и подошвы слизней в основном схо-
ден с рельефом цефалоподиума улиток. В обоих основных морфологических
типах функция рельефа одинакова. Благодаря морщинам увеличивается
поверхность тела, что особенно важно для слизней, у которых значительную
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
19'
роль играет кожное дыхание. Не случайно, что чем крупнее слизни, тем
крупнее и резче рельеф верхней стороны тела.
Окраска (см. цветную вклейку). В отличие от наземных улиток,,
поверхность тела которых обычно окрашена в черный, темно- или светло-
серые тона без всяких следов рисунка, окраска слизней более разнообразнаг
как по цветовой гамме, так и нередко за счет образования темного рисунка,
на светлом фоне. Обычно молодые слизни окрашены светлее взрослых.
Пигмент может быть сосредоточен в хроматофорах соединительной ткани
кожи или свободно лежит в виде гранул между клетками подкожной соеди-
Рис. 3. Основные типы рельефа подошвы.
I — Milacidae, II — Limacidae, III — Derocerasr IV — Mesolimax (Agriolimacidae).
нительной ткани. Так, например, у Arion ater черный пигмент находится!
в хроматофорах, а красный в межклеточных пространствах соединительной
ткани (Barr, 1927). Наиболее распространенные типы пигмента это черный,
красный и желтый. Различные сочетания их и глубина залегания в толще
покровов могут создавать многочисленные вариации окраски и рисунка.
Наиболее обычные типы окраски это черная, темно-серая, коричневая,
красноватая и охристая (серовато-желтая). Рисунок может быть пятнистым —
крупные или мелкие пятна, пятна-брызги, пятна-точки. Мелкие пятна могут
сливаться, образуя более крупные, нередко с неправильными контурами.
Слияние пятен может привести к образованию парных (1—2) цветных полос
вдоль спины (особенно вдоль киля), а на мантии — вдоль ее боковых краев
(многие Arion, Limax, Eumilax и др.). На мантии, кроме боковых полос,
может возникать и медиальная полоса или ряд пятен, обычно с менее четкими
контурами, чем вдоль краев мантии.
Подошва обычно окрашена значительно светлее верхней стороны тела.
Только боковые доли ее могут быть несколько темнее срединной за счет
сравнительно редких хроматофоров, к тому же залегающих значительно-
глубже, чем на верхней стороне тела.
Внутреннее строение (рис. 4)
Покровы. Тело слизней, как и у улиток, покрыто однослойным ци-
линдрическим эпителием. Изнутри к нему прилегают слои подкожной му-
скулатуры и соединительной ткани. Ресничный эпителий развит главным
образом на подошве, щупальцах, вокруг пневмостома и на боках тела вблизи
от последнего. В отличие от улиток слизни особенно богаты одноклеточными
слизистыми эпителиальными железами и одно- и многоклеточными субэпи-
телиальными железами. Последние открываются наружу узкими протоками,
залегающими между клетками эпителия. Среди этих желез различают три
основных типа: слизистые, белковые и известковые.
2*
20
ОБЩАЯ ЧАСТЬ
Наряду с этими наиболее многочисленными железами как улитки, так
и слизни имеют сложные и крупные специализированные железы. К их числу
Рис. 4. Внутреннее строение Tandonia budapestensis.
I — естественное положение органов; II — они же после препарирования, а — аорта (главный ствол);
ап — передняя аорта; бж — белковая железа; з — гонада; гл — глотка; гп — гермафродитный проток;
О — диафрагма;|жга — педальная железа; же — желудочек сердца; з — зоб; зк — задняя кишка; мп —
колумеллярный мускул; п — пенис; па — половой атриум; печ — печень; пж — придаточные железы;
пч — почка; пщ — пищевод; рп — половой ретрактор; рщ — ретрактор глазного щупальца; с — семя-
провод; аж — слюнная железа; си — средняя кишка; сод — семяяйцевод; ап — семяприемник; цне —
мозг; ап — эпифаллус; я — яйцевод.
принадлежат педальная железа и орган Семпера, имеющиеся у всех легочных
моллюсков. В основном они слагаются из слизистых и белковых клеток.
Педальная железа служит для выделения слизи, выстилающей путь, по
которому движется моллюск. Она открывается наружу на переднем конце
тела между головой и ногой. Это крупное мешковидное образование, чаще
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
21
всего лежащее в толще ноги, и лишь у Milacidae она располагается свободно
>в гемоцеле под органами внутренностного мешка (рис. 4, жп). Длина ее
различна, но обычно не менее х/2 длины тела (Barr, 1926; Phillips, Watson,
1930).
Строение педальной железы довольно сложное. Потолок ее имеет про-
дольные складки, одетые ресничным эпителием, а дно слагается из большого
числа одноклеточных слизистых желез. У Arion rufus эти железы выделяют
22
ОБЩАЯ ЧАСТЬ
два типа секрета (Chetail, Binot, 1967): клетки передней половины органа —
сложные мукополисахариды, а задней половины — простые мукополисаха-
риды.
Слизь педальной железы выталкивается наружу как за счет сокращений
ноги,' так и с помощью ресничек эпителия.
Орган Семпера — это крупное парное, образование неясного назначения.
Он располагается по обеим сторонам глотки и заходит в ротовые лопасти.
Основная масса его сложена железистыми клетками, окружающими спе-
циальный ганглий; выводные протоки не обнаружены.
Третьим видом специализированных желез является каудальная железа.
Она имеется у некоторых групп улиток, а среди наших слизней у Arion.
Внешне она представляет собой углубление на заднем конце тела, над ногой.
Сюда открываются многочисленные одноклеточные слизистые и белковые
железы (Herfs, 1922). Выделяемая этим органом слизь по консистенции от-
личается от слизи верхней стороны тела и педальной железы. Роль этой слизи
сигнальная — она указывает на готовность к копуляции (см. ниже).
Практически вся поверхность тела слизней продуцирует слизь, но неко-
торые участки — мантия-, район пневмостома, голова, перипедальная бо-
розда— в этом отношении играют особенно заметную роль. Слизь выделяется
как крупными специализированными органами, так и бесчисленными одно-
и многоклеточными железами покровов тела. У Arion ater только в эпителии
подошвы обнаружено 4 типа, а в эпителии верхней стороны тела 3 типа одно-
клеточных слизистых желез (Chetail, Binot, 1967; Wondrak, 1968, 1969а, b).
А глубже залегают еще разнообразные субэпителиальные слизистые железы.
Слизь представляет собой смесь различного типа секреций. Основную долю
в ней занимает вода. Так, в слизи улитки Oxychilus alliarius сухой остаток:
составляет 6.8%, в том числе 8% приходится на неорганические вещества,
77% на белки и 15% на углеводы (Lloyd, 1969).
Колебание в содержании неорганических ионов указывает на двойную-
природу слизи: часть ее является ультрафильтратом крови, а другая часть —
продуктом секреции слизистых желез. Этот вывод подтверждается обнару-
жением прямой зависимости между выделением слизи и гидростатическим,
давлением крови (Machin, 1964).
Вязкость слизи тесно зависит от содержания двухвалентных ионов.
Так, густая белая слизь, выделяемая при раздражении Deroceras reticulatum,.
очень богата солями кальция (Runham, Hunter, 1970).
У некоторых видов слизь содержит пигмент. Именно поэтому у Mala-
colimax tenellus, Limax jlavus и Arion subfuscus она имеет желтый цвет.
В жизни слизней слизь играет значительно более важную роль, чем у ули-
ток. Основное назначение ее — увлажнять покровы. Последнее необходимо
для успешного кожного дыхания и для поддержания покровов в таком со-
стоянии, чтобы они могли в свою очередь поглощать капельножидкую воду
и тем самым восстанавливать водный баланс животного.
В условиях перегрева и низкой относительной влажности выделение-
слизи и связанное с этим испарение воды способствует понижению темпера-
туры тела; тем самым на какой-то период осуществляется пассивная термо-
регуляция. Так, например, показано, что Arion ater способен при температуре
33.7° и влажности воздуха 24% поддерживать некоторое время температуру
тела на уровне 21°, т. е. значительно ниже верхней летальной границы (Hog-
hen, Kirk, 1944).
Известно, что в жаркое время дня и в засушливое лето многие слизни
прячутся в различные укрытия (под камни, в трещины почвы и др.), где-
окружают себя чехлом из вязкой слизи, и пережидают неблагоприятное время.
Очень важную роль играет слизь при движении. Вязкий секрет педальной
железы, выделяясь у переднего конца подошвы, предохраняет ее от трения,.
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
23
создавая «дорожку», по которой движется животное. В свою очередь жидкая
слизь, выделяемая покровами подошвы, способствует отрыванию животного
от вязкой «дорожки».
Помимо указанных главных функций слизи она имеет и ряд дополнитель-
ных. Стекая по поверхности тела, вокруг которого она образует как бы чехол,
она смывает посторонние твердые частицы, нейтрализует различные раз-
дражающие химические вещества, смывает споры болезнетворных грибков
и микробов. У многих видов слизь служит успешной защитой от хищников
как в силу своих ядовитых свойств, так и из-за вязкости ее, затрудняющей
действие нападающего.
Соединительная ткань. В сочетании с наружным эпителием
и подкожной мускулатурой соединительная ткань — непременный элемент
стенок тела моллюсков. Кроме того, она покрывает поверхность и входит
в состав стенок большинства органов внутренностного мешка, нередко обра-
зуя перемычки между ними, так называемые мезентериальные связки, а также
формирует стенки лакун, синусов и всего гемоцеля. Наиболее плотные
скопления соединительной ткани сосредоточены в ноге й щупальцах. Всюду
эта ткань слагается из клеточных элементов — фибробласты, хроматофоры,
вакуолизированные и слизистые клетки — и волокон. Последние химически
сходны с коллагеновыми волокнами позвоночных животных (Franc, 1968).
Мускулатура. У легочных моллюсков мускулатура тела слагается
из двух основных систем: подкожной и колумеллярной. Производными пер-
вой являются мощная и сложная мускулатура ноги и диафрагма, а второй —
половой ретрактор.
Если у улиток подкожная мускулатура сравнительно слабая, особенно
в области шеи, где она слагается из рыхлых слоев кольцевых и продольных
волокон, то у слизней на всех участках стенок тела она развита хорошо
и образует характерный кожно-мускульный мешок, слагающийся из мощных
слоев кольцевых и продольных мускульных волокон.
jy улиток задний отдел цефалоподиума в основном заполнен дорсовент-
ральной мускулатурой. В связи с размещением у большинства слизней в этой
части цефалоподиума внутренних органов дорсовентральная мускулатура
здесь исчезает. Только у Р armacellidae и у некоторых Trigonochlamydidae
эта мускулатура сохранилась, однако лишь в самом заднем конце цефало-
подиума.
Мускулатура ноги в значительной степени является продолжением му-
скулатуры стенок тела. Заходящие в ногу концы кольцевых волокон образуют
косую и поперечную мускулатуру. Продольная мускулатура этого органа
также является продолжением соответствующего слоя кожно-мускульного
мешка. Но, кроме них, нога обладает мощной дорсовентральной мускулату-
рой. Пересекаясь, все упомянутые виды мускульных волокон образуют гу-
стую и сложную объемную сеть, в которой залегают ряды кровеносных лакун.
Диафрагма слизней, как и у улиток, пронизана двумя слоями мускульных
волокон — продольными и поперечными. Если сама диафрагма — это спе-
циализированный участок верхней стенки тела, то ее мускулатура — это
дериват кожно-мускульного мешка, причем поперечная мускулатура воз-
никла за счет кольцевых волокон стенки тела.
У улиток колумеллярная мускулатура состоит из короткого общего ствола,
сзади крепящегося к столбику раковины, и его ветвей — ретракторов
(рис. 5, I): передний, задний, педальные, глоточный, щупальцевые (левый
и правый). У слизней она устроена значительно проще за счет исчезновения
переднего, заднего и педальных ретракторов (рис. 5, II). Оставшиеся ретрак-
торы сзади сливаются, образуя общий ствол, который крепится к верхней
стенке тела позади мантийного комплекса органов. Лишь у Philomycidae,
некоторых Arionidae (в частности, у Arion) и у Trigonochlamydidae ретрак-
24
ОБЩАЯ ЧАСТЬ
торы глотки и щупалец крепятся к стенке тела раздельно, не сливаясь в об-
щий ствол, и нередко в удалении от мантийного комплекса.
Ретрактор глотки впереди разделяется на две ветви, крепящиеся к глотке-
с двух сторон. В свою очередь ретракторы щупалец распадаются на ретрак-
торы губных и глазных щупалец. Кроме того, они дают несколько ветвей
к голове. Ретракторы щупалец проникают внутрь этих органов и крепятся к их
вершинам — особенность, характерная для всех Stylommatophora.
ст
гл п
гл
Рис. 5. Схема колумеллярной системы ретракторов тела улитки (/) и слизня (II).
гл — левый глоточный; а — задний; п — передний; пд — педальный; пр — половой; ст_столбик-
раковины улитки; щ — левый щупальцевый; ретракторы правой стороны глотки и правых щупалец не-
изображены; черным цветом даны мускулы, общие улитке и слизню.
Тип'ветвления колумеллярного мускула нередко варьирует даже в пре-
делах вида (Wiktor, 1973). Чаще всего от общего ствола сначала отделяется
ретрактор левых щупалец, а затем ретракторы глотки и правых щупалец.
Концевые ветви ретракторов щупалец обособляются в непосредственном
соседстве с теми органами и точками стенок тела, которые они обслуживают,
т. е. в области головы.
Несмотря на то что у большинства современных Stylommatophora, в том
числе и у многих слизней, половой ретрактор представляет собой специали-
зированный мускул, независимый от колумеллярной системы ретракторов,
по-видимому, он все же является ее производным. В пользу такой точки зре-
ния говорят достаточно многочисленные примеры, когда у некоторых улиток
он и сейчас представляет собой ветвь колумеллярного мускула (Лихарев,
1962). Среди наших слизней у Philomycidae этот мускул является ветвью рет-
рактора левых щупалец. Однако как у улиток, так и у слизней половой рет-
рактор чаще всего независим от колумеллярной мускулатуры. Обычно сзади
он крепится к диафрагме или вблизи от нее, а впереди к пенису или, реже,
к другим совокупительным органам.
Раковина и раковинная сумка. Одно из главных от-
личий слизней от улиток — это отсутствие на поверхности тела раковины..
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
25
Рудимент последней, если он сохранился, лежит в толще мантии в особой
полости — раковинной сумке. Чаще всего раковина имеет вид округло-
прямоугольной или неправильно овальной пластинки, состоящей из двух
основных частей: небольшой эмбриональной, или нуклеуса, и более крупной
дефинитивной, или спатула. Последняя сверху имеет более или менее за-
метную концентрическую исчерченность — линии нарастания. Вся верхняя
•сторона раковины покрыта тонким роговым слоем — периостракумом. Ниж-
няя сторона раковины обычно представляет собой группу кристаллов извести,
порой хаотически связанных друг с другом.
К числу немногих характерных особенностей раковины, позволяющих
различать отдельные группы слизней, относится нуклеус. Обычно это самая
высокая точка раковины (при рассматривании сбоку).
Можно различить три основных типа раковины. 1. Нуклеус лежит у зад-
него края раковины слева от продольной оси (Agriolimacidae, Limacidae,
Trigonochlamydidae). 2. Нуклеус лежит отступя от заднего края на продоль-
ной оси раковины (Milacidae). 3. В отличие от уже названных групп нуклеус
относительно очень крупный и спирально закручен (1 оборот) и лежит на
продольной оси раковины (Parmacellidae).
Процесс редукции раковины заканчивается на разных этапах. У боль-
шинства семейств она приобретает облик пластинки с ясным нуклеусом.
У некоторых нуклеус неразличим (Boettgerillidae), и раковина представляет
собой группу зерен, связанных между собой известковыми перемычками или
периостракумом. Эта связь между зернами может теряться,, и тогда отдель-
ные известковые конкреции залегают внутри раковинной сумки (Arion).
Завершают этот сравнительно-морфологический ряд Philomycidae, у которых
сохранилась крупная раковинная сумка, но без всяких следов раковины.
Размеры раковины зависят не только от возраста, но и от систематического
положения слизня. Чаще всего она достаточно велика, чтобы сверху при-
крывать органы мантийного комплекса, от которого ее отделяет нижняя
стенка раковинной сумки; задний край раковины выступает за пределы комп-
лекса (рис. 6). Только у Milacidae, Boettgerillidae и Eumilacinae (Limacidae)
раковина столь мала, что не может сверху полностью прикрыть мантийный
комплекс.
По-видимому, раковина придает известную эластичность верхней стенке
легочной полости, что очень важно для успешного засасывания и выталки-
вания воздуха во время дыхательных движений. Вероятно, той же цели слу-
жит и раковинная сумка, обычно заполненная какой-то жидкостью. Харак-
терно также^ что задний конец общего ствола колумеллярного мускула обычно
крепится к верхней стенке тела на уровне заднего края раковины. Все
это указывает на то, что у слизней рудиментарная раковина, утратив защит-
ную функцию наружной раковины улиток, частично сохранила свое зна-
чение как опорное образование.
Раковинная сумка хорошо развита даже в группах, лишенных рудимен-
тарной раковины (Philomycidae), что говорит о ее важном значении в жизни
.слизней. Вероятно, кроме придачи необходимой эластичности крыше легочной
полости она играет роль одного из депо, в котором сохраняется резерв воды.
У Parmacellidae задняя часть сумки имеет широкую связь с наружной
средой в течение всей жизни! животного. По-видимому, такая связь в виде
узкого микроскопического канала, открывающегося у заднего края мантии,
имеется и в других группах; в частности, он обнаружен у Limax maximus
и Lehmannia marginata (Tauber, 1900).
Дно раковинной сумки имеет кольцевидный валик и бороздку, выстланные
железистым цилиндрическим эпителием. Здесь происходит формирование
периостракума и известкового слоя раковины. Следовательно, как бороздка,
так и валик гомологичны таким же образованиям на крае мантии улиток.
IV
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
27
Рис. 6. Схемы мантийного комплекса органов у различных групп слизней.
1 — Meghimatium (Philomycidae), II — Arion (Arionidae); III — Tandonia (Milacidae); IV — Boett-
gerilla (Boettgerillidae); V — Limax (Limacidae); VI — Parmacella (Parmacellidae); VII — Trigonochlamys,
VIII — Selenochlamys (Trigonoclilamydidae). Слева — вид сверху, справа — вид снизу. Сердце, главный
ствол аорты и сосуды легкого зачернены, прямая кишка заштрихована в сетку, почка и мочеточник —
светлые, контуры раковины даны прерывистой линией, гр — гроздевидный орган; кл — общий ствол
колумеллярной системы ретракторов; лрщ — ретрактор левых щупалец; прщ — ретрактор правых
щупалец; ргл — ретрактор глотки; рп — половой ретрактор.
Легочная полость и мантийный комплекс о р г а-
п о в.1 Главная особенность легочной полости слизней заключается в том,
что у них она относительно меньших размеров, чему улиток. Если у послед-
них она обычно занимает большую часть нижнего оборота внутренностного
мешка, то у слизней она располагается только под задней половиной мантии.
Особенно бросается в глаза, что у этих животных она значительно короче,
чем у улиток. Меньшие размеры легочной полости не могли не сказаться на
форме и положении связанных с нею органов, т. е. на легком с дыхательной
сетью кровеносных сосудов, на сердце, почке с мочеточником и, наконец, на
прямой кишке. Подобно всем легочным моллюскам эти важнейшие для жизни
органы — так называемый мантийный комплекс — располагаются в толще
верхней стенки легкого, т. е. в мантии. Мускулистая нижняя стенка легочной
полости — диафрагма — чаще всего лишена каких-либо органов и основное
ее назначение — осуществлять циркуляцию воздуха через легочную полость.
Форма и положение отдельных частей (органов) мантийного комплекса в раз-
ных семействах слизней отличаются определенными особенностями (рис. 6)
и могут быть использованы для таксономических целей (Лихарев, Виктор,
1979а; Wiktor, Likharev, 1980).
Как и у улиток, легочная полость слизней сообщается с внешним миром
с помощью пневмостома — круглого отверстия на правом краю мантии,
в стенках которого располагается сфинктер, обеспечивающий его открывание
и закрывание. Непосредственно над пневмостомом располагаются анус
и выделительная пора.
Большая часть потолка легочной полости покрыта сложной системой
крупных и мелких складок, многократно анастомозирующих друг с другом.
В толще этих складок залегает сеть больших и малых кровеносных сосудов.
Сочетание складок и сосудов образует легкое Pulmonata.
1 Как в данном разделе, так и в остальном тексте книги, когда речь идет о положении
органов мантийного комплекса, понятия «направо», «налево», «под» или «над» употребля-
ются, исходя из истинного положения того или иного органа, а не той картины, которая
видна после вскрытия слизня и исследования этих органов с вентральной стороны.
28
ОБЩАЯ ЧАСТЬ
У улиток легкое в основном располагается перед сердцем. Размеры по-
верхности легкого, степень развития кровеносной сети и складок находятся
в тесной и прямой зависимости от размеров животного (Drozdowski, 1970).
В связи с относительно меньшими размерами, чем у улиток, легочной полости
слизней, кровеносная сеть этих животных обладает рядом особенностей.
Во-первых, в основном она располагается по одну сторону от сердца. Во-
вторых, если у улиток кровеносные сосуды легкого открываются в довольно
длинную легочную вену, в свою очередь впадающую в предсердие, то у слиз-
ней легочная вена очень короткая, так что иногда создается впечатление, что
кровь поступает в предсердие из нескольких вен. В-третьих, у крупных
слизней, кроме основной кровеносной сети, располагающейся на потолке
легочной полости, появляется дополнительная сеть на дне этой же полости,
т. е. на диафрагме (Arion ater, Limax maximus, Eumilax brandti и др.).
Установлено (Runham, Hunter, 1970), что у Pulmonata легкое обеспечивает
около одной трети общей потребности в кислороде. Остальное покрывается
за счет кожного дыхания. Так как поверхность легкого слизней относительно
меньше, чем у улиток, то кожное дыхание играет у этих животных еще более
значительную роль. В связи с этим чем крупнее слизни, тем сильнее развит
рельеф их кожи, т. е. морщины более крупные и выпуклые, а бороздки более
глубокие. Все это способствует увеличению поверхности тела, а следова-
тельно, усилению роли кожного дыхания.
У некоторых слизней, ведущих подземный образ жизни (Selenoehlamys
pallida, Troglolestes sokolovi), кровеносная сеть легкого претерпевает значи-
тельную редукцию, вплоть до полного исчезновения, и животные поглощают
кислород только через кожу. Характерно, что это мелкие слизни с особенно
сложным рельефом кожи. Например, у последнего вида мантия покрыта круп-
ными сосочками.
Как у всех моллюсков, сердце слизней (рис. 6) располагается внутри
перикардиальной сумки, которая узким реноперикардиальным отверстием
сообщается с полостью почки. Оно состоит из тонкостенного предсердия
и мускулистого желудочка. На границе между ними располагается клапан,
который регулирует поступление крови в желудочек и при сжатии его пре-
дупреждает отток крови назад в предсердие.1
У улиток сердце располагается на потолке легочной полости слева от
основания почки. Ось его обычно более или менее параллельна продольной
оси нижнего оборота внутренностного мешка. У слизней сердце занимает
иное положение. Во-первых, в связи с частичным погружением почки вглубь
внутренностного мешка оно смещается к ее переднему краю. Во-вторых,
ось сердца располагается косо к оси тела. Чаще всего она наклонена направо
под углом от 45 до 90°, а у Trigonochlamydidae этот поворот (деторсия) может
достигать еще больших размеров, так что у некоторых видов желудочек ока-
зывается перед предсердием, а сеть сосудов легкого позади сердца. Это явление
вслед за Плате (Plate, 1891) называют опистопневмонией; оно связано с по-
воротом всего мантийного комплекса направо на 180°. Больший или меньший
наклон сердца направо характерен почти всем семействам слизней. Только
у Philomycidae и Arionidae (рис. 6,1, II) ось сердца наклонена налево (торсия),
т. е. от продольной оси тела. Вероятно, такое отступление от обычной
картины коррелятивно связано с антемедиальным положением пневмостома,
в то время как у остальных семейств он располагается постмедиально или
почти медиально.
В заключение отметим, что у Рarmacellidae перед предсердием распола-
гается своеобразный гроздевидный орган неизвестного происхождения и на-
значения (рис. 6, VI, ГР).
1 Подробнее строение сердца и всей кровеносной системы изложено ниже (с. 30)..
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
39'
Выделительная система слизней, как и других легочных моллюсков,
состоит из почки и мочеточника. Последний (особенность, характерная только-
для слизней) нередко открывается наружу не непосредственно, как у улиток,
а через мочевой пузырь.
У большинства улиток почка имеет треугольную форму и лежит на по-
толке легочной полости вблизи от ее задней стенки, вершиной вперед; длина
ее заметно больше ширины.1 У слизней почка тоже располагается в задней
части потолка легочной полости, но нередко смещена за пределы последней
в толщу внутренностного мешка или под диафрагму. При этом меняется ее
форма. Вместо треугольной она становится полулунной, бобовидной или не-
правильной и вытянута поперек оси тела. Впереди обычно она имеет углуб-
ление, в котором размещается сердце (рис. 6, III—VII). Такая почка обычна.
Рис. 7. Мантия Dvroceras reticulatum (из: Runham, Hunter, 1970).
а — анус; во — выделительное отверстие; к — клапан мочеточника, м — мантия; мп — мочевой пузырь;
мч — мочеточник; мщ — мантийная щель; пн — пневмостом; р — ректум.
для большинства семейств слизней. Исключение составляют Philomycidae и
Arionidae, у которых почка кольцом окружает сердце или почти замкнута
вокруг него (рис. 6, I, II).
От почки берет начало мочеточник. Как известно, у большинства Geophila,
к которым также принадлежат и все исследованные в данной работе слизни,
мочеточник слагается из двух отделов: первичного, берущего начало у вершины
почки и тянущегося назад вдоль нее, и вторичного, простирающегося вперед
вдоль прямой кишки.
При вскрытии слизней первичный мочеточник плохо различим. Видимая
часть мочеточника (возможно, это вторичный мочеточник) обычно начинается
у заднего края почки, сближается с прямой кишкой и далее вдоль ее левой
стороны тянется вперед, где открывается наружу рядом с анусом щелевидной
выделительной порой.
У Agriolimacidae, Boettgerillidae и Limacidae мочеточник открывается на-
ружу не непосредственно, а через более или менее крупный мочевой пузырь.
Нередко на границе между ними имеется плоский клапан, который, по-
видимому, служит для предупреждения обратного попадания продуктов
экскреции в мочеточник (рис. 7). Очень часто мочевой пузырь подтянут к стенке
почки особой связкой (возможно мускул).
Почка Agriolimacidae и Milacidae (рис. 6, III) отличается от других групп
слизней присутствием крупного языковидного выступа — лобуса, который
расположен вентрально от основной части почки и вытянут направо, нередко
заканчиваясь под прямой кишкой.
Внутренние стенки почки несут многочисленные крупные складки.
Особый венозный синус входит в нее сзади иЛи со стороны вершины лобуса.
1 Речь идет об улитках с многооборотной раковиной, без всякой тенденции к ее ре-
дукции.
SO
ОБЩАЯ ЧАСТЬ
л распадается на серию мелких сосудов, пронизывающих толщу складок.
Впереди эти сосуды собираются в почечную вену, открывающуюся в пред-
сердие (Garner, 1970).
Физиологические и биохимические аспекты экскреции у слизней изучены
много слабее, чем у улиток. Наиболее точные исследования проведены на
Deroceras reticulatum(Garner, 1970; Runham, Hunter, 1970). В моче этого слизня
азот представлен главным образом пуринами, а именно мочевой кислотой
и ксантином (около 92%). Обнаруженные у Helicidae уреаза и мочевина, по-
видимому, не свойственны слизням, а аммиак занимает совершенно незна-
чительное место. Как у других наземных моллюсков, подобно насекомым
и птицам, у слизней в мочеточнике происходит реабсорбция воды и продукты
выделения выходят наружу в виде «сухой мочи».
К мантийному комплексу принадлежит также прямая кишка — ректум.
-Это последний участок задней кишки перед анусом, отличающийся не только
положением (вся остальная кишка входит в состав внутренностного мешка),
но и более узким диаметром. Он пронизывает диафрагму чаще всего на уровне
-заднего края почки и располагается на правой стороне мантийного комплекса,
прирастая к стенке тела по внешнему краю.
Все изменения, связанные с поворотом мантийного комплекса в ту или
иную сторону или смещением его к заднему концу тела, обязательно захва-
тывают и ректум. В Ьстальном эта часть кишечника не отличается от анало-
гичной у улиток.
Кровеносная система. Как у большинства Pulmonata,
кровеносная система слизней подразделяется на три основных раздела:
сердце с легочной сетью, артериальная система сосудов, венозная система
сосудов, лакун и синусов.
Сердце состоит из тонкостенного, слабо мускулизованного предсердия
и толстостенного желудочка, стенки которого сложены густой сетью мускуль-
ных волокон. Снаружи сердце одето тонким эпителием — эпикардиумом,
который является продолжением эпителия перикардиальной сумки. Внут-
ренняя поверхность сердца, как и кровеносных сосудов, лишена эпителиаль-
ной выстилки, т. е. эндотелия. В месте соединения предсердия и желудочка
имеется кольцо плотной соединительной ткани, в котором крепятся концы
мускульных волокон сердца. Здесь же располагается атрио-вентрикулярный
клапан, состоящий из двух лепестков. Другой клапан находится между же-
лудочком и основанием аорты. При систоле (сжатии) желудочка закрывается
атрио-вентрикулярный клапан, открывается клапан аорты и кровь послу-
шает в артериальную систему. Одновременно происходит заполнение кровью
предсердия через легочную вену. При диастоле (расжатии) желудочка
атрио-вентрикулярный клапан открывается, предсердие сжимается, и кровь
перегоняется в желудочек.
С аорты начинается артериальная система кровообращения (рис. 8).
Длина аорты различная: у одних она дихотомирует сразу после того, как ее
обогнет спереди кишка, у других несколько позднее. Во всех случаях она
распадается на два главных артериальных сосуда — переднюю и заднюю
-аорты. Последняя снабжает кровью печень, кишку (кроме ректума) и гонаду.
Первая, т. е. передняя, аорта снабжает кровью все остальные внутренние
органы, а также ногу и стенки тела.
Обе ветви аорты распадаются на крупные артерии, которые в свою
очередь разветвляются на более мелкие, заканчиваясь сетью капилляров.
Из капилляров кровь поступает в систему мелких лакун, расположенных
в толще соединительной ткани внутренних органов и стенок тела. С них и на-
чинается венозная система. Из мелких лакун кровь собирается во все более
крупные лакуны и синусы. К самым крупным принадлежат головной и вис-
церальный (задний) синусы, боковые продольные синусы стенок тела (по
Рис. 8. Артериальная система кровообращения Deroceras reticulatum (из: Runham, Hunter,.
1970).
a s— аорта (главный ствол); аз — задняя аорта; ап — передняя аорта; асж — артерия слюнных желе»;
асп — артерия семяприемника; бж — белковая железа; з — гонада; газ — задняя артерия гениталий;.
зап — передняя артерия гениталий; гл — глотка; гла — артерия глотки; з — зоб; зап — задняя артерия'
печени; к кишка; мп — мочевой пузырь; на — артерия ноги; п — печень; па — артерия пениса;
пап — передняя артерия печени; пч — почка; с — сердце; сж — слюнные железы; сод — семяяйцевод;.
VHC — мозг.
32
ОБЩАЯ ЧАСТЬ
одному с каждой стороны), боковые и медиальный педальные синусы ноги.
Крупные размеры висцерального синуса — характерная особенность слиз-
ней. У улиток он имеет значительно меньшие размеры. Синусы и лакуны
образуют гемоцель животного.
Из всех синусов кровь собирается в кольцевой мантийный синус, окружаю-
щий легкое. Отсюда она поступает в легочные приносящие сосуды, которые
распадаются на капиллярную дыхательную сеть, где происходит насыщение
крови кислородом. Перед этим часть венозной крови по особым сосудам и ла-
кунам проходит через почку, после чего уже попадает в дыхательную сеть
легкого. Свежая кровь собирается в сеть выносящих легочных вен, которые,
сливаясь друг с другом, открываются в предсердие одним или несколькими
«отверстиями.
Рис. 9. Основные типы челюсти слизней.
Г — одонтогнатная (Arion)., II — оксигнатная (Deroceras), III — рудиментарная хищных слизней
(Hyrcanolestes).
У слизней сердце пульсирует более интенсивно, чем у улиток (Runham,
Hunter, 1970). Так, у Deroceras agreste с массой тела 0.213 г сердце делает
90.6 ударов в минуту, а у Arion ater с массой 16.6 г — 48.8 ударов. Уста-
новлено, что с возрастом частота биения сердца снижается.
Несмотря на столь интенсивную работу сердца, трудно поверить, что
только его усилием проталкивается кровь через все тело слизня, в том числе
через две системы капилляров (в органах и в легком). Поэтому, по-видимому,
правы те авторы (Jullien, Ripplinger, 1953), которые считают, что дыхательные
движения диафрагмы, передаваясь на висцеральный синус, в свою очередь
способствуют проталкиванию крови через легкое.
Пищеварительная система. Подобно всем Pulmonata,
у большинства слизней в связи с тем, что анус располагается в передней части
лела, пищеварительный тракт имеет вид более или менее спирально закру-
ченной петли. Впереди он начинается ртом, ведущим в ротовую или пред-
глоточную полость; за нею следует глотка, в которую открывается пара слюн-
ных желез; от глотки тянется назад узкий пищевод, переходящий в зоб
и желудок; в последний открываются протоки двух долей печени; за желуд-
ком начинается кишка, которая, образуя несколько петель и извивов, от-
крывается наружу заднепроходным отверстием или анусом.
Различия между отдельными группами слизней в строении пищевари-
тельной системы связаны в первую очередь с характером питания. Как будет
видно ниже, растительноядные пблифаги по целому ряду принципиальных
.особенностей четко отличаются от хищных слизней.
Рот расположен на передней стороне головы. Сверху и снизу он прикрыт
губами (верхняя и нижняя), а по бокам — двумя ротовыми лопастями. За
ними располагается короткая ротовая или предглоточная полость. Заднюю
«стенку ее образует хитиноидная челюсть, за которой начинается глотка.
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ 33
Как у большинства Geophila, челюсть слизней имеет вид более или менее
изогнутой дуги с острым внешним (нижним) и тупым внутренним (верхним)
краем, который погружен в толщу мягких тканей ротовой полости. У расти-
тельноядных слизней встречается два типа челюсти (рис. 9): одонтогнатная —
снаружи с четкими поперечными ребрышками и крупными складками, ко-
торые нередко выступают за пределы внешнего края в виде зубчиков (многие
Arionidae и Philomycidae)-, оксигнатная — снаружи с неясной поперечной
исчерченностью или тонкими складками, а на середине внешнего края с ту-
пым треугольным выступом (Agriolimacidae, Boettgerillidae, Limacidae, Mila-
cidae, Parmacellidae).
У хищных Trigonochlamydidae челюсть либо отсутствует (Selenochlamys,
Drilolestes и др.), либо очень слабо развита, тонкая, без срединного выступа
(Trigonochlamys и др.).
Глотка представляет собой крупный мускулистый орган, впереди слабо
отграниченный от ротовой полости. Особенно крупных размеров она достигает
у хищных TrigonochlamydidaeP Системой мускулов протракторов, ретенторов,
элеваторов и тенсоров она связана с различными точками передней и нижней
(педальной) стенок тела. Кроме того, пара сливающихся за глоткой ретрак-
торов соединяет ее с основной (колумеллярной) системой ретракторов.
Стенки глотки в свою очередь имеют сложную систему мускулов, обеспечи-
вающих работу одонтофора и радулы.
Внутренние стенки глотки выстланы цилиндрическим эпителием с мощной
кутикулой. Местами эпителий имеет ресничный покров. В частности, таким
эпителием одета так называемая пищевая бороздка, которая начинается у ос-
нования челюсти и по верхней стенке глотки простирается до пищевода.
Назначение ее — направлять частицы пищи внутрь пищевода.
С брюшной стороны большую часть полости глотки занимает крупный
и высокий язык (одонтофор). В толще его залегают хрящевидные образо-
вания, создающие ему опору и вместе с мускулами управляющие работой
радулы. Внешняя поверхность одонтофора покрыта радулой, состоящей
из тонкой роговой мембраны, на которой крепятся многочисленные ряды
зубов. Сзади радула уходит в полый выступ задней части глотки — влагалище
радулы. Здесь происходит формирование новых рядов зубов, которые по-
степенно сдвигаются вперед на смену стирающимся передним рядам.
Каждый зуб радулы состоит из двух основных частей (рис. 10): базаль-
ной пластинки, плотно прилегающей к роговой мембране, которая впереди
изгибается кверху и назад и переходит в зубную пластинку (собственно зуб);
последняя заканчивается одним или несколькими зубцами. Зубы образуют
правильные поперечные и продольные ряды. В пределах продольного ряда
характер зубов существенно не меняется, если не считать, что передние зубы
всегда заметно изношены. Что касается поперечных рядов, то у большинства
растительноядных слизней всегда достаточно четко видна дифференциация
зубов на три основных группы: срединные (по одному на середине каждого
ряда), боковые и краевые. Срединный (центральный, или медиальный) зуб
всегда симметричен, а боковые (латеральные) и краевые (маргинальные) асим-
метричны, так как чем более они удалены от срединного, тем под большим
углом их зубные пластинки вытянуты в его сторону.
Срединный зуб обычно имеет три зубца, но нередко и однозубцовый.
В первом случае различают длинный мезокон (медиальный, или основной,
зубец), по сторонам которого располагаются по одному дополнительному
зубцу. У боковых зубов дополнительные зубцы расположены асимметрично
и разные по величине; их обозначают: эктокон — дополнительный зубец на
внешней (латеральной) стороне зуба; эндокон — зубец на внутренней (ме-
1 Об особенностях строения глотки этих слизней см. с. 95.
.3 И. М. Лихарев, А. Й. Виктор
34
ОБЩАЯ ЧАСТЬ
диальной) стороне зуба. У краевых зубов эндоконы отсутствуют, а число-
эктоконов, если они имеются, может быть от 1 до 5.
Если граница между срединным и боковыми зубами хорошо заметна из-за
их различий в форме, то граница между боковыми и краевыми сглажена за
счет достаточно плавного перехода от одного типа зубов к другому. Обычно-
она более или менее совпадает с боковыми продольными перегибами всей
радулы. Эти два перегиба (правый и левый) связаны с формой одонтофора
и положением хрящевой основы этого органа. Если срединные и боковые-
Л 3
Рис. 10. Основные типы зубов радулы.
I — срединный (сверху и сбоку), II — боковой, III — краевой растительноядных слизней; IV — зуб-
хищных слизней. 1 — базальная пластинка, 2 — зубная пластинка, з — мезокон, 4 — эктокон, 5 —
эндокон.
зубы располагаются на выпуклой части одонтофора, то краевые лежат на его-
боковых склонах.
У растительноядных слизней срединные и боковые зубы сравнительно-
однообразны. Различия между видами обычно сводятся к присутствию или
отсутствию дополнительных зубцов, к размерам и форме мезо-, экто- и эндо-
конов. Срединные зубы чаще всего трехзубцовые, реже однозубцовые, т. е.
только с одним, обычно мощным, мезоконом. Боковые зубы двух- или трех-
зубцовые и обычно они в направлении от оси радулы к краевым зубам
постепенно утрачивают эндокон (если он изначально присутствовал).
Более разнообразно строение краевых зубов. Исходя в первую очередь-
из их особенностей можно наметить два основных плана строения радулы
растительноядных слизней.1
1. Арионоидная радула (рис. 11, I—III). Базальные пластинки краевых
зубов в продольном направлении короткие, в поперечном Широкие. Срединный
1 Ниже мы не учитываем 4—5 самых внешних краевых зубов, так как они сохраняют
зачаточный облик и слабо дифференцированы.
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
35
зуб „чаще всего трехзубцовый; боковые и краевые двухзубцовые (мезокон
и эктокон); плавно переходя друг в друга, они различаются тем, что у боко-
вых зубные пластинки длинные, у краевых заметно короче. Радулярная
формула:
с | б к
"3 + ~2 + ~2 •
Такая радула характерна для многих Philomycidae и Arionidae.
2. Лимакоидная радула. Базальные пластинки краевых зубов в продоль-
ном направлении длинные, в поперечном узкие. Боковые и краевые зубы четко
отличаются друг от друга, хотя на стыке их могут быть зубы переходной формы.
Как правило, у краевых зубов зубная пластинка более длинная, чем у бо-
ковых. Такая радула характерна для Agriolimacidae, Boettgerillidae, Lima-
cidae, Milacidae, Parmacellidae.
В пределах данного плана строения радулы можно наметить несколько ти-
пов этого органа.
2а. Кинжаловидный тип (рис. 11, IV— FZ). Зубные пластинки краевых
зубов однозубцовые и имеют кинжало- или шиповидный облик. У немногих
видов все или отдельные краевые зубы с небольшим эктоконом. Срединный
и боковые зубы с хорошо развитыми экто- и эндоконами и остроязыковид-
ными мезоконами. Радулярная формула (в скобках указаны сравнительно
редкие варианты):
с , б , к
Т + ~2 + 1 (2) '•
Этот тип радулы наиболее обычен среди слизней. Он характерен для всех
Agriolimacidae (за исключением Deroceras osseticum), Milacidae и Parmacel-
lidae, а также для большинства Limacidae.
26. Сигмовидный тип (рис. 11, VII—IX). Зубные пластинки краевых зу-
бов 5-образно изогнутые, с несколькими (до 5) эктоконами. Срединный и бо-
ковые зубы обнаруживают тенденцию к утрате дополнительных зубцов
и к сильному развитию мезоконов. Радулярная формула:
с , б , к
3(1) + 2 (1) + 2 — 5 •
Подобную радулу имеют слизни из рода Lehmannia.
2в. Полидентикулярный тип (рис. И, X—XII). Зубные пластинки крае-
вых зубов прямые (неизогнуты S-образно!). У самых внешних краевых зубов
до 5 эктоконов; внутренние краевые зубы двухзубцовые (мезо- и эктокон).
Срединный и боковые зубы с хорошо развитыми экто- и эндоконами; мезоконы
конические или остроязыковидные. Радулярная формула:
е , б , к к
з+т + т + 3 — 5 •
Такая радула свойственна Boettgerilla pallens, Gigantomilax lederi, Limax
flavus. Сходная картина у Deroceras osseticum.
2г. Переходный тип. Внешние краевые зубы двухзубцовые; мезокон и
эктокон почти одной длины. Внутренние краевые зубы ножевидные. Средин-
ный и боковые зубы со слабо развитыми экто- и эндоконами и остроязыко-
видным мезоконом. Радулярная формула:
с б к к
3 + 3 +т + 2 •
3*
36
ОБЩАЯ ЧАСТЬ
Подобная радула отмечена у Bielzia coerulans, Turcomilax tzvetkovi,
Caspilimax keyserlingi, Malacolimax tenellus. У последнего вида иногда встре-
чаются полидентикулярные внешние краевые зубы (Flasar, 1961).
В итоге можно сделать вывод, что у слизней с лимакоидной радулой
(Limacoidea и Zonitoided) в основном этот орган устроен по кинжаловид-
Рис. 11. Зубы радул растительноядных слизней (фото Юнгблута).
I—III — Arion subfuscus: I — краевые и внешние боковые (X 500), II — краевые (Х1000), 111 — сре-
динные и боковые (х2000); IV—VI — Deroceras agreste: IV — боковые и краевые (х 1000), V — краевые
(Х1000), VI — срединные и боковые (Х2000); VII—IX— Lehmannia marginata: VII — внутренние
краевые (х2000), VIII— внешние краевые (Х1000), IX — срединные и боковые (Х2000); X—XII —
Gigantomilax lederi: X — срединные и боковые (х800), XI — боковые и внутренние краевые (х950);
XII — внешние краевые (х850).
ному типу. Лишь у некоторых Limacidae и Boettgerillidae и у одного вида
Agriolimacidae (Deroceras osseticum) встречаются другие типы радулы, ко-
торые, по-видимому, можно считать производными кинжаловидной радулы.
Вероятно, эти отклонения явились результатом адаптации к питанию ка-
кими-то определенными типами пищи.
Принципиально иной тип радулы свойствен хищным наземным моллю-
скам, в том числе и слизням Trigonochlamydidae (рис. 12). От радул расти-
тельноядных форм он в первую очередь отличается отсутствием диффе-
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
37
ренциации зубных пластинок в пределах поперечных рядов. Все зубы имеют
кинжаловидную, на конце заостренную зубную пластинку, переходящую!
в базальную, по контурам напоминающую цифру 8 (рис. 10, IV). У некоторых
родов (Parmacellilla, Troglolestes, Selenoehlamys) близ конца зубной пластинки:
Рио. 11 (продолжение).
имеется небольшой зубец, ориентированный назад (т. е. к основанию зуба)
и придающий зубу багоровидную форму (рис. 12, IV—VI).
Другая отличительная черта радулы хищников—небольшое число зубов
в поперечном ряду: от 40 (Hyrcanolestes, Drtlolestes, Selenoehlamys) до 52—64
(Trigonochlamys). В то же время у растительноядных слизней число зубов
в одном поперечном ряду значительно больше: от 93 у самых мелких видов
(например, Deroceras laeve) до 161 у крупных (например, Limax cinereoniger);
Наконец, третьей особенностью радул хищных слизней является то,
что зубы у них значительно крупнее, чем у растительноядных. Так, у Tri-
gonochlamys зубные пластинки достигают в длину до 300 мкм, а у Selenoch-
lamys — до 440 мкм, в то время как у растительноядных Deroceras laeve
38
ОБЩАЯ ЧАСТЬ
длина самых больших шиповидных краевых зубов только 26, а у крупного
Limax cinereoniger — до 48 мкм.
Перечисленные различия в форме, числе и размерах зубов у раститель-
ноядных и хищных слизней связаны с особенностями механики работы
Рис. 12. Зубы радул хищных слизней (фото Юнгблута).
I—III — Trigonoehlamys imitatrix: I — несколько поперечных рядов (х38), II — внешняя часть рядов
(Х75), III — фрагмент одного ряда у середины (Х375); IV— VI — Selenoch.la.mys pallida: IV — не-
сколько поперечных рядов (Х38), V — внешняя часть рядов (х75), VI— участок у середины ряда
(Х150).
радулы. У первых после ощупывания пищевого объекта губными щупаль-
цами и ротовыми лопастями и определения его пригодности рот раскрывается
и выдвигается вперед передняя, т. е. приподнятая, часть одонтофора. Одно-
временно зубы радулы, которые до этого прилегали к поверхности языка
зубцами внутрь глотки, приподнимаются и зубцами ориентируются вперед.
Голова слизня делает короткие толчки вперед, и радула, как терка, скребет
поверхность пищевого объекта. При этом челюсть в свою очередь прижи-
мает пищевой объект к радуле и нижним краем помогает ей размельчать его.
Частицы пищи заталкиваются в глубь глотки периодическими втягиваниями
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ 39
одонтофора и с помощью ресничек пищевой бороздки направляются к пище-
воду.
У хищных Trigonochlamydidae радула работает иначе (Шилейко, 1966).
Она выдвигается вперед сильнее, чем у растительноядных слизней. Зубы ее
раздвигаются и вонзаются в тело добычи (почвенные черви). После этого
по мере переваривания заглоченной части червя радула, не освобождая его,
медленно втягивается внутрь глотки, затягивая туда новые участки тела
жертвы. Если добыча крупная, например дождевой червь, то этот процесс
занимает много часов и может неоднократно повторяться до полного втя-
гивания червя внутрь тела слизня.
Следовательно, если у растительноядных слизней радула служит для
размельчения пищевого объекта и- подачи частиц пищи внутрь глотки,
то у хищных она выполняет роль органа захвата и заглатывания добычи
целиком или крупными кусками.
К зобу прилегает пара крупных лопастевидных слюнных желез, которые
двумя длинными протоками открываются в глотку по сторонам от начала
пищевода. Секрет их слабо изучен, но известно, что у Arion ater и Deroce-
ras reticulatum он содержит амилазу (Runham, Hunter, 1970).
В отличие от улиток, у которых пищевод обычно довольно длинный,
у слизней он заметно короче. Внутри него имеются 6 продольных складок;
на спинной стороне между ними располагается бороздка, выстланная реснич-
ным эпителием, являющаяся продолжением аналогичной бороздки глотки
(см. выше) и также служащая для транспортировки пищевых частиц (Roach,
1968). Под эпителиальной выстилкой пищевода располагаются два слоя коль-
цевой мускулатуры, обеспечивающих перистальтику этого органа.
Пищевод переходит в зоб. При вскрытии слизней этот орРан сразу бро-
сается в глаза благодаря его величине и жидкому содержимому желтого,
красного или коричневого цвета. Обычно он слабо отграничен от пищевода и,
по-видимому, является специализированным участком последнего. Он
выстлан внутри цилиндрическим эпителием, отдельные участки которого
покрыты ресничками и содержат слизистые клетки. Под эпителием распо-
лагаются два тонких слоя мышц, обеспечивающих перистальтику и продоль-
ное сокращение всего зоба, что необходимо для перемешивания жидкости
и перетирания твердых частиц пищи. Жидкость зоба содержит большое
число пищеварительных энзимов (Runham, Hunter, 1970).
За зобом располагается желудок. Внешне он не всегда четко отграничен
от зоба. Это небольшой мешковидный орган, на спинной стороне которого
позади отверстий протоков печени имеется характерный комплекс складок
(рис. 13). Этот комплекс состоит из треугольной складки, лежащей поперек
желудка между отверстиями протоков печени^ и двух продольных складок,
сходящихся к началу кишки. Все три складки образуют два желобка, на-
чинающихся от отверстий печени и сзади сходящихся в общий медиальный
желобок — тифлозоль, уходящий в глубь кишки. От каждого печеночного
отверстия радиально расходятся небольшие короткие складки. Все складки
и стенки задней части желудка выстланы цилиндрическим ресничным эпи-
телием, под которым залегают слои кольцевых мускульных волокон.
Исследования на Deroceras reticulatum показали (Walker, 1972), что твер-
дые частицы пищи размерами более 0.5 мкм не проникают внутрь печени
через ее протоки. Ресничный эпителий желудка по определенным путям
направляет такие частицы назад в сторону кишки. Однако в задней части
желудка имеются участки эпителия, реснички которого работают в обратном
направлении, т. е. толкают частицы пищи вперед. В области задней части
тифлозоля оба потока встречаются, и здесь начинается формирование фека-
лий — основное назначение тифлозоля.
40
ОБЩАЯ ЧАСТЬ
Печень состоит из двух долей, каждая из которых самостоятельным про-
током открывается в желудок. В свою очередь доли печени состоят из ло-
пастей, число которых, по-видимому, видоспецифично. Так, у Deroceras
reticulatum передняя доля состоит из 9, а задняя из 6 лопастей. Лопасти об-
разованы многочисленными ацинями, от которых отходят короткие протоки.
Последние, сливаясь друг с другом, образуют протоки лопастей, в свою оче-
редь соединяющиеся в протоки долей.
Рис. 13. Движение пищевых частиц в желудке Deroceras reticulatum. (из: Runham, Hunter,
1970).
1 — направление движения ресничек эпителия на брюшной стороне; 11 — то же на спинной (между
валиками тифлозоля началось формирование фекального шнура); 111 — поперечный разрез через тифло-
золь.
> Все эти протоки выстланы внутри ресничным эпителием, бьющим в сто-
рону желудка. Эпителий протоков и ациний окружает кольцевая мускула-
тура, обеспечивающая перистальтику стенок. Ацини состоят из четырех
типов клеток: пищеварительных, кальциевых, экскреторных и узких (Wal-
ker, 1969).
Сравнение экстракта печени с жидким содержимым зоба показало, что
они имеют много общих энзимов и что величина pH сходна или идентична.
Поэтому ряд авторов (Evans, Jones, 1962; Walker, 1969) считает, что эти
энзимы выделяются печенью. Однако нет прямых наблюдений, указываю-
щих на поступление энзимов из печени в желудок и зоб. Не исключено, что
ряд энзимов, обнаруженных в экстракте печени, служит только для внутри-
клеточного или пристеночного пищеварения. В то же время некоторые
энзимы жидкости зоба могут быть продуктами экскреции слюнных желез,
стенок зоба или его бактериальной флоры (Runham, Hunter, 1970). Печень
служит также хранилищем запасных питательных веществ (гликоген и ли-
пиды). Известную роль она играет и в выделении продуктов обмена веществ.
' Как и у раковинных форм, кишка, начинаясь от желудка, дрлает не-
сколько петель и извивов среди других органов, почти не меняясь в диа-
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
41
метре. Лишь под самый конец, переходя в ректум, она несколько, сужается
и открывается наружу анусом. Последнее колено кишки, заканчивающееся
ректумом* (у Limacidae — последние Два колена), обозначают как заднюю'
кишку. Между нею и желудком располагается средняя кишка.
Исследования на слизнях Arion ater и Deroceras reticulatum показали, что.
внутреннее строение упомянутых отделов кишки различно (Runham, Hun-
ter, 1970).1 Начальный отдел средней кишки имеет 6 внутренних продольных
Рис. 14. Изменение формы кишечника при переходе от улитки (/) к растйтельноядным
(II, III) и хищным (IV, V) слизням.
складок. Они одеты ресничным цилиндрическим эпителием, среди клеток
которого разбросаны слизистые клетки. В заднем отделе средней кишки
указанные складки исчезают, а в эпителии появляются своеобразные клетки,
содержащие гранулы протеина, — так называемые интестинальные
клетки; функция их не ясна; возможно, они выделяют оболочку, в которую
заключен фекальный шнур.
Стенки задней кишки лишены интестинальных клеток, но имеют много
слизистых клеток. Внутренняя поверхность имеет мелкие складки, которые
по гребням снабжены ресничным эпителием. Внутренние стенки ректума
имеют несколько небольших продольных складок и одну крупную на боко-
вой стенке. Вся поверхность выстлана ресничным эпителием с многочислен-
ными слизистыми клетками. В стенках всех отделов кишки под эпителием{
залегают два слоя мускульных волокон, обеспечивающих ее перистальтику.
У многих растительноядных слизней задняя кишка дает назад слепой
выступ — слепая кишка. Гистологически она сходна с прямой кишкой,,
но эпителий ее внутренних стенок лишен ресничек. Функция ее не ясна..
1 К сожалению, пока нет сопоставимых данных по гистологии кишки других слиз-
ней и легочных улиток.
42
ОБЩАЯ ЧАСТЬ
Она нередко бывает заполнена жидкостью, и в ней всегда отсутствуют фека-
лии.
Форма пищеварительного тракта (кишечника) слизней в разных группах
обнаруживает свои особенности, важные в таксономическом отношении.
У улиток кишечник построен по следующему плану (рис. 14, Z): рот—
глотка—длинный пищевод—длинный зоб—небольшой желудок—сравни-
Рис. 15. Основные типы кишечника растительноядных слизней.
I — Arion*, II — Deroceras*, III — Limax, IV — Lehmannia.
тельно короткая S-образная средняя кишка—прямая и длинная задняя
кишка, простирающаяся по верхнему наружному краю 1—2 последних
оборотов внутренностного мешка,—ректум—анус. Характерно, что у назем-
ных улиток средняя кишка короткая и слабо извита.
Обратное имеет место у растительноядных слизней, для которых обычна
длинная средняя кишка, сильно закрученная вокруг зоба. У этих слизней
можно различать два основных плана строения кишечника (рис. 15).
1. Кишечник слагается из двух петель, каждая состоящая из двух ко-
лен: первая петля образована глоткой, пищеводом, зобом и желудком (1-е
колено) и началом средней кишки (2-е колено); вторая петля слагается из
задней части средней кишки (3-е колено) и задней кишки с ректумом (4-е ко-
лено). При переходе первой петли во вторую кишка огибает спереди аорту.
Задняя кишка располагается слева от основания колумеллярного мускула.
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
43
Двухпетлевый план строения пищеварительного тракта наиболее обычен
среди растительноядных слизней. Однако он обнаруживает несколько ва-
риантов. У одних (рис. 15, II) кишка в основном располагается позади срав-
нительно короткого зоба (Agriolimacidae, Boettgerillidae, Milacidae, Par-
macellidae). Такой план строения кишечника наиболее близок к тому, что мы.
встречаем у улиток.
У других групп слизней (рис. 15, I) зоб очень длинный, и кишка пол-
ностью располагается над и под ним, т. е. закручена вокруг него (Philo-
mycidae, Arionidae).
2. Кишечник слагается из трех петель (рис. 15, III, IV). Если у пред-
шествующих групп задняя киЩка располагалась слева от колумеллярного
мускула, то у слизней Limacidae она огибает его спереди и вытягивается
сначала назад, а затем, поворачивая вперед, к анусу, образует третью петлю
кишечника. При таком плане строения кишечника самое необычное —
это огибание кишкой колумеллярного мускула и размещение ее справа от
него. Такая ситуация более нигде не встречается не только среди слизней,
но и в классе Gastropoda.
Этот план строения кишечника также может усложняться за счет боль-
шего или меньшего закручивания кишки вокруг зоба. Последнее в особен-
ности характерно для слизней, питающихся грибами и лишайниками.
Следовательно, растительноядные слизни отличаются от улиток большей
длиной кишечника как за счет общего удлинения средней кишки и ее закру-
чивания вокруг зоба, так и за счет удлинения задней кишки путем формиро-
вания третьей петли (только у Limacidae).
Совершенно иное положение у хищных слизней Trigonochlamydidae
(рис. 14, IV, У). От двухпетлевого кишечника здесь сохраняется только
намек, так как кишка сильно укорачивается. Одновременно в связи с тем,
что в пределах этого семейства имеется тенденция к смещению мантийного
комплекса к заднему концу тела, которое захватывает и анус, кишка выпрям-
ляется и извивы ее почти сглаживаются.
К сожалению, процессы пищеварения у наземных улиток из надотряда
Stylommatophora изучены очень недостаточно. Соответствующие исследования
в основном посвящены выявлению пищеварительных энзимов и выяснению,
в каких органах у моллюсков происходит внеклеточное пищеварение, а где
внутриклеточное (Owen, 1966). Более детально и точнее изучено пищеваре-
ние у растительноядных слизней (Roach, 1968; Runham, Hunter, 1970;
Walker, 1972), в особенности у Arion ater и Deroceras reticulatum.
Установлено, что каждый отдел пищеварительного тракта обладает раз-
личной мышечной активностью. Особенно сложный комплекс движений со-
вершается в глотке и зобе. В последнем это связано с необходимостью много-
кратного перемешивания пищи и перетирания частиц ее друг о друга. Жид-
кость зоба содержит большое количество пищеварительных энзимов, являю-
щихся продуктами экскреции как слюнных желез, так, вероятно, и печени.
Возможно, что некоторые энзимы выделяются стенками зоба, а также его
бактериальной флорой.
В зобе происходит не только дальнейшее измельчение пищевых частиц,
но и внеклеточное переваривание их. В первую очередь здесь усваиваются
жиры, белки и полисахариды. За счет мускулатуры зоба и желудка оба
органа многократно обмениваются содержимым. Наиболее раздробленные
пищевые частицы размерами 0.1—0.5 мкм из желудка проникают в печень.
Этому способствуют мускульные усилия желудка и зоба и перистальтика
печеночных протоков.
В ацинях печени происходит фагоцитоз пищевых частиц, причем главную
роль здесь играют пищеварительные клетки. Кальциевые клетки накапли-
44
ОБЩАЯ ЧАСТЬ
вают кальций. Непереваренные остатки выводятся из печени в виде слизи-
стого шнура за счет биения ресничного покрова протоков.
гл
Рис. 16. Нервная система Deroceras reticulatum, (из: Runham, Hunter, 1970). Мозг зачернен.
бж — белковая>келеза; г — гонада; гл — глотка; з — зоб; зк — задняя кишка; -опт. — интенстиналь-
ный нерв; кпд — кожно-педальный нерв; нп — пениальный нерв; п — пенис; пал — паллиальный нерв;
пед — педальные нервы; печ — печень; пч — почка; рпк — реноперикардиальный нерв; сж — слюнная
железа; сод — семяяйцевод.
Функция кишки достаточно сложна. С одной стороны, здесь продолжа-
ется усвоение всех пригодных элементов пищи путем абсорбции на стенках,
с другой — идет постепенное обезвоживание непереваренных остатков и фор-
мирование фекального шнура.
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
45
В связи с отсутствием совершенного механизма для перетирания пище-
вых частиц значительная часть растительных клеток (особенно лежащих
в глубине частицы) остается непереваренной и выделяется с фекалиями.
Весь процесс пищеварения — от поедания пищи до выделения экскремен-
тов — занимает у Deroceras reticulatum около 8 часов, а у Arion ater около
12 часов. Характерно, что ректум обычно не содержит фекальных масс;
вероятно, они его проходят очень быстро.
Процессы пищеварения у хищных слизней пока совершенно не изучены.
Нервная система (рис. 16). Как у всех Pulmonata, нервная
система слизней слагается из двух основных отделов: центральной нервной
системы (ЦНС, или мозг) и периферической системы нервов; последние в не-
которых органах (нога, кишечник, гениталии) образуют сложные сети —
нервные плексусы. Мозг располагается позади глотки, в основном вокруг
пищевода (рис. 17). Только в роде Selenochlamys (Trigonochlamydidae) он
окружает переднюю часть глотки (см. рис. 444). В состав мозга входят 5 пар-
ных ганглиев — церебральные, буккальные, педальные, плевральные, па-
риетальные, и один непарный абдоминальный ганглий. Ганглии связаны
друг с другом нервными перемычками — комиссурами и коннективами.
В итоге мозг слагается из трех нервных дуг, отходящих от церебральных
ганглиев: две направлены вниз и назад (педальная и висцеральная дуги),
а одна — вперед к верхней стороне глотки (буккальная дуга).
Педальная дуга образована парой педальных ганглиев, связанных друг
с другом педальной комиссуррй, а с церебральными ганглиями церебро-
педальными коннективами (левая и правая). Висцеральная дуга слагается
из парных плевральных и париетальных ганглиев и из непарного абдоми-
нального (висцерального) ганглия; последний располагается более или менее
на середине дуги. Буккальную дугу образует пара буккальных ганглиев,
связанных друг с другом буккальной комиссурой, а с церебральными гангли-
ями церебро-буккальными коннективами (левая и правая). От каждого плев-
рального ганглия отходят две коннективы: одна ^церебральному ганглию
той же стороны, другая к педальному.
Церебральные ганглии располагаются не только над пищеводом, но и над
протоками слюнных желез. Остальные ганглии находятся под ними. Между
висцеральной и педальной дугами пролегает головная артерия. На верхней
стороне педальных ганглиев (ближе к париетальным) лежит пара стато-
цистов.
Церебральные ганглии состоят из трех отделов: переднего — процереб-
рум, среднего — мезоцеребрум и заднего — метацеребрум. Эти ганглии
иннервируют голову и е'е органы — губы, ротовые лопасти, щупальца,
глотку, а также статоцисты. К каждому глазному щупальцу отходят два
нерва: толстый обонятельный и тонкий зрительный (рис. 18). Педальные
ганглии дают многочисленные нервы к ноге. Среди них нередко выделяется
пара особенно мощных медиально лежащих нервов подошвы. Кроме того,
правый ганглий посылает нерв к пенису и атриуму. Чаще всего этот нерв
проходит через толщу правого церебрального ганглия, создавая впечатление,
что именно этот ганглий иннервирует пенис.
От плевральных ганглиев нервы не отходят — особенность всех Pulmo-
nata. Париетальные ганглии (главным образом правый) иннервируют стенки
тела, мантию и легкое. Абдоминальный ганглий посылает нервы к зобу,
желудку, печени, кишке, гениталиям (кроме пениса и атриума), к почке,
сердцу, аорте и колумеллярному мускулу). Буккальные ганглии иннерви-
руют глотку, пищевод и слюнные железы.
На слизнях особенно четко проявляются следующие две тенденции, ха-
рактерные для центральной нервной системы Stylommatophora.
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
47
Рис. 17. Мозг представителей 8 семейств слизней.
I — Meghimatium bilineatum (.Philomycidae); II — Arion rufus (Arionidae)', III — Deroceras retwulatum
•(AgriolimacidaeY, IV — Boettgerilla pattens (Boettgerillidae)', V — Eumilax brandti (.Limacidae); VI — Parma-
cella ibera (Parmacellidae)', VII — Tandonia kusceri (Milacidaey, VIII — Hyrcanolestes velitaris (Trigono-
chlamydidae) (V — вид сбоку; остальные — сверху), аг — головная артерия; га — абдоминальный
ганглий; гб — буккальные ганглии; гп — педальные ганглии; гпл — плевральные ганглии; гпр — парие-
тальные ганглии; гц — церебральные ганглии; нг — головные нервы; нгл — глоточные нервы;
нин — интестинальные нервы; ним — нервы колумеллярной системы; ноб — обонятельный нерв
глазных щупалец; нп — педальные нервы; нпа, — нерв передней части мантийного комплекса; нпа2 —
нерв задней части комплекса; нож — нервы слюнных желез; нет — нервы стенок тела; н»|2 — нерв
губных щупалец; ст — статоцист.
1. Увеличение размеров церебральных ганглиев и в первую очередь
процеребрума.1 Эти парные образования, которые среди Gastropoda имеются
только у Pulmonata, наиболее высокого развития достигают в надотряде
Stylommatophora (Van Mol, 1967). Процеребрум в первую очередь обслужи-
вает обонятельную функцию обеих пар щупалец. Именно здесь залегают
нейроны, аксоны которых образуют обонятельные нервы и сливаются с ганг-
лиями вершин щупалец. Зрительные нервы слагаются из отростков ней-
ронов, залегающих в толще задних отделов церебральных ганглиев — мета-
церебрумах. /
Особенно сильного развития достигает процеребрум у Boettgerilla и
у Tandonia. Значительно слабее эти отделы развиты у Meghimatium и у Arion
(рис. 17).
2. Прогрессирующее слияние ганглиев висцеральной дуги — другая
отличительная особенность Stylommatophora. Оно может достигать раз-
личной степени. Среди наших слизней максимальное слияние имеет место
у Eumilax, Tandonia и Boettgerilla, у которых внешне границы между гангли-
ями висцеральной дуги не различимы. Меньшее слияние свойственно мозгу
Arion, Meghimatium и Deroceras.
Только некоторым хищным улиткам и слизням, в том числе Trigonochla-
mydidae (рис. 17, VIII), свойственно сильное увеличение длины церебраль-
ной комиссуры. Поэтому в состоянии покоя все окологлоточное кольцо
1 До недавнего времени эти участки церебральных ганглиев называли глазными
лопастями, что, как выяснилось, не соответствует их функции.
1
в
ft
та
9
S
о
18.
Строение глазного щупальца и клетки х.«1ииы (//) ,.„atum (и
Runham, Hunter, 1970).
пополнительная ретина; г — хрусталик; з — основная ретина; 4 — зрительный нерв; 5 — ретрак-
тор щупальца; в — пальцевидный (обонятельный) ганглий; 7 — воротничковые клетки; 8 — обонятель-
ный нерв; 9 — полость щупальца (участок гемоцеля); ю — звездчатая клетка; н — овальная клетка;
12 — микроворсинки; 13 — пигментная клетка; 14 — светочувствительная клетка.
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
49'
как бы свободно повисает на пищеводе. Удлинение комиссуры связано с ха-
рактером питания этих слизней (стр. 95), при котором крупные не измель-
ченные радулой куски добычи (например, куски дождевого червя) проходят-
через пищевод, сильно раздувая его в поперечном сечении. Благодаря длин-
ной церебральной комиссуре окологлоточное нервное кольцо не мешает
проходу пищевого комка через пищевод.
Органы чувств. В связи с тем что у слизней практически вся
поверхность тела открыта прямому воздействию внешних факторов, много-
численные чувствительные клетки разбросаны почти по всей поверхности
тела. Особенно их много на голове, шее и подошве. Однако ведущее зна-
чение имеют органы чувств, расположенные на щупальцах, как глазных,,
так и губных.
Каждое щупальце (рис. 18) представляет собой полый цилиндрический
выступ головы с несколько вздутой (особенно у глазных щупалец) вершиной,
в который заходит участок головного синуса гемоцеля. Эпителий щупалец.,
содержит многочисленные слизистые клетки. В отдельных местах под ним
располагаются своеобразные овальные и звездчатые клетки, функция кото-
рых не ясна. Внутри щупальца залегает цилиндрический ретрактор, который
дистальным концом срастается с вершиной щупальца. При сокращении этот
мускул обеспечивает погружение внутрь вершины щупальца, вплоть до
полного вворачивания и втягивания его внутрь тела. Другие мускулы,
крепящиеся у основания щупальца, управляют движением всего органа.
Каждый щупальцевый ретрактор имеет форму полой трубки, внутри которой
залегают нерв щупальца, а у глазного щупальца два нерва — обонятельный
и зрительный (рис. 18, 4, 8).
В верхнем конце каждого глазного щупальца находится щупальцевый
(пальцевидный) ганглий, у поверхности которого располага'ются тела много-
численных нейронов. Каждый нейрон посылает отростки, с одной стороны,,
к сенсорным клеткам эпителия вершины щупальца, с другой — к обонятель-
ному нерву, заканчивающемуся в процеребруме.
Щупальцевые ганглии глазных щупалец — это основные центры обоня-
ния, которое у слизней развито сравнительно хорошо. Так, Arion ater спо-
собен различать запах отдельных съедобных грибов на расстоянии 120 см
и более (Kittel, 1956; Runham, Hunter, 1970). Губные щупальца также об-
ладают свойством различать запахи, но на значительно меньшем расстоянии.
Рядом со щупальцевыми ганглиями располагаются глаза, которые во мно-
гом сходны с таковыми у улиток. Однако имеются и существенные отличия
(Hesse R., 1902; Newell, Newell, 1968). У слизней ретина содержит меньше
светочувствительных клеток, и их. расположение внутри глаза несколько
иное. Кроме того, слизни имеют так называемую дополнительную ретину,
которая не обнаружена у улиток. Принято считать, что это образование слу-
жит для восприятия инфракрасного участка спектра (Newell, Newell,
1968).
Глаз Deroceras reticulatum представляет собой полый овальный пузырек
размером 140—180 мкм, состоящий из одного слоя клеток (рис. 18). Передняя
Стенка глаза сложена из очень крупных прозрачных клеток, образующих
роговицу, отделенную от прозрачного наружного эпителия щупальца (сли-
зистая оболочка глаза) тонким слоем коллагеновых волокон. В задней части
глаза располагаются многочисленные пигментные клетки ретины, среди
которых регулярно расположены сильно вытянутые светочувствительные
клетки. Каждая такая клетка у D. reticulatum (их около 800, в то время как
у Helix около 4000) вытянута к центру глаза и образует столбик, снаружи
покрытый микроворсинками (микровилли). Сзади, т. е. к периферии глаза,,
эти клетки заканчиваются нейрофибриллами, которые, сливаясь друг с дру-
гом, образуют зрительный нерв.
4 И. М. Лихарев, А. И. Виктор
50
ОБЩАЯ ЧАСТЬ
Рис. 19. Стотолиты Arion rufus
(из: Riedel, Wiktor, 1974).
Таким образом, щупальца
В центре глаза находится хрусталик, окруженный бесцветной жидкостью.
Диаметр хрусталика около 65 мкм, фокусное расстояние 100 мкм.
В передней части глаза, асимметрично к оптической оси, в особом углуб-
лении располагается дополнительная ретина. Она значительно меньших
размеров, чем основная ретина. В ее состав входит небольшое число свето-
чувствительных (столбчатых) клеток, разделенных обычными цилиндриче-
скими клетками, но без пигмента.
Как строение глаз, так и опыты по влиянию света на поведение слизней
показывают, что эти животные не способны различать детали предметов,
в этом отношении уступая некоторым крупным улиткам из семейства Не-
licidae. Глаза слизней рассчитаны на определение интенсивности освещения.
Характер реакции слизней на свет зависит не только от интенсивности
его, но и от времени суток. Днем они ищут затемненные места, ночью же,
когда они особенно активны, предпочитают
слабое освещение мраку, но, если свет ста-
новится слишком интенсивным, активность
животных снижается или прекращается.
Опыты показали, что слизни особенно чув-
ствительны к инфракрасному участку спектра,
влияние которого стараются избежать или
ослабить (Lewis, 1967, 1969а).
Губные щупальца также обладают орга-
нами обоняния, но строение их плохо изу- .
чено. Кроме того, на поверхности этих щу-
палец располагаются сенсорные клетки, вы-
полняющие роль органов вкуса. Последнюю
функцию несут также и ротовые лопасти, од- j
нако главное их назначение — осязание, осо- 1
бенно поверхности пищевого объекта. I
слизней играют важную и разностороннюю 5
роль, и понятно, почему скорость их регенерации очень высока: у Deroceras j
reticulatum 10—12, а у Arion ater 30—60 дней (Chetail, 1963). j
Другим важным органом чувств являются статоцисты. Это парные сфе- j
рические образования, лежащие на верхней стороне педальных ганглиев,
но иннервируемые специальными нервами церебральных ганглиев !
•(рис. 17, ст). Каждый статоцист (около 200 мкм в диаметре) внутри полый,
заполнен жидкостью и содержит округлые известковые пластинки — ста-
толиты (рис. 19). Внутренняя поверхность статоциста выстлана однослой-
ным эпителием, основу которого составляют 12—14 крупных чувствительных
клеток с крупными ресничками и редкими микроворсинками. Кроме них, ,
в состав внутренней выстилки статоциста входят мелкие клетки с большим i
числом микроворсинок и редкими ресничками. От каждой крупной клетки
отходят аксоны, в совокупности слагающие нерв статоциста.
Как органы равновесия (геотаксиса) статоцисты играют важную роль
в ориентации животного, особенно при движении по наклонной поверхности
и при закручивании тела спиралью.
Половая система. В отличие от улиток, у которых гонада
{гермафродитная железа) . слагается из отдельных долек, заключенных
в толщу печени, у слизней эти дольки собраны в один более или менее ком-
пактный округлый или удлиненный орган, лежащий в задней половине тела
среди петель кишечника и лопастей печени или же заполняющий задний
конец внутренностного мешка. )
В свою очередь каждая долька гонады слагается из округлых фолликул )
(ацини), в которых формируются как мужские, так и женские гаметы. Внутри (
фолликулы выстланы герминативным эпителием или синцитием, чередую- 1
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
51
щимся с участками, занятыми питательными клетками. В связи с тем
что почти все слизни — это протерандрические животные, мужские элементы
(сперматогонии и сперматоциты) обычно располагаются ближе к центру
фолликулы, а женские (оогонии и ооциты) держатся по периферии.
Фолликулы открываются в тонкие выносящие протоки, которые, сливаясь
друг с другом, образуют общий гермафродитный проток. Последний чаще
всего простирается вперед, нередко по пути образуя извивы, особенно при
подходе к белковой железе. Здесь он дает крупный V-образный извив — та-
лон, обычно открывающийся в так называемый квадривий. Это многокамер-
ный, сложно устроенный орган, свойственный большинству легочных мол-
люсков. В нем происходит оплодотворение яйцеклеток и формирование яиц,
а также через него транзитом проходит вперед своя сперма. В квадривий
открывается проток крупной, чаще всего языковидной белковой железы,
секрет которой формирует белковую, т. е. питательную, оболочку яйца.
В тех группах (Arionidae, Milacidae), у которых яйца имеют наружную из-
вестковую оболочку, формирующая ее скорлуповая железа также впадает
в квадривий.
От квадривия простирается вперед длинный семяяйцевод, образующий
несколько петель или извивов. Он слагается из толстого фестончатого жен-
ского канала и тонкого прямого мужского канала. Внешнюю стенку муж-
ского канала образует лентовидная простата. У Limacidae передний конец
простаты может быть сильно вздутым и нередко простирается на начало семя-
провода.
Организация гениталий до разделения семяяйцевода на семяпровод
и яйцевод у всех слизней сравнительно однообразна и очень сходна с осталь-
ными Stylommatophora. С момента разделения обнаруживаются определенные
различия, особенно заметные на органах, непосредственно участвующих
в копуляции (рис. 20).
Как мужские, так и женские половые пути впадают в атриум (половая
клоака), который открывается наружу половым отверстием. Чаще всего не-
далеко от полового отверстия находится семяприемник, служащий для
принятия и более или менее длительного хранения спермы партнера, а также
для резорбции избыточной спермы (Hryniewiecka-Szyfter, R^dziniak, 1976).
Этот орган состоит из резервуара и протока. Последний у одних впадает в дис-
тальную часть женских половых путей, у других — в основание пениса,
а у третьих — непосредственно в атриум.
Семяпровод — тонкий, нередко более или менее извитой проток, который,
обособившись от семяяйцевода, вначале обычно тянется вдоль яйцевода;
вблизи от атриума он изгибается назад, простирается вдоль пениса и впадает
в него апикально или сбоку. В некоторых группах (Gigantomilax и др.) семя-
провод настолько короткий, что указанный перегиб слабо выражен. У групп
с длинным пенисом (Limax и др.) и соответственно длинным семяпроводом
пенис и дистальный участок семяпровода связаны перепонкой, снабженной
кровеносными сосудами.
У Milacidae, Parmacellidae и Trigonochlamydidae перед впадением в пенис
семяпровод заметно утолщается за счет развития мощной кольцевой му-
скулатуры и железистого эпителия, превращаясь в эпифаллус. В этом органе
формируется сперматофор — своеобразный удлиненный пакет с хитиноид-
ными стенками, в котором при копуляции передается сперма. Обычно форма
сперматофора и фактура его стенок соответствуют форме канала эпифаллуса
и рельефу его внутренних стенок (см. рис. 531—537). У большинства групп
единственный сперматофор хранится в эпифаллусе вплоть до копуляции.
Только у Trigonochlamydidae эпифаллус формирует не один, а несколько
сперматофоров, и они до копуляции находятся внутри пениса, концами при-
крепленные к особым железистым подушкам (см. рис. 396).
4*
» VZZte
Рис. 20. Основные типы строения дистальных отделов гениталий.
I — Philomycus (Philomycidae)} II — Arion (Arionidae)} III — Deroceras (Agriolimacidae)} IV—Limax
(Limacidae); V — Trigonochlamys (Trigonochlamydidae); VI — Milax (Milacidae); VII — Candaharia
(Parmacellidae). 1 — яйцевод и влагалище; 2 — семяприемник; 3 — пенис; 4 — эпифаллус; а — атриум;
рп'— половой ретрактор; ст^б — стилофор.
МОРФОЛОГИЯ С ЭЛЕМЕНТАМИ ФИЗИОЛОГИИ
53
Пенис слизней очень разнообразен по внешнему и внутреннему строению.
У одних он имеет вид длинной и стройной трубки, у других он мешковидный,
а у третьих вздут сзади или у середины. Он может быть прямым или более
или менее изогнутым, или даже спирально закрученным.
Внутренние стенки пениса (при копуляции они становятся наружными)
обычно имеют различные складки. Одна из них может быть особенно му-
скулистой и приобретать облик конического или листовидного выступа,
который сильно набухает и служит для возбуждения партнера перед копу-
ляцией {Deroceras). Такое образование называют стимулятором. Поверхность
его нередко снабжена густо расположенными мелкими складочками, а у не-
которых групп {Lytopelte, Liolytopelte) к моменту полового созревания на нем
образуется твердая известковая пластинка, вероятно, усиливающая дей-
ствие стимулятора. Другие складки пениса могут служить для направления
спермы внутрь партнера.
У многих слизней внутри заднего конца пениса располагается муску-
листая папилла — папилла пениса. Через внутренний канал ее из эпи-
фаллуса в пенис поступает сперматофор.
Снаружи пенис нередко имеет различные придатки. У одних это крупный
бичеобразный орган — бич, с довольно толстыми мускулистыми стенками,
внутри нередко со складками и сосочками {Monochroma), вероятно, служа-
щий для возбуждения партнера.1 У других {Deroceras) придатки на пенисе
мягкие, с тонкими железистыми стенками; их обозначают как пениальные
железы. Функция этих желез неясна, но, так как при копуляции они тоже
выворачиваются, не исключено, что и они играют роль в стимуляции парт-
нера.
Кроме уже упомянутых придаточных органов, на пенисе втречаются
и другие образования, особенно в роде Deroceras.
& Подавляющее большинство слизней имеют более или менее хорошо раз-
витый пенис. Только у Arioninae он полностью отсутствует и эпифаллус
непосредственно открывается в атриум.
Участок женских половых путей после разделения семяяйцевода обозна-
чают как яйцевод (или свободный яйцевод, или матка). Если в яйцевод от-
крывается семяприемник, то участок женских путей между семяприемником
и атриумом называют вагиной (влагалище). У Milacidae и Limacidae (за
исключением родов Eumilax и Aletali max) передняя часть яйцевода заметно
шире задней или даже вздутая. В этом месте находится более или менее круп-
ная папилла —- папилла яйцевода.
Влагалище и атриум в свою очередь могут иметь придаточные органы
<{Parmacellidae, Milacidae). К ним относятся вагинальные железы, распола-
гающиеся на одном из названных органов или на обоих одновременно. Кроме
-того, атриум может иметь внутренние или наружные придатки, служащие для
возбуждения партнера {Milax, Parmacella). Ту же роль выполняет так назы-
ваемая лигула, располагающаяся внутри атриума некоторых видов Arion.
Дистальные участки половых путей снабжены особыми мускулами,
«служащими для втягивания половых органов внутрь тела после копуляции.
Наиболее важным из них является половой ретрактор. Задний конец этого
: мускула крепится к стенке тела вблизи от мантийного комплекса или же
непосредственно к диафрагме. Передний конец чаще всего прикреплен к пе-
нису, если же его нет {Arioninae), то к другим совокупительным органам —
гэпифаллусу, яйцеводу, семяприемнику. Кроме полового ретрактора, нередко
1 В пользу такой интерпретации функции бича говорит тот факт, что у слизней,
которые его имеют, отсутствуют другие стимулирующие органы гениталий. Бич слизней
не имеет ничего общего с бичом Helicoidea (Шилейко, 1978) и других раковинных Geophila,
у которых он наряду с эпифаллусом служит для формирования сперматофора и при
копуляции не выворачивается.
54
ОБЩАЯ ЧАСТЬ
имеются еще 1—2 или более ретентора, связывающие концевые участки гени-
талий с передней стенкой тела.
В заключение обзора органов дистальной части половой системы (т. е.
после разделения семяяйцевода) отметим, что не все они присутствуют одно-
временно у всех слизней. Как строение этих органов, так и их комбинации
достаточно разнообразны. Однако можно наметить несколько главных типов.
Рис. 21. Продольный разрез через пе-
нис и атриум Bielzia coerulans.
an — половой атриум; п — пенис; рп — по-
ловой ретрактор; с — семяпровод; сп — семя-
приемник; я — яйцевод. Стрелками указано
направление движения спермы и яиц.
чительно крупнее, чем в предыдущих группах и принимает на себя роль
копулятивного органа (Arioninae).
Ниже мы еще специально остановимся на поведении слизней во время
копуляции (стр. 55). Как известно, у всех легочных моллюсков этот процесс
завершается обменом спермой между партнерами. Однако осуществление
этого акта и его механизм в значительной степени зависят от строения кон-
организации гениталии.
Первый тип (рис. 20, VI, VII). Жен-
ские половые пути и (или) атриум с при-
даточными органами — вагинальными
железами; у некоторых имеется атриаль-
ный стимулятор. Мужские половые пути
с эпифаллусом и папиллой пениса (Mi-
lacidae и Parmacellidae).
Второй тип (рис. 20, I, III, IV).
Женские половые пути и атриум без-
придаточных органов. Эпифаллуса нет.
Пенис чаще всего без папиллы,1 но
внутри со стимулятором и различными
складками (Agriolimacidae, Limacinae,
Eumilacinae, Boettgerillidae, Philomyci-
dae).
Третий тип (рис. 21). Женские по-
ловые пути и атриум без придаточных
органов. Семяпровод впадает в атриум
минуя пенис. Пенис превращен в огром-
ную мощную папиллу (Bielziinae).
Четвертый тип (рис. 20, У). Женские
половые пути и атриум без придаточ-
ных органов.2 Имеется эпифаллус. Пени-
альной папиллы нет; внутри пениса
имеются железистые подушки, на кото-
рых сидят сперматофоры (Trigonochla-
mydldae).
Пятый тип (рис. 20, II). Женские
половые пути и атриум без придаточных
органов. Пениса нет. Эпифаллус откры-
вается прямо в атриум. Атриум зна-
цевых участков половой системы.
При копуляции выворачивание половых органов, выдавливание в пенис
спермы или сперматофора происходит за счет притока крови в головной синус
при одновременном сжимании стенок тела вокруг заднего синуса. Выворачи-
вание захватывает только концевые участки гениталий и лишь в определенном
пределе, зависящем от мест прикрепления полового ретрактора и ретенторов
гениталий и их возможности растягиваться.
1 Исключение — Caucasolimax. (Limacidae) п Boettgerilla (Boettgerillidae), у которых
имеется пениальная папилла.
2 Исключение — Troglolestes (Trigonochlamydidae), у которого на влагалище имеется
придаток неизвестного назначения.
РАЗМНОЖЕНИЕ И ОНТОГЕНЕЗ
55
Обычно атриум выворачивается полностью, влагалище или яйцевод —
до появления входа в проток семяприемника, пенис — до выхода наружу
переднего конца пениальной папиллы или, если ее нет, до места прикрепле-
ния полового ретрактора. Обратное втягивание внутрь тела вывернутых
органов идет за счет усилия полового ретрактора и ретенторов при одновре-
менном оттоке крови из головного синуса в задний конец тела.
Механизм копуляции в значительной степени зависит от того, подается ли
сперма партнеру в свободном виде, т. е. как желеобразный комок, или она
заключена в сперма тофоры, которыми обмениваются партнеры.
В первом случае (2-й тип гениталий) сперма поступает в пенис, как за счет
давления крови в головном синусе, так и за счет перистальтики слабой
мускулатуры семяпровода. Вывернутый пенис подает ее в атриум или даже
во входное отверстие семяприемника партнера. Только у Limax s. str. сперма
партнера прилипает к переднему концу вывернутого пениса (рис. 20, I, II)
и при его втягивании внутрь тела попадает к входному отверстию семяприем-
ника.
Во втором случае копуляция идет иначе. Как известно, сперматофор
формируется в эпифаллусе. При копуляции перистальтика мощной кольце-
вой мускулатуры этого органа выталкивает сперматофор вперед. Этому
помогает мускулатура пениальной папиллы, которая к этому моменту мак-
симально выдвинута вперед. В итоге сперматофор оказывается в атриуме
партнера, откуда он поступает в его семяприемник (1-й и 4-й типы гениталий).
Остальные типы строения гениталий обеспечивают некоторые варианты
указанных двух основных способов обмена спермой при копуляции. Так,
у Bielzia (3-й тип гениталий), у которой семяпровод и пенис впадают в атриум
независимо, хотя внутри него связаны незамкнутой бороздой, пенис превращен
в мощную папиллу с крупной предпапиллярной, т. е. задней, накопительной
камерой. Этот процесс мыслится в следующей последовательности. Перисталь-
тика мощной мускулатуры семяпровода выталкивает сперму в атриум. Пенис
за счет расслабления своей мускулатуры при одновременном замыкании
сфинктера полового отверстия насасывает сперму в накопительную камеру.
При копуляции пенис несколько выдвигается вперед, так что слегка выступает
за пределы полового отверстия (Smoleiiska, 1936), а затем впрыскивает сперму
в атриум партнера.
У Boettgerilla (рис. 252) в связи со слабым развитием пениса средний учас-
ток семяпровода преобразован в толстый веретеновидный орган с мощной
мускулатурой, задача которого создать дополнительное давление для впрыс-
кивания спермы в глубь атриума и влагалища партнера.
У Arioninae (5-й тип гениталий) в связи с редукцией пениса сильно уве-
личивается атриум. При копуляции он выворачивается наружу, открывая
вход в семяприемник и сперматофор партнера, проталкивается туда за счет
перистальтики эпифаллуса.
РАЗМНОЖЕНИЕ И ОНТОГЕНЕЗ
Копуляция. У большинства слизней копуляция обычно происхо-
дит ночью, реже в сырые, пасмурные дни. Ей предшествует серия поведен-
ческих реакций, называемых брачными играми, или танцами. Продолжитель-
ность как брачных игр, так и копуляции различна у разных видов. В общей
сложности они могут занимать от трех минут {Lehmannia marginata) до суток
(некоторые Milacidae).
Брачным играм предшествует поиск готового к копуляции партнера.
В этом деле важную роль играет обоняние и слизь, которую оставляют после
себя животное,, так как подмечено, что при попадании на след партнера
слизень неоднократно задерживается, чтобы съесть оставшуюся после него
56
ОБЩАЯ ЧАСТЬ
слизь. У Arion в этом отношении особенно важную роль играет каудальная
железа, которая к началу размножения сильно вспухает и начинает выделять.
, I? кх крупные капли густой слизи (Ger-
hardt, 1940; Quick, 1946). По-ви-
димому, слизь содержит какие-то-
вещества, сигнализирующие о го-
товности к копуляции и вызываю-
щие определенные реакции у парт-
нера.
После встречи начинаются брач-
ные танцы. В наиболее полном
виде они состоят из трех фаз: пре-
следование, кружение и соедине-
ние. По-видимому, одной из глав-
ных задач брачных танцев явля-
ется синхронизация физиологии
партнеров. В это время сперма
обильно поступает в семяяйцевод.
и семяпровод, простата обогащает-
ее своим секретом, а у видов, раз-
множающихся с помощью сперма-
тофоров, в эпифаллусе заканчива-
ется их формирование. Выворачи-
вание копулятивных органов также-
начинается уже в конце брачных
танцев и обычно идет синхронно-
у обоих партнеров. В течение всех
фаз брачных танцев происходит-
взаимное ощупывание партнеров^
с помощью щупалец и поедание-
слизи как с поверхности тела, так
и с субстрата. После окончания
преследования слизни серповидно-
изгибаются, так что их половые от-
верстия прижимаются друг к другу,,
и в таком положении они начи-
нают кружиться. В это время они
выделяют большое количество-
слизи, густо покрывающей пло-
щадку, на которой происходит ко-
пуляция. К концу кружения, а у не-
которых видов и раньше, начина-
ется выворачивание атриумов,,
стимуляторов (если они имеются)
и пенисов. При этом партнеры все;
время ощупывают друг друга вы-
вернутыми участками гениталий,
особенно активны стимуляторы
и другие придатки цениса и ат-
риума.
. Затем прекращается кружение, слизни плотно прижимаются друг к другу
копуляционными органами и сильно сжимают тела (рис. 22, III, IV). При
этом происходит выталкивание и передача спермы либо в виде желеобразных
капель, или же внутри сперматофоров. В обоих случаях сперма подается
внутрь семяприемника, щелевидный вход в который нередко заметен снаружи.
РАЗМНОЖЕНИЕ И ОНТОГЕНЕЗ
57
После завершения копуляции и разъединения партнеров некоторые слизни
^{Deroceras sturanyi Kosinska, 1980) съедают слизь, оставшуюся на месте ко-
луляции.
Описанный ход брачных игр и копуляции с некоторыми вариациями обычен
для большинства видов слизней. В частности, так идет копуляция у слизней
из родов Arion, Deroceras, всех Рarmacellidae, Limax flavus, Malacolimax
tenellus, Lehmannia marginata, Bielzia coerulans (Kunkel, 1916; Gerhardt,
1933—1940; Smolenska, 1936; Жадин, 1946; Smith, 1946; Fromming, 1954;
Wiktor, 1960b; Runham, Hunter, 1970).
У Milacidae партнеры при встрече сразу прижимаются друг к другу по-
ловыми отверстиями, несколько отогнув в стороны головы. После этого они
длительное время остаются неподвижными или только очень медленно повора-
чиваются по часовой стрелке. Брачный танец вместе с копуляцией занимают
гораздо больше времени, чем у других слизней, — от 12 до 24 часов. У Milax
gagates партнеры так плотно прижимаются друг к другу, что копуляционные
органы снаружи не видны. Слизни из рода Tandonia немного более активны —
они не столь плотно прижимаются друг к другу и партнеры на короткое время
могут расходиться; при этом видны вывернутые атриумы, пениальные папиллы
и сперматофоры.
Все упомянутые слизни совершают брачные игры и копуляцию на горизон-
тальных поверхностях, т. е. на почве, верхней стороне камней и т. п., и зна-
чительно реже на наклонных поверхностях, например на стволах деревьев.
Совсем иначе ведут себя Limax maximus, L. cinereoniger и некоторые другие
виды из подрода Limax s. str. Они копулируют в подвешенном состоянии,
т. е. над землей, прицепившись к веткам деревьев или к нижней стороне
нависающих над землей скал, к потолкам гротов и т. п. (рис. 22, I, II). При
встрече эти слизни также ощупывают и облизывают друг друга. Затем они
сплетаются телами и, одновременно приклеиваясь с помощью слизи к ветке
или к камню задними концами, повисают в воздухе вниз головами. У Limax
maximus партнеры повисают на слизистом шнуре длиной 15—45 см. Сплетаясь,
слизни постепенно выворачивают пенисы, которые в свою очередь перепле-
таются, но в обратном направлении, чем туловища.
Как известно, в спокойном состоянии у этих слизней внутри пениса за-
легают две продольные складки, соединяющиеся у заднего конца (стр. 273).
При выворачивании пениса складки набухают, образуя желоб, заканчиваю-
щийся зубчатым расширением. После переплетения пенисов у основания
каждого из них, т. е. со стороны семяпровода, появляется капля спермы.
Она постепенно стекает вниз вдоль желоба, достигает расширенного конца,
после чего происходит взаимный обмен ею. Втягивание пенисов внутрь тела
идет много быстрее, чем их выворачивание, и слизни скоро расходятся. Перед
этим один из партнеров съедает слизистую нить. Копуляция вместе с танцами
продолжается более 10 часов.
У другого вида того же подрода — Limax redii, вывернутый пенис имеет
70 см в длину, а копуляция вместе с играми продолжается от 7 часов до су-
ток и более.
Нередко в поведении партнеров наблюдаются некоторые различия в ак-
тивности. Так, у Limax maximus один копулянт более активен и в этом отно-
шении напоминает самца: он начинает преследование, а после завершения
копуляции съедает слизистую нить; другой менее активен: он следует за пер-
вым на дерево или стену и сразу уходит после обмена спермой (Langlois,
1963).
Известно, что иногда у некоторых слизней (например, Arion ater) обмен
спермой не всегда обоюдный, т. е. только один из партнеров передает другому
•свою сперму. Их роли могут меняться при последующих встречах (William-
son, 1959).
58
ОБЩАЯ ЧАСТЬ
В заключение отметим, что, к сожалению, мы пока, не располагаем све-
дениями о копуляции слизней, ведущих преимущественно подземный образ
жизни (Boettgerilla, Trigonochlamydidae').
Оплодотворение и формирование яиц. После копу-
ляции сперма, попадающая в семяприемник партнера, освобождается от слизи
или оболочки сперматофора. Последняя растворяется под действием секрета,
выделяемого стенками семяприемника. В литературе пока отсутствуют точ-
ные данные о дальнейшей судьбе этой спермы у слизней. Однако нет осно-
ваний ожидать, что в этом отношении слизни отличаются от легочных,
улиток.
По-видимому, значительная часть спермы партнера проникает из семя-
приемника по семяяйцеводу в квадривий, где и происходит перекрестное-
оплодотворение. Экспериментально показано, что чужая сперма, еще находясь
в семяприемнике, стимулирует овогенез и овуляцию, т. е. поступление зре-
лых яйцеклеток в квадривий. После оплодотворения неиспользованная «чужая»
сперма растворяется секретом стенок семяприемника, а также, вероятно,
и квадривия.
Очень распространено мнение, что некоторые Arion и Deroceras, если их
изолировать до наступления половозрелости, способны к самооплодотворению
в течении ряда генераций (Maury, Reygrobellet, 1963; Franc, 1968). В пользу
такой точки зрения также говорит афаллия, т. е. редукция пениса, свойствен-
ная нередко целым популяциям Deroceras laeve (стр. 131).
Однако пока нет ни одного исследования, выполненного на цитологиче-
ском уровне, доказывающего, что в этих случаях имеет место самооплодотво-
рение. Во всяком случае можно считать бесспорным, что в основном слизни
размножаются путем перекрестного оплодотворения.
Время, проходящее между копуляцией, оплодотворением яйцеклетки
и откладкой яиц, различно у разных групп слизней; так, у Deroceras reticu-
lation на это требуется 8—10 дней, а у Arion ater несколько недель.
После оплодотворения, еще в квадривии, каждую зиготу окружает слой
питательного белка — продукта выделения белковой железы, образующего
основную массу будущего яйца. В дальнейшем, продвигаясь вперед по жен-
скому каналу семяяйцевода, окруженная белком зигота последовательно,
одевается внутренней студневидной и внешней оболочками. На границе
между белком и внутренней оболочкой возникает перивителлиновая мембрана.
Внутренняя оболочка состоит из мукополисахаридов и небольшого ко-
личества кальция, а внешняя — из полисахаридов, протеина и кальция (Car-
rick, 1938; Bayne, 1968).
У большинства слизней яйца прозрачные, но у некоторых, в частности
у Arion и Tandonia, внешний слой заполнен кристаллами углекислой извести,
что делает яйца непрозрачными.
Откладка яиц. Обычно размножение и откладка яиц более или менее
приурочены к определенным сезонам года, но нередко аномальные .погодные
условия могут изменить сроки. Так, во Франции Arion ater обычно размно-
жается осенью, но если созревание запаздывает из-за очень сухого лета, то-
большая часть популяции не откладывает яйца до будущей весны (Laviolette,
1950а).
С другой стороны, у Parmacellidae, для жизненного цикла которых обя-
зательна летняя диапауза молоди, откладка яиц и вылупление всегда приу-
рочены к маю—июню. Как показали эксперименты в лаборатории (Жадин,
1946; Увалиева, 1975), эти сроки не могут быть сильно смещены различными
комбинациями температуры и влажности. Все это указывает на то, что у та-
ких слизней период наступления яйцекладки, как и все размножение, конт-
ролируется внутренними механизмами.
РАЗМНОЖЕНИЕ И ОНТОГЕНЕЗ
. 59
Обычно слизни откладывают яйца в щели почвы, у подножья травянистых
растений, под камни и другие предметы, лежащие на земле, т. е. в убежища,
защищенные от дневного света и с достаточной влажностью (рис. 23). При
откладке яиц в почву слизни нередко расширяют выбранную ямку или щель.
Чаще всего каждый слизень откладывает яйца порциями (кладками) и не
злаков
1954).
Рис. 23. Кладка яиц De-
roceras reticulatum у при-
корневой части ---------
(из: Лихарев,
в одно место, а в несколько мест, расположенных
по соседству. Число яиц как в одной, так и во всех
кладках, по-видимому, сильно варьирует в зависи-
мости от возраста, питания, физиологического со-
стояния слизня и погодных условий. Однако общее
число яиц достаточно видоспецифично. Так, извест-
ный вредитель сельского хозяйства Deroceras reticu-
lation в общей сложности может отложить за сезон
до 500 яиц (Carrick, 1938), Arion ater до 515 (Kunkel,
1916), Limax flavus и Arion subfuscus до 340 (там же),
Candaharia rutellum до 80 (Увалиева, 1975), C. levan-
deri до 40 (Жадин, 1946), Parmacella ibera до 70
(Шилейко, 1967).
Развитие. Эмбриогенез даже в пределах од-
ной кладки не идет во всех яйцах одновременно.
Из этого следует, что процесс дробления начинается
еще в теле материнского организма.
Среди слизней наиболее полно изучено эмбрио-
нальное развитие у Deroceras reticulatum (Carrick,
1938; Runham, Hunter, 1970) и Limax maximus
(Meisenheimer, 1896—97).
Как у всех брюхоногих моллюсков, процесс дроб-
ления идет по спиральному и детерминированному
типу. Сразу после гаструляции появляется эмбрио-
нальная почка, которая накапливает кристалличе-
ские продукты выделения. Почти одновременно в пе-
реднем конце эмбриона клетки энтодермы начинают
интенсивно абсорбировать белок яйца, сильно взду-
ваются и образуют крупный головной пузырь (рис. 24).
На заднем конце зародыша разрастается так назы-
ваемый задний пузырь, или подоцист. Оба пузыря,
щаясь, прогоняют гемолимфу сквозь тело. В этом им помогает провизорное
сердце — особый сокращающийся синус. По-видимому, задний пузырь одно-
временно выполняет роль органа дыхания.
Белок поступает в тело эмбриона через бластопор, т. е. рот, и перевари-
вается клетками, выстилающими головной пузырь, почему его еще называют
эмбриональной печенью. По мере роста зародыша формируются зачатки
мантии, ноги и щупалец. За счет впячивания эктодермы спины образу-
ется желобок — раковинная железа, внутри которой формируется эмбрио-
нальная раковина. В отличие от улиток у слизней (за исключением Раг-
macellidae') раковинная железа смыкается над раковиной. Приблизительно
на той же стадии развития вследствие неравномерности роста правой и
левой стороны спинной части зародыша происходит поворот, т. е. торсия,
мантийного комплекса на 180°; однако детали этого процесса у слизней
не изучены.
Перикардий, сердце, почка и гонада — дериваты мезодермы; точнее, они
возникают из скоплений мезенхимных клеток, образовавшихся после рас-
пада мезодермального зачатка. В начале образуется перикардиальный пу-,
зырек, от которого отшнуровываются зачатки сердца и почки. Из другого
попеременно
сокра-
60
ОБЩАЯ ЧАСТЬ
скопления мезенхимных клеток формируется гонада.1 Половые пути и при-
даточные органы гениталий имеют двойственную природу. Гермафродитный
проток, белковая железа, семяяйцевод, яйцевод и семяпровод формируются
за счет впячивания мантийной полости, т. е. имеют паллиальное происхожде-
ние, а пенис и атриум — за счет впячивания прозоподиума (Миничев, Старобо-
гатов, 1971; Luchtel, 1972а, Ь).
Рис. 24. Четыре стадии эмбриогенеза Deroceras reticulatum (из: Carrick, 1938).
1 — отверстие личиночной почки; 2 — задний пузырь (подоцист); 3 — мантия; 4 — головной пузырь;
5 — глазное щупальце; 6 — губное щупальце; 7 — ротовая лопасть.
Как у всех моллюсков, нервная система и органы чувств слизней экто-
дермального происхождения и формируются из нескольких зачатков. На
голове за счет впячивания покровов зародыша образуются процеребрум,
глаза и статоцисты. Остальные участки церебральных ганглиев и другие
ганглии ЦНС возникают путем пролиферации клеток эктодермы. Передняя
часть пищеварительного тракта — глотка, пищевод и зоб, так же как и рек-
тум, — эктодермального происхождения; печень, желудок и кишка (кроме
ректума) возникают за счет энтодермы.
Легочная полость слизней, как и у всех Pulmonata, не является гомологом
мантийной полости Prosobranchia (Regondaud, 1964). Зачаток легкого появ-
ляется в форме впячивания кожных покровов еще до образования мантийной
полости. Позднее легочный мешок открывается в мантийную полость и из
края мантии образуется пневмостом. Настоящая мантийная полость сохра-
няется лишь как рудимент, в виде небольшого углубления (так называемая
супранухальная полость) у края мантии, куда открываются ректум и моче-
точник. -
1 Согласно Лухтелю (Luchtel, 1972а), у Arion circumcsriptus гонада образуется за счет
зачатка эктодермы.
РАЗМНОЖЕНИЕ И ОНТОГЕНЕЗ
61
К концу эмбриогенеза задний и головной пузыри постепенно расса-
сываются, а дефинитивное сердце и легкое приступают к своей работе.
Пищеварительная выстилка головного пузыря входит в состав дефинитивной,
печени. Перед вылуплением зародыш проглатывает остатки белка яйца.
Эмбриональное развитие заканчивается вылуплением молоди, которая
прогрызает оболочки яиц. Первые дни пищей ее служат как остатки обо-
лочки, так и нередко абортивные яйца, т. е. такие яйца, в которых не про-
исходило развитие эмбрионов. Пока не очень ясно, с чем связано присутствие
в кладке абортивных яиц — остались или они неоплодотворенными или же
были оплодотворены, но по каким-то причинам развитие эмбрионов в них не
происходило. Отмечено только, что число абортивных яиц и яиц, развитие
которых не завершилось (эмбрион погиб), увеличивается с возрастом слизней
(Kunkel, 1916; Fromming, 1954; Kosinska, 1980) и, по-видимому, связано
со старением материнского организма. Возможно, что те же причины способ-
ствуют появлению в кладках слизней уродливых и многозародышевых яиц.
(Carrick, 1938; Николаев, 1967), эмбрионы которых чаще всего гибнут еще-
до вылупления.
Постэмбриогенез и рост. Даже из одной кладки слизни
вылупляются не одновременно. При этом уже в это время они заметно раз-
личаются по размерам. Дальнейший рост также идет неравномерно.
В довольно обширной литературе анализируется не только влияние'
питания и абиотических факторов на рост слизней, но и связь между общим
ростом этих животных и формированием гениталий, созреванием половых
продуктов, наступлением зрелости и с размножением. Отмечается, что в пост-
эмбриогенезе рост слизней слагается из трех фаз: инфантильной, ювенильной,
и взрослой (Abeloos, 1942—1945; Laviolette, 1950с; Lusis, 1961; Runham,.
Laryea, 1968; Smith, 1966). Эти фазы характеризуются как по темпу роста
тела (масса, размеры), так и по состоянию гениталий (табл. 1).
Инфантильная фаза отличается высоким темпом роста животного (угло-
вой коэффициент регрессии 0.03), зачаточной гонадой и половым трактом;
только семяяйцевод начинает дифференцировку. С наступлением ювенильной
фазы темп роста падает (угловой коэффициент 0.01), а гонада начинает интен-
сивно увеличиваться в размерах и дифференцироваться на отдельные доли:
к концу фазы масса ее достигает максимума. Внутри гонады идет интенсив-
ный сперматогенез (сперматоциты — сперматиды). Кроме того, происходит
дифференциация белковой железы, а к середине фазы заканчивается форми-
рование семяяйцевода.
Взрослая фаза знаменуется остановкой роста тела. При этом завершается
формирование всех органов полового тракта. В начале этой фазы оканчива-
ется сперматогенез, и гонада вместе с ее протоками заполнена зрелыми спер-
миями. Вслед за этим начинается овогенез, а масса гонады и ее размеры умень-
шаются. Завершается созревание белковой железы, которая к началу этой
фазы начинает функционировать. Простата, которая начинает выделять секрет
еще в ювенильной фазе, заканчивает это к середине взрослой фазы, когда
начинается развитие желез женского канала семяяйцевода. К концу взрослой
фазы приурочена: копуляция и откладка яиц.
У однолетних слизней (Arion, Deroceras и др.) вслед за взрослой фазой .
наступает фаза старения. Нередко она очень коротка и заканчивается смертью
животного. Лишь при благоприятных условиях, особенно в лабораториях,
эта фаза может растянуться и даже сопровождаться ростом, тела животного-
(Kosinska, 1980 — наблюдения над Deroceras sturanyi).
У многолетних слизней после периода покоя гонада вновь повторяет свой
цикл (Limax — Abeloos, 1943, 1944).
Описанная связь между ростом и репродуктивным циклом может несколько,
варьировать в разных группах слизней, причем эти расхождения'связаны.
Изменения в половой системе в течение постэмбриогенеза Deroceras reticulatum,(по: Runham, Laryea, 1968)
Фаза роста (стадия зрелости) ювениальная | взрослая (зрелость)
«—Не дифференцирована > < X D 5 Й а & С $ ч 3 1 IX С S X D 3 X 5 ? Сб и.
0 3 1=
0 D 3
3 н Д' о m О ф t н S £Х Ф и ф ф 3 <3 <3 а 5 X 5 С с X ! X о Й о Я ф £ И к Д' ф Оч Й ф и н сб н о о С ч с а t с. с L X С. & с. С & £ ! 3 :Г й ч >s 5 а а »S & ц. J
& Е- G * С Е С 1 S с а ч < С X 1. X 3 а а Е _ ( с § ъ з S3 5 & >£ >£ Т т 3- 3- а t ь & а С Я <3 С J X 5S 3 5
с >£ >£ S а Я 5 X :Г X 3* 3* i С а Я й ч 0 5 ч Э 3 X 3 Я ч •е-Не дифференцирована > >€ t ! :Г 3 J* I Е >s ч С КС С 5 X э
3
&
о
РАЗМНОЖЕНИЕ И ОНТОГЕНЕЗ
63'
как с различиями в морфологии гениталий, так и с особенностями физиологии.
Это заметно, если сравнить Arion ater и Deroceras reticulatum. Если у первого
мужская и женская функции гениталий четко разграничены во времени, то
у второго они частично совпадают или перекрывают друг друга во взрос-
лой фазе. Именно в силу того, что Arion ater требуется больше времени на
завершение репродуктивного цикла, он дает только одно поколение в веге-
тационный сезон, а при неблагоприятных условиях размножение переносится
на весну следующего года (Chevallier, 1969, 1971). Напротив того, D. reti-
culatum способен давать в год до 2—3 генераций, т. е. 2—3 раза повторять
жизненный цикл, каждый раз завершая его размножением.
Известно много примеров, указывающих на связь роста и репродуктивного
цикла слизней с питанием и такими важными абиотическими факторами, как
влажность, температура и световой режим. Однако кроме этих факторов и пи-
тания на рост и репродуктивный цикл влияют и внутренние механизмы.
В этом отношении очень показательны эксперименты, проведенные Лавиолетом
(Laviolette, 1950, 1954b), установившим, что в крови Arion, Limax и Dero-
ceras имеются гормоны, выделяемые гонадой и контролирующие развитие-
белковой железы и полового тракта. Природа этих гормонов пока не ясна,
но влияние гонады видно из описанной выше связи главных фаз роста слиз-
ней с состоянием гениталий (табл. 1).
В свою очередь созревание гонады контролируется другими физиологи-
ческими механизмами. Экспериментально доказано, что нейросекреторная
деятельность ЦНС способствует усилению овогенеза, а какие-то клетки глаз-
ных щупалец секретируют фермент, тормозящий овогенез и способствующий
сперматогенезу (Pelluet, Lane, 1961; Wattez, 1978). По-видимому, именно-
этот эндокринный механизм устанавливает протандрию слизней и определяет-
упомянутые выше различия в сроках наступления овогенеза у A. aternD. reti-
culatum.
Жизненные циклы. Широкий круг вопросов, связанных с фено-
логией роста и размножения, для многих слизней пока еще недостаточно-
изучен или полностью неизвестен. Особенно это касается видов, обитающих
в азиатской части СССР.
Несколько лучше обстоит дело со слизнями, обитающими в европейской
части Союза. Так как большинство из них живет также в Средней и Западной
Европе, то некоторый свет на их жизненные циклы проливают многочислен-
ные зарубежные исследования. Однако в них основная информация исходит
из наблюдений, сделанных в террариумах, т. е. в условиях влажности и тем-
пературы, близких к оптимальным.1 Наблюдений непосредственно из природы
сравнительно немного и нередко они имеют фрагментарный характер. Учи-
тывая большие отличия климата стран Западной и Средней Европы от евро-
пейских областей СССР, которые не могут не сказаться на фенологии жизнен-
ных циклов слизней, использовать зарубежные данные следует с большой,
осторожностью.
Из слизней, обитающих в СССР, наиболее полно изучены жизненные
циклы у Deroceras reticulatum (Дмитриева, 1969, 1978), Gigantomilax monti-
cola armeniacus (Акрамовский, Бабаян, 1976; Арутюнова, 1978), Candaharia
levanderi (Жадин, 1946), C. rutellum (Увалиева, 1975, 1979) и Parmacella
ibera (Шилейко, 1967).
По основным особенностям жизненных циклов и длительности жизни
слизней можно разделить на однолетние и многолетние виды. Однолетний
живут 3—5 месяцев или же 12—18 месяцев и имеют только один период раз-
множения, после которого, отложив яйца, взрослые слизни гибнут. Много-
1 В основном эти сведения собраны в работах Кюнкеля (Kunkel, 1908, 1916, 19341
и Фрёмминга (Fromming, 1954).
•С>4
ОБЩАЯ ЧАСТЬ
летние живут 21/2 — 3 года и имеют два периода размножения; после второго,
отложив яйца, животные тоже вскоре умирают.
Среди однолетних слизней наблюдаются несколько типов жизненного
щикла.
Рис. 25. Жизненные циклы некоторых слизней.
I — Deroceras reticulatum", II — Arion rufus и A. subfuscus; III — Bielzia coerulans\ IV —• Limax maximus
и L. cinereoniger. Кружками обозначены периоды яйцекладки.
1. Циклы слизней Deroceras (рис. 25, Г). Эти слизни живут около 5 ме-
сяцев. Благодаря этому генерации, начавшие свою жизнь весной, успевают
закончить цикл за один вегетационный сезон. При благоприятных климати-
ческих и погодных условиях и хорошем питании эти виды могут дать вторую
и третью генерации, жизненные циклы которых заканчиваются нередко уже
в следующем вегетационном сезоне. Наглядным примером такого типа жиз-
ненного цикла служитDeroceras reticulatum. В центральных областях европей-
ской части СССР этот слизень дает за вегетационный сезон одну генерацию, на
зимовку остаются яйца. В северо-западной зоне, в частности в Ленинградской
•области, при благоприятных погодных условиях — дождливое лето, поздние
•осенние заморозки — может появиться и вторая генерация, если не всюду,
РАЗМНОЖЕНИЕ И ОНТОГЕНЕЗ
65
то в отдельных биотопах. В этом случае на зимовку остаются не только яйца,
но'и молодые слизни (Дмитриева, 1969). Наконец, в Англии с ее влажным
и теплым климатом и мягкими короткими зимами этот слизень размножается
почти круглый год (Barnes, Weil, 1944, 1945; Barnes, 1949; Bett, 1960).
По-видимому, еще более коротким жизненным циклом обладает Dero-
ceras laeve. По нашим данным он приступает к размножению в середине
второго месяца жизни. Не случайно, что этот вид — единственный слизень,
живущий в тундре Евразии и Северной Америки, т. е. в условиях очень
короткого лета.
В южных засушливых областях с континентальным климатом местные
виды Deroceras могут давать в вегетационный сезон лишь одно поколение,
. так как в течение жаркого лета их рост и созревание задерживаются из-за
необходимости скрываться в глубоких убежищах от сухости верхних слоев
почвы и наружного воздуха. Окончание роста и размножение приходятся на
осень; на зимовку обычно остаются только яйца, и лишь в благоприятные
годы или в особенно влажных биотопах может появиться вторая генерация.
В частности, это относится к популяциям D. caucasicum, обитающим в Арме-
нии (личное сообщение Н. Н. Акрамовского).
Короткий жизненный цикл слизней Deroceras тесно связан с некоторыми
особенностями физиологии этих животных. Они отличаются высоким темпом
роста и более ранним, чем у других слизней, наступлением зрелости, в част-
ности более ранним началом овогенеза (см. выше). Благодаря этому между
копуляцией, оплодотворением и откладкой яиц проходит значительно меньше
времени, чем у других слизней (например, у Arion). Кроме того, у Deroceras
эмбриональное развитие идет значительно быстрее, чем у других видов.
Так, у D. reticulatum при влажности около 100% и температуре воздуха 15°
молодь вылупляется из яиц на 11—13-й день после яйцекладки (Дмитриева,
1969).
Судя по всему, сходным жизненным циклом обладает Milax gagates
(Galangau, 1964; Focardi, Quattrini, 1972). В Италии и Франции этот слизень
становится половозрелым и приступает к копуляции в возрасте 4—5 месяцев.
Он откладывает яйца через 15—20 дней после копуляции, а затем взрослые
животные гибнут. По-видимому, за вегетационный сезон появляется две
генерации, так как установлено, что этот слизень размножается весной и
осенью. Вероятно, зимует как молодь, так и яйца.
2. Циклы слизней Arionrufus, A. lusitanicusn A. subfuscus (Abeloos, 1942—
1944; Smith, 1966; Chevallier, 1969, 1971). Эти виды живут 12—18 месяцев
(рис. 25, II). Чаще всего, а в европейской части СССР всегда, жизненный цикл,
начавшийся с вылупления из яиц осенью, через полтора года заканчивается
размножением и откладкой яиц. Поэтому обычно зимуют как молодые, так
и почти взрослые слизни. Только в таких странах, как Франция и Англия,
при благоприятных погодных условиях (в частности, при безморозной зиме)
и хорошем питании, в отдельных популяциях эти слизни завершают жиз-
ненный цикл за 12 месяцев, начав его осенью и закончив откладкой яиц осенью
следующего года. Но и в этих странах 11/2-летний жизненный цикл достаточно
, сбычен.
Столь растянутый во времени жизненный цикл этих животных в первую
очередь связан с темпом роста и развития половой системы. У Arion темп
роста значительно ниже, чем у Deroceras, и зрелость наступает позднее. Кроме
того, овогенез заметно отстает от сперматогенеза, а поэтому откладка яиц
начинается позднее и сильно растянута во времени. Следует учесть, что и
эмбриональное развитие у Arion занимает около месяца, т. е. значительно
длиннее, чем в Deroceras.
По-видимому, жизненный цикл Malacolimax tenellus во многом сходен
с упомянутыми видами Arion. В Средней Европе этот вид живет 12—13г/2
5 И. М. Лихарев, А. Й, Виктор
66
ОБЩАЯ ЧАСТЬ
месяца и достигает зрелости (копулирует) на 7—9-м месяце жизни. Установ-
лено, что часть слизней вылупляется в декабре и приступает к размножению
в сентябре следующего года; другая часть вылупляется в марте и копулирует
в .октябре того же года. На эмбриональное развитие требуется как минимум
20-24 дня (Kunkel, 1934).
Сходный жизненный цикл имеет Bielzia coerulans (Smolenska, 1936)
(рис. 25, III). Взрослые слизни приступают к размножению в первую по-
Рис. 26. Жизненный цикл Candaharia levanderi (по данным Жадина, 1945).
Римские цифры — месяцы; t — температура (наверху — воздуха, внизу — почвы); А — изменения тем-
пературы воздуха (сплошная линия) и количества осадков (прерывистая линия); Б — жизнь слизня на
открытом воздухе (II—ИГ —весенний рост, V — размножение, X—XI — осенний рост); В — жизнь
слизня в почве (I, XII — зимовка, V, VI — откладка яиц в почву и вылупление молоди, VI—IX — лет-
няя спячка молоди, гибель взрослых слизней); изменения температуры почвы (сплошная линия).
ловину лета, а откладывают яйца во второй половине, после чего вскоре уми-
рают. Вылупление молоди наступает на 17—25-й день. Молодые слизни в те-
чение осени интенсивно растут и уходят на зимовку, почти достигнув разме-
ров взрослых животных. В течение весны завершается рост и созревание..
Следовательно, благодаря тому, что у этого вида размножение целиком при-
ходится на лето в отличие от видов Arion и Malacolimax tenellus, его жизнен-
ный цикл укладывается в 12 месяцев.
3. Цикл Parmacellidae (Жадин, 1946; Шилейко, 1967; Увалиева, 1979)
(рис. 26). Жизненный цикл этих слизней настолько своеобразен, что не имеет
аналогов среди других групп наземных моллюсков; В начале лета происходит
вылупление молодых слизней, которые, не питаясь, после 1—2 дней актив-
ности находят укрытие, втягиваются внутрь крупной эмбриональной рако-
вины, замыкают вход в нее провизорной «крышечкой» и впадают в диапаузу,,
длящуюся все лето и начало осени до наступления прохладной погоды и пер-
вых дождей. После пробуждения они выползают наружу, начинают питаться
и интенсивно расти; при этом мантия довольно быстро обрастает раковину.
К началу зимы слизни уходят в укрытие на зимовку. И, наконец, весной они.
РАЗМНОЖЕНИЕ И ОНТОГЕНЕЗ
67
выползают наружу, продолжают интенсивно питаться и расти, а во второй
половине весны и в начале лета размножаются.
В связи со строгой приуроченностью размножения к определенному вре-
мени года и гибелью взрослых животных после откладки яиц1 в течение всего
года популяции состоят из слизней одного возраста. В этом также сущест-
венная особенность жизненного цикла Parmacellidae, так как у остальных
слизней (кроме Bielzia coerulans) обычно присутствие внутри популяции пред-
ставителей нескольких генераций. Различия в размерах пармацелл внутри
популяций и между популяциями вызваны неоднородностью условий
жизни.
Длительность эмбриогенеза у разных видов колеблется в следующих
пределах (при развитии в оптимальных условиях): Candaharia, levanderi —
20—25 дней, С. rutellum — 25 — 30, Parmacella ibera — 35—42'дня. Следо-
вательно, у этих слизней эмбриональное развитие занимает больше времени,
чем у Deroceras, и ближе к тем срокам, которые необходимы для Arion.
В заключение остановимся на жизненных циклах многолетних слизней.
Наши представления о них исходят из того, что мы знаем о Limax cinereoniger,
L. maximus, L. flavus и Lehmannia marginata. Необходимо подчеркнуть, что
в основном информация почерпнута из наблюдений, сделанных в лаборатор-
ных условиях, где слизни содержались при оптимальной влажности (Kunkel,
1916; Fromming, 1954). Отдельные наблюдения над размножением этих слиз-
ней, сделанные непосредственно в природе (Средняя Европа) не могут дать
четкого представления о всем цикле, тем более что всегда остается неясным
возраст слизней, т. е. наблюдался ли первый период размножения (2-й год
жизни) или второй (3-й год). Поэтому приходится довольствоваться информа-
цией о жизненных циклах этих животных, полученной из наблюдений в ла-
бораториях, когда точно известен возраст (т. е. день вылупления из яиц)
каждого слизня.
Жизненные циклы Limax maximus и L. cinereoniger очень сходны (рис. 25, IV).
Эмбриональное развитие длится минимум 18—21 день. Вылупление из яиц
происходит осенью. Первый год жизни эти животные только растут, но не
размножаются. Первая копуляция обычно происходит в июне—июле 2-го
тода жизни, т. е. на 21—22-й месяц после вылупления. Вторая копуляция
происходит на 3-м году жизни, обычно в мае—июне. Откладка яиц тянется до
двух месяцев. Всего слизни живут 23/4—3 года.
Жизненный цикл Lehmannia marginata очень сходен с предыдущим.
Главное отличие заключается в том, что эти слизни рождаются весной. Пер-
вый период размножения приходится на осень (сентябрь—ноябрь) 2-го
года жизни, а второй — на начало весны (март) следующего года. Длитель-
ность жизни 21/2 года.
Жизненный цикл Limax flavus отличается более ранним наступлением
половозрелости — первая копуляция и откладка яиц отмечены в конце 1-го
года жизни. В отношении дальнейшего размножения этого слизня мнения
расходятся. Кюнкель (Kunkel, 1916) считает, что оно повторяется еще два
раза — на 2-м и 3-м году жизни. Фрёмминг (Fromming, 1954) с ним не со-
гласен и утверждает, что оставшееся время слизень с небольшими переры-
вами откладывает яйца почти до самой смерти. Он объясняет это тем, что дан-
ный вид живет в домах — в подпольях и подвалах — и на его половом цикле
пе сказываются ни изменения погоды, ни сезоны года. Оба автора единодушны,
что слизень живет 21/2—3 года.
У многолетних слизней связь гаметогенеза и формирования полового
тракта с общим ростом не прослежена.
1 Отдельные взрослые слизни, случайно пережившие летнюю засуху, погибают осенью.
68
ОБЩАЯ ЧАСТЬ
ПИТАНИЕ
Подавляющее большинство слизней — это растительноядные полифаги
с большей или меньшей склонностью к сапро- и копрофагии или к микофагии.
Только Trigonochlamydidae являются хищными животными.1
Основой питания растительноядных слизней служат листья, стебли,
плоды, луковицы, корнеплоды и подземные клубни, зерна многих диких
и культурных растений как в свежем виде, так и в различной степени гние-
ния. Наиболее полно изучен пищевой рацион слизней, вредящих сельскому
хозяйству, — Deroceras reticulatum, Arion hortensis, Tandonia budapestensis,
Milax gagates, Candaharia levanderi, C. rutellum, Parmacella ibera и некоторых
других видов (Жадин, 1946; Barnes, 1949; Дмитриева, 1969; Увалиева,
1975, 1979; Акрамовский, Бабаян, 1976; и др.).
Наблюдения, сделанные непосредственно в полевой обстановке, и лабо-
раторные опыты показали, что эти слизни используют в пищу очень широ-
кий ассортимент растений как в свежем виде, так на различных фазах гние-
ния. Обычно отдается предпочтение сравнительно сочным и мягким частям;
растений (листовая паренхима, мякоть сочных плодов и клубней); слизни
избегают участков с жесткими покровами или с сосудисто-волокнистыми
пучками.
Выбор пищи в значительной степени зависит от характера раститель-
ности, произрастающей в непосредственной близости от убежищ, в которых
скрываются слизни на дневное время, а также от возраста животных —
взрослые слизни охотно поедают более грубую пищу, чем молодые.
Только что вылупившиеся слизни питаются остатками своих же яиц
и невылупившимися яйцами из той же кладки, а затем переходят к пита-
нию гумусом и разлагающимися остатками растений. Лишь с возрастом
свежий растительный корм занимает все большее место в их рационе.
Даже у такого в основном растительноядного животного, как Deroceras
reticulatum, в питании заметное место занимает животный белок, в первую
очередь различные тли и мелкие двукрылые (Pallant, 1969). Аналогичное
отмечено у Candaharia levanderi, которая в дополнение к растительному
рациону охотно поедает кал, гниющие остатки мяса и рыбы и даже трупы
сородичей. Еще большую долю продукты животного происхождения зани-
мают у некоторых видов Arion, Tandonia и Milax. С другой стороны, такие
виды, как Arion subfuscus, Malacolimax tenellus, Limax cinereoniger, Bielzia
coerulans, не являясь чистыми микофагами, охотно, поедают лесные шляпоч-
ные грибы и лишайники. Принято считать, что многие Philomycidae явля-
ются чистыми микофагами.
В связи с тем что активность слизней чаще всего приходится на ночи
и сумерки, они в основном и питаются в это время. Наибольшая прожор-
ливость слизней совпадает с периодом интенсивного роста, т. е. перед раз-
множением и в начале его, и резко снижается к началу копуляции и в пе-
риод яйцекладки. В это время слизни не питаются.
Количество потребляемой пищи тесно связано также с температурой
и влажностью. При влажности воздуха 90—100% у Deroceras reticulatum
наибольшая прожорливость наблюдается при температуре 17—19° и суточ-
ная прибавка массы в среднем составляет 22.7 мг (Дмитриева, 1969).
При 24—25° потребление пищи сокращается в три раза. Лабораторные опыты
показали (Pallant, 1970; Пахорукова, 1976), что у этого слизня и у близкого'
вида D. agreste при питании наиболее излюбленной пищей (салат) процент
усвоения ее очень высок и в среднем составляет 70—85% от всего съеденного
1 Другим семейством, состоящим исключительно из хищных слизней, являются южно-
африканские Chlamydephoridae. Хищники имеются также среди полуслизней (Daudebar-
diidae, Testacellidae и др.) и некоторых улиток (Streptaxidae и др.).
СРЕДА ОБИТАНИЯ
69
корма. Сходные данные получены в опытах с Arion rufus (Stern, 1969). Ха-
рактерно, что при столь высоком уровне усвоения пищи на рост идет
только 27% ассимилированной энергии. Последнее понятно, так как слизни
затрачивают много энергии на движение и регулирование водного баланса.
О питании хищных Trigonochlamydidae пока известно очень немного,
как и вообще о питании хищных наземных моллюсков. Отмечено (Шилейко,
1966), что Hyrcanolestes velitaris, обитающий в субтропических лесах Талыша
и Ленкоранской низменности, большую часть жизни проводит в лиственном
опаде среди скоплений дождевого червя Eisenia foetida, которыми в основном
и питается. Кроме того, наблюдался каннибализм и питание другими мол-
люсками.
Интенсивность питания этого хищника довольно велика. Так, летом один
слизень длиною 2 см в среднем каждые сутки съедает одного червя 4—6 см
длиной или эквивалентное количество более мелких червей. Наконец, пе-
щерный слизень Troglolestes sokolovi питается мелкими почвенными червями
энхитреидами (Левушкин, Матекин, 1965).
СЛИЗНИ И СРЕДА ОБИТАНИЯ
Как у всех животных, взаимоотношения слизней с окружающим миром
очень сложны. Они проявляются в том, что все основные физиологические
функции и поведенческие реакции, а также численность и распределение
на местности прямо или косвенно зависят от сочетания определенных внеш-
них факторов, которые для удобства анализа их влияния на слизней могут
быть подразделены на две основные группы: абиотические и биотические.
К последним относятся окружающий растительный мир и биотоп в целом,
а также хищники, паразиты и болезни слизней. К анализу этих факторов
и их роли в жизни слизней мы и переходим.
Абиотические факторы
В жизни всех наземных моллюсков, и особенно слизней, из абиотиче-
ских факторов наибольшую роль играют температура и влажность, высту-
пающие нередко в тесной взаимосвязи. Они в значительной степени опре-
деляют ход физиологических процессов, поведение и активность этих живот-
ных. Остальные факторы — осадки, свет, почва и др. — в первую очередь
сказываются на слизнях через изменения режима температуры и
влажности.
Если у улиток крупная, нередко толстостенная наружная раковина спо-
собствует защите от перегрева, отражая подобно экрану избыток солнечной
энергии, а при наступлении засухи в известной степени защищает мягкие
части тела от высыхания, то слизни лишены подобной защиты. В то же время
важная роль покровов в кожном дыхании этих животных повышает их уяз-
вимость к потере влаги. Поэтому главной защитой от высыхания и перегрева
тела является выделение слизи, которая поддерживает кожу в увлажнен-
ном состоянии. Кроме того, слизь играет важную роль в движении и в за-
щите от врагов. Однако расходование слизи сопряжено с сокращением вод-
ного баланса и с необходимостью все время его восстанавливать.
Поэтому в отличие от многих улиток (особенно ксеробионтных), которые
при угрозе перегрева и высыхания ищут защиту внутри раковины, закрывая
вход в нее эпифрагмой, слизни всегда стремятся найти убежище с подходя-
щим микроклиматом, где, сжавшись и тем самым уменьшив поверхность тела,
окружают себя чехлом из слизи и пережидают неблагоприятное время.
Более высокая подвижность слизней, чем таковая у улиток, позволяет им
быстрее найти такое укрытие.
70
ОБЩАЯ ЧАСТЬ
Температура. Влияние этого фактора сказывается как на пове-
дении, так и на ходе отдельных жизненных процессов. Исключительно важно
влияние температуры на интенсивность дыхания слизней, а следовательно,
на общем метаболизме этих животных. Специальные эксперименты показали,
что акклимация слизней к повышению температуры состоит из двух этапов
(Roy, 1963; Rising, Armitage, 1969; Runham, Hunter, 1970). Первый, срав-
нительно короткий, сопровождается сильным нарастанием скорости дыха-
ния, которая через несколько часов резко снижается до определенного
уровня, однако более высокого, чем до начала опытов. Второй этап более
растянут во времени и приводит к дальнейшему снижению скорости дыха-
ния, нередко до исходного уровня.
Отчетливо прослеживается влияние температуры на интенсивность пи-
тания и рост слизней. Так, у Deroceras reticulatum (Дмитриева, 1969) наи-
более оптимальный режим питания приходится на температуры 17—19°.
При 25° интенсивность питания и рост животного снижаются, и длительное
содержание в таких условиях приводит к гибели.
Очень велико влияние температуры на размножение слизней. Отмечено,
что низкие температуры (ниже 10°) задерживают развитие гонады, в то время
как высокие (25° и выше) ведут к уменьшению числа ооцитов или к их пол-
ному исчезновению (Lusis, 1966; Runham, Hunter, 1970; Smith, 1966).
У Deroceras reticulatum температура 17—19° наиболее благоприятна для созре-
вания половых продуктов, а 10—15° для откладки яиц; при 5° она практи-
чески останавливается (Дмитриева, 1969).
Развитие яиц и рост слизней также тесно зависят от температуры (Run-
ham, Hunter, 1970). Так, у Tandonia budapestensis при 20° эмбриональное
развитие завершается за 3 недели, а при 7.5° растягивается до 4 месяцев.
При 20° масса молодого Arion hortensis за 12 недель достигает 500 мг,
а при 5° за то же время только 50 мг.
Температура влияет на длительность жизненного цикла слизней.
Так, например, Tandonia budapestensis в Англии после короткой зимы обычно
вылупляется в начале апреля. К следующей весне он становится половозре-
лым и откладывает яйца; следовательно, весь жизненный цикл занимает
11—12 месяцев. Но после затянувшейся зимы большинство слизней запазды-
вают с вылуплением на 1—1.5 месяца и не успевают достигнуть половозре-
лой стадии к следующей весне. А так как у этого вида летом развитие гонады
приостанавливается, то запоздавшие с развитием слизни на полный цикл
затрачивают почти 2 года (Runham, Hunter, 1970).
Слизни сравнительно слабо устойчивы к отрицательным температурам
и гибнут по прошествии определенного срока пребывания в таких условиях
(Pinder, 1969). Так, среди Deroceras reticulatum, в течение 45 часов находив-
шихся при температуре —3.5°, после переноса в комнатную температуру
ожило около 50% слизней. Эту же температуру 50% Arion hortensis выдер-
живают только 28 часов. При —4.5° оба вида погибали через 12 часов.
Они же перенесли —2.5° без потерь в течение трех недель.
Е. В. Шиков (личное сообщение) проводил весной размораживание
вмерзших в лед наземных моллюсков. При этом улитки оживали, а все
слизни были мертвыми.
Если взрослые слизни не обладают достаточной устойчивостью к отри-
цательным температурам, то их яйца в этом отношении гораздо выносливее.
Так, у Deroceras reticulatum они выдерживают в течение двух суток охлаж-
дение до —11° и в течение суток —16°. Развитие яиц начинается уже при
температуре 5°, а 15° является оптимальной температурой, так как при наи-
более коротком сроке развития (11 дней) оно сопровождается 100%-ным
отрождением молоди (Дмитриева, 1969). Ниже оптимальной температуры
не только увеличивается срок развития яиц, но и уменьшается процент вы-
СРЕДА ОБИТАНИЯ
71
Q --L.1...L 1111 1—1—1....I I...I I ,|
5 10 15 20 251
Рис. 27. Зависимость продолжитель-
ности эмбрионального развития яиц
Deroceras reticulatum от температуры
(из: Дмитриева, 1968а).
в условиях 100%-ной влажности
лупления молоди; выше 15° (17—25°) происходит небольшое удлинение
инкубационного периода и заметное увеличение процента неразвившихся
и погибших яиц (рис. 27).
Температура играет решающую роль в окончании летней диапаузы
Parmacellidae. Опыты показали, что смачивание диапаузирующей молоди
Candaharia levanderi без понижения температуры не приводило к ее про-
буждению (Жадин, 1946). Оно происходит только при сочетании дождя с низ-
кой температурой воздуха (около 3.5°). Вот почему слизни не могут выйти
из диапаузы под влиянием случайного летнего дождя, после которого вновь
наступает жаркая погода, а пробуж-
даются только с установлением про-
хладного осеннего времени.
Влажность. Как у всех на-
земных моллюсков, роль влажности
в жизни слизней очень велика. Это в пер-
вую очередь связано с тем, что в теле
этих животных вода занимает очень
большую долю. Так, у Arion ater она
составляет 86.3+3.8% массы тела,
причем на долю крови приходится
36.6+10.4% (Martin е. а., 1958). Есть
основания считать, что у других
слизней тело содержит еще больше
воды.
В течение жизни слизня постоянно
происходит расходование и пополнение
водного баланса, особенно когда жи-
вотное находится в активном состоянии.
Вода тратится не только на выделение
слизи для смачивания поверхности
тела, на движение и защиту от меха-
нических раздражителей и врагов, но
также и на все основные жизненные
функции (питание, выделение, размно-
жение). Даже при содержании слизней
воздуха и без пищи у них происходят периодические колебания массы тела
в пределах 15—30%. Они связаны с изменениями водного баланса, происхо-
дящими из-за расхода воды, главным образом на движение (Barnes, Weil,
1934; Dainton, 1954а). Восстановление водного баланса происходит в пер-
вую очередь за счет поглощения воды через кожу; другим источником яв-
ляется пища.
Многие слизни способны короткое время переносить большие потери
воды. Так, Deroceras reticulatum, Limax maximus и Arion ater выдерживают
потери воды до 50% от массы тела и при возвращении к условиям 100%-ной
влажности восстанавливают свой водный баланс за несколько часов. Кюн-
кель (Kunkel, 1934) указывает, что Malacolimax tenellus после утраты за счет
высыхания 80% массы тела может быстро восстановить водный баланс
до нормы за счет поглощения воды через кожу.
Подобно взрослым слизням их яйца и эмбрионы способны кратковре-
менно выдержать значительное высыхание — до 80% (Bayne, 1968). С другой
стороны, полное погружение в воду на 4 суток слабо сказывается на ходе
развития яиц, но более длительное пребывание значительно снижает про-
цент вылупившейся молоди (Arias, Crowell, 1963).
Как правило, слизни не могут длительное время переносить влажность
ниже 90%. Вначале всякое снижение влажности вызывает кратковременное
72
ОБЩАЯ ЧАСТЬ
повышение подвижности, которое слизень использует для поиска укрытия.
Если такое по будет найдено, то слизень впадает в оцепенение и вскоре по-
гибает. Снижение подвижности связано с тем, что потеря воды вызывает
повышение осмотического давления крови, что в свою очередь влечет подав-
ление нервной активности педальных ганглиев (Kerkut, 1959). Разбавление
крови водой способствует нарастанию активности этих ганглиев и тем самым
увеличивает подвижность животных.
На рост слизней влажность оказывает влияние, аналогичное влиянию
температуры. Они не питаются, если влажность микроклимата ниже 100%.
В сухое время они прячутся в укрытия — различные щели в почве, под
камни и другие предметы на почве, под густой полог растений, и их активность
на поверхности почвы прекращается или ограничена особо влажными ме-
стами, например берегами водоемов. По-видимому, для поддержания нор-
мального водного баланса слизням важна не столь влажность воздуха,
как наличие на почве водной пленки, из которой они и черпают необходимую
влагу, впитывая ее через кожу. Другим источником воды служит пища.
Несмотря на то что всем слизням свойственны высокие требования к влаж-
ности, в этом отношении подмечены известные различия между видами.
Так, если поместить Arion ater на границе между сухой и влажной зонами,
то после короткого периода ориентации слизень направляется в наиболее
влажное место садка (Lewis, 1969а). Несколько иначе себя ведет Milax
gagates. Если разница между сырой и сухой зонами меньше 30% относитель-
ной влажности, то днем слизни собираются в сухой зоне, а ночью во влаж-
ной (Bonavita, 1967).
В итоге можно сделать вывод, что кроме высокой водопроницаемости
покровов тела слизни лишены специальных механизмов, контролирующих
их водный баланс. Отсюда их высокие требования к влажности окружающей
среды.'
, Осадки. Высокие требования слизней к влажности, особенно к влаж-
ности почвы, делает понятным значение осадков в жизни этих животных.
Не случайно, что именно в западном Закавказье, т. е. в области, где выпа-
дает наибольшее для территории СССР количество осадков (до 2500 мм за
год), обитает самая своеобразная и богатая видами фауна слизней.
Общее количество осадков и характер их распределения в течение года
наряду с другими факторами в значительной степени определяют жизненный
цикл, характер распределения на местности, численность и географическое
распространение слизней (Соколов, 1935; Гриванов, 1936а; Дмитриева,
1969). Так, сравнительная редкость и непродолжительность летних засух
на северо-западе, западе и в центре европейской части СССР способствует
тому, что обитающие здесь слизни в безморозное время года достаточно ак-
тивны.
С другой стороны, длительная летняя засуха, обычная в средиземномор-
ских областях, а также в восточном Закавказье (в том числе и на Армянском
нагорье) и в Средней Азии, не только вызывает приостановку активности
слизней на это время (Акрамовский, Бабаян, 1976; Увалиева, 1979), но
у Parmacellidae в процессе эволюции привела к появлению летней диапа-
узы, наступающей сразу после вылупления молоди из яиц. Эта диапауза
прерывается только осенью, после снижения дневных температур и первых
прохладных дождей (Жадин, 1946).
Реакция слизней на выпадение дождя в значительной степени определя-
ется температурой воздуха. Если дождю сопутствует оптимальная темпера-
тура, то слизни становятся активными и выползают на поверхность даже
в дневное время. Если же температура неблагоприятна, то слизни остаются
в укрытиях. Однако в тех случаях, когда дождь начинает затапливать их
убежища, слизни выходят наружу и спасаются от воды на возвышенных
СРЕДА ОБИТАНИЯ
73
местах. В такое время в лесу некоторые виды поднимаются вверх по стволам
деревьев и кустов, а также на траву (многие Lehmannia, Limax, Deroceras),
а на открытых местах заползают на открытые поверхности камней и отвес-
ные склоны скал.
Не меньшую роль, чем дожди, имеет снежный покров. Во-первых, он
выступает как резерв влаги для будущей весны и лета. Во-вторых, благодаря
низкой теплопроводности снег является своеобразным теплоизолятором,
предохраняющим почву от сильного охлаждения и резких колебаний тем-
пературы. Поэтому существует прямая связь между толщиной снежного
покрова и успешной зимовкой слизней и, в особенности, их яиц (Дмитриева,
1969).
Ветер. Всякое движение воздуха приводит к усилению испарения
с поверхности тела слизней, поэтому они четко реагируют на направление
и силу ветра (Kalmus, 1942; Dainton, 1954b). Очень слабый ветер обычно
вызывает лишь вытягивание щупалец в его сторону; при более сильном
слизни начинают двигаться, причем всегда по ветру. Последнее связано со
стремлением в первую очередь защитить от высыхания головной конец
и щупальца, т. е. наиболее чувствительную часть тела. Во всех случаях
слизни стараются укрыться от ветра в каком-нибудь убежище.
Ветер не только влияет на поведение слизней, но и на распределение их
на местности. Так, Шиков (19796) показал, что в поймах крупных рек Вал-
дайской возвышенности распределение популяций Deroceras agreste в зна-
чительной степени зависит от направления господствующих ветров. В от-
личии от других видов того же рода, этот слизень предпочитает наиболее
продуваемые и сухие участки речных долин.
Свет. Отношение слизней к этому фактору достаточно сложно. В пер-
вую очередь оно выражается в наличии определенного суточного ритма
активности этих животных, который в основном подчинен более четким тре-
бованиям к влажности и температуре. Повышение активности совпадает
с периодами, когда оба фактора наиболее близки к оптимуму. Это очень убе-
дительно показал Жадин (1946) на примере' Candaharia levanderi. В годы,
когда весной выпадает достаточно осадков и влажность воздуха и почвы
оптимальная, суточная активность этого слизня тесно связана с температу-
рой, особенно с минимальной. Обычно максимальный выполз слизней при-
ходится на вечерние и утренние часы, когда температура держится в преде-
лах 6—18° (рис. 28, I). В тех случаях, когда такие температуры приходятся
только на середину ночи, происходит только один массовый выполз — ночью.
Однако в прохладные и дождливые сутки активность этого слизня смеща-
ется на дневное время (рис. 28, II).
У слизней, обитающих в областях с морским климатом, где различия
между дневными и ночными температурами выражены не так резко, как
в Средней Азии, активность слизней чаще всего приходится на утренние
и вечерние часы.
По-видимому, имеется какой-то внутренний ритм, который связан с су-
точным световым режимом. Так, опыты с Arion ater показали (Lewis, 1969b),
что если содержать этих слизней в условиях постоянной освещенности или
полного м:рака, то они некоторое время сохраняют нормальную суточную
активность. Однако по истечении двух недель она постепенно нарушается
и восстанавливается лишь после того, как подопытные животные будут воз-
вращены к нормальной суточной смене дня и ночи.
Слизни особенно чувствительны к инфракрасному участку спектра, влия-
ние которого они стараются ослабить, прячась в укрытия. Как уже указы-
валось (стр. 49), рецептором инфракрасных лучей является дополнитель-
ная ретина глаза.
74
ОБЩАЯ ЧАСТЬ
То, что слизни отнюдь не индифферентны к свету, видно из общего ха-
рактера окраски многих видов, которая в значительной степени зависит
от освещенности биотопов. Подмечено, что в популяциях, населяющих от-
крытые места (поля, огороды, луга), преобладают особи с более сильным
развитием меланина в хроматофорах. Их относительно меньше в популя-
циях, живущих в затененных биотопах (леса, кустарники и т. и.), где пре-
обладают светлоокрашенные особи. Обычно у видов, проводящих в почве
большую часть жизни (Selenochlamys pallida), темный пигмент слабо развит,
а у троглобибнта Troglolestes sokolovi тело молочно-белое, т. е. лишено пиг-
мента.
I И
Рис. 28. Суточная динамика численности Candaharta levanderi в апреле 1943 г. в ясную (/)
и дождливую {Н) погоду (из: Жадин, 1946).
1 — число слизней; 2 — температура воздуха; по оси абсцисс — часы наблюдений.
Отношение к свету отнюдь не' ограничивается его влиянием на окраску
слизней. Определенное количество света необходимо для успешного созре-
вания половых продуктов (Pelluet, Henderson, 1954; Sokolove, McCrone,
1978). С другой стороны, длительное освещение высокой интенсивности
вызывает торможение гаметогенеза, а слабый свет ускоряет созревание
спермиев. Полный мрак приводит к резорбции гамет, и гонада приобретает
облик, который нормально она имеет у стареющего животного.
Почва. Химические и физические особенности почв заметно влияют
на численность и распространение слизней. Наиболее благоприятны как
для слизней, так и для улиток щелочные и нейтральные почвы. Кислые почвы
явно избегаются слизнями, хотя в этом отношении они более устойчивы,
чем улитки. Особенно большое значение имеет количество гумуса в почве.
Его роль многогранна: с одной стороны, он является источником пищи,
особенно для молоди; с другой — присутствие гумуса увеличивает влаго-
емкость почвы; наконец, обилие органики в почве способствует развитию
густого травяного покрова, который предоставляет слизням и хорошее убе-
жище, и источник пищи.
Как и все наземные моллюски, слизни избегают засоленных почв.
Пожалуй, большее значение, чем химические, имеют для слизней физи-
черцие- особенности - почв— влагоемкость, структура и' микрорельеф.
Они явно предпочитают влагоемкие и тяжелые суглинистые и глинистые
СРЕДА ОБИТАНИЯ 75
почвы и избегают водопроницаемых, легких каменистых, песчаных и су-
песчаных почв, в первую очередь из-за их непостоянной и низкой влажности.
Последняя в особенности определяет отношение слизней к почвам. Как из-
вестно, для этих животных совершенно необходимо присутствие на поверх-
ности почв водной пленки — главного источника поддержания водного ба-
ланса слизней. Только наличие такой пленки позволяет им вести активный
образ жизни, когда влажность воздуха меньше 100%. Однако на' торфяных
болотах слизни обычно не живут, за исключением Arion subfuscus, но и он
встречается здесь в небольшом количестве.
Очень большое значение в жизни слизней, особенно на открытых биото-
пах, имеет микрорельеф почвы. Участки со сглаженным рельефом, где мало
убежищ в виде ямок и щелей в почве или под камнями, неблагоприятны для
этих животных, особенно если растительный покров в таких местах редкий.
С другой стороны, обилие таких убежищ в сочетании с достаточно густым
растительным покровом (например, на полях и огородах) способствует вы-
сокой численности и быстрому расселению слизней, в частности таких
сельскохозяйственных вредителей, как Deroceras reticulatum, Candaharia
levanderi, Parmacella ibera и др.
Растительность, основные ландшафты и биотопы
Окружающий слизней растительный мир — это в первую очередь важней-
ший элемент среды обитания, а затем уже источник пищи.
Растительный покров создает под своим пологом особый микроклимат,
порой сильно отличный от макроклимата и значительно более благоприят-
ный для многих моллюсков. Растения не только защищают живущих под
ними животных от иссушающего воздействия солнца и ветра, но и благодаря
испаряющей работе листьев создают под своим пологом более высокую влаж-
ность воздуха и почвы, чем на открытых местах.
В этом отношении очень показательно сравнить, с одной стороны, широко-
лиственные леса с хорошо развитым подлеском и густым травянистым по-
кровом и, с другой стороны, расположенный по соседству сухой луг с редким
травянистым покровом. Первая ассоциация в силу благоприятного микро-
климата, создающегося под ее пологом, обилию и разнообразию экологи-
ческих ниш, как правило, отличается высоким разнообразием фауны слизней,
отдельные виды которой могут достигать высокой численности.
Иная ситуация на сухом лугу. Здесь фауна слизней значительно беднее
и численность отдельных видов ниже. Моллюски находят убежища только
там, где травяной покров образует сгущения — в прикорневой части расте-
ний, а также в отдельных углублениях микрорельефа и под камнями.
Большинство видов слизней обитает в лиственных, особенно в широко-
лиственных, и в смешанных лесах, в затененных местах с богатой и доста-
точно влажной подстилкой или под густым травяным покровом, по оврагам,
ущельям и вдоль лесных речек и ручьев. В чисто хвойных насаждениях оби-
тают лишь немногие виды (Arion subfuscus, Malacolimax tenellus) и то лишь
при достаточной влажности и наличии шляпочных грибов и лишайников,
которыми они питаются. При этом численность этих слизней здесь заметно
ниже, чем в смешанных лесах или в хвойном лесу с хорошо развитым листвен-
ным подлеском и травяным покровом.
Отношение слизней к тому или иному типу леса в значительной мере оп-
ределяется обилием и разнообразием убежищ, где животные могут укрыться
от жары и морозов. Наиболее универсальным убежищем служит лиственная
подстилка, которая для многих видов одновременно является источником
пищи или субстратом, на котором развиваются грибы, употребляемыми дан-
ными слизнями в пищу. Хвойная подстилка неблагоприятна для наземных
76
ОБЩАЯ ЧАСТЬ
моллюсков, как из-за ее большей сухости, так и из-за непригодности в пищу
(жесткость и высокое содержание смолистых веществ).
Некоторые виды лесных слизней часто встречаются на стволах деревьев
(Limax maculatus, Caspilimax keyserlingi, Lehmannia marginata). Здесь они
питаются древесными лишайниками и грибами, а также гниющей древеси-
ной. Нередко на этих же деревьях они находят укрытие в дуплах, щелях
коры и моховом покрове. Однако их нельзя считать чисто древесными оби-
тателями, так как нередко их можно встретить и в подстилке, в гнилом ва-
лежнике, под корой гниющих на земле стволов, под камнями и в
щелях скал.
Наиболее богаты слизнями участки лиственного и смешанного леса,
располагающиеся в тенистых сырых оврагах и ущельях, вблизи от водоемов
или со скалами и каменистыми осыпями. В этих осыпях среди влажных кам-
ней, чередующихся с гниющей подстилкой и слоями перегноя, особенно
много влаголюбивых улиток и слизней.
Пойменные леса и кустарники, периодически подвергающиеся паводкам,
обычно слабо заселены слизнями. И причина этому не только в прямом воз-
действии паводка на зти животные — снос и гибель от длительного пребы-
вания в воде, но и в разрушении почвенного слоя и подстилки, лишающего
слизней как убежища, так и источника пищи. Только упомянутые обита-
тели деревьев могут найти убежище в пойменном лесу. Правда, некоторые
виды попадают в пойму именно в результате паводков, т. е. путем заноса
из других мест. Однако обычно они не образуют на новом месте устойчивых
популяций (Шиков, 1977).
Сравнительно бедны слизнями дубовые леса, как чистые дубняки, так
и в смеси с другими древесными породами (липа, граб). По-видимому, это
связано с их большей сухостью, особенно подстилки. Кроме того, вероятно,
лиственный опад дуба менее пригоден в пищу моллюсков из-за высокого со-
держания в нем дубильных веществ. Поэтому не только слизни, но и боль-
шинство улиток избегают селиться в чисто дубовой подстилке.
Сравнительно бедны слизнями и другими наземными моллюсками как
в качественном, так и в количественном отношении чисто буковые насаж-
дения с бедным травяным покровом или даже вовсе без него. Вероятно, и
здесь сказывается химизм буковой подстилки.
Как для лесных видов слизней, так и для видов, населяющих открытые
пространства, наиболее благоприятны скальные участки и каменистые осыпи,
в ^особенности сложенные из известняковых и других щелочных горных
пород. Слизням благоприятен не только химизм этих субстратов, но и оби-
лие в них всевозможных щелей, углублений и других карстовых образова-
ний, в которых накапливается гумус и поддерживается свой микроклимат,
порой очень отличный от наружного и более стабильный.
По этим же причинам многие слизни, как троглоксены, нередко обитают
у входа или в начальных участках пещер. Настоящим троглобионтом среди
слизней является только Troglolestes sokolovi, живущий в одной из пещер
Абхазии на большой глубине от входа.
В отличие от лесов, в особенности горных лесов, фауна слизней открытых
пространств — лугов, степей, как горных, так и равнинных — значительно
беднее видами. На задерненных сухих и травянистых биотопах, в особен-
ности по южным склонам со слабо расчлененным рельефом и лишенным
осыпей, отдельных камней и скал, популяции слизней отличаются низкой
численностью. В европейской части СССР обычно тут обитают только неко-
торые виды рода Deroceras (D. agreste, D. reticulatum), Arion fasciatus, а на
Кавказе Gigantomilax monticola. Ёолее влажные открытые биотопы — сырые
луга — обладают аналогичной фауной, но численность популяций заметно
выше.
СРЕДА ОБИТАНИЯ
77
В биотопах с постоянной высокой влажностью, где свободная вода име-
ется на почве в течение всего вегетационного периода — заболоченные ни-
зины, заросшие густой травой берега водоемов — фауна слизней отличается
большим однообразием. Наиболее характерным видом таких мест является
широкораспространенный Deroceras laeve, который в жаркое время года ста-
новится настоящим амфибионтом, нередко обитая совместно с амфибионт-
ными водными моллюсками (например, Lymnaea truncatula). В аридных об-
ластях Кавказа и Средней Азии в разгар лета сюда нередко вытесняются
менее влаголюбивые слизни — Lytopelte maculata, некоторые Candaharia
й Deroceras.
На равнине или в горах наличие достаточного числа подходящих убежищ,
в которых слизни могут успешно зимовать, пережидать засуху или скры-
ваться на дневное время, является важнейшим моментом, определяющим
их численность, способность к расселению и даже географическое распростра-
нение. Такие убежища, как камни, щели скал и каменистые осыпи, позволяют
многим слизням жить даже в условиях горной степи и полупустынь Армян-
ского нагорья (Акрамовский, 1976) и Средней Азии. Скрываясь в них, слизни
приурочивают свою активность к наиболее влажным сезонам, а летом по
ночам или в сумерки с выпадением росы. В связи с этим полезно вспомнить,
что в условиях жаркого континентального климата с резкими перепадами
суточной температуры каменистые осыпи аккумулируют атмосферную влагу
и влаголюбивые животные могут' проживать в глубине их очень длительное
время.
Недостаток убежищ в равнинных степях и пустынях наряду с другими
неблагоприятными факторами служит причиной отсутствия слизней в ти-
пичных природных биотопах этих зон. Здесь они могут встречаться только
по поймам рек, где кроме более высокой общей влажности воздуха они на-
ходят укрытия под достаточно густым пологом растений и в подстилке.
Однако стоит только человеку начать проводить в степь и пустыню ороси-
тельные каналы и осваивать эти земли под сельскохозяйственные угодья,
как здесь появляются некоторые слизни. Последнее хорошо видно на при-
мере расселения Candaharia levanderi в Таджикистане, Parmacella ibera
на Кавказе, Deroceras caucasicum на Кавказе и в Средней Азии.
Обширные открытые пространства, занятые плантациями, полями и ого-
родами, где почва периодически подвергается глубокому разрыхлению,
а в южных областях нередко и искусственному орошению и покрыта густым
покровом культурных растений, заселяются некоторыми синантропными
слизнями (Deroceras reticulatum, D. caucasicum, D. sturanyi, Candaharia
levanderi, Parmacella ibera). Обилие зеленой пищи и убежищ во взрыхленной
почве и под пологом растений, гниющих растительных остатков, высокая
влажность почвы и воздуха — все это способствует высокой численности
я тих видов. Отсюда проистекает и вред, который они наносят многим куль-
турным растениям (с. 108).
Если избыток тепла и низкую влажность засушливых областей юга
•СССР слизням удается нейтрализовать, подгоняя свою активность и жизнен-
ный цикл к благоприятным сезонам и к ночам, а остальное время скрываясь
в хорошо защищенных убежищах, то общий недостаток тепла является для
них непреодолимой преградой, ограничивающей их возможности распростра-
нения в‘ области с холодным климатом. В тех случаях, когда нехватка тепла
достигает пессимальных пределов, поселение в жилище человека или в тепли-
цах — единственный путь преодоления этой преграды. Однако этот путь
открыт только некоторым антропохорным видам — Limax flavus, L. ma-
culatus, Lehmannia valentiana.
78
ОБЩАЯ ЧАСТЬ
Хищники, паразиты и болезни
Как и у улитки, слизни имеют довольно обширный ассортимент врагов,,
в том числе и хищников. Ими питаются многие позвоночные животные,
однако специфических «слизнеедов» среди них нет. Из млекопитающих слиз-
ней охотно поедают ежи, кроты, землеройки и некоторые мышевидные гры-
зуны; из птиц — грачи, галки, скворцы и некоторые чайки, а из домашних
птиц — куры и утки. Слизни также входят в пищевой рацион многих ля-
гушек, жаб, саламандр, ящериц и змей.
Среди беспозвоночных животных слизнями питаются многие насекомые.
Особенно их много среди жуков-жужелиц (Carabidae). Почти исключительно
наземными моллюсками, в том числе и слизнями, питаются хищные личинки
жуков-светляков (Lampyridae), а также некоторых мягкотелок (Сantharidae).
Как взрослые насекомые, так и их личинки заметно предпочитают улиток
слизням. По-видимому, это объясняется тем, что улитки не способны так
успешно защищаться от нападения с помощью слизи, как это делают взрос-
лые и здоровые слизни, порой связывая ею ноги и ротовые придатки хищ-
ников. К тому же личинки жуков, поедая улитку, нередко используют ее
раковину как убежище. Поэтому чаще жуки и их личинки нападают на мо-
лодых или ослабленных взрослых слизней.
К числу других насекомых, которые на стадии личинки питаются слиз-
нями, принадлежат некоторые серые мясные мухи (5arcophagidae), и в осо-
бенности мухи-сциомизиды (S ciomyzidae).
Хищники, нападающие на слизней, известны и среди других групп бес-
позвоночных: некоторые губоногие многоножки (Chilopodd), отдельные се-
нокосцы (Opiliones). Даже среди ресничных червей (Turbellarid) известна
одна наземная тропическая планария — Geoplana septemlineata, которая
нападает на улиток и слизней, прокалывает им кожу своей глоткой и выса-
сывает содержимое тела моллюсков.
Слизни служат хозяевами (факультативными, промежуточными или ос-
новными) для многих паразитов. Так, в пищеварительном тракте, печени или
почке некоторых слизней обнаружено несколько видов инфузорий и кокци-
дий.
Многие слизни являются промежуточными хозяевами ряда дигенети-
ческих сосальщиков (Trematoda) из семейств Dicrocoeliidaer Brachycoeliidae
и Brachilaimidae. Взрослые черви этих групп паразитируют у домашних
и диких млекопитающих и птиц.
Из ленточных червей (Cestoda) лишь немногие имеют промежуточными
хозяевами слизней. Из них в хозяйственном отношении особенно важен
Davainea proglottina — возбудитель среди домашних и диких птиц опасного'
кишечного заболевания — давайнеоза.
Среди паразитов слизней очень много разных видов круглых червей
(Nematoda). Роль слизней в жизненном цикле этих гельминтов различна.
Для тех видов нематод, взрослая фаза которых протекает в почве, в слизнях
происходит только развитие личинки (Mermithidae). По1 выходе наружу
червь достигает половой зрелости и размножается. У других круглых чер-
вей наземные моллюски, и в частности слизни, являются промежуточными
хозяевами, в которых происходит развитие личинки до инвазионной стадии,,
т. е. пригодной для заражения окончательного хозяина (млекопитающие,,
птицы).
В легком и на поверхности тела многих слизней можно встретить микро-
скопического клеща Riccardoella limacum. Он питается некоторыми клеточ-
ными элементами соединительной ткани и крови слизней. Многие аспекты
биологии этого клеща, а также его влияние на численность слизней пока>
неясны (Turk, Phillips, 1946).
СЛИЗНИ КАК ПРОДУКТ ПАРАЛЛЕЛЬНОЙ ЭВОЛЮЦИИ
79
Объектами нападения хищников и заражения паразитами служат не
только сами слизни, но и их яйца. Однако наибольший урон яйцекладкам
наземных моллюсков, особенно в теплые и влажные дни, наносят парази-
тические грибки. Они могут покрывать поверхность яиц наподобие густого
войлока, проникать внутрь и в конечном счете сильно снижать процент вы-
лупившейся молоди.
Имеются отрывочные данные о болезнях слизней, вызываемых грибами,
бактериями и, возможно, вирусами. Однако природа их пока не изучена.
Несмотря на то что хищники, паразиты и болезни должны играть важную
роль в регуляции численности слизней, особенно в природных ассоциациях,
достаточно обоснованных данных об этом пока еще нет. Тому причиной
•скрытый образ жизни большинства слизней, быстрая их перевариваемость в же-
лудках хищников, наши слабые знания о биологии паразитов и о болезнях
•слизней и, наконец, чисто методические трудности накопления соответствую-
щей информации.
Какой-либо заметной специфичности как хищников, так и паразитов
к определенным видам слизней не прослеживается. Больше того, все упомяну-
тые враги слизней с таким же успехом нападают или паразитируют в теле
многих наземных улиток. По-видимому, главную роль играют экологические
факторы, обеспечивающие контакт между слизнями и хищниками или с ин-
вазионной стадией паразита.
СЛИЗНИ КАК ПРОДУКТ ПАРАЛЛЕЛЬНОЙ ЭВОЛЮЦИИ
Известно, что слизни — это тип организации брюхоногих моллюсков, ко-
торому свойственны определенные особенности морфологии и физиологии,
отличающие этих животных от другого более обычного среди наземных Gastro-
poda типа организации — от улиток.
Между улитками и слизнями встречаются переходные формы, так называ-
емые полуслизни. У них внутренностный мешок еще не полностью погружен
>в толщу цефалоподиума, а раковина еще наружная, но в отличие от улиток
редукция ее зашла настолько далеко, что она уже не может вместить мягкие
части животного даже при сильном сокращении тела. У одних полуслизней
(например, Testacellidae, Daudebardiidae — рис. 29) наружная раковина распо-
лагается на заднем конце цефалоподиума и мантия не закрывает ее. У других
(например, некоторые Vitrinidae — рис. 30, В) раковина распола-
гается около середины цефалоподиума и частично закрыта выростами мантии.
Ведущая и характерная черта слизней, с которой связаны все остальные, —
это погружение внутренностного мешка в толщу цефалоподиума, сопровожда-
ющееся редукцией раковины и обрастанием ее мантией. Каким путем это шло,
мюжно представить на примере морфологического ряда из семейства Vitrini-
dae (рис. 30). Если у Phenacolimax annularis при сокращении животного рако-
вина полностью вмещает цефалоподиум и мантия не имеет выростов, закры-
вающих раковину, то у Vitrina pellucida при сокращении задний конец
цефалоподиума (метаподиум) уже не вмещается в раковину, а мантия имеет
языковидный выступ, который прикрывает часть раковины.
Следующий этап мы видим у полуслизня Semilimax semilimax. Объем
раковины стал еще меньше и при сокращении цефалоподиум полностью оста-
ется вне ее. Лопасть мантии более крупная и закрывает почти 2/3 верхней по-
верхности раковины. Последний этап этого ряда — Plutonia atlantica —
настоящий слизень. Раковина почти плоская и целиком закрыта мантией.
То, что редукция раковины и обрастание ее мантией шли описанным пу-
тем, подтверждается постэмбриогенезом слизней Parmacellidae (Webb et Ber-
thelot, 1835; Жадин, 1946), во время которого можно наблюдать в основном
аналогичную картину (см. рис. 469).
Рис. 29. Полуслизень Daudebardia rufa (из: Лихарев, Виктор, 19796).
р — раковина; м — мантия.
Рис. 30. Внешний облик некоторых видов Vitrinidae (из: Лихарев, Виктор, 19796).
А — Phenacolimax annularis', Б — Vitrina pellucida', В — Semilimax semilimax', Г — Plutonia atlantica.
СЛИЗНИ КАК ПРОДУКТ ПАРАЛЛЕЛЬНОЙ ЭВОЛЮЦИИ
81
Как видно из современных классификаций, внутри надотряда Stylommato-
phora слизни возникали неоднократно и нередко в далеких друг от друга
ветвях (Wenz, Zilch, 1959—1960; Taylor, Sohl, 1962; Лихарев, Виктор, 19796;.
Шилейко, 1979). Так, согласно Шилейко, который, на наш взгляд, пред-
ложил наиболее обоснованную систему основной группы указанного надот-
ряда — отряда Geophila, из 77 семейств 11 полностью или в основном состоят?
из слизней. К ним относятся: а) растительноядные — Arionidae, Oopeltidae,
Philomycidae, Parmacellidae, Milacidae, Limacidae, Agriolimacidae, Boettgeril-
lidae, Urocyclidae-, б) хишные Trigonochlamydidae, Chlamydephoridae.
Кроме того, в некоторых семействах (Bulimulidae, Vitrinidae и др.),
где большинство родов представлено улитками или полуслизнями, имеются
отдельные «слизневые» роды.
Самый факт, что в процессе эволюции Geophila группы, состоящие из слиз-
ней, возникали неоднократно и нередко независимо друг от друга, указывает
на то, что в известных условиях этот тип организации имеет преимущества
перед организацией улитки. К таким преимуществам в первую очередь отно-
сится возможность жить или укрываться в таких убежищах, как щели в почве,
скалах и каменистых осыпях, двигаться под землей по ходам, проделанным
дождевыми червями и другими почвенными беспозвоночными, т. е. находиться
в местах, куда многим улиткам с их крупным внутренностным мешком, оде-
тым в жесткую раковину, трудно или невозможно проникнуть.
Во-вторых, улитки значительную часть энергии, особенно в активном со-
стоянии, расходуют на мускульное усилие по поддержанию и управлению’
внутренностным мешком и раковиной. Слизни освобождены от такой задачи и
за счет этого отличаются большей подвижностью, что облегчает им добывание
пищи, а также поиск укрытия от неблагоприятных физических факторов и
хищников, т. е. увеличивает их биологическую активность.
В погоне за этими преимуществами, в процессе эволюции от улитки к
слизню, организация моллюсков претерпела ряд перестроек, из которых
многие свойственны всем группам слизней независимо от их происхождения и
образа жизни. Выше в разделе, посвященном морфологии и физиологии слиз-
ней, были детально рассмотрены особенности, присущие всем или большинству
слизням, отличающие их от улиток. Кратко суммируем их.
1. Внутренностный мешок погрузился в толщу цефалоподиума, в резуль-
тате чего слизни приобрели вытянутую, более или менее цилиндрическую,
внешне билатерально симметричную форму тела (рис. 31).
2. Мантия приобретает вид овального или округло-треугольного утол-
щения на верхней стороне цефалоподиума, с пневмостомом всегда на правой
стороне.
3. Покровы отличаются заметно более выраженным рельефом, нередко-
имеют рисунок или яркую окраску. Сильно развиваются эпителиальные и
субэпителиальные железы, особенно слизистые. Слизь играет исключительно
важную и многогранную роль в жизни этих животных. В частности, увлаж-
няя покровы слизня, она повышает их проницаемость как для кислорода
(кожное дыхание), так и для воды (основной путь регулирования и восста-
новления водного баланса).
4. Усиливается слой подкожной мускулатуры, что приводит к образова-
нию кожно-мускульного мешка. Происходит редукция заднего, переднего и
педальных ретракторов колумеллярной системы улитки.
5. Раковина всегда внутренняя, пластинчатая; нередко редукция ее идет-
до распада на группу известковых зерен или до полного исчезновения.
Обычно имеется раковинная сумка — полость в толще мантии, в которой
залегает рудимент раковины и которая сохраняется даже при полной редук-
ции последней.
6 И. М. Лихарев, А. И. Виктор
.82
ОБЩАЯ ЧАСТЬ
6. Легочная полость относительно меньших размеров, чем у улиток,
что привело к сокращению дыхательной поверхности легкого. В связи с этим:
а) нередко возникают вторичные дыхательные поверхности, увеличивается
значение кожного дыхания; б) укорачивается легочная вена; в) почка сдвига-
ется назад; г) сердце смещается к переднему краю почки, а ось его располага-
ется наклонно к продольной оси тела.
Рис. 31. Положение внутренностного мешка у улитки (4), растительноядного (5) и хищ-
ного (В) слизня (из: Лихарев, Виктор, 19796).
Точками обозначено легкое, черным цветом — мускулатура ноги; заштрихованы остальные органы;
р — раковина.
7. Кровеносная система отличается сильным развитием лакун и синусов
гемоцеля, особенно заднего синуса. Гемоцель служит основным резервом
воды в теле животного и одновременно гидроскелетом.
8. Для кишечника характерно: а) у растительноядных слизней — удлине-
ние его за счет кишки и зоба; б) для хищных слизней — укорачивание кишки
и сильное развитие глотки.
9. По сравнению с улитками ЦНС слизней отличается: а) возрастающей
концентрацией ганглиев, особенно на висцеральной дуге; б) относительно
крупными размерами церебральных ганглиев, в особенности за счет более
сильного развития процеребрума, что связано с возросшей ролью обоняния;
в) крупными размерами педальных ганглиев, что объясняется большей
подвижностью слизней по сравнению с улитками.
10. По-видимому, для большинства слизней характерно присутствие вну-
три глаз дополнительной ретины — образования, отсутствующего у улиток
и служащего для определения интенсивности инфракрасного участка спектра.
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ И ФИЛОГЕНИЯ
83
Столь большое сходство в строении и физиологии слизней, порой в система-
тическом отношении далеких друг от друга, говорит о том, что под влиянием
адаптации к сходным условиям — в первую очередь к жизни в тесных и уз-
ких убежищах — их эволюция в значительной степени шла параллельными
путями, что привело к созданию общего морфо-физиологического типа ор-
низации, отличающегося более высокой биологической активностью, чем
улитки.
В то же время среди слизней видны четкие различия между раститель-
ноядными и хищными животными (рис. 31). Они прослеживаются не только-
в пищеварительной системе, но и во многих особенностях строения. Если
у растительноядных слизней внутренностный мешок располагается в сильно
разросшемся метаподиуме, то у хищных слизней и полуслизней внутренности
в основном размещаются в разросшемся прозоподиуме. Последнее связано
с тем, что у хищников наблюдается тенденция к увеличению глотки и к уко-
рачиванию кишки. Внешне увеличение прозоподиума проявляется как смеще-
ние мантии и мантийного комплекса органов к заднему концу тела.
Поэтому среди слизней четко различимы две основные жизненные формы:,
растительноядная и хищная. Кроме них, можно отметить, правда, не столь,
четко выраженную жизненную форму подземных слизней. Она встречается
как среди растительноядных (Boettgerilla, Milax verrucosus), так среди хищных
форм (Selenochlamys, Troglolestes). Таким слизням свойственно узкое,
в поперечном сечении круглое тело, с очень узкой подошвой; передний конец,
обычно сужен, в результате чего половой атриум становится длинным и
узким. Не у всех, но у многих хищных подземных слизней отмечается силь-
ное смещение пневмостома назад, а в связи с этим и деторсия мантийного
комплекса, которая может достигать 180° и приводить к опистопневмонии
(рис. 6, VIII).
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ
И ФИЛОГЕНИЯ ОТДЕЛЬНЫХ СЕМЕЙСТВ СЛИЗНЕЙ
Отряд Geophila — название, восстановленное по предложению Шилейко
(1979), — включает в себя основную массу семейств (77) надотряда Stylom-
matophora. До самого последнего времени широким признанием пользовалась,
система Stylommatophora (эта группа рассматривалась как отряд внутри
подкласса Pulmonata), разработанная Пилсбри (Pilsbry, 1896, 1901) и Тиле
(Thiele, 1931). Первый автор разделил всю группу на три подотряда по
строению почки — Heterurethra, Orthurethra, Sigmurethra, а последний под-
отряд на два инфраотряда — Aulacopoda и Holopoda. Второй автор сгруппиро-
вал близкие по раковине семейства в надсемейства.
Позднее Бейкер (Baker, 1955) дополнил систему Пилсбри четвертым под-
отрядом Mesurethra и третьим инфраотрядом Holopodopes. Последующие ис-
следования в основном сводились к изменению ранга семейственных таксо-
нов: некоторые подсемейства переводились в ранг семейств, а также выделя-
лись новые семейства и подсемейства (Pilsbry, 1948; Solem, 1959; Wenz,
Zilch, 1959-1960; Taylor, Sohl, 1962).
Как убедительно показал Шилейко (1979), классйфикация Stylommato-
phora, предложенная Пилсбри, отличается формальным подходом, в значи-
чительной степени искусственна и не соответствует новым данным. Разрабо-
танная Шилейко новая классификация Geophila, дополненная нами примени-
тельно к слизням (Лихарев, Виктор, 19796), выглядит более естественной,,
так как основана не только на сравнении отдельных систем органов, но и на
выявлении общих тенденций эволюции рассматриваемых групп.
Определение положения в пределах отряда Geophila каких-либо семейств
слизней связано с известными трудностями. Так как в этом крупном отряде:
6*
84
ОБЩАЯ ЧАСТЬ
большинство групп представлены раковинными моллюсками, т. е. улитками,
естественно, что классификация в значительной степени исходит из особенно-
стей хорошо развитой наружной раковины. Это вполне оправдано, так как
раковина отражает не только общие черты строения животного, но и эволюци-
онные тенденции той или иной группы.
В этом отношении рудиментарные раковины слизней менее перспективны
.из-за крайней простоты и значительного однообразия в строении. Правда,
как это будет показано ниже, в некоторых случаях и особенности раковин слиз-
ней дают дополнительную информацию для суждения о месте этих животных
в общей системе.
Тем не менее при обсуждении систематического положения и филогении
того или иного семейства слизней приходится исходить главным образом из
особенностей строения мягких частей тела. Однако и здесь положение ослож-
.няется широким проявлением параллелизмов как во внешнем облике, так и
во внутреннем строении слизней.
В нашем дальнейшем изложении мы в основном рассматриваем следующие
нетропические семейства слизней северного полушария, которые известны нам
по личным исследованиям: Philomycidae, Arionidae (преимущественно Arioni-
nae), Agriolimacidae, Boettgerillidae, Limacidae, Trigonochlamydidae, Parma-
cellidae, Milacidae.
До сих пор как этих, так и других слизней авторы упомянутых выше
сводок по систематике Gastropoda вслед за Пилсбри относили к двум над-
семействам: Arionoidea (или Endodontoidea) и Limacoidea (или Zonitoidea).
К первому надсемейству причисляли также улиток Endodontidae, а ко вто-
рому, кроме слизней Limacidae, Milacidae, Urocyclidae, — улиток Zonitidae,
Vitrinidae, Ariophantidae, Helicarionidae и некоторые другие семейства.
Диагнозы этих двух надсемейств явно недостаточно полны и искусственны.
По существу в них только отмечались различия в строении краевых зубов ра-
дулы. Для Arionoidea считалось характерным, что у этих зубов базальная
пластинка короткая в продольном и широкая в поперечном направлениях
или почти квадратная, а внешний зубец (эктокон) всегда короче срединного
’(мезокона). С другой стороны, у Limacoidea базальная пластинка тех же зубов
длинная и узкая, с одним шиповидным зубцом или двухзубцовая; в последнем
случае внешний зубец длиннее внутреннего.
Для классификации легочных моллюсков использование признаков радулы
как единственного критерия явно недостаточно и неизбежно приводит к на-
тяжкам. Хорошо известно, что у этих животных строение радулы зависит от
характера пищи и механизма питания. Известны почти идентичные радулы
у видов из далеких друг от друга семейств (Лихарев, 1962). Поэтому в систе-
матике Pulmonata особенности радулы следует использовать с большой осто-
рожностью и только как дополнение к характеристикам, основанным на дру-
гих системах органов.
НАДСЕМЕЙСТВО ARIONOIDEA
Выше при обсуждении морфологии слизней не раз отмечалось, что два
семейства — Philomycidae и Arionidae — по строению существенно отли-
чаются от остальных. Эти различия суммированы в табл. 2. Сравнение пока-
зывает большое сходство Philomycidae и Arionidae и значительные отличия
их от остальных семейств слизней. Поэтому есть все основания объединять оба
•семейства в надсемейство Arionoidea. В то же время нет причин включать в это
•надсемейство улиток Endodontidae, как это было принято до последнего вре-
мени только на основании отдаленного сходства краевых зубов радулы.
Если не принимать во внимание те черты Endodontidae, которые свой-
ственны всем улиткам как типу организации моллюсков, то против включения
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ И ФИЛОГЕНИЯ
85
Таблица 2
Сравнение основных особенностей строения слизней Arionoidea с Limacoidea
и Zonitoidea 1
Признаки строения Arionoidea Limacoidea и Zonitoidea
Philomycidae Arionidae (Arioninae)
Задний конец тела Киль Супрапедальная бо- розда и оторочка Плавно и широко закруглен Отсутствует Отсутствуют Заострен Имеется или доста- точно ясно намечен Имеются
Положение пневмо- стома Сильно антемедиальное За редкими исклю- чениями постме- диальное
Подошва Без педальных борозд С педальными бороз- дами
Челюсть Одонтогнатная, реже оксигнатная Всегда оксигнатная
Краевые зубы раду- лы Базальные пластинки короткие и ши- рокие Базальные пластин- ки длинные и узкие
Раковина Отсутствует Распалась на зерна или цельная Цельная
Ретракторы глотки и щупалец Сзади прикрепляют ся раздельно Сзади перед прикреп- лением сливаются в общий ствол
Сердце Лежит в центре мантийного комплекса. Ось наклонена налево Лежит ближе к ле- вому краю комплек- са. Ось наклонена направо
Почка Кольцом окружает сердце или почти замкнута вокруг него Лежит позади или немного сбоку от сердца, лишь слег- ка охватывая его
Половая система Тенденция к упрощ за счет утраты стилофора у боль- шинства родов ению: за счет постепенной редукции пениса; стилофора нет Тенденция к услож- нению за счет об- разования прида- точных органов на пенисе, вагине или атриуме
1 Из-за резких отличий этих трех надсемейств от Trigonochlamydidae сравнение
с последним не проводится.
их в данное надсемейство говорит следующее: 1) наружная раковина без вся-
ких намеков на редукцию; 2) присутствие супрапедальной борозды и ото-
рочки; 3) первично примитивная половая система, т. е. без всяких придаточ-
ных органов; 4) у большинства родов челюсть совершенно иного типа, так
как она не цельная, а состоит из многих почти квадратных пластинок, т. е.
полиплакогнатная.
Анализируя черты строения, характерные для Arionoidea, можно наметить
три категории признаков.
1. С одной стороны, это черты организации, связанные с перестройкой
в процессе перехода от улитки к слизню, но свойственные только данному
надсемейству. Так, редукция раковины до полного ее исчезновения, а также
86
ОБЩАЯ ЧАСТЬ
утеря связи между ретракторами глотки и щупалец (рис. 6, I и II) ука-
зывают, что в данной группе в этих органах процесс специализации и станов-
ления морфологического типа слизень зашел дальше, чем у остальных над-
семейств. К этой же категории относятся и различия между Arionoidea и
остальными слизнями в ориентации сердца, в форме почки и ее положении
по отношению к сердцу. Но в отличие от раковины и ретракторов эти раз-
личия указывают на два независимых пути изменения мантийного комп-
лекса в процессе перестройки улитки в слизня. Следовательно, упомянутые
особенности данного надсемейства являются чертами специализации, воз-
никшими в процессе становления морфологического типа слизень.
2. С другой стороны, у Arionoidea можно различить черты, не связанные
с упомянутой перестройкой, — форма заднего конца цефалоподиума, отсут-
ствие киля, оторочки, супрапедальной и педальных борозд, только им среди:
слизней свойственный тип челюсти и форма краевых зубов радулы. Эти осо-
бенности строения в силу своей нейтральности к процессу реорганизации
улитки в слизня сохранились в данной группе от их предков, т. е. являются,
анцестральными.
3. Последняя категория признаков касается половой системы. В связи
с этим следует вспомнить, что у Geophila в двух подотрядах — Limaxina и
Helixina — половая система в процессе эволюции изменялась в направлении,
постепенного усложнения от примитивной, т. е. без придаточных органов,
эпифаллуса и папиллы пениса, к сложной, обладающей всеми или частью из
указанных органов (Лихарев, Виктор, 19796; Шилейко, 1979). Аналогичное'
наблюдается и среди слизней, за исключением Philomycidae и Arionidae..
В этих двух семействах обнаруживается обратная тенденция — упро-
щение строения копулятивного аппарата. Так, у рода Philomycus (Philo-
mycidae) имеется стилофор с известковой стрелой, которого нет у остальных
родов семейства, т. е. у Meghimatium и Pallifera. Это образование является
характернейшей чертой Helicoidea (Шилейко, 1978, 1979). В пределах послед-
них он утрачен только у Camaenidae, Oreohelicidae и у немногих видов и под-
видов из Hygromiidae. Следовательно, у высших Geophila стилофор это первич-
ное и обычное образование, а отсутствие его вторичная и сравнительно
редкая ситуация. .
Учитывая сказанное, а также то, что упомянутые выше анцестральные
признаки Arionoidea свойственны и Helicoidea, присутствие у Philomycus сти-
лофор а служит дополнительным аргументом в пользу систематической бли-
зости Arionoidea к Helicoidea. Оба надсемейства входят в инфраотряд Helixinia
подотряда Helixina (Лихарев, Виктор, 19796; Шилейко, 1979).
Среди Arionidae стилофор не встречен, и нет оснований считать, что в этом
семействе он когда-либо имелся. Тем не менее и здесь наблюдается тенденция
к упрощению строения гениталий, которая сопровождается постепенной
передачей главной роли в копуляции от мужских органов к атриуму и частич-
но к яйцеводу. Так, если в подсемействе Ariolimacinae имеется хорошо раз-
витый пенис с мощным половым ретрактором, то у другого подсемейства —
Anadeninae, особенно у рода Prophysaon, пенис слабо развит и в помощь ему
резко усиливается мускулатура эпифаллуса, который, вероятно, играет-
роль насоса, выталкивающего сперму (Pilsbry, 1948). Наконец, у Arioninae
пенис отсутствует и главную роль в копуляции играют атриум, основание-
семяприемника и в некоторой степени дистальная часть яйцевода. Именно
поэтому половой ретрактор крепится не только к эпифаллусу, но.дает также-
ветви к яйцеводу и семяприемнику, а в атриуме нередко имеется специфиче-
ский стимулирующий орган i— лигула.
То, что Philomycidae и Arionidae — близкородственные группы и заслужи-
вают объединения в одно надсемейство, известно достаточно давно (Hoffmann,.
1924; Pilsbry, 1948). Однако сближать это надсемейство с Helicoidea по сообра-
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ И ФИЛОГЕНИЯ
87
жениям, которые мы только что изложили, до сих пор не практиковались.
И главная причина этого в том, что согласно общепринятой классификации
Пилсбри (Pilsbry, 1896) Arionoidea относятся к инфраотряду Aulacopoda, a He-
licoidea — к Holopoda.
Поэтому когда Пилсбри (Pilsbry, 1948) обнаружил у рода Philomycus
стилофор с известковой стрелой, то он категорически отрицал его гомологию
с таким же органом у Helicidae. Как единственный довод в пользу своей точки
зрения он привел то соображение, что у Philomycus стрела находится
в стилофоре постоянно и не отрывается при спаривании. По-видимому,
Пилсбри было неизвестно, что среди Helicoidea также есть виды, у которых
стрела не отрывается при копуляции.
Было бы неосторожным утверждать родство Arionoidea с Helicoidea
только на основании того, что у одного из родов Philomycidae гениталии имеют
образование, очень похожее на стилофор. Но с учетом всего комплекса при-
знаков как этого семейства слизней, так и всех Arionoidea в целом, который
мы здесь рассматриваем как анцестральный, такой вывод напрашивается
сам собой.
Поскольку изученный нами материал по Arionoidea ограничивается слиз-
нями, обитающими в Европе и внетропической Азии, т. е. одним видом из рода
Meghimatium (Philomycidae) и восемью из рода Arion (Arioninae, Arionidae),
судить об эволюции внутри каждого из семейств преждевременно. Правда,
в нашем распоряжении имеются несколько экземпляров 4 видов Ariolimacinae
и Anadeninae, но их явно недостаточно для таких широких обобщений.
НАДСЕМЕЙСТВО LIMACOIDEA
Выше уже отмечалось, что до последнего времени под этим названием
объединяли всех слизней, кроме Arionidae и Philomycidae, а также несколько
семейств улиток. Указывалось, что единственным основанием для такой трак-
товки объема этой группы служила форма краевых зубов радулы, в связи
с чем мы изложили наши возражения против подобного подхода к классифика-
ции легочных моллюсков.
В состав надсемейства входят только слизни Agriolimacidae, Boettgerillidae
и Limacidae (Лихарев, Виктор, 19796). Для этих групп характерна изначально
просто устроенная половая система, которая в процессе эволюции обнаружи-
вает явную тенденцию к усложнению за счет постепенного формирования при-
даточных органов на поверхности и внутри пениса. Однако папилла пениса
и эпифаллус обычно отсутствуют. Одно из немногих исключений составляет
небольшое подсемейство Bielziinae (Limacidae), у которого весь пенис преобра-
зован в огромную папиллу. Яйцевод, вагина и атриум всегда без придаточных
органов. Иными словами, этим слизням соответствует 2-й тип гениталий
(рис. 20, III, IV) (только у Bielziinae 3-й). В связи с этим при копуляции про-
исходит выворачивание пениса и сперма передается в виде желеобразной
массы.
Надсемейство состоит из растительноядных полифагов, склонных также
питаться гниющими остатками животного происхождения. В связи с этим из-
вестный интерес представляет то, что в этой группе многие виды имеют более
или менее развитую слепую кишку, функциональное значение которой пока
не ясно. Этого органа нет в других группах слизней.
Что касается положения Limacoidea в системе Geophila, то оно далеко не
так ясно, как у Arionoidea. По-видимому, между рецентными улитками нет
близких им групп.
В отряде Geophila тенденция к усложнению строения пениса за счет обра-
зования различных придатков свойственна также подотряду Pupillina
(Шилейко, 1979). Однако для этой группы эта тенденция в первую очередь
88
ОБЩАЯ ЧАСТЬ
проявляется в формировании очень длинного й своеобразного аппендикса
пениса, которого не бывает у Limacoidea. Кроме того, Pupillina обладают
хорошо развитой высокой и узкой раковиной, сложенной из большого числа
оборотов. Среди моллюсков с такой раковиной не встречается даже намеков
на ее редукцию, не только что становления морфологического типа слизней.
Следовательно, нет никаких оснований сближать Limacoidea с Pupillina.
Слизни возникают в группах улиток с тонкостенной раковиной геликоид-
ного, зонитоидного или сукциноидного типа, т. е. с малым числом оборотов,
Рис. 32. Два представителя семейства Succineidae.
Улитка Oxyloma (А), слизень Hyalimax (Б) и его внутренняя раковина (В) (В — из: Gude, 1914).
из которых последний сильно вздутый (Succineidae, Vitrinidae, Zonitidae,
Helicarionidae). В этом отношении очень типичным примером могут служить
Succineidae (рис. 32). Хотя большинство родов этого семейства обладает на-
ружной тонкостенной яйцевидной раковиной с очень вздутым последним обо-
ротом, здесь имеется род Hyalimax, состоящий из настоящих слизней. Важно
подчеркнуть, что раковина этих слизней очень сходна с таковой у Limacoidea.
Как и у последних, она имеет вид продолговатой слабо выпуклой пластинки,
с нуклеусом у заднего края слева от продольной оси.
Конечно, речь идет только о чисто внешнем сходстве, так как Succineidae
настолько своеобразны во всем остальном, что образуют другую ветвь Stylom-
mdtophora — отряд Succinida. Однако это сходство дает возможность предпо-
лагать, что предки Limacoidea обладали сравнительно тонкостенной яйцевид-
ной раковиной с очень вздутым последним оборотом, внешне напоминавшей
раковину Succineidae.
Перейдем к рассмотрению филогенеза отдельных семейств, образующих
Limacoidea.
Agriolimacidae (рис. 33). Среди Limacoidea наиболее примитивны
Agriolimacidae, что особенно заметно на их пищеварительном тракте. Он всегда
двухпетлевый, причем кишка в основном располагается позади сравнительно
короткого зоба (с. 42, рис. 15, II). Именно такой пищеварительный тракт
наиболее близок к плану строения кишечника улиток. Семейство слагается
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ И ФИЛОГЕНИЯ
89
следующими родами: Krynickillus, Mesolimax, Megalopelte, Lytopelten Deroceras.
За исключением последнего, который образован большим числом видов, осталь-
ные имеют по 1—2 вида.
По-видимому, предки современных Agriolimacidae, как и Limacidae, об-
ладали просто устроенной половой системой, причем цилиндрический пенис
не имел наружных придатков и стимулятора внутри. В то же время, как и
Рис. 33. Схема филогении Agriolimacidae и Boettgerillidae.
у большинства Geophila, ретрактор правых щупалец перекрещивался с пени-
сом или хотя бы с его ретрактором. Подобные отношения в настоящее время
•сохранились у Mesolimax и Megalopelte.
Позднее по какой-то причине был утрачен перекрест между гениталиями и
ретрактором правых щупалец, и они расположились независимо друг от друга.
Такая ситуация сложилась у Krynickillus, Deroceras и Lytopelte. Показа-
тельно, что у одного вида — Krynickillus urbanskii могут встречаться как
особи, сохранившие перекрест, так и без него (см. рис. 213—215).
Дальнейшая эволюция монотипического рода Megalopelte пошла в сторону
разрастания мантии, которая полностью укрыла верхнюю сторону живот-
ного. Вероятно, это связано с адаптацией к полуподземному образу жизни.
У Mesolimax мантия сохранила обычный облик и размеры. Но, вероятно,
в связи с увеличением подвижности на подошве появились дополнительные
педальные бороздки (рис. 3, IV). Во внутреннем строении также произошли
существенные изменения. Если у всех остальных Agriolimacidae задняя кишка
90
ОБЩАЯ ЧАСТЬ
лежит слева от колумеллярного мускула, то в этой группе она проходит сквозь
его основание (см. рис. 241). Другая отличительная особенность этого рода —
передний конец пениса и прилегающий
стоящая из одного рода и двух видов, по
участок семяпровода окружены пе-
репончатой манжеткой (см. рис.
240). -
После утраты перекреста между
гениталиями и ретрактором пра-
вых щупалец особенно важную
роль в соответствующей ветви
Agriolimacidae сыграла тенденция
к формированию на пенисе различ-
ных наружных придатков, а внутри
него стимулятора. Разнообразие
этих органов, имеющих непосред-
ственное отношение к копуляции,
явилось результатом интенсивного'
видообразования внутри рода De-
roceras, приведшего к дифферен-
циации четырех подродов и не-
скольких десятков видов.
К Deroceras очень близок мо-
нотипический род Lytopelte, глав-
ной особенностью которого явля-
ется своеобразное строение муску-
латуры спины, приведшее к появ-
лению киля (отчетливо виден при
сокращении животного).
Среди Agriolimacidae наиболее
примитивным и генерализованным
является род Krynickittus. За ис-
ключением утраты перекреста ге-
ниталиев с ретрактором щупалец,
никаких особенно заметных черт
специализации он не обнаруживает.
Для гениталий характерно отсут-
ствие придаточных органов, в том
числе и на пенисе, который имеет
цилиндрическую или булавовид-
ную форму, а внутри систему скла-
док.
Boettgerillidae (рис. 33).
Эта небольшая группа слизней, со-
строению наиболее близка к Agrio-
limacidae.1 В то же время она обладает комплексом только ей одной прису-
щих черт: узкое цилиндрическое в поперечном сечении круглое тело, очень-
узкая подошва, дифференциация среднего участка семяпровода в муску-
листое веретеновидное тело, длинная вагина, длинный и узкий атриум, свое-
образная папилла пениса. Что касается остальных особенностей строения,,
то многие из них здесь представлены в таком сочетании, которого нет у
других семейств. Все это указывает на особое положение этой группы:
среди Limacoidea.
Эволюция Boettgerillidae шла в сторону адаптации к подземному образу
жизни, что привело к появлению многих черт специализации (с.. 83).
1 См. с. 212 — сравнение Boettgerillidae с другими семействами.
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ И ФИЛОГЕНИЯ
91
Limacidae (рис. 34). В отличие от A griolimacidae, с которыми эту
группу до сих пор объединяли в одно семейство, Limacidae отличаются силь-
ным разнообразием строения как пищеварительной системы, так и гениталий.
Оно связано не только с определенными тенденциями в эволюции этой группы,
но и с параллелизмом в формировании некоторых органов, которые свойст-
венны только данному семейству. К их числу относится, во-первых, слепая
кишка (Metalimax, Turcomilax, некоторые Gigantomilax, часть видов Limax,
и Lehmannia). Вторым проявлением параллелизма является пениальная
железа, свойственная некоторым слизням этого семейства (Lehmannia,
часть видов Turcomilax).
Предки современных Limacidae, вероятно, во многом походили на Agrio-
limacidae, которым они наиболее близки, особенно к таким примитивным ро-
щам последнего семейства, как Krynickillus и Mesolimax. Пневмостом у них
располагался постмедиально. Кроме того, они имели двухпетлевый кишечник,
который позднее несколько перестроился за счет удлинения зоба и разме-
щения кишки вокруг него. К настоящему времени подобный план строения
пищеварительного тракта сохранился только у Eumilax. Гениталии исходных
форм семейства отличались умеренно длинным цилиндрическим пенисом,
-лишенным всяких наружных придатков, а внутри имевшим систему складок.
В настоящее время среди Limacidae такие гениталии только у Metalimax.
На первых этапах филогения семейства пошла по двум направлениям.
У одних групп по неясным причинам произошло смещение пневмостома впе-
ред. Эта перестройка организации при сохранении двухпетлевого кишечника
привела к отделению от общего ствола подсемейства Eumilacinae. Однако
у рода Metalimax имеется уже намек на третью петлю кишечника.
Дальнейшая филогения этой группы пошла в сторону формирования свое-
образного пениса, который характерен для рода Eumilax (см. рис. 367).
Естественно, это было сопряжено с сильным изменением механизма популяции.
Второй род того же подсемейства — Metalimax, по-видимому, подвергся
меньшим изменениям и во многом сохранил как облик древних Eumilacinae,
так и основные особенности предковых форм семейства. В этом роде новообра-
зованием является слепая кишка, которая открывается в ректум в том месте,
где он пронизывает диафрагму, т. е. перед основанием колумеллярного му-
скула.
Филогения остальных Limacidae шла по пути удлинения кишечника за
счет формирования 3-й петли, располагающейся на верхней стороне внутрен-
ностного мешка справа от колумеллярного мускула, основание которого она
огибает. Важное адаптивное значение этого новообразования, которое нигде
не встречается за пределами данного семейства, уже отмечалось. По-види-
мому, оно связано с возросшей ролью в пищевом рационе грибов и лишай-
ников, т. е. белковой пищи.
Дальнейший филогенез семейства проходит под знаком изменений меха-
низма копуляции, что находит выражение в различных преобразованиях пе-
ниса. Особенно своеобразные тенденции проявляются в роде Bielzia.
'В то время как у остальных групп копуляция осуществляется путем вы-
ворачивания пениса наружу, в этом роде пенис, превращенный в гигантскую
папиллу, по-видимому, не выворачивается, а лишь слегка выдвигается за пре-
делы полового отверстия и как «насос» выпрыскивает сперму в атриум парт-
нера (с. 55, рис. 21). В связи с этим пенис потерял прямую связь с семяпрово-
дом, который в свою очередь сильно трансформирован.
Столь резкие отличия в строении гениталий и в механизме копуляции дают
основание выделить Bielzia в особое подсемейство Bielziinae.
В наиболее крупной ветви семейства — подсемействе Limacinae — осо-
бенно велико разнообразие в форме, величине, во внешнем и внутреннем
«строении пениса. Кроме того, при сохранении общего трехплетлевого плана
92
ОБЩАЯ ЧАСТЬ
наблюдаются некоторые вариации и в строении кишечника. Все это ука-
зывает на относительную молодость подсемейства и на то, что формообра-
зование здесь шло более интенсивно, чем в двух остальных подсемействах.
Наиболее примитивные отношения обнаруживает среднеазиатский род
Turcomilax. Они проявляются в том, что данная группа как бы объединяет
черты других родов Limacinae — Limax, Lehmannia, Gigantomilax, но у каж-
дого вида Turcomilax эти особенности сочетаются по-разному. С другой сто-
роны этот род сохранил ряд анцестральных черт организации — умеренно
длинный пенис, заднее положение правой доли печени (как у Bielziinae).
Дальнейший филогенез Limacinae в основном проходил под знаком форми-
рования различных типов копулятивного аппарата, т. е. в первую очередь
пениса. При этом наметились два главных направления: 1) укорочивание пе-
ниса — он становился короткоцилиндрическим или мешковидным, почти
круглым; внутри него сформировались различные органы стимуляции парт-
нера — Gigantomilax, Caspilimax, Caucasolimax, Malacolimax', 2) пенис
оставался достаточно длинным, а у некоторых становился очень длинным —
некоторые Lehmannia и Limax.
Одновременно во многих родах на пенисе возникали различные наружные
придатки и внутренние структуры. Так, у многих Lehmannia имеется пени-
альная железа, у некоторых Gigantomilax — бич, у Malacolimax семяпровод
перед впадением в пенис приобретает своеобразное строение. Внутри пениса
Lehmannia и Malacolimax находится крупная складка; у Limax таких складок
несколько, и одна из них имеет вид мощного гребня; у Gigantomilax и Caspili-
max внутри пениса залегает стимулятор и кольцевидный валик; у Caucasoli-
тахвнутри заднего конца пениса располагается своеобразная папилла, а семя-
приемник становится необычно мускулистым.
К сожалению, не всегда есть возможность объяснить строение копулятив-
ного аппарата определенным механизмом копуляции, хотя такая связь, есте-
ственно, всегда имеется. Только в случае с видами подрода Limax s. str.,
отличающимися особенно длинным пенисом (у некоторых видов в несколько
раз длиннее тела животного), совершенно очевидно, что столь необычные раз-
меры этого органа связаны с копуляцией в подвешенном состоянии
(рис. 22, I).
Ясна также корреляция между размерами пениса и характером перекре-
ста гениталий с ретрактором правых щупалец. У групп с достаточно длин-
ным пенисом ретрактор проходит между ним и яйцеводом, причем семяпровод
огибает ретрактор спереди. При укорачивании пениса возможны три вари-
анта: а) ретрактор щупалец перекрещивается только с половым ретракто-
ром — подроды Gigantomilax s. str., Vitrinoides (род Gigantomilax); б) ре-
трактор щупалец не перекрещивается ни с пенисом, ни с половым ретрактором,
т. е. лежит в стороне от гениталий, — подрод Monochroma (род Gigantomilax);
в) ретрактор щупалец проходит через щель между пенисом и яйцеводом,
но семяпровод огибает его сзади — роды Caucasolimax, Caspilimax и Malacoli-
max.
НАДСЕМЕЙСТВО ZONITOIDEA
Эта группа слагается из улиток Zonitidae, полуслизней Daudebardiidae
и слизней Рarmacellidae и Milacidae.
Раковина первично с низким завитком и тонкостенная (зонитоидная),
с тенденцией к редукции. Женские половые ' пути (яйцевод, вагина) или
атриум — с придаточной (вагинальной) железой. Стимулятор, если имеется,
то без известкового или хитиноидного вооружения и располагается внутри
атриума или на нем. Мужские половые пути первично с эпифаллусом. Внутри
пениса может присутствовать мощная папилла. Во многих группах при копу-
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ И ФИЛОГЕНИЯ
93-
ляции сперма передается в сперматоформах. Полифаги (нередко детритофаги
и сапрофаги) и хищные животные.
В пользу объединения перечисленных семейств в одно надсемейство
говорит не только строение гениталий, но и сравнение раковин. У большинства
Zonitidae она довольно плоская, гладкая и тонкостенная. Эти особенности ра-
ковины следует считать если не началом ее редукции, то непременной предпо-
сылкой (рис. 35). Дальнейшее уменьшение числа оборотов при одновременном
расширении последнего мы видим у полуслизней Daudebardiidae. Некоторые
авторы склонны считать последних подсемейством Zonitidae (Riedel, 1966).
Если представить дальнейший ход редукции раковины с превращением ее во
внутреннюю, то первым этапом к этому будут раковины Parmacellidae,.
Б В
Рис. 35. Этапы редукции раковины в пределах надсемейства Zonitoidea (из: Лихарев,-
Виктор, 1979).
А — Oxychilus depresses (Zonitidae); Б — Daudebardia rufa (Daudebardiidae); В — Parmacella ibera (Par-
macellidae); Г — Tandonia rustica (Milacidae).
а еще дальше пластинчатые раковины Milacidae. При этом эмбриональная
часть, т. е. нуклеус, окажется на продольной оси таких рудиментарных рако-
вин, т. е. так, как это имеет место у названных двух семейств слизней. Это
четко их отличает от раковин Limacoidea и Trigonochlamydoidea, у которых
нуклеус лежит слева от продольной оси.
До сих пор под названием Zonitoidea объединяли почти всех слизней, кроме
Arionidae и Philomycidae, а также многие семейства улиток с зонитоидной
раковиной — Vitrinidae, Helicarionidae, Ariophantidae, у которых нередко
на вагине или атриуме имеется своеобразный стимулирующий орган с извест-
ковым или хитиноидным вооружением — саркобеллум (Van Mol, 1970,1973).
, Шилейко (1979) привел достаточно убедительные возражения против такого
объединения принципиально разнородных групп.
Обе группы слизней — Parmacellidae и Milacidae — обособились от об-
щего ствола надсемейства, по-видимому, довольно рано (рис. 36). Одной из
причин этого могла быть специализация в области питания. Если у осталь-
ных Zonitoidea эволюция пошла в сторону полифагии и, в частности, сапро-
фагии (Zonitidae), которая позднее привела к хищничеству (Daudebardiidae),
то предки указанных слизней были растительноядными животными. Ве-
роятно, именно по этой причине у слизней сохранилась более примитив-
ная радула, в то время как у большинства Zonitidae она приобрела черты спе-
циализации, особенно в наиболее крупном роде Oxychilus (Riedel, 1966).
Обособление Parmacellidae и Milacidae от общего ствола надсемейства
могло быть связано с аридизацией климата Древнего Средиземноморья — цен-
тра возникновения этих групп — на рубеже палеогена и неогена.1 В этих
1 Первые находки Mi lacidae относятся к началу олигоцена, a Parmacellidae — к концу
миоцена (Стеклов, 1966).
94
ОБЩАЯ ЧАСТЬ
условиях, особенно в период летней засухи, преимущества, которые предостав-
ляет организация слизней по сравнению с улитками, особенно эволюционно
значимы.
Вероятно, уже у общих предков этих двух семейств слизней имелась мощ-
ная папилла пениса — органа, способствующего быстрой и эффективной пе-
редаче сперматофора от партнера к партнеру, а также усилились вагиналь-
ные железы. Оба образования в условиях жаркого и сухого климата имеют осо-
бенно важное значение, так как пениальная 0 й
папилла дает возможность передавать парт- =• <о
перу легко уязвимый к сухому воздуху -с о с
сперматофор быстро и почти без выступания £ $ с
наружу копулятивных органов, а вагиналь- с с 5 й
ные железы, вероятно, способствуют сма-
чиванию этих органов. ‘
Parmacellidae. Дальнейшая фи-
логения обеих групп пошла различными пу-
тями. У Parmacellidae те. связи с жизнью в осо-
бенно аридных областях (в настоящее время
в Северной Африке, восточном Закавказье,
Рис. 36. Схема филогении
Zonitoidea.
^Zonitoidea
Средней Азии) установился совершенно необычный для слизней жизненный
цикл (с. 66, рис. 26). Главное в нем — летняя диапауза, в которую впадает
молодь сразу после вылупления, и интенсивный рост молоди после пробужде-
ния. В связи с диапаузой возник ряд специальных морфологических адапта-
ций в устье эмбриональной раковины (нуклеуса) и регресс ее приостановился
(см. рис. 461, 462) (Жадин, 1946, Шилейко, 1967).
По-видимому, эти особенности жизненного цикла Parmacellidae, а также
очень крупный капюшон мантии способствуют столь эффективной защите
от высыхания, что у этих моллюсков процесс становления «слизневого»
•облика остался как бы незавершенным. Последнее выражается в том, что
нуклеус раковины сохранил следы спирали, в которую даже проникает ло-
пасть печени, а задний конец метаподиума сохранил дорсовентральную
мускулатуру.
Аридность климата сказалась и на организации двух ветвей Parma-
cellidae. Западный род — Parmacella, живущий в условиях менее засушли-
вого климата современного Средиземноморья, восточного Закавказья, се-
верного (лесного) Ирана и западного Копетдага, отличается присутствием
своеобразного стимуляторного придатка на атриуме, который при копуля-
дии выворачивается наружу. Такой стимулятор в значительно более засуш-
ливом климате Средней Азии, по-видимому, нежелателен, и поэтому другой
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ И ФИЛОГЕНИЯ
род этого семейства — Candaharia — его не имеет, что, вероятно, связано*
с его относительной примитивностью. В то же время в этом роде усложни-
лось строение вагинальной железы.
Milacidae. Филогенез этой группы в приобретении «слизневого»*
облика зашел дальше, чем у Parmacellidae. У них меньше специальных мор-
фологических и физиологических адаптаций к защите от засухи. Одной из них
можно считать густую и очень вязкую слизь, которую эти животные вы--
деляют в особенно большом количестве, когда прячутся в какое-либо убе-
жище на жаркое время. В этих случаях она образует вокруг тела как бы
кокон, в котором скрывается моллюск. Другой адаптацией, как ну Par-
macellidae, является сильное развитие придаточных желез на вагине или
атриуме. Все это указывает на то, что, вероятно, это семейство возникло*
в менее аридных районах северного Средиземноморья, где большинство ви-
дов обитает и теперь.
Из двух родов, из которых слагается это семейство — Milax и Tandonia,-
наиболее примитивной организацией обладает последняя группа, что про-
является в отсутствии стимулятора внутри атриума — новообразования,
характерного для рода Milax.
НАДСЕМЕЙСТВО TRIGONOCHLAMYDOIDEA
Единственное входящее сюда семейство — Trigonochlamydidae слагается:
из хищных слизней, ведущих полуподземный или подземный образ жизни.
Они питаются почвенными червями, главным образом дождевыми, в ходах,
которых проводят большую часть жизни.
Рис. 37. Зубы радулы Hyrcanolesies velitaris и их положение в желобе одонтофора.-
Цифры указывают положение зубов в пределах поперечного ряда радулы.
Вся организация этих моллюсков сложилась как высокая специализация
к жизни в почве и к питанию червями. В связи с этим тело их сильно вытянуто,
стройное, в поперечном сечении круглое, с очень узкой ногой — адаптации
к жизни в ходах почвенных червей. По той же причине пневмостом смещен
к заднему краю мантии, что защищает легкое от засорения частицами почвы.
Питание червями, при котором радула служит не для раздробления пищи
на мелкие частицы, как у растительноядных моллюсков, а для захвата живой
добычи и постепенного втягивания ее внутрь слизня, привело к формирова-
нию специфической для хищных наземных моллюсков (в том числе и для хищ-
ных улиток) глотки. Она отличается особенной мускулистостью стенок.
9в
ОБЩАЯ ЧАСТЬ
присутствием дополнительных мускулов, служащих для втягивания выверну-
той части глотки внутрь тела, мощным одонтофором с продольным желобом,
внутри которого залегает радула (рис. 37). Зубы радулы приобретают ха-
рактерную кинжаловидную или багоровидную форму. При выдвижении одонто-
фора навстречу добыче он раздвигается по желобу, а зубы расходятся
в стороны. Соприкоснувшись с червем, они смыкаются и вонзаются в его
тело, а затем усилием внутренних и внешних мышц глоткй и одонтофора
втягивают участок червя внутрь глотки, где начинается его переваривание.
Крупные куски червя отрываются и подаются по пищеводу в зоб. Такие мел-
кие черви, как энхитреиды, целиком заглатываются слизнем (Левушкин,
Матекин, 1965).
Несмотря на то что в связи с хищничеством у этих слизней кишка стала
очень короткой и почти утратила петли, присущие растительноядным мол-
люскам, благодаря сильному развитию глотки многие внутренние-органы
претерпели большие изменения из-за дефицита места для размещения. На ге-
ниталиях это отразилось в том, что половой атриум стал узким и длинным,
а пенис сильно сужен к атриуму. Так как через пищевод проходят крупные
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ И ФИЛОГЕНИЯ
97
куски добычи, сильно растягивающие его стенки, то расположенный вокруг
него мозг (окологлоточное нервное кольцо) обладает очень длинной цере-
бральной комиссурой и нередко не менее длинными коннективами (рис. 17,
VIII).
Увеличение размеров глотки приводит к разрастанию црозоподиума,
что внешне проявляется как смещение мантийного комплекса к заднему
концу тела, нередко сопровождающееся деторсией этого комплекса. По той же
причине происходит постепенный распад единой колумеллярной системы
ретракторов тела на отдельные мускулы, которые задними концами кре-
пятся к стенкам тела независимо друг от друга (рис. 38). Последнее понятно,
так как крупная глотка затрудняет работу ретракторов, если они сзади
сливаются в единый общий ствол.
Следовательно, вся организация этой группы слизней подчинена одной
главной эволюционной тенденции — постепенному увеличению глотки и
оснащению ее дополнительной мускулатурой. Одновременно, особенно
во внешнем облике, возникает ряд адаптаций к жизни в почве.
Среди ныне живущих семейств отряда Geophila трудно указать ближай-
ших родственников Trigonochlamydidae. Судя по их внутренней раковине,
у которой нуклеус смещен от продольной оси налево, а линии нарастания
на спатуле вытянуты направо, эти слизни, как и Limacoidea, произошли
от улиток с сукциноидной раковиной. То, что это семейство состоит из мо-
нотипических родов (всего шесть), указывает на большую древность этой
группы. Не случайно в настоящее время Trigonochlamydidae обитает только
на Кавказе и соседних горно-лесных областях Передней Азии (Понтийские
горы, Эльбурс), т. е. в Кавказской зоогеографической провинции Среди-
земноморской подобласти, малакофауна которой отличается высоким энде-
мизмом и относительным обилием монотипических родов и подродов (Ли-
харев, Раммельмейер, 1952; Лихарев, 1958; Стеклов, 1966). Поэтому есть
основание рассматривать этих слизней как реликты тропической фауны па-
леогена.
Древние Trigonochlamydidae обладали сравнительно небольшой глоткой,
которая, как у многих растительноядных моллюсков, крепилась к передней
стенке тела вокруг рта несколькими парами коротких ретенторов. В настоящее
время подобная ситуация сохранилась только у рода Trigonochlamys (рис. 39,
I). Вероятно, другой особенностью древних слизней данного семейства был
более примитивный механизм копуляции, при котором пенис сильно вы-
ворачивался, партнер раздражался стимулятором, а сперма передавалась
в капельно-жидком виде.
Таким способом в данном семействе теперь копулируют только Seleno-
chlamys. Эта группа, сохранив анцестральный способ размножения, эволю-
ционировала в сторону узкой специализации к жизни в ходах почвенных
червей и питанию ими, что проявилось в особенно сильном увеличении
глотки, которая заняла почти всю полость тела, точнее прозоподиума, так
как от метаподиума сохранился рудимент в виде очень короткого участка
тела, лежащего позади мантии. Последняя в свою очередь подверглась исклю-
чительно сильной редукции и смещению к заднему концу тела, а мантийный
комплекс органов испытал деторсию на 180°. В связи с увеличением глотки
мозг переместился вперед и охватывает не пищевод, как обычно, а переднюю
часть глотки. По той же причине колумеллярная система ретракторов пол-
ностью распалась на самостоятельные мускулы (рис. 38, IV). Одновременно
происходило уменьшение размеров тела этих слизней.
Следовательно, Selenochlamydinae — это филогенетическая ветвь семейства,
рано отделившаяся от общего ствола в результате специализации к подзем-
ному образу жизни (рис. 40).
7 И. М. Лихарев, А. И. Виктор
98
ОБЩАЯ ЧАСТЬ
Другая ветвь — Trigonochlamydinae — хотя и испытала ряд аналогичных
перестроек организации, но ведущим фактором в ее эволюции стал переход,
к размножению с помощью сперматофоров — адаптация, направленная
Рис. 39. Схема прикрепления к глотке мускулов ретенторов и протракторов у родов;
Trigonochlamydidae.
I — Trigonoehlamys', II — Drilolestes’, III — Selenochlamys\ IV — Hyrcanolestes\ V — Parmacellilla и?
Troglolestes. Места прикрепления к глотке ретенторов заштрихованы, протракторов — зачернены.
к защите спермы от высыхания. Последнее особенно важно, так как боль-
шинство видов этого подсемейства не утратило связи с внешним миром,,
хотя и проводит значительное время во влажной подстилке и верхних слоях
почвы. Переход к размножению с помощью сперматофоров сопровождался:
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ И ФИЛОГЕНИЯ
99
формированием эпифаллуса, а внутри пениса к появлению особых складок,
к которым готовые и заполненные спермой сперматофоры прикрепляются в ожи-
дании копуляции. По-видимому, только у троглобионта Troglolestes sokolovi,
в связи с жизнью в глубине влажных пещер отпала необходимость в сперма-
тофорах, что привело к ис-
чезновению упомянутых скла-
док внутри пениса, а эпифал-
лус подвергся ' заметной ре-
дукции.
Дальнейшая специализа-
ция Trigonochlamydinae про-
явилась в основном в различ-
ном оснащении глотки му-
скулами-протракторами, свя-
зывающими этот орган со
стенками тела. Вероятно, эти
мускулы производные ретен-
торов глотки. Наиболее при-
митивные отношения имеют
место у рода Hyrcanolestes,
где основания этих мышц
располагаются по обе стороны
глотки, смыкаются впереди,
:но без определенного порядка
|(рис. 39, IV). В дальнейшем
происходит упорядочивание
положения протракторов на
глотке. В родах Troglolestes
и Parmacellilla они распола-
гаются в два продольных
ряда, а в роде Drilolestes в один
ряд, т. е. аналогично как
у Selenochlamys.
Несмотря на некоторое
увеличение глотки у Trigo-
nochlamydinae, она занимает
меньше половины, а чаще
около V3 полости тела (или
внутренностного мешка). По-
этому мозг сохранил обычное
для болыпиства легочных мол-
люсков положение, т. е. рас-
полагается позади глотки, вокруг пищевода. По той же причине боль-
шая часть ретракторов тела соединяется сзади в общий ствол — колумел-
лярный мускул, который, как обычно, крепится позади мантийного комп-
лекса. У родов Troglolestes и Drilolestes все ретракторы — глоточные и
щупальцевые — сливаются сзади; у остальных родов из общей системы вы-
падают ретракторы либо глазных (Trigonochlamys), либо губных (Hyrcano-
lestes, Parmacellilla) щупалец (рис. 38).
Ярким примером адаптации к жизни в пещерах является род Troglo-
lestes. Отсутствие света способствовало утрате пигмента в покровах и ре-
дукции глаз. Так как эти слизни не полностью закапываются в почву, а лишь
погружают в нее передний конец тела в поисках червей энхитреид, то мантия
не только не подверглась редукции, но капюшон ее столь сильно развит,
что полностью закрывает шею. Тем самым он способствует очищению ее
(Л
ф
ф
0)
о
Of)
о
£
ю
ф
и
ю
Е
ю
СЬ
(Л
Ф
0)
ф
о
с
(б
и
(Л
£
£
и
о
с
о
of)
ё
(Л
ф
-0)
ф
о
Q
V)
Е
ю
£
и
о
с
ф
ф
(Л
( Trig.
,о nochlamydinae
chlamyd id ae J
nochlamydinae^
no
hlamydoidea
<•
Рис. 40. Схема филогении Trigonochlamydidae.
7*
100
ОБЩАЯ ЧАСТЬ
от частиц грунта. Вероятно, разрастание капюшона явление вторичное и:
предки этих слизней имели мантию, смещенную к заднему концу тела и с ко-
ротким капюшоном. На это указывает заднее положение мантийного ком-
плекса и пневмостома, как это имеет место у Parmacellilla, которая ведет
подземный образ жизни в ходах червей. Не случайно в обеих группах му-
скулы-протракторы одинаково располагаются на глотке.
У остальных трех родов — Trigonochlamys, Hyrcanolestes и Drilolestes —
процесс специализации к жизни в подстилке и почве не зашел так далеко.
Поэтому хотя мантийный комплекс у них и несколько смещен назад, но не
так сильно, как у Parmacellilla, Troglolestes и Selenochlamys. Спина этих
слизней еще достаточно велика, а глотка занимает от V4 до '/3 полости тела.
В итоге, принимая за основу классификацию Geophila, предложенную
Шилейко (1979), положение рассмотренных семейств слизней в общей си-
стеме отряда выглядит следующим образом:
Надотряд Stylommatophora
Отряд Geophila
Подотряд Limaxina
Надсемейство Limacoidea (Agriolimacidae, Boettgerillidae, Lima-
cidae)
Надсемейство Trigonochlamydoidea {Trigonochlamydidae)
Подотряд Helixina
Надсемейство Zonitoidea (Zonitidae, Daudebardiidae, Parmacel-
lidae, Milacidae)
Надсемейство Arionoidea (Philomycidae, Arionidae)
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
Обсуждаемые здесь семейства слизней, за исключением Philomycidae,.
распространены только во внетропических областях северного полушария,,
т. е. в Голарктике.1 В предалах этой обширной территории основная масса
видов обитает в западной части Палеарктики — т. е. в Европе, в неко-
торых средиземноморских странах Африки и Передней Азии, на Кавказе.
Лишь немногие роды характерны для горных областей Средней Азии.
Совершенно иной облик имеет распространение Philomycidae. Эта свое-
образная группа слизней населяет умеренно теплые и тропические области
Восточной Азии и Америки (рис. 41). Только один вид — Meghimatium
bilineatum, кроме восточного Китая, Корейского полуострова, Японии, Индо-
китая и некоторых островов Малайского архипелага, обитает в смешанных
и широколиственных лесах Приморского края и бассейна среднего Амура..
В силу того что раковины слизней в таксономическом отношении мало-
выразительны, говорить о времени и месте формирования тех или иных
групп, опираясь на палеонтологические находки, очень трудно. Считается,
(Wenz, 1923—1930), что Arion и Milacidae появились в начале олигоцена,
Limax — в начале миоцена, a Parmacellidae — к концу миоцена (Стеклов,.
1966). Поэтому главным источником, по которому можно судить о том, где
возникли те или иные группы слизней, является анализ их современного
распространения.
1 Речь идет только о естественном распространении слизней. Многочисленные слу-
чаи завоза отдельных видов за пределы их природных ареалов рассматриваются особо
(с. 105).
i
к
Рис. 41. Географическое распространение слизней Milacidae (Г), Parmacellidae (2) и Philomycidae (3).
102
ОБЩАЯ ЧАСТЬ
В Европе, особенно в средних, западных и южных областях этого кон-
тинента, по-видимому, было'несколько центров возникновения многих групп,
так как именно здесь обитают все или большинство видов таких родов как
Geomalacus, Arion (Arionidae), Milax, Tandonia (Milacidae — рис. 41),
Limax, Lehmannia, Malacolimax, Bielzia (Limacidae) и многие Deroceras
(Agriolimacidae)
Вероятно, одним из таких центров являлся Балканский полуостров,
где в настоящее время обитают многие виды Limax s. str. Milacidae и De-
roceras. Горы средней Европы возможно были родиной Lehmannia.
Очень важным центром формирования многих групп слизней был Кавказ
и сопредельные области Малой Азии и Ирана. И в настоящее время только
здесь обитают слизни из родов Megalopelte (Agriolimacidae), Boettgerilla
(Boettgerillidae), Gigantomilax, Caspilimax, Caucasolimax, Eumilax, Meta-
limax (Limacidae) и все Trigonochlamydidae. Обилие на Кавказе эндемич-
ных групп — показатель длительной изоляции этой территории от сосед-
них областей суши. Как известно, последнее отразилось не только на фауне
слизней, но и на других группах наземных моллюсков (Лихарев, Раммель-
мейер, 1952; Лихарев, 1957, 1958, 1962). Однако эта изоляция не всегда была
полной. На это указывает как присутствие здесь эндемичных видов, при-
надлежащих к родам, широко распространенным в других частях Средизем-
номорской подобласти (Deroceras, Krynickillus, Milax, Parmacella), так и
обитание на Кавказе Limax maculatus, живущего также в лесах Болгарии,
Румынии, Франции и, вероятно, еще в других странах Западной и Южной
Европы. Кроме того, обитание на Кавказе трансголарктического Deroceras
laeve и транспалеарктического D. agreste указывает также на существова-
ние фаунистических обменов с соседними территориями.
Подобно наземным улиткам, на слизнях Кавказа прослеживаются опре-
деленные закономерности их распространения в пределах этой горной страны.
Большинство видов приурочены к двум лесным зоогеографическим про-
винциям — Кавказской и Гирканской (Лихарев, 1958, 1962) (рис 42). Пер-
вая особенно богата эндемичными видами и родами (Megalopelte, Boettge-
rilla, Eumilax, Metalimax, Gigantomilax, Trigonochlamys, Drilolestes, Sele-
nochlamys) — всего 19 видов, из которых большая часть обитает в западной
части Закавказья, т. е. на запад от Сурамского хребта. Вторая значительно
беднее и имеет только 3 эндемичных или субэндемичных рода (Caspilimax,
Hyrcanolestes, Parmacellilla) — всего 3 вида.
Альпийская и субальпийская зоны Большого и Малого Кавказа, за исклю-
чением Caucasolimax caucasicum (обитает в альпийской зоне Большого Кав-
каза), не имеют только им присущих видов слизней. Обычно здесь обитают
те же виды, которые живут в верхней части лесной зоны, а именно некоторые
Gigantomilax — G. brunneus, G. lenkoranus, G. monticola, G. daghestanus.
На территории СССР другим центром значительного эндемизма среди
слизней является Средняя Азия, точнее Тянь-Шань и Алайская горная си-
стема. Первому характерно присутствие слизней из рода Turcomilax (Li-
macidae) (рис. 43), некоторые виды которого проникли также на Джунгар-
ский Алатау и Тарбагатайский хребет. Эта группа отсутствует на Алайской
системе хребтов и в западном Памире, где широко распространены слизни
из рода Сandaharia (Parmacellidae). Отдельные виды этого рода обитают и на
периферийных хребтах западного Тянь-Шаня.
Оба упомянутых рода представляют собой наиболее древний элемент
фауны слизней Средней Азии. В пользу такой точки зрения говорит не только
их эндемизм, но и систематическое положение в пределах соответствующего
семейства, так .как они сохранили ряд черт строения, указывающих на их
примитивность. В связи с этим невольно напрашивается аналогия с неко-
торыми среднеазиатскими родами улиток Hygromiidae (Шилейко, 1978) —
Рис. 42. Географическое распространение Caspilimax keyserlingi (7), Eumilax brandti (2),
E. intermittens (3).
Рис. 43. Географическое распространение слизней родов Turcomilax (7) и Candaharia (2).
104
ОБЩАЯ ЧАСТЬ
семейство тоже западнопалеарктического происхождения. По-видимому,
как Turcomilax, так и Candaharia — это осколки широко распространенной
древней фауны Палеарктики, существовавшей на рубеже палеогена—нео-
гена. В связи с ростом аридности климата и усилившимся процессом распада
этой территории на провинции они оказались отрезанными от западных
членов своих семейств и дали в Средней Азии ряд эндемичных видов, сохра-
нивших некоторые примитивные особенности своих предков.
Однако подобно Кавказу изоляция фауны слизней Средней Азии не была
абсолютной. С севера сюда проникли трансголарктический Deroceras laeve
и транспалеаркт'ический D. dgreste. Отражением связей с Кавказом является
присутствие Lytopelte macUlata, который населяет южные области Средней
Азии, относительно аридные биотопы предгорий Копетдага, северного Ирана
и восточного Закавказья^ Вероятно, эти фаунистические связи установились
в плейстоцене.,
Если малакофауна горного Крыма 1 отличается особым обилием энде-
мичных видов улиток,'то среди слизней эндемиков нет. Здесь наиболее ха-
рактерны слизни Tandonia cristata, Т. kaleniczenkoi, а также Limax macu-
latus. Первые два принадлежат к эвксинским видам и обитают также в запад-
ных и южных областях Причерноморья. Ареал последнего вида тяготеет
к атлантическо-средиземноморскому типу.
На территории: советской части Восточных Карпат из слизней наиболее
характерным видом является эндемик Карпат Bielzia coerulans.
Фауна слизней Русской равнины и Урала лишена эндемиков и представ-
ляет собой обедненный дериват фауны средних областей Европы, в основном
слагающийся из широко распространенных общеевропейских видов. Она
явно тяготеет к зоне смешанных и широколиственных лесов. Лишь отдель-
ные виды проникают в степную зону по пойменным и байрачным лесам —
Arion subfuscus, Deroceras laeve, D. agreste. Эти же слизни проникают в тайгу,
a D. laeve даже в тундру, но только по долинам рек.
Хотя обширные пространства Сибири еще недостаточно изучены, со-
вершенно ясно, что фауна слизней этой территории очень бедна видами.
Кроме таких широко распространенных в Голарктике или в Палеарктике
слизней, как Deroceras laeve и D. agreste, здесь обнаружены (преимущественно
в горных областях юга Сибири — Алтай, Кузнецкий Алатау, Саяны, горы
Прибайкалья и Забайкалья) два субэндемичных вида — Arion sibirlcus и
Deroceras altaicum. Первый обитает также в Приамурье и Приморье, а второй,
кроме того, на северо-западном Тянь-Шане. На северо-востоке Сибири живет
только Deroceras laeve.
Смешанные и широколиственные леса среднего Приамурья и Приморья,
в отличие от их аналогов в Европе, характеризуются очень бедной видами
и лишенной эндемиков фауной слизней. Этим же эта фауна отличается от ме-
стной фауны наземных улиток, значительно более богатой видами, среди
которых много эндемичных (Лихарев, Раммельмейер, 1952; Шилейко, 1978).
Здесь из слизней, кроме уже упомянутого Meghimatium bilineatum, обитают
Arion sibiricus, Deroceras laeve, D. agreste, D. altaicum.
Если первый вид является восточноазиатским элементом в фауне При-
амурья и Приморья, то остальные — дериваты западнопалеарктических
родов.
СИНАНТРОПНЫЕ И АНТРОПОХОРНЫЕ ВИДЫ
В процессе хозяйственной деятельности человек сознательно или невольно
разрушает или преобразует в выгодном для себя направлении природные
ландшафты или биотопы. На их месте создаются культурные — поля, ого-
1 Фауна слизней равнинной части Крыма не отличается от сопредельных степных
областей Русской равнины (см. ниже).
СИНАНТРОПНЫЕ И АНТРОПОХОРНЫЕ ВИДЫ 105
роды, плантации — или другие антропогенные ландшафты и биотопы —
вторичные леса и луга, пустоши. При этом значительная часть видов живот-
ных и растений, исходно населявших природные биотопы, уничтожается
или вытесняется с освояемой территории, а другая часть сохраняется и
нередко даже занимает господствующее положение. Эти виды обычно отли-
чаются довольно широкой экологической валентностью и, в особенности
в услових монокультурного хозяйства, их популяции могут достигать вы-
сокой численности, в то время как в природных сообществах они занимали
довольно скромное место. Такие виды принято называть синантроп-
ными.
Крайнее проявление синантропии можно видеть у животных, которые
обитают в полном отрыве от природы — в жилье человека и в таких вспо-
могательных сооружениях, как погреба, подвалы и склады, или в столь
искусственных условиях, как оранжереи и теплицы. Упомянутые проявления
синантропии встречаются и у слизней.
Как первый шаг к синантропии можно рассматривать слизней, которые
обитают как в первичных биотопах, не затронутых или почти не измененных
человеком, так и во вторичных, т. е. в биотопах, которые возникли на месте
уничтоженных человеком первичных в результате сукцессии — вторичные
леса, луга и т. п. В культурные биотопы эти виды проникают сравнительно
редко и в небольшом количестве, почему как вредители сельскохозяйствен-
ных культур они имеют третьестепенное значение. Сюда относятся Arion
subfuscus, Deroceras laeve, D. agreste, D. subagreste, D. bakurianum.
Следующий шаг в сторону синантропии — это слизни, которые явно
предпочитают антропогенные, особенно культурные, биотопы, где в благо-
приятные годы они дают вспышки численности и могут наносить существен-
ный вред многим сельскохозяйственным культурам. Они обитают и в при-
родных биотопах, как в первичных, так и во вторичных, но там их популя-
ции не отличаются столь высокой численностью, как на полях и огородах.
Среди этих слизней много серьезных вредителей сельского хозяйства (отме-
чены звездочкой): Arion fasciatus, Deroceras sturanyi*, D. reticulatum*, D. tau-
ricum*, D. caucasicum*, Lytopelte maculata, Gigantomilax monticola*, Par-
macella ibera*, Candaharia rutellum*, C. levanderi*, C. izzatullaevi, Tan-
donia cristata*, T. kaleniczenkoi.
К третьей группе принадлежат слизни, которые на территории СССР оби-
тают чаще всего непосредственно внутри домов (например, в подвалах),
во вспомогательных помещениях (погреба, продуктовые склады), в парках,
садах (особенно в ботанических садах), в оранжереях и теплицах; в ближай-
ших природных биотопах, в том числе и во вторичных, они отсутствуют.
Чаще всего их появление в таких местах связано с непроизвольным заво-
зом и поэтому их можно рассматривать как антропохорные виды.
Сюда принадлежат Boettgerilla pallens, Limax maximus, L. flavus, L. ma-
culatus, Lehmannia valentiana и, вероятно, Tandonia kusceri. Многие из них
(кроме Boettgerilla pallens) наносят существенный вред растениям и запасам
пищевых продуктов.
Некоторые антропохорные виды могут исходно обитать на территории
СССР и в природных биотопах. Однако непосредственных контактов между
«дикими» и «культурными» популяциями нет, почему и можно считать,
что последние возникли в результате завоза человеком. Так, Boettgerilla
pallens в природных условиях обитает в западной Грузии и Абхазии. В антро-
погенных условиях этот вид встречен в оранжереях в Москве и Челябинске,
в парках Львова, Сочи и Еревана, а также во многих населенных пунктах
средней и западной Европы. Umax maximus, встреченный в парках и садах
многих крупных городов европейской части СССР, в природе живет в Вос-
точных Карпатах (с. 280). L. maculatus широко распространен в лесах Крыма
106
ОБЩАЯ ЧАСТЬ
и Кавказа, но, кроме того, обнаружен в погребах и оранжереях в Ленин-
граде, Пушкине (Ленинградская область), Севастополе, Орджоникидзе
и Тбилиси.
На этих примерах достаточно четко видно, что упомянутые антропохор-
ные виды могут быть завезены человеком очень далеко от их естественного
ареала. У остальных же слизней этой группы их «родина» находится за пре-
делами СССР. Так, у Limax flavus, который обнаружен в некоторых городах
европейской части Союза (с. 283), природный ареал находится в западном
Средиземноморье; у Lehmannia valentiana, встреченной в садоводствах Ле-
нинграда, Ростова-на-Дону и Ташкента, природный ареал находится
на Пиренейском нолуострове и в соседних областях Франции; у Tando-
nia kusceri, обитающей в Ботаническом саду Одессы, — природный уареал
в Болгарии и Румынии.
Границы между упомянутыми тремя группами синантропных слизней
в известной степени условны. Сильное антропогенное воздействие на при-
роду в некоторых областях и районах, при котором даже от вторичных при-
родных ассоциаций остается очень мало, приводит к тому, что некоторые виды
первой группы сохраняются только в парках, садах и на лугах-пастбищах
(Arion subfuscus, Deroceras agreste).
Некоторые виды второй группы, явно тяготеющие к полям и огородам,
завезены далеко от их естественного ареала. Именно с этим связано, в частно-
сти, сравнительно недавнее появление Deroceras reticulatum на Кавказе и
в юго-восточном Казахстане, Deroceras caucasicum и D. sturanyi в Средней
Азии, Parmacella ibera на черноморском побережье Кавказа. По-видимому,
эти виды завезены в указанные области в последние 30—40 лет.
Очень показательно, что европейские по происхождению слизни —
Arion rufus, A. subfuscus, A. hortensis, A. intermedius, A. fasciatus, Deroce-
ras reticulatum, Milax gagates, Limax maximus, L. flavus, Lehmannia valen-
tiana — к настоящему времени завезены и укоренились во многих странах
умеренных поясов северного и южного полушария, где активно выступают
как вредители сельского хозяйства. А трансголарктический Deroceras laeve
завезен даже во многие страны тропического пояса. По-видимому, завоз
европейских слизней на другие материки и на океанические острова начался
еще во времена парусного флота. Так, из 10 видов европейских слизней,
проживающих в Северной Америке (Pilsbry, 1948), большая часть уже отме-
чалась в американской литературе 19 века: Limax flavus в 1825, г., Arion
subfuscus в 1841 г., Deroceras reticulatum и Arion fasciatus в 1851 г. Учитывая
историю колонизации Северной Америки, есть все основания предполагать,
что завоз произошел значительно раньше.
Быстрому расселению слизней путем непроизвольного завоза в первую
очередь благоприятствует значительная устойчивость яиц этих животных
к колебаниям температуры и влажности. С другой стороны, полифагия
и неприхотливость в выборе пищи также явно способствовали завозам.
И, наконец, на новые места слизни попадали с привычными для них поле-
выми и огородными культурами.
Попытаемся наметить условия, способствовавшие становлению синантро-
пии среди слизней (Лихарев, 1965). Совершенно очевидно, что основным
фактором, стимулировавшим появление синантропных животных и растений,
была активная хозяйственная деятельность человека, приведшая к сокраще-
нию первичных природных ландшафтов и биотопов, а местами к их полному
исчезновению или существенной перестройке. При этом основная масса видов
была оттеснена в уцелевшие от антропогенного воздействия резерваты.
Но некоторые, и таких было немало, не только сохранились в новых усло-
виях, но стали процветать. Последнему способствовало создание человеком
больших площадей, засеянных определенными культурными растениями
СИНАНТРОПНЫЕ И АНТР'ОПОХОРИЫЕ ВИДЫ •
107
(монокультуры), предоставившими этим видам обильную и высококалорий-
ную пищу.
Но не только непосредственное воздействие хозяйственной деятельности
людей способствовало отбору из дикой фауны видов, которых мы называем
синантропными. Определенные биологические особенности этих животных
в свою очередь сыграли в этом важную роль.
Для слизней, проживающих на полях и огородах, где периодически про-
исходит вспашка и уборка урожая, при которых резко меняется весь ком-
плекс условий жизни, большое значение имеет длительность жизненного
цикла. Только достаточно короткий жизненный цикл, в условиях умеренного
климата занимающий не более 5—7 месяцев, может позволить этим животным
уцелеть и дать потомство за период от вспашки почвы и посадки (или посева)
до уборки урожая. Не случайно на полях и огородах обитают только виды
с таким циклом — Deroceras reticulatum и другие виды того же рода, Arion
hortensis, А. fasciatus, некоторые виды Tandonia и Milax. Исключение состав-
ляют вредные Parmacellidae, жизненные циклы которых занимают около
года. Однако у них около четырех наиболее жарких месяцев молодь про-
водит в состоянии диапаузы, зарывшись глубоко в землю, и около 1—2
месяцев на зимовке, также уйдя глубоко в почву.
Причины, вызвавшие появление антропохорных видов, несколько иные.
Как правило, эти теплолюбивые южные виды, поселяясь на севере далеко
от своих природных ареалов, в непосредственной близости от человека —
в домах, погребах, парках, оранжереях, — находят здесь не только обиль-
ный корм, но, главное, благоприятный микроклимат, которого они не имели
бы, поселившись где-нибудь поблизости в природе.
Не всегда легко установить исходный, т. е. природный, ареал того или
иного вида синантропных слизней, а следовательно, его зоогеографическую
природу. В тех случаях, когда общий ареал четко подразделяется на прирдд-
ную и антропогенную части и нет оснований ожидать контактов между
популяциями, живущими в природных биотопах и таковыми из антропо-
генных биотопов, дело обстоит достаточно просто: первичным, или природ-
ным, ареалом является местность, где данный вид живет в природе. Так,
у Lehmannia valentiana первичным ареалом является Пиренейский полу-
остров и соседние области Франции, а вторичными — многочисленные
микроареалы, связанные с завозом этого вида по оранжереям и садоводствам
остальных областей Европы, Азии и на плантации Калифорнии.
Однако нередки случаи, когда в некоторых частях своего ареала синан-
тропные виды обитают как на антропогенных биотопах, так и на соседних
природных. Так обстоит дело, в частности, с Deroceras reticulatum в средней
Европе и в северо-западных и западных областях СССР. В этих случаях при-
ходится принимать во внимание распространение в природе ближайших
в систематическом отношении видов. В примере с D. reticulatum это D. agreste
и D. turcicum. Все три вида «встречаются» на юго-востоке Европы, что дает
основание рассматривать D. reticulatum как вид среднеевропейского проис-
хождения, который в силу своей склонности к синантропии широко рассе-
лился в Европе и за ее пределами.
Другой аналогичный пример дает Boettgerilla pollens. Природный ареал
находится в западном Закавказье. Это подтверждается не только самим
фактом обитания слизня в природных условиях, но и присутствием в дан-
ном регионе другого вида того же рода — В. compressa, — который известен
лишь отсюда и обитает только в природных биотопах.
108
ОБЩАЯ ЧАСТЬ
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ СЛИЗНЕЙ
В практической деятельности людей роль слизней проявляется в двух
аспектах: как передатчиков опасных гельминтозов для домашних и промыс-
ловых животных и как вредителей многих культурных растений.
Передача паразитарных инвазий. Как и многие назем-
ные улитки, некоторые слизни служат промежуточными хозяевами различным
паразитическим червям, окончательными хозяевами которых являются до-
машние и дикие млекопитающие и птицы (с. 78). При этом чаще всего
отсутствует строгая специфичность между слизнями и гельминтами: главную
роль играет образ жизни моллюсков, определяющий возможность их встречи
и заражения соответствующей фазой паразитического червя.
Это было хорошо показано Давтяном (1947, 1948, 1950) на примере
сравнительной восприимчивости при искусственном заражении различных на-
земных моллюсков Армении к легочным нематодам овец и коз — Miillerius
capillaris, Cystocaulus nigrescens и к нескольким видам Synthetocaulus. Выяс-
нено, что слизни наряду с некоторыми гигробионтными улитками являются
облигатными промежуточными хозяевами только для Miillerius capillaris
(85—100% заражения). В жизненных циклах остальных легочных нематод
главную роль играют улитки, преимущественно ксеробионтные, а слизни
лишь второстепенную роль: для Cyctocaulus как субоблигатные, факультатив-
ные или мортальные хозяева; к видам Synthetocaulus слизни абсолютно не вос-
приимчивы даже в опытах с искусственным заражением.
Многочисленные позднейшие исследования, выполненные в других регио-
нах, подтверждают ведущую роль слизней в распространении мюллериоза
среди домашних травоядных животных (Рыбак, 1961; Егоров, 1965; Трушин,
1973). В хозяйственном отношении эта роль слизней особенно важна в об-
ластях с интенсивным отгонным живоноводством (Карпаты, Кавказ, Сред-
няя Азия), где эта болезнь сильно снижает поголовье (особенно среди
ягнят) и товарность скота.
Другим серьезным паразитарным заболеванием, распространяемым при
участии слизней, является давайнеоз. Возбудитель его — ленточный червь
Davainea proglottina. Взрослая фаза обитает в кишечнике куриных, в том
числе у домашних кур, и вызывает тяжелое заболевание, сопровождающееся
сильным истощением и прогрессирующим параличом и смертью птицы.
Особенно от этой болезни страдают цыплята.
Личинки давайнии проходят развитие в теле слизней. Они заражаются
ими, поедая куриный помет или различные остатки растений и отбросы, за-
раженные яйцами червя. Если курица съедает слизня с развившейся ли-
чинкой, то при переваривании моллюска личинка освобождается, проникает
в двенадцатиперстную кишку хозяина, где прикрепляется к стенке и начинает
расти, превращаясь во взрослого паразита. В отличие от других гельминто-
зов в распространении давайнеоза участвуют только слизни, но среди них
в этом отношении строгой видоспецифичности нет.
Меньшую роль, чем наземные улитки, играют слизни в распространении
дикроцелиоза. Это заболевание крупного и мелкого рогатого скота, реже
лошадей. Возбудитель его — ланцетовидный сосальщик (JDicrocoelium lanceo-
laturri). Взрослый червь поселяется в печени указанных животных, а также
и у диких жвачных и вызывает тяжелое заболевание. Первым промежуточным
хозяином этого гельминта является наземный моллюск, вторым — некоторые
муравьи. В тех случаях, когда жизненный цикл сосальщика проходит на
сухих равнинных или горных пастбищах, в его распространении главную
роль играют наземные улитки, преимущественно ксеробионтные. Однако
в областях Европы с влажным климатом значение слизней в распространении
этого заболевания возрастает.
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
109
Слизни — вредители культурных растений.
Слизни вредят очень широкому кругу зерновых, овощных, цветочных, тех-
нических культур, а также плантациям цитрусовых и винограда. Ежегодно
во многих странах издаются специальные брошюры и бюллетени, информиру-
ющие о вредной деятельности слизней, дающие прогнозы на ближайшее
время и инструктирующие фермеров как бороться с этими вредителями.
Хотя точно подсчитать во всех странах мира убытки, которые ежегодно при-
чиняют слизни, пока не удалось, совершенно очевидно, что они очень значи-
тельны. К тому же вредные слизни отличаются от многих других вредителей
сельского хозяйства очень широким распространением. Крупные потери
от слизней несет сельское хозяйство многих стран северной и северо-запад-
ной Европы. Так, в Англии и Уэльсе в 1967 г. только по картофелю и озимой
пшенице убытки исчислялись соответственно в 645 700 и 191 700 фунтов стер-
лингов (Shotton, 1971). Считается, что ежегодно в этих областях слизни уни-
чтожают 36 тыс. тонн картофеля (Runham, Hunter, 1970).
Если же учесть, что такие вредные слизни Европы, как Deroceras reticu-
latum и Milax gagates, будучи непроизвольно завезены в США, Канаду, на юг
Австралии и в другие страны с влажным и умеренным климатом, не только
укоренились и расселились на новом месте, но и вредят там сельскому хо-
зяйству, то общие убытки от этих слизней должны быть огромны.
Нередко большие потери от слизней несет сельское хозяйство ряда об-
ластей СССР. Так, в дождливые 1922—1923 гг. в Ленинградской области
ими было уничтожено до 20 тыс. га посевов озимой пшеницы, а в Новгород-
ской области — до 32 тыс. га (Соколов, 1933). В 1957 г. в Орловской области
слизни (главным образом Deroceras reticulatum) нанесли большой ущерб
урожаю помидоров и капусты (Бодренков, 1963), а в 1962 г. в Ленинградской
области — капусте, картофелю, землянике и другим культурам (Дмитриева,
1963).
Большие убытки терпит от слизней сельское хозяйство некоторых обла-
стей Кавказа и Средней Азии, где, кроме завезенного Deroceras reticulatum
и местных видов того же рода, особенно большой вред на полях и огородах
причиняют слизни Parmacellidae — Candaharia levanderi, С. rutellum, Parma-
cella ibera (Жадин, 1946; Увалиева, 1975, 1979; Загайный, 1978).
Слизни повреждают очень широкий диапазон культур. В северо-западных,
западных и центральных областях европейской части СССР они наносят вред
многим огородным растениям — клубням и листве картофеля, белокочанной и
цветной капусте, салату, различным корнеплодам (листве и выступающим из
почвы участкам корнеплодов), рассаде и молодым всходам многих овощей,
.листьям фасоли и гороха, плодам земляники, огурцов и помидоров. Меньший
вред они наносят краснокочанной капусте, петрушке, чесноку, луку, листьям
созревающих огурцов и земляники.
Особенно ощутимый вред они причиняют озимой пшенице и р>ки, поедая
как только что высеянные зерна, так и их всходы. В меньшей степени страдают
•от слизней овес и ячмень; практически они не трогают яровую пшеницу, лен
и гречиху (Бацйлев, 1959).
В субтропиках Кавказа и на поливных землях Азербайджана, Туркмении,
республик Средней Азии и на юго-востоке Казахстана (предгорья Заилий-
•ского Алатау) слизни вредят многим овощным культурам, цитрусам, табаку
(рассаде в парниках) и декоративным растениям.
Повреждения, причиняемые слизнями, очень характерны и легко отли-
чимы от следов деятельности других сельскохозяйственных вредителей
(рис. 44). В листьях они обычно выгрызают неправильной формы дыры, остав-
ляя в целости лишь стебель листа и наиболее крупные жилки. На корнеплодах,
клубнях картофеля, плодах земляники, помидор и огурцов они выгрызают
.различной формы и размеров каверны, обычно расширяющиеся внутрь.
по
ОБЩАЯ ЧАСТЬ
У капусты они повреждают не только поверхностные и приземные листья, но-
и выгрызают глубокие углубления в кочане. В зернах злаков они выгры-
зают как зародыш, так и эндосперм.
Характерной чертой таких повреждений являются многочисленные следы
из застывшей слизи, кучки фекалий и земли. Кроме прямого вреда слизни:
Рис. 44. Повреждения, наносимые слизнями клеверу (/) и клубням картофеля (Z/).
причиняют косвенный вред, загрязняя продукты урожая и способствуя их:
загниванию, а тем самым сокращая сроки хранения.
Переползая с одного растения на другое, слизни способствуют распростра-
нению среди сельскохозяйственных культур различных грибковых и вирусных
заболеваний — пятнистость капустная, ложная мучнистая роса лимской фа-
соли, фитофтороз картофеля (Runham, Hunter, 1970; Шапиро, 1978). Эти бо-
лезни могут нанести хозяйству убытки не меньшие, а нередко и большие,,
чем прямая вредная деятельность слизней. То, что многие из них охотно
дополняют свой рацион гифами грибов, еще более способствует заражению
растений грибковыми заболеваниями.
Особенно страдают от слизней молодые всходы озимых, которые нередко-
после нападения слизней уже не оправляются и гибнут. Даже относительно
крупные всходы не всегда восстанавливаются после нападения слизней.
РЕГУЛИРОВАНИЕ ЧИСЛЕННОСТИ
111
Нередко приходится сильно поврежденные участки озимей пересеивать
или занимать под другие культуры.
Слизни вредят не только на открытом, но и на закрытом грунте, т. е. в пар-
никах и теплицах. Здесь особенно большой вред они причиняют всходам и
рассаде многих культур. Так, в парниках табаководческих хозяйств Армении
«большой вред рассаде табака причиняют Gigantomilax monticola и Deroceras
caucastcum (Акрамовский, Бабаян, 1976). Здесь они иногда уничтожают
до 25 % рассады этой ценной культуры. Аналогичное известно для некоторых
табаководческих хозяйств окрестностей Ялты, но здесь как вредители высту-
пают Deroceras tauricum и Tandonia cristata.
Существенные убытки также наносят слизни цветоводству как на откры-
том, так и на закрытом грунте (Загайный, 1978; Косоглазов, 1972). В некото-
рых цветоводствах Ленинграда, где на гидропонном грунте выращивают
ремонтантную гвоздику, заметный вред этой культуре наносит Deroceras laeve.
РЕГУЛИРОВАНИЕ ЧИСЛЕННОСТИ ВРЕДНЫХ ВИДОВ.
Полное уничтожение слизней возможно осуществить и целесообразно
«с экономической точки зрения лишь на небольшом участке, изолированном
от природных резерватов этих животных. В основном это касается парников и
теплиц. Уничтожить всех слизней, как и любого другого вредителя сельского
хозяйства на полях и огородах, практически невозможно и экономически не
оправданно. Поэтому речь может идти лишь об ограничении численности слиз-
ней в тех местах, где они потенциально опасны для будущего урожая, и в пе-
риод, когда как с биологической, так и с экономической точки зрения такая
«борьба может дать наибольший эффект.
Борьба со слизнями слагается из умелого использования и сочетания
различных агротехнических и химических мероприятий, которые следует
проводить в периоды, когда слизни наиболее уязвимы и численность их отно-
сительно невысокая.
Агротехническая борьба. Слагается из хозяйственных меро-
приятий, направленных на ограничение вредной деятельности слизней,
не связанных с непосредственным их уничтожением путем использования пе-
стицидов или репеллентов. Эти мероприятия преследуют цель: а) изменить
условия внешней среды в сторону, неблагоприятную для жизни вредителей;
«б) увеличить устойчивость культурных растений к слизням; в) попутно ме-
ханически уничтожить слизней. Естественно, что эти мероприятия не должны
проводиться в ущерб главной задаче — выращиванию хорошего урожая запла-
нированной культуры.
Одно из наиболее эффективных мероприятий — это тщательное возделы-
вание почвы перед посевом или посадкой. Разрыхление пашенного слоя трое-
кратным боронованием может привести к сильному сокращению численности
популяции как за счет уменьшения числа убежищ в почве, так и в результате
попутного непосредственного уничтожения вредителей. При этом численность
Deroceras reticulatum может сократиться на х/4—1/3 (Runham, Hunter, 1970).
Если после культивации почвы засеять поле кормовыми травами и в течение
.лета его несколько раз прокашивать, то в результате уплотнения и задернения
верхнего слоя почвы популяции слизней будут практически уничтожены.
Наибольший эффект приносят те мероприятия, при которых в почве сокра-
щается количество щелей и углублений, могущих послужить убежищами
для слизней. Однако на тяжелых почвах, где всякое улучшение их структуры
необходимо для увеличения урожайности, уплотнение поверхностного слоя
нежелательно.
Вспашка зяби и парового поля, уничтожающая сорняки, не только
лишает слизней привычных убежищ, но и подрывает их кормовую базу.
112
ОБЩАЯ ЧАСТЬ
Очистка от сорняков и разрежение культур очень важны в деле борьбы
со слизнями, так как создают лучшие условия аэрации травостоя и снижают
влажность почвы. Все это ухуюшает условия жизни слизней. Там, где воз-
можно, следует также принимать меры по снижению уровня грунтовых вод.
На полях и огородах слизни находят убежище не только в почве, но и под.
разными предметами, лежащими на ее поверхности, — камни, куски дерна,
различный мусор, кучки растений после прополки, остатки уборки урожая,
как солома, припочвенные листья некоторых овощей, ботва и т. п. Все это
следует удалить с поля или закопать достаточно глубоко в землю. Учитывая,
что пустоши, межи, кучи камней по краям поля могут служить очагами рас-
селения слизней на соседние поля и огороды, необходимо по возможности
принять меры к изоляции этих очагов или к их ликвидации.
Следует избегать посева озимых сортов пшеницы и ржи в низких и сырых
местах, в непосредственном соседстве с кустарниками, лесом и другими при-
родными резерватами слизней. Эти культуры желательно размещать вдали
от полей с клевером и капустой, которые обычно сильнее других заражены
слизнями.
Определенный эффект в борьбе со слизнями дает умелый подбор сортов тех
или иных культур, которые отличаются наибольшей устойчивостью к этим
вредителям. Так, в Англии фермеры стремятся шире использовать сорта
картофеля менее чувствительные к слизням (Runham, Hunter, 1970). Положи-
тельный эффект дает использование ранних сортов, вегетация которых начи-
нается раньше, чем слизни становятся активными, а созревание наступает до
осеннего пика численности этих вредителей.
Такие обычные удобрения, как суперфосфат и известь, действуют на
слизней как наружные яды. Поэтому их можно употреблять с двумя целями —
как удобрения и как пестициды. Однако следует учесть, что эти вещества
лишь в пылеобразном состоянии пригодны для борьбы со слизнями. Гранули-
рованный суперфосфат надо перемолоть в порошок, а известь применять в виде
свежегашеной пушонки.
Химическая борьба. Слагается из применения ядов —мол-
люскоцидов и репеллентов, т. е. веществ, отпугивающих слизней. Длительное
время химическая борьба со слизнями сводилась к использованию различ-
ных контактных неорганических веществ как в твердом, так и в жидком виде,
или же в виде фумигантов (Лихарев, 1954). Однако из-за больших норм рас-
хода этих веществ, высокой трудоемкости их использования и недостаточной
эффективности их как яды они не нашли широкого применения.
В настоящее время как в СССР, так и за рубежом, главным химическим
средством борьбы со слизнями являются различные препараты метальдегида —
твердого полимера уксусного альдегида. Высокое токсическое действие ме-
тальдегида и эффективность его использования против слизней подтвердили
многочисленные исследования и производственные испытания (Дмитриева,
1963, 1968а; Дмитриева, Шапиро, 1974, 1975, 1976; АкрамовсКий, Бабаян,
1976; Увалиева, 1979, и др.).
Действие метальдегида на слизней достаточно сложно (Thomas, 1948).
С одной стороны, так как он обладает аттрактивными, т. е. привлекающими,
свойствами, слизни его активно ищут и поедают. В этом случае он является
для них кишечным ядом. С другой стороны, этот моллюскоцид обладает силь-
ным контактным действием, так как при попадании на тело слизней вызывает
обильное выделение слизи и обезвоживание организма. В-третьих, метальде-
гид оказывает на слизней нервно-паралитическое действие, приводящее к по-
тере локомоторной активности и в конечном счете к параличу.
Физиологический механизм действия метальдегида на Deroceras reticulatum
исследован Дмитриевой и Шапиро (1975). Авторы показали, что внешние при-
знаки отравления метальдегидом проявляются через 1—2 мин после поедания
РЕГУЛИРОВАНИЕ ЧИСЛЕННОСТИ
113
отравленной приманки. Весь процесс отравления состоит из двух физиологи-
чески противоположных стадий — гиперактивации и депрессии. Продолжи-
тельпостть и глубина этих стадий зависит от дозы моллюскоцида. Гипер-
активация сопровождается повышением интенсивности дыхания и катаболиз-
мом гликогена и липидов. После этого наступает депрессия, которая выража-
ется в потере слизнями локомоторной активности, резком снижении интен-
сивности дыхания и дальнейшем катаболизме гликогена и липидов. Если
слизни приняли среднюю летальную дозу метальдегида, то вскоре после де-
прессии наступает смерть.
В СССР метальдегид выпускается в двух формах: в виде 50%-ного смачи-
вающего порошка и 5 %-ного гранулированного препарата, где роль наполни-
теля выполняют отруби грубого помола. Оба препарата предназначены для
защиты зерновых, зернобобовых, овощных, плодовых, цитрусовых, техни-
ческих, цветочных культур и винограда. Порошкообразный метальдегид,
предназначен для борьбы со слизнями путем опрыскивания водной эмульсией
(4—8 кг/га). Гранулированный препарат применяется путем рассева или
раскладывания небольшими кучками на поверхности почвы или вокруг расте-
ний, по краям поля или теплицы со стороны дневных убежищ слизней или их
природных очагов. Норма расхода при рассеивании на зерновых — 15 кг/га,
а на остальных культурах — 30—40 кг/га.
При работе с метальдегидом следует строго соблюдать определенные
меры безопасности, главным образом защищать дыхательные пути, глаза и
кожу. После его разбрызгивания или разбрасывания по полю он сравнительно
безопасен как для человека, так и для теплокровных животных. Установлено,
что для собак летальная доза этого моллюскоцида при оральном применении
составляет 600—1000 мг/кг живого веса животного, а для крыс 300—600 мг/кг.
Что очень важно, он сравнительно безвреден для большинства насекомых и
других почвенных беспозвоночных, не оказывает отрицательного воздействия
на урожайность сельскохозяйственных культур и безвреден для растений.
Иными словами, метальдегид — это пестицид очень узкого селективного
действия. В этом его значительное преимущество перед другими пестицидами,
применяемыми против слизней.
Однако при многолетнем употреблении метальдегида в одной местности
действие его на слизней ослабевает, они как бы «привыкают» к нему. Поэтому
в ряде стран, в особенности в Англии, идут поиски и испытания новых, более
эффективных моллюскоцидов, в частности из группы карбаматов (Gough,
Woods, 1954; Gould, 1962; Symonds, 1971). Так как последние значительно
более токсичны для теплокровных, чем метальдегид, и вредно сказываются
на почвенной фауне, то некоторые авторы рекомендуют их использовать для
протравливания посевного материала, особенно семян озимой пшеницы, а так-
же в качестве добавок к препаратам метальдегида.
Биологическая борьба. Биологические методы борьбы со.
слизнями пока не разработаны. Использование их естественных врагов,
в первую очередь хищников, малоперспектйвно из-за невысокой численности
последних, трудностей «привлечения» их в места, где необходимо бороться
с вредными слизнями, а также потому, что слизни не являются главным ком-
понентом в их пищевом рационе.
По-видимому, поиски биологических средств борьбы со слизнями следует
направить в сторону изучения паразитов и болезней этих животных. В этом
отношении показательным и многообещающим является исследование жиз-
ненных циклов двух инфузорий из рода Tetrahymena, проведенное Бруксом
(Brooks, 1968). Автор показал, что если Т. limacis можно считать слабо пато-
генным факультативным.паразитом моллюсков, то другой вид — Т. rostrata
является облигатным паразитом Deroceras reticulatum (возможно, и других
моллюсков), сильно патогенным как взрослым слизням, так и их яйцам..
8 И. М. Лихарев, А. Й. Виктор
114
ОБЩАЯ ЧАСТЬ
МЕТОДЫ ИССЛЕДОВАНИЯ
Данный раздел преследует цель ознакомить читателя с теми приемами и
методами изучения слизней, которые вытекают из особенностей образа жизни
и строения этих животных. Так как слизни во многом сходны с наземными
улитками, то целый ряд приемов и методов, используемых при исследованиях
с раковинными моллюсками, сохраняют свое значение и при работе со слиз-
нями (Лихарев, Раммельмейер, 1952; Jaeckel, 1953; Лихарев, 1962; Шилейко,
1978). С другой стороны, выбор методики в первую очередь определяется
задачами исследования. Так, например, изучение тех или иных гистологиче-
ских или цитологических структур требует определенной фиксации и последу-
щей обработки изучаемого объекта, часто независимо от его систематической
принадлежности. Естественно, что изложение таких специальных методик
не входит в нашу задачу.
Сбор живого материала. В связи с тем что даже самые
мелкие виды слизней крупнее многих небольших улиток, в период их ак-
тивного образа жизни обнаружить слизней на местности значительно легче.
Как уже отмечалось, в дневное время слизни находятся в различных укрыти-
тиях: на полях, огородах и лугах — в щелях почвы, под комками земли или
под различными предметами, лежащими на поверхности почвы, у подножья
травянистых растений; в лесу, кроме того, под неплотно прилегающей корой
деревьев и пней, под лежащими на земле кусками гниющих стволов и ветвей
деревьев и в лиственном опаде. Всюду, где имеются выходы горных пород
в виде отдельно лежащих камней, скал и осыпей, под камнями и в различ-
ных щелях могут скрываться слизни. Поэтому поиск и сбор слизней днем
в сухую погоду сопряжен с переворачиванием комков земли, камней и дру-
гих предметов, под которыми они могут скрываться; их собирают в глубине
травостоя, в лиственной подстилке, среди мха, под отстающей корой и т. п.
Наиболее сложен сбор слизней среди скал и осыпей. Здесь они могут пря-
таться так глубоко, что днем до них не всегда можно добраться. Присутствие
их в таких местах нередко выдают многочисленные блестящие следы, «до-
рожки», из высохшей слизи, которые слизни оставили во время ночной
активности.
Днем эффективные сборы можно сделать в туманную погоду, во время
теплого дождя или за короткий период после него, пока еще влажность
воздуха близка к 100%. Если днем в сухую погоду сбор слизней сопряжен
с некоторыми трудностями из-за скрытого образа жизни этих животных, то
значительно легче делать сборы в сумерки, особенно после выпадения росы,
и по ночам при свете электрического фонаря, когда животные выползают
из укрытий.
Сбор слизней производится вручную и с помощью пинцета.
Наибольшую сложность представляет количественный учет слизней.
В сильно пересеченной местности, изобилующей различными укрытиями, бо-
лее или менее точный учет слизней возможен только в период их активности
путем непосредственного подсчета на единицу площади. На полях и огородах,
где условия более однородны и слизни днем прячутся в глубине почвы, можно
кроме визуального подсчета производить более точный учет путем взятия
проб почвы определенной площади (обычно 0.1 или 0.25 м2) и в нескольких
повторностях. Обработка этих проб, т. е. выборка слизней и их яиц, перено-
сится в лабораторию и может быть осуществлена несколькими спосо-
бами.
Одним из них является промывка пробы водой через систему из 2—3 сит—
верхнее сито с ячейкой площадью в 5, среднее 3, нижнее 1 мм2. Наибо-
лее удобно использовать специальный набор сит, выпускаемый для агрономи-
ческих целей. После промывки каждое сито просматривают и выбирают всех
МЕТОДЫ ИССЛЕДОВАНИЯ
115
слизней и их яйца. Для большей точности можно содержимое каждого сита
опустить в раствор сульфата магния или поваренной соли с удельным, весом
1.17. В силу флотации слизни и их яйца вместе с другими органическими
частицами всплывают на поверхность.
Недостаток метода промывки почвенных проб в том, что он сопряжен
с разрушением части молодых слизней и многих яиц, которые тем самым оста-
ются неучтенными.
Выгонку слизней на поверхность почвенной пробы можно сделать путем
медленного насыщения ее водой (до 12 часов), поступающей снизу. Однако-
и этот способ не лишен своих недостатков. Слизни сильно насыщаются влагой
и увеличивают свой вес. А по весу обычно судят о возрасте и темпе роста этих
животных. Кроме того, при этом методе из учета выпадают яйца слиз-
ней.
Как сами слизни, так и их яйца нередко располагаются на различной глу-
бине, в среднем не превышая 20 см. Поскольку нередко представляется важ-
ным выяснить особенности горизонтального распределения слизней в почве,
то при промывке или разборке пробы в сухом виде работу следует вести по
слоям определенной толщины.
Собранных в природе слизней для их дальнейшего изучения желательно-
доставить живыми в лабораторию. Лишь при невозможности сделать это — от-
сутствие поблизости такой лаборатории, недостаток времени на прижизненные
наблюдения, жаркая погода, затрудняющая сохранение живых слизней —
допустима фиксация слизней сразу на месте сбора. Перед этим их необходимо
очистить от частиц почвы и сора.
Для доставки слизней в живом виде в лабораторию их следует поместить,
в чистую пластмассовую или металлическую коробку с плотно закрываю-
щейся крышкой, снабженной многочисленными мелкими отверстиями для
свободного доступа воздуха. Внутрь коробки закладывают небольшое ко-
личество влажных сочных листьев или сырого мха для поддержания необхо-
димой влажности. Полезно слегка увлажнить эту подстилку. Необходимые
сведения о месте сбора записываются на наружной поверхности коробки или
на наружной этикетке. Не рекомендуется класть такую этикетку внутрь ко-
робки, как так слизни могут ее съесть или сильно загрязнить.
Сборы с живыми слизнями до доставки в лабораторию следует тщательно
оберегать от прямого действия солнца. Время от времени надо проверять со-
стояние слизней, заменять высохшие листья свежими, очищать отверстия
крышки от слизи. Следует учесть, что в это время главная опасность для
слизней это перегрев и недостаток свежего воздуха.
Содержание слизней в неволе. В лаборатории могут
ставиться различные прижизненные наблюдения и опыты над слизнями. Для
длительного содержания их в неволе, в частности при изучении роста и раз-
множения, слизней помещают в различные садки-террариумы, тип й величина
которых определяется поставленными задачами, размерами и числом изучае-
мых экземпляров. Следует учесть, что в слишком малых садках нередко
слизни нападают и калечат друг друга.
На дно террариумов помещается слой почвы, желательно взятой из места,
где были собраны слизни. Этот слой должен быть достаточно толстый и рых-
лый, чтобы слизни могли в нем найти или сделать углубление для откладки
яиц или укрытие от дневного света. Bo-всех случаях садки должны быть за-
щищены от солнца иобеспечивать свободный доступ воздуха при высокой влаж-
ности.1 Необходимо предусмотреть периодическую чистку садков от экскре-
ментов, слизи и остатков пищи.
1 За исключением специальных опытов по влиянию влажности на слизней.
в*
lie
ОБЩАЯ ЧАСТЬ
Для питания растительноядных слизней чаще всего используют те расте-
ния, которым данный вид оказывает наибольшее предпочтение (что выясня-
ется специальными опытами), а также некоторые искусственные корма
(Kunkel, 1916; Frii mining, 1954; Дмитриева, 1969). К числу наиболее «по-
пулярных» среди растительноядных слизней кормов принадлежат листья
салата и капусты, плоды земляники и помидор, морковь, клубни картофеля
и др.
Несколько сложнее содержать в неволе хищных слизней. Для этого им
необходим такой же тип почвы и лиственной подстилки, которые для них
привычны в природе, а также корм из дождевых и других почвенных червей.
Следует признать, что техника содержания хищных слизней очень слабо
разработана.
Наркоз. При некоторых опытах, связанных с проведением микро-
операций на слизнях, необходимо их временно усыпить. Для этого разрабо-
тано несколько методов наркотизирования слизней. Одним из них является
усыпление в специальных камерах с помощью углекислого газа (СО2), на-
сыщенного парами воды (Bailey,1969). Такой наркоз действует достаточно
долго, чтобы провести короткую операцию. Другой способ связан с использо-
ванием 0.5—1%-ного водного раствора пропиленфеноксетола. Предварительно
промытых водой слизней помещают в этот раствор и через 15 мин даже круп-
ные экземпляры усыплены, и уже можно провести на них операцию. После
окончания операции слизней в течение 20 мин следует промывать водой
и оставить на влажной торфяной пластинке до тех пор, пока они не очнутся.
Фиксация. Для умерщвления и консервации слизней чаще всего
применяют 70—75%-ный этиловый спирт. Этот -метод вполне годится для
наиболее обычных исследований в области фаунистики, диагностики и систе-
матики слизней, так как позволяет сохранять их на неопределенное время и
проводить необходимые исследования по изучению внутреннего строения
этих животных. Однако фиксация спиртом непригодна для гистологических
и цитологических исследований, которые требуют специальных фиксаторов.
Если живого слизня непосредственно перенести в спирт, то он резко со-
кращается, иногда у него лопается кожа и наружу выворачивается часть
внутренностей. Но даже если это не случится, сильно сократившийся слизень
(так называемая «жесткая» фиксация) менее удобен для дальнейших анато-
мических исследований, чем слизень, сохранивший естественную форму
тела (так называемая «мягкая» фиксация).
Поэтому перед фиксацией необходимо принять меры к тому, чтобы слизень
«заснул» в расправленном состоянии. Для этого применяют несколько спо-
собов. Из них наиболее обычным и простым является помещение живых сли-
зней в достаточно просторный сосуд с холодной водой, а еще лучше с кипяче-
ной и охлажденной до комнатной температуры водой. Сосуд должен быть
закрыт так, чтобы под крышкой не оставалось воздуха. В воде слизни сильно
вытягиваются и в таком положении через некоторое время погибают. Смерть
наступает в зависимости от температуры воды, вида и размеров животного —
через несколько часов (иногда почти через сутки).
Для ускорения усыпления слизня можно добавить в воду несколько кри-
сталлов ментола или несколько капель спиртового раствора этого вещества.
Ментол расслабляет мышцы и тормозит процесс гниения, который в теплое
время года начинается сразу после смерти слизня. Для еще более быстрого
усыпления слизней рекомендуют (Runham, Isarankura, Smith, 1965) поме-
щать их в водный раствор, содержащий 0.08% нембутала и 1 % пропиленфено-
ксетола. В этом случае слизни засыпают в течение нескольких минут.
В практике авторов этой книги хорошо зарекомендовал себя метод, пред-
.ложенный им Е. В. Шиковым. Он состоит в использовании водного раствора
^кристаллического нитрата кобальта [Co(NO3)2-6H2O]. Раствор должен
МЕТОДЫ ИССЛЕДОВАНИЯ
117
иметь интенсивно розовый цвет. Живых слизнейпомещаютвсосуд с указанной
жидкостью, и через несколько минут они гибнут, полностью расслабившись.
После усыпления отмытых от слизи животных помещают в 70—75%-ный
спирт. Перед этим крупным экземплярам (более 70 мм длиной) следует ввести
внутрь тела некоторое количество спирта. Для этого лучше всего использо-
вать обычный медицинский шприц, а спирт вводить внутрь тела через перед-
нюю часть ноги. При отсутствии шприца можно надрезать кожу слизня с ле-
вой стороны, чтобы фиксатор свободно проникал внутрь тела.
Сосуд, в котором фиксируют и в дальнейшем сохраняют слизней, должен
быть достаточно просторным, чтобы слизни в нем располагались свободно
в вытянутом положении, не давили друг на друга (полезно слизней проклады-
вать слоями ваты) и чтобы объем спирта не менее чем в 3—4 раза превосходил
общий объем слизней. Во всех случаях через сутки, а потом примерно через
две недели необходимо сменить спирт и повторять эту операцию периодически,
пока спирт в сосуде со слизнями не станет бесцветным и прозрачным.
При наличии в пробе достаточного числа экземпляров одного вида реко-
мендуется часть их зафиксировать 50%-ным спиртом «жестким» способом,
т. е. без анестезии. Можно использовать для этих целей профильтрованный
старый «отработанный» спирт с последующим переводом в свежий 70%-ный
спирт. Этот материал менее удобен для препарирования, но зато на нем будет
лучше виден рельеф кожи и такие образования, как киль и мантийная бо-
роздка. Особенно такой способ рекомендуется при фиксации Trigonochlamydi-
dae. У этих слизней при «жесткой» фиксации не только лучше виден рельеф
покровов, но благодаря особенностям их внутреннего строения они обычно
приобретают форму, характерную для определенных родов. Подобный тип
фиксации пригоден и для Lytopelte maculata. При «мягкой» фиксации этот
вид внешне не отличим от многих слизней из рода Deroceras. Но при «жесткой»
фиксации на теле его появляется четкий киль, а капюшон не только сокра-
щается, но и приподнимается над шеей — внешние признаки, хорошо отли-
чающие этот вид от всех Deroceras.
Для изучения и демонстрации окраски и ее изменчивости лучше всего
фиксировать слизней в 4°/0-ном водном растворе формалина. В этом фикса-
торе лучше сохраняется природная окраска, чем в спирте. Правда, для
препарирования такой материал малопригоден, так как ткани становятся
неэластичными. При отсутствии спирта в крайней случае допустима фиксация
слизней и в формалине (в указанной концентрации). Однако при первой воз^-
можности такой материал следует перевести в спирт, который после этого
необходимо сменить еще два раза с интервалами 1.5—2 недели.
Вскрытие. Обычно в камеральную обработку входит углубленное
исследование анестезированного или зафиксированного материала, т. е. изу-
чение как внешнего облика слизней, так и их внутреннего строения.
Не обязательно вскрытие проводить на фиксированном материале. Когда
есть возможность, целесообразно изучать внутреннее строение на анестези-
рованных экземплярах, у которых ткани мягче, чем у заспиртованных, а внут-
тренние органы сохранили естественную окраску и форму. В этом случае
слизней помещают в препаровальную ванночку, заполненную физиологиче-
ским раствором (0.6—0.65%-ный водный раствор поваренной соли).
Наиболее удобно начинать вскрытие слизня, взяв его в руки, и наносить
разрезы ножницами или ланцетом. При работе с мелкими экземплярами по-
лезно проводить вскрытие под бинокуляром.
В отличие от улиток, которых обычно вскрывают с правой стороны,
слизней вскрывают с левой стороны, начиная надрез стенки тела вдоль
верхнего края ноги (по перипедальной борозде) или немного выше
(рис. 45). Разрез прокладывают от головы и немного не доводят до заднего
конца тела. Затем делают два поперечных надреза: один позади глазных
118
01ЯЦЛЯ ЧАСТЬ
щупалец, другой через задний конец на правую сторону до ноги. Все раз-
резы нужно делать оттягивая кожу кверху, чтобы не задеть внутренние-
органы.
Затем слизня спиной кверху помещают в неглубокую препаровальную
ванночку, дно которой залито смесью парафина с воском и черным сапож-
ным кремом. К дну ванночки его прикрепляют с помощью булавок, вка-
лывая их в ногу. Пинцетом отгибают направо верхнюю стенку тела, по-
путно подрезая тонкие мезентериальные тяжи, мелкие нервы и сосуды.
Отогнув верхнюю стенку тела, закрепляют ее с помощью булавок. Следует-
принять меры к тому, чтобы не оборвать ректум и аорту. При невозмож-
ности их сохранить в неповрежденном виде следует их аккуратно подрезать,.
Рис. 45. Основные разрезы для вскрытия слизней (из: Riedel, Wiktor, 1974).
так чтобы были видны концы обоих органов. После этого весь препарат
заливается 70%-ным спиртом.
Открывшаяся картина естественного расположения внутренних орга-
нов (situs viscerum) уже достаточно информативна (рис. 4). В первую оче-
редь бросается в глаза крупная буроватая печень, большая часть которой
располагается позади кишки, образуя вершину внутренностного мешка.
У одних видов это правая лопасть печени, и в этом случае меньшая левая
лопасть располагается впереди, у других видов, наоборот. Печень в значи-
тельной степени закрывает обзор многих остальных органов.
Видимые участки кишки и зоба отличаются тонкими стенками, сквозь
которые просвечивает жидкое содержимое желтоватого или зеленоватого
цвета. У самого переднего конца тела видна белая мускулистая глотка.
В передней части внутренностного мешка располагается большинство по-
ловых органов, отличающихся от печени и кишки толстыми железистыми
или мускулистыми стенками и окраской. Гонада не всегда видна снаружи,
так как может быть закрыта участками печени и кишки. Обычно хорошо
видны отдельные участки белковой железы, особенно с нижней стороны
внутренностного мешка.
На внутренней стороне отвернутой верхней стенки тела сквозь диафра-
фрагму просвечивают органы мантийного комплекса. Видны сердце (осо-
бенно желудочек), легочная сеть сосудов и желтоватая почка. У большин-
ства видов позади мантийного комплекса крепится основание мощного
мускула колумеллярная система ретракторов тела. Этот мускул уходит
вглубь внутренностного мешка, причем у видов с двухпетлевым кишечни-
ком он располагается справа от задней кишки, а у видов с трехпетлевым
кишечником кишка огибает основание мускула спереди, затем тянется назад
и наконец поворачивает вперед к мантийному комплексу, заканчиваясь-
МЕТОДЫ ИССЛЕДОВАНИЯ U9
ректумом (рис. 15). Эта третья петля всегда хорошо видна, так как распо-
лагается в одной плоскости и лежит на верхней стороне внутренностного
мешка непосредственно под покровами спины. 1
Дальнейшее исследование отдельных органов следует вести путем осво-
бождения их от различных мезентериальных связок и пленок, от мел-
ких нервов и кровеносных сосудов. Эту процедуру целесообразно начать
с отпрепарирования гениталий, сначала у переднего конца тела, освобождая
пенис, семяприемник, половой атриум, женские половые пути и переднюю
часть семяяйцевода, а затем с заднего конца. Для этого надо осторожно
раздвинуть лопасти печени и петли кишки, найти гонаду, которая отли-
чается от печени не только окраской, но более крупнозернистой структурой,
найти гермафродитный проток и постараться освободить их от всех связок.
Таким же образом освобождают белковую железу.
Затем, чтобы полностью освободить гениталии, потребуется подрезать
вблизи от основания половой ретрактор и колумеллярный мускул. Послед-
ний следует вытащить вперед. Наконец, осторожно протаскивая через
петли кишки гонаду с ее протоком и белковую железу, можно отделить
гениталии от остальных органов. Надрезав участок кожи вокруг полового
отверстия, гениталии можно полностью изолировать и изучать и хранить
отдельно от остального слизня. Однако в диагностической практике пре-
паровку гениталий обычно ограничивают освобождением и расправлением
передних участков полового аппарата, т. е. пениса, яйцевода, семяприем-
ника и атриума, не извлекая их полностью из внутренностного мешка.
После изъятия или высвобождения половых органов другие системы
внутренностного мешка становятся легко доступными для углубленного
изучения.
Строение мантийного комплекса органов имеет немаловажное значение
в исследованиях слизней. Чтобы лучше его разглядеть, следует подрезать
диафрагму слева 2 и отвернуть ее направо. Если подрезать мантийный комп-
лекс по его заднему краю, то откроется доступ в раковинную сумку и оттуда
можно извлечь рудиментарную раковину.
Остальные органы слизня исследуются общепринятыми в малакологии
методами (Большой практикум по зоологии беспозвоночных, ч. II, 1946;
Лихарев, Раммельмейер, 1952; Лихарев, 1962; Riedel, Wiktor, 1974;
Шилейко, 1978).
После изучения внутреннего строения слизня желательно все органы
по возможности поместить на свое место и возвратить верхнюю стенку
тела в исходное положение. В таком виде вскрытый слизень сохраняется
в спирту.
1 При вскрытии третья петля иногда отделяется от внутренностного мешка и приле-
гает к верхней стенке тела.
2 См. сноску на с. 27.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
Данный раздел авторы стремились изложить соответственно традициям,,
принятым в мировой литературе для сводок подобного рода, в первую оче-
редь в свете правил и рекомендаций «Международного кодекса зоологиче-
ской номенклатуры». При этом соблюдались установившиеся в серии моно-
графий «Фауна СССР» правила и стиль.
В частности, при пользовании данной книгой как определителем сле-
дует всегда учитывать, что в характеристиках каждого таксона (например,,
рода или вида) не упоминаются признаки, свойственные вышестоящим
таксонам (семейству, подсемейству, подроду). Следовательно, прежде чем*
ознакомиться с описанием того или иного рода или вида читатель должен
внимательно прочесть характеристики всех вышестоящих таксонов, к ко-
торым этот род или вид принадлежит.
УСЛОВНЫЕ СОКРАЩЕНИЯ,
ПРИНЯТЫЕ В ТЕКСТЕ И РИСУНКАХ СИСТЕМАТИЧЕСКОЙ ЧАСТИ
В описаниях при обозначении размеров: дл. пол. — длина тела при
ползании (без щупалец), дл. сок. — длина тела при сильном сокращении,,
шир. — ширина.
В рисунках радул зубы обозначены следующим образом: С — средин-
ный зуб поперечного ряда; цифры под остальными зубами означают поряд-
ковый номер от срединного зуба.
Остальные рисунки: А — аорта (общий ствол), АЖ — железы поло-
вого атриума, АП — половой атриум или атриум, Б — бич, Б Ж — бел-
ковая железа, В — вагина или влагалище, ВЖ — вагинальная (любовная)
железа, ВТ — веретеновидное тело, Г — гонада (гермафродитная железа),
ГЛ — глотка, ГП — гермафродитный проток, ЖС — желудочек сердца,.
ЗК — задняя кишка, К — кишка, КЛ — каудальная лопасть, КМ —
колумеллярный мускул, Л — лигула, МП — мочевой пузырь, МЧ —
мочеточник, ОП — отросток пениса (аппендикс), ОСП — отросток семя-
приемника, П — пенис, П1 — задний отдел пениса, П2 — передний отдел
пениса, ПВЖ — перивагинальные железы, ПЕЧ — печень, ПЖ — пени-
альная железа, ПЛ — пластинка стимулятора пениса, ПН — пневмостом,
ПО — половое отверстие, ПР — простата, ПРАП — придаток атриума,
ПЧ — почка, Р — ректум (прямая кишка), РАК — раковина, РГ — ре-
трактор глотки, РП — половой ретрактор, РТ — ретентор, РЩЛ — ре-
трактор левых щупалец, РЩП — ретрактор правых щупалец, С — семя-
провод, СЖ — слюнные железы, СК — слепая кишка, СОД — семяяйцевод
(спермовидукт), СП — семяприемник, СТ — стенка тела, СТМ — стиму-
лятор, СФ — сперматофор, ЭП — эпифаллус, Я — яйцевод.
Линейка при рисунках — масштаб: без цифр — 1 мм, с цифрами —
соответствующее значение в миллиметрах.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ
131
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ
1 (2). Мантия закрывает верхнюю сторону животного почти до самой по-
дошвы. Край мантии по всему периметру прирос к туловищу, не обра-
зуя свободной лопасти. При движении из-под мантии выступает только
голова со щупальцами и задний конец ноги. Пневмостом лежит сильно
антемедиально — расстояние между ним и передним концом тела
равно около Vg общей длины . ....................Philomycidae (с. 380).
2 (1). .Мантия закрывает лишь часть верхней стороны животного. В тех слу-
чаях, когда она закрывает всю верхнюю сторону или почти всю, то:
а) имеется крупный капюшон, т. е. свободная лопасть мантии, прикры-
вающая шею, а при сокращении и голову слизня; б) пневмостом лежит
на значительно большем удалении от переднего конца мантии.
3 (4). Задний конец ноги имеет над подошвой каудальную ямку, сверху
прикрытую трехугольной лопастью (рис. 538). Подошва не разделена
двумя продольными бороздами на три доли..............................
................................Arionidae, род Arion Fer. (с. 386).
4 (3). Задний конец ноги без каудальной ямки. Подошва двумя продоль-
ными бороздами четко разделена на три доли (рис. 2).
5 (12). Мантия с четкой дугообразной (подкововидной) бороздой, которая
своей выпуклой стороной обращена вперед. Иногда оба конца «подковы»
. сзади смыкаются и борозда приобретает овальную или яйцевидную
форму. . . . . •.
6 (9). Мантия очень крупная — у ползущего слизня длина ее около поло-
вины длины тела или больше, у сократившегося почти полностью закры-
вает верхнюю сторону животного.
7 (8). Небольшие слизни (дл. пол. 22 мм). Тело стройное, при движении
сзади булавовидно вздутое, при сокращении цилиндрическое (рис. 421).
Мантия почти полностью закрывает верхнюю сторону тела — в движе-
нии остаются открытыми лишь часть шеи, голова, а сзади кончик ноги.
Поверхность мантии покрыта крупными сосочками. Окраска тела белая.
Как у взрослых, так и у молодых слизней раковина снаружи не видна;
эмбриональная часть ее (нуклеус) без всяких следов спиральной закру-
ченное™. Слизни обитают в глубине больших пещер......................
. . . Trigonochlamydidae, род Troqlolestes Ljov. et Mat. (с. 306).
-8 (7). Взрослые слизни много крупнее. Длина мантии у ползущего живот-
ного составляет около 1/2, у сократившегося — около 2/3 общей длины.
Тело массивное, сзади суженное, при сокращении сильно вздутое, осо-
бенно в области мантии (рис. 448). Поверхность мантии зернистая,
без сосочков. Окраска от серовато-желтой до красноватой или черной,
нередко с темным рисунком. У молодых слизней снаружи всегда виден
спирально закрученный нуклеус раковины. G возрастом мантия его
обрастает и закрывает, но обычно остается щель между мантией и ки-
лем, через которую можно увидеть эту часть раковины. Слизни живут
вне пещер.......................................Parmacellidae (с. 332).
'9 (6). Мантия значительно меньших размеров — она закрывает не более
1/3 тела даже при сокращении животного.
40 (11). При движении тело червеобразно-цилиндрическое или булавовидное,
сзади плавно вздутое. У сократившихся слизней мантия лежит на се-
редине тела или даже в задней половине его. Расстояние между крайней
передней точкой тела и мантией больше или равно длине мантии.
У ползущих слизней мантия сдвинута еще сильнее назад. Капюшон
отсутствует или он узкий и слабо выражен. Подошва покрыта нерегу-
лярными прямыми поперечными бороздками...............................
..................................... Trigonochlamydidae (с. 306).
122
1. LIMACOIDEA
11 (10). При движении тело цилиндрически-веретеновидное. Как у сократив-
шихся, так и у ползущих слизней мантия лежит в передней половине1
тела и значительно ближе к голове, чем у предыдущей группы. Капюшон
более широкий. Подошва покрыта бороздами, имеющими характерную
V-образную форму (рис. 3, 7)..........................Milacidae (с. 354).
12 (5). Мантия без подкововидной борозды. Иногда от нее остаются лишь
следы в виде двух коротких и неглубоких бороздок, отходящих вперед
и назад от пневмостома, напоминающих контур летящей птицы, но ни-
когда не переходящих на левую сторону мантии (рис. 246).
13 (14). Рудимент мантии имеет вид небольшой слабо заметной круглой
«нашлепки», лежащей у самого заднего конца тела.........................
. . . Trigonochlamydidae, род Selenochlamys О. Bttg. (с. 306)..
14 (13). Значительно более крупная, овальная или яйцевидная мантия рас-
полагается в передней половине тела, и длине ее не меньше Ч3—
1/2 общей длины или закрывает всю верхнюю сторону животного..
15 (16). Тело очень стройное, узкое, червеобразно-цилиндрическое, в по-
перечном сечении круглое благодаря очень узкой подошве. Спина,
резко сужается к острому заднему концу, который при раздражении
или после фиксации сжимается с боков, приобретая форму вертикаль-
ного плавника. Киль хорошо развит и простирается на всю спину.
На мантии от пневмостома отходит вперед и назад по одной мелкой
бороздке, которые вместе напоминают контур летящей птицы................
.......................................... Boettgerillidae (с. 210).
16 (15). Тело более массивное, если стройное и узкое, то цилиндрическо-
веретеновидное и в поперечном сечении не круглое, так как подошва
более широкая. Спина, если и сужена к заднему концу, то более плавно
и слабо. Киль лишь у некоторых видов простирается на всю спину,
но выражен слабее.
17 (18). Морщины спины и боков короткие и плоские и разделены неглубо-
кими бороздками.1 На подошве прямые поперечные бороздки боковых
долей, переходя на срединную долю, V-образно изгибаются к заднему
концу тела (рис. 3, III, IV).................. . . Agriolimacidae (с. 123).
18 (17). Морщины спины и боков более длинные, выпуклые и разделены
глубокими бороздками. На подошве поперечные бороздки прямые
(рис. 3, II)........................................ Limacidae (с. 219).
1. Надсемейство LIMACOIDEA Rafinesque, 1815
Слизни с внутренней пластинчатой раковиной, нуклеус которой распо-
лагается у заднего края ее слева от продольной оси. Задний конец цефало-
подиума в различной степени заострен. Нередко имеется более или менее
четкий киль. Кроме перипедальной, имеется также супрапедальная бороздка
и оторочка. Пневмостом и мантийная щель чаще всего располагаются
постмедиально, реже антемедиально (Eumilacinae—Limacidae), но не так
сильно, как у большинства Arionoidea. Подошва всегда трехдольчатая,
т. е. с двумя продольными педальными бороздками.
Челюсть оксигнатная. Базальные пластинки краевых зубов длинные,
но узкие (рис. И, IV—XII).
Сердце лежит ближе к левому краю мантийного комплекса; ось его на-
клонена направо. Почка располагается позади или немного сбоку от
сердца, лишь слегка охватывая его (рис. 6, V).
1 Исключение — род Mesolimax Poll. (с. 204), у которого морщины выпуклые, а бо-
роздки глубокие.
I. AGRIOLIMACIDAE
123
Гениталии обнаруживают тенденцию к усложнению за счет формирова-
ния различных придаточных органов на пенисе или внутри его (стимулятор)
или же за счет сильной перестройки общего плана строения пениса (Eumi-
lax, Bielzia—Limacidae). Яйцевод, влагалище и атриум без придаточных
органов. При копуляции сперма передается в виде желеобразной массы.
Растительноядные полифаги.
В состав надсемейства входят Agriolimacidae, Boettgerillidae, и Limacidae.
Число видов около 100.
I. Семейство AGRIOLIMACIDAE Wagner, 1935
W a g n е г, 1935: 174; М a g n е, 1952 : 30 (Deroceratinae)', Н u d е с, 1972 : 84
IDerocerasinae).
Слизни небольшие или средних размеров, самые крупные взрослые
экземпляры после фиксации едва достигают 60 мм. Мантия лежит на перед-
ней половине тела; только у Megalopelte она закрывает почти всю верхнюю
сторону животного. Пневмостом располагается постмедиально или почти
медиально. Участок мантии вокруг пневмостома слегка приподнят и утол-
щен, образуя легкое вздутие или площадку, слегка отличающуюся по
окраске от соседних участков мантии. При жизни по мантии пробегают
волны сокращений в виде нежных концентрических бороздок. Боковые
доли подошвы неравномерно покрыты поперечными бороздками, которые,
переходя на срединную долю, V-образно изогнуты к заднему концу тела.
Половое отверстие располагается за правым губным щупальцем.
Раковина с эксцентрическим нуклеусом, только у Mesolimax он лежит
почти на продольной оси раковины (рис. 236).
Кишечник двухпетлевого типа, обычно почти не перекручен; кишка рас-
полагается позади сравнительно короткого зоба (рис. 15, II). Крупная
правая доля печени образует задний конец внутренностного мешка, левая
доля меньших размеров и лежит впереди.
Сердце лежит в левой передней четверти мантийного комплекса
(рис. 46); ось его почти под углом 45° наклонена направо, т. е. в сторону
продольной оси тела. Аорта длинная или средних размеров. Кровеносная
-сеть легкого располагается перед почкой и нередко также вдоль ее правой
стороны. Почка бобовидная или полулунная, лежит почти в центре мантий-
ного комплекса, и длинная ось ее располагается поперек продольной оси
тела. Снизу от почки отходит направо крупный языковидный лобус, кото-
рый простирается под мочеточник и заднюю кишку. Мочеточник берет на-
чало от заднего края почки; впереди он переходит в крупный и широкий
мочевой пузырь, который особой связкой соединен со стенкой почки. Колу-
меллярный мускул крепится позади мантийного комплекса, основание
полового ретрактора отходит от диафрагмы впереди почки и сердца.
Гениталии характерны отсутствием всяких придаточных органов на
женских половых путях. Атриум всегда небольшой. Мужские половые
пути без эпифаллуса.
Распространение. Почти вся Голарктика, однако большин-
ство родов и видов характерны для среднеземноморских и черноморских
стран.
Замечания. В данное семейство входят роды Deroceras, Krynickil-
lus, Lytopelte, Megalopelte, Mesolimax, Toxolimax. До настоящего времени
большинство авторов безоговорочно относило их к семейству Limacidae.
Только Вагнер (Wagner, 1935) считал возможным выделить род Agriolimax
(=Deroceras) в особое подсемейство Agriolimacinae. Гудец (Hudec, 1972)
ту же группу обозначил как Derocerasinae. Впервые Виктор (Wiktor, 1973)
124
I. AGRIOLIMACIDAE
обратил внимание, что не только Deroceras, но и Krynickillus, Lytopelte
и Mesolimax занимают особое место в общей системе Limacidae.
G нашей точки зрения, главным доводом в пользу выделения указанных
родов, а также Megalopelte и Toxolimax в особое семейство служит совер-
шенно иной план строения органов пищеварения, чем у Limacidae (с. 219).
Всем этим родам присущ кишечник двухпетлевого типа, который почти
не перекручен; кишка располагается позади зоба. Кроме того, правая доля
Рис. 46. Мантийный комплекс видов рода Deroceras.
Слева — вид сверху, справа — вид снизу.
печени, а не левая, как у большинства Limacidae, всегда находится сзади
и образует вершину внутренностного мешка.
Те же особенности топографии кишечника и печени свойственны и Mi-
lacidae (с. 354). Однако у последних зоб простирается несколько дальше,
а весь кишечник перекручен сильнее.
Некоторое сходство в форме пищеварительной системы у Milacidae
и Agriolimacidae можно рассматривать как параллелизм, так как по осталь-
ным системам органов, особенно гениталиям, и по внешнему виду оба се-
мейства резко отличаются друг от друга.
Из всех родов данного семейства наиболее слабо изучен Mesolimax:
авторы, как и их предшественники, располагали лишь единичными экземп-
лярами плохой сохранности или молодыми. Род Toxolimax во многом непо-
нятен (с. 208). Однако принадлежность обеих групп к этому семейству не
вызывает сомнения.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ AGRIOLIMACIDAE
1 (2). Мантия почти полностью закрывает верхнюю сторону слизня. К туло-
вищу она приросла лишь центральным участком, а по периферии сви-
сает широкой свободной складкой................4. Megalopelte Lindh.
1. DEROCERAS
125'
2 (1). Мантия располагается в передней половине тела (составляет 1/2—
1/3 общей длины животного). Сзади она плотно прирастает к туловищу
и лишь впереди образует свободную складку — капюшон.
3 (4). Каждая из боковых долей подошвы имеет по две дополнительных
продольных бороздки (рис. 3, IV). Колумеллярный мускул перед при-
креплением к верхней стенке тела раздвоен, и в развилке проходит
задняя кишка. Передний конец пениса вместе с прилегающим участком
семяпровода окружен перепончатой манжеткой (см. рис. 240)...............
.............................................5. Mesolimax Poll .
4 (3). Боковые доли подошвы без продольных бороздок. Задний конец колу-
меллярного мускула не раздвоен и располагается справа от задней
кишки. Пенис без манжетки.
5 (6). Киль четкий и крупный; при движении он занимает заднюю половину
спины, но при сокращении приобретает облик гребня и простирается
на всю спину.................................3. Lytopelte Koch et Heyn.
6 (5). Киль как легкая угловатость заднего конца тела слабо заметен и очень
короткий, особенно при движении животного.
7 (8). При жизни, а иногда и у фиксированных экземпляров на мантии видна
серповидная бороздка, которая впереди касается пневмостома, а сво-
бодным концом отогнута назад (см. рис. 208). Пенис цилиндрический
или булавовидный без стимулятора, но с различными внутренними
складками..........................................2. Krynickillus Kai
8 (7). Мантия без серповидной бороздки. Пенис мешковидный или удлинен-
ный и сложной формы, нередко разделен перетяжкой на передний
и задний отделы, снаружи с различными придаточными органами.
Внутри он имеет стимулятор и разные складки ... 1. Deroceras Raf..
1. Род DEROCERAS Rafinesquee 1820
R afinesque, 1820 : 10 (Limax, подрод Deroceras)', Gray, 1855 : 178 (Malino):
Malm, 1868 : 79 (Hydrolimax, non Haldemann, 1842); West er lund, 1894 : 163
(Arctolimax)', Hesse P., 1926 : 21 (A griolimax)', Pilsbry, 1948 : 532; Quic k,
1960 : 163 (A griolimax)', Wiktor, 1973 : 102; Дамянов, Лихарев, 1975 : 313.
Типовой вид—Limax gracilis Rafinesque, 1820 (=Limax laevis.
Muller, 1774); по монотипии.
Небольшие или средних размеров слизни (в движении длина тела не
более 60, а чаще до 45 мм), со стройным или умеренно коренастым телом,
которое при сокращении приобретает веретеновидную форму с наибольшей
шириной в области мантии. Киль как легкая угловатость заднего конца
заметен лишь при сокращении животного. Мантия обычно занимает около
1/3 длины тела или немного больше. Между мантийной щелью и средней
линией спины чаще всего 12—14, реже 10—16 рядов морщин. Пневмостом
лежит постмедиально.
Окраска одноцветная или с темным рисунком из пятнышек, которые
нередко напоминают брызги краски. Соприкасаясь, эти пятнышки могут
образовывать сетчатый рисунок, но они никогда не сливаются в правильные
цветные полосы.
Срединные и боковые' зубы радулы трехзубцовые; лишь 1—4 самых
внешних боковых зуба утрачивают эндокон и становятся двухзубцовыми,
образуя переход к шиповидным краевым зубам. Обычно число краевых
зубов в l1^—21/2 раза больше боковых.
Ректум у одних видов со слепой кишкой, у других — без нее.
Ретрактор правых щупалец не перекрещивается с пенисом или его ре-
трактором.
126
I. AGRIOLIMACIDAE
Гонада треугольная или удлиненно-языковидная, с коротким постепенно
расширяющимся протоком. Пенис сравнительно короткий, мешковидный
или удлиненный и сложный по форме. У взрослых слизней он нередко слегка
•спирально изогнут и глубокой односторонней перетяжкой разделен на
вздутый передний и удлиненный задний отделы. В переднем располага-
ется стимулятор, а в заднем — система мягких складок. Сюда открыва-
ется семяпровод. Внутренняя поверхность пениса покрыта тонкими и
правильными рядами рубчиков.
Задний отдел пениса имеет придаточные органы. У большинства видов
•здесь находится тонкостенная пениальная железа.1 Кроме того, нередко
имеется удлиненный выступ заднего отдела пениса — аппендикс, а также
другие округлые выступы или вздутия. Различные комбинации этих при-
даточных органов и их положение на пенисе имеют большое значение при
диагностике видов.
Половой ретрактор крепится к пенису 1—3 ветвями.
Яйцевод обычно короткий, трубчатый. Семяприемник узким протоком
впадает в основание пениса близ атриума.
Распространение. Большая часть северного полушария.
Особенно много видов обитает в Средиземноморской подобласти. Некоторые
развезены почти по всему свету.
Однолетние животные. Большинство видов размножаются осенью, после
чего взрослые особи гибнут. На зимовку остаются яйца, а нередко и мо-
лодь. Многие виды известны как вредители сельского хозяйства.
Замечания. 1. Большинство видов Зимрота мы смогли обстоя-
тельно изучить как благодаря знакомству с типами, так и исследовав бога-
тые сборы из «.locus typicus» и соседних районов, собранные за последние
*60 лет. Что касается остальных видов, особенно из Румынии, описанных
главным образом профессором Александру Гроссу и его коллегами, то
с ними дело обстоит несколько хуже. Правда, проф. Гроссу передал нам
в дар паратипы некоторых своих видов. Кроме того, один из нас (А. Вик-
тор), посетив Бухарест, имел кратковременную возможность познакомиться
с типами некоторых видов слизней Румынии.
Тем не менее четкого представления о многих новых румынских видах
мы не смогли составить, а некоторые из них определенно являются сино-
нимами более старых видов. Мы считаем, что в настоящее время необхо-
дима тщательная критическая сводка всей фауны слизней Румынии.2
Выполнение этой работы мы ждем от наших румынских коллег, которые,
как мы надеемся, учтут наши взгляды на некоторые виды слизней Румынии,
высказанные здесь.
А пока часть видов из рода Deroceras, описанных Зимротом, Гроссу и дру-
гими авторами, из-за неясного таксономического положения мы приводим
в разделе «Дополнение к роду Deroceras».
До последнего времени большинство авторов избегало делить данный
род на подроды. Главной причиной этому было то, что никто не располагал
материалом по большинству видов. Последняя по времени попытка раз-
деления на подроды была сделана Виктором (Wiktor, 1973). В настоящей
работе эта классификация получила дальнейшее развитие.
1 Некоторые авторы называют этот орган флагеллумом (бичом). Одако его функция
явно иная, чем у бича других семейств Stylommatophora, где он служит для образования
•сперматофоров. Поэтому название «пениальная железа», предложенное еще Зимротом
(Simroth, 1901), нам представляется более удачным.
2 Большинство видов слизней Румынии не включены в сводку Гроссу (Grossu, 1955),
так как описаны после выхода ее в свет.
1. DEROCERAS
127
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ DEROCERAS1
1 (8). Слепая кишка хорошо развита, длина ее в несколько раз больше
ширины (подрод Agriolimax Morch).
2 (5). На светлом фоне спины и мантии разбросаны темные пятнышки-
брызги, которые нередко складываются в сетевидный рисунок. Пе-
ниальная железа имеет вид группы бугорков или коротких выступов,,
тесно прижатых друг к другу, либо состоит из 1—3 длинных заострен-
ных выступов с боковыми ветвями или бугорками.
3 (4). Семяпровод впадает в пенис между двумя небольшими вздутиями
со стороны пищевода или зоба и немного сверху........................
..................................... 9. D. (A.) turcicum (Simr.).
4 (3). Семяпровод впадает в пенис со стороны правой стенки тела и немного
снизу; в этом месте на пенисе упомянутых вздутий нет.................
..................................8. D. (A.) reticulatum (Miill.).
5 (2). Тело светлое без темного рисунка из сравнительно крупных пятны-
шек, слагающихся в сеть. Иногда бывают микроскопические темные
точки. Пениальная железа имеет вид короткого прямого или изогнутого-
пальцевидного выступа или бугорка.
6 (7). Стимулятор крупный, конический, с острой вершиной.............
.........................................6. D. (A.) agreste (L.)..
7 (6). Стимулятор небольшой тупоконический или бугорковидный . . . .
..................................... 7. D. (A.) altaicum (Simr.).
8 (1). Слепой кишки нет или она имеет вид небольшого выступа либо кар-
мана задней кишки и длина ее не больше ширины.
9 (18). Стимулятор конический или полусферический, без известковой пла-
стинки (подрод Deroceras s. str.).
10 (И). Задний конец пениса с двумя длинными изогнутыми назад выступами
(рис. 65), между которыми находится пальцевидная пениальная железа
...........................................3. D. (D.) pollonerae (Simr.).
11 (10). Задний конец пениса либо с одним выступом (аппендиксом) или
с двумя, но они изогнуты вперед. Пениальная железа другой формы
и располагается иначе.
12 (13). Задний конец пениса с двумя выступами, придающими пенису вид.
молотка. Слизни одноцветные................2. D. (D.) sturanyi (Simr.).
13 (12). Задний конец пениса другой формы. Слизни с более или менее замет-
ным рисунком из темных пятен.
14 (17). На боках и спине имеются ряды белых пятнышек, которые иногда
сливаются в пойосы. Этот рисунок образован подкожными включениями
извести. Темный рисунок всегда слагается из крупных пятен. Всегда
имеется хорошо развитый пенис.
15 (16). Пениальная железа ланцетовидной формы, неветвистая и лежит на-
против аппендикса . .........................5. D. (D.) ilium (Simr.).
16 (15). Пениальная железа имеет вид многочисленных сосочков или отрост-
ков и лежит перед аппендиксом .... 4. D. (D.) osseticum (Simr.).
17 (14). Белого рисунка и подкожных включений извести нет. Темный рису-
нок образован очень мелкими пятнышками, которые особенно заметны
после фиксации, или животное одноцветно темное. Пенис нередко отсут-
ствует (афаллия) или сильно редуцирован (гемифаллия) ................
........................................1. D. (D.) laeve (Miill.).
18 (9). Стимулятор различной формы с известковой пластинкой или без нее,
но тогда он имеет форму лопатки или веера.
1 Виды, у которых таксономический ранг и положение в системе рода неясны, в дан-
ную таблицу не включены.
128
I. AGRIOLIMACIDAE
19 (26). Стимулятор с известковой пластинкой. Пениальной железы нет.
Мантия с крупным капюшоном, который при сокращении приподнят
над шеей, а иногда вывернут наружу. При движении задний конец
мантии смещен в заднюю половину тела (подрод Liolytopelte Simr.).
.20 (23). Пенис без цилиндрического аппендикса.
.21 (22). Задний конец пениса с темным пятном и двумя короткими бугорками
по бокам от семяпровода. Пластинка стимулятора имеет вид «листа
с черешком» или «ракушки»..............15. D. (L.) caucasicum (Simr.).
22 (21). Задний конец пениса без темного пятна и бугорков по бокам от семя-
провода. Пластинка стимулятора полукруглая и по прямому краю снаб-
жена зубчиками..................17. D. (L.) maldavicum (Gr. et Lupu).
.23 (20). Пенис имеет один или два цилиндрических роговидно изогнутых
аппендикса.
.24 (25). Тело почти всегда пятнистое, коричневое. Пластинка стимулятора
яйцевидная, а край ее без зубцов . . . 16. D. (L.) bureschi (Wagn.).
25 (24). Тело серовато-красное без пятен. Пластинка стимулятора круглая,
по всему краю зубчатая . . . 18. D. (L.) occidental (Gr. et Lupu).
26 (19). Стимулятор без известковой пластинки. Пениальная железа всегда
есть. Мантия со сравнительно коротким капюшоном, прилегающим
к шее и никогда не выворачивающимся наружу. При движении вся
мантия находится в передней половине тела (подрод Plathystimulus
Wiktor).
.27 (30). Пенис шаровидный, яйцевидный или грушевидный, всегда без четких
боковых вздутий или аппендикса.
.28 (29). Стимулятор светлый и имеет форму веера или листа с узким основа-
нием. Половой ретрактор двухветвистый. Задний конец пениса с тем-
ным пятном. Темные пятна на спине и мантии лежат редко, а иногда
они лишь единичные.....................10. D. (Р.) subagreste (Simr.).
.29 (28). Стимулятор темный с широким основанием и заканчивается серповид-
ным отростком (см. рис. 124). Половой ретрактор не ветвится. Задний
конец пениса без темного пятна. Темные пятна на спине и мантии лежат
очень густо, образуя плотную сеть, или так густо, что слизни кажутся
одноцветно-черными или темно-шоколадными.............................
..................................11. D. (Р.) bakurianum (Simr.).
30 (27). Пенис более вытянутый и неправильной формы из-за присутствия
на нем чётких боковых вздутий или аппендикса.
31 (32). Задний конец пениса переходит в длинный спирально закрученный
аппендикс (см. рис. 143)..................13. D. (Р.) praecox Wiktor.
32 (31). Задний конец пениса имеет различные выступы, в том числе и аппен-
дикс, но они не закручены в спираль.
33 (34). Стимулятор у основания очень широкий, но короткий (см. рис. 137).
Вид обитает в Крыму.......................12. D. (Р.) tauricum (Simr.).
-34 (33). Стимулятор у основания узкий, но длинный. Вид обитает в Карпатах
и далее на запад.......................14. D. (Р.) rodnae.Gr. et Lupu.
1. Подрод DEROCERAS s. str.
Pilsbry, 1948 : 534; Wiktor, 1973 : 106.
Тело с относительно крупной мантией, задний край которой при дви-
жении слизня немного смещен в заднюю половину тела. Капюшон занима-
мает меньше 1/2 длины мантии, при сокращении плотно прижат к шее.
Кожа тонкая, просвечивающая, с неясным рельефом из морщин и бороздок.
Слепая кишка отсутствует или имеет вид небольшого кармашка, длина
/которого не больше ширины.
1. DEROCERAS
129
Задний конец пениса с одним или двумя слепыми отростками — аппен-
диксами, а у некоторых видов, кроме того, с типичной пениальной железой.
Стимулятор полусферический или конический, с тупой или острой верши-
ной, без известковой пластинки.
Распространение. Почти все северное полушарие. Обитатели
очень влажных биотопов, нередко по берегам водоемов.
1. Deroceras (Deroceras) laeve (Miiller, 1774) (рис. 47 —54)
Muller, 1774 : 1 (Limax); Rafinesque, 1820, Ann. of Nature, 1 : 10 (Limax
gracilis)', Westerlund, 1876 : 97 (Limax hyperboreus); Simroth, 1885 : 222, 327,
T. 9, Fig. 16, 20—22 (Agriolimax laevis, part.); 1901 : 142, T. 14, Fig. 1—5 (Agriolimax);
Taylor, 1902—1907 : 121, T. 15, Fig. 5, 7 (Agriolimax)', Величковский, 1910 :
64 (Agriolimax pseudodioicus, syn. nov.); Цветков, 1940 : 388 (Agriolimax schulzi,
syn. nov.); Цветков, Матёкин, 1946:115 (Agriolimax schulzi)', Pilsbry,
1948: 539, Fig. 289—291; Лихарев, Раммельмейер, 1952:334, рис. 260
(Agriolimax)', Walden, 1957 : 349, text-fig. 1—4, T. 1, Fig. 23—27 (Limax hyperboreus
West.=L. laeuis Mull.); Quick, 1960: 172, T. 2, Fig. 19, text-figs. 10 D, I, 11 А, В, E,
F, N (Agriolimax); Лихарев, Старобогат о в, 1967 : 31; Hu dec, 1970 : 112;
Wiktor, 1973 : 106, Fig. 39, 40, 102—106, 150—158, 238, 270—273.
Locus typicus — «Fridrichsdal» — Фридриксдал у Копенгагена (Дания).
Типы неизвестны.
В следующих музеях находятся типы синонимов данного вида: 1) в есте-
ственноисторическом музее Стокгольма лектотип Limax hyperboreus We-
sterlund, 1876 (№ 1977, обозначен Вальденом); 2) в Венском естественно-
историческом музее 7 синтипов Agriolimax pseudodioicus Velitchkovsky,
1910; 3) в Зоологическом музее Московского университета типы Agriolimax
schulzi Tzvetkov, 1940.
Материал: из СССР — более 600 экз. из 159 пунктов; из других
стран — более 400 экз.
Животное очень беспокойное и подвижное. Тело цилиндрическое и строй-
ное, блестящее, мягкое, просвечивающее, при сокращении веретеновид-
ное с притупленными концами. Мантия относительно очень большая, при
жизни сзади закругленная, после фиксации нередко угловатая; длина ее,
как правило, не уступает длине спины или немного меньше. При ползании
голова и шея выступают вперед сильнее, чем у других видов того же рода.
Киль очень короткий и тупой и заметен лишь при ползании. При сокраще-
нии морщины довольно четкие и крупные — между мантийной щелью
и средней линией спины их около 10—14 рядов.
Окраска почти одноцветная, чаще всего от красновато-коричневой до
почти черной, очень редко более светлая до темно-кремовой. При фиксации
проступают темные пятнышки — меланофоры, 'которых особенно много на
мантии; на спине их меньше, и они располагаются по бороздкам. Щупальца
черные и толстые с крупными глазными вздутиями. Подошва относительно
узкая, светлее верхней стороны тела; срединная доля ее сильнее просвечи-
вает, чем боковые, и поэтому она кажется темнее.
Слизь жидкая, бесцветная.
Размеры: дл. пол. до 25, дл. мантии до 10 мм; дл. сок. 13—16,
редко до 22, дл. мантии 5.5—7.0 мм.
Раковина (рис. 48). Размеры: длина 3—4, ширина 1.5—2.0 мм.
Радула (рис. 49):
Полость тела и многие органы покрыты бурым мезентерием. Кишечник
без слепой кишки. Задняя кишка в начале вздута, но к анусу плавно су-
жается. Зоб сильно спирально изогнут и смещен вперед.
9 И. М. Лихарев, А. Й. Виктор
Рис. 47—54. Deroceras (Deroceras) laeve (Mull.) (из: Wiktor, 1973).
4 7 — внешний вид; 48 — раковина; 49 — зубы радулы; so—52 — зуфаллические гениталии; 53 — геми-
фаллические; 54 — афаллические. иыШ аа,' _
1. DEROCERAS
131
Гениталии (рис. 50—54). Очень характерной чертой этого вида
является присутствие даже в пределах одной популяции как особей с нор-
мально развитыми мужскими половыми путями (эуфалличные), так и экземп-
ляров, у которых пенис и семяпровод полностью или частично редуциро-
ваны (афалличные и гемифалличные особи). Такое положение иногда сильно
усложняет определение данного вида.
Небольшая темная шаровидная гонада обычно почти полностью закрыта
кишечником и печенью. Проток ее короткий, почти прямой и заканчивается
довольно крупным талоном. Белковая железа дисковидная, коричневая.
Женский канал семяяйцевода свинцово-серый, простата желтая.
У эуфалличных особей пенис крупный, более или менее цилиндриче-
ский, извитой или даже спирально закрученный. Он распадается на
.3—4 отдела: 1) узкий передний отдел, открывающийся в атриум; 2) очень
вздутый отдел, нередко образующий крупный выступ, в котором лежит
конический или полусферический стимулятор; 3) закрученный отдел, в зад-
ний конец которого впадает семяпровод, а немного впереди крепится по-
ловой ретрактор; 4) узкий слепой выступ нередко на конце с 2—3 бугор-
ками.
Редукция пениса проявляется в различной степени. У одних он стано-
вится коротким изогнутым цилиндром, соединяющим семяпровод с атри-
умом. У других — это небольшое вздутие на атриуме, в которое еще впа-
дает семяпровод и даже имеется слабый половой ретрактор. Чаще чем геми-
фалличные особи встречаются афалличные. У одних от пениса остается
небольшое слепое вздутие на атриуме, которое не связано с семяпроводом.
Последний же либо полностью отсутствует, либо сохраняется лишь его
задняя часть, слепо замкнутая на переднем конце.
Яйцевод цилиндрический, разной длины. У, афаллических особей он
обычно короче, но несколько вздут. Атриум цилиндрический, длинный
и тонкостенный.
Распространение. В СССР почти повсеместно. Общее рас-
пространение — Голарктика. Завезен во многие страны южного полушария.
Образ жизни. Среди всех слизней Палеарктики наиболее влаго-
любивый и холодостойкий. Обычными местообитаниями являются болота,
берега небольших водоемов, как природных, так и искусственных, где может
находиться как на почве, так и на растениях. Обитает также на влажных
лугах и в сырых лесах. Обычен в прибрежных речных биотопах тундры.
Довольно долго выдерживает пребывание под водой.
Встречается как в природных, так и в антропогенных биотопах. В обоих
случаях популяции отличаются сравнительно постоянной, но невысокой
численностью. А
Половое созревание наступает раньше, чем у других видов того же
рода. Откладка яиц идет в течение большей части теплого времени года,
а в таких странах, как Англия, практически круглый год. Яйца отклады-
ваются как по одному, так и небольшими кучками. Размеры яиц от 1.8 X
X 1.5 до 2.0 X 1.3 мм. Вылупление молоди происходит через 4—5 недель.
Молодые слизни имеют около 4 мм длины, бесцветные, просвечивающие.
Голова и щупальца светло-розовато-коричневые. Через несколько дней
пигментация усиливается, распространяясь с головы на все тело.
Зимуют как яйца, так и слизни разного возраста.
Замечания. 1. Пилсбри (Pilsbry, 1948) и Квик (Quick, 1960) от-
мечают, что многочисленные вскрытия экземпляров, собранных из разных
мест и в разные сезоны года, показали, что у афалличных особей в гонаде
всегда имеется зрелая сперма. Кроме того, хорошо развиты простата и семя-
приемник. Афалличные особи часто живут совместно с эуфалличными.
9*
132
I. AGRIOLIMACIDAE
Последнее подтверждается и на нашем материале, собранном из различ-
ных мест СССР. Никакой связи между географическим распространением
и сезонами года в появлении той или иной формы обнаружить не удалось.
По-видимому, оба автора нравы, считая, что афаллия у D. laeve аналогична
тому, что известно у Aegopinella nitens (Mich.) (Riedel, 1955).
2. В своей монографии о европейских слизнях Зимрот (Simroth, 1885)
дает описание двух форм, отнесенных им после некоторых колебаний
к D. laeve. Для одной из них особенно характерен молоткообразный пенис.
Тот же автор несколькими годами позднее (1894) описал для Балканского'
п-ова новый вид — Agriolimax sturanyi, отметив ту же особенность гени-
талий. При этом Зимрот, по-видимому, забыл, что уже имел дело со сходным
слизнем.
Ошибка Зимрота привела к тому, что до самого недавнего времени в лите-
ратуре под названием D. laeve смешивали два вида или же одной из форм
давали какое-либо новое название.
Так, Величковский (1910), обнаружив в своем материале обе формы,
название <daevei> сохранил за слизнем с молоткообразным пенисом
(=D. sturanyi Simr.), а слизню, для которого, как он заметил, характерно
присутствие афалличных и эуфалличных особей, он дал название «Agrio-
limax pseudodioicus» (=D. laeve Mull.).1 Цветков (1940) и Цветков и Матё-
кин (1946) описали для Казахстана новый вид — Agriolimax schulzi. Про-
верка их материала показала, что они имели дело с обычным D. laeve>
(Muller).
Впервые Виктор (Urbanski, Wiktor, 1968; Wiktor, 1973) четко показал
характерные особенности и различия между D. laeve (Miill.) и D. sturanyi
(Simroth) (см. ниже).
2. Deroceras (Deroceras) sturanyi (Simroth, 1894) (рис. 55—62)
Simroth, 1885 : 222, 327, T. 9, Fig. 17, 18 {Agriolimax laevis Muller, pars.); В a-
b о r, 1894 : 56, Fig. 3, 4 (4. laevis)’, Simroth, 1894a : 392, T. 19, Fig. 5, 6 {Agriolimax)’,
Величковский, 1910 : 64 {A. laevis); Rosenwald, 1926 : 238, Fig. 4, 6—8
(4. laevis); Grossu, Lupu, 1959a : 46, Fig. 6 {romanicus, syn. nov.); Walden.,.
1966 : 63; U r b a ns k i, Wiktor, 1968 : 65, Fig. 7; Wiktor J., Wiktor A.,.
1968 : 197; Gittenberger e. a., 1970 : 84, Fig. 129; H u d e c, 1970 : 109, Fig. 1;;
Wiktor, 1973 : 113, Fig. 47, 48, 159—165 , 239 , 274, 275; Д аминов, Лихарев,,.
1975 : 317, рис. 247.
Locus typicus — «Ohrid» — окрестности оз. Охрид (Балканский п-ов)..
Лектотип2 в Венском естественноисторическом музее (№ 19493b).
Материал: из СССР — 239 экз. из 31 пункта; из других стран —
более 600 экз. (в том числе и типы).
Тело менее стройное, чем у D. laeve, с выпуклой спиной и резко сужен-
ным задним концом. Мантия довольно крупная, занимает около 1/2 длины
тела; при движении задний конец ее располагается в задней половине тела.
Кожа очень тонкая и просвечивающая, со слабым рельефом; после фикса-
ции кожа теряет прозрачность, рельеф становится незаметным, но зато на
боках появляются белые тонкие нитевидные продольные полоски.
Окраска одноцветная без рисунка из пятен и полос, довольно изменчи-
вая в отношении оттенков. Чаще всего грязновато-кремовая или серовато-
коричневая. Иногда встречаются целые популяции коричневого или даже
шоколадного цвета и наряду с совсем светлыми отдельные меланистические
особи с почти черной спиной. Мантия и середина спины всегда окрашены
несколько темнее, чем бока и подошва. Слизь водянистая и бесцветная.
1 Название pseudodioicus означает «мнимораздельнополый».
? Лектотип обозначен этикеткой за подписью Г. Валдена (Н. Walden). Публикуется
впервые.
1. DEROCERAS
133
Размеры: дл. пол. до 38, дл. сок. до 20 мм.
Раковина — см. рис. 56.
Радула (рис. 57):
с , 12—146 , 1-26
3 + 3 +2
28—41к
1
X 94—102.
Кишечник (рис. 58). Слепая кишка либо полностью отсутствует, либо
представлена небольшим «кармашком». Чаще всего внутренние органы
покрыты тонким черноватым мезентерием.
Рис. 55—62. Deroceras (Deroceras) sturanyi (Simr.) (из: Wiktor, 1973).
55 — внешний вид; 56 — раковина; 57 — зубы радулы; 58 — кишечник; 59, 60 — гениталии; 61 —
вскрытый пенис; 62 — задний конец внутренностного мешка.
Гениталии (рис. 59—62). Небольшая темная гонада лежит вблизи
задней кишки или под нею. Пенис очень характерной формы. Впереди он
несколько вздут, а задний конец с двумя четкими боковыми выступами,
134
I. AGRIOLIMACIDAE
нередко отогнутыми вперед. Оба выступа отличаются друг от друга по форме
и размерам, так как один из них обычно к концу несколько сужен и длиннее
другого, что придает всему пенису молоткообразную форму. Иногда более
длинный выступ по внутреннему краю расчленен на короткие лопасти,
что указывает на его гомологию с пениальной железой. Меньший выступ
подтянут к стенке пениса нежными мускульными волокнами. Семяпровод
и полов,ой ретрактор крепятся к пенису с правой (внешней) стороны между
упомянутыми выступами. Внутри переднего отдела пениса находится круп-
ный конический стимулятор с тупой вершиной. Семяприемник с удлиненным
резервуаром и четким тонким протоком и по длине немного больше пениса.
Распространение. В СССР обнаружен в нескольких местах
Московской (Дединово, Белые Колодези), Калининской, Воронежской
(Белогорье, Подгоренского р-на), Алма-Атинской (сады, теплицы и поля
ряда предгорных совхозов — Увалиева, 1979) областей, а также в ботани-
ческих садах и на пригородных полях и огородах Ленинграда, Челябинска,
Ростова-на-Дону, Ленинабада и Хорога (Иззатуллаев, 1972, 19756; Мухит-
динов, Рахметов, 1972). В природных биотопах найден в буковых лесах
близ Ужгорода и на пойменных лугах верхней Волги.
Общее распространение этого вида изучено пока недостаточно. Тому
причиной, во-первых, то, что его длительное время, вслед за Зимротом
(Simroth, 1885), принимали за особую сезонную форму D. laeve (Muller)
(см. с. 132 наст, работы). Во-вторых, этот слизень благодаря своей синан-
тропии легко поддается завозу и приживается на новых местах, по-видимому,
не только в культурных, но и в более или менее природных биотопах.
В настоящее время большинство специалистов его хорошо различают.
Он установлен не только для Югославии, откуда был описан, но и в Бол-
гарии, Румынии (Lupu, 1971а), Венгрии, Чехословакии, Австрии, ФРГ,
Голландии, Польше и на о-ве Готланд. Одна находка (Wiktor, 1971) сделана
в Турции (к югу от Трабзона).
Образ жизни. Обитает на открытых, умеренно влажных и очень
влажных местах, особенно часто в различных антропогенных биотопах:
вторичные кустарники, сады, парки, огороды, пустоши, а также на лугах
и в^придорожных канавах, в теплицах, парниках и погребах.
Питается зелеными частями растений, плодами и овощами. Половозре-
лость наступает к середине лета или осенью. Зимуют в основном яйца, но
находки весной отдельных взрослых особей указывают на то, что и они
переживают зиму.
Замечания. 1. Внешне, особенно темноокрашенные особи данного
вида, обнаруживают некоторое сходство с D. laeve (Muller). Оно проявля-
ется в том, что у обоих видов мантия относительно длинная; при ползании
задний конец ее располагается в задней половине тела. Однако и по внеш-
нему облику оба вида имеют существенные различия: у D. laeve при фикса-
ции проступают темные пятнышки, которых не бывает у D. sturanyi.
Гораздо более четко оба вида различаются по гениталиям. У D. sturanyi
пенис благодаря двум характерным боковым выступам на заднем конце
имеет молоткообразную форму; у эуфалличных особей D. laeve пенис более
или менее цилиндрический, извитый или слегка спирально закручен; при
афаллии пенис полностью или частично редуцирован, чего никогда не бы-
вает у D. sturanyi.
2. Светлые экземпляры D. sturanyi, на первый взгляд, можно принять
за D. agreste (L.). Однако и во внешнем облике есть существенные различия
между названными видами. Если у D. sturanyi кожа очень тонкая, просве-
чивающая и со слабым рельефом, то у D. agreste она более толстая, не про-
свечивающая и с четким рельефом. Кроме того, у последнего вида мантия
относительно короче, чем у первого.
1. DEROCERAS
135
По внутреннему строению оба слизня различаются еще более четко.
У D. agreste (как и у всех видов подрода Agriolimax) всегда имеется длинная
слепая кишка, а пенис мешковидный или цилиндрический с одним неболь-
Рис. 63—66. Deroceras (Deroceras) pollonerae (Simr.) (из: Walden, 1960).
63 — внешний вид, 64 — кишечник; 65 — гениталии; 66 — дистальная часть гениталий.
шим выступом (пениальной железой) на заднем конце; у D. sturanyi слепой
кишки нет или же она имеет вид небольшого карманчика, пенис же имеет
характерный облик «молотка».
3. Deroceras (Deroceras) pollonerae (Simroth, 1889) (рис. 63—66)
Simroth, 1889 : 179, Fig. a, b, c (Agriolimax)-, P ol loner a, 1891 : 3, Fig. 2
(4. caruanae)-, Hoffmann, 1941 : 254, Fig. 9—12 (A. dubius)- Quick, 1960 : 175,
136
I. AGRIOLIMACIDAE
text-figs. 10 H, 12 A—N, Pl. 2, Fig. 17 (A. caruanae}', Walden, 1960:30, text-fig.9,
Pl. 1, Fig. 1—15 {caruanae}', A 11 e n a, 1962 : 48; Reygrobellet, 1963 : 399,
Figs, {meridionale); Che vallier, 1970 : 387 {caruanae}', Giusti, 1973 : 213, Fig.21
{pollonerai}', Дамянов, Лихарев, 1975 : 318, рис. 248 {caruanae).
Locus typicus — «Palermo» — Палермо, Сицилия (Италия).
Типы неизвестны.
Материал: из СССР — нет; из других стран — 60 экз.
Тело более коренастое, чем у D. laeve и D. sturanyi. Окраска очень
варьирует: от почти черной и темно-серой, до красновато-коричневой
и серовато-бурой, одноцветная или с рисунком из темных пятнышек. По-
дошва от серой до бурой. Слизь жидкая и бесцветная.
Размеры: дл. пол. до 30, дл. сок. до 20 мм; мантия занимает
около х/2 длины тела.
Раковина почти прямоугольная.
Радула типичная для рода. Боковых зубов 12—16, трехзубцовые; крае-
вых зубов 48—63, шиповидные; число рядов до 120.
Слепая кишка отсутствует (Quick, 1960) или представлена небольшим
кармашком (Walden, 1960) (рис. 64).
Гениталии (рис. 65—66). Гонада небольшая гроздевидная, тем-
ная. Пенис четко разделен перетяжкой на передний и задний отделы. Зад-
ний конец его несет два крючковидно согнутых друг к другу выступа, между
которыми находится крупная пениальная железа, имеющая короткий сте-
бель и 4—5 пальцевидных выступа. Рядом с железой впадает семяпровод
и крепится половой ретрактор. Внутри переднего отдела пениса находится
короткий конусовидно-шаровидный стимулятор. Резервуар семяприемника
шаровидный или слегка удлиненный.
Распространение. В природных условиях этот вид был най-
ден в Сицилии, на Мальте, островах Тосканского архипелага, в юго-восточ-
ной Франции и в Болгарии (Пирин). Завезен на Британские и Фарерские
о-ва, в северную Францию, Бельгию, Данию, Швецию, Финляндию и в
Калифорнию.
Образ жизни. В странах, куда данный вид был завезен, он оби-
тает в огородах, на полях и свалках. В северной Европе чаще всего его на-
ходят в теплицах, где нередко он наносит существенный вред.
Замечания. 1. Джюсти (Giusti, 1973), собрав большой новый
материал по данному виду, после ревизии некоторых старых сборов довольно
убедительно показал, что: a) Agriolimax caruanae Pollonera, 1891
и A. dubius Hoffmann, 1941 представляют собой младшие синонимы
D. pollonerae (Simroth, 1889); б) данному виду наиболее близок D. panormi-
tanum (Lessona et Pollonera, 1882).
2. Указание на присутствие в Румынии D. caruanae (Poll.) (см.: Grossu,
Lupu, 1965), как показала Лупу (Lupu, 1969), относятся к другому виду —
D. padisii Lupu, 1969 (см. ниже).
4.1DerocerasI(Deroceras) osseticum (Simroth, 1901), comb. nov.
(рис. 67—76)
Simroth, 1901 : 161, T. 14, Fig. 22—26 {Agriolimax}', 158, T. 15, Fig. 20—22
(A. subagreste minutus, syn. nov., pars.); 1 Лихарев, Раммельмейер, 1952 :
336, рис. 265 {Agriolimax).
Locus typicus «.Tkwibuli» — Ткибули (западная Грузия). Лектотип
(обозначен нами) находится в ЗИН АН СССР с этикеткой: «окрестности
Тквибули, Кутаисского уезда, копи Новосельского (Кавказ), май 1895 г.,
1 См. с. 162 — замечания к D. bakurianum.
Рис. 67—76. Deroceras (Deroceras) osseticum (Simr.). Окр. Батуми.
7 — внешний вид; 8—9 XI 1958; 68 — раковина; 69 — зубы радулы; 70 — кишечник; 71_________ретрак-
торы; 72 — гениталии, 20 IV 1957; 78—75 — дистальные части гениталий; 76 — вскрытый пенис
138
I. AGRIOLIMACIDAE
колл. Кисляков, определил Зимрот». Этот экземпляр изображен у Зимрота
на табл. 14, рис. 22. Он вскрыт. Длина тела 16, мантии — 7.5 мм. Характер-
ный рисунок тела в настоящее время почти исчез. Местонахождение второго
синтипа неизвестно.
Материал: из СССР — 141 экз. (в том числе и лектотип) из
46 пунктов.
Тело при сокращении веретеновидное. Мантия закрывает около 1/3 те-
ла; спереди и сзади она закругленная, наиболее широкая у заднего края.
Морщины плоские, слабо заметные. Между мантийной щелью и средней ли-
нией спины около 12—13 рядов морщин. Киль слабо развит и тупой.
Основная окраска серовато-желтая, бледно-охристая или грязно-кремо-
вая, с характерным темным рисунком. Он слагается из довольно крупных
темно-коричневых или почти черных пятен неопределенной формы и разной
величины, иногда сливающихся друг с другом. В основном темные пятна
располагаются на мантии и спине, оставляя свободными среднюю часть
спины и мантии. Нижняя часть боков и край подошвы всегда светлые, без
темных пятен. Одно особенно крупное пятно располагается над пневмостомом.
Кроме темного рисунка имеется характерный рисунок из белых пятен,
образованный подкожными включениями извести. Этих включений особенно
много на боках животного, где они образуют по одному ряду светлых пятен,
нередко сливающихся в полосы, которые чаще всего располагаются ниже
темного рисунка, но иногда (особенно в задней трети тела) частично перекрыты
темным пигментом. Аналогичная белая полоса или ряд пятен находится
на середине спины, а отдельные белые пятнышки встречаются и на мантии.
Голова и шея слегка коричневые. Подошва кремовая, без рисунка.
Размеры: дл. пол. до 40, ширина до 5, дл. мантии до 15 мм; дл. сок,
до 30 мм.
Раковина (рис. 68) широкоовальная; длина 4.2—4.8, шйрина 2.7—3.5 мм.
Радула (рис. 69) очень своеобразная- Срединный и боковые зубы с 3 зуб-
цами и удлиненной базальной пластинкой, самый внешний боковой зуб
с 2 зубцами. Краевых зубов очень много, и они двух типов: первые (т. е. ле-
жащие медиально) 70—80 зубов шиповидные; у следующих 60—70 зубов
на внешней стороне основного зубца появляются мелкие дополнительные
зубцы, число которых нарастает до 7. Такая картина наблюдалась как у эк-
земпляра из Батумского ботанического сада, так и у экземпляра с Сурам-
ского перевала. В итоге радула выглядит следующим образом:
с 166 б 144к ,
Гз + 3 1—7 X110-
Задняя кишка очень длинная, широкая и прямая, простирается назад
почти до вершины внутренностного мешка. Настоящей слепой кишки нет,
но на месте ее виден небольшой карманообразный выступ задней кишки
(рис. 70).
Гениталии (рис. 72—76). Гонада серая или светло-коричневая,
образована несколькими крупными шаровидными фолликулами, лежит
у заднего конца зоба и полностью закрыта лопастями печени и кишечником..
Белковая железа крупная, удлиненно-языковидная, коричневая. Семяяйце-
вод очень длинный. Пенис крупный, около 1/3 длины тела. Он состоит из
двух основных отделов: переднего, или собственно пениса, мешковидного
или яйцевидного и заднего, переходящего в крючковидно изогнутый, ци-
линдрический аппендикс. Оба колена аппендикса соединены перепонкой.
Собственно пенис имеет сбоку перисторазветвленную пениальную железу
(чаще всего 2—3 ветви). Нередко это целый ряд коротких или длинных
выступов, отходящих от боковой стенки переднего отдела пениса. У осно-
вания пениальной железы в пенис впадает короткий и тонкий семяпровод,
1. DEROCERAS
13ff
а с противоположной стороны к пенису подходит двух- или трехветвистый
ретрактор: одна ветвь крепится к аппендиксу у его перегиба, другая — к ос-
нованию аппендикса, а третья — к переднему отделу пениса у основания
пениальной железы. Задний конец полового ретрактора крепится к переднему
краю диафрагмы.
Внутри переднего отдела пениса располагается тупой стимулятор, а
внутри аппендикса — длинный продольный валик.
Яйцевод очень короткий и толстый. Семяприемник небольшой, с оваль-
ным резервуаром, плавно переходящим в короткий проток. Атриум слабо
выражен.
Распространение. Западная часть Северного Кавказа и запад-
ное Закавказье: на севере от горы Фишт до Теберды, на юге от Батуми до
Сурамского хребта.
Образ жизни. Обитает в широколиственных и смешанных лесах
(елово-буковых и пихтово-буковых), а также на субальпийских лугах.
Укрывается под камнями, комками почвы и в траве.
В районе Батуми копуляция и откладка яиц отмечена в апреле и мае. Зи-
муют молодь и полувзрослые особи.
Замечания. Данный вид обладает рядом очень характерных черт,
не встречающихся или редко встречающихся у других видов того же рода.
К их числу относятся: а) темный рисунок, образованный сравнительно круп-
ными пятнами, расположенными по верхней стороне таким образом, что
середина спины и нижняя часть боков остаются свободными от них; б) свое-
образный «белый» рисунок, образованный подкожными включениями из-
вести; в) необычно большое число краевых зубов радулы, из которых около
половины имеют характерные дополнительные зубчики вдоль внешней сто-
роны основного зубца; г) размеры, положение и форма задней кишки; д) ха-
рактерная форма пениса и его придатков — пениальной железы и аппендикса.
Окраска, рисунок и некоторые особенности строения пениса данного вида
обнаруживают сходство лишь с D. ilium (Simr.) (см. ниже).
5. Deroceras (Deroceras) ilium (Simroth, 1901), comb. nov. (рис. 77—83)
Simroth, 1901 : 159, T. 15, Fig. 29—32 (Agriolimax)', Лихарев, Ранне ль-
мейер, 1952 : 337, рис. 267 (Agriolimax).
Locus typicus — «Berg II in der Nahe von Wladikawkas» — гора Ил (совре-
менное название Сапицкая будка) около Орджоникидзе. Лектотип (обозна-
чен нами) и 5 паралектотипов в ЗИН АН СССР.1
Материал: из СССР — 86 экз. (вместе с типами) из 15 пунктов.
При сокращении тело веретеновидное с максимальной шириной в области
мантии, задний конец плавно снижается или же резко вертикально срезан.
Мантия закрывает 1/4—1/8 длины тела. Между мантийной щелью и средней
линией спины 12—13 рядов плоских морщин.
При жизни фоновая окраска серовато-розовая; после фиксации серовато-
желтая или грязно-охристая. Рисунок образован сравнительно крупными
темно- или красновато-коричневыми, иногда почти пурпуровыми пятныш-
ками,2 которые образуют сгущения на мантии и на спине по сторонам от
светлой срединной полоски. Кроме этой полоски, темный пигмент отсут-
ствует на боках и подошве. Голова и шея (иногда даже под капюшоном)
1 Как показала проверка, седьмой синтип — молодой Eumilax intermittens (О. Bttg.).
2 По-видимому, пигмент легко разрушается или экстрагируется при хранении
в спирте./Этим и объясняется, почему у одного из синтипов Зимрот не показал пятнышек
(табл. 15, рис. 30). Однако когда мы тщательно рассмотрели соответствующий экземпляр,
то все же обнаружили следы темного рисунка.
Рис. 77—83. Deroceras (Deroceras) ilium (Simr.).
11 — зубы радулы; 78 — кишечник; 79 — дистальная часть гениталий; 80 — они же с другой стороны;
— фрагмент пениса; 82—.стимулятор; 83— пенис молодого'экземпляра (79—82— лектотип,
Орджоникидзе, гора Ил, 16 IV 1886; 83 — Сев. Осетия, перевал Кармадон, 5 IX 1962).
1. DEROCERAS
141
покрыты более или менее интенсивным серым налетом, образованным микро-
скопическими темными точками.
Кроме темного рисунка имеется и белый рисунок, образованный под-
кожными известковыми включениями в виде пятнышек на боках и средин-
ной полоски спины.
Размеры: дл. пол. до 40, дл. сок. до 25 мм.
Раковина округло-прямоугольная. У одного экземпляра с дл. пол. 34 мм
ее размеры составляли 4x2.5 мм.
Радула (рис. 77) типичная для рода; боковых зубов 14—17, краевых —
36—42. Число рядов около 100. Пример: 1 экз. с горы Сапицкая будка
около Орджоникидзе, радулярная формула:
с 136 б 42к
3" + ~ + ~2 + “Г" Х 9' •!
Кишечник (рис. 78). Короткая карманообразная слепая кишка может
иметься, но может и отсутствовать. В отличие от D. osseticum задняя кишка
располагается косо к оси тела.
Гениталии (рис. 79—83). Небольшая черная гонада лежит за зобом
и либо совсем открыта, либо лишь слегка закрыта лопастями печени. От
нее отходит вперед довольно толстый гермафродитный проток, который,
образовав V-образный изгиб, впадает в семяяйцевод. Пенис имеет очень
сложную форму. Передний отдел его очень сильно вздут и перетяжкой
отделен от заднего отдела. Последний заканчивается аппендиксом, верхушка
которого сильно утоньшена и спирально изогнута. На границе между обоими
отделами, но позади перетяжки в пенис открывается своеобразная пениаль-
ная железа. Она имеет ланцетовидную форму, тонкие стенки, а по внутрен-
нему краю (т. е. прилегающему к пенису) волнистые очертания. Внутри
переднего отдела пениса имеется крупный конический стимулятор, заканчи-
вающийся узкой вершиной, которая обычно располагается внутри пениаль-
ной железы. В заднем отделе пениса имеется сложная система крупных
складок. Семяпровод впадает в пенис на границе между обоими его отделами
или у основания пениальной железы.
Половой ретрактор сзади крепится к диафрагме в области сердца; впе-
реди он распадается на 3 ветви, очень короткие и плотно прижатые к пенису:
одна ветвь крепится к середине аппендикса, другая к его основанию,
а третья на границе обоих отделов пениса.
Распространение. Все находки этого вида сделаны в верховьях
рек Терек,. Подкумок и восточных притоков Кубани (река Большой Зеленчук
у Архыза) и в верховьях р. Кодор (Абхазия). Следовательно, за исключе-
нием последнего местонахождения, остальные приурочены к горнолесной
зоне центральной и частично западной части Северного Кавказа.
Образ жизни. Обитает в горных широколиственных и смешанных
(елово-буковых и пихтово-буковых) лесах, по тенистым ущельям, нередко
вблизи ручьев, на траве, под камнями и в подстилке. Поднимается в горы
до высот 1700 м над ур. моря.
Замечания. 1. Данный вид несомненно близок к D. osseticum
(Simr.). Внешний облик обоих слизней очень сходен, только у D. osseticum
голова и шея либо совсем светлые, или же лишь слегка пигментированы,
в то время как у D. Шит они всегда покрыты довольно интенсивным серым
налетом. Во внутреннем строении различий больше и они выражены более
четко: а) у D. Шит задняя кишка лежит наклонно к оси тела, у D. osseticum
она вытянута вдоль тела; б) у D. Шит краевых зубов радулы около 40, т. е.
только втрое больше, чем боковых, и они без дополнительных зубчиков,
у D. osseticum краевых зубов около 145, т. е. в 8 раз больше, чем боковых,
14г
I. AGRIOLIMACIDAE
и они имеют дополнительные зубчики по внешнему краю; в) у D. Шит
пениальная железа ланцетовидной формы, не ветвящаяся и внутри нее рас-
полагается заостренная часть стимулятора, у D. osseticum пениальная
железа ветвится и стимулятор в нее не заходит. От остальных кавказских
видов Deroceras оба слизня отличаются характером темного рисунка —
сравнительно крупные пятнышки.
2. Учитывая сравнительную бедность материала по данному виду, пока
еще трудно определить его ареал и проследить изменчивость этого слизня.
Не исключено, что со временем выяснится, что D. Шит и D. osseticum
являются подвидами одного вида.
3. Гроссу и Лупу (Grossu, Lupu, 1959) указывали на присутствие этого
вида в Румынии. Позднее выяснилось, что они имели дело с Deroceras (Lio-
lytopelte) bureschi (H. Wagner) (с. 174).
gj gg 2. Подрод AGRIOLIMAX Morch, 1865
Mo rch, 1865, J. de Conchy!., 13 : 378; Wes terlund, 1894 : 163 (Chorolimaxy,
W ikt о r, 1973 : 120.
Типовой вид — Limax agrestis Linnaeus, 1758; по последующему
обозначению — Malm, 1868.
Мантия занимает не более х/3 общей длины, и при движении задний конец
ее не заходит в заднюю половину тела. Капюшон как у Deroceras s. str.
Кожа сравнительно толстая и непрозрачная, с довольно четким рельефом
из морщин и бороздок.
Слепая кишка всегда хорошо развита, и длина ее в 2 раза и более прево-
сходит ширину.
Пенис различной формы, но всегда сравнительно короткий, мешковидный,
булавовидный или овальный. Пениальная железа имеет вид одного или не-
скольких слепых отростков, снабженных боковыми веточками или пузыревид-
ными вздутиями. Аппендикса нет. Стимулятор конический, с острым кон-
цом, без известковой пластинки.
Распространение. Почти вся Палеарктика. Главным образом
мезофильные виды, живущие на открытых биотопах, но некоторые и в лесах.
6. Deroceras (Agriolimax) agreste (Linnaeus, 1758) (цв. вкл., рис. /V;
рис. И, IV—VI; рис. 84—91)
Linnaeus, 1758 : 652 (Limax); Schrenk, 1848 : 143 (Limax pallidus); К a-
leniczenko, 1851a : 224, T. 5, Fig. 3 (Krynickillus minutus); Koch, Heyne-
m a n n, 1874 : 153, T. 6, Fig. 6 (Limax Fedtschenkoni, nom. err.); Simroth, 1886a :
29, T. 1, Fig. 15, 16 (Agriolimax fedtschenkoi, nom. emend.); 1901 : 149, T. 14, Fig. 6 (Agrio-
limax agrestis var. fedtschenkoi); 155, text-fig. 8, 9, T. 16, Fig. 18—30 (Agriolimax transcau-
casicus var. coeciger, syn. nov.); 1910 : 523 (A. transcaucasicus); 1912a : 39 (A. transcaucasi-
cus); Luther, 1915 : 1, Fig. 5b (Agriolimax); Gerhardt, 1933 : 406 (Agriolimax);
Акрамовский, 1949 : 151 (A. transcaucasicus, pars.); Лихарев, Раммель-
мейер, 1952 : 332, рис. 258 (Agriolimax); Quick, 1960 : 170, text-figs. 10 B, G, 11
D, I, К, M (Agriolimax); Wiktor, 1973 : 121, Fig. 45, 46, 102—104, 107, 190—197,
241, 278, 279; Скляр, 19756:852, рис. 1—4 (transcaucasicus); Lupu, 1976:9,
Fig. 20—27 (transcaucasicus Simr. syn. agreste L.).
Terra typica — Швеция. Типы неизвестны.
Материал: из СССР — более 600 экз. из 219 пунктов; из других
стран — более 600 экз.
Тело более стройное, чем у D. reticulatum, но массивнее, чем у D. laeve.
Спина выпуклая, в поперечном сечении полукруглая. Мантия занимает
около 1/3 длины тела.
Окраска от почти белой до кремовой с легким коричневым оттенком, без
томного рисунка. Мантия и спина немного темнее боков. Подошва тожекре-
Рис. 84—91. Deroceras (Agriolimax) agreste (L.).
^4 внешний вид; 85 — раковина; 8в — зубы радулы; 87 — кишечник; 88 — слепая кишка другого
экземпляра, 89 дистальная часть гениталий; 90 — вскрытый Чтение; 91 — гениталии молодого слизня
(54, 85. 88—-91 — из: Wiktor, 1973). • --
144
I. AGRIOLIMACIDAE
мовая, ее края немного темнее середины. Слизь бесцветная, при раздражении
слизень может выделять молочно-белую и мутную слизь.
Размеры: дл. пол. до 40, дл. сок. до 35 мм.
Раковина (рис. 85).
Радула (рис. 11, IV—VI; 86). Боковых зубов около 16, краевых — около<
20; число рядов 90—100.
Гениталии (рис. 89—91). Гонада у молодых более или менее тре-
угольная, светло-кофейного цвета, у взрослых она небольшая овальная или
лопастевидная, темно-шоколадного цвета. В отличие от D. reticulatum она
меньших размеров и никогда не располагается на вершине внутренностного
мешка, а лежит приблизительно на границе между задней и средней третью
тела, вблизи слепой кишки или же под нею. У взрослых особей, т. е. с не-
большой гонадой и крупной белковой железой, пенис с перетяжкой посере-
дине. У молодых экземпляров эта перетяжка выражена слабо или совсем
отсутствует, и пенис имеет мешковидную или почти цилиндрическую форму.
Передняя часть пениса заметно вздутая, так как внутри находится крупный
конический стимулятор с рубчатой поверхностью. Пениальная железа имеет
вид небольшого пальцевидного или крючковидного, не ветвящегося придатка,,
лежащего у заднего конца пениса. Половой ретрактор крепится к пенису
приблизительно у границы его задней и средней трети. Здесь'же впадает семя-
провод. Это место нередко заметно окрашено темным пигментом. Овальный
резервуар семяприемника плавно переходит в короткий проток. Атриум
довольно длинный и цилиндрический.
Распространение.1 В СССР — почти повсеместно: на севере
от Кольского п-ова и Болыпеземельской тундры, на юг включая Кавказ и
Крым; Средняя Азия, Алтай, Саяны, Амурская область, Приморский край,,
южная часть Камчатки, Сахалин, Курильские о-ва.
Вне СССР достоверно известен из Польши (почти вся страна), Финлян-
дии, со Скандинавского п-ова, из Дании, с Британских о-вов, с о-ва Ислан-
дия, а также из Болгарии. По-видимому, широко распространен в средней
Европе. Сообщения о Wo находках в западных и южных областях Европы,
а также в северной Африке требуют тщательной проверки, в йервую очередь,
с позиций современных^взглядов на различия между D. reticulatum (Mull.)’
и D. agreste (L.).
Мы вполне присоединяемся к взглядам Валдена (Walden, 1966) о том,
что все сведения об обитании D. agreste вне Евразии относятся к D. reticu-
latum.
Образ жизни. Обитает на открытых местах, таких как луга, бо-
лота, придорожные канавы; реже в огородах и садах. Встречается на лесных
опушках и в зарослях ольхи, но никогда не углубляется внутрь леса. Укры-
вается под кусками древесины, камнями и комками почвы и в ее трещинах.
Более обычен на равнинах и низменностях, реже встречается в горах и
исключительно на открытых местах, особенно по берегам водоемов.
На северо-западе европейской части СССР откладка яиц происходит
обычно осенью и зимуют лишь яйца. В более южных областях, кроме яиц,
зимуют полувзрослые слизни, которые приступают к размножению в конце
весны—начале лета. В связи с этим осенью можно наблюдать 2—3 генера-
ции данного вида.
Каждая кладка содержит 10—20 яиц размером 2.2x2.0 или 2.5x2.25 мм.
Только что вылупившиеся слизни имеют длину 3.5 мм, тело их белое и.
просвечивающее.
1 Ввиду того что еще до недавнего времени данный вид не отличали от D. reticulatum
характеристику распространения его в СССР и Польше мы даем только на основании
анализа лично просмотренного материала.
1. DEROCERAS
145
Хозяйственное значение. В связи с тем что данный вид
длительное время путали с D. reticulatum, широко распространилось мне-
ние, что он является серьезным сельскохозяйственным вредителем. В дей-
ствительности его роль как вредителя несравненно меньшая, чем D. reticula-
tum, так как он менее тяготеет к полям и огородам, да и к тому же числен-
ность D. agreste обычно значительно ниже, чем у D. reticulatum.
Цветков и Матйкин (1946) указывают, что в Казахстане D. agreste может
играть важную роль в передаче гельминтозов, опасных для домашних живот-
ных. Последнее нуждается в проверке, так как в Казахстане, кроме дан-
ного вида, обитает также внешне сходный с ним D. altaicum (Simr.).
Замечания. 1. Длительное время почти все авторы, в том числе
и Зимрот, рассматривали данный вид как разновидность D. reticulatum.
Впервые Лутер (Luther, 1915) убедительно показал самостоятельность обоих
видов. Тем не менее и позднее многие авторы плохо отличали слабо пигмен-
тированные формы D. reticulatum от D. agreste.
Оба вида несомненно близкородственны и во многом сходны, однако при
внимательном изучении их всегда можно отличить друг от друга по следую-
щим признакам:
а) Окраска. У D. reticulatum на кремовом фоне темно-коричневый или
почти черный рисунок из пятен, которые в типичном случае образуют сет-
чатый рисунок. У слабо пигментированных экземпляров эти пятна разбросаны
редко и не образуют сеть. У D. agreste рисунка нет.
б) Гонада. У D. reticulatum она занимает задний конец внутренностного
мешка и почти не закрыта лопастями печени. У D. agreste она лежит на зна-
чительном удалении от вершины внутренностного мешка и сильно закрыта
печенью и слепой кишкой.
в) Пениальная железа. У D. reticulatum она крупная и всегда более или
менее разветвленная. У D. agreste она небольшая и имеет вид прямого или
изогнутого пальцевидного выступа.
2. В связи с нечеткостью представлений о различиях между упомяну-
тыми двумя видами и их таксономическом статусе в литературе до сих пор
нередко используются названия, которые, как мы убеждены, должны рас-
сматриваться младшими синонимами Deroceras agreste (L.).
В первую очередь это касается Agriolimax transcaucasicus Simroth, 1901.
Автор подчеркивает его сходство с A. agrestis pallidus Schrenk, 1848,1
который, как известно, является синонимом D. agreste.
Зимрот отмечает следующие основные отличия кавказского слизня от
европейского: а) слепая кишка может быть, но может и отсутствовать;
б) окраска иногда бывает серой; в) стимулятор плоский, треугольный.
По наличию или отсутствию слепой кишки Зимрот различает две разновид-
ности — Agriolimax transcaucasicus var. coeciger Simr. и A. transcaucasicus
var. simplex Simr.
Синтипы A. transcaucasicus утрачены. Однако в ЗИН АН СССР имеются
все экземпляры, определенные Зимротом как этот вид и упомянутые им
в его работах 1910 и 1912 гг. Просмотр их убедил нас в следующем. Наряду
с типичными D. agreste (или А. agrestes pallidus Schrenk в понимании Зимрота),
покровы которых полностью лишены темного пигмента и которые имеют
хорошо развитую слепую кишку (Simroth, 1901, рис. 8), за ту же форму
Зимрот принимал молодые экземпляры других кавказских видов, а именно
Deroceras caucasicum (Simroth, 1901) и Krynickillus melanocephalus Kaleni-
czenko, 1851. Для молоди этих двух видов характерна легкая серая пигмен-
тация затылка, мантии, нередко и спины. У этих экземпляров слепая кишка
либо отсутствует (К. melanocephalus), либо чуть намечена (D. caucasicum).
1 Зимрот неправильно его называет pollens Schr.
10 И. М. Лихарев, А. Й. Виктор
146
I. AGRIOLIMACIDAE
Что касается треугольного плоского стимулятора, который, по мнению
Зимрота, характерен для A. transcaucasicus, то наши материалы убедительно
показывают, что эта особенность в равной степени свойственна как молодым
экземплярам зимротовского вида (взрослыми автор не располагал, а у нас
-они имеются), так и молоди европейских D. agreste. У взрослых transcauca-
sicus гениталии ничем не отличаются от северных agreste.
В итоге изучения слизней, определенных Зимротом как Agriolimax
transcaucasicus, а также обширных сборов, сделанных на Кавказе за послед-
ние 60 лет и хранящихся в ЗИН АН СССР, ЗИН АН АрмССР (Ереван)
и в других хранилищах, мы пришли к выводу, что светлые слизни с крупной
слепой кишкой, которых Зимрот называл A. transcaucasicus var. coeciger,
ничем не отличаются от европейских экземпляров D. agreste и упомянутую
форму следует рассматривать как младший синоним данного вида. К тому же
заключению пришла Лупу (Lupu, 1976) в отношении всех сообщений о при-
сутствии в Румынии D. transcaucasicus (Grossu, Lupu, 1959, 1963; Lupu,
1972).
Аналогичная ситуация сложилась с Agriolimax fedtschenkoi (Koch et
Heynemann, 1874). Первоначальное описание этой формы очень лаконично
и касается только внешнего вида, для которого считалась характерной красно-
вато-серая окраска. Зимрот (Simroth, 1886) сначала также рассматривал
ее как отдельный вид и дал описание пениса (обычный для D. agreste). Позд-
нее тот же автор трактует этого слизня как подвид D. agreste. В то же время
Зимрот высказывает обоснованное предположение, что своеобразная окраска
^вызвана особенностями жидкости, использованной для фиксации.1 Поздней-
шие сборы не подтвердили присутствие в Средней Азии какой-то особой
формы D. agreste. Все экземпляры были достаточно типичными. Указание
Цветкова и Матёкина на присутствие в Казахстане A. fedtschenkoi как само-
-стоятельного вида обязано тому, что Цветков вслед за Зимротом применял
название agreste к той форме, которую мы рассматриваем как D. reticulatum.
'7. Deroceras'(Agriolimax)]altaicum (Simroth, 1886), comb. nov. (рис. 92—102)
Simroth, 1886a : 28, T. 1, Fig. 14 (Agrio-limaa:); 1901 : 164, Fig. 11, T. 16, Fig. 13—
17 (Agriolimax)', 1910 : 523, T. 7, Fig. 19—21 (A. buchar var. coeciger, gyn. nov.); Лиха-
рев, Раммельмейер, 1952 : 332 (Agriolimax)', Walden, 1957 : 355, text-figs.
5—9, 13—15, 18 (A. altaicus transitorius); 363 (A. sibiriensis= A. hyperboreus, sensu Simroth,
1901, non Westerlund, 1876).
Terra typica — «Altai» — Алтай. Типы неизвестны.
Материал: из СССР — 350 экз. из 57 пунктов; из других стран —
10 экз.
Форма тела как у D. agreste. Окраска светло-серая, серая, желтовато-
коричневая или охристая. Рисунка нет, если не считать нескольких бурых
пятен, которые иногда бывают на мантии.
Размеры: дл. сок. 25—30 мм.
Радула (рис. 92). Боковых зубов 15—18, краевых — 17—22; число рядов
75—89. 1—2 самых внешних боковых зуба могут быть лишены эндоконов.
Кишечник (рис. 93, 97, 101). Слепая кишка хорошо развита.
Гениталии (рис. 94—96, 98, 99, 102). Гонада, как у D. agreste,
т. е. небольшая, лежит на значительном удалении от вершины внутренно-
стного мешка и скрыта под лопастями печени. Пенис у молодых слизней
мешковидный, у взрослых удлиненный с пережимом посередине и слегка
вздутый впереди. На заднем конце его помещается пениальная железа, кото-
1 См. аналогичный случай на с. 347.
1. DEROCERAS
147
рая у одних имеет вид небольшого бугорка, а у других — короткого тупога
выступа. Семяпровод впадает в задний отдел пениса вентрально; там же,
но немного впереди крепится половой ретрактор. Внутри переднего отдела
Рис. 92—102. Deroceras (Agriolimax) altaicum (Simr.).
92 — зубы радулы, Забайкалье; 93 — кишечник, Забайкалье; 94—9в — внешний вид и внутреннее-
строение пениса лектотипа AgriolimaxbucharSina.: 97 — слепая кишка паралектотипа того же вида; 9S —
пенис, окр. Байкала, 24 VII 1966; 99 — то же, окр. Якутска; юо — ретракторы; 101 — слепая кишка;
102 — дистальная часть гениталий; то же со вскрытым пенисом (100^-102 — Камчатка, 13 IX 1931).
пениса находится небольшой тупой стимулятор, а рядом с ним продольная
складка. Резервуар семяприемника яйцевидный, плавно переходящий
в короткий проток. Атриум сравнительно широкий.
Распространение. Западный Тянь-Шань (Киргизский и Заи-
лийский Алатау), Джунгарский хребет, Алтай, Саяны, Прибайкалье и
Забайкалье, Амурская область, Приморский край, Камчатка, Сахалин,
Курильские о-ва. Вне СССР — Монголия.
10*
.148
I. AGRIOLIMACIDAE
Образ жизни. На Алтае обитает во всех вертикальных зонах от
.степной до альпийской (Увалиева, 1969). Укрывается в щелях почвы, в под-
.стилке и под камнями.
Замечания. 1. Как внешний облик, так и особенности внутреннего
.строения указывают на большую близость данного вида к D. agreste. Ареалы
„обоих видов перекрывают друг друга. Учитывая еще недостаточную изу-
ченность фауны слизней азиатской части СССР, нельзя быть окончательно
уверенными в том, что D. altaicum самостоятельный вид, а не подвид от
D. agreste.
Еще менее нам известно о внутривидовой изменчивости D. altaicum. В кол-
лекции ЗИН АН СССР имеются несколько экземпляров из южного Алтая
.(Восточно-Казахстанская область) и Бурятской АССР, у которых пениаль-
ная железа имеет намек на ветвление. Что это — новый вид или внутривидо-
вая форма D. altaicum — пока сказать трудно.
2. Вальден (Walden, 1957), просматривая старые сборы по Сибири, сде-
ланные экспедицией Норденшельда1 и определенные Вестерлундом (Wester-
lund, 1877) как D. agreste (L.), установил, что они являются D. altaicum
(Simr.), и впервые после Зимрота значительно пополнил характеристику
„этого вида рядом существенных особенностей строения. В частности, он
обратил внимание, что некоторые экземпляры отличаются более охристой
.окраской и наличием коричневых пятнышек. Это дало ему основание описать
новую форму — Agriolimax altaicus var. transitorius. Такие экземпляры име-
ются и в коллекции ЗИН АН СССР. Подобные внутривидовые отклонения
нет оснований рассматривать даже как подвиды. Поэтому название, предло-
женное Вальденом, мы считаем, младшим синонимом D. altaicum
(Simr.).:
В той же работе Вальден предложил новое название — Agriolimax
-sibiriensis — одному экземпляру из Якутска (хранится в ЗИН АН СССР),
у которого Зимрот не обнаружил ни слепой кишки, ни пениальной железы.
После просмотра этого экземпляра мы у него обнаружили как тот, так и
другой орган. Он является типичным D. altaicum, и новое название, предло-
женное Вальденом, лишь пополнило число синонимов этого вида.
3. Об «Agriolimax buchar Simroth, 1910». Этот вид был описан по мате-
риалам, собранным в нескольких пунктах западного Тянь-Шаня (Таласский
„Алатау, Киргизский хребет), а также из окрестностей озера Балх:аш.
В ЗИН АН СССР имеются 18 синтипов этого вида.2
У всех изученных им экземпляров Зимрот отметил одну общую черту:
пенис короткий, мешковидный, сзади слегка пигментированный (серый),
с короткой и простой пениальной железой и небольшим коническим стимуля-
тором. В то же время автор обнаружил и некоторые различия, которые поз-
волили ему разделить всех слизней на две группы: 1) белые и беловатые
с длинной слепой кишкой и 2) серые и коричневатые без слепой кишки.
Соответственно он разделяет новый вид на две разновидности — Agriolimax
buchar var. coeciger и A. buchar var. simplex.
Просмотр синтипов убедил нас в том, что так называемый A. buchar
var coeciger является младшим синонимом D. altaicum (Simr.). Что же ка-
сается var. simplex, то с ним вопрос остается открытым. К сожалению,
соответствующие экземпляры были плохо законсервированы (это отмечал
.еще и Зимрот). Поэтому не исключено, что такой нежный орган, как слепая
кишка, мог не сохраниться.
1 Хранятся в Риксмузеуме в Стокгольме.
2 Нами обозначен лектотип, который найден у р. Талас близ Дмитриевки 24 VII
4905 г., сборщик В. Абрамов (рис. 94—96).
1. DEROCERAS
149
8. Deroceras (Agriolimax) reticulatum (Muller, 1774) (цв. вкл., рис. VIII’,
рис. 103—107)
Muller, 1774 : 10 (Limax); Simroth, 1885 : 218, T. 9, Fig. 5—15 (Agriolimax
agrestis, pars); 1901 : 144, T. 14, Fig. 7—12 (Agriolimax agrestis, pars); Luther, 1915 : 1,
Fig. 1 В—D (Agriolimax); Wagner, 1935 : 183, Fig. 18, 19 A, B, 22 (Agriolimax agrestis
reticulatus); 1937 : 1048 (Agriolimax); Pilsbry, 1948 : 534, Fig. 287, 288; Лихарев,
Раммельмейер, 1952 : 331, рис. 256, 257 (Agriolimax); Fromming, 1954 :
214 (Agriolimax); Wiktor, 1960b : 23, Fig. 1—3; Quick, 1960 : 164, text-figs. 10 A,
F, 11 C, G, H, L, O, pl. 2, fig. 15 (Agriolimax); Дмитриева, 1969 : 802, Wiktor,
1973 : 125, Fig. 43, 44, 103, 174—189, 242, 280, 281; Дмитриева, 1975 : 40; Lupu,
4976 : 10, Fig. 1—9 (altenai, syn. nov.).
Locus typicus — «Fridrichsdal» — Фридриксдал у Копенгагена (Дания).
Типы неизвестны и, вероятно, утрачены. Описание и рисунки экземпля-
ров, собранных в типовой местности, приводит Алтена (Altena, 1969).
Материал: из СССР — более 800 экз. из 93 пунктов; из других стран —
более 2000 экз.
Тело более коренастое, чем у D. agreste, с сильно выпуклой спиной;
задний конец клиновидно сужен. Мантия занимает около 2/5 длины тела.
У взрослых слизней фон грязно-кремовый, светло-кофейный или оливково-
кремовый. Обычно имеется четкий рисунок, образованный коричневыми,
черноватыми или темно-бурыми пятнами. Чаще всего эти пятна формируют
рисунок типа неправильной сетки (отсюда и название вида «сетчатый»).
Темный пигмент в первую очередь концентрируется по бороздкам. Взрослые
особи без пятен попадаются редко, тогда как молодые долгое время лишены
их. Интенсивность окраски и плотность расположения пятен является индиви-
дуальным признаком, и в этом отношении даже внутри одной популяции
наблюдается большое разнообразие. Общее потемнение окраски обычно
происходит к концу осени. Чаще всего пятна располагаются наиболее густо
на спине и мантии. Голова и шея покрыты более мелкими пятнами, щупальца
черноватые. Подошва кремовая, а у меланистических особей коричневая,
всегда одноцветная.
Слизь бесцветная. При раздражении слизни выделяют большое количе-
ство молочно-белой слизи.
Размеры: дл. пол. до 35, реже до 45 мм; дл. сок. до 25 мм. В запад-
ных областях Европы дл. пол. до 60 мм.
Раковина — см. рис. 104.
Радула. Число боковых и краевых зубов и число рядов сильно варьируют
и зависят от возраста.
с 14-226 21—32к
т +----з--+----j--X 93-110.
Кишечник, как у D. agreste.
Гениталии (рис. 105—107). Крупная бурая гонада в мужской
фазе достигает заднего конца внутренностного мешка и лишь слабо закрыта
печенью. Позднее она немного уменьшается в размерах и смещается вперед.
Она всегда длинная, языковидная. Крупный пенис у половозрелых особей
исегда с перехватом посередине, но у молодых слизней это сужение может и
отсутствовать. Поверхность пениса белая с шелковистым блеском. Внутри
переднего отдела пениса находится крупный стройный стимулятор. Снаружи
основание стимулятора отмечено небольшой бороздковидной вогнутостью
(характерная особенность данного вида!). На заднем конце пениса сидит
крупная пениальная железа, форма которой отличается исключительно боль-
шой изменчивостью. Она может быть одностебельчатой — с лопастями
или короткими веточками по краю или двух- и трехстебельчатой, причем
шаждый стебель дает вторичные ответвления или лопасти.
Рис. 103—107. Deroceras (Agriolimax) reticulatum (Miill.) (из: Wiktor, 1973).
юз — внешний вид; 104 — раковина; юз — гениталии; юв — дистальная часть гениталий в двух по-
ложениях; 101 — 11 вариантов пениальной железы.
1. DEROCERAS
151
В задний отдел пениса справа и немного снизу впадает семяпровод,
а несколько впереди его крепится мощный половой ретрактор. Кроме ретрак-
тора, имеется довольно крупный ретентор, соединяющий передний отдел
пениса со стенкой тела, и несколько мускульных тяжей между обоими отде-
лами пениса. Семяприемник овальный или яйцевидный, с четким протоком.
Яйцевод и атриум цилиндрические, причем последний довольно короткий.
Распространение. Вид несомненно европейский, однако четкое
представление о его природном ареале составить нелегко. И не только потому,
что не всегда можно доверять определениям некоторых, особенно старых,
авторов, но и из-за сильной синантропности данного слизня, т. е. тяготения
к антропогенным и культурным биотопам.
В СССР (по материалам ЗИН АН СССР и других коллекций) он обнаружен
в Архангельской (Архангельск, Вельск), Ленинградской, Псковской, Нов-
городской, Калининской, Ярославской, Московской, Тульской, Пензен-
ской, Орловской, Курской, Саратовской (Вольск), Челябинской, Молоде-
ченской областях, в Эстонии (Таллин), Литве (Шауляйский р-н), в Татарской
АССР, на Украине (Киев, Житомир, Одесса, Ялта, Севастополь, в Карпатах —
Ивано-Франковск, Яремча), на Северном Кавказе (Майкоп, Орджоникидзе),
в Закавказье (Ереван) ,^в Казахстане (Алма-Ата) и в Средней Азии (Хамзабад,
Ферганская область). Находки на Кавказе, в Казахстане и в Средней Азии,
•безусловно, обязаны завозу.
Вне СССР — остальная Европа. Завезен в Северную Америку, Перу,
Австралию, Тасманию, Новую Зеландию и в южную Африку.
Образ жизни. Обитает на открытых биотопах, явно избегая лесов
и даже кустарников, преимущественно на глинистых почвах. Отличается
высокой экологической толерантностью и поэтому встречается не только
в природных, но и в антропогенных и особенно на культурных биотопах.
Обычен на лугах, в придорожных канавах, на полях, огородах, пустошах,
свалках, в компостах, а в городах— в парках, садах и на кладбищах. Днем
укрывается на почве под кусками древесины, камнями, комками земли и
в трещинах почвы.
Сроки размножения и число генераций зависят от климата данной мест-
ности.1 На северо-западе европейской части СССР копуляция и откладка яиц
начинается чаще всего в конце лета и в начале осени. Зимуют обычно только
яйца. При благоприятных погодных условиях (теплое и влажное лето)
размножение начинается раньше и появляется летняя генерация слизней
(обычно немногочисленная), часть которой благополучно перезимовывает
и приступает к размножению раньше, чем генерация, вылупившаяся из
перезимовавших яиц.
Откладка яиц начинается через 10 дней после копуляции. Один слизень
в несколько приемов (по 10—20 яиц) откладывает до 700 яиц. Яйца имеют
размеры 3x2.5 мм, просвечивающие, с точечными включениями извести
у поверхности.
В Англии благодаря мягкому и влажному климату в некоторые годы сли-
зень активен весь год и число генераций достигает трех.
При оптимальной температуре (15—17°) и высокой влажности вылупле-
ние молоди происходит на 11-й день. Однако в природе такие сроки касаются
только летних генераций. Обычно же яйца, отложенные осенью, остаются
в покое всю зиму и первую половину весны, пока не установится теплая
погода без ночных заморозков.
1 Подробные сведения о биологии питания и размножения данного вида приведены
у Фрёмминга (Fromming, 1954). Однако они в основном касаются Средней и Западной
Европы. Для СССР, в частности для Ленинградской области, наиболее исчерпывающие
данные сообщает Дмитриева (1969).
152
I. AGRIOLIMACIDAE
Хозяйственное значение. Основной пищей слизня служат
зеленые части растений, а также плоды и корнеплоды. Особенно предпочитает
молодые побеги и всходы различных полевых и огородных культур. Большая
прожорливость в сочетании с высокой численностью в благоприятные («слиз-
невые») годы, явное тяготение к культурным биотопам — вот причины,
почему этот слизень в определенных климатических и хозяйственных обла-
стях стал бичом полей и огородов. В СССР он чаще вредит в западных и
северо-западных областях европейской части, особенно на тяжелых глинистых
почвах. Известны случаи, когда этот слизень местами полностью истреблял
урожай капусты и всходы озимых.
Замечания. 1. В настоящей работе, как и в более ранних наших
трудах, мы придерживаемся общепринятой точки зрения, впервые обосно-
ванной Лутером (Luther, 1915), что D. agreste (L.) и D. reticulatum (Mull.) —
это самостоятельные виды, четко различающиеся как по морфологии, так
и по образу жизни. Тем не менее хотя эта точка зрения принята большинством
специалистов (Hesse Р., 1926; Ehrmann, 1933; Quick, 1960, и др.), ряд авто-
ров (Wagner, 1935; Sods, 1943) все еще рассматривает обоих слизней как
разновидности или подвиды одного вида. Другие (Reygrobellet, 1963; Lupu,
1972) название «agreste» применяют к настоящим D. reticulatum или к его
внутривидовым формам.
2. Сильная изменчивость у D. reticulatum окраски, рисунка, формы
пениса и его придатков способствовала тому, что при знакомстве с внутри-
видовыми формами этого слизня некоторые авторы принимали их за новые
виды (см. «Дополнение к роду Deroceras»). В частности, это относится к D. ge-
ticum Grossu, 1969, который следует считать синонимом данного вида.
3. До открытия таких видов, как Deroceras turcicum (Simroth, 1894),
D. rodnae Grossu et Lupu, 1965, D. praecox Wiktor, 1966, их, по-видимому,
принимали за D. reticulatum (Mull.).
9. Deroceras (Agriolimax) turcicum"; (Simroth, |1894) (рис. 108—110)
Simroth, 1894a : 392, T. 19, Fig. 2—5 {Agriolimax)', UrbaAski, Wiktor,
1968 : 169, Fig. 9 {reticulatum, pars); Grossu, 1969 : 168, Fig. 10 {wiktori)', 169, Fig. 11
(? waldeni)', Hudec, Vasatko, 1971 : 35, Fig. 22 {wiktori)', Wiktor, 1971 : 271,
Fig. 4 {wiktori)', Lupu, 1972 : 95, T. 4, Fig. 40—48.
Locus typicus — «Ochrida-See» — окрестности оз. Охрид (Балканский
п-ов). Лектотип (обозначен нами) находится в Венском естественно-истори-
ческом музее (№ 19491); здесь же хранятся 4 паралектотипа. Гениталии
лектотипа изображены у Зимрота на рис. 2—5. Голотип D. wiktori — в кол-
лекции Гроссу (№ 1147).
Материал: из СССР нет; из других стран — 14 экз. (в том числе
типы).
Тело коренастое. Мантия немного больше 1/3 длины тела. Окраска грязно-
кремовая или коричневато-светло-серая с рисунком из многочисленных
мелких коричневых пятен, формирующими неправильную сеть. Особенно
много их по сторонам от четкой светлой килевой полоски. Киль короткий.
Слизь бесцветная, при раздражении тела молочно-белая.
Размеры: дл. сок. до 32 мм, лектотип — дл. сок. 17, дл. мантии 8 мм.
Внешне животное очень походит на D. reticulatum (Muller), отличаясь лишь
меньшими размерами.
Слепая кишка крупная; ее длина в 3—10 раз больше ширины (рис. 108).
Гениталии (рис. 109, НО). Пенис мешковидный или яйцевидный,
иногда с заметным пережимом, разделяющим его на передний и задний
отделы. Внутри его находится небольшой узкоконический стимулятор, за-
остренный к концу. Пениальная железа лежит у заднего конца пениса;
1. DEROCERAS
153
она различной формы и слагается из нескольких коротких круглых или
пальцевидных выступов. Неподалеку от пениальной железы в пенис впадает
семяпровод и крепится половой ретрактор. Характерно, что семяпровод
впадает в пенис между двумя небольшими вздутиями со стороны зоба или
пищевода и немного сверху.
Распространение. Обитает в южной Румынии (Банат, Малая
Валахия, Северная Добруджа), Югославии, Болгарии, Турции (у Трабзона).
Рис. 108—110. Deroceras
70S — слепая кишка;
(Agriolimax) turcicum (Simr.) (из: Wiktor, 1971).
10S — дистальная часть гениталий; 110 — пенис.
Образ жизни. Живет в лиственных и смешанных лесах до высот
2000 м над ур. м. (в Понтийских горах Малой Азии). Укрывается под кам-
нями и кусками древесины.
Замечания. 1. По внешнему облику и внутреннему строению вид
очень близок к D. reticulatum (Muller), от которого его не всегда легко отли-
чить. Главное и наиболее постоянное отличие между этими видами — это
положение устья семяпровода, который у D. reticulatum огибает пенис и
впадает в него со стороны стенки тела и немного снизу. Пениальная железа
у обоих видов очень изменчива, но у D. turcicum она всегда небольшая,
а у D. reticulatum значительно крупнее. Наконец первый вид обитает преиму-
щественно в лесах, а второй на открытых местах.
2. Гроссу (Grossu, 1969) описал D. waldeni Grossu, 1969 по материалам
из Северной Добруджи. Различия между ним и D. turcicum очень незначи-
тельны и укладываются в границы обычной изменчивости одного вида.
3. Подрод PLATH YST1MULUS Wiktor, 1973
Wiktor, 1973 : 130.
Типовой вид — Agriolimax subagrestis Simroth, 1892; по первона-
чальному обозначению.
Слизни, внешне похожие на Agriolimax, и отличаются только некоторыми
чертами внутреннего строения.
Слепая кишка всегда имеет вид небольшого кармашка, длина которого
не больше ширины.
154
I. AGRIOLIMACIDAE
Пенис кроме хорошо развитой пениальной железы имеет боковые взду-
тия, а иногда и типичный аппендикс. Крупный стимулятор с двух сторон
сплющен, похож на лопатку или веер и всегда без известковой пластинки.
Распространение. Южные области средней и западной Европы,.
Крым и Кавказ. Мезофильные виды, преимущественно обитающие в лесах.
10. Deroceras (Plathystimulus) subagreste (Simroth, 1892) (рис. 111—115)
Simroth, 1892 : 43 {Agriolimax)', 1894b : 13 (4. agresticulus)', 12, T. 17, Fig. 5
{Agriolimax)', 1901 : 158, T. 15, Fig. 17—28 {Agriolimax)', 159, T. 15, Fig. 1—16 {A. Ananowi,.
syn. nov.); 160, Fig. 10 (A. agresticulus, syn. nov.); 1910 : 522 {Agriolimax)', 1912a : 39, 130
(Agriolimax), 42 (A. ananowi)', Лихарев, Раммельмейер, 1952 : 335 {Agrioli-
max), 336, рис. 266 (4. ananowi).
Locus typicus — «Maikop» — Майкоп, сбор Шнейдера. 2 синтипа (высох-
шие!) находятся в Естественно-историческом музее университета им. Гум-
больдта в Берлине (№ 45704).1
Материал: из СССР — более 500 экз. (в том числе типы) из;
78 пунктов.
Тело приземистое, при сокращении веретеновидное, с максимальной ши-
риной в области мантии, к заднему концу суженное и с коротким килем.
Мантия занимает менее 1/3 длины тела. Между мантийной щелью и средней
линией спины 12—13 рядов морщин.
После фиксации фон темно-кремовый или светло-кофейныйг у живых
иногда белый. Чаще всего на светлом фоне имеется более или менее сильно'
развитый темный рисунок из неправильных пятнышек или «брызг» черного
пигмента, в большинстве случаев образующих сетчатый рисунок, так как
пигмент в первую очередь распределяется по бороздкам кожи. Над.
пневмостомом всегда лежит крупное и удлиненное темное пятно.
Остальная часть мантии покрыта мелкими неправильной формы пятнышками.
Молодые слизни, а также иногда и взрослые особи без пятнышек на спине и
мантии, за исключением упомянутого пятна над пневмостомом, которое всегда
присутствует хотя бы в виде намека. Чаще же слабо пигментированные особи
имеют отдельные крупные пятна, разбросанные по спине и мантии.
Передняя часть тела (голова, шея, в том числе и под капюшоном, бока
перед мантийной щелью) окрашена иначе, чем задняя часть (спина и мантия).
Здесь темный пигмент распределен более равномерно и образует микроскопи-
ческие точки. Щупальца всегда темные, чаще всего черные. Голова и шея у ме-
ланистических особей черноватые или черные. Подошва у светлых особей
кремовая, у темных — боковые доли кофейного цвета, часто с мелкими черно-
ватыми пятнышками.
Слизь молочно-белая.
Размеры: дл. пол. до 45, ширина до 7, дл. мантии до 17 мм; дл. сок.
25—30, реже до 35, дл. мантии до 14 мм.
Раковина овальная; у экземпляра с дл. сок. 20 мм, ее размеры 5.5 X 3.6 мм.
Радула. Пример: 1 экз. из станицы Тульская; радулярная формула:
с 176 26 20к „„
3 + 3 + 2 + 1 Х 92‘
Г ениталии (рис. 114, 115). Гонада небольшая, листообразная и
удлиненная, лежит недалеко от заднего конца внутренностного мешка под
задней кишкой и сложена из очень мелких фолликулов; цвет коричневый
или черный. Пенис довольно изменчивой формы, всегда вздутый, шарооб-
разный или грушевидный, иногда с перехватом на середине. Семяпровод
। 1 Данные экземпляры не вскрыты. Между тем известно, что Зимрот вскрывал часть
синтипов. Поэтому нет полной уверенности, что берлинские экземпляры являются син-
типами.
1. DEROCERAS
155
открывается недалеко от заднего конца пениса. Еще более апикально распо-
лагается пениальная железа. Между ними на пенисе имеется темное, порой
черное пятно. У молодых слизней пениальная железа имеет вид простого
нерасчлененного выступа.1 У особей с вполне развитыми гениталиями этот
Рис. ill. Deroceras (Р lathy stimulus) subagreste (Simr.). Краснодарский край, станица
Тульская, 8 V 1924.
орган по одному из краев расчленен на мелкие пузырьки или лопасти, а у его
основания могут появляться небольшие боковые ветви. Половой ретрактор
крепится к пенису сбоку и посередине, а иногда даже к передней части
пениса. Перед этим ретрактор дает небольшую узкую ветвь, которая при-
крепляется к заднему концу пениса, по соседству с семяпроводом и пениаль.
1 Именно такие экземпляры были использованы Зимротом для первоначального опи-
сания.
156
I. AGRIOLIMACIDAE
ной железой. Стимулятор крупный, очень длинный, веерообразный (рис. 112,
115). У основания он сужен, а к свободному концу плавно расширяется и за-
Рис. 112—115. Deroceras (Р lathy stimulus} subagreste (Simr.).
112— экземпляр с вывернутым стимулятором; ИЗ — задний конец внутренностного мешка; 114 — ди-
стальная часть гениталий с двух сторон; 115 — вскрытый пенис (112 — окр. Майкопа, 6 V 1907; ИЗ-
115 — лектотип Agriolimax ananowi Simr., окр. Орджоникидзе, 1886).
кругляется, своими складками заполняя почти весь пенис. Поверхность его
имеет четкую рубчатую скульптуру. Кроме стимулятора, в заднем конце
пениса имеется несколько складок. Резервуар семяприемника различной
формы, с четким узким протоком. Атриум короткий.
1. DEROCERAS 157
Распространение. Все многочисленные находки этого вида сде-
ланы на Северном Кавказе, а также в байрачных и пойменных лесах Красно-
дарского и Ставропольского краев, в лесистых предгорьях северного Даге-
стана; лишь в районе Туапсе—Сочи он переваливает через хребты Большого’
Кавказа, и здесь ареал его частично перекрывает ареал близкородственного
вида — D. bakurianum (Simroth). Наиболее западная находка сделана у Ново-
российска, наиболее восточная — в заказнике на р. Сулак у Чоткаула.
Кроме того, как завезенный вид он обнаружен в теплицах Пушкина
(Ленинградская область), Ростова-на-Дону и в садоводствах Ташкента.
Некоторые авторы указывали на присутствие этого вида 1 в природных
биотопах Чехословакии (Babor, 1895; Hudec, 1970), Румынии (Grossu,
Lupu, 1959а), а также в Югославии (Wagner, 1935). Судя по их описаниям и
рисункам, они имели дело с другими видами. Во всяком случае эти сообщения
требуют тщательной проверки.
Образ жизни. Обитает в лесах всех типов, как горных, так и рав-
нинных. В активное время ползает по траве и почве. Укрывается под камнями,
в подстилке и среди корней трав.
Половозрелость наступает в конце весны и начале лета. Со второй поло-
вины июля встречается только молодь. К концу осени лишь некоторые экзем-
пляры достигают мужской фазы функционирования гонады. На зимовку ухо-
дят только молодые слизни.
Хозяйственное значение. Как показывают материалы, по-
лученные от быв. Северо-Кавказской станции защиты растений, этот слизень
может быть вредителем ряда огородных культур.
Замечания. 1. Данный вид был описан по нескольким молодым
экземплярам, собранным Шнейдером близ Майкопа. Позднее Зимрот (1894b,
1901) дает серию рисунков, в том числе и по новым сборам, однако опять
молодых слизней. Автор сближал этот вид с Agriolimax agrestis (L.), т. е. с De-
roceras reticulatum (Mull.) в современном понимании. С его точки зрения, кав-
казский вид отличался от европейского в первую очередь отсутствием слепой
кишки и настоящего (т. е. конического) стимулятора и слабым ветвлением пе-
ниальной железы.
В своей монографии 1901 г. Зимрот описал под названием Agriolimax
ananowi другой новый вид из-нод Владикавказа (ныне Орджоникидзе). Для
этого вида он считал характерным изменчивую окраску (от светлых экземпля-
ров без рисунка до темных с сетчатым рисунком), отсутствие слепой кишки,
сильное ветвление пениальной железы и наличие внутри пениса лопастевид-
ного стимулятора. Позднее Зимрот (1912а, с. 40) заметил, что по характеру
строения стимулятора имеются переходные формы между subagrestis и ana-
nowi.
В ЗИН АН СССР нет синтипов первого вида, но зато имеются 62 синтипа
второго вида.2 Кроме того, здесь же находятся почти все материалы, опре-
деленные Зимротом как A. subagrestis и A. ananowi и опубликованные в его
работах 1910 и 1912 гг., а также обширные сборы за последние 60 лет, опре-
деленные уже нами. Исследование этих материалов убедило нас в том, что
никакой принципиальной разницы между обоими видами Зимрота нет. Моло-
дые ananowi полностью соответствуют описанию subagrestis. Обе формы
обитают как в типовой местности subagrestis (у Майкопа), так и в типовой
местности ananowi (у Орджоникидзе). Следовательно, Agriolimax ananowi —
это младший синоним A. subagrestis.
1 Или его синонима D. ananowi (Simroth).
2 Из серии № 4 систематического каталога, содержащей 27 экз., собранных Анано-
вым и определенных Зимротом, мы обозначили лектотип. Его размеры: дл. сок. 23, дл.
мантии 10 мм. Экземпляр был вскрыт еще до нас и гениталии отпрепарированы (см. наши
рис. 113—115).
158 I. AGRIOLIMACIDAE
2. В горах к югу от Майкопа у селения Хамышки Рейбиш собрал два
маленьких слизня (дл. сок. около 10 мм), которых Зимрот (Simroth, 1894b,
1901) описал под названием Agriolimax agresticulus. Для них характерны сле-
дующие признаки: мантия с мелкими коричневыми пятнами-брызгами; ниже
средней линии спины с каждой стороны по желтовато-белой полоске, образо-
ванной подкожными включениями извести; пенис с небольшим треугольным
стимулятором и относительно крупной разветвленной пениальной железой;
слепой кишки нет.
Других находок этого вида не было. По-видимому, он описан по молодым
D. subagreste (Simr.) и, следовательно, тоже является его младшим синонимом.
3. Как показала проверка, некоторые слизни, находящиеся в настоящее
время в ЗИН АН СССР, собранные из различных мест Закавказья и опреде-
ленные Зимротом (1910, 1912а) как Agriolimax subagrestis, относятся к Dero-
ceras bakurianum (Simroth, 1912) (см. ниже).
11. Deroceras (Plathystimulus) bakurianum (Simroth, 1912), comb, nov,
(рис. 116-127)
Simroth, 1912a : 40, T. 3, Fig. 34, T. 7, Fig. 23 (Agriolimax)-, 41, T. 3, Fig. 35,
T. 7, Fig. 24 (Agriolimax schemachensis, syn. nov.); Лихарев, Раммельмейер,
1952 : 335, рис. 262 (Agriolimax).
Locus typicus — «Bakuriani» — Бакуриани (Грузия). Лектотип (обозна-
чен нами) и 4 паралектотипа в ЗИН АН СССР. За лектотип принят экземпляр,
изображенный у Зимрота (табл. 3, рис. 34) (сбор К. Сатунина, 15 VI 1908 г.
Дл. сок. 11.5, дл. мантии 5 мм).
Материал, из СССР — 97 экз. (в том числе типы) из 31 пункта.
Тело коренастое, к заднему концу клиновидно суженное, с коротким и не-
ясным килем. Мантия крупная, длина ее больше 1/3 длины тела. Между мантий-
ной щелью и средней линией спины 10—12 рядов морщин.
Окраска варьирует, особенно из-за различной степени развития темного
рисунка. У большинства особей темные пятна располагаются так густо, что
эти слизни производят впечатление одноцветно черных или темно-шоколад-
ных. У взрослых кремовый фон обычно заметен лишь на середине спины.
У молодых слизней рисунок может иметь вид сетки. Передний конец тела (го-
лова, шея под капюшоном и по бокам от него) особенно густо покрыт черными
пятнами. Мантия и бока задней части тела покрыты ими более редко, но гуще,
чем у D. subagreste. Срединная доля подошвы кремовая, боковые более темные,
нередко местами со скоплениями темных пятнышек.
Размеры: дл. сок. до 20, дл. мантии до 7 мм.
Радула (рис. 118). Пример: 1 экз. из Сочи; формула:
Кишечник. Слепая кишка (рис. 119) имеет вид неглубокого, но четкого
кармашка.
Гениталии (рис. 120—127). Листовидная, треугольная, реже удли-
ненная гонада лежит за второй петлей кишечника. У молодых она коричневая,
у взрослых шоколадная или почти черная. Пенис яйцевидный или почти
шаровидный, иногда слегка сужен посередине. В задний конец его впадает
семяпровод, здесь же крепятся ретрактор и пениальная железа. Последняя
Рис. 116, 117. Deroceras (Plathystimulus) bakurianum (Simr.).
116 — внешний вид слизня и его подошвы; 117 — экземпляр с вывернутым пенисом (окр. Батуми,
12 IV 1957).
160
I. AGRIOLIMACIDAE
имеет вид мускулистого, крючковидно изогнутого отростка, с небольшими
железистыми бугорками на поверхности. Темного пятна, характерного для
D. subagreste, здесь нет. Стенки срединной части пениса тонкие и просвечи-
вающие.
Рис. 118—122. Deroceras (Р lathy stimulus) bakurianum (Simr.),
.118 — зубы радулы; 119 — задний конец внутренностного мешка; 120, 121 — дистальная часть генита-
лий; 122 — задний конец пениса (119—122 — окр. Батуми, 12 IV 1957).
Внутри пениса находится очень характерный стимулятор (рис. 123—
127). Он имеет широкое щитовидное основание, от которого отходит слегка
•сплющенный стебель, заканчивающийся сильно сплющенным, серповидно
изогнутым отростком. Острая вершина отростка с одной стороны тонкой пере-
понкой подтянута к стеблю, благодаря чему она всегда обращена к основанию
-стимулятора. В сложенном состоянии отросток обычно обращен вперед (к ат-
риуму), реже назад. Характерной особенностью стимулятора является то, что
обычно он окрашен в темно-серый или почти черный цвет. У молодых, а также
и у взрослых особей, долго побывавших в спирте, стимулятор светлый. В зад-
ней части пениса имеется сложная система складок.
При копуляции серповидный отросток стимулятора приобретает форму
слегка сплющенной острой шапочки.
Рис. 123 —127. Deroceras (JP lathy stimulus) bakurianum (Simr.).
123 — вскрытый пенис; 124 — стимулятор в пяти положениях; 125 — пенис лектотипа Agriolimax
roseni Simr. (частично вскрыт); 126 — пенис (в трех положениях) и стимулятор лектотипа Agriolimax
schemachensis Simr.; 127 — вскрытый пенис паралектотипа Agriolimax bakurianus Simr. (.123, 124 —
окр. Батуми, 12 IV 1957; 125 — Турция, окр. Ардануча, 9 IX 1907; 126 — гора Фитдаг, близ г. Ше-
маха, 8 VI 1908; 127 — окр. Бакуриани, 4 VI 1907).
И И. М. Лихарев, А. Й. Виктор
162
I. AGRIOLIMACIDAE
Распространение. Лесные области Закавказья. Одна находка
сделана в лесах западного Дагестана у Тлярата. Отсутствует в лесах Талыша
и Ленкоранской низменности. Наиболее западные находки сделаны у Сочи
и Батуми, наиболее восточные — в Закатальском заповеднике и у горы Фитдаг
(близ Шемахи).
Образ жизни. Обитает в различных типах влажных и мезофильных
лиственных и смешанных лесов, от субтропических колхидских до горных
буковых и дубовых. По берегам горных речек проникает и в елово-пихтовые
леса.
Размножается с конца весны до начала лета. В конце лета и осенью в по-
пуляциях преобладает молодь, хотя и встречаются единичные взрослые
особи.
В некоторых районах северной Армении отмечена вредная деятельность
этого вида на огородах.
Замечания. 1. Описание этого вида, сделанное Зимротом, отлича-
ется чрезмерным лаконизмом, а изображение пениса очень схематично.
Тем не менее наличие в ЗИН АН СССР всех типов данного вида, а также
значительные сборы, сделанные после 1912 г., позволили нам существенно
пополнить первоначальное описание.
Наибольшее сходство D. bakurianum обнаруживает с D. subagreste. Однако
различия между ними достаточно четкие: а) у второго вида темные пятнышки
на мантии и спине располагаются более редко, а иногда они лишь единичные
на светлом фоне, а молодь совсем без рисунка; у первого вида, как правило,
темные пятна располагаются очень густо, даже у молодых слизней; б) у subag-
reste на заднем конце пениса имеется темное пятно, которого нет у bakurianum;
в) у subagreste ретрактор крепится к пенису двумя ветвями; у bakurianum
он не разветвляется; г) у subagreste стимулятор светлый и имеет форму веера
или листа на узком основании; у bakurianum стимулятор темный с широким
основанием, переходящим в стебель, который заканчивается узким отростком.
2. Одновременно с данным видом в той же работе Зимрот описал еще два
новых вида: Agriolimax schemachensis и A. roseni.
Все типы обоих видов находятся в ЗИН АН СССР. Нами обозначены лекто-
типы (рис. 125, 126). Описания обоих видов страдают теми же недостатками,
что и описание!), bakurianum. Однако исследование синтипов-позволило сде-
лать определенные выводы.
Что касается синтипов D. schemachensis, то они оказались типичными
D. bakurianum. Синтипы D. roseni тоже во многом сходны с D. bakurianum,
в частности, стимулятор очень близкой формы, но пениальная железа со-
вершенно иной формы—древовидно-ветвистая. Не исключено, что это
связано с обитанием в засушливых областях северо-восточной Турции (у го-
родов Ардануч и Олты).
Поэтому, отмечая несомненную близость!), roseni &D. bakurianum, вопрос
о том, являются ли они одним видом или нет, пока приходится оставить от-
тым.
3. Об Agriolimax subagrestis minutus (Simroth, 1901). Эта форма была опи-
сана по двум экземплярам, найденным Зеленским в 1882 г. у Кутаиси. Оба
синтипа находятся в ЗИН АН СССР, но, к сожалению, совершенно высохли
и непригодны для исследования, однако принадлежность их к роду Dero-
ceras не вызывает сомнения. Позднее Зимрот (1912а) упоминает еще о двух
находках этой формы: с Казбека и из Пецкарского ущелья у Цебельды (Аб-
хазия). Эти экземпляры нами исследованы, ив обоих случаях мы констатиро-
вали, что Зимрот ошибся в определении: экземпляр с Казбека оказался моло-
дым Limax maculatus (Kai.), а из Пецкарского ущелья — Deroceras osseticum
(Simroth).
1. DEROCERAS
163
Скорее всего, указанная форма является старшим синонимом Deroceras
bakurianum, но так как типы непригодны для исследования, то нет необхо-
димости обсуждать вопрос о приоритете названий.
12. Deroceras (Platystimulus) tauricum (Simroth, 1901) (рис. 128—137)
Simroth, 1885 : 224, 329, T. 9, Fig. 24, 25 (Agriolimax Dymczewiczi, non Kale-
niczenko, 1851a); 1901 : 153, Fig. 7 (Agriolimax): Лихарев, Раммельмейер,
1952 : 334, рис. 261 (Agriolimax); H u d e с, 1972 : 84, Fig. 12 (crimense); Арутю-
нова, 1975 : 107, рис. 2, А, Б, Д; Скля р, 19756 : 79, рис. (ramosum).
Terra typica — горный Крым. Типы неизвестны.
Материал: из СССР — 170 экз. из 19 пунктов.
Мантия занимает около х/3 длина тела. Морщины невысокие и разделены
мелкими бороздками. Между мантийной вырезкой и средней линией спины
12—13 рядов морщин.
Окраска образована серовато-желтым фоном и темным рисунком из пятны-
шек темно-коричневого или почти черного цвета. При сильном развитии пи-
гмента вся верхняя сторона тела, кроме боков ниже мантии, окрашена в чер-
ный цвет; лишь местами остаются более светлые промежутки. Шея, по крайней
мере открытая часть ее, черная или покрыта серым налетом. Темный пигмент
на спине и боках в первую очередь располагается по бороздкам, но при силь-
ном развитии заходит и на морщины. На мантии он образует многочисленные
пятна с нечеткими контурами, которые часто сливаются друг с другом. Чаще
всего пигмент простирается и на боковые доли подошвы, срединная доля
всегда остается светлой.
При слабом развитии пигмента спина имеет неясный серый рисунок,
а мантия — серые пятна на светло-коричневом фоне. Но даже у таких особей
боковые доли подошвы немного темнее срединной.
В пределах одной популяции встречаются как светлые, так и темные
особи, а также переходы между ними. Однако светлых слизней всегда больше,
чем темных.
Размеры: дл. сок. 30—35 мм.
Радула (рис. 128). Пример: Ялта, Никитский ботанический сад, 1экз.;
формула:
с 176 б 24к „„
3 + 3 • + 2 + 1 Х 97‘
Кишечник (рис. 135). Задняя кишка расположена косо к оси тела. Слепой
кишки нет или она представлена небольшим кармашком.
Гениталии (рис. 129—133). Гонада крупная темно-коричневая или
почти черная, нередко образует вершину внутренностного мешка; она вытя-
нута вдоль левой стороны тела и лишь слегка прикрыта лопастями печени.
Белковая железа светло-коричневая, семяяйцевод серовато-лиловый. Пенис
удлиненный; в связи с развитием вздутий и впадин он имеет неопределенную
форму, которая сильно меняется с возрастом. Задний конец пениса вытянут
в длинный крючковидный аппендикс. Кроме того, ближе к переднему концу
имеется еще другой сферический выступ. В промежутке между ним и аппен-
диксом к пенису крепится ретрактор; здесь же впадает семяпровод и Откры-
вается длинная неразветвленная пениальная железа с округлыми высту-
пами. Стимулятор крупный, очень широкий и короткий (рис. 137). Внутри
переднего сферического вздутия располагается другое небольшое мускули-
стое тельце.
На поверхности пениса и резервуара семяприемника иногда бывают чет-
кие черные пятна (рис. 131, 132, 136).
Распространение. Лесные и лесостепные районы Крыма (Юж-
ный бе^рг,, горы и предгорья).
11*
Рис. 128—137. Deroceras (Plathystimulus) tauricum (Simr.).
12S — зубы радулы: 129, 130 — дистальная часть гениталий с двух сторон; 131 — он же у ДРУГОГО
экземпляра- 132__пенис; 133 — то же вскрытый; 134 — слепая кишка; 135 — задний коней внутрен-
ностного мешка; 136 — семяприемник; 137 — стимулятор (128—130 — Никитский ботанически
сад, 23 II 1956; 131—137 — Крымский заповедник, 2 VII 1962).
1. DEROCERAS
165
Образ жизни. Обитает в лесах, парках и садах. Укрывается в под-
стилке, трещинах почвы, в гнилых пнях и под камнями.
Размножается с февраля до апреля.
Замечания. Первоначально Зимрот (1885) принял этот вид за Agrio-
limax Dymczewtczi (Kaleniczenko, 1851а), который Калениченко описал лишь
по внешнему виду. Позднее, получив дополнительный материал из Крыма,
Зимрот изменил свои взгляды на вид, описанный Калениченко.1 Поэтому
своим экземплярам он дал новое название — A. tauricus. К сожалению, типы
этого вида, по-видимому, утрачены.
Для D. tauricum наиболее характерно присутствие аппендикса и другого
округлого придатка пениса, а также боковое положение пениальной железы.
В этом отношении он обнаруживает наибольшее сходство cD. osseticum (Simr.)
и D. ilium (Simr.), от которых, в частности, отличается формой стимулятора.
13. Deroceras (Plathystimulus) praecox Wiktor, 1966 (рис. 138—143)
Wiktor, 1966 : 449, Fig. 1—10, 15, 16; H u d e c, 1967b : 351, text-figs 4—5,
T., Fig. 3; 1970 : 116, Fig. 6, 7; Vater, 1970 : 59, Fig. 2, 3; Wiktor, 1973 : 131,
Fig. 49, 50, 200—208, 243, 282, 283.
Locus typicus — «Pokrzywna» — Покшывня, восточные Судеты, Польша.
Голотип в Зоологическом музее Вроцлавского университета.
Материал: из СССР нет; из других стран — более 200 экз. (в том
числе типы).
По форме тела сходен с D. reticulatum (Muller). Мантия занимает около
1/3 длины тела. Киль слабо выражен, заметен лишь на конце тела.
Живые слизни обычно снежно-белые, иногда без рисунка, но чаще с редко .
разбросанными довольно большими пятнами красновато-коричневого или
темно-коричневого цвета. После фиксации тело становится светло-кремовым,
а пятна черновато-коричневыми. Пятнышки имеют вид брызг, которые (в отли-
чие от D. reticulatum) разбросаны неравномерно, не связаны с рельефом кожи
и не образуют сетчатого рисунка. Голова и шея светлые или коричневые, без
рисунка или покрыты мелкими крапинками. Подошва всегда белая или кремо-
вая.
Слизь бесцветная, при раздражении слизня молочно-белая.
Размеры: дл. пол. до 40 мм.
Раковина (рис. 139).
Радула (рис. 140). Боковых зубов 16—18 (трехзубцовые, но несколько са-
мых внешних двузубцовые); краевых зубов 13—23, шиповидные; чисдо рядов
90-100.
Кишечник (рис. 141).
Гениталии (рис. 142, 143). Гонада темно-коричневая, располагается
между лопастями печени вблизи от второй петли кишечника, но йена вершине
внутренностного мешка. Особенно характерен пенис. Он сравнительно боль-
шой, с широким, немного сплющенным передним отделом и значительно более
узким задним. Задний конец пениса переходит в спирально закрученный ап-
пендикс, который по внутренней стороне спирали покрыт темным пигментом,
а на внешней стороне несет пениальную железу. Последняя имеет 3—4 же-
лезистых отростка и короткий общий проток. У основания аппендикса в пе-
нис впадает семяпровод и крепится одна из ветвей полового ретрактора; дру-
гая ветвь крепится к переднему отделу пениса. Внутри переднего отдела на-
ходится большой плоский стимулятор с широким основанием и исчерченной
поверхностью. Семяприемник с круглым или овальным резервуаром и тонким
протоком.
1 См. с. 174 наст, работы,
166
I. AGRIOLIMACIDAE
Характерный облик аппендикса позволяет узнать даже молодые особи
этого вида.
Распространение. Судеты, Западные Карпаты и их предгорья
в пределах Польши и Чехословакии.
Рис. 138—143. Deroceras (Plathystimulus') praecox Wiktor (из: Wiktor, 1973).
138 — внешний вид; 139 — раковина; Ito — зубы радулы; 141 — кишечник; 142 — гениталии; 143 —
дистальная часть гениталий с двух сторон.
Образжизни. Обитает в горных и предгорных лиственных лесах
(до высоты 1200 м над ур. м.) с преобладанием липы и бука. Предпочитает
влажные биотопы, особенно тенистые ущелья и долины, где встречается
как на мху и траве, так и на подлеске.
Половая зрелость наступает с апреля по июнь; осенью большей частью
встречаются неполовозрелые особи. Этой особенностью данный вид отличается
от многих других членов того же рода, у которых половая зрелость и размно-
жение приходятся на конец лета и осень.
1. DEROCERAS
167
14. Deroceras (Plathystimulus) rodnae Grossu et Lupu, 1965 (рис. 144—148)
Grossu, Lupu, 1965a : 28, Fig. 2; Wiktor, 1966 : 449 (praecox, pars); P i n-
t e r, 1968 : 180 (praecox); К n о r r e, 1969 : 171, Fig. 1, 3; A 1 t e n a, 1970b : 67,
Fig. 1; H u d e c, 1970:113, Fig. 4, 5; Schmid, 1971:131, Fig. 1—4; 1972:343-
Wiktor, 1973 : 135, Fig. 51, 52, 209-214, 244, 284, 285.
Locus typicus — «М. Inau» — гора Инау, массив Родна, Восточные Кар-
паты, Румыния. Голотип в Естественно-историческом музее им. Гр. Антипя,
Бухарест, № 13135.
Материал: из СССР — 9 экз. из 1 пункта; из других стран — более
150 экз. (в том числе типы).
По форме тела этот вид похож на D. praecox Wiktor и D. reticulatum (Mul-
ler). Мантия занимает от 1/3 до 1/4 длины тела.
Фоновая окраска кремовая, грязно-желтая или светло-коричневая.
Изредка бывают -экземпляры, лишенные темного рисунка, но чаще всего
встречаются особи, у которых как мантия, так и спина покрыты густо распо-
ложенными мелкими темно-коричневыми или черноватыми пятнышками,
образующими сетчатый рисунок (в основном по бороздкам).1
Слизь бесцветная, но при раздражении становится молочно-белой.
Размеры: дл; пол. до 45, дл. сок. до 32 мм.
Радула (рис. 146). Сходна с D. praecox Wiktor.
Кишечник. Слепая кишка имеет вид чуть заметного морщинистого вздутия
или слабого кармашка.
Г ениталии (рис. 147,148). Гонада смещена к заднему концу внутрен-
ностного мешка, а у молодых занимает его вершину. Крупный пенис четко
разделен на широкий передний и более узкий и длинный задний отдел с неопре-
деленными вздутиями. На заднем конце пениса располагается кустистая пе-
ниальная железа с коротким стеблем и пальчатыми ветвями. Рядом с нею кре-
пится мощный половой ретрактор. Здесь же может находиться короткий вы-
ступ, вероятно, гомологичный аппендиксу, но в отличие от D. praecox Wiktor
это образование никогда не закручено спиралью, а пениальная железа рас-
полагается в стороне от него.
Семяпровод впадает в пенис почти апикально. В этом месте нередко на-
блюдается скопление темного пигмента. Внутри переднего отдела пениса на-
ходится крупный и длинный стимулятор. Он сплющен, с узким основанием и
плавно расширяется к свободному концу. Резервуар семяприемника круглый
или овальный, с тонким и длинным протоком.
Распространение. В СССР единственная находка сделана у сел.
Квасы в Раховском районе Закарпатской области. Вне СССР: Румыния
(Родна), Польша (Росточе, Краковская Юра, Восточные Карпаты), Чехосло-
вакия, ГДР, ФРГ, Австрия, Швейцария, Франция (Вогезы).
Образ жизни. Обитает в лиственных и смешанных лесах, в под-
стилке, на траве, на подлеске и стволах деревьев до высоты 2 м. Укрывается
во мху, под кусками древесины. Наиболее обычен по берегам горных речек
и на заболоченных местах, поросших злаками.
Половозрелость наступает в конце летай осенью. На зимовку уходят глав-
ным образом яйца и молодь; лишь единичные взрослые особи встречаются
весной, и после откладки яиц они вскоре погибают. В западных областях
Европы фенология размножения несколько иная (Schmid, 1971), и взрослые
слизни встречаются до середины июля, а молодь появляется уже в начале
осени.
Замечания. Данный вид имеет много общего с D. praecox Wiktor,
1966, и Виктор вначале принимал его за внутривидовую форму последнего
1 Окраска и рисунок настолько не характерны, что определение можно вести лишь
по внутреннему строению.
Рис. 144—148,. Deroceras (Р lathy stimulus) rodnae Gr. et Lupu (из: Wiktor, 1973).
144 — внешний вид; 145 — раковина; 146 — зубы радулы; 147 — гениталии; 148 — вскрытый пенис.
1. DEROCERAS
169'
вида. Позднее накопились данные, позволяющие считать обе формы двумя са-
мостоятельными викарирующими видами. Основные различия между ними
следующие.
УТ). ргаесох фоновая окраска белая (после фиксации светло-кремовая)
с рисунком из редко разбросанных крупных пятнышек; задний конец пениса
переходит в длинный спирально закрученный аппендикс, на котором сидит
пениальная железа; половой ретрактор крепится к пенису двумя ветвями —
к переднему и к заднему отделам пениса; стимулятор короче и с более ши-
роким основанием.
У D. rodnae фоновая окраска кремовая, с рисунком из мелких пятнышек,
образующих сеть; аппендикс короткий и не закручен; пениальная железа
сидит на пенисе; половой ретрактор одноветвистый — крепится к заднему
концу пениса; стимулятор более длинный и с узким основанием.
Наконец, у первого вида половозрелость наступает весной и в начале
лета, а у второго — осенью.
4. Подрод LIOLYTOPELTE Simroth, 1901
Simroth, 1901 : 174; Wiktor, 1973 : 117; Дамянов, Лихарев, 1975 :
322.
Типовой вид — Lytopelte caucasica Simroth, 1901; по последующему
обозначению — Р. Hesse, 1926.
Слизни с относительно крупной мантией, задний конец которой при дви-
жении животного немного заходит в заднюю половину тела. Капюшон круп-
ный (занимает половину мантии), при сокращении приподнят над шеей,
а иногда даже вывернут наружу. Кожа очень тонкая, просвечивающая,
почти гладкая.
Слепой кишки нет или на месте ее незначительное расширение стенки
задней кишки.
Пенис различной формы, но всегда короткий, без типичной пениальной
железы. У заднего конца он имеет различные вздутия или придатки, которые
у некоторых видов вытянуты в длинные аппендиксы. Внутри пениса находятся
различной формы продольные складки и стимулятор, на поверхности кото-
рого у взрослых слизней образуется известковая пластинка с концентриче-
скими линиями нарастания. Наличие пластинки — самая характерная черта
этой группы. Ее форма имеет большое значение при определении вида.
Р аспространение. Восточные и Южные Карпаты, Балканский
хребет, Крым и Кавказ.
Очень влаголюбивые виды, обычно живущие по берегам водоемов.
Замечания. До сих пор большинство авторов вслед за Зимротом
(Simroth, 1901) рассматривали эту группу как подрод или секцию рода
Lytopelte О. Bttg. Считалось, что основанием для такой точки зрения служит
присутствие у всех видов Liolytopelte известковой пластинки внутри пениса —
особенность, которую рассматривали как главный отличительный признак
рода Lytopelte. При этом совершенно упускалось из виду, что по остальным
особенностям организация Liolytopelte обнаруживает большое сходство с De-
roceras и сильно отличается от Lytopelte (с. 195).
15. Deroceras (Liolytopelte) caucasicum (Simroth, 1901) (рис. 149—157)
Simroth, 1901 : 151, T. 16, Fig. 1—6 (Л. dymczewiczi, non Kaleniczenko, 1851),
157, T. 16, Fig. 31—34 (Agriolimax caspius syn. nov.), 171, T. 17, Fig. 5—14 (Lytopelte),
173, T. 17, Fig. 15—23 (Lytopelte gruslna, syn. nov.); 1912a : 43, 130 (Lytopelte)’, Акрамов-
ский, 1948 : 37, рис. 1—3 (Lytopelte caucasica armenia)’, Лихарев, Раммель-
мейер, 1952 : 338 (Agriolimax caspius), 340, рис. 269—271 (Lytopelte)’, Увалиева,
1975: 44 (биология); Скляр, 1975а : 73, рис. 1, 2 (hamatum, syn. nov.); Акрамов-
с к и й, 1976 : 207, рис. 100, табл. 12, рис. 127; Арутюнова, 1978 : 990.
Рис. 149—151. Deroceras (Liolytopelte) caucasicum (Simr.). Душанбе, 30 X 1968.
149 — внешний вид; 150 — гениталии; 151 — пенис.
Рис. 152—157. Deroceras (Liolytopelte) caucasicum (Simr.). Душанбе, 30 IX 1952.
152 — зубы радулы; 153 — раковина; 154 — кишечник; 155 — ретракторы; 156 — передний конец
слизня с вывернутым пенисом; 157 — известковая пластинка стимулятора.
172
I. AGRIOLIMACIDAE
Locus typicus — «Lagodechi» — Лагодехи, в долине левого притока р. Ала-
зани (Грузия). Лектотип (обозначен нами) и 4 паралектотипа в ЗИН
АН СССР. За лектотип избран экземпляр, найденный Млокосевичем 8—12 сен-
тября 1893 г. и изображенный у Зимрота на табл. 17, рис. 5.
Материал: из СССР — более 600 экз. (в том числе типы) из 94 пункта.
Тело мягкое, водянистое. Длина мантии составляет 1/4—х/3 длины тела, и
не менее половины ее приходится на крупный капюшон. Между мантий-
ной щелью и средней линией спины 14—16 рядов морщин (у лектотипа 16).
Окраска очень варьирует — от грязно-белой, серовато-желтой или серо-
вато-розовой до темно-бурой. Мантия окрашена в тон спины, но темнее.
Щупальца, затылок и шея всегда серые или почти черные, даже под капю-
шоном. Подошва светлая.
Более темная окраска переднего конца тела, мантии и, реже, спины
обязана густому налету микроскопических точек. В отличие от Krynickillus
melanocephalus KaL, у которого тоже имеется аналогичная пигментация,
у D. caucasicum пигмент распространяется и на края мантии, оставляя свобод-
ным только валик вокруг пневмостома. Слизь бесцветная.
Характер окраски очень зависит от освещенности биотопа. В тенистых
местах популяции отличаются более светлой окраской — грязно-белой иди
серовато-желтой и более слабым развитием темного налета; на открытых ме-
стах преобладают популяции с темной окраской за счет более сильного разви-
' тия темного налета; при этом некоторые особи становятся почти черными,
особенно в области мантии.
Потемнение связано также с возрастом. Совсем молодые слизни имеют
лишь редкие темные точки в задней половине мантии и легкий серый налет
на шее, щупальцах и затылке. С возрастом темный пигмент распространя-
ется на всю мантию и даже на спину,- а передний конец тела становится очень
темным, почти как у Krynickillus melanocephalus Kai.
Размеры: дл. сок. до 40, дл. мантии до 15 мм. Длина лектотипа 27,
а мантии 9.5 мм.
Раковина (рис. 153) овальная.
Радула (рис. 152) обычная для рода. Пример: Грузия, Бакуриани, 1 экз.;
формула:
Китпечник (рис. 154). Слепая кишка имеет вид короткого и широкого
кармана. Она более отчетливо выражена у взрослых особей, чем у молоди.
Гениталии (рис. 150, 151, 156, 157). Крупная гонада вытяну-
та по левой стороне задней трети внутренностного мешка и достигает его
вершины. Окраска ее зависит от возраста: у молодых она от светло-коричне-
вой до темно-серой, у взрослых от темно-коричневой до почти черной. От го-
нады тянется вперед толстый гермафродитный проток, образуя крупные из-
вивы. Белковая железа у молодых слизней небольшая, серовато-лиловая и
лежит слева—сверху от зоба; у взрослых она крупная и коричневая и охва-
тывает зоб полукольцом снизу. Задняя половина семяяйцевода образует
несколько крупных извивов, передняя — прямая. На этом участке к семя-
яйцеводу прилегает крупная, закругленно прямоугольная светло-желтая
простата. Яйцевод, очень короткий. Семяпровод обособляется недалеко от
атриума, тянется назад и впадает в задний конец пениса. Здесь, по бокам от
семяпровода, на пенисе сидят два небольших, но четких выступа или бугорка.
Этот участок пениса интенсивно окрашен в серый или черный цвет.
Мешковидный пенис на правой стороне имеет округлый выступ или при-
даток. У молодых слизней придаток отсутствует или он меньше собственно
лениса, у полувзрослых примерно тех же размеров, что и собственно пенис,
1. DEROCERAS
173
а у взрослых значительно крупнее. В последнем случае пенис с придатком
по форме напоминают рукавицу, где роль «пальца» выполняет собственно пе-
нис. Придаток и атриум мощным мускулом-ретентором связаны с правой стен-
кой тела. Половой ретрактор сзади крепится к диафрагме в области сердца,
а спереди распадается на 2—4 ветви, из которых одни прикрепляются к соб-
ственно пенису (к заднему концу и к основанию), а другие к его придатку.
Внутри придатка находится очень крупный листовидный стимулятор,
основанием прикрепленный на границе между двумя главными отделами
пениса. В этом месте от стимулятора отходит небольшой выступ, по форме
напоминающий гриб. В его «шляпке» закладывается известковая пластинка
со шпорой. Эта-пластинка внешне сходна с округлым листом, по поверхности
которого проходят ребра-жилки, а шпора имеет вид черешка этого листа
(рис. 157). Пластинка со шпорой возникают лишь к наступлению полово-
зрелости.
Кроме упомянутого темного пятна на заднем конце, темные точки и штрихи
нередко имеются и на других местах пениса. У темноокрашенных особей не-
редко задняя часть внутренностного мешка окутана пленкой бурого мезентерия.
Распространение. В СССР — центральные и восточные об-
ласти Кавказа, главным образом в бассейнах рек Терек, Сулак, Кура,
Араке и других более мелких рек, впадающих в Каспийское море;1 кроме
того, в Крыму. Вне СССР достоверные находки этого вида пока неизвестны,
однако возможно его присутствие в восточных областях Турции и в северных
и северо-западных областях Ирана. Завезен в сады и пригородные хозяй-
ства Ташкента, Душанбе и Алма-Аты.
Образ жизни. Очень влаголюбивый слизень, живущий чаще
всего по берегам водоемов, в том числе и искусственных, на влажных лугах
и у болот в субальпийской, лесной и нижележащих зонах, в том числе степ-
ной и полупустынной, но в этом случае только в поймах рек и вдоль ороситель-
ных каналов.
Судя по коллекционным материалам ЗИН АН СССР и ЗИН АН АрмССР,
на Кавказе взрослые слизни появляются только осенью. Это позволяет пред-
полагать, что на зимовку уходит молодь, полувзрослые слизни и яйца. К ана-
логичному выводу пришел Иззатуллаев (1975) на основании наблюдений,
сделанных в Душанбе. Увалиева (1975), которая изучала питание, рост и
размножение этого вида в пригородных хозяйствах Алма-Аты (предгорья
Заилийского Алатау), пришла к несколько иным выводам. Она отмечает,
что в течение года этот слизень откладывает яйца три раза: перезимовав-
шие полувзрослые слизни — весной, перезимовавшая молодь — летом, часть
сеголеток (генерация из перезимовавших яиц) — осенью.
Хозяйственное значение. Весной и особенно осенью вредит
многим огородным и бахчевым культурам, нередко приводя к большим
потерям урожая (томатов до 37%, огурцов до 43%, клубники до 50%)
(Мухитдинов, Рахматов, 1972; Иззатуллаев, 1975а; Увалиева, 1975, 1979).
Замечания. 1. В своей монографии Зимрот (1901) в числе новых
видов дает описание следующим трем: «Agriolimax caspius», «Lytopelte cau-
casica» и «L. grusina». В ЗИН АН СССР сохранился голотип «А. caspius»
и синтипы «L. caucasica». Позднее Акрамовский (1948) описал новый подвид
«Lytopelte caucasica armenia» (голотип в ЗИН АН АрмССР, паратипы там же
и в ЗИН АН СССР).
Исходя из наших представлений об особенностях Deroceras (Liolytopelte)
caucasicum (Simroth, 1901) и изменениях, которые претерпевает окраска,
1 Единственное указание на присутствие этого вида в западном Кавказе сделано
Зимротом (1912а) на основании изучения сборов из ущелья р. Накра (Сванетия). Про-
смотр этого материала показал, что он состоит из нескольких экземпляров очень молодых
Deroceras, видовую принадлежность которых точно определить невозможно.
174
I. agriolimacidae
слепая кишка и пенис этого слизня в течение жизни одной особи, мы пришли
к выводу, что все названные формы принадлежат одному виду. «Agriolimax
caspius» описан по молодому экземпляру, у которого хотя уже имеется при-
даток пениса, но известковая пластинка еще не сформировалась. Темная
шея и затылок, серовато-розовая окраска мантии и спины, форма простаты,
темное пятно на заднем конце пениса и наличие там двух бугорков по бокам
от семяпровода — все это признаки, характерные для D. caucasicum.
Отличия между «L. caucasica», «L. grusina» и «L. armenia» сводятся к раз-
мерам придатка пениса (меньше, приблизительно равен или крупнее соб-
ственно пениса) и к наличию или отсутствию слепой кишки. Как уже указы-
валось выше, все это лишь возрастные различия.
Учитывая, что в монографии Зимрота «L. caucasica» бъш описан более
подробно и тщательно, чем «А. caspius» и «А. grusina», а также ввиду того,
что он является типовым видом подрода Liolytopelte мы сохраняем это наз-
вание, рассматривая остальные как его синонимы.
2. В той же сводке Зимрот дает характеристику Agriolimax Dymczeviczi
(Kaleniczenko, 1851). Сам Калениченко (1851а) описал лишь внешний облик
этого слизня, который, согласно его данным, обитает в Крыму. Зимрот
пользовался двумя экземплярами, собранными позднее (у Симферополя, сбор
Ю. Вагнера, 1888 г. — хранятся в ЗИН АН СССР). Характеризуя этих слиз-
ней, он, в частности, подробно остановился на внешнем и внутреннем строе-
нии пениса и дал несколько четких рисунков.
Как описание этих экземпляров, сделанное Зимротом, так и наши иссле-
дования их же убедили нас в том, что автор имел дело с Deroceras caucasicum
(Simr.). Об этом говорит форма пениса, наличие на его заднем конце темного
пятна и двух выступов по бокам от семяпровода, а также характерный при-
даток пениса со стимулятором и грибовидным тельцем. Отсутствие известковой
пластинки со шпорой можно объяснить недостаточной зрелостью изученных
экземпляров.
Однако отдавать приоритет названию dymczeviczi перед caucasicum нет
оснований, так как внутреннее строение типов первого вида (утрачены!),
по-видимому, никогда не будет известным.
3. Недавно Скляр (1975а) описал из лесной зоны Крыма новый вид
Deroceras {Liolytopelte) hamatum. Автор отмечает большое сходство этого
видя с D. caucasicum, от которого он отличается главным образом формой
известковой пластинки.
Просмотр типов (в Институте зоологии АН УССР, Киев) убедил нас,
что эти отличия возрастного порядка и описание этого вида лишь увеличило,
число синонимов D. caucasicum.
4. Выше уже указывалось, что окраска данного вида имеет много общего
с Krynickillus melanocephalus Kai. Однако это' сходство может затруднить
определение лишь молоди обоих видов, так как взрослые хорошо различа-
ются как по окраске, характеру кожи и ее рельефу, так и особенно по пенису.
Указанное сходство молоди caucasicum и melanocephalus не раз приводило
к тому, что даже такой знаток, как Зимрот, путал оба вида, а также описал
так называемый «Agriolimax transcaucasicus simplex Simroth, 1901 (см. с. 145
наст, работы).
16. Deroceras (Liolytopelte) bureschi (Wagner, 1934) (рис. 158—162)
Wagner, 1934a : 55, Fig. 7—9 [Agriolimax (Hydrolimax)]', Gross u, Lupu,
1959a : 46, Fig. 5—6 (ilium}', G г о s s u, 1964 : 84, Fig. 1 (Lytopelte herculana); Urban-
ski, Wiktor, 1968 : 62, Fig. 6 (Lytopelte)', H u d e c, 1967b : 349, Fig.l (Lytopelte
herculana)', Lupu, 1972 : 94, Fig. 4 (Lytopelte herculana)', Дамянов, Лихарев,
1975 : 323, фиг. 254.
Рис, 158—162. Deroceras (Liolytopelte) bureschi (Wagn.).
138 — раковина; 159 — зубы радулы; 160 — гениталии (из: Urbaflski, Wiktor, 1968); 161 — дистальная
часть гениталий, пенис вскрыт; 162 — пластинки стимуляторов, вид сверху (161, 162 — Болгария)
176
I. AGRIOLIMACIDAE
Locus typicus— «Hohle Mednik» — пещера Медник, ущелье p. Искыр,
в западной части Старой Планины (Болгария). Тип в Зоологическом инсти-
туте Болгарской академии наук.
Материал: из СССР нет; из других стран — более 300 экз. (в том
числе тип).
Небольшие слизни с относительно крупной мантией, задний конец
которой заходит за середину тела. Задний конец тела довольно круто
сходит на нет.
Окраска шоколадная или темно-кофейная, часто с многочисленными тем-
ными пятнышками-брызгами, но встречаются и одноцветные особи с темной
спиной или с нечеткими пятнышками на коже. Подошва всегда светло-желтая.
Очень подвижное животное, внешне напоминающее D. laeve (MiilL).
Размеры: дл. сок. 18—24 мм.
Слепой кишки нет, или же на боковой стенке задней кишки бывают не-
большие вздутия.
Гениталии (рис. 160). Небольшая, почти черная гонада лежит
на середине внутренностного мешка и частично закрыта лопастями печени
и кишкой. Семяяйцевод толстый; женская часть его окрашена в лиловый цвет.
Яйцевод и атриум короткие. Пенис неправильной формы, толстостенный.
На заднем конце его находится длинный цилиндрический аппендикс, который
по длине не уступает остальной части пениса. У основания аппендикса имеется
более или менее четкое вздутие, в которое впадает семяпровод; здесь же
крепится одна из ветвей полового ретрактора (другая крепится к основанию
аппендикса). Внутри пениса имеется широкая и крупная складка с исчерчен-
ной поверхностью (вероятно, гомолог стимулятора), на которой сидит яйце-
видная пластинка. На верхней стороне узкого конца этой пластинки нередко
имеются мелкие бугорки; остальная поверхность пластинки покрыта кон-
центрическими линиями нарастания (рис. 162).
Распространение. В СССР не обнаружен. В Румынии — Вос-
точные и'Южные Карпаты, Западные Румынские горы, низменность Олтения;
в Болгарии — западная часть Старой Планины, Витоша, Рила.
Образ жизни. Обитает в горах и предгорьях как в лиственных, так
и хвойных лесах по влажным и тенистым местам, в особенности около водое-
мов. Укрывается в подстилке, под камнями и кусками древесины. Иногда
поднимается выше верхней границы лесной зоны.
Замечание. Такие виды, как Lytopelte olteniana Grossu, 1964,
L. herculana Grossu, 1964, L. suboccidentalis Grossu et Lupu, 1965, а также
L. lotrensis Grossu, 1970, судя по первоначальным описаниям, являются
младшими синонимами Deroceras (Liolytopelte) bureschi (Wagner, 1934). Мно-
гие особенности, в которых авторы этих «видов» усмотрели их самостоятель-
ность, объясняются различием в возрасте изученных объектов (длина аппен-
дикса, наличие или отсутствие выступа на пенисе, в который впадает семяпро-
вод, форма пластинки стимулятора). Другие очень нечеткие различия,
вероятно, связаны с экологической изменчивостью.
17. Deroceras (Liolytopelte) moldavicum (Grossu et Lupu, 1961)(рис. 163—172)
Grossu, Lupu, 1961c : 28, Fig. 1—2 (Lytopelte)', Grossu, 1964 : 83 (Lyto-
pelte)-, Wiktor, 1967b : 385 (Lytopelte)', Wiktor, 1973 : 118, Fig. 41, 42, 104, 166—
173, 240, 276, 277.
Locus typicus — «Sucevi(a» — Сукевица (область Сучава, северная Ру-
мыния). Голотип находится в Естественно-историческом музее им. Гр. Антипа,
Бухарест, № 13166.
Материал: из СССР — 18 экз. из 4 пунктов; из других стран —
20 экз.
Рис. 163—172. Deroceras {Liolytopelte) moldavicum (Gr. et Lupu) (из: Wiktor, 1973).
163 — внешний вид; 164 — раковина; 165 — зубы радулы; 166 — гениталии; 167, 168 — дистальная
часть гениталий; 169, 170 — задний конец пениса; 171, 172 — стимулятор с пластинкой.
12 И. М. Лихарев, А. И. Виктор
178
I. AGRIOLIMACIDAE
При жизни тело стройное цилиндрическое, очень вытянутое, со слабым
килем, а мантия простирается назад до середины тела. После фиксации тело
.становится более массивным; из-за втягивания при фиксации переднего
цонца тела мантия как бы смещается вперед, а задний конец спины настолько
сплющен с боков, что приобретает облик вертикального плавника.
Окраска одноцветная, без пятнышек. При жизни тело фиолетово-корич-
невое, только бока и подошва немного светлее. После фиксации вся верхняя
сторона тела приобретает голубовато-стальной или синевато-фиолетовый
цвет, подошва же становится кремовой или белой. Слизь бесцветная и водя-
нистая.
Размеры; дл. пол. до 35, дл. сок. до 23 мм.
Раковина — рис. 164.
Радула (рис, 165). Боковых зубов 15—18, краевых 18—30; число рядов
92-106,
Слепой кишки нет. Обычно все внутренние органы одеты темным мезен-
терием.
Гениталии (рис. 166—172). Гонада тонкозернистая, почти черная
и располагается внутри второй петли кишечника. Белковая железа длинная.
Семяяйцевод снабжен железистыми вздутиями. Яйцевод сравнительно широ-
кий и толстостенный. Семяприемник с длинным цилиндрическим протоком
и круглым или овальным резервуаром. Атриум узкий, трубковидный.
Пенис крупный, грушевидный, широким концом обращенный к атриуму.
Задний конец пениса резко изогнут; здесь апикально впадает тонкий семя-
провод, а немного впереди него крепится половой ретрактор. Аппендикса
нет. Внутри широкой части пениса находится конусовидный стимулятор,
который к заднему концу продолжается в виде продольной складки. К бо-
ковой стенке стимулятора крепится известковая пластинка, имеющая вид
полукруга, по обрезанному краю снабженного острыми зубчиками.1
За исключением стимулятора и складки, покрытых тонкими правильными
бороздками, остальные стенки пениса гладкие.
Распространение. В СССР: Украинские Карпаты (Квасы, За-
карпатская обл.; с. Долина и Яремча, Ивано-Франковская обл.). Вне СССР:
Польша — Бещады; Румыния — Молдавские Карпаты. Общее распростра-
нение: Восточные Карпаты.
Образ жизни. Обитает в горных лиственных, преимущественно
буковых, лесах, по тенистым ущельям и берегам речек, где скопился тол-
стый слой влажной лиственной подстилки. Популяции не отличаются высо-
кой численностью. Взрослые слизни встречаются как летом, так и осенью.
Замечание. Очень возможно, что Agriolimax (Hydrolimax) hucu-
lorum Babor et Frankenberger, 1914 и Agriolimax hydrobium Wagner, 1943
являются старшими синонимами рассматриваемого вида. Однако окончатель-
ное решение принять не представляется нам возможным, так как типы этих
видов, по-видимому, утрачены, а внутреннее строение пенисов не было изу-
чено, Не было это сделано Гроссу и Лупу (Grossu, Lupu, 1957b), которые
указывают на присутствие этих видов на Молдавской возвышенности.
48. Deroceras (Liolytopelte) occidental (Grossu et Lupu, 1966), comb. nov.
(рис. 173—474)
Grossu, Lupu, 1966:25, Fig. 1—3 (Lytopelte)', Grossu, 1970c : 105,
Fig. Id, 2a (Lytopelte).
1 Нередко пластинка покрыта слизью или спермой и может быть обнаружена лишь
после тщательной препаровки.
ДОПОЛНЕНИЕ К РОДУ DEROCERAS
179
Locus typicus — «Stina de Vale» — Стына-де-Вале, горы Бихор (Западные
Румынские горы). Голотип в Естественно-историческом музее им. Гр.Антипа,
Бухарест, № 13139. (
Материал: из СССР — нет; из Румынии — 1 экз.
Окраска одноцветная серовато-красная; спина темнее, но без пятен.
Подошва желтоватая.
Рис. 173—174. Deroceras (Liolytopelte) oceidentale (Gr. et Lupu) (из: Grossu, Lupu, 1966).
173 — гениталии; 174 — вскрытый пенис.
Размеры: дл. сок. 15—17 мм. Киль отсутствует.
Гениталии. Пенис овальный, слегка удлиненный. Сзади он имеет
два выступа приблизительно равных размеров. В один из них, т. е. в задний
конец собственно пениса, апикально впадает семяпровод; у основания дру-
гого выступа — аппендикса — крепится ретрактор. Внутри пениса находится
цилиндрический стимулятор, на конце которого сидит круглая известковая
пластинка, имеющая четкие линии нарастания и зубчатые края.
Распространение. Пока известна только одна находка в Запад-
ных Румынских горах.
Образ жизни. Обнаружен в хвойных лесах на высоте свыше 1000 м
над ур. м.
Замечание. Своеобразная окраска и форма пениальной пластинки,,
по-видимому, указывают на то, что это хороший вид.
Дополнение к роду DEROCERAS
Ниже даются характеристики видов, составленные в основном только по
первоначальным описаниям, т. е. без знакомства авторов настоящей работы
с соответствующими коллекционными материалами. Исключение составляют
12*
180
I. AGRIOLIMACIDAE
3 вида, с типами которых авторы имели возможность ознакомиться,1 что,
впрочем, мало способствовало получению четкого представления об этих
слизнях.
Неполнота оригинальных описаний большинства приведенных здесь
видов в ряде случаев ставит под сомнение факт существования их в природе
и ставит вопрос: не являются ли они формами общеизвестных «старых» видов.
Deroceras (?) absoloni (Simroth, 1916)
Simroth, 1916 : 13, Fig. 3 (Agriolimax)', Gross u, Lupu, 1959a : 42, Fig. 1.
Зимрот описал этот вид по единственному экземпляру, найденному
в одной из пещер Югославии. По-видимому, наиболее характерной чертой
является отсутствие пениальной железы.
Гроссу и Лупу свои экземпляры обнаружили в Южных Карпатах у входа
в одну из пещер близ города Бая-де-Арамэ. Как по внешнему облику, так
и по гениталиям румынские экземпляры сильно отличаются от описания Зим-
рота. В частности, они имеют хорошо развитую пениальную железу.
В связи с этим мы считаем, что румынские авторы имели дело с каким-то
другим видом и пока нет оснований считать, что D. absoloni (Simr.)
.обитает в Румынии.
Deroceras (Agriolimax) callatis Grossu et Lupu, 1963 (рис. 175—176)
Gr ossu, Lupu, 1963 : 197, Fig. 1.
Locus typicus — «Comarova» — Комарова, к северу от Мангалии (Румы-
ния). Голотип в Естественно-историческом музее им. Гр. Антипа, Бухарест,
)№ 13132.
Тело одноцветное, без пятен, светло-желтое или желтовато-серое. Киль
неясный. Кожа тонкая, просвечивающая, со слабым рельефом.
Размеры: дл. сок. 20—25 мм.
Гениталии по сравнению с общей длиной тела крупные. Гонада
красновато-коричневая с толстым протоком. Пенис удлиненно-овальный,
.со слабой перетяжкой на середине. Пениальная железа имеет вид небольшой
розетки. Рядом с нею крепится к пенису короткий, но мощный ретрактор.
.Стимулятор конический, вытянутый; рядом с ним располагается неболь-
шое плоское тельце. Семяприемник овальный.
.Слепая кишка короткая.
Распространение. Известен только из района Мангалии (Се-
верная Добруджа, Румыния).
Замечание. Привлекает внимание сходство с некоторыми формами
D. turcicum (Simroth, 1894).
Deroceras (?) crimense (Simroth, 1901) (рис. 177, 178)
Simroth, 1901 : 152, T. 16, Fig. 7—12 (A griolimax)', Лихарев, Раммель-
мейе p, 1952 : 334 (A griolimax tauricus)', H u d e c, 1972 : 84, Fig. 12.
,Locus typicus—«Salgir»— долина p. Салгир (Крым). Два синтипа
в ЗИН АН СССР (высохли!).
Небольшие слизни (дл. сок. около 20 мм), которые благодаря рисунку
из темно-бурых точек и штрихов внешне сходны с D. reticulatum (Muller).
Боковые доли подошвы темные.
Слепая кишка имеет вид двух коротких выступов.
1 Deroceras crimense (Simroth, 1901), D. pentheri (Babor, 1905), D. steidachneri (Wagner
4931). . ' . . .
Рис.. 175—181. Гениталии некоторых видов Deroceras.
175—176 — D. (Agriolimax) eallatis Gr. et Lupu (из: Grossu, Lupu, 1963: 175 — гениталии, 176 — сти-
мулятор); 177—178 — D. (?) crimense .(Simr.) (из: Simroth, 1901: 177 — внешний вид, 178 — вскрытый
.пенис); 179 — D. (Agriolimax) dobrogieum Gr. (из: Grossu, 1969); 180—181 — D. (Agriolimax) forcarti
dr. et Lupu.(H3.: Grossu, Lupu, 1961: 180 — гениталии, 181 — стимулятор).
182
I. AGRIOLIMACIDAE
Гениталии (рис. 178). Пенис с заметным пережимом на середине.
На заднем конце его находится относительно крупная пениальная железа,
состоящая из общего ствола и разнообразно ветвящихся отростков. Стимуля-
тор довольно крупный, но плоский без острой верхушки. Других складок
внутри пениса не обнаружено.
Замечания. 1. Вид был описан по двум экземплярам, найденным
Кушакевичем в 1864 г. в долине р. Салгир (северные склоны Чатырдага).
К сожалению, оба экземпляра настолько высохли, что теперь они непригодны
для дополнительного исследования.
В коллекции ЗИН АН СССР имеется несколько сборов из бассейна
р. Салгир, которые мы определили как D. tauricum (Simroth, 1901).
Учитывая эти сборы, а также то, что D. crimense описан по молодым
экземплярам (небольшие размеры слизней, слабое развитие стимулятора
пениса), Лихарев и Раммельмейер (1952) считали, что данный вид является
синонимом D. tauricum. Для того чтобы решить окончательно, является ли
D. crimense самостоятельным видом или нет, требуются дополнительные ис-
следования возрастной и экологической изменчивости как D. tauricum,
так и других видов того же рода, живущих в горном Крыму, особенно
вблизи от Чатырдага.
2. Гудец (Hudec, 1972) принял за D. crimenese (Simr.) обычные D. tau-
ricum (Simr.). Это хорошо видно из описания и рисунков автора.
Deroceras (Agriolimax) dobrogicum Grossu, 1969 (рис. 179)
Grossu, 1969 : 160, Fig. 3 (dobro gicus).
Locus typicus — «БипсачДа» — Лункавица (Северная Добруджа, Румы-
ния). Голотип в коллекции Гроссу, Бухарест, № 1142.
Кожа тонкая со светло-желтым оттенком. На мантии неравномерно раз-
бросаны коричневые точки; над пневмостомом находится большое коричне-
вое пятно. За мантией имеются короткие и длинные коричневые пятнышки,
располагающиеся в основном по бокам, так что середина спины остается
светлой.
Размеры: дл. сок. до 30 мм; некоторые особи длиной 22 мм уже
половозрелые.
Раковина у голотипа почти прямоугольной формы, но у некоторых пара-
типов яйцевидная.
Слепая кишка довольно хорошо развита, длина ее в 2 раза больше ши-
рины.
Гениталии. Простата хорошо развита и частично заходит на семя-
провод. Последний впадает в пенис апикально вблизи ретрактора. Пенис
почти цилиндрический, на его заднем конце сидит двухветвистая пениаль-
ная железа. Стимулятор очень крупный (занимает почти всю полость пениса)
и напоминает остроконечную призму с продольным желобком у основания.
Распространение. Известен только из типового местонахожде-
ния в Северной Добрудже.
Образ жизни. Был найден в глубине леса и на опушке в листвен-
ной подстилке.
Замечание. К характерным признакам принадлежит темное пятно
над пневмостомом. Однако неизвестно, насколько это пятно постоянно.
Строением пениса, стимулятора и пениальной железой этот слизень напоми-
нает некоторые формы D. reticulatum (Muller) или D. turcicum (Simroth).
ДОПОЛНЕНИЕ К РОДУ DEROCERAS
183
Deroceras (Agriolimax) forcarti Grossu et Lupu, 1961 (рис. 180, 181)
Grossu, Lupu, 1961 : 21, Fig. 1—2; A 1 t e n a, 1973 : 86, Fig. 2.
Locus typicus — «Babadag» — Бабадаг, северо-восток Северной Добруджи
(Румыния). Голотип в Естественно-историческом музее им. Гр. Антипа,
Бухарест, № 13119.
Кожа тонкая. Киль очень короткий. Окраска беловато-желтая с рисунком
из мелких коричневых пятнышек, которые неравномерно разбросаны по
поверхности тела, но местами концентрируются, образуя как бы пунктирные
линии. Слизь молочно-белая.
Размеры: дл. сок. 14—17 мм.
Гениталии. Пенис мешковидный, слегка вздутый на середине,
иногда с боковыми вздутиями на заднем конце (Altena, 1973). Стимулятор
конический, с очень широким основанием и сильно исчерченной поверхностью.
Пениальная железа имеет вид небольшого яйцевидного сплющенного от-
ростка. Впереди него крепится ретрактор и впадает семяпровод. Яйцевод
заметно расширяется к атриуму. Резервуар семяприемника почти равен
пенису и имеет короткий проток.
Распространение. Северная Добруджа (Румыния) и Словения
(Югославия) (Altena, 1973).
Образ жизни. Обитает в лиственных лесах, в гнилых пнях и в под-
стилке.
Deroceras (?) jaeckeli Grossu, 1969 (рис. 182)
Grossu, 1969 : 159, Fig. 2.
Locus typicus — «Tai des Flusses Topolog» — долина реки Тополог у Тиг-
вени (Южные Карпаты, Румыния). Голотип в коллекции Гроссу, Бухарест,
.№ 1141.
Тело бледно-желтое с красноватым оттенком. Мантия и спина покрыты
редкими коричневыми пятнышками.
Размеры: дл. сок. около 22 мм.
Гениталии. Пенис яйцевидный, иногда с округлым боковым при-
датком. Внутри задней части пениса находится гофрированная продольная
складка, у основания которой лежит конический, сплющенный с боков сти-
мулятор. Его поверхность тонко исчерчена и по одной стороне имеет продоль-
ный желобок. Семяпровод открывается в задний конец пениса вблизи от
ретрактора.1
Распространение. Известен только из типовой местности, где
найден у ручья, на траве и в лиственной подстилке.
Замечание. Систематическое положение этого вида очень неясно.
Внешний облик и форма пениса, особенно присутствие бокового вздутия,
указывают на большое сходство с представителями подрода Liolytopelte.
Тот факт, что на стимуляторе не обнаружена известковая пластинка, столь
характерная для указанной группы, возможно, связан с недостаточной зре-
лостью изученных объектов, так как это образование формируется незадолго
до перехода к размножению.
Кроме того, Гроссу не обратил внимание на то, что внутреннее строение
пениса голотипа данного вида очень сходно с таковым у Agriolimax turci-
cus Simroth, 1894, найденного у озера Охрид. У этого вида также имеется
1 Гроссу отмечает, что пениальной железы нет, но на рис. 2Ь отчетливо виден неболь-
шой короткий выступ у заднего конца пениса. Кроме того, автор считает, что слепая
кишка очень короткая. Однако с этим утверждением нельзя согласиться, если руковод-
ствоваться рис.. 2f.
Рис. 182—188. Гениталии некоторых видов Deroceras.
lf.2 — D. (?) jaeckeli Gr. (из: Grossu, 1969); 183—185 — D. (Plathystimulus) padisii Lupu (из: Grossu, 1969);
186—188 — D. (Agriolimax?) pentheri (Babor) (186, 187 — дистальная часть гениталий; 188 — ре-
тракторы; синтипы, Турция, Лифос, 2—4 00 1902).
ДОПОЛНЕНИЕ К РОДУ DEROCERAS 185
крупный конический стимулятор, который располагается у основания мощ-
ной продольной складки с изрезанным задним краем (см. рис. 2а и 2d
у Гроссу и рис. 5 у Зимрота).
Deroceras (Plathystimulus) padisii Lupu, 1969 (рис. 183—185)
Grossu, Lupu, 1965 : 26, Fig. 1 (caruanae Poll.); Lupu, 1969 : 434, Fig. 2;
Grossu, 1969 : 165, Fig. 8 (transsylvanicus)', 167, Fig. 9 (pseudolytopelte)', Lupu, 1971b :
139, Fig. 1—16.
Locus typicus — «Padis» —Падиш (Западные Румынские горы). Голотип
в Естественно-историческом музее им. Гр. Антипа, Бухарест, № 13152.
Тело сзади как бы срезано и с боков сжатое. Киль короткий. Окраска
желтовато-белая с рисунком из черноватых пятнышек на мантии и боках.
Посередине спины проходит светлая килевая полоска.
Размеры: дл. сок. 18—21 мм.
Слепая кишка отсутствует.
Гениталии. У совершенно половозрелых особей пенис глубокой
вырезкой разделен на передний и задний отделы. У молодых (дл. сок. 17—
18 мм) вырезки нет и пенис имеет продолговато-овальную форму. Внутри
переднего отдела пениса находится плоский, к концу заостренный слабо
исчерченный стимулятор. На заднем конце пениса находится пениальная
железа, которая имеет вид пучка из 5—7 пальцевидных выступов, без об-
щего ствола; лишь некоторые из этих выступов дихотомируют. Здесь же
в пенис открывается семяпровод и крепится половой ретрактор.
Распространение. Западные Румынские горы.
Образ жизни. Обитает в горных лесах на высотах до 1200 м над
ур. м. Зимуют как яйца, так и полувзрослые слизни (Lupu, 1971b). В апреле—
июне происходит размножение, после чего взрослые особи гибнут. В августе
встречается только молодь. Осенью часть сеголеток успевает отложить яйца.
Замечание. Вначале этот вид был принят за западноевропейский
D. caruanae (Pollonera, 1891) (Grossu, Lupu, 1965). Позднее Лупу (Lupu,
1969) описывает его как новый для науки вид. Почти одновременно Гроссу
(Grossu, 1969) по материалам, собранным из тех же мест, описал два новых
вида — D. transsylvanicus и D. pseudolytopelte. Как убедительно показала
Лупу (Lupu, 1971b), первый вид Гроссу описан по молодым, а второй по
взрослым особям D. padisii Lupu.
Deroceras (Agriolimax?) pentheri (Babor, 1905) (рис. 186—192)
В a b о r, 1905 : 293.
Locus typicus — «Lifos» — Лифос, к югу от города Кайсери (Турция).
Синтипы (3 экз.) в Венском естественно-историческом музее, № 39449.
Тело стройное, мантия удлиненно-овальная, подошва узкая.
Верхняя сторона тела синевато-черная, молодые экземпляры черные или
коричневато-серые со светлыми пятнышками на боках. Нижняя сторона
капюшона желтовато-белая, но по краям с синими пятнышками. Боковые
доли подошвы синевато-черные.
Размеры: дл. сок. 16—23, ширина 6—7 мм.
Слепая кишка отсутствует или представлена небольшим и неглубоким
карманом.
Гениталии. Внутренностный мешок покрыт темным мезентерием.
Темно-коричневая гонада лежит около задней кишки. Ее проток желтовато-
белый, извилистый. Крупная белковая железа имеет почти прямоугольную
форму. Семяяйцевод сильно закручен; женская часть его желтовато-белая,
186
I. AGRIOLIMACIDAE
с многочисленными железистыми фестонами; мужская заметно уже и с фио
летовой простатой. Семяпровод короткий и толстый. Пенис вытянутый, почти
цилиндрический или округлый с легким поперечным пережимом. Верхняя
стенка его тонкая, просвечивающая, голубоватая; нижняя стенка толстая
Рис. 189—194. Гениталии некоторых видов Deroceras.
189—192 — D. (Agriolimax'!) pentheri (Babor) (189 — задняя часть внутренностного мешка, 190, 191 —
пенис, 192 — стимулятор (Турция, Талар у Кейзери, 19 IV 1936); 193, 194 — D. (Agriolimax) schleschi
Gr. et Lupu (из: Grossu, Lupu, 1961a): 193 — гениталии, 194 — стимулятор.
Стимулятор обращен вершиной назад; у одних экземплятров он узкий и
острый, у других — маленький и тупой, но всегда конический. Кроме стиму-
лятора, в пенисе имеется четкая складка. Пениальная железа состоит из
двух длинных ветвей с бугорчатыми краями.
Распространение. Известен только из двух мест вулканиче-
ского массива Эрджиас (Центральная Анатолия, Турция), где был найден
на высотах 1350 и 2100 м над ур. м.
ДОПОЛНЕНИЕ К РОДУ DEROCERAS
187
Deroceras (Agriolimax) ponori Lupu, 1972
Lupu, 1972 : 97, Pl. 5, Fig. 49—54.
Locus typicus — «Ponoare» — Поноаре, Падиш (Западные Румынские
горы). Голотип в Естественно-историческом музее им. Гр. Антипа, Бухарест,
.№ 13190.
Тело коренастое, светло-желтое, с редкими темными пятнышками или
штрихами, которые иногда слагаются в сетчатый рисунок. Подошва светлая.
Размеры: дл. сок. 20—30 мм.
Слепая кишка длинная, простирается назад почти на половину задней
части внутренностного мешка.
Гениталии. Пенис овальный или мешковидный, с небольшим по-'
перечным пережимом. Пениальная железа находится на его заднем конце
и состоит из пучка из 2—5 пальцевидных отростков (нередко с мелкими взду-
тиями по бокам), которые впадают в пенис независимо друг от друга. Стиму-
лятор треугольно-конический с короткой и острой вершиной.
Распространение. Западные Румынские горы.
Замечание. Очень возможно, что данный вид является одной из
многочисленных вариаций D. reticulatum (Mull.) или D. turcicum (Simroth),
которая отличается от типичной формы только пениальной железой. Если
у D. reticulatum, в тех случаях когда пениальная железа многоветвистая,
все ветви перед впадением в пенис соединяются в общий проток, то у D. po-
nori общего протока нет и они впадают в пенис независимо друг от друга.
Deroceras (Agriolimax) schleschi Grossu et Lupu, 1961 (рис. 193, 194)
Grossu, Lu pu, 1961a : 23, Fig. 3—5.
Locus typicus — «Ciucas-Gebirgsmassiv» — горный массив Чукаш, горы
Бузэу (Румыния). Голотип в Естественно-историческом музее им. Гр. Антипа,
Бухарест, № 13120.
Тело окрашено в светло-желтый цвет с темным рисунком из мелких
пятнышек и штрихов, образующих сеть как на мантии, так и на спине.
Подошва кремовая, но боковые доли ее немного темнее средней. Кожа очень
тонкая, просвечивающая.
Размеры: дл. сок. около 19 мм.
Слепая кишка хорошо развита.
Гениталии. Пенис мешковидный, с четким пережимом на середине.
Задний отдел несколько крупнее переднего. На его конце находится малень-
кая крючковидная пениальная железа, у основания которой крепится рет-
рактор и впадает семяпровод. Внутри переднего отдела пениса помещается
своеобразный стимулятор с широким основанием и узкой крючковидно
изогнутой и сплющенной верхушкой. Сзади стимулятор переходит в довольно
большую складку, другая складка меньших размеров отходит вперед к ат-
риуму.
Распространение. Известна лишь одна находка, сделанная
в горах Бузэу, вблизи водопада, где слизни ползали по камням, растениям
и влажной подстилке.
Замечание. Форма пениса, слепой кишки и стимулятора напоминает
D. forcarti Grossu et Lupu, 1961 и другие близкие виды из подрода Agrioli-
max. Требуются дополнительные исследования данного вида.
Deroceras (?) steidachneri (Wagner, 1931) (рис. 195—199)
W agner, 1931b : 196 (Agriolimax).
Locus typicus — «Samsun» — Самсун (Турция). Синтипы в Венском есте-
ственно-историческом музее,. № 16010/5.
188
I. AGRIOLIMACIDAE
Тело с очень тонкой кожей и нежными слабыми морщинами. Между
мантийной щелью и средней линией спины 11—12 рядов морщин. Мантия
овальная. Киль слабо выражен. Окраска одноцветная, без рисунка. Спина,
Рис. 195—201. Гениталии некоторых видов Deroceras.
195—199 — D. (?) steidachneri (Wagner) (синтип, Самсун, Турция: 195, 196 — дистальная часть генита-
лий, 197 — пенис, 198 — онже вскрытый, 199 — задний конец внутренностного мешка); 200 — D. (Agrio-
limax) vranceanum Gr. (из: Grossu, 1969); 201 — D. (Plathy stimulus) zilchi Gr. (из: Grossu, 1969),
бока и мантия почти черные. Шея и затылок охристые. Нижняя сторона ка-
пюшона светло-желтая. Средняя доля подошвы светло-охристая, а боковые
заметно темнее.1
1 По-видимому, в результате длительного хранения в спирте (слизни были собраны
в 1889 г.) на синтипах появились мелкие светлые пятнышки.
ДОПОЛНЕНИЕ К РОДУ DEROCERAS 189'
Размеры: дл. сок. 24—36, ширина 5—8 мм; длина мантии 9—14,
ширина 5.5—8 мм.
Радула. Срединный зуб трехзубцовый, боковые (21) тоже, но самые внеш-
ние двухзубцовые, краевые (27) шиповидные.
Слепая кишка отсутствует.
Г ениталии. Пенис мешковидный, очень мускулистый, внутри с много-
численными складками и морщинами и без типичного стимулятора — вместо
него неопределенной формы складка. Часть морщин просвечивает сквозь
стенку заднего конца пениса. Пениальная железа имеет длинный толстостен-
ный стебель и несколько коротких пузырчатых ветвей. Ретрактор короткий
и мощный; он крепится почти у середины пениса. Крупный резервуар семя-
приемника имеет довольно длинный и тонкий проток.
Распространение. Пока обнаружен только близ города Сам-
суна (черноморское побережье Турции).
Замечание. Систематическое положение этого вида пока неясно.
В пределах рода Deroceras отсутствие типичного стимулятора необычное
явление. В то же время остальные особенности этого слизня определенно
указывают на его принадлежность к данному роду.
Deroceras (Agriolimax) vranceanum Grossu, 1969 (рис. 200)
Grossu, 1969:161, Fig. 4.
Locus typicus — «Einfluss des Baches Tiganului» — долина реки Путна
у места впадения в нее ручья Тиганулу, горы Вранче, Восточные Карпаты
(Румыния). Голотип в коллекции Гроссу, Бухарест, № 1143.
Тело темно-серое, с многочисленными темно-бурыми или почти черными
пятнышками. Их так много, что слизень кажется почти черным, лишь бока
немного светлее. Кожа толстая, с очень выпуклыми морщинами.
Размеры: дл. сок. около 20 мм.
Слепая кишка хорошо развита; ее длина в 3—6 раз больше ширины.
Гениталии. Пенис почти цилиндрический, без пережима на сере-
дине. Полость его не разгорожена на два отдела и в ней помещается хорошо
развитый конический стимулятор, слегка уплощенный и с небольшим желоб-
ком у основания. Пениальная железа располагается на заднем конце пе-
ниса и имеет вид прямого или изогнутого цилиндрического выступа с много-
численными бугорками по бокам. У основания ее крепится ретрактор и впа-
дает семяпровод.
Распространение. См. типовое местонахождение.
Все типы были собраны на поляне речной террасы под кусками древесины
и на траве.
Deroceras (Plathystimulus?) zilchi Grossu, 1969 (рис. 201) i
Grossu, 1969 : 164, Fig. 6—7.
Locus typicus — Cozla — Козла в долине p. Дуная около Железных Ворот
(Южные Карпаты, Румыния). Голотип в коллекции Гроссу, Бухарест,
№ 1144.
Небольшие и стройные слизни с тонкой просвечивающей кожей. Окраска
одноцветная грязно-серая или светло-желтая, мантия немного темнее осталь-
ных участков тела. Морщины очень слабо выражены и заметны только в зад-
ней трети тела. Пневмостом лежит у самого края мантии.
Слепая кишка имеет вид небольшого кармашка.
Гениталии. Пенис почти цилиндрический, слегка сужен на середине.
У некоторых экземпляров на заднем конце пениса имеется четкий слепой
190
I. AGRIOLIMACIDAE
выступ — аппендикс. Стимулятор тонко исчерчен и имеет вид лопатки
с закругленным внешним краем и сравнительно узким основанием. В покое
расширенная часть стимулятора согнута под прямым углом к его основанию.
Пениальная железа хорошо развита в виде 3—4 нитевидных выступов
с четкими боковыми бугорками.
Распространение. Вид был обнаружен в двух местах: близ
Железных Ворот и несколько выше по Дунаю у города Молдова-Ноуэ.
Оба местонахождения находятся в Южных Карпатах.
2. Род KRYNICKILLUS Kaleniczenko, 1851
Kaleniczenko, 1851а : 220; Р i 1 s Ь г у, 1948 : 560; Wiktor, 1971а :
270; Дамянов, Лихарев, 1975 : 324.
Типовой вид — К. melanocephalus Kaleniczenko, 1851; по следую-
щему обозначению — Pilsbry, 1922.
Слизни средних размеров (при сокращении длина более 40 мм). Тело
стройное. Мантия располагается в переднем конце тела и занимает
около 1/3 общей длины; капюшон крупный (около 1/2 длины мантии), сво-
бодно нависает над шеей. При жизни, а иногда и у фиксированных
экземпляров на мантии видна серповидная бороздка, которая лежит на
правой стороне, касаясь валика пневмостома, и свободным концом изогнута
назад (рис. 209).
Радула, как у Deroceras. Слепой кишки нет или она представлена не-
большим кармашком. Пенис без разделения на передний и задний отделы,
цилиндрический или булавовидный, т. е. слегка расширен у заднего конца.
Пениальной железы, аппендикса и других наружных придатков на пенисе
нет. Стимулятор тоже отсутствует. Внутренние стенки пениса имеют раз-
личные складки, но без рубчатой скульптуры, характерной для рода
Deroceras.
Распространение. Крым, Кавказ, черноморское побережье
Малой Азии и Балканского п-ова.
Замечания. 1. Длительное время данную группу рассматривали
как секцию или подрод рода Deroceras. В пользу этого говорили значитель-
ное внешнее сходство и общий план внутреннего строения. Впервые Вик-
тор (Wiktor, 1971а) заметил, что у видов Krynickillus на мантии имеется
характерная серповидная бороздка, которой никогда не бывает у Deroceras.
Учитывая это, а также форму пениса, отсутствие на нем всяких придатков,
а внутри него стимулятора, есть основание считать обе группы самостоя-
тельными родами.
2. Пилсбри (Pilsbry, 1948) отнес в эту группу один американский
вид — Deroceras (Krynickillus) heterura Pilsbry, у которого нет ни пениальной
железы, ни стимулятора. Однако остальные особенности этого вида, как
внешний вид, так и форма пениса, в частности наличие на нем поперечной
перетяжки, указывают на принадлежность его к одному из подродов
Deroceras.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ KRYNICKILLUS
1 (2). Голова и шея черные; мантия и. спина всегда более светлые. Семя-
приемник, яйцевод и пенис почти одинаковой длины. Яйцевод значи-
тельно короче семяяйцевода.............1. К. melanocephalus Kai.
2(1). Голова и шея темные, но светлее мантии и спины. Семяприемник
и яйцевод почти вдвое длиннее пениса. Яйцевод заметно длиннее семя-
яйцевода ..............................2. К. urbanskii (Wiktor).
2. KRYNICKILLUS
191
1. Krynickillus melanocephalus Kaleniczenko, 1851 (рис. 202—207)
К aleniczenko, 1851a : 221, T. 5, Fig. 2; В oettger 0,, 1881 : 182 (Limax)’,-
1883 : 144 [Limax (Agriolimax)}', Simroth, 1885 : 224, 326, T. 9, Fig. 23 (Agriolimax)',
1886 : 30 (Agriolimax)', 1892 : 43 (Agriolimax)', К 1 i к a, 1894 : 4 (Agriolimax)', S i m-
r о t h, 1901 : 154, T. 14, Fig. 13—21 (Agriolimax)', 1910 : 521, T. 7, Fig. 18 (Agriolimax)’,-
1912a : 36 (Agriolimax)', АкрамовсКий, 1949: 151 (Agriolimax)', Лихарев,-
Раммельмейер, 1952 : 338 (Agriolimax)', Wiktor, 1971' : 269 (Deroceras).
Locus typicus — «Stauropolin» — окрестности Ставрополя. Типы не-
известны.
Материал: из СССР — более 600 экз. из 230 пунктов; из Турции
2 экз.
Тело веретеновидное, сзади плавно заостренное с тупым,- нередко вол-
нистым килем, занимающим около половины спины. Кожа сравнительно
толстая. Морщины крупные и выпуклые; между мантийной щелью и сред--
ней линией спины около 13—15 рядов морщин.
Окраска грязновато-белая, иногда голубовато-серая. Спина и мантия-
окрашены в серый цвет (мантия всегда темнее спины), а шея, голова и щу-
пальца интенсивно черные. Характерно, что пигментация шеи распростра--
няется и на часть тела, закрытую капюшоном. Серая и черная окраска
образована мелкими точковидными меланофорами, которые в первую оче-
редь располагаются по бороздкам. Правый и левый края мантии вместе'
с валиком пневмостома, боками и подошвой остаются всегда светлыми.
Размеры: дл. сок. 40—45, ширина 12, дл. мантии 20 ММ.
Радула (рис. 204). Пример: Армения, Гергары, 1 экз.; формула:
Кишечник (рис. 205). Слепой кишки нет или она имеет вид кармашка..
Задняя часть внутренностного мешка окутана черным или бурым мезен-
терием.
Гениталии (рис. 207). Небольшая черная гонада скрыта между'
лопастями печени и кишкой и лежит на значительном удалении от зад-
него конца внутренностного мешка. От нее тянется вперед узкий, почти:
прямой гермафродитный проток, который лишь вблизи белковой железы-
сильно вздут и дает несколько извивов. Белковая железа очень крупная,
бурая. Узкая лентовидная простата вытянута вдоль всего семяяйцевода.
Цилиндрический яйцевод по длине почти равен пенису или семяпроводу.
Мускулистый цилиндрический пенис прямой или слегка спирально изогнут.
Половой ретрактор и семяпровод крепятся к пенису, сильно отступя от его’
заднего конца (на границе между средней и задней третью). Внутри пенис
имеет сложную систему меандрических или кольцевидно охватывающих-
ДРУГ друга складок. Атриум очень короткий. Резервуар семяприемника
плавно переходит в тонкий проток и по длине почти равен пенису.
Распространение. В СССР: почти весь Кавказ, пойменные
и байрачные леса Ставрополья, а также горный Крым (Чатырдаг, сбор
Рошковского в 1916 г.; гора Кошкая, Симферопольский р-н, сбор Шилейко
в 1964 г.). Вне СССР: вилайет Чорох, Турция (материалы в ЗИН АН СССР),
северный Иран (Starimihlner, Edlauer, 1957). Последнее указание требует
дополнительной проверки.
Образ жизни. Обитает преимущественно в лесной и субальпий-
ской зонах, но по берегам водоемов проникает и в нижележащие аридные
зоны. Скрывается под камнями, в подстилке, в старых пнях и валежинах.
Рис. 202—207. Krynickillus melanocephalus Kai.
202 — внешний вид, Армения, Гергеры, 9 X 1922; 203 — голова и шея; 204 — зубы радулы; 205 — кишеч-
ник; 206 — ретракторы; 207 — дистальная часть гениталий (204—207 — окр. Батуми, 5 XI 1958).
2. KRYNICKILLUS
193
2. Krynickillus urbanskii (Wiktor, 1971) (рис. 208—218)
Grossu, Lupu, 1957a : 171, Fig. 1 [Deroceras (Hydrolimax) melanocephalus Ka-
leniczenko]; Urbanski, Wiktor, 1968:72, Fig. 10 (D. melanocephalum Kai.);
Lupu, 1971a : 126, Fig. 4a (D. melanocephalum); Wiktor, 1971a : 264, Fig. 2, 3
[Deroceras (Krynickillus)]; Д аминов, Лихарев, 1975 : 324, фиг. 255, 256 [De-
roceras (Krynickillus) ].
Рис. 208—215. Krynickillus urbanskii (Wiktor) (из: Wiktor, 1971a).
208, 209 — внешний вид; 210 — раковина; 211 — зубы радулы; 212 — кишечник; 213—215 — вари-
анты положения ретрактора правых щупалец по отношению к пенису и половому ретрактору.
Locus typicus — «Tai oberhalb Stadt Karlovo» — ущелье выше города
Карлово, Болгария. Голотип в Зоологическом музее Вроцлавского универ-
ситета (Польша).
Материал: из СССР нет; из других стран — 60 экз. (в том числе
типы).
Тело сравнительно стройное. Киль занимает около 1/3 спины. Кожа
тонкая, с нежным рельефом; между средней линией спины и мантийной
щелью 18 рядов морщин.
Живые слизни черно-пепельного цвета. Спина и мантия наиболее тем-
ные, а бока книзу постепенно светлеют. Голова и щупальца черноватые. Заты-
лок и подошва кремовые. Как продолжение киля посередине спины проходит
13 и, М, Лихарев, А. И, Виктор
194
I. AGRIOLIMACIDAE
более светлая (синеватая) килевая полоса. После фиксации слизни стано-
вятся коричневато-серыми. Слизь водянистая, бесцветная.
Размеры: дл. пол. около 45, мантия до 15 мм.
Раковина (рис. 210).
Рис. 216—218. Krynickillus urbanskii (Wiktor) (из: Wiktor, 1971а).
216 — гениталии; 217 — дистальные части гениталий; .218 — вскрытый пенис.
Радула (рис. 211). Пример: 1 экз. из-под Тырново (Болгария),
с 106 26 45 к
-3- + —+ —+ —Xi32-
212).
мешок окутан светлым мезентерием,
(рис. 216—218). В отличие от предыдущего вида семя-
Кишечник (рис.
Внутренностный
Гениталии
яйцевод более тонкий и значительно короче, в связи с чем яйцевод и семя-
провод очень длинные. Последний очень тонкий, тянется вперед вдоль
семяприемника и близ атриума поворачивает назад и впадает в задний
конец. пениса рядом с тонким и длинным ретрактором. Пенис небольшой
булавовидный, внутри с нежными складками. Семяприемник имеет не-
3. LYTOPELTE '
195
«большой яйцевидный резервуар й очень длинный тонкий проток, по длине
почти равный семяпроводу. Яйцевод имеет вид сильно извитой трубки,
по диаметру почти равной пенису. Ретрактор правых щупалец иногда пере-
крещивается с пенисом или его ретрактором (рис. 213—215).
Распростр ан е н и е. Обнаружен в Румынской Добрудже, в вос-
точной Болгарии; одна находка сделана в окрестностях Трабзона (Турция).
Образ жизни. Обитает в дубовых, грабовых, буковых и смешанных
лесах, а также в зарослях кустарников. Днем прячется в подстилку
•и под корой деревьев. Живет как на равнинах, так и в горах до высоты
1500 м над ур. м.
Подвижные слизни, при движении напоминающие Lehmannia.
Замечание. Вид во многом сходен с К. melanocephalus Kai., но
отличается от него меньшими размерами, более узким телом, тонкой кожей
с нежным рельефом и отсутствием темного пятна на затылке и шее. По вну-
треннему строению он отличается коротким семяяйцеводом, длинными
яйцеводом и семяпроводом и очень длинным протоком семяприемника.
Некоторое сходство в строении пениса, а именно отсутствие пениальной
железы и стимулятора, привело к тому, что данный вид некоторое время
принимали за К. melanocephalus Kai. (Grossu, Lupu, 1957a; Urbaiiski,
Wiktor, 1968; Lupu, 1971a).
3. Род LYTOPELTE 0. Boettger, 1886
Boettger 0., 1886a : 266 [Amalia (Lytopelte)]; Simroth, 1886b : 311 [Agrio-
limax (Platytoxon)], 1901 : 174 (Tropidolytopelte); H e s s e P., 1926 : 24 (Lytopelte s. str.).
Типовой вид — Amalia (Lytopelte) longicollis O. Boettger, 1886
(=Amalia maculata Koch et Heynemann, 1874); по монотипии.
Небольшие слизни (в движении длина не более 35 мм) со стройным те-
лом, которое при сокращении становится коротковеретеновидным. При
жизни киль тупой и занимает заднюю половину спины. Мантия обычно
немного менее х/2 общей длины. После жесткой фиксации киль приобретает
облик гребня, простирающегося на всю спину. При этом мантия сильно
сокращается, а капюшон приподнимается над шеей, нередко даже отгибаясь
назад. Между мантийной щелью и средней линией спины 11—13 рядов мор-
щин. Пневмостом лежит сильно постмедиально.
Окраска пестрая. Рисунок образован мелкими пятнышками в виде
штрихов или брызг. Вдоль середины спины и мантии обычно лежит светлая
полоса, образующая несколько местных расширений и окантованная сгу-
щением темного пигмента.
Срединный зуб радулы всегда трехзубцовый. Боковых зубов около 30,
из них первые 15 тоже трехзубцовые, а остальные двухзубцовые. В отли-
чие от Deroceras краевых зубов значительно меньше, чем боковых (7); все
они кинжаловидные.
Слепой кишки нет. Ретрактор правых щупалец - не перекрещивается
с пенисом или его ретрактором. Пенис мешковидный, без придаточных ор-
ганов; как и у Deroceras, неглубокой перетяжкой он разделен на передний
и задний отделы. Внутри пениса находится стимулятор с известковым
тельцем. .
Монотипический род.
Замечания. 1. Данный род во многом сходен с Deroceras. Главные
отличия: а) при сокращении тела у Lytopelte появляется мощный киль,
а капюшон приподнимается над шеей; и то, и другое не свойственно Dero-
ceras, что говорит об ином строении подкожной мускулатуры; б) совершенно
иной план строения радулы: у Deroceras почти все боковые зубы трехзуб-
цовые,^а краевых зубов значительно больше, чем боковых.
13*
196
I. AGRIOLIMACIDAE
Указанные различия между обеими группами столь существенны, что
дают нам основание присоединиться к точке зрения Гессе и Тиле (Hesse Р.,
1926; Thiele, 1931), рассматривавших их как самостоятельные роды.
В этом отношении мы расходимся во взглядах с другими авторам®
(Grossu, Lupu, 1961b; Grossu, 1970b; Altena, 1975), которые вслед за Зим-
ротом (Simroth, 1901) объединяют Lytopelte в один род с группой Liolyto-
pelte Simroth, 1901. Последнюю мы считаем подродом рода Deroceras (с. 169).
Указанные авторы основанием для объединения Lytopelte с Liolytopelte
считали наличие у обеих групп известкового тельца на стимуляторе пениса.
Однако сходные образования встречаются в разных группах Stylomma-
tophora (например, у Gastrodontidae), где они возникают совершенно незави-
симо. В то же время общий план строения Liolytopelte во многом очень-
сходен с другими подродами Deroceras.
2. В числе синонимов Lytopelte Boettger, 1886 известен Platytoxon Sim-
roth, 1886. Бётгер опубликовал описание Lytopelte в двух изданиях, при-
чем оба они вышли в 1886 г.: а) в книге G. Radde «Fauna und Flora
des sudwestlichen Caspi-Gebietes»: б) в журнале «Jahrbucher der Deutschen
Malakozoologischen Gesellschaft», t. 13, c. 241. В последней работе он ука-
зывает, что книга Радде уже вышла в свет. В том же номере того же жур-
нала на с. 311 впервые опубликовано Зимротом описание группы Platyto-
xon. Следовательно, название Lytopelte старше, чем Platytoxon.
Lytopelte maculata (Koch et Heynemann, 1874) (рис. 219—225)
Koch, Heynemann, из: Мартенс, 1874 : 2, табл. 1, рис. 4, табл. 3,
рис. 35 (Amalia); Koch, Heynemann, 1874 : 152, Т. 6, Fig. 5 (Amalia)', Boett-
ger О., 1886a : 266, T. 2, Fig. 1 [Amalia (Lytopelte) longicollis, syn. nov.); Simroth,
1886b : 311, T. 10, Fig. 1—4 [Agriolimax (Platytoxon)]; Rosen, 1892 : 123 (transcaspia,
syn. nov.); Розен, 1894 : 40 (transcaspia); Simroth, 1901 : 169, T. 17, Fig. 1—4;
1910 : 526; 1912a : 44, T. 3, Fig. 42, T. 7, Fig. 26; Лихарев, Раммельмейер,
1952 : 340; Altena, 1970a : 175, Fig. 1 (Deroceras kandaharensis, syn. nov.?); 1975 63.
[Lytopelte (Liolytopelte) kandaharensis].
Locus typicus — «prope Maracandam» — окрестности Самарканда. Типы
неизвестны.
Материал: из СССР — более 500 экз. из 68 пунктов.
Ползущее животное обладает стройным телом, с легким пережимом
позади мантии и очень подвижно. Мантия удлиненно-овальная, сзади шире,
чем спереди. Кожа тонкая с плоскими морщинами и мелкими бороздками.
Киль образован 2—3 рядами морщин.
Окраска от красновато-серой до светло-коричневой. По середине спины
и мантии тянется светлая полоска, которая имеет ясные расширения у пе-
реднего и заднего края мантии и в передней трети спины. По сторонам от
этой полосы располагается рисунок из многочисленных серых или черных
штрихов, которые постепенно редеют к ноге и к краям мантии; особенно
плотные сгущения штрихов возникают вокруг упомянутых расширений
светлой полосы и в задней половине спины по сторонам от киля.
Живые слизни очень напоминают некоторые виды Deroceras, особенно
D. laeve (Muller).
После жесткой фиксации, в результате резкого сокращения, тело при-
обретает очень характерный облик. Оно становится коротковеретеновидным
с максимальной шириной позади почти круглой мантии; капюшон припод-
нимается над шеей, а иногда даже отворачивается назад. Киль очень рез-
кий и простирается на всю спину, т. е. до мантии. Подошва остается узкой,
и поэтому в поперечном сечении тело круглое. Рисунок становится более
четким и сложен из многочисленных темно-коричневых или почти черных
пятнышек, которые образуют сгущения по сторонам от киля, вокруг круп-
Рис. 219, 220. Lytopelte maculata (Koch et Heyn.).
219 — в движении, Душанбе, 24 X 1977; 220 — после жесткой фиксации, Самарканд, 3 IV 1.929,
198
I. AGRIOLIMACIDAE
ного заднего светлого пятна мантии и менее четкого переднего пятна. Зад-
нее пятно располагается над раковинкой. Бока, шея и подошва обычно
остаются светлыми, а затылок иногда с серым налетом. Нередко централь-
ные участки светлых мантийных пятен в свою очередь заняты темным
пигментом.
Рис. 221—225. Lytopelte maculata (Koch et Heyn.).
221 — зубы радулы; 222 — кишечник; 223 — гениталии, Ферганский хребет, 3 VII 1961; 224 — ди-
стальная часть гениталий (пенис вскрыт); 225 — Ьластинка стимулятора в трех положениях.
Темный рисунок появляется уже у самых молодых слизней, четко
оконтуривая заднее пятно мантии и киль. С возрастом темного пигмента
становится больше.
Интенсивность развития рисунка зависит не только от возраста, но
и от образа жизни. У популяций с открытых мест темного пигмента больше,
и он занимает бока тела почти до самой подошвы; к тому же и фоновая
окраска темнеет. Даже киль и .заднее пятно мантии приобретают серый
налет, но тем не менее четко выделяются на фоне темной верхней стороны
тела (/. longicollis Boettger, 1886 — syn. transcaspia Rosen, 1892).
3. LYTOPELTE
199
Популяции, обитающие в лесу или в тенистых ущельях, отличаются
более светлой фоновой окраской и менее интенсивным темным рисунком
(/. typica).
Размеры: дл. пол. до 35, дл. сок. до 26 мм.
Раковина закругленно-прямоугольная; правый край ее слегка вогнут,
внизу под нуклеусом с треугольной ямкой. Размеры: длина до 3, ширина
до 1.7 мм.
Радула (рис. 221). Впервые описание радулы данного вида опубликовано
у Мартенса (1874) на основании исследования одного экземпляра из окрест-
обнаружил
его радулы:
ностей Самарканда. Изученный нами экземпляр из Талыша
большое сходство со среднеазиатским. Ниже дается формула
с 156 186 7к
т + — + — + — X 90.
Кишечник — рис. 222.
Гениталии (рис. 223—225). Гонада темно-серая или коричневая,
лежит за зобом и почти не закрыта печенью. Проток гонады толстый и дает
несколько извивов. Белковая железа светйо-коричневая, крупная, слегка
охватывает снизу зоб. Простата располагается на передней четверти семя-
яйцевод а. Яйцевод и семяпровод приблизительно одной длины; первый
плавно расширяется к атриуму. Семяприемник с овальным резервуаром
и тонким длинным протоком, открывающимся в атриум.
Пенис мешковидный, поперечной перетяжкой разделен на крупный
передний и небольшой задний отделы. Семяпровод впадает в него почти
апикально. Задний отдел пениса внутри со сложной системой нежных
складок. Передний отдел имеет несколько вздутий. Внутри него находится
крупный стимулятор, имеющий форму кольцеобразного или спирального
валика. В центре стимулятора лежит известковое тельце в виде круглой
пластинки с зубчатыми краями с крупным отростком — шпорой, — сидя-
щим в центре пластинки и острием, направленным в просвет пениса.
Известковое тельце закладывается внутри стимулятора в виде тонкой
пластинки; позднее на краю ее формируется когтевидный выступ, который
благодаря покрывающим его тканям имеет хрящевидный облик. При даль-
нейшем росте пластинки вширь этот выступ оказывается почти в центре ее.
Под конец ткани, покрывающие тельце, разрушаются, и оно оказывается
на поверхности стимулятора.
Распространение. В СССР: Средняя Азия — плодовые леса
юго-западных склонов и отрогов Ферганского хребта, долины рек Зерав-
шан (Самарканд, Пенджикент), Вахша (Душанбе), Пянджа (Куляб); запад-
ный Копетдаг; Кавказ — Ленкоранская низменность и прилегающие пред-
горья Талыша; две находки сделаны на южных склонах Большого Кавказа
в пределах Ахсуинского района АзССР (Ахсу, Падар).
Вне СССР: Афганистан (Герат, Кандагар), северный Иран (провинции
Гилян, Мазендеран).
Образ жизни. В лесных областях населяет влажные биотопы,
преимущественно тенистые ущелья и берега водоемов с обильной сырой
подстилкой, гнилыми пнями и валежинами. В безлесных засушливых об-
ластях тяготеет к берегам природных водоемов и оросительной сети, где
в дневное время скрывается под камнями, в подстилке и в трещинах почвы.
Однолетние животные. Судя по размерам особей, собранных в разное
время года, состоянию гонады, содержанию спермы в семяприемнике,
данный вид размножается в конце осени и в середине весны, т. е. дает две
генерации в год.
В засушливых субтропических областях Средней Азии и Копетдагд
активная жизнь этого слизня прерывается продолжительной летней спяч-
800
I. AGRIOLIMACIDAE
она короче, а в го-
Зимрот относил
следующие виды
кой, когда животные укрываются глубоко в почву. В лесных областях
Ленкоранской низменности и Ферганского хребта летней спячки, по-ви-
димому, нет или она бывает не каждый
год. Длительность зимней спячки раз-
лична, но в долинах
рах длиннее.
Замечание,
к данной группе
(Simroth, 1912а): L. maculata (Koch et
Heynemann, 1874), L. longicollis (Boett-
ger, 1886), L. transcaspia Rosen, 1892,
L. boettgeri Rosen, 1892. Признавая их
близость и наличие переходов между
ними, он считал, что главные отличия
относятся к форме мантии и степени
развития темного пигмента. В настоя-
щее время у нас накопился значитель-
ный материал по Lytopelte из различ-
ных мест Средней Азии, Копетдага,
Азербайджана, Афганистана и север-
ного [Ирана. В результате изучения
изменчивости окраски мы пришли к вы-
воду, что за [исключением L. boettgeri
остальные формы являются цветовыми
вариациями L. I maculata. Отличия
в форме мантии исключительно обязаны
фиксации — в более крепком фиксаторе
мантия сокращается сильнее, чем в сла-
бом.
Что касается L. boettgeri, описанной
из Копетдага, т. е. там, где также об-
наружена обычная L. maculata, то но-
вых находок похожих форм не было.
Судя по ее внешнему облику (Simroth,
1912а : т. 3, Fig. 41). не исключено, что
она описана по темным экземплярам
L. maculata, еще более потемневшим
в результате плохой консервации.
22 6
4. Род MEGALOPELTE Lindholm, 1914
Lindholm,
Раммельмейе
рев, 1957 : 1253.
г jQT иповой
. ’^•1®
1914 : 167; Лихарев,
р, 1952 : 361; Л и х а-
г jQT иповой вид — Megalopelte
simrothi, Lindholm, 1914; по монотипии.
Q Небольшие слизни (дл. пол. до 25 мм)
с очень крупной мантией, которая почти
полностью закрывает верхнюю сторону
животного (рис. 226, 227). Мантия при-
Рис. 226. Megalopelte simrothi Lindh. Росла к туловищу ЛИШЬ центральным
Окр. Батуми, 2—5 XI 1957. участком, а по периферии свисает ши-
рокой каймой. Передняя часть этой
каймы — капюшон — особенно широка (около 1/2 длины мантии). Пневмо-
стом и мантийная щель располагаются почти медиально и лишь при дви-
4. MEGALOPELTE
201
жении животного смещаются немного назад от середины правого края
мантии.
Раковина очень тонкая и хрупкая, нуклеус ее лежит у заднего края
слева от продольной оси. Небольшая мантийная полость смещена в зад-
нюю половину тела. Срединный и боковые зубы радулы с крупными мезо-
конами и отчетливым иэкто- и эндоконами. Краевые зубы кинжаловидные.
Кишка полностью располагается за зобом. Кровеносная сеть легкого раз-
Рис. 227—232. Megalopelte simrothi Lindh. Окр. Батуми, 2—5 XI 1957.
227 — в движении; 228 — подошва; 229 — зубы радулы; 230 — кишечник; 231 — ретракторы; 232 —
мантийный комплекс, слева — вид снизу, справа — вид сверху.
вита слабо. Мешковидная почка сверху полностью закрывает сердце. Моче-
вой пузырь относительно крупный и длинный (рис. 232). Длинный цилин-
дрический пенис слагается из вздутого переднего и плавно суженного
назад заднего отдела. Ретрактор правого глазного щупальца пролегает
между пенисом и яйцеводом, огибая пенис справа и снизу (рис. 231).
Монотипический род.
Замечание. Благодаря необычайно крупной мантии внешний
облик животного резко отличается от других родов семейства. Длинный
цилиндрический пенис сближает этот род с Mesolimax Pollonera, а общий
план строения кишечника и радулы, а также отсутствие придаточных орга-
нов на женских половых путях указывают на принадлежность к Agrioli-
macidae.
202
I. AGRIOLIMACIDAE
Megalopelte simrothi Lindholm, 1914 (рис. 226—235)
Рис. 233. Megalopelte simrothi Lindh. Гениталии. Окр. Батуми, 2—5 XI 1957.
Locus typicus'. «Dsanssul» — Дзансул (бассейн p. Мургут-су, вилайет
Чорох, северо-восточная Турция). Типы неизвестны.
Материал: из СССР 13 экз. из 7 пунктов; Турция 1 экз.
4. MEGALOPELTE
203
При движении тело стройное и веретеновидное (рис. 227), при сокра-
щении валикообразное (рис. 226). У ползущих животных голова и задний
конец тела лишь слегка выступают из-под мантии. Щупальца короткие
и толстые. Киль неясный и очень короткий. Кожа тонкая со слабым релье-
фом из мелких бороздок и плоских морщин, при натяжении сквозь нее
просвечивают внутренности. От основания мантии к подошве радиально
расходятся около 40 бороздок. Мантия, голова и задний конец тела черные
или темно-серые; бока под мантией светло-серые. Пигмент в основном рас-
Рис. 234, 235. Megalopelte simrothi Lindh. Гениталии.
234 — пенис с ввернутой внутрь задней частью; 235 — он же вскрытый.
полагается на морщинах, а бороздки остаются светлые. Подошва белая
(рис. 228).
Размеры: дл. пол. до 25, дл. сок. до 20 мм.
При фиксации нередко свободная кайма мантии выворачивается кверху.
Раковина округло-прямоугольная с легкими линиями нарастания.
Радула (рис. 229). Изучен 1 экз. из Батумского ботанического сада;
формула
с 146 24к
Т + ~2“ + Т-*
Кишечник (рис. 230).
Гениталии. Гонада гроздевидная, состоит из крупных фолликул
и занимает задний конец внутренностного мешка (рис. 230, 233). Семя-
яйцевод образует один крупный извив. Простата длинная, лентовидная.
Яйцевод довольно длинный и толстый.
Пенис четко подразделен на два основных отдела: передний, сзади
мешковидно вздутый и плавно сужающийся к атриуму; задний — более
узкий с многочисленными перетяжками, плавно сужающийся к заднему
концу, к которому крепится половой ретрактор, перед этим веерообразно
распадаясь на серию мелких мускулов. Тонкий семяпровод впадает в пе-
нис апикально. Внутренние стенки задней части пениса имеют рубчатую
204
I. AGRIOLIMACIDAE
поверхность. Семяприемник с овальным резервуаром, который коротким
протоком открывается в пенис близ атриума.
У некоторых вскрытых нами экземпляров задняя часть пениса (частично
с ретрактором) была ввернута внутрь переднего отдела (рис. 234, 235).
По-видимому, данные слизни готовились или уже завершили копуляцию,
при которой, вероятно, весь задний отдел пениса выворачивается наружу.
Распространение. Кроме типового местонахождения с тер-
ритории бассейна р. Мургут-су (приток р. Чорох, Турция) несколько на-
ходок сделано близ Батуми (Ботанический сад, участок колхидского леса)
и 1 экз, найден у с. Акармара близ Ткварчели (Абхазия). Кроме того, один
молодой экземпляр, принадлежащий Венскому естественно-историческому
музею, найден 9 VI 1969 на западе Малой Азии близ оз. Абант.
Образ жизни. Обитает в очень влажных местах широколиствен-
ного леса, в подстилке и под валежинами. Слизни ведут очень скрытный
образ жизни и попадаются редко и 1—3 экземплярами.
5. Род MESOLIMAX Pollonera, 1888
Pollonera, 1888 : 8; Simroth, 1892 : 42; 1894b : 12; 1901 : 133; Н е s s е Р.,
1926 : 26; Germain, 1936 : 88.
Типовой вид — М. brauni Pollonera, 1888; по монотипии.
Слизни средних размеров (дл. сок. не более 60 мм). Тело узкое, при
сокращении веретеновидное, с коротким и слабым килем на конце. Кожа
толстая, с четким рельефом. Пневмостом располагается более или менее
постмедиально. Боковые доли подошвы (рис. 3, IV), кроме обычных много-
численных поперечных бороздок, имеют по две дополнительных продоль-
ных бороздки, которые разбивают каждую долю на три почти равных по
ширине участка. Срединная доля подошвы, как у Deroceras, покрыта попе-
речными бороздками, которые V-образно изогнуты назад.
Срединный и боковые зубы радулы с хорошо развитыми мезоконами
иЧзо слабыми экто- и эндоконами. Краевые зубы шиповидные.
Следующие особенности внутреннего строения отличают данный род
от остальных Agriolimacidae.
Раковина симметричная; нуклеус лежит почти на середине ее заднего
края (рис. 236). Задний конец колумеллярного мускула вилообразно раз-
двоен, и между его ветвями проходит задняя кишка, т. е. она как бы про-
бодает основание мускула (рис. 241). Ретрактор правых щупалец обычно
перекрещивается^с пенисом. Пенис длинный и цилиндрический; передний
конец его вместе ьс прилегающим участком семяпровода окружен перепон-
чатой манжеткой (рис. 240).
Распространение. Малая Азия; о. Родос. Указания на при-
сутствие некоторых видов на Кавказе и Балканах требуют проверки.
Замечание. До недавнего времени наши знания об этом роде были
очень фрагментарны и нечетки и основывались на нескольких сообщениях
старых авторов. К тому же каждый из видов, относимых в эту группу, был
описан по одной находке одного или нескольких экземпляров, из которых
большинство были молодыми, т. е. с недоразвитыми гениталиями.1 В итоге
выходило, что для’рода Mesolimaz характерны «внешний вид и гениталии,
как у Limaz, подошва и раковина, как у Milaz, а радула и кишечник, как
у Agriolimaz» (Germain, 1936: 69; перевод с французского наш, — Л. и В.).
Каких-либо особенностей, свойственных только данному роду, авторы не
отмечали.
1 Описание одного из видов — М. reibischi Simroth, 1892 настолько неопределенно,
что нет уверенности в том, что он действительно принадлежит данному роду. Поэтому
он не включен в определительную таблицу и обсуждается особо (с. 207).
5. MESOLIMAX
205
Впервые Виктор (Wiktor, 1971а) заметил, что у двух изученных им ви-
дов Mesolimax — М. brauni и М. escherichi — передний конец пениса и при-
легающий участок семяпровода окружены особой перепончатой манжет-
кой, а задняя кишка прободает основание колумеллярного мускула. Эти
особенности не только не встречаются среди остальных Agriolimacidae,
но и у других семейств слизней. Поэтому их можно рассматривать как
главные отличительные черты данного рода.
К сожалению, осталось неизученным внутреннее строение пениса.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ MESOLIMAX
1 (2). Тело сверху серовато-охристое, мантия с двумя темными боковыми
полосами. Пенис сильно закручен, длина его около 1/2 общей длины
гениталий...........................................1. М. brauni Poll.
2 (1). Тело сверху черное, мантия без боковых полос. Пенис слегка изогнут,
длина его около х/3 общей длины гениталий . . 2. М. escherichi Simr.
1. Mesolimax brauni Pollonera, 1888 (рис. 236—241)
Pollonera, 1888 : 8, T. 2, Fig. 19—23; Simroth, 1901 : 134; Germain,
1936 : 88, Fig. 5; Wiktor, 1971a : 274, Fig. 2E, 6; F о г с a r t, 1972 : 112.
Terra typica — «Asia Minor» — Малая Азия. Типы неизвестны.
Материал: из СССР нет; из других стран — 17 экз. плохой со-
хранности.
Тело при сокращении становится веретеновидным с максимальной ши-
риной позади мантии. Задний конец заострен, с коротким и тупым килем.
Мантия занимает около 1/3 длины тела, переднюю половину ее образует
крупный капюшон. Пневмостом лежит слегка постмедиально и окружен
светлым полем.
Основная окраска серовато-охристая. Киль и середина спины заняты
узкой кремовой килевой полоской. Мантия с двумя широкими темными
боковыми полосами, окаймленными изнутри светлыми полосами. Границы
как темных, так и светлых полос мантии нечеткие, как бы размытые. Сере-
дина мантии затемнена. Задний конец тела ниже киля с темным сетчатым
рисунком. Бока к подошве постепенно светлеют. Подошва кремовая.
Размеры: дл. сок. до 53 мм.
Внешний облик этого вида во многом сходен с некоторыми цветовыми
вариациями Gigantomilax monticola (Bttg.) и G. koenigi (Simr.).
Раковина яйцевидная (рис. 236).
Радула (рис. 237, 238). Пример: 1 экз. с о. Родос (из коллекции Вен-
ского естественно-исторического музея, № 77623); формула:
с 176 26 , 27к
з + з
Экто -и эндоконы срединного и
зубы со сравнительно короткими
довольно сильно перекручен (рис.
Гениталии (рис. 240). Пенис тонкий, цилиндрический, сильно
закручен. Длина его почти равна х/2 общей длины гениталий. Семяприем-
ник открывается в яйцевод на заметном удалении от атриума.1 Ретрактор
правых щупалец перекрещивается с пенисом.
Распространение. Типовые экземпляры этого вида были най-
дены где-то в Малой Азии. Позднее (Babor, 1898) его обнаружили в Болга-
1 Согласно Полонера (Pollonera, 1888) и Жермену (Germain, 1936), семяприемник
впадает непосредственно в атриум между пенисом и яйцеводом.
-2-Т—Х101.
боковых зубов слабо развиты. Краевые
и массивными мезоконами. Кишечник
239).
Рис. 236—241. Mesolimax braunt Poll, (из: Wiktor, 1971a);
236 — раковина; 237, 238 — зубы радулы; 239 — кишечник; 240 — дистальная часть гениталий; 241 —
ретракторы и задняя кишка.
ДОПОЛНЕНИЕ К РОДУ MESOLIMAX
207
рии, однако проведенная нами проверка материалов Бабора показала,
что он имел дело с Lehmannia пу delta (Bourg.). Наконец, недавно (Wiktor,
1971а) М. brauni найден в окрестностях города Анталья (Малая Азия,
к югу от Западного Тавра) и на о. Родос (Forcart, 1972).
2. Mesolimax escherichi Simroth, 1899 (рис. 242)
Simroth, 1899 : 36; 1901 : 133, T. 13, Fig. 1—7; Germain, 1936 : 91, Fig.
7—11; Wiktor, 1971a : 276, Fig. 7.
Locus typicus — «Sultan Dagh (Permata)» — гора Султан (Малая Азия).
Типы неизвестны.
Материал: из СССР нет; из других стран — 7 экз. плохой сохран-
ности.
Верхняя сторона тела черная, особенно
боках окраска немного светлее. Шея под
Пневмостом окружен серой пло-
щадкой. Срединная доля подошвы
белая, боковые темные, у внеш-
него края совершенно черные.
Размеры: дл. сок. до 40 мм,
длина мантии до 11 мм.
Гениталии. Пенис ци-
линдрический, слегка изогнут или
извит. Длина его около 1/3 длины
гениталий или немного меньше.
Семяпровод и половой ретрактор
крепятся к пенису апикально.
Семяприемник впадает в атриум
между пенисом и яйцеводом. Рет-
рактор правых щупалец перекре-
щивается с пенисом или лежит
рядом с ним.
Распространение. Ма-
лая Азия (Турция): хребет Сул-
тан; окрестности Анкары; Запад-
середина спины и мантия; на
капюшоном серовато-желтая.
Рис. 242. Mesolimax escherichi Simr. (из: Wik-
tor, 1971а). Дистальная часть гениталий.
ный Тавр (вилайет Анталья —
у озера Авлан близ Эльмалы и у Дёшемеальты). Найден как на открытых
травянистых склонах, где прячется под камнями, так и в лесу из кедра
ливанского.
Дополнение к роду MESOLIMAX
Ниже на основании немногих литературных сведений дается характери-
стика одного вида, систематическое положение которого во многом остается
еще неясным. Он был описан по одному очень небольшому молодому
экземпляру и других находок более не было.
Mesolimax reibischi Simroth, 1892
Simroth, 1892 : 43; 1894b : 12, T. 17, Fig. 4; 1901 : 133; 1912a : 109.
Locus typicus — «Umgegend von Maikop» — окрестности Майкопа. Тип
неизвестен.
Тело стройное, сверху черное, подошва светлая, киль короткий. При
сокращении капюшон полностью закрывает шею и голову. Внешне живот-
208
I. AGRIOLIMACIDAE
ное напоминает молодых «.Limax» ananowi Simroth ( = Caucasolimax cauca-
sicum (Simr.)].
Кишечник, как в роде Deroceras, т. е. зоб располагается впереди кишки,
а левая доля печени лежит перед ним. Черная гонада находится позади
зоба. Пенис мешковидный, внутри с мощной продольной складкой. Ретрак-
тор правых щупалец перекрещивается с пенисом.
Замечание. Крайне лаконичное и несовершенное описание делает
невозможным дать четкую характеристику этому виду. В то же время
отсутствие новых находок ставит под вопрос: существует ли этот вид
в природе?
Внешнее сходство с «Limax» ananowi [syn. Caucasolimax caucasicum
(Simr.)], отмеченное даже автором вида (Simroth, 1901), может быть неслу-
чайным. Пенис М. reibischi также обнаруживает' некоторые черты сходства
с пенисом молодого С. caucasicum как по форме, так и по наличию внутри
него продольной складки и зачатка папиллы.
Против сближения М. reibischi с С. caucasicum как будто бы говорит
отсутствие у первого вида третьей петли кишечника. Однако учитывая,
что для описания использован очень молодой экземпляр, не исключено,
что эта петля столь мала, что Зимрот ее не заметил.
Окончательно решить эту загадку можно только переисследовав голо-
тип М. reibischi. Однако неизвестно, сохранился ли он.
Дополнение к семейству AGRIOLIMACIDAE
Приведенные ниже характеристики монотипического рода Toxolimax
Simroth и соответствующего вида составлены исключительно на основании
данных первоначального описания и рисунков, так как этот вид больше
ни разу не был найден. Типы, по-видимому, утрачены.
Наши предшественники, придавая большое значение таким признакам,
как присутствие рудимента (или зачатка!) мантийной борозды и известко-
вой шпоры внутри пениса, единодушно относили Toxolimax в особый под-
род рода Mesolimax Pollonera.
Ниже мы специально останавливаемся на таксономической оценке
указанных признаков и высказываем некоторые предположения, под-
твердить или опровергнуть которые можно будет лишь тогда, когда будут
найдены слизни с идентичными особенностями. В настоящее время, когда
характеристика Mesolimax пополнилась рядом существенных признаков,
несомненно, что Toxolimax нельзя считать за подрод этого рода. В то же
время принадлежность его к Agriolimacidae совершенно очевидна.
Род TOXOLIMAX Simroth; 1899, stat. nov.
Simroth, 1899 : 37; 1901 : 135; Hesse P., 1926 : 27.
Типовой вид — Mesolimax (Toxolimax) hoplites Simroth, 1899;
по монотипии.
Небольшие слизни (дл. сок. до 22 мм). Мантия занимает около х/3 общей
длины. Пневмостом лежит постмедиально. Над ним простирается нечеткая
и короткая продольная мантийная бороздка.
Раковина с эксцентрическим нуклеусом, который снизу отграничен от
спатулы четкой септой.
Кишечник двухпетлевого типа. Пенис булавовидный, сбоку с коротким
коническим выступом, внутри которого находится известковый шип или
шпора. Ретрактор правых щупалец перекрещивается с пенисом.
Монотипический род.
TOXOLIMAX
209
Над пневмостомом пролегает ман-
245
рп
Toxolimax hoplites (Simroth, 1899) (рис. 243—245)
Simroth, 1899 : 37 [Mesolimax (Toxolimax)]; 1901 : 135, T. 13, Fig. 8—15 [Me-
solimax (Toxolimax)].
Locus typicus — «Brussa» — Бурса (северо-запад Малой Азии). Типы
неизвестны.
Тело более массивное, чем у видов Mesolimax, с четким рельефом. Тупой
киль ограничен задним концом спины,
тийная борозда, которая сзади закан-
чивается у заднего края мантии, а впе-
реди немного загибается налево и плавно
сходит на нет, не достигнув продольной
оси тела, т. е. почти так же, как
у Krynickillus urbanskii (Wiktor).
Верхняя сторона тела темно-серая
или серовато-бурая, у молодых почти
черная. Пневмостом окружен светлым
полем. Подошва светло-коричневая, бо-
ковые доли у внешнего края с серым
налетом.
Размеры: дл. сок. 20—22 мм.
Раковина довольно толстая, на зад-
нем конце вздутая.
Гениталии (рис. 244, 245). Яйце-
вод очень короткий. Семяприемник не-
большой, нечетко разделен на резер-
вуар и проток и впадает в атриум. Пенис
сзади булавовидно вздут, к атриуму
плавно сужается; длина пениса около
1/Б общей длины гениталий. Семяпровод
и ретрактор крепятся к нему субапи-
кально. Сбоку пенис имеет короткий
конический выступ с заостренным кон-
цом. Этот выступ не имеет просвета.
В толще тканей его залегает своеобраз-
ная известковая шпора или шип, около
1 мм длины. В плане она имеет вид
двояковыпуклой линзы, один конец ко-
торой оттянут и заострен. В профиль
шпора очень узкая, палочковидная и
слегка изогнута. Заостренный конец
шпоры располагается в остром конце
пениального выступа.
Распространение. Един-
ственная находка сделана близ го-
рода Бурса (северо-запад Малой Азии).
Замечания. 1. Из четырех
типовых экземпляров Зимрот вскрыл
три самых крупных. Лишь один был достаточно половозрелым (в мужской
фазе). Своеобразная шпора на пенисе была обнаружена именно у этого
экземпляра. Зимрот считал, что она выполняет функцию аналогичных
пениальных образований у других моллюсков, т. е. служит для возбужде-
ния партнера по копуляции. Однако последнее маловероятно. Шпора пол-
ностью окружена тканями стенки пениса. Острый конец ее направлен
внутрь животного, а не наружу. Поэтому при копуляции и выворачивания
14 И. М. Лихарев, А. И. Виктор
203
244
Рис. 243—245. Тoxolimax hoplites (Simr.)
(из: Simroth, 1901).
243 — внешний вид; 244 — гениталий; 24S —
«шпора».
210
II. BOETTGERILLIDAE
пениса «наизнанку» острый конец шпоры будет ориентирован от партнера,
что противоречит приписываемой ей роли.
Невольно возникает подозрение, не является ли эта шпора каким-то
посторонним образованием, например цистой паразита, а выступ пениса
следствием развития этой цисты.
2. Что касается присутствия на нижней стороне раковины септы и руди-
мента спирального завитка, то такие структуры изредка встречаются у дру-
гих видов слизней из семейств Limacidae и Agriolimacidae. По-видимому,
они представляют собой аномалию или атавизм в онтогенезе отдельных
особей и не могут рассматрираться как признаки видового ранга.
3. Наличие на мантии зачатка (или рудимента) мантийной бороздки
вряд ли дает основание говорить о родстве данного вида с родом Milax,
как это делал Зимрот. Аналогичный зачаток мы встречаем не только
у Milacidae и Boettgerillidae, но и у некоторых Agriolimacidae (Krynickillus).
По-видимому, здесь мы имеем дело с проявлением параллелизма.
4. Если считать известковую шпору тератологическим образованием,
то в остальном Т. hoplites (Simroth, 1899) очень похож на Krynickillus
urbanskii (Wiktor, 1971).
Возможно, что будущие находки смогут пролить свет на поднятые здесь
вопросы.
II. Семейство BOETTGERILLIDAE Goethem, 1972
Go е t h em, 1972 : 14.
Небольшие слизни (в движении до 60 мм, дл. сок. до 30 мм), с очень строй-
ным и узким червеобразно-цилиндрическим телом, которое в поперечном
сечении почти круглое. Морщины длинные плоские, бороздки между ними
неглубокие. Голова и шея незначительно уже остального тела; щупальца
сравнительно толстые и цилиндрические.
Спина резко сужается к острому заднему концу, который при раздра-
жении сжимается с боков, образуя как бы вертикальный плавник. Киль
хорошо развит и простирается на всю спину. Мантия удлиненно-овальная,
сзади F-образно вытянута; задний конец ее достигает середины тела, хотя
вся мантия занимает около 1/3 общей длины. Капюшон немного меньше
1/2 мантии. Пневмостом лежит постмедиально и окружен небольшим взду-
тием или площадкой (как у Agriolimacidae), окрашенной светлее соседних
участков тела.
От верхнего края пневмостома вперед и назад отходит по одной мелкой
бороздке, которые вместе напоминают контур летящей птицы (рис. 246).
Обе бороздки не достигают краев мантии и не переходят на левую сторону
ее. При жизни по мантии пробегают волны сокращений в виде нежных кон-
центрических морщин.
Подошва трехдольчатая, значительно уже, чем у слизней из других се-
мейств;1 поперечные бороздки прямые. Половое отверстие лежит непосред-
ственно позади правого губного щупальца.
Раковина относительно общих размеров тела очень маленькая, закруг-
ленно-прямоугольная и лежит под задним концом мантии. Нуклеус рас-
полагается на продольной оси заметно впереди заднего края раковины и
является самой высокой точкой на ней. По периферии раковина слагается
из крупных кристаллов, иногда довольно слабо связанных с центральной
частью.
1 Исключение — некоторые Trigonochlamydidae, у которых подошва тоже очень
узкая.
II. BOETTGERILLIDAE SIT
Краевые зубы радулы кинжаловидные; челюсть оксигнатная, срединная
выпуклость ее с четкими поперечными складками (рис. 249). Кишечник двух-
петлевого типа (рис. 250). Кишка сильно сдвинута за зоб и по длине лишь
немного больше суммарной длины зоба и глотки. Левая доля печени рас-
полагается сзади, образуя вершину внутренностного мешка..
Сердце лежит в левой передней четверти мантийного комплекса (рис. 6,.
IV); ось его под углом около 45° наклонена направо, т. е. в сторону продольной
оси тела. Предсердие закрыто почкой как сверху, так и снизу. Аорта очень
короткая. Кровеносная сеть легкого слабо развита и плохо видна. Почка
закругленно-прямоугольная, без лобуса; она лежит слева от продольной
оси тела и почти параллельна ей; передний конец ее серповидно вытянут
и изогнут. Мочеточник берет начало от заднего конца почки; он длинный,
извитой, в том числе и в дорсовентральном направлении. Передний конец
мочеточника залегает внутри длинного мочевого пузыря. Анус и мочевой:
пузырь открываются наружу перед пневмостомом, Колумеллярный мускул
крепится позади мантийного комплекса, а половой ретрактор отходит от диа-
фрагмы почти в центре между почкой и мочеточником.
Гениталиям присущи две главные черты (рис. 251): 1) средний участок
семяпровода вздут и превращен в толстостенное и очень мускулистое вере-
теновидное тело (corpus fusiformis), в то время как задний участок хотя тоже
мускулистый, но более узкий и цилиндрический, а передний тонкостенный,
и очень узкий; 2) семяприемник впадает в женские пути на большом удале-
нии от атриума, благодаря чему яйцевод короткий, а влагалище длинное.
Общий план строения гениталий простой, т. е. без всяких наружных
придатков на атриуме, женских и мужских половых путях.. Внутри пениса
имеются папилла и крупные продольные складки. Половой ретрактор пе-
редним концом крепится апикально к пенису. Ретрактор правого глазного,
щупальца пересекает половые пути, огибая пенис справа и снизу, а влагалище
сверху. Размножение, вероятно, без сперматофоров.
В семейство входит один род и два вида.
Однолетние животные, ведущие в основном подземный образ жизни.
Замечание. Систематическое положение рода Boettgerilla явилось
предметом длительной дискуссии. Зимрот (Simroth, 1910, 1912а) отнес его
в семейство Limacidae, куда он причислял как Amalia (Milacidae — в нашем
понимании — с. 354), так и Parmacella (Parmacellidae'). Гессе (Hesse Р., 1926)
разделил Limacidae на два подсемейства — Limacinae и Parmacellinae. К пос-
леднему вместе с Milax, Parmacella и Aspidoporus он отнес и Boettgerilla
(с. 356). Вагнер (Wagner, 1935) выделил Milax в самостоятельное семейство,
a Boettgerilla оставил среди Parmacellinae. Виктор (Wiktor, 1973), проана-
лизировав сходство и различия между Boettgerilla, Milax и Parmacella,
пришел к выводу, что Boettgerilla принадлежит семейству Milacidae. На-
конец, Гётем (Goethem, 1972) считает, что данный род настолько отличается
от остальных двух, что заслуживает выделения в особое подсемейство или
семейство.
Указанные авторы при решении вопроса о родстве и таксономическом
ранге Boettgerilla, как правило, принимали во внимание лишь отдельные
признаки сравниваемых групп, не рассматривая весь комплекс особенностей
строения.
Так, считалось, что в пользу сближения Boettgerilla и Milax говорит:
а) сходство раковин — нуклеус-лежит медиально и отступя от заднего края;
б) наличие на мантии Boettgerilla своеобразных бороздок, которые рассматри-
вали как гомологи подкововидной борозды у Milax-, в) присутствие у обеих
групп крупного киля; г) взгляд на веретеновидное тело семяпровода Boet-
tgerilla как на гомолог эпифаллуса у Milax.
14*
212
II. boettgerillidae
В пользу объединения Boettgerilla в одно семейство с Parmacella по су-
ществу дела никаких доводов не приводилось, кроме констатации того, что
нет оснований ни тот, ни другой род объединять в одно семейство с родом
Milax.
Ниже мы приводим сравнение Boettgerilla с рядом семейств слизней 1 почти
по всему комплексу признаков, которые были использованы в данной ра-
боте для их характеристики.
Ряд черт организации Boettgerilla свойствен только этому роду и не пов-
торяется в других группах. Поэтому по ним сравнение не проводится. Вот
эти особенности: а) червеобразно-цилиндрическое, в поперечном сечении
круглое тело; б) очень узкая подошва; в) наличие веретеновидного тела на
семяпроводе; г) семяприемник открывается в женские пути очень далеко
от атриума, в связи с чем влагалище очень длинное; д) атриум очень длинный
и узкий; г) почка вытянута вдоль левой стороны мантийного комплекса.
В итоге у нижеперечисленных семейств обнаруживаются следующие
главные черты сходства и различия с Boettgerilla.
Limacinae. 2 Сходство. Мантия с крупным капюшоном. Пневмо-
стом окружен валиком. Подошва — срединные и боковые доли покрыты
прямыми поперечными бороздками. Половое отверстие лежит позади щупа-
лец. Почка без лобуса. Ретрактор правых щупалец перекрещивается с гени-
талиями. У большинства родов левая доля печени лежит сзади. Атриум
и женские половые пути без придаточных органов. Эпифаллуса нет.
Различия. Крупные слизни (есть исключения!). Мантия без бо-
роздки. Раковина с латеральным нуклеусом у заднего края. Внешние крае-
вые зубы двух- или многозубцовые. Кишечник из трех петель. Пенис без
папиллы (есть исключение!).
Agriolimacidae. Сходство. Имеются мелкие виды. Мантия с круп-
ным капюшоном. У рода Krynickillus на правой стороне мантии имеются
бороздки, сходные с таковыми у Boettgerilla. Пневмостом окружен валиком.
Половое отверстие лежит позади щупалец. Краевые зубы кинжаловидные.
Кишечник из двух петель, кишка лежит позади зоба. Атриум и женские по-
ловые пути без придаточных органов. Эпифаллуса нет.
Различия. Киль короткий и нечеткий. Раковина с латераль-
ным нуклеусом у заднего края. Подошва — поперечные бороздки на
боковых долях прямые, а на срединной У-образно изогнутые назад. Почка
имеет лобус. Ретрактор правых щупалец не перекрещивается с гениталиями.
Правая доля печени сзади. Пенис без папиллы.
Milacidae. Сходство. Длинный киль. Раковина с медиальным
нуклеусом, лежащим впереди заднего края. Краевые зубы радулы кин-
жаловидные. Кишечник из двух петель. Пенис с папиллой.
Различия. Крупные слизни (есть исключения!). Мантия с разви-
той подкововидной бороздкой и относительно меньшим капюшоном. Под зад-
ним краем мантии имеются две небольшие ямки. Вся подошва покрыта
поперечными бороздками, У-образно изогнутыми назад. Половое отверстие
сильно смещено назад и лежит на полпути между пневмостомом и щупаль-
цами. Кишка не полностью располагается за зобом. Правая доля печени
сзади. Педальная железа лежит не в толще ноги, как у остальных семейств,
а на ее внутренней поверхности. Почка имеет лобус. Влагалище или атриум
с придаточной железой. Имеется эпифаллус и сперматофоры. Ретрактор пра-
вых щупалец не перекрещивается с гениталиями.
1 Мы оставляем вне рассмотрения Philomycidae, Arionidae и Trigonochlamydidae
в_силу очевидности их резких отличий и дальности родства с Boettgerlila.
2 Подсемейства Bielziinae и Eumilacinae не рассматриваются из-за резких отличий
в строении гениталий этих групп.
BOETTGERILLA
213
Parmacellidae. Сходство. Киль простирается на всю спину. По-
ловое отверстие лежит позади щупалец. Ретрактор правых щупалец пере-
крещивается с гениталиями. Краевые зубы радулы кинжаловидные. Ки-
шечник из двух петель. Левая доля печени сзади. У некоторых видов имеется
папилла пениса.
Различия. Крупные слцзни. Мантия относительно общей длины
тела очень крупная, с развитой подкововидной бороздкой и более крупным
капюшоном. Подошва — поперечные бороздки на боковых долях прямые,
а на срединной V-образно изогнутые назад; боковые доли покрыты тонкими
продольными бороздками. Раковина со спиральным нуклеусом, у молодых
особей выступающим наружу. Вторая петля кишечника располагается над
зобом. Женские половые пути с придаточной железой. Имеется эпифаллус
и сперматофоры.
Учитывая морфологические особенности, свойственные только Boettge-
rilla, а также черты сходства и различия с перечисленными семействами,
можно сделать два основных вывода:
1. Boettgerilla не только обладает отдельными, только ей одной свойствен-
ными признаками, но и таким сочетанием остальных черт организации,
которого нет у других семейств. Поэтому есть все основания выделить этот
род в самостоятельное семейство.
2. Наибольшее сходство это семейство обнаруживает с Agriolimacidae
и наименьшее с' Milacidae и Parmacellidae.1 По-видимому, Agriolimacidae
филогенетически наиболее близки к Boettgerillidae.
^Эволюция Boettgerillidae шла в сторону адаптации к подземному образу
жизни. Это особенно ярко сказалось на форме тела, на удлинении влагалища
и атриума. В связи с этим отдельно остановимся на веретеновидном теле се-
мяпровода Boettgerilla, которое ряд авторов принимают за эпифаллус или
его производное и придают первостепенное значение в характеристике этой
группы.
Во-первых, это образование имеется не только у Boettgerilla. У северо-
американского рода Prophysaon Bland et Binney, 1873 (Anadeninae—Ario-
nidae) гениталии имеют совершенно аналогичное образование. Пилсбри
(Pilsbry, 1948 : 677) справедливо предполагает, что этот орган служит
для извержения, т. е. выталкивания, спермы. Так как у этих слизней уста-
новлены сперматофоры, то не исключено, что веретеновидное тело протал-
кивает сперму, уже заключенную в оболочку сперматофора.
Характерна коррелятивная связь между веретеновидным телом семя-
провода и некоторыми другими чертами гениталий, проявляющаяся как
у Boettgerilla, так и у Prophysaon. Она выражается в том, что это образова-
ние присутствует в тех группах слизней, у которых влагалище длинное,
а пенис сравнительно короткий и слабо развит.
|По-видимому, копуляция при длинном влагалище требует больше энергии
в проталкивании спермы в тело партнера, чем при коротком. Поэтому при
ослаблении пениса потребовался дополнительный орган, обладающий мощ-
ной мускулатурой. Если у Prophysaon такую функцию на себя принял уча-
сток эпифаллуса, то у Boettgerilla, у которых нет эпифаллуса, аналогич-
ное образование возникло за счет семяпровода.
Ц Род BOETTGERILLA Simroth, 1910
Simroth, 1910:530; 1912а : 54; Hesse Р., 1926:35; Лихарев, Р а м-
мельмейер, 1952 : 558; Wiktor, 1961 : 125; Goethem, 1972 : 1; W i k t о г,
1973 : 56.
1 Напомним, что именно с двумя последними семействами до сих пор все автор ы
сближали Boettgerilla.
214
II. BOETTGERILLIDAE
Типовой вид — В. compressa Simroth, 1910; по монотипии.
Внешний облик и внутреннее строение — см. характеристику семейства.
Ниже приводятся дополнительные признаки, на наш взгляд более свойствен-
ные родам, чем семействам.
Окраска одноцветная, без рисунка.
Срединные и боковые зубы радулы трехзубцовые, с крупными мезоконами
и четкими экто- и эндоконами.
Гонада состоит из сравнительно крупных шаровидных фолликул. Бел-
ковая железа небольшая. Талон очень маленький и представляет собой ду-
гообразно изогнутое продолжение гермафродитного протока. Задний отдел
семяпровода имеет довольно мускулистые стенки, нередко он обвивается во-
круг яйцевода. Стенки веретеновидного тела отливают шелковистым бле-
ском. Узкий и тонкостенный передний отдел семяпровода тянется вдоль
влагалища, у атриума делает изгиб и далее простирается вдоль пениса,
в который впадает апикально. Пенис почти цилиндрический, более или ме-
нее плавно сужающийся к атриуму. Последний имеет вид длинной тонкостен-
ной цилиндрической трубки.
Распространение. Западное Закавказье, Средняя Азия (Гис-
сарский хребет). Один вид встречен во многих населенных пунктах Европы.
ТАБЛИЦА ' ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ BOETTGERILLA
1 (2). Тело охристо-коричневое или кремовое. Семяприемник с шаровидным
резервуаром и длинным узким протоком . . 2. В. compressa Simroth.
2 (1). Тело светло- или голубовато-серое до пепельно-стального цвета. Семя-
приемник с сильно вытянутым в длину резервуаром и очень коротким
протоком.......................................1. В. pallens Simroth.
1. Boettgerilla pallens Simroth, 1912 (рис. 246—254)
Simroth, 1912a : 55, T. 3, Fig. 50, T. 8, Fig. 32; Л и x a p e в, Раммель-
мейер, 1952 : 558, рис. 285, 286; Wiktor, 1959 : 1, Fig. 1, 2 (yermiformis)', 1960a :
151, T. 1—3 (vermiformis); 1961 : 130, Fig. 2—5, T. 1, Fig. 4, T. 2, Fig. 5—11, T. 3, Fig. 15—
18, T. 4, Fig. 19—24, T. 5, Fig. 27—32 (vermiformis et pallens)', Schmid, 1963 : 215,
Fig. 1—4 (vermiformis)', D a x 1, 1967 : 227; Fig. 1—2 (vermiformis)-, Goethem, 1972 ;
1, Fig. 1—26, T. 1, Fig. 1—3 (yermiformis)', Лихарев, Иззатуллаев, 1972 : 55;.
Wiktor, 1973 : 58, Fig. 17, 18, 94—96, 102, 227, 249, 250; Акрамовский, 1976 :
214, рис. 103, табл. 11, рис. 122.
Locus typicus — «Gudaut» — Гудауты (Абхазия). Голотип в ЗИН АН СССР.
Материал: из СССР — 24 экз. (в том числе голотип) из 13 пунктов; из дру-
гих стран — более 260 экз.
В движении и при мягкой фиксации тело червеобразно-цилиндрическое,
но при жесткой фиксации оно в области мантии вздуто, а сзади клинообразно
сужено. Окраска светло- или голубовато-серая, или пепельно-стальная,
на спине темнее, на боках книзу становится более светлой до беловато-кре-
мового цвета. Молодые слизни белые или беловато-кремовые, реже со слабым
розовым оттенком; у особей, еще не достигших половой зрелости, спина се-
рая, а бока белые. Подошва всегда белая или кремовая. Между мантийной
щелью и килем 9—10 рядов морщин. Слизь густая, бесцветная.
Размеры: при движении длина до 60 мм; при мягкой фиксации длина
до 40 , ширина и высота до 5, длина мантии до 10, ширина подошвы до 1.8 мм.
Раковина (рис. 248) заметно крупнее и толще, чем у В. compressa. Пре-
дельные размеры: длина 1.7 , ширина 0.9, высота 0.18 мм.
Радула сходна с В. compressa, но зубы несколько меньших размеров.
В среднем формула ее следующая (Goethem, 1972):
с 66 36 15к
у + -г + ^г + — X 94.
BOETTGERILLA
215
Гениталии (рис. 251—253). Пока единственное отличие от В. com-
pressa — резервуар семяприемника сильно вытянут в длину, а проток его
очень короткий. Внутри пениса находятся 4 более или менее крупные про-
дольные складки, а в заднем конце поме-
щается длинная и узкая папилла (зани-
мает около 1/3 пениса). Она имеет кони-
ческую форму и очень острый конец, на
вершине которого открывается папилляр-
ный канал. Стенки папиллы двойные —
внутренние очень тонкие, вневтние более
толстые и мускулистые.
Распространение и образ
жизни. В СССР в природных биото-
пах этот вид обнаружен в Абхазии — близ
Гудауты и Ткварчели и по Военно-Сухум-
ской дороге у селения Амткел, а также
в западной Грузии у Лентехи, и в Верх-
ней Сванетии в долине р. Ингури, в верх-
ней подзоне леса на высоте 1750 м над
ур. моря. В антропогенных биотопах он
найден: в парках Выборга, Ленинграда,
Львова, Сочи, Орджоникидзе и Еревана;
в оранжереях Москвы и Челябинска.
Кроме того, он обнаружен в Таджики-
стане у Варзобской горной ботанической
станции (Гиссарский хребет — Лихарев,
Иззатуллаев, 1972).
Вне СССР многочисленные находки сде-
ланы на Британских о-вах, во Франции,
Бельгии, Голландии, ФРГ и ГДР, в се-
верной Швейцарии, ЧССР, Румынии,
в южной и юго-западной Польше и в оран-
жереях Хельсинки (Schmid, 1963, 1966,
1969; Goethem, 1972; Wiktor, 1973; Col-
ville e. a., 1974.)
Образ жизни. Немногие наблю-
дения за этим слизнем, сделанные при
сборе его в природных биотопах, указы-
вают, что он населяет нижнюю часть гор-
нолесной зоны. В Грузии это так назы-
ваемые «колхидские» леса с преобладанием
съедобного каштана. Здесь, как и В. com-
pressa, он ведет почти подземный об-
раз жизни, под камнями или среди осы-
пей, заросших кустами, и в лиственной
подстилке леса.
В Европе этот слизень обитает как
в оранжереях, так й на открытом грунте —
сады, парки, лесопарки, кладбища, за-
Рис. 246. Boettgerilla pallens Simr.
Внешний вид.
росшие кустами старые свалки. Кроме того, он населяет также и менее
антропогенные биотопы, преимущественно по берегам ручьев и речек, среди
кустарников (особенно в ольховниках), по заросшим горным склонам с осы-
пями.
Предпочитает тяжелые глинистые почвы. Укрывается под крупными кам-
нями, глубоко погруженными в землю, или под грудами земли. Реже ветре-
BOETTGERILLA
217
чается под досками и другими кусками древесины. Большую часть времени
проводит в почве, в которой успешно пролагает проходы.
По наблюдениям в террариуме, слизень осенью сделал 7 кладок по 1 —
4 яйца в каждой (Daxl, 1967). Яйца имеют овально-яйцевидную форму и
длину 2.1—2.9 мм, бесцветные и водянистые. При температуре 17° и насы-
щенной влажности воздуха, под сырой лиственной подстилкой вылупление
молоди произошло на 20—22-й день. В средней Европе молодые особи встре-
чаются круглый год. Это позволяет предполагать, что слизень размножается
несколько раз в году. В неволе он довольно быстро погибает. Копуляция пока
еще никем не наблюдалась. Однако внутри семяприемника не раз обнаружи-
вали комки спермиев без всяких следов остатков сперматофоров.
Из-за скрытного образа жизни этого слизня о его питании известно очень
немного. Присутствие в фекалиях кристаллов кварца указывает, что он на-
ходит пищу под землей. У пойманных на воле слизней кишечник заполнен
аморфной беловатой массой, напоминающей мицелий грибов, без всяких сле-
дов сосудистых растений. Немногие наблюдения, сделанные в террариумах,
указывают, что в этих условиях слизень предпочитает детрит, некоторые
клубни и фрукты и очень неохотно поедает зеленые части растений.
Замечания. 1. Длительное время В. pallens была известна только
по одному экземпляру, описанному Зимротом. В 1959 г. Виктор обнаружил
в Польше несколько экземпляров, которые, как ему тогда представлялось,
отличались рядом существенных особенностей от В. pallens. В связи с этим
юн описал новый вид под названием В. vermiformis Wiktor, 1959. Однако позд-
нее, ознакомившись с голотипом В. pallens, Виктор (1973) пришел к выводу,
что В. vermiformis это ее младший синоним.
После опубликования первой работы Виктора (1959) в литературе появи-
лось много новых сообщений о находках этого вида в различных странах
Европы.
2. Вслед за Виктором многие авторы считают, что В. pallens завезена
в недавнее время в Европу-с Кавказа вместе с декоративными и другими
садовыми растениями. В пользу этой точки зрения говорит не только отсут-
ствие всяких упоминаний об этом слизне в фауне Европы до 1959 г., но и
явная приуроченность большинства находок его к антропогенным биотопам
и к крупным населенным пунктам с ботаническими садами, парками и оран-
жереями.
, Однако в ряде случаев (Goethem, 1972) его обнаруживали и в природных
биотопах, сравнительно мало измененных хозяйственной деятельностью
человека. Правда, можно предположить заселение природных биотопов после
завоза этого слизня в Европу. Однако сообщение Цейслер (Zeissler, 1968)
о находке одной раковинки данного вида в отложениях плейстоцена ГДР
как будто бы говорит об исконной автохтонности его в средней Европе.
Аналогичные сомнения возникают по поводу находок этого вида на Гис-
сарском хребте (ТаджССР — Лихарев, Иззатуллаев, 1972).
По-видимому, для того чтобы решить вопрос, где он является автохтон-
ным, а где завозным, требуется более глубокое изучение его образа жизни
в различных частях современного ареала.
2. Boettgerilla compressa Simroth, 1910 (рис. 255, 256)
Simroth, 1910 : 531, Т. 7, Fig. 29—33; 1912а : 55; Л и х а ре в, Раммел ь-
мейер, 1952 : 558; Wiktor, 1961 : 128, Fig. 1, Т. 1, Fig. 1—3, Т. 3, Fig. 12-14,
Т. 5, Fig. 25, 26.
Рис. 247—254. Boettgerilla pallens Simr.
247 — мантийный комплекс, вид снизу; 248 — раковина, сверху и снизу; 249 — челюсть; 250 — кишеч-
ник; 251 —-гениталии; 252 — дистальная часть гениталий; 253 — вскрытый пенис; 254 — ретракторы
(.246, 247 — Вроцлав, IX 1977; остальные — из: Wiktor, 1973).
218
II. BOETTGERILLIDAE
Locus typicus —«Ап der oberen (Werchnaja) Zebelda» — Верхняя Цебельда
(Абхазия). Лектотип и 6 паралектотипов в ЗИН АН СССР.1 Лектотипом вы-
бран экземпляр, найденный Калишевским 21 VIII 1905 г. в Верхней Це-
бельде и изображенный у Зимрота на табл. 7, рис. 29С.
Материал: из СССР — 9 экз. жесткой фиксации (в том числе типы),
из 4 пунктов.
Тело охристо-коричневое или кремовое. Спина несколько темнее, а бока
книзу постепенно светлеют. Подошва окрашена так же. Слизь оранжевая.
Рис. 255—256. Boettgerilla compressa Simr. (Из: Wiktor, 1961).
255 — дистальная часть гениталий; 256 — гонада.
Размеры: дл. сок. до 30 мм; у экземпляра с дл. сок. 19 мм ширина
до 3, высота 4.3, ширина подошвы .1.5 мм. Между килем и мантийной щелью
9—10 рядов морщин.
Радула. Зубы несколько крупнее, чем у В. pallens.
Гениталии. Семяприемник с шаровидным резервуаром и длин-
ным узким протоком, который почти в 2 раза длиннее резервуара. Остальное —
см. описание семейства и рода.
Распространение. Типы этого вида были найдены в несколь-
ких близлежащих пунктах вдоль Военно-Сухумской дороги (Абхазия).
Кроме того, одна находка сделана в городе Кутаиси в руинах замка Баг-
рата.
Образ жизни. Обитает в зоне широколиственных лесов с преобла-
данием съедобного каштана, на известняковых почвах. По-видимому, как
и В. pallens, ведет в основном подземный образ жизни.
Замечания. Следует отметить, что в сравнении с В. pallens, данный
вид еще очень слабо изучен, а установленные на сегодня различия между
обоими видами незначительны (окраска и форма семяприемника). Как известно,
и то, и другое у слизней нередко очень варьирует. К тому же коричневая
окраска типовых экземпляров данного вида может быть связана с особенно-
стями консервации. Так как типы в настоящее время стали очень .хруп-
кими, то нами тщательно исследован лишь экземпляр, найденный в Кутаиси.
1 Из всех типов не достает только одного экземпляра, найденного у селения Ажары.
III. LIMACIDAE
219
III. Семейство LIMACIDAE Rafinesque, 1815
Rafinesque, 1815:141 (fam. Limaxia).
Слизни различных размеров (в движении от 45 до 200 мм длиной). Тело
удлиненно-цилиндрическое, при сокращении веретеновидное. Киль всегда
имеется, но различной длины — от короткого и тупого, занимающего лишь
задний конец спины, до длинного и четкого, простирающегося йа всю спину.
Мантия овальная или яйцевидная, располагается на передней половине
тела; длина ее около 1/3 общей длины, реже немного меньше половины.
Капюшон довольно крупный, при сокращении животного полностью или
почти целиком закрывает шею и голову. Пневмостом у большинства родов
располагается постмедиально или почти медиально, только у Eumilacinae —
антемедиально. Нередко, как у Agriolimacidae, участок мантии вокруг
пневмостома приподнят и утолщен, образуя легкое вздутие или площадку.
Как и у последних, при жизни по мантии пробегают волны сокращений
в виде концентрических бороздок. Морщины спины и боков более длинные
и высокие, чем у Agriolimacidae. Поперечные бороздки подошвы (рис. 3, II)
прямые и обычно располагаются нерегулярно. Половое отверстие лежит
за правым губным щупальцем.
Сердце располагается в левой передней четверти мантийного комплекса
(рис. 6, V); ось его под углом 45° или больше наклонена направо, т. е. в сторону
продольной оси тела. Кровеносная сеть легкого в основном находится перед
почкой, но нередко еще простирается вдоль правой стороны последней.
Почка более или менее фасолевидная или полулунная; она лежит у задней
половины комплекса почти поперек продольной оси тела. Лобуса нет. Мо-
четочник берет начало от заднего края почки; впереди он открывается в круп-
ный мочевой пузырь, который нередко особой связкой соединен со стенкой
почки. За исключением рода Eumilax, у остальных анус и мочевой пузырь от-
крываются наружу перед пневмостомом. Ретракторы глотки и щупалец
сливаются в единый колумеллярный мускул, прикрепляющийся к верхней
стенке тела позади мантийного комплекса.
Кишечник трехпетлевого типа (рис. 15, III, IV) и лишь у Eumilax двух-
петлевый (рис. 365). Первая петля всегда самая длинная; третья распола-
гается на верхней стороне внутренностного мешка, справа от основания ко-
лумеллярного мускула, почти по продольной оси тела. У некоторых групп
от третьей петли отходит назад более или менее длинная слепая кишка.
У большинства родов левая доля печени располагается сзади, образуя вер-
шину внутренностного мешка.
Гениталиям характерна простота строения (рис. 20, IV), проявляющаяся
з отсутствии эпифаллуса, а также придаточных органов яйцевода и атриума.
Внутри передней части яйцевода нередко имеется сфинктер. Пенис хорошо
развит, чаще всего без стимулятора и папиллы, но у некоторых видов с би-
чом или пениальной железой. Ретрактор правых щупалец у большинства
видов перекрещивается с половыми путями или с их ретрактором.
Распространение. Европа, средиземноморские области Север-
ной Африки, Кавказ, Передняя и Средняя Азия. Некоторые виды завезены
в Северную Америку и области южного полушария с умеренным климатом.
Семейство подразделяется на три подсемейства — Limacinae, Bielziinae
и Eumilacinae.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ LIMACIDAE
1 (20). Пневмостом лежит постмедиально или почти медиально. Кишечник
трехпетлевый (рис. 15, III); третья петля, в том числе и задняя кишка,
располагается справа от основания колумеллярного мускула почти .
по оси тела. rf М|
220
III. LIMACIDAE
2 (3). Семяпровод на всем протяжении вытянут вдоль яйцевода и впадает
в атриум независимо от пениса. Пенис булавовидный (Bielziinae) . . .
.............................................. 8. Bielzia Clessin.
3 (2). Семяпровод только в начальной части вытянут вдоль яйцевода, затем
отходит от него и впадает в пенис (Limacinae).
4 (7). Имеется длинная слепая кишка, которая у взрослых слизней прости-
рается назад до вершины внутренностного мешка. Эта кишка лежит
на одной линии с 6-м коленом кишечника (ректум), т. е. асимметрично
к предшествующему 5-му колену (рис. 15, IV).
5 (6). Кожа толстая. Рисунок образован многочисленными мелкими тем-
ными пятнами, которые не сливаются в продольные полосы или ряды
более крупных пятен. Семяяйцевод очень короткий, так как недалеко
от белковой железы распадается на простату (внутри с семенным кана-
лом) и длинный яйцевод. Слизь желтая или оранжевая...................
....................7. Limax Linnaeus, подрод Limacus Lehmann.
6 (5). Кожа тонкая. Рисунок обычно слагается из темных продольных полос
или рядов крупных пятен, но бывают и одноцветно черные формы.
Семяяйцевод длинный; к нему прилегает лентовидная простата, перед-
ний конец которой вздут и слегка отходит от короткого яйцевода.
Слизь бесцветная.......................... 6. Lehmannia Heynemann.
7 (4). Слепой кишки нет или она значительно короче и располагается сим-
метрично как к 5-му, так и 6-му колену кишечника (рис. 293).
8 (17). Ретрактор правых щупалец перекрещивается с половыми путями.
9 (12). Ретрактор правых щупалец перекрещивается с пенисом и семяпрово-
дом.
10 (11). Длина пениса не менее 1/2 общей длины тела. Виды обитают в Европе
............................. 7. Limax Linnaeus, подрод Limax s. str.
11 (10). Пенис значительно короче. Виды обитают в Средней Азии.........
- ...........................4. Turcomilax Simroth (часть видов). .
12 (9). Ретрактор щупалец перекрещивается только с пенисом, а семяпровод
остается вне (т. е. сзади) места перекрещивания.
13 (14). Крупные слизни (дл. сок. до 80 мм). Мантия сзади заканчивается
острым углом. Внутри пениса имеется кольцевой валик...................
............................................3. Caspilimax Hesse.
14 (13). Небольшие слизни. Мантия сзади плавно закругленная. Внутри
пениса имеется папилла или крупная складка.
15 (16). Киль отчетливый, простирается не меньше чем на 3/4 спины.
Мантия занимает около 1/2 длины тела. Пенис короткий и расчленен
на несколько отделов; внутри заднего отдела находится крупная па-
пилла. Семяприемник с очень толстыми мускулистыми стенками. Оби-
тает на Кавказе.............................2. Caucasolimax gen. nov.
16 (15). Киль короткий и нечеткий. Мантия занимает менее х/3 длины тела.
Пенис мешковидный или короткоцилиндрический, не расчленен на от-
делы; внутри он без папиллы, но с крупной складкой. Семяприемник
тонкостенный. Обитает в Европе...................5. Malacolimax Malm.
17 (8). Ретрактор правых щупалец не перекрещивается с половыми путями.
18 (19). Кишечник, особенно вторая петля, сильно перекручен. Пенис имеет
внутри кольцевой валик. Виды обитают на Кавказе.......................
.................................. 1. Gigantomilax О. Boettger.
19 (18). Кишечник не перекручен. Пенис внутри с продольными складками.
Виды обитают на Тянь-Шане.............................................
............................4. Turcomilax Simroth (часть видов).
20 (1). Пневмостом лежит антемедиально. Кишечник двухпетлевый (рис. 365),
задняя кишка располагается слева от основания колумелдярного
мускула (Eumilacinae).
1. GIGANTOMILAX
221
21 (22). Пенис короткий с длинным слепым выступом — бичом. Половой
ретрактор распадается на большое число ветвей, которые, веерообразно
расходясь, крепятся к бичу на всем его протяжении (рис. 368); слепой
кишки нет................................. 9. Eumilax О. Boettger.
22 (21). Пенис длинный без бича. Половой ретрактор не распадается на ветви
и крепится к пенису. Имеется крупная слепая кишка...............
...................................... 10. Metalimax Simroth.
1. Подсемейство LIMAGINAE Rafinesque, 1815
Пневмостом лежит постмедиально или почти медиально.
Кишечник трехпетлевый. Семяпровод впадает в пенис. Гонада распо-
лагается в задней части тела позади белковой железы. Пенис цилиндри-
ческий или мешковидный, у некоторых видов с бичом или пениальной желе-
зой.
Распространение. В пределах ареала семейства (см. выше).
В подсемейство входят 7 родов.
1. Род GIGANTOMILAX О. Boettger, 1883
Boettger, О., 1883 : 143; Simroth, 1901 : 176; Hesse Р., 1926 : 19.
Типовой вид — Amalia (Gigantomilax) lederi О. Boettger, 1883;
по монотипии.
Большинство видов средних размеров (дл. сок. до 56 мм), лишь один вид
крупный (дл. сок. до 100 мм). Киль всегда имеется, но различной высоты
и протяженности. Мантия занимает около 1/3 длины тела. Пневмостом рас-
полагается постмедиально или медиально и окружен легким валиком.
Срединный и боковые зубы радулы трехзубцовые, краевые двухзубцо-
вые; лишь самые внешние краевые зубы (зачаточные) имеют шиповидный
облик. Кишечник без слепой кишки. Первая петля почти прямая, вторая
сильно перекручена, третья заметно короче остальных, прямая. Обе доли
печени примерно равной длины и не доходят до заднего конца внутренностного
мешка.
Ось сердца наклонена под углом 45°. Аорта длинная. Почка лежит
в центре мантийного комплекса, и длинная ось ее несколько наклонена к про-
дольной оси тела. Очень длинный мочевой пузырь роговидно изогнут. Осно-
вание полового ретрактора крепится к диафрагме перед сердцем.
Гонада располагается позади кишечника, между лопастями печени, и
образует вершину внутренностного мешка. Яйцевод и семяпровод очень ко-
роткие. Семяпровод прямо простирается к пенису, не образуя пениальной
петли. Пенис мешковидный, если же цилиндрический, то не длинный. Се-
мяприемник открывается в атриум. Внутри пениса, перед впадением в атри-
ум, имеется кольцевой валик. Нередко, кроме обычного полового ретрак-
тора, пенис имеет 1—2 ретентора, соединяющие его со стенками тела.
Ретрактор правых щупалец проходит в стороне от половых путей; у не-
которых видов он перекрещивается только с половым ретрактором.
Распространение. Кавказ и сопредельные области Турции и
Ирана.
Замечания. 1. Некоторые авторы (Soos, 1924; Hesse Р., 1928;
Urbahski, Wiktor, 1968) указывают на присутствие двух видов данного рода
на Балканском п-ове. Однако судя по описаниям и рисункам, авторы имели
дело со слизнями из рода Lehmannia. В частности, такой вывод вытекает из
того, что эти виды имеют длинную слепую кишку — образование, отсутствую-
щее у настоящих Gigantomilax.
222
III. LIMACIDAE
2. Для рода Gigantomilax наиболее характерно: а) форма кишечника —
вторая петля сильно перекручена, третья короткая; б) короткий массивный
шенис с кольцевым валиком внутри; в) ретрактор правых щупалец не пере-
крещивается с гениталиями. ...
Эти особенности строения позволяют объединить в один род (в ранге
подродов.) три группы: Gigantomilax s. str., Vitrinoides Simroth, 1891 и Mono-
.chroma Simroth, 1896. .......... ................ ...
Последнюю группу до сих пор рассматривали как самостоятельный род,
•самым характерным признаком которого считалось присутствие на пенисе
•бича. Наши исследования показали, что это образование имеется также у не-
которых видов, ранее относимых то в род Gigantomilax, то в подрод Vitrinoides
'(род Limax). Это дает основание, с одной стороны, увеличить объем группы
Monochroma,.с другой — рассматривать ее как подрод от рода Gigantomilax.
3. Что касается группы Vitrinoides, то анализ организации входящих в нее
видов показал, что, во-первых, она очень далеко стоит от рода Limax, и,
.во-вторых, очень близка к роду Gigantomilax. Последнее отмечал еще Зимрот
(Simroth, 1912а). По сути дела он относил эту группу к Limax только потому,
нто у нее ретрактор правых щупалец перекрещивается с половым ретракто-
ром. Между тем, как показали наши исследования, эта особенность присуща
также и Gigantomilax lederi (Bttg.), т. е. типовому виду рода Gigantomilax.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ И ПОДВИДОВ GIGANTOMILAX
11 (2). Очень крупные слизни (дл. сок. до 100 мм), на спине с длинными
и высокими морщинами, которые расположены в ряды, почти параллель-
ные подошве. Спина с крупным килем, всегда доходящим до ман-
тии и образованным 3 рядами морщин. Подошва очень широкая и тол-
стая, выступающая за пределы контуров тела. Мантия сзади закруглен-
ная, одноцветная (изредка с темными пятнами неопределенных очер-
таний), всегда светлее спины. Внутри пениса, кроме кольцевого валика,
имеется крупный треугольный стимулятор с рубчатой поверхностью
(рис. 261) (подрод Gigantomilax s. str.)..............................
. . ............................1. G. (G.) lederi (0. Bttg.).
2 (1). Слизни меньших размеров (дл. сок. не более 70 мм), с плоскими и бо-
лее короткими морщинами. Киль, если имеется, то меньших размеров
и не всегда простирается до мантии. Подошва более узкая и тонкая.
Мантия сзади с более или менее заметным углом и окрашена в цвет
спины, с темным рисунком или без него. Пенис с кольцевым валиком
внутри, но без стимулятора.
13 (6). Вся верхняя сторона тела, в том числе и бока, окрашена в интенсивно
черный цвет (иногда киль бывает светлый, а на мантии светлые пятна
неопределенной формы). Пенис с более или менее длинным бичом
(подрод Monochroma Simr.).
• 4 (5). При сокращении капюшон полностью закрывает передний конец
тела. Бич имеет вид короткого рога, направленного назад или в сто-
рону. Пенис сравнительно короткий, мешковидно вздутый. Вид оби-
тает в Талыше ...........................6. G. (М.) lenkoranus (Simr.).
5 (4). При сокращении капюшон не полностью закрывает передний конец
тела. Бич длинный, изогнутый, направлен концом вперед. Пенис
цилиндрический и длинный. Вид обитает в Зангёзуре.....................
. _ . .. ......................... 5. G. (М.) brunneus (Simr.).
£6 (3). Окраска иная. Бока всегда светлее спины. Пенис без бича, но иногда
на нем видны 1—2 небольших выступа по бокам семяпровода (подрод
Vitrinoides Simr.).
1. GIGANTOMILAX
223
7 (8). Спина и мантия черные или темно-серые, бока темно- или светло-
пепельные с редкими светлыми круглыми пятнышками (каждое зани-
мает 2—3 морщины). Темный пигмент представлен очень мелкими
точками (видны при 20-кратном увеличении), которые концентри-
руются на морщинах, в связи с чем бороздки более светлые. Темного
рисунка нет. Третья петля кишечника занимает около х/3—х/4 длины
тела, и оба колена ее лежат почти параллельно......................
................................. 4. G. (V.) daghestanus (Simr.).
8 (7). Окраска иная, всегда с отчетливым темным рисунком на сйине и ман-
тии. Темный пигмент собран в более крупные пятнышки или точки,
которые располагаются как по морщинам, так и по бороздкам. Бока
всегда светлые. Третья петля кишечника короче, и оба колена ее рас-
полагаются под углом друг к другу.'
9 (10). Фон верхней стороны тела красновато-серый, реже серовато-желтый..
Спина и мантия имеют красновато-коричневый рисунок из двух про-
дольных полос, внутренние края которых четкие, а внешние расплыв-
чатые. Третья петля кишечника очень короткая (х/8—х/9 длины тела)'
......................................... 3. G. (V.) koenigi (Simr.).
10 (9). Спина и мантия черные или темно-бурые, бока желтые или белые.
Молодые слизни на светлом фоне имеют рисунок, образованный круп-
ными темными пятнышками, которые могут сливаться в продольные'
полосы. В этом случае внутренние края полос менее четкие, чем внеш-
ние. Третья петля кишечника занимает х/5—х/7 длины тела.
11 (12). Половой ретрактор перекрещивается с ретрактором правых щу-
палец .......................2а. G. (V.) monticola armeniacus (Simr.),
12 (11). Половой ретрактор не перекрещивается с ретрактором правых
. щупалец................2b. G. (V.) monticola monticola (О. Bttg.).
1. Подрод GIGANTOMILAX s. str.
Очень крупные слизни с длинными и высокими морщинами, располагаю-
щимися почти параллельно килю. Киль толстый, образован тремя рядами
морщин и простирается на всю спину. Задний край мантии закругленный, без;
угла. Пневмостом лежит немного постмедиально. Подошва очень широкая,
с толстым краем, на который смещена перипедальная бороздка.
Пенис мешковидный, почти круглый, без всяких придатков. Кроме по-
лового ретрактора, к нему прикрепляются два ретентора. Внутри пениса,
кроме кольцевого валика, имеется крупная треугольная складка,— ве-
роятно, стимулятор.
Ретрактор правых щупалец перекрещивается только с половым ретрак-
тором.
Распространение. Западное Закавказье и вилайет Чорох
(Турция).
1. Gigantomilax (Gigantomilax) lederi (О. Boettger, 1883) (цв. вкл., рис. V;
рис. 11, X—XII, 257-261)
Boettger, О., 1883 : 143, Т. 4, Fig. 1 [A malia (Gigantomilax)]; Р ollonera,
1887а : 5, Т. 3, Fig. 5—7; Simroth, 1901 : 178, Т. 18, Fig. 1—14, text-fig. 12; 1910г
527; 1912 а : 47, 130; Лихарев, Рамме льмейер, 1952 : 325, рис. 251—253.
Terra typica — «Swanetien» — Сванетия (Грузия).
Типы в Зенкенбергском музее (№ 155289/2). За лектотип избран экземпляр,,
изображенный у Бетгера. Его размеры см. ниже.
Материал: из СССР — 46 экз. из 29 пунктов; из Турции 3 экз. (в Ин-
ституте зоологии АН АрмССР).
Рис. 257—261. Gigantomilax (Gigantomilax) lederi (0. Bttg.).
зубы радулы; 258 — кишечник; 259 — ретракторы; 260.-гениталии; 261 — вскрытый пенис
' _ Батуми, 10 II 1959).
1. GIGANTOMILAX
225
Тело стройное, в области задней трети мантии наиболее широкое, к заднему
концу плавно, но сильно заостренное. Мантия занимает более х/3 длины тела.
Она сильно смещена вперед, немного сужается к голове и плавно закруглена
как спереди, так и сзади. Пневмостом окружен легким валиком. Вдоль спины
тянется крупный киль, образованный тремя рядами длинных морщин, из
которых средний ряд наиболее широкий. Морщины спины очень длинные
и высокие, расположены почти параллельными рядами и разделены глубо-
кими бороздками. Между мантийной щелью и килем 12—15 рядов морщин.
При сокращении крупный капюшон обычно полностью закрывает шею
и голову, и тело приобретает булавовидную форму с максимальной шириной
и высотой в области задней части мантии.
Большая часть верхней стороны тела, за исключением киля и мантии,
окрашена в синевато- или бархатисто-черный цвет; только над краем подошвы
имеется несколько белых пятен с неясными контурами. Мантия темно-
или синевато-серая, изредка с несколькими темными пятнами. Голова, щу-
пальца и затылок темно-серые или черные; бока под капюшоном белые. Бо-
ковые доли подошвы серые, а срединная белая.
Иногда встречаются экземпляры с более светлой окраской (f. kollyi
Retowski, 1889) — мантия и киль серовато-розовые, спина серовато-темно-
фиолетовая, причем бороздки окрашены темнее, чем морщины.
Окраска меняется с возрастом. После вылупления (дл. сок. 10 мм) слизни
имеют розовую окраску; через несколько дней (дл. сок. 18 мм) спина и бока
становятся серыми, а мантия светло-серая.
Размеры: дл. пол. до 150, дл. сок. до 100, шир. сок. до 20, мантия до
35 мм; лектотип: дл. сок. 57, мантия 20, спина 39, ширина 13, шир. подошвы
11 мм.
Раковина округло-прямоугольная, 10x7 мм.
Радула (рис. 11, X—Х1Ц 257). Границы между основными типами зубов
четкие. Краевые зубы с крупным мезоконом и небольшим эктоконом. Харак-
терно, что в каждом ряду у одного из средних краевых зубов эктокон рас-
падается на несколько мелких зубчиков. Ниже приводятся радулярные фор-
мулы взрослого и молодого экземпляров.
1. Взрослый экземпляр из Боржоми; дл. сок. 100 мм:
с , 396 г 22к г к , ЗЗк . .
У + У + У +
2. Молодой экземпляр из Абхазии;
4 1 2
дл. сок. 18 мм:
к Юк
Кишечник (рис. 258). Третья петля занимает меньше 1/3 длины тела.
Гениталии (рис. 260, 261). Сравнительно небольшая гонада лежит
за зобом и скрыта между лопастями печени. Ее проток вначале тонкий и пря-
мой, затем утолщается и сильно закручен. Длинный семяяйцевод дает не-
сколько крупных извивов. Впереди к нему прилегает крупная белая простата.
Яйцевод трубчатый и сравнительно тонкий, лишь на середине немного вере-
тенообразно вздут. Семяприемник с крупным резервуаром и довольно длин-
ным протоком. Вместе с яйцеводом он впадает в очень небольшой атриум,
который слабо обособлен от пениса.
Пенис очень крупный; короткий семяпровод впадает в него снизу. К пе-
нису крепятся три мускула: диафрагмальный половой ретрактор (недалеко
от семяпровода) и два ретентора, связывающие пенис со стенками тела. Внутри
пениса находится сложная система мускульных валиков и складок. Из них
одна особенно крупная с рубчатой поверхностью. В спокойном состоянии
15 И. М. Лихарев, А. Й. Виктор
226
Ш. LIMACIDAE
она сложена и несколько закручена, а при копуляции, выворачиваясь,
набухает и приобретает треугольную форму.
Распространение. В СССР — южные склоны и отроги Боль-
шого Кавказа от бассейна р. Мзымты на западе до Сурамского перевала и рай-
она Боржоми на востоке, а также Аджаро-Имеретинский и Шавшетский
хребты. Вне указанного района сделана одна находка в Тебердинском запо-
веднике. Вне СССР — вилайет Чорох (Турция).
Образ жизни. Обитает в горных широколиственных лесах пре-
имущественно нижнего пояса, в частности с примесью благородного каштана;
кроме того, живет в пихтово-буковых лесах. Днем скрывается под корой
валежин, под камнями и в щелях скал.
2. Подрод VITRINOIDES Simroth, 1891
Simroth, 1891а : 301; Hesse Р., 1926 : 13.
Типовой вид — Limax armeniacus Simroth, 1886; по монотипии.
Слизни средних размеров. Плоские и сравнительно короткие морщины
веером расходятся от заднего края мантии. Киль узкий, образован одной
длинной морщиной и чаще всего занимает не более 2/3 спины; лишь изредка
простирается на всю спину. Мантия сзади с более или менее четким углом.
Пневмостом лежит постмедиально или почти медиально. Подошва более
узкая, чем у Gigantomilax s. str., перипедальная бороздка лежит выше ее
края.
В отличие от других подродов передний конец простаты вздут. Пенис очень
короткий, мешковидный, у взрослых особей нередко как бы вдвинут внутрь
атриума. Кроме полового ретрактора, обычно имеется еще один ретентор.
На пенисе очень часто находится небольшой слепой выступ.
Как у Gigantomilax s. str., внутри пениса имеется кольцевой валик, но
треугольной складки нет.
Ретрактор правых щупалец обычно перекрещивается с половым рет-
рактором, однако у одного подвида такого перекреста нет.
Распространение. Дагестан, 'Малый Кавказ и сопредельные
области Турции и Ирана.
2а. Gigantomilax (Vitrinoides) monticola armeniacus (Simroth, 1886),
comb. nov. (рис. 262—266).
Martens, 1880 : 143 [Limax (Krynickillus) Dymczeviczi, non Kaleniczenko, 1851a];
Simroth, 1886a : 27, T. 1, Fig. 4—7 [Limax)', 1894b : 8, T. 17, Fig. 1—2 [L. primiti-
vus); 1898a : 57 (L. colchicus), 58 (L. valentini, syn. nov. et L. simplex, syn. nov.); 1901 :
77—81, T. 4, Fig. 1—7, 10—12 (L. ordubadensis nom nov., armeniacus, monticola, primitivus,
simplex, valentini); 1910 : 507 (L. monticola)', 1912a : 12, T. 5, Fig. 5 (L. incipiens pars),
12, T. 1, Fig. 6, T. 5, Fig. 6 (L. kaznakovi), 14, T. 1, Fig. 7, T. 5, Fig. 7 (L. schmidti), 17,
T. 1, Fig. 10, T. 5, Fig. 10 (L. schelkovnikovi), 18, 123, T. 2, Fig. 11, T. 6, Fig. 11 (L. vo-
ronovi, syn. nov.); Keller, 1914 : 235 [L. monticola)', Давтян, 1947 : 8, 13, 14
(L. armeniacus)', Акрамо вский, 1949 : 151 (L. armeniacus); Давтян, 1950 : 88
(L. armeniacus); Лихарев, Раммельмейер, 1952 : 315, рис. 238 (L. armeniacus);
Акрамовский, 1971a : 109; 19716 : 9; Wiktor, 1971a : 263, Fig. 1 (L. kaznakovi);
Акр амовский, 1976 : 200, рис. 97A, табл. 11, рис. 116, 117 (Vitrinoides).
Locus typicus — «Insel Sewanga im Goktscha-See und Darytschitschach» —
окрестности оз. Севан (Арм. ССР).
Местонахождение типов данного подвида неизвестно. Так же неясно,
где находятся типы следующих его синонимов, описанных Зимротом: Limax
colchicus (=L. ordubadensis , L. valentini, L. kaznakovi, L. schmidti, L. schel-
kovnikovi). В ЗИН АН СССР имеются типы Limax incipiens (лектотип и 2
паралектотипа), L. voronovi (лектотип и 25 паралектотипов), L. colchicus
Рис. 262. Gigantomilax (Vitrinoides) monticola armeniacus (Simr.). Южная Грузия 17 VII
1973.
15*
228
III. LIMACIDAE
(2 синтипа). Мы имели возможность ознакомиться с типами L. simplex (лек-
тотип и 1 паралектотип) из Зоологического музея в Хельсинки и с синти-
пом L. primitivus из Естественноисторического музея в Вене.1
Материал: из СССР более 1000 экз. из 92»пунктов; из Турции — 30 экз.
Рис. 263—266. Gigantomilax (Vitrinoides) monticola armeniacus (Simr.). Южная Грузия,
17 VII 1973.
Zes — зубы радулы; 2G4 — кишечник; 265 — гениталии; гвв — дистальная часть гениталий.
При сокращении тело животного массивное, почти цилиндрическое,
только в задней четверти довольно резко сужается и снижается. Мантия
спереди закругленная, сзади с тупым углом (возникает лишь при сильном
сокращении тела). Мантийная щель чаще всего располагается медиально,
а иногда немного смещена вперед. Пневмостом лежит медиально. Морщины
довольно крупные и резкие; между мантийной щелью и средней линией спины
их 13—16 рядов. Киль выражен различно; изредка он простирается до мантии
как заметный гребень, но чаще занимает не менее х/2 длины спины. Впереди
1 Все лектотипы обозначены впервые.
1. GIGANTOMILAX
229'
киль невысокий, в виде длинной морщины, не пересекаемой поперечными
бороздками; сзади он выступает более резко.
Окраска и рисунок очень изменчивы и зависят от возраста и характера
местообитаний. Взрослые слизни чаще всего черные или темно-бурые. По-
дошва, бока (особенно ниже мантии) и значительная часть шеи белые или кре-
мовые. Рисунок слагается из темных пятнышек, которые постепенно редеют по
направлению от спины к подошве. Вдоль затылка проходит темная полоска,
которая может распадаться на серию пятен. Концентрация пятнышек, при-
дающих темную окраску верхней стороне тела, нарастает с возрастом.
Мантия темнеет быстрее, чем спина. В связи с этим молодые животные очень
часто имеют совершенно иной облик, чем взрослые. Очень молодые слизни
нередко полностью кремовые, и только на мантии намечаются два ряда тем-
ных точек или более крупных пятен.
С возрастом такой же рисунок возникает и на спине по сторонам от свет-
лой килевой полоски. На мантии ряды пятен вскоре сливаются, образуя две
дугообразные темные полосы, которые окаймлены узким светлым промежут-
ком. Середина мантии темнеет, и нередко здесь возникают свои пятна не-
правильной формы. Полосы на спине возникают, позднее, и они не столь чет-
кие, как на мантии. К концу роста животного наступает общее потемнение
мантии и спины (сначала коричневый цвет, затем темно-бурый и, наконец,
почти черный). Характерно, что если имеется светлая килевая полоска, то
она занимает только один ряд морщин.
Размеры: дл. сок. до 47, ширина до 11, высота до 12, длина мантии
до 20 мм.
Радула (рис. 263). Границы между тремя основными типами зубов выра-
жены не очень отчетливо и зависят от возраста (как и общее число зубов).
Срединный и боковые зубы трехзубцовые, краевые с двумя зубцами, и лишь
с возрастом возникают 2—3 самых внешних краевых зуба с одним зубцом,
т. е. шиповидные. Пример. 1 экземпляр (дл. сок. 29 мм) из Сарыкамыша
(Карский вилайет Турции); формула:
с 276 19к Зк
"3 +“3” +~ +~ X117-
Кишечник (рис. 264). Третья петля короткая, обычно около х/5—х/7
длины тела, с возрастом относительно увеличивается.
Гениталии (рис. 265, 266). Крупная темная гонада лишь слабо за-
крыта печенью и занимает задний конец внутренностного мешка. Простата
очень вздута — вдвое или более шире семяяйцевода. Семяпровод и яйцевод
короткие, но толстые. Семяпровод и половой ретрактор крепятся к пенису,
отступя от его вершины. Между ними обычно имеется небольшой короткий
выступ; иногда по другую сторону от семяпровода бывает другой похожий
выступ, и оба они слегка пигментированы. Внутри пениса находится кольце-
видный валик, часто гофрированный по краям. Атриум небольшой и корот-
кий, слабо обособлен от пениса, вместе с которым он подтянут к стенке тела
несколькими пучками мышц. Семяприемник очень крупный, впадает в атриум
между пенисом и яйцеводом.
В отличие от G. (У.) monticola monticola (О. Boettger) половой ретрактор
перекрещивается с ретрактором правых щупалец.
Распространение. В СССР этот вид обитает в большей части
горных систем Малого Кавказа, т. е. в Армении, сопредельных районах
Азербайджана и южной Грузии; по-видимому, отсутствует в полупустынях
долины Аракса и в большей части Зангезурского хребта. В Грузии он найден
на Джавахетском хребте (у оз. Тумангель) и в нескольких пунктах Аджаро-
Имеретинского хребта (Годердзский перевал, курорт Бахмаро).
230.
III. LIMACIDAE
Вне СССР — Турция, вилайеты Карс (коллекция ЗИН АН СССР) и Траб-
зон (Wiktor, 1971а).
Образ жизни. В Армении этот вид обитает в степной и лугово-
степной зонах и от нижнелесной до нижнеальпийской зоны (Акрамовский,
19716). В лесах прячется под камнями, у оснований стволов и в подстилке;
на открытых местах — в трещинах скал и в осыпях. Поднимается в горы до
высоты 3000 м. В некоторых населенных пунктах Армении отмечен как вре-
дитель огородных культур и табака (Акрамовский, Бабаян, 1976).
Однолетние животные. Откладка яиц осенью, вылупление молоди весной.
Летом идет рост и созревание. К зиме взрослые особи гибнут (Акрамовский,
1976).
Замечания. 1. Выше уже излагалось, как меняются окраска и ри-
сунок в течение жизни. Однако в некоторых популяциях у взрослых слизней
наблюдается сохранение ювенильных типов окраски. Сравнительно чаще
других отклонений встречаются популяции, где преобладают особи, сохра-
нившие полосы на мантии и светлую килевую полоску (f. typtca). Очень редко,
но все же попадаются взрослые кремовые слизни с двумя рядами точек или
пятен на мантии и разрозненными точками на спине (1. kaznakovi Simr.).
Если не учитывать возрастных различий, то изменчивость половых путей
значительно меньше. Чаще всего она выражается в более слабом развитии
двух небольших слепых выступов на пенисе по бокам от семяпровода или
в отсутствии одного из них.
2. Полиморфизм окраски данного вида, с одной стороны, и фрагментар-
ность материала, которым цользовался Зимрот, — с другой, привели к тому,
что указанный автор описал как самостоятельные виды формы, которые теперь
приходится считать синонимами данного подвида (см. выше).
Прежде чем прийти к такому выводу, нами были изучены сотни экземпля-
ров из разных мест Армении и Азербайджана, хранящихся в ЗИН АрмССР
(Ереван). В частности, были просмотрены богатые серии топотипов: «Б.» агте-
niacus (полуостров Севанга на озере Севан), «А.» primitivus (Дилижан, Арме-
ния), «Б.» voronovi (гора Кяпяз у оз. Гей-гёль, АзССР).
Изучение этого материала, а также типов большинства из названных «ви-
дов» Зимрота, которых он относил к группе Vitrinoides, привело нас к выводу,
что все они представляют собой вариации окраски, рисунка и половых орга-
нов одного*таксона — Gigantomilax monticola armeniacus (Simr.). В подавляю-
щем случае различия, подмеченные Зимротом, связаны с возрастом, реже
с особенностями фиксации. Поэтому они не имеют таксономического значе-
ния. .
2Ь". Gigantomilax (Vitrinoides) monticola monticola (О. Boettger, 1881),
comb. nov.
Boettger, 0., 1881 : 180, T. 7, Fig. 6 (Limax)', Simroth, 1912a : 5, 50, 131,
T; 3, Fig. 46, T. 8, Fig. 29 (brunneus, syn. nov.); 50, T. 3, Fig. 47, T. 8, Fig. 30 (borschomensis,
syn. nov.).
Locus typicus — «auf dem Taparowan» — у оз. Паравани (южная Грузия).
Голотип в Зенкенбергском музее Франкфурта-на-Майне (№ 155348/1).
Голотип G. borschomensis Simr. и лектотип (обозначен нами) с 5 паралекто-
типами G. brunneus Simr. в ЗИН АН СССР.
Материал: из СССР — 26 экз. из 16 пунктов.
Данный подвид внешне не отличим от предыдущего, а по внутреннему
строению отличается только одной особенностью — ретрактор правых щупа-
лец не перекрещивается ни с половыми путями, ни с их ретрактором.
Расп рос транение. По-видимому, ареал этого подвида значительно
меньше ареала предыдущего подвида и ограничен районом между хребтами
1. GIGANTOMILAX
231
Джавахетским, Самсарским и прилегающей частью Триалетского хребта
(южная Грузия). Его многократно находили в окрестностях озера Паравани
у селения Тамбовка, близ озера Табацкури, соседнего с ним перевала Цхра-
цкаро и от последнего по дороге на Бакуриани.
Образ жизни. Подобен предыдущему подвиду.
Замечания. 1. Изучение типов G. borschomensis Simr. и G. brunneus
Simr., а также топотипов «Limax» monticola О. Bttg. показало их полную иден-
тичность. Если G. brunneus описан по взрослым экземплярам, то G. borscho-
mensis по единственному полувзрослому экземпляру со светлой окраской и со
слабо намеченным рисунком.
Уже Зимрот (Simroth, 1912а, с. 51) после описания обоих «видов» от-
метил, что если бы у них ретрактор пениса перекрещивался с ретрактором
правых щупалец, то их следовало бы отнести к многочисленным армянским
«Limax», т. е. к группе Vitrinoides. Он предполагает, что в этом отразилась
филогенетическая близость обоих родов — Limax и Gigantomilax.
В настоящее время нам известно, что у большинства видов Gigantomilax,
в том числе и у типового вида G. lederi (О. Bttg.), оба ретрактора перекрещи-
ваются. Поэтому не следует придавать столь большого значения тем сравни-
тельно редким случаям, когда это не имеет места. По-видимому, потеря пере-
креста между ретракторами у популяций, населяющих указанный район
южной Грузии, произошла случайно и закрепилась в результате географи-
ческой изоляции, придав этим популяциям ранг подвида.
2. Как нам сообщил Н. Н. Акрамовский, между ареалами обоих подвидов
имеется переходная зона, где они встречаются совместно, но преобладает тот
или иной подвид. Так, в окрестностях селения Тамбовка (южная Грузия)
в популяциях G. monticola у 10% особей оба ретрактора перекрещиваются,
а у 90% этого не происходит; следовательно, здесь преобладает подвид
monticola. Однако несколько восточнее, близ города Ленинакана (западная
Армения) перекрещивание ретракторов обнаружено у 97% особей и лишь
у 3% эти мускулы не перекрещиваются; следовательно, здесь преобладает
подвид armeniacus.
3. Gigantomilax (Vitrinoides) koenigi (Simroth, 1912), comb. nov. (рис. 267)
Simroth, 1912a : 10, T. 1, Fig. 4, T. 5, Fig. 4 (Limax}-, 15, T. .1, Fig. 8, T. 5, Fig. 8
(L. oltinus, syn. nov.); 16, T. 1, Fig. 9, T. 5, Fig. 9 (L. jailanus); Акрамовский,
1943 : 270 (L. valentini); 1971a : 109; 19716 : 9; 1976 : 202, рис. 97Б, табл. 11, рис. 118
(Vitrinoides).
Locus typicus — «Кег-ogly, Kreis Olty» — Кер-оглы (вилайет Эрзурум,
Турция). Типы неизвестны.
Материал: из СССР 64 экз. из 17 пунктов; из Турции и Ирака 23 экз.
Тело цилиндрическое, в последней трети к заднему концу плавно су-
женное. Подошва относительно более узкая, чем у G. monticola. Мантия за-
нимает около х/4 длины тела, спереди закругленная, сзади нередко с очень
острым углом. Пневмостом лежит сильно постмедиально. Кожа тонкая и мяг-
кая, с плоскими морщинами; между мантийной щелью и средней линией
спины 17—18 морщин. Слегка зазубренный киль (его пересекают мелкие
бороздки) занимает около а/5 спины; при резком сокращении он иногда до-
ходит до мантии.
Фон верхней стороны тела красновато-серый, реже серовато-желтый. На
мантии видны две ясные дугообразные темно- или красновато-коричневые
полосы, которые впереди могут смыкаться. По внутреннему краю они окай-
млены светлым пространством; середина мантии затемнена за счет много-
численных точек. Такой же рисунок имеют бока тела и край мантии, при-
чем нередко точки заходят на край ноги и даже на боковые доли
подошвы. На боках точки образуют сетчатый рисунок.
232
Ш. LIMACIDAE
Вдоль середины спины простирается широкая и светлая килевая полоска
(занимает 2—3 ряда морщин). По обеим сторонам ее лежат сгущения из упомя-
нутых темных точек, благодаря чему возникают как бы две спинные полосы
Рис. 267. Gigantomilax (Vitrinoides) koenigi
(Simr.). Иракский Курдистан, 17 VI 1961.
той же интенсивности и цвета,
что и мантийные. Внутренние
края спинных полос четкие,
а внешние расплывчатые. Го-
лова и шея серые или бурые.
Подошва белая. Слизь бесцвет-
ная.
Ра з м е р ы: дл. сок. до 50,
ширина и высота до 8, дл. ман-
тии до 15 мм.
Радула. Очень сходна с G. mo-
nticola armeniacus.
Кишечник. Третья петля
у взрослых слизней равна около
1/8—1/9 длины тела, а у моло-
дых относительно длиннее.
Генитали и.1 Гонада
крупная. Простата впереди
вздута более умеренно, чем
у G. monticola. Яйцевод длин-
ный и узкий, перед впадением
в атриум сильно изогнут. Семя-
приемник длинный, но неширо-
-кий, с коротким и толстым про-
током. Пенис мешковидный, ко-
роткий; длина его обычно лишь
немного больше ширины. Атриум
у молодых хорошо выражен,
тонкостенный; пенис к нему
лежит под углом. Половой рет-
рактор крепится к вершине пе-
ниса, а семяпровод несколько
ближе к атриуму. Между ними
на пенисе имеется почти ровная
площадка. Согласно Зимроту,
пенис несет небольшой слепой
выступ — цэкум, ретрактор кре-
пится отступя от заднего конца
пениса, который в этом месте
имеет темное пятно. На нашем
материале эти особенности не
обнаружены.
Распространение.
В СССР: Аджария (долина
р. Мачахелис-Цкали), Армения
(Иджеванский, Ноемберянский
и Вединский районы) и Азер-
байджан (Талыш). Вне СССР: Турция (вилайеты Чорох и Эрзурум), Ирак
(мухафаза Эрбиль).
1 В нашем распоряжении не было половозрелых экземпляров, и описание гениталий
дается с учетом данных Зимрота (Simroth, 1912а).
1. GIGANTOMILAX
233
Образ жизни. В безлесных местах живет в осыпях по берегам
ручьев; в лесу — в подстилке, в дуплах и под
По-видимому, размножается в конце августа
и в начале осени.
4. Gigantomilax (Vitrinoides) daghestanus (Simroth,
1898), comb. nov. (рис. 268)
Simroth, 1898a : 54 (Limax); 1901 : 72, T. 3, Fig.
5—6 (Limax)-, 1912a : 9, T. 5, Fig. 3 (Limax)-, 18, T. 2, Fig.
12, T. 6, Fig. 12 (L. florenskii, syn. nov.); Давтян, 1948 : 109,
113 (L. monticola)-, А к p а м о в с к и й, 1949 : 151
(L. monticola)', 19716 : 9 (florenskii); 1976 : 202, рис. 97В,
табл. 11, рис. 119 (Vitrinoides florenskii).
Locus typicus — «Gunib» — Гуниб, централь-
ный Дагестан. Тип неизвестен. Голотип Limax
florenskii в ЗИН АН СССР.
Материал: из СССР 50 экз. из 16 пунктов;
из Турции 22 экз.
При сокращении наибольшая ширина тела при-
ходится на область мантии; от нее тело сужива-
ется назад сначала постепенно, а затем быстро.
Мантия занимает не менее х/3 длины тела, сзади
с легким углом. Между мантийной щелью и се-
рединой спины около 19—20 рядов морщин, ко-
торые веерообразно расходятся от мантии. Киль
занимает около 2/3 спины; сзади он высокий и сплю-
щенный, нередко волнистый, местами пересечен
бороздками, впереди плоский, часто в виде цель-
ной длинной морщины.
Спина и мантия одноцветные, чаще всего почти
черные или темно-серые. Пигмент концентриру-
ется по морщинам, бороздки более светлые. Бока
темно- или светло-пепельные, ниже мантии еще
светлее. На верхней стороне тела, особенно на
боках, разбросаны светлые круглые пятнышки,
каждое занимающее 2—3 морщины. Иногда про-
ступает светло-желтая килевая полоска. Подошва
белая, порой с темной оторочкой вдоль внешнего
края боковых долей. С возрастом в противопо-
ложность G. monticola окраска светлеет, а пят-
нышки проступают более интенсивно.
Размеры: дл. сок. до 66, ширина до 13, дл.
мантии до 22 мм.
Радула. В отличие от G. monticola число зубов
в одном ряде и число рядов у взрослых слизней
больше. Базальная пластинка шире. У взрослых
три самых внешних краевых зуба шиповидные,
остальные всегда с 2 зубцами (мезокон и эктокон).
В течение жизни животного идет не только уве-
корой гнилых деревьев.
Рис. 268. Gigantomilax
(Vitrinoides) daghestanus
(Simr.). Горный Дагестан,
Цудахор, 28 VI 1967.
личение числа рядов, но и нарастание числа крае-
вых зубов. Последнее видно из приводимого ниже примера с двумя экземпля-
рами разного возраста из одной популяции близ Сарыкамыш (вилайет Карс,
Турция).
234.
III. LIMACIDAE
№ Дл. сок., мм Радулярная формула
< с 186 43к Зк
2 43.3 -у+ £ Н~ g Т" 1 X 150
Кишечник. Третья петля кишечника занимает около 1/4—1/3 длины тела,
и оба колена, слагающие ее, тесно прилегают друг к другу (у G. monticola
и G. koenigi петля короче, а колена располагаются под углом).
Гениталии относительно небольшие. Передняя половина простаты
Заметно вздута (в 1х/2 раза шире задней половины, т. е. слабее, чем у G. monti-
cola). Яйцевод длиннее семяпровода, в атриум впадает под углом. Семяпро-
вод широкий и короткий, впадает в пенис терминально. У молодых особей
пенис булавовидный, шаровидно вздут у заднего конца; у взрослых он может
быть округлым. Половой ретрактор крепится к пенису сбоку, почти на се-
редине его длины. Между ретрактором и семяпроводом имеется круглая
и широкая площадка, нередко слегка пигментированная. Слепой отросток
пениса (бич?) встречен лишь у отдельных экземпляров. Семяприемник
с большим, неправильной формы резервуаром и коротким, но широким про-
током. Атриум связан со стенкой тела ретентором.
Распространение. В СССР: центральный Дагестан (Гуниб, Ме-
хельта, Агвали), южная Армения (Ленинаканский, Вединский, Даралагезский
зоогеографические участки (Акрамовский, 19716)). Вне СССР: Турция (ви-
лайеты Порох, Эрзурум, Карс).
Образ жизни. Обитает в степном и лугостепном поясе, в осыпях,
там, где отдельные камни чередуются с травой и кустарником, а также под
камнями у ручьев и в трещинах скал.
Замечание. Этот вид отличается от G. monticola и G. koenigi отсут-
ствием темного рисунка на мантии и спине (в любом возрасте), а также тем,
что окраска у него с возрастом не темнеет, а светлеет. Из особенностей внутрен-
него строения наиболее характерны размеры и форма третьей петли кишеч-
ника.
3. Подрод MONOCHROMA Simroth, 1896 stat. nov.
Simroth, 1896 : 366; 1901 : 96; Germain, 1912 : 31 (JPseudarion, syn. nov.);
Hesse P., 1926 : 20.
Типовой вид — M. brunneum Simroth, 1901; по последующей моно-
типии — Simroth, 1901.
Слизни средних размеров. Морщины плоские, на спине располагаются
почти параллельно килю. Киль образован одной морщиной. Мантия с за-
метным углом на заднем крае. Пневмостом лежит постмедиально. Подошва,
как у подрода Vitrinoides.
Пенис цилиндрический или мешковидный с длинным или коротким, но
всегда четким бичом. Внутренние стенки пениса, кроме кольцевидного ва-
лика, имеют продольные складки. Ретрактор правых щупалец не перекре-
щивается ни с половыми путями, ни с их ретрактором.
Распространение. Зангезур, Талыш и северо-западные области
Ирана.
Заме ч а н и е. Как сцноним данной группы мы рассматриваем Pseud-
hrion Germain, 1912. Обоснование — см. с. 239.
1. GIGANTOMILAX
235
5. Gigantomilax (Monochroma) brun-
neus (Simroth, 1901), comb. nov.
(рис. 269—275)
Simroth, 1901 : 96, T. 6, Fig. 1 — 18
(Monochroma)', Лихарев, P ам нель-
ме й e p, 1952 : 329, рис. 254, 255 (Mono-
chroma)', Акрамовский, 19716 : 9
(Monochroma)', 1976 : 203, рис. 98, табл. 11,
рис. 120 (Monochroma).
Locus typicus — «Karabach (Kara-
goel)» — Нагорный Карабах, оз. Ka-
ра-гёль (ныне Севлич). Из синтипов
сохранился только один молодой эк-
земпляр, в свое время переданный
Зимротом в Петербургский зоологи-
ческий музей (ныне ЗИН АН СССР),
который мы обозначаем как лектотип,
так как остальные синтипы, хранив-
шиеся в Зенкенбергском музее
Франкфурта-на-Майне, не сохрани-
лись. Окраска лектотипа почти чер-
ная, лишь срединная доля подошвы
светлая; дл. сок. 21 мм.
Материал: Из СССР 70 экз.
из 21 пункта.
При сокращении тело массивное,
цилиндрическое, наиболее широкое
позади мантии, к заднему концу бо-
лее или менее резко сужается. Кожа
толстая, с длинными морщинами,
в особенности на спине. Киль различ-
ной длины; до мантии если доходит,
то в виде низкой цельной продоль-
ной морщины; сзади слегка волни-
стый. Мантия занимает около 2/5
длины тела, спереди округлая, сзади
с закругленным углом. Капюшон
не полностью закрывает передний ко-
нец тела. Между мантийной щелью и
средней линией спины 14—16, реже до
19 РЯДОВ МОРЩИН. 1*°-'
Спина и мантия одноцветные, от
черного до темно-серого цвета. Бока
и боковые доли подошвы окрашены
светлее, срединная доля белая или
светло-серая (особенно у молодых эк-
земпляров). Пигмент сосредоточен по
морщинам в виде очень мелких точек
(значительно меньших, чем у G. monti-
cola, и чуть меньших, чем у G. koenigi).
Рис. 269. Gigantomilax (Monochroma) brun-
neus (Simr.). Зангезурский хребет, 7—
9 VI 1974. Внешний вид и фрагмент по-
дошвы.
0.01
Рис. 270—273. Gigantomilax (Monochroma') brunneus (Simr.) (продолжение).
270 — зубы радулы; 271 — кишечник; 272, 273 — гениталии.
1. GIGANTOMILAX
237
Размеры: дл. сок. до 45, ширина до 12, дл. мантии до 17 мм.
Радула (рис. 270). Срединный и боковые зубы с 3 зубцами и относительно
широкими базальными пластинками. Краевые зубы вначале с 2 зубцами,
а под конец эктокон распадается на ряд очень мелких зубчиков. Пример:
1 экз. с перевала в Зангезуре; формула:
с 286 14к 9к
т + —+ —+4=6X112.
Кишечник (рис. 271). Третья петля короткая, около х/5 длины тела, и оба
колена ее располагаются друг к другу под некоторым углом.
Рис. 274, 275. Gigantomilax {Monochroma) brunneus (Simr.)
(окончание). Гениталии.
Гениталии (рис. 272—275). Передний конец простаты очень широ-
кий. Семяпровод очень короткий, но толстый. Пенис веретеновидно-цилинд-
рический, мощный; длина его в 3 раза и более превосходит ширину. Очень
короткий семяпровод впадает в пенис апикально; здесь же крепится половой
ретрактор и располагается крупный бич, который неправильно изогнут или
имеет форму рога. Внутри заднего конца бича находится мощный продольный
валик, занимающий почти всю полость. Внутренние стенки пениса и перед-
него участка бича покрыты различными (поперечными или косыми) складками.
Атриум небольшой, с тонкими стенками и слабо обособлен от пениса.
Распространение. Кроме озера Кара-гёль (современное название
озеро Севлич, в 12 км на северо-запад от Гориса, АрмССР), он найден в сле-
дующих районах Армении: Горисский, Кафанский, Мегринский. Следова-
тельно, ареал данного вида располагается в пределах горной системы Занге-
зур.
Образ жизни. В вертикальном направлении обитает от лесной зоны
до нижнеальпийского пояса (Акрамовский, 19716), в светлых лесах, лугах
и в ксерофильном редколесье, особенно вдоль ручьев, в подстилке и под ко-
рой деревьев.
238
III. LIMACIDAE .
6. Gigantomilax (Monochroma) lenkoranus Simroth, 1912, comb. nov.
(рис. 276—279) .
' Simroth, 4912a : 48, T. 3, Fig. 44, T. 7, Fig. 27; 49, T. 3, Fig. 45, T. 7,
Fig. 28 (talyschanus, syn. nov.); Лихарев, Рам мель мейер, 1952 : 327.
Locus typicus — «Berg Kyz-Kalasi (2100 m), Zuvant» — гора Кыз-Каласи,
Зуванд (Талыш, АзССР). Типы в ЗИН АН СССР; за лектотип G. lenkoranus
Рис. 276—279. Gigantomilax (Monochroma') lenkoranus Simr.
276 — кишечник; 277 — ретракторы; 278 — дистальные части гениталий, Иран, Себелан, 27 VH 1916:
279 — то же, Талыш, Орант, 22 V 1909.
нами выбран экземпляр, изображенный у Зимрота на табл. 3, фиг. 44, а за
лектотип G. talyschanus экземпляр, изображенный на той же таблице, фиг. 45.
Материал: из СССР 43 экземпляра (в том числе типы) из 15 пунктов.
При сокращении тело массивное, с наибольшей шириной за мантией.
Киль очень короткий не более 1/4 длины тела, а нередко и отсутствует. Кожа
толстая. Мантия занимает около х/3 длины тела, сзади нередко со слабым уг-
лом, поверхность ее грубозернистая. Капюшон крупный, при сокращении
полностью закрывает голову и шею. Морщины плоские, бороздки глубокие;
1. GIGANTOMILAX
239
между мантийной щелью и средней линией спины 18—20 рядов морщин.
Верхняя сторона тела интенсивно черная, иногда с синеватым оттенком,
только шея и срединная доля подошвы светлые; затылок темно-серый.
Указанная окраска обычна для популяций из открытых мест. В лесу
наряду с типичными встречаются экземпляры со светлым килем и неопреде-
ленными светлыми пятнами на мантии (f. talyschcma Simroth, 1912).
Размеры. Дл. сок. 40—50, ширина И—12, дл. мантии 15—20 мм.
Лектотип: дл. сок. 43, ширина 12, дл. мантии 15, дл. киля 9 мм.
Радула. 1. Северный Иран, Себелан, близ Ардебиля, 1 экз. (f. typica);
формула:
27к
с 336 , 28к
3 + 3 + 2—1 Х 132>
2. АзССР, Талыш, сел. Орант, 1 экз. (f. talyschana Simr.); формула:
с 326 27к
Т + "1“ + "2=Г х 121‘
Кишечник (рис. 276) сильнее закручен, чем у G. lederi (О. Bttg.). Третья
петля около х/4—1/5 длины тела (по Зимроту, не более 1/8).
Гениталии (рис. 278, 279). Крупная светло-серая треугольная го-
нада лежит позади первой петли кишечника и сверху лишь частично закрыта
лопастями печени. Ее проток утолщается и дает извивы лишь вблизи от бел-
ковой железы. Последняя образована 2—3 лопастями (как у G. lederi, в муж-
ской фазе). Семяяйцевод длинный, почти одной ширины на всем протяжении.
Простата прилегает к его переднему концу и имеет бобовидную форму.
Яйцевод короткий с рыхлыми железистыми стенками. Семяприемник с круп-
ным вытянутым резервуаром и коротким протоком, который открывается
в атриум рядом с яйцеводом. Короткий и толстый семяпровод впадает в зад-
ний конец мешковидного пениса. Рядом с семяпроводом в пенис открывается
небольшой крючковидно изогнутый конический бич.1 Половой ретрактор
крепится к пенису близко от переднего конца, задний конец его прикрепля-
ется к диафрагме под сердцем. К переднему концу пениса крепится небольшой
ретентор, связывающий его со стенкой тела. Задний конец пениса иногда
черный. Внутренние стенки складчатые и имеют 1—2 довольно крупных
валика неправильной формы.
Яркая оранжевая окраска внутренностей, которую Зимрот считал харак-
терной для данного вида, обязана каким-то особенностям фиксации использо-
ванных автором материалов. У экземпляров, собранных позднее, внутрен-
ности белые.
Распространение. В СССР все находки сделаны в горах Талыша
близ вершин наиболее высоких гор — Мараюрт, Кыз-юрды, Кыз-каласи,
Шиндан-каласи, т. е. на юго-востоке АзССР. Вне СССР — одна находка
в Иранском Азербайджане, близ города Ардебиля, на горе Себелан (в ЗИН
АН СССР).
Образ жизни. Обитает на высотах 1800—2500 м как на высокогор-
ных лугах, так и в верхнем поясе лесной зоны. Местообитания приурочены
к вершинам гор, находящихся под воздействием частых туманов, идущих
со стороны Каспийского моря. Излюбленные убежища — скалы и осыпи.
Замечание. Жермен (Germain, 1912) описал и дал рисунки несколь-
ким новым для науки видам Limacidae, найденным в северном Иране, в том
числе Pseudarion morgani (новый вид нового рода) и Malacolimax toscannei.
К сожалению, при достаточно подробном описании внешнего вида слизней
1 Зпмрот считал, что у G. talyschanus нет бича; Однако проверка типов показала,
что этот орган у них есть, но у данных'экземпляров он был ввернут внутрь пениса.
240
III. LIMACIDAE
и их половых путей автор ничего не сказал об их кишечнике и других орга-
нах.
По его словам, Pseudarion morgani внешне напоминает Arionidae. Однако
при внимательном чтении выясняется, что это сходство ограничивается только
намеком на каудальную ямку. Из рисунка же хорошо видно, что нет настоя-
щей каудальной ямки, а лишь небольшое углубление верхней стороны тела
над концом подошвы, которое нередко возникает у многих Limacidae при
«жесткой» фиксации.
По форме тела, окраске, положению пневмостома, гениталиям оба вида
Жермена очень походят на Gigantomilax lenkoranus Simr. Поэтому очень ве-
роятно, что как Pseudarion morgani, так и Malacolimax toscannei являются
синонимами данного вида.
В той же работе Жермен описал еще 4 вида Malacolimax из северного
Ирана —morgani, pollonerai, azerbaidjanensis, mesquensis. Совершенно оче-
видно, что они не принадлежат к роду Malacolimax в современном понимании.
Внешний облик и гениталии говорят о принадлежности к роду Gigantomi-
lax, но к какому из подродов — Vitrinoides или Monochroma, — пока сказать
трудно.
2. Род CAUCASOLIMAX Likharev et Wiktor, gen. nov.
Типовой вид — Limax caucasicus Simroth, 1898.
Диагноз. В отличие от других Limacidae внутри пениса имеется па-
пилла, а стенки семяприемника очень мускулистые.
Небольшие слизни (дл. пол. до 45, дл. сок. до 35 мм). Спина с отчетливым
килем, который простирается не менее чем на 3/4 спины. Мантия относительно
крупная — длина ее немного меньше 1/2 длины тела. Пневмостом лежит
постмедиально.
Срединные и боковые зубы радулы трехзубцовые, лишь несколько самых
внешних боковых с двумя зубцами. Краевые зубы однозубцовые шиповидные
или кинжаловидные.
Кишечник не закручен. Вторая и третья петли приблизительно одной
длины и сильно смещены назад. Обе доли печени не достигают вершины внут-
ренностного мешка, которую занимает гонада. Яйцевод и семяпровод ко-
роткие. Пенис тоже короткий и расчленен на несколько отделов, внутри него
находится мощная папилла. Семяприемник впадает в атриум.
В отличие от всех других родов стенки семяприемника очень толстые
и необычного- гистологического строения. Они сложены из чередующихся
слоев кольцевых и продольных мускульных волокон. Внутренняя поверхность
выстлана высоким цилиндрическим эпителием, причем концьыэпителиальных
клеток вытянуты в просвет канала, завершаясь своеобразными нитями —
филаментами.
Ретрактор правых щупалец проходит через щель между пенисом и яйце-
водом, так что семяпровод остается позади него, т. е. как у Malacolimax и
Caspilimax.
Монотипический род.
Замечание. Данному роду наиболее близки роды Gigantomilax
О. Bttg., Caspilimax Hesse и Malacolimax Malm. Всем четырем группам
свойственны короткий семяпровод и мешковидный или короткоцилиндричес-
кий пенис. По-видимому, эти особенности гениталий определили необыч-
ное положение ретрактора правых щупалец по отношению к половым путям.
В пределах семейства только у данного рода встречается пениальная
папилла и указанное выше гистологическое строение семяприемника. Эти
особенности позволяют предполагать, что копуляция у Caucasoltmax cauca-
sicus осуществляется несколько иначе, чем у других видов Limacidae.
2. CAUCASOLIMAX
241
Caucasolimax caucasicus (Simroth, 1898), comb. nov. (рис. 280—285)
Simroth, 1898a : 55 (Limax); 55 (L. ananowi, syn. nov.); 56 (L. amaloides, syn.
nov.); 1901 : 72, T. 3, Fig. 7—12 (L. ananowi et var. imereticus, syn. nov.); 75, T. 3, Fig. 13—
17 (Limax); 75, T. 3, Fig. 18—23 (L. amaloides); 1912a : 7, T. 1, Fig. 2, T. 5, Fig. 2 (L. ana-
nowi var. alticola, syn. nov.); 122, T. 10, Fig. 48 (L. svaneticus, syn. nov.); К а л и т и н а,
1958 : 159 (Limax).
Locus typicus — «Kazbek» — Казбек. В ЗИН АН СССР имеются следую-
щие типовые экземпляры:1 L. ananowi (лектотип и 3 паралектотипа — Кав-
каз, окрестности Гудаура на Военно-Грузинской дороге, сборы Ананова),
L. ananowi var. alticola (лектотип и 3 паралектотипа — гора Хочалдаг
у Лагодехи, альпийская зона 16 VI 1907, сбор Млокосевича), L. ananowi
var. imereticus (голотип — Опршети, Имеретия, VI 1877 сбор Бротеруса),
L. caucasicus (голотип — гора Майорша, долина Крестовая, у Коби, ю.-з.
Казбек, сбор Ананова), L. svaneticus (лектотип — сев. склон Латпарского
ущелья, Сванетия, 6 VIII 1910, сбор Казнакова).
Материал: из СССР более 100 экз. (в том числе типы) из 13 пунктов.
Тело стройное, к заднему концу заметно суженное. Киль иногда достигает
мантии. Мантия спереди закругленная, сзади нередко с заметной вырезкой
против киля; капюшон крупный — более х/3 длины мантии. Кожа толстая,
с крупными морщинами, разделенными довольно глубокими бороздками;
меяеду мантийной щелью и средней линией спины 14—16 рядов морщин.
Окраска слагается из микроскопических темных точек — меланофоров,
разбросанных на светлом фоне. Она варьирует от темно-бурой или почти
черной до голубовато-серой. Середина спины и мантии всегда темнее пери-
ферии и боков тела, которые могут быть белые. Рисунок отсутствует. Пнев-
мостом окружен светлым валиком. Подошва белая, к заднему концу заметно
сужена.
Размеры: дл. пол. 40—45, дл. мантии до 18 мм; дл. сок. до 35, дл.
мантии до 14 мм.
Раковина продолговато-овальная с заметным роговым слоем, имеющим
микроскопическую кольцевую исчерченность. У слизня с дл. пол. 44 мм она
имеет следующие размеры: 7x4.7 мм.
Радула. Пример: Крестовый перевал, сбор. 3. И. Калитиной (1 экз.):
с 186 36 13к
Т + УГ + ~ х 10°-
Кишечник (рис. 282). Первая петля простирается назад немного дальше,
чем остальные, которые примерно одной длины. Место перехода пятого ко-
лена в шестое иногда заметно вытянуто, образуя подобие слепой кишки.
Гениталии (рис. 283—285). Гонада образует вершину внутренност-
ного мешка и слабо закрыта другими органами. Гермафродитный про-
ток впереди очень сильно извит. Белковая железа очень крупная. Семяяй-
цевод, кроме обычных слабых извивов, образует один очень резкий и лежащий
в дорсовентральном направлении (заметен даже у молодых экземпляров).
К передней трети семяяйцевода прилегает крупная простата, переходящая
в очень короткий, но толстый семяпровод с мускулистыми стенками. Послед-
ний апикально открывается в короткий пенис, состоящий из трех отделов:
передний, средний и задний. Снаружи по бокам от семяпровода на заднем
конце пениса нередко заметны два небольших слепых выступа. Внутренние
стенки заднего отдела пениса покрыты продольными складками. В более
широком среднем отделе пениса располагается мощная папилла. Передний
отдел сужается к атриуму и нередко к остальному пенису располагается
под углом. Внутри него находится крупная складка, которая, возможно,
2
1 Лектотипы обозначены нами.
16 И. М. Лихарев, А. Й. Виктор
285
Рис. 280—285. Caucasolimax caucasicus (Simr.). Крестовый перевал, 18 VIII 1949.
280 — внешний вид; 281 — раковина; 282 — кишечник; 283 — гениталии; 284 — дистальная часть
гениталий; 285 — задний конец пениса (стрелки указывают направления движения спермы).
3. CASPILIMAX
243'
выполняет функцию стимулятора. Половой ретрактор крепится к пенису
в двух местах: к заднему и переднему отделу.
Очень короткий яйцевод впадает в атриум напротив пениса. Между ними
располагается семяприемник, который отличается своеобразным строением
(с. 240). Он неясно расчленен на резервуар и проток. Нередко задний конец,
семяприемника слегка ввернут внутрь.
Распространение. Центральная часть Большого Кавказа.
Экология. Обитает на субальпийских и альпийских лугах среди
камней и в щелях скал. Поднимается до высоты 3000 м.
Замечания. 1. В пределах одной популяции окраска слизней может
сильно варьировать. Так, на Крестовом перевале совместно обитают темно-
бурые (f. typica) и серовато-голубые особи (f. amalioides Simr.) со всеми
переходами между ними. Молодь обнаруживает те же тенденции в окраске,,
так как встречаются как синевато-черные, так и почти белые особи.
2. Подобно тому как это имело место при рассмотрении Gigantomilax
monticola (О. Bttg.), так и здесь мы вынуждены свести в синонимы (см. выше)
ряд видов Зимрота, появление которых обязано, во-первых, изменчивости
окраски и, во-вторых-, возрастным отличиям в строении гениталий. Не слу-
чайно, что из этих «видов» три — ananowi, caucasicus, amalioides — были
описаны из соседних пунктов Военно-Грузинской дороги. Для всех их Зим-
рот считал характерным наличие зачатка эпифаллуса,1 за который
он принимал задний отдел пениса, служащий преддверьем папиллы. В то же
время он почему-то не обратил внимания на очень своеобразную форму и
строение семяприемника.
3. Род CASPILIMAX Р. Hesse, 1926, stat. nov.
Hesse P., 1926 : 14,
Типовой вид — Limax key serlingi Martens, 1880; по первоначальному
обозначению.
Крупные слизни (дл. пол. до 115, дл. сок. до 80 мм). Киль короткий и ту-
пой. Мантия занимает менее х/3 длины тела, с ясным углом на заднем крае
и с крупным капюшоном. Пневмостом лежит немного постмедиально.
Срединный и боковые зубы радулы трехзубцовые, лишь самые внешние
боковые теряют эндокон и становятся двухзубцовыми. Краевые зубы двух
типов: внутренние однозубцовые, кинжаловидные (у молодых слизней могут
отсутствовать); внешние двухзубцовые за счет крупного вторичного допол-
нительного зубца.
Кишечник не перекручен и без слепой кишки. Вторая и третья петли
приблизительно одной длины, 5-е и 6-е колена прижаты и параллельны друг
к другу. Левая доля печени образует вершину внутренностного мешка.
Ось сердца наклонена под углом 45°. Аорта длинная, задний отдел кро-
веносной сети легкого отчетливо выражен. Основание полового ретрактора
крепится к диафрагме перед сердцем.
Гонада располагается позади второй петли кишечника. Яйцевод и семя-
провод короткие. Последний апикально впадает в короткоцилиндрический
пенис, внутри которого находится стимулятор и кольцевой валик. Семя-
приемник открывается в основание пениса близ атриума.
Ретрактор правых щупалец проходит через щель между пенисом и яйце-
водом, так что семяпровод остается позади его, т. е. как у Malacolimax и
Caucasolimax.
1 Непонятно, на каком основании Зимрот так оценил этот отдел гениталий, если он
ни разу не встречал сперматофора у этих слизней. А ведь эпифаллус — это орган, в кото-
ром формируется сперматофор!
16*
244
III. LIMACIDAE
Рис. 286. Caspilimax keyserlingi (Mart.)
f. tigris (0. Bttg.). Талыш, Лерик, 31 V
1974.
Монотипический род.
Замечание. До сих пор эту
группу рассматривали как подрод
рода Limax. Однако четкие различия
в форме, размерах и внутреннем
строении пениса дают основание счи-
тать обе группы самостоятельными
родами. В то же время Caspilimax
несомненно близок кавказским родам
Gigantomilax и Caucasolimax и евро-
пейскому Malacolimax.
Caspilimaxkeyserlingi (Martens, 1880),
comb. nov. (рис. 286—290)
Martens, 1880 : 153 (Limax)', В o-
e t t g e r 0., 1886a : 269, T. 2, Fig. 2—4
(Limax tigris)', 270, T. 2, Fig. 5 (Limax taly-
schanus)', 1886c : 243 (L. tigris)', 245 (L. taly-
schanus)', Simroth, 1898a : 59 (L. baeri),
1901 : 82 (L. keyserlingi, L. talyschanus)',
182, T. 19, Fig. 10—14 (Gigantomilax robus-
tus, syn. nov.); 1910a : 504, T. 6, Fig. 5—8
(L. persicus); 1912a : 19, T. 2, Fig. 13, T. 6,
Fig. 13 (L. tigris —talyschanus)', Лихарев,
P аммел ьмейер, 1952 : 314 (Limax).
Locus typicus — «Astrabad» — Act-
рабад (современное название Горган,
Иран). Голотип данного вида, так
же как голотипы Limax baeri Simroth,
1901 и Gigantomilax robustus Simroth,
1901, лектотип (обозначен нами) и
1 паралектотип Limax persicus Simroth,
1910, хранятся в ЗИН АН СССР.
В Зенкенбергском музее хранятся
слизни из коллекции О. Бетгера под
названиями Limax tigris (№ 155450)
и L. talyschanus (№ 155319). Воз-
можно, это синтипы, но полной уве-
ренности в этом у нас нет, так как
число экземпляров под первым на-
званием больше, а под вторым меньше,
чем указывал Бётгер.
Материал: из СССР 78 экз.
из 32 пунктов; из Ирана 125 экз. (в том
числе голотип).
Тело удлиненно-веретеновидное,
стройное, к заднему концу заостренное.
При сокращении слизня капюшон ман-
тии обычно свисает вперед и полностью
закрывает шею и голову. Спина с круп-
ными морщинами, которые несут мелкие
бугорки (отчетливо видны при жизни)
и разделены глубокими бороздками.
Между мантийной щелью и средней
линией спины 15—18 рядов морщин.
3. CASPILIMAX
245
На спине на сероватом или оливковом, реже грязно-желтом фоне имеется
темный рисунок из пятен, очень разнообразных как по форме, так и по вели-
чине и по положению на теле (f. tigris О. Boettger, 1886). Иногда пятна сли-
ваются в продольные полосы (f. persica Simr.), но не столь правильные и иначе
Рис. 287—290. Caspilimax keyserlingi (Mart.).
287 — кишечник; 288 — ретракторы; 289 — гениталии, Иран, Энзели, 19 III 1915; 290 — пиг.тяльняя
часть гениталий, оттуда же.
расположенные, чем у европейского Limax maximus L. и его сородичей.
Одноцветные формы (f. typica=C. keyserlingi Martens, 1880, syn. Limax
talyschanus 0. Boettger, 1886) встречаются реже пестрых. В отличие от спины
мантия чаще одноцветная, реже с небольшим числом менее четких пятен на
капюшоне. Подошва белая или светло-желтая, край ее заметно выступает за
контуры тела. Слизь бесцветная.
Размеры: дл. пол. 100—115, шир. пол. 11—15, мантия 27—30 мм,
дл. сок. 60—80 мм; голотип — дл. сок. 63, ширина 18, дл. мантии 21 мм.
Раковина яйцевидная. У экземпляра с дл. сок. 67 мм ее размеры 8x6 мм.
Радула. Пример: Иран, Энзели; формулы:
246
III. LIMACIDAE
1. Взрослый экземпляр (дл. сок. 67 мм):
с
"3
216 ЗЗк 23к
3—2+ 1 + 2 X 171.
2. Молодой экземпляр (дл. сок. 23 мм):
с 186 24к
Т+ ~Х-120.
Кишечник: см. диагноз рода (рис. 287).
Гениталии (рис. 289, 290). Узкая языковидная или треугольная,
гонада лежит на поверхности внутренностного мешка. Ее проток вначале-
тонкий и почти прямой впереди становится толстым и сильно извитым.
Белковая железа языковидная. Женский канал семяяйцевода образует круп-
ные гофры; простата длинная, широкая и лентовидная. Семяпровод впадает
в короткий, но массивный и коленообразно изогнутый пенис, к которому
прикрепляются два мускула: мощный половой ретрактор сбоку, перед пере-
гибом, и ретентор, связывающий передний конец пениса со стенкой тела.
Внутри вздутого заднего отдела пениса помещаются на одной стенке мощный
складчатый валик, на другой (напротив валика) крупный стимулятор. Узкий
передний отдел пениса имеет тонкие и гладкие стенки. Стенки яйцевода тол-
стые, железистые, внутри гладкие. Семяприемник небольшой, с коротким
протоком. Атриум слабо выражен и плотно подтянут к стенке тела многочис-
ленными мускульными волокнами.
Распространение (рис. 42, 1). В СССР наиболее обычен в лесах
Талыша и Ленкоранской низменности (АзССР); кроме того, по одной находке
сделано в лесах близ поселков Ялама и Сулут (Худатский и Исмаилинский
районы АзССР). Сообщение Зимрота (1912а) о присутствии этого вида в ок-
рестностях Боржоми связано с ошибкой в определении. Автор имел дело с мо-
лодым Eumilax brandti (Mart.) (экземпляр в ЗИН АН СССР)..
Вне СССР — лесные области северного Ирана: провинции Мазендеран
(Forcart, 1935; Starmiihlner, Edlauer, 1957), Горган (Martens, 1880; Simroth,.
1910) и Гилян (коллекция ЗИН АН СССР).
Экология. Обитает в широколиственных лесах, как равнинных,,
так и горных (буковые, дубовые, с железняком — Parrotia persica). Днем
укрывается в дуплах, под корой гнилых пней и валежин, нередко совместно-
с Limax maculatus (Kai.).
Замечание. Зимрот в ряде работ (Simroth, 1886b, 1898а, 1901,
1910, 1912а) отмечал, что, во-первых, оба вида Бётгера — Limax talyschanus
и L. tigris (Boettger О., 1886) отличаются лишь окраской и связаны переход-
ными формами; во-вторых, L. talyschanus О. Boettger, 1886 и L. keyserlingi
Martens, 1880 являются синонимами. К тому же выводу пришел и Форкарт
(Forcart, 1935).
Исследование как типового материала (см. выше), так и сборов, накопив-
шихся в ЗИН АН СССР за последние 70 лет, убедило нас в том, что С. key-
serlingi (Mart.) при большом постоянстве во внутреннем строении отличается
значительным полиморфизмом в окраске. В связи с этим такие названия,,
как Limax tigris Bttg., L. talyschanus Bttg., L. baeri Simr., L. persicus Simr.
и Gigantomilax robustus Simr., следует рассматривать как младшие синонимы;
С. keyserlingi (Mart.).
4. Род TURCOMILAX Simroth, 1901, stat. nov.
Simroth, 1901:176; 1910a : 527; 1912a : 47; Hesse P., 1926:20.
Типовой вид — Gigantomilax (Turcomilax) nanus Simroth, 1901;
по монотипии.
4. TURCOMILAX
247
Слизни средних размеров (дл. пол. до 100, дл. сок. до 55 мм). Тело вере-
теновидное, стройное, при сокращении максимальная ширина приходится на
область мантии и начало спины. Киль тупой и простирается не более чем на
заднюю треть спины. Мантия яйцевидная, нередко сзади с тупым углом;
капюшон занимает около х/2 ее длины. Пневмостом четко постмедиальный.
Окраска одноцветная (без рисунка) — от темно-бурой до черной; подошва
светлая.
Кишечник из трех петель, не перекручен; первая петля простирается
назад дальше остальных двух. У большинства видов имеется слепая кишка.
Правая доля печени лежит сзади и образует вершину внутренностного мешка.
Раковина почти полностью лежит над мантийным комплексом. Сердце
располагается в левой четверти комплекса, прижато к его краю, а ось его
наклонена направо под углом около 45° к продольной оси тела. Аорта длин-
ная. Половой ретрактор прикрепляется у левого края диафрагмы.
Гонада лежит между зобом и второй петлей кишечника. Семяяйцевод
с длинной простатой, передний конец которой обособлен от женских
половых путей. Разделение семяяйцевода на яйцевод и семяпровод происхо-
дит очень близко от атриума. Семяпровод заметно длиннее, чем у Giganto-
milax, но короче, чем у Limax. В пенис он впадает субапикально. Несколько
впереди его к пенису крепится половой ретрактор. Пенис различной формы,
но всегда длиннее, чем у Gigantomilax. Внутри него находится несколько скла-
док или валиков.
Положение ретрактора правых щупалец по отношению к гениталиям раз-
лично. 1
Распространение. Горная система Тянь-Шаня, Джунгарский
Алатау и хребет Тарбагатай (рис. 43, 2).
Род слагается из трех подродов и 4 видов.
Замечания. 1. До сих пор все авторы вслед за Зимротом (Simroth,
1901—1910а) и Гессе (Hesse Р., 1926) относили виды, которые здесь объеди-
нены в данный род, к другим родам. Так, Т. nanus (Simr.) и Т. ferganus
(Simr.) относили в подрод Turcomilax рода Gigantomilax. С другой стороны,
Т. natalianus (Mich.) рассматривали как член рода Lehmannia, а Т. turkesta-
nus (Simr.) — как Limax.
а) Указанные авторы усматривали близость среднеазиатских Т. nanus
и Т. ferganus с кавказскими видами Gigantomilax в том, что у них ретрактор
правых щупалец не перекрещивается с пенисом, а кишечник устроен по трех-
петлевому плану. Как существенные отличия, давшие основание выделить
среднеазиатские виды в особый подрод Turcomilax, рассматривался длинный
пенис и присутствие слепой кишки — у кавказских видов пенис обычно
короткий и мешковидный, а слепой кишки нет.
С нашей точки зрения, объединение обеих групп слизней в один род не
обоснованно. Трехпетлевый тип кишечника — это особенность, свойственная
всем Limacinae. Что касается положения ретрактора правых щупалец по
отношению к гениталиям, то у обеих групп оно кардинально различное. У кав-
казских Gigantomilax s. str. ретрактор щупалец перекрещивается с ретрактором
пениса, у среднеазиатских Turcomilax1 ретрактор щупалец располагается
медиально от половых путей и не перекрещивается ни с пенисом, ни с его
ретрактором. В этом последняя группа напоминает отношения, существую-
щие у кавказской группы Monochroma Simr., от которой сильно отличается
по другим признакам.
Кроме того, как Gigantomilax s. str., так и Monochroma отличаются от
Turcomilax сильно закрученным кишечником.
1 В том объеме, как понимали эту группу Зимрот и Гессе.
848
III. LIMACIDAE
б) В пользу отнесения Т. natalianus (Mich.) к европейскому роду Leh-
mannia оба автора указывали на присутствие у этого слизня слепой кишки
и бича на пенисе. Однако на этом сходство и ограничивается. Уже внешний
облик Т. natalianus сильно отличается от Lehmannia — значительно более
крупные размеры, более толстая кожа, отсутствие рисунка, характерного для
большинства Lehmannia. У последней слепая кишка всегда очень длинная
и достигает заднего конца полости тела, а главное, иначе расположена, чем
у среднеазиатского вида. У европейских видов она служит четким продол-
жением назад задней кишки (т. е. 6-го колена кишечника), располагаясь
с нею на одной линии, и всегда значительно длиннее очень короткой задней
кишки. Между тем у Т. natalianus слепая кишка чаще всего относительно
короткая и отходит назад от места перехода (перегиба) 5-го колена в 6-е,
т. е. никогда не находится на одной линии с задней кишкой.
В силу столь существенных отличий Т. natalianus от европейских видов
Lehmannia как во внешнем облике, так и во внутреннем строении нет осно-
ваний объединять их в один род.
в) В пользу причисления Т. turkestanus к роду Limax оба названных ав-
тора в первую очередь указывали на трехпетлевый план строения кишеч-
ника, перекрещивание гениталий с ретрактором правых щупалец и обособ-
ление переднего конца простаты от женских полцвых путей.
Как уже отмечалось, указанная форма кишечника присуща всем Limacinae.
Что касается упомянутых особенностей простаты и ретрактора правых щу-
палец, то и они обычны многим родам того же подсемейства.
В то же время Т. turkestanus (Simr.), а также Т. tzvetkovi sp. nov. имеют
ряд существенных отличий от европейского рода Limax. Они меньше разме-
рами, с более коротким и слабым килем, всегда одноцветно темные, т. е.
без рисунка. Во внутреннем строении они отличаются длинной аортой (у Li-
max короткая), правая доля печени располагается сзади (у Limax впереди),
пенис и семяпровод не связаны перепонкой, а внутри пениса находится язы-
ковидная складка (у Limax такая перепонка есть, а внутренние складки
пениса иной формы).
Указанные особенности названных среднеазиатских видов свойственны
остальным видам рода Turcomilax, куда мы их и относим.
2. В систематическом плане данный род наиболее близок к кавказским
родам Limacinae'. Gigantomilax О. Bttg., Caspilimax Hesse и Caucasolimax
gen. nov. В то же время Turcomilax обнаруживает по сравнению с ними ряд
примитивных особенностей — слабая закрученность кишечника, положение
правой доли печени и др.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ TURCOMILAX
1 (6). Ретрактор правых щупалец пересекается с половыми путями, огибая
пенис справа и снизу, а женские пути сверху.
2 (3). Третья петля кишечника сзади заканчивается более или менее длин-
ной слепой кишкой. На пенисе имеется крючковидный выступ — бич,
который иногда может быть ввернут внутрь пениса, но тогда снаружи
остается небольшая ямка (подрод Michaelisia subgen. nov.) .....
....................................... 3. Т. (М.) natalianus (Mich.).
3 (2). Слепой кишки нет. Бич отсутствует (подрод Taulimax subgen. nov.).
4 (5). Пенис мешковидный и короткий, прямой или слабо изогнут.........
..................................4. Т. (Т.) turkestanus (Simr.).
5 (4). Пенис цилиндрический и длинный, сильно закрученный.............
......................................5. Т. (Т.) tzvetkovi sp. nov.
6 (1). Ретрактор правых щупалец не пересекается с половыми путями и ле-
жит от них медиально (подрод Turcomilax s. str.)................. . . .
4. TURCOMILAX
249
7 (8). Слепая кишка простирается назад до вершины внутренностного мешка.
Пенис булавовидный, короткий и прямой...................................
2. Т. (Т.) ferganus (Simr.).
8 (7). Слепая кишка короче. Пенис цилин-
дрический, длинный и сильно спирально
закручен . . 1. Т. (Т.) nanus (Simr.).
1. Подрод TURCOMILAX s. str.
Семяприемник впадает в атриум между
пенисом и яйцеводом. Пенис без бича. Рет-
рактор правых щупалец располагается меди-
ально от половых путей и с ними не пере-
крещивается. Третья петля кишечника со сле-
пой кишкой.
Распростр анение. Западный
Тянь-Шань (Ферганский, Чаткальский и
Киргизский хребты).
1. Turcomilax (Turcomilax) nanus (Simroth,
1901), comb. nov. (рис. 291—295)
Simroth, 1901 : 177, ТЛ19, Fig. 15-20 [Gi-
gantomilax (Turcomilax)]', 1910a : 529, T. 7, Fig. 26
[Gigantomilax (Turcomilax) abramowi pars, syn. nov.];
530, T. 7, Fig. 27—28 [Gigantomilax (Turcomilax) pi-
schpekensis, syn. nov.]; Лихарев, Раммель-
мейер, 1952 : 328 [Gigantomilax (Turcomilax)].
Locus typicus — «Aslanbob auf dem Berge
Baubasch-ata» — Арсланбоб, на горе Бау-
баш-ата (Ферганский хребет, Киргизия).
Лектотип (см.: Simroth, 1901, табл. 19, рис.
17 и 18) и 6 паралектотипов в ЗИН АН СССР.
Там же лектотип Gigantomilax abramowi
(см.: Simroth, 1910, табл. 7, рис. 26) и 8
паралектотипов того же вида, а также лек-
тотип G. pischpekensis. Лектотипы всех трех
видов обозначены нами.
Материал: из СССР — свыше 250 экз.
(в том числе и типы) из 46 пунктов.
При ползании тело узкое и стройное, ман-
тия занимает около 1/4 общей длины. При
сокращении тело становится вздутоверете-
новидным; мантия занимает около х/3 всей
Рис. 291. ТигсотЦах (Turcomilax)
nanus (Simr.). Ферганский хребет,
Кизыл-Унгур, 7 VI 1961.
длины; капюшон полностью или почти цели-
ком закрывает шею и голову. Между мантийной щелью и средней линией
спины 14—16 рядов морщин.
Верхняя сторона тела черная, темно-бурая или темно-серая, причем бока
всегда заметно светлее спины, а иногда даже почти белые. Нередко на мантии
видны небольшие нечеткие светлые пятна, особенно многочисленные по краям.
Подошва белая, за исключением ее края, который всегда затемнен. Пигмент
концентрируется главным образом на морщинах, что особенно заметно на
боках; бороздки белые или слабо окрашены, поэтому создаётся впечатление,
что спина и бока покрыты тонкой светлой сеткой.
250
III. LIMACIDAE
Размеры: дл. пол. 70—90, ширина 7—8, дл. мантии 20—22 мм;
дл. сок. 35—45, ширина 8—10, дл. мантии 10—15 мм; лектотип: дл. сок. 21 мм.
Раковина овальная; у экземпляра с дл. пол. 90 мм ее размеры 7x4 мм.
Рис. 292—295. Turcomilax (Turcomilax) nanus (Simr.). Ферганский хребет, Кизыл-Унгур,
7 VI 1961.
292 — зубы радулы; 293 — кишечник; 294 — гениталии; 295 — пенис.
Радула (рис. 292). Срединный и боковые зубы с 3 зубцами, краевые с 2 зуб-
цами, лишь несколько самых внешних с одним зубцом — шиповидные.
Пример: 1 экземпляр с Ферганского хребта у селения Кызыл-Унгур; формула:.
с 186 23к
3 + 3 + 2 X104'
Кишечник (рис. 293). Длина слепой кишки заметно варьирует. У молодых
слизней она относительно длиннее, чем у взрослых, у которых она чаще
всего достигает заднего края второй петли; реже она простирается до вершины
внутренностного мешка.
Мощный колумеллярный мускул крепится позади диафрагмы. Ретрактор
правых щупалец ответвляется рано, в то время как ретракторы глотки и ле-
вых щупалец ответвляются позднее, т. е. ближе к голове.
Гениталии (рис. 294, 295). Гонада серая, но границы между фол-
ликулами черные. От нее отходит вперед черный сильно извитой гермафро-
4. TURCOMILAX
251
дитный проток. Белковая железа очень крупная, желтая. Яйцевод короткий,
к атриуму плавно утолщается. Семяпровод значительно длиннее яйцевода,
но короче пениса. Пенис цилиндрический, длинный, спирально закручен-
ный, внутри с двумя крупными продольными складками. Семяприемник
Рис. 296. Turcomilax (Turcomilax) ferganus
(Simr.). Чаткальский хребет, V 1973. Гени-
талии.
с овальным резервуаром и корот-
ким протоком.
Распространение. Фер-
ганский, Чаткальский, Таласский
Алатау и Киргизский хребты.
Образ жизни. Обитает
в орехово-плодовых лесах южной
Киргизии. Предпочитает сырые и
тенистые ущелья, где в дневное
время скрывается под валежником,
в дуплах, гнилых пнях и, реже,
в подстилке.
Замечание. Вид был опи-
сан по нескольким молодым экзем-
плярам, найденным известным бо-
таником Коржинским в 1895 г.
на горе Баубаш-ата у Арсланбоба
{Ферганский хребет). Сравнение
типов с многочисленными собствен-
ными сборами из того же места
показало, что все отличия перво-
начального описания от только
что приведенного чисто возраст-
ного порядка.
В 1910 г. Зимрот описал еще
три вида из группы Turcomilax.
Один из них — Gigantomilax (Т.)
jerganus (см. ниже). Другой —
Z?. (71.) abramowi — описан по сбо-
рам из трех пунктов на Александ-
ровском хребте (ныне Киргизский).
Проверка типов этого вида показала, что экземпляры из Ала-арчинской щели
(7, из них только один был вскрыт Зимротом) являются половозрелыми
Turcomilax nanus. Другие два сбора — из окрестностей озера Колтур и из
Кыгатинской щели, как показало исследование внутреннего строения, при-
надлежат к Turcomilax turkestanus (Simroth, 1898).
Третий вид — G. (Т.) pischpekensis — описан по трем очень молодым
экземплярам, найденным на Киргизском хребте у реки Суссамыр. Из них
к настоящему времени сохранился лишь один. Сравнение его с молодыми
экземплярами Т. nanus из наших сборов показало их полную идентичность.
Что касается оранжевой окраски светлых участков поверхности тела и внут-
ренних органов, которую Зимрот считал характерной для G. pischpekensis,
то она объясняется только особенностями консервации.
Следовательно, как G. (Г.) abramowi Simr., так и G. (Т.) pischpekensis
Simr., — младшие синонимы Turcomilax nanus (Simr.).
2. Turcomilax (Turcomilax) ferganus (Simroth, 1910), comb. nov. (рис. 296)
Simroth, 1910a : 528, T. 7, Fig. 22—25 [Gigantomilax (Turcomilax)].
Locus typicus — «Fergana, Bergriicken Namangan, ostlicher Abhang» —
Чаткальский хребет, восточный склон. Лектотип (обозначен нами, см.:
252
III. LIMACIDAE
Simroth, 1910a, табл. 7, рис. 22) и 1 паралектотип (см.: там же, табл. 7,
рис. 23-25) в ЗИН АН СССР.
Материал: из СССР — 10 экз. (в том числе типы) из двух пунктов.
Слизни внешне очень сходны с Т. nanus (Simr.). Мантия занимает около
х/3 длины тела. Между мантийной щелью и средней линией спины 15 рядов
морщин. Вся верхняя сторона, в том числе и голова, черные, лишь бока ниже
мантии немного светлее. Край подошвы и оторочка темные, в целом же
подошва белая.
Размеры: дл. сок. до 55, ширина 8, дл. мантии 16 мм; лектотип:
дл. сок. 33, ширина 6, дл. мантии И мм.
Кишечник. Слепая кишка очень длинная и простирается назад до вершины
внутренностного мешка.
Гщ н и т а л и и (рис. 296). Пенис короткий, но массивный, булавовид-
ный, слабо изогнут, перед атриумом резко суженный, внутри с несколькими
складками и валиками. В остальном как у Т. nanus.
Распространение. Чаткальский хребет. Кроме типов в ЗИН
АН СССР имеются только 8 экземпляров, собранных В. В. Гороховым
на южных отрогах Чаткальского хребта в мае 1973 г.
2. Подрод MICHAELISIA Likharev et Wiktor, subgen. nov.1
Типовой вид — Limax (Lehmannia) natalianus Michaelis, 1892;
обозначен здесь.
Диагноз. Третья петля кишечника со слепой кишкой. Семяприемник
открывается в передний конец пениса вблизи от атриума. Пенис с бичом,
который иногда ввернут внутрь пениса; в этом случае на месте бича видна
ямка. Ретрактор правых щупалец огибает пенис справа и снизу, а яйцевод —
сверху.
Распространение. Тянь-Шань, Джунгарский Алатау и хребет
Тарбагатай.
3. Turcomilax (Michaelisia) natalianus (Michaelis, 1892), comb. nov.
(рис. 297-302)
'Михаэлис, 1892 : 31, 3 рис. [Limax (Lehmannia)]' Simroth, 1901 : 83,
T. 5, Fig. 3—8 [Limax (Lehmannia)]', 1910a : 508, T. 6, Fig. 9, 10 [Limax (Lehmannia)]',
510, T. 6, Fig. 11, 12 [L. (Lehmannia) stummeri, syn. nov.?]; Лихарев, Раммель-
мейер, 1952 : 323 (Lehmannia)', 324 (Lehmannia stummeri)', A 1 t e n a, 1954 : 47, Fig. 1 —
3 [Gigantomilax (Turcomilax) iliensis, syn. nov.l.
Terra typica — горы восточного Казахстана2. Лектотип (обозначен нами)
и 3 паралектотипа в ЗИН АН СССР.
Материал: из СССР — 191 экз. (в том числе типы) из 25 пунктов.
Тело сравнительно стройное и мягкое. Тупой киль простирается на зад-
нюю треть спины. Мантия занимает около х/3 общей длины, при сокращении
слизня капюшон почти полностью закрывает передний конец тела. Морщины
уплощенные, а бороздки довольно глубокие. Между мантийной щелью и
средней линией спины 16—18 рядов морщин. Задний конец мантии с тупым
закругленным углом.
1 Название дано в честь Е. П. Михаэлиса (1841—1913) — известного исследователя
Алтая и Восточного Казахстана.
2 Михаэлис описал этот вид по 10 взрослым экземплярам, найденным в двух пунктах:
1) на Гасфордовом перевале (находится на одном из северо-западных отрогов Джунгар-
ского Алатау; см.: Масальский, 1913); 2) на хребте Тарбагатай в проходе Сай-асу. В ЗИН
АН СССР имеются 3 взрослых и один молодой синтип данного вида, причем на сопровож-
дающей их этикетке указаны оба местонахождения. Поэтому точно указать locus typicus
не представляется возможным.
4. TURCOMILAX
253
Все тело, за исключением подошвы, окрашено в интенсивно черный цвет,
только бока внизу иногда серые. Пигмент в основном концентрируется по
морщинам, поэтому бороздки заметно светлее, но не так сильно, как у пре-
дыдущих видов.
Размеры: дл. пол. до 100, ширина 8—9, длина мантии 25—30 мм;
дл. сок. 45—50, ширина 9—10, дл. мантии 15—20 мм; лектотип: дл. сок. 53,
Рис. 297—302. Turcomilax (Michaelisia) natalianus (Mich.).
297 — кишечник; 298, 299 — слепая кишка двух экземпляров; goo — гениталии; 301 — вскрытый пенис
другого экземпляра (внутри виден ввернутый бич); 302 — пенис паралектотипа; (297—301 — Джунгар-
ский хребет, 12—15 VI 1969).
ширина 9, дл. мантии 20 мм; между мантийной щелью и серединой спины
20 рядов морщин.
Раковина неправильно овальная, с кожистой оторочкой, особенно ши-
рокой по левому краю; ее размеры 6x4 мм..
Радула сильно походит на таковую у Deroceras. Срединный и боковые зубы
с четкими экто- и эндоконами, лишь несколько самых внешних боковых зу-
бов без эндоконов. Краевые зубы шиповидные, без дополнительных зубцов,
Пример: 1 экземпляр из Заилийского Алатау, близ Алма-Аты; формула:
с 156 56 38к
Т + “ + Т + X 12°-
254
III. LIMACIDAE
Кишечник (рис. 297). Вторая петля значительно короче первой и слабо
закручена; третья петля короче второй; слепая кишка короткая, обычно не
доходит до вершины внутренностного мешка (рис. 298, 299).
Гениталии (рис. 300—302). Гонада крупная, круглая, черная.
Лопасти печени прикрывают ее лишь частично. Гермафродитный проток
сильно извитой. Семяпровод впадает в довольно длинный цилиндрический
пенис, задний конец которого обычно отогнут вперед и короткой мембраной
подтянут к остальной части пениса. На заднем конце пениса имеется корот-
кий конический, иногда изогнутый бич с тонкими железистыми стенками.
Нередко он вйернут внутрь пениса и на его месте снаружи видна лишь ямка
(рис. 301). Яйцевод сравнительно короткий. Семяприемник с круглым или
овальным резервуаром. Атриум очень небольшой. Несколько коротких ре-
тенторов связывают нижние концы половых путей со стенкой тела. Внутри
пениса находятся 3—4 продольных валика, из них у одного задний конец
раздвоен.
Внешний облик довольно постоянный. Иногда встречаются экземпляры
немного светлее обычного. Внутреннее строение варьирует сильнее. Так,
бывают слизни без четкой слепой кишки. Значительно чаще встречаются
слизни с ввернутым внутрь пениса бичом (около 30%). В пределах одной
популяции оба отклонения имеют место наряду с типичной картиной.
Распространение. В СССР: хребты Киргизский Алатау, Заи-
лийский Алатау, Кунгей Алатау, Джунгарский Алатау и ТарбаТатайский.
Вне СССР: одна находка сделана в окрестностях города Кульджа (КНР)
(syn. Gigantomilax iliensis Altena, 1954).
Образ жизни. Обитает в горнолесной и лугово-степной зонах на
высотах 1000—3000 м над ур. м. Скрывается под камнями, в осыпях и тре-
щинах скал.
Размно жение.1 Яйца эллипсоидальной формы (6x4 мм), с неболь-
шим бугорком на каждом из концов, стекловидно-прозрачные. При комнат-
ной температуре и высокой влажности развитие продолжается 18 дней.
Только что вылупившиеся слизни бесцветные, лишь глазные щупальца
лиловые; длина тела 10—И, ширина 2, дл. мантии 4 мм. Лишь через 2—
3 недели слизни приобретают темно-коричневую окраску (при длине около
20 мм). Осенью они становятся черными. По-видимому, в Заилийском Алатау
период размножения приходится на конец весны.
Замечание. Зимрот (Simroth, 1910) по одному экземпляру, найден-
ному где-то «в окрестностях озера Балхаш», описал новый вид — Limax
(Lehmannia) stummeri. Голотип этого вида нам неизвестен.
Внешне этот слизень очень походит на типичный Г. natalianus. Во внут-
реннем строении главное отличие — третья петля кишечника более длинная
(около х/2 длины спины) и с более длинной слепой кишкой, простирающейся
до вершины внутренностного мешка. Последнее ни Зимрот, ни мы ни разу не
встречали у Т. natalianus.
Однако нам представляется, что указанные отличия от Т. natalianus,
тем более обнаруженные лишь у одного экземпляра, недостаточны для при-
знания L. stummeri за самостоятельный вид. Вероятно, это редкое внутри-
видовое отклонение Т. natalianus.
3. Подрод TAULIMAX Likharev et Wiktor, subgen. nov.2
Типовой вид — Limax turkestanus Simroth, 1898; обозначен здесь.
Диагноз. Семяприемник открывается в передний конец пениса вблизи
от атриума. Пенис без бича, внутри с языковидной складкой. Ретрактор
1 По наблюдениям, сделанным в ЗИН АН СССР за период с 6 июня по 15 июля 1963 г.
2 Таи — (тау) по-казахски «горы».
L TURCOMILAX
255
правых щупалец огибает пенис справа и снизу, а яйцевод сверху. Третья
петля кишечника без слепой кишки.
Распространение. Тянь-Шань.
4. Turcomilax (Taulimax) turkestanus (Simroth, 1898), comb. nov.
(рис. 303—308)
Simroth, 1898a : 54 (Limax)', 1901 : 70, T. 3, Fig. 1—4 (Limax)', 1910a : 502,
T. 6, Fig. 1—4 (Limax dengis, syn. nov.); Скворцов, 1940 : 371 (Heynemannia); Цвет-
ков, 1940 : 388 (Heynemannia); Лихарев, Раммельмейер, 1952 ; 322
(Limax); Соболева, 1971 : 92 (Limax).
Terra typica — «Turkestan» — Туркестан (старое название Средней Азии).
Типы неизвестны. Один синтип Limax dengis Simroth находится в Естественно-
историческом музее Стокгольма (№ 3147).
Материал: из СССР — 95 экз. из 20 пунктов.
При сокращении тело становится веретеновидно-клиновидным, плавно
сужающимся назад, с максимальной шириной в области мантии и начала
Рис. 303—305. Turcomilax (Taulimax) turkestanus (Simr.). Пржевальск.
зоз — кишечник; 304 — ретракторы; 305 — дистальная часть гениталий.
спины. Киль слабо выражен и ограничен только задним концом тела. Мантия
занимает около 1/i—1/3 общей длины, нередко сзади с заметной угловатостью;
капюшон (около х/2 мантии) закрывает бблыпую часть шеи. Морщины плос-
кие , бороздки довольно глубокие. Между мантийной щелью и средней линией
спины около 18 рядов морщин.
Окраска черная с синеватым отливом, лишь бока ниже мантии серые.
Подошва белая, за исключением внешнего края боковых долей, которые
темные. Изредка встречаются темно-бурые экземпляры или с темно-серой спи-
ной и мантией и светло-серыми боками, подошва у них кремовая.
Размеры: дл. сок. до 55, дл. мантии до 15 мм.
Радула. Срединный и боковые зубы трехзубцовые, лишь 2—3 самых внеш-
них боковых зуба без эндоконов. Внутренние краевые кинжаловидные,
внешние двухзубцовые за счет появления вторичного эктокона. Пример:
1 экземпляр из ущелья Туюк (Киргизский Алатау); формула:
с 216 26 Юк
у + -д- + -у 4- уу X около 130.
256
III. LIMACIDAE
Кишечник (рис. 303). Третья петля довольно длинная, простирается назад
почти до вершины внутренностного мешка.
Рис. 306—308. Turcomilax (Taulimax) turkestanus (Simr.). Пржевальск.
306 — гениталии; 301 — дистальная часть гениталий; 308 — вскрытый пенис.
Гениталии (рис. 305—308). Гонада черная, круглая и лежит почти
открыто среди соседних органов. Проток ее сильно извит и заканчивается
крупным талоном. Белковая железа языковидная, желтая. Семяпровод
короткий и прямой. Пенис мешковидный или бобовидный, массивный,
4. TURCOMILAX
257
сильно изогнут, а у атриума резко сужен. Полость пениса почти полностью
занята крупным языковидным тельцем, которое свободным концом обращено
назад. Поверхность тельца покрыта мелкими сосочками; в спокойном состоя-
нии оно сложено повдоль, образуя сложные складки-воланы.
Семяпровод крепится к пенису, значительно отступя от его заднего конца.
Нередко в этом месте на пенисе имеется темное пятно. Половой ретрактор
Рис.. 309, 310. Turcomilax (Taulimax) tzvetkovi Likharev et Wiktor, sp. nov. Паратип.
Заилийский Алатау, Тургенское ущелье, 12 VII 1974.
309 — гениталии; 310 — дистальная часть гениталий.
прикрепляется перед семяпроводом. Яйцевод значительно длиннее семяпро-
вода, трубчатый, впереди расширяющийся. Семяприемник с круглым ре-
зервуаром и длинным протоком. Атриум небольшой.
Распространение. Несколько сборов из окрестностей Пржеваль-
ска (Терскей-Алатау) и с Кунгей-Алатау; многочисленные находки в Заилий-
ском Алатау; одна находка на хребте Жетыжол.
Образ жизни (Цветков, 1940). Обитает в пределах горнолесной
и лугово-степной зон, среди скал и крупных камней, по обрывистым сырым
склонам и по берегам горных речек. В сухое время уходит глубоко в землю,
под камни и в трещины скал. При дождях выползает на поверхность и пере-
водит на траву.
17 И. М. Лихарев, А. И. Виктор
258
III. LIMACIDAE
5. Turcomilax (Taulimax) tzvetkovi Likharev et Wiktor, sp. nov.1
(рис. 309, 310)
Locus typicus — в 12 км от поселка Турген у Тургенского форелевод-
ческого хозяйства, Алма-Атинская область, Заилийский Алатау. Голотип
й 2 паратипа в ЗИН АН СССР (сбор Л. Я. Боркина, 12 VII 1974 г.); осталь-
ные 82 паратипа в Зоологическом музее Московского университета (сборы
Б. Н. Цветкова, Тургенское ущелье Заилийского Алатау, 11—23 VII 1937 г.).
Диагноз. По внешнему облику этот слизень неотличим от Т. turke-
stanus (Simr.). Главные отличия: пенис длинный, внутри с продольными склад-
ками.
Размеры: дл. пол. до 61, дл. мантии до 21 мм; голотип — дл. сок. 36,.
ширина 9, длина мантии 12 мм.
Кишечник как у Т. (Г.) turkestanus.
Радула:
с 216 36 15к 15к
у 4- — + — + + ~2~ X около 140
Гениталии. Главное отличие от предыдущего вида в форме и строе-
нии пениса. Он длинный (около длины тела), сильно извитый, перед атри-
умом суженный. Внутри залегают 2—3 мощные продольные складки.
Распространение. Обитает в Тургенском ущелье (восточна®
часть Заилийского Алатау) в пределах горнолесной зоны (смешанный или
хвойный лес из тяныпаньской ели), на высотах до 2900 м над ур. м.
' 5. Род MALACOLIMAX Malm, 1868
Malm, 1868 : 66; Simroth, 1891а : 302 (Microheynemannia); Hesse P.
1926 : 12.
Типовой вид — Limax tenellus Muller, 1974; по монотипии.
Небольшие слизни (дл. пол. до 50, дл. сок. до 35 мм), с очень коротким
и нечетким килем. Пневмостом лежит сильно постмедиально.
Срединный и боковые зубы радулы трехзубцовые, лишь самые внешни®
боковые иногда двухзубцовые (без эндокона). Краевые зубы двух типов:
несколько внутренних однозубцовые — шиповидные, остальные двухзуб-
цовые (за счет вторичного дополнительного зубца).
Кишечник не перекручен; вторая петля заметно короче третьей, оба
колена которой (5-е и 6-е) лежат почти параллельно друг к другу. Левая
доля печени образует вершину внутренностного мешка.
Ось сердца наклонена под углом 45°. Аорта длинная. Основание полового-
ретрактора крепится к диафрагме перед сердцем.
Гонада лежит позади или под второй петлей кишечника. Короткий се-
мяпровод, постепенно расширяясь, впадает в задний конец мешковидного-
или короткоцилиндрического пениса. Внутри пениса находится крупная
толстая складка. Семяприемник впадает в пенис вблизи от атриума.
Ретрактор правых щупалец проходит через щель между пенисом и яйце-
водом, так что семяпровод остается позади него, т. е. как у Caspilimax и
Caucasolimax.
Распространение. Большая часть Европы. Ряд старых авто-
ров (Bourguignat, 1861; Germain, 1907; Pollonera, 1909) описали несколько-
видов из Алжира, Туниса и Палестины, которые без исследования внутрен-
него строения отнесли к этой группе. Эти данные требуют проверки.
х Название дано в честь Б. Н. Цветкова (1909—1945), много сделавшего в познании!
наземных моллюсков южного Казахстана и впервые обратившего внимание на этот вид..
В его коллекции он обозначен, как «.Limax turgensis» (nom. mus.!).
5. MALACOLIMAX
259
Malacolimax tenellus (Muller, 1774) (рис. 311—318)
Muller, 1774 : 11 (Limax); Nilsson, 1822 : 11 (Limax); Schrenk, 1848 :
444 (Limax serotinus); Held, 1849, Land Moll. Bayern : 15 (Limax cereus); Dumont,
Mbrtillet, 1852, Moll. Savoie : 10 (Limax sylvaticus); Simroth, 1885 : 210,
294, T. 7, Fig. 7, T. 8, Fig. 12, 13 (Limax); Taylor, 1902—1907 : 71, T. 10, Fig. 1—4
(Limax); Лихарев, Раммельмейер, 1952 : 313 (Limax); Quick, 1960 : 180,
Fig. 13, T. 2, Fig. 16; Or ossu, 1968 : 85, Fig. 1—3 (Limax tigvenius); L u p u, 1971 :
131, Fig. 8, 9 (L. tenellus=L. tigvenius); Wiktor, 1973 : 78, Fig. 25, 26, 109—115, 233,
251, 252.
Locus typicus — «Fridrichsdal» — окрестности Копенгагена (Дания). Типы
неизвестны.
Материал: из СССР — 71 экз. из 33 пунктов; из других стран бо-
лее 350 экз.
Тело сравнительно приземистое, мягкое, с тонкой просвечивающей ко-
жей. Мантия закрывает менее 1/3 тела, сзади со слабым углом. Морщины
плоские, бороздки мелкие. Между мантийной щелью и средней линией спины
-около 16 рядов морщин.
Верхняя сторона окрашена в желтый, зеленовато- или серовато-желтый
цвет, реже оранжево-желтая, вокруг пневмостома более светлая; голова
и щупальца черные или темно-бурые. Подошва светло-желтая или кремовая.
После фиксации на мантии нередко заметен лирообразный темный рисунок
с темным пятном в центре, а на спине темные продольные полосы. Молодь
обычно окрашена светлее — кремовая или зеленовато-кремовая. После фик-
сации желтый пигмент быстро исчезает и тело становится беловато-кремо-
вым. Слизь желтая, жидкая.
Размеры, дл. пол. до 50, дл. сок. 25—35 мм.
Раковина (рис. 312) тонкая, после фиксации легко разрушается.
Радула (рис. 313). Срединный и боковые зубы с небольшими, но четкими
экто- и эндоконами и с мощным мезоконом. Краевые зубы с длинным изо-
гнутым мезоконом и довольно крупным вторичным зубцом, который посте-
пенно смещается вперед. Боковых зубов около 25, краевых около 30; число
поперечных рядов около ИО.1
Гениталии (рис. 315—318). Гонада округлая и компактная. Ее проток
тянется к семяяйцеводу почти без извивов. Белковая железа массивная,
•белая. Семяяйцевод довольно толстый и длинный, плавно изогнут. К перед-
ней трети его прилегает простата (не заходит на яйцевод). Семяпровод перед
впадением в пенис постепенно расширяется, срастается с ним и образует
ряд небольших пигментированных вздутий. Пенис короткоцилиндрический
или булавовидный, у заднего конца вздутый, иногда неглубокой перетяж-
кой разделен на два отдела: короткий передний и более длинный задний.
Внутри последнего находится крупная и толстая складка.
Распространение. В СССР — северо-западные, западные и цен-
тральные области европейской части (на север включая Карельский перешеек
и северное побережье Ладожского озера, на восток — до Казани, на юг —
Сумская область и окрестности Киева).
Вне СССР — средние, западные (до Франции), северо-западные (Бри-
танские острова), северные (Скандинавия, Финляндия до 63°30') области
Европы. Единичные находки в северных Балканах.
Образ жизни. Обитает только в лесах — как на равнине, так
и в горах. Изредка встречается в старых парках. Явно предпочитает листвен-
ные леса, однако вместе с Arion subfuscus является одним из немногих слиз-
ней, живущих и в хвойных лесах (особенно в сосняках). К извести, по-види-
1 Описание радулы сделано по материалам из СССР. Однако как число зубов, так
и их форма, по-видимому, очень варьируют (см.: Quick, 1960; Flasar, 1961).
17*
Рис. 311—318. Malacolimax tenellus (Miill.) (из: Wiktor, 1973).
311 — внешний вид; 312 — раковина; 313 — зубы радулы; 314 — кишечник; 315 — гениталии; 316
они же, дистальная часть; 317 — задний конец пениса; 318 — вскрытый пенис.
6. LEHMANNIA
261
мому, относится индифферентно, так как обычен и.на кислых почвах. Скры-
вается в подстилке, под валежинами и в гнилых пнях.
Всю жизнь проводит на поверхности почвы; на деревья не заползает.
Главной пищей служат шляпочные грибы, а также и лишайники. Мо-
лодь питается мицелием различных грибов.
Половозрелость наступает в конце лета и в начале осени (Kunkel, 1934;
Fromming, 1954). Перед копуляцией партнеры некоторое время ползают друг
за другом гуськом. Затем передний изгибается направо и животные образуют
круг. Пенисы выворачиваются очень быстро и приобретают облик массивных
синевато-белых тел, каждый с косой толстой складкой.
В северной и средней Европе яйцекладка происходит осенью. Каждый
слизень в несколько приемов откладывает до 120 яиц, которые остаются на
зимовку. Яйца слегка овальные, размером от 3.6 X 3.1 до 3.3 X 3.0 мм, светло-
янтарные, просвечивающие. В лабораторных условиях при температуре
18—25° вылупление происходит через 20—24 дня, в естественных условиях
(по данным из западной Европы) более чем через 100 дней.1
Свежевылупившиеся слизни бесцветные, только омматофоры, мантия
и хвостовой конец окрашены в лиловый цвет. Через 6—7 месяцев окраска
становится желтоватой. С наступлением половозрелости наступает общее
потемнение окраски.
По данным из западной Европы (Quick, 1960) слизень живет 11—13 ме-
сяцев и обычно погибает це позднее чем через l1^ месяца после последней
яйцекладки.
Замечание. Данному виду, по-видимому, очень близок слабо
изученный М. kostali Babor, 1900. Судя по немногим литературным источни-
кам (Ehrmann, 1933; Lozek, 1956; Wiktor, 1973), в основном опирающимся
на первоначальное описание (Babor, Kostal, 1894; Babor, 1900), М. kostali
отличается от М. tenellus меньшими размерами (дл. сок. около 28 мм), светло-
серой окраской, причем киль несколько темнее, а середина мантии, напротив,
светлее. Во внутреннем строении Бабор указывает следующие отличия:
а) краевые зубы лишь с намеком на вторичный дополнительный. зубец и
в основном шиповидные; б) пенис длиннее яйцевода. Последнее нередко можно
видеть и у настоящих М. tenellus. Что касается указанной разницы в краевых
зубах, то она может наблюдаться даже в пределах одной радулы М. tenellus.
В итоге следует признать, что видовая самостоятельность М. kostali
очень сомнительна. Однако поскольку в нашем распоряжении не было
ни типов этого вида (местонахождение их неизвестно), ни материалов из бо-
лее поздних сборов, вопрос о его систематическом положении остается для
нас открытым.
На территории СССР этот вид был обнаружен лишь в окрестностях Во-
рохты (Восточные Карпаты — Babor, Frankenberger, 1914). Вне СССР
его указывают для Верхней Австрии и Чехии (Lozek, 1956; Jaeckel, 1962).
6. Род LEHMANNIA Heynemann, 1862
Heynemann, 1862 : 211; P olloner a, 1887b : 1 (Ambigolimax); Polio-
n e r a, 1891 : 1 (Melitolimax).
Типовой вид — Limax marginatus Muller, 1774; по моноти-
пии.
Слизни средних размеров (дл. пол. до 80, дл. сок. до 40 мм), со стройным,
сзади заостренным телом. Благодаря крупному заднему венозному синусу
и влагопроницаемой коже, тело очень мягкое и скользкое. Киль различной
1 По-видимому, в СССР вылупление перезимовавших яиц происходит в конце весны—
начале лета.
262
III. LIMACIDAE
длины, слабо выпуклый; от него к мантии простирается узкая светлая полоска,
которая создает впечатление, что киль простирается на всю спину. Мантия
занимает рколо 2/3 тела. У живых слизней по ней обычно пробегают нежные
концентрические морщинки. Пневмостом лежит постмедиально.
Окраска очень разнообразная и варьирует даже внутри популяции.
.Обычно спина и мантия имеют более или менее четкий рисунок из темных
продольных полос или рядов пятен, хотя бывают и одноцветно черные
формы. Полосы лучше выражены на мантии, где нередко, смыкаясь, образуют
лирообразный рисунок. Слизь бесцветная, жидкая.
Срединный и боковые зубы радулы с крупными мезоконами и слабо раз-
витыми экто- й эндоконами. Краевые зубы двух типов: внутренние кинжало-
видные или шиповидные, внешние — S-образно изогнутые, двухзубцовые
(за счет вторичного дополнительного зубца), ближе к краю радулы — пиль-
чатые (рис. И, VII—IX).
, Кишечник не перекручен (рис. 321). Вторая петля значительно короче
первой, а третья — второй. Имеется длинная слепая кишка, расположенная
по продольной оси тела (под покровами спины), на одной линии с задней
кишкой, т. е. с 6-м коленом кишечника. Левая доля печени образует вершину
внутренностного мешка.
Ось сердца наклонена почти под углом 90°. Аорта длинная. Основание по-
лового ретрактора крепится к диафрагме слева от почки.
Гонада располагается позади второй петли кишечника или внутри пер-
вой. Семяяйцевод длинный. Вдоль него вытянулась простата, передний ко-
нец. которой обычно вздут и слегка отходит от женских половых путей.
Яйцевод и семяпровод короткие и толстые. Семяпровод впадает в пенис
субапикально, чаще всего рядом с половым ретрактором. Обычно пенис неболь-
шой, булавовидный или конический, к атриуму суженный. Однако у неко-
торых видов пенис довольно длинный и цилиндрический. Задний конец его
напротив семяпровода имеет конический или плетевидный отросток, который
обычно называют бичом.1 Внутри пениса имеется крупная продольная
складка. Семяприемник впадает в пенис у границы с атриумом. Ретрактор
правых щупалец перекрещивается с половыми Путями как у Limax.
Распространение. Большая часть Европы, кроме обла-
стей, занятых тундрой, хвойными лесами и степями.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ LEHMANNIA
1 (4). Пенис цилиндрический, извитый, в вытянутом состоянии иногда
длиннее мантии.
2 (3). Имеется бич в виде короткого конического или длинного плетено-
видного выступа...........................6. L. jaroslaviae Grossu.
3 (2). Бича нет . . . ....................7. L. nyctelia (Bourg.).
4 (1). Пенис короткий, конический или булавовидный, к атриуму сужен-
ный.
5 (6). Бич имеет форму широкой лопасти, сидящей на пенисе рядом с по-
ловым ретрактором. С противоположной стороны в пенис впадает
семяпровод............................... 5. L. valentiana (Fer.).
6 (5). Бич (если присутствует) имеет форму короткого конического или.
длинного плетевидного выступа. На пенисе он сидит с противо-
положной стороны чем семяпровод и ретрактор, которые располагаются
рядом.
7 (8). Бич впадает в середину пениса. Яйцевод на середине сильно вздут
.................................... 4. L. medioflagellata Lupu.
1 Отсутствует у L. nyctelia (Bourg.) и L. szighethyae Wiktor, 1975.'
6. LEHMANNIA
263
8 (7). Бич располагается на заднем конце пениса. Яйцевод без вздутия.
9 (12). Семяприемник в несколько раз длиннее пениса.
10 (11). Спина и бока обычно без рисунка из продольных рядов темных
пятен и полос. Мантия с двумя черными полосами с неясными конту-
рами ..............................3. L. horezia Grossu et Lupu.
11 (10). Спина и бока с темным рисунком из рядов пятен или полос. Ман-
тия с 2—3 темными полосами с четкими контурами................
........................2. L. macroflagellata Grossu et Lupu.
12 (9). Семяприемник не более чем в 1 1/2 раза длиннее, пениса.
................................. 1. L. marginata (Muller).
1. Lehmannia marginata (Muller, 1774) (рис. 319—323)
M ii 11 e г, 1774 : 10 (Limax)’, Bouchard-Chantereaux, 1838 : 28 (Limax
arborum)’, Schrenk, 1848 : 142 (Limax livonicus)’, Taylor, 1902—1907 : 89, T. 10,
Fig. 10—17 (Limax arborum)’, Лихарев, Раммельмейер, 1952 : 323, рис. 249,
250; Quick, 1960 : 194, Fig. 16, T. 2, Fig. 18, 21; Grossu, 1970c : 110, Fig. 2 (sar-
mizegetusae, syn. nov.); Lupu, 1973 : 50, Fig. 33—49 (bielzi, syn. nov.); Wiktor,
1973 : 88, Fig. 31, 32, 121—128, 234, 253, 254.
Locus typicus — «Fridrichsdal» — окрестности Копенгагена (Дания). Типы
неизвестны.
Материал: из СССР 28 экз. из 6 пунктов; из других стран более
450 экз.
Киль очень короткий и слабо выражен. Мантия сзади с отчетливым углом.
Между мантийной щелью и средней линией спины около 29 рядов морщин.
Окраска фона и особенно рисунок очень изменчивы. Фон чаще всего свет-
ло-кремовый или беловатый, а рисунок темно-серый до почти черного. Ман-
тия всегда с лирообразным рисунком, внутри которого видна еще одна не-
четкая средняя полоса или ряд пятен. Ниже светлой килевой полоски имеются
1 или 2 пары темных полос, нижняя пара нередко распадается на ряды пя-
тен. Голова и щупальца черноватые. Подошва и бока светлые.
Размеры: дл. пол. до 75, дл. сок. до 40 мм.
Радула (см. рис. 11, VII—IX, 320). Боковых зубов 10—11, краевых
около 70. '
Кишечник (рис. 321).
Гениталии (рис. 322, 323). Половые органы, как и остальные внутрен-
ности, обычно покрыты черным мезентерием. Гонада коричневая или темно-
кремовая. Белковая железа очень крупная. Пенис относительно крупный,
к заднему концу конически расширяется. Здесь в него с одной стороны впа-
дает семяпровод, с противоположной — бич. Бич имеет форму короткого
роговидного выступа (у равнинных популяций) или плетевидно вытянут,
превосходя половину длины пениса (у горных популяций). Изредка бич
представлен более или менее четким округлым выступом или совсем отсут-
ствует. Последнее чаще всего лишь внешнее впечатление, так как бич имеется,
но в результате фиксации или после копуляции он оказывается полностью
или частично ввернутым внутрь пениса. В таких случаях на наличие бича
указывает небольшая ямка на соответствующем месте пениса.
Семяприемник несколько длиннее пениса, с толстым протоком и овальным,
нередко заостренным, резервуаром. Яйцевод по длине и ширине немного
уступает пенису. Атриум небольшой.
Распространение. В СССР — Прибалтика, Карельский пере-
шеек, о. Валаам (Ладожское озеро), Полесье, Львовская, Ивано-Франков-
ская, Черновицкая и Закарпатская области. Вне СССР — почти вся Польша
(Wiktor, 1973). В коллекции ЗИН АН СССР имеются экземпляры из Швеции
(о. Лонгёрн,) Финляндии (Аландские о-ва), ФРГ (Гольштиния), с Британ-
ских о-вов (о. Мэн) и из Исландии.
264
III. LIMACIDAE
•. Как показал Форкарт (Forcart, 1966), многочисленные литературные
данные об обитании этого вида в Европе требуют специальной проверки,
так как до недавнего времени за него принимали другие виды того же рода.
Рис. 319—323. Lehmannia marginata (Miill.) (из: Wiktor, 1973).
319 — внешний вид; 320 — зубы радулы; 321 — кишечник; 322 — гениталии; 323 — дистальная часть
гениталий.
Образ жизни. На равнине и в нижних зонах гор этот вид оби-
тает только в смешанных и лиственных лесах, особенно на буке и грабе.
В хвойных лесах он живет только в горах близ верхней границы леса,
но численность его тут заметно ниже. Здесь слизень держится на гнилых
6. LEHMANNIA
265
пнях и валежинах. В субальпийской зоне он населяет каменистые и скальные
биотопы, богатые лишайниками. Во влажную погоду интенсивно ползает
по деревьям, забираясь даже в кроны.
Главной пищей служат лишайники, произрастающие на стволах деревьев
и скалах. ФрёмминГ (Fromming, 1954) отрицает это и утверждает, что слизень
питается только зелеными частями высших растений. Однако он пользо-
вался слизнями из оранжерей Берлина и, вероятно, имел дело с другим ви-
дом того же рода (возможно, L. valentiana — см. ниже).1 По той же причине
вызывают сомнения его сообщения о сроках размножения; вылупления
из яиц, росте и длительности жизни у L. marginata.
2. Lehmannia macroflagellata Grossu et Lupu, 1962 (рис. 324—328)
Grossu, Lupu, 1962 : 198, Fig. 9—11; Wiktor, 1964 : 73 (marginata)',
Hudec, Branbane c, 1965; : 271, Fig. 1, 2, 6, 10; W i k t о r, 1973 : 93, Fig. 33,
34, 129—135, 235, 257, 258.
Locus typicus — «Pietrosul» (Румыния). Голотип в Естественноистори-
ческом музее им. Гр. Антипа, Бухарест, № 13133;
Материал: из СССР нет; из других стран 90 экз., в том числе 4 па-
ратипа.
Тело более стройное, чем у предыдущего вида. Киль слабый и короткий.
Окраска очень варьирует и образована светлым или кремовым фоном и тем-
но-серым или коричневато-черным рисунком из пятен с неясными контурами.
Размеры и число пятен очень различны, в связи с чем встречаются как свет-
лые, так и темные особи. Нередко пятна сливаются, образуя более или ме-
нее четкие продольные полосы, которые особенно заметны после фиксации
слизней. Мантия всегда с 2 темными продольными полосами, которые у зад-
него края сливаются друг с другом. Нередко там же имеется третья (средин-
ная) полоса или ряд темных пятен. Киль и средняя линия спины всегда
светлые. На боках тела между темными пятнами остаются светлые округ-
лые поля — особенность, характерная только для этого вида. Подошва и
край ноги светлые, кремовые, без рисунка.
Размеры: дл. пол. до 45, дл. сок. 25—35 мм.
Радула. Боковых зубов 9—18, краевых 38—53; число поперечных рядов
134-151.
Гениталии. Гонада округлая, темно-коричневая, с почти прямым
протоком, который лишь впереди дает несколько извивов. Семяяйцевод
довольно длинный, впереди с крупной простатой. Семяпровод короткий и
толстый, к пенису крепится с брюшной стороны рядом с половым ретрактором.
Пенис очень небольшой, булавовидный или конический, с очень длинным
бичом, который к концу плавно сужается. Семяприемник с длинным и тол-
стым протоком и небольшим резервуаром. Обычно семяприемник в 2—3
раза длиннее;пениса (без бича). Яйцевод длиннее семяпровода, сзади вздут,,
к атриуму плавно сужен. Короткий атриум мускульными волокнами связан:
со стенкой тела.
Распространение. В СССР — Украинские Карпаты (Черно-
гора). Общее распространение — Судеты, Карпаты.
Образ жизни. Обитает в горных лесах, особенно в буковых,
и в субальпийской зоне. Наиболее активен после дождя. Скрывается в дуп-
лах, под корой и в щелях скал, нередко совместно с L. marginata и L. пу-
ctelia.
1 В отличие от L. valentiana L. marginata обитает только в природных биотопах..
Рис. 324—328. Lehmannia macroflagellata Gr. et. Lupu (из: Wiktor, 1973).
324 — внешний вид; 325 — раковина; 326 — зубы радулы; 321 — кишечник; 328 — гениталии.
6. LEHMANNIA:
.3. Lehmannia horezia Grossu et Lupu, 1962 (рис. 329)
Gross u, Lupu, 1962 : 196, Fig. 7.
Locus typicus — «Dorf Vaideeni» —' Вайдеени, близ Хорезу (Румыния).
Голотип в Естественноисторическом музее им. Гр. Антипа, Бухарест,
№ 13134.
Материал: из СССР — нет; из Румынии — 4 паратипа.
Тело светло-серое, по бокам еще светлее и с желтым оттенком, чаще
всего одноцветное, реже с неясными неправильными темными пятнами или
Рис. 329. Lehmannia horezia Gr. et Lupu (из: Grossu, Lupu. 1962). Гениталии.
боковыми полосами ниже киля. Серый пигмент, в основном концентрируется
по бороздкам, образуя тонкую четкую сетку. Его нет на киле и в узкой зоне
спины от киля до мантии, благодаря чему создается впечатление присут-
ствия на середине спины узкой белой полоски. На мантии имеются 2 черные
продольные полосы с неясными контурами, которые иногда смыкаются сзади,
образуя лирообразный рисунок. Подошва одноцветная, светлая, без всяких
следов темного пигмента.
Размеры: дл. пол. 70, дл. сок. до 40 мм.
Гениталиям характерен короткий, но широкий пенис с длинным
плетевидным бичом и семяприемник с толстым и очень длинным протоком,
который в несколько раз длиннее пениса.
Распространение. В СССР не обнаружена. Вне СССР —
в Румынии в бассейне р. Лункавэц (горы «Потру, Южные Карпаты).
4. Lehmannia medioflagellata Lupu, 1968 (рис. 330)
Lupu, 1968b : 39, Fig. 1—2.
Locus typicus — «Padis» — Падиш, Западные Румынские горы. Голотиге
в Естественноисторическом музее им. Гр. Антипа, Бухарест, № 13142.
268
III. LIMACIDAE
Спина1 красновато-коричневая, бока более светлые. Середина спины
со светлой и узкой килевой полоской. Мантия того же цвета, что и Спина, с чет-
кими черными боковыми полосами, которые сзади не сливаются друг с другом.
Ниже килевой полоски лежит пара черных полос. Боковые доли подошвы
темнее срединной.
Рис. 330. Lehmannia mediofiagellata Lupu (из: Lupu., 1968b). Гениталии.
Размеры: дл. пол. 60—70, дл. сок. до 30 мм.
Гениталии от предыдущих видов отличаются тем, что: а) бич располагается
не на заднем конце пениса, а на его середине; б) яйцевод на середине вздутый.
Пенис сравнительно короткий, булавовидный. Бич плетевидно сужается
к концу, изогнут. Семяприемник почти вдвое длиннее пениса, толстый,
нечетко разграничен на резервуар и проток.
Распространение. В СССР не обнаружена. Вне СССР — За-
падные Румынские горы.
5. Lehmannia valentiana (Ferussac, 1823) (рис. 331—335)
Ferussac, 1823 : 96е, Т. 8А, Fig. 5, 6 (Limax)', М a b i 11 е, 1883 : 52 (Limax
poirieri); Aliena, 1950 : 9 (Umax poirieri); Quick, 1960 : 197, Fig. 17 A, D, T. 1,
Fig.l 14 (poirieri)', Walden, 1962 : 71, Fig. 1—19, T. 1, Fig. 20—29 (Limax)', Grossu,
1970 : 112, Fig. 3 (getica); Lupu, 1973 : 53 (getica—valentiana); Wiktor, 1973 : 96,
Fig. 35, 36, 136—142, 236, 255, 256.
Terra typica — Испания? Типы неизвестны.
Материал: из СССР — 65 экз. из 5 пунктов; из других стран
27 экз.
Тело умеренно стройное, сзади ясно заостренное. Мантия покрывает
менее 1/3 длины тела. Щупальца узкие и длинные. Окраска очень варьирует.
1 Авторы настоящей работы не располагали коллекционными материалами по дан-
ному виду. Поэтому характеристика его составлена только по данным оригинального
описания.
Рис. 331—335. Lehmannia valentiana (Fer.) (из: Wiktor, 1973).
.331 — внешний вид; ЗЗГ — зубы радулы; ззз — кишечник; 334 — дистальная часть гениталий; 335 —
вскрытый пенис.
270
III. LIMACIDAE
Фон чаще всего кремовый, но бывает коричневатым и грязно-оранжевым.
Рисунок красновато- или темно-коричневый; на мантии всегда две боковые-
полосы, между которыми лежит нечеткая средняя полоса, которая может
распадаться на ряд пятен. На спине имеется пара полос, доходящих до
заднего конца тела, ниже которой может быть еще пара, но менее четких
полос или рядов пятен. Задний конец тела без рисунка или покрыт неясной
темной сетью из штрихов и пятен. Щу-
пальца бурые. Подошва одноцветная,,
кремовая, слегка просвечивающая.
Размеры: дл. пол. до 60, дл.
сок. до 46 мм.
Радула (рис. 332). Как показал Вал-
ден (Walden, 1962), с возрастом изменя-
ется форма и нарастает число боковых
и краевых зубов, а также увеличивается
число, поперечных рядов в радуле..
У взрослых слизней боковых зубов
13—16, краевых 37—50, а рядов 110—
140.
Гениталии (рис. 334, 335)..
Гонада относительно крупная, округ-
лая, светло-коричневая. Гермафродит-
ный проток длинный, сильно извитый..
Белковая железа широкая, языковид-
ная. Семяяйцевод белый, с крупной
простатой, которая впереди заметно
вздута. Пенис конический, с крупным
бичом, имеющим форму широкой лопа-
сти. Бич впадает в пенис рядом ,с ре-
трактором, т. е. с брюшной стороны
(у остальных видов того же рода он впа-
дает в пенис с противоположной сто-
роны от ретрактора). Семяпровод откры-
вается в пенис со спинной стороны. Се-
мяприемник почти той же длины, что
и пенис, или даже длиннее; он имеет
овальный резервуар. Яйцевод трубча-
тый, заметно короче пениса.
Рис. 336. Lehmannia jaroslaviae Grossu
(из: Grossu, 1967). Гениталии.
Распространение. В СССР обнаружен в садоводствах Ленин-
града, Ростова-на-Дону и Ташкента. По-видимому, природный ареал этого
вида занимает Пиренейский п-ов и частично Францию. В настоящее время
он завезен во многие страны Старого и Нового света и на острова Тихого
океана (Altena, 1950; Walden, 1962), где его нередко принимали за L. mar-
ginata (Mull.), а иногда и за Limax maximus L. В Европе вне природного
ареала известен почти из всех стран, но здесь живет только в оранжереях
и садоводствах, где питается листьями, плодами и клубнями овощных и де-
коративных культур. При этом он может приносить существенные убытки.
6. Lehmannia jaroslaviae Grossu, 1967 (рис. 336)
Grossu, 1967 : 120, Fig. 1—6; Lupu, 1973 : 49, Fig. 28—32 (vrancensis, syn. nov)^
1974 : 87 (vrancensis).
Locus typicus — «Jaroslave» — Ярославе, долина p. Тополог (Румыния).
Голотип в коллекции Гроссу (Бухарест), № 1131.
6. LEHMANNIA
271
Материал: из СССР — 4 экземпляра из одного пункта; из Румы-
нии 2 паратипа L. jaroslaviae.
Окраска верхней стороны тела либо одноцветно черная, либо с темным
рисунком на светло-сером или почти белом фоне. Рисунок очень изменчив:
при сильном развитии он представлен продольными полосами или рядами
пятен на спине и мантии, при слабом развитии от него остаются лишь следы
полос на мантии. У темных форм от киля до мантии тянется вперед светлая
полоска. Подошва одноцветная, беловато-желтая.
Размеры: дл. пол. до 80, дл. сок. 30—40 мм; голотип имеет дл.
•сок. 35 мм.
Гениталиям присущ длинный, несколько извитый цилиндриче-
ский пенис, иногда с местными вздутиями. Бич варьирует от короткого
конического до длинного плетевидного выступа. Напротив него, к заднему
концу пениса крепятся ретрактор и семяпровод. Семяприемник всегда короче
пениса.
Распространение. В СССР единственная . находка сделана
в Молдавской ССР у с. Киприаны. Вне СССР обитает в Румынии в Южных и
Восточных Карпатах, во влажных буково-еловых лесах на высотах 700—
$00 м над ур. м.
7. Lehmannia nyctelia (Bourguignat, 1861) (рис. 337—342)
В ourguigna t, 1861, Rev. Mag. Zool., ser. 2, 13, 7 : 305, T. 2, Fig. 3—4 (Limax);
Urbanski, Wiktor, 1968 : 51, Fig. 1A—C (Limax); Wiktor, 1973: 99, Fig. 29,
30, 143—149, 237, 259, 260.
Locus typicus — «Algerie». — Алжир. Типы неизвестны.
Материал: из СССР нет; из других стран более 100 экз.
Животное по форме сходно с L. marginata, но задний конец более заострен.
Киль слабо развит. Мантия закрывает менее 1/3 тела. Окраска от темно-
коричневой до грязно-серой с неясными светлыми полосами, в том числе и
•с килевой полосой. Темный пигмент образует неясный рисунок из 1—2
пар спинных полос, связанных поперечными перемычками. На мантии ана-
логичный рисунок из двух боковых (сзади сливающихся) и одной срединной
полосы. Последняя нередко распадается на ряд пятен. Бока и подошва свет-
лые. Щупальца бурые. В верхних зонах гор иногда встречаются совершенно
черные особи.
Размеры: дл. пол. до 50, дл. сок. 30—37 мм.
Раковина очень тонкая, быстро разрушающаяся в спирту (рис. 338).
Радула (рис. 339). Срединный и боковые зубы с крупным
закругленным мезоконом; экто- и эндоконы редуцированы; краевые зубы
•однозубцовые, лишь самые внешние имеют на мезоконе несколько мелких
зубчиков. Граница между боковыми и краевыми зубами нечеткая; общее
число их 73—81. Число поперечных рядов 99—124.
Кишечник (рис. 340). Слепая кишка слегка пигментирована.
Гениталии (рис. 341, 342). Гонада лежит внутри первой петли
кишечника, светло-коричневая, с длинным и узким протоком. Белковая
железа крупная, кремовая. Семяпровод толстый, короче пениса и впадает
в него слегка отступя от его заднего конца. Пенис цилиндрический изви-
тый, значительно длиннее мантии. Задний конец его заметно вздут и изо-
гнут, у места впадения семяпровода — с темным пятном. Внутри вздутия
находится крупная складка, остальная часть пениса несет внутри мелкие
продольные складки. Половой ретрактор крепится почти апикально, рядом
с семяпроводом. Семяприемник длинный, слабо дифференцирован на ре-
зервуар и проток; последний впадает в атриум у основания пениса. Яйце-
вод короткий, почти одной толщины с пенисом.
37
Рис. 337—342. Lehmannia, nyctelia (Bourg.) (из: Wiktor, 1973).
внешний вид; 338 — раковина; 339 — зубы радулы; 340 — кишечник; 341 — гениталии; 342 _
вскрытый пенис.
7. LIMAX
273
Распространение. Пока неясное, так как, во-первых, долгое
время этот вид не отличали от L. marginata, во-вторых, по-видимому, этот
слизень легко поддается завозу и поэтому пока трудно определить границы
его природного ареала. Не исключено, что первоначальное описание этого
вида было сделано по экземплярам из популяции, возникшей путем завоза
(Алжир). Судя по тому, что в природных биотопах этот вид обнаружен
в Польше, Румынии, Венгрии и Болгарии, естественный ареал ограничен
Карпатами, Балканским хребтом и сопредельными горными областями.
В СССР этот вид пока не обнаружен, но находки его возможны в Украин-
ских Карпатах.
Образ жизни. Обитает в горных широколиственных лесах, осо-
бенно в буковых. Днем скрывается в дуплах, под корой и под камнями.
Во время дождя ползает по стволам деревьев. Выше верхней границы леса
встречается чаще всего в скальных развалах.
7. Род LIMAX Linnaeus, 1758
Linnaeus, 1758 : 652; В г а г d, 1815 : 107 (Limacella); Moquin-Tandon,
1855 : 22 (Eultmaz); M alm, 1868 : 54 (H eynemannla)', Hesse P., 1926 : 9.
Типовой вид — L. maximus Linnaeus, 1758; по последующему
обозначению — Ferussac, 1819.
Крупные слизни (дл. пол. до 200, дл. сок. до 150 мм), чаще всего с довольно’
отчетливым килем, различной длины, но никогда не простирающимся на
всю спину. Мантия занимает от до 1/3 длины тела. Пневмостом лежит
сильно постмедиально.
Срединный и боковые зубы трехзубцовые, иногда лишь самые внешние
боковые теряют эндокон и становятся двухзубцовыми. Краевые зубы двух
типов: внутренние однозубцовые, кинжаловидные или шиповидные; внеш-
ние двухзубцовые за счет развития вторичного дополнительного зубца.
Кишечник не перекручен, у подрода Limacus с длинной слепой кишкой;
5-е и 6-е колена прижаты и параллельны друг к другу. Левая доля печени
образует вершину внутренностного мешка..
Ось сердца наклонена под углом 45°. Аорта короткая. Задний отдел кро-
веносной сети легкого отчетливо выражен. Основание полового ретрактора
крепится к диафрагме у заднего края почки или за пределами комплекса.
Гонада располагается позади второй петли кишечника. В отличие от осталь-
ных родов передняя часть простаты не плотно прилегает к женскому каналу
семяяйцевода, а связана с ним тонкой перепонкой. Яйцевод заметно длин-
нее, чем у Gigantomilax и близких ему групп. Четкой поперечной перетяж-
кой он разделен на более толстый передний и более узкий задний отделы.
Длинный семяпровод апикально или субапикально крепится к крупному
цилиндрическому пенису, который иногда длиннее животного. Внутри пениса
залегают несколько продольных складок, из которых одна особенно крупная
и сзади раздвоенная. Пенис и семяпровод на всем протяжении связаны тон-
кой перепонкой (отличие от остальных родов).
Ретрактор правых щупалец проходит между пенисом и семяпроводом,
с одной стороны, и яйцеводом и простатой — с другой.
Распространение. В эту группу входят около 20 видов,
большая часть которых обитает в средиземноморских областях Европы.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ LIMAX
1 (6). Третья петля кишечника простирается назад не менее чем на длины
спины. Слепой кишки нет (рис. 15, III). Слизь бесцветная (подрод
Limax s. str.).
18 И. М. Лихарев, А. й. Виктор
«74 ш. LIMACIDAE
-2 (3). Пенис очень сильно закручен, длина его не меньше длины животного.
Киль простирается на х/2 спины. . . 1. L. (Limax) cinereoniger Wolf.
3 (2). Пенис закручен слабее, длина его равна около 1/2 длины животного.
Киль простирается на х/3 спины.
4 (5). Рисунок спины слагается из продольных рядов темных пятен или
полос на светлом фоне. Семяпровод и половой ретрактор крепятся
к пенису почти апикально, т. е. к его заднему концу...................
........................................2. L. (Limax) maximus L.
5 (4). Рисунок слагается из светлых пятен на серовато-желтом или пепельно-
сером фоне. Семяпровод крепится к пенису заметно отступя от его
заднего конца...................3. ,L. (Limax) dobrogicus Gr. et Lupu.
6 (1). Третья петля кишечника очень короткая. Имеется длинная слепая
кишка (рис. 15, IV). Слизь желтая (подрод Limacus Lehmann).
7 (8). Пенис сильно извитый. Яйцевод состоит из двух отделов: заднего
узкого и переднего вздутого, толстостенного. В последний впадает
семяприемник. Верхняя сторона тела окрашена в серовато-зеленый
или оливковый цвет. На этом фоне имеется рисунок из серых пятен
с нечеткими контурами и мелкой сетки из микроскопических точек.
Этот рисунок исчезает на заметном удалении от края подошвы . . .
............................................ 4. L. (Limacus) flavus L.
8 (7). Пенис слабо извитой, С-образно загнут или прямой. Яйцевод не под-
разделен на передний и задний отделы, цилиндрический. Семяприем-
ник впадает в атриум между пенисом и яйцеводом. Фоновая окраска
желтая или серовато-желтая. Рисунок такой же, но темнее и более
четкий. Сетка из точек по бороздам простирается до края подошвы
..................................... 5. L. (Limacus) maculatus (Kai.).
1. Подрод LIMAX s. str.
Длина киля равна от 1/3 до 1/2 длины спины. Окраска верхней стороны
тела либо одноцветно темная, либо на светлом фоне имеется рисунок
из темных продольных полос или рядов пятен, а на мантии из пятен.
Третья петля кишечника обычно не- короче второй, а у некоторых видов
и длиннее; слепой кишки нет. Семяяйцевод очень длинный, яйцевод более
короткий.
Длина пениса не менее х/2 длины тела, а нередко и значительно больше.
1. Limax (Limax) cinereoniger Wolf, 1803 (рис. 343—346)
Wolf, 1803 :7; Ferussac, 1819 : 68, T. 8D, Fig. 2 (antiquorum var. a); К a-
leniczenko, 1851b : 120 (antiquorum Razoumowski); Taylor, 1902—1907 : 53,
T. 7; Лихарев, P аммел ьмейер, 1952 : 312, рис. 237; Quick, 1960 : 187,
Fig. 15 A, E, G, T. 2, Fig. 25; W i k t о r, 1973 : 68, Fig. 21, 22, 99, 230, 264, 265.
Terra typica — ФРГ и ГДР. Типы неизвестны.
Материал: из СССР — 126 экз. из 54 пунктов; из других стран
более 700 экз.
Тело стройное, к хвосту заостренное, с довольно длинным килем —
около */г длины спины. Мантия спереди закругленная, сзади угловатая,
закрывает около животного. По общему облику животное походит на L. ma-
ximus (см. ниже), но кожа более прочная и толще. Морщины более резкие,
крупнее и длиннее. Между мантийной щелью и средней линией спины 21 —
27 рядов морщин. Окраска очень изменчива, однако на востоке Европы у этого
вида большая часть тела (за исключением киля и средней доли подошвы, ко-
торые всегда светлые) окрашена в черный или темно-серый цвет; реже ниже
7. LIMAX
275
киля, после светлых промежутков, располагаются 1, 2 или 3 пары темных
продольных полос или рядов пятен. Мантия темная, одноцветная, по краю
иногда с мелкими светлыми точками. Киль чаще всего белый, реже желтый
или розовый и всегда светлее спины. Боковые доли подошвы темнее сред-
ней, но у молодых слизней иногда вся подошва светлая. Темные щупальца
покрыты черными или бурыми точками. Слизь бесцветная, довольно жид-
кая.
Окраска и рисунок очень варьируют даже в пределах одной популяции.
В частности, они зависят и от возраста животного. Молодь обычно светлая,
кремового или коричневатого цвета, с неясным рисунком или без него; на по-
дошве темный пигмент появляется к концу роста. Однако и взрослые слизни
окрашены очень разнообразно.
Рис. 343. Limax (Limax) cinereoniger Wolf (из: Taylor, 1902—1907).
Изменчивость окраски данного вида очень хорошо описана Тейлором
(Taylor, 1902—1907). Варьируют как фон, так и рисунок. Фон может быть
белым, серым, желтым и изредка розоватым. Рисунок спины слагается
из трех пар темных полос или рядов пятен. Мантия всегда одноцветно тем-
ная.
При сильном развитии темного пигмента все животное, кроме светлого
киля и срединной доли подошвы, становится черным (var. тайга Held, 1806).1
Именно эта разновидность наиболее обычна на Восточно-Европейской рав-
нине. Здесь же, но несколько реже встречаются вариации с тремя парами
полос, из которых все или часть могут распадаться на ряды пятен (var.
renardii Kaleniczenko, 1851). Еще реже встречаются экземпляры с белым
килем, светлыми боками и с двумя парами полос или рядов пятен (var.
vera Dumont et Mortillet, 1857).
Слабое развитие пигмента приводит к появлению белых или почти белых
слизней, у которых лишь боковые доли подошвы могут иметь темный налет
(var. pallescens Dumont et Mortillet, 1857). В частности, эта форма обнаружена
в Карпатах. Там же встречается другая своеобразная форма с оливково-
бурой окраской и с одной парой темных полос ниже светлого киля (var.
transsylvanica Heynemann, 1862).
Указанными цветовыми вариациями далеко не исчерпывается полимор-
физм окраски данного вида, особенно если учесть формы, обнаруженные в за-
падных и южных областях Европы. Пока нет оснований рассматривать те
или иные вариации как подвиды, так как они могут встречаться в пределах
одной популяции. Тем не менее отмечено, что сильное развитие темного пиг-
мента, приводящее к слиянию полос и к появлению одноцветно темной
окраски, более свойственно популяциям, обитающим в северных частях
ареала и в альпийской и субальпийской зонах гор. Узкие полосы, распа-
1 Номинативная вариация встречается реже и отличается тем, что не только киль,
но и лежащая перед ним узкая полоса спины до мантии светлые.
18
276
III. LIMACIDAE
дение их на ряды пятен и исчезновение некоторых полос более обычно в южных
.популяциях, обитающих в широколиственных лесах и лесостепи.
Рис. 344—346. Limax (Limax) cinereoniger Wolf (из Wiktor, 1973).
344 — раковина; 345 — зубы радулы; 346 — гениталии.
Размеры: дл. пол. 150—200 мм.
Раковина (рис. 344).
Радула (рис. 345). Срединный и боковые (20—30) зубы с небольшими до-
полнительными зубцами, причем боковые теряют сначала эктокон, а за-
7. LIMAX
277
тем и эндокон. У краевых зубов (около 50) мезокон расщеплен, так что эти
зубы вновь приобретают двухзубцовый облик. Число поперечных рядов
150-170,
Кишечник типичный для рода, только третья петля нередко простирается
до вершины внутренностного мешка, т. е. длиннее, чем у L. maximus.
Гениталии (рис. 346). Длинная и узкая пигментированная гонада
.видна сверху среди лопастей печени. Светлый гермафродитный проток вна-
чале тонкий и прямой, а затем становится толстым и извитым, особенно пе-
ред впадением в семяяйцевод. Белковая железа крупная, языковидная.
•Сравнительно длинный яйцевод перед впадением в атриум утолщается.
Небольшой семяприемник имеет овальный резервуар и короткий проток,
который впадает в атриум у основания пениса. Очень длинный цилиндри-
ческий пенис (равен или больше длины тела) образует сложные извивы.
Внутри него имеется длинная складка, которая в задней части пениса мощ-
ная и раздвоена, а в передней ординарная и, превращаясь в тонкую мембрану,
сходит к атриуму на нет. Семяпровод и половой ретрактор крепятся к пе-
нису апикально.
Распространение. В СССР — Прибалтика, южная Карелия,
западные и центральные области европейской части РСФСР (наиболее вос-
точная находка сделана вблизи Казани — Алейникова, Акрамовский,
1968), Белоруссия, северные и западные области Украины. Одна находка
сделана вблизи Кишинева. Очень обычен в Карпатах.
В европейской части СССР этот слизень обитает главным образом в сме-
шанных и лиственных лесах. На севере, т. е. в Эстонии, Ленинградской
области и в южной Карелии, в областях, где преобладают хвойные леса,
он встречается на южных склонах, сложенных из песчаников и известня-
ков (например, силурийское плато Глинт), где температурные условия бла-
гоприятствуют сохранению оазисов с лиственными породами деревьев
(Фридолин, 1934).
Вне СССР — почти вся Европа, кроме самых северных областей.
Образ жизни. Обитает в лесах вдали от населенных пунктов,
реже в старых парках. В горах средней Европы может жить также выше
лесной зоны, в каменистых развалах и щелях скал. Днем скрывается под
корой валежин, в дуплах, под камнями, ночью ползает по земле, стволам
деревьев и по поверхностям скал.
О копуляции и жизненном цикле см. выше (с. 57, 67). Эмбриональное
развитие длится 19—24 дня. Свежевылупившаяся молодь длиной 8—9 мм,
нначале бесцветная и просвечивающая, но вскоре делается непрозрачной,
а под конец черной. Длительность жизни 23/4—3 года.
Питается различными грибами и лишайниками.
Замечания. Данный вид во многом сходен с L. maximus, в связи с чем
длительное время его рассматривали лишь как разновидность последнего.
Внешне он отличается: киль длиннее; кожа грубее, толще и с более крупными
морщинами; боковые доли подошвы чаще всего темные, а мантия одноцветная
i(y L. maximus вся подошва светлая, а мантия пятнистая); щупальца покрыты
мелкими темными пятнышками (у L. maximus без них). Во внутреннем строе-
нии наш вид отличается более длинными пенисом и третьей петлей кишечника.
2. По-видимому, данному виду очень близок слабо изученный L. bielzi
Seibert, 1873 (см. о нем: Ehrmann, 1933: Lozek, 1956; Wiktor, 1973). Зейберт
и Виктор считали его самостоятельным видом, Эрманн и Ложек — подвидом
L. cinereoniger.
От последнего L. bielzi в первую очередь отличается окраской тела.
Обычно у него киль красный, спина охристо-желтая или лиловато-розовая,1
1 В литературе на немецком языке — «цвета мяса».
278
III. LIMACIDAE'
а мантия темно-красная или черная. Внутреннее строение было изучено только*
Виктором на единственном найденном им экземпляре. Серьезных отличий от-
L. cinereoniger не обнаружено. Только пенис несколько короче и тоньше.
Систематическое положение этой формы пока не ясно. Нужно иметь не
только больше материала для анатомирования, но и проследить, как поли-
морфизм окраски всей группы форм L. cinereoniger связан с изменчивостыо-
гениталий.
С территории СССР нет указаний о находках L. bielzi или сходных по ок-
раске форм L. cinereoniger. Общее распространение (Wiktor, 1973): польские
Судеты и Карпаты, Чехословакия и Австрия (северная Штирия). р’
3. В южных и юго-восточных областях Европы слизни из группы ci-
nereoniger-maximus изучены совершенно недостаточно. В связи с этим трудно*
определить наше отношение к румынскому виду L. zilchi Grossu et Lupu,
1960. В первоначальном описании указано, что по внешнему облику он напо-
минает L. maximus, хотя и имеет некоторые свои особенности. По гениталиям
он практически неотличим от L. cinereoniger.
Создается впечатление, что главным основанием для рассматривания;
L. zilchi в качестве самостоятельного вида у авторов было убеждение, что в ус-
ловиях Румынии L. cinereoniger как холодолюбивый вид обитает только в го-
рах, а на равнинах жить не может. Между тем L. zilchi был обнаружен в рав-
нинных лесах Добруджи и окрестностей Бухареста.
Позднее (Grossu, Tesio, 1971) с целью убедиться, что это разные виды, было*
поставлено биохимическое (электрофорез) исследование белков обоих видов,
для чего были использованы 10 экземпляров L. zilchi (по 5 слизней из Доб-
руджи и из-под Бухареста) и 5 экземпляров L. cinereoniger (с гор Буцеги).
В результате обнаружены как будто бы четкие различия в электрофореграм-
мах белков гемолимфы, кишки и педальной мускулатуры.
Что касается биохимических доказательств, можно отметить, что исполь-
зование только 5 экземпляров L, cinereoniger совершенно недостаточно для по-
лучения достоверных результатов. Тем более, что не доказано, что L. cinereo-
niger в понимании авторов рецензируемой работы соответствует представле-
нию об этом виде в средней Европе, откуда он был описан и где ощлучше;
изучен.
2. Limax (Limax) maximus Linnaeus, 1758 (рис. 347—350)
Linnaeus, 1758 : 652; Muller, 1774 : 5 {cinereusp, Ferussac, 1819 : 68
{antiquorum, part); Simroth, 1885 : 204, T. 8, Fig. 1—11; Taylor, 1902—1907 : 34,
T. 6; Quick, 1960:191, Fig. 15B,C, D,F, H, T. 2, Fig. 24; Лихарев, Рам-
мельмейер, 1952 : 311, рис. 236; Wiktor, 1973 : 65, Fig. 19—20, 97, 100, 229,
261, 262.
Terra typica — Швеция? Типы, неизвестны.
Материал: из СССР — 27 экз. из 18 пунктов; из других стран —
около 200 экз.
Тело стройное, к хвосту заостренное, с коротким килем, занимающим
х/3 спины. Мантия спереди закругленная, сзади угловатая; она закрывает
около х/3 Животного. Между мантийной щелью и средней линией спины 21—
26 рядов морщин.
Основная (фоновая) окраска желтовато- или пепельно-серая, или грязно-
белая; рисунок — из 2 или 3 пар полос или рядов черных пятен; мантия ис-
пещрена черными пятнами, нередко образующими меандрический рисунок;,
щупальца розовато-коричневые без темных точек; подошва белая, без ри-
сунка.
Указанные окраска и рисунок характерны как для популяций из цен-
тральных и западных областей европейской части СССР, так и для живущих,
в средней и северной Европе. В западных и южных областях Европы ветре-
7. LIMAX
279
чаются популяции меланистические (одноцветно черные, лишь подошва
светлая) или с другой фоновой окраской и рисунком.
Рис. 347—350. Limax (Limax) maximus L. (из: Wiktor, 1973).
347 — внешний вид; 348 — раковина; 349 — зубы радулы; 350 — гениталии.
Слизь бесцветная, густая.
Размеры: дл. пол. 130 (по некоторым литературным источникам
в Западной Европе до 200), дл. сок. 100 мм.
280
III. LIMACIDAE
Радула (рис. 349). Срединный и боковые зубы с небольшими экто- и эндо-
конами. Граница между боковыми и краевыми зубами слабо заметна. Краевые^
сначала шиповидные, а под конец двухзубцовые. Общее число боковых и крае-
вых зубов в каждом ряду 62—73, число рядов 138—157. Кишечник, как.
у L. cinereoniger, только третья петля короче (занимает около 1/2 спины).
Раковина (рис. 348).
Гениталии. Отличаются от L. cinereoniger сравнительно коротким
пенисом (длина его равна около половины длины тела), который нередко
несколько вздут у заднего конца (рис. 350).
Распространение. В СССР:1 ряд изолированных друг от друга
популяций в следующих населенных пунктах и вблизи от них — Калининград,
Рига, Тарту, Ленинград, Ярославль, Рославль, (Смоленской обл.), Москва,
Горький, Орел, Киев и Керчь. В природных биотопах обитает в Восточных
Карпатах.
Общее распространение. Как синантропный вид встречается,
почти во всей Европе, особенно в северных и восточных областях континента.
Естественный ареал, по-видимому, занимает горные области западной, южной
и частично средней Европы, а также Северной Африки. Завезен в Северную
Америку, Южную Африку, Австралию и Новую Зеландию.
Образ жизни. В антропогенных условиях обычен для парков и са-
дов (убежища — куски древесины, дупла, под корой и камнями), теплиц, под-
валов, погребов, подземных каналов и туннелей. В природных условиях оби-
тает в различных лиственных и смешанных лесах.
3. Limax (Limax) dbhrogicus Grossu et Lupu, 1960 (рис. 351, 352)
Grossu, Lupu, 1960:162, Fig. 5.
Locus typicus-. «in der Nahe des Klosters-Сосо?» — близ монастыря Кокос,,
северная Добруджа, Румыния. Голотип хранится в Естественноисторическом
музее им. Гр. Антипа, Бухарест, № 9500/116.
Материал: 1 экз. из Румынии.
При жизни окраска серовато-желтая; желтизну придает обильная слизь,
выделяемая покровами. После фиксации и удаления слизи верхняя сторона
тела пепельно-серая со светлыми пятнами, придающими ей мраморный облик.
Щупальца голубоватые. Мантия овальная, спереди и сзади закругленная.
Киль очень короткий, у заднего конца спины более четкий, чем впереди.
Подошва желтая, слизь ее бесцветная и вязкая.
Размеры: дл. пол. 140—150, дл. сок. 50—60 мм.
Вторая и третья петли кишечника значительно короче первой, а третья
немного длиннее второй (рис. 351).
Гонада коричневато-желтая, состоит из 3—4 почти равных долек. Белко-
вая железа довольно крупная, белая. Простата толстая, по внешнему краю
гофрированная. Яйцевод довольно длинный, вначале узкий, но ближе к ат-
риуму заметно утолщается (рис. 352). Семяпровод почти прямой и по сравне-
нию с другими видами того же подрода короче; он впадает в пенис, заметно
отступя от заднего конца последнего. Пенис толстый и цилиндрический, ко-
роче, чем у L. maximus, с 1—2 изгибами, связанными мускулистой мембраной.
Внутри него располагаются 2 продольные складки, из которых одна развита
сильнее. Половой ретрактор крепится к заднему концу пениса. Семяприемник-
с крупным овальным резервуаром и коротким протоком, открывающимся
в атриум.
1 Кроме коллекционных данных, здесь используются только те литературные источ-
ники, где нет сомнений в правильности определения.
7. LIMAX
281
Распространение. Северная Добруджа
;из двух пунктов: близ монастыря Кокос и у горы
в Румынии. Известен
Бабадаг.
Рис. 351, 352. Limax (Limax) dobrogicus Gr. et Lupu.
-351 — кишечник; 353 — дистальная часть гениталий (семяприемник поврежден). Румыния, Добруджа.
Бабадаг, 3 X 1963.
Образ жизни. Обитает во влажных и тенистых местах дубово-липо-
вого леса. Днем скрывается под корой гниющих стволов деревьев.
2. Подрод LIMACUS Lehmann, 1864
Lehmann, 1864 : 145; Malm, 1868 : 62 (Plepticolimax); С 1 e s s i n, 1884 : 62
(Simrothia); Hesse P., 1926 : 10.
Типовой вид — Limacus Breckworthianus Lehmann, 1864 (=Limax
jlavus Linnaeus, 1758, var.); по монотипии.
Киль короткий и нечеткий. Рисунок образован многочисленными мелкими
темными пятнами, которые не сливаются в продольные полосы или ряды
более крупных пятен. Третья петля кишечника очень короткая (значительно
короче второй), с длинной слепой кишкой, которая плотно прилегает к верх-
ней стенке тела и простирается назад нередко до конца полости телд (рис. 15,
IV). Семяяйцевод очень короткий, так как недалеко от белковой железы
распадается на простату (внутри с семенным каналом) и длинный яйцевод;
оба протока, тесно прилегая друг к другу, простираются вперед почти до
атриума. Пенис короче половины длины тела.
4. Limax (Limacus) flavus Linnaeus, 1758 (рис. 353—356)
Linnaeus, 1758 : 652; D rap arnaud, 1801 : 103 (variegatus); Simroth,
1885 : 214, 317, T. 7, Fig. 8, T. 8, Fig. 17—19 (variegatus); Taylor, 1902—1907 : 78,
T. 12, Fig. 5—9; Quick, 1960 : 184, Fig. 14, T. 2, Fig. 22; W i k t о r, 1973 : 75, Fig. 23,
.24, 101, 108, 232, 266, 267.
Terra typica — Швеция? Типы неизвестны.
Материал: из СССР — 21 экз. из 5 пунктов; из других стран —
75 экз.
Тело веретеновидное, в поперечном сечении почти круглое, с очень корот-
. ким и нечетким килем. Мантия закрывает не более 1/3 тела. Кожа с тонким
рельефом из относительно мелких морщин и неглубоких бороздок. Между
мантийной щелью и средней линией спины 22—23 ряда морщин.
282
III. LIMACIDAE
Основная (фоновая) окраска взрослых животных оливковая или серовато-
зеленая. Рисунок сложен из темных микроскопических точек — меланофоров,
которые, в первую очередь концентрируются по бороздкам, но на спине ме-
Рис. 353—356. Limax (Llmacus) flavus L.
353 — раковина; 354 — зубы радулы; 355 — дистальная часть гениталий, Одесса, 5 VIII 1969; 356 —
гениталии (из: Lister, 1694).
стами распространяются и на морщины, образуя серые пятна с нечеткими кон-
турами. Еще большее развитие эти пятна имеют на мантии, где они, частично
сливаясь друг с другом, формируют меандрический рисунок. Ниже спины
пятен становится все меньше, меланофоры сохраняются только по бороздкам,,
образуя сетчатый рисунок, и плавно исчезают на известном удалении от по-
7. LIMAX
283
дошвы. Непигментированные промежутки внутри рисунка имеют облик мел-
ких светлых жемчужин.
Слизь верхней стороны тела желтая, а на мантии почти оранжевая (осо-
бенно заметно при фиксации), прозрачная и жидкая. Желтоватые тона окраски
животного в значительной степени обязаны слизи, поэтому после фиксации
тело становится более тусклым, почти серым.
По сравнению с предыдущими видами того же рода окраска этого слизня
сравнительно мало варьирует. Цвет фона в значительной степени зависит от
условий обитания. Отмечено, что в достаточно влажных местах и при избытке
пищи фон окрашен в более яркие тона, чем в менее влажных местах и при го-
лодании. Кроме того, молодь обычно окрашена темнее, чем взрослые особи.
Размеры: дл. пол. до 120, дл.сок. до 70—80 мм.
Раковина — см. рис. 353.
Радула (рис. 354). Зубы в основном однозубцовые, с мощным мезоконом.
Экто- и эндоконы отсутствуют или чуть намечены у срединного и первых
боковых зубов. Боковые зубы плавно переходят в шиповидные краевые, и
четко разграничить их невозможно. Наиболее внешние краевые зубы иногда
приобретают вторичный зубец и становятся двухзубцовыми. Общее число
зубов (по одну сторону от срединного) 50—70. Число поперечных рядов зубов
130-170.
Гениталии (рис. 355, 356). Гонада белая или серая; проток ее в на-
чале прямой, а под конец образует сложные извивы. Белковая железа языко-
видная, желтая. Семяпровод сравнительно короткий; он впадает в пенис,
немного отступя от его заднего конца, рядом с половым ретрактором. Пенис
цилиндрический, толстостенный, сильно извитый. Внутри он имеет 5—6 про-
дольных складок, из которых одна особенно длинная, но низкая, а другая
хотя и короче предыдущей, но более высокая и впереди переходит в мощный
гребень.
Яйцевод состоит из двух отделов: заднего, узкого и цилиндрического, и пе-
переднего, вздутого и толстостенного, в который открывается семяприемник.
Последний слагается из короткого мускулистого протока и овального ре-
зервуара.
Распространение. В СССР обнаружен в следующих городах:
Москва, Калининград, Тарту, Саратов, Одесса и Ростов-на-Дону, где обитает
только в погребах, подвалах и оранжереях. В Ялте он встречен в плодовом
саду.
Вне СССР: природный ареал — средиземноморские страны Европы
и Передней Азии, а также на островах Средиземного моря; в частности,
в коллекции ЗИН АН СССР имеются сборы с о-вов Сардиния и Сицилия и из
окр. Бейрута (Ливан). В остальных областях Европы, а также на других
континентах и островах, куда этот слизень был завезен, он живет только в на-
селенных пунктах. В коллекциях ЗИН АН СССР и Вроцлавского музея пред-
ставлен с о-ва Тенериф (Канарские о-ва), из Чили и ЮАР (Претория).
Образ жизни. В пределах природного ареала населяет леса и влаж-
ные биотопы безлесных областей (осыпи, щели скал и т. и.).
В населенных пунктах обитает в погребах, овощехранилищах, теплицах
и непосредственно в жилых домах, выбирая такие сырые места, как подвалы,
подполья кухонь и ванн и вдоль водосточных труб.
Копуляция (Gerhardt, 1933; Fromming, 1954; Karlin, Bacon, 1961) чаще
всего происходит ночью и обычно на почве или других горизонтальных по-
верхностях. После непродолжительного брачного танца (кружение) партнеры
располагаются половыми- отверстиями друг против друга. Пенисы быстро
выворачиваются и спирально переплетаются. Как только капли спермы, за-
ключенные в слизь, показываются наружу, оба пениса быстро втягиваются
внутрь, унося с собой сперму партнера.
284
III. LIMACIDAE
Откладка яиц (чаще всего по ночам) производится порциями, по 12 штук
в кладке (всего до 138 яиц). Яйца овальные (6x4 мм), светло-янтарного цвета
с небольшими округлыми выступами по концам, придающим яйцам форму ли-
мона. Концы яиц обычно связаны слизистой нитью. Слизень помещает свои
кладки под различные предметы и в трещины почвы. Вылупление молоди
происходит через 3—6 недель. Только что вылупившиеся слизни 10—13 мм
длиной, зеленовато-желтого цвета, а щупальца у них синие.
Хозяйственное значение. Как полифаг этот слизень не’
только наносит ущерб овощам, фруктам и другим продуктам, но, питаясь-
также различными отбросами и нечистотами, способствует заражению про-
дуктов яйцами гельминтов и другими болезнетворными началами.
5. Limax (Limacus) maculatus (Kaleniczenko, 1851)K comb. nov. (цв. вкл.»
рис. Ill; рис. 357, 358)
Kaleniczenko, 1851а : 226, Т. 6, Fig. 2 (Krynickillus): М о qu in - Т an-
don, 1855 : 25, Т. 3, Fig. 3—9 (variegatus Draparnaud, 1801, pars); Boettger O.r
1881 : 186, T. 7, Fig. 7 (ecarinatus)', Simroth, 1898a : 61 (retowskii)', 1901 : 84, T. 5,
Fig. 9—16 (flavus-variegatus-ecarinatus); Лихарев, Раммельм ей e p, 1952 : 312
(flavus)- Lupu, 1970 : 63, Fig. 2—5 (grossui): H u d e c, 1972 : 84, Fig. 11 (grossui);
Chatfield, 1977 : 1, Fig. 1 (grossul).
Locus typicus — «Kuczukoiy» — Кучук-кой (современное Кипарисное, Юж-
ный берег Крыма). Типы неизвестны.
Материал: из СССР — более 700 экз. из 192 пунктов; из Болгарии —
7 экз.
Внешне слизень сходен с L. flavus. Главные отличия: а) основная окраска
(фон) более светлая — желтая или серовато-желтая; б) рисунок более четкий,,
темно-серый или почти черный; как и у предыдущего вида, он слагается из:
различных по размерам и форме темных пятен на спине и мантии и сетки из;
мелких микроскопических точек-хроматофор, но последняя простирается до>
самой подошвы, чего не бывает у L. flavus1, в) щупальца темно-серые или почти
черные. Как и у предыдущего вида, молодь всегда окрашена темнее, чем взрос-
лые слизни, нередко одноцветно черная или с единичными светлыми пятныш-
ками.
В альпийской зоне Кавказа обитают популяции, где большинство особей
имеет одноцветно черную окраску, иногда заходящую и на края подошвы
(у аг.ecarinatus О. Boettger, 1881). По внутреннему строению эти слизни не от-
личаются от обычных пестрых форм..
Размеры: дл. пол. до 120, дл. сок. до 80 мм.
Раковина, как у L. flavus.
Радула. Срединный и боковые зубы с мощными мезоконами, дополнитель-
ных зубцов нет или они рудиментарные. Краевые зубы шиповидные, лишь са-
мые внешние имеют по одному вторичному зубцу. Пример: 1 экземпляр из
Талыша; формула:
Кишечник, как у L. flavus.
Гениталии (рис. 357). В отличие от L. flavus: а) яйцевод на всем-
протяжении цилиндрический, трубчатый, без вздутия впереди, в 2 раза шире
семяпровода; б) семяприемник впадает в атриум между пенисом и яйцеводом;
в) пенис значительно короче, С-образно изогнут, если же извитый, то слабее,,
чем у предыдущего вида; г) кроме двух крупных продольных складок, внутри
заднего конца пениса имеется несколько листовидных поперечных складок,,
которых нет у L. flavus. Указанные особенности гениталий свойственны также.-
лектотипу L. ecarinatus О. Bttg. (Зенкенбергский музей, № 155346) (рис. 358)^
7. LIMAX
285'
Распространение. В СССР — в природных биотопах обитает
в Крыму и на Кавказе. Кроме того, обнаружен в погребах и оранжереях
Пушкина (Ленинградская обл.), Ленинграда, Севастополя, Орджоникидзе,
Тбилиси и Астрахани.
Рис. 357—358. Limax (Limacus) maculatus (Kai.). Дистальная часть гениталий.
357 — лектотип Limax ecarinatus О. Boettger, Кутаиси, Зенкенбергский музей, № 155346; 358 — Крым,
лес, 1 IX 1974.
Вне СССР обнаружен в Турции, Болгарии, Румынии и на Британ-
ских о-вах.
Образ жизни. В лесах на дневное время укрывается в дуплах, под
корой пней и валежин'и под камнями; в безлесных биотопах и в альпийской
зоне прячется в скальных развалах, в осыпях и в щелях скал.
В населенных пунктах обитает в погребах, овощехранилищах, в оранже-
реях, а на юге также в садах и парках.
Замечания. 1. Последние годы считалось, что в группу Limacus
входит лишь L. flavus L. Однако недавно Лупу (Lupu, 1970) обнаружила в Ру-
286
HI. LIMACIDAE
мынии другой вид той же группы, который назвала L. grossui. Позднее его
нашли в Крыму (Hudec, 1972) и на Британских о-вах (Chatfield, 1976; Evans,
1978). Наши исследования показали, что этот слизень широко распространен
на Кавказе и в Крыму.
Удалось выявить дополнительные различия между обоими видами, в том
числе и во внешнем облике.
Открытие Лупу поставило на очередь следующие вопросы: а) с каким из
двух видов имел дело Линней — автор L. flavus; б) так как у L. flavus L.
имеется много синонимов,1 вполне естественно ожидать, что один из них от-
носится к L. grossui и, следовательно, имеет приоритет над последним назва-
нием.
К сожалению, Лупу и ее последователи эти вопросы обошли молчанием.
Ниже мы пытаемся дать на них ответы.
а) Линней дал название L. jlavus со ссылкой на работу Листера (Lister,
1694). Эта редкая книга имеется в ЗИН АН СССР. Интересующий нас вид под
небиноминальным названием изображен в ней дважды: на фронтисписе
(внешний облик) и на таблице 4, рис. 3 (гениталии). Последний рисунок нам
особенно интересен (рис. 356). На нем хорошо видно, что семяприемник впа-
дает в яйцевод, т. е. как у L. flavus L. в понимании Лупу, Гудеца, Чатфильд и
нашем.
б) Изображение гениталий, сходных с L. grossui, мы обнаружили в работе
Мокен-Тандона (Moquin-Tandon, 1855). Автор приписывает их Limax variega-
ius Draparnaud, 1801 (=L. flavus L., 1758). Однако на той же таблице изобра-
жен внешний облйк этого слизня, соответствующий нашим представлениям
о L. flavus. Это противоречие, по-видимому, связано с тем, что во Франции
•обитают оба вида.
Калениченко (Kaleniczenko, 1851а) описал и дал изображение внешнего
вида Krynickillus maculatus из Крыма. Анатомией этого вида автор не за-
нимался, однако внешний облик его слизня полностью соответствует нашим
представлениям о втором виде из подрода Limacus, обнаруженном Лупу и
названном ею L. grossui.
Как показали наши исследования, этот слизень широко распространен
в лесах Крыма, где нет L. flavus. Учитывая также, что малакофауна Крыма
и Добруджи, откуда был описан L. grossui, не впервые обнаруживает большое
сходство, мы считаем, что последнее название следует рассматривать как
младший синоним Limax (Limacus) maculatus (Kaleniczenko, 1851).
2. Согласно Лупу, L. grossui отличается от L. flavus более короткой слепой
кишкой, почти прямым и более коротким пенисом, а также образом жизни.
Если L. flavus свойственна синантропия, то второй вид обитает только в ле-
сах, вдали от человеческого жилья. Во внешнем облике обоих слизней Лупу
различий не нашла.
Как показали наши исследования, размеры слепой кишки не могут слу-
жить для различения этих видов, так как у обоих ее длина нарастает с воз-
растом и под конец жизни она достигает заднего конца внутренностного
мешка.
Оба вида могут обнаруживать тенденцию к синантропии. Так, L. macu-
latus в Крыму и на Кавказе кроме природных биотопов обитает в погребах,
подвалах жилых домов, а также в парках и садах. Кроме того, он завезен
в оранжереи Ленинградской области. С другой стороны, еще старые авторы
отмечали, что L. flavus на юге Европы обитает не только близ жилья, но и в ле-
сах и лишь в более северных областях континента живет только в населенных
пунктах.
1 Наиболее полные списки синонимов см. у Тейлора (Taylor, 1902—1907) и Гессе
•(Hesse Р., 1926).
8. BIELZIA
287'
Наиболее четко оба слизня различаются по форме и внутреннему строению
пениса и по положению семяприемника. Однако при достаточной тренировке
их можно распознать и по внешнему облику, что еще подметила Чатфильд.
(Chatfield, 1976).
3. Судя по тексту и рисункам, а также по соответствующим коллекцион-
ным материалам ЗИН АН СССР, все то, что в монографии Зимрота (Simroth,
1901) написано о L. flavus, относится к L. maculatus. Это понятно, так как ав-
тор имел дело со сборами из Крыма и Кавказа, где L. flavus не обитает.
2. Подсемейство BIELZIINAE Likharev et Wiktor, subfam. nov.
Пневмостом лежит постмедиально. Кишечник трехпетлевый. В отличие-
от остальных подсемейств Limacidae сзади располагается правая доля печени,,
а не левая.
Наиболее характерно строение гениталий (рис. 361). Семяпровод пол-
ностью вытянут вдоль яйцевода, т. е. не образует петли, заканчивающейся
пенисом, как в других группах Limacidae, а непосредственно открывается
в атриум. Пенис — очень крупный и мускулистый мешковидный орган с очень,
своеобразным внутренним строением; сзади он слепо замкнут, а впереди впа-
дает в атриум (см. рис. 21). Гонада располагается рядом с белковой железой..
Монотипическая группа.
Замечание. Столь необычное для остальных Limacidae строение-
гениталий, при котором семяпровод открывается в атриум непосредственно,,
т. е. минуя пенис, давно дало основание объединять соответствующие виды
в род Bielzia Clessin, 1887 (Р. Hesse, 1926; Thiele, 1931; Wenz, Zilch, 1959—
1960; Wiktor, 1973).
Подобный план строения указывает на совершенно иной механизм ко-
пуляции (с. 55). Следовательно, морфологические и физиологические особен-
ности полового аппарата Bielzia столь резко ртличают этот род от всех других
групп Limacidae, что, с нашей точки зрения, есть все основания выделить его-
в особое подсемейство.
Принятию подобной точки зрения нашими предшественниками, вероятно,
препятствовало внешнее сходство карпатской Bielzia coerulans с некоторыми
видами Limax s. str., в особенности с L. cinereoniger, с которым она нередко
живет совместно. Однако Limacidae вообще слабо различаются по внешнему
облику. К тому же, упомянутые слизни не так уж сходны, да и род Bielzia
не ограничивается одним видом.
8. Род BIELZIA Clessin, 1887
Н az ay, 1883 : 330 (Frauenfeldia, non Egger, 1865); Clessin, 1887 : 47; Si m-
roth, 1888 : 67 (Limacopsis)', Hesse P., 1926 : 15; Лихарев, Раммель-
мейер, 1952 : 351; Wiktor, 1973 : 81.
Типовой вид — Limax coerulans M. Bielz, 1851; по монотипии..
Слизни с отчетливым килем, простирающимся на х/2 или 2/3 спины, а у не-
которых видов почти на всю спину. Мантия занимает около х/3 длины тела.
Пневмостом лежит - сильно постмедиально.
Кишечник не перекручен, без слепой кишки. Вторая и третья петли почти
одной длины.
Ось сердца наклонена почти под углом 90°. Аорта короткая. Почка очень
своеобразной формы из-за двух выступов на правой стороне ее. Мочеточник
впадает в вытянутый мочевой пузырь. Основание полового ретрактора кре-
пится к диафрагме у переднего края мантийного комплекса.
Семяяйцевод короткий, яйцевод и семяпровод длинные и подтянуты друг
к ДРУГУ многочисленными волокнами. Внутри яйцевода, недалеко от его пе-
реднего конца, находится небольшой сфинктер.
288
III. LIMACIDAE
Большая часть стенок своеобразного пениса утолщена, образуя длинное
мускулистое тело с мощной кольцевой и продольной мускулатурой, пронизан-
ное тонким продольным каналом (рис. 21). Внутри это тело плотно приросло
к внешним стенкам пениса и лишь впереди и сзади заканчивается короткими
выступами — папиллами. Перед этими папиллами располагаются две ка-
меры — передняя и задняя. Задний конец пениса утолщен за счет рыхлой
ткани, богатой лакунами. Половой ретрактор крепится к пенису позади пе-
редней папиллы,
Распространение. Один вид населяет Карпаты и некоторые
соседние горные области (см. ниже); второй обитает на о-ве Крит [5. cretica
(Simroth, 1889)]. Считается (Hesse, 1926), что еще два вида — В. moellendorffi
(Kimakowicz, 1890) и В. montenegrina (О. Boettger, 1885) — живут в Югосла-
вии (в Боснии и Черногории). Однако так как внутреннее строение их неиз-
вестно, то принадлежность к данному роду не доказана.
Замечания. В литературе существуют две точки зрения на про-
исхождение и функцию необычного органа Bielzia, который мы рассматриваем
как пенис. Зимрот (Simroth, 1885) склонен был считать его эа «мешок любовной
стрелы» («Pfeildriise») или его гомолог. Поэтому автор считал, что этот орган
при копуляции исполняет роль стимулятора. Однако такому толкованию
противоречит строение «мешка» и отсутствие в нем любовной стрелы.
Другую точку зрения высказал Коштал (Kostal, 1895). Он обнаружил,
что передний конец мускулистого тела придатка может выдаваться в атриум
в виде короткой папиллы, и заключил, что весь орган является модифици-
рованным пенисом. К этой точке зрения склоняемся и мы.
Окончательно вопрос о происхождении данного органа можно решить,
лишь проследив путь его формирования в онтогенезе. Что касается функции
его, то он,,по-видимому, играет роль насоса или шприца, сначала насасываю-
щего свою сперму, а затем подающего ее в атриум партнера. В пользу этого
говорит как строение данного органа, так и положение полового ретрактора
и ретенторов концевых участков гениталий. При этом сфинктер яйцевода за-
пирает для своей спермы вход в женские пути как в момент ее насасывания
внутрь пениса, так и при ее выпрыскивании в атриум партнера (с. 55).
Следовательно, все указанные особенности гениталий слизней этой группы
связаны с переходом к новому типу копуляции, при которой обмен спермой
происходит без значительного выдвижения наружу копулирующих органов.
Последнее отмечала еще Смоленьска (Smolehska, 1936).
Bielzia coerulans (М. Bielz, 1851) (цв. вкл., рис. Z; рис. 359—363)
В i е 1 z М., 1851 : 14 {Limax)-, Frauenfeld, 1864 : 681, Т. 20, Fig. 1—4 {Limax
sehwabi)-, H a z а у, 1883 : 100, T. 5, Fig. 1—3, T. 6, Fig. 1—4 {L. transsilvanicus Heyn.=
L. sehwabi Frauenfeld); Simroth, 1885 : 212, T. 8, Fig. 16 {Limax)-, Bakowski,
t omni cki, 1892 : 21, T. 1, Fig. 3 {Limax)-, Kostal, 1895 : 1—15, T. 19—22 {Li-
macopsis); Smolensk a, 1936 : 212, T. 30—31; Лихарев, Раммельмейер,
1952 : 352, рис. 280; Wiktor, 1973 : 82, Fig. 27, 28, 116—120, 228, 268, 269.
Locus typicus — «Dreguscher und Arpaser Gebirge» — Трансильвания (Ру-
мыния). Типы неизвестны.
Материал: из СССР — 22 экз. из 7 пунктов; из Польши — более
250 экз.
Тело стройное, к заднему концу плавно, но сильно суженное. Мантия
занимает меньше Ч3 длины тела, сзади сильно заостренная. Около х/2 мантии
приходится на капюшон. Киль простирается на Ч2 или 2/3 спины. Морщины
четкие, килеватые, длинные, на концах заостренные; между мантийной щелью
и средней линией спины 16—17 рядов морщин.
8. BIELZIA
289
Окраска очень изменчивая и зависит от возраста и физиологического со-
стояния. Взрослые животные синеватые, синевато-зеленые или фиолетовые,
без рисунка. У не вполне взрослых особей по бокам нередко заметны продоль-
ные темные полосы. У живых взрослых слизней окраска нередко перелива-
ется. Подошва синевато-черная, одноцветная или боковые доли ее более тем-
ные, чем; средняя. Слизь светло-желтая, прозрачная. Голова и щупальца
черные; затылочная складка тоже черная, но окантована светлыми бороздами.
Рис. 359—363. Bielzia coerulans (М. Bielz).
359 — внешний вид молодого слизня; зво — раковина; 361 — гениталии; звг — передняя часть яйце-
вода (вскрыта) со сфинктером; звз — начальные участки семяпровода и яйцевода (359, зво — из: Wiktor,
1973; 361—363 — Карпаты, Яремча, VIII 1969).
Синий цвет и переливы его появляются с наступлением Половой зрелости.
При консервации окраска блекнет, переливы ее исчезают и тело приобретает
синевато-серый или синевато-черный цвет.
Молодь окрашена совершенно иначе. Обычно спина оливково-бурая
•с двумя парами темно-бурых полос ниже киля, которые нередко заходят и на
мантию. Бока, киль и голова темно-желтые, подошва от серовато-зеленой до
желтой.
Размеры: дл. пол. до 100, дл. сок. до 90 мм.
Раковина (рис. 360) удлиненно- или яйцевидно-овальная; размеры около
10x5.5 мм.
Радула. Срединный и большая часть боковых зубов трехзубцовые; мезо-
коны мощные, а дополнительные зубцы небольшие; лишь несколько самых
внешних боковых зубов, утрачивая эндокон, становятся двухзубцовыми.
Большинство краевых зубов за счет эктокона двухзубцовые, только самые
внутренние трехзубцовые; самые внешние краевые зубы недоразвитые и имеют
19 И. М. Лихарев, А. Й, Виктор
290
Ш. LIMACIDAE
однозубцовый облик. Пример: 1 экземпляр из Украинских Карпат; фор-
мула:,
с . 226 56 8к 22к 5к
Т + ~з” + “2“ + ~ + ~~2~ + т X 155-
Гениталии (рис. 361—363). Гонада светло-серая или серовато-светло-
лиловая, округлая и компактная; она лежит вблизи от второй петли кишеч-
ника, рядом с передним краем языковидной белковой железы и не закрыта
соседними органами. Проток ее на всем протяжении очень сильно извитый.
К семяяйцеводу прилегает простата, имеющая плотную зернистую структуру.
Передняя часть этого органа простирается на начало семяпровода, почти пол-
ностью, как муфта, окружая его, и впадает в него несколькими протоками.
Лишь с одной стороны эта «муфта» не замкнута, образуя желоб, в котором за-
легает яйцевод. Семяпровод в начале узкий и тонкостенный; после прохожде-
ния сквозь простату становится очень толстым, мускулистым и почти прямым^
внутри он несет тонкие продольные скйадки. Пенис булавовидный, вздутый,
с мощным половым ретрактором, который крепится к верхней стенке тела у пе-
реднего края диафрагмы.
Яйцевод плавно расширяется к атриуму. В начальной части он дает корот-
кий извив и поэтому немного длиннее семяпровода. Семяприемник с длинным
овальным резервуаром и коротким протоком. Небольшой атриум, а также пе-
редние концы пениса и остальных половых путей подтянуты к стенкам тела
пучками мускулов-ретенторов.
Распространение. В СССР —Украинские Карпаты. Вне СССР —
Восточные Судеты, Западные и Восточные Карпаты и некоторые прилегаю-
щие к ним средневысотные горы (в том числе отдельные местонахождениям
в Краковско-Ченстоховской гряде) и Южные Карпаты.
Образ жизни. Обитает в умеренно влажных и очень сырых биото-
пах всех типов горных лесов, а также в вышележащих зонах. Особенно много-
числен и активен (даже днем) после обильного летнего дождя, когда интен-
сивно ползает и питается на почве. В лесу предпочитает скрываться под ва-
лежниками и в старых пнях.
Весной И первую половину лета питается лишайниками и листьями
копытня европейского, во вторую половину лета главным образом грибами и
земляникой, а при нехватке их также лишайниками и некоторыми травами..
Копуляция происходит на земле после «брачного танца» и падает на июнь—
начало июля, а откладка яиц на июль—август. Яйца округло-овальные (4х
X 5 мм) в количестве от 3 до 90 штук откладываются в виде цепочек у подножья!
трав. Вылупление молоди наступает на 17—25-й день. Молодые слизни в мо-
мент вылупления имеют длину около 6.5 мм. Слизни быстро растут и на зи-
мовку уходят почти взрослыми. После откладки яиц слизни вскоре гибнут,,
так что продолжительность жизни данного вида около одного года
(рис. 25, III).
3. Подсемейство EUMILACINAE Likharev et Wiktor, subfam. nov.
Пневмостом лежит антемедиально. Кишечник двухпетлевый (род Eumilax)
или трехпетлевый (род Metalimax). В последнем случае третья петля очень
короткая и от нее отходит назад довольно длинная слепая кишка.1 Ректум про-
низывает диафрагму заметно ближе к пневмостому, чем у других Limacidae.
Гонада полностью скрыта между лопастями печени и, как у Bielziinae, рас-
полагается вблизи от середины тела, т. е. смещена вперед и лежит на одном
уровне с белковой железой. Семяпровод впадает в пенис.
1 При вскрытии и оттягивании кверху верхней стенки тела третья петля кишечника
выпрямляется, и может создаться впечатление, что ее нет. В такую ошибку впал Зимрот,
когда утверждал, что у Metalimax кишечник двухпетлевый.
9. EUMILAX
291
Распространение. Кавказ и черноморские склоны Понтийских
гор Малой Азии.
Подсемейство слагается из родов Eumilax и Metalimax.
9. Род. EUMILAX О. Boettger, 1881
Boettger О. 1881 : 178 (род Amalia, секция Eumilax)', 1883 : 144 (род Limax,
•секция Paralimax)', Simroth, 1901 : 98 (Paralimax)', Hesse P., 1926 : 28; Л и-
харев, Раммельмейер, 1952 : 342.
. Типовой вид — Umax (Milax) Brandti Martens, 1880; по монотипии.
Крупные (дл. сок. до 150 мм) или средних размеров (дл. сок. около 35 мм)
слизни. Киль четкий и всегда простирается на всю спину. Мантия обычно
Рис. 364—367. Eumilax brandti (Mart.).
364 — раковина; 365 — кишечник; 366 — ретракторы; 367 — вывернутые при копуляции части генита-
лий (Сумбатово, 17 VII 1915).
занимает немного больше х/3 длины тела; верхняя сторона ее покрыта мел-
кими сосочками, всегда хорошо заметными при жизни, но иногда и у фикси-
рованных экземпляров. Позади мантии почти каждый ряд морщин разделя-
ется на два. Подошва шире, чем у Metalimax.
19*
.292
III. LIMACIDAE
Ось сердца наклонена направо под углом 45° или больше. Аорта средней-
длины, обычно раздваивается под почкой. Почка полулунная, лежит по-
перек оси тела и окружает сердце сзади и по бокам. Мочевой пузырь пере-
понкой связан с перикардием. Вместе с анусом он открывается наружу
немного сзади пневмостома. Половой ретрактор крепится к диафрагме
вблизи от заднего края почки.
Кишечник двухпетлевый (рис. 365). Слепой кишки нет. Семяяйцевод
очень длинный, сзади узкий, а впереди в связи с увеличением простаты
постепенно расширяющийся. Яйцевод и семяпровод короткие, последний
очень узкий.
Крупный пенис имеет очень характерную форму. Он состоит из трех
четких отделов. Передний имеет форму короткой мускулистой трубки,
к которой обычно крепится ретентор. Несколько сзади в него открывается
короткий, но толстый крючковидно изогнутый задний отдел пениса (нередко'
его обозначали как придаток пениса), у основания которого впадает семяпровод.
При копуляции эта часть пениса выворачивается и соединяется с аналогич-
ным органом партнера (рис. 370). Еще дальше пенис переходит в длинный
бичеобразный слепой замкнутый орган, постепенно сужающийся к заднему
концу. К этому отделу пениса от его заднего конца и почти на 2/3 его длины
веерообразно крепится половой ретрактор; к «вееру» прилегает крупная
ветвь половой артерии.
До сих пор вслед за Зимротом этот своеобразный отдел пениса считали
бичом. Однако у легочных моллюсков бич никогда не связан с половым
ретрактором, и внутреннее строение его иное. У данной группы внутренние
стенки этого органа (как и заднего отдела пениса) имеют многочисленные
продольные складки и сосочки. Выворачиваясь наружу при копуляции,
эта часть пениса играет роль стимулятора.
В небольшой атриум вблизи от основания пениса впадает семяприемник,
слагающийся из короткого протока и овального резервуара. Ретрактор правых
щупалец простирается под крючковидным задним отделом пениса, над
бичеобразным отделом и под семяпроводом.
Распространение. Большой Кавказ, западное Закавказье и
черноморское побережье Малой Азии.
Замечание. Бётгер (О. Boettger, 1881),. обсуждая систематическое
положение единственного в то время известного вида — Limax brandti
(Mart.), отнес его к роду Amalia (=Milax) и в новую секцию Eumilax. Позд-
нее тот же автор (1883), открыв новый вид Limax intermittens, включил его
в новую секцию Pardlimax. Зймрот (Simroth, 1901, 1910, 1912а) справед-
ливо считал Eumilax и. Paralimax синонимами, однако вопреки приоритету
он принял для этой группы последнее название — Paralimax. Гессе
(Р. Hesse, 1926) восстановил приоритет названия Eumilax, отнес крупные
виды к подроду Eumilax s. str., а более мелкие к подроду Paralimax. Подоб-
ной же точки зрения придерживались Лихарев и Раммельмейер (1952).
Наши исследования показали, что род Eumilax состоит из двух видов
слизней — крупного Е. brandti (Mart.) и средних размеров Е. intermit-
tens (О. Bttg.), а остальные виды и вариететы, описанные Бётгером и Зим-
ротом, следует рассматривать как синонимы этих двух видов. Оба слизня
очень сходны как по внешнему облику, так и по анатомии, и нет оснований
относить их в разные подроды.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ EUMILAX
1 (2). Слизни крупные — дл. сок. до 150 мм, половой зрелости они дости-
гают при дл. сок. не менее 60 мм. Между килем и мантийной щелью
число рядов морщин больше 23................1. Е. brandti (Mart.).
9. EUMILAX
293
2 (1). Слизни средних. размеров — дл. сок. около 37 мм, при этих же раз-
мерах они достигают половой зрелости. Межйу килем и мантийной
щелью число рядов морщин не более 21 . . . : . . .С ..... .
.......................................... 2. Е. intermittens (О. Bttg.).
1. Eumilax brandti (Martens, 1880) (цв. вкл., рис. W, У/Л рис. 364—371)
Martens', 1880:143 [Limax (Milax)]; Boettger О., 1881 178 [Amalia
(Eumilax)]; 1886a : 127; Pollonera, 1887 : 1, T. 3, Fig. 8—11 (Paralimax); Boet-
tger O., 1888 : 149 (P. multirugatus, syn. nov.); 1889 : 6(P. multirug atus); Retowski,
1889 : 227 [Amalia (Paralimax)]; Taube r,-, 1900 : 373, T. 9, Fig. 1—12, T. 10, Fig. 13—
24, T. 11, Fig. 25—29 (Paralimax); S i m.r о th,- 1901 : 113, T. 9a—b, T. 10, Fig. 1—4
(Paralimax); 116, T. 10, Fig. 3 (P. br. coriaceus, syn;.nov.); 117, T. 9, Fig. 1 (P. br. nubilis,
syn. nov.); 117, T. 9, Fig. 3—6, 8—Q..(P. br„ nota-tiis, syn. nov.); 117, T. 10, Fig. 4 (P. br.
liliaceus, syn. nov.); 117, T. 9, Fig, 7 (P. marmoratus, syn. nov.); 118, T. 11, Fig. 1 (P. niger,
syn. nov.); 119 (P. niger nigerrimus, syn. novi); 119, T. 11, Fig. 2—4 (P. multirugatus);
120, T. 12, Fig. 1 (P/albomaculatus, syn. nov.); 120, T. 11, Fig. 2—6 (P. salamandroides,
syn. nov.); 122, T. 11, Fig. 7 (P. gyratus, syn. nov.); 124, T. 10, Fig. 5—6 (P. gracilis, syn.
nov.); 125, T. 10, Fig. 10 (P. ochraceus, syn. nov.); 126, T. 12, Fig. 11 (P. albocarinatus,
syn. nov.); R o s e ri,- 1903: 35 (Paralimax); Simroth, 1910:516 (P. gyratus); 517,
T. 6, Fig. 14—16 (P'.kalischewskit, syn. nov.); 519, T. 6, Fig. 17 (P. nigerrimus); 520 (P. sa-
lamandroides); 1912a : 29, 125, T. 2, Fig. 19, T. 6, Fig. 18 (P. brandti, P. br. variegatus,
P. br. punctatus, syn. nov.); 31-,,125, T. 7; Fig;.11 (P. niger); 31 (P. albomaculatus); 32 (P. sa-
lamandroides); 32 (P. kalischewskii); 32, T. 3, Fig. 24, T. 7, Fig. 20 (P. albocarinatus); 124
(P. multirugatus); Лихарев, Раммельмейер, 1952 :-344 (multirugatus);
345 (ocHraceiis, niger, albocarinatus); 345, рис. 273; 346 (gracilis, kalischewskii, gyratus);
347, рис. 274 (salamandroides); 347 (albomaculatus, nigerrimus); К а л и т и н а, 1958 : 173
(niger); 174, (salamandroides); W i k t b r, 1971 : 278, Fig. 8 (Eulimax, err.).
Locus typicus; «Borshom» — Боржоми. . ‘
В нашем распоряжении не было типов данного вида, и где они находятся
неизвестно. Однако мы исследовали и обозначили лектотипы многих синонимов
и вариететов Е. brandti (Mart.) Почти все они хранятся в ЗИН АН СССР.
Сюда относятся: Paralimax br. coriaceus Simroth — 2 экз. (лектотип найден
на горе Накерала близ Тквибули — см.: Simroth,. 4.901, табл. 10, рис. 3);
Р. br. nubilis Simroth— 6 экз. (лектотип найден в том же месте — см.
табл. 9, рис. 1); Р. salamandroides Simroth — 4 экз. (лектотип найден у дер.
Куэли близ горы Гдлахой-лам — см. табл. 11-, рис. 5); Р. gyratus Simroth
(голотип найден в долине р. Лабы); Р. kalischewskii Simroth — 6 экз. (лекто-
тип найден в верховьях р. Белой — см.: Simroth, 1910, табл. 6, рис. 14).
Кроме того, исследованы :3 типовых экземпляра Р. gracilis Simroth, храня-
щиеся в Зоологическом музее Хельсинкского университета (лектотип най-
ден в Имеретии близ Опстщети — cM.:;Simroth, 1901, табл. 10, рис. 6).
Места хранения типов остальных синонимов неизвестны.
Материал: из СССР V 307 экз. из 143 пунктов; из Турции — 2 экз.
В движении и при слабом сокращении тело длинное и стройное, напоми-
нает Limax s. str. Отношение длины мантии к общей длине зависит от сокра-
щения тела. При слабом сжатий длина мантии немного меньше х/3 общей длины,
при сильном — несколько больше. Передняя часть мантии закругленная,
задняя с тупым углом, а при сокращении1 животного овальная. Мантийная
щель удалена от переднего края мантий на//3 ее длины. Киль хорошо развит
и всегда простирается до мантии, иногда плавно снижаясь к ней. Морщины
длинные и нежные. У Взрослых особей между килем и мантийной щелью
размещается 23—31 ряд морщин, а иногда до 40. У молодых слизней число
рядов меньше. Так, у одного, экземпляра с дл. сок. 45 мм было всего 20 рядов
морщин.
Окраска и рисунок очень варьируют. По-видимому, здесь проявляется
как индивидуальная, так и популяционная изменчивость. Общий фон
коричневый или черный, а у форм с рисунком кремовый или коричневый.
Рис. 368—370. Eumilax brandti (Mart.).
368 — гениталии f. l;/pica; зв9 — дистальная часть гениталий; 340 — вскрытый пенис (Аджария, до-
лина р. Мачахели-цкали, 23 VII 1973).
9. EUMILAX
295
В большинстве случаев окраска одноцветно шоколадная или черная (f. ty-
pica=multirugatus Bttg.). Иногда киль светлый (f. albocarinata Simr.=niger
Simr.=griseus Simr.). У двухцветных форм рисунок простирается как на ман-
тию, так и на спину. Чаще на мантии имеются светлые пятна (f. nubila
Simr. = notatus Simr. = ochraceus Simr.) На спине рисунок представлен пятнами
кремового цвета (f. salamandroides Simr.=gyratusSimr. = albomaculatusSimr.),
или на кремовом фоне располагаются коричневые пятна, сливающиеся
в продольные полосы (f. ochraceus Simr.=nubilus Siwe.=notatus Simr.). Как видно
даже из рисунков в работах Зимрота, между этими формами имеются раз-
личные переходы. Слизни с крупным пятнистым рисунком чаще всего оби-
тают высоко в горах. Боковые доли подошвы обычно окрашены темнее, чем
срединная; так, если боковые коричневые, то срединная кремового цвета.
Слизь бесцветная, умеренно жидкая.
Размеры: дл. пол. до 250, дл. сок. до 150, ширина до 22, дл. мантии
до 47 мм.
Радула с хорошо развитыми экто- и эндоконами на срединном и боковых
зубах и однозубцовыми кинжаловидными краевыми зубами, число последних
во много раз больше боковых. Срединный зуб размерами заметно меньше
боковых.
Кишечник — рис. 365.
Кровеносная сеть легкого состоит из двух участков: перед сердцем основ-
ной наиболее крупный, позади сердца над почкой меньшего размера задний
участок.
Гениталии построены по типичному для данного рода плану
(рис. 367—371). Главные отличия от Е. intermittens — гермафродитный
проток сильно извит, задний отдел пениса с тупым или закругленным концом.
Распространение (рис. 42, 2). В СССР — горно-лесные и
горно-луговые зоны северных и южных отрогов Большого Кавказа (от Сочи
и горы Фишт на западе до Гуниба и Закаталы на востоке), Сурамский, Триа-
летский, Аджаро-Имеретинский, Шавшетский хребты Малого Кавказа.
Вне СССР — черноморские склоны Понтийских гор Малой Азии (Retowski,
1889; Wiktor 1971) и бассейн р. Чорох. Указание об обитании этого вида
в окрестностях Анкары (Germain 1936), вероятно, ошибочно.
Образ жизни. В пределах ареала обитает во всех типах леса,
чаще всего по тенистым сырым ущельям горных рек. Выше леса живет в ка-
менистых и скальных биотопах субальпийской и альпийской зон. Активен
по ночам. Днем даже в дождливую погоду прячется в щелях скал и крупно-
блочных осыпей, а также в сырых дуплах и под валежинами.
Откладывает яйца во влажные укрытия. Яйца бесцветные, прозрачные,
овальные размером от 6 X 8до8х9 мм. Вылупление происходит на 20—45-й
день после откладки. Только что вылупившиеся слизни окрашены в розова-
тые тона и при ползании достигают 14 мм в длину. Мантия покрыта крупными
сосочками, которые после фиксации исчезают. Даже потомство из одной кладки
в первые дни жизни обнаруживает заметный полиморфизм окраски — еди-
ничные экземпляры одноцветно розовые без всяких следов темного пигмента^
большинство же имеет темный рисунок — полосы и пятна на спине и ман-
тии — различной интенсивности и формы.
Замечания. Зимрот (Simroth 1901, 1910) описал не только боль-
шое число вариететов данного вида, но и ряд других видов, которых мы рас-
сматриваем как синонимы Е. brandti (см. выше). Располагая единичными
экземплярами и не обнаружив у них существенных различий в гениталиях,
автор неоправданно большое значение придавал отличиям в окраске и ри-
сунке верхней стороны тела этих слизней.
Наши исследования значительно большего материала показали, что
окраска фона, форма и цвет рисунка отличаются большим полиморфизмом.
Рис. 371. Eumilax brandti (Mart.). Гениталии f. salamandroid.es Simr Центральная часть
Большого Кавказа, 2000 м над ур. моря, 15 VIII 1935.
9. EUMILAX
297
В то же время они не обнаруживают никаких связей с остальными особен1
ностями строения этих животных.
Согласно Зимроту, описанные им виды нередко отличаются рельефом
кожи, в частности числом рядов морщин между килем и мантийной щелью.
Наши исследования показали, что; во-первых, это число увеличивается
с возрастом слизней, во-вторых, внешне оно зависит от степени сжатия
животного при фиксации. Дело в том, что у данного вида число морщин
во втором ряду от мантии почти вдвое большем чем в первом. При сильном
сжатии слизня первый ряд слабо заметен (он как бы сминается), и при под-
счете может создаться впечатление, что морщин больше,чем на самом деле.
Исследование типов видов Зимрота подтверждает этот' вывод.
2. Eumilax intermittens (О. Boettger, 1883) (рис. 372—375)
Boettger О., 1883 : 145, Т. 4, Fig. 7 [Limax (Paralimax)]', Simroth, 1897 :
145 (Paralimax); 1901 : 126, T. 8, Fig. 5, 10—14, T. 12, Fig. 5—10 (P. raddei, syn. nov.);
127, T. 12, Fig. 5, 6, 9 (P. raddei striatus, syn. nov.); 128, T. 12, Fig. 6 (P. raddei pictus,
syn. nov.); 128, T. 12, Fig.40 (P. raddei elegans, syn. nov.); 128, T. 12, Fig. 7 (P. r. variegatus,
syn. nov.); 128, T. 12, Fig. '6 (P. r. obscurus, syn. nov.); 129, T. 8, Fig. 7, T. 12, Fig. 12,
(P. minutus, syn. nov.); 1910 : 514, T. 6," Fig. 13 (Paralimax); 1912a : 33, 127, T. 2, Fig. 25
(Paralimax); 34 (P. raddei); 34, T. 2, Fig. 27, T. 7, Fig. 21 (P. orientalis, syn. nov.); 34, T. 2,
Fig. 28, T. 7, Fig. 22 (P. griseus, syn. nov.); Лихарев, Раммельмейер, 1952 :
348—349, рис. 275—276 [E. (Paralimax) intermittens, raddei, minutus, griseus, orientalis];
Калягина, 1958 : 173, 174, (minutus, raddei).
Locus typicus — «Kutais» — Кутаиси.
В Зенкенбергском музее как синтипы данного вида (№ 25623/7) хранятся
7 высохших слизней из родов Arion и Lehmannia. Нами были исследованы и
обозначены лектотипы следующих видов и вариететов, описанные Зимротом,
которые мы рассматриваем как синонимы данного вида. Все они хранятся
в ЗИН АН СССР. Сюда относятся: Paralimax raddei Simroth, 1901 — 5 экз.
[лектотип найден на северном склоне горы Йл (ныне гора Сапицкая будка,
Орджоникидзе) — см. табл. 8, рис. 14]; Р. г. striatus Simroth — 1 экз.
(лектотип найден на горе Адай-хох на ю.-з. от Орджоникидзе — см. табл. 12,
рис. 9); Р. г. elegans Simroth — 2 экз. (лектотип найден на горе Ил — см.
табл. 12, рис. 9); Р. г. obscurus Simroth — голотип (найден на горе Фатгус
на ю.-в. от Орджоникидзе — см. табл. 12, рис. 8, 8а); Р. minutus Simroth —
голотип (найден близ дер. Куэли у горы Галахой-^лам — см. табл. 8,
рис. 7); Р. orientalis Simroth, 1912а — 4 экз. (лектотип найден на вы-
сокогорном пастбище Кем-Яйлаг, Главный хребет, быв. Нухинский уезд —
см. табл. 2, рис. 27); Р. griseus Simroth — 1 экз. (лектотип найден на горе
Хочал-даг у Лагодехи — см. табл. 2, рис. 28).
Материал: 184 экз. из 71 пункта Большого Кавказа.
Тело стройное, к заднему концу клиновидно суженное. При сокращении
слизня длина мантии немного больше х/3 общей длины. Сзади мантия слегка
сужена, но не образует тупого угла, обычного для предыдущего вида. Ман-
тийная щель относительно длиннее, чем у Е-. brandti, а пневмостом лежит
более медиально. Киль хорошо развит и простирается на всю спину. Мор1
щины спины нежные, но относительно крупнее чем у молодых экземпляров'
предыдущего вида. Между килем и мантийной щелью 17—21 ряд морщив.
Окраска слагается из желтого или желтовато-кремового фона и черного;
черновато- или темно-коричневого рисунка, форма которого сильно варьи-
рует. В типичном случае он слагается из двух темных полос, проходящих по
бокам мантии, и двух пар полос того же цвета позади мантии, т. е. на спине —-
две по сторонам от киля и две несколько ниже. Кроме того, посередине ман-
тии может пролегать третья полоса — медиальная (f. typica). Иногда рису-
нок мантии имеет неопределенные очертания и состоит из черных и желтых
298
III. LIMACIDAE
пятен (f. picta Simr.). Бока светлее, чем середина спины, так как темный
пигмент располагается по бороздкам. Темные продольные полосы спины и
мантии нередко имеют зубчатые очертания (f. raddei Simr.) и окаймлены
светлыми промежутками (f. f. striata Simr., elegans Simr.).
Рис. 372, 373. Eumilax intermittens (0. Bttg.). Цебельда, 1 X 1910.
372 — зубы радулы; 373 — кишечник.
Наряду и нередко совместно с полосатыми формами встречаются одно-
цветно черные или черновато-серые слизни (f. f. minuta Simr., obscura Simr-,
orient alls Simr.), которых следует рассматривать как меланистические формы.
Другой крайностью являются слизни со слабым развитием темного пигмента,
в связи с чем у них нет рисунка и вся верхняя сторона окрашена в светло-
серый цвет (f. grisea Simr.).
За исключением меланистических форм, у остальных киль всегда светло-
кремовый или желтоватый. Подошва светло-кремовая, боковые доли ее
чаще всего покрыты пятнышками за счет глубоколежащих черных мелано-
<форов, срединная доля подошвы всегда одноцветно светлая. Слизь бесцвет-
ная, жидкая.
Рис. 374, 375. Eumilax intermittens (0. Bttg.).
374 — гениталии f. typiea, Цебельда, 1 X 1910; 375 — дистальная часть гениталий паралектотипа f. rad-
dei Simr., гора Ил, Орджоникидзе, 1 V 1886.
300
III. LIMACIDAE
Большинство половозрелых слизней имеют следующие размеры: дл. сок.
27—38, ширина 8—9, длина' мантии 10.5—17 мм.
Радула (рис. 372). В пределах полуряда боковые зубы сначала постепенно
утрачивают эндокон, а затем и эктокон. Однозубцовых внутренних краевых
зубов немного; преобладают двухзубцовые внешние краевые зубы, у которых
имеется эктокон. Формула радулы:
с 166 56 7к 17к 5к
"3 + “3~ + “2- + “Г + “2“ + “Г*' '
Кишечник более перекручен, чем у предыдущего вида (рис. 373).
Гениталии (рис. 374, 375). От Е. brandti отличается слабо извитым
гермафродитным протоком и заостренным концом заднего отдела пениса.
Распространение. Горнолесные й горнолуговые зоны север-
ных и южных1 отрогов хребтов Большого Кавказа (от верховьев р. Мзымты
на западе, до Гуниба и Нухи на востоке) (рис. 42, 3).
Образ жизни. Обитает в различных типах леса во влажной под-
стилке, на траве и под камнями. Выше верхней границы леса живет среди
камней и во влажном мху.
Замечания. Зимрот (Simroth, 1901, 1910, 1912а), обнаружив раз-
личия в окраске и рисунке, описал ряд видов и вариететов. Однако, как
показали наши исследования, эти различия нестойкие и связаны переходными
формами, живущими нередко совместно. Согласно Зимроту, по гениталиям
отличается только Paralimax minutus Simr., описанный по единственному
экземпляру, у которого автор обнаружил перетяжку, отделяющую биче-
образную часть пениса от остального органа. Нами установлено, что такая
перетяжка возникает в результате сильного сокращения слизня при фикса-
ции. По нашим данным, есть все основания считать упомянутые выше виды
и вариететы синонимами или экологическими формами Eumilax intermittens
(Bttg.). ' !.
В Зоологическом музее Хельсинкского, университета хранится экземпляр
слизня, который Зимрот (1901, с; 124, табл. 10, рис. 8) определил как Para-
limax intermittens Bttg. Проверка /Показала, что это молодой Е. brandti
(Mart.).
10. Род METALIMAX Simroth, 1896
Simroth, 1896 : 366; 1901 : 94; 1910 : 511; 1912a : 24; H e s s e P., 1926:29;
Лихарев, Рамме льмей ер, 1952 : 349.
Типовой вид — М. elegans Simrothj 1901; но монотипии.
Слизни средних размеров (дл. сок. до 40 мм), со стройным телом. Киль
развит различно. Мантия относительно крупная (около х/3 длины тела), сза-
ди заканчивается острым углом, а впереди округлая, с крупным капюшоном.
Пневмостом лежит сильно антемедиально. Подошва узкая.
Ось сердца наклонена направо под углом 45°. Аорта очень длинная.
Почка, как у Ей/ш/аж.-Мочевой пузырь сильно вытянут к середине мантий-
ного комплекса. Кишечник трехпетлевый (рис. 378); третья петля очень ко-
роткая, с довольно крупной слепой кишкой, которая отходит от задней кишки
перед ее погружением в диафрагму и располагается справа от колумелляр-
ного мускула.
Очень длинный семяяйцевод с лентовидной простатой разделяется на
яйцевод и семяпровод вблизи от небольшого атриума. Пенис длинный, почти
цилиндрический, без придатков, внутри со складками; семяпровод впадает
в него апикально. Семяприемник открывается в яйцевод перед его впадением
в атриум.
Распространение. Западная часть Большого Кавказа.
10. METALIMAX
301
Замечание. Кроме приведенных ниже видов, к этому роду до сих
пор относили еще Metalimax mlokosieviczi Simroth, который был описан (1912а)
по одному экземпляру (голотип; хранится в ЗИН АН СССР). Этот вид на-
столько резко отличается от остальных по внешнему облику, что Зимрот
его выделил в особый подрод Metalimacoides — за крупный волнистый киль,
простирающийся на всю спину, постмедиальное положейиё пневмостома
и интенсивно черную окраску. Как голотип, так и позднее найденный второй
-экземпляр (Simroth 1912а, с. 128) были очень молодыми слизнями с зачаточ-
ными гениталиями. Внутри пениса голотипа Зимрот обнаружил небольшую
папиллу.
Новых находок не было. Не исключено, что Зимрот имел дело с молодыми
Caucasolimax caucasicus (Simroth), у которых пенис имеет папиллу. Во вся-
ком случае можно определённо сказать, что так называемый Metalimax
{Metalimacoides) mlokosieviczi Simroth к роду Metalimax никакого отношения
не имеет.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ, METALIMAX
1 (2). Тело равномерно покрыто сравнительно крупными темными пятныш-
ками, придающими слизню «леопардовый» облик. Мантия с темными
боковыми полосами. Половой, ретрактор крепится к середине пениса
.............................> г . ...... 2. M.-J elegans, Simr.
2 (1). Темные пятнышки очень мёдкиё и не видны невооруженным глазом.
По верхней стороне тела они распределены неравномерно, образуя
местные скопления или, сливаясь, дают меандрический' рисунок.
Мантия без боковых полос. Половой ретрактор крепится к заднему
концу пениса ............................ 1. М. varius (О. Bttg.).
1. Metalimax varius (Ch Boettger, 1884) (рис. 376—384)
Boettger О., 1884 147 (Paralimax)', Simroth, 1892:42 (P. reibischi)',
1894 : 10, Fig. 3 (P. reibischi); 1897 : 146 (P ar alim ax) ;A901 : 123, T. 10, Fig. 7 (P. reibischi
et varius)', 1910 : 512; 1912a : 24, T, .2, Fig. 15, T. 6, Fig. 15; 26, T, 6, Fig. 16 (reibischi);
Лихарев, P а м м e л ь м ё й е р, 1952:350, рис. 277, 278.
Locus typicus — «Psirsk» — Псырцха (Абхазия). Как нам сообщил д-р
Янссен в Зенкенбергском музее типы этого вида отсутствуют.
Материал: 202 экз. из 58 пунктов Кавказа.
Тело веретеновидно-цилиндрическое. ‘ Мантия с большим капюшоном,
который при сокращении слизня полностью закрывает щею и голову. Киль
слабый, но простирается на всю спину. Между ним и мантийной щелью
20—21 ряд морщин. Окраска слагается из серовато-желтого или красноватот
лилового фона и черного рисунка, сложенного из мелких точек (видны под
лупой). Эти точки дают продольные сгущения по обеим сторонам светлой
килевой полоски, а ниже образуют местные сгущения, постепенно исчезаю-
щие по направлению к подошве. При сильном развитии темного пигмента
из этих точек образуется меандрический рисунок, который особенно обычен
для мантии; реже здесь возникают неправильные темные боковые полосы.
Подошва всегда светлая, без темного пигмента.
При жизни окраска слизней бывает двух типов: светло-коричневая
без рисунка, который проявляется после фиксации в виде мелких точек;
темно-коричневая со светлой килевой полосой и отчетливым пятнистым ри-
сунком, который после фиксации приобретает облик меандр. Слизь вязкая,
бесцветна^.
Размер ы: дл. пол. до 60, мантии до 20 мм; дл. сок. до 35, мантии
.до 12 мм.
Рис. 376, 377. Metalimax varius (0. Bttg.).
37в — мягкая фиксация, Краснодарский край, станица Северская, 5 VIII 1972; 377 — жесткая фикса-
ция (пенис вывернут), станица Убинская, 17 IX 1970.
10. METALIMAX
303
Радула. Срединные и боковые зубы трехзубцовые, краевые однозубцо-
вые кинжало- или шиповидные.
Слепая кишка длинная, около х/5 длины тела (рис. 378, 379).
Гениталии (рис. 382—384). Гонада располагается перед белковой
железой или рядом с нею. В связи с этим длинный гермафродитный проток
сначала тянется назад, а затем отклоняется налево и перед белковой железой
сильно утолщается и образует мно-
гочисленные извивы. Семяяйцевод
дает 2—3 крупных извива. Яйце-
вод плавно сужается к атриуму;
перед ним в него впадает семяпри-
емник, состоящий из овального
резервуара и короткого протока.
Длинный семяпровод сначала тя-
нется вдоль яйцевода, затем про-
стирается назад, делает один ви-
ток вокруг пениса и впадает в его
задний конец рядом с половым
ретрактором. Последний сзади кре-
пится к верхней стенке тела у зад-
него края почки.
Пенис цилиндрический, лишь
слабо изогнут у заднего конца,
внутри он имеет крупную двух-
ветвистую продольную складку и
небольшую спиральную, лежащую
в заднем конце пениса и окружаю-
щую выход семяпровода. Атриум
очень короткий и является как бы
продолжением пениса.
Распространение. Гор-
ные и предгорные леса северо-за-
падной части Большого Кавказа:
по северным отрогам — от Ново-
российска до р. Белой, по юж-
ным — от р. Аше до р. Гумисты.
Кроме того, завезен в городские
•оранжереи Сочи и Ростова-на-
Дону (Косоглазов, 1972).
Образ жизни. Живет в
Рис. 378—380. Metalimax varius (О. Bttg.)
(продолжение).
378 — кишечник; 379 — задняя и слепая кишка;
380 — ретракторы.
лесах разного типа, от субтропических
с тиссом и самшитом до умеренно влажных дубово-грабовых, буковых и
пихтово-буковых. Днем скрывается под валежинами, в гниющей древесине,
в подстилке и под камнями.
Немногочисленные, сравнительно малоподвижные, по-видимому, одно-
летние слизни. Молодь появляется во второй половине лета. Осенью взрос-
лые экземпляры встречаются редко.
Замечания. Зимрот (Simroth 1912а, с. 27) пишет: «. : .насколько
Metalimax varius тз. М. reibischi внешне сходны, настолько они анатомически
различны. Скрещивание исключено». Несколько выше он отмечает следую-
щие основные различия между этими видами: а) у М. reibischi имеется слепая
кишка, а у М. varius ее нет; б) у М. reibischi пенис имеет вид длинного ци-
линдрического мешка, сзади заканчивающегося полукруглым слепым
выступом — цэкумом, впереди которого впадает семяпровод и крепится
ретрактор; у М. varius пенис булавовидный, без цэкума, семяпровод и рет-
рактор располагаются апикально.
304
III. LIMACIDAE
Нами исследованы те же экземпляры, с которыми имел дело Зимрот.
Проверка показала, что у всех имеется слепая кишка. Что касается цэкума,
то он отсутствует как у экземпляров, определенных как М. reibischi, так
и у определенных как М. varius. Правда, у некоторых слизней задний конец
381
Рис. 381—384. Metalimax varius (О. Bttg.) (окончание).
381 — мантийный комплекс, вид снизу (пунктиром указаны контуры раковины); 382 — гениталии;
383 — задний конец пениса; 384 — вскрытый пенис (станица Убинская, 17 IX 1970).
пениса несколько изогнут, отчего могло возникнуть впечатление, что семя-
провод и ретрактор крепятся к пенису сбоку, отступя от заднего конца.
Наконец, в пределах одной популяции встречаются экземпляры как с почти
цилиндрическим пенисом, т. е. по Зимроту М. reibischi, так и с булавовидным
пенисом, т. е. соответствующие М. varius, по представлениям того же ав-
тора.
Все это дает основание считать, что М. reibischi (Simroth, 1892) является
младшим синонимом М. varius (О. Boettger, 1884).
10. METALIMAX
305
2. Metalimax elegans Simroth, 1901 (цв. вкл., рис. IX; рис. 385—388)
Simroth, 1901 : 94, Т. 6, Fig. 1—6; 1910 : 512; 1912а : 27, Т. 2, Fig. 17; Л и-
х а р е в, Р а м м е л ь м е все р, 1952:350.
Locus typicus— «Tkwibuli» — Ткибули (западная Грузия). Голотип в
ЗИН АН СССР.
Материал: 15 экз. из ,9 пунктов Кавказа.
Рис. 385—388. Metalimax elegans Simr. Абхазия, Ткварчели, 3 IX 1952.
385.— задняя и слепая кишка; 386 — ретракторы; 387 — гениталии; 388 — дистальная часть гениталий
(вид снизу).
Тело веретеновидное, впереди более массивное, чем у М. varius. Киль
четко выражен только на заднем конце, к мантии он продолжается в виде
светлой килевой полосы. Между килем и мантийной щелью 22—23 ряда
морщин. Окраска слагается,из серовато-лилового фона и пятнистого темного
«леопардового» рисунка (пятнышки значительно крупнее, чем у М. varius).
Пятнышки на спине и боках располагаются равномерно; только вдоль ки-
левой полосы они образуют сгущения. Напротив, на мантии, сгущаясь,
они образуют две сравнительно широкие боковые полосы, которые иногда
по внутреннему краю окантованы светлыми промежутками; на середине
20 И. М. Лихарев, А. И. Виктор
306
I. TRIGONOCHLAMYDIDAE
мантии пятнышки разбросаны редко. С возрастом как мантия, так и спина
<(за исключением киля) темнеют за счет развития тонкого темно-серого
пигмента. В этом случае пятнышки остаются заметными только на боках.
Подошва светлая, почти белая.
Размеры: дл. сок. до 40 мм.
Слепая кишка относительно короче, чем у М. varius (рис. 385).
Гениталии (рис. 387, 388). Гонада располагается за серединой тела
между петлями кишечника и сверху не видна. Семяприемник с крупным
резервуаром и очень слабо заметным коротким протоком. Сквозь стенки ре-
зервуара просвечивают многочисленные продольные складки. Пенис отно-
сительно длиннее чем у М. varius. Половой ретрактор крепится к пенису
приблизительно на середине его длины, где имеется небольшой перегиб.
Внутренняя стенка пениса с мощными продольными складками.
Распространение. Южные отроги Большого Кавказа (Рачин-
ский, Згрисский и Кодорский хребты) в пределах западной Грузии и Абха-
зии (Ткибули, Цагери, Ткварчели); на северных отрогах — близ Алагира
в Северной Осетии (сбор В. Иванова, ЗИН АН СССР).
Зимрот (Simroth, 1910) сообщает о находке этого вида у р. Киша (приток
р. Белой, бассейн Кубани). Соответствующий экземпляр хранится в ЗИН
АН СССР, и проверка показала, что это молодой М. varius. Вызывает сомне-
ние другое указание того же автора (1912а)— об обитании М. elegans близ
Суреби (верховья р. Супса, Аджаро-Имеретинский хр.). Однако этого экзем-
пляра в нашем распоряжении нет.
Образ жизни. Обитает в буковых, дубовых и каштановых лесах,
под валежинами, камнями и в лиственной подстилке.
2. Надсемейство TRIGONOCHLAMYDOIDEA Р. Hesse, 1882
Слизни с внутренней раковиной, которая чаще всего имеет сосцевидный
или клювовидный нуклеус, располагающийся у заднего края слева от
продольной оси. Задний конец цефалоподиума заострен, спина с килем.
Кроме перипедальной, имеется также супрапедальная борозда и оторочка.
Пневмостом и мантийная щель сильно смещены к заднему краю мантии. По-
дошва трехдольчатая, т. е. с двумя продольными педальными бороздами.
В отличие от других надсемейств мантия обнаруживает тенденцию к смеще-
нию к заднему концу тела.
В связи с хищничеством глотка крупная или очень крупная, со своеобраз-
ной системой мускулов, крепящих ее к стенкам тела. Челюсть рудиментарная,
а у некоторых видов отсутствует. Радула без дифференциации на обычные
три типа зубов — все они кинжало- или багоровидные, с узкими базальными
пластинками, по форме напоминающими цифру 8.
Мантийный комплекс органов обнаруживает тенденцию к большей
деторсии, чем у Limacoidea и Zonitoidea.
Гениталии без придаточных органов на женских половых путях и атри-
уме (исключение род Troglolestes). Большинство видов формируют сперма-
тофоры.
Хищники.
В состав надсемейства входит одно семейство с 6 монотипическими родами.
I. Семейство TRIGONOCHLAMYDIDAE Р. Hesse, 1882
Hesse Р., 1882 : 32 (subfam. Trigonochlamydina)', Cockerell, 1891 : 225;
Hesse P., 1926:44.
При движении тело удлиненно-цилиндрическое или сзади вздутое,
а к голове плавно суженное. Подошва узкая или очень узкая, ширина ее
I. TRIGONOCHLAMYDIDAE
307
всегда значительно меньше ширины тела. При сильном сокращении на
спине появляется четкий киль, а все тело приобретает характерный для каж-
дого рода облик, который в первую очередь определяется размерами и формой
глотки. Мантия чаще всего с подкововидной бороздой. Хотя размеры и поло-
жение мантии на теле различны у разных родов, тем не менее отчетливо
прослеживается тенденция к ее уменьшению и постепенному смещению
к заднему концу тела, а пневмостома к заднему краю мантии. В связи с этим
меняется топография органов мантийного комплекса. Половое отверстие
располагается за правыми щупальцами или несколько отступя от них.
Раковина снаружи не видна, только у Рarmacellilla сквозь щель между
спиной и мантией виден нуклеус. В типичном случае небольшой сосцевидный
или клювовидный нуклеус располагается близ заднего края раковины,
немного левее ее продольной оси, напоминая наружную раковину некоторых
Ancylidae. Спатула очень тонкая, иногда слагается из отдельных известко-
вых зерен, связанных тонким периостракумом, на котором видны линии
нарастания.
Кишечник хищного типа, т. е. с мощной глоткой, специфической радулой
и короткой кишкой (рис. 403). От исходных двух петель остались лишь
1—3 легких изгиба. Ротовая полость сравнительно с другими семействами
длинная, при сокращении животного она нередко выворачивается наружу.
Глотка занимает бблыпую часть полости тела. Кроме основных глоточных
ретракторов — двух ветвей колумеллярного мускула, — она имеет не-
сколько ретенторов, соединяющих ее с передней стенкой тела, или протрак-
торов, соединяющих глотку со стенками шеи (рис. 38, 39). Влагалище ра-
дулы очень узкое и располагается на верхней стороне задней части глотки.
Мощный одонтофор (язык) сверху по продольной оси имеет глубокий желоб,
в котором располагается радула (рис. 12, 37). Она слагается из кинжало-
видных или багоровидных зубов, размеры которых плавно нарастают от
дна желоба к середине его склонов и затем также плавно уменьшаются
к верхнему краю желоба. Челюсть слабо развита, очень тонкая, гладкая,
без срединного выступа; у некоторых видов она отсутствует.
Колумеллярная система ретракторов имеет тенденцию к распаду на само-
стоятельные мускулы, которые задними концами прикрепляются к стенкам
тела независимо друг от друга и вдали от мантийного комплекса (рис. 38).
Ретрактор правого глазного щупальца перекрещивается с гениталиями
(исключение — Selenochlamys).
Для мозга характерна очень длинная церебральная комиссура, что свя-
зано с особенностями питания этих слизней (рис. 17, VIII, с. 47). Нередко
также церебро-педальные и церебро-плевральные коннективы относительно
длиннее, чем в других семействах.
Половой атриум узкий и длинный, пучками мускулов связан со стенкой
тела. Женские половые пути без придаточных органов (исключение — Troglo-
lestes). Пенис цилиндрический, в различной степени извитый, без папиллы.
Сзади он плавно переходит в эпифаллус, к которому крепится половой рет-
рактор. Внутри пениса нередко имеются различные складки; к некоторым
из них крепятся сперматофоры (у тех видов, которые ими обладают). Осно-
вание пениса окружено мускулистым чехлом. Семяпровод подтянут к осно-
ванию пениса кольцевой связкой.
Распространение. Лесные и альпийские области Кавказа,
Понтийских гор (Малая Азия) и Эльбурса (северный Иран).
Хищные слизни, ведущие подземный или полуподземный образ жизни
и питающиеся почвенными, преимущественно дождевыми червями.
По-видимому, если не все, то некоторые виды этой группы имеют мно-
голетний жизненный цикл с несколькими периодами размножения. В пользу
этого говорит то, что даже небольшие слизни, которых можно принять за
20*
308 I. TRIGONOCHLAMYDIDAE
деполовозрелую молодь, имеют полностью сформированные гениталии, а у не-
которых в семяприемнике встречен сперматофор, что указывает на имевшую
место копуляцию. В частности, это отмечено у Trigonochlamys imltatrix О.
Bttg. и у Hyrcanolestes velitaris (Mart.).
Согласно Зимроту, семейство состоит из 7 родов и 21 вида. Позднее был
описан новый монотипический род Troglolestes. Наибольшим числом видов
обладали роды Trigonochlamys, Hyrcanolestes и Pseudomilax (=Drtlolestes).
Отмечая большое сходство в строении гениталий видов одного рода, Зимрот
придавал неоправданно большое значение различиям в форме тела и в окраске.
Исследованный нами материал показывает, что даже в пределах одной
выборки при жесткой фиксации слизни могут приобретать различную форму,
которая зависит от того, насколько энергично сократился тот или иной
экземпляр. Последнее в свою очередь определяется физиологическим со-
стоянием животного: слизни, находившиеся в активном состоянии, сокра-
щаются более интенсивно, чем пребывавшие в депрессии из-за перегрева
и асфиксии.
Что касается окраски, то она зависит от возраста слизней. Как правило,
молодые окрашены светлее, нередко почти белые, а взрослые отличаются
развитием темного пигмента. В строении гениталий различия между многими
зимротовскими «видами» очень нёзначительны и, если имеются, то опять-
таки связаны с возрастом.
Согласно нашим данным, семейство Trigonochlamydidae состоит из 6 моно-
типических родов, т. е. на 2 рода меньше, чем считалось до сих пор: род
Chrysalidomilax Simr. мы считаем младшим синонимом рода Hyrcanolestes Simr.,
а род Phrixolestes Simr. синонимом Trigonochlamys О. Bttg.
Столь необычная ситуация, когда целое семейство состоит из одних
/монотипических родов, говорит о большой древности этой группы (с. 95).
В то же время не последнюю роль в становлении монотипии имел и подземный
образ жизни этих животных, при котором они очень редко появляются на
поверхности почвы, а троглобионт Troglolestes sokoloyi вообще не может
выйти за пределы своей пещеры,
Все роды данного семейства четко различаются по размерам и положению
мантии и мантийного комплекса органов, а также по размерам и строению
глотки. Особенно велики отличия у рода Selenochlamys О. Bttg., что дало
нам основание выделить его в подсемейство Seleno chlamy dinae (с, 97), четко
отличающееся по целому комплексу признаков от номинативного подсемей-
ства (рис. 40), куда относятся остальные 5 родов.
Мы не разделяем точки зрения Гессе (Р. Hesse, 1926), который предложил
подразделение данного семейства на Trigonochlamydinae и Parmacelltllinae.
По его мнению, последняя группа, куда он относил только род Рагта-
cellilla Simi., характерна тем, что «животное внешне сходно с Parmacella.
Эмбриональная раковина видна снаружи. Мантийная борозда слабо выра-
жена» (с. 54; перевод с немецкого наш., — Л. и В.).
Что касается сходства с Parmacellidae, то оно весьма относительно и огра-
ничивается тем, что в обеих группах нуклеус раковины можно рассмотреть
сквозь щель позади мантии. В остальном даже внешний облик, не говоря
о внутреннем строении, Parmacellilla очень отличен от Parmacellidae и харак-
терен для большинства Trigonochlamydidae. В пределах семейства у неко-
торых родов (Trigonochlamys, Hyrcanolestes) сосцевидный нуклеус раковины
близок по форме таковому у Parmacellilla. Что касается мантийной (подково-
видной) борозды, то и у некоторых других родов семейства она не всегда хо-
рошо выражена (Selenochlamys). Следовательно, Parmacellilla не имеет
столь крупных отличий от остальных членов семейства, чтобы заслуживать
выделения в особое подсемейство.
1: TRIGONOCHLAMYS
309
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ TRIGONOCHLAMYDIDAE
1 (2). Мантия очень большая и закрывает почти всю верхнюю сторону
животного: при движении она оставляет открытой только небольшую
часть шеи, голову и задний конец ноги; при сокращении она полностью
закрывает шею и голову . ...............4. Troglolestes Ljov. et Mat.
2 (1). Мантия значительно меньших размеров, и даже при сильном сокра-
щении животного длина её не более 1/3 общей длины тела; при сокра-
щении слизня она не закрывает шею и голову.
3 (4). Мантия лежит у заднего конца тела, длина ее в 15 раз меньше общей
длины. Глотка очень крупная и занимает почти всю полость тела . .
. ........................................6. Selenochlamys О. Bttg.
4 (3). Мантия лежит около середины или на середине тела, длина ее в 3—
5 раз меньше общей длины. Глотка занимает не более х/з полости тела.
5 (8). Передний край мантии с нешироким, но четким капюшоном, основа-
ние которого находится на уровне переднего конца подкововидной
борозды или даже немного позади нее.
!6 (7). Шея и бока ниже мантии покрыты крупными кольцевыми валиками,
придающими этой части слизня сходство с дождевым червем. Спина
покрыта сеткой борозд и полигональными морщинами. Вдоль глотки,
несколько выше периферии этого органа, тянется один ряд протракто-
ров, огибающий его сзади.......................5. Drilplestes Lindh.
7 (6). Кольцевых валиков нет. Вся верхняя сторона слизня (кроме мантии)
покрыта сеткой из глубоких борозд, очерчивающих полигональные
морщины. Глотка с двумя продольными рядами протракторов, кото-
рые впереди переходят на верхнюю сторону этого органа . . . . . .
. ..........................................2. Hyrcanolestes Simr.
8 (5). Передний край мантии с неясным и очень узким капюшоном, осно-
вание которого находится перед подкововидной бороздой.
*9 (10). При сокращении спина очень короткая, вдвое меньше длины мантии.
Между спиной и мантией имеется щель, сквозь которую виден нуклеус
раковины. Мантия покрыта крупными и плоскими полигональными
морщинами, несущими мелкие сосочки. Подкововидная борозда слабо
выражена — видна только ее правая ветвь. Раковина с крупным по-
лым нуклеусом, имеющим вид лесного ореха; внутрь его заходит
участок мантии.................................3. Parmacellilla Simr.
10 (9). При сокращении длина спины равна или больше мантии. Между
спиной и мантией нёт щели. Поверхность мантии зернистая без поли-
гональных морщин и сосочков, с четкой подкововидной бороздой.
Нуклеус очень маленький, сосцевидной или клювовидной формы
без полости внутри........................1. Trigonochlamys О. Bttg.
1. Подсемейство TRIGONOCHLAMYDINAE Р. Hesse, 1882
Мантия лежит около середины тела; длина ее в 4—5 раз меньше общей длины;
лишь у одного рода — Troglolestes Ljov. et Mat. — мантия закрывает почти
все тело. Глотка занимает не менее х/4 полости тела. Деторсия мантийного
комплекса менее 90°. Желудочек сердца находится сзади предсердия. Мозг
располагается вокруг пищевода. Ретрактор правого глазного щупальца
перекрещивается с гениталиями. Семяпровод впадает в эпифаллус апикально
или субапикально.
1. Род TRIGONOCHLAMYSJO. Boettger, 1881
Boettger О., 1881 : 173 (Pseudomi'lax, non О. Boettger, 1884), 176; Simroth,
1891b : 57; 1892a : 45; 1901 : 212, 214, 222 {Phrixolestes, syn. nov.); H e s s e P., 1926 : 48;
.Лихарев, Рам м e л ь м e й e p, 1952 : 363, 366 (Phrixolestes)-, Акрамовский,
1976 : 210.
310
I. TR1G0N0CHLAMYDIDAE
Рис. 389. Trigonochlamys imitat-
rix 0. Bttg. В движении.
Типовой вид — Trigonochlamys imitatrix О. Boettger, 1881; по1
монотипии.
При движении тело почти цилиндрическое, стройное, к концам слегка
суженное, сзади заостренное (рис. 389). Спина с четким килем. Мантия лежит
немного позади середины тела (шея длиннее спины), округло-треугольная,
с четкой подкововидной бороздой. Капюшон •
очень узкий, основание его располагается
перед подкововидной бороздой. При силь-
ном сокращении (рис. 390) тело резко взду--
вается, мантия становится короче и шире,
почти круглой, но с четким углом впереди;
отношения между длиной мантии, шеи и
спины остаются прежними; задний конец
ноги обычно слегка выступает из-под спины.
Рельеф кожи слагается из бороздок, ра-
диально расходящихся от мантии, и выпук-
лых морщин. Последние при сокращении
слизня могут распадаться на вторичные мор-
щины, менее резкие и разделенные неглубо-
кими бороздками. Между килем и передним
концом мантии 13—15 рядов морщин, между
пневмостомом и килем 4—5 рядов.
Раковина с острым выступающим кверху
нуклеусом и плоской спатулой (рис. 391, 392).
Крупная бочковидная глотка (у сократив-
шихся экземпляров длина ее около 1/2 длины
тела) сверху у переднего края имеет 6—8
ретенторов (рис. 39, /); протракторы отсут-
ствуют. Зубы радулы кинжаловидной формы
(рис. 12, Г). Челюсть тонкая, узкая, полу-
круглая.
Почка ^-образно изогнута и охватывает
сердце с правой стороны (рис. 6, VII), моче-
точник заканчивается небольшим мочевым
пузырем. Легочная сеть сосудов хорошо раз-
вита и располагается не только на потолке
легочной полости, но и на диафрагме.
Оба ретрактора глотки вместе с ретрак-
торами губных щупалец сливаются в еди-
ный колумеллярный мускул, крепящийся
за мантийным комплексом (рис. 38, III).
Ретракторы глазных щупалец не входят
в состав колумеллярной системы, и каждый
крепится к соответствующей стенке тела
у края ноги.
Семяприемник открывается в женские половые пути, поэтому влагалище
отчетливо выражено (рис. 395). Пенис крупный цилиндрический, прямой
или ^-образно изогнутый. Сзади он плавно переходит в короткий эпифаллус,
в который апикально открывается семяпровод. В этом месте к эпифаллусу
крепится половой ретрактор, другим концом прикрепляющийся к стенке
шеи или к диафрагме. Передняя половина пениса окружена мускулистым
чехлом, передний край которого срастается с пенисом, а задний остается сво-
бодным. Перед впадением в атриум пенис очень сужается.
Внутри пениса имеются 1—2 продольных валика, а в задней части не-
сколько крупных бугорков, к которым прикрепляются сперматофоры
1. TRIGONOCHLAMYS
311
фрис. 396). Последние имеют вид светло-коричневых лент, каждая с коротким
•стеблем, которым сперматофор крепится к пенису.
Монотипический род.
Рис. 390—396. Trigonochlamys imitatrix О. Bttg.
390 — при сокращении (из: Simroth, 1901); 391 — раковина сверху; 392 — она же сбоку; 393 — зубы
радулы (из: Hesse, 1882); 394 — кишечник сбоку; 395 — гениталии; 396 — вскрытый пенис с тремя
сперматофорами (394—396 — окр. Батуми, 8 IV 1957).
Замечания. В своей монографии Зимрот (Simroth, 1901) описал
новый род Phrixolestes Simr. При этом он отметил большое сходство этой
труппы с родом Trigonochlamys как во внешнем облике, так и во внутреннем
строении. Главными особенностями нового рода автор считал: тело при со-
312
I. TRIGONOCHLAMYDIDAE
кращении более стройное, покрыто более мелкими морщинами, нога не
выступает из под спины; внутри заднего конца пениса на продольном ва-
лике сидит пирамидальное известковое тельце, играющее роль стимулятора.
В распоряжении Зимрота были только два экземпляра. Один был найден
в Аджарии по дороге с перевала Хуло в Батуми; другой близ Трабзона
(Понтийские горы, Турция). Вскрыт был только первый экземпляр. Обна-
ружение у него внутри пениса известкового тельца стало главным доводом
в пользу описания нового рода и двух видов — Phr. adsharicus и Phr. роп-
ticus. ' i.
На наш взгляд, так называемый стимулятор Phr. adsharicus — это инородное
тело (возможно песчинка), случайно попавшее внутрь пениса, может быть
во время копуляции. Остальные же отличия от Trigonochlamys уклады-
ваются в обычную индивидуальную изменчивость или связаны с физиологи-
ческим состоянием слизней во время фиксации.
Следовательно,. название Phrixolestes является младшим синонимом
Trigonochlamys, а оба вида — Phr. adsharicus и Phr. ponttcus — синонимы
Tr. imitatrix.
Trigonochlamys imitatrix 0. Boettger, 1881 (рис. 389—396)
Boettger 0., 1881 : 174, T. 7, Fig. 3 (? Pseudpmilax lederi), 177,, T. 7, Fig. 5;
Hesse P., 1882 : 29, T. 2, Fig. 1; Boettger .0., 1883 140; 1886b : 126 (semiplumbeus,
syn. nov.); Retowski, 1889 : 226 (boettgeri, syn. nov.);' S imr о th, 1901 : 214—221,
T. 22, Fig. 2, T. 23, Fig. 1—6; 220, T. 22, Fig. 3, T. 23, Fig. 7-8 (boettgeri); 221, T. 22,
Fig. 1, T. 23, Fig. 9—19 (minor, syn. nov.); 222 (lederi)-, 224, T. 22, Fig. 4, T. 24, Fig. 1—15
(Phrixolestes adsharicus, syn. nov.); 225, T. 22, Fig. 5 (Phr. ponticus, syn. nov.); 1912a : 64,
T. 4, Fig. 53 (armeniaca, syn. nov.); 65, T. 4, Fig. 54, T. 8, Fig. 35 (pontica, syn. nov.); 68-
(distans, syn. nov.); Лихарев, Раммельмейер, 1952 : 364, рис. 291; 365, рис. 292
(pontica); 365 (armeniaca); 366 (distans); 366, рис. 293, 294 (Phrixolestes adsharicus); Акра
ио век ий, 1976 : 210, рис. 101А, табл. 12, рис. 129 (armeniaca).
Locus typicus; «Kutais» — Кутаиси.
Голотип хранится в Зенкенбергском музее (Франкфурт-на-Майне, ФРГ,
инв. № 155442)/ Кроме того, нами исследованы типовые экземпляры и уста-
новлены лектотипы следующих синонимов (все из ЗИН АН СССР): Tr. boet-
tgeri Retowski, 1889 — 3 экз. (лектотип, дл. сок. 30 мм); Tr. distans Simroth,
1912 — 2 экз. (лектотип, дл. сок. 17 мм); Tr. pontica Simroth, 1912 — 1 экз.
(лектотип, дл. сок. 12 мм).
Материал: из СССР — 60 экз. из 31 пункта; из Турции — 3 экз.
(типы Tr. boettgeri Retowski).
Окраска зависит от возраста слизней. Взрослые экземпляры чаще всего,
синевато-церные, только подошва светлее: боковые доли темно-серые, сре-
динная почти белая. Молодые слизни окрашены светлее — от светло-корич-
невого до серовато-бурого с рисунком из мелких черных точек, особенно-
густо лежащих на спине и более редко на боках. Точки располагаются глав-
ным образом по бороздкам и по краям мантии. Киль нередко несколько свет-
лее спины.
Размеры: голотип — дл. сок. 32, ширина 12.5, высота 14 мм; наибо-
лее крупный из измеренных экземпляров при движении имел длину 94,
ширину 10, длину шейного отдела (прозоподиум) 40, длину спины 37, длину
мантии 17 мм. После жесткой фиксации он сжался до 38 мм.
Распространение. В СССР: Грузия — лесная зона Аджаро-
Имеретинского и Шавшетского хребтов, южные склоны Рачинского хребта
(окрестности Кутаиси, Ткибули); Армения — субтропические микроучастки
Мегринского и Кафанского районов (Акрамовский, 1976 — Tr. armeniaca
Simroth); Азербайджан — внешние склоны Малого Кавказа (Ханларский,-
Шамхорский р-ны), одна находка в окрестностях Кусары (северо-восток
2. HYRCANOLESTES
313
АзССР). Вне СССР: Понтийские горы в окрестностях Ризе (Retowski,
1889 — Tr. boettgeri) и Трабзона (Simroth, 1901 — Phrixolestes ponticus).
Одна находка сделана в зимнем саду гостиницы «Ленинград» (Ленинград),
куда этот вид был завезен вместе с растениями.
Образ жизни. Обитает в субтропических лесах, во влажной лист-
венной подстилке, под камнями. и валежником. В пасмурную влажную
погоду выползает наружу.
Замечание. Целый ряд видов, описанных разными авторами
(см. выше), мы склонны рассматривать как синонимы данного вида. Все раз-
личия между ними в основном лишь в окраске, которая, как мы уже отме-
чали, меняется с возрастом. В наших сборах и сборах А. Риделя из Батум-
ского ботанического сада можно обнаружить все цветовые переходы между
перечисленными' видами и настоящими Tr. imitatrix.
2. Род HYRCANOLESTES Simroth, 1901
Boettger О., 1881 : 173 (Pseudomilax part.); Simroth, 1901 : 212, 226;
1912a : 75 (Chrysalidomilax, syn. nov.); H e s s e P., 1926:49; Лихарев, Р.аммель-
мейер, 1952 : 366; Ш и л e й к о, 19676 : 29 {Trigonochlamys)-, Акрамо вский,
1976:210.
Типовой вид — Parmacella velitaris Martens, 1880; по последую-
щему обозначению — Lindholm, 1925.
При движении тело цилиндрическое, узкое, у переднего конца закруг-
ленное, сзади — заостренное. Мантия удлиненно-овальная, длина ее около
х/4 длины тела, сзади округло-притупленная, спереди заостренная; капюшон
более широкий, чем у Trigonochlamys, но уже, чем у Drilolestes, — основание
его на уровне переднего края подкововидной борозды. Мантия несколько
смещена назад от середины тела, однако спина всегда заметно длиннее шеи.
Киль доходит до мантии, но отчетливо выражен только в задней части спины.
Подошва умеренно узкая.
При сокращении тело становится веретеновидным, т. е.< вздутым в сред-
ней части и плавно суженным к концам, или короткоцилиндрическим —
к концам резко суженным. Спина сжата с боков и с острым килем. Мантия
становится короткоовальной и лежит почти на середине тела. Шея по длине
равна спине или немного длиннее. Задний конец ноги обычно не выступает
из-под спины (рис. 397—400).
Тело покрйто цолигональными морщинамй, не столь выпуклыми, как
у Trigonochlamys, но более крупными.
Раковина очень тонкая, с чётким сосцевидным нуклеусом у заднего края
(рис. 401, 402). Спатула к краям становится очень тонкой, нередко распада-
ется на отдельные известковые зерна, связанные периостракумом.
Глотка (рис. 405) бочковидная (у сократившихся экземпляров длина
ее около х/4'длины тела), без ретенторов, но с многочисленными протракто-
рами, располагающимися в два более или менее регулярных продольных
ряда, которые впереди переходят на верхнюю сторону глотки. Основные
ретракторы глотки (т. е. ветви колумеллярной системы) крепятся к ней у ее
переднего конца. Зоб очень крупный, кишка дает два изгиба. Зубы кинжало-
видные (рис. 37), в пределах одного поперечного ряда около 40 зубов. Че-
люсть тонкая, дуговидная (рис. 9, НТ).
Мантийный комплекс сходен с Trigonochlamys, но дыхательная сеть лег-
кого ограничена только потолком легочной полости (рис. 406, 407).
Из состава колумеллярной системы ретракторов выпали ретракторы губ-
ных щупалец (отличие от Trigonochlamys), которые задними концами кре-
пятся к боковым стенкам тела (рис. 38, II). Общий ствол ретракторов глотки
и глазных щупалец сзади прикрепляется у заднего края мантийного комп-
лекса. Мозг с очень длинной церебральной комиссурой (рис. 17, VIII).
314
1. TRIGONOCIILAMYDIDAE
Пенис очень длинный, цилиндрический, сильно извитый или сзади спи-
рально скрученный (рис. 408—411). Короткий эпифаллус плавно переходит
в пенис. Семяпровод впадает в эпифаллус апикально или немного отступя от
его заднего конца. В отличие от Trigonochlamys, чехол пениса прирастает
Рис. 397, 398. Hyrcanolestes velitaris (Mart.) Окр. Ленкорани, 5 V 1964.
к последнему как передним, так и задним краями. Перед впадением в атриум
пенис сужается. От чехла отходит тонкая кольцевая связка, подтягивающая
семяпровод к пенису.
Внутри заднего конца пениса, близ входа в эпифаллус, имеется крупный
выступ, к которому крепятся 1—2 сперматофора. Передний отдел пениса
внутри имеет многочисленные мелкие бугорки.
Монотипический род.
Hyrcanolestes velitaris (Martens, 1880) (рис. 397—411)
Martens, 1880 : 154 (Parmacella)', Boettger О., 1881 : 175, T. 7, Fig. 4
(Pseudomilax bictflor, syn. nov.); 1889 : 932 (Pseudomilax)', Simroth, 1901 : 229, T. 22,
Fig. 9, T. 25, Fig. 1—11; 229, T. 22, Fig. 8, T. 26, Fig. 1—6 (valentini, syn. nov.); 230 (bi-
color)', 232 (Pseudomilax bicolor)', 1910a : 535, T. 7, Fig. 35—37 (armeniacus, syn. nov.);
1912a : 68, T. 4, Fig. 56, T. 9, Fig. 36 (kaznakovi, syn. nov.); 70, T. 4, Fig. 57 (varius, syn.
2. HYRCANOLESTES
31
nov.); 70, T. 9, Fig. 37 (obscurus, syn. nov.); 71, T. 4, Fig. 59, T. 9, Fig. 38 (Jursovi, syn.
nov.); 72, T. 4, Fig. 60, T. 9, Fig. 39 (declivis, syn. nov.); 76, T. 4, Fig. 63, T. 9, Fig. 41,
T. 10, Fig. 42 (Chrysalidomilax sphingiformis, syn. nov.); Акрамовский, 1947 : 271
(armeniacus)', Лихарев, Раммельмейер, 1952 : 368, рис. 296 (kaznakovi)',
368, рис. 297 (varius)', 369 (obscurus): 369, рис. 295, 298 (valentini)', 369 (armeniacus)', 370
(fursovi); 370, рис. 299 (declivis)', 371, рис. 300, 301 (Chrysalidomilax sphingiformis)', HI fl-
fl e й к о,- 19676 : 29, рис. 5, 6 (Trigonochlamys bicolor, Tr. sphingiformis)', Акрамов-
ский, 19716 : 9; 9 (armeniacus); 1976 : 211, рис. 101Б, 101В, табл. 12, рис. 128 (armeni-
acus).
Рис. 399—402. H.yrcanolestes velitaris (Mart.).
-399 — лектотип H. declivis Simr., гора Фитдаг, 8 V 1909; 400 — синтип Chrysalidomilax sphingiformis
Simr., окр. Ленкорани, 6 III 1904; 401 — раковина сверху; 402 — она же сбоку (399, 400 — из: Sim-
roth, 1912а).
Locus typicus; «Astrabad» — Астрабад (ныне Горган), северный Иран.
Следующий типовой материал по данному виду и его синонимам имеется
в ЗИН АН СССР (лектотипы установлены нами): Н. velitaris Martens — 2 экз.
(за лектотип избран экземпляр, изображенный у Simroth, 1901, Т. 22, Fig. 9);
Н. declivis Simroth — 2 экз. (изображение лектотипа см.: Simroth, 1912а,
Рис. 403—407. Hyrcanolestes velitaris (Mart.).
403 — кишечник сверху; 404 — он же сбоку; 405 — глотка справа; 406 — мантийный комплекс сверху;
407 — он же снизу.
2. HYRCANOLESTES
317
T. 4, Fig. 60 a, с); H. fursovi Simroth — 2 экз. (лектотип см: там же, Т. 4,
Fig. 58 b—d); Н. kaznakovi Simroth — 7 экз. (лектотип см.: там же, Т. 4,
Fig. 56 a, b); Н. obscurus Simroth (голотип); Н. varius Simroth (голотип).
Рис. 408, 409. Hyrcanolestes velitaris (Mart.).
408 — гениталии, окр. Ленкорани, 22 III 1965; 409 — передняя часть пениса
и пениальный чехол.
Типы остальных видов не сохранились, в том числе в коллекции Зен-
кенбергского музея нет типов Pseudomilax bicolor О. Bttg. и Hyrcanolestes
valentini Simr., хотя Зимрот указывал, что они хранятся там.
Материал: из СССР — 69 экз. из 32 пунктов; из Ирана — 2 экз.
Окраска чаще всего темно-серая или синевато-серая, реже почти черная
или бурая. При сокращении тела на растянутых участках боков окраска
светлеет. Глазные щупальца всегда серые или черные. Подошва белая или
желтоватая. Борозды обычно темнее морщин. Иногда на мантии и прилегаю-
318
1. TRIGONOCHLAMYDIDAE
щих к ней участках тела имеются мелкие темные пятнышки или точки.
Обычно передний конец тела светлее заднего. Молодые слизни окрашены
светлее взрослых — от светло-серых до синевато-серых.
Рис. 410, 411. Hyrcanolestes velitaris (Mart.).
410 — гениталии, окр. Ленкорани, 5 V 1964; 411 — вскрытый пенис.
Размеры. Из измеренных при жизни экземпляров наиболее круп-
ный имел: дл. пол. 70, ширину 5.5, длину мантии 17, длину шеи 23.5, длину
спины 30 мм. Он же после фиксации: дл. сок. 26.8, ширина 8, длина мантии
8 мм.
Кроме указанных в описании рода особенностей раковины, она харак
терна еще присутствием у правого заднего угла своеобразного выроста,
который чуть намечен у молодых слизней, но с возрастом постепенно уве-
личивается (рис. 401, 402). На нем даже видны кольца нарастания, анало-
3. PARMACELLILLA
319'
гичные таким же на спатуле. Он обнаружен у представителей разных по-
пуляций. Функциональное значение этого образования неясно.
Размеры раковины: у взрослого слизня при дл. сок. 28 мм длина
раковины около 3 мм.
Распространение. В СССР: леса Ленкоранской низменности
и ТалЫша, северные и восточные отроги Малого Кавказа, южные отроги
восточной части Большого Кавказа; наиболее западная находка сделана
у Боржоми; наиболее северные у Телави, Лагодехи и Закаталы. Вне СССР:
северные склоны Эльбурса (северный Иран).
Образ жизни. Обитает во влажной лесной подстилке и в верхних
слоях почвы. Наиболее обычен для лесной зоны и реже встречается в ниж-
ней части альпийской зоны. Питается дождевыми червями (Шилейко, 1966).
3. Род PARMACELLILLA Simroth, 19101
Simroth, 1910а : 536; Hesse Р., 1926 : 54.
Типовой вид — Parmacellilla filipowitschi Simroth, 1910; по мо-
нотипии.
При сокращении тело становится веретеновидным, к голове плавно,,
к заднему концу круто суженным (рис. 412). Подошва очень узкая. Мантия
располагается на задней половине тела, неправильно округлая впереди,,
с очень узким капюшоном (около 1 мм). Подкововидная борозда слабо выра-
жена. Пневмостом находится вблизи от заднего края мантии. Спина корот-
кая — длина ее меньше длины мантии, но больше чем у Selenochlamys.
Вдоль спины тянется мощный киль, который впереди как бы врезается
в мантию; в этом месте имеется щель, сквозь которую вйден нуклеус внутрен-
ней раковины. Вся верхняя сторона тела, в том числе и мантия, покрыта
крупными и плоскими полигональными морщинами, придающими слизню-
чешуйчатый облик. Каждая морщина несет мелкие сосочки, сходные с та-
ковыми на мантии Troglolestes. Затылочные бороздки широко расстав-
лены.
Нуклеус раковины очень своеобразной формы, напоминающий лесной
орех (рис. 415). В отличие от Parmacellidae, у которых нуклеус образован,
одним оборотом спирали, в данной группе эта часть раковины сохранила
лишь намек на былую спиральную закрученность — не более х/2 оборота.
Спатула плоская, не больше нуклеуса. Стенки последнего имеют характер-
ную микроструктуру, состоящую из правильных рядов мелких светлых
точек. Внутрь нуклеуса заходит участок мантии.
Задний конец тела, т. е. спина, заполнен дорсовентральной мускулату-
рой, и сюда не проникают внутренности.
Кишечник обычный для семейства, т. е. с крупным зобом и короткой,
слабо изогнутой кишкой (рис. 417). У единственного изученного экземпляра
Зимрот обнаружил довольно крупный конический слепой выступ на зобе
(до впадения протоков печени), и он был склонен его гомологизировать с кри-
сталлическим стебельком низших Prosobranchia и Bivalvia. Однако положе-
ние этого выступа скорее дает основание рассматривать его как новообра-
зование или аномалию.
Цилиндрическая глотка занимает около х/3 внутренностного мешка или
*/4 общей длины тела (рис. 39, V). От ее боков отходят два регулярных ряда.
1 Характерстика этого рода составлена на основании анализа оригинального опи-
сания и дополнительного исследования голотипа единственного вида.
320
1. TRIGONOCHLAMYDIDAE
протракторов, связывающих ее с верхней стенкой шеи. Короткая радула
плавно сужается назад и заканчивается в узком радулярном влагалище,
плотно прижатом к задней половине глотки. Зубы багоровидные (рис. 416).
Челюсть сравнительно крупная.
Мантийный комплекс органов вслед за мантией сильно смещен назад.
Кровеносная сеть легкого хорошо развита, но ограничена лишь потолком
легочной полости.
Колумеллярная система ретракторов (рис. 38, II) отличается от типичной
для других семейств слизней только тем, что ретракторы губных щупалец
не сливаются с остальными и сзади крепятся к боковым стенкам тела. Кроме
того, они посылают по одной дополнительной ветви к глотке.
Пенис цилиндрический, длинный, спирально закрученный (рис. 418,
419). Сзади апикально в него впадает семяпровод и крепится половой ретрак-
тор, который задним концом соединен с верхней стенкой тела у переднего
края мантийного комплекса. Передняя часть пениса окружена мускульным
чехлом и кольцевой связкой с семяпроводом. Внутренние стенки пениса
имеют продольные складки и бугорок, к которому крепится сперматофор.
Атриум длинный и узкий; сзади в него впадают короткий яйцевод, семя-
приемник и пенис.
Монотипический род.
Parmacellilla filipowitschi Simroth, 1910 (рис. 412—419)
Simroth, 1910а : 536, Т. 8, Fig. 38—50; Starmiihlner, Edlau er,
1957 : 471 {philtppowitscht Simr., err.).
Locus typicus: «Astrabad» — Астрабад (ныне Горган), северный Иран.
Голотип в ЗИН АН -СССР. Другой экземпляр, найденный австрийской
экспедицией в Иране и определенный как данный вид (Starmiihlner, Edlauer,
1957), согласно письму хранителя коллекции моллюсков Венского естест-
венноисторического музея д-ра О. Паджета, не сохранился.
Основная (фоновая) окраска верхней стороны тела голотипа темно-бурая
с красноватым оттенком, подошва почти оранжевая. В бороздках сосредото-
чен тонкий темный пигмент, что делает их особенно четкими. Красноватые
тона окраски тела, а также некоторых внутренних органов (печени, белковой
железы, простаты), вероятно, обязаны особенностям фиксатора; по-видимому,
исходно тело было белое с легким серым налетом.
Размеры голотипа: при дл. сок. 16.5 мм, ширина 6.5, высота 7,
длина мантии 6, длина спины 3.5, ширина подошвы 2 мм. Согласно Зимроту,
длина нуклеуса раковины около 2 мм. В настоящее время от раковины ни-
чего не сохранилось.
Распространение. Первая находка этого вида была сделана
Филиповичем 20 IV 1905 г. у г. Астрабада, вторая — австрийской экспе-
дицией (по-видимому, тоже один экземпляр) 18 XI 1949 г. близ Каспий-
ского моря между городами Чалус и Бабольсер. Следовательно, этот вид
обитает в северном Иране у подножья северных отрогов Эльбурса.
Образ жизни. Австрийский экземпляр был найден в зарослях еже-
вики и самшита среди влажного мха и подстилки. Вероятно, данный вид
ведет полуподземный образ жизни.
Замечание. Форкарт (Forcart, 1935) отмечает еще две находки
данного вида в речных выносах Прикаспийской низменности северного
Ирана. Опубликованные в той же работе фотографии этих раковин указы-
вают на то, что автор ошибся в определении и имел дело с нуклеусами ка-
ких-то Parmacellidae.
Рис. 412—419. Рarmacellilla filipowitschi Simr. Голотип.
412 — вид сбоку; 413 — вид снизу; 414 — задний конец тела сверху; 415 — нуклеус раковины снизу;
416 — зубы радулы; 417 — кишечник; 418 — гениталии; 419 — вскрытый пенис (413—418 — из: Simroth.
1910а).
21 И. М. Лихарев, А. Й. Виктор
322
I. TRIG0N0CHLAMYD1DAE
4. Род TROGLOLESTES Ljovuschkin et Matekin, 1965 1
Левушкин, Матекин, 1965 : 36.
Типовой вид — Troglolestes sokolovi Ljovuschkin et Matekin, 1965;
по монотипии.
При движении тело булавовидно-цилиндрическое, спереди плавно, сзади
более резко суженное (рис. 420). Мантия большая и закрывает почти всю
верхнюю сторону животного, оставляя открытой часть шеи, голову и зад-
ний конец ноги; при сокращении она полностью закрывает шею и голову.
Пневмостом располагается сильно постмедиально, почти у заднего конца
мантии. Капюшон крупный, занимает переднюю четверть мантии. Вся верх-
няя поверхность мантии покрыта многочисленными крупными сосочками,
которые сохраняются даже после фиксации. Подкововидная борозда выра-
жена слабо; хорошо видна ее правая часть, которая впереди поворачивает
налево и плавно сходит на нет. Половое отверстие располагается позади
и немного ниже основания правого глазного щупальца. Щупальца короткие,
особенно губные, но массивные. Глаза'рудиментарные, с недоразвитым хру-
сталиком.
Раковина сходна с таковой у Trigonochlamys (рис. 423, 424).
Глотка занимает около 1/3 общей длины тела (у сократившихся особей),,
очень мускулистая (рис. 427). Два регулярных ряда протракторов связы-
вают ее с боковыми и верхней стенками шеи. Радула с багоровидными зу-
бами (рис. 425). В центре каждого ряда располагается один маленький
шиповидный зуб без багоровидного зубца, по бокам от него еще по одному
сходному зубу, к краям багоровидный зубец становится четче и крупнее.
Челюсть отсутствует или столь слабо выражена, что осталась незамеченной.
Легочная полость очень маленькая. Дыхательная сеть кровеносных со-
судов не обнаружена. Сердце располагается слева и впереди почки, в уг-
лублении последней (рис. 432). Ось его наклонена направо, предсердие перед
желудочком.
Колумеллярная система ретракторов тела отличается примитивным-
строением (рис. 428). Общий ствол распадается на три главные ветви — пра-
вая и левая щупальцевые, а между ними глоточная. Последняя на подходе-
к глотке разветвляется на две йетви, которые крепятся к заднему концу
глотки.
Мозг — см. рис. 430.
Для половой системы характерно присутствие на влагалище, почти
в том же месте, где впадает семяприемник, особого мускулистого придаточ-
ного органа (рис. 431). Внутри него имеется полость, на дне которой поме-
щается папилла. Имеется эпифаллус, который впереди плавно переходит
в пенис, а сзади в семяпровод.2
Монотипический род.
Troglolestes sokolovi Ljovuschkin et Matekin, 1965 (рис.'420—432)
Л e в у ш к]и н,] Матекин,] 1965 : 35,
(ДО-.
Тело снежно-белое, лишь в заднем конце слегка просвечивают розоватые
внутренние органы.
Размеры: дл. пол. до 32 мм. Ц|
1 Характеристика этого рода, как и соответствующего вида, составлена только-
на основании анализа оригинального описания и рисунков. Единственный экземпляр
Tr. sokolovi, имеющийся в коллекции ЗИН АН СССР (дар П. В. Матекина), сильно по-
страдал при фиксации.
2 В тексте оригинального описания отрицается наличие эпифаллуса, но на прило-
женном рисунке он хорошо виден.
Рис. 420 432. Troglolestes sokolovi
420 — в движении; 421
Ljov. et Mat. Воронцовская пещера.
422- молодой слизень (глотка частично
глотка; 428 — ретракторы- 429 — мЛг- л т ° у’ 425 ~ зубы РаДУ-™; 42в — кишечник; 427 —
дочкой отмечен^Ридаточный =
И IU -;-о.осгальные из: Левушкин, Матекин, 1965). 3 1 4 ориг.,
21*
324
I. TRIGONOCHLAMYDIDAE
Распространение. Воронцовская пещера близ Сочи (бассейн
р. Кудепсты), в одном из глубинных залов на рыхлом сыром глинистом
грунте. Животные ползают по поверхности почвы и периодически в погоне
за энхитреидами погружают передний конец тела в толщу грунта.
5. Род DRILOLESTES Lindholm, 1925
Boettger О., 1884 : 147 (Pseudo milax, non О. Boettger, 1881); Simroth,
1892b : 52 (Pseudolimax, non 0. Boettger, 1881); 1901 : 230 (Pseudomilax), 1912a : 73 (Pse-
udomilax): Lindholm, 1925:168 (Drilolestes nom. nov..); Hesse P., 1926:51
(Pseudomilax): W e n z, 1947 : 36 (Chlamydolimax nom. nov.); Лихарев, Раммель-
мейер, 1952 : 371 (Pseudo mt lax).
Типовой вид — Pseudomilax retowskii 0. Boettger, 1884; по монотипии.
При движении тело узкоцилиндрическое и мантия располагается позади
середины, так что шея немного длиннее спины. В результате сокращения
тело слизня становится веретеновидным, сильно вздутым в области мантии
или позади нее; при этом шейный отдел заметно длиннее спинного (рис. 433).
Спина сильно сжата с боков, а посередине ее выступает резкий и высокий
киль. В целом слизень приобретает много общих черт с Hyrcanolestes'. в рель-
ефе спины сеть крупных полигональных морщин; мантия со сравнительно
широким капюшоном, основание которого располагается на уровне перед-
ней части подкововидной борозды. Во внешнем облике главное отличие от
Hyrcanolestes это своеобразный рельеф шеи и боков ниже мантии. Он слага-
ется из крупных и выпуклых кольцевидных валиков, придающих этой
части слизня сходство с дождевым червем. Поверхность этих валиков глад-
кая, так как за исключением затылочных и боковых борозд ее не пересекают
другие продольные или косые борозды.
Раковина и мантийный комплекс органов сходны с Hyrcanolestes (рис. 439,
440).
Крупная цилиндрическая глотка занимает около 1/3 длины тела. По обеим
сторонам вдоль нее, несколько выше периферии, располагается один ряд
протракторов, связывающих ее с боковыми стенками тела; из них особенно
мощны самые задние мускульные пучки (рис. 436). Основные ретракторы
глотки (ветви колумеллярной системы) крепятся к ее заднему концу. Зубы
радулы кинжаловидные, их около 40 в поперечном ряде. Размеры зубов
плавно нарастают от середины к краям радулы и лишь 2—3 самых крайних
меньше предыдущих. Челюсть не обнаружена.
Колумеллярная система ретракторов как у большинства слизней
(рис. 38, /), т. е. ретракторы обеих пар щупалец и глотки сзади сливаются
в общий ствол, который крепится к стенке тела у заднего края мантийного
комплекса. Только ретракторы губных щупалец дают дополнительную ветвь
к стенке тела.
Мозг с длинной церебральной комиссурой при сравнительно коротких
коннективах.
Гениталии (рис. 437, 438) во многом сходны с Trigonochlamys: довольно
длинное влагалище; чехол пениса передним краем прирастает к пенису,
а задний остается свободным; внутри задней части пениса несколько круп-
ных бугорков, которые с возрастом превращаются в поперечные складки.
К ним крепятся сперматофоры. Внутренние стенки передней части пениса
имеют три продольные складки.
Монотипический род.
Drilolestes retowskii (О. Boettger, 1884) (рис. 433—440)
Boettger О., 1884 : 146 (Pseudomilax): Simroth, 1901: 231, Т. 22, Fig. 7
(Pseudomilax): 232 (Ps. reibischi, syn. nov.); 232, T. 22, Fig. 6,T.26, Fig. 7—11 (Ps. ananowi,
syn. nov.); 1912a : 74 (Ps. reibischi): 75, T. 4, Fig. 62, T. 9, Fig. 40 (Ps. orientalis, syn. nov.);
5. DRILOLESTES
325
Лихарев, Раммельмейер, 1952:372 {Ps. orientalis); 372, рис.302, 303 (Ps.
ananowi); 372; 373 (Ps. reibischi); Калягина, 1958 : 165 (Ps. ananowi); Акрамов-
ский, 19716 : 9 (Hyrcanolestes orientalis); 1976 : 211, рис. 101Г, табл. 12, рис. 130 (H. ori-
entalis).
Рис. 433—435. Drilolestes retowskii (О. Bttg.). Окр. Сухуми, 20 XI 1958.
433 — внешний вид; 434, 435 — гениталии и вскрытый пенис.
Locus typicus; «Psirsk» — Псырцха, Абхазия. Где находятся и сохрани-
лись ли типы Pseudomilax retowskii О. Bttg. и Ps. reibischi Simr. — не из-
вестно. Голотип Ps. ananowi Simr. и лектотип (обозначен нами) вместе с пара-
лектотипом Ps. orientalis Simr. — в ЗИН АН СССР.
Материал: 30 экз. из 24 пунктов.
В типичном случае окраска слагается из белого или желтоватого фона,
на котором местами располагается темный пигмент, придающий соответствую-
326
I. TRIGONOCHLAMYDIDAE
щйм участкам тела светло-, темно- или свинцово-серую окраску. Как ми-
нимум в серые тона окрашена верхняя часть шеи и середина спины (вместе
с килем). При более сильном развитии пигмента не только вся мантия, но
и большая часть поверхности спины и шеи приобретают темно-серую окраску,
которая постепенно светлеет и исчезает вблизи от ноги. Совсем молодые
Рис. 436—438. Drilolestes retowskii (О. Bttg.) (продолжение).
4зв — кишечник (звездочкой отмечено положение мозга на пищеводе); 437, 438 — дистальная часть
гениталий и вскрытый пенис (436 — окр. Сухуми, 20 XI 1958; 437, 438 — Дарьяльское ущелье у сел.
Балта, 25 VI 1964).
экземпляры (дл. сок. менее 10 мм) совершенно белые. С возрастом пигмен-
тация усиливается, особенно у популяций, обитающих в альпийской зоне.
При хранении заспиртованных слизней на свету темно-серая окраска
переходит в бурую.
Между пневмостомом и передним концом тела около 25 кольцевидных
валиков-морщин.
Размеры: при дл. сок. 35 мм (наиболее крупный экземпляр из Су-
хуми) длина шеи 8, спины 11, мантии 13 мм; ширина тела 8, подошвы 3 мм;
5. DRILOLESTES
327
длина раковины 2.8 мм. Лектотип Pseudomilax orientalis Simroth, найденный
в альпийской зоне г. Хочалдаг (близ Лагодехи, сбор Млокосевича, 6 VII
1906 г.) имеет следующие размеры: дл. сок. 16, шея 4, спина 7, мантия 6 мм;
ширина тела 5.5, подошвы 1.8 мм.
Рис. 439, 440. Drilolestes retowskii (О. Bttg.) (окончание). Мантийный комплекс.
439 — вид сверху; 440 — visa снизу.
Распространение. Лесная и альпийская зоны Большого Кав-
каза и северных отрогов Малого Кавказа к востоку от Джевахетского хребта;
наиболее западная находка сделана в окрестностях Новороссийска, наиболее
восточная — у сел. Тлярата в Дагестане. В ЗИН АН СССР имеется один
экземпляр, найденный в одном плодоводческом хозяйстве Ташкента у под-
ножья кавказского фундука, с которым, вероятно, был завезен с Кавказа.
Образ жизни. В пределах указанного ареала обитает в листвен-
ной подстилке и в верхнем слое почвы. Питается дождевыми червями.
2. Подсемейство SELENOCHLAMYDINAE Likharev et Wiktor, subfam. nov.
Мантия лежит у заднего конца тела, длина ее в 15 раз меньше общей
длины. Глотка очень крупная и занимает почти всю полость тела. Мантий-
ный комплекс испытал деторсию на 180°, в связи с этим желудочек сердца
находится перед предсердием. Мозг располагается вокруг передней части
глотки. Ретрактрр правого глазного щупальца не перекрещивается с гени-
талиями. Эпифаллус отсутствует. Семяпровод впадает в пенис сбоку.
6. Род SELENOCHLAMYS О. Boettger, 1883
Boettger О., 1883 : 141; Simroth, 1891b : 57; 1892а : 47; 1892b : 54; 1901 :
212, 213;^Н esse, Р., 1926 : 53; Лихарев, Рамме ль мейер, 1952 : 373.
Типовой вид — Selenochlamys pallida О. Boettger, 1883; по моно-
типии.
328
I. TRIGONOCHLAMYDIDAE
Тело цилиндрическое, слабо суженное к концам; при сокращении оно
равномерно вздувается и напоминает личинку мухи (рис. 441). Очень малень-
кая, круглая, полулунная или фасолевидная мантия располагается у са-
мого заднего конца животного. Подкововидной борозды на ней нет или она
слегка намечена. Затылочные борозды широко расставлены, и между ними
располагаются два ряда морщин. Боковые борозды почти параллельны за-
тылочным. Глазные щупальца короткие и массивные. Подошва очень узкая.
Между мантией и задним концом ноги пролегает короткий, но резкий киль.
Раковина очень небольшая, хрупкая, округло-ромбическая 1 (рис. 443).
Крупная веретеновидная глотка занимает бблыпую часть внутренност-
ного мешка. За исключением тонкой верхней, остальные стенки глотки очень
мускулистые. От задней половины ее отходит один продольный ряд мощ-
ных протракторов (более 20), соединяющих ее с боковыми стенками тела
(рис. 444). В продольной щели мощного одонтофора располагается радула,
состоящая из багоровидных зубов (рис. 12, IV—VI). В пределах одного
поперечного ряда около 40 зубов. Челюсти нет.
Короткий пищевод довольно быстро расширяется в длинный и объеми-
стый зоб. Последний заканчивается слабо выраженным желудком, в который
открываются протоки печени. Обе доли печени почти одинаковые по форме
и размерам. От желудка отходит кишка, которая вначале дает небольшой
изгиб, а затем почти прямая и, плавно сужаясь, простирается до ануса,
т. е. до заднего конца тела.
Мантийный комплекс органов находится у заднего конца тела и занимает
площадь, значительно бблыпую, чем мантия (рис. 6, VIII). В связи с силь-
ной деторсией желудочек сердца лежит перед предсердием (опистопневмо-
ния), а относительно крупная мешковидная почка с очень коротким моче-
точником располагается слева от сердца. Легочная сеть кровеносных сосу-
дов не обнаружена.
Колумеллярная система ретракторов тела (рис. 38, IV) полностью распа-
лась на независимые друг от дуга ретракторы щупалец и глотки. Ретрактор
правых щупалец не перекрещивается с гениталиями. Церебро-педальные
и церебро-плевральные коннективы очень длинные; церебральная комис-
сура сравнительно короче, чем у других родов семейства (рис. 444).
По строению пениса этот род резко отличается от остальных Trigono-
chlamydidae (рис. 446, 447). Семяпровод впадает в него сбоку, сильно от-
ступя от его заднего конца. В связи с этим пенис сзади заканчивается длин-
ным слепым выростом, к которому апикально крепится половой ретрактор.
При вскрытии пениса видно, что передняя часть его имеет внутри три про-
дольных валика, покрытых мелкими сосочками, а задняя часть (слепой
выступ) имеет один мощный продольный гребень, который лишь сзади дает
несколько ветвей. На этом гребне располагается крупная языковидная
складка — стимулятор. Продольный разрез пениального гребня показал,
что семяпровод пронизывает его до стимулятора, у основания которого он
открывается в полость пениса. Никаких следов сперматофора не обнаружено
ни в пенисе, ни в семяприемнике, и, судя по строению пениса, положению
полового ретрактора и стимулятора, оплодотворение происходит путем выво-
рачивания пениса до появления наружу стимулятора, и сперма передается
в капельно-жидком виде.
Семяприемник с хорошо выраженным протоком, открывается в атриум
рядом с яйцеводом. Половой ретрактор задним концом крепится к верхней
стенке тела перед мантийным комплексом.
Монотипический род.
1 Присутствие раковины отмечается впервые. Ее легко обнаружить после мацерации
слизня.
Рис. 441 444. Selenochlamys pallida 0. Bttg. Военно-Сухумская дорога, 7 VII 1974.
441 - вид сверху; 442- тот же экземпляр сзади; 44а - раковина; 444 - кишечник сбоку (1-железа
Налепа, г - церебральные ганглии, з — подглоточные ганглии). железа
330
I. TRIGONOCHLAMYDIDAE
Selenochlamys pallida 0. Boettger, 1883 (рис. 441—447)
Boettger 0., 1883 : 142, T. 5, Fig. i;Simro th, 1892b : 54, T. 6, Fig. 9—16;
1912a : 62, T. 4, Fig. 52, T. 8, Fig. 34 (plumbea, syn. nov.); Лихарев, Раммель-
мейер, 1952 : 374, рис. 304 (pallida); 374 (plumbea).
Locus typicus; «Kutais» — Кутаиси. Голотип данного вида вместе с кол-
лекцией О. Бётгера, находился в Зенкенбергском музее (Франкфурт-на-
Рис. 445—447. Selenochlamys pallida О. Bttg. Военно-Сухумская дорога, 7 VII 1974.
445 — гениталии; 446 — продольный разрез пениса; 447 — продольный разрез пениса через гребень
(схематизированно).
Майне, ФРГ), но в настоящее время там его нет (сообщение хранителя кол-
лекции д-ра Янсена). Голотип синонима — S. plumbea Simr. — находится
в ЗИН АН СССР.
Материал: из СССР — 14 экз. из 9 пунктов; из Турции — 1 экз.
Тело молочно-белое, лишь голова, особенно глазные щупальца, а также
задний конец тела за мантией темно-серые. У некоторых взрослых экземпля-
ров между затылочными бороздами пролегает светло-серая полоса. Только
один экземпляр (голотип S. plumbea Simroth) имеет темно-серую окраску
верхней стороны тела, за исключением боков, где в связи с натяжением по-
кровов при фиксации окраска более светлая.
При сокращении слизня тело чаще всего равномерно вздувается, приоб-
ретая форму, напоминающую личинку мухи, но иногда (вероятно, из-за
3. ZONITOIDEA
331
переполненного зоба) средняя часть тела вздувается и слизень становится
вздуто-веретеновидным.
Мантия чаще всего круглая или овальная (ширина больше длины);
нередко при сильном сокращении киль врезается в задний край мантии
и она приобретает полулунную или фасолевидную форму. На ее поверхности
около пневмостома иногда виден остаток правого крыла подкововидной
борозды.
Размеры: при дл. сок. 22 мм ширина и высота тела около 6, длина
мантии 1.5, ширина 1.8 мм.
В толще мантии залегает небольшая (длина около 1 мм) округло-ром-
бическая раковина (рис. 443).
Распространение. Известно 10 находок этого вида. Первая
была сделана в окрестностях Кутаиси (голотип S. pallida О. Bttg.). Вторая
по времени (1 экз.) — между станицами Даховской и Каменномостской,
т. е. к югу от Майкопа (Simroth, 1892b). Третья близ города Ардануч, бас-
сейн р. Чорох, северо-восточная Турция (голотип S. plumbea Simr.). В кол-
лекциях ЗИН АН СССР и ЗИН ПАН (Варшава) имеются несколько сборов
из западной Грузии — окрестности Кутаиси, Гелати, Ткибули, Чихареши,
Лентехи; в Абхазии — окрестности Ткварчели и Амткел.
Образ жизни. Обитает в нижней подзоне широколиственных ле-
сов с преобладанием каштана и лещины, внутри известняковых осыпей,
обычно в богатой перегноем почве, скопившейся между камнями.^Ведет
подземный образ жизни и питается почвенными червями.
3. Надсемейство Z О NITOIDEA Morch, 1864
М[о rjc h, 1864, Vidensk. Meddel. Dansk Naturh. For., 20 : 5.
Улитки, полуслизни и слизни с различной степенью редукции раковины:
от наружной более или менее прижатой, в которую свободно втягиваются
мягкие части тела, до внутренней пластинчатой раковины, нуклеус которой
располагается на продольной оси, несколько отступя от заднего края. Зад-
ний конец цефалоподиума в различной степени заострен. Нередко на спине
имеется более или менее четкий киль. Кроме перипедальной имеется также
супрапедальная борозда и оторочка. Подошва чаще всего трехдольчатая,
т. е. с двумя продольными педальными бороздками.1
Челюсть чаще всего оксигнатная. Базальные пластинки краевых зубов
длинные, но узкие.
Гениталии обнаруживают тенденцию к усложнению за счет формирования
различных придаточных желез на влагалище и (или) атриуме, реже на яйце-
воде. Стимулятор, если имеется, то располагается внутри атриума или на
нем. Мужские половые пути чаще всего с эпифаллусом, реже вторично его
утратили. У большинства родов при копуляции сперма передается в сперма-
тофорах. .
У слизней этого надсемейства пневмостом и мантий-
ная щель всегда располагаются постмедиально. На мантии имеется подково-
видная бороздка. Подошва всегда трехдольчатая. Сердце находится у левого
края мантийного комплекса, ось его наклонена направо. Почка лежит по-
зади или немного сбоку от сердца (рис. 6, ИГ).
Растительноядные полифаги (нередко вместе с тем детритофаги и сапро-
фаги) и хищные животные.
В состав надсемейства входят улитки Zonitidae, полуслизниТЛамйейагс/г-
idae, слизни Parmacellidae и Milacidae. Число видов около 550.
1 Педальные бороздки утрачены у улиток Vitreinae (Zonitidae).
332
I. PARMACELLIDAE
I. Семейство PARMACELLIDAE Gray, 1860
Gray, 1860:268; Hesse P., 1926:30; Germain, 1930 : 111.
Крупные слизни (дл. пол. до 100 мм или немного больше) с массивным
телом (цв. табл., VI). Крупная мантия (длина ее около половины общей
длины) располагается на передней части тела, а у ползущих животных про-
стирается немного за середину тела; имеется крупный капюшон, длина
которого больше половины длины всей мантии; сверху она несет четкую
подкововидную борозду. Пневмостом сильно смещен назад. Наибольшая
высота и ширина тела приходится на участок, занятый задней частью мантии.
Боковые доли подошвы густо изрезаны тонкими продольными бороздками
и редкими, но более глубокими поперечными бороздками. Последние пере-
ходят на срединную долю, где они F-образно изогнуты назад. Между килем
и мантией имеется глубокая щель, сквозь которую виден задний конец
эмбриональной раковины.
При сокращении тело животного сильно вздувается в области мантии,
капюшон закрывает большую часть шеи, а задний конец тела лишь слегка
выступает из-под мантии (рис. 448).
Половое отверстие лежит за правым губным щупальцем.
Нуклеус (эмбриональная часть раковины) спирально закручен (около
1 оборота), янтарный, блестящий, почти прозрачный, у некоторых видов
с характерной микроскульптурой из спиральных рядов ямок, у остальных
гладкий или с очень тонкой спиральной исчерченностью. Спатула имеет
вид белой известковой пластинки с ясными линиями нарастания и
сверху покрыта тонким периостракумом, который в виде кожистой каймы
выступает за ее края. Граница между нуклеусом и спатулой всегда четко
выражена.
Внутренние органы располагаются под мантией и в переднем отделе
тела. Задний конец (участок с килем) кроме кожно-мускульного мешка
Снабжён системой косых дорсовентральных мускулов, т. е. по строению схо-
ден с задним отделом ноги улиток.
Срединный и боковые зубы радулы с 3 зубцами, краевые зубы шиловид-
ные, иногда с небольшими эктоконами. Влагалище радулы относительно
небольшое, и в нем располагается лишь незначительная часть терки.
Кишечник двухпетлевого типа (рис. 451). В связи с сильным смещением
пневмостома назад задняя кишка располагается позади остальных частей
кишечника и вытянута в направлении назад кверху. Глотка крупная.
Колумеллярный мускул крепится к задней стенке полости тела под
эмбриональной раковиной. Почти сразу он распадается на ретракторы глотки
и щупалец (рис. 452).
Сердце лежит в левой передней четверти мантийного комплекса (рис. 6,
VI)', ось его под углом более 45° наклонена направо, т. е. в сторону продоль-
ной оси тела. Аорта различной длины. Кровеносная сеть легкого распола-
гается справа от почки. Почка фасолевидная, вытянута вдоль левой стороны
комплекса. Впереди с вентральной стороны к ней прилегает гроздевидная
железа неизвестной функции. Лобуса нет. Мочеточник начинается от заднего
края почки; наружу он открывается рядом с анусом, перед пневмостомом.
Мочевого пузыря нет.
• Гонада занимает заднюю часть внутренностного мешка. От нее тянется
вперед длинный гермафродитный проток. Белковая железа очень крупная.
Простата состоит из двух частей: задней округлой, прилегающей к белковой
железе, и передней лентовидной, вытянутой вдоль семяяйцевода. Семя-
приемник открывается в яйцевод.
I. PARMACELLIDAE
333
Для женских половых путей характерно развитие на влагалище различ-
ных железистых образований, которые у некоторых видов очень сложно
устроены. Атриум небольшой.
Семяпровод переходит в веретенообразный эпифаллус, который в свою
очередь впадает в пенис. Последний четко подразделен на два отдела: зад-
ний — внутри с многочисленными мелкими сосочками, а иногда и с папиллой,
и передний, стенки которого внутри имеют продольные складки.
Половой ретрактор сзади крепится к диафрагме или к верхней стенке
переднего отдела тела, а впереди к Заднему концу пениса или, реже, к перед-
нему концу эпифаллуса. Ретрактор правых щупалец проходит между пени-
сом и женскими путями.
При копуляции сперма передается в сперматофорах (рис. 458).
Распространение (рис. 41,2). Ареал семейства состоит из двух
изолированных друг от друга частей: а) Северная Африка (от Александрии
до Марокко) вместе с Португалией и южной Испанией; б) восточное Закав-
казье, северный Иран, Копетдаг, Средняя Азия, Афганистан. Кроме того,
некоторые виды завезены на Черноморское побережье Кавказа, в южную
Францию и на Канарские острова.
Жизненный цикл и рост (с. 66). Отличительной особенностью
слизней этого семейства является наличие летней диапаузы и особых юве-
нильных образований на эмбриональной раковине (нуклеус), дающих воз-
можность молоди переносить летнюю засуху. Эти образования состоят из
провизорной «крышечки» и системы известковых складок в углу устья.
«Крышечка»1 является гомологом эпифрагмы и вместе со складками форми-
руется сразу после вылупления молоди из яиц. Особый клювовидный выступ
«крышечки» вместе с устьевыми складками обеспечивает ей полуподвижное
сочленение с раковиной.
Через несколько дней, после вылупления молодые слизни закапываются
в землю или прячутся в достаточно влажные убежища, закрывают устье
раковины «крышечкой» и впадают в спячку — летнюю диапаузу (рис. 26).
Пробуждение наступает лишь осенью с первыми прохладными дождями.
Молодые слизни вытягиваются из раковины, отбрасывают «крышечку», вы-
ходят на поверхность почвы и начинают питаться и расти. Одновременно
^начинается рост спатулы, и под первыми наслоениями ее исчезает устьевая
арматура нуклеуса.
Рост слизней идет в течение всей осени, а после зимовки продолжается
весной. Через несколько дней после пробуждения от летней спячки молодые
слизни становятся столь большими, что не в состоянии вместиться в нуклеус;
Особенно интенсивно растет передний отдел животного и мантия. Края ее
завертываются наружу, формируют внутри спатулу и постепенно закрывают
нуклеус. К концу роста в последнем остается толькр небольшой выступ пе-
чени.
Со второй половины весны и в начале лета слизни приступают к размно-
жению. Копуляции предшествуют «брачные игры». Партнеры возбуждают
друг друга щупальцами, а некоторые особыми стимуляторами, и, наконец,
обмениваются сперматоформами. Откладка яиц производится во влажные
укрытия или в специально выдолбленные углубления почвы.
Пармацеллы однолетние животные, и взрослые слизни обычно гибнут
вскоре после откладки яиц.
1 Вначале была обнаружена у некоторых североафриканских видов (Webb, Berthelot,
1835; Gerhardt, 1936с), позднее у Parmacella ibera из Талыша (Шилейко, 1967а), а затем
нами у среднеазиатских видов.
334
I. PARMACELLIDAE
ТАБЛИЦЫ ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ PARMACELLIDAE
[1. В основном по гениталиям
1 (2). Атриум с двумя коническими мускулистыми выступами, из которых
у большего вершина слегка закручена. У основания семяприемника
имеется небольшой шарообразный придаточный орган (род Parmacella
Cuvier, 1804). Вид обитает на Кавказе, в лесных областях северного
Ирана и на западном Копетдаге.................Parmacella ibera Eichw.
2 (1). Атриум без указанных выступов, вместо которых в него (или в перед-
нюю часть влагалища) впадает лентовидная плоская, более или менее
извитая железа, так называемая любовная железа. Семяприемник
без придаточных органов (род Candaharia God win-Austen, 1888). Виды
обитают в Средней Азии, Афганистане и северо-восточном Иране.
3 (6). Нуклеус покрыт спиральными рядами микроскопических ямок (видны
при 20—70-кратном увеличении).
4 (5). Эпифаллус впадает в пенис апикально. Половой ретрактор крепится
на границе между пенисом и эпифаллусом. Пенис без аппендикса . . .
...................................... Candaharia rutellum (Hutton),
5 (4). Эпифаллус впадает в пенис, отступя от его заднего конца. Половой
ретрактор крепится к вершине пениса. Пенис с крупным коническим
аппендиксом............................Candaharia izzatullaevi sp. nov.
6(3). Нуклеус гладкий, без указанной микроскульптуры.
7 (8). Нуклеус почти круглый. Эпифаллус впадает в пенис апикально. Внутри
заднего отдела пениса имеется папилла, пронизанная каналом эпи-
фаллуса ..................................Candaharia aethiops (West.).
8 (7). Нуклеус удлиненно-овальный Эпифаллус впадает в пенис, сильно
отступя от его заднего конца, т. е. сбоку. Пенис внутри без пациллы
..........................................Candaharia levanderi (Simr.).
j2. По раковинам!1
1 (4). Нуклеус имеет микроскульптуру из спиральных рядов ямок, которые
видны при 20—70-кратном увеличении.
2 (3). Указанная скульптура видна при 70-кратном увеличении...........
. .................................Candaharia rutellum (Hutton).
3 (2). Скульптура видна при 20-кратном увеличении.....................
...................................Candaharia izzatullaevi sp. nov.
4 (1). Нуклеус без спиральных рядов ямок, совершенно гладкий или с не-
ясными спиральными линиями.
5 (6). Нуклеус почти круглый (отношение длины к ширине равно 1.2) . . .
...........................................Candaharia aethiops (West.).
6 (5). Нуклеус удлиненно-овальный (отношение длины к ширине 1.7—1.8).
7 (8). Продольная ось нуклеуса перпендикулярна оси спатулы. Вид оби-
тает на Кавказе, в северном Иране и западном Копетдаге................
........................................ Parmacella ibera Eichw.
8 (7). Продольная ось нуклеуса располагается под заметным тупым углом
к оси спатулы. Вид обитает в Средней Азии, Афганистане и северо-
восточном Иране............................. Candaharia levanderi (Simr.).
1 Эта таблица имеет лишь вспомогательное значение и рассчитана на определение
хорошо сохранившихся пустых раковин рецентных видов. Более надежным является
определение по гениталиям.
1. PARMACELLA
335
1. Род PARMACELLA Cuvier, 1804
Cuvier, 1804 : 442; Webb, Berthelot, 1835 : Cl. V, Pl. 63 (Cryptella)',
Gray, 1855 : 58 (Drusia); Westerland, 1897 : 117 (Clathropodium)', Simroth,
1912b : 43 (Euparmacella)', Hesse P., 1926 : 39.
Типовой вид — P. olivieri Cuvier, 1804; по монотипии.
Внешний вид — см. характеристику семейства.
Влагалище очень крупное и сложной формы; чаще всего имеет вид фасоле-
видного органа с мощными мускулистыми и железистыми стенками, нижней
стороной приросшего к стенке тела. В атриум, кроме влагалища и пениса,
открывается крупный мускулистый придаток, внутри с двумя листовидными
складками — стимуляторами. В основание атриального придатка впадает
другой выступ, но меньших размеров чем сам придаток, который иногда
называют малым атриальным придатком.
Семяприемник с коротким мускулистым протоком, у основания которого
сидит небольшой шаровидный и мускулистый отросток с узким просветом.
Яйцевод очень короткий; у половозрелых особей он слабо заметен, так как
обычно скрыт разросшимся влагалищем.
Распространение. Северная Африка (от Александрии до Ма-
рокко), Португалия, южнай Испания, южная Франция, восточное Закав-
казье, Северный Иран, западный Копетдаг.
По-видимому, род объединяет около 5 видов.
Замечания. Изредка встречаются слизни, лишенные малого при-
датка атриума. Как показывают наши исследования Parmacella ibera Eich-
wald, 1841, а также указания Гессе (Hesse Р., 1885), касающиеся Р. valen-
ciennsis Webb et van Beneden, 1936, отсутствие этого органа следует рассмат-
ривать как очень редкое проявление индивидуальной изменчивости.
Указание Гамбетты (Gambetta, 1924), что оба придатка атриума при копу-
ляции выворачиваются, неточны. На самом деле выворачивается только
большой придаток; при этом расправляются и набухают оба стимулятора,
которые в покое лежат внутри этого придатка. Малый придаток остается
внутри тела слизня.
Parmacella ibera Eichwald, 18411(рис. 448—459)
Eichwald, 1841:199, Т. 38, Fig. 1—3; Simroth, 1883:1, T. 1 (olivieri
Cuvier); Boettger 0., 1886a : 271 (olivieri)', W esterlund; 1897 : 118 (Clathro-
podium vitrinaeforme)', Simroth, 1901 : 204, T. 20, Fig. 1, 2, 5, 24—27 (olivieri)', Ger-
main, 1912 : 16, Fig. 4 (simrothi); Лихарев, Раммельмейер, 1952 : 357,
рис. 283, 284 (olivieri subsp. ibera)', F or c ar t, 1959 : 39, Fig. 4.
Locus typicus — «Iberia» — Грузия (вероятно, Кахетия). Типы неиз-
вестны.
Материал: из СССР — более 500 экз. (в том числе голотип Clathro-
podium vitrinaeforme Westerlund) из 102 пунктов; из Ирана —134 экз.___
Окраска серовато-желтая до охристой, иногда почти черная. Рисунок
образован двумя боковыми полосами на мантии и несколькими пятнами.
Затылок, шея и спина темные.
Степень развития темного пигмента очень варьирует как в течение жизни,
так и в пределах популяции. Молодь обычно имеет четкий рисунок из полос
и пятен на мантии. С возрастом количество темного пигмента может увели-
чиваться, особенно при обитании на открытых местах (поля, огороды).
Особенно сильное развитие пигмента приводит к одноцветно черной окраске.
Значительно реже встречаются слизни без темного пигмента — одноцветно
серовато-желтые.
336
I. PARMACELLIDAE
Между мантийной щелью и килем 11—15 рядов морщин. Боковые доли
подошвы с нежной и густой продольной исчерченностью; срединная доля
значительно шире боковых.
Размеры: дл. сок. до 65, дл. мантии до 40, дл. спины до 12 мм.
Рис. 448—451. Parmacella, ibera Eichw. Сочи, IV 1962.
448 — внешний вид сократившегося слизня; 449 — раковина; 450 — зубы радулы; 451 — кишечник.
Раковина (рис. 449) широкоовальная. Спатула с ясными линиями нара-
стания. Нуклеус удлиненно-овальный, гладкий, блестящий, янтарного
цвета. Его продольная ось располагается под прямым углом к оси спатулы.
Длина всей раковины до 16.5, ширина до 10.5 мм; длина эмбриональной
части до 4.5, ширина до 2.5 мм.
Радула (рис. 450). У 4 экземпляров из Сочи следующая средняя формула:
с 356 56 13к
3- + — + — + ~ ХЮ2.
2
1. PARMACELLA
ЗЗТ
Гениталии (рис. 453—459). Гонада крупная, треугольная. Узкий
семяпровод четко отграничен от значительно более широкого цилиндриче-
ского эпифаллуса, в который он впадает апикально. Эпифаллус плавно рас-
ширяется по направлению к пенису; приблизительно на границе между его>
Рис. 452—454. Parmacella ibera Eichw. Сочи, IV 1962.
452 — ретракторы; 453 — гениталии; 454 — дистальные части гениталий при естественном положении
вид справа.
задней и средней третью он резко изогнут, так что оба участка 'прилегают
друг к другу, и связаны мелкими волокнами. Задний отдел пениса распола-
гается под углом как к переднему отделу, так и к эпифаллусу, от которого
внешне мало отличим. Передний отдел пениса сзади грушевидно вздут,
а к атриуму плавно сужен.
Половой ретрактор впереди крепится на границе между пенисом и эпи-
фаллусом, а сзади к верхней стенке тела перед мантийным комплексом.
Яйцевод относительно короткий и сжат с боков. Влагалище полностью
скрыто внутри очень крупной вагинальной железы, образующей несколько
крупных, складок с очень мощными стенками, внутри покрытыми многочислен-
ными сосочками. Одной стороной вагинальная железа серией мускульных
22 И. М. Лихарев, А. И. Виктор
338
1. PARMACELLIDAE
лент подтянута ко дну полости тела. В эту железу (точнее, во влагалище)
рядом с яйцеводом впадает очень крупный семяприемник, обладающий
коротким протоком. У основания протока находится небольшой шаровид-
ный отросток семяприемника.
Рис. 455—457. Parmacella ibera Eichw. Сочи, IV 1962.
— дистальные части гениталий при естественном положении, вид снизу; 456 — вскрытый пенис и
эпифаллус; 457 — части гениталий, вывернутые при копуляции.
В небольшой малозаметный атриум рядом с пенисом и влагалищем от-
крывается крупный атриальный придаток, имеющий вид конуса, вершина
которого нередко закручена. В основание этого органа впадает конический
выступ меньших размеров, чем придаток, но прямой. Атриальный придаток
лежит за глоткой поперек продольной оси тела и тесно подтянут многочислен-
ными мускулами к нижней стенке тела; его выступ располагается за половым
.отверстием вдоль подошвы.
Внутренние стенки задней и средней трети эпифаллуса покрыты мелкими
сосочками, а передней трети — неопределенно ветвящимися складками.
Задний отдел пениса имеет внутри характерную треугольную складку,
а остальная поверхность занята крупными сосочками, часть з- которых
прап
ibera Eichw. Сочи, IV 1962.
Рис. 458, 459. Parmacella
45$ __ сперматофор; 459 — гениталии молодого слизня.
22*
340
1. PARMACELLIDAE
выступает в просвет переднего отдела пениса. По-видимому, задний отдел
функционирует как папилла пениса. Передний отдел внутри с продольными
складками.
Внутри придатка атриума имеются две крупные языковидные складки,
у основания которых открывается выступ придатка. При копуляции эти
-складки выворачиваются наружу, набухают и играют роль стимуляторов.
Очень часто внутри резервуара семяприемника можно обнаружить очень
длинный сперматофор (рис. 458). Стенки его образованы красноватым хитино-
подобным веществом и имеют гофрированный участок. Большая часть сперма-
тофора закручена в спираль. Один из концов плавно нитевидно утоньшается
и заканчивается резким расширением — якорем. Характерно, что после
копуляции якорь сперматофора всегда находится внутри шаровидного от-
ростка семяприемника.
Распространение. Природный ареал включает западный Копет-
даг, а также горные и предгорные области восточного Закавказья (наиболее
западные находки у Телави и Лагодехи), как южные отроги Большого Кав-
каза, так и северные и северо-восточные отроги Малого Кавказа, побережье
Каспийского моря от Махачкалы до Я ламы, Ленкоранскую низменность и
Талышские горы. Вне СССР — горнолесные области северного Ирана (Эль-
бурс) и Южнокаспийскую низменность. Завезен на Черноморское побережье
Кавказа, где в настоящее время распространился в культурных биотопах
-от Сухуми до Сочи.
Образ жизни. Очень тепло- и влаголюбивый вид. Предпочитает
нижний пояс мезофильных горных лесов, однако легко расселяется по бере-
гам рек и оросительных каналов, далеко проникая как в предгорную,
так даже и в нагорную степь. В этих условиях обнаруживает отчетливую
синантропию.
Жизненный цикл. Наиболее полные данные по этому вопросу
-приведены у Шилейко (1967а) на основании трехлетних наблюдений, сде-
ланных в Талыше и Ленкоранской низменности.
После зимовки массовый выход почти взрослых слизней на поверхность
почвы начинается в феврале. До начала размножения идет интенсивное
питание и завершается рост слизней. Копуляция начинается в конце марта,
а откладка яиц — в начале мая и заканчивается в первых числах июня.
После откладки яиц слизни погибают. Количество яиц в одной кладке
колеблется от 6 до 70 штук. Яйца овальные, 4.5 X 3.6 мм.
Развитие яиц идет 5—6 недель. Вылупившаяся молодь имеет только
.эмбриональную раковину (нуклеус), вход в которую закрывает красноватая
роговая «крышечка», характерные ювенильные складки в субангулярной
части устья и соответствующие им выступы на «крышечке», обеспечивающие
ей полуподвижное сочленение с раковиной.
Сразу после вылупления молодые слизни закапываются в почву или
глубоко прячутся под различные предметы, после чего впадают в летнюю
диапаузу. В конце сентября с повышением влажности (дожди) и с понижением
-температуры слизни пробуждаются и начинают интенсивно питаться и
расти. С пробуждением «крышечка» отбрасывается. К декабрю слизни почти
достигают предельных размеров и уходят на короткую зимовку.
Хозяйственное значение. Благодаря интенсивному размно-
жению, высокой численности и прожорливости, особенно осенью и весной,
эти слизни наносят серьезный вред многим плодовым и овощным, а также
декоративным растениям. Особенно большие убытки они приносят на план-
тациях цитрусовых и чая Ленкоранской субтропической зоны.
Замечания. 1. Длительное время многие авторы (Simroth, 1901;
-О. Boettger, 1886а, 1889; Лихарев, Раммельмейер, 1952) рассматривали дан-
ный вид как вариетет или подвид Р. olivieri Cuvier, 1804. Кювье описал свой
1. PARMACELLA
341
вид по экземплярам, которые Оливье (Olivier, 1803) якобы нашел в Месопота-
мии (Ирак). По-видимому, эти экземпляры не сохранились. С тех пор не было
никаких указаний о проживании в Ираке как Р. olivieri, так и других видов
того же рода. Это дало основание Форкарту (Forcart, 1959) утверждать,
что типы Р. olivieri были найдены не в Месопотамии, а в Александрии (Еги-
пет), где Оливье тоже делал сборы. Как младшие синонимы Р. olivieri Фор-
карт рассматривает Р. alexandrina Ehrenberg, 1831 яР. festae Gambetta, 1924.
Кроме того, Форкарт, располагая 2 экземплярами Р. ibera, собранными
в Сухуми и посланными ему Лихаревым, не обнаружил у них малого атри-
ального придатка и сделали вывод, что в то время как Р. olivieri имеет два
придатка на атриуме, у Р. ibera этот орган в единственном числе.
Если оба экземпляра Форкарта не были аномальными,1 то остается только
предположить, что он не заметил второго (малого) придатка, который обычно
скрыт глубоко среди других органов.
Много раньше Форкарта аналогичную ошибку совершил Жермен (Ger-
main, 1912). Им описаны три новых вида из гирканских провинций северного
Ирана — Р. morgani, Р. pollonerai, Р. hyrcanensis. Судя по рисункам ге-
ниталий, эти виды отличаются от Р. ibera только отсутствием малого придатка
атриума. Между тем нами просмотрены гениталии 61 экземпляра Parmacella
из тех же районов. У всех по два атриальных придатка. По-видимому,
указанные три вида Жермена являются синонимами Р. ibera, так как по
остальным признакам они полностью соответствуют кавказской пармацелле,
иногда отличаясь лишь немного более крупными размерами: в северном Иране
длина слизней до 65, на Кавказе до 55 мм.
Судя по довольно хорошему рисунку в работе Кювье, Р. olivieri отличается
•от Р. ibera в основном только размерами и формой обоих придатков атриума.
Если у кавказской пармацеллы они резко различаются (рис. 455), то
у ,Р. olivieri они примерно одной длины, но один массивный конический,
а другой тонкий червеобразный. Наконец, если Форкарт прав и Р. festae
Gambetta является синонимом Р. olivieri Cuvier, то, судя по имеющимся
в ЗИН АН СССР экземплярам первого вида,2 нуклеус этого слизня имеет
четкую скульптуру из спиральных рядов точкообразных ямок (хорошо видны
даже при слабом увеличении). Такой скульптуры нет на нуклеусе Р. ibera
Eichw., который совершенно гладкий.
Учитывая сказанное, а также разделяя сомнение Форкарта в том, что
Р. olivieri Cuvier была описана из Месопотамии, мы считаем, что Р. ibera
Eichw. является самостоятельным видом, отличающимся от вида, описанного
Кювье, как морфологически, так и различным распространением. К сожа-
лению, Эйвальд описал Р. ibera только по раковине. Однако это не дает нам
оснований предлагать другое название кавказской пармацелле, как это
сделал Жермен (Germain, 1912; новое название — Р. simrothi).
2. Не раз путаницу в определении Р. ibera вызывали находки пустых
эмбриональных раковин. Они остаются от молодых слизней, погибших во
время летней диапаузы, когда у них еще не начала формироваться спатула.
Вестерлунд (Westerlund, 1897) такие раковины принял за новый род
и вид, который назвал Clathropodium vitrinaeforrne. Форкарт (Forcart,
1935) такие же раковины принял за остатки Рarmacellilla filipowitschi
Simroth, 1910, т. е. за представителя семейства Trigonochlamydidae.
1 Нами просмотрены гениталии сотен экземпляров Р. ibera. За одним исключением
все они имели по 2 атриальных придатка. Единственный экземпляр, у которого не было
этого органа, был найден в 1864 г. на острове Сара у Ленкорани. Мы внимательно изу-
чили все 4 экземпляра из того же сбора, что и 2 экземпляра, посланные Форкарту. Все
-они имели по два придатка.
2 Кстати, как и Форкарт, мы их получили от Р. Брандта из Киренаики (Ливия),
т. е. с «terra typica» Р. festae Gambetta.
342
I. PARMACELLIDAE
2. Род CANDAHARIA Godwin-Austen, 1888
Godwin-Austen, 1888 : 217; Simroth, 1912b : 43 (Pro parmacella, syn.
nov.); Godwin-Austen, 1914 : 314 (Kandaharia, syn. nov.)
Типовой вид — Parmacella rutellum Hutton,1849; по монотипии.
Внешний вид — см. характеристику семейства.
Влагалище всегда имеет форму цилиндрической или более или менее
вздутой трубки, окруженной слоем желез (вагинальная железа), нередко
образующим валик или муфту. Кроме семяприемника, во влагалище от-
крывается длинный более или менее извитый лентовидный орган с узким
просветом, который называют любовной железой (рис. 463).
Атриум без придатков; семяприемник без отростка, но с длинным прото-
ком. Яйцевод сравнительно длинный и всегда хорошо заметен позади вла-
галища.
Распространение (рис. 43, 2). Средняя Азия (Тянь-Шаньская,.
Алайская и Памиро-Дарвазская горные системы), горные области Афгани-
стана, северо-восточный Иран. Известно 4 вида (см. ниже).
Замечания. 1. До недавнего времени вслед за Зимротом (Simroth,
1912b) среднеазиатские виды этого семейства относили в подрод Ргорагта-
cella Simroth, 1912 рода Parmacella Cuvier, 1804 (Hesse P., 1926; Лихарев,
Раммельмейер, 1952; Wenz, Zilch, 1959—1960). В эту же группу входила
Р. kainarensis Simroth, 1912. Недавно Лихарев и Старобогатов (1967) пока-
зали, что данный вид является младшим субъективным синонимом Р. ru-
tellum Hutton, 1849 — вида, для которого Гудвин-Аустен (Godwin-Austen,
1888) установил специальный род Candaharia. Следовательно, согласно при-
оритету Candaharia является старшим синонимом Proparmacella.
2. Резкие различия в строении гениталий Parmacella и Candaharia дают
основание рассматривать их как самостоятельные роды. У Parmacella нет
любовной железы, но зато атриум и семяприемник имеют очень характерные-
придаточные органы, которые отсутствуют у Candaharia.
1. Подрод CANDAHARIA s. str.
Эпифаллус впадает в пенис апикально. На границе между ними илиг
к заднему отделу пениса крепится половой ретрактор.
1. Candaharia (Candaharia) rutellum (Hutton, 1849) (рис. 460—463)
Hutton, 1849 : 649 (Parmacella)', Simroth, 1912b : 46, T. 1, Fig. 3, 4 (P. kai-
narensis); Лихарев, Раммельмейер, 1952 : 355 (P. kainarensis); Лиха-
рев, Старобогатов, 1967 : 187, рис. 15 (Parmacella); Увалиева, 1975 : 44-
(Parmacella, биология).
Locus typicus — «Kandahar» — Кандагар (Афганистан). Типы данного-
вида и его синонима неизвестны.
Материал: из СССР — более 300 экз. из 55 пунктов; из Афганистана —
20 экз.
Верхняя сторона тела серая или серовато-желтая. На этом фоне распо-
лагается рисунок, образованный многочисленными пятнышками — «брызгами.
За счет их сгущения на мантии возникают продольные полосы, чаще всего-
вдоль левого края, реже вдоль правого, а иногда и на середине. Кроме того,
по две продольные полосы тянутся вдоль заднего конца ноги и на затылке.
Одноцветные экземпляры, т. е. без темного рисунка, встречаются редко..
Подошва окрашена в цвет фона верхней стороны тела.
Размеры: дл. сок. до 55, дл. мантии до 22, длина спины до 13 мм..
2. CANDAHARIA
343
Раковина (рис. 460—462) удлиненно-овальная. Продольная ось нуклеуса
располагается к спатуле под тупым углом. Поверхность нуклеуса покрыта
Рис. 460—463. Candaharia (Candaharia) rutellum (Hutton).
460 — раковина; 461—462 —нуклеус раковины; 463 — гениталии, Дарбазблиз Ташкента, 23 III 1914.
•спиральными рядами микроскопических ямок — точек (видны при 70-крат
лом увеличении). Длина эмбриональной раковины 2.8—3.3, ширина 2.0—2.3 мм.
Радула. Пример: степь у Дарбаза, близ Ташкента: формула:
с . 216 36 21к
3 + 3 + 2+ 1Х102,
344
I. PARMACELLIDAE
Гениталии (рис. 463). Гонада треугольная. Белковая железа
очень крупная, языковидная. К ней прилегает несколько меньших размеров
задний отдел простаты. Эпифаллус цилиндрический, в 2 раза длиннее пениса;
он образует несколько извивов и плавно переходит в задний отдел пениса,
к которому плотно подтянут многочисленными тяжами. Оба отдела пениса
внешне разделены перетяжкой. Внутри задний отдел покрыт многочислен-
ными сосочками, а передний легкими продольными складками.
Яйцевод и влагалище довольно длинные. Последнее (полностью или
только его задняя половина) муфтообразно охвачено вагинальной железой.
Овальный резервуар семяприемника подобно шляпке гриба сидит на длинном
протоке, который сильно расширяется перед влагалищем, открываясь в его
просвет несколькими крупными складками. Рядом впадают яйцевод и любов-
ная железа. Эта железа очень извита и вместе с влагалищем связана со стенкой
тела несколькими мускульными тяжами.
Распространение. В СССР: горы и предгорья Тянь-Шаня,
Алайская и Памиро-Дарвазская горные системы. Вне СССР: горные области
Афганистана.
Образ жизни. Обитает во влажных местах, особенно по берегам
природных водоемов и оросительных каналов. В неблагоприятное время
укрывается в щели почвы, под камни и у входа в пещеры. Поднимается
в горы до 3 200 м (Киргизский хребет), а вдоль оросительной сети спускается
в предгорья и в полупустыни (особенно в северных областях ареала).
В природных биотопах численность этого слизня незначительна, но в мес-
тах с поливным земледелием она может быть очень высокой.
Жизненный цикл. По данным Увалиевой (1979), в окрестностях
Алма-Аты весной (апрель—май) слизни интенсивно питаются, растут, ко-
пулируют и в июне откладывают яйца. Каждый слизень откладывает 10—
80 желтоватых яиц (4.3x3.1мм), после чего гибнет. Молодь вылупляется
на 25—30-й день, а через 3—4 дня она закапывается в почву, втягивает
тело в раковину и закрывает устье «крышечкой». Диапауза продолжается
до конца августа—начала сентября. После пробуждения сеголетки начинают
интенсивно питаться и расти вплоть до наступления холодов и ухода на
зимовку (ноябрь—март).
Хозяйственное значение. Как вредитель сельскохозяй-
ственных культур этот слизень отмечен только в предгорьях Заилийского
Алатау. Весной он в основном вредит в парниках рассаде перца, баклажанов,
и помидор. Осенью молодые слизни сильно повреждают урожаи тех же и
других культур.
Замечания. Оригинальное описание (Hutton, 1849) сделано по-
экземплярам, найденным близ Кандагара (южный Афганистан). Хуттон
дал столь краткую и общую характеристику внешнего вида этого слизня,
что она подходит почти к любому виду того же семейства. Однако у нас нет
сомнений в правильности идентификации наших материалов с видом Хут-
тона. Во-первых, в южном Афганистане до сих пор не найден другой какой-
либо вид пармацеллид. Во-вторых, в той же работе дается описание раковины
нового вида «Vitrina baccata», которая очень походит на эмбриональную ра-
ковину данного слизня. Хуттон отмечает даже точечную микроскульптуру.
Все сказанное говорит за то, что в обоих случаях автор имел дело с С. ги-
tellum.
В 1912 г. Зимрот описал новый вид — Parmacella kainarensis (сбор
А. Казнакова, 22 IV 1897 г., в Кайнаре — Гиссарский хребет). Характе-
ристика этого вида сделана по одному молодому экземпляру длиной 25 мм.
В ЗИН АН СССР имеются еще 4 экз. из того же сбора, которых Зимрот не
видел.
2. CAND AH ARIA •
345
Внимательное изучение описания этого вида, а также экземпляров из
того же сбора, где был и голотип,1 убедило нас в том, что Р. kainarensis —
младший синоним С. rutellum.
2. Candaharia (Candaharia) aethiops (Westerlund, 1896), comb. nov.
(рис. 464—467)
Westerlund, 1896 : 183 (Parmacella anthiops, nom. err.); Simroth, 1901:
202, T. 20, Fig. 13—17 (P. korschinskii var. rubra, var. anthiops West.); 1912b : 49 (P. anthi-
ops); Линдгольм, 1922 : 305 (P. aethiops, nom. emend.); Лихарев, Раммель-
мейер, 1952:354, рис. 281 (Parmacella).
Locus typicus — «Langs dem Flusse Usun-kur-ssu» — вдоль реки Узун-
кур-су, Ферганский хребет. Голотип утрачен.
Материал: из СССР — более 300 экз. (в том числе голотип Рагта-
cella korschinskii rubra Simroth) из 51 пункта.
Окраска варьирует от совершенно одноцветно черной до серовато-жел-
той с более или менее отчетливым темным рисунком. У молоди окраска
всегда светлее, а рисунок более четкий. Подошва всегда светлая. При уме-
ренном развитии темного пигмента на шее всегда имеются две четкие про-
дольные полосы. На мантии пигмент образует сгущения у левого края вплоть
до образования полосы; реже аналогичные сгущения возникают вдоль пра-
вого края мантии и на середине. Вне мантии пигмент в первую очередь
концентрируется по бороздкам.
Размеры:, дл. сок. до 45, мантии до 25, задней части до 15 мм.
Раковина (рис. 464) короткоовальная, а нуклеус почти круглый, бле-
стящий; и гладкий. Длина нуклеуса 2.7—3.1, ширина 2.2—2.5 мм.
Радула. Пример; Киргизский хребет, ущелье Аламедын, 1 экз.; формула:
с 236 66 ЗОк
Т + "У + ~2~ + Т" х но.
Г ениталии (рис. 465—467). Семяпровод тонкий, длинный и извитый.
Эпифаллус четко подразделен на два отдела: задний — веретеновидно
вздутый с толстыми мускулистыми стенками, и передний — более тонкий
и цилиндрический, впадающий в пенис апикально. Передний отдел эпифаллуса
серией мускульных лент (из них задняя наиболее крупная) подтянут
к пенису. Половой ретрактор крепится к переднему концу эпифаллуса.
Пенис резкой перетяжкой подразделен на короткий задний и длинный перед-
ний отделы. Задний имеет внутри короткую папиллу, которая является про-
должением эпифаллуса. Как сама папилла, так и стенки этого отдела пениса
покрыты мелкими сосочками. Передний отдел пениса внутри с правильными
продольными складками.
Яйцевод короткий, одной длины с шаровидно вздутым влагалищем.
Последнее, как муфтой, окружено вагинальной железой. Во влагалище впа-
дают: семяприемник с круглым или овальным резервуаром и протоком почти
той же длины и более или менее извитая любовная железа. Атриум короткий,
внутри с продольными складками. Нередко вагинальная железа закрывает
и его.
Распространение. Киргизский, Ферганский, Гиссарский, Дар-
вазский хребты, хребет Петра I; одна находка сделана у Хорога (Памир).
Одноцветные черные особи наиболее обычны в Ферганском хребте.
Образ жизни. Обитает преимущественно в горах, реже в пред-
горьях, по сырым ущельям, близ водоемов. Укрывается под камнями, в гни-
лых пнях и под валежинами.
В противоположность многим другим видам своего семейства избегает
культурных биотопов.
1 Голотип следует считать утраченным.
346
I. PARMACELLIDAE
Замечания. 1. Данный вид былописан Вестерлундом (Wester-
lund, 1896) по единственному экземпляру, найденному академиком Коржин-
I
Рис. 464, 465. Candaharia (Candaharia) aethiops (West.). Тянь-Шань, ущелье Аламедын,
14 VI 1972).
464 — раковина; 465 — гениталии.
ским на Ферганском хребте в тенистом лесу вдоль р. Узун-кур~су
14 VIII 1895 г. на высоте 2166 м. Внутреннее строение этого экземпляра
(голотип!) было изучено Зимротом (Simroth, 1901).
2. CANDAHARIA
347
В той же работе Зимрот дает описание другого экземпляра, тоже из
сборов Коржинского с Ферганского хребта, но найденного у Таран-базара.
Этот экземпляр отличается от первого только красноватой окраской. Обна-
ружив столь несущественные отличия, Зимрот справедливо считал, что оба
Рис. 466, 467. Candaharia (Candaharia) aethiops (West.). Тянь-Шань, ущелье Аламедын,
14 VI 1972.
466 — дистальная часть гениталий; 467 — продольный разрез пениса.
экземпляра принадлежат к одному виду. Однако в нарушение правила прио-
ритета названий он дал этому виду название «korschinskii п.. sp.». Соответственно
первый экземпляр он отнес к «Parmacella korschinskii anthiops West.», а вто-
рой — к «Р. korschinskii rubra var. nov.»
Мы убеждены, что, как это бывало не раз с материалами, прошедшими
исследование Зимрота, красным цветом второй экземпляр обязан особенно-
стям спирта, использованного для консервации. Поэтому придание какого-
либо таксономического значения этому признаку не имеет смысла.
348
I. PARMACELLIDAE
2. В той же работе Вестерлунда данный вид назван «Р. anthiops n. sp.»
Линдгольм (1922) справедливо указал, что в это название вкралась типо-
графская опечатка — следует читать «aethiops». Во-первых, вывод этот вы-
текает из смысла описания данного вида, так как Вестерлунд подчеркивает:
«Животное . . . целиком (даже подошва) черное . . .» (с. 183; перевод с латин-
ского наш, — Л. 'в. В.). Во-вторых, в архиве ЗИН АН СССР сохранились
«Списки материалов, посланных на определение Вестерлунду», где рукою
автора написано «Parmacella aethiops n. sp.»
2. Подрод LEVANDERIA Likharev et Wiktor, subgen. nov.1
Типовой вид — Parmacella levanderi Simroth, 1901.
Диагноз. Эпифаллус, плавно сужаясь, впадает в пенис сбоку, не-
сколько позади границы, между его передним и задним отделами. Половой
ретрактор крепится к вершине заднего конца пениса.
3. Candaharia (Levanderia) levanderi (Simroth, 1901), comb. nov.
(рис. 468—474)
Simroth, 1901 : 203, T. 20, Fig. 4, 10, 21, 22, T. 21 (Parmacella}-, 1912b : 44,.
T. 1, Fig. 1 (P. kaznakovi, syn. nov.); 45, T. 1, Fig. 2 (P. roseni, syn. nov.); Жадин,.
1946 : 15 (P. korschinskii)-, Лихарев, P аммел ьмейер, 1952 : 355 (Parmacella).
Locus typicus — «Samarkand» — Самарканд. Лектотип в Зоологическом
музее Хельсинкского университета.
Материал: из СССР — более 1000 экз. (в том числе лектотип Parma-
cella levanderi Simroth и голотипы Р. roseni Simroth и Р. kaznakovi Simroth)
из 147 пунктов; из Афганистана и Ирана — 23 экз.
Окраска чаще всего одноцветная серая или серовато-желтая; мантия
обычно окрашена немного темнее спины и боков. Иногда встречаются популя-
ции, имеющие темный рисунок на верхней стороне тела из пятен и полос.
Но чаще подобный рисунок имеет только молодь, и с возрастом он исчезает
за счет общего потемнения покровов. При сокращении на заднем краю ман-
тии появляется сильный, но закругленный угол. Подошва всегда светлее
верхней стороны-тела и одноцветная.
Размеры: дл. пол. до 100 мм, дл. сок. до 55, дл. мантии до 28,
дл. задней части до 16 мм; лектотип — дл. сок. 55 мм.
Раковина (рис. 470) овальная. Нуклеус сильно удлиненно-овальный:
его продольная ось почти перпендикулярна оси спатулы. Размеры нуклеуса:
дл. 4.0—5.2, ширина 2.3—3.1 мм.
Радула. Пример: окрестности Самарканда, 1 экз.; формула:
Гениталии (рис. 471—474). Семяпровод, плавно расширяясь, пе-
реходит в сильно извитый эпифаллус, длина которого приблизительно
равна длине семяпровода и в 2 раза больше длины пениса. Последний изгиб
эпифаллуса особыми мускульными связками плотно подтянут к пенису.
Оба отдела пениса более или менее четко разграничены перетяжкой. Эпи-
фаллус впадает в пенис сбоку, немного отступя от этой перетяжки. Слепо,
замкнутый, почти треугольный задний отдел пениса внутри покрыт много-
численными мелкими сосочками, а снаружи к его вершине крепится половой
ретрактор. Внутренняя поверхность переднего отдела пениса покрыта
продольными складками.
1 Название дано по типовому виду, который в свою очередь был назван в честь
финского зоолога К. Левандера.
Рис. 468, 4§9. Candaharia (Levanderia) levanderi (Simr.).
des — полувзрослый слизень; 469 — молодой слизень после летней диапаузы (Гиссарский хребет
Ходжа-Оби-Гарм, X 1977).
350
I. PARMACELLIDAE
Яйцевод довольно длинный и мускулистый. Он открывается рядом
с семяприемником {и любовной железой в широкое цилиндрическое влага-
лище, задняя часть которого окружена железистой муфтой. Резервуар семя-
Рис. 470—474. Candaharia (Levanderia) levanderi (Simr.).
470 —1раковина; 471 — гениталии; 472 — дистальная часть гениталий; 473 — продольный разрез
пениса и эпифаллуса; 474 — гениталии молодого слизня 471—473— Самарканд, III 1912; 474 —
Гиссарский' хребет, ущелье Кондара, 26 V 1943).
приемника обычно круглый (до тех пор, пока его не растянет сперматофор);
проток его вначале узкий и тонкостенный, но затем резко бочковидно рас-
ширяется и становится мускулистым. Внутри эта часть протока семяприем-
ника имеет много крупных продольных складок.
Любовная железа длинная и очень извитая. Атриум почти незаметен.
2. CANDAHARIA
351
Изложенный план строения женских половых путей и их придаточных
органов характерен для взрослых экземпляров, приступивших к размно-
жению. У молодых слизней (рис. 474; дл. пол. 20 мм; эмбриональная рако-
вина еще полностью открыта) влагалище без железистой муфты и немного1
длиннее яйцевода, любовная железа короткая и лишь с намеком на извивы,
семяприемник пальцевидный, т. е. без подразделения на проток и резурвуар;
имеется небольшой, но четкий атриум.
Распространение. В СССР: Алайская горная система от Зерав-
шанского хребта на западе до хребтов Алайского и Петра I на востоке; кроме
того, Чаткальский хребет. Вне СССР найдена в двух пунктах: Доши (северный
Афганистан) и Горган (северо-восточный Иран). В последнем случае, воз-
можно, завезен.
Образ жизни и жизненный цикл этого вида в условиях
Душанбе и Варзобской ботанической станции (южный склон Гиссарского
хребта) подробно изучен Жадиным (1946).1 Обитает до высот 2300 м, однако
наибольшей численности достигает в долинах, особенно в зоне поливного
земледелия.
Зимует в почве и под различными укрытиями, обычно на глубине около
2 см, т. е. ниже промерзающего слоя. После зимовки на поверхности почвы
появляется чаще всего с конца января—начала февраля, а в горах на 2 недели
позднее. С установлением теплой погоды слизни начинают интенсивно пи-
таться и расти (рис. 26).
Размножение обычно начинается в апреле, в горах немного позднее
чем в долинах. Спариванию всегда предшествует «брачный танец» — круже-
ние на избранной небольшой площадке. Через 9—11 дней после копуляции
начинается яйцекладка. Каждый слизень в несколько приемов откладывает
до 40 яиц, которые он помещает либо в специально выдолбленные в почве
ямки глубиной 2—7 см, или под камни и комки почвы. Яйца имеют диаметр
3—5 мм.
Вылупление происходит на 20—25-й день. Молодые слизни вначале
энергично ползают, но не питаются. На второй день они заклеивают устье
эмбриональной раковины эпифрагмой («крышечкой») и впадают в летнюю
диапаузу, которая длится с середины июня до середины октября. После
пробуждения, обычно вызванного первыми прохладными осенними дождями,
слизни начинают столь интенсивно питаться и расти, что через месяц раз-
растающаяся мантия почти полностью скрывает эмбриональную раковину,
а к началу зимовки (декабрь) они могут достигать 40 мм длины (в движении)
и веса 1 г.
Хозяйственное значение. Из-за большой прожорливости,
особенно весной и осенью, высокой плодовитости и явного тяготения к куль-
турным биотопам этот слизень может приносить большие убытки в теплицах
и на огородах, особенно в зоне поливного земледелия. Наряду с культурными
растениями (капуста., земляника, листья тыквы, лука и рассада других
культур) слизень охотно поедает многие дикие растения. Благодаря этому
невозделанные участки по соседству с огородами и полями являются природ-
ными резерватами вредителя.
Кроме растений эти слизни охотно едят трупы животных и кал.
Замечания. 1. К вопросу о типах С. (L.) levanderi (Simroth).
К сожалению, Зимрот (Simroth, 1901), давая описание данного вида, нигде
четко не сказал, чьи сборы он использовал. При диагнозе указано (с. 203) —
«Ташкент, Самарканд». Несколькими страницами раньше (с. 188) автор со-
общает, что получил несколько экземпляров от финского ученого Левандера.
1 Как показала проверка исходного материала, автор ошибочно определил этот вид
как Parmacella korschinskii Simroth, 1901 (—Candaharia aethiops Westerlund, 1896). На са-
мом деле он имел дело с С. levanderi (Simr.).
352
I. PARMACELLIDAE
В коллекции Зоологического музея в Хельсинки имеется один экземпляр,
вскрытый и определенный Зимротом как Parmacella levanderi и собранный
где-то у Самарканда. Его гениталии полностью соответствуют табл. 20,
рис. 21 монографии Зимрота. Этот экземпляр мы обозначаем как лектотип.
Для описания данного вида Зимрот также использовал материалы, по-
лученные им из Петербургского зоологического музея (ныне ЗИН АН СССР).
Из них один экземпляр из Самарканда погиб, но 5 экземпляров (паралекто-
типы!) из Ташкента (сбор Лидского, 1887 г.) сохранились. Зимрот их не
анатомировал.1 Наша проверка показала, что эти экземпляры принадлежат
к другому виду — Candaharia rutellum (Hutton).
Наконец, в Зоологическом музее Московского университета имеется
один экземпляр настоящей С. levanderi, определенный Зимротом (сбор. Н. Бо-
гоявленского, 1895 г., Самарканд). Возможно, что это тоже паралектотип.
2. К вопросу о систематическом положении Parmacella kaznakovi Simroth
и Р. roseni Simroth. Оба вида были описаны (Simroth, 1912b) с территории
Таджикистана и Бухарской области Узбекистана. Голотипы находятся
в ЗИН АН СССР.
Если сравнить нуклеусы раковин и гениталии обоих видов с таковыми
у Р. levanderi и учесть возрастные изменения гениталий последнего (см. выше),
то бросается в глаза большое сходство всех трех видов. Немногие отличия
Р. kaznakovi от Р. levanderi связаны с тем, что Зимрот имел дело с не вполне
взрослым экземпляром. Этим объясняется, что у Р. kaznakovi якобы: а) про-
ток семяприемника более стройный; б) любовная железа менее извита; в) ва-
гинальная железа простирается только на заднюю часть влагалища. В итоге
есть все основания считать Р. kaznakovi Simroth синонимом Candaharia
levanderi (Simroth).
Что касается Р. roseni, то главной особенностью этого вида Зимрот счи-
тал присутствие небольшого бугорка на пенисе у места впадения в него эпи-
фаллуса. Сходное образование, а иногда и два бугорка по обе стороны от
эпифаллуса мы не раз замечали даже в пределах одной популяции С. le-
vanderi. Их присутствие не представляет таксономического интереса. Здесь
эпифаллус серией мелких мышечных волокон подтянут к пенису. Эти обра-
зования возникли из-за местного сокращения указанных волокон. Тут
возможно и влияние фиксации. Следовательно, есть все основания рассмат-
ривать Р. roseni Simr. как еще один синоним Candaharia levanderi (Simroth).
4. Candaharia (Levanderia) izzatullaevi Likharev et Wiktor, sp. nov.2
(цв. вкл. рис. IP, рис. 475—479)
Материал. Голотип и 40 паратипов найдены 3. И. Иззатуллаевым
24—25 апреля 1973 г. в окрестностях Пенджикента (Таджикистан), вдоль
арыков. Кроме того, им же и еще ранее 3. В. Личковой были найдены 5 па-
ратипов в Ботаническом саду АН ТаджССР в г. Душанбе. Голотип и 42
паратипа в ЗИН АН СССР; 3 паратипа в Института зоологии и паразито-
логии АН ТаджССР. Наконец, в коллекции Б. Н. Цветкова (Зоологический
музей Московского университета) мы обнаружили 6 экз. (паратипы) этого
вида, собранных у ст. Джизак (УзССР).
Диагноз. Пенис с крупным аппендиксом.
Тело окрашено в лимонно-желтый цвет с четким темным рисунком, ко-
торый состоит из трех продольных полос на мантии и двух полос, вытянутых
вдоль киля и шеи. Кроме того, отдельные пятна различной формы и размеров
1 Один из этих экземпляров изображен на табл. 20, рис. 4 монографии Зимрота.
2 Вид назван в честь 3. И. Иззатуллаева (Душанбе), впервые обратившего наше
внимание на этот вид и передавшего нам свои сборы.
2. CANDAHARIA
353
разбросаны как по мантии, так и по бокам вдоль подошвы. Форма и локали-
зация’ отдельных компонентов рисунка варьируют даже в пределах попу-
ляции. Между килем и мантийной щелью 16 рядов морщин.
•Рис.475—479. Candaharia (Levanderia) izzatullaevi Likharev et Wiktor, sp. nov. Паратип.
Душанбе, Ботанический сад, 22 IV 1943.
-475 — раковина; 476 — дистальная часть гениталий; 477 — пенис и эпифаллус; 478 — продольный
разрез через пенис; 479 — контур одного из сосочков пениса.
Размеры: дл. сок. до 32, дл. мантии до 16, дл. задней части до
<8 мм. Голотип: дл. сок. 30, дл. мантии 16, дл. задней части 7 мм.
Раковина (рис. 475) овальная. Нуклеус удлиненно-овальный; поверх-
_23 И- М. Лихарев, А, И. Виктор
354
II. MILACIDAE
ность покрыта спиральными рядами микроскопических точек-ямок (видны
при 25-кратном увеличении). Размеры всей раковинки: длина до 10, ширина
до 6 мм; размеры нуклеуса 3.3x2.Змм.
Радула. Пример: Душанбе, Ботанический сад, 1 экз.; формула:
с 216 20к
з+— + — Х84.
Гениталии (рис. 476—479) очень сходны с таковыми у С. levanderi.
Основное отличие: задний конец пениса близ впадения в него эпифаллуса
имеет крупный конический аппендикс. Нередко, по-видимому, как следствие
копуляции аппендикс остается ввернутым внутрь пениса. В этом случае
снаружи на его месте видна ямка, а если пенис вскрыть, то аппендикс имеет
вид крупного выступа, покрытого (как и все внутренние стенки этого отдела
пениса) мелкими сосочками. По одной из сторон ввернутого аппендикса
проходит борозда, ведущая внутрь эпифаллуса.
Замечание. По гениталиям наш вид обнаруживает наибольшее
сходство с С. levanderi, отличаясь главным образом присутствием крупного
аппендикса на пенисе. Правда, как уже отмечалось выше, у levanderi на
пенисе вблизи от эпифаллуса иногда бывают 1—2 небольших выступа, однако
они никогда не достигают таких размеров, как аппендикс у нашего вида.
Для внешнего облика С. izzatullaevi характерна лимонно-желтая окраска
и точечная микроскульптура эмбриональной раковины. Как известно,,
подобная скульптура встречается еще у С. rutellum (Hutton), но последний
вид отличается совершенно иным планом строения гениталий.
II. Семейство MILACIDAE Ellis, 1926
Ellis, 1926:252; Germain, 1930:106; Cockerell, 1935 : 143; W a g-
n e r, 1935 : 189.
Слизни различных размеров (дл. пол. от 45 до 100 мм и более) с коренастым
или стройным телом. Киль всегда хорошо развит, нередко гребневидный,
у большинства видов занимает всю спину, у остальных ограничен задней
третью спины. Мантия небольшая (не более 1/3 общей длины), овальная
или яйцевидная, т. е. суженная впереди, полностью располагается на перед-
ней половине тела. Капюшон сравнительно небольшой, при сокращении
животного плотно прилегает к шее. Поверхность мантии при жизни с отчет-
ливой мелкозернистой скульптурой без всяких следов концентрических ли-
ний. Характерно присутствие здесь четкой подкововидной борозды, иногда
оба конца ее смыкаются. Под задним концом мантии по бокам от киля име-
ются две небольшие полости. Иногда в этом месте мантия имеет четкую
вырезку.
Спина и бока покрыты крупными и длинными, но слабо выпуклыми мор-
щинами, разделенными глубокими бороздками. Между мантийной щелью и
средней линией спины (или килем) чаще всего 9—12 рядов морщин. Подошва
довольно широкая, но меньше ширины тела; срединная доля ее несколько
шире боковых. Вся поверхность подошвы равномерно покрыта многочислен-
ными бороздками, которые изогнуты назад, приобретая форму латинской
буквы V. Этот рельеф более четок и регулярен, чем на срединной доле
у Agriolimacidae. Половое отверстие лежит на полпути между пневмостомом
и щупальцами. Слизь очень густая и липкая.
Раковина всегда полностью внутренняя, пластинкообразная, более или
менее овальная. Нуклеус лежит строго на продольной оси раковины, не-
сколько отступя от заднего края, и является самой выпуклой частью верхней
стороны. Граница между нуклеусом и спатулой слабо выражена.
II. MILACIDAE 355
Срединный и боковые зубы радулы трехзубцовые; лишь несколько самых
внешних боковых зубов утрачивают эндокон и становятся двухзубцовыми.
Краевые зубы кинжало- или шиповидные, иногда с небольшим эктоконом.
Кишечник двухпетлевого типа, обычно сильно перекручен. Зоб простира-
ется назад дальше чем вторая петля. Правая доля печени образует вершину
внутренностного мешка. Колумеллярный мускул крепится у заднего края
мантийного комплекса (перед ним или сзади).
Сердце лежит в левой передней четверти мантийного комплекса (рис. 6,
III), ось его под углом 45° наклонена направо, т. е. в сторону продольной
оси тела. Аорта очень длинная. Кровеносная сеть легкого располагается
перед почкой. Почка неправильно бобовидная; ее длинная ось более или
менее параллельна продольной оси тела; снизу от почки отходит направо
крупный языковидный лобус, который располагается под мочеточником и
ректумом. Мочеточник начинается от заднего конца почки; он прямой или
слабо 5-образно изогнут и вместе с анусом открывается наружу перед пневмо-
стомом. Мочевого пузыря нет или он лишь слегка намечен как расширение
переднего конца мочеточника. Между почкой и передним концом мочеточника
имеется связка.
' Педальная железа (рис. 4, ЖП) имеет вид узкого мешка, свободно лежа-
щего на внутренней поверхности ноги. Как известно, у других семейств
юна всегда погружена в толщу мускулатуры ноги.
Семяпровод впадает более или менее апикально в хорошо развитый эпи-
фаллус. Внутренние стенки последнего имеют сложные складки, между кото-
рыми формируются шипы сперматофора. Пенис различной формы, без
наружных придатков. Внутри него имеется папилла, а иногда также раз-
личные складки и сосочки. Семяприемник открывается в яйцевод. Внутри
задней части влагалища имеется венец мощных треугольных складок, не-
редко образующих вагинальную папиллу, которая может выдаваться в ат-
риум. В свою очередь атриум имеет внутри различные складки, морщины и
юосочки (род Tandonia) или стимулятор (род Milax).
Одной из характерных особенностей гениталий является присутствие
особых придаточных желез — вагинальных желез, открывающихся во
влагалище или атриум.1 Ретрактор правых щупалец не перекрещивается
с половыми путями.
Сперматофор обычно имеет удлиненно-веретеновидную форму и сужен
к обоим концам. Стенки его кожистые, медно-золотистые и покрыты рядами
шипов, которые нередко дихотомически ветвятся.
Яйца с известковой оболочкой.
Распространение (рис. 41, 7). Большинство видов обитают
в средиземноморских и черноморских странах, особенно на Балканском
полуострове. Лишь немногие населяют Британские острова, западные, се-
веро-западные и центральные области Европы.
Теплолюбивые слизни; обитают на равнинах и в нижних горизонтах
гор, как на открытых местах, так и в лесах. Многие живут в очень ксеротерм-
ных условиях — на сухих каменистых, особенно известняковых местах
с редким травяным покровом и кустарником.
Замечания. Длительное время виды этого семейства объединяли
в один род Amalia (=Milax) и относили к семейству Limacidae (Simroth,
1 Вагнер (Wagner, 1930, 1931) сообщает, что у некоторых видов этих желез нет.
Как показала проверка тех же экземпляров, с которыми имел дело автор, эти железы
у них имеются. По-видимому, эти образования развиты хорошо только у половозрелых
-слизней.
23*
356
II. MILACIDAE
1885; 1901). Эллис (Ellis, 1926), обратив внимание на большие различия
как во внешнем облике, так и в анатомии между Limax и Milax, выделил
последний род в особое семейство Milacidae. Почти одновременно с ним
Гессе (Hesse Р., 1926) счел возможным выделить роды Milax, Boettgerilla,.
Aspidoporus и Parmacella в особое подсемейство Parmacellinae, оставив
их в семействе Limacidae. Основанием обособления указанных родов слу-
жило присутствие у них подкововидной борозды на мантии, наличие эпи-
фаллуса и различных придаточных органов на влагалище и атриуме.1 *
Жермен (Germain, 1930), обратив внимание на то, что Parmacella резко;
отличается от всех Limacidae как по внешнему облику, так и по внутреннему
строению, выделил этот род в особое семейство Parmacellidae (см. с. 332).
Род Milax он оставил среди Limacidae, но в подсемействе Milacinae? Вскоре
Вагнер (Wagner, 1935) вновь возводит эту группу в ранг семейства. Так,
прежнее подсемейство Parmacellinae было поделено на два семейства (Wenz,
Zilch, 1959-1960; Wiktor, 1973).
Однако и в этой трактовке Milacidae не представляли естественной:
группы. Ряд важнейших черт организации резко отличают Boettgerilla.
как от Milax, так и от Aspidoporus. Не случайно Гётем (Goethem, 1972)<
считает, что Boettgerilla заслуживает выделения в самостоятельное подсе-
мейство или даже семейство (с. 210).
До сих пор совершенно неясным осталось положение загадочной группы
Aspidoporus Fitzinger.
В итоге, в семействе Milacidae можно оставить только род Milax в том
понимании и объеме, которое вкладывали в эту группу Эллис, Гессе и Ваг-
нер.
Последние два автора предложили свои варианты разделения этого
рода на подроды и секции. Они отмечали, что границы между подразделе-
ниями не очень четкие. По-видимому, эта нечеткость связана, во-первых,,
с недостаточно тщательным и неадекватным описанием многочисленных
видов; во-вторых, при описании видов и классификации их внутри рода:
уделялось большое внимание внешнему облику слизней и положению ва-
гинальных желез и стимулятора, но почти совсем не рассматривалась форма
и внутреннее строение эпифаллуса, пениса и других копулятивных органов.
Как известно, форма и размеры желез очень изменчивы и, в частности,,
зависят от возраста и физиологического состояния. Между тем внешнее'
и внутреннее строение пениса, эпифаллуса, влагалища и атриума отличается:
большим постоянством и позволяет не только полнее охарактеризовать все-
семейство, но и наметить различия между родами и подродами.3
Исходя из этого мы считаем, что Milacidae состоят из двух родов — Milaxr:
Gray и Tandonia Less, et Poll., характеристики которых приводятся ниже.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ MILACIDAE
1 (2). Тело сравнительно коренастое. Атриум крупный и вздутый, внутри:
с коническим или языковидным стимулятором. Придаточные железы.
(1 или 2) впадают в атриум асимметрично, у основания стимулятора:
....................................................... 1. Milax Gray.
1 Гессе (Hesse Р., 1926) признавал значительную условность объединения указанных:
родов в одно подсемейство.
3 Вопрос о систематическом положении родов Boettgerilla и Aspidoporus Жермен:
обошел молчанием. То, что Эллис уже выделил род Milax в особое семейство, по-видимому,,
он не знал.
* Впервые для характеристики этого семейства и входящих в него видов мы исполь-
зуем папиллу пениса и признаки внутреннего строения влагалища. Следует также учесть,
что внутреннее строение эпифаллуса коррелятивно связано с формой сперматофора и дает-
представление о нем даже в тех случаях, когда его нет в распоряжении исследователя-
1. MILAX
35T
2 (1). Тело сравнительно стройное. Атриум небольшой, без стимулятора.
Придаточные железы впадают во влагалище (вблизи от атриума), кото-
рое они либо окружают венцом, или же располагаются по двум сторо-
нам симметричными пучками или клубками........................... . . .
........................................2. Tandonia Less, et Poll.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ MILACIDAE
(по живым слизням)
1 (8). Крупные слизни; во время движения длина их больше 50 мм,
чаще же больше 70 мм.
2 (5). Общая окраска светлая; на этом фоне имеются многочисленные
темные пятнышки, не сливающиеся друг с другом.
3 (4). Тело почти белое или желтоватое. Темные пятнышки располага-
ются равномерно и редко, не образуя сетчатого рисунка...............
........................................ Tandonia rustica (Mill.).
4 (3). Тело розовато- или грязно-кофейное, заметно более темное на спине.
Пятнышки располагаются неравномерно — на спине их больше, к по-
дошве меньше и образуют сетчатый рисунок............................
................................... Tandonia kusceri (Н. Wagn.).
5 (2). Общая окраска темная, одноцветная или спина густо покрыта
очень мелкими (микроскопическими) темными пятнышками, сливаю-
щимися друг с другом.
6 (7). Киль кремовый или оранжевый, спина по сторонам от цего темно-
коричневая. Тело стройное, к заднему концу плавно суженное . . .
...................................... Tandonia budapestensis (Hazay).
7 (6). Киль окрашен в один цвет с пепельной или почти черной спиной
или лишь немного светлее спины. Тело коренастое, к заднему концу
резко суженное................................Milax gagates (Drap.).
8 (1). Небольшие слизни, во время движения длина их не больше 45 мм.
9 (10). Мантия с темными боковыми полосами . Milax caucasicus (Simr.).
10 (9). Мантия без боковых полос.
11 (12). Тело серовато-оливковое или пепельно-желтоватое, без рисунка
из темных пятнышек, лишь спина может быть немного темнее боков
...............................................Tandonia cristata (Kai.).
12 (И). На грязно-кремовом фоне имеется рисунок из большого числа тем-
ных пятнышек, которые, сливаясь, могут придавать спине черную
окраску, а на мантии образовывать сетчатый рисунок. Киль всегда
светлый..................................Tandonia kaleniczenkoi (Cl.).
1. Род MILAX Gray, 1855
Gray, 1855 May : 174; Moquin-Tan d on, 1855 Sept. : 19 (Amalia); M a-
b i 11 e, 1868 : 143 (Lallemantia); Lesson a, Pollonera, 1882 : 57 (Pirainea);
Hesse P., 1926 : 33 (Milax s. str., sect. Lallemantia); VI a g n e r, 1930 : 52 (Milax
s. str., sect. Lallemantia).
Типовой вид — Limax gagates Draparnaud, 1801; по последующему
обозначению — Hesse P., 1926.
Слизни небольшие или средней величины (дл. пол. не более 70 мм).
При жизни тело коренастое, плавно сужающееся к острому заднему концу.
Киль четкий и простирается на всю спину. Окраска довольно разнообраз-
ная, но обычно темная: от темно-серой до черной. Мантия с двумя боковыми
полосами неопределенной формы; внутри подковообразной борозды разбро-
саны мелкие неправильные пятнышки.
358
II. MILACIDAE
Предсердие сверху и снизу закрыто почкой. Лобус ее очень длинный.
Основание полового ретрактора крепится к диафрагме почти под центром
почки.
Атриум бочковидно вздутый, внутри него находится конический или языко-
видный спирально скрученный стимулятор. Придаточные железы несколь-
кими протоками асимметрично впадают в атриум (у основания стимулятора).
Задний конец эпифаллуса всегда шире переднего. Пенис с эпифаллусом
короче семяприемника или почти равен ему.
Распространение. Британские острова, средиземноморские
страны, Закавказье.
Данный род состоит из двух подродов: Milax s. str. и Micromilax Hesse,
1926. Единственный известный вид последнего подрода обитает на Кипре
и в Палестине.
Подрод MILAX s. str.
Единственная придаточная железа в виде компактной массы лежит под
глоткой или зобом в стороне от половых путей и впадает в атриум несколь-
кими тонкими и длинными протоками (до И штук) с мускулистыми стенками
и шелковистым блеском.
Распространение, как у рода.
Кроме приведенных ниже М. caucasicus (Simroth, 1912) и М. gagates
(Draparnaud, 1801), к данному подроду относятся М. altenai Forcart, 1972,
М. nigricans (Schulz, 1836) и М. parvulus Wiktor, 1968.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ MILAX
1 (2). Небольшие слизни — дл. пол. до 45, дл. сок. до 33 мм. За счет сильного
сгущения многочисленных темных пятнышек спина кажется почти
черной. Киль кремовый. Стимулятор небольшой, конический . . . .
....................................... 1. М. (М.) caucasicus (Simr.).
2 (1). Слизни средних размеров — дл. пол. до 70, дл. сок. до 55 мм. Верхняя
сторона тела одноцветная (без пятнышек) от пепельной до черной.
Киль окрашен в цвет спины или лишь немного светлее. Стимулятор
более крупный языковидно-конический, с одной стороны сплющен
.........................................2. М. (М.) gagates (Drap.).
1. Milax (Milax) caucasicus (Simroth, 1912) (рис. 480—486)
Simroth, 1912a : 52, T. 3, Fig. 48, T. 8, Fig. 31 {Amalia)-, Лихарев, Рам-
мельмейер, 1952 : 360, рис. 288; Ш' и л e й к о,' 19676 : 29.
Locus typicus — «Borzom» — Боржоми (Грузия). Голотип в ЗИН АН
СССР.
Материал: из СССР — 8 экз. (в том числе голотип) из 6 пунктов.
Тело умеренно коренастое, сзади плавно клинообразно суженное. Ман-
тия яйцевидная с максимальной шириной в задней трети, а к переднему
концу плавно сужается; перед килем всегда вырезана. Мантийная борозда
хорошо выражена, однако задние концы ее не сливаются друг с другом.
Пневмостом небольшой и лежит слегка постмедиально. Морщины на спине
и боках слабо выражены, между килем и мантийной щелью около 12 рядов
морщин.
Голотип после фиксации имеет черновато-фиолетовую окраску, особенно
плотную на спине и постепенно более светлую (за счет редкого расположе-
ния черных пятнышек) к подошве. У других экземпляров на грязно-кремовом
фоне разбросаны многочисленные мелкие черные пятнышки, которых особенно
Рис. 480, 481. Milax (Milax) caucasicus (Simr.). Талыш, 24 III 1965.
4S0 — внешний вид (из полового отверстия выступает стимулятор); 481 — раковина.
Рис. 482—486. Milax (Milax) caucasicus (Simr.) (окончание).
482 —зубы радулы; 483 — кишечник; 484 — ретракторы; 485 — гениталии; 486 — вскрытый пенис
(видна папилла).
1. MILAX
361
много на спине (в связи с чем она кажется почти черной) и которые постепенно
редеют к ноге. Киль всегда без черного рисунка, т. е. кремовый. Бока
ниже мантии, особенно часто с правой стороны, тоже могут оставаться свет-
лыми. Шея, голова и щупальца черноватые или совершенно черные.
Мантия с более или менее четкими боковыми полосами; они изменчивой
формы и обычно очень темные, почти черные. Середина мантии всегда свет-
лее, иногда почти кремовая, но чаще темнее, за счет развития серого или
коричневого пигмента, который может давать начало третьей, т. е. медиаль-
ной, цветной полосы. У голотипа в центре мантии имелось яркое оранжевое
пятно.
При жизни боковые доли подошвы покрыты многочисленными желтыми
пятнышками, а срединная доля серовато-синяя. После фиксации желтый пиг-
мент исчезает. Однако у большинства экземпляров подошва затемнена
мелкими черными меланофорами, которые отчетливо видны на боковых
долях и слабо просвечивают на срединной доле. Эта окраска подошвы сохра-
няется и после фиксации.
Слизь желтая.
Размеры: дл. пол. до 45 при ширине до 6 и длине мантии до 15 мм.
После фиксации дл. сок. до 33 при ширине до 7 и длине мантии до
9 мм.
Радула (рис. 482). Пример: 1 экз. из окрестностей Ленкорани: формула:
с 116 66 23к
3 + 3 + 2 + 1 Х 93‘
Ретракторы и кишечник см. рис. 483, 484.
Гениталии (рис. 485, 486). Гонада небольшая и светлая. Проток ее
почти прямой, лишь у белковой железы делает несколько извивов. Белковая
железа очень крупная. Семяяйцевод длинный и в начале образует крупную
петлю, затем почти прямой. Вдоль него тянется лентовидная простата, ко-
торая затем переходит в довольно длинный семяпровод. Последний впадает
в эпифаллус апикально, лишь с небольшим смещением от центра. Эпифаллус
слабо конический или почти цилиндрический, сильно изогнут, так что зад-
ний конец его находится ближе к атриуму, чем передний, перед впадением
в пенис заметно суженный. Пенис короткоцилиндрический, сильно раздутый,
перед атриумом резко сужается. Внутри него находится большая папилла,
а перед нею продольные складки. У границы перехода эпифаллуса в пенис
крепится мощный диафрагмальный половой ретрактор. Зимрот сообщает
еще о втором ретракторе, который прикрепляется к заднему концу эпифал-
луса. Однако при просмотре голотипа этот мускул не был обнаружен.
Яйцевод — длинная и толстая цилиндрическая трубка. У голотипа семя-
приемник с длинным протоком и небольшим заостренным резервуаром;
у других экземпляров резервуар очень большой и вздутый, а проток очень
короткий. Эти различия, по-видимому, связаны с возрастом и физиологи-
ческим состоянием изученных объектов. Влагалище короткое, но широкое,
плавно переходящее в атриум. Крупная лопастевидная придаточная железа
лежит под глоткой и пищеводом. Она открывается в атриум с помощью
пучка тонких, но толстостенных протоков. Внутри атриума, на той же
стенке, куда открываются протоки придаточной железы, располагается
небольшой конусовидный стимулятор. Поверхность его гладкая, а основание
окружено небольшим валиком.
Распространение. Голотип найден около Боржоми (Грузия).
7 экз. найдены близ Ленкорани и в ближайших горах Талыша (АзССР).
Образ жизни. Обитает в лиственных лесах и скрывается в под-
стилке, под камнями и в почве. Популяции, по-видимому, отличаются низкой
численностью.
362
II. MILACIDAE
Замечание. К числу наиболее близкородственных видов принад-
лежат Milax parvulus Wiktor, 1968, известный из Болгарии, и описанный
с о-ва Родос М. altenai Forcart, 1972. Хотя из-за недостатка материала из-
менчивость всех трех видов еще не прослежена, но анатомически они разли-
чаются очень четко.
2. Milax (Milax) gagates (Draparnaud, 1801) (рис. 487, 488)
D г ар arn aud, 1801 : 100 {Limax); 1805 : 122, Т. 9, Fig. 1—2 {Limax); Ger-
main, 1930 : 107, Fig. 46, 52 (part.); Phillips, Watson, 1930 : 72, Figs. 13,
16, 20, 21, T. 1, Fig. 6, T. 2, Figs. 4, 6, 16 17; P i 1 s b r y, 1948 : 563, Fig. 301, 302;
Quick, 1960 : 149, Fig. 8A, E, K, Fig. 9A, Pl. 1, Fig. 12; V a 1 о v i r t a, 1969 : 345,
Fig. 1.
Locus typicus — «Montpellier»? — Монпелье? (Франция). Типы неизвестны.
Материал: из СССР нет; из других стран 28 экз.
Тело сравнительно коренастое. Рельеф кожи на спине и боках очень хорошо
выражен; морщины уплощенные, но борозды между ними глубокие, особенно
продольные. Ниже киля располагаются 3—4 ряда почти параллельных
ему морщин; остальные ряды наклонные. Мантия занимает не менее х/3
длины тела, у киля с ясной вырезкой. Подкововидная борозда очень четкая,
задние концы ее не смыкаются. Между мантийной щелью и килем около
12[рядов морщин.
Окраска верхней стороны тела от пепельной до черной; спина всегда наи-
более темная, бока книзу постепенно светлеют. Киль обычно окрашен
в цвет спины, но иногда несколько светлее. Мантия с темными боковыми
полосами. Подошва всегда светлее верхней стороны тела: у серых особей
она кремовая, у черных серая. Слизь бесцветная.
Размеры: дл. пол. 50—70, дл. сок. 45—55 мм.
Раковина овальная; ее размеры 4x2.5 мм (рис. 487).
Радула:
с 176 5—66 21—22к
Т + ~ + 2 + 1
Гениталии (рис. 488). Семяпровод впадает в эпифаллус более или
менее апикально. Эпифаллус булавовидный, сильно изогнут, так что его
задний конец ориентирован почти вперед, нередко с небольшим вздутием
у заднего конца. От пениса он отграничен легкой перетяжкой, к которой
крепится ретрактор. Пенис слабоконический, сзади заметно вздутый, а впе-
реди к атриуму плавно суженный. Внутри расширенной части пениса нахо-
дится папилла.
Яйцевод — толстая и прямая трубка, всегда короче семяприемника.
Последний с шаровидным резервуаром и толстым протоком. Атриум корот-
кий. В него несколькими тонкими протоками открыватеся крупная придаточ-
ная железа. Внутри атриума, у места впадения протоков железы, находится
языковидно-конический стимулятор. Поверхность его гладкая, и лишь
у самой вершины иногда можно видеть 2—3 очень небольших сосочка.
-Сперматофор червеобразный, с рядами древовидно ветвящихся шипов.
Распространение. Британские о-ва, Франция, ФРГ, Голлан-
дия. Указания на обитание его в средиземноморских странах, возможно,
относятся к другим видам.
Легко подвергается завозу. В частности, недавно обнаружен в некоторых
теплицах западной Финляндии. (Valovirta, 1969). Кроме того, завезен и
прижился на многих континентах и островах с умеренным климатом.
Образ жизни. В областях с умеренно теплым морским климатом
обитает как в природных, так и в культурных биотопах. В первом случае
придерживается открытых мест, в частности каменистых осыпей. В антропо-
2. TANDONIA
363
генных условиях обитает в садах, парках, огородах и полях, в теплицах
и парниках.
В некоторых странах (Англия, США и др.) наносит существенный вред
полевым и огородным культурам, а также в теплицах и парниках.
487 — раковина; 488 — гениталии и атриальный стимулятор (1).
Замечания. 1. От М. caucasicus (Simr.) данный вид отличается
более крупными размерами, а также иной окраской и строением гениталий.
2. Принадлежит к наиболее крупным представителям своего рода.
Особенно близок к нему М. nigricans (Schulz in Philippi, 1836) (syn.
M. insularis Lessona et Pollonera, 1882). Последний, в частности, отличается
от М. gagates наличием на поверхности стимулятора большого числа шипо-
видных сосочков.
2. Род TANDONIA, Lessona et Pollonera, 1882
'Lessona, Pollonera, 1882 : 54; Hesse P., 1926 : 33 {Milax s. str., sect.
Tandonia)', Wagner, 1930a : 52 {Milax s. str., sect. Tandonia).
Типовой вид — Limax marginatus Draparnaud, 1801 (non Muller,
1774) =Limax rusticus Millet, 1843; по последующему обозначению — Hesse
P., 1926.
364
IL MILACIDAE
При жизни тело стройное. Слизни небольшие или средней величины
'(дл. пол. до 100 мм). Киль простирается на всю спину или ограничен только
задней частью ее. Окраска различная, одноцветная или с рисунком из тем-
ных пятнышек. У светлоокрашенных форм на мантии обычно имеются две
'боковые полосы.
Предсердие не закрыто почкой. Лобус ее умеренно длинный (см. рис. 6,
III). Половой ретрактор крепится к диафрагме позади почки.
Атриум узкий и небольшой, без стимулятора, но нередко внутри с раз-
личными складками или выступами. Придаточные железы различной формы;
они несколькими короткими протоками впадают во влагалище. Эпифаллус
обычно стройный и длинный, длиннее семяприемника. Пенис сравнительно
короткий.
Распространение. Южные Альпы и Балканский п-ов. Отдель-
ные виды обитают в западной и средней Европе, Крыму, Малой Азии и
в Северной Африке.
Подрод TANDONIA s. str.
При движении бока почти параллельные, лишь у заднего конца резко
сужаются. Киль всегда простирается на всю спину. Фоновая окраска обычно
светлая, с темным рисунком из пятнышек. При. сильном развитии рисунка
воздается впечатление общей темной окраски животного.
Придаточные железы слагаются из небольших лопастевидных или мешко-
видных долек или трубочек. Эти железы либо венцом окружают влагалище
и впадают в него у границы с атриумом, или образуют 1—2 клубка или грозди,
которые несколькими короткими протоками симметрично открываются
во влагалище.
Кроме обычного диафрагмального ретрактора, пенис нередко имеет
1—2 ретентора, связывающих его со стенкой тела.
В данную группу входят около 20 видов.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ TANDONIA
1 (4). Небольшие слизни (дл. сок. не более 30 мм). Мантия без боковых тем-
ных полос. Пенис и эпифаллус приблизительно одной длины. Пени-
альная папилла выступает в просвет переднего отдела пениса (пре-
пуциум) в виде кольцевой складки (рис. 496, 504).
2 (3). Окраска серовато-оливковая или пепельно-желтая, без рисунка,
из темных пятнышек. Семяприемник с длинным резервуаром и корот-
ким протоком. Придаточные железы имеют вид рыхлых образований
из лепестковидных фолликул. Атриум небольшой и тонкостенный
.................................... 1. Т. (Т.) cristata (Kai.).
3 (2). Окраска (фоновая) грязно-кремовая с рисунком из большого числа
темных пятнышек, которые, сливаясь, могут придавать спине черный
цвет (киль всегда светлый), а на мантии образовывать сетчатый рису-
нок. Семяприемник с шаровидным резервуаром и длинным протоком.
Придаточные железы имеют вид двух плотных образований неопре-
деленной формы. Атриум более крупный, вздутый и толстостенный
................................ 2. Т. (Т.) kaleniczenkoi (Cl.).
-4 (1). Слизни более крупные (дл. сок. больше 30 мм). Мантия с двумя боко-
выми темными полосами. Эпифаллус всегда заметно длиннее пениса.
Пениальная папилла имеет форму мощного конуса или гриба (рис. 510,
514).
5 (6). Семяпровод плавно переходит в эпифаллус (граница между ними
нечеткая). Эпифаллус образует несколько спиральных извивов . . .
......................................4. Т. (Т.) kusceri (Н. Wagn.).
2. TANDONIA
365
6 (5). Граница между семяпроводом и эпифаллусом четкая. Эпифаллус
не образует спиральных извивов.
7 (8). Тело коренастое, беловато-кремовое, с редкими темными пятнышками.
Резервуар семяприемника сильно вытянут и сзади заострен. Семя-
провод впадает точно в центр заднего конца эпифаллуса.................
.................................... 3. Т. (Т.) rustica (Mill.).
8 (7). Тело стройное, более темно окрашено, особенно спина, за счет более
сильного развития пятнышек, но киль всегда светлый. Резервуар
семяприемника слабо вытянут и сзади более закругленный. Семяпро-
вод впадает в задний конец эпифаллуса немного эксцентрично ....
................................5. Т. (Т.) budapestensis (Hazay).
1. Tandonia (Tandonia) cristata (Kaleniczenko, 1851), comb. nov.
(рис. 489—496)
Kaleniczenko, 1851a : 225, T. 6, Fig. 1 (Krynickillus); Grossu, Lupu,
1961b : 141, Fig. 10—11 (Milax cristatus nanus); Urbanski, Wiktor, 1968 : 80,
Fig. 14 (Milax cristatus nanus); Д аминов, Лихарев, 1975 : 333, рис. 263 (Milax);
Арутюнова, 1975 : 107, рис. 2В, Г, Е (Milax).
Locus typicus — «Otuz» — Отуз (совр. Щебетовка), южный Крым. Типы
неизвестны.
Материал: из СССР — 47 экз. из 12 пунктов; из других стран —
55 экз.
Небольшие слизни с узким и стройным телом. Мантия небольшая, около
•х/4 общей длины, спереди и сзади закругленная, у киля с небольшой вырезкой.
Киль четкий и острый. Рельеф кожи образован крупными и длинными мор-
щинами, разделенными глубокими бороздками. Между килем и мантийной
щелью 8—10 рядов морщин. Подкововидная борозда мантии очень четкая;
нередко оба конца ее сзади соединяются друг с другом, образуя замкнутую
окружность.
Живые слизни серовато-оливковые или пепельно-желтоватые; спина и
особенно мантия несколько темнее. После фиксации тело становится светло-
. или голубовато-пепельным и борозды особенно четкими, так как темный пиг-
мент, скрытый глубоко в коже в виде мелких зерен, размещается главным
образом вдоль них. Голова и шея черноватые, щупальца почти черные.
Подошва обычно светло-кремовая, но иногда боковые доли с серым налетом.
Молодые особи заметно светлее взрослых, иногда беловатые со слегка темной
спиной. Слизь бесцветная или желтая.
Размеры: дл. сок. 22—30, мантия 6—9 мм.
Раковина — рис. 490.
Радула (рис. 491). Боковых зубов около 9, краевых 25—38; число рядов
около 100. Пример: 1 экз., собранный у Ялты; формула:
Гениталии (рис. 493—496). Семяпровод длинный и тонкий. Он впа-
дает в задний конец эпифаллуса, но немного асимметрично. Эпифаллус
булавовидный или в виде конуса, вершиной обращенного к пенису. Обычно
он заметно короче пениса, но иногда той же длины. Чаще всего лежит на
одной оси с пенисом, реже (особенно у не вполне взрослых слизней) распо-
лагается к последнему под углом. Половой ретрактор хорошо развит. Пе-
нис цилиндрический, бочковидный или с несколькими вздутиями, толстостен-
ный и мускулистый. У переднего Конца к нему крепится ретентор, связываю-
щий его с боковой стенкой тела. Основную часть пениса занимает мощная
папилла, которая лишь передним закругленным концом выступает в корот-
Рис. 489. Tandonia (Tandonia) cristata (Kai.). Крым, Ялта, 20 II 1956.
2. TANDONIA
367
кий препуциум (рис. 495). За нею располагается вторая папилла, а между
ними узкая камера. Обе папиллы пронизаны тонким каналом, сзади пере-
ходящим в канал эпифаллуса.
Яйцевод прямой и длинный — по длине равен пенису или даже больше
и имеет вид цилиндрической трубки. Влагалище короткое с широким и мус-
Рис. 490—496. Tandonia (Tandonia) cristata (Kai.) (окончание).
490 — раковина; 491 — зубы радулы; 492 — ретракторы; 493 — дистальная часть гениталий; 49я, —
то же у другого экземпляра; 495 — вскрытый пенис; 496 — продольный разрез через эпифаллуо и пенис.
кулистым вздутием у места впадения семяприемника. Последний без чет-
кого подразделения на проток и резервуар, обычно толстый и длинный,
к заднему концу суженный и несколько заостренный. Внутренняя поверх-
ность протока семяприемника покрыта продольными складками, которые
постепенно увеличиваются в размерах по направлению к атриуму. Прида-
точные железы имеют вид плотных клубков или гроздей, состоящих из удли-
ненных фолликул. Два или три таких клубка впадают во влагалище на гра-
нице с атриумом. Последний очень короткий и внутри имеет несколько
продольных пластинчатых складок.
Распространение. В СССР обнаружен в Крыму (Севастополь,
Ялта, Симферополь), Одессе и в буковых лесах окрестностей Ужгорода
368
II. MILACIDAE
(коллекция ЗИН АН СССР). Кроме того, достоверные ' находки сделаны
в Болгарии и восточной Румынии. Указания на распространение этого вида
в Малой Азии (Retowski, 1889; Simroth, 1899, 1901) требуют проверки.
Образ жизни. В Закарпатской области, а также в Болгарии и
Румынии этот вид чаще всего встречается в буковых и смешанных лесах,
особенно на дне долин и по их тенистым склонам. По-видимому, предпочи-
тает глинистые почвы. Днем прячется под камнями и в лесной подстилке.
В Крыму и Одессе этот слизень встречен только в парках, огородах и
в теплицах, где он весной может наносить заметный вред рассаде табака и
других культур.
Замечания. 1. Долгое время все наши знания об этом виде ограни-
чивались довольно лаконичным описанием внешнего вида, данным Калени-
ченко (Kaleniczenko, 1851а). Лишь сравнительно недавно Гроссу и Лупу
(Grossu, Lupu, 1961b) обнаружили его в восточной Румынии (Кокос), и так
как их экземпляры отличались небольшими размерами (дл. сок. 14 мм),
то они пришли к выводу, что имеют дело с особым подвидом — nanus-
Gr. et Lupu. Среди наших материалов из Крыма и Болгарии встречаются
молодые Т. cristata, по внешнему облику и гениталиям идентичные указан-
ной форме. Поэтому мы рассматриваем ее как синоним данного вида.1
2. Наибольшее родство Т. cristata обнаруживает с Т. kaleniczenkoi,
с которой иногда обитает совместно. Последний вид отличается следующим:
тело более коренастое и крупнее; фиксированные экземпляры окрашены
темнее, с более отчетливыми пятнышками, образующими на мантии сетча-
тый рисунок; зубы радулы заметно крупнее, боковых зубов меньше — только 5;
семяприемник, атриум, придаточные железы и влагалище иной формы
(рис. 502); внутреннее строение пениса также несколько отличается (рис. 504).
2. Tandonia (Tandonia) kaleniczenkoi (Clessin, 1883), comb. nov.
(рис. 497-504)
Boettger 0., 1882 : 98 (Amalia Retowskii, nom. nudum); Clessin, 1883 : 39,
T. 2, Fig. 11 (Amalia kalenzkoi, nom. err.); Лихарев, Раммельмейер, 1952 :
362 (Milax)', Forcart, 1973:21 (Milax samsunensis, non Forcart, 1942).
Locus typicus— «Strateis» — Стратеиз (теперь район Ялты). Типы не-
известны.
Материал: из СССР — 7 экз. из 4 пунктов.
У фиксированных экземпляров тело коренастое, к заднему концу клино-
образно суженное. Мантия широкоовальная, занимает около г/3 длины тела.
Пневмостом лежит слегка постмедиально. Морщины крупные и выпуклые;
между мантийной щелью и килем их 9—11 рядов. Мантийная борозда слабее-
выражена, чем у Т. cristata, но достаточно четкая. Киль менее выпуклый.
Срединное поле подошвы заметно шире боковых.
Фон верхней стороны тела грязно-кремовый. Рисунок образован боль-
шим числом неправильных и различных по форме черных пятнышек. На
спине эти пятнышки обычно сливаются и придают ей черную окраску (кроме-
киля, который всегда светлый). Бока книзу постепенно светлеют, но отдель-
ные меланофоры достигают края подошвы. Мантия тоже покрыта темными
пятнышками, которые местами, сливаясь, образуют сетчатый рисунок.
Боковых полос на мантии нет. Голова и шея черноватые или совершенно-
черные. Подошва серовато-кремовая или сероватая, в толще ее как на сре-
динном, так и на боковых полях залегают отдельные черные хроматофоры,
которые слегка просвечивают сквозь ткани.
1 Рисунок гениталий Milax cristatus nanus Gr. et Lupu, опубликованный в работе
Урбанского и Виктора (Urbanski, Wiktor, 1968), сделан по молодому экземпляру.
, • eel i Кпым Никитский ботанический сад,
Рис. 497. Tandonia (Tandonia) kaleniczenko^ (Cl.). Крым, ник
24 И. М. Лихарев, А. Й. Виктор
370
II. MI LAC I DAE
Размеры: дл. сок. до 27 мм, ширина 7, длина мантии 8.5 мм.
Раковина, коллумелярный мускул и кишечник — см. рис. 498, 500,
501.
Радула (рис. 499). Зубы более крупные чем у Т. cristata. Пример: 1 экз.
из Севастополя; формула:
с 56 36к
з+ — + — X 104.
Гениталии (рис. 502—504). Гонада небольшая, слагается из мелких
светло-серых фолликул и почти целиком закрыта лопастями печени. Белко-
Рис. 498—501. Tandonia (Tandonia) kaleniczenkoi (Cl.).
498 — раковина; 499 — зубы радулы; soo — кишечник; S01 — ретракторы.
вая железа очень крупная и длинная. Длинный и тонкий семяпровод после
нескольких извивов впадает в задний конец булавовидного эпифаллуса.
Последний, плавно сужаясь, переходит в сходный по форме и размерам
пенис, который, наоборот, расширяется к атриуму. Пенис внутри имеет
мощную папиллу и продольные складки, в общем сходные с Т. cristata.
Однако задней папиллы нет, а предпапиллярная камера! крупнее
(рис. 504).
Яйцевод длинный и тонкий, в виде прямой трубки. Перед впадением
в атриум в него открывается семяприемник с шаровидным резервуаром
и тонким прямым протоком. Внутренняя поверхность протока семяприем-
ника у как у Т. cristata. Влагалище короткое, трубчатое. В него открываются
.две лепесткообразные зернистые придаточные железы с короткими и тонкими
протоками. Атриум крупный широкоовальный, боком подтянут к стенке
тела. Внутри него находятся пять продольных гофрированных скла-
док.
Распространение. В СССР: Южный берег -Крыма (Севасто-
поль, Ялта), Симферополь; вне СССР: Понтийские горы (Турция) (см. с. 380).
2. TANDONIA
371
Замечание. Клессин характеризовал данный вид лишь по внеш-
нему облику. Зимрот (Simroth, 1901), хотя, по-видимому, не располагал
материалами по роду Amalia из Крыма, учитывая описания Калениченко
Рис. 502—504. Tandonia (Tandonia) kaleniczenkoi (Cl.). Крым, Никитский ботанический
сад, 23 II 1956.
502 — гениталии; 503 — вскрытый атриум; 50i — вскрытый пенис (видна папилла; прерывистой линией
обозначен просвет задней части пениса и эпифаллуса).
(Г. cristata) я Клессина (Т. kaleniczenkoi), пришел к выводу, что здесь оби-
тают два вида. Наши исследования это подтвердили, и хотя несомненно
желательны дальнейшее изучение и сравнение обоих слизней, можно кон-
статировать, что различия между ними как во внешнем облике, так и в ана-
томии достаточно четкие.
24*
.372
II. MILACIDAE
3. Tandonia (Tandonia) rustica (Millet, 1843), comb. nov. (рис. 505—510)
Draparnaud, 1805 : 124, T. 9, Fig. 7 (Limax marginatus, non Muller, 1774);
Millet, 1843: I, T. 63, Fig. 1 (Limax)-, Leydig, 1876 : 273, T. 12, Fig. 19—21
(Limax carinatus, non Risso, 1826); Simroth, 1885 : 225, 335, T. 7, Fig. 19, T. 10,
Fig. 1—3 (Amalia marginata); Gerhardt, 1940 : 568 (Milax marginatus); For-
cart, 1942a : 118 (Milax); Fromming, 1954 : 227 (Milax); Hu d ec, 1963 : 149,
Fig. 1, T. 13, Fig. 1—3 (Milax); Lupu, 1968 : 31, Fig. 1, 6 (Milax); Wiktor, 1973 :
.51, Fig. 13—14, 87, 103, 225, 245, 246 (Milax).
Terra typica — «France» — Франция.
Материал: из СССР нет; из других стран более 200 экз.
Тело коренастое. При ползании бока становятся почти параллельными
и лишь у заднего конца тело резко сужается. Мантия занимает около 1/3 об-
щей длины, сзади широко закруглена, а у киля слегка вырезана, с четкой
подкововидной бороздой. Киль очень выпуклый.
Верхняя сторона тела почти белая. На этом фоне разбросаны мелкие
черные пятна, располагающиеся главным образом по бороздкам. На мантии
две темные боковые полосы с нечеткими границами. Киль всегда без рисунка,
голова и щупальца черноватые. Подошва светло-кремовая, без пятны-
шек.
Слизь густая, вытягивающаяся в нити, бесцетная; при раздражении сли-
зень выделяет более жидкую и слегка молочно-белую слизь.
Размеры: дл. пол. 80—100, дл. сок. до 45 мм.
Раковина овальная (рис. 506). Нуклеус сильно выдается кверху.
Радула (рис. 507). Боковых зубов 19—21, из них 1—4 самые внешние
без эктокона, т. е. двухзубцовые; остальные же, как и срединный зуб, трех-
зубцовые. Шиповидных краевых зубов 27—39. Число рядов в радуле
около 120.
Г ениталии (рис. 509, 510). Пенис и эпифаллус неясно разграничены
и плавно переходят друг в друга. Эпифаллус в 3—4 раза длиннее пениса.
Тонкий и длинный семяпровод впадает точно в задний конец эпифаллуса.
Пенис у, переднего конца резко конусовидно вздут и утолщен, после чего,
плавно сужаясь, впадает в короткий атриум. Внутри вздутия находится
мощная папилла, покрытая мелкими сосочками, придающими ей вид вино-
градной грозди.
Яйцевод и влагалище довольно толстые, трубковидные. Семяприемник
с длинным и крупным, сзади заостренным резервуаром и коротким, но тол-
стым протоком. У границы между влагалищем и атриумом открываются мно-
гочисленные желтые мелкие трубчатые придаточные железы (каждая железа
..впадает самостоятельным протоком).
Распространение. Наиболее обычен в Средней Европе — Швей-
цария, Австрия, Венгрия, ФРГ (к югу от 52° 40' с. ш.), ЧССР (только в Че-
хии), Польские Судеты. На запад достигает Пиренеев, и центральной Фран-
ции (Савойя, Юра), на юг — Корсики и Калабрии, северных областей Юго-
славии. Есть сообщения о находках этого вида в Бельгии, Голландии и
.Западных Румынских горах.
Образ жизни. Вид теплолюбивый, тяготеет к известнякам. Оби-
тает в горных лесах, особенно на тенистых склонах и в осыпях. Днем
скрывается в подстилке и под камнями. Нередко встречается в руинах
старых замков, хотя не проявляет тяготения к антропогенным биото-
пам.
Замечание. Данный вид наиболее близок к Т. budapestensis (Hazay)
и Т. kusceri (Wagn.). От обоих отличается более светлой окраской. Кроме
того, от первого вида — заостренным семяприемником, а от второго — за-
метно более коротким эпифаллусом.
SOS
0.01
аж
Рис. 505—510. Tandonia [Tandonia) rustica (Mill.).
внешний вид; SOB — раковина; 507 — зубы радулы; 508 — кишечник; 509 — гениталии' 510__
вскрытый пенис (505—509 — из: Wiktor, 1973; 510 — Польша, 14 IX 1962).
374
II. MIL ACID ЛЕ
4. Tandonia (Tandonia) kusceri (H. Wagner, 1931), comb. nov..
(рис. 511—514)
Wagner, H., 1931b : 200 (Milax)', 1931a : 72, Fig. 1, 2 (Milax)-, 1934a : 57, Fig. 11—
12 (Milax)-, Grossu, Lupu, 1961b : 137 —139, Fig. 6—9 (M. rusticus longipenis, M..
rusticus f. balcanicus), Hudec, 1964 : 187, Fig. 1 (M. bojanensis); 1965 : 11, Fig. 1 (M. bo-
janensis); 1967a : 20 (M. bojanensis); Lupu, 1968a : 32, Fig. 1—4 (M. rusticus, M. longi-
penis, M. longipenis balcanicus, M. longipenis bojanensis); Urbabski, Wiktor,
1968 : 86, Fig. 18, 19 (Milax); Osanova, 1970 : 77 (Milax); Hudec, Vasatko,.
1971 : 19, Fig. 8 (Milax); О ш а н о в а, 1972 : 147, рис. 1—2 (Milax); Д а м я н о в,.
Лихарев, 1975 : 330, рис. 261, 262, (Milax).
Locus typicus — «Svetka Petka» — Светка Петка близ Ниша (Югославия)..
Материал: из СССР — 5 экз. из 2 пунктов; из других стран — бо-
лее 200 экз.
Тело коренастое, бока почти параллельные, к заднему концу резко,
суженные. Киль в задней трети тела; он очень четкий и выступает как гре-
бень, а дальше к мантии продолжен одним рядом плоских морщин.
У живых слизней фон розовато-кофейный или серовато-розовый, но после
фиксации он становится серовато-светло-коричневым. Рисунок очень неж-
ный и слагается из мелких черных пятнышек, которые располагаются по
бороздкам; на спине их больше, а к ноге их становится меньше. Мантия
такой же окраски, как и спина, но несколько темнее, с характерным для
рода рисунком из двух нечетких боковых полос, спереди соединяющихся
в виде подковы. Кроме того, иногда еще по середине мантии проходит третья
нечеткая полоса, тоже состоящая из пятнышек. Голова и щупальца корич-
невато-черные. Киль окрашен в цвет фона, без рисунка.
Слизь очень густая и прозрачная, при раздражении на спине выделяется,
более жидкая молочно-белая слизь.
Размеры: дл. пол. до 100, дл. сок. до 65 мм.
Раковина — рис. 512.
Радула. Боковых трехзубцовых зубов 16—18, краевых шиповидных:
37—41; число рядов в радуле ИЗ—115.
Гениталии (рис. 513, 514). Длинный и тонкий семяпровод, отделив-
шись от семяяйцевода, образует несколько резких извивов, а затем, уже
почти прямой, плавно переходит в длинный цилиндрический эпифаллус.
Последний слегка расширяется по направлению к пенису, сначала прямой,
затем делает несколько извивов. Стенки эпифаллуса с мощной кольцевой:
мускулатурой, а внутри со сложной системой мелких складок. Пенис зна-
чительно шире эпифаллуса и подразделяется на более узкий задний и сильно
вздутый передний отделы. Внутри пениса находится мощная папилла,
сходная с таковой у Т. rustica. Ретрактор крепится между пенисом и эпифал-
лусом.
Яйцевод довольно длинный и толстый, плавно расширяется к атриуму.
Семяприемник с шаровидным резервуаром и длинным протоком. С яйцеводом
он сливается так близко от атриума, что влагалище почти не выражено.
Атриум очень короткий. Придаточные железы слагаются из коротких жел-
тых трубочек, в виде нескольких пучков сидящих у основания влагалища.
Распространение. В СССР этот вид обнаружен в одном из
парков и в Ботаническом саду Одессы (одна находка сделана в 1939 г.,
другая — в 1968 г. — коллекция ЗИН АН СССР). Вне СССР — большая
часть Болгарии (в том числе Южная Добруджа), Югославия (восточная Сер-
бия), Румыния (Северная Добруджа), европейская часть Турции.
Образ жизни. В Болгарии обитает на каменистых склонах, по-
крытых кустарником и лесом. Встречается и на открытых ксеротермных био-
топах, преимущественно на почвах, богатых известью. Нередко образует-
большие популяции.
, 376
II. MILACIDAE
Как указывает Ошанова (1972), яйца этого слизня имеют диаметр 4—
4.5 мм, а эмбриональное развитие продолжается 16—20 дней.
Замечания. 1. По окраске и рисунку данный вид наиболее сходен
с другим балканским слизнем — Т. serbica (Н. Wagner, 1931). Однако у по-
следнего, кроме уже упомянутых мелких темных пятнышек, на спине и бо-
ках разбросаны редкие, но крупные многоугольные темные пятна, которых
нет у Т. kusceri. По гениталиям Т. serbica отличается от Т. kusceri вздутым'
неправильной формы пенисом и более коротким эпифаллусом.
2. Очень возможно, что описанная Линдгольмом (Lindholm, 1908)
Amalia rossica Lindh. идентична T. kusceri (Н. Wagner, 1931).
В пользу такого предположения говорит не только то, что rossica была
описана по экземплярам, найденным в Одессе, т. е. там же, где обитает и
kusceri, но и значительное сходство внешнего облика обоих слизней.
К сожалению, Линдгольм не приводит сведений о внутреннем строении
своего вида. Типы же утрачены. Поэтому, хотя не исключено, что A. ros-
sica является старшим синонимом Т. kusceri, учитывая, что наши знания
о последнем гораздо полнее, мы не считаем разумным прибегать к правилу
приоритета и давать предпочтение названию, предложенному Линдголь-
мом.
5. Tandonia (Tandonia) budapestensis (Hazay, 1881), comb. nov.
(рис. 515—522)
Menegazzi, 1855 : 64, T. 2, Fig. 1—7 (Limax gagates, non Draparnaud, 1801);
Leydig, 1876 : 276, T. 12, Fig. 22 (Limax gracilis, non Rafinesque, 1820); Hazay,.
1881 : 37, T. 1, Fig. 1 (Amdlia); Kimakowicz, 1884 : 103 (A. cibiniensis); Simroth,.
1886b : 323 (A. gracilis); Phillips, Watson, 1930 : 65, Fig. 1, 4, 6, 8—11, 14,.
17—19, T. 1, Fig. 1—3, T. 2, Fig. 1, 2, 5, 8—13 (M. gracilis); Gerhardt, 1936a:
315 (M. gracilis); Boettger C., 1943 : 27 (Milax); Lozek, 1956 : 189, T. 24, Fig. 8
(M. gracilis); Quick, 1960 : 160, Fig. 8C, G, L, 9C, T. 2, Fig. 20 (Milax); Grossu,
Lupu, 1961b : 131, Fig. 1 (M. gracilis); Hu d e c, 1963 : 149, Fig., 1—2, T. 13, Fig. 2,.
3a, 4a—c (Milax); Wiktor, 1967a : 331, Fig. 5 (Milax); Urbanski, Wiktor,
1968 : 80 (Milax); Wiktor, 1971a : 262 (Milax); 1973 : 54, Fig. 15, 16, 92, 108, 226,
247, 248 (Milax); Дамянов, Лихарев, 1975:328, рис. 257 (Milax).
Locus typicus — «Budapest» — Будапешт. Типы неизвестны.
Материал: из СССР — нет; из других стран — более 50 экз.
Тело очень стройное, к заднему концу плавно суженное. .Киль отчетли-
вый. Мантия занимает около 1li длины тела. Подошва узкая.
За счет сильного развития на верхней стороне тела очень мелких темно-
коричневых пятнышек тело кажется одноцветно темным. Лишь на киле,
где пятнышек нет, и на нижней части боков, где пятнышки располагаются
редко, проступает основная, т. е. фоновая, серовато-кремовая или оран-
жевая окраска. Мантия тоже сильно испещрена темными пятнышками,
которые, сгущаясь вдоль подкововидной борозды, образуют две боковые
полосы. Голова черноватая. Подошва одноцветно коричневато-кремовая
или с легкими, почти прозрачными темными пятнышками. Слизь густая и
бесцветная, при раздражении желтоватая и жидкая.
Размеры: дл. пол. до 70, дл. сок. до 40 мм.
Раковина (рис. 517) узкая, овальная, симметричная. В отличие от
Т. rustica (Mill.) нуклеус не выдается кверху.
Радула (рис. 518). Боковых зубов 8—15, из них 2—4 самых внешних
двухзубцовые, остальные же, как и срединный зуб, трехзубцовые. Краевых
зубов 29—49, все они шиповидные. Число урядов в радуле около 94.
Гениталии (рис. 522). Гонада относительно крупная, вытянутая.
Белковая железа удлиненной формы и слагается из небольших продолго-
ватых долей. Семяпровод длинный и [тонкий; он впадает в эпифаллус
2. TANDONIA
377
апикально, но в отличие от Т. rustica немного асимметрично. Эпифаллус
толстый, цилиндрический или веретеновидный, сзади несколько расширен,
почти в два раза длиннее пениса. Пенис конический или шаровидный, в пе-
редней части заметно широкий, к атриуму суженный.
Обе придаточные железы сложены из мелких лопастей и несколькими
протоками впадают в матку близ атриума.
516
Рис. 515—519. Tandonia (Tandonia) budapestensis (Hazay) (из: Wiktor, 1973 ).
-515 — внешний вид; 516 — участок подошвы; 517 — раковина;''5Г« — зубы радулы; 519 — кишечник.
Семяприемник сзади всегда притупленный в отличие от Т. rustica. Резер-
вуар его яйцевидный или овальный, проток сравнительно короткий.
Сперматофор, за исключением одного из концов его (около 1/6 общей
длины), покрыт различной формы шипами. В начале эти шипы располага-
ются двумя рядами по одной стороне сперматофора и каждый на конце раз-
двоен. Затем рядов становится четыре, они располагаются равномерно по
периферии, и форма шипов усложняется. К противоположному концу сперма-
тофора шипы становятся мельче, и они прижаты к поверхности.
Распространение. Ареал этого вида плохо изучен. По-види-
мому, в природных биотопах он обитает в южной Европе и в некоторых
южных областях Средней Европы. Как синантропный вид, он встречается
в южной Польше, Чехии, на Британских о-вах, в Исландии и о-вах Гебриды.
Одна находка сделана недавно в антропогенных условиях близ Трабзона
(черноморское побережье Турции).
В литературе имеется только два указания о присутствии этого слизня
на территории СССР (под названием Amalia gracilis Leydig). Первое сделано
в одной из ранних работ Зимрота (Simroth, 1886b), где он говорит о присут-
378
II. MILACIDAE
ствии данного вида в Севастополе. По-видимому, автор не анатомировал со-
ответствующий сбор и ограничился лишь внешним сходством. Во всяком
случае позднее (1901) он уже не упоминает о присутствии этого вида не
только в Крыму, но и вообще в России.
Другое указание об обитании этого вида в лесных оврагах Калининград-
ской области сделано Гильбертом (Hilbert, 1909а). Вероятно и здесь имела
Рис. 520—522. Tandonia (Tandonia) budapestensis (Hazay). '
его — мантийный комплекс, вид снизу; 521 — ретракторы; 522 — гениталии.
место ошибка в определении, так как для данной территории больше не
встречается упоминаний об этом слизне.
Образ жизни. В природных биотопах этот вид обитает преиму-
щественно по каменистым осыпям, заросшим кустарниками. В антропоген-
ных условиях живет в парках, огородах и на свалках. Может встречаться
на открытых и сухих местах.
Отмечен как вредитель огородных культур.
Замечание. От остальных видов того же рода, упомянутых в на-
стоящей работе, отличается довольно характерной окраской, хотя иногда
возникают трудности с определением особей, окрашенных нетипично. По ге-
ниталиям отличается от других видов Tandonia, асимметричным впадением
семяпровода в эпифаллус, формой эпифаллуса, отсутствием ретентора на
пенисе и формой папиллы пениса.
ДОПОЛНЕНИЕ К СЕМЕЙСТВУ MILACIDAE
379
Дополнение к семейству MILACIDAE
В заключение обзора видов данного семейства дается характеристика
еще одного вида — Milax samsunensis Forcart, 1942, систематическое поло-
жение которого пока еще остается неясным. Описание было сделано по един-
ственному экземпляру. Он хранится в Базеле (Швейцария); нам не удалось
его получить для переисследования.
Такие особенности гениталий, как отсутствие придаточных желез на
влагалище и атриуме и стимулятора внутри атриума, ставят этот вид со-
вершенно особняком как от Milax, так и от Tandonia. Столь необычное для
Milacidae строение гениталий вместе с отсутствием коллекционного мате-
риала затрудняет определение нашего отношения к этому виду. Не ясно,
следует ли его выделить в особый род или же рассматривать указанные
черты гениталий как аномалию или возрастные особенности.
Milax samsunensis Forcart, 1942 (рис. 523)
Forcart, 1942b : 239, Fig. 1; 1953 : 21
Locus typicus — «Dervent Burnu» — Дервент Бурну, между Самсуном
и Чаршамба (вилайет Самсун, Турция). Голотип в Базельском естественно-
историческом музее, № 4600а.
Рис. 523. Milax samsunensis Forcart. Дистальная часть гениталий (из: Forcart, 1942b).
Тело стройное, сзади как бы обрубленное. Киль простирается на всю
спину.
Окраска фона серовато-фиолетовая.1 Верхняя сторона, за исключением
киля, покрыта темно-серыми пятнышками, которые на спине имеют вид
густо лежащих точек. На мантии эти пятнышки крупнее, но разбросаны более
редко. Только в области подкововидной борозды они заметно гуще. Подошва
1 По Форкарту, «цвета мяса».
380
I. PHILOMYCIDAE
имеет ту же фоновую окраску, но несколько светлее; рисунок на ней слага-
ется из светло-серых пятнышек.
Размеры: дл. сок. 20, ширина 4.5 мм; длина мантии 6.2, ширина 4.5 мм.
Гениталии. Тонкий семяпровод впадает в эпифаллус почти
апикально. Эпифаллус и пенис очень похожи друг на друга как по форме,,
так и по размерам: оба они короткоцилиндрические, почти шаровидные и
разделены глубокой кольцевой перетяжкой, к которой крепится половой
ретрактор. Пенис внутри с крупной и мускулистой папиллой, занимающей
почти всю полость. Короткое влагалище очень мускулистое и толстостенное;;
в атриум оно выдается в виде папиллы,1 пронизанной тонким каналом.
Яйцевод прямой, сзади довольно толстый, к переднему концу плавно сужен-
ный. Семяприемник состоит из очень крупного яйцевидного резервуара
и сравнительно длинного (длиннее пениса) протока. При слиянии с яйце-
водом узкий канал протока резко расширяется, образуя особую камеру,,
лежащую позади папиллы влагалища. Стенки этой камеры имеют складки.
Влагалище и атриум лишены придаточных желез; атриум без стимулятора.
Распространение. Известны две находки этого вида, которые
сделаны на небольших низменностях черноморского побережья Малой Азии:
одна к востоку от города Самсун (Forcart, 1942b), а другая близ города
Зонгулдак (Forcart, 1953).
Замечание. Если из-за отсутствия в нашем распоряжении голотипа
данного вида мы лишены возможности высказаться о его таксономическом:
положении, то по поводу находки у Зонгулдака наше мнение определилось
после того, как мы получили от Форкарта единственный экземпляр, най-
денный у этого города. Установлено, что как по внешнему облику, так и по
внутреннему строению этот экземпляр является Tandonia kaleniczenkot
(Clessin).
4. Надсемейство A R I О NOIDEA Gray, 1840
Полуслизни и слизни, раковина которых претерпела различную степень
редукции вплоть до полного ее исчезновения (Philomycidae). Спина в попе-
речном сечении полукруглая, а задний конец цефалоподиума плавно и ши-
роко закруглен. Имеется четкая перипедальная бороздка; супрапедальная
бороздка и оторочка неясные. Пневмостом и мантийная щель располага-
ются сильно антемедиально. Подошва без продольных педальных бороздок.2
Челюсть чаще всего одонтогнатная, реже оксигнатная. Базальные пла-
стинки краевых зубов короткие, но широкие (рис. 11, 1—Ш).
Сердце лежит почти в центре мантийного комплекса, ось его наклонена
налево. Почка кольцом окружает сердце или почти замкнута вокруг него
(рис. 6, I, II).
Гениталии обнаруживают тенденцию к вторичному упрощению за счет
редукции стилофора (Philomycidae) или постепенной утраты пениса (Arioni-
dae). Имеется эпифаллус, формирующий сперматофоры.
Растительноядные полифаги.
В состав надсемейства входят Philomycidae и Arionidae. Число видов
около 80.
I. Семейство PHILOMYCIDAE Gray, 1847
Gray, 1847 : 170 (Philomycini, subfam.); Keferstein, 1866a : 188 (Philomyci-
dae)-, 1866b : 64, T. 1; Tryon, 1884, Struct, a. Syst. Conch., 3 : 83 (Tebennophoridae)-»
Hoffmann, 1924 : 363, Fig. 1—16, T. 22, Fig. 1—19; Pilsbry, 1948:748, Fig. 402—412.
1 Форкарт о ней не пишет, но она ясно видна на рисунке.
2 Среди восточноазиатских и североамериканских Arionidae имеются роды с посг-
медиальным пневмостомом и с педальными бороздками.
I. PHILOMYCIDAE
381
Слизни небольшие или средних размеров (дл. пол. до 100, дл. сок. до
75 мм) с червеобразным, снизу сильно уплощенным телом, напоминающим
сплющенную половину продольно расщепленного цилиндра; сзади оно при-
тупленное, без киля и каудальной железы. Мантия гладкая, очень крупная,
закрывает всю верхнюю сторону и бока почти до самой подошвы, до которой
остается узкий промежуток. Почти по всему периметру край мантии сраста-
ется с телом, капюшона нет. При движении из-под мантии выступают голова
со щупальцами и задний конец ноги. Пневмостом лежит сильно антемеди-
ально, т. е. приблизительно на 1/8 длины тела от переднего конца мантии.
Половое отверстие находится непосредственно позади правых щупалец.
Подошва немного шире тела, не разделена на продольные доли. Движение-
аритмичное.
Раковины нет, но в толще мантии имеется большая полость, заполнен-
ная жидкостью.
Краевые зубы радулы двухзубцовые, базальная пластинка этих зубов
почти прямоугольная и от внутренних к внешним зубам постепенно расши-
ряется. Челюсть одонто- или оксигнатная. Кишечник двухпетлевого типа,
сильно перекручен; первая петля вместе с зобом длиннее второй и прости-
рается назад дальше. Передняя лопасть печени располагается слева между
витками кишечника; задняя лопасть меньших размеров, лежит за первой
петлей и образует вершину внутренностного мешка.
Ретракторы глотки, щупалец и головы представляют собой самостоятель-
ные мускулы, которые сзади раздельно прикрепляются к боковым стенкам
тела у подошвы (рис. 529). При этом каждый ретрактор перед прикреплением
веерообразно распадается на серию отдельных мускулов. Половой ретрак-
тор лежит косо к оси тела, так как является ветвью ретрактора левых щу-
палец; впереди он крепится к пенису.
Сердце лежит почти у середины левого края мантийного комплекса
(рис. 6, 7); ось его больше чем под углом 45° наклонена налево, т. е. от про-
дольной оси тела. Аорта сравнительно короткая и проходит сквозь почку.
Кровеносная сеть легкого в основном располагается перед почкой, но также
простирается назад по бокам от нее. Почка бобовидная; ее длинная ось слегка
наклонена к продольной оси тела; имеется небольшой лобус. Мочеточник
начинается от заднего конца почки и открывается наружу рядом с анусом,,
позади пневмостома. Мочевого пузыря нет.
Для гениталий характерны хорошо развитый пенис, длинный и обычно
очень вздутый проток семяприемника, отсутствие эпифаллуса и придаточных
желез на женских путях.1 У некоторых видов имеется лигула или стилофор
со стрелой, напоминающий стилофор Helicoidea (Шилейко, 1978).
Распространение. Природный ареал семейства имеет дизъ-
юнктивный характер (рис. 41, 5): с одной стороны, восточная и юго-восточ-
ная Азия с соседними островами (Приамурье, Приморье, Япония, Корей-
ский полуостров, восточные провинции КНР, Индокитай, Филиппины и
западная часть Зондского архипелага), с другой — юго-восточные области
Северной Америки, часть Центральной Америки и Колумбия.
Некоторые виды завезены на о. Маврикия, Гавайские о-ва и в Чили.
В семейство входят три рода: Meghimatium Hasselt, 1823; Philomycus
Rafinesque, 1820; Palltfera Morse, 1864. Первый обитает в Азии, остальные
два в Америке. Общее число видов около 20.
Внешний облик представителей всех родов очень сходен. Главные раз-
личия в строении гениталий.
х У некоторых американских видов атриум окружен железистым слоем, который,
может слегка заходить и на влагалище.
382 I. PHILOMYCIDAE
Род MEGHIMATIUM Hasselt, 1823
H a s s e 1 t, 1823, Algem. Konst-en Letterbode, Harlem, 2 : 232; Benson, 1842 :
486 {Incilaria)', Hoffmann, 1924 : 385; Jutting, 1952: 424.
Типовой вид — M. striatus Hasselt, 1823; по монотипии.
Внешнее строение — см. характеристику семейства.
Общая окраска кремовая, желтоватая или оранжевая. На этом фоне
ксегда имеется рисунок, состоящий из черноватых или коричневых пятен,
которые, сливаясь, образуют полосы или сетку. Подошва всегда светло-
кремовая или желтая.
Срединные и боковые зубы радулы с мощными, но тупыми мезоконами.
Срединные зубы без дополнительных зубцов, у боковых же имеются экто-
коны, которые у внутренних чуть намечены, но у внешних более заметны.
Еще сильнее они развиты на краевых зубах.
Распространение. Значительная часть восточной и юго-восточ-
ной Азии и соседних островов и архипелагов.
К этому роду принадлежат 4 вида: М. bilineatum (Benson, 1842), М. pi-
ctum (Stoliczka, 1873), M. striatum (Hasselt, 1823), M. tarmes (Sarasin, 1899).
На территории СССР обитает только первый вид.
Meghimatium bilineatum (Benson, 1842) (рис. 524—530)
Benson, 1842 : 486 {Incilaria)', Schrenck, 1867 : 693 {Incilaria)', Simroth,
1901:62 {Philomycus)', Hotirnunn, 1924:364, Fig. 1—7, T. 22, Fig. 1, 2, 6—12
[Philomycus {Meghimatium)]', Wachtler, 1934 : 168, Fig. 18 {Philomycus)', Лиха-
рев, Раммельмейер, 1952 : 306, рис. 234 [Philomycus {Meghimatium)]', Jut-
ting, 1952:430, Fig. 84—86; A 11 e n a, 1966:234, Fig. 1.
Locus typicus — «Chusan» — о. Чжоушань (КНР). Типы неизвестны.
Материал: из СССР — 62 экз. из 7 пунктов, из других стран —
50 экз.
Внешнее строение — см. характеристику семейства.
Окраска верхней стороны тела (мантии) слагается из желтовато-белого
или кремового фона и темно-коричневого рисунка, состоящего из полос и пя-
тен. Посередине до заднего конца тела простирается коричневая полоса
•с неправильными контурами, ограниченная немного более темными пятнами.
Сходные, но более темные полосы проходят по бокам. Их верхние края имеют
неправильные очертания и окрашены более интенсивно. В связи с этим
полосы сильнее контрастируют с вышележащими светлыми промежутками,
чем срединная полоса. Книзу боковые полосы переходят в систему мелких
пятен, доходящих до края ноги. Пневмостом окружен светлым полем. Голова
спереди и сверху черная. Подошва кремовая.
Молодые особи заметно светлее — фоновая окраска светло-серая, а ри-
сунок проще и слагается только из трех темных полос.
Размеры: дл. сок. до 50, ширина до 8 мм.
Кишечник, система ретракторов, мантийный комплекс — см. рис. 6, I,
526, 527, 529. Подробности — см. характеристику семейства и рода.
Гениталии (рис. 530). Гонада округлая, лежит позади зоба и пол-
ностью скрыта между лопастями печени. Проток ее тонкий, извилистый и
не заканчивается талоном.1 Форма белковой железы изменяется с возрастом.
Семяяйцевод сравнительно короткий и рано разделяется на длинные семя-
провод и яйцевод. Тонкий семяпровод, как обычно, тянется вперед до атриума,
затем изгибается назад и впадает в пенис почти апикально. Пенис трубковид-
яый, сзади более толстый, к атриуму плавно суженный; половой ретрактор
1 Возможно, что это особенность семейства или рода.
MEGHIMATIUM
383'
крепится к нему апикально, рядом с семяпроводом. Этот мускул пролегает
под ретрактором правого глазного щупальца, и при сильном сокращении жи-
вотного оба они могут перекрещиваться.
Рис. 524, 525. Meghimatium.
bilineatum (Bens.).
524 — взрослый слизень, Ки-
тай, Юньнань, 17 IV 1957;
525 — молодой слизень, лес
у с. Кухари, Хабаровский край,
23 IV 1971.
Яйцевод имеет вид длинной трубки, слегка сужающейся вперед. В него
(недалеко от атриума) впадает семяприемник. После их слияния образуется
короткое влагалище. Семяприемник имеет округлый резервуар и очень
длинный проток, благодаря чему весь орган длиннее яйцевода или пениса.
У половозрелых особей проток посередине веретеновидно вздут и нередко,
даже шире резервуара. У молодых экземпляров он тонкий и стройный, т. е..
384
I. PHILOMYCIDAE
не вздутый. Атриум крупный грушевидный, т. е. сзади широкий, а к поло-
вому отверстию резко суженный. Влагалище впадает в него сзади, а пенис
сбоку.
Характерно внутреннее строение некоторых половых органов. Внутри
^задней части пениса находятся многочисленные мелкие сосочки, располо-
Рис. 526—529. Meghimatium bilineatum (Bens.). Нагасаки, Япония, 1888.
526 — кишечник; 521 — почка и сердце, вид снизу; 528 — челюсть; 529 — ретракторы.
жеиные в три продольные полосы. Впереди сосочки двух полос постепенно
удлиняются и переходят в косые складки — пластинки, сосочки третьей
полосы простираются до самого атриума без изменений. Стенки вздутой
части протока семяприемника утолщены за счет мощного развития желез
и сложной системы внутренних продольных складок. Внутри атриума,
вблизи устьев пениса и влагалища, находится мощная кольцевая складка
и сплющенная лигула.
Передний конец пениса проходит через развилку, образованную ретрак-
торами правых щупалец (рис. 529).
Распространение. В СССР этот вид был найден лишь несколько
раз (все материалы в ЗИН АН СССР). Две находки сделаны к югу от Хаба-
ровска (в горах Хехцир у ж.-д. станции Корфовская и еще южнее у ж.-д.
«станции Вяземская), три сбора в окрестностях с. Петропавловка
MEGHIMATIUM
385
(около 50 км от Хабаровска вниз по Амуру) и одна находка близ бухты
Ольга (Японское море, южное Приморье). Вне СССР: Корейский п-ов, вос-
точные и южные провинции КНР (в том числе Сычуань и Юньнань — по мате-
Рис. 530. Meghimatium bilineatum (Bens.). Гениталии. Нагасаки, Япония, 1888.
риалам ЗИН АН СССР), Вьетнам, Япония и некоторые острова Восточно-
Китайского и Южно-Китайского морей (Тайвань и др.), а также о-ва Ява
и Калимантан. Завезен на Гавайские о-ва и о. Маврикий.
Образ жизни. На Дальнем Востоке СССР обитает в смешанных
и широколиственных лесах, в гнилых пнях и на грибах.
1/а25 И. М. Лихарев, А. Й. Виктор
386
II. ARIONIDAE
II. Семейство ARIONIDAE Gray, 1840
Gray, 1840 : 101; Hesse P., 1926 : 58; Pilsbry, 1948 : 660; Quick.
1960 : 119; Riedel, Wiktor, 1974 : 85.
Мантия закрывает только переднюю часть тела. Челюсть одонтогнатная.
Срединный зуб радулы с тремя четкими зубцами, боковые 1 и краевые зубы
двухзубцовые, и лишь самые внешние недоразвитые краевые зубы однозуб-
цовые. Как и у Philomycidae, в отличие от остальных семейств базальная пла-
стинка краевых зубов в поперечном направлении постепенно расширяется,,
в продольном укорачивается.
Кишечник двухпетлевого типа и сильно перекручен. Почка кольцом
окружает сердце. Для гениталий характерно наличие эпифаллуса.
Распространение. Европа, Северная Африка, Сибирь, Даль-
ний Восток, Гималаи, западные области Северной Америки, Центральная
Америка (на юг до Коста-Рики). Около 60 видов.
Замечания. 1. Приведенный выше диагноз Arionidae составлен
в основном по данным Пилсбри (Pilsbry, 1948), который дал наиболее пол-
ную характеристику семейства и входящих в него подсемейств.
Авторы настоящей работы, располагая довольно большим материалом
по европейско-сибирскому роду Arion и лишь небольшими сборами по северо-
американским и восточноазиатским Arionidae, в данное время не считают
возможным провести ревизию всего семейства и изменить или дополнить.,
его характеристику.
2. Семейство слагается из. 4 подсемейств, сильно различающихся как;
по внешнему облику, так и по внутреннему строению. Наиболее примитивны:
Binneyinae — полуслизни с крупным наружным внутренностным мешком,
закрытым раковиной, хотя и подвергшейся некоторой редукции, но еще
наружной. Они обитают на западе Северной Америки.
Остальные подсемейства объединяют настоящих слизней с внутренней
раковиной-пластинкой, которая у Arion редуцирована до группы известко-
вых зерен и кристаллов.
Обширное подсемейство Ariolimacinae (некоторые представители дости-
гают 260 мм длиной) характеризуется слиянием задних концов ретракторов
глотки и щупалец в общий мускульный ствол — особенность, указывающая
на их относительную примитивность. Эта группа обитает вдоль тихоокеан-
ского побережья Северной Америки — от Аляски до Калифорнии.
Для третьего подсемейства — Anadeninae характерно раздельное при-
крепление задних концов ретракторов глотки и щупалец, что свойственно
также и Arioninae (см. ниже), но в отличие от последних у них всегда име-
ется хорошо развитый пенис, а вторая петля кишечника длиннее первой.
Ареал этого подсемейства дизъюнктивный, так как слагается из двух изо-
лированных частей: 1) американской (приблизительно как у Ariolimacinae^
и восточноазиатской — от западных Гималаев, через южный Китай до Япо-
нии.
Наконец, четвертое подсемейство т— Arioninae отличается отсутствием
пениса, иной формой кишечника и распространением в Палеарктике.
Некоторые авторы (Cockerell, 1891; Pilsbry, 1948) склонны к данному
семейству относить род Oopelte Morch, 1867, обитающий в южной Африке,
выделяя его в особое подсемейство Oopeltinae Cockerell, 1891. Зимрот (Sim-
roth, 1910b) возражал против этого, считая целесообразным выделить эту
группу в особое семейство.
Алтена (Altena, 1966), суммируя собственные и литературные данные,
подробно рассмотрел доводы в пользу как одной, так и другой точки зре-
1 Лишь первые 1—2 боковых зуба иногда трехзубцовые.
ARION
387
иия, но воздержался от окончательного вывода. Нам представляется, что
-Oopelte заслуживает выделения в самостоятельное семейство. В дополнение
ж тому, что привел Алтена, в пользу этого говорит и внешний облик слизней—
крупная мантия, которая хотя и сдвинута к переднему концу, но закры-
вает больше половины тела животного (Altena, 1966, Т. 1—2).
Подсемейство ARIONINAE Gray, 1840
Hesse Р., 1926 : 59; Pilsbry, 1948 : 666; Quick, 1960 : 119; Wiktor,
1973 : 22; Riedel, Wiktor, 1974 : 86.
Тело относительно массивное и широкое. Пневмостом лежит антемеди-
ально (от переднего края мантии на расстоянии 1/3—1/4 ее длины). Капюшон
очень короткий и занимает х/4—1/5 мантии. Педальных бороздок нет, но на
подошве видно срединное локомоторное поле. Над задним краем подошвы
.имеется крупная каудальная ямка.
Раковина внутренняя. У рода Geomalacus. Allman, 1842 представлена
известковой пластинкой без четкого нуклеуса, а у рода Arion от нее оста-
лась группа зерен или кристаллов, лежащих в толще мантии.
Втягивание переднего конца тела обеспечивают три самостоятельных
ретрактора (рис. 539): два щупальцевых (левый и правый) и глоточный.
Места прикрепления задних концов ретракторов расставлены широко:
щупальцевые у заднего края диафрагмы, глоточный — между щупальце-
выми, но еще более назад. Правый ретрактор лежит слева от эпифаллуса
и яйцевода, не перекрещиваясь с ними.
Боковые зубы радулы плавно и незаметно переходят в краевые. Первая
петля кишечника всегда длиннее второй. Гонада располагается за кишеч-
ником. Пенис и влагалище отсутствуют; эпифаллус, яйцевод и семяприем-
ник открываются независимо друг от друга прямо в атриум. Половой ретрак-
тор впереди распадается на 2—3 ветви.
Распространение. Бблыпая часть Европы, Сибири и Даль-
него Востока СССР; несколько .видов обитают в Северной Африке.
Замечания. Основная масса видов (около 30) принадлежит к роду
Arion Ferussac, 1819. Два вида с Пиренейского п-ова и из юго-западной Ир-
ландии образуют род Geomalacus Allman, 1842. Наконец, два рода — Letour-
neuxia Bourguignat, 1866 (5 видов из Алжира и Марокко) и Ariunculus
Lessona, 1881 (около 7 видов из Восточных Альп, Пиренейского и Апеннин-
ского п-овов, о-ва Сардинии и Алжира) пока еще очень плохо изучены,
в связи с чем систематическое положение их во многом неясно.
Род ARION Ferussac, 1819
Ferussac, 1819 : 50, 53; М о quin-T an d on, 1855 : 10 (Locked)', S i m-
roth, 1901:40; 0 k 1 a n d, 1923; 4; Hesse P., 1926 : 60; P i 1 s b r y, 1948:666;
Quick, 1960 : 124, Wiktor, 1973 : 22.
Типовой вид—A. empiricorum Ferussac, 1819, part. syn. Limax
ater или L. rufus Linnaeus, 1758; по последующему обозначению — Hesse P.,
1926.
Тело несколько уплощенное. Задний конец спины мягко закруглен;
при движении из-под него выступает задний край подошвы, который
может вытягиваться, образуя треугольник. Киля нет, или он нечеткий.
Мантия овальная с зернистой поверхностью. Подошва — наиболее широкая
часть тела. Глубокая перипедальная бороздка лежит высоко над толс-
тым краем подошвы; супрапедальная бороздка нечеткая; оторочка узкая,
ло четкая и образована одним рядом длинных морщин. Имеются три заты-
25*
388
II. ARIONIDAE
лочные складки. Большинство видов имеют по паре продольных темных
полос на спине и мантии. Половое отверстие лежит вблизи от мантийной
щели.
От внутренней раковины осталась группа известковых зерен и кристал-
лов, лежащих в толще мантии. Кишечник более или менее сильно перекру-
чен. Левая печень образует вершину
Рис. 531—537. Сперматофоры различных видов
Arion (из: Riedel, Wiktor, 1974).
внутренностного мешка, внутри ко-
торой лежит гонада. Педальная
железа компактная и занимает
1/3—2/3 длины ноги.
Сердце лежит почти в цент-
ре мантийного комплекса (рис.
6, II); ось его под утлом
около 45° наклонена налево,
т. е. от продольной оси тела.
Аорта длинная. Кровеносная
сеть легкого располагается пе-
ред почкой, а у некоторых
видов, кроме того, простира-
ется назад по ее правой сто-
роне. Почка кольцевидная, с пе-
рикардием и сердцем в центре;
лобуса нет. Мочеточник очень
короткий и открывается на-
ружу рядом с анусом, почти
на одном уровне с пневмосто-
мом. Мочевого пузыря нет.
Половой ретрактор крепится
у заднего края комплекса,
а остальные ретракторы еще
более сзади.
Эпифаллус хорошо развит.
Половой ретрактор впереди
распадается на 2—3 ветви:
одна к протоку семяприемника,
вторая к яйцеводу и, наконец,
иногда бывает третья ветвь
к атриуму; сзади этот мускул
прикрепляется к диафрагме
Довольно длинный
1 задний —
531 — A. fasciatus Nilss.; 532, 533 — A. circurnscriptus
John.; 534 — A. hortensis (F6r.); 535 — A. rufus (L.);
536, 537 — A. subfuscus (Drap.).
справа от ретрактора левых щупалец (рис. 539).
и нередко очень массивный атриум состоит из двух отделов:
мускулистый, куда открываются половые пути и семяприемник; передний—
железистый, открывающийся наружу половым отверстием. У некоторых ви-
дов внутри заднего отдела атриума или в прилегающей к нему части яйце-
вода имеется особый орган — лигула, по-видимому, выполняющий функ-
цию стимулятора. Лигула (рис. 547) образована двумя волнистыми склад-
ками, концы которых могут срастаться, образуя овальный валик. Сперма-
тофор длинный, иногда с зубчатыми продольными ребрами (рис. 531 —
537).
Распространение. Большая часть равнинных и горных лесных
областей Европы, Урала, средней и восточной Сибири и Дальнего Востока
СССР. Несколько видов завезено в Северную Америку.
1 Внешне не всегда четко разграничены.
ARION
889
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ ARION
1 (6). Крупные слизни (дл. пол. 80—150, дл. сок. 50—80 мм) на спине
с четкими, крупными и высокими, иногда острыми морщинами. Тело
окрашено в один цвет, за исключением края подошвы, который у не-
которых может быть иного цвета или пестрый. Взрослые особи без
боковых темных полос на мантии и спине.1 Внутри крупного атриума
находится большая лигула языковидной или ложковидной формы
(подрод Arion s. str.).
2 (3). Яйцевод в несколько раз длиннее атриума.......................
.................................... 3. A. (A.) lusitanicus Mab.
3 (2). Яйцевод приблизительно той же длины что и атриум.
4 (5). Передний отдел атриума длиннее заднего; боковое вздутие заднего
отдела очень небольшое, так что в целом атриум почти симметричный
.............................................. 2. A. (A.) ater (L.).
5 (4). Передний отдел атриума короче или равен заднему; боковое вздутие
заднего отдела очень большое, так что атриум резко асимметричный
. . ............................................1. A. (A.) rufus (L.).
6 (1). Средние и небольшие слизни (дл. пол. 15—80, дл. сок. 10—45 мм),
на спине с менее четкими небольшими и низкими морщинами. Окраска
не одноцветная: на спине и мантии боковые полосы; у меланистических
форм, когда рисунок сливается с темной спиной, всегда нижняя часть
тела (над подошвой) остается светлой. Атриум меньших размеров
и без лигулы.
7 (12). Даже у сократившихся слизней половое отверстие располагается
впереди мантийной щели. Между щелью и средней линией спины 14—
16 рядов морщин. Резервуар семяприемника яйцевидный или заострен
на конце. (Подрод Carinarion Hesse).
8 (9). Мантия имеет мелкие темные пятна, особенно четкие после фиксации
слизня. На спине боковые полосы имеют четкие верхние границы и не-
четкие, как бы размытые, нижние. Эпифаллус без вздутия у основания
(у атриума), несколько отступя от переднего конца с четким поясом
из темного пигмента.....................6. А. (С.) circumscriptus John.
9 (8). Мантия без пятен. Нижние границы боковых полос спины всегда
четкие, но зато верхние иногда нечеткие. Эпифаллус со вздутием у ос-
нования и обычно без темной опояски.
10 (11). При жизни спина и мантия темно-пепельные с коричневатым от-
тенком; после фиксации коричневый оттенок исчезает и слизни стано-
вятся темно-серыми или почти черными. Бока светлые, без желтых или
оранжевых полос. Размеры: дл. пол. до 25, дл. сок. до 20 мм. Эпифал-
лус с небольшим вздутием на границе с атриумом. Атриум почти
прямоугольный.............................. 7. А. (С.) silvaticus Lohm.
11 (10). При жизни спина и мантия светло-серые, без коричневатого оттенка.
На боках ниже темно-серых боковых полос имеется по одной желтой
или оранжевой полоске, которые после фиксации исчезают. Размеры:
дл. пол. до 50, дл. сок. до 32 мм. Эпифаллус с крупным вздутием на гра-
нице с атриумом. Атриум конический или треугольный.....................
.................................... 8. А. (С.) fasciatus (Nilss.).
12(7). У сократившихся слизней половое отверстие располагается позади
или под мантийной щелью. Между нею и средней линией спины 17—
20 рядов морщин^ Резервуар семяприемника более или менее шаровид-
ный, без острой вершины.
13 (14). Слизни маленькие: дл. пол. до 27, дл. сок. до 10 мм. Тело массивное,
с относительно широкой подошвой и короткими щупальцами. При
1 Взрослые А. lusitanicus Mab. иногда имеют боковые полосы на спине.
390
II. ARIONIDAE
раздражении тело резко сокращается и приобретает полусферическую
форму, а на мантии и прилегающем участке спины появляются круп-
ные сосочки. Окраска бледно-желтая, беловатая, пепельная или даже
каштановая. Только на мантии иногда могут быть тонкие серые боко-
вые полосы (подрод Microarion Hesse)........................ . .
.................................10. А. (М.) intermedins (Norm.).
14 (13). Слизни крупнее, более стройные, при раздражении не способны так
сократиться, чтобы тело приняло полусферическую форму; сосочки
на мантии и спине очень мелкие. Окраска значительно более темная,
особенно у меланистических форм, у которых светлыми остаются
только бока. Боковые полосы, если они имеются, всегда значительно
более темные.
15 (16). При жизни спина темно-коричневая до почти черной, подошва
ярко-желтая до оранжевой; после фиксации спина голубовато-пепель-
ная или темно-серая, подошва кремовая. Тело сравнительно стройное.
Размеры: дл. пол. до 35, дл. сок. до 15 мм. Передний отдел яйцевода
по длине почти равен заднему. (Подрод Kobeltia Seibert) . .........
..................................... 9. А. (К.) hortensis Fer.
16 (15). В окраске преобладают оранжевые, ржавые, коричневые или черные
цвета. Подошва от светло- до серовато-желтой. Тело более массивное
и крупное. Передний отдел яйцевода в несколько раз длиннее заднего,
(подрод Mesarion Hesse).
17 (18). Рельеф спины образован сравнительно крупными и длинными мор-
щинами — между мантийной щелью и средней линией спины их 19—
20. В окраске как фона, так и боковых полос преобладают оранжевые,
ржавые или коричневые оттенки. Яйцевод и эпифаллус сравнительно
длинные................................4. А. (М.) subfuscus (Drap.).
18 (17). Рельеф спины образован более широкими и короткими морщинами —
между мантийной щелью и средней линией спины их 17—18, а иногда
14—15. В окраске преобладает черный цвет. Яйцевод и эпифаллус
сравнительно короткие..................... 5. А. (М.) sibiricus Simr.
1. Подрод ARION s. str. нЩ
Крупные слизни, которые в движении имеют длину не менее 70 мм;
бывают экземпляры длиной до 250 мм. Кожа толстая, с очень крупными
й резкими морщинами. Киля нет. Взрослые особи окрашены различно,
но всегда одноцветные (за исключением подошвы и ее края, которые могут
быть иного цвета чем спина), без рисунка из полос и пятен.
’""^Яйцевод четко разделен на передний и задний участки, на границе между
которыми крепится одна из ветвей полового ретрактора. Крупный атриум
обычно перетяжкой разделен на два отдела — передний и задний. Задний
отдел имеет боковые вздутия. Внутри его близ устья яйцевода имеется круп-
ная языковидная или ложковидная лигула. Семяприемник очень большой
с яйцевидным резервуаром.
Распространение. Входящие в эту группу 6 видов населяют
западные, северные и средние области Европы.
Arion (Arion) rufus (Linnaeus, 1758) (рис. 535, 538—543)
Linnaeus, 1758 : 652 (Limax); Ferussac, 1819 : 60, T. 1, Fig. 1—8, T. 2,
Fig. 1—3, T. 3, Fig. 1—7 (empiricorum, pars); Simroth, 1885 : 231, T. 7> Fig. 23-31,
T. 10, Fig.' 19—25, T. 11, Fig. 1—4 (empiricorum); Taylor, 1902—1907:167 (ater,
pars); 0 kl and, 1923 : 13 (ater, pars); Boettger, C., 1949 : 169, Fig. 2; W i k-
t о r, 1973 : 23, Fig. 1, 2, 53, 59—64, 81, 215; Riedel, Wiktor, 1974 : 91, Fig. 33,
34, 41, 47, 50, 120—122, 127—133.
ARION
391
Terra typica — Швеция? Типы неизвестны.
Материал: из СССР — 2 экз. из двух пунктов; из других стран —
более 200 экз.
Спина сзади несколько уплощена. При раздражении животное сильно
сокращается, принимая полусферическую форму. Пневмостом очень боль-
шой. Подошва с толстым краем.
Обычно в пределах популяции все особи окра-
шены одинаково. Чаще встречаются популяции из чер-
ных особей. Но нередки как коричневые, так и крас-
ные популяции. Черная окраска спины может соче-
таться с оранжевым или желтым цветом края подошвы.
В окраске решающее значение имеет наличие в пок-
ровах двух пигментов: черного — меланина и крас-
ного — руфина. Албонико (Albonico, 1948) показал,
что присутствие этих пигментов связано с генотипом.
Экологические факторы оказывают влияние только на
интенсивность развития меланина, количество кото-
рого определяет окончательную окраску животного:
если меланина мало, то окраска красная, умеренно —
коричневая, много — черная.
Размеры: дл. пол. до 150, дл. сок. до 80, ши-
рина до 20 мм.
Слизь бесцветная или желтая, при раздражении
молочно-белая или красная.
Радула. Боковых зубов 17—28, краевых 27—47,
число рядов 130—152.
Кишечник (рис. 540) сходен с другими видами, но
сильнее перекручен.
Гениталии (рис. 541—543). Гонада круглая,
дисковидно уплощенная. Гермафродитный проток пря-
мой, лишь впереди извитый. Белковая железа оваль-
ная, крупная. Семяяйцевод длинный закрученный.
Семяпровод тонкий, по крайней мере в 11/2 раза
длиннее эпифаллуса, в который он плавно переходит.
Яйцевод узкий, сравнительно короткий. Семяприемник
с большим шаровидным или яйцевидным резервуаром.
Эпифаллус плавно расширяется вперед, и основание
его окружено валиком. Атриум асимметричный, раз-
личной формы благодаря наличию одного большого
бокового и нескольких более мелких вздутий. Перед-
ний отдел атриума значительно уже и короче заднего,
который шире и вдвое длиннее. В заднем отделе атриума, близ устья
яйцевода, находится крупная широкояйцевидная лигула.
Распространение. В СССР достоверные находки вида были
сделаны только близ Пушкино (окрестности Москвы), Кусково (ныне район
Москвы) и в южной Карелии у озера Сандал. Совершенно очевидно, что эти
популяции возникли в результате завоза. К настоящему времени,^по-види-
мому, сохранилась только популяция в Пушкино.
Природный ареал вида в основном занимает среднюю и часть западной
Европы: северная граница начинается от устья Вислы и далее идет на запад,
включая Голландию; западная проходит где-то во Франции; южная вдоль
Альп; восточная в западной Польше и западной Чехословакии. Известен
островной ареал в Боснии (СФРЮ). Присутствие этого вида в Швеции, южнрй
Финляндии, на Британских о-вах обязано завозу.
•WTO»?
ARION
393
Образ жизни. Влаголюбивый вид, обитающий по берегам лесных
речек, на влажных лугах, близ болот, реже в парках и по придорожным
канавам, заросшим сочной травой.
Многоядный вид, однако основную долю в питании составляют растения
(листья, плоды, клубни и т. п.) и растительный перегной. Неоднократно отме-
чалось, что этот слизень поедает трупы и кал животных и даже способен
к хищничеству (иногда поедает мелких беспозвоночных, птенцов мелких
птиц, молодь грызунов).
В условиях теплого влажного климата (например, в Англии) копуляция
происходит осенью. После короткого брачного танца (кружение) партнеры
располагаются друг к другу затылками, изогнув тело полукругом
(рис. 22, 7F). Атриумы выворачиваются, а лигулы набухают и прижимаются
друг к другу. Затем происходит обмен сперматофорами и обратное ввора-
чивание гениталий. Весь процесс длится около двух часов. Через 2—4 не-
дели начинается откладка яиц.
Яйца овальные, крупные (5x4 мм), белые, кожистые. Один слизень в 2—
3 приема откладывает от 300 до 500 яиц. Вылупление происходит через 4 недели
и более. Молодь вначале белая или светло-желтая, но через несколько дней
на мантии и спине появляется пара темных полос, которые сохраняются не-
сколько месяцев до общего потемнения верхней стороны тела. Полосатая
молодь отличается от A. subfuscus (Drap.) более грубыми морщинами спины.
Длительность жизни около одного года.
Замечания. 1. Линней (Linnaeus, 1758) описал по внешним особен-
ностям два вида слизня, которых он различал только по окраске. Красного
он назвал Limax rufus, черного — Limax ater. Ферусак (Ferussac, 1819),
заметив, что обе формы связаны цветовыми переходами, решил, что они пред-
ставляют один вид, который он назвал Arion empiricorum. Лишь сравни-
тельно недавно ряд авторов (Quick, 1947; С. Boettger, 1949; Altena, 1963
и др.) показали, что в Европе обитают два близких вида, имеющие красные
и черные формы, хорошо различимые только по гениталиям, и с более или
менее разобщенными ареалами. Один из них обозначают как Arion ater
(L.), а другой — A. rufus (L.).1 Самостоятельность этих видов подтвердили
опыты по скрещиванию их и получение гибридов с промежуточными осо-
бенностями. Природные гибриды встречаются в местах, где оба вида обита-
ют совместно (Cain, Williamson, 1958; Walden, 1966).
2. В литературе XIX и начала XX века неоднократно указывалось на
присутствие в различных областях европейской части СССР слизней Limax
и Arion, относимых к видам ater L., rufus L. и empiricorum Fer. (Georgi, 1800;
Eichwald, 1830; Двигубский, 1831; Kaleniczenko, 1851b; Hensche, 1861 —
1866; Belke, 1866; Надеждин, 1868; Krol, 1878; Braun, 1883, 1884; Розен,
1905). Естественно, что авторы руководствовались лишь особенностями
внешнего вида слизней.
Зимрот (Simroth, 1901) справедливо отнесся с недоверием к этим со-
общениям, считая, что они связаны с ошибками в определениях и авторы
скорее всего имели дело с крупными экземплярами Arion subfuscus (Drap.).
Исключение, вероятно, можно сделать лишь сообщениям Двигубского и
Розена о находках A. empiricorum под Москвой, так как в коллекции ЗИН
1 Вероятно, навсегда останется загадкой, имел ли дело Линней с этими двумя ви-
дами или же с цветовыми вариациями одного из них.
Рис. 539—543. Arion {Arion) rufus (L.).
539 — ретракторы; 540 — кишечник; 541, 542 — гениталии; 543 — вскрытый атриум (539, 541 —
у оз. Сандалы, южная Карелия, 11 VIII 1920; 540, 542, 543 — из: Wiktor, 1973).
26 И. М. Лихарев, А. И. Виктор
394
II. ARIONIDAE
АН СССР имеется экземпляр A. rufus из Кусково, а в Зоологическом музее
МГУ из Пушкино.
Совершенно невероятным представляется указание Герстфельдта (Ger-
stfeldt, 1859) об обитании A. ater L. в бассейне р. Вилюй (Якутия).
2. Arion (Arion) ater (Linnaeus, 1758) (рис. 544, 545)
Linnaeus, 1758 : 652 (Limax); Peru ssac, 1819 : 60, T. 1, Fig. 1—8, T. 2,
Fig. 1—3, T. 3, Fig. 1—7 (empiricorum, pars); Taylor, 1902—1907 : 167 (pars); Q u-
i c k, 1960 : 138, Fig. 5C, 6A, B, L, T. 1, Fig. 3.
Terra typica — Щвеция? Типы неизвестны.
Материал: из СССР нет; из других стран — 18 экз.
Данный вид внешне не отличим от А. rufus, но обычно несколько меньших
размеров (дл. пол. до 130 мм). Окраска чаще всего черная, но встречаются корич-
пч
же
544,
(L.).
Рис.
ater
рп
544
545. Arion (Arion)
Норвегия, Тронд-
хейм, VI 1954.
544 — мантийный комплекс и при^
летающие органы; . 545 — генита-
лии.
невые и вариации других
цветов. Слизь бесцветная,
желтоватая или красная.
Наиболее характерно
устроены гениталии. Ат-
риум довольно симметрич-
в два раза короче переднего,
отсутствует. Лигула неболь-
ный, узкий и длинный; задний отдел его почти
Боковое вздутие атриума слабо выражено или
шая и лежит у места перехода яйцевода в атриум. Семяпровод короче, чем
у A. rufus, но зато эпифаллус длиннее. Яйцевод более широкий и впадает
в атриум в середину его заднего отдела.
Распространение. В СССР пока не обнаружен. Как и в случае
с предыдущим видом, ареал этого вида известен лишь приблизительно, так
как многие старые данные требуют проверки в свете новейших представле-
ний о структуре подрода Arion, s. str. По-видимому, природный ареал
A. ater располагается севернее A. rufus, хотя местами оба вида обитают
вместе (возможно, из-за завоза). В целом в природных биотопах A. ater
обычен на Британских и Шетландских о-вах; на севере ФРГ, в Дании, Шве-
ARION
395
ции (до 60°), Норвегии (до 70°) и на островах Балтийского моря — Гот-
ланд, Оланд и Борнхольм.
Образ жизни. Биономия во многом сходна с предыдущим видом.
Наиболее часто встречается на торфяниках и верещатниках, а также в при-
морских лесах.
3. Arion (Arion) lusitanicus Mabille, 1868 (рис. 546 , 547)
Mabille, 1868 : 134; Simroth, 1891 : 339, T. 4, Fig. 1—13, T. 5, Fig. 5,
T. 6, Fig. 1—2; A 11 e n. a, 1956 : 89, Fig. 1—6; Quick, 1960 : 135, Fig. 3G, 5A, D,
E, 6C, E, H, I; A 1 t e n a, 1971 : 183.
Locus typicus-. «sierra d’Arribada», близ Лиссабона (Португалия). Типы
неизвестны.
Материал: из СССР — 1 экз.; из других стран — 17 экз.
Внешне слизни этого вида во многом сходны с Arion rufus и A. ater.
Однако размеры их несколько меньше (дл. пол. до 100 мм). Рельеф покро-
ап
зп
Рис. 546, 547. Arion (Arion) lusitanicusMab.
Калужская обл., окр. Ферзикова, 15 VI
1931.
S46 — дистальная часть гениталий; 547 — вскры-
тый атриум.
вов еще более четкий. Окраска очень
различная. Встречаются экземпляры
одноцветно черные, темно- или зеле-
новато-пепельные, красноватые, оран-
жевые и желтые. Часто край подошвы
иного цвета чем спина. Молодые
особи обычно имеют темные продоль-
ные полосы на мантии и спине;
у взрослых мантия всегда одноцветная, а на спине иногда остается рису-
нок или его следы (после фиксации обычно исчезает).
Гениталии. В отличие от предыдущих видов яйцевод очень взду-
тый, мускулистый и длинный. Внутри него находится большая сильно
вытянутая лигула и несколько продольных складок. Эпифаллус длиннее
семяпровода, на границе с атриумом с четким кольцевым валиком. Семя-
приемник с шаровидным или яйцевидным резервуаром. Атриум очень корот-
26*
396
II. ARIONIDAE
Кий и и небольшой; заднему отделу его соответствует небольшой участок,
расположенный перед выходами яйцевода, семяприемника и эпифаллуса;
передний отдел еще меньше и слабо выражен.
В целом общий план строения гениталий напоминает таковой у A. sub-
fuscus (Drap.) (см. ниже).
Распространение. В СССР единственная находка (1 экз.)
сделана 15 VI 1931 г. Б. Н. Цветковым в Калужской области у с. Ферзиково
(39 км к востоку от Калуги) близ ручья. Экземпляр хранится в ЗИН АН СССР.
Наиболее полная сводка сведений о распространении этого вида дана
Алтеной (Altena, 1971). Из нее явствует, что природный ареал занимает
Пиренейский п-ов, южную Францию и северную Италию. В остальных
местах Европы, где этот вид был обнаружен (Британские о-ва, Швейцария,
ФРГ, Болгария и в СССР), он обитает в культурных биотопах (или вблизи
от них) небольшими локальными популяциями, возникшими в результате
завоза.
Биономия, питание и размножение очень слабо изучены. По-видимому,
слизень склонен жить в садах и питается культурными растениями.
Замечание. Многие аспекты биономии и распространения этого
Вида пока еще недостаточно изучены из-за того, что длительное время его
принимали за другие виды того же подрода.
2. Подрод MESARION Hesse, 1926
Hesse Р., 1926 : 65; Wiktor, 1973 : 29; Riedel, Wiktor, 1974 : 103.
Типовой вид — Limax subfuscus Draparnaud, 1805; по первона-
чальному обозначению.
Слизни средней величины, дл. пол. до 80 мм. Кожа умеренно толстая
с мелкими морщинами. Киля нет. Окраска от коричневой до почти черной.
При более светлой общей окраске на спине видна пара темных полос, а на
мантии лирообразный рисунок^ Правая полоса мантии почти полностью окру-
жает пневмостом. Бока окрашены несколько светлее спины. Подошва кре-
мовая.
Семяприемник с шаровидным или яйцевидным резервуаром. Яйцевод
состоит из двух участков (между ними крепится ветвь полового ретрактора),
из которых передний в несколько раз длиннее заднего. Атриум по сравнению
с остальными органами небольшой, толстый, без лигулы и не разделен пере-
тяжкой на передний и задний отделы.
К подроду относятся три вида. Два рассматриваются ниже, третий —
А. (7W.) simrothi Kunkel, 1909 — известен только из Шварцвальда (ФРГ).
4. Arion (Mesarion) subfuscus (Draparnaud, 1805) (рис. 548—551)
Draparnaud, 1805 : 125, T. 9, Fig. 8 (iimaz); Kaleniczenko, 1851b :
114, T. 4, Fig. 1 (krynickti)', Simroth, 1885 : 236, 280, T. 7, Fig. 32—35, T. 11, Fig. 5—
10 (subfuscus u. brunneus)', Taylor, 1902—1907 : 193, T. 24, Fig. 1—6; 0 kl an d,
1923 : 24, Fig. 13—21, T. 1, Fig. 2—3; Poska-Teiss, 1927 : 16, T. 1, Fig. 1, 2, 6
(esthonicusY, Лихарев, Раммельмейер, 1952:304, рис. 231; Лихарев,
1954 : 49, цв. таб., рис. 1, 2; Q u i с к, 1960 : 133, Fig. ЗН, 6D, F, G, Т. 1, Fig. 2, 9; W i к-
t о г, 1973 : 29, Fig. 3, 4, 54, 65—69, 216, 217; Riedel, Wiktor, 1974 : 103, Fig. 137,
138, 141—145.
Terra typica — Франция. Типы неизвестны.
Материал: из СССР — более 600 экз. из 256 пунктов; из других
стран — более 2000 экз.
Тело удлиненное, при рассматривании сверху с почти параллельными
Краями. Мантия занимает около х/3 длины тела, овальная, с широко за-
Рис. 548, 549. Arion (Mesarion) subfuscus (Drap.).
548 _взрослый слизень, Литва, 29 VI 1907; 549 — молодой экземпляр, окр. оз. Имандра, 6 IX 1961»
398
П. ARIONIDAE
Кругленным задним концом. Морщины спины тонкие, слабовыпуклые,
между средней линией спины и мантийной щелью 19—20 рядов морщин.
У сократившихся особей половое отверстие располагается непосредственно
за мантийной щелью или под нею.
Окраска очень варьирует даже в пределах популяции и в течение жизни
одной особи. Фон разных оттенков — от коричневого до оранжевого, чаще
ржавый или серовато-коричневый. Середина спины обычно наиболее темная,
даже темно-шоколадная, боковые полосы не различимы (/. brunnea Lehmann,
Рис. 550, 551. Arion (Mesarion) subfuscus (Drap.).
650 — гениталии (с двух сторон), Витебская обл., 20 VII 1894; 551 — кишечник (из: Wiktor, 1973).
1862). Иногда встречаются особи с четкими полосами на спине и с лирообраз-
ным рисунком на мантии (f. typica). Упомянутый рисунок наиболее обычен
для молодых особей и для экземпляров в ранней фазе половой активности.
Обе формы встречаются совместно.
Чаще рисунок на спине и мантии выражен нерезко, но тем не менее доста-
точно отчетливо. Вблизи от пневмостома от ближайшей полосы отходит
как бы клин, благодаря чему почти все отверстие окружено темным пигмен-
том (очень характерная особенность данного вида!).
После фиксации оранжевый пигмент быстро исчезает, окраска становится
более серой или черной. Какой-либо зависимости между окраской и эколо-
гическими условиями обнаружить не удалось. Подошва всегда кремовая.
У полосатых форм, как правило, слизь оранжевая или желтая, а у форм без
полос очень часто бесцветная или слабо окрашенная.
Размеры: дл. пол. 35—80, дл. сок. до 45, шир. сок. 6.5—9 мм.
Радула (рис. 11, I—III}. Переход от боковых зубов к краевым очень
нечеткий. Общее число зубов в ряде 43—70, число рядов 125—150.
Кишечник — см. рис. 551.
Гениталии (рис. 550). Относительно небольшая гонада обычно
закрыта лопастями печени. Гермафродитный проток, тонкий, близ белко-
ARION
399
вой железы, извитый. Белковая железа языковидная, обычно смещена к сред-
ней части зоба. Яйцевод длинный и мускулистый; задний участок его тонкий
и короткий, а передний вздутый и толстостенный. На границе их нередко
яйцевод перегнут. Семяприемник с длинным протоком. После копуляции,
когда резервуар заполняется спермой, он сильно увеличивается, а проток
становится более коротким. Вторая ветвь полового ретрактора прикреп-
ляется на границе между резервуаром и протоком семяприемника. Бывает
еще третья ветвь, которая прикрепляется к заднему отделу атриума. Срав-
нительно длинный и тонкий семяпровод плавно переходит в длинный и извитый
эпифаллус, который на границе с атриумом окружен заметным валиком.
Атриум небольшой, бочковидный. Задний отдел его при впадении семяпри-
емника и яйцевода слегка карманообразно вытянут. Сперматофор около 20 мм
в длину, сужается к концам, а по выпуклому краю с мелкими зуб-
чиками.
Распространение. В СССР очень широко распространен по
лесным и лесостепным областям европейской части; проникает в соседнюю
лесотундру, а местами (Кольский п-ов) и в тундру. Обычен в Карпатах и их
лесных и лесостепных предгорьях, по западным и восточным склонам среднего
и южного Урала. В пределах степной зоны обитает в байрачных и пойменных
лесах Украины. Недавно обнаружен на Северном Кавказе в лесу у г. Орджо-
никидзе (Иванов, 1978). Отсутствует в Крыму.
Вне СССР — почти вся Европа, кроме самых южных областей. Завезен
в Северную Америку и Новую Зеландию.
Образ жизни. Обитает в лиственных, смешанных и хвойных
лесах, лесотундре и в горной тундре Кольского п-ова. Кроме того, он обычен
на лесных полянах и в кустарниках. В горах поднимается значительно выше
лесной зоны. Иногда встречается на торфяных болотах. В дневное время пря-
чется под валежины, в гнилых пнях, под камни и в подстилку. В антропо-
генных биотопах обитает сравнительно редко и преимущественно в старых
парках и на кладбищах.
Излюбленной пищей служат шляпочные грибы, однако данный слизень
питается также живыми и отмершими частями растений, трупами животных
и калом позвоночных. В сырые годы может наносить некоторый ущерб по-
лям и огородам, расположенным по соседству с лесом и кустарником.
Копуляция обычно приходится на конец весны и начало лета. Откладка
яиц происходит через 3 недели после копуляции. Один слизень в несколько
приемов может отложить свыше 300 яиц.
Яйца размером 2.25x2.25 или 3.25x2.25 мм, непрозрачные, кожистые,
кремовые или желтые. Слизни откладывают их в темные места чаще всего
под различные предметы, в мох или в углубления почвы. В зависимости
от температуры вылупление происходит через 26—40 дней, а иногда через
80^—100 дней. Только что вылупившаяся молодь достигает 6 мм в длину.
Окраска ее серая или светло-коричневая, на спине темнее; щупальца лило-
вые. Через 3—4 дня появляются парные темные полосы на спине и мантии.
О жизненном цикле — см. с. 65.
Замечание. Ряд авторов придает большое таксономическое зна-
чение изменчивости окраски и рисунка, выделяя множество цветовых форм,
а некоторые из них рассматривая как подвиды или даже самостоятельные
виды. Особенно это касается двух вариаций: с боковыми полосами и без них —
A. subfuscus (Drap.) и A. brunneus Lehm.
Наши исследования показали, что существенной разницы между этими
формами нет. Внутреннее строение их идентично. Что же касается окраски,
то она изменяется в течение жизни одной особи. Поэтому молодь будет
отвечать особенностям' А. subfuscus, а взрослые — А. brunneus. Последняя
форма внешне на первый взгляд сходна с A. sibiricus Simroth (см. ниже).
400 II. ARIONIDAE
5. Arion (Mesarion) sibiricus Simroth, 1901 (рис. 552—558)
Simroth, 1901:53, T. 2, Fig. 1—6; Лихарев, Рамме льмейер,
1952 : 304 (A. subfuscus subsp. sibiricus).
Locus typicus-. «Irkutsk» — Иркутск. Лектотип (обозначен нами) и 4 па-
ралектотипа в ЗИН АН СССР. За лектотип избран один из экземпляров,
собранных Мааком близ Иркутска (см.: Simroth, 1901, табл. 2, рис. la, 1b);
Материал: из СССР — 100 экз. (в том числе и все типы) из
36 пунктов.
Спина чаще всего черная или почти черная. По бокам проходят четкие
черные полосы; между ними и почти черной спиной видны светлые зоны.
Ниже полос бока желто-коричневые, к подошве еще светлее. Обычно темный
пигмент сосредоточен по бороздкам, что особенно заметно на крае подошвы,
где он дает серию темных точек. Часть экземпляров полностью черные или
черно-бурые. В этом случае пигмент проникает даже на подошву. У осталь-
ных подошва светлая. Мантия всегда темная со слабо выраженными поло-
сами, которые лишь слегка отграничены от более светлой середины. Голова
черноватая, но шея всегда остается светлой.
Очень характерен рельеф спины. Морщины сравнительно короткие и
поэтому кажутся широкими. Между средней линией спины и мантийной
щелью обычно 17—18, изредка 14—15 рядов морщин, т. е. немного меньше,
.чем у предыдущего вида. Слизь бесцветная.
Размеры: дл. сок. 20—30, шир. сок. 7—9, дл. мантии 8—11 мм; лек-
тотип: дл. сок. 29, шир. сок. 8, дл. мантии 11 мм.
Радула (рис. 554). Боковых+краевых зубов 47, число рядов 124.
Кишечник — см. рис. 556.
Г ениталии (рис. 557, 558). Обнаруживают большое сходство с A. sub-
fuscus. Единственная разница, прослеживаемая на всем материале и, по-
видимому, не являющаяся результатом фиксации, это более короткие и
толстые яйцевод и эпифаллус. Как и у предыдущего вида, внутри яйцевода
находятся две продольные складки, которые в заднем отделе атриума иногда
образуют плоский и широкий выступ, напоминающий лигулу.
Распространение. Недостаток материала затрудняет воз-
можность дать четкую характеристику ареала данного вида. В пределах
средней и восточной Сибири он был найден: 1) в верховьях р. Она (бассейн
р. Абакан, между Абаканским хребтом и Западным Саяном, Хакасская АО) —
несколько местонахождений; 2) близ Иркутска; 3) близ Енисейска; 4) у
р. Лена ниже Турукты (Якутия). Наибольшее число находок сделано
в Приморском крае и на юге Хабаровского края (в бассейне среднего и южного
Амура); обнаружен в Южном Сахалине.
Указание Хохуткина (1961) об обитании данного вида на среднем Урале
при проверке не подтвердилось.
Образ жизни. Обитает в смешанных и лиственных лесах, в том
числе и в пойменных.
Замечание. Этот вид несомненно очень близок A. subfuscus (Drap.).
Основные отличия: 1) более темная окраска с преобладанием черного пиг-
мента; 2) более короткие и широкие морщины на спине; 3) яйцевод и эпи-
фаллус короче и толще. Ареалы обоих видов разделены обширными про-
странствами западной Сибири.
3. Подрод CARINAR1ON Р. Hesse, 1926
Hesse Р., 1926 : 65; W i k t о г, 1973 : 34; Riedel, Wiktor, 1974 : 109.
Типовой вид — Arion circumscriptus Johnston, 1828; по первона-
чальному обозначению.
Рис. 552, 553. Arion (Mesarion) sibiricus Simr.
552 - Алтай, верховья p. Ижи, 13 VI 1912; 5S3 — лектотип, Иркутск.
Рис. 554—558. Arion (Mesarion) sibiricus Simr.
554 — зубы радулы; 555 — челюсть; 556 — кишечник; 557 — гениталии, у дер. Майхэ, Приморский край.
5 VI 1927; 558 — дистальная часть гениталий, Кузнецкий Алатау, 1897.
ARION
403
Слизни средних размеров — длина не более 50 мм. Кожа умеренно тол-
стая. У молодых экземпляров посередине спины проходит киль, дости-
гающий заднего края мантии. У взрослых от него может оставаться только
след. Морщины тонкие и слабовыпуклые, между мантийной щелью и средней
линией спины 14—16 рядов морщин. У сократившихся животных половое
отверстие лежит впереди мантийной щели.
Верхняя сторона тела пепельно-серая, всегда с боковыми темными поло-
сами. На мантии правая полоса проходит над пневмостомом, не проникая ниже
его. Подошва белая или светло-кремовая. Для гениталий характерно, что
резервуар семяприемника яйцевидный или четко заостренный, а атриум длин-
ный и сплющенный, без лигулы. Яйцевод короткий и не разделен на перед-
ний и задний участки.
Распространение. В СССР встречается только в западных обла-
стях европейской части.1 Кроме того, виды этого подрода обитают почти
во всей остальной Европе, доходя на западе до Испании, на севере до Скан-
динавии и на юге до северных Балкан.
6. Arion (Carinarion) circumscriptus Johnston, 1828 (рис. 559—562)
J ohnston, 1828 : 76; Mabille, 1868 : 138 (Bourguignati)', L ohm an d er,
1937 : 97, Fig. 6—9, 14—15; Лихарев, Раммельмейер, 1952 : 303; L eh-
tinen, 1957:5; H u d e c, 1960:204, Fig. 3—4; C h e v a 1 1 i e r, 1972 : 20, Fig. 22;
Wiktor, 1973 : 34, Fig. 7, 8, 55, 70—73, 220; Riedel, Wiktor, 1974 : 110,
Fig. 146—147, 152—155, 160—161.
Terra typica — Европа. Типы неизвестны.
Материал: из СССР — 120 экз. из И пунктов; из других стран —
более 300 экз. •
Тело удлиненное, узкое и слегка дорсовентрально приплюснутое. Окраска
очень характерная. Спина голубовато-стального или пепельного цвета, иногда
темно-серая. Середина спины и мантии темная; на этом фоне заметны небольшие
и нерегулярные черные пятнышки, которые более отчетливо видны после
фиксации, особенно на мантии. Боковые полосы темно-серые, иногда почти
черные; верхние границы их четкие, тогда как нижние размытые, так как
пигмент плавно переходит на бока. Благодаря этому бока под полосами очень
темные, а к подошве постепенно светлеют. Тем не менее пигмент хотя и в малом
количестве, но доходит и до края ноги. Подошва у живых слизней голубовато-
белая, после фиксации кремовая.
Размеры: дл. пол. до 40, чаще 25, дл. сок. до 27, чаще 15—20 мм.
Радула. Боковых-)-краевых зубов 26—34, число рядов 97—118.
Кишечник — см. рис. 560.
Гениталии (рис. 561, 562). Яйцевод почти такой же толщины,
как и эпифаллус. Семяпровод длинный и тонкий. Эпифаллус короткий и тон-
кий, у основания без кольцевидного валика, но зато с четким кольцом из тем-
ного пигмента. Семяприемник обычно с сильно вытянутым резервуаром и
узким протоком, без вздутия у основания. Атриум сильно сплющен и очень
длинный, обычно длина его в два раза больше ширины.
Распространение. В СССР найден в окрестностях Ленинграда
и в Железноводске (Северный Кавказ); в последнее место, безусловно, за-
везен. Возможно, обитает в природных биотопах Прибалтики и западных
областей Белоруссии и Украины.
Вне СССР достоверные находки известны из Финляндии (Valovirta, 1967),
Швеции (Lohmander, 1937), Польши (Wiktor, 1973), Чехословакии (Hudec,
1 Здесь и ниже все сведения о распространении в СССР видов этого подрода мы даем
только по материалам коллекции ЗИН АН СССР, так как литературные данные не яв-
ляются достоверными.
404
II. ARIONIDAE
1960), Голландии (Gittenberger, Backhuys, Ripken, 1970 — «А. silvaticus»)'
и Франции (Ghevallier, 1972). Все другие сообщения о находках этого слизня
требуют проверки в свете современных знаний о различиях между видами
подрода Carinarion Hesse.
Рис. 559—562. Arion (Carinarion) circumscriptus John, (из: Wiktor, 1973).
559 — внешний вид; 560 — кишечник; 561, 5вг — гениталии.
Образ жизни. Это явственно лесной вид, предпочитающий влаж-
ные смешанные и лиственные леса, особенно ольховые. Чаще всего обитает
по долинам и оврагам, особенно там, где много лиственного опада. Охотно
прячется на дневное время в подстилку, под обломки древесины и т. п.,
но нередко и днем ползает по поверхности почвы или по подстилке. Мало-
подвижное животное и никогда не заползает не только на деревья, но даже
на пни.
В Польше обитает на равнинах, а в горах уступает место родственному
A. silvaticus.
ARION
405
Замечания. Данный вид наиболее близок к A. silvaticus Lohm.,
от которого отличается присутствием пятнышек на спине и мантии, темными
боками, а также голубоватым оттенком окраски. Для гениталий характе-
рен тонкий и короткий эпифаллус с темным кольцом на нем, отсутствие
вздутий на протоке семяприемника и на эпифаллусе, а также длинный атриум.
На существование трех самостоятельных видов, ранее относимых в один,
впервые обратил внимание Ломандер (Lohmander, 1937). Самостоятельность
A. fasciatus (Nilsson) теперь общёпризнана, но, по мнению многих авторов,
две другие формы Ломандера следует считать подвидами и называть A. cir-
cumscriptus circumscriptus Johnston и A. circumscriptus silvaticus Lohmander.
Однако наши знания морфологии, экологии и географического распростра-
нения этих трех форм — A. fasciatus Nilsson, A. circumscriptus Johnston и
A. silvaticus Lohmander — подтверждают точку зрения, что они самостоятель-
ные виды.
7. Arion (Carinarion) silvaticus Lohmander, 1937 (рис. 563—565)
Lohmander, 1937 : 94, Fig. 10—11 (circumscriptus var. silvaticus); Wiktor,
1971b : 132; 1973 : 38, Fig. 9, 10, 55, 74—76, 89, 90, 223; Riedel, Wiktor, 1974 :
114, Fig. 148—149, 156—158, 162.
Terra typica — Швеция. Типы неизвестны.
Материал: из СССР — 18 экз. из 6 пунктов; из других стран —
более 400 экз.
Тело еще более сильно сплющено, чем у предыдущего вида. Самой харак-
терной является окраска. Спина темная, тогда как бока ниже полос очень
светлые. У живых экземпляров спина и мантия темно-пепельные, со слегка
коричневым оттенком. Характерные для A. circumscriptus черные пятнышки
на спине и мантии у данного вида отсутствуют, и спина несколько более тем-
ная. Боковые полосы почти черные. Благодаря темной спине верхние границы
полос нечеткие, зато нижние хорошо выделяются на фоне светло-кремовых
боков. На первый взгляд ниже полос темный пигмент отсутствует, однако
под лупой можно заметить небольшие пятнышки, сосредоточенные по бо-
роздкам. Подошва обычно совершенно белая, но с более кремовым оттенком,
чем у предыдущего вида. Слизь бесцветная.
После фиксации коричневый оттенок окраски спины исчезает, и она ста-
новится пепельной или черноватой, а подошва кремовой. Контраст между
темной спиной и светлыми боками становится еще более четким.
Размеры: дл. пол. до 25, дл. сок. до 20 мм.
Радула. По форме зубов сходна с предыдущим видом, но они меньших
размеров и их больше. Боковых+краевых зубов 30—38, число рядов 106—126.
Гениталии (рис. 564, 565). По форме занимают промежуточное
положение между A. circumscriptus John, и A. fasciatus (Nilsson). Прежде
всего это касается размеров и формы эпифаллуса — он средней величины,
без кольца из темного пигмента и на границе с атриумом несет небольшое
вздутие. Сходное образование имеет проток семяприемника, но оно меньших
размеров, чем у A. fasciatus. Яйцевод короткий и толстый, как у A. circum-
scriptus. Наиболее характерным признаком гениталий данного вида является
атриум, который почти прямоугольной формы, сплющен и длина его лишь
немного больше ширины.
Распространение. В СССР данный вид обнаружен в Литов-
ской ССР (близ Шяуляя и Пагегяя), в окрестностях Ленинграда (Пушкин,
Можайское), у Яремчи (Ивано-Франковская обл., Карпаты). Кроме того,
известен из Финляндии (Valovirta, 1967), Швеции (Lohmander, 1937), Польши
и Чехословакии (Wiktor, 1973), Болгарии (Дамянов, Лихарев, 1975), Гол-
ландии (Gittenberger, Backhuys, Ripken, 1970), Франции (Chevallier, 1972).
406
II. ARIONIDAE
Судя по описанию и рисункам из одной работы Гроссу (Grossu, 1970d), по-
видимому, обитает и в Румынии.
Образ жизни. Биономия этого вида еще недостаточно изучена.
Как и A. circumscriptus, обитает в смешанных и лиственных лесах, особенно-
Рис. 563—565. Arion (Carinarion) silvaticus (Lohm.) (из: Wiktor, 1973).
563 — внешний вид; 564, 565 — гениталии.
в долинах и вблизи от водоемов. В Польше он живет только в горах и пред-
горьях, заменяя тут равнинного A. circumscriptus.
Замечание. Этот вид обнаруживает ряд особенностей, сближающих
его то с Л. circumscriptus, то с Л. fasciatus. По гениталиям он наиболее схо-
ден с последним видом, только атриум сплющен, тем самым напоминая
A. circumscriptus, но заметно короче. Размерами и темной окраской он схо-
ден с Л. circumscriptus, но при этом обладает рядом специфических особен-
ностей — темная спина и более светлые бока.
ARION
407
8. Arion (Carination) fasciatus (Nilsson, 1823) (рис. 566—568)
Nilsson, 1823 (non 1822) : 3 (Limaz'r, P о 1 1 о n e г a, 1887c : 19, Fig. 31, 34
(Nilssoni)-, Lohmander, 1937 : 90, Fig. 1—5, 12—13, 17; Lehtinen, 1957 : 5;
H u d e c, 1960 : 204, Fig. 1—2; W i к t о r, 1964 : 68; V a 1 о v i r t a, 1967 : 31;
Pinter, 1968:180; Grossu, 1970d : 69; Ant, 1971 : 257; W i к t о r, 1971b:
132; Wiktor, 1973:40, Fig. 11, 12, 58, 77—80, 221, 222; Riedel, W i к t о r,
1974 : 116, Fig. 150—151, 160—161, 164—168.
Locus typicus'. «Lund» — Лунд, Швеция. Типы неизвестны.
Материал: из СССР — 140 экз. из 28 пунктов; из других стран —
более 300 экз.
Слизень заметно более сплющен и шире, чем два предыдущих вида. Тело
почти всегда светлое, как бы выцветшее. Фон кремовый или желтовато-пе-
пельный. Середина спины и мантии несколько темнее, без каких-либо пят-
нышек. Боковые полосы темно-пепельные, с четкими границами как сверху,
так и снизу. У живых слизней кроме темных полос несколько ниже их за-
метны слабее выраженные узкие желтые, оранжевые или красноватые по-
лоски. При фиксации спиртом эти полоски вместе с желтоватым оттенком
фона исчезают и тело становится светло-пепельным. Подошва при жизни
кремовая, после фиксации белая. Слизь обычно бесцветная, но, как утверж-
дают некоторые авторы, иногда желтая.
Размеры: дл. пол. до 50, дл. сок. до 32 мм.
Радула.’Боковых+краевых зубов 30—35; число рядов 96—125.
Кишечник (рис. 567) сравнительно с другими видами данного рода слабо
перекручен.
Гениталии (рис. 568). Копулятивные органы по сравнению с дру-
гими видами того же подрода заметно крупнее. Яйцевод тонкий. Семяпро-
вод очень длинный, а эпифаллус необычно больших размеров: длинный,
толстый и на границе с атриумом имеет очень большое вздутие. Резервуар
семяприемника яйцевидный или клювообразно вытянут. Проток его очень
толстый, иногда почти той же ширины, что и основание резервуара. Перед
впадением в атриум проток имеет очень большое и мускулистое вздутие.
Атриум, хотя и очень варьирует, обычно имеет вид сплющенного конуса,
узким концом обращенного наружу. Сперматофор (рис. 531) сравнительно
крупный, с мелкими зубчиками цо ребрам.
Распространение. В СССР обнаружен в ряде мест' Москов-
ской, Калининской, Липецкой, Орловской, Курской, Псковской, Калуж-
ской и Ивано-Фрацковской областей, а также в окрестностях Ленинграда,
в Архангельске и Киеве.
Отчетливое тяготение вида к антропогенным биотопам не позволяет четко
определить, какие из указанных местонахождений принадлежат к природ-
ному ареалу, а какие связаны с завозом.
Вне СССР достоверные находки сделаны в Финляндии, Польше, Чехо-
словакии, Венгрии, Румынии и Болгарии. Вероятно, широко распространен
почти по всей Европе. Завезен в США (Burch, 1962).
Образ жизни. В СССР и в Польше, а также, по-видимОму, и в дру-
гих странах этот слизень оказывает явное предпочтение культурным био-
топам — огороды, парки, поля, старые кладбища и т. п. Охотно живет
во влажных местах под кустами и деревьями. Днем прячется под камни,
деревянные обломки и под другие предметы на почве. В Польше активен
в течение всего года. Встречается вместе с такими синантропными моллю-
сками, как Arion hortensis (For.), Trichia hispida (L.), Cepaea nemoralis (L.)
и др.
Замечание. По внешнему виду наиболее сходен с A. silvaticus,
но крупнее, более сплющен, светлее окрашен и имеет кроме темных боко-
Рис. 566—568. Arion (Carinarion) fasciatus Nilss. (из: Wiktor, 1973).
566 — внешний вид; 567 — кишечник; 568 — гениталии.
ARION
409
вых полос упомянутые желтые или оранжевые полоски. Гениталии отли-
чаются более крупным эпифаллусом, имеющим большое вздутие, другим
вздутием на протоке семяприемника и формой атриума.
4. Подрод KOBELT1A Seibert, 1873
Seibert, 1873, Nachr. Bl. deutsch. malak. Ges., 5 : 81; Hesse P., 1926 : 66;
Wiktor, 1973 : 42; Riedel, Wiktor, 1974 : 119.
Типовой вид — Arion hortensis Ferussac, 1819; по первоначаль-
ному обозначению.
Слизни небольшие (дл. пол. до 35 мм). Кожа умеренно толстая с мелкими
морщинами. Киля нет. Окраска темная, спина от буровато-черной до го-
лубовато-темно-серой. Боковые полосы проходят по самому краю мантии,
так что правая почти полностью окружает пневмостом. Подошва ярко-жел-
тая, оранжевая или коричневато-желтая, после фиксации кремовая. При со-
кращении тела половое отверстие располагается позади мантийной щели.
Передний участок яйцевода почти той же длины, что и задний, но не-
сколько более толстый. Резервуар семяприемника шаровидный. Атриум
небольшой, без лигулы.
К этой группе относятся 4 вида; из них 2 обитают на Пиренейском п-ове,
один в Альпах, а последний в западной и средней Европе (см. ниже).
9 . Arion (Kobeltia) hortensis Ferussac, 1819 (рис. 569—572)
Ferussac, 1819 : 65, T. 2, Fig. 4—5; Moquin-Tandon, 1855 : 14, T. 1,
Fig. 28—30 (fuscus, part.); Simroth, 1885 : 238, 276, T. 7, Fig. 42, T. 11, Fig. 16—18;
0 k 1 a и d, 1923: 40, Fig. 35—41, T. 1, Fig. 7; Q u i c k, 1960 : 130, Fig. 4A—F, К—M,
T. 1, Fig. 4; Valovirta, 1967 : 31; Grossu, 1970d : 70; Wiktor, 1973 : 43,
Fig. 5, 6, 57, 85, 86, 218, 219; Riedel, Wiktor, 1974 : 119, Fig. 139, 140, 174, 175.
Terra typica, — вероятно, Франция. Типы неизвестны.
Материал: из СССР — 1 экз.; из других стран — более 400 экз.
Тело сильно удлиненное, со слабовыпуклой спиной. Рельеф кожи тонкий,
между мантийной щелью и средней линией спины 18—20 рядов морщин.
Окраска при жизни темно-коричневая до почти черной. Молодь голубо-
вато-серая. После фиксации коричневый оттенок исчезает и спина становится
голубовато-пепельной или темно-серой. Боковые полосы спины и мантии
черные. Если верхняя граница полос всегда четкая, то нижняя неясная,
так как на боках пигмент постепенно исчезает по направлению к подошве.
При жизни иногда над боковыми полосами заметно по одной желтовато-
коричневой полоске. Слизь густая, желтая или оранжевато-желтая.
Размеры: дл. пол. до 35, дл. сок. около 15 мм.
Радула. Срединный и боковые зубы по форме походят на таковые у видов
из подрода Carinarion Hesse, краевые имеют более высокий мезокон, тогда
как эктокон рудиментарный. Боковыхф-краевых зубов 30—35, число рядов
100-120.
Кишечник (рис. 570) сравнительно немного перекручен; зоб сзади имеет
отчетливый круглый карман (возможно, желудок).
Гениталии (рис. 571, 572). У свежезафиксированных слизней
половые органы желтовато-розового цвета. Яйцевод длинный, почти равен
суммарной длине семяпровода и эпифаллуса. У места прикрепления ветви
полового ретрактора он слегка изогнут. Семяпровод сравнительно короткий.
Эпифаллус имеет вид вытянутого, несколько уплощенного посередине ко-
нуса, который на границе с атриумом имеет слабое валикообразное вздутие.
Атриум бочковидный или несколько суженный к половому отверстию.
В литературе (Quick, 1960) существует описание двух типов спермато-
форов, что дает основание подозревать таксономическую неоднородность
27 и. М. Лихарев, А. Й. Виктор
410
II. ARIONIDAE
исходного материала (рис. 534). В одном случае сперматофор (длина 6.5 мм)
имеет веретенообразную форму, почти прямой, лишь передний конец за-
гнут, гладкий, без ребер и зубчиков на них. Другой тип — сперматофор ко-
нической формы (длина 5 мм), с загнутым передним концом и широким осно-
ванием, снабженным несколькими крупными шипами; вдоль тела сперма-
Рис. 569—572. Arion (Kobeltla) hortensis Fer. (из: Wiktor, 1973).
569 — введший вид; 570 — кишечник; 571, 572 — гениталии.
тофора тянется ребро, имеющее многочисленные мелкие зубчики, ориентиро-
ванные назад.
Распространение. В пределах СССР единственная достоверная
находка (1 экз.) сделана А. А. Шилейко в 1969 г. в буковом лесу близ Ужго-
рода (Закарпатская область); Указывает ли эта находка на то, что в Закар-
патье имеется островок природного ареала или эта популяция возникла в ре-
зультате вторичного освоения естественного биотопа, пока сказать трудно.
Основной природный ареал занимает Пиренейский и Апеннинский п-ова,
Францию и Британские о-ва. Как синантропный вид обычен в средней Европе
и Скандинавии, реже в Северной Африке и Болгарии. Завезен в Северную
Америку.
Обр аз жизни. Обитает в смешанных и лиственных лесах и в те-
нистых кустарниках, как на равнинах, так и в горах. Как синантропный
ARION
411
вид обычен в садах, огородах, парках, на кладбищах, пустошах и т. п.
Часто обитает в оранжереях, парниках, во влажных хранилищах овощей и
фруктов. Особенно в большом количестве встречается в сельской местности
и в дачных поселках. Зимует в подвалах, погребах и канализационных
трубах.
Питается в основном плодами и зелеными частями растений, в связи
с чем нередко наносит существенный вред, особенно в огородах и теплицах.
Очень опасен декоративным культурам, клубнике, рассаде капусты, сель-
дерею, салату, а также некоторым бобовым.
Длительность жизни около 16 месяцев. Приступает к копуляции и откла-
дывает яйца начиная с четырехмесячного возраста и в течение большей части
года (в Западной Европе). Копуляция происходит в вечернее время. Ей
предшествуют брачные танцы — кружение, потирание радулами, слизыва-
ние слизи.
Яйца диаметром 3.5 мм. В течение жизни один слизень порционно (по 12—
80 яиц в одной кладке) откладывает около 150—200 яиц. Их развитие в за-
висимости от температуры длится от 19 дней до 7 недель.
Только что вылупившиеся слизни (длина около 4—7 мм) фиолетово-
коричневого цвета, без всяких следов киля — признак, по которому их всегда
можно отличить от молоди видов из подрода Carinarion Hesse.
5. Подрод MICROARION Р. Hesse, 1926
Hesse Р., 1926 : 66; Wiktor, 1973 : 45; Riedel, Wiktor, 1974 : 123.
Типовой вид — Limax intermedins Normand, 1852; по первона-
чальному обозначению.
Небольшие слизни (дл. пол. до 27, дл. сок. до 15 мм). Кожа умеренно тол-
стая, киля нет. Морщины короткие и очень выпуклые. При сокращении
на мантии и прилегающем участке спины морщины вздуваются в виде со-
сочков. Окраска серовато-белая, желтая или желтовато-серая. Подошва
всегда светлее верхней стороны тела. Боковых полос нет или они еле заметны.
В отличие от Mesarion, Carinarion и Kobeltia тело относительно массивное
и широкое.
Яйцевод цилиндрический, короткий, толстый и не разбит на участки.
Семяприемник с большим шаровидным резервуаром. Атриум бочковидный,
без лигулы.
К подроду принадлежат четыре вида, из которых один обитает в Порту-
галии, другой — в Штирии (Австрия), третий — в окрестностях Праги
(ЧССР), а четвертый — см. ниже.
10. Arion (Microarion) intermedins (Normand, 1852) (рис. 573—576)
Normand, 1852 : 6 {Limax)', Cl essin, 1884 : 116 {flavus)', Simroth.,
1885 : 237, 289, T. 7, Fig. 41, T. 11, Fig. 14—15 {minimus)', Taylor, 1902—1907 : 240,
Fig. 243—250, T. 24, Fig. 18—23; P i 1 s b r y, 1948 : 676, Fig. 367; Quick, 1960 : 125,
Fig. 4H, T. 1, Fig. 7; Gittenberger e. a., 1970 : 65, Fig. 63, 68; Wiktor, 1973 :
46, Fig. 37, 38, 56, 82—84, 224; Riedel, Wiktor, 1974 : 123, Fig.' 169—173.
Locus typicus'. «Valenciennes» — Валенсьенн (Франция). Типы неизвестны.
Материал: из СССР — нет; из других стран — более 200 экз.
Тело массивное, подошва относительно широкая, а щупальца короткие
и толстые. Морщины спины сравнительно высокие и короткие. При раздра-
жении тело приобретает форму полусферы.
При жизни верхняя сторона тела бледно-желтая, беловатая, пепельная
или даже каштановая. На мантии могут быть заметны очень тонкие серые
полосы, образующие рисунок в форме лиры. Щупальца и голова черноватые.
27*
412
II. ARIONIDAE
Подошва ярко-желтая. Слизь желтая. После фиксации слизни становятся
кремовыми или грязно-белыми, с более темной спиной и более четкими по-
лосами. Желтая окраска подошвы исчезает.
Размеры: дл. пол. 15—20, дл. сок. до 10 мм.
зп
Рис. 573—576. Arion (Microarion) intermedius Simr. (из: Wiktor, 1973).
573 — внешний вид; 574 — кишечник; 575, 576 — гениталии.
Радула. Форма зубов как у A. hortensis Fer. Боковых+краевых зубов
26—35, число рядов 95—120.
Кишечник (рис. 574) сравнительно мало перекручен.
. Гениталии (рис. 575, 576). Передняя часть семяяйцевода обильно
снабжена железами и широкая. Резервуар семяприемника в наполненном
состоянии достигает больших размеров. Его короткий, толстый и мускули-
стый проток посередине сильно вздут. Семяпровод тонкий и немного длин-
нее эпифаллуса. Последний представляет собой длинный конус, который
у основания имеет кольцевидный валик. Атриум неопределенной формы
или бочковидный. В местах впадения в него половых путей и семяприемника
ARION
413
образует неправильные вздутия или карманчики. Снаружи на этих вздутиях,
особенно вблизи от эпифаллуса, иногда видны мелкие выступы или сосочки,
которым внутри атриума соответствуют небольшие углубления.
Распространение. С территории СССР был указан лишь один
раз (Мельниченко, 1936 — Смоленская область). Вероятно, в основе этих
данных лежит ошибка в определении видовой принадлежности найденных
слизней. В СССР в природных биотопах этот вид не проживает. Восточная
граница его ареала проходит через западную Польшу, на севере доходя
до устья Вислы, а на юге до окрестностей Вроцлава и через западную
Чехословакию. К западу ареал охватывает почти всю западную и среднюю
Европу, Британские и Шетландские о-ва и южную часть Скандинавского п-ова.
Завезен в Северную Америку, южную Африку, Австралию, Тасманию
и Новую Зеландию.
Образ жизни. Обитает во влажных лесах, богатых подстилкой
и подлеском, особенно в буковых и ольховниках; очень редок в хвойных ле-
сах. Скрывается в подстилке, под валежником, камнями и во мху. Некото-
рые авторы (Quick, 1960) считают, что он может жить и вне леса,,однако
последнее нами никогда не отмечалось. Живет на равнинах и в нижних го-
ризонтах горных долин, не выше 900 м над ур. м.
Питается по ночам, главным образом зелеными частями растенйй и гри-
бами.
Живет до П/г лет. Половозрелость наступает на 5—9-м месяце. Один
слизень может в течение жизни отложить свыше 350 яиц. Яйца круглые,
диаметром 2—3.5 мм, беловатые, обычно откладываются под мох. Вылуп-
ление наступает через 1—4 недели.
ЛИТЕРАТУРА
Акрамовский Н. Н. Наземные моллюски
лавового плато в окрестностях гор.
Еревана. — Науч. тр. Ереван, гос.
ун-та, 1947 (1943), с. 259—273.
Акрамовский Н. Н. Новый подвид голого
слизня из южных частей Советской
Армении (Gastropoda, Limacidae). —
ДАН АрмССР, 1948, т. 8, вып. 1,
с. 37—41.
Акрамовский Н. Н. Наземные моллюски
территории селения Гнишик в Совет-
ской Армении. — В кн.: Зоол. сб. Вып. 6.
Ереван, 1949, с. 127—183.
Акрамовский Н. Н. Некоторые итоги изу-
чения современной фауны моллюсков
Армении. — В кн.: Моллюски. Пути,
методы и итоги их изучения. Л., 1971а,
с. 108—111.
Акрамовский Н. Н. Краткий каталог
современной фауны моллюсков Совет-
ской Армении. — Биол. журн. Арме-
нии, 19716, т. 24, вып. 6, с. 3—12.
Акрамовский Н. Н. Моллюски. — В кн.:
Фауна Армянской ССР. Ереван, 1976.
272 с.
Акрамовский И. Н., Бабаян Г. С. Разра-
ботка мер борьбы с горным слизнем —
вредителем табака в парниках Арме-
нии. — Изв. с.-х. наук (мин. с.-х.
АрмССР), 1976, вып. 10, с. 68—72.
Алейникова М. М., Акрамовский Н. Н.
К эколого-фаунистической характери-
стике моллюсков (преимущественно на-
земных) Среднего Поволжья. — В кн.:
Сборник кратких сообщений Казанск.
гос. ун-та, 1968. Зоология, 2,
с. 99—105.
Арутюнова Л. Д. Заметка о некоторых
наземных моллюсках южнобережья
Крыма. — Биол. журн. Армении, 1975,
т. 28, вып. 10, с. 104—109.
Арутюнова Л. Д. О годовых циклах двух
видов слизней Deroceras caucasicum
и Vitrinoides monticola (Gastropoda :
Limacidae) в условиях лабораторной
культуры. — Биол. журн. Армении,
1978, т. 31, вып. 9, с. 990—992.
Бацылев Е. Г. Борьба со слизнями. —
Сад и огород, 1959, № 6, с. 25—26.
Большой практикум по зоологии беспо-
возночных. А. В. Иванов, А. С. Мон-
чадский, Ю. И. Полянский, А. А. Стрел-
ков. Ч. II. М. 1946. 631 с.
Бодренков Г. Е. О вспышке массового
размножения слизней в Орловской об-
ласти в 1957 году. — Уч. зап. Орловск.
гос. пед. ин-та, 1963, т. 18, вып. 1,
с. 21—38.
Вашадзе В. Н. Материалы по изучению
биологии и экологии беспанцырных
моллюсков [Eumilax (Paralimax) inter-
mittens Bttg.] и меры борьбы. — Тр.
Сухум, бот. сада, 1952, вып. 7, с. 210—
212.
Величковский В. Моллюски. Очерки фауны
Валуйского уезда Воронежской губер-
нии. Харьков, 1910, вып. 6. 111 с.
Гриванов К. П. Ареал распространения
и зоны вредности слизней (Agriolimax)
в европейской части СССР. — В кн.:
Итоги науч.-исслед. раб. ВИЗР за
1935 год. Л. 1936, с. 34—37.
Давтян Э. А. Сравнительная восприимчи-
вость моллюсков к инвазированию ли-
чинками Mullerius capillaris, Cystocau-
lus nigrescens и Synthetocaulus spp. —
Тр. н.-иссл. вет. ин-та (мин. с.-х.
АрмССР), 1947, вып. 5, с. 3—20.
Давтян Э. А. Восприимчивость моллюсков
к заражению личинками некоторых
легочных нематод овец и коз. — Тр.
Ереван, зоо-вет. ин-та, 1948, т. 10,
с. 105—119.
Давтян Э. А. Динамика инвазированности
моллюсков личинками и факторы, влия-
ющие на их выхождение. — В кн.:
Зоол. сб. Вып. 7. Ереван, 1950, с. 83—
101.
Дамянов С. Г., Лихарев И. М. Сухоземни
охлюви (Gastropoda terrestria). В кн.:
Фауна на България. Т. 4. София,
1975. 425 с. (болг.).
Двигубский И. Опыт естественной истории
всех животных Российской империи
с изображением животных. Животные
мягкие и раковинные. М., 1831. 72 с.
Дмитриева Е. Ф. Эффективность и дли-
тельность действия препаратов металь-
дегида в борьбе с полевыми слизнями.—
Тр. Всес. н.-иссл. ин-та защиты расте-
ний, 1963, вып. 19, с. 260—266.
Дмитриева Е. Ф. Биология сетчатого
слизня (Deroceras reticulatum Miiller)
и меры борьбы с ним в условиях Ле-
нинградской области. — Автореф. канд.
дис., Л., 1968а. 19 с.
Дмитриева Е. Ф. Влияние температурного
фактора на скорость инкубации яиц
сетчатого слизня (Диагностика осен-
него развития и жизнеспособности). —
В кн.: Моллюски и их роль в эко-
системах. Л. 19686, с. 61—62.
ЛИТЕРАТУРА
415
Дмитриева Е. Ф. Динамика численности,
рост, питание и размножение сетчатого
слизня (Deroceras reticulatum) в Ле-
нинградской области. — Зоол. журн.,
1969, т. 48, вып. 6, с. 802—810.
Дмитриева Е. Ф. Влияние пищи на рост,
половое созревание и плодовитость сет-
чатого слизня (Deroceras reticulatum
Miill.). — Зап. Ленингр. с.-х. ин-та,
1973, т. 212, с. 93—95.
Дмитриева Е. Ф. Влияние температуры
и влажности верхнего слоя почвы
на интенсивность вылупления сетча-
того слизня (Deroceras reticulatum Mul-
ler). — В кн.: Моллюски. Их система,
эволюция и роль в природе. Л., 1975,
с. 40—43.
Дмитриева Е. Ф. «Критические» периоды
в онтогенезе сетчатого слизня (Dero-
ceras reticulatum) в связи с биоклимати-
ческими условиями Ленинградской об-
ласти. — Научн. тр. Ленингр. с.-х.
ин-та, 1978, т. 351, с. 62—64.
Дмитриева Е. Ф., Шапиро Я. С. Гранули-
рованный метальдегид — активный ток-
сикант кишечного действия. — Зап.
Ленингр. с.-х. ин-та, 1974, т. 239,
с. 110—113.
Дмитриева Е. Ф., Шапиро Я. С. Некото-
рые функциональные изменения в ор-
ганизме сетчатого слизня (Deroceras
reticulatum Muller) при отравлении
метальдегидом. — Зап. Ленингр. с.-х.
ин-та, 1975, т. 270, с. 115—118.
Дмитриева Е. Ф., Шапиро Я. С. Физиолого-
токсикологические аспекты постэмбрио-
генеза сетчатого слизня. — Научн. тр.
Ленингр. с.-х. ин-та, 1976, т. 297,
с. 91—94.
Егоров Ю. Г. Гельминтозы жвачных и меры
борьбы с ними. Минск. 1965. 140 с.
Жадин В. И. Жизненный цикл одного
среднеазиатского слизня. — Природа,
1945, № 2, с. 66—67.
Жадин В. И. К биологии Parmacella kor-
shinskii Smrth. и о мерах борьбы с ней
в Таджикистане. — Зоол. журн., 1946,
т. 25, вып. 1, с. 15—26.
Загайный А. А. Для защиты от слиз-
ней. — Защита растений, 1978, № 3,
с. 49.
Иванов В. А. К фауне слизней северных
склонов Центрального Кавказа. —
В кн.: Экология животных северных
склонов Центрального Кавказа. Орджо-
никидзе, 1978, с. 92—94.
Иззатуллаев 3. Первые находки Deroceras
sturanyi и Oxychilus kutaisianus в Тад-
жикистане. — В кн.: Вопросы зоологии
Таджикистана. Душанбе, 1972, с. 80—
86.
Иззатуллаев 3. Некоторые данные по био-
логии слизней (Mollusca, Pulmona-
ta) — вредителей сельскохозяйственных
культур Таджикистана. — Изв. АН
ТаджССР, отд. биол. наук, 1975а, № 4
(61), с. 22-24.
Иззатуллаев 3. К изучению фауны мол-
люсков окрестностей города Хорога
(Mollusca). — В кн.: Зоологический
сборник, ч. 2. Душанбе, 19756, с. 5—7.
Калениченко И. (Kaleniczenko J.). Descrip-
tion d’un nouveau genre de Limaces
de la Russie meridionale (Krynickil-
lus). — Bull. Soc. Nat. Moscou, 1851a,
v. 24, pt 1, p. 215—228.
Калениченко И. (Kaleniczenko J.). De-,
scription des limaces, que. se trouvent
dans 1’Ukraine. — Bull. Soc. Nat. Mos-
cou, 1851b, v. 24, pt 2, p. 109—126.
Калитина 3. И. К изучению зонального
распределения наземных моллюсков се-
верных склонов Центрального Кав-
каза и Восточного Предкавказья. —
Тр. Сев.-Осетин, мед. ин-та, 1958,
т. 7, вып. 1, с. 157—181.
Косоглазов А. А. Малакофауна (Gastro-
poda, Pulmonata) оранжерей юго-вос-
тока Европейской части РСФСр. —
Зоол. журн., 1972, т. 51, вып. 2,
с. 289—290.
Левушкин С. И., Матекин П. В. Biospeo-
logia sovietica XXV. Troglolestes soko-
lovi gen. n., sp. n. — первый трогло-
бионтный слизень. — Бюл. Моск, о-ва
испыт. природы. Отд-ние биол., 1965,
т. 70, вып. 3, с. 35—46.
Линдгольм В. A. (Lindholm W. A.). Mate-
rialen zur Molluskenfauna von Sud-
westrussland, Polen und der Krim. —
Зап. Новорос. о-ва естествоиспыт.,
1908, т. 31, с. 199—232.
Линдгольм В. А. Заметки о палеаркти-
ческих наземных и пресноводных мол-
люсках. — Ежегод. Зоол. муз., 1922,
т. 23, с. 304—320.
Линдгольм В. A. (Lindholm W. A.). On
a misapplied generic name for Caucasian
slugs. — Proc. Malac. Soc. Lond., 1925,
v. 16, p. 167—168.
Лихарев И. M. Слизни — вредители сель-
ского хозяйства. — М,—Л., 1954. 75 с.
Лихарев И. М. Географическое распро-
странение наземных моллюсков Кав-
каза и некоторые пути происхождения
этой фауны. — В кн.: Матер, к совещ.
по вопр. зоогеографии суши. Львов,
1957, с. 73—75.
Лихарев И. М. Наземные моллюски [Кав-
каза]. — В кн.: Животный мир СССР.
Т. 5. М., 1958, с. 464—476.
Лихарев И. М. Клаузилииды (Clausilii-
dae). — В кн.: Фауна СССР. Моллюски.
Т. 3, вып. 4. Нов. сер., № 83. М.—Л.,
1962, с. 1—317.
Лихарев И. М. Некоторые факторы, опре-
деляющие распространение синантроп-
ных наземных моллюсков. — В кн.:
Моллюски. Вопросы теоретической и
прикладной малакологии. М.—Л.,
1965, с. 48—51.
Лихарев И. М., Виктор А. Й. Мантийный
комплекс органов слизней и его таксо-
номическое значение. — В кн.: Мол-
люски, основные результаты их изу-
чения. Л. 1979а, с. 11—13.
Лихарев И. М., Виктор А. Й. Паралле-
лизмы в строении и систематическое
416
ЛИТЕРАТУРА
положение слизней надотряда Stylom-
matophora. — Тр. Зоол. ин-та АН
СССР, 19796, т. 80, с. 70—86.
Лихарев И. М., Иззатуллаев 3. О новых
завезенных видах слизней фауны Тад-
жикистана. — Докл. АН ТаджССР,
1972, т. 15, № 3, с. 66-68.
Лихарев И. М., Раммельмейер Е. С. На-
земные моллюски фауны СССР.—
В кн.: Опред. по фауне СССР. Вып. 43.
М,—Л., 1952. 511 с.
Лихарев И. М., Старобогатов Я. И. Ма-
териалы по фауне моллюсков Афгани-
стана. — Тр. Зоол. ин-та АН СССР,
1967, т. 42, с. 159—197.
Мартенс Э. Слизняки (Mollusca). — В кн.:
А. П. Федченко. Путешествие
в Туркестан. Вып. 1, т. 2, ч. 1. СПб,
1874, с. 1—64.
Масальский В. И. Туркестанский край. —
В кн.: Россия. Т. 19. 1913. 861 с.
Мельниченко А. Н. Сравнительно-фауни-
стический обзор наземных моллюсков
Жигулевских гор и Западной об-
ласти. — В кн.: К фауне Куйбышев-
ской области. М.—Куйбышев, 1936,
с. 3—20.
Миничев Ю. С., Старобогатов Я. И. Ос-
новные особенности эволюции половой
системы гастропод. — Зоол. журн.,
1971, т. 50, вып. 9, с. 1309—1322.
Миничев Ю. С., Старобогатов Я. И. К по-
строению системы эвтиневральных брю-
хоногих моллюсков. — В кн.: Мол-
люски. Их система, эволюция и роль
в природе. Л., 1975, с. 8—11.
Михаэлис Е. Описание новых и малоизвест-
ных моллюсков южного Алтая и север-
ной Джунгарии. — Зап. Акад, наук,
1892, т. 69, кн. 1, с. 31—36.
Мухитдинов А. Б., Рахматов Э. К фауне
слизней Северного Таджикистана. —
В кн.: Материалы (тез. докл.) конф,
проф.-препод, колл, естеств.-географ,
фак. Ленинабадск. пед. ин-та, Ленин-
абад, 1972, с. 25—27.
Надеждин Н. Список брюхоногих слизня-
ков (Mollusca, Gastropoda), водящихся
в окрестностях Москвы. — Изв. о-ва
любит, естествозн., антропол. и этногр.,
1869, т. 3, вып. 3, с. 177—184.
Николаев В. А. Морфо-биологические осо-
бенности яиц сетчатого слизня Deroce-
ras reticulatum Miill. — Уч. зап. Орлов,
пед. ин-та, 1967, т. 35, с. 123—128.
Ошанова Н. Голи охлюви (семейство Ario-
nidae, Limacidae и Milacidae) на Ви-
тоша. — Изв. Зоол. ин-т с музей Бълг.
АН, 1972, 35, с. 139—154 (болг.).
Пахорукова Л. В. Количественное иссле-
дование питания полевого (Deroceras
agrestis) и сетчатого (D. reticulatum)
слизней. — Зоол. журн., 1976, т. 55,
вып. 1, с. 29—31.
Розен О. В. (Rosen О.). Beitrag zur Kennt-
niss der Molluskenfauna Transcaspiens
und Chorossans. — Nachrichtenbl. dtsch.
malak. Ges., 1892, Bd 24, S. 121—126.
Розен О. В. Материалы к познанию фауны
слизняков Закаспийской области и Хо-
росана. — В кн.: Фауна Закасп. обл.
Т. 2, 1894, с. 39-57.
Розен О. В. Моллюски Окской экспеди-
ции. — Дневн. Зоол. отд-ния О-ва
любит, естествозн., антропол. и этногр.,
1905, т. 3, вып. 6, с. 79—85.
Рыбак В. Ф. Наземные моллюски окрест-
ностей с. Кончезера и их роль в рас-
пространении мюллериоза. — Тр. Ка-
рельск. фил. АН СССР, 1961, вып. 30,
с. 92-96.
Скляр И. Я. Новый для науки вид слизня
рода Deroceras (Gastropoda, Limacidae)
из Крыма. — Вестн. зоол., 1975а, № 4
с. 73—76.
Скляр И. Я. Про знахождение Deroceras
transcaucasicum Simr. на Украш! (Ga-
stropoda, Limacidae). — Допов. АН
УРСР. Сер. Б, 19756, с. 852—855.
Скляр И. Я. Новый для науки вид слизня;
рода Deroceras (Gastropoda, Limacidae)
из Крыма. — Вестн. зоол., 1975в, № 6,
с. 79-82.
Соболева Т. Н. Черный слизень (Limax
turkestanus) — промежуточный хозяин
трематод Brachylaemus spinulosus (Hoff-
man, 1899). — Тр. Ин-та зоол. [АН
КазССР], 1971, т. 31, с. 92—98.
Соколов А. М. Материалы изучения слиз-
ней и мер борьбы с ними. — Тр. Ива-
нов. с.-х. ин-та, 1935, 1, с. 110—155.
Стеклов А. А. Наземные моллюски нео-
гена Предкавказья и их стратиграфи-
ческое значение. — Тр. Геол, ин-та
АН СССР, 1966, вып. 163. 262 с.
Трушин И. Н. Роль экологических факторов
в естёственной зараженности моллюсков
личинками мюллериев. — В кн.:
Пробл. общ. и прикл. гельминтол.
М. 1973, с. 344—347.
Увалиева К. К. О распространении мол-
люсков в Восточно-Казахстанской об-
ласти. — Изв. АН КазССР, Сер. биол.,
1969, вып. 3, с. 53—59.
УвалиеваК. К. Жизненные циклы и вредо-
носность слизней Deroceras caucasi-
cum (Simroth) и Parmacella rutellum
(Hutton) в Алма-Атинской области. —
В кн.: Моллюски. Их система, эволю-
ция и роль в природе. Л., 1975, с. 44—
46.
Увалиева К. К. Слизни — вредители сель-
скохозяйственных культур юго-востока
Казахстана и меры борьбы с ними.
Алма-Ата. 1979, с. 1—59.
Фридолин В. Ю. Значение элементов рель-
ефа как оазисов-убежищ реликтовой
фауны в бассейне Финского залива,
Хибинских гор и центральной Каре-
лии. — Изв. Всес. геогр. о-ва, 1934,
в. 3, с. 294—307.
Хохуткин И. М. О распространении на-
земных моллюсков на Урале. — Зоол.
журн., 1961, т. 40, вып. 2, с. 178—183.
Цветков Б. Н. К познанию фауны мол-
люсков пастбищ Алма-Атинской об-
ласти. — Тр. Каз. научн.-исслед. вет.
ин-та, 1940, т. 3, с. 379—419.
ЛИТЕРАТУРА
417
Цветков Б. Н., Матёкин П. В. К система-
тике и экологии голого слизня — про-
межуточного хозяина легочных гель-
минтов овец и коз Южного Казах-
стана. — Изв. АН КазССР, 1946, т. 33,
с. 113—119.
Шапиро Я. С. Борьба с моллюсками. —
Защита растений, 1977, № 9, с. 48.
Шапиро Я. С. Изучение вирофорной спо-
собности четырех видов слизней. —
Научи, тр. Ленингр. с.-х. ин-та, 1978,
т. 351, с. 56—61.
Шиков Е. В. О расселении наземных мол-
люсков во время половодий. — Зоол.
журн., 1977, т. 51, вып. 3, с. 361—367.
Шиков Е. В. Наземная малакофауна
природных и антропогенных лесов и лу-
гов Валдайской возвышенности и при-
легающих низменностей. — В кн.:
Моллюски, основные результаты их изу-
чения. Л., 1979а, с. 205—207.
Шиков Е. В. Фауна наземных моллюсков
населенных пунктов Валдайской воз-
вышенности и сопредельных террито-
рий. — Зоол. журн., 19796, т. 58, № 7,
с. 969—976.
Шиков Е. В. Зависимость распределения
слизней рода Deroceras Rafinesque,
1820 в поймах крупных рек Валдайской
возвышенности от направления господ-
ствующих ветров. — Экология, 1979в,
№ 5, с. 97—99.
Шилейко А. А. К биологии почвенных
хищных слизней. — В кн.: Материалы
II Всесоюзного совещания по почвен-
ной зоологии. М., 1966, с. 155—156.
Шилейко А. А. О биологии размножения
и ювенильных адаптациях слизня
Parmacella ibera Eichw. — Зоол. журн.,
1967а, т. 46, вып. 6, с. 946—948.
Шилейко А. А. Некоторые новые и
малоизвестные моллюски Талыша. —
Научи, докл. высш, школы. Биол.
науки, 19676, № 4, с. 24—32.
Шилейко А. А. Наземные моллюски над-
семейства Helicoidea. — В кн.: Фауна
СССР. Моллюски. Т. 3, вып. 6. Нов.
сер., № 117. Л., 1978, 384 с.
Шилейко А. А. Система отряда Geophila
(=Helicida) (Gastropoda: Pulmonata). —
Тр. Зоол. ин-та, 1979, т. 80, с. 44—79.
Abeloos М. Etapes de la croissance chez
Arion rufus. — C. r. Acad. Sci. Paris,
1942, v. 215, p. 96—97.
Abeloos M. Forme de la croissance des
especes du genre Limax. — C. r. Acad.
Sci. Paris, 1943, v. 217, p. 159—161.
Abeloos M. Recherches experimentales sur
la croissance; la croissance des mollus-
ques Arionides. — Bull. biol. . France-
Belg., 1944, v. 78, N 3—4, p. 215—256.
Abeloos M. Evolution par variation du nom-
bre des phases de croissance chez les
mollusques Limaciens. — C. r. Acad.
Sci. Paris, 1945, v. 220, N 1—5, p. 62-
63.
Albonico R. Die Farbenvarieteten der gros-
sen Wegschnecken Arion empiricorum
Fer. und deren Abhangigkeit von den
Umweltsbedingungen. — Rev. Suisse
Zool., 1948, v. 55, N 25, p. 347—425.
Altena С. O. Regteren. The Limacidae
of the Canary Islands. — Zool. Verhan-
del. Leiden, 1950, v. 11, p. 1—34.
Altena C. 0. Regteren. Notes sur les limaces.
3. Sur la presence en France d’Arion
lusitanicus Mabille. — J. Conchyl.,
1956, v. 95, p. 89—99.
Altena C. 0. Regteren. Notes sur les lima-
ces, 6—7. — Basteria, 1962, v. 26,
N 3-4, p. 47-57.
Altena C. 0. Regteren. Notes sur les lima-
ces. 8. Sur I’etat de deux especes nomi-
nates de genre Arion. — Basteria, 1963,
v. 27, N 1—2, p. 1—6.
Altena C. 0. Regteren. Notes sur les lima-
ces. 10. A propos d’une petite collection
de Succineidae, Philomycidae et Lima-
cidae de File Maurice. — J. Conchyl.,
1966a, v. 105, p. 232—235.
Altena C. 0. Regteren. Notes on Land
Slugs. 11. Arionidae, Milacidae and
Limacidae from South Africa (Mollusca,
Gastropoda, Pulmonata). — Zool. Meded.,
1966b, v. 41, N 20, p. 269—298.
Altena C. 0. Regteren. Notes sur les lima-
ces. 14. Sur trois especes de Deroceras
de la Catalogue dont deux nouvelles. —
J. Conchyl., 1969, v. 107, p. 101—108,
Altena C. 0. Regteren. Notes on land slugs,
16. Deroceras from Afghanistan, inclu-
ding description of D. kandaharensis. —-
Fieldiana. Zoology, 1970a, v. 41, N 15,
p. 175—178.
Altena C. 0. Regteren. Notes sur les lima-
ces. 17. Sur deux especes de Deroceras. --
Basteria, 1970b, v. 34, N 3—4, p. 67—74,
Altena C. 0. Regteren. Neue Fundorte
von Arion lusitanicus Mabille. — Arch,
Moll., 1971, Bd 101, S. 183—185.
Altena С. O. Regteren. Notes on land slugs,
20. On two species of Deroceras from
Jugoslavia and Austria. — Basteria,
1973, v. 37, p. 85-92.
Altena C. 0. Regteren. Notes on Land
Slugs. 22: a catalogue of the genus
Lytopelte (Limacidae) and a note on
L. kandaharensis (Altena). — Nautilus,
1975, v. 89, N 2, p. 62—63.
Ant H. Das Datum Veroffentlichung von
Arion fasciatus (Nilsson). — Mitt, dtsch,
malak. Ges., 1971, Bd 2, N 19, S. 257,
Arias R. 0., Crowell H. H. A contribution
to the biology of the grey garden slug. —
Bull. S. Calif. Acad. Sci., 1963, v. 62,
N 2, p. 83—97.
Babor J. F. Uber den Cyclus der Ge-
schlechtsentwicklung der Stylommato-
phoren. — Verhandl. dtsch. zool. Ges,
Leipzig, 1894, Bd 4, S. 55—61.
Babor J. F. Doplinki k znamostem о ces-
kych slimdcich. 1. Limacidae. — VSst,
Kral. Ceske spol. nauk, Tr. math.-pfir.,
1895 (1894), v. 28, S. 1—22.
Babor J. F. Uber die von Herrn Dr. H. Re-
bel im Jahre 1896 in Ostrumelien ge-
sammelten Nacktschnecken. — Ann. na-
418
ЛИТЕРАТУРА
turhist. Hofmuseums, 1898, Bd 13,
S. 40-41...
Babor J. F. Uber die Nacktschnecken der
Grazer Umgegend. — Verb, dtsch. zool.
Ges., 1900, Bd 10, S. 148—150.
Babor J. F. Nacktschnecken. — In: Pent-
her A., Zederbauer E. Ergebnisse einer
naturwissenschaftlichen Reise zum Erd-
• schias—Dagh (Kleinasien) im Jahre
1902. — Ann. naturhist. Hofmuseums,
1905, Bd 20, S. 291-294.
Babor J. F., Frankenberger Zd. Zur Kenn-
tnis der karpathischen Weichtiere. —
Verb, zobl.-bot. Ges. Wien, 1914,
Bd 64, S. 109—122.
Babor J. F., Kostal J. Prispevky ku poz-
nam pomerft pohlavnich u nekterych
Limacidii. — Bohm. Ges. Wiss. Sitzungs-
ber., 1894 (1893), № 51. 7 p.
Bailey T. G. A new anaesthetic technique
for slugs. — Experientia, 1969, v. 25,
N H, P- 1225.
Baker H. B. Heterurethrous and Aulaco-
pod. — Nautilus, 1955, v. 68, N 4,
p. 109-112.
Bakowski J., Lomnicki A. M. Miqczaki
(Mollusca). Lwow, 1892. 234 S.
Barnes H. F. The slugs in our gardens. —
New Biology, London, 1949, N 6, p. 29—
49.
Barnes H. F., Weil J. W. Slugs in gardens:
their numbers, activities and distri-
bution. Part 1. — J. Anim. Ecol.,
1944, v. 13, p. 140—175.
Barnes H. F., Weil J. W. Slugs in gardens:
their numbers, activities and distribu-
tion. Part 2. — J. Anim. Ecol., 1945,
v. 14, p. 71—105.
Barr R. A. Some observations on the pedal
gland of Milax. — Quart. J. microscop.
Sci., 1926, N. S„ v. 70, p. 647—667.
Barr R. A. Some notes on the mucous
and skin glands of Arion ater. — Quart.
J. microscop. Sci., 1927, N. S., v. 71,
p. 503—525.
Bayne C. J. Histochemical studies on the
egg-capsules of eight gastropod mol-
luscs. — Proc, malac. Soc. Lond., 1968,
v. 38, N 3, p. 199-212.
Belke G. Notice sur 1’histoire naturelie
du district de Radomysl (gouvernement
de Kief). — Bull. Soc. Nat. Moscou,
1866, v. 39, pt 2, p. 490—571.
Benson W. H. Mollusca. In: Cantor Th.
General features of Chusan, with remarks
on the flora and fauna of. that island. —
Ann. Mag. Natur. Hist., 1842, v. 9,
p. 486—489.
Belt J. A. The breeding seasons of slugs
in gardens. — Proc. zool. Soc. Lond.,
1960, v. 135, N 4, p. 559—568.
Beyer W. N., Saari D. M. Activity and eco-
logical distribution of the slug Arion
subfuscus (Draparnaud) (Stylommato-
' phora, Arionidae). — Amer. Midland Na-
tur., 1978, v. 100, N 2-, p. 359-367.
Bielz M. Verzeichnis der Land- und Siiss-
wassermollusken Siebenbiirgens.—Ver-
1 handl: Mitth. siebenbiirg. Ver. Natur-
wiss. Hermannstadt, 1851, Bd 2, S. 14—
16, 55—59, 62-65.
BoettgerC. R. Die Nomenklatur der in Deu-
tschland vorkommenen Arten der Land-
schneckengattung Milax Gray. — Arch.
Moll., 1943, Bd 75, S. 27—29.
Boettger C. R. Zur Kenntnis der grossen
Wegschnecken (Arion s. str.) Deutsch-
lands. — Arch. Moll., 1949, Bd 78,
S. 169-186.
Boettger O. Sechstes Verzeichniss transkau-
kasischer, armenischer und nordpersi-
scher Mollusken. — Jahrb. dtsch. malak,
Ges., 1881, Bd 8, p. 167—261.
Boettger O. Nacktschnecken aus Epirus und
von den Jonischen Inseln. — Nachri-
chtenbl. dtsch. Malak. Ges., 1882,
Bd 14, S. 96—101.
Boettger O. Siebentes Verzeichniss von
Mollusken der Kaukasuslander, nach
Sendungen des Hrn Hans Leder, z. Z.
in Helenendorf bei Elisabetpol. —
Jahrb. dtsch. malak. Ges., 1883, Bd 10,
S. 135—198.
Boettger 0. Die Binnenmollusken des Ta-
lysch-Gebietes. — In: Radde G. Fauna
und Flora des siidwestlichen Caspi-
Gebietes. Leipzig, 1886a, S. 256—350.
Boettger 0. Neuntes Verzeichniss (IX)
von Mollusken der Kaukasuslander nach
Sendungen des Hrn Hans Leder, z. Z.
in Helendorf bei Elisabetpol (Trans-
kaukasien). — Jahrb. dtsch. malak. Ges.,
1886b, Bd 13, S. 121—156.
Boettger 0. Die Binnenmollusken Trans-
caspien und Chorossans. — Zool. Jahrb.,
1889, Abt. 3, Bd 4, S. 925—992.
Bonavita A. Hygrocinese chez la limace
Milax gagates Draparnaud (gasteropode
pulmone). — C. r. hebd. Seanc. Acad.
Sci. Paris, 1967, v. 265, p. 55-57.
Bouchard-Chantereaux M. Catalogue des
mollusques terrestres et fluviatiles ob-
serves dans le departement du Pas-de-
Calais. — Mem. Soc. Agr. Sci. Arts
Boulogne, ser. 2, 1838 (1837), v. 1,
p. 141—230.
Bourguignat J. F. Des limaces algerien-
nes. — Rev. Mag. zool., ser. 2, 1861,
v. 13, N 7, p. 299—306.
Brard С. P. Histoire des coquilles terrestres
et fluviatiles qui vivent aux environs
de Paris. Paris et Geneve, 1815-, p. 1—23,
1—239, 1—17.
Braun M. Zur Molluskenfauna der Ostseepro-
vinzen. — Nachrichtenbl. dtsch. malak.
Ges., 1883, Bd 15, S. 174—181.
Braun M. Beitrage zur Kenntniss der Fauna
baltica. 2. Die Land- und Susswassei-
mollusken der Ostseeprovinzen. — Arch.
Nat. Liv., Ser. 2, 1884, Bd 9, N 5,
S. 401—504.
Brooks W. M. Tetrahymenial ciliates as pa-
rasites of the grey garden slug. — Hil-
gardia, 1968, v. 39, N 8, p. 205—276.
Burch J. B. How to know the eastern'land
snails. Dubuque, Iowa, 1962. 214 p.
Cain A. J., Williamson M. N. Variation
and specific limits in the: Arion ater
ЛИТЕРАТУРА 419
aggregate. — Proc. Malac. Soc. Load.,
1958, v. 33, p. 72—86.
Carrick R. The life-history and development
of Agriolimax agrestis L. the grey field
slug. — Trans. Roy. Soc. Edinb., 1938;
v. 59, N 3, p. 563—597.
Chatfield J. E. Limax grossui Lupu 1970,
a slug new to the British Isles. — J.
Conchoid 1976, v. 29, N 1, p. 1—4.
Chetail M. Etude de la regeneration du ten-
tacule occulaire chez un Arionidae (Arion
rufus L.) et un Limacidae (Agriolimax
agrestis L.). — Arch. anat. micr. morph,
exp., 1963, v. 52, p. 129—203.
Chetail M., Binot D. Particularites histo-
chimiques de la glande et de la sole
pedieuse d’Arion rufus (Stylommato-
phora: Arionidae).—Malacologia, 1967,
v. 5, N 2, p. 269-284.
Chevallier H. Taxonomie et biologie des
grands Arion de France (Pulmonata:
Arionidae). — Malacologia, 1969, v. 9,
N 1, p.. 73—78. -
Chevallier H. Les limaces de Bretagne. —
Penn ar Bed., 1970, v. 7, N 62, p. 370—
389.
Chevallier H. Cycle biologique des grands
Arion en France (Gastropoda, Pulmo-
nata). — Atti Soc. ital. sci. natur.,
. 1971, v. 112, N 2, p, 316—320.
Chevallier H. Arionidae (Mollusca, Pulmp-
nata) des Alpes et du Jura frangais. —
i Haliotis, .1972, v. 2, N 1, p. 7—23.
Clessin S. Anhang zur Molluskenfauna der
Krim. — Malak. BL, N. F., 1883, Bd 6,
S. 37—52.
Clessin S. Deutsche Excursions-Mollusken-
Fauna. 2 Aufl. Nurnberg, 1884, S. 1—658.
Clessin S. Die Molluskenfauna Oesterreich-
: Ungarns und der Schweiz. Nurnberg,
; 1887. 858 S. .
Cockerell T. D. A.- On the geographical
distribution of slugs. — Proc. Zool. Soc.
, Lond.. 1891, p. 214—226.
.Cockerell T. D. A. African slugs. — Nau-
tilus, 1935, v. 48, p. 142—143.
Colville B., Lloyd-Evans L., Norris A.
. Boettgerilla pallens Simroth, a new
, British species. — J. Conchol., 1974,
v. 28, N 4, p. 203—208.
Cuvier G. Sur la Dolabelle, sur la Testacelle
et sur un nouveau genre de mollusques
a coquille cachee, nomee Parmacelle. —
Ann. Mus. hist, natur. Paris, 1804,
v. 5, p. 435—444.
Dainton В. H. The activity of slugs:
I. The. induction of activity by changing
temperatures. — J. exp. Biol., 1954a,
v. 31, p. 165—167.
Dainton В. H. The activity of slugs:
II. The effect of light and air currents. —
J. exp. Biol., 1954b, v. 31, p. 188—197.
Daxl R. Ein Beitrag zur Biologie von Boett-
., gerilla vermiformis Wiktor, 1959.—
Z. angewandte Zool., 1967, Bd 54,
S. 227—231.
Draparnaud J.-P.-R. Tableaux des mollus-
ques terrestres et fluviatiles de la France.
Paris, 1801, p. 1—116.
Draparnaud J.-P.-R. Histoire naturelie
des mollusques terrestres et fluviatiles
de la France. Paris, 1805, p. 1—164,
Drozdowski A. Vergleichende Studieniiber
die Lungenmorphologie bestimmter
Schneckenarten (Gastropoda, Pulmo-
nata). — Zool. polon., 1970, Bd 20,
S. 217-256.
Dumont F., Mortillet G. Histoire des mol-
lusques terrestres et d’eau douce vivants
et iossiles de la Savoie et du bassin
du Leman. — Bull. Soc. hist, natur,
Savoie, 1852, v. 2, p. 14—142.
'Ehrmann P. Mollusken. — In: Tierwelt Mit-
teleuropas. Leipzig, 1933, Bd 2; N 1,
264 S. .
Eichwald Ed. Naturhistorisehe Skizze von
Lithauen, Vblhynien und Podolien,
Wilna, 1830. 256 S.
Eichwald Ed. Fauna Caspio-Caucasia. Petro-
pol, 1841. 233 p.
Ellis R. A. British snails. Oxford, 1926,
275 p.:
Evans N. J. Limax pseudoflavus Evans:
a critical description and comparison
with related species. — Ir. Nat. J.-,
1978, v. 19, N 7, p. 231—236.
Evans W. A. L., Jones E. G. Carbohydrases
in the alimentary tract of the slug
Arion ater L. — Compar. Biochem. Phy-
siol., 1962, v. 5, N 3, p. 149—160.
Ferussac J. B. L. D’Audebard. Histoire
naturelie generale et particuliere des
mollusques terrestres. et fluviatiles. T. II,
pt. 1. Histoire naturelle pulmones sans
opercule. Paris, 1819, p. 1—96; Paris,
1823, p. 96a—96z, 96a—96X.
Flasar I. Uber die Variabilitat der Radula
bei der Nacktschnecke Limax tenellus
Miill. — Zool. Anz., 1961, Bd 167,
N 9—12, S. 381—389.
Focardi S., Quattrini R. Structure of the
reproductive apparatus and life cycle
of Milax gagates (Draparnaud) (Mol-
lusca Gastropoda Pulmonata). — Boll.
zooL, 1972, v. 39, N 1, p. 9—27.
Forcart L. Die Mollusken der nordpersischen
Provinz Masenderan und ihre tiergeo-
graphische Bedeutung. — Arch. Natur-
gesch., N. F., 1935, Bd 4, N 3, S. 404^
447.
Forcart L. Die Verbreitung der Limaciden
und Milaciden in der Schweiz. — Arch,
Moll., 1942a, Bd 74, S. 114—119.
Forcart L. Zwei neue Schnecken aus Tiir-
kei. — Arch. Moll., 1942b, Bd 74,
S. 239—242.
Forcart L. Verzeichnis der von Herrn
H. W. E. Crockewit in der Tiirkei ge-
sammelten Land- und Siisswasser-Mol-
lusken (excl. Clausiliidae). — Basteria,
1953, Bd 17, N 1—2, S. 19—28.
Forcart L. Revision de Parmacella olivieri
Cuvier et position systematique du genre
Parmacella Cuvier. — Basteria, 1959,
v. 23, N 3, p. 39—45.
Forcart L. Alpine und nordische Arten
der Gattung Lehmannia Tleynemanij
420
ЛИТЕРАТУРА
(Limacidae). — Arch. Moll., 1966, Bd 95,
S. 225-236.
Forcart L. Milacidae und Limacidae der
Insel Rhodes. — Basteria, 1972, v. 36,
N 2-5, S. 105-116.
Franc A. Sous-classe des Pulmones. —
In: Traite de zoologie. T. 5, fasc. 3,
1968, p. 325—607.
Frauenfeld G. Zoologische Miscellen, 3. —
. Verb, zool.-bot. Ges. Wien, 1864, Bd 14,
,S. 671—696.
Fromming E. Biologie der mitteleuropa-
ischen Landgastropoden. Berlin, 1954.
404 S.
Galangau V. Le cycle sexuel annuel de Mi-
lax gagates Drap. (Gasteropode pulmone)
et ses deux pontes. — Bull. Soc. zool.
France, 1964, v. 89, N 4, p. 510—513.
Garner J. H. A study of excretion in the
slug Agriolimax reticulatus. Ph. D.
thesis. 1970. University of Wales.
Georgi I. G. Geographisch-physikalische
Beschreibung des Russischen Reiches.
Th. 3, Bd 6. Konigsberg, 1800, S. 1465—
2222.
Gerhardt U. Zur Kopulation der Limaciden.
I. Mitteilung. — Z. Morph. Okol. Tiere,
1933, Bd 27, S. 401—450.
Gerhardt U. Zur Biologie der Kopulation
der Limaciden. II. Mitteilung. — Z.
Morph. Okol. Tiere, 1934, Bd 28,
S. 229—258.
Gerhardt U. Weitere Untersuchungen zur
Kopulation der Nacktschnecken. — Z.
Morph. Okol. Tiere, 1936a, Bd 30,
S. 297—332.
Gerhardt U. Weitere Untersuchungen zur
Kopulation der Stylommatophoren. —
- Z. Morph. Okol. Tiere, 1936b, Bd 31,
S. 433—442.
Gerhardt U. Zur Frage des Operculums
bei Parmacella (Gastropoda, Pulmo-
i' nata). — Zool. Anz., 1936c, Bd 113,
S. 41—44.
Gerhardt U. Weitere Untersuchungen zur
Sexualbiologie der Limaciden. — Z.
Morph. Okol. Tiere, 1937, Bd 32, S. 518—
541.
Gerhardt U. Neue biologische Untersuchun-
gen an Limaciden. — Z. Morph. Okol.
Tiere, 1939, Bd 35, S. 183—202.
Gerhardt U. Neue biologische Nacktschnec-
kenstudien. — Z. Morph. Okol. Tiere,
1940, Bd 36, S. 557—580.
Germain L. Liste des mollusques recueillis
par M. H. Gadeau de Kerville, pendant
son voyage en Khroumirie. — Bull. Mus.
hist, natur., 1907, v. 13, N 2, p. 154—158.
Germain L. Etudes sur la faune malacologi-
que terrestre et fluviatile de 1’Asie
Anterieure. 1. Parmacellidae et Lima-
cidae. — Bull, de la Delegation en
Perse, 1912, N 2, p. 4—45.
Germain L. Mollusques terrestres et flu-
viatiles (premiere partie). In: Faune
de France, v. 21. Paris, 1930. 447 p.
Germain L. Mollusques terrestres et flu-
viatiles d’Asie-Mineure. — In: Voyage
zoologique d’Henri Gadeau de Kerville
en Asie-Mineure. T. 1, pt 2. Paris,
1936. 492 p...
Gerstfeldt G. Uber Land- und Siisswasser-
Mollusken Sibiriens und des Amur-
Gebietes. — Mem. Savants etrangers,
1859, Bd 9, S. 507—548.
Gittenberger E., Backhuys W., Ripken Th.
E. J. De landslakken van Nederland.
Amsterdam, 1970. 177 p.
Giusti F. Notulae Malacologicae. XVIII.
I Molluschi terrestri e salmastri delle
Isole Eolie. — Lav. Soc. Ital. Biogeogr.,
N. S., 1973 (1972), N 3, p. 113-306.
Godwin-Austen H. H. Land and freshwater
Mollusca of India. Part VI. London,
1888, p. 207—266.
Godwin-Austen H. H. Land and freshwater
Mollusca of India. V. 2, part XII. Lon-
don, 1914, p. 311—442.
Goethem J. Contribution a 1’etude de Boett-
gerilla vermiformis Wiktor, 1959 (Mol-
lusca, Pulmonata). — Bull. Inst. Sci.
natur. Belg., ser. Biologie, 1972, v. 48,
N 14, p. 1-16.
Goethem J. L. Recherches systematiques
sur les Urocyclinae (Mollusca, Gastro-
poda, Urocyclidae). — Ann. Soc. zool.
Belg., 1977, v. 106, N 2—4, p. 123—132.
Gough H. C., Woods A. Seed dressings
for the control of wheat bulb fly. —
Nature Lond., 1954, v. 174, p. 1151—
1153.
Gould H. J. Tests with seed dressings to con-
trol grain hollowing of winter wheat
by slugs. — Plant Path., 1962, N 11,
p. 147-152.
Gray J. E. In: W. Turton. A manual of the
land and fresh-water shells of the Bri-
tish islands. 2d edit., throughly revised
and greatly enlarged by J. E. Gray.
London, 1840. 324 p.
Gray J. E. A list of genera recent Mollusca,
their synonimie and types. — Proc. Zool.
Soc. Lond., 1847, v. 15, p. 129—206.
Gray J. E. Catalogue of Pulmonata or air-
breathing Mollusca in the collection
of the British Museum. Part 1. London,
1855, IV4-192 p.
Gray J. E. On the arrangement of the land
pulmoniferous Mollusca into families. —
Ann. Mag. Natur. Hist, 3d ser., 1860,
v. 6, p. 267—269.
Grossu A. V. Gastropoda Pulmonata. In:
Fauna Republic! Populate Romine. —
Mollusca. Bucuresti, 1955, v. 3, f. 1.
518 p.
Grossu A. V. Prezen(a elementelor caucaziene
in fauna malacologica a Romaniei. —
Anal. Univ. Bucuresti, ser. biol., 1964,
An. 13, p. 82—102.
Grossu A. V. Contributions a la connaissance
du genre Lehmannia Heynemann (Ga-
stropoda, Limacidae). — J. conchyl.,
1967, v. 106, N 4, p. 119—128.
Grossu A. V. A new species of the genus
Limax in Romania. — Arch. Moll., 1968,
Bd 98, S. 85—90.
ЛИТЕРАТУРА
421
Grossu А. V. Beschreibung einiger neuer
Deroceras-Arten (Gastropoda, Limaci-
dae). — Arch. Moll., 1969, Bd 99,
S. 157—170.
Grossu A. V. Revizuirea speciilor genului
Arion Ferussac in Romania (Gastropoda,
Arionidae). — Comunic. de Zoologie,
Soc. St. Biol. R. S. R., 1970a, p. 61—74.
Grossu A. V. A comparative study to the
species of Lytopelte Boettger of Rou-
mania (fam. Limacidae, Gastr. Pulmo-
nata) and a description of a new species
L. lotrensis n. sp. — Proc, malac. Soc.
Lond., 1970b, v. 39, p. 105—110.
Grossu A. V. Zwei neue Arten der Gattung
Lehmannia Heynemann aus Ruma-
nien. — Malak. Abh. Staatl. Mus. Tierk.
Dresden, 1970c, Bd 3, N 10, S. 109—114.
Grossu A. V., Lupu D. Deroceras (Hydro-
limax) melanocephalus, eine kaukasische
Limacide in Rumanien. — Arch. Moll.,
1957a,. Bd 86, S. 171—172.
Grossu A. V., Lupu D. Contribution a la
connaissance de Limacides de la faune
roumaine. — Trav. Mus. Hist. Natur.
«Gr. Antipa», 1957b, v. 1, p. 141—146.
Grossu A. V., Lupu D. Fur Rumanien neue
Deroceras-Arten und ihre tiergeogra-
phische Bedeutung. — Arch. Moll., 1959,
Bd 88, S. 41—49.
Grossu A. V., Lupu D. Revision der Gattung
Limax aus Rumanien, nebst Beschrei-
bung einiger neuer Arten. — Arch. Moll.,
1960, Bd 89, S. 157-165.
Grossu A. V., Lupu D. Zwei neue Arten
der Gattung Deroceras Rafinesque 1820.—
Arch. Moll., 1961a, Bd 90, S. 21—25.
Grossu A. V., Lupu D. Reviziurea speciilor
genului Milax Gray 1855, existente
in Romania. — Anal. Univ. С. I. Par-
hon, ser. §t. natur., Biol., 1961b, v. 28,
an 10, p. 127—148.
Grossu A. V., Lupu D. Die Gattung Lyto-
pelte (Limacidae) in den Karpathen. —
Arch. Moll., 1961c, Bd 90, S. 27—31.
Grossu A. V., Lupu D. Zur Kenntnis der
Gattung Lehmannia Heynemann (Lima-
cidae) und deren grosse Variabilitat,
nebst Beschreibung neuer Arten. —
Arch. Moll., 1962, Bd 91, S. 191—201.
Grossu A. V., Lupu D. Variabilite du genre
Deroceras (fam. Limacidae, Gasteropo-
des terrestres) et description d’une nou-
velle espece. — Trav. Mus. Hist. Natur.
«Gr. Antipa», 1963, Bd 4, S. 193—200.
Grossu A. V., Lupu D. Especes nouvelles
du genre Deroceras (Gastropoda, Lima-
cidae) en Roumanie. — Trav. Mus. Hist.
Natur. «Gr. Antipa», 1965, v. 5, p. 25—
31.
Grossu A. V., Lupu D. Le genre Lytopelte
(Limacidae) dans les Carpathes Occiden-
tales (M. Apuseni). — Trav. Mus. Hist.
Natur. «Gr. Antipa», 1966, v. 6, p. 25—
30.
Grossu A. V., Tesio C. Etudes biochimiques
pour la taxonomie et la systematique
du genre Limax (Gastropoda Limaco-
morpha). — Atti Soc. ital. sci. natur.,
1971, v. 112, N 3, p. 289—300.
Hazay J. Die Molluskenfauna von Buda-
pest. — Malak. Bl., 1881, N. F., Bd 3,
S. 1—69.
Hazay J. Malakozoologischer Ausflug in das
Trachyt- und Kalkgebirge Ober-Un-
garns. — Malak. Bl., N. F., 1883, Bd 6,
S. 88—109.
Hazay J. Az eszaki karpatok es videkenek
Molluska Fauna]a kiilonos tekintettel
a magas Tatra tenyeszetere. — Math,
termes. Kozlem, 1884, v. 19, N 6,
S. 315-381.
Hensche A. Preussens Molluskenfauna. —
Schrift. physik-okonom. Ges. Konigs-
berg. 1861, Bd 2, S. 73—96.
Herfs A. Studien an der Hautdriisen der
Land- und Siisswassergastropoden. —
Arch, mikroscop. Anat., 1922, Bd 96,
S. 1—38.
Hesse P. Miscellen. — Jahrb. dtsch. malak.
Ges., 1882, Bd 9, S. 29-37.
Hesse P. Nacktschnecken von T anger und
Gibraltar. — Malak. BL, N. F., 1885,
Bd 7, S. 9—17.
Hesse P. Die Nacktschnecken der palSark-
tischen Region. — Abh. Arch. Moll.,
1926, Bd 2, N 1. 152 S.
Hesse P. Mazedonische Gastropoden, gesam-
melt von Herrn Prof. Dr. F. Doflein. —
Zool. Jahrb. Jena, 1928, Bd 45, S. 1—30.
Hesse R. Uber die Retina des Gastropoden-
auges. — Verhandl. dtsch. zool. Ges.,
1902, Bd 12, S. 121—125.
Heynemann F. D. Einige Mittheilungen
uber Schneckenzungen mit besonderer
Beachtung der Gattung Limax. — Ma-
lak. BL, 1862, Bd 10, S. 200-216.
Hilbert R. Die Molluskenfauna des Nord-
s&mlandischen Kiistengebiets in Le-
bensgenbssenschaften. — Nachrichtenbl.^
dtsch. malak. Ges., 1909, Bd 41, S. 35—
43.
Hoffmann H. Zur Anatomie und Systematik
der Philomyciden. — JenSische Z. Na-
turwiss., 1924, Bd 60, S. 363—396.
Hoffmann H. Uber einige Limaciden. —
Zool. Anz., 1941, Bd 136, S. 243—259.
Hogben L., Kirk R. L. Studies on tempe-
rature regulation. I. The Pulmonata
and Oligochaeta. — Proc. Roy. Soc.
Lond., ser. B, 1944, v. 132, p. 239—252.
Hryniewiecka-Szyfter S., Rgdziniak E. The
localization of the lysosomal enzymes
in the bursa copulatrix of the Helix
pomatia L. — Bull. Soc. amis sci. lettr.
Poznah, ser. D, 1976, v. 16, p. 125—134.
Hudec V. Rozdily na pohlavnich organach
plzakh Arion circumscriptus Johnst.
a Arion fasciatus (Nilss.). — Casopis
Nar. muz., odd. prirod., 1960, v. 129,
N 2, p. 204—205.
Hudec V. Poznamky к druhdm rodu Milax
Gray (Mollusca, Pulmonata) z 6SSR. —
Zool. listy, 1963, v. 12, N 2, p. 149-155.
423
ЛИТЕРАТУРА
Hiidec' V. Milax (Milax) bojanensis n. sp.
aus- Bulgarian. — Arch. Moll., 1964,
: Bd 93, S. 187—191.
Hudec V. Poznamky к anatomii nekterych
druhft plzfl u Bulharska. — Casopis Nar.
muz., odd. prirod., 1965, v. 134, N 1,
p. 11—16.
Hudec V. Poznamky к anatomii nekterych
druhfl plzfl u Bulharska. — Casopis Nar.
muz., odd. pfirod., 1967a, v. 136,
. N. 1, p. 16—24.
Hudec V. Neue Funde der Nacktschnecken
Lytopelte herculana Grossu, 1964 in Bul-
garian und Deroceras praecox Wiktor,
1966 in der Tschechoslowakei (Mol-
lusca).—Acta Soc. zool. Bohemoslov.,
1967b-, v. 31,. N -4, p. 349—354.
Hudec V. Fur die Tschechoslowakei neue
.Nacktschneckenarten (Pulmonata, Li-
. 1 macidae, Deroceras). — Biologia, Bra-
tislava, 1970, v. 25, N 2, p. 109—122.
Hudec V. Poznamky к anatomii nekterych
druhfl .plzfl z Krymu. — Casopis Nar.
muz., odd. prirod., 1972, v. 141, N 1—2,
. p. 73—91. .
Hudec V., Brabanec J.- Limax (Lehmannia)
macroflagellatus (A. Grossu — D. Lupu
- 1962) — eine neue - Nacktschnecke fiir
G. S. S. R. — Sbor. Nar. muz. Praze,
1965, Bd .21, B, N 5, S. 271—282.
Hudec V., VaSatko J. Beitrag zur Mollus-
' kenfauna Bulgarians. — Acta sci. natur.
Acad. sci. Bohemoslov., N. S., 1971,
t. 5, fasc. 2, p. 1—38.
Hutton Th. Notices of some land and fresh-
' ' water shells occuring in Afghanisthan. —
J. Asiat. Soc. Bengal, N. S;, 1849,
'• v. 18, N 2, p. 649-661.
Jaeckel S. G. A. Erganzungen und Berichti-
gungen zum rezenten und quartaren
Vorkommen der mitteleuropaischen Mol-
. lusken. — In: Zilch A., Jaeckel S. G. A.
Mollusken. Die Tierwelt Mitteleuropas.
Bd 2, N 1. Erganzung. Leipzig, 1962,
S. 25—294.
Jaeckel S. H. Praktikum der Weichtier-
kunde. Jena, 1953, S. 1—87.
Johnston G. On the Class Mollusca in Dr.
Flemming’s work on British animals,
with descriptions of some new species. —
Edinb. New Philos. J., 1828, v. 5, p. 76.
Jiillien A., Ripplinger J. Sur les correla-
tions fonctionnelles existent entre les
systeme respiratoire et circulatoire chez
1’escargot (Helix pomatia). — C. r. Soc.
Biol. Paris, 1953, v. 147, p. 826—830.
Jutting W. S. S. van Benthem. Systematic
studies on the non-marine Mollusca
of the Indo-Australien Archipelago. —
<. Treubia, 1952, v. 21, N 1—3, p. 291—435.
Kalmus H. Anemotaxis in soft-skinned
animals.— Nature London, 1942,
. v. 150, p. 524.
Karlin E. J., Bacon Ch. Courship, mating
and egg-laying behaviour in the Lima-
cidae (Mollusca). — Trans. Amer, micro-
scop. Soc., 1961, v. 80, N 4, p. 399—406.
keterstein W. Zur Anatomie von Philomy-
cus carolinensis. — Z. wiss. Zool., 1866a,
Bd 16, S. 183-189.
Keferstein W. Ueber Anatomie der Gat-
tungen Incillaria Benson und Meghima-
tium Hasselt im Vergleich mit der von
Philomycus Rafinesque. — Malak. BL,
1866b, Bd 13, S. 64—70.
Keller C. Zur Tiergeographie des Kauka-
sus. — In: Rikli M. Natur- und Kultur-
bilder aus den Kaukasuslandern und
Hocharmenien. Zurich, 1914, S. 229—
244.
Kennedy C. Y. A porphyrin pigment in the
integument of Arion ater (L.). — J.
marine biol. Assoc. U. K.,. 1959, v. 38,
N 1-2, p. 27—31.
Kerkut G. A. A neurophysiological analy-
sis of behaviour. — Proc. 15th Int.
Zool. congr., 1959, p. 845—847.
Kimakowicz M. Beitrag zur Molluskenfauna
Siebenbiirgens. — Verb. Mitt, sieben-
biirg. Ver. Naturwiss. Hermannstadt,
1884, Bd 34, S. 57-117.
Kittel R. Untersuchungen iiber den Ge-
ruchs- und Geschmackssin bei den Gat-
tungen. Arion und Limax (Mollusca:
Pulmonata). — Zool. Anz., 1956, Bd 157,
N 9—10, S. 185—195.
Klika B. Verzeichniss der von Dr. V. Vavra
wahrend seiner Kaukasus-Reise gesam-
melten Binnenconchylien. — Sitzungsb.
bohm. Ges. Wiss., math.-naturw. CL,
1894, Jhg 1893, S; 1—7.
Knorre D. Deroceras todnae Grossu et Lupu,
ein Neufund fiir die deutsche Fauna
(Gastropoda, Limacidae). — Arch. Moll.,
1969,- Bd 99, S.-171-174.
Koch C., Heynemann D. F. Neue Nacktsch-
necken aus Turkestan. — Jahrb. dtsch.
malak. Ges., 1874, Bd 1, S. 151-153.
Kosinska M. The life cycle of Deroceras
sturanyi (Simroth, 1894) (Pulmonata,
Limacidae). — Zool. Polon., 1980, v. 28,
N 2, p.
Kostal J. Prispevek ku poznani kopulacniho
apparatu u Limacopsis, coerulans Sim-
roth (Bielz). —,Vestn. Ceske spol. nauk,
1895; v. 46, p. 1—15.
Krol J. Beitrag zur Kenntniss der Mollus-
ken-Fauna Galiziens. — Verhandl. zool.-
bot. Ges. Wien, 1878, Bd 28, S. 1—10.
Kunkel K. Vermehrung und Lebensdauer
der Nacktschnecken. — Verhandl. dtsch.
zool. Ges., 1908, Bd 22, S. 153—161.
Kunkel K. Zur Biologie der Lungenschnec-
ken. Heidelberg, 1916. 440 S.
Kunkel K. Zur Biologie des Limax tenellus
Nilsson mit besonderer Beriicksichtigung
der Kopula. — Zool. Jahrb., Allg. Zool.
Physiol., 1934, Bd 53, S. 553-566.
Langlois T. H. The conjugal behaviour
of the introduced european giant slug
Limax maximus as observed on S. Bass
Island, Lake Erie. — Ohio J. Sci.,
1963, v. 65, p. 208—304.
Laviolette P. Sur un retard de la maturite
genitale observe chez Arion rufus. —
Bull. Soc. linn. Lyon, 1950a, v. 19,
- p. 52—56.
ЛИТЕРАТУРА
423
Laviolette Р. L’evolution de la glande
hermaphrodite d’Arion rufus et ses
raports avec la croissance. — C. r. Soc.
biol. Paris, 1950b, v. 144, p. 135—136.
Laviolette P. Etude cytologique et expe-
rimentale de la regeneration genitale apres
castration chez Arion rufus L. — Ann.
Sci. natur. Zool. Paris, ser. 11, 1954a,
v. 16, p. 427—535.
Laviolette P. Role de la gonade dans le de-
terminisme glandulaire du tractus geni-
tal chez quelques Gasteropodes Arionidae
et Limacidae. — Bull. biol. France-
Belg., 1954b, v. 88, p. 310—332.
Lehmann R. Neue Nacktschnecken aus
Australien. — Malak. Bl., 1864, Bd 11,
S. 145-149.
Lehtinen P. T. Siruetanalajien Arion cir-
cumscriptus Johnst. ja Arion fasciatus
Nilss. esiintyminen Suomessa. — Luon-
non Tutkija, 1957, v. 61, N 1, p. 5—12.
Lessona M., Pollonera C. Monografia dei
Limacidi italiani. Torino. 1882, 82 p.
Lewis R. D. Studies on the activity of the
slug Arion ater (L.). Ph. D. thesis,
University of Wales, 1967.
Lewis R. D. Studies on the locomotor
activity of the slug Arion ater (Linnaeus).
I. Humidity, temperature and light reac-
tions. — Malacologia, 1969a, v. 7, N 2—
3, p. 295—306.
Lewis R. D. Studies on the locomotor
activity of the slug Arion ater (Linnaeus).
II. Locomotor activity rhythms. — Ma-
lacologia, 1969b, v. 7, N 2—3, p. 307—
312.
Leydig F. Die Hautdecke und Schale der
Gastropoden, nebst einer Uebersicht der
einheimischen Limacinen. — Arch. Na-
turg., 1876, Bd 42, N 1, S. 209—292.
Linnaeus C. Systema Naturae. . . Editio
decima, reformata, 1, Holmiae, 1758.
. IV+824 p.
Lister M. Exercitatio Anatomica. Londini,
1694, XI+208 p.
Lloyd D. C. Studies on the odour of Oxychi-
lus alliarius (Pulmonata, Zonitidae). —
Malacologia, 1969, v. 9, N 1, p. 274.
Lohmander H. Uber die nordischen Formen
Arion circumscriptus Johnst. — Acta
Soc. Fauna Flora fenn., 1937, v. 60,
p. 90—112.
Lozek V. Klic ceskoslovenskych mekkystt.
Bratislava, 1956. 437 s.
Luchtel D. Gonadal development and sex
determination in Pulmonata Molluscs.
I. Arion circumscriptus. — Z. Zellforsch.
1972a, Bd 130, N 3, S. 279—301.
Luchtel D. Gonadal development and sex
determination in Pulmonate Molluscs.
II. Arion ater rufus and Deroceras reti-
culatum. — Z. Zellforsch., 1972b, Bd
130, N 3, S. 302-311.
Lupu D. Donnes nouvelles concernant la posi-
tion systematique de quelques especes
de Milax Gray, 1855, decrites anterieu-
rement, — Trav. Mus. Hist.. Natur. «Gr.
Antipa», 1968a, v. 9, p. 31—37.
Lupu D. Contributions a la connaissance
des Limacides (Gastropoda — Pulmo-
nata) des Monts Apuseni. — Trav. Mus.
Hist. Natur. «Gr. Antipa», 1968b, v. 9,
p. 39-44.
Lupu D. О noua specie a genului Deroceras
Rafinesque 1820 (Gastropoda — Lima-
cidae). — Revista Muzeelor, 1969, v. 6,
N 5, p. 434.
Lupu D. Contributions а Г etude des Lima-
cides de Roumanie. — Trav. Mus. Hist,
Natur. «Gr. Antipa», 1970, v. 10, p. 61—
71.
Lupu D. Le conspectus de la famille des
Limacides de Roumanie, avec quelques
reconsiderations systematiques basees sur
des caracteres d’anatomie comparate. —
Trav. Mus. Hist. Natur. «Gr. Antipa»,
1971a, y. 11, p. 121—137.
Lupu D. Etude sur le development de De-
roceras padisii Lupu, 1969 (=Deroceras
transsylvanicus Grossu, 1969, Deroceras
pseudolytopelte Grossu, 1969) et son
cycle biologique. — Trav. Mus. Hist.
Natur. «Gr. Antipa», 1971b, v. 11,
p. 139—147.
Lupu D. Etude anatomique comparatee
de quelques especes du genre Deroceras
Rafinesque, 1820. — Trav. Mus. Hist.
Natur. «Gr. Antipa», 1972, v. 12, p. 85—
100.
Lupu D. Etude anatomique comparative
des especes Limax cinereoniger Wolf
1803 et Limax maximus Linne 1758 et des-
cription de deux especes nouvelle du genre
Lehmannia Heynemann. — Trav. Mus.
Hist. , Natur. «Gr. Antipa», 1973, v. 13,
p. 47—66.
Lupu D. Contributions a 1’etude des gaste-
ropodes dans la region Vrancea. —
Trav. Mus. Hist. Natur. «Gr. Antipa»,
1974a, v. 14, p. 85—88.
Lupu D. La revision des re presentants de la
famille des Arionidae (Gastropoda —
Pulmonata) de Roumanie. — Trav. Mus.
Hist. Natur. «Gr. Antipa», 1974b, v. 15,
p. 31—44.
Lupu D. Deroceras agreste L. 1758 et Dero-
ceras altenai n. sp. (Gastropoda, Pul-
monata) en Roumanie. — Trav. Mus.
Hist. Natur. «Gr. Antipa», 1976, v. 17,
p. 9-13.
Lusis О. Postembrionic changes in the repro-
ductive system of the slug Arion rufus
L. — Proc. Zool. Soc. Lond., 1961,
v. 137, p. 433—468.
Lusis O. Change ,introduced in. the repro-
ductive system of Arion ater L. by va-
rying environmental conditions. — Proc.
Malac. Soc. Lond., 1966, v. 37, N 1,
p. 19—26.
.Luther A. Zuchtversuche an Ackerschnecken
(Agriolimax reticulatus Mull, und A. ag-
restis L.). — Acta Soc. Fauna, Flora
fenn., 1915, v.' 40, N 2-, p. 1—42.
Mabille. J. Des Limacieus europeens..—
J{ev. Mag. Zool., ser'. 2, 1868,, yj 20,
. p. .129-146/ ', .
424
ЛИТЕРАТУРА
Mabille J. Sur quelques especes de mollus-
ques terrestres. — Bull. Soc. philom.,
Paris, 1883, v. 7, p. 39—53.
Machin J. The evaporation of water from
Helix aspersa. 1. The nature of the eva-
porating surface. — J. exp. Biol., 1964,
v. 41, N 4, p. 759—769.
Magne A. Les Deroceratinae de la faune
girondine. — Proces verbaux Soc. sci.
phys. natur. Bordeaux, 1952 (1946—48),
. p. 30-32.
Malm A. W. Skandinaviska Land-Sniglar,
Limacina, afgebildade efter lefvande exem-
plar och beskrifna. — Goteborg vet.
Samh. Handl., 1868, Bd 10, S. 26—93.
Martens E. Aufzahlung der von Dr. Ale-
xander Brandt in Russisch-Armenien
gesammelten Mollusken. — Bull. Acad,
sci. St.-Petersb., 1880, v. 26, p. 142—158.
Martin A. W., Harrison F. M., HustonM. J.,
Stewart D. M. The blood volumes of some
representative molluscs. — J. exp. Biol.,
1958, v. 35, p. 260-279.
Maury M.-F., Reygrobellet D. Sur les di-
stinctions Specifiques chez les mollusques
Limacides du genre Deroceras. — C. r.
hebd. Seanc. Acad. Sci. Paris, 1963,
v. 257, N 1, p. 276—277.
Meisenheimer J. Entwicklungsgeschichte
von Limax maximus L. Th. I. — Z.
wiss. Zool., 1896, Bd 62, N 3, S. 415—
468.
Meisenheimer J. Entwicklungsgeschichte
von Limax maximus L. Th. II. —
Z. wiss. Zool., 1898, Bd 63, N 4, S. 573—
664.
Mengazzi L. Malacologia Veronese. Verona,
1855, 334 p.
Millet P. A. Description de plusieure espe-
ces nouvelles de mollusques de France. —
Mag. Zool. Paris (2), 1843, (Classe V),
2, mollusques, p. 1—4.
Moquin-Tandon A. Histoire naturelie des
mollusques terrestres et fluviatiles de
France, part 2. Paris, 1855. 646 p.
Muller 0. F. Vermium terrestrium et flu-
viatilium sen animalium infusorium, hel-
minthicorum et testaceorum, non mari-
norum succincta historia, 2. Havniae
et Lipsiae, 1774, 214 p.
Newell P. F., Newell G. E. The eye of the
slug, Agriolimax reticulatus (Mull.). —
Invertebr. Receptors. Symposia Zool.
Soc. Lond., 1968, N 23, p. 97—111.
Nilsson S. Historia molluscorum Sveciae
terrestrium et fluviatilum breviter de-
lineate. Lundae, 1823. XX-j-124 p.
Normand N. A. J. Description de six
limaces nouvelles observees aux environs
de Valenciennes. Valenciennes, 1852. 8 p.
Hkland F. Arionidae of Norway. — Viden-
skapsselskapets Skrifter, Christiana,
1923 (1922), N 5, S. 1—62.
Olivier G. A. Voyage dans I’Empire Otho-
mane. V. 3, Paris, 1803. 128 p.
Osanova N. Die Nacktschnecken im west-
lichen Teil des Balkan-Gebirges (Bul-
garien). — Malak. Abh. S.taatl. Mus.
Tierk. Dresden, 1970, Bd 3, N 1, S. 71—
79.
OwenG. Digestion. — In: Physiology of Mol-
lusca, v. 2. New York—London, 1966,
p. 53—96.
Pallant D. The food of the grey field slug
[Agriolimax reticulatus (Miiller)] in wo-
odland. — J. Anim. Ecol., 1969, v. 38,
N 2, p. 391—397.
Pallant D. A quantitative study of feeding
in woodland by the grey field slug
[Agriolimax reticulatus (Miiller)]. —
Proc, malac. Soc. Lond., 1970, v. 39,
N 1, p. 83-87.
Pallant D. The food of the grey field slug,
Agriolimax reticulatus (Miiller), on grass-
land.— I. Anim. Ecol., 1972, v. 41,
N 3, p. 761—769.
Pallant D. An energy budget for Agriolimax
reticulatus (Miiller) on grassland. — J.
Concho!., 1974a, v. 28, N 4, p. 243—245.
Pallant D. Assimilation in the grey field
slug, Agriolimax reticulatus (Miiller). —
Proc, malac. Soc. Lond., 1974b, v. 41,
N 2, p. 99-107.
Pelluet D., Henderson N. E. The effects
of visible radiation on the ovotestis
of the slug Deroceras reticulatum. —
Proc. Nova Scotian Inst. Sci., 1954,
v. 23, p. 420—421.
Pelluet D., Lane N. J. The relation bet-
ween neurosecretion and cell differen-
tiation in the ovotestis of slugs (Gastro-
poda: Pulmonata). — Canad. J. Zool.,
1961, v. 39, N 6, p. 789—805.
Phillips R. A., Watson H. Milax gracilis
(Leydig) in the British Isles. — J. Con-
cho!., 1930, v. 19, N 3, p. 65-93.
Pilsbry H. A. The Aulacopoda: a primary
division of the monotremate Pulmo-
nata. — Nautilus, 1896, v. 9, N 10,
p. 109—111.
Pilsbry H. A. Observations upon the nomen-
clature of slugs. — Nautilus, 1922, v. 35,
N 3, p. 77—80.
Pilsbry H. A. Land Mollusca of North
America (North of Mexico). V. II,
part 2. — Monogr. Acad, natur. Sci.
Philadelphia, N 3, 1948. XLVII+521 —
1113 p.
Pinder L. С. V. The biology and behaviour
of some slugs of economic importance
Agriolimax reticulatus, Arion hortensis
and Milax budapestensis. — Ph. D. the-
sis, University of Newcastle-upon-Tyne,
1969.
Pinter L. Tiergeographisch bedeutsame Mol-
luskenfunde in Ungarn. — Malak. Abh.
Staatl. Mus. Tierk. Dresden, 1968a,
Bd 2, N 9, S. 177—183.
Pinter L. A nyugati pilis puhatestii fau-
naja (Mollusca). — Kiilonlnyomat az Al-
lattani Kozlemenyek. Szamabol, 1968b,
v. 55, p. 1—4, 105—113.
Plate L. Studien uber opistopneumonen
Lungenschnecken. I. Daudebardia und
Testacella. — Zool. Jahrb., Abt. Anat.
Ontog., 1891, Bd 4, S. 505—630. -
ЛИТЕРАТУРА
425
Pollonera С. Sulla classificazione dei Lima-
cidi del sistema europeo. — Boll. Mus.
zool. anat. comp. Univ. Torino, 1887a,
v. 2, N 23, p. 1—6.
Pollonera C. Intorno ad alcuni Limacidi
europei poco noti. — Boll. Mus. zool.
anat. comp. Univ. Torino, 1887b, v. 2,
N 21, p. 1—4.
Pollonera C. Specie nuove о mal conoscinte
di Arion europei. — Atti Accad. Sci.
Torino, 1887c, v. 22, N 5, p. 290—313.
Pollonera C. Appunti die malacologia. —
Boll. Mus. zool. anat. comp. Univ.
Torino, 1888, v. 3, N 43, p. 1—10.
Pollonera C. Appunti di malacologia. VII.
Intorno ai Limacidi di Malta. — Boll.
Mus. zool. anat. comp. Univ. Torino,
1891, v. 6, N 99, p. 1—4.
Pollonera C. Note malacologhice. Sulla
Limacidi della Siria e della Palestine. —
Boll. Mus. zool. anat. comp. Univ.
Torino, 1909, v. 24, N 608, p. 1—8.
Poska-Teiss T. Die Nacktschnecken Est-
lands. — Arch. Moll., 1927, Bd 59,
S. 1-19.
Quick H. E. The mating proces in Arion
hortensis Ferussac and Arion subfuscus
Draparnaud. — J. conchol., 1946, v. 22,
N 8, p. 178—182.
Quick H. E. Arion ater (L.) and A. rufus
(L.) in Britian and their specific dif-
ferences. — J. conchol., 1947, v. 22,
' N 10, p. 249—261.
Quick H. E. British slugs (Pulmonata:
Testacellidae, Arionidae, Limacidae). —
Bull. Brit. Mus. (Natur. Hist.), Zool.,
1960, v. 6, N 3, p. 103—226.
Rafinesque C. S. Analyse de la nature ou
tableau de 1’univers et des corps organi-
ses. Palerme, 1815. 224 p.
Rafinesque C. S. Annals of nature or annual
synopsis of new genera and species
of animals, plants, etc., discovered in
North America. — First Annual num-
ber, Philadelphia, 1820. 16 p.
Regondaud J. Origine embrionnaire de la
cavite pulmonaire de Lymnaea stagna-
lis L. Considerations particulieres sur
la morphogenese de la commissure visce-
rale. — Bull. biol. France-Belg., 1964,
v. 48, N 2, p. 433—471.
Retowski O. Liste der von mir auf meiner
Reise von Konstantinopel nach Batum
gesammelten Binnenmollusken. — Ber.
Senkenb. naturf. Ges., 1889 (1888—1889),
S. 225—265.
Reygrobellet D. Une nouvelle espece de li-
macide Deroceras meridionale n. sp. —
Bull. Soc. zool. France, 1963, v. 88,
N 4, p. 399—402.
Riedel A. Male copulatory organs deficiency
in the Stylommatophora with a special
reference to Retinella nitens (Mich.). —
Ann. Mus. zool. Polonici, 1953, v. 15,
N 7, p. 83—100.
Riedel A. Zonitidae (excl. Daudebardiinae)
der Kaukasuslander (Gastropoda). —
Ann. zool., Warszawa, 1966, v. 24,
N 1, 303 S.
28 и. M. Лихарев, А. Й. Виктор
Riedel A., Wiktor A. Arionacea. Sslimaki
krqzalkowate i slinikowate (Gastropoda:
Stylommatophora). — In: Fauna Polski,
v. 2. Warszawa, 1974, 140 S.
Rising T. L., Armitage К. B. Acclimation
to temperature by the terrestrial gastro-
pods, Limax maximus and Philomycus
carolinianus: oxygen consumption and
temperature preference. — Comp. Bio-
chem. Physiol., 1969, v. 30, p. 1091 —
1114.
Roach D. K. Rhythmic muscular activity
in the alimentary tract of Arion ater
(L.) (Gastropoda: Pulmonata). — Comp.
Biochem. Physiol., 1968, v. 24, p. 865—
878.
Rosenwald K. Beeinflussung des Geschlechts-
wechsels von Limax laevis. — Z. in-
dukt. Abstammungs — und Vererbungs-
lehre, ,1926, Bd 43, S. 238—251.
Roy A. Etude de 1’acclimation thermique
chez la limace Arion circumscriptus. —
Canad. J. Zool., 1963, v. 41, N 4, p. 671—
698.
Runham N. W., Hunter P. J. Terrestrial
slugs. London, 1970. 184 p.
Runham N. W., Isarankura K., Smith В. J,
Methods for narcotizing and anaestheti-
zing gastropods. — Malacologia, 1965,
v. 2, N 2, p. 231—238.
Runham N. W., Laryea A. A. Studies on the
maturation of the reproductive system
of Agriolimax reticulatus (Pulmonata:
Limacidae). — Malacologia, 1968, v. 7,
N 1, p. 93—108.
Schlesch H. Die Land- und Siisswassermol-
lusken Lettlands mit Beriicksichtung
der in dem Nachbargebiet vorkommenden
Arten — Korresp. Bl. naturforsch. Ver.
Riga, 1942, Bd 64, S. 246—360.
Schmid G. Zur Verbreitung und Anatomie
der Gattung Boettgerilla. — Arch. Moll.,
1963, Bd 92, S. 215—225.
Schmid G. Weitere Funde von Boettgerilla
vermiformis. — Mitt, dtsch. malak. Ges.,
1966, Bd 1, N 8, S. 131—136.
Schmid G. Boettgerilla vermiformis auch
in Frankreich und Belgien. — Mitt,
dtsch. malak. Ges., 1969, Bd 2, N 13,
S. 20—21.
Schmid G. Deroceras rodnae, eine neue
Nacktschnecke in Siidwestdeutschland.—
Veroff. Landesstelle Naturschutz Land-
schaftspflege, В aden—Wurttemberg,
1971, Bd 39, S. 131-142.
Schmid G. Nacktschnecken aus Baden—
Wurttemberg. — Mitt, dtsch. malak.
Ges., 1972, Bd 2, N 22, S. 332-344.
Schrenck L. Reise und Forschungen im
Amur-Lande in den Jahren 1854—1856.
II. Mollusken des Amur-Landes und des
Nordjapanisehen Meeres. St.-Petersburg.
1867, S. 260—976.
Schrenk A. G. Ubersicht der Land- und
Siisswassermollusken Livlands. — Bull,
Soc. natur. Moscou, 1848, v. 21, pt 1,
p. 135—185.
Shotton F. E. Outline of the practical prob-
lem. — In: Slug research Conference,
426
ЛИТЕРАТУРА
University college. Cardiff, 1971, p. 2—3.
Simroth H. Anatomie der Parmacella
Olivieri Cuv. — Jahrb. dtsch. malak.
Ges., 1883, Bd 10, S. 1—47.
Simroth H. Versuch einer Naturgeschichte
der deutschen Nacktschnecken und ihre
europaischen Verwandten. — Z. wiss.
Zool., 1885, Bd 42, S. 203—366.
Simroth H. Weitere Mittheilungen fiber
palaearktische Nacktschnecken. — Jahrb.
dtsch. malak. Ges., 1886a, Bd 13,
S. 16-34.
Simroth H. Ueber bekannte und neue pa-
laearktische Nacktschnecken. — Jahrb.
dtsch. malak. Ges.,. 1886b, Bd 13,
S. 311—342.
Simroth H. Uber die azorisch-portugiesische
Nacktschneckenfauna und ihre Beziehun-
gen. — Zool. Anz., 1888, Bd 11, S. 66—
70, 86—90.
Simroth H. Beitrage zur Kenntniss der
Nacktschnecken. — Nachrichtenbl. dtsch.
malak. Ges., 1889, Bd 21, S. 177—186.
Simroth H. Die Nacktschnecken der portu-
giesisch-azorischen Fauna in ihrem Ver-
haltniss zu denen der palaarktischen
Region iiberhaupt. — Nova Acta Ksl.
Leop.-Carol. dtsch. Akad. Natur. Halle,
1891a, Bd 56, N 2, S. 201—423.
Simroth H. Ueber kaukasische Limaciden
und Testacelliden. — Verhandl. dtsch.
zool. Ges., 1891b, Bd 1, S. 51—58.
Simroth H. Ueber die nackten Limaciden
und Testacelliden des Kaukasus. — Sit-
zungsber. naturforsch. Ges. Leipzig,
1892a (1890—1891), S. 40—49.
Simroth H. Ueber einige Raublungenschnec-
ken des Kaukasus. — In: Festschr. fur
Leuckart. Leipzig. 1892b, S. 48—58.
Simroth H. Ueber einige von Herrn Dr.
Sturany auf der Balkanhalbinsel erbeu-
tete Nacktschnecken. — Ann. naturhist.
Hofmus., Wien, 1894a, Bd9, S. 391—394.
Simroth H. Beitrage zur Kenntnis der
kaukasisch-armenischen N acktschnecken-
fauna. — Sitzungsber. k. bomisch. Ges.
Wiss., math.-naturw. KI., 1894b (1893),
Bd 45, S. 8—23.
Simroth H. Vorlaufige Mittheilung eine
Bearbeitung der russischen Nacktschnec-
kenfauna betreffend. — Ежегод. Зоол.
муз., 1896,..т. 1, с. 355—368.
Simroth Н. Uber verschiedene Nacktschnec-
ken. — Sitzungsber. naturforsch. Ges.
Leipzig, 1897 (1895—96), Bd 22—23,
S. 140—154.
Simroth H. Uber die Gattung Limax in Russ-
land. — Ежегод. Зоол. муз., 1898a,
т. 3, с. .52—57.
Simroth Н. Uber die Gattung Parmacella. —
Ежегод. Зоол. муз., 18986, т. 3, с. 124—
135.
Simroth Н. Uber einige kleinasiatische
Nacktschnecken. — Sitzungsber. natur-
forsch. Ges. Leipzig,. 1899 (1897—98),
Bd 24—25, S. 35—38. .
Simroth H. Die Nacktschnecken des Rus-
sischen Reiches. St.-Petersburg, 1901.
321 S. .
Simroth H. Kaukasische und asiatische
Limaciden und Raublungenschnecken.—
Ежегод. Зоол. муз., 1910a, т. 15, с. 499—
560.
Simroth Н. Die Landnacktschnecken der
Deutsch Siidpolar-Expedition 1901—
1903. — Dtsch. Siidpolar-Exp., 1910b,
Bd 12, Zool. N 4, H. 3, S. 137—180.
Simroth H. Neue Beitrage zur Kenntnis der
kaukasischen Nacktschnecken. — Изв.
Кавказ, муз., 1912a, т. 6, с. 1—140.
Simroth Н. Ueber die im Friihjahr 1897 von
Herrn Kaznakov in den Gebirgen Bu-
chara’s erbeuteten Parmacellen. — Еже-
год. Зоол. муз., 19126, т. 17, с. 41—52.
Simroth Н. Ueber einige von Herrn Dr. Ab-
solon in der Herzegowina erbeutete
hohlenbewohnende Nacktschnecken. —
Nachrichtenbl. dtsch. malak. Ges., 1916,
Bd 48, S. 1—16.
Smith B. J. Maturation of the reproductive
tract of Arion ater (Pulmonata: Arioni-
dae). — Malacologia, 1966, v. 4, N 2,
p. 325—349.
Smolenska J. Observations biologiques sur
Bielzia coerulans (Bielz). — Ann. Mus.
zool. polon., 1936, v. 11, N 13, p. 241—
261.
Sokolove P., G., McCrone E. J. Reproduc-
tive maturation in the slug, Limax
maximus, and the effects of artificial
photoperiod. — J. Comp. Physiol.,' A,
1978, v. 125, N 4, p. 317—325.
Solem A. Systematics of the land and fresh-
water Mollusca of the New Hebrides. —
Fieldiana: Zoology, 1959, v. 43. 238 p.
Sods L. Mollusca. — In: Explorationes zoo-
logicae al E. Csiki in Albania peractae.
Budapest, 1924, p. 177—197.
South A. Biology and ecology of Agriolimax
reticulatus (Miill.) and other slugs:
special distribution. — J. Animal Ecol.,
1965, v. 34, N 2, p. 403—417.
Starmuhlner F., Edlauer A. Ergebnisse der
Osterreichischen Iran-Expedition 1949/
50. — Sitzunsber. Osterreich. Akad.
Wiss., Math.-naturw. KI., Abt. 1, 1957,
Bd 166, H. 9-10, S. 435—494.
Stephenson J. W., Knutson L. V. A resume
of recent studies of invertebrates asso-
ciated with slugs. — J. economic ento-
mol., 1966, v. 59, N 2, p. 356—360.
Stern G. Bilan energetique de la limace
Arion rufus (mollusque pulmone) en pe-
riode de croissance. — C. r. Acad . sci.
Paris, 1969, v. D269, N 11, p. 1015—1018.
Symonds В. V. The evaluation of new che-
micals for the control of Agriolimax
' reticulatus (Muller). — In: Slug research
Conference. University college. Cardiff,
1971, p. 44.
Szabo J., Szabo M. Lebensdauer und
Korpergrosse einiger Nacktschnecken. —
Zool. Anz., 1931, Bd 106, S. 106—111.
Tauber H. Beitrage zur Morphologie der
Stylommatophoren. — Ежегод. Зоол.
’ муз.,' 1900, т. 5, с. 373—411. ;
ЛИТЕРАТУРА
427
Taylor D. W., Sohl N. F. An outline
of gastropod classification. — Malacolo-
gia, 1962, v. 1, N 1, p. 7—32.
Taylor J. W. Monograph of the land and
freshwater Mollusca of the British Isles.
Testacellidae, Limacidae, Arionidae. Le-
eds, 1902—1907. XX-f-312 p.
Thiele J. Handbuch der systematischen
Weichtierkunde. 2. Jena, 1931, S. 461—
778.
Thomas D. C. The use of metaldehyde against
slugs. — Ann. Appl. Biol., 1948, v. 35,
p. 207—227.
Turk F. A., Phillips S. M. A monograph
of the slug mite Riccardoella limacum
(Schrank). — Proc. Zool. Soc. Lond.,
1946, v. 115, p. 448—471.
Urbanski J., Wiktor A. Beitrage zur Kenn-
tnis bulgarischer Nacktschnecken (Moll.,
Pulm.). (Systematische, zoogeographi-
sche und okologische Studien iiber die
Mollusken der Balkan—Halbinsel.
VIII). — Bull. Soc. amis Sci. lettr.
Poznan, ser. D, 1968 (1967), v. 8, p. 47—
95.
Valovirta I. List of Finnish land gastropods
and their distribution. — Ann. zool. fen-
nici, 1967, v. 4, p. 29—32.
.Valovirta I. First record of Milax gagates
- (Drap.) (Gastropoda, Milacidae) in Finn-
land. — Ann. zool. fennici, 1969, v. 6,
p. 345-347.'
Van Mol J.-J. Etude morphologique et phy-
logenetique du ganglion cerebroide des
Gasteropodes Pulmones (mollusques).—
Mem. Acad. R. Belg., coll. in-8°, ser. 2,
1967, v. 37, N 5, 168 p.
Van Mol J.-J. Revision des Urocyclidae
(Mollusca, Gastropoda, Pulmonata). Ana-
tomie, systematique, zoogeographic. —
Ann. Mus. R. Afr. Centr., 1970, v. 180.
234 p.
Van Mol J.-J. Notes anatomiques sur les
Helicarionidae (Mollusques, Gasteropo-
des, Pulmones). II. Etudes des genres
Pseudaustenia, Dyakia, Helicarion et
comprenant la description de Papuarion,
genre nouveau. Discussion sur la classi-
fication des Helicarionidae et les affi-
nites des Urocyclidae. — Ann. Soc. R.
zool. Belg.., 1973, v. 103, N 2—3, p. 209—
237.
Vater G. Lehmannia macroflagellata Grossu
et Lupu 1962 und Deroceras praecox
Wiktor 1966 im Hruby Jasenik (Altva-
tergebirge). — Casopis slezskeho Muz.,
ser. A, 1970, v. 19, p. 57—60.
Wachtler W. Der Nierenapparat der Stylom-
matophoren Lungenschnecken verglei-
chend-anatomisch betrachtet. — Zool.
Anz., 1934, Bd 105, N 7—8, S. 161—172.
Wagner J. Beitrage zur Kenntnis der Gat-
tung Milax. — Ann. Mus. Nat. Hung.,
1929, Bd 26, S. 327—338.
Wagner H. Morphologische und anatomische
Studien an Milax. — ZooL Anz., 1930a,
Bd 88, S. 39-57.
.Wagner H. Ujalb adatok a Milax-fajok
ismeretehez.—Allatt. Kozlem., 1930b,
Bd 27, N 1-2, S. 97—107.
Wagner H. Die in die Unterfamilie Parma-
cellidae gehorenden Formen des Natur-
historisches Museums in Wien. — Ann.
naturhist. Mus. Wien, 1931a, Bd 46,
S. 57—76.
Wagner H. Diagnosen neuer Limaciden aus
dem Naturhistorischen Museum Wien. —
Zool. Anz., 1931b, Bd 95, S. 194—202.
Wagner J. Die Nacktschnecken des konig-
lichen Naturhistorischen Museums in So-
fia. — Mitt. k. naturw. inst. Sofia,
1934a, Bd 7, S. 51—60.
Wagner J. Die Nacktschnecken Ungarns,
Croatiens und Dalmatiens. I. — Ann.
Mus. Nat. Hung., 1934b, Bd 28, S. 1—30.
Wagner J. Die Nacktschnecken Ungarns,
Croatiens und Dalmatiens. II. — Ann.
Mus. Nat. Hung., 1935, Bd 29, S. 169—
212.
Wagner H. Zoologische Ergebnisse der
Forschungreise N. Vasvari’s in Klein-
asien. — Matematikai es Termeszettudeo-
manyi Ertesito, 1937, Bd 56, N 3,
S. 1042—1060.
Walden H. W. Notes on some Sibirian slugs,
especially on the type of Limax hyper-
boreus Westerlund. — Arkiv zool., ser. 2,
• 1957, v. 10, N 5, S. 347—366.
Walden H. W. Om ett par for Sverige nya,
antropochora landmollusker, Limax va-
lentianus Ferussac och Deroceras caruane
(Pollonera), jamte nfigra andra, kultur-
bunda arter. — Medd. Goteborgs Mus.,
Zool. Avd., ser. B, 1960, v. 8, N 8,
p. 1—48.
Walden H. W. On the variation, nomencla-
ture, distribution and taxonomical po-
sition of Limax (Lehmannia) valentianus
Ferussac (Gastropoda, Pulmonata). — Ar-
kiv Zool., ser.’ 2, 1962, v. 15, N 3,
. S. 71—96.
Walden H. W. Einige Bemerkungen zum
Erganzungsband zu Ehrmann’s «Mol-
lusca», in «Die Tierwelt Mitteleuropas».—
Arch. Moll., 1966, Bd 95, S. 49—68.
Walker G. Studies on digestion of the slug
Agriolimax reticulatus (Muller) (Mol-
lusca, Pulmonata, Limacidae). Ph. D.
thesis, University of Wales. 1969.
Walker G. The digestive system of the
slug, Agriolimax reticulatus (Muller):
experiments on phagocytosis and nutri-
ent absorbtion.—Proc, malac. Soc. Lond.,
1972, v. 40, N 1, p. 33—43.
Wattez Chr. Influence du complexe cephali-
que (tentacule oculaires cerveau) sur
1’evolution, en culture in vitro, de go-
nades infan tiles et juveniles d’Arion
subfuscus Drap. (Gasteropode Pulmone).—
Gen. and Comp. Endocrinol., 1978,
v. 35, N 4, p. 360—374.
Webb Ph. B., Berthelot S. Sur le genre
Cryptella et description d’une nouvelle
espece (C. canariensis). — Guerin Mag.
28*
428
ЛИТЕРАТУРА
Zool., 1835, aim. V, pt 1, Cl. V, pl 63,
[8 p. — пагинация не обозн.].
Wenz W. Gastropoda extramarina tertiaria.
I—XI. — In: G. Diener (ed.). Fossilium
Catalogus, I: Animalia. Ps 17, 18, 20—
23, 32, 38, 40, 43, 46. Berlin, 1923—
1930, S. 1—3387.
Wenz W. Zur Taxonomie der Euthyneura.—
Arch. Moll., 1947, Bd 76, N 1/3, S. 36.
Wenz W., Zilch A. Euthyneura. — In:
Handbuch d. Palaozoologie. Gastropoda,
Th. 2. Berlin, 1959—1960. 834 S.
Westerlund C. A. Neue Binnenmollusken
aus Sibiriens. — Nachrichtenbl. dtsch.
malak. Ges., 1876, Bd 8, S. 97—104.
Westerlund C. A. Sibiriens Land- och Sot-
vatten-mollusker. — Svenska Vetensk.
Acad. Handl., N. F., 1877, t. 14, N 12,
111 S.
Westerlund C. A. Specilegium Malacologi-
cum. Neue Binnenconchylien aus der
Palaarktischen Region. — Nachrichtenbl.
dtsch. malak. Ges., 1894, Bd 26, S. 163—
177.
Westerlund C. A. Neue centralasiatische
Mollusken. — Ежегод. Зоол. муз., 1896,
т. 1, с. 181—198.
Westerlund С. A. Beitrage zur Mollusken-
fauna Russlands. — Ежегод. Зоол. муз.,
1897, т. 2, с. 117—143.
Wiktor A. Boettgerilla vermiformis n. sp.
(Mollusca, Pulmonata). — Comm. Poz-
nan Soc. Friends Sci. Math. Natur.,
1959, v. 4, p. 1—2.
Wiktor A. Description of Boettgerilla ver-
miformis Wiktor (Mollusca, Pulmo-
nata). — Bull. Soc. amis Sci. Lettr.
Poznan, ser. D, 1960a, v. 1, p. 151—156.
Wiktor A. Kopulacja pomrowia plamistego
Deroceras reticulatum Mull. (Mollusca,
Pulmonata). — Poznanskie Tow. przyj.
nauk., mat.-przyr., 1960b, v. 19, N 7(2),
p. 23—30.
Wiktor A. Materialen zur Kenntnis der
Gattung Boettgerilla Simroth, 1910 (Ga-
stropoda, Limacidae). — Ann. Zool.,
Warszawa, 1961, v. 19, N 4, p. 125—145.
Wiktor A. Miqczaki Ziemi Klodzkiej i got
przyleglych. Studium faunistyczno-zoo-
geograficzne. — Poznanskie Tow. przyj.
nauk., mat.-przyr., 1964, v. 29, N 1,
p. 1—129.
Wiktor A. Eine neue Nacktschnecken-Art
(Gastropoda, Limacidae) aus Polen. —
Ann. Zool., Warszawa, 1966, v. 23,
N 18, p. 449—457.
Wiktor A. Nowe dla Polski slimaki nagie
z rodzin Limacidae i Milacidae (Gastro-
poda). — Fragmenta Faunistica, 1967a,
v. 13, N 19, p. 323—335.
Wiktor A. Lytopelte (Liolytopelte) molda-
vica Grossu et Lupu (Gastropoda, Lima-
cidae) w Polsce. — Przegl. Zool., Wroc-
law, 1967b, v. 11, N 4, S. 385—388.
Wiktor A. Die von der niederlandischen
biologischen Expedition in der Tiirkei
in 1959 gesammelten Nacktschnecken
(Milacidae und Limacidae) mit Beschrei-
bung einer neuen Deroceras-Art aus
dem Balkan-Gebiet und der Tiirkei. —
Zool. Meded., 1971a, Bd 45, N 23,
S. 261—280.
Wiktor A. Wspolczesne miqczaki (Mollusca)
rezerwatu Muszkowicki Las Bukowy i
okolic. — Ochrona przyrody, Krakow,
1971b, v. 37, p. 127—134.
Wiktor A. Die Nacktschnecken Polens
(Arionidae, Milacidae, Limacidae). —
In: Monografie Fauny Polski. T. 1;
Warszawa—Krakow, 1973. 182 S.
Wiktor A., Likharev I. M. Phylogenetische
Probl erne bei Nacktschnecken aus dem
Familien Limacidae und Milacidae (Ga-
stropoda, Pulmonata). — Malacologia,
1979, v. 18, p. 123—131.
Wiktor A., Likharev I. M. The pallial
complex of Holarctic terrestrial slugs
(Pulmonata, Stylommatophora) and its
importance for classification. — Zool.
polon., 1980 (1979), v. 27, N 3, p. 421—
460.
Wiktor J., Wiktor A. Charakterystyka
fauny miqczakow polskiej czqsci Karko-
noszy ze szczegolnym uwzglqdnieniem
Karkonoskiego Parku Narodowego. —
Ochrona przyrody, Krakow, 1968, v. 33,
s. 193—214.
Williamson M. Studies on the colour and
genetics of the black slug. — Proc. Roy.
phys. Soc. Edinb., 1959, v. 27, p. 87—93.
Wolf J. Die Wiirmer. — In: Sturm’s Deut-
schlands Fauna etc. Abth. 6, H. 1—2
(nach Quick, 1960).
Wondrak G. Elektronenoptische Untersu-
chungen der Korperdecke von Arion
rufus L. (Pulmonata). — Protoplasma,
1968, N 66, S. 151—171.
Wondrak G. Elektronenoptische Untersu-
chungen der Drusen- und Pigmentzellen
aus der Korperdecke von Arion rufus L.
(Pulmonata). — Z. mikroskop. anat.
Forsch., 1969a, Bd 80, S. 17—40.
Wondrak G. Die Ultrastruktur der Sohlen-
driisenzellen von Arion rufus L. —
Malacologia, 1969b, v. 9, N 1, p. 303—
305.
Zeissler H. Boettgerilla vermiformis in Ple-
istozan von Ehringsdorf. — Arch. Moll.,
1968, Bd 98, S. 113—114.
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ МОЛЛЮСКОВ1
abramovi, Gigantomilax 249, 251
absoloni, Deroceras, Agriolimaz 8, 180
adsharicus, Phrixolestes 312
aethiops, Candaharia, Parmacella 11, 334,
345, 346*, 347*
agreste, Deroceras, Limax, Agriolimax 8,
32, 36*, 73, 76, 102, 104—107, 127,
134, 142, 143*, 148, 152
agresticulus, Agriolimaz 154, 158
Agriolimacidae 5, 8, 18, 25, 29, 33, 35,-
43, 54, 81, 84, 87—89*, 90, 91, 100,
122, 123, 208
Agriolimacinae 123
Agriolimax 8, 127, 142
Agriolimaz 125
albocarinata, Eumilax brandti f. 295
albocarinatus, Eumilax, Paralimax 293
albomaculatus, Eumilax, Paralimax 293
alexandrina, Parmacella 341
alliarius, Oxychilus 22
altaicum, Deroceras, Agriolimax 8, 104, 127,
146, 147*
altenai, Deroceras 149
altenai, Milax 358, 362
alticola, Limax ananowi 241
Amalia 357
amaloides, Caucasolimax Caucasians f. 243
amaloides, Limax 241
Ambigolimax 261
Anadeninae 86, 87, 386
ananowi, Agriolimax 154, 157
ananowi, Limax 208, 241
ananowi, Pseudomilax 324
Ancylidae 307
annularis, Phenacolimax 79, 80
anthiops, Parmacella 345
anthiops, Parmacella korschinskii 345, 347
antiquorum Fer., Limax 274, 278
antiquorum Raz., Limax 274
arborum, Limax 263
Arctolimax 125
Ariolimacinae 86, 87, 386
Arion 5, 6, 12, 14, 17—19, 22, 23, 27*,
32*, 42*, 47, 52*, 56—58, 61, 63, 65-
68, 87, 100, 102, 121, 386, 387
Arion s. str. 12, 389, 390
Arionidae 12, 14, 16—18, 23, 28, 33, 35, 43,
51, 81, 84—87, 93, 100, 121, 380, 386
Arioninae 5, 12, 53—55, 84—87, 386, 387
Arionoidea 11, 84—87, 100, 122, 380
Ariophantidae 84, 93
Ariunculus 387
armenia, Lytopelte caucasica 169, 173
armeniaca, Trigonochlamys 312
armeniacus, Gigantomilax monticola, Li-
max, Vitrinoides 9, 63, 76, 223, 226, 227*,
228*
armeniacus, Hyrcanolestes 314
Aspidoporus 356
ater, Arion, Limax 12, 14, 19, 22, 28, 32,
39, 41, 43, 45, 49, 50, 57—59, 63, 71 —
73, 387, 389, 390, 393, 394*
atlantica, Plutonia 79, 80
Aulacopoda 83, 87
azerbaidjanensis, Malacolimax 240
baccata, Vitrina 344
baeri, Limax 244
bakurianum, Deroceras, Agriolimax 8, 105,
128, 158, 159*—161*
balcanicus, Milax rusticus 374
bicolor, Pseudomilax, Trigonochlamys 314
bielzi, Lehmannia 263
bielzi, Limax 277
Bielzia 10, 55, 91, 102, 220, 287
Bielziinae 10, 54, 87, 91, 92, 220, 287
bilineatum, Meghimatium, Incilaria, Phi-
lomycus 11, 46», 100, 104, 382, 383*—
385*
Binneyinae 386
boettgeri, Lytopelte 200
boettgeri, Trigonochlamys 312
Boettgerilla 6, 9, 18, 27*, 47, 54, 55, 58,
83, 89*, 102, 213, 356
Boettgerillidae 9, 17, 18, 25, 29, 33, 35,
36, 43, 54, 81, 84, 87, 89, 90, 100, 122,
210
bojanensis, Milaz 374
borschomensis, Gigantomilax 230
Bourguignati, Arion 403
brandti, Eumilax, Limax, Amalia, Para-
limax, Eulimax 10, 14, 28, 46*, 103*,
291*, 292, 293, 294*, 296*
brauni, Mesolimax 9, 205, 206*
breckworthianus, Limacus 281
brunneus, Arion 396, 399
brunneus, Gigantomilax 230
1 Курсивом выделены синонимы, неверные определения и прочие несоответствия
с принятой в книге номенклатурой; жирным шрифтом —। названия родов и семейств
слизней; жирные цифры обозначают страницу с началом описания соответствующего
таксона; звездочкой отмечены страницы с рисунками.
430
УКАЗАТЕЛЬ
brunneus, Gigantomilax, Monochroma 9,
102, 222, 234, 235*, 236*, 237*
buchar, Agriolimax 146, 148
budapestensis, Tandonia, Amalia, Milax
11, 20*, 21*, 68, 70, 357, 365, 372,
376, 377*
Bulimulidae 81
bureschi, Deroceras, Agriolimax 8, 128,
174, 175*
callatis, Deroceras, 8, 180, 181*
Camaenidae 86
Candaharia 11, 52*, 77, 94*, 95, 102, 103*,
104, 342
Candaharia s. str. 11, 342
Carination 12, 389, 400
carinatus, Limax 372
caruanae, Deroceras, Agriolimax 135, 136
Caspilimax 9, 92, 102, 220, 243
caspius, Agriolimax 169, 173
caucasicum, Deroceras, Lytopelte 8, 65, 77,
105, 106, 111, 128, 145, 169, 170*, 171*
caucasicus, Caucasolimax, Limax 9, 208,
240, 241, 242*
Caucasians, Milax, Amalia 11, 357, 358, 359*
Caucasolimax 9, 54, 92, 102, 220, 240
cereus, Limax 259
Chlamydephoridae 68, 81
Chlamydolimax 324
Chorolimax 142
Chrysalidomilax 313
cibinensis, Amalia, 376
cinereoniger, Limax 10, 37, 56*, 57, 64*,
67, 68, 274, 275*, 276*
cinereus, Limax 278
circumscriptus, Arion 12, 59, 388*, 389,
400, 403, 404*, 406
Clathropodium 335
coeciger, Agriolimax buchar 146, 148
coeciger, Agriolimax transcaucasicus 145
cderulans, Bielzia, Limax, Limacopsis 10,
36, 54*, 57, 64*, 66-68, 104, 287, 288,
289*
colchicus, Limax 226
compressa, Boettgerilla 9,107, 214, 217, 218*
coriaceus, Paralimax brandti 293
cretica, Bielzia 288
crimense, Deroceras, Agriolimax 8, 180, 181*
crimense, Deroceras 163
cristata, Tandonia, Krynickillus, Milax 11,
104, 105, 111, 357, 364, 365*, 366* 371
Cryptella 335
daghestanus, Gigantomilax, Limax 9, 102,
223 233*
Daudebardiidae 68, 79, 92, 93, 94*, 100, 331
declivis, Hyrcanolestes 315
dengis, Limax 255
depressus, Oxychilus 93
Deroceras 4—6, 8, 19*, 32*, 42*, 47, 52*,
53, 57, 58, 61, 63—65, 73, 77, 89*, 90,
102, 117, 123, 124*, 125, 126
Deroceras s. str. 8, 127, 128
Derocerasinae 123
Derocerattnae 123
distans, Trigonochlamys 312
dobrogicum, Deroceras 8, 181*, 182
dobrogicus, Limax 10, 274, 280, 281*
Drilolestes 11, 16, 33, 37, 96*, 98*, 99*,
100, 102, 309, 324
Drusia 335
dubius, Agriolimax 135
Dymczewiczi, Limax, Agriolimax 163, 165,
169, 174, 226
ecarinatus, Limax 284, 285*
elegans, Eumilax intermittens f. 298
elegans, Metalimax 10, 300, 301, 305*
elegans, Paralimax raddei 297
empiricorum, Arion 387, 390, 393, 394
Endodontidae 84
escherichi, Mesolimax, 9, 205, 207*
esthonicus, Arion 396
Eulimax 273
Eumilacinae 10, 18, 25, 54, 91, 122, 220, 290'
Eumilax 6, 10, 19, 47, 53, 91, 102, 221, 290,
291
Euparmacella 335
fasciatus, Arion, Limax 12, 76, 105—107,
388*, 389, 405, 407, 408*
fedtschenkoi, Agriolimax 142, 146
fedtschenkoi, Agriolimax agrestis 142, 146
fedtschenkoni, Limax 142
ferganus, Turcomilax, Gigantomilax 9, 249,
251*
testae, Parmacella 341
filipowitschi, Parmacellilla 11, 319, 320,
321*, 341
flavus, Arion 411
flavus, Limax 10, 22, 35, 57, 59, 67, 77,
105, 106, 274, 281, 282*, 284-286
florenskii, Limax, Vitrinoides 233
forcarti, Deroceras 8, 181*, 183
Frauenfeldta 287
fursovi, Hyrcanolestes 315
fuscus, Arion 409
gagates (Drap.), Milax, Limax 11, 56*, 57,
65, 68, 72, 106, 109, 357, 358, 362,
363*
gagates Men., Limax 376
Gastrodontidae 196
Gastropoda 3, 13, 43, 47, 79, 84
Geomalacus 102, 387
Geophila 3, 5, 13, 14, 29, 33, 81, 83, 86,
87, 89, 97, 100
getica, Lehmannia 268
geticum, Deroceras 152
Gigantomilax 9, 51, 91, 92, 102, 220, 221
Gigantomilax s. str. 9, 92, 222, 223
gracilis Leyd., Limax, Amalia, Milax 376-
gracilis Raf., Limax 125, 129
gracilis (Simr.), Eumilax, Paralimax 293
grisea, Eumilax intermittens f. 298
griseus, Eumilax, Paralimax 297
grossui, Limax 284
grusina, Lytopelte 169, 173 _ "
gyratus, Eumilax, Paralimax 293
hamatum, Deroceras Г69, 174
Helicarionidae 84, 88, 93 ‘
УКАЗАТЕЛЬ
431
Helicidae 30, 50
Helicoidea 53, 86, 381
Helix 49
Helixina 86, 100
Helixinia 86
herculana, Lytopelte 174, 176
heterura, Deroceras 190
Heterurethra 83
Heynemannia 273
Holopoda 83, 87
Holopodopes 83
hoplites, Toxolimax, Mesolimax 9, 208, 209*
horezia, Lehmannia 10, 263, 267*
hortensis, Arion 12, 68, 70, 106, 107, 388*,
390, 409, 410*
huculorum, Agriolimax 178
Hyalimax 88
hydrobium, Agriolimax 178
Hydrolimax 125
Hygromiidae 86, 102
hyperboreus, Limax, Agriolimax 129, 146
hyrcanensis, Parmacella 341
Hyrcanolestes 10, 32*, 37, 96*, 98*, 99*,
100, 102, 309, 313
ibera, Parmacella 11, 46*, 59, 63, 67, 68, ,
75, 77, 93*, 105, 106, 109, 333, 334,
335, 336*—339*
ibera, Parmacella oliviert 335, 340
iliensis, Gigantomilax 252
ilium, Deroceras, Agriolimax 8, 127, 139,
140*
ilium, Deroceras 174
imereticus, Limax ananowi 241
imitatrix, Trigonochlamys 10, 38*, 310*,
311*, 312
Lncilaria 382
incipiens, Limax 226
intermedins, Arion, Limax, 12, 14, 106,
390, 411, 412*
intermittens, Eumilax, Limax, Paralimax
10, 103*, 139, 292, 293, 297, 298*, 299*
izzatullaevi, Candaharia .11, 105, 334, 352,
353*
jaeckeli, Deroceras 8, 183, 184*
jailanus, Limax 231
jaroslaviae, Lehmannia 10, 262, 270*
kainarensis, Parmacella 342, 344
kaleniczenkoi, Tandonia, Milax 11, 104,
105, 357, 364, 368, 369*—371*, 380
kalenzkoi, Amalia 368
kalischewskii, Eumilax, Paralimax 293
Kandaharia 342
kandahariensis, Lytopelte, Deroceras 196
kaznakovi, Gigantomilax monticola arme-
niacus f. 230
Kaznakovi, Hyrcanolestes 314
kaznakovi, Limax 226
kaznakovi, Parmacella 348, 352
keyserlingi, Caspilimax, Limax, 9, 36, 76,
103*, 243, 244*, 245*
Kobeltia 12, 390, 409
koenigi, Gigantomilax, Limax, Vitrinoides
9, 223, 231, 232*
kollyi, Gigantomilax lederi f. 225
korschinskii, Parmacella 348
kostali, Malacolimax 261
krynickii, Arion 396
Krynickillus 6, 8, 18, 89*, 90, 91, 102, 123—
125, 190
kusceri, Tandonia, Milax 11, 47*, 105, 106,
357, 364, 372, 374, 375*
laeve, Deroceras, Limax, Agriolimax 8, 14,
37, 58, 65, 77, 102, 104, 105, 111, 125,
127, 129, 130*, 132
Lallemantia 357
lederi, Gigantomilax, Amalia 9, 35, 36*,
221, 222, 223, 224*
lederi, Trigonochlamys, Pseudomilax 312
Lehmannia 4, 5, 10, 35, 42*, 73, 91, 92,
102, 220, 261
lenkoranus, Gigantomilax 9, 102, 222, 238*
Letourneuxia 387
levanderi, Candaharia, Parmacella 11, 63,
66*, 67, 68, 71, 73, 74*, 75, 77, 105,
109, 334, 348, 349*, 350*
Levanderia 11, 348
liliaceus, Paralimax brandti 293
Limacella 273
Limacidae 5, 9, 18, 19*, 25, 29, 33, 35, 36,
40, 43, 51, 53, 81, 84, 87, 89*, 91, 100,
122, 124, 219, 356
Limacinae 9, 54, 91, 92, 220, 221
Limacoidea 8, 36, 84, 85, 87-89*, 93, 97,
100, 122
Limacopsis 287
Limacus 10, 220, 274, 281
Limax 4, 5, 10, 19, 27», 42*, 51, 52*, 61,
63, 73, 91, 92, 102, 220, 273, 356
Limax s. str. 10, 55, 57, 92, 102, 273, 274
Limaxia 219
Limaxina 86, 100
Liolytopelte 8, 53, 128, 169
livonicus, Limax 263
Lochea 387
longicollis, Lytopelte, Amalia 195, 196, 200
longicollis, Lytopelte maculata f. 198
longipenis, Milax rusticus 374
lotrensis, Lytopelte 176
lusitanicus, Arion 12, 65, 389, 395*
Lytopelte 9, 53, 89*, 90, 124, 125, 169, 195
macroflagellata, Lehmannia 10, 263, 265, 266*
maculata, Lytopelte, Amalia 9, 77, 104,
105, 117, 195, 196, 197*, 198*
maculatus, Limax, Krynickillus 10, 76, 77,
102, 104, 105, 246, 274, 284, 285*
Malacolimax 10, 92, 102, 220, 258
Malino 125
marginata (Miill.), Lehmannia, Limax 10,
25, 36*, 55, 57, 67, 76, 261, 263, 264*
marginatus Drap., Limax, Amalia, Milax 372
marmoratus, Paralimax 293
maura, LimaX cinereoniger var. 275
maximus, Limax 10, 14, 25, 28, 56*, 57, 59,
64*, 67, 71, 105, 106, 273, 274, 278, 279*
medioflagellata, Lehmannia 10, 262, 267, 268*
Megalopelte 9, 16, 89*, 102, 123, 124, 200
Meghimatium 6, 11, 27*, 47, 86, 87, 381, 382
melanocephalum, Deroceras 193
432
УКАЗАТЕЛЬ
melanocephalus, Krynickillus, Limax, Agrio-
limax, Deroceras 8,145,174,190,191,192*
Melitolimax 261
meridtonale, Deroceras 136
Mesarion 12, 390, 396
Mesolimax 6, 9, 18, 19*, 89*, 91, 122—125,
201, 204, 207
mesquensis, Malacolimax 240
Mesurethra 83
Metalimacoides 301
Metalimax 10, 53, 91, 102, 221, 290, 300
Michaelisia 10, 248, 252
Microarion 12, 390, 411
Microheynemannia 258
Micromilax 358
Milacidae 5, 11, 14, 16, 18, 19*, 21, 25, 29,
33, 35, 43, 51, 53-55, 81, 84, 92—94*,
95, 100, 101*, 102, 122, 124, 331, 354
Milax 5, 6, 11, 52*, 53, 68, 94*, 95, 102,
107, 356, 357
Milax s. str. 11, 358
minimus, Arion 411
minor, Trigonochlamys 312
minuta, Eumilax intermittens f. 298
minutus, Agriolimax subagrestis 136
minutus, Eumilax, Paralimax 297
minutus, Krynickillus 142
mlokosieviczi, Metalimax 301
moellendorffi, Bielzia 288
moldavicum, Deroceras, Lytopelte 8, 128,
176, 177*
Monochroma 9, 53, 92, 222, 234
montenegrina, Bielzia 288
monticola, Gigantomilax monticola, Limax,
Vitrinoides 9, 102, 105, 111, 223, 230
monticola, Limax 233
morgani, Malacolimax 240
morgani, Parmacella 341
morgani, Pseudarion 239
multirugatus, Eumilax, Paralimax 293
nanus, Milax cristatus 365, 368
nanus, Turcomilax, Gigantomilax 9, 246,
249», 250
natalianus, Turcomilax, Limax, Lehmannia
10, 248, 252, 253*
niger, Eumilax, Paralimax 293
nigerrimus, Paralimax niger, Eumilax 293
nigricans, Milax 358
Nilssoni, Arion 407
nitens, Aegopinella 132
notatus, Paralimax brandti 293
nubila, Eumilax brandti f. 295
nubilis, Paralimax brandti 293
nyctelia, Lehmannia, Limax 10, 262, 271,
272*
obscura, Eumilax intermittens f. 298
obscurus, Hyrcanolestes 315
obscurus, Paralimax raddei 297
occidental, Deroceras, Lytopelte 8, 128,
178, 179*
ochraceus, Eumilax, Paralimax 293
ochraceus, Eumilax brandti f. 295
olivieri, Parmacella 335
olteniana, Lytopelte 176
oltinus, Limax 231
Oopelte 386, 387
Oopeltidae 81
Oopeltinae 386
orbubadensis, Limax 226
Oreohelicidae 86
orientalis, Eumilax intermittens f. 298
orientalis, Eumilax, Paralimax 297
orientalis, Hyrcanolestes 324
orientalis, Pseudomilax 324
Orthurethra 83
osseticum, Deroceras, Agriolimax 8, 35, 36,
127, 136, 137*
Oxychilus 93
Oxyloma 88*
padisii, Deroceras 8, 184*, 185
pallens, Boettgerilla 9, 35, 46*, 105, 107,
214, 215*, 216*
pallescens, Limax cinereoniger var. 275
pallida, Selenochlamys 11, 28, 38*, 74,
327, 329*, 330*
pallidus, Agriolimax agrestis, Limax 142, 145
Pallifera 86, 381
panormitanum, Deroceras 136
Paralimax 291, 292
Parmacella 5, 11, 27*, 53, 94*, 102, 335
Parmacellidae 5, 11, 16, 18, 23, 25, 28, 35,
43, 51, 53, 54, 57—59, 66, 67, 71, 72,
79, 81, 84, 92—94*, 95, 100, 101*, 107,
109, 121, 320, 331, 332, 334
Parmacellilla 11, 37, 96*, 98*, 99*, 100,
102, 309, 319
Parmacellillinae 308
parvulus, Milax 358, 362
pellucida, Vitrina 79, 80*
pentheri, Deroceras, Agriolimax 8, 184*,
185, 186*
persica, Caspilimax keyserlingi f. 245
persicus, Limax 244, 246
philippowitschi, Parmacellilla 320
Philomycidae 11, 14—18, 23—25, 28 , 29,
33, 35, 43, 54, 68, 81, 84—87, 93, 10»,
101*, 121, 380
Philomycini 380
Philomycus 52*, 86, 87, 381
Phrixolestes 309, 311
picta, Eumilax intermittens f. 298
pictum, Meghimatium 382
pictuS, Paralimax raddei 297
Pirainea 357
pischpekensis, Gigantomilax 249, 251
Plathystimulus, 8, 128, 153
Platytoxon 195, 196
Plepticolimax 281
plumbea, Selenochlamys 330
poirieri, Lehmannia, Limax 268
pollonerae, Deroceras, Agriolimax 8, 127,
135*
pollonerai, Deroceras 136
pollonerai, Malacolimax 240
pollonerai, Parmacella 341
ponori, Deroceras 8, 187
pontica, Trigonochlamys 312
ponticus, Phrixolestes 312
praecox, Deroceras 8, 128, 152, 165, 166*
primitivus, Limax 226
Proparmacella 342
Prophysaon 86
УКАЗАТЕЛЬ
433
Prosobrancjhia 60, 319
Pseudarion 234, 239
pseudodioicus, Agriolimax 129, 132
pseudolytopelte, Deroceras 185
Pseudomilax 309, 313, 324
Pulmonata 13, 27, 28, 30, 32, 45, 47, 60,
83, 84
p unctatus, Paralimax brandti 293
Pupillina 87, 88
raddei, Eumilax intermittens f. 298
raddei, Eumilax, Paralimax 297
ramosum, Deroceras 163
redii, Limax 57
reibischi, Mesolimax 9, 207
reibischi, Metalimax, Paralimax 301, 303
reibischi, Pseudomilax 324
renardii, Limax cinereoniger var. 275
reticulatum, Deroceras, Limax, Agriolimax
8, 22, 29*—31*, 39, 40*, 41, 43, 44*,
45, 46*, 48*—50, 58—60*, 62—65,
68, 70, 71*, 75—77, 105—107, 109, 112,
113, 127, 145, 149, 150*
reticulatus, Agriolimax agrestis 149
Retowskii, Amalia 368
retowskii, Drilolestes, Pseudomilax 11, 324,
325*—327*
.retowskii, Limax 284
robustus, Gigantomilax 244, 246
rodnae, Deroceras 8, 128, 152, 167, 168*
roseni, Agriolimax 162
roseni, Parmacella 348, 352
rossica, Amalia 376
rubra, Parmacella korschinskii 345, 347
rufa, Daudebardia 80*, 93*
rufus, Arion, Limax 12, 21, 46*, 50*, 56*,
64*, 65, 69, 106, 387, 388*, 389, 390,
391*, 392*
rustica, Tandonia, Limax, Milax 11, 93*,
357, 365, 372, 373*
.rutellum, Candaharia, Parmacella 11, 59, 63,
67, 68, 105, 109, 334, 342, 343*
salamandroides, Eumilax brandti f. 295
.salamandroides, Eumilax, Paralimax 293
samsunensis, Milax 11, 368, 379*
sarmizegetusae, Lehmannia 263
schelkovnikovi, Limax 226
schemachensis, Agriolimax 158, 162
schleschi, Deroceras 8, 186*, 187
schmidti, Limax 226
schulzi, Agriolimax 129, 132
schwabi, Limax 288
Selenochlamydinae 11, 97, 99*, 308, 327
Selenochlamys 11, 16, 27*, 33, 37, 45, 83,
96*, 97, 98*, 99*, 100, 102, 122, 309, 327
semilimax, Semilimax 79, 80*
semiplumbeus, Trigonochlamys 312
serbica, Tandonia 376
serotinus, Limax 259
sibiricus, Arion 12, 104, 390, 399, 400, 401*,
402*
sibiricus, Arion subfuscus 402
sibiriensis, Agriolimax 146, 148
Sigmurethra 83
silvaticus, Arion 12, 389, 405, 406*, 407
silvaticus, Arion circumscriptus 405
simplex, Agriolimax buchar 148
simplex, Agriolimax transcaucasicus 145
simplex, Limax 226
simrothi, Arion 396
simrothi, Megalopelte 9, 200*, 201*, 202
simrothi, Parmacella 335
Simrothia 281
sokolovi, Troglolestes 11, 28, 69, 74, 76,
99*, 322, 323*
sphingiformis, Hyrcanolestes, Chrysalidomi-
lax 315
steidachneri, Deroceras, Agriolimax 8, 187,
188*
Streptaxidae 68
striata, Eumilax intermittens f. 298
striatum, Meghimatium 382
striatus, Paralimax raddei 297
stummeri, Limax, Lehmannia 252, 254
sturanyi, Deroceras, Agriolimax 8, 57, 61,
77, 105, 106, 127, 132, 133*
Stylommatophora 13, 24, 45, 47, 51, 81, 83,
88, 100, 126
subagreste, Deroceras, Agriolimax 8, 105,
128, 153, 154, 155*, 156*
subfuscus, Arion, Limax 12, 22, 36*, 59, 64*,
68, 75, 104—106, 388*, 390, 393, 396,
397*, 398*, 400
suboccidentalis, Lytopelte 176
Succineidae 88
Succinida 88
svaneticus, Limax 241
sylvaticus, Limax 259
szighethyae, Lehmannia 262
talyschana, Gigantomilax lenkoranus f. 239
talyschanus, Gigantomilax 238
talyschanus, Limax 244—246
Tandonia 5, 6, 11, 27*, 47, 57, 68, 94*, 95,
102, 107, 356, 357, 363
Tandonia s. str. 11, 364
tarmes, Meghimatium 382
Taulimax 10, 248, 254
tauricum, Deroceras, Agriolimax 8, 105, 111,
128, 163, 164*
Tebennophoridae 380
tenellus, Malacolimax, Limax 10, 22, 36,
57, 65, 66, 68, 71, 75, 259, 260*
Testacellidae 68, 79
tigris, Caspilimax keyserlingi f. 245
tigris, Limax 244, 246
tigvenius, Limax 259
toscannei, Malacolimax 239
Toxolimax 9, 123, 124, 208
transcaspia, Lytopelte 196, 200
transcaucasicus, Agriolimax 142, 145
transitorius, Agriolimax altaicus 146, 148
transsilvanica, Limax cinereoniger var. 275
transsilvanicus, Limax 288
Trigonochlamydidae 5, 10, 15—18, 23, 25,
28, 33, 36, 39, 43, 47, 51, 54, 58, 68, 69,
81, 84, 85, 95—100, 102, 117, 121, 122,
306, 341
Trigonochlamydina 306
Trigonochlamydinae 10, 98, 99, 309
Trigonochlamydoidea 10, 93, 95, 99, 100, 306
Trigonochlamys 10, 16, 27*, 33, 37, 52*,
96*, 97, 98*, 99*, 100, 102, 309
Trigonochlamys 313
484
УКАЗАТЕЛЬ
Troglolestes 11, 16, 18, 37, 54, 83, 96*, 98*,
99*, 100, 121, 309, 322
Tropidolytopelte 195
truncatula, Lymnaea 77
turcicum, Deroceras, Agriolimax 8, 107, 127,
152, 153*
Turcomilax 9, 91, 92, 102, 103*, 104, 220,
246
Turcomilax s. str. 9, 248, 249
turkestanus, Turcomilax, Limax, Heyne-
mannia 10, 248, 255*, 256*
typica, Caucasolimax caucasicus f. 243
typica, Eumilax brandti f. 295
typica, Eumilax intermittens f. 297
typica, Gigantomilax lenkoranus f. 239
typica, Gigantomilax monticola armenia-
- cus f. 230
typica, Lytopelte maculata f. 199
tzvetkovi, Turcomilax 10, 36, 248, 257*, 258
valentini, Limax 226, 231
variegatus, Limax 281, 284
variegatus, Paralimax brandti 293
variegatus, Paralimax raddei 297
varius, Hyrcanolestes 314
varius, Metalimax, Paralimax 10, 301, 302*—
304*
velitaris, Hyrcanolestes, Parmacella 10, 47*,.
. 69, 95*, 313, 314*—318*
vera, Limax cinereoniger var. 275
vermiformis, Boettgerilla 216, 217
verrucosus, Milax 83
Vitreinae 331
vitrinaeformae, Clathropodiam 335, 341
Vitrinidae 79—81, 84, 88, 93
Vitrinoides 9, 92, 222, 226
voronovi, Limax 226, 230
vranceanum, Deroceras 8, 188*, 189
vrancensis, Lehmannia 270
urbanskii, Krynickillus, Deroceras 8, 89,
190, 193*, 194*, 209, 210
Urocyclidae 81, 84
waldeni, Deroceras 152
wiktori, Deroceras 152
valenciennsis, Parmacella 335
valentiana, Lehmannia, Limax 10, 77, 105—
107, 262, 268, 269*
valentini, Hyrcanolestes 314
zilchi, Deroceras 8, 188*, 189
zilchi, Limax 278
Zonitidae 84, 88, 92—94*, 100, 331
Zonitoidea 11, 36, 84, 85, 92—94*, 100, 331
ОГЛАВЛЕНИЕ
Предисловие ................................................................ 3
Систематический указатель видов ...................>....................... 8
Общая часть ....... .................................................. 13
Морфология с элементами физиологии.......................................... 13
Внешний вид............................................................ 14
Форма тела и размеры (14). — Голова (14). — Шея (14). — Мантия
(14). - Спина (17). — Нога (17). — Рельеф (17). — Окраска (19).
Внутреннее строение .................................................. 19
Покровы (19). — Соединительная ткань (23). — Мускулатура (23). —
Раковина и раковинная сумка (24). — Легочная полость и мантийный
комплекс органов (27). — Кровеносная система (30). — Пищеваритель-
ная система (32). — Нервная система (45). — Органы чувств (49). —
Половая система (50).
Размножение и онтогенез ................................................... 55
• Копуляция (55). — Оплодотворение и формирование яиц (58). — От-
кладка яиц (58). — Развитие (59). — Постэмбриогенез и рост (61). —
Жизненные циклы (63).
Питание ................................................................. 68
Слизни и среда обитания.................................................... 69
Абиотические факторы . . ... . .................................... 69
Температура (70). — Влажность (71). — Осадки (72). — Ветер (73). —
Свет (73). — Почва .(74).
Растительность, основные ландшафты и биотопы.......................... 75
Хищники, паразиты и болезни .......................................... 78
Слизни как продукт параллельной эволюции................................... 79
Систематическое положение и филогения отдельных семейств слизней........... 83
Надсемейство Arionoidea ....................................... . . 84
Надсемейство Limacoidea ............................................. 87
Agriolimacidae (88). — Boettgerillidae (90). — Limacidae (91).
Надсемейство Zonitoidea ............................................... 92
Parmacellidae (94). — Milacidae (95).
Надсемейство Trigonochlamydoidea....................................... 95
Географическое распространение ............................................. 100
Синантропные и антропохорные виды........................................... 104
Хозяйственное значение слизней ............................................. 108
Передача паразитарных инвазий (108). — Слизни — вредители куль-
. . турных. растений (109).
436
ОГЛАВЛЕНИЕ
Регулирование численности вредных видов .................................... lit
Агротехническая борьба (111). — Химическая борьба (112). — Биологи-
ческая борьба (ИЗ).
Методы исследования........................................................ 114
Сбор живого материала (114). — Содержание слизней в неволе (115). —
Наркоз (116). — Фиксация (116). —Вскрытие (117).
Систематическая часть......................................................... 120'
Условные сокращения, принятые в тексте и рисунках систематической части . . . 120'
Таблица для определения семейств.............................................. 121
1. Надсемейство Limacoidea Rafinesque, 1815 .................................. 122
I. Семейство Agriolimacidae Wagner, 1935 ............................... 123-
1. Род Deroceras Rafinesque, 1820 ................................. 125-
Дополнение к роду Deroceras...................................... 179'
2. Род Krynickillus Kaleniczenko, 1851 ............................ 190’
3. Род Lytopelte О. Boettger, 1886 ................................ 195
4. Род Megalopelte Lindholm, 1914 ................................. 2001
5. Род Mesolimax Pollonera, 1888 ................................... 204
Дополнение к роду Mesolimax....................................... 207
Дополнение к семейству Agriolimacidae............................. 208
Род Toxolimax Simroth, 1899 ...................................... 208
II. Семейство Boettgerillidae Goethem, 1972 .............................. 210
Род Boettgerilla Simroth, 1910.................................... 213
III. Семейство Limacidae Rafinesque, 1815 ................................. 219
1. Подсемейство Limacinae Rafinesque, 1815 ......................... 221
1. Род Gigantomilax 0. Boettger, 1883 ............................. 221
2. Род Caucasolimax Likharev et Wiktor, gen. nov................... 240
,3. Род Caspilimax P. Hesse, 1926 ................................. 243
4. Род Turcomilax Simroth, 1901 ............................... . 246
5. Род Malocolimax Malm, 1868 ..................................... 258
6. Род Lehmannia Heynemann, 1862 .................................. 261
7. Род Limax Linnaeus, 1758 ..................................... 273
2. Подсемейство Bielziinae Likharev et Wiktor, subfam. nov. . ......... 287
8. Род Bielzia Clessin, 1887 ...................................... 287
3. Подсемейство Eumilacinae Likharev et Wiktor, subfam. nov............ 290
9. Род Eumilax 0. Boettger, 1881 .................................. 291
10. Род Metalimax Simroth, 1896 .................................... 300
2. Надсемейство Trigonochlamydoidea P. Hesse, 1882 ........................... 306
I. Семейство Trigonochlamydidae P. Hesse, 1882 ........................... 306
1. Подсемейство Trigonochlamydinae P. Hesse, 1882 ................. 309
1. Род Trigonochlamys O. Boettger, 1881 .................... 309
2. Род Hyrcanolestes Simroth, 1901 ............................... 313
3. Род Parmacellilla Simroth, 1910 ............................... 319
4. Род Troglolestes Ljovuschkin et Matekin, 1965 ................. 322
5. Род Drilolestes Lindholm, 1925 324
2. Подсемейство Selenochlainydinae Likharev et Wiktor, subfam. nov. . . 327
6. Род Selenochlamys 0. Boettger, 1883 ............................. 327
3. Надсемейство Zonitoidea Morch, 1864 .................................... 331
I. Семейство Parmacellidae Gray, 1860 . . ................................ 332
1. Род Parmacella Cuvier, 1804 ................................. 335
2. Род Candaharia Godwin-Austen, 1888 .............................. 342
ОГЛАВЛЕНИЕ 437
II. Семейство Milacidae Ellis, 1926 .................................... 354
1. Род Milax Gray, 1855 .......................................... 357
2. Род Tandonia Lessona et Pollonera, 1882 ....................... 363
Дополнение к семейству Milacidae................................ 379
4. Надсемейство Arionoidea Gray, 1840 ..................................... 380
I. Семейство Philomycidae Gray, 1847 ................................ 380
Род Meghimatium Hasselt, 1823 .................................. 382
II. Семейство Arionidae Gray, 1840 .................................... 386
Подсемейство Arioninae Gray, 1840 ................................... 387
Род Arion Ferussac, 1819 ........................................387
Литература .....................*........................................ 414
Алфавитный указатель латинских названий моллюсков........................... 429
Илья Михайлович Лихарев, Анджей Йозефович Виктор
СЛИЗНИ.ФАУНЫ СССР И СОПРЕДЕЛЬНЫХ СТРАН
(GASTROPODA TERRESTRIA NUDA)
В серии: Фауна СССР. Моллюски. Т. III, вып. 5
Новая серия, № 122
Утверждено к печати
Зоологическим институтом Академии наук СССР
Редактор издательства И. П. Дубровская
Технический редактор Г. А. Бессонова
Корректоры Н. П. Кизим, Э. Н. Липпа, А. X. Салтанаева
ИБ М 9199
Сдано в набор 14.04.80. Подписано к печати 10.11.80. М-21032.
Формат 70Х108*/1в. Бумага типографская 2. Гарнитура
обыкновенная. Печать высокая. Печ. л. 272/2-f-l вкл. ('/а печ. л.)
Усл. печ. л. 38.67. Уч.-изд. л. 40.2. Тираж 1250.
Изд. М 7458. Тип. зак. 1186. Цена 6 р. 60 к.
Ленинградское отделение издательства «Наука»
199164, Ленинград, Б-164, Менделеевская линия, 1
Ордена Трудового Красного Знамени
Первая типография издательства «Наука»
199034, Ленинград, В-34, 9 линия, 12