






















































































































































































































































Author: Медведев Л.Н. Зайцев Ю.М.
Tags: insecta hexapoda насекомые энтомология систематика животных специальные зоологические науки биология
ISBN: 978-5-87317-615-1
Year: 2009




Text
Ю.М. Зайцев, Л.Н. Медведев
Личинки жуков-листоедов России
РОССИЙСКАЯ АКАДЕМИЯ НАУК
Институт проблем экологии и эволюции им. А.Н. Северцова
Программа Отделения биологических наук РАН
«Биологические ресурсы России: оценка состояния
и фундаментальные основы мониторинга»
Ю.М. Зайцев, Л.Н. Медведев
Личинки жуков-листоедов России
Товарищество научных изданий КМК
Москва ❖ 2009
УДК 595.768.12
ББК 28.691.89
3 17
Зайцев Ю.М., Медведев Л.Н. Личинки жуков-листоедов России. М.: Т-во научных
изданий КМК. 2009. 246 с., ил.
В работе рассматривается одно из крупнейших семейств отряда жесткокрылых, многие
из которых являются опасными вредителями растений сельского и лесного хозяйства.
Дана морфо-экологическая характеристика преимагинальных стадий жуков-листоедов
фауны России, включающая 448 известных видов, входящих в состав 122 родов и 15
подсемейств, а также таблица отличий личинок семейства листоедов от личинок других
жесткокрылых. Приведена общая морфо-экологическая характеристика 15 подсемейств
и 122 родов, составлены определительные таблицы личинок, подсемейств и родов и ви-
дов с указанием их общего распространения в регионе, с указанием трофических связей
и некоторых аспектов их биоэкологии. Определительные таблицы сопровождаются ори-
гинальными рисунками.
Работа рассчитана на энтомологов, экологов, а также специалистов в области защиты
растений.
На обложке: личинки Phyllodecta vulgatissima L. Ориг. фото О.Г. Волкова.
Издание осуществлено при финансовой поддержке
Программы Отделения биологических наук РАН
«Биологические ресурсы России:
оценка состояния и фундаментальные основы мониторинга»
ISBN 978-5-87317-615-1
© Зайцев Ю.М., Медведев Л.Н., 2009
© Т-во научных изданий КМК, 2009
© Институт проблем экологии и эволюции
им. А.Н. Северцова РАН, 2009
Содержание
Введение............................................................4
Общая часть.........................................................6
Морфология..........................................................6
Развитие и экология................................................12
Хозяйственное значение.............................................26
Сбор, хранение и препарирование личинок для определения............27
Специальная часть..................................................29
Таблица для определения подсемейств семейства Chrysomelidae........29
1. Подсемейство Donaciinae.........................................31
2. Подсемейство Megalopodinae......................................44
3. Подсемейство Zeugophorinae......................................46
4. Подсемейство Orsodacninae.......................................48
5. Подсемейство Criocerinae........................................49
6. Подсемейство Clytrinae..........................................57
7. Подсемейство Cryptocephalinae...................................72
8. Подсемейство Chlamisinae........................................87
9. Подсемейство Lamprosominae......................................88
10. Подсемейство Synetinae.........................................90
11. Подсемейство Eumolpinae........................................91
12. Подсемейство Chrysomelinae....................................102
13. Подсемейства Galerucinae и Alticinae..........................148
14. Подсемейство Hispinae.........................................205
15. Подсемейство Cassidinae.......................................209
Литература........................................................227
Алфавитный указатель латинских названий насекомых.................235
Введение
Семейство жуков-листоедов является одним из наиболее крупных среди жест-
кокрылых, насчитывает в настоящее время почти 50 000 видов в мировой фауне и
около 1 200 видов в России. Эта группа постоянно привлекала внимание исследо-
вателей, но особенно интенсивное ее изучение характерно для нескольких после-
дних десятилетий. Это объясняется в основном большой практической значимос-
тью листоедов, включающих значительное число серьезных вредителей сельскохо-
зяйственных культур и древесно-кустарниковых пород. Вместе с тем совершенно
очевидно, что успешное решение проблем борьбы с вредными формами и защиты
растений возможно лишь при условии достаточной разработанности таксономии
группы, обеспечивающей точное определение видовой принадлежности и знания
экологии и преимагинальных стадий видов.
На фоне больших успехов в изучении имагинальной стадии наши познания о
личинках значительно отстают, хотя их изучение представляет большой общебио-
логический и практический интерес. Построение естественной системы семейства
невозможно без использования личиночных признаков, дающих богатый факти-
ческий материал для понимания родственных связей отдельных таксонов и фило-
генетических связей. В практическом отношении вред, причиняемый личинками
листоедов, не уступает, а в ряде случаев даже превышает вред, приносимый други-
ми жуками. Личинки листоедов в пределах одинаковых экологических групп внеш-
не достаточно однообразны, вследствие чего не вредящие формы неоднократно
смешивались с вредителями. Поэтому определитель личинок листоедов окажется
полезным не только для энтомологов-систематиков и экологов, но и для специали-
стов по защите растений и агрономии, а также для студентов биологических и сель-
скохозяйственных факультетов.
Основоположником изучения личинок листоедов фауны СССР был Д.А. Ог-
лоблин, который, однако, из-за преждевременной гибели в 1942 г. не опубликовал
свои работы. Его материалы доработаны Л.Н. Медведевым, в 1971 г. был опублико-
ван «Определитель личинок листоедов Европейской части СССР» (Оглоблин, Мед-
ведев, 1971). Появление определителя значительно облегчило задачу изучения ли-
чинок соседних регионов, в частности Сибири и Дальнего Востока. За короткое
время появилось около двадцати работ, посвященных личинкам Сибири (Медве-
дев, 1968; Медведев, Чернов, 1969; Медведев, Зайцев, 1971, 1973, 1974, 1975, 1976,
1977, 1977а; Дубешко, Медведев, 1972, 1975; Медведев, Дубешко, 1973; Медведев,
Долгий, 1974, 1974а; Медведев, Воронова, 1976, 1977), и стало возможно составле-
ние достаточно полного определителя по этому региону (Медведев, Зайцев, 1978).
В последующие тридцать лет опубликовано достаточно много работ отечествен-
ных и зарубежных авторов, в том числе определитель личинок Центральной Евро-
пы (Steinhausen, 1994а) и Японии (Kimoto, Takizawa, 1994), так что наши знания о
личинках значительно пополнились.
Предлагаемый определитель построен по тому же принципу, что и предыду-
щие. Он содержит определительные таблицы подсемейств, родов и видов и под-
робные характеристики подсемейств и большинства родов. Видовые описания не
включались во избежание перегрузки текста, но в видовых определительных таб-
4
лицах приводятся, помимо основных, и дополнительные признаки, а также сведе-
ния о кормовых растениях, экологии и распространении.
В определитель включено 16 подсемейств (все, встречающиеся в регионе), 122
рода и 448 видов. Если учесть, что фауна листоедов региона насчитывает около
1 000 видов, то, очевидно, что видовая изученность личинок еще очень далека от
полной, особенно в подсемействе Alticinae. Тем не менее, все основные роды се-
мейства уже известны. Для таких крупных родов, как Donacia, Labidostomis,
Cryptocephalus, Chrysolina, Gonioctena, Cassida, выявлены неизвестные ранее при-
знаки и разработана новая система, позволившая наметить естественные группи-
ровки.
Вместе с тем авторы отчетливо представляют, что определение видов в отдель-
ных группах может быть затруднительно, так как выявленные видовые признаки не
всегда достаточно надежны и подчас перекрываются индивидуальной изменчивос-
тью. Однако совершенствование видовых таблиц неразрывно связано с накоплени-
ем дополнительных материалов, поиском новых морфологических признаков, даль-
нейшей разработкой методик препарирования, что является делом будущего. Авто-
ры надеются, что появление настоящей работы ускорит и облегчит последующее
изучение личинок листоедов.
5
ОБЩАЯ ЧАСТЬ
МОРФОЛОГИЯ
Тело личинок листоедов более или менее, иногда значительно удлиненное, реже
широкоовальное, прямое или С-образно изогнутое, с выпуклой дорсальной и более
плоской вентральной стороной, вальковатое или уплощенное дорсовентрально.
Окраска скрытоживущих форм белая или желтоватая, открытоживущих личинок —
более или менее темных тонов или пестрая, имеющая определенный рисунок, иногда
желтоватая, зеленоватая. Покровы тела мягкие, за исключением головной капсулы
(всегда склеротизованной, иногда очень сильно), а также во многих случаях — ног
(особенно голенелапки), вершины брюшка и склеритов тела (особенно дорсально-
го склерита переднеспинки).
Головная капсула (рис. 1, 7) небольшая, заметно уже грудных сегментов, полу-
сферическая или уплощена спереди, в большинстве случаев гипогнатическая (до
опистогнатической у Cassidinae), с хорошо развитыми лобными швами, соединяю-
щимися назади в срединный, эпикраниальный шов, в типичном случае довольно,
длинный. Иногда швы плохо заметны за счет сильной склеротизации головы (у
чехликоносцев) или недоразвиты (Cassidinae) и различимы только при специаль-
ной обработке или при рассматривании головной капсулы изнутри. У специализи-
рованных минирующих форм голова сильно уплощена дорсовентрально, эпикра-
ниальный шов сильно укорачивается вплоть до полного исчезновения, и тогда кон-
цы лобных швов доходят непосредственно до затылочного отверстия, а затылоч-
ные доли головы вытягиваются кзади.
Эпикраниальный шов отделяет друг от друга более или менее выпуклые те-
менные участки, тогда как между ветвями лобного шва расположен лоб, обычно
треугольной формы. Наличник и верхняя губа всегда развиты, более или менее по-
перечные, особенно наличник, отделены швами друг от друга и ото лба, реже на-
личник и верхняя губа сильно уменьшаются и сливаются со лбом, так что их грани-
цы почти не различимы; обыкновенно это связано с сильной склеротизацией го-
ловной капсулы. Первичная хетотаксия головы довольно постоянна, во всяком слу-
чае, в пределах каждого подсемейства; нередко появляются также дополнительные
щетинки — вторичная хетотаксия, особенно на лбу и темени.
Первичные щетинки лба нередко получают нумерацию, которая имеет безус-
ловное вспомогательное значение для описательных целей, но до сих пор базиру-
ется исключительно на топологической основе и используется в нескольких круп-
ных подсемействах (Cryptocephalinae, Chrysomelinae, Cassidinae), причем для каж-
дого подсемейства обозначения предлагались независимо, а потому они несопос-
тавимы, хотя и построены по общему принципу, с использованием буквенного и
цифрового индекса. Общеприняты четыре буквенных индекса: v — теменные, f—
лобные, с — наличниковые, 1 — губные. Цифра обозначает условный номер ще-
тинки в пределах группы (например, f2 — вторая лобная щетинка). Гомологичность
щетинок, особенно у разных подсемейств, совершенно не изучена.
Большое значение в таксономии личинок, особенно для низших систематичес-
ких категорий, имеет верхняя губа (рис. 1, 3). Здесь обозначаются передний край,
6
боковые края, передние углы (в месте перегиба от переднего к боковым краям).
Посередине переднего края часто имеется срединная выемка, очень разнообразная
по форме. Щетинки губы подразделяются на группы: маргинальные — на пере-
днем крае, дискальные — на центральной поверхности губы, щетинки срединной
выемки; реже обозначаются угловые (на передних углах) и боковые (на боковых
краях). Нижняя сторона верхней губы, переходящая в эпифаринкс, часто несет сво-
еобразную скульптуру, обычно в виде мельчайших шипиков, собранных в отдель-
ные группы или поля.
Глазки у открытоживущих форм хорошо развиты, лежат по бокам головной
капсулы несколько выше основания усиков, плоские и темно пигментированные
или светлые с более или менее выпуклой роговицей в числе 1-6. У скрытоживущих
личинок глазки обыкновенно отсутствуют.
Усики всегда короткие, расположены у передненаружных углов лобной плас-
тинки, 1-3-члениковые; предпоследний членик обычно несет крупный конусовид-
ный или округлый сенсориальный придаток, рядом с которым помещается после-
дний членик, по размерам редко превосходящий придаток, но несущий короткие
щетинки — сенсиллы, что позволяет отличить истинный членик от придатка. Иногда
последний членик усика редуцируется до щетинки.
Мандибулы средней величины, сильно хитинизированные, без молярного выс-
тупа на внутреннем крае, большей частью треугольной формы или слабо удлинен-
ные, обычно с 2-5 зубцами на вершине и с 1-2 щетинками на наружном крае, редко
с простой острой вершиной. Максиллы состоят из небольшого кардо, крупного сти-
песа, более или менее слитых друг с другом галеи и лацинии, 2-4-членикового че-
люстного щупика, сидящего на крупном пальпигере (рис. 1,2); у Donaciinae в связи с
особенностями питания галея и лациния сильно модифицированы. Нижняя губа (рис.
1,2) состоит из плохо обособленных друг от друга субментума, ментума и пременту-
ма, без настоящей лигулы. Нижнегубные щупики 1-2-члениковые, нередко окруже-
ны с боков и сзади узким дуговидным склеритом, который обозначается как менталь-
ный склерит. Максиллы отделены от нижней губы перепончатым участком и под-
вижны относительно ее (за исключением личинок-чехликоносцев). Горло отсутствует.
Исключительно важную роль в систематике личинок листоедов играют хето-
таксия и склериты грудных и брюшных сегментов. Хетотаксия развита в той или
иной степени у всех личинок, а склериты — преимущественно у открытоживущих
форм. Классификации и терминологии склеритов (в основном для подсемейств
Chrysomelinae и Galerucinae) посвящено много работ, причем одни авторы уделяли
большее внимание сравнительно-морфологической классификации, а другие огра-
ничивались чисто топологической системой. Первая абсолютно необходима для фи-
логенетических построений, но сложна для практических целей определения, ибо
гомологизация отдельных склеритов подчас вызывает значительные трудности. Вто-
рая — проста и удобна в практической работе, но затрудняет сравнительный анализ.
Для личинок, лишенных склеритов, разрабатывалась номенклатура хетотак-
сии (Donaciinae, отчасти Eumolpinae) независимо от вышеуказанных систем, но на
той же топологической основе. Это привело к значительной номенклатурной пест-
роте. Н. Патерсон (1931) положил в основу своей классификации гипотезу, соглас-
но которой каждый склерит возникает за счет хитинизации покровов вокруг осно-
вания каждой первичной щетинки, а, следовательно, в прототипе любой сегмент
7
Рис. 1. Детали строения личинок листоедов:
1 — голова: а — усик, с — наличник,/— лоб, I — верхняя губа, ш — мандибула, о — глазки,
s. е — эпикраниальный шов, s.f — лобные швы, s.fc — фронтоклипеальный шов, v — темя;
2 — нижняя губа и максилла: с — кардо, g+/ — галея и лациния, I — лигула, язычок, т —
ментум, ms — ментальный склерит, рт — прементум, р. тх — челюстные щупики, р. I —
нижнегубные щупики, s — стипес, sm — субментум; 3 — верхняя губа: а — базальный выс-
туп, б — базальные углы, в — срединная выемка, г — дискальные щетинки, д — маргиналь-
ные щетинки, е — срединные щетинки; 4 — нога: с — тазик, ch — хелониум,/ — бедро, t —
голенелапка, и — коготок, v — вертлуг; 5 — левый яйцевой зубчик среднегруди Phratora для
разрезания хориона яйца; 6 — щетинки: а — щетинковидная, б — нитевидная, в — нитевид-
ная с оплавленной вершиной, г — зазубренная, д — булавовидная, е — булавовидная на теке,
ж — щетинковидная на теке, з — шиловидная.
несет два поперечных ряда щетинок, каждая из которых расположена на неболь-
шом округлом склерите. Кроме того, этот автор полагал, что хетотаксия исключи-
тельно постоянна на протяжении всей личиночной стадии. По мере накопления
новых материалов выводы Патерсона неоднократно подвергались критике, но в
своей основе они являются верными. Однако в процессе развития у многих личи-
8
нок появляются вторичные щетинки и склериты, что затрудняет гомологизацию
при отсутствии полной серии личиночных возрастов. Особенно сложна гомологи-
зация щетинок в тех случаях, когда на одном склерите появляется большое число
щетинок. Поэтому для форм, имеющих склериты, удобнее сравнивать именно их, а
не щетинки. Но, тем не менее, в основе хетотаксии лежит единая схема, позволяю-
щая выделить гомологические группы, вне зависимости от наличия или отсутствия
склеритов.
Мы предлагаем и используем далее следующую номенклатуру (рис. 2), осно-
ванную на используемой в отечественной литературе терминологии, с добавлени-
ем удачных индексов, введенных Kimoto (1962а), которые допускают составление
формул и могут получить международное признание. Прописные буквы означают
название группы, строчные имеют следующие значения: а — передний; р — зад-
ний; i — внутренний; е — наружный; ad — вторичный спинной (дорсальный); as —
вторичный брюшной (стернальный). Номенклатура разработана для форм, имею-
щих склериты, но приложима для всех морфологических типов личинок (в после-
днем случае слово «склерит» следует заменять словом «группа» для хетотаксии или
«поле» для соответствующего участка покрова).
Первичные склериты
I. Дорсальная группа (D) I
внутренний претергальный (Dai)
внутренний посттергальный (Dpi)
наружный претергальный (Dae)
наружный посттергальный (Dpe)
II. Стигмальная группа (DL)
крыловые (на груди) (DL)
стигмальные (на брюшке) (DL)
передние (Dla)
задние (DLp)
III Эпиплевральная группа (ЕР)
передние (Ера) задние (Ерр)
IV. Гипоплевральная группа (Р)
передние (Ра)
задние (Рр)
V. Стернальная группа (S)
парастернальные (PS)
внутренние (Psi)
наружные (Pse)
стернальные (ES)
стернеллярные (SS)
Вторичные склериты
1 .Дорсальная группа (ad)
угловой (ad 1)
вставочный (ad 2)
передний (ad 2а)
задний (ad 2р)
переднекрайние (ad 3, ad 4)
интеркалярный (ad 5)
II . Стернальная rpynna(as) престерналь-
ный (as 1)
боковой (as 2)
Рис. 2. Схема расположения и номенклатура склеритов. Объяснение в тексте.
Настоящая классификация является в своей основе филогенетической, но сле-
дует иметь в виду, что точная практическая интерпретация отдельных склеритов,
особенно у личинок старших возрастов, нередко вызывает значительные затрудне-
ния в уклоняющихся от основного типа вариантах. В таких случаях нередка интер-
претация на типологической основе, правильность которой может быть проверена
только специальными исследованиями, невозможными при практическом опреде-
лении. Приведем следующий пример. При слиянии внутреннего и наружного тер-
гального склеритов объединенный склерит почти всегда выглядит поперечным. У
Phratora на груди имеется округлый претергальный склерит и поперечный пост-
тергальный. Первый находится на месте Dai и обозначается как внутренний пре-
тергальный; задний ряд принимается за Dpi+Dpe, а наружный претергальный при-
знается исчезнувшим. Топологически такая классификация верна, но филогенети-
чески кажущийся склерит Dai является в действительности слитым Dai+Dae, но
его округлая форма и расположение на месте Dai приводят к филогенетически не-
верному обозначению, которое, однако, более удобно в практическом определении.
Подобные неточности вполне допустимы и часто даже неизбежны при описаниях
личинок и составлении таблиц, если не затрагиваются филогенетические вопросы.
Для точной гомологизации склеритов аберрантных форм необходимо изуче-
ние всех возрастов, особенно молодых, и достоверное выявление первичных щети-
нок.
Сегменты тела большей частью слабо хитинизированы, мягкие; покровы час-
то имеют характерную микроскульптуру в виде мельчайших многоугольников, ши-
пиков или гранул, обычно более или менее склеротизованных; густота и располо-
жение микроскульптуры в значительной степени определяют окраску и рисунок
личинок. Переднегрудной сегмент без поперечной складчатости, в большинстве
случаев с крупным склеритом, занимающим большую часть дорсальной поверхно-
сти. Тергиты прочих сегментов (кроме последнего и предпоследнего) с 2-3 попе-
речными складками. Последние сегменты часто модифицированы: 9-й тергит не-
редко сильно склеротизован, может нести характерную скульптуру, толстые щетин-
ки, а иногда — короткие крючки или шипики на вершине, что связано с его опор-
10
ними функциями. 10-й сегмент обычно небольшой, чаще подогнут на нижнюю сто-
рону тела и выполняет функцию подталкивателя. Анальное отверстие, лежит на
10-м сегменте, имеет вид щели, реже оно трехлучевое.
Отдельные щетинки могут отличаться друг от друга по длине; если эта разница
значительная, то длинные щетинки обозначаются как макрохеты, а короткие —
микрохеты. Во многих случаях, но далеко не всегда, макрохеты представляют пер-
вичные щетинки, а микрохеты являются вторичными щетинками. Форма щетинок
очень разнообразна (рис. 1, 6). Чаще всего они щетинковидные, т.е. утончающиеся
к вершине и заостренные на ней, но могут быть также нитевидными — одинаковой
толщины по всей длине и с притупленной вершиной, пиловидными — с зазубрин-
ками по краям, булавовидными, веретеновидными, пузыревидными или с утолщен-
ной цилиндрической базальной частью — текой. Место сочленения щетинки с по-
кровом обычно хорошо заметно в виде круглого колечка, но у Cassidinae сочлене-
ния щетинок не выражены, а различия между щетинками и выростами не всегда
явственные.
Стигмы имеются на переднегрудном и 1-8-м брюшных сегментах; переднег-
рудные стигмы часто сдвинуты кзади, в область среднегруди. Иногда последняя
пара брюшных стигм редуцируется или, напротив, превращается в удлиненные,
сильно хитинизированные шипы. К стигме может примыкать стигмальная плас-
тинка. Строение стигм и стигмальных пластинок весьма разнообразно (рис. 32, 33)
и служит хорошим диагностическим признаком.
Ноги 4-члениковые, состоят из тазика, вертлуга, бедра и голенелапки (рис. 1,
4), редко граница между вертлугом и бедром неявственная и ноги выглядят 3-чле-
никовыми; коготок хорошо развит, короткий или удлиненный, часто с зубцом при
основании, у многих видов с более или менее развитым хелониумом. У некоторых
минирующих форм ноги редуцированы, на их месте остается небольшой округлый
склерит.
Личинки младших возрастов отличаются от взрослых в основном отсутствием
вторичных склеритов и вторичной хетотаксии (а также наличием первичных скле-
ритов у форм, у которых они исчезают по мере роста), более длинными щетинками,
часто с булавовидной вершиной, слабым развитием скульптурных образований,
нередко более темным телом, относительно более крупной головой и грудью и не-
большим брюшком. У личинок 1-го возраста на крыловых склеритах, особенно на
среднегрудных (иногда и на 1-м тергите брюшка), имеется хорошо заметный зуб-
чик, служащий для разрезания хориона яйца (рис. 1, 5).
Личинки листоедов могут быть отличены от личинок других жесткокрылых по
приведенной ниже краткой определительной таблице:
1(2). Ноги 5-члениковые или имеются 1-3-члениковые церки................
...................................Adephaga, Hydrophiloidea, Staphilinoidea
2(1). Ноги 3-4-члениковые или отсутствуют, церки отсутствуют или несочленен-
ные.
3(4). Мандибулы с хорошо выраженным молярным выступом, реже без него, но
тогда с простекой, максиллы с хорошо развитыми галеей и лацинией........
........................Lamellicornia, Dascilloidea, Clavicornia, Heteromera
11
4(3). Мандибулы без молы и простеки; галея и лациния обычно более или менее
слиты.
5(6). Максиллы неподвижны относительно друг друга и образуют более или менее
сплошную пластинку с нижней губой.............Malacodermata — Sternoxia
6(5). Максиллы подвижны относительно друг друга и явственно отделены от ниж-
ней губы.
7(8). 10-й сегмент брюшка с продольной бороздкой перед анусом.Teredilia
8(7). 10-й сегмент брюшка без продольной бороздки перед анусом.
9(10). Горло развито..........................Phytophaga (Cerambycidae)
10(9). Горло не развито.
11(14). Членистые ноги обычно имеются, если отсутствуют, то эпикраниальный
шов не развит. Усики 1-3-члениковые. Гипофарингеальная вилка отсутствует.
12(13). Нижнегубные щупики отсутствуют или зачаточные. Вершина мандибул про-
стая. Имеется щитовидный нижнегубной склерит. Личинки в семенах и плодах...
.................................................Phytophaga (Bruchidae)
13(12). Нижнегубные щупики 1-2-члениковые. Вершина мандибул с зубцами. Щи-
товидный нижнегубной склерит отсутствует. Личинки практически не встречаются
в семенах и плодах............................Phytophaga (Chrysomelidae)
14(11). Членистые ноги отсутствуют. Эпикраниальный шов обычно раз вит. Усики
состоят из тактильного конуса, иногда также из явственного членика. Гипофарин-
геальная вилка имеется, редко отсутствует..Rhynchophora (Curculionidae)
В определительных таблицах учтена морфология личинок старших возрастов,
поэтому определение рекомендуется проводить по личинкам второго и более стар-
шего возрастов.
РАЗВИТИЕ И ЭКОЛОГИЯ
Столь обширная, разнообразная и экологически пластическая группа, какой
являются листоеды, изучена недостаточно полно, а главное — неравномерно. В
приводимой ниже краткой характеристике особенностей развития, биологии и эко-
логии группы мы вынуждены остановиться лишь на главнейших ее моментах, без
детализации. Многие частные моменты экологии приводятся в характеристике под-
семейств. Нужно особо отметить, что экологические признаки, как показали иссле-
дования последних лет, являются для таксономических и филогенетических целей
не менее важными, чем морфологические.
Яйцо и кладка
Яйца листоедов имеют типичное для большинства представителей жесткок-
рылых строение. Форма яиц от почти круглой (Galerucella, Pyrrhalta), овально-вы-
тянутой (у большинства листоедов) до грушевидной (часть Pyrrhalta) или плоской
(Argopus), Цвет яиц различный: светло-молочный, кремовый, грязно-сиреневый,
коричнево-красный или желтый. У многих видов яйцевая оболочка гладкая, блес-
тящая и покрыта прозрачной липкой жидкостью (Chrysomelinae, Criocerinae, часть
Galerucinae), некоторые имеют плотный хорион со своеобразной скульптурой (боль-
шинство Galerucinae, Alticinae). Скульптура хориона яйца у подсемейства Alticinae
12
Рис. 3. Яйцевой чехлик (1-4) и яйцо (5-8) жуков-листоедов:
1 — Labidostomis:, 2 — Smaragdincr, 3 — Clytra; 4 — Cryptocephalus', 5 — Galeruca\ 6 —
Pyrrhalta; 7 — Luperus} 8 — Aspidomorpha.
слабо выражена и заметна лишь при достаточном увеличении. Самые мелкие яйца
(до 0,6 мм) — у Alticinae, самые крупные (до 1,8 мм) — у Chrysomelinae. Иногда у
микропиле имеются затвердевшие выделения придаточных желез самки в виде ха-
рактерного комочка. Основные типы яиц показаны на рис. 3 и 4.
Количество яиц в одной яйцекладке колеблется от 1 до 120. Они откладывают-
ся одиночно (Synetinae, Clytrinae, Cryptocephalinae) или собраны в кладки (Chry-
somelinae, Criocerinae, Cassidinae, Galerucinae), чаще всего на листьях, в любом ме-
сте их поверхности или вдоль центральной проводящей жилки (Lilioceris, Sterno-
platys fulvipes. Pyrrhalta maculicollis), В большинстве случаев яйца откладываются
на нижнюю сторону листа (у всех открытоживущих и минирующих видов), а у поч-
вообразующих — или непосредственно в почву, или на растение около его основа-
ния. Яйца Synetinae, Clytrinae, Cryptocephalinae часто не связаны с какими-либо
растениями; они свободно рассеиваются по поверхности почвы, где происходит их
дальнейшее развитие. Эмбриональное развитие яиц при оптимальных условиях
длится от 8-10 (большинство видов) до 40-45 дней (многие Clytrinae, Cryptoce-
phalinae). Различные виды и даже группы листоедов по разному защищают кладку
яиц и отдельные яйца от неблагоприятных условий. Самки Synetinae, Clytrinae, Cry-
ptocephalinae с помощью задних ног и ямки стернита брюшка покрывают яйца экс-
13
Рис. 4. Форма куколок листоедов:
1 —Labidostomis\ 2 — Chrysolincr, 3 — Phratorcr, 4 — Pyrrhaltcr, 5 — Dactylispa, 6 — Cassida.
крементами, которые образуют чехлик, получивший название яйцевого (Медведев,
1962). В таких чехликах яйца хорошо защищены от иссушения, тогда как при уда-
лении чехлика они через 2-3 дня высыхают и гибнут. Хорион этих яиц очень тон-
кий и не имеет клейкой оболочки. Яйца Criocerinae и Chrysomelinae при откладке
прикрепляются к субстрату и между собой клейкой жидкостью. Отродившиеся
молодые личинки, прорывая хорион, также покрываются ею, что позволяет им дер-
жаться на кормовом растении. Это особенно характерно для Lilioceris, Chrysolina
14
virgata. Жуки Phratora, Plagiodera и все Cassidinae покрывают отложенные яйца
жидкими прозрачными выделениями придаточных половых желез, которые, быст-
ро подсыхая на воздухе, образуют своеобразный щиток—оотеку. У Cassidinae стро-
ение оотеки и количество яиц в ней резко отличаются у различных видов и являют-
ся хорошим систематическим признаком. У большинства Galerucinae яйца, имею-
щие плотный хорион, хорошо защищены от высыхания, что позволяет им нахо-
диться в достаточно сухой среде и даже на прямом солнечном свету. Некоторые
виды маскируют кладки, покрывая их сверху экскрементами (Cassida, Argopistes}
или пенистыми образованиями (Pallasiola, Diorhabda). Colasposoma при откладке
яиц прогрызает в стебле злаков отверстие приблизительно 1,5 мм в диаметре и от-
кладывает в полость стебля яйца, после чего отверстие замазывается экскремента-
ми, что обеспечивает защиту и маскировку.
У некоторых листоедов отмечено яйцеживорождение и живорождение. При
яйцеживорождении в отложенных яйцах личинка уже сформирована, обычно про-
сматривается через хорион и выходит наружу через несколько часов или даже ми-
нут после откладки яиц. При живорождении самка отрождает личинок.
Развитие и экологические группы личинок
Число личиночных возрастов у листоедов колеблется у различных групп от
трех (наиболее часто) до четырех или пяти. Развитие длится от нескольких недель
до двух лет. Личинки последнего возраста перед окукливанием часто становятся
малоподвижными, у них утолщается тело и укорачиваются конечности — стадия
предкуколки.
Весьма разнообразные условия обитания и питания личинок позволяют выде-
лить среди них следующие экологические группы.
1. Водные формы. Живут под водой на корнях и стеблях растений, отличаются
беловатой окраской, приспособленным для сосания ротовым аппаратом и наличи-
ем дыхательных крючков. Сюда относятся все Donaciinae.
2. Почвенные формы. Живут в почве, обычно на корнях растений, обгрызая
преимущественно тонкие придаточные боковые корешки, иногда вгрызаются в глубь
корня. Отличаются беловатой окраской, отсутствием или слабым развитием глаз и
склеритов, С-образным или тонким и сильно удлиненным телом. К этой группе
относятся Eumolpinae и значительная часть Galerucinae и Alticinae. Поскольку ли-
чинки двух последних групп (особенно первая) сравнительно недавно перешли к
обитанию в почве, они сохраняют ряд признаков, свойственных открытоживущим
формам, в частности, имеют в той или иной степени развитые склериты.
3. Открытоживущие формы. Обитают на листьях, реже на стеблях растений и
составляют наиболее распространенную группу (рис. 5, 2, б). Отличаются доволь-
но разнообразной пигментной окраской, хорошо развитыми глазами, ногами и боль-
шей частью склеритами; тело прямое, более или менее уплощенное с нижней сто-
роны. Кроме того, эта экологическая группа имеет многочисленные образования
защитного характера. Сюда относятся Criocerinae, Chrysomelinae, Cassidinae, часть
Galerucinae и Alticinae.
4. Чехликоносцы. Личинки обитают в специальных личиночных чехликах, от-
крытых на переднем конце, которые строятся из частиц субстрата (обычно почвы)
и экскрементов (рис. 5, 4, 5). Личинки отличаются сильно хитинизированной го-
15
Рис. 5. Общий вид личинок листоедов:
1 — Donacicr, 2 — Zeugophorcr, 3 — Megalopus\ 4 — Labidostomis\ 5 — Cryptocephalus (в
чехлике); 6 — Chrysolincr, 7 — Gonioctena; 8 — Plagiodera (I возраст, только что вышедшая из
яйца); 9 — Galeruca.
ловной капсулой и белым туловищем с редуцированными склеритами; задняя часть
брюшка подогнута под переднюю, ноги длинные. Чехликоносцы встречаются на
листьях и стеблях растений, в подстилке, на поверхности почвы, под камнями и
16
различными укрытиями, а также в муравейниках. Однако наличие чехлика, в кото-
ром личинка проводит всю жизнь, создает настолько специфические условия, что
различие сред обитания чехликоносцев не влечет за собой и, собственно, не требу-
ет каких-либо особых адаптации; напротив, личинки даже одного вида могут пере-
ходить из одной среды обитания в другую. В качестве особой экологической под-
группы из чехликоносцев можно выделять только мирмекофилов, хотя значитель-
ная их часть является факультативными мирмекофилами. К чехликоносцам отно-
сятся Clytrinae, Cryptocephalinae, Lamprosominae и Chlamisinae, к мирмекофилам —
ряд Clytrinae (Clytra, Tituboed) и факультативно — единичные виды Cryptocephalus.
5. Минирующие формы. Личинки этой группы выгрызают мины в листьях ра-
стений (рис. 6, 2) и отличаются сильно уплощенным дорсовентрально телом, от-
сутствием или слабым развитием глаз и ног, наличием папилловидных выростов на
боках сегментов, отсутствием эпикраниального шва и некоторыми деталями стро-
ения ротового аппарата, особенно верхней губы. Сюда относятся Zeugophorinae,
Hispinae и некоторые Alticinae.
6. Внутристеблевые формы. Обитают внутри стеблей, выгрызая в них ходы,
реже углубляются в корни. Отличительной особенностью является тонкое и удли-
ненное или С-образное, слабо хитинизированное беловатое тело, короткий эпикра-
ниальный шов. К этой группе относятся некоторые Alticinae, Megalopodinae, а так-
же тропические Sagrinae.
Трофические связи и трофическая специализация являются тем фактором, ко-
торый сыграл основную роль в становлении листоедов как семейства и появлении
столь значительного видового разнообразия. Изучение кормовых растений листое-
дов началось одновременно со становлением систематики как науки.
Листоеды России связаны примерно с 50 семействами и почти с 400 видами
растений. Значительное число видов тяготеет к ивовым, березовым, сложноцвет-
ным, крестоцветным, розоцветным, осоковым, злаковым, губоцветным, бобовым,
гречишным. Следует подчеркнуть, что листоеды связаны исключительно с высши-
ми (за одним исключением) и почти полностью (свыше 99%) с покрытосеменны-
ми. Предпочтение двудольных перед однодольными характерно для всех подсе-
мейств, кроме Donaciinae и Criocerinae, в которых наблюдается обратное соотно-
шение. Связь определенных групп листоедов с крупными таксонами растений от-
ражает как исторические процессы формирования подсемейств, так и эволюцию
ротового аппарата и процесса поедания растительной ткани. Среди листоедов тра-
диционно принято выделение монофагов, олигофагов и полифагов. Однако пони-
мание объема и границ этих категорий значительно отличается у различных авто-
ров. Так, к монофагам обычно относят виды, связанные в питании с единственным
видом растения, хотя среди листоедов такие виды почти неизвестны, а относимые
к монофагам виды, как правило, связаны с несколькими очень близкими видами
растений и являются по существу узкими олигофагами. Мы полагаем, что о спект-
ре кормовых растений листоедов следует судить не по имагинальной, а по личи-
ночной стадии. Это позволяет создать значительно более четкую классификацию
трофической специализации, приводимую ниже.
1. Фитофаги. Личинка развивается исключительно за счет живой раститель-
ной ткани. Имаго — фитофаги, со спектром кормовых растений, как правило, бо-
лее широким, чем у личинки.
17
1.1. Монофаги. Личинки питаются видами растений одного рода. Имаго — мо-
нофаги или олигофаги.
1.2. Олигофаги. Личинки питаются видами растений одного семейства. Имаго —
олигофаги, реже полифаги.
1.3. Полифаги. Личинки питаются видами растений различных семейств. Има-
го — полифаги, всегда с более широким спектром по сравнению с личинками.
2. Фитосапрофаги и сапрофаги. Личинки развиваются за счет растительного
детрита или характеризуются смешанным питанием. Имаго — фитофаги с трофи-
ческим спектром более узким, чем у личинок.
Если первая группа не требует дополнительного пояснения, то на группе фито-
сапрофагов и сапрофагов следует остановиться подробнее. Сюда относится подав-
ляющее большинство личинок-чехликоносцев и некоторые виды Chrysolina и
Crosita. Ее происхождение от первой группы несомненно. Прослеживается длин-
ный ряд переходов от облигатного обитания на растениях на всем протяжении раз-
вития или только на начальных его стадиях через все более высокую долю участия
сапрофагии, при которой еще охотно используется зеленый корм (часто вне зависи-
мости от его видовой принадлежности), до облигатной сапрофагии, высшая сте-
пень которой наблюдается у мирмекофилов, вплоть до перехода к питанию пищей
животного происхождения, как это отмечено у Clytra (Skwarra, 1927). У Chrysolina
и Crosita типичной сапрофагии нет, но нередко наблюдается переход к фитосапро-
фагии при резко неблагоприятных условиях обитания. Жуки, относящиеся к этой
трофической группе, большей частью являются полифагами и очень часто антофи-
лами, значительно реже олигофагами, причем связанными почти исключительно с
древесными растениями.
Есть определенная связь между степенью специализации личинок и имаго.
Выше уже отмечалось, что у фитофагов личинки отличаются более узкой специа-
лизацией, чем имаго, а у сапрофагов наблюдается обратное соотношение. Кроме
того, для видов с личинкой, обитающей на листьях или внутри тканей растений,
преобладает моно- и олигофагия (Criocerinae, Chrysomelinae, Hispinae, Cassidinae),
а в случае обитания личинок на корнях в почве или воде доминирует олигофагия
или полифагия (Donaciinae, Eumolpinae, почвообитающие Galerucinae и Alticinae).
Факт нахождения листоедов на растении не всегда свидетельствует о том, что
это растение является кормовым. Даже в том случае, если жук питается данным
растением, ошибка вполне вероятна, ибо известно много примеров, когда при не-
благоприятных условиях, особенно в определенные периоды развития, листоеды
переходят на совершенно не свойственные им кормовые растения. Установление
кормового растения личинки обычно позволяет достоверно решить вопрос о тро-
фических связях вида. В том случае, когда вид связан со значительным числом ра-
стений, как правило, один вид или род растения оказывается наиболее предпочита-
емым. В качестве примера можно привести Cassida rubiginosa и С. vibex, питание
которых подробно изучено (Палий, Клепикова, 1957; Zwolfer, Eichhorn, 1966). У
первого вида в природе жуки отмечены на шести родах и 21 виде сложноцветных,
а личинки — на пяти родах и 12 видах, но свыше половины особей питается на
Cirsium arvense; у второго вида жуки встречаются на семи родах и 16 видах слож-
ноцветных, а личинки — на пяти родах и 11 видах, но около двух третей особей
зарегистрировано также на С. arvense. Таким образом, основное кормовое расте-
18
ние для этих видов очевидно. При выкармливании в садках, в условиях отсутствия
выбора, оба вида могут не только питаться, но даже полностью завершать развитие
на видах растений, которые не заселяются ими в природе.
Для листоедов характерно значительное число биологических форм, отражаю-
щих процессы трофической специализации и связанного с ними формообразова-
ния. Биологическими формами обычно называются популяции вида, обитающие в
пределах общего ареала и часто даже одинаковых стаций на различных кормовых
растениях, причем переход с одного кормового растения на другое затруднен или
невозможен, что свидетельствует о специфичности обмена у каждой формы. Био-
логические виды отмечены, в частности, у Lochmaea саргеае (на иве и березе),
Pyrrhalta lineola (на иве и ольхе), Bromius obscurus (на кипрее и винограде),
Galerucella grisescens (на гречишных и розоцветных). Иногда биологические фор-
мы различаются хотя бы частично и по ареалу. Так, у транспалеарктической Gale-
rucella nymphaeae кувшинковая форма охватывает весь ареал, кроме крайнего юга,
гречишная форма—весь ареал, кроме севера, а смородиновая форма является очень
массовой к востоку от Оби, встречаясь здесь совместно с двумя другими формами,
но практически отсутствует в Европе. Gonioctena pallida на протяжении всего аре-
ала связана с черемухой и рябиной, а в Предбайкалье и лесотундре Западной Сиби-
ри к ней добавляется ивовая форма.
У личинок листоедов можно выделить следующие способы питания:
1. Объедание (грубое или выборочное обгрызание, скелетирование) листьев.
Характерно для Criocerinae, Chlamisinae, Chrysomelinae, Cassidinae, частично —для
Cryptocephalinae, Galerucinae, Alticinae.
2. Минирование листьев — выедание паренхимы листа между верхним и ниж-
ним эпидермисом. Характерно для Zeugophorinae, Hispinae и некоторых Alticinae.
3. Питание внутри стеблей и черешков (Megalopodinae, некоторые Alticinae,
Hydrothassa, Prasocuris).
4. Питание внутри плодов. Отмечено только у Lochmaea crataegi и Crioceris.
Данные о питании Podagrica в плодах мальвовых, очевидно, ошибочны. У Lochmaea
переход к питанию в плодах не влечет морфологических изменений личинки.
5. Объедание корней. Свойственно Eumolpinae и некоторых Galerucinae и Alti-
cinae.
6. Высасывание соков из стеблей растений (исключительно водных). Харак-
терно для Donaciinae.
7. Питание растительным детритом. Свойственно Clytrinae, Cryptocephalinae.
Все упомянутые выше способы питания выражены очень четко, хотя в отдель-
ных случаях намечается переход от одного способа к другому, даже в пределах вида:
Так, молодые личинки Cryptocephalinae нередко скелетируют листья, а позже пере-
ходят на питание детритом в подстилке: личинки некоторых Alticinae сначала гры-
зут корни у корневой шейки, а затем переходят на внутристеблевое питание. Ли-
чинки Hydrothassa и Prasocuris встречаются как на листовой пластинке, так и в
полых черешках листьев.
Энергетика питания детально изучена всего лишь для десятка видов листоедов
(Сате, 1966; Богачева, Дубешко, 1975), в том числе для нескольких сибирских ви-
дов. Литературные данные и наши наблюдения показывают, что коэффициент при-
роста составляет величину порядка 0,1-0,2 для личинок и 0,01-0,07 — для имаго.
19
Особенно высок коэффициент у молодых личинок. Количество пищи, потреблен-
ной за полный период развития (от яйца до яйца), в 15-35 раз превышает средний
вес имаго. Если принять среднюю величину для листоедов за 25 мг, то при наличии
данных о плотности вида и весе имаго нетрудно определить порядок потребленной
ими фитомассы. Основной поправкой здесь должна быть величина фитомассы, унич-
тожаемая личинками, не завершающими свое развитие.
Жук-листоед потребляет в 2-8 раз (в среднем — в 5-6 раз) больше раститель-
ной пищи, чем личинка, но количество пищи жука примерно равно тому количе-
ству, которое потребляет потомство одной самки в личиночной стадии, ибо смерт-
ность личинок особенно высока у молодых возрастов, потребляющих ничтожно
мало пищи.
Степень усвояемости пищи значительно выше уровня 10%, как это принима-
лось в ряде работ (Гиляров, 1966), и близка к 50%. Особенно высока она у личинок,
потребляющих главным образом калорийную паренхимную ткань и оставляющих
нетронутым эпидермис.
Куколка
Куколка большей частью свободная, реже в коконе (Donaciinae, Criocerinae)
или в личиночном чехлике (у чехликоносцев). Окраска большей частью светлая:
белая, желтая, оранжевая, зеленая, иногда с темными пятнышками; перед выходом
жука куколка обычно темнеет. Морфологически куколки довольно однообразны и,
за исключением отдельных групп, различаются только в пределах крупных таксо-
нов (подсемейства, трибы). Основные типы куколок показаны на рис. 4. Развитие
длится обычно от нескольких дней до нескольких недель; зимовка в стадии кукол-
ки не характерна.
При окукливании личинки обычно в какой-то мере связаны с кормовым расте-
нием, на котором шло их развитие, они окукливаются или на нем, или где-нибудь
вблизи него. Нами выделяются следующие группы по месту окукливания.
Окукливание на кормовом растении характерно для ряда открытоживущих ли-
чинок, которые развиваются на листьях. При окукливании личинки с помощью
подталкивателя прочно прикрепляются к нижней стороне листа и линяют; у видов
родов Chrysomela. Gastrolina и Gastrolinoides куколки свободно повисают на сбро-
шенном личиночном чехлике. Молодые куколки окрашены в светлые тона, но че-
рез некоторое время темнеют и принимают покровительственную окраску.
Окукливание на листьях отмечено для Plagiodera. Gastrolinoides. Linaeidea,
Galerucella, части Oulema, Cassidinae.
Окукливание в почве типично для большинства открытообитающих и всех
почвообитающих личинок. Почвообитающие личинки мигрируют в верхние слои
почвы и окукливаются, чаще всего на глубине 2-3 см. Перед окукливанием личин-
ки строят колыбельки. Некоторые личинки Gonioctena находятся в состоянии пред-
куколки, а затем окукливаются. Куколки — от светлых до оранжевых цветов. Их
кутикула плохо защищает тело от иссушения, вынутые из почвы куколки быстро
погибают.
Окукливание в полости стеблей отмечено только у Prasocuris phellandrii'. ли-
чинки 1-го возраста скелетируют листовые пластинки, а старшие возрасты про-
20
грызают стебель и проникают в полость, где идет их дальнейшее развитие и окук-
ливание.
Окукливание в минах отмечено только у Hispinae. Перед окукливанием личин-
ка передвигается к краю мины. Часть минирующих личинок окукливается в повер-
хностном слое почвы.
Окукливание в чехликах характерно для всех личинок-чехликоносцев (Clytrinae,
Cryptocephalinae, Lamprosominae, Chlamisinae). При окукливании личинки некото-
рых видов заползают на растение и прочно прикрепляются к нему устьем, после
этого они поворачиваются головой к заднему концу чехлика и линяют; отродивши-
еся молодые жуки прогрызают чехлик и выходят наружу. Большинство чехлико-
носпев окукливаются на почве и в подстилке.
Окукливание под водой на корнях или корневищах растений в плотном перга-
ментовидном коконе наблюдается у Donaciinae.
Защитные приспособления личинок
Личинки более подвержены воздействию окружающей среды, чем имаго, по-
этому у многих из них выработались специфичные защитные приспособления. У
почвообитающих форм меньше хищников и паразитов, но они более чувствитель-
ны к такому фактору, как влажность. Липоидная оболочка кутикулы и миграция в
верхние слои при избыточной влажности обеспечивают им выживание. Личинки
Chrysomela, Plagiodera, Gastrolina, Gastrolinoides, Phratora и др. имеют кожные
железы. Испарение воды через них предохраняет, видимо, от перегрева, а, кроме
того, железы выделяют салициловую кислоту и жидкость с резким запахом бром-
бензола, отпугивающую врагов и хищников. У личинок Oulema, Lerna, Lilioceris
тело покрывается пенистой жидкостью, смешанной с экскрементами. Это предох-
раняет их не только от иссушения, но от паразитов и хищников. Личинки Gonioctena,
Paropsides на дорсальной стороне брюшка между 7-м и 8-м сегментами имеют паль-
цевидные железы, которые при малейшем раздражении выпячиваются и выделяют
жидкость с резким запахом, что, несомненно, имеет защитное значение. Обитание
личинок в минах предохраняет их от иссушения. У личинок Cassidinae экзувии при
линьке не сбрасываются, а сохраняются на хвостовых нитях и загибаются на спин-
ную сторону, образуя защитный чехлик, который часто несет и экскременты.
Личинки, обитающие во влажной среде (Cassida panzeri, С. concha), имеют
длинные, тонкие боковые выросты, что обеспечивает, видимо, большое испаре-
ние. У типичных ксерофильных видов (Cassida lineola) боковые выросты сильно
укорочены и хитинизированы, что, несомненно, связано с условиями обитания.
Это характерно и для пустынных видов (Медведев, Радзивиловская, 1970). Ли-
чинки Criocerinae, Galerucinae при раздражении выделяют изо рта или сгибов ног
капли темно-бурой или ярко-желтой жидкости, сильно изгибают тело, быстро
двигаются.
Такие личинки обычно ядовиты и не поедаются большинством птиц. Личи-
ночный чехлик у Clytrinae, Cryptocephalinae является хорошим средством защиты.
При опасности личинки быстро втягивают туловище в чехлик и закрывают устье
головой. Личинки, вынутые из чехлика, через 2-3 дня гибнут от иссушения. Защит-
ным приспособлением у личинок 1-го возраста Gonioctena является групповое оби-
тание. Днем они держатся на листьях веерообразно, на ночь сползаются в плотную
21
Рис. 6. Общий вид личинок листоедов:
1 — Psylliodes', 2 — Hispa\ 3 — Cassida piperatcr, 4 — C. concha.
группу. При одиночном содержании в садках личинки плохо питаются и чаще всего
гибнут.
Типы биологических циклов развития
Накопившийся обширный фактический материал по биологии многих видов
позволяет охарактеризовать основные типы циклов развития листоедов. В выделе-
нии биологических циклов мы берем за основу зимующую стадию и наличие или
отсутствие летней диапаузы. Количество поколений за год мы считаем второсте-
пенным признаком, так как оно, будучи тесно связанным с продолжительностью
теплого периода, очень сильно колеблется даже у отдельных видов; к тому же, в
пределах рассматриваемого региона практически все виды оказываются моноволь-
тинными.
Мы выделяем четыре основных типа с несколькими подтипами:
Тип I. Генерация однолетняя (иногда до двух или нескольких поколений в году).
Зимует имаго.
Подтип 1а. Летняя диапауза имаго отсутствует. К этому подтипу относится по-
давляющее большинство листоедов, преимущественно мезофилов и гигрофилов,
связанных с лесной зоной. Перезимовавшие жуки кладут яйца весной или в начале
лета и вскоре погибают. Развитие личинки завершается в начале или середине лета.
Вышедшее из куколки второе поколение обычно питается и осенью уходит на зи-
мовку, иногда жуки уходят на зимовку сразу после выхода из куколок, не питаясь
22
(Gonioctend); в других случаях первое поколение жуков живет до осени и встреча-
ется в конце лета вместе с жуками второго поколения {Galerucella), Представите-
лей этой группы можно разделить на весенние (Chrysomeld) и летние (Phratora,
Lochmaea, Phaedon, Aided) формы в зависимости от сроков появления личинок.
Подтип 16: имеется летняя диапауза имаго. Развитие идет по тому же плану, что
и у предыдущего, но яйцекладка и развитие личинок обычно протекают в весенний
и раннелетний период, молодые жуки в наиболее жаркий период лета диапаузиру-
ют. Диапауза либо прерывается осенью, либо переходит в зимовку. Характерно для
некоторых степных и горно-степных Cassida, в частности, С. undecimnotata.
Тип II. Генерация однолетняя. Зимуют яйца.
Подтип Па: летней диапаузы нет. Личинки появляются весной или в начале
лета, взрослые жуки — со второй половины лета, активно питаются и спаривают-
ся, яйца откладывают в конце лета, после чего погибают или частично перезимовы-
вают. Характерно для Pallasiola и некоторых Galeruca, в частности G. tanaceti и,
очевидно, G. ротопае. Сюда же предположительно относится и Entomoscelis.
Подтип Пб: имеется летняя диапауза имаго. Аналогичен предыдущему вариан-
ту, но развитие личинки сдвинуто на весенний период и начало лета, жуки нового
поколения диапаузируют в наиболее жаркий период, а осенью приступают к кладке
яиц. Значительная часть жуков также перезимовывает. Характерно для всех степ-
ных видов Galeruca и Macromonycha.
Тип III. Генерация однолетняя. Зимуют личинки.
Подтип Ша: зимует молодая личинка первого возраста. Имеется летняя диапа-
уза имаго. Перезимовавшие личинки выходят обычно ранней весной и заканчива-
ют развитие к началу лета; развитие куколки идет быстро, так что молодые жуки
появляются в начале или первой половине лета и, как правило, диапаузируют после
кратковременного питания или сразу же после выхода из куколок. Очевидно, воз-
можен также вариант без диапаузы. Спаривание и откладка яиц происходят в конце
лета, личинки питаются в течение короткого срока, а затем уходят на зимовку. Жуки
погибают осенью или частично перезимовывают. Такое развитие характерно для
большинства степных, горно-степных и петрофитных видов Chrysolina.
Подтип Шб: зимует личинка последнего возраста перед окукливанием. Летняя
диапауза отсутствует. Цикл развития в общем аналогичен варианту 1а, но развитие
личинок растянуто до конца лета, вследствие чего они уходят на зимовку, а окукли-
вание и выход жуков происходят весной следующего года. Такое развитие отмече-
но только у представителей подсемейства Zeugophorinae.
Тип IV. Генерация двухлетняя. Зимует личинка. Этот тип развития характерен
для чехликоносцев. Молодые жуки появляются весной или летом и после непро-
должительного питания приступают к спариванию и откладке яиц. Срок развития
яйца сильно варьирует, но обычно значительный, около месяца (возможно, имеет
место летняя диапауза яиц). Вышедшие личинки активно питаются и строят чех-
лик, на зимовку уходит обычно первый, реже второй возраст. В течение второго
лета продолжается развитие личинки, которое может заканчиваться к концу лета, и
на вторую зимовку идет молодой жук, не покидающий личиночного чехлика, или
зимует личинка, а окукливание и выход жука происходят уже следующей весной.
Кроме чехликоносцев, сюда же должны быть условно отнесены плохо изученные
Donaciinae, у которых развитие личинки длится более года.
23
Циклы развития многих листоедов еще неизвестны, но можно полагать, что
развитие подавляющего большинства видов палеарктической фауны должно укла-
дываться в один из перечисленных выше семи вариантов. Показательно, что зи-
мовка куколок не зарегистрирована достоверно ни у одного вида, а данные о зи-
мовке куколок, например, Criocerinae, не подтверждаются.
Абиотические факторы среды и их роль
Абиотические факторы определяют многие черты биологии и экологии листо-
едов, в частности стадиальное распределение, особенности образа жизни, тип цик-
ла развития, скорость развития отдельных стадий и степень выживаемости. Наи-
большее значение имеют такие факторы, как температура и влажность, в меньшей
степени — свет.
Скорость развития отдельных стадий связана с температурой в прямо пропорци-
ональном отношении. Количество тепла за вегетационный период определяет число
поколений в году (моно-, би- или поливольтинность). Различные стадии и возрасты
неодинаково реагируют на температурные условия, но у большинства вариации сро-
ков развития очень значительны. Так, у Aspidomorpha отношение максимального срока
развития к минимальному составляет для яиц — 3,5, для личинок 1-2-го возраста —
3,5, 3-4-го возраста — 2,3, 5-го возраста — 8,1, для куколки — 4,3.
По отношению к температуре (и климату в целом) выделяются теплолюбивые
и холодостойкие формы с рядом промежуточных градаций. По отношению к влаж-
ности можно выделить следующие группы:
Гидрофилы — формы, связанные с поверхностью водоемов, водной и прибреж-
ной растительностью; они хорошо выносят длительное погружение в воду или даже
развиваются под водой.
Гигрофилы — тяготеют к прибрежной растительности, болотам, но избегают
погружения в воду.
Мезофилы — обитатели умеренно влажных биотопов (леса, луга и т.п.). К этой
группе относится большинство видов региона.
Ксерофилы — обитатели сухих открытых пространств (степи и остепненные
участки, каменистые выходы и южные склоны сопок, пески, а на юге — пустыни).
Влажность также оказывает влияние на скорость развития предимагинальных
фаз и процент выживаемости, особенно при пониженной температуре.
По отношению к свету можно выделить светолюбивые и умеренно светолюби-
вые формы. Большинство личинок избегают прямых солнечных лучей.
Адаптация к экстремальным условиям
Большинство листоедов, обитающих в высокоширотных и высокогорных рай-
онах, адаптируются к низким температурам и короткому вегетационному периоду.
Одной из главнейших приспособительных черт является переход к яйцеживо-
рождению и живорождению, что ускоряет онтогенез, перемещая процесс развития
яиц в благоприятные условия внутри тела самки. Яйцеживорождение отмечено у
горного Chrysochloa, многих арктических и горных Chrysolina и у ряда Gonioctena.
Яйцеживорождение среди листоедов свойственно только представителям подсе-
мейства Chrysomelinae, так что они составляют основное ядро арктической и высо-
когорной фауны.
24
В условиях низких летних температур преобладают группы с открытообитаю-
щей личинкой и выпадают виды с почвообитающими личинками или чехликонос-
цами, так как для первых температура почвы недостаточна для развития, вторые,
имеющие двухлетний цикл развития, не могут выжить в зимний период.
Как известно, большинство открытообитающих личинок окукливаются в по-
чве. В условиях тундры окукливание обычно происходит на поверхности почвы,
чаще всего во мху. Chrysomela taimyrensis окукливается не на кустах ивы, а на не-
высоких дерновинках и во мху, т.е. ближе к поверхности почвы, лучше прогревае-
мой и защищенной от ветра.
Хищники, паразиты и симбионты
Листоеды имеют большое число врагов (особенно в предимагинальных стади-
ях), которые в значительной степени контролируют их численность. Весьма пол-
ный список хищников и паразитов опубликовали Жоливе и Теодорид (Jolivet, Theo-
dorides, 1952). Не перечисляя все известные виды, укажем на основные группы.
Из хищников основную роль играют насекомые. Это, в первую очередь, кло-
пы-щитники семейства Pentatomidae, особенно синий щитник Zicrona coerulea, по-
едающий личинок Galerucinae, Altica и Chrysomelinae, а также другие щитники:
Jalla, Eysarcoris, Picromerus, Arma, Rhacognathus, в меньшей степени другие хищ-
ные клопы. Из хищных жуков нужно отметить жужелиц (особенно Calosoma, Са-
rabus, Harpalus, Pterostichus, Brachinus и тесно связанную с листоедами Lebid), ста-
филинов (Staphylinus, Ocipus и др.), божьих коровок (Coccinella, Hippodamia и др.),
мертвоедов и мягкотелок. Из других, насекомых отмечались хищные мухи (Asilidae,
личинки Syrphidae), златоглазки, уховертки, муравьи, складчатокрылые осы, хищ-
ные трипсы. Клещи-краснотелки поедают яйца, пауки — взрослых жуков, редко
личинок. На почвенных личинок часто нападают нематоды.
Из позвоночных наибольшее значение имеют птицы, особенно мелкие насеко-
моядные — синицы, славки, мухоловки, скворцы, воробьи, дрозды, дятлы и др., а
также земноводные—лягушки и особенно жабы. Менее заметна роль ящериц, ежей,
землероек и кротов. Некоторые рыбы (хариус в озерах Байкал и Хубсугул) поедают
листоедов Macroplea.
Из паразитов наиболее известны перепончатокрылые, особенно Chalcididae,
Braconidae и Pteromalidae. В яйцах развиваются Tetrastichus, Anaphes, Anaphoides,
на личинках — Diosphilum, Eulophus, Schizonotus, Asecodes. Им не уступают пара-
зитические мухи (главным образом из родов Meigenia, Erinnia, Erinniopsis. Picconia,
Macquartia, Megaselia). Из других паразитов укажем грегарин, волосатиков, нема-
тод, клещей Trombidiidae (эктопаразиты имаго), а также паразитические лабульбе-
ниевые грибы. Высокую смертность жуков и личинок вызывает гриб Sporotrichus
globuliferum.
У жуков и личинок Donacia, Adoxus, Cassida обнаружены симбиотические бак-
терии, живущие в клетках придатков средней кишки, в полости мальпигиевых со-
судов и в особых карманах влагалища. При откладывании яиц симбионты попада-
ют на поверхность хориона каждого яйца. Личинки получают симбионтов, съедая
часть яйцевой оболочки. Не исключено, что бактерии-симбионты встречаются у
многих видов листоедов. Их роль еще не вполне ясна, возможно, они принимают
участие в переваривании клетчатки.
25
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
Значительное число жуков-листоедов, особенно в личиночной стадии, являют-
ся серьезными вредителями сельского и лесного хозяйства. Их экономическое зна-
чение часто недооценивается, так как наносимые листоедами повреждения редко
приводят к полной гибели растения, а причиняемый растениям физиологический
ущерб плохо поддается количественной оценке.
Характерной особенностью листоедов является узкая пищевая специализация.
Поэтому каждый вредный вид, как правило, тесно связан с повреждаемой им куль-
турой. В настоящее время для Палеарктики уже зарегистрировано несколько сотен
вредящих листоедов.
Листоеды повреждают практически все части растений, хотя и в разной степе-
ни. Наиболее распространено повреждение листьев, что приводит к уменьшению
ассимилирующей поверхности растений. Повреждения могут представлять грубое,
или сплошное, объедание, при котором остаются лишь самые толстые жилки, вы-
борочное поедание, часто сводящееся к скелетированию, и, наконец, минирование.
Оба последних типа повреждений (особенно минирование) более серьезны, ибо
при них уничтожается исключительно ассимилирующая ткань, но практически сте-
пень вреда больше зависит от численности и размеров вредителя. Повреждения
стебля (личинки стеблевых блошек) встречаются гораздо реже, при этом преобла-
дают внутристеблевые повреждения — выгрызание ходов, что нарушает проводя-
щую деятельность растения, а у молодых растений и всходов часто наблюдается
отмирание верхушечного листа.
Повреждения корней (личинки большинства блошек и Eumolpinae) наблюда-
ются весьма часто, причем личинки обычно объедают придаточные корни и корне-
вые волоски, но могут вгрызаться и в главный корень и заметно нарушают мине-
ральное питание и поступление воды. Цветы и плоды личинками практически не
повреждаются.
Степень вредоносности того или иного вида зависит в первую очередь от его
численности и способности образовывать очаги массового размножения. Именно
это является основанием для выделения первостепенных, второстепенных, третье-
степенных и случайных вредителей. Третьестепенные вредители среди листоедов
причиняют весьма незначительный вред, которым, казалось бы, можно было пре-
небречь. Однако растение может нести заметный ущерб от комплекса заселяющих
его видов, если даже причиняемый каждым в отдельности видом вред незначите-
лен. В ряде районов Сибири и Дальнего Востока некоторые достаточно серьезные
вредители являются потенциальными.
Значительное число видов листоедов, которые указывались ранее как вредите-
ли, оказались не вредящими. Причины большинства подобных ошибок — недоста-
точные экологические сведения. В настоящее время для региона зарегистрировано
около 200 вредящих видов листоедов, что составляет почти 20% общей фауны.
Большинство вредителей относится к четырем подсемействам: Cryptocephalinae,
Chrysomelinae, Galerucinae и Alticinae. Ряд подсемейств почти или совершенно не
имеют практического значения — это Donaciinae, Megalopodinae, Lamprosominae,
Hispinae и Cassidinae.
26
Интродуцированные культуры, даже если они ввезены очень давно, практи-
чески не повреждаются листоедами, исключение составляют интродуцированные
виды листоедов — Leptinotarsa decemlineata и Zygogramma suturalis. К таким куль-
турам относятся томаты, огурцы, табак, тыквенные, подсолнечник, морковь, куку-
руза, картофель. Переходы листоедов на эти культуры отмечаются иногда с близко-
родственных растений.
Основной ущерб наносится зерновым и крестоцветным культурам, свекле, пло-
довым и ягодным культурам, а на Дальнем Востоке также бобовым. Из лесных по-
род заметно повреждаются дуб, вяз, все ивовые и березовые. Из технических куль-
тур повреждаются лен, конопля, хмель. Из вредителей пастбищ известно всего 2
вида. Так. Pallasiola absinthii часто размножается в массе в степях Сибири и объе-
дает полностью полынь, оставляя только голые стебли. Массовое размножение
Theone silphoides приводит как к сильному повреждению растительности пастбищ,
так и к отравлению скота при случайном поедании ядовитых жуков и личинок,
находящихся на растении.
Детальные данные по вредящим видам листоедов приведены в ряде справоч-
ников (Balachovsky, Mesnil, 1935; Медведев, 1974).
СБОР, ХРАНЕНИЕ И ПРЕПАРИРОВАНИЕ ЛИЧИНОК ДЛЯ
ОПРЕДЕЛЕНИЯ
Личинок следует фиксировать в 70°-ном спирте, причем перед фиксацией их
следует опустить на 1-3 минуты в крутой кипяток, но не кипятить, а затем тщатель-
но промыть в чистой воде. В полевых условиях их хранят в небольших пробирках с
70°-ным спиртом. Пробирки закрывают ватными пробками и помещают в стеклян-
ную банку с притертой пробкой и слоем ваты на дне, которая также заливается
спиртом. Следует помнить, что фиксированные личинки должны свободно поме-
щаться в пробирках, в противном случае при длительном хранении тело личинок
сильно деформируется, что затрудняет их измерение. Этикетки пишутся только ту-
шью на кальке, пропарафиненной или обычной плотной бумаге. При этикетирова-
нии указывается местонахождение, дата, кормовое растение, условия сбора (напри-
мер, в почве, в муравейнике, на нижней стороне листа, в стебле и т.п.) и биотоп
(например, луг, опушка леса, полынная степь, альпийский луг и т.д.). Вместе с ли-
чинками можно фиксировать небольшие части кормовых растений, это позволяет
при камеральной обработке уточнить кормовое растение. Обычно если название
растения неизвестно, то оно берется в гербарий, причем растение и личинка отме-
чаются одним и тем же номером. При сборе мирмекофильных личинок Clytrinae
следует взять некоторое количество муравьев, чтобы в дальнейшем иметь возмож-
ность определить их. Полезно собирать также образцы повреждений, т.е. листья,
куски стеблей или корней и хранить в виде гербария. Личинок, пораженных эндо-
паразитами, следует поместить в сухие просторные пробирки для выведения пос-
ледних.
Яйца и куколки собираются и фиксируются так же, как и личинки, однако по-
мещать их в кипяток не следует. У куколок Donaciinae, имеющих очень плотный
кокон, необходимо перед фиксацией надрезать или проткнуть его иглой.
27
При определении материала наиболее удобно пользоваться бинокуляром, а так-
же микроскопом (например, при изучении строения верхней губы, лабио-максил-
лярного комплекса, стигм, усиков и т.п., требующем значительного увеличения).
При рассматривании личинок их следует помещать в воду или слабо разбавленный
глицерин, но не спирт, который быстро испаряется. Допускать высыхания личинок
не следует. Однако поскольку щетинки гораздо лучше заметны на подсохших ли-
чинках, можно в случае необходимости подсушивать личинку, но только при нали-
чии достаточного числа экземпляров. Для лучшего выявления склеритов и микро-
скульптуры тела личинок полезно выварить в 10%-ном растворе щелочи или мо-
лочной кислоты, предварительно отделив голову. Для выявления слабоокрашен-
ных щетинок или склеритов личинок помещают в раствор туши или краски. При
изготовлении постоянных препаратов используются жидкость Фора-Берлезе (ра-
створ гуммиарабика, глицерина и хлоралгидрата) и канадский бальзам, для вре-
менных препаратов берется раствор желатина или концентрированный глицерин.
Наиболее удобна жидкость Фора, так как при ее использовании не требуется пред-
варительного обезвоживания и к тому же она в значительной степени просветляет
препарат.
Иногда для точной диагностики личинок необходимо знать их прижизненную
окраску, а многие личинки, особенно Cassidinae, в спирте теряют естественную
окраску. В таких случаях перед фиксацией необходимо тщательно описать цвет
личинок или в крайнем случае фиксировать их в 4%-ном растворе формалина.
Предлагаемый ниже определитель включает определительные таблицы подсе-
мейств, родов и видов (если в роде более одного вида), а также подробные характе-
ристики подсемейств и родов. Определители для видов ориентированы, в основ-
ном, на личинок старших возрастов, в большинстве случаев определение очень
облегчается данными по кормовому растению. Отдельные виды личинок не могли
быть включены в определитель из-за неполноты и неадекватности описаний, или
их описания не содержат признаков, используемых в ключе, или если описание
известно только для первого возраста. Для родов и видов указаны только главней-
шие синонимы. Номенклатура приводится по каталогу родов листоедов (Seeno,
Wilcox, 1982) с небольшими изменениями. Распространение видов дано только для
России и в основном с использованием следующих индексов:
ЕЧ — европейская часть
СК — Северный Кавказ
СБ — Сибирь
ЗСБ — Западная Сибирь
ВСБ — Восточная Сибирь
ДВ — Дальний Восток
АР — Арктика
Сведения о более детальном распространении видов можно получить из суще-
ствующей литературе (Медведев, 1992; Медведев, Дубешко, 1992; Беньковский,
1999; Lopatin et al., 2004).
28
СПЕЦИАЛЬНАЯ ЧАСТЬ
Таблица для определения подсемейств семейства Chrysomelidae
1(2). Ноги отсутствуют, тело сильно уплощенное (рис. 5, 2). Zeugophorinae
2(1). Ноги всегда хорошо развитые.
3(6). Последняя пара брюшных стигм превращена в направленные назад острые
хитиновые шипы.
4(5). Тело вальковатое, слабо С-образно изогнутое (рис. 5, 7). Эпикраниальный шов
развит. Жевательные доли максилл превращены в хитиновые стилеты (рис. 15, 72).
Шиловидные стигмы лежат на 8-м сегменте брюшка, довольно крупные. Личинки
на подводных частях растений.................................. Donaciinae
5(4). Тело уплощенное дорсовентрально, прямое (рис. 6, 2). Эпикраниальный шов
отсутствует (рис. 78, 1). Жевательные доли максилл не превращены в хитиновые
стилеты. Шиловидные стигмы лежат на 9-м сегменте брюшка (слабо отграничен-
ном от 8-го), короткие (рис. 78, 6). Личинки минируют листья.....Hispinae
6(3). Стигмы брюшных сегментов не превращены в шипы.
7(14). Верхняя губа, наличник и лоб слиты вместе (рис. 21,7); ноги очень длинные,
коготки без пульвилл, задняя половина брюшка сверху сильно выпуклая, расши-
ренная и подогнута под переднюю (рис. 5, 4); голова с каждой стороны с 6 глазка-
ми; тело помещается в твердом открытом спереди чехлике (рис. 20). Усики 3-чле-
никовые, с сенсориальным придатком на 2-м членике; 3-й членик обычно пред-
ставлен только щетинкой.
8(13). Голова без теменного канта, ее передняя часть равномерно выпуклая (рис. 24, 7).
9(10). 2-й членик усиков с широким, полушаровидным сенсориальным придатком
на вершине (рис. 19, 5). Коготки без зубцов при основании. Голенелапка снизу с
толстыми шиловидными щетинками................................. Clytrinae
10(9). 2-й членик усиков с узким конусовидным сенсориальным придатком на вер-
шине (рис. 26, 4). Голенелапка снизу без шиловидных щетинок.
11(12). Усики короткие. Коготки с крупным зубцом при основании, сильно изогну-
тые, голенелапка снизу с 2 рядами из 7-8 расширенных на вершине щетинок с
изогнутыми концами (рис. 34, 4). Губные щупики 2-члениковые. Стипес с 1 щетин-
кой в средней части. Личиночный чехлик цилиндрический.. Chlamisinae
12(11). Усики длинные (рис. 35, 3). Коготки со слабым зубцом или без зубца (рис.
35, 9). Губные щупики 3-члениковые, стипес в средней части с продольным рядом
из 4 щетинок (рис. 35, 5). Личиночный чехлик конический.Lamprosominae
13(8). Голова с резким теменным кантом, обособляющим совершенно плоские лоб
и переднюю часть темени (рис. 28, 7). Коготки без зубца при основании. Голенелап-
ка снизу с толстыми шиловидными щетинками................Cryptocephalinae
14(7). Верхняя губа и наличник отделены друг от друга и ото лба швами (рис. 16);
ноги сравнительно короткие.
15(28). Боковые стороны сегментов тела без ветвистых отростков; 9-й сегмент брюш-
ка без длинных отростков. Челюстные щупики 3-4-члениковые.
16(17). Стерниты брюшных сегментов с парными двигательными буграми. Стигмы
с двойным отверстием (рис. 15, 3). Анальное отверстие находится на дорсальной
29
стороне тела. Голова с каждой стороны с 6 глазками. Тело покрыто слизистыми
выделениями и экскрементами...............................Criocerinae
17(16). Стерниты брюшных сегментов без двигательных бугров. Стигмы с простым
отверстием. Анальное отверстие помещается на вентральной стороне тела.
18(19). Ноги чрезвычайно короткие, особенно голенелапка. Лобные швы головы
неявственные или слабо развитые (рис. 12, 7). Голова с 3-5 маленькими глазками с
каждой стороны. Склериты на сегментах тела не развиты. Мандибулы без явствен-
ных зубцов на вершине (рис. 12,2). Личинки крупные, развиваются внутри стеблей
растений...............................................2. Megalopodinae
19(18). Ноги не укороченные, голенелапка всегда хорошо развита. Лобные швы
хорошо развиты. Вершина мандибул с явственными зубцами. Если личинки внут-
ри стеблей, то они мелкие, с 1 глазком с каждой стороны.
20(25). Коготки без хелониума (рис. 40, 6). Голова без глазков, с длинным эпикра-
ниальным швом (рис. 37). Личинки в почве, питаются корнями растений.
21(22). Вершина брюшка с урогомфами (рис. 13, 6-7). Тело удлиненное, средне-
спинка, заднеспинка и 1-8-й тергиты брюшка с многочисленными гранулами. 9-й
тергит с густыми порами (рис. 13, 4). Стигмы с двойным отверстием. Голова с 1
глазком с каждой стороны. Коготки без хелониума..........Orsodacninae
22(21). Вершина брюшка без урогомф, редко с 1-2 зубчиками. Верх тела без густых
гранул. 9-й тергит без густых пор. Стигмы с простым отверстием.
23(24). Тергиты 1-5-го брюшных сегментов с мелкими немногочисленными шипи-
ками; тергиты 9-10-го сегментов несут сильно склеротизованный щиток (рис. 12,
8). Усики 1-члениковые, на вершине членика большая конусообразная папилла (рис.
12, 6). Лоб с 4 щетинками у переднего края (рис. 12, 5). Передняя часть нижней
губы сильно выпуклая, без склеротизованной фигуры вокруг щупиков (рис. 12, 7)
..............................................................10. Synetinae
24(23). Тергиты 1-5-го брюшных сегментов без шипиков; 9-10-й сегменты — ино-
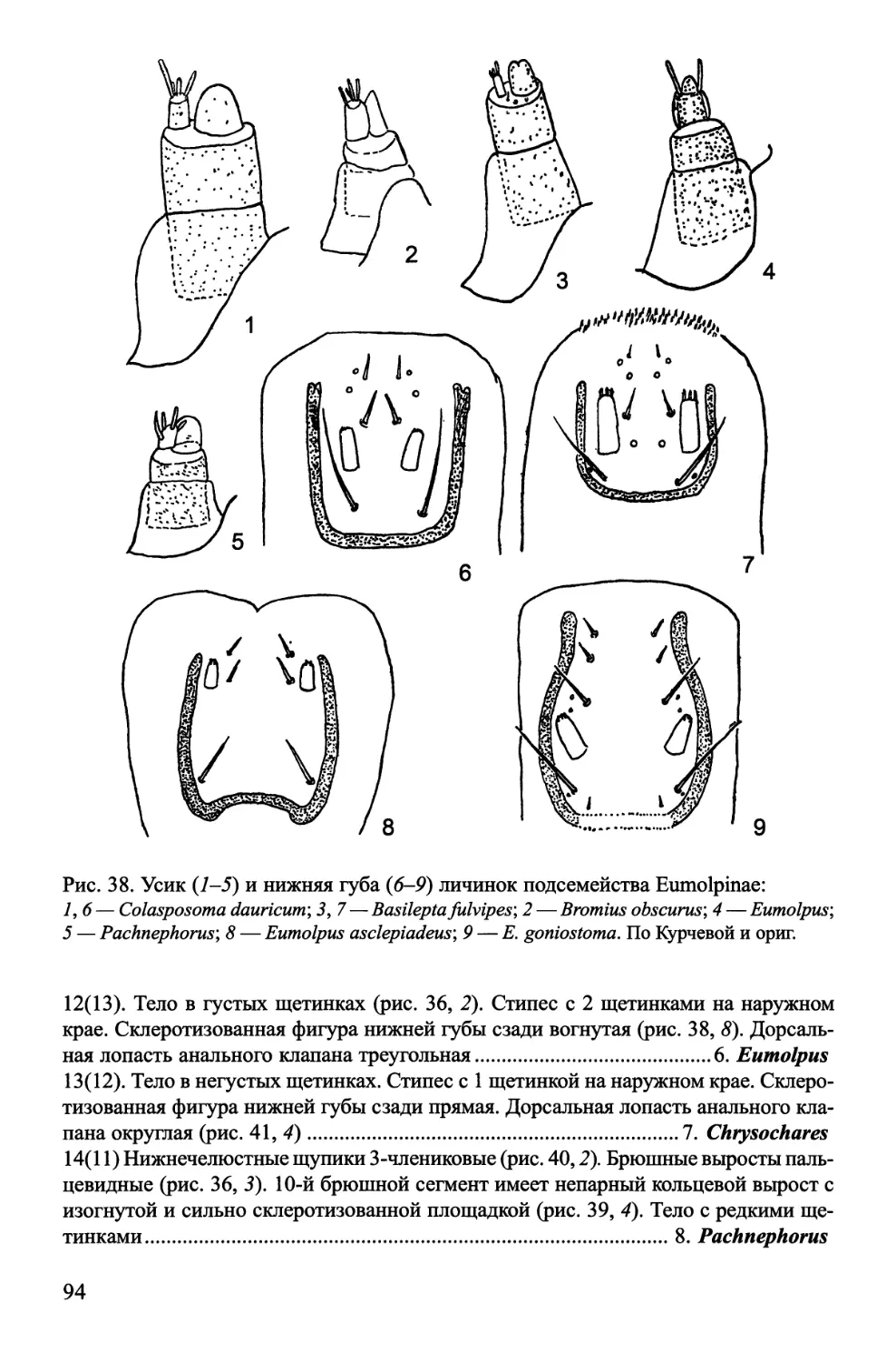
го строения. Усики 2-члениковые (рис. 38, 7), вершина 1-го членика несет большую
папиллу, почти равную расположенному рядом с ней 2-му членику. Лоб с 5 парами
щетинок (рис. 37). Передняя часть нижней губы слабо выпуклая, со склеротизован-
ной фигурой, окружающей щупики сзади и с боков (рис. 38, 6-9).
........................................................11. Eumolpinae
25(20). Коготки с хелониумом (рис. 68, 3). Голова с глазками и длинным эпикрани-
альным швом или без глазков и с очень коротким эпикраниальным швом.
26(27). Голова с каждой стороны с 6 глазками, усики 3-члениковые (рис. 43, 7)....
................................................Chrysomelinae
27(26). Голова с каждой стороны с 1 глазком или без глазков, усики 2-члениковые
(рис. 61, 7)......................................Galerucinae, Alticinae
28(15). Боковые стороны сегментов тела с длинными ветвистыми отростками; 9-й
сегмент брюшка расположен на дорсальной стороне 8-го сегмента, с двумя длин-
ными, тонкими, направленными вперед (над телом) отростками, несущими шкур-
ки личинок предыдущих возрастов и экскременты личинки (рис. 80). Тело сильно
уплощенное, с распластанными краями, спереди покрывающими всю голову (рис.
6, 3, 4). Челюстные щупики 2-члениковые...................15. Cassidinae
30
1. подсемейство donaciinae
Личинки белого цвета, с бледно-рыжевато-желтыми головой и ногами, слегка
С-образно изогнутые, с максимальной толщиной у средних брюшных сегментов,
кпереди довольно сильно суженные (рис. 5, 7). Голова (рис. 7, 3) очень маленькая,
слабо хитинизированная, прогнатическая, может почти полностью втягиваться в
переднеспинку. Эпикраниальный шов довольно длинный, верхняя губа и наличник
хорошо развиты, обособленные. Лоб пятиугольный, его форма изменчива у разных
видов, темя с несколькими щетинками. Глазков по 5 с каждой стороны, в виде тем-
ных пигментных пятен, образующих два косых ряда (3 в переднем и 2 в заднем
ряду). Усики трехчлениковые, с сенсориальными члениковидными придатками на
вершинной части 2-го членика. Наличник поперечный, с 4 щетинками. Верхняя
губа со слабо выемчатым посредине передним краем и с постоянным количеством
щетинок, которые подразделяются на 3 пары дискальных, 3 пары маргинальных,
лежащих на переднем крае, пару угловых на месте перехода переднего края в боко-
вой край и пару боковых на боковых краях. Мандибулы короткие, треугольные,
сильно хитинизированные, с 2 крупными зубцами на вершине и часто с мелкими
тупыми зубчиками на внутреннем крае, с 1 щетинкой на наружном крае. Нижнече-
люстные щупики трехчлениковые; слитые вместе галея и лациния превращены в
сильно склеротизованные узкие, удлиненные стилеты. Нижняя губа состоит из не-
склеротизованных субментума и ментума и узкого дуговидного или V-образного
склерита у основания лигулы. Нижнегубные щупики 1-члениковые.
Грудные сегменты увеличиваются в диаметре спереди назад, брюшко из 10
сегментов, из них наиболее толстые — 3-5-й сегменты; каждый сегмент с 2 или
более поперечными складками и с выпуклостями на плевральных участках. Стиг-
мы на переднегрудном сегменте и 1-7-м сегментах брюшка простые, округлые; стиг-
мы 8-го сегмента брюшка сближены друг с другом на спинной стороне сегмента и
превращены в длинные изогнутые острые шипы, направленные назад, которые обо-
значаются как дыхательные коготки (рис. 7, 7, 2). Существует также мнение, что
стигмы, в том числе и последняя их пара, не имеют функции органов дыхания, оно
обеспечивается деятельностью клеток кожного покрова. Хетотаксия сегментов тела
с хорошо выраженными первичными щетинками только у личинок первого возрас-
та. После первой линьки первичные макрохеты укорачиваются и становятся плохо
различимыми, смешиваясь с большим количеством шипиковидных, реже волоско-
видных вторичных микрохет (рис. 7,4-8). Различаются следующие основные груп-
пы щетинок (рис. 10, 70; 11,7): тергальные (передние — претергальные и задние —
посттергальные); стигмальные (на груди — крыловые) — между стигмами и тер-
гальными щетинками; эпиплевральные — непосредственно под стигмами; гипоп-
левральные — на боках сегментов под эпиплевральными; стернальные — на сред-
ней части стернитов. Ноги 3-члениковые, с небольшим коготком часто имеющим
зубец (рис. 7, 9-11). Длина тела взрослых личинок до 8-16 мм.
Самки откладывают яйца на подводные части кормовых растений у поверхно-
сти воды кучками или рядами (рис. 7,13). Кладки помещаются на стеблях, нижней
стороне листьев, в пазухах листьев или между двумя листьями. Яйца покрываются
и приклеиваются к субстрату затвердевшим в воде секретом и студнеобразной мас-
сой. Развитие яйца длится 10-14 дней. Молодая личинка прогрызает оболочку яйца
31
Рис. 7. Личинки подсемейства Donaciinae:
Дыхательный коготок личинки: 1 — внешний вид, 2 — поперечный разрез; 3 — голова; 4-8 —
форма щетинок: 4 — Macroplea mutica, 5 — Donacia versicolorea, 6 — D. cinerea, 7 —
Plateumaris affinis, 8 — P braccata; 9-11 — коготок: 9 — Donacia, 10 — Plateumaris, 11 —
Macroplea', 12 — участок корневища с коконами Donacia (верхний кокон открыт вследствие
выхода жука, черные точки — места прикрепления отпавших коконов); 13 — кладка D. to-
mentosa. По Бевингу и ориг.
и окружающую ее студнеобразную массу и обычно опускается на дно (реже про-
должает развитие во влагалище листа). Попав на дно, личинки глубоко закапыва-
ются в ил, добираются до тонких корней растений, которыми и питаются, переходя
по мере роста на более толстые корни. Как правило, личинки каждого вида разви-
ваются на одном определенном виде растения, в отличие от имаго, живущих обыч-
но на 2-3 кормовых растениях.
Добравшись до корней, личинка протыкает дыхательными коготками наруж-
ные ткани растения, проникая ими до воздухоносных тканей, откуда воздух посту-
пает через дыхательные коготки в трахеи личинки. На том месте, где воткнуты ды-
хательные коготки, личинка находится длительное время, передвигаясь вверх по
32
корням лишь по мере роста. Личинка прогрызает мандибулами отверстие в наруж-
ных покровах растения, погружает в него ротовые части, плотно затыкая ими от-
верстие, и высасывает из тканей растений соки. Личинка окружена слоем студне-
образной слизи и находится под толстым слоем донного ила, что в значительной
мере защищает ее от хищников.
Развитие личинки обычно длится более года, нередко до двух лет и только в
отдельных случаях — несколько месяцев. Взрослая личинка изготовляет под водой
тонкий пергаментовидный кокон из вещества, которое накапливается в двух специ-
ализированных мальпигиевых сосудах. Кокон прикрепляется к стеблям или кор-
ням (рис. 7,12) и соединяется с ними при помощи прогрызаемого личинкой отвер-
стия в растении, через которое кокон заполняется воздухом. Кокон светло- или тем-
но-бурый, иногда тонкий и прозрачный, но его оболочка почти непроницаема для
воздуха и жидкостей (куколки в коконах, помещенные в формалин, оставались жи-
выми 40-50 дней). Куколка свободного типа, светлая, с восковым оттенком. Для
выхода из кокона взрослый жук отгрызает его верхнюю часть. Развитие обычно
заканчивается к осени и зимует жук в коконе; в других случаях зимует личинка.
Характерной особенностью Donacia Fabricius является их симбиоз с бактерия-
ми, находящимися в двух мальпигиевых сосудах. У личинки бактерии находятся в
слепых отростках средней кишки и только перед окукливанием переходят в маль-
пигиевы сосуды. В регионе пять родов, но личинки дальневосточного рода Sominella
(с 1 видом — S. macrocnemia Fischer-Waldheim) неизвестны. Роды и виды подсе-
мейства различаются с трудом, известны в основном широко распространенные
виды; сибирские виды не изучены. Поскольку морфология родов очень близка, мы
не даем их описания, а ограничиваемся расширенной определительной таблицей.
Таблица для определения родов подсемейства Donaciinae
1(6). Лоб широкий: его ширина между боковыми углами равна длине или немного
превышает ее (рис. 8, 7), верхняя губа с более или менее явственными боковыми
углами (рис. 9, 4-13; 10, 7); на максиллах отросток лацинии длинный и тонкий, иг-
ловидный (рис. 8, 4).
2(3). Хеты тергитов груди и брюшка преимущественно короткие, конусовидные
(рис. 8, 7), среди них встречаются более или менее многочисленные длинные и
тонкие хеты на всех или только 4-7-м тергитах брюшка; 3 маргинальные хеты сбли-
жены или наружная отодвинута от двух других на расстояние, не превышающее
или не более чем вдвое превышающее диаметр ее основания (рис. 9, 4, 5, 7, 8, 10-
13; 10, 7); если наружная маргинальная хета далеко отодвинута от двух других и в
1,5 раза ближе к угловой, чем к средней маргинальной (рис. 9, 9), то серповидные
выросты 8-го сегмента брюшка очень длинные, в 2,0-2,4 раза длиннее ширины
головы........................................................1. Donacia
3(2). Все хеты тергитов груди и брюшка длинные, тонкие, оттянуты на конце в во-
лосовидное острие (рис. 8, 6); на 6-7-м или только на 7-м тергитах брюшка они
длиннее, чем на передних; наружная маргинальная хета верхней губы далеко ото-
двинута от двух других, в 1,5-2,0 раза ближе к угловой, чем к средней маргиналь-
ной (рис. 9, 6); серповидные отростки 8-го сегмента брюшка короткие, только в
1,2-1,3 раза длиннее ширины головы.
33
Рис. 8. Личинки Donaciinae:
1—3 — лоб: 1 — Donacia cinerea, 2 — Plateumaris discolor, 3 — Plateumaris weisei; 4-5 —
максилла: 4 — Donacia cinerea, 5—Plateumaris discolor, 6-7—хеты брюшных тергитов: 6—
Macroplea appendiculata, 7 — Donacia', 8-14 — левая мандибула: 8,9 — Donacia thalassina,
10—Macroplea appendiculata, 11 —Donacia clavipes, 12—D. vulgaris, 13—D. marginata, 14 —
34
4(5). Лоб с 6 щетинками; щетинки наличника не утолщенные, с каждой стороны
попарно сближены на расстояние 5 диаметров их оснований; щетинки верхней губы
неутоленные; среди тергальных хет на 6-м и 7-м сегментах брюшка встречаются
очень длинные; претергальные группы обоих сегментов развитые, на 6-м сегменте
состоят из 33-^44, на 7-м — из 8-^42 хет (рис. 11,5).......2. Macroplea
5(4). Лоб с 8 щетинками; щетинки наличника утолщенные, с каждой стороны по-
парно сближены на расстояние двух диаметров их оснований; щетинки верхней
губы утолщенные; очень длинные тергальные хеты имеются только на 7-м сегмен-
те брюшка, хеты 6-го сегмента не длиннее, чем на предыдущих сегментах; претер-
гальные группы 6-го и 7-го сегментов редуцированы, на 6-м сегменте состоят из 15,
на 7-м — из 13 хет (рис. 11,9). Передний край верхней губы со слабой широкой
выемкой между средними маргинальными хетами обеих сторон; на дне выемки
может быть маленький округлый выступ; молярный край мандибулы с 2, внутрен-
ний зубец — с 1 зубчиком. Личинки на корнях водных растений, преимущественно
на Potamogeton и Myriophillum. 1 вид — N. voronovae L. Medvedev в Забайкалье .
........................................................3. Neohaemonia
6(1). Лоб узкий: его ширина между боковыми углами меньше длины (рис. 8, 2, 3),
верхняя губа спереди обычно закруглена, без боковых углов (рис. 10, 2-4): на мак-
силлах отросток лацинии короткий и широкий (рис. 8, 5)..4. Plateumaris
1. Род Donacia Fabricius
Личинки Donacia связаны обычно с озерами, прудами, старицами, медленно
текущими реками, иногда с очень небольшими водоемами, но, так или иначе, со
свободной поверхностью воды. В регионе встречается 24 вида, известны личинки
22 наиболее обычных видов. Личинки D. clavareaui Jacobson (Narita, 1991) и D.
provosti Fairmaire (Chang, 1965) не включены, поскольку их описания не содержат
признаков, используемых в ключе.
Таблица для определения видов рода Donacia
1(20). Посттергальная группа 2-4-го сегментов брюшка узкая, образована 1-2 ряда-
ми хет с отдельными дополнительными хетами посередине и на боках, ее ширина в
3-10 раз меньше расстояния до внутренней претергальной группы (рис. И, 7, 3, 4).
2(3). Перепончатые части тазика и бедра густо покрыты шипиками микроскульпту-
ры (рис. 8, 7 7), как и нижняя сторона тела. Верхняя губа с 18 хетами, так как угло-
вые хеты удвоены (рис. 9,4), живые личинки светло-зеленые. На Butomus, Scolochloa.
ЕЧ, СК, СБ...........................................D. tomentosa Ahrens
3(2). Перепончатая часть тазика без шипиков или с немногими шипиками, бедро
без шипиков (рис. 9, 7); микроскульптура нижней стороны тела менее явственная.
Верхняя губа с 16 хетами (рис. 10, 5, 7, 5, 10). Живые личинки белые или бледно-
кремовые.
D. cinerea; 15-16—прементум и ментальный склерит: 75—D. semicuprea, 16—D. thalassina;
17—19 — нога: 77 — D. tomentosa, 18 — D. impressa, 19 — D. cinerea. ЛГ — лопасть галеи,
MC — ментальный склерит, ОЛ — отросток лацинии. По Беньковскому.
35
Рис. 9. Личинки Donaciinae:
1-3 — нога: 1 — Donacia thalassina, 2—D. vulgaris, 3—D. simplex; 4-13 — верхняя губа: 4 —
Donacia tomentosa, 5 — D. versicolorea, 6 — Macroplea appendiculala; 7 — Donacia dentata,
36
4(5). Внутренняя претергальная группа 1-4-го сегментов брюшка широкая, образо-
вана 3 рядами хет с дополнительными хетами или 4 рядами хет (рис. 11,3); на 1—4-
м сегментах брюшка наружные претергальные группы образованы 34 рядами хет,
наружные и внутренняя претергальные разделены, с 1-3 промежуточными хетами
вдоль заднего края; тергальные группы средне-, заднегруди и 1-6-го сегментов
брюшка образованы короткими одинаковыми хетами, среди которых на 5-м и 6-м
сегментах находятся единичные более длинные хеты. На Carex rostrata. Транспале-
аркт....................................................D. obscura Gyllenhal
5(4). Внутренняя претергальная группа 1—4-го сегментов брюшка узкая, образова-
на 2-3 неправильными рядами хет (рис. 11,7, 4).
6(7). Срединная интеркалярная группа хет средне- и заднегруди многочисленная,
образована 5-10 хетами (рис. 10, 70), на 1-4-м сегментах брюшка наружные и внут-
ренняя претергальные группы разделены, с отдельными промежуточными хатами,
образованы 2-3 рядами хет, посттергальная образована 1 рядом с дополнительны-
ми хетами спереди и сзади или 2 рядами хет; тергальные группы средне-, заднегру-
ди и 1-6-го сегментов брюшка образованы короткими, одинаковыми хетами, среди
которых на 5-м и 6-м сегментах находятся отдельные более длинные хеты. На Spar-
ganium angustifolium, Myriophillum apicatum. ЕЧ, СБ, ДВ.D. sparganii Ahrens
7(6). Срединная интеркалярная группа хет средне- и заднегруди отсутствует или
малочисленная, образована 1-3 хетами (рис. 10, 6, 7).
8(11). Тергальные группы средне- и заднегруди и 1-3-го сегментов брюшка кроме
обычных коротких хет с более или менее многочисленными очень длинными хетами
(в 3-4 раза длиннее коротких) (рис. 10, 6, 7), посттергальная группа 1-6-го сегмен-
тов брюшка посредине образована 1 рядом хет с отдельными дополнительными
хетами.
9(10). Все маргинальные хеты верхней губы сближены (рис. 9, 5); на средне- и зад-
негруди внутренняя посттергальная группа образована 1 рядом с отдельными до-
полнительными хетами; передняя стернальная группа более широкая, образована
посредине 3-^4, а на сторонах 2-3 (реже всюду 2) рядами хет (рис. 10, 7): ни 1-м
сегменте брюшка наружные и внутренняя претергальные группы широко разделе-
ны, на 2-^4-м сегментах — с отдельными хетами в промежутке между ними или
соединены. На Potamogeton natans, Typha lalifolia. ЕЧ, СК, СБ, ДВ.......
.....................................................D. versicolorea Brahm
10(9). Наружная маргинальная хета верхней губы немного отодвинута от 2 других
(рис. 9, 7); на средне- и заднегруди внутренняя посттергальная группа образована 2
рядами хет: передняя стернальная группа более узкая, образована 1-2 (реже 3) ря-
дами хет (рис. 10, б); на 1-4-м сегментах брюшка наружные и внутренняя претер-
гальные группы разделены, с отдельными хетами в промежутке между ними — или
соединены; наружные и внутренняя претергальная группы образованы 2 рядами
хет, посттергальная группа посредине с 1 рядом и дополнительными хетами, на
сторонах с 2 рядами хет. На Sagittaria, Alisma, Sparganium, Typha, Carex. ЕЧ, СБ ..
.........................................................D. dentata Hoppe
8 — D.fennica,9— D. crassipes, 10 — D. thialassina, 11 —D. cinerea, 12 — D. impressa; 13 —
D. bicolora. Хеты верхней губы: a — проксимальная, б — боковая, в — дистальная, г —
угловая, д — медиальная, с — маргинальные. По Беньковскому.
37
Рис. 10. Личинки Donaciinae:
1-5 — верхняя губа: 1 — Donacia marginata, 2 — Plateumaris weisei, 3 — P. discolor, 4 — P
affinis, 5 — P braccata; 6-10 — среднегрудь: 6 — Donacia dentata, 7 — D. versicolorea, 8 —
D. cinerea, 9 — D. crassipes, 10 — D. sparganii. D-D и V-V — срединные линии верхней и
38
11(8). Тергальные группы средне-, заднегруди и 1-3-го сегментов брюшка образо-
ваны короткими хетами, если среди них встречаются единичные более длинные
хеты, тогда посттергальная группа 1-6-го сегментов брюшка посредине образована
2-3 рядами хет.
12(13). Наружные и внутренняя претергальные группы 1-4-го сегментов брюшка
соединены (рис. 11, 4) на 1-4-м сегментах брюшка наружные претергальные груп-
пы образованы 2 рядами с дополнительными хетами пли 3 рядами хет; внутренняя
претергальная — 3 рядами с дополнительными хетами; посттергальная — 2-3 ря-
дами хет, ее ширина в 3-5 раз меньше расстояния до внутренней претергальной
группы; тергальные группы средне-, заднегруди и 1-6-го сегментов брюшка обра-
зованы короткими хетами, среди которых находятся отдельные длинные. На Carex,
Sparganium, Juncus, Eleocharis, Potamogeton, Alisma, Typha. Транспалеаркт.
....................................................D. aquatica Linnaeus
13(12). Наружные и внутренняя претергальные группы 1-4-го сегментов брюшка
широко разделены (иногда в промежутке между ними с отдельными хетами, сто-
ящими в 1 ряд вдоль заднего края) (рис. 11, 7).
14(15). Ментальный склерит всюду широкий, посредине почти не расширен (рис.
8, 75). На Glyceria, Carex. ЕЧ, СБ..................D. semicuprea Panzer
15(14). Ментальный склерит на концах узкий, посредине немного расширен (рис.
8, 76).
16(17). Молярный край мандибулы прямой или волнистый, но без явственных зуб-
цов или большого треугольного выступа (рис. 8, 77); на 1—4-м сегментах брюшка
наружные претергальные группы образованы 2 рядами с дополнительными хета-
ми; внутренняя претергальная — 2-3 рядами; посттергальная — 1 рядом с допол-
нительными хетами или 2 рядами хет, ее ширина в 8-9 раз меньше расстояния до
внутренней претергальной группы; тергальные группы средне-, заднегруди и 1-6-
го сегментов брюшка образованы короткими хетами, среди которых на 4—6-м сег-
ментах находятся длинные. На Phragmites, Glyceria. Транспалеаркт..........
....................................................D. clavipes Fabricius
17(16). Молярный край мандибулы с 2 зубчиками или большим треугольным выс-
тупом (рис. 8, 8, 9).
18(19). Передний край верхней губы с явственной выемкой (рис. 9, 70); на 1-4-м
сегментах брюшка наружные претергальные и внутренняя претергальная группы
образованы 2-3; посттергальная — 1 рядом с дополнительными хетами или 2 ряда-
ми хет, ее ширина в 6-16 раз меньше расстояния до внутренней претергальной груп-
пы; тергальные группы средне-, заднегруди и 1-6-го сегментов брюшка образова-
ны короткими хетами. На Eleocharis, Carex. Транспалеаркт .. D. thalassina Germar
19(18). Передний край верхней губы прямой или с очень слабой широкой выемкой
(рис. 9, 5); на 1-4-м сегментах брюшка наружные претергальные и внутренняя пре-
тергальная группы образованы 2 рядами с дополнительными хетами; посттергаль-
ная — 1 рядом с дополнительными хетами посредине и на сторонах или 2 рядами
хет, ее ширина в 5-6 раз меньше расстояния до внутренней претергальной группы;
нижнем сторон. Группы хет среднегруди: а — внутренняя претергальная, б — наружная пре-
тергальная, в — внутренняя посттергальная, г — наружная посттергальная, д — боковая
интеркалярная, е — срединная интеркалярная, ж — передняя стернальная. По Беньковскому.
39
тергальные группы средне-, заднегруди и 1-6-го сегментов брюшка образованы
короткими хетами, среди которых на 4—6-м сегментах брюшка находятся длинные
(иногда также единичные длинные хеты на 1-3-м сегментах). На Scolochloa. ЕЧ,
СБ.......................................................D. fennica Paykull
20(1). Посттергальная группа 1—4-го сегментов брюшка широкая, образована За-
рядами хет, ее ширина не более чем вдвое меньше расстояния до внутренней пре-
тергальной группы (рис. 11,5, 6).
21(22). Срединная интеркалярная группа средне- и заднегруди многочисленная,
соединяется с боковыми интеркалярными группами и образована 8-14 хетами (рис.
10, 9); на 1-4-м сегментах брюшка наружные претергальные и внутренняя претер-
гальная группы образованы 5, посттергальная — 4 рядами хет, ширина последней в
1,3-1,5 раза меньше расстояния до внутренней претергальной группы. На Nuphar,
Nymphaea. ЕЧ, СБ......................................D. crassipes Fabricius
22(21). Срединная интеркалярная группа средне- и заднегруди отсутствует или ма-
лочисленная, образована 1-^4 хетами (рис. 10, 8).
23(26). Молярный край мандибулы волнистый, иногда бываете широким выступом,
но без четких зубцов (рис. 8,14).
24(25). Передний край верхней губы прямой или с очень маленькой узкой выемкой
(рис. 9, 77); перепончатая часть тазика без шипиков микроскульптуры (рис. 8,19);
на 1—4-м сегментах брюшка наружные претергальные и внутренняя претергальная
группы разделены, с единичными хетами в промежутке между ними вдоль заднего
края, наружные образованы 3-^4, внутренняя — 4, посттергальная — 3 рядами хет,
ширина последней не более чем в 1,8 раза меньше расстояния до внутренней пре-
тергальной группы; тергальные группы средне-, заднегруди и 1-7-го сегментов
брюшка образованы короткими хетами, среди которых на 6-7-м сегментах брюшка
находятся длинные хеты (подрод Donaciella Reitter). На Typha, Carex. ЕЧ, СК, СБ
.........................................................D. cinerea Herbst
25(24). Передний край верхней губы выемчатый (рис. 9, 72). Перепончатая часть
тазика спереди с шипиками микроскульптуры (рис. 8,75); на 1-4-м сегментах брюш-
ка наружные претергальные и внутренняя претергальная группы разделены, иног-
да с отдельными хетами в промежутке между ними, или соединены узкой полоской
хет вдоль заднего края, наружные образованы 3—4, внутренняя — 3-5, посттергаль-
ная — 3 рядами хет, ширина последней в 1,7 раза меньше расстояния до внутрен-
ней претергальной группы; тергальные группы средне-, заднегруди и 1-7-го сег-
ментов брюшка образованы короткими хетами, среди которых на 6-7-м сегментах
брюшка находятся отдельные длинные хеты. На Scirpus. ЕЧ, СБ..............
........................................................D. impressa Paykull
26(23). Молярный край мандибулы с 2-3 (редко только с 1) четкими зубчиками,
иногда также имеется 1 зубчик на внутреннем зубце (рис. 8, 72, 13).
27(30). Молярный край мандибулы с 3 зубчиками, внутренний зубец с 1 зубчиком
(рис. 8,13).
28(29). Посттергальная группа 2-6-го сегментов брюшка образована 4 рядами хет
(рис. 11,5); дистальные хеты верхней губы короче медиальных (рис. 9, 73); на 1-4-
м сегментах брюшка наружные претергальные и внутренняя претергальная группы
разделены, иногда с единичными хетами в промежутке между ними, образованы 4
рядами хет, ширина посттергальной группы в 1,3 раза меньше расстояния до внут-
40
Рис. 11. Личинки Donaciinae:
1-7 — второй сегмент брюшка: 1 — Donacia thalassina, 2 — Macroplea appendiculata, 3 —
Donacia obscura, 4 — D. aquatica, 5 — D. bicolor, 6—D. marginata, 7 — Plateumaris discolor,
8,9—6-й и 7-й сегменты брюшка: 8—Macroplea appendiculata, 9—Neohaemonia voronovae.
D-D и V-V — срединные линии верхней и нижней сторон. Группы хет второго сегмента
брюшка: а — внутренняя претергальная, б — наружная претергальная, в — посттергальная,
г — стернальная, д — педальная. По Беньковскому.
41
ренней претергальной группы; перепончатая часть тазика спереди с шипиками; тер-
гальные группы средне-, заднегруди и 1-7-го сегментов брюшка образованы корот-
кими хетами, среди которых на 6-7-м сегментах брюшка находятся отдельные длин-
ные хеты. На Sparganium, Carex. ЕЧ, СБ................D. bicolora Zschach
29(28). Посттергальная группа 1-6-го сегментов брюшка образована 3 рядами хет
(рис. 11,6); дистальные и медиальные хеты верхней губы равной или почти равной
длины (рис. 10, 7); на 1-4-м сегментах брюшка наружные претергальные и внут-
ренняя претергальная группы разделены, с 2 хетами в промежутке между ними,
наружные образованы 4, внутренние — 4-5, посттергальная — 3 рядами хет, шири-
на последней в 1,8 раза меньше расстояния до внутренней претергальной группы;
перепончатая часть тазика спереди с шипиками; тергальные группы средне-, зад-
негруди и 1-7-го сегментов брюшка образованы короткими хетами, среди которых
на 5-7-м (иногда также на передних) сегментах брюшка находятся отдельные длин-
ные хеты. На Sparganium, Carex, Scolochloa. ЕЧ, СБ....D. marginata Hoppe
30(27). Молярный край мандибулы с 1-2 зубчиками, внутренний зубец с 1 зубчиком
или без него (рис. 8, 72).
31(32). Тазик с 13-22 хетами, его перепончатая часть спереди с шипиками (рис. 9,
2) на 1-4-м сегментах брюшка наружные претергальные и внутренняя претергаль-
ная группы разделены, иногда с 2 хетами в промежутке между ними, наружные
образованы 3-4, внутренние — 4 рядами хет, посттергальная группа образована 3-
4 рядами хет, ее ширина в 1,6-2,0 раза меньше расстояния до внутренней претер-
гальной группы; тергальные группы средне-, заднегруди и 1-7-го сегментов брюш-
ка образованы короткими хетами, среди которых на 5-7-м сегментах брюшка нахо-
дится несколько более длинных хет. На Typha, Sparganium, Glyceria. Транспалеаркт
......................................................D. vulgaris Zschach
32(31). Тазик с 25-28 хетами, его перепончатая часть без шипиков (рис. 9, 5); на 1-
4-м сегментах брюшка наружные претергальные и внутренняя претергальная груп-
пы разделены, с единичными хетами в промежутке между ними вдоль заднего края,
наружные образованы 3 рядами с дополнительными хетами, внутренние — 3-4,
посттергальная — 3 рядами хет, ширина последней в 1,2 раза меньше расстояния
до внутренней претергальной группы; тергальные группы средне-, заднегруди и 1-
7-го сегментов брюшка образованы короткими хетами, среди которых на 7-м сег-
менте брюшка находится несколько более длинных хет. На Sparganium. Транспале-
аркт .................................................D. simplex Fabricius
2. Род Macroplea Samouelle (Haemonia Dejean)
Личинки Macroplea живут в тех же стациях, что и Donacia, а также по берегам
морей. В регионе 4 вида, известны личинки 2 видов.
Таблица для определения видов рода Macroplea
1(2). Посттергальные группы 1-4-го сегментов брюшка широкие, образованы 2
рядами с дополнительными хетами или 3 рядами хет (рис. 11, 2); молярный край
мандибулы с 2-3 треугольными зубчиками, внутренний зубец с 1 зубчиком (рис. 8,
70), на 1-4-м сегментах брюшка наружные претергальные группы образованы 2-3
42
рядами хет, внутренняя претергальная — 2 рядами с дополнительными хетами,
посттергальная — 2-3 рядами. На Myriophyllum, Potamogeton, Carex. ЕЧ, СБ...
.....................................................М. appendiculata Panzer
2(1). Посттергальные группы 1-4-го сегментов брюшка узкие, образованы 1 рядом
с отдельными дополнительными хетами; на 1-4-м сегментах брюшка наружные и
внутренняя претергальные группы образованы 2 рядами с дополнительными хета-
ми, посттергальная — 1 рядом с дополнительными хетами. внутренняя претергаль-
ная — 2 рядами с дополнительными хетами, посттергальная — 2-3 рядами. На
Zostera, Ruppia, Potamogeton. Транспалеаркт..........М. mutica Fabricius
4. Род Plateumaris Thomson
Личинки обычно связаны с заболоченными берегами водоемов и болотами, не
покрытыми водой. В регионе 10 видов, известны личинки 6 видов. Не включены
личинки Р sericea L. и Р constricticollis Jacoby (Kanasawa, 1985) из-за несовмести-
мости описаний с ключом.
Таблица для определения видов рода Plateumaris
1(2). Наружные претергальные группы 1-6-го сегментов брюшка округлые, обра-
зованы 4-5 рядами хет; число хет в педальных и стернальный группах 4-го, 5-го и
6-го сегментов брюшка: 40 и около 50, 21 и 30, 18 и 9 соответственно; ширина
верхней губы в 1,1 раза больше длины; все маргинальные хеты находятся на рав-
ном расстоянии одна, от другой (рис. 10, 4); мандибулы в 1,3 раза шире длины. На
Carex. ЕЧ.....................................................Р. affinis Kunze
2(1). Наружные претергальные группы 1-6-го сегментов брюшка удлиненные, по-
перечные, образованы 2-3 рядами хет (рис. 11, 7).
3(4). Число хет в педальных и стернальных группах 4-го, 5-го и 6-го сегментов брюш-
ка: 60 и 75, 47 и 50, 30 и 5 соответственно; лоб в 1,4 раза длиннее ширины; наруж-
ная маргинальная хета верхней губы отодвинута от 2 других; дистальные хеты ко-
роче маргинальных (рис. 10, 5). На Phragmites. ЕЧ, СК, СБ.Р braccata Scopoli
4(3). Педальные и стернальные группы 4-го сегмента брюшка содержат меньше
хет: 34-46 и 32-40 хет соответственно, а стернальные группы 6-го сегмента — боль-
ше: 11-17 хет; лоб в 1,2-1,3 раза длиннее ширины (рис. 8, 2, 3).
5(6). Угловые хеты лба приближены к боковым углам (рис. 8, 3); ширина мандибул
в 1,2 раза меньше длины; дистальные хеты верхней губы немного длиннее марги-
нальных (рис. 10,2); число хет в педальных и стернальных группах 4-го, 5-го и 6-го
сегментов брюшка: 34-38 и 40, 34-36 и 26, 19 и 11-15 соответственно. На Carex.
Север ЕЧ, СБ, ДВ............................................Р weisei Duvivier
6(5). Угловые хеты лба отодвинуты далеко назад от боковых углов (рис. 8, 2), ман-
дибулы равной длины и ширины; дистальные хеты верхней губы короче марги-
нальных (рис. 10, 3), число хет в педальных и стернальных группах 4-го, 5-го и 6-го
сегментов брюшка: 46 и 32, 35 и 21, 24-27 и 13-17 соответственно. На Carex. ЕЧ
............................................................Р discolor Panzer
43
2. ПОДСЕМЕЙСТВО MEGALOPODINAE
Краткая характеристика подсемейства составлена по описанию и рисункам
японского вида Colobaspis japonica Baly.
Личинки удлиненные, С-образно изогнутые, белые, с более темными: головой
и последним сегментом брюшка и слегка затемненным склеритом переднеспинки
(рис. 5, 3; 13, 1-2). Голова с хорошо развитым длинным эпикраниальным швом и
неявственными или очень слабыми фронтальными швами (рис. 12, 7). Верхняя губа
и наличник обособлены ото лба, последний с 1-2 парами щетинок. Глазки мелкие,
по 3-5 с каждой стороны. Усики 4-члениковые, с папиллой на 3-м членике. Манди-
булы мощные, с тупым вершинным зубцом и зазубренностью на внутреннем крае
(рис. 12, 2). Челюстные щупики 3-члениковые, губные — 2-члениковые. Переднес-
пинка с дорсальным склеритом, прочие сегменты тела без склеритов, тергиты с
поперечным рядом щетинок (рис. 12, 4). Стигмы щелевидные. Ноги очень корот-
кие, особенно голенелапки, которые значительно уже предыдущих члеников (рис.
12, 3).
Личинки, по данным Монроза (Monros, 1954), обитают внутри стеблей расте-
ний. В регионе два рода и два вида на юге ДВ; более обычна Clytraxeloma cyanipennis
Kraatz, живущая на Maackia amurensis.
44
Рис. 13. Детали строения личинок подсемейств Megalopodinae, Zeugophorinae, Orso-
dacninae:
1-4 — общий вид: 7, 2 — Colobaspis japonica, 3 — Zeugophora annulata, 4 — Orsodacne
cerasi; 5 — верхняя губа Zeugophora turneri; 6, 7 — вершина брюшка Orsodacne'. 6 — О.
lineola, 7 — О. cerasi. По Кимото и Такизава, Коксу, Дубешко.
Рис. 12. Детали строения личинок подсемейств Megalopodinae и Synetinae:
1-4 — Megalopus'. 1 — голова, 2 — мандибула, 3 — нога, 4 — вершина брюшка; 5-9 —
Syneta betulae'. 5 — голова, 6 — усик, 7 — нижняя губа, 8 — вершина брюшка сверху, 9 — то
же, снизу. По Курчевой и Монрозу.
45
3. подсемейство zeugophorinae
Тело продолговатое, сильно уплощенное дорсовентрально, с угловидно высту-
пающими наружу боками сегментов (рис. 5, 2). Голова плоская, прогнатическая,
вдвое шире длины, позади середины округло-расширенная, без эпикраниального
шва, лобный склерит неправильно трапециевидный, доходит до затылочного от-
верстия, вследствие чего обе половины темени не соприкасаются друг с другом
(рис. 14, 7). По бокам головы имеется несколько длинных щетинок. Глазков по 2 с
каждой стороны. Усики 3-члениковые. Наличник и верхняя губа обособлены, на-
личник поперечный, без щетинок, но с порой с каждой стороны, верхняя губа попе-
речная, с округленным передним краем, усаженным плоскими, к вершине расши-
ренными хетами (рис. 14, 3). Галея и лациния нижних челюстей образуют жева-
тельную лопасть, последняя узкая, с 3 расширенными на концах плоскими хетами
на вершине и с усаженным очень мелкими шипиками внутренним краем (рис. 14,
6), челюстные щупики 3-члениковые (рис. 14, 2). Прементум, ментум и субментум
слиты вместе, нижнегубные щупики 1-члениковые, тесно придвинутые друг к дру-
гу, язычок смещен на дорсальную сторону нижней губы, при осмотре снизу незаме-
тен, так как прикрыт нижнегубными щупиками. Мандибулы с удлиненной, упло-
щенной, разделенной на 2-4 зубца вершинной частью, без щетинок на наружном
крае. Ноги отсутствуют, на их месте на вентральной стороне грудных сегментов
имеются округленные, слабо хитинизированные небольшие склериты. Тергиты и
стерниты брюшка посередине с поперечной изогнутой складкой, усаженной по зад-
нему и боковым краям очень мелкими шипиками (рис. 14, 77), помогающими дви-
жению личинки. Длинные щетинки на тергитах брюшка лишь по бокам, обычно по
2-3 с каждой стороны. Брюшко из 10 сегментов: 1-8-й сегменты несут стигмы, 10-й
сегмент очень маленький. Стигмы поперечные, с двойной щелью на внутреннем
крае. Длина взрослой личинки 5-8 мм, окраска светло-желтая, голова рыжая.
Яйца откладываются весной поодиночке на листья кормовых растений, они
продолговато-овальные, желтоватые, 0,6-0,8 мм длиной. Личинки выгрызают в
пластинке листа широкие пятнообразные мины, хорошо заметные вследствие по-
чернения отмирающей кожицы верхней стороны листа над миной. В каждой мине
находится от 1 до 8 личинок. По окончании периода питания, обычно во второй
половине лета, личинки уходят в почву; диапаузирующие личинки зимуют и окук-
ливаются весной следующего года. Куколка свободная, желтоватая. Взрослые жуки
появляются в начале лета. На яйцах и личинках паразитируют перепончатокрылые
семейства Mymaridae.
Подсемейство представлено одним родом Zeugophora Kunze с 9 видами.
Таблица для определения видов рода Zeugophora
1(6). Мандибулы с 2-3 зубцами. Передний край верхней губы с 10-14 длинными
хетами (рис. 14, 4, 5). На ивовых (подрод Zeugophora s.str.).
2(3). Мандибулы с 3 зубцами (рис. 14, 5). На тополе, осине, иве. ЕЧ, СБ.
..................................................Z. flavicollis Marsham
3(2). Мандибулы с 2 зубцами (рис. 14, 7, 9). На тополе, осине, иве.
46
4(5). Верхняя губа с 2 парами щетинок и 2 порами близ переднего края и с парой
плоских хет перед ними (рис. 13, 5). ЕЧ, СБ.................Z. turneri Power
5(4). Верхняя губа с 3 парами щетинок близ переднего края, все плоские хеты ле-
жат только на переднем крае (рис. 14, 4, 5). ЕЧ, СК, СБ (Z. subspinosa Fabricius
также ДВ).........................Z. subspinosa Fabricius, Z. scutellaris Suffrian
6(1). Мандибулы с 4 хорошо развитыми зубцами (рис. 14,10). Передний край верх-
ней губы с многочисленными (не менее 40) короткими хетами (рис. 14, 3). На бе-
ресклете (подрод Pedrillia Westwood). ДВ.........Z. annulata Baly (рис. 13, 3).
Рис. 14. Детали строения личинок рода Zeugophora.
Z. scutellaris: 1 — голова, 4 — верхняя губа, 9 — мандибула; Z. annulata: 2 — лабио-максил-
лярный комплекс, 3 — верхняя губа, 10 — мандибула; Z. flavicollis: 5 — верхняя губа, 6 —
максилла, 8 — мандибула, 11 — брюшной тергит; Z. subspinosa: 7 — мандибула.
47
4. ПОДСЕМЕЙСТВО orsodacninae
Тело удлиненное, параллелестороннее, уплощенное, белое со слабо склероти-
зованными светлобурыми головой, переднеспинкой и 9-м тергитом (рис. 13, 4).
Голова не втянута в переднеспинку, эпикраниальный шов очень короткий, лобные
швы сходятся почти у заднего края головы, щетинки хорошо развиты, многие из
них длинные. 1 глазок с каждой стороны. Фронтоклипеальный шов неявственный.
Мандибулы с 2 крупными апикальными и 3-4 небольшими субапикальными зуб-
цами, максиллы с широкой жевательной лопастью, челюстные щупики 3-членико-
вые. Нижняя губа с широким прямоугольным ментумом и более узким пременту-
мом, губные щупики 2-члениковые. Язычок прямоугольный, с 2 вершинными ще-
тинками и парой сенсилл между ними. Усики короткие, 3-члениковые. Склерит
переднеспинки со щетинками по всей поверхности. Тергиты средне-, заднегруди и
1-8-го сегментов брюшка со щетинками и многочисленными гранулами, стигмы с
двойным отверстием, 9-й брюшной тергит с густыми порами, несет на вершине
урогомфы, 10-й сегмент сдвинут вентрально. Ноги хорошо развиты, бедра доволь-
но длинные, с 4 длинными и 5 короткими щетинками, тибиотарзус с 4 длинными
щетинками, коготки тонкие и длинные, слегка изогнутые. Известны личинки 1-го
возраста, выведенные из яиц, их образ жизни не известен, возможно, на корнях. В
регионе 1 род Orsodacne Latreille, с 2 видами.
Таблица для определения видов рода Orsodacne
1(2). Урогомфы короче, 0,03-0,04 мм (рис. 13, 6). Ширина головной капсулы около
0,3 мм. ЕЧ..........................................О. lineola Panzer
2(1). Урогомфы длиннее, 0,05-0,08 мм (рис. 13, 7). Ширина головной капсулы око-
ло 0,26 мм. ЕЧ, СК, СБ.............................О. cerasi Linnaeus
48
5. ПОДСЕМЕЙСТВО criocerinae
Тело удлиненное или короткое, сверху, особенно в задней половине, выпуклое
(рис. 15, 7), у живых личинок большей частью покрыто слизистыми выделениями
желез и чехликом из жидких экскрементов. Голова с каждой стороны с 6 глазками,
эпикраниальный шов длинный, лобные швы прямые или изогнутые, темя с 8 ще-
тинками, лоб с 6-8 щетинками и 2-4 порами, наличник поперечный, в основании
сильно хитинизированный, с 4 щетинками и 2 порами (рис. 16); верхняя губа (рис.
17, 7) с глубокой треугольной выемкой на переднем крае, последний умеренно вы-
пуклый или прямой, иногда зазубрен, с 2-7 маргинальными щетинками с каждой
стороны, с 6 дискальными щетинками и 2 порами у основания. Мандибулы с 3
хорошо развитыми и 2 редуцированными зубцами, нижнечелюстные щупики 3-
члениковые, кардо нижних челюстей снаружи с 1 щетинкой, галея с 8-12 сравни-
тельно длинными щетинками, лациния редуцирована; нижнегубные щупики одно-
члениковые. Переднеспинка с эпиплевральными склеритами, несущими по 2 ще-
тинки, тергальные склериты груди и брюшка часто разбиты на большое количество
мелких вторичных склеритов (склеритовидных площадок), несущих по 1 щетинке
и расположенных поперечными рядами (рис. 18), иногда плохо различимых; ще-
тинки на склеритах короткие или отсутствуют. Стерниты 1-8-го сегментов брюшка
со сближенными парными двигательными буграми, покрытыми чрезвычайно мел-
кими шипиками, облегчающими передвижение личинки. Анальное отверстие в виде
поперечной щели, находится на дорсальной стороне тела (рис. 15, 4), что, очевид-
но, связано со способностью покрывать спинную поверхность тела полужидкими
экскрементами. Стигмы продолговатые, с двойным отверстием (рис. 15, 3). Когот-
ки с небольшим хелониумом.
Для Criocerinae характерны одинаковые условия обитания личиночной и има-
гинальной фаз. Перезимовавшие жуки откладывают весной на растения продолго-
ватые, довольно крупные (0,75-1 мм) яйца, одиночные или по нескольку штук. Раз-
витие яйца длится 12-14 дней, вышедшие личинки скелетируют листья, оставляя
нетронутой, в отличие от взрослых жуков, нижнюю пленку эпидермиса, у некото-
рых видов личинки живут внутри плодов спаржи, развитие личинки проходит за 2-
4 недели. Выросшие личинки покидают кормовое растение и уходят в почву, где
строят кокон из частиц субстрата и выделяемого личинкой пенистого, легко засты-
вающего вещества, в котором происходит окукливание. Стадия куколки длится обыч-
но 3-4 недели, молодой жук не покидает кокона до полного окрашивания, а иногда
и зимует в нем. В год обычно развивается одно поколение.
В регионе четыре рода.
Таблица для определения родов подсемейства Criocerinae
1(2). Тело сильно удлиненное, его длина более чем в 3 раза превышает максималь-
ную высоту, за серединой слабо выпуклое (рис. 15,2). Щетинки на верхней стороне
тела, особенно на тергитах брюшка, неявственные или отсутствуют. Передний край
верхней губы по бокам от срединной выемки без выемок и зазубрин и с 2 марги-
нальными щетинками с каждой стороны (рис. 17, 7). Коготки крючкообразные, при
49
Рис. 15. Общий вид (7, 2) и детали строения личинок подсемейства Criocerinae:
1 — Lilioceris] 2 — Crioceris] 3 — стигма Lilioceris merdigera] 4 — вершина брюшка Oulema
lichenis. По Хенриксену и ориг.
основании подковообразно изогнуты (рис. 17, 77). Личинки не покрывают тело эк-
скрементами. На Asparagus.................................... 1. Crioceris
2(1). Тело умеренно удлиненное, сильно выпуклое за серединой, его длина в 2-2,5
раза превышает максимальную высоту (рис. 15, 7). Щетинки верхней стороны тела
явственные. Коготки серповидные, у основания не изогнуты. Тело личинки покры-
то экскрементами.
3(4). Средне- и заднеспинка либо с крупными неразделенными посттергальными
склеритами (рис. 18, 7), либо они разбиты на многочисленные склеритовидные
площадки, не образующие правильного поперечного ряда (рис. 18, 2). Передний
край верхней губы закругленный, с широко округленными наружными углами, не
зазубренный или едва зазубрен. Лоб с 8 щетинками и 4-6 порами. На лилейных и
диоскорейных.................................................2. Lilioceris
4(3). Средне- и заднеспинка с 1 правильным поперечным рядом мелких склерито-
видных площадок, соответствующих посттергальным склеритам (рис. 18, 4).
5(6). Передний край верхней губы округленный, с широко закругленными наруж-
ными углами, несущими 6 щетинок и очень слабые плохо заметные зазубрины. Лоб
с 8 щетинками и 2 порами. На сложноцветных................ 3. Lema (s.str.)
6(5). Передний край верхней губы прямой или слабо округленный, наружные углы
образуют явственный перегиб или тупой угол и несут не более 3 щетинок.
7(8). Передний край верхней губы без зазубрин, совершенно прямой (рис. 17, 6).
Лоб с 6 щетинками и 2 порами. На пасленовых ..3. Lema (подрод Microlema Pic)
8(7). Передний край верхней губы с явственными зазубринами с каждой стороны
от срединной выемки, прямой или слабо округленный (рис. 17, 7). Лоб с 6-8 щетин-
ками и 2 порами. На злаках...................................4. Oulema
50
Рис. 16. Головные капсулы личинок подсемейства Criocerinae:
1 —Lilioceris rugata; 2 — Crioceris sp.; 3 —Lema decempunctata\ 4 — Oulema oryzae.
1. Род Crioceris Muller
Верхняя губа с 6 щетинками на диске и треугольной вырезкой на переднем
крае, без зазубрин; с 4-6 крупными переднекрайними щетинками и 3 микрохетами
по бокам вырезки (рис. 17, 7). Основание наличника сильно хитинизировано, с 4
макрохетами и 2 порами. Лоб с 8 щетинками и 2 порами. Лобные швы в срединной
части умеренно выпуклые (рис. 16, 2). Крыловые склериты среднеспинки с 2-3
плохо заметными щетинками, щетинки верха короткие, с закругленными вершина-
ми, на брюшных тергитах слабо различимые, склериты между основаниями ног
слабо выражены. Коготки крючковидно загнутые, в основании сильно расширен-
ные, образующие узкий и длинный пяточный выступ (рис. 17, 77). Тело удлинен-
ное, в задней половине незначительно утолщенное (рис. 15, 2). Личинки обычно не
покрывают тело чехликом из экскрементов, живут на листьях или внутри плодов
спаржи. Яйца темно-оливковые или зеленоватые, откладываются поодиночке или
по 3-4 на листья и плоды спаржи.
51
Рис. 17. Верхняя губа (1-10), коготок (11-13) и микроскульптура покровов тела (14,
15) личинок подсемейства Criocerinae:
1 — Crioceris sp.; 2 — Lilioceris rugata; 3 — L. scapularis\ 4 — L. lilii', 5 — L. merdigercr, 6 —
Lema decempunctatcr, 7 — Oulema oryzae; 8 — O. gallaeciana; 9 — O. dilutipes; 10 — O.
melanopus; 11 — Crioceris', 12 — Lilioceris', 13 — Oulema', 14 — O. oryzae', 15 — O, dilutipes.
52
В регионе 6 видов, преимущественно в южных районах.
Таблица для определения видов рода Crioceris
1(2). Брюшко по бокам с рядом крупных, явственных и хорошо отграниченных бу-
горков, лежащих непосредственно под стигмами. Окраска тела сероватая, оливко-
вая или зеленовато-желтая, длина тела взрослой личинки 8,5-10 мм. На листьях
Asparagus. Юг ЕЧ, СК..................................С asparagi Linnaeus
2(1). Брюшко по бокам с продольным рядом небольших, мало заметных и плохо
отграниченных бугорков. Окраска тела беловатая, длина тела взрослой личинки 8-
9,5 мм.
3(4). Голова желтая. Передний край верхней губы прямой, с узкой срединной выем-
кой и с 6 переднекрайними щетинками. В плодах спаржи. Транспалеаркт.....
................С. duodecimpunctata Linnaeus
4(3). Голова темно-бурая. Передний край верхней губы закругленный, с широкой
треугольной выемкой и с 4 переднекрайними щетинками (рис. 17, 7). На листьях
спаржи. Транспалеаркт.........................С. quatuordecimpunctata Scopoli
2. Род Lilioceris Reitter
Передний край верхней губы умеренно закруглен, с треугольной срединной
вырезкой, по бокам от которой с каждой стороны имеются 4-7 маргинальных ще-
тинок. Лоб с 8 щетинками и 4 порами (рис. 16, 7). Покров тела с очень хорошо
заметной микроскульптурой, исчезающей на складках сегментов и вокруг склери-
тов. Средне- и заднеспинка и тергиты 1-7-го сегментов брюшка с 6-8 мелкими
претергальными склеритами (рис. 18, 7), щетинки верхней стороны тела короткие,
но хорошо заметные, все тергальные склериты брюшка с 1, эпиплевральные — с
2-5 и гипоплевральные — с 3 щетинками. Коготки изогнуты, у основания не рас-
ширены, без выступа (рис. 17, 72). Склериты между основаниями ног обычно при-
сутствуют. Тело сильно выпуклое в задней половине (рис. 15, 7), покрыто сверху
экскрементами. Живут открыто на листьях лилейных, яйца оранжевые, кладутся
по 3-6 штук на нижнюю сторону листа.
В регионе 6 видов; неизвестны личинки L. ruflcollis Baly (юг ДВ, на Smilax и
Dioscorea) и L. faldermanni Guerin (СК, на Lilium).
Таблица для определения видов рода Lilioceris
1(4). Посттергальные склериты средне- и заднегруди крупные, хорошо выражен-
ные (рис. 18, 7). Микроскульптура брюшных покровов более густая.
2(3). Передний край верхней губы с 6 щетинками. Поры у основания губы раздви-
нутые (рис. 17, 2). На Dioscorea. Юг ДВ.....................L. rugata Baly
3(2). Передний край верхней губы с 14 щетинками. Поры у основания верхней губы
сближенные (рис. 17, 3). На Dioscorea. Юг ДВ.............L. scapularis Baly
4(1). Посттергальные склериты средне- и заднегруди разбиты на многочисленные
мелкие склеритовидные площадки (рис. 18, 2, 3). Микроскульптура брюшных по-
кровов более редкая.
53
Рис. 18. Грудь (1-3) и заднеспинка (4, 5) личинок родов Lilioceris и Oulema'.
1 — L. rugata; 2 — L. lilii; 3 — L. merdigera', 4 — O. dilutipes\ 5 — O. melanopus.
5(6). Передний край верхней губы с 8 щетинками; 6 дискальных щетинок верхней
губы образуют почти правильный поперечный ряд (рис. 17, 4). Непарный стерналь-
ный склерит переднегруди с 2 щетинками. Посттергальные склеритовидные пло-
щадки средне- и заднегруди (рис. 18, 2) несут 9-10 щетинок; имеются 2 претер-
гальные щетинки. Эпиплевральные и крыловые склериты груди темноокрашенные.
На лилейных (Lilium, Fritillaria, реже Convallaria, Scilla). Транспалеаркт.
..........................................................L. lilii Scopoli
6(5). Передний край верхней губы с 10 щетинками; 6 дискальных щетинок верхней
губы не образуют правильного ряда (рис. 17, 5). Непарный стернальный склерит
переднегруди с 4 щетинками. Посттергальные склеритовидные площадки средне-
54
и заднегруди (рис. 18, 3) несут около 15 щетинок; претергальные щетинки отсут-
ствуют. Эпиплевральные и крыловые склериты груди не отличимы по окраске от
тела. На лилейных (Lilium, Convallaria), иногда на луке и чесноке. Транспалеаркт
................................................... L. merdlgera Linnaeus
3. Род Lema Lacordaire
У номинативного подрода темя с 2 щетинками и 1 порой с каждой стороны,
лоб с 8 щетинками и 2 порами, наличник поперечный, с 4 щетинками и 2 порами;
основание и бока верхней губы (рис. 17, 6) умеренно закругленные, наружные углы
слабо зазубренные, с 4 щетинками, передний край с треугольной срединной выем-
кой, боковые дискальные щетинки сдвинуты к переднему краю, 2 поры у основа-
ния сближены. Границы между лбом, наличником и верхней губой четкие, щетин-
ки на склеротизованных участках не развиты. Коготки изогнуты, у основания не
расширены, без выступа. Подрод Microlema Pic характеризуется следующими при-
знаками: темя с 1 щетинкой и порой с каждой стороны, лоб с 6 щетинками и 2
порами, наличник в основании сильно хитинизированный, граница со лбом слабо
выражена (рис. 16, 3), бока и передний край верхней губы прямые, без зазубрин, с
глубокой треугольной выемкой посередине и с 2 щетинками по бокам; диск с 3
парами длинных щетинок и 2 порами (рис. 17, 6).
В регионе 4 вида, из них 2 только на юге ДВ.
Таблица для определения видов рода Lema
1(4). Передний край верхней губы округленный, с глубокой срединной выемкой и
боками (подрод Lema s.str.).
2(3). Голова, переднеспинка и ноги темнобурые. На Cirsium. Транспалеаркт.
......................................................L. cyanella Linnaeus
3(2). Голова, переднеспинка и ноги желтобурые. На Commelina. Юг ДВ.......
...................................................L. diversa Baly
4(1). Передний край верхней губы прямой, с неглубокой срединной выемкой и с 2
сенсорными щетинками по бокам (рис. 17, 6) (подрод Microlema). На пасленовых
(Lycium). Алтай, ДВ................................L. decempunctata Gebler
4. Род Oulema Gozis
Темя с каждой стороны с 2-3 щетинками и 2 порами, лобные швы умеренно
выпуклые, лоб с 6-8 щетинками и 2 порами, наличник поперечный, с 4 щетинками
и 2 порами (рис. 16, 4). Передний край верхней губы прямой, с узкой срединной
вырезкой и 2 зазубринами по бокам, несущими щетинки; поры у основания губы
расставленные (рис. 17, 7-10). Границы между лбом, наличником и верхней губой
нечеткие. Склерит переднеспинки с рядом щетинок по переднему и заднему краям.
Коготки изогнуты, у основания без расширения и выступа (рис. 17,13). Тело с мик-
роскульптурой и мелкими щетинками, сверху покрыто полужидкими экскремента-
ми. Личинки скелетируют листья злаков, выедая в них длинные узкие полосы. В
регионе 9 видов.
55
Таблица для определения видов рода Oulema
1(2). Тело с очень густой микроскульптурой и густыми щетинками, особенно на
тергитах брюшка (рис. 17, 14). Верхняя губа с кососрезанным ковнутри передним
краем и узкой глубокой вырезкой, без зазубрин. На злаках. Юг ДВ.О. oryzae
Kuwayama (О. suvorovi Jacobson)
2(1). Тело с редкой микроскульптурой и негустыми щетинками (рис. 17, 75). Верх-
няя губа с закругленным передним краем и зазубринами по бокам от срединной
выемки (рис. 17, 8-10).
3(4). Щетинки склерита переднеспинки расположены в основном по переднему
краю. Верхняя губа на диске с 8 микрохетами (рис. 17, 8). На дикорастущих, реже
культурных злаках. Повсеместно, кроме крайнего севера. Широко распространен-
ные в Палеарктике виды..................................................
.................О. gallaeciana Heyden (О. lichenis Voet), О. erichsoni Suffrian
4(3). Щетинки склерита переднеспинки расположены по переднему и заднему кра-
ям. Верхняя губа на диске с 4-6 микрохетами.
5(6). Посттергальная область средне- и заднеспинки с рядом из 10 щетинок, крыло-
вые склериты с 2 щетинками (рис. 18, 4). Верхняя губа — рис. 17, 9. На дикорасту-
щих и культурных злаках, вредит просу и сорго. Юг ДВ..О. dilutipes Fairmaire
6(5). Посттергальная область средне- и заднеспинки с рядом из 12 щетинок, крыло-
вые склериты с 3 щетинками (рис. 18, 5). Верхняя губа — рис. 17, 70. На культур-
ных и дикорастущих злаках, сильно вредит. ЕЧ, СК, СБ.О. melanopus Linnaeus
56
6. подсемейство clytrinae
Личинки с толстым белым телом, крупной головой и длинными ногами, задняя
часть брюшка подогнута под переднюю, а срединные тергиты брюшка сильно рас-
ширены, образуя своеобразный горб (рис. 5, 4). Живут в чехликах, которые над-
страиваются по мере роста (рис. 20).
Голова (рис. 19, 7) гипогнатическая, большая, широкая, сильно хитинизирована.
Эпикраниальный шов длинный, лобные швы широко расходящиеся, часто более или
менее волнистые. Темя слабо выпуклое и уплощенное, лоб сильно уплощен. Налич-
ник сливается со лбом, так что шов между ними неразличим. Лоб и темя часто с
явственной скульптурой в виде морщинок, грубых точек или вдавлений. Усики (рис.
19,5) 2-члениковые, на вершине 2-го членика имеется полушаровидный сенсориаль-
ный придаток, а также щетинка, представляющая рудимент 3-го членика; основа-
ние усиков окружено хитиновым валиком, иногда довольно высоким, в котором
может прятаться усик. У основания усиков лежат сильно выпуклые глазки, по 6 с
каждой стороны, собранные в 2 группы: 4 глазка за основанием усика и 2 под ним.
Число и расположение щетинок на голове довольно постоянно. На наличнике
имеются 2 срединные и по 3 боковые щетинки, на лбу обычно выделяется 3 ряда
щетинок — передний (обычно 6), средний (4-6) и задний (2), а также имеются 1-2
щетинки у бокового края близ основания усиков. На темени находятся 1-2 средин-
ные щетинки и 2-3 наружно-теменные; на боковой поверхности головы имеется
около 8 щетинок с каждой стороны; иногда на голове появляются многочисленные
вторичные щетинки или шипики. Форма щетинок разнообразна: булавовидная,
нитевидная, щетинковидная или пильчатая.
Верхняя губа очень маленькая, срастается с наличником (ее граница у взрос-
лых личинок плохо заметна), с 8 короткими щетинками и обычно двувыемчатым
передним краем. Мандибулы с 3-4 зубцами и 1-2 щетинками на наружной стороне.
Лабио-максиллярный комплекс (рис. 19, 4) занимает всю нижнюю поверхность го-
ловы, кардо в виде поперечной пластинки, обычно подогнутой и скрытой в складке
сочленения головы с грудью, стипес крупный и длинный, пальпигер плохо обособ-
лен, несет 4-члениковый щупик, галея срастается с пальпигером, мембрановидная
лациния недоразвита, срастается с галеей и лежит на ее верхней стороне. Субмен-
тум крупный, продолговатый, прементум небольшой, слабо поперечный, на пере-
днем крае с 2-члениковыми губными щупиками.
Грудные сегменты примерно одинаковой величины, переднегрудной тергит с 2
сильно хитинизированными склеритами, занимающими почти всю поверхность и
слабо разграниченными друг от друга по средней линии. Средне- и заднегрудь очень
мягкие, разделены 2 поперечными складками на про-, мезо- и метатергит; мезотер-
гит с 6 слабо хитинизированными склеритами (рис. 24, 2), несущими немногочис-
ленные щетинки (2 внутренние тергальные, 2 наружные тергальные и 2 крыловые);
иногда склериты частично или полностью исчезают, а поверхность покрыта густы-
ми вторичными щетинками; боковые и нижние части сегментов с единичными ко-
роткими щетинками. Ноги сильно удлиненные, их длина равна диаметру тела или
превышает его, голенелапка едва короче бедра, коготки тонкие и длинные, почти
прямые; щетинки на нижней стороне голенелапки толстые и короткие, их обычно
4, расположенных попарно.
57
Рис. 19. Детали строения личинок подсемейства Clytrinae:
1—3 — голова: 1 — Clytra atraphaxidis, 2 — Tituboea macropus, 3 — Labidostomis tridentata, I
возраст; 4-5: Smaragdina semiaurantica: 4 — лабио-максиллярный комплекс, 5 — усик; 6-
7 — голенелапка: 6 — Labidostomis, 7 — Smaragdina.
Брюшко состоит из 10 сегментов, из них 1-8-й несут стигмы; 4-9-й сегменты
выпуклые, на спинной стороне гораздо длиннее, чем на брюшной, вследствие чего
вершинная половина брюшка подогнута книзу; 10-й сегмент небольшой и отчасти
прикрыт сверху предыдущим. Строение брюшных сегментов сходно с грудными,
но склериты более мелкие и плохо заметные или вообще отсутствуют; дорсальные
щетинки длинные и крепкие, на 1-4-м сегментах направлены назад, а на подогну-
тых 5-9-м — вперед. Стигмы с широкими овальными ячеистыми стигмальными
пластинками.
Личинки 1-го возраста отличаются меньшей степенью дифференциации; у них
еще не развита скульптура головы и склерита переднеспинки, отсутствуют допол-
нительные щетинки на голове (рис. 19, 5), груди и голенелапках, имеется зубчик
для разрезания хориона яйца на наружных склеритах средне- и заднегруди.
Личиночные чехлики имеют форму расширяющегося кзади цилиндра, на вер-
шине замкнутые; их длина примерно вдвое больше ширины. Поверхность чехлика
с разнообразной скульптурой, имеющей большое систематическое значение.
Самка откладывает яйца поодиночке или небольшими группами и покрывает
их яйцевым чехликом из экскрементов, строение которого различно у разных ро-
дов. У Labidostomis Germar (рис. 3, 7) и Coptocephala Lacordaire яйца стебельчатые.
Развитие яйца длится около 15-25 дней, вышедшая из яйца личинка находится 1-2
58
дня, до полной хитинизации, в закрытом яйцевом чехлике, а затем прогрызает его
крышечку. В спокойном состоянии личинка высовывает из чехлика переднюю по-
ловину тела, но при малейшем раздражении она прячется внутрь, закрывая упло-
щенной головной капсулой входное отверстие.
По мере роста личинки яйцевой чехлик надстраивается на открытом конце и
превращается в личиночный чехлик; материалом служат экскременты и окружаю-
щий субстрат. Использование частиц минералов сильно повышает крепость чехли-
ка. Развитие идет 2 года с 2 зимовками, а число возрастов равняется трем, как и у
очень близкого подсемейства Cryptocephalinae. Перед линькой или зимовкой ли-
чинка закрывает устье крышечкой. При окукливании личинка также запечатывает
устье, одновременно прикрепляя его к какому-либо субстрату, после чего она пере-
ворачивается головой к заднему, более широкому концу чехлика. Стадия куколки
(рис. 4, 7) длится 15-20 дней. Взрослый жук прогрызает заднюю часть чехлика и
выходит наружу.
Местообитание личинок различно. Большинство живет на почве, в подстилке,
под камнями, молодые личинки Labidostomis Germar обычно встречаются на расте-
ниях, но по мере подрастания уходят в подстилку. Некоторые виды живут в мура-
вейниках (Clytra Laicharting) и приурочены к определенным хозяевам или отдают
им предпочтение. Личинки попадают в муравейник различными путями. Муравьи
затаскивают в гнезда яйца, очень похожие на семена, а иногда и личинок, или ли-
чинки заползают самостоятельно. Они не приносят какой-либо пользы муравьям,
так что это сожительство представляет комменсализм. Личинки держатся в широ-
ких галереях, забираясь подчас очень глубоко, но перед окукливанием поднимают-
ся кверху.
Питаются личинки преимущественно растительным детритом, но могут по-
едать и зеленые растения, а личинки Clytra могут питаться и животной пищей: яй-
цами, личинками и куколками муравьев.
В регионе 7 родов.
Приводимая ниже определительная таблица построена на признаках личинок
старших возрастов (за исключением особо оговоренных случаев), но включены
также и признаки личинок 1-го возраста.
Таблица для определения родов подсемейства Clytrinae
1(6). Темя в густых многочисленных щетинках.
2(5). Лоб в густых многочисленных не булавовидных щетинках. Темя и склерит
переднеспинки без ячеистой скульптуры.
3(4). Голенелапка на нижней стороне с 6-9 шиповидными щетинками (рис. 19, 6).
Щетинки головы и склерита переднеспинки умеренно густые. Личиночный чехлик
с торчащими волосковидными нитями (редко без нитей), без перетяжки перед усть-
ем. Личинка 1-го возраста: вторичные щетинки отсутствуют (рис. 23, 7), голенелап-
ка снизу с 4 щетинками; яйцевой чехлик цилиндрический, удлиненный, гладкий, с
длинной нитью на заднем полюсе (рис. 3, 7)............... 1. Labidostomis
4(3). Голенелапка на нижней стороне с 4 шиповидными щетинками. Щетинки голо-
вы и склериты переднеспинки расположены очень густо (рис. 21,3, 5), личиночный
чехлик без волосковидных нитей, с кольцевой перетяжкой перед устьем, кувшино-
59
образный (рис. 20, 2). Личинка 1-го возраста: вторичные щетинки отсутствуют;
яйцевой чехлик со спирально расположенными рядами квадратных ячеек, без нити
на заднем полюсе.........................................2. Cheilotoma
5(2). Лоб с 14 булавовидными щетинками, расположенными в 3 ряда: 6, 6, 2 (рис.
19, 2); в мелкой ячеистой скульптуре, с 2 невысокими продольными ребрышками.
Темя и склерит переднеспинки в ячеистой скульптуре, с густо расположенными
короткими не булавовидными щетинками. Средне- и заднеспинка без склеритов (за
исключением следов эпиплевральных склеритов), в довольно густых щетинках;
тергиты брюшка без склеритов и дополнительных щетинок (рис. 22, 3). Личиноч-
ный чехлик с очень тонкими стенками, сверху с поперечными невысокими мор-
щинками, у переднего края с несколькими короткими сильно сближенными про-
дольными пластинчатыми килями (рис. 20, 6)..................3. Tituboea
6(1). Темя с 6-10 щетинками (редко с короткими вторичными шипиками). Лоб и
грудные тергиты без густо расположенных дополнительных щетинок.
7(10). Темя с каждой стороны у эпикраниального шва с 1 щетинкой (рис. 24, 7). Лоб
и темя с булавовидными и простыми щетинками. Верхняя губа двувыемчатая. Тер-
гиты брюшка без явственных склеритов, с 2 поперечными рядами щетинок.
8(9). Верхняя сторона личиночного чехлнка с сильно развитыми продольными или
поперечными килями, остальная поверхность без явственной скульптуры (рис. 20,
3-5). Средне- и заднеспинка с поперечным рядом из 6 слабо развитых склеритов
(рис. 22, 4).Темя в грубой ячеистой скульптуре, часто образующей морщины. Лоб с
12 булавовидными щетинками: 6, 4, 2. Личинка 1-го возраста: лоб с 10 булавовид-
ными щетинками (4, 4, 2); склерит переднеспинки с 12 щетинками на переднем
крае и с 8 на базальной части; яйцевой чехлик в виде полураспустившегося бутона,
с 5 рядами тонких пластинок (рис. 14, 5). Личинки в муравейниках.4. Clytra
9(8). Личиночный чехлик сверху почти гладкий или с очень тонкими и нежными
штрихами и морщинками, снизу с узкой продольно-морщинистой полосой, от ко-
торой к заднему концу идут грубые морщинки. Личинка 1-го возраста: лоб (рис. 22,
2) с 12 булавовидными щетинками (6, 4, 2); вершинный членик нижнечелюстных
щупиков удлиненный; склерит переднеспинки с 12-14 щетинками на переднем крае;
средне- и заднеспинка без явственных склеритов; яйцевой чехлик со спирально
расположенными рядами квадратных ячеек......................5. Lachnaia
10(7). Темя с каждой стороны у эпикраниального шва с 2 щетинками (рис. 25, 3, 7).
Голова без явственных булавовидных, но нередко с утолщенными зазубренными
щетинками. Гребни личиночного чехлика развиты слабо. Яйцевой чехлик со спи-
рально расположенными рядами квадратных ячеек (рис. 14, 7). Личинки в под-
стилке.
11(12). Склерит переднеспинки с густой зернистостью, с булавовидными щетинка-
ми по переднему краю (рис. 25, 9). Верхняя губа с широкой трапециевидной выем-
кой, слабым срединным выступом или без него. Личиночный чехлик сверху с 3
слабыми продольными килями..............................6. Coptocephala
12(11). Склерит переднеспинки грубо морщинистый, с рядом коротких и длинных
простых щетинок по переднему краю (рис. 25, 4, 5). Верхняя губа с широкой дуго-
видной выемкой, со срединным выступом или без него. Личиночный чехлик без
явственной скульптуры....................................7. Smaragdina
60
Рис. 20. Форма чехлика личинок подсемейства Clytrinae:
1 — Labidostomis', 2 — Cheilotoma', 3 — Clytra atraphaxidis', 4 — C. quadripunctata', 5 —
C. arida, 6 — Tituboea macropus.
1. Род Labidostomis Germar
Темя и лоб в густых точках и с многочисленными, равномерно расположенны-
ми длинными простыми или короткими, слегка зазубренными щетинками (рис. 21,
2). Верхняя губа с широкой выемкой и слабым срединным выступом или без него.
Склерит переднеспинки сильно хитинизированный, с многочисленными длинны-
ми щетинками, особенно вдоль переднего края (рис. 21, 4). Средне- и заднеспинка
с 1 рядом из 6 склеритов с многочисленными щетинками. Тергиты первых сегмен-
тов брюшка с 2 поперечными рядами щетинок (2 в переднем и 4 в заднем ряду),
эпи- и гипоплевральные склериты с 1 щетинкой. Голенелапка снизу с 7-9 шиловид-
ными щетинками, сверху — с 10-12 простыми щетинками (рис. 19, 6). Коготки
длинные, простые.
61
Личинка 1-го возраста отличается отсутствием вторичных щетинок на голове
и переднеспинке (рис. 19, 3), лоб с 3 поперечными рядами щетинок (2, 4, 2), темя с
10 щетинками, причем с каждой стороны имеется только 1 внутренне-теменная ще-
тинка. Голенелапка снизу с 4 шиловидными щетинками.
Личиночные чехлики простые, со слабой скульптурой, но обычно с длинными
торчащими нитевидными образованиями (рис. 20, У), редко голые. Яйцевой чехлик
цилиндрический, удлиненный, гладкий, с длинной нитью на заднем полюсе, по-
средством которой чехлик прикрепляется к субстрату, яйца откладываются на рас-
тение, группами по 10-12 штук. Молодые личинки питаются листьями или под-
стилкой, личинки 2-го и 3-го возрастов живут в подстилке и под камнями.
В регионе 17 видов. Личинки различаются с большим трудом. Ниже приводит-
ся таблица по признакам личинок 3-го возраста.
Таблица для определения видов рода Labidostomis
1(6). Темя и лоб с многочисленными короткими и слегка зазубренными булавовид-
ными щетинками (рис. 21,2). Личиночный чехлик состоит только из экскрементов,
без нитевидных образований.
2(5). Щетинки лба булавовидные.
3(4). Передний край верхней губы почти прямой, в середине со слабым выступом.
Личиночный чехлик с характерной сетчатой скульптурой. СБ, ДВ............
.................................................L. bipunctata Mannerheim
4(3). Передний край верхней губы с широкой овальной выемкой. Личиночный чех-
лик простой. Транспалеаркт...........................L. tridentata Linnaeus
5(2). Щетинки лба зазубренные, передний край верхней губы с широкой и неглубо-
кой срединной выемкой, на дне выемки со слабым широким выступом (рис. 23, 3).
ВСБ, ДВ................................................L. chinensis Lefevre
6(1). Темя и лоб с многочисленными простыми длинными и укороченными, слегка
зазубренными щетинками (рис. 23, 4). Личиночный чехлик с мелкими частицами
почвы и длинными и короткими нитевидными образованиями.
7(12). Темя и лоб со склеротизацией в виде шипиков. Передний край верхней губы
с широкой и неглубокой срединной выемкой, иногда на дне выемки имеется не-
большой срединный выступ.
8(11). Склеротизация лба и темени густая, передний край переднеспинки с очень-
густым и спутанным рядом щетинок. Личиночный чехлик с очень густыми длин-
ными и торчащими нитевидными образованиями.
9(10) Лоб с менее густыми (около 40) щетинками (рис. 21,2). Верхняя губа двувы-
емчатая, с округленным срединным выступом. Голенелапка сверху с 8 щетинками.
Личиночный чехлик без явственной скульптуры. Юго-восток ЕЧ, СБ, ДВ......
.....................................................L. sibirica Germar
10(9). Лоб с более густыми (около 100) щетинками. Верхняя губа без срединного
выступа. Голенелапка сверху с 11-13 щетинками. Личиночный чехлик с морщини-
стой поверхностью, морщинки верхней стороны поперечные, нижней — продоль-
ные. ЕЧ, СК, СБ......................................L. longimana Linnaeus
11(8). Склеротизация лба и темени редкая, передний край переднеспинки с более
правильным рядом из редких коротких и длинных щетинок. Личиночный чехлик с
редкими и короткими нитевидными образованиями. ДВ....L. amurensis Heyden
62
Рис. 21. Лоб (1-3) и переднеспинка (4, 5) родов Labidostomis и Cheilotoma-.
1,4 — L. sibiriccr, 2 — L. bipunctatcr, 3,5 — Cheilotoma.
12(7). Темя и лоб без шиповидной склеротизации (рис.23, 5, 6). Голенелапка — рис.
23, 9, 10.
13(14). Передний край верхней губы прямой (рис. 23, 2). Переднеспинка по пере-
днему краю с 18-20 длинными простыми щетинками (рис. 23, 8). Юг ЕЧ, СК...
.......................................................L. senicula Kraatz
14(13). Передний край верхней губы с овальной выемкой (рис. 23, 7). Переднес-
пинка по переднему краю с 8-9 длинными простыми щетинками (рис. 23, 7). Юг
ЕЧ, СК, Тува...........................................L. metallica Lefevre
2. Род Cheilotoma Chevrolat (Chilotoma Redtenbacher)
Верхняя губа с неглубокой широкой вырезкой, без срединного выступа, с 8
щетинками, наличник отделен от лба явственным волнистым швом, поверхность
наличника блестящая, гладкая, с 2 щетинками посередине и с 3 на передних углах.
Лоб (рис. 21,3) почти плоский, с небольшим ямковидным вдавлением посередине,
темя умеренно и равномерно выпуклое, лоб и темя в густой пунктировке, несущей
короткие, зазубренные, но не булавовидные щетинки, на задней части лба имеются
63
Рис. 22. Личинки Clytrinae:
1,2 — Lachnaia sexpunctatcr. 1 — общий вид, 2 — голова; 3 — Tituboea macropus, средне-,
заднегрудь и 1-й тергит брюшка; 4 — Clytra quadripunctata, тергиты груди. По Фиори и ориг.
многочисленные мелкие шипики. Мандибулы с 4 тупыми зубцами. Склерит пере-
днеспинки (рис. 21, 5) мелкоморщинистый, в многочисленных щетинках, более
длинных по краям; передний край прямой. Средне- и заднеспинка с 6 плохо замет-
ными склеритами каждая, расположенными в поперечный ряд. Голенелапка снизу
с 4 толстыми шиповидными щетинками. Длина тела взрослой личинки около 6 мм.
Личиночный чехлик (рис. 20, 2) кувшинообразный, с перетяжкой перед усть-
ем; скульптура поверхности сильно сглаженная, сверху и снизу с продольной, по
бокам с поперечной исчерченностью, перед устьем часто с 1-2 поперечными реб-
рышками. Длина чехлика до 8-8,5 мм. Яйцевой чехлик со спирально расположен-
ными рядами квадратных ячеек.
Личинки в подстилке в степях. В регионе 2 вида, известна личинка Ch. musci-
formis Goeze (ЕЧ, СК, СБ).
64
Рис. 23. Личинки Labidostomis'.
1, 2 — верхняя губа: 1 — L. metallica, 2 — L. senicukr, 3-6 — лоб: 3 — L. chinensis, 4 — L.
amurensis, 5 — L. metallica, 6 — L. senicukr, 7,8 — склерит переднеспингки: 7 — L. metallica,
8 — L. senicukr, 9, 10 — голенелапка: 9 — L. metallica, 10 — L. senicula.
65
3. Род Tituboea Lacordaire (Antipa auct.)
У взрослой личинки верхняя губа со слабо двувыемчатым передним краем и с
8 короткими щетинками. Наличник отделен ото лба валикообразным возвышени-
ем, непосредственно перед ним с 2 овальными вдавлениями, его поверхность без
скульптуры, блестящая, с 2 срединными и 6 боковыми щетинками. Лоб в мелкой
ячеистой скульптуре, с 2 невысокими продольными ребрышками, поверхность лба
кнаружи от них вогнутая, между ними выпуклая в передней половине и вогнутая в
задней; 6 коротких булавовидных переднелобных щетинок, 6 срединных (из них
наружные и срединные булавовидные) и 2 заднелобные; перед срединными щетин-
ками имеются еще 3 очень короткие булавовидные щетинки (рис. 19, 2). Темя в
мелких нежных ячейках и в густых, толстых, коротких, не булавовидных щетин-
ках. Мандибулы с 4 тупыми зубцами. Склерит переднеспинки в густых мелких не-
жных ячейках и в довольно густых толстых недлинных щетинках, рассеянных по-
чти равномерно по всему склериту. Средне- и заднеспинка без явственных склери-
тов, с 2 поперечными складками, склеротизованным пятном на боковых краях (бо-
лее крупным на переднеспинке), в густо расположенных щетинках (рис. 22,3). Тер-
гиты первых брюшных сегментов с немногочисленными щетинками, собранными
в неправильный поперечный ряд. Голенелапка с 4 щетинками на нижней стороне.
Коготки сравнительно короткие, не более 1/4 длины голенелапки.
Личиночный чехлик: длина около 8 мм, стенки очень тонкие, поверхность с
концентрическими морщинками, на верхней стороне у переднего края с короткими
продольными килями, начинающимися в одной точке и расходящимися кпереди,
срединный киль очень высокий, боковые гораздо ниже (рис. 20, б).
1 вид —Т. macropus liliger. Личинки встречаются в муравейниках Plagiolepis.
Юг ЕЧ, СК.
4. Род Clytra Laicharting
У взрослой личинки передний край верхней губы с выемкой и обычно средин-
ным выступом, лоб и темя в ячеистой скульптуре. Лоб с 12 щетинками, образующи-
ми 3 поперечных ряда (6,4, 2). Темя с более или менее булавовидными щетинками;
1 внутренне-теменная щетинка с каждой стороны (рис. 24, У). Переднеспинка в
ячеистой скульптуре с многочисленными толстыми простыми и булавовидными
щетинками. Средне- и заднеспинка с 1 поперечным рядом плохо заметных склери-
тов (рис. 24, 2), внутренние тергальные склериты сильно поперечные, широко рас-
ставленные, с 1 длинной и 1 короткой щетинкой, наружные тергальные склериты с
1 длинной щетинкой, крыловые склериты с 5 щетинками на переднем крае. Терги-
ты первых сегментов брюшка без склеритов, с двумя рядами щетинок (2 в пере-
днем и 4 в заднем ряду). Голенелапка с 4 щетинками на нижней стороне. Длина
личинки до 7-8 мм.
Личинка первой стадии отличается отсутствием скульптуры на голове и пере-
днеспинке и более редкими щетинками на переднеспинке.
Личиночный чехлик с толстыми стенками и резкими продольными или попе-
речными ребрами на верхней стороне. Яйцевой чехлик в виде полураспустившего-
ся бутона, с 5 довольно правильными рядами тонких пластинок (рис. 3, 3).
66
Рис. 24. Детали строения личинок рода Clytrcr.
С. arida\ 1 — голова, 2 — грудь, 3, 4 — щетинка, 9 — верхняя губа; С. atraphaxidis', 6 —
верхняя губа; С. quadripunctatcr. 5 — щетинка, 8 — верхняя губа, 10 — переднеспинка; С.
laeviusculcr. 7 — верхняя губа, 11 — переднеспинка.
Личинки и куколки в муравейниках. В регионе 5 видов.
Таблица для определения видов рода Clytra
1(2). Темя в грубых, неодинаковых ячейках, сливающихся назади в продольные
морщинки (подрод Clytrella L. Medvedev). Щетинки лба и темени резко булавовид-
ные (рис. 19, У). Верхняя губа с очень слабым срединным выступом (рис. 24, 6).
Личиночный чехлик на верхней стороне с многочисленными тонкими поперечно-
67
дуговидными килями, обращенными выпуклостью к устью чехлика (рис. 20, 3). В
степях в гнездах Cataglyphis. Транспалеаркт..........С. atraphaxidis Pallas
2(1). Темя в нежных одинаковых морщинках.(подрод Clytra s.str.) Верхняя губа с
заметным срединным выступом. Щетинки лба и темени самое большее слабо була-
вовидные (рис. 24, 3-4). Личиночный чехлик с продольными килями. В гнездах
Formica.
3(4). Склерит переднеспинки с явственными морщинками, образующими крупные
неправильные ячейки (рис. 24,11). срединный выступ выдается из выемки верхней
губы (рис. 24, 7). ЕЧ, СК, ЗСБ.....................С. laeviuscula Ratzeburg
4(3). Склерит переднеспинки равномерно зернистый, без явственных морщинок
(рис. 24,10).
5(6). Срединный выступ не выдается из выемки верхней губы (рис. 24,8). Щетинки
лба и темени короткие, толстые, утолщаются к вершине и образуют заметную була-
ву (рис. 24, 5). Кили чехлика сверху не доходят до его переднего края, переднебоко-
вые края его сверху с прямоугольными выступами (рис. 20, 4). Транспалеаркт.
С quadripunctata Linnaeus
6(5). Срединный выступ примерно наполовину выступает из выемки верхней губы
(рис. 24, 9). Щетинки лба примерно втрое длиннее, чем у предыдущего вида, слег-
ка сужаются к вершине, образуют едва заметную булаву (рис. 24, 3, 4). Щетинки
темени практически простые (рис. 24,1). Кили чехлика сверху доходят до его пере-
днего края, переднебоковые края чехлика сверху закругленные (рис. 20, 5). СБ, ДВ
......................................С arida Weise
5. Род Lachnaia Lacordaire (Lachnaea auct.)
В регионе 1 вид — L. sexpunctata Scopoli. У личинки 1-го возраста (рис. 22,1)
передний край верхней губы двувыемчатый, с небольшим срединным выступом.
Лоб с 8 переднелобными щетинками, расположенными в 2 ряда (6, 2), 4 срединны-
ми и 2 заднелобными; 2 переднелобные щетинки 1-го ряда щетинковидные, про-
чие — булавовидные, короткие. Темя с каждой стороны с 1 внутренней булавовид-
ной и с 3 наружными, из них срединная щетинковндная, очень длинная, а осталь-
ные булавовидные (рис. 22, 2). Нижнечелюстные щупики с удлиненным вершин-
ным члеником. Склерит переднеспинки с 12-14 щетинками на переднем крае. Тер-
гиты остальных сегментов без явственных склеритов, с поперечным рядом щетинок.
Вершины бедер с 1 щетинкой на верхнем крае. Голенелапка с 4 щетинками на ниж-
ней стороне. Личиночный чехлик до 15-16 мм, почти гладкий, снизу с узкой продоль-
но-морщинистой полосой, от которой к заднему концу идут грубые морщинки. Яй-
цевой чехлик овальный, со спирально расположенными рядами квадратных ячеек.
Взрослые жуки встречаются обычно на дубе, личинки отмечены под камнями;
имеются также данные о питании личинок лишайниками и мхом на коре деревьев.
Запад ЕЧ.
6. Род Coptocephala Lacordaire
Головная капсула (рис. 25, 6) в умеренно густых равномерных точках, темя в
густых равномерных точках, с каждой стороны от теменного шва с двумя сближен-
68
ними простыми щетинками со слегка округленными вершинами. Лоб с 14 щетин-
ками (8, 4, 2), причем щетинки заднего ряда зазубренные, а переднего — слегка
уплощенные у вершины. Щетинки наличника и верхней губы простые, верхняя
губа с широкой трапециевидной выемкой. Склерит переднеспинки (рис. 25, 9) в
умеренно густых равномерных точках, с закругленными боковыми краями и с ря-
дом редких булавовидных щетинок по переднему краю и простых — по заднему.
Стигмы с крупными стигмальными пластинками. Голенелапка снизу с 4 шиповид-
ными, а сверху — с 5-6 простыми щетинками; коготок простой, длинный и прямой.
Личиночный чехлик сверху с тремя слабыми продольными килями, сходящи-
мися назади.
Личинка 1-го возраста. Трапециевидная выемка верхней губы резко выражена
(рис. 25, 8), щетинки переднеспинки простые. Яйцевой чехлик короткий, оваль-
ный, со спиральными рядами квадратных ячеек, несет на заднем полюсе нить, ко-
торой прикрепляется к субстрату.
В регионе 6 видов; известны личинки двух широко распространенных видов,
обычно связанных с полынью.
Таблица для определения видов рода Coptocephala
1(2). Срединный выступ выемки верхней губы не выражен. Щетинки заднего лоб-
ного ряда слабо зазубренные, остальные простые (рис. 25, 7). Тува, ВСЕ, ДВ.
.........................................................С. orientalis Baly
2(1). Срединный выступ выемки верхней губы выражен. Щетинки заднего и сред-
него лобного ряда зазубрены, переднего — простые (рис. 25, 6). ЕЧ, СК, СБ..
....................................................С unifasciata Scopoli
7. Род Smaragdina Chevrolat (Gynandrophthalma Lacordaire)
Лоб слабо зернистый, с тремя поперечными рядами простых или зазубренных
щетинок (6-8,4,2). Темя морщинистое, с 2 внутренними и 2 наружными простыми
щетинками; иногда голова с многочисленными вторичными короткими шиповид-
ными щетинками, резко отличающимися от длинных первичных. Верхняя губа с
дуговидной плавной выемкой, чаще без срединного выступа. Склерит переднес-
пинки сильно хитинизирован, морщинистый, на прямом переднем крае с много-
численными простыми щетинками разной длины. Средне- и заднеспинка с 1 попе-
речным рядом склеритов; внутренние тергальные склериты с 2-3 длинными и 2
короткими щетинками, наружные тергальные с 1 длинной и 1 короткой, крыловые
склериты с 5-6 щетинками, тергиты брюшных сегментов с двумя рядами щетинок
(4 в переднем, 2 в заднем). Голенелапка снизу с 4 шиповидными, сверху с 4 просты-
ми щетинками, коготок простой, длинный (рис. 19, 7).
Личиночный чехлик обычно без явственной скульптуры, с очень слабыми спин-
ными, брюшными и боковыми килями. Яйцевой чехлик короткий и широкий, оваль-
ный, со спирально расположенными рядами квадратных ячеек, разделенных резки-
ми ребрышками, без нити.
У личинки 1-го возраста передний край верхней губы с простой широкой вы-
емкой. Лоб с 3 поперечными рядами расширенных к вершине зазубренных щети-
69
нок (6 переднелобных, из них 2 простые и лежат несколько впереди от прочих, 4
срединные, 2 заднелобные). Темя с 2 внутренними и 2 наружными простыми ще-
тинками, а также несколькими щетинками около глаз и рядом коротких щетинок у
заднебоковой покатости. Склерит переднеспинки с 12-14 довольно длинными ще-
тинками на переднем крае и с 12-16 щетинками в базальной части диска.
Личинки в подстилке. В регионе 10 видов, известны личинки старших возрас-
тов двух видов. Не включены личинки S. aurita Linnaeus и S. salicina Scopoli, изве-
стные лишь по первому возрасту. Род морфологически очень близок к Coptocephala
Lacordaire.
Таблица для определения видов рода Smaragdina
1(2). Темя и передняя часть лба, помимо длинных первичных щетинок, с многочис-
ленными короткими, шиповидными вторичными щетинками (рис. 25, 75). Первич-
ные щетинки лба и темени преимущественно пиловидные. Стигмальные пластин-
ки очень крупные. Срединная выемка верхней губы без выступа. Юг ДВ.....
...............................................5. semiaurantica Fairmaire
2(1). Голова только с первичными щетинками.
3(4). Голова со слабым теменным кантом между лбом и теменем (как у Crypto-
cephalus). На ивах. ЕЧ, СК, СБ, ДВ......................5. aurita Linnaeus
4(3). Голова без теменного канта между лбом и теменем.
5(6). Переднеспинка сильно поперечная, с заостренными боковыми краями и силь-
но хитинизированным гребнем по заднему краю, несет многочисленные щетинки
по переднему (свыше 50) и заднему (свыше 60) краям (рис. 25, 5). Щетинки головы
простые. Срединная выемка верхней губы без выступа (рис. 25, 2). Стигмальные
пластинки маленькие, не превышают диаметра стигмы. В степях, жуки тяготеют к
карагане. Юг СБ.......................................S. collaris Fabricius
6(5). Переднеспинка умеренно поперечная, с широко округленными боковыми кра-
ями, без гребня по заднему краю, с правильным рядом чередующихся негустых
макро- и микрохет по переднему краю и спутанным рядом — по заднему (рис. 25,
4). Голова преимущественно с пиловидными щетинками. Срединная выемка верх-
ней губы с тупым выступом (рис. 25, 5). Стигмальные пластинки крупные, превы-
шают диаметр стигмы. Жуки на вязе, личинки в подстилке в Вязниках. Монголия,
возможно нахождение в Забайкалье..................5. mandzhura Jacobson
Рис. 25. Детали строения личинок родов Smaragdina и Coptocephala'.
S. semiaurantica'. 1 — голова; S. collaris'. 2 — передний край головы, 5 — переднеспинка;
S. mandzhura: 3 — голова, 4 — переднеспинка; С. unifasciata'. 6 — голова, 8 — верхняя губа
(1-й возраст), 9 — переднеспинка; 7 — голова С. orientalis.
70
71
7. ПОДСЕМЕЙСТВО cryptocephalinae
Личинки очень схожи с личинками Clytrinae (рис. 5, 5), скарабеоидного типа, с
толстым белым или желтоватым телом, задняя часть брюшка подогнута под пере-
днюю, а срединные тергиты брюшка сильно расширенные.
Головная капсула сильно хитинизирована, с отвесной, плоской передней час-
тью, включающей лоб и часть темени, эта уплощенная часть головной капсулы обыч-
но отделена от выпуклой части темени резким кантом, проходящим кзади от лоб-
ных швов и примерно параллельно им, иногда теменной кант не развит. Верхняя
губа, наличник и лоб слиты друг с другом, однако границы между ними более или
менее заметны. Хетотаксия головной капсулы довольно постоянна, в связи с чем
щетинки получают условные обозначения (рис. 26, 7). Верхняя губа с 6-8 просты-
ми щетинками, ее передний край двувыемчатый, часто со срединным выступом.
Наличник с 10-14 простыми щетинками и 2-4 порами. Лоб с поперечными рядами
простых или зазубренных щетинок, причем 2 краевые щетинки переднего ряда чаще
всего простые. Темя чаще с простыми щетинками. Теменной кант с рядом из 4-10
простых или зазубренных щетинок, из которых V2 чаще всего бывает длиннее про-
чих. Лоб обычно с зернистой скульптурой. Глазков по 6 с каждой стороны, из них 4
за основанием усиков, а 2 отодвинуты книзу. Усики (рис. 26, 5, 4) 3-члениковые; на
вершине широкого 2-го членика лежат небольшой 3-й членик, несущий 1 короткую
и 1 длинную щетинки, и крупный яйцевидный сенсориальный придаток. Мандибу-
лы с 4 зубцами (рис. 26, 5), лабио-максиллярный комплекс сильно развит и занима-
ет всю нижнюю поверхность головы. Галея срастается с пальпигером, мембрано-
видная; лациния редуцирована, челюстные щупики четырехчлениковые. Нижне-
губные щупики двухчлениковые. Переднеспинка всегда сильно хитинизирована, с
многочисленными короткими и длинными щетинками на краях и диске, причем
иногда переднекрайние щетинки могут быть зазубрены, эпиплевральные склериты
отсутствуют. Средне- и заднеспинка (рис. 26, 2) с поперечным рядом из 6 плохо
заметных, очень слабо хитинизированных склеритов (2 внутренних тергальных, 2
наружных тергальных и 2 крыловых), кзади от крылового лежит небольшой скле-
рит, который следует считать эпиплевральным. Тергиты груди и брюшка разделя-
ются поперечной складкой на небольшой протергит и крупную заднюю часть, пред-
ставляющую мезо- и метатергит. Тергиты брюшка с 1 поперечным рядом из 4 пло-
хо заметных склеритов, которые иногда вообще не различимы. Щетинки туловищ-
ных сегментов всегда простые, длинные и короткие (макро- и микрохеты). Стигмы
обычно с крупными стигмальными пластинками, которые более или менее редуци-
руются у степных и пустынных форм. Ноги длинные (рис. 27, 4-9) с крупными
коготками, без пульвилл, голенелапка на верхней стороне, с тонкими, на нижней с
толстыми шиповидными щетинками, иногда шиповидные щетинки появляются на
бедре. Личиночный чехлик без скульптуры или она едва намечена в виде слабых
продольных килей.
Биология Cryptocephalinae отличается от Clytrinae прежде всего отсутствием
мирмекофилии. Личинки живут в подстилке и питаются гниющими листьями, а
также на листьях растений, особенно молодые, которые позднее по мере роста пе-
реходят в подстилку. Яйцевые чехлики обычно со спирально-ячеистой скульпту-
рой, не стебельчатые. Молодые личинки появляются во второй половине лета и
72
Рис. 26. Детали строения личинок подсемейства Cryptocephalinae:
1 — голова Cryptocephalus с индексами щетинок; 2 — грудь С. janthinus', Suffrianus'. 3 —
усик, 5 — мандибула; Pachybrachis\ 4 — усик.
затем уходят на зимовку, следующей весной происходит первая линька, на вторую
зимовку уходят личинки 2-го или 3-го возраста. Молодые жуки появляются весной
или летом. Всего имеется 3 личиночных возраста. Хозяйственного значения личин-
ки Cryptocephalinae не имеют. Питание молодых личинок на листьях деревьев не
приносит сколько-нибудь заметного ущерба. Возможная положительная роль ли-
чинок Cryptocephalinae и Clytrinae в почвообразовании не выяснена, хотя они при-
нимают участие в переработке растительного детрита.
73
Рис. 27. Детали строения личинок подсемейства Cryptocephalinae:
Stylosomus-. 1 — грудь, 9 — нога; Pachybrachis hieroglyphicus'. 3 — среднегрудь, 4 — нога;
2 — среднегрудь Р scriptidorsum; нога: 5 — Cryptocephalus regalis, 6 — С. stschukini, 7 —
С. agnus, 8 — С. limbellus.
Таблица для определения родов подсемейства Cryptocephalinae
1(2). Теменной кант не развит, вертикальная, лишенная скульптуры передняя часть
головы отделена теменным перегибом от горизонтальной, густо-точечной (рис. 28,
5). Стигмы груди и брюшка в виде сильно удлиненной волнистой полоски (рис. 33,
1,2)...........................................................1. Suffrianus
74
2(1). Теменной кант развит на всем протяжении или хотя бы над основаниями уси-
ков в виде гребня (рис. 28, 2).
3(4). Голенелапка снизу с 7-9 щетинками (рис. 27, 4). Наличник отделяется ото лба
четким поперечным выступом, срединная часть наличника светлая, нехитинизиро-
ванная (рис. 28, 7). Лоб с 3 рядами щетинок (6, 4, 2) и 2 порами ... 2. Pachybrachis
4(3). Голенелапка снизу с 4-5 щетинками.
5(6). Лоб и теменной кант между V2 и V3 не имеет пор (рис. 28, 2), щетинки лба
расположены в 4 ряда (6, 2, 2, 2)........................3. Cryptocephalus
6(5). Лоб (около f2) и теменной кант (между V2 и V3) с порами (рис. 28, 4), щетинки
лба расположены в 4 ряда (4, 4, 2, 2)........................4. Stylosomus
1. Род Suffrianus Weise
Голова (рис. 28, 5) округло-вытянутая, темя в задней половине в густой зерни-
стости и с одинаковыми мелкими шипиками, теменной кант не выражен, но пере-
гиб явственный, область теменного канта с каждой стороны с рядом из 5 зазубрен-
ных щетинок и 1 порой (между V2 и V3), причем V2 в 2 раза длиннее прочих. Эпик-
раниальный шов длинный, лобные швы плавно изогнуты, отходят под тупым уг-
лом, лоб округлой формы, без зернистости, с 14 зазубренными щетинками, распо-
ложенными в 5 рядов (2, 6, 2, 2, 2), и 2 порами; щетинки наличника и верхней губы
простые, последняя с треугольным срединным выступом. Усики с коротким 3-м
члеником (рис. 26, 5). Мандибулы (рис. 26, 5) сбоку с 1 короткой и 1 длинной ще-
тинками. Переднеспинка хитинизированная, с правильным рядом из 12 коротких и
длинных щетинок по переднему и 6 — по заднему краям. Крыловые склериты с 3
макро- и 1 микрохетой, наружный тергальный — с 1 макро- и 1 микрохетой, внут-
ренний тергальный — с 1 макро- и 3 микрохетами. Стигмальные пластинки груди
сильно удлиненные, червеобразные; стигмальные отверстия в виде поперечной
щели; стигмы брюшка такого же строения, но немного меньше грудных (рис. 33, 7,
2). Голенелапка длинная, снизу с 4 короткими и крепкими, сверху с 4 длинными
щетинками, коготки слабо изогнуты.
Род включает единственный вид — 5. pumilo Suffrian, встречающийся на ивах
на юге ВС, ДВ, Сахалине, Курильских островах. Описание приводится по личинке
I возраста.
2. Род Pachybrachis Redtenbacher
Головная капсула (рис. 28, 7) сильно хитинизированная, темя слабо морщини-
стое, с редкими щетинками, теменной кант с более развитым гребнем только над
усиками, далее он сглажен, с рядом из 5 щетинок, из которых Vp V2, V3 длинные и
простые, V4 и V5 короткие и зазубренные; между V2 и V3 имеется пора, расположен-
ная ковнутри от них ближе к лобному шву, последние почти прямые, расходятся от
короткого эпикраниального шва под тупым углом. Лоб в густой зернистости, с 12
слабо булавовидными или зазубренными щетинками, расположенными в 3 ряда (6,
4, 2), и 2 порами, причем первый ряд щетинок и 2 поры лежат почти на уровне
середины шва. Наличник отделяется ото лба поперечным четким выступом (рис.
28, 7), с каждой стороны с 1 булавовидной щетинкой, в средней части хитинизиро-
75
Рис. 28. Детали строения личинок подсемейства Cryptocephalinae:
1-6 — головная капсула: 1 — Pachybrachis hieroglyphicus', 2 — Cryptocephalus sericeus} 3 —
Suffrianuspumilo; 4 — Stylosomus; 5 — Cryptocephalus limbellus\ 6 — C. agnus; 7-9 — верхняя
губа: 7 — Pachybrachis hieroglyphicus\ 8 — Cryptocephalus nitidus; 9 — C. cordiger.
ванный, светлый и гладкий, 3-й членик усика узкий и длинный, на вершине с очень
длинной простой щетинкой, папилла крупная, коническая и длиннее последнего
членика (рис. 26, 4). Верхняя губа с широким прямым передним краем, без выемки,
по бокам с каждой стороны с 9 крупными тупыми щетинками. Склерит переднес-
пинки в густой зернистости, с каждой стороны по переднему краю с двойным, а по
заднему — с одним спутанным рядом. Стигмы переднегруди с большими, овально-
вытянутыми стигмальными пластинками, отверстие стигмы расположено близ края
пластинки, примерно в 5 раз уже ее. Стигмальные пластинки сегментов брюшка
втрое шире отверстий стигмы. Голенелапка (рис. 27, 4) сильно хитинизированная,
снизу с 7-9 шипиковидными щетинками; коготок простой, острый. Личиночный
чехлик гладкий, без скульптурных образований. Личинки в подстилке.
В регионе 13 видов; известны личинки 4 видов.
76
Таблица для определения видов рода Pachybrachis
1(2). Крыловые склериты среднеспинки (рис. 27, 2) с 3 длинными и 12-14 коротки-
ми щетинками, наружные претергальные с 1 длинной и 3 короткими, внутренние с
2 длинными и 6-7 короткими щетинками; краевая щетинка первого ряда лба (fd)
длинная и простая. ЕЧ, СК, СБ, ДВ.................Р. scriptidorsum Marseul
2(1). Крыловые склериты среднеспинки с 4-6 длинными щетинками.
3(4). Крыловые склериты среднеспинки (рис. 27, 3) с 4 длинными и 5-6 короткими
щетинками, наружные претергальные с 1 длинной и 2 короткими, внутренние — с
3 длинными и 5-6 короткими щетинками. Краевая щетинка первого ряда лба (f6)
короткая, на вершине зазубрена (рис. 28, 7). ЕЧ, СК, СБ, ДВ.............
...............................................Р. hieroglyphicus Laicharting
4(3). Крыловые склериты среднеспинки с 5 длинными и 8-9 короткими щетинка-
ми, наружные претергальные склериты с 1 длинной и 4 короткими, внутренние с 2
длинными и 11-12 короткими щетинками. Краевая щетинка первого ряда лба длин-
ная и простая. Транспалеаркт, кроме севера. Указанные ниже 2 вида, очевидно, бу-
дут обьединены...................................Р. fimbriolatus Suffrian, R mendax Suffrian
3. Род Cryptocephalus Geoffroy
Один из наиболее богатых видами родов, морфологическая характеристика
которого в основном совпадает с характеристикой подсемейства. Теменной кант
явственный, с рядом зазубренных или простых щетинок вдоль него в количестве от
4-5 до 7-10, поры на лбу и темени отсутствуют. Сильно хитинизированный гре-
бень вдоль теменного канта развит, хотя бы в области усиков. Голенелапка сильно
хитинизированная, снизу с 4-5 (редко более) крепкими щетинками. Личинки боль-
шей частью живут в подстилке, под камнями, однако в некоторых случаях (особен-
но молодые личинки) могут встречаться на растительности.
В регионе около 100 видов, разделяемых на ряд подродов, не упоминаемых
ниже.
Личинки большинства видов различаются неплохо, особенно старших возрас-
тов; у 1-го возраста скульптура головы еще не развита, а ее хетотаксия впослед-
ствии заметно меняется. Существует пять определительных таблиц для этого рода:
по Западной Европе (Steinhausen, 1994), Европейской части СССР (Оглоблин, Мед-
ведев, 1971), Сибири и Дальнего Востока (Медведев, Зайцев, 1978), Монголии (Мед-
ведев, 1982), Японии (Kimoto, Takizawa, 1994). Однако, не все виды могли быть
включены в приводимый ниже определитель из-за несовместимости признаков, ис-
пользованных разными авторами и неполноты описаний; к тому же, в определителе
Штайнхаузена многие виды, очевидно, были включены по личинкам 1-го возраста.
Таблица для определения видов рода Cryptocephalus
1(24). Лобные швы сливаются с теменным кантом на уровне хеты V3 участок меж-
ду лобным швом и теменным кантом широкий (рис. 29, 1-3).
2(5). Верхняя губа с длинным узким срединным выступом, выдающимся за края
губы (рис. 29, 7). Голенелапка с 4 щетинками снизу.
77
Рис. 29. Детали строения личинок рода Cryptocephalus\
1-4 — голова: 1 — С. octomaculatus, 2 — С. elegantulus, 3 — С. labiatus, 4 — С. coryli', 5-8 —
верхняя губа: 5 — С. fulvus, 6 — С. bipunctatus, 7 — С. octomaculatus, 8 — С. coryli', 9-10 —
переднеспинка: 9 — С. hypochaeridis, 10 — С. aureolus', 11—17 — стигмы: 11 — С. pini, 12 —
С. hypochaeridis, 13 — С. fulvus, 14 — С, elegantulus, 15 — С. ocellatus, 16 — С. bipunctatus,
17 — С. coryli. По Штейнхаузену.
3(4). ЕЧ, СК. Личинка на сухих листьях тополя.........С. octomaculatus Rossi
4(3). Юг ДВ. Лоб с ячеистой структурой. Средне- и заднегрудь с бурыми бугорками
на боках..............................................С. tetradecaspilotus Baly
5(2). Верхняя губа без срединного выступа или он небольшой и не выступает за
края губы (рис. 29, 5). Сюда же относятся личинки С. confusus Suffrian и С. nigro-
fasciatus Jacoby.
6(9). Стигмальные пластинки небольшие, продолговатые, не более чем в 3 раза длин-
нее, но не шире стигмы (рис. 29,14). Хета V2 вдвое длиннее хет V345 (рис. 29, 2).
7(8). Хеты лба короткие булавовидные (рис. 29, 2). Голова черная, тергит переднес-
пинки, бедра и голенелапки темно-бурые. Жуки на деревьях, кустарниках, часто на
полыни. Транспалеаркт..............................С elegantulus Gravenhorst
8(7). Хеты f-f4 короткие, булавовидные; хеты f5-f6 длинные, остроконечные. Голо-
ва, тергит переднеспинки, бедра и голенелапки светло-бурые. Жуки на деревьях,
кустарниках. Транспалеаркт............................С. coerulescens Sahiberg
9(6). Стигмальные пластинки значительно превышают диаметр стигм.
78
10(11). Стигмальные пластинки на груди С-образные, на брюшке круглые (рис. 32,
13-14). Все хеты лба длинные и слегка зазубренные (рис. 31, 3). Связан с ивами.
Транспалеаркт.........................................С. exiguus Schneder
11(10). Стигмальные пластинки круглые (у С. fulvus Goeze, возможно, продолгова-
тые, см. тезу 15).
12(17). Стигмальные пластинки не очень большие, не более чем в 4 раза крупнее
стигмы.
13(14). Склерит переднеспинки на переднем крае среди длинных простых щетинок
также с 4-5 короткими булавовидными щетинками. Стигмальные пластинки в 2-3
раза крупнее стигм (рис. 29, 77). Личинки питались на сухих сосновых иглах. ЕЧ,
СБ, ДВ....................................................С. pini Linnaeus
14(13). Склерит переднеспинки на переднем крае без коротких булавовидных ще-
тинок. Стигмальные пластинки в 3-4 раза больше стигм. Хета V2 длинная и острая.
15(16). Хеты V3-V5 вдвое короче V2 Выемка верхней губы с небольшим выступом.
Стигмальные пластинки, по Штейнхаузену, круглые (рис. 29, 73), по нашим дан-
ным продолговатые (рис. 32, 25, 26). Голова желто-бурая тело светло-оранжевое.
ЕЧ, СК, СБ, ДВ............................................С. fulvus Goeze
16(15). Хеты Vj и V3-V5 вдвое короче V2 Выемка верхней губы без выступа. Голова
темно-бурая, тело желто-белое. Транспалеаркт..........С. bilineatus Linnaeus
17(12). Стигмальные пластинки большие, по крайней мере в 5 раз больше стигм.
18(21). Стигмальные пластинки большие, в 5-6 раз больше стигм. Хета V2 длин-
ная, вдвое длиннее прочих.
19(20). Хета Vx остроконечная, хеты V3-V5 притупленные (рис. 29, 3). Выемка вер-
хней губы с небольшим, но явственным выступом. Личинка питалась сухими лис-
тьями ивы. ЕЧ, СК, СБ, ДВ.............................С labiatus Linnaeus
20(19). Хеты V! и V3-V5 притупленные. Выемка верхней губы без выступа. Личинка
питалась сухими листьями Hypericum и Erigeron. ЕЧ, СК.С pygmaeus Fabricius
21(18). Стигмальные пластинки в 7-10 раз крупнее стигм, широко овальные (рис.
29, 75). Хета V2 длинная и острая, прочие тупые и вдвое короче.
22(23). Голова темно-бурая, лоб в густой грубой зернистости, частично собранной в
ряды. Личинки питались сухими листьями ольхи. ЕР, СК, ЗСБ ... С ocellatus Drapiez
23(22). Голова светло-бурая. Лоб в очень густой мелкой зернистости, без заметных
рядов. Личинки питались сухими листьями ивы. ЕЧ...........С rufipes Goeze
24(1). Лобные швы сливаются с теменным кантом заметно ниже хеты V3 или не
сливаются, участок между теменным кантом и лбом более или менее параллель-
ный.
25(30). Теменной кант с каждой стороны с 9-13 хетами.
26(27). Теменной кант с простыми острыми хетами одинаковой длины (рис. 28, 2).
Передний край верхней губы с широким слабым выступом. Стигмальные пластин-
ки круглые, втрое крупнее стигмы. Голенелапка снизу с 6-8 шипами. Внутренние
тергальные склериты заднеспинки с 2 макро- и 2 микрохетами, крыловой склерит с
3 макро- и 1 микрохетой. Лоб без явственной зернистости. Чехлик с продольными
ребрышками. ЕЧ, СК, СБ................................С sericeus Linnaeus
27(26). Теменной кант с простыми острыми хетами разной длины и формы. Внут-
ренние тергальные склериты заднеспинки с 2 макро- и 3 микрохетами, крыловой
склерит с 4 макро- и 2 микрохетами.
79
Рис. 30. Детали строения личинок рода Cryptocephalus'.
1-5 — С. flavipes'. 1 — голова, 2 — верхняя губа, 3 — передне- и среднегрудь, 4 — грудная
стигма, 5 — брюшная стигма; 6, 7 — передне- и среднегрудь: 6 — С. bidentulus, 7 — С.
stschukini', 8 — голова С. coerulans. По Мосейко и ориг.
80
28(29). Теменной кант с 13 парами хет, длинные хеты простые, короткие — зазуб-
рены (рис. 30, 7). Передний край верхней губы с явственной выемкой, имеющей
срединный выступ (рис. 30,2). Тергиты груди—рис. 30,3. Стигмы груди и брюшка —
рис. 30, 4-5. ЕЧ, СК, СБ, ДВ..........................С. flavipes Fabricius
29(28). Теменной кант с 10 парами коротких булавовидных и удлиненных зазубрен-
ных хет (рис. 30, 8). Передний край верхней губы с едва намеченной выемкой. Лоб
с явственной зернистостью. СБ, ДВ.....................С. coerulans Marseul
30(25). Теменной кант с каждой стороны с 4-6 хетами (рис. 28, S, 9).
31(34). Голенелапка снизу с 6-7 шипами
32(33). Голенелапка снизу с 6 шипами в 2 ряда и простой щетинкой (рис. 27, 5).
Теменной кант хорошо развит, с хетами равной длины. Темя с короткими густыми
щетинками. Хеты f6 простые, прочие зазубрены. Стигмы — рис. 32, 23, 24. СБ, ДВ
.........................................................С. regalis Gebler
33(32). Голенелапка снизу с 7 шипами (рис. 27, 7). Теменной кант развит только на
уровне глазков в виде короткого гребня, далее отсутствует, хета V2 длинная, прочие
короткие (рис.28, 5). Темя с короткими редкими щетинками. Все хеты лба простые.
Стигмальные пластинки хорошо развиты, гораздо крупнее диаметра стигм, осо-
бенно на груди (рис. 32, 7, 2). Под полынью. СБ, ДВ......С agnus Weise
34(31). Голенелапка снизу не более чем с 4 шипами (рис. 27, 6).
35(36). Голенелапка снизу только с двумя шипами. Все хеты теменного канта корот-
кие, равной длины. Стигмальные пластинки маленькие, круглые, едва вдвое круп-
нее стигмы. Выемка верхней губы с небольшим выступом, не достигающим пере-
днего края. Темя за теменным кантом с 40-50 парами жестких щетинок. Щетинки
склерита переднегруди сидят на круглых светлых основаниях. ЕЧ, СК, СБ....
......................................................С. schaefferi Schrank
36(35). Голенелапка снизу с 4 шипами.
37(42). Стигмальные пластинки груди и брюшка редуцированные или в виде очень
узкой полоски, менее диаметра стигм (рис. 32, 3, 4).
38(41). Склерит переднеспинки густо зернистый, по переднему и заднему краям со
спутанными рядами из простых заостренных хет. Верхняя губа без срединного вы-
ступа. Лоб продольный, яйцевидный, с простыми длинными хетами (рис. 31, 7).
Бедра снизу, помимо простых щетинок, с 1-3 шипами.
39(40). Темя с многочисленными длинными тонкими и короткими утолщенными
щетинками. Щетинки склерита переднеспинки у основания окружены светлыми
колечками, наружные претергальные склериты среднеспинки с 4 щетинками (рис.
30, 7). СБ, ДВ.....................................С. stschukini Faldermann
40(39). Темя с простыми щетинками разной длины. Щетинки склерита переднес-
пинки у основания без светлых колечек, наружные претергальные склериты сред-
неспинки с 5 щетинками (рис. 30, 6). Тува.............С. bidentulus Suffrian
41(38). Склерит переднеспинки морщинистый, по переднему и заднему краям со
спутанными рядами из длинных простых и коротких зазубренных хет, в основном
по переднему краю. Верхняя губа со срединным выступом, лоб слабо овальный, с
зазубренными щетинками. Стигмы — рис. 32, 5, 6. Под полынью. Степи ЕЧ, СК,
ЗСБ..............................................С. gamma Herrich-Schaffer
42(37). Стигмальные пластинки хотя бы на груди хорошо развиты.
43(44). Стигмальные пластинки на груди продолговатые, червеобразные, на брюш-
81
Рис. 31. Лоб личинок рода Cryptocephalus:
1 — stschukini; 2 — crux; 3 — exiguus; 4 —janthinus; 5 — bivulneratus; 6 — koltzei.
ке редуцированные (рис. 32, 7, 8). Лоб (рис. 31,2) слабо продольный, на вершине
притуплен, с зазубренными щетинками (кроме f6). Верхняя губа с крупным прямо-
угольным выступом, достигающим уровня переднего края (рис. 32, 2). На полыни.
СБ, ДВ...................................................С. crux Gebler
82
44(43). Стигмальные пластинки на брюшных сегментах явственные.
45(46) Стигмальные пластинки груди и брюшка резко различаются по форме, на гру-
ди круглые, на брюшке рожковидные (рис. 32, 75, 16). Темя с 36-38 парами простых
коротких и длинных щетинок, теменной кант с с 5 парами простых щетинок. Верши-
на лба прямо срезана (рис. 31,6), щетинки лба тонкие и длинные, зернистость лба
наиболее густая в центре, по бокам и у основания. Юг ДВ....С. koltzei Weise
46(45). Стигмальные пластинки груди и брюшка почти не различаются по форме,
на груди обычно крупнее.
47(68). Стигмальные пластинки груди и брюшка округлые или короткоовальные,
реже на груди слабо продолговатые, но тогда заметно шире диаметра стигм.
48(57). Стигмальные пластинки груди и брюшка очень большие, круглые, оваль-
ные или почковидные, в 5-8 раз шире диаметра стигм (рис. 33, 5, 7, 8). Голенелапка
снизу с 2 явственными и 2 более слабыми шипами.
49(50). Хеты лба длинные, утолщены и зазубренные, f4-f6 простые. Верхняя
губа без срединного выступа (1-й возраст) или с небольшим выступом. Крыловые
склериты среднегруди с 5 щетинками. Стигмы — рис. 33, 7, 8. ЕЧ, СК, СБ..
......................................................С. octacosmus Bedel
50(49). Хеты лба короткие.Верхняя губа со срединным выступом.
51(56). Срединный выступ верхней губы крупный, лежит на одном уровне с ее боко-
выми краями (рис. 29, 6). Все хеты лба примерно одинаковой длины, зазубренные.
52(55). Стигмы круглые (рис. 29,16).
53(54). Передний край переднеспинки с 1 правильным рядом щетинок.Крыловые
склериты среднегруди с 5 щетинками. Лоб и темя сплошь в явственной зернистос-
ти. ЕЧ, СК, СБ, ДВ..................................С. bipunctatus Linnaeus
54(53). Передний край переднеспинки с большим количеством микрохет (25-27 с
каждой стороны), расположенных в 2-3 сильно спутанных ряда, и с 5 макрохетами.
Крыловые склериты среднегруди с 6 щетинками. ЕЧ, СК, СБ. Известна личинка 1-
го возраста...........................................С. biguttatus Scopoli
55(52). Стигмы овальные (рис. 33, 6). Зернистость лба редкая, неправильная, обра-
зует хорошо выраженные цепочки. Крыловые склериты среднегруди с 6 щетинка-
ми. На ивах. ЕЧ, ЗСБ.......С. decemmaculatus Linnaeus (С. bothnicus Linnaeus)
56(51). Срединный выступ верхней губы небольшой, не достигает уровня боковых
краев. Зернистость лба очень редкая, равномерная, со слабо выраженными цепоч-
ками. Хеты f5 и f6 простые, в 1,5 раза длиннее прочих (рис. 31,4). Крыловые скле-
риты среднегруди с 5 щетинками. Стигмы — рис. 33, 5. Транспалеаркт......
......................................................С. janthinus Germar
57(48). Стигмальные пластинки груди и брюшка округлые или короткоовальные,не
более чем в 3 раза превышают диаметр стигм.
58(61). Стигмальные пластинки небольшие, незначительно (не более чем в 2 раза)
превышают диаметр стигм. Срединный выступ верхней губы треугольный. Лоб с
закругленной вершиной (рис. 31,5). Сибирские виды.
59(60). Все хеты лба простые (рис. 31, 5). Стигмальные пластинки — рис. 32, 77,
72. Юг СБ........................................С. bivulneratus Faldermann
60(59). Хеты f4-f6 простые, прочие зазубренные (рис.28, 6). Голенелапка снизу с 3-
4 явственными шипами (рис. 27, 8). Стигмальные пластинки — рис. 32, 77, 18. На
полыни. Юг СБ.........................................С. limbellus Sufirian
83
Рис. 32. Стигмы личинок подсемейства Cryptocephalinae:
1-26 — Cryptocephalus (нечетные — грудные, четные — брюшные): 1,2 — agnus, 3, 4 —
stschukini, 5, 6—gamma, 7,8 — crux, 9,10 — cordiger, 11,12 — bivulneratus, 13,14 — exiguus,
15,16—koltzei, 17,18 — limbellus, 19,20 — ochroloma, 21,22 — mannerheimi, 23,24 — regalis,
25, 26 — fulvus; 27 — стигма брюшка Stylosomus.
61(58). Стигмальные пластинки груди и брюшка умеренно крупные, примерно в 3-
4 раза превышают диаметр стигм (рис. 32, 9, 10). Европейские виды, частично за-
ходящие в Азию.
62(63). Срединный выступ верхней губы длинный, прямоугольный, срезан на вер-
шине, достигает переднего края (рис. 28, 8). Хеты лба булавовидные, f6 длинные и
зазубренные, зернистость лба редкая, в центре — поперечные нитеобразные обра-
зования. Передний край переднеспинки с каждой стороны с неправильным рядом
из 9 макрохет и 3-4 микрохет. Внутренние тергальные склериты на среднеспинке с
2 макрохетами и 2 микрохетами, на заднеспинке с 2 макрохетами и 4 микрохетами;
крыловые склериты средне- и заднеспинки с 4 макрохетами и 3-4 микрохетами. На
лиственных. ЕЧ, ЗСБ....................................С. nitidus Fabricius
63(62). Верхняя губа без длинного прямоугольного выступа.
64(65). Передний край переднеспинки с одним правильным рядом хет, с каждой
стороны с 4 макрохетами и 7-8 микрохетами, задний край с немногочисленными
микрохетами, остальная часть диска с единичными макрохетами. Крыловые скле-
риты среднеспинки с 4 макрохетами и 2 микрохетами. ЕЕЧ, СБ, ДВ.........
.....................................................С. nitidulus Fabricius
65(64). Передний край переднеспинки со спутанным, обычно более или менее двой-
ным рядом хет.
84
2
Рис. 33. Стигмы груди (7, 3, 5, 7) и 1-го тергита брюшка (2, 4, 8) личинок родов
Suffrianus и Cryptocephalus'.
1,2 — S. pumilo; 3,4 — С. distinguendus', 5 — С. janthinus', 6 — С. decemmaculatus', 7,8 — С.
octacosmus.
66(67). Верхняя губа без крупного выступа (рис. 28,9). Все хеты лба булавовидные.
Стигмы — рис. 32, 9,10. ЕЧ, СК, СБ, ДВ.................С. cordiger Linnaeus
67(66). Срединный выступ верхней губы треугольный, почти достигает переднего
края. Хеты f6 в 2 раза длиннее прочих, зазубренные, остальные булавовидные. Все
стигмальные пластинки округлые (рис. 33,3, 4). Зернистость лба равномерная, ред-
кая. На березах. Транспалеарктический вид........С. distinguendus Schneider
68(47). Стигмальные пластинки узкие, продолговатые, не шире или даже уже диа-
метра стигм.
69(74). Теменной кант с 5 хетами, V2 длинная, остроконечная, прочие короче, пило-
видные. Внутренние тергальные склериты с 1 макро- и 1 микрохетой. Стигмы —
рис. 32,19, 20. Чехлик с резким спинным килем.
70(71). Выемка верхней губы с большим прямоугольным выступом, достигающим
переднего края (рис. 29, 8). Лоб и темя с густой зернистостью, лобные швы нераз-
личимы (рис. 29, 4). Стигмы — рис. 29,17. ЕЧ, СК, СБ, ДВ..С. coryli Linnaeus
85
71(70). Выемка верхней губы с небольшим выступом, не достигающим переднего
края. Лоб с густой равномерной зернистостью, доходящей до теменного канта, лоб-
ные швы различимы. СБ, ДВ.
72(73). Верхняя губа со слабым срединным выступом, хеты f6 булавовидные или
зазубренные. Передний край переднеспинки с рядом из 6 макро-, 1 микрохеты и 4
пор с каждой стороны. Стигмы — рис. 32, 19, 20. Личиночный чехлик по бокам с
развитыми продольными гребнями. СБ, ДВ................С. ochroloma Gebler
73(72). Верхняя губа с развитым треугольным выступом. Хеты f6 простые или сла-
бо зазубрены. Передний край переднеспинки с рядом из 6 макро-, 6 микрохет и 2
пор с каждой стороны. Стигмы — рис. 32,21,22. Личиночный чехлик по бокам без
продольных гребней. СБ, ДВ..........................С. mannerheimi Gebler
74(69). Теменной кант с 5 недлинными хетами одинаковой длины, Выемка верхней
губы с небольшим выступом, не достигающим переднего края. Личиночный чех-
лик с продольными гребнями.
75(76). Передний край переднеспинки со спутанным двойным рядом щетинок (рис.
29, 9). Голова светлобурая с темной негустой зернистостью. Хеты лба тупые. Стиг-
мы — рис. 29,12. ЕЧ, СК, СБ.....С. hypochaeridis Linnaeus (С. cristula Dufour)
76(75). Передний край переднеспинки с простым рядом щетинок (рис. 29, 70). Го-
лова темнобурая с густой зернистостью. Хеты лба остроконечные. ЕЧ, СК, ЗСБ ..
........................................................С aureolus Sufirian
4. Род Stylosomus Suffrian
Темя и особенно лоб (рис. 28, 4) в густой зернистой скульптуре. Эпикраниаль-
ный шов длинный, темя с редкими простыми короткими щетинками, теменной кант
явственный, без гребня, с 4 короткими зазубренными и 1 очень длинной простой
щетинкой и порой между V2 и V3. Лоб с 12 зазубренными щетинками (4, 4, 2 и 2) и
2 порами, расположенными ковнутри от f2. Верхняя губа с широкой и неглубокой
выемкой и едва намеченным срединным выступом. Склерит переднеспинки хити-
низирован только в передней половине, слабо поперечный, с рядом из 5 длинных и
12-14 коротких щетинок и пор по переднему краю и с поперечным спутанным ря-
дом из 7-8 длинных и коротких щетинок по середине склерита. Склериты груди
(рис. 27, 7) и брюшка слабо выражены, на груди у переднего края имеется 4-6 мел-
ких щетинок, внутренние тергальные с 1 макро- и 3 микрохетами, наружные тер-
гальные — с 1 макро- и 1 микрохетой, крыловые склериты крупные, округлые, с 2
макро- и 3 микрохетами. Стигмальная пластинка груди крупная, округлая, отвер-
стие стигмы в 6-7 раз меньше диаметра пластинки (рис. 32, 27). Стигмальная плас-
тинка брюшных сегментов в 1,5 раза меньше грудной. Ноги (рис. 27, 9) длинные,
голенелапка снизу с 4 щетинками, утолщенными в основании и постепенно утонча-
ющимися к вершине, сверху — с длинными тонкими щетинками. Коготок тонкий,
слабо изогнут.
В регионе 3 вида на тамариске (5. tamaricis Herrich-Schaffer) и Myricaria (S'.
sinensis Lopatin). Описание рода дается по личинкам 1-го возраста S. sinensis Lopatin,
встречающегося в Туве.
86
8. ПОДСЕМЕЙСТВО chlamisinae
Представители этого подсемейства близки к Cryptocephalinae и Clytrinae, по-
вторяя в общих чертах их морфологию и отличаясь рядом второстепенных призна-
ков. В регионе 1 род — Chlamisus Rafinesque.
Голова округло-вытянутая, умеренно хитинизированная, без теменного канта,
эпикраниальный шов длинный, лобные швы слабо изогнутые, темя с 3-4 щетинка-
ми вдоль лобного шва, 1 щетинкой у теменного шва и с несколькими мелкими на
боковых частях; лоб (рис. 34, 7) с 14 щетинками, обычно расположенными в 5 ря-
дов (2, 2, 4, 4, 2). Лоб, наличник и верхняя губа сросшиеся, без швов, передний
край верхней губы сильно хитинизированный, со слабым срединным выступом и 6
плоскими длинными маргинальными щетинками с каждой стороны (рис. 34, 2).
Наличник с поперечным рядом из 2 щетинок и 4 пор. Все щетинки головы тонкие и
длинные. Усики 2-члениковые, с довольно узкой конусовидной папиллой на вто-
ром членике. Челюстные щупики 4-члениковые, губные — 2-члениковые. Стипес с
1 щетинкой в передненаружном углу и с 1 щетинкой на диске. Склерит переднес-
пинки (рис. 34, 3) без скульптуры, с тонкими щетинками и порами по переднему
краю и в задней трети. Прочие сегменты груди и брюшка с редкими тонкими длин-
ными и короткими щетинками. Вертлуги слабо обособлены от бедер, с 1 щетинкой.
Голенелапка снизу с 2 продольными рядами из 7-8 тонких длинных щетинок с рас-
ширенными концами и изогнутыми на вершине. Коготки сильно изогнутые, с круп-
ным зубцом при основании (рис. 34, 4). Личиночный чехлик цилиндрический, но
заметно расширен кпереди, с концентрическими рядами тупых бугорков.
Личинки обитают преимущественно на растениях, очень часто на стеблях.
Единственный вид рода Ch. pubiceps Chujo встречается на юге ДВ на малине,
его личинки были найдены на поверхности почвы.
Рис. 34. Детали строения личинки подсемейства Chlamisinae:
Chlamisus pubiceps: 1 — голова, 2 — верхняя губа, 3 — переднеспинка, 4 — голенелапка.
87
9. ПОДСЕМЕЙСТВО LAMPROSOMINAE
Тело скарабеоидного типа, довольно толстое, желтовато-белое, задняя часть
брюшка подогнута под переднюю, срединные тергиты брюшка расширены (рис.
35, 7, 2). Голова (рис. 35, 3) без теменного канта, лоб, наличник и верхняя губа
слиты вместе, глазков 5 с каждой стороны (2, 2, 1). Челюстные щупики 3-членико-
вые, губные — 2-члениковые, стипес продолговатый, кардо небольшой (рис. 35, 5).
Мандибулы двузубчатые, с острой вершиной (рис. 35, 6). Усики длинные, 3-члени-
ковые, с удлиненными первыми двумя члениками и небольшой конусовидной па-
пиллой на 3-м членике (рис. 35, 3, 8). Склерит переднеспинки со щетинками на
переднем крае и отчасти на диске (рис. 35, 4). Хетотаксия средне-, заднегруди и
тергита брюшка — рис. 35, 4. Стигма — рис. 35, 7. Ноги длинные, включая хорошо
обособленный вертлуг, коготки тонкие, слабо изогнутые, без зубца на основании
(рис. 35, 9). Яйцевые чехлики с различной скульптурой, иногда с крепящей нитью,
личиночные чехлики могут быть конические, часто с загнутой вершиной, но у на-
ших видов цилиндрические или кувшинообразные. Личинки питаются на стеблях
кормовых растений, с которых часто спускаются в подстилку. В регионе 2 слабо
отличающихся рода, в связи с чем описание родов не приводятся. Питание на ара-
лиевых.
Таблица для определения родов подсемейства Lamprosominae
1(2). Верхняя губа с прямым передним краем. Вершинные тергиты брюшка с до-
вольно длинными щетинками (рис. 35, 7). 1 вид, О. concolor Stum, на СК, питание
на Hedera...............................................Oomorphus Curtis
1(2). Верхняя губа с выемчатым передним краем. Вершинные тергиты брюшка без
длинных щетинок (рис. 35, 2). 1 вид, О. nigrocoeruleum Baly на юге ДВ, питание
жуков на Kalopanax, Akanthopanax....................Oomorphoides Monros
88
Рис. 35. Детали строения личинок подсемейства Lamprosominae:
Oomorphus concolor. 1 — общий вид; 3 — голова; 9 — нога; 2 — Oomorphoides cupreatus,
общий вид; 4-8 — Oomorphoides martensi‘. 4 — грудь и 1-й тергит брюшка, 5 — лабио-мак-
силлярный комплекс, 6—мандибула, 7—стигма, 8 — вершина усика. По Кимото и Такизава
и ориг.
89
10. подсемейство synetinae
Тело белое или кремоватое, со складками на тергитах всех сегментов, кроме
переднегрудного, несущего склерит, и 9-10-го сегментов брюшка. Верх покрыт
щетинками, тергиты 1-5-го брюшных сегментов с шипиками. На брюшке по бокам
3 ряда бугорков — стигмальный, эпиплевральный и гипоплевральиый; стерниты с
редкими щетинками. Лобные швы дважды вогнуто-выпуклые, эндокарина длин-
ная, лоб с 4 щетинками у переднего края (рис. 12, 5). Темя с 3 парами щетинок,
наличник с 4, верхняя губа с 6 щетинками. Глазков нет. Усики 1-члениковые, вер-
шина членика с большой конусовидной папиллой и 7 шиловидными щетинками
(рис. 12, 6). Мандибулы с 3-4 зубцами. Нижнечелюстные щупики 4-члениковые.
Сросшиеся галея и лациния несут на вершине и по внутреннему краю 9 крупных и
3 мелких щетинки. Субментум с 3 парами щетинок, ментум вытянут в длину и вздут,
с узким ментальным склеритом (рис. 12, 7). Нижнегубные щупики 1-члениковые.
9-10-й тергиты несут крупный склерит в виде треугольного щитика, хетотаксия
вершины брюшка сверху и снизу показана на рис. 12, 8, 9. Ноги 3-члениковые,
граница между вертлугом и бедром неразличима, голенелапка с 7 щетинками, ко-
готки длинные и острые, с маленькой щетинкой на нижней стороне. Длина тела
взрослой личинки до 9 мм.
Личинки живут и окукливаются в почве, обычно в оподзоленной суглинистой
и песчаной, достаточно сухой, до глубины в 70 см. Связаны с ельниками и березня-
ками, особенно на опушках и вырубках.
В регионе один род — Syneta Lacordaire с 2 видами.
Таблица для определения видов рода Syneta
1(2). Мандибулы с 4 зубцами. Темя с короткими и длинными щетинками, в задней
части с рядом очень коротких щетинок с каждой стороны (рис. 12, 5). Коготок за-
метно расширен к основанию. Вся таежная зона Палеарктики, на корнях березы....
........................................................5. betulae Fabricius
2(1). Мандибулы с 3 зубцами. Темя с короткими щетинками, в задней части только
с 1 короткой щетинкой. Коготок очень тонкий, практически не расширен к основа-
нию. Юг ДВ, Сахалин, Курилы, в хвойных лесах на корнях лиственницы.......
...........................................................S. adamsi Baly
90
11. подсемейство eumolpinae
Тело более или менее С-образное или почти прямое (рис. 36). Окраска белая
или кремовая, с желтоватой или желто-бурой головой и складчатыми покровами.
Голова (рис. 37) гипогнатическая, умеренно склеротизованная, эпикраниальный шов
довольно длинный, лобные швы дважды вогнуто-выпуклые, эндокарина на лбу раз-
витая. Темя с немногочисленными щетинками, в пределах 4-11 пар, лоб с 3-5 пара-
ми щетинок. Глазки отсутствуют. Усики 2-члениковые, 1-й членик крупный, с ши-
рокой, мягкой, несклеротизованной основной частью, в которую может втягивать-
ся более узкая и склеротизованная вершинная часть; на вершине 1-го членика нахо-
дятся небольшой второй членик, несущий на вершине несколько щетинок, и кону-
совидная сенсориальная папилла, примерно такой же величины, как и второй чле-
ник (рис. 38, 7-5). Наличник и верхняя губа хорошо развиты, обособленные, на-
личник с 4 щетинками, образующими поперечный ряду основания; верхняя губа с
6 щетинками на диске и обычно с 8 мелкими щетинками непосредственно на пере-
днем крае. Мандибулы мощные, с 2-3 зубцами и 2 щетинками на наружном крае;
сочленовый бугорок и сочленовая ямка хорошо развиты (рис. 40,4). Максиллы (рис.
40, 7) представлены небольшими кардо и стипесом, галея и лациния слиты друг с
другом и с пальпигером; на расширенном пальпигере находятся 4-члениковые (редко
3-члениковые), щупики, отличающиеся постоянной хетотаксией: 1-й и 3-й членики
с 2 щетинками, 2-й — без щетинок, 4-й — с небольшой щетинкой и с боковым
изогнутым, довольно длинным выростом, располагающимся в углублении и же-
лобке членика на его наружной стороне. Нижняя губа в виде сплошной мягкой пла-
стинки, субментум представлен базальной частью пластинки, несущей 3 пары ще-
тинок, к ментуму следует относить переднюю часть пластинки, на которой имеют-
ся 1-члениковые щупики, окруженные узкой склеротизованной фигурой, незамк-
нутой впереди, которая обозначается как ментальный склерит; у его основания рас-
положены 2 длинные, а у вершины 2 пары коротких щетинок. Переднеспинка с
более или менее явственным срединным склеритом, тергиты средне- и заднегруди
и 1-8-го сегментов брюшка с 3-4 поперечными складками и немногочисленными,
реже довольно густыми щетинками. Брюшные сегменты с 3 рядами бугорков по
бокам: над стигмами лежат стигмальные бугорки, под стигмами находятся эпип-
левральные и гипоплевральные бугорки; далее, уже на боковых частях стернитов,
находится еще ряд бугорков, которые иногда бывают вытянуты в длинные отростки
и обозначаются, по аналогии с личинками слоников, как адвентриты; все бугорки
несут щетинки или шипики. Стерниты с довольно короткими, но многочисленны-
ми щетинками. Стигмы на переднегруди и на 1-8-м сегментах брюшка кольцеоб-
разные (рис. 40, 5). 9-10-й сегменты представляют опорные образования в виде
околоанальных бугров, пластинок и шипиков, 9-й тергит с более или менее выра-
женным склеритом, морфология 10-го сегмента довольно сложная и резко отлича-
ется у разных родов; 9-й сегмент обычно охватывает 10-й, расположенный более
или менее вентрально (рис. 39,1-6). Ноги (рис. 35, 6) довольно слабые; у основа-
ния ног имеется по мозолистому бугорку, сильнее хитинизированному, чем окружа-
ющие покровы; вертлуг с 3 щетинками, бедро с 6 щетинками, голенелапка с 8 ще-
тинками, коготки острые, тонкие, слабо серпообразно изогнутые, без пульвилл.
91
Рис. 36. Общий вид личинок подсемейства Eumolpinae:
1 — Colasposoma', 2 — Eumolpus', 3 — Pachnephorus. По Курчевой и ориг.
Личинки Eumolpinae (по крайней мере, палеарктические виды) живут в почве и
питаются корнями растений, являясь, очевидно, олигофагами. Яйца откладывают-
ся в почву (у Colasposoma Motschulsky в стебли растений); окукливание в почве.
Биология и жизненный цикл изучены недостаточно полно; число личиночных воз-
растов, судя по некоторым данным, равняется трем.
В регионе 13 родов, из которых 4 встречаются только на юге ДВ; их личинки
неизвестны, но из них обычны только Cleoporus variabilis Baly на розоцветных,
прочие рода очень редки.
Таблица для определения родов подсемейства Eumolpinae
1(10). Передний край лба утолщен и склеротизован (рис. 37, 7, 2). Брюшные сег-
менты без длинных выростов на стернитах (рис. 36, 7). Мандибулы короткие и
широкие, часто с тупым зубцом на режущем крае.
2(3). Тергиты груди и брюшка у личинок старших возрастов с первичной и вторич-
ной хетотаксией, каждый тергит с 50-60 щетинками. Мандибулы без зубца на ре-
жущем крае. Бугры 10-го сегмента, помимо щетинок, с многочисленными опорны-
ми шипиками, на 9-м сегменте дорсальные бугры попарно слиты, вентральные и
боковые — с многочисленными опорными шипиками (рис. 39, 7)..............
.........................................................1. Colasposoma
3(2). Тергиты груди и брюшка только с первичной хетотаксией, каждый тергит с
10-18 щетинками. На 9-м сегменте дорсальные бугры разделенные.
92
Рис. 37. Головные капсулы личинок подсемейства Eumolpinae:
1 — Bromius obscurus; 2 — Basilepta fulvipes; 3 — Eumolpus goniostoma. По Курневой и ориг.
4(5). Лобное утолщение в виде поперечного валика с продольным гребнем, возвы-
шается несколько надо лбом и сильно над наличником (рис. 41, 7). 10-й сегмент
брюшка в виде окружающих анальное отверстие бугров, густо покрытых опорны-
ми шипиками. Склеротизованная фигура нижней губы по бокам округло выпуклая,
назади прямая или чуть вогнутая (рис. 41, 3).............2. Chloropterus
5(4). Лобное утолщение не образует валика, незначительно возвышается над на-
личником (рис. 37, 7). Бугры 10-го сегмента без опорных шипиков.
6(7). Лоб с 5 парами щетинок (рис. 37, 7). Сенсориальная папилла усика не длиннее
и не толще 2-го членика, с простой вершиной (рис. 38, 2). Передний край нижней
губы двулопастной, без шипиков; склеротизованная фигура в виде полукруга. Вен-
тральные бугры 9-го сегмента разделенные, с немногочисленными шипиками (рис.
39, 3)......................................................3. Bromius
7(6). Лоб с 3-4 парами щетинок (рис. 37, 2). Сенсориальная папилла усика почти
вдвое длиннее и шире 2-го членика (рис. 38,3). Передний край нижней губы равно-
мерно дуговидный, с многочисленными шипиками; склеротизованная фигура в виде
прямоугольника (рис. 38, 7).
8(9). Передний край верхней губы равномерно дуговидный, без толстых шипиков.
Сенсориальная папилла усика с двулопастной вершиной (рис. 38, 3). Голенелапка с
6 щетинками. Лоб с 2 длинными щетинками в задней части (рис. 37, 2). Вентраль-
ные бугры 9-го сегмента слитные, с единичными щетинками, без явственных опор-
ных шипиков (рис. 39, 2)....................................4. Basilepta
9(8). Передний край верхней губы прямой, с очень толстыми щетинками (рис. 42,2).
Сенсориальная папилла усика с конической вершиной (рис. 42, 5). Голенелапка с 4
щетинками. Лоб с 2 короткими щетинками в задней части (рис. 42, 7).5. Pagria
10(1). Передний край лба без утолщения и более сильной склеротизации (рис. 37,
3). Брюшные сегменты с более или менее длинными выростами и щетинками на
стернитах (рис. 36, 2). Мандибулы удлиненные, без тупого зубца на режущем крае.
11(14). Нижнечелюстные щупики 4-члениковые (рис. 40, 3). Брюшные выросты
сосцевидные (рис. 36, 2). 10-й брюшной сегмент образует. 3 разделенные лопасти
вокруг анального отверстия: непарную дорсальную с 2 щетинками на вершине и 2
вентральные в виде косо срезанных столбиков.
93
Рис. 38. Усик (7—5) и нижняя губа (6-9) личинок подсемейства Eumolpinae:
1,6 — Colasposoma dauricum; 3, 7 — Basilepta fulvipes', 2 — Bromius obscurus; 4 — Eumolpus',
5 — Pachnephorus', 8 — Eumolpus asclepiadeus', 9 — E. goniostoma. По Курневой и ориг.
12(13). Тело в густых щетинках (рис. 36, 2). Стипес с 2 щетинками на наружном
крае. Склеротизованная фигура нижней губы сзади вогнутая (рис. 38, <$). Дорсаль-
ная лопасть анального клапана треугольная.....................6. Eumolpus
13(12). Тело в негустых щетинках. Стипес с 1 щетинкой на наружном крае. Склеро-
тизованная фигура нижней губы сзади прямая. Дорсальная лопасть анального кла-
пана округлая (рис. 41,4)..................................7. Chrysochares
14(11) Нижнечелюстные щупики 3-члениковые (рис. 40,2). Брюшные выросты паль-
цевидные (рис. 36, 3). 10-й брюшной сегмент имеет непарный кольцевой вырост с
изогнутой и сильно склеротизованной площадкой (рис. 39, 4). Тело с редкими ще-
тинками....................................................8. Pachnephorus
94
Рис. 39. Вершина брюшка личинок подсемейства Eumolpinae:
1 — Colasposoma dauricum; 2 — Basilepta fulvipes\ 3 — Bromius obscurus; 4 — Pachnephorus
tessellatus', 5 — Eumolpus asclepiadeus\ 6 — E. goniostoma\ 7 — Chloropterus versicolor. По
Курчевой и ориг.
1. Род Colasposoma Motschulsky
Тело толстое, С-образно изогнутое, светло-кремовое, дорсальная часть покры-
та многочисленными тонкими, умеренно длинными щетинками, вентральная часть
с более толстыми щетинками, направленными кзади (рис. 36, 7). Голова блестящая,
без скульптурных образований, несколько уже переднеспинки. Эпикраниальный
шов доходит до середины головной капсулы, эндокарина и лобные швы слабо за-
метные. На темени расположено до 11 микро- и макрохет с каждой стороны, из них
2 макро- и 2 микрохеты лежат близ лобных швов, 3 микрохеты на вершине темени,
остальные по бокам. Лоб несет 4 пары микрохет (3 пары ближе к основанию, 1
пара посередине). Передняя часть лба сильно хитинизирована, с ясно выраженным
95
Рис. 40. Детали строения личинок подсемейства Eumolpinae:
Colasposoma dauricunr. 1 — максилла, 6 — нога; Eumolpus goniostomcr. 4 — мандибула, 5 —
стигма; Chrysochus'. 3 — челюстной щупик, 7 — голенелапка; Pachnephorus'. 2 — челюстной
щупик, 5 — голенелапка. По Курчевой и ориг.
возвышением, отделяющим лоб от наличника, но не выступающим над ним. На-
личник слабо хитинизирован, с 1 микрохетой с каждой стороны. 2-й членик усиков
очень маленький, несет 2 длинные и 2 короткие тупые щетинки на вершине и 1
очень мелкую на боковой части; папилла почти одинаковой длины и ширины, кол-
пачкообразная (рис. 38, 7). Мандибулы с 1 зубцом. Субментум с 3 парами хет, скле-
ротизованная фигура ментума прямоугольной формы, с закругленными задними
углами (рис. 38, 6). Нижнечелюстной щупик 4-члениковый (рис. 40, 7), 4-й членик
несет 1 микрохету и 4 короткие тупые щетинки на вершине.
Переднеспинка слабо хитинизированная, несет макро- и микрохеты, образую-
щие более или менее заметный круг на каждой половине. Тергиты брюшных сег-
ментов резко сужены посередине передними и задними валиками, хеты переднего
валика расположены в спутанный, заднего — в правильный ряд.
9-й брюшной сегмент охватывает кольцом анальный сегмент (рис. 39, 7). Сред-
ние и боковые вентральные пары 9-го сегмента явственно слитые и несут большое
количество шипиков, эпиплевральная пара бугорков несет по 3 макрохеты и по 6
шипиков, дорсальная, пара сливается между собой, несет 8 макрохет и 12 шипиков.
Анальный сегмент с трехлучевым отверстием (причем его ветви примерно одина-
ковы), дорсальная лопасть с 2 щетинками и 8 шипиками, вентральные лопасти с
многочисленными шипиками.
Ноги слабые (рис. 40, 6). Бедро с 6, голенелапка с 8 хетами, коготок слабо сер-
повидно изогнут. Длина тела до 8 мм.
При откладке яиц самка прогрызает отверстия в стеблях злаков, откладывает в
них 8-12 яиц и замазывает сверху экскрементами; выходящие личинки прогрыза-
ют ход внутри стебля и корня, а затем уходят в почву, где питаются корешками
злаков, повоя, вьюнка.
96
В регионе 1 вид — С dauricum Mannerheim (юг ВСБ, ДВ).
2. Род Chloropterus Morawitz
Тело явственно С-образное (рис. 41,7). Передняя часть лба сильно склеротизо-
вана и утолщена в виде мощного поперечного валика с продольным гребнем и мор-
щинками. Темя с 11 парами щетинок, в том числе с продольным рядом из 6 щети-
нок на задненаружных участках темени. Мандибулы широкие и короткие, с тупым
зубцом на режущем крае. Усик — рис. 41,2. Нижнечелюстные щупики 4-членико-
вые. Ментальный склерит с выпуклыми боковыми и прямой или слегка вогнутой
задней частью (рис. 41, 3). Склерит переднеспинки явственный. Тело в негустых
щетинках. Стерниты брюшка с бугорками, без длинных выростов. Голенелапка с
относительно тонкими, умеренно длинными щетинками. 10-й сегмент в виде окру-
жающих анальное отверстие бугров, густо покрытых опорными шипиками. Аналь-
ное отверстие в виде трехлучевой щели. Длина взрослых личинок до 8 мм.
В регионе 1 вид — Ch. versicolor Morawitz, встречающийся в степной зоне ЕЧ,
СК в засоленной почве.
3. Род Bromius Chevrolat (Adoxus Kirby)
Тело явственно С-образное, передний край лба склеротизован, приподнят над
наличником, но без развитого гребня (рис. 37, 7). Темя с 4 парами щетинок (не
считая щетинок на боковых частях головной капсулы). Мандибулы широкие и ко-
роткие, с тупым зубцом на режущем крае. Челюстные щупики 4-члениковые. Мен-
тальный склерит полукруглый. Склерит переднеспинки явственный. Тело в негус-
тых щетинках. Стерниты брюшка с бугорками, без длинных выростов. Голенелап-
ка с относительно тонкими недлинными щетинками. 10-й сегмент вытянут назад и
имеет вид усеченного конуса с немногочисленными шипиками на вершинной пло-
щадке (рис. 39, 3). Анальное отверстие в виде трехлучевой щели. Длина взрослой
личинки до 10 мм.
Личинки живут в почве на корнях кипрея, а на ДВ, очевидно, и на корнях ви-
нограда. В роде один вид — В. obscurus Linnaeus, распространенный повсеместно,
кроме крайнего севера и степей.
4. Basilepta Motschulsky
Тело слабо С-образно изогнутое, щетинки верха тонкие, длинные и редкие,
низа — довольно густые, более короткие, частично превращающиеся в шипики.
Лоб с 3 парами щетинок в передней половине, темя с 5 парами (рис. 37, 2).
Лобная пластинка с хитинизированным передним краем, не образующим выпукло-
го валика. Наличник несет 2 микрохеты. Мандибулы с 2 слабо заметными зубцами.
1-й членик усиков почти квадратный, с 2 порами и крохотной щетинкой, 2-й ма-
ленький, вдвое длиннее ширины, на вершине с 3 очень мелкими щетинками, па-
пилла почти вдвое толще и выше 1-го членика, с раздвоенной вершиной (рис. 38,
3). Субментум несет 3 пары хет. В основании ментума лежат 2 макрохеты, 2 сенсор-
ные поры расположены в средней части, а впереди — 2 макро- и 2 микрохеты и
97
Рис. 41. Личинки подсемейства Eumolpinae:
1—3 — Chloropterus versicolor. 1 — общий вид, 2 — усик, 3 — нижняя губа; 4 — Chrysochares
asiatica, вершина брюшка. По Курчевой и ориг.
пара сенсорных пор (рис. 38, 7). Челюстные щупики 4-члениковые. 4 пары бугров
9-го брюшного сегмента несут на себе малочисленные шипики и щетинки. Вент-
ральная пара бугров сливается между собой, несет 3 пары шипиков, боковая вент-
ральная — 4 шипика, эпиплевральная пара с 1 макро- и с 1 микрохетой, дорсальная
пара имеет макрохету и шипик (рис. 39, 2). Бедро с 5 щетинками, голенелапка с 6
щетинками, коготок тонкий, слабо изогнут. Длина тела до 4,5 мм.
В регионе 2 вида. Известна личинка В. fulvipes Motschulsky (юг СБ и ДВ), раз-
вивающаяся на корнях полыни. В. balyi Harold встречается на Сахалине и Курилах
на ольхе.
98
5. Род Pagria Lefevre (Colposcelis Dejean)
Тело личинки белое, слабо С-образно изогнуто, щетинки верха светлые, тон-
кие, снизу коричневые, крепкие и шипиковидные. Голова светло-желтая, мандибу-
лы коричневые. Головная капсула округлая, сжатая с боков, затылок с неглубокой
треугольной выемкой, которая сильно склеротизована по краям и утолщена в виде
валика (рис. 42, 7). Эпикраниальный шов едва намечен, лобные швы слабо выраже-
ны. Эндокарина длинная и светлая, доходит почти до середины головной капсулы.
Вершина лба сильно склеротизовано в виде узкой полосы, по бокам с выступаю-
щими углами. Лоб с наиболее длинными щетинками в основном по переднему краю.
Наличник в основании с двумя порами и двумя мелкими щетинками. Верхняя губа
на диске с шестью крепкими щетинками, расположенными в два ряда, боковые
щетинки первого ряда почти в 2 раза длиннее внутренних; передний край прямой,
с 12 маргинальными щетинками (рис. 42,2). Мандибулы узкие, на вершине с двумя
зубцами, с острым внутренним краем (рис. 42, 3). Нижнечелюстные щупики 4-чле-
никовые; нижнегубные —1-члениковые, ментальный склерит в виде своеобразной
фигуры (рис. 42, 5). Усики 2-члениковые, 1-й членик на вершине с конусовидной
папиллой и очень маленьким 2-м члеником (рис. 42, 6). Склерит переднеспинки
слабо хитинизирован, с длинными и короткими щетинками (рис. 42, 7), эпиплев-
ральный склерит с одной длинной щетинкой. Тергиты средне- и заднегруди с двумя
рядами склеритов, первый ряд состоит из двух мелких, слабо выраженных склери-
тов, второй — из шести склеритов, крыловые склериты с двумя длинными и двумя
короткими щетинками (рис. 42, 7). 1-8-й тергиты брюшка с тремя рядами склери-
тов, эпиплевральные склериты с одной длинной и одной короткой щетинками (рис.
42, 8). Склериты груди и брюшка крупные и соприкасаются друг с другом (рис. 42,
9). Стерниты брюшка — рис. 42,10. Анальное отверстие в виде трехлучевой щели.
Голенелапка короткая, снизу и на вершине с короткими шипиковидными щетинка-
ми, коготок слабо изогнут (рис. 42, 4).
Питание на корнях бобовых. В регионе 2 вида на юге ДВ: R consimile Baly, R
ussuriensis Moseyko & L. Medvedev. Описание составлено по личинке Р signata
Motschulsky из Вьетнама. Все данные об обитании Р signata в Палеарктике, вклю-
чая ДВ, ошибочны (Мосейко, Медведев, 2005).
6. Род Eumolpus liliger (Chrysochus Chevrolat)
Тело слабо С-образное (рис. 36, 2). Передний край лба без утолщения и более
сильной склеротизации (рис. 37, 3). Усики с умеренно крупной папиллой (рис. 38,
4). Темя с 5 парами щетинок. Мандибулы удлиненные, без тупого зубца на режу-
щем крае. Челюстные щупики 4-члениковые (рис. 40, 3). Ментальный склерит с
выпуклыми боковыми частями. Склерит переднеспинки слабо выражен. Верхняя
сторона тела в густых щетинках. 2-7-й сегменты брюшка с сосцевидными вырос-
тами по бокам стернитов, более длинными на 4-7-м сегментах. Голенелапка с отно-
сительно тонкими, очень длинными щетинками, включая и щетинку на коготке
(рис.40, 7). 10-й сегмент образует 3 отдельные лопасти вокруг анального отвер-
стия: непарную треугольную дорсальную и две вентральные в виде косо срезанных
столбиков; все три лопасти, особенно вершинные площадки вентральных, сильно
99
Рис. 42. Детали строения личинки Pagria signatcr.
1 — голова, 2 — верхняя губа, 3 — мандибула, 4 — голенелапка, 5 — нижняя губа, 6—усик,
7 — переднеспинка, 8 — 2-й тергит брюшка, 9 — среднеспинка, 10 — 2-й стернит брюшка.
склеротизованы. Анальное отверстие в виде продольной щели. Длина взрослой
личинки до 15 мм.
В регионе 3 вида, личинки которых живут на корнях растений из семейства
ластовневых (Vincetoxicum, Cynanchum, Asclepias). Неизвестна личинка К globicollis
Lefevre. С юга ДВ.
100
Таблица для определения видов рода Eumolpus
1(2). Вентральные лопасти 10-го сегмента сильно хитинизированные, разделенные
каждая поперечной бороздой посередине (рис. 39, 6). У ментального склерита вы-
ражены только боковые части (рис. 38, 9). Щетинки верха желто-золотистые. Юг
СБ...................................................Е. goniostoma Weise
2(1). Вентральные лопасти 10-го сегмента слабо хитинизированные, поперечные
борозды лопастей едва различимы (рис. 39, 5). Ментальный склерит с выпуклыми
боковыми и задней частями (рис. 38, 8). Щетинки верха бесцветные. ЕЧ, СК, юг
ЗСБ..................................................Е. asclepiadeus Pallas
7. Род Chrysochares Morawitz
Личинки очень сходны с предыдущим родом, отличаясь следующими призна-
ками: щетинки на теле гораздо более редкие; 2-й членик усика с 3 длинными ще-
тинками; стипес у наружного края с 1 щетинкой; ментальный склерит сзади пря-
мой; дорсальная лопасть анального клапана округлая (рис. 41, 4). Длина взрослых
личинок до 20 мм.
В регионе 1 вид — Ch. asiatica Pallas (юг ЕЧ, СК); жуки на ластовне, кендыре
и верблюжьей колючке, личинки отмечены в песчаной почве около корней лас-
товня.
8. Род Pachnephorus Redtenbacher
Тело очень слабо С-образно изогнутое. Передний край лба без утолщения и
более сильной склеротизации. Темя с 5 парами щетинок (не считая щетинок на
боковых частях головной капсулы). Усики с крупной папиллой, превышающей раз-
меры второго членика (рис. 38, 5). Мандибулы удлиненные, без тупого зубца на
режущем крае. Челюстные щупики 3-члениковые (рис. 40,2). Ментальный склерит
с прямыми боковыми и задней частями. Склерит переднеспинки слабо выражен.
Верхняя сторона тела с редкими щетинками и слабо выраженными поперечными
складками на тергитах. 1-8-й сегменты брюшка с длинными пальцевидными выро-
стами по бокам (рис. 36,3). Голенелапка с мощными толстыми шипами (рис. 40,8).
10-й сегмент брюшка несет на вершине фигурный опорный вырост, с сильно скле-
ротизованной изогнутой на вершине площадкой, этот вырост отделен от передней
части сегмента глубокой складкой (рис. 39, 4). Анальное отверстие в виде попереч-
ной щели. Длина взрослой личинки до 7 мм.
В регионе 5 видов; известна личинка Р. tessellatus Duftschmidt, обычного на
юге лесной, в лесостепной и степной зонах Палеарктики и связанного с песчаной
или засоленной почвой.
101
12. ПОДСЕМЕЙСТВО CHRYSOMELINAE
Тело продолговатое, но не очень удлиненное, или довольно короткое, оваль-
ное, толстое; нижняя сторона уплощена, верхняя выпуклая, иногда очень сильно
(рис. 5, 6-8). С-образная изогнутость выражена, но незначительно, особенно у ви-
дов с коротким толстым телом. Окраска довольно разнообразная, преобладают се-
рые и бурые тона, склериты обычно темнее остальной части покровов и могут кон-
трастировать с ней по окраске, у некоторых видов тело темное, с более или менее
заметным металлическим отливом (некоторые Chrysolina Motschulsky, Timarcha La-
treille), или оранжевое до яркого красноватого (Entomoscelis Chevrolat, Leptinotarsa
Stal, Zygogramma Chevrolat), щетинки верха могут быть хорошо развиты, длинные,
особенно у молодых личинок, реже тело почти голое.
Голова гипогнатическая, немного уже грудных сегментов, выпуклая и округ-
лая. Верхняя губа, наличник и лоб явственно отделены друг от друга, эпикраниаль-
ный шов хорошо развит, довольно длинный, лобные швы равномерно дуговидно
изогнутые. Глазков 6 с каждой стороны, расположенных двумя группами по 2 и 4
глазка с сильно выпуклой роговицей. Мандибулы простые, довольно крупные и
мощные, расширены к вершине, с 4-5 зубцами и обычно с 1-2 щетинками на на-
ружном крае. Максиллы с небольшим, но хорошо развитым продолговатым кардо
и широким, слабо удлиненным стипесом; жевательная лопасть явственная, челюс-
тные щупики 4-члениковые (рис. 43, 2). Нижняя губа с явственными ментумом и
прементумом, губные щупики 2-члениковые. Горло и гипофарингеальный мост от-
сутствуют. Строение ротового аппарата, за исключением формы верхней губы, от-
личается значительным постоянством в пределах подсемейства. Верхняя губа круп-
ная, поперечная, с выемкой на переднем крае, очень различной у разных родов и
иногда почти исчезающей. Наличник в виде сильно поперечной пластинки. Усики
3-члениковые. В типичном случае на голове имеется небольшое количество пер-
вичных щетинок (рис. 43, 7), отличающихся постоянным местоположением и чис-
лом (верхняя губа — 2 пары, наличник — 3 пары, лоб и темя — по 5 пар). Очень
часто появляются вторичные щетинки, иногда очень многочисленные.
Сегменты груди и брюшка с 2, реже с 1 или 3 поперечными рядами склеритов;
иногда склериты разбиваются на большое число мелких склеритовидных пласти-
нок или вообще исчезают. У молодых личинок на среднегруди хорошо заметны
зубчики для разрезания хориона яйца. У ряда родов на боках тергитов имеются
отверстия дорсальных желез; у некоторых родов, близких к Gonioctena Chevrolat,
функцию этих желез выполняют парные выпячивания в виде двух длинных отрос-
тков, лежащих на границе 7-го и 8-го тергитов и втянутых внутрь тела. Брюшко с 7
или 8 парами стигм, они круглые, с одним отверстием, без стигмальной пластинки.
Стерниты 1-8-го сегментов брюшка без двигательных бугров, лишь у некоторых
родов на 5-7-м стернитах имеются парные выросты, несущие функцию ложных
ножек. Ноги сравнительно короткие, толстые, коготки простые или с зубцом при
основании, обычно с короткими пульвиллами. Анальное отверстие лежит вентраль-
но, в центре присасывательного диска на 10-м сегменте.
Внешне личинки Chrysomelinae довольно схожи с открытоживущими личин-
ками Galerucinae и Alticinae, но легко отличаются от них числом глазков.
102
Рис. 43. Детали строения личинок подсемейства Chrysomelinae:
1 — головная капсула с индексами щетинок; 2 — лабио-максиллярный комплекс.
Подавляющее большинство личинок Chrysomelinae питается открыто на лис-
тьях кормовых растений, чаще на нижней стороне листа, лишь личинки Hydrothassa
Thomson и Prasocuris Latreille могут обитать внутри полых стеблей и черешков, где
и окукливаются, однако это свойственно главным образом личинкам старших воз-
растов, тогда как молодые личинки большей частью питаются на листьях. Яйца
откладываются группами на нижнюю сторону листа, реже внутрь стеблей и череш-
ков (Hydrothassa Thomson), они удлиненной формы, с гладким хорионом и окраше-
ны в яркие цвета (желтый, оранжевый, красный). Развитие яйца длится около неде-
ли, но у некоторых форм сроки развития яиц сильно укорачиваются, до нескольких
часов, а также наблюдаются яйцеживорождение и живорождение. Для Gastrophysa
Chevrolat отмечен партеногенез, но эти данные требуют проверки. Число личиноч-
ных стадий колеблется от 3 до 4, причем 3 личиночные стадии характерны для
форм, имеющих дорсальные железы. Окукливание обычно в почве; Chrysomela
Linnaeus, Plagiosterna Motschulsky, Gastrolina Motschulsky, Gastrolinoideis Chujo et
Kimoto и Plagiodera Erichson окукливаются на листьях, a Hydrothassa Thomson и
Prasocuris Latreille — внутри стеблей и черешков. Куколка открытого типа (рис. 4,
2-3). Развитие личинок длится от 2 до 5 недель, а куколки — 4-10 дней. В год
развивается обычно одно поколение.
Представители этого подсемейства наиболее адаптированы к суровым клима-
тическим условиям и представлены в регионе очень обильно, образуют эндемич-
ные виды и даже рода и заходят далеко на север, заселяя материковую тундру и
арктические острова. В регионе встречается 25 родов; неизвестны личинки энде-
мичного алтайского рода Oreothassa Jacobson (близкого к Hydrothassa} и централь-
но-азиатского высокогорного Cystocnemis Motschulsky, проникающего из МНР и
Казахстана в южный Алтай.
103
Таблица для определения родов подсемейства Chrysomelinae
1(2). Брюшко с 7 парами стигм на 1-7-м сегментах. Коготки без зубца у основания.
Окраска тела с более или менее ярким металлическим блеском... 1. Timarcha
2(1). Брюшко с 8 парами стигм на 1-8-м сегментах. Окраска тела обычно без метал-
лического блеска (редко у Chrysolina Motschulsky).
3(26). Крыловые склериты грудных тергитов и стигмальные склериты брюшных
тергитов с крупным отверстием желез (рис. 44, 6).
4(5). Передний край верхней губы без срединной выемки (рис. 44, 4). Первый тер-
гит брюшка (рис. 55, 7) отличается от последующих наличием претергальных скле-
ритов (очень редко они бывают также на 2-3-м брюшных тергитах).... 8. Phratora
5(5). Передний край верхней губы с явственной срединной выемкой (рис. 44, 2).
6(17). Коготки с крупным зубцом в базальной половине, эпикраниальный шов длин-
ный.
7(16). Усики короткие, 2-й членик поперечный, 3-й членик короткий, не длиннее 2-
го, самое большее вдвое длиннее ширины. На ивовых, березовых и ореховых.
8(9). Тергиты 1-2-го брюшных сегментов близ боковой части переднего края тер-
гальных склеритов с маленьким, несущим одну щетинку склеритом; тергальные
склериты с 2 щетинками; микроскульптура хитинового покрова между склеритами
сильно развита и оставляет свободной на грудных сегментах лишь срединную (свет-
лую) продольную полосу.......................................9. Plagiodera
9(8). Тергиты 1-2-го брюшных сегментов близ тергальных склеритов без следов
добавочного склерита.
10(15). Тергальные склериты груди и брюшка не более чем с 4 щетинками.
11(12). Склериты нижней стороны тела не развиты............10. Plagiosterna
12(11). Склериты нижней стороны тела развиты.
13(14). Тело умеренно удлиненное, брюшко в 1,5 раза длиннее груди. Тергальные
склериты груди с 3-4 щетинками.............................11. Chrysomela
14(13). Тело сильно удлиненное, брюшко в 2 раза длиннее груди. Тергальные скле-
риты груди с 1-2 щетинками...................................12. Gastrolina
15(10). Тергальные склериты груди с 10-15, брюшка — с 25-30 довольно длинны-
ми щетинками. Склериты нижней стороны тела не развиты...13. Gastrolinoides
16(7). Усики длинные, 2-й членик одинаковой длины и ширины, 3-й членик очень
длинный, в 3-4 раза длиннее ширины. На розоцветных.........18. Sternoplatys
17(6). Коготки без зубца при основании. Эпикраниальный шов более короткий.
18(23). Лобные швы в основной части параллельные на некотором расстоянии, да-
лее расходятся.
19(22). Переднеспинка без эпиплевральных склеритов.
20(21). Тергиты брюшных сегментов без претергальных склеритов (рис. 55, 5) ....
...........................................................15. Hydrothassa
21(20). Тергиты брюшных сегментов с рудиментами претергальных склеритов (рис.
55, 77)....................................................17. Prasocuris
22(19). Переднеспинка с небольшими, но отчетливыми (по 1 с каждой стороны)
эпиплевральными склеритами, несущими по 1 щетинке (рис. 56, 6). 1-4-й брюш-
ные сегменты с небольшими претергальными склеритами; посттергальные брюш-
ные склериты с 3 щетинками..............................16. Agrostithassa
104
Рис. 44. Детали строения личинок подсемейства Chrysomelinae:
1-4 — верхняя губа, 8-10 — коготок: 1 — Gastrophysa atrocyanea, 2 —Phaedon concinnus,
3 — G. mannerheimi, 4,9 — Phratora obtusicollis, 8 — Ph. vulgatissima, 10 — Ph. vitellinae', 5,
7—дорсальные железы и щетинки на склерите Paropsides', 6—отверстие дорсальной желе-
зы брюшного тергита Plagiosterna.
23(18). Лобные швы расходятся под углом от самого основания, не параллельные
на некотором протяжении (рис. 54,2). Посттергальные склериты брюшных сегмен-
тов с 2 щетинками.
24(25). Микроскульптура хитинового покрова тела заметна лишь на промежутках
между склеритами и не покрывает последние. Среднеспинка с 4 претергальными
склеритами (рис. 56, 5), несущими по 1 щетинке, или тергальные склериты неяс-
ные и их местоположение лишь обозначено присутствием щетинок..........
........................................................12. Gastrophysa
25(24). Микроскульптура (состоящая из бугорков или шипиков) хитинового покро-
ва тела покрывает не только промежутки, но и склериты. Среднеспинка с 2 претер-
гальными склеритами, несущими 2 щетинки (рис. 56, 2).......19. Phaedon
105
26(3). Крыловые склериты грудных сегментов и стигмальные (разделенные или
слитые) склериты брюшных сегментов без отверстий желез.
27(38). Тело, особенно в задней половине со спинной стороны, сильно выпуклое;
нижняя сторона тела уплощенная (рис. 5, 6). Склериты обычно плохо заметны или
отсутствуют.
28(29). Тело светлое, с контрастным темным рисунком; голова с округлыми крупны-
ми черными пятнами, переднеспинка с 4 пятнами, тело с 4 нерезкими темными поло-
сами (рис. 47, 7). Стигмальные склериты бугровидно выпуклые.4. Ambrostoma
29(28). Тело светлое или темное, но всегда без контрастного рисунка (не считая
темных мелких склеритовидных площадок, только у Paropsides переднеспинка рез-
ко двуцветная).
30(33). Стигмы брюшных сегментов окружены большим бугровидно приподнятым
стигмальным склеритом, несущим мелкие щетинки; прочие склериты на верхней
стороне 1-7-го сегментов брюшка отсутствуют. Тело, особенно в задней половине,
сильно выпуклое, обычно оранжевое.
31(32). Лоб с 14 недлинными щетинками, темя с 5 щетинками с каждой стороны
(рис. 45, 7). Лобные швы явственные. Верхняя губа с маленьким зубчиком на каж-
дой стороне неширокой и довольно глубокой срединной выемки. Средне- и заднес-
пинка с 2 посттергальными склеритами и со следами претергальных склеритов ..
..........................................................2. Leptinotarsa
32(31). Лоб с 10 длинными щетинками, темя с 20-24 щетинками с каждой стороны
(рис. 45, 3). Лобные швы очень слабые, плохо заметные. Верхняя губа (рис. 45, 4) с
неглубокой широкой выемкой. Средне- и заднеспинка (рис. 45, 6) без тергальных
склеритов.................................................3. Zygogramma
33(30). Стигмы брюшных сегментов окружены узким кольцевидным плоским или
слабо выпуклым склеритом, лишенным щетинок. Верхняя губа без зубчиков по краям
срединной выемки.
34(37). Тело равномерно выпуклое, среднеспинка не уже прочих сегментов. Окрас-
ка темная, реже светлая, но голова обычно темнее тела (если светлее, то верх с
явственными склеритами).
35(36). Верх обычно со склеритами или со склеритовидными площадками, если
без них, то либо лоб только с 10-14 первичными щетинками, либо стерниты брюш-
ка с явственными склеритами. Острие коготка сильно серповидно изогнутое (рис.
47, 7-11). На различных травянистых растениях...............5. Chrysolina
36(35). Верх без тергальных склеритов или склеритовидных площадок, с коротки-
ми щети нками, низ без склеритов, самое большее с мелкими склеритовидными
площадками вокруг щетинок. Окраска всегда темная, голова немного темнее тела,
с многочисленными короткими вторичными щетинками. Острие коготка слабо изог-
нуто (рис. 47, 12-14). Исключительно на полыни.................7. Crosita
37(34). Тело с седловидным перегибом в области среднеспинки, которая значительно
уже прочих сегментов. Окраска светлая, голова немного светлее тела Верх без скле-
ритов и склеритовидных площадок, с короткими щетинками. Коготки с зубцом, Лоб
с 26 щетинками (рис. 48, 7). На альпийских лугах на зонтичных и сложноцветных
.....................................6. Oreina Chevrolat (Chrysochloa Hope)
38(27). Тело co слабо и равномерно выпуклой верхней стороной (рис. 5, 7). Склери-
ты обычно явственные, резко отграниченные.
106
39(42). Головная капсула с многочисленными вторичными щетинками, не отличи-
мыми по длине от первичных щетинок (рис. 54, 3). Тергиты груди и брюшка с 3
поперечными рядами склеритов.
40(41). Второй поперечный ряд средне-, заднеспинки и брюшных тергитов вклю-
чает 6 склеритов одинаковой величины (рис. 57, 4). Лоб с 46-48 щетинками...
........................................................20. Entomoscelis
41(40). Второй поперечный ряд средне-, заднеспинки и брюшных тергитов вклю-
чает 2 склерита, которые значительно крупнее прочих (рис. 57, 5). Лоб с 20-22 ще-
тинками.....................................................21. Colaphus
42(39). Головная капсула только с первичными щетинками или темя с немногочис-
ленными вторичными щетинками. Тергиты груди и брюшка с 2 поперечными ряда-
ми склеритов (иногда склериты редуцированные). Коготки с зубцом на основании.
43(44). На границе 7-го и 8-го брюшных тергитов нет выпячивающихся желез в
виде двух пальцевидных отростков. Щетинки верха короткие, всегда гораздо коро-
че длины склеритов. Переднеспинка не бывает резко двуцветной. Внутренние тер-
гальные склериты разделенные (рис. 56, 77)................22. Apterocuris
44(43). На границе 7-го и 8-го брюшных тергитов имеется выпячивающаяся железа
(рис. 44, 5) в виде двух пальцевидных отростков (эта железа в спокойном состоя-
нии втянута внутрь и плохо заметна, для ее выворачивания следует сжать передний
конец тела личинки).
45(46). Щетинки тела чрезвычайно короткие, часто едва различимые, сидят на хи-
тинизированных бугорках или зубчиках (рис. 44, 7), на тергальных склеритах ще-
тинки гораздо короче соответствующих зубчиков. Все внутренние тергальные скле-
риты слиты попарно. Переднеспинка резко двуцветная, желтая с черно-бурой сере-
диной (рис. 57, 7)........................................ 23. Paropsides
46(45). Щетинки тела нормальные, обычно длинные. Хотя бы часть внутренних
тергальных склеритов не слита попарно. Переднеспинка не бывает резко двуцвет-
ной.
47(48). Лоб с 10 щетинками (рис. 54, 4). Внутренние претергальные склериты груд-
ного отдела часто с 3-4 щетинками, нередко слиты попарно. Питание не на бобо-
вых (кроме G. fulva Motshulsky, но тогда голова светлая с черными пятнами).
..........................................................24. Gonioctena
48(47). Лоб с дополнительными щетинками (рис. 54, 6). Внутренние претергаль-
ные склериты грудного отдела с 2 щетинками. На бобовых....25. Cercyonops
1. Род Timarcha Latreille
Тело, включая голову, зеленоватое или бурое, с металлическим отливом, в круп-
ной морщинистой скульптуре, низ без металлического блеска.
Верхняя губа со срединной выемкой, с 4-6 парами маргинальных щетинок.
Наличник с поперечным рядом из 10-16 щетинок. Лоб с 2 вдавлениями, примерно
с 20 парами коротких, плохо заметных щетинок. Темя также с короткими вторич-
ными щетинками. Переднеспинка с крупным склеритом, плохо отграниченным и
выделяющимся только сглаженной скульптурой, с мельчайшими щетинками, тер-
гиты средне- и заднегруди и 1-8-го сегментов брюшка без склеритов и щетинок
или с мельчайшими щетинками, 9-й тергит брюшка на заднем крае с щетинками;
107
Рис. 45. Детали строения личинок подсемейства Chrysomelinae:
1, 2—Leptinotarsa decemlineatcr. 1 — голова; 2 — коготок; 3-8 — Zygogramma suturalis'. 3 —
голова, 4 — верхняя губа, 5 — переднеспинка; 6 — среднеспинка, 7 — 1-й тергит брюшка,
8 — коготок.
1-7-й сегменты брюшка несут стигмы. 10-й сегмент брюшка маленький, лежит на
вентральной стороне 9-го, подталкиватель хорошо развит. Голенелапка толстая, с
обособленной нижней частью (рис. 46, 4-6), Коготки без зубца, но сильно расши-
ренные в основной части. Личинки на Galium, а также на почве и под камнями.
Окукление в почве.
В регионе 4 вида.
108
Рис. 46. Детали строения личинок рода Timarchcr.
1-3 — верхняя губа: 1 — Т. tenebricosa, 2 — Т. goettingensis, 3 — Т. hummeli; 4-6—голенелап-
ка: 4 — Т. tenebricosa, 5 — Т. goettingensis, 6 — Т. hummeli. По Штейнхаузену и Охрименко.
Таблица для определения видов рода Tim arch а
1(2). Верхняя губа с 6-8 парами щетинок на диске и с 7 парами на переднем крае
(рис. 46, 7). Голенелапка снизу с 16 щетинками (рис. 46, 4), бедро с 35-40 щетинка-
ми. Тело сверху сине-зеленое, снизу буро-желтое. На Galium. ЕЧ, СК........
.....................................................Т. tenebricosa Fabricius
2(1). Верхняя губа с 4-5 парами щетинок на диске. Голенелапка снизу не более чем
с 12 щетинками, бедро с 25-30 щетинками.
3(4). Наличник с 7-8, верхняя губа с 7 парами маргинальных щетинок (рис. 46, 2).
Голенелапка снизу с 12 щетинками (рис. 46, 5). Тело сверху буро-зеленое с зеленой
переднеспинкой. На Galium. ЕЧ.......................Т. goettingensis Linnaeus
4(3). Наличник с 5 парами щетинок, верхняя губа с 5 парами маргинальных щети-
нок (рис. 46, 3). Голенелапка снизу с 6 щетинками (рис. 46, 6). СК........
.....................................................Т. hummeli Faldermann
109
2. Род Leptinotarsa Stal
Голова (рис. 45, 7) только с первичными щетинками, верхняя губа с маленьким
зубчиком на каждой стороне срединной выемки. Переднеспинка желтоватая, толь-
ко на заднем крае буроватая, с единичными мелкими щетинками на заднем крае.
Средне- и заднеспинка с явственным крыловым склеритом, несущим короткие ще-
тинки, 2 посттергальными склеритами, несущими по 1 щетинке каждый, и со сле-
дами претергальных склеритов. Эпиплевральные грудные склериты с многочис-
ленными крепкими щетинками. Тергальные склериты 1-7-го сегментов брюшка
отсутствуют, на их место имеются лишь очень короткие отдельные щетинки. Стиг-
мы брюшных сегментов окружены бугровидно приподнятым стигмальным склери-
том; эпиплевральные склериты довольно крупные, с несколькими щетинками; вен-
троплевральные склериты гораздо меньше, с 2 щетинками. Стерниты груди и брюш-
ка с 1 непарным срединным склеритом, несущим 4-6 щетинок, но бокам от него
имеется еще 5-8 щетинок, которые на последних сегментах разбиваются на 2 груп-
пы. Тергиты 8-го и 9-го сегментов брюшка с большим непарным склеритом, несу-
щим на заднем крае крепкие щетинки. Коготки с крупным зубцом при основании
(рис. 45, 2). Окраска тела ярко-оранжевая, розоватая или желто-оранжевая.
Личинки 1-го возраста с хорошо развитыми склеритами.
На картофеле, реже на томатах, баклажанах, а также дикорастущих паслено-
вых, окукление в почве. Единственный завезенный в Европу вид — колорадский
картофельный жук — L. decemlineata Say — встречается от западных границ Рос-
сии до Приморья. Опаснейший вредитель картофеля.
3. Род Zygogramma Chevrolat
Личинка светло-желтая с темными ротовыми частями, глазками, усиками и
коготками. Голова (рис. 45, 5) слабо склеротизована, лобные швы практически не-
различимы, темя с 20-24 притупленными щетинками с каждой стороны, эпикрани-
альный шов и эндокарина четкие. Лоб широкий, гладкий, с 10 длинными щетинка-
ми. Наличник короткий, склеротизован в основании, с 6 длинными щетинками и 2
порами. Верхняя губа (рис. 45, 4) с базальными выступами, ее передний край с
неглубокой широкой выемкой, на диске с 4 длинными щетинками и 2 порами, на
переднем крае с 4 щетинками с каждой стороны и с 6 щетинками на дне выемки.
Мандибулы треугольные, с 5 зубцами, сбоку с 2 щетинками и 2 порами. Переднес-
пинка (рис. 45, 5) слабо склеротизована, по переднему и заднему краям со склеро-
тизованными участками в виде пятен, несущими короткие щетинки; эпиплевраль-
ный склерит отсутствует. Средне- и заднеспинка (рис. 45, 6) без дорсальных скле-
ритов, с 2 рядами мелких щетинок, крыловые склериты слабо склеротизованы, по-
чти не окрашены. Сегменты брюшка без склеритов, с 2 рядами мелких шипиков,
лишь на 8-м и 9-м тергитах заметны слабоокрашенные склериты с длинными про-
стыми щетинками (рис. 45, 7). Стерниты без склеритов, с неправильным рядом креп-
ких коротких щетинок. Стигмы внутри с ресничным аппаратом, стигмы груди круп-
нее прочих. Микроскульптура покровов тела в виде светлых овальных зерен. Голе-
нелапка снизу и сверху с 2 щетинками, коготок с прямоугольным зубцом при осно-
вании (рис. 45, 8).
110
Личинка 1-го возраста светло-оранжевая, склериты светло-коричневые, щетинки
длинные, светлые, зазубренные. Темя с 15-16 щетинками с каждой стороны, лоб с
10 щетинками, щетинки переднего края лба, наличника и верхней губы остроко-
нечные, незазубренные. Склерит переднеспинки слабо склеротизован, все щетин-
ки расположены по его краям. Средне- и заднеспинка (рис. 15) и брюшные тергиты
(рис. 16) с 2 рядами выпуклых отчетливых склеритов, каждый из которых с 1-2
длинными щетинками, эпиплевральные склериты с 3-4 щетинками, на 8-9-м тер-
гитах склериты сливаются в непарную пластинку. Склериты низа, кроме гипоплев-
ральных, плохо заметные, каждый с 1 щетинкой.
Питание на амброзии. В регионе 1 вид — Z. suturalis Fabricius, интродуциро-
ванный из Канады на СК и юг ДВ для борьбы с амброзией.
4. Род Ambrostoma Motschulsky
Тело (рис. 47, 7) светлое с темным рисунком, голова желтая, верхняя губа, на-
личник, передний край лба, два округлых пятна на лбу, небольшое пятно под осно-
ванием усиков, почковидное пятно за основанием усиков и два крупных косых пят-
на посередине теменных долей черные или черно-бурые. Переднеспинка белая, с 2
блестящими черными широко расставленными продольными пятнами и 2 попереч-
ными пятнами на переднем крае кнаружи от первых и с желтизной у задних углов,
остальная часть тела белая, с очень узкой серой срединной линией, 2 дорсальными
и 2 боковыми темными нерезкими полосами (дорсальные полосы представляют
участки густой микроскульптуры). Голова поперечно-овальная, с короткими ще-
тинками (до 15) по бокам теменных долей. Лоб с очень короткими первичными и
вторичными щетинками (около 60), его передний край более склеротизован и утол-
щен. Наличник узкий, со склеротизованной задней половиной, с тремя короткими
щетинками с каждой стороны, сильно сдвинутыми кнаружи. Верхняя губа с глубо-
кой срединной выемкой, 2 короткими толстыми маргинальными щетинками с каж-
дой стороны и с 4 длинными щетинками на диске. Мандибулы короткие и широкие,
почти не сужены к вершине, с 5 короткими и тупыми зубцами.
Верх с микроскульптурой, более густой и окрашенной на темных полосах; сре-
динный склерит переднеспинки с очень короткими, едва заметными редкими ще-
тинками, эпиплевральные склериты переднеспинки с короткими, но явственными
щетинками. Прочие тергиты только с крыловыми (на груди) и стигмальными (на
брюшке) склеритами; крыловые с 26-35, а стигмальные с 8-10 короткими щетин-
ками каждый. Поверхность тергитов с редкими, неравномерно разбросанными,
очень короткими щетинками, 9-й тергит едва хитинизирован, с более длинными
щетинками. Низ без склеритов и микроскульптуры, с довольно густыми, умеренно
длинными крепкими щетинками. 10-й сегмент лежит на вентральной стороне, со
склеритовидными площадками. Анальное отверстие в виде продольной щели. Го-
ленелапка, бедро и вертлуг с многочисленными щетинками, голенелапка заметно
расширена кпереди, особенно с нижней стороны под коготком. Коготки неболь-
шие, с прямоугольным зубцом при основании. Длина тела до 9 мм.
У личинки 1-го возраста тело белое, голова, переднеспинка, ноги и склериты
тела темные; лоб и темя с многочисленными щетинками. Тергиты груди и брюшка
с 2 рядами щетинок, каждая из которых лежит на отдельном склерите; на 8-9-м
111
Рис. 47. Детали строения личинок подсемейства Chrysomelinae:
1 —Ambrostoma, внешний вид; 2-4 — микроскульптура покровов: 2 — Chrysolina latimargo,
3 — Ch. bungei, 4 — Ch. graminis; 5, 6 — второй стернит брюшка, 7—14 — коготок: 5,12 —
Crosita pigra, 6,10 — Ch. koltzei, 7 — Ch. perforata, 8 — Ch. septentrionalis, 9 — Ch. bungei,
11 — Ch. polita, 13 — Crosita kowalewskyi, 14 — C. longipes.
112
тергитах они сливаются в 1 непарный склерит. Эпиплевральные склериты круп-
ные, с 7-8 щетинками, средне- и заднеспинка и 1-й тергит брюшка имеют сильно
хитинизированные бугровидные склериты.
Личинки питаются на листьях вяза; окукливание в почве. Жуки отмечались
также на боярышнике и черемухе.
1 вид — A. quadriimpressum Motschulsky на юге ВСБ и ДВ.
5. Род Chrysolina Motschulsky (Chrysomela auct.)
Тело сверху сильно выпуклое, снизу уплощенное, заметно расширено кзади.
Голова с первичными и некоторым количеством вторичных щетинок, коротких или
такой же длины, как первичные, большей частью лоб с 8-10, темя с 10-16 щетинка-
ми, наличник с 6 щетинками, верхняя губа с 4 щетинками на диске и с глубоковы-
емчатым посередине передним краем, несущим короткие щетинки. Мандибулы с 5
зубцами. Усики с хорошо развитым тактильным придатком на вершине 2-го члени-
ка. Средне-, заднеспинка и тергиты брюшка с плохо выраженными, нередко разде-
ленными на мелкие части или совершенно исчезающими склеритами; нижняя сто-
рона тела с более или менее развитыми склеритами. Личинки 1-го возраста обычно
с более длинными и густыми щетинками и часто с развитыми склеритами.
Жуки и личинки живут открыто на травянистых растениях, питаясь листьями,
а также на почве и под камнями, многие из них наиболее активны ночью, для неко-
торых видов отмечено живорождение и яйцеживорождение. 4 личиночных возрас-
та. Окукливание в почве.
В регионе около 80 видов, разделяемых на ряд подродов, не упоминаемых ниже.
Таблица для определения видов рода Chrysolina
Личинки 1-го возраста имеют хорошо развитые склериты на тергитах, которые
у личинок последующих возрастов обычно разбиваются на склеритовидные пло-
щадки или вообще исчезают. Предлагаемая ниже таблица составлена практически
для личинок старших возрастов.
1(50). Тергиты средне- и заднегруди и брюшка с явственными склеритами или скле-
ритовидными площадками.
2(7). Коготки с острым, направленным вперед зубцом, нередко выглядят расщеп-
ленными (рис. 47, 7). Голова темная. Верх с многочисленными склеритовидными
площадками, образующими до 6 неправильных поперечных рядов, и длинными
щетинками (рис. 52, 7). Микроскульптура явственная. На полыни.
3(4). Выемка верхней губы с явственным срединным зубцом, по бокам которого
имеются еще 2 мелких зубчика; края выемки также слабо зубцевидные (рис. 50, 7).
Микроскульптура верха равномерная. Юг СБ.............Ch. perforata Gebler
4(3). Выемка верхней губы без явственного срединного зубца, со слабо зубцевид-
ными краями выемки.
5(6). Выемка верхней губы широкая, неглубокая, со следами 2-3 зубчиков на осно-
вании (рис. 50, 2). Микроскульптура верха равномерно густая. АР (о-в Врангеля)
.................................................Ch. arctica L. Medvedev
113
6(5). Выемка верхней губы более узкая и глубокая, без следов зубчиков на основа-
нии, но с небольшими выемками по бокам от срединной выемки (рис. 50, 5). Мик-
роскульптура верха образует 3 темные нерезкие полосы в задней половине, разде-
ленные светлыми участками слабо окрашенной микроскульптуры. Юг СБ......
................................................Ch. aeruginosa Faldermann
7(2). Коготки с прямоугольным или тупым зубцом, либо без зубца.
8(17). Верхняя губа с крупным треугольным зубцом в срединной выемке (рис. 50,
4). Коготки без зубца. Покровы тела с микроскульптурой. Голова темная. Степные
формы, питание на полыни. Сюда же должны относиться неизвестные личинки Ch.
rufilabris Faldermann, из степей СБ.
9(12). Срединный зубец выемки верхней губы не длиннее 1/2 глубины выемки, да-
леко не доходит до переднего края. Передний край губы прямой, образует тупой
угол с краем выемки (рис. 50, 4). Верх со склеритовидными площадками и доволь-
но крупными склеритами, в основном посттергальными (рис. 52, 2). На полыни,
злаках и других растениях. Сюда же должна относиться Ch. infuscipes Weise (юг
СБ). Все следующие 3 вида очень близки, их видовая самостоятельность требует
подтверждения.
10(11). Тергиты груди и брюшка с крупными склеритами (рис. 51, 8-9), стигмы
брюшка расположены на крупных стигмальных склеритах, склериты переднеспин-
ки гладкие, без склеротизованных пятен. Юг СБ....Ch. discriminata Jacobsom
11(10). Тергиты груди и брюшка со слабо выраженными мелкими склеритами (рис.
51,10), стигмы брюшка расположены на мелких стигмальных склеритах, склериты
переднеспинки с поперечными вдавлениями и мелкими склеротизованными пят-
нами. Монголия, возможен на юге СБ................Ch. mongolensis Lopatin
12(9). Срединный зубец выемки верхней губы длинный, достигает ее переднего
края, который плавно переходит в край выемки (рис. 50, 5). Верх со сплошными
тергитами в виде 2 поперечных полос на каждом (возможно, они разбиваются на
склеритовидные площадки у старших возрастов). Известны в основном молодые
личинки.
13(16). Лоб примерно с 30 щетинками (с 10 щетинками у 1-го возраста). Склерит
переднеспинки со щетинками в основном по переднему и заднему краям.
14(15). Крыловые склериты с 8 щетинками. На полыни. Бурятия............
.............................................Ch. pusa Lopatin (1-й возраст)
15(14). Крыловые склериты с 15 щетинками. Тело с сильным темно-бронзовым от-
ливом. На полыни..................................Ch. brunnicornis Weise (Ch. koshantschikovi Jacobson)
16(13). Лоб примерно с 50 щетинками. Крыловые склериты с 14 щетинками (рис.
52, 3). Склерит переднеспинки со щетинками по всей поверхности. На полыни.
Тува. Сюда же должна относиться неизвестная личинка Ch. urjanchaica Jacobson —
вида, очень близкого к Ch. convexicollis Jacobs, и с тем же ареалом....
.....................................Ch. convexicollis Jacobson (1-й возраст)
17(8). Верхняя губа без треугольного зубца в срединной выемке.
18(19). Тергиты брюшка с 2 неправильными поперечными рядами склеритов, несу-
щих многочисленные щетинки (рис. 51, 5). Коготки без зубца на основании (рис.
51, 6). На Artemisia. ЕЧ............................Ch. carnifex Fabricius
19(18). Тергиты брюшка с 2-3 поперечными рядами склеритовидных площадок,
несущих каждая по 1 щетинке.
114
20(25). Верх с длинными первичными щетинками на склеритовидных площадках и
более короткими густыми вторичными щетинками на несклеритизованных участ-
ках покрова. Голова темная с вторичной хетотаксией. Склеритовидные площадки
каждого тергита образуют 2 поперечных ряда.
21(24). Покровы с явственной микроскульптурой. Коготки без зубца или со слабым
зубцом. На грудных тергитах 4 склеритовидные площадки в переднем и 8-9 — в
заднем ряду (рис. 52, 4), на брюшных — 8 в переднем и 7 — в заднем ряду. Пере-
дний край наличника выемчатый.
22(23). Вторичные короткие щетинки между склеритовидными площадками очень
густые, не собраны в ряды. Тело рыжебурое. На полыни. ЕЧ, СБ, ДВ........
..............................................Ch. aurichalcea Mannerheim
23(22). Вторичные короткие щетинки между склеритовидными площадками еди-
ничные, собраны в ряды. Тело желтое. На полыни. ЕЧ, СБ, ДВ..............
Ch. relucens Rosenhauer
24(21). Покровы с очень мелкой микроскульптурой, расположенной лишь на от-
дельных участках, преимущественно на сгибах. Коготки с явственным прямоуголь-
ным или тупым зубцом. Передний край наличника без выемки, На всех тергитах по
10 площадок в переднем и заднем ряду (рис. 52, 5). Вторичные короткие щетинки
между склеритовидными площадками довольно густые, не собраны в ряды. Алтай,
Тува, Саяны........................Ch. undulata Gebler
25(20). Все щетинки верха расположены на склеритах или склеритовидных пло-
щадках, обычно мало отличаются по длине.
26(39). Голова черная или черно-бурая, обычно темнее тела. Сюда же, очевидно,
относится личинка Ch. analis Linnaeus (ЕЧ).
27(30). Выемка верхней губы с полукруглым выступом на дне. Верх, особенно сред-
не- и заднеспинка, с 2 рядами четко выраженных склеритов, каждый из которых
несет по несколько щетинок, микроскульптура в виде овальных, сильно хитинизи-
рованных площадок с зубчиками на вершине. Коготки без явственного выступа или
он едва намечен. Верх с длинными и тонкими щетинками, оплавленными на вер-
шине. На сложноцветных.
28(29). Крыловые склериты среднегруди с 3 толстыми длинными и 8-9 короткими
щетинками (рис. 52, 6). Верхняя губа с неглубокой срединной выемкой, ее пере-
дний край с каждой стороны с 3 длинными шиповидными и 3 короткими щетинка-
ми (рис. 50, 6). ЕЧ, СБ............................Ch. marginata Linnaeus
29(28). Крыловые склериты среднегруди с 2 длинными и 8 более короткими щетин-
ками с оплавленными на вершине концами. Верхняя губа с глубокой срединной
выемкой, передний край губы с каждой стороны с 6 остроконечными щетинками,
из которых 3 более длинные (рис. 50, 7). Тундра и лесотундра СБ.........
.....................................................Ch. instabilis Maklin
30(27). Выемка верхней губы дуговидная, без выступа на дне. Верх с мелкими ок-
руглыми склеритовидными площадками, каждая из которых несет только 1 щетин-
ку. Коготки с крупным зубцом на основании.
31(32). Склеритовидные площадки очень мелкие, плохо очерченные, часто неяв-
ственные (рис. 53, 3). Непарный дорсальный склерит появляется с 7-го брюшного
тергита. Личинки на гречишных. СБ, ДВ.........Ch. koltzei Weise (см. тезу 79)
115
32(31). Склеритовидные площадки довольно крупные, хорошо очерченные. Непар-
ный дорсальный склерит появляется обычно с 8-го брюшного тергита. Личинки не
на гречишных.
33(34). Склеритовидные площадки тергитов многочисленные, собраны в 2-3 ряда
(до 50 штук) в передней и в 1-2 ряда (до 40 штук) в задней половине каждого терги-
та. Щетинки верха длинные, значительно превышают диаметр склеритовидных пло-
щадок. Тело темное, с более или менее явственным металлическим отливом. Дан-
ные требуют подтверждения. Питание, возможно, на Linaria. ЕЧ, СК.......
....................................................Ch. gypsophilae Kiister
34(33). Склеритовидные площадки тергитов менее многочисленные (не более 50
на каждом тергите). Щетинки короткие, не более чем вдвое превышают диаметр
склеритовидных площадок (рис. 52, 8). Тело светлое, без металлического отлива.
Микроскульптура в виде слабо хитинизированных вытянутых многоугольников,
тесно прилегающих друг к другу.
35(36). Вид с Сахалина. На среднегруди 4 площадки в переднем и 14 в заднем ряду,
на брюшных тергитах по 16 и 18 площадок в переднем и заднем ряду (рис. 60, 1).
Лоб слабо хитинизирован, особенно в передней части, с 24 щетинками (рис. 49, 3).
Микроскульптура в виде негустых выпуклых зернышек. Возможен на ДВ......
..................Ch. pirka Takizawa (возможно, подвид Ch. cavigera Sahiberg)
36(35). Тундровые виды. Склеритовидные площадки тергитов более многочислен-
ные (рис. 52, 8, 9). Микроскульптура в виде слабо хитинизированных вытянутых
многоугольников, тесно прилегающих друг к другу.
37(38). Щетинки верха примерно вдвое длиннее диаметра склеритовидных площа-
док. Лоб слабо хитинизирован, особенно в передней части, с 24 щетинками. Кры-
ловые склериты с 9 щетинками. Переднекрайние щетинки верхней губы тонкие
(рис. 50, 8). На крестоцветных. АР..Ch. cavigera Sahiberg (Ch. tolli Jacobson)
38(37). Щетинки верха мелкие, шиповидные, примерно равны диаметру склери-
товидной площадки. Лоб сильно хитинизирован, с 28 щетинками. Крыловые скле-
риты с 6 щетинками. Переднекрайние щетинки верхней губы толстые (рис. 50, 9).
На Nardosmia, Senecio, Oxytropis. АР...........Ch. subsulcata Mannerheim
39(26). Голова светлая, иногда с бурыми пятнами.
40(43). Тундровые виды. Голова иногда с бурыми пятнами, заметно светлее тела, с
вторичными длинными щетинками; лоб с 24-26 щетинками.
41(42). Склеритовидные площадки темноокрашенные. Покровы тела с явственной
микроскульптурой, разделенной промежутками (рис. 47, 2). Коготки с крупным
прямоугольным зубцом (рис. 47, 8). Щетинки тела короткие, не длиннее или едва
длиннее склеритовидных площадок. Каждый тергит примерно с 48-50 склерито-
видными площадками, образующими 2 неправильных ряда в передней (26 площа-
док) и 1 ряд в задней половине (22-24 площадки) (рис. 52, 9). Голова светлая, с
бурыми пятнами. АР.............................Ch. septentrionalis Menetries
42(41). Склеритовидные площадки бесцветные, около 35—40 на каждом тергите.
Покровы тела с неявственной, бесцветной микроскульптурой, в виде вплотную
смыкающихся многоугольников, не разделенных промежутками (рис. 47, 3). Когот-
ки со слабым зубцом (рис. 47, 9). Ар. (Новосибирские о-ва и прилегающие районы
материка)............................................Ch. bungei Jacobson
43(40). Виды не заходят в тундру.
116
Рис. 48. Головные капсулы личинок родов Oreina и Chrysolina'.
1 — Oreina basilea} 2 — Chrysolina koltzei', 3 — Ch. haemochlora; 4 — Ch. virgata; 5 — Ch.
nikolskyi} 6 — Ch. cerealis.
117
Рис. 49. Головные капсулы личинок родов Chrysolina и Oreina\
1 — Chrysolina pedestris', 2 — Ch. porphyrecr, 3 — Ch. pirka\ 4 — Oreina rugulosa. По Кимото и
Такизава, Михайлову и ориг.
44(45). Питание на Pedicularia palustris. Каждый тергит примерно с 78-80 темноок-
рашенными склеритовидными площадками, образующими 2 неправильных ряда
(42 площадки) в передней и 2 в задней половине (36-38 площадок) (рис. 53, 7).
Щетинки тела короткие, не длиннее или едва длиннее склеритовидных площадок.
Покровы тела с явственной микроскульптурой, разделенной промежутками (рис.
47, 2). Коготки с крупным прямоугольным зубцом (рис. 47, 8). Саяны, в горных
степях...................................................Ch. latimargo Weise
45(44). Питание на других растениях, в основном на Linaria, Plantago. Тергиты брюш-
ка с 2-3 спутанными рядами склеритовидных площадок. Коготки с зубцом.
46(49). Склеритовидные площадки тергитов брюшка очень мелкие и неявствен-
ные, выглядят как темные основания щетинок, эпиплевральный склерит также раз-
делен на отдельные площадки.
47(48). Верхняя губа с неглубокой выемкой. Тело бело-желтое. Все склеритовид-
ные площадки груди и брюшка очень мелкие (рис. 52, 7). На Linaria. ЕЧ, СК, СБ,
ДВ...............................................Ch. sanguinolentha Linnaeus
48(47). Верхняя губа с глубокой выемкой (рис. 51,7). Тело желто-коричневое. Скле-
ритовидные площадки тергитов груди — рис. 51, 7. На Plantago. Сюда же относит-
ся Ch. jenisseiensis Breit (ЕЧ, СК, СБ)..................Ch. limbata Fabricius
118
Рис. 50. Верхняя губа личинок рода Chrysolincr.
1 — Ch. perforata', 2 — Ch. arctica; 3 — Ch. aeruginosa; 4 — Ch. mongolensis; 5 — Ch.
convexicollis; 6— Ch. marginata; 7— Ch. instabilis; 8 — Ch. cavigera; 9 — Ch. subsulcata; 10 —
Ch. montana; 11 — Ch. geminata; 12 — Ch. polita; 13 — Ch. exanthematica; 14 — Ch. koltzei.
49(46). Склеритовидные площадки тергитов брюшка крупные, явственные, каж-
дый с одной щетинкой. Щетинки светлые. Коготки с острым зубцом. Тело желто-
коричневое. Переднеспинка с щетинками в основном по переднему и заднему кра-
ям. Склеритовидные площадки средне- и заднеспинки крупнее, чем на брюшке. На
Plantago. ЕЧ, СК..................................Ch. haemoptera Linnaeus
50(1). Тергиты средне- и заднегруди и брюшка без дорсальных склеритов и склери-
товидных площадок.
51(56). Верхняя сторона тела в длинных густых щетинках, не образующих пра-
вильных рядов, среди которых выделяются более длинные первичные и более ко-
роткие вторичные (рис. 53, 2). Голова с густыми вторичными щетинками (не менее
40 на лбу).
52(37). Лоб с 30-60 щетинками. Срединная выемка верхней губы глубокая (рис. 50,
10).
53(54). Тело грязно-серое, переднеспинка и ноги желтые, голова темно-коричне-
вая. Склерит переднеспинки с каждой стороны примерно с 20 очень длинными
119
Рис. 51. Детали строения личинок рода Chrysolina'.
1-3 — верхняя губа: 1 — Ch. limbata, 2 — Ch. varians, 3 — Ch. hyperici\ 4 — мандибула Ch.
Vartans; 5, 6 — Ch. camifex: 5 — тергит брюшка, 6 — коготок; 7, 8 — заднеспинка: 7 — Ch.
limbata, 8 — Ch. discriminata; 9, 10 — 1-й тергит брюшка: 9 — Ch. discriminata, 10 — Ch.
mongolensis.
первичными щетинками, в основном по краям. Средне- и заднеспинка с 2 попереч-
ными рядами макрохет и многочисленными микрохетами. Коготки без зубца. На
Saussurea. Алтай.........................................Ch. montana Gebler
54(53). Тело желто-белое, переднеспинка и ноги светло-желтые, голова желто-бу-
рая. Склерит переднеспинки с каждой стороны примерно с 10 умеренно длинными
щетинками, расположенными только по боковым краям. Средне- и заднеспинка
только с микрохетами. Коготки с прямоугольным зубцом. Возможен на Алтае...
........................................................Ch. dudkoi Mikhailov
55(52). Лоб примерно с 70 щетинками. Срединная выемка верхней губы неглубо-
кая. Коготки с небольшим тупым зубцом. На Aster. Юг ДВ..
.......................................................Ch. lineigera Jacobson
56(51). Верхняя сторона тела с короткими, часто едва заметными негустыми ще-
тинками.
57(64). Голова и переднеспинка светлые. Питание не на зверобое.
58(59). Щетинки на брюшных тергитах собраны в 5-7 спутанных ряда, темные.
Лоб с 30-40 щетинками. Коготки со слабым зубцом. Микроскульптура покровов
негустая. На губоцветных (Mentha, Lamium, Salvia, Galeopsis), возможно на Trollius,
Ranunculus, Plantago. Транспалеаркт..................Ch. staphylaea Linnaeus
59(58). Щетинки на брюшных тергитах собраны в 2 правильных или спутанных
ряда.
120
60(61). Коготки с крупным зубцом. Лоб с 10 длинными и 10-16 короткими щетин-
ками, щетинки верха хорошо заметные, на переднеспинке в основном по краям.
Верхняя губа с глубокой выемкой. Микроскульптура покровов густая, бесцветная,
плохо заметная, щетинки тела светлые. На губоцветных (Lamium, Galeopsis, Leo-
nurus). ЕЧ, СК, СБ...................................Ch. fastuosa Scopoli
61(60). Коготки с небольшим зубцом.
62(63). Лоб с немногочисленными щетинками. Микроскульптура покровов неяв-
ственная. На зонтичных (Chaerophyllum, Anthriscus, Aegopodium). ЕЧ....
Ch. oricalcia Miiller
63(62). Лоб с 24-30 щетинками почти одинаковой длины. Переднеспинка с щетин-
ками по всей поверхности. Микроскульптура покровов густая, явственная. На
Anthriscus. ЕЧ, СК.....................................Ch..chalcites Germar
64(57). Голова темная или бурая, обычно заметно темнее тела, если светлая, то пи-
тание только на зверобое.
65(86). Лоб только с 10-14 первичными щетинками (рис. 48, 2).
66(77). Коготки без зубца или он едва намечен. Микроскульптура явственная. Ще-
тинки верха очень короткие.
67(76). Питание только на зверобое. Срединная выемка верхней губы узкая и глу-
бокая (рис. 50, 77). Микроскульптура мелкая, не очень густая, более светлая.
68(69). Верхняя губа темная со светлым дуговидным участком близ переднего края
(рис. 51,2). Мандибула с темной базальной частью (рис. 51,4). Голова светло-ко-
ричневая. Переднеспинка с многочисленными микрохетами по всей поверхности.
Крыловые склериты с 4-5 микрохетами. Голова с первичными щетинками. ЕЧ, СК,
СБ...................................................Ch. varians Schaller
69(68). Верхняя губа сплошь темная или с небольшими светлыми пятнами.
70(73). Голова черная или темно-бурая.
71(72). Переднеспинка примерно с 20 щетинками, в основном по краям. Стерниты
брюшка с 6 щетинками каждый. 8-9-й тергиты без дорсального склерита или он
едва намечен. Юг СБ (типичная форма), юг ДВ (sbsp. ussuriensis Jacobson), Саха-
лин (sbsp. yezoensis Matsumura)..................Ch. difficilis Motschulsky
72(71). Переднеспинка примерно с 60 щетинками по всей поверхнеости. Стерниты
брюшка с 16 щетинками каждый. 8-9-й тергиты с заметным дорсальным склери-
том. ЕЧ, СК..........................................Ch. geminata Paykull
73(70). Голова коричневая или светло-коричневая.
74(75). Голова заметно темнее тела, только с первичными щетинками. Верхняя губа
темная со светлыми пятнами, как на рис. 51,3. ЕЧ, СК.Ch. hyperici Forster
75(74). Голова едва темнее светлозеленого тела. Средне- и заднеспинка с 7 щетин-
ками (5 длинных и 2 коротких) в районе крылового склерита. Голова с первичными
и вторичными щетинками. СК...........................Ch. abchasica Weise
76(67). Питание на мяте и других губоцветных; степные подвиды отмечались на
полыни. Срединная выемка верхней губы широкая, полукруглая (рис. 50, 73). Мик-
роскульптура крупная, очень густая, темная. СБ, ДВ....................
............................................Ch. exanthematica Wiedemann
77(66). Коготок с явственным зубцом.
78(85). Микроскульптура покрова явственная, темная. 7-9-й тергиты брюшка с дор-
сальным склеритом.
121
Рис. 52. Заднеспинка личинок родов Chrysolincr.
1 — Ch. arctica; 2 — Ch. mongolensis; 3 — Ch. convexicollis; 4 — Ch. aurichalcea; 5 — Ch.
undulata; 6 — Ch. marginata; 7 — Ch. sanguinolentha; 8 — Ch. cavigera; 9 — Ch. septentrionalis.
79(80). Верх co следами склеритовидных площадок вокруг оснований щетинок (рис.
53, 3). Зубец коготка прямой или притуплен (рис. 47, 10). Верхняя губа с 5 марги-
нальными щетинками с каждой стороны (рис. 50,14). На Polygonum. Сахалинский
подвид на Lamium. (см. также тезу 23). Юг СБ и ДВ.......Ch. koltzei Weise
80(79). Верх без сколько-нибудь явственных склеритовидных площадок, но иногда
со следами склеротизации у оснований щетинок. Верхняя губа с 4 парами марги-
нальных щетинок. На мяте, реже на других губоцветных.
81(82). Голова и переднеспинка коричневые, темнее светлокоричневого тела, ще-
тинки короткие, но явственные. Зубец коготка приострен (рис. 47, 77). Выемка вер-
хней губы глубокая, с округленными углами (рис. 50, 72). ЕЧ, СК, СБ, ДВ.
......................................................Ch. polita Linnaeus
82(81). Голова и тело темнобурые, нередко с металлическим отливом, с единичны-
ми, очень короткими незаметными щетинками. Зубец коготка крупный.
83(84). Выемка верхней губы с явственно тупыми углами. Микроскульптура покро-
вов более густая, темная и контрастная, особенно снизу. Зубец коготка угловатый.
ЕЧ...................................................Ch. coerulans Scriba
84(83). Выемка верхней губы с округленными углами. Микроскульптура покровов
менее густая, темная и контрастная. Зубец коготка округленный. ЕЧ, СК...
Ch. herbacea Duftschmidt (Ch. menthastri Suf&ian)
85(78). Микроскульптура покрова неявственная, бесцветная (рис. 47, 4). 8-9-й тер-
гиты брюшка с дорсальным склеритом. Наличник и переднеспинка светлее голо-
вы. Выемка верхней губы глубокая. На сложноцветных, включая полынь. ЕЧ, СК,
СБ, ДВ....Ch. graminis Linnaeus
122
& <3$ <3$
<&> <ЗЬ
<з£
Рис. 53. Заднеспинка личинок родов Chrysolina и Crosita\
1 — Chrysolina latimargo\ 2 — Ch. montana; 3 — Ch. koltzei; 4 — Ch. diversipes\ 5 — Ch.
haemochlora\ 6 — Crosita longipes', 7 — C. altaica\ 8 — C. kowalewskyi; 9 — C. pigra.
86(65). Лоб хотя бы с единичными вторичными щетинками (не менее 18). Стерни-
ты брюшка с более или менее явственными склеритами (рис. 47, 6). Личинки на
различных растениях, если на полыни, то только на Дальнем Востоке в мезофиль-
ных условиях. Микроскульптура хорошо развита.
87(88). Голова с очень густыми щетинками (на лбу свыше 100). Тергиты груди и
брюшка почти сплошь с многочисленными короткими щетинками, не образующи-
ми явственных поперечных рядов, но собраны в пре- и постдорсальные группы
(рис. 53, 4). Микроскульптура темная, мозаичная, основания щетинок темные. Ко-
готки с тупым зубцом. Питание на губоцветных (Glechoma, Linaria, Galium). ЕЧ,
СК, СБ......Ch. diversipes Bedel (Ch. violacea Muller, Ch. goettingensis Linnaeus).
88(87). Хетотаксия головы и верха негустая, лоб не более чем с 32 щетинками, тер-
гиты с короткими щетинками, собранными в 2 поперечных ряда, или щетинки от-
сутствуют.
89(94). Коготки без зубца или он едва намечен. Лоб с 30-32 щетинками (рис. 48, 5).
Микроскульптура темная, в виде отдельных многоугольников.
90(93). Дорсальная сторона тергитов без щетинок, лишь у переднего края каждого
тергита имеются 2 очень короткие щетинки (рис. 53, 5).
91(92). Тело темно-серое. Лобные швы расходятся сначала под острым, затем под
тупым углом. Темя в основном с первичными хетами (рис. 48, 3). Тергиты средне-,
заднегруди и брюшка с немногими хетами. Крыловые склериты с 5-7 хетами. На
зонтичных (Aegopodium, Conioselinum, Angelica, Heracleum, Pleurospermum). Юг
СБ от Алтая до Прибайкалья........................Ch. haemochlora Gebler
123
92(91). Тело грязно-желтое. Лобные швы сначала параллельны, затем расходятся
под почти прямым углом. Темя с более чем 60 микрохетами (рис. 49, 7). Тергиты
средне-, заднегруди и брюшка с многочисленными хетами. Крыловые склериты с
9-19 хетами. ЗСБ, Алтай.................................Ch. pedestris Gebler
93(90). Дорсальная сторона тергитов с короткими, но явственными щетинками,
образующими 2 поперечных ряда на каждом (6 в переднем и 10 в заднем ряду).
Тело желтовато-белое. На сложноцветных (Cacalia, Eupatorium, Petasites). Юг ДВ
................................................Ch. angusticollis Motschulsky
94(89). Коготки с явственным зубцом.
95(96). Лоб с 30-32 щетинками; вторичные щетинки собраны у переднего края и
вдоль лобных швов (рис. 48, 6). Микроскульптура бесцветная, очень густая, в виде
вплотную примыкающих друг к другу многоугольников. Верхняя губа с неглубо-
кой выемкой, возможно, без маргинальных щетинок. Тело светло-коричневое, час-
тично с металлическим отливом. На губоцветных (Mentha, Thymus, Sideritis). ЕЧ,
СК, СБ, ДВ..............................................Ch. cerealis Linnaeus
96(95). Признаки иные.
97(100). Виды ДВ. Лоб с 18-20 щетинками; немногочисленные вторичные щетин-
ки собраны у его переднего края (рис. 48, 4).
98(99). Лобные швы в основной части параллельные, а затем расходятся под тупым
углом (рис. 48, 4). Тело желтовато-белое. На губоцветных (Lycopus). Юг ДВ..
.....................................................Ch. virgata Motschulsky
99(98). Лобные швы расходятся от основания сначала под острым, затем под тупым
углом (рис. 48, 5). Тело серовато-бурое. На полыни. Юг ДВ, Сахалин, Курилы.
......................................................Ch. nikolskyi Jacobson
100(97). Эндемики Кавказа.
101(102). Лобные швы прямые, расходятся под тупым утлом. Лоб с 10 первичными и 16
вторичными щетинками, темя с 54—56 щетинками. Голенелапка с 4 длинными щетин-
ками сверху и 3 длинными щетинками снизу. Верхняя губа с 5 маргинальными щетин-
ками с каждой стороны,! мелкой щетинкой в выемке и 1 очень мелкой на дне. Крыло-
вой склерит переднеспинки с 10 длинными, средне- и заднеспинки с 5 щетинками.
Покровы тела слабо склеротизованы. На губоцветных. СК.....Ch. caspica Weise
102(101). Лобные швы в основании параллелесторонние, далее расходятся.
103(104). 5-й зубец мандибул заострен. Лобные швы параллельны на коротком рас-
стоянии, лоб и темя каждые с 26 щетинками примерно одинаковой длины. Верхняя
губа с 6 парами маргинальных щетинок и парой мелких внутри выемки. Крыловой
склерит переднеспинки с 12 длинными, средне- и заднеспинки с 1 длинной, 7 сред-
ними и 3 короткими щетинками. Покровы тела сильно склеротизованы. голенелап-
ка сверху и снизу с 2 щетинками. СК..........................Ch. rosti Weise
104(103). 5-й зубец мандибул косо срезан на вершине. Голенелапка с 4-5 щетинка-
ми сверху.
105(106). Голенелапка с 4 длинными и 1 короткой щетинками сверху и 2 длинными
снизу. Лобные швы параллельны на большом расстоянии, лоб с 12 первичными и
40—42 вторичными, темя с примерно 60 одинаковыми щетинками (рис. 49, 2). Кры-
ловые склериты переднеспинки с 14-15, средне- и заднеспинки с 8 щетинками. На
сложноцветных, особенно Petasites, бурачниковых, норичниковых, лютиковых, пас-
леновых. СК.......................................Ch. porphyrea Faldermann
124
106(105). Голенелапка с 3 длинными и 1 короткой щетинкками сверху и 4 длинны-
ми снизу. Лобные швы параллельны на коротком расстоянии, лоб с 8 первичными и
34-36 вторичными , темя с немногочисленными одинаковыми щетинками. Крыло-
вые склериты переднеспинки с 12 длинными, средне- и заднеспинки с 3 длинными
и 9 короткими щетинками. На пасленовых (Scopolia). СК...................
...................................................Ch. trapezicollis Bechyne
6. Род Oreina Chevrolat (Chrysochloa Hope)
Тело продолговатое, среднеспинка уже и короче передне- и заднеспинки, вслед-
ствие чего тело с явственным перехватом за переднеспинкой, которая назади силь-
но приподнята и частично прикрывает среднеспинку. 8-9-й сегменты брюшка уже
предыдущих, подвижные. Голова и переднеспинка у старших возрастов желтые или
коричневые, всегда светлее прочих сегментов. Голова (рис. 48, 7) округлая, с равно-
мерно расположенными хитинизированными пятнами. Темя в задней половине с
немногочисленными длинными и короткими щетинками, вдоль лобного шва с каж-
дой стороны с 3-4 длинными и 1-2 короткими щетинками. Лобные швы расходятся
под углом, никогда не бывают параллельными в верхней части. Лоб с 10 первичны-
ми и обычно с вторичными щетинками. Наличник узкий, с рядом из 6 щетинок.
Верхняя губа с 4 дискальными щетинками и 2-4 порами, срединная выемка треу-
гольная, широкая, с 2-4 короткими щетинками на дне, с каждой стороны от нее
имеется 5-6 маргинальных щетинок. Склерит переднеспинки слабо хитинизиро-
ван, тергиты средне-, заднегруди и брюшка с поперечной срединной складкой и со
спутанными рядами пор или щетинок; крыловые склериты со щетинками, иногда
сильно хитинизированные. У личинок со светлым телом на 7-9-м сегментах замет-
ны непарные хитинизированные склериты. Стигмы иногда окружены широким
хитинизированным кольцом. Микроскульптура покровов в виде овальных площа-
док или сливается и образует характерные только для данного рода крупные или
мелкие морщины. Коготки с прямоугольным или тупым зубцом при основании.
У личинок младших возрастов голова и переднеспинка темнее, чем верхняя
сторона тела, щетинки верха короткие или длинные.
Для данного рода характерно живорождение или яйцеживорождение. Имеется
4 возраста. Трофически имаго и личинки связаны со сложноцветными и с зонтич-
ными растениями.
Морфологически род очень близок к Chrysolina Motschulsky и Crosita Weise. В
регионе 3 вида.
Таблица для определения видов рода Oreina
1(4). Тело сверху угольно-черное или темно-бурое, блестящее; голова и переднес-
пинка светло-желтая или светло-коричневая.
2(3). Лоб с 6-8 вторичными щетинками (подрод Chrysochloa s.str.). Тело угольно-
черное, покровы без микроскульптуры, плотные, крупно морщинистые. Голова и
переднеспинка от светло-бурой или оранжевой до темно-бурой. Верхняя губа с не-
глубокой выемкой. На сложноцветных в горно-лесном поясе и на альпийских лу-
гах. Саяны..........................................О. redikorzevi Jacobson
125
3(2). Лоб с 58-60 первичными и вторичными щетинками (рис. 49, 7) (подрод
Allorinula Weise). На Centaurea. Запад ЕЧ, Урал (реликт)..............
О. coerulea Olivier (О. rugulosa Sffrian)
4(1). Тело сверху желтое, голова и переднеспинка немного светлее тела, крыловые
склериты не окрашены, стигмы не окружены темными кольцами (подрод Romalorina
Weise). Верхняя губа с глубокой выемкой. Микроскульптура тела в виде овально
вытянутых площадок. Лоб с 26 щетинками (рис. 48, 7), переднеспинка несет ще-
тинки по всей поверхности. На Heracleum, Saussurea в горно-лесном поясе и на
альпийских лугах. Юг СБ.О. basilea Gebler
7. Род Crosita Weise
Тело продолговатое, несколько расширено кзади. Голова с короткими первич-
ными и вторичными щетинками, слабо отличающимися друг от друга, на лбу около
24 щетинок, наличник с 6 щетинками, верхняя губа со срединной выемкой, несет 2
поры и 4 щетинки на диске и несколько коротких щетинок на переднем крае.
Мандибулы с 5 острыми зубцами и 2 щетинками на верхнем крае.
Склерит переднеспинки всегда хорошо развит, умеренно или сильно хитини-
зирован, с более или менее морщинистыми, очень короткими щетинками по пере-
днему и боковым краям. Средне- и заднеспинка и тергиты брюшка без явственных
склеритов или со слабо намеченными эпиплевральными склеритами, низ со слабо
выраженными склеритами. Щетинки тела всегда короткие, шиловидные, обычно
плохо заметные, редкие. Ноги довольно короткие, сильные, голенелапка снизу с 3
довольно тонкими щетинками, сверху с 3-5 щетинками. Коготки простые или с
более или менее развитым зубцом. Окраска верха бурая или темно-бурая, голова
всегда темная.
Род Crosita очень близок к Chrysolina и не всегда может быть четко отделен от
последнего, особенно от личинок, не имеющих склеритов или склеритовидных
площадок.
Личинки ведут скрытый или полускрытый образ жизни, встречаясь под курти-
нами полыни, в подстилке или в верхнем слое почвы на глубине до 5 см. Питание
только на полыни. В ночное время личинки заползают на растения для питания.
Обитают в горных районах аридной зоны на каменистых грунтах, что повело к
возникновению адаптивных приспособлений: переход к скрытному или полускрыт-
ному образу жизни, редукции склеритов и щетинок тела, усилению ног и особенно
голенелапки и коготков. Цикл развития с зимующим яйцом, весенней личинкой и
летней диапаузой имаго также адаптивен к аридным и экстрааридным условиям
существования. В регионе 4 вида.
Таблица для определения видов рода Crosita
1(2). Коготки с острым длинным зубцом, направленным вперед, так что коготок
выглядит расщепленным (рис. 47, 72). Щетинки на тергитах образуют два непра-
вильных поперечных ряда на каждом (рис. 53, 9; 47, 5). Лоб с 34-36 щетинками.
Микроскульптура явственная, темная. Тува..................С. pigra Weise
2(1). Коготки с тупым зубцом либо без зубца.
126
Рис. 54. Головная капсула (1-3, 5, 6) и лоб (4) личинок подсемейства Chrysomelinae:
1 — Sternoplatys fulvipese 2 — Gastrophysa mannerheimi; 3 — Entomoscelis orientalise 4 -
Gonioctena viminalise 5 — G. fulvae 6 — Cercyonops caraganae.
3(4). Коготки с тупым зубцом (рис. 47, 14). Щетинки верха густые, распределены
более или менее сплошь по всей поверхности тергитов (рис. 53, 6). Микроскульп-
тура явственная, темная. Лоб с 50-52 щетинками. Алтай, Тува................
.........................................................С. longipes Jacobson
4(3). Коготки без зубца или он едва намечен (рис. 47,13).
5(6). Микроскульптура тела очень густая, темная, в виде вплотную смыкающихся и
поэтому плохо заметных многоугольников. Щетинки тергитов негустые, но разбро-
саны по всей поверхности, образуя 5-6 неправильных поперечных рядов, без свет-
лых колечек у основания (рис. 53, 7). Лоб с 62-64 щетинками. Юго-восток ЕЧ, Ал-
тай ........................................................С. altaica Gebler
6(5). Микроскульптура тела умеренно густая, в виде темных многоугольников, раз-
деленных явственными промежутками. Щетинки тергитов довольно редкие, собраны
в 2 поперечных ряда, со светлыми колечками на основании (рис. 53, 8). Лоб с 30-32
щетинками. Юг Алтая и Прибайкалья......................С. kowalewskyi Gebler
8. Род Phratora Chevrolat (Phyllodecta Kirby)
Эпикраниальный шов длинный, лобные швы расходятся под прямым углом.
Лоб с 8 щетинками (отсутствует пара Q и 2 порами, наличник слабо хитинизиро-
ван, с 4 щетинками. Верхняя губа со слабо вогнутым, но не выемчатым передним
127
краем, с 4 щетинками и 2 порами на диске и 5 короткими толстыми маргинальными
щетинками с каждой стороны (рис. 44, 4). Склерит переднеспинки с 10 щетинками
на переднем и 6-10 на заднем крае, эпиплевральные склериты переднеспинки ма-
ленькие или отсутствуют (в последнем случае его местоположение определяет со-
ответствующая щетинка). Средне- и заднеспинка с 2 внутренними претергальны-
ми, несущими по 2 щетинки, и с 4 постгергальными склеритами, из них внутрен-
ние с 1, а наружные с 3 щетинками. 1-й, реже 2-3-й тергиты брюшка несут по 2
претергальных (внутренних) склерита с 1 щетинкой, иногда исчезающих; на пос-
ледующих тергитах они отсутствуют (рис. 55, 7); внутренние посттергальные скле-
риты, имеют по 3 щетинки, на всех брюшных тергитах; на 7-8-м тергитах они сли-
ты в непарный склерит; наружные посттергальные отсутствуют. Крыловые и стиг-
мальные склериты с 3 щетинками и отверстиями желез; эпиплевральные склериты
с 2 щетинками. Микроскульптура имеется у всех видов, но степень ее развития
варьирует у некоторых видов (Ph. vulgatissima Linnaeus, Ph. laticollis Suf&ian). Про-
межутки между склеритами верха могут быть светлые («светлый» тип) или с густы-
ми тесно сближенными темными пластинками микроскульптуры, так что верх выг-
лядит темным («темный» тип) или темная микроскульптура расположена по бокам
тела между тергальными и эпиплевральными склеритами, образуя вместе со скле-
ритами темные продольные полосы («полосатый» тип).
Жуки и личинки скелетируют листья ив, осины и тополей. Яйца кладутся груп-
пами до 20 штук, обычно в два ряда, одна самка откладывает 300-400 яиц. Окукли-
вание в почве. В регионе 8 видов.
Таблица для определения видов рода Phratora
1(4). Коготки с зубцом при основании (подрод Chaetocera Weise). Нижняя сторона
тела с бесцветными склеритами. Ноги светлые. Щетинки верха бело- или золотис-
то-желтые.
2(3). Коготки с крупным прямоугольным зубцом при основании (рис. 44, 8). Преоб-
ладают «светлый» и «темный» типы, реже «полосатый» тип. На различных ивах,
иногда на чозении. Вся лесная зона Палеарктики......Ph. vulgatissima Linnaeus
3(2). Коготки с длинным, узким и острым зубцом при основании (рис. 44, 9). Пре-
обладает «полосатый» тип. На иве Шверина. ВСБ, ДВ..........................
.................................................Ph. obtusicollis Motschulsky
4(1). Коготки (рис. 44,10) без зубца при основании (подрод Phratora s.str.).
5(6). Склериты нижней стороны тела бесцветные, плохо заметные. Щетинки верха
и ноги светлые. Преобладает «темный» тип, реже «светлый» или «полосатый». На
осине, тополе. Вся лесная зона Палеарктики..............Ph. laticoilis Suffrian
6(5). Склериты нижней стороны тела пигментированные, явственные, заметно тем-
нее фона.
Рис. 55. Детали строения личинок подсемейства Chrysomelinae:
1 — 1-й и 2-й тергиты брюшка Phratora laticollis', 2,4,9 — среднеспинка: 2 — Chrysomela
vigintipunctata, 4 — Ch. lapponicum, 9 — Gastrolina peltoidea', 3, 5, 8,10,11 — тергит брюш-
ка: 3 — Plagiosterna aenea, 5 — Hydrothassa marginella, 8 — Agrostithassa glabra, 10 —
Gastrolinoides japonica, 11 —Prasocuris phellandrii', 6 — 2-й тергит брюшка Plagiodera', 7 —
4-й стернит брюшка Chrysomela tremulae. По Хеннигу и ориг.
128
129
7(8). Щетинки тела черные, обычно темнее тела. Микроскульптура верха неравно-
мерная, образует отдельные темные пятна между склеритами. Переднеспинка тем-
ная, с 2 желтыми пятнами у срединной линии. Тело широкое, сужено кзади, упло-
щенное сверху; голова значительно уже груди. На ивах, чозении, осине и тополях.
Вся лесная зона Палеарктики.........................Ph. vitellinae Linnaeus
8(7). Щетинки верха светлые, не темнее или светлее тела. Микроскульптура верха
густая и равномерная, не образует отдельных пятен; верх выглядит темным. Тело
удлиненное, цилиндрическое, голова незначительно уже груди.
9(12). Переднеспинка светлая посередине
10(11). Переднеспинка черная, с широкой светлой полосой, занимающей средин-
ную треть. На ивах. Север ЕЧ, СБ, ДВ, АР............Ph. polaris Schneider
11(10). Переднеспинка светлая с темнобурыми боками. На осине, иве, чозении, то-
поле. Вся лесная зона Палеарктики.................Ph. atrovirens Cornelius
12(9). Переднеспинка черная (кроме узкой светлой срединной линии). На ивах. ЕЧ,
СК.....................................................Ph. tibialis Suf&ian
9. Род Plagiodera Erichson
Личинки темно-серого цвета, с темной головой, ногами и склеритами. Темя
впереди с каждой стороны с 4 длинными и 1 короткой щетинками, эпикраниаль-
ный шов длинный, лобные швы прямые, расходятся под тупым углом. Лоб с 6 ще-
тинками, расположенными у переднего края, щетинки и f2 отсутствуют. Налич-
ник поперечный, в основании узко хитинизирован, с рядом из 6 мелких щетинок.
Верхняя губа с узкой глубокой треугольной выемкой, с рядом из 4 длинных и 2
коротких щетинок и 4 пор на диске, передний край ее с каждой стороны с 3 широ-
кими и плоскими маргинальными щетинками, лежащими далеко от переднего края,
и 1 длинной наружной. Склерит переднеспинки разделен светлой полосой, с 10
короткими щетинками по переднему и 8 по заднему краю. Наружные претергаль-
ные склериты груди не развиты, внутренние посттергальные склериты с 2, наруж-
ные посттергальные с 3 микрохетами. Тергиты 1-6-го брюшных сегментов (рис.
55, 6) посередине с 2 сближенными тергальными склеритами, несущими по 2 мак-
рохеты; кнаружи от этих склеритов на 1-3-м брюшных сегментах находится не-
большой склерит с 1 щетинкой, стигмальные склериты с 3 микрохетами. На терги-
тах 7-8-го сегментов брюшка все склериты сливаются вместе. Эпиплевральные
склериты с 1 макро- и 2 микрохетами. Покров тела с густой темноокрашенной мик-
роскульптурой, исчезающей на середине стернитов; на груди срединная светлая
полоса также лишена микроскульптуры. Голенелапка снизу с 2, сверху с 4 щетин-
ками, коготок с прямоугольным зубцом при основании.
В России род представлен единственным видом Р. versicolorea Laicharting,
широко распространенным по всей Палеарктике. Жуки и личинки питаются листь-
ями различных ив, в основном узколистных. Окукливание на листьях.
10. Род Plagiosterna Motshulsky (Linaeidea Motschulsky)
Тело светло-желтое, верх со склеритами и 2 продольными полосами по бокам,
низ без склеритов, голова светлая, с пятнами по бокам темени и с V-образным пят-
130
ном в середине лба. Верх с микроскульптурой в виде овально-вытянутых площа-
док, наиболее густо расположенных около склеритов, несущих дорсальные желе-
зы. Эпикраниальный шов длинный, лобные швы прямые и расходятся под прямым
углом, лоб с 4 длинными и 4 короткими щетинками, расположенными в два ряда,
наличник узкий, сильно хитинизирован только в основании, с рядом из 6 коротких
щетинок; верхняя губа поперечная, по бокам основания с острыми выростами, кпе-
реди сильно сужена, с глубокой срединной выемкой, достигающей половины ее
ширины, с 3 короткими и толстыми маргинальными щетинками с каждой стороны
и с рядом из 4 длинных и 2 коротких щетинок и пор на диске. Переднеспинка свет-
лая, морщинистая, по бокам с крупными пятнами, окруженными мелкими щетин-
ками. Средне- и заднеспинка с крупными внутренними и мелкими наружными пре-
и посттергальными склеритами, которые иногда могут сливаться друг с другом.
Сегменты брюшка с одним рядом склеритов (рис. 55, 3), из которых внутренние,
начиная с 7-го сегмента, сливаются вместе. Эпиплевральные склериты мелкие, с 2-
3 щетинками. Голенелапка узкая, снизу с 2 щетинками в базальной части, коготок с
прямоугольным зубцом при основании.
Жуки и личинки питаются на ольхе; окукливание на листьях.
В России 1 вид — Р. aenea Linnaeus, широко распространенный по всей Пале-
арктике.
11. Род Chrysomela Linnaeus (Metasoma Stephens)
Тело личинок светлое, сверху и снизу с хорошо развитыми темноокрашенны-
ми склеритами, несущими мелкие щетинки, и микроскульптурой, голова и ноги
темные. Темя в задней половине с 4 щетинками, вдоль лобных швов с 4 длинными
щетинками. Эпикраниальный шов длинный, лобные швы расходятся под острым
углом. Лоб с 12 длинными щетинками, отсутствуют. Наличник широкий, хитини-
зирован по углам основания, с рядом из 6 мелких щетинок. Верхняя губа с глубокой
треугольной срединной выемкой и с 3 толстыми маргинальными щетинками с каж-
дой стороны, на диске ряд из 4 длинных толстых щетинок и 4 пор. 2-й членик уси-
ков квадратный, 3-й слабо вытянутый, с мелкими сенсорными щетинками на вер-
шине. Эпиплевральные склериты переднеспинки маленькие. Средне- и заднеспин-
ка с крупными внутренними тергальными склеритами, наружные претергальные
не развиты, наружные посттергальные у части видов крупные, у некоторых сильно
редуцированы. Тергиты брюшных сегментов с одним рядом склеритов, внутрен-
ние склериты 6-7-го сегментов сливаются в непарный склерит. Стерниты груди и
брюшка с развитыми склеритами и крепкими щетинками. Тергиты груди и брюшка
с отверстиями кожных желез, выделяющих жидкость с запахом бромбензола. Голе-
нелапка вытянутая, коготки с прямоугольным зубцом при основании.
В регионе 9 видов, связанных в питании с деревьями и кустарниками семей-
ства ивовых, лишь один вид может переходить на березу.
Таблица для определения видов рода Chrysomela
1(8). Тергальные склериты брюшка сливаются в один непарный склерит с 6-го сег-
мента.
131
2(3). Переднеспинка вся темноокрашенная, без светлого пятна посередине. На иве.
ЕЧ, СК, СБ..............................................Ch. saliceti Weise
3(2). Переднеспинка с отчетливым крупным светлым пятном посередине или свет-
лая с темными боковыми и задним краями.
4(5). Переднеспинка светлая, с бурой каймой вдоль заднего края и широкой бурой
каймой на боках. Микроскульптура покрова негустая, склериты на ее фоне резко
очерченные. На осине, тополе, редко на иве. Транспалеаркт... Ch. populi Linnaeus
5(4). Переднеспинка с прямоугольным или округлым светлым пятном посередине,
ее передний край, так же, как бока и задний край, темные.
6(7). Парастернальные склериты стернитов брюшка с 3 щетинками; гипоплевраль-
ные склериты с 6 щетинками (рис. 55, 7). На осине, редко на иве. Вся лесная зона
Палеарктики.........................................Ch. tremulae Fabricius
7(6). Парастернальные и гипоплевральные склериты стернитов брюшка с 5 щетин-
ками. На иве. Вся лесная зона Палеарктики...........Ch. cupreum Fabricius
8(1). Тергальные склериты брюшка сливаются в один непарный склерит с 7-го сег-
мента.
9(10). Средне- и заднеспинка с каждой стороны с одним наружным посттергаль-
ным склеритом (рис. 55,2). Микроскульптура покрова на стернитах густая и равно-
мерная, на тергитах заметна преимущественно около переднего и заднего краев.
На иве. Вся лесная зона Палеарктики.............Ch. vigintipunctata Scopoli
10(9). Средне- и заднеспинка с каждой стороны с разделенным на две части наруж-
ным посттергальным склеритом (рис. 55, 4). Микроскульптура покрова тергитов
густая и равномерная, на стернитах менее густая.
11(12). Микроскульптура тергитов брюшка, особенно в передней их части и посе-
редине, настолько густая, что тергальные склериты кажутся слитыми в один непар-
ный склерит. На иве и березе. Вся лесная зона Палеарктики..............
.................................................Ch. lapponicum Linnaeus
12(11). Мироскульптура тергитов брюшка менее густая, тергальные склериты рез-
ко выделяются на светлом фоне покрова и не кажутся слитыми.
13(14). Эпиплевральные склериты с 2-3 щетинками; задние парастернальные скле-
риты 2-го брюшного стернита с 2 щетинками. На ивах. Вся лесная зона Палеаркти-
ки ..................................................Ch. collaris Linnaeus
14(13). Эпиплевральные склериты с 3-4 щетинками; задние парастернальные скле-
риты 2-го брюшного стернита с 3 щетинками. На Salix arctica.
15(16). Континентальная часть Арктики от Таймыра до Чукотки............
..............................................Ch. taimyrensis L. Medvedev
16(15). Остров Врангеля.......................Ch. vrangeliana L. Medvedev
12. Род Gastrolina Motschulsky
Личинки грязно-серые, по бокам с темными продольными полосами. Тело силь-
но удлиненное, суживается кпереди и кзади от первого брюшного сегмента. Голова
темная, эпикраниальный шов длинный, лобные швы расходятся под тупым углом,
прямые, лоб с 8 длинными щетинками, расположенными в центре. Наличник с ря-
дом из 6 щетинок, сильно хитинизирован только в основании. Верхняя губа на дис-
ке с рядом из 4 щетинок, с неглубокой прямоугольной срединной выемкой и пря-
132
мым передним краем, с каждой стороны с 3 короткими и толстыми маргинальными
щетинками. Склерит переднеспинки морщинистый, с мелкими щетинками по пе-
реднему и заднему краям. Средне- и заднеспинка с 2 рядами склеритов (наружные
претергальные отсутствуют), несущих по 1-2 щетинки; крыловые склериты с мел-
кими щетинками (рис. 56, 7). Тергиты брюшка только с 2 посттергальными склери-
тами, каждый с 2-4 щетинками, начиная с 6-го сегмента они сливаются попарно.
Крыловые и стигмальные склериты с крупным отверстием желез, на 8-м тергите
брюшка они сливаются с тергальными. Эпиплевральные склериты с 4-6 коротки-
ми щетинками. Низ с хорошо развитыми склеритами, брюшные стерниты включа-
ют гипоплевральные, 2 пары парастернальных (передняя пара очень мелкая) и не-
парный стернальный. Покровы сверху с густой микроскульптурой в виде овально-
вытянутых площадок, наиболее густой около склеритов с кожными железами, сни-
зу — с гораздо более редкой. Ноги сильно хитинизированы, голенелапка снизу с 4,
сверху с 10 щетинками, коготки с крупным прямоугольным зубцом при основании.
Личинки обгрызают листья древесных пород, окукливаются на листьях (рис.
5, 3). В регионе 2 вида.
Таблица для определения видов рода Gastrolina
1(2). На среднегруди претергальный склерит гораздо крупнее (рис. 56, 7), а на зад-
негруди заметно меньше внутреннего посттергального. На маньчжурском орехе.
Юг ДВ.......................................................G. depressa Baly
2(1). На среднегруди претергальный склерит равен или едва крупнее(рис. 55, 9), а
на заднегруди заметно крупнее внутреннего посттергального. На ольхе. ВСБ, ДВ
..........................................................G. peltoidea Gebler
13. Род Gastrolinoides Chujo et Kimoto
Род близок к Gastrolina, с которым ранее и объединялся. Тело личинки умерен-
но удлиненное, светло-желтое, с многочисленными светлыми щетинками и 2 ши-
рокими продольными темными полосами по бокам груди и брюшка. Голова тем-
ная, темя спереди и по бокам с 4-5 длинными и многочисленными короткими ще-
тинками, эпикраниальный шов длинный, лобные швы расходятся под прямым уг-
лом, лоб с 20 щетинками, в основном по переднему краю, из которых 6 очень длин-
ные, наличник поперечный, хитинизирован только в основании, с рядом из 6 щети-
нок. Верхняя губа с глубокой срединной выемкой, 3 толстыми короткими и 1 мел-
кой маргинальными щетинками с каждой стороны. Переднеспинка с каждой сторо-
ны с большим коричневым пятном, по переднему и заднему краям со спутанными
рядами из многочисленных щетинок. На груди внутренние и наружные тергальные
склериты сливаются, крыловые склериты сильно хитинизированные, каждый с 1
очень длинной и 28-30 короткими щетинками. На брюшных сегментах все склери-
ты сливаются в 2 широких склерита с многочисленными щетинками (рис. 55, 70);
начиная с 5-го сегмента они сливаются в один непарный склерит. Эпиплевральные
склериты с 2 длинными и 8-10 короткими щетинками. Крыловые и стигмальные
склериты с отверстием кожных желез. Стерниты груди и брюшка слабо окрашены,
с более редкими спутанными рядами щетинок. Верх с густой микроскульптурой,
133
134
которая бугровидно приподнята или образует мелкие шипики, низ с очень светлой
микроскульптурой. Голенелапка сверху с 12-13, снизу с 2 щетинками. Коготок силь-
но хитинизирован только в базальной половине, с прямоугольным зубцом при
основании.
Жуки и личинки на разнолистной и маньчжурской лещине. В роде 1 вид G. ja-
ponica Harold, встречающийся на юге ДВ.
14. Род Gastrophysa Chevrolat (Gastroidea Hope)
Личинки грязно-серые, по бокам с каждой стороны с продольными темными
полосами. Голова темно-коричневая, эпикраниальный шов длинный, лобные швы
расходятся под тупым или острым углом (рис. 54, 2), темя с каждой стороны с 4-6
щетинками, лоб с 8 щетинками, расположенными в 2 ряда (4,4). Верхняя губа (рис.
44, /) узкая, поперечная, сильно хитинизированная, со слабой широкой или прямо-
угольной глубокой срединной выемкой, с рядом из 4 утолщенных щетинок и 2 пор
на диске; передний край губы с каждой стороны с 2-3 крепкими маргинальными
щетинками, иногда далеко отодвинутыми от переднего края. 2-й членик усиков с
конусовидной папиллой на вершине, которая значительно короче узкого и вытяну-
того 3-го членика. Переднеспинка с явственными склеритами, разделенными свет-
лой полосой, с рядом из 12 щетинок по переднему и 6 по заднему краю. Средне- и
заднеспинка (рис. 56, 8) с 2 рядами склеритов, несущих по 1 щетинке. Наружные
претергальные склериты часто разделяются на 1-2 мелких, а наружный посттер-
гальный далеко отодвинут от внутреннего, сближен с крыловым и несет 3—4 ще-
тинки. Крыловые и эпиплевральные склериты с 2 щетинками.
Тергиты брюшка с 2 рядами склеритов, на 1-6-м сегментах они разделены, а
начиная с 7-го сливаются попарно; склериты первого ряда с 1, а второго — с 2
щетинками. Стерниты груди и брюшка с явственными склеритами, которые на 7-9-
м сегментах сливаются в один непарный. Покровы тела с микроскульптурой в виде
овально-вытянутых площадок, наиболее густо расположенных около склеритов с
кожными железами. Ноги сильно хитинизированы, коготки в базальной части уз-
кие, без выступа, голенелапка снизу с 3, сверху с 2 щетинками.
Личинки на растениях семейства гречишных. В России 4 вида.
Таблица для определения видов рода Gastrophysa
1(2). Микроскульптура покрова верхней и нижней сторон тела состоит из бледно-
желтых пластинок и только на линии расположения дорсальных желез (на всех
тергитах, кроме переднеспинки) несет темно-бурые пластинки, образуя на боках
тергитов по широкой продольной темной полосе. Склериты светло-желтые, плохо
Рис. 56. Детали строения личинок подсемейства Chrysomelinae:
1,2,5,9,10—среднеспинка: 1 — Gastrolina depressa, 2—Phaedon concinnus, 5 — Gastrophysa
atrocyanea, 9 — G. viridula, 10— G. mannerheimi; 3, 4,11 — 1-й тергит брюшка: 3 —Phaedon
concinnus, 4 — Ph. cochleariae, 11 — Apterocuris sibirica’, 5 — 3-й стернит брюшка Phaedon
cochleariae', 6,7 — Sternoplatys fulvipes'. 6 — грудь, 7 — 2-й тергит брюшка. По Хеннигу и
ориг.
135
видны на светлом фоне покрова, переднеспинка светло-желтая, с несколькими бо-
лее темными мелкими пятнышками на диске. Верхняя губа с глубокой треугольной
выемкой. На Polygonum, Rumex. ЕЧ, СК, СБ...............G. polygoni Linnaeus
2(1). Микроскульптура покрова верхней стороны тела состоит только из темноок-
рашенных пластинок, вследствие чего окраска равномерно-темная, склериты и бока
переднеспинки темно-коричневые, хорошо заметные.
3(4). Переднеспинка хитинизирована только по бокам, ее щетинки окружены четки-
ми узкими хитинизированными колечками. Верхняя губа со слабой, но широкой вы-
емкой (рис. 44, 3). Средне- и заднеспинка имеют вблизи наружного претергального
склерита соответственно 2 и 1 мелких склерита с очень короткими щетинками (рис.
56,10). На Polygonum на засоленных участках. Юг СБ.....G. mannerheimi Stal
4(3). Переднеспинка сплошь хитинизированная, ее щетинки не окружены четкими
хитинизированными колечками. Верхняя губа с явственной срединной выемкой (рис.
44, /).
5(6). Тергиты тела с сильно выпуклыми склеритами, покровы в мелкой поперечной
морщинистости и с очень густой микроскульптурой, щетинки коричневые. Внутрен-
ние пре- и посттергальные склериты средне- и заднеспинки примерно одинаковой
величины (рис. 56, 8). На Rumex, Polygonum. Юг ДВ.G. atrocyanea Motschulsky
6(5). Тергиты тела с плоскими склеритами, покровы слабо морщинистые, с уме-
ренно густой, более светлой микроскульптурой, щетинки светлые. Претергальные
склериты средне- и заднеспинки значительно меньше посттергальных (рис. 56, 9).
На Polygonum, Rumex, Rheum. ЕЧ, СК, СБ....................G. viridula Degeer
15. Род Hydrothassa Thomson
Молодые личинки почти черные, взрослые темно-серые, со светлыми краями
сегментов, снизу серовато-желтые. Голова темно-коричневая, темя назади с тонки-
ми щетинками, вдоль лобного шва с каждой стороны с 3 длинными щетинками.
Эпикраниальный шов короткий, отходящие от него лобные швы на значительном
протяжении идут параллельно, далее расходятся под острым углом. Лоб с 8 щетин-
ками близ переднего края, наличник хитинизирован только в основании, с рядом из
6 щетинок. Передний край верхней губы закруглен, с неглубокой прямоугольной
срединной выемкой, с каждой стороны с 2 длинными и утолщенными и 2 коротки-
ми маргинальными щетинками; на дне выемки 2 мелкие щетинки. 1-й членик уси-
ков узкий, 2-й вытянутый, длиннее ширины, 3-й сильно вытянутый, конический, в
3 раза длиннее ширины. Переднеспинка с редкой зернистостью и с немногочислен-
ными щетинками по переднему и заднему краям. Эпиплевральные склериты пере-
днеспинки редуцированные или отсутствуют. Наружные посттергальные склериты
груди слабо выражены, с 1 микрохетой; наружные претергальные редуцированы и
представлены только щетинкой, крыловые с 2 макрохетами. На сегментах брюшка
различимы только внутренние посттергальные склериты, с 1 макро- и 1 микрохетой
(рис. 55, 5). Эпиплевральные склериты с 1 макро- и 2 микрохетами, на 7-8-м терги-
тах все склериты сливаются в один непарный склерит. Сегменты груди и брюшка с
отверстием кожных желез. Склериты стернитов груди и брюшка развиты, каждый с 1
крепкой щетинкой. Микроскульптура тела, особенно верхней стороны, в виде много-
угольников, тесно прилегающих друг к другу. Коготки без зубца при основании.
136
Личинки питаются растениями семейства лютиковых. Небольшие яйца откла-
дываются в стебли кормовых растений, личинка на листьях или, реже, в стеблях и
черешках; три личиночных возраста. Окукливание происходит в стеблях, в кото-
рые проникают выросшие личинки. В регионе 2 вида.
Таблица для определения видов рода Hydrothassa
1(2). Макрохеты переднеспинки длинные, резко отличаются от немногочисленных
коротких микрохет; претергальные склериты среднеспинки с 2 щетинками; рассто-
яние между тергальными склеритами брюшных сегментов короче ширины самих
склеритов. На Caltha, Ranunculus. ЕЧ, СБ, ДВ........Н. marginalia Linnaeus
2(1). Макрохеты переднеспинки короткие и плохо отличаются от многочисленных
микрохет, претергальные склериты среднеспинки с 1 щетинкой, расстояние между
тергальными склеритами почти вдвое превосходит ширину последних. На Caltha,
Ranunculus. ЕЧ, СБ................................Н. hannovarana Fabricius
16. Род Agrostithassa Jacobson
Близка к роду Hydrothassa (возможно, является ее подродом), но легко отлича-
ется наличием хорошо развитой пары претергальных склеритов на первых 3-4 тер-
гитах брюшка (рис. 55, 8). Переднеспинка на переднем крае с 10, на боковом с
каждой стороны с 1 и на заднем крае с 8 макрохетами, эпиплевральный склерит
небольшой, с 1 макрохетой. Внутренние претергальные склериты средне- и заднес-
пинки с 2 щетинками, внутренние посттергальные с 1, наружные посттергальные с
1 длинной и 1 короткой щетинкой. Претергальные склериты брюшных сегментов с
1 щетинкой, посттергальные (по-видимому, слитые внутренний и наружный) скле-
риты с 3 щетинками. Стернальные склериты на первых четырех сегментах узко
разделенные, начиная с 5-го сегмента постепенно сливаются, образуя один непар-
ный склерит. Микроскульптура покрова очень густая и темная, на стернитах более
светлая. Коготки без зубца.
Личинки обгрызают листья и цветы лютика. В России 1 вид, A. glabra Herbst
(ЕЧ, ЗСБ).
17. Род Prasocuris Latreille
Личинки темноокрашенные, сверху черно-зеленые, снизу желтовато-зеленые
с черной головой, переднеспинкой и тергитом 9-го сегмента брюшка. Голова со
сравнительно коротким эпикраниальным швом, лобные швы узко параллельные,
лоб и темя с первичными и несколькими вторичными щетинками, верхняя губа с
глубокой выемкой на переднем крае. Микроскульптура покрова тела на верхней
стороне очень густая, состоит из тесно сближенных, темно окрашенных многоуголь-
ников, на нижней стороне значительно более разреженная. Щетинки на склеритах
короткие, переднеспинка без эпиплевральных склеритов, на переднем крае с каж-
дой стороны с 5, на заднем с 4 макрохетами; средне- и заднеспинка с глубокой
поперечной бороздкой, с хорошо развитыми, несущими 2 щетинки, внутренними
посттергальными и несколько более мелкими наружными посттергальными скле-
137
ритами; внутренние претергальные склериты очень маленькие, с 1 щетинкой, на-
ружные почти полностью редуцированы и их местоположение можно определить
лишь по наличию короткой щетинки, плохо заметной среди микроскульптуры по-
крова. Крыловые склериты с широкими отверстиями желез. Внутренние посттер-
гальные склериты 1-6-го тергитов брюшка хорошо развиты, с 2 щетинками (одна
из них очень короткая), внутренние претергальные очень маленькие; наружные тер-
гальные отсутствуют (рис. 55,11). На поперечной складке, разделяющей тергиты, с
каждой стороны находятся по 3 небольших, более светлых, похожих на склериты
пятна, образовавшихся от слияния элементов микроскульптуры покрова; на 7-8-м
тергитах брюшка внутренние тергальные склериты, сливаясь, образуют один боль-
шой непарный склерит. Коготки без зубца на основании.
Личинки живут на листьях или в стеблях кормовых растений, обычно во влаж-
ных стациях. 3 личиночных возраста; окукливание в стеблях.
В России два вида.
Таблица для определения видов рода Prasocuris
1(2). Стерниты средне- и заднегруди с 1 непарным склеритом. На Veronica. ЕЧ, СК
.....................................................Р. junci Brahm
2(1). Стерниты средне- и заднегруди с 3 склеритами, из них передний непарный.
Личинки в стеблях и черешках листьев Cicuta, Sium, Oenanthe, реже Caltha; иногда
на листьях Caltha. Транспалеаркт.....................Р. phellandrii Linnaeus
18. Род Sternoplatys Motschulsky
Личинки желтые, с чертой головой и бурыми, сильно выпуклыми склеритами.
Темя близ эпикраниального и лобного швов с каждой стороны с 3 длинными ще-
тинками. Эпикраниальный шов длинный (рис. 54, 1). Лоб с 8 длинными щетинка-
ми, наличник отделен ото лба узкой хитинизироваиной полосой, с рядом из 6 длин-
ных щетинок. Верхняя губа с узкой глубокой треугольной выемкой, с 3 утолщенны-
ми и 1 тонкой маргинальными щетинками с каждой стороны. 3-й членик усиков
узкий и длинный. Переднеспинка по переднему краю с 10, а по заднему с 8 длин-
ными щетинками, тергиты груди с пре- и посттергальными склеритами, несущими
по 1 длинной щетинке, наружный посттергальный с 3 щетинками, крыловые и эпип-
левральные склериты с 2 длинными щетинками (рис. 56, 6). 1-й тергит брюшка с 1
или 2 рядами склеритов, прочие тергиты брюшка (рис. 56, 7) с 1 рядом склеритов,
из которых внутренние тергальные несут по 3-4 длинные щетинки и, начиная с 7
сегмента, сливаются в непарные склериты. Стерниты груди и брюшка с развитыми
склеритами. Крыловые и стигмальные склериты с отверстием кожных желез. Мик-
роскульптура покровов развита, густая, в виде светлых овальных зерен. Ноги силь-
но хитинизированные, коготки с зубцом при основании.
В регионе 3 вида, связанных с розоцветными.
Таблица для определения видов рода Sternoplatys
1(2). 1-й тергит брюшка с 1 рядом склеритов, внутренние тергалыные склериты с 4
длинными щетинками. На Filipendula. ВСБ, ДВ..........S. fulvipes Motschulsky
138
2(1). 1-й тергит брюшка с 2 рядами склеритов, внутренние посттергальные склери-
ты с 3 длинными щетинками. На Potentilla. СБ..........S. clementzi Jacobson
19. Род Phaedon Latreille
Личинка с темно-бурыми головой, ногами, пятнами на переднеспинке и скле-
ритами. Эпикраниальный шов длинный, лобные швы расходятся под острым уг-
лом. Темя вдоль лобных швов с 3 щетинками, лоб с 8 первичными щетинками, f6
отсутствует. Наличник хитинизирован в основании, с рядом из 6 коротких щетинок.
Передний край верхней губы со слабыми зазубринами и неглубокой прямоугольной
срединной выемкой, с каждой стороны с 3 толстыми длинными и 2 короткими марги-
нальными щетинками, на диске ряд из 4 щетинок и 2 пор. 2-й членик усиков продол-
говатый, вдвое длиннее ширины, 3-й членик втрое длиннее ширины. Переднеспинка
с 10 длинными щетинками на переднем и с 8 — на заднем краях, эпиплевральный
склерит переднеспинки с 1 микрохетой. Средне- и заднеспинка (рис. 56,2) с 2 рядами
пре- и посттергальных склеритов, несущих 1-2 макрохеты, причем внутренние и на-
ружные претергальные сливаются в один склерит. Крыловые склериты с 3 щетинка-
ми. Тергиты брюшка с 2 рядами склеритов, или же престигмальные склериты реду-
цируются и от них остается лишь мелкая щетинка или пора, эпиплевральные с 2
макрохетами. На 7-9-м сегментах внутренние склериты сливаются в один непарный.
Склериты низа (рис. 56, 5) развиты, каждый с 2—3 щетинками. Микроскульптура
покровов густая, заходящая и на склериты. Коготки без базального зубца.
Личинки на крестоцветных и лютиковых. 3 личиночных возраста, развивают-
ся открыто на листьях, окукливаются в почве. В регионе 5 видов.
Таблица для определения видов рода Phaedon
1(4). Претергальные склериты средне- и заднеспинки разделены на внутренние и
наружные, каждый с 1 щетинкой. Внутренние претергальные склериты на 1-2-м и
нередко на 3-6-м тергитах брюшка имеются, хотя бы с одной стороны (рис. 56, 3).
2(3). Тело со светлыми рыжеватыми щетинками. Стернальные склериты средне- и
заднегруди почти всегда неразделенные. Наружные претергальные склериты на 1-
6-м тергитах брюшка обычно отсутствуют, очень редко имеются только на одной
стороне 1-2-го тергитов. Посттергальные склериты на 1-6-м брюшных тергитах с
1 длинной и 2 короткими щетинками. Обычно на Triglochin maritima, также на
Ranunculus, а жуки еще на Cochlearia, Plantago во влажных и обычно засоленных
биотопах. ЕЧ, СБ...................................Ph. concinnus Stephens
3(2). Тело с темнобурыми щетинками. Стернальные склериты средне- и заднегруди
разделены на 3 части. Наружные претергальные склериты 1-6-й тергитах брюшка
всегда развиты, хотя бы на одной стороне. Посттергальные склериты на 1-6-м брюш-
ных тергитах с 2 длинными и 1 очень короткой щетинкой. На крестоцветных. ЕЧ,
СК, СБ.............................................Ph. armoraciae Linnaeus
4(1). Претергальные склериты средне- и заднеспинки не разделены, каждый с 2
щетинками. Внутренние претергальные склериты иногда присутствуют на правой
или левой стороне 1-го брюшного тергита (рис. 56, 4), но всегда отсутствуют на 2-
6-м тергитах.
139
5(6). Посттергальные склериты 1-6-го брюшных сегментов с 1 длинной и 2 корот-
кими щетинками. На крестоцветных. Транспалеаркт....Ph. cochleariae Fabricius
6(5). Посттергальные склериты с 4 длинными щетинками на 1-4-м брюшных сег-
ментах и с 3 длинными щетинками на 5-6-м сегментах. Данный вид, возможно,
синоним или подвид предыдущего. На крестоцветных. Юг ДВ.................
.........................................................Ph. brassicae Baly
20. Род Entomoscelis Chevrolat
Головная капсула (рис. 54, 5) с многочисленными, довольно длинными щетин-
ками, эпикраниальный шов короткий, отходящие от него лобные швы в основании
узко- или широкопараллельные, далее расходятся под острым или тупым углом,
лоб хитинизирован в верхней части, с 40—42 щетинками, наличник широкий, хити-
низирован в основании, с рядом из 4—6 щетинок и 4 пор. Верхняя губа со средин-
ной выемкой, с каждой стороны с 3 короткими и 3-4 длинными маргинальными
щетинками. Щетинки верха тела на вершине тупоокругленные или оплавленные.
Склерит переднеспинки с многочисленными длинными щетинками, разбросанны-
ми по всему диску. Средне- и заднеспинка и сегменты груди с 3 поперечными ряда-
ми склеритов, по 6 в каждом ряду, разделенных поперечными складками (рис. 57,
/). На каждом склерите от 2 до 9 щетинок. Первый ряд склеритов иногда разбит на
многочисленные склеритовидные площадки с 1 щетинкой на каждом (рис. 57, 4).
Крыловые склериты с многочисленными щетинками. Тергиты брюшка с пре- и
посттергальными склеритами, эпиплевральные и гипоплевральные склериты буг-
ровидные, с многочисленными щетинками. На последних тергитах, начиная с 7-го,
склериты начинают сливаться, образуя непарную дорсальную пластинку. Стерни-
ты груди и брюшка выражены, с короткими крепкими щетинками. 7-й стернит брюш-
ка с ложными ножками в виде сосочковидных выростов, такие же, но менее разви-
тые ложные ножки имеются на 5-6-м стернитах. Микроскульптура тела густая, в
виде мелких тупых шипиков. Коготки с прямоугольным зубцом при основании или
без него.
Окукливание в почве. В регионе 3 вида на травянистых растениях.
Таблица для определения видов рода Entomoscelis
1(4). Головная капсула сильно хитинизированная, лобные швы в основной полови-
не широко параллельные. Наличник с 6 щетинками, верхняя губа с широкой треу-
гольной или дуговидной выемкой. Средне- и заднеспинка в 1-м ряду с 6 склерита-
ми, с 2-7 щетинками каждый (рис. 57, /). Коготки без зубца. На крестоцветных и
лютиковых (Adonis). Транспалеаркт........................Е. adonidis Pallas
2(3). Склериты брюшных стернитов с более густыми щетинками: парастернальные
с 11, задние гипоплевральные с 7, эпиплевральные с 11, задние гипоплевральные с
7, эпиплевральные с 17 щетинками (рис. 59, 7). Верхняя губа с глубокой выемкой
(рис. 58, //). На крестоцветных и Glaucium...............Е. suturalis Weise
3(2). Склериты брюшных стернитов с более редкими щетинками: парастернальные
с 7, задние гипоплевральные с 4, эпиплевральные с 10 (рис. 59, 8). Верхняя губа со
слабой выемкой (рис. 58, 12).............................Е. adonidis Pallas
140
4(1). Головная капсула хитинизирована по бокам, вдоль эпикраниального и основа-
ний лобных швов, последние в основной половине узко параллельные (рис. 54, 5).
Наличник с 4 щетинками и 4 порами, верхняя губа с прямоугольной выемкой. Скле-
риты средне- и заднеспинки в 1-м ряду разбиваются на отдельные площадки, каж-
дая с 1 щетинкой (рис. 57, 4). Коготки с прямоугольным зубцом при основании. На
гречишных. Юг ВСБ и ДВ............................Е. orientalis Motschulsky
21. Род Colaphus Dahl (Colaphellus Weise)
Головная капсула с первичными и вторичными щетинками. Лобные швы рас-
ходятся под острым углом. Наличник с 6 щетинками, верхняя губа с 4 щетинками и
4 порами на диске, 3 парами маргинальных щетинок и плоской выемкой на пере-
днем крае. Склерит переднеспинки с многочисленными крепкими щетинками. Сред-
не- и заднеспинка с 3 поперечными рядами склеритов (1-й поперечный ряд не очень
четкий), несущих короткие многочисленные щетинки (рис. 57, 5); внутренние пре-
тергальные склериты среднеспинки соприкасаются или почти соприкасаются друг
с другом; второй поперечный ряд с 2 крупными склеритами. Стерниты груди с до-
вольно многочисленными щетинками. Тергиты брюшка сходны в общем с грудны-
ми, но на последних тергитах наблюдается тенденция к слиянию внутренних тер-
гальных склеритов первого и второго рядов, которые на шестом тергите сливаются
в непарный склерит, на 7-м тергите все тергальные, а на 8-9-м тергитах все склери-
ты слиты вместе. 6-7-й брюшные стерниты с ложными ножками. Промежутки между
склеритами в микроскульптуре; все щетинки короткие, нередко плохо заметны.
Коготки без зубца. Длина взрослой личинки до 8,5 мм.
Жуки и личинки питаются на крестоцветных. Окукливание в почве. В регионе
3 вида.
Таблица для определения видов рода Colaphus
1(4). Внутренние тергальные склериты груди и брюшка сильно сближенные. На
средне- и заднеспинке передний ряд представлен 3 парами вторичных склеритов
(рис. 57, 2). Вторичная хетотаксия головы более густая; на переднеспинке щетинки
разбросаны по всей ее поверхности.
2(3). Лоб и темя с многочисленными щетинками, на темени вдоль лобных швов по
10-13 щетинок. Микроскульптура очень густая, в виде плоских пластинок (рис. 57,
3). Щетинки тела темные. На различных крестоцветных. ЕЧ, СК............
................................................С. sophiae hoefti Menetries
3(2). Лоб и темя с менее густыми щетинками, на темени вдоль лобных швов по 7-8
щетинок. Микроскульптура менее густая, несет мелкие шипики (рис.57, 6). Ще-
тинки тела светлые. На различных крестоцветных, чаще на Isatis. СБ.....
........................................................С. alpinus Gebler
4(1). Внутренние тергальные склериты, особенно претергальные, явственно раз-
двинуты на груди и брюшке. На средне- и заднеспинке передний ряд представлен 2
парами четких вторичных склеритов (1-я и 3-я пара), тогда как вторая пара разбита
на мелкие склеритовидные площадки (рис. 57, 5). Вторичная хетотаксия головы
редкая, на темени вдоль лобных швов по 5-6 щетинок. Микроскульптура негустая.
141
Щетинки тела темные. На Cardamine в сырых местах в тайге. Алтай, Западный
Саян....................................................С. foveolatus Gebler
22. Род Apterocuris Jacobson
Окраска светло-серая, голова, склериты и ноги коричневые. Головная капсула
округлая, первичные щетинки длинные, тонкие и имеют следующее расположе-
ние: на темени — 10, на лбу — 10, на наличнике — 6, на верхней губе — 4. Кроме
того, на теменных участках по 3 плохо заметных вторичных щетинки, расположен-
ных в ряд, а на лбу и верхней губе — по 3 пары мелких пор. Эпикраниальный шов
короткий, лобные швы широко параллельные в основании. Передний край верхней
губы глубоко вырезан. Мандибулы короткие, с 5 зубцами и 2 щетинками. Жеватель-
ная лопасть с длинными шипами. На каждой стороне склерита переднеспинки 12
первичных и около 15 вторичных щетинок, в основном по краям. Эпиплевральные
склериты переднеспинки очень малы, плохо заметны, каждый с 1 короткой щетин-
кой. Тергиты средне- и заднеспинки с 2 рядами склеритов, несущих на претергаль-
ных и внутренних посттергальных 1 щетинку, на наружных посттергальных и кры-
ловых — 3; кроме того, на последних склеритах имеются плохо заметные вторич-
ные щетинки. На эпиплевральных, гипоплевральных и стернальных склеритах груди
по 2 щетинки. Перед стернальными склеритами имеются 2-4 маленьких склерита,
несущих по щетинке, позади них — 2 щетинки без склеритов. Тергиты брюшных
сегментов (рис. 56, 77) также с 2 рядами склеритов, имеющих по щетинке, но на
последних тергитах наблюдается тенденция к слиянию склеритов. На 7-8-м терги-
тах сливаются тергальные склериты, а на 9-м тергите все склериты слиты вместе.
Тергальные железы отсутствуют. Брюшные стерниты с 2 рядами склеритов, в пер-
вом ряду 2 небольших склерита, каждый с 1 щетинкой; второй ряд состоит из 5
склеритов, имеющих по 2-3 щетинки. 5-8-й брюшные стерниты с ложными ножка-
ми, сильно развитыми на 6-7-м стернитах. Промежутки между склеритами в мик-
роскульптуре. Коготок с тупым зубцом.
Личинки питаются на Delphinium в черневой тайге. Род включает единствен-
ный, эндемичный для Алтая вид — A. sibirica Gebler.
23. Род Paropsides Motschulsky
Тело толстое, грязно-желтое, с хорошо выраженными темно-коричневыми скле-
ритами и оранжевым низом. Голова темно-коричневая, сильно хитинизированная,
с очень короткими первичными и отдельными вторичными щетинками, темя слег-
ка морщинистое, с многочисленными щетинками; эпикраниальный шов умеренно
длинный, лобные швы расходятся под прямым углом. Лоб с 16 мелкими и 4 круп-
ными щетинками. Наличник резко поперечный, с хитинизированным основанием,
с 6 щетинками и 4 порами посередине. Верхняя губа с 4 щетинками и 2 порами на
диске, ее передний край с выемкой, достигающей 1/3 ширины губы и с 3 парами
мелких, утолщенных маргинальных щетинок. Склерит переднеспинки (рис. 57, 7)
черно-бурый, с широкими желтыми боковыми краями и с очень мелкими, редкими
щетинками. Щетинки на склеритах остальных сегментов очень короткие, практи-
чески отсутствуют и заменены короткими хитинизированными бугорками (рис. 44,
142
Рис. 57. Детали строения личинок подсемейства Chrysomelinae:
1,2, 4, 5 — среднеспинка: 1 — Entomoscelis adonidis, 4 — Е. orientalis, 2 — Colaphus alpinus,
5 — C. foveolatus', 3,6 — микроскульптура покровов: 3 — Colaphus sophiae, 6 — C. alpinus',
7 — переднеспинка и среднеспинка Paropsides.
7) . Средне- и заднеспинки (рис. 57, 7) с непарным пре- и посттергальным склери-
том, небольшим наружным претергальным и разделенным на две части крыловым,
причем крупный задний крыловой склерит, очевидно, включает и наружный пост-
тергалыный; эпиплевральный склерит крупный, 2 гипоплевральных небольшие.
Склериты брюшных сегментов аналогичны грудным, лишь задний стигмальный
обособлен от посттергального, стигма окружена группой мелких склеритов. Скле-
риты 8-9-го тергитов сливаются в одну сплошную пластинку с многочисленными
щетинками, из которых наиболее длинные расположены по краям. Между 7-м и 8-м
тергитами брюшка находится выпячивающаяся железа в виде двух пальцевидных
выростов (рис. 44, 5). Нижняя сторона тела, помимо обычных склеритов, с мелки-
ми склеритовидными площадками. Коготок тонкий, острый, серповидно загнут, с
зубцом. Длина тела взрослой личинки до 11 мм.
Жуки и личинки питаются на боярышнике, иногда переходят на грушу уссу-
рийскую. Окукливание в почве. В России 1 вид, Р. duodecimpustulatus Motschulsky,
встречающийся в ВСБ и на юге ДВ.
24. Род Gonioctena Chevrolat (Phytodecta Kirby)
Личинки видов подрода Gonioctena s.str. большей частью (кроме первой ста-
дии) грязно-коричневые, с темно-бурыми головой, склеритами тела и ногами. Эпик-
раниальный шов длинный, лобные швы расходятся под острым углом, лоб с 10
щетинками (рис. 54, 4), наличник хитинизирован в основании, с рядом из 6 щети-
нок и 2 пор, верхняя губа с широкой и глубокой треугольной вырезкой, с 3-4 марги-
нальными щетинками с каждой стороны. Переднеспинка с многочисленными ще-
тинками. Тело с хорошо развитыми, реже бесцветными и плохо заметными склери-
тами грудного и брюшного отделов и выворачивающимся наружу при раздражении
парным выпячиванием на границе 7-го и 8-го тергитов брюшка в виде двух длин-
ных отростков. Склериты средне- и заднеспинки и 1-6-го тергитов брюшка распо-
ложены двумя поперечными рядами; имеется рудимент третьего (морфологически
143
Рис. 58. Детали строения личинок подсемейства Chrysomelinae:
1 — среднеспинка Gonioctena fulva\ 2,4 — стерниты переднегруди: 2 — Gonioctena decem-
notata, 4 — G. sibiriccr, 3,5,6— 1-й тергит брюшка: 3 — Gonioctena sibirica, 5 — G. japonica,
6 — G. linnaeana; 7,10 — коготок: 7 — G. gracilicornis, 10 — G. viminalis\ 8,9 — среднеспин-
ка: 8 — G. gracilicornis, 9 — G. viminalis; 11,12 — верхняя губа: 11 — Entomoscelis suturalis',
12 — E. adonidis. По Штейнхаузену и ориг.
первого ряда) из 2 или 4 маленьких вторичных склеритов около переднего края
средне- и заднеспинки и 1-3-го тергитов брюшка. Склериты 7-9-го сегментов брюш-
ка сливаются в один непарный склерит.
Личинки подрода Goniomena зеленовато грязно-желтые, сегменты тела без ясно
выраженных склеритов, местоположение которых намечено лишь ограниченным
количеством одиночных щетинок (рис. 59, 5), переднеспинка с более или менее
длинными щетинками лишь на краях. Коготки с прямоугольным зубцом при осно-
вании.
Жуки и личинки питаются листьями деревьев и кустарников, преимуществен-
но ивовых. Окукливание в почве. В регионе 18 видов.
Таблица для определения видов рода Gonioctena
1(2). Голова светлая, с 2 крупными темными пятнами в передней части лба и задне-
боковыми частями темени (рис. 54, 5). Тергиты груди и брюшка с 2 рядами склери-
тов (рис. 58, 7), причем передний ряд представлен бесцветными, а задний — тем-
ноокрашеннымй склеритами (подрод Brachyphytodecta Chen). На леспедеце дву-
цветной. ВСЕ, ДВ......................................G. fulva Motschulsky
2(1). Голова темная или светлая, но всегда без темных пятен на лбу.
3(22). Верхняя и нижняя стороны тела с хорошо обособленными темными склери-
тами (подрод Gonioctena s.str).
4(11). Склериты верхней стороны тела почти все разделены отчетливыми проме-
жутками, рудиментарные склериты первого ряда заметны не только на 1-м тергите
брюшка, но и на части последующих (рис. 58, 3).
144
5(10). Тергальные склериты брюшка, как правило, с 1 длинной макрохетой и 2 очень
короткими микрохетами (рис. 58, 3).
6(9). Склерит переднеспинки без кожистых морщинок.
7(8). Стернит переднегруди с 2 длинными и 2 укороченными щетинками (рис. 58,
2). Склериты брюшных сегментов сильно хитинизированные. На осине, реже на
тополе, иве. Транспалеаркт........G. decemnotata Marsham (G. rufipes Degeer)
8(7). Стернит переднегруди с 2 длинными и 6 укороченными щетинками (рис. 58,
4). Склериты брюшных сегментов слабо хитинизированные. На черемухе, рябине,
боярышнике. СБ, ДВ........................................G. sibirica Weise
9(6). Склерит переднеспинки с кожистыми морщинками. На иве. Сибирь (кроме
западной), Дальний Восток. Север ЕЧ, СБ, ДВ..........G. flavicornis Suffrian
10(5). Тергальные склериты брюшка с 3 длинными щетинками каждый, причем
краевые щетинки лишь немного короче срединной (рис. 59, 7). На иве, особенно
коротконожковой. ДВ....................................G. coreana Bechyne
11(4). Склериты верхней стороны тела сильно расширены, тесно соприкасаются,
разделены узкими промежутками; внутренние посттергальные склериты на всех
тергитах брюшка слиты в непарный склерит, рудиментарные склериты первого ряда
на всех тергитах брюшка, начиная со 2-го, слиты с расположенными назади скле-
ритами.
12(15). Наружные посттергальные склериты начиная с грудных сегментов слива-
ются с непарным внутренним посттергальным (рис. 58, 6).
13(14). Верхняя сторона брюшных тергитов с 2 сплошными непарными склерита-
ми между стигмами (слитые вместе дорсальные и стигмальные, рис. 58, 5). Эпи-
плевральные склериты с 5 макро- и примерно 50 микрохетами. На ольхе. Курилы
(Кунашир).......................................G. japonica Chujo et Kimoto
14(13). Верхняя сторона передних брюшных тергитов с обособленными стигмаль-
ными склеритами, причем передний стигмальный разбит на внутреннюю и наруж-
ную части (рис. 58, 6). Эпиплевральные склериты с 2 макро- и 3—4 микрохетами.
На ивах. Вся лесная зона Палеарктики.................G. linnaeana Schrank
15(12). Наружные посттергальные склериты лишь начиная с 3-го тергита брюшка
сливаются с непарным внутренним посттергальным.
16(19). Склериты очень сильно блестящие, морщинистые, сидящие на них щетин-
ки не окружены темными участками. Эпиплевральный склерит 1-го тергита брюш-
ка с 7-8 длинными щетинками. Щетинки тела довольно толстые, хотя бы у основа-
ния, заметно сужаются к острым вершинам, короткие или средней величины, мик-
рохеты в 2-3 раза короче макрохет. Наружный посттергальный склерит груди с 3
макро- и 3—4 микрохетами.
17(18). Крыловой склерит с 5-6 макро- и 3-4 микрохетами (рис. 58, 8). Коготки с
тупым зубцом (рис. 58, 7). На ивах. ВСБ, ДВ..........G. gracilicornis Kraatz
18(17). Крыловой склерит с 7 макро- и 3 микрохетами. Коготки с прямым зубцом.
На спирее, возможны на ивах. Север ЕЧ, СБ, ДВ..........G. affinis Gyllenhal
19(16). Склериты тела слабо блестящие, неявственно морщинистые, сидящие на
них щетинки окружены темными участками, крыловой склерит с 3-4 длинными и
8-9 короткими щетинками (рис. 58,9), эпиплевральный склерит 1-го тергита брюшка
с 4-5 длинными и 5-6 короткими щетинками. Коготки с прямоугольным сглажен-
145
Рис. 59. Детали строения личинок родов Gonioctena, Cercyonops и Entomoscelis'.
1,2, 5,6— 1-й тергит брюшка: 1 — Gonioctena согеапа, 2 — G. ogloblini, 5 — G. pallida, 6 —
Cercyonops caraganae', 3-4 — среднеспинка: 3 — G. jacobsonv, 4 — G. kamiyav, 7-8 — 2-й
стернит брюшка: 7 — Entomoscelis suturalis', 8 — E. adonidis. По Штейнхаузену и ориг.
ным зубцом (рис. 58, 70). На ивах, осине. Вся лесная зона Палеарктики....
.......................................................G. viminalis Linnaeus
20(3). Нижняя сторона тела без явственных темноокрашенных склеритов.
21(28). Верхняя сторона тела с мелкой и редкой, но явственной микроскульптурой.
Щетинки верха умеренной длины или короткие; если микроскульптура плохо вы-
ражена, тогда щетинки очень короткие (подрод Gonioctena s.str.).
22(23). Верхняя и нижняя стороны тела без явственных темноокрашенных склери-
тов, с очень короткими, почти шиповидными светлыми щетинками (рис. 59, 2). На
ольхе. ВСБ, ДВ. Сюда же должна относиться связанная с ивой личинка G. sundmanni
Jacobson (север ЕЧ, СБ, ДВ)........................G. ogloblini L. Medvedev
146
23(22). Верхняя сторона тела с явственными темными склеритами.
24(27). Наружные посттергальные/склериты груди с 1-3 длинными и 3-6 коротки-
ми щетинками.
25(26). Наружные посттергальные склериты груди с 1 длинной и 3 короткими ще-
тинками. Крыловые склериты с 6 длинными и 16-18 мелкими щетинками с темно-
окрашенными вершинами (рис. 59, 3). На тополе. ВСБ, ДВ..................
........................................G. jacobsoni Ogloblin et L. Medvedev
26(25). Наружные посттергальные склериты груди с 3 длинными и 6 короткими
щетинками. Крыловые склериты с 4 длинными и 8 короткими щетинками. На ивах,
чозении. ДВ.............................G. chujoi L. Medvedev
27(24). Наружные посттергальные склериты груди без длинных щетинок, с 13-14
короткими щетинками. Крыловые склериты с 3 длинными и 10—12 мелкими свет-
лыми щетинками (рис. 59, 4). На березе. ДВ..............G. kamiyai Kimoto
28(21). Сегменты тела без явственной микроскульптуры, с длинными и довольно
густыми светлыми щетинками (рис. 59, 5). На розоцветных, реже на ивах (подрод
Goniomena Motsch.).
29(30). Задний ряд щетинок на тергитах грудного и брюшного отделов с каждой
стороны с 3 макрохетами. На рябине и черемухе. ЕЧ, СБ...................
...............................................G. quinquepunctata Fabricius
30(29). Задний ряд щетинок на тергитах грудного и брюшного отделов с каждой
стороны с 4 макрохетами. На рябине, черемухе, иве. ЕЧ, СБ .... G. pallida Linnaeus
25. Род Cercyonops Jacobson
Личинки внешне похожи на личинок Gonioctena, окрашены в грязно зеленова-
то-серый цвет, с хорошо обособленными темными склеритами. Лоб с дополнитель-
ными щетинками (рис. 54, 6). Внутренние претергальные склериты грудного отде-
ла с 2 щетинками, кнаружи от него расположены 2 склерита (первый с 1, второй с 2
щетинками), впереди от него заметны 2 рудимента склеритов с одной очень мелкой
щетинкой на каждом из них; внутренние посттергальные склериты с 3 длинными
щетинками; наружные посттергальные склериты крупные, неправильной формы, с
7 длинными щетинками, крыловые склериты с 8 щетинками. Склериты брюшных
тергитов (рис. 59, 6) имеют следующее число длинных щетинок: внутренние пре-
тергальные — 2 (расположенные, как и на грудном отделе, одна позади другой),
внутренние посттергальные — 4, наружные пре- и посттергальные и преститмаль-
ный — 3, постстигмальный — 4; близ переднего края брюшных сегментов нахо-
дятся рудименты склеритов переднего поперечного ряда (с каждой стороны по 3,
из которых два близ стигм) с очень маленькой щетинкой на каждом из них.
Жуки и личинки питаются листьями нескольких видов бобовых растений (Phaca,
Caragana, Vicia, Lathyrus). Род включает единственный вид С. caraganae Gebler,
распространенный в СБ и на северо-востоке ЕЧ.
147
13. ПОДСЕМЕЙСТВА GALERUCINAE и alticinae
Личинки обоих подсемейств морфологически очень близки и не могут быть
четко разграничены, так что ниже приводится их общая морфологическая характе-
ристика.
Личинки, живущие открыто на растениях, обычно темноокрашенные, с длин-
ными щетинками, хорошо развитыми склеритами и с 1 глазком на боках головной
капсулы позади основания усиков (рис. 5,9). Обитающие в почве или внутри расте-
ний личинки большей частью белые или желтые, с более темноокрашенными голо-
вой, ногами, переднеспинкой и тергитом 9-го сегмента брюшка, склериты осталь-
ных частей тела чаще бледноватые и нередко плохо заметные или могут вообще
отсутствовать; глазки имеются обычно только у личинок 1-го возраста, а у последу-
ющих возрастов исчезают. Тело удлиненное, никогда не бывает С-образным, иног-
да очень тонкое и длинное (рис. 6, 7). Голова небольшая, сильно хитинизирован-
ная, гипогнатическая, выпуклая (только у минирующих форм она уплощенная и
выглядит прогнатической), эпикраниальный шов длинный (большинство Galeru-
cinae), короткий (большинство Alticinae) или отсутствует (некоторые рода Alticinae),
глазки с сильно выпуклой роговицей, усики очень короткие, 1-2-члениковые, с силь-
но развитой базальной мембраной, на вершине 1-го членика находится хорошо раз-
витый сенсориальный конус. Наличник и верхняя губа хорошо развиты и обособ-
лены, реже шов между наличником и лбом не вполне явственный, верхняя губа
поперечная, с простым или выемчатым передним краем. Мандибулы с 3-5 хорошо
развитыми зубцами, нередко с пучком коротких толстых щетинок на внутреннем
крае у основания; наружный край с 2 щетинками. Максиллы с обособленными га-
леей и лацинией, причем лациния очень слабо хитинизирована, с щетинками и мел-
кими шипиками, или галея и лациния слиты вместе, рудиментарны. Челюстные
щупики короткие, обычно 3-члениковые. Нижняя губа слабо хитинизирована, мен-
тум ограничен назади дуговидным хитиновым склеритом, субментум продолгова-
то-четырехугольный, с 2-3 парами щетинок, губные щупики 2-члениковые (у ми-
нирующих Alticinae отсутствуют), язычок короткий, толстый, с немногочисленны-
ми щетинками. Эпифаринкс перепончатый, с короткими нежными волосками, ги-
пофаринкс с шиловидными щетинками, преимущественно на боках.
Переднеспинка с крупным срединным склеритом, несущим щетинки большей
частью только по краям; по бокам срединного склерита лежит довольно крупный,
более или менее треугольный эпиплевральный склерит. Средне- и заднеспинка раз-
делены поперечными бороздками на 2, а тергиты 1-8-го сегментов брюшка на 2-3
поперечных поля. Средне- и заднеспинка с 2 поперечными рядами тергальных скле-
ритов, по 4 в ряд, с хорошо развитыми крыловыми и эпиплевральными склерита-
ми, иногда с дополнительным третьим рядом вторичных склеритов. Тергиты 1-8-
го брюшных сегментов с 2-3, редко с 4 рядами склеритов. В отдельных случаях
количество тергальных склеритов может уменьшаться (обычно за счет наружных),
что наблюдается у некоторых скрытоживущих форм; реже склериты вообще исче-
зают. 9-й тергит брюшка несет округло-прямоугольный склерит с довольно длин-
ными щетинками, особенно по краям, а у некоторых Alticinae также с 1-2 зубчика-
ми или шипами на вершине. 10-й сегмент брюшка небольшой, расположен на ниж-
ней стороне 9-го сегмента и сверху не виден; выполняет функцию подталкивателя,
148
1-8-й сегменты брюшка несут стигмы с простым отверстием. У ряда форм имеют-
ся дорсальные защитные железы на груди и брюшке (от 8 до 11 пар). Ноги всегда
развиты, хотя у скрытоживущих форм они более короткие, коготки с различно раз-
витым хелониумом (рис. 68, 3, 6).
Достоверных признаков, четко разделяющих оба подсемейства, по-видимому,
нет, что еще лишний раз свидетельствует о филогенетической близости обоих под-
семейств, хотя отдельные естественные группы хорошо обособляются.
По характеру местообитания рассматриваемые подсемейства распадаются на
несколько достаточно четких групп. Личинки многих Galerucinae и Altica Geofroy
живут открыто на листьях. Они легко отличимы наличием глаз, темной окраски и
хорошо выраженных, обычно крупных склеритов и многочисленных длинных ще-
тинок.
Подавляющее большинство личинок Alticinae и некоторые Galerucinae обита-
ют в почве на корнях растений и иногда могут вгрызаться в корни, клубни или в
прикорневую часть стебля, переходя частично или полностью к внутритканевому
питанию. Личинки этой группы имеют белую или желтоватую окраску, склериты
большей частью плохо заметны, форма тела обычно червеобразная, тонкая и удли-
ненная, глаза обычно отсутствуют, эпикраниальный шов нередко укорочен, на вер-
шине сильно хитинизированного 9-го тергита брюшка часто имеются шипы, облег-
чающие передвижение в плотном субстрате.
Группа внутристеблевых личинок представлена отдельными видами из родов
Chaetocnema Stephens и Psylliodes Latreille. Судя по известным представителям, эта
группа морфологически сходна с предыдущей и, по-видимому, является ее произ-
водной. Сюда же следует отнести и некоторых Alticinae, обитающих в черешках и
мясистой толще листьев.
Наиболее специализированной является группа родов Alticinae, минирующая
листья. Для них характерно уплощенное, светлоокрашенное тело, специализиро-
ванные прогнатическая головная капсула и ротовые части, укорочение ног и редук-
ция склеритов и щетинок.
Окукливание у подавляющего большинства видов происходит в почве, обычно
в специально изготовляемой земляной ячейке, лишь некоторые Galerucinae окукли-
ваются на листьях (рис. 4, 4).
За год развивается, как правило, одно поколение. Кладка яиц обычно происхо-
дит весной или в начале лета, продуктивность самок колеблется от нескольких де-
сятков до нескольких сот яиц. Яйца откладываются в поверхностный слой почвы
или в ткани растений по 1-3 штуки или на листья группами по 6-20 штук. Развитие
яйца длится в среднем 5-8 дней. Число личиночных возрастов колеблется от 3 до 4.
Личинки Galerucinae изучены сравнительно полно и в настоящее время извес-
тны основные рода. Иное положение среди Alticinae, для которых характеристика
большинства родов, в том числе обильных видами, основывается на изучении 1-2
видов личинок. Вполне понятно, что последующее пополнение наших сведений о
личинках Alticinae может привести к пересмотру характеристик родов.
Для России известно 25 родов Galerucinae и 31 род Alticinae. Ряд родов и видов
известны только по определителю личинок Японии (Kimoto, Takizawa, 1994), со-
ставленному на японском языке, только определительные таблицы переведены на
английский. Поэтому такие рода лишь частично включены в наш определитель, а
149
Рис. 60. Схема склеритов переднеспинки, среднеспинки и 1-го тергита брюшка:
1 — Chrysolina pirka\ 2 — Fleutiauxia armata\ 3 — Aulacophora nigripennis; 4 — Sangariola
punctatostriata} 5 — Liprus punctatostriatus} 6 — Phygasia fulvipennis} 7 — Nonarthra cyanea\
8 — Hemipyxis plagioderoides\ 9 — Philopona vibex. По Кимото и Такизава.
описание родов опущены или даны очень кратко по признакам упомянутым в клю-
чах и рисункам.
Таблица для определения родов из подсемейств Galerucinae и Alticinae
1(72). Эпикраниальный шов нормальный или короткий, но всегда отчетливый; лоб-
ные швы назади сходятся вместе и соединяются с эпикраниальным швом (рис. 61,
7). Нижнегубные щупики всегда развиты. Задний край головной капсулы без удли-
ненных отростков.
2(71). Эпикраниальный шов длинный или умеренно короткий. Бока сегментов без
длинных пальцевидных отростков.
3(86). Тергиты груди и брюшка без отверстий дорсальных желез в области крыло-
вых и стигмальных склеритов.
4(21). Эпиплевральные склериты переднеспинки не менее чем с 2 макрохетами;
наружные претергальные склериты средне- и заднеспинки не менее чем с 1 макро-
150
хетой. Голова с каждой стороны с 1 глазком. Тергиты брюшных сегментов с 2 попе-
речными рядами склеритов.
5(10). Склериты верхней стороны тела бугорковидно выпуклые, с торчащими в раз-
ные стороны щетинками (рис. 62, 7).
6(9). Внутренние претергальные склериты средне- и заднеспинки разделенные.
Голенелапка короткая, коготок умеренно длинный, гораздо короче половины голе-
нелапки, с сильно развитым хелониумом.
7(8). Верхняя губа со слабо заостренными базальными углами; края срединной
выемки не выдаются вперед в виде зубчиков (рис. 61, 5). Тело черное, склериты
верха сильно или умеренно бугровидные.......................1. Galeruca
8(7). Верхняя губа с острыми базальными углами; края срединной выемки выдают-
ся вперед в виде зубчиков (рис. 61, 6). Тело пепельно-серое, склериты верха уме-
ренно бугровидные...........................................2. Pallasiola
9(6). Внутренние претергальпые склериты средне- и заднеспинки слитые (рис. 63,
7). Голенелапка удлиненная, коготок длинный, почти равен половине длины голе-
нелапки, со слабо развитым хелониумом (рис. 63, 3).............3. Theone
10(5). Склериты верхней стороны тела плоские.
11(12). Все склериты тела светлые, не отличаются по окраске от мягких покровов
или светлее их. Эпиплевральные склериты переднеспинки с 8-10 щетинками.
...........................................................4. Diorhabda
12(11). Все склериты тела темные, темнее мягких покровов или не отличаются от
них по окраске.
13(14). Верхняя губа с широкой, но неглубокой срединной выемкой. Престигмаль-
ные склериты на брюшных сегментах развиты.................... 5. Clitena
14(13). Верхняя губа без срединной выемки, ее передний край пряк£Ьй.
15(16). Щетинки тела, особенно головы, щетинковидные, с заостренной вершиной
(иногда очень короткие, но тогда в виде зубчика), темно-коричневые. Склериты низа
с характерной гребенчатой микроскульптурой (рис. 67, 7). Окукливание на расте-
нии........................................................6. Galerucella
16(15). Щетинки тела с оплавленной или округленной вершиной, светлые, если тем-
ные, то не утончающиеся к вершине и не длинные. Окукливание в почве.
17(18). Склериты крупные, тесно примыкают друг к другу, трудно различимые из-
за густой маскирующей микроскульптуры. Эпиплевральные склериты сегментов
брюшка в виде пальцевидных выростов (рис. 64, 7). Все внутренние пре- и пост-
тергальные склериты заднегруди и брюшка попарно слиты, между наружными и
внутренними претергальными склеритами груди появляются 2 (а на брюшных сег-
ментах 1) маленьких склерита с короткой тонкой щетинкой (рис. 64, 6). Передний
край верхней губы с едва намеченной срединной выемкой, над которой расположе-
но поперечное поле из очень мелких и тонких щетинок (рис. 64, 3) ...7. Apophylia
18(17). Склериты явственные, раздвинутые. Эпиплевральные склериты сегментов
брюшка не образуют выростов. Внутренние пре- и посттергальные склериты зад-
негруди не слиты попарно. Передний край верхней губы без поперечного поля мел-
ких щетинок.
19(20). Гипоплевральные склериты брюшка с 2 макро- и 1-2 микрохетами. Эпип-
левральные склериты переднеспинки с 6-11 щетинками.........8. Lochmaea
20(19). Гипоплевральные склериты брюшка с 1 макро- и 1-7 микрохетами. Эпип-
151
левральные склериты переднеспинки с 3-5 или 10-11 щетинками.9. Pyrrhalta
21(4). Эпиплевральные склериты переднеспинки с 1 макрохетой.
22 (27) Тергиты средне- и заднегруди с 3, а брюшка — с 4 рядами склеритов (рис.
67, 5).
23(24) Среднеспинка без тергальных склеритов, на заднеспинке внутренние пре-
тергальные склериты отсутствуют, прочие разделены, каждый с 1 щетинкой (рис.
66, 6). Верхняя губа (рис. 66, 2) на переднем крае с 3 парами очень толстых упло-
щенных хет...............................................13. Calomicrus
24(23) Среднеспинка с тергальными склеритами, на заднеспинке внутренние пре-
тергальные склериты имеются. Верхняя губа на переднем крае без очень толстых
уплощенных хет.
25(26). Интеркалярные склериты брюшных тергитов не слиты (рис. 67, 5). Брюш-
ные стерниты без мелких вторичных склеритов.................10. Спеогапе
26(25). Интеркалярные склериты брюшных тергитов слиты (рис. 60, 2). Брюшные
стерниты с мелкими вторичными склеритами.................11. Fleutiauxia
27(22). Тергиты средне- и заднегруди с 2 рядами склеритов (иногда имеется 2 мел-
ких вторичных склерита перед претергальными, но тогда на брюшке не более 3
рядов склеритов).
28(73). Тергиты брюшных сегментов с 3 (редко с 4) поперечными рядами склери-
тов или щетинок, если склериты не развиты (рис. 68, 5).
29 (34). Тергит 9-го сегмента брюшка почти весь покрыт частой ячеистой скульпту-
рой, с округленным вершинным краем. Склериты светлые, плохо заметные (нужна
окраска!).
30(33). Интеркалярные склериты брюшных тергитов сильно сближенные, крупные,
но не слитые.
31(32). Наружные претергальные склериты средне- и заднеспинки развиты, но без
щетинок. Личинки в почве на; корнях. (Сюда же, очевидно, относится очень близ-
кий к Luperus Geoffroy род Euluperus Weise, представленный 1 видом—Е. xanihopus
Duftschmidt, встречающимся на юге ЕЧ и СК)..................12. Luperus
32(31). Наружные претергальные склериты средне- и заднегруди развиты и несут 1
щетинку. Личинки преимущественно внутри стеблей или корней растений..
.........................................................14. Aulacophora
33(30). Интеркалярные склериты брюшных тергитов попарно слитые (рис. 69, 5)
.........................................................25. Derocrepis
34(29). Тергит 9-го сегмента брюшка без ячеистой скульптуры.
35(50). Наружные претергальные склериты средне- и заднеспинки отсутствуют или,
если развиты, без щетинок.
36(37). Склериты тела незаметные, различимы только при окрашивании. На корнях
розоцветных.................................................26. Batophila
37(36). Склериты тела хотя бы слабо различимы. На корнях розоцветных не встре-
чаются.
38(39). Тергит 9-го сегмента брюшка с 5 парами щетинок, наличник без щетинок. На-
ружные претергальные склериты средне- и заднеспинки развиты, но без щетинки. Внут-
ренние пре- и посттергальные склериты брюшка попарно слитые.27. Lythraria
39(38). Тергит 9-го сегмента брюшка не менее чем с 6 парами щетинок; наличник с
каждой стороны с 3 нормальными щетинками.
152
40(43). Внутренние пре- и посттергальные склериты средне- и заднеспинки раз-
дельные
41(42). Все внутренние склериты брюшка раздельные. Наружные претергальные
склериты средне- и заднеспинки отсутствуют (рис. 60, 4)....28. Sangariola
42(41). Внутренние претергальные склериты 1-7-го сегментов брюшка раздельные,
широко расставленные; а интеркалярные и посттергальные попарно слитые. На-
ружные претергальные склериты средне- и заднеспинки отсутствуют. Задний край
9-го тергита брюшка округленный или с одним загнутым кверху коротким острием,
склериты тела хорошо заметные..............................29. Phyllotreta
43(40). Внутренние пре- и посттергальные склериты средне- и заднеспинки попар-
но слитые.
44(45). Все внутренние склериты брюшка, включая интеркалярные, раздельные.
Наружные претергальные склериты средне- и заднеспинки отсутствуют (рис. 60,
5). Тергит 9-го сегмента брюшка хитинизирован, покрыт точками (рис. 70, 3).
...............................................................30. Liprus
45(44). Внутренние пре- и посттергальные склериты брюшка попарно слитые.
46(47). Все внутренние склериты брюшка, включая интеркалярные, попарно сли-
тые. Наружные претергальные склериты средне- и заднеспинки отсутствуют (рис.
60, б)......................................................31. Phygasia
47(46). Интеркалярные склериты брюшка раздельные. Наружные претергальные
склериты средне- и заднеспинки развиты, но без щетинки.
48(49). Наружные посттергальные склериты средне- и заднеспинки с 1 микрохе-
той,. склериты тела плохо заметные. Задний край тергитов 9-го сегмента брюшка
округленный или с 1-2 короткими загнутыми кверху остриями.32. Longitarsus
49(48). Наружные посттергальные склериты средне- и заднеспинки с 3 макрохета-
ми, склериты тела хорошо заметные. Задний край 9-го сегмента брюшка округлен-
ный .......................................................33. Crepidodera
50(35). Наружные претергальные склериты средне- и заднеспинки отчетливые, со
щетинкой (у Paraluperodes Ogloblin и Psylliodes Latreille щетинка часто очень ко-
роткая или сохраняется только пора).
51(52). Все внутренние тергальные склериты сегментов брюшка попарно сливают-
ся, образуя на середине каждого тергита 3 непарных, расположенных один за дру-
гим склерита.............................................15. Phyllobrotica
52(51). По крайней мере, интеркалярные склериты остаются разделенными; если
слиты все три пары, то вершина брюшка с 2 шиловидными остриями.
53(58). Задний край тергита 9-го сегмента брюшка с 2 шипами.
54(57). Внутренние тергальные склериты брюшка сливаются хотя бы на части сег-
ментов.
55(56). Все внутренние тергальные склериты брюшка сливаются, образуя на сере-
дине каждого тергита 3 непарных склерита ... 37. Chaetocnema (подрод Tlanomd)
56(55). Посттергальные внутренние склериты 6-8-го сегментов брюшка попарно
слитые, на 1-5-м сегментах неявственно слитые или разделенные, внутренние пре-
тергальные склериты на 1-7-м сегментах разделенные, на 8-м сегменте слиты вме-
сте (рис. 75, 4). На мальвовых.............................34. Podagrica
57(54). Все тергальные склериты брюшка разделенные.........35. Psylliodes
58(53). Задний край тергита 9-го сегмента брюшка без шипов, округленный.
153
59(70). Внутренние претергальные и посттергальные склериты брюшка попарно
слитые (рис. 67, 2), хотя бы на 6-8-м сегментах.
60(61). Посттергальные внутренние склериты 6-8-го сегментов брюшка попарно
слитые, на 1-5-м сегментах неявственно слитые или разделенные, внутренние пре-
тергальные склериты на 1-7-м сегментах разделенные, на 8-м сегменте слиты вме-
сте (рис. 75, 4), На мальвовых..............................34. Podagrica
61(60). Внутренние претергальные и посттергальные склериты брюшка попарно
слитые, начиная с первых сегментов.
62(63). Склериты тела темные, резко очерченные, хорошо заметные. Брюшные тер-
гиты с 4 рядами склеритов (рис. 67,2). Личинки крупные, живут открыто на листь-
ях растений................................................. 17. Atrachya
63(62). Склериты тела светлые, слабо очерченные, плохо заметные, образуют на
брюшке не более 3 рядов. Личинки мелкие, ведут скрытый образ жизни. Щетинка
на наружных претергальных склеритах средне- и заднеспинки очень короткая или
сохраняется только пора.
64(69). Наружные претергальные склериты средне- и заднеспинки крупные. Ли-
чинки на корнях бобовых.
65(66). Верхняя губа со слабым срединным выступом и 5 маргинальными щетинка-
ми с каждой стороны (рис. 68, 4). Склериты тела довольно заметные. 9-й тергит
брюшка с зернистой скульптурой (рис. 68, 5). Ноги без шиповидных щетинок (рис.
68, 6)......................................................18. Medythia
66(65). Верхняя губа с явственным срединным выступом (рис. 68, 7). Склериты
тела едва намеченные. 9-й тергит брюшка без зернистой скульптуры (рис. 68, 2).
Ноги с шиповидными щетинками (рис. 68, 3).
67(68). На 8-м тергите непарный посттергальный склерит не слит с прочими (рис.
68,2). Верхняя губа с 3 парами маргинальных щетинок (рис. 68,7). 9-й тергит брюш-
ка с 8 маргинальными и 4 дискальными щетинками (рис. 68,2). На корнях бобовых
..............................................19. Monolepta
68(67). На 8-м тергите внутренние и наружные посттергальные склериты сливают-
ся в одну большую непарную пластинку. Верхняя губа с 6 парами маргинальных
щетинок (рис. 76, 2). 9-й тергит брюшка с 12 маргинальными и 8 дискальными
щетинками (рис. 76, 9). На корнях пасленовых .............36. Epithrix
69(64). Наружные претергальные склериты средне- и заднеспинки небольшие. Ли-
чинки на корнях растений; на бобовых не встречаются.........35. Psylliodes
70(59). Все внутренние тергальные склериты брюшка разделенные.
71(72). На брюшных сегментах эпиплевральные склериты с 4, гипоплевральные —
с 3 щетинками...............................................16. Exosoma
72(71). На брюшных сегментах эпиплевральные и гипоплевральные склериты с 2
щетинками. Личинки в стеблях злаков...............37. Chaetocnema (s.str.)
73(28). Тергиты брюшных сегментов с 2 поперечными рядами склеритов (рис. 71,
2, 3), наружные претергальные склериты средне- и заднеспинки отсутствуют или
слабо развитые и без макрохеты.
74(75). Тело без явственных склеритов, замещенных бугорками. 9-й тергит брюшка
с простой округленной вершиной и 3 парами щетинок. Наличник с 3 парами щети-
нок ...................................................38. Hermaeophaga
75(74). Тело с явственными, реже плохо различимыми склеритами.
154
76(79). Покров тела между склеритами с густой, темноокрашенной микроскульп-
турой, склериты темноокрашенные.
77(78). Наружные посттергальные склериты средне- и заднеспинки с 2 макрохета-
ми, внутренние претергальные и посттергальные склериты сегментов брюшка по-
парно слитые (образуют на каждом тергите два непарных срединных склерита),
престигмальные склериты хорошо развитые.......................39. Altica
78(77). Наружные посттергальные склериты средне- и заднеспинки с 1 макрохе-
той, внутренние тергальные склериты (кроме слитых в один непарный склерит внут-
ренних претергальных среднеспинки) широко раздвинутые; престигмальные скле-
риты отсутствуют........................................40. Hippuriphila
79(76). Покров тела между склеритами без ясно заметной микроскульптуры, скле-
риты светлоокрашенные.
80(83). Внутренние претергальные и посттергальные склериты груди и брюшка (кро-
ме слитых посттергальных 8-го сегмента брюшка) разделенные.
81(82). Щетинки тела короткие. Мандибулы с 4 зубцами. Личинки средней величи-
ны, минируют листья гречишных..............................41. Mantura
82(81). Щетинки тела длинные, слабо булавовидные (рис. 73, 5-7). Мандибулы с 5
зубцами. Личинки очень мелкие, очевидно, открытоживущие и питаются на мхах
..........................................................42. Mniophila
83(80). Внутренние претергальные и посттергальные склериты груди и брюшка
попарно слитые.
84 (85). Тергит 9-го сегмента брюшка сплошь покрыт хитинизированной ячеистой
скульптурой, с округленным вершинным краем и 12 щетинками..43. Asiorestia
85(84). Тергит 9-го сегмента брюшка лишь в задней части немного интенсивнее
хитинизированный, без ячеистой скульптуры, с округленным или снабженным 2
короткими, загнутыми кверху остриями задним краем, с 10 щетинками.....
...........................................................44. Aphthona
86(3). Хотя бы тергиты брюшка с отверстиями дорсальных желез в области стиг-
мальных склеритов (рис. 62, 77).
87(96). Интеркалярные склериты на брюшных тергитах имеются. Грудные тергиты
с 2, брюшные — с 3 рядами склеритов.
88 (91). Дорсальные железы на грудных сегментах отсутствуют, на брюшке пред-
ставлены на 1-8-м тергитах. Эпиплевральные склериты переднеспинки с 1 щетин-
кой.
89(90). Тело и склериты темноокрашенные. Внутренние пре- и посттергальные скле-
риты брюшка попарно слитые; наружный посттергальиый не крупнее интеркаляр-
ного (рис. 62, 77)........................................ 20. Agelastica
90 (89). Тело и склериты светло-желтые. Только внутренние посттергальные скле-
риты брюшка слитые; наружный посттергальиый значительно крупнее интеркаляр-
ного (рис. 67, 4).....................................21. Morphosphaera
91(88). Дорсальные железы имеются на 1-3-м грудных и 1-8-м брюшных сегмен-
тах.
92(93). Эпиплевральные склериты переднеспинки с 1, наружные претергальные
склериты средне- и заднеспинки с 3 щетинками. Тело светлое, с желтыми головой
и склеритами..............................................22. Sermylassa
93(92). Эпиплевральные склериты переднеспинки и наружные претергальные скле-
155
риты средне- и заднеспинки с 2 щетинками.
94(95). Верхняя сторона тела с буровато-желтыми склеритами, низ без склеритов
.............................................................23. Agelasa
95(94). Верх с темно-бурыми склеритами или склеритовидными площадками вок-
руг оснований щетинок; низ со склеритами, иногда плохо заметными.......
...........................................................24. Galerucida
96(87). Интеркалярные склериты на брюшных тергитах отсутствуют, тергиты гру-
ди и брюшка с двумя рядами склеритов (рис. 60, 7). Дорсальные железы имеются
на 1-3-м грудных и 1-8-м брюшных сегментах.................45. Nonarthra
97(2). Эпикраниальный шов очень короткий. Склериты отсутствуют. Личинки жи-
вут в минах.
98(99). Бока сегментов с довольно длинными пальцевидными отростками (рис. 74,
10)........................................................46. Philopona
99(98). Бока сегментов с короткими отростками (рис. 74,12).АЛ. Hemipyxis
100(1). Эпикраниальный шов отсутствует, лобные швы оканчиваются назади на краю
затылочного отверстия, голова уплощенная с оттянутыми задними углами и глубо-
ко вырезанным верхним краем затылочного отверстия (рис. 72, 2). Нижнегубные
щупики отсутствуют. Щетинки и склериты тела редуцированы. Личинки миниру-
ют листья.
101(106). Верхняя губа на боках с бахромой из густых, длинных изогнутых хет
(рис. 61, 4).
102 (105). Бока сегментов брюшка с торчащими наружу пальцевидными отростка-
ми. Усики двучлениковые.
103(104). Верхняя губа с очень глубокой выемкой на переднем крае, без явствен-
ных щетинок на диске (рис. 61, 4)............................48. Argopus
104(103). Верхняя губа со слабой выемкой на переднем крае, с явственными ще-
тинками на диске (рис. 61,7)............................49. Sphaeroderma
105(102). Бока сегментов брюшка с небольшими округлыми выпуклостями. Усики
одночлениковые.............................................50. Pentamesa
106(101). Верхняя губа на боках без длинных изогнутых хет, со слабо выемчатым
передним краем. Бока сегментов брюшка с небольшими округлыми выпуклостями.
107(108). Микроскульптура тела в виде округлых цельных пластинок (рис. 77, 3).
Внутренний край мандибул с многочисленными длинными щетинками посередине
(рис. 77,1)..................................................51. Dibolia
108(107). Микроскульптура тела в виде сильно поперечных пластинок, вторично
разбитых на очень мелкие участки (рис. 77, 4). Внутренний край мандибул с 5-6
толстыми и короткими щетинками посередине..................52. Argopistes
1. Род Galeruca Geoffroy
Личинки обычно черные, матовые, с грязно зеленовато-серым низом. Голова с
длинным эпикраниальным швом, темя с 12 макрохетами, лоб шире длины, с 8 или
12 щетинками, наличник широкий, хитинизирован в основании, с рядом из 6 щети-
нок. Верхняя губа (рис. 61, 5) в основании с выступом, по бокам со слабо заострен-
ными углами, передний край с неглубокой срединной выемкой, с каждой стороны
от нее с 2 длинными утолщенными и 4 тонкими, иногда укороченными маргиналь-
156
Рис. 61. Детали строения личинок подсемейств Galerucinae и Alticinae:
1, 2 — голова; 1 — Pallasiola, 2 — Argopus; 3-8 — верхняя губа: 3 — Clitena; 4 —Argopus;
5 — Galeruca; 6 — Pallasiola, 7 — Sphaeroderma; 8 — Cneorane. Рис. 7 по Бионди.
ными щетинками; на диске ряд из 2 пор и 4 макрохет, причем внутренние в 2 раза
короче наружных. Мандибулы с 5 зубцами, без щетинок на внутреннем крае. Пере-
днеспинка с многочисленными длинными щетинками в основном по краям, эпип-
левральные склериты с 8-11 щетинками. Тергиты грудных и брюшных сегментов с
сильно выпуклыми, бугровидными склеритами, усаженными торчащими щетинка-
ми, с 2 рядами пре- и посттергальных склеритов, на брюшке эпиплевральные скле-
риты с 9-12, гипоплевральные с 6-8 щетинками, престигмальные склериты отсут-
ствуют (рис. 62, 7). Тергальные склериты на 8-м тергите брюшка частично слива-
ются друг с другом, тергит 9-го сегмента поперечный, с 4 щетинками вдоль пере-
днего края и с многочисленными длинными щетинками вдоль боковых и задних
краев. Микроскульптура покровов тела в виде мелких густых зернышек, щетинки
тела светло-коричневые, на вершине не заостренные. Склериты низа с редкими
щетинками. Коготки без выступа при основании. Длина тела до 12-14 мм.
157
Рис. 62. Строение 1-го тергита {1-5. 7-12} и 2-го стернита брюшка (6) личинок
подсемейства Galerucinae:
1 — Galeruca daurica; 2—Diorhabda tarsalis’, 3 — Clitena fuscipennis', 4—Galerucella grisescens;
5, 6 — G. nymphaeae', 7 — Lochmaea capreae; 8 — Pyrrhalta maculicollis', 9 — P viburni; 10 —
Exosoma lusitanica, 11 — Agelastica alni; 12 — A. coerulea.
Личинки питаются на листьях травянистых растений, обычно вместе с жука-
ми, изредка попадаются на поверхности почвы под растениями; окукливаются в
почве. Отмечено 3 личиночных возраста.
В регионе 13 видов. Известны личинки 8 видов, различающиеся с большим
трудом.
Таблица для определения видов рода Galeruca
1(2). Эпиплевральные склериты переднеспинки с 5-6, эпиплевральные склериты
брюшных сегментов — с 5, плевральные — с 4, наружные претергальные — с 2
хетами (подрод Haptoscelis Weise). Верхняя губа с каждой стороны с 2 толстыми и
2 тонкими маргинальными хетами. Тело серо-зеленое. ЕЧ, СК, ЗСБ........
................................................G. melanocephala Ponza
2(1). Эпиплевральные склериты переднеспинки с 8-11, эпиплевральные склериты
брюшных сегментов с 9-12, плевральные с 6-8, наружные претергальные — с 3-4
хетами. Верхняя губа с каждой стороны с 2 длинными утолщенными и 4 тонкими
маргинальными хетами.
3(6). Внутренние посттергальные склериты большинства брюшных сегментов по-
парно слитые.
158
4(5). Тело желтобелое с желтой головой. Переднеспинка с 3 поперечными рядами
щетинок. 9-й тергит брюшка с 2 дискальными и 6 маргинальными щетинками (под-
род Emarhopa Weise). На вьюнке. Юг ЕЧ......................G. rufa Germar
5(4). Тело черное или чернобурое. Все внутренние посттергальные склериты брюш-
ных сегментов попарно слитые со 2-го тергита, образуют непарный склерит, несу-
щий 12 щетинок. Наружные претергальные склериты отодвинуты от внутренних
на расстояние, равное их диаметру, и несут по 4 щетинки. Склерит переднеспинки
по переднему краю со спутанным двойным рядом щетинок (подрод Galeruca s.str.).
Микроскульптура тела очень густая и темная, щетинки тела желто-бурые. На Thalic-
trum, Aconitum. ЕЧ, СК, ЗСБ............................G. laticollis Sahiberg
6(3). Внутренние посттергальные склериты передних брюшных сегментов разде-
ленные, наружные претергальные склериты с 3-4 щетинками (подрод Galeruca s.str.).
7(10). Склерит переднеспинки по переднему краю с правильным рядом из череду-
ющихся коротких и длинных щетинок. Внутренние посттергальные склериты раз-
делены на 1-4-м или 1-5-м сегментах брюшка.
8(9). Внутренние претергальные склериты сегментов брюшка отодвинуты друг от
друга на расстояние, заметно превышающее их ширину, внутренние посттергаль-
ные склериты с 4 щетинками. Лоб с 8 щетинками. На полыни. ЕЧ, СК, СБ....
........................................G. interrupta circumdata Duftschmidt
9(8). Внутренние претергальные склериты сегментов брюшка отодвинуты друг от
друга на расстояние, меньшее их ширины, внутренние посттергальные склериты с
6 щетинками. Лоб с 12 щетинками. На полыни. СБ, ДВ......G. daurica Joannis
10(7). Склерит переднеспинки по переднему краю со спутанными рядами из чере-
дующихся коротких и длинных щетинок. Внутренние посттергальные склериты
разделены на 1-7-м сегментах брюшка.
11(14). Наружные претергальные склериты брюшка с 3 (редко 2-4) щетинками.
12(13). Наружные претергальные склериты сегментов брюшка очень маленькие,
отодвинуты от внутренних на расстояние в 5 раз большее их ширины. На Knautia,
Succisa, Centaurea, Cirsium. ЕЧ, СК, СБ................G. ротопае Scopoli
13(12). Наружные претергальные склериты сегментов брюшка небольшие, но бо-
лее крупные, чем у G. ротопае, отодвинуты от внутренних на расстояние в 3 раза
большее их ширины. Полифаг, чаще на сложноцветных. Повсеместно в Палеаркти-
ке, на север до тундры.................................G. tanaceti Linnaeus
14(11). Наружные претергальные склериты брюшка с 6 щетинками. На Thalictrum.
Юг СБ, ДВ..................................................G. weisei Reitter
2. Род Pallasiola Jacobson
Очень близка к роду Galeruca Geoffroy, но от последней легко отличается сле-
дующими признаками: лоб с 10 длинными щетинками (рис. 61, 7), верхняя губа
при основании с острыми углами, загнутыми ковнутри, передний край с неглубо-
кой выемкой, края которой образуют зубцевидные выступы (рис. 61, 6), щетинки
верха более темные и более закругленные на вершине. Окраска живых личинок
пепельно-серая. Длина взрослой личинки до 9 мм.
Жуки и личинки на листьях полыни, окукливание в почве.
В роде 1 вид — R absinthii Pallas, встречающийся на юге СБ и ДВ.
159
3
1
Рис. 63. Детали строения личинок Galerucinae:
1-3 — Theone silphoides: 1 — среднеспинка, 2 — 1-й тергит брюшка, 3 — голенелапка; 4 —
Pyrrhalta luteola, 4-й тергит брюшка.
3. Род Theone Gistl
Личинка черно-бурая^ склериты тела бугорковидно выпуклые, буровато-жел-
тые, щетинки длинные, рыжевато-желтые. Переднеспинка сильно поперечная, с
более темно пигментированной серединой диска, на переднем крае с 28-36 щетин-
ками (из них 12-14 очень длинные), расположенными группой близ срединной по-
лоски и в виде неправильного ряда вдоль края; базальный край с 20-22 щетинками,
боковые края с 6 щетинками. Эпиплевральные склериты переднеспинки с 11-12
щетинками, стигмальные склериты с 1 длинной и 4 короткими щетинками. Внут-
ренние претергальные склериты средне- и заднеспинки (рис. 63, 7) слиты попарно,
с 6 щетинками с каждой стороны, внутренние посттергальные склериты разделен-
ные, с 5 щетинками; наружные претергальные с 3-5 щетинками, в 3-4 раза меньше
наружных посттергальных, последние с 8-9 щетинками; крыловые склериты с 12
щетинками. На 1-7-м тергитах брюшка (рис. 63,2) внутренние претергальные скле-
риты разделенные, с 5-7 щетинками, внутренние посттергальные слитые, с 4 ще-
тинками с каждой стороны; наружные претергальные с 3, наружные посттергаль-
ные с 3-4, постстигмальные с 8-10 щетинками. Внутренние тергальные склериты
8-го тергита брюшка слитые попарно, с каждой стороны с 4 щетинками на претер-
гальном и 5 щетинками на посттергальном, соединенном с наружными посттер-
гальными. Тергит 9-го сегмента брюшка сильно поперечный, с широко дугообраз-
ным вершинным краем, с 4 щетинками близ переднего края и с 14 щетинками с
каждой стороны вдоль краев и частично на диске. Голенелапка довольно длинная, с
крупным длинным коготком, равным примерно половине длины голенелапки, и с
редуцированным хелониумом, не покрывающим коготка (рис. 63, 3).
Один вид этого рода — Th. silphoides Dalman встречается на юго-востоке ЕЧ.
Личинки живут на листьях полыни; окукление в почве. Заметно повреждают паст-
бищные угодья, при массовом размножении могут являться причиной отравления
скота.
160
4. Род Diorhabda Weise
Личинки этого рода очень сходны с Pyrrhalta. отличаясь, прежде всего, светло-
окрашенными склеритами; окраска тела от сплошь желтой до темной за счет силь-
ного развития микроскульптуры. Тело сильно удлиненное, склерит переднеспинки
резко поперечный, длина которого примерно в 2,5-3 раза меньше ширины; щетин-
ки расположены помимо переднего и заднего краев также на середине диска, в том
числе и вдоль срединной линии. Эпиплевральные склериты переднеспинки с 8-10,
брюшка с 7-8 щетинками (рис. 62, 2). Склериты средне- и заднеспинки обычно с
неявственными границами. Наружные претергальные склериты брюшных терги-
тов с 2 щетинками.
Жуки и личинки живут на листьях; 3 личиночных возраста, окукливание в по-
чве.
В регионе 3 вида, чаще в полупустынной зоне. Личинки на растениях, окукле-
ние в почве.
Таблица для определения видов рода Diorhabda
1(2). Тело без микроскульптуры, светлое, со светлыми склеритами (молодые ли-
чинки с темными склеритами). Коготки с крупным зубцом и большим хелониумом.
Верхняя губа с едва вогнутым передним краем и слабым базальным выступом.
Внутренние посттергальные склериты брюшных сегментов (начиная со 2-го) сли-
ваются попарно (рис. 60, 3). Забайкалье, на солодке.......D. tarsalis Weise
2(1). Тело с явственной микроскульптурой, образующей темный фон или рисунок.
Коготки без явственного зубца, с небольшим хелониумом. Внутренние посттергаль-
ные склериты 1-5-го брюшных сегментов раздельные (рис. 60, 7, 2).
3(4). Внутренние тергальпые склериты на среднеспинке слитые, на заднеспинке
узко разделенные, с 3 щетинками; наружные претергальные склериты средне- и
заднеспинки небольшие, с 1 щетинкой. Внутренние пре- и посттергальные склери-
ты 1-7-го сегментов брюшка с 3 щетинками. На верблюжьей колючке. Юг ЕЧ, СК
.....................................................D. persica Faldermann
4(3). Внутренние тергальные склериты средне- и заднеспинки раздельные, с 5 ще-
тинками; наружные претергальные склериты средне- и заднеспинки более круп-
ные, с 4 щетинками. На 1-7-м сегментах брюшка внутренние претергальные скле-
риты с 5, внутренние посттергальные с 4—5 щетинками. На гребенщике. Юг ЕЧ, СК
........................................................D. elongata Brulle
5. Род Clitena Baly
Голова, ноги и склериты тела темноокрашенные. Темя с редкими мелкими
щетинками назади и с рядом из 4 длинных щетинок с каждой стороны вдоль лоб-
ных швов. Эпикраниальный шов короткий. Лоб с 6 первичными щетинками, рас-
положенными в 2 ряда (4, 2), и 2 порами, наличник поперечный, с рядом из 6 мел-
ких щетинок и 2 пор, причем наружные щетинки крупнее прочих. Верхняя губа
(рис. 61, 3) сильно хитинизированная, в основании с сильным выступом, на пере-
днем крае с широкой неглубокой выемкой, по бокам от нее с каждой стороны с 5-6
161
маргинальными щетинками, из которых 2 наружные длинные и толстые; диск с 4
длинными щетинками и 2 порами. Переднеспинка морщинистая, с редкой зернис-
тостью, с 10 длинными и 10 короткими щетинками по переднему и 6 длинными по
заднему краю; эпиплевральный склерит с 5 щетинками. Тергиты груди и брюшка с
2 рядами склеритов. Внутренние претергальные склериты средне- и заднегруди с 3
щетинками (1 макро- и 2 микрохеты), наружные претергальные с 2 микрохетами,
каждый посттергальный склерит с 1 макрохетой. Крыловые склериты с 3 макро- и
3-4 микрохетами, на всех брюшных сегментах внутренние тергальные склериты
сливаются попарно и имеют соответственно 2 макро- и 4 микрохеты и 2 макро- и 2
микрохеты. Наружные претергальные с 1 макро- и 1 микрохетой, посттергальные —
с 1 макрохетой, пре- и посттергальные склериты развиты, каждый с 1 макро- и 1
микрохетой, эпиплевральные с 1 макро- и 5 микрохетами, гипоплевральные с 1
макро- и 4 микрохетами (рис. 62, 3). Стерниты груди и брюшка развиты, с редкими
щетинками. Голенелапка толстая и короткая, снизу с 2, сверху с 3 щетинками. Кого-
ток длинный, изогнут, с прямоугольным выступом при основании, хелониум силь-
но развит и прикрывает полностью коготок.
Жуки и личинки на листьях клена, окукливание в почве. В регионе 1 вид — С.
fuscipennis Jacoby на юге ДВ.
6. Род Galerucella Crotch
Очень близок к роду Pyrrhalta, отличаясь достоверно только по комплексу при-
знаков. Голова, ноги и склериты тела темноокрашенные. Щетинки тела всегда тем-
ноокрашенные, тонкие, постепенно утончаются к вершине, склериты тела могут
быть сильно расширенные, морщинистые, с нерезкими границами и укороченны-
ми щетинками или же явственные, обособленные, с длинными щетинками, превы-
шающими в 2-3 раза ширину склерита. Внутренние претергальные склериты брюш-
ка сливаются в один непарный склерит или разделенные. Эпиплевральные склери-
ты груди и брюшка с 2—4 щетинками, гипоплевральные с 1 макро- и 1-2 микрохета-
ми, престигмальные склериты развиты. Микроскульптура верха тела между скле-
ритами неявственная, развита только на самих склеритах в виде мелких густых
зернышек, образующих мелкие шипики. Склериты низа слабо хитинизированы,
состоят из овально-вытянутых пластинок с 4—6 мелкими зубчиками (рис. 62, 6; 67,
7), с утолщенными и укороченными щетинками.
Окукливание на листьях. В регионе 3 вида на травянистых растениях, обычно
водных или прибрежных, реже на кустарниках.
Таблица для определения видов рода Galerucella
1(2). Внутренние претергальные склериты 1-8-го сегментов брюшка разделены по
средней линии (рис. 62, 4). Гипоплевральные склериты с 3 щетинками. Склериты
тела четкие, не морщинистые, разделены значительными промежутками. Щетинки
верха в 2-3 раза длиннее склерита. На Polygonum, Lysimachia, Hydrocharis, Rumex,
реже на травянистых розоцветных. Вся лесная зона Палеарктики............
......................................................G. grisescens loannis
2(1). Внутренние претергальные склериты 1-8-го сегментов брюшка сливаются в 1
162
непарный (рис. 62, 5). Гипоплевральные склериты с 2 щетинками. Склериты тела
нечеткие, морщинистые, плохо отделены друг от друга, щетинки верха короткие
(за исключением эпиплевральных и гипоплевральных).
3(4). Эпиплевральные склериты переднеспинки с 4 щетинками. Щетинки склери-
тов 8-го тергита брюшка длинные. На Brasenia и других гигрофильных растениях.
Юг ДВ............................................G. nipponensis Laboissiere
4(3). Эпиплевральные склериты переднеспинки с 2 щетинками. Щетинки склери-
тов 8-го тергита брюшка короткие. На кувшинковых, Lysimachia, Potamogeton,
Polygonum, Rumex, Sagittaria, Potentilla, местами на смородине. Широко распрост-
ранена по всей Палеарктике, на север до тундры.....G. nymphaeae Linnaeus
7. Род Apophylia Thomson
Тело стройное, немного дорсовентрально уплощенное, темно-бурое; голова и
ноги темно-коричневые; низ плоский, светло-желтый. Склериты крупные, тесно
примыкают друг к другу, трудно различимые из-за густой маскирующей микро-
скульптуры покровов тела в виде густых зерен; щетинки верха светлые, очень ко-
роткие, со слабо булавовидными или округленными вершинами, каждая щетинка
расположена на небольшом возвышении склерита. Эпиплевральные склериты сег-
ментов брюшка в виде пальцевидных выростов, которые придают личинкам силь-
но уплощенную форму, склериты верха плоские (рис. 64, 7).
Головная капсула округлая (рис. 64, 2), затылок с широкой треугольной выем-
кой, темя в задней половине гладкое, без щетинок, эпикраниальный шов короткий.
Лобные швы вначале прямые и отходят под острым углом, далее слабо изогнуты и
доходят до основания усиков. Лоб узкотреугольный, в середине с 2 слабыми косы-
ми вдавлениями и 6 длинными остроконечными щетинками, расположенными в 2
ряда (4, 2), вдоль границы с наличником узко склеротизован, наличник узкий, с
рядом из 6 коротких щетинок. Верхняя губа (рис. 64, 3) умеренно склеротизован-
ная, узкая, основание в середине с треугольным выступом, по бокам без острых
выступов, передний край почти прямой, со слабой и широкой, едва намеченной
срединной выемкой, над которой расположено поперечное поле из очень мелких и
тонких щетинок, по бокам переднего края с каждой стороны с 2 длинными, сбли-
женными и 4 тонкими, более укороченными маргинальными щетинками, на диске
8 пор и 4 дискальные щетинки, расположенные косым рядом, причем наружные
щетинки длиннее внутренних. Мандибулы с 5 зубцами, на внутреннем крае с пуч-
ком из 3-4 тонких щетинок, сбоку с 2 порами и 2 щетинками (рис. 64, 4). Переднес-
пинка поперечная, с двумя квадратными склеритами, разделенными узкой светлой
полосой, склериты с отдельными ямками и небольшими бугорками, с более длин-
ными щетинками по переднему краю и редкими на диске; эпиплевральный склерит
овально-вытянутый, выпуклый, примыкает к боковому краю переднеспинки, с 4
щетинками (рис. 64, 5). Склериты груди и брюшка с двумя рядами склеритов, каж-
дый из которых несет по 1-2 толстых и по 2-3 очень тонких и коротких щетинки
(рис. 64, б). Все внутренние пре- и посттергальные склериты заднегруди и брюшка
попарно слиты. Между наружными и внутренними претергальными склеритами
груди появляются 2 (а на брюшных сегментах 1) маленьких склерита с короткой
щетинкой. Крыловые склериты с 3 длинными щетинками. Эпиплевральные скле-
163
Рис. 64. Детали строения личинки Apophylicr.
1 — общий вид, 2 — голова, 3 — верхняя губа, 4 — мандибула, 5 — переднеспинка, 6 — 1-й
тергит брюшка, 7 — 9-й тергит брюшка, 8 — 3-й стернит брюшка, 9 — нога.
риты сегментов брюшка с 2 длинными щетинками. На 9-м тергите брюшка все
склериты сливаются в непарную широкую пластинку (рис. 64, 7), сильно нависают
над подталкивателем. Стигмы круглые, с тонким склеротизованным кольцом. Стер-
ниты груди и брюшка развиты, но слабо склеротизованы, с крепкими длинными
щетинками (рис. 64, 8). Ноги сильно хитинизированные, коготки слабо изогнуты,
острые, в основании подковообразно расширены и с конусовидным хелониумом
(рис. 64, 9).
164
Длина тела личинки 3-го возраста 7,5 мм.
В регионе 2 вида на юге ДВ, их личинки неизвестны. Описание рода составле-
но по личинке Apophylia furcigera Chujo из Вьетнама.
8. Род Lochmaea Weise
Голова, ноги и склериты тела темноокрашенные. Темя назади в редких щетин-
ках, в передней половине с рядом из 3—4 щетинок вдоль лобных швов. Эпикрани-
альный шов длинный, лобные швы расходятся под острым углом, лоб с 6 щетинка-
ми (4, 2), наличник поперечный, с рядом из 6 коротких щетинок. Верхняя губа в
основании с сильным выступом, хитинизирована только в основной половине, с
округленным передним краем, без срединной выемки, по бокам с 2 толстыми и 4
короткими маргинальными щетинками с каждой стороны. Переднеспинка с 7-9
щетинками по переднему и 4 по заднему краю, эпиплевральные склериты с 6-10
щетинками. Внутренние претергальные склериты средне- и заднегруди с 1-3, а
брюшка с 1-2 щетинками; на всех сегментах брюшка (рис. 62, 7) внутренние пре- и
посттергальные склериты сливаются в один непарный склерит, эпиплевральные
склериты с 3—4 щетинками, гипоплевральные с 2 длинными и 1 короткой, престиг-
мальные склериты развиты или отсутствуют. Стерниты груди и брюшка с развиты-
ми склеритами, с 1-2 щетинками на каждом. Голенелапка удлиненная, коготки с
тупым зубцом.
Жуки и личинки живут на листьях деревьев и кустарников, реже на вереске.
Имеются три личиночные стадии, окукливание в почве. В регионе 3 вида.
Таблица для определения видов рода Lochmaea
1(2). Брюшные сегменты с престигмальными склеритами. Гипоплевральные скле-
риты брюшных сегментов с 2 длинными щетинками. В завязях и молодых плодах
терновника, боярышника и яблони. Вся лесная зона Палеарктики............
........................................................L. crataegi Forster
2(1). Брюшные сегменты без престигмальных склеритов (рис. 62, 6). Гипоплевраль-
ные склериты брюшных сегментов с 3 хетами (2 макро- и 1 микрохета).
3(4). Внутренние претергальные склериты с каждой стороны с 3 длинными щетин-
ками. Темя с 5 парами длинных щетинок. На вереске, лесная зона. Европа. Указа-
ния для Сибири нуждаются в уточнении.................L. suturalis Thomson
4(3). Внутренние претергальные склериты с каждой стороны только с 1 длинной
щетинкой (рис. 62, 7). Темя с 6 парами длинных щетинок. На иве и березе. Вся
лесная зона Палеарктики, заходит в тундру..............L. capreae Linnaeus
9. Род Pyrrhalta loannis (Galerucella Crotch)
Голова, ноги и склериты тела темноокрашенные. Голова умеренно хитинизи-
рованная, назади в мелких редких щетинках, вдоль лобных швов с 2 длинными
щетинками. Эпикраниальный шов короткий, лоб с 6 щетинками (4, 2), на границе с
наличником хитинизирован в виде валика, наличник с 6 мелкими щетинками. Пе-
редний край верхней губы прямой, слабо изрезан или с едва намеченной вырезкой,
165
с каждой стороны с 2 толстыми длинными и 4 тонкими короткими маргинальными
щетинками. Склерит переднеспинки слабо хитинизирован, с редкими рядами ще-
тинок по переднему и заднему краям или с многочисленными щетинками по всему
диску; эпиплевральный склерит переднеспинки с 1-2 длинными и 1-10 короткими
щетинками. Внутренние претергальные склериты груди и брюшка с 1-3 длинными
и 1-6 короткими щетинками. Престигмальные склериты брюшных сегментов раз-
виты или отсутствуют, эпиплевральные с 2-4 длинными и 1-5 короткими, гипоп-
левральные с 1 длинной и 1-5 короткими щетинками. Щетинки верха с притуплен-
ными или оплавленными вершинами, никогда не бывают заостренными на конце,
обычно светлые. Стерниты груди и брюшка явственные, их микроскульптура в виде
овальных редких зернышек. Голенелапка короткая, коготки со сглаженным зубцом
при основании.
Жуки и личинки на листьях древесных и травянистых растений, имеются 3
личиночных возраста, окукливание в почве. В регионе 9 видов; неизвестна личин-
ка Р semifulva Jacoby, живущая на плодовых; описание личинки Р flavescens Weise,
развивающейся в бутонах липы на юге ДВ, очень краткое и неудовлетворительное
(Синчилина, 1962). Выделяется ряд подродов, иногда рассматриваемых как рода.
Таблица для определения видов рода Pyrrhalta
1(6). Внутренние претергальные склериты брюшных тергитов с 5-8 парами круп-
ных щетинок; между внутренними и наружными претергальными склеритами име-
ется по 1 изолированной щетинке (рис. 62, 8).
2(3). Внутренние претергальные склериты брюшка с 5 длинными щетинками (рис.
63, 4). Тело желтое, у взрослых личинок вдоль боков с каждой стороны с продоль-
ной темной полосой. На листьях ильмовых. ЕЧ, СК, ЗСБ.....Р luteola Mtiller
3(2). Внутренние претергальные склериты брюшка с 1-2 длинными щетинками.
Виды ДВ.
4(5). Эпиплевральные склериты переднеспинки с 2 длинными и 8 короткими, внут-
ренние претергальные склериты брюшка с 2 длинными и 3 короткими щетинками.
Короткие щетинки верха тела темно-коричневые. На вязе. Юг ДВ...........
....................................................Р. aenescens Fairmaire
5(4). Эпиплевральные склериты переднеспинки с 1 длинной и 10 короткими ще-
тинками, внутренние претергальные склериты брюшка с 3 длинными и 4 коротки-
ми щетинками (рис. 62, 8). Короткие щетинки верха тела светло-коричневые. На
вязе. Юг ДВ............................................Р. maculicollis Motschulsky
6(1). Внутренние тергальные склериты брюшных сегментов с 1 парой крупных
щетинок; без изолированной щетинки между внутренними и наружными претер-
гальными склеритами (рис. 62, 9).
7(8). Брюшные сегменты с престигмальными склеритами (рис. 62,9). Эпиплевраль-
ные склериты переднеспинки с 2 щетинками и порой. Верхняя губа с едва намечен-
ной срединной выемкой. Наружные претергальные склериты груди с очень малень-
кой щетинкой. На калине. ЕЧ, СК, СБ, ДВ..................R viburni Paykull
8(7). Брюшные сегменты без престигмальных склеритов.
9(10). Наружный тергальный склерит средне- и заднеспинки развит, с длинной
щетинкой. Стерниты брюшка темно-коричневые. Внутренние тергальные склери-
166
ты груди с 2 длинными щетинками и 2 порами, эпиплевральные склериты с 4 ще-
тинками. На ивах и ольхе. Вся лесная зона Палеарктики.В lineola Fabricius
10(9). Наружный претергальный склерит средне- и заднегруди редуцирован, с по-
рой. Стерниты брюшка светлые. Внутренние тергальные склериты груди с 1, эпип-
левральные склериты с 1-2 длинными щетинками.
11(12). Эпиплевральный склерит брюшка с 2 длинными и 2 очень короткими ще-
тинками. Тело желтое, с темно-бурыми склеритами. На Lythrum. Широко распрос-
транена по всей Палеарктике.......................В calmariensis Linnaeus
12(11). Эпиплевральный склерит брюшка с 1-2 щетинками. Тело светло-желтое со
светло-бурыми склеритами
13(14). Эпиплевральный склерит брюшка с 1 длинной и 1 короткой щетинками.
Голова сильно хитинизирована, с нерасширенными лобными швами. Тело светлое,
желтозеленое, со светло-бурыми склеритами. На травянистых розоцветных. Вся
лесная зона Палеарктики...............................В tenella Linnaeus
14(13). Эпиплевральный склерит брюшка с 1 длинной щетинкой. Голова слабо скле-
ротизована, с сильно расширенными лобными швами. Тело желтое, с темно-буры-
ми склеритами. На Cirsium, Lythrum. ЕЧ, СБ, редкий вид.В. pusilia Dufthchmidt
10. Род Cneorane Baly
Личинка светло-желтая, с темной головой, ногами и склеритами. Темя с каж-
дой стороны с 10 щетинками, расположенными в основном вдоль лобных швов.
Эпикраниальный шов короткий. Лоб гладкий, с 8 щетинками, расположенными в 3
ряда (2, 2, 4), и 2 порами. Наличник узкопоперечный, с рядом из 6 коротких щети-
нок. Верхняя губа (рис. 61,5) поперечная, ее основание в середине с сильным выс-
тупом, на диске ряд из 4 длинных тонких щетинок и 2 пор; передний край с широ-
ким срединным выступом, усаженным густыми шиловидными щетинками, с каж-
дой стороны от выступа расположены 4 утолщенных и 2 тонких маргинальных
щетинки. Мандибулы треугольные, усики 1-члениковые, сенсориальный конус уз-
кий и вытянутый (так что усики выглядят двучлениковыми). Переднеспинка почти
квадратная, со светлой узкой полосой, разделяющей склериты, по переднему краю
с каждой стороны с рядом из 4 длинных и 6 коротких щетинок, а по заднему — с 1
длинной и 3 короткими щетинками. Средне- и заднеспинка (рис. 67, 3) с 3 рядами
склеритов; вторичные склериты первого ряда мелкие и с короткими щетинками,
склериты 2-го и 3-го рядов с длинными тонкими щетинками, причем внутренние
тергальные попарно сливаются; наружные посттергальные с 4 длинными, крыло-
вые склериты с 4 длинными и 2 короткими щетинками. Стигма с округлым стиг-
мальным отверстием и с овально-вытянутой стигмальной пластинкой. Все эпип-
левральные склериты груди с 1 щетинкой. 1-8-й тергиты брюшка (рис. 67, 5) с 4
рядами склеритов; склериты первого ряда (2 на первом тергите и 4 на прочих) мел-
кие и не имеют щетинок, второй и четвертый ряд состоят из 5 склеритов с длинны-
ми щетинками; внутренние тергальные склериты попарно слившиеся; третий ряд
состоит из 2 раздвинутых интеркалярных склеритов. Стигмальные склериты ма-
ленькие, эпиплевральные склериты с 4 длинными щетинками. На 9-м тергите брюш-
ка все склериты сливаются в сплошной, почти квадратный склерит с рядом длин-
ных щетинок по краям и с 4 в середине. Склериты низа хорошо выражены. Микро-
167
Рис. 65. Детали строения личинок Galerucinae:
1-3 —Aulacophora nigripennis: 1 — общий вид, 2 — голова, 3 — 9-й тергит брюшка; 4-6 —
Fleutiauxia armata\ 4—общий вид, 5—голова, 6 — 9-й тергит брюшка. По Кимото и Такизава.
скульптура редкая, в виде небольших овальных зернышек, по краям самих склери-
тов в виде неправильных многоугольников (рис. 67, 7). На 9-м тергите брюшка мик-
роскульптура у основания сегмента в виде вытянутых шестиугольников, тесно при-
легающих друг к другу и с вытянутыми вершинами, которые образуют своеобраз-
ные мелкие шипики, направленные в одну сторону. Ноги длинные, голенелапка уз-
кая и длинная, снизу с 8, сбоку с 4 и сверху с 3 щетинками. Коготки простые, ост-
рые, с небольшим хелониумом и щетинкой у основания. Единственная личинка
была найдена в листовом опаде под Lespedeza. Во Вьетнаме нами были найдены
личинки вида этого рода в цветках кустарниковых бобовых. Указание на нахожде-
ние личинок в почве на корнях Lespedeza ошибочны.
В регионе 1 вид — С. violaceipennis rufipes Weise, на юге ДВ.
11. Род Fleutiauxia Laboissiere
Тело тонкое С-образно изогнутое светло-желтое, слабо хитинизированы (рис.
65, 4). Голова с немногочисленными щетинками (рис. 65, 5). Схема склеритов
груди и брюшка — рис. 60, 2, средне-, заднеспинка и брюшко с двумя мелкими
вторичными склеритами с передней стороны у переднего края, так что на груди
три ряда склеритов, на брюшке — четыре ряда (рис. 60, 2). Интеркалярные скле-
риты брюшных тергитов слиты, стерниты с мелкими вторичными склеритами.
Вершинный тергит брюшка слабо хитинизирован без ячеистой скульптуры, с не-
многочисленными вторичными щетинками (рис. 65, 6). Длина тела взрослой ли-
чинки свыше 5 мм.
Личинки в почве, жуки питаются на Lespedeza (Kimoto, Takizawa, 1994). В Рос-
сии один вид — Е armata Baly на юге ДВ. Нами имаго данного вида на юге Амур-
ской области после выхода из мест зимовки неоднократно отмечались при интен-
сивном питании на листьях Atractylodes ovata, питание на Lespedeza требует под-
тверждения.
168
12. Род Luperus Geoffrey
Окраска охряно-желтая, с рыжеватой головой, ногами и тергитом 9-го сегмен-
та брюшка. Голова немного длиннее своей ширины, с коротким эпикраниальным
швом, без глазков, лоб не длиннее своей ширины, с 3 макрохетами и 3 мелкими
порами, темя с 6 макрохетами, усики двучлениковые, мандибулы с 3 хорошо разви-
тыми острыми зубцами (остальные 2 зубца редуцированы), на внутреннем крае
недалеко от основания с пучком толстых коротких щетинок; нижнечелюстные щу-
пики трехчлениковые, нижнегубные — двучлениковые, кардо нижних челюстей
без щетинок. Переднеспинка с 8 (4 длинные и 4 короткие) щетинками на переднем
и с 6 (4 длинные и 2 короткие) на заднем крае, эпиплевральные склериты переднес-
пинки маленькие, с одной макрохетой, наружные претергальные склериты средне-
и заднеспинки без щетинок, внутренние тергальные склериты попарно слиты, кры-
ловые склериты с 1 макро- и 2 микрохетами. Тергиты 1-8-го сегментов брюшка с 3
поперечными рядами склеритов, внутренние тергальные склериты попарно сли-
тые, промежуточные склериты сближенные, сильно поперечные, постстигмальные
крупные, тергит 9-го сегмента брюшка с ячеистой скульптурой и закругленным вер-
шинным краем, стернит 9-го сегмента брюшка с поперечным склеритом, несущим
4 длинных щетинки. Покров тела между склеритами мелкозернистый, без микро-
скульптуры.
Жуки питаются листьями древесных пород (ольха, береза, ива, реже сосна и
ель), личинки живут и окукливаются в почве, питаясь, очевидно, корнями травяни-
стых растений.
В регионе 8 видов, известны личинки 1-го возраста 4 видов.
Таблица для определения видов рода Luperus
1(2). Мандибулы с 4 зубцами, 9-й тергит брюшка с 6 краевыми щетинками (рис. 66,
10) ЕЧ, ЗСБ...............................L. luperus Sulzer (L. lyperus Bedel)
2(1). Мандибулы с 2-3 зубцами. 9-й тергит брюшка с 8-12 щетинками.
3(4). Мандибулы с 2 зубцами. 9-й тергит брюшка (рис. 66,11) с 8 щетинками (4 на
диске, расположены попарно, 2 на боковых краях). Вся лесная зона Палеарктики
...................................................L. longicornis Fabricius
4(3). Мандибулы с 3 зубцами, 9-й тергит брюшка (рис. 66, 12) с 12 щетинками (8
длинных краевых и 4 коротких на диске). Вся лесная зона Палеарктики.....
......................................................L. flavipes Linnaeus
13. Род Calomicrus Stephens
Тело удлиненное, светло-желтое, со светло-коричневой головой. Головная кап-
сула (рис. 66, 1) с коротким эпикраниальным швом. Лобные швы расходятся под
острым углом. Усики (рис. 66, 3) одночлениковые, с одной щетинкой и тремя шипи-
ками. Глазки отсутствуют. По бокам головы располагаются по четыре макрохеты,
перед наличником — две микрохеты. На темени щетинки располагаются в два ряда
по 4 микрохеты в каждом. Мандибулы (рис. 66, 4) с тремя зубцами и пучком корот-
ких щетинок на внутреннем крае. Верхняя губа (рис. 66, 2) в центре переднего края
169
Рис. 66. Детали строения личинок Calomicrus и Luperus'.
1-8 — Calomicrus minutus: 1 — голова, 2 — верхняя губа, 3 — усик, 4 — мандибула, 5 —
голенелапка, 6 — тергиты груди, 1-го и 4-го тергитов брюшка, 7 — 8-й тергит брюшка; 8 —
9-й тергит брюшка; 9 — С. pinicola, 9-й тергит брюшка; 10-12—Luperus, 9-й тергит брюшка:
10 — L. longicornis, 11 —L. luperus, 12 — L.flavipes. По Дубешко и Штейнхаузену.
с пучком острых шипиков и с двумя типами маргинальных щетинок, одни заост-
ренные макрохеты расположены с верхней стороны полукругом близ вершины,
другие шесть очень толстых уплощенных хет — с нижней стороны, 4 дискальные
щетинки располагаются в два ряда.
Нижнечелюстные щупики трехчлениковые. На галее имеется десять мелких
сенсорных щетинок, на лацинии — две макрохеты. Стипес с внешней стороны не-
сет одну макрохету, субментум две, ментум — две макро- и две микрохеты. Нижне-
губные щупики одночлениковые. Переднеспинка (8,4) с крупным склеритом, несу-
щим по 4 макрохеты по переднему и заднему краям, ее эпиплевральный склерит с
2 щетинками (требует проверки, может быть 1). Все тергиты, начиная со средне-
спинки, разделены поперечной складкой. На средне- и заднеспинке различаются
два ряда макрохет, на 1-3-м брюшных тергитах четко различаются уже три ряда
(рис. 66, 6). Среднеспинка без тергальных склеритов, на заднеспинке внутренние
претергальные склериты отсутствуют, прочие разделены, каждый с 1 щетинкой.
Крыловые склериты средне- и заднеспинки с одной макро- и одной микрохетами.
8-й тергит брюшка с частично слитыми склеритами (рис. 66, 8), 9-й тергит с круп-
170
ным, хорошо склеротизованным склеритом с ячеистой скульптурой, закругленным
вершинным краем, 4-8 маргинальными и 4 дискальными макрохетами (рис. 66, 8).
Голенелапка (рис. 66, 5) с пятью щетинками, коготки с крупным хелониумом.
Характеристика рода составлена по неполному и, очевидно, неточному описа-
нию личинки 1-го возраста С. minutus loannis (Дубешко, 1982). В России 5 видов.
Таблица для определения видов рода Calomicrus
1(2). 9-й тергит с крупным, хорошо склеротизованным склеритом с ячеистой скуль-
птурой, 8 маргинальными и 4 дискальными макрохетами (рис. 66, 8). Личинки на
корнях Pulsatilla на юге ВСЕ и ДВ.......................С. minutus loannis
2(1). 9-й тергит с продолговатыми светлобурыми пятнами, без ячеистой скульпту-
ры, с 4 маргинальными и 4 дискальными макрохетами (рис. 66, 9). ЕЧ, на корнях
травянистых растений в сосняках....................С. pinicola Duftschmidt
14. Род Aulacophora Lacordaire
Тело узкое, цилиндрическое, кремово-желтое (рис. 65, 7). Головная капсула —
рис. 65, 2. Все тергальные склериты груди с 1 щетинкой, внутренние пре- и пост-
тергальные склериты попарно слитые (рис. 60, 5). Тергиты брюшка с 3 поперечны-
ми рядами склеритов, несущими по 1 щетинке, внутренние пре- и посттергальные
попарно слиты, интеркалярные не слиты, но очень крупные, поперечные и сильно
сближенные. На последних тергитах брюшка наружные тергальные склериты сли-
ваются с внутренними. Хитинизированная площадка 9-го тергита брюшка с ячеис-
той структурой, с 4 щетинками вдоль закругленного заднего края (рис. 65, 3).
Жуки питаются на тыквенных, личинка в почве. В регионе 1 вид —A. nigripennis
Motschulsky, встречающийся на юге ДВ. Характеристика рода составлена частично
по личинке южнопалеарктического вида A. foveicollis Lucas.
15. Род Phyllobrotica Chevrolat
Окраска личинки белая, голова рыжевато-бурая, наличник, лобные швы, края
головной капсулы и усиковых впадин черно-бурые, глазков нет, эпикраниальный
шов очень короткий, темя с 6 макрохетами, лоб немного длиннее своей ширины, с
6 макрохетами, усики двучлениковые, мандибулы с 5 зубцами. Переднеспинка силь-
но поперечная, с 10 щетинками на переднем крае, с 4 — на заднем и 4 дискальными
щетинками на вдавленной части диска. Склериты грудных и брюшных сегментов
плохо ограниченные, с короткими щетинками. Тергальные склериты средне- и зад-
неспинки небольшие, с 1 щетинкой на каждом. Склериты брюшных сегментов
расположены на каждом тергите 3 поперечными рядами, внутренние претергаль-
ные склериты слиты в непарный склерит. Наружные претергальные склериты
слиты с престигмальными в один несущий 2 щетинки склерит; интеркалярные,
постстигмальные и наружные посттергальные склериты с 1 щетинкой, внутрен-
ние посттергальные склериты попарно сливаются, образуя 1 непарный склерит с
2 щетинками. Тергит 9-го сегмента брюшка с 8 маргинальными и 8 дискальными
щетинками.
171
Личинки живут в почве на корнях Scutellaria, обычно на сырых лугах и болоти-
стых лесных полянах. Окукливание в почве.
В регионе 3 вида. Известна личинка Ph. quadrimaculata Fabricius, встречаю-
щаяся в ЕЧ и СБ.
16. Род Exosoma Jacobson
Эпикраниальный шов нормальный, не укороченный. Срединный склерит пе-
реднеспинки с немногочисленными короткими щетинками по краям, эпиплевраль-
ные склериты с 1 щетинкой каждый. Средне- и заднеспинка с 2 претергальными и
2 посттергальными склеритами, несущими по 1 щетинке каждый, крыловые скле-
риты с 3, эпиплевральные склериты с 1 щетинкой, брюшные тергиты (рис. 62,10) с
3 поперечными рядами склеритов: 2 пары претергальных, 1 пара интеркалярных, 2
пары посттергальных; все тергальные склериты несут по 1 щетинке; престигмаль-
ные склериты с 1 щетинкой, постстигмальные — с 2 щетинками, эпиплевральные —
с 4, гипоплевральные — с 3 щетинками.
Жуки питаются на Hemerocalis. Приводимое выше описание рода сделано по
личинке западноевропейского вида Е. lusitanica Linnaeus.
В регионе 1 вид — Е. collare Hummel (ЕЧ, СК, ЗСБ), личинка которого неизве-
стна.
17. Род Atrachya Dejean (Luperodes Motschulsky)
Окраска зеленовато-серая, с темно-бурыми головой, переднеспинкой, склери-
тами тела и 9-м тергитом брюшка. Темя в задней половине с очень мелкими щетин-
ками или порами, в передней вдоль лобных швов с каждой стороны с 2 крупными
щетинками. Эпикраниальный шов короткий. Лоб с 6 длинными щетинками и 2 по-
рами. Наличник узко хитинизирован в основании, с рядом из 6 щетинок и 2 пор.
Верхняя губа в основании с выступом, на диске с рядом из 2 пор и 4 щетинок,
причем наружные вдвое длиннее внутренних; ее передний край прямой, с каждой
стороны с 6 маргинальными щетинками. Склерит переднеспинки с правильным
рядом из 12 щетинок по переднему и со спутанным из 10 по заднему краю; эпип-
левральный склерит с 1 щетинкой. Средне- и заднеспинка с 2 рядами склеритов,
внутренние тергальные склериты сливаются в непарные, каждый из которых имеет
соответственно по 2 и 4 щетинки; наружные претергальные с 1, а наружные пост-
тергальные с 3 щетинками, крыловые склериты с 7 щетинками, из которых 3 очень
мелкие. 1-8-й тергиты брюшка (рис. 67,2) с 4 рядами склеритов, первый ряд состо-
ит из 2 очень мелких вторичных склеритов с едва заметной щетинкой на каждом
(на последних сегментах они едва различимы), второй и четвертый ряд с 5 склери-
тами, так как внутренние сливаются в 1 непарный; третий ряд включает 2 интерка-
лярных склерита. Эпи- и гипоплевральные склериты с 3 щетинками. На 9-м сег-
менте все склериты сливаются в 1 непарный. Стерниты груди и брюшка с темноок-
рашенными склеритами, каждый с 2 щетинками. Ноги сильно хитинизированные,
коготки при основании имеют сглаженный зубец. Микроскульптура тела в виде
негустых овальных площадок, она заходит на склериты в виде тесно примыкаю-
щих друг к другу многоугольных площадок. Длина тела до 10 мм.
172
Рис. 67. Детали строения личинок подсемейства Galerucinae:
1 — микроскульптура стернитов Galerucella nymphaeae\ 2, 4 — 1-й тергит брюшка: 2 —
Atrachya menethriesi, 4 — Morphosphaera japonica, Cneorane violaceipennis'. 3 — заднеспин-
ка, 5 — 2-й тергит брюшка, 7—микроскульптура 9-го тергита брюшка; Galerucidaflavipennis'.
6 — 1-й тергит брюшка, 8 — заднеспинка.
Личинки на листьях бобовых, особенно сои; жуки — полифаги, но предпочи-
тают бобовые. Окукливание в почве. Серьезный вредитель. В регионе 1 вид — А.
menethriesi Falderman на юге ДВ.
18. Род Medythia Jacoby (JParaluperodes Ogloblin)
Личинки светло-желтые, со светло-бурой головой, переднеспинкой, 9-м терги-
том брюшка и ногами; склериты тела слабоокрашенные. Темя с 5-6 длинными и 4-
173
5 короткими щетинками. Эпикраниальный шов короткий, лоб с 6 длинными и 4
короткими щетинками, наличник слабо хитинизирован, с 6 очень мелкими щетин-
ками. Верхняя губа (рис. 68, 4) поперечная, на диске с рядом из 4 щетинок и 2 пор,
ее передний край слабо зазубрен, без срединной выемки, в середине с очень мелки-
ми густыми щетинками, по бокам с каждой стороны с 5 толстыми коническими
маргинальными щетинками. Усики 2-члениковые, с крупной конусовидной папил-
лой. Мандибулы с 3 зубцами. Склерит переднеспинки с рядом из 4 щетинок по
переднему и 2 — по заднему краям, эпиплевральный склерит небольшой, с 1 ще-
тинкой. Средне- и заднеспинка с 2 рядами склеритов, внутренние тергальные сли-
ваются в непарные с 2 щетинками каждый, наружные претергальные с очень мел-
кой плохо различимой щетинкой, посттергальные с 1 щетинкой, крыловые склери-
ты с 2 длинными и 4-5 очень мелкими щетинками. 1-7-й сегменты брюшка с 3
поперечными рядами склеритов, внутренние склериты первого и третьего рядов
сливаются в непарные склериты с 2 щетинками, второй ряд состоит из 2 широко
расставленных интеркалярных склеритов, каждый с 1 мелкой щетинкой, эпиплев-
ральные и гипоплевральные склериты с 2 щетинками. На 8-м тергите склериты
третьего ряда слиты в один, с 6 длинными щетинками; на 9-м тергите все склериты
слиты, с 12 крупными щетинками по краям (рис. 68, 5) и 6 мелкими на диске, пос-
ледний имеет очень густую зернистость и сливается с третьим рядом 8-го тергита.
Стерниты груди и брюшка слабоокрашенные. Голенелапка толстая, щетинки тон-
кие, заостренные, коготки- острые, слабо изогнутые, без выступа при основании
(рис. 68, б). Микроскульптура покровов тела редкая, в виде светлых овальных зе-
рен, заходит только на края склеритов. Длина тела до 5 мм.
Жуки питаются на листьях и плодах, а личинки — в корневых клубеньках сои.
В регионе 1 вид — М. suturalis nigrobilineatus Motschulsky на юге ДВ.
19. Род Monolepta Erichson
Личинки этого рода очень близки к роду Paraluperodes, отличаясь немногими
признаками. Верхняя губа с более явственным срединным выступом и 3 марги-
нальными щетинками с каждой стороны, а наружные дискальные щетинки при-
мерно вдвое длиннее внутренних (рис. 68, 7). Щетинки верхней стороны остроко-
нечные. Склериты тела едва намеченные, слабо хитинизированные; на 8-м тергите
брюшка посттергальные и эпиплевральные склериты не слиты с непарным пост-
тергальным склеритом (рис. 68, 2). 9-й тергит брюшка без зернистой скульптуры, с
8 маргинальными и 4 дискальными щетинками. Голенелапка сверху и сбоку с ко-
роткими толстыми, почти шиловидными щетинками (рис. 68, 3).
Личинки в почве на корнях бобовых, окукливание в почве. В регионе 10 видов,
из них 9 на юге ДВ. Известна личинка М. quadriguttata Motschulsky.
20. Род Agelastica Chevrolat
Тело личинки узкое, вытянутое, темно-бурое, голова и ноги темные. Эпикра-
ниальный шов длинный, темя у лобных швов с 3 щетинками с каждой стороны,
позади глаз с 5-6 щетинками; лоб с 6 длинными и 2 короткими щетинками и 2
порами. Наличник хитинизирован только по бокам основания, с рядом из 4 щети-
нок и 2 пор. Верхняя губа узкая, хитинизированная только в основании, передний
174
Рис. 68. Детали строения личинок родов Monolepta и Medythia\
Monolepta quadrimaculata\ 1 — верхняя губа, 2 — 8-й и 9-й тергиты брюшка, 3 — нога;
Medythia suturalis'. 4 — верхняя губа, 5 — 8-й и 9-й тергиты брюшка, 6 — нога.
край закругленный, без срединной выемки, с каждой стороны с 5 толстыми корот-
кими и 1 длинной тонкой маргинальными щетинками; диск с 4 длинными щетинка-
ми и 2 порами. Переднеспинка и тергиты средне- и заднегруди со светлой средин-
ной полосой, склерит переднеспинки с редкими щетинками по краям, эпиплевраль-
ный склерит с 1 маленькой щетинкой. Средне- и заднегрудь с 2 рядами склеритов,
внутренние претергальные склериты с 2 длинными и 2-3 короткими щетинками,
внутренние посттергальные с 2 длинными и 2-4 короткими, наружные претергаль-
ные с 5-6 короткими, наружные посттергальные с 2 длинными щетинками. Крыло-
вые склериты с 5 длинными и 6-7 мелкими щетинками. Сегменты брюшка (рис.
62, 77) с 3 рядами склеритов, внутренние тергальные сливаются в один непарный
склерит, каждый с 2 длинными и 3-8 мелкими щетинками, интеркалярные склери-
ты широко раздвинутые, с 1 щетинкой. Стигмальные склериты 1-8-го сегментов
брюшка с крупным отверстием кожных желез, 3-4 длинными и 3-5 короткими ще-
тинками. Эпиплевральные склериты бугровидно приподнятые, с 8-9 короткими и
175
длинными щетинками, гипоплевральные склериты с 2 длинными и 3-6 короткими
щетинками. На 9-м тергите все склериты сливаются в один непарный, несущий 24
маргинальных и 8-10 дискальных щетинок. На стернитах груди имеются ряды из
мелких склеритов, каждый с 1 мелкой щетинкой. Микроскульптура покровов тела
в виде вытянутых многоугольных площадок, тесно примыкающих друг к другу. Го-
ленелапка короткая, коготки в основании узкие, серповидно загнуты, без выступа.
Длина тела до 12 мм.
Молодые личинки скелетируют, а старших возрастов прогрызают в листьях
ольхи сквозные овальные отверстия. 3 личиночных возраста; окукливание в почве.
В регионе 2 вида.
Таблица для определения видов рода Agelastica
1(2). Наружные претергальные склериты брюшных сегментов редуцированы или
отсутствуют, на их месте только 1 мелкая щетинка. Гипоплевральные склериты с 2
длинными и 3-4 короткими щетинками (рис. 62, 72). На ольхе. ДВ..........
..........................................................A. coerulea Baly
2(1). Наружные претергальные склериты брюшных сегментов развиты, разбиты на
2 мелких склерита, каждый с 1 щетинкой. Гипоплевральные склериты с 2 длинны-
ми и 7-8 более короткими щетинками (рис. 62, 77). На ольхе. ЕЧ, СК, ЗСБ.
..........................................................A. alni Linnaeus
21. Род Morphosphaera Baly
Голова и ноги темно-бурые, тело, включая склериты, желтовато-белое. Эпик-
раниальный шов короткий, лоб с 4 щетинками у переднего края, темя с 8 длинными
щетинками, в том числе по 3 вдоль лобных швов. Глазки развиты. Мандибулы с 4
зубцами. Верхняя губа с 4 дискальными щетинками и широко закругленным пере-
дним краем, без срединной выемки. Эпиплевральные склериты переднеспинки с 1
щетинкой. Тергиты средне- и заднегруди с 2 рядами склеритов; все тергальные скле-
риты обособленные, но внутренние сильно сближенные; крыловой склерит с 3 мак-
ро- и 2-3 микрохетами. 1-8-й брюшные тергиты с 3 рядами склеритов и с дорсаль-
ными железами на стигмальных склеритах; внутренние посттергальные слиты в
непарный склерит, наружные претергальные отсутствуют, интеркалярные широко
расставленные, с 1 щетинкой, стигмальные разделены на передний и задний, с же-
лезой между ними, эпиплевральные с 4 макро- и 1 микрохетой, гипоплевральные
склериты с 2 макро- и 2 микрохетами (рис. 67, 4). Склериты низа явственные. Ко-
готки без явственного зубца, с крупным хелониумом. Личинка этого вида отличает-
ся слабо заметными склеритами тела и практически не выраженным срединным
склеритом переднеспинки. Длина тела до 13 мм.
Личинки развиваются в Японии на листьях фикусов; кормовое растение в Рос-
сии не выявлено. В регионе 1 вид — М. japonica Homstedt. На юге ДВ.
22. Род Sermylassa Reitter
Тело желтовато-белое, с желтыми склеритами и головой; края головкой капсу-
лы, наличник и ноги темно-коричневые. Эпикраниальный шов довольно короткий,
176
примерно в 4 раза короче длины лба по средней линии. Верхняя губа со срединной
выемкой на переднем крае. Мандибулы с 4 хорошо развитыми и 1 редуцированным
зубцом. Переднеспинка со спутанным рядом коротких щетинок вдоль краев, с 1-2
дискальными щетинками с каждой стороны; эпиплевральные склериты небольшие,
округленные, с 1 щетинкой; стигмальные склериты переднегруди с 1 щетинкой.
Внутренние претергальные склериты средне- и заднеспинки слитные, с 2 щетинка-
ми с каждой стороны; наружные претергальные склериты такой же величины, как
один внутренний, с 3 щетинками; посттергальные склериты имеют то же распо-
ложение и количество щетинок, как и претергальные; крыловые склериты с 5 ще-
тинками. 1-8-й тергиты брюшка с 3 рядами склеритов, из которых внутренние пре-
тергальные и посттергальные склериты попарно слитые.
Жуки и личинки питаются на подмареннике. Окукливание в почве. В роде 1
вид — 5. halensis Linnaeus, встречающийся в ЕЧ и на юге ЗСБ.
23. Род Agelasa Motschulsky
Тело желтое, с темно-бурой головой и более светлыми склеритами. Эпикрани-
альный шов короткий. Лоб с 8 макро- и 8 микрохетами, темя с 4 макро- и 1 микро-
хетой с каждой стороны. Верхняя губа с глубокой срединной выемкой, мандибулы
с 5 зубцами. Все сегменты груди и 1-9-й сегменты брюшка с парой защитных же-
лез, открывающихся в виде щелей в области крыловых и стигмальных склеритов.
Переднеспинка с 10 щетинками на переднем крае, 6 на заднем и 6 на диске, эпип-
левральный склерит с 2 щетинками, железа лежит между срединными и эпиплев-
ральными склеритами. Внутренние тергальные склериты средне- и заднеспинки
узко разделенные или с тенденцией к соединению, с 3—4 щетинками каждый, на-
ружные претергальные — с 1, посттергальные с 3 щетинками, крыловые — с 4,
эпиплевральные разделены на 2 мелких склерита, каждый с 1 щетинкой. Тергиты
брюшка с 3 рядами склеритов, внутренние пре- и посттергальные попарно слитые,
соответственно с 3—4 и 2 парами щетинок; наружные претергальные с 2, посттер-
гальные, престигмальные и интеркалярные с 1, постстигмальные с 2, эпиплевраль-
ные с 4 щетинками. 9-й тергит с 2 поперечными рядами щетинок (на середине дис-
ка и на заднем крае), с 10 в переднем и 18-20 в заднем ряду. Склериты нижней
стороны тела не развиты. Коготок тонкий, сильно изогнут, с большим хелониумом.
Длина тела взрослой личинки до 10 мм.
Личинки питаются листьями Actinidia. 3 личиночных возраста. Окукливание в
почве. В регионе 1 вид — A. nigriceps Motschulsky, на юге ДВ.
24. Род Galerucida Motschulsky
Тело светло-желтое, голова, ноги, склериты или склеритовидные площадки
вокруг оснований щетинок темно-бурые. Эпикраниальный шов короткий, лоб с 6
длинными щетинками, темя с 4 щетинками с каждой стороны. Верхняя губа со
срединной выемкой. Сегменты груди и 1-9-й сегменты брюшка несут железы. Пе-
реднеспинка на переднем крае с 14, на заднем с 8, на боках с 2 щетинками; эпип-
левральные склериты с 2 щетинками, имеется также маленький вторичный скле-
рит у передненаружного края срединного склерита. Внутренние пре- и посттер-
177
гальные склериты средне- и заднегруди (рис. 67, 8) попарно слитые, с 3—4 парами
щетинок, наружные претергальные с 1, посттергальные с 3 щетинками, постстиг-
мальные с 2 щетинками. На 1-7-м сегментах брюшка (рис. 67, 6) все внутренние
пре- и посттергальные склериты попарно слитые, с 3—4 парами щетинок, наружные
тергальные и интеркалярные с 1 щетинкой, пре- и постстигмальные с 3, эпиплев-
ральные с 6 щетинками. Длина тела до 16 мм. Личинки обитают на, листьях, име-
ются 3 личиночных возраста. Окукливание в почве. 3 вида на юге ДВ; известны
личинки двух обычных видов.
Таблица для определения видов рода Galerucida
1(2). Склериты тела светлые, плохо заметные (особенно у старших возрастов), на
верхней стороне с темными склеритовидными площадками вокруг оснований ще-
тинок. Вторичный склерит перед внутренним претергальным склеритом отсутствует.
Тело до 9 мм. На винограде. Юг ДВ....................G. flavipennis Solsky
2(1). Склериты тела явственные, хитинизированные. Перед внутренними претер-
гальными склеритами средне- и заднегруди имеется небольшой вторичный скле-
рит. Тело до 16 мм. На Polygonum. Юг ДВ...........G. bifasciata Motschulsky
25. Род Derocrepis Weise
Тело белое, тонкое и длинное, голова, склерит переднеспинки, ноги, после-
дний склерит брюшка светло-желтый, склериты крупные, не окрашены, слабо очер-
ченные и трудно различимые, щетинки длинные, редкие и светло-желтые.
Головная капсула (рис. 69, 7) слабо склеротизованная, овально-вытянутая, с
параллельными боковыми сторонами. Эпикраниальный шов короткий, лобные швы
слабо выражены, прямые, затылок с треугольной выемкой, эндокарина намечена
только в вершинной части, темя в задней части гладкое, без щетинок, с группой
пор, по бокам голова с длинными щетинками. Лоб гладкий, с 6 длинными щетинка-
ми и 4 порами, по границе с наличником более склеротизован и утолщен, наличник
узкий, по бокам с 2 порами. Верхняя губа (рис. 69,2) узкая, передний край прямой,
со слабым вершинным выступом, несущим мелкие щетинки, 4 дискальные щетин-
ки основного ряда и 2 поры расположены косым рядом, причем наружные в 2 раза
длиннее внутренних; 2 дискальные щетинки вершинного ряда короче предыдущих
и расположены вблизи переднего края губы, по углам переднего края расположено
по 5 маргинальных щетинок, причем одна из них длинная и плоская. Усики одно-
члениковые (рис. 69, 3), на вершине с очень крупной сосцевидной папиллой и од-
ной щетинкой. Мандибулы с 3 зубцами. Переднеспинка с 2 склеритами, слабо отде-
ленными друг от друга, каждый по переднему краю с 4, а по заднему с 2 длинными
остроконечными щетинками, эпиплевральный склерит переднеспинки с одной
щетинкой (рис. 69, 4). Склериты тергитов груди и брюшка крупные, на средне- и
заднегруди с 2, а на брюшке с 3 рядами склеритов. Внутренние пре- и посттергаль-
ные склериты средне- и заднегруди попарно слиты (рис. 69, 4), каждый с 2 щетин-
ками, наружные претергальные с одной короткой щетинкой, наружные пост- тер-
гальные с одной длинной и одной короткой щетинками. Крыловые склериты с 3
длинными и одной короткой щетинками. Все внутренние склериты сегментов брюш-
178
Рис. 69. Детали строения личинки Derocrepis rufipes'.
1 — голова; 2 — верхняя губа; 3 — усик; 4 — передне- и среднеспинка; 5 — 4-й тергит
брюшка; 6 — 4-й стернит брюшка; 7 — 9-й тергит брюшка; 8 — голенелапка.
ка попарно слиты (рис. 69, 5), с 2 длинными щетинками; эпиплевральный склерит
с 1 длинной и 2 короткими щетинками. На 8-м сегменте внутренние и наружные
посттергальные склериты сливаются в одну большую непарную пластинку, несу-
щую 4 длинные щетинки. 9-й тергит брюшка слабо склеротизованный, с явствен-
ной ячеистой микроскульптурой и рядом длинных щетинок у основания и верши-
ны (рис. 69, 7). Стерниты груди и брюшка слабо склеротизованы 69, 6). Ноги корот-
кие, коготки узкие, хелониум хорошо развит; голенелапка с крепкими шипиковид-
ными щетинками (рис. 69, 8). Длина тела личинки последнего возраста до 10 мм.
В России 2 вида, известна личинка Derocrepis rufipes Linnaeus (ЕЧ, СК, СБ).
Питание на бобовых.
26. Род Batophila Foudras
Тело тонкое и длинное, белое, голова, склерит переднеспинки и 9-й тергит
брюшка светложелтые, склериты тела различимы только при специальной обра-
ботке и окраске.
Эпикраниальный шов длинный, около трети длины лобного шва. Эпиплевраль-
ный склерит переднеспинки с 1 щетинкой. Наружные претергалые склериты сред-
не-и заднеспинки отсутствуют. Тергиты брюшка с тремя рядами склеритов, без от-
179
верстия желез, 9-й тергит с простой округленной вершиной и 6 парами длинных
щетинок примерно одинаковой длинны, из них 3 пары на диске (2,2, 2) и 3 пары по
краям.
В регионе 3 вида на розоцветных, известна личинка В. rubi Paykull (ЕЧ, ЕС,
ЗСБ), живущая на корнях малины и очевидно других розоцветных.
27. Род Lythraria Bedel
Личинка бледно-желтая, с рыже-желтыми головой и ногами. Глазков нет; эпик-
раниальный шов короткий, лоб шире своей длины, с 6 макрохетами, темя с 8 мак-
рохетами, усики двучлениковые, с небольшим сенсориальным придатком на вер-
шине 1-го членика, наличник без щетинок, верхняя губа со слабо хитинизирован-
ным передним краем, усаженным короткими щетинками; мандибулы с 5 зубцами,
нижнечелюстные щупики трехчлениковые, нижнегубные двучлениковые. Передне-
спинка на переднем крае с 10, на заднем с 6 макрохетами; средне- и заднеспинка со
слитыми попарно внутренними тергальными склеритами, наружные претергаль-
ные склериты без щетинок, наружные посттергальные склериты с 1 макрохетой (и с
зубчиком для разрезания хориона яйца), крыловые склериты с 2 макрохетами. Терги-
ты 1-7-го брюшных сегментов (рис. 71, 4) с 3 поперечными рядами склеритов; внут-
ренние тергальные склериты попарно слитые, прочие все с 1 макрохетой, тергит 9-го
сегмента брюшка с 10 макрохетами, стернит с широким склеритом, с 1 макрохетой,
коготки с большим хелониумом. Описание составлено по личинке 1-го возраста.
Жуки встречаются на сырых лугах и болотах на Lysimachia и Astragalus.
Единственный вид рода — L. salicariae Paykull (ЕЧ, СК, СБ).
28. Род Sangariola Jacobson
Тело светлое с темными головой, склеритом переднеспинки и 9-го тергита
брюшка, узкое и длинное (рис. 74, 7). Голова с недлинным эпикраниальныим швом,
лоб с 4 макрохетами у переднего края, наличник с 4 хетами (рис. 74, 2). Эпиплев-
ральный склерит переднеспинки с 1 щетинкой. Склериты последующих тергитов
неявственные, практически не хитинизированные. Средне- и заднегрудь с 2 рядами
склеритов, внутренние пре- и посттергальные склериты не слитые попарно, наруж-
ные претергальные отсутствуют, брюшные тергиты с 3 рядами склеритов, внутрен-
ние и интеркалярные склериты не слиты попарно (рис. 60, 4). 9-й тергит брюшка
умеренно хитинизирован, с округленной вершиной и с 4 парами щетинок, его по-
верхность редко пунктирована посередине (рис. 74, 3).
Личинки в почве внутри корней, питание жуков отмечено на лилейных. 1 вид,
5. punctatostriata Motschulsky.
29. Род Phyllotreta Chevrolat
Личинки желтые или желтоватые, удлиненные с темноокрашенными головой,
ногами и нередко буроватыми склеритами. Голова у минирующих видов (Ph.
nemorum) широкая, у прочих слегка удлиненная; лоб с 6 щетинками, темя с 3-5
макрохетами, эпикраниальный шов короткий, глазков нет; усики 2-члениковые, с
180
очень маленьким 2-м члеником и сравнительно крупным сенсориальным конусом;
мандибулы с 5 отчетливыми зубцами; нижнечелюстные щупики 3-члениковые, ниж-
негубные 2-члениковые. Средне- и заднеспинка без наружных претергальных скле-
ритов (исключение — Ph. vittula). Тергиты сегментов брюшка (рис. 71, б) с 3 попе-
речными рядами склеритов, промежуточные склериты всегда разделены широким
промежутком; тергит 9-го сегмента брюшка с округленным или снабженным од-
ним загнутым кверху коротким острием задним краем (рис. 72,14). Коготки с хоро-
шо развитым хелониумом.
Почти все виды питаются крестоцветными и резедовыми растениями, един-
ственным исключением является Ph. vittula Redt., которая развивается на злаках.
Большая часть видов, особенно в имагинальной стадии, сильно вредит культурным
растениям, уничтожая всходы; личинки питаются главным образом придаточными
корнями, реже внутренними частями стеблей или минируют листья. Окукливание в
почве.
В регионе около 35 видов; известны личинки только широко распространен-
ных видов.
Таблица для определения видов рода Phyllotreta
1(16). Вершина 9-го тергита брюшка более или менее широко закругленная, без
загнутого кверху острия. Наружные претергальные склериты средне- и заднеспин-
ки отсутствуют (рис. 71,5).
2(3). Внутренн ие посттергальные склериты 1-7-го тергитов брюшка широко рас-
ставленные (рис. 71, 6), на 8-м тергите сближенные и слиты с каждой стороны с
наружными посттергальными склеритами. Голова сравнительно узкая, лоб замет-
но длиннее своей ширины. Тергит 9-го сегмента брюшка с широко закругленным
выпуклым вершинным краем, уплощенный (рис. 72, 8). Окраска желтовато-белая,
голова бурая, со светлыми швами и передним краем лба, переднеспинка, ноги, скле-
риты тела и 9-й тергит брюшка светло буровато-рыжие. Личинки выгрызают ходы
в черешках листьев хрена. ЕЧ, СК, СБ................Ph. armoraciae Linnaeus
3(2). Внутренние посттергальные склериты 1-7-го тергитов брюшка попарно сли-
тые, образуют один непарный, несущий с каждой стороны 1 щетинку и 1 пору скле-
рит; на 8-м тергите все 4 посттергальных склерита слиты вместе.
4(5). Голова сравнительно широкая, лоб заметно шире своей длины. Тергит 9-го
сегмента брюшка (рис. 72, 9) с широко округленным задним краем, без вдавления.
Окраска светло-желтая, голова, переднеспинка и ноги темно-бурые, склериты тела
и 9-й тергит брюшка светло-бурые, отчетливо заметные даже при слабом увеличе-
нии. Минируют листья крестоцветных, из культурных крестоцветных повреждают
преимущественно корнеплодные. ЕЧ, СК, СБ, ДВ.........Ph. nemorum Linnaeus
5(4). Голова сравнительно узкая, лоб заметно длиннее своей ширины. Окраска бе-
лая или желтовато-белая, голова, переднеспинка и 9-й тергит брюшка светло-бу-
рые, склериты слабо пигментированы и плохо заметные.
6(7). 9-й тергит брюшка с явственным вдавлением и равномерно округленным зад-
ним краем, буроватый (рис. 72,10). Переднеспинка светлая. Щетинки на склеритах
тела сравнительно короткие. На корнях крестоцветных. ЕЧ, СК, СБ, ДВ......
....................................................Ph. undulata Kutschera
181
7(6). 9-й тергит брюшка без вдавления.
8(9). Переднеспинка светлая. 9-й тергит брюшка с толстыми щетинками и широко
закругленным задним краем (рис. 72, 77). На корнях крестоцветных. ЕЧ, СК, СБ,
ДВ.....................................................Ph. nigripes Fabricius
9(8). Переднеспинка темно-окрашенная.
10(13). 9-й тергит брюшка с довольно короткими тонкими щетинками и округлен-
ным задним краем.
11(12). 9-й тергит брюшка длиннее ширины, с узко округленным задним краем (рис.
72, 76). На корнях резеды и других крестоцветных. СК .... Ph. nodicornis Marsham
12(11). 9-й тергит брюшка шире длины, с широко округленным задним краем. На
корнях крестоцветных. ЕЧ, СК, СБ, ДВ......................................
....................................Ph. striolata Fabricius (Ph.vittata Fabricius)
13(10). 9-й тергит брюшка с длинными толстыми щетинками и широко
закругленным или притупленным задним краем.
14(15) 9-й тергит брюшка с очень толстыми щетинками и сильно притупленным,
почти обрубленным задним краем (рис. 72, 72). На корнях крестоцветных. ЕЧ, СК,
СБ..................................................Ph. tetrastigma Cornelius
15(14). 9-й тергит брюшка с умеренно толстыми щетинками и широко закруглен-
ным задним краем (рис. 72, 73). На корнях крестоцветных. ЕЧ, СК, СБ, ДВ...
.........................................................Ph. ochripes Curtis
16(1). Вершина 9-го тергита брюшка с узким, загнутым кверху зубцом (рис. 72,14).
17(18). Средне- и заднеспинка без наружных претергальных склеритов. На корнях
крестоцветных. Первый вид — транспалеаркт, второй — ЕЧ, СК, ЗСБ...........
........................................Ph. atra Fabricius, Pr. cruciferae Goeze
18(17). Средне- и заднеспинка с наружными претергальными склеритами. На кор-
нях крестоцветных и, возможно, злаков. Повсеместно в Палеарктике..........
....................................................Ph. vittula Redtenbacher
30. Род Liprus Motschulsky
Тело светлое с бурой головой, склеритом переднеспинки и 9-го тергита брюш-
ка, узкое и удлиненное (рис. 70, 7). Голова с умеренно длинным эпикраниальным
швом, лоб с 6 макрохетами в передней части, наличник с 4 хетами (рис. 70, 2).
Эпиплевральный склерит переднеспинки с 1 щетинкой. Склериты последующих
тергитов слабо хитинизированы, плохо различимы. Средне- и заднегрудь с 2 ряда-
ми склеритов, внутренние пре- и посттергальные склериты попарно слиты, наруж-
ные претергальные отсутствуют (рис. 60, 5), 9-й тергит брюшка сильно хитинизи-
рован, покрыт густой и мелкой пунктировкой, с 5 парами щетинок (рис. 70, 3).
Личинки обитают в почве на корнях Hosta и других лилейных. В России на юге
ДВ 1 вид, L. punctatostriatus Motschulsky.
31. Род Phygasia Dejean
Тело светлое с темной головой, узкое и удлиненное (рис. 74, 4). Голова с до-
вольно коротким эпикраниальным швом, лоб с 8 макрохетами в передней части,
наличник с 4 хетами (рис. 74, 5). Эпиплевральные склериты переднеспинки с 1
182
Рис. 70. Детали строения личинок подсемейства Alticinae:
1-3 — Liprus punctatostriatus'. 1 — общий вид, 2 — голова, 3 — 9-й тергит брюшка; 4-7 — 9-
й тергит брюшка Longitarsus*. 4 — L. linnei, 5 — L. melanocephalus, 6 — L. anchusae, 7 — L.
pratensis. По Кимото и Такизава, Бионди, Штейнхаузену.
щетинкой. Склериты последующих тергитов плохо различимы. Средне- и заднег-
рудь с 2 рядами склеритов, внутренние пре- и посттергальные склериты попарно
слиты, наружные претергальные отсутствуют (рис. 60, б). 9-й тергит брюшка уме-
ренно хитинизирован, покрыт пунктировкой, с округленной вершиной, 2 парами
длинных и 3 парами коротких щетинок (рис. 74, б).
Личинки обитают в почве. Жуки отмечены на черемухе и Cynanchus. В России
1 вид, Ph. fulvipennis Baly на юге ДВ.
32. Род Longitarsus Latreille
Личинки тонкие, сильно удлиненные, белые, с темноокрашенными головой,
ногами, пятнами на переднеспинке, тело их может укорачиваться и удлиняться бла-
годаря наличию сильно развитых межсегментных перепонок. Голова направлена
183
Рис. 71. Детали строения личинок подсемейства Alticinae:
Altica oleracecr. 1 — среднеспинка, 2 — 1-й тергит брюшка; Phyllotreta armoraciae'. 5 — задне-
спинка, 6 — 3-й тергит брюшка; 3 — 1-й тергит брюшка Hippuriphila modeeri', 4, 8, 9 — 2-й
тергит брюшка: 4 — Lythraria salicariae, 5 —Aphthona nonstriata, 9 — Longitarsus parvulus',
7,10, — 9-й тергит брюшка: 7 — L. jacobaeae, 10 — A. nonstriata.
184
вперед, уплощенная, ее затылочное отверстие лежит преимущественно на вентраль-
ной стороне, эпикраниальный шов короткий, лоб длиннее своей ширины, с 4 длин-
ными щетинками на переднем утолщенном крае, с 2 микрохетами и 4 мелкими
порами вдоль середины, теменные микрохеты по 1 с каждой стороны, глазки отсут-
ствуют, усики двучлениковые, с сильно развитой базальной мембраной, их 1-й чле-
ник короткий, широкий, на вершинной пленчатой части с группой сенсориальных
придатков, 2-й членик округло-конический; мандибулы с 5 зубцами, из которых 2-й
и 3-й (считая от внутреннего края) крупные, прочие мелкие. Переднеспинка с 8
макрохетами на переднем крае и с 6 на заднем, эпиплевральный склерит малень-
кий, с 1 щетинкой, внутренние тергальные склериты средне- и заднеспинки и 1-7-й
тергиты брюшных сегментов (рис. 71, 9) попарно слитые, с каждой стороны с 1
щетинкой, наружные претергальные склериты средне- и заднеспинки без щетинок,
тергиты брюшных сегментов с 3 поперечными рядами склеритов; промежуточные
склериты далеко раздвинутые, тергит 9-го сегмента брюшка на вершине округлен-
ный или с 1-2 загнутыми кверху короткими хитинизированными остриями.
Биология личинок плохо изучена, личинки известны лишь у немногих евро-
пейских видов. Жуки питаются листьями различных травянистых растений, личинки
грызут корни тех же растений; у ряда видов зимуют личинки. Окукливание в почве.
В регионе около 50 видов.
Таблица для определения видов рода Longitarsus
1(8). Вершинный край 9-го тергита брюшка с 1-2 короткими, загнутыми кверху
остриями.
2(3). Вершинный край 9-го тергита брюшка с 1 острием (рис. 70, 4). Личинки вбу-
равливаются в мясистые корни Symphytum. ЕЧ, СК.......L. linnaei Duftschmidt
3(2). Вершинный край 9-го тергита брюшка с 2 остриями (рис. 70, 5).
4(5). Диск 9-го тергита брюшка светлый. На корнях Echium. ЕЧ, СК... L. echii Koch
5(4). Диск 9-го тергита брюшка сплошь или хотя бы в вершинной половине черно-
бурый.
6(7). Переднеспинка с 4 темными пятнами. На корнях или в основании черешков
подорожника. ЕЧ, СК, СБ..........................L. melanocephalus Degeer
7(6). Переднеспинка с 5 неправильными темными полосками. На корнях лютико-
вых, также подорожника и Rhinantus. ЕЧ, СК, СБ...........L. luridus Scopoli
8(1). Вершинный край 9-го тергита брюшка без шиовидных отростков, округлен-
ный.
9(10). Диск 9-го тергита брюшка темно-бурый, с очень слабым поперечным вдавле-
нием перед вершиной (рис. 71, 7). На корнях Senecio. ЕЧ, СК, СБ.........
....................................................L. jacobaeae Wathouse
10(9). Диск 9-го тергита сплошь или в большей части светлый.
11(14). Склерит переднеспинки с многочисленными буроватыми пятнами.
12(13). Диск 9-го тергита с точечной зернистостью, без бурых краев. На корнях
губоцветных. ВСБ.................................L. transbaicalicus Ogloblin
13(12). Диск 9-го тергита с бурыми краями. Тело коренастое, в 5-6 раз длиннее
ширины. На корнях коровяка. ЕЧ, СК, СБ.................L. tabidus Fabricius
14(11). Склерит переднеспинки без явственных пятен.
185
15(26). Диск 9-го тергита в задней половине с о мелкими темными пятнышками
или с 1-2 ямками перед утолщенным задним краем.
16(19). Диск 9-го тергита в задней половине с мелкими темными пятнышками и
затемненным задним краем.
17(18). Задняя половина 9-го тергита темнее передней, светлобурая. На корнях мяты
........................................................L. lycopi Foudras
18(17). Задняя половина 9-го тергита не темнее передней. На корнях Ballota. ЕЧ,
СК......................................................L. ballotae Marsham
19(16). 9-й тергит с 2 небольшими ямками у вершинного края (рис. 70, б), без мел-
ких темных пятнышек в задней половине.
20(21). Тело узкое и длинное, примерно в 14 раз длиннее ширины. На корнях бурач-
никовых. ЕЧ, СК, СБ.....................................L. anchusae Paykull
21(20) Тело менее узкое и длинное, примерно в 5-9 раз длиннее ширины.
22(25). 9-й тергит с 1 большой плоской ямкой перед утолщенным светлобурым зад-
ним краем (рис. 70, 7).
23(24). На корнях Tussilago. Тело примерно в 9 раз длиннее ширины. ЕЧ, СК, СБ,
ДВ.................................................L. suturellus Duftschmidt
24(23) На корнях Plantago.Teno примерно в 5-6 раз длиннее ширины. ЕЧ, СК, ЗСБ
........................................................L. pratensis Panzer
25(22). 9-й тергит с 2 овальными плоскими ямками перед утолщенным светлобу-
рым задним краем. На корнях Plantago. ЕЧ, СК, СБ, ДВ....
.............................................L. lewisi Baly (£. scutellaris Rey)
26(15). 9-й тергит без вдавлений и затемненных участков у вершинного края.
27(28). На корнях бурачниковых, особенно Echium. ЕЧ, СК, СБ..............
........................................................L. exoletus Fabricius
28(27). На корнях льна, также Boraginaceae, особенно Pulmonaria. Вредит. ЕЧ, СК,
СБ......................................................L. parvulus Paykull
33. Род Crepidodera Chevrolat (Chalcoides Foudras)
Голова с очень коротким эпикраниальным швом, с каждой стороны с 5 темен-
ными и 4 лобными макрохетами, лоб заметно длиннее своей ширины; передний
край верхней губы без вырезки; мандибулы с 5 зубцами и с небольшим пучком
коротких щетинок на внутреннем крае, кардо нижних челюстей с короткой щетин-
кой на наружном крае. Переднеспинка с каждой стороны с 5 макрохетами на пере-
днем крае и с 3 макрохетами на заднем, эпиплевральные склериты переднеспинки
маленькие, с 1 макрохетой. Внутренние склериты средне- и заднеспинки попарно
слитые, с каждой стороны с 1 макрохетой, наружные претергальные склериты без
щетинок, наружные посттергальные с 2 щетинками (и с хитиновым зубцом для
разрезания хориона яйца), крыловые склериты с 3-4 щетинками. Тергиты 1-7-го
брюшных сегментов с 3 поперечными рядами склеритов, внутренние претергаль-
ные склериты попарно слитые (рис. 75, S); промежуточные, наружные тергальные
и престигмальные склериты с 1, постстигмальные с 2 щетинками; эпиплевральные
и гипоплевральные склериты с 2 щетинками.
В регионе встречаются 7 видов. Жуки питаются листьями ивы, осины и топо-
ля, личинки на корнях.
186
Таблица для определения видов рода Crepidodera
1(4). Крыловые склериты средне-и заднегруди с 3 щетинками. Тело светло-желтое
до желто-белого.
2(3). Стигмы бурые, резко отличаются от светлого тела. Тергиты брюшка шире длин-
ны (рис. 75, 8). Щетинки тела бурые. Транспалеаркт.........G aurata Marsham
3(2). Стигмы желтые, едва различимые на фоне тела. Тергиты брюшка длиннее
ширины (рис. 75, 9). Щетинки тела светло-желтые. Транспалеаркт......С. plutus
Latreille
4(1). Крыловые склериты средне-и заднегруди с 4 щетинками. ЕЧ, СК, СБ.......
.......................................................С. fulvicornis Fabricius
34. Род Podagrica Foudras
Тело удлиненное, без боковых выступов на брюшных сегментах. Голова с до-
вольно длинным эпикраниальным швом, длина лобной пластинки немного больше
ширины, вдоль срединной линии лба 2 пары макрохет и 2 пары пор, и у наружных
углов еще 1 пара макрохет; темя с 5 парами микрохет. Верхняя губа без выемки, с
пучком из 6-7 щетинок с каждой стороны переднего края и с 2 щетинками у осно-
вания (рис. 75, 7). Склерит переднеспинки с 6 макрохетами на переднем крае и с 6
на заднем, эпиплевральные склериты переднеспинки маленькие, с 1 макрохетой.
Средне- и заднеспинка с разделенными внутренними и наружными тергальными
склеритами, наружный претергальный склерит явственный, с 1 щетинкой. Брюш-
ные тергиты (рис. 75, 4) с 3 поперечными рядами склеритов; внутренние претер-
гальные склериты на 1-7-м сегментах разделенные, на 8-м слитые; внутренние по-
сттергальные на 6-8-м сегментах попарно слитые (на 8-м вместе с наружными по-
сттергальными), на 1-5-м разделенные (часто на 3-5-м неявственно слитые). 9-й
тергит с округленной вершиной и с вдавлеииен посередине, несет 14 макрохет (4,
4, 4, 2), у Р fuscicornis Linnaeus с 2 шипами на вершине.
Хенриксен (Henriksen, 1927) в описании личинки Р fuscicornis Linnaeus упо-
минает об отсутствии склеритов, которые в действительности хорошо различимы
после специальной обработки.
В регионе 3 вида на мальвовых (Malva, Althaea); личинки живут сначала на
мягкой коре корневой шейки, позже в стеблях. По данным Палия (1962), личинки Р
fuscicornis Linnaeus живут в соплодиях Althaea.
Таблица для определения видов рода Podagrica
1(2). 9-й тергит брюшка с парой шипов на заднем крае (рис. 75, 2). ЕЧ, СК...
.......................................................R fuscicornis Linnaeus
2(1). 9-й тергит брюшка без шипов.
3(4). Внутренние пре- и посттергальные склериты на всех сегментах попарно сли-
ты (рис. 75, 3). ЕЧ, СК........................................R malvae liliger
4(3). Внутренние претергальные склериты на груди и 1-7-м сегментах брюшка раз-
дельные, на 8-м сегменте попарно слиты (рис. 75, 4). ЕЧ, СК.................
......................................................R mcnthricsi Faldermann
187
3
Рис. 72. Детали строения личинок подсемейства Alticinae:
1,2 — голова: 1 —Aphthona euphorbiae, 2 — Dibolicr, 3 — мандибула Argopus bicolor, 4,5 —
заднеспинка: 4 — Mantura rustica, 5 — Asiorestia ferruginecr, 6 — 4-й тергит брюшка
Chaetocnema tibialis', Psylliodes chrysocephakr. 7 — заднеспинка и 1-й тергит брюшка, 15 —
9-й тергит брюшка; 8-14, 16 — 9-й тергит брюшка Phyllotreta: 8 — Ph. armoraciae, 9 — Ph.
nemorum, 10 — Ph. undulata, 11 —Ph. nigripes, 12 — Ph. tetrastigma, 13 — Ph. ochripes, 14 —
Ph. atra, 16 — Ph. nodicornis.
188
35. Род Psylliodes Latreille
Личинки желтовато-белые, удлиненные, с небольшой уплощенной, направлен-
ной вперед головой; голова, ноги и часто тергит 9-го сегмента брюшка темно-бу-
рые или рыжеватые, склериты тела у молодых личинок немного затемнены, у более
взрослых слабо заметные. Эпикраниальный шов короткий; темя с 10 макрохетами
и небольшим количеством микрохет; лоб с 6 макрохетами и 2-4 микрохетами; уси-
ки 2-члениковые, с сенсориальным придатком на вершине 1-го членика; наличник
с 6 короткими щетинками; глазки у взрослых личинок отсутствуют; мандибулы с 5
зубцами. Нижнечелюстные щупики 3-члениковые, нижнегубные щупики очень ко-
роткие, 2-члениковые. Переднеспинка с 10 макрохетами на переднем и с 6 на зад-
нем краях; эпиплевральные склериты маленькие, с 1 щетинкой. Средне- и заднес-
пинка (рис. 72, 7) с небольшими наружными претергальными склеритами, с чрез-
вычайно короткой щетинкой на них, иногда лишь с порой; внутренние тергальные
склериты попарно слитые. Тергиты брюшных сегментов (рис. 72, 7) с 3 поперечны-
ми рядами склеритов; внутренние тергальные склериты свободные или попарно
слитые, интеркалярные склериты широко расставленные. Тергит 9-го сегмента
брюшка с округленным или снабженным 2 короткими остриями задним краем (рис.
72,16). Коготки с большим хелониумом.
Личинки живут в почве, питаясь корнями растений и нередко вгрызаясь в них,
особенно у корневой шейки, или прокладывают ходы в стеблях растений. Окукли-
вание в почве. В регионе около 30 видов.
Таблица для определения видов рода Psylliodes
1(8). Вершинный край 9-го тергита брюшка простой, округленный. Внутренние
тергальные склериты 1-7-го тергитов брюшка попарно слитые. Сюда же относится
Р angusticollis Baly с юга ДВ, живущий на пасленовых.
2(3). Тергит 9-го сегмента брюшка с 10 (4, 4, 2) длинными щетинками; непарный
претергальный склерит 1-7-го тергитов брюшка не шире посттергального. Личин-
ки питаются корнями конопли, хмеля и крапивы и нередко вгрызаются в подзем-
ную часть стебля. Широко распространена по всей Палеарктике, кроме севера....
..........................................................R attenuata Koch
3(2). Тергит 9-го сегмента брюшка с 14 (4,4,4,2) короткими щетинками, непарный
претергальный склерит 1-7-го тергитов брюшка заметно шире посттергального.
4(5). Тергит 9-го сегмента брюшка с короткими щетинками. Молодые личинки пи-
таются мелкими придаточными корешками картофеля, взрослые личинки нередко
вгрызаются в корни. ЕЧ, СК, ЗСБ...........................R affinis Paykull
5(4). Тергит 9-го сегмента брюшка с длинными щетинками. Личинки минируют в
основании стеблей Solanum. ЕЧ, СК, СБ.
6(7). Тергиты с явственно различимыми склеритами. Тело желтое. В стеблях пасле-
новых. ЕЧ, СК, ЗСБ......................................R dulcamarae Koch
7(6). Тергиты с едва различимыми склеритами. Тело светлое, беложелтое. В стеб-
лях Carduus, Cirsium. ЕЧ, СК, СБ, ДВ....................R chalcomera liliger
8(1). Вершинный край 9-го тергита брюшка с 2 загнутыми кверху короткими хити-
низированными остриями (рис. 72, 75).
189
9(10). Тергальные склериты груди и брюшка неявственные. Тергит 9-го сегмента
брюшка не шире длины, с 1 парой дискальных щетинок. На крестоцветных. Саха-
лин, Курилы.............................................Р. punctifrons Baly
10(9). Тергальные склериты груди и брюшка хорошо различимы. Все три пары внут-
ренних тергальных склеритов 1-8-го тергитов брюшка не слиты попарно друг с
другом (рис. 72, 7), за исключением посттергальных на 8-м тергите. Лоб и темя с
немногочисленными микрохетами, наружные претергальные склериты средне- и
заднеспинки с микрохетой, эпиплевральные склериты брюшных сегментов с 1 мак-
ро- и 1 микрохетой. Тергит 9-го сегмента брюшка заметно шире длины, с 12-14
щетинками, в том числе с 2 парами дискальных. Личинки выгрызают ходы в стеб-
лях крестоцветных.
11(12). Голова, переднеспинка, склериты тела и 9-й тергит брюшка светлые, желто-
ватые, особенно у старших возрастов. Размеры меньше, ширина головной капсулы
личинок 1-3-го возрастов соответственно 0,18-0,22, 0,25-0,29 и 0,32-0,4 мм. ЕЧ,
СК, СБ......................................................Р. cuprea Koch
12(11). Голова, переднеспинка, склериты тела и 9-й тергит брюшка темные, бурые.
Размеры крупнее, ширина головной капсулы личинок 1-3-го возрастов соответствен-
но 0,25-0,33, 0,36-0,42 и 0,49-0,61 мм. ЕЧ, СК, СБ.Р. chrysocephala Linnaeus
36. Род Epithrix Foudras
Описание рода, данное ранее (Оглоблин, Медведев, 1971; Медведев, Зайцев,
1978) и составленное по кратким и недостаточно подробным описаниям американ-
ских видов, оказалось неточным. Ниже мы приводим его переописание на основе
морфологии личинки Epithrix pubescens Koch.
Тело белое, тонкое и длинное; голова, склерит переднеспинки, последний скле-
рит брюшка и ноги светло-желтые, склериты тела крупные, но слабо очерченные и
трудно различимые, щетинки длинные, остроконечные, светло-желтые.
Головная капсула (рис. 76, 7) слабо склеротизованная, овально-вытянутая, с
параллельными боковыми сторонами. Эпикраниальный шов короткий, лобные швы
едва намечены, прямые, затылок с треугольной выемкой, темя в задней части толь-
ко с порами, в передней части с длинными щетинками. Лоб с 6 длинными щетинка-
ми и 2 порами, на границе с наличником склеротизован в виде узкой полосы, на-
личник с 1 мелкой щетинкой и рядом из 4 пор. Верхняя губа (рис. 76, 2) в середине
переднего края со слабым выступом, несущим мелкие густые щетинки; на диске с 4
длинными щетинками и 2 порами.
Усики одночлениковые (рис. 76, б), на вершине с крупной конусовидной папил-
лой. Глазки не развиты. Мандибулы (рис. 76,5), с 4 зубцами, сбоку с порой и 2 щетин-
ками, с внутренней стороны с 2 толстыми щетинками. Лабио-максиллярный комп-
лекс слабо склеротизованный, нижнегубные щупики одночлениковые, максилляр-
ные трехчлениковые. Переднеспинка с гладкими склеритами, слабо отделенными друг
от друга, каждый по переднему краю с рядом из 4 длинных и 2 коротких щетинок, ее
эпиплевральный склерит с 1 щетинкой (рис. 76,3). Склериты тергитов груди и брюшка
крупные, плотно примыкают друг к другу, на средне- и заднегруди с 2, на брюшке с 3
рядами. Внутренние пре- и посттергальные склериты груди попарно слиты, с 2 длин-
ными щетинками, наружные претергальные — с 1 короткой, наружные посттергаль-
ные с 1 длинной и 1 короткой, а крыловые с 3 длинными и 1 короткой щетинками
190
(рис. 76, 3). На брюшке внутренние склериты попарно слиты (рис. 76, 4), каждый с
2 длинными щетинками, интеркалярные склериты отделены друг от друга, наруж-
ные пре- и посттергальные склериты с одной щетинкой, эпиплевральный с одной
длинной и 2 укороченными щетинками. На 8-м сегменте внутренние и наружные
посттергальные склериты сливаются в одну большую непарную пластинку. 9-й тер-
гит брюшка овально-вытянутый, слабо склеротизован только на вершине, по краю
с 12 щетинками (рис. 76, 9). Стерниты груди и брюшка развиты, но слабо склероти-
зованы, расположение склеритов на стернитах брюшка показано на рисунке 76, 7.
Микроскульптура покровов тела развита слабо. Голенелапка с короткими ши-
пиковидными щетинками (рис. 76, 8).
37. Род Chaetocnema Stephens
Личинки Chaetocnema s.str. белого цвета, с темно-бурыми головой, ногами,
переднеспинкой и 9-м тергитом брюшка, склериты сегментов светло-бурые. Голова
без глазков, со светлыми швами, длинным эпикраниальным швом; темя с 8 макро-
хетами; лоб с 6 макрохетами, немного шире своей длины; усики 2-члениковые, с
небольшим сенсориальным придатком на вершине 1-го членика. Мандибулы с 5
зубцами и пучком из 3 коротких и толстых хет на внутреннем крае близ основания.
Переднеспинка с 6 макрохетами на переднем и с 6 на заднем крае; эпиплевральные
склериты крупные, с 1 макрохетой. Средне- и заднеспинка с наружными претер-
гальными склеритами; крыловые склериты с 3 щетинками. Тергиты брюшных сег-
ментов с 3 поперечными рядами склеритов; внутренние тергальные склериты 1-6-
го тергитов брюшных сегментов разделенные, эпиплевральные и гипоплевраль-
ные склериты с 2 щетинками; тергальные склериты с 1 щетинкой, тергит 9-го сег-
мента брюшка с округленным вершинным краем. Коготки с большим хелониумом.
Личинки подрода Tlanoma отличаются от номинального подрода прежде всего
наличием на заднем крае 9-го тергита брюшка 2 загнутых кверху шипиков; кроме
того, эпикраниальный шов короче, лоб с 8 макрохетами, внутренние тергальные
склериты 1-8-го тергитов брюшка слиты попарно (рис. 72,6), эпиплевральные скле-
риты с 3 щетинками.
Личинки живут в почве на корнях растений или внутри стеблей. В регионе
около 30 видов.
Таблица для определения видов рода Chaetocnema
1(4). Вершинный край 9-го тергита брюшка простой, округленный. Внутренние
тергальные склериты 1-6-го тергитов брюшка не слиты друг с другом (подрод
Chaetocnema s.str.). Личинки выгрызают ходы внутри молодых стеблей пшеницы,
ржи, ячменя, овса и дикорастущих злаков. Длина личинки до 5 мм.
2(3). Внутренние претергальные и посттергальные склериты средне- и заднеспин-
ки попарно слиты; внутренние посттергальные склериты тергитов брюшка почти
вдвое крупнее интеркалярных склеритов. Вся Палеарктика, кроме севера...
....................................................Ch. hortensis Geof&oy
3(2). Внутренние пре- и посттергальные склериты средне- и заднеспинки разделен-
ные; внутренние посттергальные склериты тергитов брюшка такой же величины,
как и интеркалярные склериты. ЕЧ, СК, СБ, ДВ........Ch. aridula Gyllenhal
191
Рис. 73. Детали строения личинок родов Hermaeophaga и Mniophila:
1-3 — Hermaeophaga: 1 — голова, 2 — тергиты груди и 1-го тергита брюшка, 3 — вершина
брюшка; 4-7 — Mniophila: 4 — голова, 5 — тергиты груди, 6 — 1-й тергит брюшка, 7 —
вершина брюшка. По Коксу и Вельх.
4(1). Вершинный край 9-го тергита брюшка с 2 загнутыми кверху короткими ост-
риями. Все три пары внутренних склеритов 1-8-го тергитов брюшка слиты попар-
но (рис. 72, 6). Лоб и темя без микрохет, наружные претергальные склериты сред-
не- и заднегруди с макрохетой, эпиплевральные склериты брюшных сегментов с 3
макрохетами, 9-й тергит брюшка с 14 щетинками (подрод Tlanoma Motschulsky).
Личинки на корнях маревых. ЕЧ, СК, СБ...................Ch. tibialis liliger
192
38. Род Hermaeophaga Foudras
Тело узкое и длинное, светлое. Голова (рис. 73, 7) овальная, эпикраниальный
шов умеренно длинный, лобные швы расходятся под прямым углом. Верхняя губа
без выемки, с 2 парами дискальных хет и группой микрохет на переднем крае. На-
личник с 3 парами хет, лоб с 4 парами хет разной длины, темя с 6 парами длинных
хет, из них 2 за усиками. Усики 2-члениковые с крупной папиллой, глазки отсут-
ствуют. Мандибулы с 5 зубцами, но у взрослой личинки зубцы 3 и 4 слиты, образуя
единый широкий зубец. Максиллы и нижняя губа типичного для Alticinae строе-
ния, челюстной щупик 3-члениковый. Переднеспинка с 5 парами длинных щети-
нок вдоль переднего края и с 3 парами за серединой. Средне-, заднеспинка и брюш-
ные тергиты (рис. 73, 2) без склеритов, но с соответствующими им бугорками по-
стоянной формы, образующими 2 поперечных ряда на каждом тергите, бугорки в
каждом ряду выглядят слитыми и несут единичные длинные щетинки. 9-й тергит
полукруглый, с 1 длинной и 2 короткими щетинками на каждой стороне и полу-
круглой ямковидной структурой с каждой стороны перед вершиной (рис. 73,3). Ноги
типичного строения, пульвиллы большие. Длина взрослой личинки около 6 мм.
Известна личинка Н. mercurialis Fabricius, живущая на корнях Mercurialis и
вгрызающаяся внутрь корней. ЕЧ, СК. В России 1 вид.
39. Род Altica Geoffroy (Haltica Fabricius)
Личинки темноокрашенные, с хорошо развитыми склеритами, несущими до-
вольно длинные щетинки. Голова с каждой стороны с 1 глазком; эпикраниальный
шов короткий, темя с 10, лоб с 6 макрохетами, усики двучлениковые, с крупным
сенсориальным придатком на наружной стороне вершинной части 1-го членика,
верхняя губа с 4 дискальными щетинками, слабо хитинизированным передним кра-
ем, на котором с каждой стороны находится по 2 более крупных, ланцетовидных,
слегка изогнутых хеты и 2-3 маленьких; мандибулы с 5 зубцами; кардо нижних
челюстей снаружи с небольшой щетинкой. Тергиты грудных и брюшных сегмен-
тов с двумя поперечными рядами склеритов (рис. 71,7, 2), большинство из которых
несут по 1 макрохете; передний край переднеспинки с 10, задний с 4 и боковые
края с 1 макрохетой; покров тела между склеритами с довольно густой микроскуль-
птурой. Коготки с хорошо развитым хелониумом. 9-й тергит брюшка без зубцов.
Жуки и личинки питаются листьями различных травянистых, реже древесных
растений, окукливание происходит в поверхностном слое почвы.
В регионе около 17 видов. Биология и личинки изучены плохо, главным обра-
зом вследствие большой трудности определения видов.
Таблица для определения видов рода Altica
1(10). Эпиплевральные склериты брюшных сегментов с 2 длинными щетинками и
1 микрохетой или порой.
2(3). Микрохета эпиплевральных склеритов очень короткая, часто едва различи-
мая. На Epilobium. ЕЧ.............................A. impressicollis Reiche
3(2). Микрохета эпиплевральных склеритов длинная, явственная.
193
4(7). Хелониум всегда белый. Верхняя губа короче, светлая только на переднем крае
(рис. 75, 5). Щетинки верха очень светлые, беложелтые, просвечивающие.
5(6). Расстояние между внутренними и наружными посттергальными склеритами
7-го брюшного тергита очень малое, так что склериты почти касаются друг друга.
Тело более склеротизовано, темнобурое, ноги бурые. На Epilobium. ЕЧ, СК..
.............................................................A. lythri Aube
6(5). Расстояние между внутренними и наружными посттергальными склеритами
7-го брюшного тергита шире, хотя бы на ширину наружного склерита. Ноги бурые.
На винограде. Юг ЕЧ, СК..........................................A. ampelophaga Guerin
7(4). Хелониум бурый или каштаново-бурый с желтым краем. Верхняя губа длин-
нее, в значительной части светлая. Щетинки темнее, светло-рыжие или коричнева-
тые.
8(9). Светлая часть верхней губы рыже-желтая, поперечная, охватывает переднюю
половину (рис. 75, 7). На березе. ЕЧ....................A. aenescens Weise
9(8). Светлая часть верхней губы беловатая, треугольная, охватывает значительную
часть губы (рис. 75, 6). На дубе. ЕЧ, СК.............A. quercetorum Foudras
10(1). Эпиплевральные склериты брюшных сегментов с 2 длинными щетинками,
но без микрохеты и поры.
11(18). Крыловые склериты груди с 2-3 порами, несущими микрохеты. На лещине.
ЕЧ, СК.................................................A. brevicollis Foudras
12(11). Крыловые склериты груди с 1 порой, несущей микрохету. Следующие виды
почти не различимы морфологически.
13(16). Стернальный склерит брюшных стернитов поперечный, почти вдвое шире
длины.
14(15). Микроскульптура верха густая, темнобурая, тело выглядит темным. На
Myricaria, Tamarix, иногда на Hippophae и иве. ЕЧ, СК, СБ .... Л. tamaricis Schrank
15(14). Микроскульптура верха менее густая, светлобурая, тело выглядит светлобу-
рым.
16(17). Расстояние между наружными пре- и посттергальными склеритами брюш-
ных тергитов вдвое больше поперечника склерита. Питание на Carduus. ЕЧ, СК,
СБ, ДВ.................................................A. carduorum Guerin
17(16). Расстояние между наружными пре- и посттергальными склеритами брюш-
ных тергитов примерно равно поперечнику склерита. Питание на Oenotheraceae,
Polygonaceae. Транспалеаркт............................A. oleracea Linnaeus
18(11). Стернальный склерит брюшных стернитов слабо поперечный, в 1,5 раза
шире длины. На Sanguisorba. ЕЧ, СК, СБ...................................
.................................A. helianthemi Allard (A. pusilia Duftschmidt)
40. Род Hippuriphila Foudras
Окраска грязновато-серая, голова, ноги и склериты темно-бурые. Лоб шире
своей длины, с 6 макрохетами, темя с 6 макрохетами; эпикраниальный шов доволь-
но короткий, наличник с 6 щетинками, верхняя губа с очень слабо хитинизирован-
ным передним краем, усаженным небольшим количеством коротких щетинок, ман-
дибулы с 5 зубцами, их наружный край с очень короткой щетинкой близ основания,
внутренний край недалеко от основания с пучком из трех коротких толстых хет,
194
нижнечелюстные шипики трехчлениковые, нижнегубные двучлениковые, кардо
нижних челюстей снаружи с очень короткой щетинкой. Передний край переднес-
пинки с 8, задний с 6 макрохетами, внутренние претергальные склериты слитые,
наружные отсутствуют, крыловые склериты средне- и заднеспинки с 2 макрохета-
ми, прочие тергальные склериты с 1 макрохетой. Тергиты 1-7-го сегментов брюш-
ка (рис. 71, 3) с 2 поперечными рядами склеритов, между обоими рядами среди
микроскульптуры находятся 2 очень мелкие щетинки (рудименты интеркалярных
склеритов); расстояние между претергальными склеритами не менее чем втрое шире
промежутка между внутренними посттергальными склеритами, тергит 9-го сегмента
брюшка с 9 маргинальными и 6 дискальными щетинками. Покров тела между скле-
ритами на тергитах с частой, на стернитах с более редкой микроскульптурой, со-
стоящей из буроватых щитков разной величины и формы.
В регионе 2 вида, обычна Н. modeeri Linnaeus, встречающаяся по всей лесной
зоне ЕЧ и СБ.
41. Род Mantura Stephens
Приводимая ниже краткая характеристика рода составлена по описаниям аме-
риканских видов.
Голова с коротким эпикраниальным швом, мандибулы с 4 зубцами, нижнегуб-
ные щупики имеются, подбородок с 2 парами щетинок. Склерит переднеспинки с
немногочисленными щетинками по краям, эпиплевральный склерит переднеспин-
ки с 1 щетинкой. На средне- и заднеспинке (рис. 72, 4) наружные претергальные
склериты отсутствуют, внутренние тергальные раздельные, с 1 щетинкой, наруж-
ные посттергальные с 1, крыловые с 3 щетинками. Брюшные тергиты с 2 попереч-
ными рядами склеритов, которые не сливаются друг с другом, за исключением внут-
ренних посттергальных 8-го сегмента; все тергальные склериты несут по 1 щетин-
ке. 9-й тергит с округленным наружным краем, без шипиков, по крайней мере, с 8
щетинками, из них 2 дискальных.
Виды этого рода связаны главным образом с гречишными. Личинки минируют
листья. В регионе 3 вида, обычна М. rustica Linnaeus на Polygonum и Rumex (ЕЧ,
СК, СБ, ДВ).
42. Род Mniophila Stephens
Личинки светло-желтые с желтыми склеритами, несущими на верхней сторо-
не длинные, более или менее булавовидные щетинки. Голова (рис. 73, 4) с каждой
стороны с одним глазком, эпикраниальный шов умеренно длинный, темя с 8 макро-
и 2 микрохетами, лоб с 6 микрохетами, усики двухчлениковые с большим коничес-
ким сенсориальным придатком на первом членике. Верхняя губа с 4 дискальными
щетинками, ее передний край с 2 парами микрохет между ними. Мандибулы с 5
зубцами и с пучком щетинок у основания разной длины и толщины. Кардо и стипес
несут по 1 щетинке. Склерит переднеспинки с 8 парами щетинок. Средне-, заднег-
рудь и тергиты брюшка с двумя поперечными рядами склеритов. На груди (рис. 73,
5) наружные претергальные склериты с 1 щетинкой, наружные посттергальные сли-
ты с крыловыми, с 2 щетинками, эпиплевральные передний и задний имеют по 1
195
щетинке. На тергитах брюшка (рис. 73, б) наружные претергальные склериты с 1
щетинкой, наружные посттергальные слиты с задними стигмальными, начиная с 8-
го тергита, склериты сливаются (рис. 73, 7). Покровы между склеритами без мик-
роскульптуры. Коготки с хорошо развитым хелониумом. Личинки очень мелкие,
длина менее 1,5 мм у первого возраста.
Известна только личинка первого возраста, которая, предположительно, откры-
тоживущая и питается на мхах, так же как и жуки. В России 1 вид — М. muscorum
Koch на СК.
43. Род Asiorestia Jacobson (Crepidodera Stephens)
Личинка желтовато-белая, с темно-бурыми головой, ногами, склеритами груд-
ных и брюшных сегментов и тергитом 9-го сегмента брюшка. Эпикраниальный
шов по длине равен одной трети длины головы, лоб не длиннее своей ширины, с 6
макрохетами, темя с 10 макрохетами, глазки имеются лишь у личинки 1-го возрас-
та; усики двучлениковые, мандибулы с 5 зубцами, нижнечелюстные щупики трех-
члениковые, нижнегубные двучлениковые. Переднеспинка с 10 щетинками на пе-
реднем и с 6 на заднем крае. Внутренние тергальные склериты средне- и заднес-
пинки и 1-7-го сегментов брюшка попарно слитые, в виде непарных сильно попе-
речных склеритов; наружные претергальные склериты средне- и заднеспинки без
щетинок; наружные посттергальные склериты средне- и заднеспинки с 1 макро- и 2
микрохетами, крыловые склериты с 2 макро- и 1 микрохетой (рис. 72, 5). Наруж-
ные тергальные и стигмальные склериты 1-7-го сегментов брюшка с 1, эпиплев-
ральные и гипоплевральные склериты с 2 щетинками, тергит 9-го сегмента с 8 мар-
гинальными и 4 дискальными щетинками, с ячеистой скульптурой.
Жуки встречаются на сырых лугах, болотах, а также на посевах злаков, пита-
ются на злаках, реже на бобовых, щавелях, сложноцветных, крапиве. Основными
кормовыми растениями личинок являются, по-видимому, злаковые. Молодые ли-
чинки вбуравливаются в молодые стебли или в проростки злака и выгрызают в них
продольный ход. Личинки зимуют и окукливаются весной следующего года; неред-
ко вредят посевам культурных злаков.
В регионе окодо 10 видов. Известна личинка широко распространенного в ЕЧ,
СК, СБ. вида A. ferruginea Scopoli.
44. Род Aphthona Chevrolat
Личинки белые, с темными головой, ногами, переднеспинкой и тергитом 9-го
сегмента брюшка, задний край которого округленный или с 2 загнутыми кверху
короткими остриями (рис. 71, 70). Тело сильно удлиненное, тонкое. Лоб с 6 макро-
хетами, почти одинаковой длины и ширины, эпикраниальный шов короткий (рис.
72, 7), нижнегубные щупики двучлениковые; подбородок с 4 щетинками, кардо
нижних челюстей с 1 короткой щетинкой снаружи.
Переднеспинка на переднем крае с 8, на заднем с 6 макрохетами. Склериты
тела прозрачные, плохо заметные, эпиплевральный склерит переднеспинки с 1
щетинкой, наружные претергальные склериты средне- и заднеспинки отсутствуют,
внутренние пре- и посттергальные склериты на всех сегментах попарно слитые
196
(рис. 71, 8), тергит 9-го сегмента брюшка с 4 дискальными и 8 краевыми щетинка-
ми (рис. 71, 70). Ноги сравнительно короткие, хелониум немного длиннее коготка.
Личинки питаются теми же растениями, на которых встречаются жуки, либо
минируя листья, либо подгрызая тонкие придаточные корни. Окукливание в почве.
В регионе около 40 видов. Личинки известны только для двух европейских видов.
Таблица для определения видов рода Aphthona
1(4). Задний край тергита 9-го сегмента брюшка с 2 широко расставленными, заг-
нутыми кверху хитиновыми остриями (рис. 71,10); передний край переднеспинки
с каждой стороны и крыловые склериты грудных сегментов с 1 микрохетой.
2(3). Переднеспинка бурая со светлыми пятнами по бокам. Теменной шов корот-
кий, около 1/4 лобного шва. Тергит 9-го сегмента брюшка с очень короткими ост-
риями. На Iris, Anabasis. ЕЧ, СК, СБ, ДВ..............A. semicyanea Allard
3(2). Переднеспинка одноцветно бурая. Теменной шов длинный, около 1/2 лобного
шва. Тергит 9-го сегмента брюшка с более длинными остриями. Личинки миниру-
ют толстую базальную часть листьев Iris; на болотах. ЕЧ, СК, ЗСБ.......
...................................A. nonstriata Goeze (A. coerulea Geoffrey)
4(1). Задний край тергита 9-го сегмента брюшка закругленный, без хитиновых от-
ростков; передний край переднеспинки без микрохет; крыловые склериты грудных
сегментов с 2 микрохетами.
3(4). Передний край тергита переднеспинки с каждой стороны с одной микрохетой
между 3-й и 4-й макрохетами. Крыловые склериты средне- и заднегруди с 2 макро-
хетами и 1 микрохетой (рис. 75, 77). На молочае. ЕЧ, СК.A. cyparissiae Koch
4(3). Передний край тергита переднеспини без микрохеты между 3-й и 4-й макро-
хетами.
5(6). Крыловые склериты средне- и заднегруди с 2 макрохетами и 2 микрохетами
(рис. 75, 10). Личинки живут в почве, питаются придаточными корнями льна; вре-
дят посевам, отмечен также на молочае. ЕЧ, СК, СБ...A. euphorbiae Schrank
6(5). Крыловые склериты средне- и заднегруди с макрохетой и 1 микрохетой. На
молочае. ЕЧ, СК....................................A. venustula Kustschera
45. Род Nonarthra Baly
Тело продолговатое, умеренно широкое, светлое с темными головой, переднес-
пинкой, склеритами и 9-м тергитом (рис. 74, 7). Голова с коротким, но явственным
эпикраниальным швом, темя с 2 парами, лоб с 3 парами длинных щетинок (рис. 74,
8) склерит переднеспинки с длинными щетинками в передней половине, ее эпип-
левральный склерит с 2 щетинками. Средне- и заднеспинка с 2 рядами склеритов,
внутренние тергальные склериты не слитые, наружные претергальные склериты
развиты (рис. 60, 7). Брюшные тергиты с 2 рядами склеритов, несущих по 1 щетин-
ке, внутренние пре- и посттергальные склериты попарно слитые, наружные пре-
тергальные отсутствуют, наружные посттергальные с очень длинной щетинкой.
Грудные и брюшные сегменты с отверстиями защитных желез. 9-й тергит довольно
короткий (рис. 74, 9), с 6 парами щетинок.
В России 1 вид — N. cyanea Baly на юге ДВ, жуки обычно на цветах, личинки
питаются на мхах.
197
Рис. 74. Детали строения личинок подсемейства Alticinae:
1—3 — Sangariola punctatostriata'. 1 — общий вид, 2 — голова; 3 — 9-й тергит брюшка; 4-6 —
Phygasia fulvipennis\ 4 — общий вид, 5 — голова, 6 — 9-й тергит брюшка; 7-9 — Nonarthra
суапеа: 7 — общий вид, 8 — голова, 9 — 9-й тергит брюшка; 10—11 — Philopona vibex 10 —
общий вид, 11 — голова; 12-13 — Hemipyxis plagioderoides*. 12 — общий вид, 13 — голова;
14 — Pentamesa, общий вид. По Кимото и Такизава, Лопатину и Дашевской.
46. Род Philopona Erichson
Тело уплощено дорсовентрально, светлое, без склеритов. Эпикраниальный шов
короткий, лоб с 6 щетинками (2,4), наличник без щетинок, верхняя губа с 5 щетин-
ками (рис. 74, 77). Глазки отсутствуют. Бока всех грудных и брюшных сегментов
вытянуты в тонкие пальцевидные отростки, оканчивающиеся каждый 1-3 толсты-
ми щетинками. Тергиты груди и брюшка с 6 щетинками каждый; 2 в переднем и 4 в
заднем ряду (рис. 60, 9). Личинки минируют листья.
198
Краткая характеристика рода составлена по описаниям американских видов.
Единственный в регионе вид — Ph. vibex Erichson встречается на юге ДВ, был
отмечен на подорожнике.
47. Род Hemipyxis Dejean (Sebaethe Baly)
Тело продолговатое, умеренно широкое, светлое с темной головой (рис. 74,
72). Голова с коротким эпикраниальным швом и немногочисленными короткими
щетинками (рис. 74, 73). Переднеспика с короткими щетинками по краям, сидя-
щими на небольших округлых склеритовидных площадках, ее эпиплевральные
склериты с одной щетинкой. Все сегменты тела с боковыми выростами. Средне-
и заднеспинка с 2 рядами небольших не слитых склеритов; наружные претер-
гальные склериты отсутствуют (рис. 60, 8). Брюшные тергиты с 2 рядами терги-
тов, внутренние претергальные попарно слиты, наружные претергальные отсут-
ствуют, обе пары постергальных склеритов раздельные. Все склериты несут по 1
короткой щетинке. Последний тергит брюшка своеобразной формы, благодаря
выростам (рис. 74, 72).
В регионе 1 вид — Н. plagioderoides Motschulsky на юге ДВ. Жуки отмечались
на Veronica, Clerodendron, Lamium.
48. Род Argopus Fischer
Личинки белые, с рыжими головой, переднеспинкой, переднегрудью и нога-
ми; тело уплощенное, с очень короткими, почти незаметными щетинками; склери-
ты обособлены преимущественно на переднеспинке. Голова уплощенная, с направ-
ленным вперед ротовым отверстием, с очень глубоко овально вырезанным верх-
ним краем затылочного отверстия и соответственно сильно оттянутыми узкими зад-
ними отростками головной капсулы (рис. 61,2); лоб немного шире своей длины, с
6-8 мелкими порами (с очень короткими щетинками), лобные швы не сходящиеся,
в задней половине почти параллельные, оканчиваются на краю затылочного отвер-
стия, эпикраниальный шов отсутствует. Усики двучлениковые, 1-й членик их почти
вдвое длиннее своей ширины, 2-й очень короткий. Верхняя губа (рис. 61,4) широ-
кая, с глубоко и широко выемчатым передним краем, с округленными боковыми
краями, усаженными очень длинными, частыми, изогнутыми хетами, частично за-
нимающими и передний край. Мандибулы с 3 крупными и 1 маленьким зубцами,
внутренний их край близ основания с короткими щетинками (рис. 72, 3). Нижне-
губные щупики отсутствуют. Средне- и заднеспинка с поперечной бороздкой, с 4
претергальными и 4 посттергальными очень короткими щетинками, крыловые скле-
риты отчетливые, с 4 очень короткими щетинками. Брюшные сегменты на боках с
довольно длинными пальцевидными отростками, их тергиты с 4 претергальными и
6 посттергальными очень короткими щетинками. Покров тела с очень мелкой бу-
горчатой микроскульптурой.
Личинки минируют листья, окукливание в почве. Род связан с ломоносом
(Clematis), в регионе 6 видов.
199
Рис. 75. Детали строения личинок подсемейства Alticinae:
1-4 — Podagrica'. 1 — верхняя губа Р menetriesi, 2 — вершина брюшка Р fuscicomis, 3 — 1-й
тергит брюшка Р malvae, 4 — 7-8-й тергиты брюшка Р menetriesi', 5-7—верхняя губа Alticcr.
5—A. lythri, 6—A. quercetorum, 7—A. aenescens; 8-9— 1-й тергит брюшка: 8 — Crepidodera
aurata', 9—С. plutus; 10—11 — заднеспинка Aphthona’. 10—A. euphorbiae; 11 —A. cyparissiae.
По Штейнхаузену, Бартковской и Вархаловскому и ориг.
Определительная таблица видов рода Argopus
2(1). Сегменты брюшка с длинными пальцевидными выростами по бокам. На
Clematis, Pulsatilla. ЕЧ, СК...................A. bicolor Fischer-Waldheim
1(2). Сегменты брюшка с небольшими треугольными выступами по бокам. На
Cirsium. Юг ДВ, Сахалин, Курилы................A. punctipennis Motschulsky
3(4). Пальцевидные выросты округлены на вершине. На Clematis, Pulsatilla. ЕЧ,
СБ, ДВ................................A. nigritarsis Gebler (A. clypeatus Baly)
49. Род Sphaeroderma Stephens
Личинки чрезвычайно сходны с родом Sphaeroderma, отличаясь только фор-
мой и хетотаксией верхней губы (рис. 61, 7), хотя эти различия установлены по
единичным видам и вплследствии могут не подтвердиться. Личинки минируют
листья, окукливание в почве. Питание на Cirsium и Carduus. В регионе 5 видов, из
них 3 на юге ДВ.
200
о
о
Рис. 76. Детали строения личинки Epithrix pubescens\
1 — голова; 2 — верхняя губа; 3 — передне- и среднеспинка; 4 — 4-й тергит брюшка; 5 —
мандибула; 6 — усик; 7 — 4-й стернит брюшка; 8 — голенелапка; 9 — 9-й тергит брюшка.
Определительная таблица видов рода Sphaeroderma
1(2). Виды ЕЧ и СК. Сегменты брюшка с небольшими выступами по бокам. На
сложноцветных. Среднеспинка без склеритов..................................
.......................................5. testacea Fabricius, 5. rubidum Graels
2(1). Виды юга ДВ.
3(4). Среднеспинка с 2 парами склеритов, среднегрудь с сильно хитинизирован-
ным непарным склеритом. Сегменты брюшка с небольшими выступами по бокам
..........................................................S. unicolor Kimoto
201
4(3). Среднеспинка без склеритов, среднегрудь без непарного склерита. Сегменты
брюшка с треугольными выростами по бокам...................5. balyi Jacoby
50. Род Pentamesa Harold
Тело уплощенное, с короткими немногочисленными щетинками (рис. 74, 14).
Голова уплощена, прогнатическая, с очень глубоко вырезанным верхним краем за-
тылочного отверстия и оттянутыми задними отростками, лоб доходит до затылоч-
ного отверстия, с 6 короткими щетинками, лобные швы не сходящиеся, эпикрани-
альный шов отсутствует. Усики одночлениковые. Верхняя губа широкая, глубоко и
широко выемчатая на переднем крае, усажена по краям очень длинными и густыми
хетами, ее диск с 8 щетинками (рис. 77, 2). Мандибулы с 3 крупными зубцами и
группой хет снизу близ основания. Нижнечелюстные щупики 3-члениковые, ниж-
негубные щупики отсутствуют. Тергиты средне- и заднегруди разделены попереч-
ной бороздкой, передняя часть с 2 щетинками, задняя — с 3 с каждой стороны.
Брюшные тергиты разделены двумя складками на 3 части, каждая соответственно с
3, 1, 3 парами щетинок, на боках с небольшими выпуклостями.
Личинки минируют листья. В регионе 1 вид, Р duodecimpunctata Motschulsky
(юг СБ и ДВ), связанный с Clematis. Известна личинка среднеазиатского вида Р
kondarensis Lopatin.
51. Род Dibolia Latreille
Личинки белые, с темными ногами, головой, переднеспинкой и переднегру-
дью. Голова с каждой стороны с 1 глазком, уплощенная, с направленным вперед
ротовым отверстием, с глубоко угловидно выемчатым задним краем затылка (рис.
72, 2) и удлиненными задними частями головной капсулы. Лобные швы прямые,
сходятся под тупым углом на краю затылочного отверстия, эпикраниальный шов
отсутствует. Темя в передней части с мелкими щетинками; лоб заметно шире дли-
ны, с 4-6 очень короткими щетинками. Наличник узкий, сильно хитинизирован-
ный, сросшийся со лбом, с 1 щетинкой на боковых углах и 2 в середине. Усики 2-
члениковые, с очень коротким 2-м члеником, несущим щетинки. Верхняя губа по-
перечная, со слабо выемчатым и слабо хитинизированным передним краем, несу-
щим мелкие густые щетинки. Мандибулы с 4 хорошо развитыми зубцами, на внут-
реннем изогнутом крае у основания с рядом длинных тонких щетинок (рис. 77,1).
Лабио-максиллярный комплекс сильно хитинизированный, нижнечелюстные щу-
пики 3-члениковые, нижнегубные редуцированы. Склерит переднеспинки разде-
лен светлой полосой, каждый с рядом из 6 щетинок по переднему и 4 по заднему
краю. Средне- и заднеспинка с поперечной бороздкой, претергальная часть с 2,
посттергальная с 6, крыловые склериты с 3 длинными и 1 мелкой щетинками. Брюш-
ные сегменты на боках с небольшими округлыми выпуклостями, их тергиты разде-
лены поперечной бороздой, с 4 претергальными и 6 посттергальными короткими
щетинками. Ноги коренастые, сильно хитинизированные, коготки небольшие, в
основании сильно изогнуты. Стернит груди сильно хитинизирован, с 4 щетинками
и продольным хитинизированным килем. Микроскульптура в виде округлых слабо
хитинизированных площадок (рис. 77, 3).
202
Рис. 77. Детали строения личинок подсемейства Alticinae:
1 — мандибула Dibolia metallica', 2 — верхняя губа Pentamesa', 3-4 — микроскульптура по-
кровов: 3 — Dibolia metallica, 4 — Argopistes', 5-7 — 9-й тергит брюшка Dibolia'. 5 — D.
depressiuscula', 6 — D. rugulosa; 7 — D. metallica', 8-11 — 4-й стернит брюшка Dibolia'. 8 —
D. depressiuscula', 9 — D. rugulosa', 10 — D. femoralis, 11 —D. metallica. По Штейнхаузену,
Лопатину и Дашевской и ориг.
Личинки минируют листья губоцветных и бурачниковых. Окукливание в по-
чве. В регионе 10 видов.
Определительная таблица видов рода Dibolia
1 (2). Тело желто-белое, пластинковидная скульптура покровов едва различима. Тер-
гит переднегруди в середине светлобурый. 9-й тергит брюшка с округленной вер-
шиной и 6 парами щетинок. На Marrubium. ЕЧ, СК, ЗСБ........D. cynoglossi Koch
2(1). Тело темно-желтое до светло-бурого, пластинковидная скульптура покровов
хорошо различима, тергиты иногда с поперечными полосами, без скульптуры.
3(4). 9-й тергит брюшка с притупленной вершиной (рис. 77, 5). 3 пары щетинок на
брюшном стерпите образуют ломаную линию (рис. 77, S). На Galeopsis. ЕЧ, СК, СБ
.....................................................D. depressiuscula Letzner
203
4(3). 9-й тергит брюшка округлен, начиная от основания.
5(6). Округлость 9-го тергита брюшка в форме полукруга (рис. 77, 6); 3 пары его
щетинок лежат на одной дуговидной линии (рис. 77, 9). На Stachys. ЕЧ, СК, ЗСБ
....................................................D. rugulosa Redtenbacher
6(5). Округлость 9-го тергита брюшка уплощена по бокам (рис. 77, 7).
7(8). Вентральные щетинки стернита брюшка образуют ломаную линию (рис. 77,
70). На Salvia. ЕЧ, СК..............................D.femoralis Redtenbacher
8(7). Вентральные щетинки стернита брюшка образуют дуговидную линию (рис.
77, 77). На Salvia. ЕЧ, СК......D, metallica Motschulsky (D. schillingi Letzner)
52. Род Argopistes Motshulsky
Личинки очень схожи с родом Dibolia, отличия сводятся к следующим призна-
кам: голова и ноги слабее хитинизированы, верхняя губа с прямым основанием,
внутренний край мандибул прямой, с 5-6 короткими утолщенными щетинками,
тергит и стернит переднегруди слабо хитинизированы, микроскульптура тела в виде
узких поперечных площадок, которые в свою очередь разбиты на продольные уча-
стки (рис. 77, 4).
Личинки минируют листья амурской сирени. В регионе 2 вида на юге ДВ; из-
вестна личинка наиболее обычного вида — A. biplagiatus Motschulsky.
204
14. ПОДСЕМЕЙСТВО HISPINAE
Тело удлиненное, уплощенное дорсовентрально (рис. 11,2), иногда очень силь-
но, так что верхняя и нижняя стороны совершенно плоские. Окраска беловатая или
желтоватая, с более темными головой, переднеспинкой и последним тергитом брюш-
ка, реже сплошь светлая. Голова небольшая, сильно втянута в переднеспинку, про-
гнатическая, вытянута назади в два отростка, глубоко вдающихся в переднеспинку;
эпикраниальный шов не развит, вследствие чего лобный склерит доходит до заты-
лочного отверстия; лобные швы явственные (рис. 78, 7). Глазков 4-6 пар, чаще все-
го 5 пар. Усики 3-члениковые. Наличник и верхняя губа небольшие или средней
величины. Мандибулы треугольные, довольно короткие, с 1-3 зубцами на вершине
и острым режущим внутренним краем. Челюстные щупики 2-члениковые; губные
щупики 1-члениковые.
Сегменты тела в более или менее явственной микроскульптуре, с поперечной
бороздкой посередине на верхней и нижней сторонах; эти борозды обычно не до-
ходят до боковых краев и ограничены спереди и сзади выпуклыми валиками, несу-
щими двигательную функцию. Хетотаксия развита слабо. Ноги (рис. 78, 3) корот-
кие и слабые, 3-члениковые (тазик, бедро и голенелапка), основания ног сильно
раздвинутые, коготки с парными пульвиллами; иногда ноги полностью отсутству-
ют, заметны лишь склериты — рудименты тазиков.
Брюшко состоит из 16 сегментов, но 8-9-й сегменты сливаются друг с другом
и граница между ними нередко почти неразличима, а 10-й сегмент очень малень-
кий; по бокам сегментов имеются выросты, иногда весьма длинные, пальцевид-
ные, иногда более или менее склеротизованные (рис. 78, 5).
Склериты на сегментах тела отсутствуют, кроме сплошных склеритов на пере-
днеспинке. переднегруди, последнем тергите брюшка, особенно в его задней час-
ти, соответствующей 9-му тергиту, реже на последнем стерните брюшка.
Стигмы на переднегруди и 1-7-м сегментах брюшка простые, кольцевидные;
стигмы 8-го сегмента брюшка лежат дорсально, обычно у заднего края тергита,
перемещаясь на 9-й тергит, большей частью в виде направленных кверху хитини-
зированных шипов (рис. 78, 6), реже простые.
Личинки минируют листья различных растений или, у некоторых тропических
форм, обитают в пазухах и влагалищах листьев, между прилегающими друг к другу
листьями или внутри стеблей. У минирующих форм яйца откладываются в ткани
листьев. В одной мине может быть несколько личинок. Имеются три личиночных
возраста. Окукливание в мине.
Куколки отличаются своеобразной морфологией (рис. 4, 5) и хорошо отлича-
ются у разных видов.
В регионе 5 родов, из них 4 только на юге ДВ. Известны личинки двух родов.
Таблица для определения родов подсемейства Hispinae
1(2). Голова и склериты тела темноокрашенные, резко отличаются от несклероти-
зованных участков. Микроскульптура явственная (рис. 78,2). Тело узкое, более или
менее цилиндрическое. Боковые лопасти брюшных сегментов без отростков (рис.
205
Рис. 78. Детали строения личинок подсемейства Hispinae:
Hispa atra: 1 — голова, 2 — микроскульптура покровов, 3 — нога, 4—верхняя губа; Dactylispa
anguloscr. 5 — боковой отросток брюшка, 6 — 8-й и 9-й тергиты брюшка.
11,2). Шипы 9-го тергита брюшка тонкие, отодвинуты от заднего края и не выда-
ются за него.....................................................1. Hispa
2(1). Голова и склериты тела светлоокрашенные, не отличаются по окраске от не-
склеротизованных участков. Микроскульптура неявственная. Тело широкое, упло-
щенное дорсовентрально. Боковые лопасти брюшных сегментов с тонкими отрос-
тками (рис. 78, 5). Шипы 9-го тергита брюшка крупные, лежат непосредственно на
заднем крае и выдаются за него (рис. 78, 6)..................2. Dactylispa
1. Род Hispa Linnaeus (Hispella Chapuis)
Тело цилиндрическое, несколько уплощено дорсовентрально (рис. 11, 2). Ок-
раска кремово-белая, голова, склериты переднеспинки, переднегруди и 9-го терги-
та брюшка темно-бурые. Наличник не отделен ото лба. Глазков по 4 с каждой сто-
роны в виде небольших пигментированных пятен, 2 передние глазка выпуклые,
задние плоские. Щетинки на голове короткие, единичные, по бокам головы с каж-
дой стороны 5-6 более длинных щетинок (рис. 78, 7). Передний край верхней губы
с дуговидной срединной выемкой, с более длинными и густыми щетинками на пе-
редних углах (рис. 78, 4). Переднеспинка с крупным склеритом, средне- и заднес-
206
пинка и 1-8-й тергиты брюшка с поперечными бороздками, не доходящими до кра-
ев и ограниченными спереди и сзади выпуклыми двигательными валиками, на 7-8-
м тергитах эти бороздки очень слабые. 1-7-й сегменты брюшка с крупными про-
стыми стигмами и выступающими по бокам двигательными буграми. 9-й тергит
уплощен, слабо отделен от 8-го, с крупным склеритом, с 2 поперечными вдавлени-
ями у основания и большим подковообразным вдавлением посередине; вершина
тергита с 2 щетинками; на боковых краях тергита помещаются стигмы, вытянутые
в тонкие длинные зубцы, направленные назад и вверх. Переднегрудь с крупным
склеритом, на средне- и заднегруди двигательные валики широко прерванные по-
середине, 1-7-й стерниты брюшка с двигательными валиками, более короткими,
чем на тергитах. 9-й стернит брюшка не склеротизован, с 4 щетинками у переднего
края. 10-й сегмент в виде двух поперечных валиков, ограничивающих анальное
отверстие: Голенелапка короче и тоньше бедра, оканчивается двумя большими пуль-
виллами и тонким коготком, не выступающим за пульвиллы (рис. 78, 3). Сегменты
тела в явственной микроскульптуре (рис.78, 2). Щетинки туловища немногочис-
ленные, короткие, в основном на боках тергитов и на склерите переднеспинки. Длина
тела взрослой личинки до 6-6,5 мм.
Личинки младших возрастов отличаются только более светлой окраской и рез-
че отграниченными склеритами.
Минируют листья Роа и Agropyron, в каждой мине находится по одной ли-
чинке, ход прокладывается от вершины листа к основанию, мины узкие, длиной
до 5 см.
В роде один вид — Н. atra Linnaeus, распространенный в ЕЧ, СК и на юге СБ
до Байкала.
2. Род Dactylispa Weise
Тело одноцветно желтоватое, склеротизованные участки не отличаются по ок-
раске от несклеротизованных покровов, контур тела продолговато-овальный, дор-
совентральное уплощение сильное. Голова типичного для минирующих Hispinae
строения, но отличается слабой склеротизацией и светлой окраской; задняя поло-
вина головы в мелкой поперечной морщинистости. Переднеспинка с крупным, плохо
ограниченным, светлоокрашенным склеритом, покрытым поперечными нежными
морщинками; средне- и заднеспинка, xaKyHispa Linnaeus, 1-8-й тергиты брюшка с
двигательными бороздками, хорошо развитыми на всех тергитах, кроме 8-го. Боко-
вые края сегментов брюшка выдаются в стороны в виде округлых лопастей, круп-
ных на 1-5-м и слабых, вплоть до полного исчезновения, на 6-8-м сегментах; каж-
дая лопасть оканчивается тонким отростком почти одинаковой длины и ширины,
несущим короткую щетинку; кпереди от отростка находится еще одна щетинка (рис.
78, 5). 9-й тергит не отграничен от 8-го, хитинизирован сильнее предыдущих, одна-
ко без явственного склерита, с 2 поперечными вдавлениями, соединенными по бо-
кам. Стигмы переднегруди и 1-7-го сегментов брюшка выпуклые, в виде коротких
трубочек, стигмы 9-го сегмента в виде крепких шипов, направленных вверх и назад
(рис. 78, 6). Переднегрудь с довольно четко отграниченным светлоокрашенным
продольно-морщинистым склеритом, на средне- и заднегруди двигательные бороз-
дки не выражены, но заменяются 2^4 вдавлениями или следами срединного скле-
207
рита. Брюшные стерниты без склеритов, двигательные бороздки на 1-7-м стерни-
тах. Взрослая личинка до 5 мм длины.
Личинки и куколки в минах листьев полыни, мины обширные, занимают по-
чти всю площадь листа. В минах обычно по 1-3 личинки.
В нашей фауне известно 4 вида, все с юга ДВ, известны личинки 2 видов.
Таблица для определения видов рода Dactylispa
1(2). Стигмальные шипы 9-го тергита брюшка широкие и изогнутые кнаружи. Бо-
ковые выросты 7-го сегмента брюшка умеренно толстые. На полыни. Юг ДВ.
......................................................D. angulosa Solsky
2(1). Стигмальные шипы 9-го тергита брюшка тонкие и прямые. Боковые выросты
7-го сегмента брюшка тонкие. Юг ДВ, Сахалин.............D. masoni Gestro
208
15. ПОДСЕМЕЙСТВО cassidinae
Живые личинки желто-зеленые, зеленые или светло-коричневые, с широким,
овальным, сильно уплощенным дорсовентрально телом, с плоской нижней и слабо
выпуклой верхней стороной (рис. 11, 3, 4). Голова опистогнатическая, прикрыта
переднеспинкой и не видна сверху, уплощенно-шаровидная или поперечная. Эпик-
раниальный шов большей частью плохо заметен (различим в виде утолщения на
нижней поверхности темени), лобные швы обычно недоразвитые, их местополо-
жение можно определить лишь по распределению пигмента и микроскульптуры на
лбу (рис. 79, 7). Наличник с рядом из 6 мелких щетинок. С каждой стороны головы
имеется по 5 пигментированных глазков, с сильно выпуклой роговицей. Усики 3-
члениковые, первый членик широкий и короткий, уплощенный, 2-й очень малень-
кий. Верхняя губа поперечная, на диске с рядом из 4 крепких щетинок, с закруглен-
ными боками и со срединной выемкой на переднем крае, иногда слабо развитой;
дно выемки бывает прямое или слабо выпуклое, обычно с 4-6 длинными и тонки-
ми или короткими и утолщенными щетинками; передний край с каждой стороны от
выемки с 3-6 маргинальными щетинками, которые могут быть тонкие с острыми
вершинами или плоские и толстые с округленными вершинами. Мандибулы корот-
кие и широкие, с 5-6 зубцами (рис. 79, 3). Лабио-максиллярный комплекс слабо
хитинизирован, кардо нижних челюстей слито со стипесом (рис. 79, 2), нижнече-
люстные щупики 2-члениковые, нижнегубные 1-члениковые (рис. 79,2). Переднес-
пинка очень крупная, без срединного склерита, иногда с мелкими склеротизован-
ными пятнышками или темными щетинками, образующими своеобразный рису-
нок, с 4 отростками с каждой стороны, из них 2 отростка находятся на переднем
крае и обозначаются как переднекрайние (наружный и внутренний); они могут быть
в основании отделены друг от друга или слиты вместе. На боковом крае находятся
боковой и стигмальный отростки, у основания последнего расположена стигма груди.
Средне- и заднеспинка имеют с каждой стороны по 2 отростка (передний и зад-
ний), не сросшихся в основании; они или одинаковой длины, или же передний не-
сколько короче заднего. 1-8-й сегменты брюшка несут с каждой стороны по 1 боко-
вому отростку, чаще всего на 1-5-м сегментах они постепенно укорачиваются кза-
ди, а на 6-8-м — заметно длиннее предыдущих. Боковые отростки имеют выросты,
обозначаемые как вторичные ответвления, их количество варьирует у различных
видов. Вторичные ответвления оканчиваются теками в виде острой иглы, вытяну-
той шишки или пузырька. Длина и форма боковых отростков различны, на первых
сегментах брюшка они могут достигать от 1/5 до 1/2 ширины сегмента, могут быть
тонкими, вытянутыми, или же толстыми, с расширенными основаниями. 9-й сег-
мент брюшка расположен на дорсальной стороне 8-го, с 2 отростками (хвостовыми
нитями). Они обычно загибаются на спинную сторону и вытягиваются вдоль тела,
на них сохраняются экзувии и экскременты, образуя личиночный чехлик (маска, по
Steinhausen, 1950). Выделяются 4 типа чехликов: из экзувиев, компактный из экск-
рементов, составной из экскрементов и из экскрементов в виде длинных нитей.
Хвостовые нити могут быть сильно хитинизированными и тонкими, достигая по-
ловины длины тела, или же толстыми и короткими, сросшимися или свободными в
основании. За единичным исключением, они лишены вторичных ответвлений, но
209
Рис. 79. Детали строения личинок подсемейства Cassidinae:
1 — голова Cassida prasincr, С. pallidicollis'. 2 — лабио-максиллярный комплекс, 3 — манди-
була, 10 — вторичные ответвления; 4 — нога С. nebuloscp 5-8 — боковые отростки; 5 —
Thlaspida, 6 — Pilemo stoma, 7 — C. ferruginea, 8 — C. panzerir, 9,11,12 — вторичные ответв-
ления: 9 — С. berolinensis, 11 — С. amurensis, 12 — С. parvula.
210
часто имеют у основания хорошо развитые шипики или щетинки, сидящие на рас-
ширенном основании — теке. Ноги (рис. 79, 4) короткие и толстые, с очень корот-
кой голенелапкой, коготки маленькие, сильно изогнутые, с прямоугольным высту-
пом при основании или без него.
Яйца откладываются в мае-июле группами, реже поодиночке на нижней сто-
роне листьев и покрываются специальной оотекой, представляющей обычно плен-
чатую капсулу или отдельные пластинки, скрепленные на одном конце, реже сплош-
ной комочек, в котором заключены яйца. Оотека образуется за счет быстро засты-
вающих выделений придаточных желез. Число яиц в оотеке обычно 2-9, но у неко-
торых видов может достигать 80; сверху оотека нередко покрывается экскремента-
ми. Яйца продолговатые, заостренные на обоих концах, уплощены дорсовентраль-
но. Развитие яиц длится 4-6 дней. Вышедшие молодые личинки питаются на лис-
тьях, обыкновенно на их нижней стороне, в отдельных случаях на цветках и стеб-
лях. Число личиночных возрастов у разных видов колеблется от 4 до 5. При линьке
личиночные шкурки сдвигаются назад и часто остаются на хвостовых нитях (рис.
80, 4), так что в таком случае для определения возраста личинки достаточно сосчи-
тать число шкурок. Характерной особенностью личинок является их способность
покрывать тело сверху экскрементами, которые помещаются на загнутых на спин-
ную сторону хвостовых нитях вместе с личиночными шкурками младших возрас-
тов (рис. 80, 7-5). Защитная роль такого личиночного чехлика очевидна. Таким же
чехликом и личиночными шкурками покрыта куколка. Личиночная стадия длится
12-20 дней. Как в личиночной, так и в имагинальной стадии Cassidinae при опти-
мальных условиях являются монофагами или узкими олигофагами. Окукливание
происходит на растениях, обычно на нижней стороне листа, к которому куколка
прикрепляется задним концом тела. Куколки, как и личинки, имеют боковые выро-
сты и хвостовые нити (рис. 4, 6), причем куколки разных видов резко отличаются
друг от друга, гораздо более, чем жуки, в отличие от остальных подсемейств листо-
едов (кроме Hispinae), у которых куколки очень однообразны. Все развитие проте-
кает за 4-5 недель. В течение лета развивается 1 поколение, в отдельных случаях,
особенно в южных районах, возможно 2 поколения. Зимуют жуки в подстилке. В
Палеарктике практически все виды связаны с травянистыми растениями.
В регионе 7 родов.
Таблица для определения родов подсемейства Cassidinae
1(2). Боковые отростки на груди не развиты, на брюшке очень короткие, прижаты к
телу и мало заметны. Тело удлиненное, мокрицеобразное, сильно сужено кзади, в
передней половине сильно выпуклое (рис. 81,7).................1. Ischyronota
2(1). Боковые отростки груди и брюшка хорошо развитые, явственные. Тело упло-
щенное или умеренно выпуклое.
3(8). Коготки при основании с широким или узким прямоугольным выступом (рис.
82,10, 77).
4(5). Верхняя губа со слабой, едва намеченной срединной выемкой и с 5 маргиналь-
ными щетинками с каждой стороны (рис. 83, 7). Боковые отростки сегментов с 5-6
вторичными ответвлениями с каждой стороны, которые приблизительно вдвое ко-
роче самих отростков; 2-й членик усиков короткий (рис. 82, 7).2. Hypocassida
211
Рис. 80. Хвостовые чехлики личинок подсемейства Cassidinae:
1 — нитевидный из экскрементов; 2 — компактный из экскрементов; 3 — составной i
крементов; 4 — из экзувиев. По Штейнхаузену.
212
5(4). Верхняя губа с глубокой треугольной срединной выемкой и с 5-6 маргиналь-
ными щетинками с каждой стороны. Боковые отростки сегментов с 7-14 вторич-
ными ответвлениями с каждой стороны.
6(7). Верхняя губа с 5 маргинальными щетинками с каждой стороны (рис. 83, 2).
Вторичные ответвления боковых отростков (в количестве 7-9) тонкие, очень длин-
ные, на вершине с длинными шиловидными теками (рис. 79, 6). 2-й членик усиков
длинный (рис. 82, 2)......................................3. Pilemostoma
7(6). Верхняя губа с 6 маргинальными щетинками с каждой стороны (рис. 83, 3).
Боковые отростки сегментов с 10-14 недлинными вторичными ответвлениями с
каждой стороны, на вершине с округло-вытянутыми теками (рис. 79, 5)....
............................................................4. Thlaspida
8(3). Коготки при основании без выступа или же он сильно сглажен (рис. 82, 72).
9(12). Стигмы 1-го брюшного сегмента не отличаются по размерам и хитинизации
от последующих. Верхняя губа со срединной выемкой и с 3-5 маргинальными ще-
тинками с каждой стороны примерно одинакового строения. Хвостовые нити ли-
чинок 1-го возраста без вторичных ответвлений (кроме С. flaveola Thunberg, С. azurea
Fabricius). Чехлик разнообразного строения.
10(11). Голова с многочисленными треугольными спинулами посередине. Верхняя
губа с каждой стороны с 4 шиловидными маргинальными щетинками и с глубокой
прямоугольной срединной выемкой, несущей 7-8 коротких щетинок (рис. 81, 5).
Хвостовые нити тонкие, в основании сближены, но не сросшиеся (рис. 81,2), снизу
с редкими щетинками. Коготки со сглаженным зубцом. Личиночный чехлик треу-
гольный. Только юг ДВ.....................................5. Glyphocassis
11(10). Голова без многочисленных треугольных спинул посередине. Комплекс при-
знаков иной...................................................6. Cassida
12(9). Стигмы 1-го брюшного сегмента гораздо крупнее и более хитинизированы,
чем последующие. Верхняя губа со срединной выемкой и с 4 маргинальными ще-
тинками с каждой стороны, из которых две внутренние толще наружных (рис. 84,8,
9). Хвостовые нити личинок 1-го возраста с многочисленными длинными вторич-
ными ответвлениями (рис. 85, 7). Чехлик из экзувиев.....7. Aspidomorpha
1. Род Ischyronota Weise
Тело удлиненное, мокрицеобразное, сильно сужается кзади, в передней поло-
вине сильно выпуклое (рис. 81, 7). Верхняя губа (рис. 81,4) без выемки на пере-
днем крае, 6 срединных маргинальных щетинок равномерно распределены. Боко-
вые отростки на груди не развиты, на брюшке отсутствуют или очень короткие,
особенно на первых сегментах, плохо различимые, направлены назад и более или
менее прижаты к телу, вторичные ответвления не развитые или едва заметные (на
последних сегментах брюшка). Хвостовые нити раздвинутые или сросшиеся у ос-
нования, довольно длинные, без щетинок, несут экзувий. Верхняя сторона тела без
склеротизованного рисунка, с короткими щетинками. Коготки с небольшим зубцом
при основании.
В регионе 3 вида, встречающихся на юго-востоке ЕЧ, СК и в Туве на солянках;
более обычна L desertorum Gebler.
213
Рис. 81. Детали строения личинок подсемейства Cassidinae:
1,4 — Ischyronota desertorunv. 1 — общий вид, 4 — верхняя губа; 2, 3, 5 — Glyphocassis
spilotar. 2 — общий вид, 3 — мандибула, 5 — верхняя губа; 6 — Cassida versicolor, верхняя
губа; 7 — Cassida canaliculata, верхняя губа. По Штейнхаузену, Чо и Ли и ориг.
2. Род Hypocassida Weise
Тело личинки овально-вытянутое, с наибольшей шириной в области первых
брюшных сегментов. Верхняя губа (рис. 83, 7) со слабой прямоугольной срединной
214
выемкой, на дне которой имеются 6-8 микрохет, и с 5 маргинальными щетинками с
каждой стороны, 2-й членик усиков короткий (рис. 82, 7). Переднеспинка сплошь
усеяна мелкими зернышками, со склеротизованным рисунком в виде 2 неправиль-
ных колец или склеритов, разделенных светлой полосой. Боковые отростки пере-
днеспинки одинаковой длины, составляют около половины ее ширины, с 5-6 вто-
ричными ответвлениями; переднекрайние отростки сросшиеся в основании. Пере-
дние боковые отростки средне- и заднегруди заметно уже в основании и короче
задних, боковые отростки сегментов брюшка примерно одинаковой величины.
Щетинки верха сильно редуцированы, плохо заметные. Хвостовые нити сильно
хитинизированные, в основании сближены, без щетинок, тонкие и длинные, дости-
гают половины длины тела, несут компактный чехлик из экскрементов. Подталки-
ватель короткий, его второй членик треугольный, анальное отверстие шелевидное.
Коготки с прямоугольным зубцом при основании. На полевом вьюнке, отмечался
как случайный вредитель свеклы.
В России 1 вид — Н. subferruginea Schrank, встречающийся в ЕЧ, СК, СБ.
3. Род Pilemostoma Desbrocher
Тело личинки сильно вытянутое, верх с длинными щетинками, которые не ко-
роче 1/2 вторичных ответвлений. Верхняя губа с глубокой узкой треугольной выем-
кой, по бокам которой расположено по 2 микрохеты, с каждой стороны от выемки с
4 маргинальными щетинками (рис. 83, 2). 2-й членик усика длинный, слабо кони-
ческий, с крупным сенсорным придатком на вершине (рис. 82, 2). Боковые отрост-
ки тонкие, расположены на увеличенных боковых выростах тела, с каждой сторо-
ны с 7-14 вторичными ответвлениями (рис. 79,6). Передние боковые отростки сред-
не- и заднеспинки тоньше и почти на 1/2 короче задних. Боковые отростки 1-5-го
сегментов брюшка постепенно уменьшаются кзади, на 6-8-м сегментах вдвое длин-
нее, чем на 5-м. Хвостовые нити слабо хитинизированные, в основании не сросши-
еся и без щетинок, длинные и тонкие. Коготки с прямоугольным зубцом при осно-
вании (рис. 82, 70). На Senecio, Pulicaria. 1 вид — R fastuosa Schaller (ЕЧ, СК, СБ).
4. Род Thlaspida Weise
Тело широкоовальное, с длинными боковыми отростками, несущими 10-14
недлинных вторичных ответвлений с каждой стороны и оканчивающимися вытя-
нутыми шишковидными теками (рис. 79, 5). Верхняя губа с глубокой округло-треу-
гольной срединной выемкой и с 6 маргинальными щетинками с каждой стороны,
из которых 4 толстые и торчащие и 1 мелкая тупая расположены на самом крае
губы, а 1 длинная и шиловидная значительно отодвинута от переднего края (рис.
83, 3). Мандибулы с 6 зубцами, из которых 2 срединных наиболее длинные и широ-
кие. Переднеспинка с 2 крупными хитинизированными пятнами в виде двух кру-
гов, окруженных рядом из крупных хет с темными основаниями, все боковые отро-
стки примерно одинаковой длины. На средне- и заднеспинке передние отростки
заметно короче задних; боковые отростки 1-5-го сегментов брюшка заметно укора-
чиваются кзади, на 6-м и 8-м сегментах они длиннее, чем на пятом, но заметно
короче, чем на 7-м. Хвостовые нити длинные и тонкие, широко расставленные, на
215
Рис. 82. Детали строения личинок подсемейства Cassidinae:
7,3-5 — усик: 1 —Hypocassida, 3 — Cassida undecimnotata, 4 — С. amurensis, 5 — С. prasina\
6-9 — переднеспинка: 6 — С. stigmatica, 7 — С. lineola, 8 — С. mandli, 9 — С. mongolica\
Pilemostoma-. 2 — усик, 10 — коготок; 11-13 — коготок: 11 — Thlaspida, 12 — С. nebulosa,
13 —Aspidomorpha difformis\ 14 — вершина брюшка С. тиггаеа.
вершине с мелкими шипиками, в основании не сросшиеся, снизу и с боков с хета-
ми. Прочие сегменты тела с явственными поперечными рядами из крупных хет и
многочисленными мелкими шипиками. Коготки тонкие, серповидные, их основа-
216
ние вдвое уже длины коготка, с прямоугольным выступом (рис. 82, 77). Хвостовые
нити с компактным чехликом из экскрементов в виде отдельных, плотно прилегаю-
щих друг к другу нитей.
В России 1 вид — Th. lewisii Baly на юге ДВ; ее личинка неизвестна. Описание
составлено по личинке Th. biramosa Boheman из Вьетнама и Th. lewisii Baly.
5. Род Glyphocassis Spaeth
Тело овально-вытянутое, светло-коричневое, с редкими короткими светло-ко-
ричневыми щетинками, теки булавовидные (рис. 81, 2). Микроскульптура покро-
вов тела в виде мелких светлых овальных зерен, более густая на межсегментных
складках. Боковые отростки довольно короткие и конические, к вершине постепен-
но утончаются, с каждой стороны с 7-8 вторичными шиловидными ответвления-
ми, в основании сильно расширены, особенно на брюшных сегментах. Голова без
явственных лобных швов, с многочисленными треугольными спинулами посере-
дине. Верхняя губа широкая (рис. 81, 5), в основании слабовыпуклая, по бокам
закругленная, по переднему краю склеротизованная в виде узкой полосы, передний
край прямо срезан ковнутри, с глубокой прямоугольной срединной выемкой, на дне
ее имеется 7-8 коротких щетинок, по бокам с каждой стороны с 4 шиловидными
маргинальными щетинками; четыре дискальные щетинки основного ряда располо-
жены косым рядом, две щетинки вершинного ряда очень короткие и отодвинуты от
переднего края верхней губы. Мандибулы с 6 зубцами и острым режущим внутрен-
ним краем (рис. 81,5). Переднеспинка слабо поперечная, с двумя слабо склероти-
зованными пятнами и с группой мелких, слабозаметных щетинок в центре диска.
Переднекрайние боковые отростки расположены на общем основании, наружные
заметно уже внутренних. Боковые и стигмальные отростки одинакового строения.
Задние отростки средне- и заднегруди более расширены в основании и заметно
длиннее передних. Боковые отростки 1-5-го сегментов брюшка постепенно умень-
шаются кзади, 6-й несколько длиннее предыдущих, 7-й и 8-й равны и заметно длин-
нее предыдущих. Граница между сегментами груди и брюшка с очень плохо разли-
чимыми спутанными рядами светло-коричневых щетинок. Хвостовые нити скле-
ротизованные, тонкие, в основании сближены, но не сросшиеся, снизу с редкими
щетинками, в 2,5 раза длиннее 7-го бокового отростка, к вершине утончаются. Ли-
чиночный чехлик плоский и рыхлый, треугольной формы, с овальными отверстия-
ми. Стигмы светло-коричневые, на груди более крупные, чем на брюшке. Коготки
со сглаженным зубцом. Личинка 1-го возраста на наружной стороне хвостовых ни-
тей без ответвлений.
В России 1 вид — Glyphocassis spilota Gorham, встречается на юге ДВ, питает-
ся на Ipomoea.
6. Род Cassida Linnaeus
Один из наиболее крупных родов данного подсемейства, морфологическая ха-
рактеристика которого в общем близка к характеристике подсемейства; от родов
Hypocassida Weise, Pilemostoma Desbrocher и Thlaspida Weise отличается отсутствием
явственного прямоугольного выступа при основании коготков, от Aspidomorpha Норе
217
Рис. 83. Верхняя губа личинок подсемейства Cassidinae:
1 —Hypocassida; 2 — Pilemo stoma; 3 — Thlaspida; 4 — Cassida flaveola; 5 — C. pallidicollis;
6 — C. scraphina; 7 — C. panzeri; 8 — C. vittata; 9 — C. concha; 10 — C. parvula.
примерно одинаковыми размерами стигм груди и брюшка, а от Glyphocassis Spaeth
в основном комплексом признаков.
В регионе 43 вида. Из видов, не включенных в определительную таблицу (ли-
чинки неизвестны или недостаточно описаны) С. hemisphaerica Herbst — на Silene
и Dianthus, С. subreticulata Suffrian — на Lychnis и Saponaria, несколько видов с юга
ДВ — С. fuscorufa Motschulsky (на полыни), С. spaethi Weise (на полыни, пижме),
С. velaris Weise (на мари). Часть этих видов условно введена в таблицу по аналогии
с близкородственными формами.
Таблица для определения видов рода Cassida
1(4). Хвостовые нити толстые, короткие, несут многочисленные вторичные ответв-
ления и мало отличаются от боковых отростков (рис. 85,3). Верхняя губа с 3 марги-
нальными щетинками с каждой стороны (рис. 83,4). Стигмы черные. Чехлик отсут-
ствует. На гвоздичных.
2(3). Верх пятнистый, грязно-желтый. Боковые отростки с черными основаниями,
к вершинам светлее. На видах подсем. Alsinoideae (Stellaria, Cerastium, Malachium,
Honckenya). Транспалеаркт, обычно на лугах.............С flaveola Thunberg
3(2). Верх однотонно грязно-желто-зеленый. Боковые отростки темные. На видах
подсем. Silenoideae (Silene, Saponaria). ЕЧ, СБ. Тяготеет к известковым склонам,
очень редка............................С. azurea Fabricius (С. ornata Creutzer)
218
4(1). Хвостовые нити тонкие и длинные, гораздо длиннее боковых
отростков, без вторичных ответвлений (рис. 85, 4).
5(10). Отростки переднеспинки (особенно передние) очень короткие, конические,
не более чем вдвое длиннее ширины, резко отличаются друг от друга по величине;
вторичные ответвления гораздо короче поперечника отростков (рис. 82, 6). Верх-
няя губа с 5 маргинальными щетинками с каждой стороны.
6(7). Переднекрайние внутренние отростки переднеспинки заметно длиннее наруж-
ных, сильно сближены (рис. 82, 6). Все отростки короткоконические, с очень корот-
кими вторичными ответвлениями. Переднеспинка без рисунка. Хвостовые нити
длинные, не сросшиеся в основании, тонкие. Чехлик из экзувиев. На Tanacetum.
ЕЧ, СК, СБ. Чаще в сухих местах.......................С. stigmatica Suffrian
7(6). Переднекрайние внутренние отростки переднеспинки гораздо короче наруж-
ных, широко раздвинутые. Хвостовые нити короткие, с толстыми сросшимися ос-
нованиями.
8(9). Отростки переднеспинки очень короткие, наружные переднекрайние менее
чем вдвое длиннее внутренних; вторичные ответвления развиты слабо (рис. 82, 7).
Рисунок переднеспинки в виде 2 незамкнутых кругов и общей складки, параллель-
ной основанию. Тело удлиненное, зеленое, со светлым рисунком. Чехлик из экск-
рементов, компактный, часто отпадает у последних возрастов. На полыни, преиму-
щественно веничной. ЕЧ, СК, СБ, ДВ. Тяготеет к степям и сухим местам....
.........................................................С lineola Creutzer
9(8). Отростки переднеспинки гораздо длиннее, наружные переднекрайние более
чем вдвое длиннее внутренних; вторичные ответвления многочисленные (рис. 82,
8). Рисунок переднеспинки в виде 2 кругов. Тело удлиненное, коричневое. Чехлик
из экскрементов, компактный. На полыни, особенно на красночерешковой. Юг ДВ.
В сухих местах...........................................С. mandli Spaeth
10(5). Отростки переднеспинки длинные, более или менее линейные, с хорошо раз-
витыми вторичными ответвлениями, которые обычно не короче поперечника отро-
стков (рис. 82, 9).
11(12). Верхняя губа с прямым передним краем, без срединной выемки, с 6 корот-
кими срединными и с 3 краевыми щетинками с каждой стороны (рис. 81,6). Хвос-
товые нити без выроста с наружной стороны основания и без хет на трубчатых
основаниях. На вьюнковых (подрод Taiwania Spaeth). Юг ДВ................
....................................................С. versicolor Boheman
12(11). Верхняя губа с дуговидным передним краем,и с явственной срединной вы-
емкой.
13(32). Верхняя губа с 3 краевыми щетинками с каждой стороны (рис. 83, 5).
14(15). Хвостовые нити с явственным выростом с наружной стороны у основания,
без хет в основании, широко расставленные (рис. 85, 2). Все боковые отростки не-
сут по 5-7 длинных вторичных ответвлений с каждой стороны. Окраска изумруд-
но-зеленая, без явственного рисунка. Тело широкоовальное. Чехлик из экзувиев.
На мари белой, лебеде, свекле, щирицевых. Юг ДВ. На лугах...............
...........................................С. piperata Норе (С. japana Baly)
15(14). Хвостовые нити без выроста с наружной стороны основания.
16(21). Хвостовые нити с многочисленными хетами, сидящими на столбчато удли-
ненных основаниях в базальной трети или половине (рис. 85, 5). На маревых.
219
17(18). Вторичные ответвления боковых отростков оканчиваются короткими пу-
зырьковидными хетами (рис. 79, 9). Переднеспинка со склеротизованным рисун-
ком. Темя и лоб склеротизованы. Чехлик из экзувиев. На лебеде. ЕЧ, СК, СБ, ДВ. В
степях и остепненных стациях........................С berolinensis Suffrian
18(17). Вторичные ответвления боковых отростков оканчиваются более или менее
длинными цилиндрическими хетами, иногда расширенными на вершине (рис. 79,
10). Переднеспинка без рисунка.
19(20). Стигмы переднегруди узко трапециевидные, втрое уже основания бокового
отростка. Хвостовые нити темные, с густой мелкой зернистостью, с 8-10 толстыми
хетами на столбчатых основаниях в базальной половине (рис. 85, 4). Чехлик из
экзувиев. На мари. Юг СБ, ДВ. В степях и на остепненных склонах..........
....................................................С. pallidicollis Boheman
20(19). Стигмы переднегруди широко трапециевидные, немного уже основания бо-
кового отростка. Хвостовые нити светлые, со слабой зернистостью, с 18-20 тонки-
ми хетами на столбчатых основаниях, заходящими в вершинную половину (рис.
85, 6). Чехлик из экскрементов, составной. На мари, лебеде, свекле и щирицевых.
Транспалеаркт, в открытых мезофильных стациях.......С nebulosa Linnaeus
21(16). Хвостовые нити без хет на трубчатых основаниях или они единичные и
короткие у самого основания.
22(23). Хеты в срединной выемке верхней губы очень толстые, конические, плотно
прилегающие друг к другу (рис. 83, 6). Хвостовые нити без хет. Тело удлиненно-
овальное. Питание на Convolvulus и Clematis (подрод Alledoya Hincks).....
......................С. scraphina Menetries (СК), С. vespertina Boheman (ДВ)
23(22). Хеты в срединной выемке верхней губы умеренно утолщенные, с заострен-
ными вершинами.
24(27). Хвостовые нити в базальной части с единичными хетами на трубчатых ос-
нованиях. Срединная выемка верхней губы прямоугольная (рис. 83, 9). Тело округ-
лое или эллиптическое. Боковые отростки очень длинные (рис. 11, 4). На гвоздич-
ных (подрод Mionycha Wse.).
25(26). Тело желтое, с полосой белых пятен от заднего края переднеспинки до 6-го
сегмента брюшка. Чехлик из экзувиев, иногда с остатками экскрементов. На Silene,
реже на Spergula, Saponaria. Она отличается бледно-желтым телом с просвечиваю-
щими покровами, более крупными размерами и наличием составного чехлика из
экскрементов. Юг ЕЧ и СБ, СК. (К этому виду очень близка недостаточно описан-
ная С subreticulata Suffrian — ЕЧ, СК, СБ, ДВ, также живущая на гвоздичных) ..
...................................................С. margaritacea Schaller
26(25). Тело одноцветно-желтое. На Lychnis. Юг ДВ. На лугах ... С. concha Solsky
27(24). Хвостовые нити без хет на трубчатых основаниях.
28(29). Срединная выемка верхней губы глубокая. Хвостовые нити без экскремен-
тов. Анальное отверстие V-образное или неявствеино 3-лучевое. Оба членика под-
талкивателя цилиндрические. Рисунок переднеспинки в виде двух неправильных
окружностей, изолированных друг от друга. На Matricaria. СК..............
........................................................С. inquinata Brulle
29(28). Срединная выемка верхней губы неглубокая, округлая (рис. 83, 7). Тело про-
долговато-овальное, Чехлик из экскрементов, компактный. На сложноцветных.
30(31). Боковые отростки удлиненные, слабо сужены к вершине; их вторичные от-
220
Рис. 84. Верхняя губа личинок рода Cassida и Aspidomorpha'.
1 — С. тиггаеа, 2 — С. viridis\ 3 — С. amurensis; 4 — С. undecimnotata\ 5 — С. denticollis\ 6 —
С. rubiginosa', 7 — С. sanguinosa\ 8 — A. transparipennis; 9 — A. diffbrmis.
ветвления тонкие и длинные, превышают ширину отростка (рис. 79, 8). Тело более
широкое. На Cirsium, Arctium, Scorzonera, Mulgedium. Транспалеаркт, обычно на
лугах.........................................................С. panzeri Weise
31(30). Боковые отростки короткие, явственно конические, с очень короткими вто-
ричными ответвлениями, которые гораздо меньше ширины отростков (рис.79, 7).
Тело более узкое. На Pulicaria. ЕЧ, СК, СБ.................С ferruginea Goeze
32(13). Верхняя губа с 4-5 маргинальными щетинками с каждой стороны.
33(38). Хвостовые нити у основания сросшиеся на заметном протяжении (рис. 85,
7) и утолщенные, довольно короткие (около 1/3-1/4 длины тела). Верхняя губа с 4
маргинальными щетинками с каждой стороны и резкой трапециевидной выемкой,
основание которой прямое. 8-й тергит брюшка без крепких длинных щетинок. Тело
узкое, яйцевидное, суженное кзади. Боковые отростки довольно короткие (около 1/4
ширины тела), вторичные ответвления оканчиваются пузыревидной хетой (рис. 79,
72). На маревых и гвоздичных (подрод Cassidulella Strand).
34(35). Ноги и боковые отростки темные, стигмы черные, со светлым кольцом, хво-
стовые нити бурые. Передний край верхней губы с трапециевидной выемкой (су-
жается к основанию) (рис. 83, 5). Тело голубовато-зеленое, с узкой темной средин-
ной полоской и прилегающей к ней полосой со светлыми пятнами. Чехлик из экск-
рементов, компактный, небольшой. На Spergula, свекле. ЕЧ, СК, СБ, ДВ.........
..............................................................С. vittata Villers
35(34). Ноги, боковые отростки и стигмы светлые. Передний край верхней губы с
обратно-трапециевидной выемкой (расширяется к основанию; рис. 83, 70).
36(37). Хвостовые нити бурые или черные в основании, к вершине светлее. Тело
светло-зеленое, с узкой темной полоской вдоль срединной линии и широкой поло-
221
сой желто-белых пятен. Переднеспинка без рисунка. Чехлик из экскрементов, со-
ставной. На гвоздичных (Silene, Spergula, Stellaria), также на свекле. Транспале-
аркт, встречается в сухих местах.........................С. nobilis Linnaeus
37(36). Хвостовые нити светлые, не несут чехлика. Переднеспинка со слабым ри-
сунком. На лебеде, Suaeda и других маревых. Транспалеаркт, связан со степями и
сухими стациями..........................................С. parvula Boheman
38(33). Хвостовые нити не сросшиеся и не утолщенные у основания. Верхняя губа
с 5 маргинальными щетинками с каждой стороны (часто 1-2 внутренние щетинки
мелкие и плохо заметные). Сюда должна относиться также личинка С. velaris Weise
с юга ДВ; жуки на мари.
39(44). 8-й тергит брюшка с крепкими длинными хетами, гораздо более крупными,
чем обычные хеты спинной стороны (рис. 82,14). Стигмы светлые. Вторичные от-
ветвления боковых отростков оканчиваются щетинковидными хетами. Переднес-
пинка без рисунка.
40(43). Выемка верхней губы неглубокая, округлая, без явственных внутренних уг-
лов (рис. 84, 7). Подталкиватель длинный, цилиндрический, не короче половины
боковых отростков 8-го сегмента. Хвостовые нити длинные, около 1/2 длины тела.
Чехлик из экскрементов в виде длинных нитей (рис. 80, 7).
41(42). Вторичные хеты боковых отростков на конце кнопковидно утолщены. Вы-
емка верхней губы очень слабая (рис. 81, 7). На Salvia (подрод Lordiconia Reitter)
ЕЧ, СК, ЗСБ.......................................С. canaliculata Laicharting
42(41). Вторичные хеты боковых отростков на конце заострены. Выемка верхней
губы полукруглая (рис. 84, 7).Тело светло-зеленое, с желто-зелеными боковыми
отростками и с 3 продольными темными полосами. На сложноцветных (Inula,
Pulicaria). Транспалеаркт, идет на север до лесотундры. В сырых местах....
.......................................................С murraea Linnaeus
43(40). Выемка верхней губы глубокая, с дуговидно выступающим основанием и
явственными внутренними углами (рис. 84,2). Подталкиватель короткий, треуголь-
ный, не длиннее трети боковых отростков 8-го сегмента (подрод Odontionycha Weise).
Тело светло-зеленое, посередине с темной полоской, окаймленной светлыми поло-
сами. Чехлик из экскрементов, компактный. На губоцветных (Mentha, Lycopus,
Stachys, Salvia, Melissa, Nepeta, Galeopsis). Транспалеаркт, в лесах и на сырых лугах
.........................................................С viridis Linnaeus
44(39). 8-й тергит брюшка без длинных хет, отличающихся от обычных хет спин-
ной стороны.
45(46). Срединная выемка верхней губы округленно-трапециевидная, с прямым
основанием и без явственных внутренних углов; нижняя сторона губы с явствен-
ной зубчатой микроскульптурой посередине и по бокам. Внутренние маргиналь-
ные щетинки губы незначительно короче наружных (рис. 84,3). Усик немного длин-
нее ширины у основания, конический (рис. 82, 4). Вторичные ответвления боковых
отростков оканчиваются длинными гладкими шиловидными хетами (рис. 79, 77).
Подталкиватель цилиндрический, длинный (подрод Ташата Spaeth). Переднеспин-
ка без рисунка, стигмы светлые. Боковые отростки 7-8-го сегментов брюшка в 1,5
раза длиннее предыдущих. Хвостовые нити длинные, до половины длины тела;
боковые отростки составляют около половины ширины тела. Чехлик из экскремен-
тов, составной. На вьюнковых (Calystegia). Юг ДВ.........С. amurensis Kraatz
222
46(45). Срединная выемка верхней губы глубокая, с дуговидно выступающим ос-
нованием и резкими, острыми внутренними углами; нижняя сторона губы по бо-
кам без зубчатой микроскульптуры. Вторичные ответвления боковых отростков окан-
чиваются короткими овальными или коническими зазубренными хетами. На слож-
ноцветных.
47(48). Усики колпачковидные, не длиннее ширины (рис. 82, 3). Маргинальные
щетинки верхней губы (рис. 84, 4) одинаковой длины, толстые (подрод Lordicassis
Reitter). Боковые отростки в 7-8 раз короче ширины тела; хвостовые нити в 3-3,5
раза короче длины тела. Переднеспинка с явственным рисунком. Стигмы свет-
лые. Боковые отростки 7-8-го брюшных сегментов немного длиннее предыду-
щих. Чехлик из экскрементов, компактный. На Carduus. Алтай................
...................................................С. undecimnotata Gebler
48(47). Усики цилиндрические, заметно длиннее ширины (рис. 82, 5). Маргиналь-
ные щетинки верхней губы обычно тонкие, 1-2 внутренние гораздо короче наруж-
ных, часто плохо заметные (рис. 84, 5). Длина боковых отростков не менее 1/5-1/3
ширины тела.
49(58). Стигмы светлые.
50(53). Выемка верхней губы глубокая и более узкая; срединные щетинки очень
короткие, далеко не доходят до переднего края выемки, часто плохо заметные (рис.
84, 5). Отростки 7-8-го сегментов брюшка значительно длиннее предыдущих. Тело
светло-зеленое.
51(52). Переднеспинка без явственного склеротизованного рисунка. Боковые отро-
стки длинные, до 0,4-0,5 ширины тела. Вершины вторичных отростков с рашпиле-
видной скульптурой. Подталкиватель цилиндрический. Голова с неявственными
темными пятнами, хвостовые нити черные, блестящие, со светлыми вершинами.
Чехлик из экзувиев. На Achillea. Оба вида — ЕЧ, СК, СБ. Обычны на лугах...
....................................С. prasina liliger, С. sanguinolentha Muller
52(51). Переднеспинка с явственным склеротизованным рисунком. Боковые отрос-
тки более короткие, около 1/4 ширины тела. Вершины вторичных отростков без
рашпилевидной скульптуры. Подталкиватель треугольный. Голова с темными чет-
кими пятнами, хвостовые нити бурые. Чехлик из экскрементов, компактный. На
Achillea, Artemisia, Tanacetum. ЕЧ, СК, СБ, ДВ........С. denticollis Suffrian
53(50). Выемка верхней губы широкая, менее глубокая; срединные щетинки длин-
ные, хорошо заметные, достигают или немного не достигают переднего края выем-
ки (рис. 84, б). Переднеспинка с более или менее явственным рисунком.
54(57). Боковые отростки средне- и заднеспинки примерно одинаковой длины.
55(56). Боковые отростки брюшка примерно одинаковой длины. Подталкиватель
длинный (около 1/2 длины отростков 8-го сегмента), цилиндрический (рис. 85, 8).
Рисунок переднеспинки слабо выражен. Тело продолговато-овальное, длина боко-
вых отростков около 1/2 ширины тела, длина хвостовых нитей около 1/4 длины
тела. На Rhaponticum. ВСБ, в степях и других сухих стациях................
......................................................С mongolica Boheman
56(55). Боковые отростки 7-8-го сегментов брюшка гораздо длиннее предыдущих.
Подталкиватель очень короткий, треугольный (рис. 85, 9). Рисунок переднеспинки
явственный. Тело коротко-овальное, длина боковых отростков около 1/3 ширины
тела, длина хвостовых нитей не менее 1/2 длины тела. На сложноцветных подсем.
223
Рис. 85. Хвостовые нити (7-7) и подталкиватель (5, 9) личинок родов Cassida и
Aspidomorpha'.
1 — A. transparipennis, I возраст; 2 — С. piperata\ 3 — С. flaveola\ 4 — С. pallidicollis\ 5 — С.
berolinensis\ 6 — С. nebulosa', 7 — С. parvula\ 8 — С. mongolica\ 9 — С. vibex.
224
Cynareae (Carduus, Cirsium, Arctium, Centaurea, Serratula, Sonchus, Silybium, Onopor-
don). Транспалеаркт. Обычно на лугах.......................С. vibex Linnaeus
57(54). Боковые отростки средне- и заднеспинки различной длины: передние за-
метно короче и тоньше задних. Подталкиватель длинный, цилиндрический. Пере-
днеспинка с явственным рисунком. Тело продолговато-овальное, длина боковых
отростков около 1/3 ширины тела, длина хвостовых нитей около 1/3 длины тела. На
Centaurea. ЕЧ, СК.......................................С. pannonica Suffrian
58(49). Стигмы черные. Боковые края тела и основания боковых выростов темные.
59(60). Срединная выемка верхней губы (рис. 84, 6) трапециевидная (сужается к
основанию). Подталкиватель длинный, до половины длины отростков 8-го сегмен-
та, цилиндрический или конический. Переднеспинка с тонкими булавовидными
светлыми хетами. Боковые отростки тонкие, без мелких шипиков. Только передние
боковые отростки с темными основаниями. Тело яйцевидное, серо-зеленое. На слож-
ноцветных подсем. Cynareae (Carduus, Cirsium, Arctium, Centaurea, Onopordon, Sily-
Ьшп).Транспалеаркт, идет довольно далеко на север. На лугах................
.........................................................С. rubiginosa Muller
60(59). Срединная выемка верхней губы (рис. 84, 7) обратнотрапециевидная (рас-
ширяется к основанию). Подталкиватель очень короткий, треугольный. Переднес-
пинка с толстыми зубцевидными темными хетами. Боковые отростки более тол-
стые, с мелкими шипиками, особенно у основания. Все боковые отростки с тем-
ными основаниями. Тело более узкое, эллипсовидное, темно-буро-зеленое. На
Tanacetum, Achillea. ЕЧ, СК, СБ. На сырых лугах, болотах.
.....................................................С. sanguinosa Suffrian
7. Род Aspidomorpha Hope
Тело овально-вытянутое, наибольшая ширина в области первых брюшных сег-
ментов. Верхняя губа с широкой срединной выемкой, на дне которой расположено
6 тонких щетинок, и с 4 маргинальными щетинками с каждой стороны, из которых
2 внутренние значительно толще наружных (рис. 84, 8, 9). Глазков по 5 с каждой
стороны головы, мандибулы слабо хитинизированы, с 6 зубцами. Коготки при ос-
новании без явственного зубца (рис. 82, 13), Боковые отростки длинные и тонкие,
примерно равны половине ширины тела, с каждой стороны с 5-6 вторичными от-
ветвлениями. Переднекрайние отростки переднеспинки одинаковой длины, на сред-
не- и заднеспинке передние заметно короче задних; на 1—4-м сегментах брюшка
они постепенно уменьшаются кзади, на 5-м немного длиннее, чем на 4-м, на 6-8-м
вдвое длиннее, чем на 4-м. Переднеспинка в середине с продольным полем из тон-
ких и светлых хет, ее склерит по бокам с рисунком в виде 2 коричневых пятен.
Стигмы 1-го сегмента брюшка примерно в 2 раза крупнее последующих и окруже-
ны широким хитинизированным кольцом. Хвостовые нити не сросшиеся при осно-
вании, длинные и тонкие, с чехликом из экзувиев. Стерниты брюшка с рядами из
длинных хет.
Личинки 1-го возраста имеют на наружной стороне хвостовых нитей много-
численные длинные ответвления (рис. 85, 7), исчезающие у более старших возра-
стов.
В России 2 вида на юге ДВ. Жуки и личинки питаются на Calystegia.
225
Таблица для определения видов рода Aspidomorpha
1(2). Передний край верхней губы (не считая срединной выемки) прямой (рис. 84,
9). Переднеспинка с 2 слабо хитинизированными пятнами у задних углов, в середи-
не со слабым продольным полем из светлых хет, боковые отростки без мелких ши-
пиков .....................................A. transparipennis Motschulsky
2(1). Передний край верхней губы явственно вогнут (рис. 84, 8). Переднеспинка с 2
четкими хитинизированными пятнами у задних углов, в середине с продольным
полем из светлых густых и мелких хет, боковые отростки с густыми хетами.
................................................A, difformis Motschulsky
226
Литература
Бабенко З.С. 1952. Кувшинковый листоед (Galerucella nymphaeae L.) как вредитель смороди-
ны в условиях Томской области // Бюлл. Сиб. бот. сада. Вып. 3. С. 36-39.
Беньковский А.О. 1995. Морфология преимагинальных стадий жука-листоеда Galeruca
melanocephala Ponza (Coleoptera, Chrysomelidae) // Энтомологическое обозрение. T. 74.
Вып. 1. С. 15-18.
Беньковский А.О. 1999. Определитель жуков-листоедов европейской части России и евро-
пейских стран ближнего зарубежья. М. 204 с.
Беньковский А.О. 2001. Сем. Chrysomelidae. Подсемейство Donaciinae (листоеды - радужни-
цы), личинки // Определитель пресноводных беспозвоночных России и сопредельных
территорий. СПб.: Наука. Т. 5. С. 359-366.
Богачева И.А., Дубешко Л.Н. 1975. Экология и энергетика питания листоеда Gonioctenapallida
L. в тайге и лесотундре Сибири // Экология. № 3. С. 94-96.
Богданов-Катьков Н.Н. 1922. Хреновый листоед, или бабануха. Пг. 82 с.
Бровдий В.М. 1973. Жуки-листоеды Galerucinae // Фауна Украины. Киев. Т. 19. Вып. 7. 194 с.
Васильев Р.А. 1964.Фауна и экология листоедов Хасанского района Приморского края и био-
логия наиболее вредных видов в лесном хозяйстве. Автореф. канд. дис. М.
Гиляров М.С. 1966. Изучение беспозвоночных животных как компонента биоценоза // Про-
грамма и методика биогеоценологических исследований. М.: Наука. С. 163-194.
Гиляров М.С, Курчева Г.Ф. 1956. Особенности строения личинки листогрыза Chloropterus
versicolor F. Мог., обусловленные обитанием в почве // Зоол. журн. Т. 35. Вып. 3. С. 395-
399.
Гиляров М.С., Медведев Л.Н. 1964. Семейство Chrysomelidae — Листоеды // Определитель
обитающих в почве личинок насекомых. М.: Наука. С. 507-530.
Гусев Г.В. 1955. Соевый листоед как вредитель сельскохозяйственных культур на Сахалине //
Зоол. журн. Т. 34. Вып. 4. С. 806-809.
Джолова Н.Г. 1965. Насекомые — вредители овощных культур в Прибайкалье. М.: Наука.
Дубешко Л.Н. 1982. К познанию личинок листоедов Южной Сибири и Монголии // Зоологи-
ческие исследования в МНР. М. С. 152-157.
Дубешко Л.Н., Медведев Л.Н. 1972. Новый вид листоеда (Coleoptera, Chrysomelidae) — вре-
дитель ольхи в Прибайкалье // Экология вредных и полезных насекомых. Воронеж. С.
79-87.
Дубешко Л.Н., Медведев Л.Н. 1975. К познанию личинок листоедов (Coleoptera, Chrysomelidae)
фауны Монголии // Насекомые Монголии. Л.: Наука. Т. 3. С. 234-243.
Зайцев Ю.М. 1997. Морфология личинки Oomorphoides martensi (Coleoptera, Chrysomelidae)
из юго-восточной Азии // Зоол. журн. Т. 76. № 3. С. 371-374.
Зайцев Ю.М, Медведев Л.Н. 1983. Личинки листоедо-щитоносок трибы Aspidomorphini
(Chrysomelidae, Cassidinae) из Вьетнама // Фауна и экология животных Вьетнама. М.:
Наука. С. 130-145.
Зайцев Ю.М., Павлов С.И. 1986. Морфо-экологическая характеристика личинок рода Donacia
Поволжья // Экология животных Поволжья и Приуралья. Куйбышев. С. 44-50.
Зайцев Ю.М, Охрименко Н.В. 1989. Неизвестные личинки жуков-листоедов рода Chrysolina
(Coleoptera, Chrysomelidae) с Кавказа // Вести, зоол. № 2. С. 72-74.
Криволуцкая Г.О., Медведев Л.Н. 1966. Фауна жуков-листоедов (Coleoptera, Chrysomelidae)
Курильских островов // Энтомофауна лесов Курильских островов, п-ова Камчатки, Ма-
гаданской области. М.-Л.: Наука. С. 25-33.
Курчева Г.Ф. 1958. Почвообитающие личинки листогрызов подсемейства Eumolpinae
(Coleoptera, Chrysomelidae), наиболее распространенные в европейской части СССР //
Acta Soc. Entomol. Cechosloveniae. T. 55. No. 4. P. 383-393.
227
Курчева Г.Ф. 1967. Личинки Eumolpinae и Syneta betulae F. (Coleoptera, Chrysomelidae) // Эн-
томол. обозр. Т. 46. Вып. 1. С. 222-233.
Лопатин И.К., Дашевская Н.И. 1978. Описание личинки Pentamesa kondarensis Lop.
(Coleoptera, Chrysomelidae) // Фауна и экология животных Таджикистана. Душанбе: ТГУ.
С. 44-49.
Матис Э.Г. 1970. Описание личинок и куколок некоторых малоизвестных видов и имаго од-
ного нового вида (Coleoptera, Cassidinae) // Энтомологические исследования в Кирги-
зии. Фрунзе: Илим. С. 23-33.
Медведев Л.Н. 1962. Систематика и биология личинок Clytrinae (Coleoptera, Chrysomelidae) //
Зоол. журн. Т. 41. Вып. 9. С. 1334-1344.
Медведев Л.Н. 1968. О личинках Hispinae (Coleoptera, Chrysomelidae) фауны СССР // Зоол.
журн. Т. 47. Вып. 1. С. 79-84.
Медведев Л.Н. 1972. Фауна листоедов (Coleoptera, Chrysomelidae) Сахалина // Экология вред-
ных и полезных насекомых. Воронеж. С. 96-107.
Медведев Л.Н. 1974. Сем. Chrysomelidae — листоеды // Насекомые и клещи — вредители
сельскохозяйственных культур. Л.: Наука. Т. 2. С. 157-196.
Медведев Л.Н. 1981. Колорадский картофельный жук. Положение в системе, эволюция, мор-
фология имаго и личинки // Колорадский картофельный жук. М.: Наука. С. 27-54.
Медведев Л.Н. 1982. Листоеды МНР. Определитель. М.: Наука. 304 с.
Медведев Л.Н., 1992. Сем. Chrysomelidae —листоеды. Определитель насекомых Дальнего
Востока СССР. СПб.: Наука.Т. 3. Ч. 2. С. 533-602.
Медведев Л.Н., Воронова Н.В. 1976. Описание личинки Ambrostoma (Chrysomelidae) и рас-
пространение рода Ambrostoma в Монголии // Насекомые Монголии. Л.: Наука. Вып. 4.
С. 237-240.
Медведев Л.Н., Воронова Н.В. 1977. Обзор видов рода Diorhabda фауны МНР // Раститель-
ный и животный мир МНР. Л.: Наука. С. 238-244.
Медведев Л.Н., Долгин М.М. 1974. Ранее неизвестные личинки листоедов с Алтая. Сообщ 1
// Зоол. журн. Т. 53. Вып. 4. С. 642-645.
Медведев Л.Н., Долгин М.М. 1974а. Ранее неизвестные личинки листоедов с Алтая. Сообщ.
2 // Зоол. журн. Т. 53. Вып. 6. С. 942-945.
Медведев Л.Н., Дубешко Л.Н. 1973. К познанию личинок жуков-листоедов Прибайкалья //
Фауна и экология насекомых Восточной Сибири и Дальнего Востока. Иркутск. С. 171—
180.
Медведев Л. Н., Дубешко Л.Н. 1992. Определитель листоедов Сибири. Иркутск: Изд-во Ир-
кутск. ун-та. 220 с.
Медведев Л.Н., Зайцев Ю.М. 1971. Новые личинки Eumolpinae (Coleoptera, Chrysomelidae) с
Дальнего Востока // Зоол. журн. Т. 50. Вып. 7. С. 1026-1033.
Медведев Л.Н., Зайцев Ю.М. 1973. Личинки листоедов Восточной Сибири и Верхнего При-
амурья // Энтомофауна Советского Дальнего Востока. Владивосток. С. 153-165.
Медведев Л.Н., Зайцев Ю.М. 1974. Личинки листоедов-чехликоносцев (Coleoptera,
Chrysomelidae, Camptosoma) из Монголии // Насекомые Монголии. Л.: Наука. Вып. 2. С.
203-210.
Медведев Л.Н., Зайцев Ю.М. 1975. Описание личинки листоеда Paropsides duodecimpustulata
Gebl. // Насекомые средней тайги Якутии. Якутск. С. 49-54.
Медведев Л.Н., Зайцев Ю.М. 1976. Обзор личинок рода Crosita Motsch. (Col., Chrysomelidae)
// Зоол. журн. Т. 56. Вып. 1. С. 52-59.
Медведев Л.Н., Зайцев Ю.М. 1977. Обзор личинок рода Cryptocephalus (Coleoptera,
Chrysomelidae) из МНР // Растительный и животный мир МНР. Л.: Наука. С. 223-236.
Медведев Л.Н., Зайцев Ю.М. 1977а. Материалы к познанию личинок листоедов (Coleoptera,
Chrysomelidae) Монголии // Насекомые Монголии. Л.: Наука. Вып. 5. С. 353-371.
228
Медведев Л.Н., Зайцев Ю.М. 19776. Морфология личинки рода Cneorane (Coleoptera, Chry-
somelidae) Дальнего Востока // Фауна и экология насекомых Восточной Сибири и Даль-
него Востока. Иркутск: Изд-во Иркутск, гос. ун-та. С. 122-130.
Медведев Л.Н., Зайцев Ю.М. 1978. Обзор личинок рода Chrysochloa (Coleoptera, Chry-
somelidae) фауны СССР // Зоол. журн. Т. 57. Вып. 7. С. 1024-1032.
Медведев Л.Н., Зайцев Ю.М. 1978а. Личинки жуков-листоедов Сибири и Дальнего Востока.
М.: Наука. 182 с.
Медведев Л.Н., Зайцев Ю.М. 1980. Новые данные о личинках листоедов (Coleoptera, Chryso-
melidae) Монголии // Насекомые Монголии. Л.: Наука. Вып. 7. С. 97-106.
Медведев Л.Н., Ковалев О.В. 1983. Теоретические основы интродукции амброзиевых листо-
едов рода Zygogramma Chevr. (Coleoptera, Chrysomelidae) в СССР для биологической
борьбы с амброзией // Энтомол. обозр. Т. 67. Вып. 1. С. 17-32.
Медведев Л.Н., Оглоблин Д.А. 1965. Обзор личинок Cryptocephalinae (Coleoptera, Chrysome-
lidae) лесной зоны европейской части СССР // Зоол. журн. Т. 44. Вып. 7. С. 1018-1027.
Медведев Л.Н., Радзивиловская М.А. 1970. К познанию личинки Ischyronota elevata Rtt. (Co-
leoptera, Chrysomelidae, Cassidinae) из Средней Азии // Паразитофауна растений и жи-
вотных Узбекистана. Ташкент: Изд. Ин-та зоол. и паразитол. АН Уз. ССР. С. 199-204.
Медведев Л.Н., Самодерженков Е.В. 1986. Жизненные формы листоедов (Coleoptera, Chrys-
omelidae) // Экология жизненных форм почвенных и наземных членистоногих. Тр. МГПИ
им. В.И. Ленина. М. С. 91-102.
Медведев Л.Н., Чернов Ю.И. 1969. Новый вид листоеда рода Chrysomela (Coleoptera, Chiyso-
melidae)—важный компонент биоценозов Таймыра // Зоол. журн. Т. 48. Вып. 4. С. 532-537.
Мищенко А.И. 1940. Насекомые — вредители полевых и овощных культур Дальнего Восто-
ка. Хабаровск.
Мосейко А.Г., Медведев Л.Н. 2005. К систематике рода Pagria Lefevre 1884 с замечаниями
по роду Rhyparida Baly 1861 // Энтомол. обозр. Т. 84. Вып. 1. С. 108-124.
Оглоблин Д.А. 1927. Описание личинки Chaetocnema breviuscula Fald. (Coleoptera, Chryso-
melidae) // Защита растений. T. 4. № 1. С. 1-5.
Оглоблин Д.А. 1936. Листоеды, Galerucinae // Фауна СССР. Насекомые жесткокрылые. Т. 26.
М.-Л.: Изд-во АН СССР. 455 с.
Оглоблин Д.А., Медведев Л.Н. 1965. Обзор личинок Cryptocephalinae лесной зоны европей-
ской части СССР // Зоол. журн. Т. 44. Вып. 7. С. 1018-1027.
Оглоблин Д.А., Медведев Л.Н. 1971. Личинки жуков-листоедов (Coleoptera, Chrysomelidae)
европейской части СССР. Л.: Наука. 123 с.
Охрименко Н.В. 1993. Неизвестная личинка жука-листоеда рода Chrysolina (Coleoptera,
Chrysomelidae) с Кавказа // Вести, зоол. Вып. 3. С. 77-79.
Охрименко Н.В. 1995. Описание личинок двух видов жуков-листоедов рода Chrysolina (Co-
leoptera, Chrysomelidae) с Кавказа // Вести, зоол. Вып. 5-6. С. 13-17.
Охрименко Н.В. 1997. Преимагинальные стадии, распространение и экология жука листоеда
Timarcha hummeli (Coleoptera, Chrysomelidae) // Вести, зоол. Т. 31. Вып. 4. С. 82-84.
Палий В.Ф., Клепикова П.И. 1957. Экология щитоносок Центрально-черноземной полосы
РСФСР с описанием куколок наиболее распространенных видов // Энтомол. обозр. Т.
36. Вып. 1. С. 75-95.
Палий В.Ф. (при участии Клепиковой П.И.). 1959. Описание личинок жуков-щитоносок
(Cassidinae) европейской части СССР // Энтомол. обозр. Т. 38. Вып. 4. С. 805-818.
Палий В.Ф. 1961. Фауна вредных земляных блошек СССР. Фрунзе: Изд-во АН Кирг. ССР.
Палий В.Ф. 1962. Распространение, экология и биология земляных блошек фауны СССР.
Фрунзе: Изд-во АН Кирг. ССР. 118 с.
Самойлов Т.П. 1936. К экологии жуков-листоедов и слоников в заповеднике Горно-Таежной
станции Д.-В. филиала АН СССР // Тр. Горно-Таежной станции Д.-В. филиала АН СССР.
Т. 1. Владивосток. С. 239-264.
229
Синчилина Е.М. 1962. Массовые вредители бутонов лип в Приморье // Сообщения ДВ фили-
ала Сибирского отделения АН СССР. Владивосток. Вып. 16. С. 111-118.
Томилова В.П., Нарушевич Т.С. 1965. Кувшинковый листоед — вредитель смородины на
Байкале // Изв. Вост.-Сиб. отдела Геогр. о-ва СССР. Т. 64.
Энгельгард В., Мищенко А.И. 1931. Насекомые — вредители риса в Дальневосточном крае.
Хабаровск: Сельхозгиз. 69 с.
Яковлев Б. 1960. Колорадский картофельный жук. Рига.
Anderson W.H. 1938. Description of the larvae of Chaetocnema denticulata (liliger) and Chaetoc-
nema pulicaria Melsheimer (Coleoptera: Chrysomelidae) // Proc. Entomol. Soc. Wash. Vol.
40. No. 6. P. 161-169.
Balachovsky A., Mesnil L. 1935. Les insectes nuisibles aux plantes cultivees. Paris. 1137 p.
Bartkowska J., Warchalowski A. 1978. Studien an Blattkafer-Larven. I. Gattung Haltica Koch
(Coleoptera, Chrysomelidae) // Polskie Pismo Entomologiczne. T. 48. P. 583-591.
Bienkowski A. 1995. Morphology of larva of Phaedon concinnus Stephens and its distinction from
larvae of allied species (Insecta: Coleoptera: Chrysomelidae) // Entomologische Abhandlun-
gen, Staatliches Museum fur Tierkunde Dresden. Bd. 56. Nr. 9. S. 171-176.
Biondi M., De Nardis G. 1998. Descrizione della larva di primo stadio di Longitarsus minimus e L.
pratensis И Fragmenta entomologica. Roma. T. 30. Fasc. 2. P. 261-270.
Biondi M., De Nardis G. 1999. Morphological description of the first-instar larva of Derocrepis
sodalis (Coleoptera, Chrysomelidae, Alticinae) // Ital. J. Zool. Vol. 66. P. 393-397.
Blunck H. 1932. Zur Kenntnis der Lebensgewohnheiten und der Metamorphose getreidebewohnender
Halticinen // Z. angew. Entomol. Bd. 19. S. 358-394.
Bokermann WN. 1964. A. Notes on the biology of Poropleura cuprea KI. (Coleoptera, Chry-
somelidae, Chlamisinae) // Ann. Acad, brasil. Cienc. T. 36. No. 2. P. 165-172.
Borner C., Blunck H. 1919. Larven der Kafergattung Phyllotreta II Illustr. Landwirtsch. Ztg. Nr.
75-76. S. 1-4.
Boving A.G. 1910. Natural history of the larvae of Donaciinae // Int. Rev. Hydrobiol. Hydrogr.,
Biol. Suppl. I ser. Vol. 3. P. 1-108.
Boving A.G. 1927. Descriptions of larvae of the genera Diabrotica and Phyllobrotica, with a dis-
cussion of the taxonomic validity of the subfamilies Galerucinae and Halticinae // Proc. Ento-
mol. Soc. Wash. Vol. 29. No. 9. P. 193-205.
Boving A.G. 1929. Beetle larvae of the subfamily Galerucinae // Proc. Nat. Mus. U. S. Vol.75. No.
2. P. 1-43.
Boving A.G., Craighead F.C. 1931. An illustrated synopsis of the principal larval forms of the
Coleoptera // Entomol. amer. N.S. P. 1-351.
Came P.B. 1966. Growth and food consumption during the larval stages of Paropsis atomaria
(Coleoptera, Chrysomelidae) // Entomol. exp. et appl. T. 9. P. 105-112.
Carpenter G.H. 1966. A new cabbage-eating larva, Psylliodes chrysocephala Lin. // J. Econ. Biol.
(Entomol.). Vol. 1. P. 152-156.
Chang Y. 1965. On the external anatomy and the caudal claws of the larva of Donacia provosti I I
Acta Entom. Sinica. Vol. 14. No. 6. P. 613-616.
Chen S.H. 1935. Recherches sur les Chrysomelinae de la Chine et du Tonkin. Tesis. Paris: Fac. Sci.
Univ. 22 p.
Cho H.W, Lee J.E. 2006. Immature stages of Glyphocassis spilota (Gorham) (Coleoptera Chry-
somelidae: Cassidinae) from Korea // Entomological Research. Vol. 36. P. 42-44.
Chuan Chien S. 1940. Chaetotaxy in Chrysomelid larvae // Sinensia. Vol. 11. P. 549-572.
Cox M.L. 1981. Notes on the biology of Orsodacne Latreille with a subfamily key to the larvae of
the British Chrysomelidae (Coleoptera) // Entomol. Gaz. Vol. 32. P. 123-135.
Cox M.L. 1982. Larvae of the British genera of chrysomeline beetles (Coleoptera Chrysomelidae)
// Systematic Entomology. Vol. 7. P. 297-310.
230
Cox M.L. 1991. The larvae of British Phaedon (Coleoptera: Chrysomelidae: Chrysomelinae) I I
Entomol. Gaz. Vol. 42. P. 267-280.
Dobson R.M. 1960. The immature stages of the flea beetles Psylliodes cuprea (Koch) and Psyl-
liodes chrysocephala (L.) (Col., Chrysomelidae) II The Entomologist’s Monthly Magazine.
Vol. 46. P. 1-4.
Doguet S. 1994. Faune de France. France et regions limitrophes. 80. Coleoptera: Chrysomelidae 2.
Alticinae. Paris. 694 pp.
Donisthorpe A. 1902. The life history of Clytra quadripunctata L. И Trans. Entomol. Soc. London.
P. 11-24.
Emden F.L., van. 1967. The taxonomic significance of the characters of immature insects I I Annu.
Rev. Entomol. Vol. 2. P. 91-106.
Fiori G. 1948. Contributi alia conoscenza morfologica ed etologica dei Coleotteri. II. Lachnaea
italica Wse. // Boll. 1st. Entomol. Univ. Stud. Bologna. T. 17. P. 188-195.
Fiori G., 1952. Contributi alia conoscenza morfologica ed etologica dei Coleotteri V. Coptocepha-
la kiisteri Kraatz e Cryptocephalus frenatus Laich. I I Boll. 1st. Entomol. Univ. Stud. Bologna.
T. 18. P. 182-196.
Fiori G. 1957. Contributi alia conoscenza morfologica ed etologica dei Coleotteri VI. La larva del
Crisomelide Clitrine Tituboea bigultata 01.1 I Ann. Fac. Agron. Univ. Sassari, Studi Sassaresi.
Ser. 3. T. 5. P. 88-96.
Gomandt H., 1955. Die Kafergattung Phyllodecta Kirby H Dtsch. Entomol. Z., N.F. Bd. 2. H. 1-2.
S. 1-100.
Grandi G. 1932. Morfologia ed etologia comparata dei Insetti a regime specializzato II. La morfo-
logia delle larve minatrici di due Coleotteri delle sottofamiglia degli Alticini I I Mem. R. Ac-
cad. Sci. 1st. Bologna. Ser. 8. T. 9. P. 3-10.
Grandi G. 1935. Morfologia ed etologia comparata dei Insetti a regime specializzato VI. La morfo-
logia delle larve minatrici degli Hispini dei genus Hispa L. e Hispella Chap. (Coleoptera-
Chrysomelidae) И Mem. R. Accad. Sci. 1st. Bologna. Ser. 9. T. 13. P. 3-10.
Grandi G. 1938a. Morfologia ed etologia comparata dei Insetti a regime specializzato XV. La mor-
fologia e l”etologia delle larve di tre Coleotteri delle famiglia dei Crisomelidi e dei Curcu-
lionidi I I Boll. 1st. Entomol. Univ. Stud. Bologna. T. 11. P. 1-16.
Grandi G. 1938b. Morfologia ed etologia comparata dei insetti a regime specializzato XIV. La
morfologia e T’etologia delle larve di due Coleotteri Crisomelidi delle tribu degli Alticini I I
Mem. R. Accad. Sci. 1st. Bologna. Ser. 9. T. 5. P. 165-170.
Gressitt J.L. 1962. The tortoise beetles of China I I Proc. Calif. Acad. Sci. Vol.. 27. No. 17. P. 433-
592.
Gressitt J.L., Kimoto S. 1963. The Chrysomelidae of China and Korea, IIИ Рас. Ins. Monogr. IB.
Hennig W. 1938. Ubersicht fiber die Larven der wichtigsten deutschen Chrysomelinen I I Arb. phys-
iol. angew. Entomol. Bd. 5. H. 2. S. 85-136.
Henriksen K. 1927. Larveme И V. Hansen. Biller 7. Bladbiller og Bonnebiller. Danmarks Fauna.
Copenhagen. Bd. 31. S. 290-376.
Hiller E. 1975. Zur Kenntnis des Schwarzbraunen Kiefemblattkafers, Luperus pinicola (Duft.).
(Coleoptera, Chrysomelidae) I IZ. angew. Entomol. Bd. 79. S. 396-421.
Jolivet P. 1950. Les parasites, predateurs et phoretiques des Chrysomeloidea de la faune franco-
beige I I Bull. Inst. Sci. nat. Belg. T. 26. Fasc. 34. P. 4.
Jolivet P. 1952. Quelques donnees sur la myrmecophilie des Clytrides I I Bull. Inst. Sci. nat. Belg. T.
28. Fasc. 8. P. 1-12.
Jolivet P, Theodorides J. 1952a. Les parasites, phoretiques et predateurs des Chrysomeloidea I I
Bull. Inst. Sci. nat. Belg. T. 28. Fasc. 20. P. 1-19.
Kanervo V. 1946. Tutkimuksia lepan lehtikuoriaisen, Melasoma aenea L., luontaisista vihollisista
И Ann. zool. «Vanamo». Vol. 12. No. 3. P. 1-206.
231
Kimoto S. 1952. A phylogenetic consideration of Chrysomelinae based on immature stages of
Japanese species И J. Fac. AgfL Kyushu Univ. Vol. 12. No. 2. P. 57-88.
Kimoto S. 1957. Biological studies on earlier stages of Japanese Chrysomelinae (Coleoptera, Chry-
somelidae) I I Sci. Bull. Fac. Agr., Kyushu Univ. Vol. 16. No. 1. P. 41-57.
Kimoto S. 1962. Description., of immature stages of Japanese Chrysomelinae, belonging to a ge-
neric group Chrysolina, Gonioctena, Phola and Phaedon II J. Fac. Agr. Kyushu Univ. Vol. 12.
No. 2. P. 89-104.
Kimoto S. 1962a. Description of immature stages of Japanese Chrysomelinae, belonging to a ge-
neric group Chrysomela I I J. Fac. Agr. Kyushu Univ. Vol. 12. No. 2. P. 105-116.
Kimoto S., Takizawa H. 1994. Leaf beetles (Chrysomelidae) of Japan. Tokai University Press. 539
pp.
Klausnitzer B. 1970. Die Larve von Phytodecta quinquepunctata (F.) (Col. Chrysomelidae) I I En-
tomol. Nachr. Ber. Bd. 3. S. 34-36.
Kurosa K. 1959. Illustrated insect larvae of Japan. Tokyo.
Lipp H. 1935. Lebensweise von Metasoma aenea L. in der Mark I I Dtsch. Entomol. Z. S. 1-64.
Lopatin I.K., Aleksandrovich O.R., Konstantinov A.S. 2004. Check list of leaf-beetles (Coleoptera,
Chrysomelidae) of the eastern Europe. Olsztyn: Wydawnictwo Mantis. 343 p.
Maim J.S., Crowson R.A. 1981. The systematic position of Orsodacne Latr. and Syneta Lac. (Co-
leoptera, Chrysomelidae), in relation to characters of larvae, internal anatomy and tarsal ves-
titure I I J. Nat. Hist. Vol. 15. P. 727-749.
Manolache C.J., Dobreanu E., Manolache F. 1939. Recherches morphologiques sur 1’Altise de la
Mauve (Podagrica fuscicornis L.) I I Verb. VII. Int. Kongr. Entomol. Bd. 4. S. 2544-2560.
Manolache C.J., Dobreanu E., Manolache F. 1943. Observati morfologice si biologice a supra in-
sectei Podagrica malvae (liliger) // Bui. Soc. Nat. Romania. T. 17. P. 27-66.
Marshall J.E. 1979. The larvae of the British species of Chrysolina (Chrysomelidae) I I Syst. Ento-
mol. Vol. 4. No. 4. P. 409-417.
Marshall I.E. 1980. A key to some larvae of the British Galerucinae and Alticinae (Coleoptera:
Chrysomelidae) I I Entomol. Gaz. Vol. 31. P. 275-283.
Medvedev L.N., Matys E.G. 1975. Neue palaarktische Schildkaferlarven der Gattung Cassida L. //
Dtsch. Entomol. Z., N.F. Bd. 22. H. 1-3. S. 137-143.
Monros F. 1949. Descripcion de las metamorfosis de «Lamposoma chorisiae» Monros 6 consid-
eraciones taxonomicas sobre «Lamprosominae» (Coleoptera, Chrysomelidae) // Acta zool. T.
7. p. 449-466.
Monros F. 1954. Megalopus jacobyi, nueva plaga de Solanaceas en el noroeste Argentine, con
notas sobre biologia 6 taxonomia de Megalopinae (Coleoptera, Chrysomelidae) // Rev. Agr.
noroeste argent. T. 1. No. 2. P. 167-179.
Narita Y. 1991. Description of the larva of Donacia clavareaui (Coleoptera Chrysomelidae) //
Elytra. T. 19. P. 21-23.
Newton H.C.F. 1934. On the biology of Psylliodes hyoscyami (L.), the Henbane flea beetle, with
descriptions of the larval stage // Ann. Appl. Biol. Vol. 21. P. 153-161.
Ohno M. 1956. Note on a Chrysomelid-beetle, Gastrophysa atrocyanea Motsch. (Coleoptera Chry-
somelidae) in Japan П Insect. Ecology. Vol. 5. No. 13. P. 147-157.
Ohno M. 1958. On the genus Paropsides Motsch. of Japan I I Kontyu. Vol. 26. No. 1. P. 33-40.
Okhrimenko N.V. 1992. The larva of the leaf beetle Chrysolina rosti kubanensis L. Medvedev and
Okhrimenko, 1991, from the Caucasus (Coleoptera Chrysomelidae) I I Russian Entomol. J.
Vol. 1. No. 1. P. 63-65.
Osborne J. A. 1880. On the eggs and larvae of some Chrysomela and other (allied) species of Phy-
tophaga // Entomol. Month. Mag. Vol. 18. P. 150-154.
Paterson N.F. 1930. The bionomics and morphology of the early stages of Paraphaedon tumidulus
Germ. H Proc. Zool. Soc. London. P. 627-676.
232
Paterson N. 1931. Studies on the Chrysomelidae. Pt II. The bionomics and comparative morpholo-
gy of the early stages of certain Chrysomelidae (Coleoptera, Phyt.) // Proc. Zool. Soc. London.
P. 879-949.
Peterson A. 1961. Larvae of Insects. Pt 2. Columbus, Ohio. 416 p.
Reineck G. 1913. Beitrag zur Lebensweise von Cryptocephalus janthinus Germ. // Dtsch. Ento-
mol. Z. H. 2. S. 163-168.
Reineck G. 1929. Beitrag zur Lebens- und Entwicklungsweise von Coleopteren H Z. wiss. Inst.
Biol. Bd. 24. S. 220-226.
Rhynehart J.G. 1922. On the life history and bionomics of the flax flea-beetle Longitarsus parvulus
(Payk.) with descriptions of the hitherto unknown larval and pupal stages // Sci. Proc. R.
Dublin Soc. Vol. 16. P. 497-541.
Sanderson E.D. 1902. Notes upon the structure and classification of Chrysomelid larvae П Proc.
Entomol. Soc. Wash. P. 21-31.
Seeno T.N., Wilcox J. A. 1982. Leaf beetle genera (Coleoptera: Chrysomelidae) // Entomography.
Vol. 1. P. 1-221.
Shaheen A.A. 1973. Morphology of adult and immature stages of Aulacophora foveicollis Lucas //
Agric. Res. Rev. Vol. 51. No. 1. P. 79-90.
Smirnov N.N. 1960. Nutrition of Galerucella nymphaeae L. — mass consumer of water-lily H
Hydrobiologia. Vol. 15. No. 3. P. 208-224.
Skwarra E. 1927. Uber die Emahrungsweise der Larven von Clytra quadripunctata L. // Zool. Anz.
Bd. 71. H. 3/4. S. 83-96.
Steinhausen W.R. 1950. Vergleichende Morphologic, Biologic und Okologie der Entwicklungs-
Stadien der in Niedersachsen heimischen Schildkafer (Cassidinae, Chrysomelidae) und deren
Bedeutung fiir die Landwirtschaft. Braunschweig. 67 S.
Steinhausen W.R. 1965. Vergleichende Morphologic des Labrum von Blattkaferlarven (Coleoptera
Chrysomelidae) // Dtsch Entomol. Z., N.F. Bd. 13. H. 4/5. S. 313-322.
Steinhausen W.R. 1978. Bestimmungstabelle fur die Larven der Chrysomelidae (partim) H B. Klaus-
nitzer. Ordnung Coleoptera Larven. Berlin. S. 336-343.
Steinhausen W.R. 1982. Status of Alticinae Larvae Research (Coleoptera: Chrysomelidae) H Spix-
iana, Suppl. 7. P. 39-42.
Steinhausen W.R. 1994a. 116. Familie: Chrysomelidae // B. Klausnitzer: Die Larven der Kaefer
Mitteleuropas. Bd. 2. Krefeld: Goecke & Evers. S. 231-314.
Steinhausen W.R. 1994b. Larvae of Palearctic Timarcha Latreille // Jolivet, Cox & Petitpierre (ed.).
Novel aspects of the biology of Chrysomelidae. Kluver Acad. Publ. P. 119-125.
Steinhausen W.R. 1995. Neue Blattkafer-Larven (Coleoptera: Chrysomelidae) // Entomol. Nachr.
Ber. Bd. 39. H. 1/2. S. 65-67.
Steinhausen W.R. 1996. Status of West Palearctic leaf beetle larvae research П Jolivet & Cox (ed.).
Chrysomelidae biology. Vol. 3. General studies. Amsterdam: SPB Academic Publishing. P.
65-91.
Steinhausen W.R. 2004. Neue Westpalaearktische Blattkafer-Larven und -Puppen (Coleoptera,
Chrysomelidae) П Entomol. Bl. Bd. 100. H. 2. S. 91-103.
Swietojanska J. 2005. Comparative description of first instar larvae of Cassida nobilis Linnaeus,
1758 and Cassida vittata Villers, 1789 (Coleoptera: Chrysomelidae, Cassidinae) // Genus.
Vol. 16. No. 1. P. 49-68.
Takizawa H. 1971. On the larvae of the genus Chrysolina Motsch. in Japan (Coleoptera, Chry-
somelidae) II Entomol. Rev. Japan. Vol. 23. No. 2. P. 102-109.
Takizawa H. 1972. Descriptions of larvae of glanduliferous group of Galerucinae in Japan, with
notes on subdivisions of the subfamily // Insecta Matsumurana, Suppl. 10. P. 1-22.
Takizawa H. 1976. Larvae of the genus Gonioctena Chevrolat (Coleoptera Chrysomelidae): de-
scription of Japanese species and the implication of larval characters for the phylogeny H
Kontyu, Tokyo. Vol. 44. No. 4. P. 444-468.
233
Takizawa H. 1989. Notes on larvae of the subfamily Chrysomelinae (Coleoptera Chrysomelidae),
part 11 I Kanagawa-Chuho, Yokogama. No. 90. P. 243-256.
Takizawa H. 2005. Supra-generic subdivisions of the subfamily Alticinae based on larval charac-
ters, with descriptions of larvae of Hispaniolan species (Coleoptera Chrysomelidae) I I Insecta
Matsumurana, new ser. Vol. 62. P. 187-206.
Tolg F. 1913. Psylliodes attenuata Koch, derHopfen- oderHanf-Erdfloh. I. Teil: Morphologic und
Biologic der Praimaginalstadien I I Verh. Zool.-Bot. Ges. Wien. Bd. 63. S. 1-25.
Urban C. 1913. Beitrage zur Lebensgeschichte der Kafer. 1. Luperus saxonicus Gmel. 2. Liiperus
niger Goeze H Entomol. Bl. Bd. 9. H. 3/4. S. 16-19, 57-58.
Urban C. 1922. Zur Biologie der Zeugophora flavicollis Mrsh. (Col. Chrysom.) I I Dtsch. Entomol.
Z. Bd. 17. S. 405-408.
Urban C. 1932. Galeruca melanocephala Ponza // Entomol. Bl. Biol. Syst. Kafer. Bd. 28. S. 5-11.
Van Emden H.F. 1962. Key to the species of British Cassidinae larvae (Col. Chrysomelidae) H
Entomol. Mon. Mag. Vbl.98. P. 33-36.
Vig K. 1989. The morphology of Phyllotreta larvae (Col. Chrysomelidae) and determination keys
to larvae of more common species П Noveny-vedelen. Vol.. 25. No. 9. P. 412-419.
Vig K. 1991. Morphological description of immature stages of horse radish flea-beetle, Phyllotreta
annoraciae Koch (Coleoptera, Chrysomelidae) П G. Agric. Vol. 3. No. 1. P. 51-62.
Welch R.C. 1972. The biology of Hermaeophaga mercurialis F. (Coleoptera, Chrysomelidae) П
Entomol. Gaz. Vol. 23. No. 3. P. 153-166.
Zwolfer H., Eichhorn O. 1966. The host ranges of Cassida spp., attacking Cynareae (Compositae)
in Europe IIZ. angew. Entomol. Bd. 58. H. 4. S. 384-397.
234
Алфавитный указатель латинских названий
насекомых
abchasica, Chrysolina 121
absinthii, Pallasiola 27, 159
adamsi, Syneta 90
Adephaga 11
adonidis, Entomoscelis 140, 143, 144, 146
aenea, Plagiostema 131
aenescens, Altica 194, 200
aenescens, Pyrrhalta 166
aeruginosa, Chrysolina 114,119
affinis, Plateumaris 32, 38, 43
affinis, Psylliodes 189
Agelasa 156, 177
A. nigriceps 177
Agelastica 155, 174
A. alni 158, 176
A. coerulea 158, 176
agnus, Cryptocephalus 74, 76, 81, 84
Agrostithassa 104, 137
A. glabra 128, 137
Alledoya 220
Allorinula 126
alpinus, Colaphus 141, 143
altaica, Crosita 123, 127
alni, Agelastica 158,176
Altica 23, 25, 149, 155, 193
A. ampelophaga 194
A. aenescens 194, 200
A. brevicollis 194
A. carduorum 194
A. helianthemi 194
A.impressicollis 193
A. lythri 194,200
A. oleracea 184,194
A. pusilia 194
A. quercetorum 200
A. tamaricus 194
Alticinae 3, 5, 12, 13, 15, 17, 18, 19, 26, 30,
102, 148, 149, 150, 157, 183, 184, 188, 198
Ambrostoma 106, 111, 112
A. quadriimpressum 113
ampelophaga, Altica 194
amurensis, Cassida 210, 216, 221, 222, 224
amurensis nigrilabris, Labidostomis 62, 65
analis, Chrysolina 115
Anaphes 25
Anaphoides 25
anchusae, Longitarsus 183,186
angulosa, Dactylispa 206, 208
angusticollis, Chrysolina 124
angusticollis, Psylliodes 189
annulata, Zeugophora 45, 47
Antipa 58, 66
Aphthona 155, 196
A. coerulea 197
A. cyparissiae 200
A. euphorbiae 188, 197, 200
A. nonstriata 184,196
A. semicyanea 196
A. venustula 197
Apophylia 151, 163, 164
A. furcigera 165
appendiculata, Macroplea 34, 36, 41, 43
Apterocuris 107, 142
A. sibirica 135, 142
aquatica, Donacia 39, 41
arctica, Chrysolina 113,119,122
Argopistes 15, 156, 203, 204
A. biplagiatus 204
Argopus 12, 156, 157, 199
A. bicolor 188,200
A. clypeatus 200
A.nigritarsis 200
A.punctipennis 200
arida, Clytra 61, 67, 68
aridula, Chaetocnema 191
Arma 25
armata, Fleutiauxia 150,168
armoraciae, Phaedon 139
armoraciae, Phyllotreta 181,184,188
asclepiadeus, Eumolpus 94, 95
Asecoides 25
asiatica, Chrysochares 101
Asilidae 25
Asiorestia 155, 196
A. ferruginea 188, 196
asparagi, Crioceris 53
Aspidomorpha 13, 24, 213, 221, 224, 225
A. difformis 216,221,226
A. transparipennis 221, 224, 226
atra, Hispa 206, 207
atra, Phyllotreta 182,188
Atrachya 154, 172
A. menethriesi 173
atraphaxidis, Clytra 58, 61, 67, 68
atrocyanea, Gastrophysa 105, 135, 136
atrovirens, Phratora 130, 189, 152, 168, 171
attenuata, Psylliodes 171
Aulacophora 152, 168, 171
235
A. foveicollis 171
A. nigripennis 150, 168, 171
aurata, Crepidodera 187,200
aureolus, Cryptocephalus 78, 86
aurichalcea, Chrysolina 115,122
aurita, Smaragdina 70
azurea, Cassida 218
ballotae, Longitarsus 186
balyi, Basilepta 98
balyi, Sphaeroderma 202
basilea, Oreina 117,126
Basilepta 93, 97
B. balyi 98
B. fiilvipes 94, 95, 98
Batophila 152, 179
B.rubi 180
berolinensis, Cassida 210, 220, 224
betulae, Syneta 45, 90
bicolor, Argopus 188, 200
bicolora, Donacia 37,41,42
bidentulus, Cryptocephalus 80, 81
bifasciata, Galerucida 178
biguttatus, Cryptocephalus 83
bilineatus, Cryptocephalus 79
biplagiatus, Argopistes 204
bipunctata, Labidostomis 62, 63
bipunctatus, Cryptocephalus 78, 83
biramosa, Thlaspida 217
bivulneratus, Cryptocephalus 82,83,84
bothnicus, Cryptocephalus 83
braccata, Plateumaris 33, 38, 43
Brachinus 25
Brachyphytodecta 144
Braconidae 25
brevicollis, Altica 194
Bromius 93, 97
B. obscurus 19, 93, 94, 95, 97
Bruchidae 12
brunnicomis, Chrysolina 114
bungei, Chrysolina 112,116
calmariensis, Pyrrhalta 167
Calomicrus 152, 169, 170
C. minutus 171
C. pinicola 171
Calosoma 25
canaliculata, Cassida 214, 222
capreae, Lochmaea 19, 165
Carabus 25
caraganae, Cercyonops 146, 147
carduorum, Altica 194
camifex, Chrysolina 114,120
caspica, Chrysolina 124
Cassida 14, 15, 23, 25, 217, 218, 221
C. amurensis 210, 216, 221, 222, 224
C. azurea 218
C. berolinensis 210, 220, 224
C. canaliculata 214, 222
C. concha 21, 22, 218, 220
C. denticollis 221, 223
C. ferruginea 210
C. flaveola 218, 224
C. fuscorufa 218
C. hemisphaerica 218
C. inquinata 220
C.japana 219
C. lineola 21,216,219
C.mandli 216,219
C. margaritacea 220
C. mongolica 216, 223, 224
C. murraea 116,221,222
C. nebulosa 210, 216, 220, 224
C. nobilis 222
C. omata 218
C. pallidicollis 210,218,220,224
C. pannonica 225
C. panzeri 21, 210, 218
C. parvula 210, 210, 222, 224
C. piperata 22, 219, 224
C. prasina 210, 216, 223
C. rubiginosa 18,221,225
C. sanguinolentha 223, 225
C. sanguinosa 221
C. scraphina 218
C. spaethi 218
C. stigmatica 216, 219
C. subreticulata 218, 220
C. undecimnotata 23, 221, 223
C. velaris 218,222
C. versicolor 214, 219
C. vespertina 220
C.vibex 18,224,225
C. viridis 221,222
C. vittata 218
Cassidinae 3, 6, 10, 13, 15, 18, 19, 20, 21, 26,
30, 209,210,211,212,214,216
Cassidulella 221
Cataglyphis 68
cavigera, Chrysolina 116,119,122
Cerambycidae 12
Cercyonops 107, 146, 147
C. caraganae 146, 147
cerasi, Orsodacne 45, 48
cerealis, Chrysolina 117,124
Chaetocera 128
coerulea, Aphtona 197
Chaetocnema 149, 153, 154, 194
236
Ch. aridula 191
Ch. hortensis 191
Ch. tibialis 188,192
Chalcididae 25
chalcomera, Psylliodes 189
Chalcoides 186
Cheilotoma 60, 61, 63
Ch. musciformis 64
Chilotoma 63
chinensis, Labidostomis 62, 65
Chlamisinae 3, 17, 19, 21, 29, 87
Chlamisus 87
Ch. pubiceps 87
Chloropterus 93, 97
Ch. versicolor 95, 97, 98
chrysocephala, Psylliodes 188,190
Chrysochares 101
Ch. asiatica 98, 101
Chrysochloa 106, 125
Chrysochus 96, 99
Chrysolina 5, 14, 16,18, 23, 24, 102, 104, 106,
113, 117, 118, 119, 120, 122, 123, 125, 126
Ch. abchasica 121
Ch. aeruginosa 114,119
Ch. angusticollis 124
Ch. analis 115
Ch. arctica 113, 119, 122
Ch. aurichalcea 115, 122
Ch. brunnicomis 114
Ch.bungei 112,116
Ch. camifex 114, 120
Ch. caspica 124
Ch. cavigera 116, 119, 122
Ch. cerealis 117,124
Ch. chalcites 121
Ch. coerulans 122
Ch. convexicollis 114, 119, 122
Ch. difficilis 121
Ch. discriminate 114,120
Ch. diversipes 123
Ch. dudkoi 120
Ch. exanthematica 119, 121
Ch. fastuosa 121
Ch. geminata 119, 121
Ch. goettingensis 123
Ch. graminis 112, 122
Ch. gypsophilae 116
Ch. haemochlora 117,123
Ch. haemoptera 119
Ch. herbalcea 122
Ch. hyperici 120, 121
Ch. infiiscipes 114
Ch. instabilis 115,119
Ch. jenisseiensis 118
Ch. koltzei 112, 115, 117, 119, 122, 123
Ch. kozhantchikovi 114
Ch. latimargo 112, 118, 123
Ch. limbata 118, 120
Ch. lineigera 120
Ch. marginata 115, 119, 122
Ch. menthastri 122
Ch. mongolensis 114, 119, 120, 122
Ch. montana 119, 120, 123
Ch. nikolskyi 117,124
Ch. oricalcia 121
Ch. pedestrid 118,124
Ch. perforata 112, 113, 119
Ch. pirka 116, 118, 150
Ch, polita 112, 119, 122
Ch. porphyrea 118,124
Ch. pusa 114
Ch. relucens 115
Ch. rosti 124
Ch. rufilabris 114
Ch. sanguinolentha 118, 122
Ch. septentrionalis 112, 116, 122
Ch. staphylea 120
Ch. subsulcata 114, 119
Ch. tolli 116
Ch. trapezicollis 125
Ch. undulata 115,122
Ch. uijanchaica 113
Ch. ussuriensis 121
Ch. varians 120, 121
Ch. violacea 123
Ch. virgata 115, 117, 124
Ch. yezoensis 121
Chrysomela 20, 21, 23, 103, 104, 113, 131
Ch. collaris 132
Ch. cupreum 132
Ch. lapponicum 128, 132
Ch. populi 132
Ch. saliceti 132
Ch. taimyrensis 25, 132
Ch. tremulae 128, 132
Ch. vigintipunctata 128, 132
Ch. vrangeliana 132
Chrysomelidae 3, 12
Chrysomelinae 3, 6, 12, 13, 14, 15, 18, 19, 24,
25, 26, 30, 102, 103, 104, 105, 108, 112, 127,
128, 135, 143, 144
chujoi, Gonioctena 147
cinerea, Donacia 32, 34, 35, 37, 38, 40
Clavicomia 11
clavipes, Donacia 34, 39
clavareaui, Donacia 35
237
clementzi, Sternoplatys 139
Cleoporus 92
C. variabilis 92
clypeatus, Argopus 200
Clitena 151, 157, 158, 161
C. fuscipennis 162
Clytra 13, 17, 18, 59, 60, 66
C. arida 61, 67, 68
C. atraphaxidis 58, 61, 67, 68
C. laeviuscula 67, 68
C. quadripunctata 61, 64, 67, 68
Clytraxeloma 44
C. cyanipennis 44
Clytrella 67
Clytrinae 3, 13, 17, 21, 29, 57, 58, 59, 61, 64,
72, 73, 87
Cneorane 152, 157, 167
C. violaceipennis rufipes 168, 173
Coccinella 25
cochleariae, Phaedon 135, 140
coerulans, Cryptocephalus 80, 81
coerulea, Agelastica 158, 176
coerulea, Aphthona 197
coerulea, Oreina 126
coerulea, Zicrona 25
coerulescens, Cryptocephalus 78
Colaphellus 140
Colaphus 107, 141, 143
C. alpinus 141, 143
C. foveoiatus 142, 143
C. sophiae hoefti 141
Colasposoma 92
C. dauricum 94, 95, 96, 97
collare, Exosoma 172
collaris, Chrysomela 132
collaris, Smaragdina 70
Colobaspis 44
Colobaspis japonica 44
concha, Cassida 21, 22, 218, 220
concinnus, Phaedon 105, 135, 139
concolor, Oomorphus 88, 89
confusus, Criphtocephalus 78
consimile, Pagria 99
constricticollis, Plateumaris 43
convexicollis, Chrysolina 114, 119, 122
Coptocephala 58, 60, 68, 69, 70
C. orientalis 69, 70
C. unifasciata 69, 70
cordiger, Cryptocephalus 76, 84, 85
coreana, Gonioctena 145, 146
coryli, Cryptocephalus 78, 85
crassipes, Donacia 37, 38, 40
crataegi, Lochmaea 165
Crepidodera 153, 186, 187, 196
C. aurata 187, 200
C. fulvicomis 187
C. plutus 187
Criocerinae 3, 12, 13, 14, 15, 17, 18, 19, 20, 21,
24, 30, 49, 50,51,52
Crioceris 50,51,52,53
C. asparagi 53
C. duodecimpunctata 53
C. quatuordecimpunctata 53
cristula, Cryptocephalus 86
Crosita 18, 106, 123, 125, 126
C. altaica 123, 127
C. kowalewskyi 112, 123, 127
C. longipes 112,123,127
C.pigra 112, 123, 126
cruciferae, Phyllotreta 182
crux, Cryptocephalus 82, 84
Cryptocephalinae 3, 6, 13, 17, 19, 21, 26, 29,
72, 73, 74, 76, 84, 87
Cryptocephalus 5, 13, 16, 73, 75, 77, 78, 80, 82,
84, 85
C. agnus 74, 76, 81, 84
C. aureolus 78, 86
C. bidentulus 80, 81
C. biguttatus 83
C. bilineatus 79
C. bipunctatus 78, 83
C. bothnicus 83
C. bivulneratus 82, 83, 84
C. coerulans 80, 81
C. coerulescens 78
C. confusus 78
C. cordiger 76, 84, 85
C. coryli 78, 85
C. cristula 86
C. crux 85, 82
C. decemmaculatus 83, 85
C. distinguendus 85
C. elegantulus 78
C. exiguus 79, 82, 84
C. flavipes 80, 81
C. fulvus 78, 79, 84
C. gamma 81, 84
C. hypochaeridis 78, 86
C. janthinus 73, 82, 83, 85
C. koltzei 82, 83, 84
C. labiatus 78, 79
C. limbellus 74, 76, 83, 84
C. mannerheimi 84, 86
C. nitidulus 76, 84
C. nitidus 84
C. ocellatus 78, 79
238
C. ochroloma 84, 86
С. octacosmus 83, 85
С. octomaculatus 78
С. pini 78, 79
С. pygmaeus 79
С. regalis 74,81,84
С. rufipes 79
С. schaefferi 81
С.sericeus 76, 79
С. stschukini 74, 80, 81, 82, 84
С. tetradecaspilotus 78
cuprea, Psylliodes 190
cupreatus, Oomorphoides 89
martensi, Oomorphoides 89
cupreum, Chrysomela 132
Curculionidae 12
cyanea, Nonarthra 150, 194, 198
cyanella, Lema 55
cyanipennis, Clytraxeloma 44
cynoglossi, Dibolia 203
cyparissiae, Aphthona 197, 200
Cystocnemis 103
Dactylispa 114,202,206,207,208
D. angulosa 206, 208
D. masoni 208
Dascilloidea 11
daurica, Galeruca 158, 159
dauricum, Colasposoma 94, 95, 96, 97
decemlineata, Leptinotarsa 27, 108, 109
decemmaculatus, Cryptocephalus 83, 85
decemnotata, Gonioctena 144, 145
decempunctata, Lema 52, 55
dentata, Donacia 36, 37, 38
denticollis, Cassida 222, 223
depressa, Gastrolina 133, 135
depressiuscula, Dibolia 203
Derocrepis 152, 178
D. rufipes 179
desertorum, Ischyronota 213, 214, 215
Dibolia 156, 188,202
D. cynoglossi 203
D. depressiuscula 203
D. femoralis 203, 204
D. metallica 203
D. rugulosa 203, 204
D. schilling! 204
difficilis, Chrysolina 221
difformis, Aspidomorpha 216, 221, 226
dilutipes, Oulema 52, 54, 56
Diorhabda 15, 151, 158
D. elongata 161
D. persica 161
D. tarsalis 161
Diosphilum 25
discolor, Plateumaris 34, 38, 41, 43
discriminata, Chrysolina 114,120
distinguendus, Cryptocephalus 85
diversa, Lema 55
diversipes, Chrysolina 123
Donacia 5, 16, 25, 32, 33, 35
D. aquatica 39, 41
D. bicolora 37, 41, 42
D. cinerea 32, 34, 35, 37, 38, 40
D. clavareaui 35
D. clavipes 34, 39
D. crassipcs 37, 38, 40
D. dentata 36, 37, 38
D. fennica 37, 40
D. impressa 35, 37, 40
D. marginata 34, 38, 41, 42
D. obscura 37, 41
D.provosti 35
D. semicuprea 35, 39
D. simplex 36, 42
D. sparganii 37, 38
D. thalassina 34, 35, 36, 37, 39, 41
D. tomentosa 32, 35, 36
D. versicolorea 32, 36, 37, 38
D. vulgaris 34, 36, 42
Donaciella 40
Donaciinae 3, 7, 15, 17, 18, 20, 21, 23, 26, 27,
29,31,32, 33, 34, 38
dudkoi, Chrysolina 120
dulcamarae, Psylliodes 189
duodecimpunctata, Crioceris 53
duodecimpunctata, Pentamesa 202
duodecimpustulatus, Paropsides 143
echii, Longitarsus 185
elegantulus, Cryptocephalus 78
elongata, Diorhabda 161
Emarhopa 159
Entomoscelis 23, 102, 107, 140, 146
E. adonidis 140, 143, 144, 146
E. orientals 127, 141, 143
E. suturalis 140, 144, 146
Epithrix 154, 190
E. pubescens 190, 201
erichsoni, Oulema 56
Erinnia 25
Erinniopsis 25
Eulophus 25
Euluperus 152
E. xaniopus 152
Eumolpinae 3, 7, 15, 18, 26, 30, 91, 92, 94, 95,
96, 98
Eumolpus 92, 93, 94, 99, 101
239
E. asclepiadeus 95, 101
Е. globicollis 100
Е. goniostoma 95, 96, 101
euphorbiae, Aphthona 188,197,200
exanthematica, Chrysolina 119, 121
exiguus, Cryptocephalus 79, 82, 84
exoletus, Longitarsus 186
Exosoma 154, 172
E. collare 172
E. lusitanica 158, 172
Eysarcoris 25
faldermanni, Lilioceris 53
fastuosa, Chrysolina 121
fastuosa, Pilemostoma 215
femoralis, Dibolia 203, 204
fennica, Donacia 37, 40
ferruginea, Cassida 210
ferruginea, Asiorestia 188,196
fimbriolatus, Pchybrachis 77
flaveola, Cassida 218,224
flavescens, Pyrrhalta 166
flavicollis, Zeugophora 46, 47
flavicomis, Gonioctena 145
flavipennis, Galerucida 173, 178 -
flavipes, Luperus 169,170
Fleutiauxia 152, 168
F. armata 150, 168
Formica 68
foveicollis, Aulacophora 171
foveolatus, Colaphus 142, 143
fulva, Gonioctena 127, 144
fulvicomis, Crepidodera 187
fulvipennis, Phygasia 150,183,198
fulvipes, Basilepta 94, 95, 98
fulvipes, Stemoplatys 13, 127, 135, 138
fulvus, Cryptocephalus 78, 79, 84
furcigera, Apophylia 165
fuscicomis, Podagrica 187,200
fuscipennis, Clitena 162
fuscorufa, Cassida 218
Galeruca 13, 16, 23, 151, 156, 157, 158
G. daurica 158, 159
G. interrupta circumdata 159
G. laticollis 159
G. melanocephala 158
G. pomonae 23, 159
G.rufa 159
G. tanaceti 23, 159
G. weisei 159
Galerucella 12, 20, 23, 151, 162, 165
G. grisescens 19, 158, 162
G. nipponensis 163
G. nymphaeae 19, 158, 163, 173
Galerucida 156, 177
G. bifasciata 178
G. flavipennis 173, 178
Galerucinae 3, 12, 13, 15, 18, 19, 21, 25, 26, 30,
102, 148, 149, 150, 157, 158, 160, 168, 173
gallaeciana, Oulema 52
gamma, Cryptocephalus 81,84
Gastroidea 135
Gastrolina 20, 21, 103, 104, 132, 133
G. depressa 133, 135
G. peltoidea 128, 133
Gastrolinoides 20, 21, 103, 104
G.japonica 128, 135
Gastrophy sa 103, 105, 135
G. atrocyanea 105, 135, 136
G. mannerheimi 105, 127, 135, 136
G. polygon! 136
G. viridula 135, 136
geminata, Chrysolina 119,121
glabra, Agrostitassa 128,137
Glyphocassis 213,217,218
G. spilota 214, 217
globicollis, Eumolpus 100
goettingensis, Timarcha 109
Gonioctena 5, 16, 20, 21, 23, 24, 102, 107, 143,
144, 146
G. affmis 145
G. chujoi 147
G. coreana 146
G. decemnotata 144, 145
G. flavicomis 145
G. fulva 127, 144
G. gracilicomis 144, 145
G. jacobsoni 146, 147
G.japonica 144, 145
G. kamiyai 146, 147
G. linnaeanus 144, 145
G. ogloblini 146
G. pallidus 19, 146, 147
G. quinquepunctatus 147
G. rufipes 145
G. sibirica 144, 145
G. sundmanni 146
G. viminalis 127, 144, 146
Goniomena 144
goniostoma, Eumolpus 94, 95
gracilicomis, Gonioctena 144, 145
graminis, Chrysolina 112,122
grisescens, Galerucella 19, 158, 162
Gynandrophthalma 69
gypsophilae, Chrysolina 116
haemochlora, Chrysolina 117,123
Haemonia 42
240
haemoptera, Chrysolina 119
halensis, Sermylassa 177
Haptoscelis 158
Haltica 193
hannoverana, Hydrothassa 137
Haptoscelis 158
Harpalus 25
helianthemi, Altica 194
Hemipyxis 156, 199
H. plagioderoides 150, 198, 199
hemisphaerica, Cassida 218
herbacea, Chrysolina 122
Hermaeophaga 154, 192
H. mercurialis 193
Heteromera 11
hieroglyphicus, Pachybrachis 74, 76
Hippodamia 25
Hippuriphila 155, 194
H. modeeri 184, 195
Hispa 22, 206
H. atra 206, 207
Hispella 206
Hispinae 3, 17, 18, 19, 21, 26, 205
hortensis, Chaetocnema 191
hummeli, Timarcha 109
Hydrophiloidea 11
Hydrothassa 19, 103, 104, 136, 137
H. hannoverana 137
H. marginella 128, 137
hyperici, Chrysolina 120, 121
Hypocassida 211,214,216,217,218
H. subferruginea 215
hypochaeridis, Cryptocephalus 78, 86
impressa, Donacia 35, 37, 40
impressicollis, Altica 193
infiiscipes, Chrysolina 114
instabilis, Chrysolina 115, 119
inquinata, Cassida 220
interrupta circumdata, Galeruca 159
Ischyronota 211,213
I. desertorum 213, 214, 215
jacobaeae, Longitarsus 184,185
jacobsoni, Gonioctena 146, 147
Jalla 25
janthinus, Cryptocephalus 83
japana, Cassida 219
japonica, Colobaspis 44
japonica, Gastrolinoides 128, 135
japonica, Gonioctena 144, 145
japonica, Morphosphaera 173, 176
jenisseinsis, Chrysolina 118
junci, Prasocuris 138
kamiyaj, Gonioctena 146, 147
koltzei, Chrysolina 112, 115, 117, 119, 122, 123
koltzei, Cryptocephalus 82, 83, 84
kowalewskyi, Crosita 112, 123, 127
kozhantchikovi, Chrysolina 114
labiatus, Cryptocephalus 78, 79
Labidostomis 5, 13, 14, 16, 58, 59, 61, 62, 63,
65
L. amurensis nigrilabris 62, 65
L. .bipunctata 62, 63
L. chinensis 62, 65
L. longimana 62
L. metallica 63, 65
L. senicula 63, 65
L. sibirica 62, 63
L. tridentata 58, 62
Lachnaia 60, 68
L. sexpunctata 64, 68
laeviuscula, Clytra 67, 68
Lamellicomia 11
Lamprosominae 3, 17, 21, 26, 29, 88, 89
lapponicum, Chrysomela 128, 132
laticollis, Galeruca 159
laticollis, Phratora 128
latimargo, Chrysolina 112, 118, 123
Lebia 25
Lerna 21, 50, 52, 55
L. cyanella 55
L. decempunctata 52, 55
L. diversa 55
Leptinotarsa 102, 106, 110
L. decemlineata 27, 108, 110
lewisi, Longitarsus 186
lewisii, Thlaspida 217
lichenis, Oulema 50, 56
lilii, Lilioceris 52, 54
Lilioceris 13, 14, 21, 50, 51, 52, 53, 54
L. faldermanni 53
L. lilii 52, 54
L. merdigera 50, 52, 54, 55
L. ruficollis 53
L. rugata 51, 52, 53, 54
L. scapularis 52, 53
limbata, Chrysolina 118,120
limbellus, Cryptocephalus 74, 76, 83, 84
Linaeidea 130
lineigera, Chrysolina 120
lineola, Cassida 21, 216, 219
lineola, Orsodacne 45, 48
lineola, Pyrralta 19,167
linnaeanus, Gonioctena 144, 145
linnei, Longitarsus 183,185
Liprus 153, 182
L. punctatostriatus 182, 183
241
Lochmaea 23, 151, 158, 165
L. capreae 19, 165
L. crataegi 165
L. suturalis 165
longicomis, Luperus 169, 170
longimana, Labidostomis 62
longipes, Crosita 112, 123, 127
Longitarsus 153, 183, 185
L. anchusae 183, 186
L. ballotae 186
L. echii 185
L. exoletus 186
L. jacobaeae 184, 185
L. lewisi 186
L. linnei 183, 185
L. luridus 185
L. lycopi 186
L. melanocephalus 183,185
L. parvulus 186
L. pratensis 183,186
L.scutellaris 186
L. suturellus 186
L. tabidus 185
L. transbaicalicus 185
Lordicassis 223
Lordiconia 222
Luperodes 172
Luperus 13, 152, 169, 170
L. flavipes 169, 170
L. longicomis 169, 170
L. luperus 169, 170
L. lyperus 169
L. minutus 171
luperus, Luperus 169, 170
luridus, Longitarsus 185
lusitanica, Exosoma 158, 172
luteola, Pyrrhalta 160, 166
lycopi, Longitarsus 186
Lythraria 152, 180
L. salicariae 180, 184
lythri, Altica 194, 200
Macquartia 25
macrocnemia, Sominella 33
Macromonycha 23
Macroplea 25, 32, 35, 42
M. appendiculata 34, 36, 42, 43
M. mutica 32, 43
macropus, Tituboea 58, 61, 64
maculicollis, Pyrrhalta 13, 158, 166
Malacodermata 12
malvae, Podagrica 187
mandli, Cassida 216,219
mandzhura, Smaragdina 70
mannerheimi, Cryptocephalus 84, 86
mannerheimi, Gastrophysa 105, 127, 135, 136
Mantura 155, 195
M. rustica 188, 195
margaritacea, Cassida 220
marginata, Chrysolina 115,119,122
marginata, Donacia 34,38,41,42
marginella. Hydrothassa 128, 137
Medythia 154, 173, 175
M. suturalis nigrobilineatus 174
Megalopodinae 3, 17, 26, 30, 45
Megalopus 16, 45
Megaselia 25
Meigenia 25
melanocephala, Galeruca 158, 185
melanocephalus, Longitarsus 183
melanopus, Oulema 52, 54, 56
Melasoma 131
Mendax, Pachybrachis 77
menetriesi, Atrachya 173
menetriesi, Podagrica 187,200
menthastri, Chrysolina 122
mercurialis, Hermaeophaga 193
merdigera, Lilioceris 50, 52, 54, 55
metallica, Dibolia 203
metallica, Labidostomis 63, 65
Microlema 50, 55
minutus, Calomicrus 171
minutus, Luperus 171
Mionycha 220
Mniophila 155, 192, 195
M. muscorum 196
modeeri, Hippuriphila 184,185
mongolensis, Chrysolina 114, 119, 120, 122
mongolica, Cassida 26, 223, 224
Monolepta 154, 174, 175
M. quadriguttata 174, 175
montana, Chrysolina 119, 120, 123
Morphosphaera 155, 176
M. japonica 173, 176
murraea, Cassida 216,221,222
musciformis, Chilotoma 64
muscorum, Mniophila 196
mutica, Macroplea 32, 43
Mymaridae 46
nebulosa, Cassida 210, 216, 220, 224
nemorum, Phyllotreta 181,188
Neohaemonia 35
N. voronovae 35, 41
nigriceps, Agelasa 177
nigripennis, Aulacophora 150, 168, 171
nigripes, Phyllotreta 182,188
nigritarsis, Argopus 200
242
nigrocoeruleus, Oomorphoides 88
nikolskyi, Chrysolina 117,124
nipponensis, Galerucella 163
nitidulus, Cryptocephalus 76, 84
nitidus, Cryptocephalus 84
nobilis, Cassida 222
nodicomis, Phyllotreta 182,188
Nonarthra 156, 197
N. cyanea 150, 197, 198
nonstriata, Aphthona 184, 197
nymphaeae, Galerucella 19, 158, 163, 173
obscura, Donacia 37, 41
obscurus, Bromius 19, 33, 34, 95, 97
obtusicollis, Phratora 105, 128
ocellatus, Cryptocephalus 78, 79
ochripes, Phyllotreta 182,188
ochroloma, Cryptocephalus 84, 86
Ocipus 25
octacosmus, Cryptocephalus 83, 85
octomaculatus, Cryptocephalus 78
Odontionycha 222
ogloblini, Gonioctena 146
oleracea, Altica 184,194
Oomorphoides 88
O. cupreatus 89
O. martensi 89
O. nigrocoeruleum 88
Oomorphus 88
O. concolor 88, 89
Oreina 24, 106, 117, 125
O.basilea 117, 126
O. coerulea 126
O. redikortzevi 125
O. rugulosa 118, 126
Oreothassa 103
orientalis, Coptocephala 69, 70
orientalis, Entomoscelis 127, 141, 143
Orsodacne 45, 48
O. cerasi 45, 48
O. lineola 45, 48
Orsodacninae 3, 30, 45, 48
Oulema 20, 21, 50, 52, 54, 55
O. dilutipes 52, 54, 56
O. erichsoni 56
O.gallaeciana 52, 56
O. lichenis 50, 56
O. melanopus 52, 54, 56
O. oryzae 51, 52, 56
O. suvorovi 56
Pachnephorus 92, 94, 96, 101
P. tessellatus 95, 101
Pachybrachis 73, 75, 76
P.fimbriolatus 77
P. hieroglyphicus 74, 77
P.mendax 77
P. scriptidorsum 74, 76
Pagria 93, 99
P. consimile 99
P. signata 99, 100
P. ussuriensis 99
Pallasiola 15, 23, 151, 157, 159
P. absinthii 27, 159
pallidicollis, Cassida 210, 218, 220, 224
pallidus, Gonioctena 19, 146, 147
pannotica, Cassida 225
panzeri, Cassida 21, 210, 218
Paraluperodes 153, 173, 174
Paropsides 21, 105, 107, 142, 143
P. duodecimpustulatus 143
parvula, Cassida 210, 218, 222, 224
parvulus, Longitarsus 186
pedestris, Chrysolina 118,124
Pedrillia 47
peltoidea, Gastrolina 128,133
Pentamesa 156,198,202,203
P.duodecimpunctata 202
P. kondarensis, 202
Pentatomidae 25
perforata, Chrysolina 112, 113, 119
persica, Diorhabda 161
Phaedon 23, 105, 135, 139
Ph. armoraciae 139
Ph. brassicae 140
Ph. cochleariae 135, 140
Ph. concinnus 105, 135, 139
phellandrii, Prasocuris 20, 128, 138
Philopona 156, 198,
Ph. vibex 150, 198, 199
Phratora 10, 14, 15, 21, 23, 104, 127, 128
Ph. atrovirens 130
Ph. laticollis 128
Ph. obtusicollis 105, 128
Ph. polaris 130
Ph. tibialis 130
Ph. vitellinae 105, 130
Ph. vulgatissima 105, 128
Phygasia 153, 182
Ph. fulvipennis 150, 183, 198
Phyllobrotica 153, 171
Ph. quadrimaculata 172
Phyllodecta 127
Phyllotreta 153, 180, 181, 188
Ph. armoraciae 181, 184, 188
Ph. atra 182, 188
Ph. cruciferae 182
Ph. nigripes 181,188
243
Ph. nodicomis 182, 188
Ph. ochripes 182, 188
Ph. striolata 182
Ph. tetrastigma 182, 188
Ph. undulata 181, 188
Ph. vittata 182
Ph. vittula 182
Phytodecta 143
Phytophaga 12
Picconia 25
Picromerus 25
pigra, Crosita 112, 123, 126
Pilemostoma 210, 213, 215, 216, 217, 218
P. fastuosa 215
pini, Cryptocephalus 78, 79
pinicola, Calomicrus 171
piperata, Cassida 22, 219, 224
parka, Chrysolina 116,118,150
Plagiodera 15, 16, 20, 21, 103, 104, 128, 130
P. versicolorea 130
plagioderoides, Hemipyxis 150, 198, 199
Plagiolepis 66
Plagiostema 20, 103, 104, 105, 128, 130
P. aenea 131
Plateumaris 32, 35, 43
P. affinis 32, 38, 43
P. braccata 32, 38, 43
P. constricticollis 43
P. discolor 34, 38, 41, 43
P. sericea 43
P. weisei 34, 38, 43
plutus, Crepidodera 187
Podagrica 153, 154, 187,200
P. fuscicornis 187, 200
P. malvae 187
P. menethriesi 187, 200
polaris, Phratora 130
polita, Chrysolina 112,119,122
polygoni, Gastrophysa 136
pomonae, Galeruca, 23, 159
populi, Chrysomela 132
porphyrea, Chrysolina 118, 124
prasina, Cassida 210, 216, 223
Prasocuris 19, 103, 104, 137, 138
P.junci 138
P. phellandrii 20, 128, 138
pratensis, Longitarsus 183,186
provosti, Donacia 35
Psylliodes 22, 149, 153, 154, 189
P. affinis 189
P. angusticollis 189
P. attenuata 189
P. chalcomera 189
P. chrysocephala 188, 190
P. cuprea 190
P. dulcamarae 189
P. punctifrons 190
Pteromalidae 25
Pterostichus 25
pubescens, Epithrix 190, 201
pubiceps, Chlamisus 87
pumilio, Suffiianus 75, 76, 85
punctatostriata, Sangariola 150, 180, 198
punctatostriatus, Liprus 182,183
punctifrons, Psylliodes 190
punctipennis, Argopus 200
pusa, Chrysolina 114
pusilia, Altica 194
pygmaeus, Cryptocephalus 79
Pyrrhalta 12, 13, 14, 152, 161, 165, 166
P. aenescens 166
P. calmariensis 167
P. flavescens 166
P. lineola 19, 167
P. luteola 160, 166
P. maculicollis 13, 158, 166
P. pusilia 167
P. semifulva 166
P. tenella 167
P. vibumi 158
quadriguttata, Monolepta 174, 175
quadriimpressum, Ambrostoma 113
quadrimaculata, Phyllobrotica 172
quadripunctata, Clytra 61, 64, 67, 68
quatuordecimpunctata, Crioceris 53
quercetorum, Altica 200
quinquepunctatus, Gonioctena 147
redikortzevi, Oreina 125
regalis, Cryptocephalus 74, 81, 84
relucens, Chrysolina 115
Rhacognathus 25
Rhynchophora 12
rosti, Chrysolina 124
rubi, Batophila 180
rubidum, Sphaeroderma 201
rubiginosa, Cassida 18, 221, 225
rufa, Galeruca 159
ruficollis, Lilioceris 53
rufilabris, Chrysolina 114
rufipes, Cryptocephalus 79
rufipes, Derocrepis 179
rufipes, Gonioctena 145
rugata, Lilioceris 51,52,53,54
rugulosa, Dibolia 203, 204
rugulosa, Oreina 118, 126
rustica, Mantura 188, 195
244
Sagrinae 17
salicariae, Lythraria 180,184
saliceti, Chrysomela 134
salicina, Smaragdina 70
Sangariola 153, 180
S. punctatostriata 150, 180, 198
sanguinolentha, Cassida 223, 225
sanguinolentha, Chrysolina 118,122
sanguinosa, Cassida 221
scapularis, Lilioceris 52, 53
schaefferi, Cryptocephalus 81
schilling!, Dibolia 204
Schizonotus 25
scraphina, Cassida 218
scriptidorsum, Pachybrachis 74, 76
scutellaris, Zeugophora 47
Sebaethe 199
semiaurantica, Smaragdina 58, 70
semicuprea, Donacia 35, 39
semicyanea, Aphthona 196
semifulva, Pyrrhalta 166
senicula, Labidostomis 63, 65
septentrionalis, Chrysolina 112, 116, 122
sericea, Plateumaris 43
sericeus, Cryptocephalus 76, 79
Sermylassa 155, 156
S. halensis 177
sexpunctata, Lachnaia 64, 68
sibirica, Apterocuris 135, 142
sibirica, Gonioctena 144, 145
sibirica, Labidostomis 62, 63
signata, Pagria 99, 100
silphoides, Theone 27, 160
simplex, Donacia 36, 42
sinensis, Stylosomus 86
Smaragdina 13, 58, 60, 69, 70
S. aurita 70
S. collaris 70
S. mandzhura 70
S. salicina 70
S. semiaurantica 58, 70
Sominella 33
S. macrocnemia 33
spaethi, Cassida 218
sophiae hoefti, Colaphus 141
sparganii, Donacia 37, 38
Sphaeroderma 156,157,200
S. balyi 202
S. rubidum 201
S. testacea 201
S. unicolor 201
spilota, Glyphocassis 214,217
Staphylinoidea 11
Staphylinus 25
staphylea, Chrysolina 120
Stemoplatys 104, 138
S. clementzi 139
S. fulvipes 13, 127, 135, 138
Stemoxia 12
stigmatica, Cassida 216, 219
stschukini, Cryptocephalus 74, 80, 81, 82, 84
Stylosomus 74, 75, 76, 84, 86
S. sinensis 86
S. tamaricis 86
subferruginea, Hypocassida 215
subreticulata, Cassida 218,220
subspinosa, Zeugophora 47
subsulcata, Chrysolina 116,119
Suffrianus 73, 74, 85
S. pumilio 75, 76, 85
sundmanni, Gonioctena 146
suturalis, Entomoscelis 140, 144, 146
suturalis, Lochmaea 165
suturalis nigrobilineatus, Medythia 174
suturalis, Zygogramma 27, 108, 111
suturellus, Longitarsus 186
suvorovi, Oulema 56
Syneta 90
S. adamsi 90
S. betulae 45, 90
Synetinae 3, 13, 30, 45, 90
Syrphidae 25
tabidus, Longitarsus 185
taimyrensis, Chrysomela 25, 132
Taiwania 219,222
tamaricis, Altica 194
tanaceti, Galeruca 23, 159
tarsalis, Diorhabda 161
tnbricosa, Timarcha 109
tenella, Pyrrhalta 167
Teredilia 12
tessellatus, Pachnephorus 95, 101
testacea, Sphaeroderma 201
tetradecaspilotus, Cryptocephalus 78
Tetrastichus 25
tetrastigma, Phyllotreta 182,188
thalassina, Donacia 34, 35, 36, 37, 39, 41
Theone 151, 160
Th. silphoides 27, 160
Thlaspida 210, 213, 215, 216, 217, 218
Th. biramoisa 217
Th. lewisii 217
tibialis, Chaetocnema 188, 192
tibialis, Phratora 130
Timarcha 102, 104, 107, 109
T. goettingensis 109
245
T. hummeli 109
Т. tenebricosa 109
Tituboea 17, 58, 60, 66
T. macropus 66
Tlanoma 153, 193
tolli, Chrysolina 116
tomentosa, Donacia 32, 35, 36
transbaicalicus, Longitarsus 185
transparipennis, Aspidomorpha 221, 224, 226
trapezicollis, Chrysolina 125
tremulae, Chrysomela 128, 132
tridentata, Labidostomis 58, 62
Trombididae 25
tumeri, Zeugophora 45, 47
undecimnotata, Cassida 23, 221, 223
undulata, Chrysolina 115,122
undulata, Phyllotreta 181, 188
unicolor, Sphaeroderma 201
unifasciata, Coptocephala 69, 70
urjanchaica, Chrysolina 114
ussuriensis, Chrysolina 121
ussuriensis, Pagria 99
variabilis, Cleoporus 92
varians, Chrysolina 121
velaris, Cassida 218,222
venustula, Aphthona 197
versicolor, Cassida 214, 219
versicolor, Chloropterus 95, 97, 98
versicolorea, Donacia 32, 36, 38
versicolorea, Plagiodera 130
vespertina, Cassida 220
vibex, Cassida 18,224,225
vibex, Philopona 150, 198, 199
vibumi, Pyrrhalta 158
vigintipunctata, Chrysomela 128, 132
viminalis, Gonioctena 127, 144, 146
violacea, Chrysolina 123
violaceipennis rufipes, Cneorane 168, 173
virgata, Crinoline 15, 117, 124
viridis, Cassida 221, 222
viridula, Gastrophysa 135, 136
vitellinae, Phratora 105, 130
vittata, Cassida 218
vittata, Phyllotreta 182
vittula, Phyllotreta 182
voronovae, Neohaemonia 35, 41
vrangeliana, Chrysomela 132
vulgaris, Donacia 34, 36, 42
vulgatissima, Phratora 105, 128
weisei, Galeruca 159
weisei, Plateumaris 34, 38, 43
xanihopus, Euluperus 152
yezoensis, Chrysolina 121
Zeugophora 16, 46, 47
Z. annulata 45, 47
Z. flavicollis 46, 47
Z. scutellaris 47
Z. subspinosa 47
Z. tumeri 45, 47
Zeugophorinae 3, 17, 19, 23, 29, 45, 46
Zicrona coerulea 25
Zygogramma 102, 106, 110
Z. suturalis 27, 108, 111
246
Научное издание
ЗАЙЦЕВ Юрий Михайлович,
МЕДВЕДЕВ Лев Никандрович
ЛИЧИНКИ ЖУКОВ-ЛИСТОЕДОВ РОССИИ
Москва: Товарищество научных изданий КМК. 2009. 246 с., ил.
при участии ИП Михайлова К.Г.
Главный редактор издательства К.Г. Михайлов
Верстка. К.Г. Михайлов
Для заявок:
123100, Москва, а/я 16, Издательство КМК
эл. почта: mikhailov2000@gmail.com
http ://avtor-kmk.ru
Отпечатано в ГУП ППП «Типография «Наука» АИЦ РАН
121099 Москва, Шубинский пер., 6.
Подписано в печать 11.11.2009. Заказ № 1921
Формат 70x100/16. Объем 15,5 печ. л. Бум. офсетная. Тираж 800 экз.