










































































































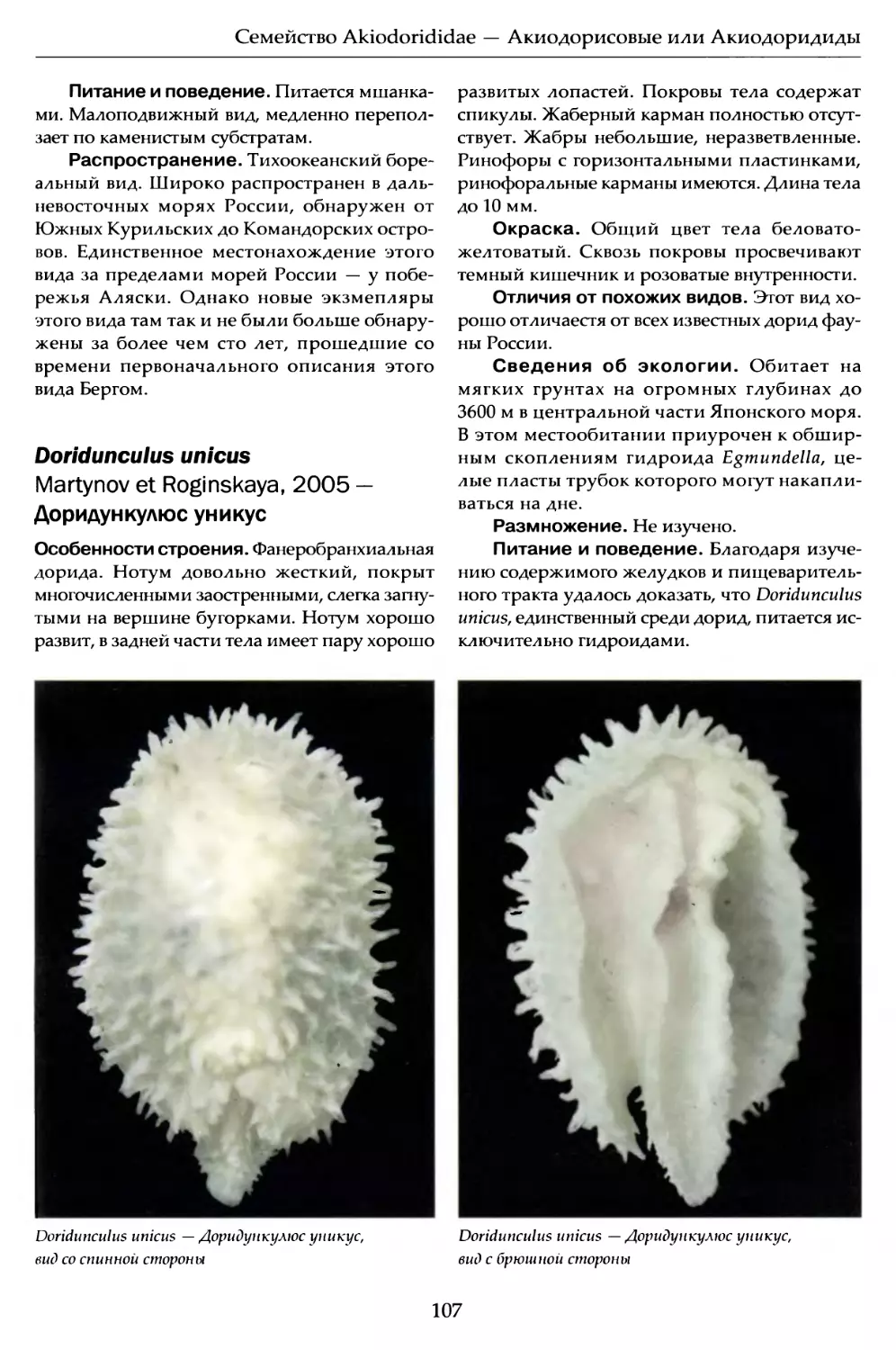











































































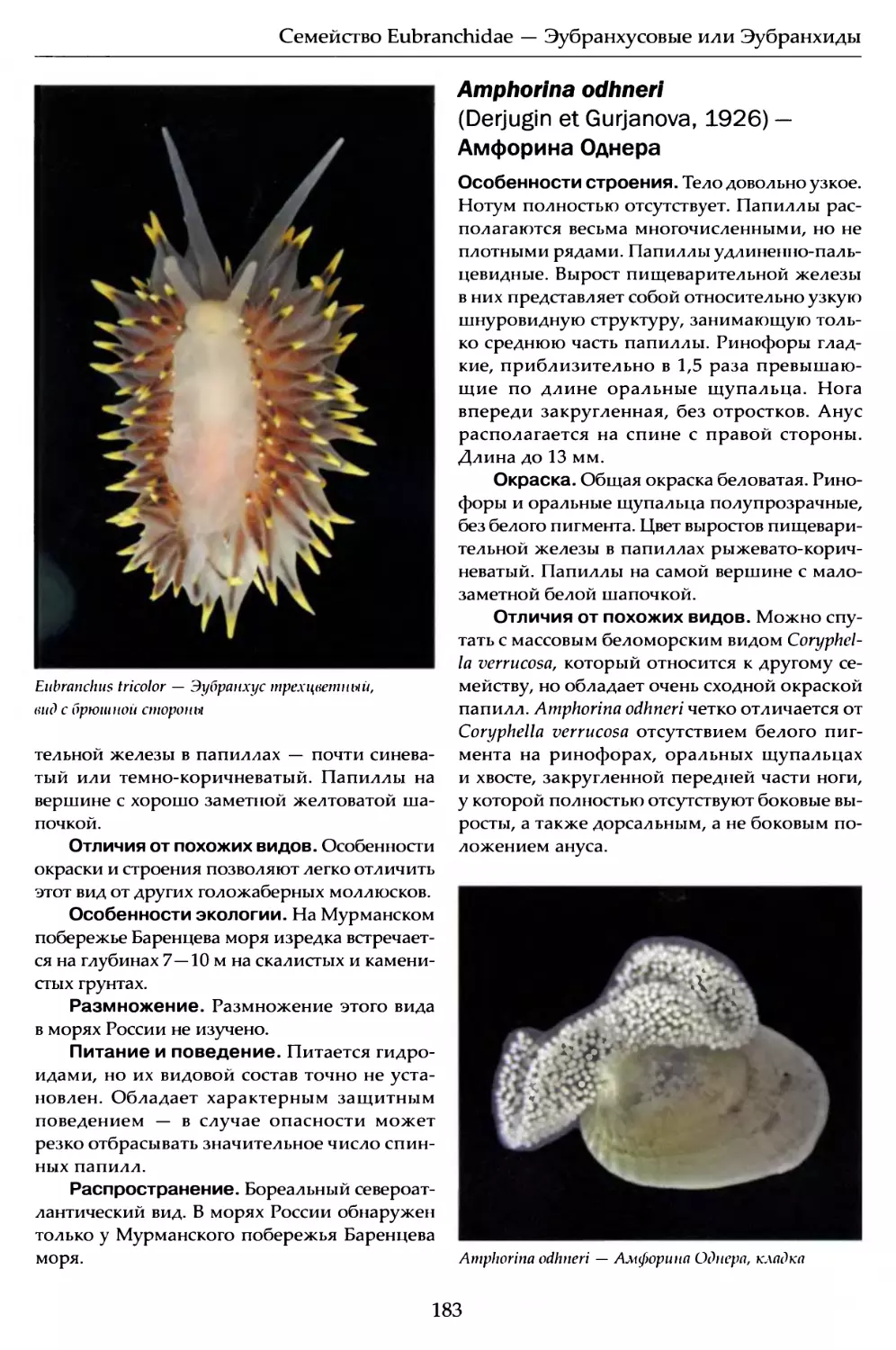

















































Author: Мартынов А.В. Коршунова Т.А.
Tags: общая экология биоценология гидробиология биогеография зоология моллюски
ISBN: 978-5-93457-358-5
Year: 2011




Text
Александр Мартынов • Татьяна Коршунова
Заднежаберные
моллюски
морей России
ПЛЮС
цэаггэон
ЗООЛОГИЧЕСКИЙ МУЗЕЙ
МОСКОВСКОГО ГОСУДАРСТВЕННОГО
УНИВЕРСИТЕТА ИМЕНИ М. В. ЛОМОНОСОВА
Александр Мартынов • Татьяна Коршунова
Заднежаберные
МОЛЛЮСКИ
морей России
АТЛАС-ОПРЕДЕЛИТЕЛЬ
с обзором биологии
Москва
Издательство «Фитон+»
УДК 574(084)
ББК 28.082
М29
Рекомендовано к печати редакционным советом
Научно-исследовательского Зоологического музея
Московского государственного университета имени М. В. Ломоносова
Рецензент:
М. В. Калякин — доктор биологических наук, директор Зоологического музея МГУ имени
М. В. Ломоносова
Мартынов А. В., Коршунова Т. А.
М 29 Заднежаберные моллюски морей России. Атлас-определитель с обзором
биологии / Александр Мартынов, Татьяна Коршунова. — М.: ЗАО «Фитон+»,
2011. - 232 с.: ил.
ISBN 978-5-93457-358-5
Это первый иллюстрированный атлас-определитель заднежаберных моллюсков морей Рос-
сии — удивительной группы морских животных, которая включает и хорошо известных дайверам
голожаберных моллюсков. Особую уникальность атласу придает тот факт, что он основан на много-
летней работе авторов — профессиональных биологов.
Книга дает полную картину разнообразия обитающих у нас видов. Знакомит со строением,
размножением и привычками, а также единственной в своем роде эволюционной историей этих
необычных существ, привлекающих своими неземными формами и иррациональной окраской.
Атлас богато иллюстрирован авторскими цветными фотографиями, сделанными практически во
всех морях России — от берегов Баренцева и Белого морей до таких экзотических уголков, как воды
Камчатки и Курильских островов.
Книга адресована не только ученым, но и широкому кругу читателей — любителям подводного
плавания, преподавателям, студентам, словом, всем, кто интересуется природой.
Martynov A., Korshunova Т.
М 35 Opisthobranch Molluscs of the Seas of Russia. A colour guide to their taxonomy
and biology / A. Martynov, T. Korshunova. — M.: Fiton+, 2011. — 232 p.
The new title opens for readers amazing world of remarkable group of marine animals, who forgot her-
itage of their own ancestors and discarded one of the most characteristic feature of the phylum Mollusca —
shell. Such important loss led then to the dramatic evolutionary changes shaped this super-attractive for
both amateurs and scientists marine group that well known under the names of sea slugs, nudibranchs or
opisthobranch molluscs. During last decade numerous colour guides on the nudibranchs have appeared but
present book is the first colour atlas of the Russian sea slugs ever published and covers all vast Russian seas
from freezing Arctic to the subtropical Black Sea and southern Japan Sea. The extraterrestrial shapes and
psychedelic colours of the nudibranchs are itself enough reasons to stop on this book but numerous high
quality colour photographs of 103 species of the Russian opisthobranchs are only one of the many levels of
the new title. The authors, professional biologists, specialists on the opisthobranchs molluscs, already have
described 16 new species, 7 new genera and a new family of the Opisthobranchia from the Russian Seas,
personally collected and studied their animals, including SCUBA diving in sometimes severe conditions
of the Arctic Barents Sea and North Pacific Kamchatka region, unfold for readers convoluted evolutionary
history of the sea slugs.
УДК 574(084)
ББК 28.082
Охраняется ГК РФ, часть 4. Воспроизведение всей книги илилюбой ее части запрещается
без письменного разрешения издательства. Любые попытки нарушения закона будут
преследоваться в судебном порядке.
ISBN 978-5-93457-358-5
© ЗАО «Фитон+», 2011
Заднежаберные
МОЛЛЮСКИ
морей России
АТЛАС-ОПРЕДЕЛИТЕЛЬ
с обзором биологии
СОДЕРЖАНИЕ
Предисловие....................................................6
От авторов.....................................................9
Вступление ................................................... 10
БИОЛОГИЯ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ
ЗАДНЕЖАБЕРНЫХ И ГОЛОЖАБЕРНЫХ МОЛЛЮСКОВ
Кто же такие голожаберные моллюски
и почему еще существует выражение «заднежаберные моллюски»... 13
СТРОЕНИЕ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
Введение в морфологию заднежаберных моллюсков:
что нужно обязательно знать, чтобы лучше понять эту книгу ... 17
ПОВЕДЕНИЕ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
Удивительные привычки удивительных моллюсков .................30
ЖИЗНЕННЫЙ ЦИКЛ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
Нужен ли партнер гермафродиту и зачем детям раковина? ........35
АТЛАС ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
МОРЕЙ РОССИИ
Как устроен атлас ..........................................41
О том, что необходимо знать читателю о названиях животных,
чтобы пользоваться атласом..................................43
ЭВОЛЮЦИЯ, ШАГ ПЕРВЫЙ. ВСЁ ЕЩЁ С РАКОВИНОЙ
Отряд Cephalaspidea — Цефаласпиды ..........................46
Семейство Acteonidae — Актеониды .......................46
Семейство Cylichnidae — Цилихниды.......................48
Семейство Philinidae — Филиниды.........................50
Семейство Aglajidae — Аглаиды...........................53
Семейство Retusidae — Ретузиды..........................55
Семейство Volvulellidae — Вольвулеллиды ................61
Семейство Atyidae — Атииды..............................62
Отряд Anaspidea — Анаспиды...................................63
Семейство Akeridae — Акериды ...........................63
Семейство Aplysiidae — Аплизииды или Морские зайцы .....67
ЭВОЛЮЦИЯ, ШАГ ВТОРОЙ. ПРОРЫВ НАВЕРХ
Отряд Thecosomata — Раковинные крылоногие моллюски..........71
Семейство Limacinidae — Лимациновые......................71
Отряд Gymnosomata — Безраковинные крылоногие моллюски ......73
Семейство Clionidae — Клионовые..........................73
4
ЭВОЛЮЦИЯ, ШАГ ТРЕТИЙ. БЕГСТВО ПОД ЗЕМЛЮ
Отряд Acochlidiida — Акохлидииды .....................................76
Семейство Parhedylidae — Пархедилиды .............................77
Семейство Microhedylidae — Микрохедилиды..........................79
Семейство Asperspinidae — Асперспиниды ...........................79
ЭВОЛЮЦИЯ, ШАГ ЧЕТВЁРТЫЙ. ПЕРВАЯ ПОПЫТКА
Отряд Sacoglossa — Сакоглоссы ........................................81
Семейство Limapontiidae — Лимапонциевые...........................81
Семейство Hermaeidae — Гермаеиды .................................87
ЭВОЛЮЦИЯ, ШАГ ПЯТЫЙ. ГЕНИАЛЬНЫЙ НЕУДАЧНИК
Отряд Notaspidea — Нотаспиды..........................................89
Семейство Pleurobranchidae — Плеуробранхусовые....................89
Семейство Pleurobranchaeidae — Плеуробранхеивые ..................98
ЭВОЛЮЦИЯ, ШАГ ШЕСТОЙ. ВЕРШИНЫ ЭВОЛЮЦИИ
Отряд Doridacea — Голожаберные моллюски дориды ...................... 99
Семейство Chromodorididae — Хромодорисовые ......................102
Семейство Akiodorididae — Акиодорисовые......................... 105
Семейство Polyceridae — Полицеровые .............................108
Семейство Vayssiereidae — Весьеровые............................ 116
Семейство Anculidae — Анкуловые................................. 118
Семейство Dorididae — Дорисовые ................................ 120
Семейство Discodorididae — Дискодорисовые ...................... 122
Семейство Onchidorididae — Онхидорисовые ........................126
Семейство Goniodorididae — Гониодорисовые....................... 144
Отряд Nudibranchia — Голожаберные моллюски........................... 146
Подотряд Arminacea — Арминацеа ...................................... 147
Семейство Doridoxidae — Доридоксовые ............................ 147
Семейство Dironidae — Дироновые ................................. 148
Подотряд Dendronotacea — Дендронотацеа .............................. 150
Семейство Tritoniidae — Тритониевые............................. 150
Семейство Dendronotidae — Дендронотусовые........................152
Семейство Dotoidae — Дотоиды.................................... 158
Семейство Embletoniidae — Эмблетониевые ........................ 161
Подотряд Aeolidacea — Эолидацеи......................................163
Семейство Flabellinidae — Флабеллиновые .........................163
Семейство Pseudovermidae — Псевдовермисовые......................180
Семейство Eubranchidae — Эубранхусовые...........................182
Семейство Tergipedidae — Тергипесовые........................... 193
Семейство Facelinidae — Фацелиновые ............................ 215
Семейство Aeolidiidae — Эолидовые ...............................216
НЕОБХОДИМОЕ ПОСЛЕСЛОВИЕ................................................221
Некоторые важные термины.............................................224
Указатель латинских названий ........................................226
Избранные работы авторов.............................................229
5
Предисловие
Наверняка вы слышали слово «биоразнообразие»,
которым ученые обозначают всю совокупность
живых существ на нашей планете, но вряд ли все-
рьез пытались представить себе его настоящие
масштабы. Нам кажется, что мы неплохо пред-
ставляем себе разнообразие млекопитающих, жи-
вущих на нашей планете, хотя и забываем при
этом о том, что нам мало знакомы мелкие зверь-
ки — грызуны, насекомоядные или летучие мыши,
составляющие на самом деле подавляющее боль-
шинство видов млекопитающих. Кто-то неплохо
знает птиц, кто-то увлекается бабочками, но
все-таки я рискну предположить, что почти ни-
кто из наших читателей не представляет себе
истинные масштабы разнообразия жизни на Зем-
ле. А ведь мы знаем еще далеко не о всех видах жи-
вотных, растений и грибов, населяющих нашу
планету с незапамятных времен. Каждый год уче-
ные описывают новые и новые виды животных,
в основном, конечно, мелких и малозаметных. Об-
щий счет идет уже на миллионы видов, живые
существа обнаруживаются повсюду, и, конечно,
очень многие из них населяют моря и океаны —
колыбель жизни на Земле.
Эта книга познакомит нас с малой толикой
громадного разнообразия живых форм. Но посвяще-
на она, в отличие от многих подобных изданий, не
птицам, бабочкам, динозаврам или крупным мле-
копитающим, а совершенно таинственной для
подавляющего большинства читателей группе жи-
вых существ — заднежаберным моллюскам. Что
такое моллюск — мы еще более-менее представ-
ляем. Кто-то помнит из школьных уроков даже
о том, что есть на свете брюхоногие моллюски —
те самые, у которых на брюшной стороне тела
действительно имеется мускулистая нога, обе-
спечивающая их передвижение, и двустворчатые
моллюски — в соответствии с названием дей-
ствительно укрытые двумя почти симметрич-
ными створками. Более эрудированная публика
вспомнит и о головоногих моллюсках. Это тоже
«говорящее» название, ноги у осьминога и его род-
ственников растут прямо из «головы» (берем это
слово в кавычки, так как использовать в данном
случае этот термин приходится с некоторыми
оговорками). Но я почти уверен, что кроме био-
логов и, может быть, некоторых дайверов, о задне-
жаберных моллюсках почти никто и почти ни-
чего не знает. А оказывается, что не где-то
в таинственных джунглях или недоступных мор-
ских глубинах, а буквально у берегов нашей страны
тихо и незаметно обитают разнообразные по
окраске и строению существа со своими привыч-
ками, пристрастиями, требованию к местам
обитания, солености, температуре, корму...
Не будем забегать вперед и пересказывать
то, о чем вы узнаете из этой книги. Прежде чем
начать знакомиться с ней, давайте обратим
внимание не только на то, что нам открывает-
ся совершенно неизвестный ранее мир мелких
морских беспозвоночных животных. Давайте от-
метим для себя, что проникнуть так глубоко
в мир этих братьев наших меньших нам удается
только потому, что нашлись ученые, готовые не
только сами исследовать жизнь, строение
и историю этих существ, но и познакомить
с ней нас, несведущих. Материалы для этой кни-
ги были получены в результате многолетних
кропотливых поисков, находок, разочарований,
сомнений и в конце концов открытий. Для этого
авторы использовали акваланги и хитрые прибо-
ры, позволяющие вылавливать интересующих их
животных на морском дне, сложнейшие микро-
скопы и бинокуляры — и коллекции многих от-
ечественных и зарубежных зоологических музеев,
горы научной литературы — и специальные ка-
меры, позволяющие запечатлеть портреты глав-
ных героев книги и случаи из их сложной, напря-
женной и не всегда понятной нам жизни. Я всегда
полагал, что если ученый хорошо знаком с пред-
метом своих исследований, то он сможет хорошо
рассказать об этом и заинтересовать этим пред-
метом даже тех, кто раньше был очень далек от
данной темы. Очень надеюсь, что так и полу-
чится.
Еще одна интрига этой книги заключается
в том, что у нас в руках — не просто атлас-опре-
делитель. Да, в нем собраны сведения о внешнем
виде, биологии и распространении заднежаберных
моллюсков, и в этом отношении книга — и атлас
тоже. Однако авторы попытались пойти дальше
и рассказать нам еще и про эволюцию обсуждае-
6
мых животных. На всякий случай напомню, что
пока все сомнения в том, что эволюция действи-
тельно привела к существованию на Земле громад-
ного числа разнообразных живых существ, выгля-
дят вполне ненаучно. Для настоящих биологов
посмотреть на то, как реально происходят эво-
люционные изменения животных, или хотя бы
проследить ее этапы на примере конкретных
групп — мечта, которая сбывается далеко не всег-
да. В книге о заднежаберных моллюсках нам обе-
щают показать, как она происходит, и расска-
зать о том, как можно расшифровать историю
группы родственных организмов, постепенно вы-
рабатывавших различные, порой весьма замысло-
ватые адаптации, т.е. приспособления к конкрет-
ным условиям существования. Это трудная
задача, но даже если кому-то покажется, что она
решена в данной книге не в полной мере — вы все-
таки уже и сами включитесь в увлекательное ис-
следование. Последний реверанс мы сделаем в адрес
тех читателей, которым покажется, что книга
все-таки получилась слишком научной и поэтому
малопонятной. Известно немало случаев, когда
ученые, пытаясь рассказывать о сложных вопро-
сах, упрощают их настолько, что в изложении
уже почти не остается научных фактов. Мне ка-
жется, что такие примеры можно считать не-
уважением к читателю. В нашей книге авторы
и редактор постарались разделить сугубо научную
и научно-популярную части текста. И можно на-
деяться, что читатель не испугается или, как
говорили раньше, не убоится сложных вопросов,
а напротив — постарается расширить свой кру-
гозор, вникнув во все описанные здесь закоулки на-
учного исследования.
Пожелаю читателю удачи и надеюсь сам то-
же получить удовольствие от книги, задуманной
и сделанной в рамках серии о животных, выпу-
скаемых издательством «Фитон+» совместно
с Научно-исследовательским Зоологическим музе-
ем Московского государственного университета
имени М. В. Ломоносова
Директор Зоологического музея
МГУ имени М. В. Ломоносова,
доктор биологических наук
М. В. Калякин
Colga minichevi — Колга Миничева
8
От авторов
Уважаемый читатель, эта книга откроет перед
Вами окно в завораживающий мир заднежаберных
моллюсков, основу которых составляют знамени-
тые голожаберники, полностью лишенные ракови-
ны, хорошо известные как ныряльщикам-дайверам,
так и просто любителям природы. Вся представ-
ленная здесь информация основывается на большой
и серьезной научной работе, которую авторы кни-
ги ведут более двух десятилетий. Работе, специ-
ально посвященной этим, наверное самым удиви-
тельным моллюскам морей России. Тот раздел
систематики, который изучает заднежаберных
моллюсков, очень развит в современной науке. По-
мимо обширного корпуса научных работ имеется
огромный слой популярной литературы — от
статей в журналах до толстенных атласов. Де-
сятки, а возможно уже и сотни цветных книг на
английском. Есть популярные атласы на норвеж-
ском, корейском, японском и других языках. Но
нет на русском. Это совершенно несправедливо
хотя бы уже потому, что первый голожаберный
моллюск из морей России — Tritonia tetraquetra
был описан российским ученым Палласом факти-
чески во времена Линнея — в 1788 году!
Но, к счастью, за два последних десятилетия
был совершен настоящий прорыв в наших знаниях
о голожаберных моллюсках морей России. Особенно
парадоксально это выглядит в свете того, что
именно два этих десятилетия стали для россий-
ской науки подлинным временем испытаний и вы-
живания. Поэтому, представляя этот первый рос-
сийский атлас заднежаберных моллюсков морей
России, основанный исключительно на собствен-
ных научных исследованиях и оригинальных фото-
графиях, мы испытываем подлинное чувство гордо-
сти. Начавшись на берегах одного из самых
отдаленных морей России — Японского, наши ис-
следования были распространены на большинство
других российских морей. Авторы лично побывали
как на студеных побережьях Баренцева и Белого
морей, так и на почти субтропических берегах Чер-
ного моря. Не остались без внимания такие экзо-
тические уголки, как Курильские острова и побере-
жье Камчатки. Был исследован даже единственный
известный голожаберный моллюск Балтийского
моря, в котором вода чуть солонее речной. Во всех
этих районах авторы не только собирали свои объ-
екты в приливно-отливной зоне (на так называемой
литорали), но и проводили работы с применением
лекговодолазного снаряжения — аквалангов.
Подобный подход позволил неизмеримо рас-
ширить потенциал исследования. Единственное
место, где мы не смогли пока побывать лично, —
глубокая Арктика (к которой относятся такие
моря, как Карское, Лаптевых, Восточно-Сибир-
ское и др.). Заменой им стали почти арктические
Баренцево и Белое моря, на глубинах которых оби-
тает настоящая арктическая фауна. Но это не
означает, что фауна голожаберных моллюсков
Арктики осталась неисследованной. Напротив,
в нашем распоряжении были обширные материа-
лы, собранные многочисленными русскими и со-
ветскими экспедициями в течение всего XX сто-
летия, начиная с ранних исследований «В помощь
поморам русского севера», стартовавших еще
в 1898 году. В результате заднежаберные моллю-
ски Арктики оказались на сегодняшний день
очень хорошо изученными. Одним из важных ре-
зультатов обработки материалов, накопившихся
в ведущих российских зоологических институтах
и музеях почти за 150 лет, но остававшихся
вплоть до недавнего времени не исследованными,
стало обнаружение нами уникального нового рода,
который был назван в честь Мурманска и всего
Мурманского побережья — Murmania. Научное
описание этого нового рода и вида, поднятого с глу-
бин еще начале XX века, было опубликовано лишь
в 2006 году! Подобное — не редкость в система-
тике. Собранные десятки, а иногда и более сотни
лет назад, зафиксированные особым образом эк-
земпляры ожидают в коллекциях своего часа, когда
ими займется один из редких специалистов по
данной группе организмов.
Результат многолетней работы — перед вами,
и включает не только атлас, который содержит
более 100 великолепно иллюстрированных видов
заднежаберных моллюсков, но и увлекательное пу-
тешествие сквозь удивительные эволюционные
преобразования, которые претерпел «скромный»
раковинный предок на своем пути превращений
в подлинно иррациональные формы и краски голо-
жаберных моллюсков.
9
Вступление
Несмотря на триумфальное шествие
идеи эволюции, ставшей достоянием
науки более полутора веков назад,
вплоть до настоящего времени в биологии сохра-
няется несколько парадоксальная ситуация —
даже профессиональные систематики продол-
жают выделять виды так же, как это делали и во
времена Линнея, т. е. вне всякой связи с историей
их формирования. А идея эволюции в система-
тике выделена фактически в специальную об-
ласть современной биологии — так называемую
филогенетическую систематику, метод построе-
ния формальных древовидных схем, своего рода
«генеалогий» организмов. Анализ же эволюци-
онного аспекта становления вида до сих пор
представляют как исключительно особый, теоре-
тический уровень науки, а не естественную, инте-
гральную часть систематики, каковой она и явля-
ется на самом деле. Вот почему нам кажется так
важно продемонстрировать некие всем будто бы
знакомые и почти тривиальные утверждения
(например, о важности эволюции) на нетриви-
альном, свежем материале. И та группа морских
организмов — заднежаберные моллюски, кото-
рой мы профессионально занимаемся более
20 лет, предоставляет поистине уникальную воз-
можность для этого. Пожалуй, ни в одной дру-
гой группе живых существ нет такого сочетания
фантастического эволюционного «букета» раз-
ных форм, «застывших» на различных эволюци-
онных «стадиях» с невероятной эстетической
привлекательностью составляющих ее видов.
Это обстоятельство и определило несколько
необычное построение нашей книги в сравнении
со многими другими подобными изданиями.
Мы всячески старались избежать ситуации, ког-
да читатель вместо системы знаний, пусть даже
упрощенной и ориентированной на неспециа-
листа, получает хаос бесчисленных цветных изо-
бражений, число которых со временем только
возрастает. Выбранный нами путь в известной
степени комбинирует традиционную системати-
ку с эволюционным подходом, но не в сторону
его формализации, подобно современной фило-
генетике, а делает упор именно на содержатель-
ный аспект преобразования формы организмов,
который может быть не менее увлекательным.
Нам было интересно показать, как одна сложная
система организации (например, раковинный
моллюск) превратилась в другую, например
в моллюска без раковины, а не просто констати-
ровать факт, что раковинные и безраковинные
группы на каком-то уровне ответвились от обще-
го эволюционного древа. Два столпа, на которых
держится предлагаемый здесь подхол достаточ-
но традиционны: морфология (т. е. строение
организмов) и онтогенез (т. е. анализ жизненных
циклов). И старая традиционная наука, взятая
под несколько необычным углом зрения, вполне
возможно еще сыграет свою не последнюю роль
и для сверхсовременной молекулярной биоло-
гии, протянув ей путеводную нить — нить систе-
матической иерархии. И само собой, ничто не
препятствует использованию этой книги по
«прямому назначению», т. е. как атласа, посколь-
ку именно цветной атлас заднежаберных моллю-
сков России и является его основой.
Одновременно сам атлас был написан с це-
лью именно серьезного знакомства с фауной
заднежаберных моллюсков морей России. К со-
жалению, даже при таком подходе невозможно
в сколько-нибудь популярном формате предста-
вить всех заднежаберных моллюсков фауны Рос-
сии — подобная работа является самостоятель-
ной задачей, скорее научной, чем популярной
(желающие ознакомиться со специализирован-
ной научной литературой, посвященной изуче-
нию фауны заднежаберных моллюсков России,
а также их систематике и эволюции могут найти
список научных работ авторов в приложении).
Прежде всего, есть некоторое число крайне ред-
ких видов, для которых нет не только прижиз-
ненного, но даже удовлетворительного изобра-
жения в фиксированном состоянии. Кроме того,
целый ряд видов представлен в виде «сомни-
тельных» указаний, никогда не подтвержденных
после их первой публикации. Особо следует
сказать о раковинных заднежаберных отряда
Cephalaspidea. Эти зарывающиеся формы часто
бывают труднодоступны, а кроме того имеют
в своем составе целый ряд очень сходных видов,
различение которых может составить головную
10
боль даже для специалиста. По этой причине
при отборе представителей этой группы учи-
тывалась хотя бы минимальная их доступность
для наблюдения, а также их особый системати-
ческий или эволюционный интерес. В резуль-
тате из более чем 50 видов цефаласпид фауны
России в атласе представлены лишь 15. Но это
не означает, что по атласу нельзя составить
впечатление о составе цефаласпид морей Рос-
сии. Напротив, все основные группы в нем
представлены. В тексте при сравнении видов
также нередко даются указания на те виды, ко-
торые не вошли непосредственно в атлас. В то
же время все небольшие группы заднежабер-
ных, вроде акохлидиид и птеропод, имеют
100 % репрезентативность. Наконец, основные
группы заднежаберных моллюсков, наиболее
богатые видами, как в мировой фауне, так
и в фауне морей России, — дориды и другие
голожаберные моллюски имеют представи-
тельство примерно от 70 до 85 %. Это, безуслов-
но, высокие цифры, и практически любой вид
голожаберников, собранных в пределах морей
России и сопредельных акваторий, сможет
быть отнесен, как правило, к определенному
виду или реже к близко родственным группам.
Статистика по отдельным группам заднежа-
берных в результате имеет следующий вид:
• Отряд Cephalaspidea (Цефаласпиды) —
около 30 % (15 видов)
• Отряд Anaspidea (Анаспиды) —
100 % (3 вида)
• Группа Pteropoda (Крылоногие
моллюски) —
100 % (2 вида, не считая заносных)
• Отряд Acochlidiida (Акохлидииды) —
100 % (3 вида)
• Отряд Sacoglossa (Сакоглоссы) —
около 80 % (6 видов)
• Отряд Notaspidea (Нотаспиды) —
100 % (3 вида)
• Отряд Doridacea (Голожаберные
моллюски-дориды) —
около 70 % (27 видов)
• Отряд Nudibranchia (собственно
голожаберные моллюски) —
около 85 % (44 вида)
Всего, таким образом, в атлас включено
103 вида заднежаберных моллюсков, из которых
к полностью безраковинным относится 81 вид.
За немногими исключениями, перечисленными ниже, все фотографии заднежаберных мол-
люсков сделаны непосредственно авторами данной книги, Т. А. Коршуновой и А. В. Мартыновым.
Фотографии следующих видов были предоставлены:
• Akera bullata и Coryphella nobilis — О. В. Савинкиным
(Институт проблем экологии и эволюции РАН, г. Москва),
• Akiodoris lutescens — Н. П. Санамян (Камчатский филиал тихоокеанского института
географии ДВО РАН, г. Петропавловск-Камчатский),
• Aplysia juliana и A. parvula — М. В. Переладовым (Всероссийский научно-
исследовательский институт рыбного хозяйства и океанографии, г. Москва),
• Coryphella polaris — Н. А. Червяковой
(Биологический факультет МГУ им. М. В. Ломоносова, г. Москва),
• Calycidoris guentheri и Coryphella salmonacea — Bjorn Gulliksen
(University of Tromso, Норвегия),
• Dendronotus robustus — Erling Svensen (Egeroya, Норвегия),
и используются с их разрешения.
Авторы благодарят К. Э. Санамян и Н. П. Санамян (Камчатский филиал тихоокеанского
института географии ДВО РАН, г. Петропавловск-Камчатский), Б. И. Сиренко (Зоологический
институт РАН, г. С.-Петербург), О. В. Савинкина (Институт проблем экологии и эволюции
РАН, г. Москва) и В. В. Гульбина (Институт биологии моря ДВО РАН, г. Владивосток) за по-
мощь во время полевых работ и изучения коллекций, сотрудников межкафедральной лабо-
ратории электронной микроскопии Биологического факультета МГУ имени М. В. Ломоносо-
ва (г. Москва), а также Michael Schrodl (Баварская государственная зоологическая коллекция,
Мюнхен, Германия) за многолетнее продуктивное сотрудничество.
11
БИОЛОГИЯ
ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ
ГОЛОЖАБЕРНЫХ И ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
Кто же такие голожаберные моллюски
и почему еще существует выражение «заднежаберные моллюски»
Как уже было указано, данная книга — цвет-
ной атлас, в котором изображения представ-
ляют самостоятельную ценность и которые
читатель может изучать в известном смысле
отдельно от текста. Однако не следует забы-
вать, что одновременно данная работа являет-
ся еще и введением в систематику голожабер-
ных моллюсков. Поэтому наилучший эффект
будет достигнут, если читатель возьмет на се-
бя некоторый труд ознакомиться с общим
строением этих удивительных животных
и с их привычками. Это не так трудно, как,
возможно, кажется на первый взгляд. Особен-
но если помнить про «путеводную нить» всей
биологии — систематику, и важнейший ее
принцип — иерархию. Последовательно «сво-
рачивая» данные о тысячах отдельных видов
во все более общие, но значительно менее
многочисленные понятия — категории, мож-
но гораздо легче выучить этот универсальный
язык жизни, который затем может быть при-
менен к любым частным случаям — отдель-
ным видам. Помимо ключевого принципа
иерархии, необходимо будет усвоить еще
и некоторое количество названий на латин-
ском языке. В XVIII веке латынь еще была
основным языком всей науки, поэтому и «отец
систематики» Карл Линней «раздал» живот-
ным и растениям названия именно на этом
языке. И хотя во времена Линнея была описа-
на лишь очень незначительная часть живого
мира, известного нам сегодня, правила описа-
ния видов, заложенные «отцом систематики»,
сохранились и по сей день, ведь «эффект осно-
вателя» очень силен в любой области челове-
ческой деятельности. По той же причине мы
унаследовали латинский язык как основной
язык научного описания живого мира. И даже
если названия видов взяты из других языков
(например, древнегреческого или русского),
они записаны латинским алфавитом и им
приданы соответствующие окончания. На-
пример, хорошо известное видовое название
лососевой рыбы кижуча, на латыни будет зву-
чать почти также — Oncorhynchus kisutch. По-
скольку слово кижуч было заимствовано из
бесписьменного тогда языка одного из наро-
дов Камчатки. Сходным образом, русское на-
звание лисицы-корсака было латинизировано
и стало основой для его латинского имени,
т. е. официального научного названия Vulpes
corsac. Но эти примеры — скорее исключения.
Большинство научных названий организмов
являются родными латинскими словами или
латинизированными древнегреческими. Для
названий иногда используются и даже ничего
не значащие, но красиво звучащие сочетания.
Конечно, никто не будет требовать от вас зна-
ния весьма сложной грамматики латинского
языка. Научные названия видов и групп систе-
матической иерархии — не более чем свое-
образные ярлыки-метки, которые мы «накле-
иваем» на них, чтобы уметь их различать. Но
выучив небольшое число латинских названий,
вы фактически приобщитесь к тому великому
языку, который в совершенстве знали и Нью-
тон, и Ломоносов, и Линней.
Преимущества же знания не только назва-
ний видов, но и других групп систематической
иерархии заключаются как раз в том, что, даже
не зная определенное название того или иного
вида, мы можем точно очертить круг форм,
13
БИОЛОГИЯ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
к которым он относится. Для этого необходи-
мо понять, что моллюск является представите-
лем семейства, например, хромодоридид (на
латыни это будет Chromodorididae). К этому
семейству относятся сотни видов и десятки ро-
дов голожаберных моллюсков, обитающих
в тропических водах — таких, как Красное мо-
ре или побережье Вьетнама. Поскольку с опре-
деленными оговорками можно признать, что
легче определить признаки семейства, то
встретив под водой какого-нибудь хромодори-
са (Chromodoris) или глоссодориса (Glossodoris),
мы можем пока уверенно сказать, что это
хромодоридида, а выяснение деталей оста-
вить на потом. Более того, если мы находим-
ся у берегов российских морей, и обнаружим
там хромодоридиду, то можно будет с уверен-
ностью сказать, что это один из видов рода
Cadlina (кадлина), поскольку других предста-
вителей семейства Chromodorididae в морях
России не обитает. По крайней мере, пока не
обнаружено.
Какое же место в систематической иерар-
хии занимают голожаберные моллюски? Не-
сколько упрощая, их положение можно ука-
зать следующим образом:
Царство Animalia (животные). К этой
наиболее общей группе относятся организмы,
в большинстве случаев не способные к само-
стоятельному синтезу органических веществ
с использованием энергии солнечного света
(как это имеет место у растений). Другая харак-
терная черта животных — способность к актив-
ному движению (даже у сидячих, неподвиж-
ных на первый взгляд организмов). К царству
животных, таким образом, относится и микро-
скопическая амеба, и многотонный гренланд-
ский кит, и похожие на растения колонии ги-
дроидов и мшанок. В свою очередь в царство
Animalia входит (помимо одноклеточных), око-
ло 30 типов многоклеточных животных, таких,
например, как губки (Porifera), кишечнопо-
лостные (Cnidaria) или хордовые (Chordata).
Голожаберники относятся к типу моллюсков
(Mollusca) — обширной группе морских, пре-
сноводных и наземных животных, насчитываю-
щей не менее 100 000 видов, т. е. понятие
Mollusca «сворачивает», включает в себя, раз-
нообразие почти сотни тысяч видов, тысяч ро-
дов, сотен семейств и отрядов и около десятка
классов. Вместо того чтобы бесконечно пере-
числять все эти тысячи видов, мы просто ука-
зываем, что это — моллюски. Очень удобно
и практично. Итак:
Тип Mollusca (моллюски). К этому ти-
пу относятся на первый взгляд очень разные
по внешнему строению животные: головоно-
гие (кальмары, осьминоги), брюхоногие мол-
люски (улитки) и почти червеобразные, мало-
известные широкой публике соленогастры.
Тем не менее все эти различные организмы
объединяет уникальный набор признаков: на-
личие в глотке радулярного аппарата (особым
образом устроенных зубов моллюсков) и ман-
тийного комплекса органов. Мантия — это
своеобразная секреторная складка вокруг тела
моллюска, неизвестная в таком виде у других
типов животных. В большинстве случаев ман-
тия секретирует раковину — твердый внеш-
ний скелет моллюсков. Также мантия отграни-
чивает особую — мантийную — полость,
имеющуюся только у моллюсков. Внутри ман-
тийной полости находятся жабры и несколько
специализированных желез; там же открыва-
ется задняя кишка, т. е. анальное отверстие
и половое отверстие.
Класс Gastropoda (гастроподы или
брюхоногие моллюски). К этому классу от-
носятся виды, у которых имеются хорошо вы-
раженная, как правило, спиральнозакручен-
ная твердая раковина и ползательная
подошва — нога. В большинстве случаев для
них характерна также обособленная мантий-
ная полость с одной или двумя жабрами (кте-
нидиями) внутри. Возникновение современ-
ных брюхоногих моллюсков было обусловлено
специфическим процессом, который произо-
шел у их древних предков. Раковина, вместе
с внутренними органами и мантийной поло-
стью, по до сих пор не вполне ясным причи-
нам развернулась относительно ноги на 180
градусов. В результате все современные гастро-
поды оказались перекрученными и органы
мантийной полости открываются у них не так,
как было у предков: отверстие мантийной по-
лости, жабры и анальное отверстие направле-
ны вперед, и находятся фактически над голо-
вой, тогда как раньше они «смотрели» назад.
Сообразуясь с понятием перекрученности,
этот процесс назвали торсией (от лат. torsio —
вращение, скручивание).
14
Систематическое положение голожаберных и заднежаберных моллюсков
Гидроидный полип,
представитель кишечнополостных (Cnidaria)
Представител ъ
типа мшанок (Bryozoa)
И вот своеобразный «реванш» за искаже-
ние предковой морфологии осуществила как
раз та самая группа, которой посвящена дан-
ная книга, а именно,
Подкласс Opisthobranchia (заднежа-
берные брюхоногие моллюски или просто
заднежаберные моллюски). У них мантий-
ная полость начала постепенно «сползать на-
зад», передвинувшись сначала в правое боко-
вое, а затем и почти в заднее положение.
Мантийная полость, естественно, потянула за
собой и жабру, и анальное отверстие. Процесс
этот обозначили как деторсия, т. е. обратная
торсия. Деторсия не привела заднежаберных
назад к своим предкам. Вместо этого, волею
эволюционной судьбы, сформировалась, на-
верное, одна из самых удивительных групп жи-
вотного мира. Попытаться дать этой группе
определение — отдельная и очень серьезная
задача. Деторсия, конечно, не обусловила все
многообразие заднежаберных, но стала своего
рода «спусковым крючком», триггером даль-
нейших радикальных преобразований органи-
зации данной группы, основным трендом ко-
торой стала редукция. Дело в том, что
заднежаберные в первую очередь характеризу-
ются тем, что они утратили в сравнении с ра-
ковинными предками, и только потом тем, что
они приобрели. У многих представителей это-
го подкласса нет (или очень сильно мо-
дифицировано) того, что составляет основу
любого брюхоногого моллюска: раковины,
мантийной полости, настоящей жабры. Но
и приобрели они много. Научились употре-
блять в пищу невкусные, а порой просто опас-
ные или ядовитые организмы, такие как ки-
шечнополостные, губки или мшанки. Список
«изобретений» заднежаберных можно продол-
жать очень долго (это и заимствование чужих
стрекательных клеток для собственной защи-
ты, и использование хлоропластов водорослей
для получения дополнительной энергии, и ряд
других). Пожалуй, самым выдающимся и фан-
тастическим по степени эволюционного успе-
ха их «проектом» стали голожаберные моллю-
ски (Nudibranchia). Именно по этой причине
говорить об этой группе бессмысленно без
долгого и обстоятельного разговора о том, кто
же такие заднежаберные моллюски, какие под-
группы включают и как соотносятся с собствен-
но голожаберными моллюсками.
15
БИОЛОГИЯ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
Строение голожаберных моллюсков группы Aeolidacea
16
Строение заднежаберных моллюсков
СТРОЕНИЕ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
Введение в морфологию заднежаберных моллюсков:
что нужно обязательно знать, чтобы лучше понять эту книгу
Мы выяснили, что Opisthobranchia и Nudi-
branchia относятся к типу Mollusca (моллюсков),
и соответственно обладают целым комплексом
признаков, присущих моллюскам в целом (ман-
тийная полость, жабры, радулярный аппарат).
В то же время такой характернейший признак
(как всего типа моллюсков, так и класса брюхо-
ногих — улиток), как раковина, во многих груп-
пах заднежаберных частично редуцируется или
полностью исчезает. Процесс редукции ракови-
ны протекал независимо в различных таксонах,
и это привело к появлению внешне очень сход-
ных, но имеющих различное эволюционное про-
исхождение отрядов заднежаберных моллюсков,
в конечном счете происходящих от одного пред-
ка. Из наиболее ярких примеров внешне необы-
чайно сходных, но имеющих серьезные внутрен-
ние отличия, доказывающих их независимое
происхождение, следует, конечно, указать пред-
ставителей отряда сакоглосс (Sacoglossa) и голо-
жаберных моллюсков из группы эолид (Aeolida-
сеа). И в том и другом случае сформировался
полностью безраковинный моллюск, лишенный
настоящей жабры. «Освободившееся» простран-
ство на спинной стороне тела было занято осо-
быми удлиненными неразветвленными выроста-
ми — папиллами, нередко располагающимися
в особого рода пучках.
Получившиеся в итоге организмы нередко
столь сходны внешне, что неспециалисты с боль-
шой вероятностью отнесут их к одной группе.
Такие явления конвергенции (или параллельного
развития) весьма обычны в животном мире. До-
статочно вспомнить, что дельфины очень сходны
с рыбами и также имеют плавники, с помощью
которых они передвигаются в водной среде. В та-
ких случаях уже упомянутая систематическая
иерархия и «завернутая» в нее информация о де-
талях морфологического строения является той
самой путеводной нитью, которая позволит из-
бежать ошибочного сближения этих двух таксо-
нов. Уже Аристотелю было известно, что дель-
фины, в отличие от рыб, дышат легкими
и кормят своих детенышей молоком. Таким об-
разом они относятся к млекопитающим, т. е.
классу Mammalia, как сказал бы современный
систематик. Из этого также следует, что дельфи-
ны вторично перешли к обитанию в водной сре-
де, поскольку все имеющиеся данные свидетель-
ствуют, что предки первых млекопитающих
были исключительно наземными животными.
Точно также и в случае независимого происхо-
ждения отрядов Sacoglossa и Nudibranchia, у нас
имеется достоверная информация, что две эти
группы произошли независимо от различных
древних заднежаберных моллюсков, у которых
еще была раковина. Это не абстрактное, теорети-
ческое знание. В группу сакоглосс входят не-
сколько семейств, у которых еще имеется вполне
развитая раковина, сходная с таковой у настоя-
щих раковинных Opisthobranchia из отряда це-
фаласпид. И раковинные сакоглоссы значитель-
но отличаются от современных раковинных
представителей отряда нотаспид (Notaspidea),
наиболее близких к настоящим голожаберным
моллюскам по целому ряду внутренних призна-
ков. А в группу анаспид, куда входят хорошо из-
вестные тропические «морские зайцы» (Aplysia,
аплизии), у которых раковина тоненькая, внутрен-
няя, завернута под мантией, входят одновременно
и любители холодных вод акеры (Akerа), еще об-
ладающие вполне развитой наружной ракови-
ной, хотя уже очень тонкой, как и у аплизий.
Все приведенные выше факты преследуют
очень важную цель, а именно — продемонстри-
ровать, что в основе практически любой группы
безраковинных заднежаберных моллюсков ле-
жат раковинные предки, причем, как правило,
до сих пор обитающие в современных морях.
Поэтому вся та сложная эволюционная мозаика
из модификаций мантии, жабры, радулярного
аппарата (зубов) и других органов, наложенная
на независимую редукцию (исчезновение) рако-
вины, и привела к появлению целого «букета»
групп, которые составляют нынешних Opistho-
branchia. Понимание того, что за этими столь
эстетически прекрасными и в сущности столь
похожими моллюсками стоят сложные процес-
сы их исторического формирования, несомнен-
но придает особое измерение этой книге. Мы не
просто разглядываем красивые изображения,
а делаем это с большим внутренним смыслом:
постигаем обычно скрытую от нас эволюцион-
ную историю группы.
17
БИОЛОГИЯ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
Итак, вышеупомянутый «букет заднежа-
берных моллюсков» состоит из следующих
отрядов:
• Отряд Cephalaspidea — Цефаласпиды
• Отряд Anaspidea (=Aplysiomorpha) —
Анаспиды или морские зайцы
• Отряд Thecosomata — Текосоматы
или раковинные крылоногие моллюски
• Отряд Gymnosomata — Гимносоматы или
безраковинные крылоногие моллюски
• Отряд Sacoglossa — Сакоглоссы
• Отряд Acochlidiacea — Акохлидииды
• Отряд Notaspidea (=Pleurobranchoidea) —
Нотаспиды
• Отряд Doridacea (=Anthobranchia) —
Голожаберные моллюски-дориды
• Отряд Nudibranchia — Голожаберные
моллюски
Обобщенные изображения каждого из
указанных отрядов заднежаберных моллюсков
читатель найдет на стр. 18-19. Характеристика
каждого из этих отрядов будет приведена да-
лее, непосредственно перед описанием видов
CEPHALASPIDEA
SACOGLOSSA
головной
щит
лопасти
головного
щита
раковина
ANASPIDEA
ринофоры
висцеральный
мешок
18
Строение заднежаберных моллюсков
NOTASPIDEA
NUDIBRANCHIA
DORIDACEA
ВСКРЫТАЯ ДОРИДА
Cryptobranchia Phanerobranchia
оральный рот ринофоры
глаз
гонады
желудок
кишечник
сердце
половая
система
центральная
нервная
система
пище-
варительная
железа
глотка
вскрытый
нотум
анус
ПОЛОВАЯ СИСТЕМА ДОРИДЫ
19
БИОЛОГИЯ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
Большой отряд дорид делится на две основные группы:
• криптобранхиальные дориды (Cryptobranchia), т. е. такие, у которых жабры втягиваются в особую полость
(слева криптобранхиальная дорида Cadlina laevis — вверху жабры вытянуты, внизу жабры втянуты в полость);
• фанеробранхиальные дориды (Phanerobranchia), т. е. такие, у которых жаберная полость отсутствует
(справа фанеробранхиалъная дорида Acanthodoris pilosa — вверху жабры вытянуты, внизу жабры сокращены,
жаберная полость отсутствует). Моря России — подлинное царство ф)анеробранхиалъных дорид, тогда как
видов криптобранхий известно в нашей фауне совсем немного
Согласно распространенной точке зрения, фанеробранхиальные дориды эволюционно предшествовали
криптобранхиальным. Открытие в водах Камчатки уникального переходного звена Onchimira cavifera
Martynov et al., 2009, доказало, что все произошло с точностью до наоборот: сначала возникли более сложные
криптобранхии, а фанеробранхиальные дориды произошли от них путем редукции
20
Строение заднежаберных моллюсков
из этих групп, обитающих в морях России. Для
некоторых из этих групп даны не только наи-
более «типичные» представители (в системати-
ке, из-за огромного разнообразия таксонов,
слово «типичный» надо использовать с боль-
шой осторожностью), но и различные виды,
характеризующие большие подгруппы внутри
того или иного отряда. Так, отряд голожабер-
ных моллюсков-дорид, Doridacea, широко
представленный в морях России, включает две
традиционные группы — криптобранхиаль-
ных дорид (Cryptobranchia), у которых имеется
особая полость вокруг жабр, а радула состоит
из значительного числа рядов зубов, и фанеро-
бранхиальных дорид (Phanerobranchia), у них
полость вокруг жабр отсутствует, а радула со-
стоит из небольшого числа рядов зубов. Но
природа очень любит «нарушать» те рамки,
которыми человек пытается ее ограничить.
В подтверждение этого совсем недавно у берегов
Камчатки нами была сделана сенсационная на-
ходка — был обнаружен новый род (и вид), ко-
торый сочетал в себе признаки двух больших
групп дорид, т. е. Cryptobranchia и Phanero-
branchia. Новый вид так и был назван — Onchi-
mira cavifera, что в вольном переводе означает
«удивительная онхидоридида, несущая жабер-
ную полость». У этого нового рода имеется жа-
берная полость, как и у криптобранхиальных
дорид, но другие признаки внутреннего строе-
ния в точности соответствуют таковым
фанеробранхиальных дорид из семейства
Onchidorididae. Подобные явления, когда в при-
роде можно обнаружить реальные, а не гипоте-
тические переходные звенья между двумя боль-
шими группами организмов, являются гораздо
более частым феноменом среди морских бес-
позвоночных, чем это можно было бы себе пред-
ставить. И обусловлены они фактом эволюции.
Подробнее об этом будет рассказано ниже.
И представители всех без исключения от-
рядов заднежаберных моллюсков обнаружены
в морях России! Разнообразие их конечно да-
леко не столь велико, как в тропиках, но если
пытливый читатель задастся целью на личном
опыте (например при погружениях с аквалан-
гом) познать все разнообразие Opisthobranchia,
то он будет вознагражден.
Любая группа организмов характеризуется
как сходством, так и отличиями в сравнении
с ближайшими группами организмов. Задне-
жаберные моллюски обладают целым рядом
общих признаков внешнего и внутреннего
строения, которые, однако, имеют свой опреде-
ленный вариант в каждой конкретной группе
и которые будут упоминаться в кратких очер-
ках, сопровождающих описание видов. Отме-
тим среди этих признаков наиболее существен-
ные. Для удобства термины, описывающие
строение (морфологию) заднежаберных мол-
люсков, сгруппированы последовательно от об-
ласти головы к задней части тела, а после этого
к органам внутреннего строения:
Органы головного отдела:
Головной щит — кожная складка, покры-
вающая головную лопасть у раковинных зад-
нежаберных моллюсков отряда цефаласпид
(Cephalaspidea). Головной щит выделяет
обильную слизь, способствующую скольже-
нию, что позволяет моллюскам лучше пере-
двигаться в грунте.
Оральный парус — широкая полукруглая
или трапециевидная складка вокруг рта у ряда
заднежаберных (нотаспиды, некоторые дори-
ды) и голожаберных моллюсков.
Оральные щупальца — пара околоротовых
щупалец разнообразного строения. Основной
орган осязания. У нотаспид семейства плеуро-
бранхид (Pleurobranchidae) оральные щупальца
имеют вид свернутых трубок, соединенных ши-
рокой перепонкой; такой оральный аппарат
иногда называют парусом. У дорид оральные
щупальца варьируют от слабо обособленных
лопастей с боковыми щелями до цельных паль-
цевидных выростов, а иногда полностью отсут-
ствуют. У семейств Tritoniidae, Dendronotidae,
некоторых арминид (Arminidae) оральные щу-
пальца отсутствуют, а у большой группы голо-
жаберных моллюсков Aeolidacea, они напротив,
почти всегда хорошо развиты.
Ринофоры (в дословном переводе «несущие
нос») — пара щупальцевидных органов химиче-
ского чувства, расположенных в передней части
тела. Иногда их еще называют щупальцами вто-
рой пары. В отличие от млекопитающих, для
которых зрение является основным или, по край-
ней мере, не менее важным, чем обоняние, ис-
точником информации об окружающем мире,
для многих морских обитателей мир состоит
главным образом из запахов, а глаза в лучшем
случае способны лишь различать свет и тень.
Заднежаберные моллюски — не исключение. По
запахам, т. е. ничтожному количеству молекул
21
БИОЛОГИЯ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
Оральный парус и ротовое отверстие представителя отряда нотаспид Berthella californica крупным планом
Dendronotus dalli. Поверхность тела голожаберных моллюсков дендронотид несет множество разветвленных
отростков, особенно мощных в околоротовой области
22
Строение заднежаберных моллюсков
Для нотаспид характерны свернутые в трубку
ринофоры (хорошо заметна продольная щель).
Ринофоры — хемосенсорный орган, основной
инструмент, с помощью которого заднежаберные
моллюски ориентируются в пространстве
Голожаберные моллюски из группы Dendronotacea
(как Dendronotus frondosus) также обладают
цельными пластинчатыми ринофорами,
но они дополнительно окружены мощными
разветвленными отростками
Для голожаберных моллюсков-дорид типичны цельные
ринофоры, покрытые многочисленными пластинками,
как например у акантодориса (Acanthodoris pilosa)
Многие голожаберники-эолидацеи (Aeolidacea),
как например Cuthonella marisalbi, обладают
простыми гладкими ринофорами
23
БИОЛОГИЯ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
тех или иных веществ в воде, они находят и еду,
и партнера для размножения. Ринофоры чрезвы-
чайно разнообразны по форме — от свернутых
в трубку, «ухообразных» у заднежаберных мол-
люсков отрядов анаспид (Anaspidea) и нотаспид
(Notaspidea) до сложных органов, состоящих из
стебля, окруженного разветвленными отростка-
ми у голожаберных моллюсков из семейства
тритониид (Tritoniidae) или покрытого плотны-
ми рядами пластинок у большинства голожа-
берных моллюсков из отряда дорид (Doridacea)
и др. Часто представляют просто пару гладких
щупалец. Обычно способны втягиваться в осо-
бые ринофоральные карманы.
Органы висцерального мешка:
Висцеральным мешком называют массив-
ную заднюю часть тела многих заднежаберных
моллюсков, содержащую большинство вну-
тренних органов, мантийную полость, ракови-
ну и жабру. У многих сакоглосс, всех нотаспид
и голожаберных моллюсков висцеральный ме-
шок сливается с головным отделом.
Мантия и мантийная полость — специ-
фический орган, имеющийся только у моллю-
сков. Представляет собой особого рода кожную
складку, которая секретирует твердый наруж-
ный скелет — многослойную раковину. Ман-
тийная складка образует также мантийную
полость — один из важнейших органов моллю-
сков. В мантийной полости располагается одна
или две жабры, там же находятся выводные про-
токи почек, анальное и половое отверстия.
У большинства брюхоногих моллюсков мантий-
ная полость открывается в передней части тела
над головой. Жабра в этом случае также направ-
лена вперед. Поэтому одно из традиционных
названий этой большой группы (подкласса)
брюхоногих — переднежаберные (по латыни
Prosobranchia). В данной же книге идет речь
о заднежаберных брюхоногих моллюсках
(Opisthobranchia). Традиционно им также при-
дается статус подкласса, но в последние два
десятилетия в связи с коренной ломкой пред-
ставлений о систематической иерархии, рас-
пространением так называемой филогенетиче-
ской и молекулярной таксономии, системы
могут выглядеть гораздо сложнее простой дилем-
мы переднежаберные/заднежаберные. Почему
заднежаберные? Дело в том, что мантийная по-
лость у большинства представителей этой группы
развернута в сравнении с переднежаберными
примерно на 90° вправо. В результате мантийная
полость оказывается сбоку, а не над головой, и жа-
бра «смотрит» в заднебоковом направлении.
У различных представителей заднежаберных
моллюсков процесс смещения мантийной поло-
сти или жабры может идти еще дальше, в резуль-
тате чего эти органы оказываются действительно
на самом заднем конце тела. У тех групп заднежа-
берных моллюсков, у которых раковина подвер-
глась редукции (или исчезла), вполне ожидаемо
редуцируется или сильно видоизменяется ман-
тия, которая и ответственна за синтез раковины.
Так, у анаспид, у которых имеется тоненькая вну-
тренняя раковина, мантия и мантийная полость
еще сохраняются, хотя и в уменьшенном виде,
а вот у голожаберных моллюсков, у которых рако-
вина полностью исчезает во взрослом состоянии,
мантия сильно трансформируется, превращаясь,
например, в кожный щит (нотум), покрываю-
щий спину у нотаспид и дорид, а мантийная по-
лость и вовсе исчезает, жабра при этом может
прикрепляться непосредственно к боковой стенке
тела (у нотаспид).
Настоящая жабра (ктенидий) — называ-
ется так потому, что сохраняет сходство с жа-
берным аппаратом других моллюсков и имеет
с ним общее происхождение. Напротив, вто-
ричные жабры, присущие большинству голо-
жаберных моллюсков, — это простые выросты,
имеющие, в отличие от настоящих жабр, иную
схему кровоснабжения, но выполняющие та-
кую же (дыхательную) функцию. Кольцеобраз-
ные жабры представителей отряда дорид явля-
ются настоящими жабрами, но только сильно
видоизмененными. Один непарный ктенидий
свойственен большинству отрядов заднежабер-
ных моллюсков — цефаласпидам, анаспидам,
нотаспидам, архаичным семействам сакоглосс.
У акохлидиид, большинства сакоглосс и у всех
представителей отряда голожаберных моллю-
сков (Nudibranchia) настоящий ктенидий пол-
ностью отсутствует.
Раковина — раковина является характер-
ным признаком типа Mollusca, но ее отсутствие
вовсе не означает принадлежность того или
иного вида к другому типу животных, посколь-
ку у самых различных классов, относящихся
к этому типу, раковина может уменьшаться
в размерах или даже полностью исчезать. Сре-
ди заднежаберных моллюсков более или менее
развитая наружная раковина имеется только
у представителей отряда Cephalaspidea и неко-
24
Строение заднежаберных моллюсков
Основной орган дыхания у заднежаберных моллюсков — жабра. Она может располагаться с правой стороны
под кожной складкой (нотумом), как у нотаспиды Berthella californica, перемещаться на спинную сторону,
как у большинства дорид, или реже располагаться на брюшной стороне
торых архаичных семейств из отряда сакоглосс
(Sacoglossa). У видов из отрядов Anaspidea
и Notaspidea раковина в большинстве случаев
внутренняя, и находится под мантией. В этом
случае раковина представляет собой обычно
овальную вогнутую пластинку разного размера
с бортиком по краю и рудиментарным (т. е.
сильно уменьшенным) не выступающим завит-
ком, расположенным в левом заднем углу рако-
вины. Наконец, все без исключения голожабер-
ные моллюски из обоих отрядов, т. е. Doridacea
и Nudibranchia во взрослом состоянии полно-
стью лишены раковины. Однако нотум, как про-
изводное мантии, вполне естественно сохраняет
секреторную активность и у безраковинных
групп. Так, многие голожаберные моллюски из
отряда дорид несут внутри нотума множество
иголочек-спикул, которые вместе образуют
ажурную сеть, служащую для лучшей защиты
мягкого тела этих моллюсков. Также лишены
раковины представители отрядов Acochlidiida
и большинство семейств отряда Sacoglossa. Но
даже в этом случае раковина полностью не ис-
чезает из жизненного цикла заднежаберных
Редкий случай брюшного расположения жабр
у особой группы дорид — корамбид (Corambe obscura)
25
БИОЛОГИЯ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
Среди голожаберных моллюсков только дориды
обладают настоящими жабрами, тогда как
для остальных групп характерны так называемые
вторичные жабры, которые могут выглядеть как
множество разветвленных пучков вдоль боков тела
(как у дендронотацеи Tritonia tetraquetra), или в виде
неразветвленных папилл, как у группы Aeolidacea
Мощные жабры архидориса (Archidoris pseudoargus)
крупным планом, способные втягиваться в особую
полость. Архидорис — один из самых крупных видов
дорид российских морей, своему недавнему появлению
у побережья Баренцева моря обязан происходящему
в течение
последних десяти лет потеплению Арктики
Обычное для дорид спинное расположение жабр
(Onchidoris bilamellata). Каждая жабра онхидориса
представляет собой неразветвленный вырост,
несущий особые пластинки
У целого ряда голожаберных моллюсков-дорид вдоль
края имеются специальные защитные образования,
отпугивающие хищников, как эти лимонно-желтые
железы у Cadlina laevis
У колги (Colga minichevi) каждая жабра еще дополни-
тельно ветвится, в результате чего жаберный аппарат
таких дорид напоминает небольшой кустарник
Спинная сторона многих дорид (например у Cadlina
laevis) несет мощный кожный щит (нотум),
окружающий тело со всех сторон
26
Строение заднежаберных моллюсков
моллюсков. Подобно большинству голожабер-
ных моллюсков, сакоглоссы и акохлидииды со-
храняют крошечную, но нормально развитую
раковину на личиночной стадии, что является
одним из лучших подтверждений происхожде-
ния безраковинных заднежаберных от раковинных
предков и реальности эволюции.
Нотум и связанные с ним структуры:
Нотум — кожный щит, покрывающий
дорсально (т. е. со спины) тело голожаберных
моллюсков. Нотум — это видоизмененная ман-
тия. У многих голожаберных моллюсков нотум
имеет тенденцию к полной или частичной ре-
дукции. В некоторых случаях нотум имеет под-
кожные железы, секретирующие защитные
вещества. Поверхность нотума у некоторых го-
ложаберных моллюсков отряда дорид покрыта
особыми образованиями — кариофиллидиями.
Кариофиллидий представляет собой микро-
скопический цилиндрический вырост, на конце
которого располагается розетка длинных спи-
кул. Они многочисленны и из-за их очень мел-
ких размеров поверхность нотума кажется
гладкой или бархатистой. Считается, что ка-
риофиллидии играют роль механорецептора,
т. е. своеобразного органа осязания.
Вторичные жабры — различные респира-
торные структуры, появляющиеся у голожабер-
ных моллюсков после редукции (исчезновения)
настоящего ктенидия. Они могут располагаться
на обратной стороне нотума плотными рядами
в виде листовидных пластинок (у дорид из се-
мейства филлидиид, Phyllidiidae), продольных
складок, папилл (у голожаберных моллюсков
сем. Arminidae), древовидных структур по кра-
ям нотума у голожаберных моллюсков Tritonii-
dae и Dendronotidae.
Жаберный карман — полость в задней ча-
сти нотума у многих дорид, куда втягиваются
жабры (см. стр. 20).
Железы нотума («мантийные» железы) —
многоклеточные подкожные железы у неко-
торых криптобранхиальных дорид, главным
образом хромодоридид, секретирующие за-
щитные вещества.
Папиллы (цераты) — спинные выросты
главным образом у голожаберных моллюсков
традиционных подотрядов эолидацей (Aeoli-
dacea) и арминацей (Arminacea). У всех эоли-
дацей и у арминацей из семейства янолид
(Janolidae) в папиллы проникает вырост пище-
варительной железы (дивертикулюм). У пода-
вляющего большинства эолидацей вырост пи-
щеварительной железы внутри вершинной
части папиллы формирует особый мешок —
книдосак, где накапливаются попадающие
с пищей стрекательные клетки (нематоцисты)
кишечнополостных, которые в дальнейшем ис-
пользуются для защиты моллюсков.
У всех представителей голожаберных моллюсков группы Aeolidacea, как например у Trinchesia pustulata, спинная
сторона покрыта папиллами, где хранятся «похищенные» у книдарий стрекательные клетки
27
БИОЛОГИЯ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
Спинные выросты-папиллы эолидацей служат
не только для дыхания, но и как важный орган защиты:
на их вершинах хранятся готовые к использованию
стрекательные (жгучие) клетки. Хорошо заметен узкий
вырост пищеварительной железы у Eubranchus tricolor,
проходящий сквозь всю папиллу и оканчивающийся
особым мешочком (книдосаком). Для этого вида
характерно особое защитное поведение — в случае
опасности он резко отбрасывает часть папилл
У эолидацеи Trinchesia viridis на вершинах папилл
белого пигмента нет, поэтому мешочек-книдосак,
содержащий чужие стрекательные клетки,
хорошо заметен. Расположение спинных папилл —
важный систематический признак в этой группе.
Папиллы могут покрывать спину многочисленными
неправильными рядами или принимать регулярный
вид, формируя правильные, хорошо разделенные
друг от друга ряды
Независимое возникновение сходных
буккальных помп в семействах
Akiodorididae (В-С) и Onchidorididae (D-E)
Спикулы — микроскопические известко-
вые иголочки прямой или слегка изогнутой
формы, расположенные в подкожном слое но-
тума или мантии. Наиболее характерны для
отрядов акохлидиид (Acochlidiida) и дорид
(Doridacea). Спикулы служат для укрепления
мягкого нотума, могут располагаться рыхло
или формировать густую плотную сеть. В по-
следнем случае, как например, у некоторых
видов семейства онхидоридид (Onchidoridi-
dae), нотум приобретает жесткость, сравнимую
со свойствами цельной раковины.
Органы пищеварительной системы:
Буккальная помпа (глоточный насос) —
особый мускулистый вырост глотки, характер-
ный для ряда семейств дорид, не имеющих
жаберной полости (так называемых фанеро-
бранхиальных дорид). Буккальная помпа по-
могает высасывать содержимое из мшанок,
асцидий и некоторых других колониальных
животных, которыми питаются дориды. Бук-
кальные помпы у различных семейств дорид
имеют разный план строения и возникли в хо-
де эволюции независимо на основе верхней
стороны глотки, исходно способной к сократи-
тельным движениям. Можно различить, по
меньшей мере, два основных типа буккальных
помп: первый, при котором помпа формиру-
ется на основе почти всей верхней стороны
28
Строение заднежаберных моллюсков
глотки, при этом концентрированных муску-
лов не образуется. Такой тип глотки можно
найти у семейств Akiodorididae и Anculidae.
Второй тип глотки характерен для семейств
Onchidorididae и Goniodorididae. В этом случае
буккальная помпа формируется только на
основе самой передние части глотки, в резуль-
тате чего она оказывается опоясанной мощным
лентовидным мускулом, который называется
периферическим. Буккальная помпа является
очень важным признаком, помогающим опре-
делить систематическое положение тех или
иных видов и направление их эволюции.
Радула — специфический орган, свой-
ственный исключительно моллюскам. Радула
занимает большую часть внутреннего объема
глотки и у различных таксонов состоит из раз-
ного числа твердых хитиноидных зубцов, рас-
положенных на эластичной подложке пра-
вильными поперечными рядами. Это один из
важнейших органов моллюсков, с помощью
которого осуществляется захват и первичная
переработка пищи. То или иное изменение
жизненной стратегии, например переход от
хищничества к растительной диете, ведет, как
правило, к значительным изменениям в строе-
нии зубов радулы. Поэтому радула является
одним из важнейших систематических при-
знаков (см. стр. 34), позволяющих различать
как отдельные виды, так и таксоны более высо-
кого ранга: роды, семейства и т. д.
Жевательный желудок — особый отдел,
расположенный между пищеводом и желези-
стым желудком у многих цефаласпид, всех ана-
спид и крылоногих моллюсков. Жевательный
желудок предназначен для дополнительной
механической переработки твердой или гру-
бой пищи, такой как раковины моллюсков или
слоевища водорослей и содержит разное чис-
ло особых жевательных пластинок — от трех
(у многих цефаласпид) до значительного числа
(у анаспид и крылоногих моллюсков).
Пищеварительная железа — крупный ор-
ган пищеварительной системы, внутри клеток
которого осуществляется процесс пищеваре-
ния. Следует отметить, что в отличие от млеко-
питающих, у которых процесс пищеварения
осуществляется в основном в полости кишечни-
ка, у моллюсков пищеварение преимуществен-
но внутриклеточное. Именно по этой причине
пищеварительная железа (иногда неверно на-
зываемая печенью) играет у заднежаберных
моллюсков очень важную роль. Форма и строе-
ние пищеварительной железы имеет большое
значение в систематике многих заднежаберных
моллюсков. У раковинных цефаласпид пищева-
рительная железа располагается внутри завитка
раковины и частично повторяет его спиральную
форму. У безраковинных заднежаберных мол-
люсков висцеральный мешок сливается с голов-
ным отделом и пищеварительная железа рас-
прямляется, становясь мощным компактным
органом у нотаспид и дорид. В ходе дальней-
шей эволюции пищеварительная железа из
компактного органа превращается в разветвлен-
ный, состоящий подчас из значительного числа
ветвей, комплекс. Подобное строение пищева-
рительной железы независимо появилось у двух
отрядов заднежаберных моллюсков — Sacoglos-
sa и Nudibranchia. Особой сложности развет-
вленная пищеварительная железа достигает
у голожаберных моллюсков группы Aeolidacea.
У них даже принято выделять переднюю и за-
днюю части пищеварительной железы.
Этим перечнем строение заднежаберных
моллюсков, разумеется, не исчерпывается. Для
того чтобы подробно описать все вариации
в строении нервной, кровеносной или пищева-
рительной систем понадобится множество от-
дельных томов. Но даже это небольшое пере-
числение некоторых «узловых» признаков
поможет читателю несколько приподняться
над простым разглядыванием картинок и по-
нять, что за каждым таксоном стоят очень
сложные морфологические сети, обеспечиваю-
щие функционирование того или иного вида.
Участок радулы представителя рода Cadlina,
электрон 1 ю-м икроскопи ческое изображен не
29
БИОЛОГИЯ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
ПОВЕДЕНИЕ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
Удивительные привычки удивительных моллюсков
Заднежаберные моллюски «создали» свой осо-
бый мир: с необычной едой, несъедобной для
большинства животных — губками, кишечно-
полостными, мшанками, с необычными при-
вычками — похищением чужих клеток и даже
молекул, культивированием в собственном ор-
ганизме одноклеточных зеленых водорослей.
В этом мире есть и крошечные (3-5 мм) весьеры
(Vayssierea), сверлильщики червей-спирорбид,
и полностью оторвавшиеся от дна планктон-
ные рыбообразные филлирое (Phyllirhoe),
и ушедшие в пространство между песчинками,
почти превратившиеся в червей — псевдовер-
месы (Pseudovermis). Хорошо знакомый многим
ныряльщикам «испанский танцор» — гекса-
бранхус (Hexabranchus) — гигант, достигающий
иногда полуметра, ползает по рифу, как обыч-
ный брюхоногий моллюск, но может неожи-
данно взмыть над поверхностью дна в вибри-
рующем танце на своих странных «крыльях»,
являющихся видоизменением края мантии-
нотума. Наконец, большая группа древних зад-
нежаберных моллюсков — цефаласпид, у ко-
торых еще сохраняется наружная раковина,
зарылись в песок и ил, где они стали передви-
гаться с помощью особого разрастания голов-
ной лопасти — головного щита и питаться
«чем придется» — мелкими фораминифера-
ми, моллюсками или червями.
Пищевые пристрастия, а также стратегии
защиты голожаберных моллюсков примерно
соответствуют двум их основным группам — от-
рядам дорид (Doridacea) и собственно голожа-
берным моллюскам (Nudibranchia). Дориды, за
немногими исключениями, питаются губками,
или реже мшанками. Поскольку и губки
и мшанки — колониальные неподвижные жи-
вотные, хищниками дорид можно назвать лишь
условно. Они пасутся на жертве, как коровы на
лугу. Но есть среди дорид и настоящие хищни-
ки — виды из семейства гимнодоридиды (Gym-
nodorididae) нападают и целиком проглатыва-
ют некоторых голожаберных моллюсков-эолид.
Упоминавшаяся крошка весьера — тоже дори-
да, но странная и вторично лишенная жабр. Ее
можно встретить на мелководье в заливе Петра
Великого Японского моря. Нет жабр и у отно-
сительно крупных тропических филлидиид.
Однако во всем остальном они — несомненные
дориды. Вот только способ питания у них не-
типичен даже для моллюсков. Радулы — зубов
моллюсков — у филлидиид (Phyllidiidae) нет,
но они умудряются поедать губок так же, как
и их зубастые родственники. Вместо того, чтобы
отрывать зубами кусочки губки, филлидии сна-
чала частично «на месте» обрабатывают участок
губки ферментами слюнных желез, и лишь по-
том всасывают уже полупереваренную пищу
мускулистой глоткой. Такое вот «следующее по-
коление» пищеварительного аппарата. Губки —
весьма пренеприятные, хотя и неподвижные,
создания секретируют множество ядовитых ве-
ществ. И дориды-губкоеды, особенно в тропи-
ках, в том числе упомянутые филлидии, ис-
пользуют вещества из съеденных губок для
последующего синтеза собственных, также ядо-
витых для морских животных-хищников, ве-
ществ (отсюда упомянутая в начале главы мета-
фора о «похищении» молекул). Для человека
голожаберные моллюски не опасны, но пытать-
ся их отведать не следует.
Большинство голожаберных моллюсков из
отряда Nudibranchia внешне гораздо более раз-
нообразны, чем дориды. Тритонии — обычно
крупные, внешне неуклюжие создания. А вот
мелкие их потомки эолиды очень изящны...
Почти все представители этого отряда свою
жизнь связали с книдариями (кишечнополост-
ными) — мягкими кораллами, актиниями, ги-
дроидными полипами. И эти голожаберники
уже больше хищники, чем дориды. Тем не ме-
нее многие эолиды также пасутся на непо-
движных, условно беззащитных колониях ги-
дроидных полипов. Вот только проблем с их
поеданием гораздо больше — у гидроидов жгу-
чие стрекательные клетки. Похожие клетки
имеют на вооружении и смертельные для чело-
века австралийские кубомедузы. Но победа над
полипами далась голожаберным моллюскам не
сразу. У видов рода тритония, или родственной
им тропической марионии еще нет никаких
особых приспособлений для хранения опасных
стрекательных клеток — книдоцист. Вероятно
в слизи, выделяемой этими моллюсками, осо-
бенно железами передней части ноги, содер-
жатся вещества, изолирующие чувствительный
30
Поведение заднежаберных моллюсков
Пищевые пристрастия заднежаберных моллюсков
очень разнообразны. На этом фото, например,
запечатлен уникальный момент нападения хищного
крылоного моллюска Clione limacina (морского ангела)
на мирного поедателя планктона Limacina helicina
(морского черта)
В пищевом поведении эолидацей (например, Nudibranchus
rupium) возможны различные стратегии: откусывание
отдельного полипа целиком вместе с оболочкой-текой
и высасывание содержимого полипа или высверливание
отверстий в стволиках колонии. Полипы и их оболочки
при этом остаются не тронутыми
Один из самых массовых видов Aeolidacea прибрежных вод Камчатки Himatina trophina
может поедать не только гидроидов, но даже ракообразных
31
БИОЛОГИЯ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
эпителий пищеварительного тракта от ожогов
стрекательных клеток. Да и сам эпителий голо-
жаберных моллюсков особенный. В различных
семействах голожаберных моллюсков незави-
симо появляются спинные выросты — папил-
лы, на вершинах которых в особых мешочках
(называемых книдосаки) накапливаются жи-
вые стрекательные клетки съеденных полипов.
У особой группы — семейства арминид вместо
спинных папилл по бокам тела формируются
особые пузырьки, где также накапливаются
стрекательные клетки. В тропических морях из
этого семейства обычен род армина (Armina).
Пищеварительная железа многих голожабер-
ников, в первую очередь у эолид (иногда не-
верно называемая печенью) не в виде цельного
органа, как у большинства моллюсков, а в виде
системы разветвленных каналов, заходящих
в папиллы, по которым реснички эпителия
гонят стрекательные клетки на их вершину. Где
стрекательные клетки используются для защи-
ты... уже не полипа, а самого голожаберного
моллюска. Этот парадокс вошел во все учебни-
ки. В то же время у некоторых арминацей
(Arminacea) — таких как живущая у нас в Япон-
ском море окрашенная в дивный апельсино-
вый цвет дирона папиллы уже есть, а мешочков-
книдосаков на их вершине, куда поступают
стрекательные клетки, еще нет. Также как и нет
разветвленной пищеварительной железы. Но
даже эолиды, особенно их молодь, порой, весь-
ма осторожно пробуют полип своими щупаль-
цами, а лишь потом приступают к трапезе.
А виды рода дото и вовсе не осмеливаются тро-
гать самих гидроидных полипов, и сверлят ра-
дулой аккуратные дырочки в стеблях колоний,
высасывая частично их содержимое — там ни-
каких стрекательных клеток нет. Эти приме-
ры — доказательство того, что было множество
эволюционных «экспериментов», прежде чем
сформировались эолиды, вершина удивитель-
ной приспособленности к суровым книдариям.
И осколки этих экспериментов все еще вполне
успешно ползают в разных морях.
Те из голожаберных моллюсков, которые
наиболее хорошо приспособлены к поеданию
книдарий, поступают со своими жертвами бес-
церемонно. Крупные беломорские и японо-
морские эолидии «натягивают» собственную
голову на не менее крупную актинию, и могут
в таком состоянии пребывать часами, посте-
пенно разрушая актинию челюстями и раду-
Aeolidia papillosa на заиленной литорали Белого моря
32
Поведение заднежаберных моллюсков
Участок радулы представителя рода Boreoberthella. Здесь и на стр. 34 электронно-микроскопические изображения
лой. Похожим образом действует корифелла
изящная на Баренцевом море, но только не
с актинией, а с отдельным маленьким полипом
колонии гидроидов. И времени у нее на это
уходит куда меньше — она попросту перекусы-
вает стебелек, соединяющий полипа с колони-
ей, и проглатывает несчастного. А дендронотус
ирис с тихоокеанского побережья Северной
Америки отправляется прямо в «пещеру», где
скрывается его жертва — цериантария (род-
ственник актиний), на которую он нападает.
Цериантария буквально засасывает дендроно-
туса в свою трубку, откуда он возвращается
уже одинокий и...сытый. Беломорские и даль-
невосточные дендронотусы не отличаются
столь экзотическими привычками. Иногда
эолиды нападают друг на друга. Мы наблю-
дали однажды, как две корифеллы из залива
Петра Великого, скользя с внутренней стороны
поверхностной водной пленки, пытались уку-
сить Друг друга челюстями за кончики хвостов,
как играющие щенки. Только вряд ли игры ко-
рифелл были столь безобидны. Довольно круп-
ная паракорифелла, посаженная в сосуд с дю-
жиной юных корифелл, через несколько дней
оказалась совершенно лысой — дерзкая моло-
дежь объела все ее папиллы.
Каждому из перечисленных пищевых
предпочтений, сформировавшихся в той или
иной группе, обычно соответствует весьма
определенный тип строения радулы. Так, у со-
скребателей-губкоедов дорид и нотаспид, раду-
ла состоит из большого числа сходно устроен-
ных зубов (см. стр. 29, здесь и далее фотографии
радул сделаны с помощью электронного ми-
кроскопа для наших оригинальных исследова-
ний), тогда как голожаберники-эолиды, поеда-
ющие отдельных гидроидов полипов обладают
мощным срединным зубом, окруженным лишь
двумя боковыми зубами (см. стр. 34, такой тип
радулы называется трисериалъныи) или они
вовсе отсутствуют (см. стр. 34, унисериалъный
тип радулы). У тех дорид, которые перешли от
питания губками к более дифференцированно-
му поеданию мшанок, число рядов радулярных
зубов уменьшилось, а зубы стали более разно-
образны по форме, как например у семейства
онхидоридид (там же). Травоядные аплизии,
пасущиеся на крупных бурых и красных водорос-
лях, также обладают очень мощным челюстным
33
БИОЛОГИЯ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
Трисериалъная радула Pseudovermis (Aeolidacea)
и радулярным аппаратом для того, чтобы
справляться с этой грубой пищей. Напротив,
также «сидящие» на растительной диете сако-
глоссы научились использовать совершенно
иную пищевую стратегию — они не заглатыва-
ют водорослевую массу целиком, а прокалыва-
ют в стенке водоросли крошечную дырочку,
после чего высасывают ее внутреннее содержи-
мое (включая функциональные хлоропласты,
которые затем перемещаются в папиллы сако-
глосс и служат для дополнительного полу-
чения энергии). Вполне естественно, что для
такой ювелирной работы необходима совер-
шенно особая радула, которая присуща только
сакоглоссам. В каждом ряду остается всего
один зуб радулы, который становится очень
узким, ножевидным, идеально подходящим
для прокалывания стенок водорослей.
Унисериальная радула Murmania (Aeolidacea)
Продольный срез через глотку представителя рода Aplysia
Участок радулы
представителя рода Onchimira (Doridacea)
Участок радулы
представителя рода Placida (Sacoglossa)
34
Жизненный цикл заднежаберных моллюсков
ЖИЗНЕННЫЙ ЦИКЛ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
Жизненный цикл заднежаберных моллюсков:
нужен ли партнер гермафродиту и зачем детям раковина?
За редчайшими исключениями все представи-
тели огромной группы заднежаберных моллю-
сков (а их насчитывается несколько тысяч видов)
являются гермафродитами. Каждая особь лю-
бого из видов заднежаберных моллюсков имеет
одновременно и мужские, и женские органы
размножения. Благодаря такой комбинации
репродуктивный аппарат заднежаберных,
и особенно голожаберных моллюсков, является
одним из самых сложных среди морских бес-
позвоночных. Половая система заднежаберных
моллюсков состоит из двух основных отделов —
гонады (половой железы), где образуются гаме-
ты (половые клетки), и целого комплекса про-
токов, резервуаров и желез, проходя через
которые женская половая клетка последова-
тельно оплодотворяется, покрывается оболоч-
ками, и наконец, вместе со множеством других
оплодотворенных половых клеток заключается
в общую сложную оболочку — кладку.
Свои кладки голожаберные моллюски при-
крепляют к твердым субстратам — к обратной
стороне камней, скалам, водорослям, гидрои-
дам и пр. Общей особенностью кладок боль-
шинства голожаберных моллюсков является их
спиральная форма. Количество витков спира-
ли, детали ее строения в большинстве случаев
видоспецифичны — т. е. каждый вид отклады-
вают кладку определенной формы, найдя кото-
рую в природе, можно затем идентифициро-
вать и «хозяина», ее отложившего. Некоторые
виды откладывают очень коротенькие, прижа-
тые к субстрату кладки, выглядящие как сверну-
тый в спираль шнур из 1,5-2 оборотов (напри-
мер у мелких видов семейства эубранхид из
группы Aeolidacea), или мощные кладки в виде
высоко приподнятых над субстратом лент из
более чем 5 оборотов спирали (каку многих нотас-
пид и дорид). Яйца внутри кладки уже оплодо-
творены и находятся в процессе развития.
Многие представители заднежаберных моллюсков
обладают очень сложными копулятивными аппаратами,
как например у Onchidoris muricata, снабженными
причудливыми выростами. Строение копулятивного
аппарата — один из важнейших систематических
признаков. В различных группах заднежаберных
моллюсков копулятивные аппараты могут нести
еще более причудливые шипы, папиллы, стилеты
Все заднежаберные моллюски - гермафродиты,
т. е. обладают одновременно как мужскими,
так и женскими половыми протоками, которые
открываются с правой стороны тела, под кожной
складкой-нотумом. По этой причине во время
копуляции пара особей (как изображенные дориды
Onchidoris muricata) располагается в такой вот
характерной позе
35
БИОЛОГИЯ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
На первый взгляд гермафродитам для
оплодотворения партнеры не требуются. Это
тем не менее заблуждение. Дело в том, что лю-
бой половой процесс, в отличие от бесполого
размножения, всегда предполагает рекомбина-
цию, своего рода перемешивание мужского
и женского генетического материала. С помо-
щью этого процесса поддерживается опреде-
ленный уровень разнообразия (полиморфиз-
ма) в популяции, что в целом способствует ее
выживанию. Хорошо известно, что любая по-
рода может придти в упадок, если скрещива-
ния будут только близкородственными. Чтобы
избежать подобного эффекта, заднежаберные
моллюски, хотя и сочетают в одной особи
и мужскую и женскую, тем не менее обладают
мощной системой защиты от самооплодотво-
рения и потенциального «ухудшения» свойств
вида. Но это не исключает того, что, в некото-
рых случаях самооплодотворение все-таки
происходит.
Каждое яйцо кладки в процессе развития
претерпевает ряд последовательных делений
(которые биологи называют дроблением), в ре-
зультате чего из одной-единственной оплодот-
воренной яйцеклетки (зиготы) образуется
сложный многоклеточный зародыш. И у всех
заднежаберных моллюсков, как и у большин-
ства других морских беспозвоночных, итогом
развития яйца является не образование юве-
нильной (молодой) формы, близкой по строе-
Новорожденные голожаберные моллюски, как правило,
совсем не похожи на своих родителей: для того чтобы
завершить свой жизненный цикл, они должны пройти
через особую планктонную стадию личинки-велигера,
снабженную раковиной и специальными ресничками
нию к половозрелым особям, а образование
той или иной стадии личинки, резко отличной
по строению от взрослых форм.
Личинка, как правило, обладает специфи-
ческими приспособлениями, которые позво-
ляют ей существовать некоторое время в во-
дной толще — разного рода лопастями,
покрытыми длинными ресничками. У всех
брюхоногих моллюсков (к которым относятся
и заднежаберные моллюски) подобная реснич-
ная лопасть личинок называется парусом, или
по-другому, велюмом. Отсюда наиболее распро-
страненный термин, обозначающий подобную
личинку, — велигер. У подавляющего большин-
ства заднежаберных моллюсков из кладки непо-
средственно выходит свободноплавающая
личинка-велигер, которая немедленно «отправ-
ляется» в планктон, где проводит как минимум
несколько дней, а чаще — несколько недель. За
время, проведенное в планктоне, личинка может
быть унесена течениями на значительное рас-
стояние — сотен, а порой и тысяч километров от
места, где она вылупилась из кладки. Личинка
таким образом может выполнять очень важную
расселителъную функцию, которой во многом
лишена взрослая, относительно малоподвижная
стадия заднежаберных моллюсков. Свободно-
плавающие личинки заднежаберных моллюсков
могут быть двух типов — те, которые питаются
во время нахождения в планктоне (в этом случае
они называются планктотрофнъгми), и те, кото-
рые существуют исключительно за счет заранее
запасенных питательных веществ (такие личин-
ки называются лецитотрофными). Помимо рес-
ничного паруса характернейшей чертой свобод-
ноплавающих личинок заднежаберных
моллюсков является наличие раковины. При
этом даже у полностью лишенных каких-либо
следов раковины на взрослой стадии голожабер-
ных моллюсков и сакоглосс на стадии личинки
имеется хорошо развитая тонкостенная ракови-
на. Почему это происходит?
Дело в том, что столь привычные для нас
особенности размножения млекопитающих
в реальности — лишь один из того значитель-
ного числа вариантов, который можно об-
наружить в живой природе. Новорожденная
морская свинка принципиально сходна со
взрослой особью, хотя ее пропорции еще да-
леки от половозрелого животного. Чего никак
нельзя сказать о заднежаберных моллюсках.
И тот причудливый путь, который проходит
36
Жизненный цикл заднежаберных моллюсков
Жизненный цикл большинства заднежаберных
моллюсков включает донную половозрелую стадию
и планктонную личинку, снабженную ресничной
лопастью — парусом, и заключенную в тонкостенную
спиральнозавитую или овальную раковинку
большинство как моллюсков, так и морских
беспозвоночных весьма недвусмысленно напо-
минает нам факт, что любой организм не суще-
ствует только как взрослая форма, а в виде
совершенно определенного жизненного или
онтогенетического цикла.
«Силы притяжения» между различными
стадиями жизненного цикла столь сильны, что
любой признак, однажды в нем возникнув, не
исчезает затем «насовсем» и внезапно, а надолго
остается его составной частью. Даже если та или
иная черта строения, в общем, особой необходи-
мости животным не несет. Таковы жаберные
щели, появляющиеся на ранних стадиях разви-
тия любого млекопитающего, а затем полно-
стью исчезающие. Такова рыбообразная стадия
головастика, имеющаяся в жизненном цикле
большинства амфибий. Такова раковина, имею-
щаяся у личинок большинства во взрослом со-
стоянии безраковинных голожаберных моллю-
сков. В чем причина широкого распространения
подобных явлений? Часть из них можно объяс-
нить адаптацией личинок к существенно отлич-
ным условиям их обитания в сравнении со
взрослыми особями. Вполне возможно, что на-
личие раковины чуть лучше защищает их от воз-
можных хищников, как если бы такой оболочки
не было вовсе. Но это только часть объяснения.
Несомненно, что жаберные щели, даже руди-
ментарные, не нужны зародышу млекопитаю-
щих, который все необходимое, включая кисло-
род, получает через плаценту матери. Но
у дальних предков млекопитающих — асцидий,
ланцетников и хрящевых рыб, жаберные щели
выполняли и выполняют важнейшую для орга-
низма функцию дыхания в водной среде. И в он-
тогенетическом цикле млекопитающих сохра-
нились следы прошлых состояний этого
признака, характерных для взрослых особей, но
только на ранних эмбриональных стадиях. Это
явление также может быть названо рекапитуля-
цией признаков. Так, у целого ряда голожабер-
ных моллюсков развитие вторично укорачивает-
ся, а из кладки выходит не свободноплавающая
личинка, а ползающая молодь, больше похожая
на взрослых голожаберных моллюсков. Тем не
менее подобное укорачивание не происходит
внезапно, не оставляя следов в онтогенетическом
цикле. Свободноплавающая стадия велигера
«перемещается» внутрь оболочки яйца, где не-
редко можно различить даже зачатки раковины,
уже совершенно не нужной для защиты.
Человек после рождения не говорит боль-
ше года. Совершенно очевидно, что если бы
Homo sapiens был создан по заранее четко за-
данному плану, рождался бы сразу разговари-
вающим как Цицерон и половозрелым. Вместо
этого любой человек должен «ждать» более
десяти лет, чтобы достичь репродуктивного
периода, и именно в процессе этого растянуто-
го созревания (т. е. жизненного цикла) форми-
руется чудесный орган, выделяющий человека
из ряда других млекопитающих, — наш мозг.
Формируется та сложная биологическая и со-
циальная сеть, именуемая языком, основным
регулятором которой и является мозг. При
этом совершенно неопровержимым является
тот факт, что вне круга людей язык у ребенка
никогда не разовьется. Многочисленные исто-
рии с детьми, которые «воспитывались» жи-
вотными и не говорят, являются несомненным
тому подтверждением. Т. е. человеческий язык
(«человечность» как таковая) является вовсе не
целью жизненного цикла Homo sapiens, а хруп-
кой, нестабильной «верхушкой», надстройкой
этого цикла, которая при несоблюдении ряда
условий никогда не разовьется. В то же время
отсутствие языка не помешает организму ис-
полнять функционирование других черт чело-
веческого онтогенеза, сформировавшихся в от-
деленные геологические эпохи, и составляющие
основу морфологии и физиологии класса мле-
копитающих. Иначе говоря, бессмысленно об-
суждать происхождение и эволюцию любой
37
БИОЛОГИЯ ЗАДНЕЖАБЕРНЫХ МОЛЛЮСКОВ
половозрелой взрослой формы вне ее онтоге-
нетического цикла, нередко весьма сложного.
Ряд стадий этого цикла, унаследованных от
далеких предков, являются более консерва-
тивными (как жаберные щели), другие (как
например, человеческий язык) — совсем не-
давно приобретенными и оттого еще оконча-
тельно не «зафиксировавшиеся» в онтогенети-
ческом цикле.
Именно благодаря подобному консер-
ватизму онтогенетических циклов живых ор-
ганизмов и возможно существование науки
систематики. Подобный консерватизм позво-
ляет вычленять наиболее общие, предковые
черты жизненных циклов, которые можно со-
поставить с организацией типов и классов,
и более частные, производные черты много-
численных потомков, объединенных ныне
в семейства, роды, виды. Именно прежде всего
такому консерватизму организации, а не ис-
ключительно адаптациями и объясняется
присутствие раковины у личинок безраковин-
ных голожаберных моллюсков. Однако, как
уже говорилось, онтогенетический цикл лю-
бого живого существа характеризуется одно-
временно устойчивостью (включая целый ряд
признаков, унаследованных от далеких и близ-
ких предков), так и изменчивостью, некоторой
нестабильностью и неоднозначностью оконча-
тельной взрослой формы каждого вновь по-
являющегося из яйца организма. Именно бла-
годаря этой второй составляющей каждого
жизненного цикла стала возможной эволю-
ция организмов.
Прямая связь между онтогенетическим
циклом любого организма и эволюцией осо-
бенно хорошо прослеживается на примере
особого процесса, который биологи сейчас на-
зывают педоморфозом, а раньше именовали
неотенией. Педоморфоз — это эволюционный
процесс, при котором те стадии жизненного
цикла, которые у предков появлялись только
у ювенильных особей (очень молодых экзем-
пляров, «детей»), а у взрослых затем полностью
исчезали, у потомков сохраняются и на взрос-
лых, половозрелых стадиях. Прекрасным при-
мером процесса педоморфоза являются целый
ряд видов из группы хвостатых амфибий, куда
относятся такие всем хорошо известные живот-
ные, как тритоны и саламандры. Наиболее зна-
менитая педоморфная, неотеническая форма
хвостатых амфибий — это аксолотль, который
является фактически «головастиком-перерост-
ком» южноамериканской хвостатой амфибии
амбистомы. Как и любой нормальный голова-
стик, аксолотль имеет хорошо развитые на-
ружные жабры, которые у взрослых форм, как
хвостатых, так и безхвостых амфибий, полно-
стью исчезают. Но аксолотли половозрелы!
Они являются фактически взрослой формой,
но с личиночными признаками, т. е. с наруж-
ными жабрами. Аксолотли все еще способны
при той или иной комбинации внешних усло-
вий превращаться в «окончательную», безжа-
берную взрослую форму — амбистому. Но
целый ряд других хвостатых амфибий полно-
стью утерял способность к превращению в без-
жаберную форму. Поэтому у взрослых стадий
таких видов амфибий, как сирены и протеи,
«навсегда застыли» детские черты других зем-
новодных — наружные жабры. Подобные фак-
ты поэтому недвусмысленно связывают жиз-
ненный цикл (индивидуальное развитие)
и эволюцию (историческое развитие).
Как оказалось, подобные сдвиги онтогене-
тического, жизненного цикла в ту или иную
сторону (т. е. в сторону более взрослой или
более ювенильной формы) — очень распро-
странены среди самых различных организмов.
В частности, нами впервые было показано, что
«загадочное» семейство голожаберных мол-
люсков корамбид (Corambidae), которое мно-
гие полагали наиболее архаичной группой
дорид из-за брюшного положения их жабр
и анального отверстия, на самом деле таковым
не является, а все его «странные» признаки
хорошо укладываются в личиночную морфо-
логию целого ряда «нормальных» семейств со
спинными жабрами. Как выяснилось, все без
исключения дориды проходят в своем разви-
тии стадию, на которой анус еще находится на
брюшной стороне, и только затем перемеща-
ется на спину, где одновременно начинают
формироваться жабры. А у корамбид анус
остался на брюшной стороне и у взрослых
особей. И жабры соответственно развились
также на брюшной, а не на спинной стороне
(см. стр. 39). Получилась самая настоящая эво-
люционная загадка, решить которую оказа-
лось возможно только после изучения онтоге-
неза дорид и обнаружения в морях России
переходного звена — нового рода и вида Loy
meyeni Martynov, 1994, обладающего одновре-
менно личиночными признаками в строении
38
Жизненный цикл заднежаберных моллюсков
Доказательства процесса педоморфоза у голожаберных моллюсков-дорид: сравнение взрослых педоморфтых
таксонов (В, D, Corambe obscura) со взрослыми (Af Cadlina laevis) и ранними ювенильными особями (С, Е,
Cadlina laevis) обычных (не педоморфных) видов. D, Е - электронно-микроскопические изображения
жабр и их спинным расположением, харак-
терным для большинства других видов семей-
ства онхидоридид (см. ниже).
К настоящему времени накопилось очень
много свидетельств, что подобные сдвиги жиз-
ненных циклов и являются основным механиз-
мом эволюции. И как бы это не ущемляло че-
ловеческую гордость, взрослый Homo sapiens
демонстрирует целый ряд признаков (пропор-
ции черепа, форма ушной раковины и др.)/
которые гораздо более сходны с детенышами
шимпанзе, чем с их взрослыми особями. Ре-
грессивный на первый взгляд «сброс» взрослых
обезьяньих черт позволил человеку развить
совершенно новые качества «давно известных»
органов млекопитающих — передних конеч-
ностей и мозга, именно этим обеспечив себе
небывалый прогресс. Сходным образом, педо-
морфные голожаберные моллюски корамби-
ды, помимо личиночных черт, приобрели со-
вершенно новые признаки, не известные не
только у других голожаберников, но среди
остальных моллюсков, например особую за-
щитную, периодически отслаивающуюся мяг-
кую кутикулу. Именно благодаря регрессивно-
му недоразвитию раковины образовались
столь прогрессивные по многим другим при-
знакам «бесконечно разнообразные» заднежа-
берные моллюски. Именно подобным образом,
используя два этих основных компонента —
прогресс одних признаков и регресс других,
эволюция и ткет свое величественное полотно,
над разгадкой бесчисленных деталей которого
ученые бьются уже не одно столетие.
39
АТЛАС
ЗАДНЕЖАБЕРНЫХ
МОЛЛЮСКОВ
морей России
В этой части книги авторы знакомят
читателей с разнообразными и интерес-
нейшими обитателями морей России —
заднежаберными моллюсками. Очерки
содержат поистине уникальную инфор-
мацию об их внешнем облике, строении
и образе жизни. Яркие описания дополне-
ны редчайшими фотографиями, сделан-
ными в сложных условиях арктических
и дальневосточных морей России.
Переходя от отряда к отряду, от группы
к группе, вы сможете воочию убедиться
в грандиозности идеи эволюции и прикос-
нуться к таинству превращения вполне
рядовой улитки с раковиной в эфемерное
чудо природы — голожаберного моллюска.
Именно поэтому Атлас станет для вас
еще и путеводителем по важнейшему
разделу биологии — теории эволюции.
КАК УСТРОЕН АТЛАС
Информация о каждом виде организова в дан-
ной работе как небольшой очерк. В каждом из
очерков есть ряд стандартных разделов, поэто-
му вы можете легко сравнивать между собой
представленные здесь виды. Итак, каждое опи-
сание состоит из следующих разделов:
Особенности строения. В этом разде-
ле кратко указаны признаки данного вида, ко-
торые помогут вам отличить его от ближай-
ших родственников. При описании признаков
будет использован ряд необходимых терминов.
Наиболее важные из них перечислены в пре-
дыдущей главе.
Окраска. В отличие от многих других
организмов, большинство голожаберных мол-
люсков ярко окрашены. При этом особенности
окраски часто видоспецифичны, т. е. уникаль-
ны для данного вида. Сопоставив особенности
строения и окраски, как на изображениях, так
и в тексте описания, вы составите наиболее
полное представление о конкретном виде зад-
нежаберного моллюска и сможете сравнить
его с любым другим видом.
Отличия от похожих видов. Благодаря
этому разделу, читатель сможет легче сориен-
тироваться на какие признаки обратить внима-
ние при определении того или иного вида
и с какими близкородственными или сходны-
ми видами его можно спутать.
Особенности экологии. Экология —
это прежде всего наука, о взаимоотношении
организмов друг с другом и со средой. По-
этому не бывает «плохой экологии», как обыч-
но пишут газетчики, а нужно говорить о «пло-
хих» или «неудовлетворительных» условиях
среды, в которой обитает тот или иной вид жи-
вых организмов (например, человек). В дан-
ной книге в разделе «особенности экологии»
будут указаны главным образом физические
характеристики экосистемы, в которой обита-
ет тот или иной заднежаберный моллюск, т. е.
грунты (скальные, каменистые, песчаные, или-
стые), на которых живет конкретный вид, глу-
бина его обитания, в некоторых особых случа-
ях — соленость воды и пр. Биологические
особенности (которые также являются частью
науки экологии), такие как питание и раз-
множение, вынесены для удобства в отдельные
разделы.
Размножение. В данном разделе при-
ведены краткие сведения о сроках размноже-
ния вида, особенностях строения его кладки
и личинки. В отношении устройства личинок
возможны два основных типа развития. В боль-
шинстве случаев из яйца выходит микроскопи-
ческая личинка, имеющая раковинку и особый
ресничный парус — велюм (за что она и полу-
чила название — велигер). Подобная личинка
почти не имеет запасов желтка, поэтому, на-
ходясь в планктоне, вынуждена питаться —
еще более мелкими микроскопическими водо-
рослями. Спустя некоторое время (обычно не
более нескольких недель) подросшая личинка
опускается на дно, где претерпевает процесс
метаморфоза (т. е. превращения). При этом ли-
чиночные черты, такие как раковинка, исчеза-
ют или сильно видоизменяются, и личинка
превращается в донную молодь. Напротив,
при прямом развитии из яйца сразу выходит
ползающая молодь, только очень маленькая
и лишенная многих признаков взрослых мол-
люсков, которая приступает к питанию непо-
средственно пищей, свойственной взрослым,
на дне, а не в планктоне. Между двумя этими
основными типами развития заднежаберных
моллюсков есть несколько промежуточных.
Так, в некоторых случаях личинка является
планктонной, но при этом не питается, как
и при прямом развитии. Происходит это по-
тому, что в яйце имеется значительный запас
питательных веществ. Такие непитающиеся
планктонные личинки называются лецито-
трофными.
Питание и поведение. Эти особеннос-
ти у каждого вида также являются предметом
самого серьезного изучения наукой экологи-
ей. В данной книге мы конечно не будем при-
водить сложные расчеты метаболизма, но
кратко перечислим наиболее характерные
особенности питания и поведения того или
иного вида.
Распространение. Это также очень
важная характеристика любого вида. Особен-
ностями распространения организмов на зем-
ном шаре занимается особая наука биогеогра-
фия. Чтобы лучше понимать этот раздел,
необходимо будет запомнить несколько на
первый взгляд причудливых, но по своей сути
41
довольно простых терминов. Эти выражения
входят в арсенал любого биогеографа, изучаю-
щего распространение организмов в морской
среде. Итак, в зависимости от области Мирово-
го океана, в которой распространен тот или
иной вид, он получает ту или иную характери-
стику. Арктические виды, как это очевидно
следует уже из названия, обитают почти ис-
ключительно в пределах Северного Ледовитого
океана (иногда его еще называют Арктиче-
ским). Бореальные виды обитают южнее —
в умеренных, т. е. тоже холодных, но не ледя-
ных водах Северного полушария Земли.
Слово бореальный происходит от названия се-
верного ветра — Борея. К бореальным водам
относятся северные части как Атлантического,
так и Тихого океанов. Поэтому виды, которые
обитают как в северных частях Атлантики, так
и в северных частях Тихого океана, но отсут-
ствуют при этом в настоящей Арктике (т. е.
между Баренцевым и Чукотским морями) на-
зываются амфибореалъными. Иногда выделя-
ют еще амфиатлантические виды, т. е. такие
виды, которые обитают как по европейскому,
так и по американскому побережью северной
Атлантики, и амфипацифические виды, кото-
рые можно встретить в северной части Тихого
океана как вдоль российского, так и вдоль аме-
риканского берега.
В летний период бореальные воды могут
прогреваться до значительных, почти субтро-
пических температур в + 20-25° С. Такие усло-
вия, например, наблюдаются в нашем заливе
Петра Великого, что в Японском море. Зимой
там царит Арктика — часть бухт замерзает,
а температура может опускаться вплоть до на-
стоящих арктических значений - 2° С. Летом,
напротив, вода может прогреваться до + 25° С.
Даже в почти арктическом Белом море, в не-
которых бухтах температура может достигать
летом + 20° С! Такие особенности физических
параметров среды, т. е. морской воды, во
многом будут определять и биологические
особенности той или иной экосистемы. Не
удивительно поэтому, что зимой в заливе
Петра Великого можно встретить самого на-
стоящего арктического голожаберного мол-
люска хлямиллю (Chlamylla atypica, см. ниже),
а летом, напротив, тропическую гониодори-
деллу (Goniodoridella savignyi) (т. е. такой вид,
который в норме распространен лишь в са-
мых тепловодных частях океана, где никогда
не бывает сезонного понижения темпера-
туры). В российской части Японского моря
летом в отдельные, наиболее теплые годы
отмечены заходы таких теплолюбивых живот-
ных, как морские змеи, рыба-луна и каракати-
ца. Поэтому единственное место в России, где,
если повезет, можно встретить самых настоя-
щих тропических «морских зайцев» — апли-
зий — это как раз некоторые районы Япон-
ского моря.
Многие из видов, обитающих в Черном
море, также хорошо адаптированы к подоб-
ной двойственности, поскольку летом морская
вода там может достигать почти тропических
значений в + 28° С, тогда как зимой опускаться
до + 7° С. Поэтому в Черном море обитает це-
лый ряд настоящих субтропических видов,
вселившихся туда из Средиземного моря, но
способных переживать сезонное охлаждение
воды зимой. Благодаря такой двойственности
в сезонных вариациях условий обитания, це-
лый ряд видов может получить и двойные
эпитеты с точки зрения биогеографии. Так,
упомянутый голожаберник Chlamylla atypica,
распространенный как в Арктике, так и в уме-
ренных водах Атлантического и Тихого океа-
нов, с точки зрения биогеографии будет име-
новаться как арктическо-бореальный. По
этой причине наука биогеография обладает
сложной, весьма разветвленной системой тер-
минов. Далеко не все из них необходимы для
целей настоящей книги. Из некоторых допол-
нительных терминов необходимо знать такие,
например, выражения как циркумполярный,
т. е. распространенный по всей Арктике,
и циркумтропический, т. е. распространен-
ный по всему тропическому поясу Мирового
океана. Наконец, для глубоководных живот-
ных, обитающих в океане глубже 500-1000 м
обычные «правила» биогеографии не приме-
нимы. Дело в том, что на больших глубинах
температура почти одинакова, и весьма низ-
кая — от 0° С и обычно не выше + 4° С. Поэто-
му в какой бы биогеографической области,
выделенной на основе распределения мелко-
водных животных — бореальной или тропи-
ческой, не обитал глубоководный организм,
это будет иметь минимальное значение, по-
скольку почти идентичные особенности среды
на больших глубинах потенциально позволят
расселиться ему почти по всему Мировому
океану.
42
О ТОМ, ЧТО НЕОБХОДИМО ЗНАТЬ ЧИТАТЕЛЮ
О НАЗВАНИЯХ ЖИВОТНЫХ,
ЧТОБЫ ПОЛЬЗОВАТЬСЯ АТЛАСОМ
Для того чтобы лучше понять не только эту
книгу, но и любой атлас морских беспозвоноч-
ных (как, впрочем, и иных организмов), с мак-
симумом картинок и минимумом текста, кни-
гу, в которой благодаря этому неизмеримо
возрастает значение (и доля) названий, читате-
лю надо иметь базовые представления о том,
как именно присваиваются научные названия
организмам, что они означают и какие особые
приемы используются при их образовании.
Любое научное название практически любого
живого организма всегда состоит из двух слов,
записанных латинским алфавитом, — имени
рода и видового эпитета. Это основной прин-
цип биологической номенклатуры (области
науки, которая изучает названия), и известен
он как принцип биноминальной, т. е. двойной,
состоящей из двух названий, номенклатуры.
В наиболее развитом и последовательном виде
этот принцип был применен Карлом Линне-
ем, хотя в той или иной форме, принцип двух
названий был известен и до Линнея.
Подавляющее большинство линнеевских
родов (т. е. тех родов, которые установил Карл
Линней в своей классической «Системе приро-
ды», двенадцать прижизненных изданий кото-
рой выходили с 1735 по 1767 годы) были впо-
следствии разделены на целый ряд новых родов.
Процесс выделения новых родов (так же как
и видов, семейств и даже иногда новых отрядов
и классов) продолжается и в наше время.
Помимо родового и видового названий,
любое настоящее научное название всегда
включает в себя и имя автора, впервые описав-
шего конкретный вид, которое указывается
сразу после видового эпитета. В ряде случаев
имя автора не упоминают, например, в эколо-
гических и тому подобных публикациях. Но
любая серьезная систематическая сводка всегда
включает в себя имя автора и год, в который он
впервые опубликовал новое название. Такой
же формат записи используется при упомина-
нии таксонов более высокого ранга, чем вид —
родов, семейств, и даже типов.
В зоологии имена некоторых авторов при-
нято приводить в сокращенном виде: до одной
буквы сокращены имена таких классических ав-
торов, как Линней (латинское Linnaeus,
сокращенный вариант — L.) и Фабрициус
(Fabricius, R). Все остальные имена обычно при-
водятся полностью. Любое имя автора таксона,
каким бы причудливым алфавитом оно не за-
писывалось на его родине, в научном названии
фиксируется только латинскими буквами. Это
относится и к русским фамилиям, которые так-
же записываются латиницей. В случае, если на-
звание вида остается в комбинации с тем же
родом, в котором оно было исходно предложе-
но, имя автора указывается без скобок, в случае
если впоследствии вид был помещен в другой
род, т. е. возникла новая комбинация родового
и видового названия, имя автора заключается
в скобки. Так, например, широко распростра-
ненный в Белом море голожаберный моллюск-
дорида Cadlina laevis (Linnaeus, 1762) был перво-
Памятник Карлу Линнею в г. Уппсала
43
начально описан Линнеем в составе рода Doris.
Значительно позднее, огромный сборный лин-
неевский род Doris был разделен на значитель-
ное число морфологически хорошо обоснован-
ных родов (многие из которых, в свою очередь,
стали основами для новых семейств). Одним из
таких родов и стала Cadlina, выделенная круп-
ным датским специалистом по голожаберным
моллюскам Бергом спустя более чем столетие
после Линнея, в 1878 году. Поэтому в новой ком-
бинации Cadlina laevis, имя Линнея берется те-
перь в круглые скобки.
Помимо основного названия многие виды
имеют еще и дополнительные названия, назы-
ваемые синонимами. Дело в том, что в «досете-
вой период» развития человечества, коммуни-
кация между учеными не только из разных
стран, но даже из разных городов одной страны
была нередко весьма затруднена. В таких усло-
виях почти неизбежна ситуация, когда разные
ученые описывали один и тот же вид под раз-
ными названиями. Такие названия считаются
теперь синонимами и приводятся под основ-
ным, действительным или, как говорят систе-
матики, валидным (от франц, valide — важный,
действующий) названием. В данной книге по-
добные названия снабжены специальным ука-
занием — «синонимы». Другим источником
синонимов является широкая изменчивость
особенно массовых, обычных видов. В случае
некоторых широко распространенных видов
число синонимичных названий может дости-
гать сотен! В данном атласе мы, конечно, не
будем приводить все, а это подчас десятки
и сотни синонимов, но одно или два когда-то
популярных названия для некоторых видов бу-
дет указано. Это поможет читателю правильно
соотнести описанные здесь виды с видами,
приводимыми в популярной литературе или
даже в других сводках, так как там они могут
быть упомянуты под другим названием, сино-
нимом. Предпочтение, какое название считать
действительным, какое — синонимом, опреде-
ляется принципом приоритета. Чем раньше
тот или иной ученый опубликовал описание
того или иного вида и сопроводил его опреде-
ленным названием, тот видовой эпитет и будет
признан валидным. Но из этого правила име-
ется и ряд исключений, регулируемых кодек-
сом зоологической номенклатуры.
Кроме того, необходимо иметь представле-
ние о так называемой открытой номенклатуре.
К методу открытой номенклатуры систематики
прибегают, когда по тем или иным причинам
невозможно точно определить тот или иной
вид, род или даже семейство. В этом случае на-
звание может выглядеть следующим образом:
например, Cadlina sp. — если точно не известен
вид, в данном случае, рода Cadlina, Chromodoridi-
dae gen. sp. (или gen. indet.) — если точно не из-
вестен род, в данном случае, семейства
Chromodorididae, и т. д. Кроме того, могут упо-
требляться такие конструкции как Cadlina spp. —
в этом случае подразумевается, что речь идет
о нескольких близких трудноразличимых ви-
дах, в данном случае рода Cadlina.
Особо следует сказать о соотношении ла-
тинских научных названий и «обычных» назва-
ний животных, иногда называемых «народны-
ми». Для некоторых групп организмов, таких
как растения, млекопитающие (звери) или
птицы, существует богатая традиция «народ-
ных» названий. В этом случае, практически
каждый вид, даже весьма редкий, имеет свое
уникальное название не только на латинском,
но и на родном для систематика языке, напри-
мер на русском. Ситуация осложняется, когда
мы от общеизвестных организмов переходим,
например, к морским беспозвоночным. Более
или менее устоявшаяся система параллельных
названий на латинском и родном языке суще-
ствует только в странах, имеющих свободный
выход к морским побережьям, население кото-
рых хотя бы в прошлом повседневно соприка-
салось с морской фауной и в дополнение к это-
му имеет еще особую культурную традицию,
своего рода очень дифференцированный взгляд
на окружающую природу, как, например,
в Японии. Этого безусловно нельзя сказать
о многих других странах мира, где доминируют
скорее обобщенные подходы к названиям орга-
низмов, не имеющих прямого практического
значения. Мы скорее скажем «червяк», «жу-
чок», «улитка», чем задумаемся о конкретном
названии того или иного организма.
Вполне естественно, что по мере развития
наших контактов с подводными обитателями
традиция «раздавать» обычные названия начи-
нает создаваться заново прямо «на лету», и как
правило, с помощью прямого перевода латин-
ского названия на тот или иной язык. Подобный
прямой перевод далеко не всегда приводит
к приемлемым, благозвучным комбинациям.
В России ситуация осложняется еще и тем, что
44
бум переводной литературы и сетевых изданий
не дал возможности ни переводчикам, ни поль-
зователям задуматься о необходимости аккурат-
ного подхода к переводу названий. В результате
в русский язык потоком хлынули разного рода
«лунные ракушки», «морские слизни» и «нуди-
бранчи», прямая калька даже не с латыни, а с ан-
глийского языка, вызывающие недоумение у спе-
циалистов и приводящие к путанице. Поэтому
в отношении морских беспозвоночных мы при-
держиваемся мнения, что желательно запоми-
нать их латинское название. В конце концов чи-
татель будет вознагражден тем, что он произносит
эти названия так же как и профессионалы-систе-
матики! К сожалению, для языков, которые не
используют латинский алфавит, а к ним отно-
сится и русский язык, подобный подход вызыва-
ет известные трудности. Поэтому в данной книге
рядом с латинским названием приводится его
транслитерация па кириллице или перевод. Во
всех случаях родовое название никогда не пере-
водилось. Многие самые старые родовые назва-
ния голожаберных моллюсков основывались,
например, на именах различных богов и их мно-
гочисленных родственников, например Doris
(Дорис, дочь Океана), Tritonia (Тритон, сын По-
сейдона), Actaeon (Актеон, сын пастуха, прогне-
вавший Артемиду) и т.д. Последующие поколе-
ния систематиков, выделяя новые роды,
использовали названия старых родов как основу,
в результате чего на свет появились разнообраз-
ные Archidoris, Pseudotritonia или Japonactaeon.
После подобных «многослойных» процедур из-
влечь хоть каплю смысла и благозвучия из ито-
говых названий (Древняя Дорис, Фальшивый
Тритон, Актеон из Японии!!!) представляется
отнюдь не тривиальной задачей даже для та-
лантливого филолога. Один из родов, выделен-
ный классиком систематики голожаберных
моллюсков XIX века, датчанином Рудольфом
Бергом, красиво звучащая Rostanga (обитающая
и в фауне России), имеет, например, почти
идентичное название со шведским городком
Rostanga. Но есть ли тут какая-то связь так види-
мо и останется загадкой — Берг не предоставил
нам комментариев по этому поводу. Что же ка-
сается видовых эпитетов, то в большинстве слу-
чаев для них также приводилась лишь трансли-
терация. В небольшом числе случаев, когда
латинское название можно было перевести на
русский однозначно и благозвучно, такой пере-
вод был представлен, например, Cylichna alba —
Цилихна белая. В случае, если видовое название
на русском языке написано с заглавной буквы,
это означает, что вид назван в честь конкретного
человека, как правило специалиста в области
систематики. Что же касается непереведенных
названий, то поскольку латинские и древнегре-
ческие словари сейчас широко доступны в Сети,
любознательный читатель, желающий выяс-
нить перевод того или иного названия (если они
конечно не основаны на именах собственных
или случайных комбинациях букв), всегда смо-
жет сделать это самостоятельно.
Баренцево море. Литоральная ванна
45
• Эволюция, шаг первый •
ВСЁ ЕЩЁ С РАКОВИНОЙ
ОТРЯД CEPHALASPIDEA — ЦЕФАЛАСПИДЫ
Это наиболее древняя и архаичная группа заднежаберных моллюсков, которая наглядно демон-
стрирует реальность эволюции на примере редукции внешнего скелета — раковины. И хотя
основная цель данной книги — рассказ о моллюсках без раковины, а у цефаласпид, а также род-
ственных им морских зайцев, напротив, раковина еще есть, весь обширный корпус морфологи-
ческих, систематических, физиологических и молекулярных данных свидетельствует в пользу
того, что именно эти раковинные моллюски — цефаласпиды — являются отдаленными пред-
ками безраковинных голожаберных моллюсков, а не наоборот. Причем потеряли голожаберные
моллюски раковину не в одночасье. Этому предшествовал длительный процесс ее постепенной
редукции, в результате которой мы можем наблюдать среди заднежаберных моллюсков целый
«букет» разнообразных промежуточных форм, дающий нам возможность проследить все или
почти все этапы эволюции.
У цефаласпид имеется хорошо развитая, обычно цилиндрическая или веретеновидная
раковина, однако с почти полностью погруженным или лишь частично выступающим за-
витком. «Полноценный» завиток (т. е. обороты раковины) имеется лишь у немногих цефал-
аспид, прежде всего у семейства Acteonidae. В то же время у некоторых семейств, таких как
Aglajidae и Gastropteridae, которые по основному плану строения, несомненно, еще принад-
лежат к отряду цефаласпид, раковина стала внутренней и подверглась значительной редук-
ции. Т. е. эти семейства демонстрируют начальные этапы того пути, который в конце концов
привел к появлению полностью безраковинных, на первый взгляд столь необычных, голожа-
берных моллюсков.
У всех цефаласпид имеется головной щит, который помогает им частично зарываться
и скользить в верхнем слое ила или мелкого песка. Нога обычно выражена хуже, чем у других
брюхоногих, часто треугольной формы из-за того, что по бокам формируются особые складки —
параподии.
СЕМЕЙСТВО ACTEONIDAE —
АКТЕОНОВЫЕ ИЛИ АКТЕОНИДЫ
Среди древних цефаласпид семейство актеонид
одно из наиболее архаичных. У него отсутству-
ют еще многие признаки, свойственные «на-
стоящим» цефаласпидам, например головной
копулятивный аппарат и цельный головной
щит. По этой причине в некоторых системах
это семейство даже выделяют в отдельный от-
ряд Acteonida, чтобы подчеркнуть его отличия
от остальных цефаласпид. У актеонид имеются
также и дополнительные оральные лопасти, на-
поминающие оральные щупальца морских
зайцев-анаспид и некоторых голожаберных
моллюсков. Раковина актеонид, как правило,
высокая, башенковидная, напоминающая ско-
рее раковину брюхоногих моллюсков, а не боль-
шинства цефаласпид. Также, в отличие от мно-
гих (но не всех цефаласпид), у актеонид
полностью отсутствует особый отдел пищева-
рительной системы — жевательный желудок
с особыми твердыми пластинками. Копулятив-
ный аппарат актеонид, в отличие от большин-
ства цефаласпид, располагается внутри мантий-
ной полости рядом с женским отверстием, что
делает актеонид сходными с нотаспидами и го-
ложаберными моллюсками, сильно отличаю-
щимися от этого семейства по внешним при-
знакам. Большинство видов актеонид обитает
в тропических и субтропических морях. В фау-
не России пока обнаружен только один вид.
46
Семейство Acteonidae — Актеоновые или Актеониды
Japonactaeon nipponensis
(Yamakawa, 1911) —
Японактеон ниппонский
Особенности строения. Имеется хорошо раз-
витая, овальная наружная раковина с развитым
завитком. Головной щит мощный, впереди за-
кругленный, в задней части, перед раковиной,
формирует пару широких удлиненных лопа-
стей. Нога короткая, без заметных параподий.
Высота раковины до 7 мм.
Окраска. Раковина обладает очень харак-
терной окраской — многочисленные узкие
светлые полосы по черному фону. Головной
щит также темного цвета.
Отличия от похожих видов. Крупная вы-
сокая раковина черного цвета с хорошо разви-
тым завитком в сочетании с темным головным
щитом позволяет легко отличить этот вид от
любых других заднежаберных моллюсков фау-
ны России.
Особенности экологии. Обитает на мяг-
ких, песчаных и илисто-песчаных грунтах. От-
мечен от литорали до глубины 1,5-2 м.
Размножение. Достоверных данных нет.
Вероятно, подобно другим цефа л аспидам, от-
кладывает слизистые мешкообразные кладки
на поверхности грунта.
Питание и поведение. Может почти пол-
ностью зарываться в грунт, так что над поверх-
ностью остается только верхняя часть ракови-
ны. С помощью головного щита, который
выделяет обильную слизь, способствующую
скольжению, моллюск может быстро передви-
гаться в грунте. Питается различными червя-
ми, которых извлекает из толщи грунта с по-
мощью мощной радулы.
Распространение. Субтропическо-боре-
альный, тихоокеанский вид. В морях России
этот примечательный вид обитает только в за-
ливе Петра Великого Японского моря. Кроме
того, отмечен у Японских островов.
Japonactaeon nipponensis — Японактеон ниппонский, вид со спинной стороны
47
Отряд Cephalaspidea — Цефаласпиды
СЕМЕЙСТВО CYLICHNIDAE —
ЦИЛИХНОВЫЕ ИЛИ ЦИЛИХНИДЫ
Большое семейство, обладающее всеми при-
знаками «типичных» цефаласпид. Наружная
раковина, как правило, характерной для це-
фаласпид «свиткообразной» формы с сильно
погруженным завитком (но есть и представи-
тели с выступающим завитком раковины).
Головной щит с короткими задними лопастя-
ми. Копулятивный аппарат головной. Хоро-
шо развитый жевательный желудок с тремя
(у большинства видов) мощными твердыми
пластинками. Жевательные пластинки свет-
лые, с простой гладкой или слабо складчатой
поверхностью. Радула имеется. Цилихни-
ды — характерный компонент донных сооб-
ществ холодноводных морей и широко пред-
ставлены в фауне России. Встречаются также
и в тропических морях.
Acteocina matusimana
(Nomura, 1939) —
Актеоцина матусимана
Особенности строения. Имеется хорошо
развитая, овально-цилиндрическая тонко-
стенная наружная раковина. Завиток рако-
вины выступает, но небольшой. Головной
щит маленький, впереди простой, закру-
гленный, в задней части, перед раковиной,
формирует пару коротких лопастей. Нога
короткая, без заметных параподий. Высота
раковины до 5 мм.
Окраска. Цвет раковины варьирует от
почти белого до желтоватого или коричне-
ватого. Головная лопасть беловатая, пла-
стинки жевательного желудка светлые, поч-
ти не заметны (у видов из рода Retusa (см.
ниже) пластинки могут просвечивать сквозь
раковину).
Отличия от похожих видов. Acteocina
matusimana можно отличить от очень похо-
жей на нее по строению раковины Retusa in-
stabilis (см. семейство Retusidae) благодаря
заостренному краю первого оборота (над
устьем раковины). У Retusa instabilis этот край
закругленный. Кроме того, сквозь переднюю
часть раковины Retusa instabilis у живых эк-
земпляров просвечивают три темные пла-
стинки жевательного желудка. У Acteocina
matusimana эти пластинки светлые и почти не
заметны сквозь раковину.
Особенности экологии. Обитает на мяг-
ких, песчаных и илисто-песчаных грунтах. От-
мечен от литорали до глубины 1,5-2 м.
Размножение. Не изучено. Вероятно, по-
добно другим цефаласпидам, откладывает сли-
зистые мешкообразные кладки на поверхности
грунта.
Питание и поведение. Может почти
полностью зарываться в грунт, так что над
поверхностью остается только верхняя часть
раковины. С помощью головного щита, кото-
рый выделяет обильную слизь, способствую-
щую скольжению, моллюск может быстро
передвигаться в грунте. Питается форамини-
ферами, которых после заглатывания из-
мельчает тремя особыми мощными пластин-
ками, располагающимися в жевательном
желудке.
Распространение. Субтропическо-боре-
альный тихоокеанский вид. Известен из залива
Петра Великого Японского моря, отмечен
у Японских островов.
Acteocina matusimana — Актеоцина матусимана,
вид со спинной стороны
48
Семейство Cylichnidae — Цилихновые или Цилихниды
Cylichna alba
(Brown, 1827) —
Цилихна белая
Синонимы:
Cylichna corticata (Beck in Moller, 1842)
Cylichna consobrina (Gould, 1859)
Cylichna arctica (Minlchev, 1977)
Особенности строения. Имеется хорошо раз-
витая почти цилиндрическая наружная рако-
вина с толстыми стенками. Завиток раковины
полностью погруженный и не выступает. Го-
ловной щит удлиненный, впереди простой, за-
кругленный, в задней части, перед раковиной,
формирует пару коротких лопастей. Нога ко-
роткая, без заметных параподий. Высота рако-
вины до 15 мм.
Окраска. Цвет раковины варьирует от
почти белой (чаще у глубоководных экземпля-
ров) до желтоватой и оранжево-коричневой.
Головная лопасть беловатая, в средней части
просвечивает розоватое пятно — жевательный
желудок.
Отличия от похожих видов. Можно спу-
тать с некоторыми видами из рода Retusa, ряд
из которых также обладает почти цилиндри-
ческой раковиной с погруженным завитком.
Cylichna alba отличается от них более крупными
размерами и крепкой раковиной.
Особенности экологии. Обитает на мяг-
ких, песчаных и илисто-песчаных грунтах. От-
мечена от литорали (у Мурманского побере-
жья Баренцева моря) до больших глубин
в 2000 м.
Размножение. Мало изучено. Вероятно,
подобно другим цефаласпидам, откладывает
слизистые мешкообразные кладки на поверх-
ности грунта.
Питание и поведение. Может почти пол-
ностью зарываться в грунт, так что над поверх-
ностью остаются только верхняя часть ракови-
ны. С помощью головного щита, который
выделяет обильную слизь, способствующую
скольжению, моллюск может быстро передви-
гаться в грунте. Питается фораминиферами
и возможно мелкими моллюсками, которых
после заглатывания измельчает тремя особы-
ми мощными пластинками, располагающими-
ся в жевательном желудке.
Распространение. Арктическо-бореаль-
ный вид. Является одним из самых массовых
Cylichna alba — Цилихна белая,
вид со спинной стороны
видов Северных и Дальневосточных морей Рос-
сии, отмечен повсеместно от Баренцева до
Японского моря. В Европе отмечен по крайней
мере до севера Великобритании. Известен так-
же с восточного побережья США.
Cylichna occulta
(Mighels et Adams, 1842) —
Цилихна оккульта
Особенности строения. Имеется хорошо раз-
витая толстостенная овальная наружная рако-
вина. Завиток раковины полностью погружен-
ный и не выступает. Головной щит короткий,
впереди простой, закругленный, а в задней ча-
сти, перед раковиной, формирует пару корот-
ких лопастей. Нога без заметных параподий.
Высота раковины до 12 мм.
Окраска. Раковина неравномерно покры-
та желтовато-коричневатым пигментом, остав-
ляющим не пигментированные беловатые об-
ласти значительного размера. Головная лопасть
беловатая, в задней части слегка желтоватая.
49
Отряд Cephalaspidea — Цефаласпиды
Отличия от похожих видов. От Cylichna
alba хорошо отличается овальной, почти бо-
чонкообразной, а не цилиндрической ракови-
ной. В Арктических морях обитает два близких
к Cylichna occulta вида: Cylcihna densistriata (Leche,
1878) и Cylichna scalpta (Reeve, 1855). Первый от-
личается от Cylcihna occulta почти шаровидной
раковиной, тогда как второй, напротив, почти
прямоугольной формой с более развернутым
устьем. Схожие виды из рода Retusa имеют бо-
лее мелкую и тонкую раковину.
Особенности экологии. Обитает на мяг-
ких, песчаных и илисто-песчаных грунтах.
Отмечена от нижнего горизонта литорали
(в Белом море) до глубин в 200-300 м.
Размножение. Почти ничего не известно
о размножении этого вида. Вероятно, подобно
другим цефаласпидам, откладывает слизистые
мешкообразные кладки на поверхности грунта.
Питание и поведение. Может почти пол-
ностью зарываться в грунт, так что над поверх-
ностью остается только верхняя часть ракови-
ны. С помощью головного щита, который
выделяет обильную слизь, способствующую
Cylichna occulta — Цилихна оккулъта,
вид со спинной стороны
скольжению, моллюск может быстро передви-
гаться в грунте. Питается фораминиферами
и возможно мелкими моллюсками, которых
после заглатывания измельчает тремя особы-
ми мощными пластинками, располагающими-
ся в жевательном желудке.
Распространение. Арктическо-бореаль-
ный вид. Широко распространен в Северных
и Дальневосточных морях России, отмечен по-
всеместно от Баренцева до Японского моря.
В Европе отмечен по крайней мере до севера
Великобритании. Известен также с восточного
побережья США.
СЕМЕЙСТВО PHILINIDAE —
ФИЛИНИДЫ
У большинства представителей этого семей-
ства раковина превращается в тонкую вогну-
тую пластинку, скрытую под кожей. Филини-
ды в эволюционном смысле — продолжение
тенденций, которые наметились уже у цилих-
нид. Подобно цилихнидам, большинство фи-
линид обладает тремя мощными светлыми
жевательными пластинками с гладкой или
чуть складчатой поверхностью. Радулярный
аппарат имеется, также весьма сходен с тако-
вым у цилихнид. Головной щит у филинид еще
более разрастается. Раковина, напротив, суще-
ственно редуцируется в сравнении с цилихни-
дами. Развитая наружная раковина имеется
лишь у небольшого числа филинид. Как и ци-
лихниды, представители семейства Philinidae
широко распространены в арктических и уме-
ренных морях, и представлены в фауне России
целым рядом родов и видов. Имеются и тро-
пические представители.
Phlllne argentata
Gould, 1859 -
Филине серебристая
Особенности строения. Раковина тонкая,
внутренняя, просвечивает сквозь покрываю-
щую ее мантию. Крупные зубчатые отростки
в задней части раковины отсутствуют. Голов-
ной щит относительно короткий, овальный,
закруглен в передней и задней частях. Нога
с хорошо развитыми параподиями. Длина те-
ла до 15 мм.
50
Семейство Philinidae — Филиниды
Philine argentata — Филине серебристая,
вид со спинной стороны
Окраска. Тело полупрозрачное, просвечи-
вающие внутренности беловатые (кроме ко-
ричневатой пищеварительной железы). По
поверхности тела могут быть разбросаны мел-
кие белые пигментные крапинки.
Отличия от похожих видов. В морях Рос-
сии обитает целый ряд видов из семейства фи-
линид. Здесь представлены только три наибо-
лее обычных на мелководье Японского моря
вида. В этом районе Philine argentata можно спу-
тать с двумя другими видами — Philine scalpta
и Yokoyamaia ornatissima. Philine scalpta отличает-
ся от Philine argentata более крупными размера-
ми и мощной удлиненной, а не овальной го-
ловной лопастью. Кроме того, Philine scalpta
обитает в очень мелководных хорошо прогре-
ваемых бухтах от литорали до глубины обычно
не более 5 м, тогда как Philine argentata, напро-
тив, предпочитает более холодноводные участ-
ки и глубины обычно более 10 м. Yokoyamaia
ornatissima отличается от Philine argentata харак-
терной суженой, клиновидной головной лопа-
стью и характерным задним краем раковины,
несущим мощные зубцы. Все эти виды также
имеют целый ряд важных отличий по внутрен-
ним признакам и по строению раковины.
Особенности экологии. Обитает на мяг-
ких, песчаных и илисто-песчаных грунтах. От-
мечена на мелководье (5 м), но более обычен
несколько глубже, от 10-20 до 100-200 м.
Размножение. Почти ничего не известно
о размножении этого вида. Вероятно, подобно
другим цефаласпидам, откладывает слизистые
мешкообразные кладки на поверхности грунта.
Питание и поведение. Может почти пол-
ностью зарываться в грунт, так что над поверх-
ностью остаются только верхняя часть тела.
С помощью головного щита, который выделя-
ет обильную слизь, способствующую скольже-
нию, моллюск может быстро передвигаться
в грунте. Питается мелкими моллюсками, ко-
торых после заглатывания измельчает тремя
особыми мощными пластинками, располагаю-
щимися в жевательном желудке.
Распространение. Бореальный тихооке-
анский вид. Широко распространен в Япон-
ском море и у Японских островов.
Philine scalpta
A. Adams, 1862 —
Филине скальпта
Особенности строения. Раковина тонкая,
внутренняя, просвечивает сквозь покрываю-
щую ее мантию. Крупные зубчатые отростки
в задней части раковины отсутствуют. Голов-
ной щит очень мощный, удлиненный, слегка
приострен или закруглен в передней части
и закруглен в задней части. Нога с хорошо раз-
витыми параподиями. Длина тела до 25 мм,
при высоте раковины до 15 мм.
Окраска. Тело полупрозрачное, просвечи-
вающие внутренности беловатые (кроме
коричневато-розоватой пищеварительной же-
лезы). По поверхности тела могут быть разбро-
саны мелкие белые пигментные крапинки.
Отличия от похожих видов. От Philine ar-
gentata отличается мощной удлиненной голов-
ной лопастью и предпочтительным обитанием
на самых мелководных участках (литораль —
5 м). Внешне схожая Yokoyamaia ornatissima от-
личается от Philine scalpta характерной суженой,
почти клиновидной головной лопастью и ха-
рактерным задним краем раковины, несущим
мощные зубцы.
51
Отряд Cephalaspidea — Цефаласпиды
Philine scalpta — Филине скалътпа,
вид со спинной стороны
Особенности экологии. Обитает на мяг-
ких, песчаных и илисто-песчаных грунтах.
Предпочитает очень мелководные участки
с хорошо прогреваемой водой, от литорали до
глубины обычно не более 5 м.
Размножение. Размножение отмечено
в августе. Крупные мешкообразные слизистые
кладки прикрепляются к поверхности грунта
с помощью короткого узкого стебелька.
Питание и поведение. Может почти
полностью зарываться в грунт, так что над
поверхностью остаются только верхняя часть
тела. С помощью головного щита, который
выделяет обильную слизь, способствующую
скольжению, моллюск может быстро пере-
двигаться в грунте. Питается мелкими мол-
люсками, которых после заглатывания из-
мельчает тремя особыми мощными плас-
тинками, располагающимися в жевательном
желудке.
Распространение. Субтропическо-боре-
альный вид. Отмечен в заливе Петра Великого
Японского моря, у Японских островов и побе-
режья Кореи.
Yokoyamaia ornatissima
(Yokoyama, 1927) —
Йокоямайа украшенная
Особенности строения. Раковина тонкая,
внутренняя, просвечивает сквозь покрываю-
щую ее мантию. Имеются характерные круп-
ные зубчатые отростки в задней части ракови-
ны. Головной щит удлиненный, клиновидный,
заметно приострен в передней и закруглен
в задней части. Нога с хорошо развитыми па-
раподиями. Длина тела до 10 мм.
Окраска. Тело полупрозрачное, просвечи-
вающие внутренности розоватые.
Отличия от похожих видов. Yokoyamaia
ornatissima хорошо отличается от других видов
филинид характерной суженой, клиновидной
головной лопастью и особым задним краем
раковины, несущим мощные зубцы.
Особенности экологии. Обитает на мяг-
ких, песчаных и илисто-песчаных грунтах. От-
мечен от глубин 5-10 м до 50 м.
Размножение. Не изучено.
Питание и поведение. Может почти пол-
ностью зарываться в грунт, так что над поверх-
ностью остается только верхняя часть тела.
Семейство Aglajidae — Аглаиды
Yokoyamaia ornatissima — Йокоямайа украшенная,
вид со спинной стороны
С помощью головного щита, который выделя-
ет обильную слизь, способствующую скольже-
нию, моллюск может быстро передвигаться
в грунте. Питание не изучено, вероятно в раци-
он могут входит фораминиферы.
Распространение. Бореальный тихооке-
анский вид, распространен в Японском море
и у Японских островов.
СЕМЕЙСТВО AGLAJIDAE —
АГЛАИДЫ
Необычное семейство цефаласпид, по форме
напоминающих скорее настоящих голожа-
берных моллюсков. Раковина у аглаид пред-
ставляет собой крошечную рудиментарную
пластинку и снаружи не заметна. Радула и ру-
диментарные пластинки жевательного желуд-
ка имеются только у одного рода Odontoglaja,
тогда как у всех остальных представителей
они полностью исчезли. Виды из этого семей-
ства, как правило, обладают яркой окраской,
что также более характерно для голожабер-
ных моллюсков, чем для цефаласпид. Тем не
менее общее разделение тела аглаид на отде-
лы, когда все еще можно выделить типичные
для цефаласпид мощный головной щит, ман-
тию и боковую жабру внутри мантийной по-
лости, в сочетании с характерными для цефа-
ласпид признаками внутреннего строения,
несомненно свидетельствуют, что семейство
Aglajidae сформировалось в результате вто-
ричного перехода цефаласпид от зарывающе-
гося образа к жизни к обитанию на поверх-
ности грунта. Большинство аглаид обитают
в тропических и субтропических морях, явля-
ясь в том числе характерным компонентом
сообщества коралловых рифов. Поэтому не
удивительно, что в преимущественно холод-
новодных морях России обитает лишь два
представителя из этого семейства.
Melanochlamys diomedea
(Bergh, 1893) -
Меланохлямис диомедеа
Особенности строения. Тело удлиненное, по-
хожее на таковое безраковинных моллюсков.
Можно четко выделить два отдела — более ко-
роткий овальный головной щит без дополни-
тельных отростков и больший по размерам
висцеральный отдел тела, покрытый сверху
мантией и содержащий внутренние органы.
Раковина рудиментарная, внутренняя, снару-
жи незаметна. Нога с хорошо развитыми пара-
подиями. Длина тела до 15 мм.
Окраска. По сероватому полупрозрачно-
му фону разбросаны многочисленные мелкие
крапинки и пятнышки, которые, частично
сливаясь, образуют характерную черновато-се-
роватую или иногда коричневатую общую
окраску тела.
Отличия от похожих видов. От единствен-
ного другого вида из семейства аглаид, обнару-
женного в морях России, Philinopsis gigliolii, за-
метно отличается однотонной окраской,
состоящей только из сероватых крапинок,
и полностью лишенной ярких синих полос
и оранжевых пятен.
Особенности экологии. Обитает на мяг-
ких, песчаных и илисто-песчаных грунтах. В от-
личие от большинства аглаид предпочитает
более глубокие участки с менее прогретой во-
дой. Отмечен в основном на глубинах 15-80 м,
изредка встречается на литорали.
53
Отряд Cephalaspidea — Цефаласпиды
Melanochlamys diomedea — Меланохлямис диомедеа,
вид со спинной стороны
Размножение. Размножение не изучено.
Питание и поведение. Может частично
зарываться в грунт. Отмечено питание нема-
тодами.
Распространение. Тихоокеанский боре-
альный вид. Широко распространен в Японском
море, отмечен у Южных курильских островов
и у побережья Хоккайдо. По тихоокеанскому
побережью Северной Америки обитает от Але-
утских островов до Калифорнии.
Philinopsis gigliolii
(Tapparone-Canefri, 1874) —
Филинопсис Гиглиоли
Особенности строения. Тело удлиненное,
массивное, высокое. Четко выделяется два при-
мерно равных по размерам отдела — мощный
удлиненный головной щит без дополнитель-
ных отростков и висцеральный отдел тела, по-
крытый сверху мантией. Раковина рудимен-
тарная, внутренняя, снаружи незаметна. Нога
с хорошо развитыми параподиями. Один из
самых крупных видов цефаласпид фауны Рос-
сии, длина тела достигает 30 мм.
Окраска. Окраска включает желто-оран-
жевые пятна небольшого размера, густо раз-
бросанные по темно-коричневому, почти чер-
ному фону. Края ноги, головного щита
и мантии оторочены яркими желтовато-оран-
жевыми, черными и синими полосками.
Отличия от похожих видов. От единствен-
ного другого вида из семейства аглаид, обнару-
женного в морях России, Melanochlamys diome-
dea, данный вид заметно отличается более
разнообразной окраской, включающую яркие
синие полосы и оранжевые пятна, и высоким
массивным телом.
Особенности экологии. Обитает на мяг-
ких, песчаных и илисто-песчаных грунтах. Явно
предпочитает мелководья с хорошо прогревае-
мой водой, на глубинах от 1 до 10 м.
Размножение. Размножение не изучено.
Питание и поведение. Может частично
зарываться в грунт. Хищный вид. Питается
другими, более мелкими раковинными це-
фаласпидами, а возможно и другими мол-
люсками.
Philinopsis gigliolii — Филинопсис Гиглиоли,
вид со спинной стороны
54
Семейство Retusidae — Ретузиды
Распространение. Тихоокеанский суб-
тропическо-бореальный вид. В морях России
обнаружен только в заливе Петра Великого
Японского моря. Отмечен также в водах Япо-
нии и Китая.
СЕМЕЙСТВО RETUSIDAE —
РЕТУЗИДЫ
Весьма специализированное семейство, состоя-
щее почти исключительно из мелких представи-
телей. Сохраняя общий «план строения» цефа-
ласпид (небольшой головной щит с короткими
задними лопастями, головной копулятивный
аппарат и жевательный желудок с тремя пла-
стинками), ретузиды полностью утеряли такой
характерный орган большинства моллюсков
(включая заднежаберных), как радулу. Пластин-
ки жевательного желудка имеют у ретузид спе-
цифическое строение — их жевательная поверх-
ность покрыта крупными и мелкими бугорками.
Особенности расположения и число бугорков
специфичны для каждого вида. Кроме того, же-
вательные пластинки ретузид имеют характер-
ную темно-коричневую, почти черную окраску.
Ретузиды широко распространены как в холод-
ных и умеренных, так и в субтропических морях.
В фауне России обитает целый ряд видов этого
семейства, их систематика до сих остается пред-
метом исследования.
Retusa instabilis
Minichev, 1971 —
Ретуза нестабильная
Особенности строения. Имеется хорошо раз-
витая, овальная или овально-цилиндрическая
тонкостенная наружная раковина. Степень вы-
раженности завитка раковины варьирует от
выступающего до погруженного. Головной
щит маленький, впереди простой, закруглен-
ный, а в задней части, перед раковиной, фор-
мирует пару коротких лопастей. Нога корот-
кая, без заметных параподий. Высота раковины
до 4 мм.
Окраска. Цвет раковины варьирует от поч-
ти белого до желтоватого или коричневатого.
Головная лопасть беловатая, в средней части
просвечивают три темные пластинки жеватель-
ного желудка.
Retusa instabilis — Ретуза нестабильная,
вид со спинной стороны
Отличия от похожих видов. Систематика
рода Retusa до сих пор полностью не устоялась.
Не случайно виды из этого рода нередко награж-
дались такими видовыми эпитетами, как «неста-
бильная» (instabilis) или «изменчивая» (variabilis).
Дело в том, что раковина у видов рода Retusa,
в частности степень погружения завитка в рако-
вину, может варьировать в довольно широких
пределах, тогда как признаки внутреннего строе-
ния являются значительно более стабильными.
Поэтому идентификация ряда видов может
встретить определенные затруднения. Retusa in-
stabilis сходна по форме раковины с Retusa perten-
uis, но существенно отличается по строению пла-
стинок жевательного желудка и копулятивному
аппарату. Лучше всего при различении этих ви-
дов использовать экологический и географиче-
ский критерий — Retusa instabilis обитает исклю-
чительно в заливе Петра Великого Японского
моря, в самых мелководных участках, от литора-
ли до глубины около 2 м. Напротив, Retusa perten-
uis обитает повсеместно в морях России, но в за-
ливе Петра Великого встречается обычно глубже
50 м, и никогда — на литорали. В то же время
55
Отряд Cephalaspidea — Цефаласпиды
Retusa instabilis — Ретуза нестабильная, изменчивость раковины
в северных частях ареала, например в Баренце-
вом море, Retusa pertenuis на литорали обитать
может, a Retusa instabilis там, напротив, отсут-
ствует. Ситуация усугубляется еще и тем, что
совместно с Retusa instabilis в заливе Петра Вели-
кого может встречаться представитель другого
семейства — цилихнид (Cylichnidae) — Acteocina
matusimana. Эти два вида имеют очень разное
внутреннее строение, но крайне похожи друг на
друга по форме раковины. Acteocina matusimana
можно отличить благодаря заостренному
краю первого оборота (над устьем раковины).
У Retusa instabilis этот край закругленный. Кро-
ме того, сквозь переднюю часть раковины Retusa
instabilis у живых экземпляров просвечивают
три темные пластинки жевательного желудка.
У Acteocina matusimana эти пластинки светлые
и почти не заметны сквозь раковину.
Особенности экологии. Обитает на мяг-
ких, песчаных и илисто-песчаных грунтах. От-
мечен от литорали до глубины 1,5-2 м.
Размножение. Ничего не известно о раз-
множении этого вида. Вероятно, подобно дру-
гим цефаласпидам, откладывает слизистые меш-
кообразные кладки на поверхности грунта.
Питание и поведение. Может почти пол-
ностью зарываться в грунт, так что над поверх-
ностью остается только верхняя часть ракови-
ны. С помощью головного щита, который
выделяет обильную слизь, способствующую
скольжению, моллюск может быстро передви-
гаться в грунте. Питается фораминиферами,
которых после заглатывания измельчает тремя
мощными пластинками, располагающимися
в жевательном желудке.
Распространение. Бореальный тихооке-
анский вид. Достоверно известен только из за-
лива Петра Великого Японского моря, но несо-
мненно обитает также и у Японских островов.
Retusa pertenuis
(Mighels, 1843) -
Ретуза тонкая
Особенности строения. Имеется хорошо раз-
витая, овальная или овально-цилиндрическая
тонкостенная наружная раковина. Завиток сла-
бо выступает. Головной щит маленький, впере-
ди простой, закругленный, в задней части, пе-
56
Семейство Retusidae — Ретузиды
Retusa per tenuis — Рету за тонкая,
вид с брюшной стороны
ред раковиной, формирует пару коротких
лопастей. Нога короткая, без заметных парапо-
дий. Высота раковины до 3 мм.
Окраска. Цвет раковины варьирует от
почти белого до желтоватого.
Отличия от похожих видов. Retusa perte-
nuis характеризуется слабовыступающим, но
все еще заметным завитком раковины. Это
вид является самым обычным видом из рода
Retusa в Северных и Дальневосточных морях
России. Тем не менее в том или ином отдель-
ном регионе наряду с этим видом могут оби-
тать и другие виды ретузид, которых нередко
трудно отличить между собой по признакам
раковины, в то время как они очень хорошо
различаются по внутреннему строению. В не-
которых случаях (но далеко не во всех!), с раз-
умной осторожностью, можно использовать
географический и экологический критерий
для различения видов. Так, Retusa pertenuis мо-
жет обитать на литорали Баренцева моря, но
никогда не встречается на литорали и мелко-
водье (2-5 м) залива Петра Великого Япон-
ского моря, где, напротив, обычной является
Retusa instabilis.
Особенности экологии. Обитает на мяг-
ких, песчаных и илисто-песчаных грунтах. От-
мечена от литорали (на Мурманском побере-
жье Баренцева моря) до глубины 500 м.
Размножение. Ничего не известно о раз-
множении этого вида. Вероятно, подобно
другим цефаласпидам, откладывает слизи-
стые мешкообразные кладки на поверхности
грунта.
Питание и поведение. Может почти
полностью зарываться в грунт, так что над
поверхностью остается только верхняя часть
раковины. С помощью головного щита, кото-
рый выделяет обильную слизь, способствую-
щую скольжению, моллюск может быстро
передвигаться в грунте. Питается форамини-
ферами, которых после заглатывания из-
мельчает тремя особыми мощными пластин-
ками, располагающимися в жевательном
желудке.
Распространение. Бореально-арктичес-
кий вид. Широко распространен во всех морях
России, от Баренцева до Японского.
Retusa truncatula
(Bruguiere, 1792) —
Ретуза усеченная
Особенности строения. Имеется хорошо
развитая тонкостенная наружная раковина
овально-цилиндрической формы, характер-
но расширенной в нижней части. Осевые
складки на раковине хорошо выражены. За-
виток полностью погруженный. Головной
щит маленький, впереди простой, закру-
гленный, в задней части, перед раковиной,
формирует пару коротких лопастей. Нога
короткая, без заметных параподий. Высота
раковины до 4 мм.
Окраска. Цвет раковины варьирует от
почти белого до желтоватого.
Отличия от похожих видов. От Retusa
variabilis, другого широко распространенного
вида черноморских ретузид, Retusa truncatula
достаточно уверенно отличается характерным
расширением в нижней части раковины и на-
личием на раковине резких осевых (вертикаль-
ных) складок. В других морях России этот вид
полностью отсутствует.
Особенности экологии. Обитает на мяг-
ких, песчаных и илисто-песчаных грунтах.
57
Отряд Cephalaspidea — Цефаласпиды
Retusa truncatula — Ретуза усеченная, раковина со стороны устья и сверху
Retusa truncatula — Ретуза усеченная,
раковина со стороны устья, изображение получено
с помощью сканирующего электронного микроскопа
По крайней мере в прибрежных водах Кры-
ма, взрослые экземпляры этого вида отсут-
ствуют на самом мелководье, обитая глубже
20-30 м.
Размножение. Вероятно, подобно другим
цефаласпидам, откладывает слизистые меш-
кообразные кладки на поверхности грунта.
Ювенильные особи этого вида изображены на
стр. 59.
Питание и поведение. Может почти
полностью зарываться в грунт, так что над
поверхностью остаются только верхняя часть
раковины. С помощью головного щита, кото-
рый выделяет обильную слизь, способствую-
щую скольжению, моллюск может быстро
передвигаться в грунте. Питается форамини-
ферами, которых после заглатывания из-
мельчает тремя особыми мощными пластин-
ками, располагающимися в жевательном
желудке.
Распространение. Атлантический боре-
ально-субтропический вид. Широко распро-
странен вдоль берегов Европы от побережья
Испании до Норвегии и в Средиземном мо-
ре. Обычен по всей северной части Черного
моря.
58
Семейство Retusidae — Ретузиды
Retusa truncatula — Ретуза усеченная, ювенильные особи; вид со спинной и брюшной стороны
Retusa variabilis
(Milaschevitsch, 1912) —
Ретуза изменчивая
Особенности строения. Имеется хорошо
развитая тонкостенная наружная раковина
овальной формы, не расширенная в нижней
части. Осевые складки на раковине не выраже-
ны, имеется только спиральная исчерчен-
ность. Завиток полностью погруженный. Го-
ловной щит маленький, впереди простой,
закругленный, а в задней части, перед ракови-
ной, формирует пару коротких лопастей. Но-
га короткая, без заметных параподий. Высота
раковины до 4 мм.
Окраска. Цвет раковины варьирует от
почти белого до желтоватого.
Отличия от похожих видов. Retusa vari-
abilis входит в число двух наиболее обычных
видов ретузид, известных из Черного моря.
От Retusa truncatula, другого широко распро-
страненного вида черноморских ретузид,
Retusa variabilis достаточно уверенно отлича-
ется овальной (а не овально-цилиндрической)
формой раковины без расширения и отсут-
ствием резких осевых (вертикальных) скла-
док на раковине. В других морях России этот
вид полностью отсутствует. Следует пом-
нить, что другие виды ретузид с полностью
погруженным завитком, не включенные
в данную книгу Retusa minima (Yamakawa,
1911) и Retusa succincta (A. Adams, 1862), яв-
ляются довольно обычными в Японском мо-
ре и весьма сходны с Retusa variabilis по фор-
ме раковины.
Особенности экологии. Обитает на
мягких, песчаных и илисто-песчаных грун-
тах. По крайней мере, в прибрежных водах
Крыма, взрослые экземпляры этого вида от-
сутствуют на самом мелководье, обитая глуб-
же 20-30 м.
Размножение. Ничего не известно о раз-
множении этого вида. Вероятно, подобно
другим цефаласпидам, откладывает слизи-
стые мешкообразные кладки на поверхности
грунта.
Питание и поведение. Может почти пол-
ностью зарываться в грунт, так что над поверх-
ностью остаются только верхняя часть ракови-
ны. С помощью головного щита, который
выделяет обильную слизь, способствующую
скольжению, моллюск может быстро передви-
59
Отряд Cephalaspidea — Цефаласпиды
Retusa variabilis — Ретуза изменчивая, раковина со стороны устья и сверху
Retusa variabilis — Ретуза изменчивая,
раковина со стороны устья, изображение получено
с помощью сканирующего электронного микроскопа
Пищевой объект многих представителей
семейства ретузид: раковинные простейшие
из группы фораминифер
гаться в грунте. Питается фораминиферами,
которых после заглатывания измельчает тремя
особыми мощными пластинками, располагаю-
щимися в жевательном желудке.
Распространение. Биогеографическая
природа этого вида не ясна. Широко распро-
странен по всей северной части Черного моря,
но из других морей не известен. Возможно, Re-
tusa variabilis является эндемиком Черного мо-
ря, но происходит от одного из близких среди-
земноморских видов, например таких, как
Retusa umbilicata.
60
Семейство Volvulellidae — Вольвулеллиды
СЕМЕЙСТВО VOLVULELLIDAE —
ВОЛЬВУЛЕЛЛИДЫ
Одно из наиболее специализированных се-
мейств цефаласпид, еще сохранившее раз-
витую наружную раковину. Также имеется
рудиментарный головной щит и головной ко-
пулятивный аппарат. Радула и жевательные
пластинки полностью редуцированы. Но ха-
рактеристика вольвулеллид не исчерпывается
этими исключительно регрессивными черта-
ми. В глотке у этого семейства, лишенной
обычной радулы, развился довольно сложный
хоботок, отсутствующий у других цефаласпид.
Этот признак несомненно можно рассматри-
вать как прогрессивный относительно других
групп этого отряда. Как именно функциониру-
ет этот хоботок при питании до сих пор не из-
вестно. К этому семейству относится един-
ственный род Volvulella, который тем не менее
представлен довольно значительным числом
видов, распространенных главным образом,
в субтропических и тропических морях. В фау-
не России известен только один вид.
Volvulella sculpturata
Minichev, 1971 —
Вольвулелла скульптурированная
Особенности строения. Имеется хорошо
развитая тонкостенная наружная раковина ха-
рактерной узковеретеновидной формы, резко
заостренной с обоих концов. Завиток полно-
стью погруженный. Головной щит очень ма-
ленький, простой, закругленный. Нога корот-
кая, без заметных параподий. Высота раковины
до 6 мм.
Окраска. Цвет раковины и тела белова-
тый, за исключением верхней части, где имеет-
ся довольно яркое оранжевое пятно. У более
мелких экземпляров подобное пятно может
отсутствовать.
Отличия от похожих видов. Благодаря
особой форме раковины четко отличается от
большинства видов цефаласпид фауны морей
России.
Особенности экологии. Обитает на мяг-
ких, песчаных и илисто-песчаных грунтах на
глубинах от 8 до 50 м.
Размножение. Ничего не известно о размно-
жении этого вида. Вероятно, подобно другим це-
Volvulella sculpturata — Вольвулелла
скульптурированная, вид со спинной стороны
фаласпидам, откладывает слизистые мешко-
образные кладки на поверхности грунта.
Питание и поведение. Может почти
полностью зарываться в грунт, так что над по-
верхностью остается только верхняя часть ра-
ковины. С помощью головного щита, который
выделяет обильную слизь, способствующую
скольжению, моллюск может быстро пере-
двигаться в грунте. Объект питания не изве-
стен, но особенности строения пищеваритель-
ной системы семейства вольвулеллид резко
отличают его от других цефаласпид. Подобно
ретузидам, у них полностью отсутствует раду-
ла, но кроме этого полностью редуцировались
еще и пластинки жевательного желудка. Глот-
ка у вольвулелл модифицировалась в особый
мускулистый хобот. Все эти черты крайней
специализации пищеварительной системы,
несомненно, свидетельствуют и об особом
способе питания и пищевом объекте Volvulella
sculpturata.
Распространение. По-видимому, боре-
альный тихоокеанский вид. Известен пока
только из северо-западной части Японского
моря.
61
Отряд Cephalaspidea — Цефаласпиды
СЕМЕЙСТВО ATYIDAE —
АТИИДЫ
Это семейство в целом обладает всеми
чертами, характерными для большинства
цефаласпид: свиткообразной наружной ра-
ковиной, головным щитом, головным копу-
лятивным аппаратом, радулой и тремя пла-
стинками жевательного желудка. Пластинки
атиид характеризуются рифленой, гребен-
чатой жевательной поверхностью, что отли-
чает их от пластинок всех предшествующих
семейств цефаласпид. Атииды обитают поч-
ти исключительно в тропических и субтро-
пических морях, поэтому вполне законо-
мерно, что в фауне России представители
этого семейства обнаружены исключитель-
но в южных районах дальневосточных мо-
рей, прежде всего в Японском море, где вода
летом прогревается почти до субтропиче-
ских значений.
Cylichnatys angusta — Цилихнапшс узкий,
вид со спинной стороны
Cylichnatys angusta
(Gould, 1859) -
Цилихнатис узкий
Синонимы:
Cylichnatys incisula (Yokoyama, 1928)
sensu Golikov, Scarlato, 1967
non Yokoyama, 1928
Особенности строения. Раковина очень
тонкая, овальная. Завиток раковины не вы-
ступает. Головной щит короткий, впереди
и в задней части простой, закругленный. Но-
га короткая, по бокам формирует хорошо
выраженные параподии. Высота раковины
до 6 мм.
Окраска. Раковина полностью прозрач-
ная, сквозь нее у живых особей просвечивает
тело моллюска, покрытое многочисленными
темно-коричневыми крапинками и пятнышка-
ми. Сходную окраску, но с меньшим числом
полос темно-коричневого пигмента имеет го-
ловная лопасть.
Отличия от похожих видов. От всех цефа-
ласпид фауны России хорошо отличается
очень тонкой прозрачной раковиной, сквозь
которую просвечивает темное пигментирован-
ное тело.
Особенности экологии. Обитает на
мягких, песчаных и илисто-песчаных грун-
тах. Отмечен на небольших глубинах (от 3-5
до 30 м), главным образом в заливе Петра
Великого Японского моря, где довольно
обычен.
Размножение. Ничего не известно о раз-
множении этого вида. Вероятно, подобно
другим цефаласпидам, откладывает слизи-
стые мешкообразные кладки на поверхности
грунта.
Питание и поведение. Может почти
полностью зарываться в грунт, так что над по-
верхностью остается только верхняя часть ра-
ковины. С помощью головного щита, который
выделяет обильную слизь, способствующую
скольжению, моллюск может быстро пере-
двигаться в грунте. Питание не изучено, воз-
можно питается фораминиферами и мелки-
ми моллюсками.
Распространение. Бореально-субтропи-
ческий вид. В морях России известен только
в Японском море. Широко распространен в во-
дах Японии.
62
Семейство Akeridae — Акериды или Акеровые
ОТРЯД ANASPIDEA — АНАСПИДЫ
Данный отряд по особенностям внутреннего и внешнего строения фактически является одним
из вариантов «плана строения» отряда цефаласпид. Так же, как и цефаласпиды, тело анаспид
достаточно четко разделено на два отдела — головную лопасть и висцеральный мешок, так же,
как и у цефаласпид, у них имеется головной копулятивный аппарат. Внутри мантийной полости
имеется хорошо развитая единственная жабра — настоящий ктенидий. Наиболее архаичные
анаспиды из семейства акерид (Akeridae), представленные и в фауне России, даже имеют тонкую
наружную раковину и по общему облику еще ближе стоят к настоящим цефаласпидам.
В отличие от акерид все остальные анаспиды имеют лишь внутреннюю пластинкообразную
раковину, почти незаметную снаружи. Тем не менее анаспиды в целом имеют комплекс свое-
образных черт, которые позволяют их выделять как хорошо обособленную группу. У большин-
ства из них есть хорошо развитые, свернутые в трубку ринофоры и оральные щупальца (но эти
органы отсутствуют у акерид). Параподии (боковые края ноги) у анаспид особенно хорошо вы-
ражены, формируя мощные боковые лопасти. Наконец, у всех анаспид, даже цефласпидоподоб-
ных акерид, имеется мощно развитый жевательный желудок с большим числом жевательных
пластинок разного размера (а не только трех, как у цефаласпид).
Наиболее известными представителями отряда Anaspidea являются знаменитые «морские
зайцы», аплизии, названные так из-за крупных свернутых в трубку ринофоров, отдаленно напо-
минающих длинные уши. Аплизии, как и большинство анаспид, обитают преимущественно
в тропических морях. Но некоторые виды сумели проникнуть и в моря России. Многие виды
способны к кратковременному плаванию, с использованием крыловидных параподий. Все без
исключения анаспиды — растительноядные моллюски.
СЕМЕЙСТВО AKERIDAE —
АКЕРИДЫ ИЛИ АКЕРОВЫЕ
Наиболее архаичное семейство анаспид, еще со-
храняющее целый ряд признаков, связывающих
их с отрядом цефаласпид. К таким признакам
следует отнести тонкую наружную раковину
свитковидной формы, простой головной щит
без свернутых в трубку ринофоров и оральных
щупалец и мощную треугольную ногу, боковые
края которой, параподии — еще не срослись
с мантией, как у аплизий. Но наличие в жева-
тельном желудке большого числа пластинок
указывает на их родство с более «типичными»
анаспидами — аплизиями. Благодаря указан-
ным особенностям строения, семейство акерид
систематики могли относить как к отряду цефа-
ласпид, так и к анаспидам. Подобная двойствен-
ность легко объяснима с эволюционной точки
зрения — акериды с одной стороны еще сохра-
няют признаки цефаласпид, а с другой — уже
приобрели признаки более «высокоорганизо-
ванных» анаспид и крылоногих моллюсков.
К семейству акерид относится единственный
род Акега, в который входит небольшое число
видов, обитающих как в умеренных, так и в тро-
пических морях. В фауне России один вид.
Akera bullata
MOIIer, 1776 -
Акера буллата
Особенности строения. Имеется хорошо раз-
витая тонкостенная овальная наружная ракови-
на. Завиток раковины полностью погруженный
и не выступает. Головной щит мощный, очень
сильно удлинен. Обособленные ринофоры
и оральные щупальца отсутствуют. Нога очень
широкая, треугольная, с мощными боковыми
лопастями-параподиями. Длина тела может
достигать значительных размеров, до 60 мм.
Окраска. Раковина блестящая, нерав-
номерно покрыта желтовато-коричневатым
пигментом. Головная лопасть и параподии
характерной мозаичной окраски, состоящей
из множества светлых пятен на коричневом
фоне.
Отличия от похожих видов. Особенности
строения данного вида позволяют легко отли-
чить его от других заднежаберных моллюсков
фауны России.
Особенности экологии. Обитает на мел-
ководье, от литорали до глубины около 7 м.
Размножение. В морях России не отмече-
но. В европейских водах откладывает крупные
63
Отряд Anaspidea — Анаспиды
Akera bullata — Акера буллата,
вид со спинной стороны
Akera bullata — Акера буллата,
вид с брюшной стороны
слизистые мешкообразные кладки. Развитие
со стадией свободно плавающей личинки-
велигера.
Питание и поведение. Очень активный,
подвижный вид, с разнообразным поведени-
ем. Может активно передвигаться по различ-
ным субстратам, предпочтение отдается пес-
чаным и илистым грунтам. Так же, как все
анаспиды, акера — растительноядный орга-
низм, и поэтому предпочитает скопления
различных водорослей, например зеленой во-
доросли Enteromorpha, напоминающей по
форме длинные узкие ленточки с неровными
краями. Наконец, Akera bullata способна к на-
стоящему, хотя и кратковременному плава-
нию, что даже удалось запечатлеть на одной
из фотографий (стр. 65). Плавание акеры от-
даленно напоминает таковое крылоногих
моллюсков, которые произошли от форм, по-
хожих на этот род (см. ниже).
Распространение. Атлантический боре-
альный вид. Широко распространен в морях
Европы от Норвегии до Средиземного моря.
В фауне России отмечен только по Мурман-
скому побережью Баренцева моря. Два пе-
риода, когда этот вид был достоверно обнару-
жен в Баренцевом море (вторая половина
1920-х годов и 2007-2009) уверенно совпадают
с периодами максимального потепления вод
данного региона.
Появление Akera bullata всегда сопровожда-
ют не отдельные, случайно обнаруженные эк-
земпляры этого вида, а целые его «стаи», со-
стоящие из десятков и даже сотен особей.
Наблюдение за скоплениями акер может доста-
вить подлинное удовольствие ныряльщику.
64
Семейство Akeridae — Акериды или Акеровые
Плывущая Akera bullata (Акера буллата), вид с брюшной стороны
65
Отряд Anaspidea — Анаспиды
Следует отметить, что периодиче-
ские потепления Арктики и вхо-
дящего в нее Баренцева моря —
закономерное явление, не
обязательно связанное с деятельно-
стью человека. Оно зависит от мно-
гих факторов, в том числе и от того,
насколько близко к Мурманскому
побережью Баренцева моря подхо-
дит одна из последних «веточек»
мощного теплого течения Гольф-
стрим — Нордкапского течения. Это
течение несет с собой множество
личинок самых различных беспозво-
ночных, включая и голожаберных
моллюсков. Насколько успешным
будет их оседание из планктона на
Мурманском побережье Баренцева
моря, зависит от благоприятной
летней температуры моря. Так, на-
чиная примерно с 2000 года, на
Мурманском побережье наблюдает-
ся совершенно очевидное потепле-
ние, и августовская температура
морской воды на поверхности мо-
жет достигать +12°С. Напротив,
обычная многолетняя температура
воды в августе для этого региона со-
ставляет лишь +8" С. Четыре граду-
са — очень существенная величина
для морских организмов, поэтому
начиная с 2000 года мы обнаружили
в этом регионе целых десять видов
заднежаберных моллюсков, кото-
рые никогда прежде в морях России
не регистрировались.
Так, у Мурманского побережья Ба-
ренцева моря было отмечено пер-
вое за всю историю наблюдений
появление крупного эффектного го-
ложаберного моллюска-дориды
Archidoris pseudoargus ярко-желтого
цвета. К таким видам и относится
Akera bullata. Но в отличие от других
новых для фауны России видов, так-
же найденных на Мурманском побе-
режье в бухте Дальнезеленецкой,
в течение 2000-2010 годов, акеру
в этом регионе уже находили. Прои-
зошло это чуть западнее, в Кольском
заливе (на берегу которого, в част-
ности располагается прославлен-
ный город Мурманск), но почти сто-
летие назад, во второй половине
1920-х годов. И именно для этого
периода было также отмечено зна-
чительное потепление моря.
Akern bullata — Акера буллита
66
Семейство Aplysiidae — Аплизииды или Морские зайцы
Aplysia juliana (Аплазия юлиана) у острова Монерон среди морских ежей
СЕМЕЙСТВО APLYSIIDAE —
АПЛИЗИИДЫ
ИЛИ МОРСКИЕ ЗАЙЦЫ
К этому семейству относятся наиболее «ти-
пичные» анаспиды — знаменитые тропиче-
ские морские зайцы. Аплизииды обладают
характерной формой тела, включающей
мощный висцеральный отдел с хорошо вы-
раженными боковыми складками, сросши-
мися с мантией, и таким же крупным голов-
ным отделом, вытянутым в виде длинной
«шеи» и несущим по паре больших, сверну-
тых в трубку ринофоров и оральных щупа-
лец. Раковина аплизиид представляет собой
вогнутую пластинку, обросшую мантией.
Снаружи раковина почти не заметна. Под
мантией и раковиной располагается крупная
жабра. Задние края мантии часто вытянуты
в форме трубки, называемой сифоном, кото-
рый принимает участие в обеспечении при-
тока к жабре свежей воды. Аплизии обладают
мощной радулой, состоящей из значительно
числа поперечных рядов зубов, с централь-
ным зубом, и такими же хорошо развитыми
челюстями (см. стр. 34). Подобное сильно раз-
витое «вооружение» глотки помогает аплизи-
ям справляться с весьма грубой растительной
пищей — водорослями, основным элементом
их рациона.
Как уже отмечалось, аплизииды — это ис-
ключительно тепловодные организмы, рас-
пространенные в тропических и субтропиче-
ских морях, что накладывает существенные
ограничения на возможности появления апли-
зий в умеренных водах. Поэтому в морях Рос-
сии аплизии зарегистрированы лишь в двух
районах, которые географически очень близки
к обычному для аплизиид ареалу обитания
и одновременно характеризуются высокими
летними температурами морской воды благо-
даря гидрологическим особенностям (в част-
ности, теплым течениям), что позволяет апди-
зиям выжить. Например, в северной части
Японского моря тропические аплизии полза-
ют среди типичных северных морских ежей
Strongylocentrotusl Не исключено, что некото-
рые особи обитающих там апдизий способны
даже пережить неблагоприятное зимнее по-
нижение температуры. В противном случае,
один из обнаруженных там видов, Aplysia ju-
liana, вряд ди успел бы дорасти до весьма вну-
шительных размеров — около 10-15 см — за
короткий летний период.
67
Отряд Anaspidea — Анаспиды
Aplysia juliana
Quoy et Gaimard, 1832 —
Аплизия Юлиана
Особенности строения. Имеет характерный
«аплизийный» внешний облик. Отличитель-
ной чертой внешнего строения данного вида
является особая присоска, которую формирует
задняя часть ноги. Длина тела может достигать
значительных размеров, до 200 мм.
Окраска. По коричневатому или зелено-
ватому фону разбросаны белые крапинки
и пятнышки разного размера.
Отличия от похожих видов. Особенности
строения данного вида позволяют легко отли-
чить его от других заднежаберных моллюсков
фауны России. Можно спутать только с другим
видам аплизий, Aplysia parvula, также обнару-
женным в Японском море, от которого Aplysia
juliana хорошо отличается более крупными
размерами и особенностями окраски (в част-
ности, отсутствием черной каймы по краю па-
раподий).
Особенности экологии. Обитает на мел-
ководье, на глубине около 3-5 м, на скалистом
и каменистом грунте. Как и большинство ви-
дов аплизий, данный вид способен секретиро-
вать защитную «чернильную» жидкость, по-
добно осьминогам и каракатицам. Но
в отличие от других видов, «чернильная» жид-
кость Aplysia juliana не пурпурного, а молочно-
белого цвета.
Размножение. В морях России не отмече-
но. В тропических водах откладывает крупные
слизистые кладки в виде шнуров. Развитие
со стадией свободно плавающей личинки-
велигера.
| Процесс размножения у аплизий характе-
ризуется некоторыми любопытными чер-
тами. Поскольку копулятивный аппарат апли-
зий, как и у цефаласпид, находится на голове,
а женское генитальное отверстие — далеко
позади, в висцеральном мешке рядом с жа-
брой, то эта особенность позволяет аплизиям
при копуляции подчас выстраиваться в целые
цепочки особей, в которых каждая предыду-
щая особь оплодотворяет последующую.
Питание и поведение. Очень активный,
подвижный вид. Может активно передвигаться
по различным субстратам. Aplysia juliana — ти-
пичный растительноядный организм, в состав
диеты которого входят бурые и красные водо-
росли.
Распространение. Циркумтропический
вид. Отмечен во многих тепловодных районах,
включая Индийский океан и Карибское море.
В фауне России обнаружен только в водах
у острова Монерон.
Aplysia parvula
Guilding in Morch, 1863 —
Аплизия парвуля
Особенности строения. Имеет характерный
«аплизийный» внешний облик. В отличие от
предыдущего вида задний край сросшихся па-
раподий очень низкий, не образует широкий
«воротник» вокруг сифона. Это один из самых
мелких видов аплизий, достигающий в длину
не более 30 мм.
В водах Японского моря, побли-
зости от побережья острова Са-
халин, находится крошечный, но
уникальный остров Монерон.
Уникальность его заключается
в том, что хотя вблизи от него лет-
ние температуры морской воды
ниже, чем в заливе Петра Велико-
го, но непосредственно вокруг
острова они остаются высокими.
Происходит это благодаря одной
из веточек теплого Цусимского
течения, которое постоянно
«бьет» непосредственно в остров
Монерон, «подогревая» воду во-
круг него. Теплое течение обе-
спечивает и постоянный приток
личинок богатейшей фауны со-
седней Японии, где число видов
заднежаберных моллюсков в разы
превышает таковую России. Сре-
ди тех немногих видов, которые
благодаря теплому течению смог-
ли не только достичь Монерона,
но осесть в его водах, и успешно
дорасти до взрослой стадии,
в первую очередь необходимо на-
звать два вида аплизий. Зта уди-
вительная «игра природы» позво-
ляет нам наслаждаться оазисом
тропической фауны в сердце
весьма холодной северной части
Японского моря.
68
Семейство Aplysiidae — Аплизииды или Морские зайцы
Aplysia parvula — Аплизия парвуля, вид со спинной стороны
Окраска. Общая окраска тела обычно корич-
невато-красноватая, но встречаются и зеленоватые
экземпляры. Окраска может быть однородной,
или она дополняется разбросанными белыми
пятнышками. Другой характерной особенностью
этого вида является темная полоса, идущая по
краю параподий, нередко прерывистая.
Отличия от похожих видов. Особенности
строения данного вида позволяют легко отли-
чить его от других заднежаберных моллюсков
фауны России, включая другой вид аплизий,
Aplysia juliana, благодаря мелким размерам
и особенностям окраски (в частности, наличию
черной каймы по краю параподий).
Особенности экологии. Обитает на мелко-
водье, на глубине около 3-5 м, на скалистом и ка-
менистом грунте. Обнаружен также в сообще-
стве обрастания искусственных сооружений —
причалов, различных плавучих конструкций
и т. п.
Размножение. В морях России не отмече-
но. В тропических водах откладывает крупные
слизистые кладки в виде шнуров. Развитие
со стадией свободно плавающей личинки-
велигера.
Питание и поведение. Очень активный,
подвижный вид. Может активно передвигаться
по различным субстратам, предпочтение от-
дается песчаным и илистым грунтам. Aplysia
parvula — типичный растительноядный орга-
низм, в состав рациона которого входят бурые
и красные водоросли.
Распространение. Циркумтропический
вид, заходящий в умеренные воды. Отмечен
во многих тепловодных районах, включая Ин-
дийский океан и Карибское море, но также
и в бореальных водах Европы. В фауне России
обнаружен только в двух районах Японского
моря — водах у острова Монерон и в заливе
Петра Великого.
69
• Эволюция, шаг второй •
ПРОРЫВ НАВЕРХ
ГРУППА ПЛАНКТОННЫХ КРЫЛОНОГИХ
МОЛЛЮСКОВ-ПТЕРОПОД
(традиционный отряд Pteropoda)
Наблюдая в тропиках за огромными травоядными аплизиями (так называемыми «морскими
зайцами», см. выше), медленно ползающими по дну и достигающими гигантских размеров в 1 м
(т. е. «величиной почти с собаку»), очень трудно себе представить, что они являются ближайши-
ми родственниками небольших планктонных моллюсков, поражающих своей элегантной фор-
мой и необычными привычками. Эти обитатели толщи воды также относятся к заднежаберным
моллюскам, но к особой группе — крылоногих моллюсков или птеропод (от лат. pteros — крыло,
poda — нога). Между тем все полученные наукой данные о строении крылоногих моллюсков
(включая сравнение ДНК) однозначно свидетельствуют в пользу их родства с анаспидами, в том
числе и с неуклюжими аплизиями. Боковые края ноги анаспид, правда скорее более близких по
строению к современному роду акера, чем к аплизиям, разрослись, а остальная часть ползатель-
ной подошвы, напротив, уменьшилась, сформировав уникальный плавательный аппарат, напо-
минающий крылья — т. е. именно то, за что птеропод и назвали крылоногими моллюсками.
Преобразование организации (зеленые стрелки) донной цефаласпиды (внизу)
в планктонного крылоногого моллюска без раковины (наверху) минуя несколько промежуточных стадий.
Обратный ход эволюции (красные стрелки) невозможен
70
Семейство Limacinidae — Лимациновые
У раковинных заднежаберных моллюсков,
даже у типично донных, нога имеет треуголь-
ную форму за счет особых лопастей по бокам,
называемых параподиями. Благодаря этому
среди них довольно много форм, которые спо-
собны с большей легкостью, чем другие гастро-
поды, использовать для плавания модифици-
рованные крыдья-параподии. Такие, на первый
взгляд, неуклюжие крупные существа как мор-
ские зайцы — аплизии, оказывается, иногда
весьма неплохо плавают. Представители семей-
ства Gastropteridae, относящиеся к раковинным
цефаласпидам, уже наполовину донные, напо-
ловину пелагические моллюски. И наконец,
в продолжение этой тенденции, появились
крылоногие моллюски — птероподы, полно-
стью лишенные связи с дном и ведущие весьма
активный пелагический образ жизни. В то вре-
мя как многие текосоматы — мирные собирате-
| Погрузившись в ледяные воды Белого моря
практически в любое время года, но предпо-
чтительнее летом, ныряльщик без труда встре-
тит представителей крылоногих моллюсков,
получивших романтическое имя морских анге-
лов, чей внешний вид действительно отмечен
вполне неземными чертами. Но морские анге-
лы имеют совсем не ангельский нрав! В каче-
стве объекта питания они используют других
крылоногих моллюсков, помельче и не таких
проворных — лимацин. За свою темную окра-
ску они получили название... морских чертей.
ди планктона, гимносоматы — специализиро-
ванные хищники и охотятся главным образом
па своих раковинных сородичей, а также на
морских стрелок-хетогнат и медуз.
Во многих старых учебниках все еще
можно встретить название отряда
Pteropoda в качестве общего для всей
группы крылоногих моллюсков. Но уже
достаточно давно за существенную
разницу в строении птеропод разде-
лили на два независимых отряда—ра-
ковинных крылоногих моллюсков —
текосомат (Thecosomata, от дргреч.:
лека—оболочка, раковина; сома—те-
ло) и безраковинных гимносомат
(Gymnosomata, от дргреч.: гимнов —
обнаженный). Парадоксальным об-
разом, в самое недавнее время
на очередном витке изучения
птеропод снова заговорили о необхо-
димости рассматривать, как и раньше,
обе группы в рамках единого отряда
Pteropoda. Такой «маятник мнений» —
достаточно обычная вещь в таксоно-
мии. Несмотря на подобные колеба-
ния, всегд а есть наиболее устоявшаяся
система, которой придерживается
большинство специалистов.
ОТРЯД THECOSOMATA —
РАКОВИННЫЕ КРЫЛОНОГИЕ МОЛЛЮСКИ
Как вытекает уже из названия этого отряда, относящиеся к этой группе птероподы всегда об-
ладают тонкой наружной раковиной. У некоторых текосомат раковины несут причудливые вы-
росты, каку видов из родов Cavolinia и Diacria. Имеется либо пара крыльев, либо одно широкое
полукруглое крыло, каку видов рода Corolla. Радула присутствует, особые крючья отсутствуют.
Раковинные крылоногие моллюски питаются фитопланктоном. После гибели текосомат их
раковинки опускаются на дно, и в некоторых районах океана грунт почти целиком состоит из
раковин птеропод. К этому отряду относится 8 семейств, из которых в морях России известно
лишь одно.
СЕМЕЙСТВО LIMACINIDAE —
ЛИМАЦИНОВЫЕ
Раковина у этого семейства спирально завитая,
простого строения, без выростов. Имеется пара
хорошо выраженных крыльев. К этому семей-
ству относится единственный род Limacina,
представители которого широко распростра-
нены в умеренных и арктических морях север-
ного полушария.
71
Отряд Thecosomata —Раковинные крылоногие моллюски
Limacina helicina
(Phipps, 1774) -
Лимацина или Морской чертик
Особенности строения. Limacina helicina —
представитель крылоногих моллюсков (Pte-
ropoda). Помимо Limacina и СНопе, к птеропо-
дам относятся еще более ста видов, но живут
они в основном в планктоне теплых морей.
Собственно под названием «крылоногие мол-
люски» скрываются две различные группы
(хотя и родственные друг другу) — текосома-
ты (Thecosomata), или раковинные крылоно-
гие (к ним принадлежит лимацина) и гимно-
соматы (Gymnosomata), безраковинные
крылоногие (сюда относится клион). Тело
Limacina helicina — Лимацина или Морской чертик
Limacina helicina заключено в прозрачную
хрупкую раковинку, спирально завитую
в одной плоскости, так что завиток выдается
очень слабо. Раковина левозавитая. На по-
верхности раковины хорошо заметна осевая
скульптура в виде многочисленных узких,
приподнятых ребер. Характерной чертой
строения лимацины является пара крупных,
расширенных дистально крыльев, в расправ-
ленном состоянии по размеру превышающих
раковину. Длина до 8 мм.
Окраска. Общая окраска черновато-
коричневатая. Просвечивающее сквозь раковину
тело более интенсивного темного оттенка, тогда
как крылья более светлые, коричневатые.
Отличия от похожих видов. В теплые
годы в Белое море заходит другой вид рода
Limacina — Limacina retroversa, который хорошо
отличается от Limacina helicina высокой башен-
ковидной раковиной. Обычно Limacina retro-
versa в Белом море не встречается.
Особенности экологии. Наиболее харак-
терной чертой биологии Limacina helicina, по
крайней мере, в Белом море, является ярко вы-
раженная сезонность появления этого вида.
Практически отсутствуя в планктоне в течение
большей части года, в конце мая — первой по-
ловине июня тысячи особей Limacina helicina
«внезапно» появляются близ поверхности во-
ды. Уже в июле лимацины также внезапно ис-
чезают. Лимацины, подобно клионам (СНопе
limacina), способны совершать вертикальные
миграции в толще воды.
Размножение. У лимацин описано слож-
ное размножение с переопределением пола.
Сроки размножения в Белом море приходятся
на июнь.
Питание и поведение. Столь же необыч-
но, как и их внешний облик, питание лимацин.
Они продуцируют особую ловчую сеть из сли-
зи, размер которой значительно превышает
размеры животного. В эту сеть лимацина ло-
вит различных мелких планктонных животных,
в том числе бактерий и ракообразных. После
того как сеть заполняется, лимацина съедает
ее вместе с пойманными организмами. Стиль
плавания лимацины — быстрый и немного
«дерганный», поскольку взмахи крыльев очень
частые. Основной враг лимацины — крылоно-
гий моллюск клион, плавает гораздо быстрее
ее, и способы защиты от этого хищника у Lima-
cina helicina чисто пассивные.
72
Семейство Clionidae — Клионовые
Limacina helicina — Лимацина или Морской чертик (слева) служит пищей
для Clione limacina — Морского ангела или клиона (справа)
Распространение. Арктическо-бореаль-
ный вид. В Арктике отмечен в Баренцевом,
Карском и Чукотском морях. В северной Ат-
лантике по европейскому побережью на юге
доходит до Бискайского залива, по амери-
канскому — до мыса Кейп-Код. Некоторые
систематики выделяют несколько отдельных
подвидов Limacina helicina, обитающих в се-
верной части Тихого океана. В Белом море,
по-видимому, повсеместно.
ОТРЯД GYMNOSOMATA — БЕЗРАКОВИННЫЕ
КРЫЛОНОГИЕ МОЛЛЮСКИ
У всех представителей этой группы раковина полностью отсутствует. Обычно есть пара хорошо раз-
витых крыльев. Радула имеется, некоторые виды обладают также и челюстями. Около рта у ряда
представителей располагаются два кармана, содержащие особые крючья, которые способны выво-
рачиваться и удерживать жертву. Гимносоматы — хищники, нападающие на раковинных крылоногих
моллюсков. К этому отряду относится 7 семейств, из которых в морях России известно лишь одно.
СЕМЕЙСТВО CLIONIDAE —
КЛИОНОВЫЕ
Представители этого семейства обладают особы-
ми выворачивающимися щупальцами (так назы-
ваемыми буккальными конусами) и мощными
крючьями, которые в неактивном состоянии по-
мещаются внутри особых карманов, открываю-
щимися по бокам от ротового отверстия. Радула
имеется, челюсти обычно отсутствуют. К этому
семейству относится небольшое число родов, из-
вестных как в умеренных, так и тропических во-
дах Мирового океана, но особенно широко рас-
пространенных в арктических морях.
73
Отряд Gymnosomata — Безраковинные крылоногие моллюски
Clione limacina — Морской ангел или клион
74
Семейство Clionidae — Клионовые
Cllone limacina
(Phipps, 1774) -
Морской ангел или клион
Особенности строения. Тело клионов удли-
ненное, крупная голова с двумя щупальцами.
Рот окружен особыми выворачивающимися
щупальцами, которые отсутствуют у других
заднежаберных моллюсков. В средней части
тела располагается пара плавательных кры-
льев. Это видоизмененные боковые края ноги
(параподии), хорошо развитые у заднежабер-
ных моллюсков отряда Anaspidea, которые
и являются предками крылоногих моллюсков.
Между крыльями располагается рудимент
средней части ноги, свернутый в виде треу-
гольной складки. Морские ангелы могут до-
стигать в длину 5 см.
Окраска. Голова и кончик хвоста окра-
шены в розовато-оранжевый цвет. Сквозь
почти прозрачное тело просвечивают вну-
тренние органы — оранжево-красные нерв-
ные узлы, пищеварительная железа и органы
размножения.
Особенности экологии. Морской ан-
гел — исключительно планктонный организм.
В зависимости от условий среды и обилия пи-
щи клионы совершают в толще воды верти-
кальные миграции, то опускаясь почти к само-
му дну, то поднимаясь почти к поверхности.
В тихую погоду на Белом море с лодки нередко
можно наблюдать клионов, плывущих недале-
ко от поверхности воды.
Размножение. Клионы — гермафродиты,
как и почти все заднежаберные моллюски, но
только у них размножение приобретает не-
обычные черты. У клионов созревает сначала
мужская часть половой системы, и два физио-
логических (т. е. временных) самца обменива-
ются спермой, которая поступает в особый
резервуар в основании копулятивного органа.
После этого дозревает и женская часть, и особи
снова, вторично копулируют, причем сперма
партнера поступает уже в другой, женский се-
мяприемник.
Питание и поведение. Принято счи-
тать, что клионы питаются строго опреде-
ленной пищей (такой способ питания назы-
вают монофагией). И хотя имеются старые
данные, что в условиях аквариума клионы
могут поедать, например, мясо северного
оленя, все последующие наблюдения свиде-
тельствуют исключительно в пользу моно-
фагии. Всю свою жизнь морские ангелы пи-
таются другими крылоногими моллюсками,
главным образом представителями рода
Limacina, и прежде всего Limacina helicina (см.
выше). Уже начиная с ювенильной стадии
маленькие клионы нападают на личинок ли-
мацин. Клионы плавают гораздо проворнее
своих жертв, поэтому догнать лимацину им
не составляет труда. Вокруг рта морского ан-
гела расположены особые выворачивающие-
ся щупальца, в основании которых прячутся
мощные крючья. Когда клион настигает ли-
мацину, он мгновенно выбрасывает щупаль-
ца, удерживая ими жертву, а крючья вонзает
в ее тело (см. стр. 73). Лимацина пытается
уплыть и избежать гибели, но ее слабые кры-
лья не способны справиться с мощным захва-
том клиона. Плавая вместе с жертвой, иногда
в течение нескольких часов, морской ангел
извлекает тело лимацины из раковины с по-
мощью крючьев и поедает.
Распространение. Арктическо-бореаль-
ный, циркумполярный вид (т. е. обитающий
как в Северном Ледовитом океане, так и в се-
верных частях Тихого и Атлантического океа-
нов, см. также словарь). Обычен в Белом мо-
ре. В северной Атлантике повсеместно, по
европейскому побережью на юге доходит до
Британских островов, по американскому по-
бережью — до Северной Каролины. Север-
ная часть Тихого океана. Антарктический вид
Clione antarctica иногда считают подвидом
Clione limacina.
| В массовых количествах лимацины появля-
ются в Белом море на очень короткий пери-
од, в июне-июле, после чего исчезают совер-
шенно, тогда как клионы встречаются в разном
количестве на протяжении всего года. Поэтому
до сих пор остается неясным, чем питаются
клионы большую часть года, когда лимацин
практически нет. Возможно они разыскивают
отдельных лимацин, а возможно все-таки пере-
ходят на питание другими объектами. Недавно
было выдвинуто предположение о способности
клиона к исключительно эффективному запаса-
нию и экономному расходу питательных ве-
ществ, позволяющей ему голодать многие ме-
сяцы в ожидании появления лимацин.
75
• Эволюция, шаг третий •
БЕГСТВО ПОД ЗЕМЛЮ
ОТРЯД ACOCHLIDIIDA (=ACOCHLIDIACEA) —
АКОХЛИДИИДЫ
Пожалуй, одна из самых необычных групп заднежаберных моллюсков, во внешнем строении которой
остается очень мало «следов» их раковинных предков. Тело акохлидиид четко разделено на два от-
дела: мощный висцеральный мешок и удлиненную узкую головную лопасть. Голова несет одну или
две пары простых щупалец: оральные щупальца и ринофоры. Раковина у акохлидиид полностью
отсутствует. Вместо этого висцеральный мешок у многих видов содержит в покровах большое коли-
чество особых известковых иголочек, называемых спикулами. Покров из спикул отчасти заменяет
раковину, выполняя функцию защитного чехла. Нога акохлидиид настолько редуцирована, что прак-
тически не принимает участие в локомоции, а мощный висцеральный мешок, можно сказать, «во-
лочится» позади животного, если моллюск передвигается по твердой поверхности. Однако, в той
необычной среде, в которой обитают акохлидииды — а это пространство между песчинками в толще
грунта, называемое интерсгициаль, этот кажущийся недостаток превращается в преимущество.
Поскольку среда интерстициали подразумевает наличие некоторого узкого пространства,
то частицы грунта, которые эту среду формируют, должны лежать рыхло, оставляя своего рода
щели между отдельными песчинками. Именно по этой причине более знакомые тонкозерни-
стые пески и илы, размер частиц которых весьма мал, плохо подходят на роль среды для интер-
стициали. Наилучшими же для нее будут грубозернистые пески, с большим размером состав-
ляющих их частиц, которые населены в том числе и акохлидиидами. И их червеобразная форма
тела, с длинным висцеральным мешком, обременительным на твердых поверхностях, стала иде-
альной при передвижении в узких пространствах.
Акохлидииды, помимо «странностей» внешнего строения, характеризуются целым рядом других
необычных черт. Так, среди них встречаются раздельнополые виды, в то время как остальные задне-
жаберные моллюски в подавляющем своем большинстве гермафродитны. Помимо этого, акохли-
дииды на фоне всех заднежаберных выделяются и тем, что несколько видов среди них сумели освоить
настоящие пресные водоемы, а не только приустьевые участки моря с пониженной соленостью. Все-
го известно более полусотни видов акохлидиид, объединяемых в несколько семейств.
Обычному ныряльщику, дайверу,
вряд ли часто приходит в голову,
что все те широко рекламируемые
красоты коралловых рифов или
скалистых ландшафтов, покрытые
несметным количеством морских
ежей и голотурий, занимают лишь
ничтожную часть от общей площади
дна Мирового океана. Большую же
его часть занимают совершенно
«скучные» мягкие грунты — пески,
илы и им подобные, заселенные бо-
гатейшей фауной, в состав которой
входят почти все известные типы
беспозвоночных. Среди обитателей
мягких грунтов есть и гигантские
черви-эхиуриды, и чрезвычайно
многочисленные многощетинковые
черви средних размеров, двуствор-
чатые моллюски и заднежаберные
моллюски-цефаласпиды того же
среднего калибра. И наконец, есть
организмы, масштаб которых таков,
что отдельная песчинка для них
примерно то же, что для человека
жилой дом небольших размеров.
Вот эти организмы принято назы-
вать интерстициальными. Акох-
лидииды, о которых идет здесь
речь, относятся как раз к этой эко-
логической группе. Они нашли
свою нишу и весьма успешно в ней
существуют.
76
Семейство Parhedylidae — Пархедилиды
| Группа акохлидиид является особой и для
истории российской науки. Дело в том, что
честь открытия мелких интерстициальных акох-
лидиид (крупные пресноводные акохлидииды
были открыты ранее) принадлежит выдающему-
ся российскому морфологу и эмбриологу Алек-
сандру Онуфриевичу Ковалевскому. Свои от-
крытия Ковалевский сделал на берегах Черного
моря, работая на знаменитой Севастопольской
биологической станции. Для того чтобы обнару-
жить акохлидиид, Ковалевский разработал про-
стой, но очень эффективный метод исследова-
ния грунта, который применяется до сих пор.
А грунт из Черного моря, в котором и были обна-
ружены уникальные, ни на что не похожие мол-
люски, был добыт специально для Ковалевского
военным кораблем при содействии вице-
адмирала С. П. Тыртова. Эта на первый взгляд
незначительная деталь прекрасно иллюстрирует
то уважительное отношение к науке, которое су-
ществовало в прежней России и которое посте-
пенно возрождается и в наше время. Детальное,
ставшее классическим, описание двух новых
видов акохлидиид из Черного моря Ковалевский
опубликовал на заре XX века, в 1901 году. Пара-
доксальным образом в советское время это от-
крытие было предано забвению, и за минувшее
столетие никто не опубликовал новых находок
акохлидиид из Черного моря. К большому сожа-
лению, в мире сейчас редко вспоминают об от-
крытиях Ковалевского, хотя акохлидиидами за-
нимаются довольно активно.
За прошедшие несколько лет мы приложили
немало усилий для того, чтобы исправить это
положение. Нами были подробно с примене-
нием легководолазной техники исследованы
грубозернистые пески вокруг Севастополя, и,
в конце концов, было подтверждено суще-
ствование в Черном море обоих видов акох-
лидиид, описанных Ковалевским!
СЕМЕЙСТВО PARHEDYLIDAE —
ПАРХЕДИЛИДЫ
Представители этого семейства обладают двумя
парами хорошо выраженных длинных щупа-
лец. Висцеральный мешок удлиненный. Глаза
имеются. Морские формы. К этому семейству
относится небольшое число родов, обитающих
в субтропических и тропических водах.
Parhedyle tyrtowii
(Kowa levsky, 1900) —
П архед иле Тыртова
Особенности строения. Висцеральный ме-
шок мощный, широкий и длинный, изогну-
тый. Имеется две пары длинных щупалец —
ринофоров и оральных щупалец. Спикулы не
заметны. Длина до 3 мм.
Окраска. Тело почти однородно молочное
белое, другие элементы окраски практически
не выделяются.
Отличия от похожих видов. От остальных
акохлидиид, известных в фауне России (вклю-
чая другой вид, достоверно известный из Чер-
ного моря, Pontohedyle milaschewitschi), хорошо
отличается мощным, загнутым висцеральным
мешком, молочно-белой однородной окраской
и двумя парами длинных щупалец.
Особенности экологии. Обитает в грубо-
зернистом песке в сублиторали.
Размножение. Не изучено. По-видимому,
откладывает маленькие сплюснутые кладки,
прикрепляя их к поверхности частиц грунта.
Питание и поведение. Активный, под-
вижный вид. Передвигается между частицами
грунта. Питание не изучено.
Распространение. По данным Ковалевско-
го, вид в начале XX века был довольно обычен
в Черном море в районе Севастополя. Но на
основании наших материалов, полученных лишь
в самое недавнее время, вид является очень ред-
ким и после долгих и утомительных поисков был
обнаружен единственный экземпляр (!). Данный
вид назван Ковалевским в честь вице-адмирала
русского флота С. П. Тыртова.
Parhedyle tyrtowii — Пархедиле Тыртова,
вид с правой стороны
Отряд Acochlidiida (=Acochlidiacea) — Акохлидииды
Parhedyle tyrtowii — Пархедиле Тыртова,
вид со спинной стороны
Parhedyle tyrtowii — Пархедиле Тыртова, головной
отдел частично втянут в висцеральный мешок
Pontohedyle milaschewitschi —
Pontohedyle milaschewitschi —
Понтохедиле Милошевича, вид со спинной стороны
Понтохедиле Милошевича, вид со спинной стороны
78
Семейство Asperspinidae — Асперспиниды
СЕМЕЙСТВО MICROHEDYLIDAE —
МИКРОХЕДИЛИДЫ
Виды из этого семейства обладают лишь одной
парой уплощенных оральных щупалец. Висце-
ральный мешок удлиненный. Глаза имеются.
Морские формы. К этому семейству относится
небольшое число родов, обитающих в суб-
тропических и тропических водах.
Pontohedyle milaschewitschi
(Kowalewsky, 1901) —
Понтохедиле Милошевича
Особенности строения. Висцеральный ме-
шок мощный, широкий и длинный, изогну-
тый. Имеется только одна пара уплощенных
оральных щупалец, формирующая харак-
терный молотообразный облик передней
части головы. Спикулы не заметны. Длина
тела до 3 мм.
Окраска. Тело полупрозрачно-белова-
тое. Сквозь висцеральный мешок просвечи-
вают зеленовато-коричневатые внутренности.
На голове располагаются небольшие белые
пятнышки.
Pontohedyle milaschewitschi —
Понтохедиле Милошевича, вид с правой стороны
Отличия от похожих видов. От других
акохлидиид, известных в фауне России, хоро-
шо отличается наличием только одной пары
головных отростков — уплощенных оральных
щупалец.
Особенности экологии. Обитает в грубо-
зернистом песке в сублиторали.
Размножение. Не изучено. По-видимому,
откладывает маленькие сплюснутые кладки,
прикрепляя их к поверхности частиц грунта.
Питание и поведение. Активный, по-
движный вид. Может активно передвигаться
между частицами грунта. Питание вида не
изучено.
Распространение. Данный вид относи-
тельно нередок в Черном море в районе Севасто-
поля, в то время как в других районах отмечен
не был. Кроме того, вид широко распространен
в Средиземном море, обнаружен и в теплово-
дных районах Атлантического океана. Вид на-
зван А. О. Ковалевским в честь К. О. Милаше-
вича, крупного отечественного специалиста по
раковинным моллюскам, работавшего в нача-
ле XX века.
СЕМЕЙСТВО ASPERSPINIDAE —
АСПЕРСПИНИДЫ
Представители данного семейства обладают
двумя парами коротких щупалец. Висцераль-
ный мешок укороченный. Глаза у большинства
видов отсутствуют. Морские формы. К этому
семейству относится лишь два рода. Большая
часть видов обитает в умеренных и субтропи-
ческих водах.
Asperspina murmanica
(Kudinskaja et Minichev, 1978) —
Асперспина мурманская
Особенности строения. Висцеральный ме-
шок относительно узкий и короткий. Имеется
две пары коротких щупалец — ринофоров
и оральных щупалец. Сквозь мешок заметен
довольно плотный слой спикул. Длина тела
до 3 мм.
Окраска. Тело моллюска полупрозрач-
но-беловатое. Сквозь висцеральный мешок
просвечивает розовато-красная пищевари-
тельная железа.
79
Отряд Acochlidiida (=Acochlidiacea) — Акохлидииды
Отличия от похожих видов. От других
акохлидиид, известных в фауне России, хоро-
шо отличается сочетанием относительно ко-
роткого висцерального мешка, окраски и на-
личия двух пар коротких щупалец. Кроме того,
на Мурманском побережье Баренцева моря,
другие акохлидииды, кроме Asperspina тиг-
manica, отсутствуют.
Особенности экологии. Обитает в грубо-
зернистом песке на литорали.
Размножение. Не изучено. По-видимому,
откладывает маленькие сплюснутые кладки,
прикрепляя их к поверхности частиц грунта.
Питание и поведение. Активный, подвиж-
ный вид. Может активно передвигаться между
частицами грунта. Питание не изучено.
Распространение. Данный вид впервые
обнаружен на Мурманском побережье Барен-
цева моря, и до сих пор нигде за его пределами
не найден.
Asperspina murmanica — Асперспина мурманская,
вид со спин нои стороны
Asperspina murmanica — Асперспина мурманская,
вид с брюшной стороны
Asperspina murmanica — Асперспина мурманская, вид с правой стороны
80
• Эволюция, шаг четвёртый •
ПЕРВАЯ ПОПЫТКА
ОТРЯД SACOGLOSSA — САКОГЛОССЫ
Представители этого отряда, несмотря на внешнее сходство с голожаберными моллюсками, воз-
никли от них независимо, от архаичных раковинных заднежаберных моллюсков, демонстрируя
яркий пример параллельной эволюции. Под термином параллельная эволюция в биологии
подразумевается независимое возникновение сходных признаков у не родственных напрямую
групп. В отличие от голожаберных моллюсков, основная специализация сакоглосс — питание
содержимым клеток водорослей-макрофитов, преимущественно зеленых и красных. Для этого
у сакоглосс радула видоизменилась в набор узких кинжаловидных зубов, а дорсальная часть глот-
ки превратилась в помпу, высасывающую сквозь отверстия, проделанные радулой, содержимое
водорослей.
От голожаберных моллюсков — эолид, с которыми можно спутать сакоглосс, они хорошо
отличаются наличием только одной пары щупалец — ринофоров, тогда как у эолид еще име-
ются и оральные щупальца.
| Подобно голожаберным моллюскам-
эолидам, использующим чужие стрека-
тельные клетки, сакоглоссы используют для
своих целей хлоропласты водорослей, попа-
дающих в их организм вместе с едой. Здесь
опять имеет место случай удивительного па-
раллелизма с голожаберными моллюсками —
пищеварительная железа сакоглосс состоит
из разветвленной системы каналов, дающих
отростки в спинные папиллы. Внутри папилл,
в этих выростах и накапливаются хлоропласты,
которые могут использоваться сакоглоссами
как дополнительный источник энергии (хлоро-
пласты при этом сохраняют способность фото-
синтезировать внутри чужого организма!).
СЕМЕЙСТВО LIMAPONTIIDAE —
ЛИМАПОНЦИЕВЫЕ
Одно из самых крупных семейств безрако-
винных сакоглосс. Большинство представи-
телей несет на спинной поверхности разное
количество удлиненных папилл. Внутрь
каждой папиллы заходит по выросту пище-
варительной железы, и кроме того, отростки
одной из желез репродуктивного аппарата
в виде тонких беловатых веточек. Имеется
небольшое число видов, которые полностью
лишены спинных папилл, но сохраняют об-
щие черты внутреннего строения с теми ви-
дами, у которых папиллы хорошо развиты.
Подобные лишенные папилл представите-
ли, относящиеся к роду Limapontia, широко
распространены в литоральных «лужах» Ба-
ренцева моря: отмечены также и для Белого
моря.
Alderia modesta
(Loven, 1844) -
Алдерия скромная
Особенности строения. Тело довольно ши-
рокое. Большая часть спины свободная, по
краям располагаются короткие и длинные
пальцевидные папиллы. На голове имеется
пара коротких треугольных ринофоров.
Оральные щупальца отсутствуют. Длина до
10 мм.
Окраска. Общая окраска тела пестрая —
оливковая, зеленоватая, коричневатая. Выро-
сты пищеварительной железы в папиллах
очень светлые, и на фоне пестрого тела почти
не заметны.
81
Отряд Sacoglossa — Сакоглоссы
Alderia modesta — Алдерия скромная,
вид со спинной стороны
Alderia modesta — Алдерия скромная,
вид с брюшной стороны
Отличия от похожих видов. От другой,
обычной в Белом и Баренцевом морях сако-
глоссы — Limapontia senestra данный вид хоро-
шо отличается широким телом и наличием
папилл на дорсальной стороне.
Особенности экологии. Alderia modesta
тесно связана с зеленой нитчатой водорослью
Vaucheria. В эстуариях достигает значительной
численности — более 1000 экз/м2, причем
плотность поселений этого вида, зарегистри-
рованная в северной Норвегии, достигала
9 000 экз/м2. В Белом море, возможно, являет-
ся довольно обычным видом в опресненных
Alderia modesta — Алдерия скромная, кладка
приустьевых участках, но до настоящего вре-
мени имеются лишь отдельные находки. При
нормальной солености обычно удается найти
лишь немногочисленные экземпляры Alderia
modesta.
Размножение. На Баренцевом море от-
мечено в августе. Кладки в виде удлиненных
вздутых шнуров моллюски откладывают на зе-
леные нитчатые водоросли. Из кладок могут
выходить велигеры двух типов — плавающие
или ползающие.
Питание и поведение. Питается содер-
жимым клеток нитчатой водоросли Vaucheria.
Подвижный вид.
Распространение. Амфибореальный вид.
Распространен в северной Атлантике от Нор-
вегии до Франции, известен вдоль Мурманско-
го побережья Баренцева моря, по тихоокеан-
скому побережью Северной Америки, изредка
обнаруживается в заливе Петра Великого
Японского моря. В Белом море достоверно об-
наружен пока в эстуарии реки Малая Кудьма
в г. Северодвинск, а также у Соловецких остро-
вов, но возможно распространен в эстуариях
повсеместно.
82
Семейство Limapontiidae — Лимапонциевые
Limapontia senestra — Лимапонция сенестра,
вид со спинной и с правой стороны
Limapontia senestra
(Quatrefages, 1844) —
Лимапонция сенестра
Синоним:
Limapontia cocksi (Alder et Hancock, 1848)
Особенности строения. Тело довольно узкое,
достаточно высокое и полностью лишено каких-
либо выростов-папилл. Ринофоры у взрослых
особей пальцевидные, у молодых — треуголь-
ные, очень короткие. Отверстие копулятивного
аппарата расположено на голове, позади пра-
вого глаза. За ринофорами хорошо заметны
черные глаза. Длина до 6 мм.
Окраска. Общая окраска тела от темно-
коричневой до почти черной. Область вокруг
глаз и внешняя поверхность ринофоров ли-
шена пигмента и образует пару характерных
пятен на голове. В задней части спины вокруг
ануса также имеется небольшое беловатое
пятнышко.
Отличия от похожих видов. Близкий вид
Limapontia depressa, живет исключительно на
илистой или песчаной литорали в Баренцевом
море, и в Белом море не обнаружен.
Особенности экологии. Живет главным
образом в литоральных лужах, на каменистой
и скалистой литорали, где нередко можно
обнаружить тысячи экземпляров Limapontia
senestra.
Размножение. В Белом море размноже-
ние отмечено в июле-августе. Откладывают
кладки в виде шаровидных полупрозрачных
мешочков на зеленых нитчатых водорослях.
Недавно отложенные кладки содержат жел-
тые яйца, в то время как кладки с готовыми
Limapontia senestra — Лимапонция сенестра,
кладка на пищевом объекте — зеленой водоросли
83
Отряд Sacoglossa — Сакоглоссы
к вылуплению зародышами имеют буровато-
черный цвет. На свет они появляются уже
полностью сформированные, отличаясь от
взрослых в основном размерами (т. е. разви-
тие прямое).
Питание и поведение. Высасывают содер-
жимое зеленых нитчатых водорослей.
Чрезвычайно подвижный вид, активно
ползающий среди зеленых нитчатых водо-
рослей. Интересно брачное поведение этого
вида: готовые к копуляции особи приближа-
ются к друг другу с вывернутыми копулятив-
ными аппаратами, и резким движением пе-
ниального стилета впрыскивают сперму
партнеру под кожу в специальное вздутие на
правой стороне тела.
Распространение. Атлантический боре-
альный вид. Распространен в северной Атлан-
тике от Норвегии до северной Испании, а так-
же на Мурманском побережье Баренцева моря.
Отмечен в Белом море. Близкий вид Limapontia
capitate был указан из Черного моря, но изобра-
жений черноморских особей в нашем распоря-
жении пока нет.
Ercolania boodleae — Эрколания Сюдлея,
вид со спи и нои стороны
Ercolania boodleae
(Baba, 1938) -
Эрколания бодлея
Особенности строения. Тело довольно узкое.
Большая часть спины покрыта длинными вере-
теновидными папиллами. На голове распола-
гается пара длинных, свернутых в трубку рино-
форов. Оральные щупальца отсутствуют.
Длина до 15 мм.
Окраска. Общая окраска почти черная,
включая спинные папиллы. Вырост пищевари-
тельной железы в папиллах плохо заметен
сквозь темный фон. На вершине каждой па-
пиллы находится яркое оранжевое пятно.
Отличия от похожих видов. По особен-
ностям окраски и строения данный вид хо-
рошо отличается от всех сакоглосс фауны
России.
Особенности экологии. Данный вид
обитает преимущественно на своеобразной
зеленой водоросли хетоморфа — Chaetomorpha
linum, представляющей собой длинные нити,
состоящие из многочисленных коротких взду-
тий, разделенных желобками, напоминаю-
щих четки.
Размножение. Кладки в виде удлиненных
вздутых шнуров, нередко завитых в виде непра-
вильных спиралей, моллюски откладывают на
поверхность камней и на скопления зеленых
водорослей.
Питание и поведение. Питается содер-
жимым клеток зеленых водорослей. Подвиж-
ный вид.
Распространение. По-видимому, боре-
ально-субтропический вид. Довольно часто
встречается в заливе Петра Великого Японско-
го моря, обнаружен в заливе Анива (Южный
Сахалин). Отмечен в водах Японии.
Ercolania viridis
(A. Costa, 1866) —
Эрколания зеленая
Особенности строения. Тело довольно узкое.
Большая часть спины покрыта длинными вере-
теновидными папиллами. На голове распола-
гается пара длинных ринофоров. Оральные
щупальца отсутствуют. Длина до 6 мм.
Окраска. Общая окраска тела варьирует
от беловатой до темной, почти черной, вклю-
84
Семейство Limapontiidae — Лимапонциевые
Ercolania viridis — Эрколания зеленая, со сшитой и с брюшной стороны
Ercolania viridis — Эрколания зеленая, кладка
Ercolania viridis — Эрколания зеленая,
ювенильная особь со сннннон стороны
Ercolania viridis — Эрколания зеленая,
пищевой объект — зеленая нитчатая водоросль
85
Отряд Sacoglossa — Сакоглоссы
чая спинные папиллы. Найденные нами осо-
би были довольно мелкими, и все со светлым
фоном окраски, но у некоторых особей па-
пиллы были покрыты темным пигментом.
Вырост пищеварительной железы в папиллах
зеленоватый или красноватый.
Отличия от похожих видов. По особен-
ностям окраски и строения данный вид хо-
рошо отличается от других сакоглосс, из-
вестных в Черном море, где из безраковинных
сакоглосс был ранее отмечен только один
вид — Calliopaea bellula, у которого полно-
стью отсутствует зеленая или черная пиг-
ментация.
Особенности экологии. Данный вид
можно обнаружить среди различных водорос-
лей, в том числе зеленых.
Размножение. Кладки в виде удлиненных
вздутых шнуров, нередко завитых неправиль-
ными спиралями, моллюски откладывают на
скопления зеленых водорослей.
Питание и поведение. Питается содер-
жимым клеток зеленых водорослей. Подвиж-
ный вид.
Распространение. Атлантический суб-
тропическо-бореальный вид. Широко распро-
странен по Европейскому побережью, отмечен
у Восточного побережья США и в Карибском
бассейне. Встречается у Крымского побережья
Черного моря.
Placida dendritica
(Alder et Hancock, 1843) —
Плацида дендритика
Особенности строения. Тело умеренно ши-
рокое. Большая часть спины покрыта длинны-
ми веретеновидными папиллами. На голове
располагается пара длинных, свернутых в труб-
ку ринофоров. Оральные щупальца отсутству-
ют. Длина до 10 мм.
Окраска. Общая окраска беловатая, на го-
лову и ринофоры заходят многочисленные вет-
ви пищеварительной железы темно-зеленого
или реже желтовато-коричневатого цвета.
Вырост пищеварительной железы в папиллах
сходного цвета. На поверхности папилл раз-
бросаны многочисленные мелкие белые пят-
нышки и крапинки.
Отличия от похожих видов. По особен-
ностям окраски и строения данный вид хоро-
Placida dendritica — Плацида дендритика
(летняя форма), вид со спинной и брюшной стороны
86
Семейство Hermaeidae — Гермаеиды
Placida dendritica — Плацида дендритика
(зимняя форма), вид со спинной стороны
шо отличается от большинства сакоглосс
фауны России. Единственный вид, с которым
его можно спутать, это Hermaea vancouverensis,
от которой Placida dendritica отличается мощ-
ными разветвленными ветвями пищевари-
тельной железы, просвечивающей сквозь
спинную сторону. Кроме того, Placida dendri-
tica связана с зелеными водорослями, такими
как ульва (Ulva) или кодиум (Codium), тогда
как Hermaea vancouverensis обитает на специа-
лизированной красной водоросли родомела
(Rhodomela).
Особенности экологии. Обитает от лито-
рали до глубины 2-5 м, на различных зеленых
водорослях, прежде всего на крупных слоеви-
щах ульвы.
Размножение. Кладки в виде удлинен-
ных вздутых шнуров, нередко завитых в виде
неправильных спиралей, моллюски отклады-
вают на поверхность слоевищ зеленых водо-
рослей.
Питание и поведение. Питается содер-
жимым клеток зеленых водорослей. Подвиж-
ный вид.
Распространение. Амфибореальный
вид. По Европейскому побережью доходит до
Средиземного моря, отмечен на Восточном
побережье США. В Тихом океане обитает по
американскому побережью от Британской
Колумбии (Канада) до Калифорнии. В морях
России обнаружен в Белом и Баренцевом мо-
рях (где очень редок) и в Японском море (где
встречается чаще).
СЕМЕЙСТВО HERMAEIDAE —
ГЕРМАЕИДЫ
Близкие по внешнему строению к предше-
ствующему семейству гермаеиды отличают-
ся от него в основном лишь тонкими дета-
лями репродуктивного аппарата. Так же,
как и лимапонциевые, это семейство вклю-
чает значительное число видов, покрытых
папиллами и обладающих значительным
внешним сходством с голожаберными мол-
люсками. В морях России известен только
один вид.
Hermaea vancouverensis
(O’Donoghue, 1924) —
Гермаеа ванкуверская
Возможный синоним:
Stiliger akkeshiensis Baba, 1935
Особенности строения. Тело относительно
узкое. Большая часть спины свободная, по
краям располагаются довольно длинные ве-
ретеновидные папиллы. На голове распола-
гается пара длинных, свернутых в трубку ри-
нофоров. Оральные щупальца отсутствуют.
Длина до 5 мм.
Окраска. Общая окраска беловатая, на го-
лове имеется характерное треугольное пятно,
очерченное двумя тонкими линиями. Вырост
пищеварительной железы в папиллах темно-
зеленый.
Отличия от похожих видов. Можно спу-
тать с другим видом сакоглосс, Placida dendritica,
у которого также имеются сходные зеленова-
тые папиллы. Hermaea vancouverensis заметно
отличается от Placida dendritica менее развет-
вленной пищеварительной железой, просвечи-
вающей сквозь спинную сторону. Кроме того,
87
Отряд Sacoglossa — Сакоглоссы
Placida dendritica связана с зелеными водорос-
лями, тогда как Hermaea vancouverensis обитает
на красной водоросли родомеле (Rhodomela).
Развитие со стадией свободно плавающей
личинки-велигера.
Особенности экологии. Hermaea vancouve-
rensis тесно связана с красной водорослью Rho-
domela. Эта водоросль похожа на небольшие
кустики и очень широко распространена на
литорали Дальневосточных морей России. Her-
maea vancouverensis также обнаружена только на
литорали.
Размножение. Кладки в виде удлиненных
вздутых шнуров моллюски откладывают на
красную водоросль Rhodomela.
Питание и поведение. Питается содер-
жимым клеток красной водоросли Rhodomela.
Подвижный вид.
Распространение. Тихоокеанский боре-
альный вид. Широко распространен вдоль аме-
риканского побережья от Аляски до Калифор-
нии. В морях России обнаружен только у двух
самых крупных Южных Курильских остро-
вов — Кунашира и Итурупа.
Hermaea vancouverensis — Гермаеа ванкуверская,
вид со спинной стороны
Белое море. Литораль
88
• Эволюция, шаг пятый •
ГЕНИАЛЬНЫЙ НЕУДАЧНИК
ОТРЯД NOTASPIDEA (= PLEUROBRANCHOIDEA) —
НОТАСПИДЫ
Этот отряд — подлинная реликтовая группа в современной фауне. Если бы волею эволюционных
судеб нотаспиды полностью вымерли, происхождение голожаберных моллюсков оставалось бы для
нас если не полной, то очень трудноразрешимой загадкой. Дело в том, что по особенностям внутрен-
него строения, прежде всего по строению репродуктивного аппарата, нотаспиды практически не от-
личаются от голожаберных моллюсков. Некоторые особенности внешнего строения, в частности на-
личие у нотаспид мощного спинного кожного щита, покрывающего все тело вместо настоящей мантии,
идентичны таковым у голожаберных моллюсков группы дорид. На основании этих и некоторых дру-
гих подобных фактов было даже предложено либо включить нотаспид в отряд голожаберных моллю-
сков (Nudibranchia), либо объединить Notaspidea и Nudibranchia в новую группу Nudipleura.
В то же время у нотаспид еще остается целый ряд признаков внешнего и внутреннего строе-
ния, объединяющих их с раковинными заднежаберными моллюсками. Прежде всего, это наличие
у нотаспид внутренней пластинковидной рудиментарной раковины, скрытой под кожей. Кроме
этого, у них имеется одна крупная жабра, располагающаяся, как и у цефаласпид, с правой сто-
роны тела. Но мантийная полость у нотаспид, в отличие от цефаласпид, уже полностью исчезла,
и жабра прикрепляется непосредственно к боковой стенке тела. Таким образом, нотаспиды яв-
ляются важным эволюционным связующим звеном между безраковинными голожаберными
моллюсками и одной из древних групп раковинных заднежаберных моллюсков — цефаласпида-
ми. Не исключено, что предковой группой для нотаспид, а значит, и всех «бесконечно разно-
образных» голожаберных моллюсков были актеониды, еще обладающие хорошо развитой
башенковидной раковиной (см. выше). У актеонид, в отличие от большинства цефаласпид,
копулятивный аппарат располагается не на голове, а рядом с женским генитальным отверсти-
ем, поблизости от жабры, т. е. там же, где и у современных нотаспид.
К нотаспидам относится всего лишь около ста видов. Обеспечив своим возникновением всю
последующую бурную эволюцию голожаберных моллюсков (число которых, по самым скром-
ным подсчетам, превышает три тысячи видов), они остались в современной фауне крошечной
реликтовой группой. Гениальным неудачником, без которого не появились бы последующие
«вершины эволюции».
Нотаспиды обитают как в умеренных, так и в тропических водах, имеются также антаркти-
ческие и глубоководные представители. В фауне России обнаружены представители трех видов,
из обоих семейств нотаспид.
СЕМЕЙСТВО PLEUROBRANCHIDAE —
ПЛЕУРОБРАНХУСОВЫЕ
ИЛИ ПЛЕУРОБРАНХИДЫ
Крупнейшее семейство нотаспид. Характеризу-
ется ринофорами, объединенными основания-
ми и впереди соединенными с оральным пару-
сом небольшого размера. Всегда имеется
довольно крупная внутренняя раковина. Нотум
хорошо развит, накрывает ноту, ринофоры
и единственную крупную жабру. У ряда видов
на жабре располагаются особые бугорки. Челю-
сти хорошо развиты, состоят из многочислен-
ных удлиненных крестообразных микроскопи-
89
Отряд Notaspidea (= Pleurobranchoidea) — Нотаспиды
Ж
22
£3
Japonacteon nipponensis
Tylodina fungina_______
Bathydoris clavigera
Bathydoris spiralis____
Cadlina laevis
Archidoris pseudoargus
Goniodoris castanea
Doridoxa ingolfiana
Tochuina gigantea
Goniaeolis atypica
61 791 Pleurobranchus areolatus — Pleurobranchus membranaceus Pleurobranchus grandis Pleurobranchus testudinarius
66
52
22
Boreoberthella augusta
Bathyberthella antarctica
Bathyberthella orcadenis
Bathyberthella tomasi
Bathyberthella zelandiae
— Berthella californica
Berthella canariensis
Berthella martensi
Berthella platei
Berthella plumula
Berthella sideralis
Berthella stellata
Pleurehdera haraldi
Berthellina citrina
1&rth£lliBfLe&WLr4sl_
Tomthompsonia antarctica
Pleurobranchella nicobarica
Pleurobranchaea californica
Euselenops luniceps
os
RM
Z
я
S
’5
Пример современного представления эволюционного процесса
с использованием формальных компьютерных методов: эволюционное дерево отряда Notaspidea
и положение на нем нового рода Boreoberthella (отмечено красной стрелкой).
Глубоководный недавно открытый род нотаспид Boreoberthella (красная стрелка)
является ближайшим родственником мелководного тропического рода Pleurobranchus (синяя стрелка).
Из работы Martynov & Schrodl, опубликованной в 2008 году
90
Семейство Pleurobranchidae — Плеуробранхусовые или Плеуробранхиды
ческих элементов. Радула мощная, состоит из
значительного числа рядов зубов. Каждый ряд,
в свою очередь, может насчитывать у некоторых
видов до нескольких сотен зубов.
В тропиках из этого семейства довольно
обычны крупные, приметные моллюски из ро-
да Pleurobranchus, в бореальных водах чаще
можно встретить представителей рода Berthella,
а в водах Антарктиды обитает особый род Bathy-
berthella. Наконец, совсем недавно, в северной
части Тихого океана нами был обнаружен но-
вый глубоководный род и вид, который был на-
зван Boreoberthella augusta. В результате эволюци-
онного анализа этого рода оказалось, что он
наиболее вероятно является ближайшим род-
ственником тропического рода Pleurobranchus,
хотя и значительно отличается от него по внеш-
ним признакам. Этот новый род — первый глу-
боководный представитель нотаспид, обнару-
женный в северной части Тихого океана.
Формирование настоящих голожа-
берных моллюсков обусловлено
всего несколькими основными эво-
люционными процессами, проте-
кавшими внутри предковой группы
нотаспид. Среди таких процессов
нужно в первую очередь выделить
следующие: 1. Отделение ринофо-
ров от орального паруса. 2. Пре-
вращение ринофоров из трубко-
видных в цельные. 3. Полная
редукция раковины. 4. Модифика-
ция единственной боковой жаб-
ры — ее перемещение на спинную
сторону или полная редукция.
Именно указанные выше преобра-
зования основных внешних органов
нотаспид и привели к формирова-
нию голожаберных моллюсков.
Какие доказательства можно приве-
сти в пользу реальности подобного
хода эволюции? Неоценимую по-
мощь здесь нам может оказать уже
подробно разобранный выше ана-
лиз онтогенетических (или жиз-
ненных) циклов той или иной группы.
Ринофоральный аппарат, преобра-
зования которого занимали ключе-
вое место при формировании голо-
жаберных моллюсков, еще весьма
архаичен у нотаспид и состоит из
двух свернутых в трубку пластинок,
соединенных с оральным парусом,
напоминая в целом видоизменен-
ный головной щит цефаласпид. за-
дние края которого «загнулись»
и сместились вперед, образовав
трубчатые ринофоры (стр. 92А,
электронно-микроскопическое изо-
бражение). У голожаберных моллю-
сков, напротив, ринофоры цельные
и полностью отделены от орального
паруса (стр. 92C-D). Другая харак-
терная особенность ринофоров но-
таспид — то, что они объединены
своими основаниями, сильно сбли-
жены, тогда как, например, у всех
дорид ринофоры всегда значитель-
но разделены. При этом, как все
классические эволюционные иссле-
дования. так и самый современный
анализ с использованием молеку-
лярных данных. всегда представляет
голожаберных моллюсков (включая
дорид) в качестве потомков нота-
спид, а не наоборот. В этом случае
в онтогенетическом цикле дорид
должны были сохраняться те или
иные следы взрослых черт строения
их предков — нотаспид. И исследо-
вания индивидуального развития
дорид, в том числе специально про-
веденные нами с использованием
сканирующего электронного микро-
скопа, однозначно свидетельствуют,
что подобные следы — реально су-
ществующий факт. Так взрослые до-
риды, например, широко распро-
страненная в Белом море Cadlina
laevis, во взрослом сотоянии имеют
по два полностью замкнутых рино-
форальных кармана (в каждом рас-
полагается по ринофору), не соеди-
ненных друг с другом и с оральным
парусом (стр. 92С). В то же время
ранние ювенильные стадии этого
же вида (Cadlina laevis) демонстри-
руют принципиально иное устрой-
ство ринофорального аппарата:
ринофоры сближены своими осно-
ваниями. карманов не имеют, впе-
реди смыкаются с небольшим
оральным парусом (стр. 92 В). Та-
ким образом, строение рифоноров
у личинок дорид почти идентично
таковому у взрослых нотаспид! За
тем исключением, что трубковид-
ная. складчатая форма ринофоров
нотаспид уже не проявляется у до-
рид даже на личиночных стадиях
и полностью «стерлась» из их эво-
люционной истории. Но основной
принцип строения ринофоров ли-
чинок дорид и взрослых нотаспид
практически одинаков.
Это не что иное, как рекапитуля-
ция (появление признаков пред-
ков на одной из ранних стадий
жизненного цикла потомков),
мнимое отсутствие которой не
раз ставилось «в вину» основате-
лю онтогенетического метода, вы-
дающемуся немецкому биологу
Эрнсту Геккелю. Не меньше вни-
мания взаимодействию онтогене-
за и эволюции уделяла и отече-
ственная школа эволюционной
морфологии, блестящим пред-
ставителем которой был Алексей
Николаевич Северцов. Но в совре-
менной науке, где предпочтение
зачастую отдается формальному
компьютерному анализу, для срав-
нительного изучения жизненных ►
91
Отряд Notaspidea (= Pleurobranchoidea) — Нотаспиды
Изображения головного отдела заднежаберных моллюсков,
полученные с помощью сканирующего электронного микроскопа
циклов остается все меньше
и меньше места. А ведь современ-
ная техническая база неизмеримо
превышает те возможности, кото-
рые были у Геккеля и Северцова.
Рекапитуляции между тем реаль-
но существуют! И данные, полу-
ченные нами при изучении голо-
жаберных моллюсков, являются
одним из лучших тому доказа-
тельств. Все эти факты неоспори-
мо свидетельствуют, что тщатель-
ный анализ сходств и различий
целых жизненных циклов близко-
родственных групп, а не только их
половозрелых форм — один из
наилучших способов реального,
а не формального, «теоретическо-
го» изучения эволюции.
Ринофоральный аппарат нотаспид
и голожаберных моллюсков под- ►
92
Семейство Pleurobranchidae — Плеуробранхусовые или Плеуробранхиды
брасывает и другие эволюцион-
ные «сюрпризы», которые обора-
чиваются трудноразрешимыми
проблемами в систематике. Все
вышесказанное о сходстве рино-
форальных аппаратов нотаспид
и дорид относится только к одно-
му (правда наиболее богатому
родами и видами) из двух ныне
существующих семейств нота-
спид — Pleurobranchidae. Но есть
и второе семейство, пусть читате-
ля не смутит его очень сходное
написание с предыдущим — так
иногда бывает в систематике. На-
зывается другое семейство Pleu-
robranchaeidae, и его ринофо-
ральный аппарат сходен не со
взрослыми доридами и не с их ли-
чинками, а со взрослыми предста-
вителями совершенно другой
группы голожаберных моллюсков,
куда входят такие крупные при-
метные виды, кактритония и ден-
дронотус. Дело в том, что и у три-
тоний (стр. 92 F), и у некоторых
архаичных арминацей из родов
Heterodoris и Doridoxa, и у нота-
спид из семейства Pleurobran-
chaeidae (стр. 92 Е) ринофо-
ральный аппарат представлен
чрезвычайно широко разросшим-
ся оральным парусом, который
сместил когда-то единые ринофо-
ры и «развел» их далеко по боко-
вым сторонам головы. У дорид
ринофоры также разведены дале-
ко по бокам головы, но механизм,
который к этому привел, был прин-
ципиально иной, чем у тритоний
и плеуробранхаеид. Тогда как
у последних ринофоры лишь вы-
теснялись на периферию головы
разрастающимся оральным пару-
сом, у дорид, когда-то единые ри-
нофоры, напротив, окружались
передним краем нотума, а не
орального паруса. Впоследствии
края нотума полностью замкну-
лись и, образовав ринофоральные
карманы, также разошлись на пе-
риферию нотума, а оральный па-
рус так и остался небольшим, а те-
перь еще и полностью скрытым
под передним краем нотума.
Принимая во внимание все изло-
женные факты о существенно раз-
личных механизмах образования
цельных ринофор у дорид и ден-
дронотацей, мы полагаем крайне
преждевременным утверждения
о том, что все голожаберные мол-
люски имеют монофилетическое
(т. е. единое) происхождение, и их
необходимо считать одним отря-
дом Nudibranchia. Напротив, при-
веденные факты свидетельствуют
скорее в пользу полифилети-
ческого (т.е. различного, неодно-
кратного) происхождения с одной
стороны дорид (традиционный
подотряд Doridacea), а с другой
строны, остальных голожаберных
моллюсков, относящихся к тра-
диционным подотрядам Dendrono-
tacea, Arminacea и Aeolidacea, от
различных нотаспид. Т. е. в пер-
вом случае для дорид предковой
группой нотаспид будут Pleuro-
branchidae, а для остальных голо-
жаберных моллюсков — нотаспиды
из семейства Pleurobranchaeidae.
Это определяет наше отрицатель-
ное отношение и к объединению
Nudibranchia и Notaspidea в группу
Nudipleura.
Нотаспид лучше рассматривать
как архаичную, предковую по от-
ношению к голожаберным мол-
люскам группу, обладающую
существенно отличным от голо-
жаберников онтогенетическим
циклом. Различные представите-
ли этого архаичного, еще единого
и не стабилизировавшегося цик-
ла нотаспид в дальнейшем дали
различных голожаберных мол-
люсков. По этой причине мы со-
храняем три отдельных отряда:
нотаспид (Notaspidea), дорид
(Doridacea или Anthobranchia)
и голожаберных моллюсков
(Nudibranchia, объединяет в свою
очередь три традиционных под-
отряда — Dendronotacea, Armina-
cea и Aeolidacea). Также допусти-
мо неоднократно упоминавшееся
в данной книге выражение «голо-
жаберные моллюски-дориды»,
поскольку все эти группы имеют
близкое строение и происхожде-
ние и традиционно объединялись
в рамках одного отряда Nudi-
branchia.
Boreoberthella augusta
Martynov et Schrodl, 2008 —
Бореобертелла августа
Особенности строения. Тело удлиненно-
овальное, нотум относительно широкий, впе-
реди без вырезки. Ринофоры в виде свернутых
в трубку пластинок, сближены своими осно-
ваниями. Оральный парус широкий, трапе-
циевидный, с несколько оттянутыми и утол-
щенными краями. Единственная жабра,
располагающаяся под нотумом с правой сто-
роны тела, состоит из многочисленных жабер-
ных пластинок. Обособленные бугорки на ра-
хисе ктенидия отсутствуют. В области середины
ктенидия открывается анальное отверстие.
Имеется внутренняя раковина — она крупная,
очень тонкая и полностью или частично лише-
на твердой кальциевой основы. Копулятивный
орган не втяжной, в виде широкой лопасти
располагается впереди от жабры. Длина до
40 мм.
93
Отряд Notaspidea (= Pleurobranchoidea) — Нотаспиды
Boreoberthella augusta — Бореобертелла августа, вскрытый фиксированный экземпляр со спинной стороны
Boreoberthella augusta — Бореобертелла августа, фиксированный экземпляр, вид с правой стороны
94
Семейство Pleurobranchidae — Плеуробранхусовые или Плеуробранхиды
Boreoberthella angusta — Боуеобеутелла августа,
фиксированный экземпляр, вид со спинной стороны
Boreoberthella augusta — Бореобертелла августа,
фиксированный экземпляр, вид с брюшной стороны
Окраска. Прижизненная окраска не из-
вестна. Вероятно, как многие глубоководные
виды, имеют однородную светло-розоватую
или желтоватую окраску.
Отличия от похожих видов. Похожих ви-
дов из отряда нотаспид, обитающих на глуби-
нах свыше 200 м, в морях России нет.
Особенности экологии. Обитает на пес-
чаных грунтах с примесью камней. Обнаружен
на глубинах от 150 до 540 м.
Размножение. Не известно.
Питание и поведение. Питается губ-
ками.
Распространение. Глубоководный вид.
Судя по имеющимся у нас данным, широко рас-
пространен в верхней батиали северной части
Тихого океана от Японского моря до Командор-
ских островов. За пределами российских вод
пока не обнаружен. Возможно обитает также
в районах Тихого океана, примыкающих к Але-
утским островам.
Berthella californica
(Dall, 1900) -
Бертелла калифорнийская
Особенности строения. Тело удлиненно-
овальное, нотум относительно широкий, впе-
реди без вырезки, скульптура покровов в ви-
де небольших бугорков, разбросанных по
нотуму. Ринофоры в виде свернутых в трубку
пластинок, сближены своими основаниями.
Оральный парус широкий, трапециевидный,
с несколько оттянутыми и утолщенными
краями. Единственная жабра, располагаю-
щаяся под нотумом с правой стороны тела,
состоит из многочисленных жаберных пла-
стинок. Обособленные бугорки на рахисе кте-
нидия отсутствуют, однако в месте присоеди-
нения каждой жаберной пластинки к центру
ктенидия имеется утолщение. В области се-
редины ктенидия открывается анальное от-
верстие. Имеется внутренняя раковина — она
95
Отряд Notaspidea (= Pleurobranchoidea) — Нотаспиды
Berthella californica — Бертелла калифорнийская,
вид со спинной стороны
Berthella californica — Бертелла калифюрнийская,
вид с брюшной стороны
Berthella californica — Бертелла калифюрнийская, вид с правой стороны
96
Семейство Pleurobranchidae — Плеуробранхусовые или Плеуробранхиды
Berthella californica — Бертелла калифорнийская
крупная, тонкая, вогнутая, накрывает значи-
тельную часть внутренностей. Ее скульптура
в виде концентрических угловатых линий.
Копулятивный орган втяжной, узкий, кони-
ческий. Длина до 40 мм.
Окраска. Основная окраска тела белова-
тая. По ноту му и его обратной стороне, краям
ноги и оральному парусу разбросаны разного
размера ярко-белые пятнышки и точки, со-
стоящие из микроскопических кристаллов,
концентрирующихся преимущественно вну-
три бугорков. По всему краю нотума проходит
тонкая белая линия.
Отличия от похожих видов. Похожих ви-
дов из отряда нотаспид, обитающих на мелко-
водье, в морях России нет. От голожаберных
моллюсков Berthella californica отличается свер-
нутыми в трубку ринофорами и единственной
настоящей жаброй, располагающейся под но-
тумом с правой стороны, а не на его спинной
стороне.
Особенности экологии. Обитает на каме-
нистых и скалистых грунтах, на мелководье, от
литорали, до 20-30 м.
Размножение. Кладка представляет со-
бой широкую завитую ленту белого цвета
длиной до 9 см. При температуре около 20° С
яйца в кладке развивались около 2 недель,
затем из них вышли велигеры с турбоспираль-
ной раковинкой.
Berthella californica — Бертелла калифюрнийская,
кладка
97
Отряд Notaspidea (= Pleurobranchoidea) — Нотаспиды
Питание и поведение. Питается бесске-
летными губками. Малоподвижный вид, часто
обнаруживается на субстрате — камнях, лами-
нарии вне всякой связи с пищевым объектом.
Распространение. Широко распростра-
ненный тихоокеанский бореальный вид. По
российскому побережью отмечен в Японском
море, у тихоокеанского побережья Камчатки
и Командорских островов, по американско-
му — от Аляски до Калифорнии.
СЕМЕЙСТВО
PLEUROBRANCHAEIDAE —
ПЛЕУРОБРАНХЕИВЫЕ
Небольшое семейство нотаспид. Характери-
зуется ринофорами, далеко разнесенными
по сторонам от обширного широкого ораль-
ного паруса. Раковина полностью отсутству-
ет. Нотум сильно редуцирован, почти не на-
крывает ногу и единственную крупную
жабру. Челюсти хорошо развиты, состоят из
большого числа удлиненных многоугольных
микроскопических элементов. Радула мощ-
ная, состоит из множества рядов зубов. Есть
виды, у которых в каждом ряду, в свою оче-
редь, может насчитываться до нескольких
сотен зубов.
В тропических и субтропических морях из
этого семейства довольно обычны крупные
представители родов Pleurobranchaea и Euse-
lenops, в батиали Индийского океана обитает
специфический глубоководный род Pleuro-
branchella. Представители семейства Pleurobran-
chaeidae, известны из морей России только по
одному ювенильному экземпляру, обнаружен-
ному в водах у острова Монерон. Причиной
этого, так же, как и в случае с аплизиями, несо-
мненно, является существующее там теплое
течение. В нашем распоряжении пока нет при-
жизненного изображения представителя этого
семейства, поэтому его фотография в атласе не
приводится.
Pleurobranchaea japonlca
(Thiele, 1925) -
Плевробранхея японская
Особенности строения. Нотум гладкий, он
практически полностью слит с боками тела
и задней дорсальной поверхностью ноги, за ис-
ключением правой задней боковой стороны
тела, где он прикрывает основание жабры. Ри-
нофоры довольно крупные относительно тела,
в виде неправильных трубок с боковой попе-
речной щелью. Полукруглый фронтальный
парус гладкий по краю. По краям парус несет
треугольные лопасти с боковой щелью. Задняя
дорсальная часть ноги очень массивная, ника-
ких следов шпоры (небольшого отростка верх-
ней части ноги, характерного для большинства
других более южных плевробранхей) нет. Глот-
ка сильно вывернута. Жабра крупная, изогну-
тая, практически не прикрыта краем нотума.
Состоит из 23 широких треугольных жаберных
пластинок.
Окраска. Информация о прижизненной
окраске единственного ювенильного экземпля-
ра, собранного у берегов острова Монерон, от-
сутствует. Окраска крупных живых особей из
вод Японии включает серовато-коричневатый
фон, на который «наброшена» густая сеточка
из более темных линий.
Отличия от похожих видов. Похожих ви-
дов из отряда нотаспид, обитающих на мелко-
водье, в морях России нет.
Размножение. В морях России не изуче-
но, в водах Японии данный вид откладывает
кладки в виде толстого шнура. Развитие со ста-
дией свободноплавающей личинки-велигера
с наружной раковинкой.
Особенности экологии. Преимуществен-
но мелководный вид, но может спускаться до
значительной глубины (400 м). Этот вид обыч-
но обитает на илистых или песчаных грунтах
на спокойных мелководьях. В водах у острова
Монерон обнаружен на каменистом грунте
с примесью песка.
Питание и поведение. Сведения по пита-
нию отсутствуют. Возможно, питается акти-
ниями, как ряд других видов плевробранхей.
Малоподвижный вид.
Распространение. В водах Японии из-
вестен с побережья всех островов, кроме Хок-
кайдо, по их тихоокеанской и япономорской
сторонам. Отмечен в Восточно-Китайском
и Желтом морях. В морях России обнаружен
только в водах у острова Монерон. Не исклю-
чено, что в дальнейшем, кроме ювенильных
экземпляров, в водах у острова Монерон
будут обнаружены и половозрелые особи
этого вида.
98
• Эволюция, шаг шестой •
ВЕРШИНЫ ЭВОЛЮЦИИ
ОТРЯД DORIDACEA (= ANTHOBRANCHIA) —
ГОЛОЖАБЕРНЫЕ МОЛЛЮСКИ-ДОРИДЫ
Дориды — одна из крупнейших групп голожаберных моллюсков. В морях России она занимает
особое место, и ее представителей можно найти повсеместно. Характерной чертой этой группы
является расположение жабры, имеющей вид кольца, в задней части спинной стороны.
Эволюционные преобразования дорид:
удлинение или укорочение жизненного цикла
относительно предков — нотаспид
КРИПТОБРАНХИАЛЬНЫЕ
ДОРИДЫ
Жабры
развива-
ются на
спинной
стороне
0
е
НОТАСПИДЫ
ЖИЗНЕННЫЙ ЦИКЛ
Ринофоры
остаются
сближен-
ными
Жабра
развивается
на брюшной
Развивается
жаберная
полость
стороне
сбоку
Ринофоры
расходятся
в сторону
Анус пере-
ФАНЕРОБРАНХИАЛЬНЫЕ
ДОРИДЫ
Ринофоры
сближены
Жабры еще
не появились
Анус
на брюшной
стороне
сбоку
мещается
на спинную
сторону
Жабры появляются
на спинной стороне
Жаберная полость
не развивается
Анус
Жабры
развиваются
на брюшной
стороне
КОРАМБИДЫ
остается
на брюшной
стороне
О О
Анус на
брюшной
стороне
в задней
части —
ЭВОЛЮЦИЯ
Модель эволюции дорид, представленная в виде перестроек жизненных (онтогенетических) циклов
99
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
Cadlina laevis — Кадлина гладкая
Acanthodoris pilosa — Акантодорис шиповатый
Corambe obscura — Корамбе темная
Polycera quadrilineata — Полицера четырехполосая
Дориды являются одной из ключевых групп как тропических коралловых рифов, так и хо-
лодных скальных мелководий северной части Тихого океана (например, российской Камчатки).
Насчитывается более 10 семейств, сотни родов и более 2000 видов дорид. В морях России обита-
ют не менее 40 видов этой группы, и большинство как массовых, так и редких представителей
приведено ниже. Криптобранхиальные дориды включают в свою диету главным образом губок,
в то время как фанеробранхиальные питаются преимущественно мшанками. Отсутствие жабер-
ного кармана (или, по крайней мере, ясное направление на его редукцию), таким образом, во
многом коррелирует с наличием узкой радулы и питанием мшанками.
100
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
Основные формы строения жаберного аппарата дорид,
изображение получено с помощью сканирующего электронного микроскопа
101
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
Особенности строения отряда до-
рид проще всего понять исходя из
предковой для них морфологии от-
ряда нотаспид из семейства плев-
робранхид (Pleurobranchidae, см.
выше). В процессе превращения
организации плевробранхид в та-
ковую дорид единственная боковая
жабра плевробранхид была окру-
жена разросшимися складками
нотума, а затем переместилась на
спинную сторону моллюска. Склад-
ки нотума со временем срослись,
сформировав таким образом свое-
образную вторичную жаберную по-
лость, иначе называемую жабер-
ным карманом. Единственная
жабра в узкой округлой полости
кармана изогнулась, сформировав,
таким образом, жаберное кольцо.
Параллельно процессу формиро-
вания жаберного кармана проис-
ходил процесс окружения трубча-
тых ринофоров плевробранхид
передними складками нотума,
формирование из них ринофораль-
ных карманов, их отделение друг от
друга, замыкание и расхождение
по бокам передней части тела.
В результате этих двух основных
эволюционных поцессов сформи-
ровались криптобранхиальные
дориды, т.е. дориды, обладающие
хорошо развитой жаберной поло-
стью. У группы криптобранхиаль-
ных дорид (Cryptobranchia) жабры
способны целиком втягиваться
в жаберный карман (стр. 101 А, В).
Все остальные разнообразные
модификации жаберного аппарата
дорид, главным образом редукци-
онные, являются производными
(стр. 101C-F) от основной предко-
вой, криптобранхиальной органи-
зации. Это важнейшее положение
эволюционной истории дорид, ко-
торое уверенно объясняет все бес-
численные на первый взгляд от-
дельные варианты. Среди основных
производных от криптобранхиаль-
ной можно выделить следующие
типы организации дорид:
1) фанеробранхиальные — жа-
берная полость полностью реду-
цирована, жабры остаются на
спинной стороне и прикрепля-
ются непосредственно к наруж-
ным покровам (стр. 101 С, D);
2) гипобранхиальные — жаберная
полость полностью редуцирована,
жабры располагаются на брюш-
ной стороне под складкой нотума
в задней части (стр. 101Е, F);
3) абранхиальные — жабры и жа-
берная полость полностью реду-
цированы.
Среди трех этих основных типов ор-
ганизации фанеробранхиальная
является наиболее распространен-
ной и ее приобретали независимо
самые различные семейства до-
рид, в то время как гипобранхиаль-
ная и абранхиальная свойственна
лишь небольшому числу специали-
зированных групп.
Классические систематики, рабо-
тавшие с доридами, полагали, что
среди них существуют две большие
группы, которые соответствуют двум
их основным типам организации:
Cryptobranchia (криптобранхи-
альные дориды, в переводе —
скрытожаберные) и Phanerobran-
chia (фанеробранхиальные, или
открытожаберные дориды). Тогда
как сложное строение жаберной
полости дорид, устроенной сход-
ным образом у самых различных
ее представителей, предполагает
единственное происхождение
группы Cryptobranchia, весь имею-
щийся к настоящему времени кор-
пус фактов свидетельствует, что
фанеробранхиальные дориды —
лишь регрессивный уровень орга-
низации, достигнутый независимо
различными семействами дорид.
Единая по происхождению (моно-
филетическая) группа Cryptobran-
chia, таким образом, представляет
собой предковый для всех дорид
уровень организации. По этой при-
чине Cryptobranchia и Phanero-
branchia не используются здесь
в качестве основных систематиче-
ских подгрупп, на которых подраз-
деляет дорид традиционная систе-
матика. Фанеробранхиальные
семейства мозаично разбросаны
среди тех семейств криптобран-
хий, от которых они происходили.
Чтобы обозначить этот факт макси-
мально ясным образом, перед
описанием строения каждого вида
указывается к какому типу органи-
зации — крипто- или фанеробран-
хиальному он относится.
СЕМЕЙСТВО CHROMODORIDIDAE —
ХРОМОДОРИСОВЫЕ
ИЛИ ХРОМОДОРИДИДЫ
Обширное семейство исключительно крип-
тобранхиальных дорид, чрезвычайно разно-
образное в суб тропических и тропических
водах, в морях России представлено только
одним родом Cadlina. Тепловодные предста-
вители семейства обычно ярко и разнообраз-
но окрашены, в элементы окраски входят
пятна, полоски и г. д. Среди харашерных для
фонических морей родов, так или иначе зна-
комых любому ныряльщику, можно назвать
хромодорисов (Chroniodoris), глоссодорисов
(Glossodoris) и целый ряд других — Hypselo-
doris, Ceratosonia и ир. Неродственные виды
могут иметь сходную окраску и принадле-
102
Семейство Chromodorididae — Хромодорисовые или Хромодоридиды
жать к определенной так называемой цвето-
вой группе. Тело хромодоридид обычно удли-
ненное, с высокими боковыми сторонами
тела, так что края нотума значительно при-
подняты и не прикрывают края ноги. Нотум
по краю нередко содержит различным обра-
зом расположенные подкожные железы, се-
кретирующие ихтиотоксины (т. е. ядовитые
для рыб вещества). Жабры у многих родов
с уменьшенными в размерах пластинками,
отчего они приобретают характерный узко-
конический вид. Ринофоры обычные для
многих дорид, с многочисленными пластин-
ками. Оральные щупальца у большинства
пальцевидные, у Cadlina — треугольные с бо-
ковой бороздой. Глотка содержит хорошо
развитые челюсти, состоящие из отдельных
элементов. Радула характеризуется наличи-
ем центрального зуба, который может быть
хорошо развитым и зазубренным или глад-
ким, рудиментарным. Латеральные (боковые)
зубы радулы всегда в той или иной степени
зазубрены, их форма и расположение зубчи-
ков очень разнообразна и характеризует от-
дельные роды. Тропические хромодоридиды
питаются главным образом безспикульными
губками из семейства Aplysiellidae. Кладка
хромодоридид обычно в виде широкой фес-
тончатой спирали.
Cadlina laevis
(L„ 1767) -
Кадлина гладкая
Особенности строения. Криптобранхиаль-
ная дорида. Cadlina laevis — единственный
представитель обширного тропического се-
мейства Chromodorididae, проникшего в Арк-
тику. В покровах имеются спикулы, однако
нотум довольно мягкий. Характерно наличие
жаберного кармана, куда жабры способны втя-
гиваться целиком. Ринофоры с горизонтальны-
ми пластинками, ринофоральные карманы
имеются. Длина до 30 мм.
Окраска. Общая окраска тела беловатая,
характерные многочисленные желтые подкож-
ные железы располагаются вдоль боковых кра-
ев нотума. У некоторых экземпляров эти желе-
зы могут быть почти белые.
Отличия от похожих видов. От других до-
рид, обитающих в северных морях России, от-
личается наличием желтых желез (не пятен),
просвечивающих по краю нотума, и наличием
жаберного кармана. Кроме того, от Adalaria
proxima и Onchidoris muricata также отличается
заметно более крупными размерами и относи-
тельно мягким нотумом.
Особенности экологии. Встречается под
камнями, довольно обычен на пластинах лами-
нарии на глубине 5-10 м. В массовых количествах
отмечен зимой на литорали Белого моря.
Размножение. В Белом море размно-
жение отмечено с марта по июнь. Судя по
очень низким предпочитаемым температу-
рам нереста и распространению, является
настоящим арктическим видом. Кладка
в виде широкой спирально закрученной
ленты беловатого или желтоватого цвета.
Развитие прямое, из кладки выходит ползаю-
щая молодь.
Питание и поведение. Питается бесске-
летными губками. Малоподвижный вид, ча-
сто обнаруживается на субстрате — камнях,
ламинарии вне всякой связи с пищевым объ-
ектом.
Распространение. Арктическо-бореаль-
ный вид. Распространен в северной Атлантике
от Норвегии до Северной Испании, у Гренлан-
дии, на восточном побережье США, на Мур-
манском побережье Баренцева моря. В Аркти-
ке известен из морей Карского и Лаптевых,
а также из центрального полярного бассейна.
Достоверных данных о наличии этого вида
в дальневосточных морях нет (там обитает не-
сколько похожих видов из рода Cadlina, окон-
чательное систематическое положение кото-
рых пока не установлено). В Белом море
обнаружен повсеместно.
Cadlina laevis — Кадлина гладкая, кладка
103
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
Cadlina laevis — Кадлина гладкая, вид со спинной (слева) и брюшной (справа) стороны
Cadlina laevis — Кадлина гладкая
104
Семейство Akiodorididae — Акиодорисовые или Акиодоридиды
Ранняя ювенильная радула Cadlina laevis
Взрослая радула Cadlina laevis
Взрослая радула Doridunculus unicus
Взрослая радула Doridunculus unicus
СЕМЕЙСТВО AKIODORIDIDAE —
АКИОДОРИСОВЫЕ
ИЛИ АКИОДОРИДИДЫ
Небольшое с точки зрения видового состава, но
очень важное с точки зрения эволюционной
истории семейство фанеробранхиальных до-
рид. Долгое время полагали, что эта лишь
группа родов семейства онхидоридид (см. ни-
же). Однако детальный анализ целого ряда
признаков внутреннего строения, прежде всего
радулы и буккальной помпы, ясно продемон-
стрировал, что это группа резко отлична от
всех онхидоридид и заслуживает выделения
в самостоятельное семейство.
В пределах морей России известны два ро-
да акиодорид, один из них был описан совсем
недавно российскими исследователями изуни-
Недавно предложенная новая мо-
дель эволюции дорид хорошо объ-
ясняет факт существования неза-
висимого семейства акиодоридид.
Согласно новым представлениям,
криптобранхиальные дориды (до-
риды с хорошо развитой жаберной
полостью) являются эволюционно
первичными, тогда как фанеро-
бранхиальные дориды (дориды
без жаберного кармана) образо-
вались как вторичные, редукцион-
ные варианты.
Дело в том, что по строению раду-
лы акиодоридиды (особенно ан-
тарктический ponArmodoris) очень
похожи на криптобранхиальный
род Cadlina, но наличие у кадлины
жаберной полости и ее отстугствие
у акиодоридид, приводили к тому,
что как традиционная систематика,
так и «современная» филогене-
тическая систематика помещали
кадлину и акиодоридид в две со-
вершенно разные, неродственные
группы дорид. Согласно же новым
представлениям, столь разная на
первый взгляд внешняя морфоло-
гия достаточно легко может быть
достигнута путем простого недо-
развития жаберной полости в про-
цессе онтогенеза.
Известно всего пять родов акиодо-
ридид. Хотя акиодоридиды обна-
ружены в самых различных райо-
нах Мирового океана, в своем
распространении они связаны ли-
бо с большими глубинами, либо
с холодными водами.
105
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
кального местообитания — глубоководных
котловин Японского моря, глубина в которых
достигала 3600 м! В этом экстремальном ме-
стообитании были обнаружены значительные
скопления нового вида Doridunculus unicus, ко-
торый, как выяснилось, единственный из всей
обширной группы дорид, перешел на питание
гидроидными полипами. Гидроидная диета
широко распространена среди других голожа-
берных моллюсков, но это был первый дока-
занный случай питания гидроидными полипа-
ми представителя группы дорид.
Akiodorls lutescens
Bergh, 1880 —
Акиодорис желтоватый
Особенности строения. Фанеробранхиальная
дорида. Нотум мягкий, покрыт многочислен-
ными шаровидными бугорками. Каждый бо-
лее крупный бугорок окружен несколькими
мелкими. Нотум хорошо развит. Покровы тела
содержат спикулы. Жаберный карман полно-
стью отсутствует. Жабры крупные, разветвлен-
ные. Ринофоры с горизонтальными пластинка-
ми, ринофоральные карманы имеются. Длина
тела до 40 мм.
Окраска. Общий цвет тела беловатый, под
жабрами располагаются более яркие желтова-
тые железы.
Отличия от похожих видов. Можно
спутать с некоторыми крупными белыми он-
хидоридидами, например, Adalaria slavi. В от-
личие от нее Akiodoris lutescens лишен харак-
терной пигментации в виде многочисленных
мелких разбросанных пятнышек. Кроме то-
го, спинные бугорки Adalaria slavi более одно-
родные по размерам и имеют характерную
лампообразную форму, тогда как бугорки
Akiodoris lutescens резко подразделяются на
крупные и мелкие, и более округлые по фор-
ме. Некоторым сходством с ним обладает
Onchidoris bilaniellata, который хорошо отли-
чается наличием многочисленных пятен ин-
тенсивного коричневого цвета.
Сведения об экологии. Обитает на скали-
стых и каменистых грунтах, обычно на глуби-
нах 20-40 м, но может спускаться и до 700 м.
Размножение. Не изучено.
Akiodoris lutescens — Акиодорис желтоватый, Akiodoris lutescens — Акиодорис желтоватый,
вид со спишюй стороны фиксированi1ый экземпляр со спитюй cmopoiiы
106
Семейство Akiodorididae — Акиодорисовые или Акиодоридиды
Питание и поведение. Питается мшанка-
ми. Малоподвижный вид, медленно перепол-
зает по каменистым субстратам.
Распространение. Тихоокеанский боре-
альный вид. Широко распространен в даль-
невосточных морях России, обнаружен от
Южных Курильских до Командорских остро-
вов. Единственное местонахождение этого
вида за пределами морей России — у побе-
режья Аляски. Однако новые экзмепляры
этого вида там так и не были больше обнару-
жены за более чем сто лет, прошедшие со
времени первоначального описания этого
вида Бергом.
Doridunculus unicus
Martynov et Roginskaya, 2005 —
Доридункулюс уникус
Особенности строения. Фанеробранхиальная
дорида. Нотум довольно жесткий, покрыт
многочисленными заостренными, слегка запя-
тыми на вершине бугорками. Нотум хорошо
развит, в задней части тела имеет пару хорошо
развитых лопастей. Покровы тела содержат
спикулы. Жаберный карман полностью отсут-
ствует. Жабры небольшие, неразветвленные.
Ринофоры с горизонтальными пластинками,
ринофоральные карманы имеются. Длина тела
до 10 мм.
Окраска. Общий цвет тела беловато-
желтоватый. Сквозь покровы просвечивают
темный кишечник и розоватые внутренности.
Отличия от похожих видов. Этот вид хо-
рошо отличаестя от всех известных дорид фау-
ны России.
Сведения об экологии. Обитает на
мягких грунтах на огромных глубинах до
3600 м в центральной части Японского моря.
В этом местообитании приурочен к обшир-
ным скоплениям гидроида Egntundella, це-
лые пласты трубок которого могут накапли-
ваться на дне.
Размножение. Не изучено.
Питание и поведение. Благодаря изуче-
нию содержимого желудков и пищеваритель-
ного тракта удалось доказать, что Doridunculus
unicus, единственный среди дорид, питается ис-
ключительно гидроидами.
Doridunculus unicus — Доридункулюс уникус, Doridunculus unicus — Доридункулюс уникус,
вид со спинной стороны
вид с брюшной стороны
107
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
Пищевой объект Doridunculus unicus —
колония гидроидных полипов Oplorhiza
Распространение. Абиссальный глубоко-
водный вид. Известен только из глубоководных
частей Японского моря.
СЕМЕЙСТВО POLYCERIDAE —
ПОЛИЦЕРОВЫЕ
ИЛИ ПОЛИЦЕРИДЫ
Обширное семейство фанеробранхиальных до-
рид. Полицериды продолжают дальнейшую
тенденцию на редукцию нотума, которая на-
метилась уже у акиодоридид. Тело полицерид
обычно удлиненное, нотум практически исчез
или превратился в разного рода отростки по
краям спины. Жаберный карман полностью
редуцирован. У части родов, таких как Nem-
brotha и Roboastra, тело совершенно гладкое, ли-
шенное отростков, тогда как представители
родов Triopha, Colga и Polycera имеют разное ко-
личество отростков на нотуме, особенно по его
краю. Ринофоры покрыты пластинками, ряд
представителей имеет ринофоральные карма-
ны, но у некоторых родов (Palio, Polycera) даже
они полностью редуцируются. Оральные щу-
пальца обычно — лишь простые треугольные
складки по бокам у рта со щелью наверху.
Окраска очень разнообразная: так, например,
бореальный род Palio окрашен в неприметные
желтовато-зеленоватые тона, а тропические
Nembrotha несут многочисленные яркие (оран-
жевые, синие) полосы по светлому или тем-
ному фону. В отличие от акиодоридид бук-
кальная пома у полицерид не развилась.
У большинства представителей имеются хоро-
шо развитые монолитные челюсти неправиль-
но-треугольной или четырехугольной формы
с крыловидным расширением наверху. Как
и у большинства фанеробранхиальных дорид,
радула сильно редуцирована, и состоит из не-
большого числа зубов в поперечном ряду. Вто-
рой латеральнй зуб характерной формы. Цен-
тральный зуб, если есть, обычно в виде простой
пластинки.
Полицериды питаются, главным образом,
мшанками, причем, если представители рода
Colga, имеющие очень крупные зубы и хоро-
шо выраженный центральный зуб, откусыва-
ют части кустистых колоний мшанок, то по-
лицеры, у которых зубы мельче и центральный
зуб отсутствует, — питаются как кустистыми,
так и корковыми мшанками. Виды из рода
Nembrotha могут пожирать других голожабер-
ных моллюсков, например, эолидаций. Поли-
цериды распространены как в тропических
и субтропических, так и в умеренных водах.
Широко представлены полицериды и в фауне
России. Всего в семействе насчитывается более
10 родов.
Colga mlnlchevl
Martynov et Baranets, 2002 —
Колга Миничева
Особенности строения. Фанеробранхиальная
дорида. Нотум почти полностью редуцирован,
за исключением фронтального паруса. Спин-
ная сторона густо покрыта многочисленными
продольными рядами (до 8) удлиненных от-
ростков. Вдоль границы спины и боков с каж-
дой стороны расположено значительное число
сходных длинных узких отростков. Жабры
мощные, разветвленные. Жаберный карман от-
сутствует. Ринофоры с большим числом гори-
зонтальных пластинок. Ринофоральные карма-
ны отсутствуют. Тело довольно мягкое. Длина
до 50 мм.
108
Семейство Polyceridae — Полицеровые или Полицериды
Окраска. Общая окраска беловатая или
желтоватая. Кончики спинных и боковых вы-
ростов и жабр покрыты ярким белым пиг-
ментом.
Отличия от похожих видов. Форма тела,
окраска и значительные размеры позволяют
отличить данный вид от большинства других
фанеробранхиальных дорид фауны России.
Однако Colga minichevi — не единственный вид
рода Colga, обитающий в морях России. Для их
различения от Colga minichevi удобно использо-
вать географический критерий и батиметриче-
ский критерий (т. е. особенности распределе-
ния по глубинам). В Баренцевом море, на
глубинах от 18 до 440 м (обычно глубже 100 м)
обитает исключительно Colga villosa. Этот вид
полностью отсутствует в дальневосточных мо-
рях России, где на глубинах, обычно свыше
100 м, обитает еще один вид из рода Colga —
Colga pacifica. Напротив, Colga minichevi обитает
в дальневосточных морях обычно на очень не-
больших глубинах в 5-15 м (отдельные экзем-
пляры обнаружены вплоть до 50 м). Кроме
того, Colga pacifica хорошо отличается от Colga
minichevi наличием всего 1-3 рядов отростков
на спинной стороне, тогда как у последней их
всегда достоверно больше.
Особенности экологии. Судя по нашим
данным, является одним из наиболее массовых
видов, обитающих на каменистых и скалистых
грунтах на небольших глубинах 5-15 м вдоль
Курильских островов и тихоокеанского побе-
режья Камчатки.
Размножение. До сих пор ничего не из-
вестно о размножении этого вида.
Питание и поведение. Питается боль-
шим набором различных видов мшанок, у ко-
торых имеются разветвленные колонии. Загла-
тывает их целиком или отрывает от них
большие куски с помощью мощных челюстей
и радулы. Умеренно подвижный вид.
Распространение. Научное описание этого
вида было сделано лишь в 2002 году российскими
учеными. Тихоокеанский широко распростра-
ненный бореальный вид. Обитает, по-видимому,
во всех дальневосточных морях России, но осо-
бенно обычен у Курил и Камчатки. За пределами
морей России до сих пор не отмечен.
Colga minichevi — Колга Миничева, Colga minichevi — Колга Миничева,
вид со спинной стороны вид с брюшной стороны
109
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
Colga minichevi — Колга Миничева
по
Семейство Polyceridae — Полицеровые или Полицериды
Palio dubia
(М. Sars, 1829) -
Палио сомнительный
Особенности строения. Фанеробранхиальная
дорида. Тело более узкое, чем у многих других
дорид, удлиненное. Вся поверхность спины
и боков покрыта редкими довольно мелкими
бугорками. Нотум практически полностью ре-
дуцирован, за исключением фронтального па-
руса. Жаберный карман отсутствует. Позади
жабр выделяются мясистые постжаберные ло-
пасти. Ринофоры с горизонтальными пластин-
ками. Ринофоральные карманы отсутствуют.
Тело мягкое, спикулы отсутствуют. Длина до
15 мм.
Окраска. Общая окраска светло-сероватая,
оливковая, серо-зеленоватая, пластинки рино-
форов светло-коричневые. Спинные бугорки
покрыты довольно ярким желтым пигментом.
Выросты позади жабр белого цвета.
Отличия от похожих видов. Можно спу-
тать с другой беломорской доридой, обладаю-
щей относительно узким удлиненным телом,
Ancula gibbosa, от которой отличается сероватой
или оливковой окраской тела, отсутствием от-
ростков вокруг ринофоров и очень короткими
отростками позади жабр.
Особенности экологии. Обитает на лито-
рали и верхней сублиторали под камнями, на
фукусах, а также встречается на пластинах ла-
минарии. На литорали Белого моря особенно
многочисленен осенью.
Размножение. В Белом море размноже-
ние отмечено с мая по июль. Кладка в виде до-
вольно узкой розоватой ленты, закрученной
в спираль из 1,5-2 оборотов. Развитие со стади-
ей велигера.
Питание и поведение. Питается корко-
выми мшанками, в том числе, вероятно Еи-
cratea loricata. Малоподвижный вид. Обладает
необычным для голожаберных моллюсков
(но характерным для сакоглосс) репродук-
тивным поведением. Копулятивный аппа-
рат, вооруженный многочисленными мелки-
ми шипиками, действует как игла шприца,
которым партнеры вводят сперму под кожу
друг другу, а не в одно из женских половых
Palio dubia — Полно сомнительный,
вид со спинной стороны
Palio dubia — Полно сомн и тельный,
вид с брюшной стороны
111
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
Polio dubia — Полно сомнительный
112
Семейство Polyceridae — Полицеровые или Полицериды
отверстий, как это делает подавляющее чис-
ло других голожаберных моллюсков. По
этой причине вагинальное отверстие у этого
вида полностью редуцировалось.
Распространение. Атлантический боре-
альный вид. Распространен в северной Атлан-
тике от Норвегии до Франции, на восточном
побережье США, на Мурманском побережье
Баренцева моря и в Белом море. В дальнево-
сточных морях обитает, по-видимому, близ-
кий вид.
Polycera quadrilineata
(Muller, 1776) -
Полицера четырехполосая
Особенности строения. Фанеробранхиальная
дорида. Тело более узкое, чем у многих других
дорид, удлиненное. Поверхность спины и бо-
ков гладкая. Нотум практически полностью
редуцирован, за исключением фронтального
паруса. Жаберный карман отсутствует. Позади
жабр имеется пара удлиненных постжаберных
выростов. Ринофоры с горизонтальными пла-
стинками. Ринофоральные карманы отсут-
ствуют. Тело мягкое, спикулы отсутствуют.
Длина тела ювенильных баренцевоморских
экземпляров около 6 мм. В других частях аре-
ала взрослые экземпляры этого вида могут
достигать 40 мм.
Окраска. Общая окраска беловатая. Спин-
ные и боковые бугорки покрыты довольно яр-
ким желтым, иногда почти оранжевым пиг-
ментом. У крупных особей ярко-желтые точки
на спине замещаются полосками. Выросты по-
зади жабр белого цвета, кончик также покрыт
желтым пигментом.
Отличия от похожих видов. Можно
спутать с другой фанеробранхиальной ши-
роко распространенной в морях России до-
ридой, обладающей относительно узким
удлиненным телом, Ancula gibbosa, от кото-
рой отличается только наличием пары от-
ростков после жабр. Наиболее сходна с очень
редкой в морях России, также фанеробран-
хиальной доридой Goniodoridella savignyi, от
которой отличается гладкими ринофорами,
Polycera quadrilineata — Полицера четырехполосая, Polycera quadrilineata — Полицера четырехполосая,
вид со спинной стороны вид с брюшной стороны
113
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
Polycera quadrilineata — Полицера четырехполосая
лишенными пластинок и наличием больше-
го числа отростков на фронтальном парусе.
Кроме того, для их различения можно ис-
пользовать географический критерий — го-
ниодориделла известна только из Японского
моря, тогда как четырехполосая полицера
известна только из Баренцева моря.
Особенности экологии. В Баренцевом
море найдены только ювенильные экземпляры
на корковых мшанках, обрастающих некото-
рые красные водоросли, на глубине 1-2 м.
Размножение. В Баренцевом море раз-
множение не отмечено. За пределами морей
России, в водах Британских островов, полице-
ры откладывают кладку в виде довольно узкой
беловатой ленты, закрученной в спираль из
нескольких оборотов. Развитие со стадией ве-
лигера.
Питание и поведение. Питается корковы-
ми мшанками. Подвижный вид.
Распространение. Атлантический боре-
альный вид. Распространен в северной Ат-
лантике от Норвегии до Франции, на восточ-
ном побережье США, на Мурманском
побережье Баренцева моря. Появление юве-
нильных экземпляров этого вида в Баренце-
вом море также связано с процессом потеп-
ления Арктики.
114
Семейство Polyceridae — Полицеровые или Полицериды
Triopha catalinae
(Cooper, 1863) —
Триофа каталинская
Синоним:
Triopha pacifica (Volodchenko, 1941)
Особенности строения. Фанеробранхиальная
дорида. Тело удлиненное, очень широкое. Но-
тум полностью редуцирован, вдоль границы
спины и боков с каждой стороны расположено
по одному ряду коротких булавовидных от-
ростков, по 6 с каждой стороны. Имеется ши-
рокий фронтальный парус, покрытый по краю
сходными отростками. Ринофоральные карма-
ны высокие. Ринофоры покрыты многочислен-
ными плотно расположенными пластинками.
Вся спина покрыта округлыми бляшками. Жа-
берный карман отсутствует. Жабры мощные,
разветвленные. Может достигать очень боль-
ших размеров, вплоть до 80 мм в длину.
Окраска. Общая окраска сероватая или
беловатая. Боковые отростки спины, а также
бляшки и жабры окрашены в красный цвет.
Отличия от похожих видов. Форма тела,
окраска и значительные размеры позволяют
с легкостью отличить данный вид от любых
других фанеробранхиальных дорид фауны
России.
Особенности экологии. Обитает от лито-
рали до глубины 70 м, наиболее часто встреча-
ется в прибрежной зоне на глубинах 7-20 м.
Размножение. Кладка представляет со-
бой довольно широкую белую ленту, закручен-
ную в спираль из нескольких оборотов. Разви-
тие со стадией личинки-велигера.
Питание и поведение. Питается боль-
шим набором различных видов мшанок, у ко-
торых имеются разветвленные колонии. Загла-
тывает их целиком или отрывает от них
большие куски с помощью мощных челюстей
и радулы. Умеренно подвижный вид.
Распространение. Тихоокеанский бо-
реальный вид. Обитает, по-видимому, во
всех дальневосточных морях России, и далее
по американскому побережью повсеместно
отмечен от Алеутских островов до Кали-
форнии.
Triopha catalinae — Триофа каталинская,
вид со спин нои стороны
Triopha catalinae — Триофа каталинская,
вид с брюшной стороны
115
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
СЕМЕЙСТВО VAYSSIEREIDAE —
ВЕСЬЕРОВЫЕ ИЛИ ВЕСЬЕРИДЫ
Семейство весьерид резко отличается по
внешнему строению от большинства других
дорид. Тело у взрослых экземпляров достига-
ет в длину лишь 3-6 мм, обычно оранжевато-
красного цвета, удлиненное, изящное, высо-
кое, полностью лишено каких-либо выростов
за исключением пары гладких конических ри-
нофор, не имеющих выраженных ринофо-
ральных карманов. Оральные щупальца от-
сутствуют, имеются лишь малозаметные
валики по бокам рта. Анус смещен из задней
части тела в среднюю и немного вправо. Нога
узкая. Радула с небольшим числом сильно за-
зубренных зубов в каждом ряду. Строго гово-
ря, весьериды достигают, наверное, пределов
в редукции исходной сложной морфологии
криптобранхиальных дорид.
Очень специализированно и питание ви-
дов этого семейства. Весьериды питаются, раз-
мягчая трубки полихет из семейства спи-
рорбид (Spirorbidae), живущих в спирально
завитых известковых домиках, секретом ораль-
ных желез и просверливая в трубках отверстия
радулой, а затем высасывая оттуда червя, что
уникально для дорид.
Крайняя необычность внешнего строения
этого семейства (червеобразная форма тела
без каких-либо следов жабр) заставляли ис-
следователей то сближать весьерид с сако-
глоссами из рода лимапонтия, или даже вы-
делять в самостоятельный отряд Okadaiida.
Судя по строению радулы, весьериды могут
быть потомками широко распространенного
тропического семейства гимнодоридид
(Gymnodorididae), среди которых имеются
как крупные виды, так и мелкие формы
с уменьшенным размером жабр (имеются
указания на нахождение безжаберных незре-
лых гимнодоридид), и от которых они могли
произойти путем педоморфоза.
Для весьерид известны мелкие уплощенные
округлой формы кладки с незначительным ко-
личеством крупных, богатых желтком яиц, из
которых выходит ползающая червеобразная
молодь. В семейство входит единственный род.
Vayssierea elevens (Весъера изящная) на пищевом объекте — многощетинковых червях спирорбидах
116
Семейство Vayssiereidae — Весьеровые или Весьериды
Vayssierea elegans
(Baba, 1930) -
Весьера изящная
Синонимы:
Okadaia elegans Baba, 1930
Okadaia tecticardia Slavoshevskaya, 1971
Особенности строения. Фанеробранхиальная
дорида (вернее будет определить ее как абранхи-
альную, т. е. безжаберную, поскольку жабры
полностью отсутствуют). Тело очень узкое, почти
червеобразное. Поверхность спины и боков глад-
кая. Нотум полностью редуцирован. Какие-либо
выросты на теле, за исключением пары простых
гладких ринофоров отсутствуют. Ринофораль-
ные карманы также отсутствуют. Тело мягкое,
спикул нет. Одна из самых мелких дорид, длина
взрослых экземпляров не превышает 6 мм.
Окраска. Тело красного или краснова-
то-оранжевого цвета. Реже встречаются свет-
ло-желтые экземпляры. На спине рядом
с анальным отверстием иногда просвечивает
небольшой красноватый ренальный орган
и массивная коричневатая печень. Ринофоры
на конце лишены окраски.
Отличия от похожих видов. Сходных ви-
дов в фауне России нет. Красновато-оранжевой
окраской обладает еще и другая дорида Ros-
tanga alisae, которая значительно отличается от
Vayssierea elegans широким телом, нормально
развитым нотумом, наличием жабр и жабер-
ного кармана.
Особенности экологии. Обитает от лито-
рали до глубины 5 м и является самым массо-
вым видом голожаберных моллюсков в заливе
Петра Великого во все сезоны года. Селится
преимущественно под камнями, но может
встречаться и в ризоидах бурых водорослей.
Размножение. Размножение в заливе Пет-
ра Великого происходит с конца мая по сентябрь.
Кладка маленькая, уплощенная, по форме ва-
рьирует от округлой до бобовидной, прикрепля-
ется обычно к камням. В строме кладки обычно
находится около десяти яиц красного цвета.
Питание и поведение. Обладает одним
из самых необычных среди голожаберных мол-
люсков пищевым поведением. Совместными
Vayssierea elegans — Весьера изящная,
вид со спи иной стороны
Vayssierea elegans — Весьера изящная,
вид с брюшной стороны
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
действиями радулы и ферментов оральных же-
лез высверливает отверстия в известковых
трубках сидячих многощетинковых червей из
семейства спирорбид, а затем высасывает их
содержимое. По этой причине Vayssierea elegans
обычно можно обнаружить поблизости от ха-
рактерных спиралевидных трубок спирорбид.
Распространение. Поскольку внешне все
виды этого семейства крайне схожи, точное рас-
пространение родов и видов не совсем ясно, но
можно утверждать, что в водах Японии, у южных
Курильских островов и в заливе Петра Великого
Японского моря обитает Vayssierea elegans (Baba,
1930), а в водах Австралии и Новой Каледонии —
Vayssierea caledonica Risbec, 1928. Наконец, согласно
альтернативному мнению, все мелкие безжабер-
ные дориды из рода Vayssierea, относятся к одному
виду, первоначально описанному из Китая —
Vayssierea felis (Collingwood, 1881).
СЕМЕЙСТВО ANCULIDAE —
АНКУЛОВЫЕ ИЛИ АНКУЛИДЫ
Семейство фанеробранхиальных дорид. Руди-
ментарный жаберный карман имеется только
у одного рода Diaphorodoris, тогда как все
остальные входящие в семейство группы ха-
рактеризуются полным отсутствием жаберной
полости. Нередко это семейство считают сино-
нимом гониодоридид, однако, они четко от-
личаются строением буккальной помпы. Тело
обычно высокое, гладкое, часто покрыто силь-
но удлиненными отростками разного размера.
У рода Trapania имеются два мощных отростка
позади жабр, тогда как у Ancula имеется зна-
чительное количество подобных отростков.
Жабры слаборазветвленные. Тело высокое
(как, например, у рода Ancula), либо сильно
прижатое, так что нога и спина практически
сливаются (как у рода Hopkinsia). Голова несет
обычно пару щупалец и в большинстве случа-
ев не прикрыта нотумом. Буккальная помпа
представляет собой полушаровидную струк-
туру, сильно выступающую из большей части
дорсальной стороны глотки, не имеющую раз-
витого периферического мускула. Радула с не-
большим числом сильно зазубренных зубов
в каждом ряду. Распространены анкулиды
главным образом в тропических и субтропи-
ческих водах. В морях России отмечено только
два рода.
Ancula gibbosa
(Risso, 1818) -
Анкула гребенчатая
Синоним:
Ancula cristata Alder, 1841
Особенности строения. Фанеробранхиальная
дорида. Тело более узкое, чем у многих других
дорид, удлиненное, мягкое, но спикулы в по-
кровах присутствуют. Нотум полностью реду-
цирован. Спинная поверхность гладкая. Жа-
берный карман отсутствует. Жабры окружены
10-15 крупными пальцевидными отростками.
Ринофоры с горизонтальными пластинками,
в их основании с каждой стороны имеется по
паре отростков, ринофоральные карманы от-
сутствуют. Кроме того, пара щупальцевидных
выростов располагается на голове. Длина бело-
морских особей до 16 мм.
Окраска. Общая окраска тела беловатая,
кончики жабр, ринофоров и всех выростов на
теле покрыты желтоватым или оранжевым
пигментом, который может отсутствовать у не-
которых экземпляров.
Отличия от похожих видов. Можно спу-
тать с другой беломорской доридой, обладаю-
щей относительно узким удлиненным телом,
Palio dubia, от которого отличается белой окра-
Ап си la gibbosa — Анкула гребенчатая
118
Семейство Anculidae — Анкуловые или Анкулиды
Ancula gibbosa — Анкула гребенчатая,
вид со спинной стороны
Ancula gibbosa — Анкула гребенчатая,
ювенильная особь со спинной стороны
ской тела и длинными выростами вокруг рино-
фор и жабр.
Особенности экологии. Обитает под кам-
нями, на скалах, обычен на литорали, встреча-
ется в верхней сублиторали. Мелких особей
этого вида довольно регулярно обнаруживали
в нижнем горизонте литорали на беломорской
биологической станции «Мыс Картеш». В от-
дельные годы в местах с хорошими гидродина-
мическими условиями (например на литорали
острова Харлов в Баренцевом море) особи этого
вида и их кладки сплошной коркой покрывали
камни и скалы. Численность вида от года к году
подвержена значительным колебаниям: в пер-
вой половине 1970-х годов Ancula gibbosa была
одним из самых массовых литоральных видов
в губе Дальне-Зеленецкой Баренцева моря, тог-
да как в конце 1990-х не было найдено ни одного
экземпляра. Похожий феномен отмечен для
этого вида и в водах Британских островов.
Размножение. В Белом море размноже-
ние отмечено в июле-августе. Кладка в виде
узкой, слегка загнутой, беловатой ленты. Раз-
витие со стадией велигера.
Питание и поведение. По-видимому, пи-
тается кустистой мшанкой Eucratea loricata.
Распространение. Амфибореальный
вид — широко распространен в Северной Ат-
лантике, известен из Баренцева и Белого мо-
рей, по тихоокеанскому побережью Северной
Америки, от Аляски до Калифорнии (нередко
американские особи рассматриваются как са-
мостоятельный вид Ancula pacifica (MacFarland,
1905), изредка встречается и в дальневосточных
морях России, известен из Средиземного моря.
Обитает главным образом, на литорали, спу-
скаясь до глубины 5-10 м.
Goniodoridella savignyi
Pruvot-Fol, 1933 —
Гониодориделла Савиньи
Особенности строения. Фанеробранхиаль-
ная дорида. Тело более узкое, чем у многих
других дорид, удлиненное. Поверхность
спины и боков гладкая. Нотум практически
полностью редуцирован, за исключением
119
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
фронтального паруса. Жаберный карман от-
сутствует. Позади жабр имеется пара удлинен-
ных постжаберных выростов. Ринофоры глад-
кие, без пластинок. Ринофоральные карманы
отсутствуют. Тело мягкое, спикулы отсутству-
ют. Длина тела взрослых полововозрелых эк-
земпляров около 3-6 мм.
Окраска. Общая окраска беловатая. Спин-
ные и боковые бугорки покрыты довольно яр-
ким желтым пигментом. Выросты позади жабр
белого цвета, кончик также покрыт желтым
пигментом.
Отличия от похожих видов. Наиболее
сходна с другой очень редкой в морях Рос-
сии, также фанеробранхиальной доридой
Polycera quadrilineata, от которой отличается
гладкими ринофорами, лишенными пласти-
нок и наличием только двух отростков на
фронтальном парусе. Кроме того, для их
различения можно использовать географи-
ческий критерий — гониодориделла извест-
на только из Японского моря, тогда как четы-
рехполосая полицера известна только из
Баренцева моря.
Особенности экологии. Единичные эк-
земпляры обнаружены под мелкими камнями,
поросшими мшанками, на глубине 2—3 м.
Размножение. Не изучено.
Питание и поведение. Питается мшанка-
ми. Подвижный вид.
Рас п ростран ен ие. Т роп ическо-субтроп и-
ческий вид, заходящий в низкобореальные об-
ласти. В морях России обнаружен только в за-
ливе Петра Великого Японского моря.
СЕМЕЙСТВО DORIDIDAE —
ДОРИСОВЫЕ ИЛИ ДОРИДИДЫ
Исторически дориды начались с линнеевского
рода Doris, который вскоре превратился в це-
лое семейство Dorididae, а затем и в отряд/
подотряд Doridacea. Глубокие исторические
корни этого семейства привели к тому, что
оно до сих пор наименее четко определено как
группа. В разные исторические периоды его то
разбивали на почти десяток самостоятельных
семейств, то, наоборот, объединяли в него поч-
Goniodoridella savignyi — Гониодориделла Савинъи,
Goniodoridella savignyi — Гониодориделла Савинъи,
вид со сн ин нои стороны
вид с брюшной стороны
120
Семейство Dorididae — Дорисовые или Доридиды
Криптобранхиальный уровень орга-
низации оказался значительно бо-
лее консервативным, чем фанеро-
бранхиальный. Именно поэтому
среди криптобранхий так много
«унылых» родов, различить которые
может только специалист. Фауна
морей России — это в принципе
подлинное царство именно фане-
робранхиальных, а не криптобран-
хиальных дорид. Так, из общего
списка дорид фауны России
31 вид — это фанеробранхиальные
дориды, тогда как лишь 8 относится
к криптобранхиальным. Сходные
пропорции дает и фауна дорид Се-
верной Атлантики. А вот в тропиках
соотношение это перевернуто
с точностью до наоборот — огром-
ное число видов криптобранхий,
и относительно небольшое — фа-
неробранхиальных дорид. Это на-
водит на интересные эволюцион-
ные выводы, в частности, такой, что
более сложный аппарат для втяги-
вания жабр, каковым является жа-
берная полость, тратит и избыточ-
ное количество энергии, при
недостатке которой, в более суро-
вых условиях севера, он будет
«сброшен» из жизненного цикла
в первую очередь. Жабры в конце
концов, можно не менее успешно
просто сократить, не прибегая при
этом к разного рода «переуслож-
ненным» полостям.
ти все, что возможно, валили в кучу «все что
нужно и не нужно». Поэтому мы несколько
необычным образом определим это семейство
от противного — доридиды это то, что остает-
ся после выделения из них по крайней мере
двух более-менее признанных криптобранхи-
альных семейств — хромодоридид (см. выше)
и дискодоридид (см. ниже). Т. е. у доридид ра-
дула, как правило, состоит из гладких, а не за-
зубренных зубов, в репродуктивной системе во
многих случаях отсутствует копулятивный ап-
парат в виде цельной конической папиллы
и обыкновенно нет кариофиллидий (также см.
ниже) на нотуме. Все это, конечно, может ва-
рьировать в широких пределах (включая по-
явление зазубренных зубов радулы у некото-
рых родов), существуют и разного рода
аномалии в репродуктивном аппарате, размы-
вая и без того нечеткую картину. В остальном
в отношении скульптуры нотума, жабр и т. д.
и т. п. доридиды могут выглядеть внешне очень
по-разному.
В морях России из этой группы известно
лишь небольшое число видов, из которых здесь
приводится лишь один.
Archidoris pseudoargus
(Rapp, 1827) -
Архидорис псевдоаргус
Особенности строения. Криптобранхиаль-
ная дорида. Нотум мягкий, покрыт многочис-
ленными шаровидными бугорками. Каждый
более крупный бугорок окружен несколькими
мелкими. Нотум хорошо развит. Покровы тела
содержат спикулы. Жаберный карман хорошо
развит. Жабры крупные, разветвленные. Рино-
форы с горизонтальными пластинками, рино-
форальные карманы имеются. Одна из самых
крупных дорид фауны России, длина тела
до 12 см.
Окраска. Типичная окраска лимонно-
желтая, но встречаются экземпляры со слабо
выраженными сероватыми или коричневаты-
ми пятнами.
Отличия от похожих видов. Благодаря
своим размерам и особенностям морфологии
вид легко отличается от других представителей
дорид. Некоторым сходством с ним обладает
Onchidoris bilmnellata, который хорошо отлича-
ется наличием многочисленных пятен интен-
сивного коричневого цвета, полным отсутстви-
ем жаберной полости, перазветвлепными (хотя
и крупными жабрами).
Archidoris pseudoargus — Архидорис исевдоаргус,
кладка
121
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
Archidoris pseudoargus — Архидорис псевдоаргус,
Archidoris pseudoargus — Архидорис псевдоаргус,
вид со спинной стороны
вид с брюшной стороны
Особенности экологии. Обитает на ска-
листых и каменистых грунтах, на глубинах
7-20 м.
Размножение. Кладка в виде широкой
фестончатой спиральной ленты, прикреплен-
ной к субстрату ребром, белого или желтова-
того цвета. Развитие со стадией свободнопла-
вающей личинки-велигера.
Питание и поведение. Питается, как мно-
гие другие криптобранхиальные дориды, губ-
ками. Малоподвижный вид, медленно пере-
ползает по каменистым субстратам.
Распространение. Бореальный северо-
атлантический вид. Широко распространен
в северной Атлантике. В водах России известен
только с Мурманского побережья Баренцева
моря. Появление этого вида в Баренцевом мо-
ре было впервые зарегистрировано только
в 2000 году. Связано он с процессом потепле-
ния Арктики, которое наблюдается в послед-
ние 10 лет. В результате этого средняя темпе-
ратура поверхности моря в августе повысилась
с +8° С до +12° С. Этого на первый взгляд не-
значительного повышения оказалось достаточ-
но для массового оседания личинок этого вида
на Мурманском побережье Баренцева моря.
Ранее ареал этого вида располагался более чем
на тысячу километров юго-западнее и никогда
не охватывал российскую часть Баренцева мо-
ря. Развитие личинок до взрослых половозре-
лых особей стало возможно благодаря более
благоприятным температурам, и стало не еди-
ничным, а массовым явлением. Поэтому в на-
стоящее время дайверы могут наслаждаться
зрелищем процветающей популяции этого
крупного голожаберного моллюска в пределах
Баренцева моря.
СЕМЕЙСТВО DISCODORIDIDAE —
ДИСКОДОРИСОВЫЕ
ИЛИ ДИСКОДОРИДИДЫ
Одно из крупнейших семейств криптобранхи-
альных дорид. Наиболее характерной чертой
этого семейства является присутствие особого
рода микроскопических выростов гготума —
так называемых кариофиллидий. Каждый ка-
122
Семейство Discodorididae — Дискодорисовые или Дискодоридиды
риофиллидий имеет сложное строение и со-
стоит из основания и нескольких спикул
(иголочек) на вершине, расположенных в фор-
ме звездочки или цветка (от латинского слова
caryophyllaceous — гвоздика). В основании ка-
риофиллидий располагается чувствительное
нервное окончание, и они выполняют роль
своеобразного рецептора механических раз-
дражений. Таких кариофиллидий на поверх-
ности нотума дискодоридид тысячи, из-за чего
спинная сторона у многих представителей это-
го семейства имеет характерный «войлочный»
вид. Окраска очень разнообразна, но преобла-
дают неяркие тона. Ринофоры с пластинками.
Оральные щупальца обыкновенно в виде цель-
ных пальцевидных выростов. Копулятивный
аппарат, как правило, цельный, конический.
В ряде случаев имеются различные дополни-
тельные органы.
Дискодоридиды широко распространены
в тропических, субтропических и бореальных
водах. В водах России отмечены представители
двух родов.
из-под нотума, светло-коричневатые, почти сли-
ваются с общим тоном нотума. Такие пятна более
заметны в направлении к краям нотума. Второй
вид пятен — наиболее характерная особенность
Diaulula sandiegensis. Они обычно темно-коричне-
вые. Форма, размеры и количество пятен варьи-
рует значительно, но пятна располагаются всегда
отдельно друг от друга, и никогда не бывают
очень крупными. Каждая особь имеет свой уни-
кальный рисунок пятен.
Отличия от похожих видов. Благодаря
характерной окраске — редким темно-корич-
невым пятнам, располагающимся на кремо-
вом, светло-коричневом фоне, Diaulula sandie-
gensis хорошо отличается от любых других
видов криптобранхиальных дорид, обитающих
в пределах морей России.
Особенности экологии. Обитает на ска-
листых и каменистых грунтах, от литорали до
глубины в среднем 10-20 м, изредка до 72 м.
Детом в заливе Петра Великого это одна из
наиболее обычных литоральных дорид, «пасу-
щаяся» на 1убках Adocia cinerea и Haliclona per-
molis. Осенью этот вид можно заметить даже
Diaulula sandiegensis
(Cooper, 1863) —
Диаулула сандиегенская
Особенности строения. Криптобранхиаль-
ная дорида. Тело овальное, утолщенное, до-
вольно жесткое. Нотум широкий, утончен по
краям, сильно нависает над ногой. Вся без ис-
ключения поверхность нотума покрыта много-
численными плотно расположенными карио-
филлидиями, которые различимы только при
использовании оптики, без которой нотум
производит впечатление гладкой поверхности.
Ринофоральные карманы имеются. Пластинки
ринофоров скошенные, плотно прилегают
друг к дру1у. Мощные разветвленные жабры
располагаются почти правильным кольцом
с анусом в центре. Посторальные лопасти в виде
двух широких лопастей, охватывающих всю
переднюю часть ноги и смыкающихся в сере-
дине подо ртом с образованием выемки.
Оральные щупальца длинные, пальцевидные.
Длина до 50 мм, изредка до 80 мм.
Окраска. Общая окраска нотума — светло-
кремовая с желтоватым оттенком. Два сорта пя-
тен разбросаны по всему нотуму. Первые — более
мелкие, очень тусклые, как будто просвечивают
Diaulula sandiegensis — Диаулула сандиегенская,
вид со спинной стороны
123
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
в бухтах, примыкающих к центру г. Влади-
востока на урезе воды среди плит набережной.
Реже встречается зимой и весной.
Размножение. Кладка в виде широкой
фестончатой спиральной ленты, прикреплен-
ной к субстрату ребром, белого или желтова-
того цвета. Развитие со стадией свободнопла-
вающей личинки-велигера. Кладки этого вида
полностью отсутствуют на литорали во все се-
зоны года, но в июле на литорали появляется
молодь. Возможно Diaulula sandiegensis в наших
водах размножается в верхней сублиторали
весной — в начале лета.
Питание и поведение. Питается, как
и многие другие криптобранхиальные дориды,
губками. Малоподвижный вид, медленно пере-
ползает по каменистым субстратам.
Распространение. Тихоокеанский боре-
альный вид. На российском побережье досто-
верно отмечен в Японском море и у Командор-
ских островов. По американскому побережью
распространен от Аляски до Калифорнии.
Rostanga alisae
Martynov, 2003 —
Ростанга Алисы
Особенности строения. Криптобранхиаль-
ная дорида. Тело овальное. Нотум широкий,
утолщен по краям, сильно нависает над ногой.
Вся без исключения поверхность нотума по-
крыта многочисленными плотно расположен-
ными кариофиллидиями. Ринофоральные
карманы имеются. Пластинки ринофоров
расположены почти вертикально, немного-
численные. Жабры располагаются правиль-
ным кольцом с анусом в центре, слаборазвет-
вленные. Оральные щупальца длинные,
пальцевидные. Длина до 12 мм.
Окраска. Общая окраска нотума оранже-
вато-красноватая. Оттянутый кончик стебля
ринофор беловатый или беловато-оранже-
ватый. Ринофоральные пластинки оранжева-
то-красноватые. Жабры оранжево-беловатые.
Между ринофорами просвечивает беловатое
пятно.
Отличия от похожих видов. От всех крип-
тобранхиальных дорид, отмеченных в морях
России, хорошо отличается оранжево-красно-
ватой общей окраской. Единственная другая
дорида фауны России, обладающая сходным
Rostanga alisae — Ростанга Алисы,
вид со спинной стороны
цветом тела — Vayssierea elegans (см. выше), су-
щественно отличается от Rostanga alisae узкой,
почти червеобразной формой тела, полным
отсутствием нотума и жабр.
Сведения об экологии. Данный вид обна-
ружен на глубине 1-4 м, почти исключительно
на красной губке Ophlitaspongia pennata, покры-
вающей камни, скалы или друзы крупной ми-
дии Crenornytilus grayanus. Rostanga alisae доволь-
но редкий вид.
Размножение. Отмечено в августе. Кладка
в виде довольно узкой фестончатой спираль-
ной ленты, прикрепленной к субстрату реб-
ром, розоватого цвета. Развитие со стадией
свободноплавающей личинки-велигера.
Питание и поведение. Питается, как
и мггогие другие криптобранхиальные дориды,
губками. Малоподвижный вид, медленно пере-
ползает по каменистым субстратам.
Распространение. Научное описание этого
вида дагго одним из авторов данной книги лишь
в 2003 году. Известен исключительно из россий-
ской части Япоггского моря. Возможно обитает
и у берегов японского острова Хоккайдо.
124
Семейство Discodorididae — Дискодорисовые или Дискодоридиды
Эволюционная последовательность превращений форл1 организации заднежаберных моллюсков,
допускающая только прямое преобразование форлия (зеленые стрелки)
и не допускающая обратное (красные стрелки) — от раковины до мягкого тела
Наименее противоречиво объяснить
причину столь большого внутренне-
го сходства и столь большого внеш-
него несходства родов Onchimira
и Corambe можно лишь с помощью
адекватной эволюционной моде-
ли, которая исходила бы из реально
существующих в природе процес-
сов, а не абстрактных сценариев.
И такая модель есть. Ее основа —
сравнительный анализ онтогене-
за, жизненных циклов. Достаточно
подробно преимущества такого под-
хода были обсуждены выше. Имен-
но в онтогенезе криптобранхиальных
дорид, а значит, и у криптобранхи-
альной онхимиры имеется ранняя
стадия, на которой анальное отвер-
стие занимает брюшное, а не спин-
ное положение, характерное для
взрослой стадии. И именно у взрос-
лых, половозрелых корамбид, столь
сходных по внутренним признакам
с «нормальными» доридами (в том
числе и с онхимирой) анус оказыва-
ется брюшным, а значит, и жабры,
которые всегда развиваюся побли-
зости от него, тоже брюшные.
Наиболее простым, столь любимым
учеными «экономным» объяснением
(вспомните «бритву Оккама») будет
как раз то, что у корамбид нормаль-
ные взрослые внешние признаки
«просто недоразвились», в то время
как внутренние развились вполне
согласно «плану строения» осталь-
ных онхидоридид, породив в резуль-
тате эту систематическую загадку,
которая не дает покоя исследовате-
лям вот уже почти 150 лет. В процес-
се решения этой головоломки вы-
яснилось, что корамбиды совсем не
древняя, архаичная группа, а наобо-
рот, одна из «конечных», наиболее
сильно «уклонившихся» от исходной
основы групп онхидоридид.
Сходным образом, присутствие
рудименатарной жаберной поло-
сти на ранних стадиях развития
видов из рода Onchidoris, которые
во взрослом состоянии никогда не
имеют даже следа жаберной поло-
сти, четко указывает, что сначала
были криптобранхиальные, слож-
ные дориды, а только потом воз-
никли фанеробранхии, как упро-
щенный их вариант. И реальное,
а не мифическое существование
«удивительной онхидоридиды, не-
сущей жаберную полость» — On-
chimira cavifera в водах Камчатки
и указывает нам на реально суще-
ствующее недостающее эволю-
ционное звено между крипто-
бранхиями и фанеробранхиями.
И ставит «с головы на ноги» и всю
современную систему дорид, уже
почти «потерявшуюся» за фор-
мальными компьютерными фило-
генетическими деревьями.
Поэтому мы рады приветствовать
читателей в их начинающемся путе-
шествии по семейству онхидоридид,
которое наиболее разнообразно
именно в морях России!
125
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
СЕМЕЙСТВО ONCHIDORIDIDAE —
ОНХИДОРИСОВЫЕ
ИЛИ ОНХИДОРИДИДЫ
Это семейство, наверное, лучший из извест-
ных нам примеров доказанной эволюционной
преемственности между на первый взгляд
столь различными типами внешней организа-
ции, как криптобранхиальная Onchimira и ги-
побранхиальная, ни на что не похожая Со-
rambe. И не удивительно, что еще не так давно
все систематики выделяли Corambe в самостоя-
тельное семейство Corambidae (корамбиды)
и ставили их в основание системы дорид, как
будто бы они были самой древней и архаич-
ной группой. Любопытно, что подобный
взгляд на систему дорид во многом «по инер-
ции» сохраняется до сих пор. Тем не менее
два этих столь различных рода — Onchimira
и Corambe — объединены в одно семейство на
основании столь глубокого сходства в дета-
лях строения чрезвычайно сложно устроен-
ных внутренних органов — буккальной пом-
пы, радулы и репродуктивного аппарата, что
сомневаться в их ближайшем родстве просто
невозможно.
Как уже говорилось, буккальная помпа
и малозубая дифференцированная радула
нужна онхидоридидам для того, чтобы наибо-
лее эффективно «вскрывать» всеми упомяну-
тыми инструментами, как консервным ножом,
твердые колонии мшанок — их основного пи-
щевого объекта. К настоящему времени в мо-
рях России известно семь родов и более десяти
видов из этого семейства.
Onchimira cavifera
Martynov, Korshunova,
Sanamyan et Sanamayn, 2009 —
Онхимира кавифера или
«удивительная онхидоридида,
несущая жаберную полость»
Особенности строения. Криптобранхиаль-
пая дорида. Единственный представитель
Onchimira cavifera — Онхимира кавифера, Onchimira cavifera — Онхимира кавифера,
вид со спинной стороны вид с брюшной стороны
126
Семейство Onchidorididae — Онхидорисовые или Онхидоридиды
Onchimira cavifera — Онхимира кавифера, хорошо заметна развитая полость вокруг жабр
семейства онхидоридид, имеющий хорошо
развитую, полнофункциональную жаберную
полость, в которую жабры способны втяги-
ваться целиком. Эта особенность связывает
большинство видов онхидоридид, полно-
стью лишенных жаберной полости (группа
Phanerobranchia, фанеробранхиальные дори-
ды) с остальными доридами, которые, на-
против, обладают хорошо развитой жабер-
ной полостью (группа Cryptobranchia,
криптобранхиальные дориды). Иначе гово-
ря, онхимира является самым настоящим
переходным звеном в эволюции голожабер-
ных моллюсков. Нотум у онхимиры хорошо
развит, почти гладкий или покрыт редкими
очень короткими, узкими цилиндрическими
бугорками. Покровы тела мягкие и содержат
рыхлый каркас из спикул. Жабры неразвет-
вленные. Ринофоры с горизонтальными пла-
стинками, ринофоральные карманы имеют-
ся. Оральный парус этого вида имеет
характерную трапециевидную форму. Дли-
на до 26 мм.
Окраска. Общая окраска тела сероватая.
Жабры и ринофоры более светлые. Пигмент-
ные пятна отсутствуют.
Отличия от похожих видов. Комбина-
ция окраски, строения покровов, формы
жабр и орального паруса позволяют уверен-
но отличить онхимиру от любых других как
фанеробранхиальных, так и криптобранхи-
альных дорид.
Особенности экологии. Основное мес-
тообитание данного вида — обросшие кор-
ковыми мшанками крупные валуны на глу-
бине 15-25 м. Здесь Onchimira cavifera обитает
нередко совместно с другими видами из се-
мейства онхидорид, такими как Adalaria olgae
и Adalaria slavi.
Размножение. Пока ничего не известно
о размножении этого вида.
Питание и поведение. Вероятно, питает-
ся корковыми мшанками. Малоподвижная
форма.
Распространение. Данный вид был впер-
вые обнаружен только в 2008 году, и официаль-
но зарегистрирован только в 2009. В настоящее
время достоверно Onchimira cavifera известна
только из прикамчатских вод Тихого океана.
По всей вероятности, ареал этого вида является
более обширным, и включает, по меньшей ме-
ре, Курильские и Командорские острова.
127
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
Calycidoris guentheri
Abraham, 1876 —
Калицидорис Гюнтера
Особенности строения. Криптобранхиаль-
ная дорида. Среди всего семейства онхидори-
дид, только у родов Onchimira и Calycidoris
еще имеется жаберная полость. Однако в от-
личие от полностью функциональной жабер-
ной полости онхимиры, калицидорис имеет
значительно редуцированную полость, не
способную полностью сокращаться над жа-
брами. Тело у Calycidoris guentheri содержит
мощный каркас из спикул. Нотум развит, по-
крыт многочисленными жесткими кониче-
скими или цилиндрическими короткими
выростами. Жабры неразветвленные. Рино-
форы с горизонтальными пластинками,
ринофоральные карманы имеются. Длина
до 30 мм.
Окраска. Окраска желтоватая или бело-
ватая. Заметные пигментные пятна отсут-
ствуют.
Отличия от похожих видов. Комбина-
ция окраски, строения покровов, формы
жабр и орального паруса позволяют уверен-
но отличить калицидориса от любых других
как фанеробранхиальных, так и криптобран-
хиальных дорид.
Особенности экологии. Данный вид
обитает, по всей видимости, почти исключи-
тельно на мягких — т. е. илистых и песча-
ных — грунтах. Обнаружен на глубинах от
9 до 270 м.
Размножение. До сих пор ничего не из-
вестно о размножении этого вида, хотя он яв-
ляется одним из самых массовых в пределах
своего ареала. Вероятно, откладывает широ-
кие спиральные кладки беловатого или жел-
товатого цвета.
Питание и поведение. Объект питания
достоверно не известен. Данный вид по-
видимому, медленно передвигается по мягким
грунтам.
Распространение. Арктический вид,
широко распространенный почти по всем
морям Северного Ледовитого океана от Ба-
ренцева моря до Чукотского. Заходит также
в северную часть Берингова моря, условия
в которой почти не отличаются от арктиче-
ских. В Белом море тем не менее не обнару-
жен.
Calycidoris guentheri — Калицидорис Гюнтера,
вид со спинной стороны
Acanthodoris pilosa
(Abildgaard in MOIIer, 1789) —
Акантодорис шиповатый
Особенности строения. Фанеробранхиальная
дорида. Тело мягкое, хотя покровы содержат
разбросанные спикулы. Нотум хорошо развит,
обычно покрыт коническими короткими вы-
ростами. Жаберный карман отсутствует. Жаб-
ры крупные, много ветвистые, что нетипично
для семейства онхидоридид. Ринофоры с гори-
зонтальными пластинками, ринофоральные
карманы имеются. Длина до 30 мм.
Окраска. Окраска чрезвычайно вариабель-
на, от снежно-белой до совершенно черной.
Обычны сероватые, коричневатые и желтова-
тые особи. Встречаются особи и с ярко-желты-
ми ринофоральными пластинками. Внутрен-
нее строение у разноокрашенных экземпляров
не демонстрирует межвидовых отличий.
Отличия от похожих видов. Светлоокра-
шенные экземпляры Acanthodoris pilosa можно
спутать с другими доридами, окрашенными
в белый цвет. От Cadlina laevis отличается от-
сутствием желтых подкожных желез, отсут-
128
Семейство Onchidorididae — Онхидорисовые или Онхидоридиды
Acanthodoris pilosa — Акантодорис шиповатый, Acanthodoris pilosa — Акантодорис шиповатый,
вид со спинной стороны вид с брюшной стороны
Acanthodoris pilosa (Акантодорис шиповатый) в типичном местообитании — на бурой водоросли Аскофиллюм
129
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
Вариации окраски Acanthodoris pilosa
(Акантодорис ш и повит ы й)
ствием жаберного кармана и, как правило,
хорошо выраженными коническими выроста-
ми нотума, от Adalaria proximo и Onchidoris mu-
ricata отличается мягким телом и коническими
мягкими, а не жесткими грибовидными или
копьевидными бугорками нотума.
Особенности экологии. Типичный оби-
татель беломорской литорали, особенно харак-
терен для зарослей бурой водоросли Ascophyl-
lum nodosum. На веточки этой же водоросли
Acanthodoris pilosa нередко откладывает свои
кладки. Кроме того, встречается на камнях
Acanthodoris pilosa — Акантодорис шиповатый,
кладка
и других твердых субстратах. В Белом море из-
вестен главным образом на литорали, в то вре-
мя как в районе Шпицбергена обнаружен на
глубинах около 300 м. Значительно реже встре-
чается в дальневосточных морях России.
Размножение. В Белом море размноже-
ние отмечено с июня по сентябрь. Кладка
в форме широкой белой, прикрепленной
к субстрату узким ребром слегка изогнутой
или спирально завитой ленты. Развитие со ста-
дией велигера.
Питание и поведение. Питается корковы-
ми мшанками, как и почти все виды семейства
Onchidorididae, взламывая отдельные зооиды
мшанок зубами радулы и высасывая содержи-
мое специальным органом — буккальной пом-
пой, в которую превращена передняя дорсаль-
ная часть глотки (характерный признак всего
семейства онхидоридид). В Белом море этот
вид питается мшанкой Flustrella hispida, которая
часто обрастает веточки Ascophyllum nodosum.
Распространение. Амфибореальный
вид — широко распространен в Северной Ат-
лантике и по тихоокеанскому побережью Се-
верной Америки от Аляски до Калифорнии,
отмечен и в дальневосточных морях России,
возможно, обитает в Средиземном море.
Adalaria proxima
(Alder et Hancock, 1854) —
Адалярия проксима
Особенности строения. Фанеробранхиаль-
ная дорида. Нотум покрыт многочисленными
довольно широкими, но приостренными, поч-
ти копьевидными бугорками. В центре спины
крупные бугорки более-менее равномерно че-
редуются с мелкими. Покровы тела жесткие,
содержат каркас из многочисленных перекре-
щивающихся спикул. Нотум умеренно развит.
Жаберный карман отсутствует. Ринофоры
с горизонтальными пластинками, ринофо-
ральные карманы имеются. Длина беломор-
ских особей до 20 мм.
Окраска. Общая окраска тела беловатая,
иногда с желтоватым оттенком.
Отличия от похожих видов. Очень легко
спутать с Onchidoris muricata, с которым часто оби-
тает совместно. Скорее всего, по этой причине
достоверные находки Adalaria proxima в Белом мо-
ре относятся лишь к самому недавнему времени.
130
Семейство Onchidorididae — Онхидорисовые или Онхидоридиды
Adalaria proxima — Адалярия проксима,
вид со спинной стороны
Adalaria proxima — Адалярия проксима,
вид с брюшной стороны
Отличается от него приостренными копьевид-
ными, а не закругленными булавовидными бу-
горками. Следует иметь в виду, что при сокраще-
нии бугорков у Adalaria proxima они становятся
округлыми, и это может затруднить идентифи-
кацию. Лучше всего эти виды различаются по
зубам радулы. От других беломорских дорид,
имеющих белую окраску, отличается жестким,
пронизанным спикулами нотумом.
Особенности экологии. В Белом море
основной биотоп данного вида — обросшие
корковыми мшанками талломы (слоевища)
ламинарии на глубине 5-10 м. Здесь Adalaria
proxima обитает нередко совместно с близким
видом Onchidoris muricata. Вероятно встречается
также под камнями на литорали.
Размножение. Размножение на Белом море
отмечено нами в конце мая — начале июня. Клад-
ка в виде узкой ленты из 2-3 оборотов. Развитие
со стадией не питающегося велигера.
Питание и поведение. Питается корковы-
ми мшанками, включая Membranipora, Cribrilina
annulata и Cauloramphus sp. Малоподвижная
форма. При питании взламывает зооиды мша-
нок радулой и высасывает их содержимое бук-
кальной помпой.
Распространение. Амфибореальный
вид — широко распространен в Северной Ат-
лантике и встречается по тихоокеанскому по-
бережью Северной Америки. В водах России
известен также с Мурманского побережья Ба-
ренцева моря, возможно обитает в Японском
море. Указания из других Арктических морей,
а также из Охотского моря, относятся к близ-
кому виду.
Adalaria proxima — Адалярия проксима, кладка
131
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
Adalaria olgae
Martynov, Korshunova,
Sanamyan et Sanamayn, 2009 —
Адалярия Ольги
Особенности строения. Фанеробранхиальная
дорида. Нотум покрыт многочисленными уме-
ренно узкими, закругленными бугорками.
В центре спины крупные бугорки более-менее
равномерно чередуются с мелкими. Покровы
тела довольно мягкие, хотя и содержат жесткий
каркас из многочисленных перекрещивающих-
ся спикул. Нотум хорошо развит. Жаберный
карман отсутствует. Ринофоры с горизонталь-
ными пластинками, ринофоральные карманы
имеются. Длина до 15 мм.
Окраска. Общая окраска тела яркая ли-
монно-желтая, иногда даже с зеленоватым
оттенком.
Отличия от похожих видов. От всех онхи-
доридид, имеющих желтоватый цвет тела, дан-
ный вид отличается яркой лимонно-желтой
окраской.
Особенности экологии. Основное место-
обитание данного вида — обросшие корковы-
ми мшанками крупные валуны на глубине
15-25 м. Здесь Adalaria olgae обитает нередко со-
вместно с другими видами из семейства онхи-
дорид, такими как Adalaria slavi и Onchimira
cavifera.
Размножение. Пока ничего не известно
о размножении этого вида. Вероятно, от-
кладывает спиральные кладки в виде узкой
ленты.
Питание и поведение. Скорее всего, пи-
тается корковыми мшанками. Малоподвижная
форма.
Распространение. Данный вид был
впервые обнаружен только в 2008 году
и официально зарегистрирован в 2009. В на-
стоящее время достоверно Adalaria olgae из-
вестна из прикамчатских вод Тихого океана.
По всей видимости, ареал этого вида явля-
ется более обширным и включает, по мень-
шей мере, Курильские и Командорские
острова.
Adalaria olgae — Адалярия Ольги, Adalaria olgae — Адалярия Ольги,
вид со спинной стороны вид с брюшной стороны
132
Семейство Onchidorididae — Онхидорисовые или Онхидоридиды
Adalaria olgae — Адалярия Ольги
133
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
Adalaria slavi
Martynov, Korshunova,
Sanamyam et Sanamayn, 2009 —
Адалярия Славы
Особенности строения. Фанеробранхиаль-
ная дорида. Нотум покрыт многочисленными
умеренно узкими, лампообразными закру-
гленными бугорками. В центре спины круп-
ные бугорки более-менее равномерно череду-
ются с мелкими. Покровы тела довольно
мягкие, хотя и содержат жесткий каркас из
многочисленных перекрещивающихся спи-
кул. Нотум хорошо развит. Жаберный карман
отсутствует. Ринофоры с горизонтальными
пластинками, ринофоральные карманы име-
ются. Длина до 26 мм.
Окраска. Общая окраска тела снежно-
белая. По нотуму разбросано большое число
очень мелких белых пигментных пятнышек.
Отличия от похожих видов. Многие, даже
очень сильно отличающиеся по внутренним
признакам виды из семейства онхидоридид
могут обладать сходным внешним габитусом
Adalaria slavi — Адалярия Славы
и, что особенно вводит в заблуждение, такой
же белой окраской. Этот вид, например, доста-
точно легко спутать с Adalaria proxima. Харак-
терным признаком, хорошо отличающим
Adalaria slavi — Адалярия Славы,
вид со спи иной стороны
Adalaria slavi — Адалярия Славы,
вид с брюшной стороны
134
Семейство Onchidorididae — Онхидорисовые или Онхидоридиды
Adalaria slavi от Adalaria proxima и других белых
онхидоридид, является обязательное наличие
многочисленных мелких белых пятнышек, так
называемой пигментной «пыли».
Особенности экологии. Основное ме-
стообитание данного вида — обросшие кор-
ковыми мшанками крупные валуны на глу-
бине 15-25 м. Здесь Adalaria slavi обитает
нередко совместно с другими видами из се-
мейства онхидорид, такими как Adalaria olgae
и Onchimira cavifera.
Размножение. Пока ничего не известно
о размножении этого вида.
Питание и поведение. Вероятно, питает-
ся корковыми мшанками. Малоподвижная
форма.
Распространение. Данный вид был впер-
вые обнаружен только в 2008 году и официаль-
но зарегистрирован в 2009. В настоящее время
достоверно Adalaria slavi известна лишь из при-
камчатских вод Тихого океана. По всей види-
мости, ареал этого вида является более обшир-
ным и включает, по меньшей мере, Курильские
и Командорские острова.
Adalaria jannae
Millen, 1987 -
Адалярия Яны
Особенности строения. Фанеробранхиальная
дорида. Нотум покрыт многочисленными до-
вольно узкими закругленными бугорками. Бо-
лее крупные папиллы располагаются по краям
нотума, центр спины занимают в основном
мелкие папиллы, последние также есть на са-
мом краю нотума. Нотум умеренно редуциро-
ван. Покровы тела содержат жесткий каркас из
многочисленных перекрещивающихся спикул.
Жаберный карман отсутствует. Ринофоры с го-
ризонтальными пластинками, ринофоральные
карманы имеются. Длина обычно не более
10 мм, изредка до 15 мм.
Окраска. Общая окраска тела полупро-
зрачно-беловатая, иногда с желтоватым оттен-
ком. За жаберным кольцом располагается ха-
рактерная для этого вида крупная ярко-белая
железа.
Отличия от похожих видов. Очень трудно
отличить Adalaria jannae от Onchidoris muricata
Adalaria jannae — Адалярия Яны,
вид со спинной стороны
Adalaria jannae — Адалярия Яны,
вид с брюшной стороны
135
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
или Onchidoris тасгоротра. У Adalaria jannae
вдоль средней части спины имеется ряд, со-
стоящий только из мелких бугорков, тогда как
у обоих видов мелких беловатых онхидорисов
они чередуются с более крупными. Кроме то-
го, Adalaria jannae имеет очень характерный
признак — выступающую округлую или оваль-
ную крупную ярко-белую железу, располо-
женную за жабрами на краю нотума. Напро-
тив, у Onchidoris muricata и Onchidoris тасгоротра
эта железа почти не выступает, уплощена, фак-
тически распластана по краю нотума и гораздо
хуже заметна. Лучше всего эти виды различа-
ются по зубам радулы.
Сведения об экологии. Обитает от литора-
ли до глубины 13 м, обычно под небольшими
камнями, обросшими корковыми мшанками.
Размножение. Размножение особей из
тихоокеанских вод зарегистрировано с июля
по ноябрь, в заливе Петра Великого — с марта
по июль. Кладка в виде толстого шнура, бело-
ватая, полупрозрачная. При температуре
+8° С кладка развивается 11-13 дней. Развитие
со стадией личинки-велигера.
Питание и поведение. В водах залива
Петра Великого питается корковыми мшанка-
ми Cribrilina annulata. При питании взламывает
фронтальную мембрану корковых мшанок ра-
дулой и высасывает содержимое зооидов бук-
кальной помпой.
Распространение. Тихоокеанский боре-
альный вид. Широко распространен в северной
части Тихого океана от залива Петра Великого
по российскому побережью на север включая
Курильские острова и Камчатку до Калифор-
нии по американскому берегу на юге.
Onchidoris muricata
(М й Iler, 1776) -
Онхидорис муриката
Особенности строения. Фанеробранхиальная
дорида. Нотум покрыт многочисленными бу-
лавовидными закругленными бугорками.
В средней части спины мелкие бугорки преоб-
ладают над крупными. Нотум умеренно реду-
цирован. Покровы тела содержат жесткий кар-
Onchidoris muricata — Онхидорис муриката, Onchidoris muricata — Онхидорис муриката,
вид со спинной стороны вид с брюшной стороны
136
Семейство Onchidorididae — Онхидорисовые или Онхидоридиды
Onchidoris muricata (Онхидорис муриката) на пищевом объекте — корковой мшанке
Кладка Onchidoris muricata (Онхидорис муриката) на пищевом объекте — корковой мшанке
137
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
кас из многочисленных перекрещивающихся
спикул. Жаберный карман отсутствует. Рино-
форы с горизонтальными пластинками, рино-
форальные карманы имеются. Длина обычно
не более 10 мм, изредка до 15 мм.
Окраска. Общая окраска тела беловатая,
нередко с желтоватым оттенком.
Отличия от похожих видов. Очень легко
спутать с Adalaria proximo, от которой отличается
булавовидными закругленными, а не приострен-
ными копьевидными бугорками нотума. Не ме-
нее трудно отличить Onchidoris muricata от Adalar-
ia jannae. Эта проблема может возникнуть, только
если речь идет о тихоокеанских особях, посколь-
ку ни в Белом, ни в Баренцевом морях Adalaria
jannae не обитает. В отличие от Onchidoris muricata,
Adalaria jannae вдоль средней части спины имеет
ряд только мелких бугорков, тогда как у On-
chidoris muricata они чередуются с более крупны-
ми. Кроме того, Adalaria jannae имеет очень харак-
терный признак — выступающую округлую или
овальную крупную ярко-белую железу, располо-
женную за жабрами на краю нотума. Напротив,
у Onchidoris muricata эта железа почти не выступа-
ет, уплощена, фактически распластана по краю
нотума и гораздо хуже заметна. Лучше всего эти
виды различаются по зубам радулы.
Сведения об экологии. В Белом море вид
обычен на пластинах ламинарии, покрытых кор-
ковыми мшанками, на глубине 5-10 м. Встреча-
ется также на литорали под камнями.
Размножение. В Баренцевом море много-
численны копулирующие особи этого вида
и их кладки в виде узких лент из 1,5-2 оборотов
встречаются в июле и августе. В Белом море
размножение отмечено с мая по август вклю-
чительно. Развитие со стадией велигера.
Питание и поведение. На Белом море пи-
тается главным образом корковой мшанкой
Membranipora membranacea, возможно в диету
включены также Cribrilina annulata и Cauloramphus
sp. Малоподвижная форма. При питании взла-
мывает зооиды корковых мшанок радулой и вы-
сасывает их содержимое буккальной помпой.
Распространение. Амфибореальный вид —
широко распространен в Северной Атлантике
и встречается по тихоокеанскому побережью Се-
верной Америки. В водах России известен вдоль
Мурманского побережья Баренцева моря, из Бе-
лого моря, прикамчатских вод и у Командорских
островов. По всей видимости, распространен на
мелководье этих районов повсеместно.
Onchidoris тасгоротра
Martynov, Korshunova,
Sanamyam et Sanamayn, 2009 —
Онхидорис макропомпа
Особенности строения. Фанеробранхиальная
дорида. Нотум покрыт многочисленными була-
вовидными закругленными бугорками. В сред-
ней части спины мелкие бугорки преобладают
над крупными. Нотум умеренно редуцирован.
Покровы тела содержат жесткий каркас из мно-
гочисленных перекрещивающихся спикул. Жа-
берный карман отсутствует. Ринофоры с гори-
зонтальными пластинками, ринофоральные
карманы имеются. Длина до 15 мм.
Окраска. Общая окраска тела беловатая.
Отличия от похожих видов. Очень легко
спутать с Onchidoris muricata, от которого надеж-
но отличается только внутренними признака-
ми. Поэтому, с известной долей осторожности,
для различения этих видов можно использовать
географический критерий. В Белом и Баренце-
вом морях обитает только один вид из рода
Onchidoris, имеющий беловатую или светло-
желтоватую окраску без пятен, поэтому найден-
ных там мелких онхидорисов (предварительно
отличив их от также сходной Adalaria proxima),
можно уверенно определять как Onchidoris mu-
ricata. Напротив, у побережий Камчатки и Ко-
мандорских островов обитают как Onchidoris
muricata, так и Onchidoris тасгоротра. И хотя по-
следний вид является в российской части Тихо-
го океана гораздо более обычным, в данном
случае встреченные экземпляры лучше опреде-
лять как Onchidoris sp. (так называемая откры-
тая номенклатура, указывается только род,
вид не известен), а более точное определение
предоставить специалистам. Для отличия ти-
хоокеанских онхидорисов от Adalaria jannae см.
очерк об Onchidoris muricata.
Сведения об экологии. Основное место-
обитание данного вида — обросшие корковы-
ми мшанками мелкие камни на глубине
6-15 м.
Размножение. Пока ничего не известно
о размножении этого вида.
Питание и поведение. Питается корковы-
ми мшанками. Малоподвижная форма.
Распространение. Данный вид был впер-
вые обнаружен только в 2008 году, и официаль-
но зарегистрирован в 2009. В настоящее время
Onchidoris тасгоротра достоверно известен из
138
Семейство Onchidorididae — Онхидорисовые или Онхидоридиды
Onchidoris тасгоротра — Онхидорис макропомпа, Onchidoris тасгоротра — Онхидорис макропомпа,
вид со спинной стороны вид с брюшной стороны
Onchidoris тасгоротра — Онхидорис макропомпа
139
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
прикамчатских вод Тихого океана и Командор-
ских островов. По всей видимости, ареал этого
вида является более обширным и включает, по
меньшей мере, Курильские острова.
Onchidoris bilamellata
(Linnaeus, 1767) —
Онхидорис биламеллата
Особенности строения. Фанеробранхиальная
дорида. Нотум мягкий, покрыт многочислен-
ными булавовидными закругленными бугор-
ками. В средней части спины мелкие бугорки
преобладают над крупными. Нотум умеренно
редуцирован. Покровы тела содержат спику-
лы. Жаберный карман отсутствует. Жабры
у этого вида особенно крупные. Ринофоры
с горизонтальными пластинками, ринофо-
ральные карманы имеются. Длина до 40 мм.
Окраска. Общая окраска тела состоит из
многочисленных сливающихся коричневых пя-
тен разного размера, разбросанных по желто-
ватому фону.
Отличия от похожих видов. Этот круп-
ный и уникальный по особенностям морфоло-
гии (с мощными, по краям спирально завиты-
Onchidoris bilamellata — Онхидорис биламеллата,
вид со спинной стороны
Onchidoris bilamellata — Онхидорис биламеллата,
вид с брюшной стороны
Onchidoris bilamellata — Онхидорис биламеллата,
вид со спинной стороны
140
Семейство Onchidorididae — Онхидорисовые или Онхидоридиды
ми жабрами) вид легко отличается как от
других представителей рода Onchidoris, так и от
других дорид.
Сведения об экологии. Обитает на скали-
стых и каменистых грунтах, как правило на ли-
торали, но может опускаться до глубины 50 м.
Размножение. Кладка в виде широкой
фестончатой спиральной ленты, прикреплен-
ной к субстрату ребром, белого или желтова-
того цвета. Развитие кладок беломорских эк-
земпляров до свободноплавающих велигеров
продолжалось около 12 суток.
Питание и поведение. Данный вид обла-
дает не только уникальным для рода Onchidoris
строением, но и крайне необычными для всех
дорид пищевыми пристрастиями. Зубы раду-
лы у этого вида сильно удлинены, и, подобно
консервному ножу, могут вскрывать крупные
домики усоногих раков-балянусов, в изобилии
обитающих на литорали, а также в обраста-
нии искусственных конструкций и сооруже-
ний — буев, причальных стенок, судов. По
этой причине Onchidoris bilamellata особенно
часто встречается именно в обрастаниях (вклю-
чая корпуса судов), а также на литорали.
Распространение. Амфибореальный
вид — широко распространен в Северной Ат-
лантике и встречается по тихоокеанскому по-
бережью Северной Америки. В водах России
известен с Мурманского побережья Баренцева
моря, из Белого моря, прикамчатских вод
и у Командорских островов. Относительно ред-
кий вид, но местами может образовывать ло-
кальные скопления.
Кладка Onchidoris bilamellata (Онхидорис биламеллата)
на скальном субстрате среди пищевого объекта —
балянусов
Loy meyenl
Martynov, 1994 —
Лой Мейена
Особенности строения. Криптобранхиаль-
ная дорида с ювенильной жаберной поло-
стью. Тело неправильно-овальное, уплощен-
ное, асимметричное. Поверхность нотума
гладкая. Ринофоральные карманы имеются.
Ринофоры гладкие, конические. Задние ло-
пасти нотума полностью срастаются своими
нижними краями, верхними же — только на
небольшом заднем участке, образуя белова-
тый узкий шов. После образования шва верх-
ние края лопастей постепенно расходятся,
образуя своеобразный жаберный карман,
дно которого образовано сросшимися ниж-
ними краями лопастей нотума. Жаберный
карман каплевидный и несколько смещен на
левую сторону тела. В расширенной части
кармана располагаются три небольшие од-
новетвистые жабры. Передний край ноги
утолщенный, не раздвоенный, посередине
имеет небольшую выемку. Оральные щу-
пальца с треугольными лопастями. Длина до
6,5 мм.
Окраска. Периферическая часть нотума
беловатая, полупрозрачная. Срединное взду-
тие розовое, сквозь него просвечивают белова-
тые внутренности. Вся средняя часть между
вздутием и периферией нотума, от ринофо-
ральных карманов до жаберного отверстия, за-
нята коричневым пигментом, представленным
в виде небольших, часто сливающихся пятны-
шек, крапинок, штрихов.
Отличия от похожих видов. От всех дру-
гих дорид, известных в фауне России, отлича-
ется сросшимися асимметричными задними
лопастями нотума, образующими каплевид-
ный жаберный карман.
Сведения об экологии. В отличие от
большинства дорид, этот вид обитает исклю-
чительно в специфическом биотопе — на пес-
чаных грунтах, на глубине 40-100 м.
Размножение. Пока ничего не известно
о размножении этого вида.
Питание и поведение. Не изучено. Воз-
можно, питается кольчатыми червями.
Распространение. В настоящее время до-
стоверно известен только из залива Петра Ве-
ликого Японского моря и Охотского моря. По
всей вероятности, этот вид достаточно широко
141
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
Loy meyeni — Лой Мейена,
вид со спинной стороны, хорошо заметна
уникальная жаберная полость ювенильного типа
Loy meyeni — Лой Мейена,
вид с брюшной стороны, заметен шов
от слияния краев жаберной полости
распространен на песчаных грунтах в дальне-
восточных морях России. Вид назван в честь
выдающегося российского теоретика-эволю-
циониста Сергея Викторовича Мейена.
Corambe obscura
(Verrill, 1870) -
Корамбе темная
Особенности строения. Гипобранхиальная
дорида. Тело почти правильно округлое, упло-
щенное, симметричное. Поверхность нотума
гладкая. Ринофоральные карманы имеются.
Ринофоры покрыты небольшим числом про-
дольных складок. Задние лопасти нотума от-
сутствуют. Жабры целиком смещены на брюш-
ную сторону, и прикрыты задним краем
нотума. Жабры имеют особую пластинчатую
форму. Передний край ноги в середине со зна-
чительной выемкой. Оральные щупальца тре-
угольные. Длина до 6 мм.
Окраска. Нотум со спинной стороны по-
крыт сложным узором из многочисленных сли-
вающихся точек, пятнышек и штрихов черного,
коричневого, желтоватого и белого цветов.
Отличия от похожих видов. От всех дру-
гих дорид, известных в фауне России, отлича-
ется вентральным (брюшным) положением
жабр и особого рода пестрой окраской.
Сведения об экологии. Нередко обита-
ет в сильно опресненных, хорошо прогре-
ваемых эстуариях рек, но еще в зоне действия
морской среды. Населяет различные суб-
страты, покрытые корковыми мшанками, —
небольшие камни, друзы мидий Mytilus gal-
loprovincialis, буи и другие искусственные
конструкции.
Размножение. Этот вид, как и вся группа
корамбид, обладает необычной скоростью раз-
множения и созревания. Кладки в виде узкого
шнура, завитого в спираль из нескольких оборо-
тов, усеивают субстрат в большом количестве. Из
кладки выходит свободноплавающая личинка-
велигер, которая очень быстро оседает на суб-
страт и превращается в маленького моллюска.
Ювенильные особи тоже растут чрезвычайно бы-
стро, достигают стадии половозрелости меньше
чем за несколько недель и приступают, в свою
очередь, к откладыванию кладок. Продолжи-
тельность жизни у корамбе не более месяца.
Столь стремительное половое созревание
142
Семейство Onchidorididae — Онхидорисовые или Онхидоридиды
Corambe obscura — Корамбе темная,
вид со спинной стороны
Corambe obscura — Корамбе темная,
вид с брюшной стороны
Corambe obscura (Корамбе темная) на пищевом объекте — корковой мшанке
143
Отряд Doridacea (= Anthobranchia) — Голожаберные моллюски-дориды
Corambe obscura — Корамбе темная, кладка
Распространение. В морях России об-
наружен только в Черном море. Предполага-
ется, что Corambe obscura является здесь занос-
ным видом — его личинки были занесены
в Черное море из Атлантического океана
с балластными водами судов, и найдя в Чер-
ном море подходящие условия, успешно его
заселили. Новый вселенец в некоторых райо-
нах Черного моря, например в Севастополь-
ской бухте, может образовывать значитель-
ные скопления. В бассейне Атлантического
океана данный вид отмечен по американско-
му побережью в Чесапикском заливе, Флори-
де, Саргассовом море, Мексиканском заливе.
В водах Бразилии обитает очень близкий вид,
который, возможно, является лишь одной из
форм Corambe obscura.
характерно для большей части видов группы
корамбид (Corambidae), которой раньше при-
давали статус семейства. Сейчас выяснено, что
корамбиды лишь одна из подгрупп семейства
онхидоридид, с которыми их объединяет целый
ряд уникальных признаков внутреннего строе-
ния. Высокий темп созревания привел к тому,
что не все органы их предков развиваются до
нормального состояния. Подобный феномен
называется педоморфозом. У корамбид, относи-
тельно их предков — видов из рода Adalaria, це-
лый ряд признаков остается на уровне ранних
ювенильных особей — например вентральное
(брюшное) положение анального отверстия
и недоразвитые пластинки на ринофорах.
Питание и поведение. Питается исклю-
чительно корковыми мшанками, быстро пере-
двигаясь по субстрату.
СЕМЕЙСТВО GONIODORIDIDAE —
ГОНИОДОРИСОВЫЕ
ИЛИ ГОНИОДОРИДИДЫ
Гониодоридиды продолжают тенденцию на
редукцию и видоизменение нотума, наметив-
шуюся у онхидоридид. По строению внутрен-
них органов — радулы и буккальной помпы,
гониодоридиды почти не отличаются от онхи-
доридид. Нотум может быть либо гладкий (как
у рода Goniodoris), либо чаще снабжен разного
рода отростками по краю и па спине (каку Оке-
nia). Гониодоридиды — относительно неболь-
шое семейство, обитающее как в тропических,
так и в умеренных водах. В морях России от-
мечено пока лишь два вида, но только один из
них известен достоверно.
Случаи педоморфоза, когда взрос-
лые особи сохраняют признаки ли-
чиночных стадий своих предковых
форм, широко распространены
в природе. Более того, это один из
основных способов эволюции.
В наиболее развитой форме он ха-
рактерен для некоторых хвостатых
амфибий — тритонов и амбистом.
У амбистом, например, личинка
с наружными жабрами может долгое
время не превращаться в безжабер-
ную форму, и она становится поло-
возрелой, фактически так и не до-
стигнув окончательной взрослой
нормальной стадии.
Явление педоморфоза может силь-
но «смущать» систематика, и в исто-
рии этой науки были нередки слу-
чаи, когда близкородственные
таксоны, один из которых включал
нормальные, полностью дифферен-
цированные формы, а другой — пе-
доморфные, относили к разным,
неродственным семействам. Про-
исходило это потому, что система-
тики далеко не всегда знали, какие
стадии онтогенеза (индивидуально-
го развития) проходит тот или иной
вид, и не могли соотнести половоз-
релые педоморфные организмы
с этими стадиями. Поэтому так важ-
но исследовать не только взрослые
формы, но и весь онтогенетический
цикл того или иного вида, от яйца до
взрослого состояния.
144
Семейство Goniodorididae — Гониодорисовые или Гониодоридиды
Okenia distincta
Baba, 1940 —
Окения особенная
Особенности строения. Фанеробранхиальная
дорида. Тело удлиненное, несколько расширен-
ное в средней части. По краю имеется кайма с 6-8
удлиненными коническими отростками. Сход-
ные выросты, в числе 5-6, располагаются на спи-
не. Жабры неразветвленные. Ринофоры довольно
длинные, практически гладкие. Впереди от рино-
фор располагаются два удлиненно-конических
отростка. За ними на переднем крае головы на-
ходятся два коротко конических оральных щу-
пальца. Длина взрослых особей — до 10 мм.
Окраска. Общая окраска тела полупро-
зрачно-беловатая, с разбросанными светло-
коричневыми пятнышками.
Отличия от похожих видов. Форма тела,
окраска и значительные размеры позволяют
отличить данный вид от большинства других
фанеробранхиальных дорид фауны России.
Особенности экологии. Данный вид весь-
ма обычен в заливе Петра Великого Японского
моря на литорали и в сублиторали на глубине
до 5 м, под камнями, покрытыми некоторыми
видами мшанок.
Размножение. Кладка в виде узкого шну-
ра белого цвета, завернутого в спираль из одно-
го оборота. Развитие со стадией свободнопла-
вающей личинки-велигера.
Питание и поведение. Питается кусти-
стыми мшанками из рода Amathia sp. Малопод-
вижный вид.
Okenia distincta — Окения особенная,
вид со спинной стороны
Распространение. Низкобореальный ти-
хоокеанский вид. Известен только из залива
Петра Великого Японского моря и прибреж-
ных вод острова Хоккайдо (Япония).
Okenia distincta (Окения особенная), только что отложившая кладку
145
Отряд Nudibranchia — Голожаберные моллюски
Баренцево море. Закат
ОТРЯД NUDIBRANCHIA — ГОЛОЖАБЕРНЫЕ МОЛЛЮСКИ
На протяжении всей книги неоднократно упоминались выражения вроде «бесконечно разно-
образные голожаберные моллюски» или «самая обширная по объему группа заднежаберных».
И в самом деле, этот отряд, даже после того, как его состав был ограничен тремя традиционны-
ми подотрядами — Dendronotacea, Arminacea и Aeolidacea (т. е. даже после того, как дориды —
Doridacea — были вынесены за его пределы и стали рассматриваться как самостоятельная груп-
па в ранге отряда), все равно остается очень значительным по числу видов и одновременно
чрезвычайно разнообразным по строению. Поэтому, вместо того чтобы долго перечислять все
многообразие признаков входящих в отряд Nudibranchia видов, более рациональным будет ука-
зать лишь несколько наиболее важных, наиболее общих характеристик. И самым естественным
с точки зрения биологии образом это можно сделать в рамках теории эволюционных превра-
щений организмов. Итак. В основании настоящих голожаберных моллюсков лежит предковая
группа, жизненный цикл которой с одной стороны характеризовался наличием взрослой формы
с целым рядом регрессивных черт, прежде всего полной редукцией настоящей жабры (ктенидия)
и раковины, с другой — прогрессивным развитием головного отдела, с мощным разрастанием
орального паруса и формированием цельных ринофоров, несущих разного рода выросты или
пластинки. Подобная форма имела также умеренно развитый нотум, а ее пищеварительная же-
леза была цельным, компактным органом. Наконец, голожаберники сформировали еще один
важный прогрессивный признак — мощные цельные челюсти. Именно совокупность этих новых
черт, в особенности наличие широкого орального паруса и мощных челюстей, позволила голо-
жаберным моллюскам в конечном счете «освоить» одну из самых проблематичных — из-за на-
личия стрекательных клеток — для питания групп беспозвоночных — книдарий, более широко
известных под названием кишечнополостные. Из реально существующих ныне видов максималь-
но близко к такой предковой форме стоят некоторые представители традиционного подотряда
Arminacea, прежде всего виды родов Doridoxa и Heterodoris, обитающие и в морях России (см.
ниже). «Типичные» арминацеи, относящиеся к семейству Arminidae, похожи по общему плану
строения на доридоксу, но заметно отличаются от нее сближенными, а не разделенными рино-
форами — признаком, возникшим среди арминид несомненно вторично.
Последующая эволюция этой исходной предковой формы голожаберных моллюсков за-
ключалась с одной стороны в дальнейшей редукции нотума, превращении его боковых краев
либо в кустистые разветвленные вторичные жабры — так образовался подотряд дендронотацей
(Dendronotacea), либо в формировании на спинной стороне значительного числа особых удли-
ненных выростов — папилл, характерных главным образом для подотряда эолидацей (Aeolidacea).
Параллельно преобразованию внешних признаков шла модификация пищеварительной желе-
зы, которая из цельного компактного органа постепенно превратилась в мощную систему раз-
146
Семейство Doridoxidae — Доридоксовые или Доридоксиды
ветвленных каналов и трубок, имеющихся уже у некоторых дендронотацей (Dendronotus) и ар-
минацей (Janolus), но максимального развития достигающую у подотряда эолидацей (Aeolidacea).
Процесс эволюционного формирования «типичной» эолиды, у которой система взаимодействия
между пищеварительной железой и спинными папиллами достигла своего совершенства, не
был «удачным выстрелом с одного раза». Именно поэтому, как свидетельство множества не реа-
лизовавшихся до конца попыток, среди голожаберных моллюсков существует целый ряд групп,
своеобразных «эволюционных тупиков», у которых, например, спинные папиллы имеются, но
выросты пищеварительной железы в них еще не заходят (как у рода Dirona), или ветви этой же-
лезы уже заходят в папиллы, но особый мешочек, называемый книдосаком, в котором накапли-
ваются стрекательные клетки, полученные вместе с едой, еще не сформировался (как у рода
Janolus). И только у настоящих Aeolidacea, к которым относится много различных семейств, вся
эта система представлена в полном виде — имеется и разветвленная пищеварительная железа,
заходящая в папиллы, и мешочек-книдосак на их вершинах.
Голожаберные моллюски — своеобразная вершина эволюции заднежаберных моллюсков,
хотя биологи в наше время и предпочитают не выделять «вершин», а мыслить «деревьями»,
представляя эволюцию как некий непрерывный процесс со множеством отдельных «вершинок».
И тем не менее! Голожаберные моллюски, освоившие всевозможные местообитания — от пес-
чаного дна до обрастания судов, все мыслимые глубины — от литорали до абиссальной «бездны»
в 3000-4000 метров, и практически неуязвимые для хищников благодаря «воровству» чужих стре-
кательных клеток, поистине могут быть названы «вершиной» подкласса Opisthobranchia.
ПОДОТРЯД ARMINACEA — АРМИНАЦЕА
Традиционная группа голожаберных моллюсков, которая всегда вызывала большие споры сре-
ди специалистов и в настоящее время в исходном виде понимается редко. Дело в том, что этот
подотряд состоит из целого веера форм, представляющих собой разные эволюционные стадии
развития голожаберных моллюсков, и четкие границы между этими стадиями отсутствуют, что
затрудняет выделение отдельных групп. Происходит это из-за постепенного процесса транс-
формации исходной предковой формы, напоминающей доридоксу и тритонию, в многочислен-
ных, более «совершенных» потомков, все более и более становящихся похожими на настоящих
эолидацей, но осуществляющих это неоднократно и независимыми путями. Поэтому, чтобы
лучше понять разнообразие арминацей (как, впрочем, и многих других крупных групп задне-
жаберных) и направления эволюции, по которым они развивались, следует обращать внимание
на характеристики отдельных семейств.
Большинство «типичных» арминацей, относящихся к семейству арминид (Arminidae), оби-
тает на мелководье тропических морей и в морях России не известны.
СЕМЕЙСТВО DORIDOXIDAE —
ДОРИДОКСОВЫЕ
ИЛИ ДОРИДОКСИДЫ
Одно из наиболее архаичных семейств не только
подотряда арминацей, но и всего отряда голожа-
берных моллюсков. Доридоксовые еще сохраня-
ют широкий оральный парус с расставленными
по сторонам от него ринофорами. Причем рино-
форы располагаются в ринофоральных карма-
нах, имеющих форму свернутой трубки, не пол-
ностью замкнутой с внешней стороны. По этому
признаку доридоксовые очень сходны с предста-
вителями семейства Pleurobranchaeidae отряда
нотаспид. Весьма реалистичным выглядит сцена-
рий, в соответствии с которым цельные ринофо-
ры голожаберных моллюсков возникли внутри
трубчатых ринофоров нотаспид, которые пре-
вратились, таким образом, в ринофоральные
карманы. Доридоксы имеют хорошо развитые
челюсти и радулу с обособленным центральным
зубом и большим числом боковых рядов зубов.
К доридоксовым относится только один род
Doridoxa и два мало исследованных вида, обитаю-
щих в северной части Атлантического океана
и в Арктике, нередко на больших глубинах.
147
Отряд Nudibranchia — Голожаберные моллюски
Doridoxa walteri — Доридокса Вальтера,
фиксированный экземпляр, вид со спинной стороны
Doridoxa walteri — Доридокса Вальтера,
фиксированный экземпляр, вид с брюшной стороны
Doridoxa walteri
(Krause, 1892) -
Доридокса Вальтера
Синоним:
Dermatobranchus walteri Krause, 1892
Особенности строения. Тело довольно широ-
кое, удлиненно-клиновидное, впереди находит-
ся пара разнесенных ринофоральных карманов.
Перед ними берет начало оральный парус,
а сзади — гладкий редуцированный край ноту-
ма, не имеющий отростков. Ринофоры несут
пластинки. Оральный парус трапециевидный,
без выемки. Поверхность спины покрыта низки-
ми бугорками. Анус в виде крупного округлого
отверстия на боку тела. Длина тела до 40 мм.
Окраска. Прижизненная окраска тела до-
стоверно не известна. Вероятней всего, общий
цвет беловатый или слегка желтоватый, с про-
свечивающими более темными внутренностя-
ми и розоватой пищеварительной железой.
Отличия от похожих видов. По строению
тела хорошо отличается от других голожабер-
ных моллюсков фауны России.
Особенности экологии. Обитает на мяг-
ких грунтах, возможно с примесью камней, на
глубинах от 42 до 227 м.
Размножение. Не изучено.
Питание и поведение. Не изучено. По-
видимому, передвигается по мягким грунтам
и питается одной из групп кишечнополостных.
Распространение. Достоверно отмечен в мо-
рях Баренцевом, Карском и Лаптевых. Вероятно,
арктический вид, нахождение которого возможно
в сопредельных частях северной Атлантики.
СЕМЕЙСТВО DIRONIDAE —
ДИРОНОВЫЕ
ИЛИ ДИРОНИДЫ
Небольшое семейство голожаберных моллю-
сков. Тело широкое, впереди имеет широкий
волнистый оральный парус. Спинная часть по-
крыта крупными листовидными или вздутыми
гладкими папиллами (Dirona albolineata (Eliot,
1905) — дирона белополосая), или мелкими со-
сочками (JDirona picta (MacFarland, 1905) — диро-
на раскрашенная). Ринофоры с длинными сте-
блями, несущими в верхней трети характерные
пластинки. Анус открывается с правой стороны
в задней части тела. Имеются мощные челюсти.
Радула только с двумя продольными рядами
зубов. Центральный зуб квадратный с выступаю-
щим пальцевидным отростком, второй лате-
ральный — мощный, крыловидно изогнутый.
В отличие от большинства других голожабер-
148
Семейство Dironidae — Дироновые или Дирониды
ных моллюсков из групп Arminacea и Aeolidacea,
дирониды обладают широким спектром пита-
ния, включающим не только гидроидов и мша-
нок, но даже мелких брюхоногих моллюсков.
Включает единственный род дирона (Diro-
na). В водах России обитает один вид.
Dirona pellucida
Volodchenko, 1941 —
Дирона полупрозрачная
Синоним:
Dirona aurantia Hurst, 1966
Особенности строения. Тело высокое, удли-
ненное, по форме приближается к овально-
прямоугольному. Спина покрыта папиллами,
более густо расположенными по краю, и более
редкими в средней части спины. Папиллы круп-
ные, веретеновидные. Ринофоры несут пластин-
ки, расположение которых трудно различимо
вследствие фиксации. Оральные щупальца от-
сутствуют, вместо них имеется довольно широ-
кий оральный парус, волнистый по краю. Нога
широкая, впереди утолщена, но не раздвоена.
Анус располагается по краю правой стороны
далеко позади, почти на заднем конце тела.
Крупный вид, длина может достигать 80 мм.
Окраска. Данный вид обладает характерной
окраской — ярко-оранжевой, с небольшими яр-
кими белыми пятнами, покрывающими спинную
сторону, фронтальный парус, папиллы. Кончик
папилл также окрашен белым пигментом.
Отличия от похожих видов. По строению
тела и окраске хорошо отличается от всех голожа-
берных моллюсков, обитающих в морях России.
Особенности экологии. В дальневосточ-
ных морях встречается относительно редко, от
литорали до глубины 20 м на каменистых
и скалистых грунтах.
Размножение. Кладка в виде длинного
спирально завитого, уложенного фестонами
шнура розоватого цвета. Развитие со стадией
свободноплавающей личинки-велигера.
Питание и поведение. Умеренно подвиж-
ный вид. Диета состоит преимущественно из
гидроидов.
Распространение. Тихоокеанский бореаль-
ный вид. Обитает во всех дальневосточных морях
России от Командорских островов на севере до
Японского моря на юге. Широко распространен
также и в прибрежных водах Северной Америки,
от Аляски до штата Вашингтон.
Dirona pellucida — Дирона полупрозрачная,
вид со спиннои стороны
Dirona pellucida — Дирона полупрозрачная,
вид с брюшной стороны
149
Отряд Nudibranchia — Голожаберные моллюски
ПОДОТРЯД DENDRONOTACEA — ДЕНДРОНОТАЦЕА
Для этой традиционной подгруппы голожаберных моллюсков характерно наличие нескольких
пар разветвленных отростков вдоль краев спины (иногда их число достигает значительной
величины). Некоторые группы, к которым, например, относятся обитающие в Баренцевом
море представители рода Doto, вместо древовидных отростков несут на спине шишковидные
выросты. Пищеварительная железа у ряда групп цельная, в то время как у других разветвлен-
ная. Подавляющее большинство дендронотацей лишены книдосаков, но у видов рода Нап-
cockia на вершинах разветвленных выростов формируется несколько своеобразных мешочков,
в которых также накапливаются стрекательные клетки. У всех дендронотацей хорошо раз-
виты челюсти и за редким исключением радула с обособленным центральным зубом и боль-
шим числом боковых рядов зубов. Так же как и Aeolidacea, жизненный цикл различных пред-
ставителей Dendronotacea тесно связан с книдариями. Крупные дальневосточные Tritonia
tetraquetra питаются морскими перьями и так называемыми мягкими кораллами — особой
группой кишечнополостных из отряда альционарий (Alcionaria), тогда как более мелкие
представители, такие как виды родов Doto и Dendronotus, обитают среди колоний гидроидных
полипов и питаются ими.
СЕМЕЙСТВО TRITONIIDAE — ных ринофоральных карманов. От них впере-
ТРИТОНИЕВЫЕ ИЛИ ТРИТОНИИДЫ
Одно из центральных семейств подотряда ден-
дронотацей. Виды, входящие в это семейство, как
правило, обладают массивным телом и хорошо
развитым оральным парусом. Все тритонииды
обладают крупными челюстями и широкой ра-
дулой с большим числом боковых рядов зубов
и центральным зубом. Пищеварительная железа
в этом семействе еще сохраняет предковую фор-
му — в виде цельного, компактного, не развет-
вленного органа. Питаются тритонииды, глав-
ным образом, альционариями.
В морях России отмечено три вида, из
которых здесь приводится один наиболее
массовый.
Tritonia tetraquetra
(Pallas, 1788) -
Тритон ия тихоокеанская
Синонимы:
Tritonia diomedea Bergh, 1894
Tritonia primorjensis Minichev, 1971
Non Tochuina tetraquetra sensu Bergh, 1879
Особенности строения. Особи этого вида до-
стигают крупных для голожаберных моллю-
сков размеров — до 300 мм в длину и мотут
иметь вес до 1,7 кг. Тело массивное, удлиненно
клиновидное, впереди имеется пара разнесен-
ди берет начало оральный парус, а сзади — ре-
дуцированный край нотума, на котором
располагаются вторичные жабры в виде дре-
вовидных выростов. Оральный парус трапе-
циевидный, по переднему краю он двулопаст-
ной с выемкой по середине и выростами по
краю. Ринофоры несут около 20 коротких вер-
тикальных двуветвистых отростков, окружаю-
щих гладкий стебель. Поверхность спины по-
крывают широкие бугорки, формирующие
сеть низких гребней. Вторичные жабры пред-
ставляют собой невысокие древовидно-раз-
ветвленные кустики с толстым стеблем. Анус
в виде крупного округлого отверстия, располо-
женного на боку тела под краем спины. На
границе первой и второй трети тела распола-
гается область генитальных отверстий, окру-
женных утолщенным бортиком.
Окраска. Общая окраска тела от розова-
той до ярко-оранжевой. Узкая белая линия
окантовывает ноту, сходная линия тянется по
переднему краю фронтального паруса. Мало-
заметная прерывистая белая линия может
иметься на дорсолатеральном крае тела между
жабрами. Выросты орального паруса и жабры
от ярких оранжево-красных до почти белых.
Ринофоральные выросты бледно-желтые или
коричневатые. Стебель ринофоров с белым
кончиком.
Отличия от похожих видов. По строению
тела и окраске хорошо отличается от других
голожаберных моллюсков фауны России.
150
Семейство Tritoniidae — Тритониевые или Тритонииды
Tritonia tetraquetra — Тритония тихоокеанская,
Tritonia tetraquetra — Тритония тихоокеанская,
вид со спинной стороны
вид с брюшной стороны
Tritonia tetraquetra — Тритония тихоокеанская
151
Отряд Nudibranchia — Голожаберные моллюски
| Tritonia tetraquetra — важный объект нейро-
физиологии. У этого вида отдельные нерв-
ные клетки могут достигать в диаметре 1 мм,
что облегчает манипуляции с ними в сравне-
нии с нейронами других объектов. На основе
моделей, построенных багодаря изучению
функционирования нервной системы трито-
нии, делаются, с определенными допущения-
ми выводы и о работе человеческого мозга.
Особенности экологии. Один из самых
массовых сублиторальных видов. Широко рас-
пространен на скалистых, каменистых и мяг-
ких грунтах. Может встречаться уже начиная
от глубины 2 м, но обычно обитает глубже, от
10-15 м. В Японском море обитает еще глубже,
ниже 30-40 м, и, по-видимому, на мягких, или-
стых и песчаных грунтах. Максимальная глуби-
на нахождения — 640 м. Особенности биоло-
гии этого крупного вида в морях России
в целом изучены недостаточно.
Размножение. Кладка в виде спирально
завитого шнура зеленоватого цвета, развитие
со свободно плавающим велигером.
Питание и поведение. Питается своеоб-
разными колониальными кишечнополостны-
ми из отряда морских перьев, например в во-
дах Северной Америки отмечено питание на
Ptilosarcus gurneyi, Virgularia spp., Stylatula elon-
gata. Причем, в зависимости от вида морских
перьев, окраска и размеры взрослых особей
тритонии, поедающих их, могут различаться.
Малоподвижный вид.
Распространение. Тихоокеанский боре-
альный вид. Широко распространен во всех
дальневосточных морях России, от Берингова
на севере, до Японского на юге, и в прибреж-
ных водах Северной Америки, от Алеутских
островов по меньшей мере до штата Калифор-
ния, более южные указания нуждаются в под-
тверждении.
СЕМЕЙСТВО DENDRONOTIDAE —
ДЕНДРОНОТУСОВЫЕ
ИЛИ ДЕНДРОНОТИДЫ
По наличию мощных кустистых выростов
вдоль боковых краев данное семейство напо-
минает семейство тритониид, от которого хо-
рошо отличается узким телом, небольшим
оральным парусом, несущим длинные развет-
вленные выросты, а также частично развет-
вленной пищеварительной железой, отростки
которой могут даже заходить в спинные раз-
ветвленные отростки. Радула характеризуется
мощным центральным зубом и значительным
числом боковых зубов. К этому семейству от-
носится лишь один род Dendronotus, центр раз-
нообразия которого приходится на северную
часть Тихого океана.
Dendronotus frondosus
(Ascani us, 1774) —
Дендронотус фрондозус
Особенности строения. Тело узкое, ораль-
ный парус относительно узкий, с небольшим
количеством разветвленных отростков. Вдоль
спины располагается в среднем 4-6 пар (у очень
крупных экземпляров их число может дости-
гать 10) характерных отростков, довольно длин-
ных и разветвленных. Веточки пищеваритель-
ной железы могут проникать в эти отростки,
а также в ринофоральные карманы, но книдо-
саков там не образуют. Карманы ринофоров
имеют латеральную папиллу. Строение спин-
ных отростков и выростов фронтального пару-
са ювенильных особей значительно отличается
от такового взрослых экземпляров. Так, особи
длиной около 1 мм имеют неразветвленные
выросты как на спине, так и на парусе, тогда
как у экземпляров длиной более 3 мм спинные
отростки дихотомически ветвятся, но вторич-
ные выросты очень короткие, так что создается
впечатление почти гладких отростков. Длина
тела обычно до 50 мм, но может достигать
100 мм и более.
Окраска. Общая окраска тела обычно пе-
страя — по беловатому фону разбросано мно-
жество коричневатых, розоватых, красноватых
пятен, полосок, которые иногда формируют
своеобразный узор. Кроме того, множество
снежно-белых и золотистых крапинок разбро-
сано по всему телу, включая жабры. Часто эти
крапинки располагаются внутри мелких бугор-
ков. У некоторых особей в жабры проникают
коричневатые просвечивающие веточки пище-
варительной железы. Наряду с пестрыми, встре-
чаются розовые и темно-красные, почти одно-
родные особи, также нередко покрытые
152
Семейство Dendronotidae — Дендронотусовые или Дендронотиды
золотистой «пылью». Наконец, как на Баренце-
вом, так и на Белом морях, часто можно обна-
ружить полупрозрачно-белых дендронотусов.
Верхние участки жабр у таких экземпляров
всегда полупрозрачные и бывают либо такого
же цвета, как и остальное тело, либо белыми
или золотисто-белыми за счет кончиков просве-
чивающей изнутри пищеварительной железы.
Однородно белые особи D. frondosus могут копу-
лировать с типичными, пестрыми экземпляра-
ми. Встречаются и переходные по окраске особи
между «пестрой» и «белой» формами.
Отличия от похожих видов. Однородно
белые и розоватые вариететы можно легко спу-
тать с Dendronotus dalli. Необходимо отметить,
что вероятней всего все указания тихоокеанско-
го вида Dendronotus dalli для северной Атланти-
ки и Белого моря основаны на неверном опре-
делении слабоокрашенных вариететов
Dendronotus frondosus. Достоверных указаний
Dendronotus dalli из Белого и Баренцева морей
на сегодняшний день нет. Dendronotus frondosus
четко отличается от Dendronotus dalli отсутстви-
ем непрозрачного белого пигмента на поверх-
ности кончиков жабр и выростов фронтально-
го паруса; вместо этого сквозь полупрозрачные
жабры изнутри просвечивают беловатые и/
или золотистые кончики ветвей пищевари-
тельной железы. Эти два вида также различа-
ются строением радулы. От второго, широко
распространенного в Белом море вида рода
Dendronotus — Dendronotus robustus, хорошо от-
личается как экологией — последний обитает
исключительно на мягких грунтах на глубине
свыше 10 м, так и морфологией — Dendronotus
robustus, оправдывая свое название, характери-
зуется массивным широким телом и очень ши-
роким оральным парусом.
Особенности экологии. В Баренцевом
и Белом морях является одним из главных ком-
понентов в гидроидном обрастании различных
твердых субстратов от литорали до глубины
10-15 м. Массовые скопления образует на ис-
кусственных конструкциях, например, мидие-
вых коллекторах. Весьма обычен также в обра-
стании гидроидных полипов Obelia longissima
Dendronotus frondosus — Дендронотус ф)рондозус,
вид со спинной стороны
Dendronotus frondosus — Дендронотус фрондозус,
вид с брюшной стороны
153
Отряд Nudibranchia — Голожаберные моллюски
Dendronotus frondosus (Дендронотус фрондозус) на пищевом объекте — колонии гидроидных полипов обелии
(обелии длиннейшей) на пластинах ламина-
рии (Laminaria saccharin а).
Размножение. Период размножения в Бе-
лом море — с конца мая по начало октября.
Кладка в виде узкого, извитого спирального
шнура, беловатого или светло-розового цвета.
В августе в Баренцевом море на литорали
встречается множество молоди этого вида. Раз-
витие со стадией личинки-велигера.
Dendronotus frondosus — Дендронотус фрондозус,
кладка
Питание и поведение. В Белом море
основным компонентом диеты этого вида яв-
ляется гидроид Obelia longissima. Кроме того,
отмечено питание гидроидом Diphasia falax. Ча-
сто обитает непосредственно на пищевом объ-
екте — колонии гидроидов, обкусывая гидро-
теки с помощью мощных челюстей и радулы.
Одну крупную колонию обелии может атако-
вать несколько десятков особей Dendronotus
frondosus одновременно. Может плавать корот-
кое время с помощью боковых волнообразных
изгибаний тела. Имеются старые данные о по-
едании Dendronotus frondosus в Британских во-
дах треской и камбалой, а также морской со-
бачкой Blennius pholis (редкий случай для
малосъедобных или ядовитых голожаберных
моллюсков), впоследствии, впрочем, не под-
твержденные. Для Белого моря случаи поеда-
ния Dendronotus frondosus рыбами и другими
хищниками также никогда не были докумен-
тированы. Хотя разветвленные спинные выро-
сты дендронотуса можно трактовать как крип-
тическую, защитную адаптацию, широкое
распространение красноватых и коричневатых
154
Семейство Dendronotidae — Дендронотусовые или Дендронотиды
| Один из видов рода Dendronotus, обитаю-
щий по тихоокенскому побережью Север-
ной Америки, Dendronotus iris (Cooper, 1863),
приобрел способность плавать с помощью
боковых изгибаний тела настолько хорошо,
что этот вид нападает на одиночных ане-
мон — цериантарий (Ceriantharia), обитаю-
щих в высоких трубках, подплывая к ним со
стороны венчика щупалец.
особей Dendronotus frondosus, хорошо заметных
на фоне гидроидов, свидетельствует в пользу
почти исключительно дыхательной функции
дорсальных отростков, иногда называемых так-
же вторичными жабрами.
Распространение. Амфибореальный вид,
обитающий в северной Атлантике и в северной
части Тихого океана, проникающий по азиат-
скому побережью на юг вплоть до Желтого мо-
ря, хотя уже в Японском море значительно
менее обилен, чем в Белом. Границы распро-
странения в Арктике точно не установлены, обы-
чен в Баренцевом море. Обитает, по-видимому,
во всех дальневосточных морях России. В Белом
море распространен повсеместно.
Dendronotus dalli
Bergh, 1879 —
Дендронотус Долла
Non Dendronotus dalli sensu Roginskaya, 1987
Особенности строения. Тело по строению
и размерам близко к описанному выше Den-
dronotus frondosus.
Окраска. Общая окраска тела однород-
ная — розоватая, беловатая, желтоватая. Са-
мой характерной чертой данного вида являют-
ся полоски белого непрозрачного пигмента,
которые покрывают сверху кончики развет-
вленных отростков на спине, оральном парусе,
ринофоральных карманах.
Отличия от похожих видов. Однородно бе-
лые и розоватые вариететы Dendronotus frondosus
можно легко спутать с Dendronotus dalli. Тем не ме-
нее отличить одного от другого можно благодаря
наличию у первого белых пигментных полосок,
покрывающих поверхность разветвленных выро-
стов. Напротив, имеющийся у некоторых одно-
родно окрашенных особей Dendronotus frondosus
белый пигмент на вершинах спинных выростов
всегда располагается внутри отростков, а не по-
крывает их снаружи, как у Dendronotus dalli.
Dendronotus dalli — Дендронотус Долла, Dendronotus dalli — Дендронотус Долла,
вид с правой стороны вид с брюшной стороны
155
Отряд Nudibranchia — Голожаберные моллюски
Dendronotus dalli — Дендронотус Долла
156
Семейство Dendronotidae — Дендронотусовые или Дендронотиды
Dendronotus dalli — Дендронотус Долла, кладка
Особенности экологии. Достаточно обы-
чен в гидроидном обрастании твердых субстра-
тов на глубинах 7-15 м, но может достигать
глубины более 100 м.
Размножение. В водах Камчатки размно-
жение отмечено в августе. Кладка в виде узкого
извитого спирального шнура, беловатого или
светло-розового цвета. Развитие со стадией
личинки-велигера.
Питание и поведение. В целом сходно
с таковым Dendronotus frondosus. Отмечено пи-
тание различными гидроидами.
Распространение. Тихоокеанский широ-
ко распространенный вид. В морях России,
по-видимому, является достаточно обычным
видом в прибрежных водах Камчатки, Коман-
дорских островов, Берингова моря, Северных
Курильских островов, но уже в Японском мо-
ре достоверно не отмечен. Широко распро-
странен также в прибрежных водах Северной
Америки, от Берингова пролива до штата Ва-
шингтон.
Dendronotus robustus
Verrill, 1870 -
Дендронотус крепкий
Особенности строения. Тело широкое, мас-
сивное, оральный парус широкий, с многочис-
ленными отростками. Вдоль спины располага-
ется в среднем 6-7 пар довольно коротких
разветвленных отростков. Карманы ринофоров
имеют латеральную папиллу. Особи из Белого
моря достигают 90 мм.
Окраска. Общая окраска тела коричнева-
то-красноватая, с разбросанными многочис-
ленными мелкими белыми пятнышками.
Кончики разветвленных спинных выростов
и отростков фронтального паруса покрыты
сверху белым пигментом.
Отличия от похожих видов. От другого
беломорского представителя рода Dendrono-
tus — Dendronotus frondosus, хорошо отличается
широким массивным телом, широким фрон-
тальным парусом и более короткими спинны-
ми отростками.
Особенности экологии. Dendronotus
robustus приспособлен к жизни на мягких, или-
стых и песчаных грунтах. Судя по имеющимся
данным, в Белом море, в отличие от Баренцева
моря, где он обнаружен на глубинах свыше
100 м, обитает уже с глубины 10-15 м. Макси-
мальная известная глубина обитания этого
вида в Белом море — 125 м. Впервые был
описан из района Новой Англии (США),
с литорали, но большая часть последующих
находок этого вида лежит на глубинах свыше
ста метров.
Размножение. Ничего не известно о раз-
множении этого вида, несмотря на, по-
видимому, достаточную обычность его на
доступных глубинах в Белом море. Особи,
собранные в районе Массачусетса в июле,
были половозрелые.
Питание и поведение. В отличие от дру-
гих видов рода Dendronotus, характеризуется
необычной диетой, включающей полихет,
детрит — попутно захваченный несъедобный
материал, и лишь небольшую часть гидроид-
ных колоний. Желудки особей из Двинского
залива Белого моря содержали множество
клубков полихет из семейства Oweniidae,
фрагменты их трубок, а, кроме того, фрагмен-
ты ракообразных, детрит, склеенные комки
песка, и даже — у одной особи — многочис-
ленные надкрылья жуков. В то же время фраг-
менты колоний гидроидов были представле-
ны в незначительном количестве. Широкое
тело позволяет данному виду успешно пере-
двигаться по поверхности ила и песка. Судя
по спектру пищевых предпочтений, Dendro-
notus robustus, по крайней мере в ряде районов
северной Атлантики и Субарктики, несомнен-
но предпочитает полихет, но может кроме
того безвыборочно захватывать с помощью
широкого фронтального паруса различных
беспозвоночных и попутный несъедобный
материал.
157
Отряд Nudibranchia — Голожаберные моллюски
Dendronotus robustus (Дендронотус крепкий) в естественном местообитании — на илисто-песчаном грунте
Распространение. Циркумполярный вид,
заходящий в северные части северной Атланти-
ки и северной части Тихого океана, на юге, воз-
можно, доходящий до Японского моря. В Ба-
ренцевом море обитает на глубинах свыше ста
метров, в то время как в Белом море, где обна-
ружен в Двинском и Кандалакшском заливах,
может подниматься значительно выше.
ид входит единственный, но очень обильный
по числу видов род Doto, широко представ-
ленный как в тропических, так и в умеренных
водах. Все без исключения дотоиды — мелкие
формы, обитающие среди гидроидов и пита-
ющиеся ими.
В морях России встречается, по меньшей
мере, два вида.
СЕМЕЙСТВО DOTOIDAE —
ДОТОИДЫ
Своеобразное семейство, характеризующееся
значительным числом редукционных черт,
если за точку отсчета взять организацию три-
тониид и дендронотид. Вторичные развет-
вленные жабры преобразуются в цельные
шишковидные отростки по краям спины,
оральный парус сильно уменьшается в разме-
рах и теряет характерную для дендронотацей
форму, ринофоры превращаются в простой
гладкий стебель, но сохраняют при этом ри-
нофоральные карманы. Во внутреннем строе-
нии также имеются существенные черты ре-
дукции — в радуле остается только один
крупный центральный зуб. В семейство дото-
Doto coronate
(Gmelin, 1791) -
Дото короната
Особенности строения. Тело узкое, слабо су-
живается к заднему концу. Спинная сторона
несет с каждого бока по одному ряду крупных
булавовидных отростков, покрытых, в свою
очередь, большим числом мелких конических
бугорков, из-за чего они несколько напомина-
ют по форме шишку. Оральные щупальца от-
сутствуют, вместо них имеется очень малень-
кий полукруглый оральный парус. Гладкие
конические ринофоры способны втягиваться
внутрь крупных воронковидных ринофораль-
ных карманов. Передний край ноги без боко-
вых отростков. Длина тела до 10 мм.
158
Семейство Dotoidae — Дотоиды
Doto coronata — Дото короната,
вид со спинной стороны
Doto coronata — Дото короната,
вид с брюшной стороны
Группа особей Doto coronata (Дото короната), демонстрирующая размах изменчивости
159
Отряд Nudibranchia — Голожаберные моллюски
Окраска. На спинной стороне по белова-
тому фону разбросано большое число сливаю-
щихся мелких пятен и крапинок, цвет которых
может варьировать в довольно широких преде-
лах — от светло-розоватого до темно-фиолето-
вого и темно-коричневого. Большинство из
бугорков на спинных отростках также марки-
рованы одним более темным пятном.
Отличия от похожих видов. Виды из рода
Doto обладают очень характерным габитусом,
который трудно спутать с таковым других ви-
дов голожаберных моллюсков. Помимо Doto
coronata, в Баренцевом море обитает только
еще один вид из этого рода — Doto fragilis, ко-
торый отличается от первого вида почти одно-
родной светло-коричневатой окраской и за-
метно более широким телом.
Особенности экологии. Наиболее значи-
тельной численности данный вид достигает
в сообществе обрастания плавучих искусствен-
ных сооружений — понтонов, буев и пр. В этом
биотопе Doto coronata обитает на колониях ги-
дроидных полипов Obelia longissima, покрываю-
щих широкие слоевища бурой водоросли ла-
минарии. Был обнаружен также под камнями
в нижнем горизонте литорали. Может также
спускаться до глубины 10-15 м.
Размножение. В Баренцевом море раз-
множение отмечено с конца июля по начало
сентября. Кладка представляет собой довольно
узкую розоватую ленту, часто завитую в виде
неправильных спиралей. Развитие со стадией
свободноплавающей личинки-велигера.
Питание и поведение. Медленно пере-
двигается среди столонов гидроидных поли-
пов, высверливает радулой в них отверстия
и высасывает их содержимое.
Doto coronata — Дото короната,
кладка
Распространение. Атлантический боре-
альный вид. В морях России обнаружен только
вдоль Мурманского побережья Баренцева
моря. По европейскому побережью на юг от-
мечен вплоть до Испании.
Doto fragilis
(Forbes, 1838) -
Дото хрупкий
Особенности строения. Тело относительно
широкое, слабо суживается к заднему концу.
Спинная сторона несет по бокам по одному
ряду крупных булавовидных отростков, покры-
тых, в свою очередь, большим числом мелких
конических бугорков, из-за чего они несколько
напоминают по форме шишку. Оральные щу-
пальца отсутствуют, вместо них имеется очень
маленький полукруглый фронтальный парус.
Гладкие конические ринофоры способны втя-
гиваться внутрь крупных воронковидных рино-
форальных карманов. Передний край ноги без
боковых отростков. Длина тела до 6 мм.
Doto fragilis — Дото хрупкий,
вид со спинной стороны
160
Семейство Embletoniidae — Эмблетониевые или Эмблетонииды
Doto fragilis — Дото хрупкий,
вид с брюшной стороны
Окраска. Общий цвет тела однородный,
светло-коричневый, кремовый. Бугорки на
спинных выростах, края ринофоральных кар-
манов и фронтального паруса маркированы
мелкими белыми крапинками.
Отличия от похожих видов. От другого
баренцевоморского вида из рода Doto, Doto
coronata хорошо отличается почти однород-
ной светло-коричневатой окраской и заметно
более широким телом.
Особенности экологии. Обитает на ка-
менистых и скалистых субстратах на глубине
около 7-10 м.
Размножение. В морях России не изу-
чено.
Питание и поведение. Малоподвижный
вид. Питается гидроидными полипами.
Распространение. Атлантический бо-
реальный вид. В морях России обнаружен
только вдоль Мурманского побережья Ба-
ренцева моря. По европейскому побережью
на юг отмечен до атлантического побережья
Испании.
СЕМЕЙСТВО EMBLETONIIDAE —
ЭМБЛЕТОНИЕВЫЕ
ИЛИ ЭМБЛЕТОНИИДЫ
Крайне специализированное семейство, воз-
можно продолжающее редукционные тенден-
ции предшествующего семейства дотоид. Так-
же как и у дотоид, у эмблетониид имеется
небольшое число папилл еще более простого
строения и маленький оральный парус. Ри-
нофоры простые гладкие, без карманов. Во
внутреннем строении между дотоидами
и эмблетониидами тем не менее имеются
весьма существенные различия.
К этому семейству относится лишь один
род Embletonia с небольшим числом видов,
один из которых был совсем недавно обнару-
жен нами в интерстициали на Мурманском
побережье Баренцева моря. Склонность к оби-
танию в интерстициали была отмечена для
этого вида и в других частях ареала.
Embletonia pulchra
(Alder et Hancock, 1844) —
Эмблетония прелестная
Особенности строения. Тело очень узкое,
почти червеобразное. Спинная сторона несет
с каждого бока по одному ряду крупных, удли-
ненных, гладких отростков. Оральные щупаль-
ца отсутствуют, вместо них имеется маленький
двулопастной фронтальный парус. Гладкие ко-
нические ринофоры лишены ринофоральных
карманов. Передний край ноги без боковых от-
ростков. Длина тела до 7,5 мм.
Окраска. Общий цвет тела однородный,
светло-коричневый, кремовый. Мелкие белые
крапинки разбросаны по спине и папиллам.
Отличия от похожих видов. Сходных видов
в Баренцевом море нет. В Черном и Японском мо-
рях обитает внешне сходная Tenellia adspersa, отли-
чающаяся от Embletonia pulchra закругленным, а не
двулопастным фронтальным парусом
Особенности экологии. В отличие от
большинства других голожаберных моллюсков,
обитает в очень необычной среде — в интерсти-
циали, особом пространстве между частицами
крупнозернистого песка и битой ракушки. На
Баренцевом море в подобном местообитании
Embletonia pulchra обнаружена на литорали.
Размножение. В морях России не изучено.
161
Отряд Nudibranchia — Голожаберные моллюски
Embletonia pulchra — Эмблепюния прелестная,
вид со спинной стороны
Питание и поведение. Подвижный вид.
Питание не изучено.
Распространение. Атлантический боре-
альный вид. В морях России обнаружен только
вдоль Мурманского побережья Баренцева мо-
ря. По европейскому побережью на юг отмечен
вплоть до Средиземного моря включительно.
| Долгое время полагали, что представители
рода Embletonia несут на вершинах папилл
мешочки для хранения стрекательных клеток
(книдосаки), подобно видам из подотряда
Aeolidacea. Считалось также, что в отличие от по-
следних, эмблетонииды могут нести сразу не-
сколько мешочков — книдосаков на вершине
каждой папиллы, выглядящих как подушечки.
Новейшие исследования наших коллег из Гер-
мании между тем продемонстрировали, что ви-
ды из рода Embletonia действительно могут на-
капливать стрекательные клетки на вершинах
папилл, но привычного для подотряда эолида-
цей книдосака при этом не образуется.
Embletonia pulchra — Эмблепюния прелестная,
Embletonia pulchra — Эмблепюния прелестная,
детали строения спинной папиллы
вид с брюшной стороны
162
Семейство Flabellinidae — Флабеллиновые или Флабеллиниды
ПОДОТРЯД AEOLIDACEA — ЭОЛ ИДАЦЕИ
Все последующие виды относятся к другой большой подгруппе голожаберных моллюсков, тра-
диционному подотряду Aeolidacea, или эолидам. Подавляющее большинство эолид выработали
очень специфический тип защиты. Пищеварительная железа трансформируется у них из цель-
ного органа в систему разветвленных трубок, выросты которых проникают в спинные папиллы,
формируя в дистальной части папиллы особый небольшой мешочек — книдосак. В книдосаке
накапливаются неразряженные стрекательные клетки — нематоцисты, поступающие в пищева-
рительную железу эолид вместе с их основной пищей — различными книдариями. Таким об-
разом, стрекательные клетки книдарий используются для защиты чужого организма — голожа-
берного моллюска. Подобная адаптация возникла не сразу. В целом ряде ныне существующих
видов можно проследить практически все этапы постепенного становления этого сложного за-
щитного приспособления. Так, есть группы, уже имеющие папиллы, но с цельной пищевари-
тельной железой; у других форм пищеварительная железа проникает в спинные выросты, но
книдосаки еще не формируются (как у дендронотусов); и, наконец, группа эолид — характери-
зуется полным набором органов для защитного использования нематоцистов. Голожаберные
моллюски имеют очень долгую историю приспособления к питанию книдариями. Так, архаич-
ные Pleurobranchaea, еще сохраняющие настоящую жабру — ктенидий на правой стороне тела,
уже могут поедать актиний. Последующая эволюция одного из главных стволов голожаберных
моллюсков была во многом направлена на появление узкоспециализированных на питании кни-
дариями таксонов и привела, в конечном счете, к появлению эолид.
В пределах подотряда эолидацей совершенно отчетливо выражена эволюционная тенден-
ция к упрощению организации и сокращению числа исходно значительного числа рядов па-
пилл, неоднократно возникающая в разных семействах. Эта тенденция сопровождалась (а в не-
котором смысле и была следствием) все большей «ювенилизацией» взрослой половозрелой
формы, т. е. образованием педоморфных видов. В крайних случаях, как и в случае с корамбида-
ми из отряда дорид (см. стр. 144), это приводит к появлению фактически половозрелых особей
с ярко выраженными признаками ранних ювенильных экземпляров. Среди эолидацей к по-
добной ювенильной морфологии приближается ряд мелких видов из семейства тергипедид,
один из которых — Tenellia adspersa во взрослом состоянии обладает всеми признаками ранних
ювенильных особей — наличие маленького орального паруса вместо оральных щупалец в со-
четании с небольшим числом папилл и чрезвычайно коротким жизненным циклом.
СЕМЕЙСТВО FLABELLINIDAE —
ФЛАБЕЛЛИНОВЫЕ
ИЛИ ФЛАБЕЛЛИНИДЫ
Обширное семейство эолидацей, еще сохра-
няющее ряд архаичных черт, объединяющих
их с подотрядом арминацей. К таким призна-
кам относится широкий край нотума, имею-
щийся у некоторых видов, а также боковое по-
ложение анального отверстия. Радула с двумя
дополнительными боковыми зубами, которые
могут быть рассмотрены как остатки многочис-
ленных рядов зубов, имеющихся у арминацей.
У некоторых флабеллинид сохраняется (или
восстанавливается заново) совсем древний при-
знак невтяжного копулятивного аппарата, ха-
рактерный для раковинных актеонид и ряда
нотаспид.
Флабеллиниды особенно разнообразны
в умеренных и арктических водах (включая
фауну России), но имеются и тропические
представители.
Chlamylla atyplca
(Bergh, 1899) —
Хлямилля нетипичная
Особенности строения. Тело широкое. Па-
пиллы образуют вдоль каждой из сторон спи-
ны сплошной покров, без видимых переры-
вов. Нотум хорошо развит, формируя
выступающие боковые края спины. Сильно
разветвленные веточки пищеварительной же-
лезы под кожей выглядят как две сплошные
полосы по краям спины. Пищеварительный
163
Отряд Nudibranchia — Голожаберные моллюски
Chlamylla atypica — Хлямилля нетипичная, вид елевой стороны
Chlamylla atypica — Хлямилля нетипичная,
вид со спинной стороны
вырост в папиллах в виде узловатого шнура,
чаще занимает лишь среднее положение в па-
пилле, но может заполнять и больший объем.
Ринофоры крупные, конические, у большин-
ства особей длиннее оральных щупалец при-
близительно в 1,5 раза. Отростки переднего
края ноги отсутствуют. Анус открывается
с правой стороны в середине тела под закраи-
ной (краями нотума). Область половых отвер-
стий окружена комплексом складок. Впереди
ануса, под закраиной, начинается волнистая
кожная складка, которая переходит в непра-
вильно треугольный по форме воротничок пе-
ниса. На переднем конце этого воротничка на-
ходится отверстие пениса. Длина до 50 мм.
Окраска. Основной цвет тела беловато-
желтоватый. Имеются две вариации окраски
пищеварительной железы (включая выросты
в папиллах) — оранжевая и розовая. В районе
перикардия, а также на ринофорах и ораль-
ных щупальцах разбросаны белые пятнышки
и крапинки. На верхней части папилл распо-
лагается широкое, но прерывистое белое
кольцо.
Отличия от похожих видов. Данный вид
хорошо отличается от Coryphella nobilis отсут-
ствием передних боковых отростков у ноги,
а также наличием мощной складки-воротника
вокруг наружного пениального отверстия,
продолжающейся довольно длинной про-
дольной складкой в сторону анального отвер-
164
Семейство Flabellinidae — Флабеллиновые или Флабеллиниды
стия, хорошо заметной, если рассматривать
моллюска с правой боковой стороны. От Со-
ryphella salmonacea хорошо отличается про-
стой, а не в форме листа клевера, формой
головы. Наконец, еще один вид широкотелых
флабеллинид — Coryphella polaris, четко отли-
чается от Chlamylla atypica яркой желтоватой
общей окраской тела и очень длинными ри-
нофорами, которые почти в 3-4 раза длиннее
оральных щупалец.
Особенности экологии. В Белом море
обнаружен на мягких грунтах, на глубинах
15-20 м. Возможно, является довольно обыч-
ным видом, но редко указывался из-за недо-
статочной изученности беломорских субли-
торальных голожаберных моллюсков.
Примечательно географическое распростра-
нение этого вида в зависимости от темпера-
турных условий. Судя по предпочитаемым
температурам нереста (от -2° с до +5° С),
Chlamylla atypica является настоящим арктиче-
ским видом. В южной части ареала, в заливе
Петра Великого Японского моря, Chlamylla
atypica выходит для размножения на литораль
исключительно в зимний период, когда темпе-
ратуры достигают предельно низких, арктиче-
ских значений. В летний период, когда темпе-
ратуры в заливе Петра Великого могут
подниматься выше +20° С, Chlamylla atypica на
литорали полностью отсутствует. В условиях
Белого моря этот вид может найти подходя-
щие низкие температуры круглогодично на
относительно небольшой глубине. Интересно
отметить, что на литорали этот вид в Белом
море не встречается. Наиболее удивительным
является распространение этого вида на юг
вплоть до Восточно-Корейского залива (Север-
ная Корея). На мелководье в этом заливе тем-
пература моря столь высока, что позволяет
существовать там субтропической фауне (со-
гласно нашим данным, там обитает настоя-
щий субтропический вид Homoidoris japonica),
в то время как на глубинах свыше 100 м темпе-
ратура остается низкой, что фактически дает
возможность проникать в субтропики таким
арктическим видам, как Chlamylla atypica.
Размножение. Репродуктивные особен-
ности этого вида известны только для Япон-
ского моря, где размножение на литорали
происходит с конца декабря по март включи-
тельно. Кладка в виде широкой ленты белова-
того или розоватого цвета, завитой в спираль.
Развитие со стадией личинки-велигера. В каж-
дой яйцевой капсуле содержится до 10 заро-
дышей, что не характерно для других голожа-
берных моллюсков.
Питание и поведение. Пищевые предпо-
чтения этого вида в Белом море не известны.
В Японском море на литорали питается оди-
ночными атекатными гидроидами. Может
обитать как на мягких, так и на твердых грун-
тах. В период размножения может образовы-
вать на литорали массовые скопления.
Распространение. Арктическо-бореаль-
ный вид. Гренландия (Дэвисов пролив), Барен-
цево и Чукотское моря (вероятно, распростра-
нена в Арктике шире), все дальневосточные
моря России; на юге доходит до Восточно-
Корейского залива (Северная Корея), где опу-
скается на глубины свыше 100 м. В Белом море
известен из Кандалакшского залива.
Paracoryphella islandica
(Odhner, 1937) -
Паракорифелла исландская
Особенности строения. Тело широкое, не-
сколько суженное к заднему концу. Нотум по
краю хорошо выражен, нависает над боками
тела. Папиллы многочисленными неправиль-
ными рядами покрывают спину с обоих краев,
оставляя свободной довольно широкую сред-
нюю ее часть. Папиллы удлиненно-пальце-
видные. Вырост печени в них в виде шнура,
занимающего большую часть папиллы. Ветви
пищеварительной железы сильно разветвлен-
ные. Ринофоры небольшие, конические, у всех
просмотренных особей меньше крупных
оральных щупалец в 1,5 раза. Край ноги обра-
зует впереди короткие треугольные отростки.
Анус открывается около середины тела под
краем нотума. Характерной особенностью дан-
ного вида является невтяжной копулятивный
орган, располагающийся в передней части те-
ла справа. Длина до 30 мм.
Окраска. Основная окраска покровов
оранжевая. По всей спине, основаниям папилл
и немного по ним самим разбросаны мелкие
белые пятнышки. Ринофоры и оральные щу-
пальца на концах присыпаны белой пылью.
Цвет ветвей печени и выростов ее в папиллах
светло-коричневый. У оснований ринофор на-
ходятся черные глаза. За ними просвечивает
165
Отряд Nudibranchia — Голожаберные моллюски
Paracoryphella islandica — Паракорифелла исландская,
вид со спинной стороны
Paracoryphella islandica — Паракориф)елла исландская,
вид с брюшной стороны
розовый овальный желудок, вытянутый попе-
речно и продолжающийся в розовую кишку,
которую можно проследить под закраиной.
Отличия от похожих видов. Можно спу-
тать с другими видами из семейства флабелли-
нид, которые обладают широким телом, таки-
ми как Chlamylla atypica, Coryphella polaris
и Coryphella nobilis или Coryphella trophina. По-
мимо деталей окраски, Paracoryphella islandica
хорошо отличается от всех этих видов коротки-
ми ринофорами, которые в 1,5-2 раза короче
мощных оральных щупалец.
Особенности экологии. Встречается от
литорали до глубины 3-5 м на каменистых
грунтах.
Размножение. Особенности размноже-
ния этого вида впервые были изучены в зали-
ве Петра Великого Японского моря. Размно-
жение происходит с декабря по апрель при
температуре от -1,5° С до +7° С. Кладка в виде
узкого шнура белого или розоватого цвета, за-
крученного против часовой стрелки в спираль
из 4-5 оборотов. В каждой яйцевой капсуле,
размеры которых варьировали от 115x97 до
139x100 мкм, находилось от одного до двух за-
родышей. Развитие при температуре +4° С
шло около месяца, затем из кладки вышли
свободноплавающие велигеры с турбоспи-
ральной раковинкой.
Питание и поведение. Питается атекат-
ными гидроидами Hataia parva. Малоподвиж-
ный вид.
Распространение. Своим видовым на-
званием — исландская — данный вид обязан
тому обстоятельству, что его впервые обнару-
жили и описали у берегов Исландии. Инте-
ресно, что, несмотря на это, Paracoryphella islan-
dica у берегов «острова вулканов» до сих пор
не была повторно найдена, а большинство до-
стоверных находок происходит из морей Рос-
сии. Данный вид, по всей вероятности, может
быть назван амфибореальным. В настоящее
время он отмечен из Баренцева, Белого морей,
прибрежных вод Камчатки и в Японском мо-
ре, а также, вероятно, распространен и в водах
Аляски.
166
Семейство Flabellinidae — Флабеллиновые или Флабеллиниды
Coryphella polaris
Volodchenko, 1946 —
Корифелла полярная
Особенности строения. Тело широкое. Па-
пиллы образуют вдоль каждой из сторон спи-
ны сплошной покров, без видимых перерывов.
Нотум хорошо развит, формируя выступаю-
щие боковые края спины. Вырост пищевари-
тельной железы в папилле очень тонкий, ните-
видный. Ринофоры необычайно крупные,
конические, приблизительно в 3 раза длиннее
оральных щупалец. Отростки переднего края
ноги отсутствуют. Анус открывается с правой
стороны, в середине тела под закраиной (края-
ми нотума). Пениальный воротничок отсут-
ствует. Длина до 40 мм.
Окраска. Основной цвет тела (включая па-
пиллы) ярко-желтый. Оральные щупальца
и ринофоры покрыты на вершинах белым пиг-
ментом. На вершинах папилл располагается
широкое белое кольцо (заходящее на самый
кончик папиллы), хорошо заметное на общем
желтом фоне. Окраска пищеварительного вы-
роста в папиллах маскируется желтой окра-
ской внешнего чехла папиллы.
Отличия от похожих видов. От других ви-
дов беломорских флабеллинид с широким те-
лом хорошо отличается яркой желтой общей
окраской, мощными длинными ринофорами
и очень тонкими выростами пищеварительной
железы в папиллах.
Особенности экологии. Почти ничего не
известно о биологии этого чрезвычайно редко-
го вида. За всю историю его изучения собраны
всего лишь несколько экземпляров. Благодаря
новым данным по Белому морю, удалось вы-
яснить, что он может обитать на каменистом
заиленном грунте.
Размножение. Не изучено.
Питание и поведение. Возможно, питает-
ся крупными атекатными гидроидами. Мало-
подвижный вид.
Распространение. Арктическо-бореаль-
ный вид. В Арктике — Баренцево, Карское, Во-
сточно-Сибирское моря, в Тихом океане —
Охотское море, прибрежные воды Южных
Курильских островов.
Coryphella polaris (Корифелла полярная) в естественном местообитании — на заиленном каменистом грунте
167
Отряд Nudibranchia — Голожаберные моллюски
Coryphella salmonacea
(Couthouy, 1838) —
Корифелла лососевая
Особенности строения. Тело широкое. Па-
пиллы образуют вдоль каждой из сторон спи-
ны сплошной покров, без видимых перерывов.
Нотум умеренно развит, формируя выступаю-
щие боковые края спины. Пищеварительный
вырост в папиллах занимает значительный объ-
ем. Ринофоры крупные, конические, длиннее
оральных щупалец приблизительно в 1,5 раза.
Голова у многих, но, по-видимому, не у всех
особей, в форме трилистника — характерная
черта данного вида. Имеются короткие тре-
угольные отростки переднего края ноги. Анус
открывается с правой стороны, в середине тела
под закраиной (краями нотума). Область по-
ловых отверстий без заметных складок. Длина
до 30 мм.
Окраска. Необычное название вида связа-
но с нежно-оранжевой, «лососевой окраской»
выростов пищеварительной железы в папил-
лах, а также ринофоров и оральных щупалец.
Общая окраска тела более светлая.
Отличия от похожих видов. От всех видов
широкотелых флабеллинид, обитающих в Ар-
ктическом бассейне, Coryphella salmonacea отли-
чается формой головы в виде листа клевера.
Особенности экологии. По-видимому,
предпочитает мягкие грунты с примесью камней.
В распределении этого вида по глубинам имеется
примечательная особенность: в высокой Арктике,
у Земли Франца Иосифа, может обитать на са-
мом мелководье, около 2 м глубины, тогда как
южнее вид заметно тяготеет к более глубоким
местообитаниям — так на Мурманском побере-
жье Баренцева моря, по-видимому, мельче, чем
на глубине 50 м не обитает. Отмечен до 340 м.
Размножение. В морях России не изучено.
Питание и поведение. Может питаться
роющими актиниями и гидроидами. Умерен-
но подвижный вид.
Распространение. Арктический вид. Оби-
тает в Баренцевом, Белом, Карском, Лаптевых
и Восточно-Сибирском морях. В дальневосточ-
ных морях на больших глубинах, обычно ниже
100 м, обитает очень близкий вид Coryphella
japonica Volodchenko, 1941, достоверно отличаю-
щийся по строению копулятивного аппарата.
Coryphella salmonacea (Корифелла лососевая) в естественном местообитании — на скалистом грунте
168
Семейство Flabellinidae — Флабеллиновые или Флабеллиниды
Coryphella sahnonacea — Корифелла лососевая
Coryphella nobllls
(Verrill, 1880) -
Корифелла благородная
Особенности строения. Тело широкое. Па-
пиллы образуют вдоль каждой из сторон спи-
ны сплошной покров, без видимых перерывов.
Нотум хорошо выражен, заметно выступает
над боками тела, вдоль него располагается
часть папилл. Вырост пищеварительной желе-
зы в папиллах в виде тонкого шнура, занимаю-
щего только середину папиллы. Папиллы лан-
цетовидные, довольно широкие. Ринофоры
в мелких складках и морщинках, примерно
в 1,5 раза длиннее оральных щупалец. Края
ноги образуют впереди короткие отростки.
Анус открывается сбоку с правой стороны тела,
в его передней половине. Длина до 30 мм.
Окраска. Общая окраска тела беловатая
или слегка желтоватая. Дистальная половина
оральных щупалец покрыта редкими белыми
крапинками, которые концентрируются при
сокращении щупалец, иногда формируя дор-
сально сверху тонкую белую полоску. Верхняя
четверть ринофор покрыта сходной белой
«пылью». На хвосте короткая широкая белая
полоска. Цвет ветвей пищеварительной железы
и ее выростов в папиллах оранжеватый. Па-
пилла на своей вершине с широким белым
колечком, самый кончик папиллы, как прави-
ло, остается полупрозрачным.
Отличия от похожих видов. От обычной
в Белом море Coryphella verrucosa данный вид
хорошо отличается заметно более широким
телом и непрерывными рядами папилл. Co-
ryphella nobilis можно спутать с другими широ-
котелыми флабеллинидами, также известны-
ми в Белом море, прежде всего с Chlamylla
atypica, которая, по-видимому, также не редка
на глубинах 15-25 м в Кандалакшском заливе.
Этот вид хорошо отличается от Coryphella nobilis
отсутствием передних боковых отростков у но-
ги, а также наличием мощной складки-ворот-
ника вокруг наружного пениального отверстия,
продолжающейся в сторону анального отвер-
стия и хорошо заметной с правой боковой сто-
роны. Другой вид, с которым Coryphella nobilis
можно было бы легко спутать, — Coryphella sal-
топасеа, но достоверные современные находки
последнего вида в Белом море отсутствуют.
169
Отряд Nudibranchia — Голожаберные моллюски
Coryphella nobilis — Корифелла благородная
Coryphella nobilis — Корифелла благородная, вид со спинной стороны
170
Семейство Flabellinidae — Флабеллиновые или Флабеллиниды
В отличие от Coryphella nobilis Coryphella salmona-
cea обладает более широким телом, более
плотно покрытым папиллами. Кроме того,
они достовено различаются по характерной
форме головы — у Coryphella salmonacea с вен-
тральной стороны голова напоминает клевер-
ный лист, и также по почти полному отсут-
ствию боковых отростков на ноге у этого вида.
Наконец, еще один вид широкотелых флабел-
линид — Coryphella polaris, четко отличается
яркой желтоватой общей окраской тела
и очень длинными ринофорами, которые поч-
ти в 3-4 раза длиннее оральных щупалец.
Особенности экологии. По меньшей ме-
ре в условиях Белого моря относительно ши-
рокое тело Coryphella nobilis позволяет этому
виду успешно существовать на мягких грунтах,
избегая таким образом конкуренции с самым
массовым беломорским видом корифелл, Co-
ryphella verrucosa, обитающим на твердых грун-
тах (или в обрастании), в более мелких водах.
В Белом море приурочен к сублиторали,
вплоть до максимальных глубин, от 25 до 250 м
и, вероятно, мягким грунтам, хотя, возможно,
встречается и на каменистых грунтах, как это
имеет место у берегов Новой Англии. Судя по
батиметрическому распространению этого ви-
да в Белом море, Coryphella nobilis предпочитает
низкие температуры.
Размножение. Особенности размноже-
ния в Белом море не известны. У берегов Новой
Англии размножение зарегистрировано с мая
по август, кладка в виде узкой волнистой лен-
ты. Развитие со стадией личинки-велигера.
Питание и поведение. До настоящего
времени особенности питания Coryphella nobilis
в Белом море были неизвестны. Возможно,
в диету данного вида входят крупные гидроид-
ные полипы Tubularia и Sertularia cupressina. На
глубинах свыше 50 м, где тубулярия отсутству-
ет, Coryphella nobilis вероятно питается роющи-
ми актиниями. Учитывая крупный размер ги-
дрантов тубулярий, Coryphella nobilis скорее
всего не может заглатывать их целиком, пере-
тирая ножку, как поступают другие виды кори-
фелл с гидроидами, у которых гидранты мель-
че, а обкусывает их постепенно, начиная
с щупалец.
Распространение. Северная Атлантика,
по европейскому побережью — у Исландии,
Фарерских островов и северной Норвегии, Грен-
ландии; недавнее указание из вод Ирландии
нуждается в подтверждении; по американскому
побережью — у Новой Англии; в Арктике — Ба-
ренцево, Белое, Карское и Восточно-Сибирское
моря; в Тихом океане — во всех дальневосточ-
ных морях России.
Coryphella gracilis
(Alder et Hancock, 1844) —
Корифелла изящная
Особенности строения. Тело узкое. Папиллы
распадаются на несколько неотчетливых групп.
Нотум умеренно выражен, немного выступает
над боками тела. Папиллы удлиненно-
пальцевидные, довольно тонкие. Вырост пище-
варительной железы в них в виде толстого
шнура, занимающего почти всю папиллу. Ри-
нофоры гладкие, примерно в 1,5 раза короче
длинных оральных щупалец или почти равны
им. Края ноги образуют впереди довольно
длинные отростки. Длина до 25 мм.
Окраска. Общая окраска тела полупро-
зрачно-беловатая. На оральных щупальцах
Coryphella gracilis — Корифелла изящная,
вид со спинной стороны
171
Отряд Nudibranchia — Голожаберные моллюски
Coryphella gracilis — Корифелла изящная,
вид с брюшной стороны
сверху обычно имеется белая полоска. Разроз-
ненные белые крапинки заметны с внутренней
стороны верхней трети ринофоров. На хвосте
слабо выраженная белая полоска, которая
может отсутствовать. В передней части тела
у крупных особей отчетливо просвечивает не-
сколько витков семяпровода. Цвет ветвей пи-
щеварительной железы и ее выростов оранже-
ватый. Папилла на своей вершине с довольно
узким белым прерывистым колечком, самый
кончик папиллы остается полупрозрачным.
Отличия от похожих видов. Легко спутать
с обычным беломорским видом корифелл,
Coryphella verrucosa. Заметно отличается от по-
следнего более узким, грациозным телом
с меньшим количеством папилл (у экземпля-
ров сопоставимых размеров), передним краем
головы с постоянной, отчетливой выемкой,
более длинными передними отростками но-
ги и отчетливо просвечивающими сквозь
переднюю часть спинной стороны, за рино-
форами, несколькими петлями семяпровода.
Последний признак, особенно у крупных эк-
земпляров, позволяет безошибочно отличить
Coryphella gracilis от Coryphella verrucosa. У по-
следней семяпровод очень короткий, не об-
разует подобных петель и практически не
просвечивает дорсально.
Особенности экологии. Обитает на каме-
нистых и скалистых грунтах. В Баренцевом море
весьма обычен, а на литорали даже значительно
преобладает по численности над обычной в Бе-
лом море Coryphella verrucosa; предпочитает рас-
щелины в скалах, поросшие гидроидами из
рода эудендриум (Eudendrium). На Белом море
более редок, немногочисленные находки также
приурочены к литорали, где этот вид находили
совместно с Coryphella verrucosa.
Размножение. В Баренцевом море раз-
множение отмечено в августе. На литорали
в это время обнаружена многочисленная мо-
лодь. Кладка в виде узкого шнура беловатого
или розоватого цвета, завитого в спираль. Раз-
витие со стадией личинки-велигера.
Питание и поведение. В Баренцевом мо-
ре вид четко приурочен к колониям атекатного
гидроида Eudendrium annulatum, которым
и питается. В водах Британии, а также Новой
Англии, Coryphella gracilis также предпочитает
питаться гидроидами из рода Eudendrium, хо-
тя особи, найденные на глубине свыше 25 м,
поедали других гидроидов.
Обычно с одной достаточно крупной коло-
нии Coryphella annulatum можно снять несколько
десятков как крупных, так и ювенильных экзем-
пляров Coryphella gracilis, постоянно обитающих
среди веточек гидроида. На литорали под кам-
нями можно обнаружить мелких особей Co-
ryphella gracilis, без какой-либо видимой связи
с колониями Eudendrium. Пищевое поведение
заключается в нападении на отдельные полипы
колонии, незащищенные у эудендриума твер-
дой текой, при этом корифелла, широко рас-
крыв ротовое отверстие, фактически натягивает
свою голову на полип, а затем, совместными
движениями челюстей и радулы, откусывает его
у ножки и постепенно проглатывает, продол-
жая перетирать полип радулой.
Coryphella gracilis — Корифелла изящная,
кладка
172
Семейство Flabellinidae — Флабеллиновые или Флабеллиниды
Распространение. Амфиатлантический
вид; Северная Атлантика — по европейскому
побережью от Норвегии до Франции (имеется
одно старое указание из Испании) и по амери-
канскому побережью от Ньюфаундленда до
Новой Англии. В водах России обитает по
Мурманскому побережью Баренцева моря
и Белом море. В дальневосточных морях Рос-
сии обитает вид-двойник, Coryphella amabilis
(Hirano et Kuzirian, 1991).
Coryphella amabilis
(Hirano et Kuzirian, 1991) —
Корифелла амабилис
Особенности строения. Тело моллюска тон-
кое и изящное. Оральные щупальца пальце-
видные, закругленные на конце и отогнуты на-
зад. Ринофоры гладкие, примерно в 1,5 раза
длиннее оральных щупалец. С каждой сторо-
ны спины вдоль ее края имеется узкая закраи-
на (край рудиментарного нотума), которая
тянется до самого конца тела. На закраине рас-
полагаются три группы папилл — одна перед-
няя и две задние. Группы состоят из 1-8 папилл.
Папиллы пальцевидные или веретеновидные,
с приостренными или закругленными верши-
нами. Голова впереди ровная. Нога довольно
узкая, впереди имеет по бокам очень короткие
треугольные отростки. Анус открывается во
второй половине тела, недалеко от его середи-
ны, под первой папиллой 2-й группы папилл.
Длина до 10 мм.
Окраска. Общая окраска тела беловатая.
Цвет печеночного выроста в папиллах светло-
кирпичный или розовато-красноватый. На верх-
них частях папилл разбросаны снежно-белые
пятнышки, у части папилл эти крапинки могут
формировать полукольцо. Сквозь спину просве-
чивают: беловатые ганглии с черными глазами,
петли семяпровода, прозрачный перикардий
с сердцем, розоватая гонада. Оральные щупаль-
ца и ринофоры несут сверху полоску белых кра-
пинок. На голове белый пигмент отсутствует.
Отличия от похожих видов. Легко спутать
с обычным видом дальневосточных корифелл,
Coryphella athadona. Заметно отличается от послед-
него более узким, грациозным телом с меньшим
количеством папилл (у экземпляров сопостави-
мых размеров), передним краем головы с посто-
янной, отчетливой выемкой, более длинными
Coryphella amabilis — Корифелла амабилис,
вид со спинной стороны
передними отростками ноги и отчетливо просве-
чивающими сквозь переднюю часть спинной
стороны, за ринофорами, несколькими петлями
семяпровода. Последний признак, особенно
у крупных экземпляров, позволяет безошибочно
отличить Coryphella amabilis от Coryphella athadona.
У последней семяпровод очень короткий, не
образует подобных петель и практически не
просвечивает дорсально.
Особенности экологии. Встречается
крайне редко в заливе Петра Великого в зимне-
весенний период на литорали под камнями
среди мелких особей Coryphella athadona. Раз-
множение. Кладки в форме тонких волнистых
розоватых спиралей. Развитие со стадией
личинки-велигера.
Питание и поведение. Возможно, питает-
ся гидроидом Haitaia parva.
Распространение. Тихоокеанский боре-
альный вид. Отмечен в заливе Петра Великого
Японского моря, у Шантарских и Командор-
ских островов, а также в водах у острова Хок-
кайдо (Япония). В Баренцевом и Белом морях
обитает вид-двойник, Coryphella gracilis.
173
Отряд Nudibranchia — Голожаберные моллюски
Coryphella verrucosa
(М. Sars, 1829) -
Корифелла бугорчатая
Особенности строения. Тело узкое. Нотум
полностью отсутствует. Папиллы отчетливо рас-
падаются на две основные группы — переднюю
и заднюю, с промежутком между ними. Задняя
группа, в свою очередь, распадается еще на не-
сколько менее отчетливых групп. Папиллы
удлиненно-пальцевидные, умеренно толстые.
Вырост пищеварительной железы в них в виде
толстого шнура, занимающего почти всю папил-
лу. Необходимо отметить, что в южной Норве-
гии встречаются особи Coryphella verrucosa с очень
короткими папиллами, отчего весь облик живот-
ного становится совершенно иным. Именно на
подобных экземплярах было основано первона-
чальное описание Coryphella verrucosa, и они от-
ветственны за название этого вида — бугорчатая.
Несмотря на то что классический короткопа-
пильный вариетет не отмечен в водах России,
обнаруженные в мае 2007 г. беломорские особи
отчасти ему соответствуют. Ринофоры выглядят
почти гладкими, хотя они мотут быть покрыты
небольшими бугорками и морщинками, подли-
не примерно равны оральным щупальцам. Края
ноги образуют впереди довольно длинные от-
ростки. Анус открывается в первой половине
тела. Длина до 38 мм.
Окраска. Общая окраска тела беловатая. На
оральных щупальцах сверху обычно имеется тон-
кая белая полоска. Похожая полоска может быть
и с внутренней стороны ринофоров, но очень ча-
сто она полностью отсутствует. На хвосте замет-
ная длинная белая полоска. Цвет ветвей пищева-
рительной железы и ее выростов в папиллах очень
изменчив, встречаются особи с оранжеватыми,
розовато-сероватыми, коричневато-красными,
коричневатыми отростками. Папилла с узким бе-
лым колечком на вершине, самый кончик папил-
лы, как правило, остается полупрозрачным.
Отличия от похожих видов. Легко спутать
с другим, редким в Белом море видом корифелл,
Coryphella gracilis. Заметно отличается от послед-
него более широким, массивным телом с боль-
шим количеством папилл (у экземпляров сопо-
ставимых размеров), передним краем головы без
Coryphella verrucosa — Корифелла бугорчатая,
вид со спинной стороны
Coryphella verrucosa — Корифелла бугорчатая,
вид с брюшной стороны
174
Семейство Flabellinidae — Флабеллиновые или Флабеллиниды
Coryphella verrucosa — Корифелла бугорчатая
175
Отряд Nudibranchia — Голожаберные моллюски
Coryphella verrucosa — Корифелла бугорчатая,
кладка
постоянной, отчетливой выемки (хотя времен-
ные выемки могут образовываться при движе-
нии или питании моллюска), более короткими
передними отростками ноги и не просвечиваю-
щим сквозь переднюю часть дорсальной сторо-
ны семяпроводом. Последний признак, особенно
у крупных экземпляров, позволяет безошибочно
отличить Coryphella verrucosa от Coryphella gracilis.
У последней семяпровод довольно длинный, со-
стоит из двух петель и заметно просвечивает
сквозь спинную сторону.
Особенности экологии. Пожалуй, самый
массовый вид беломорских голожаберных
моллюсков. Встречается на камнях, скалах,
пластинах ламинарии, от литорали до глуби-
ны 10-15 м. Особенно массовые скопления на
Белом море образует среди гидроидного обра-
стания мидиевой культуры. В Баренцевом мо-
ре, напротив, довольно редок на литорали,
становясь массовым на глубинах более 7 м.
Размножение. Размножение в Белом мо-
ре отмечено с мая по август включительно,
в Баренцевом отмечено в августе. Кладка в виде
прямого узкого шнура, завитого в спираль, бе-
ловатого или розоватого цвета. Развитие со
стадией личинки-велигера.
Питание и поведение. В Белом море
обычным пищевым объектом этого вида явля-
ются колонии гидроидных полипов Obelia long-
issima. Кроме того, зимой на Белом море, а так-
же в условиях эксперимента Coryphella verrucosa
предпочитала гидроидов Clava multicornis и Со-
ryne loveni. В водах Британских островов Co-
ryphella verrucosa обычгго питается крупными
гидроидами Tubularia indivisa, а в водах Новой
Англии может даже переходить на питание
сложными асцидиями. На Белом море тем не
менее подобные пищевые объекты для Coryphel-
la verrucosa не документированы.
Несмотря на отчетливое предпочтение
данным видом колоний гидроидных полипов
Obelia longissima, часто встречается вне всякой
связи с пищевым объектом, на заиленных, ли-
шенных гидроидов пластинах ламинарии, на
нижней поверхности камней. Как и другие
узкотелые эолиды, Coryphella verrucosa довольно
подвижна и приспособлена к жизни на веточ-
ках гидроидов. Нападает на отдельные полипы
обелии, откусывает их вместе с теками у сте-
белька, пустые теки выбрасываются вместе
с фекальными массами. По-видимому, изредка
может нападать на особей своего вида.
Близкий вид, обитающий в дальневосточных
морях, Coryphella athadona, значительно более
агрессивен. Нередко нападая на особей своего ви-
да, даже близких по размерам, он, по всей види-
мости, полностью вытеснил Coryphella verrucosa
с литорали в сублитораль (в отличие от Белого
и Баренцева морей, в дальневосточных морях
Coryphella verrucosa доходит до глубины 67 м, прак-
тически полностью отсутствуя на литорали).
Распространение. Амфибореальный вид —
широко распространен в северной Атлантике
и встречается по тихоокеанскому побережью Се-
верной Америки. По азиатскому побережью на
юге доходит до Японского моря. В водах России
обитает по Мурманскому побережью Баренцева
моря и в Белом море, где весьма обычен, во всех
дальневосточных морях — довольно редок.
Coryphella athadona
Bergh, 1875 -
Корифелла атадона
Особенности строения. Тело удлиненное, от-
носительно узкое. Дорсальные папиллы рас-
полагаются группами. Наиболее хорошо вы-
ражены две первые группы, соответствующие
передней части пищеварительной железы.
Остальные группы папилл, относящиеся
к ее задней части, выражены менее явно, одна-
ко можно проследить еще 3-4 пучка. Нотум по
боковым краям тела имеется, но слабый и пре-
рывистый — в промежутке между папиллами
передней и задней печени она отсутствует.
176
Семейство Flabellinidae — Флабеллиновые или Флабеллиниды
Coryphella athadona — Корифелла атадона,
вид со спинной стороны
Coryphella athadona — Корифелла атадона,
вид с брюшной стороны
Папиллы веретеновидные или удлиненно-паль-
цевидные, с дивертикулом пищеварительной
железы, который занимает ее большую часть.
Ринофоры равны по длине оральным щупаль-
цам или немного длиннее их. Щупальцевидные
выросты переднего края ноги отсутствуют.
Окраска. Общая окраска тела беловато-жел-
товатая. Начинаясь из одной точки примерно на
середине головы, две белые линии, составленные
из точек, расходятся под углом и достигают кон-
чиков оральных щупалец. Этот признак очень
характерен для Coryphella athadona и проявляется
в большей или в меньшей степени у всех живых
особей. Белый пигмент также образует довольно
широкую полосу на хвостике и — у некоторой
части особей — прерывистую линию вдоль сере-
дины спины. Цвет выростов пищеварительной
железы в папиллах широко варьирует от темно-
вишневого, малинового и кирпичного до желто-
ватого и светло коричневого.
Отличия от похожих видов. Легко спутать
с другим, более редким в дальневосточных мо-
рях видом корифелл, Coryphella verrucosa. Хоро-
шо отличается от последнего простым закру-
гленным передним краем ноги без выростов
и характерным расположением белых пигмент-
ных линий на голове — в виде треугольника.
Особенности экологии. Самый массовый
вид этого семейства в Дальневосточных морях
России. Обитает под камнями, на скалах и валу-
нах, среди друз крупной мидии Crenomytilus graya-
nus, на слоевищах и в ризоидах водорослей, в со-
ставе обрастания буев, и даже среди гидроидов,
Coryphella athadona (Корифелла атадона) на пищевом
объекте — колонии гидроидных полипов
177
Отряд Nudibranchia — Голожаберные моллюски
Coryphella athadona — Корифелла атадона
Coryphella athadona — Корифелла атадона,
кладка
покрывающих камчатского краба Paralithodes кат-
chatica. Отмечен от литорали до глубины 15-25 м.
Массовые скопления образует летом в литораль-
ных «лужах» на тихоокеанском побережье Кам-
чатки. Может дрейфовать с оторванными водо-
рослями на поверхности моря.
Размножение. В заливе Петра Великого
приходится на май-июнь, в более холодных во-
дах (например на Камчатке), смещается на июль
и август. Кладка в виде тонкого беловато-розова-
того шнура, завитого в спираль из нескольких
оборотов, часто в виде фестонов. Развитие со ста-
дией свободноплавающей личинки-велигера.
Питание и поведение. Активный, под-
вижный вид. Предпочитает колонии гидроид-
ных полипов Obelia longissima. Вид в целом более
агрессивен, чем другие флабеллиниды, как по
отношению к особям своего вида, так и пред-
ставителям других видов флабеллинид.
Распространение. Тихоокеанский боре-
альный вид. Обитает в дальневосточных морях
повсеместно, от Японского моря до Командор-
ских островов включительно.
178
Семейство Flabellinidae — Флабеллиновые или Флабеллиниды
Hlmatlna trophina
(Bergh, 1894) -
Химатина трофина
Особенности строения. Тело широкое. Па-
пиллы образуют вдоль каждой из сторон спи-
ны сплошной покров, без видимых перерывов.
Нотум хорошо выражен, заметно выступает
над боками тела. Папиллы ланцетовидные, до-
вольно широкие; вырост пищеварительной
железы в них в виде тонкого шнура, занимаю-
щего только ее середину. Ринофоры в мелких
складках и морщинках, примерно в 1,5 раза
длиннее оральных щупалец. Края ноги образу-
ют впереди короткие отростки. Анус открыва-
ется сбоку с правой стороны тела, в его перед-
ней половине. Длина до 40 мм.
Окраска. Общая окраска тела беловатая или
слегка желтоватая. Цвет ветвей пищеварительной
железы и ее выростов в папиллах оранжеватый.
Отличия от похожих видов. От других
видов, обитающих на мелководье дальнево-
сточных морей России, данный вид хорошо
отличается заметно более широким телом
и непрерывными рядами папилл.
Himatina trophina — Химатина трофина,
кладка
Особенности экологии. Обитает на ска-
листых и каменистых грунтах. По тихоокеан-
скому побережью, в водах юго-восточной Кам-
чатки, является одним из самых массовых
видов. Отмечен от литорали до глубины 115 м,
Himatina trophina — Химатина трофина,
вид со спинной стороны
Himatina trophina — Химатина трофина,
вид с брюшной стороны
179
Отряд Nudibranchia — Голожаберные моллюски
Himatina trophina — Химатина трофина
но отчетливо предпочитает наиболее мелкие
участки от 6 до 12 м.
Размножение. Кладка представляет со-
бой изогнутый тонкий спиральный шнур. Раз-
витие со стадией личинки-велигера.
Питание и поведение. Активный, под-
вижный, хищный вид. Видимо, предпочитает
гидроидов, но может поедать полихет, мелких
голожаберных моллюсков, а также ракообраз-
ных-амфипод из рода Caprella («морских ко-
зочек»).
Распространение. Тихоокеанский боре-
альный вид. Распространен в морях России
в Беринговом море, у Командорских островов,
в Охотском море, у Северных Курильских
островов, в заливе Петра Великого Японского
моря и у Восточной Камчатки. В Тихом океане
по американскому побережью от Аляски до
Орегона.
СЕМЕЙСТВО PSEUDOVERMIDAE —
ПСЕВДОВЕРМИСОВЫЕ
ИЛИ ПСЕВДОВЕРМИДЫ
Это семейство можно назвать «акохлидиидами
голожаберных моллюсков», поскольку это
единственная группа, обитающая исключитель-
но в интерстициали, пространстве между части-
цами грунта. «Pseudovermidae» можно переве-
сти как «псевдочерви», но скорее — по форме
и манере передвижения — они напоминают
маленьких змеек. Несмотря на множество ре-
дукционных признаков, псевдовермиды еще
сохраняют некоторые из типичных черт, харак-
терных для эолидацей, в частности, наличие не-
больших папилл, содержащих книдосак. При-
чем имеются как виды с довольно развитыми
папиллами, так и со слабовыраженными, оста-
ющимися лишь в виде крошечных бугорков.
180
Семейство Pseudovermidae — Псевдовермисовые или Псевдовермиды
| Отдельно следует сказать об авторе этого
вида — Софье Переяславцевой. Женщины
в науке XIX века были все еще большой редко-
стью, что уж говорить о той должности, которую
занимала долгие годы Переяславцева — пост
директора Севастопольской биологической
станции. Софья Переяславцева стала одной из
первых, кто всерьез стал изучать фауну и систе-
матику моллюсков Черного моря. Далеко не все
ей удалось на этом пути, но то, что удалось, за-
служивает глубокого уважения.
Pseudovermis paradoxus
Perejaslavtseva, 1891 —
Псевдовермис парадоксальный
Особенности строения. Тело имеет змеевид-
ную форму. Нога очень сильно редуцирована,
моллюск фактически ползает на брюшной сто-
роне тела. Спинные папиллы сохраняют кни-
досак, но сами редуцировались до крошечных
бугорков-сосочков. Голова несет притупленную
удлиненную лопасть, в которую превращен
оральный парус, оральные щупальца отсут-
ствуют. Спикулы не заметны. Длина до 6 мм.
Окраска. Тело полупрозрачное, серовато-
желтоватое. Сквозь покровы просвечивают
зеленовато-коричневатые ветви пищевари-
тельной железы.
Отличия от похожих видов. Резко отли-
чается, благодаря своей ярко выраженной
червеобразной форме, от других голожабер-
ных моллюсков. От обитающих с ними сов-
местно акохлидиид отличается отсутствием
разделения тела на головную лопасть и висце-
ральный мешок.
Особенности экологии. Обитает в грубо-
зернистом песке в сублиторали.
Размножение. По-видимому, откладыва-
ет маленькие шаровидные кладки, прикрепляя
их к частицам грунта. Вместе со взрослыми
особями нередко можно обнаружить значи-
тельное число ювенильных экземпляров, кото-
рые значительно короче взрослых и несут лишь
одну-две пары рудиментарных сосочковидных
папилл.
Pseudovennis paradoxus — Псевдовермис
парадоксальный, вид со спинной стороны
Pseudovennis paradoxus — Псевдовермис
парадоксальный, вид со спинной стороны
181
Отряд Nudibranchia — Голожаберные моллюски
Pseudovernits paradoxus (Псевдовермис парадоксальный)
свернутый в кольцо
Питание и поведение. Активный, под-
вижный вид. Может активно передвигаться
между частицами грунта. Питание не изучено.
Возможно, питается мелкими кольчатыми
червями, которые в изобилии были обнаруже-
ны вместе с псевдовермисами.
Распространение. Эндемик Черного
моря. Довольно обычный вид на Крымском
побережье Черного моря в районе Сева-
стополя.
СЕМЕЙСТВО EUBRANCHIDAE —
ЭУБРАНХУСОВЫЕ
ИЛИ ЭУБРАНХИДЫ
Своеобразное семейство мелких эолидацей.
Большинство видов обладает ограниченным
числом крупных папилл. Эубранхиды не-
сколько неожиданным образом комбинируют
черты строения двух крупных семейств эоли-
дацей: радулы, состоящей из трех рядов зубов
(признак семейства флабеллинид) и особой
дополнительной железы копулятивного аппа-
рата (признак семейства тергипедид). Детали
строения радулярных зубов, однако, суще-
ственно отличаются от флабеллинид. Особен-
ности внешнего строения, в частности поло-
жение ануса между ветвями правой и задней
пищеварительной железы, соответствуют та-
ковому тергипедид.
Достаточно широко распространено как
в тропических, так и в бореальных водах. Очень
разнообразен видовой состав и в морях России:
здесь представлено 7 видов из 5 родов.
Eubranchus tricolor
Forbes, 1838 —
Эубранхус трехцветный
Non Eubranchus tricolor sensu Roginskaya, 1962
Особенности строения. Один из самых
крупных и красивых видов данного семей-
ства. Тело широкое. Нотум полностью отсут-
ствует. Папиллы располагаются многочис-
ленными, плотными рядами. Папиллы
в средней части широкие, к вершинам резко
сужаются, образуя характерный оттянутый
кончик. Вырост пищеварительной железы
в них в виде довольно узкого шнура, зани-
мающего только среднюю часть папиллы.
Ринофоры гладкие, мощные, примерно
в 2,5-3 раза длиннее оральных щупалец. Нога
впереди закругленная, без отростков. Анус
располагается на спине с правой стороны.
Длина до 30 мм.
Окраска. Общая окраска беловатая. Рино-
форы и оральные щупальца полупрозрачные,
без белого пигмента. Цвет выростов пищевари-
Eubranchus tricolor — Эубранхус трехцветный,
вид со спинной стороны
182
Семейство Eubranchidae — Эубранхусовые или Эубранхиды
Eubranchus tricolor — Эубранхус трехцветный,
вид с брюшной стороны
тельной железы в папиллах — почти синева-
тый или темно-коричневатый. Папиллы на
вершине с хорошо заметной желтоватой ша-
почкой.
Отличия от похожих видов. Особенности
окраски и строения позволяют легко отличить
этот вид от других голожаберных моллюсков.
Особенности экологии. На Мурманском
побережье Баренцева моря изредка встречает-
ся на глубинах 7—10 м на скалистых и камени-
стых грунтах.
Размножение. Размножение этого вида
в морях России не изучено.
Питание и поведение. Питается гидро-
идами, но их видовой состав точно не уста-
новлен. Обладает характерным защитным
поведением — в случае опасности может
резко отбрасывать значительное число спин-
ных папилл.
Распространение. Бореальный североат-
лантический вид. В морях России обнаружен
только у Мурманского побережья Баренцева
моря.
Amphorina odhneri
(Derjugin et Gurjanova, 1926) —
Амфорина Однера
Особенности строения. Тело довольно узкое.
Нотум полностью отсутствует. Папиллы рас-
полагаются весьма многочисленными, но не
плотными рядами. Папиллы удлиненно-паль-
цевидные. Вырост пищеварительной железы
в них представляет собой относительно узкую
шнуровидную структуру, занимающую толь-
ко среднюю часть папиллы. Ринофоры глад-
кие, приблизительно в 1,5 раза превышаю-
щие по длине оральные щупальца. Нога
впереди закругленная, без отростков. Анус
располагается на спине с правой стороны.
Длина до 13 мм.
Окраска. Общая окраска беловатая. Рино-
форы и оральные щупальца полупрозрачные,
без белого пигмента. Цвет выростов пищевари-
тельной железы в папиллах рыжевато-корич-
неватый. Папиллы на самой вершине с мало-
заметной белой шапочкой.
Отличия от похожих видов. Можно спу-
тать с массовым беломорским видом Coryphel-
la verrucosa, который относится к другому се-
мейству, но обладает очень сходной окраской
папилл. Amphorina odhneri четко отличается от
Coryphella verrucosa отсутствием белого пиг-
мента на ринофорах, оральных щупальцах
и хвосте, закругленной передней части ноги,
у которой полностью отсутствуют боковые вы-
росты, а также дорсальным, а не боковым по-
ложением ануса.
Amphorina odhneri — Амфорина Однера, кладка
183
Отряд Nudibranchia — Голожаберные моллюски
Atnphorina odhneri — Амфорина Однера,
вид со спинной стороны
Atnphorina odhneri — Амфорина Однера,
вид с брюшной стороны
Atnphorina odhneri (Амфорина Однера) в естественном местообитании — зарослях гидроидов и мшанок
184
Семейство Eubranchidae — Эубранхусовые или Эубранхиды
Особенности экологии. По нашим на-
блюдениям, вид довольно обычен в сублито-
ральной зоне некоторых районов Белого мо-
ря. В других местах, например на Мурманском
побережье Баренцева моря изредка встреча-
ется на каменистой литорали. У Новой Земли
(в проливе Маточкин Шар) найден в большом
количестве в зарослях гидроидов и мшанок,
на глубинах свыше 20 м.
Размножение. Размножение в Белом мо-
ре отмечено с апреля по июль. Кладка в виде
довольно широкой ленты розового цвета, свер-
нутая в кольцо. Развитие со стадией личинки-
велигера.
Питание и поведение. Питается гидрои-
дами, но их видовой состав точно не установ-
лен; возможно, это атекатные гидроиды из
рода Согупе. Умеренно подвижная форма, оби-
тающая как под камнями, так и в зарослях ги-
дроидов.
Распространение. Бореальный североат-
лантический вид. Известен только из Баренце-
ва и Белого морей.
Amphorlna horii
(Baba, 1960) -
Амфорина Хори
Особенности строения. Тело удлиненное,
но относительно широкое. Ринофоры паль-
цевидные тонкие, в 1,5-2 раза длиннее ораль-
ных щупалец. Папиллы длинные узковерете-
новидные. Дивертикул пищеварительной
железы занимает преимущественно среднее
положение в папилле. Кончик папиллы за-
острен. Нога впереди немного расширена
и закруглена без каких-либо выростов. Дли-
на до 6 мм.
Окраска. На голове и ринофорах имеется
пара широких продольных полосок ржавого
или темно-оранжевого пигмента. Сходный
оранжево-ржавый пигмент в виде мелких кра-
пинок разбросан по большей части спины
и внешнему краю ноги. На спине, под основа-
ниями ринофор имеется крупное треугольное
оранжево-ржавое пятно. Вырост пищевари-
тельной железы в папиллах зеленоватый или
серовато-зеленоватый, в основании с черными
крапинками.
Отличия от похожих видов. Можно спу-
тать с некоторыми мелкими видами из другого
Спинные выросты-папиллы Amphorina odhneri
(Амфорина Однера)
Amphorina horii (Амфорина Хори)
с кладкой
185
Отряд Nudibranchia — Голожаберные моллюски
семейства Tergipedidae, также обитающих
в Японском море, например Trinchesia pupillae.
От этого вида A. horii хорошо отличается на-
личием оранжевых полосок на голове, ораль-
ных щупальцах и ринофорах.
Особенности экологии. Обнаружен на
самом мелководье, не глубже 1,5 м в составе
гидроидного обрастания бурых водорослей
коккофоры и саргассума.
Размножение. Размножение в морях Рос-
сии не регистрировалось.
Питание и поведение. Умеренно подвиж-
ная форма, которую в основном можно наблю-
дать в зарослях гидроидов.
Распространение. По-видимому, приа-
зиатский субтропическо-бореальный вид.
В морях России обнаружен только в заливе
Петра Великого Японского моря. В водах Япо-
нии, напротив, распространен широко: залив
Сагами, Внутреннее море Сето, япономорское
побережье острова Хонсю и др.
Nudlbranchus rupium
(Muller, 1842) -
Нудибранхус рупиум
Синоним:
Eubranchus exiguus sensu Roginskaya,
non Alder et Hancock
Особенности строения. Тело узкое, изящное.
Нотум полностью отсутствует. Папиллы распо-
лагаются немногочисленными, очень редкими
рядами. Папиллы довольно длинные, пальцевид-
ные. Срединного вздутия и соответствующего
ему выроста пищеварительной железы нет. Ри-
нофоры гладкие, примерно в 1,5-2 раза длиннее
оральных щупалец. Нога впереди закругленная,
без отростков. Анус располагается на спине с пра-
вой стороны. Длина до 12 мм.
Окраска. Общая окраска тела сероватая.
По телу, ринофорам, оральным щупальцам
и папиллам разбросаны многочисленные, мел-
Nudibranchus rupium — Нудибранхус рупиум,
вид со спи in юн стороны
Nudibranchus rupium — Нудибранхус рупиум,
вид с брюшной стороны
186
Семейство Eubranchidae — Эубранхусовые или Эубранхиды
Nudibranchus rupium — Нудибранхус рупиум, Nudibranchus г upturn — Нудибранхус рупиум,
вид со спинной стороны
вид с брюшной стороны
кие, почти не сливающиеся коричневато-оран-
жеватые пятнышки. Только у некоторых особей
из Японского моря, и редко — из Белого, эти
пятна полностью отсутствуют, и тело равномер-
но покрыто серовато-оливковой «пылью». На
ринофорах и оральных щупальцах, кроме того,
разбросаны мелкие белые крапинки, а в сред-
ней части, как правило, имеется довольно ши-
рокое коричневатое колечко. Кончики ринофо-
ров и оральных щупалец полупрозрачные. Цвет
выростов пищеварительной железы в папиллах
зеленоватый, серовато-зеленоватый, травяно-зе-
леный. В середине папиллы может быть непол-
ное колечко из белых точек. У вершины папилл,
как правило, довольно узкое коричневатое коль-
цо. Кончик папиллы с размытой голубоватой
шапочкой или почти прозрачный. Кроме того,
сквозь спинную часть тела, как правило, просве-
чивает ствол пищеварительной железы также
зеленоватого цвета.
Отличия от похожих видов. По Мурман-
скому побережью Баренцева моря обитает
очень близкий вид Nudibranchus exiguus, кото-
рый отличается от Nudibranchus rupium более
крупными коричневыми пятнами на спинной
стороне, которые могут иногда формировать
две прерывистые полосы вдоль спины, а также
цветом пищеварительной железы, сероватой,
розоватой или почти бесцветной у Nudibran-
chus exiguus, и зеленоватой у Nudibranchus ru-
pium (у взрослых экземпляров), удлиненной
формой папилл и структурой зубов радулы.
Форма кладок также заметно отличается у этих
двух видов: у Nudibranchus rupium кладки в виде
шнура, часто завернутого в полукольцо, тогда
как у Nudibranchus exiguus они в форме малень-
ких овальных или округлых мешочков. Еще
один похожий вид — Tergipes tergipes, хорошо
отличается от Nudibranchus rupium очень редки-
ми папиллами (только по одной папилле
в каждом ряду, тогда как у Nudibranchus rupium
в каждом ряду несколько папилл) и отчетли-
выми красновато-оранжевыми полосками на
голове. Более существенно эти два вида отли-
чаются анатомически, являясь представителя-
ми различных семейств.
187
Отряд Nudibranchia — Голожаберные моллюски
Nudibranchus rupium (Нудибранхус рупиум) в естественном местообитании — колонии гидроидных полипов обелии
Ювенильная особь Nudibranchus rupium (Нудибранхус рупиум) атакует отдельный полип обелии
188
Семейство Eubranchidae — Эубранхусовые или Эубранхиды
Nudibranchus rupium (Нудибранхус рупиум) в уникальном местообитании — гидроидном обрастании на крабе-пауке
Особенности экологии. В Баренцевом
и Белом морях наиболее обычен среди обраста-
ния гидроидов из семейства Obeliidae на лами-
нарии, растущей на искусственных плавучих
конструкциях, а также гидроидного обрастания
на канатах и в искусственной мидиевой культу-
ре, от поверхности до глубины около 3-5 м, где
может образовывать массовые скопления.
Размножение. На Белом море размноже-
ние отмечено с мая по сентябрь включительно.
Взрослый экземпляр Nudibranchus rupium
(Нудибранхус рупиум) с кладками
Кладка от изогнутой овальной до кольцеобраз-
ной. Развитие со стадией личинки-велигера.
Питание и поведение. Основным пище-
вым объектом данного вида на Белом море яв-
ляется гидроид Obelia longissima. На одной до-
статочно крупной колонии Obelia longissima
в мае можно обнаружить сотни взрослых
и ювенильных Nudibranchus rupium. Вся жизнь
данного вида, после оседания личинок из план-
ктона, протекает на колонии обелии. Мелкие
размеры и изящная форма Nudibranchus rupium,
позволяют ему легко двигаться по веточкам
гидроида. Питается нудибранхус выедая тело
отдельных гидрантов обелии, оставляя в отли-
чие от корифелл теку нетронутой. Кладки от-
кладываются также на веточки гидроида.
Распространение. Амфибореальный
вид. Северная Атлантика по европейскому по-
бережью от Норвегии до Нидерландов (юж-
ная граница ареала точно не определена),
Гренландия и по американскому побере-
жью — отмечен в водах Массачусетса; в водах
России — в Баренцевом и Белом морях. Север-
ная Пацифика — по американскому побере-
жью от Аляски до Орегона, по российскому
побережью — во всех дальневосточных морях
включая Японское.
189
Отряд Nudibranchia — Голожаберные моллюски
Nudibranchus exiguus
(Alder et Hancock, 1842) —
Нудибранхус маленький
Особенности строения. Тело узкое, изящное.
Нотум полностью отсутствует. Папиллы рас-
полагаются немногочисленными, очень редки-
ми рядами. Папиллы довольно короткие, ха-
рактерно вздутые, угловато расширенные
к середине. Ринофоры гладкие, примерно
в 1,5-2 раза длиннее оральных щупалец. Нога
впереди закругленная, без отростков. Анус рас-
полагается на спине с правой стороны. Длина
до 8 мм.
Окраска. На спинной стороне располага-
ются довольно крупные коричневые пятна.
Отличия от похожих видов. От близкого
вида Nudibranchus rupium, с которым Nudibran-
chus exiguus может обитать совместно, отлича-
ется более крупными коричневыми пятнами
на спинной стороне, которые могут иногда
формировать две прерывистые полосы вдоль
спины, а также цветом пищеварительной же-
лезы, сероватой, розоватой или почти бесцвет-
ной у Nudibranchus exiguus, и зеленоватой
у Nudibranchus rupium (у взрослых экземпля-
ров), более короткими вздутыми папиллами
и структурой зубов радулы. Форма кладок так-
же заметно отличается у этих двух видов:
у Nudibranchus rupium кладки в виде шнура, ча-
сто завернутого в полукольцо, тогда как у Nudi-
branchus exiguus они в форме маленьких оваль-
ных или округлых мешочков.
Особенности экологии. В Баренцевом
море изредка встречается среди гидроидного
обрастания гидроидов из семейства Obeliidae
на ламинарии, растущей на искусственных
плавучих конструкциях, а также гидроидного
обрастания на канатах и в искусственной ми-
диевой культуре, от поверхности до глубины
около 3-5 м.
Размножение. В Баренцевом море раз-
множение зарегистрировано в августе. Кладка
в виде маленьких сфер или дисков. Развитие со
стадией личинки-велигера.
Питание и поведение. Основным пище-
вым объектом данного вида является гидроид
Obelia longissima. Поведение сходно с предыду-
Nudibranchus exiguus — Нудибранхус маленький,
вид со спинной стороны
Nudibranchus exiguus — Нудибранхус маленький,
вид с брюшной стороны
190
Семейство Eubranchidae — Эубранхусовые или Эубранхиды
Nudibranchus exiguus — Нудибранхус маленький,
кладка
щим видом. Кладки откладываются также на
веточки гидроида.
Распространение. Бореальный атланти-
ческий вид. Северная Атлантика по европей-
скому побережью от Норвегии до Северной
Испании, по американскому побережью — от-
мечен в водах Массачусетса. В России обнару-
жен только в Баренцевом море.
Aenlgmastyletus alexeil
Martynov, 1998 —
Энигмастилетус алексея
Особенности строения. Тело узкое, изящное.
Нотум полностью отсутствует. Папиллы рас-
полагаются немногочисленными, очень редки-
ми рядами. Папиллы довольно длинные, не-
правильно веретеновидные, с неравномерным
вздутием недалеко от своей вершины. Имеется
срединное вздутие и соответствующий ему вы-
рост пищеварительной железы. Ринофоры
гладкие, примерно в 2-2,5 раза длиннее ораль-
ных щупалец. Передняя часть ноги несет удли-
ненные отростки, которые по размеру лишь
чуть меньше оральных щупалец. Анус распо-
лагается на спине с правой стороны. Длина до
14 мм.
Окраска. Общая окраска тела полупро-
зрачно-сероватая. Спина, бока и частично па-
пиллы покрыты овальными, удлиненными
или округлыми, отделенными друг от друга
оранжево- и темно-коричневыми пятнышка-
ми. Кроме коричневого пигмента по телу, па-
пиллам, ринофорам и особенно по перикар-
дию разбросаны снежно-белые крапинки.
Окраска выроста пищеварительной железы
внутри папиллы сероватая, серовато-коричне-
вая, серовато-зеленоватая. На внешней поверх-
ности папиллы имеется субапикальная нерав-
номерная полоска коричневого пигмента
и кольцо из белых точек на вздутии папиллы.
Отличия от похожих видов. От других
представителей семейства эубранхид, обнару-
женных в Японском море (таких как Nudibran-
chus rupium и Leostyletus pseudomisakiensis), отли-
чается длинными отростками переднего края
и характерной окраской, состоящих из отдель-
ных, не сливающихся с друг другом и фоном
коричневатых пятен.
Особенности экологии. В заливе Петра
Великого обитает на глубинах 1,5-4 м и приу-
рочен к верхушечным частям листьев морских
трав Zoster a marina, Zoster a asiatica и Zoster a caes-
pitosa, покрытых различными гидроидами. От-
дельные особи способны дрейфовать вместе
с оторванными листьями Zostera в приповерх-
ностных водах и достигать литорали.
Aenigmastyletus alexeii — Энигмастилетус алексея,
вид со спинной стороны
191
Отряд Nudibranchia — Голожаберные моллюски
Размножение. Размножение зарегистри-
ровано в августе и сентябре. Кладка представ-
ляет собой прямую или слегка завернутую
широкую ленту, прикрепленную к субстрату
очень узким ребром (в ребро укладывается
2-4 слоя яиц). Цвет кладки беловато-полупро-
зрачный. Длина колеблется от 3 до 20 мм при
средней высоте 2 мм. В процессе развития за-
родышей кладка сильно разбухает и начинает
напоминать свежеотложенную кладку близко-
го вида Leostyletus pseudomisakiensis. При темпе-
ратуре +26...+28° С развитие зародышей дли-
лось 4-5 дней, после чего из кладки выходили
свободно плавающие велигеры с яйцевидной
личиночной раковиной.
Питание и поведение. Этот и следующий
вид обладают очень необычным половым по-
ведением. Копулятивный аппарат состоит из
нескольких твердых образований сложной фор-
мы — так называемых стилетов. Стилеты спо-
собны к резким быстрым колебательным дви-
жениям. Из всего семейства эубранхид такой
системой обладает лишь небольшая группа ро-
дов. Предполагается, что подобная система по-
явилась в ответ на особый образ жизни этого
вида — на верхушках морских трав и водорос-
лей, подверженных сильному волнению.
Распространение. Обнаружен пока толь-
ко в заливе Петра Великого Японского моря.
Несомненно, однако, что данный вид имеет го-
раздо более широкое распространение, охва-
тывающее по меньшей мере воды вокруг Япон-
ских островов, а возможно, и простирающееся
дальше на юг.
Leostyletus pseudomisakiensis
Martynov, 1998 —
Леостилетус псевдомисакенский
Особенности строения. Общий габитус данно-
го вида сходен с Aenigmastyletus alexeii, но тело
более изящное, ринофоры более тонкие и длин-
ные, могут в три раза превышать длину ораль-
ных щупалец. Строение папилл идентично
Aenigmastyletus alexeii. Отдельные папиллы (обыч-
но самая внутренняя из них) ряда правой перед-
ней пищеварительной железы могут по длине
более чем в 2 раза превышать ринфоры. Перед-
няя часть ноги несет короткие треугольные от-
ростки, по размерам не превышающие 1/4 дли-
ны оральных щупалец. Длина тела до 13 мм.
Leostyletus pseudomisakiensis — Леостилетус
псевдомисакенский, вид со спинной стороны
Окраска. Спина, бока и частично папиллы
покрыты мелкими штрихами и пятнышками
светло-коричневого цвета. У крупных экземпля-
ров на спине и боках эти пятнышки могут прак-
тически полностью сливаться, образуя однород-
ный коричнево-оранжевый фон, а пятнышки на
папиллах напоминают таковые Aenigmastyletus
alexeii. У мелких экземпляров пятнышки слива-
ются не полностью, а на папиллы практически
не заходят. Кроме этого, по телу, папиллам
и ринофорам разбросаны снежно-белые кра-
пинки. На ринофорах, в верхней их половине
можно различить одно-два неправильных ко-
ричневатых колечка. Выросты пищеваритель-
ной железы в папиллах в основании коричнева-
тые, далее сероватые, серовато-зеленоватые.
Отличия от похожих видов. От других
представителей семейства эубранхид, обнару-
женных в Японском море (таких как Nudibran-
chus rupium и Aenigmastyletus alexii), отличается
очень короткими отростками переднего края
и характерной окраской, состоящей у половоз-
релых экземпляров из сливающихся с друг
другом и фоном коричневатых пятен.
192
Семейство Tergipedidae — Тергипесовые или Тергипедиды
Leostyletus pseudomisakiensis — Аеостилетус
псевдомисакенский, вид с брюшной стороны
Особенности экологии. В заливе Петра
Великого обитает на глубинах 3-5 м и, по-ви-
димому, строго приурочен к верхушечным
частям листьев морской травы Zostera asiatica,
покрытых различными гидроидами. В этом
биотопе Leostyletus pseudomisakiensis может
обитать совместно с Aenigmastyletus alexeii.
Размножение. Размножение зарегистри-
ровано в сентябре. Кладка представляет прямой
или слегка изогнутый шнур, прикрепленный
к субстрату широким основанием, куда уклады-
вается более 10 слоев яиц. Цвет кладки белова-
тый или кремовый. При температуре +26...+280 С
развитие зародышей длилось 4-5 дней, после
чего из кладки выходили свободноплавающие
велигеры с яйцевидной раковиной.
Питание и поведение. Этот вид, как
и предыдущий, обладает очень необычным по-
ловым поведением и сложной системой стиле-
тов, которая существенно отличается у Leostyle-
tus pseudomisakiensis от Aenigmastyletus alexeii.
Распространение. Обнаружен пока толь-
ко в заливе Петра Великого Японского моря.
Несомненно, однако, что данный вид имеет го-
раздо более широкое распространение, охва-
тывающее по меньшей мере воды вокруг Япон-
ских островов, а возможно, и простирающееся
дальше на юг.
СЕМЕЙСТВО TERGIPEDIDAE —
ТЕРГИПЕСОВЫЕ
ИЛИ ТЕРГИПЕДИДЫ
Одно из самых разнообразных семейств голо-
жаберных моллюсков группы эолид, представ-
ленных в морях России. В состав этого семей-
ства входят как крупные глубоководные виды,
обладающие широким телом, покрытым зна-
чительным числом папилл, как, например,
Murmania antiqua, так и мелкие мелководные
представители из родов Trinchesia и Tergipes,
имеющие лишь редкие немногочисленные
спинные папиллы. Характерной внешней чер-
той этого семейства, позволяющей хорошо от-
личать его от семейства Flabellinidae, является
положение анального отверстия среди папилл
правой стороны в средней части спины, а не на
боковой стороне тела, также справа, вне пря-
мой связи с папиллами. Кроме того, имеется
целый ряд внутренних признаков, резко отли-
чающих тергипедид от других семейств эолид,
в частности комбинация наличия дополни-
тельной железы копулятивного аппарата и ра-
дулы с единственным зубом в каждом попереч-
ном ряду.
Научное описание Murmania anti-
qua — этого уникального вида — бы-
ло опубликовано лишь несколько лет
назад российскими исследователя-
ми. Обычно открывают лишь новые
виды, но в данном случае отличия
в строении были столь велики, что
был выделен также и самостоятель-
ный новый род, который был назван
в честь прославленного российского
города Мурманска и Мурманской
области, в прилежащих к которым
частям Баренцева моря этот новый
таксон и был обнаружен. Мурмания
обладает целым рядом древних, ар-
хаических черт, не свойственных
большинству представителей се-
мейства тергипедид.
193
Отряд Nudibranchia — Голожаберные моллюски
Murmania antiqua — Мурмания древняя,
фиксированный экземпляр, вид со спинной стороны
Murmania antiqua — Мурмания древняя,
фиксированным экземпляр, вид с левой стороны
Murmania antiqua
Martynov, 2006 —
Мурмания древняя
Особенности строения. На дорсальной ча-
сти головы имеется треугольная складка,
вклинивающаяся между ринофорами. Тело
высокое, по бокам — закраина с закруглен-
ным краем. Ветви пищеварительной железы
очень многочисленные, разветвленные, без
прямого соответствия рядам папилл. Преа-
нальных рядов папилл более 20, постанальные
не редуцированы. Нет заметной границы
между передней и задней частями пищевари-
тельной железы. Анус располагается около
середины спины, несколько сдвинут назад.
Длина тела до 50 мм.
Окраска. Прижизненная окраска не из-
вестна.
Отличия от похожих видов. Особенности
внешнего и внутреннего четко отличают этот
вид от большинства других видов группы эо-
лид. Некоторое поверхностное сходство имеет-
ся только с представителями совершенно
другого семейства Aeolidiidae, широко распро-
страненными в морях России на мелководье
(например, Aeolidia papillosa). От этих видов
Murmania antiqua хорошо отличается особенно-
стями расположения рядов спинных папилл,
наличием мощной треугольной складки в об-
ласти головы и многими другими чертами.
Особенности экологии. Умеренно глубо-
ководный вид. Обитает на глубинах от 50 до
350 м, по-видимому, на мягких, илисто-песча-
ных грунтах, возможно с примесью камней.
Размножение. Не изучено.
Питание и поведение. Не изучено. Судя
по всему, питается крупными атекатными ги-
дроидами и актиниями, медленно передвига-
ясь по грунту.
Распространение. Известен пока только
из южных частей Баренцева моря.
194
Семейство Tergipedidae — Тергипесовые или Тергипедиды
Cuthonella abyssicola
Bergh, 1884 —
Кутонелла глубоководная
Особенности строения. Тело широкое, мае-
сивное, покрыто многочисленными рядами
веретеновидных и пальцевидных папилл. Пе-
редняя часть закруглена и лишена боковых от-
ростков. Длина до 40 мм.
Окраска. Общая окраска тела беловатая.
У половозрелых особей сквозь спинную сторо-
ну просвечивает ярко-розовая гонада. Окраска
выроста пищеварительной железы внутри па-
пиллы красноватая. Кончик папиллы светлый,
с просвечивающим книдосаком.
Отличия от похожих видов. От других эо-
лид с широким телом (например из семейства
Flabellinidae, см. выше) отличается строением ря-
дов папилл, покрывающих как спинную, так
и боковую стороны тела, закругленным перед-
ним краем ноги, лишенным даже коротких от-
ростков и положением ануса, открывающегося
с правой стороны среди папилл, а не непосред-
ственно на боковой стороне, лишенной папилл.
Cuthonella abyssicola — Кутонелла глубоководная,
фиксированный экземпляр, вид со спинной стороны
От других представителей рода Cuthonella, широ-
ко распространенных на мелководных участках
моря, отличается крупными размерами и значи-
тельно большим числом спинных папилл.
Особенности экологии. Глубоководный
вид. Обитает на глубинах от 50 м до более чем
1000 м, по-видимому, на мягких илисто-песчаных
грунтах, возможно с примесью камней.
Размножение. Не изучено.
Питание и поведение. Судя по всему, пи-
тается крупными атекатными гидроидами,
медленно передвигаясь по грунту.
Распространение. К глубоководным ви-
дам не применимы обычные биогеографиче-
ские термины, в том числе и использованные
в данной книге. Так, та или иная биогеографи-
ческая область характеризуется вполне опреде-
ленным температурным режимом. В то же вре-
мя температура на больших глубинах как
в тропическом поясе, так и в бореальном всегда
почти одинаковая и очень низкая, порядка
+2...+4° С или даже ниже, так что глубоководные
виды могут иметь всесветное распространение.
Данный вид отмечен в северной Атлантике и не-
которых районах Арктики, в частности, в Барен-
цевом море и море Лаптевых.
Cuthonella hiemalls
(Roginskaya, 1987) —
Кутонелла зимняя
Особенности строения. Тело довольно узкое.
Нотум полностью отсутствует. Папиллы рас-
полагаются довольно многочисленными (13-15),
плотными рядами. Папиллы удлиненно-
пальцевидные. Вырост пищеварительной желе-
зы в них в виде довольно толстого шнура, зани-
мающего большую часть объема папиллы.
Гладкие конические ринофоры немного короче
оральных щупалец. Длина до 26 мм.
Окраска. Общая окраска тела беловато-
желтоватая. У половозрелых особей сквозь
спинную сторону просвечивает ярко-розовая
гонада. Верхняя половина ринофоров и кончи-
ки оральных щупалец присыпаны белой пы-
лью. Окраска печеночного выроста внутри па-
пиллы темно-бежевая в темно-коричневых
пятнах. Кончик папиллы светлый, с просвечи-
вающим книдосаком.
Отличия от похожих видов. От большин-
ства беломорских эолид отличается темно-
195
Отряд Nudibranchia — Голожаберные моллюски
Ранние ювенильные экземпляры Cuthonella hiemalis (Кутонелла зимняя) только чпю вышедшие из кладки
коричневой окраской пищеварительной железы
в папиллах. Данный вид очень легко спутать с
близкими видами Cuthonella marisalbi и Cuthonella
concinna. От них Cuthonella hiemalis отличается на-
личием светлых вершин папилл, без белой ша-
почки, несколько большим числом рядов папилл,
а также анатомическими признаками. Кроме то-
го, взрослые, крупные экземпляры Cuthonella hie-
malis встречаются главным образом зимой и ран-
ней весной, тогда как крупные особи Cuthonella
marisalbi и Cuthonella concinna встречаются почти
исключительно летом. Помимо указанных видов,
Cuthonella hiemalis можно спутать с некоторыми
экземплярами Cuthonella verrucosa, имеющими
коричневую окраску папилл, от которых Cutho-
nella hiemalis хорошо отличается закругленными
передними краями ноги, более уплощенным те-
лом, папиллами, расположенными как на спин-
ной, так и на боковой сторонах тела, а также от-
сутствием белой полосы на хвостике.
Особенности экологии. Обитает главным
образом на литорали в зарослях бурых водорос-
лей Ascophyllum nodosum и Fucus serratus, а также
под камнями. В Белом море Cuthonella hiemalis
обнаружена пока только в районе Беломорской
биологической станции МГУ (Еремеевский по-
рог), где, по-видимому, весьма обычна.
Размножение. В Белом море размноже-
ние отмечено с января по май, таким образом
этот вид нерестится при максимально низких,
арктических температурах (наиболее благо-
приятная температура для раннего эмбриоге-
неза от -1,6 до +3,0° С). Несмотря на это, в Арк-
тике Cuthonella hiemalis никогда не была
Cuthonella hiemalis — Кутонелла зимняя,
кладка
196
Семейство Tergipedidae — Тергипесовые или Тергипедиды
зарегистрирована. Приверженность к предель-
но низким температурам нереста возможно
сохранилась у Cuthonella hiemalis от предкового
вида, сходного с близкой к ней Cuthonella abys-
sicola, которая сохранила архаичные черты
в своем строении и одновременно привержен-
ность к местообитаниям с очень низкой темпе-
ратурой — на больших глубинах (1000-1500 м)
в северной Атлантике или на меньших глуби-
нах, но в высокой Арктике. Очень близкий
к Cuthonella hiemalis вид, дальневосточная Cutho-
nella soboli, также размножается в зимний пери-
од. Кладка Cuthonella hiemalis представляет со-
бой кирпично-красный шнур, закрученный
в спираль. В каждой яйцевой капсуле по одно-
му зародышу. Развитие прямое, из кладки вы-
ходит ползающая молодь без раковины.
Питание и поведение. В весенне-летний
период отмечено питание атекатными гидрои-
дами Clava multicornis и Coryne loveni, зимой —
Dynamena pumila. Достаточно малоподвижные
эолиды, обитающие на фукоидах. Во время
отлива прячутся в гуще водорослей или под
камнями. В отличие от корифелл целиком за-
глатывающих гидрантов, Cuthonella hiemalis вы-
сасывает содержимое гидроидных полипов
через небольшое отверстие, которое прогрыза-
ет радулой и челюстями, оставляя внешнюю
оболочку гидранта пустой.
Распространение. Атлантический боре-
альный вид. Известен главным образом из
Белого моря, менее достоверные находки —
в Баренцевом.
Cuthonella soboli
Martynov, 1992 —
Кутонелла Соболь
Особенности строения. Тело удлиненное, не-
сколько расширенное в средней части и сужа-
ющееся в задней. Гладкие конические ринофо-
ры с приостренной, но несколько усеченной
вершиной по толщине равны оральным щу-
пальцам, но немного длиннее их. Пальцевид-
ные или веретеновидные, заостренные на вер-
шины папиллы покрывают спинную сторону
тела косыми поперечными рядами, которые
впереди оканчиваются на уровне ринофор.
Общее число рядов папилл у половозрелых
особей по обеим сторонам тела 11-15. Число
преанальных рядов папилл у половозрелых
Cuthonella soboli — Кутонелла Соболь,
вид со спинной стороны
моллюсков обычно 6. Передняя часть ноги не-
сколько расширена с малозаметной выемкой
посередине, но обособленные отростки отсут-
ствуют. Длина до 30 мм.
Окраска. Общая окраска тела беловато-
желтоватая. Передняя часть головы и спины
желтоватые. Сквозь большую часть спины и но-
ги просвечивает охряно-желтая гонада с ярко-
красными скоплениями зрелых спермиев. На
краях ринофоров, оральных щупалец и головы
концентрируется оранжево-охряный пигмент,
верхняя треть ринофор присыпана белой пы-
лью, отдельные белые точки встречаются на
оральных щупальцах. Окраска пищеваритель-
ного выроста внутри папиллы табачно-бурая.
Кончик папиллы полупрозрачный, сквозь него
просвечивает книдосак. Поверхность папиллы
на вершине и ниже состоит из очень маленьких
матовых железок.
Отличия от похожих видов. От большин-
ства эолид, обитающих в дальневосточных мо-
рях России отличается уплощенным телом
и темно-коричневой окраской пищеваритель-
ной железы в папиллах. Очень похожий вид из
197
Отряд Nudibranchia — Голожаберные моллюски
Cuthonella soboli — Кутонелла Соболь,
изменчивость окраски
этого же рода, Cuthonella hiemalis обитает толь-
ко в Белом и Баренцевом морях. Помимо ука-
занного вида Cuthonella soboli можно спутать
с некоторыми экземплярами Coryphella athado-
na, имеющих коричневую окраску папилл, от
которых Cuthonella soboli хорошо отличается
более уплощенным телом, папиллами, рас-
положенными как на спинной, так и на боко-
вой сторонах тела, а также отсутствием белых
полос в виде треугольника на голове.
Особенности экологии. Обитает от лито-
рали до глубины нескольких метров. Полово-
зрелые особи были найдены в основном под
камнями, молодь — на различных водорослях,
например на Coccophora. Взрослые моллюски
начинают встречаться на литорали с конца де-
кабря по начало мая.
Размножение. Размножение начинается
в январе, пик его приходится на апрель — на-
чало мая. Кладка представляет собой спираль-
ный шнур из 2-4 оборотов, завитый против ча-
совой стрелки. Окраска кладки варьирует от
светло-розовой до охряно-оранжевой. Развитие
длится от 1 до 1,5 месяца. Из кладки выходят сво-
бодно плавающие велигеры, имеющие яйцевид-
ную раковинку, довольно большую ногу с кры-
шечкой и относительно небольшой парус. Судя
по тому, что в некоторых капсулах еще до вы-
хода велигеров у зародышей начинает разру-
шаться личиночная раковинка, вполне вероятно,
что развитие в некоторых случаях может быть
прямым. Планктонный период жизни велиге-
ров, судя по всему, укорочен, приблизительно
через неделю можно наблюдать осевших личи-
нок без раковинки и дорсальных выростов с ро-
зоватыми, довольно большими глазами.
Питание и поведение. Питаются, судя по
всему, атекатными гидроидами. Достаточно
малоподвижные эолиды.
Распространение. Тихоокеанский бореаль-
ный вид. Обитает во всех дальневосточных морях
России, от Японского моря до Командорских
островов. За пределами России, по-видимому,
может встречаться у берегов острова Хоккайдо.
Cuthonella concinna
(Alder et Hancock, 1843) —
Кутонелла изящная
Особенности строения. Тело довольно узкое.
Нотум отсутствует. Папиллы располагаются до-
вольно многочисленными (10-11), плотными ря-
дами. Папиллы удлиненно-пальцевидные. Вы-
рост пищеварительной железы в них в виде
довольно толстого шнура, занимающего боль-
шую часть объема папиллы. Гладкие конические
ринофоры немного длиннее оральных щупалец.
Передний край ноги со слабо выраженными бо-
ковыми лопастями. Длина до 18 мм.
Окраска. Общая окраска тела беловато-
желтоватая. Сквозь спинную сторону поло-
возрелых особей просвечивает розоватая го-
нада. Верхняя половина ринофоров и кончики
оральных щупалец присыпаны белой пылью.
Окраска печеночного выроста внутри папил-
лы темно-коричневая. Кончик папиллы с бе-
лой шапочкой.
Отличия от похожих видов. От большин-
ства беломорских и баренцевоморских эолид от-
личается темно-коричневой окраской пищевари-
тельной железы в папиллах. Данный вид очень
легко спутать с близким видом Cuthonella hiemalis.
От последнего Cuthonella concinna отличается на-
личием белой шапочки на вершинах папилл
(у Cuthonella hiemalis вершина папилл светлая,
198
Семейство Tergipedidae — Тергипесовые или Тергипедиды
Cuthonella concinna — Кутонелла изящная,
вид со спинной стороны
Cuthonella concinna — Кутонелла изящная,
вид с брюшной стороны
полупрозрачная), несколько меньшим числом
рядов папилл, а также анатомическими призна-
ками. Кроме того, взрослые, крупные экземпля-
ры Cuthonella hiemalis встречаются главным обра-
зом зимой и ранней весной, тогда как крупные
особи Cuthonella concinna встречаются летом. Cu-
thonella concinna можно также спутать с некоторы-
ми экземплярами Coryphella verrucosa, имеющи-
ми коричневую окраску папилл, от которых
Cuthonella concinna хорошо отличается закруглен-
ными (хотя и несколько оттянутыми латерально)
передними краями ноги, без боковых отростков,
более уплощенным телом, папиллами, располо-
женными как на спинной, так и на боковой сто-
ронах тела, а также отсутствием белой полосы на
хвостике. См. также сравнение этого вида с Cutho-
nella marisalbi в следующем очерке.
Особенности экологии. На литорали
Мурманского побережья Баренцева моря пред-
почитает селиться в среднем и даже в верхнем
горизонте литорали, остающемся во время от-
лива без воды максимально долгое время.
Размножение. В Баренцевом море раз-
множение отмечено в августе. Кладка пред-
ставляет собой довольно толстый розоватый
шнур, закрученный в спираль. В каждой яйце-
вой капсуле по одному зародышу. Развитие со
стадией личинки-велигера.
Cuthonella concinna — Кутонелла изящная,
кладка
199
Отряд Nudibranchia — Голожаберные моллюски
Питание и поведение. По-видимому, пи-
тается гидроидами Sertularia sp.
Достаточно малоподвижные эолиды.
Пищевое поведение, возможно, сходно с пре-
дыдущим видом.
Распространение. Амфибореальный вид.
Северная Атлантика по европейскому побере-
жью от Норвегии до Франции, западное побе-
режье Гренландии, Исландия и по американ-
скому побережью — Новая Англия. Северная
Пацифика — по американскому побережью от
Аляски до Орегона. В водах России достоверно
известен только в Баренцевом и Белом морях.
Cuthonella marisalbi
(Roginskaya, 1963) —
Кутонелла беломорская
Особенности строения. Тело довольно узкое.
Нотум отсутствует. Папиллы располагаются до-
вольно многочисленными (10-11), плотными
рядами. Папиллы удлиненно-пальцевидные.
Вырост пищеварительной железы в них в виде
довольно толстого шнура, занимающего боль-
шую часть объема папиллы. Гладкие кониче-
ские ринофоры немного длиннее оральных щу-
палец. Передний край ноги с мощными
боковыми лопастями. Длина до 20 мм.
Окраска. Общая окраска тела беловато-
желтоватая. Сквозь спинную сторону полово-
зрелых особей просвечивает розоватая гонада.
Верхняя половина ринофоров и кончики ораль-
ных щупалец присыпаны белой пылью. Окрас-
Cuthonella marisalbi — Кутонелла беломорская,
кладка
ка выроста пищеварительной железы внутри
папиллы сероватая, светло-коричневая. Кончик
папиллы с широкой белой шапочкой.
Отличия от похожих видов. Данный вид
очень близок к Cuthonella concinna, и вопрос
о его синонимии с этим видом остается откры-
тым. По внешним признакам Cuthonella mari-
salbi отличается от Cuthonella concinna более
длинными и стройными папиллами, отчетли-
во выраженными мощными боковыми лопас-
тями ноги и сероватыми, светло-коричневаты-
ми выростами пищеварительной железы
в папиллах. Хотя оба вида отмечены в Белом
море, но именно в этом регионе Cuthonella
marisalbi является локально массовым видом,
тогда как Cuthonella concinna, напротив, обычна
в верхнем горизонте литорали Мурманского
побережья Баренцева моря.
Особенности экологии. Встречается на
бурой водоросли Fucus serratus, обросшей ги-
дроидами. Обитает главным образом на лито-
рали. Особенно массовые скопления данный
вид образует на Еремеевском пороге близ Бе-
ломорской биологической станции МГУ. Из-
редка отмечен вплоть до глубины 10 м.
Размножение. В Белом море размножение
зафиксированно с конца мая по начало сентября.
Кладка представляет собой довольно толстый
розоватый шнур, закрученный в спираль. В каж-
дой яйцевой капсуле по одному зародышу. Раз-
витие со стадией личинки-велигера.
Питание и поведение. По-видимому, пи-
тается гидроидами Diphasia falax и Sertularia sp.
Достаточно малоподвижные эолиды. Пищевое
поведение, возможно, сходно с таковым у пре-
дыдущего вида.
Распространение. Достоверно известен
только из Белого моря. Должен обитать также, по
меньшей мере, в соседнем Баренцевом море.
Catriona gymnota
(Couthouy, 1838)
Катриона гимнота
Особенности строения. Тело довольно узкое.
Нотум полностью отсутствует. Папиллы рас-
полагаются довольно многочисленными (около
10) плотными рядами. Папиллы удлиненно-
пальцевидные. Вырост пищеварительной желе-
зы в них в виде довольно толстого шнура, зани-
мающего большую часть объема папиллы.
200
Семейство Tergipedidae — Тергипесовые или Тергипедиды
Cuthonella marisalbi (Кутонелла беломорская) на пищевом объекте — колонии гидроидных полипов
Cuthonella marisalbi — Кутонелла беломорская,
вид со спинной стороны
Cuthonella marisalbi — Кутонелла беломорская,
вид с брюшной стороны
201
Catriona gymnota — Катриона гимнота,
вид со спинной стороны
Гладкие конические ринофоры близки по раз-
мерам оральным щупальцам. Длина до 9 мм.
Окраска. Характерными особенностями
данного вида является сочетание оранжевых
ринофор, ярко-розовых выростов пищевари-
тельной железы в папиллах и широких колец
белого пигмента у вершин папилл.
Отличия от похожих видов. От всех видов
из родов Cuthonella и Trinchesia, отмеченных в фа-
уне России, Catriona gymnota хорошо отличается
уникальными особенностями своей окраски.
Особенности экологии. В морях России
обнаружена на скалистом и каменистом грун-
тах на глубине 7-10 м.
Размножение. В морях России не изучено.
Питание и поведение. Малоподвижный
вид. Питается гидроидами.
Распространение. Атлантический боре-
альный вид. Северная Атлантика по европей-
скому побережью от Норвегии до Франции,
а по американскому побережью отмечен в во-
дах Новой Англии. В водах России обнаружен
только вдоль Мурманского побережья Барен-
цева моря.
Catriona gymnota — Катриона гимнота,
вид с брюшной стороны
Trinchesia ornata
(Baba, 1937) -
Тринчесия украшенная
Особенности строения. Тело почти цилин-
дрическое, несколько суживающееся к заднему
концу и переходящее в маленький хвостик. Го-
лова равна по ширине ноге. Ринофоры пальце-
видно-конические, гладкие. Оральные щупаль-
ца пальцевидные, несколько вздутые на конце,
чуть длиннее и тоньше ринофор. Нога впереди
с мощными коническо-треугольными отростка-
ми, длина которых составляет примерно поло-
вину длины оральных щупалец. Насчитывается
4-5 ветвей передней правой пищеварительной
железы, соответствующие рядам папилл. Анус
располагается перед первой папиллой первого
ряда правой задней пищеварительной железы.
Длина до 20 мм.
Окраска. Общая окраска тела охристая,
бледно-оранжевая. Выросты пищеварительной
железы внутри папилл серовато-зеленоватые.
Передняя часть чехла папиллы покрыта по
всей длине лазоревым, ярко-голубым пигмен-
том. Ближе к вершине папиллы этот пигмент
переходит в неровное снежно-белое субапи-
202
Семейство Tergipedidae — Тергипесовые или Тергипедиды
Trinchesia ornata (Тринчесия украшенная) в процессе откладывания кладки
кальное кольцо, а внизу папиллы лазоревая
полоска сильно утончается.
Отличия от похожих видов. Можно спу-
тать с другим видом этого рода, Trinchesia viridis,
обычным в Белом море, но встречающимся
также и в Японском, причем иногда совместно
с Trinchesia ornata. Данный вид отличается от
Trinchesia viridis характерной ярко-голубой
Trinchesia ornata — Тринчесия украшенная,
вид со спинной стороны
окраской внешнего чехла спинных папилл
и хорошо выраженными отростками передне-
го края ноги.
Особенности экологии. В заливе Петра
Великого достаточно редкий вид, обитает
в основном на литорали, иногда опускаясь до
2 м, на камнях, поросших гидроидами из се-
мейства Sertullariidae.
Размножение. Кладка представляет собой
очень узкую ленту, прикрепленную к субстрату
ребром, завитую против часовой стрелки в спи-
раль из 2-4 оборотов. Цвет ее бледно-желтый.
Развитие со стадией личинки-велигера.
Питание и поведение. Питается гидрои-
дами из семейства Sertulariidae. Передвигается
по поверхности камней и по коротким веточ-
кам сертуляреллы.
Расппространение. По-видимому, при-
азиатский субтропическо-бореальный вид.
В морях России обнаружен только в заливе
Петра Великого Японского моря. В водах Япо-
нии, напротив, распространен широко: залив
Сагами, залив Осака, Внутреннее море Сето;
япономорское побережье острова Садо, залив
Тояма и др.
Trinchesia viridis
(Forbes, 1840) -
Тринчесия зеленая
Особенности строения. Тело довольно узкое.
Нотум полностью отсутствует. Папиллы удли-
ненно-пальцевидные, располагаются доволь-
но многочисленными, плотными рядами.
203
Отряд Nudibranchia — Голожаберные моллюски
Trinchesia viridis — Тринчесия зеленая
Trinchesia viridis — Тринчесия зеленая,
вид со спинной стороны
Trinchesia viridis — Тринчесия зеленая,
вид с брюшной стороны
204
Семейство Tergipedidae — Тергипесовые или Тергипедиды
Насчитывается 4-5 ветвей передней правой
пищеварительной железы, соответствующие
рядам папилл. Гладкие конические ринофоры
немного длиннее оральных щупалец. Длина
до 20 мм.
Окраска. Тело беловатое с желтоватым от-
тенком. Ринофоры и оральные щупальца при-
мерно на две трети покрыты белым пигмен-
том. Цвет выростов пищеварительной железы
в папиллах от темно-зеленого до зеленовато-
коричневатого. Неправильные снежно-белые
пятна разбросаны в основном по передней гра-
нице папилл. Книдосак хорошо выделяется,
желтовато-беловатый, вершина папилл про-
зрачная, иногда со слабым оранжевым пигмен-
том. На хвостике белая полоска.
Отличия от похожих видов. От всех дру-
гих видов из рода Trinchesia, отмеченных в мо-
рях России (включая близкий вид Trinchesia
pustulata), хорошо отличается зеленоватой или
зеленовато-коричневатой окраской выроста
пищеварительной железы в папиллах.
Особенности ЭКОЛОГИИ. Достаточно
обычный в Баренцевом море вид, в Белом море
более редок, был обнаружен только в районе
Ругозерской губы Кандалакшского залива.
Обитает главным образом под камнями на ли-
торали и приурочен к гидроидам семейства
Sertulariidae.
Размножение. Размножение в Белом мо-
ре отмечено в июле. Кладка в виде узкой ленты
белого цвета, завитой в спираль. Количество
яиц в кладке достигает 2000. Развитие со стади-
ей личинки-велигера.
Питание и поведение. Питается гидроида-
ми из семейства Sertulariidae, в Баренцевом мо-
Trinchesia viridis — Тринчесия зеленая, кладка
ре — Sertularella rugosa (L., 1758). На Белом море
возможно также в диету входят гидроиды рода
Sertularella. Передвигается по поверхности кам-
ней и по коротким веточкам сертуляреллы.
Распространение. Амфибореальный вид.
Северная Атлантика — по европейскому по-
бережью от Норвегии до Франции, западное
побережье Гренландии, Исландия и по амери-
канскому побережью — Новая Англия. Север-
ная Пацифика — по американскому побере-
жью от Аляски до Орегона, по российскому
побережью — во всех дальневосточных морях
включая Японское.
Trinchesia perca
(Er. Marcus, 1958) —
Тринчесиа перка
Особенности строения. Тело узкое. Нотум
полностью отсутствует. Папиллы удлиненно-
пальцевидные, располагаются довольно мно-
гочисленными, но неплотными рядами. На-
считывается 2-3 ветви передней правой
пищеварительной железы, соответствующие
рядам папилл. Гладкие пальцевидные рино-
форы, почти равны или немного короче ораль-
ных щупалец. Длина до 10 мм.
Окраска. Общая окраска желтоватая, по-
лупрозрачная. Характерной особенностью
окраски данного вида является диффузное пят-
но серовато-коричневатого пигмента на голове
и в передней части спины. Другой характерной
чертой данного вида являются коричневатые
каналы пищеварительной железы, отчетливо
просвечивающие сквозь спинную сторону.
Между ветвями пищеварительной железы
у зрелых особей хорошо заметны кремовые
фолликулы гонады.
Отличия от похожих видов. Данный вид
по особенностям формы тела и окраски хорошо
отличается от всех представителей тергипедид,
отмеченных как в Черном море, так и в других
морях России.
Особенности экологии. Trinchesia perca
обитает в кутовых, распресненных частях моря
поблизости от эстуариев рек, под камнями, на
которых можно встретить и другую богатую
фауну — полихет, губок, мидий и других бес-
позвоночных.
Размножение. Размножение отмечено
в августе. Кладка бобовидная, розоватая.
205
Отряд Nudibranchia — Голожаберные моллюски
Trinchesia perca — Тринчесиа перка,
вид со спинной стороны
Trinchesia perca — Тринчесиа перка,
вид с брюшной стороны
Trinchesia perca (Тринчесиа перка) атакует актинию Диадумене
206
Семейство Tergipedidae — Тергипесовые или Тергипедиды
Trinchesia регса — Тринчесиа перка, кладка
Развитие со стадией свободноплавающей ли-
чинки-велигера.
Питание и поведение. В отличие от боль-
шинства других видов семейства тергипедид,
Trinchesia регса питается не гидроидными поли-
пами, а мелкими актиниями, которые также
относятся к типу книдарий. Довольно подвиж-
ный вид, обитающий на твердых поверхностях,
обычно под камнями.
Распространение. Встречается точечно
в различных тропических и субтропических
районах Мирового океана.
Trinchesia pustulata
(Alder et Hancock, 1854) —
Тринчесия пузырчатая
Особенности строения. Тело узкое. Нотум
полностью отсутствует. Папиллы удлинен-
но-пальцевидные, располагаются довольно
многочисленными, но неплотными рядами.
Насчитывается 4 ветви передней правой пе-
чени, соответствующие рядам папилл. Глад-
кие пальцевидные ринофоры почти равны
или немного короче оральных щупалец.
Длина до 15 мм.
Окраска. Общая окраска тела беловатая.
Ринофоры, оральные щупальца и спинные па-
пиллы в верхней части несут редкие мелкие
белые крапинки. Выросты пищеварительной
железы и ветви пищеварительной железы,
просвечивающие дорсально сквозь покровы,
желтоватые, оранжеватые, изредка светло-
коричневые с желтоватым оттенком. Вершина
папилл прозрачная.
Отличия от похожих видов. По форме
тела и расположению папилл Trinchesia pus-
tulata напоминает Trinchesia viridis, но хорошо
отличается от этого вида желтоватым или
По своему статусу Trinchesia регса
существенно отличается от боль-
шинства других заднежаберных
моллюсков фауны России и со-
предельных стран. Данный вид яв-
ляется видом-вселенцем, т. е.
«чужеродным» видом для нашей
фауны, ареал которого еще недав-
но не охватывал моря России.
Многие виды-вселенцы, например,
растений или насекомых представ-
ляют собой серьезную проблему
для карантинных служб любой
страны, поскольку потенциально
они могут серьезно нарушить эко-
логический баланс той или иной
территории или акватории.
К счастью, в случае с Trinchesia
регса никакой угрозы экосистемам
Черного моря (кроме эстетического
удовольствия ныряльщиков) этот
голожаберный моллюск не несет.
В эстуарий Севастопольской бухты
данный вид попал, скорее всего,
с одним из многочисленных судов,
причем, вероятно, не во взрослом
состоянии, а в виде личинок, зака-
чанных в балластную систему судна
вместе с забортной водой за тыся-
чи километров от Черного моря.
В районе Севастополя личинки
нашли подходящие условия и смог-
ли вырасти во взрослых моллюсков,
которые, в свою очередь, приступи-
ли к размножению в совершенно
новой для них обстановке.
Для Trinchesia регса и раньше была
отмечена способность «к переме-
не мест». Причем до такой степени,
что вопрос о настоящей родине
этого вида до сих пор является
предметом дебатов. Предполага-
ется, что данный вид первоначаль-
но обитал в районе мощных эсту-
арных систем побережья Бразилии
и других тропических районов Ат-
лантического океана. Кроме того,
данный вид обнаружен в таких «не-
ожиданных» местах, как солонова-
тое озеро в районе Сан-Франциско,
в Чесапикском заливе, что на вос-
точном побережье США, и даже на
Гавайских островах — т. е. ареал
этого вида, пусть и точечно, охва-
тывает почти весь мир.
В Черном море данный вид был об-
наружен нами впервые в 2007 году.
Причем Trinchesia регса оказался
выловленным вместе с его пище-
вым объектом, мелкой актинией
Diadumene lineata, которая тоже не
была ранее указана для Черного
моря. Судя по всему, хищник
и жертва перемещаются при помо-
щи человека по Мировому океану
одновременно
207
Отряд Nudibranchia — Голожаберные моллюски
Trinchesia pustulata — Тринчесия пузырчатая,
вид со спинной стороны
Trinchesia pustulata — Тринчесия пузырчатая,
вид с брюшной стороны
оранжеватым цветом папилл (у Trinchesia viridis
папиллы зеленого или зеленовато-коричнева-
того цвета).
Сведения об экологии. В Баренцевом
море обитает, главным образом, на камени-
стой литорали, где предпочитает литораль-
ные ванночки с прогретой водой; изредка
Trinchesia pustulata — Тринчесия пузырчатая,
кладка
обнаруживается в обрастании ламинарий на
глубине до 2 м.
Размножение. Размножение отмечено
в августе. Кладка бобовидная, беловатая. Раз-
витие прямое, из кладки выходит червеобраз-
ная ползающая молодь без раковины.
Питание и поведение. На Баренцевом
море этот вид был обнаружен поблизости от
большого числа различных гидроидов, но
питание ими показано не было. Довольно
подвижный вид, обитающий на твердых по-
верхностях, обычно под камнями.
Распространение. Амфибореальный
вид. Северная Атлантика по европейскому
побережью от Норвегии до Франции,
Гренландии, Исландия и по американскому
побережью — Новая Шотландия и Новая
Англия. Северная Пацифика — по амери-
канскому побережью — Британская Колум-
бия. Возможно, тихоокеанские экземпляры
относятся к близкому виду. В морях России
этот вид отмечен только в Баренцевом и Бе-
лом морях.
208
Семейство Tergipedidae — Тергипесовые или Тергипедиды
Trinchesia pupillae — Тринчесия пупилла,
вид со спинной стороны
Trinchesia pupillae
(Baba, 1961) -
Тринчесия пупилла
Особенности строения. Тело узкое. Нотум
полностью отсутствует. Ринофоры почти пря-
мые, приостренные на конце, толще пальце-
видных оральных щупалец и длиннее их при-
мерно в 2 раза. Насчитывается лишь две ветви
передней правой пищеварительной железы,
соответствующие рядам папилл. Нога впереди
с прямым краем, передние углы закругленны
и слегка оттянуты в стороны. Анус располага-
ется перед первым рядом задней правой пече-
ни, между первой и второй папиллами. Длина
до 6 мм.
Окраска. Тело беловатое. Спина, голова,
ринофоры, оральные щупальца и папиллы
присыпаны бело-золотистой пылью. Ринофоры
и оральные щупальца примерно в середине не-
сут оранжево-красные колечки. Вырост пище-
варительной железы в папиллах коричневый
с сероватым оттенком. Ветви пищеварительной
железы, просвечивающие дорсально сквозь по-
кровы, «состоят» из черно-коричневых точек.
Вершина папилл полностью прозрачная.
Отличия от похожих видов. По форме
тела и расположению папилл Trinchesia pupillae
Trinchesia pupillae — Тринчесия пупилла, вид с правой стороны
209
Отряд Nudibranchia — Голожаберные моллюски
хорошо отличается от всех известных видов
этого семейства, обитающих в заливе Петра
Великого Японского моря.
Сведения об экологии. В заливе Петра
Великого встречается довольно редко, главным
образом в гидроидном обрастании бурой во-
доросли коккофоры (Cocophora langsdorphii).
Размножение. Размножение зарегистри-
ровано в начале сентября. Кладка бобовидная,
полупрозрачно-беловатая, прикреплена
к субстрату в средней части. Кладка заключает
в себе около ста беловатых яиц. Развитие про-
текает около недели, и из кладки выходят сво-
бодноплавающие велигеры с яйцевидной ли-
чиночной раковинкой.
Питание и поведение. Питается различ-
ными гидроидами. Довольно подвижный вид,
обитающий на твердых поверхностях, обычно
под камнями.
Распространение. По-видимому, при-
азиатский субтропическо-бореальный вид.
В морях России обнаружен только в заливе Пе-
тра Великого Японского моря. В водах Японии,
напротив, распространен широко: тихоокеан-
ское побережье острова Хонсю, залив Осака,
залив Цуруга; Внутреннее море Сето; японо-
морское побережье: залив Тояма.
Tenellia adspersa
(Nordmann, 1845) —
Тенеллия искристая
Особенности строения. Тело удлиненное.
Голова несет впереди по бокам вместо ораль-
ных щупалец маленький фронтальный па-
рус. Ринофоры гладкие, длинные, почти ци-
линдрические. Папиллы немногочисленные.
По форме они в основном неправильно паль-
цевидные, только самые крупные близки
к веретеновидной форме. Вырост пищевари-
тельной железы занимает большую часть
объема папиллы. Спина несет по обеим сто-
ронам около двух продольных рядов папилл.
Насчитывается лишь две ветви передней пра-
вой пищеварительной железы, соответствую-
щие рядам папилл. Анус открывается в меж-
печеночном пространстве ближе к переднему
ряду передней пищеварительной железы.
Длина тела до 4 мм.
Окраска. По беловатому, полупрозрачному
фону разбросаны коричневатые или черноватые
Tenellia adspersa — Тенеллия искристая,
вид со спинной стороны
Tenellia adspersa — Тенеллия искристая,
увеличенная передняя часть тела
210
Семейство Tergipedidae — Тергипесовые или Тергипедиды
Tenellia adspersa — Тенеллия искристая
211
Отряд Nudibranchia — Голожаберные моллюски
крапинки и пятнышки. Папиллы в сходных, но
немногочисленных крапинках. Ринофоры
и оральный парус полупрозрачные.
Отличия от похожих видов. Комбинация
небольшого закругленного орального паруса
без срединной выемки, более чем один про-
дольный ряд папилл и сероватая окраска пи-
щеварительной железы в папиллах позволяют
хорошо отличать данный вид от других голо-
жаберных моллюсков с небольшим числом
спинных папилл, таких как Embletonia pulchra
и Tergipes tergipes.
Сведения об экологии. Обитает примуще-
ственно в мелководных распресненных районах
Азовского,Черного и Балтийского морей.
Размножение. Замечательной особенно-
стью Tenellia adspersa является ее крайне короткий
жизненный цикл — моллюски становятся поло-
возрелыми спустя всего лишь две недели после
выхода из яйцевой оболочки. Подобный жизнен-
ный цикл отмечен лишь для немногих других
голожаберных моллюсков. Кладка в виде неболь-
ших мешочков, белых или бледно-розовых. Клад-
ки могут быть двух типов — с более мелкими
яйцами, которые развиваются быстрее, но с об-
разованием типичной плавающей личинки-ве-
лигера, или с более крупными яйцами, которые
развиваются медленнее, с образованием ползаю-
щей личинки, вскоре теряющей раковинку.
Питание и поведение. Питается широ-
ким спектром гидроидов, включая один из
наиболее обычных в морях России гидроид
Obelia longissima и характерную для Черного
и Азовского морей Cordylophora caspia. Доволь-
но подвижный вид, активно передвигающийся
по веточками гидроидов.
Распространение. По-видимому, атлан-
тический бореально-субтропический вид. Ве-
роятно, расселился вместе с судами в воды
Японии и Бразилии, а также тихоокеанского
побережья США. В Атлантике известен от Сре-
диземного моря до Норвегии, в Америке —
вдоль восточного побережья США. В морях
России и сопредельных стран в массовом коли-
честве был обнаружен в Азовском и Каспий-
ском морях и единично — в Японском море.
Единственный вид голожаберных моллюсков,
обнаруженный в сильно опресненном Фин-
ском заливе Балтийского моря.
Terglpes terglpes
Forsskal, 1775 —
Тергипес тергипес
Особенности строения. Тело удлиненное. Го-
лова несет впереди короткие оральные щупаль-
ца. Ринофоры гладкие, длинные, конические.
Папиллы немногочисленные, веретеновидные.
Вырост пищеварительной железы занимает
большую часть объема папиллы. Спина несет
по обеим сторонам лишь по одному ряду про-
дольных рядов папилл. Анус открывается
в межпеченочном пространстве ближе к перед-
нему ряду передней пищеварительной железы.
Длина тела до 8 мм.
Окраска. Общая окраска сероватая, полу-
прозрачная. Сквозь спинную сторону просве-
Tenellia adspersa характеризуется
собранием редких или уникальных
особенностей. Обитает на мелково-
дье, от литорали до глубины 8 м.
Обнаружен на различных субстра-
тах, покрытых гидроидами, — водо-
рослях, камнях; обычен в обраста-
нии искусственных сооружений.
Tenellia adspersa являет уникальный
для голожаберных моллюсков при-
мер эвригалинности, т. е. способ-
ности к обитанию в воде с различ-
ной соленостью. В экспериментах
с азовоморскими особями моллю-
ски выдерживали изменения соле-
ности от 3 до 40 промилле, те. от
почти пресной до сверхсоленой во-
ды. Тем не менее вид в целом явно
предпочитает районы с понижен-
ной соленностью. Так, за двенад-
цать лет изучения Nudibranchia за-
лива Петра Великого нами была
обнаружена единственная особь
этого вида, тогда как на экспери-
ментальных панелях, выставленных
в Азовском море в районе г. Жданов
(ныне Мариуполь), на одной коло-
нии гидроида Peregonimus megas
весом 100 г советскими исследова-
телями в 1960-х годах было найдено
более 7000 половозрелых особей.
Кроме того, сотни особей были об-
наружены в гидроидном обраста-
нии нефтяных платформ в Каспий-
ском море. В целом этот вид можно
рассматривать как единственного
голожаберного моллюска, способ-
ного обитать в практически пресных
водах: в верхнем эстуарии реки
Темзы Tenellia adspersa была най-
дена вместе с дафниями и пресно-
водными коловратками.
212
Семейство Tergipedidae — Тергипесовые или Тергипедиды
Tergipes tergipes, носящий необыч-
ное двойное название с одинаковым
написанием рода и вида (такое до-
пускается в систематике), ставит
своеобразную «точку» в длинном
эволюционном ряду регресса от ис-
ходно весьма сложной организации
наиболее архаичных тергипедид,
близких к Murmania antiqua, до сверх-
упрощенного строения Tergipes ter-
* * *
gipes. У этого вида с каждой стороны
тела остается лишь по одному ряду
папилл, а в передней пищеваритель-
ной железе осталось лишь по одной
ветви и одной папилле. Дальше упро-
щаться почти некуда. Морфологиче-
ский регресс тем не менее способ-
ствовал прогрессу экологическому:
изящная миниатюрная форма терги-
песа наилучшим образом способ-
ствует его обитанию на отдельных
тоненьких веточках колоний гидро-
идных полипов рода Obelia. Напро-
тив, широкотелые, сложные Murma-
nia antiqua и Cuthonella abyssicola
лучше приспособлены к обитанию на
мягких грунтах и питанию более круп-
ной пищей — крупными атекатными
Tubularia - п одоб н ы м и гидроидам и,
а возможно, и актиниями.
чивает коричневатый центральный канал пи-
щеварительной железы, такого же цвета
выросты пищеварительной железы в папил-
лах. Характерным признаком этого вида явля-
ются также оранжево-ржавые полоски пиг-
мента на голове в основании ринофоров
и оральных щупалец.
Отличия от похожих видов. Наличие ко-
ротких, но хорошо выраженных оральных щу-
палец в сочетании с особенностями окраски
позволяет надежно отличить Tergipes tergipes от
других видов с небольшим числом рядов па-
пилл — Embletonia pulchra и Tenellia adspersa.
Сведения об экологии. Обитает на мел-
ководье, от литорали до глубины 8 м. Обнару-
жен на различных субстратах, покрытых ги-
дроидами, — водорослях, камнях, обычен
в обрастании искусственных сооружений.
Размножение. В Баренцевом море раз-
множение зарегистрировано в августе. Кладка
Tergipes tergipes — Тергипес тергипес,
вид со си и и нои стороны
Tergipes tergipes — Тергипес тергипес,
вид с брюшной стороны
213
Отряд Nudibranchia — Голожаберные моллюски
Tergipes tergipes (Тергипес тпергипес) в естественном местообитании
на пищевом объекте — колонии гидроидных полипов обелии
в виде маленького мешочка беловатого цвета.
Развитие со стадией свободноплавающей ли-
чинки-велигера.
Питание и поведение. Питается широким
спектром гидроидов, включая один из наиболее
обычных в морях России гидроид Obelia longis-
sima. Довольно подвижный вид, активно пере-
двигающийся по веточками гидроидов.
Распространение. Атлантический боре-
альный вид. В Атлантике известен от Среди-
земного моря до Норвегии, а также вдоль вос-
точного побережья США. В морях России
обнаружен только у Мурманского побережья
Баренцева моря.
Tergipes tergipes — Тергипес тергипес, кладка
214
Семейство Facelinidae — Фацелиновые или Фацелиниды
СЕМЕЙСТВО FACELINIDAE —
ФАЦЕЛИНОВЫЕ
ИЛИ ФАЦЕЛИНИДЫ
Из этого обширного, преимущественно те-
пловодного семейства в морях России к на-
стоящему времени обнаружено лишь два
вида (в книге будет представлен только один
массовый вид). Характерной чертой этого
семейства во внешнем строении является по-
ложение анального отверстия — между ря-
дами задней части пищеварительной желе-
зы, а не в промежутке между ее передней
и задней частями, как у предшествующего
семейства тергипедид. Для внутреннего
строения характерны радула с единственным
зубом в каждом поперечном ряду и отсут-
ствие особой железы копулятивного органа,
присутствующей у тергипедид. Среди фаце-
линовых много видов с достаточно необыч-
ным для голожаберных моллюсков поведе-
нием и пищевыми пристрастиями. Так,
целый ряд видов хищничает на других видах
голожаберников из подотряда Aeolidacea.
Hermissenda crassicornis
(Eschscholtz, 1831) —
Хермиссенда толстоусая
Особенности строения. Тело умеренно ши-
рокое, изящное. Голова широкая с крупными
коническими заостренными на конце ораль-
ными щупальцами, которые располагаются
перпендикулярно продольной оси тела
и в 2-3 раза превышают по длине короткие
конические ринофоры. Папиллы многочис-
ленные, образуют неотчетливые группы. Па-
пиллы пальцевидные и веретеновидные. Нога
впереди с характерными угловатыми отрост-
ками. Анус располагается между 2 и 3 группа-
ми папилл. Длина тела до 40 мм.
Окраска. Общая окраска тела беловатая,
чуть отступя до переднего края начинается
оранжевая линия, которая проходит через
всю середину головы, сильно утончается меж-
ду ринофорами, а далее, за глазами, снова
становится равной по ширине головному от-
резку, затем снова исчезает и последний раз
появляется в районе перикардия. У самого
края обоих оральных щупалец начинается
Hermissenda crassicornis — Хермиссенда толстоусая,
вид со спинной стороны
Hermissenda crassicornis — Хермиссенда толстоусая,
увеличенная передняя часть тела
215
Отряд Nudibranchia — Голожаберные моллюски
тонкая голубоватая линия, которая проходит
по всей внешней поверхности и раздваивается
у их основания; одно ответвление проходит
между ринофорами, далее линии от обоих
щупалец несколько расширяются соответ-
ственно расширению оранжевой линии, а за-
тем, сузившись, сопровождают последнюю на
всем ее протяжении и далее, когда оранжевая
линия уже заканчивается, доходят до конца
тела. На каждой боковой поверхности головы
также имеются по два коротких толстых оран-
жевых мазка.
Отличия от похожих видов. Сочетание
формы тела и окраски хорошо отличает дан-
ный вид от других голожаберных мол-
люсков.
Особенности экологии. В заливе Петра
Великого Японского моря обитает от лито-
рали (редко) до глубины 3-5 м, где был обна-
ружен среди трубок полихеты Bispira sp.,
покрытых в незначительной степени различ-
ными гидроидами. В бухтах Японского моря
на север от залива Петра Великого, где вода
летом прохладнее на 2-3 градуса, вид, кажет-
ся, встречается чаще; в бухте Туманной был
найден в литоральной ванне. В сборах с пла-
стин Laminaria japonica на ламинариевых
плантациях в заливе Опричник близ поселка
Каменка среди огромного множества
Coryphella athadona встречались изредка и Нег-
missenda crassicornis. Ламинарии были покры-
ты гидроидным обрастанием, состоящим
главным образом из Obelia longissima. В этом
биотопе Hermissenda crassicornis, несомненно,
может выступать как активный хищник: в ее
желудках были найдены челюсти и радулы
Coryphella athadona.
Размножение. Размножение в заливе
Петра Великого зарегистрировано в августе
и сентябре. Моллюски откладывали кладки
в виде тонких извитых шнуров розоватого цве-
та, закрученных против часовой стрелки
в спираль из значительного числа оборотов.
Развитие со стадией свободноплавающей
личинки-велигера.
Питание и поведение. На тихоокеан-
ском побережье Северной Америки для это-
го вида характерна разнообразнейшая дие-
та — различные гидроиды, сцифоидные
медузы, застрявшие на мелководье, морские
перья, мелкие ракообразные и, наконец,
другие голожаберные моллюски. Для вида
отмечен каннибализм. Hermissenda crassi-
cornis — один из немногих видов Nudibranchia,
который сам является постоянным объектом
питания, в данном случае для цефаласпиды
Navanax inermis, а также фанеробранхиаль-
ной дориды из рода Nembrotha. Было описано
нахождение этого вида в желудке рыб из се-
мейства керчаковых.
Распространение. Тихоокеанский боре-
альный вид. Широко распространен по амери-
канскому побережью от Аляски до Мексики.
В водах России отмечен в Японском море
и у Южных Курильских островов.
СЕМЕЙСТВО AEOLIDIIDAE —
ЭОЛИДОВЫЕ ИЛИ ЭОЛИДИДЫ
Небольшое специализированное семейство.
Внешний вид может быть достаточно разно-
образен, многие тропические представители
имеют гораздо более узкое тело, чем харак-
терные для морей России представители ро-
да Aeolidia. Всех их объединяет значительное
сходство во внутреннем строении: радула
унисериальная, единственный зуб в ряду —
гребневидный, очень широкий. Подобная
морфология радулы связана с пищевой спе-
циализацией эолид: в их диету входят почти
исключительно крупные кишечнополостные
актинии («морские анемоны»), и чтобы спра-
виться с такой массивной пищей, эолидидам
и понадобилась мощная радула. Подобно
предшествующему семейству фацелинид,
анус эолид располагается, как правило, сре-
ди ветвей задней пищеварительной железы,
а для копулятивного аппарата характерно от-
сутствие специализированной дополнитель-
ной железы.
Но несмотря на то, что это семейство не
является самым богатым по числу видов или
разнообразию признаков, именно оно волею
исторических судеб стало основой для имено-
вания всей обширной группы голожаберных
моллюсков подотряда Aeolidacea.
В водах России обитают представители
двух родов — Aeolidia и Cerberilla. В атласе
представлены два вида из рода Aeolidia —
один наиболее массовый и мелководный,
второй — более редкий, обитающий на зна-
чительных глубинах в северной части Тихого
океана.
216
Семейство Aeolidiidae — Эолидовые или Эолидиды
Aeolidia herculea
(Bergh, 1894) -
Эолидия геркулея
Синоним:
Aeolidia grandis (Volodchenko, 1941)
Aeolidia farallonensis Gosliner et Behrens, 1996
Особенности строения. Тело широкое, покрыто
многочисленными уплощенными папиллами,
расположенными скошенными в переднем на-
правлении рядами. Анальное отверстие на боко-
вой поверхности тела, справа, под рядами задней
пищеварительной железы. Длина тела до 30 мм.
Окраска. Прижизненная окраска не из-
вестна.
Отличия от похожих видов. Единственный
глубоководный вид рода Aeolidia, заметно отли-
чающийся как от других голожаберных моллю-
сков, так и от мелководной Aeolidia papillosa, в част-
ности, по положению ануса на боковой стороне
тела (признак эволюционно древний), а не между
ветвями пищеварительной железы.
Особенности экологии. Обитает на мяг-
ких грунтах, возможно с примесью камней.
Обнаружен на значительных глубинах, от 418
до 1491 м в северной части Тихого океана.
Размножение. Не изучено.
Питание и поведение. Не изучено. Веро-
ятно, питается актиниями.
Распространение. Глубоководный вид.
Aeolidia papillosa
(Linnaeus, 1761) —
Эолидия папильчатая
Особенности строения. Тело широкое, по-
крыто многочисленными уплощенными па-
пиллами, расположенными скошенными в пе-
реднем направлении рядами. Гладкие
конические ринофоры равны или немного ко-
роче оральных щупалец. Анальное отверстие
расположено на спинной стороне справа, меж-
ду рядами задней пищеварительной железы.
Длина тела до 120 мм.
Окраска. Наиболее типичная окраска —
серовато-коричневатая, состоит из многочислен-
ных мелких точек и крапинок, разбросанных по
телу и папиллам. Кроме того, встречаются
и красноватые, желтоватые, зеленоватые экзем-
пляры. Оральные щупальца покрыты белым
пигментом. На голове, перед ринофорами, осо-
бенно у мелких экземпляров, располагается ха-
рактерное белое треугольное пятно.
Отличия от похожих видов. От других
мелководных голожаберных моллюсков подо-
тряда Aeolidacea хорошо отличается широким
телом и характерной особенностью, свойствен-
ной всем видам рода Aeolidia — ветви пищева-
рительной железы, несущие папиллы, скоше-
ны в направлении от центра спины вперед, а не
от центра спины назад, как у подавляющего
большинства других эолидацей.
Aeolidia herculea — Эолидия геркулея, фиксированный экземпляр, вид с правой стороны,
хорошо заметны скошенные в переднем направлении ветви пищеварительной железы
217
Отряд Nudibranchia — Голожаберные моллюски
Aeolidia papillosa — Эолидия папилъчатпая,
вид со спинной стороны
Aeolidia papillosa — Эолидия папилъчатпая,
вид с брюшной стороны
Особенности экологии. Aeolidia papillosa
один из наиболее известных голожаберных
моллюсков нашей фауны (хотя и не самых
массовых). Обитает на каменистых, скалистых,
а иногда и на песчаных субстратах. Обнару-
жен от литорали до глубины 153 м.
Размножение. Размножение в Белом мо-
ре с середины июля до конца сентября. Кладки
Aeolidia papillosa — Эолидия папилъчатпая, кладка
имеют вид спиральных шнуров, извитых в виде
фестонов. В самых крупных кладках количество
яиц может достигать 1 млн.
Питание и поведение. В Белом море пи-
таются актиниями Metridium senile, Tealea felina
и Bunodactys Stella. Молодь, растущая на свет-
лых коричневых или оранжевых актиниях, вы-
растает во взрослых особей со светлым, корич-
невым или оранжевым содержимым папилл.
В заливе Петра Великого Aeolidia papillosa пое-
дают актиний Cnidopus japonica. Эолидия не-
обычайно широко растягивает глотку, охваты-
вая половину актинии, и, видимо, совместными
действиями челюстей и радулы скребет ее, по-
сле чего может бросить недоеденной.
Распространение. Амфибореальный
вид. Широко распространен в Белом, Баренце-
вом и всех дальневосточных морях. В северной
Атлантике по европейскому побережью оби-
тает от Норвегии до Франции, отмечен в Ис-
ландии, а по американскому побережью ука-
зан с Баффиновой Земли и до Новой Англии.
В Тихом океане по американскому побере-
жью известен от Аляски до Калифорнийского
залива.
218
Семейство Aeolidiidae — Эолидовые или Эолидиды
Беломорские особи Aeolidia papillosa (Эолидия папилъчатая) в естественном местообитании —
илисто-песчаном грунте среди ракушек и камней
219
Баренцево море, губа Ярнышная. Прилив
Баренцево море, губа Ярнышная. Отлив
220
Необходимое послесловие
ЭВОЛЮЦИЯ И СИСТЕМАТИКА:
ВЗГЛЯД СО СТОРОНЫ ГОЛОЖАБЕРНЫХ МОЛЛЮСКОВ
Заднежаберные моллюски, в силу своей необы-
чайной эволюционной пластичности, т. е. спо-
собности завоевывать новые типы местообита-
ний и существенно менять исходный предковый
облик, являются одной из лучших модельных
групп, на которой особенно удобно проиллю-
стрировать такие важнейшие и наиболее об-
щие понятия биологии, как эволюция и систе-
матика.
2009 год был особым — годом двойного
юбилея Дарвина и его книги. Поэтому помимо
прошедших закономерных и многочисленных
юбилейных статей и конференций, представ-
ляется небезынтересным взглянуть не только
на сам доказанный факт эволюции, на ставшие
классическими постулаты синтетической тео-
рии эволюции, но и на другие немаловажные
компоненты эволюционной теории, которые
далеко не всегда получают должное освеще-
ние. Непревзойденную роль в становлении
дарвиновской теории сыграла классическая
систематика, и тот факт, что сам Дарвин был
прежде всего натуралистом и систематиком,
а потом уже эволюционистом. Несмотря на
это, о связях систематики и эволюции говорят
не часто, точнее ровно в том объеме, который
подчеркивает, что именно процесс эволюции
ответственен за появление бесконечного раз-
нообразия живых форм.
В качестве специальной проблемы для эво-
люционной теории Дарвин выделял отсутствие
или редкость переходных разновидностей. Это
было вполне естественно для науки того вре-
мени, когда самые знаменитые «потерянные
звенья» еще не были открыты или не были ин-
терпретированы с точки зрения эволюции.
Именно отсутствие подобных звеньев нередко
ставится в упрек креационистами в адрес тео-
рии эволюции. Поэтому так важно было от-
крытие кистеперых рыб для понимания ран-
них этапов эволюции наземных позвоночных.
Обнаружение переходных таксонов позвоноч-
ных продолжается и сейчас — достаточно на-
звать тиктаалика, открытого лишь в 2004 году
ископаемого организма, сочетающего черты
кистеперых рыб и земноводных.
Значительный интерес к поиску переход-
ных групп и эволюционным исследованиям
позвоночных животных хорошо объясним ши-
роким распространением знаний об их морфо-
логии и классификации. В то же время, несмо-
тря на значительное число опубликованных
работ по эволюции и систематике самых раз-
личных беспозвоночных животных, подчас да-
же самые сенсационные открытия остаются
уделом обсуждения лишь узкой группы спе-
циалистов. Это происходит прежде всего из-за
слабой информированности ученых других
специальностей о разнообразии и строении
беспозвоночных. Между тем, среди последних
«потерянных звеньев» значительно больше,
и там иногда можно наблюдать почти «вжи-
вую» как шла эволюция, оставившая следы
в современной фауне в виде отдельных родов
и видов. Кроме того, огромная доля нередко
формальных, основанных на статистических
алгоритмах эволюционных деревьев, характер-
ная для современного этапа развития зоологии,
подчас оставляет мало места для классических
эволюционно-морфологических исследований
и тесно связанного с ними понятия о переход-
ных группах. В этом отношении классическая
систематика и морфология может все еще
очень много дать для изучения эволюции.
Одной из групп беспозвоночных, на кото-
рой особенно удобно и наглядно изучать эво-
люцию, являются брюхоногие заднежаберные
моллюски — главная тема данной книги.
Основным эволюционным процессом, кото-
рый доминировал среди заднежаберных, была
редукция (т. е. потеря) раковины. Это привело
к появлению среди Opisthobranchia целого вее-
ра независимо возникших безраковинных
групп высокого ранга (отрядов). Одной из та-
ких групп, наиболее эволюционно успешной,
морфологически разнообразной и обильной
221
по числу видов, являются голожаберные мол-
люски. В свою очередь, среди голожаберных
моллюсков выделяется большая группа дорид
(Doridacea или Anthobranchia), насчитывающая
около 2000 видов. Особая привлекательность до-
рид как модельной группы для изучения эво-
люции заключается в их сходном плане строе-
ния, допускающего тем не менее некоторую
степень эволюционной пластичности. Пода-
вляющее большинство представителей этой
группы имеет развитый кожный спинной
щит — нотум, полностью покрывающий вну-
тренние органы и часто накрывающий ногу,
в передней части которого располагается пара
органов химического чувства — ринофоров,
а в задней части располагается самый характер-
ный внешний орган дорид — розетка дорсаль-
ных жабр. Криптобранхиальные (Crypto-
branchia), или скрытожаберные дориды имеют
вокруг жабр особую полость — жаберный кар-
ман, в который специальные мышцы могут
полностью втянуть жабры и сомкнуть наружное
отверстие кармана. Для криптобранхий также
характерен нормально развитый нотум, полно-
стью накрывающий ногу. В задней части нотум
никогда не бывает расщеплен на лопасти, но
иногда сильно редуцирован и оттянут. Фанеро-
бранхиальные (Phanerobamchia) или открыто-
жаберные дориды полностью лишены жабер-
ной полости. Жабры при этом прикрепляются
прямо к задней части нотума и также при по-
мощи особых мышц способны резко сокра-
щаться. Сами жабры у обеих групп устроены
сходным образом. Также принципиально сход-
но устроены основные системы органов, напри-
мер пищеварительная и половая системы.
В ходе экспедиционных работ у тихооке-
анского побережья Камчатки (фауна Nudi-
branchia которой вплоть до наших исследова-
ний была очень слабо изученной) в августе
2008 года были обнаружены несколько десят-
ков экземпляров неизвестного представителя
семейства онхидоридид (Onchidorididae) об-
ладающего развитым жаберным карманом.
Дальнейшее изучение данных особей под-
твердило, что жабры способны полностью
втягиваться в полость, а ее края могут полно-
стью сокращаться над втянутыми жабрами.
В то же время, по строению радулы и глотки
обнаруженные особи являются типичными
представителями семейства Onchidorididae,
т.е. имеется специфический глоточный орган
онхидоридид — буккальная помпа и узкая ра-
дула (зубы моллюсков).
Обнаружение в водах Камчатки нового ро-
да и вида, который получил название Onchimira
cavifera (которое можно перевести как «удиви-
тельная онхидоридида, несущая жаберную по-
лость»), обладающего с одной стороны настоя-
щим жаберным карманом криптобранхиального
типа, а с другой стороны специализированной
глоткой и радулой, типичными для фанеробран-
хиального семейства Onchidorididae (представи-
тели которого в большинстве своем полностью
лишены жаберного кармана), является серьез-
ным аргументом в пользу вторичного происхо-
ждения большинства фанеробранхиальных до-
рид путем редукции жаберного кармана.
Onchimira cavifera является своеобразным
вектором, который помогает определить пра-
вильное направление эволюции: если специа-
лизированные черты в строении пищевари-
тельной системы, которыми обладают все
представители онхидоридид, не известны за
пределами этого семейства, а напротив, нали-
чие жаберной полости — характерная черта
большинства других дорид, то онхимира, об-
ладающая одновременно и жаберным карма-
ном, и радулой, и буккальной помпой (особым
выростом глотки) безоговорочно онхидори-
дидного типа — очевидно указывает, что новый
род стоит в основании Onchidorididae, и что
жаберная полость затем редуцировалась
у остальных онхидоридид.
Особая важность открытия онхимиры за-
ключается в том, что этот таксон является самым
настоящим потерянным звеном — организмом,
который связывает две большие таксономиче-
ские группы с различным строением. Именно
потерянных звеньев часто не хватало Дарвину,
чтобы эволюционная теория выглядела более
реалистичной. После обнаружения Onchimira
cavifera у берегов Камчатки, а ранее еще и Loy
meyeni Martynov, 1994 в Японском море, можно
построить эволюционную картину развития
этой группы в Тихом океане, которая будет
основана не только на теоретически постулиро-
ванных направлениях эволюции, а на совершен-
но реальных родах и видах. И можно проследить
все стадии той редукции и модификации, кото-
рую претерпела исходная организация крипто-
бранхиальных дорид от Onchimira cavifera до
Adalaria jannae, Loy meyeni и, наконец до абер-
рантных (т. е. на сильно отличающихся от
222
«обычных» дорид) педоморфных видов из рода
Corambe, жабры которых вторично перемести-
лись вентрально (т. е. на брюшную сторону)
в заднюю часть нотума, как это исходно наблю-
дается у личинок всех дорид, и у взрослых особей
древних моллюсков моноплакофор и хитонов.
Эта история еще раз подчеркивает необычайную
устойчивость на первый взгляд хрупкого жиз-
ненного цикла. Чтобы «разрушить» исходный
более сложный цикл криптобранхиальных до-
рид и сформировать упрощенный, педоморф-
ный цикл корамбид, потребовался целый ряд
эволюционных стадий, которые остались в со-
временной фауне в виде соврешенно определен-
ных родов и видов — онхимиры, адалярии, лоя,
причем в той же географической области, где
и происходило, судя по всему, их формирова-
ние — в северной части Тихого океана.
И наконец. Суммируя все сказанное, хоте-
лось бы еще раз обратить внимание читателя,
особенно если он не является профессиональ-
ным биологом, на ту мысль, которую мы всяче-
ски старались до него донести. Ведь если от-
влечься от тривиального взгляда на окружающий
нас мир как на «неизменный» и «постоянный»,
то, внимательно читая эту книгу, можно вдруг
осознать всю хрупкость границ между, казалось
бы, столь бесконечно различными раковинны-
ми актеонидами и похожими на мягкую поду-
шечку крошечными голожаберниками корам-
бидами. И как, последовательно анализируя все
промежуточные этапы, можно вдруг понять
пути столь грандиозного эволюционного про-
цесса преобразования «простой» улитки в это
иррационально красивое чудо природы, голо-
жаберного моллюска.
Авторы будут признательны, если дайве-
ры, любители или профессиональные биоло-
ги, обнаружив в морях России интересного за-
днежаберного моллюска, систематическое
положение которого они не смогли опреде-
лить, используя эту книгу (или в процессе
идентификации выяснилось, что это редкий
вид), свяжутся с нами, а возможно даже сфото-
графируют найденный экземпляр, затем за-
фиксируют его и передадут авторам. Ин-
струкции, как это можно сделать, будут
предоставлены в каждом конкретном случае
специально. Российская наука только выиграет
от этого. Адрес, по которому с нами можно
связаться, — martynov@zmmu.msu.ru.
Баренцево море
223
НЕКОТОРЫЕ ВАЖНЫЕ ТЕРМИНЫ
Абранхиальные дориды — группа голожаберных мол-
люсков отряда дорид, не имеющая систематическо-
го статуса, у которой жабры и жаберная полость
полностью редуцированы.
Амфибореальный вид — бореальный вид, распростра-
ненный одновременно в северных частях как Атлан-
тического, так и Тихого океанов.
Бореальный вид — вид, распространенный в северных
частях Атлантического и/или Тихого океанов и от-
сутствующий в большинстве морей Северного Ле-
довитого (Арктического океана), и адаптированный
к обитанию при средней летней температуре в диа-
пазоне около +10...+20°С. Хорошо переносит зим-
нее охлаждение вплоть до 0° С.
Велигер — планктонная свободноплавающая личинка,
имеющаяся у большинства брюхоногих моллюсков,
включая группу заднежаберных (Opisthobranchia).
Тело велигера одето в тонкую личиночную раковин-
ку и впереди снабжено лопастью, покрытой длин-
ными ресничками, — называемой велюмом или
парусом. Велигер имеет также хорошо развитые
органы равновесия, необходимые для ориентации
в планктонной среде — статоцисты. Многие личин-
ки-велигеры питаются, находясь в планктоне, и име-
ют развитую пищеварительную систему. Такой тип
велигера называется планктотрофным. Другие, на-
против, не питаются, и существуют за счет нако-
пленных запасов желтка. Такой тип велигера назы-
вается лецитотрофным.
Гермафродитизм/гермафродиты — совмещение
в одном организме как женских, так и мужских
структур полового аппарата. Подавляющее боль-
шинство заднежаберных моллюсков — герма-
фродиты.
Гипо бранхиальные дориды — не имеющая системати-
ческого статуса группа голожаберных моллюсков
отряда дорид, у которых жаберная полость полно-
стью редуцирована, а жабры располагаются на
брюшной стороне под складкой нотума в задней
части.
Жевательный желудок — особый отдел, расположен-
ный между пищеводом и железистым желудком
у многих цефаласпид, всех анаспид и крылоногих
моллюсков. Жевательный желудок предназначен
для дополнительной механической переработки
твердой или грубой пищи, такой как раковины
моллюсков или слоевища водорослей и содержит
разное число особых жевательных пластинок — от
трех (у многих цефаласпид) до значительного числа
(у анаспид и крылоногих моллюсков).
Интерстициаль/интерстициальный — пространство
между частичками грунта, и мелкие организмы его
населяющие (например, заднежаберные моллюски
группы акохлидиид).
Книдосак — особый мешочек, образованный разрас-
танием выроста пищеварительной железы внутри
вершинной части спинных выростов-папилл боль-
шинства голожаберных моллюсков подотряда
Aeolidcaea и некоторых видов из подотрядов
Arminacea и Dendronotacea, в котором накаплива-
ются попадающие с пищей стрекательные клетки
(нематоцисты) кишечнополостных.
Конвергенция — эволюционный процесс, близкий по
смыслу к понятию параллелизм. В обоих случаях
достаточно удаленно родственные группы приоб-
ретают независимое друг от друга сходство. Поня-
тие конвергенции обычно используют при харак-
теристике сходства действительно очень удаленно
родственных групп, например рыбообразной фор-
мы ихтиозавров (рептилии) и дельфинов (млеко-
питающие). Конвергенция и параллелизм могут
быть обусловлены различными причинами: на-
пример наличием в жизненном цикле даже уда-
ленно родственных групп сходной ювенильной
стадии, которая при тех или иных условиях может
стать полноценным признаком взрослых стадий.
В другом случае конвергенция может быть напря-
мую не связана с общими онтогенетическими при-
знаками. В этом случае сходство появляется дей-
ствительно «заново». К таковому можно отнести
разнообразные случаи мимикрии, например меж-
ду тропическими голожаберными моллюсками
и плоскими червями.
Копулятивный аппарат — мужской совокупитель-
ный орган. Имеет чрезвычайно разнообразное
строение у представителей различных групп, мо-
жет быть снабжен дополнительными твердыми
стилетами или железами. Строение копулятивно-
го аппарата нередко является важным признаком
в систематике самых различных групп заднежа-
берных моллюсков.
Криптобранхиальные дориды — монофилетическая
группа отряда дорид, обладающая развитой жабер-
ной полостью (жаберным карманом), куда жабры
способны полностью втягиваться.
Нематоцисты — то же, что стрекательные клетки.
Параллелизм — эволюционный процесс, близкий по
смыслу к понятию конвергенция. В обоих случаях
достаточно удаленно родственные группы приоб-
ретают независимое друг от друга сходство. Поня-
224
тие параллелизма обычно используют при характе-
ристике сходства относительно более родственных
групп, например, говорят о параллельной редук-
ции раковины среди различных отрядов (например,
голожаберных моллюсков и сакоглосс) одного под-
класса заднежаберных моллюсков. Параллелизмы
в эволюции, как правило, связаны с общей онтоге-
нетической базой. Причем они могут быть как ре-
грессивного (как в случае редукции раковины), так
и прогрессивного свойства (в случае, например,
сходного добавления стадий при разрастании дор-
сальной стороны глотки и независимого формиро-
вания нового прогрессивного признака — буккаль-
ной помпы).
Параподии — боковые треугольные выросты ноги у ря-
да цефаласпид и анаспид. У птеропод параподии
превращаются в крыловидные отростки, с помо-
щью которых они плавают в водной толще.
Педоморфоз (неотения) — широко распространенное
явление регрессивной перестройки жизненного
(онтогенетического) цикла живых организмов, при
котором те или иные признаки, ранее появляю-
щиеся только на ранних (ювенильных) стадиях, а за-
тем исчезающие, напротив, сохраняются и на взрос-
лых стадиях.
Прямое развитие — особый модифицированный
тип онтогенетического цикла, при котором сво-
бодноплавающая планктонная личинка не разви-
вается, а из кладки выходит непосредственно дон-
ная ползающая молодь. Между прямым
и планктонным развитием есть целый ряд про-
межуточных случаев, при которых, например,
имеется хорошо развитая личинка с личиночной
раковиной, но не выходящей в планктон, а разви-
вающейся внутри яйцевых оболочек. В случае же
глубоко модифицированного прямого развития
личиночная раковинка может развиваться лишь
в очень слабой степени.
Редукция — эволюционный процесс уменьшения сте-
пени развитости той или иной структуры живого
организма вплоть до полного ее исчезновения.
У заднежаберных моллюсков наиболее ярко выра-
женной является редукция раковины.
Ринофоральные карманы — обычно замкнутая или
реже полузамкнутая полость, куда ринофоры спо-
собны полностью втягиваться. У многих голожабер-
ных моллюсков ринфоральные карманы полностью
редуцируются.
Стрекательные клетки (нематоцисты) — особые
защитные клетки, свойственные только представи-
телям типа книдарий (более известных под назва-
нием кишечнополостных). Каждая такая клетка
содержит внутри плотно скрученный жгут, а на по-
верхности — чувствительный волосок. При каса-
нии чувствительного волоска внутренний жгут
резко распрямляется и вонзается в тело жертвы,
куда поступает также и небольшая порция токси-
на. Каждая клетка имеет, конечно, микроскопиче-
ские размеры, но в случае выстреливания тысяч
подобных клеток жертве может быть нанесен су-
щественный урон. Известно например, что австра-
лийские кубомедузы смертельны даже для челове-
ка. Обычно не смертельная, но имеющая яд такой
силы, что он способен приводить к кратковремен-
ным неприятным вегетативным эффектам, неболь-
шая гидроидная медуза-крестовик (Gonionemus
vertens), обитает в водах и нашего залива Петра Ве-
ликого, что в Японском море. Целый ряд голожа-
берных моллюсков научился успешно использо-
вать это весьма грозное и эффективное оружие
в своих целях. Не выстрелившие стрекательные
клетки, после поедания голожаберниками их хозя-
ев — книдарий — транспортируются по каналам
пищеварительной железы к вершинам папилл, где
накапливаются в особых мешочках-книдосаках,
а затем используются для собственной защиты
моллюсков.
Ювенилизация — термин, по смыслу близкий к поня-
тию педоморфоз. При ювенилизации, признаки
бывшие у предков только на ранних стадиях онто-
генетического цикла, у потомков появляются на
взрослых, половозрелых стадиях жизненного цик-
ла. Иногда понятие ювенилизации использовать
удобнее, чем термин педоморфоз, поскольку явля-
ясь более общим, с его помощью может быть оха-
рактеризована различная степень проявления юве-
нильных признаков на взрослых стадиях, даже
относительно небольшая. При педоморфозе обыч-
но речь идет о ярко выраженной ранней ювениль-
ной морфологии на взрослых стадиях онтогенети-
ческого цикла.
Ювенильный — признак, относящийся к одной из не-
половозрелых, как правило, достаточно ранних ста-
дий жизненного цикла.
Циркумтропический вид — тропический вид, отмечен-
ный в водах Мирового Океана повсеместно (т. е. как
в тропических водах Индийского и Тихого океанов,
так и Атлантического океана).
Фанеробранхиальные дориды — не имеющая система-
тического статуса группа голожаберных моллюсков
отряда дорид, у которых жаберная полость полно-
стью редуцирована, жабры при этом остаются на
спинной стороне и прикрепляются непосредствен-
но к наружным покровам.
225
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИИ
Acanthodoris pilosa ... 128,129
Acochlidiida ... 76-80
Acochlidiacea ... 76-80
Actaeon ... 45
Acteocina matusimana ... 48
Acteonida ... 46
Acteonidae ... 46
Adalaria jannae ... 135
Adalaria olgae ... 132
Adalaria proxima ... 130,131
Adalaria slavi ... 134
Adocia cinerea ... 123
Aenigmastyletus alexeii ... 191
Aeolidacea ... 163-219
Aeolidia farallonensis (syn.) ... 217
Aeolidia grandis (syn.) ... 217
Aeolidia herculea ... 217
Aeolidia papillosa ... 217, 218
Aeolidiidae ... 216-219
Aglajidae ... 53
Akera bullata ... 63,64
Akeridae ... 63-66
Akiodoris lutescens ... 106
Akiodorididae ... 105-107
Alderia modesta ... 81, 82
Amathia sp. ... 145
Amphorina horii ... 185, 186
Amphorina odhneri ... 183
Anaspidea ... 63-69
Ancula cristata (syn.) ... 118
Ancula gibbosa ... 118
Ancula pacifica ... 119
Anculidae ... 118,119
Animalia ... 14
Anthobranchia ... 99-144
Aplysia juliana ... 67, 68
Aplysia parvula ... 68, 69
Aplysiellidae ... 103
Aplysiidae ... 67-69
Archidoris ... 121
Archidoris pseudoargus ... 121
Armina ... 32
Arminacea ... 147-149
Arminidae ... 146
Armodoris ... 105
Ascophyllum nodosum ... 130, 196
Asperspina murmanica ... 79, 80
Asperspinidae ... 79
Atyidae ... 62
В
Bathyberthella ... 91
Berthella ... 91
Berthella californica ... 95, 96
Boreoberthella ... 33, 90
Boreoberthella augusta ... 93, 94
Bryozoa ... 15
Cadlina laevis ... 103-105
Calycidoris guentheri ... 128
Calliopaea bellula ... 86
Caprella ... 180
Catriona gymnota ... 200, 202
Cauloramphus sp. ... 131, 138
Cavolinia ... 71
Cephalaspidea ... 46-62
Ceratosoma ... 102
Cerberilla ... 216
Chaetomorpha linum ... 84
Chlamylla atypica ... 163,164
Chordata ... 14
Chromodorididae ... 102-104
Chromodoris ... 102
Clava multicornis ... 176,197
Clione antarctica ... 75
Clione limacina ... 72, 73
Clionidae ... 73-75
Cnidaria ... 14,15
Cnidopus japonica ... 219
Cocophora langsdorphi ... 210
Codium ... 87
Colga minichevi ... 108,109
Colga pacifica ... 109
Colga villosa ... 109
Corambe obscura ... 142,143
Corambidae ... 38, 126, 144
Cordylophora caspia ... 212
Coryne loveni ... 176,197
Coryphella athadona ... 177
Coryphella amabilis ... 173
Coryphellagracilis ... 171
Coryphella japonica ... 168
Coryphella nobilis ... 169,170
Coryphella polaris ... 167
Coryphella salmonacea ... 168
Coryphella verrucosa ... 174-176
Crenomytilus grayanus ... 124,178
Cribrilina
annulata ... 131,136,138
Cryptobranchia ... 102
Cuthonella abyssicola ... 195
Cuthonella concinna ... 198,199
Cuthonella hiemalis ... 195,196
Cuthonella marisalbi ... 200
Cuthonella soboli ... 197
Cylichna alba ... 49, 50
Cylichna arctica (syn.) ... 49
Cylichna consobrina (syn.) ... 49
Cylichna corticata (syn.) ... 49
Cylcihna densistriata ... 50
Cylichna occulta ... 49, 50
Cylichna scalpta ... 50
Cylichnatys angusta ... 62
Cylichnatys incisula (syn.) ... 62
Cylichnidae ... 48, 49
D
Dendronotacea ... 150-162
Dendronotidae ... 152-158
Dendronotus dalli ... 155
Dendronotus frondosus ... 152,153
Dendronotus robustus ... 157,158
Dermatobranchus
walteri (syn.) ... 148
Diacria ... 71
Diaphorodoris ... 118
Diaulula sandiegensis ... 123,124
Dirona aurantia ... 149
Dirona pellucida ... 149
Dironidae ... 148-151
Diphasia falax ... 154, 200
Discodorididae ... 122-125
Doridacea ... 99-146
Dorididae ... 120,121
Doridoxa walteri ... 148
Doridoxidae ... 147
Doridunculus unicus ... 107
Doris ...44, 45,120
Doto coronata ... 158,159
Doto fragilis ... 160
Dotoidae ... 158, 159
226
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Diadumene lineata ... 207
Dynamena pumila ... 197
E
Egmundella ... 107
Embletonia pulchra ... 161,162
Embletoniidae ... 161
Enteromorpha ... 64
Ercolania boodleae ... 84
Ercolania viridis ... 84, 85
Eubranchidae ... 185-191
Eubranchus exiguus ... 186
Eubranchus tricolor ... 182,183
Eucratea loricata ... Ill, 119
Eudendrium ... 172
Eudendrium annulatum ... 172
Euselenops ... 98
F
Facelinidae ... 215
Flabellinidae ... 163-179
Flustrella hispida ... 130
Fucus serratus ... 196, 200
G
Gastropoda ... 14
Gastropteridae ... 46, 71
Glossodoris ... 14,102
Goniodoridella savignyi ... 119,120
Goniodorididae ... 144, 145
Goniodoris ... 144
Gymnodorididae ... 30, 116
Gymnosomata ... 71-74
H
Haliclona permolis ... 123
Hancockia ... 150
Hataiaparva ... 166
Hexabranchus ... 30
Hermaea vancouverensis ... 87, 88
Hermaeidae ... 87
Hermissenda crassicornis ... 215, 216
Heterodoris ... 93,146
Himatina trophina ... 31,179,180
Homo sapiens ... 37, 39
Homoidoris japonica ... 165
Hopkinsia ... 118
Hypselodoris ... 102
J
Janolidae ... 27
Janolus ... 147
Japonactaeon ... 45
Japonactaeon nipponensis ... 47
Laminaria japonica ... 216
Laminaria saccaharina ... 154
Leostyletus
pseudomisakiensis ... 192,193
Limacina ... 71, 75
Limacina helicina ... 72
Limacina retroversa ... 72
Limacinidae ... 71
Limapontia ... 81
Limapontia capitata ... 84
Limapontia cocksi (syn.) ... 83
Limapontia depressa ... 83
Limapontia senestra ... 83
Limapontiidae ... 81-85
Loy meyeni ... 141,142
M
Mammalia ... 17
Melanochlamys diomedea ... 53, 54
Membranipora ... 131
Membranipora membranacea ... 138
Metridium senile ... 219
Microhedylidae ... 79
Mollusca ... 14,17,24
Murmania antiqua ... 194
Mytilus galloprovincialis ... 142
N
Navanax inermis ... 216
Nembrotha ... 108,216
Notaspidea ... 89-98
Nudibranchia ... 146-218
Nudibranchus exiguus ... 190
Nudibranchus rupium ... 186
Nudipleura ... 89, 93
О
Obelia longissima ... 153, 154, 160, 176,
178, 189, 190,212-216
Okadaia elegans (syn.) ... 117
Okadaia tecticardia (syn.) ... 117
Okadaiida ... 116
Okenia ... 144
Okenia distincta ... 145
Onchidorididae ... 126-143
Onchidoris bilamellata ... 140
Onchidoris
тасгоротра ... 138,139
Onchidoris muricata ... 136
Onchimira cavifera ... 126
Oncorhynchus kisutch ... 13
Opisthobranchia ... 15, 221
Ophlitaspongia pennata ... 124
Oweniidae ... 157
p
Palio ... 108
Palio dubia ... 111,112
Paracoryphella
islandica ... 165,166
Paralithodes kamchatica ... 178
Parhedyle tyrtowii ... 77, 78
Parhedylidae ... 77
Peregonimus megas ... 212
Phanerobranchia ... 20, 21, 102
Philine argentata ... 50, 51
Philine scalpta ... 51, 52
Philinidae ... 50, 51
Philinopsis gigliolii ... 54
Phyllidiidae ... 27, 30
Phyllirhoe ... 30
Placida dendritica ... 86
Pleurobranchaea japonica ... 98
Pleurobranchaeidae ... 98
Pleurobranchella ... 98
Pleurobranchidae ... 89-97
Pleurobranchoidea ... 89-97
Pleurobranchus ... 90,91
Polycera ... 108
Polycera quadrilineata ... 113
Polyceridae ... 108-115
Pontohedyle
milaschewitschi ... 78, 79
Porifera ... 14
Pseudotritonia ... 45
Pseudovermis ... 30, 34
Pseudovermidae ... 180, 181
Pseudovermis
paradoxus ... 181, 182
Pteropoda ... 70-72
Ptilosarcus gurneyi ... 152
227
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
Retusa instabilis ... 55
Retusa minima ... 59
Retusa per tenuis ... 56
Retusa succincta ... 59
Retusa truncatula ... 57, 58
Retusa umbilicata ... 60
Retusa variabilis ... 59, 60
Retusidae ... 55-60
Rhodomela ... 87, 88
Roboastra ... 108
Rostanga ... 45
Rostanga alisae ... 117, 124
Sacoglossa ... 81-88
Sertularella rugosa ... 205
Sertularia sp. ... 200
Sertularia cupressina ... 171
Sertulariidae ... 203-205
Spirorbidae ... 116
Stiliger akkeshiensis (syn.) ... 87
Strongylocentrotus ... 67
Stylatula elongata ... 152
Tealeafelina ... 219
Tenellia adspersa ... 210
Tergipedidae ... 193-213
Tergipes ... 193
Tergipes tergipes ... 212,213
Thecosomata ... 71, 72
Tochuina tetraquetra ... 150
Trapania ... 118
Trinchesia ornata ... 202, 203
Trinchesia perca ... 205, 206
Trinchesia pupillae ... 186, 209
Trinchesia pustulata ... 207, 208
Trinchesia viridis ... 203, 204
Triopha catalinae ... 115
Triopha pacifica (syn.) ... 115
Tritonia ... 45
Tritonia diomedea (syn.) ... 150
Tritonia primorjensis (syn.) ... 150
Tritonia tetraquetra ... 150,151
Tritoniidae ... 150
Tubularia ... 171,213
Tubularia indivisa ... 176
u
Ulva ...87
V
Vaucheria ... 82
Vayssierea ... 30, 118
Vayssierea caledonica ... 118
Vayssierea elegans ... 116,117
Vayssiereafelis ... 118
Vayssiereidae ... 116,117
Virgularia spp. ... 152
Volvulella ... 61
Volvulella sculpturata ...61
Volvulellidae ... 61
Vulpes corsac ... 13
Yokoyamaia ornatissima ... 52, 53
z
Zostera ... 191
Zostera asiatica ... 191, 193
Zostara caespitosa ... 191
Zostera marina ... 191
Примечание:
Синонимы имеют пометку (syn.);
жирным шрифтом выделены номера страниц, на которых есть иллюстрации.
228
ИЗБРАННЫЕ РАБОТЫ АВТОРОВ
А. Публикации по систематике и фауне заднежаберных моллюсков:
1. Мартынов А. В., 1992. Новый вид голожаберных
моллюсков из Японского моря с замечаниями
о роде Cuthonella (Gastropoda, Opisthobranchia)
И зоологический журнал. Т. 71, № 12. С. 18-24.
2. Мартынов А. В., 1994. Материалы к ревизии голо-
жаберных моллюсков семейства Corambidae
(Gastropoda, Opisthobranchia) Сообщение 1: Си-
стематика // Зоологический журнал. Т. 73, № 10.
С. 1-15.
3. Мартынов А. В., 1997. Subclassis Opisthobranchia.
Список видов животных, растений и грибов лито-
рали дальневосточных морей России / Сост.
О. Г. Кусакин, М. Б. Иванова, А. П. Цурпало и др.).
Владивосток: Дальнаука. С. 77-80.
4. Мартынов А. В., 1997. Заднежаберные моллюски
Командорских островов с замечаниями по фауне
Дальневосточных морей России. Донная фауна
и флора Командорских островов / Ред. А. В. Ржав-
ский. Владивосток: Дальнаука. 230-241 с.
5. Мартынов А. В., 1998. Заднежаберные моллюски
(Opisthobracnhia) семейства Eubranchidae: таксо-
номическая структура и два новых вида из Япон-
ского моря Ц Зоологический журнал. Т. 77, № 7.
С. 763-777.
6. Мартынов А. В., 1998. Subclassis Opisthobranchia
И А. В. Адрианов, О. Г. Кусакин. Таксономический
каталог биоты залива Петра Великого Японского
моря. Владивосток: Дальнаука. С. 204-209.
7. Мартынов А. В., Чабан Е. М., 1998. Aplysia parvula
и A. juliana из прибрежных вод острова Моне-
рон — первая находка представителей Aplysiidae
(Opisthobranchia, Anaspidea) в фауне России
И Ruthenica. Vol. 8, № 1. d. 17-28.
8. Токмакова Н. П., Петрова О. М., Мартынов А. В.,
1999. Сравнительная гистологическая характери-
стика эпителия пищеварительной железы неко-
торых видов заднежаберных моллюсков // Биоло-
гия моря. Т. 25, № 1. С. 51-55.
9. Мартынов А. В., 1999. Голожаберные моллюски
северо-западной части Японского моря (с замеча-
ниями об отряде Nudibranchia): Автореф. дис......
канд. биол. наук. СПб.: Зоологич. ин-т РАН. 24 с.
10. Martynov А. V., 2001. Order Nudibranchia 11 In: List of
species of free-living invertebrates of Eurasian Arctic
Seas and adjacent deep waters / Eds. В. I. Sirenko et al.
St. Petesrbrug: Zoological Institute RAS. P. 109-110.
11. Мартынов A. B., 2002. Два новых вида рода Trinche-
sia Ihering, 1879 из залива Петра Великого Япон-
ского моря (Nudibranchia, Tergipedidae), с замеча-
ниями о таксономии семейства // Ruthenica. Т. 12,
№ 1. С. 45-54.
12. Мартынов А. В., Баранец О. Н., 2002. Ревизия рода
Colga Bergh (Opisthobranchia, Polyceridae), с опи-
санием нового вида из северной части Тихого
океана // Ruthenica. Т. 12, № 1. С. 23^-3.
13. Мартынов А. В., Редькин Д. В., 2002. Новый для
фауны России бореальный голожаберный мол-
люск Archidoris pseudoargus (Nudibranchia, Dorid-
idae) из губы Дальне-Зеленецкой Баренцева моря
И Биологические основы устойчивого развития
морских экосистем. Мурманск. С. 198-200.
14. Martynov А. V., 2003. A new species of the genus Ros-
tanga Bergh, 1879 (Mollusca: Opisthobranchia) from
the Peter tne Great Bay, the Japan Sea, with a discus-
sion on the genus Boreodoris Odhner, 1939 // Ru-
thenica. Vol. 13, № 2. P. 141-147.
15. Martynov A. V., Roginskaya I. S., 2005. A new species of
the genus Doridunculus G.O. Sars, 1878 (Mollusca,
Nudibranchia): hydroid-feeding dorid from the abys-
sal depths of the Sea of Japan // Ruthenica. Vol. 14,
№ 2. E 135-145.
16. Martynov A. V., 2006. Nudipleura Ц In book: Yu. I. Kan-
tor, A. V. Sysoev. Marine and brackish water Gastro-
poda of Russia and adjacent countries: an illustrated
catalogue. Moscow: KMK Scientific Press Ltd.
P. 267-294.
17. Martynov A. V., 2006. Sacoglossa 11 In book: Yu. I. Kan-
tor, A. V. Sysoev. Marine and brackish water Gastro-
poda of Russia and adjacent countries: an illustrated
catalogue. Moscow: KMK Scientific Press Ltd.
P. 265-267.
18. Martynov A. V., Korshunova T. A., Savinkin О. V., 2006.
Shallow-water opisthobranch molluscs of the Mur-
man coast of the Barents Sea, with new distribution-
al data and remarks on biology 11 Ruthenica. Vol. 16.
№ 1-2. P. 59-72.
19. Martynov A. V., Korshunova T. A, Grintsov V. A., 2007.
Opisthobranch molluscs of the Northern Black Sea.
I. Short history of studies and the first record of
a non-indigenous nudibranch species Trinchesia
perca (Er. Marcus, 1958) (Nudibranchia: Tergipedi-
dae) // Ruthenica. Vol. 17, № 1-2. P. 43-54.
20. Мартынов А. В., Коршунова T. А., Санамян H. П.,
Санамян К. Э., 2008. Голожаберные моллюски
прибрежных вод Камчатки: недооцененное раз-
нообразие Северо-Западной Пацифики // Мате-
риалы IX Международной научной конференции
«Сохранение биоразнообразия Камчатки и при-
легающих морей», г. Петропавловск-Камчатский.
Сборник статей / Отв. ред. А. М. Токранов. Кам-
чатский филиал Тихоокеанского института гео-
графии ДВО РАН. С. 85-88.
21. Мартынов А. В., Кортунова Т. А., Санамян Н. П.,
Санамян К. Э., 2009. Заднежаберные моллюски
(Gastropoda: Opisthobranchia) прибрежных вод
острова Старичков. Биота острова Старичкова
и прилегающей к нему акватории Авачинского
залива // Труды Камчатского филиала Тихооке-
анского ин-та географии ДВО РАН. Вып. 6.
С. 227-240.
22. Neusser Т. Р, Martynov А. V., Schrodl М., 2009. Heart-
less and primitive? Computer-based 3D reconstruc-
tion of the polar acochlidian gastropod Asperspina
murmanica Ц Acta Zoologica. Vol. 90. P. 228-245.
23. Martynov A. V., Korshunova T. A., Sanamyan N. P, Sa-
namyan К. E., 2009. Description of the first crypto-
branch onchidoridid Onchimira cavifera gen. et sp.
nov. and of three new species of the genera Adalaria
and Onchidoris (Nudibranchia: Onchidorididae)
from Kamchatka waters // Zootaxa. № 2159. P. 1^13.
229
Б. Публикации по эволюционной истории в контексте систематики
заднежаберных моллюсков:
1. Мартынов А.В., 1994. Материалы к ревизии голо-
жаберных моллюсков семейства Corambidae
(Gastropoda, Opisthobranchia). Сообщение 2: Про-
исхождение корамбид Ц Зоологический журнал.
Т. 73, №11. С. 36-43.
2. Martynov A.V., 1995. Materials for the revision of the
nudibranch family Corambidae (Gastropoda,
Opisthobranchia). Communication 2. Origin of the
Corambidae // Hydrobiological Journal. Vol. 31, № 7.
C.59-66.
3. Мартынов A. B., 1998. Забытый антарктический
род Guyvalvoria и его родственник из Баренцева
моря — представители особой линии в семействе
Tergipediaae // Материалы XIII (4) Всероссийской
малакологической конференции. СПб.: Зоологич.
ин-т РАН.
4. Martynov А. V., 1999. Buccal pumps, gills pockets and
new understanding of suctorial pnanerobranchial
dorids Ц Systematic, Phylogeny and Biology of
Opisthobranch Molluscs (2nd International Work-
shop of Malacology) Menfi, Italy. June 10-14 1999.
P. 13-14.
5. Мартынов A. B., 2000. О таксономическом положе-
нии рода Echinocorambe Valdes et Bouchet, 1998
(Gastropoda: Nudibranchia) Ц Материалы XIV (5)
Всероссийской малакологической конференци.
СПб.: Зоологич. ин-т РАН. С. 50-52.
6. Millen S. V., Martynov А. V., 2005. Redescriptions of
the nudibranch genera Akiodoris Bergh, 1879 and
Armodoris Minicnev, 1972 with description of a new
species of Akiodoris and description of new family
Akiodorididae Ц Proceedings of the California Aca-
demy of Sciences. Vol. 56, № 1. P. 1-22.
7. Martynov A. V, 2006. Archaic Tergipedidae of the
Arctic and Antarctic: Murmania antiqua gen. et sp.
nov. from the Barents Sea and a revision of the genus
Guyvalvoria Vayssiere with the descriptions of two
new species // Ruthenica. Vol. 16, № 1-2. P. 73-88.
8. Martynov A. V., Schrodl M., 2008. The new Arctic
side-gilled sea slug genus Boreoberthella (Gastrop-
oda, Opisthobranchia): Pleurobranchoidean system-
atics and evolution revisited // Polar Biology. Vol. 32.
P. 53-70.
9. Мартынов А. В., Коршунова T. А., Санамян H. П.,
Санамян К. Э., 2009. Революционная находка: об-
наружение переходного звена в эволюции голо-
жаберных моллюсков у берегов Камчатки. В книге:
Сохранение биоразнообразия Камчатки и приле-
гающих морей // Материалы X Международной
научной конференции, посвященной 300-летию
со дня рождения Г. В. Стеллера. Петропавловск-
Камчатский: Камчатпресс. С. 175-183.
В. Теоретические работы по онтогенезу и эволюции:
1. Мартынов А. В., 2009. От онтогенеза к эволюции:
систематика в ожидании смены парадигмы.
Эволюция и систематика: Ламарк и Дарвин
в современных исследованиях // Со. трудов Зоо-
логического музея МГУ / Ред. А. В. Свиридов,
А. И. Шаталкин. Т. 50. С. 145-229.
2. Martynov А. V., 2010. A new concept of ontogenetic
systematics: Toward the synthesis of taxonomy, on-
togeny, and phylogenetics. Molecular Phylogenetics,
Molpny-2 // Second Moscow International Conferen-
ce on Molecular Phylogenetics, Moscow State Uni-
versity. 18-21 May 2010. P. 55-56.
Г. Работы по истории зоологических коллекций:
1. Martynov А. V, 2002. The shell collection of 2. Мартынов А. В., Слепкова H. В., Долголенко M. A.,
J. H. Chemnitz in the Zoological Institute RAN // Ru- 2003. О возможном происхождении трёх гравиро-
thenica. Vol. 12, № 1. Р. 1-18. ванных наутилусов из ранних коллекций Петров-
ской Кунсткамеры // Ювелирное искусство и ма-
териальная культура. СПб.: Государственный
Эрмитаж. С. 75-81.
Д. Научно-популярные статьи по заднежаберным моллюскам:
1. Мартынов А. В., 2006. Ускользающая красота 3. Мартынов А. В., 2007. Знакомьтесь: морской ангел
И Нептун - 21 Век. № 3. С. 104-109. Ц Нептун - 21 Век. № 1. С. 103-104.
2. Мартынов А. В., 2006. Десять тысяч лет под водой
И Нептун - 21 Век. № 6. С. 108-113.
Александр Владимирович Мартынов
Татьяна Алексеевна Коршунова
ЗАДНЕЖАБЕРНЫЕ МОЛЛЮСКИ
МОРЕЙ РОССИИ
Атлас-определитель с обзором биологии
Фотографии Т. А. Коршуновой, А. В. Мартынова
В книге также использованы фотографии
О. В. Савинкина, Н. П. Санамян, М. В. Переладова, Н. А. Червяковой,
Bjorn Gulliksen, Erling Svensen
Редактор В. В. Джанаева
Корректор Л. И. Боршова
Главный художник М. В. Суханова
Верстка и подготовка к печати М. Н. Синельниковой
ЗАО «Фитон+» — www.phytonflowers.ru
Изд. лиц. ЛР № 066624 от 26.05.99
Гигиенический сертификат № 77.99.2953.П.5535.9.99 от 13.09.99
Налоговая льгота —
общероссийский классификатор продукции ОК 005-93, том 2
95 3000 — книги и брошюры
Отдел оптовых продаж — тел. (499) 256-25-75, salesC^fiton-knigi.ru
Розничная продажа — интернет-магазин www.plantarya.ru
Формат 70хЮ0*/1б- Гарнитура Newton. Бумага мелованная.
Усл. печ. л. 18,85. Тираж 800 экз. Заказ N« Р-474.
Отпечатано в типографии филиала ОАО «ТАТМЕДИА»
«ПИК «Идел-Пресс». 420066, г. Казань, ул. Декабристов, 2.
Мартынов Александр Владимирович — научный
сотрудник Зоологического Музея МГУ имени
М. В. Ломоносова, кандидат биологических наук.
Участник многочисленных экспедиций по исследова-
нию морей России. Основная область научных ин-
тересов —морфология, систематика и эволюционная
история морских беспозвоночных. Автор несколь-
ких новых родов и видов глубоководных офиур, со-
бранных в разных районах Мирового Океана на
глубинах 5-6 тысяч метров. Обнаружил в морях
России и описал целый ряд новых таксонов задне-
жаберных моллюсков. Полученные им новые микро-
морфологические и онтогенетические данные
позволили пересмотреть систематику голожаберных
моллюсков и иглокожих из класса офиур.
Коршунова Татьяна Алексеевна — старший на-
учный сотрудник Института Высшей Нервной Дея-
тельности и Нейрофизиологии РАН, кандидат
биологических наук. Основная область научных
интересов — клеточные и молекулярные меха-
низмы обучения и памяти. В качестве объекта ис-
следования использует животных с простой
нервной системой — моллюсков. Активный участ-
ник многочисленных экспедиций по изучению
морей России, включая погружения в сложных
условиях Северных и Дальневосточных морей.
Автор большинства фотографий, использованных
в настоящем Атласе.