















































































































































































































































































































































































































Author: Бриттон Г.
Tags: материальные основы жизни биохимия молекулярная биология биофизика общая биохимия биология физиология
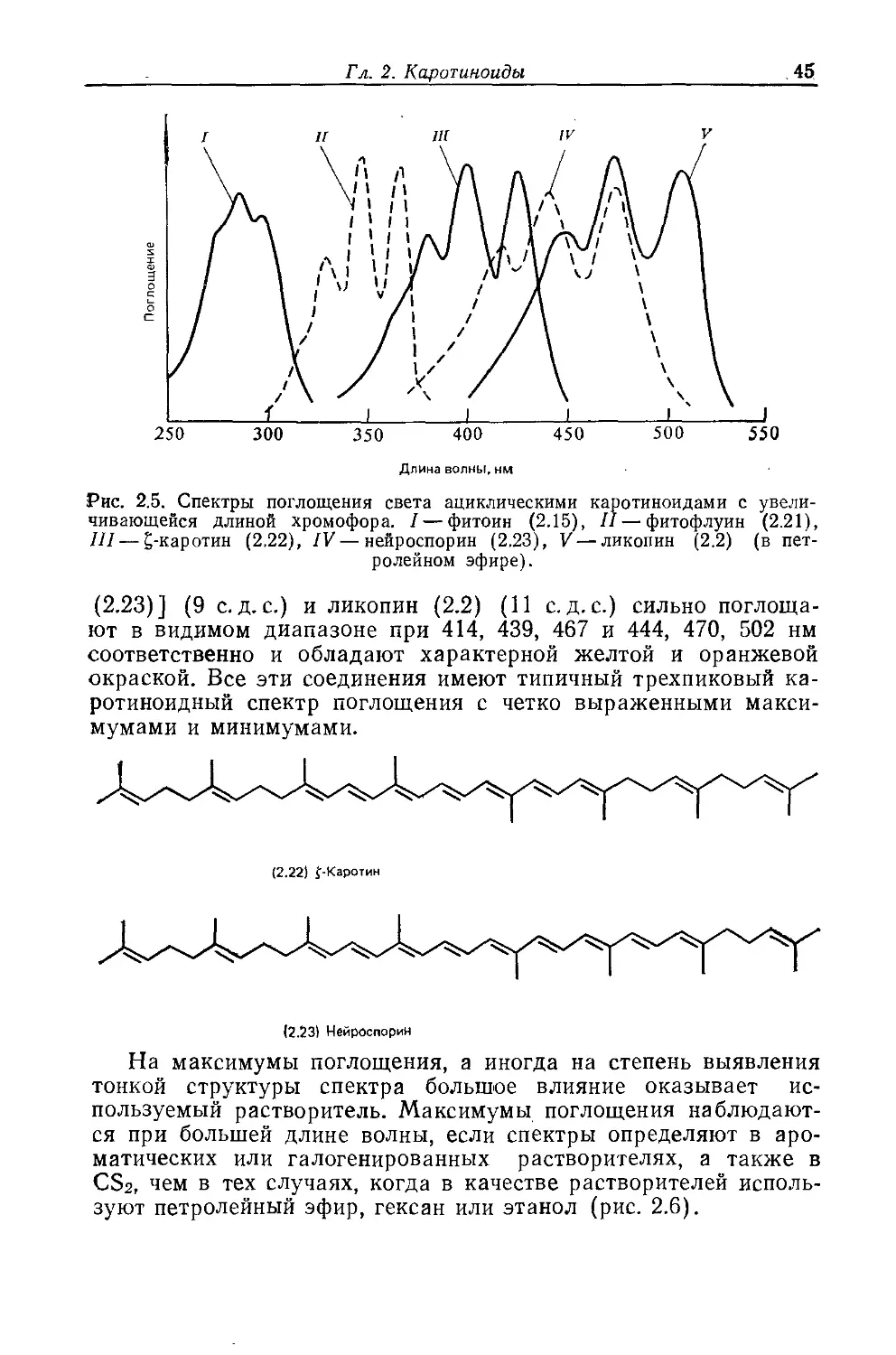
Year: 1986




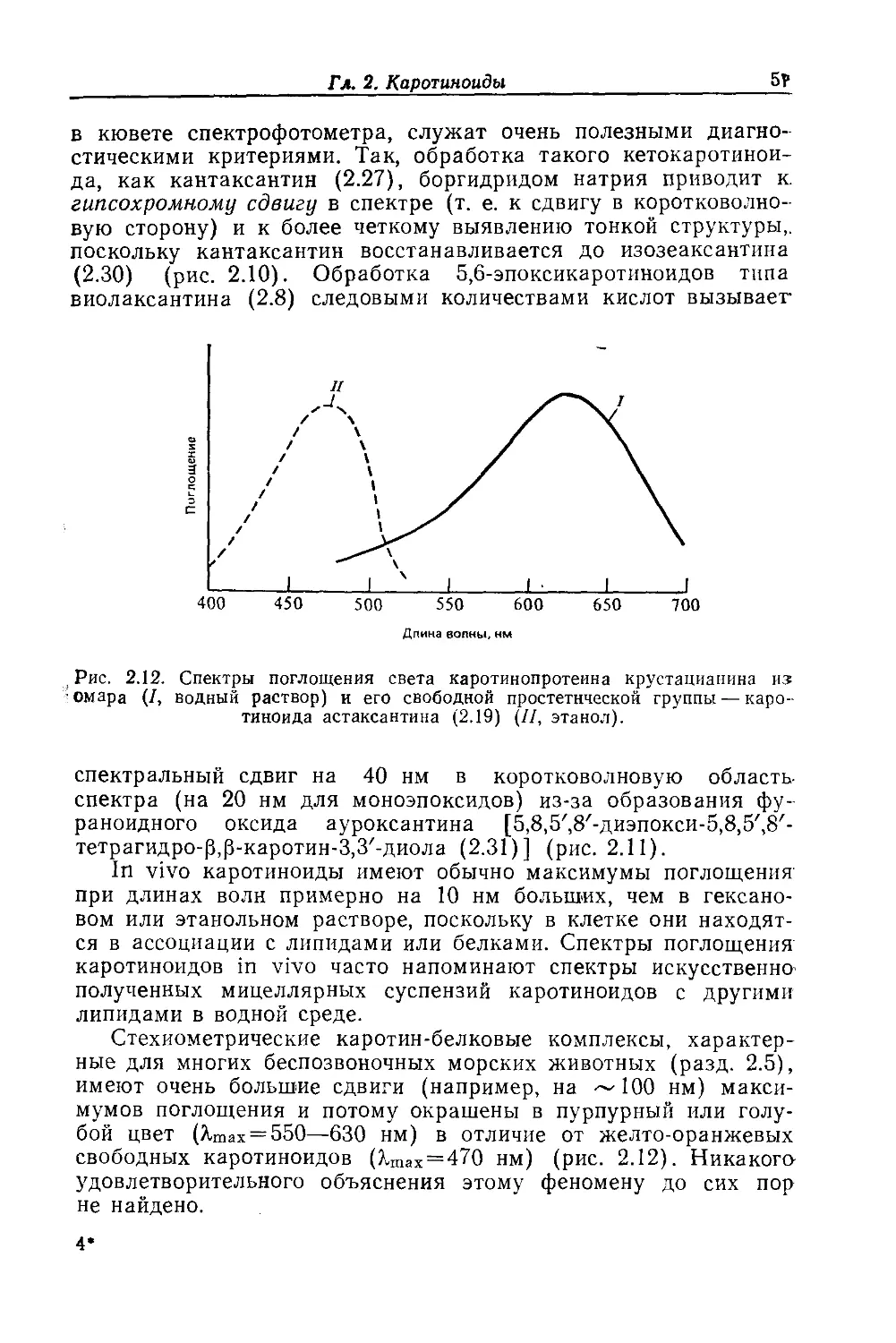
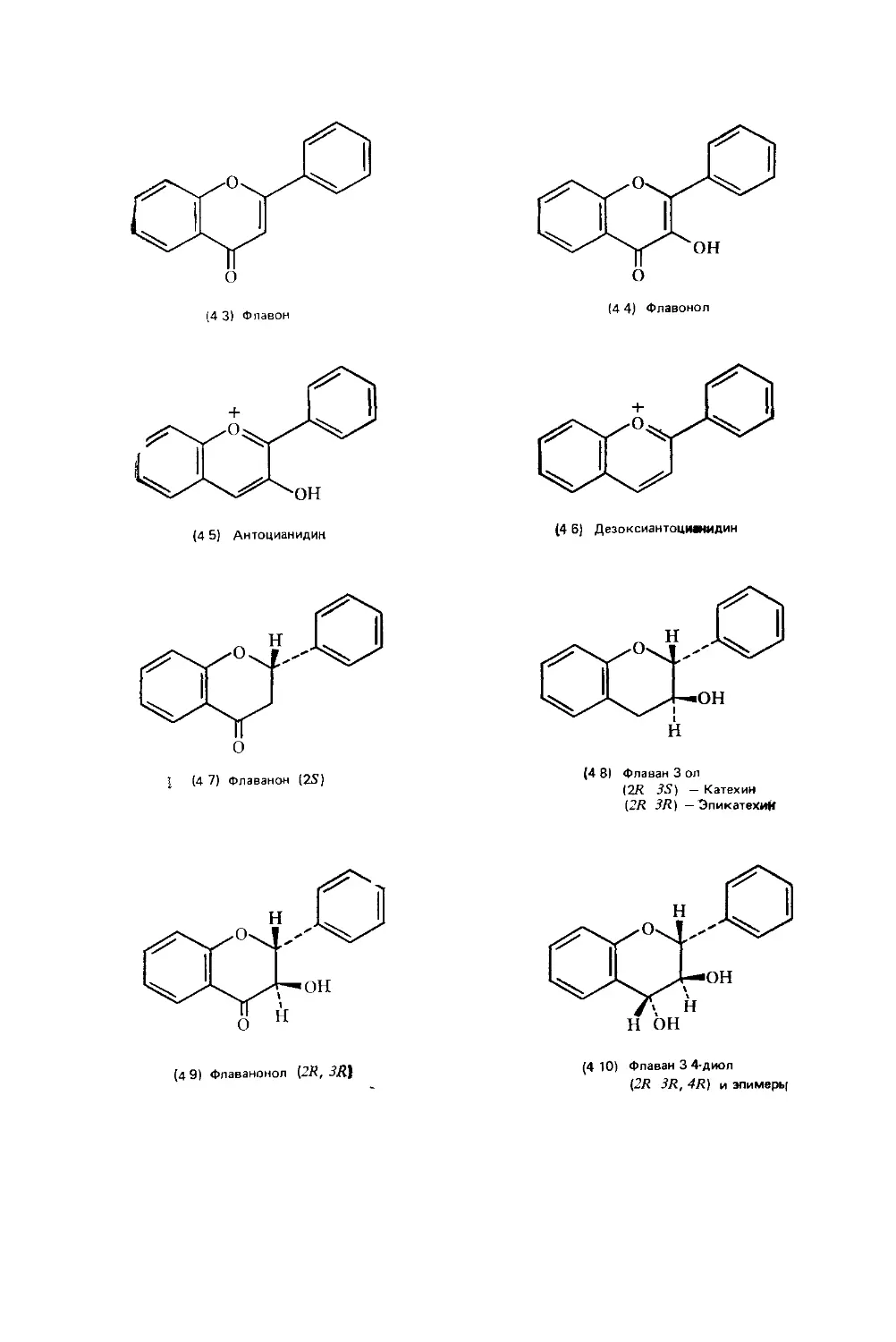
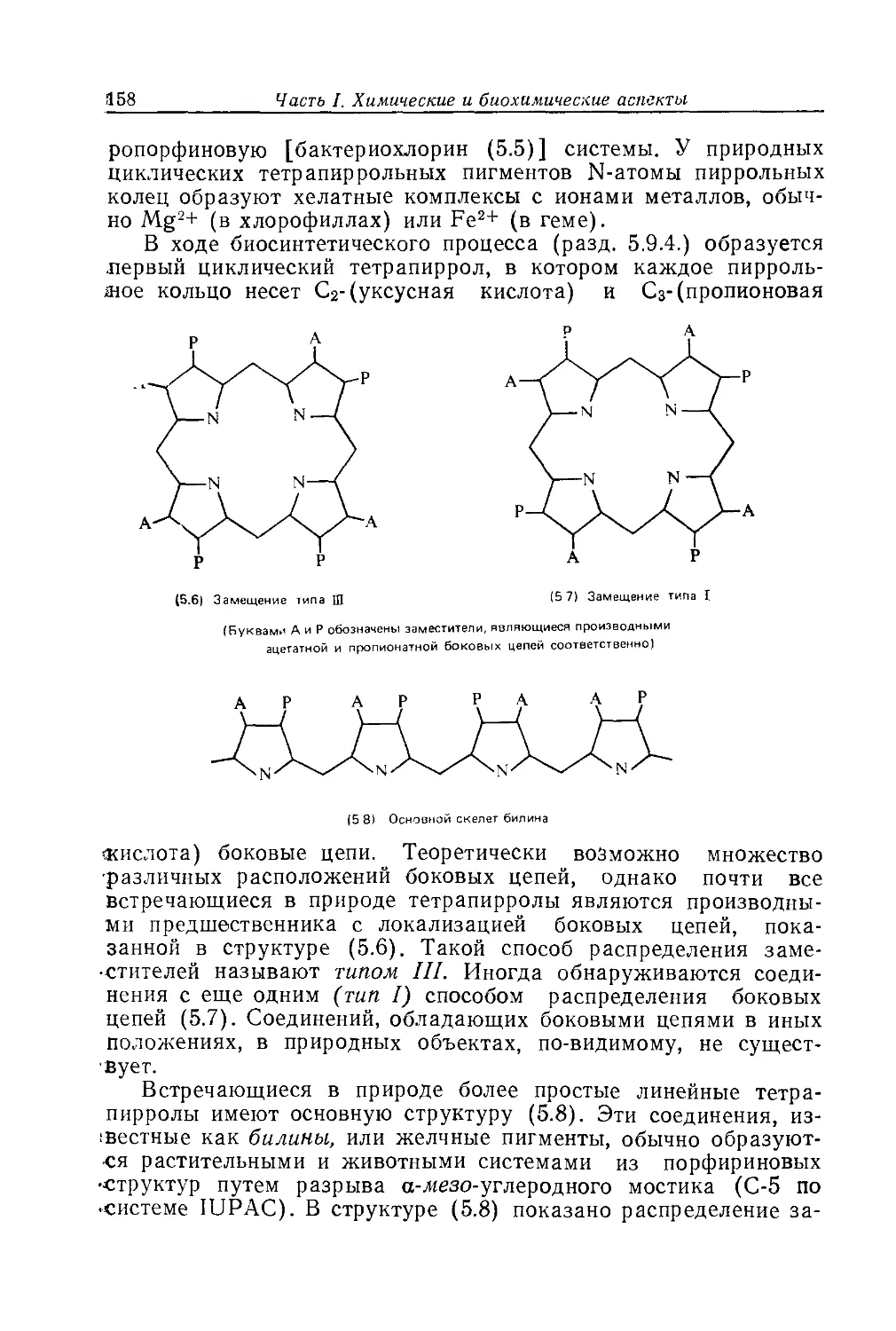
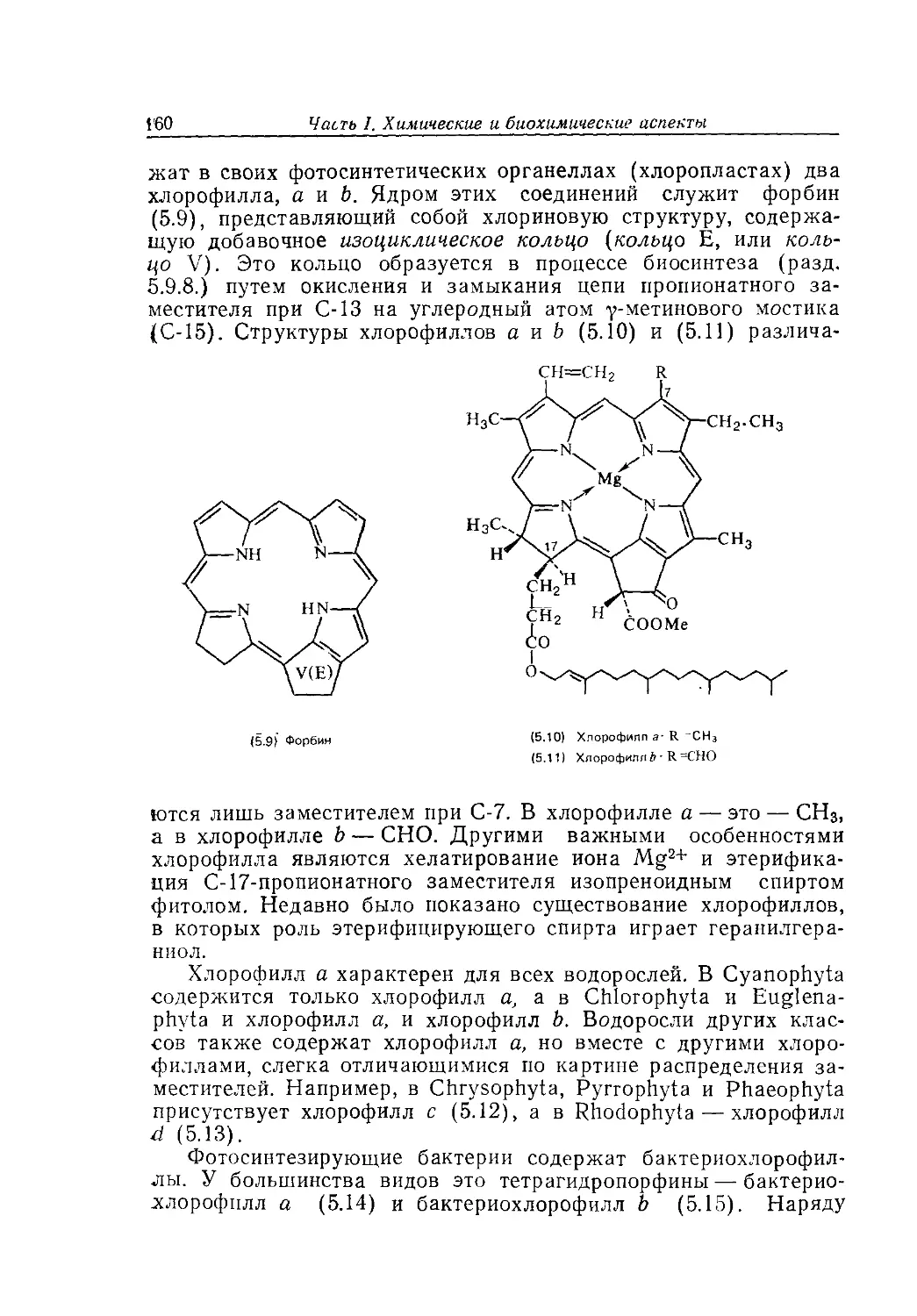
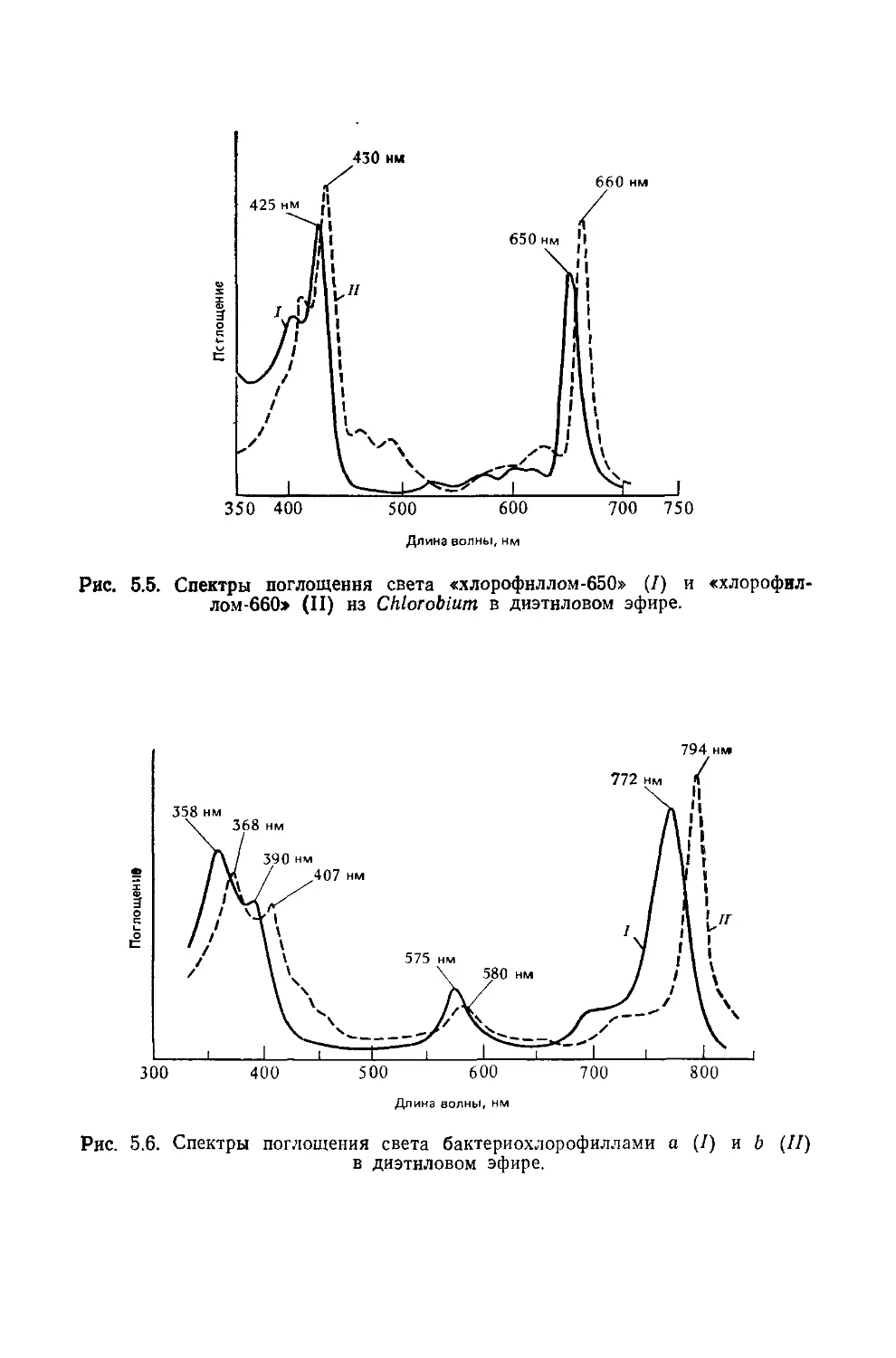
Text
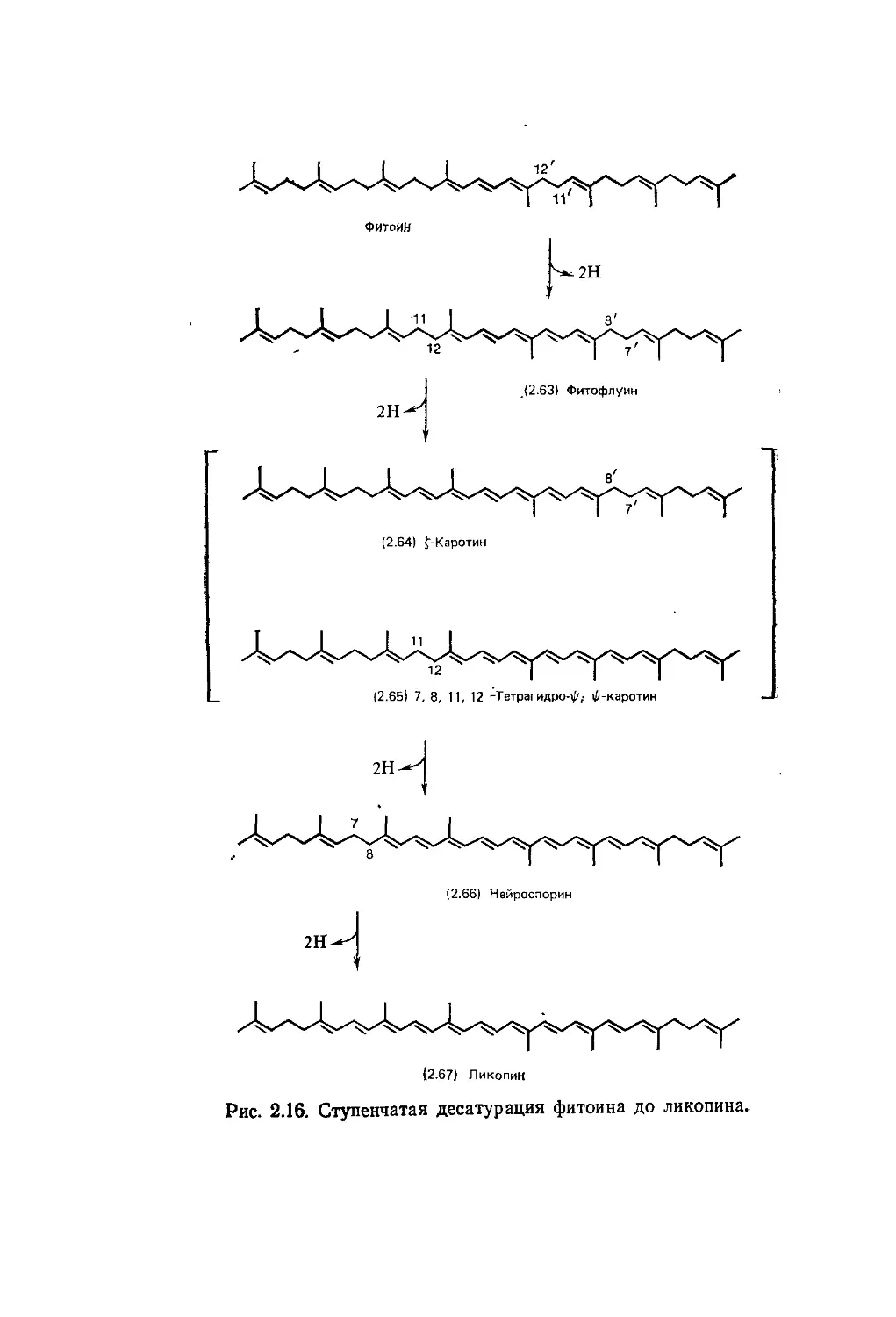
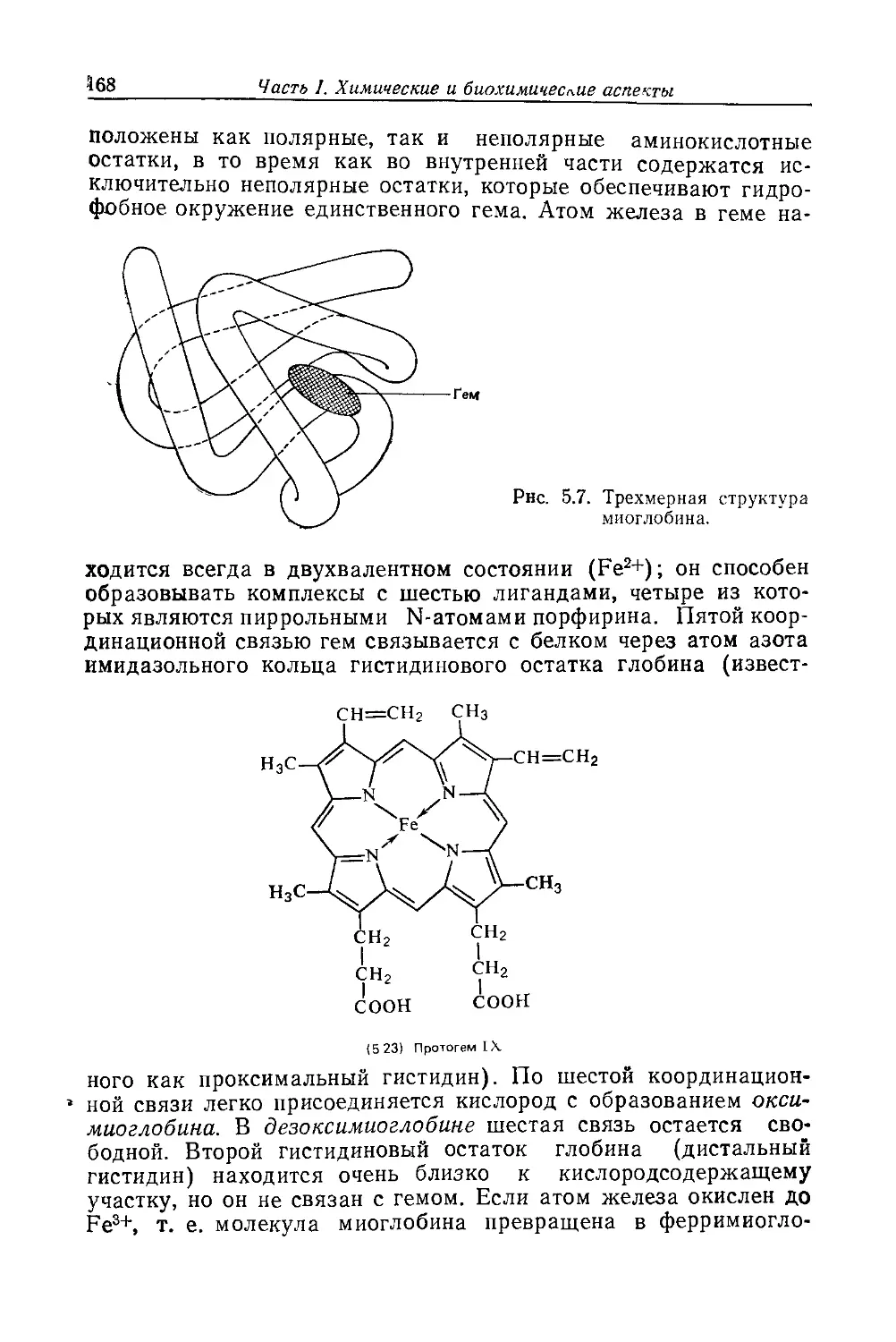
Биохимия природных пигментов
The Biochemistry
of Natural Pigments
G. BRITTON
Cambridge University Press
Cambridge New Rochelle
London New York
Cambridge University Press
Cambridge
London New York New Rochelle
Melbourne Sydney
Г.Бриттон
БИОХИМИЯ
ПРИРОДНЫХ
> ПИГМЕНТОВ
Перевод с английского
канд. биол. наук В. Д. ЦЫДЕНДАМБАЕВА
под редакцией
проф. М. Н. ЗАПРОМЕТОВА
Москва ,,Мир“ 1986
ББК 28.072
Б87
УДК 577.117.3
Бриттон Г.
Б87 Биохимия природных пигментов: Пер. с англ. — М.:
Мир, 1986. — 422 с., ил.
Книга английского автора — первая биохимическая монография, охватываю-
щая все краски живой природы. В доступной, но строго научной форме она зна-
комит с химией и биохимией пигментов, их распространением в природе (от бак-
терий до млекопитающих), многообразием функций.
Предназначена для биохимиков и физиологов, зоологов и ботаников, для
студентов и преподавателей биологических вузов.
Б
2007020000—261
041/01/—86
143—86, ч. 1
ББК 28.072
Редакция литературы по биологии
Cambridge University Press 1983
This book was originally published in the
English language by Cambridge University
Press of Cambridge, England.
© перевод на русский язык, «Мир», 1986
Предисловие редактора перевода
Предлагаемая вниманию читателя книга Г. Бриттона «Биохимия
природных пигментов» уникальна по замыслу. Автор — известный специа-
лист в области химии и биохимии каротиноидов — взял на себя труд п сме-
лость собрать воедино накопленные к настоящему времени биохимией, хи-
мией и биологией сведения о природных пигментах. Если говорить точнее,
то это первая монография, охватывающая (преимущественно с биохими-
ческих позиций) все краски живой природы, даже те, которые обусловлены
прежде всего не наличием специфических пигментов, а интерференцией све-
та в тонких слоях полимерных пленок (например, крылья бабочек и птиц,
чешуя рыб).
Для осуществления своего замысла автору пришлось пользоваться ли-
тературой по оптике и молекулярной спектроскопии, органической химии и
молекулярной биологии, биохимии и физиологии человека и животных, на-
секомых и микроорганизмов, высших и низших растений, а также по таким
более узко специализированным разделам, как фотобиология и фотосинтез,
биолюминесценция и хемилюминесценция, химия и фотохимия процессов зре-
ния, поведение животных и экологические аспекты животного и раститель-
ного мира.
Книга Г. Бриттона написана простым и ясным языком. Она содержит
минимум необходимого материала и поэтому легко читается, временами
приближаясь к стилю научно-популярного изложения. При всем этом книга
глубоко научна, и для читателя, заинтересующегося каким-либо вопросом
более детально, снабжена в конце каждого раздела достаточно обстоятель-
ной библиографией, включающей как монографии, так и обзоры и публика-
ции последних конференций и симпозиумов.
Можно ли объединить все разнообразие красок живой природы в рам-
ках общей биохимической проблемы? После знакомства с книгой Г. Брит-
тона становится ясным, что на этот вопрос следует дать отрицательный от-
вет. Действительно, шесть рассматриваемых в книге крупных групп природ-
ных пигментов—-каротиноиды, хиноны, О-гетероциклические пигменты (глав-
ным образом флавоноиды), тетрапирролы (гем, хлорофиллы, билииы, кобал-
амины и др.), прочие N-гетероциклические пигменты (флавины, феназины,
феноксазины, беталаины и др.) и меланины — во многом, а в некоторых
случаях и полностью отличаются друг от друга как по биогенетическим
путям, так и по функциям и распространению в природе. В соответствии
с этим в книге рассматриваются такие разнообразные и крупные проблемы,
как биохимия зрения (и, в частности, цветового зрения), биохимия фотосин-
теза у бактерий, прокариотических сине-зеленых водорослей, эукариотиче-
6
Предисловие редактора перевода
ских водорослей и высших растений, вопросы фотобиологии (фитохром и его
участие в процессах роста и морфогенеза растений, в фототропизмах и в
фототаксисе водорослей и бактерий; изменение окраски животных в зави-
симости от стадий развития, внешних условий или стрессовых воздействий)
и экологические аспекты пигментации у растений и животных.
При таком широком размахе в некоторых случаях совершенно естест-
венна неполнота илн ограниченность информации о функциях той или иной
группы рассматриваемых соединений.
Так, например, в книге Г. Бриттона не приведены сведения о влиянии
флавоноидов на процессы роста, развития и репродукции растений, об их
способности регулировать процесс окислительного фосфорилирования, об их
антимутагенной, Р-витаминной и других видах биологической активности, а
также о путях их метаболизма в организме животных и человека.
Можно не согласиться и с тем, что автор вычленяет хиноны в особую
группу природных пигментов, отрывая их от исходных фенольных соедине-
ний (гидрохинонов), хотя он сам признает большую гетерогенность пред-
ставителей этой группы, их малое участие в окраске наружных покровов
или тканей и отсутствие общей биологической функции.
Однако если учесть, что основной целью автора было описание лишь
пигментных функций тех или иных веществ, то эти пробелы вполне могут
быть оправданы.
Особая ценность книги Г. Бриттона состоит в том, что на фоне стреми-
тельно возрастающего объема научной информации и все более узкой спе-
циализации исследователей она дает возможность со сравнительно неболь-
шой затратой времени ознакомиться с современным состоянием наших зна-
ний в области химии, биологии и биохимии природных пигментов, имеющих
столь важное значение в жизни человека и животных, высших и низших
растений, грибов и микроорганизмов.
М. Н. Запрометов
Предисловие
В то время когда я пишу эти строки, листья на деревьях в моем
саду меняют свою окраску с летней зеленой на желтые, красные и коричневые
цвета осени, розы и другие цветы все еще расцвечены яркими красками, а
большинство болотных птиц на берегу уже нарядились в свои зимние на-
ряды. Подобные простые явления служат хорошим примером как распрост-
ранения окраски, так и ее разнообразия в живой природе; но они настоль-
ко привычны, что обычно мы считаем их чем-то само собой разумеющимся.
У некоторых животных окраска и характер ее распределения имеют боль-
шое значение для маскировки, позволяющей не привлекать внимания хищ-
ников. У других животных ее функция состоит в оповещении о своем при-
сутствии потенциального брачного партнера. Яркие цвета окрашенных цвет-
ков и плодов важны для привлечения опыляющих и разносящих семена жи-
вотных. Глаз человека радуют насыщенные или нежные оттенки всевозмож-
ных садовых и полевых цветов и краски ярко окрашенных плодов в при-
родных условиях или в витринах магазинов. Нет ничего удивительного в
том, что ученые уже давно заинтересовались природой окраски растений и
животных и изучают механизмы, лежащие в основе ее образования и про-
явления. Сейчас уже хорошо известно, что существуют два принципиально
различных механизма формирования окраски в природе: физические или оп-
тические явления, связанные со структурой клеток и тканей, обусловлива-
ют структурную окраску, а присутствие поглощающих свет веществ — пиг-
ментов — обусловливает пигментную окраску. Настоящая книга посвящена
биохимии природных пигментов — молекул, создающих множество красок в
живом мире. Однако роль многих пигментов не ограничивается тем, что они
придают окраску; способность поглощать видимый свет делает их полезны-
ми и в других отношениях; они необходимы для таких жизненно важных
процессов, как, например, улавливание света при фотосинтезе, восприятие
света и различение цветов в процессе зрения, а также многие другие опо-
средованные светом реакции и регуляторные механизмы. Все эти вопросы,
касающиеся роли природных пигментов, и будут рассмотрены в данной
книге.
Она разделена на две части. В первой части описаны основные особен-
ности химии и биохимии главных групп природных пигментов, вторая часть
посвящена биологическим аспектам и охватывает основные функции пиг-
ментов в природе. В книге используется описательный подход и основное
внимание уделяется главным свойствам и принципам функционирования
пигментов. Такое описание не может быть исчерпывающим, поскольку его
ограничивает объем глав, иначе каждая глава разрослась бы до серии в
8
Предисловие
несколько томов. Цель же настоящей книги заключается в том, чтобы дать
общую картину, привлечь внимание к наиболее интересным проблемам, при-
вить вкус к их изучению и снабдить читателя ключевыми ссылками. Я с
большой тщательностью подошел как к отбору освещаемых в книге тем,
так и к определению степени детальности их изложения. Читатели могут
ие согласиться с моим выбором, либо могут подумать, что я неоправданно
расставил акценты или неудачно распределил материал по главам, однако
я вижу предмет, описываемый в данной книге, именно таким, как я его
представил. Написание и подготовка этой книги были связаны с новыми
ощущениями, часто приятными, иногда расстраивающими и отнимающими
время и внимание, которые, возможно, требовались для другого. В ходе
подготовки, однако, я много прочел и много узнал о природных пигментах.
Это было очень плодотворно, и я надеюсь, что мне удалось донести до чи-
тателя хотя бы часть добытых мною знаний и того огромного интереса, кото-
рый возбуждает во мне данный предмет.
Наконец — и это мне доставляет большое удовольствие — я должен вы-
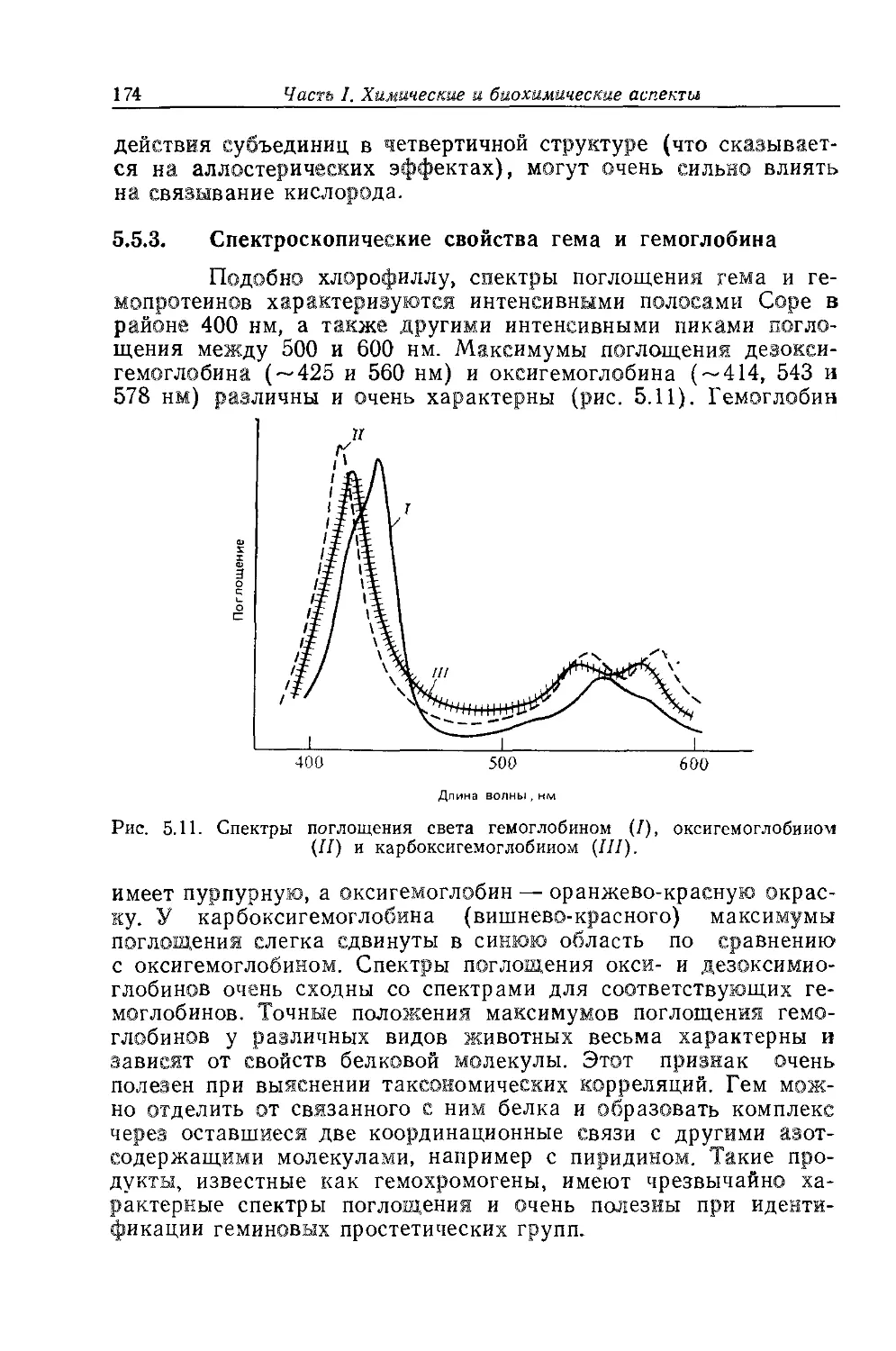
разить свою благодарность очень многим людям. Прежде всего я хотел бы
поблагодарить своих родителей за их жертвы и поддержку в продолжение
всех лет моего обучения: я счастливо провел эти годы, изучая мир при-
родных пигментов. Приношу также мою благодарность доктору Э. Хэсламу
и профессору Т. В. Гудвину, которые поощряли меня в моей работе и у
которых я научился столь многому. Я признателен сотрудникам моей ис-
следовательской группы за снисходительность, проявленную ими на протя-
жении этих лет, когда я посвящал книге то время и внимание, на которые
с полным правом могли рассчитывать они. Я также хотел бы поблагодарить
доктора Эрнста Кирквуда, миссис Марион Жуве и других сотрудников
«Кэмбридж Юниверсити Пресс» за их труд по превращению моего машино-
писного текста в книгу.
Я особенно благодарен членам моей семьи, которые терпеливо мири-
лись с тем, что я тратил время и энергию на подготовку книги и часто не
уделял им должного внимания. Моя жена Пат вынесла все это со стойкостью
и терпением. Более того, она очень помогла мне, превратив мою несовер-
шенную и местами неразборчивую рукопись в безупречный машинописный
текст. Мои дети — Ребекка и Джонатан — временами были лишены общения
со мной и того отцовского руководства, которое необходимо в их возрасте
и которое я сам хотел бы осуществить. Это им я посвящаю настоящую кни-
гу в надежде на то, что они смогут получить столько же удовольствия, сколь-
ко получил я, соприкоснувшись с миром живой природы, где цвет играет
такую большую роль.
Г. Бриттон
Час ib 1
Химические
и биохимические аспекты
Глава 1
Свет и цвет
1.1. Введение
1.1.1. Электромагнитное излучение Солнца
Все процессы жизнедеятельности на Земле в конечном
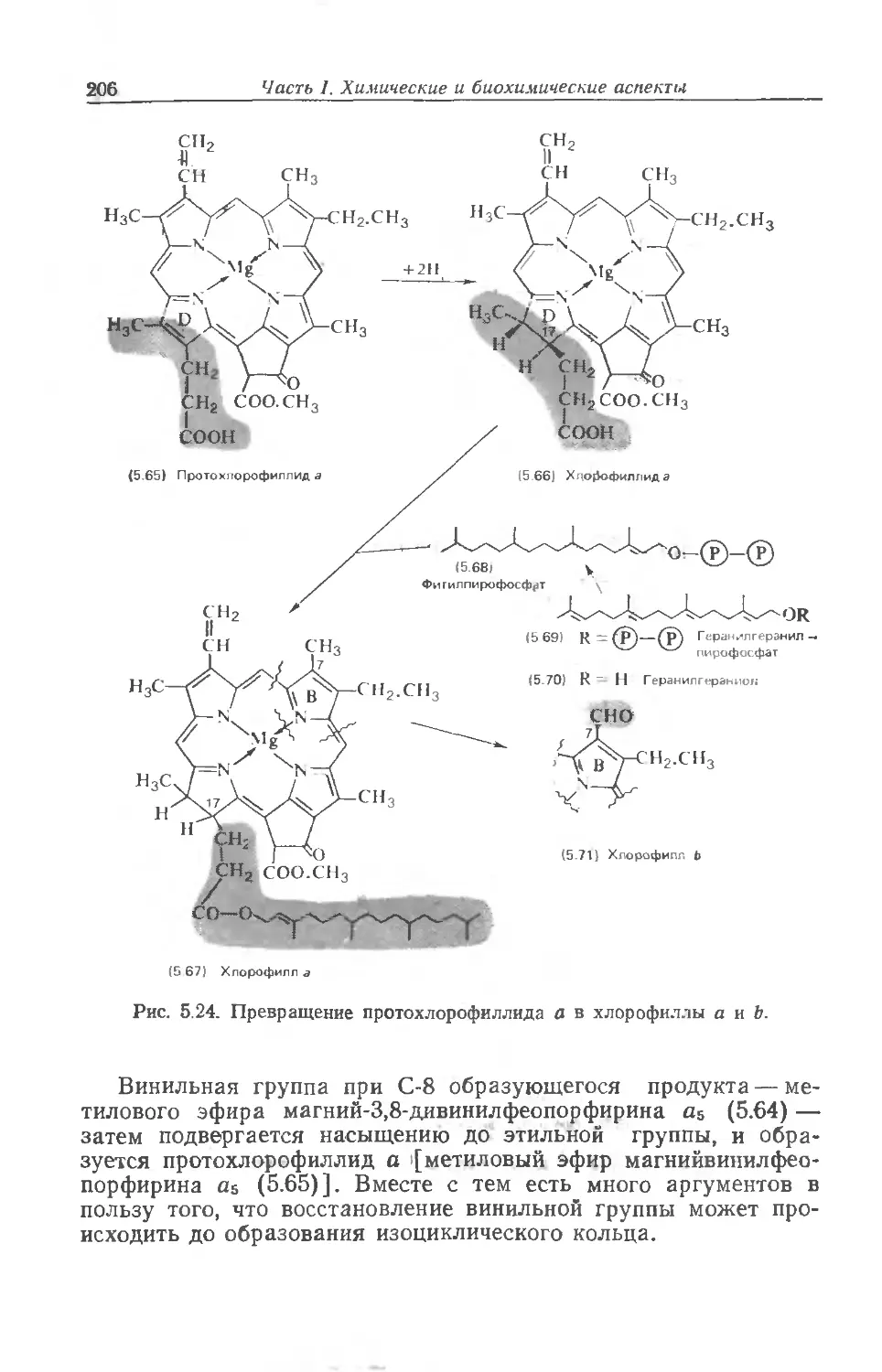
итоге зависят от той части огромных ресурсов солнечной энер-
гии, которая достигает поверхности нашей планеты. Солнце ис-
пускает широкий спектр электромагнитных излучений, от длин-
новолнового инфракрасного (ИК.) и радиочастотного до корот-
коволнового ультрафиолетового (УФ) и у-лучей. (рис. 1.1).
Однако земная атмосфера эффективно отфильтровывает боль-
шую часть этого излучения, особенно обладающие высокой
энергией и губительные для тканей живых организмов УФ-,
рентгеновские и у-лучи.
1.1.2. Видимый свет
Среди разных видов излучений, которые достигают
поверхности Земли, с наибольшей легкостью пронизывают ат-
мосферу лучи с длинами волн 380—750 нм (1 нм = 10-9 м).
Этот диапазон длин волн, называемый видимым светом, имеет
первостепенное значение для поддержания жизни. Животные,
в том числе и человек, обладают очень сложными фоторецептор-
ными системами для обнаружения видимого света, а также для
точного различения света с разной длиной волны в процессе
цветового зрения. Окраска и способность приобретать окраску
стали, таким образом, очень важными в мире живого.
Видимый свет — это тот диапазон световой энергии, кото-
рый используется растениями и микроорганизмами в процессе
фотосинтеза. С помощью фотосинтеза атмосферная двуокись
углерода фиксируется в такой химической форме, которая ис-
пользуется не только самими растениями, но и служит первич-
ным источником пищи для всего живого мира. Различные фо-
торецепторы регистрируют также вариации в количестве до-
Часть I. Химические и биохимические аспекты
ступного видимого света, например различия в длине дня и
ночи, и это лежит в основе чрезвычайно важных механизмов,
регулирующих рост и развитие. Для того чтобы быть окрашен-
ными, различать цвета и осуществлять такие процессы, как
рецепция света, фотосинтез и фоторегуляция, живые организмы
должны обладать специфическими механизмами, позволяющи-
ми им поглощать свет в видимой части спектра. Поэтому мо-
лекулы природных пигментов, или биохромов, обладающие осо-
бым свойством поглощать свет с длинами волн 380—750 нм,
Рнс. 1.1. Спектр электромагнитного излучения.
имеют фундаментальное значение. Целью настоящей книги яв-
ляется обзор основных химических и биохимических свойств
природных пигментов, а также по возможности полное описа-
ние их функционирования на молекулярном уровне.
1.2. Цвет и его восприятие
1.2.1. Цвет
Одновременное восприятие всего диапазона 380—
750 нм создает у человека ощущение белого света. Другие жи-
вотные способны воспринимать излучение с длинами волн, вы-
ходящими за пределы этого диапазона. Например, пчелы «ви-
дят» УФ-лучи, которые невидимы для человека.
Ощущение цвета создается в том случае, если мы воспри-
нимаем лучи только части видимого диапазона. Белый свет
представляет собой совокупность электромагнитных излучений
с диапазоном длин волн 380—750 нм. Если его разделить пу-
тем пропускания через призму, то образуется ряд полос, каж-
дая из которых охватывает гораздо более узкий диапазон длин
волн. Мы видим эти лучи как ряд цветов, в частности красный,
оранжевый, желтый, зеленый, голубой, синий и фиолетовый
Гл. 1. Свет и цвет
11
цвета радуги, которые образуются в результате преломления
солнечного света на капельках воды. Ощущение каждого от-
дельного цвета связано с длиной волны светового луча данно-
го диапазона; например ощущение желтого цвета мы получаем
от света с длиной волны ~ 580 нм. Ощущения, которые люди
с нормальным цветовым зрением отождествляют с определен-
ными длинами волн, перечислены на рис. 1.2. Можно также
III'
Фиоле—Синий-Зеленый—Желтый тОранже--Крас-
товый ; вый ный;
Г ! I I
X • I 1
Комплементарный \ • [ i |
Цвет (---}Желтый“Оранже--Крас- Пурпур- Синий-[—
।вый» ный ный।
Длина волны^нм 400 500 600 700 800
Рис. 1.2. Видимый участок спектра. Показаны цвета, которые люди с нор-
мальным цветовым зрением идентифицируют как свет с определенной дли-
ной волны, а также комплементарные (цвета, полученные «вычитанием» от-
дельных длин волн) цвета, наблюдающиеся в тех случаях, когда свет оп-
ределенного цвета, или диапазона длин волн, «вычитается» из спектра бе-
лого света.
получить ощущение определенного цвета при смешивании све-
та с разными длинами волн, например желтый цвет может
быть получен при смешивании красного и зеленого света.
Вместе с тем ощущение цвета можно получить путем вычи-
тания из совокупности лучей, составляющих белый свет, до-
вольно узкой полосы (в диапазоне длин волн 20—30 нм).
В этом случае виден цвет, комплементарный цвету отсутствую-
щей полосы длин волн. Таким образом, если белый свет про-
пустить через фильтр или вещество, которое поглощает, напри-
мер, синий свет, т. е. свет в диапазоне длин волн 480+30 нм,
то выходящий луч будет иметь цвет, комплементарный синему,
т. е. желтый. Комплементарные, или «вычитаемые», цвета, на-
блюдающиеся после исключения света определенной длины
волны из белого света, также перечислены на рис. 1.2.
1.2.2. Восприятие света и различение цветов
Приведенные выше рассуждения о свете и цвете часто
используют при объяснении нашей способности «видеть цвет».
Хотя различение и описание цветов в значительной степени
субъективны, в их основе должен лежать фундаментальный
физиологический механизм, который не только позволял бы об-
12
Часть I. Химические и биохимические аспекты
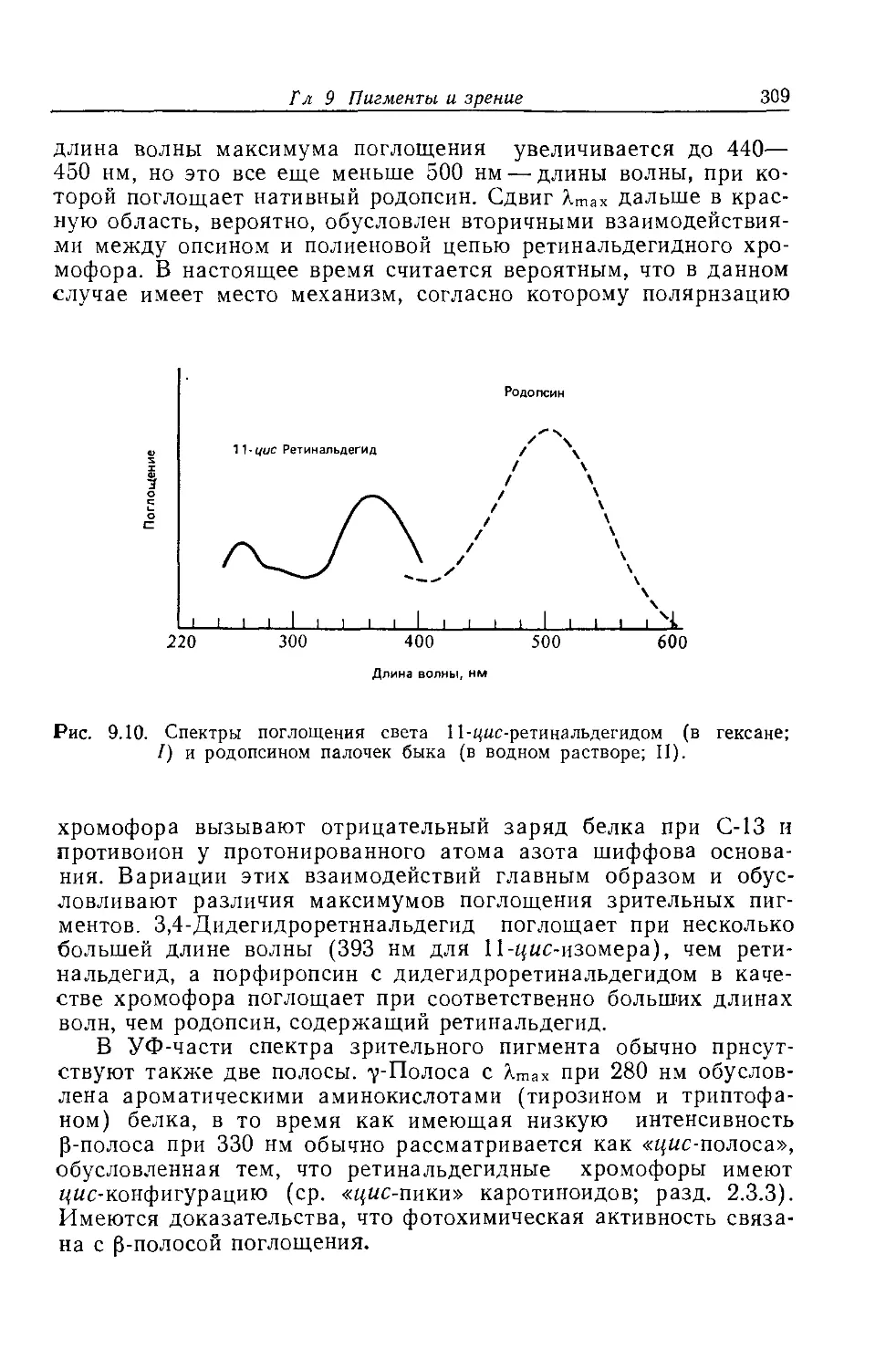
иаруживать электромагнитное излучение в диапазоне длин
волн 380—750 нм, но был бы также ответственным за различ-
ные ощущения, возникающие в ответ на излучение определен-
ной длины волны внутри этого диапазона. Насколько нам из-
вестно, растения и микроорганизмы зрения не имеют, но и они
способны использовать энергию света определенных длин волн,
например для фотосинтеза (гл. 10), и способны двигаться или
расти по направлению к источнику света либо от него (гл. 11).
Однако лишь в животном царстве механизмы восприятия све-
та и различения цветов развились в точный и чувствительный
аппарат зрения, который мы знаем и которым пользуемся.
1.2.3. Глаз и цветовое зрение
У животных органом восприятия света и различения
цветов служит глаз. У человека и многих животных в сетчатке
глаза имеются специфические клетки: палочки и колбочки, ко-
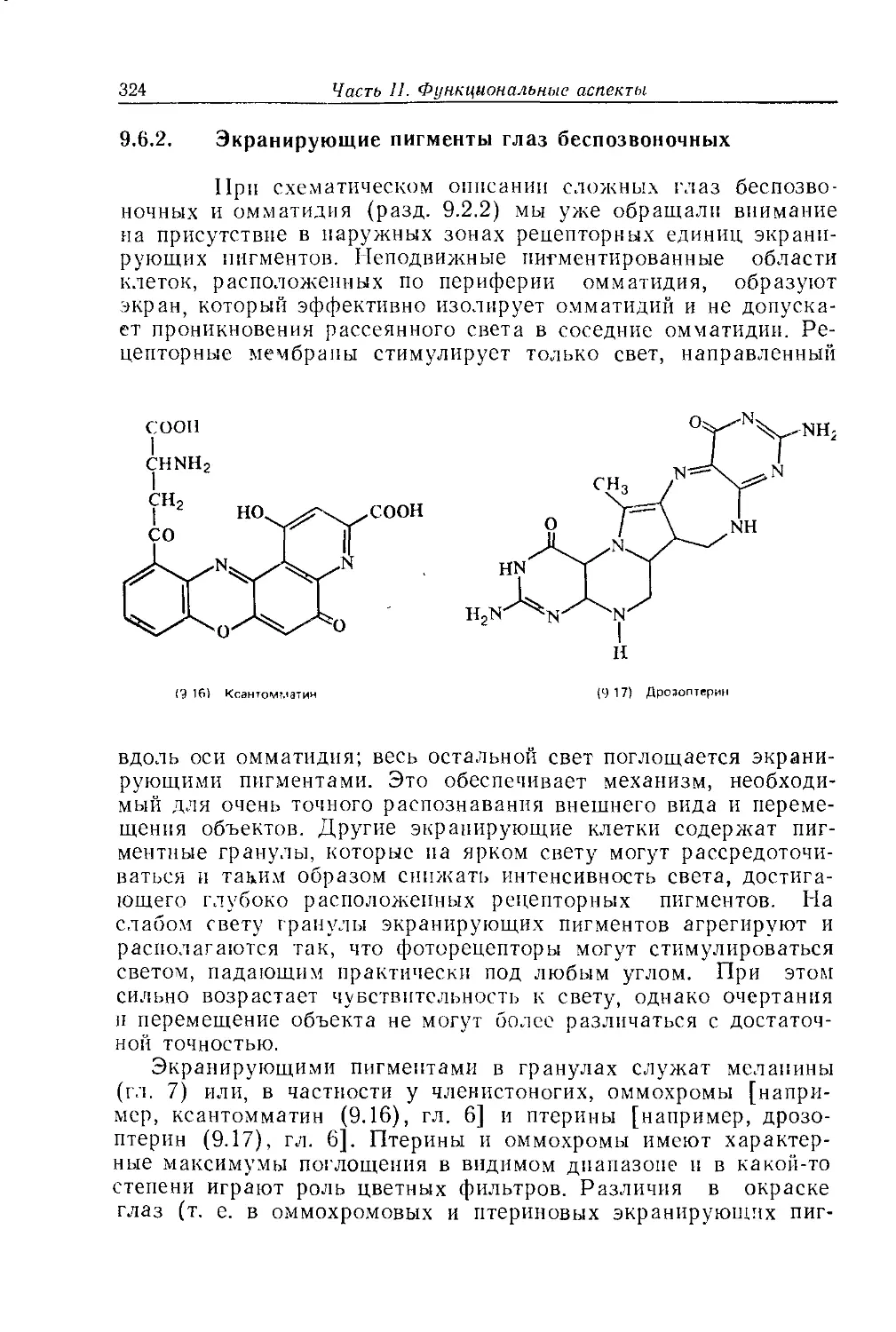
торые содержат фоторецепторы, или зрительные пигменты. Па-
лочки в сетчатке млекопитающих ответственны за восприятие
света низкой интенсивности. Они содержат пигменты, скотопси-
ны, которые чувствительны к свету очень низкой интенсивности.
Максимум чувствительности человеческого скотопсина, родоп-
сина, расположен приблизительно при 520 нм; вместе с тем
глаз воспринимает свет в достаточно широком диапазоне длин
волн, близких к этому значению. Поглощение света соответст-
вующих длин волн зрительным пигментом приводит к генери-
рованию электрического и нервного импульса. Свет разных
длин волн вызывает в скотопсине одну и ту же реакцию. Вме-
сте с тем колбочки содержат ряд зрительных пигментов, фо-
топсинов, которые чувствительны к свету определенной длины
волны. У человека имеются три таких пигмента, чувствитель-
ные к синему, зеленому и красному свету. Эти три фоторецеп-
тора охватывают почти весь диапазон видимого спектра и обес-
печивают механизм различения цветов, достаточно чувстви-
тельный для ощущения очень тонких различий в окраске, ее
оттенках и нюансах. Данная трихроматическая система и вхо-
дящие в нее пигменты более подробно описаны в гл. 9. В со-
временном цветном телевидении также используется трихрома-
тическая система, в которой любой цвет или оттенок получают
путем сложения красного, зеленого и синего цвета.
1.3. Цвет у живых организмов
В то время как на все живые организмы или ткани па-
дает белый свет, например солнечный, отражается от них и по-
падает на глаз наблюдателя только часть видимого диапазона,
Гл. 1. Свет и цвет
13
т. е. эти организмы и ткани выглядят окрашенными. Окраска
может быть обусловлена физической природой поверхности
ткани (структурная окраска) или же присутствием особых хи-
мических соединений (пигментов, или биохромов), которые спе-
цифически поглощают видимый свет некоторых длин волн.
1.3.1. Структурная окраска
В животном царстве есть много примеров, когда на-
блюдаемая окраска является результатом таких оптических
феноменов, как рассеяние, интерференция или дифракция све-
та содержащимися в тканях микроскопическими структурами.
Такую окраску называют структурной. Структурная окраска —
весьма обширная и важная тема исследований, но подробное
описание разных видов структурной окраски и оптических яв-
лений, которые их вызывают, выходит за рамки этой книги. По-
этому ниже мы охарактеризуем эти явления лишь вкратце.
1.3.2. Светорассеяние; синева Тиндаля
Очень маленькие частицы, диаметр которых меньше
длины волны красного или желтого света, могут отражать или
рассеивать коротковолновые компоненты белого света сильнее,
чем длинноволновые. Наиболее простой пример этого явления —
голубизна неба. Мельчайшие частички пыли или других обра-
зований в атмосфере рассеивают падающий белый свет таким
образом, что свет, отражающийся по направлению к поверхно-
сти Земли, содержит больше коротковолновых (синих и фиоле-
товых), чем длинноволновых (красных и желтых), лучей, и по-
этому мы видим небо голубым. Этот процесс часто называют
рассеянием Рэлея или Тиндаля, а образующийся цвет известен
как синева Тиндаля.
В большинстве случаев нерадужная (непереливчатая) си-
няя окраска у животных представляет собой синеву Тиндаля.
Так, голубой цвет глаз у людей обусловлен рассеянием белого
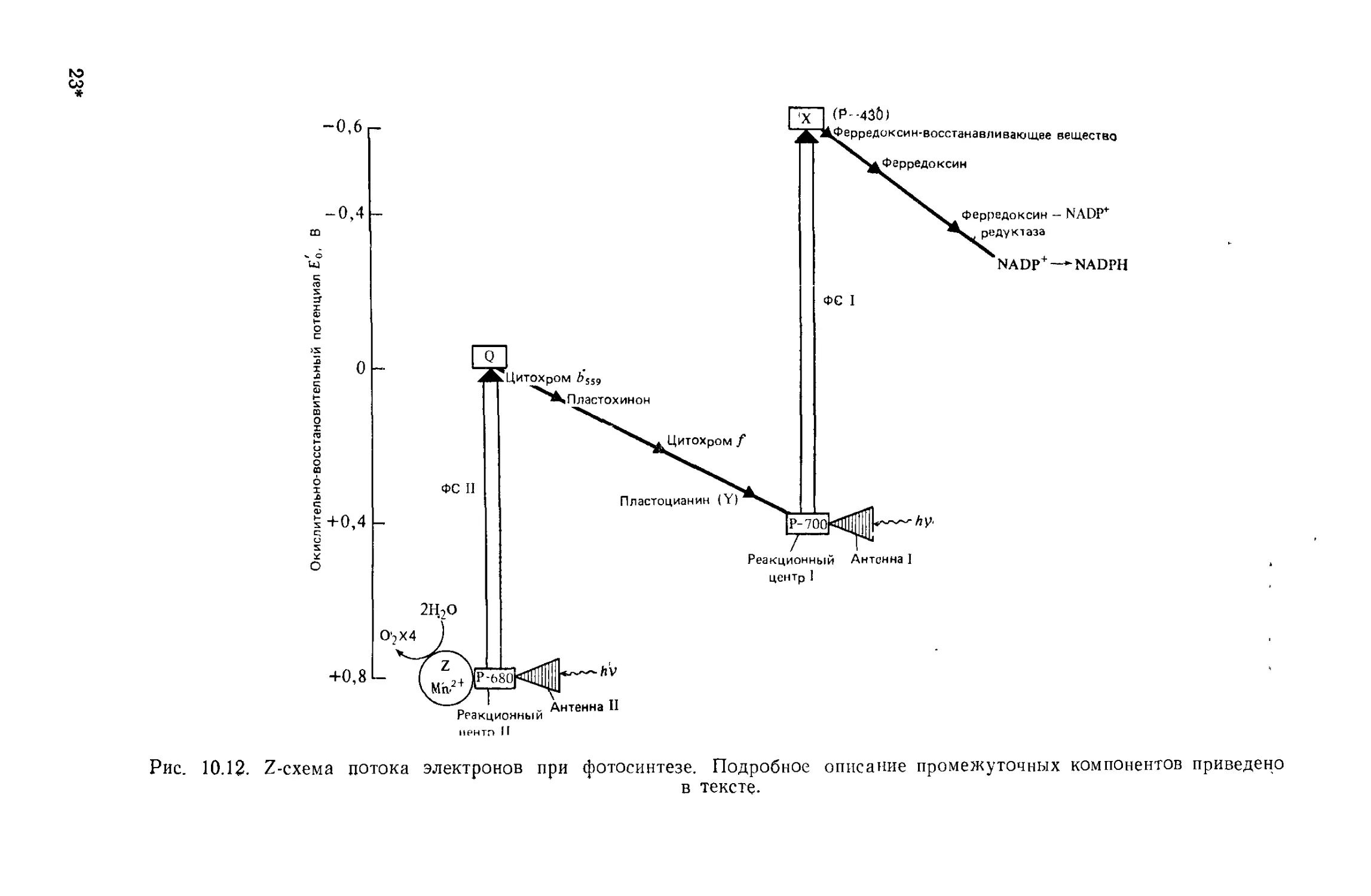
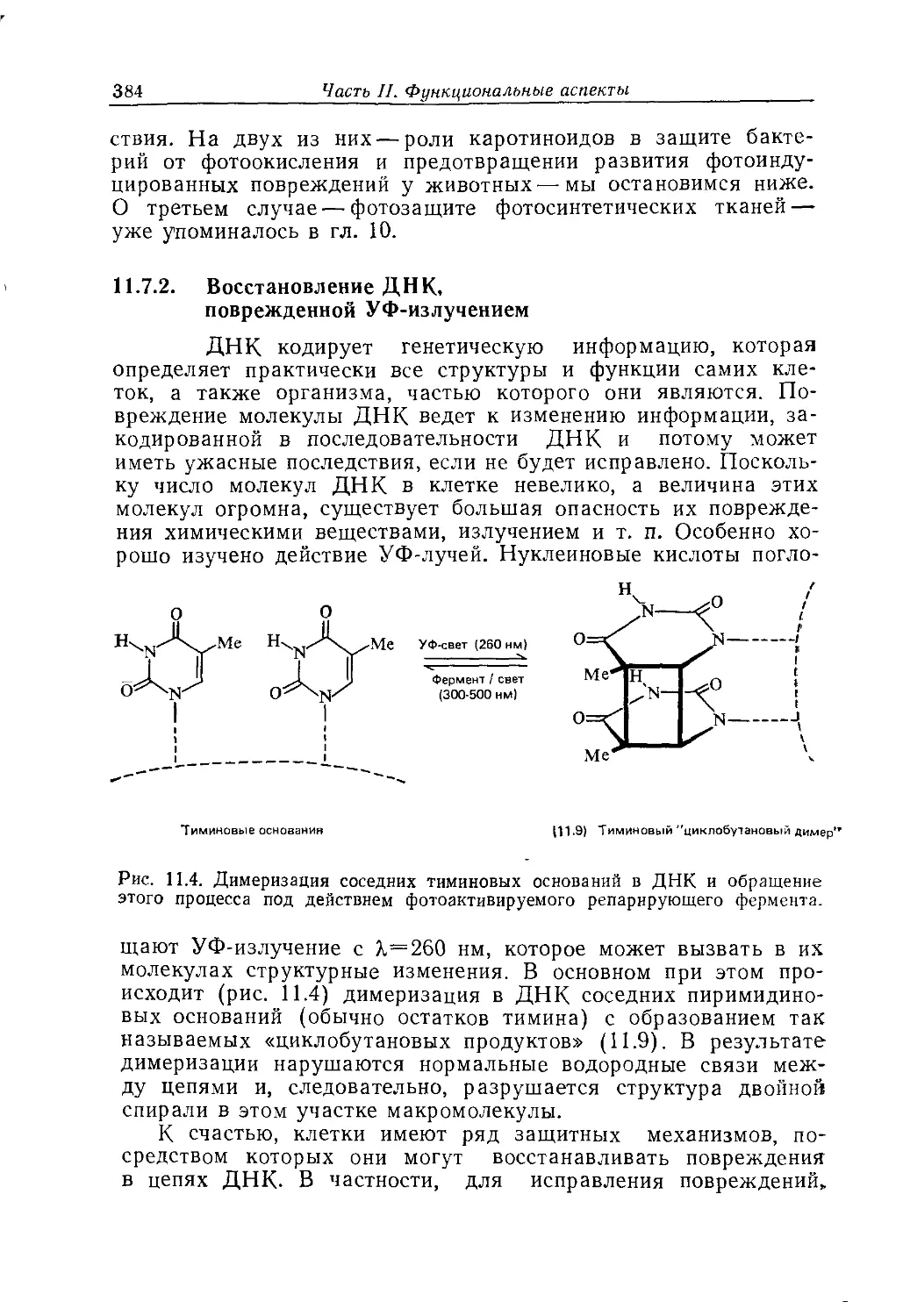
света мельчайшими белковыми частицами радужной оболочки.
У многих птиц с голубым оперенением, например у лазоревок,
волнистого попугайчика и других попугаев, внутри кератино-
вых бородок пера содержатся светорассеивающие частицы в
форме мельчайших наполненных воздухом пластинок.
В пользу структурного происхождения синевы Тиндаля сви-
детельствует тот факт, что до сих пор из тканей не удалось вы-
делить ни одного синего пигмента, а также то, что синяя окра-
ска не выявляется при просматривании тканей в проходящем
белом свете. Ткани выглядят матовыми без радужной окраски,
14
Часть 1. Химические и биохимические аспекты
причем при рассматривании практически под любым углом они
окрашены одинаково.
Зеленая окраска, особенно у перьев, часто обусловлена на-
ложением синего цвета структурного происхождения и желтого
цвета пигмента.
1.3.3. Радужная окраска
К числу наиболее ярких визуальных эффектов, наблю-
даемых в природе, относится сверкающая радужная структур-
ная окраска, часто встречающаяся в животном царстве, в част-
ности у птиц, насекомых и рыб. Для радужной переливчатой
окраски характерно то, что наблюдаемые цвета изменяются в
зависимости от угла зрения. Этот эффект обусловлен двумя оп-
тическими явлениями — интерференцией и дифракцией.
Интерференция. Явление интерференции, возможно, наи-
лучшим образом известно на примере тонкой пленки масла на
поверхности воды. Свет, отраженный от нижней поверхности
(границы масло — вода) пленки, проходит несколько большее
расстояние, чем свет, отраженный от верхней поверхности —
границы масло — воздух. Когда различие между пройденными
расстояниями эквивалентно половине длины волны света, два
световых луча, отраженные от верхней и нижней поверхностей,
находятся в противофазе и гасят друг друга. Таким образом,
в наблюдаемом отраженном свете отсутствует свет отдельных
длин волн, в результате чего отраженный луч приобретает ок-
раску. При более остром угле зрения расстояние, которое про-
ходит луч между верхней и нижней поверхностями, больше.
Следовательно, в другой части спектра (при большей длине
волны) произойдет интерференция, и потому здесь будет на-
блюдаться иная окраска.
У животных известно много примеров окраски, образую-
щейся в результате интерференции. У многих насекомых тон-
кой пленкой, обусловливающей появление ряда интерферирую-
щих цветов при рассматривании под разными углами, служит
просвечивающее крыло. Многие бабочки имеют на поверхно-
сти чешуек крыльев пластинки с мельчайшими воздушными
пузырьками между ними. Расстояние между пластинками при-
мерно постоянно, п поэтому в довольно широком диапазоне уг-
лов зрения окраска почти не меняется.
Окраска, обусловленная интерференцией, обычно встреча-
ется у птиц, например у самца павлина. Уплощенные крючочки
на бородках пера пластинчатых структур, играющих роль интер-
ферирующей пленки, повернуты таким образом, что их плос-
кая поверхность обращена в сторону наблюдателя. Во многих
случаях яркость переливающихся цветов усиливается в при-
Гл. 1. Свет и цвет
15
сутствии подстилающей черной поверхности (меланин), кото-
рая поглощает весь остальной свет. Благодаря такой подстил-
ке оперение птиц часто имеет металлический блеск.
Дифракция. Довольно близкий к описанному выше эффекту
радужного окрашивания дает дифракция, хотя она менее ха-
рактерна для природных тканей, чем интерференция. Искусст-
венные дифракционные решетки, используемые в некоторых
оптических приборах, состоят из серии очень близко и на оди-
наковом расстоянии друг от друга расположенных параллель-
ных линий, нанесенных на отшлифованную поверхность. При-
мером радужной окраски, образующейся на природных пла-
стинчатых структурах, которые ведут себя как дифракционные
решетки, является перламутровая окраска раковин моллюсков.
1.3.4. Структурная белизна
Понятие структурной белизны можно проиллюстриро-
вать на примере снега, который своим блестящим белым цве-
том обязан отражению падающего белого света от поверхности
бесчисленных мелких кристалликов. Сходный эффект дает от-
ражение света другими твердыми или жидкими частицами ли-
бо поверхностями, содержащимися в среде с иным показате-
лем преломления. Частицы не должны быть слишком малы,
чтобы не происходило различного рассеяния лучей с разной
длиной волны (тиндалевское рассеяние). Приведем несколько
примеров структурной белизны в природе — белые волосы (от-
ражение от пузырьков воздуха, заключенных в прозрачное
твердое вещество), белые перья (отражение от множества ма-
леньких бесцветных крючочков на бородке пера), молоко (от-
ражение от капелек в эмульсии, состоящей из двух жидкостей
с разными показателями преломления), белые бабочки (отра-
жение от пронизанных жилками и сетчатых, покрытых чешуй-
ками поверхностей), а также белые и серебристые рыбы (от-
ражение от кристаллов гуанина).
1.3.5. Химическая окраска, обусловленная природными
пигментами
Разнообразие природной окраски является большей
частью результатом преимущественного поглощения света не-
которых длин волн химическими веществами, содержащимися в
ткани. Химические вещества, ответственные за это поглощение,
являются природными пигментами. Именно биохимии этих
природных пигментов, или биохромов, посвящена данная кни-
га. Практически все природные пигменты представляют собой
органические вещества, но они значительно отличаются от пиг-
16
Часть I. Химические и биохимические аспекты
ментов, выпускаемых промышленностью и широко используе-
мых в качестве красок для рисования и красителей. Однако,
прежде чем приступить к подробному описанию различных
групп природных пигментов, необходимо рассмотреть в общем
виде те свойства молекул, которые обусловливают поглощение
видимого света.
1.4. Поглощение света
1.4.1. Энергия и длина волны
Когда атом (или молекула) поглощает квант энергии
электромагнитного излучения, он переходит из одного энерге-
тического состояния в другое. Электрон может занимать толь-
ко определенные дискретные энергетические уровни, и погло-
щение кванта света возможно только в том случае, когда энер-
гия поглощаемого излучения Е равна разности энергий между
двумя этими уровнями. Однако, согласно «правилам отбора»,
не все такие переходы «разрешены»; некоторые из них являют-
ся «запрещенными». Будет ли квант поглощен, строго зависит
от его энергии, а следовательно, от длины волны излучения,
так как
Е = hv,
где Е — энергия кванта, h — постоянная Планка, a v — частота
излучения, связанная с длиной волны % соотношением
v = с/Х,
в котором с — скорость света. Таким образом, между энергией
и длиной волны имеет место обратная зависимость, или, други-
ми словами, чем больше разность энергий двух энергетических
состояний, тем короче длина волны излучения, необходимого
для осуществления перехода между ними. Энергия УФ- и ви-
димого излучения достаточно велика для осуществления элект-
ронных переходов, т. е. кванты этих видов излучения способны
вызывать переход электронов из более низкого в более высо-
кое энергетическое состояние. Кроме того, каждое энергетиче-
ское состояние электронов даже в наиболее простых двухатом-
ных молекулах характеризуется различными колебательными и
вращательными энергетическими уровнями. Разности колеба-
тельных и вращательных энергий значительно меньше, чем
электронных, поэтому они соответствуют квантам ИК-излуче-
ния.
Гл. 1. Свет и цвет
17
1.4.2. Поглощение излучения атомами
При поглощении атомом кванта света его электрон пе-
реходит из основного состояния в возбужденное, причем энер-
гия кванта точно соответствует разности энергий между двумя
энергетическими состояниями. Поскольку существует однознач-
ное соответствие между энергией, частотой и длиной волны из-
лучения, при простом переходе электрона с одного уровня на
другой поглощенное излучение или свет может быть только-
какой-то одной длины волны и в спектре поглощения будет на-
блюдаться единственная линия.
1.4.3. Поглощение излучения молекулами
В случае молекул ситуация не так проста. Даже двух-
атомные молекулы велики по сравнению с атомами, и их уже
нельзя рассматривать как жесткие частицы. В них происходят
молекулярное вращение и колебания ядер, причем энергия
вращательного и колебательного движения также квантуется
(рис. 1.3). Таким образом, любой электрон в молекуле в ос-
новном состоянии может находиться на нескольких колеба-
тельных энергетических уровнях, причем для каждого из по-
следних возможно несколько вращательных энергетических
уровней. То же самое справедливо и для электрона в возбуж-
денном состоянии. И хотя разницы колебательных и враща-
тельных энергий малы по сравнению с разницей электронных
энергий, при обсуждении электронных переходов их необходи-
мо учитывать. Следовательно, для осуществления электронного-
перехода энергия кванта не обязательно должна иметь одно-
строго определенное значение; она должна соответствовать
разности между основным и возбужденными состояниями для
различных колебательных и вращательных уровней. Возбужде-
ние электронов может сопровождаться колебательным и вра-
щательным возбуждением молекул. В итоге при электронном
возбуждении энергия поглощается в некотором диапазоне длин
волн излучения, и поэтому для молекул спектроскопические
линии поглощения расширяются до полос поглощения с цент-
ром, соответствующим длине волны максимального поглощения
(Хтах); обычно ширина полосы составляет 50—100 нм. Как
правило, невозможно достичь достаточно хорошего разрешения
полос поглощения, по которому можно было бы восстановить
тонкую структуру колебательных и вращательных уровней.
Переход электрона в возбужденное состояние на более вы-
сокий энергетический уровень происходит чрезвычайно быстро
('---1015 с), тогда как более тяжелые атомные ядра не могут
в течение этого времени сколько-нибудь заметно переместиться.
2—84
18
Часть I. Химические и биохимические аспекты
Е2-^^ /Е2---------------------
Второе электронное
возбужденное состояние
2
•Ет
0>
Вращательные
уровни
'Электронный
переход
V3
Колебательные
уровни энергии N?
•Первое электронное
возбужденное состояние
Некоторые
из возможных
электронных
переходов
Ео
Vo
к. Вращательные
уровни
•Ro|
го
Основное электронное
состояние
1
Рис. 1.3. Диаграмма,
ные и вращательные)
ронные переходы для
иллюстрирующая электронные (а также колебатель-
энергетические уровни и некоторые возможные элект-
атома (X) и двухатомной нли многоатомной молеку-
лы (Б).
Следовательно, во время электронных переходов положения
ядер, или межъядерные расстояния, практически не меняются
(принцип Франка — Кондона).
Молекулярные колебания имеют более или менее гармони-
ческий характер, так что колеблющиеся ядра дольше всего на-
ходятся в крайних положениях, где движение наиболее мед-
ленное. Таким образом, электронные переходы наиболее веро-
ятны в том случае, когда межъядерные расстояния максималь-
Гл. 1. Свет и цвет
19*
ны или минимальны, и, следовательно, вероятность поглощения
кванта с энергией, соответствующей этим положениям, боль-
ше, чем в других случаях. В результате наиболее интенсивным
будет поглощение света, состоящего именно из таких квантов.
В общем положение полосы поглощения Хщах определяется
энергией, необходимой для наиболее вероятных электронных
переходов, а интенсивность поглощения отражает вероятность,
происходящего перехода.
1.4.4. Абсорбционные свойства некоторых простых молекул
В простой молекуле способен возбуждаться любой
электрон. Требуемая для этого энергия зависит прежде всего
от типа занимаемой им орбитали (рис. 1.4). Так, для возбуж-
дения электрона на о-орбитали необходима наиболее высокая
энергия. Следовательно, труднее всего возбудить электроны в
Рис. 1.4. Пять электронных состояний, характерных для органических моле-
кул, и наиболее обычные электронные переходы, которые могут происходить
в результате поглощения ими УФ- илн видимого света.
таких простых органических молекулах, как насыщенные угле-
водороды. Единственными доступными электронами у них явля-
ются электроны о-связей, и для перехода электрона о-связи на
антисвязывающую (разрыхляющую) п*-орбиталь необходимо
поглощение большого количества энергии. Таким образом, в
простой молекуле, такой, как молекула метана (СН4), для воз-
буждения электронов требуется энергия ~600 кДж/моль. Это
2*
20
Часть I. Химические и биохимические аспекты
количество энергии может дать электромагнитное излучение в
УФ-диапазоне, в случае метана (газ) с длиной волны ~122 нм.
Углерод-углеродная двойная связь С —С может рассматри-
ваться как л-связь. Переход электрона со связывающей л-ор-
бнтали на разрыхляющую л*-орбиталь достигается значитель-
но легче, чем переход о—мт*. Меньшее количество энергии,
которое необходимо в этом случае, может быть обеспечено
УФ-излучением несколько большей длины волны (162 нм для
этилена, СН2 = СН2).
Когда гетероатом, такой, как атом кислорода или азота, со-
держится в насыщенной или ненасыщенной молекуле, его не-
связывающий неспаренный р-электрон (обозначается п) может
быть переведен на о*-разрыхляющую орбиталь, п—>-о*-Пере-
ходы требуют еще меньшей энергии и могут достигаться при
поглощении энергии более длинноволнового электромагнитного
излучения (183 нм для метанола, СН3ОН).
Переход п—>л* в молекулах, содержащих группу С = О,
требует наименьшего количества энергии. Так, ацетон,
(СН3)2С = О, для перехода п—>л* имеет максимум поглоще-
ния при 280 нм.
1.4.5. Абсорбционные свойства сопряженных полиенов
Так же как и для простых примеров, описанных выше,
в случае сложных молекул чем легче происходит возбуждение
электрона, тем меньшее количество энергии требуется для это-
го и тем большую длину волны может иметь свет, способный
осуществить переход электрона в возбужденное состояние. Это
утверждение хорошо иллюстрируется простым качественным
сравнением светопоглощающих свойств полиенов с сопряжен-
ными связями.
Если вместо одной двойной связи в молекуле содержится
ряд сопряженных двойных связей, можно считать, что л-элект-
роны этих двойных связей делокализованы по всей системе со-
пряженных связей. По мере увеличения длины такой системы
возрастает и степень ее стабилизации в результате резонансной
делокализации, причем для максимальной резонансной стаби-
лизации система сопряженных двойных связей должна быть
плоской. В возбужденном состоянии такая стабилизация даже
больше, чем в основном, так что при увеличении длины систе-
мы двойных связей разница энергий двух состояний становится
меньше. Таким образом, в сопряженной системе электронное
возбуждение (л—>-л*-переход) достигается значительно легче,
чем в случае изолированной двойной связи. При удлинении со-
пряженной системы двойных связей требуемая для возбужде-
ния энергия становится меньше и в конце концов может быть
Гл. 1. Свет и цвет
21
достигнута стадия, на которой возбуждение происходит при
поглощении видимого света. При этом часть спектра белого
света поглощается, а остальная проходит насквозь или отра-
жается, так что соединение выглядит окрашенным. По мере
роста цепи с сопряженными двойными связями длина волны
максимального поглощения увеличивается и наблюдаемая окра-
ска изменяется: от желтой к оранжевой,, затем к красной и
фиолетовой. Этот эффект прекрасно иллюстрируется на приме-
ре каротиноидной группы природных пигментов (гл. 2).
Структуры, ответственные за поглощение света, называют
хромофорами. В случае упомянутых выше полиенов хромофо-
ром является система сопряженных двойных связей. Другие
функциональные группы или заместители в молекуле могут
изменять положение (главным образом в сторону увеличения
длины волны) максимума поглощения; такие группы называ-
ются ауксохромами. Они могут либо увеличивать длину хро-
мофора, либо облегчать электронные переходы, повышая ста-
бильность возбужденного состояния.
1.4.6. Поглощение света некоторыми
группами природных пигментов
Каротиноиды (гл. 2) являются полиенами, которые
имеют хромофор, представляющий собой протяженную систе-
му сопряженных двойных связей. Другие группы природных
пигментов обязаны своей окраской другим хромофорам.
В большинстве случаев в их образовании участвует сопряжен-
ная или ароматическая л-электронная система, в которой при-
сутствуют добавочные электрон-донорные или электрон-акцеп-
торные группы; особенно важны атомы азота и кислорода.
Разделение зарядов, характерное для молекул этого типа, мо-
жет вносить значительный вклад в общую резонансную струк-
туру, что приводит к высокой степени стабилизации, особенно
в возбужденном состоянии. Поэтому необходимая для возбуж-
дения энергия невелика, и может происходить поглощение ви-
димого света. Этот случай хорошо иллюстрируют хиноны и
аналогичные им системы, которые служат основой многих
природных пигментов. Наиболее существенной особенностью
других классов соединений является вклад в гетероароматиче-
скую систему электронов атомов кислорода и азота, не участ-
вующих в образовании связей.
В частности, интенсивное поглощение достигается, когда ди-
польный момент молекулы в основном состоянии отличается
от дипольного момента молекулы в возбужденном состоянии,
т. е. возбуждение приводит к перераспределению заряда. Чем
больше перераспределение заряда, или дипольный момент пере-
22
Часть I. Химические и биохимические аспекты
хода, тем больше интенсивность полосы поглощения света. Ди-
польные моменты перехода представляют собой векторы. Для
сложных молекул возможно существование нескольких элект-
ронных возбужденных состояний с различными дипольными мо-
ментами перехода и с различной пространственной ориента-
цией.
Когда хромофоры пигментов расположены близко друг к.
другу, между ними могут возникать электронные (экситонные)
взаимодействия благодаря сопряжению дипольного момента
перехода одной молекулы с соответствующими моментами пе-
рехода других сходных или идентичных молекул. Это приводит
к образованию различных возбужденных электронных энерге-
тических уровней и к расщеплению полос поглощения. На-
блюдается также аналогичное влияние экситонов на полосы
кругового дихроизма. Обнаружение экситонного расщепления
может дать полезную информацию об относительной ориента-
ции хромофорных молекул, например об упорядоченном распо-
ложении их в стопках.
Электронные и структурные особенности, обусловливающие
светопоглощающие свойства главных групп природных пиг-
ментов, будут рассмотрены в соответствующих главах.
1.4.7. Триплетные состояния, флуоресценция
и фосфоресценция
Когда молекулы пигментов поглощают энергию света,
они активируются и переходят в более высокоэнергетическое
возбужденное электронное состояние. Такое возбужденное со-
стояние с электронами на разрыхляющих орбиталях неста-
бильно, и после отдачи избытка энергии электроны возвраща-
ются в основное состояние. Это может осуществляться несколь-
кими путями (рис. 1.5). Прежде всего по мере потери возбуж-
денной молекулой избытка колебательной энергии происходит
быстрая (10~12 с) колебательная релаксация, в результате че-
го молекула возвращается на более низкий колебательный уро-
вень. Существует несколько способов релаксации колебатель-
но-возбужденных состояний. Простейшим из них является пе-
реход в основное электронное состояние либо путем испускания
излучения в процессе флуоресценции, либо путем передачи
энергии возбуждения какой-то другой молекуле, которая в
свою очередь становится электронновозбужденной. Поскольку
колебательная энергия уже потеряна, энергия флуоресценции
или передаваемая энергия всегда меньше, чем поглощенная.
Поэтому длина волны флуоресценции больше, чем у поглощен-
ного излучения. Аналогичным образом при передаче энергия
может поглощаться только молекулой, имеющей максимум по-
Гл. 1. Свет и цвет
23
Синглетные уровни
Триплетные уровни
Рис. 1.5. Некоторые пути, по которым возможна потеря энергии молекулы
в первом возбужденном синглетном состоянии. Показаны колебательная ре-
лаксация, флуоресценция, межсистемный переход и фосфоресценция.
глощения при большей длине волны, чем у первоначально воз-
бужденной молекулы, например с каротиноида (Хтах = 450 нм)
на хлорофилл (Хтах = 680 нм) при фотосинтезе.
Второй возможностью является переход в более низко-
энергетическое триплетное состояние. В возбужденных состоя-
ниях, являющихся результатом поглощения света, все элект-
роны обычно спарены, и такие состояния называются синглет-
ными. В триплетном состоянии два электрона не спарены, и по-
тому такое состояние обычно более стабильно, чем соответст-
вующее синглетное (правило Хунда). Следовательно, переход
из низшего синглетного возбужденного состояния (Si) в три-
плетное состояние’ (Т1) энергетически выгоден, хотя вероят-
ность его мала и он происходит довольно медленно. Однако ес-
ли синглетное состояние живет достаточно долго, то синглет-
24
Часть I. Химические и биохимические аспекты
триплетный переход (3]—>Л), обычно называемый межсистем-
ным, может происходить у значительного числа возбужденных
синглетных молекул. Триплетные состояния обычно гораздо бо-
лее долгоживущие, чем исходные синглетные возбужденные
состояния.
В некоторых случаях триплетное состояние, подобно воз-
бужденному синглетному, возвращается в основное состояние
(So) путем перехода, сопровождающегося излучением энергии
(Т]—*-30), правда, вероятность этого процесса мала. Посколь-
ку изменение энергии при таком переходе невелико, испускае-
мый свет имеет значительно большую длину волны как по
сравнению с поглощенным, так и по сравнению со светом, ис-
пускаемым в виде флуоресценции; такое выделение энергии
называется фосфоресценцией. Поскольку переход 1\—>30 име-
ет низкую вероятность и триплетное состояние может быть
долгоживущим, фосфоресценция может продолжаться в тече-
ние нескольких секунд, т. е. значительно дольше, чем флуорес-
ценция, которая обыкновенно протекает в течение 10~9—10 7 с
после исходного поглощения света.
В биологических системах потеря избытка энергии в ре-
зультате процессов, не сопровождающихся испусканием излу-
чения, имеет особое значение. Избыток энергии может быть
перенесен как от молекул в синглетном возбужденном состоя-
нии 3], так и от молекул в более долгоживущем триплетном
состоянии Т\ на другие близко расположенные молекулы и мо-
жет использоваться для осуществления химических реакций.
Именно такие процессы, не сопровождающиеся излучением или
испусканием света, обусловливают превращение поглощенного
света в химическую энергию при фотосинтезе, а также множе-
ство других важных реакций в биологических тканях (гл. 9—11).
1.5. Спектроскопические методы исследования
природных пигментов
Из приведенного выше краткого обсуждения ясно, что
при изучении природных пигментов поглощение света имеет
фундаментальное значение. Спектроскопия электронного по-
глощения, с помощью которой регистрируют поглощение УФ-
и видимого света, является основным спектроскопическим ме-
тодом, применяющимся как для выявления свойств пигментов,
так и для их количественного анализа. Однако специфические
свойства пигментов в отношении поглощения света позволяют
исследовать их и другими методами, главным образом резо-
нансной рамановской спектроскопией и методом кругового ди-
хпоизма. Так же как и при изучении других органических мо-
Гл. 1. Свет и цвет
25
лекул, чрезвычайно полезными могут оказаться и методы инф-
ракрасной (ПК) спектроскопии, ядерного магнитного резонанса
(ЯМР) и масс-спектрометрии.
1.5.1. Спектроскопия поглощения видимого света
Природные пигменты по приведенному выше опреде-
лению поглощают свет в видимом диапазоне спектра электро-
магнитного излучения, т. е. между длинами волн 380 и 750 нм.
Поэтому их спектр поглощения видимого света имеет по край-
ней мере один максимум поглощения при длине волны (Хтах),
характерной для хромофора молекулы пигмента. Это свойство,
а также общая картина спектра дают полную информацию о
молекулярной структуре и обычно используются при первых
попытках идентифицировать пигмент. Положение Хтах сильно
зависит от используемого растворителя, а у некоторых групп
пигментов и от величины pH. На спектры поглощения пигмен-
тов in vivo часто влияет ближайшее микроокружение моле-
кулы.
Подробно свойства спектров поглощения света различными
группами пигментов, а также некоторыми индивидуальными
пигментами будут приведены в соответствующих главах этой
книги.
При исследовании функционирования пигментов применяют
очень сложные модификации основного спектроскопического
метода измерения поглощения света. Такие модификации по-
зволяют изучать очень быстро протекающие процессы (в тече-
ние пико- или наносекунд). Исследуемую систему периодически
освещают короткими интенсивными вспышками света и затем
регистрируют изменения в спектре поглощения. Подобные ме-
тоды позволили получить очень ценную информацию при ис-
следовании первичных реакций фотосинтеза.
1.5.2. Количественный спектрофотометрический анализ
Спектры поглощения света чрезвычайно ценны также
для точного, чувствительного и воспроизводимого количествен-
ного анализа пигментов. Интенсивность полосы поглощения
при какой-либо длине волны регистрируют экспериментально
как абсорбцию, экстинкцию, поглощение, или оптическую плот-
ность раствора. Она прямо пропорциональна как концентрации
пигмента в растворе, так и расстоянию, проходимому светом
через раствор (законы Ламберта — Бэра).
При таком анализе используются два коэффициента. Коэф-
фициент молярного поглощения (или экстинкции) определяет-
ся как поглощение света 1 М раствором вещества при длине
26
Часть I. Химические и биохимические аспекты
светового пути через раствор 1 см. Коэффициент удельного по-
глощения (или экстинкции) А1см1% или (Eicm1 )—это погло-
щение света 1%-ным (вес/объем) раствором вещества при дли-
не светового пути 1 см. Для большинства групп природных
пигментов существуют справочные таблицы этих коэффициен-
тов.
Зная стандартный коэффициент поглощения, концентрацию
любого раствора можно определить по величине его поглоще-
ния. Так, количество х г пигмента в у мл раствора находят по
формуле
х = (Аху)/(А>%1см-100),
где А — измеренная величина поглощения света раствором.
1.5.3. Дифференциальные спектры
Два образца одного и того же вещества, находящиеся
в разных условиях, могут иметь несколько разные максимумы
поглощения, а также несколько разные интенсивности погло-
щения, но эта разница столь мала, что ее трудно заметить при
изучении этих спектров по отдельности. Однако вариации в
этих величинах гораздо легче уловить, если один из образцов
использовать в качестве стандарта, против которого снимается
спектр другого образца. Получаемые при этом дифференциаль-
ные спектры являются очень высокочувствительным средством,
с помощью которого обнаруживают небольшие изменения в
светопоглощающих свойствах. Например, дифференциальные
спектры «свет — темнота», в которых сравнивают поглощение
света освещенным образцом и образцом, содержащимся в
темноте, оказались чрезвычайно ценными при выявлении не-
значительных изменений этой величины, которые имеют место
при освещении фотосинтезирующих тканей или частиц. Диффе-
ренциальные спектры окисленных и восстановленных форм бы-
ли использованы для получения информации об участии цито-
хромов в цепи переноса электронов и об окислительно-восста-
новительном состоянии отдельных цитохромов в определенных
условиях. С помощью этого основного метода и многих его
изощренных модификаций мы узнали очень много нового о фи-
зических состояниях пигментов и их функционировании в фото-
синтезе и транспорте электронов.
1.5.4. Рамановская спектроскопия
С помощью рамановской спектроскопии регистрируют
изменения колебательной и вращательной энергии, причем для
возбуждения таких изменений используют поглощение УФ- и
Гл. 1. Свет и цвет
27
видимого света. Когда монохроматический свет проходит через
образец, некоторое количество света рассеивается под прямым
углом к падающему лучу. При изучении этого процесса было
обнаружено, что длина волны рассеянного света отличается от
длины волны падающего света, поскольку в колебательной и
вращательной энергии молекул за то короткое время, в про-
должение которого фотоны падающего света «улавливаются»
молекулами образца, происходят определенные изменения.
В силу этого величины длин волн соответствуют поглощению
или испусканию ИК-излучения. Получаемый при этом спектр
содержит ряд полос поглощения (рамановских линий), каждая
из которых соответствует отдельному типу колебаний в моле-
куле, например колебания валентной связи С = С обусловлива-
ют полосу поглощения при 1500 см"1. Важно, однако, что
ИК-поглощение и рамановские спектры никогда не бывают
идентичными. Различия между ними могут давать ценную ин-
формацию о молекулярной симметрии, так как симметричная
молекула или колебание не дает полосы в ИК-спектре погло-
щения, но может давать сильную рамановскую полосу.
При работе с пигментами обычно применяется резонансная
рамановская спектроскопия. Когда длина волны падающего,
или возбуждающего, света приближается к той, при которой
происходит максимальное поглощение света образцом, улав-
ливание кванта света становится гораздо более вероятным. По-
этому рассеяние света значительно увеличивается, а интенсив-
ность рамановских линий сильно возрастает. В образце, содер-
жащем смесь соединений, резонансное усиление регистрируется
только для тех рамановских линий, которые обусловлены ко-
лебаниями молекул, возбужденных падающим УФ- или види-
мым светом. При этом получают информацию об определенных
молекулах, поглощающих свет; другие молекулы, которые не
поглощают свет возбуждающей длины волны, не дают резо-
нансно-усиленных рамановских линий. Резонансный раманов-
ский метод, таким образом, особенно ценен для исследования
пигментов in situ. Пигменты, находящиеся в панцире, коже и
т. п., могут быть обнаружены и количественно изучены с по-
мощью этого метода без предварительной экстракции из ткани
и очистки от примесей.
Другой областью применения этого, метода является иссле-
дование взаимодействий пигмент (хромофор) — белок, по-
скольку в этом случае метод позволяет получить избирательную
информацию о хромофорной молекуле без разрушения комп-
лекса. В частности, можно обнаружить переход хромофорной
молекулы в основное состояние, который происходит при свя-
зывании с белком или в результате других изменений в ее мик-
роокружении. Таким образом можно изучать свойства хромо-
28
Часть I. Химические и биохимические аспекты
форов в их природном окружении. Этот подход оказался чрез-
вычайно полезным при изучении связывания ретинальдегида с
опсином в зрительных пигментах (гл. 9).
1.5.5. Линейный дихроизм
В предыдущих разделах все рассуждения о поглоще-
нии света относились к естественному неполяризованному све-
ту. Однако не менее ценную информацию можно извлечь также
из феноменов, связанных с поглощением поляризованного
света.
Согласно электромагнитной теории, световая волна состо-
ит из электрических и магнитных векторных компонентов, ко-
торые находятся под прямыми углами друг к другу и к на-
правлению распространения волны. Частота колебаний явля-
ется частотой излучения. Свет, испускаемый природным ис-
точником или обычной лампой накаливания, неполяризован.
Однако если его пропустить через поляризатор, то пройдет
лишь свет с определенной ориентацией электрических и маг-
нитных векторов. Пигмент, у которого хромофорные группы
расположены беспорядочно, будет поглощать свет определен-
ной длины волны независимо от того, поляризован свет или
нет. Если же благодаря упорядоченной ориентации хромофо-
ров в природной структуре имеет место асимметрия, то по-
глощение будет зависеть от плоскости поляризации луча света.
Существуют две взаимно перпендикулярные плоскости поляри-
зации, характеризующиеся соответственно максимальным и ми-
нимальным поглощением, для которых можно получить ди-
хроичное отношение. Этот феномен лежит в основе линейного
дихроизма. Исследования с помощью линейного дихроизма
оказались очень полезными при изучении ориентации пигмент-
ных хромофоров в упорядоченных биологических структурах,
особенно в фотосинтетических пигмент-белковых комплексах.
1.5.6. Круговой дихроизм
Родственный линейному дихроизму круговой дихроизм
(КД) позволяет не только определять асимметрию, уже суще-
ствующую в молекуле пигмента, но и обнаруживать асиммет-
рию, индуцированную в хромофоре, например при связывании
пигмента с белком. Если совместить два плоскополяризованных
луча с плоскостями поляризации под прямым углом друг к дру-
гу, имеющих одну и ту же длину волны и амплитуду, но
различающихся по фазе на л/2 (или на Зл/2, для того чтобы
вызвать вращение в обратном направлении), то образующийся
в результате световой луч приобретет круговую поляризацию
с направлением вращения плоскости поляризации вправо или
Гл. 1. Свет и цвет
29
влево. Асимметричный, или оптически активный, пигмент по-
глощает право- или левополяризованный свет неодинаково.
Эффект кругового дихроизма изменяется с изменением длины
волны, так что может быть получен спектр КД. Круговой ди-
хроизм наблюдается лишь в тех. областях спектра, в которых
имеются полосы поглощения. В некотором роде сходную ин-
формацию об оптической активности молекул пигментов дает
метод дисперсии оптического вращения (ДОВ), который позво-
ляет измерить изменения вращения плоскости поляризации
света при изменении длины его волны. Эти методы особенно
полезны в органической химии при определении относительной
и абсолютной конфигурации хиральных молекул.
Круговой дихроизм, однако, используют не только при оп-
ределении оптической активности хиральных молекул. Его ус-
пешно применяют при выявлении асимметрии, индуцируемой в-
результате включения обычно симметричной или нехиральной
молекулы в организованную структуру, например при асиммет-
ричном связывании с белком. Индуцированная асимметрия мо-
жет приводить к различному поглощению право- и левовраща-
ющего поляризованного света, т. е. в результате КД можно на-
блюдать в области главных полос поглощения хромофора. При
этом удается выявлять различные формы пигмента in situ и по-
лучать сведения об искажениях конформации молекулы хромо-
фора, вызванных связыванием с другими молекулами или обу-
словленных структурной организацией.
1.6. Значение окраски в природе
Многие природные пигменты принимают участие в
важных метаболических или физиологических процессах. Осо-
бенно детально изучено функционирование хлорофилла и дру-
гих пигментов в фотосинтезе и роль гемоглобина как перенос-
чика кислорода. Во многих случаях, однако, единственной из-
вестной функцией пигмента является то, что он придает окра-
ску организму или той его части, которая содержит данный
пигмент.
В растительном царстве ярко окрашенные цветки и плоды,,
контрастно выделяющиеся на общем фоне зеленой окраски
листвы, привлекают внимание насекомых и других животных.
Благодаря этому растения извлекают для себя пользу при
опылении и распространении семян. В животном же царстве
окраска, обусловленная пигментами или структурными особен-
ностями, может играть как сигнальную роль, так и маскиро-
вочную.
Более подробно значение окраски в природе мы обсудим в
гл. 8.
30
Часть I. Химические и биохимические аспекты
1.7. Заключение и комментарии
Любые попытки объяснить то многообразие способов,
>с помощью которых живые организмы реагируют на свет или
используют его, требуют понимания механизмов взаимодейст-
вия между светом и молекулами. Молекулы, обладающие осо-
бым свойством поглощать свет в видимой части спектра, явля-
ются природными пигментами. Без пигментов и без упомянутых
выше взаимодействий та жизнь, какой мы ее знаем, была бы
невозможна. Цвет в природе играет большую роль. Эта глава
«мела своей целью дать очень краткое описание физических
основ цвета, а также физических и химических способов, с по-
мощью которых может достигаться и обнаруживаться окраска
в природе.
Можно сравнительно просто определить, какую природу —
химическую (т. е. обусловленную пигментом) или физическую
(обусловленную структурой) — имеет данный цветовой эф-
фект. Идентификация и характеристика пигмента обычно яв-
ляется стандартной задачей в органической химии. В последу-
ющих главах первой части этой книги приведены основные хи-
мические свойства наиболее крупных групп природных пигмен-
тов. Гораздо более сложной является проблема взаимодействия
молекул пигмента с их ближайшим микроокружением, напри-
ме с белками в мембранах. Применение сложных современных
физико-химических методов, таких, как резонансная раманов-
ская спектроскопия, линейный и круговой дихроизм и ядерный
магнитный резонанс, позволяет решить эту проблему, а также
получить информацию о молекулярных изменениях, которые
претерпевают некоторые пигменты при их функционировании.
Вторая часть этой книги представляет собой обзор функций
природных пигментов как в роли окрашивающих агентов, так
я в роли участников гораздо более сложных процессов, таких,
как фотосинтез, зрение и другие фотореакции, которые могут
протекать за время порядка пикосекунд.
Если об окраске, обусловленной пигментами, известно очень
много, то примеры детального изучения окраски, обусловлен-
ной структурой, немногочисленны. Вместе с тем систематиче-
ское исследование микроскопических структур, участвующих в
образовании структурной окраски, должно быть очень увлека-
тельно и информативно.
1.8. Рекомендуемая литература
Большинство читателей книг по биохимии, подобных
этой, были бы удовлетворены общим описанием светопоглоща-
ющих свойств молекул, которое можно найти в учебниках по
Гл. 1. Свет и цвет
31
органической химии и в монографиях по фотобиологии, напри-
мер в двухтомнике Клейтона (Clayton, 1971). Не исключено,,
однако, что найдутся и такие, которые захотят более глубоко
понять физические процессы и теоретические концепции, позво-
ляющие более строго определять и интерпретировать эти све-
топоглощающие свойства. Им следует познакомиться с теорети-
ческой работой Маррелла (Murrell, 1963).
Есть много книг и обзоров, которые посвящены широкому
кругу проблем химии, биохимии и биологии природных пигмен-
тов в целом и отдельных групп пигментов. В эту книгу вклю-
чен материал лишь из небольшого их числа, прежде всего наи-
более всеобъемлющие работы последних лет, в которых в свою
очередь приведены ссылки на более старые издания. Если мы
обратимся к некоторым старым работам, читающимся с огром-
ным интересом, то мы как бы вновь пройдем тот путь, по ко-
торому шло развитие исследований природных пигментов. Мно-
гие из первых работ по растительным пигментам были выпол-
нены химиками, однако работы по окраске животных явились-
продолжением описательных работ классических натуралистов.
Две статьи Мак-Мунна (MacMunn, 1883 и 1890) и книга Нью-
бигина (Newbigin, 1898) содержат обширные обзоры ранних ра-
бот по природной окраске у животных. Рост количества знаний
о химии пигментов животных позволил Дэнису Фоксу (Denis
Fox, 1953) написать исчерпывающую монографию, посвящен-
ную окраске животных. Эта книга недавно (в 1976 г.) была пе-
реработана; к ней были добавлены приложения, и она стала
очень полезным источником информации по распространению-
пигментов у животных. В третьей книге этого же автора, рас-
считанной на неспециалистов (Fox, 1979), читатель найдет
увлекательное описание окраски в природе. Очень популярная
книга Фокса и Веверса (Fox, Vevers, 1960) также дает инте-
ресное, широкое описание предмета. Очень много информации
о физиологических аспектах «зоохромов» можно почерпнуть из-
основательного труда Нидхама (Needham, 1974). Более огра-
ниченной по охвату, но несомненно содержащей полезный ма-
териал является монография Вюйома (Vuillaume, 1969) о пиг-
ментах у беспозвоночных животных.
Особенно полезным источником информации о пигментах
растений служит двухтомник под редакцией Гудвина (Goodwin„
1976), где содержится подробное описание химии, распростра-
нения, биосинтеза и функций главных классов растительных
пигментов, а также имеются чрезвычайно ценные главы об экс-
периментальных методах, используемых для их изучения. Вто-
рое издание книги под редакцией Цзигана (Czygan, 1980) так-
же в общем посвящено пигментам растений. Другие книги и
статьи, посвященные более специальным аспектам, например
32
Часть 1. Химические и биохимические аспекты
каждой отдельной группе пигментов, будут отмечены в после-
дующих главах.
Окраска, обусловленная структурой, изучена не столь хоро-
шо, как окраска, обусловленная пигментами. В книгах Фокса
(Fox, 1976) и Фокса и Веверса (Fox, Vevers, 1960) в общих
чертах даны главные характеристики структурной окраски и
•описаны физические явления, которые ее вызывают. Моногра-
фия Симона (Simon, 1971) также содержит введение в эту про-
блему; ей же посвящены несколько подробных статей, вышед-
ших в последнее время, например статья Хаксли (Huxley,
1975). Однако для правильного понимания физических явле-
ний, которые при этом затрагиваются, читателю следует вос-
пользоваться учебником по физике или по оптике.
В настоящее время спектроскопические методы представ-
ляют собой существенную часть обучения химиков-органиков;
этим методам посвящен ряд учебников. К примеру, учебник
Банвелла (Banwell, 1972) включает разделы по электронному
поглощению и рамановской спектроскопии. Библиотечка по ор-
ганической химии должна включать и другие полезные книги.
Для более детального ознакомления с методом кругового ди-
хроизма следует обратиться к монографии Снацке (Snatzke,
1967), а с методом резонансной рамановской спектроскопии
биологических молекул — к обзорам Варшела (Warshel, 1977)
и Кэри (Сагеу, 1978). Юнге (Junge, 1976) дает превосходное
описание кинетической флеш-спектроскопии, главным образом
в применении к фотосинтезу. Подробное описание спектроско-
пических свойств различных групп пигментов можно найти в
работах, цитируемых в последующих главах; в них же рас-
сматривается применение спектроскопических методов для изу-
чения пигментов.
1.9. Дополнительная литература
Banwell С. N. (1972). Fundamentals of molecular spectroscopy, 2nd edition,
London, McGraw-Hill.
Carey P. R. (1978). Resonance Raman spectroscopy. In: Biochemistry and bio-
logy, Quart. Rev. Biophys., 11, 309.
Clayton R. K. (1971). Light and living matter, vols 1 and 2, New York,
McGraw-Hill.
Czygan F.-C. (ed.) (1980). Pigments in plants, 2nd Edition, Stuttgart and New
York, Qustav Fischer.
Fox D. L. (1953). Animal biochromes and structural colours, Cambridge Uni-
versity Press.
Fox D. L. (1976). Animal biochromes and structural colors, 2nd edition, Ber-
keley, Los Angeles and London, University of California Press.
Fox D. L. (1979). Biochromy: natural coloration of living things, Berkeley, Los
Angeles and London, University of California Press.
Fox H. M., Vevers G. (1960). The nature of animal colours, London, Sidgwick
and Jackson.
Гл. 1. Свет и цвет
33
Goodwin Т. W. (ed.) (1976). Chemistry and biochemistry of plant pigments,
2nd edition, vols 1 and 2, London, New York and San Francisco, Academic
Press.
Huxley J. (1975). The basis of structural colour variation in two species of
Papilio, J. Entomol., 50A, 9.
Junge IF. (1976). Flash kinetic spectrophotometry in the study of plant pigments.
In: Chemistry and biochemistry of plant pigments, 2nd edition, vol. 2, ed.
T. W. Goodwin, p. 233, London, New York and San Francisco, Academic
Press.
MacMunn C. A. (1883). Studies on animal chromatology, Proc. Birmingham
Nat. Hist. Soc., 3, 351.
MacMunn C. A. (1890). Contribution to animal chromatology, Quart. J. Microsc.
Sci., 30, 51.
Murrell J. N. (1963). The theory of the electronic spectra of organic molecules,
London, Mathuen.
Needham A. E. (1974). The significance of zoochromes, Berlin, Heidelberg and
New York, Springer-Verlag.
Newbigin M. I. (1898). Colour in nature. London, John Murray.
Simon H. (1971). The splendor of iridescence: structural colors in the animal
world, New York, Dodd, Mead.
Snatzke G. (ed.) (1967). Optical rotatory dispersion and circular dichroism in
organic chemistry, London, Heyden.
Vulllaume M. (1969). Les pigments des invertebres, Paris, Masson.
Warshel A. (1977). Interpretation of resonance Raman spectra of biological mole-
cules, Ann. Rev. Biophys. Bioeng., 6, 273.
3—84
Глава 2
Каротиноиды
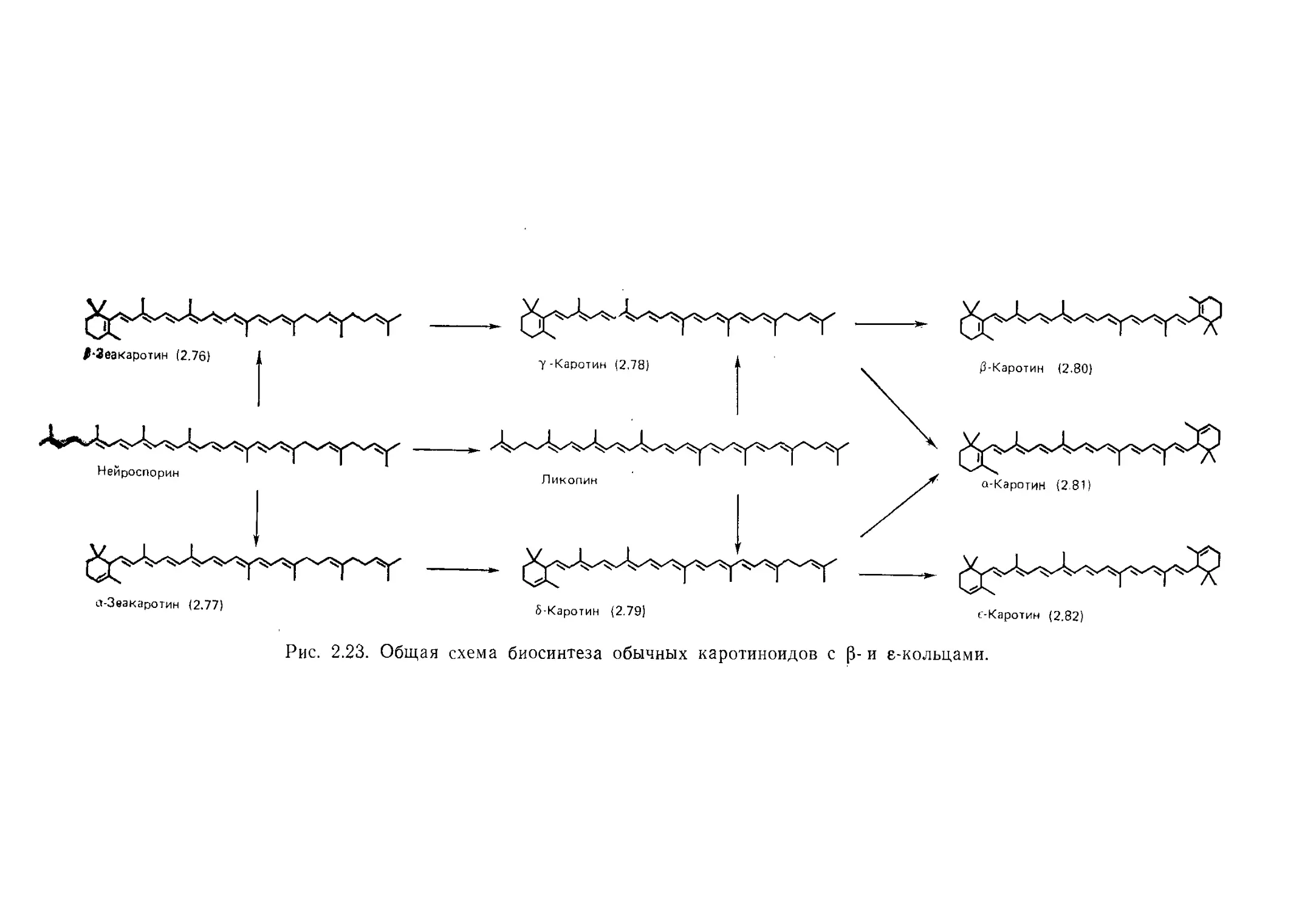
2.1. Введение
Из всех классов природных пигментов каротиноиды,
по-видимому, наиболее широко распространены и, несомненно,
принадлежат к числу наиболее важных соединений. Они обна-
ружены у всех представителей растительного царства как в фо-
тосинтезирующих, так и в нефотосинтезирующих тканях, а
также часто встречаются у микроорганизмов. Они полностью
или частично обусловливают окраску многих животных, осо-
бенно птиц, рыб и насекомых. Каротиноиды и их производные
имеют большое значение для животных, поскольку они являют-
ся основой зрительных пигментов, ответственных за восприятие
света и различение цветов.
2.2. Структура и номенклатура
2.2.1. Основная структура
Практически все каротиноиды представляют собой
или тетратерпены, т. е. С^-соединения, углеродный скелет ко-
торых построен из восьми Cs-изопреновых фрагментов (2.1),
или их производные. Основная структура молекулы симметрич-
на и состоит из двух Сго-половин; примером такого соединения
может служить ликопин (2.2) — красный пигмент томатов.
С
I
С (S
(2.1) Изопреновая единица
(2.2) Ликопин
Гл. 2. Каротиноиды
35
Основная структура иногда бывает модифицирована: на од-
ном или на обоих концах молекулы может присутствовать ше-
стичленное (или иногда пятичленное) кольцо, как, например,
у р-каротина (2.3). Последний является оранжевым пигментом
корней моркови и обычно рассматривается как «прародитель»
для всей группы каротиноидов.
даротиноидные углеводороды известны под названием ка-
ротинов. Все их производные с кислородсодержащими функ-
циональными группами именуются ксантофиллами. В кароти-
ноидах обнаруживается большинство обычных кислородсодер-
жащих функциональных групп, например гидрокси-, метокси-,
эпокси-, кето-, альдегидная и карбоксильная группы; при этом
соответствующие группы могут быть этерифицированы или
гликозилированы. Заместители обычно локализованы у Сэ-кон-
цевых групп (см. ниже). До настоящего времени в природе не
найдены каротиноиды, включающие азот-, серу- или галогенсо-
держащие заместители.
2.2.2. Номенклатура
Идентифицировано и охарактеризовано около 500 при-
родных каротиноидов. Многие из них, особенно наиболее важ-
ные и известные уже в течение многих лет, носят общеприня-
тые тривиальные названия. Обычно такие названия происходят
от биологического источника, из которого эти каротиноиды бы-
ли впервые выделены. В последние годы для внесения ясности
в случайную и часто запутанную тривиальную номенклатуру
была введена новая полусистематическая номенклатура.
В этой книге хорошо известные каротиноиды обычно будут
называться своими тривиальными названиями, но в соответст-
вии с современной практикой при первом упоминании каждому
каротиноиду будет дано также полусистематическое наимено-
вание.
2.2.3. Правила IUPAC для номенклатуры каротиноидов
Все специальные названия каротиноидов основаны на
названии «каротин», соответствующем структуре и порядку
нумерации углеродных атомов, показанным на рис. 2.1, на
3*
36
Часть I. Химические и биохимические аспекты
котором также приведен ряд возможных структур концевой
Cg-группы. Наименование того или иного каротиноидного угле-
водорода составляется путем добавления в виде приставки к
слову «каротин» двух греческих букв, характеризующих две
концевые Cg-группировки. Греческие приставки помещают в ал-
фавитном порядке: р (бета), у (гамма), е (эпсилон), х (каппа),
Ф (Фи), X (хи), ф (пси).
Типы С9 концевых групп
Рис. 2.1. Основная структура каротиноидов; типы С9-концевых группы и схема
нумерации согласно правилам IUPAC.
Группировки заместителей обозначаются приставкой или
суффиксом согласно обычным правилам органической химии.
Система нумерации показана на рис. 2.1. Если две конце-
вые группы различны, то не помеченные штрихом номера да-
ются атомам С той группы, которая обозначена (греческой
буквой) в названии соединения первой. Формула должна быть
изображена таким образом, чтобы С-атомы, обозначенные без
штрихов, были расположены с левой стороны.
2.2.4. Некоторые примеры структуры каротиноидов
и их номенклатуры
Каротины. Наиболее известный из всех каротиноидов
р-каротин (2.3) обозначается по системе ШРАС как р,р-каро-
тин. Его изомер а-каротин (2.4) стал называться р,е-каротином,
Гл. 2. Каротиноиды
37
а ациклический каротиноид ликопин (2.2) получил название
ф,ф-каротина.
Каротиноидные углеводороды, отличающиеся от исходных
каротинов уровнем восстановления, являются важными био-
синтетическими промежуточными продуктами. Их название об-
разуется добавлением к исходному названию приставки «де-
гидро» или «гидро» (с соответствующим коэффициентом), а
также указанием номеров атомов углерода, к которым при-
соединены или у которых отняты водородные атомы. Примером
может служить р-зеакаротин (2.5), имеющий полуспстематиче-
ское название 7',8'-дигидро-р,г|:-каротин.
(2.5) З-Зеакаротин
Ксантофиллы. Все кислородсодержащие производные каро-
тиноидных углеводородов в настоящее время называют ксанто-
филлами, хотя в прошлом название «ксантофилл» использова-
лось только для обозначения каротинолов (каротиноидов, со-
держащих спиртовые группы) и даже одного-едпнственного со-
единения — лютеина (см. ниже).
В этом разделе будет приведено лишь несколько примеров
строения молекулы и номенклатуры некоторых наиболее важ-
ных из сотен встречающихся в природе ксантофиллов. Из всех
ксантофиллов наиболее важными и наиболее распространенны-
ми янляются каротинолы. Самые широкораспространениые и
известные из них — это пигменты листьев — лютеин (2.6) и зе-
аксантин (2.7), которые являются дигидроксипроизводными
а-каротина и p-каротина соответственно и называются по но-
вой системе р,8-каротин,-3,3'-диолом и р,р-каротин-3,3'-диолом.
Известны также ксантофиллы с гидроксигруппами в других по-
ложениях молекулы, например при С-2 и С-4 в кольце, а ацик-
лические соединения с третичными гидроксигруппами при С-1
широко распространены у некоторых бактерий (разд. 2.4.4).
38
Часть I. Химические и биохимические аспекты
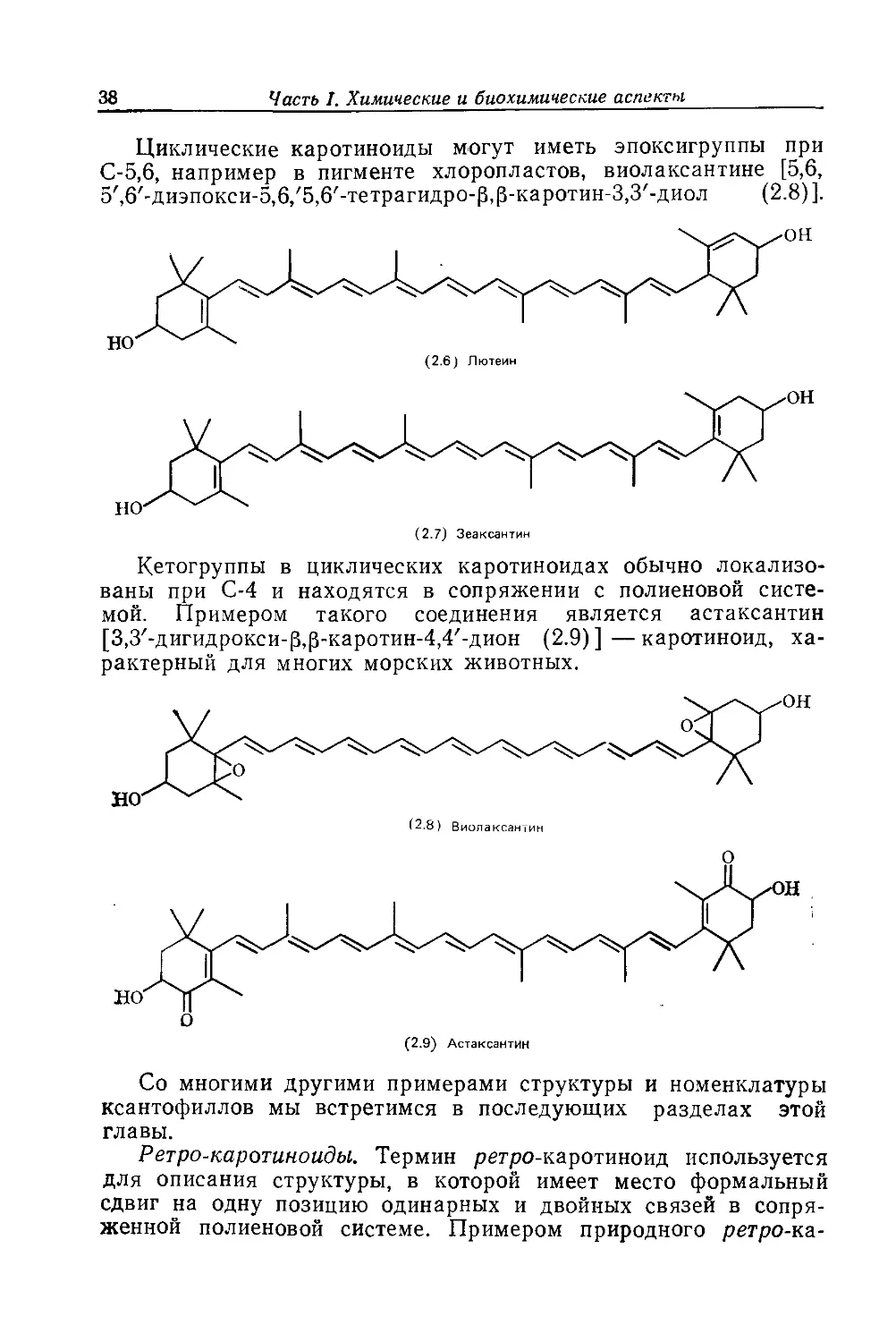
Циклические каротиноиды могут иметь эпоксигруппы при
С-5,6, например в пигменте хлоропластов, виолаксантине [5,6,
5',6'-диэпокси-5,6/5,6'-тетрагидро-|3,|3-каротин-3,3'-диол (2.8)].
Кетогруппы в циклических каротиноидах обычно локализо-
ваны при С-4 и находятся в сопряжении с полиеновой систе-
мой. Примером такого соединения является астаксантин
[3,3'-дигидрокси-|3,р-каротин-4,4'-дион (2.9)]—каротиноид, ха-
рактерный для многих морских животных.
(2.9) Астаксантин
Со многими другими примерами структуры и номенклатуры
ксантофиллов мы встретимся в последующих разделах этой
главы.
Ретро-каротиноиды. Термин ретро-каротиноид используется
для описания структуры, в которой имеет место формальный
сдвиг на одну позицию одинарных и двойных связей в сопря-
женной полиеновой системе. Примером природного ретро-ка-
Гл. 2. Каротиноиды
39
ротиноида может служить родоксантин [4',5'-дидегидро-4,5/-
ретро-р,р-каротин-3,3'-дион (2.10)], который придает красный
цвет присемянникам («ягодам») тисса.
Примечание. Согласно новой номенклатуре, в паре цифр,
предшествующей приставке «ретро», первая цифра означает-
атом углерода, который формально потерял протон, а вторая —
углеродный атом, который его приобрел.
Гомо-, апо- и нор-каротиноиды. Хотя большинство кароти-
ноидов представляют собой С40-соединения, некоторые могут
иметь больше или меньше, чем 40, атомов углерода. С45 и С50-
каротиноиды, которые найдены у некоторых бактерий, часто
называют высшими или гомо-каротиноидами. Они представля-
ют собой нормальную С40-каротиноидную молекулу с одним или
двумя Сэ-заместителями при С-2, С-2'. Они могут быть цикли-
ческими или ациклическими, как, например, декапреноксантин
[2,2'-бис(4-гидрокси-3-метилбут-2-енил)-е,е-каротин (2.11)] и
бактериоруберин [2,2'-бис (З-гидрокси-З-метилбутил) -3,4,3',4'-
тетрадегидро-1,2,1',2'-тетрагидро-ф,ф-каротин-1, l'-диол (2.12) ].
Каротиноиды с меньшим, чем 40, числом атомов углерода
подразделяются на две категории. Молекулы, в которых С40-уг-
СН2ОН
40
Часть I. Химические и биохимические аспекты.
леродный скелет укорочен формальным удалением фрагментов
с одного или обоих концов, называются апо-(или диапо-) каро-
тиноидами. Например, Сзо-соединение З’Цитраурин [3-гидрокси-
8'-апо-р-каротин-8'-аль (2.13)]. Этот и близкие к нему апо-ка-
ротиноиды обусловливают окраску апельсинов и плодов других
цитрусовых.
(2.13) (З-Цитраурин
Нор-каротиноид — это такой каротиноид, в котором отсут-
ствует один или небольшая группа атомов С не на концах мо-
лекулы, а в ее внутренней части. Например, актиния (Actinia
equina) содержит заметное количество пурпурного пигмента
актиниоэритрина, который лишен С-2 и С-2' и поэтому назы-
вается 3,3'-дигидрокси-2,2'-динор-р,р-каротин-4,4'-дион-3,3'-ди-
ацилатом (2.14).
2.2.5. Стереохимия
Геометрическая изомерия. Полиеновая система каро-
тиноидов предоставляет широкий простор для существования
большого числа геометрических (цис-транс) -изомеров. Так, ли-
копин (2.2)—симметричная молекула с И сопряженными
двойными связями — теоретически способен существовать в
1056 формах, а для несимметричных каротиноидов возможно
наличие еще большего числа изомеров. К счастью, для практи-
ческой реализации многих потенциальных изомеров существу-
ют пространственные препятствия, и поэтому они образуются
с трудом. Тем не менее имеется 72 возможных стерически «не-
запрещенных» изомера ликопина (о которых не следует забы-
вать) (рис. 2.2). По соглашению термины цис- и транс- в при-
менении к каротиноидам используются для обозначения относи-
тельного расположения тех заместителей при двойной связи,
которые образуют часть главной цепи углеродных атомов.
Гл. 2. Каротиноиды
41
В природе большинство каротиноидов существует исключи-
тельно или главным образом в полностью транс- (полностью £)
форме. Имеется, однако, несколько важных исключений. Фито-
ин (7,8,11,12,7',8\11\12'-октагидро-ф,ф-каротин), обычно рас-
сматриваемый в качестве первого С^-углеводородного проме-
жуточного продукта биосинтеза каротиноидов (разд. 2.6.3), как
Затрудненные Незатрудненные
Рис. 2.2. Стерически «затрудненные» и стерически «незатрудненные» цис-
двойные связи.
правило, выделяют в виде 15-цис(2)-изомера (2.15). Возмож-
но, что самым замечательным zpc-каротиноидом является про-
ликопин — поли-^ис-изомер ликопина, встречающийся в приро-
де у ряда растений. Структура проликопина недавно была ус-
тановлена; было показано, что он представляет собой 7,9,7',9'-
тетра^ис-ликопин (2.16).
В общем же следовые количества zpc-изомеров, присутству-
ющие в экстрактах природных объектов, обычно рассматрива-
ются как артефакты, образующиеся в результате стерических
изменений природных полностью тра«с-каротиноидов.
(2.16) Проликопин
42
Часть I. Химические и биохимические аспекты
Одинарные связи, входящие в состав полиенового хромофо-
ра, имеют конформацию s-транс. В циклических каротиноидах
связь С-6,7 обычно принимает з-^ис-конформацию как в крис-
таллическом состоянии, так и в растворе. Другие детали кон-
формации каротиноидов будут обсуждаться позже в связи с
их светопоглощающими свойствами (разд. 2.3.3).
Абсолютная конфигурация. Многие органические молекулы
являются хиральными. Это значит, что они существуют только
в виде одного из двух возможных энантиомеров — стереоизоме-
(2.18) (ЗЯ)-З-Гидрокси-/^-кольцо
(2. 1 7) (6Я) е Кольцо
(2.19) (ЗЯ)-Концевая группа (2.20} (3S)-Концевая группа
астаксантина (2.9) астаксантина (2.9)
ров, несовместимых со своим зеркальным отражением. Наибо-
лее распространенный тип хиральности наблюдается при нали-
чии асимметрического атома углерода, который теоретически
может существовать в двух пространственных конфигурациях.
Хиральность часто наблюдается у каротиноидов, и обычно она
обусловлена присутствием асимметрического атома углерода в
молекуле. Примером хиральных центров является атом С-6 в
s-кольце каротиноидов (2.17) и атом С-3 в обычных цикличе-
ских каротин-3-олах (2.18).
Абсолютная конфигурация многих хиральных каротиноидов
была определена главным образом с помощью оптических мето-
дов [дисперсии оптического вращения (ДОВ), кругового ди-
хроизма (КД)] и ядерного магнитного резонанса (ЯМР). Не-
давно было обнаружено, что у разных живых организмов
встречаются различные оптические изомеры каротиноидов. Так
дрожжи Phaffia rhodozyma образуют (ЗЯ, 3'R)-астаксантин
(2.19), в то время как омар накапливает преимущественно
(3S, 3'5)-изомер (2.20) наряду с (ЗЯ, 3'R)- и мезо-(ЗР, 3'S)-
формами в меньших количествах.
Гл. 2. Каротиноиды
43
2.3. Свойства
2.3.1. Общие физические свойства
Каротиноиды являются липидами. Они растворимы в
органических растворителях и могут быть экстрагированы из
природных объектов полярными растворителями, такими, как
ацетон и спирты. Даже ксантофиллы с четырьмя и более гидр-
оксигруппами в молекуле практически нерастворимы в воде.
Однако они становятся растворимыми после гликозилирования
или образования комплексов с белками. In vivo каротиноиды^
обычно локализованы в липофильных, гидрофобных районах
клетки, таких, как липидные глобулы, кристаллические струк-
туры и мембраны (в последних они находятся в комплексе с
белками).
За исключением таких промежуточных продуктов в биосин- ,
тезе, как фитоин, фитофлуин и обычно ^-каротин, каротиноиды
при комнатной температуре представляют собой твердые веще-
ства. Большинство из них может быть получено в кристалли-
ческом виде при использовании соответствующих смесей рас-
творителей.
2.3.2. Стабильность
Выделенные каротиноиды как без растворителя, так
и в растворе очень чувствительны к свету и теплу (которые
вызывают чмс-трамс-изомеризацию), к кислотам (которые так-
Рис. 2.3. Катализируемая кислотой изомеризация 5,6-эпоксикаротиноида в
5,8-фур аноидный оксид.
же вызывают цмс-трамс-изомеризацию и особенно превращение
5,6-эпоксидов в 5,8-фураноидные оксиды; рис. 2.3) и в некото-
рых случаях к основаниям (которые вызывают аутоокисление
З-гидрокси-4-кето-концевых групп, как, например, у астаксанти-
на; рис. 2.4). Система сопряженных двойных связей делает ка-
ротиноиды чрезвычайно подверженными окислительному обес-
цвечиванию кислородом воздуха.
44
Часть I. Химические и биохимические аспекты
In vivo каротиноиды обычно являются составной частью
структуры клетки или органеллы и находятся в ассоциации
или с белками, или с другими клеточными липидами. В этих
условиях каротиноиды стабилизированы и защищены от пере-
численных выше вредных влияний.
Рис. 2.4. Окисление (З-гидрокси-4-кето) -концевой группы каротиноидов.
In vitro каротинопротеины, особенно астаксантин-белковые
комплексы беспозвоночных (разд. 2.5), также значительно бо-
лее стабильны, чем свободные каротиноиды.
2.3.3. Поглощение света
Хромофор, ответственный за поглощение каротиноида-
ми видимого света, представляет собой систему сопряженных
двойных связей (гл. 1). С увеличением длины полиеновой
л-электронной системы стабильность первого возбужденного со-
стояния повышается; при этом электронное возбуждение про-
исходит легче, оно требует меньше энергии и осуществляется
светом с большей длиной волны.
Такой возрастающий батохромный эффект (сдвиг в длинно-
волновую сторону) может быть проиллюстрирован спектрами
поглощения биосинтетического ряда (разд. 2.6.4) ациклических
(2.21) Фитофлуин
каротиноидов с возрастающей длиной хромофора (рис. 2.5).
Так, фитоин (2.15) [3 сопряженные двойные связи (с. д. с.)] и
фитофлуин [7,8,11,12,7',8'-гексагидро-ф,ф-каротин (2.21)]
(5 с. д. с.), имеющие Хтах при 275, 285, 296 и 331, 348, 367 нм
соответственно, бесцветны; g-каротин [7,8,7/,8/-тетрагидро-ф,ф-
каротин (2.22)] (7 с.д. с.) имеет слабое поглощение в голубом
участке спектра (Хтах = 378, 400 и 425 нм) и потому выглядит
слегка желтоватым, а нейроспорин [7,8-дигидро-ф,ф-каротин
Гл. 2. Каротиноиды
.45
Длина волны, нм
Рис. 2.5. Спектры поглощения света ациклическими каротиноидами с увели-
чивающейся длиной хромофора. I— фитоин (2.15), II — фитофлуин (2.21),
III — ^-каротин (2.22), IV—нейроспорин (2.23), V — ликопин (2.2) (в пет-
ролейном эфире).
(2.23)] (9 с.д.с.) и ликопин (2.2) (И с.д.с.) сильно поглоща-
ют в видимом диапазоне при 414, 439, 467 и 444, 470, 502 нм
соответственно и обладают характерной желтой и оранжевой
окраской. Все эти соединения имеют типичный трехпиковый ка-
ротиноидный спектр поглощения с четко выраженными макси-
мумами и минимумами.
(2.23) Нейроспорин
На максимумы поглощения, а иногда на степень выявления
тонкой структуры спектра большое влияние оказывает ис-
пользуемый растворитель. Максимумы поглощения наблюдают-
ся при большей длине волны, если спектры определяют в аро-
матических или галогенированных растворителях, а также в
CS2, чем в тех случаях, когда в качестве растворителей исполь-
зуют петролейный эфир, гексан или этанол (рис. 2.6).
46
Часть 1. Химические и биохимические аспекты
При циклизации ликопина в е,е-каротин (2.24) или у,у-ка-
ротин (2.25) из сопряженной системы удаляются две двойные
связи, в результате чего спектры поглощения этих циклических
каротиноидов очень сходны со спектром нейроспорина. В слу-
чае каротиноидов, имеющих p-кольцо, таких, как (3-каротин,
Длина волны, нм
Рис. 2.6. Влияние растворителя на спектр поглощения света типичного каро-
тиноида ликопина (2.2). I — в петролейиом эфире или этаноле; II—в хло-
роформе или бензоле, III — в сероуглероде.
двойные связи внутри кольца сопряжены со связями главной
полиеновой цепи. Однако из-за пространственных ограничений
между метильными заместителями в кольце и главной полиено-
вой цепью молекула изгибается относительно одинарной связи
С-6,7 таким образом, что л-орбиталь 5,6-двойной связи кольца
не лежит в одной плоскости с л-электронной системой полиена.
Наблюдающееся при этом уменьшение степени перекрывания
{2.25) у} у -Кароти!
Гл. 2. Каротиноиды
47
350 400 450 500 550
Длина волны, нм
Рис. 2.7. Влияние р-колец на спектры поглощения света каротиноидами.
I— р-каротин (2.3), II— у-каротии (2.78) и III — ликопин (2.2) (в петро-
лейном эфире).
л-орбиталей означает, что эффективная длина хромофора мень-
ше, чем полная длина системы из И сопряженных двойных
связей, и, следовательно, максимум поглощения имеет место
при более короткой длине волны, чем в случае ликопина. Изгиб
молекулы приводит также к ослаблению тонкой структуры,
т. е. к менее выраженным максимумам и минимумам в спект-
ре. На рис. 2.7. сравниваются спектры ликопина, у-каротина
(Р,ф-каротина) и р-каротина.
(2.26) Эхиненон
(2.27) КзНтаксантин
48
Часть I. Химические и биохимические аспекты
Влияние на тонкую структуру спектров еще более заметно
у каротиноидов, содержащих в сопряжении с полиеновой сис-
темой карбонильную группу. Сопряженная группа С = О эф-
фективно удлиняет хромофор так, что максимум поглощения
сдвигается в более длинноволновую область, однако при этом
Длина волны, нм
Рис. 2.8. Влияние сопряженных карбонильных групп на спектры поглощения
света каротиноидами (в петролейном эфире). I— (3-каротин (2.3), II— эхи-
ненон (2.26) (одна группа С = О), III—кантаксантин (2.27; две группы
С=О).
почти полностью утрачивается тонкая структура спектра. На
рис. 2.8 сравниваются спектры p-каротина, эхиненона [р,р-ка-
ротин-4-она (2.26)] и кантаксантина [|р,р-каротин-4,4'-диона
(2.27)].
Другие заместители, например ОН-группа, обычно оказыва-
ют незначительное влияние на спектр поглощения либо не ока-
Рис. 2.9. Сравнение спектров
ностью транс-ликопина (/)
Длина волны, нм
поглощения света ( в петролейном эфире) пол-
н поли-цнс-изомера проликопина (2.16; II).
Гл. 2, Каротиноиды 49
зывают его вовсе. Так, спектры (3-каротина, (3-криптоксантина
[Р,(3-каротин-3-ола (2.28)], изокриптоксантина [р,(3-каротин-4-
ола (2.29)], зеаксантина (2.7) и изозеаксантина [р,(3-каротин-
4,4'-диола (2.30)] практически идентичны как по положению
^-тах, так и по форме.
Полоса поглощения цис-пзомеров менее интенсивна и сдви-
нута в более коротковолновую область, чем у соответствующих
полностью транс-соединений (рис. 2.9). Это обусловлено иска-
4—«4
so
Часть 1. Химические и биохимические аспекты
Рис. 2.10. Спектроскопическая проба на сопряжение кетогрупп у каротинои-
дов. Спектры полощеиия света сняты в этаноле. I — кантаксантин (Рф-ка-
ротин-4,4'-дион; 2,27); II — изозеаксантин (2.30), продукт восстановления
кантаксантииа под действием NaBH<.
350 400 450 500
Длина волны,нм
Рис. 2.11. Спектроскопическая проба на 5,6-эпоксигруппы в циклических каро-
тиноидах. Спектры поглощения света сняты в этаноле. I — диэпоксикароти-
яноид виолаксантии (2.8); II — ауроксаитии (2.31), фураноидный оксид, обра-
зующийся в результате добавления капли разбавленной НС1.
_1
550
жением расположенной в одной плоскости полиеновой системы
из-за поворота части молекулы вокруг одинарной связи, при-
мыкающей к ^«с-двойной связи; при этом уменьшаются стери-
ческие препятствия между двумя атомами водорода («неза-
трудненная щ/с-конфигурация») или между водородным ато-
мом и метильной группой («затрудненная ^«с-конфигурация»)
(рис. 2.2).
Сдвиги максимума поглощения, происходящие в результате
химических реакций, протекающих в очень маленьком объеме
Гл. 2. Каротиноиды
5р
в кювете спектрофотометра, служат очень полезными диагно-
стическими критериями. Так, обработка такого кетокаротинои-
да, как кантаксантин (2.27), боргидридом натрия приводит к.
гипсохромному сдвигу в спектре (т. е. к сдвигу в коротковолно-
вую сторону) и к более четкому выявлению тонкой структуры,,
поскольку кантаксантин восстанавливается до изозеаксантина
(2.30) (рис. 2.10). Обработка 5,6-эпоксикаротиноидов типа
виолаксантина (2.8) следовыми количествами кислот вызывает-
. Рис. 2.12. Спектры поглощения света каротинопротеина крустацианина из
омара (/, водный раствор) и его свободной простетнческой группы — каро-
тиноида астаксантина (2.19) (II, этанол).
спектральный сдвиг на 40 нм в коротковолновую область-
спектра (на 20 нм для моноэпоксидов) из-за образования фу-
раноидного оксида ауроксантина [б.в.б'.в'-диэпоксн-бДб'Х-
тетрагидро-0,0-каротин-3,3'-диола (2.31)] (рис. 2.11).
In vivo каротиноиды имеют обычно максимумы поглощения
при длинах волн примерно на 10 нм больших, чем в гексано-
вом или этанольном растворе, поскольку в клетке они находят-
ся в ассоциации с липидами или белками. Спектры поглощения
каротиноидов in vivo часто напоминают спектры искусственно-
полученных мицеллярных суспензий каротиноидов с другими
липидами в водной среде.
Стехиометрические каротин-белковые комплексы, характер-
ные для многих беспозвоночных морских животных (разд. 2.5),
имеют очень большие сдвиги (например, на ~ 100 нм) макси-
мумов поглощения и потому окрашены в пурпурный или голу-
бой цвет (Хтах = 550—630 нм) в отличие от желто-оранжевых
свободных каротиноидов (Z.max = 470 нм) (рис. 2.12). Никакого
удовлетворительного объяснения этому феномену до сих пор
не найдено.
4»
52 .. Часть I. Химические и биохимические аспекты
2.4. Распространение в природе
Каротиноиды представляют собой наиболее широко
распространенный класс природных пигментов. Они обнаруже-
ны повсеместно в растительном царстве, а также обычно
встречаются у животных и микроорганизмов.
2.4.1. Высшие растения
Фотосинтезирующие ткани. Все зеленые ткани высших
растений содержат одни и те же главные каротиноиды, которые
локализованы (возможно, исключительно) в хлоропластах.
К ним принадлежат 0-каротин (2.3), лютеин (2.6), виолаксан-
тин (2.8) и неоксантин [б'.б'-эпокси-б^-дидегидро-ЗДб'.б'-тет-
рагидро-0,0-каротин-3,5,3'-триол (2..32)]. Они часто содержатся
в хлоропластах вместе с меньшими количествами а-каротина
(2.4), зеаксантина (2.7), 0-криптоксантина (2.28) и антераксан-
тина [5,6-эпокси-5,6-дигидро-0,0-каротин-3,3'-диола (2.33)]. Ка-
(2.33) Антераксантин
ротиноиды, вероятно, локализованы в гранах хлоропластов в
виде хромопротеинов. Некоторые, если не все, хлоропластные
каротиноиды могут присутствовать in vivo в различных формах
в разных участках хлоропласта. Значение этого факта и воз-
можные функции каротиноидов в хлоропластах будут обсуж-
даться в гл. 10.
Нефотосинтезирующие ткани. Многие желтые цветки и оран-
жево-красные плоды обязаны своей окраской каротиноидам, ко-
торые обычно локализованы в хромопластных структурах. Жел-
тые цветки, например у нарцисса и одуванчика, часто содер-
Гл 2. Каротиноиды
53
жат большие количества каротиноидных эпоксидов, таких, как
виолаксантин (2.8), в то время как оранжевые (например, у аб-
рикоса) и красные (например, у томата) плоды часто окраше-
ны соответственно 0-каротином и ликопином или их простыми
гидроксипроизводными. В некоторых случаях, однако, пигмен-
тация обусловлена присутствием каротиноидов, которые ветре-
(2,36) Капсорубин
чаются только у одного или у очень небольшого числа видов.
Так, цветки эшшольции калифорнийской (Eschscholtzia califor-
nica) содержат ретро-каротиноид эшшольцксантин (4',5'-диде-
гидро-4,5/-ретро-0,0-каротин-3,3'-диол (2.34)], присемянники
(«ягоды») тисса (Taxus baccata) содержат ретро-каротиноид-
ный кетон родоксантин (2.10), а знакомая нам окраска крас-
ного перца (Capsicum аппишп) обусловлена циклопентановыми
каротиноидами капсантином [3,3'-дигидрокси-0,х-каротин-6/-
оном (2.35)] и капсорубином [З.З'-дигидрокси-х.х-каротин-
6,6'-дионом (2.36)].
В цветках и плодах, а также в стареющих осенних листьях
гидроксикаротиноиды обычно этерифицированы жирными кис-
лотами (например, пальмитиновой кислотой).
51
Часть I. Химические и биохимические аспекты
Хотя в корнях каротиноиды обычно не встречаются, в кор-
нях моркови (Daucus carota) содержится большое количество
Р- и а-каротина, и именно от моркови этот класс пигментов и
получил свое название.
2.4.2. Водоросли
Каротиноиды обычно содержатся в хлоропластах во-
дорослей, в том числе и у обычных морских водорослей. Меж-
ду хлоропластными каротиноидами разных классов водорослей
наблюдаются значительные качественные различия, и поэтому
-ОСОСН3
(2.37) Фукоксантин
состав каротиноидов пытаются использовать для решения не-
которых проблем в систематике и эволюции водорослей. Каро-
тиноиды иногда присутствуют также вне хлоропласта, напри-
мер в светочувствительном «глазке» Euglena (гл. 11) ив реп-
родуктивных участках у колониальных видов, таких, как Ulva —
зеленого «морского салата», обычно обнаруживаемого в при-
брежных водах или в мелководье между скалами. Кетокароти-
ноиды, в том числе и астаксантин (2.9), могут накапливаться
вне хлоропласта у некоторых зеленых водорослей (Chlorophyta)
при неблагоприятных условиях культивирования, главным об-
разом при недостатке минеральных веществ или азота.
О -рамнсзз
(2.38) Миксоксантофилл
Состав каротиноидов в хлоропластах зеленых водорослей
обычно сходен с их составом в хлоропластах высших расте-
ний, что предполагает явную эволюционную связь. Большинст-
во других классов водорослей образуют ацетиленовые или ал-
леновые каротиноиды. Годовой природный синтез фукоксантина
[5,6-эпокси-3,3/,5/-тригидрокси-6/,7/-дидегидро-5,6,7,8,5',6/ - гекса-
Гл. 2. Каротиноиды
55
гидро-р,р-каротин-8-он-3'-ацетата (2.37)]—характерного каро-
тиноида бурых морских водорослей (Phaeophyta)—оценивает-
ся в несколько миллионов тонн.
Примитивные прокариотические сине-зеленые водоросли,
или бактерии (Cyanophyta, или Cyanobacteria), синтезируют
fi-каротин и несколько его простых гидрокси- и кетопроизвод-
ных, но многие виды также накапливают миксоксантофилл
[2'- (p-L-рамнопиранозилокси) -3',4'-дидегидро-1',2'-дигидро-р,ф-
каротин-3,1'-диол (2.38)] и другие гликозиды каротиноидов, ко-
торые более типичны для нефотосинтезирующих бактерий.
2.4.3. Грибы
Каротиноиды широко распространены в природе, но у
грибов они встречаются лишь у некоторых видов. Blakeslea tri-
spora, например, синтезирует в мицелии p-каротин в таких ко-
личествах, что возможно его производство путем ферментации.
(2.39) Торулародин
Большинство каротиногенных грибов накапливают только каро-
тины, особенно p-каротин и у-каротин; ксантофиллы являются
редкостью. Очень немногие из макроскопических грибов, съе-
добных и ядовитых, окрашены каротиноидами. К ним относит-
ся съедобный гриб лисичка (Cantharellus cibarius), который
окрашен в желтый цвет кантаксантином (2.27). Красные дрож-
жи (Rhodotorula spp.) образуют характерную для них кароти-
ноидную кислоту торулародин [З'Л'-дидегидро-р.ф-каротин-Ш'-
карбоновую кислоту (2.39)].
2.4.4. Бактерии
Если даже кратковременно экспонировать агаровую
пластинку на открытом воздухе, то можно увидеть рост на ней
микробных колоний, в том числе многих желтых, оранжевых
и красных бактерий и дрожжей. Пигменты, обусловливающие
их окраску, во многих случаях являются каротиноидами. И, хо-
тя распространение каротиноидов у бактерий систематически
не изучалось, все же можно сделать некоторые обобщения.
Н ефотосинтезирующие бактерии. Каротиноиды, найденные у
представителей многих классов и семейств нефотосинтезирую-
щих бактерий, обычно локализованы в клеточных мембранах
56
Часть I. Химические и биохимические аспекты
или в клеточной стенке. Часто биосинтез каротиноидов инду-
цируется светом. Многие виды накапливают простые С40-каро-
тиноиды, такие, как p-каротин, у-каротин и их производные.
Некоторые бактерии синтезируют С45- и С50-структуры. Так, ха-
рактерным каротиноидом галофильных Halobacteria являр^ч
(2.40) Сардинаксантин
(2.41) "К.п. 450”
(2.42) "Диапонейроспорин”
(2.43) Слирилпоксантин
(2.44) Гидро ксисфероидин
(2.45) Хлоробактин
О Me
(2.4в) Окенен
Гл. 2. Каротиноиды
57
ациклический С50-пигмент бактериоруберин (2.12), в то время
как некоторые виды Flavobacterium, Sarcina и Corinebacterium,
в том числе некоторые обычные загрязнители воздуха и расти-
тельные и животные патогены, содержат циклические С50-каро-
тиноиды, такие, как декапреноксантин (2.11), сарцинаксантин
[2,2'-бис-(4-гидрокси-3-метилбут-2-енил)-у,у-каротин (2.40)] и
«К- п. 450» [2,2'-бис-(4-гидрокси-3-метилбут-2-енил)-р,р-каротин
(2.41)]. Недавно у некоторых патогенных видов Streptococcus
it Staphylococcus был обнаружен ряд С30-тритерпеноидных ка-
ротиноидов. И, хотя они названы «диапокаротиноидами», эти
соединения, например диапонейроспорин (2.42), являются ис-
тинными тритерпеноидами, образующимися из фарнезилпиро-
фосфата через С30-аналог фитоина (разд. 2.6.3). У нефотосин-
тезирующих бактерий обычно встречаются также гликозиды
Сзо-, С4о- и Сбо-каротиноидов.
Фотосинтезирующие бактерии. Каротиноиды обнаружены у
всех фотосинтезирующих бактерий, у которых они являются
важными компонентами фотосинтетического аппарата (гл. 10).
Пурпурные несерные бактерии (Rhodospirillaceae) характери-
зуются присутствием ациклических пигментов с третичными
гидрокси- или метоксигруппами при С-1 и С-1', примером чему
служат спириллоксантин [1,Г-диметокси-3,4,3',4'-тетрадегидро-
1,2,1',2'-тетрагидро-ф,ф-каротин (2.43)] из Rhodospirillum rub-
rum. и гидроксисфероидин [1'-метокси-3',4'-дидегидро-1,2,7,8,1',
2'-гексагидро-ф,ф-каротин-1-ол (2.44)] из Rhodopseudomonas
sphaeroides. В пурпурных и зеленых серных бактериях (Chro-
matiaceae и Chlorobiaceae) обычно содержатся ароматические
каротиноиды, такие, как хлоробактин [ф,ф-каротин (2.45)] и
юкенон [|1'-метокси-1',2'-дигидро-%,ф-каротин-4'-он (2.46)].
2.4.5. Животные
Каротиноиды обусловливают окраску наружных покро-
вов у многих животных, принадлежащих ко всем классам, кро-
ме млекопитающих.
Позвоночные; млекопитающие. Хотя каротиноиды чрезвы-
чайно важны для млекопитающих как предшественники вита-
мина А и часто присутствуют в небольших количествах в пече-
ни и жировых тканях, не известно ни одного случая, когда бы
эти соединения в сколько-нибудь заметной степени участвова-
ли в формировании нормальной окраски наружных покровов у
какого-либо вида млекопитающих. Правда, известны случаи,
когда люди, в рацион питания которых входили в чрезмерно
большом количестве морковь или апельсины, обладали столь
высокой подкожной концентрацией каротиноидов, что их кожа,
58
Часть I. Химические и биохимические аспекты
особенно на ладонях и на ступнях, приобретала оранжевый от-
тенок.
Птицы. Самая обычная желтая окраска яичного желтка
свидетельствует об универсальной важности каротиноидов для
птиц. Кроме того, не является необычным окрашивание у них
каротиноидами кожи и особенно оперения. Желто-красные
перья у многих видов птиц пигментированы каротиноидами.
В качестве наиболее известного примера можно привести ро-
зово-красноватую окраску перьев фламинго, обусловленную
кетокаротиноидами, главным образом кантаксантином (2.27).
(2.47) Д, у Каротин
У многих птиц зеленая окраска перьев является результатом
комбинации основной голубой структурной окраски и фоновой
желтой, обусловленной каротиноидами.
Рыбы, амфибии и рептилии. Каротиноиды могут окрашивать
кожу (например, у золотой рыбки) или мышцы (например, у
лосося) рыб. У рыб чаще других встречаются такие каротинои-
ды, как астаксантин и его сложные эфиры. Обусловленная ка-
ротиноидами желтая окраска иногда встречается у амфибий и
рептилий.
(2.48) Реньератин
Беспозвоночные; насекомые. Каротиноиды ответственны за
окраску некоторых (но далеко не всех) желтых, оранжевых и
красных насекомых (см. также гл. 3 и 6). Хорошо известными
примерами являются саранча Schistocerca (p-каротин), коло-
радский жук Leptinotarsa decemlineata (кантаксантин) и божья
коровка Coccinella septempunctata, содержащая большое число
каротинов, главным образом редко встречающийся 0,у-каротин
(2.47) и родственные ему соединения, содержащие у-кольцо.
Морские беспозвоночные. Обусловленная каротиноидами
пигментация особенно характерна и важна почти для всех клас-
сов морских беспозвоночных. Типичными каротиноидами для
них являются кетосоединения, такие, как кантаксантин и аста-
ксантин. Иногда обнаруживают и другие необычные каротино-
Гл. 2. Каротиноиды.
59
иды. У широко размножившихся в тропических морях окрашен-
ных в разные цвета губок (Porifera) преобладают арильные
каротиноиды, такие, как реньератин [0,%-каротин (2.48)]. Ак-
тиния (Actinia equina) — обычный обитатель скальных вод; ок-
рашена в пурпурный цвет нор-каротиноидным эфиром актинио-
эритрином (2.14).
У многих морских беспозвоночных преобладающий кароти-
ноид присутствует не в свободной форме, а в виде стехиомет-
рического комплекса с белком (разд. 2.5).
Каротиноиды и каротинопротеины чаще всего содержатся в
эпидермисе или в панцире беспозвоночных животных, а также
(иногда в высоких концентрациях) в репродуктивных органах
и яйцах, хотя значение их присутствия в последних остается
пока неизвестным.
2.5. Каротинопротеины
У морских беспозвоночных кетокаротиноиды обычно
присутствуют в виде стабильных водорастворимых каротино-
протеиновых комплексов. В этих комплексах каротиноиды в
стехиометрических соотношениях связаны с белками — липо-
протеинами или гликопротеинами. В некоторых случаях каро-
тинопротеины могут прочно связываться со структурным мате-
риалом наружных покровов тела, хитином или карбонатом
кальция. Некоторые каротинопротеины имеют сложную субъ-
единичную структуру, примером чему служит пигмент панциря
омара крустацианин, в то время как другие, в частности фио-
летовый каротинопротеин морской звезды Asterias rubens, зна-
чительно более просты.
Образование каротинопротеинового комплекса обычно при-
водит к значительному батохромному сдвигу в спектре погло-
щения (рис. 2.12), и потому эти комплексы часто имеют пур-
пурный, голубой или зеленый цвет в отличие от желтой или
оранжевой окраски свободных каротиноидов. По-видимому,
важную роль в спектральном сдвиге играют кетогруппы каро-
тиноидных простетических групп, обычно представленных кан-
таксантином или астаксантином, однако природа каротинопро-
теинового связывания пока остается неясной. Показано лишь,
что во взаимодействии каротиноидов с белком не участвуют
ковалентные связи, поскольку свободный каротиноид легко вы-
свобождается при денатурации белка нагреванием или органи-
ческими растворителями, причем иногда этот процесс обратим.
Вероятно, наиболее известным примером окраски, обу-
словленной каротинопротеиновым комплексом, является омар
Homarus vulgaris. Его панцирь имеет серо-голубой цвет благо-
даря присутствию каротинопротеина крустацианина. При варке
60
Часть I. Химические и биохимические аспекты
каротинопротеин денатурирует и выделяется свободный каро-
тиноид астаксантин, а также продукт его аутоокисления аста-
цин. Поэтому вареный омар выглядит красным.
Хотя каротинопротеиновые комплексы легко денатурируют
при избытке тепла или органических растворителей, в водных
растворах они значительно более стабильны по отношению к
свету или кислороду, чем свободные каротиноиды.
2.6. Биосинтез
2.6.1. Введение
Было показано, что каротиноиды синтезируются толь-
ко в растениях и микроорганизмах. В организме многих живот-
ных могут накапливаться и даже модифицироваться каротино-
иды, полученные с пищей (разд. 2.8), однако биосинтез каро-
тиноидов de novo никогда не был продемонстрирован однозначно
ни в одной из животных систем. Биосинтетический путь, ко-
торый будет обсуждаться ниже, представляет собой некое обоб-
щение, основанное на исследованиях многих каротиногенных
систем, главным образом листьев и хлоропластов, плодов и
хромопластов томата, грибов и бактерий, а также полученных
из них ферментных препаратов.
Каротиноиды являются тетратерпенами, и их биосинтез
происходит по нормальному изопреноидному пути, дающему
начало также и другим важным природным продуктам, таким,
как каучук, стероиды, содержащиеся во многих эфирных мас-
лах моно-, сескви- и дитерпены, а кроме того, боковым цепям
хинонов, участвующих в переносе электронов.
Путь биосинтеза каротиноидов может быть разбит на не-
сколько стадий: 1) образование Сго-промежуточного продукта
геранилгеранилпирофосфата; 2) образование фитоина — пер-
вого Сад-каротипа; 3) ряд реакций десатурации; 4) циклизация
и связанные с ней реакции с участием двойной связи С-1,2;
5) окончательные модификации.
Первая стадия образования геранилгеранилпирофосфата
является общей для биосинтеза всех изопреноидов, в то время
как остальные характерны только для биосинтеза каротинои-
дов.
2.6.2. Образование геранилгеранилпирофосфата
Большинство экспериментальных данных, касающихся
начальных стадий изопреноидного пути биосинтеза, получены
на примере образования холестерина, а не каротиноидов. Од-
нако нет оснований считать, что составляющие его реакции для
этих классов соединений различаются.
СН3. СО. S. СоА СН3 ОН
к ~ООС 'С' г
2ХCH, . СО.SCoA----ч—*-CH3.CO.CH2CO.SCoA——sr-----•- \ s' X
'3 Л Л СН2 сн2
CoASH CoASH /9 ел\ гляг с' л
(2.49) Ацетоацетил-СоА (2.50) ГМГ-СоА
2 NADPH
/1
®—Аденозин
(2.51) Мевалоновая кислота (МВК|;
(2.53) МВК-5-пирофосфат (2.52) МВК-5-фосфат
| Ангидродекарбоксилаза
СН3
I z-x х-х ИППФ-изомераза
сн2о-$ЧЭ ...............
СН2 V/C\
н н
(2.54) Изопентенилпирофосфат (ИППФ)
ГМ Г-СоА-редуктаза.
(2.55) Диметилаллилпирофосфат
(ДМАПФ)
(2.58) Гераниягеранилпирофосфат (ГГПФ)
Рис. 2.13. Образование геранилгеранилпирофосфата (ГГПФ) по основному
пути биосинтеза изопреноидов из ацетил-СоА.
<62
Часть I. Химические и биохимические аспекты
Первым общим предшественником изопреноидов является
.ацетат в виде ацетил-СоА. Биосинтетический путь от ацетил-
'СоА до геранилгеранилпирофосфата (ГГПФ) представлен на
рис. 2.13. Ацетил-СоА (3 молекулы) превращается через ацето-
.ацетил-СоА (2.49) в З-гидрокси-З-метилглутарил-СоА [ГМГ-
СоА (2.50)]. Данные об участии в этом пути в качестве проме-
жуточного продукта малонил-СоА, как в случае биосинтеза
жирных кислот, противоречивы.
ГМГ-СоА подвергается восстановлению в две стадии до ме-
валоновой кислоты {МВД (2.51)]. Эта ГМГ-СоА-редуктазная
^реакция является главной контрольной точкой биосинтеза хо-
лестерина; справедливо ли это для каротиногенеза, еще пред-
стоит выяснить. МВК представляет собой первое соединение,
которое в биосинтезе изопреноидов используется исключитель-
но в качестве промежуточного продукта. Это соединение, по-
разному меченное радиоактивными (3Н и 14С) и стабильными
(2Н и 13С) изотопами, было синтезировано и широко применя-
лось в качестве субстрата при изучении биосинтеза каротинои-
дов.
На следующих стадиях МВК дважды подвергается фосфо-
рилированию с образованием 5-фосфата (2.52) и 5-пирофосфа-
та мевалоновой кислоты (2.53) при участии киназ и АТР. За-
тем МВК-5-пирофосфат декарбоксилируется с образованием
-«изопреновой единицы» — изопентенилпирофосфата [ИППФ
(2.54)]. Изомераза катализирует обратимую изомеризацию
ИППФ и диметилаллилпирофосфата [ДМАПФ (2.55)]. Эти две
молекулы являются первыми субстратами пренилтрансфераз-
ных ферментов, которые катализируют образование изопрено-
идных цепей. ДМАПФ функционирует в качестве «затравоч-
ной» молекулы, которая конденсируется с молекулой ИППФ с
«образованием Сю-промежуточного продукта геранилпирофосфа-
та [ГПФ (2.56)], являющегося предшественником монотерпе-
нов. Последовательное присоединение еще двух молекул ИППФ
.дает Cis-фарнезилпирофосфат [ФПФ (2.57)]—предшествен-
ник сесквитерпенов, стероидов и тритерпенов, а также
С20-ГТПФ (2.58). Процесс удлинения цепи может продолжать-
ся с образованием длинноцепочечных полипренолов, или же
ГГПФ может использоваться для синтеза С2о-дитерпенов
(в том числе фитола — боковой цепи хлорофилла) или Сад-ка-
ротиноидов.
2.6.3. Образование фитоина
В первой стадии биосинтетического процесса, которая
.свойственна исключительно каротиноидам, принимают участие
щве молекулы ГГПФ и образуется первый С40-каротиноидный
Рис. 2-14. Возможный механизм образования фитоииа.
64
Часть I. Химические и биохимические аспекты
промежуточный продукт. Этим промежуточным продуктом яв-
ляется фитоин (2.61), а не ликоперсин (7,8,11,12,15,7',8',11',12',
15'-декагидро-ф,ф-каротин, 2.62)—Сад-аналог стероидного
предшественника сквалена, как предполагалось первоначально.
В биосинтезе фитоина участвует Сад-циклопропановый проме-
жуточный продукт — префитоинпирофосфат [ПФПФ (2.59)].
Возможные механизмы образования ПФПФ из ГГПФ и фитои-
на из ПФПФ приведены на рис. 2.14. Последний промежуточ-
ный продукт в этой цепи — ион карбония (2.60) — стабилизи-
руется в результате потери протона, что приводит к образова-
нию фитоина. Последнее более вероятно, чем формальное при-
соединение Н_ от NADPH, которое должно было бы привести
к образованию ликоперсина.
Биосинтез фитоина из МВК, ИППФ, ГГПФ и ПФПФ был
продемонстрирован с помощью многочисленных неочищенных
ферментных препаратов, выделенных, например, из хлоропла-
стов, хромопластов томата, грибов и бактерий.
Сзо-Тритерпеновые каротиноиды, характерные для некото-
рых бактерий (разд. 2.4.4), вероятно, синтезируются аналогич-
ным путем из ФПФ и Сзо-прескваленпирофосфата.
Стереохимия. Фитоин, образуемый большинством каротино-
генных систем, в том числе высшими растениями, по-видимому,
является 15-гщс-изомером (2.15). Образование этого изомера
сопровождается потерей 1 -про-S-водородного атома в каждой
молекуле ГГПФ, а у некоторых бактерий полностью транс-фи-
тоин (2.61) синтезируется непосредственно в результате потери
1-npo-S-водородного атома у одной молекулы ГГПФ и 1-npo-R-
водородного атома у другой (рис. 2.15).
2.6.4. Десатурация
При образовании окрашенных каротиноидов из фитои-
на прежде всего происходят четыре последовательные десату-
рации, каждая из которых приводит к введению двойной связи
и дальнейшему удлинению полиенового хромофора на две со-
пряженные двойные связи (рис. 2.16). Промежуточными про-
дуктами в этой последовательности реакций являются фито-
•флуин (2.63), ^-каротин (2.64) и нейроспорин (2.66), а конеч-
ным продуктом — ликопин (2.67). У многих микроорганизмов
^-каротин целиком или частично замещен его несимметричным
изомером 7,8,11,12-тетрагидро-ф,ф-каротином (2.65). Превра-
щение в ликопин и другие каротиноиды меченых фитоина, фи-
тофлуина и ^-каротина было достигнуто с помощью бесклеточ-
ных препаратов из пластид томата и из бактерий.
У многих микроорганизмов процесс десатурации ингибиру-
ются дифениламином, а у высших растений — некоторыми гер-
СП
I
32
Рис. 2.15. Стереохимия отщепления водорода при биосинтезе полностью транс- и 15-^ис-фитоииа.
(2.65) 7, 8, 11, 12 -Тетрагидро-0,« 0-каротин
2Н^
(2.67) Ликопин
Рис. 2.16. Ступенчатая десатурация фитоина до ликопина..
Гл. 2. Каротиноиды
G7
бицидами; при этом происходит накопление фитоина. При по-
следующем удалении ингибитора происходит образование бо-
лее ненасыщенных каротиноидов, по-видимому, за счет накопив-
шегося фитоина.
Почти все природные каротиноиды имеют полностью транс-
конфигурацию, и, следовательно, в тканях, где образуется
15-1{МС-фитоин, на одной из стадий процесса десатурации долж-
на происходить изомеризация. Показано, что эта изомеризация
в различных системах может происходить на разных стадиях.
Предполагают, что у зеленых водорослей имеет место изомери-
Рис. 2.17. Стереохимия потери водорода в реакции десатурации. Символы
Нав, Has, H5R и H5S указывают, что эти водородные атомы происходят из
2-npo-R-, 2-npo-S-, 5-npo-R- и 5-npo-S-водородных атомов мевалоната соот-
ветственно.
зация ^-каротина, у томатов — фитофлуина и у некоторых бак-
терий и грибов — фитоина. У бактерий, синтезирующих только
транс-фитоин, обнаружены только транс-изомеры каротинои-
дов.
Механизм реакций десатурации еще не выяснен, однако экс-
перименты со стереоспецифически меченой 3Н при С-2 или С-5
МВК свидетельствуют о том, что введение каждой двойной свя-
зи происходит путем транс-элиминирования водорода (рис.
2.17). Были представлены доказательства, согласно которым
необходимыми кофакторами этого процесса в пластидах выс-
ших растений служат NADP+ и FAD. У бактерий предполага-
ют участие в нем системы переноса электронов.
2.6.5. Последующие реакции; общие соображения
Последовательность реакций десатурации отнюдь не
приводит к образованию полностью сопряженного пентадекае-
на ЗДЗ'Д'-тетрагидро-ф.ф-каротина (2,68). Она останавлива-
ется на стадии ликопина, в котором остается насыщенной
связь С-3,4. Двойные связи С-1,2 остаются изолированными и
не участвуют в формировании главного полиенового хромофо-
ра. Однако у большинства каротиногенных систем ликопин не
является конечным продуктом, а служит лишь промежуточным
звеном в биосинтезе главных нормально образующихся каро-
5*
68
Часть 1. Химические и биохимические аспекты
тиноидов. В частности, ликопин может подвергаться различным
модификациям по изолированной связи С-1,2 с образованием •
как ряда характерных для фотосинтезирующих бактерий ацик-
(2.68) 3, 4, 4-Тетрагидроликопин
(2.69) Родопин
лических каротиноидов, так и многих более широко известных
моноциклических и бициклических каротиноидов, типичных для
растений.
2.6.6. Последующие реакции; биосинтез ациклических
каротиноидов
Из ряда дополнительных реакций, которые могут про-
исходить по двойной связи С-1,2, простейшие наблюдаются в
ходе биосинтеза ациклических каротиноидов. Наиболее ярким
примером является присоединение воды с образованием 1-гидр-
окси- и 1-метоксикаротиноидов, характерных для фотосинтези-
рующих и некоторых других бактерий. В случае гидратации
ликопина по двойной связи С-1,2 образуется родопин ф1,2-ди-
Рис. 2.18. Механизм образования 1-гидрокси-1,2-дигидрокаротиноидов.
гидро-ф,ф-каротин-1-ол (2.69) ]—главный каротиноид пур-
пурной бактерии Rhodomicrobium vanniellii. Возможный меха-
низм этой реакции приведен на рис. 2.18.
У фотосинтезирующих бактерий за такой гидратацией обыч-.
но следует метилирование (с помощью S-аденозилметионина)
третичной гидроксильной группы и десатурация связи С-3,4.
Гл. 2. Каротиноиды
69
Предложенные к настоящему времени схемы биосинтеза сферо-
идина '[ 1-метокси-3,4-дидегидро-1,2,7',8'-тетрагидро-ф,ф-кароти-
на (2.70)] и спириллоксантина (2.43) —главных каротиноидов
Rhodopseudomonas sphaeroides и Rhodospirillutn rubrum. — соот-
ветственно представлены на рис. 2.19.
Однако С-1-гидроксилирование и О-метилирование могут
происходить на более ранних стадиях десатурации; у этих бак-
терий при определенных условиях культивирования были обна-
ружены гидрокси- и метоксипроизводные фитоина, фитофлуина
и 1,2,7,8-тетрагидро-ф,ф-каротина.
Описанные биосинтетические реакции протекают в анаэроб-
ных условиях. Поступление О2 в культуру R. sphaeroides вы-
зывает быстрое превращение желтого сфероидина в его крас-
ное кетопроизводное сфероиденон [1-метокси-3,4-дидегидро-
1,2,7',8'-тетрагидро-ф,ф-каротин-2-он (2.71)] (рис. 2.20).
Галобактерии (Halobacteria), приспособившиеся к жизни в
условиях сильного засоления, содержат ациклические С45- и
Cso-каротиноиды, такие, как бактериоруберин (2.12). Предпо-
лагают, что они образуются в реакции, аналогичной гидратации
по связи С-1,2 (рис. 2.18), с участием инициирующих электро-
фильных агентов — С5-фрагментов (рис. 2.21).
2.6.7. Циклизация
Жесткость сопряженной полиеновой системы кароти-
ноидов предотвращает интенсивную циклизацию по типу, ко-
торый имеет место у ди- и тритерпеноидов. Циклизация у ка-
ротиноидов ограничивается образованием единственного шести-
членного кольца на одном или на обоих концах молекулы ацик-
лического предшественника. Циклизация каротиноидных про-
межуточных продуктов может рассматриваться как дополни-
тельный процесс, начинающийся протонной атакой по С-2 у
концевой двойной связи С-1,2. Далее, как показано на рис. 2.22,
образуется ион карбония (2.72), который может стабилизиро-
ваться путем потери протона у С-6, С-4 или С-18; при этом
формируются соответственно р-кольцо (2.73), е-кольцо (2.74)
или в редких случаях у-кольцо (2.75). Кольца разных типов не
способны превращаться друг в друга.
Общая схема биосинтеза обычных каротиноидов, содержа-
щих р- и е-кольца, приведена на рис. 2.23. Согласно этой схеме,
существуют две главные точки, в которых может происходить
циклизация. Если в норме десатурация завершается до начала
циклизации, то непосредственным предшественником моноцик-
лических у-каротина (2.78) и 6-каротина [е,ф-каротина (2.79)],
а следовательно, и дициклических р-каротина (2.80), а-кароти-
на (2.81) и е-каротина (2.82) является ликопин. Если же цик-
•Спириллоксантин (2.43)
Гл. 2. Каротиноиды
71
лизация происходит до окончания десатурации, то ключевыми
промежуточными продуктами являются нейроспорин, р- и
а-зеакаротин [Т'.в'-дигидро-б,^-каротин (2.76) и 7',8'-дигидро-
е,ф-каротин (2.77)]. В любом случае циклизация происходит в
Сфероидин
(2.71) Сфероиденон
Рис. 2.20. Аэробное превращение сфероидина в сфероиденон.
каротиноидной «полумолекуле», которая достигла уровня нена-
сыщенности ликопина; концевые группы с одинарной связью
С-7,8 циклизоваться не могут.
Рнс. 2.21. Биосинтез ациклических С45- и Сзо-каротинондов.
Многие превращения, показанные на рис. 2.23, были проде-
монстрированы в опытах с ферментными системами из бакте-
рий и высших растений (из пластид томатов), причем у бакте-
Рис. 2.19. Предполагаемые схемы биосинтеза сфероидина и спириллоксан-
тииа у фотосинтезирующих бактерий.
Рис. 2.22. Общий механизм циклизации каротиноидов, приводящий к обра-
зованию трех типов колец — 0, у и е.
рии это удалось после исключения из реакционной смеси инги-
битора циклизации — никотина.
Стереохимия. Эксперименты с применением стабильных изо-
топов внесли ясность в стереохимию сгибания молекулы, Н+-
атаки и циклизации при формировании 0-кольца (рис. 2.24).
Несмотря на то что установлены хиральность при С-6, а также
стереохимия водородной атаки при С-2 и потери водорода при
С-4 (рис. 2.25), стереохимия образования е-кольца все еще ос-
тается не ясной. Полученные к настоящему времени результаты
свидетельствуют о том, что при биосинтезе 0- и е-колец стерео-
химия сгибания молекулы может фундаментально различаться.
Циклические С50-каротиноиды. У некоторых нефотосинтезиру-
ющих бактерий циклизация инициируется электрофильными
Сб-фрагментами (рис. 2.26), что приводит к образованию С45-
и Сбо-каротиноидов, имеющих при атоме С-2 0-, g- или у-кольца
Сб-заместители, например «К. п. 450» (2.41), декапреноксанти-
на (2.11) и сарцинаксантина (2.40).
Стереохимия циклизации при образовании этих «высших
каротиноидов» отличается от установленной для С4о-соеди-
нений.
/•Зеакаротин (2.76)
ct-Зеакаротин (2.77)
5-Каротин (2.79)
Рис. 2.23. Общая схема биосинтеза обычных каротиноидов с |3- и е-кольцами.
(3-Каротин (2.80)
а-КаротиН (2.81)
оКаротин (2.82)
74
Часть I. Химические и биохимические аспекты
Н Н
Рис. 2.25. Известные к настоящему времени детали стереохимии образова-
ния е-кольца.
2.6.8. Окончательные модификации
В предыдущих разделах описаны главные реакции, по-
средством которых синтезируются основные структуры ацикли-
ческих и циклических каротиноидов. Индивидуальные кароти-
ноиды образуются в результате последующих модификаций. Не-
которые из этих модификаций, и прежде всего включение кис-
лородсодержащих функциональных групп, происходят повсе-
местно. Другие, по-видимому, уникальны и характерны для
биосинтеза единственного каротиноида, встречающегося лишь
у одного вида или у группы видов. Диапазон структурных мо-
дификаций циклических каротиноидов шире, чем у соединений
ациклического ряда. Главные структурные модификации ацик-
лических каротиноидов, в частности у фотосинтезирующих бак-
терий, были приведены ранее (разд. 2.6.6).
У циклических каротиноидов чаще всего в качестве допол-
нительной группы встречается ОН-группа. Особенно часто гидр-
оксилирование имеет место при С-3, однако встречаются также
и 2-гидрокси- и 4-гидроксикаротиноиды. Последние, как прави-
ло, окисляются далее до 4-кетокаротиноидов, таких, как кан-
таксантин (2.27) (рис. 2.27). Гидроксильные группы иногда на-
ходятся в других положениях молекулы, например при С-19.
Из процессов гидроксилирования изучено только введение
гидроксила в положение С-3. У растений и бактерий (3/?,3'/?)-
зеаксантин (р^-каротин-ЗД'-диол (2.83)] образуется путем
76
Часть I Химические и биохимические аспект и
(2 27) Кантаксантин
Рис 2 27 Образование кетока-
ротиноида кантаксантина.
Рис 2 28. Стереохимия введения 3-гидроксигруппы зеаксантина. Н5Я и H5s
были исходно 5-npo-R- и 5-иро-5-водородными атомами мевалоната.
гидроксилирования p-каротина. Источником ОН-группы служит
молекулярный О2, и ее присоединение катализируется оксида-
зой со смешанной функцией (стереохимия реакции показана на
рис. 2.28). Лютеин (2.6)—главный ксантофилл листьев — ве-
роятно, образуется аналогичным образом из а-каротина. Хи-
ральность при С-З' в данном случае противоположна хирально-
сти при С-З, а также хиральности у зеаксантина, и потому оче-
видно, что стереохимия гидроксилирования у них различна.
Хлоропластные ксантофиллы виолаксантин (2.8) и неоксан-
тин (2.32) содержат 5,6-эпоксигруппы. Описано ферментатив-
Гл. 2. Каротиноиды
77
ное эпоксидирование зеаксантина до виолаксантина. Эпоксиды
[или близкие к ним перекисные структуры (2.84)] могут быть
важными промежуточными продуктами при различных модифи-
кациях каротиноидов. На рис. 2.29 показаны возможные меха-
низмы образования: а) алленовой концевой группы неоксанти-
на, б) ацетиленовых концевых групп, таких, как у каротинои-
да водорослей аллоксантина [7,8,7',8'-тетрадегидро-р,р-каротин-
3,3'-диола (2.85)], в) циклопентанового кольца капсантина
(2.35) и капсорубина (2.36) и г) ретро-каротиноида эшшольц-
ксантина (2.34). Хотя все эти схемы выглядят очень правдопо-
добными, практически нет биохимических доказательств, кото-
рые бы их подтверждали.
2.6.9. Биосинтез арилкаротиноидов
Обнаружение у бактерий нескольких каротиноидов, со-
держащих арильные концевые группы, представляет большой
интерес, поскольку это свидетельствует в пользу существования
еще одного, ранее неизвестного, пути биосинтеза ароматичес-
кого кольца из мевалоната, а не по шикиматному пути или из
ацетата с помощью поликетидного механизма (гл. 3).
Образование 1,2,5-триметилфенильной концевой группы, та-
кой, как у хлоробактина (2.45), включает простую миграцию
одной из метильных групп при С-1 в положение С-2. Гораздо
более сложная перегруппировка необходима для биосинтеза
1,2,3-триметилфенильной концевой группы, содержащейся в
океноне (2.46), Несмотря на то что пока не получено подтверж-
дающих доказательств, предполагают, что в такой перегруппи-
ровке участвует промежуточный продукт, имеющий структуру
призмы Ладенбурга (рис. 2.30).
78
Часть I. Химические и биохимические аспекты
2.7. Регуляция и контроль биосинтеза каротиноидов н
2.7.1. Грибы и бактерии
Генетический контроль. При изучении биосинтеза ка-
ротиноидов широко использовались мутанты плесневого гриба
Phycomyces blakesleeanus. Существуют три главные группы му-
тантов — мутанты, накапливающие ликопин; мутанты, накап-
ливающие фитоин; и мутанты, не способные синтезировать ка-
ротиноиды. Обширные комплементационные исследования по-
казали, что в нормальном каротиногенезе участвуют только три
гена, обозначаемые как car R, саг В и саг А, и что каждая из
трех групп мутантов соответствует мутации в одном из этих
генов. Циклизация (ликопина?) осуществляется продуктом ге-
на car R; две копии этого продукта, т. е. два циклазных фер-
мента в едином ферментном комплексе, участвуют в образова-
нии p-каротина. Аналогичным образом четыре копии продукта
гена саг В функционируют в дегидрогеназном комплексе, ко-
торый осуществляет четыре последовательные реакции десату-
рации, необходимые для превращения фитоина в ликопин.
Предполагают, что биосинтез в целом происходит на мульти-
ферментном комплексе, включающем десатуразы и циклазы.
Свет. Свет стимулирует дополнительный синтез каротинои-
дов у многих грибов и бактерий, которые в темноте обычно об-
разуют умеренное их количество. У многих других грибов и
бактерий, однако, синтез каротиноидов в темноте происходит в
очень ограниченной степени или не происходит вовсе, но может
начаться в ответ на одновременное кратковременное воздейст-
вие света и кислорода. После освещения перед началом каро-
тиногенеза обычно наблюдается индукционный период (около
4 ч), необходимый для синтеза ферментов. Подобный механизм
Рис. 2.29. Возможные механизмы образования различных структур ксаитО'
ф ил лов из 5,6-эпоксида или 5,6-перекиси (начало см, на с, 78),
80
Часть I Химические и биохимические аспекты
фотовозбуждения специфического биосинтеза обеспечивает на-
личие каротиноидов только тогда, когда они необходимы для
защиты организма от вредного действия избытка света и кис-
лорода.
В настоящее время выяснено, на какую стадию образова-
ния каротиноидов влияет свет. У Mycobacterium при освещении
усиливается образование ГГПФ, а образование ПФПФ целиком
зависит от освещения. У гриба Neurospora crassa ферменты,
катализирующие как образование, так и десатурацию фитоина,
по-видимому, являются фотоиндуцируемыми.
Рис. 2.30. Предполагаемый механизм биосинтеза 1,2,3-триметилфенильной
концевой группы
У фотосинтезирующих бактерий свет необходим как для
синтеза каротиноидов, так и для образования других компонен-
тов фотосинтетических мембран (гл. 10).
Условия культивирования. У многих грибов и бактерий ко-
личественный и качественный состав каротиноидов изменяется
при изменении условий культивирования. На количество и со-
став образующихся каротиноидов сильно влияют природа ис-
точников углерода и азота, отношение углерод/азот, доступ-
ность минеральных солей, витаминов и ростовых факторов, сте-
пень аэрации, pH среды и температура.
Химический контроль. Известно, что многие вещества сти-
мулируют или подавляют синтез каротиноидов или вызывают
качественные изменения в их составе у микроорганизмов. Не-
которые из таких соединений, например дифениламин и нико-
тин, широко использовались в исследованиях биосинтеза каро-
тиноидов (разд. 2.6.4 и 2.6.5).
Гл 2 Каротиноиды
84
Интересный пример химического контроля каротиногенеза
в природных биологических системах обнаружен у некоторых
гетероталлических грибов, в частности у Blakeslea trispora.
При раздельном выращивании ( + )- и (—)-штаммы не образу-
ют каротиноиды в заметных количествах, тогда как в смешан-
ных культурах происходит интенсивный синтез р-каротина.
Этот синтез индуцируется триспоровой кислотой (2.86) — гор-
0
(2 86) Триспор я I кислота
моном, который образуется под действием комбинации фермен-
тов из двух штаммов. Триспоровая кислота представляет собой
метаболит p-каротина и ее главная функция заключается в
стимулировании споруляции и репродукции. Не исключено, что
стимуляция каротиногенеза является частью механизма, обес-
печивающего усиление образования триспоровой кислоты.
2.7.2. Растения и водоросли
Генетический контроль. Из генетических доказательств
следует, что в зеленых тканях высших растений и у водорослей
реакции десатурации и циклизации находятся под непосредст-
венным ядерным контролем. Так, были выделены мутанты ку-
курузы (Zea mays) и зеленой водоросли Scenedesmus obliquus,
накапливающие фитоин, ((-каротин или ликопин.
Наиболее важные генетические исследования проведены на
томатах. Были сделаны некоторые предположения относитель-
но участков действия генов, которые контролируют биосинтез
каротиноидов в плодах томата. Некоторые гены действуют до
стадии образования фитоина и контролируют количество пред-
шественников, вступающих в процесс биосинтеза каротинои-
дов. Другие гены контролируют соответственно десатурацию
фитоина, циклизацию ликопина до p-каротина, приводящую к
образованию е-, а не p-кольца, или стереохимию конечного про-
дукта. Так, «теневые» (ghost) мутанты томата имеют «белые»
плоды, содержащие в большом количестве фитоин; плоды ли-
нии «высокий р» («high-р») накапливают в высоких концентра-
ьиях p-каротин вместо накапливаемого в нормальных условиях
ликопина; дельта-штамм в качестве главного пигмента содер-
жит 6-каротин; а у плодов томата сорта «танжерин» вместо
6—84
Часть I. Химические и биохимические аспекты
«2
---—-----—--------------------------------------- /-------
полностью трамс-ликопина в качестве главного карбтиноида
присутствует проликопин.
Синтез каротиноидов в хлоропластах. В фотосинтезирующих
тканях каротиноиды локализованы в хлоропластах; весьма ве-
роятно, что они и синтезируются в этих органеллах. Этиолиро-
ванные проростки и выращенные в темноте культуры Euglena
gracilis синтезировали лишь небольшие количества каротинои-
дов, главным образом ксантофиллов. В ответ на кратковремен-
ное освещение нормальные хлоропластные каротиноиды синте-
зируются по мере образования функционирующих хлороплас-
тов. Полагают, что действие света опосредовано фитохромом.
Каротиноиды представляют собой неотъемлемую часть самой
•структуры хлоропластов, и регуляция их синтеза тесно взаи-
мосвязана с синтезом хлорофилла и других компонентов хлоро-
пластов (гл. 10).
Многие водоросли образуют хлоропласты, а следовательно,
и нормальные хлоропластные каротиноиды даже при росте в
темноте.
Синтез каротиноидов в созревающих плодах. У многих пло-
дов созревание сопровождается интенсивным синтезом кароти-
ноидов по мере замены хлоропластов на хромопласты и изме-
нения окраски с зеленой на красную. Образующиеся в ходе со-
зревания плодов или уже-после их сбора каротиноиды часто
отсутствуют в исходных хлоропластах, например ликопин в то-
матах и капсантин в перце. Свет обычно не оказывает сущест-
венного влияния на процессы созревания или накопления ка-
ротиноидов, тогда как температура является важным факто-
ром этих процессов.
Химический контроль. Известно, что многие вещества сти-
мулируют или ингибируют синтез каротиноидов. В их число
входят некоторые гербициды, например Sandoz 6706 [4-хлор-
5-(диметиламино) - 2(а,а,а-трифтор-л-толил) -3(2Н) - пиридази-
нон], который может ингибировать синтез каротиноидов (бло-
кируя десатурацию фитоина) и таким образом предотвращать
правильное развитие хлоропластов.
2.8. Метаболизм каротиноидов у животных
2.8.1. Интактные каротиноиды
Хотя многие животные используют каротиноиды для
окраски, полагают, что они не способны синтезировать эти со-
единения и получают их в необходимых количествах с пищей.
Например, фламинго, живущие в зоопарке, теряют свою харак-
терную привлекательную розовую окраску при недостатке ке-
токаротиноидов, которые в природных условиях они получают,
Гл. 2. Каротиноиды
питаясь \ крошечными рачками. Многие животные (птицы,
а также Насекомые и другие беспозвоночные) могут, однако, мо-
дифицировать структуру поступающих с пищей каротиноидов,
в частности путем введения кетогрупп в положение С-4. Было
показано, что некоторые животные — главным образом птицы
и беспозвоночные — превращают р-каротин (2.3) и зеаксантин
(2.7) в кантаксантин (2.27) и астаксантин (2.9) соответственно^
Рис. 2.31. Образование нор-каротиноида актиниоэритрииа.
Интересной и необычной модификацией является образова-
ние у актиний «ор-каротиноида актиниоэритрииа (2.14) из:
сложных эфиров астаксантина с помощью процесса, включаю-
щего превращение шестичленного кольца в пятичленное. Пред-
ложенный механизм (рис. 2.31) включает бензильнокислотную
перегруппировку промежуточного трикетосоединения.
У ряда животных были обнаружены каротиноиды, которые-
не могли присутствовать в пище или образовываться из при-
сутствующих в пище с помощью какого-либо известного мета-
болического процесса. Например, божья коровка Coccinella
septempunctata содержит необычные каротиноиды с у-кольцом,
которые, вероятно, образуются с помощью микробных симби-
онтов. Происхождение (6S, 6'S) -е-каротина, присутствующего в:
окрашенных жировых каплях сетчатки некоторых птиц (гл. 9),.
остается загадкой.
2.8.2. Образование ретинальдегида
и зрительных пигментов
У многих животных, в том числе у человека, наиболее-
важными продуктами метаболизма каротиноидов являются ви-
тамины А [ретинол (2.87) и 3,4-дидегидроретинол (2.88) и со-
6*
84
Часть I Химические и биохимические аспекты.
ответствующие альдегиды — ретинальдегид (2.89) и 3,4-диде-
гидроретинальдегид (2.90)]. Ретинальдегид (ретиналь, рети-
нин) образуется в слизистой кишечника в результате окисли-
тельного расщепления на две половины молекулы р-каротина
(рис. 2.32). Этот процесс катализируется р-каротин-15,^'-ди-
оксигеназой и идет через образование промежуточного перекис-
ного соединения (2.91). Ретинальдегид и ретинол легко пре-
вращаются друг в друга в присутствии NAD(H) или NADP(H)
алкогольдегидрогеназами, содержащимися в различных тканях,
(2 89) Ретинальдегид (2 90) 3 4 Дидегидроретинальдегид
главным образом (у млекопитающих) в печени и в сетчатке
глаза. В крови витамин А транспортируется в виде комплекса
с липопротеином (ретинолсвязывающим белком), а запасы эфи-
ров ретинола хранятся в печени.
Процесс зрения зависит от группы фоточувствительных пиг-
ментов, которые локализованы в сетчатке глаза. Эти зритель-
ные пигменты представляют собой комплексы опсинов (глико-
липопротеинов) и 11-1|пс-ретинальдегида или 11-цпс-дегидро-
ретинальдегида. Более детально эти комплексы и их функции
в процессе зрения обсуждаются в гл. 9.
2.9. Функции каротиноидов
Основные функции каротиноидов в биологических тка-
нях обусловлены главным образом их способностью поглощать
свет. Роль каротиноидов в фотосинтезе, защите от вредного
действия света, фототропизме и фоторецепции, а также их
вклад в окраску содержащих их тканей обсуждаются во вто-
рой части данной книги.
Других функций у каротиноидов не обнаружено. Содержа-
ние каротиноидов в высоких концентрациях в репродуктивных
Гл 2 Каротиноиды
85
тканях грибов, водорослей, растений и животных указывает на
возможную роль и участие этих соединений в процессах раз-
множения, однако четкие данные по этому поводу отсутствуют.
2.10. Каротиноиды как пищевые красители
Уже в течение многих лет каротиноиды используют в
качестве красителей в пищевой промышленности. Их присут-
ствие во многих природных пищевых продуктах делает их иде-
Рис 2 32 Образование ретинальдегида путем окислительного расщепления
Р-каротина.
ально пригодными для этой цели. В пищевые жиры, особенно
в маргарин, добавляют p-каротин, благодаря чему организм
получает дополнительное количество необходимого для него
витамина А. Водорастворимые или по крайней мере дисперги-
руемые в воде производные p-каротина, кантаксантин и апо-
каротиноиды, применяются для окрашивания напитков и дру-
гих пищевых продуктов. Если число каротиноидов, которые мо-
гут быть использованы, и особенно диапазон красок, который
может быть получен, увеличатся, то применение каротиноидов
в пищевой промышленности станет еще более широким.
86
Часть I Химические и биохимические аспекты
2.11. Использование каротиноидов в медицине
В медицине каротиноиды используются главным обра-
зом для профилактики или лечения авитаминоза А. В этих слу-
чаях рекомендуется пищевой рацион, богатый каротиноидами,
или специальные препараты. Вместе с тем, как недавно пока-
зано, при некоторых кожных заболеваниях, которые обостря-
ются под действием солнечного излучения, каротиноиды игра-
ют защитную роль. В частности, большие дозы p-каротина зна-
чительно смя! чают симптомы эритропоэтической порфирии.
Для этой болезни характерно нарушение метаболизма порфи-
ринов, в результате чего они накапливаются в организме и на-
чинают играть роль фотосенсибилизаторов. У таких больных
под влиянием солнечного света появляется зуд, жжение и отеч-
ность. В настоящее время рассматривается также возможность
использования каротиноидов, главным образом p-каротина и
кантаксантина, а также ретиноидов (производных витамина А)
для защиты от некоторых видов рака кожи, вызываемого дей-
ствием УФ-излучения или химических веществ.
2.12. Другие полиеновые пигменты
Светопоглощающие свойства каротиноидов обусловле-
ны главным образом наличием в них сопряженной полиеновой
системы. Поэтому не удивительно, что спектры поглощения со-
пряженных полиенов некаротиноидной природы очень сходны
(2 92) Кортизалин
соон
(2 93) Валлемиг. А
со спектрами собственно каротиноидов, причем настолько
сходны, что те немногие сопряженные некаротиноидные полие-
ны, которые сейчас известны, считали сначала каротиноидами.
Все некаротиноидные полиеновые пигменты имеют грибное
или бактериальное происхождение. Плодовые тела гриба Corti-
ciutn salicinum содержат красный пигмент кортизалин (2.92),
а другой гриб, Wallemia sebi, обычно обнаруживаемый на внут-
Гл. 2. Каротиноиды
87
ренней стороне пчелиных ульев, содержит пиррольные полиены,
например валлемию А (2.93). Из некоторых бактерий были по-
лучены арильные полиены, причем оказалось, что они присутст-
вуют в них наряду с каротиноидами. Скользящие бактерии ти-
па Cytophaga— Flexibacterium содержат флексирубины —
сложные эфиры со структурой, представленной формулой
(2.94), и их хлорпроизводные. Некоторые морские виды Xantho-
monas содержат сложные эфиры бромированных арильных по-
лиенов, например ксантомонадин I (2.95).
Биосинтез этих полиенов не изучался. По-видимому, они не-
изопреноидного происхождения и, скорее всего, синтезируются
по поликетидному пути (гл. 3). Информация о распростране-
нии, локализации внутри клетки и функциях какого-либо из
этих пигментов отсутствует.
2.13. Заключение и комментарии
Каротиноиды представляют собой классический при-
мер группы природных пигментов. Все члены этой группы об-
ладают очень близкой структурой, основанной на сопряженном
полиеновом хромофоре, который и обусловливает их светопо-
глощающие свойства. Они дают возможность наблюдать чет-
кую корреляцию между максимумом поглощения и длиной
хромофора. Каротиноиды чрезвычайно широко распространены
у живых организмов и принимают участие во всех фотофунк-
циях, обычно связанных с природными пигментами. Именно по
этим причинам (а не потому, что автор лучше знаком с дан-
ным классом соединений) каротиноиды были рассмотрены бо-
83
Часть I Химические и биохимически- аспекты
лее детально, чем будут рассматриваться некоторые другие
группы пигментов.
Каротиноиды предоставляют химику-органику большие воз-
можности проявить свое мастерство в установлении структуры
молекул. Новые соединения открывают довольно часто, осо-
бенно у микроорганизмов и морских животных. В большинстве
случаев это сравнительно небольшие модификации основной
структуры, однако всегда существует вероятность обнаружить
ранее неизвестный тип кольца или открыть азот-, серу- или га-
логенсодержащие каротиноиды. Необходимо также помнить,
что полная характеристика каротиноида должна включать оп-
ределение его стереохимии, и в первую очередь хиральности,
поскольку в настоящее время известно, что «один и тот же»
каротиноид может существовать в различных хиральных фор-
мах. Искусство химика проверяется также по способности раз-
рабатывать новые методы синтеза, особенно стереоконтроли-
руемого синтеза полиеновой цепи, и вводить хиральные конце-
вые группы.
Возможно, самые трудные проблемы в области изучения
каротиноидов связаны с их биохимией. Главные пути биосин-
теза и последовательности реакций _\же хорошо известны, и
предстоит определить лишь стереохимию и механизм некоторых
из этих реакций. Однако особого внимания требуют исследо-
вания на ферментном уровне, не исключено, что ферменты ор-
ганизованы в связанные с мембранами комплексы, и это мо-
жет создавать значительные методические трудности. До тех
пор пока не будет достигнут прогресс в этой области, невоз-
можно выяснить детали механизмов регуляции и контроля био-
синтеза каротиноидов (особенно фоторегуляции). Что касается
вопроса о происхождении каротиноидов у животных, то здесь
не исключены сюрпризы; старая идея о том, что животные по-
лучают каротиноиды только с пищей, сейчас кажется несостоя-
тельной. Животные способны модифицировать попавшие с пи-
щей каротиноиды различными путями, в том числе с помощью
стереохимических превращений. Исследование последних очень
перспективно, несмотря на то что метаболические превращения
обычно происходят медленно и в связи с этим их трудно обна-
ружить
Ассоциация каротиноидов с белками, в основном с образо-
ванием синих комплексов у морских беспозвоночных, — об-
ласть, в которой можно ожидать прогресса уже в ближайшем
будущем. Будучи интересными сами по себе, эти комплексы
служат очень полезными моделями взаимодействия белка с не-
большими липидными молекулами, и полученные при этом ре-
зультаты будут представлять несомненную ценность для мно-
гих разделов биохимии. Микроокружение и белковые комплек-
Гл 2 Каротиноиды
89
сы каротиноидов в разных тканях, особенно в фотосинтетиче-
ских мембранах, — также важная область исследования. Полу-
ченные здесь результаты несомненно будут способствовать луч-
шему пониманию механизма фотосинтеза. Существует широкий
простор для проверки оригинальных идей и исследований функ-
ций и механизма действия каротиноидов у различных видов
живых организмов и в самых разных тканях.
Использование каротиноидов в качестве пищевых добавок,
красителей и провитаминов А уже происходит в больших мас-
штабах и, вполне вероятно, будет расширяться. Эго связано с
}величением спроса на природные красители, которые часто
предпочитают чисто синтетическим красящим веществам, а
также с тем, что постепенно налаживается промышленное про-
изводство каротиноидов. В будущем не исключено открытие
новых функций каротиноидов у животных, в том числе и у че-
ловека. Каротиноиды применяются и будут применяться в даль-
нейшем в медицине в их собственной роли, а не только в ка-
честве предшественника витамина А.
2.14. Рекомендуемая литература
В качестве стандартного справочника может быть ис-
пользована обширная монография под редакцией Ислера (Is-
ler, 1971), поскольку она дает очень подробный обзор по хи-
мии, спектроскопии, биосинтезу, функциям и промышленному
использованию каротиноидов. Эта книга очень хорошо допол-
няется двухтомной «Сравнительной биохимией каротиноидов»
Гудвина (Goodwin, 1980), для которой характерен более биоло-
гический и биохимический подход. Первый том посвящен рас-
тительному (и микробному), а второй животному царству. «Хи-
мия и биохимия растительных пигментов» (под ред Гудвина;
Goodwin, 1976) содержит хотя и не всеобъемлющие, но чрез-
вычайно полезные статьи по химии (Moss, Weedon), распрост-
ранению в природе (Goodwin) и биосинтезу (Britton) кароти-
ноидов. Более старые работы Фокса (Fox, 1953), Цехмейстера
(Zechmeister, 1962), Каррера и Юкера (Karrer, Jucker, 1950)
содержат информацию, которая также может быть полезной.
Химия и биохимия каротиноидов ежегодно освещается ис-
черпывающими литературными обзорами в серии «Терпеноиды
и стероиды», издаваемой в рамках серии «Chemical Society’s
Specialist Periodical Reports» (например, Britton, 1979), и в
выходящих раз в три года публикациях трудов Международ-
ного симпозиума по каротиноидам (1966, 1969, 1972, 1975, 1978,
1981) (см. Plenary and Session Lectures, 1967, 1969, 1973, 1976,
1979, 1982).
90
Часть I. Химические и биохимические аспекты
Каротинопротеины специально обсуждаются в обзорах Чиз-
мана и др. (Cheesman et al., 1967), Загальского (Zagalsky,
1976) и Бриттона (Симпозиум по каротиноидам, Britton, 1981);
много статей по этому вопросу собрано в книге Ли (Lee, 1977).
Обзор использования каротиноидов в медицине был пред-
ставлен Мэтьюз-Ротом, (Mathews-Roth, 1981) в трудах Симпо-
зиума по каротиноидам, а возможная роль каротиноидов
в предотвращении рака обсуждается в работе Пето и др. (Peto
et al., 1981). Методы, полезные для работы с каротиноидами,
описаны в ряде обзоров (Britton, Goodwin, 1971; Davies, 1976,
Liaaen-Jensen, Jensen; 1965; Liaaen-Jensen, 1971).
2.15. Дополнительная литература
Britton G. (1976). Biosynthesis of carotenoids. In: Chemistry and biochemistry
of plant pigments, 2nd edition, vol. 1, ed. T. W. Goodwin, p. 262, London,
New York and San Francisco, Academic Press.
Britton G. (1979). Carotenoids and polyterpenoids. In: Specialist Periodical
Reports: Terpenoids and steroids, vol. 9, ed. J. R. Hanson, p. 218, London,
The Chemical Society.
Britton G-, Goodwin T. W. (1981). Biosynthesis of carotenoids, Methods Enzy-
mol., 180, 654.
Cheesman D. F., Lee IF. L., Zagalsky P. F. (1967). Carotenoproteins in inver-
tebrates, Biol. Rev., 42, 131.
Davies В. H. (1976). Carotenoids. In: Chemistry and biochemistry of plant pig-
ments, 2nd edition, vol. 2, ed. T. W. Goodwin, p. 38, London, New York
and San Francisco, Academic Press.
Fox D. L. (1953). Animal biochromes and structural colours. Cambridge Uni-
versity Press.
Goodwin T. IF. (1952). The comparative biochemistry of carotenoids, London,
Chapman and Hall.
Goodwin T. IF. (1976). Distribution of carotenoids. In: Chemistry and bio-
chemistry of plant pigments, 2nd edition, vol. 1, ed. T. W. Goodwin, p. 225,
London, New York and San Francisco, Academic Press.
Goodwin T. IF. (1980). The comparative biochemistry of the carotenoids, 2nd
Edition, vol. 1, London, Chapman and Hall. (Vol. 2, in press.)
Isler O. (ed.) (1971). Carotenoids, Basel and Stuttgart, Birkhauser-Verlag.
Karrer P„ Jucker E. (1950). Carotenoids (trans. E. A. Braude), Amsterdam,
Elsevier.
Lee IF. L. (1977). Carotenoproteins in animal coloration, Stroudsberg, USA,
Dowden, Hutchinson and Ross.
Liaaen-Jensen S. (1971). Isolation, reactions. In: Carotenoids, ed. O. Isler,
p. 61, Basel and Stuttgart, Birkhauser-Verlag.
Liaaen-Jensen S., Jensen A. (1965). Recent Progress in carotenoid chemistry,
Prog. Chem. Fats other Lipids, 8, 129.
Moss G. P., Weedon В. C. L. (1976). Chemistry of the carotenoids. In: Chemistry
and biochemistry of plant pigments, 2nd edition, vol. 1, ed. T. W. Goodwin,
p. 149, London, New York and San Fancisco, Academic Press.
Peto R., Doll R„ Buckley J. E., Sporn M. B. (1981). Can dietary bets-carotene
materially reduce human cancer rates? Nature, 290, 201.
Plenary and Session Lectures, First International Symposium on Carotenoids
other than vitamin A 1966(1967), Pure Applied Chem., 14, 227. Also published
as Carotenoids other than vitamin A-L, London, Butterworth.
Гл. 2. Каротиноиды
91
Plenary and Session Lectures. Second International Symposium on Carotenoids
other than vitamin A (1969), Pure Applied Chem., 20, 365. Also published
as Carotenoids other than vitamin A-II, London, Butterworth.
Plenary and Session Lectures, Third International Symposium on Carotenoids
other than vitamin A, 1972 (1973), Pure Applied Chem., 35, 1. Also publi-
shed as Carotenoids other than vitamin A-III, London, Butterworth.
Plenary and Session Lectures, Fourth International Symposium on Carotenoids,
1975 (1976), Pure Applied Chem., 47, 97. Also published as Carotenoids-4
(Berne, 1975), ed. В. C. L. Weedon. Oxford: Pergamon.
Plenary and Session Lectures, Fifth International Symposium on Carotenoids,
1978 (1979), Pure Applied Chem., 51, 436—675, 857—887. Also published
as Carotenoids-5 (Madison, 1978), ed. T. W. Goodwin, Oxford, Pergamon.
Plenary and Session Lectures, Sixth International Symposium on Carotenoids
1981. Published (1982) as Carotenoid chemistry and biochemistry, eds.
G. Britton and T. W. Goodwin, Oxford, Pergamon.
Zagalsky P. F. (1976). Carotenoid-protein complexes, Pure Applied Chem., 47,
103.
Zechmeister L. (1962). Cis-trans isomeric carotenoids, vitamin A and aryl-
polyenes, Vienna, Springer.
Глава 3
Хиноны
3.1. Введение
Хиноны представляют собой большую и довольно гете-
рогенную группу соединений. Их окраска варьирует от бледно-
желтой через оранжевую, красную, пурпурную и коричневую
до почти черной. Они являются важными пигментами у ряда
грибов, лишайников и некоторых групп беспозвоночных жи-
вотных. Хиноны широко распространены также у высших рас-
тений, но, как правило, содержатся в тканях, которые обычно
не видны, например в коре, ядровой древесине и корнях. Они
редко вносят заметный вклад в наружную окраску высших рас-
тений. Некоторые хиноны, однако, являлись основой важней-
ших красителей еще в античные времена, например хны и ма-
рены.
Некоторые изопренилированные хиноны, например убихи-
нон, менахинон, филлохинон и пластохинон, представляют со-
бой чрезвычайно важные биологические молекулы. Те или
иные из них можно обнаружить во всех живых тканях, но они
не являются пигментами и потому подробно рассматриваться
не будут.
3.2. Строение
Основная хиноновая структура представляет собой не-
насыщенный циклический дикетон, являющийся производным
моноциклического или полициклического ароматического угле-
водорода. Хиноновые структуры формально могут быть полу-
чены окислением соответствующих дигидроксифенолов. Так,
простейшими примерами служат орто- пли 1,2-бензохинон (3.1)
и пара- или 1,4-бензохинон (3.2)—производные бензола, ко-
торые могут рассматриваться как продукты окисления дигидр-
оксифенолов пирокатехина (3.3) и гидрохинона (3.4) соответ-
ственно.
Хиноны могут происходить от различных ароматических
углеводородов и соответственно получают свои названия и по-
рядок нумерации углеродных атомов от исходного углеводоро-
да. Так, показанные ниже производные нафталина (3.5) и (3.6)
называются соответственно 1,2- и 1,4-нафтохиноном. Из нес-
Гл. 3. Хиноны
93
кольких возможных антрахиноновых структур в природных
пигментах часто встречается только 9,10-хиноновая структура
(3.7), хотя иногда обнаруживаются и производные, содержа-
щие 1,2-хиноновую структуру.
(3.4) Хинол
(3 2) 1,4-Бензохинон
(3.6) 1,4 Нафтохинон
(3.7) 9,10-Ан+рахннон
(3.8) 2,6-Нафтохинон
Хиноновая система не обязательно ограничена одним коль-
цом. Две кетогруппы могут находиться в разных кольцах, но
они должны быть сопряжены с соответствующей л-электронной
системой для того, чтобы образовалась растянутая хиноновая
структура. Одним из простейших примеров такой структуры
служит 2,6-нафтохинон (3.8). Примеры сложных структур,
включающих растянутые хиноновые системы, можно встретить
среди пигментов тлей (афинов; разд. 3.4.4).
Наиболее широко распространенными и имеющими наи-
большее значение как пигменты являются 1,4-нафтохиноны и
9,10-антрахиноны, однако в последующем обзоре нам встре-
тится и несколько других хиноновых структур.
В природных хиноновых пигментах основной хиноновый
скелет обычно содержит замещающие группы; наиболее часто
•94
Часть I. Химические и биохимические аспекты
встречаются среди них метильные, гидрокси- и метоксигруппы.
Фенольные гидроксигруппы могут быть гликозилированы (осо-
бенно у высших растений). Некоторые из более сложных хино-
нов имеют длинные боковые цепи, которые в ряде случаев мо-
гут складываться в добавочные кольца. Другие сложные хино-
ны могут рассматриваться как димеры обычных простых наф-
тохиноновых и антрахиноновых структур.
Структура некоторых из наиболее важных встречающихся
в природе хинонов приведена в последующих разделах этой
главы.
3.3. Свойства
3.3.1. Химические свойства
Хиноны представляют собой скорее сопряженные цик-
лические дикетоны, чем ароматические системы. Однако они в
значительной степени стабилизируются за счет образования ре-
зонансных заряженных ароматических структур (рис. 3.1). Как
травило, 1,2-хиноны менее стабильны, а потому более реакци-
онноспособны, чем 1,4-хиноны или растянутые хиноновые сис-
темы.
О О о+
Рис. 3.1. Вклад заряженных ароматических структур в стабилизацию хино-
нов.
Наиболее характерной химической реакцией для хинонов
является обратимое восстановление до соответствующего фе-
нола. Так, 1,2-бензохинон (3.1) и пирокатехин (3.3) или 1,4-
бензохинон (3.2) и гидрохинон (3.4) легко превращаются друг
в друга. Относительно малозамещенные хиноны, особенно 1,2-
хиноны, чрезвычайно подвержены полимеризации, протекаю-
щей по свободнорадикальному механизму.
Многие природные хиноны имеют фенольные ОН-группы и
как следствие слегка кислую реакцию. Благодаря этому они
способны к ионизации и образованию солей в щелочной среде.
Это свойство особенно ярко выражено в случае гидроксибензо-
гхинона (3.9) и 2-гидрокси-1,4-нафтохинона (3.10).
Гл. 3. Хиноны
95
3.3.2. Физические свойства
Почти все природные хиноны представляют собой твер-
дые вещества и легко кристаллизуются. Большинство из них
хорошо растворимы в органических растворителях, тогда как
гликозиды и некоторые карбоновые кислоты способны раство-
ряться в воде. Хиноны, которые одновременно являются фено-
лами или карбоновыми кислотами, растворимы в щелочных
водных растворах.
Высшие полициклические хиноны ведут себя подобно по-
лимерам и довольно трудно растворяются в водной или орга-
нической среде.
3.3.3. Поглощение света
Детальное теоретическое объяснение светопоглощаю-
щих свойств хинонов лежит вне круга тем, затронутых в этой
книге. Однако краткое описание спектров поглощения хинонов-
и обсуждение факторов, которые влияют на их максимум пог-
лощения, существенны для оценки значения этих соединений
как пигментов.
Спектры поглощения прародительских хинонов приведены
на рис. 3.2. Наиболее простой спектр — спектр 1,4-бензохино-
на — имеет интенсивную полосу поглощения (Z) при ~ 240 нм,
полосу «электроннотранспортного перехода» средней интенсив-
(3.9) Гидроксибензохинон
(3.10) 2-Гидрокси-1,4-нафтохииоИ
ности (ZZ) при ~285 нм и лишь слабое хиноидное п—»-л*-пог-
лощение в видимом диапазоне при ~434 нм (полоса III). Длят
нестабильного 1,2-бензохннона максимумы поглощения нахо-
дятся при значительно более высоких значениях длин волн:
для полос II и III при 375 и 586 нм соответственно. Хиноидные'
п—>л*-переходы «запрещены», и поэтому полоса поглощения
III имеет низкую интенсивность. Хотя хиноны поглощают свет
в видимой области спектра, это поглощение слишком мало,,
чтобы они выглядели окрашенными. Правда, последнее возмож-
но при очень высоких концентрациях хинонов.
96
Часть I. Химические и биохимические аспекты
Спектры нафтохинонов, антрахинонов и высших хинонов
отличаются значительно большей сложностью благодаря при-
сутствию в них полос поглощения, обусловленных бензеноид-
ными переходами в дополнение к хиноидному поглощению.
Главные полосы поглощения 1,4-нафтохинона локализованы
при 245, 257 и 335 нм, а у антрахинона — при 243, 263, 332 и
•405 нм (рис. 3.2).
Рис. 3.2. Главные полосы поглощения в электронных спектрах 1,2-бензохи-
нона (Л), 1,4-бензохинона (Б), 1,4-нафтохинона (В) и антрахинона (Г). По
оси ординат отложены приблизительные величины поглощения (логарифми-
ческая шкала) для каждого соединения, поэтому сравнивать между собой
относительные интенсивности полос поглощения различных соединении не
следует.
Заместители очень сильно влияют на спектры хинонов.
Правда, при введении заместителя в молекулу 1,4-бензохино-
на полосы / и III почти не изменяются, но полоса II претерпе-
вает значительный «красный сдвиг» (в порядке усиления влия-
ния заместителя) на 27 нм для СН3, 69 нм для ОСН3 и 81 нм
для ОН. Влияние второго заместителя менее значительно.
В результате замещения полоса поглощения II может оказать-
ся сдвинутой в видимый диапазон спектра и соединение станет
окрашенным.
В случае нафтохинонов (рис. 3.3) замещение в хиноновом
кольце оказывает слабое действие на положение максимумов
поглощения, в то время как замещение в ароматическом коль-
це, особенно ОН-группами, вызывает батохромный эффект и
Гл. 3. Хиноны
97
Рис. 3.3. Спектры поглощения 1,4-нафтохинона (/), 5-гидрокси-1,4-нафтохи-
нона (II) и 5,8-дигидрокси-1,4-нафтохинона (III). Величины поглощения (ло-
гарифмическая шкала) даны приблизительно, и для различных соединений
сравнивать их не следует.
некоторые УФ-полосы поглощения сдвигаются в видимый диа-
пазон. 77ери-гидроксилирование( например, в положениях 5 и
8 у 1,4-нафтохинона) дает наиболее ощутимый эффект, бензе-
ноидная полоса при этом сдвигается почти на 100 нм [напри-
(3,11а) Н; 5-Гидрокси-1,4-нафтохинон
(3,11b) R=OH, 5,8 дигидрокси-1,4-нафтохиноН
мер, у 5-гидрокси-1,4-нафтохинона (3.11а) с 335 до 429 нм].
Нафтазарины (3.11b) с двумя пери-гидроксигруппами облада-
ют сильным многополосным бензеноидным поглощением при
525 нм. Эта структура образует основу спинохромов — интен-
сивно окрашенных пигментов, найденных у иглокожих (разд.
3.4.2).
7—84
98 Часть I. Химические и биохимические аспекты
Сходным образом у антрахинонов наблюдается сложное
бензеноидное поглощение. У них на максимум поглощения до-
минирующее влияние оказывает присутствие ОН- и алкокси-
групп, особенно в случае наличия нескольких заместителей.
Влияние «-гидроксилирования более значительно, чем Р-гидро-
ксилирования (за исключением тех случаев, когда р-гидрок-
сильная группа находится в положении, соседнем к «-гидрок-
сильной) .
В щелочной среде гидроксипроизводные хинонов способны
ионизироваться, что приводит к значительным батохромным
сдвигам. Такие «щелочные сдвиги» имеют большое диагности-
ческое значение, особенно в рядах нафтохинонов и антрахино-
нов. Так, например, анион 2-гидрокси-1,4-нафтохинона (3.12)
имеет оранжевую окраску (ХШах = 459 нм). Сходным образом
анион 5-гидрокси-1,4-нафтохинона окрашен в фиолетовый цвет
(%тах = 538 нм), а у 2,4-, 5,6- и 5,8-дигидрокси-1,4-нафтохино-
нов — в синий (/.max=650, 571 и 655 нм соответственно). Дру-
гим полезным диагностическим свойством является то, что вос-
становление до гидрохинонов (при помощи обработки NaBH4)
приводит к образованию ароматической системы, спектр кото-
рой относится к бензеноидному типу.
Спектры поглощения хинонов обычно измеряют в этаноле;
в тех же случаях, когда используются другие растворители, на-
пример хлороформ, значения 7imax несколько отличаются от
стандартных. Таблицы максимумов поглощения природных -и
модельных хинонов приведены в монографии Томсона (Thom-
son, 1971), в которой также обсуждаются их ИК-, ЯМР- и
масс-спектры.
3.4. Распространение в природе
Окрашенные хиноны обнаружены главным образом у
высших растений, грибов и бактерий, а в животном царстве —
у членистоногих и иглокожих. Хиноны очень легко образуются
окислением соответствующих гидрохинонов. Во многих случа-
Гл. 3. Хиноны
99
ях выделяемые хиноны могут быть артефактами, возникаю-
щими вследствие процедуры экстракции или высушивания рас-
тений или растительных тканей на воздухе.
3.4.1. Бензохиноны
Из природных источников, главным образом из грибов
и различных тканей высших растений, было выделено около
100 бензохинонов, из которых почти все являются 1,4-бензохи-
нонами. К бензохинонам принадлежат и такие важные в биоло-
гическом отношении молекулы, как пластохинон (3.13) и уби-
хинон (3.14). Образцы убихинонов обычно получают в виде
смесей молекул с различной длиной изопреноидной цепи (изо-
(3,13) Пластохинон
(3 14) Убихинон-п
пренилогов). В определенной ткани или у того или иного вида
обычно преобладает один из изопренилогов. Наиболее часто
встречаются убихинон-9 и убихинон-10 с девятью и десятью
изопреновыми остатками соответственно. В целом бензохино-
ны не являются сильно окрашенными соединениями и не вно-
сят сколько-нибудь ощутимого вклада в природную окраску
организмов.
3.4.2. Нафтохиноны
Природные нафтохиноны включают витамины К, фил-
лохинон (3.15) и менахинон (3.16) —широко распространенные
и важные в биологическом отношении молекулы, которые, од-
нако, не являются пигментами. Эти вещества имеют одно и то
же нафтохиноновое ядро, но различные изопреноидные боковые
цепи. Филлохинон (витамин Ki) из высших растений имеет фи-
тольную боковую цепь, тогда как бактериальные менахиноны
(витамин Кг), подобно убихинонам, различаются длиной боко-
вой цепи. Чаще всего встречаются соединения с шестью-де-
вятью изопреновыми единицами. Распространение других наф-
тохинонов в растениях систематически не изучалось. 1,4-Наф-
тохиноны время от времени обнаруживают в различных тканях
высших растений — в листьях, цветках, плодах, корнях, коре
7*
100
Часть I Химические и биохимические аспекты
и древесине. Среди наиболее известных примеров можно на-
звать юглон (3.17) и его производные из зеленых частей грец-
кого ореха (Juglans regia), а также лавсон (3 18) из лосонии,
или хенны (Lawsonia alba). 1,4-Нафтохиноны иногда обнару-
живаются также в грибах, например моллисин (3.19)—жел-
тый пигмент из культур Mollisia fallens Другие нафтохиноны
(1,2-, 1,5- и 2,6 ) в природе встречаются редко.
(3 17) Юглон
(3 16) Менахинон п
(3 18) Лавсон
(3 19) Моллисин
Спинохромы. В животном царстве около 20 близких по
строению сильно окрашенных красных, пурпурных или синих
нафтохиноновых пигментов было найдено у иглокожих, глав-
ным образом у морских ежей, а также у морских звезд и офи-
ур. Эти соединения, известные как спинохромы или эхинохро-
мы, были впервые выделены из кальцинированных частей жи-
вотных, например из игл панциря морских ежей Однако они
также содержатся в полостной жидкости, яйцах и внутренних
органах. Большинство видов синтезируют смесь из шести и бо-
лее пигментов, которые содержатся в иглах и в панцирях глав-
ным образом в виде кальциевых и магниевых солей, а в дру-
Гл 3 Хиноны
10]
гих тканях, возможно, в виде комплексов с белками В струк-
турном отношении эти пигменты иглокожих представляют со-
бой высокозамещенные и сильно оксигенированные производ-
ные юглона или нафтазаринов
Широкое использование тривиальных названий спинохро-
мов часто приводит к недоразумениям, поскольку все главные
соединения имеют по нескольку синонимов. Названия спино-
R1 R2 R3
]3 20) Спинохром А ОН И СОМе 3 ацетил 2 7 дигидроксинафтазарин
(3 21) Спинохром В н н ОН 2 3 7 тригидроксиюглон
(3 22) Спинохром С он он СОМс 3 ацетил 2 6 7 три идроксинафтазарин
]3 23) Спинохром D он н ОН 2 3 6 тригид оксинафтазарин
(3 24) Спинохром Е он он ОН 2 3 6 7 тетрагидроксинафтазарин
(3 25) Эхинохром А он СН2СН3 ОН 6 этил-2 3 7 тригидроксинафтазарин
Рис 3 4 Структура и номенклатура некоторых спинохромовых и эхинохро-
мовых пигментов
хромы А—Е и эхинохром А присвоены в настоящее время ше-
сти главным членам этой группы веществ, однако лучше при-
менять полусистематическую номенклатуру, согласно которой
пигменты называют как замещенные юглоны и нафтазарины.
Структуры (3 20—3 25) и полусистематические названия этих
главных соединений приведены на рис 3 4
3.4.3. Антрахиноны
Антрахиноны являются самой большой группой при-
родных хинонов В растениях, грибах и лишайниках найдено
почти 200 представителей этой группы Наиболее широко рас-
пространенным антрахиноном является, вероятно, эмодин
(3 26), который был выделен из плесневых и высших грибов,
лишайников, цветковых растений и насекомых. Однако в све-
жесобранных растительных тканях содержится очень мало эмо-
дина или он вообще отсутствует. В большинстве случаев (если
не во всех) при разрушении или высушивании тканей в собст-
102
Часть I. Химические и биохимические аспекты
венно эмодин легко превращаются содержащиеся в тканях in
vivo гликозиды эмодина или восстановленные (гидрохиноновые)
формы этих гликозидов. Наиболее известным антрахиноном
высших растений является, по-видимому, ализарин (3.27) —
главный пигмент марены, Rubio, tinctorium, получаемый из из-
мельченных корней растения. Ализарин был, вероятно, самым
важным красителем в античные времена.
Антрахиноны животных. Некоторые насекомые отряда Сос-
cidae окрашены антрахинонами. Большинство этих пигментов
представляют собой карбоновые кислоты, и некоторые из них,
главным образом С-глюкозид карминовой кислоты (3.28) —
(3 26) Эмодин
(3.27) Ализарин
карминово-красное красящее вещество кошенили — и красная
кермезиновая кислота (3.29), используются в качестве краси-
телей уже в течение многих веков.
Недавно было обнаружено, что окраска группы австралий-
ских криноид (морских лилий) обусловлена красными или пур-
пурными антрахинонами, такими, как родоптилометрин (3.30),
и сходными пигментами, имеющими в положении 4 антрахино-
новой системы колец длинную боковую цепь. Эти вещества
весьма сходны с некоторыми антрахиноновыми метаболитами
грибов. Галлахром — пигмент некоторых многощетинковых
червей — интересен как пример редкой 1,2-антрахиноновой
структуры (3.31).
Антрациклиноны. Антрациклиноны из культур некоторых
стрептомицетов явно близки к тетрациклиновым антибиотикам.
Структурно, однако, они могут рассматриваться как замещен-
ные антрахиноны, например аклавинон (3.32).
3.4.4. Высшие хиноны
Среди высших или более сложных хинонов интересны
главным образом афины, изучавшиеся наиболее интенсивно.
Афины являются производными протоафинов, которые содер-
жатся в гемолимфе различных тлей. Совершенно очевидно,
Гл. 3. Хиноны
103
что протоафины, например (3.33), являются производными
нафтохинонов. После гибели насекомого протоафин превра-
щается в стабильный конечный продукт красного цвета эритро-
афин (3.34) через менее стабильные промежуточные соедине-
ния, такие, как желтый ксантоафин (3.35) и оранжевый хризо-
(3 29) Кермезиновая кислота
(3 28) Карминовая кислота
(3.30) Родоптилометрин
(3 31) Галлахром
(3 32) Аклавинон
афин (3.36). Разные виды тлей синтезируют разные прото-,
ксанто-, хризо- и эритроафиновые структуры с различной сте-
реохимией. Приведенные выше афины представляют собой
пигменты, впервые выделенные из Aphis fabae (бобовой тли).
Все они являются высшими хинонами, в молекулах которых
две карбонильные группировки разделены значительным рас-
стоянием в сложной полициклической сопряженной струк-
туре.
Другими интересными сложными хинонами являются гипе-
рицин (3.37), содержащийся в растениях рода Hypericum
(разд. 3.7.2), и несколько соединений, известных как фринге-
(3.37) Гиперицин
(3.38) Фрингелит
Гл. 3. Хиноны
105
литы, например (3.38), который был найден в ископаемых
криноидах юрского периода. Гиперицин и фрингелиты пред-
ставляют собой димеры антрахинонов.
3.5. Вклад в окраску
Изопренилированные хиноны убихинон, пластохинон
и т. п., которые обнаруживаются у всех высших растений,
не принимают участия в формировании окраски этих растений.
Другие хиноны, особенно нафтохиноны и антрахиноны, чрез-
вычайно широко распространены в различных тканях высших
растений, но и они редко вносят существенный вклад в их
окраску. В то же время они часто обусловливают окраску,-
обычно желтую, оранжевую или коричневую, плесневых и выс-
ших грибов, а также лишайников.
Только в животном царстве были обнаружены наиболее'
характерные примеры окрашивания хинонами. Так, спинохро-
мы и эхинохромы придают пурпурную, синюю или зеленую'
окраску большинству морских ежей; красиво окрашенные де-
коративные представители некоторых видов морских ежей ча-
сто выставляются дома. Антрахиноновые кислоты обусловли-
вают красную окраску некоторых щитовок и приготавливаемо-
го из них пищевого красителя кошенили. Протоафины, содер-
жащиеся в гемолимфе многих видов тлей, при биологических
значениях pH ионизированы и образуют фиолетово-красные
анионы, которые главным образом и обусловливают темную
окраску гемолимфы. После гибели насекомых ферменты гемо-
лимфы превращают протоафины в стабильные окрашенные
продукты — афины, которые и придают посмертную окраску
насекомым. Афины характерны для темноокрашенных видов
тлей, тогда как хорошо известная зеленая тля, не содержащая
в гемолимфе афинов, содержит близкие им соединения — водо-
растворимые гликозиды афанины, имеющие зеленую окраску.
3.6. Биосинтез
3.6.1. Введение
При биосинтезе пигментов любого класса для созда-
ния основной структуры используется один главный путь1.
Различные индивидуальные соединения образуются затем с
помощью последующих модификаций основной структуры.
1 В биосинтезе флавоноидных пигментов (разд. 4) всегда используются
два главных биогенетических пути (шикиматный и поликетидный). —-Прим,
ред.
106
Часть I. Химические и биохимические аспекты
В случае же хинонов этого не происходит: они представляют
собой биосинтетически гетерогенную группу веществ. Как вы-
ражение крайнего случая, различные организмы могут синте-
зировать один и тот же хинон с помощью различных биосин-
тетических путей.
Большинство хинонов образуются по одному из двух глав-
ных путей. В некоторых случаях образование углеродного ске-
лета идет по третьему главному пути. Эти же три главных
пути обеспечивают образование всех основных групп хинонов,
т. е. бензохинонов, нафтохинонов, антрахинонов и высших
хинонов.
Число индивидуальных хинонов, для которых был бы ис-
следован биосинтез, очень невелико. Во многих случаях путь
биосинтеза хинона был просто выведен исходя из структурных
соображений. И потому положение вещей в этой области Том-
сон образно назвал «кучкой фактов, поддерживающих гору
спекуляций» (Thomson, 1971).
3.6.2. Общие соображения
Хиноны представляют собой окисленные ароматические
соединения, и потому не удивительно, что они могут синтезиро-
ваться посредством главных путей, с помощью которых обычно
образуются ароматические соединения. Этими путями являются
поликетидный путь из ацетата и малоната и шикиматный путь
биосинтеза. В случае некоторых нафтохинонов и антрахинонов
синтез идет по третьему главному пути — изопреноидному.
К сожалению, наши знания биосинтеза хинонов еще слишком
фрагментарны для того, чтобы сделать даже самые общие
выводы о регулирующих этот процесс факторах.
Поэтому в данном разделе будет представлено только са-
мое общее описание важнейших реакций основных биосинте-
тических путей. Вместе с тем здесь будут приведены некоторые
из наиболее важных и хорошо документированных примеров.
3.6.3. Поликетидный путь
Возможно, наиболее часто используется для биосинте-
за хиноновых пигментов поликетидный путь. Этот процесс, до-
вольно близкий к ступенчатому удлинению цепи при биосинтезе
жирных кислот, приведен на рис. 3.5. На первой стадии конден-
сация ацетил-СоА и малонил-СоА дает ацетоацетил-СоА. В от-
личие от биосинтеза жирных кислот образующаяся кетогруппа
на этой стадии не восстанавливается. Вместо этого процесс
удлинения цепи продолжается путем добавления С2-фрагментов
из малонил-СоА до тех пор, пока не достигается требуемая
Гл. 3. Хиноны
107
„SCoA
ч
сн3—со
ООС
Ацетил СоА
CH2.CO.S Со А
Малонил СоА
СН3.СО.СН2-СО
SCo А О0С s Со А ООС
СН2. СО ( С1
Ацетоацетил
2 Малонил СоА
' СН3.СО.СН2.СО-СН2-СО.СН2.СО.S Со А
Рис. 3 5 Главный поликетидный путь биосинтеза фенолов и хинонов.
длина цепи. Такая система из чередующихся СО- и СН2-групп
называется поликетидной системой.
Когда молекула поликетида соответствующим образом из-
гибается, может происходить циклизация, протекающая путем
удаления молекулы воды от соответствующим образом располо-
женных СО- и СН2-групп, что и приводит к образованию аро-
матической или хиноновой молекулы.
Картина распределения кислородсодержащих функций че-
рез один углеродный атом часто может служить отличительной
особенностью молекул, образовавшихся по этому пути, хотя
добавление или удаление .замещающих групп часто маскирует
тот факт, что данное вещество было синтезировано по полике-
тидному пути. Образование хинона по поликетидному пути
можно показать с помощью опытов по включению ацетата
(или малоната), меченного радиоактивными или стабильными
изотопами, а способ изгиба поликетидной молекулы можно
выяснить, изучая продукты расщепления меченой молекулы.
108
Часть I. Химические и биохимические аспекты
(3 39)
(3 40) Орселлиновая кислота
(3 41) Спинулозин
Рис. 3.6. Поликетидиый механизм биосинтеза бензохинона спинулозина.
<3 42)
(3 43) Яваницин
.Рис. 3.7. Образование нафтохинона яваницина из гептаацетилполикетида.
Бензохиноны. Простым примером биосинтеза соединений
бензохиноновой группы служит синтез грибного метаболита
спинулозина (3.41), который образуется из тетраацетатного
поликетида (3.39) через орселлиновую кислоту (3.40) (рис.
,3.6).
Нафтохиноны. Для нафтохиноновой группы хинонов в экс-
периментах по включению меченного ацетата или малоната
культурой плесневого гриба Fusarium javanicum было установ-
лено, что яваницин (3.43) образуется из гептаацетильного поли-
кетида (3.42), как это показано на рис. 3.7. Необычно проис-
ходящее при этом восстановление терминальной карбоксильной
труппы до метильной. Менее простым примером служит обра-
зование моллисина (3.19)—другого грибного метаболита, про-
Гл. 3. Хиноны
109
дуцируемого культурой Mollisia caesia. Здесь биосинтез опять-
таки происходит из ацетата и малоната, но в этом случае об-
разуются две поликетидные цепи — одна из трех, а другая из
четырех Сг-фрагментов, а не одна более длинная цепь.
(3 26) Эмодин
Рис. 3.8. Биосинтез антрахинона эмодииа из октаацетилполикетида.
Антрахиноны. Вероятно, многие, а возможно, и большинство
антрахинонов, особенно антрахиноны типа эмодина (3.26), так-
же синтезируются по поликетидному механизму. Считается,
что сам эмодин образуется путем соответствующего изгиба и
конденсации октаацетилполикетидной цепи, как показано на
рис. 3.8. Введение или удаление замещающих групп на позд-
0MS) л«м>«ммм* immdrD
110
Часть 1. Химические и биохимические аспекты
них стадиях биосинтеза может затем давать начало ряду при-
родных эмодинподобных хинонов. Для объяснения биосинтеза
других антрахинонов, таких, как пигмент лишайников солори-
новая кислота (3.44) и хинон насекомых лаккаиновая кисло-
та D (3.45), необходимы альтернативные способы изгиба поли-
кетидной цепи.
(3 46) б Пирромицинон
Рис. 3.9. Образование антрациклинона е-пирромициноиа из поликетида.
Высшие хиноны. Многие их хинонов с более крупными мо-
лекулами, особенно хиноны, образуемые грибами, также, по-
видимому, синтезируются по ацетатно-малонатному пути. Изо-
топные исследования показали, что антрациклинон е-пирроми-
цинон (3.46) образуется из девяти ацетатных (малонатных)
фрагментов и одного пропионатного остатка, который служит
в качестве затравки и на котором формируется вся цепь (рис.
3.9).
3.6.4. Шикиматный путь
Другой главный путь биосинтеза ароматических со-
единений и хинонов связан с промежуточным образованием
шикимовой кислоты (3.47). Это путь, по которому синтезируют-
ся важные, хотя и неокрашенные клеточные компоненты —
убихинон, менахинон, филлохинон и пластохинон. Фактически
основные детали этого пути биосинтеза хинонов выяснены в
результате исследований образования убихинона и менахино-
Гл. 3. Хиноны
111
МеПбопизм
’углеводов
соон
I
с=о
I
— сн,
I 2 -
но—с—н
н—с—он
н-с—он
сн2о—®
(3 49) 5 Дегидрохинная
кислота
5 ДегидрошиКймошЯЙ
кислота
(3 47) Шикимовая кислота
(3 50) 5 Фосфошикимовая
(Енолпируватныи эфир)
кислота
(3 52* Хоризмовая кислота
(3 53) Префеновая кислота
Рис. 3.10. Начальные стадии шикиматного пути биосинтеза фенольных соеди-
нений и хинонов.
на. Многие окрашенные хиноны, и в первую очередь Мафтохи-
ноны, также синтезируются по этому пути. С шикиматом мы
еще встретимся как с промежуточным иродуктом прж биосин-
тезе флавоноидных пигментов (гл. 4).
Подробно начальные стадии шикиматного пути приведены
на рис. 3.10. Предшественники из общего метаболического
112
Часть I Химические
и
*ские аспекты
(3 55) Гомогентизиновая кислота
Рис 311 Биосинтез бензохиноновогб кольца убихинона Для введения пре
нильной боковой цепи и метильного и метоксильных заместите чей возможны
несколько альтернативных последовательностей реакций Другие бензохи-
ноны могут синтезироваться сходным образом из шикимата через п гидро-
ксибензоат или гомогентизат
фонда углеводов дают начало С7-сахару, а именно 7-фосфату
З-дезокси-О-арабиногептулозоновой кислоты [ФДАГК (3 48)],
который затем подвергается циклизации с образованием 5-де-
гидрохинной кислоты (3.49) Последняя превращается в шики-
мовую кислоту (3 47) и далее в 5-фосфошикимовую кислоту
(3 50) Присоединение следующего С3-фрагмента [в виде фос-
фоенолпирувата (3 51)] к 5-фосфошикимовой кислоте дает
хоризмовую кислоту (352), которая после внутримолекулярной
перегруппировки превращается в префеновую кислоту (3 53)
Эти чрезвычайно важные промежуточные соединения являются
предшественниками ароматических аминокислот триптофана,
фенилаланина и тирозина, флавоноидных пигментов и многих
hooc.ch(nh2).ch2-ch2-cooh
(3 56) Глутаминовая кислота
Юг тон
Рис 3 12 Вероятные пути биосинтеза нафтохинонов в том числе менахчно-
на, филлохинона, лавсона и юглона, из шикимата ТПФ — тиаминпирофосфат.
8—84
314
Часть I Химические и биохимические аспекты
природных продуктов с ароматической структурой Природные
хиноны образуются как через префеновую кислоту, так и из бо-
лее ранних предшественников, таких, как шикимовая или хо-
ризмовая кислоты.
Бензохиноны. Бензохиноновое кольцо убихинона образуется
из шикимата через хоризмат (3.52) и п-гидроксибензоат (3 54)
(рис 3 11) Возможно, что некоторые другие природные бензо-
хиноны синтезируются из шикимата либо вышеуказанным пу-
тем через /г-гидроксибензоат, либо через гомогентизат (3.55)
или даже через С6-С3-коричные кислоты (гл. 4). Однако экспе-
риментальные данные для подтверждения этой гипотезы отсут-
ствуют
Нафтохиноны. Роль шикимата в биосинтезе хинонов наилуч-
шим образом изучена на примере нафтохинонов. Выяснены
-основные детали пути образования менахинона, филлохинона,
лавсона (3 18) и юглона (3 17) (рис 3 12). В опытах с введе-
нием меченых предшественников было показано, что шикимат
включается в молекулу хинона в виде интактного Су-фрагмен-
та, образующего бензеноидное (А) кольцо и одну из хиноновых
карбонильных групп. Три остальных углеродных атома проис-
ходят из некарбоксильных углеродных атомов глутамата (3 56)
или р-кетоглутарата (3 57). Ключевым промежуточным соеди-
нением является, по-видимому, о-сукцинилбензоат (3.58), обра-
зующийся в результате присоединения к шикимату тиаминпи-
рофосфатного производного полуальдегида янтарной кислоты
(образовавшегося из глутамата или p-кетоглутарата). Конеч-
ные стадии, во время которых о-сукцинилбензоат превращается
в лавсон, юглон и родственные хиноны, пока не выяснены.
Считается, что собственно 1,4-нафтохинон (3.59) и, возможно,
также 1,4-нафтогидрохинон (3 60) являются промежуточными
продуктами биосинтетического пути, ведущего к образованию
логлона В биосинтезе менахинона участвуют нафтилкарбоновая
кислота (3.61) и 2-метилнафтохинон [менадион (3.62)].
3.6.5. Мевалонатный путь
Путь через мевалоновую кислоту, очевидно, исполь-
зуется для получения полиизопреноидных боковых цепей уби-
хинона, менахинона и т. п Известны также примеры исполь-
зования коротких изопреноидных заместителей (С5 или Сю) в
промежуточных продуктах для введения недостающих углерод-
ных атомов в систему колец нафтохинонов или антрахинонов.
Образующиеся при этом нафтохиноны и антрахиноны являют-
ся, таким образом, с биосинтетической точки зрения замещен-
лыми бензохинонами и нафтохинонами соответственно.
00
Шикигизт ---------
(3 47)
(3 64) п Гидроксифенилпируват
о
(3 63) Химафилин
Рис 3.13 Биосинтез иафтохииоиа химафилина при говместном участии шикиматиого и мевалонатного путей.
116
Часть I. Химические и биохимические аспекты
Нафтохиноны. Мевалонатный путь биосинтеза нафтохино-
нов используется растениями сем. Pyrolaceae. Именно по этому
пути синтезируется химафилин (3 63). Хиноновое кольцо (В)
этого соединения образуется, как и у бензохинонов, обычным
путем из шикимата. Недостающие же атомы бензеноидного
кольца А и метильный заместитель образуются из мевалоната.
Вероятный путь биосинтеза химафилина приведен на рис. 3.13.
Он включает перегруппировку п-гидроксифенилпирувата (3.64)
в гомогентизат (3.55), который затем превращается в гомоар-
(3 68) Алканнин
Рис. 3.14. Образование пренилироваииого нафтохинона алканнина нз л-гид-
роксибеизоата (шикиматный путь) и геранилпирофосфата (из мевалоната)
«бутин (3.65). Пренилирование последнего дает замещенный
гидрохинон (3.66), который циклизуется в гидрохиноновую
форму химафилина (3 67). Последняя окисляется далее в соб-
ственно химафилин. В настоящее время выделены (в виде
глюкозидов) почти все промежуточные продукты этого предпо-
лагаемого биосинтетического пути.
В случае алканнина (3.68), полученного из корней предста-
вителей сем. Boraginaceae, кольцо А образуется из шикимата
через /г-гидроксибензоат, а остальные десять углеродных ато-
мов, включая атомы хинонового кольца (кольца В), предостав-
ляются двумя молекулами мевалоната. Биосинтез (рис. 3.14)
включает в первую очередь пренилирование п-гидроксибензоа-
та геранилпирофосфатом с образованием промежуточного про-
дукта (3.69). Затем следует окислительная циклизация и вве-
дение кислородсодержащих групп.
Гл 3 Хиноны
117
Антрахиноны. Биосинтез антрахинонов в растениях сем.
Rubiaceae, Bignoniaceae и Verbenaceae, по-видимому, довольно
близок к синтезу алканина. Ализарин (3.27) биосинтетически
представляет собой замещенный нафтохинон. В него включает-
ся метка и из шикимата, и из мевалоната. Кольца А и В об-
разуются из шикимата через о-сукцинилбензоат, как это было
описано выше для биосинтеза юглона (рис. 3.12), в то время
как кольцо С образуется из мевалоната. Одна метильная груп-
па пренильной боковой цепи в ходе образования ализарина
теряется, однако у других близких антрахинонов она сохра-
няется (иногда в окисленной форме).
3.6.6. Фенольная конденсация
Фенольная конденсация представляет собой биосинте-
тический процесс, характерный для организмов, синтезирующих
фенольные соединения и хиноны. В ходе этого процесса проис-
ходит димеризация по свободнорадикальному механизму и об-
разуется связь между весьма реакционноспособными группами
мономеров. Многие из встречающихся в природе крупных хи-
нонов имеют структуру, свидетельствующую о том, что они
являются димерами или другими продуктами конденсации наф-
тохинонов или антрахинонов. Иногда в ткани, где они нахо-
дятся, присутствуют также и мономеры. Общеизвестным при-
мером подобной ситуации служит одновременное присутствие
7-метилюглона (3.70) и нескольких его димеров (3.71—3.73)
у видов Drosera и Diospyros, в том числе и в древесине эбено-
вого дерева, где некоторые из них являются предшественника-
ми черного пигмента.
В тех или иных случаях может происходить несколько по-
следовательных фенольных конденсаций. Так, гиперицин яв-
ляется диантрахиноном, в котором мономеры соединены в трех
местах.
Фенольная конденсация происходит также при образовании
протоафинов у тлей и при посмертной трансформации прото-
афинов в ксантоафины, однако химия этих процессов гораздо
сложнее, чем до сих пор рассмотренных.
Образование полимеров (меланинов) с помощью той же
фенольной конденсации хиноидных мономеров будет описано
в гл. 7.
3.6.7. Общая оценка
Сложность биосинтеза хинонов заключается в том,
что он может происходить по совершенно различным альтер-
нативным путям. Грибы, по-видимому, образуют хиноны в
основном с помощью поликетидного пути, тогда как у высших
118
Часть I Химические и биохимические аспекты
растений они обычно синтезируются несколькими путями, в том
числе и шикиматным, нередко в комбинации с изопреноидным;
в последнем с участием мевалоната формируется часть систе-
мы колец нафтохинонов и антрахинонов.
Хиноновые пигменты животных, например спинохромы и
эхинохромы иглокожих и антрахиноны насекомых и криноид»
по-видимому, не поступают в организм с пищей. У этих живот-
ных происходит самостоятельный биосинтез хинонов, и в из-
ученных случаях было показано, что он происходит по полике-
тидному механизму. Не исключено, что эти хиноны образуются
микроорганизмами-симбионтами; действительно, эта возмож-
ность в некоторых случаях кажется весьма вероятной, напри-
мер в случае биосинтеза пигмента насекомых — карминовой
кислоты (3.28).
3.7. Функции и биологическое действие
У хиноновых пигментов нет какой-либо одной общей
функции. Они не играют важной роли в качестве пигментов
в фотосинтезе или фоторецепторов в других процессах. Они
Гл 3 Хиноны
119
редко имеют первостепенное значение в качестве пигментов,
придающих внешнюю окраску. Обычно хиноновые пигменты
вносят лишь незначительный вклад в окраску организмов и
тканей, в которых они образуются и накапливаются.
3.7.1. Окрашивание
Известно несколько примеров окрашивания (обычно
в желтый, оранжевый или коричневый цвет) грибов и лишай-
ников нафтохинонами и антрахинонами. Ярко-красная, пурпур-
ная и даже синяя или зеленая окраска некоторых беспозво-
ночных животных, например морских ежей и ряда близких
к ним форм, некоторых криноид (морских лилий), червецов и
тлей—наиболее известные примеры проявления пигментных
свойств хинонов. Обычно хиноны не являются пигментами выс-
ших растений, однако у немногих видов они ответственны за
желтую окраску.
3.7.2. Функции, не связанные с поглощением света
Многие хиноны обладают важными биологическими
функциями, которые не зависят от их светопоглощающих
свойств. Даже в тех случаях, когда соединение может погло-
щать свет в видимой части спектра и, следовательно, окраше-
но, это свойство не используется при функционировании моле-
кул данного вещества.
Перенос электронов. Возможно, наиболее существенную
роль играют изопренилированные молекулы убихинона и мена-
хинона, которые являются дыхательными коферментами в си-
стемах переноса электронов у животных, растений и микроор-
ганизмов, а также близкий к ним по строению хинон — пласто-
хинон фотосинтетических систем переноса электронов в хлоро-
пластах (гл. 10). Важным свойством в данном случае являет-
ся легкость и обратимость восстановления хинонов через семи-
хиноновый радикал до гидрохинонов.
Биологическое действие хинонов, синтезируемых одними
организмами, на другие организмы. Многие хиноны обладают
разрушающим действием, и поэтому они могут синтезироваться
организмами в целях обеспечения безопасности или защиты.
Простые бензохиноны — жгучие, токсичные и едкие — исполь-
зуются некоторыми насекомыми, например жуком-бомбарди-
ром, в качестве защитных выделений, которые могут «выстре-
ливаться» нагретыми до высокой температуры прямо в напа-
дающего Некоторые грибные нафтохиноны и антрациклиноны
обладают антибактериальными или противовирусными свой-
ствами и поэтому обеспечивают соответствующую защиту про-
120
Часть I Химические и биохимические аспекты
дуцирующего их организма. Простой нафтохинон юглон (3.17),
синтезируемый грецким орехом, токсичен для многих других
растений.
По крайней мере в одном случае функции хинонов обеспечи-
вают скорее нападение, а не защиту. Так, фитопатогенный
плесневый гриб Fusarium martii образует несколько нафтохино-
нов Как было показано, один из них, мартицин (3 74), вызы-
вает увядание растения-хозяина и является, таким образом,
важным компонентом атакующего механизма патогена.
Известно, что некоторые хиноны оказывают раздражающее,
токсическое или разъедающее действие на ткани человека и
(3 74) Мартицин
других млекопитающих. Наблюдающаяся у многих людей
аллергическая реакция при контакте с популярным комнатным
растением Primula obconica вызывается бензохиноном прими-
ном (3.75). Некоторые нафтохиноны, содержащиеся в древе-
сине, могут вызвать раздражение, чихание и экзему у плотни-
ков и столяров, которым приходится дышать воздухом, содер-
жащим древесную пыль.
Сложный высший хинон гиперицин (3 37) и близкие к нему
соединения обусловливают токсичность растений рода Hyperi-
cum (зверобой) для животных. Гиперицин, по-видимому, дей-
ствует на животных как фотосенсибилизатор, вызывающий фо-
тоокисление светочувствительных белков. Такое действие мо-
жет быть причиной воспалений, отеков, а в наиболее тяжелых
случаях и гибели животного.
3.7.3. Спинохромы и эхинохромы
Относительно функций спинохромов и эхинохромов у
иглокожих было сделано много предположений. К, настоящему
времени большая часть из них либо была опровергнута, либо,
в лучшем случае, не подтвердилась. Эти соединения обладают
алгистатическими свойствами и могут быть полезны животным
как вещества, подавляющие рост паразитических сине-зеленых
водорослей.
Гл- 3. Хиноны
121
3.8. Использование в промышленности и медицине
3.8.1. Красители
В античные времена некоторые хиноновые пигменты
очень ценились как красители. Растения, служившие источни-
ками этих пигментов, часто выращивали на больших полях и
плантациях для снабжения сырьем красильщиков. Вероятно,
наиболее широко распространенной из античных красок была
марена, получаемая из измельченных корней Rubia tinctorum,
которую применяли древние персы, египтяне, греки и римляне
Красящим веществом марены является смесь антрахинонов,
•среди которых преобладает ализарин (3.27), который служит
он о
(3 76) Мориндон
основой стабильных красных красок. Хотя ализарин сейчас
практически вышел из употребления как краситель, он все еще
находит применение при изучении роста костей.
Хна — желто-коричневый препарат из листьев Lawsonia
alba в течение долгого времени использовалась в Африке и на
Востоке для окрашивания и в косметических целях. Следы хны
до сих пор обнаруживают на ногтях египетских мумий. Кра-
сящим веществом ее является нафтохинон лавсон (3 18), кото-
рый легко экстрагируется из листьев L. alba водным раствором
карбоната натрия.
В качестве примеров можно привести также экстракты, со-
держащие нафтохинон алканин (3 68) и его эпимер шиконин,
которые использовались римлянами и древними японцами, и
корни моринды, применявшиеся в Индии, на Яве и в Новой
Зеландии, красящим веществом которых являются антрахино-
ны, подобные мориндону (3.76).
Наконец, необходимо упомянуть о хиноновых красителях,
приготовляемых из насекомых. Вероятно, наиболее известным
античным красителем этого типа является кармин, который
получали из огромного числа червецов Kermococcus ilicis. Его
красящую основу составляет антрахинон кермезиновая кисло-
та (3.29). Сходное соединение карминовая кислота (3.28) из
другого насекомого — Dactylopius coccus — является активным
началом кошенили, впервые использованной, вероятно, инками
в Перу, и до сих пор в небольших количествах применяющей-
122
Часть I. Химические и биохимические аспекты
ся для окрашивания пищевых продуктов, а также гистологиче-
ских препаратов.
С развитием современной синтетической химии крупномас-
штабное производство и использование природных хиноновых
красителей сильно сократилось; лишь некоторые из них ис-
пользуются до сих пор в небольших количествах в качестве
красителей для пищевых продуктов, вин и косметики.
3.8.2. Использование в медицине
Еще в древнейшие времена препараты хинонов приме-
няли в медицине в качестве слабительных. Нафтохиноны и ан-
трахиноны ответственны за хорошо известное и широко исполь-
зуемое послабляющее действие александрийского листа, кру-
шины и ревеня.
I । 1 1 1
Me ОАс ОН ОН Me
(3 77) Стрептоварицин А (стереохимия не показана)
В последнее время у некоторых природных хинонов обна-
ружена противовирусная, антибактериальная и фунгицидная
активность, причем число таких хинонов растет. В некоторых
случаях их можно использовать в качестве антибиотиков при
лечении инфекционных заболеваний. Так, стрептоварицины,
например (3.77), представляющие собой оранжевые макролид-
ные нафтохиноны, проявляют заметную активность in vivo
против Mycobacterium tuberculosis. Антрациклиноны также об-
ладают антибиотической активностью, подобной активности
тетрациклинов, которых они напоминают по структуре.
3.9. Заключение и комментарии
Каротиноиды, описанные в предыдущей главе, пред-
ставляют собой однородную группу пигментов и являются
производными одного и того же основного углеродного скелета.
Гл. 3. Хиноны
123
Для хинонов наблюдается существенно иная картина. Эта
группа включает соединения с различными углеродными ске-
летами и системами колец, обладающие, однако, одним общим
свойством — циклической ендионовой, или хиноновой, структу-
рой. Сколько-нибудь исчерпывающий систематический обзор
состава хинонов природных тканей в настоящее время отсут-
ствует, однако совершенно очевидно, что химикам-органикам
еще предстоит открыть и установить структуру значительного
числа разнообразных новых хинонов. Возможно, при этом
будут решены некоторые сложные стереохимические проблемы.
Имеется также ряд неисследованных возможностей и в
проблеме биосинтеза хинонов. В настоящее время выяснено,
что они синтезируются по двум (иногда трем) главным био-
синтетическим путям, однако пока детали этих путей изучены
на слишком малом числе примеров. Необходимо отметить, что
в данном случае мы имеем интересную ситуацию, когда два
•совершенно различных пути используются для биосинтеза
очень сходных и даже одних и тех же соединений. Это позво-
ляет разрабатывать вероятные пути и механизмы биосинтеза
индивидуальных природных хинонов. Существует также широ-
кий простор для экспериментального выяснения биосинтетиче-
ских путей, ведущих к тем или иным соединениям, и изучения
принимающих в них участие ферментных систем, которые,
за очень редкими исключениями, остаются совершенно неис-
следованными. Пока нет никаких данных, касающихся регуля-
ции биосинтеза хинонов. К проблемам, созревшим для биохи-
мического изучения, можно отнести также локализацию хино-
новых пигментов внутри клетки, возможную связь хинонов
с белками или другими веществами и функции хинонов в
тканях.
Чем больше становится известно о действии, оказываемом
(некоторыми хинонами на живые ткани, и чем яснее установлен
механизм этого действия, тем больший интерес вызывают хи-
ноны как лекарственные средства, и это в свою очередь стиму-
лирует поиски новых природных типов и структур ХИНОНОВ.
3.10. Рекомендуемая литература
В качестве дополнительного источника сведений о при-
родных хинонах следует воспользоваться всеобъемлющей
монографией Томсона (Thomson, 1971), которая дает исчерпы-
вающий обзор химии всех природных хинонов, в том числе
пигментов, которые были открыты к 1970 г. Она также содер-
жит живое описание первых работ по установлению структуры
некоторых соединений классическими методами и заниматель-
ную историю использования некоторых важнейших природных
124
Часть I Химические и биохимические аспекты
хиноновых красителей В этой же книге и в более позднем
обзоре того же автора затронут также биосинтез хинонов
(Thomson, 1976а) Детали биосинтеза ароматических соеди-
нений (в том числе и хинонов) по шикиматному пути обсуж-
даются в обзоре Хэслама (Haslam, 1979) Два тома пр. химии
хиноидных соединений (Patai, 1974) содержат важную инфор-
мацию о многих природных хинонах Полезные сведения из
ранних работ по химии и биохимии хинонов можно найти в
более старой монографии Мортона (Morton, 1965) С экспери-
ментальными методами, используемыми в исследованиях хино-
нов, можно познакомиться в обзоре Томсона (Thomson, 1976).
Подробности методов в применении к тем или иным индиви-
дуальным хинонам обсуждаются в работах, на которые ссы-
лается в своей монографии Томсон (Thomson, 1971)
3.11. Дополнительная литература
Haslam Е (1979) Shikimic acid metabolites In Comprehensive organic cne-
mistry, vol 5, ed E Haslam, p 1167, Oxford and London, Pergamon
Morton R A (ed) (1965) Biochemistry of quinones, London, Academic Press.
Patai S (ed) (1974) The chemistry of the quinonoid compounds, vols 1 and 2,
London and New York, Wiley
Thomson R H (1971) Naturally occurring quinones, 2nd edition London and
New York, Academic Press
Thomson R H (1976a) Quinones, nature, distribution and biosynthesis In:
Chemistry and biochemistry of plant pigments, 2nd edition, vol 1, ed.
T W Goodwin, p 527, London, New York and San Francisco, Academic
Press
Thomson R H (1976) Isolation and identification of quinones In Che-
mistry and biochemistry of plant pigments, 2nd edition, vol. 2, ed.
T W. Qoodwm, p 207, London, New York and San Francisco, Academic
Press
Глава 4
О-Гетероциклические пигменты — флавоноиды
4.1. Введение
Флавоноиды синтезируются почти исключительно выс-
шими растениями В их число входят антоцианы, ответствен-
ные за наиболее яркие цвета, в которые окрашены растения,—-
интенсивно красные, пурпурные и синие цветки и плоды. Бла-
годаря своей яркой окраске они контрастно выделяются на
фоне зеленой листвы и тем самым привлекают внимание чело-
века и животных Другие флавоноиды также могут вносить
свой вклад в формирование окраски растений, хотя на первый
взгляд этот вклад и неочевиден.
4.2. Структура и номенклатура
4.2.1. Структурные классы
Все природные флавоноиды являются О-гетероцикли-
ческими соединениями, структурную основу которых составляет
трициклическая молекула флавона [2-фенилхромона, или 2-фе-
нилбензо-у-пирона (4 1)] или флавана [2-фенилбензопирана
(42)]. Таким образом, основная флавоноидная структура со-
стоит из двух бензольных колец, обозначаемых А и В, соеди-
ненных Сз-фрагментом, который вместе с кислородным атомом
образует у-пироновое кольцо Порядок нумерации атомов в.
такой структуре показан в формуле (4.1). Атомам бензопиро-
новой системе присвоены номера без штрихов, в то время;
как кольцо В рассматривается как заместитель и его атомам;
даны номера со штрихами
(4 1) Флавон
(4 2) Флаван
(4 4) Флавонол
(4 5) Антоцианидин
(4 8) Флаван 3 ол
(2R 3S) — Катехин
{2R 3R} — ЭпикатехиЙ
(4 9) Флаванонол [2R, 3R}
(4 10) Флаван 3 4-диол
(2R 3R, 4R) и эпимеру
Гл. 4. О-Гетероциклические пигменты — флавоноиды
127
(4.12) Халкон
(4.13) Аурон
Рис. 4.1. Основные скелеты главных классов флавоноидов.
Связующий С3-фрагмент, который образует гетероцикличе-
ское кольцо, определяет класс, к которому принадлежит тот
или иной флавоноид. Этот фрагмент может быть представлен1
несколькими состояниями, которые имеют различную степень
окисления. Каждое из этих состояний соответствует отдельному
классу флавоноидов, как это показано на рис. 4.1. Наиболее
важными из этих классов являются флавоны (4.3), их 3-гидро-
ксипроизводные— флавонолы (4.4) и антоцианидины (4.5),
в которых гетероциклическое кольцо при выделении их из.
кислых растворов находится в форме соли флавилия. Реже
встречаются 3-дезоксиантоцианидины (4.6). Производными
флавана являются флаваноны (4.7), флаван-3-олы, или катехи-
ны (4.8), флаванонолы, или дигидрофлавонолы (4.9), и фла-
ван-3, 4-диолы, или проантоцианидины (4.10). На рис. 4.1 пока-
заны также некоторые классы веществ, родственные флаво-
ноидам,— изофлавоны (4.11), халконы (4.12) и ауроны (4.13).
Хотя эти последние и не обладают 2-фенилхромоновым скеле-
том и потому, строго говоря, не являются флавоноидами, они
тем не менее столь близки к флавоноидам химически и био-
синтетически, что их всегда включают в группу флавоноидов.
Отметим, что, если различные классы истинно флавоноидных
соединений обычно именуют производными флавона или фла-
вана, для одной из групп сохранено старое название антоциа-
f28
Часть I Химические и биохимические аспекты
нидины; тривиальные названия часто используются и для не-
которых других групп, например для катехинов. Другие ста-
рые термины, такие, как антоксантин, антохлор и химохром,
в настоящее время используются редко.
4.2.2. Характер замещения в кольце (рис. 4.2)
Внутри каждого класса флавоноиды характеризуются
числом и положением заместителей в ароматических кольцах.
‘Обычно эти заместители являются ОН-группами, которые мо-
тут быть метилированными или гликозилированными. Локали-
₽ИС. 4 2. Наиболее обычное расположение заместителей в молекулах флаво-
* '* ’ ноидов.
зация некоторых ОН-групп является следствием общего био-
синтетического пути (разд. 4.6). У большинства флавоноидов
кольцо А имеет ОН-группы либо при С-7, либо при С-5 и С-7.
Эти ОН-группы редко метилируются. Кольцо В практически
всегда гидроксилировано при С-4'1 и обычно также при С-З' и
С-5'; в этих последних двух положениях ОН-группы часто ме-
тилированы
•4.2.3. Гликозилирование
In vivo флавоноиды существуют в большинстве слу-
чаев, а возможно, и всегда в виде гликозидов. У тех флаво-
ноидов, которые обладают С-З-гидроксигруппой, в особен-
1 Известны флавоноиды, не гидроксилированные у атома С 4' кольца В,
-Хотя они более редко встречаются в природе. — Прим ред.
Гл 4 О Гетероциклические пигменты — флавоноиды 129
ности у антоцианидинов и флавонолов, это положение являет-
ся преимущественным для гликозилирования, однако гликозид-
ные остатки при С-7, С-4' или С-5 также встречаются очень
часто. Не является необычным присутствие сахаров более чем
в одном положении флавоноидной молекулы, а также гликози-
лирование последней ди- или трисахаридами. Природные пиг-
менты обычно представляют собой р-гликозиды с D-глюкозой
в качестве наиболее широко распространенного моносахарида,
однако L-рамноза и D-галактоза также встречаются часто,
тогда как другие сахара — реже. В некоторых случаях сахар
может быть ацилирован фенолкарбоновой кислотой, обычно
гидроксикоричной.
Нет ничего необычного и в присутствии в экстракте свобод-
ного флавоноида (агликона). Однако свободные флавоноиды
отсутствуют в ткани in vivo; они быстро образуются в ходе
экстракции под действием гликозидазных ферментов, которые
в ряде случаев продолжают функционировать даже в присут-
ствии высоких концентраций органических растворителей.
4.2.4. Некоторые важные примеры
Наиболее важными флавоноидными пигментами расте-
ний in vivo являются антоцианы (антоцианы представляют со-
бой гликозиды, соответствующие им агликоны носят название
Общая структура антоцианидинов
(4 14) R1 =R2 =Н Пеларгонидин
(4 15) R1 =ОН , R2 =Н Цианидин
(4 16) R1 =R2 ОН Дельфинидин
Общая структура флавонолов
(4 17) R1 = R2 =Н Кемпферол
(4 18) R1 =ОН , R2 =Н Кверцетин
(4 19) R1 = R2 ОН Мирицетин
антоцианидинов). Среди них наиболее широко распространены,
особенно в лепестках цветков, пеларгонидин (4 14), цианидин
(4.15) и дельфинидин (4 16). Флавонолы с ОН-группами в со-
ответствующих положениях в кольцах А и В также широко
распространены — это кемпферол (4.17), кверцетин (4.18) и
Мирицетин (4.19).
9—84
130
Часть I. Химические и биохимические аспекты
Число индивидуальных флавоноидных агликонов велико;
оно включает около 300 флавонов и флавонолов, 50 халконов,
20 антоцианидинов. Число же их возможных гликозидов и
ацилированных гликозидов поистине огромно. В этой книге
будут рассмотрены лишь немногие из флавоноидов. Однако
представленная ниже информация относится ко всем предста-
вителям данного флавоноидного класса.
4.2.5. Номенклатура
Как видно даже на примере упоминавшихся выше
соединений, среди названий флавоноидов преобладают три-
виальные, обычно являющиеся производными от названия
того растения, из которого данное соединение было впервые
выделено или в котором оно обычно содержится, например
дельфинидин был выделен из дельфиниума. Это приводит к
затруднениям и недоразумениям, поскольку даже очень близ-
кородственные соединения, например разные гликозиды одного
и того же агликона могут иметь совершенно несходные наи-
менования. Как выражение крайнего случая иногда для одного
и того же соединения используют два совершенно различных
названия.
4.3. Свойства
4.3.1. Химические свойства
Химические свойства флавоноидов уже давно интен-
сивно исследовались методами классической органической хи-
мии. С их помощью было разработано несколько главных пу-
тей синтеза флавоноидов. Флавоноиды обычно вступают в реак-
ции, характерные для их замещающих групп, например гидро-
ксильных. Связующий Сз-фрагмент гетероциклического кольца
может подвергаться восстановлению или окислению; при этом
возможны ограниченные превращения флавоноидов одного
класса в флавоноиды другого. Щелочное расщепление, часто
требующее жестких условий, приводит к разрыву флавоноид*
ной молекулы на два фрагмента, содержащие бензольные
кольца. Эта реакция может оказаться полезной для установле-
ния распределения заместителей в кольце, правда, подобную
информацию сейчас обычно получают спектроскопическими ме-
тодами (УФ- и ЯМР-спектроскопией, масс-спектрометрией).
4.3.2. Физические свойства
Природные флавоноидные гликозиды сравнительно не-
плохо растворимы в воде и могут экстрагироваться ею или
(как это обычно и делают) водными растворами спиртов из
Гл. 4. О-Гетероциклические пигменты — флавоноиды 131
растительных тканей. Антоцианы несут положительный заряд,
и их экстракция требует умеренно кислых условий. Обыч-
но флавоноиды более стабильны в отношении света, умерен-
ного нагревания и изменений pH, чем большинство других
пигментов1.
Очистка. Разделение и очистка флавоноидов легко дости-
гаются с помощью хроматографии на бумаге или тонкослойной
хроматографии. Для быстрого получения результатов при опре-
делении в растительных тканях флавоноидов обычно применяют
двумерную хроматографию. Даже бесцветные соединения легко
обнаруживаются при освещении хроматограмм УФ-светом,
особенно в присутствии паров аммиака, когда наблюдается
характерная флуоресценция.
4.3.3. Поглощение света
Как уже отмечалось в гл. 1, длина волны света, по-
глощенного молекулой, является функцией той степени лег-
кости, с которой происходит переход электронов на более вы-
сокие энергетические уровни. Чем длиннее сопряженная цепь
или хромофор молекулы и чем больше число участвующих в
нем функциональных групп или ауксохромов, тем выше сте-
пень стабилизации возбужденного состояния и тем легче оно
достигается, т. е. тем меньшая энергия требуется для возбуж-
дения и тем большей длины свет поглощается. Это утвержде-
ние очень хорошо иллюстрируется на примере флавоноидов.
Производные флавана (рис. 4.3). У гидроксилированных
производных флавана, например у флаван-3-олов (4.8) и фла-
ван-3,4-диолов (4.10), одинарная связь С-3,4 эффективно раз-
деляет два бензольных кольца. У этих соединений, таким об-
разом, переходы происходят только в изолированных хромо-
форах бензольных колец, и они поглощают только в УФ-диа-
пазоне спектра при 275—280 нм, как и соответствующие про-
стые фенолы. Флаваноны (4.7) и изофлавоны (4.11) обладают
кольцом А, сопряженным с С-4-карбонильной группой, и пото-
му имеют такой же максимум поглощения, как гидроксиацето-
фенон.
Производные флавона (рис. 4.4). У флавонов (4.3) и флаво-
нолов (4,4) имеет место сопряжение между С-4-карбонильной
группой и кольцом В, и потому спектры имеют две интенсив-
ные, хорошо разделенные полосы поглощения. Полоса I в длин-
1 Флавоноиды некоторых классов весьма неустойчивы к действию солнеч-
ного света, а также в условиях щелочных значений pH. Это относится преж-
де всего к катехинам (4.8), проантоцианидинам, флаван-3,4-диолам (4.10),
а также к антоцианидинам (4.5) и их гликозидам антоцианам. — Прим ред.
9*
132
Часть I. Химические и биохимические аспекты
Рис. 4.3. Спектры поглощения УФ света этанольными растворами производ-
ных флавана: флаван-3,4-диола (/) и флаванона (II).
Рис. 4.4. Спектры поглощения света этанольными растворами флавона (А)
и флавонола (5).
новолновой части связана с хромофором, состоящим из коль-
ца В, сопряженного с карбонильной группой, а более коротко-
волновая полоса II — с кольцом А. В случае флавонолов ви-
нильная ОН-группа при С-3 влияет на подвижность электрона.
Полоса I смещается в более длинноволновую область, чем у
соответствующих флавонов, и поглощение в синем диапазоне
Гл. 4. О-Гетероциклические пигменты — флавоноиды
133
Рис. 4 5 Спектры поглощения света этанольными растворами халкона (7)
и аурона (77)
Рис 4 6 Спектры поглощения света антоцианидином 7 — при pH 1,0 преоб-
ладает соль флавилия (4 20); 77 — при pH 4,0 преобладает псевдоосновная
форма (4 21), 777 — при pH 7,5 преобладает форма ангидрооснования (4 22).
спектра может оказаться достаточным для того, чтобы флаво-
нол приобрел желтую окраску
Халконы и ауроны (рис 4 5) Большая копланарность, воз-
можная у халконов (4 12), а еще в большей степени у ауронов
(4 13), облегчает возбуждение электронов и, таким образом,
134
Часть I. Химические и биохимические аспекты
способствует сдвигу Лтах в длинноволновую часть спектра. По-
этому халконы желтые, а ауроны оранжевые.
Антоцианидины (рис. 4 6). У антоцианидинов и их гликози-
дов, обычно выделяемых в кислом растворе в виде флавилие-
вых солей (4.20), электрон гетероциклического атома кислоро-
(4 20) Соль флавилия (цианцдин)
да принимает участие в образовании л-связей в гетероарома-
тическом кольце, так что хромофором становится вся молеку-
ла. Поэтому из всех флавоноидов антоцианидины поглощают
свет с наибольшей длиной волны и окрашены в оранжевый,
красный, пурпурный или синий цвет. Однако по мере повыше-
(4 22) Хиноидное основание
Гл 4 О-Гетероциклические пигменты — флавоноиды 135
ния pH раствора антоцианидинов до нейтрального, образуется
все больше и больше бесцветных псевдоосновных форм [2-гид-
роксихроменов (4.21)]. При величинах pH выше 7 в неболь-
шом количестве образуются хиноидные основания (4.22), по-
глощающие в более длинноволновой части спектра и окрашен-
ные в синий цвет; в водных растворах эти основания неустой-
чивы Однако они являются антоциановой формой, которая,,
видимо, стабильна in vivo и содержится главным образом в-
лепестках цветков, которые часто имеют синий цвет более глу-
бокого тона, чем это можно было бы предполагать по спектрам
Рис 4 7 Влияние замещающих ОН-групп на спектры поглощения света ан-
тоцианами Спектры (в этаноле+HCl) I — гликозидов пеларгонидина (4 14);
II — цианидина (4 15), III — дельфинидина (4 16)
поглощения выделенных антоцианов (в форме флавилиевых
солей).
Влияние заместителей У флавоноидов всех классов ОН-за-
местители вносят несвязывающие электроны, чем увеличивают
степень делокализации, стабилизируют возбужденное состояние
и тем самым облегчают возбуждение электронов. Влияние
увеличения степени гидроксилирования на светопоглощающие
свойства хорошо видно на примере спектров трех обычных
антоцианидинов (рис. 4.7) Пеларгонидин (4 14), цианидин
(4 15) и дельфинидин (4 16) с одной, двумя и тремя ОН-груп-
пами в кольце В соответственно имеют оранжевую, красную
и пурпурную окраску. Сходный эффект может быть обнаружен
и у флавоноидов других классов; например, флавонолы кемп-
ферол (4.17), кверцетин (4 18) и мирицетин (4 19) имеют
taiax (в этаноле) при 368, 374 и 478 нм соответственно ОН-
группы в других положениях молекулы не дают такого эффек-
136
Часть I. Химические и биохимические аспекта
та. Метилирование, гликозилирование или ацилирование ОН-
групп обычно уменьшает или вообще исключает этот батохром-
ный эффект.
Влияние pH и хелатирования. О влиянии изменений величи-
ны pH на ионизацию и спектры поглощения антоцианидинов
мы уже упоминали. Все классы флавоноидов представляют
собой фенолог, и поэтому их спектры заметно изменяются при
высоких pH (выше pH 9), когда происходит ионизация феноль-
ных ОН-групп, а также при образовании хелатов с ионами
металлов. (В случае антоцианидинов и их гликозидов при вы-
соких значениях pH наблюдается необратимый распад этих
соединений.) При этом их максимумы поглощения сдвигаются
в более длинноволновую часть спектра и многие флавоноиды
приобретают яркую окраску. Ионизация флавонолов и род-
ственных им соединений, вызываемая выдерживанием в парах
аммиака, часто используется для обнаружения их на хромато-
граммах.
4.4. Распространение
4.4.1. У животных
Сообщений о присутствии флавоноидов у животных
очень мало. Флавоны и флавонолы широко, хотя отнюдь не
повсеместно, обнаруживаются у личинок и взрослых особей
отряда Lepidoptera, особенно у белых и кремовых бабочек.
Эти соединения насекомые, вероятно, получают из растений,
которыми они питаются. Так, например, гусеница Eupithecia
oblongata может быть окрашена в синий или в красный цвет
в зависимости от антоциана, содержащегося в цветках, кото-
рыми она питается. Такая способность принимать окраску,
подобную окраске окружающей среды, явно полезна насеко-
мому в целях маскировки. Флавоноиды и флавоноидоподобные
соединения были идентифицированы также в тканях виноград-
ной улитки Helix pomatia (флавон?), у морского гидроида
Sertularella и пресноводного жука-плавунца Dytiscus (аурон).
Присутствие флавоноида у названного последним вида (хищ-
ника) является неожиданным и необъяснимым.
4.4.2. У микроорганизмов
Синтез флавоноида у микроорганизмов был убеди-
тельно показан лишь недавно. Плесневый гриб Aspergillus сап-
didus продуцирует флавонин (4.23), очевидно, по тому же са-
мому биосинтетическому пути, который используется высшими
растениями.
Гл. 4. О-Гетероциклические пигменты — флавоноиды
137
4.4.3. У высших растений
Флавоноиды являются почти исключительно пигмен-
тами высших растений. Они могут синтезироваться и накапли-
ваться во всех растительных тканях — в листьях, древесине,
корнях, плодах, семенах и во всех частях цветков, главным
образом в лепестках. Природные гликозиды растворимы в воде
и обнаруживаются обычно в клеточном соке или в вакуолях.
В недавних сообщениях высказано предположение, что не-
большие количества флавоноидов могут присутствовать в хло--
ропластах высших растений.
Были проведены широкие систематические исследования со-
става флавоноидов в высших растениях различных видов и
сортов. Для получения фингерпринтов, позволяющих быстро
определять состав флавоноидов у того или иного вида расте-
ния, использовали двумерную хроматографию на бумаге. Было
показано, что состав флавоноидов является чрезвычайно важ-
ным таксономическим признаком как при установлении родства
между видами, принадлежащими к одному семейству, так и
при выявлении различий между близкородственными видами.
В этом отношении может быть очень характерным присутствие
флавоноидов тех или иных классов и распределение замести-
телей у индивидуальных соединений.
4.5. Участие в формировании окраски растений и
4.5.1. Антоцианы
Из всех флавоноидов именно антоцианы вносят наи-
больший вклад в (формирование окраски растений. Эти соеди-
нения ярко окрашены в оранжевый, красный, пурпурный или
синий цвет и обусловливают окраску почти всех красно-синих
цветков. Известный пример — красная роза, за окраску кото-
рой ответственны производные цианидина. Была установлена
четкая корреляция между окраской цветков и структурой анто-
цианов, которые в них содержатся. В ходе систематического
138
Часть I. Химические и биохимические аспены
исследования было обнаружено, что большинство оранжевых
цветков содержат пеларгонидин, красно-вишневых — цианидин,
а пурпурно-синих — дельфинидин.
Антоциановая окраска характерна для многих красных
плодов, таких, как земляника, малина, вишня, яблоки, в кото-
рых наличие антоцианов является признаком зрелости. Боль-
шинство «черных» плодов, например ежевика, черный вино-
град, на самом деле окрашены в очень глубокий красный или
пурпурный цвет благодаря присутствию антоциана в чрезвы-
чайно высоких концентрациях. Это утверждение прекрасно ил-
люстрируется тем фактом, что черный виноград дает красное
вино, в котором содержание антоцианов уже значительно ни-
же. Окрашенными благодаря присутствию антоцианов могут
быть и другие части растений, такие, как листья (краснокочан-
ная капуста) или стебли (ревень).
Антоцианы часто образуются в большом количестве в моло-
дых побегах и листьях, которые поэтому приобретают красную
окраску в отличие от зеленой у зрелых листьев. Общеизвест-
ным примером служит темно-красная окраска стеблей и
листьев у первых весенних побегов розы. В некоторых случаях
красный антоциан сохраняется до зрелости, обусловливая
красную окраску листвы некоторых декоративных видов. Крас-
ный цвет осенних листьев также может быть следствием уси-
ленного Синтеза антоцианов. Происходящее осенью разруше-
ние хлорофилла делает антоциановую окраску более заметной.
Окраска, обусловленная антоцианами, особенно цветков и
плодов, в которых доминируют хиноидные формы пигментов
(4.22), может зависеть от таких факторов, как pH, образова-
ние хелатных комплексов с металлами и копигментация. Влия-
ние изменений pH на длину волны поглощаемого антоцианами
света уже обсуждалось (разд. 4.3.3). Антоцианы, особенно те,
которые содержат орто-ДиГидроксизамёстители в кольце В,
способны образовывать хелатные комплексы с ионами метал-
лов, ЧТО обычно приводит К сдвигу Zmax В ДЛИННОВОЛНОВУЮ
сторону, т. е. их синяя окраска становится более темной. Хо-
рошо известен пример гортензии Hydrangea macrophylla. У это-
го растения один и тот же антоциан дельфин-идин-3-глюкозид
присутствует в чашелистиках как розовых, так и синих цвет-
•йЪв, оДн&кб в сйних цйетках он образует комплексы с ионами
Р*^МОния и молибдена. Обогащение почвы соответствующими
Ион аки металлов приводит к появлению синих цветков на кус-
Имевшем ранее розовые Цветки. И наоборот, дефицит этих
сблей может вызвать превращение синих цветков в розовые.
ь Многие садоводческие программы предусматривают выве-
дёнйё “разновидностей цветков с «лучшей», т. е. более интен-
ЙгЬнбЙ’ Или редкой окраской или необычным ее распределе-
Гл. 4. О-Гетероциклические пигменты — флавоноиды
139
нием. Говоря языком биохимии, цель заключается в достиже-
нии качественных или количественных изменений состава ан-
тоцианов. В качестве примера можно назвать попытки создать
голубую розу. В этом случае цель заключалась в том, чтобы
создать голубую окраску с помощью либо соответствующей
копигментации, либо с помощью замены цианидина на дель-
финидин. Последней замены достичь чрезвычайно трудно, по-
скольку роза в нормальных условиях не обладает необходи-
мыми ферментами для осуществления добавочного гидроксили-
рования.
4.5.2. Другие флавоноиды
Вклад в окраску растений других классов флавонои-'
дов обычно менее очевиден, правда халконы, и особенно ауро-
ны, иногда ответственны за окраску желтых цветков, например
у львиного зева Antirrhinum majus.
Флавоны и флавонолы практически не поглощают свет ви-
димой области спектра, но тем не менее ответственны за бе-
лую и кремовую окраску цветков, которые без них были бы
почти прозрачными. Необходимо также отметить, что флавоны
и флавонолы поглощают свет в области ближнего УФ, «види-
мого» для пчел и других насекомых. Хотя эти соединения
и цветки, содержащие их, кажутся нам белыми или бес-
цветными, они выглядят «окрашенными» для пчел и таким
образом помогают привлекать насекомых. Флавоны и флавоно-
лы практически всегда содержатся в листьях, хотя из-за при-
сутствия хлорофилла они не вносят никакого вклада в их
окраску.
Мономерные флаваны, флаваноны и их гидроксипроизвод-
ные непосредственно не обусловливают окраску растений.
Вместе с тем димеры, олигомеры и полимеры, главным обра-
зом катехинов и проантоцианидинов, придают коричневую окра-
ску осенним и сухим листьям, а также темной ядровой древе-
сине многих растений. Подобные соединения («конденсирован-
ные дубильные вещества») обусловливают также коричневатую
окраску чая1.
1 Золотисто-вишневый цвет настоя хорошего черного чая обусловлен ди-
мерными продуктами (теафлавинами и теарубигинами), образующимися из
катехинов в процессе ферментации листьев чайного растения. Коричневатая
же окраска настоя свидетельствует о «переферментации». Продукты, обуслов-
ливающие цвет чая, лишь весьма условно могут быть причислены к конденси-
рованным дубильным веществам, поскольку их молекулярная масса слишком
мала. — Прим. ред.
140
Часть I Химические и биохимические аспекты
4.5.3. Совместное окрашивание (копигментация)
Будучи бесцветными или почти бесцветными, гидро-
ксифлаваны, флавоны и флавонолы вносят важный вклад в
окраску многих цветков путем копигментации. Эти соединения
часто присутствуют в цветках вместе с антоцианами и обра-
зуют с ними комплексы, которые поглощают свет более ин-
тенсивно и при больших длинах волн, чем одни только анто-
цианы. Многие синие цветки обязаны своей окраской присут-
ствию именно таких комплексов, причем некоторые из них бы-
ли недавно выделены. Например, пигмент цветков синего ири-
са содержит хиноидное основание ацилированного дельфини-
дин-3,5-диглюкозида, стабилизированное путем копигментации
с С-гликозилфлавонами. В других случаях копигментационный
комплекс может быть стабилизирован ионами металлов, таки-
ми, как Mg2+. Вместе с тем у некоторых синих цветков, на-
пример у Lobelia, окраска обусловлена не копигментацией и
не комплексообразованием с металлами. У этих цветков синее
хиноидное основание само по себе стабилизировано путем
взаимодействия с кофеильными группами ацилированного са-
харного заместителя.
Другие аспекты окрашивания растительных тканей флаво-
ноидами и другими пигментами будут обсуждаться в гл. 8.
4.6. Биосинтез ,
4.6.1. Введение
Биосинтез флавоноидов осуществляется в очень боль-
ших масштабах. Показано, что у высших растений на него
расходуется почти 2% всего углерода, фиксируемого при фото-
синтезе. В общих чертах биосинтез флавоноидов изучен доста-
точно хорошо, однако многие детали, главным образом касаю-
щиеся ферментов, еще ждут своего выяснения. Удобнее всего
рассматривать биосинтетический путь по стадиям: 1) образо-
вание основного Сб-С3-Сб-скелета, включая два главных пути
биосинтеза фенольных соединений — поликетидный и шикимат-
ный, 2) пути, по которым флавоноиды различных классов об-
разуются из основного Сб-С3-Сб-предшественника, и возможные
взаимопревращения между флавоноидами различных классов,
3) окончательные модификации, такие, как гидроксилирование,
метилирование и гликозилирование, которые дают начало мно-
гим индивидуальным флавоноидам внутри каждого класса.
Гл 4 О-Гетероциклические пигменты — флавоноиды
141
4.6.2. Образование основного С6-С3-С6-скелета
Уже давно было показано, что кольцо А флавоноидной
молекулы происходит из ацетатных фрагментов, в то время
как кольцо В и связывающие оба бензольных кольца три угле-
родных атома составляют фенилпропаноидный остаток, проис-
ходящий из шикимата. Подробно шикиматный путь, приводя-
щий к образованию фенилаланина, был рассмотрен в гл. 3.
В биосинтезе флавоноидов фундаментальное значение имеет
реакция, катализируемая ферментом фенилаланин-аммиак-
лиазой (ФАЛ), в ходе которой фенилаланин (4.24) дезамини-
руется (рис. 4.8) с образованием транс-коричной кислоты (4 25).
(4 24) R= Н Фенилаланин
(4 26) R- ОН Тирозин
(4 25) R-H транс Коричная кислота
(4 27) R-OH п Кумаровая кислота
Рис 4.8. Дезаминирование феиилаланииа и тирозина до транс-коричной и
n-кумаровой кислот соответственно.
В некоторых случаях сходным образом тирозин (4.26) дез-
аминируется до n-кумаровой кислоты (4.27), однако послед-
няя обычно образуется путем гидроксилирования коричной
кислоты с помощью 4-гидроксилазы коричной кислоты — окси-
дазы со смешанной функцией, которая использует NADPH и
молекулярный кислород. В ходе этой реакции происходит ин-
тересная внутримолекулярная миграция водорода, так назы-
ваемый NIH-сдвиг1 (рис. 4.9).
Образовавшиеся коричные кислоты затем активируются в
реакции с коферментом А (рис. 4.10), аналогичной синтезу
ацетил-СоА из ацетата. Циннамоил-СоА (4.28) служит затрав-
кой в поликетидном процессе (разд. 3.6.3), в ходе которо-
го он конденсируется с тремя молекулами малоната, ак-
тивированных СоА, или сходного тиоэфира. В результате цик-
лизации поликетидной системы (4.29) образуется гидроксили-
1 Термином «NIH-сдвиг» принято обозначать внутримолекулярную мигра-
цию атома водорода в бензольном ядре (арильном фрагменте) в процессе
монооксигеназного гидроксилирования из пара-положения (С-4) в лета-поло-
жение (С-З). Такая миграция (сдвиг) была впервые обнаружена в Нацио-
нальном институте здоровья, Бетесда, США (National Institute of Health —
сокращенно NIH) при исследовании лара-гидроксилнрования L-фенилаланина
под действием оксидазы со смешанной функцией (монооксигеназы) в печени
животных. — Прим. ред.
Рис. 4 9. Предполагаемый механизм гидроксилирования коричной кислоты
Гл. 4. О-Гетероциклические пигменты — флавонсиды
143
Циииамат -}- АТР ->- Циниамоил-АМР-j-РР,
Циннгмоил-АМР+CoASH------> Циннамоил-S.СоА + АМР
Рнс. 4.10. Образование циннамоил-СоА-тиоэфиров.
1 z ' (4 30) Халкон
Рис. 4.11. Происхождение С6-Сз-Сб-скелета флавоноидов: механизм образо-
вания халконов.
рованное кольцо А и завершается формирование Сб-Сз-С^ске-
лета халкона (4.30) (рис. 4.11). Образование халкона изщ-ку-
мароил-СоА и малонил-СоА под действием ферментов из куль-
туры клеток петрушки (Petroselinum hortense) было показано
экспериментально.
4.6.3. Образование флавоноидов различный классов
Сейчас уже хорошо известно, что халконы играют
центральную роль в биосинтезе флавоноидов всех других
классов. Это показано на рис. 4.12, который суммирует приня-
тые в настоящее время взгляды на возможные биогенетические
взаимопревращения между флавоноидами различных классов.
Однако, прежде чем многие из этих возможных взаимопревра-
щений можно будет рассматривать как доказанные, необходи-
Рис. 4.12. Общая схема биосинтеза флавоноидов иллюстрирующая установленные (->) и предполагаемые (--------------*)
биогенетические взаимосвязи между флавоноидами различных классов.
Гл 4 О Гетероциклические пигменты — флавоноиГи
145
мо проделать большую работу по выявлению соответствующих
ферментов.
Флаваноны. В растворах происходит спонтанная изомери-
зация халконов в соответствующие флаваноны. Вместе с тем
в природе эта реакция катализируется халкон-изомеразой, и ее
продуктом является (2S)-флаванон (4.7). In vivo процесс легко
обратим, и поэтому очень трудно определить, халкон ли или со-
ответствующий флаванон служит непосредственным предшест-
венником флавоноидов других классов. Доступные в настоящее
время доказательства, полученные главным образом в работах
с суспензионными культурами клеток петрушки и других видов
растений, свидетельствуют в пользу того, что прямыми пред-
шественниками являются флаваноны. Этот вопрос может быть
разрешен однозначно лишь при исследовании ферментативных
систем, в которых не функционирует халкон-флаванон-изомера-
за.
Ауроны. Ауроны могут быть легко получены из халконов хи-
мическим окислением. Было показано также их образование
m vivo с помощью ферментативного окисления халконов, од-
нако механизм этой реакции неясен.
Флавоны. Считается, что флавоны образуются скорее путем
окисления соответствующих флаванонов, чем дегидратацией
флаванонолов, однако механизм реакции в этом случае также,
не установлен.
Флаванонолы. Показано прямое ферментативное образова-
ние этих соединений из флаванонов. В культурах Haplopappus
gracilis за него, по-видимому, ответственна микросомальная,
оксидаза со смешанной функцией. Происходящее при этом
гидроксилирование — очень важный процесс, поскольку не ис-
ключено, что флаванонолы служат промежуточными продуктами
при биосинтезе флавоноидов других классов (содержащих 3-
гидроксигруппу).
Флавонолы. Химическое окисление флаванонолов до флаво-
нолов довольно легко себе представить. Подобное ферментатив-
ное дегидрирование почти наверняка происходит в биосинтезе-
флавонолов, хотя прямые доказательства этого до сих пор от-
сутствуют.
Антоцианидины. Превращение флаванонолов в антоцианиди-
ны было показано у нескольких видов растений и подтверждено
в экспериментах с суспензионными культурами клеток Однако,
механизм этого превращения неизвестен. Степень окисления
этих соединений формально одна и та же, и поэтому был пред-
ложен гипотетический путь, не включающий реакций окисления
и восстановления (рис. 4.13). Биосинтез того небольшого числа
3-дезоксиантоцианидинов, которые встречаются в природе^
остается пока загадкой.
10—84
Рис. 4.13. Предполагаемый механизм биосинтеза антоцианидинов.
Гл 4 О-Гетероциклические пигменты — флавоноиды
147
Катехины и флаван-3,4-диолы (проантоцианидины). Эти
производные флавана имеют более низкую степень окисления,
чем халконы. Об их биосинтезе мало что известно. Чтобы объ-
яснить существующие между ними взаимосвязи (рис. 4.14),
предложена лишь гипотетическая схема, предусматривающая
восстановление халкона-флаванона до флавена (4.31).
Изофлавоны. Эти соединения образуются из халконов в про-
цессе, включающем миграцию кольца В.
4.6.4. Завершающие модификации
Присутствие ОН-групп при С-5, С-7 и С-4' у большин-
ства флавоноидов обусловлено основным путем их биосинтеза.
Индивидуальные флавоноиды образуются в результате удаления
одной или нескольких ОН-групп, дальнейшего гидроксилирова-
ния, О- или С-метилирования или гликозилирования. Эти мо-
дификации обычно происходят на последней стадии биосинтеза,
т. е. уже после образования основного Сб-С3-Сб-скелета, хотя
удаление кислорода у С-5, возможно, происходит на стадии
поликетида.
Показано ферментативное гидроксилирование флавоноид-
ного кольца В при С-З' или С-5'. Вместе с тем не исключено,
что перед включением в молекулу флавоноида м-кумаровая кис-
лота гидроксилируется до кофейной кислоты (4.32). Добавоч-
ное гидроксилирование может происходить практически при
любой степени окисления флавоноидной молекулы.
(4 32) Кофейная кислота
В реакциях О-метилирования в качестве донора метильных
групп используется S-аденозилметионин, причем ферменты, ка-
тализирующие эти реакции, чрезвычайно специфичны как по
отношению к месту метилирования, так и по отношению
к классу метилируемого соединения. В нормальных условиях
метилтрансферазы флавоноидов катализируют метилирование
только на уровне флавонов и флавонолов и не катализируют
метилирование антоцианидинов.
Что касается довольно простого процесса — О-гликозилиро-
вания, то источником моносахаридных фрагментов в нем явля-
ются активированные UDP-производные.
10*
Рис, 4,14. Предполагаемый механизм биосинтеза производных флавана (замещающие ОН-группы в кольцевых не пока-
заны).
Гл. 4. О-Гетероциклические пигменты — флавоноиды.
149
Механизмы С-метилирования и С-гликозилирования мало
изучены. По-видимому, в этих процессах используются те же
самые доноры, что и при 0-замещении, т. е. S-аденозилметио-
нин и UDP-caxapa.
4.6.5. Окислительная полимеризация флавоноидов
Бесцветные гидроксилированные флаваны (катехины,
флаван-3,4-диолы) могут давать начало окрашенным продуктам
в результате процессов окислительной димеризации и полимери-
(4 33) Теафлавин
он
(Галлоил)
зации. Хорошо известный коричневато-красный цвет чая обус-
ловлен продуктами такого рода, в частности теафлавином (4.33).
4.6.6. Генетический контроль биосинтеза флавоноидов
Наследуемость состава антоцианов в цветках послужила
основой классических генетических экспериментов Менделя, а
строгий генетический контроль биосинтеза флавоноидов был изу-
чен более подробно, чем у какого-либо другого класса веществ
растительного происхождения. Описаны многие различия от-
дельных генов, которые приводят к значительным качественным
и количественным вариациям состава флавоноидов. Многие
биохимические эффекты, например, общий биосинтез флавоно-
идов (т. е. наличие окрашенных или альбиносных фенотипов),
накопление флавоноидов определенных классов или индивиду-
альных соединений, структурные модификации, такие, как гид-
роксилирование, метилирование или гликозилирование, и рас-
пределение флавоноидов в различных органах растений, корре-
150
Часть I. Химические и биохимические аспекты
лируют со специфическими генетическими факторами. Генети-
ческие исследования помогли установить пути биосинтеза фла-
воноидов, а также заложили основу многих обширных программ
селекции, осуществленных в целях получения новых сортов куль-
турных растений.
4.6.7. Другие факторы, влияющие на биосинтез
флавоноидов
На биосинтез флавоноидов и его регуляцию оказыва-
ют влияние многие внутренние факторы и факторы окружаю-
щей среды. К наиболее важным из них относятся свет и стрес-
совые условия, такие, как ранение или инфекция. Однако в
большинстве случаев их влияние изучено на физиологическом,
а не биохимическом уровне.
Свет. Наиболее широко исследовалось влияние света. Обыч-
но свет стимулирует синтез флавоноидов, особенно антоцианов,
влияя главным образом на активность участвующих в этом
процессе ферментов. По своей реакции на индукцию светом эти
ферменты подразделяются на две группы. ФАЛ и ферменты,
которые превращают коричную кислоту в n-кумароил-СоА,
индуцируются значительно быстрее, чем ферменты, катализи-
рующие более поздние биосинтетические реакции и образующие
вторую группу. Синтез ферментов de novo начинается после-
индукции светом, причем главной регуляторной точкой является
ФАЛ. У многих видов показано участие в этом процессе фито-
хрома, однако могут также функционировать и другие фоторе-
цепторы, например флавин или флавопротеин. Интересно отме-
тить, что регулируется, по-видимому, только образование кольца
В (шикиматный путь), тогда как синтез кольца А (поликетид-
ный путь) не подвержен регуляции.
Повреждение или инфекция. Синтез флавоноидов в зеленых
растениях часто усиливается после механических повреждений
или заражения патогенными организмами. Наблюдаемое увели-
чение содержания флавоноидов может, однако, просто отражать
общее возрастание активности фенилаланин—аммиак-лиазь»
вызванное необходимостью образования фенольных фитоале?
синов и связанное с повышением устойчивости растения к з •
болеванию.
Избыточное образование зараженными тканями аитоцианог
легко заметно, например в случае поражения грибом листьер
персика и миндаля, которое выражается в курчавости листьев.
Пораженные листья приобретают вид ярких оранжево-красных
стручков или плодов. Другой пример — яблоки. Незрелые пло
ды, пораженные личинками насекомых, обычно синтезируют
повышенное количество антоцианов и преждевременно выгля-
Гл 4. О-Гетероциклические пигменты — флавоношты 151
дят почти зрелыми. По этому признаку их можно легко отли-
чить на дереве от здоровых плодов.
Возраст. Образование флавоноидов в зеленых тканях расте-
ний, по-видимому, зависит также от таких факторов, как воз-
раст и общая стадия развития. В молодых тканях содержится
больше флавоноидов, чем в зрелых. В частности, антоцианы
часто образуются в больших количествах в новых побегах и в
листьях; в результате последние могут иметь красную окраску
в отличие от зеленой у зрелых листьев.
4.7. Метаболизм флавоноидов у животных
Все травоядные животные поглощают с пищей большое
количество флавоноидов. Однако они редко утилизируются жи-
вотными для пигментации или других целей (разд. 4.4.1). В подав-
ляющем же большинстве случаев флавоноиды, подобно другим
чужеродным фенольным соединениям, либо превращаются в
сульфаты или глюкуронаты и выделяются, либо расщепляются
до фенолкарбоновых кислот (рис. 4.15,Л). Сообщения о каком-
либо вредном действии флавоноидов на животных отсутствуют.
4.8. Разрушение флавоноидов микроорганизмами
Работ, посвященных разрушению флавоноидов микро-
организмами, известно очень мало. Показано лишь, что некото-
рые грибы могут расщеплять С3-связующий фрагмент флавоно-
идной молекулы и продуцировать таким образом простые фено-
лы из колец А и В (рис. 4.15,5). Вполне вероятно также, что
те почвенные микроорганизмы, которые способны окислять и
расщеплять ароматические кольца простых фенолов, могут
сходным образом разрушать и флавоноиды.
4.9. Функции флавоноидов в растениях
Главная функция флавоноидов в растениях состоит в
пигментации тканей, в которых они синтезируются и накаплива-
ются (гл. 8). Не исключено, что по крайней мере некоторые
флавоноиды могут выполнять другие важные функции. Сильное
поглощение флавонами, флавонолами и антоцианами света в
УФ-диапазоне позволило предположить, что они могут защи-
щать ткани от вредного действия УФ-излучения.
Отмечены также и другие формы защитного действия фла-
воноидов. Было высказано мнение, например, что флавоноиды,
содержащиеся в листьях, могут отпугивать насекомых и таким
образом в течение долгого времени предохранять растение от
повреждений. Вместе с другими растительными фенолами фла-
воноиды также, по-видимому, участвуют в формировании устой1-
чивости растений к болезням или инфекции.
152
Часть 1. Химические и биохимические аспекты
А Животные
Гомопротокатеховая кислота
Б Микроорганизмы
+С0
Протокатеховая кислота
Рис. 4.15. Метаболизм гликозидов кверцетина, например рутина, у живот-
ных (4) и у микроорганизмов (5). Gly — гликозил.
Наконец, в последнее время флавоноиды обнаружены в хло-
ропластах, причем они были найдены у всех видов, в которых
их искали. Это позволяет предполагать, что флавоноиды могут
иметь важную, но пока неизвестную функцию в этой органелле.
4.10. Использование антоцианов в качестве
пищевых красителей
Применение антоцианов в качестве красящих добавок
к пище ограничено использованием экстрактов из сильно пигмен-
тированных плодов, таких, как черный виноград и черника. Эти
Гл. 4. О-Гетероциклические пигменты — флавоноиды.153
экстракты представляют собой многокомпонентные смеси, вклю-
чающие много различных водорастворимых веществ. Чистые
препараты антоцианов пока не используются в качестве пищевых
красителей, поскольку при значениях pH выше 4 их окраска не-
стабильна. Однако в настоящее время разрабатываются методы,
с помощью которых антоцианидины в форме солей флавилия
или хиноидных оснований можно стабилизировать в диапазоне
pH, характерном для пищевых продуктов и напитков. Так, на-
пример, при этом используют самоассоциацию молекул в форме
катионов в высоких концентрациях, благодаря которой полу-
чают интенсивную и стабильную окраску. Помогает также ко-
пигментация, особенно с присоединением ацетальдегида, что да-
ет сильноокрашенные и стабильные пигменты даже при pH 6.
4.11. Заключение и комментарии
После гетерогенных хинонов в этой главе мы рассмот-
рели группу соединений, имеющих один и тот же углеродный
скелет — флавоноиды. Различная степень окисления обусловли-
вает существование разных классов флавоноидов, каждый из
которых характеризуется собственными светопоглощающими
свойствами. Соединения одного из классов — антоцианы (воз-
можно, в хиноноидной форме) придают цветкам и плодам наи-
более яркую окраску. Они вызывают особый интерес у садово-
дов, и изменение их состава лежит в основе многих программ
по выведению растений с новой необычной окраской цветков.
Состав флавоноидов, в том числе и множества их гликозид-
ных производных, служит и будет продолжать служить полез-
ным признаком для выявления таксономических корреляций.
В систематических исследованиях, вероятно, будут обнаружены
все новые типы флавоноидов, структуру которых предстоит
расшифровать химикам-органикам. Многое еще предстоит сде-
лать и для выяснения биосинтеза флавоноидов. Прямые дока-
зательства предложенных ферментативных взаимопревращений
различных классов флавоноидов во многих случаях все еще от-
сутствуют, механизмы многих реакций до конца не выяснены, а
подробных исследований, посвященных катализирующим их
ферментам, почти нет. В последнем случае исключение состав-
ляют работы с суспензионными культурами клеток некоторых
растений (в частности, петрушки). Хотя физиологические фак-
торы и факторы окружающей среды (например, свет), которые
регулируют биосинтез флавоноидов, в целом выявлены, меха-
низмы, регулирующие состав флавоноидов и их раздельный
биосинтез, особенно антоцианов, в различным образом окра-
шенных участках цветков и других растительных тканей, почти
неизвестны. Их выяснение имеет особый интерес для садово-
154
Часть I. Химические и биохимические аспекты
дов, поскольку позволит лучше понять механизм копигментации
и другие феномены, ответственные за изменение основной окрас-
ки, обусловленной антоцианами in vivo. Именно при изучении
флавоноидов очень полезным может оказаться метод культуры
растительных тканей, особенно в случае их биосинтеза и регу-
ляции, а также механизмов обогащения окраски.
Вместе с тем многие проблемы остаются нерешенными. Если
небольшие количества флавоноидов содержатся в растительных
хлоропластах, то какова их функция в этих органеллах? Дей-
ствительно ли они отсутствуют у водорослей, хотя присутству-
ют и играют какую-то роль в хлоропластах растений? Многие
животные, в том числе человек, потребляют большие количест-
ва флавоноидов с пищей. Каким образом они метаболизиру-
ются? Оказывают ли они какое-либо вредное действие? Ис-
пользуются ли они в организме животных ?!
По мере роста требований к применению пищевых добавок
интерес к флавоноидам (особенно к антоцианам) как к про-
мышленным пищевым красителям природного происхождения
несомненно будет увеличиваться. Поэтому вопросы, связанные
с их метаболической судьбой в организме животных, и в част-
ности человека, приобретают еще большую важность. Со вре-
менем будут найдены методы промышленного синтеза некото-
рых флавоноидов или их крупномасштабного биологического
производства, возможно, с использованием культур тканей. Осо-
бое значение приобретут и способы стабилизации окраски анто-
цианов в пищевых препаратах.
4.12. Рекомендуемая литература
Наиболее фундаментальной книгой о флавоноидах явля-
ется большой том под редакцией Харборна, Мэйбри и Мэйбри
(Harborne, Mabry, Mabry, 1975). В нем содержится исчерпываю-
щая информация по всем аспектам химии и биохимии этих соеди-
нений. Остается также полезной более старая книга под редак-
цией Гейссмана (Geissman, 1962) по химии флавоноидов. Более
биохимический или систематический подход выбран Харборном
(Harborne, 1967) в его «Сравнительной биохимии флавоноидов».
1 Если функции флавоноидов в хлоропластах высших растений мало ис-
следованы, то об их метаболизме в организме животных и человека, а также
о биологической активности известно довольно много. Показано, что флаво-
ноиды совершенно не токсичны. Они в значительных количествах содержатся
в большинстве съедобных плодов и ягод и обладают Р-витаминной актив-
ностью Довольно детально изучен и метаболизм флавоноидов многих классов
(см., например, монографию R. R. Schelin «Mammalian metabolism of plant
xenobiotics», Acad Press, London — New York—San Francisco, 1978). — Прим-
ped.
Гл 4 О-Гетероциклические пигменты — флавоноиды 155
Другая книга под редакцией Харборна (НагЬогпе, 1964) по
биохимии фенольных соединений в общем также содержит по-
лезный материал. Две недавно вышедшие обзорные статьи
Суэйна (Swain, 1976а) и Уонга (Wong, 1976) посвящены соот-
ветственно природе и свойствам флавоноидов и их биосинтезу.
Функции флавоноидов в растениях обсуждаются Харборном
(НагЬогпе, 1976). Статья Хэслама (Haslam, 1979) о шикимат-
ном пути биосинтеза включает описание биосинтеза флавонои-
дов, причем механизмы реакций в ней рассматриваются на-
столько подробно, насколько это позволяют экспериментальные
данные. В статье Гризебаха (Grisebach, 1980) приведена имею-
щаяся на сегодняшний день информация по избранным аспек-
там биосинтеза флавоноидов. Совсем новый обзор Тимберлэй-
ка (Timberlake, 1980) позволяет вникнуть в проблемы, связан-
ные с использованием антоцианов в качестве пищевых краси-
телей.
Методы, обычно применяющиеся при исследовании биохи-
мии флавоноидов, обсуждаются Суэйном (Swain, 1976b), а бо-
лее детально они описаны еще в одной книге Харборна (Наг-
Ьогпе, 1973).
4.13. Дополнительная литература
Geissman Т. A. (ed.) (1962). The chemistry of flavonoid compounds, New York,
Macmillan.
Grisebach H. (1980). Recent developments in flavonoid biosynthesis. In: Pig-
ments in plants, 2nd edition, ed. F.-C. Czygan, p. 187, Stuttgart and New
York, Gustav-Fischer.
.Harborne J. B. (ed.) (1964). Biochemistry of phenolic compounds, London and
New York, Academic Press.
Harborne J. B. (1967). Comparative biochemistry of the flavonoids, London
and New York, Academic Press.
Harborne J. B. (1973). Phytochemical methods. London, Chapman and Hall.
Harborne J. B. (i976). Functions of flavonoids in plants. In- Chemistry and
biochemistry of plant pigments, 2nd edition, vol. 1, ed. T. W. Goodwin,
p. 736, London, New York and San Francisco, Academic Press.
Harborne J. В, Mabry T. J., Mabry H. (eds.) (1975). The Flavonoids, London,
Chapman and Hall.
Haslam E. (1979). Shikimic acid metabolites In- Comprehensive organic che-
mistry, vol. 5, ed. E Haslam, p. 1167, Oxford and London, Pergamon
Swain T. (i976a). Nature and properties of flavonoids. In: Chemistry and bio-
chemistry of plant pigments, 2nd edition, vol. 1, ed. T. W. Goodwin, p. 425,
London, New York and San Francisco, Academic Press.
Swain T. (1976b). Flavonoids. In: Chemistry and biochemistry of plant pig-
ments, 2nd edition, vol. 2, ed. T. W. Goodwin, p. 166, London, New York
and San Francisco, Academic Press.
Timberlake C. F. (1980). Anthocyanins—occurrence, extraction and chemistry,
Food Chem , 5, 69.
Wong E. (1976). Biosynthesis of flavonoids. In: Chemistry and biochemistry of
plant pigments, 2nd edition, vol. 1, ed T. W. Goodwin, p. 464. London.
New York and San Francisco, Academic Press.
Глава 5
Тетрапирролы
5.1. Введение
N-Гетероциклическое соединение пиррол (5.1) яв-
ляется очень стабильной гетероароматической системой, хотя
простые монопирролы встречаются в природе нечасто (см., од-
нако, пигменты wallemla). Ди- и трипирролы также редки, хо-
тя красный пигмент бактериального происхождения продигио-
зин, как стало теперь известно, представляет собой линейный
трипиррол (5.2). Вместе с тем циклические тетрапиррольные
природные пигменты широко распространены. К ним относятся
такие важные вещества, как хлорофилл — зеленый светоулав-
ливающий пигмент растений и гем, составляющий основу транс-
портирующих кислород красных пигментов крови. Родственны-
ми им соединениями являются также билины — линейные тет-
н
(5.1) Пиррол
ОМе R1 R2
(5.2) Продигиозин; R1, R2, R3 = Н или алкил
рапирролы, или тетрапирролы с открытой цепью. Эта группа
включает желчные пигменты животных, фикобилины — вспомо-
гательные фотосинтетические пигменты некоторых водорослей,
а также хромофор фоторегуляторного пигмента растений — фи-
тохрома.
5.2. Общие структурные свойства
Наиболее важными природными тетрапиррольными пиг-
ментами являются порфирины, содержащие «суперкольцо», или
макроцикл, в котором четыре пиррольных остатка связаны одно-
углеродными мостиками. Основным скелетным каркасом явля-
ется порфин, структура которого и общепринятая нумерация
атомов по Фишеру приведены на рис. 5.1. Четыре пиррольных
кольца обозначаются А, В, С, D или I, II, III, IV, а связываю-
щие метиновые мостики, или лгезо-углеродные атомы, — буква-
ми а, р, у, б. Периферические углеродные атомы входящих
Гл. 5. Тетрапирролы
157
Рис. 5.1. Основная структура н схема нумерации
по Фишеру для порфиновой системы колец.
в макроцикл пиррольных колец получили номера от 1 до 8.
У различных природных пигментов эти углеродные атомы несут
добавочные боковые цепи (см. ниже). В 1960 г. комиссия
IUPAC рекомендовала новую номенклатуру и систему нумера-
ции атомов молекулы порфина (рис. 5.2). Хотя широкоизвест-
Рис. 5.2. Система нумерации ШРАС для порфина.
(5 5) Тетрагидропорфин
(бактериохлорин!
ная система обозначения атомов и колец по Фишеру до сих
пор сохранила свою популярность, в этой книге мы будем при-
держиваться системы, рекомендованной IUPAC.
Основная порфиновая структура (5.3) присутствует в геме,
в то время как хлорофиллы и бактериохлорофиллы содержат
соответственно дигидропорфиновую [хлорин (5.4)] и тетрагид-
358
Часть I. Химические и биохимические аспекты
ропорфиновую [бактериохлорин (5.5)] системы. У природных
циклических тетрапиррольных пигментов N-атомы пиррольных
колец образуют хелатные комплексы с ионами металлов, обыч-
но Mg2+ (в хлорофиллах) или Fe2+ (в геме).
В ходе биосинтетического процесса (разд. 5.9.4.) образуется
.первый циклический тетрапиррол, в котором каждое пирроль-
гное кольцо несет С2-(уксусная кислота) и Сз-(пропионовая
(5.6) Замещение типа Ш <57> Замещение типа I
(Буквами А и Р обозначены заместители, являющиеся производными
ацетатной и пропионатной боковых цепей соответственно)
(5 8) Основной скелет билина
жислота) боковые цепи. Теоретически возможно множество
•различных расположений боковых цепей, однако почти все
встречающиеся в природе тетрапирролы являются производны-
ми предшественника с локализацией боковых цепей, пока-
занной в структуре (5.6). Такой способ распределения заме-
стителей называют типом III. Иногда обнаруживаются соеди-
нения с еще одним (тип I) способом распределения боковых
цепей (5.7). Соединений, обладающих боковыми цепями в иных
положениях, в природных объектах, по-видимому, не сущест-
Вует.
Встречающиеся в природе более простые линейные тетра-
пирролы имеют основную структуру (5.8). Эти соединения, из-
вестные как билины, или желчные пигменты, обычно образуют-
ся растительными и животными системами из порфириновых
•структур путем разрыва а-лезо-углеродного мостика (С-5 по
•системе IUPAC). В структуре (5.8) показано распределение за-
Гл. 5. Тетрапирролы
159’
мещающих боковых цепей билина, образовавшегося в резуль-
тате а-расщепления порфирина типа III.
5.3. Общие светопоглощающие свойства тетрапирролов
Тетрапиррольный макроцикл представляет собой плос-
кую высокосопряженную систему. Делокализация электрона-
распространяется по всему макроциклу, что придает ему в зна-
чительной степени ароматический характер. В такой ситуации
возбуждение электрона происходит очень легко. Перераспре-
деление заряда, сопровождающее возбуждение электрона, не-
изотропно, что приводит к появлению нескольких дипольных
моментов, которые в свою очередь обусловливают возникнове-
ние ряда интенсивных полос поглощения в большинстве случа-
ев в диапазоне 470—700 нм, т. е. полос интенсивного красно-
го, пурпурного или зеленого цвета. Очень интенсивная полоса
Соре, которая находится приблизительно при 400 нм, обуслов-
лена симметричным расположением четырех пиррольных N-
атомов; она чрезвычайно характерна для тетрапиррольного
макроцикла.
Протяженность и поляризация сопряженного хромофора, а
следовательно, и положение максимумов поглощения зависят от
степени гидрирования. На спектр поглощения влияет также-
расположение заместителей в пиррольных кольцах. Эти эффек-
ты имеют большое диагностическое значение при идентифика-
ции структурных типов. Ионизация в кислых или щелочных ус-
ловиях также вызывает изменения в спектрах.
Линейные тетрапирролы имеют более простые спектры по-
глощения в видимом диапазоне. Характерное для них увели-
чение степени поляризации приводит к сдвигу наиболее длин-
новолновой полосы поглощения в еще более длинноволновую-
область, а полоса Соре, характерная для макроциклических
структур, естественно, отсутствует. По мере насыщения двойных
связей, особенно при л/езо-углеродных атомах, у линейных тет-
рапирролов уменьшается длина сопряженного хромофора, а сле-
довательно, и длина волны максимума поглощения.
В следующих разделах этой главы будут рассмотрены неко-
торые из наиболее важных природных тетрапирролов и во мно-
гих случаях приведены подробные спектры поглощения.
5.4. Хлорофиллы
5.4.1. Структура и распространение
Вклад, вносимый хлорофиллами в окраску раститель-
ных тканей, проявляется во всеобщем распространении зелениз
Во время вегетации. Все зеленые ткани высших растений содер-
1'60
Часть I. Химические и биохимические аспекты
жат в своих фотосинтетических органеллах (хлоропластах) два
хлорофилла, а и Ь. Ядром этих соединений служит форбин
(5.9), представляющий собой хлориновую структуру, содержа-
щую добавочное изоциклическое кольцо (кольцо Е, или коль-
цо V). Это кольцо образуется в процессе биосинтеза (разд.
5.9.8.) путем окисления и замыкания цепи пропионатного за-
местителя при С-13 на углеродный атом у-метикового мостика
(С-15). Структуры хлорофиллов а и b (5.10) и (5.11) различи-
(5.10) Хлорофилл a- R СН3
(5.11) Хлорофилл b- R =€НО
ются лишь заместителем при С-7. В хлорофилле а — это — СНз,
а в хлорофилле b — СНО. Другими важными особенностями
хлорофилла являются хелатирование иона Mg2+ и этерифика-
ция С-17-пропионатного заместителя изопреноидным спиртом
фитолом. Недавно было показано существование хлорофиллов,
в которых роль этерифицирующего спирта играет геранилгера-
ниол.
Хлорофилл а характерен для всех водорослей. В Cyanophyta
-содержится только хлорофилл а, а в Chlorophyta и Euglena-
phyta и хлорофилл а, и хлорофилл Ь. Водоросли других клас-
сов также содержат хлорофилл а, но вместе с другими хлоро-
филлами, слегка отличающимися по картине распределения за-
местителей. Например, в Chrysophyta, Pyrrophyta и Phaeophyta
присутствует хлорофилл с (5.12), а в Rhodophyta— хлорофилл
d (5.13).
Фотосинтезирующие бактерии содержат бактериохлорофил-
лы. У большинства видов это тетрагидропорфины — бактерио-
хлорофилл а (5.14) и бактериохлорофилл Ъ (5.15). Наряду
Гл. 5. Тетрапирролы
161
с фитолом в качестве спиртов в состав молекулы этих бакте-
риохлорофиллов входят фарнезол (5.16) и геранилгераниол
(5.17). Серные бактерии Chlorobium содержат ряд хлоробиум-
хлорофиллов (бактериохлорофиллы end), которые являются
дигидропорфинами (5.18).
Хлорофиллы локализованы в хлоропластах высших расте-
ний и водорослей и в более простом фотосинтетическом аппа-
(5.12) Хлорофилле
(5.13) Хлорофилл t?
рате прокариотических сине-зеленых водорослей и фотосинте-
зирующих бактерий. In vivo, было обнаружено несколько
спектрофотометрически различающихся форм хлорофиллов, что
указывает на возможные различия в их функционировании
в процессе фотосинтеза (гл. 10). Вероятно, спектрофотометри-
ческие различия обусловлены микроокружением молекул хло-
рофилла, например ассоциацией их с белками или специальной
упаковкой.
5.4.2. Общие свойства
Хлорофиллы представляют собой сложные эфиры; они
растворимы в большинстве органических растворителей. Из
тканей, в которых они содержатся, их можно экстрагировать
полярными органическими растворителями, в частности ацето-
ном и спиртами. Хлорофиллы не стабильны и легко разруша-
ются при действии света, Ог, тепла, кислот и щелочей. В раст-
воре, даже при комнатной температуре, хлорофиллы а и b под-
вергаются изомеризации до близких по структуре хлорофиллов
а' и Ь', которые, вероятно, являются эпимерами (по СООМе
в кольце Е) собственно хлорофиллов. В присутствии кислот
11—84
162
Часть I. Химические и биохимические аспекты
удаляется ион магния и в результате образуется феофитин
(5.19). Это происходит настолько легко, что феофитин в доволь-
но больших количествах обычно обнаруживается как артефакт
на хроматограммах растительных экстрактов. Более жесткая
обработка кислотой приводит к отщеплению этерифицирующего
спирта (фитола и т. п.), в результате чего образуется водораст-
{5 14) Бактериохлорофилл а
(К = фарнезил или Геранилгеранил)
(5 15)Бактериохлорофилп в
(5.16) Фарнезол
r1=ch2.ch3, сн2.сн2.сн3
или СН2.СН(СН3)2
r2= СН3 или сн2.сн3
R3=H,CH3mhCH2-CH3
(5 18) Хлорофиллы СЫогоЫит (бактериохлорофиллы с и d)
Гл. 5. Тетрапирролы
163
воримый феофорбид (5.20). Феофорбиды и их метиловые эфи-
ры, а также хлорофиллид (5.21) являются продуктами щелоч-
ного гидролиза хлорофиллов в отсутствие кислорода. Поэтому
омыление служит полезным способом разрушения хлорофилла
и применяется в целях облегчения работы с другими липидами
растений. Водорастворимые продукты разрушения хлорофилла
эффективно удаляются промыванием экстракта водой.
Некоторые из этих продуктов широко используются в иссле-
дованиях биосинтеза хлорофилла с применением радиоактивных
(5 19) феофитин (Я=Фитил)
(5 20) Феофорбид (R=H)
(14С и 3Н) и стабильных (13С) изотопов. Продукты разрушения
хлорофилла относительно стабильны, и работать с ними легко.
Кроме того, их легко очистить и получить для них ЯМР- и
другие спектры, которые значительно более просто интерпрети-
ровать, чем соответствующие спектры собственно хлорофиллов.
5.4.3. Спектроскопические свойства
Спектры поглощения хлорофиллов а и b в диэтиловом
эфире приведены на рис. 5.3. Полосы Соре расположены при
430 и 455 нм соответственно, а наибольшие длины волн а-полос
поглощения составляют 662 и 641 нм соответственно. Главные
свойства спектров хлорофиллов с и d (рис. 5.4), а такжё хло-
робиум-хлорофиллов (рис. 5.5) сходны со свойствами спектров
хлорофиллов а и Ь, однако максимумы поглощения их спектров
различаются.
Бактериохлорофиллы а и b (рис. 5.6) также дают полосы
Соре приблизительно при 400 нм (358 и 368 нм соответственно),
однако характерная для них более сильная поляризация тетра-
11*
Длина волны, нм
Рис, 5.3. Спектры поглощения света хлорофиллом а (/) и хлорофиллом b (II)
в диэтнловом эфире.
Рис. 5.4. Спектры поглощения света хлорофиллом с (/) и хлорофиллом d (II)
в диэтнловом эфире.
Длина волны, нм
Рис. 5.5. Спектры поглощения света «хлорофиллом-650» (/) и «хлорофил-
лом-660» (II) из Chlorobium в диэтнловом эфире.
Поглощение
Длина волны, нм
Рис. 5.6. Спектры поглощения света бактериохлорофиллами а (7) и b (II)
в диэтнловом эфире.
166
Часть I. Химические и биохимические аспекты
гидропорфинового хромофора вызывает появление a-полос по-
глощения при значительно больших, чем у хлорофиллов, дли-
нах волн (772 и 794 нм соответственно), находящихся в инфра-
красной области спектра. Поэтому бактериохлорофиллы не
имеют интенсивной окраски.
5.4.4. Хлорофилл и его производные у животных
Свободного хлорофилла в животных тканях обычно
нет; его можно обнаружить лишь в кишечнике травоядных. Не-
которые виды животных, в частности водные беспозвоночные,
содержат симбиотические одноклеточные водоросли. Интерес-
ным примером представляется одна из тропических асцидий,
которая, как было обнаружено, содержит неизвестный ранее
тип прокариотических зеленых водорослей Prochloron, в насто-
ящее время рассматриваемый многими исследователями как
«недостающее звено» в эволюционной цепи высших растений и
(5 21) Хлорофиллид
(5 22] Филлоэритрин
их хлоропластов. Другие животные способны сохранять и на-
капливать функционирующие хлоропласты из растительного
материала, поступающего с пищей. Одним из примеров может
служить кишечнодышащий моллюск Elysia virldis, который спо-
собен накапливать до 108 хлоропластов (в одном организме).
Источником этих хлоропластов является сифоновая водоросль
Codium fragile, причем скорость фотосинтеза в хлоропластах,
содержащихся в животном, сравнима со скоростью этого про-
цесса в интактных водорослях. Очевидно, все такие животные
должны содержать хлорофилл и поэтому окрашены обычно в
зеленый цвет.
Гл. 5. Тетрапирролы
167
Продукты распада хлорофиллов, включая феофорбиды, хло-
рины и филлоэритрин (5.22), in vivo в растениях встречаются
редко (хотя они и являются распространенными артефактами).
В то же время ряд животных, в частности некоторые многоще-
тинковые черви и эхиуриды, используют их в качестве пигмен-
тов наружных покровов. Эти соединения могут образовываться
в животных из получаемого с пищей хлорофилла или поступать
непосредственно с распадающимися растительными остатками,
которыми питаются животные. У травоядных или всеядных, в
том числе у человека, главным продуктом распада хлорофилла
является филлоэритрин, который содержится в пищеваритель-
ном тракте и в желчи.
5.5. Гем и гемопротеины
Простетическая группа наиболее известных животных
пигментов — белка крови гемоглобина и мышечного белка мио-
глобина, так же как хлорофилл, является порфирином. Гемо-
глобин и миоглобин являются гемопротеинами, т. е. белками,
содержащими в качестве простетической группы порфирин с
хелатированным ионом железа, или гем. К гемопротеинам от-
носятся также цитохромы и некоторые ферменты, такие, как
пероксидаза и каталаза. В функционирующих гемоглобине н
миоглобине железо находится в восстановленной форме, Fe2+;
если же в их молекулах присутствует окисленная Ге3+-форма,
то они неактивны. Что касается цитохромов, то их нормальное
функционирование зависит от легкости взаимопревращения
окисленной и восстановленной форм.
5.5.1. Гемоглобин и миоглобин: структура и свойства
Жизненно важным пигментом крови, переносящим
кислород у большинства животных, в том числе у млекопитаю-
щих, является гемоглобин — гемопротеин, который в качестве
простетической группы содержит протогем (5.23), представляю-
щий собой Fe-хелатный комплекс протопорфирина IX. Мышцы
содержат структурно и функционально сходный с ним пигмент —
миоглобин. Эти два белка были первыми белками, трехмерная
структура которых была установлена с помощью рентгенострук-
турного анализа. Миоглобин имеет единственную полипептид-
ную цепь, состоящую из 153 аминокислотных остатков (мол.
масса 17 800). Его трехмерная структура показана на рис. 5.7.
Пептидная цепь миоглобина свернута таким образом, что его
молекула очень компактна. Около трех четвертей цепи имеет
структуру а-спирали, в которую входят восемь различных спи-
рализованных сегментов. С наружной стороны молекулы рас-
168
Часть I. Химические и биохимические аспекты
положены как полярные, так и неполярные аминокислотные
остатки, в то время как во внутренней части содержатся ис-
ключительно неполярные остатки, которые обеспечивают гидро-
фобное окружение единственного гема. Атом железа в геме на-
Рнс. 5.7. Трехмерная структура
миоглобина.
ходится всегда в двухвалентном состоянии (Fe2+); он способен
образовывать комплексы с шестью лигандами, четыре из кото-
рых являются пиррольными N-атомами порфирина. Пятой коор-
динационной связью гем связывается с белком через атом азота
имидазольного кольца гистидинового остатка глобина (извест-
ного как проксимальный гистидин). По шестой координацион-
ной связи легко присоединяется кислород с образованием окси-
миоглобина. В дезоксимиоглобине шестая связь остается сво-
бодной. Второй гистидиновый остаток глобина (дистальный
гистидин) находится очень близко к кислородсодержащему
участку, но он не связан с гемом. Если атом железа окислен до
Fe3+, т. е. молекула миоглобина превращена в ферримиогло-
Гл. 5. Тетрапирролы
169
бин, то последний способен присоединять по шестой координа-
ционной связи лишь воду, а не кислород и потому не может
быть переносчиком кислорода.
Белковая структура гемоглобина более сложна. Гемоглобин
млекопитающих (в том числе человека) имеет мол. массу 64 500
и состоит из четырех полипептидных цепей, каждая из которых
содержит свой собственный гем. Преобладающая форма гемо-
глобина у взрослых людей •— гемоглобин А — имеет две пары
полипептидных цепей: a-цепей, каждая из которых состоит из
141 аминокислотного остатка, и 0-цепей — по 146 остатков
в каждой. В содержащемся в небольшом количестве у взрос-
лых людей гемоглобине А2 и в зародышевом гемоглобине F 0-
цепи замещены другими полипептидами. Несмотря иа то что
аминокислотные последовательности полипептидных цепей ге-
моглобина и миоглобина в значительной степени различаются,
трехмерные структуры их чрезвычайно сходны и гемы в моле-
кулах того и другого занимают гидрофобные полости внутри
свернутых полипептидных цепей. Проксимальный и дистальный
остатки гистидина входят в число девяти аминокислот, которые
одинаковы во всех миоглобинах и гемоглобинах у ряда изучен-
ных в этом отношении видов животных.
Четыре полипептидные цепи гемоглобина ассоциированы в
примерно тетраэдрическую структуру и образуют почти сфери-
ческую молекулу. Каждая из a-цепей контактирует с двумя 0-
цепями, в то время как между двумя a-цепями или между дву-
мя 0-цепями взаимодействие почти отсутствует. Каждый из че-
тырех гемов гемоглобина способен присоединить одну молекулу
кислорода. Кислородсодержащая форма гемоглобина называ-
ется оксигемоглобином (5.24 b), а форма, не содержащая кис-
лорода,— дезоксигемоглобином (5.24 а). Когда дезоксигемо-
глобин поглощает кислород, в его трехмерной структуре про-
исходит ряд изменений, главным образом перемещение атома
Fe2+ в плоскость системы колец гема (см. ниже и рис. 5.8).
Как и в случае миоглобина, окисление Fe2+ до Fe3+ приводит
к образованию неактивной формы гемоглобина — метгемоглоби-
на, которая не способна присоединять молекулярный кислород.
5.5.2. Гемоглобин и миоглобин: функционирование
при переносе кислорода
Способность обратимо образовывать комплексы с кис-
лородом обусловливает жизненно важную роль гемоглобина
как переносчика кислорода у животных. У млекопитающих ге-
моглобин содержится в красных кровяных клетках (эритроци-
тах) и отвечает за перенос кислорода из легких по артериям,
артериолам и капиллярам в различные ткани тела. Он также
170
Часть I. Химические и биохимические аспекты
t Пппги-пгти гольца гема
Г лобин
(5 24а) Двэоксигемоглобин
О2
(5.24b) Оксигемоглобин
Рнс. 5.8. Связывание кислорода гемом в гемоглобине.
помогает переносить двуокись углерода по обратному пути от
тканей в легкие. Способность артериальной крови переносить
кислород в присутствии гемоглобина в 70 раз выше, чем в его
отсутствие. Сродство к кислороду у миоглобина значительно
больше, чем у гемоглобина, поэтому миоглобин может прини-
мать кислород от гемоглобина для использования или сохране-
ния его в мышечных клетках.
Детали механизма связывания кислорода гемоглобином
сложны, но хорошо изучены. Будучи чрезвычайно важным с
физиологической точки зрения, этот процесс служит прекрас-
ной иллюстрацией аллостерических взаимодействий и регуля-
ции. Характерные особенности связывания кислорода гемогло-
бином могут быть суммированы следующим образом:
1. Кривая связывания — освобождения кислорода гемогло-
бином имеет сигмоидную форму (тогда как форма соответст-
вующей кривой для миоглобина гиперболическая). Это свиде-
Гл. 5. Тетрапирролы
171
тельствует о том, что связывание кислорода гемом — коопера-
тивный процесс, т. е. связывание кислорода одним гемом об-
легчает его связывание другими гемами.
2. Сродство гемоглобина к кислороду зависит от величины
pH и содержания СОг.
3. Органические фосфаты, особенно 2,3-дифосфоглицерат
(ДФГ), также оказывают влияние на сродство гемоглобина
к кислороду.
Кооперативное связывание кислорода. Когда молекула гемо-
глобина диссоциирована на составляющие ее полипептидные
цепи, поведение последних очень сходно с поведением миогло-
бина: их кривые освобождения кислорода имеют гиперболиче-
Oj—НЬ в активно метаболизирующей ткани, ^2
главным образом в мышцах
<ЗО2
СО V “ ~ 1 ‘
2 В альвеолярных капиллярах легких
н+
н+
Рис. 5.9. Уравнение, суммирующее эффект Бора. НЬ — гемоглобин.
скую форму, на их кислородсвязывающие свойства не влияют
СОг, ДФГ или величина pH. Аллостерические свойства гемо-
глобина обусловлены взаимодействием четырех субъединиц его
молекулы. При связывании кислорода прежде всего происходит
перемещение атома железа гема. Как уже отмечалось ранее,
в дезоксигемоглобине атом железа в геме вынесен (на 0,75А)
за пределы плоскости порфиринового макроцикла. Связывание
кислорода приводит к тому, что атом железа перемещается в
плоскость гема оксигемоглобина, а проксимальный гистидин
приближается к кольцу гема (рис. 5.8). Такое перемещение вы-
зывает последующие небольшие изменения третичной структу-
ры субъединицы, в частности изменяется положение тирозино-
вого остатка и смежной С-концевой аминокислоты. В результа-
те исчезают некоторые взаимодействия между субъединицами;
следовательно, четвертичная структура дестабилизируется. При
этом изменяется конформация других субъединиц, что облегча-
ет связывание ими кислорода. И наоборот, освобождение кис-
лорода одним из гемов приводит к изменениям конформации и
взаимодействий между субъединицами, что облегчает освобож-
дение кислорода другими гемами.
Влияние СО? и величины. pH (эффект Бора) (рис. 5.9).
С понижением pH освобождение Ог гемоглобином облегчается.
То же происходит при повышении концентрации СОг. Это чрез-
вычайно важно с физиологической точки зрения, так как в тка-
!72 Часть 1. Химические и биохимические аспекты
нях с быстро протекающим обменом веществ, таких, как мы-
шечная ткань, образуется много СОа и кислот. Высокие уровни
СОа и Н+ стимулируют освобождение Оа из гемоглобина, и та-
ким образом удовлетворяется потребность в большом количест-
ве кислорода в метаболически активных тканях. После освобож-
дения кислорода дезоксигемоглобин присоединяет Н+ и СО?.
Для альвеолярных капилляров легких характерна более высо-
кая концентрация кислорода, и там по мере связывания дезо-
ксигемоглобином кислорода происходит освобождение Н+ и СОа.
Структурные превращения, которые претерпевает при этом мо-
лекула гемоглобина, были подробно выяснены. При переходе
от оксигемоглобина к дезоксигемоглобину в результате кон-
формационных изменений карбоксилсодержащие аминокислот-
НЬ—ДФГ+ 4 О2 нь (О2)4+ ДФГ
Рис. 5.10. Уравнение, отражающее действие дифосфоглицерата (ДФГ) на
гемоглобин (НЬ).
ные остатки приближаются к некоторым гистидиновым остаткам
и концевым NHa-группам. С изменением локального заряда
микроокружения повышается величина рК кислого остатка и
таким образом возрастает его сродство к Н+ СО? также зна-
чительно легче связывается с дезоксигемоглобином, чем с ок-
сигемоглобином. Она связывается с концевой NHz-группой каж-
дой цепи с образованием карбаминопроизводных.
Влияние ДФГ. У человека ДФГ снижает сродство гемогло-
бина к кислороду в 26 раз. Это очень важно с физиологической
точки зрения, поскольку в отсутствие этого механизма гемо-
глобину было бы трудно освобождать много кислорода в ка-
пиллярах тканей. Такое действие ДФГ обусловлено его способ-
ностью связываться с дезоксигемоглобнном, но не с оксигемо-
глобином. Одна молекула ДФГ связывается с гемоглобиновым
тетрамером, располагаясь в центральном пространстве в непо-
средственной близости ко всем четырем субъединицам. Связы-
вание ДФГ и О2 — взаимоисключающие процессы. В ходе ок-
сигенации конформационные изменения приводят к значитель-
ному уменьшению центрального пространства в гемоглобино-
вом тетрамере и молекула ДФГ вытесняется. Однако при этом
необходимо нарушение ДФГ-белкового взаимодействия, что
затрудняет связывание кислорода гемоглобином. Для поглоще-
ния ДФГ в свою очередь требуется расщепление связи гемо-
глобин—О?, так что ДФГ облегчает высвобождение кислорода.
Функционирование ДФГ в упрощенном виде представлено урав-
нением на рис. 5.10.
Гл. 5. Тетрапирролы
173
Влияние других веществ. Сродство гемоглобина к окиси уг-
лерода гораздо больше, чем к кислороду; следовательно, СО
может вытеснять кислород из оксигемоглобина. Образующийся
при этом карбоксигемоглобин не способен служить переносчи-
ком кислорода, и поэтому окись углерода является весьма эф-
фективным адом. Карбоксигемоглобин имеет вишнево-красную
окраску, характерную для цвета лица людей, отравившихся
окисью углерода, что позволяет легко диагностировать отрав-
ление. Функционирование гемоглобина могут серьезно нару-
шать различные лекарственные препараты. Известно, что про-
дукты метаболизма ацетанилида, фенацетина и некоторых дру-
гих лекарственных препаратов индуцируют окисление гемогло-
бина до Ре3+-формы (метгемоглобина), приводя к серьезному
снижению кислороднереносящей способности крови.
Гемоглобин плода. Так же как и у большинства других жи-
вотных, у человека на разных стадиях развития организма
имеются различные типы гемоглобина в крови. Гемоглобин
плода и гемоглобин взрослого человека различаются по спект-
рам поглощения света и электрофоретическим свойствам.
В крови зародыша на ранних стадиях его развития присутству-
ет гемоглобин третьего типа. Зародышевый гемоглобин F обла-
дает более высоким сродством к кислороду, чем гемоглобин А
взрослых людей. Благодаря этому возможен оптимальный пе-
ренос кислорода от гемоглобина А матери к гемоглобину F
плода. Более высокое сродство гемоглобина F к кислороду
подтверждается также тем, что он связывает ДФГ менее проч-
но, чем гемоглобин А.
Генетические дефекты в структуре и функционировании ге-
моглобина. Существует много генетических вариаций человече-
ского гемоглобина. Наиболее известная из них найдена при
«серповидно-клеточной анемии» — мутации одного гена, кото-
торая в гомозиготном состоянии вызывает деформацию эритро-
цитов с образованием клеток, имеющих форму серпа. Гемогло-
бин S таких серповидных клеток отличается от нормального
гемоглобина лишь одним аминокислотным остатком в р-цепях.
В нем происходит замена полярной глутаминовой кислоты на
неполярную аминокислоту валин, что приводит к очень сильно-
му снижению растворимости дезоксигемоглобина S, хотя раст-
воримость оксигемоглобина при этом остается нормальной.
Дезоксигемоглобин S образует волокнистый осадок, который
вызывает деформацию и разрушение эритроцитов и как следст-
вие — хроническую гемолитическую анемию.
В настоящее время известно более 100 мутантных гемогло-
бинов. Некоторые из замен являются безвредными «поверхност-
ными» заменами, тогда как другие, которые затрагивают кис-
лородсвязывающие участки, третичную структуру или взаимо-
174
Часть I. Химические и биохимические аспекты
действия субъединиц в четвертичной структуре (что сказывает-
ся на аллостерических эффектах), могут очень сильно влиять
на связывание кислорода.
5.5.3. Спектроскопические свойства гема и гемоглобина
Подобно хлорофиллу, спектры поглощения гема и ге-
мопротеинов характеризуются интенсивными полосами Соре в
районе 400 нм, а также другими интенсивными пиками погло-
щения между 500 и 600 нм. Максимумы поглощения дезокси-
гемоглобина ( — 425 и 560 нм) и оксигемоглобина ( — 414, 543 и
578 нм) различны и очень характерны (рис. 5.11). Гемоглобин
Длина волны , нм
Рис. 5.11. Спектры поглощения света гемоглобином (/), оксигемоглобином
(II) и карбоксигемоглобином (III).
имеет пурпурную, а оксигемоглобин — оранжево-красную окрас-
ку. У карбоксигемоглобина (вишнево-красного) максимумы
поглощения слегка сдвинуты в синюю область по сравнению
с оксигемоглобином. Спектры поглощения окси- и дезоксимио-
глобинов очень сходны со спектрами для соответствующих ге-
моглобинов. Точные положения максимумов поглощения гемо-
глобинов у различных видов животных весьма характерны и
зависят от свойств белковой молекулы. Этот признак очень
полезен при выяснении таксономических корреляций. Гем мож-
но отделить от связанного с ним белка и образовать комплекс
через оставшиеся две координационные связи с другими азот-
содержащими молекулами, например с пиридином. Такие про-
дукты, известные как гемохромогены, имеют чрезвычайно ха-
рактерные спектры поглощения и очень полезны при иденти-
фикации геминовых простетических групп.
Гл. 5. Тетрапирролы
175
5.5.4. Распространение гемоглобина
Гемоглобин является пигментом крови практически у
всех позвоночных, а также у многих беспозвоночных животных.
Однако он не обнаружен у Porifera, Coelenterata, Rotifera,
Sipuncula, Polyzoa, Brachiopoda, Onychophora, Tardigrada,
Chaetognata и Protochordata и довольно редок у Arthropoda.
У позвоночных гемоглобин локализован в эритроцитах. У бес-
позвоночных он может содержаться в специальных частицах,
но чаще присутствует в растворенном виде в крови или гемо-
лимфе. Гемоглобин встречается почти во всех тканях тела.
Размеры гемоглобинового комплекса варьируют в очень ши-
роких пределах. Как уже упоминалось, гемоглобин млекопита-
ющих имеет тетрамерную форму с мол. массой 64 500. У других
представителей животного царства были найдены его мономер-
ные и димерные формы (с мол. массой 17 000 и 34 000 соответ-
ственно), в то время как у многих беспозвоночных сложные мо-
лекулы значительно крупнее и могут иметь мол. массу до
3 000 000.
Состав и последовательность аминокислот в гемоглобинах
различных видов животных сильно варьируют, что приводит
к различиям в максимумах поглощения света, растворимости,
изоэлектрической точке, сродстве к кислороду и устойчивости
изолированных пигментов к кислотам, щелочам и нагреванию.
Даже среди млекопитающих имеются значительные различия
в стабильности и форме кристаллов выделенных гемоглобинов.
Эти структурные различия ограничиваются белковой частью
молекулы, тогда как все формы гемоглобинов содержат одну и
ту же протогемовую простетическую группу.
Наряду с различиями в гемоглобине между семействами,
родами, видами или даже подвидами могут также существо-
вать различия в гемоглобине у какого-либо индивида в разные
периоды жизни или даже в одно и то же время. Как упомина-
лось выше, человек имеет различные типы гемоглобина в крови
в разные периоды жизни. Та же ситуация встречается и у дру-
гих видов; например, гемоглобины цыпленка и курицы или го-
ловастика и лягушки различны, причем во всех случаях пиг-
мент молодого организма имеет более высокое сродство к кис-
лороду.
У многих животных имеется более чем один гемоглобиновый
компонент в крови; эти компоненты в некоторых случаях вы-
полняют разные функции. Два гемоглобиновых компонента ти-
хоокеанского лосося, Onchorhynchus keta, сильно различаются
между собой по сродству к кислороду, этот факт может объяс-
няться необходимостью для этого вида проводить часть жизни
в соленой воде, а часть — в пресной.
176
Часть I. Химические и биохимические аспекты
Хотя гемоглобин обычно считается продуктом чисто живот-
ного происхождения, одна его форма — леггемоглобин, была
обнаружена у бобовых растений. Его присутствие ограничено
клетками корневых клубеньков, содержащих симбиотические
азотфиксирующие бактерии (Rhizobiutn spp.). Гемопротеины со
свойствами гемоглобинов обнаружены также у некоторых гри-
бов и простейших.
5.5.5. Вклад гемоглобина в окраску животных
Гемоглобин эритроцитов в капиллярных кровеносных
сосудах кожи придает розоватый оттенок коже людей «белой»
расы. У большинства других позвоночных эта окраска скрыта
волосами, перьями, чешуей или завуалирована другими кожны-
ми пигментами. Гемоглобином обусловлена розовая окраска
языка, внутренней части ушей и кончика носа у многих млеко-
питающих. Некоторые специфические ткани или участки тела
могут окрашиваться гемоглобином в красный цвет и играть при
этом роль предупреждающих сигналов, или признаков опреде-
ленного пола (гл. 8), например голая шея у некоторых грифов,
бородка у индюка и других птиц, а также ягодицы у павиана.
Среди беспозвоночных обусловленная гемоглобином окраска
наблюдается у многих многощетинковых и однощетинковых
кольчатых червей (например, у «мотыля»), а также при неко-
торых условиях у ветвистоусых и листоногих Crustaceae. Одна-
ко гемоглобин вносит свой вклад в окраску лишь небольшого
числа животных, позвоночных и беспозвоночных.
5.5.6. Хлорокруорин
Близкородственным гемоглобину является другой ге-
мопротеин — хлорокруорин, или хлорогемоглобин, который слу-
жит переносчиком кислорода в зеленой крови очень ограничен-
ной группы многощетинковых червей. Простетическая группа
хлорокруорина— хлорокруорогем (5.25)—отличается от про-
тогема тем, что в нем вместо винильной группы при С-3 нахо-
дится формильная, т. е. альдегидная, группа. Во всех других
отношениях, а именно образованием комплекса хлорокруороге-
ма с глобиновым белком, а также сродством такого комплекса
к кислороду хлорокруорин сходен с гемоглобином. В спектре
поглощения оксихлорокруорина полоса Соре наблюдается при
430 нм, а а- fJ-полосы поглощения — при 604 и 558 нм, т. е. при
больших длинах волн, чем у оксигемоглобина. Окси- и дезокси-
формы хлорокруорина по окраске почти не различаются, одна-
ко для этого пигмента характерен сильный дихроизм, и его
Гл. 5. Тетрапирролы
177
окраска при разбавлении меняется от красной до зеленой. Лишь
у очень небольшого числа видов хлорокруорин придает телу
животного зеленоватую окраску.
5.5.7. Гемоцианин, гемоэритрин и гемованадии
Эти соединения служат пигментами крови или дыха-
тельными пигментами у очень небольшого числа беспозвоноч-
ных. Они представляют собой металлопротеины и включены в
этот раздел условно, так как, несмотря на названия, не явля-
ются ни производными гема, ни производными порфирина. На-
пример, гемоцианины брюхоногих моллюсков, таких, как улитка
Helix pomatia, представляют собой гигантские белки (мол.
масса 9> 10е). Их функциональной единицей, которая связыва-
ет одну молекулу Ог, служит пара атомов меди, окруженная
компактно свернутым полипептидом с мол. массой 50 000. Ог
семи до девяти таких функциональных единиц составляют фраг-
мент с мол. массой 4—5*105, а молекула гемоцианина включа-
ет 20 таких фрагментов.
5.5.8. Цитохромы электронтранспортной цепи
Цитохромы представляют собой группу небольших ге-
мопротеинов, у которых в отличие от гемоглобина и миоглоби-
на входящий в состав их гема атом железа легко подвергается
обратимому окислению и восстановлению. Это свойство придает
им чрезвычайно важное биологическое значение в переносе
электронов. Цитохромы содержат все животные, растения и
аэробные микроорганизмы. К настоящему времени выявлено и
довольно детально изучено большое число цитохромов. На ос-
12—84
178
Часть 1. Химические и биохимические аспекты
новании природы содержащейся в них простетической группы
и способа ее связи с белками их можно разделить на четыре
главные группы — цитохромы а, Ъ, с и d.
Простетическая группа цитохрома b представляет собой
протогем (5.23), как и у гемоглобина. Гемогруппа цитохрома а,
обозначаемая как гем а (5.26), отличается от протогема заме-
ной метильной группы при С-18 на СНО-группу и модификаци-
ей винильной группы при С-З путем присоединения к ней Ci5-
(фарнезил) изопреноидной цепи. Название «цитохром d* приме-
няется к цитохромам с дигидропорфириновой (хлориновой)-
простетической группой, содержащей железо (5.27); боковые
цепи заместителей у этой простетической группы могут варьи-
ровать. Группа цитохрома с включает все цитохромы, у кото-
(5.27) Простетическая группа цитохрома d
(заместители R1, R2 и R3 достоверно не выяснены)
Гл. 5. Тетрапирролы
\7<3>
рых боковые цепи гема связаны с белком ковалентными тио-
эфирными связями, например (5.28).
Индивидуальные цитохромы внутри этих групп обозначают
нижними индексами, например цитохром д6, или в их название
входит длина волны a-полосы в спектре поглощения, например
цитохром д-550.
Количественный вклад цитохромов в суммарную концентра-
цию тетрапирролов у организмов, содержащих хлорофилл или
гемоглобин, незначителен, однако они жизненно необходимы
для функционирования этих организмов, В митохондриях эука-
риотических клеток они являются основой высокоорганизован-
ной электронтранспортной цепи (рис. 5.12), которая использу-
ется для аэробного окисления восстановленных коферментов
Рис. 5.12. Роль цитохромов в митохондриальной электронтранспортной си-
стеме.
12
a so
Часть I. Химические и биохимические аспекты
(например, NADH), образующихся в ходе окислительного рас-
щепления питательных веществ. С электронтранспортной цепью
связан процесс окислительного фосфорилирования, в котором
генерируется большая часть АТР клеток. В цепи участвуют
несколько цитохромов, и гемовое железо каждого из них пре-
терпевает окисление и восстановление по мере прохождения
Фис. 5 13. Механизм функционирования цитохрома Р450 при окислении суб-
страта RH до ROH оксидазой со смешанной функцией.
’электрона по всей цепи вплоть до молекулярного кислорода.
•Сходным образом цитохромы функционируют в фотосинтети-
ческом переносе электронов. Без электронтранспортных систем
•синтез АТР, необходимого для протекания биохимических реак-
ций, был бы невозможен. Роль цитохромов и других пигментов
в этих процессах, особенно в фотосинтетическом транспорте
электронов, рассмотрена в гл. 10.
Цитохромы жизненно необходимы для функционирования
клеток, но они не вносят никакого вклада в окраску организ-
мов.
5.5.9. Цитохром Р450
Цитохром Р450 представляет собой гемопротеин, кото-
рый широко используется при биологическом окислении, ката-
лизируемом оксидазами со смешанной функцией. Р450 является
щитохромом типа b с протогемом IX в качестве простетической
.группы. В результате реакции с окисью углерода Ее2+дает ста-
Гл. 5. Тетрапирролы
181
бильный продукт с Хтах = 450 нм, от чего и произошло его наз-
вание «Р450».
Оксидазы со смешанной функцией катализируют введение
одного атома молекулы кислорода в органическую молекулу
RH с образованием окисленного продукта ROH. Второй атом
кислорода восстанавливается до воды. Второй субстрат [кофер-
мент, обычно NAD(P)H] используется при этом в качестве до-
нора электронов. Вся система представляет собой небольшую
электронтранспортную цепь, включающую флавопротеин и ци-
тохром Р450, который принимает электроны от восстановленного
флавина в две одноэлектронные стадии и передает эти элект-
роны на молекулярный кислород. Субстрат RH в ходе реакции,
по-видимому, связывается с цитохромом Р450. Возможный меха-
низм этой реакции приведен на рис. 5.13. Характерно, что такое
гидроксилирование протекает с сохранением конфигурации.
Примерами реакций, катализируемых оксидазами со смешанной
функцией, могут служить гидроксилирование стероидов в мик-
росомах печени, а также гидроксилирование лекарственных
препаратов (детоксикация). Индукция цитохрома Р45о происхо-
дит под влиянием многих чужеродных органических соединений.
5.5.10. Другие гемопротеиновые ферменты
Помимо цитохромов гемовая простетическая группа
необходима также некоторым другим ферментам для их ката-
литического действия. В число таких гемопротеинов входят пе-
роксидазы и каталазы из различных растительных и живот-
ных источников. У этих ферментов порфирин обычно представ-
лен протогемом. Например, пероксидаза хрена с мол. массой
44 000 содержит одну гемовую группу, которая катализирует
окисление фенольных соединений с помощью Н2О2. Каталаза
(из печени быка) имеет мол. массу 248 000 и содержит четыре
гемогруппы. Этот фермент катализирует расщепление Н2О2 до
воды с чрезвычайно высокой скоростью.
5.6. Свободные порфирины у животных
Свободные порфирины—протопорфирин (5.29), уро-
порфирин (5.30) и копропорфирин (5.31)—обычно присутству-
ют в моче и фекалиях животных. Довольно часто они встреча-
ются также в различных тканях животных, однако, как пра-
вило, в количествах, недостаточных для придания окраски этим
тканям. У беспозвоночных свободные порфирины и гемы, а так-
же их белковые конъюгаты встречаются спорадически, обычны-
ми они являются лишь у червей и моллюсков. Наиболее извес-
тен пример окраски наружных покровов у дождевого червя:
182
Часть I. Химические и биохимические аспекты
пурпурно-багровый цвет передне-дорзальной поверхности его
тела обусловлен протопорфирином. Раковины некоторых мол-
люсков содержат очень большие количества уропорфирина, ча-
сто изомера уропорфирина I (5.32). Свободные порфирины чрез-
вычайно редко встречаются или вообще не обнаружены у про-
стейших, кишечнополостных, членистоногих и протохордовых.
НООС.Н2С.Н2С СН2.СН2-СООН
(5 29) Протопорфирин
(5 30) Уропорфирин Ш
У позвоночных, особенно у млекопитающих, свободные пор-
фирины редки, но они иногда встречаются в некоторых внут-
ренних тканях. Имеется, однако, много примеров присутствия
протопорфиринов в яичной скорлупе некоторых птиц, благода-
ря чему их скорлупа может быть окрашена в коричневый цвет
и иметь сложный рисунок окраски. Порфирины легко связыва-
ются с минеральными веществами, и обусловленная ими окрас-
ка яичной скорлупы, как правило, удивительно устойчива. Нет
также ничего необычного в присутствии в оперении птиц коп-
ропорфирина III, причем иногда в больших количествах. Заме-
чательная ярко-красная окраска перьев бананоеда обусловлена
присутствием хелатного комплекса уропорфирина III с медью.
Этот пигмент хорошо растворим в воде, хотя, к счастью, лишь
Гл. 5. Тетрапирролы
183
при щелочной реакции среды, так что птице не угрожает потеря
окраски в ее природной среде обитания — во влажных тропи-
ческих лесах.
Подводя итог, следует отметить, однако, что в общем вклад
свободных порфиринов в окраску животных невелик.
(5 31) Копропорфирин III
(5 32) Уропорфирин!
5.7. Витамин Bi2
Витамин В12 играет важную роль в организме млеко-
питающих, в том числе человека. Дефицит этого витамина ле-
жит в основе патогенеза заболевания, именуемого перни-
циозной анемией. Он является также незаменимым ростовым
фактором для некоторых микроорганизмов. Структурно вита-
мин Ви (цианокобаламин) представляет собой циклический
тетрапиррол с катионом Со+ в качестве хелатируемого иона
металла (5.33). В случае витамина В12 макроцикл является не
стандартным порфирином, а представляет собой коррин, в кото-
184
Часть I. Химические и биохимические аспекты
ром б-метиновый мостик (С-20) отсутствует, и в результате
между кольцами А и D имеется непосредственная ковалентная
связь. В то же время а- и 0-метиновые мостики (С-5, С-15) не-
сут добавочные метильные группы и вся система целиком нахо-
дится в гораздо более восстановленном состоянии, чем в слу-
чае порфиринов. Важное значение имеет также присутствие в
ней нескольких ацетамидных и пропионамидных заместителей.
Наиболее необычным свойством молекулы является содержа-
ние диметилбензимидазолрибозилфосфата, связанного через
фосфатную группу с боковой цепью кольца D и через атом
азота имидазольного кольца (координационной связью) с ато-
мом кобальта. Диметилбензимидазольная группа может быть,
заменена на аденин.
(5 33) Витамин В12
Цианокобаламин Х-СК
он он
Гл. 5. Тетрапирролы
185
Обычно витамин В12 выделяют из тканей в растворе, содер-
жащем ионы CN-, которые играют роль шестого лиганда Со+,
но комплекс витамина с CN не активен in vivo. Метаболически
активная форма (В^-кофермент) вместо CN содержит адено-
зин, непосредственно связанный с Со+ через С-5 рибозного
остатка. Кофермент участвует в реакциях перегруппировки, та-
ких, как перегруппировка метилмалонил-СоА в сукцинил-СоА.
Витамин В12 синтезируется исключительно микроорганизма-
ми: ни животные, ни растения его не образуют. Животные по-
лучают свою дозу В12 главным образом от микроорганизмов ки-
шечника. В медицинской промышленности витамин В12 получа-
ют из микробных культур.
In vivo витамин В]2 присутствует в столь малых количест-
вах, что его темно-красная окраска не сказывается на внеш-
нем виде микроорганизмов.
5.8. Линейные тетрапирролы — билины
5.8.1. Введение
Билины распространены широко, хотя и нерегулярно, у
представителей как растительного, так и животного царства.
У животных они представляют собой продукты катаболизма
гема. У растений и водорослей они также являются производ-
ными порфиринов; у некоторых водорослей они образуются в
•больших количествах в виде белковых конъюгатов, которые
играют важную роль в фотосинтезе. В структурном отношении
билины представляют собой линейные (или открытые) тетра-
пирролы— производные структуры (5.8). Углеродные мостики
(а, Ь, с), связывающие пиррольные кольца, могут быть как на-
сыщенными (—СН2—), так и ненасыщенными (—СН = ). Если
все три углеродных мостика насыщены, то молекула представ-
ляет собой билан, если же один мостик ненасыщен, то — билен.
Билидиены и билитриены имеют соответственно два и три не-
насыщенных (—СН = )-мостика. Таким образом, среди струк-
тур, представленных ниже, уробилиноген (5.37) служит приме-
ром билана, уробилин (5.39) — b-билена, билирубин (5.35) — а,
с-билидиена, а биливердин (5.34) — а, Ь, с-билитриена. Все эти
соединения, как показано, существуют преимущественно в ди-
кетоформе.
5.8.2. Билины животных (желчные пигменты)
Структура и образование. У млекопитающих разруше-
ние гема гемоглобина и других гемопротеинов приводит к обра-
зованию открытых тетрапирролов, издавна известных как желч-
(5 34) Биливердин IX а
он
(5 37) Уробилиноген
(5.38) Стеркобилин
Гл. 5. Тетрапирролы
187
ные пигменты. Этот катаболический процесс происходит глав-
ным образом в ретикулоэндотелиальных клетках селезенки,
печени, костного мозга и в меньшей степени почек. Источником
главной массы гемов, катаболизирующихся по этому пути, явля-
ется гемоглобин стареющих эритроцитов; аналогичным обра-
зом утилизируются также гемовые простетические группы дру-
гих белков, например гемсодержащих ферментов.
В ходе образования билинов происходит ферментативное
расщепление порфириновой системы гема по а-метиновому мо-
стику (С-5), что приводит к образованию первого открытого
соон соон
(5 39) Уробилин
тетрапиррола — билитриена биливердина 1Ха (5.34). Это зеле-
ный желчный пигмент, который окрашивает фекалии большин-
ства птиц и амфибий. У большей части млекопитающих, в том
числе и у человека, центральная (—СН = )-группа восстанавли-
вается до (—СН2—)-группы с образованием оранжевого били-
диена билирубина (5.35). Он превращается в диглюкуронид
(5.36), который проходит через кишечник. Бактерии кишечни-
ка осуществляют дальнейшее восстановление последнего с об-
разованием бесцветных продуктов, таких, как билан уробили-
ноген (5.37). Наряду с восстановлением (—СН = )-мостиков
происходит также восстановление винильных боковых цепей до
этильных, что приводит к образованию «л*езо»-структур. Эти
продукты на воздухе окисляются до желто-коричневых биленов
стеркобилина (5.38) и уробилина (5.39), которые главным об-
разом и придают окраску фекалиям и моче. Более подробно
эти процессы рассматриваются ниже при обсуждении биосинте-
за тетрапирролов (разд. 5.9.14).
Свойства. Биливердин и билирубин представляют собой кис-
лоты и поэтому растворимы в водных растворах едких щело-
чей. Их соли с большинством ионов нещелочных металлов в
воде нерастворимы; кальциевая соль билирубина является глав-
ным компонентом желчных камней. Благодаря раскрытию пор-
фиринового макроцикла пиррольные кольца и углеродные ато-
мы метиновых мостиков становятся более доступными для хи-
188
Часть I. Химические и биохимические аспекты
мической атаки, и, следовательно, более реакционноспособными.
Для билинов характерно интенсивное поглощение света в ви-
димой области, однако полосы Соре они не дают. Насыщение
по метиновым мостикам укорачивает хромофор, и поэтому мак-
симумы поглощения их спектров наблюдаются при более корот-
ких длинах волн. Так, у биливердина %гаах = 680 нм (в кислой
среде), у билирубина %таХ = 450 нм, а у уробилина Хтах = 490нм.
Биланы, такие, как уробилиноген, в видимом диапазоне спект-
ра не поглощают.
Распространение. Хотя образование билинов из молекул ге-
ма наиболее интенсивно изучалось у млекопитающих, этот про-
соон соон
(5.40) Биливердин IX у
цесс широко распространен у всех представителей животного
царства. Билины были найдены у большинства главных групп
животных. Они служат пигментами покровов тела многих бес-
позвоночных, особенно червей (например, Nereis diversicolor,
обычного ярко-зеленого морского червя прибрежных скалистых
банок) и насекомых. Хорошо известная зеленая окраска крови
и покровов тела многих кузнечиков, гусениц и т. п. обусловлена
билинами биливердинового типа, а вовсе не хлорофиллом, как
думали первоначально. Общим для этих беспозвоночных явля-
ется присутствие глаукобилина (мезобиливердина), у которого
винильные боковые цепи восстановлены до этильных групп.
У позвоночных билины хорошо известны как пигменты,
обусловливающие окраску желчи и фекалий, но они редко
придают окраску каким-либо поверхностным тканям. У чело-
века, однако, такая ситуация известна — при заболевании жел-
тухой кожа принимает желтоватый оттенок. Пигмент, ответст-
венный за этот симптом, представляет собой билирубин, кото-
рый присутствует в крови в значительных количествах. Другой
известный пример такого рода у позвоночных — синяя или зе-
леная окраска яичной скорлупы у некоторых птиц, которая
обусловлена биливердином. К таким птицам относятся лесная
завирушка (Prunella modularis), а также некоторые домашние
утки (известны «голубые» утиные яйца). У некоторых насеко-
Гл. 5. Тетрапирролы
189
мых расщепление гема происходит при у-лгезо-углеродном ато-
ме и в результате образуются билины у-ряда. Так, биливердин!
1Ху (5.40) найден в покровах тела гусеницы белой бабочки-
капустницы (Pieris brassicae).
Функции. Несмотря на интенсивное изучение билинов, осо-
бенно у позвоночных, их участия в функционировании какого,-
либо органа не обнаружено.
5.8.3. Фикобилины и фикобилипротеины
Распространение в природе, свойства и структура..
В растительном царстве билины присутствуют в виде белковых,
конъюгатов (фикобилипротеинов) у водорослей трех типов —
Rhodophyta (красные водоросли), Cyanophyta (сине-зеленые-
водоросли) и Cryptophyta (криптомонады). Билипротеины водо-
рослей представляют собой кислые водорастворимые глобуляр-
ные белки. Первоначально предполагали, что они обладают
большой молекулярной массой, но, как теперь выяснилось, их
молекула состоит из небольших субъединиц (мол. масса
— 20 000). Фикобилиновые простетические группы прочно, т. е.
ковалентно, присоединены к белку и могут быть удалены лишь,
после чрезвычайно жесткого химического воздействия. Это»
очень усложняет выяснение структуры фикобилинов. Фикоби-
липротеины агрегируют с образованием частиц, называемых
'фикобилисомами (разд. 10.8.2 и рис. 10.17). Последние играют
важную вспомогательную роль при улавливании света в про-
цессе фотосинтеза.
В водорослях преобладают билипротеины двух главных
групп — красные фикоэритрины и синие фикоцианины. Боль-
шинство видов водорослей содержит как фикоэритрин, так и.
один или несколько фикоцианинов, среди которых обычно пре-
обладает какой-либо один билипротеин. Красные водоросли в>
основном содержат фикоэритрин, а сине-зеленые — фикоциа-
нин, хотя имеются представители Cyanophyta, у которых преоб-
ладает красный фикоэритрин. Относительные количества при-
сутствующих фикоцианина и фикоэритрина зависят от спект-
рального состава света, например освещение зеленым светом»
благоприятствует синтезу поглощающего в зеленом диапазоне-
фикоэритрина, в то время как на красном свету преобладает
синтез фикоцианина. В дополнение к этим двум фикобилипро-
теинам в ядре фикобилисомной структуры содержится неболь-
шое количество аллофикоцианина.
Спектроскопически можно различить несколько типов фико-
эритринов и фикоцианинов. Типичные спектры поглощения пиг-
ментов, выделенных из одной водоросли, показаны на рис. 5.14.
Хотя все фикоэритрины, выделенные из водорослей, обладают
190
Часть I. Химические и биохимические аспекты
характерным главным максимумом поглощения при 560—
570 нм (максимумы поглощения пигментов из криптомонад мо-
гут слегка выходить за пределы этого диапазона), спектры по-
глощения фикоэритринов из Cyanophyta, Rhodophyta и Crypto-
phyta, а также фикоэритринов, выделенных из разных видов,
могут различаться. То же относится и к фикоцианинам (Л,тах =
610—620 нм). Аллофикоцианин, который в фикобилисоме при-
Рис. 5.14. Спектры поглощения света фикоцианином (/), фикоэритрином (II)
л аллофикоцианином (III) из сине-зеленой водоросли Chlorogloea. Спектры
в водном растворе, pH 7,0.
нимает энергию от фикоэритрина и фикоцианина, поглощает
даже при еще большей длине волны (650 нм).
Хромофорами простетических групп фикоцианина и фико-
эритрина являются фикоцианобилин и фикоэритробилин соот-
ветственно. Однако ситуация не всегда столь проста; некото-
рые билипротеины содержат два различных хромофора, напри-
мер фикоцианин из Rhodophyta включает как фикоцианобилин,
так и фикоэритробилин.
Выяснение структуры этих фикобилиновых хромофоров
оказалось чрезвычайно сложным. Сейчас уже показано, что,
подобно желчным пигментам животных, все они являются
IXa-изомерами и образуются путем разрыва а-метинового мос-
тика порфирина. В старых работах обработка фикобилипротеи-
нов кипящим метанолом приводила к высвобождению модифи-
цированных хромофоров «фикоэритробилина-690» [фикобили-
вердина (5.41)] и «фикоэритробилина-590» [фиковиолина
(5.42а)]. Истинные хромофоры — фикоцианобилин и фикоэри-
Гл. 5. Тетрапирролы
191
тробилин — строятся из них путем образования тиоэфирной
связи между этилиденовой группировкой кольца А и цистеино-
вым остатком белка (рис. 5.15). Можно привести еще один
пример. Предполагают, что фикоуробилин из фикоэритрина
Rhodophyta образуется из хромофора (5.43) в результате об-
разования тиоэфирных связей с заместителями в кольцах А и
D. (Для удобства на рисунках хромофоры представлены в виде
линейных структур без учета их истинной природной конфор-
мации.)
соон соон
(5 41) Фикоэритробилин-690 = Фикобиливердин
СООН COOR
(5 42а) R =Н; Фикоэритробилин 590 = Фиковиолин
(5 42b) R=CH3, Аплизиовиолин
(5 43) Фикоуробилин
<92
Часть I. Химические и биохимические аспекты
Остаток цистеина
Предполагаемое дополнительное
связывание фикоуробилина
Рис. "515. Предполагаемое ковалентное связывание с белком фикоэритро-
билина, фикоцианобилина и фикоуробилина.
Распространение и функции в растениях. В царстве расте-
ний распространение фикобилинов ограничивается тремя клас-
сами водорослей. Концентрация билипротеинов у водорослей
обычно высока — для фикоцианина зарегистрированы значения,
достигающие 24% сухой массы водорослей. Столь высокие
концентрации билипротеинов обусловливают характерную го-
лубую и красную окраску большинства представителей Cyanop-
hyta и Rhodophyta. У этих водорослей билипротеиновые пиг-
менты присутствуют в виде агрегатов (билисом) в фотосинте-
тических структурах, где они играют важную роль в улавли-
®ании света. Их функционирование в фотосинтезе будет обсуж-
даться в гл. 10.
Гл. 5. Тетрапирролы
193
Дальний красный свет 730 нм
Красный свет 660 нм
Рис. 5 16 Предполагаемая модель комплекса тетрапиррол — белок в Рефор-
ме фитохрома и механизм его превращения в Ptr-форму.
Распространение у животных. Имеется много сообщений о
присутствии билипротеинов, таких, как фикоцианин и фикоэри-
трин у беспозвоночных животных, в частности моллюсков, при-
чем некоторые из них используют эти пигменты для формиро-
вания окраски тела и раковин. Хорошо изученным примером
служит аплизиовиолин (5.42b) — метиловый эфир фикоэритро-
билина — в заметных количествах присутствующий в пурпур-
ном оборонительном секрете моллюска Aplysia. Известно так-
13-64
194
Часть I. Химические и биохимические аспекты
же окрашивание фикобилинами некоторых рыб, например го-
лубого губана. Рыбы получают фикобилины из водорослей, ко-
торыми они питаются.
5.8.4. Фитохром
Все высшие растения содержат фитохром — сине-зе-
леный фотохромный пигмент, который контролирует большое
число разнообразных метаболических процессов, а также раз-
личные стадии развития организмов. Фитохром представляет
собой белок с мол. массой 120 000. В качестве хромофора про-
стетической группы он содержит линейный тетрапиррол, или
билин. Фитохром существует и функционирует в двух фор-
мах— в форме Рг, которая имеет максимум поглощения в
красном диапазоне спектра (660 нм), и в форме Pfr с макси-
мумом поглощения в дальней красной области (730 нм). Эти
две формы легко переходят друг в друга; при поглощении
красного света форма Рг превращается в форму Ргг, которая в
свою очередь дает форму Рг в результате поглощения дальнего
красного света.
соон соон
(5.44) Хромофор фитохрома (Рг)
Детали структуры белка и ультраструктуры фитохрома пол-
ностью не установлены; структура белка у разных видов ра-
стений, по-видимому, различна. Тетрапиррольный хромофор,
который может быть выделен (5.44), оказался сходным с фико-
билинами водорослей. Точно характер связи хромофора с бел-
ком еще не известен, но ясно, что эта связь отличается от той,
которая имеется у фикобилипротеинов. На рис. 5.16 показаны
предполагаемое связывание тетрапиррол — белок у Реформы
фитохрома и механизм ее превращения в Реформу.
Более подробно функционирование фитохрома у растений
обсуждается в гл. И.
Гл. 5. Тетрапирролы
195
5.9. Биосинтез и метаболизм тетрапирролов
5.9.1. Введение
Насколько сейчас известно, все живые организмы ис-
пользуют для образования порфиринов один и тот же меха-
низм независимо от того, гем или хлорофилл является конеч-
ным продуктом. Как у животных, так и у растений линейные
билины образуются из порфириновых предшественников.
Удобнее всего рассматривать биосинтез порфиринов по ста-
диям: а) образование б-аминолевулиновой кислоты (АЛК)’,
б) образование монопиррола, порфобилиногена: в) образова-
ние уропорфириногена — первого тетрапиррольного макроцик-
ла; г) модификация путем введения боковых цепей, приводя-
щая к образованию протопорфириногена; д) дегидрирование
макроцикла, приводящее к образованию протопорфирина IX;
е) образование хелатного комплекса с ионом металла, приво-
дящее к гему или предшественнику хлорофилла — магнийсодер-
жащему протопорфирину IX; ж) последующие модификации,
ведущие к образованию хлорофилла.
Стадии от (а) до (е) являются общими при биосинтезе
гема и хлорофилла.
5.9.2. Образование б-аминолевулиновой кислоты
б-Аминолевулиновая кислота [АЛК (5.45)] может
синтезироваться двумя различными путями. В случае основного
пути у животных и, возможно, у бактерий используется фер-
мент АЛК-синтетаза (б-аминолевулинат-синтаза), который ка-
тализирует образование АЛК из сукцинил-СоА и глицина.
Реакция протекает с участием пиридоксальфосфата через об-
разование промежуточного шиффова основания (рис. 5.17).
Такие промежуточные продукты, как, например, а-амино-0-ке-
тоадипиновая кислота, остаются связанными в виде шиффовых
оснований (5.46), а перед отделением АЛК от пиридоксаль-
фосфата происходит декарбоксилирование. Была выяснена
стереохимия этого процесса. 2-про-5-Водородный атом глицина
сохраняется в ходе всего процесса; это указывает на то, что
при такой последовательности реакций в одном случае проис-
ходит сохранение, а в другом — инверсия конфигурации.
У водорослей и высших зеленых растений большая часть,
если не вся, АЛК образуется путем переаминирования (рис.
5.18). АЛК-трансаминаза (аминолевулинат — аминотрансфера-
за) катализирует перенос аминогруппы от аминокислоты, на-
пример аланина, на у, б-дикетовалериановую кислоту [а-кето-
13*
El н
с—соон
NHj Гикцнн
HOOC сн2.сн2—c
II
Сукцинил-СоА О
SCoA[h]
-I A
И с.сн
nCOO-
CHj.COOH
Пиридоксальфосфат
^c—соон
H N
CoASH
И н
/С\
„ / ХСО.СН2.СН2.СООН
rljN
(5.45) 5- Аминолевулиновая кислота
Пиридок сальфосфат
глицина и сукцинил-СоА.
Рис. 5.17. 6-Аминолевулинатсинтетазный путь образования 6-АЛК из
Гл. 5. Тетрапирролы
197
глутаровый альдегид (5.49)], которая в свою очередь образует-
ся из L-глутамата (5.47) в результате переаминирования до
а-кетоглутарата (5.48) и восстановления последнего.
5.9.3. Образование порфобилиногена
Асимметричная кнорровская конденсация двух моле-
кул АЛК происходит в ходе процесса (рис. 5.19), который
включает альдольную конденсацию, удаление воды и образо-
сн3
соон
Аланин
с=о
I
соон
Пируват
Рис. 5.18. Образование б-АЛК путем переаминирования.
вание шиффова основания. Эту реакцию катализирует один
фермент — АЛК-дегидратаза (порфобилиногенсинтаза), и ее
продуктом является порфобилиноген [ПБГ 5.50)]. На послед-
ней стадии происходит потеря водорода при С-2 пиррольного
кольца, причем эта реакция является стереоспецифичной, по-
скольку отщепляется атом, происходящий из про-/?-водородного
атома дри С-5 АЛК.
5.9.4. Образование первого тетрапиррола —
уропорфириногена III
Это наиболее интересная и наиболее сложная часть
всего биосинтетического пути. Тетрапиррольный макроцикл об-
разуется из четырех идентичных монопиррольных фрагментов
в форме ПБГ. Простая конденсация по типу «голова-к-хвосту»
четырех молекул ПБГ привела бы к образованию тетрапирро-
А. Суммарный процесс
соон
СООН
к
Сг12
соон
сн2
сн2
соон сн2
-I I
сн2
‘ СН2
Сн2
н
(5 50) Порфобилиноген
nh2
(ПБ1 )
Б Возможный механизм
соон
ООН
соон
сн2
фермент---NH2
сн2
сн2
сн2
сн2
НС
Фермент---N=C
сн2
Н2О
сн2
сн2
nh2
nh2
nh2
СООН
f
соон сн2
ООН
соон
сн2
сн2
н+ с=
сн2
сн2
сн2
1 л
с-4)н
фермент—N—С
сн2
Н20
Фермент—N= С
сн2
сн2
1
nh2
nh2 н н
nh2
соон
соон сн2
сн2 сн2
н \==/
Фермент—N-ттС
н+ СН2 V н
nh2
(5 50) ПБГ
Рис 5 19 Образование порфобилиногена- возможный механизм, включаю-
щий образование шиффова основания между АЛК и лизиновым остатком
АЛК-дегидратазы
Гл. 5 Тетрапирролы
199
ла уропорфириногена I (5.51). Обычно же первым циклическим
тетрапирролом, который может быть выделен в качестве про-
межуточного продукта пути биосинтеза хлорофилла и гема,
является другой изомер — уропорфириноген III (5.52), в кото-
ром положения боковых цепей ацетатного и пропионатного
заместителей у одного из пиррольных колец формально изме-
нены на обратные.
Образование уропорфириногена III из четырех молекул
ПБГ — сложный процесс, который все еще продолжают интен-
сивно изучать. В нем участвуют два фермента —ПБГ-дезами-
наза (уропорфириноген I — синтаза) и уропорфириноген Ш-ко-
синтетаза, — которые, по-видимому, функционируют совместно
а не строго последовательно. При денатурации косинтетазы,
например нагреванием до 55—60 °C, первый фермент остается
активным, и в результате реакции образуется уропорфирино-
ген I. Однако он уже не может изомеризоваться до уропорфи-
риногена III. Вероятная последовательность происходящих со-
бытий приведена на рис. 5.20. Четыре молекулы ПБГ соби-
раются последовательно, начиная с кольца А, в линейный би-
лан (5.53). Эта реакция протекает в присутствии ПБГ-дезами-
назы, которая затем катализирует образование ключевого
промежуточного продукта — гидроксипроизводного этого била-
на (5 54). Последний быстро циклизуется косинтетазой с обра-
зованием уропорфириногена III. В этой реакции может прини-
мать участие стабилизируемый ферментом промежуточный про-
дукт—метиленпирроленин (5.55). В ходе циклизации происхо-
дит внутримолекулярная перегруппировка в кольце D, вероят-
но, по механизму, ответственному за образование промежуточ-
ного продукта спиро-типа (5.56). В отсутствие косинтетазы ли-
нейный тетрапиррол может легко циклизоваться до уропорфи-
риногена I.
5.9.5. Превращение в протопорфириноген (рис. 5.21)
Следующей стадией биосинтетического пути является
последовательное декарбоксилирование четырех ацетатных бо-
ковых цепей уропорфириногена до четырех метильных групп.
На этой стадии пропионатные боковые цепи не изменяются.
Уропорфириноген I может таким образом превращаться в
копропорфириноген I (5.57). Однако обычно в процессе био-
синтеза из уропорфириногена III образуется копропорфирино-
ген III (5 58). Удаление всех четырех карбоксильных групп
катализируется одним ферментом — уропорфириноген Ш-де-
карбоксилазой. Промежуточные продукты в нормальных усло-
виях, по-видимому, не отщепляются от фермента, однако не-
большие количества таких продуктов были получены из фека-
(5.Б1) Уропорфириноген 1
(5.52) Уропорфириноген Ш
Рис 5 20 Предполагаемый механизм образования уропорфириногеиа III (А=
=--СП2СООН; Р = —СН2СН2.СООН).
Гл. 5. Тетрапирролы
20!
лий крыс. Имеющиеся доказательства подтверждают следую-
щую последовательность декарбоксилирования колец: D, А, В
и С. Была также определена стереохимия процесса декарбокси-
лирования: он протекает с сохранением конфигурации (рис.
5.21, Б).
Две пропионатные боковые цепи копропорфириногена III
(в кольце А, а затем в кольце В) далее модифицируются в
винильные группы с помощью копропорфириногеноксидазы,
для функционирования которой необходим молекулярный кис-
лород. Копропорфириноген I такой модификации не подвер-
гается. Промежуточными продуктами модификации копропор-
фириногена III могут быть соединения с гидроксилированны-
ми пропионатными боковыми цепями, а ее конечным продук-
том является протопорфириноген IX (5.59). Установлено так-
же, что при этом наблюдается транс-отщепление водорода
(рис. 5.21, Б).
5.9.6. Дегидрирование до протопорфирина IX
Прежде чем произойдет образование хелатного ком-
плекса с металлом, порфириногенный макроцикл в результате
удаления шести водородных атомов должен быть превращен в
сопряженный окрашенный порфирин (рис. 5.22, А). Последова-
тельность их удаления неизвестна. Продукт этой реакции—
протопорфирин IX (5.60) является последним промежуточным
продуктом, общим для биосинтеза хлорофилла и гема.
5.9.7. Образование хелатного комплекса с металлом
Биосинтез гема. Гем (протогем; 5.61) образуется из
протопорфирина IX и Ре2+-иона под действием фермента фер-
рохелатазы (рис. 5.22, Б). Обычно этот фермент прочно связан
с клеточными частицами. Его можно обнаружить в митохонд-
риях животных клеток, в хлоропластах растений и в мембранах
фотосинтезирующих бактерий. В некоторых случаях, однако,
в клетках присутствует также и растворимая феррохелатаза.
Очищенная феррохелатаза способна вводить в протопорфири-
ны (которыми могут быть не только протопорфирины IX), по-
мимо ионов Fe2+, ионы других металлов, например Zn2+ и
Со2+. Некоторую роль в этом процессе играет, по-видимому,
липид (вероятно, фосфолипид).
Биосинтез хлорофилла. Хотя считают, что введение иона
магния при биосинтезе хлорофилла также происходит на ста-
дии протопорфирина IX (рис. 5.22, В), на практике чрезвычай-
но трудно разделить образование хелатного комплекса с ме-
таллом от метилирования пропионатной боковой цепи при С-13.
Изомеры I
НООС HjC HjC CH, соон ноос
Уропорфириноген I
(5 57) Копропорфириноген |
Уропорфириноген III
^/(5 58) Копропорфириноген III
снон сн, соон
(5 59) Протопорфириноген । х
Гл. 5. Тетрапирролы
203
Б
1) Декарбоксилирование ( стадия 1 )
СН2.СН2.СООН
2) Элиминирование ( стадия 2
Включение ОН с сохранением
Рис. 5.21. Превращение уропорфириногена в копропорфирииоген и затем в
протопорфнрииогеи IX. А. Путь превращения. Б. Стереохимия.
Однако была показана этерификация магнийпротопорфирина
IX (5.62) до метилового эфира (5.63) с участием S-аденозилме-
тионина. Это подтверждает идею о том, что в нормальных
условиях хелатирование предшествует метилированию. Что
касается фермента, ответственного за введение иона Mg2+ в
молекулу протопорфирина IX, то здесь пока получены лишь
предварительные результаты.
5.9.8. Последующие стадии биосинтеза хлорофилла а
Образование изоциклического кольца (кольца Е) и
протохлорофиллида а. Метилированная пропионатная боковая
цепь в положении 13 кольца С метилмагнийпротопорфири-
на IX (5.63) используется для образования изоциклического
кольца (кольца Е) с помощью последовательности реакций,
показанной на рис. 5.23. Были идентифицированы промежуточ-
ные продукты этой последовательности реакций, протекающих
по типу р-окисления.
Протопорфириноген IX
<5 61) Гем (протогем)
(5 62) Магнийпротопорфирин IX
(5 63) Метил'магнийпротопорфирин IX
Рис. 5 22 А. Превращение протопорфнриногена IX в протопорфирин IX
Б. Хелатирование Fe2+ с образованием гема В Введение магния с образо-
ванием метилмагнийпротопорфирина IX
(5.65) Протохлорофил л ид а
Рис. 5.23. Образование изоциклического кольца и протохлорофиллида а.
206
Часть 1. Химические и биохимические аспекта
(5 67) Хлорофилла
Рис. 5.24. Превращение протохлорофиллида а в хлорофиллы а и Ь.
Винильная группа при С-8 образующегося продукта — ме-
тилового эфира магний-3,8-дивинилфеопорфирина as (5.64) —
затем подвергается насыщению до этильной группы, и обра-
зуется протохлорофиллид а {метиловый эфир магнийвинилфео-
порфирина as (5.65)]. Вместе с тем есть много аргументов в
пользу того, что восстановление винильной группы может про-
исходить до образования изоциклического кольца.
Гл. 5. Тетрапирролы
207
Образование хлорофилла а. Превращение протохлоро-
филлида а в хлорофилл а включает только две реакции — гид-
рирование кольца с образованием дигидропорфиринового (хло-
ринового) макроцикла и этерификацию последнего Сго-изопре-
ноидным спиртом фитолом (рис. 5.24). Хотя эти реакции ка-
жутся простыми, в целом образование хлорофилла а из про-
тохлорофиллида— процесс чрезвычайно сложный.
Сначала происходит восстановление кольца D транс-присо-
единением двух водородных атомов, что приводит к образова-
нию хлорофиллида а (5.66). У некоторых растений и водорос-
лей, например у Chlorella, которая может синтезировать хлоро-
филл в темноте, эта реакция является простой катализируемой
ферментом темновой реакцией. Большинству растений, однако,
для синтеза хлорофилла необходим свет. Кроме того, процесс
насыщения кольца D, по-видимому, является фотопревраще-
нием, причем не свободного протохлорофиллида а, а протохло-
рофиллид-белкового комплекса, известного как протохлоро-
филлндный галохром, который содержит одну молекулу прото-
хлорофиллида на молекулу белка. Полученные данные ука-
зывают на то, что голохромный белок представляет собой фо-
тофермент протохлорофиллидредуктазу. Образование комплек-
са протохлорофиллид—фермент — NADPH происходит в тем-
ноте, однако для восстановления кольца D протохлорофиллида
необходим свет.
Завершающей стадией биосинтеза хлорофилла а (5.67) яв-
ляется этерификация остатка пропионовой кислоты при С-17
(кольцо D) хлорофиллида а фитолом. Хорошо известен фер-
мент хлорофиллаза, который (в присутствии 30%-кого ацето-
на!) гидролизует или осуществляет переэтерификацию фитиль-
ной сложноэфирной группы хлорофилла. После многолетних
споров в настоящее время установлено, что хлорофиллаза,
действующая в обратном направлении, не принимает участия
в нормальном биосинтетическом пути. В качестве донора фи-
тильной группы идентифицирован фитилпирофосфат (5.68), об-
разующийся из геранилгеранилпирофосфата (5.69). Недавно у
высших растений были обнаружены разновидности хлорофил-
ла, содержащие геранилгераниол (5.70) и ди-, а также тетра-
гидрогеранилгераниол; это позволяет предположить, что в не-
которых случаях этерифицирующим спиртом может служить
геранилгераниол, который, вероятно, восстанавливается в фитол
уже после присоединения к порфирину.
5.9.9. Образование хлорофилла b
Скорее всего, хлорофилл b (5.71) образуется из хло-
рофилла а путем окисления метильной группы при С-7 до аль-
дегидной. Пока ничего не известно о механизме этого предпо-
208
Часть I Химические и биохимические аспекты
лагаемого процесса. Альтернативные предположения, соглас-
но которым эти два хлорофилла синтезируются различными
или расходящимися путями или хлорофилл а образуется из
хлорофилла Ь, были отвергнуты.
5.9.10. Образование хлорофиллов end
О биосинтезе хлорофиллов с и d ничего не известно.
На основании их структуры можно предположить, что хлоро-
филлы Ci (5.72) и с2 (5.73) образуются из промежуточных про-
дуктов пути биосинтеза хлорофилла а, а хлорофилл d (5.74) —
из самого хлорофилла а (рис. 5.25).
5.9.11. Образование бактериохлорофиллов
Были получены мутантные штаммы бактерий (RJiodop-
seudomonas spp.), накапливающие протохлорофиллид а,
хлорофиллид а и другие близкие к ним соединения, у которых
винильная группа в кольце А модифицирована в гидроксиэтиль-
ную или ацетильную. Как видно на биосинтетической схеме,
приведенной на рис. 5.26, модификация боковой цепи при С-3
происходит перед гидрированием кольца В, приводящем к об-
разованию бактериохлорофиллида а (5.75). Этерификация фи-
толом или близким к нему спиртом, таким, как фарнезол, яв-
ляется заключительной стадией образования бактериохлоро-
филла а (5.76). Бактериохлорофилл b (5.77) образуется, по-
видимому, путем дегидрирования бактериохлорофилла а.
5.9.12. Образование хлоробиум-хлорофиллов
(бактериохлорофиллов cud)
Относительно биосинтеза нескольких хлорофиллов
Chlorobium известно очень мало. По-видимому, они образуют-
ся из гидроксиэтильного промежуточного продукта (5.78) био-
синтеза бактериохлорофиллов и дополнительные алкильные
заместители при С-8, С-12 и С-20 вводятся позже путем при-
соединения к винильным группам; донором при этом служит
S-аденозилметионин.
5.9.13. Биосинтез витамина Bi2
Тип III расположения боковых цепей в витамине В12
указывает на фундаментальное сходство его биосинтеза с био-
синтезом гема и хлорофилла. Корриновое кольцо происходит
от АЛК и ПБГ, и начальные стадии его биосинтеза идентичны
стадиям образования порфирина. Уропорфириноген III вклю-
чается в корриновую систему без какой-либо фрагментации
Гл 5 Тетрапирролы
20»
(5 72) Хлорофилл с, (R ( П2 СИ,)
(5 64) (R -СП -СП;) Метилмагнии-
3,8 дивинипфеопорфирин as
{ReCH2 СПз) Протохлорофиллида
(5 73) Хлорофилл с2 (К -( 11 - Cl I ,}
л. (5 74) Хлорофилл^
(5 67) Хлорофилл a
Рис. 5.25. Возможные пути образования хлорофиллов end.
или перегруппировки четырех колец. Более поздние стадии
биосинтеза витамина В12 в настоящее время интенсивно из-
учаются.
5.9.14. Образование билинов
У животных билины образуются в результате катабо-
лического распада гема, главным образом гема гемоглобина,
в результате окислительного элиминирования одного из угле-
14—84
(5 78)
{5.77| Бактериохлорофилл b
фарн«ип или геранилгерзнилХ
Рис. 5.26. Образование бактериохлорофиллов и хлоробиумхлорофиллов.
Гл- 5. Тетрапирролы
211
родных атомов метиновых мостиков и потери хелатированного
атома железа. Расщепление происходит преимущественно в
ретикулоэндотелиальных клетках селезенки, печени, костного'
мозга и в меньшей степени почек. Хотя механизм этого процес-
са еще до конца не ясен, известно, что два вводимых кетоновых,
или лактамных, атома кислорода происходят из двух различ-
ных молекул О2 и что в этом процессе участвуют цитохром-
связанная гем-оксигеназа, NADP+ и аскорбиновая кислота.
Метиновый углерод освобождается в виде окиси углерода, же-
лезо транспортируется к месту хранения общих запасов желе-
за в организме, а глобин расщепляется до аминокислот, по-
ступающих в общий метаболический фонд. У млекопитающих
происходит разрыв исключительно а-метинового мостика (С-5)
с образованием биливердина 1Ха (5.79). Расположенный в
центре молекулы биливердина углеродный атом метиновой
группы затем восстанавливается до СН2 под действием редук-
тазы [с кофактором NAD(P)H], которая присутствует в пече-
ни и селезенке. Образовавшийся таким путем билирубин (5.80)
транспортируется в виде альбуминового комплекса в печень,
где происходит его связывание с глюкуроновой кислотой и
другими сахарами, которое катализируется микросомным фер-
ментом. Водорастворимый конъюгат поступает в желчь и за-
тем в кишечник. В кишечнике билирубиновые конъюгаты гид-
ролизуются p-глюкуронидазой, после чего претерпевают даль-
нейшие превращения, главным образом восстановление с по-
мощью кишечных бактерий. Эти превращения включают гидри-
рование винильных боковых цепей, последовательное восстанов-
ление двух остающихся метиновых мостиков и ступенчатое вос-
становление некоторых (двух) двойных связей пиррольных ко-
лец с образованием d- и /-уробилиногенов (5.81, 5.82) и /-стер-
кобилиногена (5.83). Билиногены окисляются на воздухе и дают
d- и /-уробилин (5.84, 5.85) и /-стеркобилин (5.86) —желто-ко-
ричневые пигменты, которые окрашивают мочу и фекалии.
Предполагаемая суммарная схема образования билинов у
млекопитающих приведена на рис. 5.27. К сожалению, пока
нет данных, касающихся образования билинов у низших жи-
вотных.
У растений. У водорослей, так же как и у животных, окис-
лительный разрыв порфиринового кольца происходит практи-
чески только по углероду а-метинового мостика, который те-
ряется в виде окиси углерода. Источником билинов у водорос-
лей является, скорее всего, гем, а не хлорофилл, и механизм
его распада, вероятно, сходен с механизмом распада у живот-
ных. Детали заключительных стадий распада, в результате
которого образуются фикобилины с характерной структурой,
пока не ясны. Для образования билипротеинов у водорослей; и
14*
(5 80) Билирубин
CH, CH,
I I
соон COOH
(5 79) Биливердин IX a
соон COOH
Б
Гемоглобин эритроцитоз
Ретикуло-
эндотелиальная
система
Печень
Кишечник
рис 5 27,А
Билирубин
В плазме крови в виде
Э альбуминового комплекса
Билирубин
Г люкуроновая
кислота и т п.
Конъюгаты билирубина
3 Желчь
Конъюгаты билирубина
Глюкуронидаза
Билирубин
Разрушение
микроорганиз-
мами и
аутоокисление
рис 5 27, В
Уробилин, стеркобилин
Рис. 5 27. Схема образова-
ния билинов из гема у мле-
копитающих. А. (с, 212).
Распад гема до билирубина
в ретикулоэндотелиальных
тканях Б. Конъюгация би-
лирубина с альбумином и его
транспорт в кишечник. В.
(с. 214). Распад билирубина,
осуществляемый микроорга-
низмами и его аутоокисле-
ние в кишечнике.
(5.83) / -Стеркобилиноген
(5.86) / - Стеркобилин
(5.84) (R=CH=CH2) : d-Уробилин
(5.85) (R=CH2CH3) : / -Уробилин
Гл. 5. Тетрапирролы
215
включения их в фикобилисомы необходим одновременный син-
тез белка de novo. До сих пор не известно, происходит ли ана-
логичным путем биосинтез билиновой простетической группы
фитохрома.
5.9.15. Факторы, контролирующие синтез
и накопление тетрапирролов
Внешние факторы — влияние окружающей среды и
условий питания. На синтез тетрапирролов могут влиять раз-
личные внешние факторы (например, окружающая среда или
питание). Наиболее известным из факторов окружающей среды
является освещение, необходимое для биосинтеза хлорофилла
или бактериохлорофилла в большинстве фотосинтезирующих
систем. Например, у Rhodopseudomonas sphaeroides бактерио-
хлорофилл синтезируется только в анаэробных условиях на
свету, хотя синтез гема (цитохромов) у этой бактерии проис-
ходит в темноте в присутствии кислорода воздуха. Активность
АЛК-синтетазы на свету значительно выше, чем в темноте. Хо-
рошо известно, что большинство высших растений при выдер-
живании в темноте становятся этиолированными, т. е. они не
способны синтезировать нормальные хлоропласты или хлоро-
пластные пигменты, особенно хлорофиллы. Однако при пере-
носе на свет этиолированные проростки образуют большое
количество хлорофилла за период около 48 ч и приобретают
нормальную зеленую окраску. Такой синтез хлорофилла проис-
ходит по стадиям. Очень быстро, даже после кратковременного
освещения, небольшое количество протохлорофиллида а (ас-
социированного с протохлорофиллидголохромным белком), со-
держащегося в этиолированных тканях, превращается в хлоро-
•филлид а, который впоследствии более медленно этерифици-
руется до хлорофилла а. По этому пути образуется небольшое
количество хлорофилла в течение первых двух часов позелене-
ния. Затем наступает индукционный период, длящийся в тече-
ние нескольких часов, за которым следует быстрый синтез
•de novo гораздо больших количеств хлорофилла. Во время
индукционного периода происходит интенсивный синтез фер-
ментов и значительно возрастает активность АЛК-синтетазы.
Образование хлорофилла представляет собой лишь часть го-
раздо более сложного процесса формирования хлоропластов,
причем факторы, регулирующие образование различных хлоро-
пластных компонентов, точно скоординированы (гл. 10).
Хлороз, т. е. недостаток хлорофилла, является общим сим-
птомом многих болезней растений. Понижение содержания
хлорофилла в тканях может быть обусловлено нехваткой пи-
тательных веществ, особенно азота, железа и магния. Трудно
216
Часть I Химические и биохимические аспекты
сказать, влияют ли эти факторы только на синтез хлорофилла
или на формирование хлоропластов в целом.
У большинства красных и сине-зеленых водорослей на син-
тез и концентрацию билипротеинов влияют такие факторы, как
интенсивность и спектральный состав падающего света. Эти
же факторы определяют, синтез какого из них, фикоцианина
или фикоэритрина, будет преобладающим. При освещении
красным светом преимущественно образуется поглощающий
красные лучи фикоцианин, в то время как зеленый свет благо-
приятствует образованию поглощающего в этой области фико-
эритрина.
У животных синтезы двух компонентов гемоглобина — гема
и глобина — по-видимому, тесно скоординированы. В ответ на
понижение уровня кислорода в окружающей среде происходит
возрастание количества гемоглобина (до 20% У человека).
Это наблюдается, например, на больших высотах над уровнем
моря. При этом происходит также усиление синтеза аллостери-
ческого эффектора — 2,3-дифосфоглицерата, — что облегчает и
делает более эффективной «разгрузку» кислорода из оксигемо-
глобина. Эти феномены привлекли внимание широкой публики
во время проведения Олимпийских игр 1968 г. в Мехико. Атле-
ты из Кении и Эфиопии, привычные к условиям большой вы-
соты над уровнем моря, выиграли большинство медалей в беге
на средние и длинные дистанции. Спортсмены из других
стран, не успевшие в должной мере акклиматизироваться, не
показали высоких результатов, поскольку у них, по-видимому,
уровни гемоглобина и 2,3-дифосфоглицерата недостаточны для
того, чтобы мышцы работали эффективно в течение длитель-
ного времени в разреженной атмосфере высокогорья.
Весьма вероятно, что во многих рассмотренных выше слу-
чаях первой реакцией на внешний фактор является изменение
количества или активности АЛК-синтетазы.
Внутренние факторы — биохимический контроль. В биосин-
тезе циклических тетрапирролов — гема, хлорофиллов и кор-
ринов — у всех живых организмов (животных, растений и
микроорганизмов) главные участки биохимического контроля
находятся около АЛК и ее ключевых ферментов — АЛК-синте-
тазы и АЛК-дегидразы. Механизм действия этого контроля в
индивидуальных случаях пока изучен недостаточно. Множе-
ство экспериментальных данных свидетельствует в пользу того,
что у разных организмов и в разных тканях могут функциони-
ровать несколько различных механизмов. Тот факт, что АЛК-
синтетаза обычно является нерастворимым ферментом мито-
хондрий, в то время как почти все другие ферменты биосинте-
за порфиринов растворимы и находятся в цитоплазме, позво-
ляет предположить, что в контролирующем механизме опреде-
Гл 5 Тетрапирролы
217
ленную роль играет проницаемость митохондриальной мембра-
ны для АЛК. В большинстве случаев, однако, регулируется
активность или концентрация АЛК-синтетазы или фермента,
активирующего АЛК-синтетазу. По-видимому, наиболее важ-
ной и распространенной является регуляция синтеза или ак-
тивности фермента конечным продуктом.
В тех случаях, когда организм или ткань синтезирует раз-
личные тетрапирролы, например бактериохлорофилл и гем,
каждый из биосинтетических путей, вероятно, контролируется
отдельно.
5.10. Нарушения метаболизма порфиринов
5.10.1. Порфирии
В нормальных условиях у животных синтез и распад
порфиринов контролируются очень четко. У человека обнару-
жена группа расстройств — врожденных или приобретенных
нарушений ферментов, участвующих в биосинтезе гема,—
для которых характерно присутствие в различных тканях орга-
низма свободных порфиринов и (или) их предшественников.
Такие нарушения, называемые порфирией, клинически обнару-
живаются по повышенному выделению этих соединений.
При эритропоэтической порфирии наблюдается недостаток
уропорфириноген Ш-косинтетазы и накопление больших коли-
честв уропорфириногена I, обусловливающее винно-красное
окрашивание мочи и интенсивную флуоресценцию (в красной
области спектра) зубного дентина и других тканей в УФ-свете.
При острой перемежающейся порфирии поражается печень.
В ней накапливаются порфирины и их предшественники, осо-
бенно АЛК и порфобилиноген, которые обнаруживаются также
в моче. Во время перемежающихся приступов больной испы-
тывает острую боль в брюшной полости. Эти две главные
группы порфирий включают несколько типов порфирий, раз-
личающихся природой накапливающихся порфиринов или их
предшественников, типом наследования, а также клиническими
свойствами и симптомами.
Причиной психического расстройства английского короля
Георга III, по-видимому, была именно острая перемежающаяся
порфирия (пестрая порфирия). Каждому его приступу пред-
шествовала острая боль в брюшной полости, а в бюллетенях
о состоянии здоровья отмечалось, что его моча имела характер-
ный красный цвет. Сообщалось также о наличии симптомов,
известных сейчас как симптомы порфирии, у его родственников
по восходящей и нисходящей линии
При различных типах порфирии в моче и фекалиях могуг
накапливаться уро- и копропорфирины (5 30, 5 31), уро- и ко-
218 Часть I. Химические и биохимические аспекты
пропорфириногены (изомеры I и III) (5.51, 5.52, 5.57, 5.58),
протопорфирин IX (5.60), АЛК (5.45) и ПБГ (5.50). Их на-
копление обусловлено главным образом дефектами в регуля-
ции активности АЛК-синтетазы. Порфирию можно вызвать
экспериментально введением различных лекарственных препа-
ратов, в том числе барбитуратов.
Свободный порфирин в тканях тела может действовать как
сенсибилизатор опасных фотоокислительных процессов, поэто-
му многим больным порфирией присуща повышенная чувстви-
тельность к свету и они страдают от действия попадающего
на кожу солнечного света. Интересно, что в некоторых случаях
от вредных эффектов фотосенсибилизации защищает р-каро-
тин. Это напоминает защитное действие каротиноидов при фо-
тоокислении кислородом в синглетном состоянии—процесса,
часто сенсибилизируемого порфиринами у бактерий и растений
(гл. 10 и 11).
5.10.2. Нарушения образования билинов
Желтуха. Патологические нарушения, связанные с из-
бытком билинов, называют желтухой. Они могут быть резуль-
татом либо избыточного образования, либо пониженной дегра-
дации билинов или же нарушения их выведения. В случае
желтухи у человека избыток билирубина (5.80) придает ин-
тенсивную желтую окраску плазме крови и тканям организма.
Билирубин переходит из крови в межтканевое пространство и
интенсивно поглощается соединительными тканями.
Имеется несколько причин возникновения желтухи у взрос-
лых. В норме билирубин переходит из ретикулоэндотелиальной
системы в печень, где он связывается с глюкуроновой кислотой
и экскретируется в желчь. Если распад гемоглобина в значи-
тельной степени усиливается, печень не справляется с удале-
нием билирубина из крови (ретенционная, или паренхиматоз-
ная, желтуха). Чаще желтуха возникает в результате закупор-
ки желчных путей или некроза печени, что ведет к регургита-
ции компонентов желчи в кровоток (регургитационная, или
обтурационная, желтуха). В этом случае в крови и в тканях
обнаруживается повышенное содержание связанного билиру-
бина.
Преходящая и безвредная желтуха довольно часто наблю-
дается у новорожденных. В печени новорожденных не хватает
ферментов, ответственных за связывание билирубина; послед-
ний же присутствует у них в больших количествах из-за рас-
щепления избытка эритроцитов во время жизни зародыша в
матке. В результате накапливается свободный билирубин. Эту
безопасную и легкую желтуху необходимо отличать от стой-
Гл 5 Тетрапирролы
219
кой и опасной желтухи, сопровождающей гемолитические бо-
лезни новорожденных. В этом случае билирубин может изби-
рательно поглощаться тканями мозга, вызывая его стойкое по-
вреждение.
Желтуха бывает не только у человека и млекопитающих.
Зеленая «желтуха», вызванная биливердином (5.79), описана
У щук.
Образование синяков. Временная черно-синяя и желтая
•окраска синяков появляется у человека, когда кровь попадает
в межтканевые пространства, где гемоглобин разрушается до
таких продуктов, как биливердин (зеленый), уробилин (оран-
жевый) и биливиолины (фиолетово-синие).
5.11. Функции тетрапиррольных пигментов
Некоторые функции тетрапирролов, не связанные с их
светоплоглощающими свойствами, мы уже отмечали (особен-
но кислородпереносящую функцию гемоглобина). Однако
большинство биологических функций этих пигментов связано
с их светопоглощающими свойствами. Так, например, они при-
дают окраску тканям, участвуют в фотосинтезе и служат фо-
торецепторами. Этим темам будут посвящены гл. 10 и 11.
5.12. Заключение и комментарии
Тетрапиррольные пигменты, представляющие собой
группу соединений со столь жизненно важными биологически-
ми функциями, изучались чрезвычайно интенсивно. Поэтому о
механизмах их образования и функционирования известно
больше, чем в случае какой-либо другой группы пигментов.
Установление трехмерных структур миоглобина и гемоглобина
и механизма, с помощью которого гемоглобин функционирует
в транспорте кислорода, представляет собой один из классиче-
ских образцов научного исследования. Во многом ясным стал
также путь, по которому хлорофиллл используется как глав-
ный светособирающий пигмент в фотосинтезе (гл. 10). Основ-
ные аспекты биосинтеза порфиринов (и коррина), в том числе
его детали и стереохимия, изучены в очень элегантных опытах
с помощью классических радиоизотопных методов и усовер-
шенствованных методов введения и анализа 13С- и 2Н-метки.
Желающий изучать биосинтез порфиринов не смог бы сделать
ничего лучшего, чем прежде всего внимательно прочитать эти
работы. Тем не менее даже при таком положении вещей ход
некоторых биосинтетических превращений до сих пор полностью
не установлен. Для изучения образования бактериохлорофиллов,
необычных хлорофиллов с и d из водорослей, модифицирован-
220
Часть I. Химические и биохимические аспекты
ных гемов некоторых цитохромов и т. д., имеются широкие
возможности. Необходимо завершить также структурные ис-
следования этих соединений. Нуждаются в детальных химиче-
ских исследованиях свободные порфирины, найденные у низ-
ших животных. До конца не выяснены взаимодействия между
хлорофиллами и белками в специфических комплексах в фо-
тосинтетическом аппарате и ориентация хлорофиллов внутри
этих комплексов. Факторы, которые регулируют образование
этих комплексов и биосинтез самих молекул хлорофилла
(и других порфиринов), — это та область исследований, где
биохимиков ждут открытия.
Свойства линейных тетрапирролов, или билинов, совершен-
но отличны от свойств макроциклических порфиринов, но опять-
таки и здесь существует много захватывающих проблем. Эти
соединения, известные как желчные пигменты, образуются в
организме животных, в том числе и человека, путем разруше-
ния гема. Механизмы распада и превращений с образованием
продуктов, которые в конце концов выделяются, заслуживают
повторного исследования с применением современных физико-
химических методов. К проблемам, заслуживающим внимания,
относятся также образование билинов в синяках, при желтухе
и при формировании окраски скорлупы птичьих яиц. Широкое
поле исследований предоставляют механизмы отложения и
связывания билинов и свободных порфиринов с минеральными
веществами в яичной скорлупе и факторы, которые регулируют
сложный характер окрашивания скорлупы у разных видов.
Растительные билины (фикобилины) у сине-зеленых и красных
водорослей содержатся в фикобилисомах и функционируют
как вспомогательные пигменты в фотосинтезе. Многие вопросы,
касающиеся этих пигментов, такие, как способ их поглощения
и связывания с белком, остаются пока без ответа. Наиболее
интересным растительным билином является фитохром — «не-
уловимое» вещество, которое служит посредником во многих
световых реакциях у представителей растительного мира. Пол-
ная картина фитохромных фотоциклов, когда она в конце
концов будет целиком выяснена на молекулярном уровне, по-
видимому, будет столь же впечатляющей, как и картина цик-
лов превращения зрительного пигмента млекопитающих.
5.13. Рекомендуемая литература
Из множества книг, посвященных тетрапиррольным
пигментам, выделяется монография под редакцией Смита
(Smith, 1975). В ней исчерпывающе рассмотрена химия этих
соединений, а также представлено множество данных об их
биосинтезе. Хорошим источником информации может служить
Гл. 5, Тетрапирролы
221
обширное семитомное издание под редакцией Долфина (Dolp-
hin, 1978), правда, к настоящему времени вышли только два
тома. Полезным остается сборник докладов, представленных
на симпозиуме по порфиринам и родственным им соединениям
(Goodwin, 1968). Отдельным аспектам тетрапиррольных пиг-
ментов посвящен ряд книг и статей. Например, Джексон (Jack-
son, 1976) приводит данные по химии и распространению хло-
рофиллов, а Вернон и Сили (Vernon, Seely, 1966) обсуждают
все аспекты химии и биохимии хлорофилла. Информацию по
порфиринам у животных читатель может получить в специаль-
ных статьях Кеннеди (Kennedy, 1969, 1976), а также Риминг-
тона и Кеннеди (Rimington, Kennedy, 1962); он найдет ее так-
же в соответствующих разделах книг по пигментам животных
Фокса (Fox, 1976), Фокса и Веверса (Fox, Vevers, 1960) и
Нидхэма (Needham, 1974). Из многих публикаций, посвящен-
ных билинам как у животных, так и у растений, можно поре-
комендовать следующие: Bouchier, Billing, 1967; Hudson,
Smith, 1975; Lathe, 1972; O’Carra, O’hEocha, 1976; With, 1968;
Rudiger, 1980. Химия и биохимия фитохрома описана Смитом
и Кендриком (Smith, Kendrick, 1976), а данные о цитохроме
Р450 собраны в книге под редакцией Купера, Розенталя, Снай-
дера и Уитмера (Cooper, Rosenthal, Snyder and Witmer, 1975).
Что касается биосинтеза порфиринов, то его общую карти-
ну с акцентом на механизмы реакций и стереохимию дает
статья Ахтара и Джордана (Akhtar, Jordan, 1979), в то время
как биохимические и биологические аспекты рассмотрены в
работах Богорада (Bogorad, 1976) и Шнейдера (Schneider,
1980). Появившаяся совсем недавно статья Баттерсби и др..
(Battersby et al., 1980) разъясняет ряд моментов, бывших ра-
нее сомнительными. Информацию о биосинтезе порфиринов и
билинов можно получить из фундаментальных книг, цитиро-
ванных выше, главным образом из монографии под редакцией
Смита (Smith, 1975). И наконец, следует упомянуть о двух
специальных статьях, читающихся с большим интересом, —
это обзор Перутца (Perutz, 1970), в котором объясняется ме-
ханизм кооперативного связывания гемоглобина с кислородом,
и статья Тшуди и Шмида (Tschudy, Schmid, 1972), посвященная
болезням, входящим в группу порфирий.
5.14. Дополнительная литература
Akhtar М., Jordan Р. М. (1979). Porphyrin, chlorophyll and corrin biosynthesis.
In: Comprehensive organic chemistry, vol. 5, ed. E. Haslam, p. 1121, Ox-
ford, Pergamon.
Battersby A. R., Fookes C. J. R., Matcham G. W. J., McDonald E. (1980). Bio-
synthesis of the pigments of life: formation of the macrocycle, Nature,
285, 17.
222
Часть I. Химические и биохимические аспекты
Bogorad L. (1976). Chlorophyll biosynthesis. In: Chemistry and biochemistry of
plant pigments, 2nd edition, vol. 1, ed. T. W. Goodwin, p. 64, London, New
York and San Francisco, Academic Press.
Bouchier I. A. D., Billing В. H. (eds ) (1967). Bilirubin metabolism, Oxford,
Blackwell.
Cooper D. F., Rosenthal O., Snyder R., Witmer C. (eds.) (1975). Cytochromes
Pisa and b5, New York, Plenum.
Dolphin D. (ed.) (1978). The porhyrins, vols. 1 and 2 (7 vols projected), New
York, Academic Press.
Box D. L. (1976). Animal biochromes and structural colours, 2nd edition, Berke-
ley, Los Angeles and London, University of California Press.
Box H. M„ Vevers G. (1960). The nature of animal colours, London, Sidgwick
and Jackson.
Goodwin T. W. (ed.) (1968). Porphyrins and related compounds, London and
New York, Academic Press.
Hudson M. B., Smith, К. M. (1975). Bile pigments, Chem. Soc. Revs., 4, 363.
Backson A. H. (1976). Structure, properties and distribution of chlorophylls. In:
Chemistry and biochemistry of plant pigments, 2nd edition, vol. 1, ed.
T. W. Goodwin, p. 1, London, New York and San Francisco, Academic
Press.
Kennedy G. F. (1969). Pigments of Annelida, Echiuroidea, Sipunculoidea, Priapu-
loidea and Phoronidea, Chem. Zool., 4, 311.
Kennedy G. F. (1976). Survey of avian eggshell pigments, Comp. Biochem.
Physiol., B55, 117.
Lathe G. H. (1972). The degradation of haem by mammals and its excretion
as conjugated bilirubins, Essays in biochemistry, 8, p. 107, London and New
York, The Biochemical Society—-Academic Press.
Needham A. E. (1974). The significance of zoochromes, Berlin, Heidelberg and
New York, Springer-Verlag.
O’Carra P., OhEocha C. (1976). Algal biliprotein and phycobilins. In: Chemistry
and biochemistry of plant pigments, 2nd edition, vol 1, ed. T. W. Goodwin,
p. 328, London, New York and San Francisco, Academic Press.
Perutz M. B. (1970). Stereochemistry of cooperative effects of haemoglobin,
Nature, 228, 726.
.Rimington C„ Kennedy G. Y. (1962). Porphyrins. In: Comparative biochemistry,
vol. 4, eds. M. Florkin and H. S. Mason, p. 557, New York and London,
Academic Press.
Rudiger W. (1980). Plant biliproteins. In: Pigments in plants, 2nd edition, ed.
F.-C. Czygan, p. 314, Stuttgart and New York, Gustav Fischer.
Schneider H. (1980). Chlorophyll biosynthesis. Enzymes and regulation of enzy-
me activities In: Pigments in plants, 2nd edition, ed. F.-C. Czygan, p. 237,
Stuttgart and New York, Gustav Fischer.
Smith H., Kendrick R. E. (1976). The structure and properties of phytochrome.
In- Chemistry and biochemistry of plant pigments, 2nd edition, vol. 1,
ed. T. W. Goodwin, p. 378, New York, London and San Francisco, Academic
Press.
Smith К- M. (ed.) (1975). Porphyrins and metalloporphyrins, Amsterdam, Else-
vier.
Tschudy D. P., Schmid R. (1972). The porphyrias. In: The metabolic basis of
inherited diseases, 3rd edition, eds. J. B. Stanbury, J. B. Wyngaarden and
D. S. Predrickson, p. 1087. New York, McGraw-Hill.
Vernon L. P„ Seely G. R. (eds.) (1966). The chlorophylls. New York and Lon-
don, Academic Press.
'With T. K- (1968). Bile pigments. (Trans. J. P. Kennedy), New York and London,
Academic Press.
Глава 6
Другие неполимерные N-гетероциклические
пигменты
6.1. Введение
Как уже отмечалось в предыдущей главе, простая си-
стема азотсодержащего гетероароматического кольца пиррола
лежит в основе многих чрезвычайно важных природных пигмен-
тов. В этой главе мы познакомимся с другими азотсодержащи-
ми гетероциклическими системами, которые также являются
основой пигментов некоторых классов. У большинства этих
пигментов скелетами молекул служат конденсированные ди-,
три- или олигоциклические гетероароматические системы колец,
а также их частично восстановленные производные. У этих
соединений возбуждение электрона обычно протекает относи-
тельно легко, особенно когда молекула содержит длинный со-
пряженный хромофор или несколько заместителей. Они могут
иметь желтую, красную, и пурпурную или синюю окраску.
Обычно полосы поглощения л—>л*-перехода присутствуют у
них в диапазоне длин волн, близком к диапазону у соответст-
вующего карбоцикла, а дополнительные п—>л*-переходы обу-
словливают важное, хотя и менее интенсивное поглощение (за-
прещенные переходы) при больших длинах волн.
Из нескольких групп пигментов, которые будут описаны,,
чрезвычайно важными веществами, синтезируемыми всеми
живыми организмами, являются пурины и птеридины, однако-
они функционируют в качестве пигментов лишь у небольшого-
числа животных. Близкий к ним рибофлавин синтезируется
только растениями и микроорганизмами. Он играет чрезвы-
чайно важную роль у животных в качестве витамина, но редко
служит пигментом. Оммохромы являются продуктами исклю-
чительно животного происхождения (членистоногих). Фенази-
новая группа пигментов синтезируется только бактериями, ко-
торые образуют также другие азотсодержащие пигменты. Бета-
лаины, не содержащие систем конденсированных колец, обра-
зуются исключительно в растениях.
В этой главе будут описаны последовательно основные ха-
рактеристики каждой из групп пигментов: структура, распро-
странение, свойства, функции и биосинтез.
224
Часть I Химические и биохимические аспекты
6.2. Пурины, птерины и флавины
6.2.1. Введение
Вероятно, наиболее важными простыми (т. е. неполи-
мерными) N-гетероцнклическими пигментами в животном цар-
стве после тетрапирролов являются птерины. Общепринято
рассматривать одновременно с ними очень близкие к птеринам
пурины и флавины. Хотя пурины и не поглощают видимый
свет, тем не менее они ответственны за структурную белую и
другие структурные окраски у многих животных (гл. 1). Фла-
вины вносят незначительный вклад во внешнюю окраску орга-
низмов, однако рибофлавин является важной фоторецепторной
молекулой, и мы его рассмотрим в гл. 11.
В последующих разделах вначале будут описаны основные
кольцевые системы скелета молекул, а затем по очереди будут
рассмотрены распространение в природе, свойства и биологи-
ческое значение каждой группы пигментов. Наконец, будет
описан биосинтетический путь, дающий начало всем трем
группам пигментов.
6.2.2. Основные типы кольцевых систем
Скелеты пуриновых, птеридиновых и флавиновых
молекул содержат по четыре гетероциклических атома азота,
как это показано на рнс. 6.1. Скелет пуриновых молекул (6.1)
Пурин (6.1}
Рис 6.1. Основной скелет молекул пурина, птеридина и изоаллоксазина.
Показаны различные системы нумерации, применяющиеся для пигментов
различных классов.
имеет шестичленное пиримидиновое кольцо и пятичленную ими-
дазольную систему. Птеридин (6.2) довольно близок по струк-
туре к пурину, но содержит шестичленную пиразиновую систе-
му вместо имидазольной. Изоаллоксазиновый скелет (6.3), ле-
жащий в основе молекулы флавина, представляет собой заме-
щенный (бензо) птеридин. Как видно на рис. 6.1, для этих трех
групп используются различные системы нумерации атомов, так
Гл. 6 Другие неполимерные пигменты
225
что во избежание путаницы следует обращаться с ними крайне
осторожно, например при сравнении характера распределения
заместителей у представителей различных групп.
6.2.3. Пурины
Распространение в природе. Пурины аденин (6.4) и
гуанин (6.5) встречаются у всех организмов, будучи компонен-
тами нуклеиновых кислот и нуклеотидов. Гуанин является так-
же одним из пуринов, участвующих в формировании и распре-
делении окраски у животных, мочевая кислота (6.6) также
чрезвычайно широко распространена, тогда как ксантин (6.7)
и изогуанин (6.8) встречаются реже. Эти пурины не погло-
щают видимый свет, но для них характерно сильное поглоще-
ние в УФ-свете, и поэтому некоторые животные, главным об-
разом насекомые, могут их видеть. Пурины вносят важный
вклад в окрашивание животных благодаря своему участию в
формировании структурной окраски, особенно белого и сереб-
(6 4) Аденин
(6 5) Гуанин
(6 7) Ксантин
(6 8) Изогуанин
ристого оттенков. В частности, гуанин накапливается в боль-
ших количествах в некоторых тканях в микрокристаллической
или гранулярной форме. Его кристаллы или частицы, будучи
должным образом ориентированы в тканях, отражают весь па-
дающий свет и таким образом придают ткани белую или се-
ребристую окраску, как это нам хорошо знакомо на примере
чешуи рыб. В коже и чешуе рыб гуанин содержится в спе-
циальных клетках, известных как гуанофоры или иридофоры,
причем отражающие свойства гуаниновых кристаллов изме-
няются в ответ на изменения уровня фонового освещения
15-84
226
Часть / Химические и биохимические аспекты
(гл. 8). В некоторых случаях, как, например, у золотой рыбки,
гуанофоры «выложены» снизу каротиноидами, что придает ей
золотистый цвет. Гуанин и другие пурины формируют также
структурную белую и серебристую окраску у некоторых амфи-
бий и беспозвоночных. Тиндалевское рассеяние света мельчай-
шими частицами гуанина в совокупности с черной меланиновой
окраской подстилающих тканей дает синюю окраску. Пурины
не оказывают влияния на окраску или внешний вид каких-
либо растений или микроорганизмов.
6.2.4. Птерины
Птерины впервые были открыты как пигменты крыльев
бабочек, однако сейчас они стали известны в качестве окра-
шенных компонентов многих других насекомых, а также рако-
образных, амфибий и рептилий. Развернутое химическое на-
звание птерина — 2-амино-4-гидроксиптеридин (6.9). Число из-
(6 9) Птерин (2 амино-4 гидроксиптеридин)
вестных пигментов животных с птерином в качестве основного
скелета сейчас уже велико. Индивидуальные соединения раз-
личаются главным образом природой заместителей при С-6 и
С-7, а также степенью окисления атомов азота в положениях 5
и 8. У Lepidoptera (бабочки) птерины встречаются преимуще-
ственно, хотя и не исключительно, у Pieridae. К наиболее из-
вестным птеринам бабочек относятся белый лейкоптерин (6.10),
который содержат белые бабочка-капустница (Pieris brassicae)
и бабочка-репница (Р.гарае), хризоптерин (6.11), желтый пиг-
мент бабочки-лимонницы (Gonepteryx rhamni), и красный эри-
троптерин (6.12), присутствующий у бабочки с оранжевой
окраской — зорьки Euchloe cardamin.es. Птерины содержатся
в кристаллической форме в чешуйках крыльев, а желтые, оран-
жевые и красные соединения придают соответствующие цвета
сложному рисунку на крыльях бабочек. Несмотря на то что
лейкоптерин является характерным птерином белых бабочек,
он не является белым пигментом; белый цвет их крыльев имеет
структурное происхождение.
Впервые птерины были обнаружены у Lepidoptera, но поз-
же было показано, что они являются обычными пигментами у
Гл. 6. Другие неполимерные пигменты
227
Hymenoptera; ксантоптерин (6.13), например, придает желтую
окраску обыкновенной осе (Vespula vulgaris). Птерины, однако,
в нормальных условиях не содержатся в наружных покровах
тела видов Diptera — даже двукрылые сирфиды (журчалки),
по внешнему виду напоминающие ос (мимикрия), не исполь-
(6 10) Лейкоптерии
(6 12) Эритроптерин
(6.13) Ксантоптерин
зуют их для формирования желтой окраски. Вместе с тем пте-
рины могут присутствовать у них в глазах. Например, глаза
плодовой мушки Drosophila melanogaster содержат смесь пте-
ринов, в том числе димерный дрозоптерин (6.14). Птерины
обнаружены в наружных покровах тела и в глазах у ракооб-
разных и других членистоногих, однако у последних они из-
учались гораздо менее интенсивно, чем у насекомых.
Сейчас известно, что желтая, оранжевая и красная окраска
у рыб, а также у амфибий и рептилий, таких, как лягушки,
<614) Дрозоптерин
15'
228
Часть I. Химические и биохимические аспекты
жабы, саламандры и змеи, обусловлена птеринами, которые
локализованы (в гранулярной форме) в ксантофорных и эри-
трофорных клетках (гл. 8). Наиболее распространенными пте-
риновыми пигментами у этих позвоночных являются сепиапте-
рин (6.15) и димеры дрозоптеринового типа.
Иногда в глазах позвоночных птерины служат для отраже-
ния света, эту же роль они очень часто играют у насекомых и
других членистоногих (гл. 9).
У других представителей животного царства, а также у
растений и микроорганизмов окрашивание птеринами (или
(6.17) Биоптерин
другими птеридинами) не встречается, хотя некоторые из них
важны в других отношениях. Производное птеридина — фолие-
вая кислота (6.16), которая синтезируется микроорганизма-
ми,— является незаменимым витамином для человека и боль-
шинства животных. Биоптерин (6.17) также имеет существен-
ное биологическое значение, например, как кофактор при гид-
роксилировании фенилаланина с образованием тирозина.
Общие свойства. Птерины (а также встречающиеся в приро-
де пурины) представляют собой амфотерные молекулы со
слабыми кислотными и основными свойствами. Они плохо ра-
створимы в воде, хорошо растворяются в разбавленных кис-
лотах и щелочах, умеренно растворимы в полярных органиче-
ских растворителях и не растворяются в неполярных. Боль-
шинство птеринов являются потенциальными окислительно-вос-
становительными агентами, однако при физиологических пар-
циальных давлениях кислорода они полностью окислены.
Птеридины с кислородсодержащими заместителями, в том
числе и птерины, могут существовать как в кето-, или хиноид-
ной форме (6.18), так и в виде енола (6.19). Птерины в целом
отличаются фотолабильностью.
Гл. 6. Другие неполимерные пигменты
229
Светопоглощающие свойства. В спектре поглощения птери-
на обычно присутствуют три (иногда два) максимума, но, как
правило, в видимой области спектра расположен только один
из них (рис. 6.2). Влияние заместителей видно из сравнения
максимумов поглощения (в щелочном растворе) лейкоптерина
он
(6.18) Кето-или хиноидная форма (6.19) Енольная форма
^птерина птерина
(6.10) (240, 285 и 340 нм), ксантоптерина (6.13) (255 и
391 нм), хризоптерина (6.11) (252 и 385 нм) и эритроптерина
(6.12) (240, 310 и 475 нм). Как можно было ожидать, прини-
мая во внимание амфотерные свойства молекул птеринов, их
максимумы поглощения зависят от величины pH. Сдвиги мак-
Длина волны, нм
Рис. 6.2. Спектры поглощения света дрозоптерином (6.14) в щелочном рас-
творе (/) н в кислом растворе (II).
симумов обычно не превышают ~40 нм в диапазоне pH от
1 до 13.
Окраска, присущая различным птеринам в живых тканях,
обычно обусловлена поглощением света в более длинноволно-
вой области спектра, чем это можно было бы ожидать на
основании их максимумов поглощения. Например, ксантопте-
рин с Хтах=390 нм, имеющий слабо-желтую окраску in vitro,
230
Часть I. Химические и биохимические аспекты
дает оранжевый цвет in situ. Наблюдаемый здесь батохромный
эффект может быть обусловлен конъюгацией с белками или
сильной стабилизацией кетохиноидной таутомерной формы пиг-
мента (6.18).
Большинство птеринов флуоресцируют в УФ-свете, причем
их полосы испускания сдвинуты на 150—200 нм в длинновол-
(6 20) Рибофлавин
(6 21) восстановленный рибофлавин
(6 22) Полухиноновый радикал
новую область по сравнению с соответствующими полосами
поглощения. In situ при связывании пигмента с белками про-
исходит тушение такой флуоресценции.
6.2.5. Флавины
В основе флавиновых молекул лежит изоаллоксазино-
вая структура, которая представляет собой птеридин (не пте-
рин), конденсированный с бензольным кольцом. Флавиновая
структура имеет метильные заместители в положениях 6 и 7
этого бензольного кольца, а также заместитель, присоединен-
ный к гетероциклическому атому азота в положении 9. У един-
ственного широко распространенного в природе флавина — ри-
бофлавина или рибитилфлавина [6,7-диметил-9-рибитилизоал-
локсазина (6.20) ] — последний заместитель представляет собой
сахароспирт рибит.
Распространение. Рибофлавин имеет важное биологическое
значение, поскольку он является функциональной частью фла-
вопротеинов и двух коферментов — флавинмононуклеотида
(рибофлавинфосфата, FMN) и флавинадениндинуклеотида,
FAD, которые используются во многих окислительно-восстано-
Гл. 6. Другие неполимерные пигменты 23t
вительных реакциях. Восстановленные формы этих кофермен-
тов— FMNH2 и FADH2 — содержат восстановленный рибофла-
вин (6.21). Флавины и флавиновые коферменты часто встре-
чаются в виде прочно связанных простетических групп у мно-
гих ферментов, главным образом ферментов дыхательной цепи.
Рибофлавин в качестве главного компонента этих коферментов
и ферментов встречается у всех живых организмов. Считается,
что животные не способны его синтезировать и должны полу-
чать с пищей, т. е. он представляет собой витамин (В2). Расте-
ния и микроорганизмы синтезируют рибофлавин, который тре-
буется животным. Образование витамина В2 дрожжами ис-
пользуется в промышленных масштабах уже на протяжении
многих лет.
Есть сообщения об эндогенном синтезе рибофлавина у не-
которых насекомых, например у таракана Periplaneta, однако,
скорее всего, у них за этот синтез ответственны микробы-эндо-
симбионты.
Несмотря на повсеместное распространение в природе, ри-
бофлавин редко участвует в формировании внешней окраски
живых организмов и никогда — у высших растений. Микроор-
ганизмы, используемые для промышленного получения рибо-
флавина, могут окрашиваться им в желтый цвет, однако обыч-
но они представляют собой искусственно полученные мутант-
ные штаммы, для которых желтая окраска никакого значения
не имеет. Иногда рибофлавин вносит свой вклад в желтую
окраску у беспозвоночных, таких, как пиявки и черви, и может
быть главным желтым пигментом наружных покровов голоту-
рий Holothuria forskali.
Рибофлавин обычно накапливается в сетчатке глаза у по-
звоночных. Хорошим примером тому служат такие животные,
как галаго (Galago), у которого золотисто-желтые кристаллы
рибофлавина составляют прозрачный отражающий слой, вы-
стилающий сзади сетчатку. В глазах членистоногих рибофла-
вин в отличие от птеринов не обнаружен.
Общие свойства. Присутствие рибита, содержащего много
гидроксильных групп, делает рибофлавин хорошо растворимым
в воде. Флавиннуклеотиды, содержащие, кроме того, фосфат-
ную, а в случае FAD и сахарную группы, еще лучше раствори-
мы в воде. Свободный рибофлавин растворим в полярных орга-
нических растворителях, таких, как ацетон или спирты, но не-
растворим в хлороформе. Он легко восстанавливается, а его
восстановленная форма вновь легко окисляется. Это свойство
используется при биологическом функционировании флавино-
вых коферментов и в электротранспортной цепи. (гл. 10).
Восстановление происходит в две одноэлектронные стадии, т. е.
через образование полухинонового радикала (6.22).
232
Часть I. Химические и биохимические аспекты
Светопоглощающие свойства. Рибофлавин имеет максимумы
поглощения света при 223, 267, 373, 445 и 475 нм (рис. 6.3), и
поэтому его раствор имеет ярко-желтый цвет. Сходство его
спектра со спектром p-каротина (гл. 2) приводило ко множе-
ству недоразумений относительно того, какой из этих пигмен-
тов, содержащихся в очень малых количествах, является истин-
ным первичным рецептором в ряде фотореакций (гл. И).
УФ-излучение индуцирует сильную флуоресценцию рибо-
флавина в диапазоне 520—565 нм. В присутствии флавинну-
200 300 400 500 550
Длина волны, нм
Рис. 6.3. Спектр поглощения рибофлавина при pH 7.0.
клеотидов и других ароматических структур, в том числе
остатков ароматических аминокислот в белках, происходит ту-
шение этой флуоресценции
6.2.6. Биосинтез пуринов, птеринов и рибофлавина
Биосинтез всех этих трех групп соединений происходит
по одному и тому же основному пути; как птерины, так и ри-
бофлавин синтезируются через пуриновые промежуточные про-
дукты. Поэтому в приводимом ниже описании сначала пред-
ставлен путь образования пурина гуанина, а затем описаны
реакции, посредством которых гуанин (в виде гуанозин-9-три-
фосфата, GTP) может превращаться либо в птерин, либо в
рибофлавин.
Образование гуанина. Пурины гуанин (6.5) и аденин (6.4)
входят в состав нуклеиновых кислот всех живых организмов.
Поэтому их биосинтез был изучен очень подробно, и путь, по
которому происходит образование пуриновой системы колец
из небольших фрагментов, очень хорошо известен. Все живые
организмы синтезируют свои пурины по одному и тому же
пути (рис. 6.4).
Гл. 6, Другие неполимерные пигменты 233
Сборка пуриновой системы колец происходит на молекуле
рибозо-5-фосфата (6.23). Сначала последний активируется в
результате превращения в а-5-фосфорибозил-1-пирофосфат
(6.24), при участии киназы и АТР. Затем пирофосфатная
группа (6.24) заменяется аминогруппой и образуется 5-фосфо-
рибозил-1-амин (6.26). Источником аминогруппы является
амидная NH2-rpynna глутамина (6.25). Этот азот составит пер-
вую часть пуриновой системы колец. В ходе этой реакции про-
исходит инверсия конфигурации при С-1 рибозы. Появляющая-
ся таким образом p-конфигурация затем сохраняется на протя-
жении всего биосинтетического пути.
Затем к аминогруппе при С-1 фосфорибозиламина с по-
мощью амидной связи присоединяется глицин (6.27). Для про-
текания этой реакции, продукт которой известен как глицина-
мидрибонуклеотид (6.28), необходим АТР. Следующим этапом
является перенос формильной (СНО) группы с кофермента
метенил-№-10-тетрагидрофолиевой кислоты (интересно, что
сам этот птерин образуется по этому же пути) на свободную
аминогруппу (6.28) в присутствии трансформилазы; при этом
образуется формилглицинамидрибонуклеотид (6.29).
На этой стадии все атомы, которые войдут в состав имида-
зольного кольца пуринового ядра, уже присоединены к фосфо-
рибозе. Однако до того, как замкнется имидазольное кольцо,
происходит перенос еще одного атома азота — опять от глута-
мина. Кислород глицинамидного фрагмента амида (6.29) за-
мещается на NH-группу, которая в конечном итоге войдет в
состав пиримидинового кольца пурина. И лишь у продукта
этой реакции — a-N-формилглицинамидинрибонуклеотида (6.30)
в результате процесса дегидратации, при участии АТР, проис-
ходит замыкание кольца. Имидазольное ядро образовавшегося
таким образом 5-аминоимидазолрибонуклеотида (6.31) затем
карбоксилируется путем присоединения СО2 (при участии био-
тина) с образованием 4-карбокси-5-аминоимидазолрибонуклео-
тида (6.32).
Второй атом азота пиримидиновое кольцо получает от аспа-
рагиновой кислоты (6.33). Вначале аспартат присоединяется
амидной связью к карбоксильной группе имидазола. Затем
полученное сукцинокарбоксамидное производное (6.34) расщеп-
ляется и в составе имидазолкарбоксамидного продукта (6.35)
остается лишь аминогруппа аспартата. Последний углеродный
атом пуринового ядра дает формил-№°-тетрагидрофолиевая
кислота; появляющийся при этом 5-формамидоимидазол-4-кар-
боксамидрибонуклеотид (6.36) замыкается в кольцо с образо-
ванием первого пуринового продукта — инозиновой кислтты
[IMP (6.37)].
(6.25) Глутамин
©-о.сн^о н АГр ®-О.СДо н
(СН2)2
CH.NH
СООН
(р)-о.сн2О Г\н^|
ОН он
hWo
AMP /—X
он он
СООН
(6.23) Рибозо-5-
фосфат
(6.24) о-5-Фосфорибозил-
1 -пирофосфат
(СН2)2
ch.nh2
СООН
,ДннД
н у у Н
ОН он
(6.26) б-ФосфорибоЗИЛ/
1 -амин
Глутаминовая
кислота
ho]oc.ch2.nH2|
(6.27) Глицин
hVVoH
(р)-о.сн2О nh.co.ch2.nh.|cho|
Днн/\
н у ун
он он
Метен ил-N5'1 °-ТГФ
©—о.сн2 0 nh|co.ch2.~nh^
Дннд,
н у у н
он он
(6.29) о -14-ФормилглиЦинамид-
рибонуклеотид
(6.28) Глицинамидрибонуклеотид-
н
I
(6.25)
Н2С СИ Глутамин
I II -------------
^С. О
NH
। Ух
Рибоза—Q9
СН
II
о
NH
I У
Рибоза—QP)
А
АТР НС
II СН
,С /
H2N^ N
Рибоза—(?)
(6.29)
(6.30) P-N-Формилглицинамидин -
рибонуклеотид
(6.31) 5-Аминоимидазол-
рибонуклеотид
(6.33) Аспарагиновая кислота Биотин
О
II
NHf с--<хн
II СН -*-у
Фумаровая
jajiS л| кислота
Рибоза—(Р)
соон
I
СН2 О
I II
CH—NH-C- N
I С" \\
СООН II СН
Н.Х
соон
I
сн2
CHNH2
I
соон
|со2|
ООС k
—
II /СН
H2NxC^N
Рибоза—(Р)
(6.35) 5-Аминоимидазол- (6.34) Б-Аминоимидазол-4-N-
4*карбоксамидрибонуклеотид су кцино карбоксамидрибонуклеотид
(6.32) 4-Карбокси-5-аминоимидазол
рибонуклеотид
Рис. 6.4. Главные особенности пути биосинтеза гуанииа и дргих пуринов.
Гл. 6. Другие неполимерные пигменты
235
о
h2n
Формил —N
СН ТГФ
H2N^'n'
Рибоза - (F)
О
1° _ >С N
__ H3N "У^СИ
Жк >-'Nz
NH I Z-X
Рибоза — (г)
"-N
I
Рибоза-(
(6.35)
(6.36) 5 Формамидопмидазол - (6.37) IMP
4 - карбоксамид -
рибонуклеотид
Гуанин
О
1 м <б-25)
С"-^ Глутамин
Рибоза- (р)
Рибоза—(Р)
(6.39) GMP
(6.38) Ксантиловая кислота
(начало см. на с. 234).
Гуаниновая структура в виде гуанинмонофосфата [GMP
(6.39)] образуется из IM.P в результате двухстадийного про-
цесса. Сначала в положение С-2 пуринового ядра под дей-
ствием ПАО+-дегидрогеназы вводится кислород и образуется
ксантиловая кислота (6.38). Затем введенный кислород заме-
щается аминогруппой, происходящей из глутамина. Собственно
гуанин освобождается путем расщепления нуклеотида GM.P.
Образование птеринов. Птериновое производное — фолиевая
кислота (6.16)—является витамином для многих животных,
в том числе для человека, и должна поступать с пищей или
от кишечных микроорганизмов. Поэтому биосинтез фолиевой
кислоты у бактерий, в частности у Escherichia coli, исследован
очень подробно. По этому же пути у животных синтезируются
птерины, которые затем используются ими для формирования
окраски.
Биосинтез птеринов является продолжением пуринового
биосинтетического пути. Гуаниннуклеотид [GM.P (6.39)] под-
вергается дальнейшему фосфорилированию до трифосфата
[GTP (6.40)], 9-рибозотрифосфатная группа GTP использует-
ся для увеличения имидазольного кольца до пиразинового.
Подробно последовательность реакций показана на рис. 6.5.
(Примечание. Важно помнить, что схемы нумерации, исполь-
зуемые для пуринового и птеринового рядов, различны; см.
разд. 6.2.2.) Сначала расщепляется имидазольное кольцо GTP
0 в "гЛа- т . "? г см₽((.з9>—I12lANA-v h2NAnA СН ^СН.СН2О-®-®-@ 'СН—сн 1 1 ОН ОН (6 40) GTP о он ОН „ он он В II и II 0 /А. /СН—СН-СН2ОН .СИ —СН—СН2О-®-®-© П jTYY л 1°У - 1Г h2n'AnA'xnA h2n-i4n/41 н Н (6.42) Дигидронеоптерин (6.41) 7.8-Дигидронеоптеринтрифосфат Рис. 6.5. Механизм увеличения кольца при синтезе птеринов из пуринов. я'2 —-СН.СП2О-®-®-® ГН-СНОН-СПОН NH 1 2 0 ОН ОН Чг с—СН-СН—СН2О-®-®~® сн2 1
Гл. 6. Другие неполимерные пигменты 237
и теряется атом С-8. Впоследствии место двух недостающих
атомов пиразинового кольца, т. е. атомов птерина С-6 и С-7,
займут углеродные атомы 1 и 2 трифосфорилированной рибо-
зы. 2-Амино- и 4-гидрокси-(или кето-)заместители находятся
на своем месте, так что первый птеридиновый продукт уже
представляет собой птерин, а именно 7,8-дигидронеоптеринтри-
фосфат (6.41). Последний дефосфорилируется с образованием
собственно 7,8-дигидронеоптерина (6.42).
7,8-Дигидронеоптерин является ключевым соединением, ко-
торое дает начало (по различным разветвляющимся путям)
фолиевой кислоте, биоптерину и многим птериновым пигмен-
там животных. До сих пор в отношении последующих разветв-
лений главного биосинтетического пути и взаимопревращений
образующихся продуктов существует некоторая неопределен-
ность. Те разветвления, которые, по последним данным, ка-
жутся наиболее вероятными, приведены на рис. 6.6. Главные
особенности, отраженные на этой схеме, таковы: 1) С-6-боко-
вые цепи подвергаются постепенному укорочению и модифика-
ции, а в положение С-7 вводятся новые заместители; 2) на
различных стадиях в ходе модификации боковых цепей 7,8-ди-
гидроптериновая структура дегидрируется до более обычных
полностью ароматических птеринов. Все природные птерины и
дигидроптерины могут быть получены путем различных пре-
вращений, однако детали биохимии большинства предполагае-
мых реакций еще полностью не исследованы.
Весьма вероятно, что димеризация, с помощью которой об-
разуются птерины, подобные дрозоптерину (6.14), происходит
на поздних стадиях биосинтеза, но опять-таки детальные ис-
следования этого процесса не проводились.
Биосинтез рибофлавина (рис. 6.7). Биосинтез рибофлавина
у растений и особенно у микроорганизмов также включает
разрыв имидазольного кольца GTP и потерю атома С-8. По-
следующие реакции, однако, отличаются от реакций в случае
биосинтеза птеринов. Рибозотрифосфатный заместитель у про-
межуточного 6-гидрокси-2,4,5-триаминопиримидинового произ-
водного (6.43) замещается рибитильной группой (а возможно,
превращается в нее), и 2-аминозаместитель рибитилпиримиди-
нового продукта (6.44) модифицируется в гидроксильную груп-
пу. Затем к промежуточному дигидроксидиаминопиримидиново-
му производному (6.45) присоединяется С4-фрагмент (проис-
хождение его неизвестно) с образованием важного промежуточ-
ного продукта 6,7-диметил-8-рибитиллюмазина (6.46).
После этого рибофлавин образуется из двух молекул диме-
тилрибитиллюмазина под действием рибофлавинсинтазы.
В этой реакции С4-остаток, состоящий из углеродных атомов
6 и 7 и связанных с ними метильных заместителей, переносится
СН2ОН
о
с нон
СН-Ю-(Р)
h2n
снон
Дигидронеоптерин-
фосфат
снон
о
СН2Ск
I ДЕ
сно-^ч>
HN
N.
о
h2n
снон
сн3
снон
н Дигидронеоптерин-
2’3’ циклофосфат
с=о
h2n
н
Дигидроптерин
h2n
h2n
н
Дигидронеоптерин
N
сн3
снон
снон
h2n
Дигидробиоптерин Н
Птерин
Дрозоптерин
(6 14)
Н Сепиалтепин
Рис. 6.6 Возможные биосинтетические взаимосвязи между птеринами.
о
о
GTP (6 40)
(6.47) Рибофлавин (=6 20)
4 Рибитиламино 5 амино 2,6*
дигидроксипиримидин
Рнс. 6.7. Схема пути биосинтеза
рибофлавина из GTP (рибитнл
= СН.СНОН СНОН СНОН СН2ОН).
240
Часть I. Химические и биохимические аспекты
с одной молекулы люмазина на другую для достраивания ди-
метилбензольного кольца рибофлавина (6.47).
Получены мутантные штаммы дрожжей Sacckaromyces се-
revisiae с нарушенным синтезом рибофлавина, у которых на-
капливаются предполагаемые промежуточные продукты био-
синтеза рибофлавина.
6.2.7. Факторы, контролирующие биосинтез пуринов,
птеринов и флавина
Несмотря на то что биосинтетические пути этих соеди-
нений тесно взаимосвязаны, факторы, контролирующие их об-
разование, различаются. В случае синтеза рибофлавина у мик-
роорганизмов хорошо изучен генетический контроль и проде-
монстрирована регуляция через обратную связь конечным про-
дуктом.
На синтез и отложение пуринов, и особенно птеринов,
влияют многие факторы. Обычно эти вещества локализованы
в специфических пигментных клетках, ксантофорах и эритро-
форах, которые не только ответственны за окраску наружных
покровов тела животных, но и контролируют изменения окрас-
ки, происходящие в ответ на изменения факторов окружающей
среды, таких, как цвет поверхности, на которой они находятся.
Известен гормональный контроль пигментации, главным обра-
зом в ходе созревания и развития организма. Помимо действия
на синтез птеринов факторы, контролирующие пигментацию,
действуют также на биосинтез каротиноидов и меланинов. Ре-
гуляция окраски, характер ее распределения у животных и
механизмы изменения окраски будут обсуждаться в гл. 8.
6.3. Феназины
6.3.1. Структура
Дибензопиразиновая структура (6.48) феназина имеет
внешнее сходство с изоаллоксазиновым скелетом флавинов.
Феназины представляют собой небольшую группу, состоящую
примерно из 30 зачастую ярко окрашенных пигментов, обра-
зуемых исключительно бактериями. Как правило, в качестве
заместителей в положениях 1 и 6 присутствуют гидрокси- и
карбоксигруппы, а к азоту пиразинового кольца присоединен
кислород или метильная группа. Наиболее известными природ-
ными феназинами являются пиоцианин (6.49) и феназин-N-OK-
сидиодинин (6.50). Другими примерами служат оксихлорора-
фин (6.51) и аэругинозин В (6.52), содержащие в качестве
заместителей карбоксамидную и сульфоновую группы соответ-
ственно.
Гл. 6. Другие неполимерные пигменты
241
6.3.2. Свойства
Многие феназины, особенно карбоновые кислоты, за-
метно растворимы в воде и накапливаются в культуральной
среде. Часто они образуются в столь высоких концентрациях,
что осаждаются в виде плотного осадка. Большинство
встречающихся в природе феназинов имеет лишь ограничен-
(6 48) Основной дибенэопирэзиновый
скелет феназинов
(6 49) Пиоцианин (6 50) Иодинин
(6.51) Оксихлорорафин
ную растворимость в органических растворителях, хотя многие
из них можно экстрагировать хлороформом из подкисленных
водных растворов.
Обычно феназины интенсивно окрашены и позволяют полу-
чить широкое разнообразие оттенков. Они характеризуются не-
сколькими полосами поглощения в ультрафиолете и по крайней
мере одной главной полосой в видимой области (400—600 нм),
которой феназины обязаны своей окраской (рис. 6.8). Боль-
шинство феназинов желтые (Хтах=400—450 нм), однако
иодинин — пурпурный (Хтах=530 нм), а пиоцианин--------СИНИЙ
(Хтах=695 нм). Оксихлорорафин и его дигидропроизводное
существуют в форме зеленого л-комплекса.
16—84
242
Часть 1, Химические и биохимические аспекты
6.3.3. Распространение
Распространение феназинов ограничено несколькими
видами бактерий; Pseudomonas, Streptomyces, Brevibacterium
и Nocardia. Иногда, однако, обусловленная феназинами окрас-
ка может встретиться и в тканях животных. Так, Pseudomonas
является обычным микробным паразитом кожи человека и дру-
Рис. 6.8. Спектр поглощения света иодииина (6 50) в хлороформе
тих животных; пиоцианин, синтезируемый этим организмом,
может в некоторых случаях окрашивать гной зараженных им
ран в синий цвет. Сине-зеленая окраска, иногда обнаруживае-
мая в шерсти овец, также обусловлена пиоцианином из Р.аеги-
g inosa.
6.3.4. Биосинтез
Биосинтез феназинов наиболее интенсивно изучался
на примере Pseudomonas aeruginosa и Р. phenazinium, синте-
зирующих в основном пиоцианин (6.49) и иодинин (6.50) со-
ответственно. Главные особенности этого биосинтетического
пути были выявлены в опытах по включению меченых предше-
ственников, а также при изучении накапливающихся проме-
жуточных продуктов у мутантных штаммов. В то же время
детали большинства индивидуальных реакций и катализирую-
щие их ферменты до сих пор не установлены.
Биосинтез феназинов представляет собой еще одну ветвь
главного шикиматного пути биосинтеза ароматических соеди-
нений (гл. 3 и 4). Предполагаемый путь образования фенази-
нового ядра показан на рис. 6.9. Шикимовая кислота (6.3)
весьма эффективно включается в феназины. Происхождение
атомов азота феназиновой системы пока не известно, однако
Гл. 6. Другие неполимерные пигменты
243
СООН
{6 56) Антраниловая кислотг
СООН
(6 57) Феназин 1, 6 дикарбоновая кислота
Рис. 6 9. Возможные пути образования основной феназиновой структуры из
шикимовой кислоты
предполагается участие в ее образовании азотзамещенного ши-
киматного промежуточного продукта (еще не идентифицирован-
ного), такого, как (6.54). Может быть также использован
альтернативный путь через хоризмовую кислоту (6.55) и ее
азотсодержащие производные. Очевидным кандидатом в пред-
шественники ранее считалась антраниловая кислота (6.56), од-
нако сейчас известно, что она в феназины не включается.
Каким бы ни был механизм образования феназин-1,6-ди-
карбоновой кислоты (6.57), в настоящее время ясно, что она
является первым феназиновым продуктом и предшественником
других феназинов, синтезирующихся по ответвляющимся и
альтернативным путям. Некоторые из этих путей показаны на
рис. 6.10. Считается, что ОН-группы при С-1 и С-6 вводятся
прямым окислительным замещением карбоксильных групп,
в то время как ОН-группы в других положениях появляются
в результате обычного ароматического гидроксилирования.
N-Окисление, приводящее к образованию N-оксидной структу-
ры иодинина, представляет собой ферментативный процесс,
16*
(6.57) Феназин-1, 6-
1, 6-Дигидроксифеназин
дикарбоновая кислота
1 -Г идро ксифеназин
(6.50) Иодинин
1. 8* Дигидроксифеназин
Рис. 6.10. Некоторые взаимопревращения феназинов, в том числе пути биосинтеза пиоцианина (6.49) и иодинина (6.50),
Гл. 6. Другие неполимерные пигменты
245
причем роль субстрата в нем могут играть разные феназины.
N-Метильный заместитель пиоцианина образуется обычным
образом из S-аденозилметионина.
Синтез феназинов бактериями в большой мере зависит от
состава культуральной среды и от условий культивирования,
однако детали контролирующих и регулирующих этот процесс
механизмов остаются невыясненными.
6.3.5. Биологическая активность
Феназины, синтезируемые одним видом, могут оказы-
вать влияние на другие виды и их ткани, причем это влияние
осуществляется различными путями. Они были первыми бак-
териальными продуктами, для которых была показана анти-
биотическая активность против других микроорганизмов. Бак-
териостатические свойства иодинина и пиоцианина сейчас хо-
рошо известны. По-видимому, они обусловлены взаимодей-
ствием феназинов с ДНК (преимущественно интеркаляция
плоской ароматической системы колец в молекулу ДНК). Есть
сообщения, что феназинди-М-оксиды обладают канцеростатиче-
ской активностью.
(6 58) феноксазиновая система колец
Некоторое биологическое значение могут иметь также окис-
лительно-восстановительные свойства феназинов. Пиоцианин
подавляет активность сукцинатдегидрогеназы мозга, но в то
же время активирует некоторые другие дегидрогеназы. Сооб-
щалось об активном поглощении и концентрировании фенази-
нов тканями животных, однако пока не известно, имеет ли
какое-либо физиологическое значение этот факт, или он слу-
чаен.
6.4. Феноксазины
Феноксазиновая циклическая система (6.58) в струк-
турном отношении очень сходна с феназиновой. Наиболее из-
вестными феноксазиновыми пигментами являются оммохромы.
246
Часть I. Химические и биохимические аспекты
6.4.1. Оммохромы
Структура. Эти окрашенные в темные цвета вещества
впервые были экстрагированы из омматидий сложных глаз
членистоногих и сначала считались меланинами. Они подраз-
деляются на две главные подгруппы — омматины и оммины,
представляющие собой соответственно димеры и олигомеры
производных кинуренина (6.59); были выделены и другие под-
группы, однако полностью они пока не охарактеризованы.
Наиболее обычным оммохромом является желтый ксантомма-
тин (6.60), который легко восстанавливается до окрашенного
соон
(6 59) Кинуренин
(6 60) Ксантомматин
(6 61) Дигидро ксантомматин
(6 62) -Оммин А
Гл 6 Другие неполимерные пигменты
247
в красный цвет дигидроксантомматина (6.61). Эти соединения
служат примерами основной омматиновой структуры, состоя-
щей из феноксазиновой циклической системы с двумя аспар-
татными боковыми цепями, одна из которых циклизована и
образует четвертое кольцо. Вместе с тем оммины представляют
собой более крупные недиализующиеся молекулы, содержащие
серу. Структура относительно простого тримерного оммина,
оммина А, представлена формулой (6.62).
Рис. 6 11. Спектры поглощения (в слабокислом метаноле) ксантомматина (Z)
и дигидроксантомматина (И).
В глазах беспозвоночных оммохромы находятся в гранулах
в виде комплексов с белками.
Светопоглощающие свойства. Оммохромы сильно поглощают
в ультрафиолете, а также имеют характерные полосы поглоще-
ния в видимом диапазоне при ~440—500 нм (рис. 6.11), при-
чем их максимумы поглощения зависят от pH. In vivo как ом-
матины, так и оммины выглядят темноокрашенными; они мо-
гут быть желто-коричневыми, пурпурными или почти черными.
Из-за этого их часто принимали за меланины.
Распространение. Оммохромы представляют собой харак-
терные пигменты глаз насекомых и других членистоногих. Они
функционируют в глазах не как фоторецепторы, а как защит-
ные пигменты, которые предохраняют фоторецепторы от по-
вреждения рассеянным светом (гл. 9). Оммохромы широко
распространены в наружных покровах членистоногих и других
беспозвоночных, главным образом у головоногих, а также най-
дены в яйцах и различных тканях некоторых червей. Предпо-
H2N. /-COOH
CH
I
CH2
c=o
NH.CHO
(6.63) Триптофан
(6.64) Формилкинуренин . (6.59) Кинуренин
(6.62) Оммин А
(6 65)3 Гидроксикинуренин
(6.66) Феноксазиновый промежуточный
продукт
H2N /СООН
сн
Рис. 6.12. Путь биосинтеза ксантомматина
(6.60) Ксантомматин
Гл. 6. Другие неполимерные пигменты
249
лагалось, что они встречаются и у некоторых позвоночных, но
это не было доказано.
Биосинтез. Много информации о биосинтезе оммохромов
было получено из подробных генетических исследований, про-
веденных на плодовой мушке Drosophila melanogaster, дикий
тип которой в качестве коричневого глазного пигмента содер-
жит ксантомматин. Известно много мутантов Drosophila,
имеющих аномальную окраску глаз из-за повреждений пути
биосинтеза оммохрома, в результате чего накапливаются про-
межуточные соединения этого процесса.
Путь биосинтеза ксантомматина приведен на рис. 6.12. Не-
смотря на то что важнейшие особенности этого пути были уста-
новлены, детали отдельных реакций в большинстве случаев еще
не вполне понятны. Биосинтез начинается с размыкания кольца
триптофана (6.63), в результате чего образуется формилкинуре-
нин (6.64), который содержит аспартатную боковую цепь.
Спонтанное деформилирование последнего дает собственно ки-
нуренин (6.59), который гидроксилируется до 3-гидроксикину-
ренина (6.65). Это соединение подвергается окислительной ди-
меризации, приводящей к образованию оксазинового мостика
омматиновой структуры (6.66). Затем в результате циклизации
аспартатной боковой цепи с замыканием на замещающую ами-
ногруппу второй молекулы гидроксикинуренина образуется чет-
вертое кольцо. Оммины также синтезируются из 3-гидроксики-
нуренина, но не через промежуточную омматиновую структуру.
Родственными оммохромам в структурном отношении явля-
ются папилиохромы — белые и желтые соединения, найденные
только в крыльях бабочек сем. Papilionidae. Эти пигменты так-
же представляют собой производные кинуренина, возможно,
в виде комплексов с не содержащими азота хинонами. Собст-
венно кинуренин и гидроксикинуренин также иногда служат
желтыми пигментами.
Функции. Функции оммохромов ограничены их участием в
формировании внешней окраски и защитной ролью в глазу бес-
позвоночных.
6.4.2. Микробные феноксазины. Актиномицины
Феноксазиноновый хромофор (6.67) содержится также в
молекулах нескольких микробных пигментов, многие из которых
имеют важные антибиотические свойства. Наиболее известны
из них актиномицины (6.68) — антибиотики красного цвета, син-
тезируемые плесневым грибом Streptomyces. Весьма вероятно,
что биосинтез этих микробных пигментов включает конденса-
цию 3-гидроксиантраниловой кислоты (6.69) или ее произвол-
250
Часть I. Химические и биохимические аспекты
(6.67) Феноксазиночсвый Хромофор
(6.68) Актиномицины (R,К'-пентапептидные цепи) (6.69) З-Гидроксиантраниловая кислота
ных с помощью процесса, сходного с процессом образования
ксантомматина из 3-гидроксикинуренина.
6.5. Беталаины
6.5.1. Введение
Беталаины являются исключительно растительными
пигментами. Они имеют очень ограниченное распространение и
хорошо известны по интенсивной красно-пурпурной окраске кор-
неплодов столовой (красной) свеклы. Они растворимы в воде
и на первый взгляд сходны с антоцианами; одновременно пиг-
менты этих двух групп в одном объекте не встречаются. При-
мерно до 1960 г. практически ничего не было известно о химии
или биохимии беталаинов, однако сейчас показано, что суще-
ствуют две главные группы этих пигментов — красно-фиолето-
вые бетацианины и желтые бетаксантины.
6.5.2. Структура
Бетацианины. За одним исключением, в основе всех
встречающихся в природе бетацианинов лежат только два аг-
ликона— бетанидин (6.70) и его С-15-эпимер изобетанидин
(6.71). Единственным исключением является 2-декарбоксибета-
нидин (6.72), к настоящему времени обнаруженный в цветках
только одного вида растений (Carpobrotus acinaciformis). Основ-
ная структура бетацианинов состоит из двух N-гетероцикличе-
ских систем—дигидроиндольной и дигидропиридиновой, свя-
занных между собой двухуглеродным мостиком.
В большинстве случаев природные бетацианины представля-
ют собой гликозиды, у которых к ОН-группе в положении С-5
Гл. 6, Другие неполимерные пигменты
251
или С-6 (но никогда к обеим) дигидроиндольного кольца при-
соединен моно- или дисахарид. Чаще всего такими сахаридами
являются глюкоза или глюкуроновая кислота. Остатки сахаров
могут быть ацилированы, обычно коричными кислотами.
Бетаксантины. У бетаксантинов сохраняются дигидропири-
диновое кольцо и связующий С2-мостик, однако дигидроиндоль-
ное кольцо заменено аминокислотой или аминогруппой, напри-
мер пролином в индикаксантине (6.73). Другие представители
(6.70) Бетанидин (R' = R‘=COOH; R3=H)
(6.71) Изооетанидин (R1 =R, = COOH ; R2-Н)
(6.72) Карбоксибетанидин (К'= R -П ; R2=COO11)
этой группы содержат вместо пролина гидроксипролин, аспара-
гиновую кислоту, глутаминовую кислоту или глутамин, метио-
нин, дигидроксифенилаланин, а также моно- или дигидроксифе-
нилэтиламин. Во всех случаях связывание с общей для всех них
частью молекулы происходит через N-атом аминокислоты. Схе-
мы нумерации атомов вызывают много недоразумений. Для
разных бетаксантиновых агликонов используются разные схе-
мы, поскольку нумерация начинается с аминного азота амино-
кислоты или аминного остатка. Нумерация бетаксантинов от-
личается от нумерации бетацианинов.
6.5.3. Свойства
Как агликоны, так и гликозиды представляют собой во-
дорастворимые ионизируемые (кислые) соединения. Они оптиче-
ски активны, а для бетанидина была установлена абсолютная
конфигурация при С-2 и С-15. Эпимеризация при С-15 проис-
ходит очень легко, возможно, даже спонтанно. Бетаксантины
имеют желтую окраску с максимумом поглощения света при
~480 нм, в то время как бетацианины обладают значительно
252
Часть I. Химические и биохимические аспекты
более протяженной сопряженной системой и поглощают при
больших длинах волн (534—554 нм), а потому окрашены в
красно-фнолетовые цвета. Химия беталаинов подробно не изу-
чалась, однако хотелось бы обратить внимание на одно их
свойство —это относительную легкость взаимопревращений раз-
личных беталаинов. Так, например, в присутствии L-пролина и
аммиака бетанин (бетанидин-5-О-|3-В-глюкозид) легко превра-
щается в бетаксантин индикаксантин (6.73).
(6 73) Индикаксантин
6.5.4. Распространение
Ядовитый гриб Amanita muscarla (мухомор) содержит
один фиолетовый и несколько желтых пигментов, которые были
идентифицированы как беталаины, например мускааурин I
(6.74). Кроме этого гриба беталаины обнаружены только у
высших растений, причем лишь у представителей отдельных се-
мейств порядка Centrospermae, в том числе и у многих экзоти-
ческих растений, таких, как кактусы. Чаще всего они содер-
жатся в цветках, но могут присутствовать и в листьях, плодах
или корнях; наиболее известным примером беталаинов служит
красновато-пурпурный пигмент корнеплодов столовой свеклы
(Beta vulgaris). Окраска последних обусловлена главным обра-
разом присутствием бетанина (бетанидин-5-О-р-П-глюкопирано-
зида). Пока не обнаружено примеров, когда бетацианины и ан-
тоцианы встречались бы у одних и тех же видов, правда, дру-
гие флавоноиды могут присутствовать наряду с бетацианинами.
6.5.5. Биосинтез
О биосинтезе беталаинов имеется мало четкой информа-
ции, хотя известно, что ароматические аминокислоты, такие, как
тирозин и дигидроксифенилаланин [ДОФА (6.75)], включаются
Гл, 6. Другие неполимерные пигменты
253
как в дигидроиндольное, так и в дигидропиридиновое кольцо
бетанидина. Полагают, что биосинтетический путь (рис. 6.13)
включает разрыв дигидроксифенильного кольца ДОФА, за ко-
торым следует рециклизация с образованием беталамовой кис-
лоты (6.76). Дигидроиндольное кольцо [циклоДОФА (6.77)
образуется путем циклизации ДОФА] затем присоединяется
через атом азота к альдегидной группе беталамовой кислоты с
образованием бетанидина. Когда таким же образом к бетала-
но
(6.74) Мускааурин I
мовой кислоте присоединяются другие аминокислоты или ами-
ны, образуются бетаксантины.
Что касается гликозилирования, то сведения относительно-
того, когда оно происходит, на ранних (например, на стадии
циклоДОФА) или на поздних (например, на стадии бетаниди-
на) стадиях биосинтеза, противоречивы. Изобетанидин образу-
ется эпимеризацией бетанидина. Известно, что такая эпимери-
зация происходит in vitro очень легко, даже спонтанно, однако-
in vivo она представляет собой контролируемый процесс, ката-
лизируемый ферментом. Об энзимологии и механизмах реак-
ций, протекающих в ходе биосинтеза беталаинов, почти ничего
не известно.
По-видимому, у некоторых видов для синтеза беталаинов
абсолютно необходим свет. У других видов образование пиг-
ментов происходит и в темноте, но при освещении белым све-
том интенсивность синтеза возрастает. Весьма вероятно, что
биосинтез беталаинов находится под прямым ядерным генети-
ческим контролем.
Рис. 6.13. Предлагаемый путь биосинтеза бетанидина.
Гл. 6. Другие неполимерные пигменты 255-
6.5.6. Функции и использование
Физиологические функции беталаинов в растениях не из-
вестны. Считают, что они играют ту же роль, что и антоцианы,
которых они замещают, так что в плодах и цветках они могут
служить для привлечения насекомых, птиц и т. д. в целях рас-
пространения семян и опыления (гл. 8). Широкая известность
бетанина как природного пигмента корнеплодов столовой свек-
лы способствовала росту интереса к возможному использова-
нию этого соединения или содержащих его экстрактов в каче-
стве промышленных пищевых красителей.
6.6. Другие N-гетероциклические пигменты
Азотсодержащие гетероциклические пигменты, не при-
надлежащие ни к одной из уже описанных главных групп, бо-
лее или менее часто обнаруживаются только у микроорганиз-
мов, в частности у бактерий, хотя некоторые алкалоиды выс-
ших растений, например берберин (6.78), окрашены и, следо-
вательно, могут рассматриваться как растительные пигменты.
Интенсивно окрашенные азотсодержащие бактериальные-
продукты включают производные индола, например виолацеин>
(6.79) из Chromobacterium violaceum, пирролы, например три-
пиррол продигиозин (6.80) из Serratia spp., и димерные произ-
водные пиридина, например индигоидин («бактериальный инди-
го») из Pseudomonas indigofera (6.81). Светопоглощающие свой-
ства последнего обусловлены диаза-о-дифенохиноновой структу-
рой. Биосинтез и функциональное значение этих соединений в
большинстве случаев не исследовались.
Обычно думают, что индиго является растительным пигмен-
том, однако растительный индиго, использовавшийся в качестве
красителя для материи и для раскраски тела (вайда) древни-
ми, является артефактом. Сок растений Indigofera tinctoria и
Isatis tinctoria содержит индикан — бесцветный глюкозид 3-гид-
роксииндола (6.82). Пигмент же индиго (6.83) образуется толь-
ко после его гидролиза и окисления.
Сходная ситуация имеет место и в случае 6,6-диброминдиго
(6.84), использовавшегося древними римлянами и другими на-
родами в качестве краски, называемой тирским пурпуром.
В данном случае это также артефакт, образующийся в резуль-
тате фотохимического окисления животного продукта. Природ-
ный же предшественник, синтезируемый различными морскими-
моллюсками, такими, как Мигех и Nycella, недавно был иден-
тифицирован как тиривердин (6.85).
о
(6 60> Продигиозин
(6 65) Тиривердин
(6 84> 6,6' -Диброминдиго
/Тирский пурпур)
Гл. 6, Другие неполимерные пигменты 257
6.7. Заключение и комментарии
Пигменты, описанные в этой главе, не принадлежат все
к какой-либо одной группе, однако для удобства они собраны
вместе на основании их общего структурного свойства — присут-
ствия по крайней мере одного азотсодержащего кольца. Однако
некоторые классы пигментов имеют явное структурное и функ-
циональное сходство; пурины, птерины и флавины, например,
обладают сходной системой гетероароматических колец, а так-
же подобными свойствами. Рассмотренные в данной главе пиг-
менты предоставляют широкое поле для дальнейших химичес-
ких исследований. Поскольку исчерпывающего систематическо-
го поиска новых N-гетероциклических пигментов не проводи-
лось, можно не сомневаться в том, что такие пигменты еще бу-
дут открыты; не исключено даже, что будут обнаружены со-
вершенно новые группы пигментов. Среди них могут встретить-
ся сложные структуры с интересной стереохимией, трудно под-
дающиеся исследованию методами синтеза. Здесь много рабо-
ты и биохимику. В общих чертах биосинтетические пути для
большинства групп уже выяснены, однако подробности меха-
низмов отдельных реакций и катализирующие их ферменты все
еще не установлены. Мало известно также о регуляции путей
биосинтеза. N-Гетероциклические пигменты, как правило, не ис-
пользуются широко в качестве пищевых красителей, хотя в на-
стоящее время интенсивно окрашенные водорастворимые бета-
лаины вызывают интерес в этом отношении.
Подводя итог, следует отметить, что изучение N-гетероцик-
лических веществ остается одной из наиболее заброшенных об-
ластей изучения пигментов, хотя именно они предоставляют ши-
рокое поле для интересных исследований.
6.8. Рекомендуемая литература
РазнообразныеN-гетероциклические пигменты, описан-
ные в этой главе, напоминают разрозненную коллекцию; поэтому
не удивительно, что не существует книги или обзора, которые
охватили бы все группы этих пигментов. Основные книги о пиг-
ментах животных (Fox, Vevers, 1960; Needham, 1974; Fox,
1976) и пигментах растений (Goodwin, 1976) наряду со сведе-
ниями о главных пигментах содержат информацию и о боль-
шинстве из этих азотсодержащих пигментов. Изучающим дан-
ный вопрос необходимо обратиться к более специализирован-
ным книгам или статьям, в которых сконцентрирована инфор-
мация о пигментах определенных групп.
В случае пуринов и птеринов особенно полезны статьи Бра-
уна (Brown, 1971), Циглера (Ziegler, 1961, 1965) и Циглера и
Хармсена (Ziegler, Harmsen, 1969), а также два последних то-
17—84
258
Часть 1. Химические и биохимические аспекты
ма из серии трудов симпозиумов (Pfleiderer, 1975; Kisliuk,
Brown, 1979). Ряд статей, в частности Плота идр. (Plant, 1974),
посвящены рибофлавину и его биосинтезу. Последние достиже-
ния методологии в области птеринов и флавинов включены в
два тома «Methods in enzymology» (McCormick, Wright, 1971,
1980). Оммохромам повезло меньше. В обзоре Линцена (Lin-
zen, 1974) приведены подробности биосинтетических исследова-
ний и ссылки на более ранние работы по химии и биохимии
этих соединений. Что касается микробных феназинов, то статьи
Лейзингера и Марграфа (Leizinger, Margraff, 1979), а также
Ингрэма и Блэквуда (Ingram, Blackwood, 1970) освещают их
в общих чертах и с акцентом на прикладную микробиологию
соответственно. Глава, написанная Пиаттелли (Piattelli, 1976)
в книге Гудвина о растительных пигментах (Goodwin, 1976), со-
держит информацию по химии и биохимии беталаинов. Крат-
кое описание некоторых других азотсодержащих пигментов
входит в статью Томсона (Thomson, 1976) в этой же книге.
6.9. Дополнительная литература
Brown G. М. (1971). The biosynthesis of pteridines, Adv. Enzymol., 35, 35.
Fox D. L. (1976). Animal Biochromes and structural color, 2nd edition. Ber-
keley, Los Angeles and London, University of California Press.
Fox H. M., Vevers G. (1960). The nature of animal colours, London.
Goodwin T. W. (ed.) (1976). Chemistry and biochemistry of plant pigments,
2nd edition, vol. 1. London, New York and San Francisco, Academic Press.
Ingram I. M„ Blackwood A. C. (1970). Microbial production of phenazines, Adv.
Appl. Microbiol., 13, 267.
Kisliuk R. L., Brown G. M. (eds.) (1979). Chemistry and biology of pteridines,
New York, Elsevier-North Holland.
Leisinger T., Margraff R. (1979). Secondary metabolites of the fluorescent
Pseudomonads, Microbiol. Rev., 43, 422.
Linzen B. (1974). The tryptophan-ommochrome pathway in insects, Adv. Insect.
Physiol., 10, 117.
McCormick D. B., Wright L. D. (eds.) (1971). Methods in enzymology, vol. 18B,
New York, London and San Francisco, Academic Press.
McCormick D. B., Wright L. D. (eds.) (1980). Methods in enzymology, vol. 66,
New York, London and San Francisco, Academic Press.
Needham A. E. (1974). The significance of zoochromes.
Pfleiderer W. (ed.) (1975). Chemistry and biology of pteridines, Berlin.
Piattelli M. (1976). Betalains. In: Chemistry and biochemistry of plant pigments
2nd edition, vol. 1, ed. T. W. Goodwin, p. 560, London, New York and San
Francisco, Academic Press.
Plaut G. W. E., Smith С. M., Alworth W. L. (1974). Biosynthesis of water-
soluble vitamins, Ann. Rev. Biochem., 43, 899.
Thomson R. H. (1976). Miscellaneous pigments. In: Chemistry and biochemistry
of plant pigments, 2nd edition, vol. 1, ed. T. W. Goodwin, p. 597, London,
New York and San Francisco, Academic Press.
Ziegler I. (1961). Genetic aspects of ommochrome and pterin pigments, Adv.
Genet., 10, 349.
Ziegler I. (1965). Pterine als Wirkstoffe und Pigments, Ergebn. Physiol., 56, 1.
Ziegler I., Harmsen R. (1969). The biology of pterins in insects , Adp. Insect.
Physiol., 6, 139.
Глава 7
Меланины
7.1. Введение
Черная окраска широко распространена у живых орга-
низмов . Термин «меланин» обычно применяют для описания не-
растворимых полимерных пигментов, ответственных за большин-
ство природных окрасок черного цвета. Его используют также
в настоящее время для описания сходных полимерных веществ,
которые обусловливают природную коричневую, красную и жел-
тую окраску, главным образом перьев птиц и волос. Меланины
являются единственными пигментами, широко используемыми
для окрашивания среди млекопитающих, в том числе человека.
Термин «меланин» не несет информации о химической струк-
туре пигментов, за исключением той, что они представляют со-
бой полимеры с относительно высокой молекулярной массой.
Были предприняты попытки различить и охарактеризовать от-
дельные классы меланиновых пигментов. Так, черные пигмен-
ты животных обычно называют э у меланинами, в то время как
желто-коричневые разновидности известны под названием фео-
меланинов. Имеющие некоторое сходство с ними не содержа-
щие азота черные пигменты растений часто называют алломе-
ланинами.
Знания о химии и биохимии меланинов в значительной сте-
пени ограничены из-за больших технических трудностей, возни-
кающих при изучении этих пигментов. Меланины обычно нерас-
творимы ни в одном из растворителей, и поэтому их трудно
выделить и очистить. Действительно, в случае меланинов почти
невозможно узнать, имеем ли мы дело с чистым веществом в
том смысле, что в пробе содержится только один вид молекул.
Какие-либо диагностические пробы, которые могли бы оказать-
ся полезными для меланинов, а также общие методы специфи-
ческого расщепления меланиновой молекулы на специфические
фрагменты или субъединицы отсутствуют. Невозможно даже
доказать идентичность двух образцов меланинов. Связывание с
белком, которое в нормальных условиях имеет место по край-
ней мере в случае меланинов животных, делает выделение и
изучение этих пигментов еще более трудным.
На фоне упомянутых экспериментальных трудностей и сле-
дует рассматривать современное состояние знаний о биохимии
меланинов.
17*
260
Часть I. Химические и биохимические аспекты
7.2. Химия
7.2.1. Общая характеристика
При исследовании природных веществ установление хи-
мической структуры часто помогает предположить способ био-
синтеза изучаемого соединения. В случае меланинов мы имеем
дело в значительной степени с обратной ситуацией, именно био-
химические исследования позволили сделать первые предполо-
жения относительно химической структуры данной группы ве-
ществ.
Сейчас известно, что меланины являются полимерами хино-
идных соединений. Классическая структура меланина — это
длинноцепочечный полимер, состоящий из индолил-5,6-хиноно-
вых (7.1) единиц (рис. 7.1). Ситуация, однако, далеко не так
Рис.7.1. Фрагмент структуры меланина как длинноцепочечного полимера ин-
долил-5,6-хинона (7.1).
проста. Во-первых, любой образец меланина почти наверняка
состоит из смеси макромолекулярных видов. Во-вторых, мела-
нины редко являются гомополимерами, обычно их молекулы со-
стоят из различных мономерных единиц. Кроме того, разные
меланины, например из различных живых организмов, построе-
ны из разных мономерных единиц. Ситуация еще более ослож-
няется возможностью разветвления полимерной цепи и пере-
крестного связывания между цепями.
Гл. 7. Меланины
261
Несмотря на сложность проблемы, биохимические и хими-
ческие исследования по глубокому расщеплению позволили вы-
яснить главные структурные особенности различных классов
меланинов. Необходимо, однако, отдавать себе отчет в том, что
ни в одном случае мы не имеем возможности даже приблизить-
ся к полному установлению структуры. Ниже мы обсудим глав-
ные структурные особенности и свойства прежде всего пигмен-
тов наиболее широко изучавшейся группы — эумеланинов, а за-
тем меланинов других главных групп.
7.2.2. Эумеланины
Распространение. Название «эумеланины» применяют к
черным азотсодержащим полимерным пигментам, обычно живот-
ного происхождения. Однако эумеланины синтезируются также
некоторыми растительными тканями. Различают несколько ти-
пов эумеланинов, выделенных из ряда источников. Каждый тип
эумеланинов имеет характерный набор элементов и харак-
терный состав продуктов расщепления в жестких условиях:
при кипячении с 6М НС1 или при нагревании со щелочью до
300 °C. Наиболее интенсивно изучен сепиомеланин — темно-ко-
ричневый пигмент защитных чернил каракатицы (Sepia offici-
nalis) и меланин из меланом млекопитающих. В меньшей сте-
пени охарактеризованы меланины, содержащиеся в черных во-
лосах, перьях и коже, а также в сосудистой оболочке глаза и в
некоторых внутренних органах. Некоторые черные пигменты
растений, в том числе те, которые образуются в результате по-
вреждений клубней картофеля и бананов, также могут принад-
лежать к эумеланинам. Сходные пигменты (ДОФА-меланины)
образуются из дигидроксифенилаланина [ДОФА (7.2)] под
действием растительных и грибных ферментов.
Структура. Классическая идея (рис. 7.1) о том, что эумела-
нин представляет собой линейный полимер индолил-5,6-хинона
(7.1), в сущности верна, однако чрезвычайно упрощенно отража-
ет положение вещей. Действительно, главными мономерами яв-
ляются индолил-5,6-хинон и его восстановленные производные;
кроме того, в полимере может присутствовать небольшое число
пиррольных мономеров (7.3), образующихся, по-видимому, из
индольных предшественников. В меланинах меланомы могут
также содержаться ДОФА и ДОФА-хиноновые (7.4) фрагмен-
ты. Исследования с помощью электронного парамагнитного ре-
зонанса (ЭПР) указывают на присутствие небольшого количе-
ства свободных радикалов, возможно, полухиноновых структур.
In vivo с эумеланином могут быть связаны тяжелые металлы,
особенно медь, цинк или железо. Степень перекрестного связы-
262
Часть I Химические и биохимические аспекты
вания меланиновых цепей не определяли, но, вероятно, она не-
велика.
Фрагмент предполагаемой структуры сепиомеланина пока-
зан на рис. 7.2. Структура других эумеланинов, вероятно, напо-
минает эту, однако она может отличаться от нее числом моно-
мерных субъединиц и частотой их встречаемости. Природные
полимеры представляют собой длинноцепочечные макромолеку-
лы. In vivo они связаны с белком, возможно, через сульфгид-
рильную группу цистеина.
(7 2) Дигидроксифенилаланин
(7 3)
(7 4) Дофа хинон
Свойства. Эумеланины практически нерастворимы в воде и
в органических растворителях. Они чрезвычайно инертны и ста-
бильны и подвергаются изменениям лишь в результате наибо-
лее жестких химических воздействий. Вместе с тем эумелани-
ны могут обесцвечиваться при длительном выдерживании на
воздухе на ярком солнечном свету или, особенно эффективно,
при продолжительном окислении перекисью водорода. Обесцве-
чивание человеческих волос, используемое «перекисными блон-
динками», хорошо известно.
Как и следовало ожидать, эумеланины, имеющие черный
цвет, поглощают свет во всем видимом диапазоне. Поглощение
происходит более интенсивно при более коротких длинах волн,
и, возможно, это не является простым следствием высокосопря-
женной системы.
С помощью дифракции рентгеновских лучей твердых образ-
цов эумеланинов животных была показана упаковка в стопки
индольных и других ароматических фрагментов с образованием
л-комплекса. Упакованные в стопки фрагменты могут быть уда-
ленными друг от друга членами одной полимерной цепи или
же членами разных полимерных цепей.
Гл. 7. Меланины
263
Рис 7 2 Фрагмент предполагаемой структуры сепиомеланина. Частота встре-
чаемости остатков пиррольных мономеров умышленно сильно преувеличена,
чтобы проиллюстрировать несколько различных возможностей связывания
В действительности эти мономеры составляют лишь очень небольшую часть
всех мономеров.
7.2.3. Феомеланины
Распространение. Феомеланины имеют желтую, красную
или коричневую окраску. Некоторые авторы применяют назва-
ние «феомеланин» только к желтым или светло-желтоватым
пигментам и называют красные вещества «эритромеланинами».
В этой книге все эти пигменты мы будем называть «феомела-
нинами». Распространение феомеланинов, по-видимому, огра-
ничивается волосами и перьями, а, возможно, также и веснуш-
264
Часть I. Химические и биохимические аспекты
ками, которые у людей часто сопутствуют рыжим волосам.
Лучше других изучены пигменты рыжих волос человека и крас-
ных перьев некоторых линий домашних кур. Содержание фео-
меланинов в красных перьях обычно значительно выше, чем в
рыжих волосах.
Рис. 7.3 Фрагмент предполагаемой структуры галлофеомеланина.
Структура. Наиболее интенсивно изучались красные пигмен-
ты куриных перьев, особенно у красных родайлендов. С ними,
по-видимому, сходны пигменты рыжих волос человека и других
животных, например орангутана. Эти красные пигменты извест-
ны под общим названием галлофеомеланины и представляют со-
бой сложные серусодержащие макромолекулы, образованные из
дофахинона и цистеина. Главные единицы, из которых постро-
ен полимер, no-видимому, происходят от бензотиазола (7.5);
фрагмент возможной структуры такого полимера показан на
рис. 7.3. При обработке волос или перьев кислотой освобожда-
ется небольшое количество димерных пигментов — трихосидери-
(7 ь) Бензотиазол
(7 6) Трихосидерин
Гл. 7. Меланины
265
нов (7.6). Являются ли эти пигменты природными продуктами
или артефактами, остается неизвестным.
Свойства. Феомеланины отличаются от большинства эумела-
нинов растворимостью в разбавленных щелочах. Вместе с тем
у них есть и общие с эумеланинами свойства — это нераствори-
мость в других растворителях и химическая стабильность.
Феомеланины и трихосидерины интенсивно поглощают свет
в видимом диапазоне (500—550 нм), что и обусловливает их
красноватую окраску.
7.2.4 Алломеланины
Распространение. Черные пигменты, известные как алло-
меланины, являются, по-видимому, главным образом продуктами
высших растений (они составляют важную часть черных защит-
ных покровов многих зрелых семян) и грибов; у последних они
обнаруживаются в спороносном слое и в спорах съедобных гри-
бов и в черных гифах плесневых грибов, таких, как Phycomyces.
Структура. Алломеланины грибов изучались наиболее интен-
сивно, однако их структура еще далека от выяснения. Харак-
терной особенностью алломеланинов является то, что они со-
держат мало или вообще не содержат азота и потому не могут
быть полимерами индола. Многие из них, по-видимому, пред-
ставляют собой полимеры простых фенолов, таких, как пиро-
катехин (7.7), и их хинонов и рассматриваются как пирокате-
хиновые меланины. Имеющиеся доказательства свидетельству-
ют в пользу того, что между пирокатехиновыми фрагментами
на различных стадиях окисления может происходить далеко
идущая конденсация с образованием полимерных структур,
в которых мономерные единицы соединены С—С- и С—О—С-
связями, как, например, показано во фрагменте структуры
(7.8). Пирокатехин, по-видимому, является главным продуктом
расщепления растительных алломеланинов. Алломеланины гри-
бов Aspergillus niger и Daldinia concentrica, вероятно, содержат
периленовые мономеры, такие, как (7.9), представляющие со-
бой производные 1,8-дигидроксинафталина (7.10).
Свойства. Алломеланины, так же как меланины других
групп, нерастворимы ни в каких растворителях и химически
стабильны. Они обычно поглощают свет во всем видимом диа-
пазоне, однако иногда наблюдаются максимумы поглощения,
главным образом при ~450 нм.
7.2.5. Склеротииы
Склеротины представляют собой полимерные вещест-
ва, наиболее характерные для экзоскелета членистоногих. Они
ответственны за затвердение и связанное с ним потемнение ку-
тикулы у многих видов насекомых.
266
Часть 1 Химические и биохимические аспекты
Химия склеротиков и биохимия процесса склеротизации изу-
чены мало, однако некоторые их особенности уже известны.
Склеротин не простой продукт полимеризации хинонов, как ме-
ланин. Это белок, который способен сополимеризоваться с об-
разованными при участии ферментов хинонами, подобными
дофахинону, а также с другими близкими к тирозину молеку-
лами. В сополимеризации могут участвовать тирозиновые ос-
татки белка. Усиленный синтез меланина обычно следует по-
сле завершения процесса затвердения кутикулы при склероти-
зации.
7.3. Распространение меланинов
7.3.1. У животных
За исключением гемоглобина крови, меланины являют-
ся единственными пигментами, которые синтезируются и широко
используются млекопитающими. Они также представляют со-
бой обычные пигменты оперения птиц. Черная кожа, волосы,
шерсть и перья окрашены эумеланинами. Желтый, рыжий и ко-
ричневый цвет волос и шерсти обусловлены феомеланинами,
которые служат также пигментами в коричневых, а в некоторых
случаях в желтых и красных перьях. Пигментами желтых и
красных перьев могут быть и каротиноиды (гл. 2). Феомела-
нины найдены только у млекопитающих и у птиц, тогда как
эумеланины широко встречаются также у рыб и беспозвоноч-
ных. К наиболее хорошо известным примерам относятся чер-
ная окраска многих насекомых и других членистоногих (эуме-
ланины присутствуют вместе со склеротином). Однако в неко-
торых случаях темная окраска у ряда насекомых и других чле-
нистоногих вызывается не меланинами, как думали раньше,
а оммохромами (гл. 6).
Наряду с их значением в качестве черных, коричневых и
красных пигментов меланиновые частицы или слои обычно от-
ветственны за структурную окраску, обусловленную физически-
ми или структурными причинами (гл. 1).
Меланины обычно рассматриваются как пигменты наружных
покровов, однако их находят также и в других тканях. В ра-
дужной оболочке глаза содержится меланин, который ответ-
ствен за их окраску как непосредственно, в случае карих и чер-
ных глаз, так и опосредованно, благодаря светорассеянию кол-
лоидных частиц, обусловливающему голубой цвет глаз. Функ-
ционирование меланинов в качестве защитных пигментов в гла-
зах некоторых видов животных будет рассмотрено в гл. 9. Внут-
ренние ткани, такие, как substantia nigra мозга млекопитаю-
Гл. 7. Меланины
267
щих, также часто содержат значительные количества меланино-
вых пигментов. Избыточное образование меланинов служит ха-
рактерным признаком некоторых опухолей человека — меланом.
Сепиомеланин секретируется и испускается в качестве за-
щитного чернильного облака многими головоногими моллюска-
17 9)
(7 10) 1 8 Дигидроксинафталин
ми, например осьминогом или кальмаром. Собранные чернила
используются художниками как темно-коричневая «сепия».
Устойчивость меланинов делает сепию идеальным по долговеч-
ности материалом, хорошо сохраняющим цвет.
Внутриклеточная локализация. В наружных покровах тела
меланины обычно присутствуют в специфических клетках, из-
вестных как меланоциты или меланофоры. Внутри этих клеток
меланины могут быть локализованы в специфических органел-
лах меланосомах и в ассоциации с белком в виде гранул, обыч-
но около 1 мкм длиной, которые имеют различную форму у
268
Часть I. Химические и биохимическиеаспег.ты
разных видов. Структуры и факторы, которые влияют на мор-
фологию и физиологию этих видов, играют чрезвычайно важ-
ную роль в механизмах изменения окраски у животных и бу-
дут описаны более подробно в гл. 8.
7.3.2. У растений и микроорганизмов
Черные пятна, полосы и другие метки часто можно уви-
деть на листьях, цветках и других органах высших растений,
где они могут служить указателями направления для опыляю-
щих их насекомых. Черные оболочки семян и стручков также
очень обычны, однако здесь может быть более важной инерт-
ность материала, чем его черный цвет. Как правило, пигменты,
обусловливающие такую черную окраску, не исследуют, а при-
нимают без доказательства, что они являются соединениями
меланинового типа. В растениях встречаются индольные эуме-
ланины. Они могут синтезироваться при повреждении тканей,
например в плодах банана и в клубнях картофеля. Однако наи-
более характерными растительными меланинами являются эу-
меланины, содержащие мало азота, или не содержащие его
совсем. Алломеланины встречаются во многих окрашенных
в темный цвет оболочках семян, где они содержатся в пигмен-
тированных эпидермальных клетках. Алломеланины являются
обычными пигментами спор или гимениальной пластинки у плес-
невых и других грибов. Некоторые бактерии синтезируют чер-
ные пигменты, которые, по-видимому, принадлежат к мелани-
новому типу, однако они не были охарактеризованы. Феомела-
нины в растительном царстве не обнаружены.
7.4. Биосинтез
7.4.1. Введение
Предшественником эумеланинов и феомеланинов являет-
ся тирозин. Удалось достичь включения тирозина в эумеланин во-
лос, перьев и в чернила головоногих моллюсков, а также тиро-
зина и цистеина в феомеланин. При биосинтезе алломеланинов
используются другие фенолы, главным образом пирокатехин
(7.7). Ферментативное превращение тирозина в производные
индола активно изучалось, и теперь очевидно, что индольные
молекулы полимеризуются с образованием меланинов. Для ме-
ханизмов такой полимеризации были предложены правдоподоб-
ные схемы, однако ни одна из них не подтверждена экспери-
ментально.
Гл. 7. Меланины
269
7.4.2. Биосинтез эумеланинов
Ферментативное окисление тирозина. Ферментативное
образование пигментов меланинового типа из тирозина было
впервые продемонстрировано с препаратом грибно! о фермента,
получившего название тирозиназы. Сходное окисление тирозина
ферментом из мучного червя (Tenebrio molitor) и аналогичны-
ми препаратами из картофеля и грибов интенсивно исследовал
Рэпер. Его работы, продолженные позже Мэзоном, привели к
появлению классической гипотетической! схемы меланогенеза
Рэпера — Мэзона (рис. 7.4). Согласно этой схеме, образование
меланина происходит практически в три стадии. Сначала тиро-
зин (7.11) последовательно окисляется до ДОФА (7.12) и
дофахинона (7.13), а последний циклизуется с образованием
окрашенного в красный цвет дофахрома (7.14). На второй
стадии дофахром превращается в бесцветную 5,6-дигидрокси-
индол-2-карбоновую кислоту (7.15), а затем в собственно 5,6-
дигидроксииндол (7.16). Наконец, окисление последнего до ин-
долил-5,6-хинона (7.17) с последующей полимеризацией дает
меланиновую макромолекулу. Сходный процесс рассматривал-
ся для образования эумеланина млекопитающими и птицами.
Фермент тирозиназа представляет собой полифенолоксидазу
(о-дифенол: О2 оксидоредуктазу) с ионом Си2+ в качестве про-
стетической группы. Она, по-видимому, имеет две различные
активности — аэробно окисляет тирозин до ДОФА и затем
•окисляет сам ДОФА. Остальные стадии образования мелани-
нов могут происходить неферментативным путем.
Схема Рэпера — Мэзона, по-видимому, в основном верна, од-
нако более современные работы неизбежно привели к ее рас-
ширению и модификации. Меланины, полученные ферментатив-
ным путем из разных субстратов, например из тирозина, ДОФА,
дофамина (7.18) и 5,6-дигидроксииндола, выглядят сходными,
однако имеют разные свойства. Сейчас также предполагают,
что небольшое количество индольных мономеров разрушается
до пиррольных фрагментов (рис. 7.5).
Полимеризация. О процессе полимеризации, с помощью ко-
торого меланиновая макромолекула строится из мономерных
единиц, практически ничего не известно. В молекуле хинона, по-
добного индолил-5,6-хинону (7.17), атомы углерода в положе-
ниях 4 и 7 обладают высокой реакционной способностью. Свя-
зывание мономеров по этим положениям через атом азота, ато-
мы углерода 2 и 3 гетероциклического кольца происходит до-
вольно легко (рис. 7.6). Такая же легкость присуща сополи-
меризации с молекулами других мономеров, таких, как дофа-
хинон. Наличие столь большого числа реакционноспособных
атомов в молекулах мономеров обеспечивает возможность пе-
(7,16) 5,6-Дигидроксииндол
(7 17) Цндолил-5,6-хинрн
Рис. 7.4. Схема меланогенеза Рэпера — Мэзона.
Гл. 7. Меланины
271
рекрестного связывания. Не все остатки мономеров остаются в
форме хинонов. Многие из них присутствуют в виде соответст-
вующих фенолов или частично восстановленных форм. В неко-
торых случаях процесс полимеризации может протекать на бел-
ковом матриксе.
ch2.ch2.nh2
(7 18) Дофамин
Модельная структура эумеланина (сепиомеланина) на рис.
7.2 представляет собой как раз такую структуру, образование
которой, по-видимому, происходит в ходе описанных выше про-
цессов.
7.4.3. Биосинтез феомеланинов
Феомеланины образуются по модифицированному эуме-
ланиновому пути. Ферментативное окисление тирозина дает
опять-таки дофахинон (7.13). Однако в этом случае последний
взаимодействует с серусодержащей аминокислотой цистеином
Рис. 7.5. Деградация индольных мономеров до пиррольных единиц.
(7.19), вероятно, с образованием бензотиазолового промежуточ-
ного продукта, такого, как (7.20). Димеризация или полимери-
зация мономерных молекул может затем привести соответствен-
но к образованию молекул трихосидеринового (7.6) типа или
феомеланинов. Было продемонстрировано включение меченых
тирозина и цистеина в феомеланин, а красно-коричневый фео-
меланиноподобный пигмент был получен путем окисления ДОФА
(7.2) грибной полифенолоксидазой в присутствии цистеина. Пред-
212
Часть I Химические и биохимические аспекты
Рис. / 9 Механизм окислительной полимеризации индолил-5,6-хинона.
полагаемый механизм, показанный на рис. 7.7, является, одна-
ко, чисто спекулятивным и не был подтвержден эксперимен-
тально.
7.4.4. Биосинтез алломеланинов
Растения и грибы синтезируют много веществ фенольной
природы. Окисление этих веществ полифенолоксидазой дает во
многих случаях хиноны, которые легко полимеризуются до чер-
ных безазотистых веществ алломеланинового типа. В результа-
те ферментативного окисления пирокатехина (7.7) образуется
не только о-бензохинон, но и такие продукты, как гидрокси-/г-
бензохинон (7.21). Природные пирокатехиновые алломеланины,
по-видимому, образуются путем полимеризации хиноновых мо-
лекул, подобных упомянутым выше, с образованием разветв-
ленных структур, таких, как (7.8).
Нафтоловые алломеланины, синтезируемые Aspergillus ni-
ger и другими грибами, по-видимому, образуются аналогичным
путем из 1,8-дигидроксинафталина (7.10) (поликетидный про-
дукт) через хиноновый и периленовый промежуточные продук-
ты, такие, как (7.9).
Рис. 7 7. Предполагаемый механизм биосинтеза феометанииов и трихосидеринов из дофахинона и ццстецна
274 Часть I, Химические и биохимические аспекты
7.4.5. Общие выводы
Выяснены две общие особенности биосинтеза меланинов:
1) начальные стадии включают ферментативное окисление фе-
нольных или полифенольных предшественников с образовани-
ем хинонов, и 2) высокореакционноспособные хиноны легко
подвергаются полимеризации или сополимеризации, приводя-
щей к образованию окрашенных меланиновых макромолекул.
Создается впечатление, что делавшиеся до сих пор попытки
классифицировать меланиновые вещества на эумеланины, фео-
о
о
(7.2V Гидракси-П-бензохинон
меланины и алломеланины и установить биосинтетические пути
этих трех групп позволяют получить лишь весьма упрощенную
картину. Большинство (если не все) живых тканей обладают
ферментами полифенолоксидазного типа. Все живые ткани со-
держат фенольные соединения, часто (особенно у растений)
весьма разнообразные. В результате окисления любого из этих
соединений полифенолоксидазой — либо его одного, либо в ком-
бинации с другими фенольными или нефенольными веществами
(например, цистеином) — происходит образование хинонов, спо-
собных полимеризоваться с образованием макромолекулярных
продуктов со свойствами меланинов.
Таким образом, один и тот же основной биосинтетический
процесс приводит к образованию огромного числа продуктов,
которые могут выглядеть сходными, но иметь совершенно раз-
ную структуру. Различия возникают благодаря разнообразию
в степени окисления мономерных предшественников и су-
ществованию различных последовательностей и механизмов по-
лимеризации, сополимеризации и перекрестного связывания.
Детали этих механизмов все еще не установлены ни для одно-
го индивидуального природного меланина. Не исключено, что
еще будут идентифицированы новые структурные типы мелани-
нов, молекулы которых построены из пока не открытых моно-
мерных единиц.
Гл. 7. Меланины
275
7.5. Факторы, влияющие на меланогенез
7.5.1. У растений и грибов
Контроль меланогенеза у растений и грибов подробно не*
изучался. Вместе с тем известно, что у растений синтез мела-
нина в темных пятнах и характер расположения этих пятен,
обусловливающий общий внешний вид листьев и цветков, на-
ходится под генетическим контролем. Меланины могут синте-
зироваться в ответ на механическое повреждение тканей, напри-
мер у плодов после удара появляются бурые и черные пятна.
У ряда грибов наблюдается корреляция между синтезом мела-
нинов и спорообразованием, однако значение этого явления по-
ка не известно. Это может быть связано с образованием проч-
ной инертной устойчивой оболочки спор, а не с их окрашива-
нием, как в случае оболочек семян растений.
7.5.2. У животных
У животных меланины обычно содержатся в особых ор-
ганеллах и специфических клетках, так что, скорее всего, регу-
ляторный механизм контролирует процесс дифференциации и
образования этих клеток и органелл, а не биосинтез меланинов
непосредственно. Конечно, в основном синтез и отложение ме-
ланинов, обусловливающие внешнюю окраску или характер ее
распределения у животного, находятся под генетическим конт-
ролем. Так, люди негроидной расы имеют черную кожу вне за-
висимости от окружающей среды. Однако факторы среды и се-
зонные изменения также могут быть чрезвычайно важными.
Известно, например, несколько примеров стимуляции мелано-
генеза низкими температурами или такими факторами, как
влажность и роение (у пчел).
Беспозвоночные и пойкилотермные позвоночные. У этих жи-
вотных изменения окраски как быстрые, так и медленные весь-
ма обычны и, как правило, происходят с участием меланинов и
меланинсодержащих клеток. Однако рассматривать в этом слу-
чае меланины изолированно нельзя, поскольку в окрашивании
клеток участвуют также другие пигменты и другие клетки или
типы органелл. Мы обсудим изменения окраски и механизмы
таких изменений в гл. 8.
Млекопитающие и птицы. Меланиновая пигментация у мле-
копитающих и птиц находится под прямым генетическим конт-
ролем. Тип изменения окраски у них отличается от того, кото-
рый обычен для беспозвоночных, амфибий, рептилий и рыб,
18*
276
Часть I. Химические и биохимические аспекты
правда, у некоторых видов наблюдаются сезонные вариации
окраски.
Широко изучалась пигментация человеческой кожи. Пока-
зано, что в коже происходит быстрое обновление меланинов:
они теряются при слущивании эпидермиса и затем вновь синте-
зируются. У представителей темных рас меланина синтезирует-
ся и накапливается значительно больше, чем у представи-
телей так называемой белой расы. Меланоциты обнаруживают-
ся в коже человеческого зародыша, принадлежащего к негро-
идной расе, уже на одиннадцатой неделе развития.
У людей белой расы хорошо заметен рост содержания ме-
ланина в коже в результате экспозиции на солнечном свету
при загорании. Загар формируется в две стадии. Сначала, сра-
зу же после облучения солнечным светом, синтезируется неко-
торое количество меланина в результате фотоокисления пред-
шественников и быстро образуется «заслон», защищающий от
солнечных ожогов. Этот быстрый загар возникает в основном
под действием света с длиной волны 350 нм. Главный и стой-
кий загар развивается спустя два дня после экспозиции на
солнце и достигает максимума примерно через семь дней. Ме-
ланиновые гранулы образуются в эпидермальных меланоцитах,
а затем они распространяются в кератиноциты (клетки кожи).
Стойкий загар (как и болезненный ожог кожи) наиболее эф-
фективно вызывается светом с длиной волны ~300 нм. На ос-
нове биохимических процессов, обусловливающих загар, вырос-
ла целая индустрия летнего отдыха, однако механизм этих про-
цессов все еще до конца не ясен.
На синтез меланина в коже человека влияет гормон гипо-
физа— меланоцитстимулирующий гормон (МСГ). У женщин
белой расы избыточный синтез меланинов часто происходит в
некоторых участках кожи во время беременности, что отража-
ет обычно повышение активности гипофиза. Меланинстимулиру-
ющие гормоны стимулируют меланогенез и распределение мела-
нинов и у других позвоночных и беспозвоночных животных. Из-
вестно, что меланогенез у многих низших животных стимулиру-
ется светом, но этот эффект явно отсутствует в случае темных
перьев птиц или волос млекопитающих.
Сезонные вариации. Хорошо известно, что оперение многих
птиц в брачный период отличается от зимнего оперения. Эти
различия часто обусловлены вариациями синтеза меланинов
(эумеланинов и феомеланинов) в новых перьях, которые растут
после линьки. Вероятно, происходящие при этом процессы на-
ходятся как под гормональным, так и под генетическим контро-
лем. Некоторые млекопитающие и птицы, живущие в Арктике,
летом имеют темную окраску, а зимой полностью или частично
белую, т. е. они извлекают максимальную пользу из подобной
Гл 7. Меланины
277
маскировки во все времена года. Факторами, вызывающими се-
зонную линьку и контролирующими меланогенез в растущих
волосах и перьях, могут быть температура и продолжитель-
ность дня.
7.5.3. Аномальный меланогенез
Альбинизм. Альбиносные представители млекопитаю-
щих и птиц являются рецессивными гомозиготами, у которых в
коже, глазах, а также в волосах или перьях отсутствует меланин,
и потому они не окрашены в отличие от темного доминантного
фенотипа. Такие животные имеют меланоциты, в которых со-
держатся только бесцветные тени меланиновых гранул, посколь-
ку они лишены тирозиназы (полифенолоксидазы) и потому не
ch(oh).ch2.nh сн3
(7 22) Адреналин
способны превращать тирозин в меланин. Отмечена необычная
восприимчивость животных-альбиносов к болезням, хотя не из-
вестно, связано ли это с неспособностью образовывать меланин.
Болезни человека, связанные с нарушением метаболизма ме-
ланина. У человека и других животных многие опухоли имеют
темную окраску. В таких опухолях (меланомах) синтезируют-
ся и накапливаются избыточные количества меланина. Много
меланина содержится также в родинках и родимых пятнах.
Усиление и ослабление меланогенеза наблюдается при ряде
заболеваний, в том числе болезни Паркинсона, аддисоновой бо-
лезни, а также при витилиго. Однако изменения меланогенеза
являются при этих заболеваниях, по-видимому, вторичными.
В некоторых случаях источником мономеров для меланиновых
молекул может служить адреналин (7.22) или его предшествен-
ники, используемые в нормальных условиях для образования
адреналина.
7.6. Функции
Единственная функция меланинов растений состоит в
формировании общей окраски растений или характера ее распре-
деления. Что касается очевидной корреляции между синтезом
меланинов и споруляцией у различных грибов, то ее значение
пока не известно.
278
Часть I. Химические и биохимические аспекты
Две главные функции меланинов у животных, а именно их
роль как пигментов наружных покровов и как светозащитных
пигментов в глазах и коже, обсуждаются в гл. 8 и 11.
7.7. Заключение и комментарии
Из всех групп природных пигментов, рассмотренных в
этой книге, меланины несомненно являются наиболее трудными
для изучения. Выделение, очистка и химическое разрушение ме-
ланинов представляют такую проблему, что их пришлось оха-
рактеризовать не по какому-либо химическому свойству, а про-
сто как группу темных, чрезвычайно стабильных полимеров. По-
этому в области структуры и биосинтеза меланинов перед ув-
леченными и настойчивыми исследователями структуры — ши-
рокое поле деятельности. Мы еще очень далеки как от возмож-
ности определить структуру какого-либо меланина, так и от
полного понимания его биосинтеза. В этой связи хотелось бы
отметить, что меланины из многих объектов вообще не изуча-
лись. В детальной разработке нуждается еще один вопрос —
белковое окружение меланинов in vivo.
По-видимому, у меланинов нет других функций, кроме пиг-
ментной, но даже в этом случае они заслуживают внимания как
единственные пигменты, ответственные за окраску тканей у че-
ловека. Желание миллионов представителей белых рас увели-
чить содержание меланина в коже при загорании на солнечном
свету привело к созданию целой индустрии летнего отдыха.
Необходимо помнить, что в основе расовых конфликтов и апар-
теида лежат всего лишь различия в содержании кожных мела-
нинов. По этой причине ни одна другая группа природных пиг-
ментов не имеет столь далеко идущих социальных последствий.
Как ни трудны химические проблемы, связанные с меланинами,
разрешить их, вероятно, все-таки легче, чем проблемы социаль-
ные.
7.8. Рекомендуемая литература
Для дополнительного чтения о меланинах, особенно об их
химии, распространении и биосинтезе, можно порекомендовать
четыре книги и обзорные статьи, написанные Томсоном (Thom-
son, 1962), Николаусом (Nicolaus, 1968), Сваном (Swan, 1974)
и Блойсом (Blois, 1978). Хотя они и были написаны в течение
последних 20 лет, на протяжении которых число изученных
меланинов все возрастало, и была накоплена огромная инфор-
мация; фундаментальных изменений в области меланинов за
это время не произошло. Каждая из этих статей содержит об-
ширный список оригинальных работ, который может быть ис-
Гл. 7. Меланины
279
пользован для получения детальной информации по любому
частному вопросу. Наиболее подробные сведения о меланинсо-
держащих клетках и их функционировании при изменении окрас-
ки животных, а также о гормональной регуляции синтеза и рас-
пространения меланинов можно найти в специальной моногра-
фии (Bagnara, Hadley, 1973).
7.9. Дополнительная литература
Bagnara J. Т., Hadley М. Е. (1973). Chromatophores and color change: the
comparative physiology of animal pigmentation, Englewood Cliffs, New
Jersey, Prentice Hall.
Blois M. S. (1978). Melanins. In: Photochemistry and photobiology reviews,
vol. 3, ed. К. C. Smith, p. 115, New York, Plenum.
Nicolaus R. A. (1968). Melanins, Paris, Hermann.
Swan G. A. (1974). Structure, chemistry and biosynthesis of the melanins,
Fortschr. Chem. Org. Naturst., 31, 521.
Thomson R. H. (1962). Melanins. In: Comparative biochemistry, vol. 3, eds.
M. Florkin and H. S. Mason, p. 727, New York and London, Academic
Press.
Часть II
Функциональные аспекты
Глава 8
Значение окраски в природе
8.1. Введение
Наиболее очевидная и фундаментальная функция пиг-
ментов в живых организмах состоит в том, что они придают тка-
ням окраску. В этой главе сделана попытка объяснить, почему
важно это свойство и как оно используется животными и расте-
ниями.
Мы распознаем принадлежность животных или растений к
тому или иному виду в первую очередь визуально. Вероятно,
первыми регистрируемыми зрительными впечатлениями являют-
ся общие размеры и форма объекта, однако чрезвычайно важ-
ны также его окраска и характер ее распределения. Последнее
обычно используется как своего рода «тонкая настройка» в це-
лях различения видов, близких по форме и размерам, но в не-
которых случаях окраска и ее распределение могут быть наи-
более явными характерными признаками. Большинство видов
животных реагируют на такие визуальные сигналы и использу-
ют свои зрительные возможности как средство для распознава-
ния пищи, врагов или брачных партнеров. Параллельно с этим
у большинства животных и многих растений развилась способ-
ность к окрашиванию со своим типом распределения окраски.
Эта способность используется животными и растениями для то-
го, чтобы оповестить об их присутствии или скрыть его. Окрас-
ка и характер ее распределения изменчивы, причем изменения
могут быть связаны с временем года, стадиями развития или
быстро появляться в ответ на изменение условий окружающей
среды.
Все эти аспекты будут рассмотрены в данной главе, основ-
ная цель которой — охарактеризовать значение и важность
окраски в животном царстве. Однако и для многих растений
важно быть окрашенными либо иметь окрашенные органы
или ткани (такие, как цветки и плоды) для привлечения
Гл. 8. Значение окраски в природе
281
тех животных-переносчиков, которые помогают распростране-
нию видов растений посредством непроизвольного их опыления
или распространения семян. Будут рассмотрены современные
виды, образовавшиеся в результате миллионов лет эволюции и
отбора по тем признакам, которые давали хотя бы незначи-
тельное преимущество для выживания индивида или вида в це-
лом. Какие-либо предположения об эволюции пигментации об-
суждаться не будут. Цель данного раздела заключается лишь
в демонстрации путей, с помощью которых окраска и характер
ее распределения используются с выгодой для себя теми жи-
вотными и растениями, которые существуют сегодня.
8.2. Окраска и характер ее распределения у животных
Окраска внешних покровов помогает животным укры-
ваться или маскироваться (покровительственная окраска), а
также служит для опознавания или предупреждения (предуп-
реждающая окраска). При этом используется как структурная
окраска, так и окраска, обусловленная пигментами (любого
класса).
8.2.1. Покровительственная окраска
Цель покровительственной окраски заключается в мас-
кировке, т. е. в том, чтобы сделать животное как можно более
незаметным в его природных условиях обитания. Покровитель-
ственные цвета, имеющие большое значение для животных, ис-
пользуются ими для того, чтобы избежать обнаружения хищ-
ником, но они также используются и хищниками, чтобы не быть
замеченными будущей жертвой и тем облегчить ее поимку.
Покровительственная окраска может быть имитацией фоновой
окраски, или она может быть очень сложной, исчерченной по-
лосами, искажающей форму и делающей животное незаметным.
Некоторые животные приобретают более яркую или более тем-
ную окраску лишь с помощью сочетаний различных оттенков
серого цвета, в то время как другие способны воссоздавать в
деталях окраску поверхности, на которой находятся, а третьи
даже могут копировать окраску пестрых поверхностей. В не-
которых случаях то, что на расстоянии выглядит однотипным,
в действительности может быть окрашено в яркие цвета, раз-
личимые лишь с близкого расстояния. Необходимо также пом-
нить, что внешний вид животного нельзя рассматривать без
учета ее природной среды обитания. Так, тигр или зебра очень
заметны в зоопарке, но окраска из чередующихся полос может
быть чрезвычайно эффективной для маскировки этих живот-
ных в среде, окружающей их в естественных условиях.
282
Часть 11. Функциональные аспекты
Животные, перемещающиеся по изменяющейся поверхности,
не могут быть хорошо замаскированы все время. Некоторым ви-
дам, например крабу Carcinus, присущ полиморфизм окраски.
Известны его разные формы, отличающиеся друг от друга по
окраске, соответствующей поверхности определенного типа.
Краб целенаправленно ищет такую поверхность и проводит
там большую часть времени.
Многочисленные опыты подтвердили значение покровитель-
ственной окраски для выживания видов. Если насекомые од-
ного и того же вида, но принадлежащие к двум по-разному
окрашенным формам обитают на одноцветной поверхности, то
те из них, чья окраска контрастирует с поверхностью, с гораздо
большей вероятностью станут жертвами хищников, чем те,
окраска которых соответствует окраске поверхности.
8.2.2. Предупреждающая окраска
Предупреждающая, или сем этическая, окраска обычно
состоит из бросающихся в глаза больших контрастных ярких
пятен, имеющих четкие границы. Они узнаются и интерпрети-
руются видящим их животным как предостережение или, на-
оборот, как приглашение в зависимости от предшествующего
жизненного опыта.
Отпугивающая окраска. Об окраске и характере ее распре-
деления говорят, что они отпугивающие, если они предупреж-
дают того, кто их видит, об опасности или неприятном воз-
действии. Хорошо известным примером такой предупреждаю-
щей окраски служит окраска ос (например, Vespula vulgaris),
чьи контрастные желтые и черные полосы вызывают страх да-
же у людей. Бросающаяся в глаза окраска у других насекомых,
например красно-черная окраска божьих коровок (Coccinella
spp.), служит предупреждением потенциальному хищнику о
том, что ее обладатели несомненно невкусны. Таким животным
нет нужды прятаться или пытаться избежать встречи с хищ-
ником. Хищники, руководствуясь своим предшествующим опы-
том, не преследуют их.
Опознавательная окраска. Яркая привлекающая внимание
окраска (и характер ее распределения), которая служит для
привлечения других животных, известна как опознавательная.
В большинстве случаев она предназначена для особей своего
вида, но противоположного пола и является частью сексуально-
аттрактивного демонстрационного поведения. Обычно ярко окра-
шен только самец и только он активен в демонстрационном по-
ведении, однако в некоторых случаях особи обоих полов сход-
ны и активны как те, так и другие. Помимо того что сексуаль-
ное демонстрационное поведение имеет сигнальную функцию,
Гл. 8. Значение окраски в природе
283
благодаря ему, по-видимому, ускоряется созревание гонад и го-
товность к спариванию, а также увеличивается плодовитость.
Развитие сексуально-демонстрационной окраски, как правило,
является сезонным и, вероятно, находится под гормональным
контролем.
Часто сексуально-демонстрационная окраска вынуждена
вступать в компромисс с требованиями маскировки. Яркие цве-
та и вызывающий характер их распределения необходимы жи-
вотному только в период ухаживания или во время угрожаю-
щего поведения по отношению к сопернику, в остальное же вре-
мя они должны быть замаскированы.
Использование привлекающей окраски и характера ее рас-
пределения в несексуальных целях менее обычно, однако в не-
которых случаях с ее помощью привлекается добыча. Некото-
рые глубоководные рыбы, например удильщики, обычно хорошо
маскирующиеся, имеют ярко окрашенную и часто светящуюся
«приманку» для привлечения добычи. Сходной цели служат и
щупальца актиний.
Псевдосематическая окраска. Псевдосематическая окраска
имитирует предупреждающую (сематическую) окраску другого
вида. В большинстве случаев встречается псевдоотпугивающая
окраска, ведущая к тому, что особи безвредного и «вкусного»
вида не атакуются и не поедаются потому, что они внешне на-
поминают особей неродственного опасного или «невкусного»
вида. Например, многие журчалки приобрели осоподобную
желто-черную окраску, а некоторые тараканы точно имитиру-
ют божьих коровок.
8.3. Пигментные клетки животных — хроматофоры
Во внешних покровах животных, особенно беспозвоноч-
ных и пойкилотермных (холоднокровных) позвоночных, пигмен-
ты локализованы в специальных клетках. В наиболее ценной мо-
нографии о пигментных клетках животных и изменениях окрас-
ки последних (Bagnara, Hadley, 1973) для описания этих окра-
шенных клеток рекомендуется использовать термин «хромато-
фор». Такая номенклатура будет принята и в настоящей книге,
несмотря на то, что другие авторы используют и альтернатив-
ные термины, например «хроматоциты». Различают несколько
видов хроматофоров, в первую очередь по цвету, за который
они ответственны.
8.3.1. Меланофоры
Черные пигментные клетки, содержащие меланин (гл.
7), известны как меланофоры, правда, в случае млекопитающих
и птиц такие клетки чаще называют меланоцитами. У позвоноч-
284
Часть 11. Функциональные аспекты
ных существуют по крайней мере два типа меланофорных кле-
ток, которые отличаются по локализации, внешнему виду и ре-
акции на гормоны. Почти у всех пойкилотермных животных
чаще всего встречаются дермальные меланофоры — очень круп-
ные клетки до 0,5 мм в диаметре, участвующие в быстрых из-
менениях окраски. Эпидермальные меланофоры в отличие от
дермальных в быстрых изменениях окраски играют незначи-
тельную роль. Они представляют собой тонкие удлиненные
клетки, которые обычны для рептилий и амфибий, но редко
встречаются у рыб. Сходные эпидермальные клетки (меланоци-
ты) обусловливают окрашивание кожи меланином у млекопита-
ющих, а также пигментацию меланинами, в том числе и феоме-
ланином, волос и перьев.
Фундаментальным свойством всех меланофоров является их
способность синтезировать свой меланиновый пигмент. По край-
ней мере у позвоночных, синтез меланина внутри меланофора
включает отложение меланиновых полимеров на матриксе скру-
ченных в спираль белковых нитей. Такой матрикс называется’
премеланосомой. Премеланосома содержит фермент тирозина-
зу (полифенолоксидазу), которая и катализирует синтез мела-
нина (гл. 7). Сформировавшиеся субклеточные органеллы назы-
вают меланосомами, или меланиновыми гранулами, причем аг-
регация и распределение этих частиц играют большую роль при
изменении окраски животных (разд. 8.4.2).
Судя по немногочисленным исследованиям меланинсодержа-
щих клеток и органелл у беспозвоночных животных, они, по-ви-
димому, напоминают аналогичные клетки и органеллы позво-
ночных.
8.3.2. Ксантофоры и эритрофоры
Меланофоры обычно окрашены в черный цвет, хотя не-
которые из них, содержащие феомеланин, могут иметь желтую
или оранжево-красную окраску. Большинство желтых, оранже-
вых и красных клеток наружных покровов принадлежат к раз-
ным типам клеток, известным как ксантофоры (желтые) или
эритрофоры (красные). Преобладающие пигменты в этих клет-
ках— каротиноиды (гл. 2), содержащиеся в липидных каплях
или в виде каротиноидных пузырьков. Яркую окраску многим
пойкилотермным позвоночным, таким, как лягушки и жабы,
придают птерины (гл. 6), как сами по себе, так и в комбинации
с каротиноидами. Птерины локализованы в специализированных
небольших органеллах, называемых птериносомами, которые
распределены по всей цитоплазме. Даже у видов, которые окра-
шены главным образом каротиноидами, в развивающихся ксан-
тофорах и эритрофорах сначала синтезируются и становятся
Гл 8 Значение окраски в природе
285,
видимыми птерины, тогда как каротиноиды, которые должны
быть получены из пищи, выявляются лишь позднее.
Птериновые пигменты глаз Drosophila локализованы в ор-
ганеллах, удивительно сходных с птериносомами позвоночных.
8.3.3. Иридофоры
У беспозвоночных и пойкилотермных позвоночных
очень распространены иридофоры (иногда их называют гуано-
форами), представляющие собой клетки наружных покровов.
Несмотря на то что иридофоры не являются в прямом смысле
пигментсодержащими клетками, подобными меланофорам,
ксантофорам и эритрофорам, они тем не менее играют важную
роль в формировании окраски и внешнего вида животных. Ири-
дофоры содержат органеллы, которые ориентированы таким
образом, что эффективно отражают свет, и таким путем форми-
руют основу многих радужных, или «металлических», цветов
структурного происхождения. Главными «пигментами» иридо-
форов служат пурины — чаще всего гуанин (гл. 6), —которые
имеют белую окраску или вообще бесцветны, но, будучи орга-
низованы в виде пачек или бляшек, отражают все или некото-
рые лучи видимого света и таким образом в большинстве слу-
чаев обусловливают серебристый и золотистый металлический
блеск, хорошо известный на примере чешуи рыб.
8.3.4. Ассоциации хроматофорных клеток
Функциональная единица хроматофоров дермы. В дер-
мальном слое кожи позвоночных, которые могут быстро и пол-
ностью изменять свою окраску, обычно присутствуют хроматофо-
ры трех типов, причем они имеют специфическую локализацию.
Ксантофоры (или эритрофоры) расположены сверху, иридофо-
ры— непосредственно под ними, а дермальные меланофоры
формируют базальный слой. Как правило, число клеток этих
трех типов различно. Такая ассоциация клеток трех типов (рис.
8.1) составляет функциональную единицу хроматофоров дермы,
благодаря которой и обеспечиваются быстрые изменения окрас-
ки. Поглощающие свет меланофоры и отражающие свет иридо-
форы ответственны за темные и светлые участки соответствен-
но, в то время как ксантофорам и эритрофорам отводится глав-
ным образом роль желтого фильтра. Иридофоры могут также
участвовать в формировании структурной голубой окраски. Бы-
стрые изменения окраски осуществляются благодаря изменени-
ям вклада, вносимого в ее формирование клетками разных ти-
пов, составляющими функциональную единицу (разд. 8.4).
286
Часть 11 Функциональные аспекты
Эпидермальная меланиновая единица. Эпидермальные мела-
нофоры позвоночных определяют цвет наружных покровов в
ассоциации с другими клетками. Меланиновый пигмент пере-
носится из места его образования — меланофора — в соседнюю
или окружающие его клетки. В целом все меланизированные
клетки можно рассматривать как эпидермальную меланиновую
единицу. Способность клеток переносить пигменты особенно
.важна для млекопитающих и птиц, поскольку она позволяет
Фис. 81. Схематическое изображение функциональной единицы хроматофо
ров дермы.
меланинам транспортироваться из собственно меланофоров в
эпидермальные клетки (из которых образуются специализиро-
ванные структуры — волосы, перья и клюв) и откладываться
там.
8.3.5. Эмбриональное развитие хроматофоров
Развитие пигментсодержащих клеток и характер об-
разования пигментов у позвоночных подробно изучены Местом
образования хроматофоров у всех классов позвоночных явля-
ется нервный гребень. Например, у земноводных здесь образу-
ются все три основных типа пигментированных клеток — мела-
нофоры, иридофоры и ксантофоры. Наряду с хроматофорами
нервный гребень может давать начало клеткам многих других
типов, однако факторы, определяющие тип образующихся кле-
ток, пока не известны. Развитие клеток по пути именно хрома-
тофора определяется очень рано, еще до образования нервно-
го валика, и появление хроматофоров разных типов происходит
в определенной последовательности. Вначале дифференцируют-
ся меланофоры дермы, а уже затем ксантофоры и иридофоры.
Гл 8 Значение окраски в природе 287
Эпидермальные меланофоры появляются сравнительно поздно,
в некоторых случаях лишь незадолго до метаморфоза.
В ходе эмбриогенеза пигментные клетки должны переме-
щаться на большие расстояния от нервного гребня до наруж-
ных покровов, которым они придают окраску. Миграция хро-
матобластов (развивающихся затем в хроматофоры) была про-
демонстрирована экспериментально на примере лягушки Rana
pipiens В основном пигментные клетки перемещаются в форме
хроматобластов, причем их перемещение происходит еще до
того, как в них синтезируется большая часть пигмента. Вместе
с тем наблюдали и некоторые ограниченные перемещения диф-
ференцированных меланофоров.
Характер распределения окраски обусловлен миграцией хро-
матобластов в генетически предопределенные участки. Белые
пятна у некоторых видов или альбиносных мутантов наблюда-
ются, как правило, потому, что хроматобласты не способны
дифференцироваться в хроматофоры, хотя они и переместились
в белые участки. Сходным образом у голубых мутантов или у
нормальных зеленых лягушек ксантобласты (частично диффе-
ренцированные ксантофоры) локализованы в соответствующих
им местах, однако они не способны образовывать желтый пиг-
мент.
Природа пигментов, образуемых дифференцированными
хроматофорами, генетически детерминирована, в то время как
процесс дифференциации клеток и синтез пигмента могут регу-
лироваться гормональными факторами.
8.4. Изменение окраски у животных
Некоторые животные сохраняют одну и ту же окраску
и одно и то же ее распределение на протяжении всей своей
жизни. За исключением незначительных модификаций (таких,
как солнечный загар и потеря пигментов волос), это справедли-
во для человека и многих других млекопитающих. Единствен-
ный пигмент, который должен у них синтезироваться, — это
пигмент, требующийся для возмещения потерь, связанных ли-
бо с износом и повреждением пигментированных тканей, либо с
линькой.
У многих животных, однако, внешний вид меняется как на
разных стадиях развития, так и в разное время года. Как пра-
вило, эти изменения (по крайней мере частично) связаны с из-
менениями окраски или характера ее распределения в наруж-
ных покровах. Есть также группа животных, в основном это
беспозвоночные и пойкилотермные позвоночные, которые спо-
собны очень быстро изменять свою окраску и ее рисунок в ог-
288
Часть II. Функциональные аспекты
вет на стрессовые воздействия или изменения окружающей
среды.
Эти изменения окраски обусловлены либо изменениями ко-
личества содержащегося пигмента в результате его синтеза или
разрушения, либо изменениями эффективности, с которой про-
является его окраска.
8.4.1. Изменения окраски, обусловленные синтезом
или разрушением пигмента
Такой тип изменения окраски называют по-разному —
морфологическим, морфогенетическим или хромогенным. К не-
му принадлежат изменения (или перераспределение) окраски,
связанные с линькой оперения или шерсти и заменой их со-
ответственно перьями или волосами другого цвета. Сюда же
относятся и сезонные изменения, связанные с маскировкой (на-
пример, белая окраска зимой) или с размножением (приобре-
тение брачной окраски).
К этому же типу принадлежат и некоторые изменения
окраски, обусловленные факторами окружающей среды. Обыч-
ным примером таких изменений служит адаптация к цвету
поверхности. Так, у животных, которых содержат на темной по-
верхности, синтезируется больше меланина и образуется боль-
ше меланосом, тогда как у животных, оказавшихся на более
светлой поверхности, наблюдается потеря меланина. Общее чис-
ло пигментных клеток в коже адаптирующегося животного мо-
жет увеличиваться или уменьшаться; при этом может происхо-
дить как пролиферация уже существующих меланофоров, так
и меланизация еще недифференцированных меланобластов.
Аналогичные изменения отмечены и для хроматофоров других
типов. Так, у рыб светлая поверхность обусловливает усиление
образования «пигмента» иридофоров гуанина и одновременно
распад меланина и, наоборот, образование меланина, происхо-
дящее на темном фоне, сопровождается исчезновением гуанина.
Желтый пигмент ксантофоров ведет себя так же, как и мела-
нин, т. е. его синтез усиливается в ответ на потемнение фона.
8.4.2. Физиологические изменения окраски
Физиологические, или хромомоторные, изменения
окраски обычно происходят значительно быстрее, чем в только
что описанном случае, поскольку они не обусловлены синтезом
или распадом пигментов. В этом случае мобилизация уже су-
ществующих пигментированных органелл внутри клетки опре-
деляет эффективность проявления различных пигментов и, сле-
довательно, общую окраску, приобретаемую животным. Обыч-
Гл. 8. Значение окраски в природе
289
но такие реакции протекают легко и обратимо. Они характер-
ны для пресмыкающихся, земноводных, рыб и многих беспоз-
воночных, но не встречаются у млекопитающих или птиц.
Обычный механизм. У всех животных, за исключением голо-
воногих и крылоногих моллюсков, изменения окраски происхо-
дят в результате перемещения пигментных органелл в хромато-
форе (рис. 8.2). Например, в меланофорах меланиновые гра-
Меланофорные клетки
агрегированы
Светлый фон
.Светлая окраска
Меланиновые гранулы
Темный фон
Темная окраска
Меланиновые гранулы
Диспергированы-
Рис. 8.2. Обычный механизм физиологических изменений окраски (посвет-
ление — потемнение) путем агрегации и диспергирования меланиновых гра-
нул (меланосом) внутри меланофора.
нулы (или меланосомы) могут распределяться по периферии
клетки, где они эффективно поглощают свет, обусловливая тем
самым темную окраску, или же могут собираться в небольшом
объеме в центре клетки, благодаря чему поглощается лишь не-
значительное количество падающего света. Таким образом,
диспергирование и агрегация меланосом обеспечивает быстрое
потемнение или посветление кожи. Функциональная единица
хроматофоров дермы (разд. 8.3.4) содержит не только мелано-
форы, но также иридофоры и ксантофоры (или эритрофоры),
в которых пигментные частицы или гранулы также могут быть
диспергированы или собраны в одном месте, внося больший или
меньший вклад в поглощение света и окраску. Благодаря этому
животное может быстро и обратимо менять окраску в широком
диапазоне цветов и довольно точно имитировать окраску по-
верхности, на фоне которой оно находится. Именно таким пу-
тем достигается то замечательное разнообразие окраски, кото-
рое характерно для хамелеона.
Механизм изменения окраски у головоногих. Головоногие
моллюски, например кальмар и осьминог, а также некоторые
крылоногие моллюски используют иной механизм изменения
19—84
290
Часть II Функциональные аспекты
окраски (рис. 8.3). Эти животные имеют структуры, представля-
ющие собой крошечные органы, которые состоят из клеток пя-
ти различных типов, включая центральный хромофор. Сокраще-
ние радиальных мышечных волокон вызывает растяжение хро-
мофора (до размеров, в 7 раз превышающих исходный диа-
метр), что сопровождается рассредоточением его пигментных
гранул. Хроматофоры могут быть темно-коричневыми, красны-
ми и желтыми [темные хроматофоры содержат оммохром (гл.
6), а не меланин], и окраска кожи в данный момент времени
А Б
Рис 8 3. Механизм физиологических изменений окраски у головоногих мол-
люсков А Светлая окраска Меланиновые гранулы агрегированы путем сжа-
тия центрального хроматофора в небольшом объеме Б. Темная окраска.
Меланиновые гранулы диспергированы в результате сокращения радиальных
мышечных волокон, вызывающего сильное растяжение центрального хрома-
тофора.
зависит от того, в какой степени расширены хроматофоры всех
этих типов. У животных с такими хроматофорными органами
изменения окраски происходят чрезвычайно быстро (менее чем
за 1с).
8.4.3. Факторы, контролирующие изменения окраски
Все изменения окраски, связанные с приспособлением
к цвету фона, независимо от того, происходит ли при этом из-
менение концентрации пигментов или используется механизм аг-
регации-диспергирования пигментных гранул, регулируются, по-
видимому, одними и теми же факторами. Главным регулирую-
щим фактором скорее всего служит альбедо, т. е. соотношение
между количеством света, отраженного от фоновой поверхности,
и интенсивностью прямого падающего света. На изменение
окраски могут оказывать влияние и другие факторы окружаю-
щей среды. Так, низкие температуры или повышенная влаж-
ность часто приводят к усилению пигментации как путем син-
теза (меланин, птерин, оммохром), так и посредством усиления
Гл 8 Значение окраски, в природе 291
поглощения, транспорта и отложения (экзогенные каротинои-
ды). У некоторых видов (водных и наземных) позвоночных и
беспозвоночных обнаружено существование суточных ритмов
изменения окраски.
Часто синтезу пигментов в ходе длительной адаптации к
фоновой окраске предшествуют более быстрые физиологические
изменения цвета, осуществляющиеся путем агрегации или дис-
пергирования пигментных органелл; возможно, синтез пигмен-
тов является необходимым следствием таких изменений.
8.4.4. Механизмы регуляции изменения окраски
Изменения окраски обоих типов могут регулироваться
гормональными механизмами или нервной системой. Гормон ги-
пофиза, известный как меланоцитстимулирующий гормон
(МСГ), или интермедин, регулирует синтез меланина и диспер-
гирование меланосом. МСГ оказывает влияние и на более ярко-
окрашенные пигментные клетки — иридофоры, ксантофоры и
эритрофоры. МСГ представляет собой пептид, аминокислотный
состав которого у разных видов животных несколько различает-
ся. Все к настоящему времени охарактеризованные образцы
(8 1 ) Мелатонин
(8 2) Адреналин (R-CH3)
(8 3 ) Норадреналин (R = Н)
(8 4 ) Тироксин
19*
292
Часть II. Функциональные аспекты
МСГ позвоночных включают активный гептапептид: —Met—
Glu—His—Phe—Arg—Trp—Gly—.
Другой гормон — мелатонин (8.1), секретируемый эпифи-
зом, — регулирует реакцию на свет или альбедо у многих ви-
дов, ингибируя процесс потемнения. Адреналин (8.2) и норад-
реналин (8.3), действующие как нейрогормоны, и тироксин
(8.4), действующий как инициатор морфологической диффе-
ренцировки, также могут влиять на пигментацию и изменение
окраски. В качестве вторичного посредника в механизме дей-
ствия МСГ и адреналина при изменении окраски у позвоноч-
ных, вероятно, участвует сАМР (3',5'-циклический аденозннмо-
нофосфат). Большое значение для функционирования этих гор-
монов имеют тиоловые группы белков.
Считают, что физиологические изменения окраски у рако-
образных опосредованы несколькими гормонами. Различают от-
дельные гормоны, вызывающие диспергирование или концент-
рирование красного, черного и белого пигментов. Они, по-види-
мому, являются пептидами и синтезируются в глазных стебель-
ках, где локализованы фоторецепторные пигменты, улавливаю-
щие изменение фоновой окраски.
Изучение биохимии гормонов, участвующих в механизмах
окраски у животных, только начинается.
8.5. Окраска у растений
Представители растительного царства окрашены пре-
имущественно в зеленый цвет. Зеленый цвет, во всем своем раз-
нообразии оттенков, услаждает глаз человека, но можно не со-
мневаться, что и он стал бы монотонным, если бы зеленый фон
не оживлялся всплесками других ярких и контрастных цветов.
Ярко окрашенные цветки и плоды невольно притягивают глаз.
Однако значение их окраски, по-видимому, не только в этом,
но имеет гораздо более фундаментальную основу. Зеленая
окраска растений и хлорофилл, который ее обусловливает, иг-
рают чрезвычайно важную роль в процессе фотосинтеза, под-
держивающего существование любого растения. Значение же
контрастирующей окраски цветков и плодов заключается в том,
что она облегчает распространение и выживание вида.
Цветки привлекают внимание не только человека, но и мно-
гих других более мелких животных, которые оказывают расте-
ниям огромную услугу, выполняя роль переносчиков пыльцы.
Пчелы, вероятно, в этом отношении изучены лучше, чем любые
другие переносящие пыльцу животные. Пчелы способны разли-
чать четыре основных «цвета», включая ультрафиолет в диапа-
зоне 340—380 нм. Свет красных длин волн они не видят, пред-
почитая синие, желтые или поглощающие в УФ-свете (белые)
Гл. 8. Значение окраски в природе
293
цветки. Другие животные, служащие переносчиками пыльцы,
имеют иные цветковые «привязанности». Колибри, например,
являющиеся важными опылителями в тропиках, предпочитают
красные и оранжевые цвета. Некоторые опылители проявляют
предпочтение к палевой окраске; ночные бабочки, жуки и
(иногда) летучие мыши больше любят белые (ультрафиолето-
вые), а дневные бабочки —не очень яркие цвета, такие, как
розовый или розовато-лиловый.
Яркая, бросающаяся в глаза окраска плодов привлекает пи-
тающихся плодами животных, в первую очередь птиц, которые
потом разносят неперевариваемые семена на очень большие
расстояния.
По-видимому, эволюция у растений была направлена на от-
бор пигментов, придающих цветкам и плодам окраску, которая
привлекала бы животных — главных опылителей и разносчиков
семян. Так, виды растений, опыляемые преимущественно пчела-
ми, содержат в цветках бесцветные флавоноиды или синие ан-
тоцианы, в то время как тропические виды, опыляемые главным
образом колибри, имеют красные цветки, содержащие красные
антоцианы.
Необходимо подчеркнуть важную роль в качестве пигментов
растений веществ, обладающих сильным поглощением в ближ-
нем ультрафиолете. В отличие от глаз человека глаза многих
других животных, преимущественно насекомых, чувствительны
к УФ-свету и, следовательно, видят соединения, поглощающие
его.
Вещества, поглощающие видимый и УФ-свет, имеют значе-
ние при формировании «рисунка» цветков. В ходе эволюции вы-
работалось несколько очень сложных типов рисунков, многие из
которых приспособлены к особенностям насекомых-опылителей.
Так, многие цветки, например у наперстянки (Digitalis purpu-
rea), имеют «медовые линии» или подобные им метки, которые
указывают пчелам и другим насекомым путь к нектару. С этой
же целью могут использоваться и высокие концентрации УФ-по-
глощающего «пигмента», особенно в основаниях лепестков. Ин-
тенсивное УФ-поглощение служит для насекомых указателем
их цели — центра цветка, содержащего нектар и пыльцу. Ри-
сунок цветка формируется благодаря как локальному усилению
образования пигмента в определенных участках лепестков, так
и наложению второго пигмента на главный. Формирование ри-
сунка цветков находится под строгим и сложным генетическим
контролем.
Окраску и рисунок почти у всех цветков обусловливают две
группы пигментов — каротиноиды (гл. 2) и флавоноиды (гл. 4).
Красная, пурпурная и синяя окраска практически всегда обус-
ловлена антоцианами. Белые и кремовые цветки содержат
294
Часть II. Функциональные аспекты
большое количество флавонов и флавонолов, которые являются
УФ-поглощающими пигментами, видимыми для насекомых.
Желтая или оранжевая окраска обычно вызвана каротиноида-
ми, но в некоторых случаях, целиком или частично, она обу-
словлена флавоноидами классов халконов и ауронов, или же
бетаксантинами (гл. 6). Смеси водорастворимых флавоноидных
пигментов, например желтого и красного, дают, как и следова-
ло ожидать, оранжевый цвет. Вместе с тем сочетание пурпурно-
го водорастворимого антоциана и желтого липофильного каро-
тиноида создает обычно коричневую окраску, например у цвет-
ков лакфиоли (Cheiranthus cheiril}.
Окраска плодов, особенно красная, пурпурная и синяя, обыч-
но обусловлена антоцианами, хотя в некоторых случаях (на-
пример, у томата и апельсина) она имеет каротиноидную при-
роду.
Растительные пигменты как бы оповещают животных о на-
личии цветка или плода. Цель такого оповещения почти всегда
заключается в привлечении полезных животных, а не в пред-
упреждении неблагоприятного воздействия. Таким образом,
у растений роль окраски, обусловленной пигментами, значи-
тельно более проста, чем у животных. Однако имеется несколь-
ко отдельных случаев, в которых экологические факторы могут
быть чрезвычайно сложными.
8.6. Заключение и комментарии
Окраска и характер ее распределения, особенно у жи-
вотных, имеют большое значение в поведенческом и экологиче-
ском аспектах, однако они могут оказаться малоинтересными в
аспекте биохимическом, если не считать, конечно, идентифика-
ции участвующих в их образовании пигментов. Вместе с тем су-
ществует ряд биохимических вопросов, достойных внимания, ко-
торые все еще остаются без ответа. Особый интерес вызывают
вопросы, касающиеся механизмов регуляции синтеза пигментов
и отложения их в тканях, а также изменения окраски. В каче-
стве примера можно назвать механизмы, с помощью которых
факторы окружающей среды (такие, как свет и температура),
вероятно, при участии гормонов вызывают быстрые или мед-
ленные изменения окраски, или пути, по которым реализуется
генетический контроль распределения окраски. В настоящее
время биохимические аспекты этих явлений еще чрезвычайно
трудны для изучения, но они несомненно привлекут значитель-
но большее внимание исследователей в ближайшем будущем.
В растительном царстве синтез пигментов является лишь
частью грандиозных морфологических изменений, которые про-
исходят по мере развития и созревания цветков и плодов,
Гл. 8. Значение окраски в природе
295
а также частью трансформации хлоропластов в хромопласта,
сопровождающей эти изменения. Механизмы регуляции и конт-
роля происходящего при этом синтеза пигментов — увлекатель-
ная область исследования для биохимика.
8.7. Рекомендуемая литература
Много примеров, иллюстрирующих значение окраски и
характера ее распределения у представителей животного цар-
ства, приведены в книгах общего характера (Broughton, 1964;
Fox, 1976, 1979; Fox, Vevers, 1960; Needham, 1974). Эта же те-
ма более подробно и широко обсуждается в сохраняющей свое
значение работе Котта (Cott, 1940). Пигментным клеткам жи-
вотных и измерениям окраски посвящено много книг и статей.
Среди них монографии Бэгнэра и Хэдли (Bagnara, Hadley,
1973), в которой приводится чрезвычайно полезное общее опи-
сание данной области исследования, и более специальные пуб-
ликации Делла Порта и Мюльбока (Della Porta, Miihlbock,
1966) о меланоцитах, Фингермана (Fingerman, 1970) о хрома-
тофорах, а также Новалеса (Novales, 1969) и Рили (Riley,
1972) о регуляции изменений окраски, дополняющих более ста-
рую книгу Паркера (Parker, 1948). В еще более специальном
труде Сирла (Searle, 1968) приведен обзор генетических дан-
ных по окраске покровов тела у млекопитающих.
Общий обзор значения окраски у растений опубликован
Харборном (Harborne, 1976). Много интересной информации об
экологическом значении окраски цветков и опылении можно
извлечь из книги Фэгри и Ван-дер-Пила (Faegri, Van der Pijl,
1971).
8.8. Дополнительная литература
Bagnara J. T., Hadley M. E. (1973). Chromatophores and color change: the com-
parative physiology of animal pigmentation, Englewood Cliffs, New Jersey,
Prentice-Hall.
Broughton W. B. (ed.) (1964). Colour and life, London, Institute of Biology.
Cott H. B. (1940). Adaptive coloration in animals, London and New York, Ox-
ford University Press.
Della Porta G., Miihlbock 0. (eds.) (1966). Structure and control of the mela-
nocyte, New York, Springer-Verlag.
Faegri K.., Van der Pijl L. (1971). The principles of pollination ecology, 2nd
edition, Oxford, Pergamon.
Fingerman M. (1970). Comparative physiology: chromatiphores, Ann. Rev. Phy-
siol , 32, 345
Fox D. L. (1976). Animal biochromes and structural colors, 2nd edition, Berke-
ley, Los Angeles and London, University of California Press.
Fox D. L. (1979). Biochromy: natural coloration of living things, Berkeley, Los
Angeles and London: University of California Press.
296 Часть II. Функциональные аспекты
Fox Н. М., Vevers G. (1960). The nature of animal colours, London, Sidgwick
and Jackson
Harborne I B. (1976). Functions of flavonoids in plants. In: Chemistry and bio-
chemistry of plant pigments, 2nd edition, vol. 1, ed T. W. Goodwin, p. 736.
London, New York and San Francisco, Academic Press.
Needham A. E. (1974). The significance of zoochromes, Berlin, Heidelberg and
New York, Springer-Verlag.
Novales R R. (ed ) (1969). Cellular aspects of the control of color changes,
Amer. Zool, 9, 427.
Parker G. H. (1948). Animal color changes and their neurohumors, Cambridge
University Press.
Riley V. (ed ) (1972). Pigmentation: its genesis and control, New York, Apple-
ton-Century-Crofts
Searle A. G (1968). Comparative genetics of coat colour in mammals, New York,
Academic Press.
Глава 9
Пигменты и зрение
9.1. Введение
Первая часть этой книги посвящена биохимии природ-
ных пигментов различных классов, которые придают окраску
содержащим их тканям. Далее (в гл. 8) подчеркивалась важ-
ность свойства быть окрашенным как для выживания индиви-
да, так и для распространения вида. Ясно, что все это имеет
значение лишь в том случае, если окраску и характер ее рас-
пределения могут увидеть и распознать различные животные.
Другими словами, животные должны обладать способностью об-
наруживать свет, а также различать свет разных длин волн.
С этой целью у них развились фоторецепторные органы — гла-
за, в которых центральную роль играют поглощающие свет
фоторецепторы, или зрительные пигменты. В дополнение к соб-
ственно фоторецепторным пигментам часто используются дру-
гие пигменты, играющие вспомогательную роль. В связи с этим
в книге о природных пигментах нельзя не остановиться на
процессах фоторецепции и зрения. И не только потому, что фо-
торецепторные молекулы интересны сами по себе, но также и
потому, что большинство других природных пигментов были бы
не нужны и никогда ие появились, если бы такого механизма
различения цветов не существовало.
9.2. Глаз
Большинство живых организмов способны реагировать
на свет, потому что они обладают какими-либо фоторецептор-
ными клетками, органеллами или молекулами. Но лишь в жи-
вотном царстве эта способность реагировать на свет использу-
ется наиболее эффективно в процессе зрения. Термин «зрение»
означает не просто обнаружение света, но также восприятие
положения, формы и перемещения в пространстве объекта, а во
многих случаях и различение цветов. Для истинного зрения не-
обходим аппарат, с помощью которого в рецепторных клетках
происходило бы формирование истинного изображения; для это-
го и развились фоторецепторные органы, или глаза. Существу-
ют два основных типа глаз; глаза одного типа свойственны поз-
воночным, а другого — некоторым беспозвоночным.
Сосудистая оболочка
Радужная
оболочка
Роговица
.Ресничные
мышцы
Стекловидное
тело
Желтое пятно
Хрусталик /
/ Сетчатка
Центральная ямка
Зрительный нерв
Рис, 9.1. Глаз позвоночных. Схематически показаны структурные особеннос-
ти типичного глаза позвоночных (например, человека).
Стекловидное
тело
Свет из
хрусталика
Ри^. 9.?. Схематическое изображение структуры сетчатки позвоночных (мас-
штаб не соблюдается).
Гл 9 Пигменты и зрение
299
9.2.1. Глаз позвоночных
Глаз позвоночных (рис. 9.1) представляет собой пре-
ломляющий орган с одной большой линзой — хрусталиком. Свет
попадает в глаз через роговицу и радужную оболочку и фоку-
сируется хрусталиком на чувствительной поверхности — сетчат-
ке. Наиболее чувствительная область сетчатки, находящаяся
точно в фокусе, называется центральной ямкой; она может быть
защищена пигментированным экранирующим слоем — желтым
пятном.
Пигментный эпителий/
Наружная пограничная
сосудистая обогочка
мембраны и пигменты
Рис. 9 3. Схематическое изображение фоторецептора позвоночных
(палочки).
Структура фоточувствительной сетчатки более подробно по-
казана на рис. 9.2. Сетчатка содержит ряд плотноупакованных
фоторецепторных клеток; свет достигает их, пройдя через сеть
нервных клеток. Сетчатка включает рецепторные клетки двух
типов — палочки, которые в сетчатке человека имеют размер
~28 мкм в длину и 1,5 мкм в диаметре, и суживающиеся к
концу более короткие колбочки. Некоторые животные имеют
палочки нескольких типов; существуют и двойные колбочки.
Число фоторецепторных клеток в сетчатке огромно. Так, напри-
мер, число палочек в сетчатке глаза крысы оценивается по край-
ней мере в 15 миллионов.
Фоторецепторные молекулы (или зрительные пигменты) ло-
кализованы в наружных сегментах рецепторных клеток (рис.
9.3), соприкасающихся с пигментным эпителием. Последний со-
держит гранулы пигмента, поглощающие рассеянный свет, и у
некоторых видов отражающий слой — зеркальце (tapetum). Пиг-
ментный эпителий непосредственно участвует в регенерации зри-
тельных пигментов (разд. 9.4,3). Оси рецепторных клеток на-
правлены в сторону хрусталика, благодаря чему достигается
300
Часть II. Функциональные аспекты
максимум поглощения света. Зрительные пигменты локализова-
ны в стопках мембранных дисков (разд. 9.2.3), расположенных
перпендикулярно оси клетки.
9.2.2. Глаз беспозвоночных
У беспозвоночных наблюдается большое разнообразие
типов глаз. Их глаза сильно отличаются от глаз позвоночных.
Главные свойства глаз беспозвоночных хорошо выявляются на
примере сложных глаз членистоногих (рис. 9.4). Глаза членис-
Свет
Рис. 9.4. Сложный глаз беспозвоночных. Схема, иллюстрирующая свойства
фасеточного глаза членистоногих.
тоногих состоят из большого числа (до ~ 10 000) крошечных
отдельных фоторецепторных единиц, называемых омматидиями
(или фасетками). Каждый омматидий (рис. 9.5,Л) содержит
собственный хрусталик, фокусирующий часть поля зрения на
связанные с ним рецепторы. Свет проходит через хрусталик в
кристаллический конус, который направляет свет в трубчатые
ретикулярные клетки (клетки сетчатки), формирующие струк-
туру рецептора. В сердцевине омматидия расположенные рядом
концы ретикулярных клеток образуют рецепторную структу-
ру— рабдом — которая содержит зрительный пигмент и явля-
ется, таким образом, аналогом наружного слоя сетчатки поз-
воночных. В одном и том же омматидии могут содержаться
зрительные пигменты, имеющие разные максимумы поглощения
света. Несущие пигмент мембраны формируются не в располо-
Гл. 9. Пигменты и зрение
30 J
женных стопками плоских слоях, как это имеет место в глазах
позвоночных, а в плотноупакованных трубочках, называемых
микроворсинками. Часто микроворсинки присутствуют в виде
двух перпендикулярных друг другу структур, благодаря кото-
рым глаз способен различать по-разному поляризованный свет
(рис. 9.5, Б).
А ’
Хрусталик
Базальная
мембрана
Пучок пигментных клеток
Свет
Б
Кристаллический
конус
Ретикулярные клетки
Микро ворсинки
Пучок
пигментных
клеток
Ретикулярные
клетки
Рис. 9.5. А. Схема, иллюстрирующая главные структурные особенности ом-
матидия. Б. Сечение, показывающее взаимно перпендикулярное расположе-
ние микроворсинок в соседних рабдомах.
Рабдом
Ретикулярные клетки, объединенные в пучки, содержат гра-
нулы экранирующего пигмента. При ярком освещении эти гра-
нулы могут перемещаться в омматидий, ограничивая таким об-
разом количество света, которое может достигать рецепторов.
Длинные тонкие клетки, расположенные по периферии оммати-
дия, содержат дополнительный экранирующий пигмент, кото-
рый предотвращает проникновение отклонившегося света в дру-
гие омматидии.
Глаз головоногих моллюсков. Глаз некоторых головоногих
моллюсков, таких, как осьминог или кальмар, скорее напоми-
нает глаз позвоночных, чем сложные глаза, обычно встречаю-
щиеся у беспозвоночных. В общем он проявляет свойства глаза
того и другого типа, но в действительности представляет собой
рабдомерный глаз особого типа (рис. 9.6). Хотя он, так же как
и глаз позвоночных, имеет один большой хрусталик, формиру-
ющий изображение на рецепторном слое, его рецепторный слой
состоит из рабдомов, которые повернуты «лицевой стороной» к
свету и выстланы нервной тканью.
302
Часть II. Функциональные аспекты
9.2.3. Локализация зрительных пигментов
в рецепторных мембранах
Структура и расположение мембран наружного сегмен-
та палочек (рис. 9.3) и локализация зрительных пигментов
внутри этих структур в настоящее время хорошо известны, хо-
тя в этом отношении были изучены лишь несколько видов жи-
вотных. Гораздо меньше внимания уделялось рецепторным
мембранам колбочек позвоночных и микроворсинкам сложных
Рис. 9.6. Некоторые структурные особенности глаза головоногих моллюсков.
глаз беспозвоночных. Однако, по-видимому, рецепторы всех ти-
пов построены в соответствии с одним основным принципом —
принципом ориентирования молекул зрительных пигментов в
упорядоченной структуре мембраны.
Наружные сегменты палочек сетчатки позвоночных интен-
сивно иследовались с помощью дифракции рентгеновских лучей,
электронной микроскопии и других современных методов. В ре-
зультате было показано, что они содержат стопки мембранных
дисков (рис. 9.7). Эти диски представляют собой структуры, со-
стоящие из двух слоев глобулярного белка (в основном это
зрительный пигмент родопсин) и слоя липидов (главным обра-
зом фосфолипидов) между ними. Родопсин составляет большую
долю (~85°/о) мембранного белка. Молекулы зрительного пиг-
мента ориентированы в рецепторной мембране таким образом,
что поглощение света, проходящего вдоль оси палочки, макси-
мально. Была предложена модель, согласно которой молекулы
зрительного пигмента могут латерально перемещаться в мемб-
ране и вращаться вокруг оси, перпендикулярной поверхности
мембраны, причем любые другие перемещения исключены. По-
Гл. 9. Пигменты и зрение
303
Рис. 9 7. Структура фоторецеп-
торных мембран наружного сег-
мента палочек позвоночных. А.
Локализация мембранных дисков
в виде стопки в наружном сег-
менте палочки. 5. Модель ульт-
раструктуры мембранного диска.
лученные к настоящему времени данные подтверждают эту мо-
дель.
Измерения величин удельного поглощения показали, что кон-
центрации пигментов в палочках, колбочках и мембранах у
большого числа видов животных сходны между собой. Обычно
получали величины порядка 108—109 молекул зрительного пиг-
мента на сегмент или рабдом.
9.3. Зрительные пигменты
Внутри рецепторных мембран находится поглощающий
свет фоточувствительный пигмент, который играет основную
роль в первичном улавливании света. Обычно у животных име-
ется несколько зрительных пигментов (у человека, например,
четыре); причем в палочках и колбочках обнаруживаются раз-
ные пигменты. Каждый индивидуальный зрительный пигмент
характеризуется своей величиной Хтах. Эти величины для раз-
ных зрительных пигментов находятся в диапазоне между 345 и
620 нм, что обеспечивает максимальную чувствительность гла-
за к свету в этом диапазоне. Все известные зрительные пигмен-
ты (а их довольно много) имеют очень сходную структуру. Мо-
лекула любого пигмента представляет собой липопротеин, свя-
занный с небольшим хромофором. Во всем животном царстве
найдены лишь две очень сходные хромофорные группы. Неболь-
шие различия в структуре и конформации липопротеинов (оп-
синов) лежат в основе значительных вариаций величин ?.тах.
304 Часть II. Функциональные аспекты
9.3.1. Хромофор
Хромофорами зрительных пигментов служат ретиналь-
дегид [(9.1), известный также как ретиналь или ретинен] и
3,4-дидегидроретинальдегид [(9.2), использована схема нумера-
ции, принятая для каротиноидов]. Зрительные пигменты, вклю-
чающие ретинальдегидный хромофор, встречаются почти у всех
представителей животного царства, в то время как распростра-
нение 3,4-дидегидроретинальдегида ограничивается рядом прес-
новодных рыб и некоторыми видами земноводных. По этим хро-
мофорам различают два семейства зрительных пигментов — ро-
допсины (содержащие ретинальдегид) и порфиропсины (содер-
жащие дидегидроретинальдегид).
Если сравнить молекулы ретинальдегида и пигмента расте-
ний р-каротина [(9.3), гл. 2], то сразу бросается в глаза их
структурное сходство. Животные не способны синтезировать ре-
тинальдегид de novo и должны образовывать его из р-кароти-
на и родственных ему каротиноидов, получаемых с пищей, или
из витамина А [ретинола (9.4)]. В кишечнике млекопитающих
фермент с оксигеназной активностью расщепляет p-каротин на
две молекулы ретинальдегида (рис. 9.8). Ретинальдегид восста-
навливается до ретинола и сохраняется в печени в виде слож-
ных эфиров (главным образом пальмитинового). Для транспор-
та ретинола в сетчатку глаза используется специфический ли-
попротеин —- ретинолсвязывающий белок. В сетчатке ретинол
может превращаться в ретинальдегид и включаться в состав
зрительных пигментов. Для образования родопсина необходим
«стерически затрудненный» 11-ц«с-изомер ретинальдегида (9.5).
Гл. 9. Пигменты и зрение
305
КИШЕЧНИК
Ретинальдегид
Ретиниловые эфиры
Рис. 9.8. Схема образования ретинальдегида путем расщепления р-каротииаг
ферментом с оксигеназной активностью из кишечника млекопитающих.
9.3.2. Белок
Белок, с которым связывается ретинальдегид, известен
под названием опсина. Получить олеиновый белок, свободный
от липидов и углеводов, довольно трудно. Это обстоятельство,
а также нестабильность очищенного свободного белка затрудня-
ет его изучение. Термин «опсин» иногда применяют ко всему
20—84
306
Часть II Функциональные аспекты
комплексу (белка, липидов и углеводов) в целом, а иногда
лишь к его белковой части.
Все изученные к настоящему времени опсины, которые бы-
ли выделены из сетчатки многих видов животных, представля-
ют собой небольшие белки с мол. массой ~ 30 000—40 000. Для
опсинов, выделенных из палочек некоторых видов животных,
был определен аминокислотный состав (но не последователь-
ность аминокислот). Углеводная часть комплекса, состоящая из
одного (или нескольких) остатка глюкозамина и маннозы, проч-
но связана с аспарагиновым остатком молекулы белка. С бел-
ком ассоциировано также значительное количество липидов,
главным образом фосфатидилхолин и фосфатидилэтаноламин.
Вопрос о том, связаны ли эти фосфолипиды со зрительным пиг-
ментом, составляя часть его молекулы, или они просто являют-
ся загрязнениями, попавшими из липидной области рецепторной
мембраны, остается открытым.
9.3.3. Связывание ретинальдегида с опсином
Почти все исследования по связыванию ретинальдегида
.и опсина были проведены с родопсином, выделенным из пало-
чек сетчатки позвоночных, главным образом на примере круп-
ного рогатого скота. По-видимому, другие зрительные пигмен-
ты в основном сходны с этим родопсином. Необходимо рассмот-
реть два вида связывания — первичное связывание, с помощью
которого ретинальдегид ковалентно присоединяется к функцио-
нальной группе опспна, и вторичное, обусловленное нековалент-
ными взаимодействиями.
Ретинальдегид ковалентно связывается с белком путем обра-
зования шиффова основания (или альдимина) между его аль-
дегидной группой и аминогруппой опсина. По общепринятому
представлению в этом процессе участвует е-аминогруппа лизи-
нового остатка (рис. 9.9). Однако имеются некоторые экспери-
ментальные данные, свидетельствующие в пользу альтернатив-
ной гипотезы, согласно которой шиффово основание, по крайней
мере на некоторых стадиях процесса зрения, может образовы-
ваться при участии аминогруппы этаноламинового остатка фос-
•фатидилэтаноламина.
Чрезвычайно важны также нековалентные взаимодействия
между ретинальдегидом и опсином, особенно взаимодействия
между боковыми группами аминокислот и л-электронной си-
стемой полиена. Подобные взаимодействия определяют конфор-
мацию хромофора, а также вызывают поляризацию л-электрон-
ной системы. Именно эти эффекты и обусловливают незначи-
тельные вариации величины Хтах у разных зрительных пигмен-
тов.
Гл. 9. Пигменты и зрение
307
С '
Nil [
H2N—(СН2)4—СН Опои»
Остаток лизина
Альдимин^или
шиффово основание
Рис. 9.9. Ковалентное связывание 11-ч«с-ретинальдегида с опсином путем
образования шиффова основания (или альдимина) с е-аминогруппой лизино-
вого остатка.
Конформация, принимаемая опсином в результате различ-
ных нековалентных взаимодействий, позволяет связываться с
ним лишь небольшому числу изомеров ретинальдегида и его
аналогов. В природном родопсине с белком связан только
11-г|«с-ретинальдегид (по-видимому, может связываться также
его искаженный 6-5-ч«с-изомер). Результаты, полученные с по-
мощью резонансной рамановской спектроскопии и кругового
дихроизма, свидетельствуют против считавшегося ранее вероят-
ным связывания 12-$-ц«с-изомера (9.6). Полностью тра«с-ре-
тинальдегид не связывается с опсином, и изомеризация связан-
ного 11-цис-ретинальдегида в полностью транс-изомер в процес-
се зрительного цикла (разд. 9.4) приводит к отщеплению рети-
нальдегида от опсина.
9.3.4. Свойства
Стабильность. Хранящиеся в темноте экстракты зри-
тельных пигментов очень стабильны. Опсин, освобожденный из
пигментного комплекса, гораздо менее стабилен и легко разру-
201
'308
Часть II. Функциональные аспекты
шается как in vitro, так и in vivo. Его стабильность восстанав-
ливается после инкубации с 11-г|«с-ретинальдегидом. (При ин-
кубации с 9-ч«с-ретинальдегидом также образуется стабильный,
хотя и не встречающийся в природе комплекс — изородопсин.)
Родопсин стабилен в широком диапазоне pH (~5—10), тогда
(9 6) Изогнутая 6-з-цис, 12-з-4/ис-конформация
11 Qt/c-ретинальдегида
как свободный опсин сохраняет нативность в значительно бо-
лее узком диапазоне величин pH.
Поглощение света. Спектры поглощения света 11-цис-рети-
нальдегида и родопсина из палочек сетчатки быка приведены
на рис. 9.10. Для того чтобы объяснить, каким образом связы-
вание с опсином вызывает столь значительный сдвиг максиму-
ма поглощения света, была проделана огромная работа; сво-
бодный 11-г|«с-ретинальдегид поглощает при ~375 нм, а зри-
тельные пигменты — при значительно больших длинах волн, на-
пример бычий родопсин — при ~500 нм. Этот феномен нельзя
объяснить лишь одним образованием шиффова основания, по-
скольку простые N-ретинилиденальдимины (9.7) поглощают в
диапазоне 360—380 нм. При протонировании альдимина (9.8)
Гл 9 Пигменты и зрение
309
длина волны максимума поглощения увеличивается до 440—
450 нм, но это все еще меньше 500 нм —длины волны, при ко-
торой поглощает нативный родопсин. Сдвиг Zmax дальше в крас-
ную область, вероятно, обусловлен вторичными взаимодействия-
ми между опсином и полиеновой цепью ретинальдегидного хро-
мофора. В настоящее время считается вероятным, что в данном
случае имеет место механизм, согласно которому поляризацию
Рис. 9.10. Спектры поглощения света 11-1{ис-ретинальдегидом (в гексане;
/) и родопсином палочек быка (в водном растворе; II).
хромофора вызывают отрицательный заряд белка при С-13 и
противоион у протонированного атома азота шиффова основа-
ния. Вариации этих взаимодействий главным образом и обус-
ловливают различия максимумов поглощения зрительных пиг-
ментов. 3,4-Дидегидроретннальдегид поглощает при несколько
большей длине волны (393 нм для 11-г|«с-изомера), чем рети-
нальдегид, а порфиропсин с дидегидроретинальдегидом в каче-
стве хромофора поглощает при соответственно больших длинах
волн, чем родопсин, содержащий ретинальдегид.
В УФ-части спектра зрительного пигмента обычно присут-
ствуют также две полосы. у-Полоса с Zmax при 280 нм обуслов-
лена ароматическими аминокислотами (тирозином и триптофа-
ном) белка, в то время как имеющая низкую интенсивность
p-полоса при 330 нм обычно рассматривается как «цис-полоса»,
обусловленная тем, что ретинальдегидные хромофоры имеют
цпс-конфигурацию (ср. «цис-ппкп» каротиноидов; разд. 2.3.3).
Имеются доказательства, что фотохимическая активность связа-
на с p-полосой поглощения.
310 Часть 11. Функциональные аспекты
9.4. Функционирование зрительных пигментов;
циклы превращений
В сетчатке позвоночных палочки ответственны за ско-
топическое, или «ночное», зрение, т. е. улавливание света низ-
кой интенсивности. Это весьма чувствительный процесс, однако
он не включает никакого механизма различения цветов и по-
тому является монохроматическим или почти монохроматичес-
ким. При более высоких интенсивностях освещения имеет мес-
то фотопическое зрение, за которое ответственны колбочки. По-
следние содержат ряд зрительных пигментов (иногда называе-
мых фотопсинами в отличие от скотопсинов палочек), которые
максимально реагируют на свет разных длин волн, благодаря
чему возможно различение цветов. К настоящему времени ис-
следован лишь механизм скотопического зрения в палочках сет-
чатки позвоночных; однако выявленные главные особенности
этого механизма свойственны и фотопическому зрению в кол-
бочках, и зрительному процессу в глазах беспозвоночных.
В связи с этим в данной книге будут подробно освещены функ-
ционирование родопсина и некоторые аспекты цветового зрения.
9.4.1. Обесцвечивание родопсина
Вопрос о том, что происходит, когда фотон света по-
глощается зрительным пигментом родопсином, весьма сложен
и до сих пор до конца не выяснен. Экспериментальные иссле-
дования затруднены тем, что ряд стадий этого процесса проте-
кает за очень короткое время (порядка миллисекунд-пикосе-
кунд). В приведенном ниже описании суммированы некоторые
современные взгляды на функционирование родопсина, однако
по большинству проблем фотохимии нет единой точки зрения.
После достаточно долгого пребывания в темноте сетчатка
адаптируется к ней и молекулы зрительных пигментов находят-
ся в полностью регенерированной форме (см. ниже), т. е. хро-
мофор 11-цис-ретинальдегид связан с белком опсином в специ-
фической в-з-цис, 12-х-тра«с-конформации. Молекулы пигмен-
тов в рецепторной мембране ведут себя как самостоятельные
функциональные единицы. Попадающий в глаз свет падает на
рецепторные клетки сетчатки и поглощается молекулами зри-
тельного пигмента. Затем происходит обесцвечивание пурпурно-
го зрительного пигмента в ходе серии изменений, которую мы
все еще не в состоянии описать на молекулярном уровне. Вме-
сте с тем в общих чертах стадии процесса уже выяснены (рис.
9.11).
Первым событием является поглощение фотона света 11-цис-
ретинальдегидным хромофором родопсина. Это приводит к
Батородопсин
(полностью транс)
Дальнейшие конформационные
изменения
Депротонироваиие
Рис. 9.11. Молекулярные превращения в процессе обесцвечивания родоп-
сина.
312
Часть II. Функциональные аспекты
Родопсин (Х1Пак498нм)
Гипсородопсин (430 нм)
Батородопсин (543 нм; первоначально назывался прелюмироДопсином)
Люмиродопсин (497 нм)
Метародопсин I (478 нм)
Метародопсин II (380 нм)
Метародопсин HI (465 нм) = Парародопсин
транс-К-Ретинилиденолсин (440 365 нм)
Н*
траяс-Ретинальдегид (370 нм) + Опсин
Рис. 9.12. Предполагаемая последовательность промежуточных продуктов
при обесцвечивании родопсина.
электронному возбуждению, за которым следуют конформаци-
онные изменения и изомеризация до полностью трпнс-ретиналь-
дегидной структуры. Последовательные изменения формы рети-
нальдегидного хромофора сопровождаются рядом изменений
конформации опсина. В результате по мере образования пол-
ностью транс-И-ретинилиденопсина специфические взаимодей-
ствия между полиеновой системой и белком постепенно сходят
на нет и в конце концов исчезают. В комплексе белка с хро-
мофором интактной остается только ковалентная непротониро-
ванная альдиминовая связь, которая затем легко и необратимо,
расщепляется с образованием свободного полностью транс-ре-
тинальдегида и опсина.
Многие промежуточные продукты последовательности обес-
цвечивания родопсина были идентифицированы с помощью
спектроскопических методов. Полученные результаты дают
Гл. 9. Пигменты и зрение
313
представление о происходящих превращениях (рис. 9.12). Од-
нако пока не удалось установить соответствия между различ-
ными промежуточными продуктами, выявленными по характер-
ным для них спектральным свойствам, и теми или иными кон-
фигурациями и конформациями комплекса ретинальдегид — оп-
син.
Свет, по-видимому, необходим только для превращения ро-
допсина в батородопсин, вероятно, лишь для образования очень
короткоживущего (3 пс) промежуточного продукта—пребато-
родопсина. В альтернативном пути или в ответвлении главного
пути превращений принимает участие гипсородопсин. Последу-
ющие превращения могут протекать и в темноте. Веские дока-
зательства, полученные с помощью резонансной рамановской
спектроскопии и кругового дихроизма, указывают на то, что изо-
меризация 11-цис-хромофора в полностью транс-форму происхо-
дит в ходе фотоконверсии родопсина в батородопсин, вероятно,
через стадию пребатородопсина (достаточно четко установлено
наличие в батородопсине изогнутого 11-транс-хромофора). В по-
следующих продуктах — люмиродопсине и метародопсине — ре-
тинальдегидный хромофор, несомненно, содержится уже в пол-
ностью транс-форме, однако взаимодействия между полиеновой
цепью и опсином еще в значительной степени сохраняются. Ме-
тародопсин I и все предшествующие промежуточные продукты
являются, по-видимому, протонированными альдиминами. Де-
протонирование, сопровождающееся гипсохромным сдвигом,
происходит на стадии превращения метародопсина I в метаро-
допсин II. На этой же стадии происходит высвобождение ней-
ромедиатора, инициируемое изменением конформации белка.
Неожиданным и необъяснимым является батохромный сдвиг,
происходящий при образовании парародопсина (или метаро-
допсина III) из метародопсина II на пути N-ретинилиденопсину.
Совершенно очевидно, что для выяснения процесса обес-
цвечивания родопсина на молекулярном уровне необходимо за-
тратить еще немалые усилия.
9.4.2. Нервный импульс
Поглощение света молекулой зрительного пигмента,
приводящее к электронному возбуждению и последующим пре-
вращениям (результатом которых является обесцвечивание пиг-
мента), тесно связано с возникновением нервного импульса,
передаваемого в мозг. Исходное поглощение фотона света про-
исходит практически мгновенно, в то время как последующие
превращения молекул протекают в течение более длительного
времени. Известно, что нервные импульсы возникают через
несколько миллисекунд после поглощения света, и в настоящее
314
Часть II. Функциональные аспекты
время уже ясно, что генерация импульса происходит при пере-
ходе метародопсина I в метародопсин II. Мы не в состоянии
пока ответить на вопросы, каким образом фотовозбуждение
единственной молекулы пигмента может вызвать прохождение
тока через синапс на отдаленном конце рецепторной клетки и
как благодаря ничтожно малой энергии всего лишь одного фо-
тона могут осуществляться значительные перемещения заряда,
необходимые для генерации нервного импульса. Считают, что
молекулы пигментов каким-то образом контролируют ионные
каналы через внутренние мембраны рецепторных клеток. Фо-
товозбуждение молекулы пигмента вызывает изменение ее кон-
фигурации или заряда. В результате этого открывается канал
и возникает поток ионов, который вызывает поляризацию клет-
ки и таким образом стимулирует синапс. Затем ионные насосы
должны получить энергию для восстановления нормального рас-
пределения ионов. Такой механизм может объяснить необходи-
мое усиление и быстроту реакции на относительно большом
расстоянии.
9.4.3. Регенерация родопсина
Процесс фотообесцвечивания приводит к расщеплению
родопсина на составляющие компоненты — опсин и ретинальде-
гид. Ретинальдегид, высвободившийся в виде полностью транс-
изомера, прежде чем вновь связаться с опсином, должен превра-
титься в 11-циоформу. Однако такое превращение не простая
изомеризация. Ряд возможных реакций, протекающих при этом,
суммирован на рис. 9.13, причем все они проверены в экспери-
менте.
Ферментативное восстановление тдшноретинальдегида до
траноретинола, катализируемое алкогольдегидрогеназой, про-
исходит в рецепторной мембране, и для него необходим NADH.
Ретинол транспортируется из рецепторной клетки в пигмент-
ный эпителий, где он может этерифицироваться жирными кис-
лотами, главным образом пальмитиновой и стеариновой. Рети-
нол и ретиниловые эфиры, образовавшиеся из зрительных пиг-
ментов вместе с запасными, сохраняющимися в пигментном эпи-
телии, образуют внутриклеточный фонд. Изомеризация ретино-
ла, ретинальдегида и ретиниловых эфиров до цис-11-формы
продемонстрирована в эксперименте. Расщепление циоретини-
ловых эфиров (где происходит этот процесс, пока не известно)
дает 11-циоретинол, который может быть снова окислен в ре-
цепторе до альдегида. Полагают, что рекомбинация 11-цмс-ре-
тинальдегида с опсином происходит спонтанно.
Разные виды животных могут использовать различные пути
регенерации родопсина. Так, у крыс наибольшее значение име-
Гл. 9. Пигменты и зрение
315
ет прямая реизомеризация полностью транс-ретинальдегида
в 11-ццс-ретинальдегид, в то время как у крупного рогатого
скота главным процессом считается более окружной путь —
через ретинол и ретиниловые эфиры.
Регенерация родопсина через ретинол и ретиниловые эфиры
протекает довольно медленно по сравнению с процессом фото-
обесцвечивания; она может занимать несколько минут. Поэтому
данным путем родопсин способен синтезироваться в количест-
Полностью транс ретинальдегид ч • - 11 цмс ретинальдегид
Полностью транс ретинол
1Т цис ретинол
Эфиры полностью транс ретинола
Эфиры Т Т цис ретинола
Рис. 9.13. Возможные пути превращения полностью грамс-ретинальдегида в
11-цыс-ретинальдегид в процессе регенерации родопсина.
вах, достаточных для осуществления зрительного цикла лишь
при низких интенсивностях света. При дневном свете, когда
интенсивность освещения высока, родопсин будет оставаться
практически полностью обесцвеченным, так как скорость его
регенерации значительно ниже, чем скорость обесцвечивания.
В отличие от этого фотопсины колбочек, которыми опосредова-
но дневное цветовое зрение, должны регенерироваться очень
быстро, поскольку они постоянно функционируют при высоких
интенсивностях света. Предполагают, что в данном случае дей-
ствует механизм фоторегенерации, в котором изомеризация
полностью транс-ретинальдегида катализируется светом.
Путь регенерации родопсина через фонды ретинола и рети-
ниловых эфиров в пигментном эпителии обеспечивает эффектив-
ное обновление ретинальдегидной части всех зрительных пиг-
ментов палочек и колбочек. Потери ретинальдегида возмещают-
ся из запасов в пигментном эпителии. Эти запасы в свою оче-
редь по мере надобности восполняются путем транспорта рети-
316
Часть II. Функциональные аспекты
нола и его эфиров из обширного фонда, сохраняющегося в пе-
чени. В экстремальных случаях недостатка витамина А (рети-
нола) запасы в печени истощаются, и, хотя снабжение сетчат-
ки поддерживается до последней возможности, в конце концов
может образоваться его дефицит. В результате организму не
удается восполнять потери ретинальдегида и, следовательно,
регенерировать зрительный пигмент. Наступает слепота, внача-
ле обратимая, но при длительном дефиците ретинола переходя-
щая в постоянную из-за денатурации опсина, нестабильного в
свободном состоянии. Существует мнение, что морковь помога-
ет видеть «в темноте» (т. е. при низких интенсивностях света);
по-видимому, оно небезосновательно, поскольку морковь слу-
жит непревзойденным источником провитамина А-—(3-кароти-
на и таким образом помогает предотвратить недостаточность
витамина А и вызываемую этой недостаточностью слепоту.
Помимо родопсина были исследованы также синтез и обмен
других компонентов рецепторной мембраны, а именно белка
(опсина), фосфолипидов и углеводов.
9.4.4. Взаимопревращения ретинола
и 3,4-дидегидроретинола
У некоторых видов земноводных и рыб ретинол может
превращаться в 3,4-дидегидроретинол (или наоборот), в резуль-
тате чего пигмент в рецепторе изменяется с родопсина на пор-
фиропсин. Опсин в рецепторе может соединяться с 11-цпс-изо-
мером как ретинальдегида, так и 3,4-дидегидроретинальдегида,
и, следовательно, тип образующегося зрительного пигмента за-
висит от того, какой хромофор присоединен к опсину. По-види-
мому, должны существовать механизмы, с помощью которых
пигментный эпителий может осуществлять взаимопревращения
этих двух ретинолов, а также выбирать «правильный» ретинол,
необходимый в данном конкретном случае.
У других видов, в частности у пресноводных рыб, ретинол и
3,4-дидегидроретинол, по-видимому, образуются из разных ка-
ротиноидов. Например, предшественником ретинола служит р-
каротин, в то время как 3,4-дидегидроретинол образуется из
ксантофиллов, таких, как лютеин (9.9) и, возможно, астаксан-
тин (9.10). Механизм этих превращений, и в первую очередь
удаления кислородсодержащих заместителей в кольце, пока не
установлен.
Замена синтеза порфиропсина на синтез родопсина (или на-
оборот) часто связана со стадиями развития животного. На-
пример, взрослые амфибии обычно содержат исключительно
(или преимущественно) родопсины, в то время как головастики
используют порфиропсины. В других случаях, например у рыб.
Гл. 9. Пигменты и зрение
317
такие изменения могут иметь сезонный характер и зависеть от
уровня освещенности или же они могут происходить в ответ на
переход из соленой морской воды в пресноводную среду обита-
ния.
9.4.5. Циклы превращений зрительных пигментов
у беспозвоночных
Было показано, что зрительные пигменты многих бес-
позвоночных, главным образом ракообразных и головоногих
моллюсков, сходны со зрительными пигментами позвоночных и
(9.10) Астаксантин
принадлежат к родопсиновому типу. В принципе те и другие
имеют также сходные циклы превращений, однако в деталях
последние различаются. Так, у беспозвоночных превращения,,
вызываемые освещением, не приводят к высвобождению ретин-
альдегида и опсина, а заканчиваются на стадии метародопси-
на, в котором траноретинальдегид остается присоединенным к
опсину. Метародопсин может находиться в протонированной
(кислой, Хтах—500 нм) или в непротонированной (щелочной,.
Агпах^380 нм) форме. Максимумы поглощения этих метародоп-
синов, например у кальмара, находятся при больших длинах
волн, чем у соответствующих N-ретинилиденопсинов позвоноч-
ных, что указывает на сохранение взаимодействия между хро-
мофорной группой и белком. Исследования с помощью круго-
вого дихроизма показали, что белковые молекулы метародоп-
сина и исходного родопсина почти не различаются по степени
спирализации. Однако родопсин в отличие от метародопсина
проявляет индуцированный круговой дихроизм, что указывает
на изогнутость углеводородной цепи ретинальдегидного хромо-
фора.
3 i 8 Часть II. Функциональные аспекты
Даже в том случае, когда в глазу содержатся несколько
зрительных пигментов с различными величинами %тах, метаро-
допсины, образующиеся из разных форм родопсинов, имеют
очень сходные максимумы поглощения. По-видимому, в ходе
образования метародопсина вторичные опсин-хромофорные
взаимодействия, которые обусловливают различные светопогло-
щающие свойства зрительных пигментов, исчезают.
Результаты спектроскопических исследований, проведенных
на глазах и на сетчатке многих видов беспозвоночных, позво-
ляют предположить, что существует очень много механизмов
фотообесцвечивания, которые пока не известны.
Фоторегенерация. У беспозвоночных регенерация зритель-
ного пигмента из метародопсина в нормальных условиях осу-
дцествляется с помощью механизма, принципиально отличаю-
щегося от соответствующего механизма позвоночных. Относи-
тельно стабильный метародопсин превращается непосредствен-
но в исходный пигмент под действием света. Фоторегенерации
подвергается как кислая, так и щелочная формы метародопси-
на. Промежуточный продукт процесса регенерации, названный
«Рз8о», был обнаружен у некоторых видов кальмара и осьмино-
га. Полагают, что этот продукт образуется, когда под действием
•света транс-ретинальдегидный хромофор кислого метародопсп-
на изомеризуется в 11-цис-форму. Затем в темноте быстро про-
исходят небольшие конформационные изменения, которые необ-
ходимы для завершения вторичных хромофор-белковых взаимо-
.действий родопсина.
В некоторых случаях регенерация может осуществляться и
биохимическим путем, однако в нормальных условиях гораздо
большее значение имеет фотоиндуцируемый процесс. По-види-
мому, у некоторых насекомых уровень зрительных пигментов
.днем контролируется фоторегенерацией, а ночью для этой цели
используется биохимическая регенерация.
Другой путь был описан для фоторегенерации родопсинов в
глазах некоторых головоногих. В качестве фотокатализатора у
них in vivo служит, по-видимому, ретинохром — второй фото-
чувствительный пигмент, который локализован в слое, подсти-
лающем рецепторы. Идентичность фотокатализатора и ретино-
хрома и механизм функционирования последнего пока не уста-
новлены.
‘9.5. Некоторые аспекты цветового зрения
Родопсин в палочках сетчатки представляет собой пиг-
мент, обусловливающий зрение при низких интенсивностях све-
та. Его максимум поглощения света находится примерно при
•500 нм, но с его помощью глаз способен лишь обнаруживать
Гл. 9. Пигменты и зрение
319*
свет, но не способен различать цвета. У большинства позвоноч-
ных, в том числе и у человека, за цветовое зрение ответствен-
ны другие рецепторные клетки сетчатки — колбочки, и в них
для этой цели используется другая группа зрительных пигмен-
тов — фотопсины.
9.5.1. Цветовое зрение у человека
По вполне понятным причинам наиболее подробно
изучено цветовое зрение у человека. В данном случае это три-
хроматический процесс, за который ответственны рецепторы
трех цветов, чувствительные к разным частям видимого спект-
ра. Эти цветовые рецепторы (колбочки) наиболее многочислен-
ны в сетчатке, в области центральной ямки, которая в связи с
этим наиболее цветочувствительна. Каждый из трех различных
колбочковых рецепторов содержит свой зрительный пигмент,
который и определяет его спектральную чувствительность. У че-
ловека эти три пигмента имеют значения А,тах при 440, 535 и
575 нм и, следовательно, чувствительны соответственно к сине-
му, зеленому и красному свету. Различные формы цветовой
слепоты у человека обычно обусловлены отсутствием одного или
нескольких из этих рецепторных пигментов, поскольку человек
теряет способность реагировать на свет, который поглощается
этим пигментом. Например, человек, лишенный пигмента с
Хтах = 575 нм (поглощающего красные лучи), видит только си-
ние и зеленые цвета и не чувствителен к свету более длинных
волн.
При низких интенсивностях света (например, при свете лу-
ны) человек не различает цвета, поскольку чувствительность,
колбочек составляет лишь около одной тысячной светочувстви-
тельности палочек. Однако концентрация пигментов в колбоч-
ках и палочках почти одинакова, и фоточувствительность изо-
лированного пигмента колбочек, по-видимому, не ниже чувст-
вительности родопсина палочек.
В структурном отношении пигменты колбочек сходны с ро-
допсином палочек и представляют собой комплексы 11-цис-
ретинальдегидного хромофора с опсином. Различия в их мак-
симумах поглощения света обусловлены различиями в конфор-
мации опсина и во вторичном связывании хромофора. Пигмен-
ты колбочек выделить гораздо труднее, чем родопсин палочек,
и потому детали их структуры и промежуточные продукты соот-
ветствующих циклов превращения выяснены менее подробно.
Полагают, что механизмы фотообесцвечивания, регенерации;
пигмента и формирования нервного импульса, которые имеют
место при колбочковом зрении, в принципе сходны с соответ-
320
Часть 11. Функциональные аспекты
ствующими механизмами в фоторецепторах палочек, однако
пока еще мало экспериментальных данных, подтверждающих
это мнение.
'9.5.2. Цветовое зрение у животных
Отнюдь не все виды животных обладают механизмами,
позволяющими различать цвета, и имеют колбочковые фоторе-
цепторные пигменты; считается, что многие млекопитающие, в
том числе крупный рогатый скот и овцы, цветовым зрением не
обладают. Вместе с тем многие позвоночные и беспозвоночные
способны видеть и различать цвета. Они используют ряд фото-
рецепторов, максимально чувствительных к свету разных длин
волн, и, по-видимому, обладают трихроматической системой,
подобной той, которая присуща человеческому глазу. Так, зо-
лотая рыбка (Carassius auratus) имеет колбочки трех типов и
три зрительных пигмента с величинами Хтах при 455, 530
и 625 нм соответственно. У нее, как и у многих пресноводных
рыб, пигменты представлены порфиропсинами, т. е. имеют в ка-
честве хромофора 3,4-дидегидроретинальдегид.
У многих земноводных ситуация намного сложнее. Лягушки,
как правило, имеют палочки двух типов, называемые «красны-
ми» и «зелеными» палочками, которые поглощают соответст-
венно зеленые и синие лучи. Кроме того, они имеют колбочки
разных типов, в том числе двойные колбочки, которые содер-
жат два или три разных пигмента, чувствительные в разных
участках спектра. У взрослых амфибий это родопсин с 11-цис-
ретинальдегидом в качестве хромофора; у головастиков пигмен-
ты очень сходны с пигментами взрослых особей, но содержат
11-цнс-3,4-дидегидроретинальдегид (порфиропсины).
Цветовым зрением обладают и некоторые беспозвоночные
животные. У пчел, например, имеются четыре пигмента с макси-
мумами поглощения в диапазонах 300—340, 400—480, 480—500
и 500—650 нм. Пигмент с максимальной чувствительностью
в диапазоне 300—340 нм позволяет насекомым видеть длинно-
волновые УФ-лучи как особый цвет. К сожалению, подробного
изучения биохимии цветового зрения у насекомых не проводи-
лось.
В прошлом для зрительных пигментов, которые были либо
выделены из животных различных видов, либо просто обнару-
жены спектроскопически, часто использовались тривиальные
названия. Так, пигменты, имевшие желтую, синюю или фиоле-
товую окраску, называли соответственно хризопсином, циано-
псином и йодопсином, а термин родопсин применяли для крас-
ных и пурпурных пигментов независимо от их происхождения.
Эта терминология неудобна, поскольку при ее использовании
Гл. 9. Пигменты и зрение
321
нельзя получить представления ни о структуре, ни о возмож-
ных взаимосвязях этих пигментов, выделенных из разных видов
животных.
9.6. Вспомогательные пигменты в процессе зрения
9.6.1. Позвоночные
Меланины. Меланины содержатся в различных тканях
глаза позвоночных. Сосудистая оболочка и пигментный эпите-
лий, расположенные на задней стороне глаза, содержат мела-
ниновые гранулы, использующиеся в качестве экранирующих
пигментов, которые поглощают отклонившийся свет всех длин
волн. Меланин, выделенный из сосудистой оболочки и пигмент-
ного эпителия некоторых млекопитающих, представляет собой
индольный эумеланин (гл. 7), связанный с белком.
У многих млекопитающих, в том числе и у человека, мела-
нины содержатся также в задней части радужной оболочки и
формируют экран, который не позволяет видеть красный цвет
крови в капиллярах. Этот красный цвет можно видеть в глазах
животных-альбиносов, которые лишены меланинового слоя. Ка-
рие и желтые глаза окрашены меланиновыми гранулами стромы
радужной оболочки, в то время как голубой цвет глаз у челове-
ка и у некоторых других животных обусловлен рассеянием све-
та мельчайшими частицами белка или меланина в радужной
оболочке. Меланин, содержащийся в радужной оболочке, в
процессе зрения, вероятно, роли не играет.
Зеркальный слой (tapetum lucidum). В сетчатке некоторых
животных, которые ведут преимущественно дневной образ
жизни, но хорошо видят и в сумерках, под рецепторным слоем
лежит отражающий слой. Этот слой у одних видов находится
перед слоем пигментного эпителия, у других за ним, а у треть-
их является его частью. Функции такого отражающего слоя,
или «зеркальца», заключаются в возвращении света, который
не был поглощен фоторецептором при первом прохождении че-
рез сетчатку. Благодаря этому повышается чувствительность
глаза. Зеркальный слой обычно состоит из упорядоченных
кристаллов, которые действуют как 01ражатели и обусловлива-
ют сияние глаз. Обычно кристаллы состоят из птеридинов и
пуринов, таких, как гуанин [(9.11), гл. 6]. У некоторых живот-
ных, например, у кошки, лемура, галаго, зеркальный слой име-
ет желтый цвет, поскольку он состоит из кристаллов рибофла-
вина (9.12). Это позволяет повысить чувствительность глаза к
голубому цвету, так как рибофлавин поглощает синий и УФ-
свет с длиной волны 450 нм и ниже и испускает его в виде
21—84
322
Часть !!. Функциональные аспекты
флуоресценции при 520 нм. Свет с длиной волны 520 нм более
эффективно поглощается родопсином в рецепторах.
Желтое пятно (macula lutea). У человека и у других прима
тов, ведущих дневной образ жизни, в центре сетчатки есть об-
ласть высокой остроты зрения — область центральной ямки. Она
в минимальной степени покрыта нервной тканью, а от стекло-
видного тела отделена желтым фильтрующим слоем, называе-
мым «желтым пятном». По-видимому, его функцией является
поглощение некоторой части синего света с длиной волны
450 нм. При этом селективно снижается чувствительность кол-
бочковых рецепторов к синему свету, что способствует умень-
(9 12) Рибофлавин
шению размывания изображения из-за коротковолновой хрома
тической аберрации хрусталика. На основании спектра погло-
щения можно предположить, что пигмент желтого пятна чело-
века является каротиноидом, однако точно он пока не охарак-
теризован.
Светофильтры из масляных капель. Прежде чем свет дости-
гает фоторецепторной мембраны в сетчатке глаза, он должен
пройти через внутренний сегмент рецепторной клетки. У неко-
торых пресмыкающихся и птиц эти сегменты содержат окра-
шенные масляные капли. Диаметр капель обычно такой же,
как у фоточувствительных наружных сегментов, так что весь
свет, прежде чем достигнуть зрительного пигмента, должен
пройти через них. У цыплят в сетчатке содержатся рецепторные
клетки шести типов, и масляные капли присутствуют в кол-
бочках пяти типов. Идентифицированы красные, оранжево-
желтые, лимонно-желтые и даже бесцветные, но поглощающие
в УФ-свете масляные капли. Каждому из морфологических
типов колбочек присущи свои капли.
Было обнаружено, что окрашенные масляные капли содер-
жат в чрезвычайно высоких (почти 1 М) концентрациях сво-
бодные каротиноиды. У индейки в красных, желтых и почти
бесцветных каплях идентифицированы соответственно астаксан-
тин (9.10), лютеин (9.9) и галлоксантин — С27-апо-каротиноид
[Ю'-апо-р-каротин-ЗДО'-диол (9.13)], по-видимому являющийся
производным лютеина или зеаксантина (9.14).
Гл. 9. Пигменты и зрение
323
Интересно отметить, что каротин желтых капель с Хтах =
= 440 нм, который ранее называли «сарциненом», оказался очень
редким (63, 6zS)-e, е-каротином (9.15). Происхождение астак-
сантина, и особенно данной изомерной формы е-каротина, ос-
тается загадкой, поскольку эти пигменты, по всей вероятности,
не содержатся в пище птиц (разд. 2.8.1).
(9 15) (6S 6 S) с, с К тотин
Каким бы ни было происхождение этих пигментов, ясно,
что масляные капли в сетчатке глаза птиц улучшают различе-
ние цветов при цветовом зрении. Различным образом окрашен-
ные капли поглощают свет разных длин волн, так что рецептор-
ной мембраны достигает тоже свет разных длин волн, где он и
поглощается зрительным пигментом. Благодаря этому обеспе-
чивается механизм различения цветов, который позволяет силь-
но сузить диапазон длин волн света, достигающего рецептора,
п надежно разделить диапазоны чувствительности разных рецеп-
торных клеток.
Сходные окрашенные масляные капли содержатся в сет-
чатке глаз некоторых рептилий. Пигменты таких капель у раз-
личных видов черепах имеют спектры, типичные для каротино-
идов, однако с химической точки зрения они пока не охаракте-
ризованы. Считают, что в данном случае функционирует меха-
низм различения цветов, сходный с механизмом, существующим
У птиц.
211
324
Часть 11. Функциональные аспекты
9.6.2. Экранирующие пигменты глаз беспозвоночных
При схематическом описании сложных глаз беспозво-
ночных и омматидия (разд. 9.2.2) мы уже обращали внимание
на присутствие в наружных зонах рецепторных единиц экрани-
рующих пигментов. Неподвижные пигментированные области
клеток, расположенных по периферии омматидия, образуют
экран, который эффективно изолирует омматидий и не допуска-
ет проникновения рассеянного света в соседние омматидии. Ре-
цепторные мембраны стимулирует только свет, направленный
(9 16) Ксантомматин
(9 17) Дрозолтерин
вдоль оси омматидия; весь остальной свет поглощается экрани-
рующими пигментами. Это обеспечивает механизм, необходи-
мый для очень точного распознавания внешнего вида и переме-
щения объектов. Другие экранирующие клетки содержат пиг-
ментные гранулы, которые па ярком свету могут рассредоточи-
ваться и таким образом снижать интенсивность света, достига-
ющего глубоко расположенных рецепторных пигментов. На
слабом свету гранулы экранирующих пигментов агрегируют и
располагаются так, что фоторецепторы могут стимулироваться
светом, падающим практически под любым углом. При этом
сильно возрастает чувствительность к свету, однако очертания
п перемещение объекта не могут более различаться с достаточ-
ной точностью.
Экранирующими пигментами в гранулах служат меланины
(гл. 7) или, в частности у членистоногих, оммохромы [напри-
мер, ксантомматин (9.16), гл. 6] и птерины [например, дрозо-
птерин (9.17), гл. 6]. Птерины и оммохромы имеют характер-
ные максимумы поглощения в видимом диапазоне н в какой-то
степени играют роль цветных фильтров. Различия в окраске
глаз (т. е. в оммохромовых и птериновых экранирующих пиг-
Гл. 9. Пигменты и зрение
325
ментах) у мутантных штаммов Drosophila лежат в основе мно-
гих фундаментальных генетических исследований, которые бы-
ли проведены с этой мушкой.
9.7. Заключение и комментарии
В животном царстве встречается много разных струк-
тур глаза, однако, насколько это известно, всем им присущи
одни и те же механизмы зрения и сходные ретинальальдегид-
белковые пигменты. Ряд усовершенствований основного меха-
низма (например, наличие пигментов с различными величинами
Хтах в качестве цветных фильтров) обеспечивает оптимальную эф-
фективность зрения при слабом свете или различение цветов при
обычном освещении, что необходимо животному в природных
условиях обитания (например, на земле или в воде). Вызывает
удивление поразительное сходство зрительных пигментов и
циклов их превращения. Ретинальдегид-опсиновый комплекс
представляется идеальным для целей улавливания света. Пред-
полагают, что системы зрительных пигментов возникали совер-
шенно независимо друг от друга по крайней мере трижды в хо-
де эволюции животного мира.
Поскольку современные биохимические и физико-химические
методы развиваются очень быстро, можно ожидать, что вскоре
накопится обширная информация о циклах превращений зри-
тельных пигментов и их промежуточных продуктах, а также об
опсин-хромофорных взаимодействиях, особенно для родопсина
палочек. Пройдет, однако, еще немало времени, прежде чем
станут известны все детали структуры некоторых короткоживу-
щих промежуточных продуктов, что позволит оценить значение
небольших изменений конформации, взаимодействий белок-—
хромофор и особенностей поглощения света. Следует также вы-
яснить механизм генерации нервного импульса в ответ на погло-
щение фотона зрительным пигментом. Даже после того как мы
ответим на некоторые вопросы о функционировании родопсина
у тех немногих видов, которые наиболее подробно изучены
(человек, крыса, крупный рогатый скот), предстоит огромная
работа по изучению биохимии цветового зрения у млекопитаю-
щих, а также зрительных пигментов и циклов их превращений
у других животных.
9.8. Рекомендуемая литература
Зрение чрезвычайно важно для эффективного функцио-
нирования практически всех животных, в том числе и человека.
Поэтому процесс зрения довольно подробно описан в учебни-
ках по биохимии и физиологии. Число обзорных статей, посвя-
22—84
326
Часть II. Функциональные аспекты
щенных широкому набору тем, связанных со зрением, огромно.
Для исчерпывающего ознакомления со всеми аспектами зре-
ния читателю следует рекомендовать серию томов, озаглавлен-
ную «Глаз», под редакцией Дэвсона (Davson, 1977). В частно-
сти, для биохимиков наиболее полезен один из томов — «Фо-
тобиология зрения» (Knowles, Dartnall, 1977), который содер-
жит обширный список ссылок на более ранние работы. В моле-
кулярной биологии и биофизике зрения идеи быстро меняются,
и нет ничего необычного в том, что здесь существует много про-
тиворечивых мнений. Различные теории, касающиеся про-
цесса зрения, подробно обсуждаются в обзорной статье Отто-
лени (Ottolenghi, 1980).
9.9. Дополнительная литература
Davson Н. (ed.) (1977). The eye, 2nd edition, 5 volumes, New York, London
and San Francisco, Academic Press.
Knowles A., Dartnail H. J. A. (1977). The photobiology of vision (The eye, 2nd
edition, ed. H. Davson, vol. 2B), New York, London and San Francisco,
Academic Press.
Ottolenghi M. (1980). The photochemistry of rhodopsins, Adv. Photochem.,
12, 97.
Глава 10
Фотосинтез
10.1. Введение
Фотосинтез — это процесс, с помощью которого зеле-
ные растения, водоросли и некоторые бактерии способны пере-
водить энергию солнечного света в химическую форму, доступ-
ную для биосинтеза клеточных компонентов. Полученная таким
путем химическая энергия используется и запасается растения-
ми и, кроме того, служит (через пищевые цепи) первичным ис-
точником энергии для нефотосинтезирующих организмов, в
первую очередь для животных. Фотосинтез растений является
также источником необходимого для жизни кислорода. Поэто-
му вся жизнь на нашей планете зависит от фотосинтеза.
В фотосинтезе высших растений и водорослей (рис. 10.1)
энергия света поглощается и используется для расщепления
молекул воды. Этот простой процесс (световая реакция) при-
водит к выделению кислорода и к образованию восстановитель-
ных эквивалентов, которые затем используются в последова-
тельности темновых реакций для фиксации двуокиси углерода
в доступной форме углеводов. Углеводы могут утилизироваться
как энергетические запасы или как источник углерода для
синтеза всех других молекул, в которых нуждается растение.
В ходе фотосинтеза происходит образование АТР по сопря-
женному механизму фотофосфорилирования.
Как можно видеть из рис. 10.1, в расщеплении двух моле-
кул воды участвуют четыре кванта (фотона) света; при этом
выделяется одна молекула кислорода и образуются четыре вос-
становительных эквивалента. Еще четыре кванта обеспечивают
энергией перенос этих четырех восстановительных эквивалентов
с образованием в конечном итоге NADPH, который и использу-
ется вместе с АТР для восстановления молекулы СО2 и по-
следующего образования углеводов. Таким образом, для вос-
становления одной молекулы СО2 и выделения одной молекулы
О2 необходимо минимум восемь квантов (фотонов).
Некоторые зеленые и пурпурные бактерии, которые в нор-
ме живут в анаэробных условиях, например в иле и в стоячей
воде, также фототрофны и способны восстанавливать СО2 до
углеводов, правда, в ходе фотосинтеза О2 у них не выделяет-
ся. Эти бактерии не способны использовать энергию света для
22*
328
Часть II. Функциональные аспекты
расщепления воды; они используют другие доноры водорода
(электрона) Н2А (рис. 10.2). Три главные группы фотосинте-
зирующих бактерий используют разные доноры электрона при
фотосинтезе. Пурпурные несерные бактерии Rhodospirillaceae
(прежде называвшиеся Athiorhodaceae), например Rhodospi-
rillurn rubrum, в нормальных условиях утилизируют простые
А 2Н2О———О2+4Н++4е~
4Л v
Б 4Н++ 4e“+2NADP+-----2NADPH
В 2NADPH+CO2 +2Н+ АТР » [СН2О] +2NADP++H2O
Рис. 10.1. Суммарный процесс фотосинтеза у растений. А, Б. Стадии, в ко-
торых используется энергия света, или световые реакции. В. Последующие
темновые реакции фиксации углерода.
органические молекулы, в то время как зеленые серные бакте-
рии (Chlorobiaceae, прежде называвшиеся Chlorobacteriaceae)
например Chlorobium spp., как правило, используют неоргани-
ческие серусодержащие соединения (например, H2S) или водо-
род. Пурпурные серные бактерии, принадлежащие к Chromatia-
сеае (прежде называвшиеся Thiorhodaceae), например Chrorna-
tium spp., могут утилизировать как органические молекулы,
так и неорганические серусодержащие соединения.
hv
СО2 + 2Н2А ---> [СН2О] + 2А + Н2О.
Рис. 10.2. Суммарный процесс бактериального фотосинтеза.
Детали синтеза углеводов и механизмов фотофосфорилиро-
вания лежат за пределами настоящей книги. Однако мы оста-
новимся здесь на роли в этих процессах пигментов, поскольку
они имеют фундаментальное значение в улавливании и утили-
зации энергии света. Светособирающая роль хлорофилла в фо-
тосинтезе— вероятно, наиболее яркий пример специфических
биологических фотофункций природного пигмента. Функциони-
рование каротиноидов и фикобилинов в качестве вспомогатель-
ных пигментов также прямо связано с их светопоглощающими
свойствами. Другие окрашенные молекулы, в том числе цито-
хромы и флавопротеины, участвуют в фотосинтезе как часть
электронтранспортных систем; способность этих соединений
поглощать видимый свет не имеет отношения к их функциони-
рованию. Ниже будут освещены вопросы о том, как поглощаю-
щие свет пигменты расположены в фотосинтетическом аппара-
Гл. 10. Фотосинтез
329
те высших растений, водорослей и фотосинтезирующих бактерий
и как эти пигменты функционируют в процессе утилизации
энергии света.
10.2. Фотосинтетический аппарат эукариот: хлоропласты
10.2.1. Морфология хлоропластов
В фотосинтезирующих клетках активные пигменты рас-
положены внутри ламеллярных мембран в виде функциональ-
но-организованных единиц. У фотосинтезирующих эукариот
(высших растений и большинства водорослей) несущие пигмент
мембраны заключены в специфических органеллах — хлоропла-
стах. У высших растений морфологические различия между
хлоропластами невелики, в то время как у водорослей форма и
размеры хлоропластов значительно варьируют. Chlorella, на-
пример, имеет единственный чашевидный хлоропласт, тогда
как хлоропласты некоторых видов Spirogyra представляют со-
бой длинные, спирально закрученные образования, лежащие
вдоль всей клетки.
Обобщенная структура хлоропласта высших растений пока-
зана на рис. 10.3,Л. Клетки листьев растений могут содержать
несколько сотен таких хлоропластов — эллиптической или лин-
зообразной формы — длиной около 3—10 мкм. Хлоропласт
состоит из двойной наружной мембраны (или оболочки), заклю-
чающей в себе матрикс — строму, которая содержит внутрен-
ние фотосинтетические мебраны.
10.2.2. Оболочка хлоропласта
Оболочка хлоропласта представляет собой непрерыв-
ную двойную мембрану, которая функционирует как селектив-
ный барьер при транспорте метаболитов внутрь органеллы или
из нее. Полагают, что внутренняя мембрана может играть
некоторую роль в формировании новых внутренних ламелл.
У некоторых видов растений к внутренней мембране оболочки
хлоропласта прилегает протяженная система трубочек и пу-
зырьков. Эта система, известная как периферический ретикулум,
по-видимому, характерна для растений, обладающих Сгпутем
фиксации углерода (разд. 10.6), но иногда она обнаруживается
и у некоторых С3-растений, в частности в стрессовых условиях
окружающей среды. Оболочка хлоропластов хлорофилла не
содержит, однако в ней присутствуют каротиноиды, а именно
зеаксантин (10.1), антераксантин (10.2) и виолаксантин (10.3),
которые с помощью ферментов могут превращаться друг в дру-
га. В последнее время появляется все больше данных, свиде-
330
Часть II. Функциональные аспекты
Б
Рис. 10 3. А. Обобщенная схема хлоропласта высших растений. Б. Увели-
ченное изображение граны тилакоида, иллюстрирующее главные структур-
ные особенности тилакоидных мембран
тельствующих о том, что оболочка является важным участком
синтеза хлоропластных веществ, в первую очередь галактоли-
пидов, пренилхинонов и, возможно, некоторых каротиноидов.
10.2.3. Строма
Внутренний белковый матрикс хлоропласта известен
под названием строма. Наряду с фотосинтетическими мембра-
нами ламелл (см. ниже) с помощью электронной микроскопии
в строме обнаружены и другие структуры. К ним относятся
рибосомы и тяжи ДНК, которые участвуют в хлоропластной
саморегуляции и репликации, зерна запасного полисахарида
крахмала, осмиофильные глобулы (иначе, пластоглобулы),
Гл 10 Фотосинтез
331
которые, возможно, служат экстраламеллярными фондами мем-
бранных липидов, и пиреноиды (у некоторых водорослей) или
сходные с ними стромальные центры (у высших растений). Точ-
ные функции последних двух небольших электроноплотных
структур неизвестны, однако показано, что большую часть их
составляет белок и что они обладают рибулозодифосфат-карбо-
ксилазной активностью
(10 3) Виолаксантик
10.2.4. Фотосинтетическая ламеллярная система мембран
Фотосинтетические пигменты содержатся в протяжен-
ной системе внутренних мембран хлоропласта, которые являют-
ся местом, где происходит фотосинтез С помощью электронной
микроскопии в хлоропласте удалось различить группы замкну-
тых мешкообразных дисков, названных тилакоидами., которые
расположены один над другим, образуя грану (рис. 10 3,5).
Внутренний объем, ограниченный мембраной одного тилакоида,
известен как локус (loculus; от лат. — ящичек с перегородками,
полочками, ларчик). Концы тилакоидов, находящиеся в кон-
такте со стромой, называют краями (margins), а участки, где
два тилакоида плотно прилегают друг к другу, — перегородка-
ми (partition). Стопки гран связаны между собой мембранами,
которые называют одиночными тилакоидами, или ламеллами
стромы.
332
Часть II. Функциональные аспекты
10.2.5. Ультраструктура тилакоидной мембраны
хлоропласта
Структура мембраны как тилакоидов гран, так и тила-
коидов стромы (одиночных тилакоидов) была подробно изучена
с помощью электронной микроскопии и иммунологических ме-
тодов. Много информации получено также и о химическом со-
ставе этих мембран.
Их ультраструктура схематически изображена на рис. 10.4.
Как и большинство биологических мембран, тилакоиды состоят
Рис. 10.4. Схематическое изображение ультраструктуры мембран тилакои-
дов, собранных в граны, и мембран одиночных тилакоидов.
главным образом из белков и липидов, содержащихся пример-
но в равных количествах. Структура мембраны асимметрична, ее
липидный матрикс состоит из двух слоев, причем наружный слой
толще внутреннего. В липидный матрикс погружено множество
белковых частиц, которые видны как на наружной, так и на
внутренней поверхности тилакоида. Они видны также по обе
стороны от гидрофильной центральной зоны внутри мембраны
на срезе препарата, приготовленного методом замораживания —
скалывания. Эти частицы могут совершенно свободно переме-
щаться в жидком липидном матриксе. Их содержится до 6000
на 1 мкм2, и они занимают примерно 70 и 50% площади тила-
коидов гран и тилакоидов стромы соответственно. Фотохимиче-
ская активность, электронный транспорт и активность метабо-
лических ферментов ассоциированы с частицами разных типов,
и катализируемые ими реакции протекают в направлении, пер-
пендикулярном к мембране.
Между тилакоидами стромы и тилакоидами гран имеются
не только структурные, но и функциональные различия. Как бу-
дет показано ниже, фотосинтез включает две фотохимические
реакции, которые протекают в комплексах, известных как фо-
Гл. JO. Фотосинтез
333
тосистемы I и II (ФС I и ФС II). Комплексы фотосистем и
транспорта электронов локализованы в мембранной структуре
со специфическими, хотя и не обязательно регулярными, интер-
валами в виде групп глобул. Подробно локализация фотосин-
тетических пигментов в комплексах фотосистем будет описана
ниже (разд. 10.4.2). Здесь же следует рассмотреть процесс фо-
тосинтеза в целом. Это поможет лучше оценить значение лока-
лизации пигментов в специфических участках или в комплек-
сах и ту роль, которую пигменты играют в фотосинтезе.
10.3. Фотосинтез у растений; общая схема
Система тилакоидных мембран хлоропласта превраща-
ет энергию света в форму, которая может быть использована
для осуществления химических реакций. Целиком процесс фо-
тосинтеза был схематически представлен на рис. 10.1. В при-
водимом ниже обсуждении фотосинтеза рассматриваются три
стадии. Первая стадия представляет собой световую реакцию —
первичный процесс, с помощью которого энергия света погло-
щается светособирающими пигментами и переносится на фото-
химические реакционные центры. На второй стадии поглощен-
ная энергия света используется для осуществления транспорта
электронов от воды до NADP+. В ходе электронного транспор-
та устанавливается градиент заряда, или концентрации прото-
нов, через функциональные везикулы мембраны. Третья стадия
представляет собой путь, по которому NADPH, образованный
электронтранспортной системой, и АТР, генерируемый за счет
различий электрохимического потенциала протонного градиен-
та, используются для фиксации СО2 и синтеза углеводов. Хотя
в целях упрощения процесс фотосинтеза разбит на три стадии,
необходимо помнить, что поглощение света, транспорт электро-
нов и генерация электрохимического градиента в действитель-
ности очень тесно сопряжены.
10.4. Поглощение света — первичный процесс фотосинтеза
10.4.1. Фотосинтетическая единица
Типичный тилакоидный диск, например из зрелого
листа шпината, содержит около 200 фотосинтетических элект-
ронтранспортных цепей и до 100 000 молекул хлорофилла. Каж-
дая электронтранспортная цепь способна совершать один пол-
ный оборот за 15 мс (или около того), тогда как одна молеку-
ла хлорофилла не может так быстро поглощать световой квант
даже при высокой интенсивности света. Было рассчитано, что
при высвобождении одной молекулы О2 и восстановлении од-
334
Часть 11 Функциональные аспекты
нои молекулы СО2 в процесс может вовлекаться до 2500 моле-
кул хлорофилла. Исходя из этого расчета, была разработана
концепция фотосинтетической единицы, в которой происходят
первичные фотохимические реакции фотосинтеза в так называ-
емом реакционном центре, причем каждый реакционный центр
связан со сравнительно большим числом пигментных молекул,
поглощающих фотон и передающих ему энергию. Любая фо-
тосинтетическая электронтранспортная цепь включает фотоси-
Свет
Рис. 10.5. Диаграмма, иллюстрирующая механизм действия фотосинтетиче-
ской единицы.
стемы I и II, которые осуществляют две различные фотохими-
ческие реакции, причем та и другая системы имеют свой собст-
венный реакционный центр. В настоящее время известно: для
того чтобы выделилась одна молекула О2 и восстановилась одна
молекула СО2, через обе фотосистемы должно пройти по четыре
электрона (т. е. всего необходимо восемь фотонов). Отсюда
следует, что для одной фотохимической реакции требуется около
300 молекул хлорофилла. На самом деле этот расчет сильно уп-
рощен, так как в нем не учитываются влияние на данную величину
факторов окружающей среды и межвидовые различия. Однако,
если даже цифра в 300 молекул хлорофилла не точна, незави-
симо от этого концепция фотосинтетической единицы, по-види-
мому, верна. В настоящее время считают, что фотосинтетиче-
ская единица для каждой фотосистемы представляет собой
комплекс, в котором основная часть молекул хлорофилла обра-
зует антенну из светопоглощающих пигментов. Эта антенна
собирает энергию света и передает ее на специфический хлоро-
филловый димер в реакционном центре, в котором она исполь-
Гл. 10. Фотосинтез
335
зуется для функционирования фотосинтетической электронтранс-
портной системы (рис. 10.5). По последним данным, антенна
ФС I содержит 120 молекул хлорофилла а, а антенна ФС II —
лишь 60. Наряду с этими двумя фотосистемами обычно присут-
ствует третий светособирающий пигментный комплекс (ССК
или LHCP; от англ, light-harvesting chlorophyll protein), свя-
занный с ФС II. В результате большой работы, проведенной
в последние годы, удалось многое узнать об организации этих
трех фотосинтетических единиц (или частиц) в тилакоидных
мембранах. Было показано, что они распределены в мембране
неравномерно. Согласно одной из моделей, частицы ФС I рас-
пределены как по одиночным, так и по собранным в граны ти-
лакоидам, в то время как большие агрегаты, включающие ФС II
и ССК, содержатся лишь в тилакоидах гран. Другая модель
предполагает очень низкое содержание единиц ФС I в гранах,
соприкасающихся друг с другом, где локализованы главным
образом единицы ФС II и ССК. Вместе с этим допускается при-
сутствие единиц ФС I в концевых гранах и в наружных тилако-
идах. Согласно этой новейшей модели, системы ФС I и ФС II
пространственно разделены, а связь между ними осуществля-
ет «челнок» восстановительных эквивалентов. Роль челнока
может играть очень большой фонд пластохинонов.
10.4.2. Светособирающие пигменты
Из обработанных детергентами хлоропластов можно
выделить три светособирающие пигментные системы в форме
комплексов хлорофилл — белок. Так, ФС I можно разделить на
комплекс хлорофилл а — белок, который содержит около 120
молекул хлорофилла а (10.4), входящих в состав антенны, и
реакционный центр Р-700 (см. ниже, разд. 10.4.3). В состав
ФС II входит комплекс хлорофилл а — белок, содержащий ре-
акционный центр Р-680 и около 60 молекул хлорофилла а. Ни
в ФС I, ни в ФС II хлорофилл b (10.5) не содержится. В обе
фотосистемы включено некоторое количество р-каротина (10.6),
хотя не известно, является ли он частью пигментов антенны
или входит в состав реакционных центров.
ССК можно выделить в виде комплекса хлорофилл а/Ь —
белок и расщепить его на три компонента хлорофилл — белок.
ССК, по-видимому, является единственным комплексом, кото-
рый содержит хлорофилл Ь, функционирующий как вспомога-
тельный пигмент. В его состав входят в значительных количест-
вах каротиноиды, которые представлены главным образом ксан-
тофиллами [преимущественно лютеином (10.7)]. Роль ССК, тес-
но связанного с ФС II, заключается в увеличении светособира-
ющей способности ФС II, в частности путем расширения спект-
336
Часть 11 Функциональные аспекты
рального диапазона света, который может быть использован
этой фотосистемой. Исследования, проведенные с проростками
гороха, позволили предположить, что существуют светопогло-
щающие частицы размером 160А, которые состоят из ядра
ФСП — хлорофилл а-—белок, окруженного четырьмя едини-
(10 4) Хлорофилл a R - СНз
(10 7) Лютеин
цами ССК (рис 10.6). Рассматривается также возможность
использования энергии света, улавливаемой ССК, для осущест-
вления реакции ФС1.
В любой светособирающей системе спектроскопическими
методами можно выявить несколько разных форм хлорофилла
а со слегка различающимися А,тах. Антенны обеих фотосистем
содержат хлорофиллы п-660, п-670, п-678 и п-685 (названы по
длинноволновому максимуму поглощения), но наряду с ними
в ФС I имеются формы с максимумами поглощения 690, 705
и 720 нм. Различия в максимумах поглощения, по-видимому,
Гл 10 Фотосинтез
337
объясняются различиями в молекулярном окружении молекул
хлорофилла, в частности ассоциацией их с белками или взаимо-
действием с соседними молекулами хлорофилла. Например,
сдвиг максимума поглощения хлорофилла a in vivo в красную
область спектра до 720 нм обусловлен молекулярной агрега-
цией. Однако недавно было высказано предположение, что су-
ществуют четыре химически различные формы обоих хлоро-
филлов (а и Ь).
Кванты, поглощенные пигментами антенны, переходят от
молекулы к молекуле путем резонансного переноса энергии;
Рис. 10.6. Модель предполагаемой свето-
улавливающей частицы размером 160 А из
проростков гороха
при этом энергия собирается в реакционном центре. Эффектив-
ность процесса переноса энергии высока, а энергетические по-
тери в результате испускания флуоресценции или нерациональ-
ной фотохимии невелики. Перенос энергии через всю систему
антенны происходит очень быстро, протекая за пикосекунды.
Резонансный перенос энергии идет в направлении пигментов,
поглощающих при больших длинах волн, так что кванты, по-
глощенные вспомогательными пигментами в ССК, т. е кароти-
ноидами (400—500 нм) и хлорофиллом b (640—650 нм), а так-
же формами хлорофилла а, поглощающими в более коротко-
волновой области, могут передаваться на хлорофиллы, погло-
щающие при больших длинах волн, и в конце концов на реак-
ционный центр (рис. 10.7). Таким образом используется боль-
шая часть спектра солнечного света.
Защитный механизм против избыточного поглощения света.
Размеры антенны в хлоропластах позволяют транспорту элект-
ронов идти с полной скоростью даже в облачный день. Очевид-
но, в условиях яркого освещения (на ярком солнечном свету)
может поглощаться гораздо больше квантов, чем способен ис-
пользовать реакционный центр. Избыток энергии возбужденно-
го хлорофилла может удаляться различными путями. Один из них
включает внутрисистемный переход с образованием более дол-
338
Часть II. Функциональные аспекты
Рис. 10.7. Диаграмма, иллюстрирующая прохождение энергии возбуждения
через различные формы пигментов в светособирающей антенне фотосистем
I и II.
гоживущей, но все еще высокоэнергетической формы 3Хл
в триплетном состоянии. От этого «триплетного» хлорофилла
избыток энергии может передаваться на находящийся в основ-
ном состоянии молекулярный кислород, переводя его в синглет-
ное состояние 'Ог- Этот чрезвычайно реакционноспособный оки-
слитель может окислить любую подходящую акцепторную мо-
лекулу, в том числе и сам хлорофилл, и тем самым вызвать
сильные, а возможно, и летальные повреждения фотосинтети-
ческих мембран. Считают, что одной из главных функций каро-
тиноидов в антенне является защитная. Они предотвращают
подобные повреждения путем вмешательства в эту последова-
тельность событий, либо преимущественно реагируя с окисля-
Гл 10. Фотосинтез
339
* Хл Фотосинтез
например Хл)
Рис. 10 8. Потеря энергии возбуждения хлорофиллом (Хл), ведущая к об-
разованию синглетного кислорода (Юг), и три механизма, в которых каро-
тиноиды (КАР) могут функционировать как защитные агенты против опас-
ного или ведущего к летальному исходу окисления, вызываемого 1О2. 1. КАР
в основном функционирует как субстрат для окисления. 2. Переход энергии
возбуждения с 3Хл на КАР. 3. Переход энергии возбуждения с ’О2 на КАР.
ющим синглетным кислородом, либо принимая на себя энер-
гию возбуждения «триплетного» хлорофилла или «синглетного»
кислорода (рис. 10.8).
10.4.3. Фотохимические реакционные центры
Антенна и пигменты ССК участвуют в поглощении
света, в результате чего происходит возбуждение электронов, и
передают энергию возбуждения от молекулы к молекуле, пока
она не достигнет реакционного центра. В реакционном центре
эта энергия возбуждения превращается в химически утилизи-
руемую форму. Активным пигментом в реакционных центрах
обеих фотосистем служит специфический димер хлорофилла а,
который подвергается обратимому окислению.
Реакционный центр I. Пигмент реакционного центра фото-
системы I характеризуется изменениями поглощения света,
главным образом при 700 нм, которые выявляются в спектре
после освещения хлоропластов вспышками света. Он известен
как хлорофилл аг> или пигмент Р-700, и представляет собой
340
Часть И. Функциональные аспекты
специфический комплекс белка с димером, состоящим из двух
молекул хлорофилла а. Структура этого димера, или «особой
пары», по-видимому, чрезвычайно важна для функционирования
центра, но полностью она еще не установлена. Известно, одна-
ко, что тетрапиррольные кольца мономеров лежат примерно
в плоскости тилакоидной мембраны. Были предложены модели
димера, в которых молекулы воды взаимодействуют с атомом
Рис. 10.9. Модель возможных взаимодействий между водой и двумя моле-
кулами хлорофилла в димере Р-700 (хлорофилле as).
магния, находящимся в центре одной из молекул хлорофилла,
и карбонильной группой кольца V второй молекулы (рис. 10.9).
Энергия поглощенного антенной света переносится в реак-
ционный центр, и хлорофилл ai подвергается электронному воз-
буждению. Этот процесс занимает менее 30 пс. В течение по-
следующих 20 нс хлорофилл щ окисляется, вероятно, с образо-
ванием катион-радикала (Хл-Хл) + -, в котором неспаренный
электрон делокализован по всей системе обоих порфириновых
колец. Электрон, отданный хлорофиллом аг в ходе такого окис-
ления, эффективно переносится на первичный акцептор X (Р-
430). Хлорофилл «I возвращается в свое нормальное состояние
в результате получения электрона от первичного донора Y. Та-
ким образом, реакционный центр ФС I осуществляет эту часть
фотосинтетического транспорта электронов путем эффективной
передачи электронов от Y на X, т. е. с помощью процесса, ко-
торый не может происходить самопроизвольно (рис. 10.10,Л).
Гл. 10. Фотосинтез
3411
Рис. 10.10. Первичные реакции в фотосинтетических реакционных центрах.
А. Фотосистема I. Б. Фотосистема 11.
Реакционный центр II. Пигмент реакционного центра II
представляет собой также комплекс хлорофилла с белком, со-
держащий димер хлорофилла а, известный как хлорофилл «и,
или Р-680. Хотя иной характер поглощения света этим пигмен-
том указывает на то, что молекулы хлорофилла а находятся
здесь в другом молекулярном окружении или по-иному ориен-
тированы, чем в случае пигмента Р-700, процессы поглощения
света и окисления, происходящие в реакционном центре II,
сходны с аналогичными процессами в реакционном центре I.
Здесь также энергия электронного возбуждения передается
с хлорофилла антенны на хлорофилл ап, который подвергается
возбуждению с последующим окислением до катион-ратикала
и делокализацией неспаренного электрона. В этом случае элект-
рон передается на первичный акцептор электрона фотосистемы
II Q (Х-320). Затем катион-радикал хлорофилла «и восстанав-
ливается, получая электрон от донора Z. Таким образом, фото-
система II эффективно переносит электроны от Z на Q (рис.
10.10).
23—84
342
Часть II. Функциональные аспекты
10.5. Вторичные события; система фотосинтетического
транспорта электронов
В эукариотических клетках в процессе окислительного
распада питательных веществ образуются восстановительные
нуклеотиды, главным образом NADH. Для их окисления моле-
кулярным кислородом используется митохондриальная электрон-
транспортная система (рис. 10.11,А). Это термодинамически
выгодный процесс, поскольку окислительно-восстановительный
Рис. 10.11. А. Окисление NADH молекулярным О2 в митохондриях. Б. Вос-
становление NADP+ до NADPH водой и выделение молекулярного О2 при
фотосинтезе.
потенциал пары NAD+/NADH ( — 0,32 В) более отрицателен,
чем потенциал пары О2/Н2О ( + 0,82), или, другими словами,
молекулярный кислород является более сильным окислителем,
чем NAD". Ясно, что обратный процесс, в котором NAD+ (пли
NADP*-) восстанавливается до NAD(P)H с помощью воды, а
вода в свою очередь окисляется до молекулярного кислорода
(рис. 10.11,5), энергетически невыгоден и не может происхо-
дить спонтанно. В связи с этим становится очевидной чрезвы-
чайная важность фотосинтетического транспорта электронов.
Для восстановления NAD+ необходимо поступление энергии, и
для этой цели используется энергия света, улавливаемая в пер-
вичных реакциях фотосинтеза.
10.5.1. Z-схема
Из множества различных схем, предложенных для
транспорта электронов при фотосинтезе, большинству извест-
ных данных удовлетворяет лишь зигзагообразная Z-схема (рис.
Рис. 10.12. Z-схема потока электронов при фотосинтезе. Подробное описание промежуточных компонентов приведено
в тексте.
344 Часть II. Функциональные аспекты
10.12) , иллюстрирующая, каким образом свет обеспечивает
энергией две различные фотореакции. Собранная энергия света
используется для образования восстанавливающих агентов, до-
статочно сильных, чтобы передавать электроны (т. е. для вос-
становления) на ряд промежуточных компонентов электрон-
транспортной последовательности.
10.5.2. Фотосистемы I и II
Свет, поглощенный фотосистемой I, поставляет энер-
гию для переноса электронов от донора Y (окислительно-восста-
новительный потенциал +0,4 В) через хлорофилл «г (Р-700)
реакционного центра на акцептор X (окислительно-восстанови-
тельный потенциал ~ —0,6В), т. е. против градиента потенциа-
ла в +1,0 В. Восстановленная форма X затем передает элект-
роны (восстанавливает) на NADP+ (окислительно-восстанови-
тельный потенциал —0,32 В) через ферредоксин (белок, содер-
жащий железо и серу) с помощью флавопротеинового фермен-
та.
Донор электронов Y для фотосистемы I расположен в конце
последовательности компонентов электронтранспортной цепи —
после пластохинона, цитохрома f и пластоцианина. Он получает
электроны, передающиеся через эти промежуточные компонен-
ты, от соединения Q (окислительно-восстановительный потен-
циал ~0В). Q является акцептором электронов в фотосистеме
II и получает их через Р-680 от донора Z. Как уже отмечалось,
восстановительный акцептор Q передает электроны фотосисте-
ме I. Окисленный донор Z является достаточно сильным окис-
лителем, способным окислить воду, что приводит к выделению
кислорода. Таким образом, энергия для расщепления воды по-
‘Ставляется светом, улавливаемым фотосистемой II.
Общая картина такова, что при переносе электронов от до-
нора на акцептор фотосистемой I образуется сильный восста-
новитель, который может восстанавливать NADP+ до NADPH,
и слабый окислитель, который тем не менее способен получать
электроны от пластоцианина. Вместе с тем перенос электронов
с донора на акцептор фотосистемой II приводит к образованию
сильного окислителя (окисленный донор), который может
осуществлять окисление воды и выделение кислорода. Образую-
щийся при этом восстановитель (восстановленный акцептор),
хотя и более слабый, чем в фотосистеме I, достаточно силен
для передачи электронов через последовательность компонентов
пластохинон — цитохром f — пластоцианин.
Гл. 10. Фотосинтез
345
10.5.3. Природа первичных акцепторов и доноров
Компоненты электронтранспортной системы, которые
отдают хлорофиллам или получают от них электроны, были
идентифицированы главным образом по спектроскопическим
данным, однако в химическом отношении они были охарактери-
зованы далеко не во всех случаях.
Фотосистема I. Первичный акцептор X, который получает
электроны от Р-700 (хлорофилла ai), был идентифицирован по
изменениям поглощения света. Его назвали Р-430 и считают, что
он является белком, содержащим железо и серу. Донор Y
представляет собой медьсодержащий белок пластоцианин, ко-
торый поставляет электроны для восстановления окисленного
катион-радикала хлорофилла а\.
Фотосистема II. Первичный акцептор Q фотосистемы II, для
которого характерны изменения поглощения света при 320 нм,
по-видимому, является прочносвязанной формой пластохинона
(10.8). Эта форма отличается от основной формы пластохино-
на — одного из последующих компонентов в цепи переноса
электрона. С акцептором Q тесно связан неидентифицированный
компонент С-550, имеющий максимум поглощения при 550 нм.
Донор Z, который поставляет электроны для восстановления
хлорофилла Оц (Р-680), также неидентифицирован. Предпола-
гают, что он прочно связан с марганцем или цитохромом £559.
10.5.4. Природа других компонентов фотосинтетической
электронтранспортной системы
Ряд компонентов фотосинтетической системы транс-
порта электронов представляют собой молекулы, которые погло-
щают свет видимых длин волн и поэтому являются окрашен-
ными пигментами. Так, цитохромы, подобные цитохромам мито-
хондриальных систем транспорта электронов, — это железо-
(гем)содержащие белки, поглощающие свет примерно при
560 нм и потому окрашенные в красный цвет. (Общие свойства
гемов и цитохромов были описаны в гл. 5.) Флавопротеиновый
346
Часть II. Функциональные аспекты
фермент ферредоксин — ЫАВР+-оксидоредуктаза окрашен
в желтый цвет, поскольку ь качестве простетической группы
содержит рибофлавин (гл. 6). Пластоцианин представляет со-
бой медьсодержащий белок, и его интенсивная синяя окраска
обусловлена, вероятно, поглощением в результате переноса
заряда медью. Неокрашенный важный компонент системы —-
пластохинон (10.8) —представляет собой бензохинон с длинной
изопреноидной боковой цепью (свойства бензохинонов были
описаны в гл. 3).
Рис. 10.13. Взаимосвязь некоторых фотосинтетических электронтранспортных
цепей через большой фонд пластохинона (PQ). Цит — цитохром; ПЦ — пла-
стоцианин; Q — хинон.
Пластохинон выполняет в системе переноса электронов не-
сколько специфических функций (рис. 10.13). Его значительно
больше, чем других компонентов цепи, и он служит электрон-
ным «буфером», который обеспечивает гладкое функциониро-
вание цепи даже при сильных колебаниях в распределении
квантов света между двумя фотосистемами. Он способен так-
же связывать между собой несколько электронтранспортных
цепей и таким образом повышать надежность системы. Напри-
мер, если какой-либо реакционный центр II не функционирует,
то пластохинон может обеспечить работу связанного с ним ре-
акционного центра I за счет электронов, поступающих из дру-
гого реакционного центра II. В результате реакционный центр I
не будет испытывать недостатка в электронах. Другая возмож-
ная роль пластохинона упоминалась ранее (разд. 10.4.1), когда
рассматривалось распределение фотосистем в тилакоидах. Из-
за пространственного разделения разных фотосистем необходим
механизм, обеспечивающий поток электронов между ними, и
предполагают, что в этом механизме главную роль играет
пластохинон.
Вместе с тем следует отметить, что способность всех этих
соединений поглощать видимый свет не имеет отношения к их
функционированию при фотосинтезе, и в первичном поглощении
света они не участвуют.
Гл. 10. Фотосинтез
347
10.6. Темновые реакции
Хотя подробное изложение этой темы и не входит в
задачу настоящей книги, краткое описание основных ее аспек-
тов необходимо для получения полной картины фотосинтеза.
10.6.1. Фотофосфорилирование
В настоящее время принято считать, что фотосинтети-
ческая электронтранспортная цепь расположена перпендикуляр-
но тилакоидной мембране и дважды пересекает ее (рис. 10.14).
В продолжение одного оборота цепи каждый реакционный
Рис. 10.14. Расположение фотосинтетической электронтранспортной цепи в
тилакоидной мембране.
центр переносит один электрон с внутренней стороны мембра-
ны на наружную. Одновременно на внутренней стороне мембра-
ны в результате расщепления воды и окисления дигидропласто-
хинона освобождаются протоны, Таким образом, создается гра-
диент электрохимического потенциала с положительным заря-
дом (или низким pH) на внутренней стороне мембраны и с от-
рицательным зарядом (или высоким pH) на ее наружной сто-
роне. Согласно хемиосмотической гипотезе Митчелла, энергия
этого градиента служит движущей силой синтеза АТР из ADP
при помощи ATP-азы, связанной с мембраной. Когда работают
обе фотосистемы, поток электронов является нециклическим, и
говорят, что образование АТР происходит путем нециклического
фотофосфорилирования. Если же функционирует только одна
фотосистема (обычно ФС I), наблюдается циклический поток
электронов, и говорят, что образование АТР происходит путем
циклического фотофосфорилирования.
A CH2OH
c=o
н—с—он
н—с—он
I
СН2О— (Р)
СН2О-©
с=о
н—с—он
I
Н—С—он
I
СН2О-(р)
сн2о-®
-оос—с—он
с=о
Н—С—он
I
СН2О-®
соо"
—»-Н—С—ОН
СН2О-®
12 хЗ-Фосфоглицерат
Gx Рибулозо-
5-фосфат
6* Рибулозо-
1,5-дифосфат
Регенерация рибулозо-5-
фосфата (см. Б)
СН2ОН
с=о
ДигидроксиаЦетонфосфат
НО—С—н
СОО—®
Н— С —он
СН2О-®
12* 1,3-Дифосфоглицерат
I
н—с—он
СН2О-®
12 х Глицеральдегид-
'3-фосфат
хЗ
СН2О-®
с=о
НО—с—н
Х1
Глюкоза или ’
крахмал
швия
Прибавление 1 мо-
лекулы гексозьы
У///ШШ//Ш
Н—С—ОН
н—С—он
Н—С— он
Н—он
СН2О-®
ЗхФруктозо-6-фосфат
3 хФруктозо-1,6-дмфосфат
Рис. 10.15. Схема пути фотосинтетической фиксации углерода и синтеза углеводов по Кальвину — Бенсону. А. Основные
реакции, в которых фиксируется СО2 и используются NADPH и АТР, образующиеся фотосинтетической цепью транспорта
электронов и путем фосфорилирования. Суммарное уравнение: 6 СО2 + 18 АТР + 12 NADPH —> Фруктозо-6-фосфат ф-
+ 18 ADP+12 NADP++17 Pi.
сно
Н-С —он
44“лицеральдегид -
7 3-фосфат
х 2
СН2ОН
н—с—он
сн.он
CH2OH
СН2О—©
сно
ЗхФруктозо-
6- фосфат
Н3О~®
Ксилулозо-
5 фосфат
н —с —он
X 4
с Н2ОН
СН2О-©
Н—С—ОН
н—('—он
I л-4
сн2о-@
Эритроза -
4-фосфат
CHjOH
2 к Дигидрокси-
“’’'ацетон фосфат
сн2о— ©
с=о
но—с—н
н—с—он
х2 H-C-OH
н-с-он
сн2о— ©
Седугептулозо -
7-фосфат
СН2О—®
Седугептулозо -
1. 7- бисфосфат
I 6*Рибулозо!-
I ’5-фосфат ' I
(jHO
Н-С—ОН
н—С—ОН
I
СН2О-©
Рибозо-5-фосфат
СН2О-® <Uo Н—<!—ОН н—<!:—он <^н2о-(р) СН2О-® СН2О-®
||-он со2 ООС—<!?—он нч
(j—он н—с—он -_!> но^ 9=0 н-с—он
СН2О—(Р) ^н2о—(р)
Рибулозе -1,5-бисфосфат (Энедиол) Промежуточная
Р —оксокислота
СН2О-<₽)
‘ООС—С—он
— н
соон
I
н—с—он
СН2О-(Р)
2 X 3-Фосфоглицерат
Рис. 10.15 (продолжение) Б. Промежуточные продукты при регенерации ри-
булозо-1, 5-дифосфата из глицеральдегид-3-фосфата, дигидроксиацетоифосфа-
та и фруктозо-6-фосфата. В. Реакция фиксации СО2, катализируемая рибуло-
зо-1,5-дифосфат-карбоксилазой (карбоксидисмугазой).
350
Часть II. Функциональные аспекты
10.6.2. Фиксация углерода
Химическая энергия улавливается двумя путями — в
форме восстановительного кофактора NADPH и в форме АТР,
образующегося в результате сопряженного фосфорилирования
(так же как и в случае митохондриального транспорта электро-
нов). NADP и АТР поставляют восстановительные эквивален-
Рис. 10.16. Путь фотосинтетической фиксации СО2 по Хэтчу — Слэ-
ку (С4-путь).
ты и химическую энергию для фиксации углерода, с помощью
которых СО2 восстанавливается до углеводов. Хотя у разных
фотосинтезирующих организмов в образовании углеводов участ-
вуют различные метаболические последовательности, по-види-
мому, в хлоропластах всех высших растений реакции протека-
ют в основном по циклу Кальвина — Бенсона, приведенному на
рис. 10.15.
Наряду с циклом Кальвина — Бенсона (Сз-путем) необхо-
димо отметить и цикл Хэтча — Слэка (С^путь; рис. 10.16),
который встречается у некоторых тропических растений, глав-
ным образом травянистых, и позволяет осуществлять синтез
углеводов из СО2 при низких концентрациях последней и с
очень небольшими потерями воды. Особенностью С4-растений
является совместное функционирование клеток двух типов —
Гл. 10. Фотосинтез
351
клеток мезофилла и клеток обкладки сосудистого пучка. Фик-
сация СОа у них происходит в клетках мезофилла в виде мала-
та; последний затем транспортируется в клетки обкладки сосу-
дистого пучка, где он подвергается декарбоксилированию. Вы-
делившаяся СО2 фиксируется в реакции с рибулозо-1,5-бисфо-
сфатом по обычному пути Кальвина — Бенсона.
(10.9.) Хлорофилл с
10.7. Фотосинтез у эукариотических водорослей
Процесс фотосинтеза у зеленых водорослей (Chloro-
phyceae) очень напоминает аналогичный процесс в хлоропла-
стах высших растений; многие детали фотосинтеза были выяс-
нены при исследовании зеленых водорослей, и в первую оче-
редь Chlorella, Scenedesmus и Chlamydomonas. В целом фото-
синтез у всех эукариотических водорослей, по-видимому, сходен
с фотосинтезом у высших растений. У них также в ходе этого
процесса выделяется кислород, используются две фотосистемы
и функционирует Z-схема транспорта электронов. Однако раз-
ные классы водорослей различаются по составу фотосинтети-
ческих пигментов ССК. Одни водоросли, например из сем.
Chlorophyceae и Euglenophyceae, так же как и высшие расте-
ния, содержат хлорофиллы а и Ь, а в качестве вспомогательных
пигментов каротиноиды. У других водорослей хлорофилл b
отсутствует и заменен хлорофиллами end (10.9; 10.10), кото-
рые служат вспомогательными пигментами у Dinophyceae, Chry-
sophyceae, Bacillariophyceae, Xanthophyceae, Phaeophyceae и у
некоторых представителей Rhodophyceae. Наблюдаются также
значительные вариации в составе каротиноидов (гл. 2). Напри-
мер, бурые морские водоросли (Phaeophyceae, Chrysophyceae,
Bacillariophyceae), доля которых в мировой фотосинтетической
352
Часть II. Функциональные аспекты
активности весьма высока, в качестве главного каротиноида
содержат фукоксантин (10.11). При разрушении тилакоидов
бурых водорослей детергентами выявляются три пигмент-бел-
ковых комплекса: комплекс Р-700 — хлорофилл а — белок,
сходный с аналогичным комплексом у высших растений, и два
(10 11 ) фукоксантин
(10 13 ) Фикоэритрин
ССК — один комплекс фукоксантин — хлорофилл afct — белок,,
а другой комплекс виолаксантин — хлорофилл alc-tlc?. — белок.
Морские динофлагелляты содержат водорастворимый ССК, ко-
торый представляет собой комплекс перидинин (10.12)—хло-
рофилл а — белок. Фукоксантин и перидинин являются важны-
ми вспомогательными пигментами, переносящими энергию на
хлорофилл а с высокой эффективностью (70%).
В красных водорослях (Rhodophyceae), в основном содер-
жащих только хлорофилл а, присутствуют в больших количест-
вах фикобилины — комплексы между белками и линейными
тетрапирролами, подобными фикоэритрину (10.13; гл. 5),—
которые эффективно переносят энергию возбуждения на хлоро-
филл а в фотосистеме II. Фикобилины присутствуют в клетках
водорослей в виде белковых комплексов в частицах, называе-
Гл 10. Фотосинтез
353’.
мых фикобилисомами, которые присоединены к ламеллам хло-
ропластов (разд. 10.8).
Эукариотические водоросли живут в различных местооби-
таниях и характеризуются большой вариабельностью показате-
лей роста, поэтому нет ничего удивительного в том, что у не-
которых видов наряду с различиями в пигментации обнаружи-
ваются и некоторые различия в составе переносчиков электро-
нов и в их последовательности в электронтранспортной цепи.
Большое разнообразие наблюдается также и в путях метабо-
лизма углерода. Подробно в этом отношении было изучено
лишь несколько видов водорослей; поэтому не исключено, что
многие отклонения от фотосинтеза по типу высших растений
остаются пока неоткрытыми.
10.8. Фотосинтез у прокариотических
сине-зеленых водорослей
10.8.1. Общие особенности
Сине-зеленые водоросли (Cyanophyta или Cyanobac-
teria) представляют собой единственную большую группу про-
кариот, которые способны к фотосинтезу с выделением кисло-
рода, сходному с фотосинтезом у высших растений. Однако
тилакоидные мембраны у них находятся не в хлоропластах, а
распределены по всей цитоплазме клетки, преимущественно на
ее периферии. Фотосинтетические пигменты сине-зеленых водо-
о
(10 14 ) Эхиненон
рослей заметно отличаются от хлоропластных пигментов расте-
ний и водорослей. Подробно было изучено лишь несколько ви-
дов сине-зеленых водорослей, и тем не менее удалось устано-
вить главные особенности их фотосинтеза. У этих водорослей
также обнаружены две фотосистемы и электронтранспортная
цепь Z-типа, а их ФС I, по-видимому, сходна с аналогичной
системой у эукариот. Однако состав компонентов их фотосистем,,
а также переносчики и их последовательность в системе транс-
порта электронов отличаются от аналогичных параметров у
высших растений. В общем пигменты, фотосистемы и электрон-
транспортные компоненты у них, по-видимому, гораздо менее
прочно соединены с мембранными структурами, чем у фотосин-
.'354
Часть II. Функциональные аспекты
тезирующих эукариот. Были охарактеризованы пигменты сине-
зеленых водорослей. Они содержат единственный хлорофилл —
^хлорофилл а, а в качестве вспомогательных пигментов, связан-
ных с фотосистемой II, — фикобилины, локализованные в фико-
билисомах (разд. 10.8.2). Из каротиноидов с фотосистемой I
ассоциированы p-каротин и, возможно, эхиненон (10.4), а с фо-
тосистемой II — различные ксантофиллы.
Так же как у других организмов в разных условиях роста и
окружающей среды у сине-зеленых водорослей наблюдаются
.значительные изменения в морфологии, пигментации и деталях
механизма фотосинтеза.
10.8.2. Фикобилисомы
Красные водоросли и сине-зеленые водоросли (или
бактерии) отличаются от всех других фотосинтезирующих орга-
низмов тем, что в качестве вспомогательных светоулавливающих
пигментов они используют фикобилипротеины (гл. 5), которые
локализованы в специфических структурах, называемых фико-
,билисомами. Последние представляют собой макромолекулярные
Рис. 10.17. Модель структуры фикобилисомы красных водорослей.
агрегаты мол. массой до 20-106, упорядоченно расположенные
со стороны стромы вдоль тилакоидной мембраны. Эти структу-
ры, по-видимому, очень сходны у всех изученных в данном
отношении организмов. Центральное ядро фикобилисомы, ко-
торое примыкает к тилакоиду, составляет аллофикоцианин —
фикобилипротеин, обладающий поглощением (и испусканием
флуоресценции) в длинноволновой области спектра. От этого
ядра в направлении стромы радиально расходятся упакованные
в стопки палочки, содержащие фикоцианин (внутренние палоч-
ки) и фикоэритрин (наружные палочки) (рис. 10.17). В фико-
билисоме присутствуют также другие, неокрашенные белки,
Гл. 10. Фотосинтез
355-
которые, по-видимому, играют роль связующих компонентов,
поддерживающих ее структуру.
Фикобилисома представляет собой светособирающий комп-
лекс, в котором происходит перенос энергии с фикоэритрина
через фикоцианин на аллофикоцианиновое ядро. На ФС II мо-
жет переноситься около 95% уловленной энергии, что предпо-
лагает тесную пространственную взаимосвязь между фикобили-
сомами и ФС II. По-видимому, каждая фикобилисома обслу-
живает несколько реакционных центров ФС II. ФС I у сине-
зеленых водорослей, по всей видимости, сходна с аналогичной
системой высших растений; она не использует энергии света,
улавливаемого фикобилисомами.
10.9. Бактериальный фотосинтез
Свет в качестве первичного источника энергии могут
использовать три главные группы бактерий — зеленые и пур-
пурные серные бактерии (Chlorobiaceae и Chromatiaceae) н
пурпурные несерные бактерии (Rhodospirillaceae). Как прави-
ло, фотосинтезирующие бактерии имеют клеточную стенку,
сходную со стенкой других грамотрицательных бактерий, одна-
ко под этой стенкой у них расположена толстая клеточная мем-
брана. Мембрана фотосинтезирующих бактерий может много-
кратно изгибаться и впячиваться, образуя ламеллярные мем-
бранные структуры или везикулы внутри клетки. Эти структу-
ры несут фотосинтетический аппарат, который значительно от-
личается от соответствующего аппарата хлоропластов растений.
В деталях биохимия фотосинтетических процессов у бактерий
и растений также различна. Препараты фотосинтетических
мембран можно получить из разрушенных клеток в виде ча-
стиц или пузырьков, называемых хроматофорами (ср. с хрома-
тофорами животных, гл. 8), но, как правило, интактные клет-
ки, за исключением клеток зеленых серных бактерий (Chloro-
biaceae), не содержат таких дискретных структур. Зеленые сер-
ные бактерии (например, Chlorobium) отличаются от других
фотосинтезирующих бактерий тем, что лишены внутренних
мембран; вместе с тем они обладают уплощенными сигарооб-
разными структурами, называемыми хлоросомами, которые при-
соединены к плазматической мембране. Хлоросомы служат
функционально организованными светособирающими антеннами
для реакционных центров, которые локализованы в плазмати-
ческой мембране. Перенос энергии в хлоросоме происходит С’
бактериохлорофилла с (1000—1500 молекул) через бактерио-
хлорофилл а (возможно, 100 молекул в комплексе с белком)
на бактериохлорофилл с реакционного центра.
356
Часть II. Функциональные аспекты
Бактериальный фотосинтез во многом отличается от фото-
синтеза у растений. Во-первых, бактерии — единственные из
-фотосинтезирующих организмов — не способны использовать в
качестве конечного восстановителя воду. Они используют дру-
гие восстановители, которыми могут быть органические моле-
кулы или неорганические соединения серы, и, следовательно,
ИО 15) Бактериохлорофилл а
(R = фарнезил или Геранил геранил )
(10.16 ) Бактеоиохлооофилл b
•бактерии не выделяют кислород. Во-вторых, фиксация и мета-
болизм углерода у бактерий происходят не в цикле Кальвина —
Бенсона, а иным путем. В-третьих, аппарат первичного улавли-
вания света и переноса электронов у них совершенно отличен
от наблюдающихся в растительных клетках; в частности, у них
протекает только одна световая реакция, правда, она во мно-
гом сходна с реакцией в фотосистеме I растений.
н3со
оснэ
(10 17) Спириллоксантин
Вместе с тем механизм улавливания света у бактерий очень
сходен с соответствующим механизмом у растений, хотя фото-
синтетические единицы у первых меньше. Так же как в хло-
ропластах, свет поглощается пигментами антенны, энергия воз-
буждения быстро передается на реакционный центр и исполь-
зуется в качестве движущей силы в транспорте электронов.
Главным фотоактивным пигментом является бактериохлоро-
филл (БХл), в большинстве случаев бактериохлорофилл а
(10.15), а в некоторых случаях (например, у Rhodopseudomo-
Гл. 10. Фотосинтез
357
nets sphaeroides)—бактериохлорофилл b (10.16). Каротиноиды
в фотосинтезирующих бактериях представлены характерными
для них ациклическими метокси- или арил-каротиноидами (гл.
2), например спириллоксантином (10.17) у Rhodaspirillum rub-
rum. Собирающая свет антенна передает энергию возбуждения
на БХл реакционного центра, который поглощает при 870—
875 нм у Rhodospirillaceae и при 890 нм у Chromatiaceae. БХл
реакционного центра, Р-870 или Р-875 аналогичны Р-700, или
хлорофиллу ai, ФС1 высших растений. Бактериальный реак-
(10.18.) бактериофеофитин а
ционный центр содержит три полипептида (с мол. массой 28 000,
32 000 и 35 000), ассоциированных с четырьмя молекулами бак-
териохлорофилла, две молекулы бактериофеофнтина (10.18),
одну молекулу убихинона (10.19) и негемовое железо. В не-
большом количестве в препаратах реакционного центра обна-
руживаются также каротиноиды, входящие в состав главным
образом светособирающего комплекса антенны.
В тесной связи с реакционным центром находится главная
светособирающая антенна, которая у Rhodospirillaceae погло-
щает при 875 нм. Этот комплекс включает два полипептида,
а также бактериохлорофилл и каротиноиды в соотношении 1 : 1.
Соотношение БХл: реакционные центры остается постоянным и
составляет около 25: 1. У некоторых видов, например у Rhodo-
spirillum rubrum, имеется только один ССК, тогда как у других
организмов, и в первую очередь у Rhodopseudomonas spp., со-
держится и второй подобный комплекс, поглощающий при бо-
лее коротких длинах волн (Хтах = 800 и 850 нм). Он включает
два пептида, две формы бактериохлорофилла (две молекулы
БХл-850 и одну молекулу БХл-800) и каротиноид (одну моле-
кулу). Отношение числа этих комплексов к числу реакционных
358 Часть II. Функциональные аспекты
центров варьирует в зависимости от условий окружающей
среды от 10 до 100 молекул БХл-800-850 на один реакционный
центр (разд. 10.11).
В первичной световой реакции энергия возбуждения перено-
сится на Р-870 и один электрон передается от бактериохлоро-
филла «особой пары» па акцептор. Окисленный Р-870 в свою
очередь получает электрон от молекулы донора. Химическая
природа этого донора и первичного акцептора пока неизвестна.
Электронтранспортпые цепи у разных видов бактерий раз-
личаются в деталях. Наиболее подробно их изучали у Rhodo-
spirillum rubrum, у нескольс.их видов Rhodopseudomonas
(R. sphaeroides, R. capsulata, R. palustris) из Rhodospirillaceae
и у некоторых видов Chromatium и Chlorobium из классов сер-
ных бактерий. У всех этих бактерий в цепи переноса электро-
нов участвует несколько цитохромов, тогда как плаетоцианин,
по-видимому, ие используется. Те функции, которые пластоциа-
нин выполняет у растений, у бактерий может выполнять убихи-
нон. Подробное обсуждение фотосинтетического электронного
транспорта у бактерий не входит в задачу данной книги.
10.10. Синтез пигментов в связи
с развитием хлоропластов
10.10.1. Формирование хлоропластов высших растений
из этиопластов
Структурные изменения. Биохимические и структурные
изменения, происходящие в ходе формирования хлоропластов,
в основном изучали на зеленеющих этиолированных пророст-
ках. Семядоли и листья проростков покрытосеменных, прора-
стающих и растущих в темноте, отличаются от семядолей и
листьев проростков, растущих на свету, по форме (меньшие по
размерам, удлиненные) и окраске (они желтые). В этиолиро-
ванных тканях хлоропласты не образуются, а пропластиды раз-
виваются в этиопласты (около 3—5 мкм в диаметре). Характер-
ной структурной особенностью этиопластов является наличие
проламеллярных тел — квазикристаллических трехмерных труб-
чатых структур, из которых обычно выходят немногочисленные
ламеллы. После освещения выросших в темноте проростков из
этиопластных мембран образуются хлоропласты. В ходе этого
процесса кристаллическое проламеллярное тело теряет свою
структурную регулярность, а затем полностью превращается в
пластинки перфорированных мембран, которые и дают начало
тилакоидам. В недавно опубликованных исследованиях выска-
зывается предположение, что протилакоидные мембраны, сое-
диненные с трубочками проламеллярного тела, могут играть
Гл. 10. Фотосинтез
359
чрезвычайно важную роль в синтезе компонентов развивающих-
ся фотосинтетических мембран, в том числе хлорофилла и ка-
ротиноидных пигментов.
Характер развития фотосинтетической активности во време-
ни заметно зависит от вида, возраста и условий роста растения.
В течение первых минут после освещения обычно появляется
небольшая фотосинтетическая активность, которая быстро уси-
ливается в продолжение первых двух часов, тогда как весь
процесс формирования хлоропластов из этиопластов заверша-
ется за 48 ч.
Формирование активных фотосистем. Рост тилакоидной
мембраны и развитие функционирующего фотосинтетического
аппарата в ходе дифференциации этиопласта в хлоропласт —
многоступенчатый процесс, который включает не только биосин-
тез структурных и функциональных компонентов, но также и
интеграцию и сборку этих компонентов в функциональные еди-
ницы. На разных стадиях развития мембран можно выделить
тилакоиды, содержащие ФС I- и ФС П-единицы. Сначала фор-
мируются ядра ФС I и ФС II, включающие реакционные цент-
ры, а затем простые (мономерные?) формы ССК. Дифференциа-
ция первичных тилакоидов в тилакоиды стромы и гран проис-
ходит по мере синтеза ССК; в ходе такой дифференциации
размер ФС I- и ФСН-единиц увеличивается, а в процессе даль-
нейшего развития пигмент-белковые комплексы постепенно ор-
ганизуются в большие надмолекулярные структуры полностью
развитых хлоропластов.
Регуляция развития хлоропластов светом. Окончательное
состояние тилакоидных мембран зависит от условий окружаю-
щей среды, главным образом от освещения. Хлоропласты, раз-
вивавшиеся при высоких интенсивностях освещения, имеют
относительно небольшие, но высокоэффективные ФС I и ФСП,
тогда как ССК У них редуцированы. При более низких интен-
сивностях освещения, при которых поглощение света должно
быть по возможности максимально эффективным, большое зна-
чение имеет синтез ССК, связанный с организацией тилакоидов
в граны. Имеются сообщения, что первичными регуляторами
развития хлоропластов служат красный свет и фитохромная си-
стема. Однако, согласно другим сообщениям, важную роль в
данном случае играет синий свет и пока еще неизвестный фо-
торецептор.
Образование фотосинтетических пигментов. Путь биосинте-
за, по которому образуется хлорофилл, описан в гл. 5. Этиоли-
рованные проростки не содержат хлорофилла, но содержат в
небольшом количестве протохлорофиллид (10.20), связанный
с белком и локализованный в проламеллярных телах. Результа-
ты спектроскопических исследований указывают на существо-
360
Часть II. Функциональные аспекты
вание трех форм протохлорофиллида с максимумами поглоще-
ния 628, 637 и 650 нм. Эти формы различаются степенью агре-
гации или способом связывания с белком. После освещения
проростков, выросших в темноте, с помощью спектроскопиче-
ских методов был обнаружен ряд промежуточных продуктов
(Хтах = 676, 678, 682 и 672 нм), которые образуются по мере
превращения протохлорофиллида в хлорофилл а. Сначала хло-
рофилл образуется из протохлорофиллида очень быстро, одна-
ко затем наблюдается короткий индукционный период, после
Ц0.20.) Протохлорофиллид а
которого более медленно синтезируется большая часть хлоро-
филла. Предполагают, что скорость синтеза хлорофилла регу-
лируется фитохромом. Хотя ССК быстро становятся фотохими-
чески активными, вначале их размеры невелики по сравнению
с хлоропластами зеленых листьев. Главная часть хлорофилла,
синтезирующаяся в продолжение нескольких часов после ин-
дукционного периода, служит в основном для увеличения раз-
мера светсобирающих антенн.
Этиолированные проростки обычно не содержат каротинов,
ио в них присутствуют в небольших количествах ксантофиллы
и компоненты электронтранспортной цепи, в том числе цито-
хромы, пластоцианин и пластохинон. На свету параллельно с
образованием главной части хлорофилла происходит интенсив-
ный синтез хлоропластных каротиноидов (гл. 2) и компонен-
тов фотосинтетической электронтранспортной цепи, а также
включение этих молекул в тилакоиды. Синтезы компонентов
хлоропласта в высокой степени взаимозависимы и тесно связа-
ны генетически. Если развитие хлоропластов протекает нор-
мально, на необходимом уровне должно поддерживаться и их
Гл. 10 Фотосинтез
361
снабжение структурными и функциональными компонентами.
Прекращение поступления одного из компонентов, например, в
результате действия ингибитора, приведет к предотвращению
развития хлоропластов и может вызвать блокирование синтеза
других компонентов (разд. 10.10.4).
10.10.2. Развитие хлоропластов в тканях растений,
выросших на свету
Этиолированные проростки, зеленеющие на свету, пред-
ставляют собой удобную синхронную систему, на примере ко-
торой можно изучать развитие хлоропластов, однако в ходе
нормального образования хлоропластов в тканях растений, рас-
тущих на свету, процесс позеленения не происходит. В нормаль-
ных условиях в меристематических тканях развиваются не-
большие пропластиды, которые в конечном итоге и становятся
хлоропластами. Такое превращение происходит без промежу-
точного образования этиопластов. Развитие хлоропластов из
пропластид в растениях, растущих на свету, изучать довольно
трудно, и поэтому пока нет практически никакой информации
о синтезе пигментов и его регуляции в подобных системах.
У растений разных видов или растущих в условиях разной ос-
вещенности отмечены вариации в размерах ССК и в составе
входящих в него пигментов.
В ряде работ показано, что синтез и распад (turnover) хло-
рофиллов и каротиноидных пигментов продолжается и в зре-
лых, функционирующих хлоропластах, однако для более де-
тальной характеристики этих процессов необходимы дальней-
шие исследования.
10.10.3. Развитие хлоропластов у водорослей
Исследований, посвященных развитию хлоропластов у
водорослей, известно немного. Чрезвычайно удобным объектом
оказалась Euglena gracilis, поскольку она образует нормальные
хлоропласты только на свету. В клетках же, растущих в темно-
те, содержатся пропластиды (структуры, сходные с этиопласта-
ми растений), которые на свету превращаются в функциониру-
ющие хлоропласты. Первые 12 ч освещения представляют со-
бой индукционный период, в ходе которого происходит перенос
энергии, небольших молекул, восстановительных эквивалентов
и, наконец, закодированных в ядре белков в развивающуюся
пластиду, в частности из митохондрий, где происходит индуци-
руемый светом распад запасного углевода парамилона. Между
12 и ~96 ч сама пластида обладает высокой активностью, и в
это время в ней образуется большинство хлоропластных ком-
24— S4
362
Часть II. Функциональные аспекты
понентов, в том числе хлорофиллы и каротиноидные пигменты.
Функционирование пластидного и непластидного компартмен-
тов клетки, по-видимому, координируется двумя разными фо-
торецепторами. Один из них, вероятно, представляет собой си-
не-красный фоторецептор, сходный с протохлорофиллидом.
Между растущей в темноте Euglena и молодыми этиолирован-
ными высшими растениями имеется большое сходство в харак-
тере синтеза протохлорофиллида, в организации пропластид и
в фотопревращениях, с которых начинается развитие пластид.
Зеленые водоросли, особенно Chlorella, Scenedesmus и
Chlamydomonas, изучались весьма широко. При росте на свету
они имеют хлоропласты, сходные с хлоропластами высших
растений, и содержат сходный набор фотосинтетических пигмен-
(10.21) ("Каротин
тов. Эти же пигменты, однако, содержатся и в культурах, ра-
стущих в темноте. При переносе культуры на свет не происхо-
дит массированного синтеза ни одного из хлоропластных ком-
понентов, однако свет может быть необходим для фиксирова-
ния пигментов в фотосинтетических мембранах или частицах.
Было получено несколько мутантных штаммов водорослей,
у которых при выращивании в темноте состав пигментов зна-
чительно отличается от состава у дикого штамма; у них может
полностью отсутствовать хлорофилл, а биосинтез каротиноидов
может быть блокирован на одной из ранних стадий, например
на стадии ^-каротина (10.21). При освещении клеток некоторых
из этих штаммов происходит нормальное образование хлоро-
пластов, причем данный процесс в некоторых отношениях схо-
ден с позеленением этиопластов. Это делает такие штаммы
очень удобным объектом для изучения структурных изменений
и превращений пигментов.
Успешно выращивают в темноте лишь несколько видов сине-
зеленых водорослей (Cyanophyceae, или Cyanobacteria); в опы-
тах с одним из них (Chlorogloea fritschii) были получены ин-
тересные результаты. У этого организма хлорофилл и кароти-
ноидные пигменты синтезируются в темноте, но в фотосинте-
тические мембраны при этом не встраиваются. Для того чтобы
это произошло, необходим свет.
Красные (Rhodophyceae) и сине-зеленые водоросли исполь-
зуют в качестве вспомогательных пигментов фикобилины. К со-
жалению, синтез этих соединений, так же как и образование
Гл. 10. Фотосинтез
363
фотосинтетических мембран, и особенно фикобилисом, подроб-
но не изучался.
По-видимому, по крайней мере у некоторых видов, соотно-
шение образующихся фикоцианина (Хтах^620 нм) и фикоэри-
трина (2vmaxc^565 нм) в фикобилинах в большой степени опре-
деляется спектральным составом падающего света. Так, на зе-
леном свету стимулируется синтез поглощающего в зеленой
области красного пигмента фикоэритрина, в то время как на
красном свету преобладает образование поглощающего красные
лучи синего пигмента фикоцианина. Это обеспечивает макси-
мальное поглощение доступного света данными вспомогатель-
ными пигментами.
10.10.4. Общая регуляция образования хлоропластов
и синтеза пигментов
По-видимому, должен существовать общий механизм,
который регулирует образование хлоропласта в целом. Как
осуществляются при этом тонкие взаимодействия компонентов
и их контроль, не известно, однако были обнаружены тесные
генетические взаимосвязи между ними. Должны синтезировать-
ся все компоненты, и все они должны быть доступны для вклю-
чения в тилакоидные мембраны. В противном случае синтети-
ческие процессы подавляются. Например, действие некоторых
гербицидов заключается в подавлении биосинтеза каротинои-
дов. Если этиолированные проростки или культуры водорослей
Euglena, выращенные в темноте, обработать такими гербици-
дами, то нормальные каротиноиды хлоропластов не образуются
и, следовательно, не включаются в фотосинтетические мембра-
ны. В результате не синтезируются и другие компоненты хло-
ропластов, в том числе хлорофилл, и, следовательно, не про-
исходит развития хлоропласта в целом. Даже если это было бы
и не так, то подавление образования каротиноидов привело бы
к тому, что весь синтезированный хлорофилл и зарождающие-
ся фотосинтетические мембраны оказались бы без защиты от
фотоокисления (разд. 10.4.2) и разрушались бы. Поэтому гер-
бициды, подавляющие биосинтез каротиноидов в растениях,
очень эффективны.
10.11. Развитие фотосинтетического аппарата
у фотосинтезирующих бактерий
Многие фотосинтезирующие бактерии, в том числе
Rhodopseudomonas capsulata, R. palustris и R. sphaeroides,
способны расти как в анаэробных условиях на свету, так и в
аэробных условиях в темноте. Интенсивность освещения и пар-
24*
364
Часть II. Функциональные аспекты
циальное давление кислорода являются главными факторами,
управляющими развитием внутренних мембран и обеспечиваю-
щими оптимальное использование источника энергии. Так, в
темноте аэробные условия вызывают увеличение количества
синтезирующихся компонентов дыхательной электронтранспорт-
ной цепи и включение их в мембраны. Вместе с тем клетки, ра-
стущие на свету в бескислородной среде, синтезируют фотосин-
тетические мембраны и пигменты, причем максимального уров-
ня синтез достигает при низких интенсивностях освещения,
когда необходима наибольшая эффективность поглощения све-
та.
Главным фактором, регулирующим развитие фотосинтети-
ческих мембран и синтез пигментов, по-видимому, является
парциальное давление кислорода. Если оно выше определенно-
го уровня, дыхание может происходить с достаточной эффек-
тивностью, но образования фотосинтетических мембран или син-
теза пигментов при этом не наблюдается. Низкое парциальное
давление кислорода стимулирует образование фотосинтетиче-
ского аппарата и пигментов, в первую очередь реакционных
центров и главного комплекса светособирающей антенны Р-875.
В ответ на изменение интенсивности освещения изменяется и
состав пигментов. Так, у Rhodopseudomonas spp., свет низкой
интенсивности стимулирует синтез бактериохлорофилла и ка-
ротиноидов, поскольку происходит формирование вторичного
комплекса светособирающей антенны Р-800-850. Свет высокой
интенсивности подавляет формирование этого комплекса, и в ре-
зультате содержание пигментов снижается. В случае Rhodospi-
rillum rubrum, которая не содержит антенны Р-800-850, содер-
жание пигмента главной светособирающей антенны Р-875 регу-
лируется интенсивностью освещения. О том, как протекают и
регулируются процессы, в ходе которых фотосинтетические пиг-
менты образуются и включаются в мембраны, известно немного.
Гены, контролирующие синтез хлорофилла и каротиноидов, а
также, возможно, развитие активного фотосинтетического ап-
парата в целом, локализованы в хромосоме (но не в плазмиде)
и расположены очень близко друг к другу. В кодировании фо-
тосинтетического аппарата может участвовать одна большая
генетическая единица.
10.12. Дальнейшая судьба фотосинтетических пигментов
в ходе разрушения хлоропластов
Фотосинтезирующие ткани высших растений могут пре-
кращать свою фотохимическую активность по разным причинам
Наиболее известны две из них — старение листьев осенью и со-
Гл. 10. Фотосинтез
365
зревание плодов. В этих случаях хлоропласты прекращают
функционировать и либо разрушаются, либо превращаются в
хромопласты.
10.12.1. Распад хлоропластов в листьях
У большинства деревьев осенью листья изменяют свою
окраску с зеленой на желтую, красную или коричневую и опа-
дают с ветвей. Этот процесс включает распад хлоропластов и
разрушение хлорофилла. Подробно механизм разрушения хло-
рофилла пока не изучен. Известно лишь, что на ранних стади-
ях его распада теряются фитол и атом магния, в результате
чего образуется феофорбид (10.22). Порфириновая система ко-
лец затем расщепляется с образованием бесцветных соединений,
имеющих сравнительно небольшую молекулярную массу.
Хлоропластные каротиноиды целиком не теряются, о чем
свидетельствует желтая окраска старых листьев. |3-Каротин в
заметной степени окисляется через эпоксиды и апо-каротинали,
а ксантофиллы этерифицируются жирными кислотами. Ярко-
красная окраска некоторых осенних листьев обусловлена ин-
тенсивным синтезом в ходе старения антоцианов (гл. 4). Этот
процесс, однако, к распаду хлоропластов непосредственного
отношения не имеет.
10.12.2. Созревание плодов
Незрелые плоды имеют обычно зеленую окраску и со-
держат функционирующие хлоропласты. Во многих случаях,
когда плоды созревают, хлоропласты превращаются в нефото-
синтезирующие хромопласты. По мере потери хлоропластами
фотосинтетической активности разрушается и хлорофилл. Иног-
да разрушаются и хлоропластные каротиноиды, но часто вме-
сто них образуется гораздо большее количество других каро-
тиноидов, которые и придают окраску зрелым плодам. Хорошо
известным примером такой замены служат плоды томата, кото-
рые приобретают красную окраску в результате интенсивного
синтеза ликопина (10.23).
10.13. Заключение и комментарии
Процесс фотосинтеза чрезвычайно важен для жизни на
нашей планете, поскольку он осуществляет утилизацию имею-
щейся в изобилии солнечной энергии в химически доступную
форму путем фиксации СОг с образованием углеводов. Пигмен-
ты, и в первую очередь хлорофилл, играют главную роль в фо-
тосинтезе, и поэтому данный процесс еще долго будет оставить-
366
Часть II. Функциональные аспекты
ся основной областью исследования пигментов. В настоящее
время много внимания уделяется механизму и регуляции син-
теза пигментов, включению их в фотосинтетические мембраны,
ориентации молекул пигментов в фотосинтетическом аппарате
и молекулярным превращениям, которые происходят в течение
очень кратковременных первичных фотореакций. Более подроб-
ная информация о механизмах фотосинтеза позволит создать
простые модельные системы, которые могут быть использова-
соон
(10.22) Феофорбид
(10.23) Ликопин
ны для улавливания и утилизации солнечной энергии. По ме-
ре того как пища и традиционные источники энергии в виде
полезных ископаемых будут становиться все менее доступными,
возникает задача увеличения продуктивности фотосинтеза. Оп-
ределенные успехи в этом направлении уже достигнуты. Так,
в Бразилии такие высокоурожайные культурные растения, как
сахарный тростник, используются для получения сахара, из ко-
торого затем производят этанол, добавляемый в горючее для
двигателей внутреннего сгорания. Фиксированный посредством
фотосинтеза углерод, вероятно, заменит каменный уголь и
нефть в качестве главного источника сырья для химической
промышленности. Одноклеточные водоросли уже широко при-
меняются в качестве богатых витаминами и минеральными ве-
ществами питательных кормовых добавок для животных. Ин-
тенсивно изучаются с целью получения источников водорода
Гл. 10. Фотосинтез
367
системы, содержащие хлоропласты, гидрогеназы и другие ка-
тализаторы. Совершенно очевидно, что в основе успешного осу-
ществления подобных программ должно лежать детальное зна-
ние механизма поглощения света фотосинтетическими пигмен-
тами.
10.14. Рекомендуемая литература
Литература, посвященная фотосинтезу, огромна; число
оригинальных статей, публикуемых ежегодно, насчитывает ты-
сячи наименований. Обзорные статьи и книги также имеются в
изобилии, так что здесь могут быть упомянуты лишь некоторые
из них. Для неспециалистов могут оказаться полезными такие
источники вводного характера, как книги Рабиновича и Го-
винджи (Rabinowitch, Govindjee, 1969) и Грегори (Gregory,
1977), Более подробное общее описание читатель найдет в очень
авторитетной книге под редакцией Говинджи (Gowindjee, 1975).
Чрезвычайно полезными и всеобъемлющими являются пятый и
шестой тома из серии «Encyclopaedia of plant physiology» под
редакцией Требста и Эврона (Trebst, Avron, 1977) и Гиббса и
Латцко (Gibbs, Latzko, 1979) соответственно. Недавно появив-
шаяся серия книг под редакцией Барбера включает тома, по-
священные хлоропластам (т. 1, 1976), фотосинтезу (т. 2, 1977)
и модельным системам, связанным с фотосинтезом и продуци-
рованием энергии (т. 3, 1979). Все аспекты проблем, связанных
с хлоропластами растений и водорослей, включая их структу-
ру, ультраструктуру и развитие, охватывает обширная моно-
графия Кирка п Тилни-Бэссета (Kirk, Tilney-Bassett, 1978). Де-
тали развития хлоропластов и фотосинтетического аппарата
служат предметом другой книги — под редакцией Акоюноглу и
Арджироуди-Акоюноглу (Akoyunoglou, Argyroudi-Akoyunoglou,
1978). Фотосинтез и фотосинтетические пигменты бактерий под-
робно обсуждаются в монографии по фотосинтезирующим бак-
териям (Clayton, Sistrom, 1978). Доклады на проходящих раз
в три года Международных конгрессах по фотосинтезу (Metz-
пег, 1969; Forti et al., 1972; Avron, 1975; Hall et al., 1978; Akoyu-
noglou, 1981) дают представление о текущей работе в сотнях
лабораторий по всему миру, изучающих фотосинтез. Многие
методы общего характера, наиболее полезные при исследова-
нии фотосинтеза, описаны в двух томах «Methods in enzymo-
logy» (San Pietro, 1971, 1972).
10.15. Дополнительная литература
Akoyunoglou G. (ed.) (1981). Proceedings of the 5th International Congress on
Photosynthesis, Philadelphia, Balaban International Science Services.
Akoyunoglou G., Argyroudi-Akoyunoglou J. H. (eds.) (1978). Chloroplast deve-
lopment, Amsterdam, Elsevier.
368
Часть II. Функциональные аспекты
Avron М. (ed ) (1975). Proceedings of the 3rd International Congress on Pho-
tosynthesis, Amsterdam, Oxford and New York, Elsevier.
Barber J. (ed.) (1976). The intact chloroplast. (Topics in photosynthesis, vol. 1),
Amsterdam, Oxford and New York, Elsevier-North-Holland.
Barber J. (ed.) (1977). Primary processes of photosynthesis. (Topics in photo-
synthesis, vol. 2), Amsterdam, Oxford and New York, Elsevier-North Hol-
land.
Barber I. (ed.) (1979). Topics in relation to model systems (Topics in photosyn-
thesis, vol. 3), Amsterdam, Oxford and New York, Elsevier-North Hol-
land.
Clayton R K., Sistrom W. R. (eds.) (1978). The photosynthetic bacteria, New
York, Plenum.
Fortt G., Avron M.. Melandri A. (eds.) (1972). Proceedings of the 2nd Interna-
tional Congress on Photosynthesis Research, The Hague, Junk N. V.
Gibbs M„ Latzko E. (eds.) (1979). Photosynthesis II: Photosynthetic carbon
metabolism and related processes. (Encyclopaedia of plant physiology,
vol. 6), Berlin, Heidelberg and New York, Springer-Verlag.
Govindjee (ed.) (1975). Bioenergetics of Photosynthesis, New York, San Fran-
cisco and London, Academic Press.
Gregory R. P. F. (1977). Biochemistry of photosynthesis, 2nd edition. Chichester,
New York, Brisbane and Toronto, Wiley.
Hall D. 0, Coombs I., Goodwin T. W. (eds.) (1978). Proceedings of the Fourth
International Congress on Photosynthesis, London, The Biochemical Society.
Kirk J. T. 0., Tilney-Bassett R. A. E. (1978). The plastids: their chemistry, struc-
ture, growth and inheritance, 2nd edition, Amsterdam, New York and Oxford,
Elsevier-North Holland.
Metzner H. (ed.) (1969). Progress in photosynthesis research, Tubingen, Laupp.
Rabinowitch E., Govindjee (1969). Photosynthesis. New York, London, Sydney
and Toronto, Wiley.
San Pietro A. (ed ) (1971). Photosynthesis, Part A. (Methods in enzymology,
vol. 23), New York and London, Academic Press.
San Pietro A. (ed.) (1972). Photosynthesis and nitrogen fixation, Part B.
(Methods in enzymology, vol. 24), New York and London, Academic
Press.
Trebst A., Avron M. (eds.) (1977). Photosynthesis I: Photosynthetic electron
transport and photophosphorylation (Encyclopaedia of plant physiology,
vol. 5), Berlin, Heidelberg and New York, Springer-Verlag.
Глава 11
Другие фотофункции природных пигментов
11.1. Введение
Предыдущие главы второй части настоящей книги
были посвящены основным аспектам функционирования при-
родных пигментов — окрашиванию тканей, процессу зрения и
фотосинтезу. Однако существует много других функций, свя-
занных со светопоглощающими свойствами пигментных моле-
кул, правда, эти функции не так широко распространены, не
имеют столь фундаментального значения и не так хорошо
изучены, как рассмотренные в гл. 8—10. Особенности некото-
рых из этих фотофункций пигментов будут описаны в данной
главе.
11.2. Фитохром
11.2.1. Введение
Многие аспекты роста и морфологии растений регули-
руются светом. Большинство (если не все) реакций растений
на свет опосредованы одним чрезвычайно важным фоторецеп-
торным пигментом — фитохромом, который содержится в клет-
ках в очень небольших количествах. В структурном отношении
фитохром представляет собой белок, имеющий в качестве про-
стетической группы (и хромофора) линейный тетрапиррол (би-
лин). Детали структуры и свойств фитохрома приведены в
гл. 5 (рис. 5.16).
Существуют взаимно переходящие друг в друга формы
фитохрома. Одна из них—Рг — поглощает красный свет
Umax=660 нм) и превращается при этом в другую форму —
Pfr. Форма Pfr поглощает свет в дальней красной части спект-
ра (Хтах=730 нм) и в результате превращается опять в Рефор-
му. Эти две формы функционируют как переключающий меха-
низм. В большинстве случаев активной формой, или «включа-
телем», который инициирует физиологические процессы, служит
Pfr-форма. Так, когда красный свет с длиной волны ~ 660 нм
поглощается Реформой, образуется Pfr-форма, что стимулирует
многие фотореакции, например развитие цветков. Однако, если
вслед за освещением красным светом следует вспышка даль-
370 Часть II Функциональные аспекты
него красного света с длиной волны ~730 нм, то Реформа
превращается в неактивную форму Рг, и поэтому светозависи-
мый процесс будет предотвращен.
Что касается того, будет или нет протекать светозависимая
реакция, то это определяется спектром света, которым осве-
щали растение последний раз. Так, после последовательного
освещения дальним красным, красным, дальним красным и
снова красным светом фитохром останется в Pfr-форме и бу-
дет, следовательно, инициировать ответную реакцию. Вместе
с тем после освещения красным, дальним красным, красным и
опять дальним красным фотореакция не инициируется, посколь-
ку последним в данной цепи было освещение дальним красным
светом и фитохром остался в неактивной Рг-форме. Для того
чтобы фотореакция произошла, необходимо освещение крас-
ным светом в течение очень короткого промежутка времени
(порядка нескольких секунд или даже менее) хотя бы части
растения (например, одного листа). Действие красного света
в некоторых случаях может быть сведено на нет обработкой
дальним красным светом даже по прохождении значительного
времени (2—3 ч).
11.2.2. Распространение и локализация
Фитохромные системы обнаруживаются во всем расти-
тельном царстве: у высших растений, мхов, папоротников и
водорослей, правда, более-менее подробно они изучены только
у высших растений. У многих высших растений фитохром со-
средоточен в некоторых специфических тканях. Например,
в этиолированных проростках овса высокие концентрации фи-
тохрома обнаружены в паренхиме и в эпидермальных клетках,
расположенных несколько ниже (на 0,1 —1,5 мм) верхушки
колеоптиля, в то время как в самом ее кончике пигмент от-
сутствует. Внутри клетки фитохром (в Реформе) связан с
ядерной оболочкой, а также содержится в таких органеллах,
как митохондрии, амилопласты, этиопласты и хлоропласты.
По-видимому, Pfr-форма более прочно, чем Реформа, ассоции-
рована с мембранными структурами, в которых, как предпола-
гают, находятся дискретные рецепторные участки. Не исклю-
чено также наличие внутри клетки различных фондов фито-
хрома. Протекание фотореакции обусловлено, возможно, лишь
небольшим количеством прочно связанной Pfr-формы фитохро-
ма, в то время как большая часть фитохрома в Реформе
остается в свободном виде (не связанной с рецепторными уча-
стками) и в инициировании фотореакции непосредственно не
участвует.
Гл, И. Другие фотофункции природных пигментов 371
11.2.3. Примеры процессов, контролируемых фитохромом
Из многих фотореакций, опосредованных фитохромом,
лучше всего изучены, вероятно, инициация цветения, прораста-
ние семян и позеленение этиолированных тканей. В первом
случае очень кратковременное освещение даже части растения
(одного листа) светом требуемой длины волны инициирует
реакцию, для завершения которой необходимо несколько не-
дель. Ясно, что при этом происходит экспрессия новой генети-
ческой информации. В природных условиях начало цветения
определяется длиной дня, или, говоря более точно, продолжи-
тельностью темнового периода. Так, у растений короткого дня
цветение начинается в условиях длинной ночи и короткого
дня, в то время как для растений длинного дня необходимы
прямо противоположные условия — длинный световой и корот-
кий темновой период. В обоих случаях фитохром является фо-
торецептором, который опосредует реакцию.
Прорастание семян в большинстве случаев стимулируется
образующимся на красном свету Pfr-фитохромом, однако в не-
которых случаях свет может подавлять этот процесс.
Кратковременное освещение красным светом этиолирован-
ных проростков влечет за собой глубокие изменения. Послед-
ние выражаются в том, что увеличиваются размеры листьев,
ткани приобретают зеленую окраску (обусловленную синтезом
хлорофилла и других хлоропластных компонентов), а запасы
крахмала интенсивно утилизируются до начала активного фо-
тосинтеза.
У различных растений фитохромная система используется
также как регулятор состояния покоя и старения, роста корней,
движения листьев и общего поддержания внешнего вида в про-
цессе роста.
11.2.4. Способ функционирования фитохрома
Было предпринято много попыток объяснить действие
фитохрома и предлагались различные модели его функциони-
рования. Ясно одно: первичным событием является поглощение
света, вызывающее возбуждение фитохрома, в результате чего
происходит превращение Реформы в Pfr-форму и наоборот
(рис. 5.16). Изменения конформации фитохромного белка при
этом, по-видимому, невелики, однако различия в прочности
связывания между фитохромом и мембранными структурами
для двух форм могут быть значительными.
В недавних исследованиях с применением низких темпера-
тур и флеш-фотолиза обнаружены промежуточные формы меж-
ду Р1г и Рг с различными максимумами поглощения. На осно-
372
Часть II. Функциональные аспекты
вании полученных результатов была предложена схема, описы-
вающая их взаимопревращения (рис. 11.1). Эту циклическую
схему пока еще нельзя интерпретировать на молекулярном
уровне. Показано, что среди ее компонентов встречаются цис-
изомеры билинового хромофора [например, (11.1)], и предпо-
Рис. 11.1. Цикл взаимопревращений двух форм фитохрома (приводится с
учетом обнаруженных к настоящему времени промежуточных соединений).
Вероятно, имеют место также и альтернативные параллельные пути пре-
вращения. Нижние индексы у прописной буквы Р соответствуют максимумам
поглощения света промежуточными формами фитохрома. Ры и Рх — слабо
поглощающие формы. Реакции, помеченные буквой «т», являются темно-
выми.
лагают, что наряду со структурными изменениями, показанны-
ми на рис. 5.16, происходят и изменения конформации белка.
В какой-то степени взаимопревращения двух форм фитохрома
могут происходить и в темноте. При этом Реформа образуется
непосредственно из Ри-формы, в то время как Pfr-форма, ве-
роятно, образуется из промежуточного продукта Peso, а не из
Рг-формы.
Прорастание семян очень часто происходит в темноте, так
что регуляция этого процесса фоторецепторным фитохромом на
Гл. It. Другие фотофункции природных пигментов
373
первый взгляд кажется парадоксальной. Вероятно, это можно
объяснить тем, что фитохром «консервируется» в Pfr-форме во
время созревания семян. Затем в соответствующее время семе-
на поглощают воду, ткани оводняются и Pfr-фитохром начи-
нает активно функционировать.
Полагают, что действие фитохрома опосредуется вещества-
ми с гормональной активностью, такими, как цикло-АМР
А Короткий день
День Вечерние сумерки Ночь Утренняя заря День
Солнечный сеет Красный, дальний красный - 3 1 Красный: дальний красный -0,7:1 Темнота Красный: дальний красный - 0,7:1 Красный: дальний красный - 3.1
Б Длинный день
День
Вечерние
сумерки
Ночь
Утренняя
заря
День
Солнечный свет
Красный; дальний красный
- 3 1
Красный;
дальний
красный
- 0,7:1
Темнота
Красный:
дальний
красный
- 0.7:1
Красный:
дальний красный
/^>3:1
Преобладает Pfr-
‘Преобладает РГ'
Преобладает Pfr
Рис. 11.2. Преобладающие формы фитохрома при разной длине дня.
А. Короткий день/длинная ночь. Б. Длинный день/короткая ночь
(сАМР), ацетилхолин или различные ростовые вещества расте-
ний. В разных тканях, однако, существует много различных
реакций, регулируемых фитохромной системой. Некоторые из
них могут протекать очень быстро (за несколько секунд или
даже быстрее), другие же — значительно медленнее (за не-
сколько часов, дней или даже недель). По-видимому, фитохром
осуществляет свое действие с помощью нескольких разных
механизмов. К таким механизмам можно отнести влияние на
1) ионные потоки, 2) биоэлектрические потенциалы, 3) прони-
цаемость мембран или 4) активность дыхания. Однако все они
являются гипотетическими, и в настоящее время данных,
подтверждающих какой-либо из них, далеко не достаточно.
Детали действия фитохрома были установлены главным
образом при изучении влияния на него красного и дальнего
красного света, но нельзя забывать, что в природных условиях
374 Часть II. Функциональные аспекты
растения либо находятся в темноте, либо освещаются белым
светом. Свет с длиной волны между ~500 и 700 нм обеспечи-
вает равновесие, при котором преобладает Pfr-форма (~80%
всего фитохрома), в то время как свет с длинами волн вне
этого диапазона приводит к равновесию, в котором преобла-
дает Pr-фитохром. Поэтому большое значение имеют измене-
ния спектрального состава падающего света, происходящие в
течение дня. Так, на протяжении большей части дня соотноше-
ние энергий красных и дальних красных лучей в солнечном
свете составляет ~3: 1, что благоприятствует превращению
Рг—>Pfr. Однако в сумерки и в темноте это соотношение сни-
жается до 0,7: 1 (т. е. преобладает дальний красный свет) и
фитохромное равновесие сдвигается в сторону Реформы. Та-
ким образом, в условиях длинного дня в продолжение большего
периода времени в избытке будет содержаться Pfr-форма, в то
время как при коротком дне (и длинной ночи) более длитель-
ное время будет доминировать образующийся в сумерках
Рг-фитохром (рис. 11.2). Этим можно объяснить хорошо изве-
стное влияние продолжительности дня на процессы роста у
растений. Подобные эффекты, а также другие возможные су-
точные ритмы должны приниматься во внимание при оценке
действия фитохрома в природной среде обитания растений.
11.3. Фототаксис
Фототаксисом называют индуцированное светом на-
правленное движение или изменение движения либо всего ор-
ганизма, либо отдельных клеток или органелл. Индуцирован-
ное движение обычно зависит от направления падающего света
и может быть «положительным» (направленным к свету) или
«отрицательным» (от света). Интенсивность освещения может
также влиять на скорость движения; такое явление называется
фотокинезом.
Многочисленные примеры фототаксиса были обнаружены у
водорослей, динофлагеллят, грибов и бактерий; описано также
зависимое от света движение хлоропластов в клетках водорос-
лей. Предполагают, что у разных организмов в фотореакциях
принимают участие сразу несколько пигментов или групп пиг-
ментов, действие которых обусловлено их спектрами действия.
В число таких пигментов входят хлорофилл, бактериохлоро-
филл, каротиноиды, билипротеины, фитохром и рибофлавин.
К сожалению, более подробная их идентификация пока не про-
водилась.
Фототаксис изучали у многих видов, в частности у одно-
клеточных водорослей и динофлагеллят, но наиболее распро-
страненным объектом была Euglena gracilis. При постоянном
Гл. 11, Другие фотофункции природных пигментов 375
однородном освещении клеток или в темноте никакой фото-
реакции не наблюдается. Фотостимуляция Euglena происходит
в ответ на изменение интенсивности освещения выше или ниже
некоторого порогового значения. Нормальная реакция на свет
у Euglena — положительная, т. е. клетки движутся в направ-
лении источника света (интенсивность освещения при этом
возрастает) так, чтобы достигалось наибольшее образование
энергии в результате фотосинтеза. Однако свет очень высокой
Рис. 11.3. Схематическое изображение клетки Euglena gracilis (длина 40—
50 мкм).
интенсивности, который может быть опасным, вызывает отри-
цательный фототропный эффект.— наблюдается характерное
движение клеток от источника света.
На рис. 11.3 схематически изображена клетка Euglena.
С фотодвигательной реакцией связаны две области клетки —
область собственно фоторецептора (парафлагеллярное тело), а
также стигма («глазок»). Фоторецептор получает максимальное
количество света в том случае, если клетка ориентирована та-
ким образом, что падающий свет направлен прямо вниз в от-
верстие, или так называемую «глотку». Чтобы достичь этого,
и происходит соответствующее движение, или ориентация кле-
ток. Первичный фоторецептор в парафлагеллярном теле со-
стоит из упорядоченных флавиновых молекул. Механизм фото-
реакции и путь, с помощью которого в результате поглощения
света происходит двигательная реакция, пока еще не известны.
Стигма также играет важную, хотя и не главную роль в
фототактической реакции. Она содержит высокие концентра-
ции каротиноидов, которые служат экранирующими пигмента-
ми. Поскольку движение Euglena имеет сложный характер и
376
Часть II. Функциональные аспекты
включает вращение, пигментированная стигма периодически
заслоняет фоторецептор от света. Считают, что это регулирует
выпрямление и движение жгутика и таким путем контролирует-
ся направление движения.
11.4. Фототропизм
Фототропизм — это индуцируемый светом рост или об-
разование изгибов у растений и грибов, происходящие обычно
в направлении источника света (положительный фототропизм),
а иногда и от него (отрицательный фототропизм). Очевидно,
что и в этом случае происходит поглощение света фоторецеп-
торным пигментом. Спектры действия, определенные для мно-
гих фототропных реакций, имеют максимумы при 450—460 нм
и очень напоминают спектры поглощения р-каротина (11.2) и
рибофлавина (И.З). После продолжительных дискуссий в на-
стоящее время пришли к тому, что рибофлавин в большей сте-
пени, чем p-каротин, удовлетворяет требованиям фоторецептор-
ного пигмента, и почти несомненно именно он используется в
качестве фоторецепторного компонента.
(112) 3 Каротин
(113) Рибофлавин
В молодых тканях высших растений фототропные реакции
влияют на характер роста. Свет поглощается рибофлавином,
в результате чего происходит либо разрушение ауксина (индо-
лил-3-уксусной кислоты) на свету, либо его удаление из осве-
щенной области. В результате затененная сторона растения
имеет возможность больше удлиняться и побег или стебель
изгибаются в направлении света. В зрелых тканях растений
фототропные реакции обусловлены изменениями тургора под
действием света.
Гл. 11. Другие фотофункции природных пигментов 377
При перемещении или росте грибных гиф в направлении
света фоторецепторным пигментом также служит рибофлавин,
тогда как ауксин в этом не участвует. В некоторых недавно
опубликованных сообщениях высказывается предположение о-
том, что первичный рибофлавиновый фоторецептор образует
комплекс с белком и с антенной из каротиноидных молекул,
которая повышает эффективность улавливания света.
11.5. Бактериородопсин
Некоторые галофильные бактерии способны использо-
вать энергию света для образования АТР с помощью процесса,
который не похож на фотосинтез у растений или бактерий.
В частности, H.halobium используют для образования АТР
обычное аэробное дыхание, если имеется в достаточном коли-
честве необходимый для этого кислород. В условиях же не-
хватки кислорода в клеточной мембране этих бактерий появ-
ляются специфические пурпурные образования, так называемые
заплаты (patches). Пигмент, обусловливающий их пурпурную-
окраску, представляет собой белок бактериородопсин. Послед-
ний служит фоторецептором в процессе превращения энергии
света в протонный градиент, который в свою очередь является
движущей силой синтеза АТР с помощью хемиосмотического
механизма. Фоточувствительная пурпурная мембрана состоит
из липопротеинового матрикса, причем с помощью дифракции
рентгеновских лучей показано, что молекулы бактериородопси-
на расположены в этой мембране в виде жесткой двумерной
решетки.
Бактериородопсин, представляющий собой липопротеин с
мол. массой 26 000, сходен со зрительными пигментами глаза.
В качестве простетической группы он содержит ретинальдегид,
связанный в виде протонированного шиффова основания (или
альдимина) с е-аминогруппой лизинового остатка белка (11.4).
Адаптированная к темноте фотореактивная пурпурная форма
бактериодопсина имеет Хтах —560 нм и, по-видимому, состоит
из смеси двух видов, включающих полностью транс- и 13-цис-
ретинальдегидные хромофоры, которые поглощают соответ-
ственно при 570 и 550 нм. Эти виды содержатся в соотношении
1 : 1 и принимают участие в двух различных, хотя и необяза-
тельно независимых, фотоциклах. Один из них, для транс-фор-
мы, изучен лучше. Его главная особенность заключается в том,
что поглощение света приводит к обесцвечиванию хромофора
и образованию формы с Zmax = 410 нм (М4ю), в которой альди-
мин больше не протонирован (11.5). Ее хромофор, видимо,
имеет 13-цис-конформацию; вполне вероятны и другие конфор-
мационные изменения, касающиеся одинарных или двойных
25-84
378
Часть II. Функциональные аспекты
связей. Было также показано, что виды бактериородопсина,
поглощающие при 625, 610 и 550 нм, представляют собой ко-
роткоживущие (10-12—10-6) промежуточные продукту при об-
разовании М4ю. Предполагают, что между ними и видами, по-
глощающими при 520 и 640 нм, возможны взаимные переходы.
Образование М4ю из транс-бактериородопсина происходит с
потерей протона протонированным альдимином, тогда как для
регенерации бактериородопсина из М4ю необходимо поглоще-
ние протона. Эти-то направленные чередующиеся потеря и по-
глощение протонов и создают протонный градиен!'.
(11 4) Протонлров<1чмп^ форм. альдмммновои связи ретинальдегида с лизином
(115) Henpoiончрованная ферма альдиминовои связи ретинальдегида с лизином
В настоящее время даже приблизительно невозможно опре-
делить последовательность событий в фотоцикле 13-цис-бакте-
риородопсина; известно лишь, что при этом несомненно проис-
ходит г{пс-транс-изомеризация. Вероятно также, что существуют
точки, перекрестно связывающие два цикла, однако более по-
дробные данные об этом отсутствуют.
Между фотоциклами бактериородопсина и зрительного пиг-
мента родопсина (гл. 9) существуют два фундаментальных
различия. Во-первых, в случае бактериородопсина ретинальде-
гидный хромофор никогда не отделяется от белка и альдими-
новая связь остается все время интактной. Во-вторых, кинети-
ка фотоцикла молекулы транс-бактериородопсина зависит от
того, происходят ли фотоциклические изменения и у ближай-
ших соседних с ней молекул.
Образование АТР в бактериородопсиновых циклах служит
одним из лучших примеров функционирования хемиосмотиче-
Гл 11 Другие фотофункции природных пигментов
379
ского механизма. Существенной особенностью действия бакте-
риородопсина является установление протонного градиента.
Свет индуцирует депротонирование протонированной альдими-
новой группы бактериородопсина, и образующийся при этом
обесцвеченный пигмент впоследствии вновь протонируется в
ходе темновой реакции Выделение и поглощение протонов
происходит в строго определенном направлении: потеря про-
тона происходит с наружной, а поглощение протона — с внут-
ренней стороны мембраны. Таким образом, через мембрану
устанавливается градиент протонов и заряда, который и ис-
пользуется в качестве движущей силы для синтеза АТР из
ADP, катализируемого АТРазой (ср. с фотосинтетическим фос-
форилированием, гл. 10).
Функционирование бактериальной пурпурной мембраны
вызывает неослабевающий интерес. Это связано с тем, что в
его основе лежит относительно простой механизм улавливания
энергии света без участия хлорофилла, а рецепторный пигмент,
бактериородопсин, сходен со зрительными пигментами живот-
ных.
11.6. Внеглазные фоторецепторы у животных
11.6.1. Введение
В настоящее время не вызывает сомнений, что у мно-
гих животных сетчатка глаза является не единственной свето-
чувствительной тканью. Внеглазные фоторецепторы обнаруже-
ны к настоящему времени у многих видов как позвоночных,
так и беспозвоночных животных. Эти фоторецепторы не позво-
ляют животному «видеть», как это происходит при истинном
зрении, когда животное способно воспринимать образ, а также
быстро распознавать форму, положение и перемещение объек-
та в пространстве. Однако они принимают участие в опосредо-
вании долговременных эффектов, которые зависят от изменений
общей интенсивности освещения. Примерами процессов, кото-
рые регулируются светом, детектируемым внеглазными рецеп-
торами, могут служить поддержание суточных ритмов и ритмов
с более длинными периодами (лунных) изменения окраски в
ответ на изменения освещенности фона (посветление или потем-
нение кожи) и изменения сроков метаморфоза (влияние на
диапаузу у некоторых насекомых). Тот факт, что реакция на
интенсивность освещения осуществляется не с помощью глаз,
а каким-то иным путем, можно подтвердить тем, что эта реак-
ция не подавляется и не ослабляется у ослепленных животных.
Фоточувствительные ткани могут быть локализованы в специ-
фических органах, таких, как глазки (стигмы) и эпифиз, либо
25*
380
Часть II. Функциональные аспекты
.фоточувствительность может проявлять наружный покров в
целом. Ниже мы приведем несколько примеров, однако следует
отметить, что экспериментальных данных очень мало и ни в
одном из исследованных случаев фоторецепторный пигмент не
был убедительно идентифицирован. Скорее всего, существуют
различные механизмы фотореакций, и маловероятно, чтобы
они представляли собой сложные регенерационные циклы, по-
добные описанному в гл. 9 для родопсина.
11.6.2. Медиальный глазок беспозвоночных
Одним из наиболее высокоразвитых примитивных фо-
торецепторов является так называемый медиальный глазок
многих членистоногих. Так, например, мечехвост Limulus ро-
lyphemus обладает парой таких рецепторов, которые располо-
жены на средней линии тела. Хотя эти органы представляют
собой примитивные глазки, они снабжены хрусталиковыми
структурами.
Чувствительность таких глазков на порядок ниже, чем у
обычного (сложного) глаза, однако длины волн, при которых
чувствительность максимальна, у них сходны (520—530 нм).
Глазок имеет второй рецепторный пигмент с высокой чувстви-
тельностью к УФ-свету (370 нм). Считают, что пигменты глаз-
иков принадлежат к родопсиновому типу.
11.6.3. Эпифиз и подобные ему ткани
Было показано, что у многих видов рыб и земновод-
ных эпифиз или близко примыкающий к нему участок проме-
жуточного мозга чувствительны к свету и оказывают влияние
на регуляцию меланофорных реакций в механизмах изменения
-окраски. Во многих случаях эпифиз ответствен за реакцию по-
бледнения кожи; животные, у которых он удален, остаются
темными. В отсутствие света реакция эпифиза состоит в том,
что он выделяет гормон, опосредующий диспергирование ме-
ланина, в результате чего происходит потемнение тканей. Свет,
уловленный эпифизом, подавляет эту реакцию, и происходит
посветление тела.
Фоторецепторные структуры были найдены в эпифизе зем-
новодных. Аналогичные структуры были обнаружены в род-
ственных органах рептилий — в так называемом «лобном орга-
не» и в «теменном глазу». Рецепторные клетки этих структур,
по крайней мере внешне, сходны с фоторецепторами сетчатки
и связаны с мозгом нервами. Природа пигментов в фоторецеп-
торных структурах пока не выяснена.
Гл. 11. Другие фотофункции природных пигментов 381
В чем-то сходна с ними гардерова железа у новорожденных
грызунов, которая, как считают, является внеглазным фоторе-
цептором, контролирующим их циркадианные ритмы. Эта же-
леза наряду со значительным количеством протопорфирина IX
(11.7) содержит необычный, с тремя карбоксильными группа-
ми, порфирин (11.6). Напрашивается предположение, что эти
порфирины представляют собой фоторецепторные пигменты, но
оно пока не проверено.
11.6.4. Наружные покровы животных как фоторецептор
Известно, что у многих животных фоточувствительны
наружные покровы, а иногда и более глубоко лежащие ткани.
Особенно чувствительными могут быть некоторые специфиче-
ские области тела, например сифон у двустворчатых моллюсков
и асцидий. Явление фоточувствительности наружных покровов
может быть продемонстрировано на животных, у которых уда-
лены глаза, эпифиз и глазки, но которые все еще сохраняют
способность к фотореакции; она исчезает лишь в том случае,
(11 6) Гардеропорфирин
(117) Протопорфирин IX
382
Часть II. Функциональные аспекты
если поверхность тела покрыть непрозрачной краской. Обычно
в наружных покровах таких животных в значительном количе-
стве содержатся каротиноиды, а иногда присутствуют и пиг-
менты других классов. Однако специфические фоторецептор-
ные клетки у них, как правило, отсутствуют, и это затрудняет
локализацию и идентификацию фоторецепторных пигментов.
Тот факт, что такой пигмент должен содержаться лишь в не-
значительном количестве, делает его идентификацию чрезвы-
чайно трудной, поскольку он легко может быть замаскирован
гораздо большими количествами главных пигментов наружных
покровов. Исследования поведения животных и спектров дей-
ствия пигментов позволили в некоторых случаях найти ключ
(11.8) ЭхинохромА
к установлению того, к какому классу пигментов принадлежит
данное соединение.
Явление фоточувствительности наружных покровов обнару-
жено у отдельных представителей всех главных систематиче-
ских групп беспозвоночных. По-видимому, оно часто встречает-
ся у водных животных, а у насекомых фоточувствительность
играет важную роль в регуляции циркадианных ритмов.
Большинство доказательств (хотя и не очень строгих) сви-
детельствует в пользу того, что рецепторными веществами
дермы являются каротиноиды или ретинальдегид-белковые
комплексы. Зарегистрированные спектры действия, например
максимумы чувствительности фоторецепторных пигментов не-
которых двустворчатых моллюсков, усоногих раков и рыб, ча-
сто находятся в том же диапазоне, что и в случае родопсин —
порфиропсина.
У некоторых морских звезд, голотурий и насекомых, а так-
же у многих червей активными пигментами могут быть порфи-
рины. Предполагают, что и рибофлавин может обладать полез-
ной фотосенсибилизирующей функцией в наружных покровах
голотурий и в покровах спинной стороны ракообразных.
В некоторых случаях в качестве фоторецепторов функцио-
нируют нервные клетки. Хорошо известный пример — гениталь-
ный ганглий у моллюска Aplysia. Нервные клетки этого ган-
глия содержат каротиноид и гемопротеин и поэтому очень чув-
Гл. И. Другие фотофункции природных пигментов
383
ствительны к свету именно тех длин волн, которые максималь-
но поглощают эти пигменты. У других животных фоторецепто-
рами могут служить пигменты других классов. Непосредствен-
ная фоточувствительность нервного тяжа у Branchiostoma при-
писывается наличию в нем меланина, а у иглокожего Diadema
и у морской лилии Antedon наблюдаемые спектры действия
указывают на присутствие хинона эхинохромового типа (11.8).
Эти примитивные фотореакции можно разделить на две
главные группы: на явные трансформации и на более медленно
протекающие реакции, подобные наблюдаемым при циркадиан-
ных циклах. В обоих случаях механизмы их действия пока не
выяснены.
11.7. Фотозащита
11.7.1. Введение
До сих пор обсуждался вопрос о том, как свет с по-
мощью различных механизмов используется живыми организ-
мами с пользой для них, но не следует упускать из виду тот
факт, что световая энергия может также наносить повреждения
живым тканям. При поглощении света природным пигментом
•его молекула переходит в нестабильное и короткоживущее
возбужденное состояние с повышенным уровнем энергии. Этот
.избыток энергии обычно используется в организме; например,
•он обеспечивает синтез АТР (в фотосинтетическом транспорте
электронов) или образование нервного импульса, вызывающего
ответную реакцию. Примерами такого использования света слу-
жат различные фотофункции природных пигментов, рассмот-
ренные в данной книге.
Однако энергия, уловленная в возбужденных молекулах,
может также вызывать повреждения, осуществляя нежелатель-
ные химические реакции, которые могут привести к разруше-
нию живых тканей. Различные природные пигменты (особенно
порфирины) могли бы служить сенсибилизаторами для ката-
лизируемых светом повреждений, особенно в присутствии кис-
лорода, если бы не было соответствующего защитного механиз-
ма. Вредное действие на ткани может оказывать не только
видимый свет. Ультрафиолет высокой интенсивности погло-
щается белками и нуклеиновыми кислотами и потенциально
способен повреждать эти жизненно важные молекулы.
Одна из наиболее существенных функций многих природ-
ных пигментов, главным образом каротиноидов и меланинов,
заключается в защите жизненно важных тканей от фотодина-
мического и в первую очередь фотоокислительного поврежде-
ния. Подробно описаны три случая такого фотозащитного дей-
384
Часть II. Функциональные аспекты
ствия. На двух из них — роли каротиноидов в защите бакте-
рий от фотоокисления и предотвращении развития фотоинду-
цированных повреждений у животных—-мы остановимся ниже.
О третьем случае — фотозащите фотосинтетических тканей —
уже упоминалось в гл. 10.
11.7.2. Восстановление ДНК,
поврежденной УФ-излучением
ДНК кодирует генетическую информацию, которая
определяет практически все структуры и функции самих кле-
ток, а также организма, частью которого они являются. По-
вреждение молекулы ДНК ведет к изменению информации, за-
кодированной в последовательности ДНК и потому может
иметь ужасные последствия, если не будет исправлено. Посколь-
ку число молекул ДНК в клетке невелико, а величина этих
молекул огромна, существует большая опасность их поврежде-
ния химическими веществами, излучением и т. п. Особенно хо-
рошо изучено действие УФ-лучей. Нуклеиновые кислоты погло-
УФ-свет (260 нм)
Фермент / свет
(300-500 нм)
Тиминовые основания
Рис. 11.4. Димеризация соседних тиминовых оснований в ДНК и обращение
этого процесса под действием фотоактивируемого репарнрующего фермента.
(11.9) Тиминовый "циклобутановый димер
щают УФ-излучение с Х=260 нм, которое может вызвать в их
молекулах структурные изменения. В основном при этом про-
исходит (рис. 11.4) димеризация в ДНК соседних пиримидино-
вых оснований (обычно остатков тимина) с образованием так
называемых «циклобутановых продуктов» (11.9). В результате
димеризации нарушаются нормальные водородные связи меж-
ду цепями и, следовательно, разрушается структура двойной
спирали в этом участке макромолекулы.
К счастью, клетки имеют ряд защитных механизмов, по-
средством которых они могут восстанавливать повреждения
в цепях ДНК. В частности, для исправления повреждений,
Гл. 11. Другие фотофункции природных пигментов 385
вызываемых различными воздействиями, в том числе и УФ-из-
.лучением, используется механизм репарации путем выщепле-
ния, в ходе которого небольшой участок поврежденной цепи
ДНК удаляется и замещается новосинтезированным неповреж-
денным участком. Многие клетки, главным образом бакте-
риальные (но также и клетки человека) используют для репа-
рации ДНК механизм фотореактивации, с помощью которого
повреждение может быть исправлено без расщепления молеку-
лы ДНК. При этом с молекулой ДНК, содержащей пиримиди-
новый димер, связывается фермент. В результате поглощения
света (300—500 нм) комплексом фермент —ДНК фермент ак-
тивируется и восстанавливает поврежденный участок молеку-
лы, расщепляя димеры с образованием нормальных пиримиди-
новых оснований.
11.7.3. Защита от фотоокисления у бактерий
Известно много случаев, когда бескаротиноидные му-
танты бактерий (в норме образующих каротиноиды) гибнут в
результате совместного действия света и кислорода, в то время
как окрашенные организмы, принадлежащие к дикому типу,
в тех же условиях повреждений не получают. Защитное дей-
ствие каротиноидов было продемонстрировано на примере как
фотосинтезирующих, так и нефотосинтезирующих бактерий.
Фотозащитным действием обладают лишь те каротиноиды,
которые поглощают в видимом диапазоне (>400 нм), т. е. для
такого действия необходим хромофор, содержащий по крайней
мере восемь сопряженных двойных связей. Возможно, что в
некоторых случаях каротиноиды в клеточной оболочке осуще-
ствляют защиту клетки, отфильтровывая свет опасных длин
волн, однако обычно они более непосредственно участвуют в
процессе фотосенсибилизации.
Фотодинамическая гибель бактерий наблюдается при на-
личии света, кислорода, а также эндогенного или экзогенного
фотосенсибилизатора. Свет поглощается сенсибилизирующим
пигментом, который подвергается межсистемным превращениям
и переходит в более долгоживущее, но все еще высокоэнерге-
тическое триплетное состояние. Пигмент в таком состоянии
способен передавать избыток энергии молекулярному кислоро-
ду, переводя его в высокореакционноспособное синглетное со-
стояние (‘О2). Образовавшийся синглетный кислород может
окислить любую подходящую акцепторную молекулу, напри-
мер молекулу ненасыщенной жирной кислоты. При этом обра-
зуются перекиси, которые могут вызвать обширные, а иногда
и летальные повреждения чувствительных внутриклеточных
процессов.
386
Часть II. Функциональные аспекты
Каротиноид может нарушить эту последовательность собы-
тий, выступая в роли либо предпочитаемого субстрата окисле-
ния, либо акцептора избытка энергии триплетного сенсибили-
затора или Ю2.
Образующийся при этом триплетный каротиноид распадает-
ся, не вызывая нарушений. Подобный защитный механизм
очень сходен с описанными механизмами защиты фотосинтети-
ческих мембран (разд. 10.4.2), показанными на рис. 10.8.
Защита от фотоокисления, вероятно, является главной функ-
цией каротиноидов у нефотосинтезирующих микроорганизмов.
В организмах некоторых примитивных животных для защиты
от фотоокисления используются сходные механизмы.
Возможно, у некоторых организмов и в некоторых тканях
аналогичную фотозащитную функцию могут выполнять другие
поглощающие свет соединения, обладающие антиокислитель-
ным действием (например, хиноны), однако сколько-нибудь
подробно в этом отношении изучены пока только каротиноиды.
Синглетный кислород, образующийся не в фотохимических,
а в каких-то других процессах, также может быть опасен для
бактерий. Предполагают, что фагоцитоз бактерий полиморфно-
ядерными лейкоцитами животных происходит с участием об-
разующегося биохимическим путем ’О2. В этом случае кароти-
ногенные бактерии оказываются более защищенными от воз-
действия *О2, чем бескаротиноидные.
11.7.4. Фотозащита у животных
У животных защита от облучения светом обычно обес-
печивается экранирующим слоем пигмента, который либо по-
глощает свет всех длин волн, либо отфильтровывает особенно
опасные лучи. Для целей экранирования наилучшими являют-
ся, очевидно, темные пигменты, такие, как меланины, посколь-
ку они сильно поглощают во всем диапазоне видимого и УФ-
света. Наиболее известным примером фотозащитной реакции
служит пигментация кожи человека. Так называемая «белая
кожа» у человека почти прозрачна, однако кератин рогового
слоя поглощает значительную часть УФ-лучей. В ответ на про-
должительное освещение солнечным светом в процессе форми-
рования солнечного загара усиливается образование кератина
и особенно меланина. Загоревшая белая кожа пропускает лишь
5% УФ-лучей с длиной волны 300 нм, тогда как незагоревшая—
25%. У людей с темной кожей почти весь УФ-свет поглощается
значительными количествами меланина в коже. Этим обеспечи-
вается адекватная защита от высоких доз лучистой энергии,
характерных для тех областей земного шара, где живут тем-
ные расы.
Гл. 11. Другие фотофункции природных пигментов 387
Защитные меланины встречаются и у низших животных.
Черный слизень (Arion ater) накапливает в наружном покрове
меланин в количествах, пропорциональных количеству фотоди-
намически свободного порфирина. Сходные примеры корреля-
ции между содержанием меланина и свободных порфиринов
были описаны у морских животных.
Свободные порфирины, будучи фотосенсибилизаторами, мо-
гут оказывать вредное (даже губительное) действие на млеко-
питающих, в том числе и на человека. Они могут накапливать-
ся при нарушении метаболизма гема, поскольку являются про-
межуточными продуктами этого процесса. Для больных пор-
фирией (разд. 5.10.1) характерна чрезвычайная фоточувстви-
тельность; они совершенно не выдерживают освещения солнеч-
ным светом. У них отсутствует эндогенный экранирующий пиг-
мент для защиты от действия света. Если больному порфирией
ввести p-каротин, то он будет откладываться в тканях кожи и
поглощать свет опасных длин волн. Такой искусственный пиг-
ментный экран предотвращает поглощение света порфириновым
фотосенсибилизатором.
Считают, что у низших животных в качестве фильтрующих
или экранирующих пигментов функционируют не каротиноиды,
а соединения других классов, например нафтохиноны у игло-
кожих. Каротиноиды — широко распространенные пигменты
наружных покровов — являются антиоксидантами и в то же
время активно поглощают свет. Вполне вероятно, что у живот-
ных они осуществляют защиту от фотоокисления так же, как
они делают это у бактерий (разд. 11.7.3). Высокие концентра-
ции каротиноидов в яйцах животных, живущих на суше и на
мелководье, могут отражать эту фотозащитную роль.
11.8. Биолюминесценция
11.8.1. Введение
Предшествующие части этой главы, да и всей книги,
были посвящены поглощению света молекулами различных
пигментов. В заключение следует хотя бы кратко остановиться
на обратном процессе — биолюминесценции, — в котором био-
химическая энергия используется живыми организмами для
генерирования видимого света.
11.8.2 Распространение биолюминесценции
Биолюминесценция была обнаружена у морских бак-
терий и динофлагеллят, а также у небольшого числа видов
грибов. Это явление, однако, лучше изучено у представителей
388 Часть II. Функциональные аспекты
животного царства. Наиболее известный пример — светляк
(и его личинка) — наземное насекомое, которое ночью для
привлечения партнера использует вспышки или непрерывно ис-
пускаемый свет. Однако наиболее часто биолюминесценция
встречается у морских животных: рыб, ракообразных, моллюс-
ков, кольчатых червей и кишечнополостных. Биолюминесцен-
ция некоторых светящихся животных, главным образом рыб и
некоторых головоногих моллюсков, обусловлена их симбиозом
с колониями испускающих свет бактерий. У других животных
люминесценция является их собственной особенностью и не за-
висит от симбионтов.
Испускающие свет бактерии принадлежат к двум родам:
все симбиотические виды — к роду Photobacterium, а все сво-
бодноживущие — к роду Berteckea. У животных, которые ис-
пользуют Photobacterium, микроорганизмы содержатся в спе-
циальных светящихся органах. Наиболее удивительным приме-
ром такого органа служит, вероятно, «приманка» морского чер-
та. Поддерживание существования бактериальных симбионтов
в таких органах выдвигает свои проблемы. Распространение
бактерий внутри тела животного должно ограничиваться свето-
носным органом. Кроме того, они должны передаваться без
загрязнений бактериями других видов в светоносные органы
особей последующих поколений. Бактерии люминесцируют не-
прерывно, так что животным необходим какой-то механизм,
контролирующий испускание света этим органом. Такой конт-
роль может осуществляться с помощью либо 'перемещения са-
мого органа, либо хроматофоров, поглощающих испускаемый
свет, либо различных механизмов типа «жалюзи». Люминесцен-
ция у животных, которые не используют бактерий, также про-
исходит лишь в специальных светоносных органах. У таких
животных число светоносных органов обычно больше, чем у
симбиотических видов, и они, как правило, имеют характерный
рисунок расположения.
11.8.3. Механизмы
Обычно явление биолюминесценции объясняют на при-
мере окисления субстрата (люциферина) ферментом (люцифе-
разой), в результате которого на одной из стадий этого про-
цесса происходит испускание фотона. В недавних исследованиях
были выяснены детали реакций, протекающих у некоторых
организмов.
Конечно, было бы идеально разделить примеры биолюми-
несценции на сравнительно небольшое число типов и разра-
ботать классификацию по типу реакций. Однако в настоящее
Гл. 11. Другие фотофункции природных пигментов
389-
время фрагментарность наших знаний о биолюминесценции не
позволяет сделать это.
Для непрерывного испускания света бактериям необходим
фермент (названный бактериальной люциферазой), длинноце-
почечный алифатический альдегид или родственное ему соеди-
нение в качестве субстрата (так называемого люциферина),
молекулярный кислород и восстановленный флавинмононуклео-
тид (FMNH2). Полагают, что свет испускается в виде флуорес-
ценции возбужденным окисленным флавиннуклеотидом, обра-
зующимся в ходе реакции.
Лучше других изучена биолюминесценция светляка. Снача-
ла люциферин [лампирин (11.10)] реагирует с АТР с образо-
ванием связанного с ферментом люциферин-АМР-производного,
которое затем окисляется до оксилюциферина (11.11) молеку-
лярным кислородом в реакции, протекающей в несколько ста-
дий. Фермент при этом претерпевает ряд значительных кон-
формационных изменений, и в результате на одной из стадий,
происходит испускание света.
Механизмы испускания света другими организмами в прин-
ципе сходны и различаются лишь степенью ассоциации или
сложности люциферин-люциферазной системы. Так, у гидроид-
ной медузы Aequorea люцифераза и люциферин [коэлентеразин
(11.12)] остаются прочно связанными в виде стабильного фо-
(11 10) Лампирин (люциферин светляка)
(11 11) Дегидролампирин (оксилюциферин))
‘J2) Коэлентеразин
(11 13) Коэлентеразин
(связанная с белком форма в
стабильном комплексе — экворине)
390
Часть II. Функциональные аспекты
тобелкового комплекса (11.13). У кораллового полипа — Ren.il-
1а — тот же самый люциферин (11.12) сохраняется в виде суль-
фатного производного и высвобождается в результате удаления
сульфатной группы при действии 3',5'-дифосфоаденозина и
фермента люциферинсульфокиназы. В присутствии молекуляр-
ного кислорода неустойчивый люциферин окисляется до окси-
.люциферина и происходит испускание света.
(11.15) Люциферин Latia
Небольшой ракушковый рачок Cypridina образует свой лю-
циферин (11.14) и люциферазу в разных железах. Оба компо-
нента выпускаются струей в морскую воду, где происходит
люминесцентная реакция. Некоторые рыбы, очевидно, для обес-
печения себя люциферином и люциферазой используют Cypri-
dina в пищу.
Хотя биолюминесценция водных животных обычно связана
с морской средой, хорошо известна люминесценция и у одного
пресноводного вида — брюхоногого моллюска Latia. Люцифе-
рин Latia (11.15) очень сходен с каротиноидными пигментами
(гл. 2).
Окраска (т. е. длина волны) испускаемого света в разных
случаях различна; например, динофлагеллятам присуща флуо-
Гл. И. Другие фотофункции природных пигментов 391
ресценция красного цвета, a Renilla и Aequorea — сине-зелено-
го. На эту окраску в значительной степени влияет конформа-
ция фермента, которая в свою очередь подвержена влиянию
таких факторов, как температура, величина pH, давление, кон-
центрация солей и АТР.
11.8.4. Функции биолюминесценции
Цели, которым служит биолюминесценция у свободно-
живущих бактерий, пока не ясны. Гораздо легче понять воз-
можную пользу, извлекаемую из биолюминесценции животны-
ми. Люминесценция, как собственная, так и обусловленная
бактериями-симбионтами, используется животными, живущими
в темноте, с теми же целями, что и окраска животными, жи-
вущими в мире света, т. е. для привлечения жертв или брачных
партнеров, для предупреждения (предостережения), а у неко-
торых рыб для маскировки путем искажения темного силуэта,
который, если смотреть снизу, хорошо виден на светлом фоне.
11.9. Заключение и комментарии
В первой части настоящей книги были описаны основ-
ные характеристики главных групп природных пигментов.
В предыдущих главах второй части обсуждались наиболее из-
вестные и понятные биологические функции этих пигментов, а
именно окрашивание, улавливание света и распознавание цве-
та (зрение), а также улавливание энергии света в фотосинтезе.
В этой последней главе объединены некоторые другие аспекты
фотобиологии, описаны процессы, в которых природные пиг-
менты играют важную роль. Здесь рассмотрены фоторецепто-
ры, такие, как фитохром и флавины, которыми обладают расте-
ния и микроорганизмы, а также бактериородопсин, используе-
мый для образования АТР у Halobacteria.
Один из разделов посвящен встречающимся у животных,,
но пока неидентифицированным внеглазным фоторецепторам.
Кратко рассмотрены также защитные функции пигментов, ко-
торые помогают живым организмам избегать вредного действия
облучения УФ- и видимым светом. Хотя эти функции пигментов
и не вызывают столь сильного интереса, как перечисленные
выше, они чрезвычайно важны для выживания организмов. На-
конец, некоторое внимание было уделено и биолюминесценции,
с помощью которой живые организмы используют химическую-
энергию для испускания света.
В области идентификации фоторецепторов, а также изуче-
ния на молекулярном уровне изменений, происходящих в ходе
функционирования этих фоторецепторов и наблюдаемых при
392 Часть II. Функциональные аспекты
этом реакций, для биохимиков существует широчайшее поле
деятельности. Такая работа чрезвычайно трудна, поскольку
фоторецепторы обычно присутствуют в тканях в крайне малых
количествах, а изменения, по которым можно судить о реакции,
могут быть очень незначительными. Эти проблемы представ-
ляют собой вызов мастерству биохимика. Наряду с явлениями,
описанными в этой главе, по-видимому, существует много дру-
гих форм или примеров взаимодействия между светом и живы-
ми организмами (через природные пигменты), которые пока
еще не открыты и могут служить новыми объектами исследо-
вания.
11.10. Рекомендуемая литература
Как и следовало ожидать, к главе, охватывающей
столь широкое разнообразие тем и направлений, можно дать
очень много ссылок. Приводимый ниже список ни в коей мере
не может считаться исчерпывающим. Все входящие в его со-
став литературные источники очень информативны, однако в
хорошей библиотеке читатель найдет другие книги и обзоры
литературы по этим же проблемам, которые также окажутся
полезными.
Несколько книг, посвященных фотобиологии, включают боль-
шинство или даже все темы, затронутые в этой главе. Клэйтон
(Clayton, 1971) дает ценное вводное описание, а в книгах Ка-
стеллани (Castellani, 1977), Чекуччи и Вила (Checucci, Weal,
1973), Смита (Smith, 1977) и Уолкена (Wolken, 1975) освеще-
ны подробно различные аспекты фотобиологии. Ежегодная се-
рия «Photochemical and photobiological Reviews» (Smith, 1976—
1979) содержит самые современные обзорные статьи по ряду
проблем фотобиологии.
Для получения информации по фототаксису и фототропизму
читателю следует обратиться к книгам Клэйтона (Clayton,
1971) и Уолкена (Wolken, 1975) и к статье Хэнда (Hand, 1977).
Много публикаций посвящено фитохрому. В некоторых из них
рассмотрены и биохимические аспекты проблемы, в частности
в книге Митракоса и Шропшира (Mitrakos, Shropshire, 1972),
обзорах Ли (Lee, 1977), Пратта (Pratt, 1979), Сэттера и Гэл-
стона (Satter, Galston, 1976), Шропшира (Shropshire, 1977), а
также Смита и Кендрика (Smith, Kendrick, 1976). В работах
о бактериородопсине и пурпурных мембранах Halobacteria ча-
ще всего приводят ссылки на книгу под редакцией Каплана и
Гинзбурга (Caplan, Ginzburg, 1978) и обзор Оттолени (Otto-
lenghi, 1980), содержащий наиболее современные взгляды на
механизм фотоциклов бактериородопсина.
,Гл- 11- Другие фотофункции природных пигментов 393
Фотозащитное действие пигментов, и в первую очередь
каротиноидов, описано Кринским (Krinsky, 1971), а информа-
цию о фотоповреждениях и фоторепарации ДНК можно полу-
чить из книги Ванга (Wang, 1976) и из недавно вышедшего
обзора Лемана и Бриджеса (Lehmann, Bridges, 1977). Не-
сколько книг и обзоров (например, Millott, 1968; Eakin, 1974;
Menaker, 1977; Bennett, 1979; Yoshida, 1979) посвящены вне-
глазным фоторецепторам; в них подчеркивается отсутствие
биохимических сведений по этому предмету. И наконец, био-
люминесценция рассматривается в обзорах Гастингса (Has-
tings, 1968), Джонсона и Ханеды (Johnson, Haneda, 1966) и
Уарда (Ward, 1979), а также в недавно вышедшей книге под
редакцией Герринга (Herring, 1978). Соответствующие методы
собраны в томе «Methods in enzymology» (De Luca, 1978).
11.11. Дополнительная литература
Bennett Al. F. (1979). Extraocular light receptors and circadian rhythms. In:
Handbook of sensory physiology, vol. VII/6A. Vision in invertebrates, ed.
H. Autrum, p. 641, Heidelberg, Berlin and New York, Springer-Verlag.
Caplan S. R., Ginzburg M. (eds.) (1978). Energetics and structure of halophilic
microorganisms, New York, Elsevier-North Holland.
Castellani A. (ed.) (1977). Research in photobiology, New York and London,
Plenum.
Checucci A., Weal R. A. (eds.) (1973). Primary molecular events in photobiology,
Amsterdam, Elsevier.
Clayton R. K. (1971). Light and living matter, vol. 2, The biological part, New
York, McGraw-Hill.
De Luca M. (ed.) (1978). Bioluminescence and chemiluminescence (Methods in
enzymology, vol. 57), New York, Academic Press.
Eakin R. M. (1974). The third eye, Berkeley, University of California Press.
Hand W. G. (1977) Photomovement. In: The sciences of photobiology, ed.
К. C. Smith, p. 313, New York and London, Plenum.
Hastings J. W. (1968). Bioluminescence, Ann. Rev. Biochem., 37, 597.
Herring F. J. (ed.) (1978). Bioluminescence in action, London, New York and
San Francisco, Academic Press.
Johnson F. H, Haneda Y. (eds.) (1966). Bioluminescence in progress. New
Jersey, Princeton University Press.
Krinsky N. I. (1971). Function. In: Carotenoids, ed. O. Isler, p. 669, Basel and
Stuttgart, Birkhauser. _ v c
Lee J. (1977). Bioluminescence. In: The science of photobiology, ed. К. C. Smith,
p. 371, New York and London, Plenum. .
Lehmann A. R„ Bridges B. A. (1977). DNA repair, Essays. In: Biochemistry,
13, 71, London, The Biochemical Society-Academic Press. h„tnhi„Uov
Menaker M. (1977). Extraretinal photoreception. In: The science of photobiology,
ed. К. C. Smith, p. 227, New York and London, Plenum.
Millott N. (1968). The dermal light sense, Symp Zool. Soc. London,
Mitrakos K„ Shropshire W„ Jr. (eds.) (1972). Phytochrome, London and New
York, Academic Press. n , .
OHolenghi M. (1980). The photochemistry of rhodopsins, Adv. Photocnem.,
12, 97. ...
Pratt L. H. (1979). Phytochrome. In: Photochemical and photobiological reviews,
vol. 4, ed. К. C. Smith, p. 59, New York, Plenum.
26—84
394 Часть II. Функциональные аспекты
Satter R. L„ Galston A. W. (1976). The physiological functions of phytochrome.
In: Chemistry and biochemistry of plant pigments, 2nd edition, vol. 1, ed.
T. W. Goodwin, p. 681, London, New York and San Francisco, Academic
Press
Shropshire W„ Jr. (1977). Photomorphogenesis In: The science of photobiology,
ed. К. C. Smith, p. 281, New York and London, Plenum.
Smith H , Kendrick R. E. (1976). The structure and properties of phytochrome.
In: Chemistry and biochemistry of plant pigments, 2nd edition, vol. 1, ed.
T. W. Goodwin, p. 378, London, New York and San Francisco, Academic
Press.
Smith К. C. (ed.) (1976—1979). Photochemical and photobiological reviews,,
vols 1—4, New York, Plenum.
Smith К- C. (ed.) (1977). The science of photobiology, New York and London,
Plenum.
Wang S. У. (ed.) (1976). Photochemistry and photobiology of nucleic acids,
vol. 2, Biology, New York, Academic Press.
Ward W. W. (1979). Energetics of bioluminescence. In: Photochemical and Pho-
tobiological Reviews, vol 4, ed К C Smith, p. 1, New York, Plenum.
Wolken J. J (1975). Photoprocesses, photoreceptors and evolution, New York,
San Francisco and London, Academic Press.
Yoshida M. (1979). Extraocular photoreception. In Handbook of sensory phy-
siology, vol. VII/6A, Vision in invertebrates, Heidelberg, Berlin and New
York, Springer-Verlag.
Задачи
1. Считают, что в большинстве случаев белая «окраска» по
происхождению является структурной. Приведите несколько
примеров белой окраски и выскажите ваши предположения
относительно того, какими структурными элементами и физи-
ческими явлениями она может быть обусловлена.
2. Определите батохромный, гипсохромный и гипохромный
эффекты в терминах энергетических изменений и вероятностей
переходов.
3. Для количественного определения веществ в биохимии
часто используют простой колориметр, с помощью которого
измеряют количество света, прошедшего через окрашенный
раствор. Между источником белого света и образцом обычно
помещают простой светофильтр. Объясните, почему получен-
ные результаты будут наиболее точными, если используют
-фильтр, имеющий окраску, комплементарную окраске изучаемо-
го образца?
4. Образец соединения М (0,026 мг) растворили в 10,0 мл
соответствующего растворителя. Коэффициент удельной экс-
тинкции Micm1”7” для соединения М равен 1600. Рассчитайте:
1) поглощение 10 мл раствора в 1-см кювете; 2) поглощение
10 мл раствора в 3-см кювете; 3) поглощение 3 мл раствора в
1-см кювете; 4) поглощение в 3-см кювете объемом 10 мл ра-
<твора, приготовленного разбавлением 3 мл исходного раство-
ра до 10 мл.
5. Экстракт листьев в водном 80%-ном ацетоне имеет по-
глощение 0,50 и 1,00 при 645 и 663 нм соответственно (в 1-см
кювете). Рассчитайте концентрации (в мг/л) хлорофиллов а и
b в экстракте, если для хлорофилла а А1см1%=820 и 170 (при
663 и 645 нм соответственно), а для хлорофилла b AiCM1% =
100 и 450 (при 663 и 645 нм соответственно).
Упрощенный метод расчета концентраций хлорофиллов
включает использование уравнений:
Суммарный хлорофилл (мг/л) =20,2 A64s+8,02 А663
Хлорофилл а (мг/л) = 12,7 А663—2,69 А645
Хлорофилл в (мг/л) =22,9 А645—4,68 А66з,
где А645 и А663 — зарегистрированные величины поглощения при
26*
396
Задачи
645 и 663 нм соответственно. Даст ли применение этих уравне-
ний в данном случае точные значения концентрации хлорофил-
лов?
6. а) Пигмент X (3,43 мг) в гексановом растворе (500 мл)
после пятикратного разбавления имеет при 450 нм поглощение
0,346. Рассчитайте его коэффициент молярной экстинкции
(мол. масса пигмента равна 536).
б) Растворитель выпарили и полученную суспензию X инку-
бировали в аэробных условиях с препаратом кишечного фер-
мента в присутствии NADPH. После инкубации полученные
продукты экстрагировали гексаном (800 мл). Гексановый ра-
створ обладал поглощением при 450 нм (0,577) и имел новый
максимум поглощения (А = 0,272) при 366 нм. Рассчитайте об-
щее количество непрореагировавшего X и образовавшегося про-
дукта У в растворе (в мг и мкмоль), если коэффициент удель-
ной экстинкции А1см1% продукта Y (Хтах=366 нм) составляет
1740, а коэффициент молярной экстинкции 49 500. Сделайте
свои заключения о ферментативной реакции. (Внимание} Учти-
те, что X не поглощает при 366 нм, а У не поглощает при
450 нм.)
в) Очищенный продукт реакции У (1,25 мг) при инкубации
со специфическим белком образует (с количественным выхо-
дом) водорастворимый комплекс, имеющий Хтах = 500 нм. Об-
щий объем водного раствора составил 100 мл, а наблюдаемое
поглощение 1,76. Рассчитайте молекулярную массу белка, если
А1см1% (при 500 нм) для комплекса составляет 9,8 при эквимо-
лярном соотношении У: белок. (Внимание! Во всех случаях
использовались 1-см кюветы общим объемом 3 мл.)
7. В приводимой ниже таблице содержатся коэффициенты
миллимолярной экстинкции окисленной (еох) и восстановлен-
ной (еге<1) форм цитохрома с. Постройте дифференциальный
спектр (восстановленной формы минус окисленной) по этим
данным. Стоит ли пытаться различить окисленную и восстанов-
ленную формы лишь по спектрам поглощения этих форм?
Л, нм £ох £red К ®ох £r?d л £ох £red
370 28,0 14,0 440 20,0 8,0 520 8,0 13,5
380 34,5 20,0 450 13,0 4,5 530 9,0 9,0
390 45,5 32,0 460 9,5 2,0 540 9,5 8,0
400 73,0 53,0 470 6,5 1,0 550 8,0 24,0
410 105,0 110,0 480 5,5 1,5 560 6,5 2,0
415 88,0 128,0 490 5,5 2,0 570 5,5 1,5
420 60,9 94,0 500 6,0 4,0 580 4,5 1,0
430 ! 32,0 32,0 510 6,5 8,0 590 2,0 0,5
I
Задачи
397
8. Установите приблизительное положение главных макси-
мумов поглощения света для следующих каротиноидов: родо-
пина, е-каротина, 4-кето-у-каротина, 5,6-эпокси-5,6-дигидро-
ф,ф-каротина, у-каротин-5,6-эпоксида, зеаксантина, 4-кето-р-
зеакаротина, лороксантина (|3,е-каротин-3,19,3'-триола) и
Р,|3-каротин-2,2'-диона. Известно, что нейроспорин, ликопин,
у-каротин, p-каротин и эхиненон имеют Хтах при 440, 470, 460,
450 и 458 нм соответственно. (Структура и полусистематическая
номенклатура каротиноидов приведены в гл. 2.)
9. Объясните следующие наблюдения:
1) Разбавленный раствор хлорофилла в диэтиловом эфире
имеет зеленую окраску, в то время как концентрированный
раствор может выглядеть красным.
2) В УФ-свете хлорофилл имеет красную окраску.
3) Если при освещении солнечным светом смотреть на де-
рево через фильтр, прозрачный для узкого диапазона длин
волн красного света, то листья выглядят розовыми на почти
черном фоне.
10. Открыт новый вид животного, которое имеет синюю
окраску. Как вы определите, имеет ли окраска структурное
происхождение, или же она обусловлена пигментом. Если вер-
но последнее, то как можно идентифицировать класс, к кото-
рому принадлежит данный пигмент?
11. Каротиногенные бактерии инкубировали с [2-14С,
(5R)-5-3HJ -мевалоновой кислотой. Меченые каротиноиды —
фитоин, p-каротин и зеаксантин — экстрагировали, очищали, а
затем определяли их радиоактивность. Получили следующие
соотношения 14С : 3Н для:
[2-14С, (5/?) -5-3Н!] -мевалоновой кислоты 1:1
фитоина 1:1
Р-каротина 1:0,5
(3R, 3'R)-зеаксантина 1:0,25
Какие можно сделать выводы о последовательности реакций
при биосинтезе?
С1Ц он
/ 1 н
ноос ^.с
Чн', сщ ^он
। 2-,4С-(5А J-5-3Н, ] Мевалоновая кислота
12. Биосинтез нафтохинона моллисина происходит из двух
поликетидов, состоящих из трех и четырех Сг-фрагментов соот-
ветственно. Нарисуйте схему образования моллисина по этому
398
Задачи
биосинтетическому пути и объясните, как этот путь можно
экспериментально отличить от пути, в котором участвует толь-
ко одна гептакетидная цепь.
Предложите также правдоподобный способ изгибания поли-
кетидной цепи, который мог бы иметь место при биосинтезе
антрахинона солориновой кислоты (пигмента лишайников) и
лаккаиновой кислоты D (пигмента насекомых).
о
Лаккаиновая кислота D
13. Считают, что антрахинон ализарин (3.27) синтезируется
в результате комбинации шикиматного и мевалонатного путей
через пренилированный промежуточный продукт
Предложите механизм образования ализарина по этому пути,
а также по альтернативному пути либо только с помощью по-
ликетидного механизма, либо с помощью комбинации его с
шикиматным путем. Позволят ли исследования с меченым аце-
татом установить, в результате какого из альтернативных пу-
тей биосинтеза образовался ализарин?
14. Из лепестков обычного садового цветка, имеющих кре-
мовую окраску, выделили значительное количество водораство-
римого «пигмента», который можно пометить инкубацией с
14С-ацетатом или |4С-фенилаланином. Выделенное соединение
Задачи.
399
имело мол. массу 480, а значение его Хтах (в метаноле), рав-
ное 365 нм, сдвигалось до 425 нм в присутствии щелочи. Обра-
ботка этого соединения диазометаном приводила к образова-
нию его пентаметилового эфира, в то время как при действии
ацетангидрида в пиридине получали его нонаацетильное про-
изводное. В результате гидролиза пигмента р-глюкозидазой
получили продукт с брутто-формулой С15Н10О8, максимум по-
глощения которого был лишь слегка сдвинут по сравнению с
максимумом исходного пигмента (Хтах=378 нм). После обра-
ботки диазометаном этот продукт опять давал пентаметиловый
эфир, однако его ацетилирование в данном случае приводило
к образованию гексаацетильного производного. На основании
этих данных и биосинтетических соображений воссоздайте
структуру исходного пигмента и объясните, как она может
быть подтверждена.
15. Колонка для гель-фильтрации (Сефадекс G-25) была
уравновешена 0,05 М К-фосфатным буфером, pH 7,0. В колон-
ку внесли свежеприготовленный раствор дитионита натрия,
который проник в гель. Затем добавили смесь гемоглобина и
феррицианида калия, и после ее впитывания было продолжено
элюирование фосфатным буфером, pH 7,0. Пока смесь гемогло-
бин— феррицианид не достигла зоны дитионита, она имела
коричневую окраску. Однако вышедшее из зоны дитионита ве-
щество было пурпурным и постепенно по мере продвижения к
нижней части колонки и при переходе в элюат становилось
алым. Объясните эти наблюдения.
16. Опишите, какое действие на сродство гемоглобина А к
кислороду окажут следующие изменения и почему: 1) 4-крат-
ное увеличение парциального давления СОг’, 2) 4-кратное уве-
личение концентрации 2,3-дифосфогдицерата (ДФГ); 3) сни-
жение pH с 7,4 до 7,2; 4) диссоциация молекулы гемоглобина
на мономерные субъединицы.
17. Одноклеточная водоросль может расти в отсутствие
фотосинтеза на глюкозе в качестве источника углерода. В этих
условиях глюкоза метаболизируется до ацетата, который и ис-
пользуется для биосинтеза клеточных компонентов. Если клет-
ки культивировать в условиях отсутствия фотосинтеза, в каче-
стве единственного источника воды использовать оксид дейте-
рия (2Н2О), а в качестве источника углерода — обычную
[!Н]-глюкозу, то распределение ‘Н и 2Н в образующихся мо-
лекулах каротина и хлорофилла будет строго определенным.
Если же культуру перенести в условия, при которых возмо-
жен фотосинтез (свет+СОг в качестве источника углерода),
то как со временем будет изменяться характер распределения
метки в пигментах? (Источником воды остается оксид дейте-
рия.)
400
Задачи
18. Каким будет распределение метки при биосинтезе фико-
билинового хромофора [например, (5.41)] из [5-14С]-АЛК
{6-аминолевулиновой кислоты). Можно ли, пользуясь таким
распределением метки, установить, какой из двух альтернатив-
ных предшественников — гем или хлорофилл — был истинным?
19. У крысы расщепление гема происходит с образованием
биливердина, который в свою очередь превращается в билиру-
бин. В ходе расщепления в молекулу вводятся две кислород-
содержащие группы (кетогруппы). Были предложены три аль-
тернативных механизма, по которым может протекать эта реак-
ция: 1) оба вводимых атома кислорода происходят из одной
молекулы О2; 2) оба атома кислорода происходят из молеку-
лярного О2, но из двух разных молекул; 3) один атом кислоро-
да происходит из молекулярного кислорода, а другой — из воды.
Молекулярная масса нормального (16О) билирубина со-
ставляет 584. Предскажите для каждого механизма характер
распределения метки и молекулярную массу образующегося
билирубина, если реакция протекает с 18О2 (100%) и Н216О
(100%). Если расщепление гема происходит в атмосфере, со-
держащей смесь кислорода 18О2 (20%) и кислорода 16О2 (80%),
то можно ли с помощью соотношения между содержанием мо-
лекулярных видов билирубина с мол. массами 584, 586 и 588
(определенного путем масс-спектрометрии) установить, по ка-
кому из этих трех механизмов происходит расщепление гема?
20. Дрозоптерин (6.14) представляет собой димерный пте-
рин. Из какой мономерной формы (или форм) может образо-
ваться его молекула? Предложите возможный механизм диме-
ризации. Спектр поглощения дрозоптерина был определен в
нейтральной среде, в 0,1 М НС1 и в 0,1 М NaOH. В нейтраль-
ном растворе максимумы поглощения наблюдались при 265 и
485 нм. Как будут отличаться от спектра в нейтральном раст-
воре спектры в кислом и щелочном растворах?
21. Бактерия Pseudomonas phenazinium образует и секрети-
рует в культуральную среду большое количество феназинового
пигмента иодинина (6.50). Были получены два мутантных
штамма этой бактерии, один из которых (F11) накапливает
6-гидроксифеназин-1-карбоновую кислоту (см. рис. 6.10), а
другой (13Z) не образует никакого пигмента. Культуры этих
двух штаммов посеяли на агар взаимно перпендикулярными
полосами. При проверке агара через 7 дней обнаружили, что
вокруг точек пересечения культур этих двух штаммов происхо-
дит образование иодинина. Объясните этот результат.
22. Модель части структуры сепиомеланина показана на
рис. 7.2. Приведите структуру молекул мономеров, из которых
она была получена. Предложите альтернативную частичную
Задачи
401
структуру молекулы меланина, сконструированную из этих
мономеров, и предскажите схему его биосинтеза.
23. Гусеницы бражника питаются листьями тополя (Popu-
lus spp.) и ивы (Salix spp.). Полевые исследования показали,
что гусеницы, обнаруживаемые на беловатой нижней стороне
листьев тополя, имеют серую окраску, в то время как гусени-
цы, встречающиеся на ярко-зеленых листьях ивы, имеют зеле-
ную окраску. Была начата исследовательская работа по из-
учению экологических факторов и факторов питания, которые
определяют и поддерживавают такой диморфизм окраски.
Среди запланированных опытов были: 1) анализ состава и
определение содержания пигментов у гусениц обоих типов и
в листьях обоих видов, и 2) изучение влияния на диморфизм
окраски света и темноты (при выведении гусениц на листьях
различных видов растений). Какую полезную информацию да-
дут эти опыты? Предложите возможные объяснения диморфиз-
ма окраски и разработайте другие эксперименты для проверки
этих гипотез.
24. Объясните, каким образом трихроматическая система,
основанная на трех фоторецепторных пигментах, имеющих
максимумы поглощения при разных длинах волн, может обес-
печить чувствительный механизм различения цветов?
25. Сетчатка глаза птиц содержит несколько фоторецепто-
ров, включающих зрительные пигменты, и ряд интенсивно, но
по-разному окрашенных масляных капель, которые действуют
как светофильтры; свет достигает рецептора только после про-
хождения через масляную каплю. Рассмотрим систему из двух
фоторецепторов, поглощающих в диапазоне 400—600 нм
(Хтах=500 нм) и 470—670 нм (Zmax = 570 нм), и две масляные
капли, поглощающие при 400—500 нм (Zmax = 450 нм) и 420—
520 нм (Хтах==470 нм) соответственно. Какое действие окажет
каждый из этих фильтров на поглощение света каждым фото-
рецептором и каким образом это скажется на чувствительности
различения оттенков? (Предполагается, что все спектры имеют
симметричную форму.)
26. Солнечный свет может проникать в чистые воды океана
на значительную глубину. Тем не менее при этом происходит
некоторое поглощение света, и с возрастанием глубины диапа-
зон непоглощенных волн становится все уже. На глубине
1000 м остается очень узкая полоса длин волн с центром при
470 нм, которую все еще можно увидеть невооруженным гла-
зом. В более мутных прибрежных водах свет виден лишь до
глубины 50 м, глубже всего проникают волны в диапазоне
~570 нм. Каковы должны быть принципы организации пиг-
ментов и фоторецепторных систем, которые были бы наиболее
выгодны рыбам и другим животным; а) обитающим на разных,
402
Ответы
но всегда примерно одних и тех же глубинах океанских и при-
брежных вод, б) обитающим в водах океана, но способным
более свободно перемещаться между поверхностью и морским
дном, и в) свободно перемещающимся на всех глубинах как
прибрежных, так и океанских вод.
27. Прокариотические сине-зеленые водоросли (цианобак-
терии) являются простейшими фотосинтезирующими организ-
мами, выделяющими кислород. Они не содержат хлоропластов,
но имеют более или менее развитые внутренние фотосинтетиче-
ские мембраны, включающие хлорофилл а, каротиноиды и фи-
кобилины. Опишите все изменения в морфологии, ультраструк-
туре и составе пигментов, которые можно ожидать при пере-
носе цианобактерий из: 1) условий низкой освещенности в
условия высокой освещенности и высокого содержания кислоро-
да, 2) условий высокой освещенности в условия низкой осве-
щенности, 3) условий освещения белым светом в условия осве-
щения зеленым или красным светом.
28. Объясните следующие явления:
а) Осенью листья изменяют свою окраску с зеленой на
желтую, красную или коричневую.
б) Дающая много цветков роза «Маскарад» имеет зеленые
бутоны, которые распускаются в желтые цветки, приобретаю-
щие позже красную окраску.
в) Листья зеленых в нормальных условиях растений стано-
вятся желтыми, если растение содержится несколько дней в
темноте, или если оно испытывает недостаток магния или же-
леза.
29. Спектр действия фототропной реакции гриба очень схо-
ден со спектром поглощения как (З-каротина, так и рибофла-
вина. Предложите экспериментальный подход, с помощью ко-
торого можно было бы определить, какой из этих пигментов
является фоторецептором.
30. Для индуцирования цветения растениям длинного дня
необходимы условия с длинным днем и короткой ночью, тогда
как растения короткого дня цветут только в условиях короткого
дня и длинной ночи. Объясните, как один и тот же фоторецеп-
тор, фитохром, может индуцировать цветение у растений обоих
типов.
Ответы
1. См. разд. 1.3.4. Структурная белая окраска обусловлена
обычно светом, отраженным от мельчайших кристалликов, ча-
стиц или капелек.
2. Определения этих выражений, правда не в терминах ве-
роятностей переходов и изменений энергии, приведены в гл.1.
Ответы
403
Изменения энергии при переходе определяют длину волны
максимума поглощения, а вероятность перехода определяет
интенсивность поглощения.
3. Фотоячейка колориметра детектирует проходящий свет
во всем видимом диапазоне. Выбранный фильтр пропустит
лучи лишь тех длин волн, в диапазоне которых поглощает об-
разец, так что небольшие различия в концентрации приведут
к значительным различиям в поглощении.
4. 1) 0,416, 2) 1,248, 3) 0,416, 4) 0,374. Вспомните законы
Ламберта — Бэра: поглощение света веществом пропорциональ-
но концентрации вещества и длине пути света, проходящего
через его раствор.
5. Полученные с помощью упрощенных уравнений концент-
рации хлорофиллов а и b равны соответственно 11,35 и
6,77 мг/л. Эти значения идентичны полученным с применением
более сложных уравнений:
АвМ = (А1%1см для Хл а при 645 нм • Концентрация Хл а)-)-
-/-(А1'“1см ДЛЯ Хл b при 645 нм-Концентрация Хл Ь)
А66з= (А,%! см для Хл а при 663 нм• Концентрация Хл a)-f-
+ (А1%1См Для Хл b при 663 нм • Концентрация Хл Ь)
6. а) 135-103.
б) Экстрагировано 1,83 мг X (3,415 мкмоль) и 1,25 мг У
(4,40 мкмоль). Отсюда мол. масса У равна 284 в соответствии
с реакцией Х-\-О2—^2У (т. е. У=1/2Х-|-О).
в) 180 мг комплекса составляют 4,4 мкмоль, отсюда
мол. масса равна 41 000.
7. В качестве ординаты откладывают величины (еох—еге<1)
(они могут быть положительными или отрицательными), а в
качестве абсциссы — длину волны. Окисленная и восстановлен-
ная формы цитохрома имеют величины Хшах соответственно при
~410 и ~415 нм, поэтому различить эти формы или устано-
вить состав их смеси лишь на основании спектров поглощения
практически невозможно.
8. Величины Zmax находятся при 470, 440, 468, 455, 455, 450г
438, 445 и 450 нм соответственно. Нейроспорин — хромофор с
девятью сопряженными внециклическими двойными связями —
имеет А.тах = 440 нм; при сопряжении каждой дополнительной
внециклической двойной связи с хромофором А.тах увеличивает-
ся на 15 нм. (3-Кольцо имеет одну двойную связь в сопряжении
с ациклическим полиеном, благодаря которой А.тах увеличивает-
ся лишь на 5 нм. При сопряжении с хромофором кетогруппы
при С-4 Zmax увеличивается на 8 нм. Эпоксидирование двойной
связи нарушает сопряжение в этой точке; другие заместители,
404
Ответы
например гидроксильная или несопряженная кетогруппа, на
поглощение света хромофором не влияют. Двойная связь g-коль-
ца не сопряжена, она не вносит вклада в формирование хромо-
фора и не влияет на величину его Zmax-
9. Ключом к объяснению этих наблюдений является то, что
хлорофилл не только сильно поглощает свет в нескольких
участках видимого спектра, но также обладает очень сильной
красной флуоресценцией.
10. Структурный голубей цвет воспринимается как голубой
лишь в отраженном, но не в проходящем свете. Окраска мо-
жет зависеть также от угла падения света и от угла наблюде-
ния. При погружении животного в воду или в другой раство-
ритель окраска структурного происхождения, вероятно, исчез-
нет, а после испарения растворителя восстановигся. Если
окраска обусловлена пигментом, то последний можно экстраги-
ровать из тканей водой или органическими растворителями.
Информацию о хромофоре может дать резонансная раманов-
ская спектроскопия. Изучение растворимости, физико-химиче-
ских и спектроскопических свойств выделенного пигмента (см.
соответствующие главы данной книги) позволит идентифици-
ровать класс, к которому принадлежит это соединение.
11. См. в гл. 2 общий путь биосинтеза каротиноидов. Фи-
тоин образуется из восьми молекул мевалоновой кислоты (МК),
и поэтому образовавшаяся молекула фитоина сохраняет все
восемь меченых углеродных атомов и восемь атомов трития из
субстрата с двойной меткой. Образование фитоина, таким об-
разом, происходит стереоспецифически; водородный атом, те-
ряющийся из положения С-1 геранилгеранилпирофосфата в хо-
де образования С-15, ^'-двойной связи фитоина, должен быть
немеченым Н-атомом из положения С-5 МК- В p-каротине со-
храняется восемь 14С-атомов, но лишь четыре атома трития.
Тритиевые атомы должны стереоспецифически теряться в ходе
десатурации. Сходным образом должен теряться 3Н-замести-
тель из положения С-З обоих колец p-каротина при его гидро-
ксилировании до зеаксантина.
12. Наиболее вероятные способы образования этих соеди-
нений в результате изгибания молекул соответствующих поли-
кетидов приведены ниже:
Ответы
405
Моллисин
Наилучший способ проверки подобных схем состоит в исполь-
зовании меченого [ 1,2-13С2]-ацетата. Если этот 13С-субстрат
будет включаться в образующийся продукт с достаточной эф-
фективностью (не менее 1% включения), то атомы углерода,
источником которых является интактная молекула ацетата,
будут обладать 13С-13С-спин-спиновым взаимодействием при
ЯМР-спектроскопии. Если же уровень включения не превышает
10%, между другими углеродными атомами заметного спин-
спинового взаимодействия не будет происходить. Таким обра-
зом можно установить положение ацетатных фрагментов, об-
разующих поликетидную цепь, в молекуле конечного продукта.
13. Этот промежуточный продукт образуется в результате
присоединения Cs-изопренового фрагмента к о-сукцинилбензоа-
ту или к нафтол/нафтохиноновому промежуточному соедине-
нию (см. рис. 3.12). Четыре из пяти углеродных атомов этого
изопренового фрагмента будут затем использованы для образо-
вания кольца С ализарина. Метод включения [13Сг]-ацетата,
упомянутый в ответе на вопрос № 12, позволяет различить
большинство возможных альтернативных механизмов — любая
часть молекулы, происходящая от немеченого ацетата, не бу-
дет содержать метки.
14. Включение меченого ацетата и фенилаланина в состав
этого водорастворимого пигмента позволяет предположить, что
он принадлежит к классу флавоноидов. Другие его характери-
стики соответствуют структуре p-D-глюкозида мирицетина
(4.19). Обработка диазометаном приводит к метилированию
только фенольных ОН-групп, в то время как ацетилируются и
все первичные, и все вторичные ОН-группы.
406
Ответы
15. Феррицианид окисляет гемоглобин до метгемоглобина,
обладающего бурой окраской. Колонка будет задерживать
небольшие молекулы и ионы, например дитионит и феррициа-
нид, в значительно большей степени, чем крупные молекулы
гемоглобина. В процессе хроматографирования метгемоглобин,
быстрее двигающийся в колонке, достигнет зоны дитионита и
восстановится в гемоглобин. Последний в ходе дальнейшего
элюирования окисляется кислородом воздуха, растворенным в.
элюирующем буфере, с образованием оксигемоглобина.
16. Влияние СО2, Н+ и ДФГ на сродство гемоглобина к
кислороду и на кооперативность функционирования ассоцииро-
ванных гемоглобиновых субъединиц описаны в гл. 5 (разд.
5.5.2).
17. Образующийся из [*Н]-глюкозы ацетат содержит 'Н в
метильной группе (у С-2). Когда этот ацетат далее метаболи-
зируется в среде с 1 2 3Н2О, при восстановлении С-1 карбоксиль-
ной группы ацетата к нему будет присоединяться только дей-
терий. Таким образом, молекулы каротина и хлорофилла у тех
углеродных атомов, которые произошли из атомов С-1 ацетата,
будут содержать только дейтерий, но сохранят 'Н у атомов
углерода, источником которых был атом С-2 ацетата. В усло-
виях фотосинтеза, когда источником углерода служит СО2,
а единственным доступным водородом в среде является дейте-
рий из 2Н2О, во всех положениях синтезирующихся молекул
будет содержаться только дейтерий.
18. Схемы превращения АЛК в гем и хлорофилл, а также
схема образования фикобилиновой структуры из порфиринов
приведены в гл. 5. Из этих схем следует, что распределение
метки в фикобилине будет одинаковым независимо от того, гем
или хлорофилл были промежуточными продуктами его биосин-
теза.
19. Если в системе присутствует смесь кислорода 18О2 и
кислорода 16О2, то в случае механизма 1 оба кислородных ато-
ма билирубина будут происходить из одной и той же молеку-
лы, будь то 18О2 или 16О2. При биосинтезе билирубина по ме-
ханизму 2 оба кислородных атома могут поставляться как
18О2, так и 16О2. В случае же использования механизма 3 моле-
кула О2(либо 18О2, либо 16О2) поставляет лишь один из двух
атомов кислорода. Если смесь состоит из 20% 18О2 и 80% 16О2,
то синтезируемый по этим трем механизмам билирубин будет
иметь следующий состав:
1) 584 (80%), 586 (0%) и 588 (20%);
2) 584 (64%), 586 (32%) и 588 (4%);
3) 584 (80%), 586 (20%) и 588 (0%).
Ответы
407
Этот состав легко установить с помощью масс-спектромет-
рии.; Если используемый в опыте кислород был представлен
лишЬ 18О2-видом, то билирубин, образовавшийся по механиз-
му 3, содержал бы лишь один меченый 18О-атом и имел бы
мол. массу 586, в то время как синтез по механизмам 1 и 2
привел бы к образованию лишь одного молекулярного вида
билирубина с мол. массой 588. Следовательно, с помощью од-
ной масс-спектрометрии нельзя установить, какой из этих ме-
ханизмов был использован для синтеза билирубина.
20. Вероятным мономерным предшественником при синтезе
дрозоптерина является сепиаптерин (6.15), Сз-боковая цепь
которого может использоваться для формирования связываю-
щего мономеры пятичленного кольца. Спектры поглощения
дрозоптерина в кислой и щелочной среде см. на рис. 6.2, а его
свойства описаны в разд. 6.2.4. Для объяснения изменений в
спектрах следует принять во внимание способность амфотер-
ных молекул птеринов к ионизации.
21. У обоих этих штаммов содержится неполный набор
ферментов, катализирующих образование феназинов. Проме-
жуточный продукт, накапливаемый штаммом F11, может пре-
вращаться в иодинин штаммом 13Z.
22. Ниже приведены главные мономерные структуры, ве-
роятно участвующие в синтезе меланинов. Путем окислитель-
ной конденсации полифенольных структур из них может обра-
зоваться много различных меланиновых структур.
ноос^____
// \\ и другие пирролкарбоновые
х—СООН кислоты
МООС,
н
ио- хинон этого соединения
23. Следует принять во внимание общую стратегию покро-
вительственной окраски, описанную в гл. 8. Опыт 1 покажет,
действительно ли гусеница поглощает и запасает различные
пигменты, содержащиеся в листьях разных видов растений.
Однако не исключено, что различия в окраске листьев обуслов-
лены особенностями структуры, а не различиями в составе
пигментов. Опыт 2 позволит выяснить, реагируют ли гусеницы
на окраску фоновой поверхности при участии какого-либо фо-
торецепторного механизма, или приспособление к фоновой
408
Ответы
окраске происходит в ответ на какой-то другой раздражитель,
например на какое-то вещество, содержащееся в листьях од-
ного вида, но отсутствующее в листьях другого. Не следует
также упускать из виду вероятность того, что по-разному окра-
шенные формы могут представлять собой две генетически раз-
личающиеся популяции.
24. Трихроматическая система зрительных пигментов в прин-
ципе сходна с трихроматической системой, использующейся в
цветном телевидении или в цветной фотографии. Свет трех раз-
ных длин волн (соответствующий трем основным цветам)
можно смешать в бесчисленном числе комбинаций цвета и ин-
тенсивности и получить любой необходимый цвет или его от-
тенок любой интенсивности. Сходным образом, если диапазон
поглощения трех зрительных пигментов охватывает практиче-
ски весь видимый участок спектра и максимумы поглощения
этих пигментов четко разграничены, свет любого цвета (или с
любым распределением длин волн) можно разделить на три
первичных компонента, которые будут детектироваться этими
тремя пигментами.
25. Постройте графики спектров поглощения для фоторе-
цепторов в присутствии и в отсутствие масляных капель. Имей-
те при этом в виду, что капли интенсивно окрашены и значи-
тельная часть света с длинами волн в диапазоне их поглощения
не достигнет фоторецептора. Из графика станет ясно, каким
образом с помощью этих капель достигается получение более
узких и лучше разделенных спектров поглощения зрительных
пигментов.
26. Несмотря на неизбежные межвидовые различия, эволю-
ция животных, вероятно, будет происходить в направлении ис-
пользования фоторецепторов, которые наиболее эффективно
поглощают свет, проникающий на большую глубину в усло-
виях их обитания. Для окрашивания же будут использоваться
пигменты, максимально поглощающие либо максимально про-
пускающие свет в этом оптимальном диапазоне длин волн.
Свободное перемещение животных между различными среда-
ми обитания предполагает необходимость использования ими
фоторецепторов для различных условий освещения; при этом
они могут использовать более сложные комбинации пигментов.
27. 1) В этих условиях отпадает необходимость очень эф-
фективного улавливания света и, следовательно, будет доста-
точно меньшего количества вспомогательных светособирающих
пигментов. Однако при этом возникает необходимость более
эффективной защиты фотосинтетического аппарата от фото-
окисления, так что можно ожидать усиления синтеза каротина.
2) В этих условиях улавливание света должно стать макси-
мально эффективным за счет синтеза большего количества
Ответы
40 9
светособирающих пигментов; наблюдаемый эффект будет со-
провождаться увеличением размеров фикобилисом.
3) В результате такого переноса будет синтезироваться
больше фикобилинов, поглощающих соответственно в зеленой
или в красной области спектра. Этим будет достигаться мак-
симальное использование доступного света.
28. а) По мере старения листьев происходит разрушение
хлоропластов и их зеленого пигмента хлорофилла и становятся
видны содержащиеся в них желтые каротиноиды. Затем начи-
нают синтезироваться окрашенные в красный цвет антоцианы
и, наконец, окислительная конденсация фенольных соединений
приводит к образованию полимерных соединений коричневого
цвета.
б) Наблюдаемая окраска обусловлена последовательно зе-
леным хлорофиллом, желтым каротиноидом и, наконец, крас-
ным антоцианом.
в) В отсутствие света хлоропласты разрушаются и проис-
ходит распад хлорофилла. Растения становятся желтыми и
вместо хлоропластов содержат этиопласты, окраску которым
придают каротиноиды. Восстановление зеленой окраски и по-
вторное образование хлорофилла и хлоропластов происходит
при освещении растений. Дефицит железа или магния вызывает
подавление синтеза хлорофилла (разд. 5.9.7 и 5.9.15).
29. Это сделать очень трудно. Такой фоторецептор должен
содержаться в очень небольших количествах вместе с другими
неактивными пигментами, концентрация которых гораздо боль-
ше. Вероятно, для этой цели можно использовать сложный
метод спектроскопического исследования быстрых процессов.
Не исключено, однако, что единственный путь, с помощью ко-
торого можно было бы установить природу действующего фото-
рецепторного пигмента, — это его выделение и химическая иден-
тификация.
30. Рассмотрите свойства фитохрома, описанные в разд. 11.2
и иллюстрируемые рис. 11.2. Помните, что активной формой
фитохрома является, по-видимому, Pfr-форма, тогда как Рг-
форма играет лишь пассивную роль.
27—84
Предметный указатель
Абсорбция 25
Агликон 129
Адаптация к высоте 216
S-Аденозилметионин 68, 147, 149,
203, 208, 245
Адреналин 277, 292
Азотсодержащие пигменты 254—257
Аклавннон 102, 103
Актиномицин 249, 250
Актнноэритрин 40, 59, 83
Александрийский лист 122
Ализарин 102, 117, 121
Алканнин 116, 121
АЛК см. 6-Аминолевулиновая кис-
лота
АЛК-дегидратаза (порфибилиноген-
синтаза) 195—198, 216
АЛК-сннтетаза 195, 196, 215—218
АЛК-трансаминаза (аминолевули-
нат — аминотрансфераза) 196
Аллоксантин 77
Алломеланины 259, 265, 268, 272
Аллофикоцианин 189, 190, 354
Альбедо 290
Альбинизм, альбиносы 277, 287, 321
5-Аминоимидазолрибонуклеотид 233,
234
Ампнолевулинат — аминотрансфера-
за см. АЛК-трансаминаза
б-Аминолевулиновая кислота (АЛК)
195—197, 208, 215, 216
Ангидрородовнбрин 70
Антенны (светособирающие пигмен-
ты) 334—339, 364
Антераксантин 52, 329, 331
Антибиотики 249
Антибиотические свойства пигментов
119—120, 122, 249
Антоксантин 128
Антохлор 128
Антоцианидины 126—129, 133—136,
144—146
Антоцианы 125, 129, 137—139, 149—
154, 252, 365
Антрахиноны 93, 96, 98, 101 —102,
105, 109—110, 114, 118, 119, 121,
122
Антрациклиноны 102, НО, 122
Аплизиовиолин 191, 193
Апо-каротиноиды 39—40, 85, 365
Арильные полиены 87
Астаксантин 38, 42, 44, 51, 54, 58—
60, 83, 316, 317, 323
Астацин 60
Ауксохромы 21
Ауроксантин 49, 50
Ауроны 127, 133—134, 139, 145, 294
Афины 93, 102, 103, 105, 117
Аэругннозин 240, 241
Бактериальный синтез 327—ь328,
355—358, 367
Бактериородопсин 377—380
Бактериоруберин 39, 57, 69
Бактериофеофитин 357
Бактериохлорин 157, 158
Бактериохлорофиллид а 208, 210
Бактериохлорофиллы 157, 160—162,
165, 166, 208, 210, 219, 355—358,
364, 374
Батородопсин 312, 313
Батохромный эффект 44, 59
Бензотиазол 264
Бензохиноны 92—96, 99, 106, 108,
114, 116, 119—120, 272, 274
Берберин 255, 256
Бетаксантины 250—251, 253, 294
Беталаины 250—255, 257, 258
Беталамовая кислота 253, 254
Бетанидин 251—254
Бетанин (бетанидин-5-О-Р-глюкопи-
ранозид) 252
Бетацианины 250—255
Биливердины 185—188, 211, 212, 219
Билины см Желчные пигменты
Билипротеины 374
Билирубин 185—188, 211, 212, 214,
218
Биолюминесценция 387—392
Биоптерин 228, 238
Биохромы 10, 13, 15—16
Бора эффект 171 —172
Валлемия А 86, 87
Видимый свет 9—11
Виолаксантин 38, 52, 53, 76, 329, 331
Внолаксантин — хлорофилл a/ci/cz —
белок, комплекс 352
Виолацеин 255, 256
Витамин А (ретинол, 3,4-дидегид-
роретинол) 83—85, 89, 304, 305,
315, 316
— В[2 183—185, 208—209
Галлахром 102, 103
Галлоксантин 322, 323
Галлофеомеланин 264
Галобактерии 69
Гардерова железа 381
Гардеропорфирин 381
Гем 156, 158, 167, 170, 171, 174, 185,
209, 212, 213, 217, 382, 387
— биосинтез 201, 204, 215
Гемованадин 177
Гемоглобин 29, 167—177, 187, 216,
219
Предметный указатель
41k
— А 169, 173
— А2 169
— F 169, 173
— S 173
Гемолимфа 105, 175
Гемопротеины 167, 382. См. также
Гемоглобин, Миоглобин
Гемоцианин 177
Гемоэритрин 177
Геранилгеранилпирофосфат (ГГПФ)
60—62, 80, 206, 207
Геранилгераниол 161, 162, 206, 207
Геранилпирофосфат 61, 62, 116
Гербициды 64, 67, 82, 363
З-Гидроксиантраниловая кислота 249,
250
Гидроксибензоат 116
Гидрокси-п-бензохинон 272, 274
ЗТидроксииндол 255, 256
рТидрокси-р-метилглутарил-СоА
(ГМГ-СоА) 61, 62
Гидроксинафтохиноны 95—98
Гидроксисфероидин 56, 57, 70
2-Гидроксихромены 134, 135
Гидрохинон 92—94, 115, 116
Гиперицин 103, 104, 120
Гипсородопсин 312, 313
Гипсохромный сдвиг 51
Глаз беспозвоночных 300—302
— позвоночных 297—300
Глаукобилин (мезобиливердин) 188
Гликозиды флавоноидов 128—129
Глицин 233, 234
Глицинамидрибонуклеотид 233, 234
Голохромный белок (протохлорофил-
лидредуктаза) 207, 213
Гомоарбутин 115, 116
Гомогентизиновая кислота 112, 114,
115
Гомо-каротиноиды 39
Гомопротокатеховая кислота 152
Гуанин 15, 225, 226, 285, 288, 321,
322
— биосинтез 232—237
Гуанинмонофосфат 235
Гуаниннуклеотид 235
Гуанофоры см. Иридофоры
3-Дезоксиантоцианидин 126, 127, 145
Дезоксигемоглобин 168—169, 173—
175
Дезоксимиоглобин 168
Декапреноксантин 39, 57, 74
2-Декарбоксибетанидин 250, 251
Дельфинидин 129, 135, 138—140
«Диапонейроспорин» 56, 57
Дибензопиразин 240. 241
27*
6,6-Диброминдиго 255, 256
Дигидроксантомматин 246, 247
1,8-Дигидроксинафталин 265, 267, 272
Дигндрокси-1,4-нафтохиноны 97
7,8-Дигидронеоптерин 236—238
Дигидропорфирины 157, 161, 178, 207
3,4-Дидегидроретинальдегид 84, 304,
309, 316, 320
3,4-Дидегидроретинол (витамин А2)
83, 84, 316
Диметилаллилпирофосфат (ДМАПФ)
62
6,7-Диметил-8-рибитиллюмазин 237,
239
Дисперсия оптического вращения
(ДОВ) 29, 42
3,5-Дифосфоаденозин 390
2,3-Дифосфоглицерат (ДФГ) 171 —
173, 216
Дифракция 15
ДОФА (дигидроксифенилаланин)
252—254, 261, 262, 269—271
ДОФА-меланин 261
Дофамин 269, 271
Дофахинон 261, 262, 264, 269—271,
273
Дофахром 269, 270
Древесина, кора 92, 99—100, 117, 120,
137 139
Дрозоптерин 227, 237, 238, 324
ДФГ см. 2,3-Дифосфоглицерат
Желтое пятно 298, 299, 322
Желчные пигменты (билины) 156,
158, 185—194, 209, 211—215, 218—
221, 372
Желчный камень 187
Загар 276, 287, 386
а-Зеакаротин 71, 73
Р-Зеакаротин 37, 71, 73
Зеаксантин 38, 49, 52, 76, 83, 322,.
329, 331
Зеркальный слой (tapetum lucidum)
321____322
Зрение 11—13, 30, 34, 227, 228, 247,
249, 266, 285, 297—326
Зрительные пигменты (фоторецептор-
ные молекулы) 299, 302—303, 310—
318
— циклы 310—318, 325
Изоаллоксазин 224, 230, 240
Изобетанидин 250, 251
Изогуанин 225
Изозеаксантин 49, 50
Изокриптоксантин 49
412
Предметный указатель
Изоксантоптерин 238
Изопентенилпирофосфат 62
Изопреновая единица 34
Изопреновые заместители 114
Изопреноидный путь 61, 106, 118
Изопсин 320
Изородопсин 308
Изофлавоны 127, 131, 144, 147
Индиго 255, 256
Инднгоиднн 255, 256
Индигоксантин 252
Индикан (3-гидроксииндоглюкозид)
255, 256
Индолил-5,6-хинон 260, 261, 269, 270,
272
5,6-Индол-2-карбоновая кислота 269,
270
Инозиновая кислота 233, 235
Интерференция 14—15
Инфракрасная спектроскопия (ИК)
25, 98
Иодннии 240—245
Иодофоры 225, 285, 286, 291
Кантаксантин 47, 48, 50, 51, 55, 58,
76, 83, 85, 86
Капсантин 53, 77
Капсорубин 53
Карбоксигемоглобин 173—174
Карминовая кислота 102, 103, 118,
121
«-Каротян 36, 37, 52, 54, 69, 73, 76
Р-Каротин 35—37, 46—49, 52—58, 69,
73, 76, 81, 83, 85, 86, 218, 304, 305,
335, 336, 354, 365, 376
р,р-Каротин 76
Р,у-Каротин 58
у-Каротин 47, 55, 56, 69, 73
у,у-Каротин 46
6-Каротин 69, 73, 81
Е-Каротин 69, 73, 83, 322
Е,Е-Каротин 46, 323
^-Каротин 43—45, 64, 66, 81, 362
Каротиноиды 21, 23, 34—90, 122, 226,
240, 284, 293, 294, 304, 309, 322,
328, 329, 338, 339, 351, 357—365,
374—377, 382, 383, 385—387
Сз0-Каротиноиды 40, 57, 62, 64
С40-Каротиноиды 39—41
С45,Сзо-Каротиноиды 39, 56, 57, 69,
71, 72, 74, 75
Каротинопротеин 44, 51, 59—60, 90
Каталаза 167, 181
Катехины 139, 146, 149
Кверцетин 129, 135, 152
Кемпферол 129, 135
Кератин 386
Кермезнновая кислота 102, 103, 121
а-Кетоглутарат 197
Кинуренин 246—249
Колебательная релаксация 22, 23
Коэлентеразнн 389
Комплементарные цвета 11
Конденсированные дубильные веще-
ства 139
Копигментация 140, 154
Копропорфириноген (III) 199—203
Коричные кислоты 114, 139, 141, 142,
150
Коррнн 183
Кортнзалнн 86
Кофейная кислота 147
Кошениль 102, 105, 121
Коэффициент молярного поглощения
25
— удельного поглощения 26
«К. п. 450» 56, 57, 74
Красители 92, 121—122, 255, 256
Красное внно 138
В-Криптоксантин 49
Круговой дихроизм (КД) 22, 24, 28—
30, 32, 42, 307, 313, 317
Крустацианины 51, 59
Ксантнловая кислота 235
Ксантин 225
Ксантоафнн 103, 104, 117
Ксантобласты 287
Ксантомматин 246—249, 324
Ксантомонаднн I 87
Ксантоптернн 227, 229, 238
Ксантофиллы 35, 37—38, 335, 354,
360, 365
Ксантофоры 240, 284—286, 291
n-Кумаровая кислота 141, 147
Культуры клеток растений (петруш-
ки) 143, 145, 153, 154
Курчавость листьев 150
Лавсон 100, ИЗ, 114, 121
Лаккаиновая кислота D 109
Ламберта—Бэра законы 25
Лампирнн 389
Леггемоглобин 176
Лейкоптерин 227, 229, 238
Ликоперсин 63, 64
Ликопин 34, 37, 41, 44—48, 53, 64—
71, 73, 78, 81, 82, 365, 366
Линейный дихроизм 28, 30
Листья, побеги 138, 139, 150
Люмнродопсин 312, 313
Лютеин 37, 38, 52, 316, 317, 322, 335,
336
Люцифераза 388—390
Предметный указатель
413
Люциферин 388—390
Магнийпротопорфирин IX 203, 204
Мартицин 120
Масляные капли 322—323
Масс-спектрометрия 25, 98, 130
Мевалонат см. АТевалоновая кислота
Мевалонатный путь 114—117
Мевалоновая кислота, мевалонат
(МВК) 61, 62, 67, 76, 77, 114—118
Медиальный глазок 380
Межсистемный переход 23, 24, 385
Меланиновые гранулы (меланосомы)
267, 276, 277, 284, 289, 290, 291
Меланины 226, 240, 259—279, 283—
284, 286, 290, 321, 324, 386, 387
— биосинтез 268—274
Меланогенез 268—273
— аномальный 277
Меланома 261, 267, 277
Меланосомы см. Меланиновые гра-
нулы
Меланофоры 283—286, 291
Меланоцитстимулирующий гормон
(МСГ) 276, 291—292
Меланоциты 267, 276, 283—284, 295,
386
Мелатонин 291, 292
Менадион 113, 114
Менахинон 92, 99, 100, ПО, 114, 119
Метародопсииы 312—314, 317
Метгемоглобин 169, 173
Метилмагнийпротопорфирин IX 203—
205
7-Метилюглон 117, 118
Микроокружение пигментов 25, 27,
30, 161, 225
Миксоксантофилл 54, 55
Миоглобин 167—174, 219
Мирицетин 129, 135
Моллисин 100
Мориндон 121
Мочевая кислота 225
Мускааурин 252, 253
Нафтазарииы 101
Нафтохиноны 93—94, 96—101, 105,
106, 108—109, 114—122
Нейроспорин 44, 45, 64, 66, 70, 71,
73
Неоксантин 52, 76, 77
Норадреналин 292
Нор-каротиноиды 39—40, 83
Окенон 56, 57, 77
Окраска (в природе) 29, 31—32, 220,
281—283. См. также Структурная
окраска
— влияние гормонов 291—292
— изменение у животных 287—292
— наружных покровов 57—59, 175,
181, 188, 226, 240, 265—267, 278,
295
— оперения 182
— раковины моллюсков 182, 193
— суточные ритмы 291
— у растений 292—294
— яичной скорлупы 182, 220
Оксигемоглобин 168—174, 176
Оксидазы со смешанной функцией
76, 181
Оксилюциферин 390
Оксихлорорафин 240, 241
Омматины 246—249
Оммины 246—248
Оммохромы 223, 246—249, 258, 266,
290, 324
Опсины 28, 84, 305—319, 325
Оптическая плотность 25
Отравление окисью углерода 173
Папилиохромы 249
Пеларгонидин 129, 135, 138
Перенос электронов (электронный
транспорт, электронтранспортная
система) 119, 177—180
— — при фотосинтезе 327, 328, 332—
335, 337, 342—348, 352, 353, 355—
358, 360, 383
Перидинин 352
Перламутр 15
Пероксидаза 167, 181
Пигмент — белок, комплексы 28, 30,
352, 360
Пиоцианин 240—242, 244, 245
Пирокатехин 92—94, 265, 267, 268,
272
е-Пирромнцинон ПО
Пищевые красители 85, 89, 105 121,
152—154, 257
— цепи 327
Пластоглобулины 330
Пластохиноны 92, 99, 105, ПО, 119,
335, 343—346
Пластоцианин 344—347, 358, 360
Поглощение света 16—24
Покровительственная окраска 281 —
282
Покровные ткани как фоторецептор
381—383
Поликетидный путь 77, 87, 106—110,
117, 140, 141, 150
Полифенолоксидаза см. Тирозиназа
Полухиноиовый радикал 230, 231
414
Предметный указатель
Порфии 157
Порфирии 217—218
Порфирины 86, 156—183, 195—221,
365, 381, 382, 387
Порфиропсины 304, 316, 320, 382
Порфобилиноген (ПБГ) 195, 197—
199, 208, 217
Пребатородопсии 313
Предупреждающая окраска 281—283
Префеновая кислота 111,112
Префитоиипирофосфат (ПФПФ) 63,
64, 80
Примни 120
Природные пигменты 10, 15—16
Провитамин А 89, 316
Продигиозии 156, 255, 256
Проламеллярные тела 358, 359
Проликопин 41, 48, 82
Пропластиды 358, 361
Протилакоидные мембраны 358—359
Протоафииы 102—105, 117
Протогем 167, 168
Протокатеховая кислота 152
Протопорфирин I 201
Протопорфирии IX 167, 182, 195, 201,
381
Протопорфирииогеи IX 195, 199,
201—204
Протохлорофиллид а (магнийвинил-
феопорфириновый эфир) 203, 205—
209, 359—360, 362
Птеридины 223, 224, 228
Птерииосомы 284, 285
Птерины 224, 226—230, 232—238, 240,
257, 258, 285, 290
Пурины 223—226, 232—240, 258
Рамановская спектроскопия 24, 26—
28, 30, 32, 307, 313
Реакционные центры 333—346, 355—
359, 364
Рентгеноструктурный анализ 167
Реньератии 58, 59
Ретикуло-эидотелиальиая система
187, 213
Ретинальдегид (ретииаль, ретинен)
28, 83—85, 304, 310—321, 325, 378,
382
Ретииол (витамин Ai) 83—84, 86,
304, 305, 314, 315
Ретинолсвязывающий белок 84, 305
Ретииохром 318
Ретро-каротиноиды 38—39, 53, 77
Рибофлавин (витамин В2) 223, 224,
232—240 258 321 322, 374—376,
382
Родовибрии 70
Родоксантин 39, 53
Родопин 68, 70
Родопсин 12, 302, 314—316, 320, 325
Родоптилометрин 102, 103
Рэлея рассеяние 13
Сарциноксантии 56, 57, 74
Световые реакции 327
Светорассеяние 13—14
Светособирающий комплекс 335—337,
360
Сепиаптерин 227, 228, 238
Сепиомелаиии 261—263, 267, 271
Серповидно-клеточная анемия 173
Сетчатка 12, 83, 84, 231, 298, 299,
319, 321, 322
Сигнальная окраска 29, 175, 391. См.
также Предупреждающая окраска
Симбиоз 83, 118, 187, 231, 388
Синглетные состояния 23—24, 338,
385
Склеротины 265—266
Скотопическое («ночное») зрение 310
Скотопсины 12, 310
Сложный глаз 300—301
Солорииовая кислота 109, НО
Соре полосы 159
Спектроскопия 24—28
Спинохромы 97, 100—101, 118, 120
Спинулозии 108
Спириллоксаитин 56, 57, 69, 70, 356,
357
Стеркобилин 186, 187, 211, 214
Стеркобилиноген 211, 214
Стигма 375
Стрептоварицин А 122
Строма (хлоропластов) 329—331
Структурная белизна 15, 224
— окраска 13, 30, 32, 224, 266, 281,
285—286
о-Сукцинилбензоат 113, 114, 117
Сфероидеион 69, 71
Сфероидин 69—71
Z-Схема (транспорта электронов)
342—344, 351, 353
Таксономия 137, 153, 166, 175
Теафлавины 139, 149
Темновые реакции 327, 328, 346—349
Тетрагидропорфирии (бактериохло-
рии) 157, 160, 163, 166
3,4,3',4'-Тетрадегидроликопии 67, 68
Тетрапирролы 156—222
Тетратерпеиы 34
Тиамиипирофосфат 113
Тилакоидная мембрана 330—332, 340,
346, 347, 360, 363
Предметный указатель
415
----бурых водорослей 352
----сине-зеленых водорослей 353
353
Тилакоидные граны 52, 330—332, 335,
359
Тиминовые основания 384
Тиндаля синева 13—15
Тиривердин 255, 256
Тирозин 141, 268—270, 309
Тирозиназа (полифеиолоксидаза) 269,
271, 274, 277, 284
Тироксин 291, 292
Тирский пурпур 255
Торулародин 55
Триплетные состояния 22—24, 338,
386
Триптофан 248, 249, 309
Триспоровая кислота 81
Трихосидерииы 264—265, 271
Убихинон 92, 99, 105, 110, 112, 113,
119, 357, 358
Уробилиноген 186—188, 211, 214
Уробилины 187, 188, 211, 214
Уропорфирин 181 —183
Уропорфириноген 'III) 197—200, 203
Уропорфириногеиы 197, 199—201,
208, 217—218
Феназин-1,6-дикарбоиовая кислота
243
Феназины 223, 240—245, 258
Фенилаланин 141
Фенилаланин-аммиак-лиаза (ФАЛ)
141, 150
Феноксазины 245—249
Фенольная конденсация 117
Феомеланины 222, 263—266, 268,
271—273
Феофитин 162, 163
Феофорбиды 163, 167, 365, 366
Ферредоксин, 344 347
Ферримиоглобии 168—169
Феррохелатаза 201
Фикобилины и фикобилипротеины
156, 192, 211, 220, 328, 352—354,
362, 363
Фикобилисомы 189, 192, 211, 220,
353—355, 363
Фикоуробилин 191, 192
Фикоцианин 189 190, 192, 216, 354,
363
Фикоцианобилин 190, 192
Фикоэритрин 189—191, 193, 216, 352,
354, 363
Фикоэритробилин (фикобиливердии,
фиковиолин) 190—194
Фиксация углерода 350—351
Филлохинои 92, 99, 100, 110, ИЗ,
114
Филлоэритрин 166, 167
Фитилпирофосфат 206, 207
Фитоалексины 150
Фитоин 41, 43, 45, 57, 60, 62—67 78,
80—82
Фитол 62, 160, 162, 207, 208, 365
Фитофлуии 43—45, 64, 66, 69
Фитохром 83, 150, 156, 193, 194 215,
220, 221, 359, 360, 369—374, 391
Ф лаван-3,4-диолы (проантоцианиди-
ны) 126, 127, 132, 144, 147, 148
Флаванонолы (дигидрофлавонолы)
126, 127, 130, 144—146
Флаван-З-олы (катехины) 126—128,
131, 144, 148, 149
Флаваноны 126, 127, 131, 132, 139,
144, 145, 148
Флаваны 125, 131, 132, 144, 147,
149
Флавилиевые соли 133—135
Флавины и флавопротеины 150, 181,
230—240, 257, 258, 328 344, 375,
389, 391
Флавоиин 136, 137
Флавоноиды 111, 112, 125—155,
293—294
Флавонолы 126, 127, 129, 132, 133,
135, 136, 139, 140, 144, 145, 151,
294
Флавоны 125—127, 130—132, 136,
139, 144, 145, 147, 151, 294
Флексирубины 87
Флешспектроскопия 32
Флеш-фотолиз 371
Флороглюциикарбоиовая кислота 152
Флуоресценция 22—24, 131, 230,
232, 321—322, 337, 389
Фолиевая кислота 228, 235
Форбин 160
Формилглицинамидрибоиуклеотид
233, 234
Формилкииурении 248, 249
Формил-№°-тетрагидрофолиевая кис-
лота 233
Фосфоресценция 22—24
Фотодинамическое повреждение 384—
386
Фотозащита 79, 80, 84—86, 384—386
Фотопическое зрение 310
Фотопсииы 12, 310, 315, 319
Фоторегеиерация 318
Фоторецепция 12, 84, 299—301
Фотосеисибилизация 120
Фотосинтез 9, 23—26, 29, 30, 32, 84,
416
Предметный указатель
118, 140, 156, 160, 166, 185, 189,
219, 220, 326—368
Фотосинтезирующие бактерии 57, 68,
76, 80, 160, 161, 201, 327—329,
355—358, 363—364, 385—386
— единицы 333—335, 356
Фотосистемы I и II (ФС1 и ФСП)
330, 332—347, 354—357, 359
Фототаксис 374—376, 392
Фототропизм 84, 376—377, 392
Фотофосфорилирование 327, 328,
347—349
Фоточувствятельность и фотозащита
120, 380—384
Фрингелиты 104, 105
Фукоксантин 54—55, 352
Фукоксантин — хлорофилл а/сг —
белок, комплекс 352
«Халконовая форма» 134
Халконы 127, 133—134, 139, 143, 144,
147, 294
Химафилин 115, 116
Химохром 128
Хиноидное основание 134, 135
Хинол 92—94
Хиноны 92—124
Хлорины 157, .160, 167, 178, 207
Хлоробактин 56, 57, 77
Хлоробиум-хлорофиллы 208
Хлорозы 215
Хлорокруорин 176—177
Хлорокруорогем 177
Хлороксантин 70
Хлоропласты 38, 52, 54, 60, 64, 76,
82 152, 154, 160, 166—167, 201,
329—333, 358—365, 370, 374
Хлоросомы 355
Хлорофилл а 159—165, 206—209,
215, 334—337, 340, 341, 351—352
— а' 161
— щ (Р = 700) 335, 338—340, 343—
345, 352, 357
— ап (Р = 680) 335, 338, 341, 343,
345
— alb — белок, комплекс 335, 338
— b 160—163, 207—208, 335—338,
351
— Ь' 161
Хлорофиллаза 207
Хлорофилл — белок, комплексы 335—
337
Хлорофиллид а 163, 166, 206. 207,
210
Хлорофиллы 29, 62, 82, 138, 156—
167, 195—217, 219, 220, 221, 329
— биосинтез 195—210, 334—341
— сине-зеленых водорослей 354
— cad 160—164, 208, 209, 219, 351
Хна 92, 121
Хоризмовая кислота 111, 112, 243
Хризоафин 103, 104
Хризопсин 320
Хризоптерин 226, 227, 229, 238
Хроматобласты 287
Хроматофоры 283—287, 295, 355
Хромопласты 60, 64, 67, 71, 294, 365
Хромофоры 21
Хунда правило 23
Цвет 10—И
Цветки и плоды 29, 52, 53, 99 125,
129, 137—140, 149—153, 268, 280,
292____295 371
Цветовое зрение 11 —12, 318—321,
325
Цианидин 135, 139
Цианокобаламин 184
Цианопсины 320
Циркадианные ритмы 381—383
Цитохромы 167, 177—181, 215, 328
Р-Цитраурин 40
Червецы 119
Шикиматный путь 110—116, 118, 124,
140, 141, 150
Шикимовая кислота (шикимат) ПО—
113, 115, 117
Шиконин 121
Экситонные взаимодействия 22
Экстинкция 25
Электронтранспортная цепь см Пе-
ренос электронов
Электрохимические градиенты 333,
347, 377—379
Эмодин 101, 102, 106
Эпифиз 380—381
Эритроафин 103, 104
Эритроптерин 227, 229, 238
Эритрофоры 240, 284 — 286, 291
Эхиненои 47, 48, 353, 354
Эхинохромы 100, 101, 382, 383
Эшшольцксантин 53, 77, 79
Юглоны 100, 101, 113, 114, 117, 120
Яваницин 108
Ядерный магнитный резонанс (ЯМР)
25, 42, 98, 130, 163
Яичный желток 58
ЯМР см. Ядерный магнитный резо-
нанс
Указатель латинских названий
Actinia equina 59
Aequorea 389
Amantia muscaria 252
Antedon 383
Antirrhinum majus 139
Aphis fabae 103
Aplysia 193, 382
Arion ater 387
Arthropoda 175
Aspergillus candidus 136
— niger 265, 272
Asterias rubens 59
Bacillariophycea 351
Beneckea 388
Beta vulgaris 252
Bignoniaceae 117
Blakeslea trispora 55, 81
Boraginaceae 116
Brachiopoda 175
Branchiostoma 383
Brevibacterium 242
Cantharellus cibarius 55
Capsicum annuum 53
Carassius auratus 320
Carcinus 282
Carpobrotus acinaciformis 250
Centrospermae 252
Chaetognatha 175
Cheiranthus cheirii 294
Chlamydomonas 351
Chlorella 207, 329, 351, 362
•Chlorobacteriaceae 328
Chlorobiaceae 57, 328, 355
Chlorobium 161, 208, 355, 358
Chlorogloea 190
— fritschit 362
Chlorophyceae 351
Chlorophyta 160
Chromatiaceae 57, 355
Chromatium 328, 358
Chromobacterium violaceum 255
Chrysophyceae 351
Coccinella septempunctata 58
Codium fragile 166
Coelenterata 175
Corticium salicinum 86
Corynebacterium 57
Cryptophyta 189, 190
Cyanobacteria 55
Cyanophyta 55, 189, 190
Cypridina 390
Cytophaga 87
Dactylopius coccus 121
Daldinia concentrica 265
Daucus carota 54
Diadema 383
Digitalis purpurea 293
Dinophyceae 351
Diospyros 117
Diptera 227
Drosera 117
Drosophila melanogaster 227, 249, 325
Dytiscus 136
Elysia viridis 166
Escherichia coli 235
Eschscholtzia californica 53
Euchloe cardamines 226
Euglena 54, 362, 363, 375
— gracilis 82, 361, 374
Euglenophyceae 351
Eupithecia oblongata 136
Flavobacterium 57
Flexibacterium 87
Fusarium javanicum 108
— martii 120
Galago 231
Gonepteryx rhamni 226
Halobacteria 56, 69, 391
Haplopappus gracilis 145
Helix pomatia 136, 177
Holothuria forskali 231
Homarus vulgaris 59
Hydrangea macrophylla 138
Hymenoptera 227
Hypericum 103, 120
indigofera tinctoria 255
isatis tinctoria 255
Juglans regia 100
Kermococcus ilicis 121
Latia 390
Lawsonia alba 100, 121
Lepidoptera 226
Leptinotarsa decemlineata 58
Limulus polyphemus 380
Lobelia 140
Mollisia caesia 109
•— fallens 100
Myrex 255
418
Указатель латинских названий
Mycobacterium 80
— tuberculosis 122
Nereis diversicolor 188
Neurospora crassa 80
Nocardia 242
Nucella 255
Onchorhynchus keta 175
Onychophora 175
Papilionidae 249
Periplaneta 231
Petroselinum Hortense 143
Phaeophyceae 351
Phaeophyta 160
Phaffia rhodozyma 42
Photobacterium. 388
Phycomyces blakesleeanus 78, 265
Pieridae 226
Pieris brassicae 189, 226
— rapae 226
Polyzoa 175
Porifera 59, 175
Primula obconica 120
Prochloron 166
Protochordata 175
Prunella modularis 188
Pseudomonas aeruginosa 242
— indtgofera 255
— phenazinium 242
Pyrrophyta 160
Лапа pipiens 287
Renilla 391
Rhizobium spp. 175
Rhodomicrobium vanniellii 68
Rhodophyta 160, 189, 190
Rhodopseudomonas 57, 69, 208, 357,
364
— capsulata 356, 363
— palustris 358, 363
•— sphaerotdes 69, 215, 356—358, 363
Rhodospirillaceae 57
Rhodospirillum rubrum 57, 69, 328, 357,.
358, 364
Rhodotorula spp 55
Rotifera 175
Rubta tinctorum 102, 121
Rubiacea 117
Saccharomyces cerevisiae 240
Sarcina 57
Scenedesmus 351, 362
— obliquus 81
Schistocerca 58
Septia officinalis 261
Serratia 255
Sertularella 136
Sipunculata 175
Spirogyra 329
Staphylococcus 57
Streptococcus 57
Spreptomyces 249
Tardigrada 175
Taxus baccata 53
Tenebrio molitor 269
Ulva 54
Vespula vulgaris 227
Wallemia sebi 86
Xanthomonas 87
Xanthophyceae 351
Zea mays 81
ОГЛАВЛЕНИЕ
Предисловие редактора перевода .................................... 5
Предисловие........................................................ 7
Часть I. Химические и биохимические аспекты
Глава 1. Свет и цвет.............................................. 9
1.1 Введение................................................. 9
1.2. Цвет и его восприятие....................................10
1.3 Цвет у живых организмов...................................12
1.4. Поглощение света.........................................16
1.5. Спектроскопические методы исследования природных пиг-
ментов 24
1.6. Значение окраски в природе...............................29
1.7. Заключение и комментарии.................................30
1.8. Рекомендуемая литература..................................30
1.9. Дополнительная литература................................32
Глава 2. Каротиноиды............................................. 34
2.1. Введение.................................................34
2 2. Структура и номенклатура............................. 34
2.3. Свойства.................................................43
2.4. Распространение в природе................................52
2.5. Каротинопротеины.........................................59
2.6. Биосинтез................................................60
2.7. Регуляция и контроль биосинтеза каротиноидов .... 78
2.8. Метаболизм каротиноидов у животных...................... 82
2.9. Функции каротиноидов.................................... 84
2.10. Каротиноиды как пищевые красители ....... 85
2.11. Использование каротиноидов в медицине....................86
2.12. Другие полиеновые пигменты...............................86
2.13. Заключение и комментарии.................................87
2.14. Рекомендуемая литература.................................89
2.15. Дополнительная литература................................90
Глава 3. Хиноны...................................................92
3.1. Введение.................................................92
3.2. Строение.................................................92
3.3. Свойства.................................................94
3.4. Распространение в природе................................98
3 5. Вклад в окраску......................................105
420
Оглавление
3.6. Биосинтез...............................................105-
3 7. Функции и биологическое действие..............US'
3.8. Использование в промышленности и медицине .... 121
3.9. Заключение н комментарии............................ ... 122
3.10 Рекомендуемая литература........................... .... 123
3.11. Дополнительная литература.............................. .124
Глава 4. О-Гетероциклические пигменты—флавоноиды...................125
4.1. Введение.................................................125
4.2. Структура и номенклатура.................................125
4.3. Свойства.................................................130
4.4. Распростраиеиие..........................................136
4.5. Участие в формировании окраски растений..................137
4.6. Биосинтез................................................140'
4.7. Метаболизм флавоноидов у животных.........................151
4 8. Разрушение флавоноидов микроорганизмами.......151
4.9. Функции флавоноидов в растениях..........................151
4.10. Использование антоцианов в качестве пищевых красителей 152
4.11. Заключение н комментарии.................................153
4 12. Рекомендуемая литература..............................154
4.13. Дополнительная литература................................155
Глава 5. Тетрапирролы.............................................156
5.1. Введение.................................................156
5.2 Общие структурные свойства...............................156
5.3. Общие светопоглощающие свойства тетрапирролов . . . 159
5.4. Хлорофиллы...............................................159
5.5. Гем н гемопротенны.......................................167
5.6. Свободные порфирины у животных...........................181
5.7. Витамин В|2..............................................183
5.8. Линейные тетрапирролы — билины...........................185
5 9. Биосинтез и метаболизм тетрапирролов................195
5.10. Нарушения метаболизма порфиринов.........................217
5.11. Функции тетрапиррольных пигментов....................... 219
5.12. Заключение и комментарии.................................219
5.13. Рекомендуемая литература.................................220
5.14. Дополнительная литература................................221
Глава 6. Другие неполимерные N-гетероциклические пигменты 223
6.1. Введение.................................................223
6.2. Пурины, птерины и флавины................................224
6.3. Феназины.................................................240
6.4. Феноксазнны..............................................245
6.5. Беталаины.............................................. 250!
6.6. Другие N-гетероцнклические пигменты......................255
6.7. Заключение и комментарии.................................257
6.8. Рекомендуемая литература.................................257
6.9. Дополнительная литература................................258
Глава 7. Меланины...........................................259
7.1. Введение.............................................259
7.2. Химия.............................................. 260'
Оглавление 421
7.3. Распространение меланинов .... , 266
7.4. Биосинтез.......................................’ ’ 26В
7.5. Факторы, влияющие на меланогенез . . •...............275
7.6. Функции............................................’ ‘ 277
7.7. Заключение и комментарии 278
7.8. Рекомендуемая литература..............................278
7.9. Дополнительная литература..............................279
Часть II. Функциональные аспекты
Глава 8. Значение окраски в природе...................щ . ... 280
8.1. Введение.............................................280
8.2. Окраска и характер ее распределения у животных . „ . 281
8.3. Пигментные клетки животных — хроматофоры .... 283
8.4. Изменение окраски у животных.........................287
8.5. Окраска у растений...................................292
8.6. Заключение и комментарии.............................294
8.7. Рекомендуемая литература.............................295
8.8. Дополнительная литература....................... 295
Глава 9, Пигменты и зрение......................................297
9.1. Введение..................................................297
9.2. Глаз......................................................297
9 3. Зрительные пигменты..................................303
9.4. Функционирование зрительных пигментов; циклы превраще-
9.5. Некоторые аспекты цветового зрения.............318-
9.6. Вспомогательные пигменты в процессе зрения .... 321
9.7. Заключение и комментарии..................................325
9.8. Рекомендуемая литература...........325
9.9. Дополнительная литература...........326
Глава 10. Фотосинтез..................................................327
10.1. Введение................................................
10.2. Фотосинтетический аппарат эукариот: хлоропласты .
10.3. Фотосинтез у растений; общая схема......................
10.4. Поглощение света — первичный процесс фотосинтеза
10 5. Вторичные события; система фотосинтетического транспорта
электронов ...................................................
10 6. Темновые реакции.................;...................
10.7. Фотосинтез у эукариотических водорослей .
10.8, Фотосинтез у прокариотических сине-зеленых водорослей
10.9. Бактериальный фотосинтез................................
10.10. Синтез пигментов в связи с развитием хлоропластов
10 11. Развитие фотосинтетического аппарата у фотосинтезирую-
щих бактерий .................................................
10.12. Дальнейшая судьба фотосинтетических пигментов в ходе
разрушения хлоропластов.......................................
10.13. Заключение и комментарии...............................
10.14. Рекомендуемая литература
10.15. Дополнительная литература..............................
327
329
333
333
342
347
351
353
355
358
363
364
365
367
367
422 ___________________________Оглавление_____________________________
Глава 11. Другие фотофункции природных пигментов................369
11.1 Введение............................................... 369
11.2 . Фитохром.............................................369
11.3 Фототаксис..............................................374
11 4 Фототропизм.........................................376
11.5 . Бактериородопсии......................................377
11.6 . Виеглазиые фоторецепторы у животных...................379
11.7 . Фотозащита............................................383
11 8. Биолюминесценция....................................387
119 Заключение и комментарии................................391
11 10 Рекомендуемая литература............................392
11.11. Дополнительная литература..............................393
Задачи............................................................395
«Ответы...........................................................402
Предметный указатель..............................................410
^Указатель латинских названий . . 417
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги, ее оформле-
нии, качестве перевода и другие просим присылать
по адресу:
129820. Москва, И-110, ГСП
1-й Рижский пер., д. 2,
издательство «Мир»
Монография
Г. Бриттон
БИОХИМИЯ ПРИРОДНЫХ ПИГМЕНТОВ
Научный редактор Т. И. Пономарева
Мл. научн. редактор 3. В. Соллертинская
Художник В. П. Груздев
Художественный редактор Л. М. Кузнецова
Технический редактор Н. И. Манохина
Корректор Н. В. Андреева
ИБ № 5609
Сдано в набор 15.01.86. Подписано к печати 04.07.86.
Формат 6ОХ90’/1б. Бумага типографская № 1.
Печать высокая. Гарнитура литературная.
Объем 13,25 бум. л. Усл. печ. л. 26,50.
Усл. кр.-отт. 26,50. Уч.-изД. л. 27,12. Изд. № 4/4189.
Тираж 3.050 экз. Зак. 84. Цена 3 р. 90 к.
ИЗДАТЕЛЬСТВО «МИР».
129820, ГСП, Москва, И-110, 1-й Рижский пер., 2.
Московская типография № П Союзполиграфпрома
при Государственном комитете СССР по делам
издательств, полиграфии и книжной торговли.
113105, Москва, Нагатинская ул., д. 1.
Отсканировал Семенюченко Владимир
chem_vova@mail.univ.kiev.ua;
vova2002@mail.ru