






















































































































































































































































Text
ПАУКООБРАЗНЫЕ
том
VI
выл.
1
Издательство Акаде/лии Наук СССР
Москва - Ленинград
INSTITUT ZOOLOGIQUE DE L’ACADEMIE DES SCIENCES DE L’URSS
NOUVELLE SERIE n's 28
FAUNE DE L’URSS
Sous la direction de S, A. SERNOV, de VAcademic Redige par D; A. OGLOBLIN
ARACHNOIDEA
Vol. VI, n° 1
ACARIENS TYROGLYPHOIDES
par
A. A. ZACHVATKIN
EDITION DE 1’ACADEMIE DES SCIENCES DE L’URSS MOSCOU • 1941 • LENINGRAD
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ АКАДЕМИИ НАУК СССР
НОВАЯ СЕРИЯ Ха 28
ФАУНА СССР
Главный редактор акад. С. А. ЗЕРНОВ Редактор Я- А. ОГЛОБЛИН
ПАУКООБРАЗНЫЕ
Т. VI, вьш. 1
А. А. ЗАХВАТКИН
ТИРОГЛИФОИДНЫЕ КЛЕЩИ (TYROGLYPHOIDEA)
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР МОСКВА • 1941 • ЛЕНЙНГРАД
Ответственный редактор директор Зоологического института акад. О. А. Зернов
Редактор издания С. С. Смирнов
редактор издательства Н. И. Ш а р а п о в.
Тираж 1 000. Подписано к печати 18 декабря 1940 г. М 41923. Печатных листов зозд. Авт. листов 33.66. В печ. листе 46080 тип. зи. Заказ Л? 284. Цепа 31 руб. 50 коп.
Набрано во 2-й типографии ОГИЗ’а РСФСР треста «ПолиграФкиига» «Печатный Двор» им. А. 31. Горького. Ленинград, Гатчинская, 26.
Отпечатана в Ремесленном училище № 71 ПолиграФпромышлеиности. Гомеровский, 24.
Переплетно-брошировочиые работы производились в Тиио-литограФии Академии Наук.
ПРЕДИСЛОВИЕ
Огромный хозяйственный вред, приносимый амбарными клещами, ставит перед научно-исследовательскими учреждениями нашей страны вопрос о всемерном развертывании и форсировании работ по их изучению. Выполнение этих работ, однако, в чрезвычайной степени тормозится исключительно слабой разработанностью систематики тех групп клещей, к которым относятся виды, вредящие запасам сельскохозяйственных продуктов, и полным отсутствием сводок по этим группам. В связи с этим, представилось своеврсмеппым включить в план издания «Фауна СССР» такую сводку; ее выполнение было поручено мне.
В основу предлагаемой обработки семейств Tyroglyphidae, Saprogly-phidae и Glycyphagidae легли материалы, скопившиеся в моих руках в течение предшествовавших 3 лет работы по систематике и фаунистике амбарных клещей, и обширные специальные сборы, произведенные мною и Е. М. Булановой в энтомологических и терологических коллекциях Зоологического института (ЗИН) Академии Наук, Московского зоологического музея и частном собрании Н. Н. Филиппова. Выла использована и акарологическая коллекция ЗИН, содержавшая всего несколько десятков препаратов клещей названных семейств, и все другие доступные мне материалы — в общей сложности около 3000 препаратов.
При мопографировапии группы пришлось полностью переработать — практически, создать заново — всю ее систематику, начиная от самых общих приемов различения и характеристики родов и видов и кончая ее классификацией. Эта перестройка явилась, во-первых, результатом детализации систематического исследования, с привлечением большого числа признаков, остававшихся ранее неиспользованными (например: хэтотаксия тела и йог, строение первой нимфальиой стадии, особенности онтогенеза ряда признаков и др.); во-вторых, опа явилась неизбежным следствием применения к обработке даипой группы организмов принципов естественной системы, строящейся на учете суммарного (интегрального) сходства классифицируемых объектов и тщательном анализе характера связей между' ее признаками, — неизбежным потому, что имевшаяся система Tyroglyphoidea, созданная Oudemans’oM, опиралась иа произвольно и случайно выбранные единичные признаки, которым столь же произвольно придавалось значение «руководящих». В связи с этим, общий стиль пред
VI
ПРЕДИСЛОВИЕ
лагаемых здесь описаний существенно разнится от применявшихся ранее стандартов, в очень многих случаях изменен калибр таксономических категорий и внутри них произведены многочисленные перемещения. Во избежание вызываемой этими переделками опасности нарушения преемственности между предшествующей литературой и данной работой, особое внимание было обращено па проработку сипопимики и расшифровку описаний прежних авторов в свете вновь полученных результатов и модернизованной систематической методики.
В виду все еще крайпс слабой изучеппостп фауны Tyrogl.yphoidea detri-licolae нашего Союза (стоящего теперь в этом отношении все же впереди других стран), в настоящую работу включены и все описанные из Палеарктики формы, еще не найденные у пас; практика последних лет показала, что большинство из них со временем обнаруживается и в СССР; кроме того, этим полнее удовлетворяется задача работы — служить основой для дальнейших работ по систематике и фаунистике дайной группы клещей.
Предлагаемая работа была выполнена в Зоологическом институте Московского Государственного университета в 1935—1936 гг.; все рисунки п перерисовки' исполнены мною, резюме переведено М. А. Языковой. Осуществлению этой работы содействовали многие лица, которым я приношу свою самую искреннюю благодарность.
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ
I. Сем. Tyroglyphidae
1. Подсем. Туго gly phinae
1. ТРИВА TYROGLYPHINI
1. Род Tyroglyphus Latr.
Стр.
I, Т, fiH'huu) (,L.)............................................................... 86
Bl Т> iyroph«gol(|i'B( up, ...................................................... 90
a. ТРИВА TYROl’IIAOINI
2, Род Alcuroglyphus, gon. n.
I. A. ovniuB (Tr.) ............................................................... 92
1!, A. slculus (F. ot R.) ........................................................ 94
3. Род Kusinia, gen. n.
J, K. laevis (Duj.)................................................................ 95
4. Род Tyrolichus Ouds.
1. T. casei (Ouds.) ........................................................... 97
2, T. lini (Ouds.)................................................................ 98
5. Род Tyrophagus Ouds.
:l. T. noxius A. Z................................................................ 103
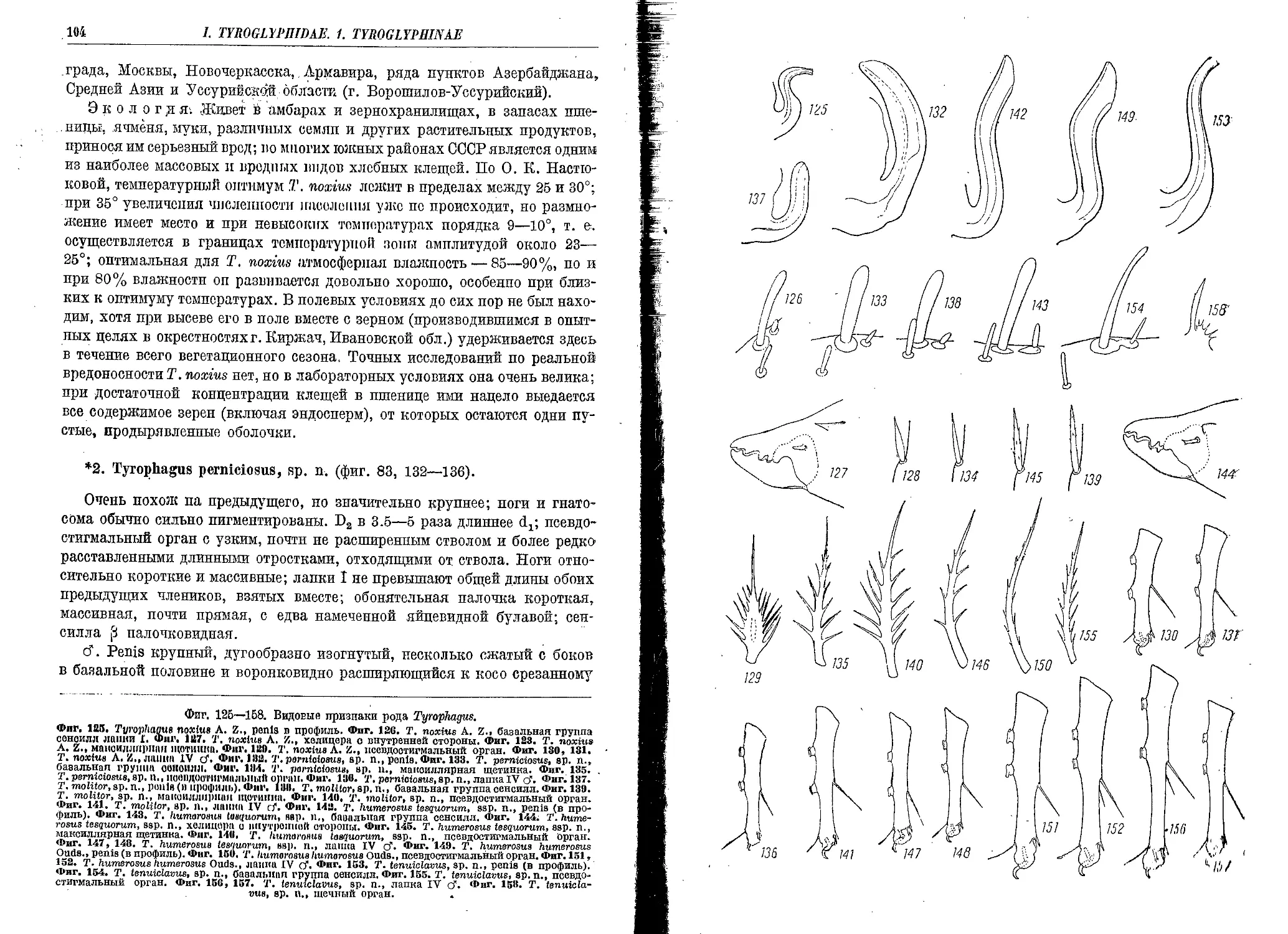
2, T. poiniciosus, sp. n........................................................ 104
8. T. molitoi, sp. n............................................................. 106
4. T. humeiosus Ouds............................................................. 106
5. T. tenuiclavus, sp. n......................................................... 109
6. T. longior (G-erv.) .......................................................... 109
6. Род Forcellinia Ouds.
1. F. fungivora (Ouds.) ......................................................... 114
2. F. diamosa, sp. n............................................................. 116
3. F. gigantonympha (Vtz.) ..................................................... 116
4. F. wasmanni (Mon.)........................................................... 117
7. Род Scatoglyphus Beil.
1. S. polytrematus Beil ......................................................... 121
2. Подсем." Rhiz о gly phi n a e
3. ТРИБА ACOTYLEDONINI
8. Род Acotyledon Ouds.
1. A. rhizoglyphoides~(A. Z.) .............'..................................... 135
2. A. sokolovi, sp. n. ............................. !........................... 137
3. A. paradox^ Ouds..'. . .............. . . ?.................................. 138
VIII
ТИРОГЛИФОИДВЫЕ КЛЕЩИ
4. A. agilis (Сап.)......................................................................140
5. A. michaeli (Ouds.) ................................................................ 140
6. A. redikorzevi (A. Z.) ...............................................................142
7. A. albcrti (A. Z.)................................................................. 146
8. A. krameri (Berl.) .............................................................. 147
9. A. leytaudi (Ouds.).................................................................. 148
10. A. batsylevi, sp. n................................................................. 148
11. A. volgini, sp. n................................................................... 150
12. A. tshemyshevi, sp. n............................................................ 152
13. A. ourynympha (Ouds.) .............................................................. 153
9. Рол Caloglyplvud Berl.
1. C. rodionovi A. Z.................................................................. 159
2. C. tshemyshevi, sp. n............................................................ 163
3. C. mandzhur, sp. n................................................................. 163
4. C. spinitarsus (Herm.) :.......................................................... 166
5. 0. polyphyllae, sp. n............................................................. 168
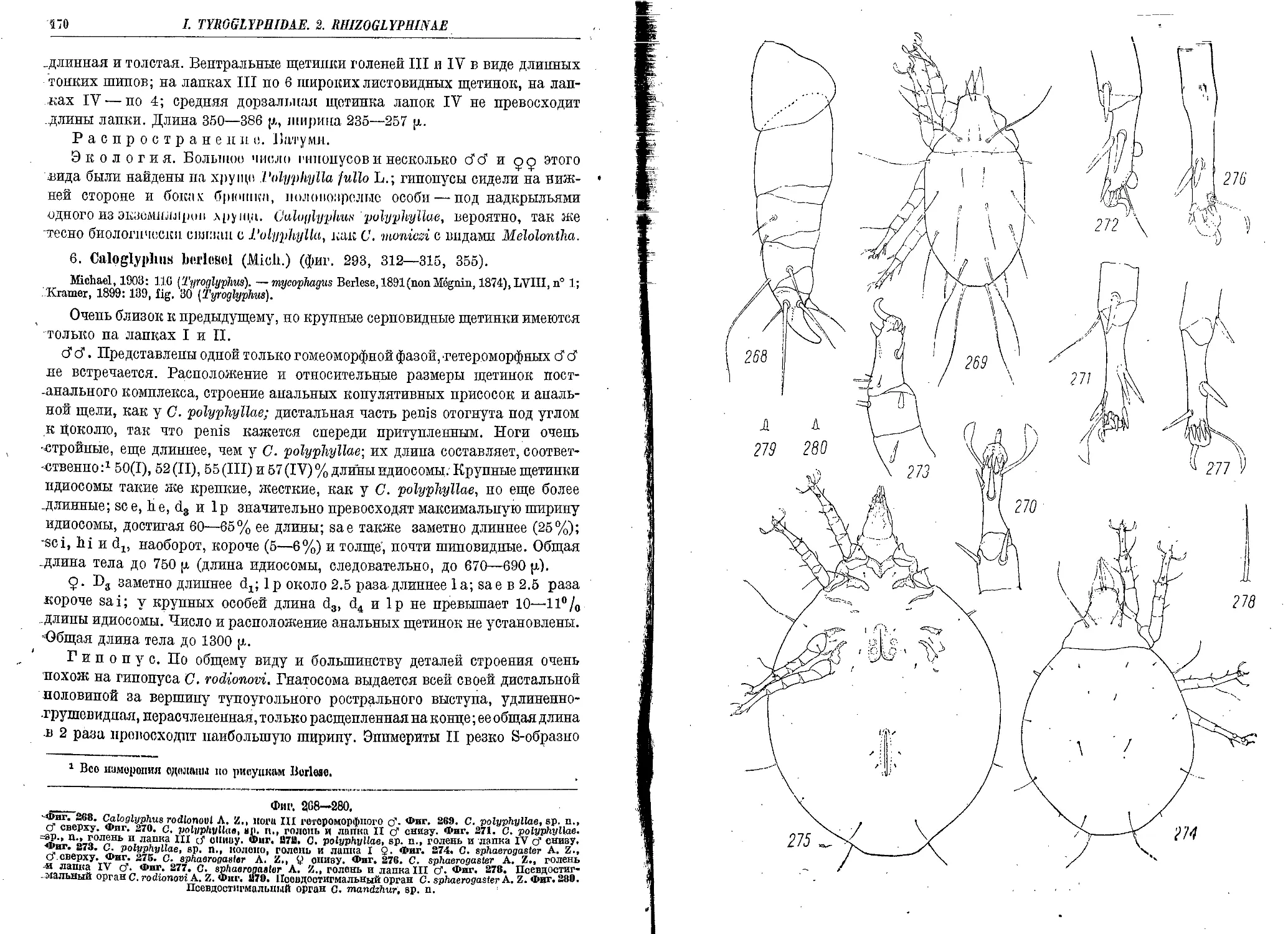
6. C. berlesei (Mich.)........................................................... 170
7. 0. moniezi A. Z.................................................................... 172
8. 0. sphaerogaster A. Z.............................................................. 173
9. C. mycophagus (M6gn.).............................................................. 176
10. 0. geotruporum, sp. n............................................................. 178
4. ТРИБА RIIIZOGLYPHINI
10. Род Rhteoglyphus Clap.
1. R. echinopus (F. ct R.) .............................................................. 182
11. Род Heteroglyphus FoA
1. H. vitium Foa......................................................................... 186
12. Род Histiogaster Berl.
1. H, bacchus, sp. n..................................................................... 190
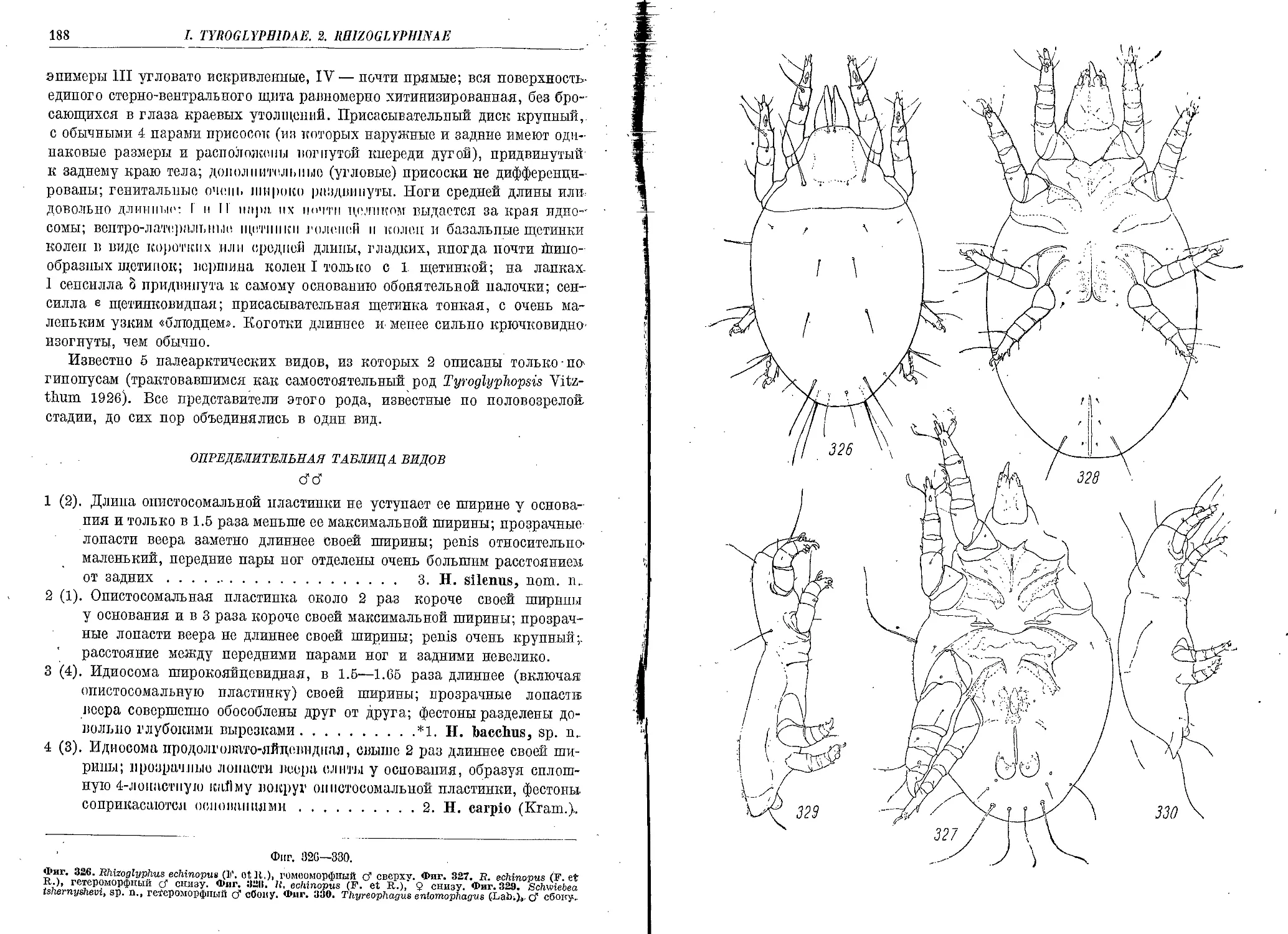
2. H. carpio (Kram.).................................................................. 192
3. H. silenus, nom. n................................................................ 192
4. H. filippovi, sp. n............................................................... 194
5. H. ocellatus (Vtz.)................................................................ 196
13. Род Schwiebea, Ouds.
1. S. tshemyshevi, sp. ................................................................ 201
2. S. ipidis Vtz....................................................................... 202
3. S. cavernicola Vtz.................................................................. 203
4. S. talpa Ouds....................................................................... 203
5. S. pachydenna, sp. ................................................................. 204
6. S. xossica, sp. n....................................................................205
14. Род Monieziella Berl.
1. M. corticalis (Mich.) '................................ .'............................ 207
2. M. berlesiana, nom. nov............................................................... 207
3. M. magna Berl......................................................................... 208
15. Род Thyreophagus Rond.
1. T. entomophagus (Lab.)......................'......................................... 209
16. Род Laclcerbaueria, gen. n.
1. L. cribratissima, sp. n............................................................... 214
17. Род Robinisca, gen. n.
1. R. macrocnemis, sp. n................................................................. 216
2. R. mycolicha (Ouds.) ............................................................... 217
3. R. tarsispina (Ouds.)............................................................ . 218
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ IX
18. Род Viedebcmtiia Ouds.
1. V. schmitzi Ouds....................................................................... 218
19. Род TroupectMia, gen. n.
1. T. nova (Ouds.)....................................................................... 220
2. T. crabronis, sp. n................................................................... 222
3. T. (?) mycoboia (Ouds.)................................................................ 222
20. Род Rodionovia, gen. n.
1. R. inopinata (Vtz.).................................................................... 233
21. Род Garsaultia Ouds.
1. G. testudo Ouds................................................................ . . . 224
22. Род Sancassania Ouds.
1. S. chelone Ouds........................................................................ 225
23. Род Schuleea, gen. n.
1. S. pamirensis, sp. n.................................................................. 226
2. S. caucasica, sp. n................................................................... 227
24. Род Froriepia Vtz.
1. F. vimariensis Vtz..................................................................... 228
II. Сем. Saproglyphidae
1. Род Ponioppidania Ouds.
1. P. littoralis (Halb.).................................................................. 235
2. Род Saproglyphus Berl.
1. S. neglectus Berl...................................................................... 238
3. Род Winlersehmidtia Ouds.
1. W. hamadryas (Vtz.).................................................................... 240
4. Род Calvo Ha Ouds.
1. C. tuberculata, sp. n.................................................................. 248
2. C. heterocoma (Mich.).................................................................. 248
3. C. romanovae, sp. n.................................................................... 252
4. C. hebeclinii (Sicher)..................................................................252
5. 0. kamenskii, sp. n.................................................................... 254
6. C. elisabethae, sp. n.................................................................. 254
7. C. reticulata, sp. n................................................................... 254
8. C. kneissli Krausse................................................................... 255
9. 0. thraca Vtz......................................................................... 255
10. 0. calliphorae Vtz................................................................... 256
11. C. bulgarica Storkau................................................................. 257
12. 0. circumspectans Vtz................................................................ 258
13. C. hagensis Ouds..................................................................... 259
14. 0. fusiformis, sp. n................................................................. 259
15. 0. striata (Vtz.).................................................................... 260
16. C. nataliae, sp. n................................................................... 262
17. 0. elliptica, sp. n.................................................................. 262
18. C. (?) zacheri Ouds.................................................................. 263
5. Род Ensliniella Vtz.
1. E. parasitica Vtz...................................................................... 266
2. E. kostylevi, sp. n.............................................................. . 267
X
ТИР0Г.1ИФ0ИДНЫЕ ЫЕЩИ
6. Род Nanaams Ouds.
1. N. minutus (Ouds.) ..................................................... 269
7. Род Vidia Ouds.
1. V. gussakovskii, sp. n...................................................270
2. V. popovi, sp. n....................................................... 272
3. V. lineata Ouds......................................................... 273
4. V. undulata Ouds........................................................ 274
III. Сем. Glycyphagidac
1. Подсем. Chortoglyphinae
1. Род Chortoglyphus Beil.
1. Ch. arcuatus (Tr.)...................................................... 286
2. Род Blomia Ouds.
1. B. thori A. Z........................................................... 288
2. B. tiibodas (Ouds.)..................................................... 290
3. B. kulagini A. Z........................................................ 291
2. Подсем. Glycyphaginae
3. Род Glycyphagus Her.
1. G. destructor (Schrk.) Ouds............................................. 296
2. G. pilosus Ouds......................................................... 297
3. G. fustifer Ouds....................................................... 298
4. G. michaeli Ouds.........................................................300
5. G. burchanensis Ouds.................................................... 301
6. G. cadaverum (Schrk.) Ouds............................................. 302
7. G. domesticus (Deg.).................................................... 302
8. G. ornatus Kram......................................................... 304
4. Род Cometacarus A. Z.
1. 0. smimovi, sp. ........................................................ 308
2. 0. oudemansi A. Z. ......................................................309
3, 0. setosus (0. L. Koch)..................................................309
3. Подсем. Aero glyphinae
б. Род Aeroglyphus, gen. n.
1. A. peregrinans (Berl.) ...................................................312
4. Подсем. Ctenoglyphinae
6. Род Diamesoglypkus, gen. n.
1. D. intermedins (Can.)...................... . ....................... 314
7. Род Ctenoglyphus Berl.
1. C. plumiger (C. L. Koch)................................................ 318
2. C. canestrinii (Arm.)................................................... 320
3. 0. palmifer (F. et R.) . . .................................. 320
4. C. multifoliatus, nom. n................................................ 321
8. Род Nycterigtyphus, gen. n.
1. N. pterophorus (Berl.).................................................. 322
5. Подсем. Labidophorinae
9. Род Xenocastor, gen. n.
1. X. fedjushini, sp. n................................................... 330
СИСТЕИА.ТНЧЕСКИН УКАЗАТЕЛЬ Х(
10. Род Labidophorus Kram.
1. L. sciuriuus (C. L. Koch)......................................................... 335
2. L. desmanae, sp. n................................................................ 337
3. L. talpae Kram.................................................................... 330
4. L. soricis Ouds.................................................................. 341,
5. L. mogerae, sp. n................................................................ 341
11. Род Gohieria Ouds.
1. G. fusca (Ouds.) ................................................................ Bil-
ls. Род Xenoryctes, gen. n.
1. X. krameri (Mich.)................................................................. 347
2. X. heptneri, sp. n................................................................. 349
13. Род Orycteroxenus, gen. n.
1. 0. dispar (Mich.)-................................................................ 352
14. Род Talpacarus. gen. n.
1. T. platygaster (Mich.)............................................................ 354
15. Род Fusacarus Mich.
I. F. laminipes Mich......V.......................................................... 357
16. Род Myacarus, gen. n.
1. M. arvicolae (Duj.)............................................................... 358
2. M. hypudaei (0. L. Koch).......................................................... 360
6. Подсем. Carpoglyphin a e
17. Род Hyadesia M6gn.
1. И. fusca (Lohm.).................................................................. 364
2. H. algivorans (Mich.) .............................................................366
18. Род Carpoglyphus Rob.
1. C. lactis (L.).................................'.................................. 367
19. Род Hericia Can.
1. H. hericia (Rob.)'................................................................ 372
2. H. georgei Mich................................................................. 373
7. Подсем. Chaetodactylinae
20. Род Senneriia Ouds.
1. S. cerambycina (Scop.) . ..........................................................382
2. S. gargantua, sp. n............................................................... 385
3. S. dissimilis, sp. n............................................................. 385
4. S. cantabrica, sp. n............................................................. 385
5. S. alfkeni (Ouds.)................................................................ 386
6. S. horrida (Vtz.)................................................................. 387
7. S. zhelochovtzevi, sp. n.......................................................... 388
8. S. potanini, sp. n.................................................................390
9. S. oudemansi, sp. n.............................................................. 390
21. Род Chaeta dactylus Rond.
I, Ch. osmiae (L. Duf.).............................................................. 394
2. Oh. reaumuri (Ouds.)...............................................................805
3. Ch. birulai. sp. n............................................................... 390
XII
ТИРОГЛИФОНДИЫЕ КЛЕЩИ
4. Ch. clavigei Ouds................................................................... 396
5. Ch. anthidii (Ouds.) ................................................................. 398
6. Ch. dementievi, sp. n................................................................. 399
22. Род Jlorsiia Ouds.
I. H. ornata (Ouds.) .................................................................... 400
2, H. helenae (Ouds.).................................................................... 401
23. Род Tortonia Ouds.
1. T. intermedia (Ouds.) ................................................................ 403
2. T. fluctuata Ouds..................................................................... 403
3. T. smitsvanburgsti Ouds............................................................... 404
4. ’ T. dogaressa, sp. n...............................'................................. 404
24. Род Neohorstia, gen. n.
1. N. mamillata, sp. n................................................................... 406
25. Род Cerophagvs Ouds.
1. O. granulatus (Duj.).................................................................. 407
26. Род Cerophagopsis, gen. n.
1. C. skorikovi, sp. n................................................................. 410
27. Род Sennertiowyx, gen. n.
J..S. manicati (Giard).................................................................... 411
ОБЩАЯ ЧАСТЬ
Семейства клещей Tyroglyphidae, Saproglyphidae и Glycyphagidae, рассматриваемые в настоящей работе, относятся к обширной серии Acaridiae подотряда Sarcoptiformes n образуют в своей совокупности провизорную группировку: Tyroglyphoidea detriticolae, соответствующую, по своему составу, прежнелгу сем. Tyrogliphidae auct. без группы Anoetidae. Представители этого комплекса характеризуются следующими основными признаками, комбинация которых отлипает их от других, смежных группировок серии Acaridiae. Ротовые части грызущего типа; хелицеры 2-члё-нистые, клешневидные, с хорошо развитым подвижным пальцем и крупнозубчатыми режущими краями; педипальпы без пластиночных разращений и бичевидных вершинных щетинок. Кожа гладкая, зернистая или шиповатая, лишь крайне редко с тонкой и более или менее параллельной штриховкой. Генитальное (яйцевыводное) отверстие $ в виде продольной щели, расположенное вентрально и совершенно обособленное от анальной щели; у большинства видов имеются генитальные щупальцы; вентральные кольце-, видные органы отсутствуют. Голени (по крайней мере двух передних пар ног) дорзально с крупной вершинной щетинкой; амбулакры состоят из более или менее хорошо развитого перепончатого praetarsus и непарного крючковидного коготка. Преимущественно сапробиотические формы, ведущие в половозрелой и нимфальных стадиях свободный образ жизни, ио широко йспрльзующие эпизоические способы расселения, осуществляемого обычно особой гипопальной стадией’ резко отличающейся по своей организации от других стадий развития.
ВНЕШНЕЕ ОТРОЕНИЕ ПОЛОВОЗРЕЛОЙ СТАДИИ
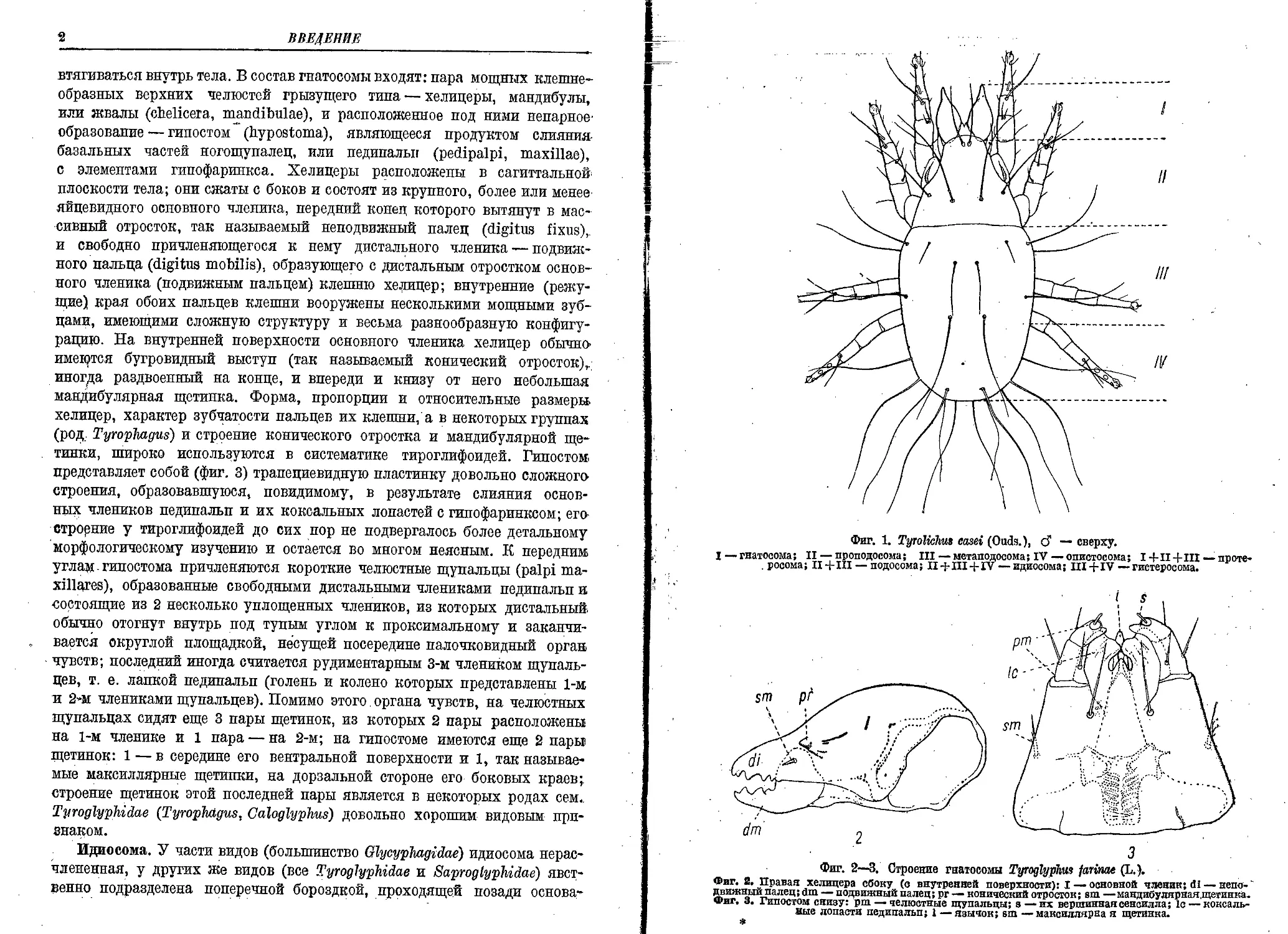
Тело тироглифоидей состоит, как и у других клещей, из двух основных отделов: комплекса ротовых частей—гнатосомы, и тела собственно, или идиосомы, несущего все 4 пары ходильных конечностей (фиг. 1).
Гнатосома (фиг. 2). Гнатосома относительно невелика, компактна и, в подавляющем большинстве случаев, свободна; лишь у очень немногих форм (часть Labidoplwnnac) она целиком скрыта в особом углублении, под передним краем идиосомы (выступающим тогда вперед в виде капюшона) — камеростоме; она обладает значительной подвижностью и способна отчасти
1 Фауна СССР, Захватили—284
2
ВВЕДЕНИЕ
втягиваться внутрь тела. В состав гнатосомы входят: пара мощных клешнеобразных верхних челюстей грызущего типа — хелицеры, мандибулы, или жвалы (chelicera, mandibulae), и расположенное под ними непарное’ образование — гипостом (hypostoma), являющееся продуктом слияния базальных частей ногощупалец, или педипальп (pedipalpi, maxillae), с элементами гипофаринкса. Хелицеры расположены в сагиттальной плоскости тела; они сжаты с боков и состоят из крупного, более или менее яйцевидного основного членика, передний конец которого вытянут в массивный отросток, так называемый неподвижный палец (digitus fixus), и свободно причленяющегося к нему дистального членика — подвижного пальца (digitus mobilis), образующего с дистальным отростком основного членика (подвижным пальцем) клешню хелицер; внутренние (режущие) края обоих пальцев клешни вооружены несколькими мощными зубцами, имеющими сложную структуру и весьма разнообразную конфигурацию. На внутренней поверхности основного членика хелицер обычно’ имещтся бугровидный выступ (так называемый конический отросток),, иногда раздвоенный на конце, и впереди и книзу от него небольшая мандибулярная щетинка. Форма, пропорции и относительные размеры, хелицер, характер зубчатости пальцев их клешни, а в некоторых группах (род. Tyrophagus) и строение конического отростка и мандибулярной щетинки, широко используются в систематике тироглифоидей. Гипостом представляет собой (фиг. 3) трапециевидную пластинку довольно сложного строения, образовавшуюся, невидимому, в результате слияния основных члеников педипальп и их коксальных лопастей с гипофаринксом; его строение у тироглифоидей до сих пор не подвергалось более детальному морфологическому изучению и остается во многом неясным. К передним углам гипостома причленяются короткие челюстные щупальцы (palpi ma-xillares), образованные свободными дистальными члениками педипальп и состоящие из 2 несколько уплощенных члеников, из которых дистальный обычно отогнут внутрь под тупым углом к проксимальному и заканчивается округлой площадкой, несущей посередине палочковидный орган чувств; последний иногда считается рудиментарным 3-м члеником щупальцев, т. е. лапкой педипальп (голень и колено которых представлены 1-м и 2-м члениками щупальцев). Помимо этого , органа чувств, на челюстных щупальцах сидят еще 3 пары щетинок, из которых 2 пары расположены на 1-м членике и 1 пара — на 2-м; на гипостоме имеются еще 2 пары щетинок: 1 — в середине его вентральной поверхности и 1, так называемые максиллярные щетинки, на дорзальной стороне его боковых краев; строение щетинок этой последней пары является в некоторых родах сем., Tyroglyphidae (Tyrophagus, Caloglyphus) довольно хорошим видовым признаком.
Идиосома. У части видов (больпшнство Glycyphagidae) идиосома нерас-члененная, у других Же видов (все Tyroglyphidae и Saproglyphidae) явственно подразделена поперечной бороздкой, проходящей позади основа-
Фиг. 1. Tyrolichus easel (Ouds.), d — сверху.
I — гнатосома; II — проподосома ; ill — метаподосома; IV — опистосома; I + II + III — проте-. росома; II+III — подосома; II III+IV — идиосома; III + IV — гистеросома.
Фиг. 2—3. Строение гнатосомы Tyroglypkus farinae (L.).
Фиг. й. Правая хелицера сбоку (о внутренней поверхности): I — основной членик; 61 — непс-' движный палец; dm — подвижный палец; рг — конический отросток; вт —мандибулярная,щетинка. Фиг. 3. Гипостом снизу: pm — челюстные щупальцы; в—их вершинная сенсилла; 1с — коксаль-ные лопасти педипальп; 1 — язычок; sm —максиллярна я щетинка.
4
ВВЕДЕНИЕ
ния ног II, на приблизительно конический, суживающийся кпереди, передний отдел — проподосому (propodosoma), несущую комплекс ротовых частей (гнатосому) и обе передние пары ходильных ног (I и II), и больший по своим размерам задний отдел — гистеросому (hysterosoma), несущую 2 задние пары ног (III и IV). Дальнейшего подразделения гистеросомы на метаподосому и лишенную конечностей опистосому (брюшко) у тирогли-фоидей не наблюдается. Не наблюдается у них также и каких-либо непосредственных следов сегментации в виде поперечных бороздок или швов (кроме уже отмеченной границы между проподосомой и гистеросомой) или метамерно расположенных щитков, пятен, или скульптурного орнамента, так что следы таковой могут быть усмотрены лишь в характере распределения щетинок спинной поверхности, имеющих более или менее ясно выраженную тенденцию располагаться в поперечные ряды. Основываясь на этом последнем признаке и на положении конечностей, можно считать, что тело половозрелых тироглифоидей состоит из 13 слившихся сегментов: б протеросомальных (acron, 2 сегмента, несущие ротовые конечности, и 2 сегмента, несущие передние пары ходильных ног) и 8 гистеросомальпых (2 сегмента, несущие задние пары ходильных ног, и 6 брюшных — опистосомальных). Это представление, впрочем, нуждается еще в подтверждении сравнительно-анатомическими и эмбриологическими данными.
Общая форма идиосомы весьма изменчива. Отношение ее длины к ширине колеблется у разных -видов от 1.2 : 1 (некоторые Sennertia) до 3— 3.4 : 1 (виды рода Monieziellay, она то пузыревидно вздута, то валькова-тая, то более или менее сильно уплощена в дорзо-вентральном направлении; типичной можно все же считать продолговато-овальную форму с отношением длины к ширине около 1.7—1.8 : 1. У живых особей спинная поверхность идиосомы лишь редко бывает равномерно выпуклой; обычно ее рельеф более расчленен, благодаря наличию 2 продольных цервикальных вдавлений на проподосоме (или соответствующей ей передней части нерасчлененной идиосомы) и парных, часто S-образно изогнутых продольных бороздок (рахидиальных бороздок) на гистеросоме, разбивающих поверхность последней на более широкую медианную (осевую) часть и 2 боковых (плевральных) поля; степень вдавленности этих бороздок и выпуклости разделяемых ими полей не только различна у разных видов, но сильно меняется даже у одной.и той же особи в зависимости от ее физиологического состояния (например сглаживается у раздутых яйцами 99, становится резче при дегидратации организма вследствие пребывания в более сухой среде и т/д.). На препаратах эти бороздки обычно становятся совершенно незаметными, и намечаемое ими продольное разделение идиосомы на 3 части бесследно исчезает. 1 *
г' 1 В некоторых -своих работах 1926—1932 гг. Oudemans придавал наличию рахидиальных бороздок Значение'важного-видового признака, найдя их случайно сохранившимися на 'некоторых препаратах/
ВНЕШНЕЕ СТРОЕНИЕ
»
Кожные покровы, У подавляющего большинства тироглифоидей кожа имеет мягкую, перепончатую консистенцию, бесцветна и очень тонка; у всех Tyroglyphidae и Saproglyphidae она, как правило, совершенно гладкая («зеркально-гладкая»), что придает живым клещам этих семейств; очень характерный стеклянный блеск; у Glycyphagidae кожа обычно покрыта тонкой зернистой или шиповатой микроскульптурой, и ее поверхность бывает совершенно гладкой лишь у видов с более толстыми, хитииизи-рованнымп кожными покровами (Chortoglyphus, Fusacarus и др.); параллельная штриховка кожи наблюдается лишь как исключение (у Aeroglyphus, Calvolia hebeclinii, С- tuberculata, сГсГ Win terschmid tia) и крайне тонка, имея совершенно другой характер, чем у паразитических Acaridiae. У некоторых Glycyphagidae кожа более или менее сильно уплотнена (хитинизи-рована), образуя в крайних случаях сплошной жесткий панцырь (con-scutum), одевающий всю или почти всю идиосому (Fusacarus, Chortoglyphus, Xenoryctes, Gohieria); в этих случаях опа также более или менее интенсивно окрашена в бурые или рыжеватые тона. У форм с мягкой, перепончатой кожей нередко имеют место уплотнение и склеротизация отдельных ее участков, приводящие к образованию на этих местах более или менее явственных щитов. Чаще всего такого рода щит имеется на спинной поверхности проподосомы; это — проподосомальный щит (scutum propodosoma-tale), вероятно, гомологичный propeltidium других паукообразных. У Туго-glyphidae и Saproglyphidae он имеет форму широкого параллелограма или. геральдического щита и занимает бблыпую часть поверхности проподосомы впереди лопаточных щетинок и практически не пигментирован (его очертания явственно видны лишь на хорошо просветленных препаратах); у тех Glycyphagidae, где он вообще имеется, проподосомальный щит более разнообразен по своей форме, всегда занимает меньшую площадь, и у рода Glycyphagus (подрода Glycyphagus s. str.) представлен узким продольным теменным килем (crista metopica), сильно напоминающим одноименное образование многих Trombidiiformes— Parasitengona. При утолщении и уплотнении кожи (например у некоторых Schwiebea из Tyroglyphidae) проподосомальный щит становится неотличимым по своей консистенции от окружающей его кожи, теряя, таким образом, свою самостоятельность. Второй спинной щит, опистосомальный (scutum opisthosomatale) имеется только у d d некоторых Rhizoglyphini (Schwiebea, Monieziella, Histiogaster, Thyreophagus), у которых он занимает заднюю половину гистеросомы и принимает участие в образовании характерной для этих форм свободной опи-стосомальной пластинки. Помимо этих щитов, сходные, но менее постоянные щитовидные образования встречаются нередко вокруг элементов коксо-стернального скелета и над основаниями передних ног; у 99 некоторых Tyroglyphidae, кроме того, иногда есть еще небольшой генитальный щит — округлая, слабо хитинизированная площадка, в центре которой открывается копулятивное отверстие.
Латинки идиосомы (фиг. 4, §7, 41, 46). На спинной и боковых поверх-
6
ВВЕДЕНИЕ
ностях идиосомы помещается от 7 до 20 пар симметрично расположенных щетинок,.образующих 6 групп: 1) теменные щетинки (setae verticales— v е, v i), имеющиеся в числе 1 или 2 пар (отсутствуют только у рода Monie-eiella) и расположенные ближе к переднему концу идиосомы, чем щетинки других групп; 2) лопаточные (s. scapulares — sc), имеющиеся также в числе
Ну
од
Р
sai
ps —
сх-
г
Ае х
Н (a
"gl
1р
see
Фиг. 4. ForeeUinia fungivora, (Oud«.), С? сбоку (дистальные членики ног. не изобраг жены).
' Расположение щитов, щетинок, желез и т. д. Щетинки:
1‘ vi — внутренние теменные; . ve — наружные , теменные;
' все — наружные лопаточные; eel—внутренние лопаточные; hi — внутренние
. плечевые;., he — наружные 1 плечевые; eh — подплече-
. . вые; di — 1-я пара спин-' вых; da —2-я пара епйн-
. вых; ds -^- 3-я пара спин- ... ных; di — 4-Я’пара спинных; . ,1а —- передние .боковые; 1р — задние боковые; вае — наружные . .крестцовые; sai—внутренние крестцовые; сх — коксальные; а — 1-я пара анальных; Pi— 1-я пара постанальных; Рз—2-я пара постанальных'; Рз—3-япара постанальных;
S —проподосомальный щит; ‘ , , ch—хитиновые образования
' Над основанием ног I; J —
. дачный орган; ре — псевдо- . стигма льный орган; о — вы-
. водное отверстие коксаль-- вых желез (« псевдостигма»);
gl -г- жировые железы; og — . их выводное отверстие; р — поровидные органы; С — хе-
; ‘лидера; Ну — гипостом; Р— яроподосома; Н—гистеросома.
1 йЛй'2 пар, образующих дорзальный- поперечный ряд у заднего края про-подосомы (или на соответствующем ему местей-у форм с нерасчлененной идиосомой); 3) группа плечевых щетинок-(s. humerales — h e, hi), состоя-» щая из 1—3 пар щетинок, расположенных в области «плеч»— у перед-»
них углов гистеросомы или» на соответствующих им плечевых выпуклостях, прячем 1 или 2 Пары из этих щетинок занимают маргинальное положение; 4) группа из 4 пар,спинных щетинок»(srdorsales. 1—4, dx—d4), образующих парный ряд вдоль средней линии спины; б) 1^-3 пары боковых щетинок (s, .laterales — L а), ^расположённых Ио краям тела или вблизи них, взад-
ВНЕШНЕЕ СТРОЕНИЕ
лей половине идиосомы, и 6) 1—2 пары крестцовых щетинок (s. sacralds — -ва), группирующихся на заднем конце идиосомы. Положение большинства этих щетинок изменчиво в значительных, но строго определенных пределах, достаточно ясных для того, чтобы в каждом отдельном случае можно было без особого труда определить принадлежность каждой пары к той или ииой из перечисленных групп и провести их гомологизацию. Размеры щетинок, в общем, чрезвычайно изменчивы: имеется вся цепь переходов •от крайне мелких, заметных лишь при самом сильном увеличении микрохэт,
77
Фиг. 6—17. Различные типы щетинок идиосомы.
5 — волосовидная; 6—щетинковидная; 7 — игольчатая; 8—шиловидная; б—лентовидная; 10 — листовидная; 11—• зазубренная; 12—тонкоперистая; 13 — пушистая; 14 — крупноперистая; 15 — гребенчатая; 16 — двусторонне-гребенчатая; 17 — опахаловидная.
размером в 1—2% длины идиосомы, до длиннейших макрохэт, превышаю-1 щих идиосому в 2 — 3.5 раза; при этом степень изменчивости в длине неодинакова у разных пар щетинок; некоторые из них (например передняя пара теменных или наружные лопаточные) имеют довольно константные размеры, тогда как у других они колеблются в исключительно широких пределах. В соответствии с этим, крайне изменчивы и соотношения их! 5»щмеров, но, как правило, последние возрастают по направлению к заднему концу идиосомы. В сильнейшей степени изменчивы и строение и форма самих щетинок, которые бывают следующих типов (фиг. 5—17), связанных между собой всеми мыслимыми переходами: 1) волосовидные, характеризующиеся своей особой тонкостью и гибкостью, не утолщающиеся к OOUO-
3 ВВЕДЕНИЕ
ванию;' 2) простые, нормального «среднего» типа щетинки, отличающиеся от волосовидных большей толщиной и жесткостью и более или менее заметным возрастанием своего диаметра к основанию; 3) игольчатые, очень, жесткие, массивные, постепенно заостряющиеся к концу, прямые щетинки, обычно небольшие по своим размерам; 4) шипбвидные, имеющие более-или менее коническую форму и обычно малые размеры; 5) уплощенные-или лентовидные, для которых характерна их сплющенность в дорзо-вентральном направлении при относительно небольшой ширине, и 6) листовидные или чешуевидные, имеющие форму тонких, более или менее широких пластинок очень разнообразной конфигурации. Щетинки первых 3 типов, бывают то гладкими, то вооруженными очень изменчивыми по своим размерам и числу вторичными отростками, в зависимости от длины и густоты расположения которых различают следующие типы их строения:
1) зазубренные — при очень малых размерах и сильной разреженности, вторичных отростков щетинки;
2) тонкоперистые —при малой длине и толщине усаживающих их ресничек и их относительно небольшой густоте;
3) пушистые — когда эти реснички расположены очень густо, в виде-более или менее короткого и тонкого сплошного опушения;
4) крупноперистые, у которых вторичные отростки относительно велики,, но тонки и гибки, и образуют бородки по обеим сторонам щетинки;
5) гребенчатые, имеющпе крепкие отростки, расположенные на одной только стороне щетинки;
6) двусторонне-гребенчатые, у которых вторичные отростки, столь же-крепкие, как у щетинок предыдущего типа, располагаются по обеим сторонам ствола щетинки.
При уплощенной форме щетинок они бывают либо крупноперистыми, либо пушистыми; чешуевидные щетинки всегда более или менее густо, пушисты, а шиповидные —всегда гладкие. В большинстве случаев щетинки спинной стороны идиосомы торчат перпендикулярно или под острым, углом к ее поверхности, и их размерами определяется, соответственно, очень разнообразный у разных видов общий «габарит» клеща, находя--щийся в известной связи с условиями обитания, так как щетинки несут, невидимому, осязательную функцию, играя роль органов -ориентации! в размерах свободных, доступных прохождению клещей пространств между частицами обитаемого ими субстрата. Очень важную биологическую роль-играют эти щетинки и в определении удельной поверхности тела и, соответственно, парусности клещей, сильнейшим образом отражающейся на их рассе лительной способности (см. стр. 24). Строение, размеры и положение-описанных щетинок дают огромное число признаков первостепенной важности и широко используются в систематике группы; при исчислении их длины, последняя обычно приводится в процентах к общей длине идиосомы. Кожное вооружение брюшной поверхности идиосомы гораздо беднее, чем спинное вооружение, и состоит обычно из коротких, однотипных,, более или.
ВНЕШНЕЕ СТРОЕНИЕ &>
менее волосовидных щетинок: 2 пар коксальных щетинок (s. coxales,—ox), расположенных, соответственно, на коксальных полях I и III, 1 подплоче-иой пары (s. subhumerales — sh), находящейся книзу от плечевых, 3 nup, мелких генитальных щетинок (s. genitales), группирующихся вокруг гепи-тальпого отверстия, и анального комплекса щетинок, часто состоящего ии а разделенных между собой групп’—преапальлой (s, praeanales — рта), сложенной 1—2 парами щетинок, и постанальиой (s. postanales — р), образованной 1—5 парами щетинок. Число, расположение и строение коксальных, подплечевых и генитальных щетинок очень постоянны во всей группе; в противоположность этому, щетинки апальпого комплекса довольно изменчивы как по числу, так и по своему размещению (разнящимся, у обоих полов) и дают ряд хороших систематических признаков.
Чашевидные органы. На гистеросоме расположены 4 пары поровидных образований, из которых 1 находится вблизи плечевых щетинок, 1 — вблизи передней пары боковых щетинок, 1 — у заднего' конца тела и 1—по сторонам анального отверстия. Эти образования, представляют собою органы чувств, гомологичные щелевидным органам других паукообразных.
Щечные органы («Klemm organe»). У Tyroglyphidae и многих Sapro-glyphidae на боковых поверхностях проподосомы расположены своеобразные кутикулярные отростки, так называемые щечные органы (фиг. 4). В большинстве случаев они имеют вид тонкой линейной пластинки, обращенной вершиной вперед или косо книзу и внутрь и обхватывающей с боков основание гнатосомы; у Uhisoglyphinae щечные отростки простые,, нередко покрытые тонким вторичным опушением, тогда как у многих Ту-roglyphinae они имеют более сложную форму: у Tyroglyphus farinae (L.), например, они представлены веером тончайше рассеченных лоцастинок (фиг. 111, 115, 117, 118).
Псевдостигмальный орган. Над основаниями ног I, на дорзальной поверхности проподосомы (resp. идиосомы), помещается псевдостигмальный орган (фиг. 4, ps), представляющий собой небольшую, более или менее-модифицированную щетинку, очень изменчивую по своей форме; у некоторых видов он имеет простую щетинковидную или палочковидную форму,: у других — более или менее утолщен и перист по краям; наибольшей Сложности достигает строение этого органа у Glycyphaginae, где он имеет вид: сильно ветвистого кустика. Псевдостигмальный орган тироглифоидей не-гомологичен одноименным образованиям у Oribatei и Trombidiiformes.
Железы. У основания псевдостигмального органа (непосредственно1 пад основаниями ног I) у многих видов помещается маленькое щеле-видное отверстие, окруженное эллиптическим хитиновым кольцом, так называемая псевдостигма (фиг. 4, о). Многие авторы принимали эту щель за настоящую стигму и пытались даже доказать, что от нее отходит внутрь тела трахейный ствол, но исследованиями Э. Г. Беккера, было установлено, что она представляет собой не что иное, как вывод
10
ВВЕДЕНИЕ
ное отверстие. коксальных желез (см. анатомический раздел). Сходное строение имеют выводные отверстия жировых желез (фиг. 4, gl), открывающиеся на гистеросоме между передней и задней парами боковых щетинок; сами эти железы имеют вид крупных мешковидных образований, расположенных по бокам гистеросомы, в ее задней половине, и наполненных сильно преломляющим свет секретом (часто приобретающим на препаратах темную окраску).
Анальное отверстие (anus — а) имеет вид более или менее длинной продольной щели, расположенной вентрально, обычно вблизи заднего края гистеросомы, но иногда и довольно сильно отодвинутой от него; у Tyroglyphidae положение ее отлично у сГ и ф, но в других группах оно, в большинстве случаев, идентично у обоих полов.
Основную часть наружного полового аппарата с? составляет копулятивный орган—penis (р); он расположен в генитальной щели (ag), в •особом углублении, частично прикрытом нависающими над ним складками кожи, помещающемся в пространстве между основаниями ног III и IV, между ногами IV или более или менее далеко позади них и лишь в исключительных случаях—между основаниями ног I (род Chortoglyphus).
Penis состоит из обычно очень сложного по своей архитектуре цоколя, снабженного целой системой аподем, к которым прикрепляются мышцы, управляющие его движением, и сильно изменчивой по своим размерам и •форме дистальной части — стилета; помимо того, иногда имеются еще 1 или несколько опорных склеритов, расположенных вокруг penis, и иногда более -крупная хитиновая площадка — эпиандрий (фиг, 48, ера), находящаяся у переднего края генитального отверстия. По сторонам penis, внутри генитальной камеры, помещаются обычно 2 пары разнообразных по своей форме генитальных щупальцев (pg), часто способных далеко выпячиваться из генитального отверстия, являющиеся органами чувств (хеморецепто-рами). С копулятивным аппаратом С функционально связаны специальные органы фиксации, имеющиеся у многих групп. У всех Tyroglyphidae ‘(кроме рода Scatoglyphus) и родов Pontoppidania (Saproglyphidae) и Chortoglyphus (Glycyphagidae) этот прикрепительный аппарат состоит из пары более или менее крупных анальных копулятивных присосок (da), расположенных по бокам анальной щели, и 2 (у Pontoppidania — 1) пар тарзальных копулятивных присосок (dt), сидящих на внешней поверхности лапок IV — по 2 на каждой. У сШ подсем. Labidophorinae и, пови-димойу, некоторых Saproglyphidae (Nanacarus) роль прикрепительных аппаратов играют более или менее резко модифицированные ноги III и IV, а в некоторых других случаях эту функцию выполняют соответственно -расширенные praetarsi пог I и II.
Наружный половой аппарат 9 представлен далеко разобщенными друг от друга генитальным и копулятивным отверстиями. Генитальное, или яйцевыводное, отверстие (фиг. 26, g, фиг. 47, ge) имеет вид продольной-щели, расположенной обычно в пространстве между эпимерами III и IV, но иногда
ВНЕШНЕЕ СТРОЕНИЕ Ц
я значительно дальше кпереди (начинаясь тогда непосредственно попади эпимер I) или, в очень редких случаях, еще дальше назад. У большинства форм генитальное отверстие прикрыто парой более или менее сильно развитых генитальных клапанов, обычно смыкающихся по средней линии в своей передней части и постепенно расходящихся назад; у некоторых Glycyphagidae имеется еще 3-й непарный клапан, замыкающий генитальное отверстие сзади; под этими клапанами помещаются 2 пары генитальных щупальцев, имеющих такое же строение, как у с?. У большинства представителей семейств Saproglyphidae и Glycyphagidae передний край генитального отверстия снабжен склеритом полулунной формы — эпиги-нием, отсутствующим у Tyroglyphidae-, иногда эпигиний соединен с стер-нумом или с особым хитиновым циркумгенитальиым кольцом, окружающим яйцевыводное отверстие, или с эпимерами ног III. Копулятивное •отверстие (ас) занимает обычно терминальное положение, • располагаясь позади заднего конца анальной щели; у части форм оно представлено малозаметной округлой порой, но у других его края сильно разрастаются и образуют цилиндрическую или коническую копулятивную трубку (так называемую bursa copulatrix), направленную назад или косо вниз (фиг. 46, Ъс).
Коксо-стернальный скелет» Ноги состоят, как обычно, из 6 члеников, но их базальные членики—тазики (coxae) сильно модифицированы и неподвижно сращены с идиосомой, образуя характерный для Acaridiae коксо-стернальный скелет; последний составлен из узких, более или менее линейных склеритов — эпимер и эпимеритов, из которых первые обычно снабжены погруженными в тело пластинчатыми аподемами, служащими для прикрепления некоторых из мышц, приводящих в движение ноги, и соединены своими внешними концами с сочленовными ямками (ацетабу-лами) ног. Обычно коксо-стернальный скелет состоит из следующих элементов: 1) эпимер I пары ног (ер I), в большинстве случаев сливающихся друг с другом по средней линии тела, образуя непарный склерит, стернум (st), имеющий форму букв V или Y; 2) эпимер II пары ног (ер II), в подавляющем большинстве случаев свободных; 3) эпимеритов II пары ног (et II), нередко совершенно обособленных от основания ног II; 4) эпимер III пары (ер III) и Б) эпимер IV пары ног (ер IV), связанных своими внешними концами с передним краем сочленовных ямок (ацетабул) соответствующих пар ног и, в большинстве случаев, с свободными внутренними Концами; иногда развиты еще и эпимериты III и IV пар ног, отходящие от заднего края их ацетабул. У некоторых форм промежутки, заключенные между эпимерами (носящие название коксальных полей и нумеруемые по очерчивающим мх спереди эпимерам), целиком более или менее плотно хитииизировапы, образуя такие же коксальные щиты (фиг. 551), как у многих клещей других1 подотрядов (например многих Trombidiiformes)-, этими случаями, гд<г «пиморы образуют утолщенный передний, а эпимериты — задний край щитов, ясно намечаются исходные точки и пути редукции тазика и коион
12
ВВЕДЕНИЕ
ного его распадения на только что названные элементы (хотя, в данной случае, скорее имеет место вторичное образование коксальных щитов в результате уплотнения поверхности соответствующих коксальных полей).
Ноги. Вследствие модификации тазиков, свободная часть ногц состоит-только из 5 члеников: 1) вертлуга (trochanter), 2) бедра (femur), 3) колена,, или коленного членика (genu, patella), 4) голени (tibia) и 5) лапки (tarsus);, на конце лапок помещается амбулакральный аппарат, состоящий из болев или мепее сильно развитого перепончатого praetarsus и непарного, обычно крючковидного коготка (unguium); строение амбулакр дает хорошие признаки для характеристики крупных таксономических категорий; у одних групп коготок сидячий(фиг. 18—19)ипричлеляется непосредственно к концу лапки при помощи особых соединительных склеритов (с), образующих, с коготком дикондильное сочленение, и тогда praetarsus имеет вид болев или менее крупного мясистого чехла, одевающего базальную часть коготка;, у других коготок стебельчатый (фиг. 20—24), т. е. помещается на дистальном конце более длинного в этих случаях praetarsus и сам не связан с лапкой; при этом он нередко сильно редуцирован и заметен лишь с трудом (а у рода Nanacarus даже полностью отсутствует), но в некоторых случаях достигает больших размеров, и praetarsus кажется тогда коротким стебельком, его поддерживающим. Пропорции ног и отдельных их члеников^ сильно изменчивы, в соответствии с чем можно наметить несколько основных типов их строения: копательный — наиболее ярко выраженный у родов Rhizoglyphus и Schtviebea, характеризующийся необыкновенной мощностью передних пар ног, снабженных крупными коническими шипами, при относительно слабом развитии задних; ходильный — пользующийся наиболее широким распространением среди тироглйфоидей и отвечающий типичной для них средней степени длины и стройности ног, и бегательный тип, характеризующийся крайним удлинением лапок и выраженный в наиболее резкой форме у рода Glycyphagus. Сильно изменчива и форма члеников, хотя в типе они все имеют форму несколько сжатого с боков цилиндра, за исключением заметно суживающейся к концу лапки. По общему плану строения ноги I и II очень сходны между собой и существенно отличаются от таковых задних пар, строение которых также почти идентично; исключения из этого правила имеют место в случаях резкой гипертрофии ног I (у def рода Tyroglyphus) или ног III (у гетероморфных сГсГ Rhizo-glyphinae), о которых еще будет речь ниже.
Вооружение ног состоит из довольно большого числа кожных органов чувств — щетинок и других сенсилл, часто более или менее модифицированных, причем каждая из пар ног имеет в этом отношении свои специфические особепиости. Вооружение ног I таково: на вертлугах и бед.-рах — по 1 вентральной щетинке; на коленах—по 2 базальных щетинки, из которых одна расположсца дорзально и одна у наружного края членика, и по 2 вершинных, расположенных одна рядом с другой посередине дистального края колена, на его дорзальной поверхности; одна из этих
ВНЕШНЕЕ СТРОЕНИЕ
1JI
щетинок представлена обычно более или менее короткой, притупленной -«обонятельной» палочкой (фиг. 93, 456), другая имеет нормальное строение, но у некоторых форм (fihortogtyphinae) отсутствует; голени вооружены жаждая 3 щетинками, из которых одна очень крупная, всегда гладкая— дорзальная вершинная, и 2 вентральных. Вооружение лапок I значительно
Фиг. 18—24. Типы строения амбулакр,
-Фиг. 18. Histiogaster bacchus, sp. п. Фиг. 19. Aleuroglyphus cvatus (Troup.). Фиг. 20. Carpoglyphus laetis (L.). Фиг. 21. Xenocastor fedjushini, sp. n. Фиг. 22. Glycyphagus destructor (Schr.). Фиг. 23. Sennertia cantabrica, sp. n. Фиг. 24. нога I Hyadesia fusca (Lohm.) (no Lohmann). Сокращения!
ta—лапка; p—praetarsus; u —коготок; c — соединительные склериты.
богаче и сложнее, чем таковое предыдущих члеников: оно состоит, в типе, 'ИЗ трех групп сенсилл — базальной, медиальной и апикальной; базаль-1ШЯ группа состоит из расположенной дорзально у основания лапки так называемой обонятельной палочки (а), имеющей более или менее булавовидную форму, дополнительного к ней штифтика Р, расположенного у 00 основания на той же сочленовной подушечке (отсутствует у ряда видов), 41 двух сидящих кнаружи от обонятельной палочки сенсилл т и 8, из кото»
14
ВВЕДЕНИЕ
рых 1-я обычно палочковидна; медиальная группа сенсилл расположена в среднем поясе лапки, но иногда сдвигается ближе к ее дистальному концу; она составлена 3 или 4 щетинками, 3 из которых сидят вентрально в виде треугольника, а одна (г) — дорзально или на внутренней поверхности лапки; у некоторых форм сенсилла г сдвинута к обонятельной палочке, располагаясь непосредственно впереди нее, и тогда обычно имеет шиповидную форму (фиг. 196, 198, 199); апикальная группа сенсилл состоит из 3—4 дорзальных вершинных щетинок, в большинстве случаев волосовидных, и венца из 3—5 вентральных шипиков (у Tyroglyphus — только 1), окружающих снизу основание амбулакры;, у многих родов сем. Tyroglyphidae имеется еще дорзальный вершинный шип, расположенный кнаружи от основания коготка. Вооружение ног II во всем сходно с таковым I пары, с той только разницей, что вершина колен имеет здесь только 1 (а не 2) сенсиллу, гомологичную обонятельной палочке колен I и сходную с нею по строению, и что на лапках II отсутствуют сенсиллы ₽ и 8. III пара ног вооружена 1 вентральной щетинкой на вертлугах, коленях и голенях и 1 дорзальной — на коленях и голенях; лапки лишены базальной группы сенсилл и имеют только 2 вентральных щетинки в среднем поясе и 1—2 вершинных дорзальных щетинки, но тот же набор апикальных шипов, как и передние-лапки. Вооружение ног IV сходно с таковым ног III, но здесь отсутствуют вентральные щетинки на вертлугах и коленях и дорзальная щетинка колен, зато на бедрах имеется по 1 вентральной щетинке. Сенсиллы, составляющие вооружение ног, очень разнообразны по своему строению; их число (на лапках), расположение некоторых из них, например базальных щетинок колен, е и др., и их строение дают ряд признаков первостепенного систематического значения.
Вторичные отличия между полами чрезвычайно разнообразны и проявляются в самых различных признаках и с самой различной степенью резкости. 'Как правило, d меньше Q и имеет более длинные и стройные ноги и более длинные щетинки тела, ио и это правило допускает некоторые исключения. В некоторых группах наблюдается своеобразный диформизм dd, представленный здесь двумя фазами: более близкой по своему строению к 99 гомеоморфной фазой и гетероморфной фазой, характеризующейся сильным утолщением III пары ног и гипертрофированным развитием вторичных половых признаков.
Фиг. 25-27.
Фиг. 25. Tyrolgyphue farinae (L.), с? снизу: m—хелицеры; h—гипостом; pg—генитальные щупальцы; Р — penis; а — анальная щель; d— анальные копулятивные присоски; ат — амбулакра; ta — лапка; t — голень; g — колено; f — бедро; tr — вертлуг; о —тазик (эпимер). Фиг. 26-Т. farinae (L.), 9 снизу: а — анальная щель; g— генитальное (яйцевыводное) отверстие; ва — анальные щетинки. Фиг. 27. T. farinae (L.), Q сверху: G— гнатосома;Р—проподосома; Н—гистеросома; щетинки: vi — внутренние теменные, уе — наружные теменные, sei— внутренние лопаточные, все — наружные лопаточные, hl — внутренние плечевые, he — наружные. плечевые, di — 1-я пара спинных, ds — 2-я пара спипных, da — 3-я пара спинных, da — 4-япара спинных, 1а — передние боковые, 1р — задние боковые, вае — наружные крестцовые, sal — внутренние крестцовые, ра—постанальные.
I
16
ВВЕДЕНИЕ
КРАТКИЙ ОЧЕРК ВНУТРЕННЕГО СТРОЕНИЯ
Анатомическому исследованию подверглись до сих пор лишь очень немногие из представителей группы Tyrogly phoidea: Tyrophagus longior (Gerv.) (Nalepa, 1884), Carpoglyphus lactis (L.) (Nalepa, 1885, и Lonnfors, 1930), Hericia hericia (Rob.) и Talpacarus platygaster (Mich.) (Michael, 1901). -Этими исследованиями был выяснен в общих чертах план их внутреннего строения, но они явно недостаточны для суждения о несомненно существующих анатомических различиях между отдельными входящими сюда группировками менее высокого таксономического ранга (семействами, родами).
Эндоскелет. В противоположность многим другим группам клещей, эндостернит у тироглифоидей сильно редуцирован; он представлен здесь маленькой продольной пластинкой, лежащей под задним отрезком пищевода (у Talpacarus), или совершенно ничтожным по своим размерам склеритом, расположенным под желудком и над задним концом подглоточного ганглия (у Carpoglyphus)1, к нему прикрепляются несколько пар мышц, в том числе musculus depressor вертлугов. Слабое развитие эндостернита, очевидно, компенсируется наличием коксо-стернального (эпимерального) скелета, большинство элементов которого имеет хорошо развитые аподемы, служащие для прикрепления многих из мышц, приводящих в движение ноги и гнатосому.
Мускулатура состоит, как и у большинства других клещей, из шести -основных групп мышц. Главными компонентами первой из них — группы мышц, приводящих в движение ротовые части, — являются две системы мощных продольных мышц — ретракторов, при помощи которых гнатосома, в целом, и хелицеры втягиваются внутрь тела; эта группа мышц заполняет почти все пространство между мозгом, желудком и наружными покровами и прикрепляется дорзальио в области проподосомальногощита. К этой же группе относятся и меньшие по своим размерам мыпщы, придающие гнатосоме вращательное движение, и сгибатели гнатосомы, позволяющие ей отгибаться вниз почти перпендикулярно. Вторую группу составляют мощные дорзо-вентральные мышцы (m. dorsoventrales), точки прикрепления которых намечены продольными вдавлениями на спинной поверхности тела (рахидиальными бороздками); сокращение этих мышц вызывает подтягивание брюшной поверхности тела к спинной, т. е. его уплощение. Укорочение тела (т. е. его сокращение в продольном направлении) производится третьей группой мышц — 4 парами продольных мышц, идущих вдоль стопок тела от плечевых выпуклостей до уровня III пары ног (эти мышцы имеются, невидимому, но у всех представителей группы). К четвертой группе относятся многочисленные мышцы, приводящие в движение ноги; их совокупность может быть, в свою очередь, подразделена на две группы: а) мышцы, приводящие в движение ногу в целом, и б) мышцы, управляющие движением отдельных частей (resp. члеников) ног. Первые
ВНУТРЕННЕЕ СТРОЕНИЕ
17
»i:i них отходят пастью от дорзальной стенки тела (разгибатели вертлуги, ш. levator trochanteris), пастью от эндостернита (сгибатели вертлуга, ли. depressor trochanter is), пастью же от стернума и эпимер II (мышцы, вращающие вертлуг и. двигающие его. вперед и назад); прикрепляются они все к проксимальному краю вертлуга.- Вторая группа ножных мышц представлена сгибателями и' разгибателями бедра (ш. flexor femoris и m. extensor femoris), колена, голени и лапки с коготком. Сгибатель каждого «членика тянется от его основания до дорзальной стенки 2-го за ним членика (например сгибатель лапки — от колена, голени — от бедра и т. д.), более короткий разгибатель — от членика, непосредственно предшествующего ему; таким образом каждый членик ноги содержит отрезки трех мускулов. Пятую и шестую группы мышц составляет мускулатура пищеварительного тракта ‘и половых органов, рассматриваемых ниже, в связи •с соответствующими системами.
- Нервная система крайне концентрирована, образуя единственную крупную ганглиозную массу (синганглий).— мозг, пронизанный пищеводом; составляющие его элементы, надглоточный и подглоточный ганглии и парные окологлоточные коммиссуры, совершенно слиты друг с другом. Число отходящих от него нервов не может считаться окончательно установленным.
Пищеварительный канал тироглифоидей состоит из 5 отделв: глотки; пищевода, желудка, или средней кишки, толстой кишки и задней кишки, открывающейся заднепрохЬдным отверстием на вентральной поверхности -опистосомальной области тела. Глотка (pharynx), начинающаяся в глубине ротовой полости и составляющая ее, непосредственное продолжение, представляет собой уплощенную эластичную трубку, играющую роль насоса, всасывающего пищу из ротовой полости и нагнетающего ее в пищевод; эта функция выполняется ею благодаря наличию двух систем мышц: б—6 пар мощных радиальных расширителей глотки (m. dilatatores pha-ryngis), идущих от середины ее верхней стенки к окружающему хитиновому скелету-и растягивающих глотку при сокращении, и нескольких поперечных замыкателей глотки (ш. constrictores pharyngis), натянутых между приподнятыми боковыми краями ее верхней желобообразной стенки и вызывающих своим сокращением (комбинированно с эластичностью самой стенки) ее прогибание вниз и замыкание просвета глотки. Кзади глотка переходит в цилиндрический пищевод (oesophagus), образуя на границе с ним перегиб, очевидно препятствующий обратному попаданию пищи в глотку; пройдя сквозь широкое отверстие в глоточном нервном кольце, пищевод впадает в объемистый, более или менее треугольный желудок (ventriculus), снабженный парой боковых слепых отростков (caeca). Желудок отделен перехватом от шаровидной или эллипсоидальной толстой кишки (colon), последняя, в свою очередь, отделена перетяжкой от посте-мопцо расширяющейся. кзади, сжатой с боков прямой кишки (rebtum), поставляющей ее дистальную часть.
2 Фауна СССР, Захватили—284
18
ВВЕДЕНИЕ
На границе между двумя этими отделами впадает пара обычно довольно длинных, более или менее расширяющихся к концу трубчатых выделительных органов; это — мальпигиевы сосуды, размеры п форма, которых различны у разных видов; у некоторых форм (например у рода Talpacarusf они совершенно отсутствуют.
Железистая система. G пищеварительным аппаратом связаны и некоторые из имеющихся у тироглифоидей желез: сюда относятся железистые-участки внутренней стенки желудка и его слепых придатков и мелкие железистые образования в толще эпителия задней кишки; последние выделяют, невидимому, слизистый секрет, обволакивающий фекальные массы тончайшей плёнкой (в известной мере аналогичной перитрофическрй мембране, образуемой насекомыми). Более крупные размеры имеют две. другие группы.желез, не связанные с пищеварительным трактом: коксальные и жировые железы, протоки которых открываются на наружной поверхности тела, в соответствии с чем о них была уже речь при описании внешнего строения тироглифоидей. Жировые железы расположены, как уже указывалось, в гистеросоме, позади основания ног IV; они имеют вид: овальных, мешковидных, выстланных низким эпителием образований,, обычно наполненных желтоватым, маслянистым, сильно преломляющим свет секретом; эти железы являются, повидимому, смазочными железами: можно думать, что тонкий жировой слой, покрывающий все тело большинства тироглифоидей и, вероятно, препятствующий его смачиванию, обра^-зуется выделяемым ими секретом. '
Органы дыхания у тироглифоидей, как и у большинства других Аса-ridiae, отсутствуют, и дыхание у них происходит через кожу; в соответствии с этим, они лишены и стигм [так называемая «псевдостигмальная» щель, имеющаяся у многих видов над основанием ног I и многими авторами принимавшаяся за настоящую, функционирующую стигму (например Oude-mans, 1932), есть на самом деле выводное отверстие коксальных желез]..
У 99 Gohieria fusca (Ouds.) (Labidophorinae) недавно обнаружены генитальные трахеи, очевидно аналогичные таковым рода Су la (Bdellidae)-и око'логенитальным воздушным камерам некоторых -других Trombidii-formes и также лишенные спиральной «нити»; несут ли они дыхательную* функцию, — неизвестно. Нет у тироглифоидей и кровеносной системы.
Мужской половой аппарат .состоит из парных, более или менее грушевидных семенников (у Carpoglyphus скорее эллипсоидальных), постепенно переходящих спереди *- без сколько-нибудь ясного отграничения — в длинные семепроводы, соединяющиеся спереди в короткий семеизверга-тельиый канал; этот канал принимает также выводные протоки чрезвычайно крупных, разнообразных по своей форме придаточных желез;, имеющихся в число 1 (у Tyroglyphidae) или 2 (у Glycyphagidae)-, в последнем случае они сильно разнятся между собой по своему строению и форме;. Строение совокупительного органа (penis) и окружающих его хитиновых частей было уже рассмотрено выше; здесь нужно только отметить,
РАЗВИТИЕ . 1»
что последние служат местом прикрепления многих мускулов, управляющих движением penis. Заслуживают еще упоминания мышцы-ретракторы генитальных щупальцев, идущие к ним от аподем эпимер IV.
В состав женской половой системы входят: так называемая совокупительная сумка (bursa eopulatrix) — трубка, в сущности играющая роль-ипгипы; канал, соединяющий ее с семеприемником; семеприемник; парные яичники и яйцеводы, соединяющиеся дистально в широкий общий яйцевод, и ряд структур, связанных с яйцевыводным отверстием. Совокупительная сумка представляет собой округлое, поровидное отверстие,, расположенное на аборальном полюсе тела и служащее, как уже указывалось, копулятивным отверстием (у ряда Glycyphagidae его края более или менее сильно разрастаются, образуя уже описанную копулятивную трубку); оно ведет в узкий канал с эластичными стенками, впадающий в пузыревидный, довольно крупный семеприемник; этот последний, в свою очередь, связан с яичниками посредством пары коротких каналов, идущих к их аборальному полюсу. Яичники имеют эллипсоидальную или грушевидную форму; от их оральных полюсов отходят длинные яйцеводы, соединяющиеся затем в короткий, очень широкий общий яйцевод (называемый обычно вагиной), снабженный мускулистыми стенками и складчатой хитиновой выстилкой (имеющей в оптическом разрезе несколько зубчатые очертания); «вагина» открывается вентрально крупным генитальным (яйцевыводным) отверстием, строение которого было уже описано.
ИСТОРИЯ РАЗВИТИЯ; СТРОЕНИЕ НЕПОЛОВОЗРЕЛЫХ СТАДИЙ
Все Tyrogly phoidea detriticolae, как правило, яйцекладущи. У некоторых видов, правда, иногда имеет место и факультативное живорождение или яйцеживорождение (Tyroglyphus farinae L., некоторые Tyropha-gus, Glycyphagus destructor Schrk.), но эти случаи у них крайне редки и представляют скорее физиологическую аномалию, чем специфическую для этих видов биологическую черту; характер таковой живорождение приобретает только у некоторых видов Caloglyphus {С. moniesi A. Z., С. poly-phyllae A.Z.'), у которых оно существует наряду с откладкой яиц, и, невидимому, у Xenocaslor и Carpoglyphus (Trouessart, 189,7); при этом у Caloglyphus moniezi развитие молоди в теле самки может итти вплоть до образования протонимф или даже гипопусов.
Постэмбриональное развитие представляет собой эпиморфоз, усложг пенный факультативным образованием особой расселительной или покоящейся стадии, так называемого гипопуса. .Как и у других клещей, функции роста и доимагинального питания здесь полностью осуществляются стадиями, близкими по своей организации и общему облику к полоноиро-л oil, т. с. личинкой и нимфами. Гипопальная же стадия представляет собой чното провизорное образование с неимагинальным типом строения, 1Ю участвующее в выполнении вегетативных функций и подготовке дефинитивного состояния организма н служащее только расселению вида,.
н
20
ВВЕДЕНИЕ
. Личинка (<фиг. 28—30, 33—35, 44—45, 49—51) характеризуется, прежде всего, наличием только 3 пар ног (ноги IV еще совершенно не развиты и представлены только подкожными скоплениями клеток, аналогичными имагинальным дискам насекомых) и особых провизорных образований — грудных палочек («Bruststiele»), расположенных на нижней поверхности проподосомы, кнаружи от коксальных щетинок!; эти органы, гомологичные так называемым «Urstiginen» эонимф многих представителей подотряда Trombidiiformes, весьма разнообразны по своей форме, но обычно имеют вид подвижных, булавовидно утолщающихся к концу, полых цилиндров; их функция остается неизвестной. В строении идиосомы характерно более слабое, чем у следующих стадий, развитие гистеросомы (очевидно, стоящее в связи с отсутствием задней пары ног), мало превосходящей по своей массе проподосому в тех случаях, когда они разделены, и отсутствие 4-й пары спинных и задних боковых щетинок; sa i, в большинстве случаев, очень длинны, играя роль- хвостовых pseudocerci, свойственных первой стадии, развития большинства наземных членистоногих. Из других хэтологических особенностей личинок (рассматриваемых более детально в специальной части этой работы) необходимо упомянуть отсутствие у них генитальных, аданальных и анальных щетинок (развита только 1 пара постанальных), отсутствие щетинок на вертлугах и отличия в положении некоторых из щетинок идиосомы, частью обусловленные стереометрическими моментами, частью же не находящиеся в видимой связи с формой тела и ее последующими изменениями (например нередко имеющие место в течение онтогенеза перемещения задних теменных щетинок, занимающих тогда у личинок положение, отличное от дефинитивного); кроме того, у личинок иногда не развиты и некоторые из других шар щетинок, но в этих- случаях они отсутствуют также у протонимф или даже во всех стадиях, включая взрослую, так что этот признак не входит в число специфических возрастных особенностей личинки. Такими особенностями является, помимо перечисленных, еще полное отсутствие у этой стадии зачатков наружных половых органов и отсутствие сенсиллы 8 и средней вершинной щетинки лапок I.
Первая нимфальная стадия, протонимфа (фиг. 31, 52—53), имеет уже 4 хорошо развитые пары ног, лишена каких бы то ни было провизорных органов и еЩе более приближается по своему строению к половозрелой. Специфические возрастные признаки этой стадии заключаются в наличии зачаточного генитального отверстия, снабженного 1 парой генитальных щупальцев, 1 пары генитальных щетинок и 3 пар аданальных щетинок
Фиг. 28—40. Пимфальпые стадии Tyroglyphidae.
Фиг. 28. Tyroglyphus farinae (Ь,), личинка сверху. Фиг. 29. Тоже, снизу. Фиг. 30. Т. farinae (L.), грудная палочка личинки. Фиг. 31. Т. farinae (L.), протонимфа снизу. Фиг. 32. Т. farinae (L.), телеонимфа снизу. Фиг. 33. Aleuroglyphus ouatus (Тг.), личинка сверху. Фиг. 34. То же, снизу. Фиг. 35. A. ooatus (Тг.), грудная палочка личинки. Фиг. 36. Tyrophagus noxiits A.Z ., личинка снизу. Фиг. 37. ТО же, сверху. Фиг. 38. Aleuroglyphus ovatus (Тг.), грудная палочка. Фиг. 39. Tyrolichus casei (Ouds.), грудная палочка личинки. Фиг. 40. Caloglyphus mandzhur, sp. п., грудная палочка личинки.
22
ВВЕДЕНИЕ
(в тех случаях, когда они, вообще, есть) и в отсутствии щетинок на вертлугах I — III и на 4 базальных члениках ног IV, а также средней .вершинной щетинки лапок I; на спинной поверхности развит, как правило, весь ассортимент щетинок, имеющийся у взрослых животных, в том числе d4 и 1 р, но размеры отдельных щетинок и их взаимоотношения обычно в той или иной мере отличны от окончательных; отлично иногда и положение некоторых из щетинок (например задних теменных) и пропорции конечностей, но во'всем остальном протонимфы имеют уже совершенно имагинальное строение.
Стадия телеонимфы (фиг. 32, 54—55) отличается от половозрелой только зачаточным состоянием наружного полового аппарата, представленного у обоих полов небольшой генитальной щелью с 2 парами щупальцев по бокам и отсутствием анальных щетинок. Ее отличия от протонимфы заключаются, соответственно, в наличии 2 пар генитальных щупальцев, полного набора генитальных щетинок, вентральной щетинки на вертлугах I — III, средней вершинной — на лапках I и всех недостававших, ранее элементов вооружения ног IV. Таким образом морфологические изменения, испытываемые Tyroglyphoidea в процессе постэмбрионального развития, в общем, невелики. В некоторых случаях нимфы всех возрастов обнаруживают более значительные отличия от взрослых, причем эти различияz могут происходить как вследствие проявления у взрослых гипертелических «надставок», так и в результате развития у нимфы особых провизорных состояний тех или иных дефинитивных структур; наиболее ярким примером такого рода могут служить нимфы Xenoryctes, ; имеющие чешуевидно расширенные, густопушистые щетинки и совершенно своеобразное строение кожи; к этому же результату могут привести и ретардации в развитии определенных признаков у нимф (например внутренних лопаточных и плечевых, передних боковых и трех передних пар спинных щетинок у Caloglyphus) или явления имагинальной редукции (редукции внутренних теменных щетинок у взрослых Ctenoglyphus palmif&r и генитальных щупальцев у Chortoglyphus и т. д.). Половая дифференцировка нимф у тироглифоидей выражена слабо; в немногих случаях (например у Carpoglyphus') она вполне явственна у телеонимф, но, насколько известно, не может быть прослежена до более ранних нимфальных возрастов; этот вопрос, впрочем, еще недостаточно разработан и требует более детального выяснения. Во всяком случае, вторичные половые различия, в том числе и самые своеобразные гиперморфозы сГсГ, -как правило, у нимф даже не намечены.
Гипопальные стадии — специфический атрибут свободноживущих Acaridiae (т. е. Tyroglyphoidea detrilicolae -J- Anoetidae): аналогичные им стадии имеются еще только у помпогих эпизоических представителей той же серии, а именно у некоторых видов Falculiferidae и Dermoglyphidae. Как уже указано, гипопальные стадии суть чисто провизорные образования, свободные от выполнения собствсшю-онтогенетических функций и резко
Фиг. 41—45. Saproglyphus neglectus Berl.
•Фиг. 41. 9 сверху; обозначения щетинок те же, Что на фиг. 27. Фиг. 42. $> снизу: ер — ЗЧШ'НППИ 1 а — анальные щетинки; ра — постаНальныё щетийкц. Фиг. 43. сГ снизу. Фиг. 44. Личинка оперк у.
Фиг. 45. То же, снизу.
24
ВВЕДЕНИЕ
отличные по своей организации от нимфальных стадий и половозрелой стадии; они лишены функционирующего ротового аппарата, имеют сильноредуцированную пищеварительную систему и существуют только за счет-использования резервных веществ, - накопленных предшествующими ним-фальными стадиями. У всех видов, способных к образованию гипопусов, эти стадии вклиниваются между первым и вторым нимфальными возрастами! (т. е. между стадиями протонимфы и'телеонимфы), причем ио своим возрастным признакам—наличию двух пар генитальных щупальцев и щетинок на; вертлугах I—III, а также числу генитальных и анальных щетинок1 — они, идентичны с телеонимфой. Гипопусы бывают двух типов — расселитель-иые и покоящиеся. Расселительные гипопусы служат исключительно распространению вида, и у всех тех форм, у которых они имеются, эта жизненная функция почти целиком осуществляется ими одними, играющими,, таким образом, специфическую роль диаспор. Все гипопусы используют пассивные способы расселения, прикрепляясь к различным животным, служащим им переносчиками, т. е., более точно, способ форезии (Lesne, 1897; «эпизоический способ» Sernander’a). В соответствии с этим у них. развит чрезвычайно богатый арсенал всевозможных органов фиксации, служащих как для схватывания переносчика, так и для прочного удержания на его поверхности, и вся их организация находится в теснейшей: зависимости от типа используемых ими переносчиков. Таковыми служат, .с одной стороны, насекомые и другие наземные членистоногие, с другой — мелкие млекопитающие (насекомоядные и грызуны), в соответствии с чем. вся совокупность расселительных гипопусов распадается на две основ-ные группы — энтомохорных и терохорнух гипопусов.
Общие всем расселительным гипопусам признаки таковы: идиосома более или менее сильно уплощена в дорзо-вентральном направлении,, с плоской или даже вогнутой брюшной и более выпуклой спинной поверхностью, в типе щитовидная, всегда, (за исключением только рода Senner-tia) явственно подразделенная на проподосому и гистеросому. Кожные-покровы обычно уплотненные и часто окрашенные в различные оттенки бурого или желтого цветов, с разнообразной (редко вполне отсутствующей), скульптурой. Гнатосома сильно редуцирована, представлена непарным:
. 1 Которым, по всем данным, гомологичны присоски или другие элементы анального прикрепительного аппарата.
Фиг. 46. Glycyphagus destructor (Schrk.), g сверху.
О—гнатосома; I—диооома; be — копулятивная трубка; щетинки: va — передние теменные,. у Р — задние теменные, sol — внутренние лопаточные, все — наружные лопаточные, h — плечевые, dt — 1-я,пара спинных, da — 2-я пара спинных, d8 — 3-я пара спинных, d4 — 4-я пара спинных, h — 1-я пара боковых, 1ц — 2-я пара боковых, 1 я — 3-я пара боковых, зае— наружные крестцовые, aal — внутренние крестцовые.
Фиг. 47—48. Glycyphagus destructor (Schrk.).
Фиг. 47. д снизу: G—гнатосома; 1 — идиосома; ер—эпигиний;§е — генитальяое(яйцевыводное> отверстие; а — анальная щель; щетинки: all — подплечевые, рга — прэанальные, ра — пост— , анальные; ноги: ta — лапка, ti—голень, g — колено, Г —бедро,.t — вертлуг, с —тазик (эпимеры),.
Фиг. 48. сГ снизу (ноги не изображены): ера—опиандрий; рга — прэанальные щетинки.
•36
ВВЕДЕНИЕ
-образованием, напоминающим так называемый tritosternum клещей группы Gamasides и несущим лишь осязательные функции, или же вовсе отсутствует. На -нижней стороне тела, позади генитального отверстия, помещается мощный прикрепительный аппарат, строение которого различно в разных группах гипопусов и служит основой для их различения. Ноги I и II (а иногда и III) обычно развиты значительно сильнее, чем задние, ж более или менее значительно выдаются за края тела; ноги III и IV почти целиком или целиком спрятаны под телом или, во всяком случае, значительно меньше выдаются за его края, чем передние; на ногах I—III у большинства видов имеются листовидно расширенные щетинки, лапки IV очень часто оканчиваются 1—4 весьма длинными парусными щетинками и в этих •случаях обычно лишены амбулакр.
Энтомохорный тип гипопусов — наиболее обычная и распространенная •среди Tyroglyphoidea гипопальная форма; гипопусы этого типа характеризуются (фиг. 57—63) наличием присасывательного аппарата, состоящего из особого присасывательного диска, расположенного вентрально в задней части тела и несущего 3—4 пары округлых мясистых присосок, и обычно 1—2 пары стернальных (или коксальных) прцсосок; этот аппарат, действующий по пневматическому принципу, обеспечивает прикрепление гипопуса к гладким поверхностям тела насекомых, используемых им для расселения; этой же цели служат й многочисленные другие органы ‘фиксации, имеющиеся у них, но играющие, невидимому, подчиненную роль: присасывательные щетинки, расширенные на конце в виде блюдца, листовидные волоски на ногах и пр.; у некоторых представителей данного чипа, переносимых сильно опушенными перепончатокрылыми, развиты •еще специальные приспособления для прикрепления к волоскам переносчика, заключающиеся в особом устройстве амбулакр, снабженных мощными винтообразно закрученными коготками и иногда приобретающих характер щипцов. В остальном строение гипопусов этого типа чрезвычайно разнообразно. .
Расселительных гипопусов терохорного типа образуют- только представители подсем. Labidophorinae (сем. Glycyphagidae). Наиболее характерная особенность гипопусов этого типа (фиг. 64—-65) заключается в наличии у них чрезвычайно своеобразного аппарата, служащего для прикрепления к волосам млекопитающих; этот аппарат состоит из пары подвижных клапанов с более или менее ребристой внутренней поверхностью и 2 пар также поперечноребристых зажимов, находящихся под ними, и образует своего рода тиски, в которые прочно зажимается волос; никаких присосок у гипопусов терохорного типа пет, но в остальном они, в общем, довольно сходны с эптомохорными.
Фиг. 49—Ов.-Нимфальные стадии Glycyphagidae.
"Фиг. 49. Glyeyphagus destructor (Schrk.), личинка сверху. Фиг. 50. То же, снизу. Фиг. 51. Личинка Gl. ornalug Kram., сверху. Фиг. 52. Протонимфа Gl. destructor (Schrk.), общий вид (снизу). Фиг. 53. То же, зачатки наружных гениталий. Фиг. 54. Телеонимфа Gl. destructor(Schrk.), общий вид(снизу). Фиг. 55. То же, зачатки наружных гениталий. Фиг. 56. Личинка Chortoglyphv.s arcuatus (Тг.).
28
ВВЕДЕНИЕ
В противоположность расселительным гипопусам, служащим средством . миграции клещей и охвата ими новых местообитаний, покоящиеся гипопальные стадии служат преимущественно сохранению вида в данном местообитании, его переживанию на месте при неблагоприятно складывающихся, здесь экологических условиях. В соответствии с этим, покоящиеся гипопусы характеризуются чрезвычайно высокой устойчивостью к действию-внешних условий .и способны проводить в состоянии диапаузы очень длительное время (измеряемое месяцами и даже годами).
Покоящиеся формы гипопусов имеются у небольшого числа видов: у Tyroglyphus farinae L., у некоторых видов рода Ghae to dactylus и у рода Glycyphagus. У Tyroglyphus farinae эта гипопальная форма близка по своей организации к расселительной, от которой отличается очень слабой подвижностью, некоторой редукцией ног, присасывательного аппарата, кожной системы органов чувств и гнатосомы; еще большее сходство она. обнаруживает с гипопусами типа Acotyledon, стоящими как бы на грани между расселительными и покоящимися типами и также характеризующимися редукцией присасывательного аппарата (стр. 24). У родов Ghaeto^ dactylus и Glycyphagus имеет место далеко идущая редукция локомоторных, прикрепительных и осязательных органов, доходящая до их полного выпадения у Gl. domesticus Deg., у которого покоящийся гипопус представлен йтаровидным, нерасчлененным телом, лишенным конечностей и весьма напоминающим цисту Tardigrada или простейших; это сходство усугубляется еще тем, что гипопус у этих родов остается заключенным в уплотняющемся экзувии протонимфы, играющем роль защитной оболочки. У Tyroglyphus farinae и Chaetodactylus покоящиеся гипопусы образуются наряду с расселительными, но у Glycyphagus последние совершенно отсутствуют.
Причины, ведущие к образованию гипопусов, различны у разных видов и для разных типов гипопальных стадий. У некоторых из изученных в этом отношении видов оно происходит регулярно у части особей каждой попу-’ляции (и каждого поколения), вне зависимости от внешних условий и носит, так сказать, характер «страховки»; в этих случаях протонимфы, дающие таких детерминированных гипопусов, нередко несколько отличаются по своему облику (например большей шириной тела) от других, не дающих гипопусов или образующих их только под давлением неблагоприятных для дальнейшего развития экологических условий; таковыми могут быть пересыхание субстрата, исчерпание корма или ухудшение его пищевых свойств, а, может быть, и температурные условия. У других видов образование гипопуса наблюдается только при неблагоприятном сочетании факторов среды, а при оптимальном состоянии оно вовсе не имеет места. У третьих.
Фиг. 57—61. Расселитсльпые гипопусы Tyroglyphidae и Saproglyphidae.
Фиг. 57. Tyroglyphusfarinae (L.), сверху. Фиг. 58. Тоже, снизу. Фиг. 59. То же, сбоку. Фиг. 69. Calvolia elliptica, ер. п., сверху. Фиг. 01. То же, снизу. Сокращения: gn — гнатосома; st — ster num; ер — эпимеры II; et — эпимериты II; ex I—IV — коксальиые поля I—IV; da — присасывательный диск; Р— проподосома; Н — гистеросома.
30
ВВЕДЕНИЕ
наконец, гипопальные стадии являются обязательными для всех особей или, по крайней мере, приобретают обязательный характер в определенные, ритмически наступающие моменты (например у ряда видов, живущих, в гнездах и строго подчиненных биологическому ритму хозяина). Необходимо еще указать, что у форм, обладающих способностью к образованию^ двух типов гипопусов — расселительных и покоящихся, причины, определяющие переход их в тот или другой тип, могут быть совершенно различны.
Признаки неполовозрелых стадий Tyroglyphoidea широко используются в систематике этой группы; особенно много внимания уделялось рассели-тельным гипопусам, по которым, описано до 40% всех относящихся сюда, видов, причем целые группы (Chaelodactylinae, Saproglyphidae) известны почти что только по ним. В результате этого, в данной группе возникла,, помимо основной систематики, базирующейся на признаках взрослых форм,, п особая «гипопальная». До последнего времени считалось, что признаки гипопусов находятся в тесной зависимости от систематического положения тех форм, к которым они относятся, и что, следовательно, можно на основании строения гипопуса судить об их близости или, другими словами, что система, построенная на основании гипопальных стадий, совпадает-« имагинальной; на основе этого допущения некоторые имагинальные роды, (например Caloglyphus и Cosmoglyphus) даже различались только по признакам их гипопусов. При монографическом исследовании группы и более детальном изучении ее представителей выяснилось, однако, что дело обстоит далеко не так; было установлено, что если в некоторых группах и имеет, действительно, место параллелизм в обеих системах, то в других, гипопальные группировки не соответствуют каким-либо имагинальным, и наоборот, четко очерченные имагинальные группы не совпадают с гипопальными (например во всех подсемействах и трибах сем. Tyroglyphidae,. в роде Acotyledon и др.); здесь, таким образом, наблюдается та же картина,, как в отношении личиночной системы Holometabola к имагинальной, и гипопальным группировкам следует, соответственно, придавать лишь провизорное, временное значение. В противоположность этому, признаки нимфальных стадий всегда укладываются в рамки имагинальной системы,, поддерживают ее и во многих случаях дают ценные дополнительные указания на систематическую близость соответствующих половозрелых форм..
ПОЛОЖЕНИЕ ГРУППЫ В СИСТЕМЕ ACARIDIAE И ЕЕ КЛАССИФИКАЦИЯ
До 20-х годов текущего столетия, ознаменовавшихся в акарологии полной перестройкой системы подотряда Sarcoptiformes, свободноживущие тироглифоидеи составляли, совместно с группой Anoetidae, сем. Tyrogly-
Фиг. 62—65. Рассслителытые гипопусы Glycyphagidae.
Фиг. 62. Энтомохорный гипопус Sennerlia horrida (Vtz.) сверху. Фиг. 63. Тоже, снизу. Фнг. 61.
Терохориый гипопус Labtdophorus sciurinus (Koch) сверху. Фиг. 65. То же, снизу» F — прикрепительный аппарат.
12
ВВЕДЕНИЕ
phidae—по одним авторам и подсем. Tyroglyphinae семейства Sdrcoptidae— по другим. Это семейство (resp. подсемейство) отличалось от смежных с ним группировок почти исключительно биологическими признаками — свободным образом жизни его представителей, противопоставлявшим его другим, паразитическим и эпизоическим группам. Морфологические признаки занимали в его диагнозе подчиненное положение и носили преимущественно негативный характер (отсутствие параллельной штриховки на поверхности кожи; нестебельчатые praetarsi). Благодаря этому, в сем. Tyroglyphidae оказались объединенными столь чуждые между собою группы, как сво-•бодноживущие Tyroglyphoidea, с одной стороны, и Anoetidae — с другой, и совершенно не были отображены бросающиеся в глаза связи между первой из этих групп и многими паразитическими или эпизоическими формами, отошедшими в другие семейства (подсемейства Analginae, Sarcopti-пае и др.). '
В принятой современными авторами Классификационной схеме Oude--mans’a (разработанной им в 1923—1927 гг. и впоследствии лишь слегка видоизмененной Vitzthum’oM и отчасти Sig Thor’ом), члены рассматриваемой 'здесь группы никак не объединены друг с другом и входят, на правах 15 автономных семейств, в когорту Diacotricha, к которой относятся также семейства Anoetidae, Canestriniidae и целый ряд эпизоических и паразитических групп: Acaridae (== Sarcoptidae), Listrophoridae, Eustathiidae и Analgesidae, Falculiferidae, Syringobiidae н Dermogly phidae. Эта первая когорта противопоставляется второй — Anacdtricha, составленной остальными группами эпизоических и паразитирующих на позвоночных Acaridiae (Avenzoaridae, Proctophyllodidae, Psoralgidae, Epidermoptidae, Psoroptidae, Heteropsori-dae, Laminiosioptidae') ц связанными с насекомыми Linobiidae nHemisarco-ptidae. Двумя этими когортами и исчерпывается состав суперкогорты (серии) Acaridiae, составляющей, совместно с серией Oribatei, подотряд Sar-coptiformes.
Различие между обеими когортами (надсемействами) — Diacotricha и Anacotricha— заключается в одном только признаке, подчеркиваемом •«самими их названиями: наличии или отсутствии внутренних (иначе передних) теменных щетинок (setae verticales interiores); Высокое систематическое значение, придаваемое этому признаку, оказывается, однако, при более трезвой оценке, произвольным и совершенно необоснованным; в самом деле, он не может претендовать на роль индикатора, так как не имеет коррелятивных связей с каким-либо достаточно крупным комплексом призна-
Фиг. 66—79. Покоящиеся гипопусы и гипопальпыо камеры различных Tyroglyphoidea.
Фиг. 66. Покоящийся гипопус Turoglyphwi farlnae (L.) сверху. Фиг. 67. То же, снизу. Фиг. 68. То же, нога I снизу. Фиг. (И). То же, нога IV снизу (по Schulze). Фиг. 70. Покоящийся гипоцус Glyeyphagus destructor (Schrk.) сверху. Фиг. 71. То же, снизу (рисунок с объекта, заключенного я жидкость Фора-Берлезе). Фиг. 711. То жо, рисунок с живого объекта. Фиг. 73. То же, нога I сверху. Фиг. 74. Покоящийся гипопус (II. miehaeli Ouds. снизу (по Michapl’ro). Фиг. 7S. Гипопапь-ная камера Gl. destructor (Schrk.). Фиг. 76. То же, Gl. miehaeli Ouds. (по М1сЬаеГю). Фиг. 77. То же, Gl. domesticus (Deg.). Фиг. 78. То же, Chaetodactylus osmiae (Duf.). Фиг. 79, Покоящийся гипопус Ch. osmiae (Dut.) снизу (по Trouessart’y).
II Фауна СССР, Захваткип—284
3i
ВВЕДЕНИЯ
ков, устанавливающим черты сходства (т. е. систематической близости) между несущими его формами. С другой стороны, этот признак далеко не имеет абсолютного (универсального) значения, так как среди Diacotricha существует ряд форм и даже целых групп, где s. verticales inter!ores сильно редуцированы (Xenocastor) или даже совершенно отсутствуют (Monieziella, все или почти все Listrophoridae и др.); больше того, наличие или отсутствие этих щетинок подвержено в отдельных группах географической и даже индивидуальной (альтернативной) изменчивости. Отсюда следует, что разграничение этих когорт совершенно искусственно и все составляющие их группировки сливаются на практике в так и остающееся нерасчлененным обширное и многообразное целое — серию Acaridiae.
Таким образом, в этой классификации группа Tyroglyphoidea перестала существовать, как таковая, распавшись на большое число мелких, якобы самостоятельных «семейств», и положение ее представителей в системе Acaridiae остается неясным, поскольку такой системы, вообще, не оказывается.
Это положение явилось естественным результатом сугубо формалистического подхода Oudemans’a и следовавших за ним авторов к построению акарологической системы. В ее основу были положены единичные, произвольно выбранные признаки, которым придавалось значение руководящих, без анализа и учета характера их связей и вне зависимости от суммарного сходства между трактуемыми формами; богатейшие фонды признаков, долженствующие быть использованными для правильного понимания сходственных отношений между ними, оставались незатронутыми систематическим исследованием, и сам выбор «руководящих» признаков постоянно менялся, что влекло за собой постоянные переделки системы и перемещения отдельных форм из одной группы в другую; кроме того, не учитывался и масштаб систематических различий, что имело следствием явную переоценку таксономического уровня отдельных группировок и их чрезмерное дробление. Все эти недостатки современной акарологической системы являются непреодолимыми в настоящий момент препятствиями для правильного суждения о действительных соотношениях между отдельными группами клещей и, в частности, для окончательной фиксации положения в пей группы Tyroglyphoidea detriticolae. Не имея возможности проработать всю обширную серию Acaridiae, я вынужден был ограничиться лишь упорядочением систематики рассматриваемой здесь группы, оставляя в стороне выяснение се связей с соседними группировками. Проведенная в этом направлении работа привела, во-первых, к реабилитации надсемейственного объединения Tyroglyphoidea detriticolae, общие признаки которого приведены на стр. 1; во-вторых, было установлено, что все многообразие свободноживущих Acaridiae, распределяемых Oudemans’oM в 21 «семейство», естественно образует только четыре группировки семейственного масштаба, за которыми, соответственно, закрепляются названия: Варго-glyphidae, Tyroglyphidae, Glycyphagidae и Anoetidae, из которых три первых
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ »»
систематически близки между собой и к ряду эпизоических и паразитических групп, а четвертая стоит в системе особняком и должна быть выделена в особое надсем. Anoetoidea. В принимаемом здесь объеме сем. Туго-glyphidae соответствует 5 «семействам» системы Oudemans’a: Tyroglyphidao Ouds. 1932, Caloglyphidae Ouds. 1932, Tyrophagidae Ouds. 1924, PJiizogly-, phidae Ouds. 1923 и Forcelliniidae Ouds. 1924; сем. Saproglyphidae, в моем понимании, равно «семействам» Saproglyphidae Ouds. 1923, Winterschmid-liidae Ouds. 1923, Ensliniellidae Vtz. 1926 (nec 1931), Czenspinskiidae Ouds. 1927, Nanacaridae Ouds. 1924, Pontoppidaniidae Ouds. 1924; к сем. Glycypha-gidae я отношу представителей принятых Oudemans’oM семейств Glycypha-gidae, Chortoglyphidae, Carpoglyphidae, Lentungulidae и часть Ensliniellidae Vtz. 1931 (роды Horstia и Tor tonin'). Для меня несомненно, что соответствующая обработка других Acaridiae, да и большинства других групп клещей, основанная на принципах естественной биологической системы, приведет к такому же резкому сокращению .числа принятых здесь семейств.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ
Группа Tyroglyphoidea detriticolae известна еще слишком недостаточно, чтобы можно было иметь какие-либо суждения о характере распределения ее представителей по зоогеографическим областям; статистика видов, описанных в каждой из них, выражает пока только степень изученности данной области, но никак не численность населяющих ее форм, и потому не может, служить этой цели; равным образом, нехватает данных и для установления приуроченности той или иной группы форм к определенной зоогеографической области. В большинстве случаев остаются еще невыясненными и-ареалы отдельных видов, намечаемые пока только единичными точками, подвергшимися более или менее интенсивному акарологическому изучению. Сейчас можно только сказать, что рассматриваемая группа в целом имеет космополитическое распространение и богато представлена в умеренных и тропических поясах обоих полушарий, заходя отдельными видами как в арктическую, так и в антарктическую зоны. Из Палеарктики известно около 180 из общего числа в приблизительно 250 описанных до сих пор видов.
Существующие данные о распространении синантропных видов тиро-глифоидей также еще далеко недостаточны для окончательного суждения об ого характере. Они позволяют пока только прийти к выводу, что распространение большинства из них (за исключением только Tyroglyphus fari-
L. и Rhieoglyphus echinopus F. et В.) не является космополитным, А ограничено более узкими и достаточно определенными рамками. Это обстоя-твльотио особенно бросается в глаза при сопоставлении списков амбарных НЛОЩой Европы и США [в США отсутствуют, например, представители родов Glycyphagus, Aleuroglyphus, Thyreophagus, Histiogastcr, Gohioria It др., но имеется ряд амбарных видов (Tyrophagus amcricanus Bnks.., ♦
36
ВВЕДЕНИЕ
«Т». armipes Bilks.), отсутствующих в Европе], но даже и отдельные европейские страны представляют в этом отношении значительные различия. Так, например, Caloglyphus rodionovi A. Z., широко распространенный на континенте, не встречается, невидимому, в Англии; Tyrophagus longior Gerv. (= infestans Berl.) приурочен к Западной Европе и у нас в СССР заменен другими видами и т. д. Распределение отдельных видов амбарного комплекса тироглифоидей по территории СССР также неодинаково, причем некоторые из них оказываются приуроченными к более северным районам (Glycyphagus michaeli Ouds.), другие — к южным (Tyrophagus noxius A. Z., Т. perniciosus A. Z.). Еще более резко выражена закономерность в распространении этих же видов в полевой обстановке, так что каждый из изучавшихся в этом отношении районов обладает своим, специфическим набором хлебных видов, встречающихся в поле в местных условиях. Так, в степной полосе Башкирии (Стерлитамакский зерносовхоз), в поле, были найдены: Tyroglyphus farinae L., Tyrophagus humerosus tesquorum A. Z., Caloglyphus rodionovi A. Z., Acotyledon sokolovi A. Z. и 3 вида Glycyphagus: Gl. destructor Schrk., Gl. michaeli Ouds. и Gl. domesticus Deg. В степях Предкавказья (Ворошиловский зерносовхоз, Благодарненского р-на) встречались Tyroglyphus farinae L., Tyrophagus perniciosus A. Z., T. humerosus tesquorum A. Z., Galoglyphus rodionovi A. Z., Acotyledon batsylevi A. Z., Glycyphagus destructor . Schrk. и Gl. domesticus Deg., в Ивановской же области (окрестности г. Кир-жач) полевая фауна хлебных клещей складывалась только из Tyroglyphus farinae L., Acotyledon sokolovi A. Z., Glycyphagus destructor Schrk., Gl. michaeli Ouds., Gl. cadaverum Schrk. и Chortoglyphus arcuatus Tr.
Для более детальной обрисовки ареалов синантропного и полевого распространения этих видов, к сожалению, еще нехватает данных.
ЭКОЛОГИЯ
Характерными для Tyroglyphoidea detriticolae биоэкологическими признаками можно считать, в самой общей форме, следующие.
а) сапробный образ жизни во всех стадиях развития;
: б) гигрофильность (связанную с малой валентностью в отношении к влажности) половозрелой и вегетативных стадий, их анемофобию и относительную эвритсрмпость;
в) малую способность всех стадий к активному расселению и широкое использование ими различных пассивных способов такового;
г) уже отмоченную выше способность к образованию специальных рас-с.елительных и покоящихся стадий: (гипопусов).
Общие условия обитания. Подавляющее большинство свободножи-вущих Tyroglyphoidea живот в мертвых, преимущественно гниющих органических материалах (главным образом растительного происхождения), служащих им одновременно и кормовым субстратом или, по крайней мере, питательной средой для микроорганизмов, составляющих
ЭКОЛОГИЯ jf
иногда их пищу. Занимая всевозможные типы местообитаний, создаваемых^ скоплениями таких субстратов в природе и человеческом хо:ш11-стве, экологический ареал этой группы еще расширяется в сторону охвата им и некоторых видов живых растительных материалов — луковиц, корнеплодов и других подземных частей высшнх растений (а в некоторых, редких, случаях даже и их надземных частей), скоплений их семян, а также грибов и водорослей. Самыми общими условиями обитания тироглифоидей в этих субстратах являются их достаточно высокая влажность, слабая аэрация и защищенность от прямого действия солнечных лучей. Значение влажности субстрата в создании необходимых для тироглифоидей условий обитания чрезвычайно велико и многосторонне и далеко не исчерпывается ее ролью, как источника снабжения водой организма при питании. В самом деле, не меньшее значение, чем «влажность корма», имеет здесь и влажность воздуха в толще субстрата и местах его залегания, так как ею обусловлена, прежде всего, сама возможность дыхания через поверхность кожи, свойственного всем представителям группы; связанная с этим типом дыхания нежность и тонкость кожных покровов, также характерная для большинства из них, и высокое процентное содержание свободной воды в их теле (по Н. Schulze, например, общее содержание воды в теле Caloglyphus rodionovi A. Z. достигает 83.03%) — все это ставит самую возможность их существования в зависимость от достаточного насыщения воздуха влагой. Для большинства видов, подвергшихся экспериментальному изучению, минимальный процент относительной влажности воздуха, при котором вообще возможно их существование, лежит выше 60%, а оптимальные для них условия создаются при 80'—90%, а иногда и 100% его влажности. Для Tyroglyphus farinae L., например, минимальной является влажность воздуха 60%, а оптимальной — от 75 до 80%; для Tyrophagus noxius A. Z. — соответственно 70 и 90%; для Aleuroglyphus ovatus Тг. приемлемый минимум атмосферной влажности лежит выше 75%, а оптимум — около 90%; оптимальная для видов родов Rhisoglyphus и Caloglyphus влажность воздуха близка к 100% (минимальная для С. rodionovi составляет окойо 90%) и т. д. В силу тесной зависимости, существующей между влажностью воздуха и таковой субстрата, 1 указанным нормам атмосферной влажности соответствуют и совершенно определенные минимальные и оптимальные для тироглифоидей влажности обитаемых ими субстратов (различные при разной температуре и для разных видов субстрата и опре
1 Влажность большинства субстратов, обитаемых тироглифоидеями, находится, в результате их гигроскопичности, в тесном соответствии с таковой окружающего воздуха и в постоянном вшш-модействии с нею, так что каждому показателю атмосферной влажности соответствует онредо» лонное (особое для каждого вида субстрата) процентное содержание воды в субстрате (поглощающем ее из влажной атмосферы до установления соответствующего равновесия или отдающем избыток содержащейся в нем воды прилегающим слоям воздуха). Это подвижное равновесно между влажностями субстрата и окружающей его атмосферы приводит к их вааимпоП регулировке, причем специфическая влагоемкость субстрата и быстрота поглощения (или отдачи) ин цоды, а также имеющаяся температурная обстановка придают различные цифровые »upiUllO)lll)l устанавливающимся здесь соответствиям.
38
ВВЕДЕНИЕ
деляемые их специфической влагоемкостыо). Так, Tyroglyphus farinae L. может развиваться в муке лишь при ее влажности не ниже 12.5%, в зерне (пшеница) — не ниже 13.4%, а оптимальная для него влажность этих продуктов равна, соответственно, около 15—16% и 18—19%; минимальной для Caloglyphus rodionovi A. Z. является влажность субстрата около 22% и т. д.
Роль ряда других факторов в создании необходимых для тироглифоидей условий обитания, в основном, сводится также к определению соответствующего режима влажности; в этой плоскости действуют как защищенность их местообитаний от действия ветра и прямых солнечных лучей, так и характер строения занимаемых ими субстратов, поддерживающие слабую аэрацию и пониженную испаряющую способность воздуха в обитаемой ими среде.
' В отношении к температуре клещи данной группы, в целом, менее требовательны, чем к влажности, и видовые различия в этом отношении вариируют в гораздо более широких пределах. Так, нулевая точка развития Tyroglyphus farinae L. лежит несколько ниже 8°; она равна: приблизительно 9° для Tyrophagus noxius A. Z., 16° дляCaloglyphus rodionovi A. Z., 20—21° для Aleuroglyphus ovatus Тг.; оптимум развития приходится: назову 20—22° для Tyroglyphus farinae, около 25° ДЛЯ Tyrophagus noxius, 31—32° для Caloglyphus rodionovi, 35° для Aleuroglyphus ovatus. Если учесть, что эти данные относятся к представителям только одного ценоза — амбарного, т. е. к группе в той или иной мере экологически близких между собой форм, то можно предположить, что границы общей термической зоны развития совокупности представителей группы Tyroglyphoidea окажутся еще значительно шире.
В отношении к кормовым свойствам субстрата у Tyroglyphoidea наблюдаются самые разнообразные картины. Большое число видов характеризуется крайней многоядностью и использует безразлично как растительные, так и животные субстраты самого различного рода (многие амбарные виды); у других наблюдаются известные ограничения в выборе субстрата, третьи, наконец, являются узко специализированными монофагами. Необходимо-, впрочем, отметить, что мы не располагаем еще достаточно точными данными для суждения о реально имеющих здесь место отношениях, так как, с одной сторопы, чрезвычайно трудно разъединить кормовые свойства субстрата от других, его особенностей (определяющих возможность обитания в нем клещей) и рассматривать их вне связи с последними, а с другой, здесь, вероятно, часто имеет место питание клещей не самим субстратом, а произрастающей на пом микрофлорой. Для немногих видов удалось показать прямую зависимость в выборе субстрата от его химизма, например от содержания в нем определенных кислот. Так, Carpoglyphus lactis L. живет только в веществах, содержащих молочную, янтарную или уксусную кислоты; аналогичная связь с наличием в субстрате уксусной кислоты явственно намечается у видов рода Histiogaster Berl. Возможно также ограничение
экология
uo
набора используемых кормовых субстратов требованиями к определен» ному структурному состоянию вещества, например порошковидному у Thyreophagus и Gohieria.
Распределение по отдельным типам местообитаний. Обширный экологический ареал группы Tyroglyphoidea detriticolae складывается из следующих 16 основных типов местообитаний, каждый из которых характеризуется определенными различиями в составе населяющего его «тироглифоценоза».
1. Запасы сельскохозяйственных продуктов в амбарах, складах и жилых помещениях. Чрезвычайно обширный комплекс местообитаний этого типа может быть расчленен па 3 основных подтипа таковых, существенно разнящихся между собой по связанному с ними набору видов. А) Запасы зерна и семян различных культурных растений и продуктов их переработки (мука, жмыхи и т. п.)-служат местообитанием многочисленным амбарным видам тироглифоидей, наиболее характерными из которых являются следующие: Tyroglyphus farinae L., Tyrophagus noxius A. Z., T. perniciosus A. Z., T. americanus Bnks., Tyrolichus casei Ouds., T. Uni Ouds., Aleuroglyphus ovatus Tr., Caloglyphus rodionovi A. Z., Thyreophagus entomophagus Lab., Chortoglyphus arcualus Tr., все виды родов Blomia и Glyeyphagus, Cometacarus smirnovi A. Z., Gohieria fusca Ouds. и «Glyeyphagus» robustus Bnks. Вблыпая часть этих видов характеризуется многоядностыо, но среди них есть и олигофаги и формы, отдающие явное предпочтение какому-либо одному виду субстрата (например Thyreophagus, Gohieria). Значительное большинство перечисленных видов более или менее широко распространено и в полевых условиях, но некоторые из них известны пока только как синантропы (Tyrophagus noxius A. Z., Tyrolichus lini Ouds., виды родов Blomia, Thyreophagus, Gohieria и др.); эти последние виды обычно характеризуются в биологическом отношении своей термофильностыо и слабым расселитель-ным потенциалом (отсутствие гипопальных стадий, слабая подвижность и т. д.). Местами, в состав амбарного комплекса тироглифоидей включаются отдельные виды, основной экологической базой которых являются различные полевые местообитания, например Tyrophagus humerosus Ouds., Т. tenuiclavus A. Z., многие виды рода Acotyledon, некоторые Calvolia и др. Запасы сухих фруктов, овощей, лекарственных трав и других высушенных растительных материалов, а также сухие вещества животного происхождения населены представителями того же амбарного комплекса, из которых наиболее обычны здесь виды Glyeyphagus. Здесь встречается и связанный преимущественно с фруктовыми продуктами Carpoglyphus lactis L., находимый также в прокисшем варенье, старых кондитерских изделиях, в випо и разлагающихся молочных продуктах. Несколько особняком стоит фауна сыра, в которую входят, помимо нескольких общеамбарных видон, как Tyroglyphus farinae, Tyrolichus casei, еще и ряд более строго приуроченных к этому субстрату видов: Tyrophagus longior Gerv. (=infestans Berl,), Aeoty* ledon sironiformis F. et R., A. terminalis Bnks. и Calvolia sac.hvri OlldH*
40
ВВЕДЕНИЕ
Б) Луковицы огородных и декоративных культур, корнеплоды и клубни обладают своим специфическим набором приуроченных к ним видов Tyroglyphoidea, из которых наиболее важны Bhizoglyphus echinopus F. et R., некоторые виды Caloglyphus (C. sphaerogaster A. Z., C. mandshur A. Z.) и Tyrophagus tenuiclavus A. Z.; на этих же субстратах встречаются иногда и наиболее многоядные из амбарных видов — Tyroglyphus farinae, Tyrophagus noxius, Caloglyphus rodionovi и др. В) Сено в сеновалах и конюшнях имеет своеобразное население тироглифоидей, складывающееся как из наиболее распространенных и эвритопиых «амбарных» видов (Tyroglyphus farinae, Tyrophagus sp. sp., большинство видов Glycyphagus, Chortoglyphus'), так и из экологически узко локализованных здесь представителей подсем. Ctenogly-phinae-, сюда заходят также и некоторые из видов, характерных для следующего типа местообитаний.
2. Скопления мертвых растительных остатков на полях широко используются тироглифоидеями в качестве местообитаний. Особенно большое значение в этом отношении имеют более крупные из таких скоплений, образуемые в результате хозяйственной деятельности человека — стога сена, ометы и кучи соломы, скопления различных отходов от очистки зерна и т. п., причем это значение возрастает пропорционально долговечности этих объектов, т. е. продолжительности их пребывания в поле. Среди тироглифоидей, населяющих эти местообитания, доминирующую роль играют хлебные клещи — Tyroglyphus farinae, виды родов Tyrophagus, Glycyphagus и др., по здесь обитают также и многие не-амбардые виды родов Acotyledon (A. redikorzevi A. Z., A. batsylevi A. Z., A. sokolovi A. Z. и др.), Tyrophagus humerosus Ouds., Т. tenuiclavus A. Z., некоторые Calvolia и Forcellinia. Сходную, но менее разнообразную фауну имеют и более мелкие скопления мертвых растительных остатков (например стерня, войлок, образующийся в результате отмирания вегетативных частей травянистых растений на задерненных участках), но только в том случае, если в них создаются и более или менее длительно поддерживаются условия влажности, необходимые для обитания клещей.
3. Почва полей. В поверхностных слоях почвы культурных полей нередко встречаются тироглифоидеи, существующие здесь за счет падалицы, непроросших зерен или других мелких объектов растительного происхождения, диффузио залегающих в почве (Tyroglyphus farinae L., Т. perniciosus A. Z., Glycyphagus sp. sp.) или же за счет подземных частей живых растений (Tyrophagus humerosus Ouds., Rhizoglyphus). Данных о населении почвы целины пока почти нет.
4. М е р т в а я лес к а я иоде т и л к а. В местообитания этого . дина также заходят отдельные амбарные виды Tyroglyphoidea (Tyroglyphus farinae, Glycyphagus destructor, Gl. michaeli'), по основную массу их населения составляют более стенотопные формы, из которых наиболее характерны следующие: Acotyledon Ishernyshevi A. Z., Forcellinia diamesa A. Z., Caloglyphus ishernyshevi A. Z., Schwiebea pachyderma A. Z., а также
ЭКОМ ГИЯ и
II еще не найденные у пас Sancassania chelone Ouds., Acotyledon eurynyni pha Ouds., Troupeauia mycobora Ouds., Schwiebea talpa Ouds., Calvolia hagmxix Ouds. Здесь найдены и менее строго приуроченные к этим местообитаниям Tyroglyphus tyrophagoides A. Z., Tyrophagus humerosus Silvester A. Z., Acotyledon solwlovi k. Z., Caloglyphus tshernyshevi A. Z. и весьма эвритоппый Rhizoglyphus echinopus F. et R.
5. Гниющая древесина. В более или менее разложившейся » древесине — в старых ппях, древесной трухе, скоплениях буровой муки
I в ходах точащих деревья насекомых и под кусками отстающей коры — оби-
тает весьма разнообразная и богатая фауна тироглифоидей: Caloglyphus tshernyshevi A. Z., ряд видов Schwiebea (Schw. rossica A. Z., Schw. ipidis Vtz., Schw. tshernyshevi A. Z.), Histiogaster carpio Kram., виды родов Troupeauia, Rodionovia и многочисленные представители сем. Saproglyphidae (Saprogly-phus, Winterschmidtia, много Calvolia)-, часть последних в той или иной мере биологически связана с короедами и. другими ксилофагамн из насекомых, служащими переносчиками их гипопусам; к этой же группировке клещей примыкают и виды, живущие в гнездах многочисленных перепончатокрылых, гнездящихся в дереве (более детально рассматриваемые-ниже).
6. Вытекающий сок деревьев и пропитанные им участки коры населены крайне оригинальными видами рода. Hericia Сап., ведущими, соответственно, полуводный образ жизни в этой жидкой среде.
7. Корни и другие подземные части цветковых растений (в поле) служат местообитанием крайне многоядному Rhizoglyphus echinopus F. et R. и некоторым видам родов Caloglyphus (С. hete~ romorphus Felt, C. phylloxerae Riley), Tyrophagus и Monieziella (? M. minuta Dem.).
8. Зеленые части живых растений используются тироглифоидеями лишь как исключение; здесь встречаются только Tyrophagus humerosus Ouds., живущий преимущественно на проростках зерновых культур (и изредка даже вредящий им) и на листьях различных тыквенных растений, пораженных паутинными клещиками, и некоторые виды Calvolia (С. hebeclinii Sich. и более эвритопная С. heterocoma Mich.).
9. Грибы. Плодовые тела шляпочных грибов и трутовиков обладают своим, довольно специфическим набором видов, состоящим из Caloglyphus туcophagus Megn., С. berlesei Mich., Histiogaster silenus k. Z., Acotyledon alberti A. Z., Forcellinia fungivora Ouds. и F. (?) lintneri Osb. и Saproglyphus neglectus Berl. Значительно большее число видов существует, несомненно, за счет низших грибов, например различных плесеней, развивающихся на обитаемых ими субстратах.
10. Скопления морских водорослей на прибрежных, камнях и сваях, в полосе прибоя, населены также весьма оригинальными видами литоральных родов Hyadesia и Pontoppidania — единственных
№
ВВЕДЕНИЕ
морских представителей’дайной группы (если не считать загадочную Аса-ridina balaenarum van Bened., найденную на китах).
11. Мхи и лишайники. Среди мхов и под лишайниками найдены отдельные виды Tyroglyphoidea (Tyrophagus dimidiatus В erm., многие экзотические Caloglyphus, Robinisca, Monieziella magna Berl., cRhi-soglyphus» occurens Berl., «Rhizoglyphus» algidus Berl. и Calvolia heterocoma Mich.), нов этом типе местообитаний они представлены несравненно слабее, чем господствующие здесь Oribatei; в общем, эти местообитания еще очень мало изучались.
12. Гнезда одиночных и общественных пчел и о с населены чрезвычайно большим количеством видов клещей группы Tyroglyphoidea, играющих здесь роль комменсалов и, в большинстве случаев, очень строго приуроченных к какому-либо одному определенному роду или даже виду хозяина; особенно много таких видов биологически «вязано с пчелами и осами, гнездящимися в дереве. Установленные до настоящего времени случаи этого рода могут быть представлены в виде следующего списка:
Хозяева
Сем. Apidae
Frosopis conformis Forst.
Xylocopa violaeea L.
» valga F.
» punctilabris Mor.
» varentzovi Mor.
/ » przewalskii Mor.
» turaniea Mor.
» nitidiventris Sm.
» kalinavskii Rad. (cireumvolans Sm.)
» dissimilis Lep.
.» combinate Rits.
» ordinaris Sm.
Xoptorfhosoma nigrita F.
» caftra L. Platynopoda tenuiscapa Westw.
Chelostoma jlorisomnis L.
•Osmia nifa L.
» leohli Duclco
» dimidiate Mor.
» tricornis F.
» panzeri Mor.
» leucogastra Mor.
» emarginate Lep.
г> leeana Kirby
» solskyi Mor.
» nigrivmtris Zett.
Zithurgus denlipes Sm.
Комменсалы
Vidia undulate Ouds.
Sennertia cerambyeina (Scop.)
Sennertia cerambyeina (Scop.), S. gargantua A. Z.
Sennertia gargantua A. Z.
Sennertia cerambyeina (Scop.)
Sennertia cerambyeina (Scop.) Sennertia cerambyeina (Scop.) Sennertia cerambyeina (Scop.) Sennertia alfkeni (Ouds.), Horstia ornate (Ouds.) Sennertia horrida (Vtz.), S. dissimilis A. Z.,
S. oudemansi A. Z., Horstia helenae (Ouds.) Sennertia bijilis (Can.), Horstia trifilis Con. Horstia puleherrima (Vtz.)
Ceroglyphus monstrosus Vtz., «Glyeyphagus» geni-eutetus Vtz., Sennertia morstetti Vtz., S. per-turbans Vtz., S.. simplex (Trag.), Horstia glabra Vtz.
Sennertia eaffra Vtz.
Sennertia greeni (Ouds.), S. koptorthosomae (Ouds.), S. hipposideros (Ouds.)
Chaetodactylus birulai A. Z.
Chaetodactylus osmiae (Duf.)
Chaetodactylus osmiae (Duf.) Chaetodactylus osmiae (Duf.) Chaetodactylus osmiae (Duf.), Ch. elaviger
Chaetodactylus riaumuri (Ouds.) Chaetodactylus riaumuri (Ouds.) Chaetodactylus riaumuri (Ouds.) Tortonia intermedia (Ouds.) Tortonia intermedia (Ouds.) Tortonia intermedia (Ouds.) Chaetodactylus ludwigi (Trouess.)
экология
И'
Megachile doderleini Fr.
» bombycina Rad.
» ligniseea KI.
Megachile sp. sp.. div.
Anthidium 7-dentatum Latr.
» stricticum F.
» latieeps Mor.
» pubeseens Mor.
» manicatum L.
» oblongatum Latr.
» cingulatum Latr.
» jlorentinum F.
» diadema Latr.
Bombus sp. sp. div.
Cerophagopsis skorikovi A. Z.
Chaetodactylus dementjevi A. Z., Vidia linrnta Ouds.
Vidia lineata Ouds., Chaetodactylus dementjevi
Vidia lineata Ouds.
Tortonia dogaressa A. Z.
Chaetodactylus anthidii (Ouds.), Tortonia smits-vanburgsti Ouds.
Neohorstia mamillata A. Z.
Neohorstia mamillata A. Z.
Sennertionyx manicati (Giard)
Sennertionyx manicati (Giard)
Sennertionyx manicati (Giard) Sennertionyx manicati (Giard) Sennertionyx manicati (Giard)
Kuzinia laevis (Duj.), Cerophagus granulatus (Duj.)
Сем. Sphegidae
Cr'abro jossorius L.
» impressus Sim.
» vagus L.
» solenius L.
* elypeatus L.
» afatius Pnz.
>» wollmanni Kohl.
Vidia gussaJiovslcii A. Z., Laekerbaiieria eribra-tissima A. Z.
Vidia gussakovskrii A. Z., V. popovi A. Z.
Vidia gussakovskii A. Z.
Vidia gussakovskii A. Z.
Vid„a popovi A. Z.
Vidia popovi A. Z.
Vidia popovi A. Z.
Сем. Vespidae
djidynerus (Lwnotus'j floricola Sauss. » » rossii Lep.
Follistes gallicus L.
Ensliniella parasitica Vtz.
Ensliniella kostylevi A. Z. «Carpoglyphus» vagdbundus Berl.
13. Муравейники. Среди палеарктических Tyroglyphoidea мирмекофилами являются лишь Forcellinia wasmanni Mon., F. gigantonympha Vtz., Froriepia vimariensis Vtz. и, вероятно, также Calvolia bulgarica Storkan и Acotyledon krameri Berl.; из экзотических видов к этой биологической группировке относятся Acotyledon oudemansi A. Z., «Tyroglyphus» javensis Ouds., «Т.» australasiae Ouds. и «Т.» vandergooti Ouds. и весьма оригинальные «Myrmoglyphus» (= Acotyledon part.): M. triscissus Vtz. и AT. bipilis Vtz.
14. Гнезда термитов. До настоящего времени известен только 1 термитофильный вид — Acotyledon feytaudi Ouds., живущий в гнездах Reticuli ter mes lucifugus Rossi.
15. Гнезда грызунов и насекомоядных. Мертвая растительная подстилка и запасы в норах мелких млекопитающих обильно населены тироглифоидеями. В состав очень богатой фауны клещей гнезд крота входят: Fusacarus laminipes Mich., Talpacarus platygastcr M'ioh., Orycteroxenus dispar Mich., Xenoryctes krameri Mich, и X. heplneri A. Z. и известные лишь по гипопусам Labidophorus talpae Kram. и Myacarus hypu-daei C. L. Koch, а также Tyroglyphus farinae L., Aleuroglyphus ovahiH Robinisca mycolicha Ouds., Viedebannttia schmitsi Ouds. и Glyoyphagut
a
ВВЕДЕНИЕ
ornatus Kram. Многие из перечисленных видов встречаются также и в гнездах землероек, где встречается еще и Labidophorus soricis Ouds., тогда как другие виды рода Labidophorus Kram. приурочены к гнездам выхухоли (L. desmanae A. Z.) и дальневосточного крота (L. mogerae A. Z.); В гнездах различных полевок живут виды родов Myacarus, Labidophorus soricis Ouds., Tyroglyphus farinae L., Forcellinia fungivora Ouds. и некоторые Tyrophagus, Aleuroglyphus ovatus Tr. и Glyeyphagus ornatus Kram.; в гнездах сусликов найден пока только Tyroglyphus farinae L.; наконец, гнезда белок служат местообитанием для Labidophorus sciurinus С. L. Koch и Calvolia heterocoma Mich. Постоянное нахождение многих из амбарных видов (и в том числе Tyroglyphus farinae) в гнездовой подстилке и запасах в норах грызунов мне кажется весьма симптоматичным и подтверждающим высказанное 3. С. Родионовым предположение о нидикольном происхождении амбарной группировки тироглифоидей (вернее, его основного ядра).
16. Пещеры. Пещерных видов известно среди тироглифоидей всего 3: «Tyroglyphus» (Caloglyphus ?) spelaeus Berl., Stereoglyphus hemisphaericus Berl. и Schwiebea cavernicola Vtz.; последняя была найдена в мелких скоплениях воды.
Способы расселения. Большинство тироглифоидей — малоподвижные, подчас неповоротливые и медленные в своих движениях формы, малоспособные к самостоятельному преодолению больших пространств. Скорость активного передвижения половозрелых особей колеблемся, у разных видов и в зависимости от рода субстрата, от 4—5 мм (6.2 — у Caloglyphus rodionovi A. Z.; От 6.2 до 7.5 — у Chortoglyphus) до 30—40 мм (Tyrophagus noxius A. Z.) в минуту (Козулина, 1936); только виды рода Glyeyphagus — лучшие бегуны во всей группе — достигают значительных скоростей порядка 40—65 (Gl. destructor) и даже до 150 мм (Gl. michaeli) в минуту. При этом, проходимый ими путь лишь редко бывает прямолинейным, так что полезное перемещение клеща (т. е. вычисленное по прямой) оказывается еще значительно меньшим указанных цифр (2.9—4.4 мм в минуту для Chortoglyphus1, 23.7—26 мм — для Tyrophagus noxius1, 41.7—59.0 мм —для Glyeyphagus michaeli и т. д.). Возможности активного расселения представителей этой группы еще сокращаются и практически ставятся в относительно очень узкие рамки их малой экологической валентностью (главным образом в отношении к влажности), позволяющей им передвигаться лишь в совершенно определен ной микроклиматической обстановке; последняя же редко бывает однородной на более или менее значительном протяжении. Между тем, общее жизненное значение рассслитсльпой функции приобретает в этой группе исключительную важность (именно в связи с стеиовалентностью половозрелой и вегетативных стадий со представителей) благодаря прерывистому характеру залегания большинства используемых ими местообитаний, разобщенных непреодолимыми для них пространствами; кроме того, неустойчивость режима влажцости во многих занимаемых ими местообитаниях также вызывает необходимость в постоянных перебросках клещевого насе
ЭЕОМГИЯ
45
ления с одной их точки в другую, в зависимости от микроклиматических колебаний в различных участках биотопа (например в различных слоях скоплений растительных остатков). Намечаемое, таким образом, противоречие между малой способностью тироглифоидей к активному (самостоятельному) расселению и биологической необходимостью в большом рассе-лительном потенциале разрешается чрезвычайно широким использованием ими различных пассивных способов расселения.
Расселение при помощи ветра (анемохория) занимает среди этих способов видное место. Его биологическое значение, с очевидностью, явствует из наличия у очень многих Tyroglyphoidea специальных приспособлений, ведущих к увеличений) удельной поверхности тела и заключающихся в большой (иногда огромной) длине щетинок идиосомы, их перистости или других модификациях (листовидной или опахаловидной форме и т. п.), способствующих повышению его парусности. Этот способ расселения используется, повидимому, всеми стадиями развития тироглифоидей, включая личинок (у большинства которых сильно развиты «хвостовые» щетинки, выполняющие здесь роль парусных pseudocerci, столь универсально распространенных у первой личиночной стадии наземных насекомых) и расселительных гипопусов; у последних приспособлением к переносу ветром могут считаться длинные парусные щетинки задней пары ног, имеющиеся у большинства видов. Возможность расселения ветром покоящихся гипопусов рода Glyeyphagus, остающихся заключенными в высокопарусиом нимфаль-ном экзувии, также вполне реальна (Megnin, 1880).
Еще большим распространением, чем анемохория, пользуются среди тироглифоидей различные способы расселения при помощи других животных, т. е. явления зоохории; первенствующую роль играет здесь эпизои-ческий способ, или форезия, заключающийся в прикреплении клещей к поверхности тела животных-переносчиков и последующих странствованиях на нем до попадания «пассажира» в благоприятную для дальнейшего развития и размножения экологическую обстановку. У тироглифоидей форезия осуществляется преимущественно расселительными гипопусами — особой стадией развития, специально приспособленной к выполнению этой функции. Она богато вооружена разнообразными органами фиксации, построена обычно по так называемому «типу сопротивления» («Trutztypus»), обеспечивающему ее сохранность от механических повреждений, и обладает повышенной, по сравнению с другими стадиями, экологической валентностью (что обеспечивает ее переживание при неизбежных сменах обстановки в проезжаемом ею пути). В качестве переносчиков гипопусами используются либо мелкие млекопитающие (грызуны и насекомоядные — всеми имеющими гипопусов Labidophorinae), либо насекомые и другие пазом шло членистоногие (всеми остальными группами тироглифоидей). При этом .некоторые виды пользуются лишь каким-либо одним определенным видом (или группой близких видов) переносчика, с которым они находятся в болоо тесных биологических отношениях (например в качество комменсалов),
46
ВВЕДЕНИЕ
тогда как другие прикрепляются безразлично ко всем движущимся объектам, с которыми приходят в соприкосновение. Ясное представление о наблюдаемых здесь отношениях может дать следующая таблица,1 в которой све-. дены зарегистрированные до сего времени случаи форезии расселительных гипопусов Tyroglyphoidea.
Вид клеща
Сем. Туго glyph! daе
Tyroglyphus farinae (L.)
Kuzinia laevis (Duj.
Forcellinia wasmanni (Mon.)
» gigantonympha (Vtz.) Acotyledon paradoxa Ouds.
» krameri (Berl.)
Caloglyphus rodionovi A. Z.
» moniezi A. Z.
» polyphyllae A. Z.
» spinitarsus Herm.
» geotruporum A.. Z.
» berlesei (Mieh.)
» mycophagus (Megn.)
Rhizoglyphus echinopus (F. et R.)
Histiogaster ocellatus (Vtz.)
» filippovi A. Z. Monieziella corticalis (Mich.)
» javensis (Ouds.) Lackerbaiieria cribratissima A. Z. Troupeauia nova (Ouds.)
» crabronicola A. Z.
» novisimilis (Vtz.)
Переносчики
Мучной хрущак {Tensbrio molitor L.)
Различные мелкие стафилины и другие сапроб-
ные жуки
Бронзовка Osmoderma eremita Scop.
Шмель Bombus terrestris (L.)
Блохи: Pulex irritans L., Hystrichopsylla talpae Curt., Ctenopsylla segnis Schonnch., Cte-nophthalmus breviatus W. et J., C. pol-lexXI. eiJ.,Ceratophyllus tesquorum W.
Суслик Citellus sp.
Полевка Microtus sp. :
Шмели (все виды рода Bombus s. 1.)
Пчелы: Xylocopa pavlovskyi Popov
Osmia przewalskii F. Mor.
Муравьи: Camponotus ligniperdus Latr., For- t mica sanguinea Latr., Lasius tuligi- | nosus Latr. (
Муравей Camponotus sp. i
Летучая мышь
Муравьи ,j
Различные клещп из сем. Parasitidae j
Мучной хрущак (Tenebrio molitor L.) ’
Амбарная огневка {Ephestia kuhniella Zell.) 1
(гусеницы)
Майские жуки (Melolontha sp. sp.)
Мраморный хрущ {Polyphylla fullo L.) Бронзовка Osmoderma eremita Scop.
Жужелица Carabus nemoralis 0. F. Miill.
Навозник Trypocopris vernalis L.
Мраморный хрущ {Polyphylla fullo L.)
Жук носорог {Oryctes nasicomis L.)
Различные многоножки, мокрицы и мелкпо жуки
? Сверчок Liogryllus eampestris (L.)
Ближе не определенный жук сем. Rhizopha-gidae
Мухи: Merodon equestris F., Eumerus sp., Sea-topse sp.
Короед Xyleborus pfeili Ratz.
Короед Ips duplicatus Sahib.
Навозник Trypocopris vernalis L.
Жук сем. Brenthidae Trachelizus bisulcatus Fl
Oca Crdbro fossorius L.
Слоник Hylobius abietis L.
Oca Crabro solenius L.
Осы: Crabro dives Lep., C. spinieollis H. S.
Жуки сем. Brenthidae: Baryrhynchus latirostris- | Gyll., Prophthalmus bourgeoisi Power. Hypomiolispa exarata Desbr., Caenorhynchodes serrirostris F., Rhy-phidorhynchus longimanus F.
1 В данном списке приведены как установленные автором, так и указанные ранее в литературе факты.
экология
41
Rodionovia inopinata (Vtz.) Schulzea pamirensis A. Z.
» caucasica A. Z. Frorwpia vimariensis Vtz. «Tyroglyphus» vethi Ouds.
» submarinus Vtz.
» maegillavryi Ouds.
» mayaorum Vtz.
Слоник Cossonus linearis L.
Пчела Megachile grombzevskii F. Mor.
Пчела Prosopis sp.
Муравей Lasius fuliginosus Latr.
Навозники: Phanaeus sp., Heliocopris sp.
Морская змея Platurus colubrinus L.
Ближе не определенный таракан
Жуки сем. Brenthidae: Trachelizus bisulcatus F., Caenorhynchodes serrirostris F., Hormocerus. reticulatus F.
Жук Brenthus deplanatus Gyll.
Сем. Saprcglyphidae
Ensliniella parasitica Vtz.
» kostylevi A. Z. Calvolia heteroeoma (Mich.) » elisdbethae A. Z.
» reticulata A. Z.
» thraea Vtz.
» ealliphorae^Vtz.
» bulgarica Storkan
» striata Vtz.
» fusijormis A. Z.
» elliptica A. Z.
» nataliae A. Z.
» circumspectans Vtz.
Vidia lineata Ouds.
» undulata Ouds.
» squamata Ouds.
» gussdkovslcii A. Z.
» popovi A. Z,
Oca Odynerus (Lionotus) florieola Sauss.
» » » rossii Lep.
Белка (Sciurus vulgaris L.)
Слепушонка (Ellobius talpinus Pall.)
Прометеева мышь (Prometheomys shaposhnikovi:
Sat.)
Пчела Osmia dalmatina F. Mor.
Ближе не определенная «мелкая металлически-, зеленая мушка» *
Синяя мясная муха Calliphora vomitoria L.
Муравьи
Короеды: Taphrorhychus sp., Piiyogenes lepi~-dus Wichm.
Blastophagus minor Ratz.
Hylesinus fraaini Pnz.
Ips acuminatus Eichh. ?
Ips stebbingi Strohm.
Пчелы: Megachile pilicerusTAor.,Megachile sp. sp._ div.
Пчела Prosopis conformis Forst.
Крыса Rattus 'jerdoni Bly th.
Осы: Crabro fossorius L., C. impressus Sm.,_ C. vagus F., C. solenius L.
Осы: Crabro clypeatus L., C. alatus Pnz., C._
wollmanni Kohl., C. impressus Sm.
Сем. Glycyphagidae
Подсем. Labidophorinae
Labidophorus seiurinus (Koch) » desmanae A. Z.
» mogerae A. Z.
» talpae Kram.
» soricis Ouds. •
Xenoryctes krameri (Mich.) » heptneri A. Z.
Myacarus hypudaei (Koch)
arvicolae Duj.
Белка (Sciurus vulgaris L.)
Выхухоль (Desmana moschata L.)
Дальневосточный крот (Mogera robusta Nehr.),
Крот (Talpa europaea L.)
Соня Museardinus avellanarius L.
Землеройки: Sorex araneus L., Crocidura sp.,.
Neomys fodiens L.
Крот (Talpa europaea L.)
Водяная крыса (Arvieola amphibius L.)
Полевки: Microtus oeconomus Pall., Evotomys-glareolus Schr.
Мышь-малютка (Micromys minutus L.)
Крот (Talpa europaea L.)
Крот (Talpa europaea L.)
Полевки: Microtus arvalis L., Evotomys glu* reolus Schr.
Крот (Talpa europaea L.)
Землеройка (Crocidura lasiura Thos.)
Полевки: Microtus subterrancus Solyn., Af, «rid» lis L., Evotomys glareohu L.
48
ВВЕДЕНИЕ
Водяная крыса (Arvicola amphMus L.) Хомяк (Criceius cricetus fuscidorsis Arg.) Хомячок (Cricetulus migratorius Pall.) Степная пеструшка (Lagurus lagurus Pall.) Бурундук (Eutamias asiaticus orientalis Bonn.) Крот (Talpa europaea L.) Землеройка (Sorex araneus L.)
Подсей. Chae todac tylinae
.Sennertia cerambycina (Scop.)
» dissimilis A. Z.
» cantabrica A. Z.
» gargantua A. Z.
» alfkeni (Ouds.)
.» horrida (Vtz.)
» zhelochovtsevi A.. Z.
» potanini A. Z.
.» oudemansi A; Z.
» bifilis (Can.)
» simplex (Tragh.)
» morstatti Vtz.
» perturbans Vtz.
» caffra Vtz.
» hipposideros (Ouds.)
» koptorthosomae (Ouas.) tChaetodacbylus osmiae (DuL)
» rdaumuri (Ouds.)
» birulai A. Z.
» claviger Ouds.
» megachilarum k. Z.
» ludwigi (Trouess.)
» anthidii (Ouds.)
Morstia ornata (Ouds.)
» helenae (Ouds.)
» irifilis (Can.)
» pulcherrima (Vtz.)
» glabra (Vtz.)
Nebhorslia mamillata A. Z.
Tortonia intermedia (Ouds.)
» fluclmta Ouds.
» smitsvanburgsli Ouds.
» dogaressa A. Z. Eennertionyx mdnicati (Giard)
Xlerophagopsis skorikovi A. Z. Eerophagus granulatus (Duj.)
Пчелы: Xylocopa violacea L., X. valga Gerst., X. varenlzovi Mor., X. nitidiventris Sm., X. przewalskii F. Mor., Osmia princeps F. Mor., 0. maeroglossa Gerst., Megachile argentata F., Anthidium ehristofi F. Mor.
Oca Polochnm repandum Spin.
Пчела Xylocopa dissimilis Lep.
» » cantabrica Lep.
» » valga Gerst., punetilabris
F. Mor.
» » kalinovskii Rad., circumvolans
Sm.
» » dissimilis Lep.
» » olivieri Lep.
» » pictifrons Sm.
» » dissimilis Lep.
» » combinata Rits.
» Koptorfhosoma rngrita F.
» » » »
» » » »
» » oaf fra L.
» Platynopoda tenuiscapa Westw.
» » » »
» Osmia rufa L., 0. tricomis F., 0. kotili Ducke, 0. dimidiata F. Mor.
» » panzeri F. Mor., 0. leucogastra
F. Mor., 0. emarginala Lep.
» Chelostoma florisomnis L.
» Osmia tricomis F.
» Majachile bombycina Rad., M. ligniseca Kg.
Oca Pseudovespa germanica L.
Пчела Lifhurgus dentipes Sm.
» Anthidium stricticum F.
» Xylocopa circumvolans Sm.
» » dissimilis Lep.
» Platynopoda tenuiscapa Westw.
» Xylocopa combinata Rits.
» » ordinaria Sm.
» K.optorthosoma nigrita F.
Пчелы: Anthidium pubescens F. Mor., A. laticeps F. Mor.
Пчелы: Osmia leeana Kirby, 0. solskyiF. Mor., 0, nigriventris (Zett.) Nyl., Stelis phae-optera Latr.
Шмель liomlms proteus Gerst.
Пчела Anthidium stietieum F.
» » 7-dentatum F.
Пчелы: Anthidium manicatum L., A. florenti-num F. ,A. cingulatum Latr., A. diadema F., A. oblongatum Latr.
» Megachile dbderleini Fr.
Шмели: Bornbus lapidarius L., B. terrestris L., B. argillaceus Latr.
экология
48
Довольно широким распространением среди тироглифоидей пользуется также зоохорное расселение половозрелых особей клещей, т. е. имагипаль-иая форезия. Известные случаи такого рода сведены в следующую таблицу:
Вид клеща
Сем. Tyroglyphidae
Tyroglyphus farinae (L.)
Переносчики
Tyrophagus «puireseentiae» Sohr. » noxius A. Z.
Tyrolichus casei (Ouds.)
Povelsenia neotropica (Ouds.)
Ebertia australis (Ouds.) Caloglyphus polyphyllae A. Z.
Olafsenia trifolium (Ouds.)
Сем. Saproglyphidae
Nanacarus minutus (Ouds.) .
Кошка (Felis domestica L.) Водяная крыса (Arvicola amphibius L.) Мышь (Mus museulus L.) Крыса (Epimys norvegicus Berk.) Скворец (Sturnus vulgaris L.) Голубь (Columba livia L.) Воробей (Passer domesticus L.) Летучая мышь Myotis dasycneme Boie Крыса (Epimys norvegicus Berk.) Мышь (Mus musculus L.) Воробей (Passer domesticus L.) Мышь (Mus musculus L.) Навозник Phanaeus sp.
» » »
Мраморный хрущ (Polyphylla fullo L.) Навозник Heliocopris bucephalus F.
Нетопырь (Eptesicus serotinus Schreb.) Землеройка (Sorex vulgaris L.).
Пчела Platynopoda tenuiscapa Westw.
Сем. Glycyphagidae
•Glyeyphagus destructor (Schrk.)
» domesticus Deg.
Aeroglyphus peregrinans (Berl.)
Nycteriglyphus pterophorus (Berl.)
Labidophorus sciurinus (Koch)
» desmanae A. Z.
Xenocastor fedjushini A. Z.
Крыса (Epimys norvegicus Berk.)
Мышь (Mus musculus L.)
Воробей (Passer domesticus L.)
Дупель (Capella solitaria Gr.)
Голубь (Columba livia L.)
Скворец (Sturnus vulgaris L.)
Дрозд (Turdus musicus L.)
Пчелы: Xylocopa violacea L., X. valga Gerst.
Шмель Bombus lapidarius L.
Летучая мышь Nyctalus noctula L.
Белка (Sciurus vulgaris L.)
Выхухоль (Besmana moschata L.)
Бобр (Castor fiber 'vistulanus Matsch.)
У последних 2 из перечисленных видов их симфористические отношения к хозяину стоят на грани к постоянному обитанию на нем, т. е. эпизоиче-скому образу жизни, являющемуся факультативным у видов рода Labi-' dophorus, а может быть, нормальным (облигатным) у Xenocastor.
Возможность расселения тироглифоидей эндозоическим способом была установлена лишь в самое последнее время (Буланова, 1936); оказалось, что яйца всех испытанных в этом отношении амбарных видов (Tyroglyphus farinae L., Tyrophagus noxius A. Z., Caloglyphus rodionovi A. Z.), xii.pnif.TO-ризующиеся вообще особой жизнестойкостью, а в некоторой море и покоящиеся гипопусы Glyeyphagus destructor gjphrk., могут проходить ЛМОО'ГВ
4 Фауна СССР, Захватили—284
50
ВВЕДЕНИЕ
с пищей через кишечный тракт мышей и выделяться с их экскрементами^ не теряя при этом своей жизнеспособности; что же касается других стадий,, то они неизменно погибали и подвергались перевариванию.
Огромную и многостороннюю роль в рассолении тироглифоидей играет-человек. Вследствие обитания в основных продуктах питания (зерно, мука), фураже, семенном материале и т. д., они касаются многих сторон хозяйственной деятельности человека и широко распространяются им вместе с зараженными продуктами, а также и непосредственно через тару, сельскохозяйственный и транспортный инвентарь, домашних животных и т. д.
Удельный вес каждого из этих способов расселения в общем рассели-тельном потенциале различных видов весьма изменчив. У очень многих видов главным или даже единственным способом является «гипопальная» форезия, и они в самой слабой степени пользуются другими способами, соответственно теряющими свое значение или даже полностью выпадающими; это имеет место у практически всех нидикольных видов (все Chaetodactylinae, Ldbidophorinae, многие Saproglyphidae и некоторые Tyrogly-phidae: Kuzinia, Forcellinia wasmanni и др.), у биологически связанных с пластинчатоусымп жуками видов рода Caloglyphus и многих, особенно-толстых и малоподвижных видов, вроде Rhizoglyphus и др. У других видов основным способом расселения служит «имагинальнЗя» форезия (роды Nycteriglyphus, Aeroglyphus), у третьих — анемохория (род Ctenoglyphus) или ее комбинация с самостоятельным передвижением (Cometacarus и, невидимому, роды Glycyphagus и Tyrophagusy некоторые виды используют, по видимому, все перечисленные способы (например Tyroglyphus farinaeR наконец, существует ряд синантропных форм, расселенпе которых, невидимому, почти целиком осуществляется человеком (Thyreophagus, Gohieria fusca, виды родов Aleuroglyphus, Blomia и Chortoglyphus').
Размножение. Имеющиеся в литературе данные о плодовитости тироглифоидей очень немногочисленны и касаются исключительно амбарных видов. Согласно этим данным, общее число яиц, откладываемых одной самкой,, составляет у Tyroglyphus farinae L. 20—30 шт., у Tyrolichus casei Ouds. И Rhizoglyphus echinopus Fum. et Rob. — около 100, у Caloglyphus rodionovi A. Z. — до 621, у Gohieria fusca Ouds. — 11—24, a у Glycyphagus destructor Schrk. всего только 8—10 шт.; таким образом, различия в объеме яйцепродукции, наблюдаемые между отдельными представителями этой группы, очень велики. В очень сильной степени вариирует яйцепродук-ция и у одного и того же вида, в зависимости от условий его обитания (например с 3 до 621 япца у Caloglyphus rodionovi).
Продолжительность индивидуального развития также крайне изменчива и в сильнейшей степени зависит от внешних условий, главным некоторых здесь является температура. Так, при оптимальной влажности, весь цикл развития Caloglyphus rodionovi (от яйца до половозрелой стадии) занимает, в зависимости от температуры, от 5 до 28 дней (а именно: при-16.5—18°—28 дней; 24° — 11 ДПОЙ; 24.7°—10 дней; 25.5° — 9 дней;..
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ ПГ
29,5° — 6 дней; 30,8° — 5 дней); у Rhizoglyphus echinopus — от 0 до 27 дней и т. д.
В соответствии с большой изменчивостью его слагаемых, очень разнообразен и общий потенциал размножения различных видов Tyroglyphoidea. Прилагаемая таблица с цифрами интенсивности размножения нескольких из амбарных видов Tyroglyphidae, составленная но работе О. К. Настюко-вой и показывающая среднее число особей к одной исходной через 1 месяц после закладки культур, дает некоторое, хотя и неполное, представление о межвидовых различиях этого потенциала, а также о масштабе его изменчивости у одного и того же вида, в зависимости от внешних условий.
АтМ|.СФераая влажность
Температура в С° 80О/о 00%
Tyroglyphus farinas L. Aleuroglyphus ovatus Тг. Tyrophagus no-anus A. Z. Tyroglyphus fa-rinae L. Aleuroglyphus ovatus Tr. Tyrophagus noxius A. Z.
7— 8 1.28 7.69
10-11.5 — 0 0.64—1.0 — 0.23 2.16
13 35.25 — —— 35.89 — 1
15—15.5 — 0.14—0.31 5.7 — 0.14— 0.31 16.6
22—22.5 43.58 0.77—1.4 17.5 173.07 1.93— 4.11 60.7
25—25.5 58.33 2.31—2.83 40.0 57.69 4.8 — 8.5 243.0'
30 2.58 6.37 39.8 — 22.87—25.4 190.0
35 0.0 7.57—7.64 0.0 0.0 35.04—52.08 9.9
Главную роль в определении потенциала переживания тироглифоидей играют, помимо уже рассмотренных физических факторов среды, многочисленные хищники, из них особую i-ажность имеют хищные клещи сем. Cheyletidae (в первую очередь виды Cheyletus, затем роды Acaropsis, Chele-tomorpha, Cheletophyes и др.), являющиеся специализированными потребителями тироглифоидей и сопутствующие им в большинстве занимаемых ими местообитаний; энергично истребляют их и другие хищные клещи (представители семейств Laelaptidae, Parasitidae и др.), а иногда и хищные личинки некоторых галлиц (например Arthrocnodax sp.).
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
Большое хозяйственное значение группы Tyroglyphoidea detriticolae определяется, прежде всего, тем, что к ней относятся все амбарные виды клещей, вредящие запасам сельскохозяйственных продуктов. Нет, практически, ни одного вида таких продуктов, который не подвергался бы в той или иной мере нападению этих клещей, но наибольший экономический эффект приобретает их деятельность как массовых вредителей зерна и продуктов его переработки, маслосемян, луковиц огородных и декоративных культур и клубнеплодов.
При нормальной влажности зерна (до 17%) клещами поирождиотой преимущественно, если не исключительно, зародыш, особая lipillUlviuvruJIli’
*
S2
ВВЕДЕПИЕ
ность которого для клещей единодушно отмечается всеми авторами; клещи проникают в него, главным образом, через трещинки и разрывы оболочки зародышевого конца зерна и выедают в нем полости — камеры питания, в которых уже происходит их дальнейшее размножение и развитие. С увеличением численности поселившейся в зерне колонии клещей (число их в одном зерне достигает иногда 200 особей) быстро увеличивается и объем выгрызаемой ими в зародыше полости, пока он пе оказывается съеденным целиком; после этого клещи обычно эмигрируют в соседние зерна или же переходят к питанию эндоспермом. Повреждения последнего наблюдаются, главным образом, при повышенной влажности зерна, вызывающей его размягчение, набухание и появление разрывов оболочки, или же при наличии механических повреждений, облегчающих доступ к нему клещей; в таких случаях нередко имеет место полное уничтожение содержимого зерна, от которого остается тогда только продырявленная оболочка, набитая клещами, их трупами, линочными шкурками и экскрементами. Объем причиняемого таким образом вреда определяется многими причинами: плотностью •клещевого населения, продолжительностью их разрушительной деятельности, состоянием зерна и условиями его хранения. По имеющимся данным, в производственных (складских) условиях число поврежденных клещами верей, даже при сравнительно небольшой их концентрации (I—II степени заражения1), достигает после нескольких месяцев хранения 30—32%; при III степени заражения зерна и большой длительности пребывания •в нем клещей, в отдельных, наиболее пораженных, гнездах все 100% зерен •иногда оказываются в той или иной мере поврежденными ими.
Не меньшее значение, чем непосредственное, повреждение зерна (выедание), имеет, вероятно, косвенный вред, наносимый зерну клещами в результате их обитания в нем. Эта сторона вредной деятельности, объем которой трудно поддается учету, заключается в засорении зерна, повышении его влажности, стимулировании самосогревания и заражении бактериозами и грибными заболеваниями. Такой эффект жизнедеятельности клещей обусловливается, прежде всего, заполнением межзерновых пространств массой клещей, их шкурок и экскрементов (вызывающим склеивание зерна в комки и ухудшающим аэрацию во всей его толще), а также и тем, что они являются переносчиками различных грибных и бактериальных болезней зерна, плесеней и т. д. и способствуют их распространению по всей его толще; кроме того, считается, что экскремецты клещей служат питательным субстратом для различных бактерий и что их скопления являются, таким образом, очагами заражения ими зерна бактериозами; наконец, ость указания, что повреждения клощой активизируют жизненные про-
1 I степенью заражения зерна клещами считается наличие их в числе, не превышающем 20 экз., в отсеве из килограммовой пробы зерна; 11 отененью — если в отсеве их больше 20, но они не образуют сплошной движущейся массы; 111 степенью—присутствие клещейвтаком количестве, что в отсеве они нс разбегаются, а образуют сплошные, движущиеся комки («войлочные массы»).
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
цессы в зерне (его дыхание, ферментативные процессы и т.д.) и тем самым усиливают (или даже вызывают) его самоувлажиение и согревание.
Результаты разрушительной деятельности клещей в зерне весьма многообразны. Прежде всего, она сказывается в постепенном снижении его всхожести, очевидно соответствующем степени и объему повреждения ими зародыша. При большой концентрации клещей (III степень зараженности) падение всхожести зерна в зараженной партии идет очень быстро; так, по Соколову, сильно зараженная клещами пшеница снизила за 30 дней (в лабораторных условиях) свою всхожесть па 64%; рожь за 67 дней — на 51% ит. д.; темп ее снижения также сильпо зависит от исходной влажности зерна; по Родионову, за 60 дней зерно (пшеница) 17% влажности потеряло 13—15% всхожести, тогда как при 14.8% влажности — лишь 6—8%. Несомненно, большую роль играет здесь и температура, определяющая интенсивность размножения и жизнедеятельности, а следовательно, и вредной деятельности клещей. Помимо снижения всхожести, зараженное клещами зерно, в известной мере, снижает свой абсолютный вес (в опытах Соколова, например, пшеница, зараженная во II сильной степени, через месяц потеряла, в среднем, 5% своего веса) и испытывает ряд изменений в своем химическом составе; так, пшеница, зараженная клещами во II сильной степени, через месяц снизила содержание жира с 1.71—1.84 до 1.3— 1.52%, белка с 11.87—16.42 до 10—10.62% и крахмала с 60.14 до 56.4%; одновременно с этим она увеличила свою влажность на 2% (с 11 до 13%).. Мука из зараженного зерна обладает повышенной влажностью, гигроскопичностью и кислотностью, при пониженном содержании клейковины; она имеет затхлый или даже гнилостный запах и горький привкус; заметно, ухудшаются и ее хлебопекарные свойства (Соколов и др.). Наиболее серьезными из вредителей зерна являются мучной (Tyroglyphus farinae L.) и удлиненные клещи (Tyrophagus noxius A. Z. и Т. perniciosus A. Z.). Характер вреда, причиняемого клещами муке и другим продуктам переработки зерна, также многообразен; помимо поедания самой муки, они вызывают ее само-увлажнение, засорение и заражение микроорганизмами, быстро приводя ее к порче и снижая ее пищевые и хлебопекарные свойства; при длительном обитании клещей в муке она слеживается, приобретает темную окраску, специфический, неприятный запах и становится непригодной it употреблению; мука грубого помола нередко выедается клещами вся, кроме остающихся нетронутыми отрубей. Муке особенно вредят Aleuroglyphus ovatus Tr., Tyreophagus, Gohieria и мучной клещ. Повреждения маслосемян имеют, в общем, такой же характер, как повреждения зерна, вариируя в зависимости от культуры и состояния семян; их производят те же виды, которые вредят зерну, плюс Tyrolichus casei Ouds., Caloglyphus rodionovi A. Z. и др.
Очень большие убытки наносят клещи этой группы и многим другим пищевым продуктам — сухим фруктам и овощам, сыру и т. д.; так, но ПОД' счетам американских авторов, сыроваренная промышленность США теряет ежегодно до 3 млн. долл, от повреждения сыра клещами.
ВВЕДЕНИЕ
Чрезвычайно серьезное экономическое значение имеют тироглифоидеи и как вредители овощного и цветоводческого хозяйств. Первую роль здесь играет луковичный клещ (Rhizoglyphus echinopus F. et R.), нападающий на луковицы всевозможных декоративных и ряда огородных культур/ клубни картофеля, свеклу и другие корнеплоды как в грунте, так и, особенно, при их храпении. Повреждения, наносимые этим видом луковицам декоративных растений (гиацинтов, тюльпанов, нарциссов, лилий, ирисов и т. д.), заключаются, прежде всего, в том, что он прогрызает между их чешуями ходы, идущие глубоко внутрь луковицы, затрагивая при этом и зачатки цветоноса и листьев, которые нередко выедаются начисто; при сильной степени заражения луковиц полости, выгрызаемые клещами в их толще, достигают значительных размеров; при вскрытии луковиц они оказываются заполненными буроватой или черной трухой, состоящей из экскрементов клещей, их линочпых шкурок, своего рода буровой муки и т. п., а также самими клещами во всех стадиях развития. Механическое повреждение тканей, производимое клещами, влечет за собой отмирание отдельных участков луковицы как непосредственно в результате самого повреждения, так и вследствие заселения пораненных поверхностей болезнетворными и гнилостными грибками и бактериями (споры которых в широкой мере разносятся клещами при их переходах из одной пораженной луковицы в другую). Поврежденная луковица вскоре начинает размягчаться и гнить, заселяясь богатой сапрофитной флорой, а также нематодами, личинками различных двукрылых и т. д. При хранении луковиц в условиях, благоприятствующих размножению клещей (при достаточно высокой температуре), зараженные ими объекты успевают совершенно разрушиться в течение периода хранения; если же развертывание их вредной деятельности тормозится хранением при низкой температуре и происходит позднее, уже в период выгонки луковиц или высадки их в грунт, то различные болезненные симптомы сказываются на вырастающем растении У «Lili нт Harris!», например, в этих случаях наблюдается болезненное состояние, известное под названием «basal rot» (Ogilvie, 1929); Scalia наблюдал у зараженных экземпляров Lilium tigrinum пятнистость, а затем и полное отсыхание листьев и всего растения, несмотря на обильную поливку; еще чаще пораженное растение долго не выказывает признаков болезни, но затем внезапно гибнет.
Степень вреда, наносимого Rhizoglyphus echinopus луковицам различных декоративных культур, неодинакова. Особенно сильно страдают от него нарциссы и различные виды лилий; что определяется относительной рыхлостью луковиц этих растений, т. о. наличием значительных промежутков между их чешуями, облегчающих клещам доступ внутрь луковиц. Тюльпаны же и, особенно, гиацинты, луковичные чешуи которых плотно прилегают друг к другу, менее подвержены нападению этого вредителя.
Представление об объеме вредной деятельности R. echinopus на декоративных луковичных культурах могут дать следующие примеры. Так, по
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
55
Woods’y, от 20 до 60% всего урожая луковиц, получаемых с обширных плантаций Lilium longiflorum var. eximium (более известной садоводам под названием Lilium Harrisi) на Бермудских островах, гибнет (при выгонке) в результате деятельности этого клеща. G-arman (1823), осмотревший около 1 млн. луковиц, ввезенных в США, указывает, что средняя зараженность их Rhizoglyphus’от равнялась 16—20%. Hodson, проанализировавший 500 образцов луковиц нарцисса (содержавших по 6—20 луковиц каждый) из Голландии и различных местностей Великобритании, нашел, что 85% из них были в большей или меньшей степени заражены луковичным клещом. В литературе нередки указания и на случаи 100% гибели целых больших партий луковиц в результате их повреждения Rhizoglyphus echinopus.
Вред, наносимый этим же видом луку, также очень велик и имеет сходный характер. По Попову (1929), проникновение клещей впутрь луковиц идет здесь со стороны неопробковевшего донца, так как они оказываются не в состоянии преодолеть их пергаментовидпую оболочку. На первых этапах повреждения донце измочаливается, теряет связь с самой луковицей и затем легко отваливается, обнажая и внутренние части; клещи тогда проникают в промежутки между чешуями; последние разъединяются, буреют и начинают гнить, и через 1%—2 месяца луковица оказывается совершенно разрушенной. Особенно быстро гибнет репчатый лук, имеющий более мясистые чешуи и содержащий больше воды. Насколько велик масштаб вреда, наносимого этим клещом луку, видно из данных того же автора, согласно которым в Татарской АССР в 1925—1926 гг. за зиму погибло до 40—50% всего запаса луковиц, хранившегося во многих хозяйствах.
По своему характеру, повреждения, причиняемые луковичным клещом клубням картофеля и георгинов, довольно сходны с теми, которые он наносит луковицам. По АрреГю и BOrner’y (1905) и Zacher’y (1927), они обычно имеют вид неправильных ходов, идущих вглубь клубня от пораженных участков поверхности и заполненных рыхлой коричневатой мукой, кишащей клещами всех стадий. Картофель подвергается нападению как в поле, так и при его хранении на складах. Особенное предпочтение клещи оказывают гниющим клубням, но проедают ходы и в совершенно здоровых тканях; цо данным тех же авторов, сильнее повреждаются следующие сорта картофеля: Richters Imperator, желтый столовый картофель, Irene и Sophie.
Необходимо отметить, что некоторыми авторами (например Maurizio, 1906) высказывались сомнения относительно первичного характера заражения Rhizoglyphus’от картофеля, т. е. того, нападает ли он на вполне здоровые клубни; этп сомнения, вероятно, основывались на том факте, что заражение ими картофеля часто комбинируется с его заражением картофельной нематодой Heterodera rostochiensis Woll. (Zimmermann, 1920) и другими вредителями. Последующие данные Zacher’a, однако, решили этот вопрос в положительном смысле, с тем только уточнением, что заражению подвергаются предпочтительно уже механически поврежденные (пораненные) клубни.
56
ВВЕДЕНИЕ
Этот же вид, вместе с некоторыми другими (Heteroglyphus vitium Foa, «Rhizoglyphus» minor Dem. и др.), наносит ощутимый ущерб культуре винограда, повреждая его корни. По данным Mangin’a и Viala (1900, 1902), Silvestri (1902), Petri (1928) и других авторов, нападению этого клеща подвергаются в первую очередь ослабленные п больные кусты винограда, но затем он переходит и на вполне здоровые. Обычно он встречается здесь вместе с филокссрон, локализуясь преимущественно на вздутиях и разращенияхкорней (образовавшихся в результате ее деятельности), но встречается также на мелких нелепых корешках и выгрызает в них ходы, постепенно распространяющиеся внутрь здоровой ткани и вызывающие ее омертвение. Эти повреждения сильно отражаются па всем развитии куста. На первых этапах заражения нарушается гармоничность его роста; ствол развивается неправильно; листья остаются мелкими; общая ассимилирующая поверхность куста сокращается; кисти уже в первый год заражения не дозревают, а в дальнейшем все больше и больше мельчают. При прогрессирующем заражении ходы клещей все дальше углубляются в толщу корней, й в случаях, когда они достигают их сердцевины, куст отмирает; обычно-это происходит на 3—5-й год его заражения клещом.
Поражения, наносимые Rnizoglyphus’от корням винограда, в некоторых случаях способствуют также его повреждению другими вредителями; так, Petri (1929) считает, что они позволяют филоксере достичь внутренних слоев коры главного корня, чем усугубляется эффект наносимых ею повреждений. По Дементьеву (1913) Rhizoglyphus echinopus (ошибочно описываемый им под названием «В. самсаsicus») является также одним из возбудителей хлороза винограда на Кавказе.
Некоторую роль играют тироглифоидеи и как вредители полеводства: известно много случаев повреждения тем же видом Rhizoglyphus подземных частей культурных злаков (пшеница, овес: Reuter, 1901; Tullgren, 1916), и есть отдельные указания на случаи серьезного повреждения видами Tyrophagus (группы humerosus') сеянцев пшеницы (Schoevers, 1926).
Наконец, значительный вред причиняется представителями того же семейства (Forcellinia fungivora Ouds., F. (?) lintneri Osb. и видами рода Acotyledon) шампиньонам в парниковых хозяйствах. Следует еще отметить-отдельные случаи экономически ощутимого ущерба, наносимого Histiogaster bacchus A. Z. производству виноградного уксуса и Carpoglyphus; lactis L. некоторым сортам вин.
Другая сторона вредной деятельности тироглифоидей заключается в их патогенном значении для человека и домашних животных. Они могут вызвать, при проглатывании их вместе, с пищей, острые желудочно-кишечные заболевания, а при попадании в дыхательные пути — катарральные и астматические явления (Родионов, 1936). Кроме того, некоторые представители этой группы (например Rhizoglyphus echinopus F. et R.-) могут быть, причиной появления своеобразных дерматитов у лиц, постоянно приходящих в соприкосновение с продуктами, зараженными этими клещами (Па
МЕРЫ БОРЬБЫ
вловский и Штейн, 1930). Наконец, известны случаи эидопаразитизма 2 видов — Histiogaster spermaticus Trouess. и Carpoglyphus alienus Bnks. в мочеполовой системе человека.
МЕРЫ БОРЬБЫ
В настоящей главе изложены лишь общие принципы борьбы с тиро-глифоидными клещами применительно к наиболее важным из многочисленных продуктов, подвергающихся их нападению. Более детально эти вопросы освещены в ряде специальных работ и, особенно, в обстоятельной сводке П. К. Чернышева. 1
1. Зерно. Совокупность мероприятий но защите зерна от хлебных клещей (т. е. амбарных тироглифоидей) может быть расчленена на следующие 3 группы: 1) собственно профилактические мероприятия, направленные к предупреждению заражения зерна клещами; 2) гигиенические (оздоровительные) мероприятия, препятствующие размножению клещей в зерне при его хранении, и 3) истребительные, которые, в свою очередь, могут быть подразделены на мероприятия разового действия (например газовая дезинсекция) и более длительно действующие (пересыпание зерна сухими акарицидами) или же, как это чаще делается, на физико-механические и химические.
Вследствие широкого распространения хлебных клещей в полевых условиях (по крайней мере, в большинстве зерновых районов) и многообразия: используемых ими путей и способов расселения, защита зерна от заражения ими должна начинаться еще задолго до его поступления на хранение в элеваторы и склады. Она должна осуществляться во всех звеньях уборочного процесса и на всех этапах пути, проходимого зерном от косовицы до ссыпки на хранение, а также охватывать все объекты, приходящие в соприкосновение с ним. При уборке комбайнами, первым таким мероприятием должна быть тщательная очистка машины от,оседающих на различных ее частях пыли и растительных остатков. Особое значение имеет такая очистка перед началом уборки, так как даже незначительные скопления детрита (в бункерах, на сетках и т. д.), оставшегося здесь с прошлого уборочного сезона, могут служить местообитанием хлебных клещей и, следовательно, источником заражения ими зерна нового урожая. В полной мере то же относится и к транспортным средствам, обслуживающим комбайн — автотранспорту, бестаркам и пр., принимающим зерно из бункеров комбайна: следует периодически производить, тщательную их очистку, следить за тем, чтобы, например, в щелях кузова не скоплялись зерно пли частицы отходов.
1 Клещи, вредящие запасам сельскохозяйственных продуктов в ССОР. < )o.iii>itojixoiiriiu, 1938. При составлении настоящей главы автор широко пользовался копоультациоП II. It. ’lop-пышева и 3. С. Родионова, любезно предоставивших в его распоряжение ряд «шшх 01ЦПШ11>ну-бликованных данных и просмотревших ее в рукописи.
'38
ВВЕДЕНИЕ
При пользовании токами и полевыми перевалочными пунктами, большое внимание должно быть уделено поддержанию их в чистоте; сами площадки следует менять'достаточно часто (так как неизбежно скопляющиеся па них просыпь, отходы и т. д. более или менее быстро заселяются клещами) и, при выборе и устройстве новых, руководствоваться рядом условий, затрудняющих их заселение клещами из окружающей обстановки. Такие площадки должны быть сухими, хорошо очищенными от растительности и ее остатков (стерни, половы и пр.). В случае, необходимости в укрывании насыпей зерна соломой требуется собл юденпс некоторых предосторожностей, вытекающих из частой зараженности ее клещами, а именно — брать для этой цели по возможности свежую и сухую солому, избегая старую, долго лежавшую в поле. Само собой разумеется, что длительное пребывание насыпей зерна под открытым небом увеличивает шансы заражения клещами пропорционально времени его нахождения в поле. 1
Уборка простыми сельскохозяйственными машинами с последующими -обмолотом и очисткой зерна представляет, в силу своей расчлененности и большой длительности, больше уязвимых точек в смысле возможности заражения зерна клещами, чем комбайновая уборка. Заражение скошенного хлеба в валках, снопах и копнах, хоть и мало реально, в виду эфемерности существования этих объектов, по все же возможно при определенных условиях (повышенной влажности). Более возможно и чаще наблюдалось на практике заселение клещами скирд, точнее их нижних, припоч-вепных слоев. Опасность заражения их также возрастает в прямом соответствии с временем стояния скирды, а также зависит и от выбора и подготовки площадки, на которой опа построена: на новых, сухих, хорошо очищенных -площадках, достаточно полно изолирующих скирду, эта опасность значительно ниже, чем при сложении скирд на стерню или дерн и на более влажные (например пониженные) участки поля или, особенно, на старые, уже да: но эксплоатируемые тока. Определенное значение имеет здесь и посещаемость скирды мышами, являющимися переносчиками многих видов хлебных клещей.
При обмолоте и очистке зерна внимание должно быть обращено на предшествующее обеззараживание и содержание в чистоте молотилок, веялок и других машин, при плохом уходе часто оказывающихся сильно зараженными клещами. Необходимо также следить за правильностью регулировки барабанов молотилки при ее работе, не допуская увеличения боя верпа, так как от целости зерновых оболочек сильно зависит степень его поражаемости клещами при дальнейшем хранении. Борьба за чистоту токов, достаточно частая их смена и очистка также имеют крупнейшее профилактическое значение. Известно, что при долгой эксплоатации тока нередко становятся настоящими рассадниками клещей, сплошь заселяющих
1 При длительном лежании зерна в поле возможно также его заражение клещами, вносимыми посещающими зерно грызунами и насекомыми.
МЕРЫ БОРЬБЫ
-5»
устилающий их слой отходов и, на значительную глубину, перемешанную с ними почву; необходимо также следить за чистотой транспортного инвентаря, тары и т. д. Вообще говоря, сжатые сроки уборки и обработки урожая в сочетании с постоянной заботой о чистоте всех объектов, приходящих в соприкосновение с ним, могут свести до минимума опасность заражения зёрна клещами на его пути с поля до зернохранилища.
Важнейший комплекс профилактических мероприятий связан с подготовкой зернохранилищ к приему нового урожая. Особое значение, в смысле 'предупреждения его заражения клещами, имеет здесь тщательная очистка складских помещений (и их подполий, где таковые имеются) от мусора, сметок и пыли, их просушка и химическое обеззараживание, а также принятие мер к изоляции хранилищ от занесения в них клещей извне. Для обеззараживания зернохранилищ обычно применяется влажная дезинсекция, заключающаяся в опрыскивании помещения раствором каустической соды, минерально-масляными эмульсиями, известью и т. д : в некоторых случаях прибегают и к газовой дезинсекции (преимущественно хлорпикрином). Мероприятия по охране складских помещений от попадания в них клещей извне включают очистку прилегающей к складу территории от растительных остатков и т. п., защиту от грызунов и птиц, недопущение помещения в склады зараженной тары и других предметов, могущих содержать клешей. Эти же меры должны соблюдаться и в дальнейшем, уже при хранении зерна.
Общее направление профилактических мероприятий, имеющих целью .препятствовать размножению клещей в зерне при его храпении, определяется подчеркнутыми выше (стр. 36) особенностями биологии этих вредителей: их гигро- и термофильностью, ансмофобией. связью с развитием сапробной микрофлоры. Ясно, что положительный эффект может быть достигнут здесь всяким мероприятием, ведущим к поддержанию в запш-дцаемых продуктах низкой температуры и низкой влажности, достаточной вентиляции и малой активности микробиологических процессов. Такими мероприятиями является, по существу, вся система хозяйственно-оперативных работ складской сети, направленных к оздоровлению зерна и созданию оптимальных условий его хранения. Целью всех этих работ является: а) максимальное охлаждение зерна, б) доведение его до сухого состояния; в) устранение факторов, вызывающих самоувлажнение и самосогревание — засоренности, колебаний температуры, недостатка вентиляции, доступа сырого воздуха и т. д. Здесь, таким образом, имеет место полное совпадение задач борьбы с хлебными клещами с таковыми правильного храпения зерна: всякая операция, ведущая к оздоровлению зерна, — сушка, очистка, охлаждение, активная вентиляция — является вместе с тем основной и первоочередной мерой борьбы с клещами, снижающей возможности их массового, экономически ощутимого размножения в продукте. Рациопал ышл конструкция зернохранилищ, подчиненная тем же общим задачам, ивллетол также немаловажным звеном в цепи факторов, создающих требуемые уоло-
60
ВВЕДЕНИЕ
вия хранения зерна; тем самым, она играет определенную роль и в его защите от вредной деятельности хлебных клещей.
Практически все операции по оздоровлению зерна имеют и, в известной: мере, побочный, истребительный эффект, т. с. являются комплексными. Так, зерносушение (как солнечное, так и термическое) не только снижает" влажность зерна (тем самым затрудняя развитие в нем клещей), но и приводит к гибели значительного процента клещей в результате комбинированного воздействия высокой температуры и дегидратации; при перелопачивании, имеющем целью снижение температуры в толще зерна, клещи гибнут и от непосредственного механического воздействия; при очистке зерна на различного типа машинах, большая часть клещей отделяется от зерна потоком ветра от вентилятора., часть же механически раздавливается' движущимся зерном и т. д. Благодаря этому, указанные операции применяются с успехом и как прямые истребительные меры борьбы против: хлебных клещей. Они также легли в основу ряда специально истребительных мероприятий, представляющих собой ту или иную модификацию этих, по существу, гигиенических операций. Большое внимание привлек к себе, например, механический метод борьбы: были предложены различные типы-специально сконструированных для уничтожения клещей барабанов «клещебоек» и других машин, но положительный эффект их работы обычно снижается тем, что они сильно бьют зерно; между тем известно, что битое, вообще механически поврежденное зерно особенно охотно заражается клещами. Это обстоятельство необходимо учитывать и при подработках зерна на зернопультах, сортировках, транспортерах и пр., тщательной регулировкой машин не допуская увеличения боя зерна.
Был также разработан специальный режим термической сушки, дающий повышенный истребительный эффект; но его применение на практике ограничивается тем обстоятельством, что быстрое и резкое повышение температуры зерна (составляющее его основу) отрицательно отзывается на нем: понижается всхожесть, ухудшаются хлебопекарные свойства.
Из химических мер борьбы с хлебными клещами наибольшее применение имеет фумигация зерна и складских помещений хлорпикрином. Главны» преимуществом этого метода является быстрый и наглядный эффект, получаемый при пользовании им и обусловленный высокой токсичностью хлорпикрина для питающихся стадий клещей. Это преимущество хлорпикрина сильно снижается рядом присущих ему серьезных недостатков, а именно:: большой устойчивостью к этому фумиганту яиц клещей (100% умерщвление' которых достигается лишь при очень высоких концентрациях яда и длительных экспозициях, например, 126 г ла 1 м8 в течение 7 суток, не применимых в производственных условиях) и отчасти их покоящихся гипопусов, приводящей к быстрому восстановлению заражения продукта клещами после прекращения действия яда (иногда уже через 8—16 дней после газации зараженность вновь начинает возрастать); снижением всхожести зерна при газации (особенно резком при его повышенной влажности),
МЕРЫ БОРЬБЫ
трудностью дегазации, а также и опасностью работ § столь сильно действующим отравляющим веществом. Вследствие этого применение хлорпикрина может быть допущено лишь в особых случаях и никак не может претендовать на ту ведущую роль, которую ему иногда придают. Газирование хлорпикрином посевного зерна даже вообще воспрещено. Кроме хлорпикрина, для газирования зерна применяется иногда сероуглерод. Он быстро проникает в толщу насыпей, не снижает всхожести зерна и легко из него испаряется. Вместе с тем CS2 весьма огнеопасен, сильпо взрывчат, и при работах с ним было немало крупных аварий, вследствие чего применение его сильно ограничивается; в больших городах оно воспрещено вообще. В последнее время для борьбы с клещами были предложены еще некоторые другие отравляющие вещества, например сероводород и циан-плав (цианистый кальций), но перспективы их применения еще неясны.
Особую группу химических мер борьбы с клещами составляет применение порошковидных, сухих акарицидов, примешиваемых в различных пропорциях к зерну. По механизму своего действия часть этих веществ (вивианит, мел, суперфосфат, тальк) относится к контактным ядам, часть же (нафталин, парадихлорбензол и др.) — к фумигантам; общим свойством тех и других является длительность их действия, так как, уничтожая уже имевшихся в продукте клещей, они, вместе с тем, предохраняют его от нового заражения в течение более или менее продолжительного срока. Применение сухих акарицидов, однако, еще не вошло в широкую оперативную практику и сильно ограничивается недостаточной разработанностью этого, метода. Наибольшее распространение получил нафталин, но и он применяется пока только для дезинсекции посевного материала, так как продовольственным продуктам он сообщает резкий неприятный запах.
Следует еще упомянуть о принципиальной возможности использования я борьбе с амбарными клещами ультракоротких волн, но и этот метод еще не вышел из стадии опытной разработки. Неясны также и перспективы применения здесь биологического метода: наличие группы специализированных хищников, истребляющих тироглифоидей (и обладающих к тому же большей, чем эти последние, экологической валентностью), а именно хищных клещей сем. Cheyletidae, как будто говорит о возможности опытной работы и в этом направлении. Однако полученные результаты не дают повода для особого оптимизма, так как положительный эффект получается лишь при определенных численных соотношениях между хищными и амбарными клещами при оптимальных температурах и зависит также от ряда других, иногда трудно контролируемых условий.
Удельный вес каждого из перечисленных методов борьбы с клещами в общей системе мероприятий по защите зерна от этих вроднтолоП, .разумеется, неодинаков. Вряд ли подлежит сомнению, что во псяк'оП рациональной системе этих мероприятий ведущая роль должна при надлежать профилактическим, методам борьбы, т. е. мерам, направленным к созданию и длительному поддержанию в охраняемых объектах такой шеологичеокой
62
ВВЕДЕНИЕ
обстановки, при которой всякое экономически ощутимое размножение-вредителя в них было бы исключено. Это положение приобретает сугубое значение в отношении к клещам, вредящим запасам, в виду особо длительного пребывания объектов в сфере действия вредителей, характеризующихся необычно быстрым размножением и многочисленностью следующих друг за другом поколений; при этих обстоятельствах истребительные мероприятия разового действия, даже самые эффективные, соответственно оттесняются на второй план.
Благодаря полному совпадению задач борьбы с клещами с таковыми, правильного хранения и оздоровления .зерна молено быть уверенным, что-приведение всей складской сети в соответствие с требованиями, предъявляемыми ей рациональными методами хранения зерна, и неуклонное соблюдение всех правил его профилактики и гигиены — наиболее верный и хозяйственно-эффективный путь к победе над амбарными клещами.
2. Мука и другие продукты переработки зерна. Борьба с клещами в муке, манной крупе и других-продуктах переработки зерна представляет? большие трудности и, по существу, еще совершенно не разработана. В связи с этим рекомендуется только сокращение сроков хранения этих продуктов при особо тщательном соблюдении всех рассмотренных профилактических мероприятий.
3. Плодоовощи. Борьба с клещами на клубне- и корнеплодах также-строится, в основном, из профилактических и гигиенических мероприятий, И здесь применение многих из этих мероприятий должно сопутствовать, уборке урожая, начиная с первых ее этапов, так как ими в значительной-степепи определяется дальнейшее состояние объектов и подверженность, их заражению клещами. Своевременная уборка и вывоз урожая- с поля,, принятие мер к предупреждению механических повреждений (картофеля, овощей) при копке (а в дальнейшем, при перевозках), достаточная полнота, послеуборочной досушки — являются начальными звеньями в цепи этих, мероприятий. Закладке продукта на хранение должны предшествовать, с одной стороны, соответствующая подготовка хранилища (очистка и обеззараживание пустых помещений, подполий, прилетающей к складу территории), с другой — предварительная подработка самого продукта: тщательная его переборка с удалением больных и поврежденных луковиц,, клубней и т. п. и рассортировка по назначению и сортам.
В виду большого процентного содержания воды в большинстве корне-и клубнеплодов, особое значение при их хранении приобретают создание в складских помещонияк опредолсиной температурной обстановки и правильная организация вентиляции. Для храпения большинства овощей наиболее благоприятна температура порядка +1—2°, устраняющая возможность размножения в лих клещей, так что основной задачей здесь, является неуклонное поддержание температуры помещений на этом уровне.1
1 Для лука-севка, хранящегося при -|-18—20° С, особое значение имеет тщательная подсушка перед поступлением его в ложку. 1
СБОР И КОНСЕРВИРОПКА 63
Помимо этого, необходимо производить периодическую очистку и обеззараживание мест, где производится переборка продуктов, и уничтожать, остатки от переборки, мусор, гнилье и т. п. (предварительно обеззараживая их известковым молоком); пол складов полезно посыпать от времени до времени негашеной известью в целях предупреждения заноса клещей извне и расползания их по территории хранилища. Сильно зараженные партии продуктов, при необходимости их временного сохранения, должны тщательно изолироваться от здоровых.
Исхребительные меры борьбы с клещами применительно к плодоовощам еще недостаточно разработаны. Довольно широкое практическое применение они имеют только в отношении к луку, чесноку и, в особенности, к декоративным луковичным культурам, как известно, особенно сильно страдающим от повреждения клещами. Довольно хорошие результаты дает, например, погружение луковиц в воду, нагретую до 42—50°, на 3— 10 минут; при тщательном проведении этого способа дезинсекции, клещи гибнут и луковицы не теряют своих посадочных качеств. По данным английских и американских авторов, с успехом применяется также погружение луковиц в подогретый раствор никотин-сульфата или 2% раствор формалина. Рекомендуется также окуривание луковиц сероуглеродом и парадихлорбензолом; по некоторым данным перспективно и окуривание овощей парами сернистого ангидрида.
4. Сухофрукты. Среди мер, принимаемых для сохранения сушеных фруктов, наибольшее значение имеет окуривание сернистым ангидридом непосредственно перед их сушкой. Этим не только улучшаются цвет, блеск и другие товарные свойства продукта, но и предупреждается его заражение клещами. Хранение сухофруктов, 'имеющих повышенную влажность, увеличивает, как и в других аналогичных случаях, опасность их заражения клещами.
Кроме перечисленных выше физиологических методов, несомненно в будущем следует ожидать применения и биологического метода в борьбе с амбарными клещами, на изучение которого следует обратить серьезное внимание.
СБОР И KOHCEPBHFOBKA КЛЕЩЕЙ
Приведенный на стр. 39—44 обзор местообитаний тироглифоидей дает общее представление о том, где следует собирать клещей этой группы. При производстве таких сборов приходится пользоваться разнообразной методикой, меняющейся в зависимости от рода объекта, подвергающегося обследованию. Наиболее универсальный из методов сбора заключается в просеивании соответствующих субстратов па мелких ситах, с последующим экстрагированием клещей из полученного отсева. Просеивание лучше всего производить па 2—3 секциях обычных хлебных сит (с крайними, и средним по диаметру отверстиями); при этом субстраты, состоящие nit крупных отдельностей (например: сено, мертвые листья), iipoAiiapuTojibiin
«4
ВВЕДЕНИЕ
измельчаются ножницами, а слишком влажным дают слегка подсохнуть. Выборку клещей из отсева можно производить вручную, просматривая иго небольшими порциями под бинокуляром (или штативной лупой с достаточным увеличением) и вынимая обнаруживаемых клещей увлажненным концом препаровальной иглы, или же, экстрагировать их автоматическим способом па аппаратах Берлезе или Тулльгрена. Действие этих аппаратов основано на стремлении клещей уйти из высыхающего субстрата; в схеме они состоят из воронки с надетой на. ее трубку пробирки с водой (или консервирующей жидкостью) и сита из тонкой металлической сетки, надеваемого на раструб воронки; исследуемый субстрат насыпается тонким слоем на сито, воронка затемняется (папример вставляется в вырез какого-либо ящика), и вся система выставляется па солнце или подогревается сверху электрической лампочкой в течение нескольких часов — до полной выгонки клещей из субстрата и перехода их в пробирку. Просеивание субстратов дополняется непосредственным осмотром (с помощью лупы или бинокуляра) объектов, не могущих быть обработанными на ситах, а также .сбором клещей и, главным образом, их гипопусов с насекомых и мелких млекопитающих. Последний может производиться как с свежих, так и с коллекционных материалов: насекомые обследуются непосредственным осмотром, а млекопитающие — вычесыванием и вытряхиванием над листом черной бумаги или же, в случае работы с коллекционными материалами, фильтрацией спирта из банок, в которых опи хранятся, с последующим просмотром осадка под бинокуляром.
Собранные клещи переносятся в консервирующую жидкость или непосредственно заделываются в постоянные препараты. Из консервирующих жидкостей наиболее пригодна жидкость Oudemans’a, состоящая из 87 объемных Пастей 70° спирта, 5 частей глицерина и 8 частей крепкой уксусной кислоты, но в крайнем случае может быть использован 70° спирт. Гораздо удобнее непосредственная заделка, живых или фиксированных клещей в постоянные микроскопические препараты. Для этой цели лучше всего пользоваться жидкостью Фора-Берлезе, состоящей из следующих
веществ: Дистиллированной воды........................................... 50 вес. ч.
Хлорал-гидрата................................... 200 » »
Глицерина.......................................... 20 » »
Гуммиарабика сухого................................. 30 » »
Гуммиарабик может быть заменен вишневым клеем, по никак не головым конторским клеем, оказывающимся непригодным для составления этой смеси; рекомендуется еще прибавить к ней 5 весовых частей хлорида кокаина, но практика показывает, что можно прекрасно обойтись и без него. Для изготовления этой смеси следует, прежде всего, растворить в указанном количество дистиллированной воды гуммиарабик (или вишневый клей); затем к полученной жидкости добавляют глицерин и хлоралгидрат, после чего ее помещают в тепло (можно в термостат), в герметически накрытой банке, и дают постоять дня 2; полученная смесь затем фильтруется
ЛИТЕРАТУРА
65
через стеклянную вату, и после этого она готова к употреблению. Изготовление препаратов при пользовании жидкостью Фора-'Верлсзе чрезвычайно просто: клещей помещают в небольшую каплю этой жидкости, нанесенной на предметное стекло, прикрывают покровным, затем несколько раз слегка подогревают препарат па пламени спиртовой горелки до достаточного просветления объектов — и препарат готов; ему остается еще только подсохнуть, для чего его следует хранить в горизонтальном положении (в папках'). Жидкость Фора-Берлезе— наилучшая из существующих смесей для заделки клещей в препараты; единственным ее недостатком может считаться то, что она слишком сильно просветляет листовидные щетинки ног, становящиеся иногда трудно заметными, но этого легко избежать при некотором навыке, «дозируя» степень нагревания препарата. При невозможности пользоваться этой жидкостью приходится заделывать клещей в глицерин-желатин, предварительно просветлив их в молочной кислоте. Обычная энтомологическая техника приготовления микроскопических препаратов абсолютно не применима к препаровке тироглифоидных клещей.
ОСНОВНАЯ ЛИТЕРАТУРА
Систематика, морфология и’ анатомия
В er 1es е А., 1882—1896, 1898, Acari, Miriapodi е Scorpion! Italian!. Padova, Firenze.", В erl.es e A.,1897. Aeari, Myriapod! e Scorpion! Italian!. Ordo Cryptostigmata. Firenze. Oanestrini G., 1888. Tiroglifidi, studio critico. Padova.
Oanestrini G., 1888 (a). Prospetto dell’Acarofanna Italiana, vol. III. Padova.
Kramer P., 1899. Tyroglyphinae. In: Oanestrini & Kramer. Demodicidae und Sar-coptidae. Das Tierreich, Lief. 7: 132—150.
Michael A. D., 1901, 1903. British Tyroglyphidae, vol. I—II. London.
N a 1 e p a A., 1885,1886. Die Anatomic der Tyrqglyphen, I—II. Sitzber. d. Akad. Wise. Wien, XC, Abt. I: 197—228; XCII, Abt. I: 116—167.
Oudemans A. C., 1897—1932. Многочисленные статьи в голландских журналах: Entomol. Вег., Tijdschr. voor Entomol., Tijdschr. Ned. Dierk. Ver.
VitzthumH., 1912—1928. Ряд статей в жури.: Arch. Naturg., Zool. Jahrb., Zool. Anz. и др. "Fitzthum H., 1931. Acarina. In: W. Kiikenthal, Handbuch der Zoologie.
[Zachvatkin А. А.] Захваткин A. A., 1937. Исследования по систематике клещей сем. Tyroglyphidae. Уч. Зап. Моск. Гос. унив., III: 169—202.
Биология, экология и хозяйственное значение
[D е v el В. L. et U s h a t i n s k a j a R. S.] Д e в e лл Б. Л. и У ш а т и н с к а я Р. С., 1935.
Какой частью зерна питаются амбарные клещи. На фронте с.-х. загот., 1935, № 9: 42—45, Hase А., 1929. Zur pathologisch-parasitologischen und epidemiologisch-hygienischen Bedeutung der Milben, insbesondere der Tyroglyphinae usw. Ztschr. Parasitenk.: 765—821.
Maurizio A., 1905. Studien fiber Milben, die in Futter- und Nahrungsmitteln leben, Landw. Jahrb. d. Schweiz, XIX: 735—758.
Maurizio A., 1906. Zur Lebensweise der Milben der Familie Tyroglyphidae in Futter- und Nahrungsmitteln. Cbl. Bakter.,Parasitenk. u. Infektionskr., Abt. 2, XV:606—623,723—737.
Michael A. D., 1901. British Tyroglyphidae, vol. I: 126—183.
NewsteadR. & Duvall H. M., 1918—1919. Bionomic, Morphological and Economie Re-
port on the Acarids of Stored Grain and Flour, pt. I. Rep. Grain Pests (War) Com., no. 2,4. NewsteadR. & Morri s, H. M., 1921. Id., pt.- II. Ibid., no. 6: 7—15.
[Rodionov Z. S.] Родионов 3. 0., 1936. Вред от хлебных клещей. Вопросы питании, 1936, № 5: 111—118.
[R о m а п о v a W. Р. J Р о м а и о в а В. П., 1936. К характеру повреждения верпа амбарными • клещами. Защита растений, 1936, № 8: 20—38.
(Shesterikova М. N. ] Шестериков а М. Н., 1932. Клещи, встроча|<>|цноон и масло-ссмепах, их биология и повреждения, наносимые ими. Тр. Инет, нищ, промышл, НКСнаба, I, вып. 8.
И Фауне СССР, Захваткин—284
66
ВВЕДЕНИЕ
Schulze Н., 1924. Ueber die Biologie von Tyroglyphus mycophagus (M6gn.), zugleich ein B'ei-trag zur Hypopusfrage. Ztschr. Morphol. u. Oekol. d. Tiere, II: 1—57.
Schulze H., 1924 (a). Zur Kenntnis der Dauerformen (Hypopi) der Mehlmilbe Tyroglyphus farinae L. Cbl. Bakteriol., Parasitenk. u. Infektionskr., Abt. 2, LX: 536—549.
[T s h e r n у s h e v P. К. ] Чернышев П. К., 1938. Клещи, вредящие запасам сельскохозяйственных продуктов в ССОР. Сольколхозгиз.
Zacher F., 1927, Die Vorrats-, Speicher- und Materialschadlinge und ihre Bekampfung.
Z a c h e r E., 1933. Haltung und Ziichtung von Vorratsschadlingen. Abderhalden’s Handb. d. biol.
Arbeitsmeth., Abt. IX, Toil 7: 4.
[Z a c h v a t k i n A. A.] 3 a x в а т к и n A. A., 1936. О распространении хлебных клещей в полевых условиях. Зоол. жури., XV, 4: 698—719.
Список сокращений, принятых в тексте
Отрезки л а и к и IV cW Tyroglyphidae
ab — расстояние между основанием лапки и проксимальным краем первой (проксимальной), тарзальной копулятивной присоски.
cd — расстояние между обеими тарзальными копулятивными присосками.
ef — расстояние между дистальным краем второй (дистальной) тарзальной копулятивной присоски и копцом лапки.
Щетинки идиосомы'
ап — анальные щетинки — setae anales. d — спинные щетинки — setae dorsales. dj—1-я пара спинных щетинок — setae dorsales primae. d2 — 2-я пара спинных щетинок — setae dorsales secundae. d3 — 3-я пара спинных щетинок — setae dorsales tertiae. d4 — 4-я пара спинных щетинок — setae dorsales quartae. h—плечевые щетинки — setae humerales.
h e —наружные плечевые щетипки — setae humerales exteriores.
hi — внутренние плечевые щетипки — setae humerales interiores.
1 —боковые щетинки — setae lateral es.
sh — подплечевые щетинки — setae subhumerales.
la — передние поясничные щетинки — setae laterales (s. lumbales) anteriores. Ip — задние поясничные щетинки — setae lumbales posteriores.
Ij — 1-я пара боковых щетинок — setae laterales primae 12 — 2-я пара боковых щетинок — setae laterales secundae 13 — 3-я пара боковых щетинок — setae laterales tertiae 14 — 4-я пара 16 ' P
•' только у Glycyphagidae.
щетинок — setae laterales quartae
5-я пара боковых щетинок — setae laterales quintae
постанальные щетинки — setae postanales.
р, — 1-я пара постанальпых щетинок — setae postanales primae.
p2 — 2-я пара постанальных щетинок — setae postanales secundae.
ps — 3-я пара постанальных щетинок — setae postanales tertiae.
pra — прэанальные щетинки — setae praeanales.
sa — крестцовые щетинки — setae sacrales.
sa e — наружные крестцовые щетинки — setae sacrales exteriores.
sa i — внутренние крестцовые щетинки — setae sacrales interiores.
sc — лопаточные щетинки — setae scapulares.
sc о — наружные лопаточные щетинки — setae scapulares exteriores.
sc i — внутренние лопаточные" щетинки — setae scapulares interiores.
v — темсппые щетинки — setae verticales.
v a — передние теменные щетинки — setae verticales anteriores • 1 Qlwwhaaidae v p — задние темсппые щетинки — setae verticales posteriores J y У JP У v e — наружные теменные щетинки — setae verticales exteriores ' ™
v i — внутренние темонпые щетинки — setae verticales interiores
боковых
}у Tyroglyphidae и Saproglyphidae*
Сенсиллы лапок
a — главная «обонятельная» палочка.
f — добавочный «обонятельный» штифтик (famulus).
7 — проксимальная сенсилла базальной группы.
6 — дистальная сенсилла базальной группы.
е—дорзальная сенсилла медиальной группы-
СПЕЦИАЛЬНАЯ ЧАСТЬ
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА СЕМЕЙСТВ
А. Взрослые, животные
1(2). Коготки сидячие, причленяющиеся к концу лапки посредством пары особых соединительных склеритов; praetarsi мясистые, подушковидные, окружающие в виде чехла основание коготка или редуцированные; с? с хорошо развитыми анальными и 2 парами тарзальных копулятивных присосок (отсутствуют только у рода Scatoglyphus Berl.); 2 без эпигиния; генитальные щупальцы пальцевидные; кожа гладкая, тонкая, перепончатой консистенции, бесцветная; идиосома явственно разделена поперечной бороздкой на проподосому и гистеросому;. щетинки идиосомы никогда: не бывают гребенчатыми или листовидными................................1. Tyroglyphidae.
2 (1). Коготки стебельчатые, сидящие 'па ^самом конце продолговатого praetarsus, не соединенные при помощи склеритов с концом лапки; с? без анальных и тарзальных копулятивных присосок (если они есть, то генитальное отверстие расположено между основаниями ног I, и кожа сильно уплотнена),'или тарзальных присосок только 1 пара; Q часто с эпигинием; генитальные щупальцы обычно дисковидные.
3 (4). Идиосома всегда явственно разделена на проподосому и гистеросому; кожа гладкая или крайне тонко штрихованная, нежная, перепончатой консистенции; все щетипки идиосомы и ног гладкие, волосовидные ................................... 2. Saproglyphidae*
4 (3). Идиосома нерасчлененная, если же с поперечной бороздкой, то усаживающие ее щетинки листовидны или двусторонне-гребенчаты, или кожа имеет плотную консистенцию и более или менее интенсивно окрашена; кожа зернистая, шиповатая или густо штрихованная, редко гладкая и тогда сильно уплотненная; щетинки идиосомы и йог очень разнообразного строения, но почти никогда не бывают волосовидными..........................3. Glycypluigldue.
В. Гипопусы
1 (2). Нижняя поверхность тела с прикрепительным аппаратом, сооФол« щим из пары подвижных клапанов и 2 расположенных под ним Ий|> *
68
i. TiRGGLiPHIDAE
поперечнорёбристых зажимов и приспособленным для схватывания и удержания волос млекопитающих, к которым прикрепляются гипопусы; спинная поверхность тела уплощенная, гладкая.............. .
........................3. Glycyphagidae (подсем. Labidophorinae).
2 (1). Нижняя поверхность тела с присасывательным диском — округлой пластинкой, несущей 3—5 пар мясистых присосок; спинная поверхность более или менее сильно выпуклая, если же уплощенная, то в густой параллельной штриховке.
3 (6). Praetarsi длинные, более или мепее стебельчатые, несущие па самом конце коготок.
4 (5). Коксальные поля III и IV замкнутые, смыкающиеся по средней линии тела, разделенные здесь только продольным линейным склеритом; у. большинства видов на проподосоме имеется пара полушаро-видпо выступающих глаз...............................2. Sapro glyph! dae.
5 (4). Коксальные поля III и IV открытые, разъединенные по средней линии тела очень широким промежутком; глаз нет.........................
. . . . 3. Glycyphagidae (подсем. Chaetodaclylinae и род Hericia).
6 (3). Praetarsi отсутствуют; коготок, таким образом, сидячий.
7 (8). Гнатосома хорошо развита, подвижная; стернальные щетинки почти всегда превращены в присоски; гистеросомальный щит налёгает передним краем на основание проподосомального щита; поверхность обоих щитов тонкопористая или усеяна округлыми ямками; генитальные щупальцы о закругленным концом ... 1. Tyro glyph! dae.
8 (7). Гнатосома более или менее редуцирована, неподвижно сращена с идиосомой или совсем отсутствует (представлена тогда только 1— 2 парами тонких щетинок — арист); стернальные Щетинки не превращены в присоски; гистеросомальный щит не налегает на проподосо-мальный; поверхность обоих щитов с более или менее сетчатым орнаментом из тонких вдавленных линий или усеяна вытянутыми в длину ямками; концы генитальных щупальцев вытянуты в тонкое острие . . . .....................3. Glycyphagidae (часть подсем. Chaetodactylinae).
I. Сем. TYROGLYPHIDAE i
Tyroglyphidae Ouds. 1932 +' Caloglyphidae Ouds. 1932+ TyropHagidae Ouds. 1924 + For-eelliniidao Ouds. 1934 + Rhizoglyphidae Ouds. 1923.
Характеристика семейства. Идиосома всегда явственно подразделена поперечной перетяжкой, проходящей позади основания ног II, на проподосому и гистеросому, тучная, сильно выпуклая как с брюш-
1 К этому же сомоПстну отнооятон следующие недостаточно описанные виды: Rlvizoglyphutt minor Dumontjov 1913 (Кавказ) » Irowmwli Berloso 1897 (Франция)
» eulamus Berloso 1921 (Италия)
» gtvmanious Berloso 1921 (Германия)
» оссигепч Borloso 1921 (Италия)
1 » algidue Borloso 1921 (Италия).
ХАРАКТЕРИСТИКА. СЕМЕЙСТНА
69
ной, так и со спинной стороны, в поперечном сечении широко-эллиптическая или почти круглая; у живых особей на спинной поверхности обычпо-наблюдаются продольные мускульные вдавления (соответствующие местам прикрепления крупных дорзо-вентральных мышц и мощных ретракторов хелицер), становящиеся незаметными у фиксированных экземпляров и на препаратах. Прижизненная окраска идиосомы беловатая, полупрозрачная, с характерным стеклянным блеском, иногда с легким сероватым, бурым и зеленоватым оттенком и расплывчатыми пятнами, определяемыми темным, цветом содержимого кишечника. Кожа тонкая, мягкой, перепончатой консистенции, бесцветная и прозрачная, в подавляющем большинстве случаев совершенно гладкая (только у с? с? Caloglyphus mandzhur очень тонкоморщинистая); проподосомальный щит всегда хорошо развит, занимающий всю площадь спинной поверхности проподосомы между передними теменными щетинками и лопаточными, широкий, слабо пигментированный или почти бесцветный; у некоторых видов рода Schwiebea, он разделен продольной полоской мягкой, перепончатой кожи на 2 узких, более или менее самостоятельных щита (фиг. 340), а у Schwiebea pachyderma неявстве-неп, в связи с общим утолщением всей кожи. У бб1 многих Rhizoglyphinae задняя часть спинной поверхности гистеросомы покрыта более или менее плотным и соответственно пигментированным опистосомальным щитом (фиг. 330, 327, 340), иногда образующим пластинчатые или сложные вееровидные разращения; кроме того, у многих форм кожа вокруг основания ног I и эпимер в той или иной мере уплотнена, образуя неправильно очерченные хитинизированные участки (фиг. 257, 328).
Вооружение спинной и боковых поверхностей идиосомы (фиг. 4, 27} складывается из 7—14 пар гладких, тонко- и редкоперистых и лишь очень-редко — густоперистых (Kuzinia; Forcellinia wasmanni) щетинок, расположение большинства которых очень постоянно у всех представителей семейства. На проподосоме помещаются следующие из них: 1) внутренние теменные (v 1), расположенные медиально на переднем ее краю, всегда тесно сближенные основаниями и мало изменчивые по размерам (равные 10—17% длины идиосомы); эти щетинки отсутствуют только у рода Monieziella; 2) наружные теменные (v е), положение и размеры которых, напротив, очень изменчивы и имеют большое таксономическое значение (фиг. 80— 86): они сидят то у середины боковых краев проподосомального щита и представлены тогда тонкими гладкими микрохэтами (у Rhizoglyphinae), то-ближе к его переднему краю, но тоже дорзально, и тогда также представлены микрохэтами (Tyroglyphus, Forcellinia), то более или менее латс-ралыю, кнаружи от передних углов проподосомального щита и почти на одной поперечной линии с передними теменными; в последнем случив они имеют значительную длину, явственно перисты и более или моноо дугообразно огибают с боков гнатосому (Tyrophagus, Tyrolichus) или торчат прямо вперед (Aleuroglyphus, Kuzinia)- у многих Rhizoglyphini V о «о-воринчпго отсутствуют; 3) наружные, лопаточные (sc с), расположенный НО-
i. T^ar.VPftWAE
юади проподосомального щита, на небольшом расстоянии от заднего края проподосомы; положение их очень константно,-и у всех без исключения видов они представлены макрохэтами; 4) внутренние лопаточные (sc i), расположенные кнутри от наружных и образующие с ними прямой или дугообразный поперечный ряд (в зависимости от того, помещаются ли они на одной линии с sc е, впереди или позади них); размеры их крайне изменчивы, и они нередко (у большинства Rhisoglyphini) совершенно отсутствуют. 'На гистеросоме располагаются следующий пары щетинок: б) наружные плечевые (й е), хорошо развитые у всех представителей семейства и расположенные латерально, на боковой поверхности передних углов гистеросомы; 6) внутренние плечевые (h i), сидящие, как уже показывает их название, кнутри от ,(й е), более дорзальпо; размеры щетинок этой пары очень изменчивы, но в большинстве случаев они уступают таковым наружных плечевых, а у большей части' Rhizoglyphini совершенно отсутствуют;. 7—10) 4 пары спинных щетинок (dx—d4), образующие 2 продольных ряда вдоль средней линии тела, из которых передние (d4) расставлены много шире, чем следующие за ними (d2); d3 раздвинуты приблизительно также широко, как dx, d4 вновь более сближены между собой; расстояния между отдельными парами спинных щетинок, измеренные в продольном направлении, подвержены некоторой изменчивости, обусловленной пропорциями •тела и степенью растяжения кожи спины (изменяющейся в зависимости ют степени «упитанности» особи или ее раздутости половыми продуктами); у сШрода Schwiebea d4 иногда сильно1 смещены латерально; длина спинных щетинок очень изменчива, ио, как правило, они постепенно увеличиваются в размерах кзади, и d4 почти всегда короче других; у Rhieoglyphini d4—d2, а иногда d3 сильно редуцированы" (род Rhizogtyphus') или отсутствуют (остальные роды); 11) передние боковые (1 а), сидящие наискось кнаружи :и назад от щетинок 2-й спинной пары (d2), близ боковых краев гистеросомы; они обычно невелики, иногда отсутствуют; 12) задние боковые (1 р), расположенные еще более латерально, чем передние боковые, и дальше назад (обычно па уровне середины расстояния .между d3 и d4); они имеются у всех видов и иногда достигают значительной длины; 13) наружные крестцовые (sac), сидящие латерально на месте перехода боковых краев идиосомы в задний (иногда они более или менее сильно смещены на брюшную поверхность тела); и, наконец, 14) внутренние крестцовые щетинки (sa i), помещающиеся терминально, на заднем краю гистеросомы; эти щетинки имеются, у всех представителей семейства и принадлежат к числу самых крупных из щетинок идиосомы. Вооружение нижней поверхности идиосомы состоит
Фиг. 80-89.
•Фиг. 80—85. Положенно теменных щетинок у различных Tyroglyphidae. Фиг. 80. Aleuroglyphus ovatus (Tr.): ve—темеппыо наружные щетинки; vl—теменные внутренние щетинки. Фиг. 81.
Kuzinia laevis (Duj.) (протопимте). Фиг. (12. У'огсвШта fungivora’ (Ouds.). Фиг. 83. Tyrophagus jterniciosus, sp. n. Фиг. 84. Ту rolichuH casei (ОШ1н). Фиг. 115. Acotyledon sp. Фиг. 86. Голень и лапка IV СУ Tyrophagus tenuiclamis, sp. n.: do-г-тпреальные копулятивные присоски; ab, cd,.ef — принятые обозначения отдельных отрепков ланки. Фиг; 87. Дистальная часть ноги IV (колено, голень и лапка) О' Rhizoglyphus echinopus (Fum. et Hob.). Фиг. 88. Анальная щель, анальные копулятивные присоски и щетинки сГ Tyroglyphus farinae (L.). Фиг, 80. То же, Tyrophagus humerosus Ouds.
12
I. 1YR0GLYPHIDAE
из пары подплечевых щетинок (sh), стандартного ассортимента коксаль-ных и генитальных щетинок и комплекса анальных, число и расположение-которых различно у обоих полов (ср. стр. 73).
Коксо-стернальный скелет очень хорошо развит. Эпимеры I неизменно слиты в мощный V-образный стернум (st), все остальные (ер II—IV) свободные, с широкими пластинчатыми аподемами, погруженными внутрь тела (только у $ Schwiebea rossica эпимеры III и IV соединены своими внутренними концами); эпимсриты П (cpt) обычно совершенно обособлены от других элементов коксо-стернального скелета и имеют вид узких линейных склеритов, расположенных поперечно на границе между проподосомой и гистеросомой и этим подчеркивающих их разделение. Длина и пропорции ног весьма изменчивы, но строение их, в общем, довольно однообразно (за исключением случаев гетероморфизма d'd'). Базальные членики пог более или менее цилиндрические, лапки обычно несколько суживаются к концу. Коготки среднего размера или крупные, сидячие, причленяющиеся к концу лапки посредством особых склеритов (art) и окруженные у основания широким мясистым, prae tarsus (рг); у большинства Rhizoglyphincie praetarsus более или менее редуцирован и одевает в виде чехла только самое основание особенно крупного у них коготка. Вооружение 4 базальных члеников ног состоит из типичного для группы в целом ассор’тимента щетинок, нередко превращенных в более или менее мощные шипы, но никогда не бывающих густоперистыми или гребенчатыми; вентральные щетинки вертлугов и бедер всегда гладкие, базальные щетинки колен никогда не достигают значительных размеров. Вооружение лапок I (фиг. 90—106, 202—224), имеющее большое систематическое значение, складывается, как правило, из следующих элементов: главной обонятельной палочки (а), обычно более или менее булавовидно утолщенной к концу; сидящего у ее основания (на той же сочленовной подушечке) штифтика £1, имеющегося у всех родов, кроме Tyroglyphus', базальной палочки расположенной кнаружи и назад от а; щетинки о, сидящей впереди т, на уровне основания а или заметно дистальнее; щетинки г, расположенной на внутренней поверхности лапки, впереди и внутрь от обонятельной палочки или непосредственно впереди нее, и имеющей тогда форму конического шипа; 4 дорзальных вершинных щетинок, из которых наружная нередко превращена в шин (иногда редуцирована); 3 очень разнообразных по своему строению, крупных вентральных щетинок, ' расположенных обычно в среднем поясе лапки, из которых 1 сидит медиально и ближе-к основанию лапки, чем обе другие, имеющие более латеральное положение; 5 вентральных вершинных шиииков, иногда достигающих значительной величины, но нередко также частично редуцированных, так что их остается только 3 (Tyrophallus') или далее 1 (Tyroglyphus). У Rhizogly-phini имеют место значительные уклонения.от этой схемы, выражающиеся в выпадении многих из элементов вооружения лапок I — сенсилл е, ог а иногда и одной из вентральных. Лапки III и IV дорзально с 3 вершин
ХАРАКТЕРИСТИКА СЕМЕЙСШ 73
ными щетинками и нередко 1 шипом, вентрально с 2 медиальными щетинками и 3—5 (у Tyroglyphus одним) вершинными шипиками.
3. Генитальное отверстие расположено в пространстве между основаниями ног III и IV или между ногами IV, или же позади их основания; pen is с более или менее трубчатой дистальной частью (стилетом) и массивным, обычно очень сложным по своей архитектуре цоколем; структурно обособленных от penis склеритов вокруг него нет; эпиандрий отсутствует; генитальные щупальцы всегда хорошо развитые, пальцевидные, с цилиндрической или конической базальной частью и четко отграниченным от нее, более или менее полушаровидным вершинным куполом. Апальпая щель более или менее сильно отодвинута от заднего края гистеросомы и тесно придвинута к генитальной; по сторонам, у ее заднего конца, у подавляющего большинства видов расположена пара относительно крупных анальных копулятивных присосок (de), строение которых довольно разнообразно. У одних форм (Tyroglyphus, многие Rhizoglyphini) они имеют вид хитинового кольца,, окруженного рамкой очень тонкой и растяжимой кожи, и в состоянии эрекции приобретают правильную цилиндрическую форму; у других форм анальные копулятивные присоски представлены полушаровидными, слабо-очерченными по контуру и обычно не хитинизированными подушками, в состоянии эрекции лишь увеличивающими свою выпуклость; при таком их строении они снабжены хорошо развитым хитиновым эндоскелетом, состоящим из имеющего форму скобки соединительного склерита, огибающего сзади края присосок и конец анальной щели; у рода Histiogaster анальные присоски целиком сильно хитинизированы и имеют очень своеобразное, уклоняющееся от нормы устройство. Анальных щетинок, как правило, только 1 пара, так как 2-я пара превращена в анальные присоски (у Scatoglyphus их 2 пары); они расположены у большинства видов на выпуклости анальных присосок, впереди их центральной поры и только у Tyroglyphus и Aleuroglyphus siculus они помещаются на значительном расстоянии впереди присосок. У рода Scatoglyphus анальные присоски отсутствуют. Позади анальной щели находятся 3 пары постанальных щетинок. (рх—р3), размеры и положение которых очень изменчивы; у рода Histiogaster их только 1 пара — рт (обе остальные совершенно метаморфизованы, образуя листовидные прозрачные лопасти по периферии опистосомальной пластинки). На внутренней поверхности лапок IV помещается по паре тарзальных копулятивных присосок более или менее блюдцевидной формы,, положение которых очень изменчиво.
2- Генитальное отверстие всегда расположено в пространстве между эпимерами III и IV, относительно небольшое; нет ни эпи-гипия, ни хитинового циркумгенитального кольца; клапаны генитального отверстия всегда хорошо развиты и имеют более или меццо сильно уплотненные, хитинизированные края, смыкающиеся по средней линии тела на значительном протяжении и затем расходящиеся дугообразно в стороны и назад (часто образуя довольно янстпеппый тупой
'74
1. TT110GLYPI11DAE
угол в.т'очке, откуда начинается их расхождение); генитальные щупальцы хорошо развитые, пальцевидные, такого же строения, как у <3. Копулятивное отверстие обычно хорошо заметное, но не образующее выдающейся над поверхностью тела трубки; у многих видов кожа вокруг него несколько уплотнена. Анальная щель всегда отделена от генитальной .довольно широким промежутком, но нередко также более или мепее сильно отодвинута и от заднего края тела, помещаясь тогда приблизительно посередине расстояния между ними (у большинства Acotyledonini)-, около--апальных щетинок в большинстве случаев 7 пар, из которых 3 пары анальных обычно представлены микрохэтами и располагаются вдоль краев anus по всей его длине, 3 пары более крупных адаиалытых окружают их снаружи кольцом, а 1 пара — постапальпых — представлена макрохэтами (часто очень крупными); у Rhizoglyphini анальный комплекс щетинок испытывает такое же сокращение, как и другие группы щетинок, и их остается всего 1 или 2 пары.
Вторичные различия между полами у Tyroglyphidae весьма разнообразны - как по объему затрагиваемых ими признаков, так и по своей резкости.
.Наиболее велики они в подсем. Rhizoglyphinae (в частности у Rhizoglyphini'), у которых, кроме того, наблюдается еще резко выраженный диморфизм сГ сТ, представленных здесь двумя фазами — гомеоморфной, относительно более близкой по своему строению к £, и гетероморфной, характеризующейся резкой модификацией и утолщением нот III и гипертрофированным развитием всех вторичных половых признаков.
Нимфальные стадии, в общем, сильно напоминают взрослых, отличаясь от них, помимо общих возрастных признаков, только некоторыми деталями хэтотаксии, или в иных, чем у взрослых, соотношениях их размеров (например у Tyrophagus), или же в уклоняющемся от дефинитивного положения v е (у Aleuroglyphus и Tyrolichus). Личинки с хорошо развитыми, трубчатыми «(только у Thyreophagusпуговковидными) грудными палочками, обычно снабженными более или менее явственно обособленным полушар обидным вершинным куполом, напоминающим аналогичное образование у генитальных щупальцев прото- и телеонимф и взрослых.
Расселительные гипопусы Tyroglyphidae (фиг. 67—59) относятся все к эптомохорному типу. Гнатосома (gn) всегда хорошо развита, подвижно сочленена с идиосомой посредством мягкой сочленовной перепонки и функционирует, невидимому, в роли органа осязания; опа обычно состоит из более крупного базального членика, очень изменчивого по своей форме и пропорциям, и пары более или менее обособленных от него дистальных
Фиг. 90—106.
-Фиг. 90. Tyroglyphus farlnao(l,.), лапка I Q сверху. Фиг. 9t. То же, снизу. Фиг. 92. Т. farinae (L.), конец лапки I о' сбоку. Фиг. 03. Т. farinae (L.), колено 1 Фиг. 94. Aleuroglyphus ovatus (Тг.), лапка I q сверху. Фиг. 95. То же, ониау. Фиг. 9(1. A. oualus (Тг.), конец папки I cf снизу. Фиг. 97. Kuzinia laevis (Duj.), протопимфа, лапка I сверху. Фиг. Oil. То же, снизу. Фиг. 99. Tyrolichus easel (Ouds.), лапка I 9 сверху. Фиг. 100. То же, скипу. Фиг. 101. Tyrophagus tenuiclavus, sp. n.. лапка о' сверху. Фиг. 102. То же, онипу. Фиг. 103. Т. tenuiclavus, sp. п., колено I. Фиг. 104. For-•nsellinia fungivora (Ouds.), лапка I 9 сверху. Фиг. IOS. To же, снизу. Фиг. 106. F. fungivora (Ouds.), конец лапки I О'. Вукооккыс обозначения см. в тексте, стр. 72.
16
Л TYROGLYPHIDA.E
члеников, несущих на конце по крепкой щетинковидной аристе (аг); у некоторых форм гнатосома совершенно не расчленена, но в таких случаях ее конец обычно более или менее глубоко расщеплен; помимо арист, на гнатосоме имеется 2 пары мелких щетинок: одна — на дорзальной поверхности дистальных члеников, другая — па боковых поверхностях базального. Идиос'ома целиком одета более или менее плотным и хитинизи-рованным панцирем, особенно жестким на сиипной поверхности, эллиптической или яйцевидной формы, с широко закругленным задним краем; спинная поверхность Идиосомы сильно выпуклая, брюшная же уплощена и обычно способна сильно прогибаться, подтягиваясь кверху мощной дорзо-вентральной мускулатурой; края идиосомы обычно уплощены. Про-подосома (рг) всегда четко обособлена от гистеросомы и сочленена с нею-подвижно, обладая способностью заметно .отгибаться кверху (под тупым углом к вентральной поверхности гистеросомы); при этих движениях задняя часть ее спинного щита глубоко уходит, под налегающий на нее передний край гистеросомы, с которым он образует по бокам подобие шарниров; форма проподосомального щита очень изменчива, но у большинства, видов приближается к треугольной; ростральный выступ обычно хорошо развит, но иногда (Robinisca, Froriepia, Viedebanttia) совершенно отсутствует, и щит имеет тогда дугообразный передний край; размеры его также очень изменчивы. Боковые края гистеросомального щита загнуты на брюшную поверхность в виде более или менее широких эпиплевр, играющих, повидимому, известную роль в прикреплении гипопуса к переносчику (представляя собою периферическое кольцо той «присоски», какой становится само тело гипопуса при прогибании его вентральной поверхности). Поверхность спинных щитов в большинстве случаев гладкая (тонкопористая), иногда в рассеянной ямчатой пунктировке, но без сетчатого рисунка и продольной или концентрической штриховки; только очень узкий участок вдоль переднего края гистеросомального щита покрыт тонкой поперечной штриховкой (иногда этот участок, по Oudemans’y, имеет перепончатую консистенцию). Все щетинки спинной поверхности обычно представлены' однотипными, тонкими, подчас трудно заметными микрохэтами,. и лишь у немногих родов (Tyroglyphus, Lackerbaueria) достигают значительных размеров, но в этом случае волосовидны; иногда лопаточные крепче-остальных и более или менее искривлены (некоторые Acotyledon)-, расположение щетинок довольно изменчиво, но, в виду трудности их обнаружения, мало используется в систематике гипопусов. Пссвдостигмальный орган чаще хорошо развит, щетинковидный. Глаза отсутствуют в подавляющем большинство случаев; только у Monieziella они хорошо развиты, и представлены полушаровидными линзами, расположенными по бокам-проподосомы (над основаниями ног I) и снабженными темнопигментиро-ванными ретинами; у Histiogaster имеются только последние, образующие-пару сближенных между собой оптических пятен на конце рострального-выступа, и нет оформленных линз. Пщкняя поверхность идиосомы обычно
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
п
уплотнена, образуя ряд более или менее хитинизироваппых щитов: стернальный — составленный коксальными полями I и II, вентральный — образованный таковыми ног III и IV, и генитальный — расположенный позади вентрального и нередко слитый с ним в одно целое; плевральные поля, окружающие с боков эти щиты, обычно также более или менее хити-низированы. Строение стернального щита довольно изменчиво; у одних видов эпимеры II не доходят до его заднего края, свободные, и тогда щит образует одно перасчлепенное целое, и коксальные поля I и II остаются открытыми; у других эпимеры II тянутся до заднего края и здесь соединяются тонкой линией с эпимеритами II, замыкая, таким образом, коксаль-•ные поля II, приобретающие тогда характер более или менее прямоугольных боковых парных щитов, обособленных от треугольного непарного среднего, составленного коксальными полями I; последние обычно открыты (если стернум не доходит до их заднего конца), но иногда замкнуты (у Forcellinia) ; у Froriepia стернум сливается с концами эпимер II и эпимерами III; боковые края стернального щита (обрисованные более или менее явственными эпимеритами II) часто образуют с его задним краем явственные боковые утлы, так что он приобретает трапециевидные очертания, но у других форм боковые углы сильно сглажены или даже не намечены вовсе и щит обрисован кзади дугообразной линией. Вентральный щит в большинстве случаев отделен от стернального поперечной полоской мягкой, перепончатой кожи, но у рода Histiogaster они не разделены вовсе и образуют иерасчлененное целое, а у Froriepia и Tyroglyphus разделены только эпимерами III; передний край вентрального щита, образованный эпимерами III, параллелен заднему краю стернального щита; в тех случаях, когда коксаль-ные поля III замкнуты и более или менее широко раздвинуты (например у Schulze а), он иногда прерван посередине, и тогда сам вентральный щит .распадается, аналогично стернальному, на составляющие его отдельности — парные коксальные поля III и IV; вдоль щита обычно тянется перепончатая продольная линия — вентральный шов, разграничивающий коксальные поля III и IV по средней линии на более или менее значительном протяжении; эпимеры IV в большинстве случаев имеют диагональное направление и нередко доходят концами до вентрального шва, замыкая, таким образом, коксальные поля III и IV. Генитальный щит значительно длиннее своей ширины и отделен от вентрального прямым или дугообразным поперечным швом, иногда очень неявственным или даже совершенно отсутствующим. Коксальные волоски почти всегда заменены присосками, расположенными, соответственно, на коксальных полях I и III; одна из пар гопи-тальных щетинок расположена в точке стыка эпимер IV и вентрального шва, и 2 другие — по сторонам генитальной щели; из последних impysitiliui превращена в генитальные присоски (dg), а внутренняя нс bjwhkimoiioiih. Генитальная щель всегда вполне явственна, с парой пальцевидных гони*’ тальпых щупальцев; у некоторых видов Acotyledon она слита о анплышИ (У других видов обычно плохо заметной) в длинную ано-гоинтплi.ny 10 щоль.
78
/. TYROGlYPHIDAE
Присасывательный диск (da) обычно крупный, с 4—5 парами хорошо развитых присосок и более или менее сильно хитинизированной поверхностью,.', окруженный кольцом очень эластичной мягкой кожи, позволяющей ему довольно сильно выпячиваться над брюшной поверхностью тела; свободной перепончатой каймы вокруг него пот, как пет и связанных- с ним эндо-скелетных образований. У части видов рода Acotyledon (paradoxa, michaeli, rhizoglyphoides) присасывательный аппарат более или менее сильно редуцирован; в крайних случаях on представлен только рудиментами 1—2 пар присосок на еле намеченном, пехитинизированном присасывательном диске.
Все 4 пары ног с хорошо развитыми, слабо изогнутыми коготками, но без всяких следов перепончатого praetarsus; иногда коготки очень длинны, а у рода Froriepia имеют даже почти щетинковидную форму. Передние пары ног (I и II) всегда развиты значительно сильнее, чем задние, которые у большинства форм почти целиком скрыты под идиосомой и выдаются за ее края только концами лапок; вооружение их 4 проксимальных члеников идентично по числу и расположению составляющих его элементов с таковым половозрелой стадии, с тем только отличием, что на вершине колен I имеется лишь 1 дорзальная щетинка (только у Schulzea их 2); строение же, размеры и форма этих элементов очень изменчивы и обычно сильно отличаются от наблюдаемых у нимф и'взрослых. Вооружение лапок I и II складывается из тех же элементов, как и у взрослых; обонятельная палочка обычно длиннее и стройнее, сенсиллы у, 8, а часто и е нередко имеют палочковидную форму; паружпая и внутренняя вентральные и 1—2 из вершинных часто листовидно уплощены, а средняя вершинная почти всегда расширена на конце в овальное блюдце и несет присасывательную функцию. Вооружение лапок III и IV состоит из значительно большего числа щетинок, чем у взрослых; многие из этих щетинок часто бывают листовидными, а 1—2 нередко гипертрофированы, достигая х/2—4/5 длины идиосомы («парусные щетинки»). Размеры гипопусов колеблятся в широких пределах — от 150 до 475 р-
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ПОДСЕМЕЙСТВ
Д (2). V е расположены латерально, у передних углов проподосомального щита, довольно длинные, тонко- и густоперистые, или же немного дальше назад (приблизительно на 4/4 расстояния между линией, проведенной через основания vi и лопаточными щетинками), и тогда, представленные микрохэтами; щетинки спинной поверхности идиосомы в большинстве случаев длинные, тонко- и редкоперистые; все щетинки пог обычно тонкие и гибкие; коготки небольшие, praetar-sus крупный, подушковидпый, одевающий большую половину коготка ............................................1. Tyroglyphinae.
2 (1). Уев виде очень мелких, гладких микрохэт, расположенных дор-зально близ середины боковых краев проподосомального щита, при-
ТАБЛИЦА ГИПОПУСОВ
близительно на одинаковом расстоянии от v i и лопаточных щетинок, или же совершенно отсутствуют; щетинки спинной поверхности идиосомы гладкие, часто более или менее утолщающиеся к основанию; щетинки ног обычно более или менее утолщенные, часто шиповидныс; коготки ног I и II крупные, praetarsus в большинстве случаев одевает-в виде чехла только основание коготка........2. Rhizoglyphinae.
ОПРЕ ЦЕЛИТЕЛЬНАЯ ТАБЛИЦА ГИПОПУСОВ
1 (2), Коготки необычайно длинные и тонкие: на ногах I они равны по. длине всей ноге, щетинковидны и почти не отличаются по своему строению и размеру от крупных щетинок лапок; присасывательные щетинки лапок I и II также непомерно длинны; эпимеры II соединены концами с стернумом . . . ........................24. Froriepia Vtz.
2 (1). Коготки нормального размера и строения, всегда много короче ла-цок; длина присасывательных щетинок не превосходит половины, длины ног I и II; эпимеры II не соединены концами с стернумом.
3 (4). На боковых краях проподосомы (над основанием пог I) находится пара крупных, полушаровидных глаз..............14. Monieziella Berl.
4 (3). Глаза отсутствуют или расположены па вершине рострального выступа и лишены сильно выпуклых линз.
б (36). Присасывательный диск хорошо развитый, более или менее плотно-хитинизированный, несущий 4 или б пар вполне развитых присосок.
6 (35). Присасывательный диск округлый или понеречноэллиптический, не длиннее своей ширины; наружные присоски всегда расположены кпереди от задних; присасывательные щетинки лапок I и II расширены на конце.
7 (8). Щетинки проподосомального щита (sc) очень длинные, волосовидные, равные 1/5—всей длины идиосомы; коксальных присосок нет;, лапки IV на конце с 1 очень длинной парусной щетинкой, превосходящей 1/2 Длины тела, и второй более короткой......................
.......................................... 1. Tyrogiyphus Latr.
8 (7). Sc короткие, не превышающие 5—8% длины идиосомы.
9 (10). Гнатосома значительно короче своей ширины у основания, трапециевидная; коксальные поля III замкнутые, широко раздвинутые; генитальный щит не обособлен от вентрального. На шмелях и в их гнездах...........................................3. Kuzinia, gen. n.
]0 (9). Гнатосома иной формы, всегда заметно длиннее своей ширины у основания; если коксальные поля III широко раздвинуты и при этом замкнуты, то генитальный щит отделен от вентрального болоо пли менее явственным поперечным швом.
11 (34). Вершина колен I только с 1 дорзальной щетинкой.
12 (13). Эпимеры III и IV свободные, далеко не достигающие сродной линии тела; вентральный и стернальный щиты по отграничены друг
30
I. TYROGLYPHIDAJ-
от друга и от плевральных полей, образуя с ними одно нсраспленен-ное целое; ростральный выступ па конце с парой близко придвинутых друг к другу глаз, представленных скоплениями темного пигмента, со слабо дифференцированными, мало выступающими линзами или вовсе без них ............................12. Histiogaster Berl.
13 (12). Эпимеры III сливаются или, во всяком случае, близко сдвинуты концами, очерчивая, таким образом, передний край вентрального щита, или же он ясно обрисован каким-либо иным, образом; коксаль-пыс поля Ш обычно замкнутые; стернальный щит явственно отграничен от плевральных полей; глаз нет.
14 (27). Обе пары коксальных присосок хорошо развиты, нормального строения.
15 (16). Лапки IV с 2 длинными парусными щетинками, из которых 1 почти равна длине всего тела; спинные щиты покрыты очень грубой и густой ямчатой скульптурой; коксальные присоски расположены на территории очень явственных краевых утолщений коксальных полей; наружная вентральная щетинка голеней I и II и базальная и наружная щетинки колен (тех же пар ног) длинные, гибкие, явственно перистые.....................................16. Lackerbaiieria, gen. n.
16 (15). Лапки IV без длинных парусных щелинок; поверхность спинных щитов гладкая (тонкопористая) или в более рассеянной и мелкой ямчатой пунктировке; краевые утолщения коксальных полей малозаметные или отсутствуют, коксальные присоски расположены вне занятой ими территории; наружные щетинки голеней и колен I и II гладкие, более или менее толстые, обычно шиповидные; базальная щетинка колен I и II гладкая, короткая и толстая.
17 (18). Присасывательный диск маленький, удаленный от заднего края тела на расстояние, сильно превышающее его длину; наружные края коксальных полей III в большинстве случаев длинные, прямые, лапки III и IV длинные и стройные, на препаратах обычно отгибающиеся под углом к остальной части ноги . . .....................
...........................................8. Acotyledon Ouds.
18 (17). Присасывательный диск крупный; расстояние между его задним краем и задним краем тела значительно меньше его длины; наружные края коксальных полей III обычно короткие, более или менее выпуклые: лапки-III и IV короче и массивнее, па препаратах обычно пе отгибающиеся под углом к предыдущим члепикам ноги.
19 (20). Коксальные поля II тесно смыкаются сейчас же позади короткого стернума, так что между их внутренними краями остается только чрезвычайно узкий продольный промежуток; поперечный диаметр присасывательного диска умещается в ширине идиосомы около 1.5 раза; присасывательная щетинка лапок II представлена очень широким листовидным образованием. Очень мелкая форма, длина 156 ц........................................ 20. Rodionovia, gen. в.
ТАБЛИЦА ГИПОГГУСОН 81
20 (19). Коксальные поля II не смыкающиеся позади стернума, разделен- . пые широким промежутком, часто открытые; поперечный диаметр присасывательного диска умещается больше 2 раз в ширине идиосомы. Длина не менее 200 р..
21 (22). Стернум и эпимеры II доходят до заднего края стернального щита, вполне разделяя и замыкая коксальные поля I и II; наружные присоски сильно сдвинуты кпереди от центральных ..................
............................ 6. Forcellinia Owls.
22 (21). Стернум не доходит до заднего края стернального щита, оставляя коксальные поля I более или менее широко открытыми; наружные присоски присасывательного диска расположены на одной поперечной линии с центральными или чуть впереди нее, или немного позади.
23 (24). Гнатосома длинная и узкая, с параллельными краями, не расчлененная на конце; передний край проподосомального щита дугообразный, идущий параллельно переднему краю гистеросомы и не образующий рострального выступа; vi отсутствуют; ноги I целиком видимые сверху................................17. R)binisca, gen. n.
24 (23). Гнатосома другой формы, если же длинная и узкая, то ветви ее развилки обособлены в виде самостоятельных цилиндрических члеников; проподосомальный щит приблизительно треугольный, образующий явственный ростральный выступ; если же его передний край дугообразный, то щит имеет полулунную форму, выступает вперед в виде козырька и закрывает почти целиком ноги I и II; v i хорошо развитые, но иногда смещенные на вентральную поверхность щита. .
25 (26). Сенсилла е лапок I и II и вершинная дорзальная щетинка колен I палочковидные, притупленные или даже несколько вздутые на конце; ростральный выступ густо пунктированный .......................
......................................10. Rhizoglyphus Clap.
.26 (25). Сенсилла е лапок I и II и вершинная дорзальная щетинка колен I в виде более или менее жестких, утончающихся к концу щетинок; ростральный выступ непунктированный............9. Caloglyphus Berl.
27 (14). Передние коксальные присоски совершенно редуцированы или заменены волосками, задние — нормально развитые или также заменены волосками, или редуцированные.
28 (31). Задние коксальные присоски хорошо развитые, нормального строения.
29 (30). Передние коксальные присоски заменены волосками; идиосома очень широкая, почти круглая; проподосомальный щит очень короткий ......................................8. Acotyledon OudH.
30 (29). Передние коксальные присоски совершенно редуцированы; идиосома продолговатая, в 1.5 раза длиннее своей ширины; длина про-подосомального щита равна г/4 длины гистеросомалыюго ....... ..................... .......................13. Sehwlobon Ои<1в.
в Фауна СССР. Захваткин—284
82
1. TYROGIAPHIDAE 1. TYROGLYPHINAE
31 (28). Задние коксальные присоски заменены волосками или редуцированы.
32 (33). Обе пары коксальных присосок заменены довольно длинными, крепкими щетинками; коксальные поля III и IV довольно широко-раздвинуты; гнатосома с лиственно обособленными дистальными члениками .....................................9. Caloglyphus Berl.
33 (32). Обе пары коксальных присосок заменены крайне мелкими, подчас трудно заметными микрохэтами, или задние редуцированы; коксальные поля III и IV смыкаются по сродней линии тела; гнатосома не-расчлененная, только слегка расщепленная па конце..............
................................... 19. Troupcauia, gen. n.
34 (11). Вершина колец I с 2 дорзальными щетинками, коксальные поля II и III замкнутые, широко раздвинутые; все коксальные поля с явственными краевыми утолщениями; наружные присоски присасывательного диска расположены значительно впереди центральных ............................................23. Sehulzea, gen. n.
35 (6). Присасывательный диск узкий, сильно вытянутый в длину, языко-видный, почти в 2 раза длиннее своей ширины; наружные и задние присоски расположены на одной поперечной линии, очень маленькие;, присасывательные щетинки лапок I и II волосовидные, не расширенные на конце ............................8. Acotyledon Ouds. (pt.).
36 (5). Присасывательный диск совершенно редуцированный, такой же-мягкой, перепончатой консистенции, как и окружающие его поля,, несущий только 2 пары сильно ‘редуцированных, плохо заметных присосок ........................8. Acotyledcn (pt.).
Недостаточно описанные роды Garsaultia Ouds., Sancassania Ouds. и Viedebanttia Ouds. не смогли быть помещены в таблицу.
1. Подсем. Tyroglyphinae
V е расположены латерально, у передних углов проподосомального’ щита и приблизительно на одной поперечной линии с v i или очень немного позади нее, и тогда — довольно длинные, тонко- и густоперистые; или же—более дорзально и несколько дальше назад (но не более, чем на. У4 расстояния между линией, проведенной через основания v i и лопаточными щетинками), и тогда — мелкие (равные х/4—*/8 длины v 1), гладкие или тонко зазубренные; sc i длиннее sc е или равны им, редко в 2—3 раза, короче; щетинки спинной поверхности идиосомы в большинстве случаев длинные, гибкие, тонко- и редкоперистые, никогда не утолщенные у основания; число их (14 пар) строго постоянно и никогда не испытывает сокращения; щетинки задней половины гистеросомы обычно очень длинны, радиально расходятся в стороны, кверху и назад и образуют вееровидный., «шлейф». Ноги, как правило, довольно стройные, менее изменчивые по своей длине и пропорциям, чем у подсем. Rhizoglyphinae, и слабее воору-
ХАРАКТЕРИСТИКА ПОДСЕМЕМСТНА
о о
О О’
жспные; все щетинки голеней и колен и средние вентральные липок тонкие и гибкие, более или менее зазубренные или редко- и тоикош'.рпстыо. Вершина лапок то с небольшим коническим дорзальным шипом (у родов Туго-lichus иForcellinia), то без него, с 1—5 вентральными шипами; сомой крупной из вершинных щетинок лапок I и II является 2-я внутрь от дорзального вершинного шипа (или его рудимента), которая всегда волосовидна; штифтик р, помещающийся впереди обонятельной палочки, то хорошо развит (у Tyrophagini), то совершенно отсутствует (у Tyroglyphini)-, сен силлы о и е всегда хорошо развиты, s у всех видов щетинковидная, расположенная на значительном расстоянии кпереди и внутрь от обонятельной палочки. Коготки небольшие; praetarsus крупный, подушковидный, одевающий большую половину коготка.
Морфологические различия между полами невелики у всех родов, кроме рода Tyroglyphus Latr.; у с? никогда не наблюдается никаких модификаций гистеросомы, ни сколько-нибудь значительных отличий от 9 в форме и пропорциях тела; совершенно отсутствует также и диморфизм <5 <5, которые здесь всегда представлены одной гомеоморфной фазой. Лостанальных щетинок у сГ всегда 3 пары, и они. все'имеют значительные размеры; тарзальные копулятивные присоски расположены обе в проксимальной половине лапок IV или на примерно одинаковом расстоянии от их основания и конца; вершинная дорзальная щетинка голеней IV имеет нормальное строение. У 9 3 пары анальных микрохэт, часто довольно длинных, 3 пары более крупных аданальных щетинок, 1 пара очень длинных постанальных щетинок (общее число элементов анального комплекса — 7 пар щетинок — остается у всех видов неизменным); анальная щель обычно сдвинута к заднему краю гистеросомы. Вторичные половые различия очень незначительны и заключаются только в несколько большей относительной длине щетинок идиосомы, а иногда в отдельных деталях вентрального вооружения конца лапок; только у рода Tyroglyphus эти- различия резко выражены. — У ним-фальных стадий не наблюдается ретардаций в развитии щетинок спинной поверхности (v е, sc i, h i, dx—d2, 1 а), но нередко в течение онтогенеза имеют, место значительные перемещения v е (которые в этих случаях у личинок расположены дальше назад, чем> у взрослых, и постепенно передви» гаются кпереди). Расселительных гипопусов образуют лишь немногие из представителей подсемейства (Tyroglyphus, Kuzinia и Forcellinia), что, очевидно, находится в связи с большой расселительной потенцией других стадий (обусловленной как их подвижностью, так и высокой парусностью)' и их широкой полифагией у большинства видов.
Подсем. Tyroglyphinae соответствует по своему составу установленным Oudemans’oM семействам Tyroglyphidae Ouds. 1932, Tyrophagidae Ouds. j 1)24 и Forcelliniidae Ouds. 1924, с добавлением некоторых форм из ого сом. (!и-loglyphidae [Aleuroglyphus ovatus Troup. 1879 и Mycetoglyphus P'onwlliniu)' fungivora Ouds. 1932]; возможно, что в него войдет также м JiiloillWHWipiCTM-цеское «семейство» Ebertiidae 1924 (род Ebertia Ouds. 1924). I) ii|)JlllllMtl<)Ml.lX
*
84
1. TfROGLYPHIDAK. 1. TYROGLYPOINAE
мною рамках это подсемейство состоит из 35—40 видов, из которых немного больше 20 описаны из Палеарктики; они распределяются по 8—9 родам, образующим 2 трибы: Tyroglyphini и Tyrophagini.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Верхняя поверхность члеником пог I и II с рельефным орнаментом в виде вильчатых или iqwmnrtpwnto разветвленных килей; все щетипки спинной поверхности утолщенные, короткие, густопушистые; с? без анальных и тарзальных копулятивных присосок, с 2 парами анальных микрохэт; анальное отверстие 9 сильно удалено от заднего края тела ................................................7. Scateglyphus Вег!..
2 (1). Членики всех ног гладкие, без килей; щетинки тела другого строения; d с хорошо развитыми анальными и тарзальными копулятивными присосками, только с 1 парой анальных микрохэт; анальная щель 9 придвинута к заднему краю тела.
3 (4). Половой диморфизм резко выражен; ноги 1yd утолщены, с крупным коническим зубцом на вентральной поверхности бедер, у 9 нормаль--ные; внутренняя вершинная щетинка колен I очень длинная, в 6—10 раз длиннее наружной, имеющей вид прямой, притупленной на конце палочки; вершина лапок с 1 вентральным шипом, без дорзального; сенсилла [3 отсутствует. (1. Триба Tyroglyphini)....................
...........................................1. Tyrogiyphus Latr.
4 (3). Вторичные половые различия слабо выражены: ноги I нормального строения у обоих полов; обе вершинные щетинки колен I приблизительно одинаковых размеров и формы; вершина лапок с 3—5 вентральными шипами и нередко с 1 дорзальным; сенсилла р хорошо развита. (2. Триба Tyrophagini)
5 (8). Конец лапок с довольно крупным коническим дорзальным шипом и 5 хорошо развитыми вентральными шипами.
6 (7). V е очень длинные (равные ®/4 длины v i), тонко- и густоперистые, расположенные латерально, немного позади поперечной линии, проведенной через основания vi; они дугообразно огибают с боков гна-тосому и направлены вершинами косо книзу и внутрь; псевдостигма л ьный орган ланцетовидный, густоперистый по краям; у помещается па одном уровне с обонятельной палочкой; у d тарзальные копулятивные присоски расположены на одинаковом расстоянии от основания и вершины лапки.................................4. Tyrolichus Otids.
7 (6). V е представлены микрохэтами (в 4—6 раз короче v i), гладкие или слабо зазубренные, расположенные дорзально значительно позади поперечной линии, проведенной через основание v i; псевдостигмаль-ный орган щетинковидный; помещается значительно дистальнее, чем обонятельная палочки; у d обе тарзальные копулятивные присоски расположены в основной половине лапки .... 6. Forcellinia Ouds.
1. TYROGIYPHUS
85
8 (б). Конец лапок обычно только с 3 вентральными шинами;1 ve длинные, тонкоперистые, располс женпые латерально, приблизи'юльио на одной поперечной линии с v i.
9 (10). L а в виде ! руппых щетинок, во много раз длиннее dx; da сильно смещены кпереди, очень длинные —- не уступающие в размерах ds; голени и - лапки 1 и II с тупым продольным килем. В гнездах шмелей .........................................3. Kuzinia, gen. n.
10 (!). L а представлены микро: этами приблизительно такого же размера как dx; d2 не смещены кпереди, короче d3; голени и лапки без киля.
11 (12). Ноги длинные и стройные; вершинные шипы лапок мелкие, прямые; S расположена значительно дистал: нее, чем обонятельная палочка; v е длинные, дугообразно огибающие с боков гиатосому, направленные вершинами книзу и внутрь..............................
......................................... 5. Tyrcphagus.Ouds.
12 (11). Ноги относительно короткие, массивные; тершинные шипы лапок крючковидно изогнутые, крупные; S распол<жепа на одном уровне с обонятельной палочкой; ve прямые или слабо дугло'раз-
ные.........................................2. Aleuniglyphus, gen. n.
1. ТРИБА TYROGLYPHINI Захваткин, 1937:179. —Tgrogtyphidae Oudemans, 1932, Entomol. Вег., VIII, n° 183:36.
Половой диморфизм резко выражен: у <5 ноги более или менее сильно веретеновидно утолщены и снабжены крупным коническим зубцом на вентральной поверхности бедер и несколькими небольшими бугорками па таковой колен. Вооружение конца лапок состоит только из 1 вентрального шипа, но у d на лапках I, непосредственно позади него, расположена еще чешуевидно расширенная у основания щетинка, налегающая на его основание; сенсилла р (штифтик впереди обонятельной палочки) отсутствует; внутренняя вершинная щетинка колен I очень длинная, в 6—10 раз длиннее наружной, имеющей вид прямой, притупленной на конце палочки. Анальные копулятивные присоски d круглые, с плотно хитинизированным периферическим кольцом, окруженные кольцом очень мягкой и растяжимой кожи, в состоянии эрекции правильной цилиндрической формы; анальная пара щетинок далеко отодвинута кпереди от присосок.
Известен только 1 род с 3 видами.
1. Род TYROGIYPHUS LATR.
Latreille, 1796; Precis d. caract. е&пёг. d. Inseetes:18b; Oudemans, 1924, Entomol. Ber., VI: 260. — Aleurdbius Canestrini, 1888:7; 1888a:367; Berlese, 1898:XXXV, n° 12; Kramer, 1899:187; Michael, 1903:71.
Тип рода: Acarus farinae L. 1758.
V e в 5—6 раз короче v i, в виде гладких прямых микрохэт, расположенные почти дорзально, значительно позади линии, проведенной череп
• Рудименты дорзального и 4-го и 5-го вентральных шипов, имеющие вид itpidlno мелких толстых щетинок, могут быть обнаружены только с иммерсией.
86
i. TYROGLYPHIDAE 1. TYROGLYPBINAE
основание v i (приблизительно на %—% расстояния между этой линией и лопаточными щетинками); щечные органы сложные, состоящие из нескольких более или менее ветвистых отростков, sci одинаковой длины, с sc е или значительно короче их; hi много короче he; d15 d2 и 1 а короткие, размеры остальных различны у разных видов. Ноги довольно длинные и стройные (за исключением пог I id); сенсилла3 лапок I расположена в проксимальной четверти лапки, па одно!! высоте с обонятельной палочкой.
. Pi расположены кнаружи от р«, почти маргинально, образуя с р2 и Рз дугообразный поперечный ряд, тянущийся параллельно заднему краю идиосомы; обе тарзальные копулятивные присоски расположены в проксимальной половине лапки (IV).
2 - Анальные и адаиальные щетинки представлены микрохэтами и только постанальпые — крупными щетинками. Положение vеу личинок и нимф не отличается от дефинитивного; у одного из видов имеются гипопальные стадии—как расселительная, так и покоящаяся.
Известны 3 вида, из которых один (Т. africanus Ouds. 1910) требует более детального изучения.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Sc i в 2—3 раза короче sc е; h i, dx, d2 и 1 а представлены микрохэтами, все остальные — крупными, явственно перистыми щетинками, из которых многие достигают и даже превосходят половину длины идиосомы и образуют такой яге пышный «шлейф», как,у видов родов Tyrophagus, Tyrolichus и др.; псевдостигмальный орган тонкий, длинный, щетинковидный, очень слабо перистый; лапки Т сГ вентрально посередине с бугровидной выпуклостью..................*2. Т. tyrophagoides, sp. в.
2 (1). Sc i одинаковой длины с sc ё; все щетинки идиосомы, кроме sc е, h е, sa i и постанальных (у с? р3), очень короткие (у <3 равны 8—20, у Б—15% длины идиосомы), гладкие; на заднем конце тела, таким
образом, только 2 пары относительно длинных (30—40% длины идиосомы) щетинок — sa i и ра; псевдостигмальный орган короткий, густоперистый, обычно луковицеобразно утолщенный у основания; лапки I сГ без бугра на вентральной поверхности . . *1. Т. farinae (L.).
*1. Tyroglyphus farinae (L.). — Мучной клещ (фиг. 2 и 3, 25—32, 57—59, 66—69, 88, 90—93, 113—120).
Linn6,1758, Syst. Nat., cd. X, vol. I, pt. 2:616; 0. L. Koch, 1841, Dtsch. Crust., XXXII,fig. 21, 22 (Acarus); Gervais, 1844, in: Walckonaer, Hist. Nat. Ins. Apt., Ill: 262; Berlese, 1884: XIV, n° 9; Oudemans, 1928, Entomol. Bor., VI:260 (7’«rog!ypte); Cancstrini, 1888:26, t. II; 1888a:399, t. XXX, fig. 1; Miohiwl, 1908:73, t. XXIX (Aleurobiue). — Biro Robin, 1860, Bull. Soc. Nat. Moscou, XXXIII:184, t. VIII; Trouponu, 1878:101, (Altwobius). —farris Oudemans, 1905, Entomol. Ber., II, n° 26:20 LdleuroMus); 1918, Arch. i. Naturg., 79, II. 10:45, t. XVII, fig. 19—23, u. t. XVIII, fig. 1—16 (Tyroglyphus).
Биология, n p о д о ii о о и о n т i>. Michael, 1903:78—79; Maurizio, 1905, 1906; Newstead et Duvall, 1921; Zachor, 1927,1938; Романова и Ильинская, 1935, Защ. Раст., 2:77; Романова, 1936, Защ. Раст., 8:20.
Sc i одинаковой длины с sc о, по превышающие 25% длины идиосомы; большинство щетинок спинной поверхности гистеросомы, а именно h i,
1. TYROGLYPHUS
87
все спинные (dx—d4), обе пары 1 и sa е, короткие, длиной в (J—длины идиосомы; h i, dj и 1 а короче других, примерно одинаковой длины («- 12%); d2—d4 и 1 р имеют также почти одинаковые размеры, но несколько длиннее предыдущих; крупными щетинками представлены только й о, ни i и одна из пар постанальных, так что на заднем конце идиосомы их только 4; все щетинки гистеросомы почти гладкие (с единичными, крайне редко расставленными ресничками). Щечные органы с 2—4 прямыми линейными отростками и тонко бахромчато рассеченным основанием. Псевдостигмальный орган довольно короткий, обычно более или менее вздутый у основания, густоперистый. Генитальные щупальцы слабо суженные у основания. Пропорции ног довольно изменчивы, вентральный шип лапок обычно довольно крупный.
d1. Лапки I более или менее сильно конически расширяющиеся к основанию, без бугра на вентральной поверхности; голени I усеченно конические; Pj едва короче р2 и не более чем в Зраза короче р3; анальные щетинки не утолщены. Длина 320—430
q. Лапки I не более чем в 4 раза, лапки IV до 6 раз длиннее своего диаметра посередине; вентральные щетинки лапок IV расположены на большом расстоянии друг от друга, одна значительно дистальнее другой. Длина 350—670 [л.
Личинка. Все щетинки спинной поверхности идиосомы (в том числе и sc) представлены микрохэтами; постанальные длиннее всех других, но и они не превышают 25 % длины идиосомы; грудные палочки стройные, слабо утолщающиеся к концу, с слабо развитым вершинным куполом.
Расселительный гипопус. Беловатый, ноги с легким розовым оттенком. Идиосома в .1.3—1.5 раза длиннее своей максимальной ширины (приходящейся на уровень переднего края гистеросомы), с тупоугольным передним краем, слабо выпуклыми (иногда почти прямыми) боковыми краями и полукругло очерченным задним концом; спинная поверхность умеренно выпуклая, покрытая резкой и густой ямчатой скульптурой. Щетинки спинной поверхности длиннее, чем у гипопусов всех других представителей семейства, тонкие, волосовидные; особенно длинны sc, из которых внутренние (sc i) достигают 25—27%, а наружные — от 12 до 20% длины идиосомы; щетинки спинной поверхности гистеросомы имеют в длину 5—10 % длины идиосомы; d3 длиннее других, sa i очень короткие; расположение щетинок уклоняется от нормы в том, что d2 сильно смещены вперед, a dx раздвинуты в стороны, образуя с d2 прямой поперечный ряд. Проподосомальный щит приблизительно вдвое короче гистеросомального, треугольный, с закругленным на конце ростральным выступом, отделенным пологими прогибами от слабо выпуклых передне-боковых краев щита; v i довольно длинные; глаза отсутствуют. Гнатосома целиком скрыта под ростральным выступом, в 1,5 раза длиннее своей ширины, с очень короткими, слабо обособленными дистальными члениками. Стернальный щит хорошо очерченный с боков мощными эпимеритами II, срастающимися концами о шт-
88 1. TYR0GLYPB1DAE. 1. TYROGLYPBINAE
мерами III; стериум и эпимеры II свободные; коксальные поля III полуоткрытые; эпимеры IV с угловато загибающимися кпереди концами; генитальный щит неясно обособлен от вентрального; вентральный шов заметен только в области коксалыплх полей IV; краевые утолщения довольно хорошо развиты. Коксальные присоски отсутствуют (имеются рудиментарные коксальные щетинки); генитальные расположены ближе к внешним краям щита, чем к генитальной щели. Присасывательный диск маленький, попергчноэллилтичсской формы; периферические присоски немного меньше центральных, боковые образуют с последними прямой поперечный ряд; дополнительные присоски недоразвиты; расстояние между задним краем присасывательного диска и задним краем тела почти равно длине диска. Ноги I и II средней длины (30—35% длины идиосомы), умеренно толстые, почти целиком видимые сверху; вентральные щетинки бедер очень длинные, вентро-латеральные голеней и колен тонкие, гладкие, довольно короткие; колени I с 1 (короткой) вершинной щетинкой; лапки приблизительно втрое длиннее своего диаметра у основания, не длиннее голени и колена, взятых вместе; обонятельная палочка очень длинная и тонкая, резко искривленная перед своим слегка вздутым концом; 7 и е щетинковидные; средняя вентральная щетинка волосовидная, обе боковые и 2 из ’вершинных узколанцетовидные. Длина ног III составляет ок"Ло 25, ног IV — около 2.% длины идиосомы; вентральные щетинки голеней длинные; лапки с 4 ланцетт идиыми и несколькими довольно длинными простыми щетинками; лапки IV па конце с очеьь длинной парусной щетинкой. Дли.,а 2ОС—260 |л, ширина 1чи—180 [л.
Покоящийся гипопус. Идиосома широко-эллиптическая, с полукругло закругленным передним краем, лишенным рострального выступа; спинная поверхность довольно сильно выпуклая, гладкая, с очень короткими и тонкими, трудно заметными щетинками (часть их, повидимому, атрофирована). Гнатосома сильно редуцирована, представлена парой тупых бородавочек, без арист. Строение коксо-стернального скелета, в общем, сходно с таковым у расселителыюго гипопуса. Присасывательный диск рудиментарный, еле намеченный, с 3 парами редуцированных, видимо не-функционирующих присосок; наружные периферические и генитальные присоски совершенно отсутствуют. Ноги короткие и толстые; ноги I выдаются за края тела концом колена, голенью и лапкой, ноги II — только лапкой, III и IV целиком спрятанные под телом; все усаживающие их щетинки представлены микрохэтами, листовидные и парусные щетинки отсутствуют;
Фиг. 107—120.
Фиг. 107. Tyrogiyphus tyrophagoidesi *р. П., (? сверху. Фиг. 101). Т. tyrophagoides, sp. п.( нога I (голень и лапка) с? в профиль. Фиг, 100. 7’. (prop/iapoidoe, ер. п., псевдостигмальный орган. Фиг. 110. Т tyrophagoides, sp. п., отровон овцой ив крупных щетинок гистеросомы. Фиг. 111. т. tyrophagoides. sp. п., щечные отростки. Фиг. 112. 7'. tyrophagoides, sp. п., мандибул > виде внутренней поверхности. Фиг. 113. Т. farinae (I..) f. typloa, йоги I (голень лапка) д. Фиг. 114. Т farinae (Ь.> f. typica, псевдостигмальный орган. «Риг. ПП. 7’. farinae (L.) f. typica, щечные отростки. Фиг. 116. Т. farinae (L.) f. typica, мандибула. Фиг. 117—ПО. Т. farmae (L.) f. /arris Ouds., различная форма -щечных отростков. Фиг. ПО. т. farinae (!..) f. /arris Ouds. псевдостигмальный орган. Фиг. 120. Tyrogiyphus farinae (I..) f. /arris Ouds., мандибула.
-90
I. TYROGLYPBIDAE. 1. TYROGLYPBINAE
обонятельная палочка имеет такую же форму, как у расселительного гипопуса. Длина около 200 у, ширина 160 р.
Распространение. Космополитический синатропный вид. В полевых условиях найден в Германии, Голландии и многих пунктах лесной и степной полосы СССР.
Экология. Tyroglyphus Jarinae (L.) один из наиболее многоядных видов амбарных тироглифоидей, живущий в самых разнообразных пищевых продуктах как. растительного, так и животного происхождения, в сене, соломе и других растительных, остатках; особенно известен как постоянный вредитель зерна и продуктов его переработки. По сравнению с другими амбарными видами характеризуется большей экологической валентностью и меньшой требовательностью к теплу и влажности; оптимальными для него являются температуры порядка 22—26° при атмосферной влажности в 80—90% (соответствующей 16—17% влажности зерна и 17— 18% муки), по он хорошо развивается и при температуре в 7—8° и влажности воздуха в 70%. В полевых условиях распространен шире, чем другие амбарные виды, причем очень обычен в гнездах мышевидных грызунов, сусликов и крота. Переносчиками гипопусов служат, в зависимости от характера местообитания и его энтомофауны, самые различные насекомые: амбарные вредители, шмели, бронзовки (Osmoderma eremita Scop.), стафилиниды и другие обитатели мертвых растительных остатков, блохи (Geratophyllus tesguorum Wagn., Ctenophthalmus breviatus Wagn. et Joff, G. teres J. et Arg., Neopsylla setosa Wagn., Pulex irritans L., Hystrichopsylla segnis Schonch. и др.); мелкие млекопитающие и птицы также служат гипопусам и половозрелой форме в качестве переносчиков.
Изменчивость Т. farinae очень велика и охватывает большое число признаков: степень утолщения ног I у <5, интенсивность окраски конечностей, относительные размеры щетинок идиосомы, строение псевдостигмального органа, щечных отростков, хелицер и т. д. В отдельных популяциях (преимущественно населяющих полевые местообитания вида) преобладают особи с бледными, едва пигментированными конечностями, почти не утолщенными у d ногами I, слабо перистым псевдостйгмальным органом и относительно длинными щетинками идиосомы; такие особи были описаны в свое время Oudemans’oM в качестве самостоятельного вида Т. farris Ouds. Этим формам, однако, нельзя придавать таксономического значения, так как они связаны с типичной полной цепью переходов во всех признаках и все эти признаки очень непостоянны.
I
*2. Tyroglyphus tyrophagoidcs, sp. в. (фиг. 107—112).
Мелкий вид, габитуальпо напоминающий представителей рода Tyrophagus,-, особенно велик,о это сходство у $ Sci в 1.8—3.2 раза короче sc е, длина которых равна 50—-(10% длины идиосомы; из щетинок спинной поверхности гистеросомы малые размеры (4—13% длины идиосомы) имеют только h i, dj, d2 и 1 а; все остальные длиною свыше 40 (до 75)% длины
1. TYROGLYPHUS
91
идиосомы, топко, но явственно, перистые (пористость особенно ннметпа н проксимальных 2/з их длины), совершенно сходные с таковыми Туп-phagus и образующие такой же, как у них, пышный, вееровидный шлейф. Щечные органы представлены 2 линейными отростками, из которых верхний резко изогнут книзу, с несколькими тончайшими зубчиками между их основаниями и такими же на концах. Псевдостигмальный орган длинный, ш,етпп-ковидный, слегка дугообразно искривленный в базальной трети, очень слабо перистый. Генитальные щупальцы сильно конически суживающиеся к основанию.
d. Лапки I почти цилиндрические, вентрально с бугровидной выпуклостью близ середины; макрохэты тела более густоперисты, чем у <?; р4 в 4 раза короче р3 и более чем в 2 раза короче р2; анальные щетинки шиповидные. Длина 290 у.
$. Лапки I около 6 раз, лапки IV приблизительно в 8 раз длиннее своего диаметра посередине. Длина 390—430 ц.
Распространение. Описывается по 1 d и 1? из Агрызского р-на Татарской АССР и 2 99 из Нерубаевского лесного промышленного хозяйства, Одесской обл.
Экология, Экземпляры, найденные в Татарской АССР, были собраны в поверхностном слое почвы (невидимому, перемешанной с растительными остатками) на току вблизи колхозного семенного амбара; экземпляры из Одесской области были собраны в мертвой лесной подстилке в широколиственных лесах.
У татарской q щетинки идиосомы заметно короче, чему украинских; так, длина d4 и d2 составляет у нее 4—5, 1 а 8, sc i 17, d4 и sc е 47% длины идиосомы, тогда как у украинских d4 и d2 имеют в длину 9—12,1 а 11—12, sc i 25—33, sc е 55—57 и d4 62—73% длины идиосомы; псевдостигмальный орган у последних также значительно длиннее. Возможно, что мы здесь .имеем дело с 2 различными подвидами, но имеющийся у меня материал явно .недостаточен для решения этого вопроса.
2. ТРИБА TYROPHAGINI
Захваткин, 1937:179; TyrophagidaeForcelliniidae Oudemans 1923, 1927.
Половой диморфизм практически отсутствует; ноги I у cf не видоизменены. Вооружение конца лапок состоит из 3—5 вентральных и нередко 1 дорзального шипов; сенсилла р всегда хорошо развита; внутренняя вершинная щетинка' колен I немного длиннее наружной. Анальные копулятивные присоски d округлые или яйцевидные, окруженные неполным и менее явственным перепончатым кольцом, в состоянии эрекции более или мопсе молушаровидные (у р. Scatoglyphus они отсутствуй.т); анальная пара що-тниок почти всегда тесно придвинута к присоскам, располагаясь обычно па их выпуклости.
В пределах Палеарктики эта триба представлена 6 родами приблиин-тольнос 20 видами; из внепалеарктических родов к лей должен быть отно-
92 I. TYROGLYPHIDAE. 1. TYROGLYPHINAE
сен род Povelsenia Ouds. 1924, а возможно, также роды Ceroglyphus Vtz.. 1919 и Ebertia Ouds. 1924; кроме того, сюда, несомненно, относятся многие из североамериканских видов tjroglyphus»: Т. americanus Banks 1906, Т. lintneri Osb., Т. breviceps Banks 1906, T. cocciphilus Banks 1906 и др..
2. Род ALEUJlOGl'.TPnUS, gen. n.
Захваткин, 1936:24 (nom. nudum).
Тип рода: Tyroglyphus ovalus Troup. J 879.
Хелицеры очень крупные и массивные, особенно у сГ, у которых их длина умещается в таковой идиосомы только 3.5—4 раза. V е расположены лате-рально, почти на одной поперечной линии с v i, в 1.5—4 раза короче последних, прямые, слабо зазубренные или тоикоперистые; размеры других щетинок идиосомы различны у разных видов, но некоторые из них всегда представлены макрохэтами; степень перистости щетинок также различна, но они никогда не бывают густоперистыми. Псевдостигмальный орган густоперистый. Ноги относительно короткие и толстые; концы лапок с 3 вентральными шинами, но без дорзального (он представлен короткой, толстоватой щетинкой).
<5. I 2 образуют с Pj дугообразно вогнутый кпереди ряд, расположенный на значительном расстоянии от заднего края идиосомы; р3 сидят почти маргинально; тарзальные копулятивные присоски помещаются на приблизительно одинаковом расстоянии от основания и конца лапок IV.
Оба вида, составляющие род Aleuroglyphus, известны только из Европы.
[ ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Sc i в 2—2.5 раза короче sc е; h i, все d и обе пары 1 представлены, очень короткими (4—10% длины идиосомы), редкопериетыми щетинками; более или менее крупные размеры имеют из щетинок гистеросомы только h е, sa е, sa i и 2 пары постанальных, образующие вместе редкий недлинный «шлейф»; анальные щетинки <5 помещаются на самой выпуклости яйцевидных, слабо очерченных спереди присосок .
.•.........................................’1. A. cvatus (Troup.).
2 (1). Обе пары sc одинаковой длины; Голыпинство щетинок гистеросомы (ho, hi, dB, d4, 1 p, обе пары sa и 1 пара постанальных) очень длинные (40—100% длины идиосом ), образующие сзади пышный радиальный «шлейф»; dj и d8 также сравнительно велики (< коло 20% длины идиосомы), анальные щетинки cf сильно отодвинуты от круглых, резко очерченных присосок....................."2. A. Siculus (F. et R.).
*1. Aleuroglyphus ovr.lus (Troupoau) (фиг. 19, 33—35, 80, 94—96, 122).
Troupean, 1878, Bull. Soc. 3ct. d’Angore, VI—VII, faec. 11:106. fig. 9—11; Kramer, 1899:140-(Tyroglyphus): Oudemans, 1932, Entomol. Her., VIII. n° 183: 368 (Caloglyphus).
Экология. Настюкова. 1938 (in Utt.).
2. ALEUROGLYPIirS
!)it
Идиосома беловатая, гнатосома и ноги насыщенного виппо-краспого <цвета, темнее, чем у других видов. Наружные края хелицер chjii.ho выпуклые. Уев 1.5—2 раза короче v i, очень тонкогребенчатые; sc i в 2—-2.5 раза короче see, длина которых составляет около 22—25% длины идиосомы; hi, dx—d3 и 1 а очень короткие (4—7% длины идиосомы), редкоперистыо, обычно с раздвоенным концом и несколько закрученными на конце отростками (ресничками); такое же строение имеют и несколько болоо длинные 1 р (9—10% длины идиосомы) и еще немного превышающие их но длине d4 (13—15%); из щетинок гистеросомы макрохэтами представлены только h е (20—27%), sa е (23—24%), sa i (40—58%), 2 пары постанальных, из которых 4 последние пары образуют своей совокупностью редкий, недлинный «шлейф»; все они тонко- и редкоперисты. Псевдостигмальный орган крупный, пластинчато уплощенный, широколанцетовидной формы, по краям с бахромой из многочисленных (около 20 с каждой стороны) игольчатых отростков. Лапки I в 2.2—2.7, лапки IV в 2.5—3 раза длиннее своего диаметра у основания, почти цилиндрические; оба крайних вентральных шипа лапок крючковидно искривлены и увеличены в размерах, особенно сильно на ногах I и II; вершинная дорзальная щетинка голеней IV короче лапки.
(5. Р2 на % длиннее pj и в 2 раза короче р3, длина которых составляет около 4j% длины идиосомы; penis почти прямой, постепенно заостряющийся к концу, почти в 3 раза длиннее своей ширины у заднего края цоколя; анальные щетинки помещаются на самой выпуклости яйцевидных, неясно очерченных спереди анальных копулятивных присосок; оба крайних вентральных вершинных шипа лапок I и II сильно утолщены (еще крупнее, чем у ф), причем внутренний значительно больше наружного. Длина 480— 550 |i.
ф. Постанальные макрохэты длиною в 25—32% длины идиосомы, анальная щель простирается вплоть до заднего края гистеросомы; оба крайних вентральных вершинных шипа лапок I и II одинаковой длины, менее толстые, чем у . Длина 580—670 pt. У личинок все щетинки спинной поверхности идиосомы очень коротки (самые крупные из них - sc е и 11 е — не превышают 10—15% длины идиосомы); р длиннее всех других, но и они не превышают 35—36% длины идиосомы; грудные палочки короткие, правильной цилиндрической формы, с усеченным концом, едва больше чем в 2 раза длиннее своего диаметра.
Распространение. Описан во Франции, затем найден в Голландии и СССР; у нас пока известен только из Москвы и ее ближайших окрестностей (с. Волховское) и из Киева.
Экология. Живет в муке; в условиях опыта хорошо размножается также в ядре подсолнуха и сильно увлажненном зерне; в полевых условиях найден в норах полевок (3. С. Родионов) и крота (Oudemans). По данным О. К. Настюковой, A. oyatus— высоко термо- и гигрофильный вид. находящий оптимальные условия для размножения при температуре в 35" и отно
94
I. TYROGLYPHIDAE. 1. TYROGLYPHINAE
сительной влажности воздуха в 90% (что соответствует 15-—17.7% влажности субстрата); минимальная температура, при которой вообще возможно его размножение, равна 22°, минимальная атмосферная влажность — 80%, верхние границы той и другой но установлены, но, по Смарагдовой, этот вид обладает соответственно высокой жаростойкостью. Размножаясь в массе, при соответствующих условиях, /I. oval,us приносит иногда ощутимый вред..
*2. Aleuroglyplnis slculiiH (Hum. el Hob.) (фиг. 123).
Fumouze et Robin, 1.807, Jouni. Aunt. 1’hyniol., IV, n° (i: 685, t. 24; Kramer, 1899:141’ (Tgroglyphus).
Окраска гнатосомы и ног менее интенсивна, чем у предыдущего вида; наружные края хелицер менее выпуклые, благодаря чему они имеют более-коническую форму. Ve в 4 раза короче vi, довольно явственно перистые; обе пары sc приблизительно одинаковой длины (около 45—50% длины идиосомы); 1 а очень короткие; d2длиною около 20%, h е и hi, примерно, равны, по длине sc; d3, d4, 1 р, обе пары sa и постанальные очень длинные (60— 100% длины идиосомы), образующие сзади животного очень пышный вееровидный шлейф; все эти щетинки почти гладкие; Псевдостигмальный орган, длиннее v е, коленчато изогнутый и зубчатый. Лапки I почти в 3 раза, лапки IV около 4 раз длиннее своего диаметра у основания, почти цилиндрические; крайние кнаружи вентральные вершинные шипы лапок незначительно крупнее среднего; вершинная дорзальная щетинка голеней IV длиннее лапки.
<5. Р2 короче рх и почти в 3 раза короче р3, длина которых составляет около 90% длины идиосомы; penis треугольный, несколько притупленный’ на конце, едва длиннее, своей ширины у основания цоколя; анальные копулятивные присоски крупные, сходные по строению с таковыми Tyroglyphus-, анальные щетинки отделены от них большим расстоянием; вершинные вентральные шипы лапок I и II не гипертрофированы. Общая длина тела (включая гнатосому) 230—340 р.
9. Постанальные макрохэты длиною в 60—70% длины идиосомы; задний конец анальной щели отделен значительным промежутком от заднего-края тола. Общая длина тела 250—650 р. — Личинка и другие стадии-развития детально не описаны.
Распространение. Сицилия, Азербайджан. Найден описавшими его авторами в сухой шпанской мушке (происходившей из Сицилии)-вместе с клещом Thyroophagus ontomophagus (Lab.) и единичными экземплярами Glycyphagus. Недавно найден в Азербайджане, в пыли колхозного амбара.
3. Род KUZINIA, gon. п.
Тип рода: Hypopus lacvis Duj. 1849.
Близок к роду Ateuroglyphus, от которого отличается сильно смещенными кпереди d2, расположенными почти на одной поперечной линии,
3. KUZINIA
95
с Щ, очень крупными 1а, наличием продольного ребра на голопях и лапках I и II и прямыми вершинными шипами лапок.
Гипопус. Идиосома обратнояйцевидная, с уплощенным, языковидпо выступающим задним концом и очень широкими, пластинчатыми боковыми краями, от степени распластанности которых сильно вариируст но ширина; спинная поверхность гладкая (крайне мелкопористая), умеренно выпуклая. Щетинки спинной поверхности все представлены микрохэтамл; они образуют 2 почти правильных парамедианных продольных ряда щетинок; sa i находятся на большом расстоянии от заднего края тела (что стоит в связи с его своеобразным устройством). Выводные отверстия жировых желез расположены на территории боковых разращений гистеросомы, приблизительно на одном уровне с 1 а. Глаз нет. Гнатосома нерасчлененная, в виде низкого, трапециевидного образования, ширина которого у основания, намного превышает длину. Стернальный щит четко очерченный; его длинные, чуть вогнутые боковые края образуют с слабо выпуклым задним краем, закругленные, но вполне явственные углы; эпимеры II свободные, почти прямые; так же как и стернум, они на значительное расстояние не доходят до заднего края стернального щита; коксальные поля III совершении замкнутые, широко раздвинутые вклинивающимися между ними коксаль-ними полями IV (последние, таким образом, спереди открыты и принимают участие в образовании четкого, прямого переднего края вентрального щита); они слиты между собою и с генитальным щитом в единое целое; поверхность, обоих щитов (стернального и вентрального) равномерно плотно хитинизи-рована; краевые утолщения узкие, лучше заметные на вентральном щите., Коксальные присоски слабо развитые. Присасывательный диск крупный,, несущий 4 пары присосок; дополнительных присосок нет. Ноги средней длины, почти целиком выступающие за края тела; вентральные щетинки, вертлугов и бедер I и II длинные; вентро-латеральные колен и голеней: (1и11) гладкие, толстоватые, но не имеющие вида шипов; дорзо-базальная щетинка колен короткая; вершина колен I с 1 щетинкой. Лапки I и II довольно короткие; на лапках I палочковидная сенсилла 7 расположена, более проксимально, чем главная обонятельная палочка; сенсилла 3-отсутствует; е в виде толстой прямой щетинки; присасывательная щетинка слаборазвитая; две из вершинных и наружные вентральные — крупные, листовидные; на лапках III и IV по 1 довольно длинной парусной щетинке.
*1. Kuzinia laevis (Duj.) (фиг. 81, 97, 98, 124, 159—163).
Dujardin, 1849, Ann. Sci. Nat., (3), XII: 244, t. II, fig. 11—13 (Hypopus).—fucorum Oudemans, 1902, Entomol. Вег., I, n°3:17; 1904, Tijdschr. Entomol., XLVI:18, t. 3, fig. 38—40-(Tyroglyphus). — lundbecki Tragardh, 1905, Fauna Aictica, IV:66 (Tyroglyphus).—echinoyus Vitzthum, 1912, Ztschr. f. wiss. Insektenbiol.: 180, fig. 11 (Rhizoglyphus).
Идиосома сильно пузыревидно вздутая, резко суживающаяся к заднему концу. Sc i несколько длиннее sc е; все щетинки спинной поверх-пости гистеросомы, кроме Щ, представлены макрохэтами длиною в 40— 70% длины идиосомы; все они явственно перистые. Псоидостигмил 1.111.1II
Э6
I. TYROGLYPHIDAE. /. TYROGLYPIHNAE
•орган очень длинный, усаженный по краям многочисленными ресничками. Щечные органы простые, шиловидные, очень нежной консистенции, с шероховатой поьерхкбетыо.
d. Penis клювовидный, с очень крупными ба;альпыми отростками цоколя. Анальные копулятивные присоски такого же типа как у Aleuroglyphus. Длина 480—ООО [А,
ф. Аданальныо и задняя пара анальных щетинок длинные, волосовидные; анальная щель заметно отодвинута от заднего края тела. Длина 520—700 р,.
Личинки характеризуются таким же положением v е, как у личинок Aleuroglyphus и Tyrolichus, и длинными опахаловидными щетинками спинной поверхности тела. У протонимф v е уже более сдвинуты кпереди, а все щетинки спинной поверхности становятся относительно очень короткими (20—23% длины идиосомы), густо перистыми и совершенно однотипными (кроме dx). Строение телеонимф уже близко к дефинитивному.
Гипопус. Более или менее интенсивного буровато-желтого цвета. Проподосомальный щит около 5 раз короче гистеросомы, серповидный, •с тупоугольно выдающимся вперед ростральным выступом; sc расположены слегка вогнутой кпереди дугой; sa хорошо развиты; передний край гистеросомального щита дугообразно выпуклый. Генитальные присоски крупные. Присасывательный диск поперечноэллиптический, в густой ямчатой пунктировке; вентральные присоски очень крупные, боковые едва больше передних, расположенные несколько впереди центральных (образуя с ними вогнутый кпереди поперечный ряд); задние присоски мясистые, чуть больше •боковых. Лапки I и II с 3 крупными и 1 мелкой ланцетовидными щетинками, лапки III с 6, IV — с 4 такими же щетинками; присасывательная щетинка лапок I и II едва выдается за конец коготка, на лапках I — с очень маленьким блюдцем, на лапках II — с булавовидно утолщенным, искривленным концом. Длина 300—390 р..
Распространение. ВсяТоларктика, на север до Гренландии; южная граница точно не установлена, но, вероятно, совпадает с таковой рода Bombus s. lat.
Экология. Живет в гнездах шмелей; гипопусы встречаются в изобилии на всех палеарктических видах рода Bombus s. 1.; изредка попадается также на некоторых других крупных пчелах (Xylocopa pavlovskyi Popov, •Osmia przcwalslcii F. Mor.).
4. Род TYR0LIC1IUS OUDS.
Oudemans, 1923, Mntomol. Ног., VI: 260.
Тип рода: Tyroglyphus oasci Ouds. 1910.
V e расположены латсралыю, чуть позади поперечной линии, провег денной через основания v i, длинные, густо- и тонкоперистые; они более .или менее явственно дугообразно изогнуты, огибая с боков гнатосому,
4. TYROLICHUS
97
и направлены концами вниз и несколько внутрь; sc i одншпсовой длины с sc е или даже длиннее их; обе пары плечевых представлены макрохэтами, так же как и все другие щетинки спинной поверхности гистеросомы, за исключением dx или dx и 1 a; d2 средней длины; все эти щетинки оченьтонко-и редкоперистые или почти гладкие. Псевдостигмальный орган ланцетовидный, густоперистый. Ноги длинные, довольно массивные (значительно толще, чем у Tyrophagus)-, конец лапок с 5 хорошо развитыми, крючковидно изогнутыми вентральными типами и 1 (такого же размера) дорзальным; сенсилла 8 лапок I расположена в проксимальной четверти лапки, на одной высоте с обонятельной палочкой; вершинная щетинка голеней I и II нормального размера.
d. Р2 и р3 расположены маргинально, рх — несколько отступя от заднего края тела; тарзальные копулятивные присоски помещаются на одинаковом расстоянии от основания и конца лапок IV.
9. Из щетинок анального комплекса 3 пары (постанальные и 2 пары аданальных) представлены макрохэтами, остальные 4 пары — очень тонкими и длинными волосовидными щетинками.
У личинок ve расположены много дальше назад, чем у половозрелой стадии, и имеют вид гладких микрохэт; грудные палочки относительно короткие, едва утолщенные в дистальной половине; у протонимф v е приобретают близкие к дефинитивным: относительные размеры, но положение их еще сходно с наблюдающимся у личинок, и только у телеонимф они сдвигаются кпереди до имагинальной нормы.—Гипопальной стадии, невидимому, не образуют.1
Известны 2 вида, относимых Oudemans’oM к 2 различным родам.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). L а представлены макрохэтами, не уступающими по размерам плечевым; на спинной поверхности гистеросомы, таким образом, только 1 пара микрохэт (dj...............................*1. Т. easel (Ouds.).
2 (1). L а представлены такими же микрохэтами, как dx. . 2. Т. lini(Ouds).
*1. Tyrolichus easel (Ouds.). — Сырный клещ (фиг. 1, 39, 84, 99, 100).
Oudemans, 1910, Entomol. Вег., Ill, n° 53:74; 1913, Arch. f. Naturg., LXXIX, H. 4, 10:62 (Tyroglyphus)-, Entomol. Вег., VI: 250 (Tyrolichus).—siro Canestrini, 1888:402, t. 31, fig. 2; Michael, 1903:117, t. XXXV—XXXVI, fig. 11—20 (Tyroglyphus).
Идиосома продолговатоовальная, с едва намеченными, сильно сглаженными плечевыми выпуклостями; v е расположены немного позади поперечной линии, проведенной через основания v i, приблизительно на х/4 короче последних; sc расположены слабо вогнутой кпереди дугой, внутренние в 1.3—1.5 раза длиннее наружных; из щетинок спинной поверхности гистеросомы только dT представлены микрохэтами (длиною около 5—7% длины
1 Приписываемые Т. casei (Ouds.) гипопусы, описанные Vitzhum’oM (1926), нороптио, пи-лиются таковыми Т. farinae (L.).
7 Фауна СССР, Захваткмп—284
98
I. TYROGLYPHIDAE. 1. TYROGLYPHINAE
идиосомы); d2 в 2.5—3 раза длиннее dy, 1 а в .4—6 раз длиннее dx, представленные, как и все остальные пары щетипок, макрохэтами; длина последних колеблется в пределах 30—60% длины идиосомы; все они редко, но довольно явственно перистые. Псевдостигмальный орган веретеновидпо утолщенный у основания, с 9—12 парами тонких игольчатых отростков. Щечные органы состоят каждый из 1 линейного отростка с прямо срезанным концом, угловатым выступом вначале дистальной половины и 2—3 тонкими зубчиками у основания. Лапки I около 3 раз длиннее своего диаметра у основания; обонятельная палочка прямая, почти цилиндрическая, чуть вздутая в своей дистальной четверти.
<5. Penis клювовидный, почти прямой, иостеиеппо суживающийся к своему приострениому коип.у, приблизительно в 3 раза длиннее своей ширины у основания цоколя. Анальные копулятивные присоски округлые, с сплошным хитинизировапным периферическим кольцом, непосредственно впереди которого сидят топкие анальные щетинки; расстояние между тарзальными копулятивными присосками немного превышает их диаметр. Длина 360— 480 |л.
2- Задние края генитальных клапанов образуют более резкий, чем обычно, почти прямой угол с внутренними. Длина 420—725 р.. У личинок d2 в 5 раз длиннее dx и достигают-40% длины идиосомы; грудные палочки в 3 раза длиннее своего максимального диаметра, с прямо усеченным концом.
Распространение. Известен в Голландии, Англии, Италии, Германии и СССР. У нас найден в Москве, в Ивановской обл., в целом ряде пунктов Смоленской обл., на Украине (Одесская обл.) и в Восточной Сибири (Иркутск).
Экология. Синантропный вид, живущий на сыре, в зерне, в семенах льна, в старых медовых сотах, на мертвых насекомых (например в коллекциях); Michael находил его на сухой спорынье. Недавно найден П. К. Чернышевым и в полевых условиях, под корою старых пней. Биология и экономическое значение Т. casei (Ouds.) еще неясны; известно, что в некоторых департаментах Франции и в Тюрингии он специально разводится и применяется в кустарном сыроваренном производстве для придачи сыру особого аромата или фальсификации его внешнего вида.
2. Tyrollchus llnl (Ouds.).
Oudcmtuw, 1928, Rntiwnol, Her., VI:807 (Ityrdborus).
Недостаточно известный вид, отличающийся от предыдущего меньшими размерами (длина d1 280 |л), малой величиной 1 а, представленных здесь такими же микрохэтами, как dx, и несколько более сдвинутыми кпереди v е (располагающимися на. одной поперечной линии с vi).
Распространение. Германия. Найден всего 1 раз, в семенах льна.
5. mOPHAGUS
99
5. Род TIROPHAGUS OUDS.
Oudemans, 1923, Entomol. Ber., VI:260.
Тип рода: Tyroglyphus putrescenliae (Schrank 1781?) Ouds. 1905.
Хелицеры нормального размера, их длина составляет не свыше а/г> длины проподосомы. V е расположены латерально, па одной поперечной липни с v i, длинные, тонкоперистые, дугообразно огибающие с боков гнатосому, направленные концами книзу и внутрь; sc i всегда заметно длиннее see: из щетинок спипной поверхности гистеросомы микрохэтами представлены только dx, 1 а и иногда d2 (в других случаях они имеют среднюю длину); все остальные — очень длинные (50—100% длины идиосомы), редко- и тонкоперистые, образующие сзади длинный и пышный шлейф. Строение псевдостигмальпого органа изменчиво; щечные органы с 1 линейным отростком и более или менее развитой тонкозубчатой лопастью у основания. Ноги длинные и стройные; конец лапок без дорзального шипа, с 3 небольшими, прямыми вентральными шипами (оба крайние кнаружи — 4-й и 5-й — вентральные шипы и дорзальный шип рудиментарны и заметны лишь при самых сильных увеличениях); сенсилла 5 лапок I находится значительно впереди (более дистально) основания обонятельной палочки; вершинная щетинка голеней I и II нормального размера.
cf. Р2 и р3 представлены макрохэтами и образуют дугообразный поперечный ряд вдоль заднего края гистеросомы; pj короткие, расположенные много впереди них; анальные копулятивные присоски косые, более или менее яйцевидные, слабо очерченные спереди (без периферического хити-низированного кольца); положение тарзальных присосок очень изменчиво.
ф. Постанальные и задние пары аданальных щетинок представлены макрохэтами.
Личинка. Положение теи их относительные размеры, как уполово-зрелойстадии; sc емного короче sc i, в виде микрохэт; столь же малую длину имеют и sa е; sa i длиннее всех других щетинок тела, играющие роль «хвостовых»; р а много короче sa i; грудные.палочки длинные, стройные, булавовидно утолщенные к концу. — Гипопальных стадий виды этого рода не образуют.
Космополитический род. Систематика рода Tyrophagus находилась до настоящего времени в хаотическом состоянии. В то время как одни авторы (Michael, Berlese, Vitzthum) считали всех его представителей относящимися к одному, сильно изменчивому, виду, называемому ими «Т. longiorQ&rv.» или «Т. dimidiatus Henn.», другие (Oudemans, 1926) пытались расчленить этот комплекс на ряд более «мелких» видов (Г. humcrosus, Т. viviparus и др.), но эти попытки, к сожалению, не увенчались успехом. Причиной этому явились как неудачный выбор признаков, положенных в основу различения отдельных форм, так и действительные трудности в нащупывании таковых, обусловленные отличным от ccMoiicTiieiuioil нормы характером различий между видами и локализацией большинства стих
X 100 1. TYROGLYPHIDAE. 1. TYROGLYPHINAE
признаков в системах, обычно не подвергающихся детальному изучению (гнатосома, обонятельный аппарат передних лапок, копулятивный орган , пропорции отдельных члеников пог). В связи с этим возникают большие трудности в сопоставлении существующих в литературе видовых названий с формами, различаемыми в настоящей работе, и мне пришлось оставить многие из этих названий неиспользованными. Так, я не нашел возможным фиксировать за какой-либо из этих форм старейшее наименование «putre-scentiae Schrk. 1781», так как в описании Schrank’a нельзя найти никаких указаний, позволяющих сделать это хоть сколько-нибудь обоснованно, и откровенно говоря, в нем и сопровождающих его рисунках даже вообще очень трудно узнать представителя рода Tyrophagus', в представлении же Oudemans’a Т. putresccnliae есть синоним Т. longior Gerv., и он объединял под этим Названием всех вообще Tyrophagus’oB, пока совсем не отказался от него в пользу названия Т. dimidiatusYA&cm. 1804, которое также долго применялось им для обозначения всех представителей рода. Для меня несомненно, что видовое название Негшанн’а должно быть отнесено к одному из «полевых» видов Tyrophagus группы Т. humerosus, но к какому именно, нельзя сказать без изучения соответствующего топотипического материала из Эльзаса (из мха). Т. infestans Berl. 1881 (пес Oudemans 1926) я рассматриваю как синоним Т. longior Gerv., каковым является, по всей вероятности, и Т. viviparus Ouds. 1926; что же касается Т. infestans Ouds. 1926 (пес Berlese 1881), то это, по всей вероятности, не что иное, как форма, несколько произвольно называемая мною Т. humerosus Ouds.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (4). D2 в 2.5—4 раза длиннее dx; ноги относительно короткие, менее стройные; лапки I не длиннее голени и колена, взятых вместе; тарзальные копулятивные присоски <5 расположены приблизительно на одинаковом расстоянии от основания и конца лапок IV.
2 (3). D2 в 2.5 —3 раза длиннее dp, penis очень маленький, резко S-образно искривленный; псевдостигмальный орган ланцетовидный, с длинными, сильно отклоненными отростками, отходящими только от его расширенной основной половины; конический отросток внутренней поверхности хелицер расположен значительно более проксимально, чем мандибулярная щетинка, вытянутый в длинное и тонкое острие; максилляр-пая щетинка ворстсновидно вытянута; сенсиллар лапок I яйцевидная. Длина: сГ 280—350 |Л, $ 320—415 р..............*1. Т. noxius A. Z.
3 (2). D2 в 3.5—4 раза длиннее df, penis очень крупный, дугообразно изогнутый; псевдостигмальный орган с узким стволом и более редко расставленными отростками, отходящими от него на протяжении 2/3 его длины; конический отросток хелицер менее заостренный, близко придвинутый к мандибулярной щетинке; максиллярная щетинка и сенсилла р слабо утолщены. Крупнее: длина с? 450—540 р, $ 550— 700 р.....................................*2. Т. perniciosus, sp. n.
Фиг. 121—124.
Фиг. 121. Tyrophagus noxius A. Z., 9. Фиг. 122. Ateuroglyphus ovatus (Tr.)r c9. Фиг. 123. Л, lion-Jus (F. et R.), 9 (no Fumouze et Robin). Фиг. 184. Kuzinia laevis (Duj.)> протонимфА.
102
I. TYROGLYPHIDAE. 1. TYROGLYPHINAE
4 (1). Da равны по длине или же пемиого (не свыше чем в 1.6 раза) длиннее их; ноги длинные и стройные; лапки I часто длиннее голени и колена, взятых вместе; расположение тарзальных копулятивных присосок изменчиво.
5 (6). Ноги очень длинные и стройные; лапки I почти в 1.5 раза длиннее, лапки TV в 1.45—1.6 раза длинное соответствующих голени и колена, вместе взятых; максиллярния щетинка довольно длинная, неутолщенная; обе вентральные щетинки лапок IV расположены в дистальной трети лапки; идиосома, относительно узкая, почти в 2 раза длиннее своей ширины..................................6. Т. longior (Gerv.).
6 (5). Ноги менее длинные; лапки I по более чем в 1,15, лапки IV не более чем в 1.3 раза длиннее соответствующих голени и колен, взятых вместе; максиллярная щетинка короткая, утолщенная; вентральные щетинки лапок IV помещаются близ середины лапки; идиосома более широкая.
7 (8). D2 в 1.3—1.6 раза длиннее df, лапки I в 1.04—1.16, лапки IV в 1.11— 1.3 раза длиннее голени и колена соответствующей пары ног, взятых вместе; penis тонкий, цилиндрический, постепенно суживающийся к очень тонкому, прямо усеченному концу, по своей форме очень напоминающий носик кофейника; обе тарзальные копулятивные присоски расположены в проксимальной половине лапки; обонятельная палочка лапок I и II очень стройная, длинная, заметно утоньшающаяся к концу; сенсилла далеко отставлена от ее основания........................
.....................................*5. Т. teriuiclavus, sp. n.
8 (7). D2 равны по длине dx; лапки I не длиннее голени и колена I, взятых вместе; лапки IV не более чем в 1.08 (дг)—1.23 (д) раза длиннее голени и колена IV; penis другой формы; расположение тарзальных копулятивных присосок изменчиво; обонятельная палочка лапок I и II короче и толще, расширяющаяся к концу; сенсилла р лапок I придвинута к самому ее основанию.
9 (14). Penis дважды коленчато изогнутый, тонкостенный, воронковидно расширяющийся к косо срезанному концу, крупный; ствол псевдостиг-мального органа нерасширенный, отростки короткие, отходящие от него на большей части его длины; сенсилла р лапок I невздутая . . . ........................................... *4. Т. humeresus Ouds.
10 (11). Дистальный перегиб penis слабый; лапки IV короче, составляющие 0.94—1.07 (д') или 0.98—1.10 (д) длины голени и колена IV, взятых вместо; расположение тарзальных копулятивных присосок очень изменчиво; отрезок of равен 0.75—1.75 отрезка аЪ; последний в 1.5— 3 раза больше расстояния между присосками. Мелкая форма: <5 288— 370 р, 350—405 р.................*4с. Т. humeresus tesquorum, ssp. n.
11 (10). Дистальный перегиб penis более резкий; лапки IV длиннее, составляющие 1.03—1.08 (д') или 1.03—1.23 (<j) длины голени и колена IV, взятых вместе; отрезок ef в 1.5—2.5 раза длиннее ab.
12 (13). Величина: д' 370—465 р, д 470—770 р; отрезок ef лапок IV в 1.45—
5, TYROPHAGUS 103
2.5 раза больше отрезка ab; последний больше расстояния между присосками в 1.25—2.8раза . . . *4а.. Т. humeresusЬитегонин (OikIh.), 13 (12). Величина: d 280—380 ц, £ 380—475 [л; отрезрЩ ef в 2—2,.5 раза больше отрезка ab; последний превосходит расстояние между при-соскамив1.25—1.35раза...........* 4b. Т. humeresus Silvester, няр. И,
.14 (9). Penis короткий, дугообразный илп почти прямой, толстостенный, с закругленным концом; ствол псевдостигмального органа ланцетовидно расширенный, отростки длинные, отходящие только от основных 3/5 его длины; сенсилла лапок I яйцевидно утолщенная. Крупный вид: длина <5 515—520 р.; £ неизвестна . . . *3. Т. molitor, sp. n.
*1. Tyrophagus noxius A. Z. (фиг. 36—38, 122, 125—131).
Захваткин, 1935:25. —Irillleri Волощук, 1936, Защита Раст., 8:155.
Мелкий вид с бледными, почти непигментированными" конечностями, Конический отросток внутренней поверхности хелицер расположен на значительном расстоянии позади (проксимальнее) мандибулярной щетИнки и вытянут в длинное, тонкое острие; максиллярная щетинка острая, гладкая, веретеновидно утолщенная. D2 в 2.5—3 раза длиннее dx и 1 а; псовдо-стигмальный орган пластинчато расширенный, ланцетовидный, с длинными, сильно отклоненными отростками, имеющимися только в его расширенной проксимальной части; вершина вытянута в длинную гладкую ость. Ноги относительно короткие; лапки I не превышают общей длины голени и колена I; обонятельная палочка лапок I и II довольно длинная, почти цилиндрическая, очень слабо вздутая к концу, слегка дугообразно искривленная; сенсилла лапок I яйцевидно утолщенная; вентральные щетинки .лапок IV расположены около середины членика; хитинизированных участков вокруг эпимер нет.
<5. Penis очень мелкий, очень резко S-образно искривленный; его проксимальная часть сжата с боков и пластинчато расширена, так что вентральный край образует выступающий тупой угол; дистальная часть почти цилиндрическая, постепенно суживающаяся к крайне тонкому концу. .Лапки IV немного короче голени и колена, вместе взятых; тарзальные копулятивные присоски расположены на почти одинаковом расстоянии от основания и конца лапки, т. е. отрезки ab и ef почти равны между собой; расстояние между присосками обычно равно их расстоянию от основания и конца лапки. Длина 280—350 р..
2- Лапки IV приблизительно равны по длине голени и колену, взятым вместе. Длина 320—415 р..
Личинка. Sc е в 2 раза короче sc i; обе пары h почти одинаково!! длины; d2 почти в 2 раза длиннее d1 и 1 a; d3 заметно короче sc i и болов чем в 2 раза короче sa i, заметно уступающих в длине идиосоме; на о только л 1.5 раза длиннее 1 а; грудные палочки длиною около 18-- 14 |л.
Распространение. Достоверно известен пока только ио Jioiuiip
104
1. TYROGLYPHIDAE. 1. TYROGLYPHINAE
града, Москвы, Новочеркасска,, Армавира, ряда пунктов Азербайджана, Средней Азии и Уссурийский области (г. Ворошилов-Уссурийский).
Экология; Живет в амбарах и зернохранилищах, в запасах пшеницы, ячменя, муки, различных семяп и других растительных продуктов, принося им серьезный вред; во многих южных районах СССР является одним из наиболее массовых и вредных видов хлебных клещей. По О. К. Настю-ковой, температурный оптимум Т. noxius лежит в пределах между 25 и 30°; при 35° увеличения численности населения уже по происходит, но размножение имеет место и при невысоких температурах порядка 9—10°, т. е. осуществляется в границах температурной зовы амплитудой около 23— 25°; оптимальная для Т. noxius атмосферная влажность — 85—90%, по и при 80% влажности оп развивается довольно хорошо, особенно при близких к оптимуму температурах. В полевых условиях до сих пор не был находим, хотя при высеве его в поле вместе с зерном (производившимся в опытных целях в окрестностях г. Киржач, Ивановской обл.) удерживается здесь в течение всего вегетационного сезона. Точных исследований по реальной вредоносности Т. noxius нет, но в лабораторных условиях она очень велика; при достаточной концентрации клещей в пшенице ими нацело выедается все содержимое зерен (включая эндосперм), от которых остаются одни пустые, продырявленные оболочки.
*2. Tyrophagus perniciosus, яр. в. (фиг. 83, 132—136).
Очень похож па предыдущего, но значительно крупнее; ноги и гнатосома обычно сильно пигментированы. Da в З-5—5 Раза Длиннее dx; псевдостигмальный орган с узким, почти не расширенным стволом и более редко1 расставленными длинными отростками, отходящими от ствола. Ноги относительно короткие и массивные; лапки I не превышают общей длины обоих предыдущих члеников, взятых вместе; обонятельная палочка короткая, массивная, почти прямая, с едва намеченной яйцевидной булавой; сенсилла палочковидная.
с?. Penis крупный, дугообразно изогнутый, несколько сжатый с боков в базальной половине и воронковидно расширяющийся к косо срезанному
Фиг. 126—168. Видовые признаки рода Tyrophagus.
Фиг. 125. Tyrophaqut noxius A. Z., penis в профиль. Фиг. 126. Т. noxius A. Z., базальная группа сенсилл лапки I. Фиг. 127. Т. noxius A. Z., хелицера о внутренней стороны. Фиг. 123. Т. noxius A. Z., макоиллпрпал щотикка. Фиг. 120. Т. noxius A. Z., псевдостигмальный орган. Фиг. 130, 131. Т. noxius A. Z., лайка IV сГ. Фиг. 132. Т. perniciosus, ер. п., penis. Фиг. 133. Т. perniciosus, ер. п., базальная группа ООНОИЛЛ. Фиг. 134. Т. perniciosus, ер. п., макоиллярная щетинка. Фиг. 135. Т. perniciosus, вр.п.,поевдоотт'МалЫ1ый орган. Фиг. 136. Т. perniciosus,ер. п., лапкаIV О'. Фиг.137. Т. molitor, ер. п.,рои1е(» профиль). Фиг. 130. Т, шоШог.вр. п., базальная группа сенсилл. Фиг. 130. Т. molitor, ер. п., макоиллярпан щетинка. Фиг. 140, Т. molitor, ер. п., псевдостигмальный орган. Фиг. 141. Т. molitor, ер. п., лапка IV сТ. Фиг. 142. Т. humerosus tesquorum, ssp. n., penie (в профиль). Фиг. 143. Т. humerosus tesquorum, вер. П,, базальная группа сенсилл. Фиг. 144; Т. humerosus tesquorum, вер. п., хелицера о инутроппой стороны. Фиг. 145. Т. humerosus tesquorum, вер. п., макоиллярная щетинка. Фиг. 140. Т. humerosus tesquorum, ssp. п., псевдостигмальный орган. Фиг. 147, 148. Т. humerosus tesquorum, евр. я., лапка IV d'. Фиг. 140. Т. humerosus humerosus Ouds., penis (в профиль). Фиг. 150. Т. humerosus humerosus Ouds., псевдостигмальный орган. Фиг. 151, 152. Т. humerosus humerosus Ouds., ланка IV О'. Фиг. 153. Т. tenuiclavus, sp. n., penis (в профиль). Фиг. 154. т. tenuiclavus, sp. п., базальная группа сенсилл. Фиг. 155. Т. tenuiclavus, sp. п., псевдостигмальный орган. Фиг. 15G, 157. Т. tenuiclavus, sp. п., лапка IV О'. Фиг. 158. Т. tenuicla-vus, sp. п., щечный орган. .
106
I. TYROGLYPBIDAE. 1. TYROGLYPH1NAE
концу. Пропорции лапок IV и расположение тарзальных присосок, как у Т. noxius. Длина 450—500 ;л.
9. Лапки IV приблизительно равны но длине голени и колену, взятым вместе. Длипа 550—700 р..
Распространение. Достоверно известен только из СССР; найден в Костроме, Барнауле и очень многих пунктах Краснодарского и Орджоникидзе! ского краев, где является самым распространенным и массовым из видов рода Tyrophagus и широко распространен в полевых условиях.
Экология. Живет в зерне (преимущественно пленчатых культур — •овса и ячменя), различных продуктах его переработки, маслосеменах и, вероятно, во многих других растительных веществах в амбарах и на складах, но также и в полевых условиях, где обитает в почве и в скоплениях самых разнообразных мертвых растительных остатков- (Миненкова, 1934; Захваткии, 1936). Специальных лабораторных работ по экологии Т. perniciosus нет.
Вредоносность. По данным В. П. Романовой (1936), повреждения, наносимые этим видом (фигурирующим в ее работе под названием Т. putre-. scentiae Schrank) зерну (пшенице), имеют такой же характер, как и производимые мучным клещом (Tyroglyphus farinae L.). Т. perniciosus очень близок (как и предыдущий вид) к вредящему зерну в США Т. americanus Banks 1906 и к описанному из Цейлона Т. caslellanii Hirst 1915; очень возможно, что он даже окажется идентичным с последним видом.
. *3. Tyrophagus inolitor, sp. п. (фиг. 137—141).
Сходен с Т. perniciosus, от которого отличается малыми размерами d2, строением penis и некоторыми деталями вооружения лапок I. D2 такой же длины, как dt и 1 а; псевдостигмальный орган заметно ланцетовидно расширенный, с длинными отростками в основной половине и вершиной, вытянутой в длинную, гладкую ость. Ноги довольно короткие; лапки I короче голепи и колена, вместе взятых. Обонятельная палочка довольно короткая и толстая, почти цилиндрическая, очень слабо изогнутая; Р яйцевидно утолщенная.
сГ. Penis небольшого размера, слабо S-образно изогнутый, с толстыми, сильно хити визированными «щенками и тупым закругленным концом. Лапки IV несколько короче или равны по длине двум предыдущим членикам ноги, взятым вместо; расположение присосок, как у Т. perniciosus. Длина 515—520 |л. — неизвестна.
Распространение. Вид описывается по 2 С<5 из Саратова. Найден во дворе мельницы, в мусоре под деревянным настилом.
*4. Tyrophagus huinerosus Ouds. (фиг. 89, 142—152).
Oudemans, 1923, Entomol. Bor., V.
Конический бугор внутренней поверхности хелицер массивный, тупой или слегка приостренный, близко придвинутый к мандибулярной щетинке;
5. TYROPHAGUS
107
максиллярная щетинка толстая, но не веретено видная, явственно iiniiyrt|M'ii-пая. И2 такой же длины, как dx и 1 а (короткие); псевдостигмальныП оргии с немногочисленными короткими отростками и цилиндрическим, iiopnciiiH-ренным стволом. Ноги средней длины: лапки I несколько короче или рпины по длине двум предыдущим членикам, вместе взятым; обонятельная палочки, почти прямая, довольно массивная, слегка утолщающаяся к концу; сенсилла р лапок I палочковидная; эпимеры часто окружены неправильной формы хитиновыми образованиями.
с?. Penis крупный, тонкостенный, дважды колепчато изогнутый; дистальное колено его расширяется в виде воронки к своему косо срезанному концу. Отношение длины лапок IV к таковой голени и колена, взятых вместе, колеблется в пределах от 0.94 : 1 до 1.08 : 1; расположение присосок гораздо более изменчиво, чем у других видов Tyrophagus. Длина 280—466 р.
2. Длина лапок IV равна 0.98—1.22 общей длины голени и колена IV, т. е. также значительно более изменчива, чем у других видов. Длина 350—770 р.
Личинка. Sc е превосходят х/2 длины sc i; h е в 2 раза короче h i; длина dlf d2 и la составляет около % длины, sc i; d3 очень длинные, в 2 раза длиннее sc i; sa i в 1.75 раза длиннее d3, значительно происходящие длину идиосомы; sa е более чем в 2 раза длиннее 1 а; грудные палочки длиною около 18 р.
Распространение. Известен пока из Голландии, Германип, СССР: Ленинградской, Московской, Ивановской и Одесской обл., Башкирии, Краснодарского и Орджоникидзевского краев и из Западной Сибири. Т. humerosus образует 3 географических расы.
*4а. Tyrophagus huinerosus humerosus (Ouds.) A. Z. (фиг. 89, 149—152).
Крупная форма: d1 370—465, £ 470—770 p. Псевдостигмальный орган тонкий, щетинковидный, заметно изогнутый S-образно; перистость его такого же типа, как и перистость крупных щетинок гистеросомы — очень короткая и редкая. Ноги обычно пигментированы, эпимеры часто окружены неправильной формы хитиновыми утолщениями.
d. Дистальный перегиб penis резкий. Отношение длины лапок IV к голени и колену IV, взятым вместе, колеблется в незначительных пределах: 1.03—1.07 : 1; отрезок ef в 1.5—2.5 раза больше отрезка ab; последний превосходит расстояние между присосками в 1.25—2.8 раза. Длина 370— 465 р.
2- Отношение длины лапок IV к длине голени п колена IV, взятых вместе, равно 1.03—1.22 : 1. Длина 470—770 р.
Распространение. Голландия, Германия, СССР. Инпостон мне из большого количества пунктов Ивановской, Московской, Килин ты с кой и Ленинградской обл. и из окрестностей Новосибирска.
Экология. Распространен почти исключительно в половой обстановке; живет в сене, соломе и других мертвых растительных остатних (чисто
108
I. TYROGLYPHIDAE. 1. TYROGLYPHINAE
встречается в скирдах и снопах), в почве, на подземных частях живых растений, а также на их надземных частях; нередко встречается на проростках культурных злаков, нанося им иногда ощутимый вред, и на листьях огурцов и других тыквсппых, пораженных паутинными клещиками (Tetra-nychidae).
*4b. Tyrophagus humcrosus Silvester, ssp. n.
Близок к типичной форме, во значительно мельче. Псевдостигмальный орган тонкий, щетипковидный.
d. Дистальный перегиб penis резкий; его дистальное «колено» несколько' длиннее, чем у типичной формы. Отношение длины лапок IV к общей длине' голени и колена, взятых вместе, равно 1.04—1.08 :1; расположение присосок очень постоянно; отрезок ef в 2—2.2 раза больше отрезка ab; последний. в 1.26—1.35 раза больше расстояния между присосками. Длина 280—380 р.
. Отношение длины лапок IV к общей длине соответствующих голени, и колена колеблется в пределах между 1.05 : 1 и 1.25: 1. Длина 380—475 р..
Распространение. Известен пока только из одного пункта: Нерубаевское лесное промысловое хозяйство, Одесской обл. Живет в мертвой растительной подстилке дубовых лесов.
*4с. Tyrophagus humerosus tesquorum, ssp. н. (фиг. 142—148).
По размерам близок к предыдущему, от которого отличается строением псевдостигмальпого органа и penis, несколько более короткими ногами и совершенно исключительной изменчивостью в расположении тарзальных, копулятивных присосок d.
Псевдостигмальный орган прямой, с относительно длинными, довольно-сильно отклоненными от ствола отростками.
d. Дистальный перегиб penis менее резкий, чем у других форм Т. humerosus. Отношение длины лапок IV к общей длине голени и колена. IV равно. 0.94—1.07 : 1, расположение присосок необычайно изменчиво: они то придвинуты к проксимальному концу ланки, как у других форм Т. humerosus, то расположены почти таким же образом, как у видов типа Т. noxius.. Длина 288—370 [г.
9. Отношение длины лапок IV к длине голени и колена IV, взятых вместе, равно 0,08—1.11 : 1. Длина 350—465 |л.
Р а с п р о с т р а и о и и о. Известен из Башкирии (много местонахождений) и с Сонорного Кавказа (окрестности г. Ворошиловска).
Эколог и я. Так же широко распространен в полевой обстановке,.. как типичная форма, по встречается и в норах грызунов и нередко заходит-в сельские зернохранилища и ссыппункты. В южных районах, где он. обитает в поле совместно с Т. perniciosus, последний замещает его в боль-' шинстве местообитаний, и он встречается почти исключительно в биотопах, остающихся вне экологического ареала Т. perniciosus — на подземных частях вегетирующих растений и на их надземных частях.
5. TYROPHAGUS 109
*5. Tyrophagus tenuiclavus, sp. и. (фиг. 86, 101—103, 103 I OB),
Довольно крупный вид, близкий к Т. humerosus, но хорошо отл и 41110111,11 llc.ii от него более длинными ногами, строением penis и рядом других признак,он. D3 в 1.4—1.6 раза длиннее dx; ствол псевдостигмального органа перасши।юн • пый, отростки средней длины, все почти одинакового размера (за исключением самых дистальных). Ноги длинные (лапкп I в 1.04—1.16 раза длиннее двух предыдущих члеников поги, вместе взятых), обычно слабо пигментированные; обонятельная наличка очень тонкая и длинная, заметно утоньшающаяся к концу; длю га се превосходит диаметр лапки у ее середпны.
d. Penis среднего размера, стройный, несильно S-образно изогнутый, .постепенно суживающийся к очень тонкому, прямоусеченному концу; по своей форме он очень напоминает носик кофейника. Лапки IV в 1.10— "1.26 раза длиннее голени и колена IV, взятых вместе; обе присоски распо-.ложены в проксимальной половине лапки. Длина 330—535 |л.
2- Лапки IV в 1.18—1.3 раза длиннее двух предыдущих члеников поги, вместе взятых, очень стройные. Длина 530—670 ц.
Распространение. Достоверно известен пока лишь из пределов СССР, где найден в следующих пунктах: Москва и ее окрестности, Ленинградская, Калининская, Ивановская обл., Северный Кавказ (г. 'Моздок) .и Камчатка (окрестности Петропавловска).
Экология. Встречается преимущественно на гниющих овощах как на складах, так и в огородах; 3. G. Родионов находил его (иа Северном Кавказе) в большом количестве на гниющих яблоках (падалице) в фруктовых садах; встречается иногда также в скирдах и других скоплениях мертвых растительных остатков. *
6. Tyrophagus longior Gerv.
Gervais,- 1844, in: Walckenaer, Hist. Nat. Ins. Apt., 111:262, t. 35, fig 5; Fumouze et Robin, 1867, Journ. Anat. Physiol., IV, n° 6:582, t. 25 (Tyroglyphus). — infestans Berlese, 1884: XIV, n° 6; Canestrini, 1888b: 405, t. 32, fig. 1 (Tyroglyphus).
Мне в натуре неизвестен. Отличается от других видов группы Т. humerosus более узкой, вытянутой в длину идиосомой и очень Длинными и стройными ногами. Максиллярная щетинка длиннее, чем у других видов, тонкая и гладкая. В2 короткие, одинаковой длины с dj и 1 а. Псевдостигмальный орган с щетинковидным, совершенно не расширенным стволом и многочисленными довольно длинными отростками, усаживающими его почти до самой вершины и слабо отклоненными от ствола. Ноги очень длинные и топкие; лапки I у обоих полов приблизительно в 1.5 раза, лапки IV в 1.45— 1.6 раза длиннее соответствующих голени и колена, взятых вместе; обо вентральные щетинки лапок IV расположены в дистальной трети лавки; обонятельная палочка почти цилиндрическая (едва утолщенная к nniqiyr.non-иому концу), довольно длинная.
d. Penis небольйюй, явственно S-образно изогнут и несколько утоиь-инается к концу; обе тарзальные копулятивные присоски рпеполоЖены и про
110
I. TYROGLYPHIDAE. 1. TYROGLYPHLNAE
ксимальной трети лапки, т. е. много базальнее, чем у других видов. Общая, длина тела: <5 170—4.60 р. г.о Fumouze et Robin, до 350 р. — по Berlese, около 550 [г — по Canestrini. Длина ср 200—*750 р,.
Распространение. Западная Европа: Англия, Франция, Италия.
Экология. Синантроп и i.i ii вид, живущий преимущественно на старом сыре; Fumouze et Robin находили его еще на сухих нарывниках; поставленные ими опыты перенесения 7'. longior в гпиющую муку дали отрицательные результаты (клещи погибли через 2 дня), что говорит о меньшей, по сравнению с другими видами, экологической пластичности Т. longior.
Под именем Т. longior Gerv. до сих пор объединяются все сииаптроппые виды рода Tyrophagus, которых, как показывает настоящая обработка, существует несколько.
6. Род FORCEEEINIA OUDS.
Oudemans, 1924, Entomol. Вег., VI: 231.—Mycetogtyphus Oudemans, 1932, ibid., VIII, n° 183:358 (part.).
Тип рода: Tyroglyphus ivasmanni Moniez 1892.
Хелицеры нормального размера. Уев 5—7 раз короче v i, в виде гладких или слегка зазубренных прямых микрохэт, расположенных почти дорзальпо, значительно позади поперечной линии, проведенной через основания v i (приблизительно па 1/4 расстояния между этой линией и лопаточными щетинками); обе пары sc одинаковой длины или внутренние длиннее наружных; все щетинки гистеросомы средней длины однотипные и тогда (так же как и sc и v i) уплощенные, густоперистые по краям и оканчивающиеся своеобразной овальной пластинкой, или же dx, d2 и 1 а представлены гладкими микрохэтами, а остальные — едва перистыми макрохэтами обычного строения, или же размеры и строение щетинок занимают промежуточное положение между этими двумя крайними типами. Псевдостигмальный орган щетинковидный, иногда слегка зазубренный; щечные отростки простые, палочковидные, с прямо усеченным концом и угловатым выступом в дистальной половине. Ноги довольно длинные и стройные; конец лапок с хорошо развитым коническим дорзальным шипом и 5 вентральными;, сенсилла у лапок I расположена значительно дистальнее, чем обонятельная, палочка; вершинная щетинка голеней I и II нормального размера.
с?. Ра представлены небольшими щетинками (не превышающими больше чем в 2 раза длину рх) и расположены па значительном расстоянии от заднего края тела, намечая вместо с рх фигуру перевернутой трапеции (как у боль-
Фиг. 159—169.
Фиг. 159. Kuzinia laevis (Duj.), гипопус, общий пид снизу. Фиг. 1вО. К. laevis (Duj.), гипопус, сверху. Фиг. 161. К. laevis (Duj.), гипопус, лапка 1 сверху. Фиг. 162. То же, снизу. Фиг. 163. К. laevis (Duj.), гипопус, лапка [II спину. Фиг. 164. Forcelliniafungivora (Ouds.), О. сверху. Фиг. 165.
F. fungivora (Ouds.), penis. Фиг. 166. F. waemanni (Moniez), <J сверху. Фиг. 167. F. 'wasmanni (Moniez), penis. Фиг. 168. F. wasmanni (Moniez), одна из щетинок идиосомы. Фиг. 169. F. dia-mesa, sp. n., конец одной из щетинок идиосомы. Буквенные обозначения те же, что и на фиг. 27;.
112
I. TYROGLYPHIDAE. 1. TYROGLYPIUIYAE
шинства Acolyledonini}, p3 длинные, сидящие маргинально; анальные копулятивные присоски такого же строения, как у Tyyophagus, без периферического хитпнизированного кольца; обе пары тарзальных присосок помещаются в проксимальной половине лапки IV. J
Макрохэтами представлена только одна из пар постапальиых щети- I цок. J
Л п ч п и к а. Положение v е и их относительные размеры пе отли- I чаются от дефинитивных; «хвостовыми» щетинками являютсяsa i, превосходящие по своей длине все остальные щетшгки тела; грудпыс палочки длинные и стройные, булавовидно утолщающиеся к концу.
Гипопус известен у 3 видов. Идиосома с гладкой, очень сильно выпуклой спинной поверхностью и пластинчатыми, подогнутыми вентрально .краями; все усаживающие ее щетинки крайне мелки. Проподосомальный щит ср слабо намеченным ростральным выступом или без такового, со слабо .дугообразным почти прямым передним краем; v i мелкие или отсутствуют. Выводные отверстия жировых желез расположены дорзально, непосредственно позади плечевых щетинок, или на том же уровне вентрально. Глаз ; и псевдостигмального органа пет. Гнатосома средней длины или длинная • ;
Цв 2—3.5 раза длиннее своей ширины), нерасчлененная. Стернальный щит четко очерченный, с длинными боковыми краями, образующими с задним краем явственный угол. Стерпум и эпимеры II доходят до его заднего края, замыкая, таким образом, коксальные поля I и II; наружные края коксаль-иых полей I отделены от эпимер II узкими полосками мягкой кожи, так что •стернальный щит оказывается разделенным на 3 самостоятельных щита .'(непарный средний, составленный коксальными полями I, и 2 боковых — жоксальными полями II); коксальные поля III замкнутые, разделенные по средней линии тела узким вентральным швом; эпимеры IV соединяются с вентральным швом далеко позади переднего края вентрального щита; генитальный щит явственно обособлен от вентрального дугообразным или .волнистым швом"; вся поверхность брюшных щитов равномерно плотно хитинизирована, без особенно заметных краевых утолщений. Коксальные •присоски хорошо развиты; присасывательный диск округлый; наружные присоски расположены значительно впереди центральных. Ноги I и II довольно длинные, почти целиком выступающие за края идиосомы; веи--тральпые щетинки вертлугов и бедер короткие, вентро-латеральные ;
•щетинки колен и голеней утолщенные в виде шипов, гладкие; базаль- ,
пая щетинка колон очень короткая, вершина колеи I дорзально только •с 1 щетинкой; лапки стройные; у щетипковидная, в виде S-образно искривленной, довольно крепкой щетинки; присасывательная щетинка лапок!
•с крупным или средней величины блюдцем, па лапках II лишь слегка утолщена к концу; средняя вентральная щетинка шиповидная, наружная и i внутренняя вентральные и вершинные щетинки простые или слегка ланцетовидно расширенные; лапки III и IV с крепкими (отчастизазубренными) .щетинками, без парусных макро хэт.
в. FORCELLINIA
113
К роду Forcellinia Ouds. относятся 4 палеарктических вида; очень возможно, что к нему относится п североамериканский Tyroglyphus lintneri Osborn 1876, известный как вредитель грибов.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОР
1 (2). Dp d2 и I а в виде тонких гладких микрохэт одинакового размера (длиною в 4—7% длины идиосомы), все остальные щетинки спинпой поверхности идиосомы очень длинные (40—60% длпны идиосомы), нормального строения (волосовидные, постепенно утончающиеся к концу), очень редко- и тонкоперистые; sc i заметно длиннее sc е;
с необычайно длинным и тонким penis. Габитуально очень похож па виды рода Tyrophagus группы humerosus Ouds..............
....................................*1. F. fungivora (Ouds.).
2 (1). Dx уплощенные, расширяющиеся к концу, густоперистые по краям, длиною около 15% длины идиосомы; d2 и 1 а приблизительно вдвое длиннее dt; все остальные щетинки спинной поверхности средней длины, уплощенные, расширяющиеся к вершине, оканчивающейся овальной или ланцетовидной прозрачной пластинкой, и густоперистые по краям; или же более длинные и редкоперистые, с очень узкой вершинной пластинкой; обе пары sc одинаковой длины; penis нормальных размеров, клювовидный.
3 (4). Все щетинки спинной поверхности идиосомы однотипные, уплощенные, расширяющиеся к концу, образующему овальную или широколанцетовидную прозрачную пластинку, относительно короткие; у d рх и р2 имеют почти одинаковую длину. Живет исключительно в муравейниках. . . ...........................*4. F. wasmanni (Mon.).
4 (3). лопатчато расширенные, густоперистые, все остальные тонкие, довольно редко- и тонкоперистые, оканчивающиеся узкой ланцетовидной пластинкой; размеры их очень разнообразны, колеблясь в пределах от 13 до 45% длины идиосомы; у J р2 почти в 2 раза длиннее рг В мертвой лесной подстилке..................*2. F. diamesa, sp. n.
Гипопусы
1 (2). Гнатосома выдается за передний край идиосомы всей своей дистальной половиной, длинная (приблизительно втрое длиннее своей ширины), с параллельными сторонами; проподосомальный щит серповидный, с небольшим, ио вполне явственным треугольным ростральным выступом, несущим на конце пару мелких щетинок (v i), не нависающий в виде козырька над основаниями ног I—II; окраска коричпевато-розовая.................................*4. F. wasmanni (Mon.).
2 (1). Гнатосома короткая (не более чем в 2 раза длиннее своей ширины у основания), целиком спрятанная (так же как и проксимальные членики пог I и II) под нависающим в виде козырька проподосомальный.
8 Фауна СССР, Захватшш 284
114
I. TYROGLYPffIDAE. 1. TYROGLYPHINAE
щитом; передний край последнего полти прямой (едва дугообразно’ выпуклый), без каких-либо намеков на ростральный выступ; v i отсутствуют; окраска охрово-бурая.
3 (4). Мелкая форма (длина около 350 ;i); коксальные поля II несколько' суживаются кзади, так что их наружные края имеют косое направление и заметно сходятся кзади; задний край вентрального щита волнистый; периферические присоски немного меньше центральных. ..............................................*1. F. fungivora (Ouds.).
4 (3). Гигантская по своим размерам форма (длина 460 у); коксальные-поля II заметно расширяются кзади, так что их наружные края почти строго параллельны друг другу (и, соответственно, средней линии тела); задний край вентрального щита дугообразный; периферические-присоски много меньше центральных . . 3. F. gigantonympha (Vtz.).
*1. Forcellinia fungivora (Ouds.) (фиг. 4, 82, 104—106, 164, 165, 173, 174).
Oudemans, 1932, Entomol. Вег., VIII, n° 183 : 360 (Myeetogtyplius).
Габитуально и по многим деталям строения очень похож на Tyrophagus humerosus Ouds. V е гладкие; sc i на 1/5—г/4 длиннее sc е, длиною около 40% длины идиосомы; h i почти в 1.5 раза короче h е; dn d2 и 1 а представлены гладкими микрохэтами, приблизительно одинакового размера (4—7 % длины идиосомы), все остальные щетинки спинной поверхности гистеросомы — тонкими редкоперистыми (как и sc и vi) макрохэтами, длиною в 35—60% длины идиосомы, имеющими совершенно такое же строение, как у видов рода Tyrophagus, и также образующими пышный радиальный «шлейф»; самые длинные из них d4, sa i и d3. Псевдостигмальный орган гладкий, щетинковидный.
с?. Р2 в 1.5 раза длиннее р4 п почти в 3 раза короче р3: длина последних составляет около 40%, длины идиосомы; дистальная часть (стилет) penis вытянута в тонкую, необычайно длинную трубку (длиннее лапки + голень IV); вентральные вершинные шипы лапок I нормальные. Длина 410—490 [л.
Постанальпые щетинки гладкие, длиною около 30% длины идиосомы. Длина 520—600 р.
Г и и о и у с. Охристо-бурый. Идиосома имеет форму усеченного спереди эллипсиса, приблизительно в 1.25 раза длиннее своей максимальной ширины, приходящейся значительно поза ди середины, сзади широко закругленная. Проподосомальпый щит в 6 раз короче гистеросомы, выступающий
Фиг. 170—179.
Фиг. 170. Forcellinia wasmanni (Montez), гипопус сверху. Фиг. 171. Тоже, снизу. Фиг. 172. F. v;as-таппг (Moniez), гипопус, голень п липка IV снизу. Фиг. 173. F. fungivora (Ouds.), гипопус снизу. Фиг. 174. F. fungivora (Ouds.), гипопус, ланка I сверху. Фиг. 175. Robinisca mycolicha (Ouds.), гипопус снизу. Фиг. 170. R. mycolicha (Oude.), гипопус, голень и лапка IV снизу. Фиг. 177. R. macrocnemis, sp. п., гипопус снизу. Фиг. 17(1. 11. macrocnemis, sp. п., гипопус, лапка I сверху. Фиг. 170.
R. macrocnemis, ер. п., гипопус, голень и лапка IV снизу.
ш»
116 I. TYROGLYPHIDAE. 1. TYROGLYPHINAE
вперед в виде козырька, с почти прямым (едва выпуклым) передним краем и закругленными передне-боковыми углами; нет никаких следов рострального выступа; v i отсутствуют. Гпатосома в 2 раза длиннее своей ширины, немного суживающаяся it своему расщепленному концу, целиком скрытая под проподосомальным щитом. Коксалыплс поля II заметно суживаются кзади, с конвергирующими назад наружными краями; задний край вентрального щита волнистый. Длина, присасывательного диска едва уступает его ширине, расстояние между задним краем присасывательного диска и задним краем тела, значительно уступает длине диска. Вентро-латеральные шипы голеней и колеи довольно короткие; наружная и внутренняя вентральные щетинки лапок так же листовидно расширены, как внутренняя вершинная; присасывательная щетинка лапок I с толстым стебельком и очень крупным, неотогнутым продолговатым блюдцем. Лапки IV с 2 ланцетовидно расширенными щетинками; их базальная щетинка гладкая, среднедорзальная короче лапки. Длина 250 у, ширина около 200 |л.
Распространение. Достоверно известна только из Англии и ряда пунктов северной и средней полосы Европейской части СССР: окрестности Ленинграда, г. Киржач Ивановской обл., Стерлитамакский и Чекмагушевский р-ны Башкирской АССР.
*2. Forcellinia diamesa, sp. н. (фиг. 169).
Занимает промежуточное положение между типично «тирофагоидной» F, fungivora (Ouds.) и крайне оригинальной мирмекофильной F. wasmanni (Mon.). V е гладкие; обе Пары sc почти одинаковой длины (около 27% длины идиосомы); h i немного длиннее 11 е; dx длиною около 15% длины идиосомы, лопатчато расширяющиеся к концу, густоперистые по краям; d2 и 1 а в 2—2.5 раза длиннее dx и приблизительно равны по длине d3, d4, 1 р и h i (30—35% длины идиосомы); sa е короче других щетинок гистеросомы (13%); sai длиннее всех остальных (около 45% длины идиосомы); все крупные-щетинки спинной поверхности гистеросомы довольно редко- и тонкоперистые, гибкие, неутолщенные, оканчивающиеся узкой ланцетовидной прозрачной пластинкой; v i и sc такого же строения, но без апикальных пластинок. Псевдостигмальный орган слегка зазубренный.
О'. Penis клювовидный, такого же размера, как у F. ivasmanni-, па лапках 1 внутренние вентральные шипы крупнее остальных, направлены косо внутрь и соприкасаются концами. Длина 550 |л.
Р а с и р о с т р а и о и и о. Вид описывается по 1 сГ, найденному в Кунцевском р-не, Московской обл. (дер. Шульгине). Найден в мертвой подстилке (из более или менее перегнивших дубовых листьев) в лиственном лесу.
3. Forcellinia gigantonynipha (Viz.) (фиг. 180, 181).
Vitzthum, 1919, Arch. f. Natiirg., LXXXIV, 6:34, fig. 9 (Tyroglyphus).
Известен только гипопус, чрезвычайно похожий на гипопуса F. fungi-тота (Ouds.), от которого он отличается только своими огромными разме-
в. FORCELLINIA
1 i7
рами и следующими деталями строения: коксальныеполя II несколько рас» ширяются кзади, в соответствии с нем боковые края стернального щита почти параллельны друг другу (и средней линии тела); задний край центрального щита дугообразный; присасывательный диск заметно шире длины, отставленный па большее расстояние от заднего края тела; лшрифо-рические присоски мельче; вентро-латеральные шипы колен и голеней I и II заметно длиннее, так же как и средняя дорзальная щетинка лапок III и IV; присасывательная щетинка с более длинным и топким стебельком и маленьким овальным блюдцем. Все остальные признаки, как у гипопуса F. fungivora Ouds. Длина 460 р., ширина 315 р.
Распространение. Описан из Германии (окрестности Лейпцига). Гипопус найдеп па муравье рода Camponotus (вид не указан).
Мне кажется очень вероятным, что этот вид идентичен с Garsaultia tesludo Ouds. 1916, но недостаточность описания Oudemans’а (ср. стр. 224) не позволяет решить сейчас окончательно Этот вопрос.
*4. Forcellinia wasmanni (Moniez) (фиг. 166—168, 170—172).
Moniez, 1892, Rev. biol. Nord France, IV:387; Berlese, 1897:108; Kramer, 1899:140; Michael,. 1903:131, t. XXXVIII (Tyroglyphus); Oudemans, 1924, Entomol. Ber., VI:231 (Forcellinia). Non Oudemans, 1913, Arch. f. Naturg., LXXIX, 10:170—173, t. XVIII, f. 17—18.
Биология. Wasmann, 1897, Zool. Anz., XX:170—173, 346—360.
V e тонко зазубренные; все остальные щетинки спинной поверхности идиосомы однотипные, сильно уплощенные, расширяющиеся к концу, образующему овальную или широколанцетовидную прозрачную пластинку с явственной срединной жилкой, и густоперистые по хсраям; обе пары sc одинаковой длины (20—25% длины идиосомы); hi заметно длиннее he, ио несколько короче sc; dx короче всех других щетинок спинной поверхности гистеросомы, длина которых колеблется в пределах 18—30% длины идиосомы (длиннее всех d3 и sa i); d2, d4 и Ip имеют почти равные размеры. Лсевдостигмальный орган слегка зазубренный.
d. Р2 едва длиннее рх и почти в 3 раза короче густоперистых р3, длина, которых составляет около 25—28% длины идиосомы; penis клювовидный, нормального размера; вентральные вершинные шипы лапок I не модифицированы. Длина 350—500 р.
Постанальпые щетинки гладкие, приблизительно равны по длине sai. Длина 530—700 р.
Гипопус. Коричпевато-розовый, Идиосома правильной яйцевидной формы, около 1.5 раза длиннее своей максимальной ширины, приходящейся несколько впереди середины, сильно суживающаяся кзади. Проподосомаль-иый щит серповидный, не нависающий в виде козырька над основаниями передних ног, с небольшим, но вполне явственным треугольным ростральным выступом, несущим на конце пару мелких щетинок. Гпатосома. длинная (в 3—3.5 раза длиннее своей ширины), с почти параллельными сторонами, выдающаяся всей своей дистальной половиной за передний край идиосомы. Коксальпые поля II прямоугольные, с немного сходящимися 1шади бокс*
118
I. TYROGLYPHIDAE. 1. TYROGLYPIIINAE
выми краями, задний край вентрального щита волнистый. Коксальные присоски крупнее, чем у других видов; длина присасывательного диска равна его ширине, кзади он заметно суживается; периферические присоски много мельче центральных; расстояние между задним краем присасывательного диска и задним краем тела составляет около 1/2 ег0 длины. Ноги I выдаются за края тела почти всем бедром и 3 следующими за ним члениками, ноги II — начиная с середины колен; вентро-латеральные шипы колен и голеней средней длины; наружная и внутренняя вентральные щетинки лапок волосовидные или едва расширенные; присасывательная щетинка лапок I с широким, но коротким блюдцем, отогнутым под прямым углом к ее стебельку. Лапки IV с 1 ланцетовидно расширенной вершинной щетинкой и зазубренной базальной щетинкой. Длина 210—260 р, ширина 160— 210 (и..
Распространение. Голландия, Германия, СССР; у нас найден пока только в окрестностях Москвы (Кунцевский р-н) и в Калининской обл. (Великолукский р-н).
Экология. Строго мирмекофильный вид, живущий в гнездах Сатро-notus ligniperdus L., Formica sanguinea L., Lasius fuliginosus L. и, вероятно, ряда других видов муравьев. Встречается в влажных частях гнезд, где питается трупами муравьев, их личинок и куколок и другими органическими остатками. Переносчиками гипопусов служат сами муравьи (иногда и их сожители), часто настолько густо облепляемые ими (число гипопусов на одном муравье иногда исчисляется тысячами), что они теряют возможность двигаться и питаться и погибают; массовое появление гипопусов F. wasmanni, наблюдаемое обычно при высыхании гнезда, может таким образом привести к гибели всей колонии муравьев.
7. Род SCATOGLYPHUS BERL.
Berlese, 1913, Redia, IX:104.
Тип рода: Scatoglyphus polytremalus Berl. 1913.
Хелицеры довольно стройные, по своим размерам не уклоняющиеся от нормы. V е раза в 2 короче vi, расположенные дорзально; немного позади линии, проведенной через основание последних, сходящиеся и почти соприкасающиеся своими концами; они имеют такое строение, как и все остальные щетинки идиосомы (кроме анальных и, может быть, стернальных), т. е. сильно утолит,сны и густо опушены. Строение щечпых органов неизвестно. Sc i много короче прямых, расширяющихся к концу sc с; h е и h I одинакового размера — короткие, приблизительно такой же длины, как dx—d3 и 1 a; d1 менее широко расставлены, чем у других родов, образуя с следующими парами d. более правильный (парный) продольный ряд; d4 сильно
Фиг. 180—18G.
Фиг. 180. Forcellinia giganlonympha (Viz.), гипопус сверху. Фпг. 181. То же, снизу. Фиг. 182. Acotyledon mlgini,Bp. п., гипопус сипну. Фиг. 183. A. volgini, sp. п., проподосомальный щит сверху. Фиг. 184. A. volgini, sp. п., гипопус, конец колена, голень и лапка I сверху. Фиг. 185. A. vol-gini, sp. п., гипопус, лапка IV снизу. Фиг, 18(1. A. tshernyshevi, sp. п., гипопус, общий вид снизу.
120
I. TYROGLYPHIDAE. 1. TYR0GLYPH1NAE
отодвинуты назад и занимают терминальное положение; соответственным же образом сдвинуты назад и 1 р и sa с, a sa i полностью смещены на брюшную поверхность тела, где сидят па вершине особой складки (или выпуклости) и почти целиком закрыты сверху ого нависающим над ними задним краем. Коксальные поля III (а может быть, и I и II) щитовидные. Ноги умеренно стройные, недлинные; поверхность члеников ног I и II (кроме вертлугов) украшена ветвистыми, У-образиыми или крестообразными килями; небольшие кили есть в на голенях LJ1 и IV, по они отсутствуют на других члениках этих пар пог; базальные щетинки колен I—III и вентральная щетинка голеней III ланцетовидно утолщенные, пушистые; некоторые из щетинок лапок I и II (невидимому 8, е и вентральные) также опушены
Фиг. 187—191. Scatogtyphus polytrematiis Beil. (по Berlese).
Фиг. 187. сГ, общий вид сверху. Фиг. 188. сГ, гистеросома, вид снизу. Фиг. 189. Q, гистеросома, вид снизу. Фиг. 190. Конец лапки. Фиг. 191. Одна из щетинок идиосомы.
и много длиннее обычного; детали апикального вооружения лапок не вполне ясны, но здесь, во всяком случае, нет массивных конических шипиков. Половой диморфизм очень невелик.
d. Пот пи анальных, ни тарзальных копулятивных присосок; обе пары анальных щетинок представлены топкими гладкими микрохэтами. Поста-нальпыо щетинки па рисунке Bcrloso по изображены: может быть, они сильно-редуцированы или отсутствуют?
$. Генитальное отверстие очень крупное, с прямолинейным задним краем. Анальная щель близко придвинута к генитальной и, соответственно, сильноудалена от заднего края тола ; ни со клапанах сидят 5 пар тонких, гладких и длинных щетинок, из которых задняя длиннее остальных; непосредственно-позади нее находится округлая (хитпиизированная) площадка, в центре которой расположено копулятивное отверстие, а с боков — пара коротких,.
7. SCA TOGLYPHUN
121
ланцетовидно утолщенных постанальных щетинок; эта площадки тицжо отделена значительным промежутком от заднего края тела.Пополов/)’ зрелые стадии неизвестны.
1. Scatoglyphus polytrematus Berl. (фиг. 187-—191).
Berlese, 1913, Redia, IX:104, t. П, fig. 19.
Беловатый, прозрачный, овальный; щетинки тела толстые, довольно' короткие, перистые; из них 6 задних довольно велики (превосходят длину лапок). Членики пог украшены ветвистыми складками или морщинами. Опина снабжена короткими щетинками такой же формы, как вышеуказанные. О' до 310 [а длины, 190 р. ширины; £ до 340 р. длины, 120 [л ширины..
Распространение. Флоренция, редок. Живет в сухом курином помете вместе с другими тироглифидами.
2. Подсем. Rhizoglyphinae
Уев виде мелких, гладких микрохэт, расположенных дорзально, близ-; середины боковых краев проподосомальиого щита, приблизительно на одинаковом расстоянии от v i и лопаточных щетипок, или же совершенно отсутствуют; sc i пе менее чем в 2 раза короче sc е или отсутствуют. Щетинки идиосомы всегда гладкие, 1 в большинстве случаев короткие; основание их часто конически или луковицеобразно утолщено; обычны случаи сильного обеднения хэтотаксии, выражающегося в полном выпадении целого ряда пар, щетипок — v е, sc 1, h i, sh, dx—d2 и иногда d3,1 а и одной из пар крестцовых; щетинки задней половины гистеросомы никогда пе достигают тех размеров, которые они имеют у большинства Tyroglyphinae, и не образуют своей совокупностью пышного вееровидного «шлейфа», столь типичного для последних. Ноги очень вариируют по своей длине и пропорциям; вершина, лапок всегда с хорошо развитым, более или менее коническим дорзальным шипом и 5 вентральными (только у рода Thyreophagus вентральных вершинных шипов 2 + 1 рудиментарный); самой крупной из вершинных щетинок лапок I и II является расположенная непосредственно у внутреннего края дорзального вершинного шипа; следующая за ней по направлению к внутреннему краю лапки щетипка короткая, более или менее утолщенная и часто превращенная в узкую, сжатую с боков пластинку, дугообразно изогнутую книзу; впереди главной обонятельной палочки а всегда находится дополнительная сенсилла р, расположенная на той же сочленовной подушечке;, коготки крупные и массивные, особенно на двух передних парах ног; ргас-tarsus в подавляющем большинстве случаев редуцированный, иногда в очень, сильной степени; медиальные щетинки лапок, латеро-вентральные голепоП и колен, а иногда еще и базальная щетинка колен I и II часто пропр/нцоцы в более или менее мощные, конические шипы; щетинки ног никогда ио ом вают зазубренными или перистыми.
1 У Acotylcdonini концы v i и иногда некоторых других щетинок <wi'l(n iiniiy/ipi'iio.
73.22
1. TYllOGLYPHIDAE. 2. RHIZOGLYPHINXE
Половой диморфизм выражен в большинстве случаев резко; у одних форм он сказывается в большей длине пог и щетинок идиосомы у d, у других — в наличии у с? опистосомального щита, одевающего заднюю часть спинной и боковых (а иногда и вентральной) поверхностей гистеросомы п образующего различные, частью очень причудливой формы, пластинчатые разращения, в деталях вооружения пог, в общей форме тела и т. д. Постанальные ще-тинки d нередко представлены крайне мелкими микрохэтамп, а иногда модифицированы; тарзальные копулятивные, присоски почти всегда смещены к самому концу лапки. У 1 пара постаиальных макрохэт пО—3 пары -анальных и столько же аданальных мпкрохэт; положение анальной щели -сильно вариирует: иногда опа удалена от заднего края тела на такое же .расстояние, как от генитальной тцели, в других же случаях настолько • смещена назад, что простирается до перехода задней поверхности гистеросомы в спинную.
У многих представителей подсем.. PMzoglyphinae наблюдается очень своеобразный диморфизм dd. В этих случаях они представлены двумя резко . различными фазами — гомеоморфной и гетероморфной. У гомеоморфных dd ноги III имеют нормальные размеры и строение, сходные с таковыми ног IV или соответствующей пары ног qq; щетинки идиосомы и ноги относительно короче. У гетероморфных же особей ноги III резко гипертрофированы и видоизменены: они сильно вздуты, их лапки укорочены и обычно слиты с сильно увеличенным в размерах когтем; одна из вентральных щетинок лапок при -этом также гипертрофирована, имеет вид крупного шипа и образует вместе с противопоставленным ей когтем своего рода клешню; praetarsus совершенно отсутствует; в соответствии с гипертрофией свободных члеников пог III сильно увеличены и их коксальные элементы. Кроме того, у гетероморфных dd, щетинки идиосомы обычно длинпее и толще, чему гомеоморфных, и ноги I—II и IV значительно длиннее и стройнее, чем у последних; нередко имеют место и различия в общей форме идиосомы, иногда испытывающей у гетероморфных особей более или менее бросающееся в глаза компенсаторное уменьшение. Способность к образованию гетероморфной фазы d d встречается почти у всех родов данного подсемейства, но обычно присуща лишь • отдельным видам каждого рода; у одних видов гетероморфная фaзadd встречаемся очень редко, у. других она более обычна, и в отдельных колониях ею представлены все или почти все dd; у третьих, наконец, опа сильно пре-• обладает над гомеоморфной вплоть до полного выпадения последней, наблю-..дающогося у ряда видов Nehwielwa.
Способность к образованию рассол отельных гипопусов присуща, вероятно, почти всем видам, что находится в полном соответствии, с малой подвижностью и слабой парусностью других стадий.
К подсем. Rhizoglypkinao отпоситсй свыше 60 видов, из которых больше 40 -входит в состав палеарктической фауны; опп естественно группируются ..в 2 трибы: Acotyledonini л llhizoglypkini.
ТАБЛИЦА РОДОВ
123
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОП сМ
1 (4). V е представлены микрохэтамп, но всегда вполне явственны; нс i. h i, sh, d15 d2 и 1 а хорошо развиты; вершинная щетпика голепсП I V нормального строения; впереди обонятельной палочкп нет пиши (г шс> тинковпдна и расположена значительно впереди и кнутри от лес); задний копец тела пемодифпцпрованпып, без оппстосомальпой пла-стппкп. (1. Триба Acotyledonini.')
2 (3). Наружная вентральная и внутренняя вершинная щетппкп лапок I и II (а иногда еще и лапок III п IV) серповидные, дугообразно расходятся в стороны и дистально расширены в полулунную прозрачную пластинку; вентральные щетинки голеней шиловидные; обе тарзальные копулятивные присоски расположены в дистальной половине лапки (IV). Крупные формы (600—1200 ;л) . . . . 9. Caloglyphus Berl.
3 (2). Наружная вентральная и все вершинные щетинки лапок I—II простые, волосовидные, не расширенные на конце (только у гетероморфных dd A. batsylevi наружная вершинная щетинка расширена в овальную пластинку); вентральные щетинки голеней пе превращены в шипы; тарзальные копулятивные присоски расположены на приблизительно одинаковом расстоянии от основания и вершины лапки IV. Более мелкие формы (350—600 р.)..................8. Acotyledon Ouds.
4 (1). V е отсутствуют; sc i, hi, dx—d2 (а часто также и d3, 1 а и sai) в подавляющем большинстве случаев отсутствуют; если же они есть, то представлены микрохэтамп, и непосредственно впереди обонятельной палочки находится крупный конический шип (гомологичен г); вершинная щетинка голеней IV превращена в полый конический шип; задний конец тела в большинстве случаев сильно модифицирован, образуя хптииизированную горизонтальную опистосомальную пластинку. (2. Триба lihizoglypliini.)
Ъ (6). Ноги IV сильно утолщены; задний конец тела немодифицированиый, без опистосомальной пластинки ...... 11. Heteroglyphus Foa.
6 (5). Ноги IV не утолщены; если же они слегка утолщены (у Histiogasler'), то имеется хорошо развитая опистосомальная пластинка, песущая 4 расположенные веером прозрачных лопасти.
7 (8). Опистосомальиая пластипка отсутствует, задний конец тела не модифицирован; рх и р2 длинные; sc i, hi, d2 п d2 имеются, хотя обычно они и очень мелки; 2 пары sa (sa е и sai); анальные копулятивные присоски без хитинового кольца; наружная п внутренняя вентральные и внутренняя вершинная щетинкв/лапок I и II несколько листовидно расширены на конце................'. . . . 10. RhizoglyphiiH (Пар.
8 (7). Опистосомальная пластинка хорошо развита; рр а обычно также и |»а представлены микрохэтамп, или р2 и р3 отсутствуют; нс j, h i, dt и da, а иногда еще и d3 и 1 а, совершенно отсутствуют; I пари на, (на, i).
124 I. TYROGLYPUIDAE. 2. R11IZ0GLYPHINAE
9(10). Опистосомальная пластинка, резко суживающаяся к основанию, с фестончатым свободным краем, несущим 4 крупные, прозрачные листовидные лопасти, расположенные веером. Ноги IV заметно утолщены; анальные копулятивные присоски с крупным, массивным хитиновым-кольцом или целиком хнтиннзнровапные; вентральные и вершинные-щетинки лапок нерасширенные. Присоски целиком сильпо хитини-зированные, очень рельефные, с ситовидной (пронизанной многочисленными отверстиями) вентральной выпуклостью и кинжаловидно утолщенными щетинками; постанальных щетинок только 1 пара (рх). ................................................12. His tio gas ter Berl.
10 (9). Опистосомальная пластинка приблизительно полукруглая, расширяющаяся к основанию, целыюкрайпая, без каких бы то ни было придатков; ноги короткие, обычно массивные; ноги III несколько короче и тоньше ног IV или одинакового размера с ними, или же ноги III сильно утолщены и модифицированы (у гетероморфных d1 d1); анальные копулятивные присоски хитииизированы только по окружности, умеренно выпуклые, не ситовидные, с простой, неуплощенной щетинкой;. 3 пары р (р, — р3).
11 (12). Непосредственно впереди обонятельной палочки находится массивный конический шип; наружные вентральные щетинки голеней и-колен I и II и две из вентральных щетинок лапок представлены более-или менее толстыми, коническими шипиками; praetarsus совершенно* отсутствует; опистосомальная пластинка расположена на уровне дорзальной поверхности гистеросомы, составляя ее непосредственное продолжение; нижняя сторона этой пластинки скошена кпереди и книзу таким образом, что расположенные на ней анальные копулятивные присоски обращены косо назад (смотреть в профиль); сМ представлены одной только гетероморфной фазой, или она, во всяком случае, сильно преобладает численно над гомеоморфной.....................
.......................................... 13. Schwiebea Ouds.
12 (11). Шипа впереди обонятельной палочки нет; все щетинки голеней и-колен и 2 вентральные щетинки лапок тонкие и гибкие, более или мепее волосовидные; praetarsus хорошо развитый; с? с? представлены, только гомеоморфной фазой.
13 (14). V i отсутствуют; 1 а хорошо развитые, довольно длинные; р2 длинные, волосовидные; see и h-е короткие (12—20 % длины идиосомы) ;ргае-tarsiiH представлен 1 или 2 вертикальными полукруглыми пластинками, прикрывающими с боков, как шоры, основание коготка; строение-опистосомалыюй властлшсн, как у tfchwicbea . . 14. Monieziella Berl.
"14 (13). V i хорошо развиты; 1 а. отсутствуют; р2 представлены крошечными микрохэтами; sc епй с длинные (33—40 % длины идиосомы); praetarsus-в виде мясистой подушечки, одевающий большую половину коготка (как у Tyroglyphinae); опистосомальная пластинка расположена на. уровне вентральной поверхности тела; спинная поверхность гистеро-
ТАБЛИЦА РОЛОН
Г25
сомы сзади редко понижается, образуя над пластинкой крутой скцт (смотреть в профиль)....................15. ThyreophuguH Itoud.
99
1 (4). V е представлены вполне явственными микрохэтамп; sc i, h i, d, n da хорошо развиты; впереди обонятельной палочки нет шипа (г щетинковидная, расположенная значительно впереди и косо кнутри от нее); апальпая щель обычно расположена в середине опистосомы, на большом расстоянии от заднего конца тела; даже в тех случаях, когда опа больше сдвинута назад, расстояние между ее задним концом и задним краем тела не уступает х/2 расстояния от ее переднего конца до генитальной щели. (1. Триба Acotyledonini.)
:2 (3). Наружная вентральная и внутренняя вершинная щетинки лапок 1 и II (а частью также и II и IV) серповидные, крупные, дугообразно расходящиеся в стороны и расширенные дистально в полулунную или ланцетовидную прозрачную пластинку; вентральные щетинки голеней в виде массивных конических шипов; praetarsus небольшой, чехло-образно одевающий только основание коготка. Крупные, иногда гигантских размеров формы (650—2000 у.) .... 9. Caloglyphus Berl.
-3 (2). Наружная вентральная и все вершинные щетинки лапок простые, волосовидные, не расширенные на конце; вентральные щетинки голеней не шиповидные; praetarsus сильнее развитый, подушковидный, одевающий всю основную половину коготка. Более мелкие формы (450—750 у.).....................................8. Acotyledon Ouds.
-4 (1). V е отсутствуют; sc 1, h 1, dx и d2 отсутствуют, если же представлены микрохэтамп, то тогда непосредственно впереди обонятельной палочки находится крупный- конический шип (род Rhizoglyphus); апальпая щель сдвинута назад, простираясь до заднего края тела; нередко даже ее клапаны слегка выдаются позади последнего; анальных микро-хэт только 1 пара, или они совершенно отсутствуют. (2. Триба ВЫго-glyphini.)
'В (6). Sc i, hi, dx и d2 представлены микрохэтамп; псевдостигмальпый орган хорошо развитый, щетинковидный; ноги очень массивные, вооруженные многочисленными крепкими шипами. Крупная (до 1000 у), очень «тучная» форма.........................10. Rhizoglyphus Clap.
45 (5). Sc i, hi,.псевдостигмальпый орган, dx и d2, а часто также и d3 и 1 a, совершенно отсутствуют. Более мелкие формы (280—630 у.).
7 (10). Непосредственно впереди обонятельной палочки находится крупный конический шип (г); только 1 пара крестцовых щетинок (sa i); анальные микрохэты совершенно отсутствуют; praetarsus сильно редуцирован или отсутствует.
•8 (9). Sc е, he, d4 и sa i очень длинные (равные 30—65% длины идиосомы); d3 длиною в 15—40% длины идиосомы; ноги довольно длинные и стройные; наружная и внутренняя вентральные щетинки лапок Т и II длин-
126 I. TYROGLYPHIDYE. 2. RHIZOGLYPHINAE
пые, волосовидные; вершина колеи I дорзально с 2 щетинками; эпимеры III и IV не соединены друг с другом . ... 12. Histiogaster Berl.
9 (8). See, he, d4 и sa i короткие (равные 7—25% длины идиосомы); cl3 отсутствуют пли представлены микрохэтами; ноги очень короткие' (равные 14—25% длины идиосомы), массивные; внутренняя вентральная щетинка лапок I и II шниовпдпая, наружная — волосовидная;, вершина колен I с 1 дорвал ьпой щетинкой, еслпжеих2, тоэпимерыШ п IV слиты концами друг с другом.................13. Schwiebea Ouds.
10 (7). Впереди обонятельной палочки нет пиша; крестцовых щетинок 2 пары (sa с и sa i); имеется 1 пара, анальных микрохэт; praetarsus-хорошо развит; идиосома сильно вытянутая в длину (в 2—3.4 раза, длиннее своей ширины), вальковатая.
11 (12). V е отсутствуют; 1 а хорошо развиты; все щетинки идиосомы короткие, жестковатые; praetarsus представлен 1 или 2 полукруглыми вертикальными пластинками, прикрывающими с боков, как шоры, основную половину коготка; идиосома крайне узкая, в 3—3.4 раза длиннее-своей ширины ....................................14. Monieziella Berl.
12 (11). V е хорошо развиты; 1 а отсутствуют; все щетинки идиосомы очень, длинные и тонкие, волосовидные; praetarsus в виде мясистой подушечки,- одевающей большую половину коготка (такого же строения, как у Tyroglyphinae)-, идиосома в 2—2.2 раза длиннее своей ширины ..............................................15. Thyreophagus Rond.
3. ТРИБА ACOTYLEDONTNI
Вооружение спинной и боковых поверхностей идиосомы не редуцировано, складывается из полного набора—15 пар щетинок; v е представлены микрохэтами, но всегда вполне развитые. Длина и пропорции ног очень-разнообразны; praetarsus развит слабее, чем у Tyroglyphinae, но всегда хорошо заметный. Вершина лапок всегда с хорошо развитым дорсальным шипом и 5 вентральными; на лапках I развиты обе пары базальных сенсилл; они сидят на небольшом расстоянии друг от друга в проксимальной трети лапки; сенсилла г всегда представлена тонкой (не шиловидной) щетинкой и. расположена значительно впереди обонятельной палочки, одна из вершинных щетипок (2-я внутрь от дорзального вершинного шипа) утолщена, ноне модифицирована. Половой диморфизм выражен у большинства форм, но ле особенно резок; он заключается обычно в том, что у d щетинки идиосомы несколько длиннее, чем. у 9, и ноги более стройные и длинные; опистосо-мального щита у d пот, вершинная щетинки голеней IV нормального строения. .У d анальные копулятивные присоски подушковидные, лишенные-кольцевой хитиновой рамки; 3 лары хороню развитых постанальпых.щетинок; тарзальные копулятивные присоски расположены на одной продольной линии. У 9 анальное отверстие в большинстве случаев расположено в середине опистосомы, на одинаковом расстоянии от генитального (яйцевывод-
8. ACOTYLEDON
127
иого) отверстия и от заднего края тела; постанальных щетинок у них 1, пара,, анальных микрохэт 3—6 пар.
К трибе Acotyledonini я отношу 2 рода: Caloglyphus Bell. 1923 п Acotyledon Ouds. 1902, объединяющие 22 палеарктических и около десятка американских п ппдо-малапскпх видов. К этой же группе, вероятно, относятся и род Achropodophorus Rosas Costa 1929.
Я. Род ACOTYLEDON OUDS.
Oudemans, 1903, Tijdsclir. Entoinol., XLV:147. —Eberhardia Oudemans, 1994, Entomol. Ber., VI:230; Захватит 1937:187.—Cosmoglyphus Oudemans, 1932, Entoinol. Вег., VIII,. n° 183:358. —Myrmogtyplius Vitzthum, 1935, Zool. Anz., 0X11:1.
Тип рода: Acotyledon paradoxa Ouds. 1903 (гипопус).
Мелкие или среднего размера формы: с? 350—600 р, £ 450—750 р. Форма и пропорции идиосомы весьма разнообразны; также изменчивы и длина и пропорции ног. Наружная вентральная и все вершинные щетинки лапок Г и II простые, волосовидные, пе расширенные на конце (исключение составляют только гетероморфные с?<5 A. batsylcvi, у которых одна, из вершинных щетинок лапок I и II расширена па конце в овальную пластинку); внутренняя и средняя вентральные представлены то шипами, то-простыми щетпнкамп.
с?. Тарзальные копулятивные присоски расположены приблизительно • на одинаковом расстоянии от основания и вершины лапки.
ф. Анальная щель иногда (у A. rhizoglyplioides, A. krameri и др.) приближена к заднему краю тела, по в большинстве случаев значительно отодвинута от него; анальных микрохэт 6 пар.
Половой диморфизм в большинстве случаев выражен довольно слабо. ’
Личинки с хорошо развитыми ve, sc i, hi, 1 а и dr— d2, чем они легко, отличаются от таковых рода Caloglyphus, у которых все эти щетинки находятся в зачаточном состоянии.
Гипопусы двух резко разнящихся между собой типов, считавшихся самостоятельными родами (Acotyledon Ouds. и Cosmoglyphus Ouds.), ио не соответствующих каким-либо определенным группировкам имагиналигой системы. Гипопусы типа Acotyledon характеризуются следующими признаками. Идиосома овальная, с тупыми, пеуплощенными краями, слабо хитинизиро-ванная; спинная поверхность равномерно выпуклая, усаженная относительно длинными, тонкими волосовидными щетинками; h е на одной поперечной линии с h i. Проподосомальпый щит треугольный, занимающий по менее х/4 спинном поверхности идиосомы; его края не образуют пластинчатых расширений. Стериальыи щит плохо очерченный; задний край его лишь слабо намечен тонкой, иехитннизированной складкой кожи; стсриум и эпимеры II короткие, свободные, далеко не достигающие заднего itpiui стернального щита,, дугообразно изогнутые; коксальные поля 1II замкнутые, довольно широко раздвинутые; коксальные поля IV разделены «ужинающимся кзади промежутком, слабо намеченные; гопптальиыП щит на обо-
128
I. TYROGLYPHIDAE. 2. RIIIZOGLYPHINAE
юоблен; поверхность всех щитов брюшной стороны идиосомы слабо хитини-зированная (почти перепончатой консистенции'), с совершенно не выраженными краевыми утолщениями. Коксальные присоски слабо развитые пли отсутствуют; присасывательный диск мягкий, перепончатой консистенции, сильно вытянутый в длину, языковидный или же сильпо редуцированный; присоски на нем в количество 1—4 пар, слабо очерченные, более или менее рудиментарные. Ноги средней длины, довольно массивные, все однотипные; ноги I целиком видимые сверху, ноги 11 видны, начиная с вершины бедер; .щетинки голспей и колеи игольчатые; лапки с 3—4 слабо ланцетовидными щетинками, передние без присасывательной щетинки (замененной длинным простым волоском).
Гипопусы типа Cosmoylyphus очень напоминают таковых рода For-'cellinia Ouds. Идиосома широкоэллинтическая пли почти круглая, с пластинчато уплощенными краями, в большинстве случаев сильно хитинизи-рованиая и окрашенная; все щетинки спинной поверхности гистеросомы представлены микрохэтамп, sc крепкие, более или менее искривленные; .11 е расположены значительно позади li 1. Проподосомальный щит очень короткий (в 8—30 раз короче гистеросомы), пластинчатый, часто с крыло-видными разращениями по бокам. Вентральная поверхность идиосомы сильно хитинизирована; стернальный щит очень четко очерченный, с длинными, почти прямыми боковыми краями и дугообразно вогнутым задним краем, отделенным от вентрального щита тонкой, перепончатой линией; стернум и эпимеры II топкие, очень длинные (лишь немного не доходят до заднего края щита), почти прямые и очень слабо сходящиеся кзади; вентральный щит явственно обособлен от генитального, с дугообразно выпуклым передним краем, длинными прямыми боковыми краями (превосходящими длину их ацетабулярного края) и волнистым задним краем; коксальные .поля III полуоткрытые; эпимеры III длинные, сходятся кпереди под острым углом и оканчиваются у вентральных щетинок, на некотором расстоянии от вентрального шва и переднего края щита; коксальные поля IV узкие, открытые, разделенные только спереди. Коксальные присоски хорошо развиты ( у A. ewynympha передние заменены щетинками); присасывательный диск округлый или поперечноэллиптический, сильно хитинизированный, с б парами хорошо развитых присосок, очень сильно удаленный от заднего края •тела. Ноги I и II довольно массивные, частично спрятанные под пластинчатыми краями проиодосомального щита; паружпыс вентральные щетинки голеней и колец превращены в крупные, кипжаловидные шипы (у A. tsher-nyshevi они тоньше, чем у других видов); лавки более или менее уплощены, с 4—5 закрученными на конце листовидными щетинками и хорошо развитой присасывательной щетинкой. Ноги 111 и IV очень стройные, с длинными тонкими лапками (па препаратах почти всегда отгибающимися под прямым углом к проксимальным членикам ноги), вооруженными 3 — 5 листовидно расширенными и более или мепео закрученными на конце щелинками.
8. ACOTYLEDON
li»O
К роду Acotyledon я отношу 12 палеарктических видов; к ному отнооятся также тропические A. dampfi Ouds. 1928, A. leefmansi Ouds. 1927, /I. oudtt-nbansi A. Z. 1937 и A. sironiformis F. et R. 1868.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
dd*
1 (2). Лостанальные щетинки расположены в 2 продольных ряда; расстояние между щетинками передней (рх) и средней пар (р2) одинаково — их основаниями намечается квадрат; расстояние между задним концом анальной щели и задним краем тела превосходит больше чем в 2 раза ее длину; ироподосомаигнатосома очень узкие.. 5. A. michaeli (Ouds.).
2 (1). Расположение постанальных щетинок иное: рх всегда расставлены значительно шире, чем р2; расстояние между задним концом анальной щели и задним краем тела не более чем в 1.5 раза превосходит ее длину; пропорции гнатосомы и проподосомы нормальные.
3 (8). Sc 1 в 5—10 раз короче sc е; расстояние между sc 1 превышает больше чем в 2 раза расстояние между каждой из щетинок этой пары и sc е; псевдостигмальный орган щетинковидный, гладкий или почти гладкий; вентральные щетинки лапок не утолщены в виде шипов; обонятельная палочка лапок I и II с булавовидным, вздутым концом; 1 р не менее чем вдвое длиннее 1 а.
4 (5). Sc i в 10 раз короче sc е; они отстоят друг от друга на расстояние в 6 раз большее, чем от sc е; все щетинки гистеросомы чрезвычайно короткие: длина he составляет не больше 15, d4, 1 р и sa i 5—7% длины идиосомы; ноги короткие и очень массивные; лапки I и II менее чем в 2 раза длиннее своего диаметра посередине.....................
.............................. *1. A. rhizoglyphoides (A. Z.).
5 (4). Sc i в 5—6.5 раза короче sc е; расстояние между ними не больше чем в 4-—4.3 раза шире промежутка, отделяющего каждую из них от se е; некоторые из щетинок гистеросомы довольно длинные: h е составляет 20—30,d4,lpHsa i от 16 до 18% длины идиосомы; ноги более длинные и стройные; длина лапок I и II превосходит больше чем в 3 раза их диаметр посередине.
( 7) 6; Р2 расположены на том же уровне, как рх, образуя с ними правильный поперечный ряд; h е длинные, составляют около 30 % длины идиосомы; ноги стройные.....................................4. A. agilis (Can.).
7 (6). расположены далеко позади plt почти на том же уровне, как р„, образуя с последними несколько дугообразно выгнутый кпереди ряд; he короче, составляют около 20% длины идиосомы; ноги массив-пые...........................................*2. A. sokolovl, нр, и.
8 (3). Sc i в 2—3 раза короче see; расстояние между sc i iipoiii.iinaoT но более чем в 1.5 раза расстояние между каждой из щетинок этой пиры
9 «Лауна СССР, Захватит!—284
130
I. lYROGLYPHIDAE. 2. RHIZOGLYPHINAE
й see; псевдостигмальпый орган перистый; обонятельная палочка' лапок I и II не образующая явственно обособленной булавы.
9 (10). Идиосома сильно вытянутая в длину,почти цилиндрическая,в2.1— 2.2 раза длиннее своей ширины; вентральные щетинки лапок довольно тонкие, игольчатые; 1 р в 3 раза длиннее 1 а. . . . *8. A.krameri(Berl.).
10 (9). Идиосома пе болео чем в 1.9 раза длиннее своей ширины; средняя и. внутренняя вентральные щетинки лапок I и II превращены в довольно массивные конические шипы; 1 р не более чем в 1.5 раза длиннее 1 а.
11 (12). Щетипки идиосомы (особенно более короткие из них) прямые, крепкие, почти игольчатые; р2 расположены на одной линии с р3..........
......................................... 7. A. alberti (A. Z.)..
12 (11). Щетипки идиосомы более или менее гибкие, не игольчатые.
13 (14). Щетипки идиосомы длинные, все очень тонкие, волосовидные;, у гомеоморфных особей длина d3 и 1 р составляет около 22,1 а 16—17% длины идиосомы; р2 расположены на одной линии с р3, в 3 раза длиннее Рр.псевдостигмальный орган утолщенный, широколапцетовидный,. с несколькими (4—6) щетинковидными отростками на своей дорзальной поверхности; у гетероморфных особей одна из вершинных щетинок: лапок I и II расширена на конце в овальную, прозрачную пластинку .................... . . ...................*10. A. batsylevi, sp. n.
14 (13). Щетинки идиосомы короче: длина d3 и 1 р не превышает 16—17, 1а—9 % длины идиосомы; некоторые из них (d1? d2, la, а иногда еще d3, d4,1 р и h i) притупленные на конце, при небольших увеличениях кажущиеся обломанными (при большом увеличении видно, что конец их шпателевидно уплощен и закруглен); р2 расположены значительно-впереди р3, намечая вместе с ними трапециевидную фигуру, не более чем вдвое длиннее рх; псевдостигмальпый орган перистый только по-краям; гетероморфная фаза неизвестна и, вероятно, отсутствует ... . . . ...................................* 6. A. redikorzevi (A. Z.).
15 (16). Щетинки спинной поверхности идиосомы короткие, длина sc i составляет около 10, dx и d2 6.5—7.5,1 р 14—15, sa е 14, sa i 26—27%. длины идиосомы; псевдостигмальный орган шире, более утолщенный .........................* 6а. A. redikorzevi redikorzevi (A. Z.).
16 (15). Щетинки идиосомы длинные: длина sc i составляет 13—14, d1nd2 9—10,1 р около 17, sa е 23—24, sa i до 32% длины идиосомы; псевдо-стигма льиый орган почти цилиндрический............................
.........................*61). A. redikorzevi borealis, ssp. n.
Фиг. 102—203. Лапки I Tyroglyphidae.
Фиг. 192. Acotyledon redikorzevi. (Л. Z.), лапка I 9 сверху. Фиг. 193. To же, снизу. Фиг. 194.
Caloglyphus rodionovi A. Z., лапин 1 гетероморфного o’ сверху. Фиг. 195. Тоже, снизу. Фиг. 19S.
Rhizoglyphus echinopus (F. et II.), ланка I гетероморфного O’ сверху. Фиг. 197. То же, снизу,-Фиг. 198. Histiogaster bacchus, ер. К., лапка 1 cf сверху. Фиг. 199. Schwiebea rossica, sp. п„ лапка I Q сверху. Фиг. 200. То же, сливу. Фиг. 201. Thyreophagus entomophagus (Lab.), лапка I § сверху.
Фиг. 202. Th. entomophagus (Lab.), лапка I сливу. Фиг. 203. Th. entomophagus (Lab.), конец лапки L 2 снизу. Буквенные оОовпачения см. в тексте, стр'. 66.
132 I. TYROGLYPHIDAE. 2. RIIIZOGLYPHINAE
99
1(8). Sc i в 6—10 раз короче sc с; расстояние между обеими щетинками этой пары превышает более чем в 2 раза расстояние между каждой из них и sc е; вентральные щетинки лапок I и II не утолщены в виде шипов; псевдостнгмал ьпып орган щетинковидный, 1 р более чем в 2 раза длиннее 1 а.
2 (3). Sc 1 около 10 раз короче sc е; расстояние между ними в 6 раз больше, чем между каждой из них и sc е; все щетинки гистеросомы очень короткие: длина d3 и составляет б—0,1 р и sa i около 7, h е не больше 15%
длины идиосомы; ноги короткие, очень массивные: лапки I и II не более чем в 2 раза, лапки IV около 3 раз длиннее своего диаметра у середины; анальная щель сильно сдвинута кзади — расстояние между ее задним концом и задним краем тела в 2.5 раза меньше, чем расстояние от ее переднего конца до заднего края генитальной щели.........
. . ............................ *1. A. rhizoglyphoides (A. Z.).
3 (2). Sc i в 6—7 раз короче sc е; расстояние между ними превышает не более чем в 4—4.3 раза промежуток между каждой из них и sc е; часть щетинок гистеросомы значительно длиннее; ноги длиннее и Стройнее, лапки I и II не менее чем в 3 раза длиннее своего диаметра посередине, расстояние между задним концом анальной щели и задним краем тела составляет больше половины промежутка между ее передним концом и задним краем генитальной щели.
4 (5). D2 в 3 раза длиннее dx, равные половине длины d3; последняя составляет около 25—27 % длины идиосомы; расстояние между генитальной щелью и анальной равно приблизительно расстоянию между задним концом последней и задним краем тела; постанальная пара щетинок расположена далеко позади анальной щели . . 5. A. michaeli (Ouds).
5 (4). D2 равны по длине dx и в 5 раз короче d3, или же, когда последние также представлены микрохэтами, одинакового с ними размера; расстояние между генитальной щелью и анальной в 1.7 раза больше расстояния между задним концом последней и задним краем тела; постанальные щетинки расположены чуть позади заднего конца анальной щели.
6 (7). D3 представлены микрохэтами, так ясе как и dj и d2; длина h е и d4 составляет около 30, sa г около 25% длины идиосомы..................
............................................ 4. A. agilis (Сан.).
7 (6). 1)3в 5 раз длинное da, равные приблизительно 15% длины идиосомы; такую же, примерно, длину (17—18%) имеют d4 и sai; he длиною в 20% идиосомы................................*2. A. sokolovi, sp. n.
8 (1). Sc i в 2—3 раза короче нее; расстояние между обеими'щетинками этой пары превышает расстояние между sc еикаждой из них не больше чем в 1.5 раза; вентральные щетинки лапок I и II по большей части пеплообразные; псевдостигмальпый орган перистый.
8. ACOTYLEDON
133
!) (10). Анальная щель близко придвинута к заднему краю толп.; идиосома сильно вытянута в длину, в 2.1—2.2 раза длиннее своей ширины; центральные щетинки лапок I и II игольчатые; 1 р в 3 раза длинно I п, . . . ......................................... *8. A. krameri (Bin,'].).
10 (9). Анальная щель расположена на приблизительно одинаковом рас-стоянии от заднего края тела и от генитальной щели; идиосома не более чем в 1.9 раза длиннее своей ширины; вентральные щетинки лапок I и II шиповидпые; 1 р не более чем в 1.5 раза длиннее 1 а.
11 (12). Щетинки идиосомы прямые, игольчатые; d3 короткие, такой же длины, как d2 (около 6% длины идиосомы); d4B 2—2.5 раза длиннее d2 ........................................... 7. A. alberti (A. Z.).
12 (11). Щетинки идиосомы более или менее гибкие, неигольчатые; d3 в 1.5 раза длиннее d2, одинакового размера с d4 (12—25% длины идиосомы).
13 (14). Щетинки идиосомы длинные, очень тонкие, волосовидные; длина d3 и d4 составляет 24—25, 1 р и sa i около 21—23, sa е 17—18% длины идиосомы; sc i немного более чем в 2 раза короче sc е; псевдостигмаль-ный орган уплощенный, широколанцетовидный, с несколькими щетинковидными отростками на своей дорзальной поверхности; средняя и внутренняя вентральные щетинки лапок I и II в виде тонких, длинных шипов.................................... * 10. A. batsylevi, sp. и.
14 (13). Щетинки идиосомы короче: длина d3 и d4 не превышает 15—16, 1 р и sa е 10—12, sa i 18% длины идиосомы; sc i в з раза короче sc е; часть щетинок спинной поверхности (dx, d2, 1 а, а иногда также и d3, d4,1 рий i) с притупленными концами; псевдостигмальный органуже, перистый только по краям; средняя и внутренняя вентральные щетинки лапок I и II в виде довольно коротких и массивных конических шипов....................................* 6. A. redikorzevi (A. Z.).
15 (16) Щетинки спинной поверхности идиосомы короткие: длина v i составляет 11—12, see 25—27, h е около 20, d2 7—7.5, d3 12—13, la 7.5—8% длины идиосомы; псевдостигмальный орган более или менее пластинчатый.................*6 a. A. redikorzevi redikorzevi (A. Z.).
16 (15). Щетинки спинной поверхности идиосомы длиннее: длина v 1 равна 7.5—14, see 30—33, he около 25, d2 10.5—11, d3 1.5—20, la 11% длины идиосомы; псевдостигмальный орган почти цилиндрический ................................* 6. b. A.redikorzevi borealis, ssp. n.
Гипопусы
1 (6). Проподосомальный щит приблизительно треугольный, с тупыми неуплощенными краями, занимающий не менее J/4 спинной понорх-ности идиосомы; присасывательный диск мягкий, перепончатой консистенции, сильно редуцированный и несущий тогда 1—2 нары рудиментарных присосок, или же он вытянут в длину, узкий, лныконпд-пый, с 4 парами также более илп менее редуцированных присосок;
134 I. TYROGLYPHIbAE. 2. RIIIZOGLYPIIIA'AE
коксальные поля I п II слабо хитинизированные, не образующие хоть сколько-нибудь ясно очерченного стернального щита; коксальные . поля III замкнутые, заметно раздвинутые; область генитального щита не отграничена ни от гаиссальных полей IV, ни от присасывательного-диска.
2 (3). Присасывательный диск с 4 парами присосок (расположенными в таком порядке : 2, 2, 4), узкий, языковидный, сильно вытянутый в длину; присоски неродной и центральной нар довольно крупные, но плохо очерченные, задние - все одинакового размера, мелкие, более рельефные; коксальные присоски более или мспее явственные................
.................................* 1 A. rhizoglyphoides (A. Z.).
3 (2). Присасывательный диск совершенно рудиментарный, с 2 парами „ более или менее редуцированных присосок; коксальные присоски отсутствуют.
4 (5). Ано-генитальная щель очень длинная, по бокам (позади середины) с парой довольно хорошо развитых присосок; 2-я, совершенно зачаточная, пара присосок находится немного позади первой и представлена только их сдвоенными центральными порами, расположёнными совсем близко друг от друга; эпимерйты II совершенно не выраженные; стернум значительно не доходит до концов сильно дугообразно искривленных эпимер II.............................* 3,. A. paradoxa Ouds.
5 (4). Генитальная щель короткая, не соединенная с анальной; по бокам ее присосок пет, и обе пары их расположены значительно позади нее; присоски передней пары гораздо сильнее раздвинуты, чем задние, и одинаковой с ними формы и размера, эпимериты II довольно явственные; стернум длинный, заходящий так же далеко назад, как почти прямые эпимеры II.........................5. A. michaeli Ouds.
Q (1). Проподосомальный щит иной формы, с листовидно уплощенными краями, прикрывающими сверху основание или более или менее значительную часть ног I и II; длина его не превышает х/8 общей длины, идиосомы;-присасывательный диск нормального строения, сильно хити-низированный, округлый или поперечно-эллиптический, с 5 парами хорошо развитых присосок; коксальные поля I и II равномерно сильно хитипизированные, образующие Очень четко очерченный, плотный стернальный щит; коксальные поля III открытые, если же замкнутые, то смыкаются по сродней линии тела или же, наоборот, очень широко раздвинуты; генитальный щит очень ясно обособленный как от вентрального щита, так и от присасывательного диска.
7 (8). Периферические щетинки гистеросомы (по крайней мере h i, й е, sal и р а) толстые, шиловидные..................7. A. alberti (A. Z.).
8 (7). Периферические щетинки гистеросомы тонкие, гибкие, волосовидные.
9 (12). Передний край цроподосомального щита почти прямо усеченный, слабо волнистый, с слегка дугообразно выпуклой средней лопастью и еле намеченными боковыми вырезами; ростральный выступ смещен
8. ACOTYLEDON 1.13
на его вентральную поверхность и целиком скрыт им или отоутотпуот, гнатосома с параллельными краями или слабо суживающаяся it концу.
10 (11). Крупная (380 у), томно окрашенная форма; проподосомильиыН щит чрезвычайно короткий, около 35 раз короче гистсросомпл!.-ного; вентральный щит много длиннее стернального, с длинным вентральным швом; коксальные поля III открытые; ноги I и II короткие и массивные...............................11. A. yolgini, sp. n.
1 (10). Мелкая (245—285 p), бледно окрашенная форма; длина проподосо-мального щита составляет около 1[8 длины гистеросомального; вентральный щит много короче стернального, без продольного (вентрального) шва; коксальные поля III замкнутые, очень широко раздвину- тые; ноги I и II длинные и тонкие: ноги I выступают за передний край шроподосомального щита, начиная с вершины бедер, ноги II—начиная с вершины колен; наружные шипы голеней и колен I и II небольшие, тонкие . . . .............................*12. A. tshernyshevi, sp. и.
12 (9). Передний край проподосомального щита глубоко дугообразно вырезанный по обеим сторонам треугольно выдающегося вперед рострального выступа, на вершине которого сидят хорошо развитые теменные щетинки; вентро-латеральные шипы голеней и колен I и II широкие, ножевидные; гнатосома грушевидная, не более чем в 2 раза длиннее своей ширины, глубоко расщепленная на конце; коксальные поля III открытые, эпимеры IV свободные.
13 (14). Ростральный выступ короче почти прямоугольных (закругленных лишь на вершине) боковых лопастей проподосомального щита (т. е. не достигает линии, проведенной через их вершины), довольно широкий; расстояние между задниц краем присасывательного диска и задним краем идиосомы равно длине стернального щита........................
..........................................* 8. A. kraineri (Berl.).
14 (13). Ростральный выступ длиннее широко закругленных, почти полукруглых боковых лопастей проподосомального щита (т. е. выдается за линию, проведенную через их вершины), вытянутый в тонкий отросток; расстояние между задним краем присасывательного диска и задним •краем идиосомы уступает длине стернального щита .......................
..............t.........................* 6. A. redikorzevi (A. Z.).
*1. Acotyledon rhizoglyphoides (A. Z.) (фиг. 204—209, 242).
Захватили, 1937:191, рис. 35—42 (Eberhardia).
По общему виду, пропорциям тела и конечностей, малой длине щетинок "гистеросомы и положению анальной щели у § напоминает некоторых прод-•ставителей трибы .Rhisoglyphini, например Schwiebea или Rhisoglyphutt, Идиосома сильно удлиненная, вальковатая, в 1.8—2 раза длиппоо ouooll максимальной ширины; боковые края ее, начиная от h е и до 1 р, почти параллельные, задний конец широко закругленный. Sc i в 10 ран короче нс о, •^близко придвинутые к их основанию, так что расстояние между но 1 пршюо*
136
1. rYROGIA'PHIDAE. 2. RHIZOGIAPHINAE
ходит около 6 раз промежуток между каждой из щетинок этой пары и sc е;. последние много длиннее всех остальных щетинок идиосомы, составляют около 25% ее длины; щетинки гистеросомы чрезвычайно короткие, тонкие, довольно гибкие; размеры их, выраженные в процентах к общей длине идиосомы, в среднем, таковы: h о 14—15; hi, dx, d2, 1 а и sa е 2.4—2.5; d3, d4, 1 р и sa i 5—6.5; ощутимых различий между полами в длине щетинок не-наблюдается; sc е и более крупные из щетинок гистеросомы слегка утолщенные к основанию. .Псо.вдостигмальный орган щетипковидный, гладкий, слегка S-образно изогнутый. Ноги относительно очень короткие и массивные, составляющие, в среднем, 28, 26, 21 и 24% длины идиосомы; лапки 1 и И в 2 раза, лапки IV в 2.8—3 раза длиннее своего диаметра близ середины; вентральные щетинки ланок прямые, жестковатые, не утолщенные в виде-шипов; обонятельная палочка лапок I и II стройная, с почти шаровидной, хорошо обособленной булавой.
d. Р2 расположены на одном уровне с р3, образуя вместе с ними прямой поперечный ряд; расстояние между рх приблизительно равно расстоянию между р3; анальная щель отделена от заднего края тела промежутком, немного превосходящим ее длину; дистальная часть penis короткая, слабо-искривленная. Длина 360—653 р,.
9. Постаиальные щетинки расположены на одном уровне с задним концом анальной щели; последняя сильно сдвинута назад, так что расстояние между ее передним концом и генитальной щелью в 2.5 раза больше промежутка, остающегося между ее задним концом и задним краем тела. Длина 535—755 [л.
Гипопус. Типа Acotyledon. Окраска беловатая с более или менее сильным буроватым оттенком. Щетинки спинной поверхности тонкие, волосовидные, довольно короткие. Передне-боковые края проподосомального-щита заметно вогнутые; ростральный выступ закругленный. Стернум короче половины проподосомы; эпимеры II направлены друг к другу под прямым углом, слегка дугообразные, с узкими аподемами; эпимериты II слабо развитые, но все же довольно явственные, намечающие трапециевидные очертания стернального щита; коксальные поля III длиннее своей ширины, их сочленовные края (обращенные, к вертлугам) дугообразно вогнутые. Коксальные присоски редуцированные, в виде несколько хитинизиро-ванпых бляшек; генитальные присоски мелкие, но вполне явственные;, врисасьна'юльпын диск очень слабо хитипизиро] апиый, сильно вытянутый в длину; присоски ла нем расположены в 3 продольных ряда: 2, 2, 4; передние и центральные присоски довольно крупные, с явственными срединными кольцевидными порами и редуцированными периферическими частями; боковые и задние периферические присоски мелкие, тесно сближенные между собой и расположенные в прямой поперечный ряд; они рельефнее и сильнее хитинизированы, чем остальные; генитальная щель нормального размера, хорошо отделенная от анальной. Сенсилла 8 лапок I щетинковидная, внутренняя вентральная щетинка голеней I и II и вентральные:
8. ACOTYLEDON
11.7
лапок IV довольно длинные, игольчатые. Длина 210—250 ширина iy;i ’
145 [л.
Распространение. Известен только из пределов СССР: Моск ли, Кострома.
Экология. Живет в старом, более или менее заплесневевшем зерно (пшеница, лен) в амбарах (особенно их подпольях).
* 2. Acotyledon sokolovi, sp. п. (фиг. 210—215).
Sc i в 6—6.4 раза короче sc е; расстояние между обеими щетинками' этой пары в 4.5 раза больше промежутка, отделяющего каждую из них от sc е; последние много длиннее всех остальных щетинок идиосомы, составляющие около 30—31% ее длины; щетинки гистеросомы, в общем, короткие; hi, d.!, d2 и 1 а представлены микрохэтамп, длиною в 2.5—3.3%;. длина h е составляет 20, d3 15, d4 17—20, 1 р 15—16, sa е 5—6, sa i 17—18% длины идиосомы; здесь характерны очень малые размеры h i, dx, d2, 1 a, и sa e и относительно большая длина 1 р, превосходящих 1 а в 5—5.5 раза; более крупные из щетинок крепкие, резко луковицеобразно утолщенные у основания, более мелкие — очень тонкие, гибкие. Псевдостигмальный орган щетинковидный. Ноги средней длины; все вентральные щетинки лапок довольно длинные и тонкие, гибкие, не шипообразные; обонятельная палочка лапок I и II толстая, с более или менее хорошо обособленной яйцевидной булавой.
с?. Гомеоморфная фаза. Различий в размерах усаживающих идиосому щетинок между d и $ нет. Р2 расставлены, примерно, так же широко, как р3;р2 расположены почти на одной линии с р3 и составляют вместе с ними слабо дугообразно выгнутый кпереди поперечный ряд; они расставлены значительно шире, чем у других видов, так что расстояние между ними умещается в расстояние между рх чуть больше 2 раз. Анальная щель, отстоит от заднего края тела на расстояние в 1.5—1.75 раза больше, чем ее длина. Ноги несколько короче, чем у 9. Длина 600—655 р.
9. Идиосома в задней своей части шире, чем у Jjy сильно раздутых, яйцами особей она имеет грушевидную форму; расстояние между генитальной щелью и анальной почти в 2 раза больше расстояния между задним, концом последней и задним краем тела. Ноги несколько длиннее, чем у d" лапки IV заметно длиннее голени и колена, взятых вместе. Длина 550—740 ;х.
Гипопус. Очень сходен с гипопусом A. rhizoglyphoides, но значительно крупнее (длина 300 — 350 [г, ширина 210—295 р.) и с болео-узким, тонко приостренным ростральным выступом.
Распространение. Известен мне из Ленинграда и его окрестностей, окрестностей Москвы, из Ивановской обл., Чекмагушепского р-на Башкирской AGGP и Барнаула.
Экология. Живет в разнообразных растительных остатних: и скирдах ржи, на зерновой просыпи в подпольях зернохранилищ и и сильно влажных семенах, в подстилке из отмерших листьев в лесах (и этом последнем
138
I. TYROGLYPHIDkE. 2. RHIZOGLYPHINAE
-субстрате найден мною под Москвой, но отсутствует в очень больших акарологических сборах из него, произведенных П. К. Чернышевым на Украине).
* 3. Acotyledon paradoxa Ouds. (фиг. 216—220, 239—241).
Oudemans, 1902, Entomol. Bor., 11'14; Tijdsch. Entoinol., XLV:147, t. 12, fig. 48—49.
Известны только протонимфа и гипопус.
И р о т о п и м ф а. В высшей степени похожа па протонимфу A. sokolovi, отличаясь от псе целым рядом топких признаков: 1) лапки длиннее и стройнее: их длина превышает диаметр посередине в 3.1—3.2 раза; 2) вентральные щетинки большинства члеников ног песколысо длиннее и значительно толще, прямые, игольчатые, частью почти шиповидные (например вентральные щетинки лапок III и IV); 3) средняя вентральная щетинка лапок I распо--ложена значительно ближе к основанию лапки, чем к заднему краю среднего вершинного шипа; расстояние между нею и боковыми вентральными щетинками довольно велико (больше диаметра лапки); 4) обонятельная палочка лапок I и II стройнее; 5) вершинная щетинка голеней III очень длинная, вдвое длиннее лапок III; 6) генитальные щупальцы крупнее, сильно расширяющиеся к концу; диаметр их полушаровидной вершины около 9 р..
От протонимф также очень близкого вида A. rhizoglyphoides (A. Z.) отличается гораздо более длинными щетинками идиосомы и ног, более длинными и стройными ногами (у протонимф A. rhizoglypholdes лапки I менее чем вдвое .длиннее своего диаметра), яйцевидной формой булавы обонятельной палочки лапок I и II (у A. rhizoglyphoides она почти круглая), расположением sc и более крупными, сильнее расширенными генитальными щупальцами (диаметр их у A. rhizoglyphoides 3.5 [J-, форма — почти цилиндрическая).
Гипопус. Типа Acotyledon. Окраска беловатая. Передне-боковые края проподосомального щита прямые или очень слабо вогнутые; ростральный выступ тупой; задний край гистеросомы широко закругленный; щетинки спинной поверхности довольно длинные, очень тонкие, волосовидные. Стернум длиннее половины проподосомы; эпимеры II направлены друг к другу под тупым углом, сильно изогнутые, с очень широкими аподемами; эпимериты II неявствеиные; проподосома (стернальный щит) отграничена сзади дугообразной поперечной линией (складкой кожи); коксальныеполя III короткие; их свободные (обращенные к основанию ног) края волнистые, так же хсак и у коксальных полей IV. Стернальные присоски отсутствуют,
Фиг. 204-—22G.
Фиг. 204. Acotyladonrhlzoglynhoidos (Л. Z.), o' сверху. Фиг. 205. A. rhizoglypholdes (A. Z.), Q снизу. Фиг. 206. A. rhizoglypholdos(A. Ноги .1 (,> сверху. Фиг. 207. Л. rhizoglypholdes (A. Z-), псевдостигмальный орган. Фиг. 200. Л, rhizotllypholdos (Л. Z.), лапка IV сГ. Фиг. 209. A. rhizoglyphoides
УА. Z.), генитальное шуиильцо Иротоиимфы, Фиг. 210. Л. sokolovi, sp. п., сГ сверху. Фиг. 211. A. sokolovi, sp. п., лапка I О сверху. Фиг. 212. Л. sokolovi, вр. п., лапка I протонимфы сверху. Фиг. 213. То же, снизу. Фиг. 214. Л. sokolovi, нр, п., липка и вершинная щетинка голени IV протонимфы. Фиг. 215. A. sokolovi, sp. п., генитальное щупальце протонимфы. Фиг. 216. A. paradoxa Ouds., протонимфа, лапка I сверху. Фиг. 217. То же, спину. Фиг. 218. A. paradoxa Ouds., папка IV сбоку. Фиг. 219. A. paradoxa Ouds., вершиппаи щетинка голени IV. Фиг. 220. A. paradoxa Ouds., генитальное щупальце. Фиг. 221. A. redikorzevi (Л. Z.), 9 сверху. Фиг. 222. A. redikorzevi (A. Z.), псевдостигмальный орган. Фиг. 223. Aeotytedon batsylevi, sp. n., Q сверху. Фиг. 224. A. batsy-utevi, sp. n., псевдостигмальный орган. Фиг. 225. A. batsylevi, sp. п., лапка I гетероморфного o' сверху. Фиг. 226. A. batsylevi, sp. п., лапка I о сверху.
140
I. TYROGLYPHIDAE. 2. RfUZOGLYPHINAE
так же как и генитальные; присасывательный диск представлен мягкой,, перепончатой подушкой, не отграниченной от вентро-генитального щита, но-довольно ясно очерченной сзади и с боков тонкой складкой кожи; на ней расположены только 2 пары сильно редуцированных присосок, из которых передние (соответствующие .передним периферическим присоскам гипопусов, других типов) хорошо различимы, а задние (гомологичные центральным присоскам) представлены только сдвоенными центральными порами и: заметны лишь на. хорошо просветленных препаратах; обе эти пары присосок тесно придвинуты к длинной, узкой апо-гепитальпой щели. Строение и. вооружение ног, как у A. rliKoolyphoides, но сенсилла s лапок I и II крупнее, шиповидная, а внутренняя: нейтральная щетинка голеней тех же пар ног и базальная. щетипка лапок IV короче и толще и вентральные щетинки бедер I значительно длиннее. Длина 244—285 р, ширина 160—195 [л.
Распространение. Описан из России без более точного указания района и местности; мне,известен пока только из Иркутска.
Экология. Гипопусы, послужившие Oudemans’y для описания видаг были собраны с ближе не определенной летучей мыши; иркутские экземпляры — гипопусы и линяющие протонимфы — собраны в большом числе, в зерне (пшеница) в амбаре.
4. Acotyledon agilis (Сап.) (фиг. 227, 228).
Canestrini, 1888:30, t. I, fig. 11—13; 1888a:408, t. 33, fig. 2; Kramer, 1899:139 (Tyrogtyph'us');: Захваткин 1937: 193 (Eberhardia).
Мне в натуре неизвестен. Судя по описаниям и рисункам Canestrini, крайне близок к A. sokolovi; отличается от него только тем, что у d р2 расположены па одном уровне с рп образуя с ними прямой поперечный ряд,, и что d3 представлены микрохэтами (на рис. 2а Canestrini они не изображены,, как и несомненно представленные микрохэтами dx, d2 и 1 а); кроме того,, ноги, невидимому, значительно длиннее и стройнее, чем у A. sokolovi-, по-рис. 2 той же работы их длина у d составляет 36, 32, 30 и 32% длины идиосомы; наконец, дистальная часть penis изображена более резко искривленной, чем это имеет место у A. sokolovi. Все остальные признаки, как у A. sokolovi. Длина 9 около 650 [л, ширина 390 [л; размеры d не указаны.
Гипопус неизвестен. Личинки размером около 200X110 |л, с длинными,, стройными, булавовидно утолщенными к концу грудными палочками; постапалышо щетинки у лих длиннее sa i, которые очень коротки. Яйцо-широко-эллипсоидальное, как у A. rhisoolyphoidos и других близких видов,, размером около 110 X 80 [л.
Распростри и о и и о. Малостей только из Италии. Живет в разлагающихся растительных веществах.
5. Acotyledon michaoll (Ouds.) (фиг. 229—233, 353).
Oudemans, 1924, Entomol. Bor., VI: 267 (Tyroglyphvs).— agilis Michael, 1903: 95, t. XXXII,. XXVI, fig. 1—7 (Rhizoglyphus); OudemujiH, 1924, Entomol. Ber., VL230 (Eberhardia)-, 1932, ibid., Vlll: 358 (Coswwglyphus).—michiwli Захнаткнп, 1937 :188 (Eberhardia).
8. ACOTYLEDON
Тип рода Eberhardia Ouds. 1923. Мне в натуре неизвестен; на ооиоиании оригинального описания и многочисленных рисунков Michael’я может быть •охарактеризован следующим образом.
О'. Гетероморфная фаза. Идиосома узкая, сильно удлинон-лая: отношение ее длины к ширине равно 2.2 : 1; наибольшая се ширины приходится на уровне 1 а, откуда она постепенно суживается до sa о; задний конец слегка усеченный. Гнатосома также очень узкая, сильно вытянутая в длину. Sc i в 3 раза короче sc е, расставленные (судя по рис. 1 табл. XXXII Michael’n) на одинаковое расстояние друг от друга и от sc е; длина последних составляет свыше 40% длины идиосомы; большинство щетинок гистеросомы очень длинные, крепкие, вероятно несколько утолщенные у основания; размеры их в процентах к длине идиосомы (вычисленные -но тому же рисунку Michael’a) таковы: h i, d2, 1 а и sa е около 14—16; h е, d3 и 1 р 40; d4 околю 60; sa 1 48; dx на рисунке не изображена и, невидимому, представлена микрохэтами. Псевдостигмальный орган не изображен и в тексте не упомянут. Анальная щель сильно отодвинута от заднего края тела, от которого отделена промежутком, превосходящим больше чем вдвое ее длину; постанальные щетинки расположены в 2 продольных ряда; расстояние между щетинками передней (рх) и средней (р2) постапальных пар одинаково их основаниями намечается квадрат; щетинки обеих этих пар -очень короткие, шиловидные, раз в 10 короче р3; дистальная часть penis вытянута в очень тонкое, прямое острие. Ноги чрезвычайно длинные и тонкие (кроме, разумеется, модифицированной III пары), составляющие (по рис. 2 табл. XXXII), соответственно, около 42, 40, 38 и 52 % длины идиосомы; лапки I и II около 5 раз, лапки IV около 7 раз длиннее своего диаметра посередине (рис. 3, 4, 6 табл. XXVI); вентральные щетинки лапок крепкие, прямые, игольчатые, но не в виде «настоящих» конических шипов, что особо подчеркивается автором; расположение тарзальных копулятивных присосок не указано, Michael даже отрицает их наличие. Средние размеры {«наиболее обычный размер в найденной мной колонии»): длина идиосомы около 570 [л, ширина — 280 р..
Гомеоморфная фаза Michael’eM, к сожалению, не описывается и на его таблицах не изображена; он только отмечает, что она существует наряду с гетероморфной.
9. Габитуально резко отличается от С. Идиосома гораздо шире, около 1.65 раза длиннее своей наибольшей ширины, с сильно дугообразно выпуклыми на всем протяжении боковыми краями; ее задний край почти прямой. Гнатосома нормальной ширины. Sc i в 6 раз короче sc е; расстояние между ними втрое больше промежутка между каждой из них и sc о; последние значительно длиннее всех других щетинок идиосомы, составляя около 36% ее длины; щетинки гистеросомы умеренной длины, крепкие, зкостк,опитые; размеры их в процентах к общей длине идиосомы(вычисло||пыо по рис. 3 табл. XXXII Michael’n) следующие: hi 7.5; he около 25; d, -1Ji; da 13.5; d3 27; d4.21.5; 1 p 23 (1 а на рисунке не изображены: вероятно, очень мелки»
142
I. ТУ1Ю(ИУРН11)АЕ. 2. /UIIZOGLYPHINAE
и тонкие) ; sa е 6; sa i 21. Анальная щель отодвинута от генитальной на такое-же расстояние, как от заднего края тела; постанальные щетинки находятся; далеко позади конца анальной щели. Поги умеренной длины; размеры их, по рис. 4 той же таблицы, составляют 39, 36, 30 и 33% длины идиосомы; пропорции лапок, примерно, как у A. redikorzevi; вентральные щетинки лапок более или менее игольчатые. Средняя длина идиосомы 530 [г, ширина 320 р..
Гипопус. Типа Acotyledon. Окраска беловатая с легким буроватым; оттенком, особенно па. ногах и по краям тела. Передне-боковые края пропо-досомального щита заметно вогнутые; ростральный выступ узкий, треугольный, с очень острой вершиной. Сториум длинный, простирающийся назад, почти так же далеко, как эпиморы П; последние широко расставлены концами и направлены под очень острым углом к продольной оси тела; эпиме-риты II явственные, почти параллельные эпимерам II; коксальные поля III,. как у A. rhizoglyphoides. Генитальная щель короткая; окологенитальные-присоски не изображены (отсутствуют?); оформленного присасывательного' диска нет, на его месте расположены 2 широко разобщенные пары мелких присосок, из которых передние очень далеко раздвинуты в стороны (соответствуют наружным периферическим), а задние значительно сближены друг с другом (гомологичны задним периферическим); помимо того, кнаружи от-присосок передней пары, почти у самых краев тела, помещается по 1 овальному образованию, строение которого неясно: Michael’ем они трактуются, как 3-я пара присосок, но их своеобразное положение говорит против такого, толкования. Размеры и пропорции ног, как у A. paradoxa и A. rhizoglyphoides-, длинных щетинок на лапках нет. В остальном сходен с двумя только-что названными видами. Длина 260 [л, ширина 160 [л.
Распространение. Известен только из Англии. Найден на. гниющих стеблях капусты.
*6. Acotyledon redikorzevi (A. Z.) (фиг. 192, 193, 221, 222, 243—248);
Захваткин, 1937:192, рис. 33—34, 43—49 (Eberhardia).
Идиосома в 1.7—1.9 раза длиннее своей максимальной ширины. Sc i. в 2.2—3 раза короче sc е; расстояние между ними только в 1.5 раза болыпе-промежутка, отделяющего каждую из них от sc е; длина последних составляет 25—33% длины идиосомы, щетинки гистеросомы умеренной длины, довольно тонкие и гибкие, не игольчатые; размеры их, выраженные в процентах к общей длине идиосомы, таковы: he 20—25; hi 5—8; dx 5—9.5; d2 7—10.5; d3 12—20; d4 12—22; 1 a 7.5—11; 1 p 9—17; sa c 9—24; sa i 12—32. Таким образом d2 почти одинакового размера с (1Х, <13 не больше чем вдвое.
Фиг. 227—238,
Фиг. 227. Acotyledon agilis (Сап.), (Г спину (по Cnnestrlni). Фиг. 22В. A. agilis (Сап.), папка IV Cf (по Ganestrini). Фиг. 22». A. mlohunli (Ouds.), гетероморфный Cf сверху (по Micbael’K*). Фиг. 230.. То же, снизу (по MichaelTo). Фиг. 231. A. mla/iaoli (Ouds.), penis (по Michael’io). Фиг. 232. A. mi-chaeli (Ouds.), Q сверху (no Michael’io). Фиг. 233. То же, спиау (по Michael’io). Фиг. 234. A. lira-men (Berl.), сГ снизу (по Berlese). Фиг. 23В. Д. )i.runieri (Вот!.), У -сверху (по Berlese). Фиг. 286. То же, снизу (по Berlese). Фиг. 237. A. hramori. (Вог).), ланка IV o’ (по Berlese). Фиг. 238. A. lira-meri (Berl.), penis (по Berlese).
'144
I. TYROGLYPHIME. 2. RHIZOGLYPHINYE
длиннее d2, Ip в 1.2—1.6 раза больше 1 а; часть более коротких щетинок спинной поверхности гистеросомы имеет шпателевидно уплощенный, широко закругленный конец; при небольшом увеличении они кажутся как бы обломанными. Псбвдостигмальпый орган ланцетовидный, сильно перистый по краям. Ноги длинные и стройные; на лапках I и II средняя и внутренняя вентральные щетинки представлены довольно толстыми, коническими шинами; вентральные щетинки ланок П.Т и IV также шиповидпые; обонятельная палочка лапок Г и II слабо булавовидно утолщенная, в общем дочти ^цилиндрическая.
3. Гомеоморфная фаза. Идиосома позади 1 р слабо суживающаяся; щетинки спиппой поверхности обычно несколько длиннее, чем у $; наиболее ощутимы различия в длине 1 р, за е и за i. Постанальные щетинки тонкие, волосовидные, рг расставлены так же широко, как р3; расстояние между р2 в З.б раза меньше и они расположены приблизительно посередине . между рг и р3; длина их превосходит длину рх в 1.7—2,2 раза. Анальная щель отстоит от заднего края тела на расстоянии, превосходящем около 1.5 раза ее длину; penis с слабо изогнутой дистальной частью. Ноги значительно длиннее, чем у 9: длина их составляет, соответственно, 36—39, 33—37, -36—37 и 38—44% длины идиосомы; лапки IV заметно короче двух предыдущих члеников ноги, взятых вместе; отрезок ab в 1.5 раза больше cd и ef, .которые равны между собой. Длина 430—516 р..
Гетероморфная ф а з а 3 неизвестна: в обеих исследованных мною колониях этого вида 33 были представлены только гомеоморф-щыми особями...
9. Идиосома сильно суживающаяся позади 1 р; боковые края ее от точек прикрепления 1 р до уровня sa е почти прямые, заметно сходящиеся кзади, дцетинки спинной поверхности немного короче; постанальные расположены .значительно позади конца анальной щели; расстояние между генитальной щелью и анальной меньше, чем расстояние между задним концом последней и задним краем тела. Длина ног составляет соответственно 30—33, 29—32, 27—32 и 32—38% длины идиосомы; лапки IV равны по длине голени и колену, взятым вместе. Длина 530—775 [л.
Гипопус. Типа Cosmoglyphus. Окраска в препаратах интенсивно ржаво-коричневая, у живых особей темнокоричневая; поверхность идиосомы очень блестящая, как бы лакированная. Идиосома очень широко овальная, иногда далее почти круглая; отношение ее длины к ширине колеблется в зависимости от степени расплпстаппости терпев, между 5 : 4 и 11 : 10 -гистеросома с дисковидно уплощенными краями; щетинки спинной поверхности длиною, в сродном, с голень I, более или менее дугообразно изогнутые. Передний край ироподосомальиого щита глубоко дугообразно вырезанный по обоим сторонам треугольно выдающегося вперед рострального выступа, на вершине которого сидят хорошо развитые v i; его крыловидные боковые лопасти более или мопсе широко закруглены; ростральный .выступ вытянут в тонкий отросток и заметно выдается концом за линию,
8. ACOTYLEDON tin
проведенную через вершины боковых лопастей. Гнатосома. ri>voiminainui, не более чем вдвое длиннее своей ширины; ее вершина глубоко piicniytiлиий, образуя пару обособленных дистальных члеников; аристы дугооОршиш изгибающиеся кнаружи. Эпимеры II не достигают заднего края стерииль-ного щита и пе соединены с эпимеритами II; коксальные поля III открытью; эпимеры IV свободные, довольно резко дугообразно изогнутые. Расстояние между задним краем присасывательного диска и задним краем идиосомы меньше длины стернального щита; периферические присоски близко придвинуты к центральным; передние несколько меньше центральных и значительно крупнее задних и боковых, размеры которых почти одинаковы; боковые присоски находятся на одной поперечной линии с срединными «порами» центральных присосок. Horn довольно длинные и стройные; ноги I выдаются за передний край вырезов проподосомального щита вершиной бедер и 3 дистальными члениками, ноги II — за вершины его боковых лопастей концом колен, голенью и лапкой; лапки I и II заметно уплощенные в дорзо-вен-тральном направлении. Вентральные щетинки вертлугов I средней длины; наружные вентро-латеральные шипы колен и голеней I и II очень крупные, уплощенные, кинжаловидные, несколько искривленные; на лапках I: сенсилла о в виде длинной, крепкой щетинки, сидящей проксимально на той же высоте, как обонятельная палочка; такой же, но еще более длинной щетинкой представлена и сенсилла г, но она расположена значительно дистальнее; все предвершинные сенсиллы щетинковидные; две из вершинных листовидно расширены и слегка закруглены па конце; присасывательная щетинка с длинным, тонким стебельком и узким, хорошо обособленным блюдцем; лапки II с 2 такими же листовидно расширенными щетинками; на лапках III и IV по 4 яйцевидно расширенных щетинки, не считая расширенной на конце в овальную пластинку дорзальной вершинной щетинки. Длина 252—308 [г, ширина 205—262 р..
Распространение. Известен пока только из 3 пунктов GGGP: Стерлитамакского зерносовхоза (Вашк. ACGP), Кировской обл. и г. Осташкова (Калининской обл.). Значительные различия, существующие между калининскими и башкирскими экземплярами этого вида, заставляют меня считать их представителями 2 разных подвидов: A. redikorzevi redikorzevi A. Z. и A. redikorzevi borealis, ssp. и.
Экология. Живет в сильно влажном прорастающем зерне и ско-4 плениях различных отходов от его очистки на токах, , перевалках и вокруг зернохранилищ; в естественных скоплениях растительных остатков пока еще не был находим, и его распространение в полевых условиях остается ощо невыясненным.
*6а. Acotyledon redikorzevi redikorzevi (A. Z.).
Щетинки идиосомы более короткие, что легко видеть при сопостпнлоиии их средних размеров у обоих подвидов (в процентах к длине идиосомы):
10 Фауна СССР, Захватная —281
146
I. TYROGLYPIIIDAE. 2. RHIZOGLYPHINAE
A. r. redikorzevi...........
A. r. borealis..............
A. r. redikorzevi ..........
A. r. borealis . ...........
9
v 1 sc e sc l h e li i di d2 d3 d4 I a 1 p sa e sa i 12.6 27 10 21.5 7 6.6 7.5 14.5 20' 8.5 14 14.5 27.5
14.4 30 13.5 25 8 9.5 9.5 16 22 10 17 24 32
11.5 26.5 8.5 22.5 6 5.5 7 12.5 13.5 7.7 9.5 9.8 16.2:
13.7 31.511.7 25 8 8 10.7 17.5 16 11 12 12.5 19
Псевдостигмальный орган довольно широкий, пластинчато уплощенный. Ноги короче, чем у следующего подвида: длина их у с? составляет, соответственно, 36, 33—34, 32—33 и 38% длины идиосомы, уд — 30, 29, 27, 32%.
Гипопус. Окраска болев томная, ржаво-коричневая. Боковые лопасти проподосомальиого щита широко закругленные, с дугообразно выпуклым наружным краем, почти полукруглые; средняя длина 10 промеренных особей 263 [г.
Распространение. Окрестности г. Стерлитамака (Башк. АССР).
*6b. Acotyledon redikorzevi borealis, ssp. n.
Щетинки идиосомы заметно длиннее; псевдостигмальный орган слабо уплощенный, почти цилиндрический. Ноги значительно длиннее, чем у типичной формы: у С длина их составляет, соответственно, 39, 37, 36.5 и 44%.. длины идиосомы, у — 33, 32, 31 и 38%.
Гипопус. Окраска более светлая, ржаво-охристая; боковые лопасти проподосомальиого щита с пря:мым наружным краем, почти прямоугольные,, закругленные лишь на вершине. Длина идиосомы 272—308 ft, в среднем (для 10 особей) 290 ft.
Распространение. Известен только из г. Осташкова (Калининской обл.).
7. Acotyledon alberti (A. Z.) (фиг. 354).
Очень сходен с предыдущим видом, от которого отличается только некоторыми особенностями хэтотаксии, а именно: щетинки идиосомы (особенно более короткие из них) толще, прямые, крепкие, почти игольчатые, концы их, видимо, приострены, а не уплощены; размеры щетинок почти точно такие же, как у A. redikorzevi redikorzevi (может быть, чуть короче), но d3 короче (У 9 “^ус? в 1.5 раза длиннее) ;усГ р2 расположены на одной линии с р,, образуя с ними правильный поперечный ряд. Размеры и все остальные признаки, как у A. redikorzevi..
Г и п о и у с. Типа Cosmoglyphus. Очень похож на гипопуса A. redikorzevi, от которого отличается лишь следующими признаками: окраска светлее (довольно светлая жолтовато-хитипово-бурая, слегка прозрачная; ноги розовые или слабого винного цвета); боковые вырезы переднего края про-подосомальпого ш,ита мопсе глубокие; периферические щетинки идиосомы жесткие, шиповидныо («short spines»); передние присоски присасывательного диска значительно меньше центральных и немного уступают в размерах задним; боковые присоски расположены на одной поперечной линии с срединными порами центральных присосок; наружные вентролатеральные шипы голеней и колеп I и II, видимо, шире, напоминающие по своей форме
8. ACOTYLEDON
147
лавровый лист («laurel-leaf-shaped»). Длина, в среднем, 200 |Л, ширппп 21 о р„ (По Michael’ю.)
Распространение. Достоверно известен пока только из Л игл ни. Живет па гниющих грибах; был найден также, ноне в столь большом число, на гниющих стеблях капусты и других гниющих растениях.
* 8. Acotyledon krameri (Berl.) (фиг. 234—238, 249, 250).
Berlese, 1881, Atti В. 1st. Veneto Sc., Lett, e Arti, ser. V, vol. VIII:13; Kramer, 1899:140 (Tyrogtyphus); Oudemans, 1932, Entomol. Ber., VIII:358 (Cosmoglyphus).
Идиосома узкая, вытянутая в длину, в 2.1—2.2 раза длиннее своей максимальной ширины, сильно суживающаяся позади Ip. Sc i в 2 раза короче sc е; расстояние между sc i менее чем в 1.5 раза больше расстояния между каждой из них и sc е ; длина последних равна приблизительно 40%. длины идиосомы;’щетинки гистеросомы умеренной длины, тонкие и гибкие, приблизительно одинаковой длины у обоих полов; размеры их, в процентах длины идиосомы: h е 25; h i 5; dx 5; d2 7; d3 9—10; d416; 1 a 6; 1 p 18; sa e 16; sa i около 30; d2, таким образом, лишь немного длиннее dx и мало уступают в размерах d3; 1 а в 3 раза короче 1 р. Псевдостигмальный орган густопсрп-стый, не утолщенный. Ноги длинные и стройные; вентральные щетинки лапок довольно тонкие, не шиповидные; обонятельная палочка с очень, слабо булавовидно утолщенным концом.
d. Известна только гомеоморфная фаза. Рх расставлены шире, чем р3; положение р2 не установлено. Расстояние между задним концом анальной щели и задним краем тела почти в 2 раза больше ее длины; дистальная часть penis прямая. Ноги несколько длиннее, чем у 9; лапки IV довольно короткие; отрезок ab заметно меньше cd, но много больше ef. Длина около 450 р,.
9. Постанальные щетинки расположены немного позади анальной щели; последняя очень коротка и сильно сдвинута назад, так что расстояние между ее передним копцом и генитальной щелью в 8 раз больше промежутка, отделяющего ее от заднего края тела. Длина ног составляет приблизительно' 40 (I), 36 (II), 34 (III) и 38—40 (IV)% длины идиосомы; лапки IV заметно длиппее голени и колена, вместе взятых. Длина около 630 р.
Гипопус. Типа Cosmoglyphus. Чрезвычайно похож на гипопуса A. redikorzevi, от которого отличается только следующими признаками: идиосома более вытянутая в длину, при самой полной распластанности боковых краев, ее ширина составляет не более 4/5 длины; края гистеросомы совершенно плоские, обособленные от ее центральной выпуклости; ростральный выступ шире (не вытянутый в «посик», треугольный) и короче; он значительно не достает концом до линии, проведенной через вершины боковых лопастей проподосомальиого щита; эти последние почти прямоуголш1Ы(\ закругленные лишь на вершине, с прямыми наружными краями; piiiiiiii.iioh‘ стериума глубже, а сам он длиннее; стернальный и вентральный щиты ужо, в соответствип с более вытянутой формой тела; расстоянии между задним краем присасывательного диска и задним краем тела бол|лно, риппоо длине
148
I. TVROGLVPniDAE, ?, RHIZOGLYPIUNAP
стернального щита; передние периферические присоски значительно меньше .центральных и не на много крупнее задних и боковых; последние несколько сдвинуты кпереди от линии, проведенной через срединные поры центральных присосок; ноги короче; ноги Т выдающиеся за передний край вырезов проподосомального щита только частью колена, голенью и лапкой, ноги III выдаются за вершины его боковых лопастей только дистальной третью голени п лапкой; присасывательный волосок лапок 1 и II несколько шире; на лапках III и JV только по 3 листовидно расширенных щетинки, не считая вершинного волоска; ноги III и IV тоньше, особенно их бедра. Размеры единственного, имеющегося у меня, экземпляра: длина 2G7 р., ширина 205 р; по Berlese, длина около 300 р.
A. krameri близок к A. redikorzevi и A. alberti, от которых хорошо отличается более вытянутой в длину идиосомой, неутолщенными в виде шипов вентральными щетинками лапок и положением анальной щели д.
Распространение. Достоверно известен только из Италии и Украины (Нерубаевское лесное промышленное хозяйство, Одесской обл.). Живет в разлагающихся растительных веществах; гипопусы используют в качестве переносчиков муравьев, различных мелких жуков и клещей.
9. Acotyledon feytaudi (Ouds.).
Oudemans, 1928, Entomol. Вег., VII: 313 (Calogtyphus)-, 1932, ibid., VIII: 358(Cosmoglgphus).
Известен только гипопус, описываемый Oudemans’ом следующим образом: «У feytaudi спинные волоски по меньшей мере в 3 раза длиннее и более-гибки (slapper), чем у krameri, и дорзальный поперечный ряд состоит не из 4, а из 6 волосков (между наружными и внутренними, впереди жировых желез, прибавляется по 1). Ноги у feytaudi короче и толще, чем у krameri-, при вытянутых ногах у krameri видна (при рассматривании сверху) половина бедер I, у feytaudi — только дистальная половина колен I. У krameri линия, - отделяющая 4 тазика I-J-II от 4 тазиков III-J-IV, слабо выгнута кпереди; у feytaudi — сильнее. У krameri присасывательная пластинка приблизительно четыреху гольная; у feytaudi—приблизительно шестиугольная». Если учесть, что «Calogtyphus krameri» Ouds. не идентичен с A. krameri (Berl. 1891), а очень близок (если .не идентичен) к A. redikorzevi, то окажется, что большинство признаков, характеризующих A. feytaudi Ouds., как.раз совпадает с теми, какие отличают от A. redikorzevi настоящего A. krameri, и различия между последним и Л. feytaudi остаются неясными; таким различием может считаться только большая длина щетинок у A. feytaudi, так как у A. krameri :и A. redikorzevi они практически одного размера.
Распростри и о н и о. Описан ио материалу из Испании и Алжира. Найден в гнездах термита lletivulitermes lucifugus Rossi.
*10. Acotyledon batsylcvl, sp. в. (фиг.- 223—226).
Sc i в 2 раза короче sc с; расстояние между обеими щетинками этой пары :в 1.5 раза больше промежутка, отделяющего каждую из них от sc е; длина
8. ACOTYLEDON I*»
последних составляет 30—35% длины идиосомы; вес щетиики сиютципимМ довольно длийные, очень тонкие и гибкие, волосовидные; размеры их и н|нВ центах к общей длине идиосомы таковы: h е 25—30; h i и dx 10—12; <la 19,5 18; d3 22,5—28; d4 22.5—31; la 16—18; Ip '21—26; sa e 17—31; ini 23 — 41; d2, таким образом, заметно длиннее dx и менее чем вдвое короче d3;lpnl.3—1.5 раза длиннее 1 а. Псевдостигмальный орган пластинчатый, широколанцетовидный, сильно перистый по краям и, кроме того, с несколькими (4—6) щетипковидлыми отростками ла своей дорзальной поверхности. Ноги длинные и стройные; па лапках I и II средняя nt-внутренняя вентральные щетинки шиповидные, как и вентральные лапок. Ши IV; обонятельная палочка передних лапок очень слабо булавовидно-утолщена.
сГ. Щетинки передней постанальной пары (рх) расставлены несколько-шире, чем р3; р2 находятся на одном уровне с р3, образуя с ними прямой-поперечный ряд; расстояние между р2 в 4 раза меньше расстояния между pv Анальная щель отделена от заднего края тела промежутком, в 1.5 раза превосходящим ее длину. Тарзальные присоски расположены более дистально, чему A. redilcorzevi.
Гомеоморфная фаза. Все вершинные щетинки лапок I и II простые, не расширенные на конце.
Гетероморфная фаза. Щетинки идиосомы, как обычно,. Длиннее, чем у гомеоморфных особей, это видно из следующей таблички,, в которой сопоставлены их средние размеры у обеих фаз (в процентах к длине идиосомы):
vi see sei lie hi (lt <12 <13 d( la Ip sa e sai
Гомеоморфные...... 14.6 30 14.5 28 12 12 12.5 22.5 22.5 16 22 22.5 87
Гетероморфные..... 17 35 16 30 12.5 12 18 27 31 18 26 30.5 40-
Ноги также длиннее и тоньше, чем у гомеоморфных особей (47, 44, 'ЗГ и 48% длины идиосомы); шипы и волоски на них крупнее; внутренняя вершинная щетинка лапок I и II дистально расширена в овальную прозрачную пластинку; модифицированные ноги III только с 1 шипом на. вентральной поверхности крючковидных лапок; отдельные щетинки неутолщены, гибкие. Длина 437—550 у.
ф. Большинство щетинок идиосомы такой же длины, как у гомеоморфных О'; заметно короче sa е (17 % длины идиосомы) и sa i (около 23%); постанальные отодвинуты далеко назад от анальной щели; расстояние между генитальной щелью и анальной меньше, чем между задним концом последней,, задним краем тела. Длина ног составляет, в среднем, 36, 34, 33.5 и Я7°% длины идиосомы. Лапки IV заметно длиннее двух предыдущих членпкон. ноги, взятых вместе, и в 6 раз превосходят свой диаметр посередине. Длина 600 у, ширина 340 р..
Гипопус. Во всем сходен с гипопусом A. redilcmw'!, но переднем несколько крупнее (288—310 р.) и окрашен.светлее.
150
I. TYROGLYPHIDAE. 2. RIllZOGLYPHlYAE
Распространение. Найден пока только 1 раз, в окрестностях т. Ворошиловска на Северном Кавказе. Живет в скоплениях сильно увлажненных, разлагающихся растительных остатков (отходы от обмолота и провеивания пшеницы).
*11. Acotyledon vol^ini, нр. и. (фиг. I№—]<%).
Известен только гипопус, характорпзующпйся своими большими размерами, крайне у пороченным пронодосомальным щитом и вооружением пог. Окраска темпокорнчпевая. Идиосома, удя и пен но-п йцевидпоп формы, спереди почти прямо усеченная; отношение ее, длины к ширине равно 1.45 : 1; спинная поверхность сильно и равномерно выпуклая, гладкая; боковые края гистеросомы пластинчатые, но более узкие, чем у видов типа A. krameri-, щетинки ее спинной поверхности короткие, прямые. Проподосомальный щит пластинчатый, чрезвычайно короткий; длина его видимой части составляет только г/35 длины гистеросомалы-юго щита; передний край щита слабо волнистый, с чуть намеченными боковыми вырезами и слегка выпуклой средней частью; ростральный выступ такой же формы, как у A. redikorzevi, и также несущий на конце пару хорошо развитых теменных щетинок, но смещенный на вентральную поверхность щита и целиком прикрытый сверху его пластинчатым передним краем; лопаточные щетинки крепкие, sc е прямые, с резко отогнутым под углом концом; sc i, как обычно, дугообразно загибающиеся назад. Гнатосома целиком скрытая под нависающим краем проподосомаль-ного щита, крупная, вДвое длиннее своей наибольшей ширины, начиная от середины равномерно суживающаяся кпереди; вершина ее, расщеплена на х/4 всей ее длины; аристы длинные, почти прямые. Эпимеры II доходят до заднего края стернального щита, по не соединены с прямыми эпимери-чами II особыми хитиновыми перемычками (как это имеет место у Forcel-linia); вентральный щит очень большой, значительно превосходящий по своей длине стернальный щит; коксальные поля III и IV открытые; эпимеры IV такого же строения, как у A. redikorzevi, но шире раздвинутые концами; вентральный шов простирается вдоль всего щита, до его заднего края; последний имеет такую же форму, как у A. redikorzevi и A. krameri. Расстояние между задним краем присасывательного диска и задним краем тела заметно уступает длине стернального щита; расположение присосок и их относительные размеры, как у A. krameri. Ноги короткие и массивные; за передний край проподосомальпого щита выдаются только самый конец колен, голени и лапки пог I и лапки йог I.I., Вентральные щетинки вертлугов I имеют
Фиг. 28!)—.253. Гипопусы рода Acotyledon Ouds.
Фиг. 239. A. paradoxa Ouds., общий иПД сверху. Фиг. 210. То же, снизу. Фиг. 241. A. paradoxa Ouds., лапка I сверху. Фиг. 242. A, rlilioglupholdoe (А. Z.), задняя часть гистеросомы с присасывательным диском, снизу. Фиг. 243. A. redlhorzovi (A. Z.), общий вид сверху. Фиг. 244. A. redi-korzevl (A. Z.), проподосомальный щит сверху. Фиг. 245. A. redikorzevi (A. Z.), гнатосома, вид снизу. Фиг. 246. A. redikorzevi (A. Z.), общий ЛИДСИИВУ. Фиг. 247. A. redikorzevi (A. Z.), нога! сверху. Фиг. 248. A. redikorzevi (A. Z.), лапка IV снизу. Фиг. 240. A. krameri (Berl.), общий вид снизу. Фиг. 250. A. krameri (Berl.), проподосомальный щит сверху. Фиг. 251. A. tshemyshevi, sp. п., проподосомальный щит сверху. Фиг. 252. A. tshemyshevi, sp. п., нога I сверху. Фиг. 253. A. tshemyshevi, sp. п., гнатосома.
152
I. TYROGLYPRIDAE. 2. RRIZOGLYPHINAE
форму ланцетовидных уплощенных шипов; Наружные вентро-латеральные шипы колен и голеней 1и11 ножсвпдные, приблизительно такой же формы, как у A. redikorzevi; лапки I и II сильно уплощенные в дорзо-вентральном направлении: их вентральные щетинки все расположены на наружном ребре лапки, образуя здесь своего рода щетку: средняя вентральная щетипка в виде крепкого шипа, обо боковые и 3 из вершинных щетинок сильно листовидно расширены; их пластинки имеют удлиненно-овальную форму и явственную срединную жилку. Ноги 11Г и 1V гораздо массивнее, чему A. redikorzevi. На ланках Ш и IV сильно листовидно расширены по 4 щетинки. Длина идиосомы 380 р., ширина 265 р..
Раскроет р а п с и и о. Вид описывается по экземплярам из Чек-магушевского р-па (Вашкпр. АССР); местообитание неизвестно.
*12, Acotyledon tshernyshevi, sp. п. (фиг. 186, 251—253).
Гипопус. Типа Cosmoglyphus. Окраска очень бледная, беловатая, с легким охристым оттенком. Идиосома широкояйцевидная, гладкая, равномерно и сильно выпуклая, усаживающие ее щетинки очень короткие, прямые, но тонкие; края гистеросомы не уплощены и не обособлены от ее центральной части; проподосомальный щит с прямо усеченным, слабо волнистым передним краем, не образующим хоть сколько-нибудь намеченного рострального выступа; длина его видимой части по средней линии составляет околоке длины гистеросомальпого щита; v i отсутствуют. Гнатосома в 2.5 раза длиннее своей ширины, с почти параллельными краями, наполовину выдающаяся за передний край идиосомы; вершина ее чуть раздвоена; аристы слабо конвергирующие, слегка дугообразно изогнутые, причем выпуклости описываемых ими дуг обращены кнаружи. Эпимеры II не достигают заднего края стернального щита и не соединены хитиновой перемычкой с эпимеритами II; коксальные поля II, таким образом, открытые; коксальные поля III почти замкнутые, очень широко раздвинутые; эпимеры IV сильно дугообразно искривленные; продольного вентрального шва нет. Расстояние между задним краем присасывательного диска и задним краем идиосомы меньше длины стернального щита; периферические присоски значительно отодвинуты от центральных; передние в несколько раз крупнее задних и боковых, имеющих одинаковые размеры. Ноги стройные; лапки I и II цилиндрические, не уплощенные. Вентральные щетинки вертлугов! очень короткие, вентро-латеральные шипы колеи и голеней I и II тонкие, почти прямые, небольшого размера;, на лапках I средняя вентральная щетинка превращена в шип; предвершинные и вершинные щетппкп, кроме внутренней, топкие, прямые, не расширенные на конце; внутренняя вершинная расширена в широколанцетовидную-пластинку; присасывательная щетинка толстая, постепенно расширяющаяся к крупному удлиненному «блюдцу». Па лапках II внутренняя вершинная имеет такую же форму, как па лапках I, по присасывательная щетинка гораздо меньше и ее блюдце почти не обособлено от стебелька. Лапки III и IV
8. ACOTYLEDON. 9. CALOGLYPHUS
1ПЗ'
c 5 листовидно расширенными щетинками. Длина 245—285 р., ширина, 200 -240 |Л.
Распространение. Известен только из Украины (Нерубаенскпп. лссопромхоз, Одесской обл.). Гипопусы найдены в мертвой лесной подстилке в небольшом числе экземпляров.
' К роду Acotyledon относится, вероятно, также известный лишь по гипопусам и недостаточно описанный:
13. Acotyledon eurynyniplia (Ouds.).
Oademans, 1912, Entomol. Ber., 111:189 (Tyroglyphus).
Гипопус. Название eurynympha дано потому, что нимфа, а именно-дейтонпмфа или гипопальная форма, необычно широка, почти круглая. В остальном это — настоящий Tyroglyphus. Ноги снабжены крупными шипами, много более крепкими, чем у Hypopus spinitarsus Herm. Протеро-торакс (= проподосомальный щит) очень короткий и между ногами I треугольно вытянув кпереди. 4 волоска на протеротораксе и 10 пар на гисте--ротораксе (= гистеросомальпом. щите) суть очень короткие толстые щетинки. Нет парусных щетинок («sleepharen»); ланцетовидные волоски почти незаметные; коготки крепкие, имеющие форму крюка («bootliukvormig»). В гниющих листьях, Гаага, Д-р Эверте, 1 экз. (По Oudemans’y.)
По Vitzthum’y, это единственная из странствующих нимф Tyrogly-phidae, снабженная волоском на тазиках!, тогда как иа тазиках III имеются присоски; тазики I и III у всех других видов вооружены одинаково. Последний признак позволит, надо думать, легко узнать Т. eurynympha, если она вновь попадет в руки исследователей; так как о нем нет речи в оригинальном описании Oudemans’a, можно предположить, что Vitzthum имел в своем распоряжении собственноручный рисунок Oudemans’a, сделанный им с типа Т. eurynympha и оставшийся, к сожалению, неопубликованным..
9. Род CALOGLYPHUS BERL.
Berlese, 1923, Redia, XV: 262 (sbg.); Oademans, 1932, Entomol. Ber., VIII: 357.
Тип рода: Tyroglyphus berlesei Michael 1903.
Крупные,-иногда гигантского размера формы: 6*500—1100 р, q 700— 2300 у. Идиосома с умеренно выступающими плечевыми буграми; боковые края ее от плеч до точек прикрепления 1 р более или менее выпуклые, дальше назад, до уровня sa i, равномерно суживающиеся, задний край очень слабо выпуклый, прямой или даже слегка вогнутый. Наружная вентральная и одна из вершинных щетинок лапок I и II увеличены в размерах, дугообразно искривлены и в дистальной половине сильно расширены, в виде полулунной или округлой (иногда неправильно очерченной) npoapii'i-пой пластинки; внутренняя и средняя вентральные щетинки тех же линек и вентральные щетинки лапок III и IV (иногда только 1 из.лих) нргн|ннцеиы в более илн менее мощные шипы; вентральные щетинки голеней и колон также более или менее шиповидные.
154
I. TYROGLYPHIDAE. 2. RKIZOGLYPHINAE
d. Тарзальные копулятивные присоски смещены к самому концу лапки.
§. Анальная щель всегда сильно удалена от заднего края тела; анальных микрохэт 3—6 пар. Половой диморфизм в большинстве случаев очень резкий.
У личинок и пимфи.1П.Н1.1х стадий ряд щетинок передней половины идиосомы v е, sc j, hi, nIi, <lj —- <1а и la сильно редуцированы. Отроение гипопусов чрезвычайно изменчиво.
К роду Cdloilhivh'ii^ относите,я, помимо описываемых здесь 10 палеарктических видов, еще ряд зп’зотпческих: Г. iiwmirsi Ouds. 1926, С. brasiliensis Ouds. 192'1, Ti/rof/li/phus wiii'ii,icvl<tl,un Vtz. 1924, T. ' spelaeus Berl. 1923, а также североамериканские 7’. hclcromorphus Felt 1896, T. armipes Banks ,1906 и др.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ cW
1 (6). Гетероморфные d'd.
.2 (3). Псевдостигмальный орган хорошо развитый, щетинковидный, длиною с диаметр голени I; кожа гладкая; р2 во много раз длиннее рг волосовидные ...................................... * 1. С. гоdionovi А. Z.
3 (2). Псевдостигмальный орган, в виде крошечной палочки (длиною в 5— 8 ре), заметный лишь при больших увеличениях.
- 4 (5). Кожа гистеросомы явственно, хотя и тонко продольноморщинистая; псевдостигмальный орган заметно расширяющийся к основанию, почти конический; р2 во много раз длиннее рн волосовидные; анальные копулятивные присоски нормального размера. . . 3. С. mand.zhur,sp. п.
• А (4). Кожа совершенно гладкая; псевдостигмальный орган цилиндрический; р2 короткие; анальные копулятивные присоски необычайно маленькие..................................*4. С. spinitarsus (Негш.).
• '6 (1)'. Гомеоморфные с? <5.
.7 (12). Анальные копулятивные присоски эллиптические, сильно хити-низированные, расположенные косо; их задние края выдаются назад значительно дальше конца анальной щели; длина последней немного превышает длину присосок; щетинки идиосомы толстые, очень жесткие. Относительно мелкие формы, не превышающие 730 р. в длину.
с'8 (9). Лапки IV с 2 крупными серповидными щетинками; d2 и d3 почти одинакового размера, обе значительно короче 1 р п более чем вдвое короче d4........................................* 7. С. moniezi A. Z.
- 9 (8). Лапки IV только с 1 крупной серповидной щетинкой или без таковых; d3 в 2.5—3.5 раза длиннее d2, почти одинакового размера с d4nlpили длиннее последних.
. 10 (п). Крупные серповидные щетинки имеются на всех лапках; дистальная часть penis тонкая, прямая; длина sc е, h е и 1 р не превышает 40—45 %, sa е 15% длины идиосомы.......................*5. С. polyphyllae, sp. n.
11 (10). На лапках III и IV пет крупных серповидных щетинок; дистальная часть penis отогнута иод углом к его цоколю, так что в вентральном
9. CALOGLYPHUS
I ii.'i
аспекте on кажется тупым;' длина sc e, h e и I p достигает <ki lift %, sa e 25% длины идиосомы ................6. С. bcrksel (Mich,),
12 (7). Анальные копулятивные присоски округлые или яйцевидные, слабо хитипизироваппые; линия, проведенная через их задние кран, проходит заметно впереди заднего конца анальной щели, последняя в 2—3 раза длиннее присосок; щетинки идиосомы обычно довольно тонкие и гибкие. Размеры изменчивы, часто очень крупные (до 1100 р.).
13 (16). Р2 во много раз длиннее р1( волосовидные, далеко выдающиеся за задний край идиосомы; щетинки гистеросомы длиппые.
14 (15). Кожа гистеросомы явственно (хотя и топко) продольно морщипи-, стая; псевдостигмальный орган в виде очень маленькой (6—8 р.) конической палочки; задний край проподосомального щита выемчатый; расстояние между р2 в 3.5 раза меньше расстояния между рх . . . ............................................ 3. С. mandzhur, sp. n.
15 (14). Кожа совершенно гладкая; псевдостигмальный орган хорошо развитый, щетинковидный, по длине близок к диаметру голени I; задний крап продосомалыюго щита без выемки, прямой или слабо выпуклый; расстояние между р2 только в 2—2.3 раза меньше расстояния между рх.............................* 1. С. rodionovi A. Z.
16 (13). Р2 такого же размера, как рх, короткие; щетинки гистеросомы короче.
17 (18). Обонятельная палочка лапок I и II топкая, цилиндрическая; идиосома сильно пузыревидно вздутая, в 1.5 раза длиннее своей максимальной ширины; гипостом слабо суживающийся кпереди, с резко S-образными боковыми краями..............* 8. С. sphaerogaster A. Z.
18 .(17). Обонятельная палочка лапок I и II массивная, сильно булавовидно утолщенная к концу; идиосома довольно стройная, почти в 2 раза длиннее своей максимальной ширины, приходящейся чуть позади плеч; гнатосома очень узкая; гипостом сильно суживающийся кпереди, с почти прямыми боковыми краями ..........................
................................... 9. С. mycophagus (Megn.)
99
1 (2),. Обонятельная палочка лапок I и II массивная, сильно булавовидно вздутая к концу; гнатосома очень узкая; гипостом сильно суживающийся кпереди, с почти прямыми боковыми краями.....................
.......................... ........ 9. С. mycophagus (Megn.).
2 (1). Обонятельная палочка лапок I и II тонкая, цилиндрическая; пропорции гнатосомы нормальные; гипостом слабо суживающийся кпереди, с более или менее резко выгнутыми боковыми краями.
: 3 (4). Пластинки серповидных щетинок лапок очень широкие; шюндос.тиг-мальный орган совершенно отсутствует. . * 5. С. polyphylhw, нр. и.
4 (3). Пластинки серповидных щетинок лапок уже, ланцотопидпыщ имеется псевдостигмальный орган (часто он очепь мелкий).
136 I. IYROGLYPHIDAE. 2. RHIZOGLYPHINAE
5 (6). Псевдостигмальный орган хорошо развитый, щетинковидный, длиною1 ' равный диаметру лапки I; sa е очень короткие, в 8—9 раз короче sa i; 1 р в 2—3.5 раза длиннее 1 а; анальных микрохэт 6 пар, из которых. 4 задние расположены ла почти одинаковом расстоянии друг от друга и образуют с каждой сторопы анальной Щ?ли по косому дугообразному ряду................................... *1. С. rodionovi A. Z.
6 (5). Псевдостигмальнын орган очень маленький (4—8 р.), палочковидный (хорошо заметный лили, при больших увеличениях); анальных микрохэт 3—(> пар, расположенно их иное.
7 (8). Анальных микрохэт только 3 лары; пластинки серповидных щетинок ланок очень узкие.......................* 4. С. spinitarsus (Herm.).
8 (7). Анальных микрохэт 4—6 пар, пластинки серповидных щетинок лапок ле очень узкие.
9 (10). Ноги очень стройные; длина d3, d4 и 1 р равна 16—21% длины идиосомы; анальных микрохэт 4 пары, из которых 2 расположены у переднего конца анальной щели, 1 — у середины ее длины и 1— на середине расстояния между средней парой и задним концом anus; генитальные щупальцы, atrium и края анальной щели ржаво-рыжие ........................................* 8. С. sphaerogaster A. Z.
10 (9). Ноги массивные; длина d3, d4 и 1 р не превышает 8—11% длины идиосомы; анальных микрохэт 6 пар, из которых 2 расположены у переднего конца анальной щели, 1.—у середины и 3 тесно сгруппированы у ее заднего конца; генитальные щупальцы, atrium и края, анальной щели пепигмеитированные..............3. С. mandzhur, sp. m
Гипопусы
1 (2). Коксальные присоски заменены щетинками; дистальные членики гнатосомы широко раздвинутые, длинные (равные по длине ее квадратному основному членику), 2-членистые; коксальные поля III и IVразъединены широким продольным .промежутком........................
...................................9. С. mycophagus (Megn.).
2 (1). Коксальные присоски нормального, строения; дистальные членики гнатосомы тесно сближенные между собой, короткие (всегда значительно короче основного членика), 1-члепистые или вовсе не обособленные от основного членика; коксальные поля III и IV разъединены узким продольным промежутком.
3 (4). Ноги I и Ц чрезвычайно короткие и толстые, конические, почти целиком спрятанные под нависающим в виде козырька серповидным проподосомальным щитом; за его полукруглый передний край выдаются. только самые кончики лапок I и коготок и окружающие его листовидные и присасывательная щетинка лапок II; v i расположены па вентральной поверхности пластинчатого проподосомальногощита,.
0. CALOGLYPBUS 1»7
значительно отступя от его переднего края. Крупная форма (-ЮО--420 |л), находимая на бронзовке Osmoderma eremita Scop . .......
........................................4. С. spinitarsus (lleriii.).
4 (3). Ноги I и II выступающие, по крайней мере, 3 дистальными члениками за края проподосомального щита, стройные; проподосомальпый щит более или менее треугольный, с выступающим вперед вершинным углом; v i расположены на вершине этого угла или непосредственно под его краем.
5 (12). Гнатосома целиком спрятанная под вершинным (ростральным) выступом проподосомального щита, за край которого выдается только дистальная часть ее арист.
«6 (7). Длина проподосомального щита составляет немного меньше */2 длины гистеросомального; гнатосома яйцевидная, ветви ее развилка очень короткие, не отчлененные от основного членика; внутренняя вентральная щетинка голеней II много больше наружной; эпимеры II сходящиеся под тупым углом; стернальный щит обрисован сзади дугообразной линией. Мелкая (200—235 р.) беловатая форма, встречаемая в массе под надкрыльями навозника Geotrupes stercorosus Forst................
....................................* 10. С. geotruporum, sp. в.
7 (6). Длина проподосомального щита составляет не более х/4 длины гистеросомального; ветви развилка гнатосомы длинные, явственно обособленные от ее основного членика; внутренняя вентральная щетинка (шип) голеней II короче наружной; эпимеры II сходятся под острым углом и обычно несколько отогнуты концами кнаружи; стернальный щит обрисован сзади более или менее трапециевидной линией. Более крупные виды.
£ (9). Проподосомальпый щит равен г/6 длины гистеросомального; ноги I и II короткие, очень массивные, выдающиеся за передний край идиосомы только коленом, голенью и лапкой; лапки очень коротки, значительно короче соответствующих колена и голени, взятых вместе, и только в 1.6—2 раза длиннее своего диаметра у основания; наружные вентральные шипы голеней I и II одинаковой длины с лапками; передне-боковые края проподосомального щита равномерно слабо выпуклые на всем своем протяжении, не образующие прогибов по сторонам рострального выступа; последний, таким образом, не обособлен. Длина 350—390 р. На мраморном хруще (Polyphylla fullo L.). . . ................. ..........................* 5. С. polyphyllae, sp. n.
:£) (8). Проподосомальпый щит в 4.5—5 раз корочегистеросомального; ноги [ иП стройные; их лапки равны по длине или длиннее соответствующих колена и голени, взятых вместе, и не менее чемвЗ раза длиннее споого диаметра у основания; наружные вентральные шипы голеней .!. и 11 много короче лапок; передне-боковые края проподосомального щита, явственно вогнутые по сторонам хорошо обособленного, прпостргн-шого рострального выступа.
158
J. TYROGLYPHIDAE. 2. RHIZOGLYPniNAE
10 (11). Ноги II выступают за края проподосомы только дистальной половиной колена, голенью и лапкой; внутренние вентральные шипы голеней I и II много тоньше наружных; боковые края гнат ос омы прямые, параллельные друг другу; ростральный выступ мепее обособлен от-проподосомальиого щита; длина вентрального щита не превосходит его максимальной ширины; ширина присасывательного диска умещается в паиболi.iiii'ii ширине тела меньше 3 раз. Длина 225—390 р. Под падкрыльямп ,майского жука (Melolontha sp. sp.)................
...........................................*7. С. moniezi A.Z.
11 (10). Ноги Г[ выступают за, края проподосомы дистальной половиной бедра, всем коленом, голенью и лапкой; внутренние вептральные-шипы голеней. I и II почти одинаковой мощности с наружными; боковые края гнатосомы выпуклые; ростральный выступ очень явственно обособленный; длина вентрального щита значительно превосходит его максимальную ширину; ширина присасывательного диска умещается 4 раза в наибольшей ширине тела. Длина 400—440 р. Переносчики неизвестны...........................3. С. mandzhur, sp. в.
12 (5). Гнатосома выдается за вершину рострального выступа вершиной своего развилка или всей своей дистальной половиной.
13 (14). Ветви развилка гнатосомы не обособлены в виде самостоятельных члеников; гнатосома в целом удлиненно-грушевидная, выступающая за передний край проподосомы всей своей дистальной половиной; боковые края стернального щита резко S-образно изогнутые; задние-ирисоски присасывательного диска немного меньше передних; вентральные щетинки голеней Д и II не шиповидные. Под надкрыльями жука-носорога (Oryctes nasicornis L.) и мраморного хруща (Polyphylla fullo L.)..................................... 6. С. berlesei (Mich.).
14 (13). Ветки развилка гнатосомы явственно обособлены в виде самостоятельных члеников; основной членик ее с параллельными краями; боковые края стернального щита почти прямые; задние присоски присасывательного диска много меньше передних; вентральные щетинки голеней I и II шиповидные.
15 (16). Гнатосома выдается за вершину рострального выступа дистальной, четвертью или третью своего основного членика и всем развилком (дистальными члепиками); ее основной членик в 3—3.5 раза длиннее1 своей ширины; длина проподосомальиого щита умещается в длине-гистеросомалыюго (f—8 раз; внутренние вентральные шипы голеней I и II немного короче наружных; наружная вентральная щетинка, колен И шиновцдиая; ностаиальпые щетинки короче голени II . . ..............................................* 1. 0. то dieno vi A. Z.
16 (15). Гнатосома выдается за вершину рострального выступа только концом своих дистальных члеников (развилка); основной членик ее в 2 раза длиннее своей ширины; длина проподосомальиого щита умещается в длине гистсросомал иного 4.5—4.7 раза; внутренние вентраль
9. CALOGLYPHUS
ПИ)
ные шипы голеней I п II в 2—3 раза короче наружных; iiupy,'dfiia.u вентральная щетинка-колен II волосовидная; постаиальные щетинки длинные, в 2 раза длиннее голени II ... * 2. С. tshernyshovi, нр. и. г
В таблицу не включены неизвестные ине в натуре и недостаточно полно описанные qq С. berlesei Mich, и С. moniezi A. Z.
* 1. Caloglyphus rodionovi A. Z. (фиг. 194, 195, 254—259, 262, 2G3, 268, 278, 288, 289, 295, 297, 298).
Захваткпп, 1935:22, рис. 4, 22—24; 1937:182, рис. 1—13 и 25—30. —mycophagus Schulze,. 1924, Ztschr. Morphol. u. Oekol. d. Tiere, 11:1—57, 17 figs. (Tyroglyphus) (non Megnin, 1874).
Биология. Schulze, 1924, Ztschr. Morph, u. Oekol. d. Tiere, 11:1—57; Шестерпкова, 1932, Tp. H.-п. пнет. пищ. промышл., I, вып. 8; Zacher, 1927, Vorrats-Speicher- u. Materialschad-linge: 269.
- Псевдостигмальный орган хорошо развитый, щетинковидный, длиною-в 30—40 р.; дорзальные щетинки гипостома довольно длинные, неутолщенные; задний край проподосомальиого щита прямой или более или менее выпуклый; боковые поверхности проподосомы впереди вертлугов I иехити-низироваиные; хитипизироваиных участков вокруг стерпума и эпимер нет.. Серповидные щетинки лапок I и II с относительно узкой ланцетовидном пластинкой; на лапках III одна из них имеет такие же размеры, вторая гораздо уже и длиннее; лапки IV с 1 узкой серповидной щетинкой. Степень полового диморфизма очень изменчива в связи с крайней изменчивостью сГсГ-;. обычно он выражен очень резко. Кожа у обоих полов гладкая.
сШ диморфны; гетероморфная фаза встречается гораздо реже гомеоморфной. Кожа совершенно гладкая, топкая. Р2 длинные, волосовидные,, в 3—5 раз длиннее рх; расстояние между ними в 2—2.3 раза меньше расстояния между рх. Анальные копулятивные присоски яйцевидные, слабо-хитинизированные, резко очерченные сзади соединительным склеритом,, в соответствии с чем линия, проведенная через их задние края, проходит несколько выше заднего конца анальной щели; длина последней в 2—2.5 раза, больше длины присосок; дистальная часть penis прямая, очень тонкая.
Гомеоморфная фаза. Длина ног очень изменчива, колеблясь.. в пределах 33—36 (I), 31—40 (II), 33—42 (III) и 37—47 (IV) % длины идиосомы; их относительная длина, таким образом, то приближается к таковой q ,. то к таковой гетероморфных особей; толщина пог, пропорции отдельных члеников и форма шипов вариируют в таких же широких пределах. Задняя (внутренняя) вентральная щетипка лапок III только у очень крупных особей (порядка 1000 р.) превращена в шип; в подавляющем большинстве случаев опа пе модифицирована-. Щетинки идиосомы довольно тонкие и гибкие; длина их также изменчива, и у промеренных мною особей колебалась в следующих пределах (в процентах к длине, идиосомы); v i 12—15; sc е 32 40; sc i 7—11; h е 25—43; hi 5—9; Щ4—6; da8—13; d3 18—40; d4 .20 06;-la 7—11; Ip 22—35; sa e 5—11; sa i 34—66. Длина 585—1000 p,.
Гетероморфная фаза. Ноги I, II и IV длинные и crpolliii.ic; их длина составляет, соответственно, 38—45 (I), 40—18 (II), 30 Io (III
-IfiO
I. TYROGLYPJIIDAE. 2. RHIZOGLYPHINAE
n 42—53 (IV)% длины идиосомы; щипы пог длинные, ножевидпые. Щетинки идиосомы много толще и, в среднем, длиннее, чем у гомеоморфных особей, .жесткие, сильно луковицеобразно утолщенные у основания; длина их, в процентах к длине идиосомы: v j. 1.1.—14; sc е 35—40; sc i 11; he 31—35; hi 8—9; dx 6—7; d2 21—22; <l3 41—45; d4 45—50; la 11—12; Ip 33—40; ,sa e 10—15; sa i 44—52. Длина 705—950 р..
ф. Ноги короче и массивпео, чем у большинства <5сГ, шипы на них также короче и массивнее; их пропорции, одпако, довольно изменчивы; щетинки идиосомы тонкие и гибкие; <12 в 1.5-.2 раза длиннее Щи в 2—3 раза
короче d3; 1 )> в 2—3.5 раза, длинное 1 a; sac очень короткие, в 8—9 раз короче sai; у крупных особен длина d8 и d4 равняется 18—20, 1 р около 15% длины идиосомы. Анальных микрохэт 6 пар, из которых четыре задание расположены па почти одинаковом расстоянии друг от друга и образуют с каждой стороны анальной щели по косому дугообразному ряду, а две, как обычно, сидят у ее переднего конца. Генитальные щупальцы и края анальной щели непигментированные. Длина 510—1240 р., ширина 255— -860 р,. Яйца размером от 110X80 до 210X125 |л. Хорион неутолщенный и непигментированный.
Личинка. Грудные палочки относительно стройные, длиною в 20 р., умеренно булавовидно вздутые; их диаметр в расширенной части не превышает 5 р., вершинный купол полушаровидный.
Гипопус. Окраска более или менее интенсивно желтовато-розовая, : конечности темнее. Идиосома в 1.5 раза длиннее своей ширины, эллиптическая; покровы плотные, довольно сильно хитипизироваппые. Пропода-'Сомальный щит в 6—8 р'аз короче гистеросомального щита, приблизительно 'треугольный; ростральный выступ умеренно выдающийся вперед, тупоугольный, с закругленной вершиной; передне-боковые края щита чуть во--гнутые по сторонам рострального выступа, далее кнаружи слабо выпуклые; .•sc выстроены в слабо дугообразный, выгнутый кпереди ряд; v i на вершине рострального выступа; гистеросомальный щит с очень слабо дугообразно выпуклым, почти прямым передним краем; поперечная штриховка у переднего края слабо намеченная; вся остальная поверхность щита очень тонкопористая, без более грубой ямчатой пунктировки; щетинки гистеросомы очень короткие; sa i расположены значительно выше постанальных, заметно кнутри от них; р а много короче голени II. Гнатосома узкая и длинная, выдающаяся за вершину рострального выступа концом основного членика и всем развилком; основной членик с прямыми, параллельными друг другу боковыми краями, в 3—3.5 раза, длиннее своей ширины; дистальные членики цилиндрические, тесно сближенные между собой. Стернальный щит
Фиг. 254—261.
-Фиг. 254. Caloglyphus rodionovi Л. Z., гетероморфный с? сверху. Фиг. 255. С. rodionovi A. Z., нога I гетероморфного rf. Фиг. 256. С. rodionovi Л. Z., лапка IV гетероморфного Фиг. 257. С. rodionovi, Z., гомеоморфный cf сверху. Фиг. 258. С. rodionovi A. Z., 9 снизу. Фиг. 259. С. rodio-..novi A. Z., нога I Q. Фиг. 280. С. mandzhur, sp. п., гетероморфный О сверху. Фиг. 261. С. mandzhur, sp. п., гомеоморфный cf снизу.
11 Фауна СССР, Захватшш —284
162
I. TYROGLYPHIDAE. 2. RHIZOGLYPHINAE
хорошо очерченный с боков слабо S-образно изогнутыми эпимеритами II. без оформленного заднего края; коксальные поляII замкнутые, эпимеры II, сходящиеся под острым углом; коксальные поля III и IV хорошо очерченные, разъединенные по средней линии узким продольным промежутком; эпимеры III дугообразные; генитальный щит явственно обособленный; коксальные и генитальные присоски, а также и наружные и задние из расположенных на присасывательном -диско мясистые, грибовидные; генитальные присоски отстоят дальше от небольшой генитальной щели, чем от наружных краев генитального щита; наружные и задние периферические присоски почти в 2 раза меньше центральных; передние почти такого же размера, как последние; ширина присасывательного диска умещается в ширине идиосомы несколько меньше 3 раз; генитальные щупальцы с тупой, закругленной вершиной. Ноги длинные и стройные; средняя длина их, выраженная в процентах к длине идиосомы, составляет, соответственно, 40, 36, 32 и 32; ноги I выдаются за края проподосомы, начиная с середины бедер, ноги II — с середины колен; голени заметно длиннее своего диаметра; длина лапок I и II в б—7 раз превосходит их диаметр посередине и обычно больше общей длины голени и колена, вместе взятых; наружные щетинки колен шиповидные; вентро-латеральные щетинки голеней в виде умеренно толстых, чуть искривленных шипов, причем внутренние лишь немного уступают в размерах наружным; последние в 2—2.5 раза короче'лапок; на лапках I и II по 5 листовидных щетинок, из которых 2 много меньше остальных 3; пластинки их узкие, ланцетовидные, слабо загнутые на конце, без продольной жилки; присасывательная щетинка с крупным, но узким блюдцем; обонятельная палочка прямая, очень длинная и тонкая, не сильно булавовидно утолщенная; В расположена много впереди у и представлена очень мелкой простой щетинкой; е в виде крепкой, слабо изогнутой щетинки, расположенной далеко впереди основания обонятельной палочки, но ниже В. На лапках III по 4, на лапках IV по 3 коротких ланцетовидных щетинки; средняя дорзальная щетинка лапок IV в 1.5 раза длиннее лапки. Длина 252—340 [х, ширина 170—220 |х.
Распространение. Достоверно известен только из СССР (Москва,. Ленинград, Ярославль, Барнаул, ряд пунктов Северного Кавказа, Башкирия) и Германии.
Экология. Синантропный, амбарный вид, живущий в влажном зерне (преимущественно па просыпях в подпольях зернохранилищ), маслосеменах, отрубях, муке и иногда на гпиющем луке и других корнеплодах. В полевых условиях встречается преимущественно в гниющей соломе и. других отходах в попосродствошюй близости от жилья и, невидимому, только в более южных районах. Высоко гигрофильный и термофильный вид. Успешно развивается при влажности субстрата не ниже 22—25, но ине выше 80%. Температурный минимум развития равен 16.5°, оптимальная температура 29—30° С; при оптимальной температуре и влажности инкубационный период длится всего 1%—2 дня, постэмбриональное развитие — от 5 до
9. CALOGLYPHUS
1 Ihf
9 дней. Вследствие быстроты развития и большой плодовитости (один ф откладывает, при благоприятных условиях, до 621 яйца) увеличение численности С. rodionovi идет в подходящих условиях очень быстро, и он обычно встречается огромными колониями. Переносчиками гипопусов служат мучной хрущак (Tenebrio molitor L.) и другие амбарные насекомые. По Schulze, этот вид нередко нападает на живых насекомых (Tenebrio и др.) и их личинок и наносит иногда значительный ущерб культурам мучных червей.. Продуктам вредит лишь при условии их повышенной влажности.
*2. Caloglyphus tshernyshevi, sp. п. (фиг. 264, 299. BOO).
Известен только гипопус. Очень близок к гипопусу С. rodionovi, отличаясь от него следующими признаками: проподосомальпый щит заметно длиннее, составляя немного менее 1/i длины гистеросомального щита; постанальные щетинки много длиннее, вдвое превосходящие длину голени II, гибкие, волосовидные; гнатосома значительно короче и выдается только концом своего развилка (дистальных члеников) за вершину рострального выступа,; основной членик ее только в 2 раза длиннее своей ширины;' коксальные поля II открытые; коксальные, генитальные, задние и боковые присоски менее мясистые, чем у С. rodionovi. Ноги имеют приблизительно такие же пропорции, как у С. rodionovi-, наружная щетинка колен I тонкая, волосовидная; внутренние вентральные щетинки голеней I и II очень маленькие, в 2—3 раза короче наружных; обонятельная палочка лапок I и II несколько короче и более резко булавовидно утолщена к концу; 8 такой же длины, как у; 4 листовидные щетинки на лапках I, 3 из них крупного размера и только одна, более короткая, средняя дорзальная щетинка лапок IV много короче. Длина 225—350 р., ширина 175—240 у.
Распространение. В лесной подстилке и под корой пней в широколиственных лесах Одесской обл. (Нерубаевский лесопромхоз).
3. Caloglyphus mandzhur, sp. п. (фиг. 40, 260, 261, 265, 280, 287, 301, 302).
. Псевдостигмальный орган очень маленький (6—8 у), в виде прямой, заметно расширяющейся к основанию палочки. Проподосомальпый щит-е более или менее глубоко выемчатым задним краем; боковые поверхности проподосомы впереди вертлугов I и неправильные участки вокруг стернума. и эпимер плотно хитинизированы, но лишь слабо окрашены. Серповидные-щетинки лапок I—III, как у С. rodionovi, но несколько более приостреиы; на лапках IV по 1 серповидной щетинке такой же формы, как на передних, погах.
<5<5 диморфны, причем гетероморфные особи многочисленнее гомеоморфных. Кожа плотнее, чем у других видов, особенно на гистеросоме, тонко, и густо продольноморщинистая (смотреть при большом увеличении), нередко, окрашенная в буроватый цвет. Р2 длинные, волосовидные, в З.б—б раз длиннее рх; расстояние между р2 в 3.5 раза меньше такового между рг,
*
164
I. TYROGEYPllIDAE. 2. RRIZOGLYPHINAE
Анальные копулятивные присоски округлые, слабо хитинизированные, четко очерченные сзади соединительным склеритом, так что линия, Проведенная через их задние края, проходит несколько впереди заднего конца анальной щели. Дистальная часть penis прямая, очень тонка'я.
Гоме оморфпая ф а в а. Ноги довольно длинные и стройные; -они всегда значительно дл и и ное. и тоньше, чем у задняя (внутренняя) вентральная щетин на ланок III по превращена в шип. Щетинки идиосомы толще и крепче, чем у (7. iodionw-i', длина их чрезвычайно изменчива, но, в среднем, опа. значительно больше, чем у С. rodionovi. Длина 690—965 ре, ширина 510 [Л.
Гетеро м. о р ф н а я: ф аза. Крупнее. Ноги чрезвычайно длинные и стройные. Длина 920—1025 н, ширина 530—600 р.
ф. Кожа совершенно гладкая, всегда бесцветная, тоньше, чем у <d; логи значительно короче и массивнее. В2 в 1.1—1.70 раза длиннее Щ и не больше чем в 1.7 раза короче d3; 1 р в 2—3.5 раза длиннее la; sa е относительно длинные, только в 2—2.6 раза короче sa I; 6 пар анальных микрохэт, из которых две помещаются у переднего конца анальной щели, одна у ее середины и три тесно сгруппированы у ее заднего конца. Генитальные щупальцы и края анальной щели непигментированные. Длина 860—1400 р., ширина до 860 р.. Яйца размером от 175X113 до 220X140 р..
Личинка характеризуется очень крупными (23—24 р.), сильно булавовидно утолщенными грудными палочками; их вздутая дистальная часть •очень напомипает-жолудь в его чашечке.
Гипопус. Светлобуровато-желтый. Идиосома эллиптическая, в 1.6 раза длиннее своей ширины; кожные покровы плотные, сильно хитинизированные. Проподосомальный щит в 4 раза короче гистёросомального, с узким, треугольным, сильно выдающимся вперед ростральным выступом, явственно отграниченным от дугообразно выпуклых на всем их протяжении'переднебоковых краев щита; vi сидят на самой вершине рострального выступа; sc относительно длинные и тонкие, как и все другие щетинки спинной поверхности тела. Гистеросомальный щит со слабо выпуклым передним краем; поперечная штриховка у переднего края слабо намеченная; вся остальная поверхность щита гладкая, мелкопористая, без более грубой ямчатой скульптуру. Гнатосома целиком спрятана под ростральным выступом, за вершину которого выдастся только дистальная половина арист; основной членик бочеиковидпый, с заметно выпуклыми боковыми краями, чуть длиннее своей ширины. Кокоальиыо поля Ш хорошо очерченные, разделенные .довольно широким продольным промежутком; вентральный щит, в целом, .значительно длинное снооЛ ширины. Коксальные присоски очень мелкие, нормального строения. Присасывательный диск небольшой (его диаметр
Фиг. 262—267.
-Фиг. 262. Caloglyphus rodionovi Л.. Z., runoiryc сверху. Фиг. 263. То же, снизу. Фиг. 264. С. tsher-nyshevi, sp. п., гипопус, передний часть тела с ногами I и II снизу. Фиг. 265. С. mandzhur, sp. n., гипопус снизу. Фиг. 266. С. geolruporum, sp. п., гипопус сверху. Фиг. 267. То же, снизу.
166
I. TYROGLYPHIDAE. 2. RHIZOGLYPHINAE
умещается в ширине тела 4 раза); наружные и задние присоски много меньше передних. Ноги I и II умеренной длины, не очень толстые, выдающиеся за края проподосомы, начиная с середины бедер; лапки I довольно длинные, около 2.5 раза длиннее своего диаметра у основания и немного уступают длине голени и колена, взятых вместе; вентро-латеральные шипы голеней и колен довольно длиппые, прямые, почти цилиндрические (не уплощенные); внутренние только немного уступают в размерах наружным; листовидные щетинки лапок плотной консистенции, с ланцетовидной пластинкой, оттянутой дистально в тонкое острие; только одна из них заметно меньше остальных; обонятельная палочка прямая, довольно массивная, слабо булавовидная. Вентральные щетинки голеней III и IV шиповидные; на лапках по 4 довольно широких ланцетовидных щетинок; на лапках III, кроме того, обе основные вентральные несколько ланцетовидно расширены; средняя дорзальная щетинка лапок IV почти в 2 раза длиннее лапки. Длина 339— 440 р, ширина 260—275 р.
Распространение. Маньчжурия: Харбин.
Экология. 'Найден в огромном количестве в гниющих головках чеснока, ввезенного из Харбина. Многие из 2$ содержали большое количество яиц (до 45 штук), в других же можно было видеть вполне сформированных, уже вылупившихся из яиц личинок и пустые оболочки яиц; таким образом, факультативное, живорождение так же свойственно этому виду, как С. moniezi.
* 4. Caloglyphus spinitarsus (Henn.) (фиг. 281, 282, 306).
Hermann, 1804, M&n. Apt6rol.:85, t. VI, fig. & (Acarus); Oudemans, 1924, Entomol. Ber., VI: 323, fig. (Hypopus')-, 1932, ibid., VIII:358, fig. (Caloglyphus).
d. Гетероморфная фаза. Длина 715 p; ширина у плеч 370 р; боковые края тела идут до 1 е почти параллельно; задний край закругленный. Все волоски длиннее, чем у q. Тонких продольных линий на спине я не заметил, возможно вследствие раздутия, вызванного молочной кислотой или хлорал-гидратом. Генитальное отверстие сходно с таковым нимфы III; его длина 85 р. Расстояние между отверстием и anus 18 р; anus — 100 р; расстояние последнего от заднего края 200 р. Каждый анальный клапан несет 1 маленькую щетинку. Анальные присоски необычно маленькие. Позади anus 3 пары коротких и 1 пара длинных щетинок. Ноги длинные; моги Ш утолщенные; лапки IV е 2 присосками, расположенными весьма дистально, что не наблюдается пи у одного другого вида. Если при рассматривании генитального отверстия опускать тубус глубже, можно заметить цоколь penis, узкий в породней части, у середины внезапно расширяющийся, кзади становящийся еще шире; сам penis, расположенный в самой задней части, очень мал. Ланцетовидные волоски. лапок I и II шире, чем у у которой они чрезвычайно узки. Maxillicoxae значительно уже, чем у q. Гомеоморфная фаза неизвестна.
£. Длина без гнатосомы 711 р; ширина в плечах 311 р, у жировых желез —
9. CALOGLYPHUS
167
370 p; края тела между плечами и жировыми железами слегка нагнуты; задний край тупой, даже слегка вогнутый (типично для Caloglyphun). Плечевые волоски сидят скорее вентрально, чем дорзально. L и 4 крестцовый длинные: sa е субмаргинальны, длиннее прочих, равны по длине логам I;' sei, h и d слегка длиннее, чем ширина лапок; на спинке те ясс тонкие линии, как у нимфы III. Генитальное отверстие маленькое (100 р.), расстояние между ним и anus 95 р; anus — 122 р; расстояние между последним и задним краем 155 р. Генитальные клапаны каждый с 2, анальные — каждый с 3 чрезвычайно мелкими щетинками. Позади anus, субмаргинально, те же 2 пары щетинок, что и у нимфы III. Копулятивное отверстие вентральное. Обращают на себя внимание короткие ноги. (Oudemans, 1932.)
Некоторым дополнением к этому описанию может служить помещенная в той же работе Oudemans’a (Acarol. Aanteek., СХН) заметка: «Tracheen bij Acaridiae», где изображен псевдостигмальный орган С. spinitarsus (называемый Oudemans’oM «супракоксальиым волоском»), имеющий вид очень короткой, прямой, закругленной на конце палочки. Совокупность зтих признаков позволяет считать С. spinitarsus близким к (7. sphaerogaster и особенно к С. mandzhur. От первого вида он сразу же отличается стройным телосложением, резким половым диморфизмом (выраженным, как обычно, в размерах щетинок, длине ног и в пропорциях гнатосомы), гетероморфизмом d* d*; от С. mandshur его отличают «необычно маленькие» анальные копулятивные присоски, короткие постанальные щетинки (рг и р2) и наличие всего 3 пар анальных микрохэт у q.
Гипопус. Хорошо отличается от всех других видов крупными размерами, чрезвычайно короткими и толстыми ногами и своеобразной формой гистеросомы и проподосомального щита. Окраска охристо-бурая. Идиосома в 1.35 раза длиннее своей ширины; гистеросома очень правильной эллиптической формы, в 13 раз длиннее узкого, пластинчатого проподосомального щита, выдающегося вперед в виде полулунного козырька. Кожные покровы очень плотные, сильно хитинизированные; спинная поверхность лишена ямчатой скульптуры; все усаживающие ее щетинки чрезвычайно короткие. Sc расположены в дугообразный ряд, идущий параллельно переднему краю гистеросомы; vi смещены на вентральную поверхность проподосомального щита и сидят значительно отступя от его переднего края. Передний край гистеросомального щита резко дугообразно выпуклый (чем отчасти и определяется правильность его формы), е неясной поперечной штриховкой; dx и hi смещены на его боковые края, he — на вентральную поверхность; sai расположены несколько кнутри от ра; последние относительно длинные. Гцатосома смещена очень далеко назад, так что она вся, вместе с принтами, спрятана под крышевидно нависающим проподосомальным щитом; ее основной членик чуть длиннее своей ширины, с параллельными боковыми краями; дистальные членики хорошо обособленные, в 1.5 раза короче основного. Стернальный щит сзади слабо очерчен; его боковые кран болео
168
I. TYROGLYPHIDAE. 2. RHIZOGLYPHINAE
явственно намечены прямыми эпимер)muni II; концы эпимер II слегка, отогнуты кнаружи; коксальные поля III слегка раздвинутые, слабо обрисованные; коксальные поля IV не разделены по средней линии тела и еле' заметно обособлены от генитального щита. Коксальные присоски нормальные. Присасывательный диск очень крупный, его присоски широко расставленные; соотношения пх разморен, примерно, таковы же, как у С. poly-phyllae, но дополнительные присоски гораздо больше. Ноги1! и II чрезвычайно короткие и толстые (около 20% длины идиосомы), конические, почти целиком спрятанные под толом; за передний край лронодосомального щита . выдаются только самые кончики лапок 1 и коготок и окружающие его листовидные и присасывательная щетинки ланок II; лапки короткие, в 2 раза длиннее своего диаметра у основания; голени, колени и бедра очень низкие, кольцевидные; вентральные шипы голеней и наружные шипы колен толстые, но короткие; листовидные щетинки лапок с очень широкой, сильно закрученной пластинкой, имеющей явственную срединную жилку, все крупные; присасывательные щетинки длиннее лапки, с узким блюдцем; обонятельная палочка длинная, несколько искривленная, с слабо булавовидноутолщенным концом; у расположена несколько выше основания обонятельной палочки; г представлена толстой, жесткой щетинкой, находится посередине длины лапки. Ноги III и IV приблизительно такой же длины, как передние; вентральные щетинки голеней мало утолщенные; листовидных щетинок по 5 на каждой лапке; дорзальная щетинка лапок IV короче самой лапки. Длина 400—420 р, ширина 200—310 р.
Распространение. Франция, Германия; СССР (окрестности Киева); указание для Англии недостоверно.
.Экология. Гипопусы встречаются преимущественно на бронзовке-Osmoderma eremita Scop., но были находимы также на жужелице Carabus. nemoralis О. F. М. (Vitzthum, 1931).
* б. Caloglyphus polyphyllae, sp. п. (фиг. 269—273, 283, 284, 294, 304, 305).
Псевдостигмальный орган отсутствует; дорзальные щетинки гипостома, короткие, толстые, конически расширяющиеся к основанию. Границы, проподосомальиого щита неясные; хитипизироваиных участков перед, вертлугами I и вокруг эпймер нет. Серповидные щетинки всех лапок с очень широкой овальной пластинкой. Кожа у обоих полов гладкая. Половой, диморфизм выражен очень резко.
d1. Износит только гомеоморфная фаза. Р2 короткие, приблизительно, такого же размера., как pt; промежуток между р2 около 2 раз меньше расстояния между р1. Анальные копулятивные присоски крупные, эллиптические, очень сильно хитипиэиронапные и рельефные, расположенные косо; их задние края заходят зшщитсЛьйо дальше назад, чем анальная щель; дистальная часть penis прямая, очепь.узкая. Ноги очень длинные, довольно толстые; I пара заметно короче других; все щетинки колен и голеней и
9. CALOGLYPHUS
Kill
вентральные щетинки всех лапок (в том числе обе центра,иьпыи щетинни лапок III) представлены очень длинными ножевидиыми шипами; нонтрпль пые вершинные шипы лапок необычайно длинные и стройные (ое.оГинин» центральные), лапкп сильно конусовидно расширяющиеся к ociioiuinnio, перед серединой резко суженные и вновь расширенные к концу. Крупные щетипки идиосомы длинные, толстые, очепь жесткие, сильно утолщенный к основанию, по своему строению сходные с таковыми гетероморфной фазы С. rodionovi-, более мелкие щетинки (sci, hi, dx и la) тоже довольно крепкие. Длина 560—640 р..
9. Ноги короткие и толстые; шипы колон, голеней и лапок толстые, прямые. Щетинки идиосомы жесткие, короткие; их размеры, а также и число-it расположение анальных микрохэт не смогли быть установлены из-за. очень плохой сохранности имеющихся 29- Длииа 70°—1000р..
Гипопус. Окраска охристо-(хитиново-)желтоватая. Идиосома в. 1.5 раза длиннее своей ширины, эллиптическая; кожные покровы плотные, сильно хитинизированные. Проподосомальный щит в 6 раз короче гистеро-сомального щита; передний край его прямоугольный, с закругленной вершиной и слегка выпуклыми на всем протяжении сторонами; sc расположены трапецией; vi смещены на вентральную поверхность щита и сидят немного-отступя от его переднего края, на вершине расположенной здесь продольной выпуклости; гистеросомальпый щит с резко дугообразно выпуклым передним краем, серповидный, четко отграниченный сзади; передний участок его поверхности покрыт очень резкой поперечной штриховкой (составленной тонкими килями), вся остальная поверхность щита покрыта рассеянными поровидными ямками; щетинки гистеросомы довольно тонкие. Гнатосома целиком спрятанная под нависающим передним краем пропо-досомального щита, за вершину которого выдается только дистальная половина ее арист; основной членик с параллельными сторонами, в 1.5 раза, длиннее своей ширины. Стернальный щит хорошо очерченный, со слегка, вогнутыми боковыми и задним краем; коксальные поля II, таким образом, открытые; коксальные поля III хорошо очерченные, слегка раздвинутые; коксальные поля IV еле обрисованные и не отделенные от генитального щита; коксальные присоски среднего размера, пуговицеобразные; присасывательный диск небольшой (егоширина умещается в максимальной ширине идиосомы свыше 3 раз); периферические присоски в 1.5 раза меньше центральных; наружные присоски несколько смещены назад; генитальные щупальцы с конически прпостренной вершиной. Ноги I и II чрезвычайно массивные, короткие (28 и 25% длины идиосомы), выдающиеся за края проподосомы 3 дистальными члениками; лапки короткие, в 1.7 — 2 раза длиннее своего-диаметра у основания; наружные вентральные шипы голеней очень крупные, ножевидные, заметно искривленные, достигающие длины лапок; нпруж'-пые шипы колен много короче соответствующих шипов гололвй; .'Iистопид-пые щетинки лапок с очень широкой, овальной, закрученной nil- конце пластинкой, с явственной срединной жилкой; присасц1ППте,||Ы|Ц)1 щетникп
470
I. TYRGGLYPHIDAE. 2. RHIZOGLYPHINAE
-длинная и толстая. Вентральные щетинки голеней III в IV в виде длинных тонких шипов; на лапках III по 6 широких листовидных щетинок, на лапках IV — по 4; средняя дорзальная щетинка лапок IV не превосходит .длины лапки. Длина 350—386 ширина 235—257 р..
Распространение. Батуми.
Экология. Большое число гипопусов и несколько б* о” и ^9 этого вида были найдены па хруще Volyphylla jullo L.; гипопусы сидели на ниж- • ней стороне и боках брюшка, половозрелые особи — под надкрыльями одного из экземпляров лруищ. Caloylyphun polyphyllae, вероятно, так же тесно биологически связан с J.'olyphylla, как С. monicsi с видами Melolontha.
6. Caloglyphus berlesei (Mich.) (фиг. 293, 312—315, 355).
Michael, 1903: 110 IPyroqlyphus). —mycophagus Berlese, 1891 (nonMegnin, 1874),LVlII,n0 1; Kramer, 1899:139, fig. 30 (Tyroglyphvs).
Очень близок к предыдущему, но крупные серповидные щетинки имеются только па лапках I и II.
с? с?. Представлены одной только гомеоморфной фазой, .гетероморфных d d не встречается. Расположение и относительные размеры щетинок пост--анального комплекса, строение анальных копулятивных присосок и анальной щели, как у С. polyphyllae; дистальная часть penis отогнута под углом к цоколю, так что penis кажется спереди притупленным. Ноги очень -стройные, еще длиннее, чем у С. polyphyllae-, их длина составляет, соответственно i1 50(1), 52 (II), 55 (III) и 57 (IV) % длины идиосомы.- Крупные щетинки идиосомы такие же крепкие, жесткие, как у С. polyphyllae, но еще более .длинные; see, he, d3 и 1р значительно превосходят максимальную ширину идиосомы, достигая 60—65% ее длины; sae также заметно длиннее (25%); sci, hi и dx, наоборот, короче (5—6%) и толще', почти шиповидные. Общая .длина тела до 750 р. (длина идиосомы, следовательно, до 670—690 р).
$ П3 заметно длиннее dx; 1р около 2.5 раза длиннее la; sae в 2.5 раза короче sai; у крупных особей длина d3, d4 и 1р не превышает 10—11% „длины идиосомы. Число и расположение анальных щетинок не установлены. •Общая длина тела до 1300 р,.
Гипопус. По общему виду и большинству деталей строения очень похож на гипопуса С. rodionovi. Гнатосома выдается всей своей дистальной половиной за вершину тупоугольного рострального выступа, удлиненно-грушевидная, перасчлененная, только расщепленная на конце; ее общая длина В 2 раза превосходит наибольшую ширину. Эппмериты II резко S-образно
1 Все измерения сделаны но рисункам Bcrleao.
____ Фиг, 908—280,
••Фиг. 268. Caloglyphus rodionovi Л. И., Нога Ш гетероморфного о”. Фиг. 269. С. polyphyllae, ер. п., СТ сверху. Фиг. 270. С. polyphyllae, ар. п., голень и лапка II О’ снизу. Фиг. 271. С. polyphyllae.
~eP-> п., голень и лапка III спину. Фиг. 272. С. polyphyllae, ер. п., голень и лапка IV сГ снизу. Фиг. 873. с. polyphyllae, ер. п., колено, голень и лапка I Q. Фиг. 274. О. sphaerogaster A. Z., С; , сверху. Фиг. 275. С. sphaerogaster A. Z., (,> снизу. Фиг. 276. С. sphaerogaster A. Z., голень •и лапка IV o’- Фиг. 277. О. sphaerogaster A. Z., голень и лапка III О’. Фиг. 278. Псевдостигмальный органе, rodionovi A. Z. Фиг, 270. Псевдостигмальный орган С. sphaerogaster A. Z. Фиг.280.
Псевдостигмальный орган о. mandzhur, ар. п.
172
I. TYROGLYPHIDAE. 2. REIZOGLYPHINAE
изогнуты; коксальные поля III очень четко очерченные, с длинными, сильно» скошенными передне-боковыми краями, образующими явственный тупой угол с прямым передним краем. Задние и боковые присоски присасывательного диска почти одинакового размера с центральными присосками, кольцевидные, правильной округлой формы. Ноги I и II относительно длинпые-и стройные, но все же несколько короче, чем у С. rodionovi (33 и 30% длины идиосомы); вооружены; их слабее, чем у всех других видов, вентро-латеральные щетинки голеней и колеи ио имеют вида шипов; все щетинки лапок IV короче самой лапки.
Р а с п р о с т р а, и е и и. е. Пока известен только из Италии.
Эколог и я. Живет па грибах и гпиющих клубпцх картофеля. Гипопусы были находимы ла дорзальной поверхности брюшка (под крыльями)-жука-носорога (Oryctes nasicornis L.) и мраморного хруща (Polyphylla fullo L.).
* 7. Caloglyphus moniezi A. Z. (фиг. 285, 303).
Захваткин, 1937 :185, рис. 31. —mycophagus Moniez, 1892 (non Megnin, 1874), Mem. Soc. Zool. Fr., V:584, 8 figg.; Oudemans, 1907, Entomol. Bor., 1:223; Vitzthum, 1924, Arch. Naturg., XC, 10 : 60, fig. 42 (Tyrogtyphus).
Недостаточно полно описанный вид, несомненно очень близкий к С. poly-phyllae и С. berlesei и отличающийся от них преимущественно строением гипопуса. В половозрелой стадии отличается от С. polyphyllae наличием 2 серповидных щетинок на лапках IV и тем, что крупные щетинки идиосомы на конце тонко зазубрены, а от С. berlesei — крупными размерами серповидных щетинок лапок III и IV. Псевдостигмальный орган, дорзальные щетинки гипостома, расположение и размеры постанальных щетинок d и число анальных микрохэт д остаются неизвестными, так как у Moniez. они не описаны и не изображены. Половой диморфизм резко выраженный; d много мельче Q. •
d. Как и у С. polyphyllae, представлен только гомеоморфной фазой. Строение копулятивного аппарата точно такое же, как у названного вида. Щетинки идиосомы жесткие; размеры их, насколько можно судить, мало-изменчивы; d2 и d3 почти одинаковой длины, обе значительно короче 1 р и более чем вдвое короче d4. Общая длина тела до 810 р.; длина идиосомы,, таким образом,Не превышает 720—730 р.
9. Характерно наличие 2 форм самок: нормальных по своим размерам (общей длиной в 1150—1250 р) яйцекладущих, окрашенных в беловатый цвет и содержащих большее или меньшее число яиц (до 35), и гигантских, живородящих особей, достнппощих в длину 2000 или даже 2262 р, окрашенных в тусклый сероватый или кирпично-красный цвет и почти неспособных, к самостоятельному передвижению из-за непомерной раздутости гистеросомы; внутри таких самок находятся, помимо яиц на разных стадиях эмбрионального развития, еще личинки и. протонимфы, иногда даже последние содержат уже вполне сформировавшихся гипопусов. Яйца длиною от 153-до 180 р, диаметром в 99—117 р, Личинка точно не описана.
9. CALOGLYPHUS
ИИ
Гипопус. Очень похож на гипопуса С. polyphyHue. Oitpuoim Пилим бледная, хитинизация спинных щитов слабее. Про1юдо<н>мл,иhlihlll ЩНФ .длиннее, равный, примерно, Vs Длины гистеросомального; передне Oohip вне края его впереди чуть прогнутые, в соответствии с чем выступ несколько уже и резче выступает вперед; sc расположены с.нш><> выгнутой кпереди дугой; передний край гистеросомального щита почти прямой, его скульптура значительно нежнее; составляющие ее поперечные липип образованы рядами мельчайших бугорков, менее правильны и местами прерываются; щетинки гистеросомы несколько крепче, жестковатые; sa i ближе придвинуты к постанальным и лишь немного уступают им в длине, поровидные ямки щита разбросаны реже и, в связи с его мепыпей плотностью и более слабой пигментацией, не так бросаются в глаза. Гнатосома и коксальные щиты такого же строения, как у С. polyphyllae. Стернальные присоски несколько крупнее, с явственным кольцевидным краевым утолщением; присасывательный диск больше: его ширина умещается в максимальной ширине идиосомы около 2.5 раза; присоски па нем также крупные, особенно периферические и дополнительные; генитальные щупальцы с широко закругленной, куполовидной вершиной. Ноги I и II несколько длиннее (30 и 28%. длины идиосомы) и много стройнее; лапки около 3 раз .длиннее своего диаметра у основания и почти в 4 раза длиннее диаметра у середины; голени I заметно длиннее своего диаметра; наружные вентральные шипы голеней I и II значительно короче и тоньше, чем у С. polyphyllae (обычно они не превосходят х/2 длины лапок); листовидные щетинки лапок I и II уже, ланцетовидные, более постепенно расширяющиеся к середине и менее резко закрученные на конце; обонятельная палочка длиннее и тоньше, почти прямая; присасывательная щетинка короче (она почти не заходит за вершину листовидных щетинок), с более крупным блюдцем; г расположена значительно более дистально. Ноги III и IV также заметно стройнее, чему С. polyphyllae, щетинки на них тоньше, листовидные волоски уже, ланцетовидные. Длина 300—362 р. (по Vitzthum’y, 1924, длина 224— 392 р, ширина 190—210 р).
Распространение. Известен из Франции, Голландии, Германии и СССР (окрестности Москвы: Волховское; Смоленск).
Э к о л о г и я. С. moniezi биологически тесно связан с майскими жуками (Melolonlha), служащими при жизни переносчиками гипопусов, а после смерти, в гпиющем состоянии, кормовым субстратом для других стадий. Более мелкие qq яйцекладущи, тогда как крупные живородящи, причем развитие молоди в их теле идет иногда вплоть до образования протоинмф и даже гипопусов.
* 8. Caloglyphus sphaerogaster A. Z. (фиг. 274—277, 279, 28И, 'JIMI), Захва/гкин, 1937:186, рис. 15—24.
Хорошо отличается от всех других видов, кроме С. wop/ш/щ.ч, полным -отсутствием полового диморфизма: пропорции тела п пог, вооруженно
П1
L TYROGLYPHIDAE. 2. Rlll/.OGLYPHLNAE
последних, относительные размеры щетинок идиосомы — практически одинаковы у обоих полов; мало отличаются и их абсолютные размеры. Гистеросома пузыревидно вздутая, резко суживающаяся кзади от точки прикрепления ее до sa i, с узким, слегка закругленным задним краем; ее ширина умещается в общей длине идиосомы только 1.3—1.5 раза. Псевдостигмальный орган представлен очень маленькой (3—5р.), цилиндрической, слегка приостренной к концу палочкой; дорзальные щетинки гипостома короткие, толстые, почти конические. Задний край проподосомальиого щита прямой или слабо выпуклы!!; боновые поверхности проподосомы вокруг вертлугов I и широкие, неправильной формы, участки вокруг стернума и эпимер сильна хитипизлровапы и окрашены в ржаво-рыжий цвет. Щетинки идиосомы короткие, довольно топкие и гибкие, более крупные из них сильно луковицеобразно утолщены у основания; размеры щетинок (в процентах к длине-идиосомы) у обоих полов таковы: v i 9—10; sc е 30—33; sc i 7—8; h e 20— 25; 11 i 8—4; Щ 2.5—4; d2 5—6; d3 12—21; d4 16—20; 1 a 3—5; 1 p 13—17; sa e 8—5; sai 15—20. Ноги сильно пигментированные, стройные и длинные, в силу крайней раздутости тела кажущиеся непропорционально тонкими; длина их составляет у d 32—34 (I и II), 30—32 (III) и 33—37 (IV) %. длины идиосомы; у £, в связи с еще большей раздутостью идиосомы, они оказываются на 3—4% короче, но имеют в точности такие же пропорции, как у с?. Серповидные щетинки ланок I и II е умеренно широкой пластинкой, па лапках III и IV несколько уже; иа лапках IV их 2; обонятельная палочка лапок I и II топкая, цилиндрическая.
сГсГ представлены только гомеоморфной фазой. Р2 представлены тонкими микрохэтами, одинакового размера с рл; расстояние между ними К 2 рана меньше расстояния между рг Анальные копулятивные присоски яйцевидные, слабо хитинизированные, резко очерченные сзади соединительным склеритом, спереди плохо обрисованные; линия, проведенная-через их задние края, проходит несколько впереди заднего конпа анальной щели; длина последней почти в 3 раза больше длины присосок; дистальная часть penis прямая, очень тонкая. Длина 900—1000 р., ширина 600—690 р.
Анальных микрохэт всего 4 пары; из них 2 помещаются у переднего конца анальной щели, 1 — у середины и 1 — иа половине расстояния между этой сродной парой и задним концом anus. Генитальные щупальцы и края, анальной щели ржаво-рыжпо. Длина 1180—1225 [л, ширина 775—930 р..
Яйца о очень толстым хорионом, окрашенным в коричневый цвет; размеры их 180—100 X 76—100 |i; число яиц, одновременно созревающих: в теле £, доходит до 25. — Гипопус и другие стадии развития неизвестны..
Распространены о. Лайдон только 1 раз. в окрестностях Москвы. Небольшая колония этого вида была паПдона в гниющем луке, сильно зара-
Фш-. 281.-285.
. Фиг. 281. Calogluphus spinitarsua (1 lerni.), rnuoiiyo сверху. Фиг. 282. To же, снизу. Фиг. 283..
С. polyphyllae, sp. п., гипопус оворху, Фиг. ШИ. то же, снизу. Фиг. 285. С. moniezi A. Z., гипопус опиву.
176 I. Ti'ROGLYPHIDAE. 2. REIZ0GLYPH1NAE
.женном Bhizoglyplius echinopus, видами Anoetus и сопуствующими им хшц-.лыми Laelaptidae.
9. Caloglyplius mycopliagus (Megn.) (фиг. 290—292, 316, 356, 357).
Megnin, 1874, Journ. Anat. Physiol., X:235, t. VII—IX (Tyroglyphus).
Недостаточно известный вид, несомненно очень близкий к предыдущему и характеризующийся также совершенно не выраженным половым диморфизмом; большинство признаков — длинные н стройные ноги, короткие .щетинки идиосомы, строение копулятивного аппарата <у, расположение и размеры постанальных щетинок, сильная пигментация генитальных щупаль-щев и краев анальной щели — общи обоим видам, и они различаются между собой только тем, что у С. mycophagus гистеросома менее шарообразно раздута (длина идиосомы у <5 почти в 2 раза, у q в 1.6 раза больше ширины), гнатосома уже, обонятельная палочка лапок I и II сильно булавовидно .вздутая, a penis крючковидно, изогнут и притуплен на конце. Общая длина тела <5 (включая гнатосому) 800—950 р., ширина 350—400 р.; длина q 1000— 1250 р., ширина 500—600 р.. Яйца размером около 150 X 100 р.. — Личинка •с крупными, умеренно вздутыми на конце грудными палочками.
Гипопус. Окраска бледнорозоватая, конечности довольно темного винного цвета. Идиосома широкая, только в 1.35 раз длиннее своей ширины. Проподосомальнып щит в 10 раз короче гистеросомального, с тупоугольно выдающимся вперед ростральным выступом и очень явственно вогнутыми на большом протяжении передне-боковыми краями, что придает ему своеобразную, не свойственную другим видам рода конфигурацию; расположение sc не установлено; vi сидят на вершине рострального выступа; h е, -sa i и р длинные, особенно последние; на рис. 2 табл. ГК Megnin’a (воспроизводимом здесь) все они изображены имеющими такую же длину, как голень и колено I, вместе взятые. Гнатосома выдается за вершину рострального выступа дистальной половиной своего развилка; основной .членик квадратный, с вогнутым посередине передним краем; ветви развилка длинные (почти такой же длины, как основной членик'), широко расставленные, 2-членистые, явственно обособленные от основного членика. Стернальный щит слабо хитипизированный, очерченный дугообразной, прерванной посередине линией; стерпум и эпимеры II мощные, последние с резко отогнутыми кнаружи концами; коксальпыс поля III очень четко очерченные, разъединенные между собой, так же как и относительно хорошо обрисованные коксаль--иые поля IV, широким продольным промежутком; генитальный щит явственно отграниченный от коксальпых полей IV; стернальные присоски заме-
Фнг. 286-296.
-Фиг. 28В. Caloglyphus sphacrogrmliir Л. анальная щель 9 и окружающие ее щетинки. Фиг. 287. "То же, С. mandzHur, sp. п. Фиг. 288 —288. 'ГО же, С. rodionovi A. Z. Фиг. 290. С. mycophagus (Megn.), 9 снизу (по Megnin) Фиг. 291. С. mycopliagus (Megn.), ноги I и II и гнатосома снизу (по .Megnin’y). Фиг. 292. с- mycopliagus (MT.gn.), копулятивный аппарат О’, направо вверху — penis в профиль (по Milgnin.). Фиг. 298. С. berlesei (Mich.), 9, очертания тела (сверху) (по Berlese).
-Фиг. 294. С. polyphyllae, sp. n., auani.itaii щель cf с копулятпвнымп прпсосками и постанальными щетинками. Фиг. 295. То же, С. rodionovi A. Z. Фиг. 296. То же, С. sphaerogaster A. Z.
12 Фауна СССР, Захваткин—284
178
I. TYROGLYPHIDAE. 2. RHIZOGLYPHINAE
иены довольно длинными крепкими щетинками; генитальные присоски.-близко придвинуты к генитальной щели; передние присоски присасывательного диска заметно мельче боковых и задних, которые лишь немного уступают в размерах центральным; боковые расположены несколько впереди центральных. Ноги довольно длинные и стройные; длина их, выраженная, в процентах длины идиосомы, составляет, соответственно, 40, 35, 21 и 30; ноги I и II почти целиком выступающие за передний край проподосомаль-кого щита; их лапкп около Я раз длиннее своего диаметра посередине; вентро-латеральные щетинкп колеи и голеней топкие, не утолщенные в виде шипов; бросаются в глаза большие размеры сильно булавовидно утолщающейся к концу, несколько искривленной обонятельной палочки лапок I и II и резкий серповидный изгиб коготков. Длина 300—330 р, ширина 220— 240 р.
Распространение. Достоверно известен только из Франции.
Экология. Живет на грибах, преимущественно па ножках плодовых тел, выгрызая здесь пещерки, на дне которых встречается многочисленными колониями. Гипопусы попадаются иногда па свежих грибах,, но более многочисленны на разложившихся, уже подсыхающих после предшествовавшего периода разжижения; они встречаются особенно часто па. различных мелких жуках, многоножках и паукообразных, служащих: им переносчиками. 99 яйцекладущи; число откладываемых ими яиц не-установлено, но оно, вероятно, довольно велико, так как в теле самок могут одновременно созревать до 12 яиц. По наблюдениям Megnin самки копулируют только в стадии телеопимфы.
Мне уже приходилось указывать (1937), что под именем Tyroglypkus или С. mycophagus до сих пор смешивались почти что все виды рода Caloglyphus и даже некоторые Acotyledon (A. alberti A. Z., A. krameri auct.).. Между тем, при достаточно внимательном отношении к очень хорошим для. того времени описанию и рисункам Megnin, легко убедиться, что рассматриваемый им вид во всех стадиях своего развития хорошо отличается, от многочисленных форм, описывавшихся под тем же названием всеми позднейшими авторами.
* 10. Caloglyphus geotruporum, sp. п. (фиг. 266, 267, 307, 308).
Hypopus «К» Dujardin, 1849, Ann. Sci. Nat., Zool., (3), XII: 263.
Известен только гипопус, хорошо отличающийся от таковых других; видов Caloglyphus своими малыми размерами и беловатой окраской. Идиосома шврокоовалышя, в ',1.4 раза, длиннее своей максимальной ширины. Проподосомальпый щит треугольный, с слегка выпуклыми переднебоковыми краями и параболически закругленным ростральным выступом, немного более чем в 2 раза корова гпстеросомальпого щита;-У1 сидят на вершине рострального выступа; гпстсросомальпый щит с совершенно прямым передним краем; поперечная штриховка в .его передней части очень нежная,, остальная поверхность гладкая, бесструктурная. Гнатосома целиком скры
9. CALOGLYPHUS
1711
тая под ростральным выступом, за края которого выдаются только он npiwi»i, яйцевидная, нерасчлененпая, только слегка расщепленная на конце. Отер-пальный щит неоформленный, мягкой, перепончатой консистенции; энпми" риты II почти поперечные, еле заметные, слабо намечающие дугообразную границу между проподо-и гистеросомой; эпимеры II, сходящиеся.под тупым углом, слабо дугообразные, с очень широкими аподемами; коксальные поля III хорошо очерченные, разъединенные узким продольным промежутком; коксальные поля IV еле дифференцированные; коксальные и гениталь-пыеприсоски мелкие,пормальпогостроения, присасывательный диск довольно крупный; задние я боковые присоски мелкие (диаметр их в 3 раза меньше диаметра центральных), расположенные дугой. Ноги короткие, довольно массивные; передние 2 пары приблизительно одинаковой длины; лапки короткие, меньше чем в 2 раза длиннее своего диаметра; вентральные щетинки бедер короткие; вентро-латеральные щетинки голеней и колен /игольчатые, причем внутренние вентральные щетинки голеней заметно (особенно на ногах II) длиннее наружных; па лапках по 4 очепь широких, сильно скрученных, листовидных щетинки с явственной сродней жилкой; присасывательная щетинка средней длины; па лапках III по G, па лапках IV по 4 листовидных щетинки, имеющих такую же форму, как и па передних ногах; вентральные щетинки лапок IV короткие, толстые, прямые. Длина 200—235 у, ширина 144—170 р..
Распространение. Известен пока из СССР, 'Франции и Германии. Гипопусы встречаются в большом числе под надкрыльями навозника Geotrupes st&rcorosus Forst., с которым этот вид, вероятно, также связан в своей биологии, как С. moniezi с майскими жуками.
4. ТРИБА RHIZOGLYPHINI
Вооружение идиосомы сильно редуцировано: v е, sc i, clx—d2 и la в подавляющем большинстве случаев совершенно отсутствуют, и только у lihizoglypl'ius они представлены очень мелкими, часто трудно заметными микрохэтами; у большинства форм отсутствуют также d3 и sh, а у некоторых (Histiogaster, Schwiebea) еще и sa е; в таких случаях вооружение спинной поверхности идиосомы складывается всего из 6—7 пар щетинок. Ноги обычно короткие, массивные. Praetarsus в большинстве случаев сильно редуцированный, коготки обычно очень крупные и массивные (особенно на ногах I п II). На лапках I только 1 базальная сенсилла (8) или их пет совсем; сенсилла г расположена непосредственно впереди обонятельной палочки и обычно представлена коническим шипом; средняя вершинная щетинка р более или менее утолщена и модифицирована и обычно имеет над узкой, изогнутой книзу пластинки. Половой диморфизм почти у нсох ферм, чрезвычайно резкий: yds большинстве случаев (за исключенном родов Rhizoglyphus и Heterogtyphus) задний конец тела (опистосома) сильно модифицирован, образуя различного рода пластинчатые разращенii.ii, и огодорыпль-ная поверхность склеротизирована и окрашена, создания, таким образом, *
180
I. TYROGLYPHIDAE. 2. RHIZOGLYPHINAE
более или менее явственно обособленный от окружающей мягкой кожи опистосомальный щит; вершинная щетинка голеней IV превращена в полый конический шип, ноги IV иногда более или менее сильно утолщены; кроме того, у многих форм общая конфигурация тела у d резко отлична от таковой 9. У 9 строение заднего конца тела не уклоняется от обычной нормы, описто-сомальный щит отсутствует, вершин пая щетипка голеней IV не модифицирована и ноги IV никогда, не бывают утолщенными. Копулятивный аппарат п anus у J обычно снят,но смещены назад; анальные копулятивные присоски в большинстве случаев плотно хитининпровапы по всей окружности; поставил иных щетинок 1, 2 .или: 3 (Bhisoglyphus) пары, и они часто представлены микрохэтами; тарзальные копулятивные присоски сдвинуты к самому концу членика, крупные, обычно расположенные наискось друг от друга. У 9 anus сильно смещен назад, занимая иногда почти терминальное положение; аданальных щетинок всего 1 пара. — У нимфальных стадий v е, sc i, hi, sh, d,—d3 и 1 а всегда отсутствуют.
В трибу Bhizoglyphini, очерчиваемую совокупностью перечисленных выше признаков, входят около 15 палеарктических видов, распределяющихся в следующие 6 родов: Bhizoglyphus Clap. 1869, Heteroglyphus Foa 1913, Schwiebea Cuds. 1916, Histiogaster Berl. 1883, Monieziella Berl. 1897 и Thyreo-phagus Rond. 1874. Как видно из этого списка, моя триба Bhizoglyphini не соответствует по содержанию и объему пи сем. Bhizoglyphidae Ouds. 1923, ни какой-либо другой из установленных Oudemans’ом групп, так как составляющие ее члены были размещены этим автором в 2 различных семействах: Caloglyphidae Cuds. 1932 (роды Histiogaster и Monieziella) и Bhizoglyphidae Ouds. 1923 (роды Bhizoglyphus и Schwiebea)-, с другой стороны, я не включил в данную группу роды Sancassania Ouds. 1916 и Garsaultia Ouds. 1916 (помещаемые Oudemans’ом рядом с родом Bhizoglyphus), известные только по гипопальной стадии, не дающей, как известно, возможности правильно судить о систематической принадлежности соответствующих имагинальных форм.
10. Род BHIZOGLYPHUS CLAP.
Clapardde, 1869, Ztschr. wiss. Zool., XVIH:506; Canestrini, 1888:8; 1888a:357; Berlese, 1896: LXXIX, n° 5; Kramer, 1899:143; Michael, 1903:80; Berlese, 1897:109.—Coepophagus AKgnin, 1880, Lus parasites: 144.—Hypopus Oudemans, 1901, Tijdschr. Ned. Dierk. Ver., (2), VII:2.
Тип рода: ll'yroglyphus echinopus Fum. et Rob. 1868.
Вторичные половые различия очень невелики. Ve отсутствуют; sc i, hpi, sh, dx—da 11 I 11 представлены микрохэтами (иногда отсутствуют),
Фиг. 297—311. Детали crpoiniint и вооружении пог гипопусов родов Caloglyphus иRhizogtyphus. Фиг. 297. Caloglyphus rodlononi Л. Z., .пинки и голень I сверху. Фиг. 298. С. radio novi A. Z., лапка IV снизу. Фиг. 299. О, luhernyshMl, нр. п., голень и лапка I сверху. Фиг. 300. С. tsher-nyshevi, sp. п., лапка IV снизу. Фиг. 301. mandzhur, sp. п., голень и лапка I сверху. Фиг. 302. С. mandzhur, sp. п., голень и ланка IV снизу. Фиг. 303. С. moniezi A. Z., лапка и голень I сверху. Фиг. 304. С. polyphyllae, нр. о., го,иеш, и лапка I сверху. Фиг. 305. С. polyphyllae, sp. n., лапка IV снизу. Фиг. 306. С. splnilamiu (Попп.), голень и лапка I сверху. Фиг. 307. С. geotru-porum, sp.-n., голень и лапка! сверху. Фиг. 308. С. geolruporum, sp. п., лапка IV снизу. Фиг. 309. Rhizoglyphus echinopus (F. et R.), голень и ланка I сверху. Фиг. 310. R. echinopus (F. et R.), то же, снизу. Фиг. 311. it. echinopus (F. et R.), лапка IV снизу.
182 I. TYROGLVPHJDAE. 2. RHIZGGLYPHINAE
остальные — довольно короткими и тонкими щетинками; обе пары крест- |
цовых (sa е н sa i) хорошо развиты; есть псевдостигмальный орган. Эпи- ]
меры III и IV свободные. Ноги короткие и массивные, вооруженные много- ij
численными крепкими шипами; такими шипами представлены наружные ]
вентральные (или непарная нонтралъшш на ногах III и IV) щетинки голе- ' ней и колен, средняя вентральная ланок и сенсилла г; последняя распо- i
ложена непосредственно впереди обонятельной палочки; 8 помещается '
кнаружи и несколько впереди от лшитдцой; средняя вершинная щетинка лапок прямая, папрапленная косо шиш; вершинные шипы (дорзальный и все 5 вентральных) очень крупные. Коготки очень крупные и массивные; praetarsus сильно редуцирован.
<3. Задний конец идиосомы не модифицирован, без всяких следов описто-сомальной пластинки; р2 короткие, р2 и р3 длинные, выстроенные в дугообразный поперечный ряд. Имеется гетероморфная фаза, но она сильно уступает в численности гомеоморфной. ,
ф. Помимо 1 пары довольно крупных постанальных щетинок, имеются
еще 3 пары аданальных микрохэт.—У нимф sci, h i, d1—d2 и 1 а отсутствуют; личинки с крупными, булавовидными грудными палочками и более длинными, чем у других стадцй, d3 и 1 а. Есть гипопальная стадия. ;
В роде Rhizoglyphus я с уверенностью могу поместить только 1 палеарктический вид—В. echinopus (F. et В.). Большое число видов этого рода описано из Северной Америки и тропической Азии, по они все требуют критического и более детального изучения.*
* 1. Rhizaglyphus echinopus (Fum. et. Rob.) — Корневой, луковичный или картофельный клещ (фиг. 87, 196—198, 309—311, 326—328, 337, 338).
Fiimouzc et Robin, 1868, Journ. Anat. Physiol., V:287, t. XX—XXI; Kramer, 1876, Arch. Natiirg., XLII, 1:43 (Tyrogtyphus); Murray, 1874, Econ. Entomol. :257; Berlese, 1897:109; Kramer, 1899:143; Michael, 1903:84, t. XXX—XXXI (Rhizogtyphus); Megnin, 1880, Les parasites: 144 {Coepophagus). — dujardini Claparede, 1869, Ztschr. wiss. Zool., XVIII.-505, t. XXXV(Hypopus). — robini Claparede, 1859, op. cit. :506, t. XXXV, fig. 7—11; Michael, 1885, Journ. R. Mier. Soo., ser. II, vol. V:19, t. Ill, fig. 15—17 (Rhizoglyphus). — mUgniniHaller, 1880, Ztschr. wiss. Zool., XXXIV: 273, t. IX, fig. 16, et X, fig. 1; Berlese, 1884, A. M. S;, XIV, n° 7 (Tyroglyphus). —spinitMSUS Canestrini, 1888:31; 1888a:413, t. XXXIII, fig. 1 [Rhizoglyphus'). —hyacinthi Boisduval, 1867, Entomol. horticole:86 (nom. mid.) (Aearus); Banks, 1906; Garman, 1921; Connecticut Agr. Expt. Sta., Bull. 225. {Rhizoglyphus).
Биология, вредоносность. Michael, 1885, Journ. R. Mier. Soc., ser. II, vol. V:19; 1993:92—96; Mangin et Viala, 1892, C. R. Acad. Sci. Paris, CXXXIV-.151—153; Garman, 1921, op, oit.; Hodson, 1928, Bull. Entomol. Res., XIX:187—200; Попов 1929, Изв. Паз. инет, с. х. и лесов., № 5: 1—29.
Очень «тучная», модщ|11пая в своих движениях форма. Размеры, относительная длина ног, пропорции ланок и длина щетипок очень изменчивы. Идиосома приблизительно в .1.6 раза длиннее своей ширины, с слабо намеченными (на препаратах) плечевыми выпуклостями, равномерно суживающаяся к своему закругленному заднему концу. See длиннее и крепче всех других щетинок идиосомы; sc i, в 10—15 раз короче sc е, представлены тонкими микрохэтами, иногда совершенно отсутствуют; hi, cLx, da и 1 а также в виде микрохэт, длиною в 1—3% длины идиосомы (иногда отсут-
10. BIHZOtilA'HIUS INll
'ствуют); d3 в 2.5—4 раза длиннее d2 и в 2.3—3 раза короче <l4’( ii(wiipiiiliu приблизительно равны по длине sa i; sa е в 1.7—2.5 раза, короче на ||ne»i-достигмальный орган щетинковидный, гладкий, приблизительно рнанмН по длине d3. Внутренняя вентральная щетинка голеней и базал ьиня ih,otiiiiiiii ,колен I и II игольчатые, наружные вентральные щетинки колен, и голгнеП в виде коротких толстых шипов; наружная и внутренняя вентральная и внутренняя вершинная щетинки лапок I и II несколько листовидно риг.-'ширены па конце; обонятельная палочка прямая, почти цилиндрическая ‘(слегка суживающаяся перед едва обозначенной булавой); дорзальная вершинная щетинка лапок III и IV длинная (в 2 или больше раз длиннее лапки).
dd. Диморфны; гетероморфные особи у нас очень редки.1 Рх представ-.лены микрохэтами, р2 в 5—6 раз длиннее рх и приблизительно в 1.5 раза короче р3. Анальные копулятивные присоски обычного строения, без хитинизированного периферического кольца; penis продолговатотрапециевидный, с короткой дистальной частью, совершенно не выдающейся за передние края цоколя.
Гомеоморфная ф а з a. Sc i, h. i и d4 отсутствует чаще, чем у 9; .длина щетинок идиосомы, в среднем, больше, чему £, несколькоменсоизменчива и обычно колеблется в следующих пределах: v i 11—17; sc с 23—32; h е 20—25; d4—d2 и 1 a 1.5—3; d3 8—10; d4 27—32; 1 p 11—30; sa e 6—20; sai 15—30; pt 3—4; p2 16—25; p3 23—35% длины идиосомы; характерна относительно большая длина, которую достигают d3, d4 и особенно 1р. ьОтносительная длина ног очень константна: они имеют в длину около 30 .(I—II) и 24—26 (III и IV)% длины идиосомы. Длина 380—725 р.
Гетероморфная фаза. Относительные размеры ног I п II и щетинок идиосомы меньше отличаются от таковых гомеоморфной фазы, чем у Caloglyphus. Ноги III и.IV значительно длиннее (приблизительно равны по длине передним); вентральные и внутренняя вершинная щетинки лапок I и II серповидно расширенные на конце, с волнистой, сильно закрученной пластинкой; вентральная щетинка голеней модифицированных ног III .в виде крепкого конического шипа; хитинизированные участки вокруг эпимер развиты значительно сильнее, особенно на коксальных полях III. .Длина 600—780 р..
Q. Щетинки идиосомы короче, чему d, их относительные размеры более изменчивы и, в общем, обратно пропорциональны размерам тела; у крупных особей, длиною около 1000 р, длина щетинок составляет (в процентах к длине идиосомы): vi 9—11; sc е 17—20; 11 е 15—17; h i, d15 d2 и 1 а 1.5--.2; d3 5—7; d4 12—16; 1 p 9—10; sa e 4—8; sai 10—15; p a 16—17; у мелких (530—600 p) размеры щетинок ближе к таковым <d, а именно: v 1
.15—16; see 28—32; he 23—32; hi, dx, d2 и 1 a 2—3; d3 6—8; <l4 «О > УГ»; dp 12—15; sae 6—14; sai 20—27; 20—25% длины идиосом!,i. !>тому iito
1 Мне онп попались только в одной пробе из ФерганскоИ долини,
184
I. mOGLUPBlDAE. 2. RUIZOGIA'PBINAE
принципу подчиняется и относительная длина ног, которая у крупных 99 составляет 21—24 (I и II) и 16—17 (III и IV)% длины идиосомы, а у мелких, соответственно, 28—30 (I и II) и 20—22 (III и IV)% длины идиосомы. Длина 530—1100 р..
Л и ч и п к а. В3и 1а приблизительно равны по длине хвостовым щетинкам (sa i), т. е. относительно много длиннее, чем у половозрелой стадии; грудные палочки около 3.5 раза длиннее своего наибольшего диаметра,, с маленьким, почти шаровидным вершинным куполом.
Г и п о п у с. Габитуалыю и ноболыпинсгну морфологических признаков-очень похож на гипопусов рода Caloglyphus (особенно С. moniezi и С. polyphyllae), от которых отличается полукругло выдающимся вперед пластинчатым ростральным выступом, покрытым сверху рассеянной пунктировкой. Более или менее интенсивного буроватого цвета. Идиосома эллиптическая, в 1.5—1.6 раза длиннее своей ширины; кожные покровы плотные, сильно хитинизированные. Проподосомальный щит в 3—3.5 раза короче гистеросомы, с широким, полукруглым, сильно выдающимся вперед ростральным выступом, отграниченным явственными прогибами от закругленных передне-боковых углов щита; ростральный выступ в резкой рассеянной пунктировке, вся остальная поверхность обоих щитов (кроме обычной штрихованной полоски .по переднему краю гистеросомального щита> гладкая; все щетинки спинной поверхности очень короткие. Гнатосома целиком спрятана под ростральным выступом, за который выдается только-дистальная половина арист; -ее базальный членик около 1.5 раза длиннее своей ширины у основания, едва заметно суживающийся к своему более или менее глубоко расщепленному концу; дистальные членики тесно сближенные, в 2.5—3 раза короче базального. Стернальный щит четко очерченный, с прямым задним краем и несколько вогнутыми боковыми; стернум и эпимеры II далеко не доходят до его-заднего края, коксальные поля I и II открытые; коксальные поля III довольно хорошо очерченные; генитальный щит явственно обособлен от коксальных полей IV. Коксальные присоски, мелкие, нормального строения. Присасывательный диск средней величины,, совершенно такого же строения, как у рода.Caloglyphus-, все периферические-присоски приблизительно одинаковой длины. Ноги I и II короткие, массивные; более изменчивые, чем обычно, по своим пропорциям; лапки в 2—-2.5 раза длиннее своего диаметра у основания; вентро-латеральные шипы, голеней и колон прямые, умеренных размеров; лапки с 3 крупными и 2 ма-
Фиг. 812—325.
Ф(иг. 312. Caloglyphus fterteiioi (Mich.), o' оиерху (ио Berlese). Фиг. 313. То же, снизу (no Berlesei. Фиг. 314. С. berlesei (Mich.), копуллтиппый ппппрпт cf (по Berlese). Фиг. 315. С. berlesei (Mich.), серповидная щетинка лапки Г (no Horlese). Фиг. 31(1. С. mpcophasus (Megn.), с?’снизу(поMegnin’у),. Фиг. 317. Schvriebea ipidis Viz., if спину (no Vltzthum’y). Фиг. 318. s. ipidis Vtz., 9 сверху (no Vltzthum’y). Фиг. 319. Monieiiolla oorticalis (Mich.), снизу (no Michael’to). Фиг. 320. M. cor-ticalis (Mich.), Q снизу (no Mlehacl’io). Фиг. 321. To me, сверху (no Michael’ю). Фиг. 322. M. cor-ticalis (Mich.), лапка I снизу (no Mlchaol'io). Фиг. 323. M. berlesiana, nom. n., о"снизу (no Berlese). Фиг. 324. M. berlesiana, nom. n., снизу (no Berlese). Фиг. 325. M. berlesiana, nom. n.,.
лапка l опоку (no Berlese)..
186
I. TYROGLYPIIIDAE. 2. RHIZOGLYPHINAE
ленькими листовидными щетинками, дугообразно изогнутые на конце; средняя вентральная щетинка ланок в виде короткого, притупленного шипа; присасывательная щетинка с узким блюдцем; обонятельная палочка почти прямая, слегка утолщающаяся к. концу; о отсутствует; г в виде толстой, притупленной на конце щетинки, Вентральные щетинки голеней III и IV шиповидные; вершина ланок с 4 листовидно расширенными щетинками; па лапках III, кроме того, такую же форму имеет одна из базальных (другая шиловидная); дорзальная щетинка, лапок IV не длиннее самой .лапки. Длина, 250—370 р.
Р а с и р о с т р а. и е п и о. Космополит; северная граница распространения в Палеарктике не установлена.
Эколог и я. Живет в луковицах лука, лилий, нарциссов, гиацинтов, -тюльпанов, орхидей и других луковичных культур, в клубнях картофеля и георгинов, в свекле и других овощах, на корнях винограда, пшеницы, овса и целого ряда другие культурных растений как в условиях хранения, так и в поле; найдеп также на опавших с дерева фруктах, в лесной подстилке, на грибах и т. п.; иногда встречается в зернохранилищах, в зерне повышенной влажности. Крайне влаголюбивый вид, хорошо развивающийся даже в совершенно разжиженных объектах. Оптимальная температура лежит в пределах между 23—26°, но размножение не прекращается и при температуре воздуха (в помещении) в 6—10°. Продолжительность индивидуального развития составляет при 18.3—23.9° 17—27 дней, при 20—26.7° — от 9 „до 13 дней. Переносчиками гипопусов служат преимущественно двукрылые, развивающиеся в тех же объектах, как и клещи: виды Scatopse, Phorbia, Clwrtophila, луковичная муха (Eumerus), некоторые мелкие жуки.
R. echinopus известен как чрезвычайно серьезный вредитель декоративных луковичных растений, лука, картофеля и других корне- и клубнеплодов; его деятельность особенно интенсивна в условиях храпения этих -объектов и нередко причиняет здесь огромный хозяйственный ущерб. 11
11. Род HETERO GLYPH US FOA
Foil, 1909, Atti Accad. Lincei Roma, Rendic. (5), 18, II: 663.
Извостод только 1 вид.
1. Iloturoglyphus vitium Кой.
Foil, 190(1, Atti, Aooud, Lined Ronin, Rendic. (5), 18, 'll: 653.
Малоизвестная форма, намоченная Кой в 1909 г. в ее биологической работе о клещах, вредящих корням винограда, по так и оставшаяся не описанной более детально, Куда по отрывочным указаниям, разбросанным . в тексте этой статьи, /Z. viliMvi, габитуальпо очень сходен с Rhizoglyphus .echinopus (F. et R.), вместо с которым он живет; dd1 легко отличаются от •таковых Bliizoglypkus своими утолщенными ногами IV, тогда как этих двух родов различаются лишь с трудом; по Foa их главное различие заклю-
/2. H1STI0GASTER
W
чается в том, что ноги III и IV имеют у Heteroglyphus одинаковую длину, в то время как у Rhizoglyphus ноги III заметно короче задних.
Распространение. Италия. Живет па корнях винограда, .совместно с Rhizoglyphus echinopus (F. ct R.).
12. Род HISTIOGASTER BERL.
Berlese, 1883, Riv. period, lav. R. Acad. Padova, XXXVIII: 45; 1897: 100. —Tyrogtyphopsis 'Vitzthum, 1926, Zool. Jahrb., Syst., XLII: 476.
Тип рода: Histiogaster silenus, nom. ,n.
Пропорции идиосомы изменчивы. V e, sc i, hi, dx, d2, 1 а и sa e отсутствуют; все остальные (sc e, h e, d3, d4, Ip и sai) представлены макрохэта-:мп (длиною в 25—60% длины идиосомы). Ноги средней длины или довольно длинные, относительно стройные; средняя вентральная щетинка лапок в виде массивного конического шипа; таким же шипом представлена ;и г, расположенная непосредственно впереди обонятельной палочки; средняя вершинная щетинка лапок I и II S-образно изогнутая, наружная— умеренной длины. Коготки крупные, массивные; praetarsus совершенно редуцирован. Морфологические различия между полами очень резки.
cW представлены только гомеоморфной фазой. Задний конец идиосомы образует горизонтальную, сильно хитинизированную опистосомальную пластинку, резко суживающуюся к основанию; свободный край этой пластинки образует 4 полукруглых фестона, несущих 4 крупные, совершенно прозрачные, листовидные лопасти, расположенные веером; анальные присоски целиком сильно хитинизировапы, скошенные, с своеобразной хитиновой решеткой в центре; рх в виде микрохэт, р2 и р3 отсутствуют (им, невидимому, гомологичны описанные листовидные лопасти). Ноги IV заметно утолщены.
9$. Задний конец идиосомы не модифицирован, ноги IV не утолщены; только I пара анальных щетинок — постанальные. Личинки с дефинитивной хэтотаксией и мелкими веретеновидными грудными палочками.
Гипопус. Идиосома яйцевидная, с гладкой (крайне тонкопористой), умеренно выпуклой спинной поверхностью; все ее щетинки представлены микрохэтами (кроме иногда очень длинных р a); sc расположены трапецией, •sa i — несколько кнаружи от постанальных. Проподосомальный щит в 2.5 раза короче гистеросомы, с округлым ростральным выступом. Пара хорошо развитых (по лишенных четко оформленных линз), раздельных глаз на вершине рострального выступа. Псевдостигмальный орган отсутствует. Выводные отверстия жировых .желез расположены латерально, приблизительно на уровне 1 а.Гнатосома в 1.5—2 раза длиннее своей ширины, па конце вильчатая. Эпимериты II идут в продольном направлении до пару энного края ацетабул III или слабо намечены, вследствие чего сторннл1>11ы11 щит совершенно не обособлен и образует с вентральным одно iiopiic’iJioiion-лос целое; эпимеры всех пар свободные, широко ра.ггг»<>д1111<М1И1>н> концами;
188
I. TYROGLYPBIDAE. 2. RHIZOGLYPHINAE
эпимеры III угловато искривленные, IV — почти прямые; вся поверхность-единого стерно-вентрального щита равномерно хитинизированная, без бросающихся в глаза краевых утолщений. Присасывательный диск крупный, с обычными 4 парами присосок (из которых наружные и задние имеют одинаковые размеры и расположены вогнутой кпереди дугой), придвинутый к заднему краю тела; дополнительные (угловые) присоски не дифференцированы; генитальные очень широко раздвинуты. Ноги средней длины или довольно длинные: Г и П пара их почти целиком выдастся за края идиосомы; вептро-ла.тералт>ныо щетинкн голеней и колен и базальпые щетинки колен в виде коротких или средней длины, гладких, иногда почти йшпо-образпых щетинок; вершина колен I только с 1 щетинкой; на лапках-1 сенсилла 8 придвинута к самому основанию обонятельной палочки; сенсилла s щетинковидпая; присасывательная щетинка тонкая, с очень маленьким узким «блюдцем». Коготки длиннее и-менее сильно крючковидно-изогнуты, чем обычно.
Известно 5 палеарктических видов, из которых 2 описаны только-ио-гипопусам (трактовавшимся как самостоятельный род Tyroglyplwpsis Vitz-thum 1926). Все представители этого рода, известные по половозрелой, стадии, до сих пор объединялись в один вид.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РИДОВ
1 (2). Длина опистосомальной пластинки не уступает ее ширине у основания и только в 1.6 раза меньше ее максимальной ширины; прозрачные лопасти веера заметно длиннее своей ширины; penis относительномаленький, передние пары ног отделены очень большим расстоянием от задних..................................... 3. Н. silenus, nom. n..
2 (1). Опистосомальная пластинка около 2 раз короче своей ширины у основания и в 3 раза короче своей максимальной ширины; прозрачные лопасти веера не длиннее своей ширины; penis очень крупный;, расстояние между передними парами ног и задними невелико.
3 (4). Идиосома широкояйцевидная, в 1.5—1.65 раза длиннее (включая опистосомальную пластинку) своей ширины; прозрачные лопасти веера совершепно обособлены друг от друга; фестоны разделены довольно глубокими вырезками....................*1. И. bacchus, sp. n..
4 (3). Идиосома продолговато-яйцевидная, свыше 2 раз длиннее своей ширины; прозрачные лопасти веера слиты у основания, образуя сплошную 4-лоиастную кайму вокруг опистосомальной пластинки, фестоны соприкасаются основаниями......................2. Н. carpio (Kram.),
Фиг. 326—330.
Фиг. 326. Rhizoglyphus echinopus (If. of, Ji,.), гомеоморфный cf сверху. Фиг. 327. В. echinopus (F. et R.), гетероморфный cf снизу. Фиг. 3211. К, ochinopus (F. et R.)> $ снизу. Фиг. 329. Schwiebea tshernyshevi, sp. n., гетероморфный сГ сбоку. Фиг. 330. Thyreophagus entomophagiis (Lath^cf сбоку.
190
I. TYROGLYPHIDAE. 2. RHIZOGLYPHINAE
Гипопусы
1 (2). Гнатосома в 1.5 раза длиннее своей ширины у основания, продол-говато-трапециевидпая; эпимериты II далеко пе доходят до ацетабул-' III; от конца последних отходит тонкий волнистый отросток, направленный вперед; р а очень длинные (около 25% длины идиосомы),, ноги III и IV длиною около 28—30% длины идиосомы, с очень длинными парусными щетин ками; передние присоски много крупнее задних и боковых................................*1. Н. bacchus, sp. п.
2 (1). Гнатосома в 2—3.5 раза длиннее своей ширины, с параллельными; сторонами; эпимериты II соединены концами с наружным краем ацетабул III; эпимеры III без волнистого отростка; р а очень короткие-(не свыше 10% длины идиосомы); ноги 1Й и IV длиною около 22—24% , длины идиосомы; парусные щетинки лапок III и IV короткие;' передние присоски одинакового размера с задними и боковыми или: меньше их.
3 (4). Вентро-латеральные щетинки голеней и колен! и II тонкие, пе превращенные в шипы; дорзальная вершинная щетинка колен I очень, крупная, приближающаяся по размерам к таковой голеней; концы, эпимер II загибаются кнаружи; коксальные присоски довольно крупные; глаза расположены в продольной оси тела; их внутренние края, параллельны на всем протяжении...............*4. Н. iilippovi^ sp. n.
4 (3). Вентро-латеральйые щетинки голеней и колеи I и II в виде коротких, довольно толстых шипов; дорзальная вершинная щетинка колен, мелкая; концы эпимер II ие отогнуты кнаружи, сходящиеся; коксальные присоски очень мелкие; глаза расположенные косо, почти соприкасающиеся передними концами и широко расходящиеся кзади . . ............................................. 5. Н. ocellatus (Vtz.).
* 1. Histiogaster bacchus, sp. n. (фиг. 18, 198, 331—333, 339).
carpio Zacher, 1927 : 270 (non Kramer 1881).
Идиосома умеренно длинная, у сГ значительно шире, чем у д; задние-пары пог отделены от передних очень небольшим пространством, так что-эпимеры III и эпимериты II более сближены между собой, чем последние с эпимерами II; ноги относительно длинные; наружная вентральная щетинка голеней I и II игольчатая. В8 короче всех других щетинок спинной, поверхности гистеросомы.
с?. Идиосома широкояйцевидиая, в 1.5—1.65 раза длиннее своей ширины, с сильно закругляющимися кзади боковыми краями. Опистосомальная. пластинка сильно поперечная, почти в 2 раза короче своей ширины у основания и около 3 раз короче спорП максимальной ширины; ее фестоны разделены широкими промежутками, в которых край самой пластинки довольно глубоко вырезан; прозрачные лопасти веера совершенно обособлены друг от друга и широко раздвинуты основаниями, короче своей
12. IIIS'HOGASTIiR
НН
длииы, заметно расширяющиеся от основания к широко :шкруглепшimу концу; дорзальный скат опистосомальной пластинки покрыт пепpnии.ив-ной крупноячеистой скульптурой; penis очень крупный, зал ими к иш II большую часть пространства между основаниями пог IV; ширила его цоколя у основания превосходит длину; анальные щетинки кишкаловидпыг, расположенные впереди центральной части присосок. Эпимеры II очень мощные, загибающиеся концами кнаружи и почти касающиеся ими тоже, очень мощных эпнмеритов II, так что коксальные поля II оказываются почти замкнутыми; очень сильно развиты и эпимеры IV. Кнутри от вершинного шипа голеней IV находится небольшой бугорок. Общая длина тела 420—515 [л; длина идиосомы (с опистосомальной пластинкой) 370-—400 р, ширина 230—270 р.
д. Идиосома в 1.8—1.9 раза длиннее своей ширины, заметно суживающаяся кзади, с более или менее выпуклыми краями. Эпимеры довольно мощные; ноги несколько короче, чем у о’; анальная щель довольно длинная.
Гипопус. Светлобуроватый. Наибольшая ширина идиосомы приходится на уровень плеч; идиосома суживается кзади сильнее, чем у других видов. Щетинки спинной поверхности довольно короткие, топкие, р а необычно длинные, волосовидные. Глаза в виде пары овальных продольных выпуклостей, расположенных на верхней поверхности рострального выступа; опи разделены очень узким продольным промежутком; их роговицы, пронизаны многочисленными порами. Гнатосома в 1.5 раза длиннее своей ширины у основания, к которому она заметно расширяется; дистальные членики довольно явственно обособленные, короткие. Эпимериты II очень-короткие, доходящие только до половины расстояния между ацета-булами II и III; стернум короткий, концы эпимер II пе отргнуты кнаружи; от концов эпимер III отходит вперед по тонкому, волнисто изогнутому отростку. Коксальные присоски слабо развиты, заметны с трудом (особенно задние); присасывательный диск окружен широкой перепончатой каймой; передние присоски значительно крупнее задних и боковых. Лапки I и II очень стройные, почти цилиндрические (в 4—4.5 раза длиннее своего диаметра у основания), заметно длиннее обоих предыдущих члеников ноги, взятых вместе; обонятельная палочка и сенсилла 8 длинные, очень стройные, вентральные щетинки сильно сдвинуты к концу лапки, длинные вершинные щетинки также очень длинны, нерасширенные. Ноги III и IV длиннее, чем у других видов; лапки заметно длиннее голени и колена, взятых вместе, с очень длинными парусными щетинками (из которых сидящие па лапках IV достигают свыше 70% длины идиосомы). Длина около 270 р, ширина 180 р.
Распространение. Достоверно известен пока только из (•<VI* и Германии; материалом для описания вида послужили экземпляры пи Предкавказья (совхоз Абрау-дюрсо).
Экология. У нас найден в дубовых бочках из-под iiiiiin; по Zneliec'y встречается в большом количестве в употребляемых при изготовлении.
192
I. TYllOGLYPHIDAE. 2. JUIIZOGLYPIIINAE
уксуса дубовых стружках и может оказывать такое влияние на процессы образования уксуса, что должен рассматриваться как вредитель.
2. Histiogaster carpio (Krain.).
Kramer, 1881, Zool. Anz., IV: Ш9; Arch. Niiturg,, XLVIII, 1: 183, t. ХШ, fig. 5—10 (Tyro-glyphus).
Очень близок к предыдущему, по отличается от пего более узкой идиосомой (особенно у о), более короткими ногами, размерами щетинок идиосомы и строением оппстосомал1,по1) пластинки (!.
сГ. Идиосома продолговато-яйцевидная, конически суживающаяся кзади, начиная от точек прикрепления 1 р, около 2.2 раза длиннее своей ширины (длина идиосомы собственно, без опистосомальной пластинки, в 2 раза больше ее ширины); длина щетинок спинной поверхности идиосомы, в процентах к ее длине (по рис. 6 Kramer’a): v i около 18; sc е 30; h е 24; d3, d4, 1 p и sa e около 35. Опистосомальная пластинка такой же формы, как у Н. bacchus, но прозрачные лопасти ее веера слиты друг с другом -у основания, образуя, таким образом, монолитную 4-лопастную кайму; доколь penis приблизительно такой же длины, как его ширина у основания. Эпимеры нормального строения; эпимериты II параллельны эпимерам II. Общая „длина тела (включая гнатосому и опистосомальную пластинку с ее выростами) 450 р; ширина идиосомы 200 р.
$. Идиосома в 2—2.1 раза длиннее своей ширины, с почти параллельными боковыми краями и широко закругленным задним концом; длина щетинок спинной поверхности близка к таковой q Н. bacchus, но d3 (как и у <5) длиннее; по рис. 5 Kramer’a, размеры их, в процентах к длине идиосомы, таковы: sc е 36; h е 30; d3 40; d4 35; 1 р и sa е 36—38. Анальная щель длинная. Общая длина тела (включая гнатосому) 520 р; длина идиосомы около 500. р, ширина 250 р. Гипопус неизвестен.
Распространение. Описан из Германии (точное местонахождение не указано). Найден в большом числе на куске гниющего дерева.
3. Histiogaster silenus, nom. и.
carpio Berlese, 1883, Riv. period, lav. R. Acad. Padova. XXXVIII: 45, t. I; 1884, fasc. XIV, 5; Canestrini, 1888a: 395, t. 35, fig. 2.
Эиачитслыго мелг.че, чем предыдущий. Идиосома у обоих полов сильно вытянута в длину, валысоватая; задние пары ног отделены, в связи с этим, .очень большим пространством от передних, так что расстояние между эпимеритами И л эпимерами IIГ во много раз больше, чем между первыми и эпимерами II. Ноги относительно короткие, наружная вентральная щетинка голеней I И .11 1П11ПОВИД1ШЯ.
Фис. 331—336,
(Фиг. 331. Histiogaster bacchus, sp. n,, o' сверху. Фиг. 332. To же, снизу. Фиг. 333. Н. bacchus, ,sp. n., 9 сверху. Фиг. 334. Thyreophague entomophagus (Lab.), cC сверху. Фиг. 335. Th. entomophagus (Lab.), задний конец тела о’ снизу. Фиг. 336. Th. entomophagus (Lab.), 9 снизу.
194
I. TYROGLYPHIDAE. 2. RHIZOGLYPHINAE
Идиосома. около 2.2—2.3 раза длиннее своей ширины (считая ее? длину вместе с опистосомальной пластинкой, без которой она чуть меньше 2 раз превышает ширину), с почти параллельными боковыми краями, постепенно закругляющимися к основанию опистосомальной пластинки. Sc е, 1 р, d4 и sa е длиной около 50% длины идиосомы. Длина опистосомальной пластинки чуть превышает ее ширину у основания и только в 1.5 раза уступает ее максимальной ширине; фестоны соприкасаются основаниями; прозрачные лопасти веера совершенно обособлены друг от друга, но соприкасаются основаниями и радиально расходятся концами; длина их приблизительно в 1.5 раза больше ширины; они не суживаются к основанию и параболически закруглены на конце. Penis небольшой, занимающий незначительную часть пространства между основаниями ног IV; ширина его цоколя у основания значительно уступает длине; анальные щетинки кинжаловидные, расположенные на одном уровне с центральной частью присосок. Эпимеры нормального строения и размера. Длина ног составляет (по рисунку Berlese): 33 (I и II), 30 (III) и 28 (IV) % длины идиосомы. 06.1 ая длина тела (включая гнатосому и опистосомальную пластинку) 300
9. Идиосома немного больше чем в 2 раза длиннее своей ширины, со; слабо выпуклыми боковыми краями, заметно суживающаяся к почти прямому заднему краю; sc е, 1 р и sa е длиною около 40—45% длины идиосомы; anus очень короткий. Общая длина тела 350 [л.
Неполовозрелые стадии неизвестны.
Распространение. Италия. Найден на гниющих трутовиках (Polyporus gelsorum) и на гниющих стволах виноградной лозы.
* 4. Histiogaster filippovi, sp. п. (фиг. 340, 341).
Известен только гипопус. Довольно интенсивного буровато-желтого цвета. Наибольшая ширина идиосомы приходится значительно позади плеч, задний край широко закругленный. Щетинки спинной поверхности очень мелкие, трудно заметные; постанальные щетинки едва достигают 5%. длины идиосомы. Глаза в виде пары овальных продольных выпуклостей, расположенных на спинной поверхности рострального выступа и разделенных довольно широким ,продольным промежутком; роговицы пронизаны многочисленными порами, их внутренние края параллельные на всем протяжении. Гиатосома в 2 раза длиннее своей ширины, с параллельными сторонами, довольно глубоко расщепленная па конце; эпимериты II доходят до наружного края ацетабул Ш; стсрпум длинный; концы эпимер II резко загибаются кнаружи; эпимеры Ш без волнистого отростка; нет ника-
Фиг. 837—343.
Фиг. 337. Rhizoglyphus eohinopus (Г. ot II.), гипопус сверху. Фиг. 338. То же, снизу. Фиг. 339.-Histiogaster bacchua, sp. п., гипопус спину. Фиг. 340. И. filippovi, sp. п., гипопус сверху. Фиг. 341.. Тоже, снизу. Фиг. 342. Schwebea Wiernyahevi, sp. п., гипопус снизу (ноги не изображены). Фиг. 343_ S. Ishernyshevi, sp. п., гипопус, нога I сверху.
196
I. TYROGLYPHIDAE. 2. RHIZOGLYPHINAE
ких следов продольного вентрального шва. Коксальные присоски крупные, особенно стернальные; присасывательный диск окружен очень явственной, широкой перепончатой каймой; периферические присоски все почти одинакового размера. Лапки I и II довольно массивные, постепенно суживающиеся к концу, чуть короче голени и колена, взятых вместе; обонятельная палочка и сенсилла 8 довольно короткие и массивные; последняя расположена заметно впереди осповапия первой. Ноги I и II довольно короткие, выдающиеся за края тела только лапками; последние не длиннее голени и колена, вместе взятых; вентральные щетинки голеней и колен III и голеней и бедер IV топкие и короткие, не утолщенные в виде шипов: ближайшая к основанию дорзальная щетинка лапок III длиною около 15, лапок IV — около 25% длины идиосомы; все щетинки лапок волосовидные, не расширенные на конце. Длина 242 ji, ширина 183 р..
Распространение. Известен только из окрестностей Владивостока. Найден на короеде Ips subelongatus Motsch.
5. Histiogaster ocellatus (Vtz.).
Vitzthum, 1928, Zool. Jahrb., Syst., LIT: 177, fig. 35—36 (Tyroglyphopsis).
Гипопус. Мне в натуре не известен. Судя по описанию и рисункам Vitzthum’a, очень близок к Н. filippovi, от которого отличается только в следующих признаках. Мельче. Окраска слабожелтоватая. Идиосома несколько шире. Щетинки спинной поверхности несколько длиннее и, несмотря на их тонкость, легче заметны, чём это обычно у странствующих нимф (= гипопусов), постанальные щетинки длиною около 10% длины идиосомы. Глаза описываются Vitzthum’oM следующим образом: «...под передним выступом notocephale (= проподосомального щита) лежит с каждой стороны по овальному, темнокаштаново-бурому светочувствительному органу. Они свободно выдаются своей передней третью, тогда как остальная часть прикрыта сверху notocephale. Я могу придать этим необычным органам только значение глаз, хотя настоящей бесцветной роговицы над этими ретинами не удается обнаружить»; таким образом они, по своему строению, сильно отличаются от таковых описанных выше видов; на рис. 35 Vitzthum’a видно, что эти глаза расположены косо: они почти соприкасаются спереди и широко дивергируют кзади. Гнатосома стройнее, раза в 3.5 длиннее своей ширины, с параллельными сторонами, чуть расширенная на конце; концы эпимер II сходящиеся, не отогнутые кнаружи; продольный вентральный шов слегка намечен. Коксальные присосоки очень мелкие; вокруг присасывательного диска нельзя заметить перепончатого края; периферические присоски мельче, чем у предыдущего вида; передние из них меньше всех остальных. Вершинная щетинка колен I на рисунке не изображена, и о ней ни словом но упоминается в тексте; вероятно, она очень мала (так как если бы опа имела такие размеры, как у Н. filippovi, она, несомненно, бросилась бы исследователю в глаза); вентро-латеральные щетинки голеней I и II представлены шинами умеренной мощности; такими
13. SCHWIEBEA
ну/
же, но гораздо более короткими шинами представлены и иарушиш литоральные щетинки колен I и II. Лапки I несколько короче и маейимнйн’, сенсилла 8 расположена (судя по рисунку) на той же высоте, как обонятельная палочка; вершинные и предвершинные щетинки (их всего по Я) лапок I и II дистально с незначительным листовидным расширением. Вентральные щетинки голеней и колеи III и голеней и бедер IV шиловидные; щетинки лапок III и IV длиннее, чем у Н. filippovi, причем по 3 из них несут на конце такие же небольшие листовидные расширения, как соответствующие щетинки передних пар лапок; самые крупные из щетинок лапок III и IV — одна из дорзальных — не превышают более чем в 2 раза длину лапки и листовидно расширены на конце. Длина идиосомы 176 р,, ширина 142 р.
Распространение. Описан по единственному экземпляру из местности Brout-Vernet (Франция). Гипопус найден на короеде Xyleborus pfeili Ratz.
13. Род SCHWIEBEA OUDS.
Oudemans, 1916, Entomol. Вег., IV: 264.
Тип рода: Schwiebea talpa Ouds. 1916.
, V e, sc i, псевдостигмальный орган, h i, dt и d3 и в большинстве случаев
также d3 и 1 а отсутствуют; отсутствует также одна из пар крестцовых (sa е). Эпимеры III и IV сходящиеся, а иногда даже сливающиеся своими внутренними концами. Ноги короткие и массивные, мощно вооруженные; вентральные щетинки голеней, базальная и наружная щетинки колен, внутренняя и средняя щетинки и е лапок I и II представлены крепкими коническими шипами; 8 тесно придвинута к основанию обонятельной па-; лочки, s расположена непосредственно впереди нее; средняя вершинная лапок I и II в виде резко дугообразно изогнутой книзу линейной пластинки; наружная вентральная щетинка волосовидная; в связи с сильным укоро-’ чением лапок, их вентральные щетинки (resp. шипы) близко придвинуты
, к вершинным, которые имеются в числе 5 вентральных и очень крупного
дорзального. Коготки очень крупные и массивные (особенно на ногах I k и II); prae tarsus совершенно редуцированный. Половой диморфизм крайне h резкий.
I d1. Идиосома довольно широкая, заметно суживающаяся кзади; зад-
I ний конец тела уплощен в полукруглую, горизонтальную, более или менее | сильно хитинизированную опистосомальиую пластинку, дорзальная по-| верхность которой составляет прямое продолжение спинной поверхности I гистеросомы и расположена в одной плоскости с нею; вентральная иопорх-
г ность этой пластинки более или менее вогнута и скошена вперед и iiiip.i,
так что анальные копулятивные присоски оказываются обращенными кою I книзу и назад й конструкция в целом напоминает (особенно ври рнсомп-|, тривании животного в профиль) аналогичное видоизменение конца тола Е у dd жуков-лубоедов (Eccoptogaster)', на спинпой поверхности щшстосо»
198 I. TYROGLYPHIBAE. 2. RHIZOGLYPHINAE
мальной пластинки помещается только 1 пара щетинок — d4, в большинстве случаев сдвинутая к самым краям пластинки (только у S. vitethumi d4 расположены в ее центральной части); р4 и р2 представлены крошечными микрохэтами. У большинства видов сГ d1 представлены только гетероморфной фазой, но у S', tshernyshevi встречаются в небольшом числе и гомеоморфные особи.
$. Идиосома обычно узко и длиннее, чем у <3, вальковатая; опистосо-мальпая пластинка отсутствует, задний конец тела не модифицирован; имеется только 1 пара анальных щетинок.
Личинка с крупными, булавовидными грудными палочками. Гипопальная стадия известна только у одного из видов; ее признаки, следовательно, трудно вводить в родовую характеристику.
К роду Schwiebea Ouds. относятся 6 палеарктических видов, 1 ориентальный (S. italica Ouds. 1923 с Явы) и 2 или 3 неарктических (Monie-ziella xylophaga Banks 1912, M. ulmi Jacot 1936 и, вероятно, M. brevipes Banks 1906). Большинство его представителей экологически связано с лесом, обитая под корой пней и в устилающей лесную почву мертвой подстилке из более или менее перегнивших листьев; 8. xylophaga живет в отмирающих стеблях люцерны.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ ‘
сГсГ
1 (2). Вершина колен I с 2 щетинками; вентральные шипы лапки, голени и колена III у гетероморфных особей тонкие, слабо развитые; лапки I и II очень короткие; р3 расположены близ заднего края опистосо-мальной пластинки, на одной поперечной линии с задней парой постанальных микрохэт (Ра). (Подрод Megninietta Jacot) . . ..............
.....................................* 6. S. (Megn.) rossica, sp. n.
2 (1). Вершина колен I только с 1 щетинкой, имеющей зато большую длину; вентральные шипы лапки, голени и колена III у гетероморфных особей очень толстые, с широко закругленным концом; лапки I и II длинные; р3 расположены на боковых краях вентральной поверхности опистосомальной пластинки, значительно впереди уровня задней пары постаиальпых микрохэт (ра). (Подрод Schwiebea s. str.)
3 (4). 1)4 расположены в центральной части опистосомальнойпластинки;
d3 и 1 а имеются налицо и представлены микрохэтами; sa е в 3 раза
1 В таблицу по пошли сГ (Т 8. paohydtirma, остающиеся неизвестными, и недостаточно описанная q 8. talpa.
Фиг. 844-ЗБ2.
Фиг. 344. Schwiebea tshernyshevi, up. п., гетероморфный o' сверху. Фиг. 345. То же, снизу. Фиг. 346. S. tshernyshevi, ер, п., гомеоморфный снизу. Фиг. 347. S. rossica, ер. п., задний конец тела С? снизу. Фиг. 348. S. pachyderma, ер. n., о, общий вид сверху. Фиг. 349. То же, снизу. Фиг. 359. 8. pachyderma, ер. п., нога I сверху. Фиг. 351. S. rossica, sp. n., д снизу. Фиг. 352. S. ipidis Vtz., гетероморфный О'сверху (по Vitzthum’y).
200
I. TYROGLYPHIDAE. 2. RHIZOGLYPBINAE
короче p3 и более чем в 2 раза короче 1р. Длина 383—600 р, . . . . !
..................................... 2. S. (s. str.) ipidis Vtz.
4 (3). D4 расположены на самом краю опистосомальной пластинки; dj.
и1 а отсутствуют; sa е несколько длиннеер3 и более чем в 2 раза длин- -нее 1р. Длина 210—365 [J.......*1. S. (s. str.) tshernyshevi, sp.n. • ’
99
1 (2). Эпимеры III слиты своими внутренними концами с эпимерами IV; - 1
вершина колеи I с 2 щетинками; наружная вентральная щетинка голеней I и II отсутствует; идиосома сильно вытянутая в длину, в 2.3—2.4 раза длиннее своей ширины; боковые края гистеросомы прямые, параллельные друг другу на большом протяжении. Длина 280—360 [J.. (Подрод Mdgninietta Jacot). . *6. S. (Megn.) rossica, sp. n.
2 (1). Эпимеры III и IV свободные, не связанные друг с другом; вершина колен I дорзально только с 1 щетинкой; обе вентральные щетинки голеней I и II хорошо развиты; идиосома менее вытянутая в длину, в 1.65 — 2 раза длиннее своей ширины; боковые края гистеросомы более .или менее выпуклые. (Подрод Schwiebea s. str.)
3 (4). Максимальная ширина идиосомы умещается в ее длине 1.66—1.7 раза; задний конец тела слегка приостренный; кожа толстая, плотная, окрашенная в слабый буроватый тон; проподосомальный щит необособленный; щетинки гистеросомы очень тонкие, гибкие, волосовидные; ноги III и IV незначительно короче и тоньше передних двух пар; обонятельная палочка лапок I и II прямая, постепенно утолщающаяся к концу. Длина 295—370 р...................................
...............................* 5. S. (s. str.) pachyderma, sp. n.
4 (3). Максимальная ширина идиосомы умещается в ее длине около 2 раз; задний конец тела широко закругленный; кожа нормальной, перепончатой консистенции, бесцветная; проподосомальный щит более или менее явственный; щетинки гистеросомы толще, более жесткие; ноги III и IV много короче и тоньше ног I и II; обонятельная палочка лапок I и II сильно искривленная, с булавовидно вздутым концом.
б (8). L а и d3 представлены микрохэтами, но вполне явственны.
6 (7). Длина 590—630 р..................... 2. S. (s.str.) ipidis Vtz.
7 (6). Длина 365—465 р..................3. S. (s.str.) • cavermcola Vtz.
8 (5). Lan dgCOBcpiucnuo отсутствуют . . *1. S. (s.str). tshernyshevi, sp. n.
1. Подрод Schwiebea s. str.
Вершина колена I с 1 дорзальной щетинкой. Тело 9 умеренно удлиненное, идиосома в 1.65—2 раза длиннее своей ширины: расстояние между основанием ног II и III невелико; эпимеры III и IV свободные, не соединенные концами.
13. SCHWIEBEA
IUI
* 1. Schwiebea (s. str.) tshernyshevi, sp. п. (фиг. 329, .’М2
d1. Гетероморфная фаза. Гистеросома сильно расширенпая в плечах, от уровня которых она постепенно и довольно равномерно суши вается кзади до уступов, намечающих основание опистосомальной пластинки, не образуя внезапного сужения позади основания ног III; наибольшая ее ширина умещается в общей длине идиосомы около 1.7 раза. D3 и 1 а совершенно отсутствуют; d4 смещены к самому краю тела; р3 расположены у боковых краев опистосомальной пластинки, почти на одной продольной линии с sa е и значительно впереди поперечной линии, проведенной через основания задней пары постанальпых микрохэт (р2); sa е лишь немного уступают в ра мерах р3 и раза в 2 длиннее 1 р; р4 ближе придвинуты к анальным присоскам, чем sa е. Вентральный шип лапок и наружные боковые шипы голеней и колен ног III очень массивные,, с широко закругленной вершиной. Длина 210—365 ширина 123—215 р..
Гомеоморфная фаза. Идиосома заметно уже, чем у гетероморфных сГ <5. Ноги короче и массивнее; эпимеры и вертлуги III развиты лишь немного слабее, чем у гетероморфных d'd'. Длина единственного экземпляра, имевшегося в моем распоряжении, 330 р., ширина 165 р..
$. Длина идиосомы превосходит около 2 раз ее наибольшую ширину; боковые края гистеросомы от плеч до места прикрепления 1 р очень слабо-выпуклые, почти прямые; задний конец тела широко закругленный. Кожа нормальной консистенции, мягкая, тонкая, перепончатая; несколько уплотнены и хитинизированы лишь коксальные поля I и II; проподосомальный щит явственный, хорошо отграниченный. Длина sc е составляет около 25% длины идиосомы; щетинки гистеросомы средней длины, жесткие, довольно толстые; d3 и 1 а отсутствуют. Ноги III и IV далеко уступают в размерах ногам I и II; эпимеры IV на значительном протяжении почти параллельны продольной оси тела; их концы отделены от концов эпимер III довольно значительным промежутком. Вертлуги I и II весьма мощные; вершинная щетинка колен I в виде крепкой щетинки, уступающей в длине вершинной щетинке голеней I лишь раза в 2; обонятельная палочка сильно-дугообразно искривленная, булавовидно утолщенная па конце. Генитальная щель помещается в пространстве, заключенном между эпимерами IV. Длина 360—465 ;л, ширина 180—240 ц,.
. Г и п о п у с. Напоминает гипопусов рода Robinisca и в еще большей степени гипопуса Caloglyphus tshernyshevi. Идиосома обратно-яйцевидная, в 1.5 раза длиннее своей ширины; спинная поверхность умеренно выпуклая, гладкая; все усаживающие ее щетинки очень короткие. Проподосомалы1ы11 щит в 4—4.2 раза короче гистеросомы, с округлым, слабо iiaMo'ioiiniiiM ростральным выступом; глаза и псевдостигмальный орган отоутотпуют, Гнатосома заметно выступающая за передний край проподосомы, и У plum длиннее своей ширины у основания; дистально она расщоплопн ни '/< длины, причем ветви развилка не обособлены в видо отдельных 4.iintiiiic.<ni,
202 I. TYROGLYPHIDAE. 2. RHIZOGLYPHINAE
Стернальный щит равномерно плотно хитинизированный, с совершенно прямым задним и прямыми, заметно сходящимися кзади боковыми краями; •стернум простирается лишь немного за середину щита; эпимеры II почти прямые, сходящиеся кзади под углом около 45°, сближенные концами. Передний край вентрального щита прямой, боковые края короткие, прямые, образующие с передним почти прямой угол; задний край S-образно выгнутый. Генитальный щит вполне обособленный; его передний край, как обычно, повторяет изгиб заднего края центрального щита, от которого он отделен довольно широким промежутком мягкой кожи. Присоски коксальпых нолей I редуцированы; сидящие па коксальпых полях III присоски хорошо ^развиты. Присасывательный диск обычного строения, придвинутый к заднему краю тела; все периферические присоски сходны по размерам, дополнительные слабо развиты. Ноги I и II длинные, довольно стройные, почти целиком видимые сверху; вентральные щетинки вертлугов и бедер короткие; вентро-латеральные щетинки голеней и колен представлены довольно крупными, но стройными, ножевидными шипами; колени I с 1 (довольно длинной) вершинной щетинкой и очень короткой проксимальной; присасывательный волосок листовидно расширенный на конце; средняя вентральная щетинка шиповидная, обе боковые и все вершинные тонкие, волосовидные. Парусных щетинок на лапках III и IV нет. Длина 205—230 jr, ширина 145—155 [г.
Описан по 2 экз., оставшимся заключенными в шкурках протонимф, 'чем была документально установлена их видовая принадлежность.
Распространение. Известен пока только из одного пункта: Нерубаевское лесное промышленное хозяйство, Одесской обл.
Экология. Живет в дубовых лесах, под корой пней.
2. Schwiebea (s. str.) ipidis Vtz. (фиг. 317, 318, 352).
talpn Vitzthum, 1923 (non Oudemans 1916), Arch. Naturg., LXXXIX, 2: 151, fig. Б0—55. — ipidis Vitzthum, 1932, Zool. Jahrb., Syst., LXIII: 697.
Очень близок к предыдущему, но отличается от него значительно более крупными размерами и некоторыми деталями хэтотаксии.
d. Гетероморфная фаза. Наибольшая ширина тела приходится не па уровень плеч, как у S. tshernyshevi, а значительно дальше назад, близ середины длины гистеросомы;' общая конфигурация тела, в соответствии с этим, иная, чем у гетероморфных d d 8. tshernyshevi-, d3 и 1 а имеются налицо и представлены очень мелкими микрохэтами. Щетинки 4-й спинной мары (d4) расположены в центральной части опистосо-мального щита, на значительном расстоянии от его боковых краев; sa е в 3 раза короче р8 и более чем в 2 раза короче 1 р. По бокам анальной щели расположены 2 пары мелких округлых образований, по форме и размерам напоминающих анальные поры. Во всех остальных признаках сходен с гетероморфными dd 8. tshernyshevi. Длина 383—600 р, ширина 260— .267 р.. Гомеоморфная фаза неизвестна.
13. schwiebea ana
9. Отличается от таковой S. tshernyshevi только более крупными размерами и наличием d3 и 1 а, представленных микрохэтами. Длила 500- -600 р, ширина на уровне плеч 235—260 р. — Гипопус и другие неполовозрелые стадии неизвестны.
Распространение. Известен пока только из Германии. Живет в маточных ходах короедов Ips laricis F. и Hylastes palliatus Gy 11.
3. Schwiebea (s. str.) cavernicola Vtz.
Vitzthum, 1932, Zool. Jahrb., Syst., LXIII: 697.
Известен только по самкам. Крайне близок к В. ipidis Vtz., от которого отличается только меньшими размерами (длина 365—465 р), меньшей длиной щетинок идиосомы и лапок, более сдвинутой назад гнатосомой и неясным проподосомальным щитом.
Распространение. Найден в пещерах в Югославии (в скоплениях воды).
4. Schwiebea (s. str.) talpa Ouds.
Oudemans, 1916, Entomol. Вег., IV: 265.
Описан по одной 9, найденной в окрестностях Вонна, в гниющих листьях. Оригинальное описание Oudemans’a, в переводе с голландского, таково:
Schwiebea nov. gen. относится к Tyroglyphinae, напоминает также Туго-glyphus’a, но имеет спинной щит, как Suidasia Ouds. и Herida Сап. Он отличается от обоих родов короткими толстыми ногами и наличием всего 2 щетинок на задней половине проподосомы. Вертлуги I также с сильно хитинизированной спинной поверхностью... В. talpa nov. sp. напоминает крота в миниатюре, имеет 333 у. в длину; на всех лапках по 3 или 4 коротких шипа; короткие ноги с этими копательными когтями («graatnagels») говорят о роющем образе жизни, как у Acarus (Sarcoptes).
Приведенный диагноз может быть несколько дополнен указаниями Vitzthum’a, располагавшего собственноручным рисунком Oudemans’a и имевшего, таким образом, возможность сравнивать с ним описанных им Schwiebea ipidis Vtz. Согласно этим указаниям, аутентичная В. talpa во всех деталях идентична с В. ipidis Vtz., отличаясь только тем, что у нее проподосомальный щит простирается назад дальше уровня лопаточных щетинок и что все более крупные щетинки гистеросомы, хотя и изогнуты, но более крепки. Все эти данные, однако, недостаточны для того, чтобы по ним можно было составить себе ясное представление о В. talpa и ее отношениях к описываемым здесь формам. Малые размеры Oudemans’oncicoro вида (9 333 р) и его местообитание (гниющие листья) наводят на мысль, что он может оказаться идентичным с формой, описанной здесь под iiniiiin,-нием В. pachyderma, но против этого говорит отсутствие у послодпой хоть сколько-нибудь ясно намеченного проподосомального щита и утолщений хитина на верхней поверхности вертлугов I и тонкость со щетинок. О том,
204
I. TYROGLYPHIDAE. 2. RHIZOGLYPHINAE
что (S', talpa не идентична с видом Vitzthum’a, писал еще сам Oudemans, а вероятность ее идентичности с S. tshernyshevi (более близкой к ней по-размерам) мне кажется очень малой, так как местообитания их совершенно-различны. Таким образом S. talpa Ouds. остается пока не расшифрованной.
* 5. Schwiebea (s. str.) pachyderma, sp. n. (фиг. 348—350).
Q. По общей формо тела и пропорциям напоминает маленького Rhi-zoglyphus. Идиосома относительно очень широкая, ее максимальная ширина умещается в длине только 1.05—1.7 раза; боковые края гистеросомы за. плечами слегка стянутые, затем умеренно дугообразно выпуклые до точки прикрепления 1 р; задний конец тела слегка приостренпый. Кожа очень плотная, толще, чем у других видов Schwiebea и даже всех других известных мне видов Tyroglyphoidea, окрашенная в слабый, но явственный буроватый тон; сколько-нибудь ясно отграниченного проподосомального щита нет. Sc е длиннее и толще всех прочих щетинок идиосомы: длина их составляет около 20—22% длины последней; все щетинки гистеросомы очень тонкие, гибкие, волосообразные; самые длинные из них h е, достигающие 12—15% длины идиосомы; 1 а и d3 сильно смещены назад, мелкие (3—4.5%), но все же) значительно крупнее, чем у 8. ipidis; d4 несколько длиннее 1р и sa, имеющих почти одинаковые размеры (около 9% длины идиосомы; р а длиннее, почти равные h е. Характерна относительно большая длина ног III и IV, лишь слабо уступающих в этом отношении передним. Эпимеры III дугообразные, ориентированные по поперечной оси тела, и почти соприкасающиеся концами с концами сильно скошенных назад, почти прямых эпимер IV; вертлуги I и II особенно мощные; вершинная сенсилла колен I и II в виде небольшой, притупленной на конце щетинки; обонятельная палочка прямая, постепенно утолщающаяся к закругленному концу; лапки III и IV относительно стройные; последние имеют длину, в 2.5 раза превышающую их диаметр, и заметно длиннее голени и колена, взятых вместе. Генитальная щель расположена целиком в пространстве между эпимерами IV; задние края генитальных клапанов расходятся под почти прямым углом; генитальные щупальцы мелкие. Длина 295—370 [г, ширина 175—220 [г. <3 неизвестен.
Распространение. Вид известен пока только из 2 пунктов СССР: Винницкой обл. УССР и окрестностей Москвы (Шульгино, Кунцевского р-иа). Из Виппицкой обл. происходит почти весь материал, послуживший для описания вида. Живет исключительно в мертвой лесной подстилке дубовых (а может быть, и других широколиственных) лесов.
2. Подрод Megninietta Jacot 1936
Jacot, 1936, Ann. Entomol. Soc. Amor., XXIX: 631.
Вершина колена I с 2 дорзальными щетинками. Тело $ очень сильна вытянутое в длину, узкое; ширина идиосомы умещается в ее длине 2.3—
13. SCHWIEBEA.
2.4 раза; расстояние между основаниями ног II и III очень велико; нни* меры III сливаются с концами эпимер IV.
*6. Schwiebea (Megninietta) rossica, sp. и. (фиг. 199, 200, 347, 351).
сГ. Гетероморфная фаза. Вся передняя половина гистеросомы до уровня задних краев вертлугов III равномерно расширенная, с параллельными краями; затем внезапно стягивается, образуя здесь ясно •обозначенные уступы, за которыми уже постепенно суживается кзади до •основания опистосомальной пластинки. Г»3 и 1 а совершенно отсутствуют, d4 смещены к самому краю тела; р3 расположены у заднего края опистосомальной пластинки, на одной поперечной линии с р2, значительно ближе •к средней линии тела, чем sae; последние несколько длиннее р3; р4 находятся значительно дальше от копулятивных присосок, чем от sa е, образуя с ними и р2 правильный дугообразный ряд. Вентральный шип лапок и наружные боковые шипы голеней и колен ног III тонкие, слабо развитые. Длина 235—310 [л, ширина 100—170 ;л.
Гомеоморфная фаза 3 неизвестна.
Q. Идиосома сильно вытянутая в длину, что придает самкам большое габитуальное сходство с таковыми Monieziella; отношение ее длины к ширине равно 2.3—2.4 : 1; боковые края прямые, почти строго-параллельные друг другу на всем протяжении; задний конец тела широко закругленный. Кожа нормальной консистенции, мягкая, тонкая, бесцветная; коксальные поля I и II нехитинизированные; проподосомальный щит явственный, четко отграниченный от окружающей его мягкой кожи, разделенный продольной полоской мягкой, перепончатой кожи на 2 узких самостоятельных щита. Длина sc е не превышает 7—8% длины идиосомы; щетинки гистеросомы очень короткие, тонкие, но жестковатые; длина he равна 6—7, 1 р, d4 и sa е 4—5, р а около 6% длины идиосомы; dg и 1 а отсутствуют. Все ноги очень коротки. Эпимеры III слиты концами с эпимерами IV, образуя с каждой стороны пару изящных хитиновых конструкций; обонятельная палочка резко дугообразно искривленная, путовковидно вздутая на конце; диаметр голеней и коленных члеников и бедер I—II уступает их длине; наружные вентральные щетинки голеней I и II отсутствуют. Генитальная щель простирается вперед до уровня передних краев эпимер III; задние края генитальных клапанов расходятся под очень тупым углом; генитальные щупальцы относительно мелкие. Длина 280—360 р.; ширина 110— 360 р.; ширина 110—150 р.
Личинка (как и $, протонимфа и телеонимфа) с сильно вытянутой в длину идиосомой; грудные палочки стройные, длиною около 9 |х, при диаметре в расширенной части в 2.1 р.. — Гипопус неизвестен.
Распространение. В окрестностях Москвы (дор. Шульгине, Кунцевского р-на) и в Великолукском р-не Калининской обл. Жннет более или менее крупными колониями под корой березовых пней.
206'
1. TYROGEYPHIDAE. 2. RHIZOGLYPEINXE
14. Род MONIEZIELLA BERL.
Berlese, 1897: 107.
Тип рода: Monieziella, berlesiana, nom. n.
Очень близок к предыдущему. V i отсутствуют, так же как и sc i, h i и 3 передние пары d (dj—da); 1 а длинные, такой же, примерно, длины, как 1 р. У Q развиты обе пары sa, представленные относительно длинными щетинками, и 1 пара аданальп ых. Строение опистосомы сГ такое же, как у Schwiebea. Эпимеры III и IV параллельные друг другу на всем протяжении, свободные. Вооружение лапок слабое,, чем у предыдущего рода, и скорее напоминает таковое рода Т hyrcophagus-, дорзальный вершинный шип тонкий, вентральные — слабо развитые; е пе имеет формы конического ши-паи, вероятно, редуцирована; наружная и внутренняя вентральные щетинки представлены топкими щетинками. Коготки слабее, чем у Schwiebea-, ргае-tarsus значительно более крупный, образующий 1 или 2 полукруглые вертикальные пластинки, прикрывающие с боков основание коготка наподобие шор. Тело у обоих полов сильно вытянутое в длину; максимальная ширина идиосомы умещаемся в ее длине ус? 2.3 — 2.6 раза, у $ 3 — 3.4 раза. Гетероморфной фазы у сГ неизвестно. — Нимфальные стадии не описаны.
Гипопус весьма напоминает таковых рода Histiogaster, но имеет более вытянутое в длину тело и пару хорошо развитых глаз, расположенных на боковых краях проподосомы (над основаниями ног I) и состоящих каждый из полушаровидио выступающей прозрачной линзы, окаймленной снизу скоплением темного пигмента.
К роду Monieziella я отношу 2 европейских вида, до сих пор постоянно смешивавшихся друг с другом: М. corticalis Mich. 1885 и М. berlesiana, nom. n. (М. mali Berlese 1897, non Schimer 1868, M. corticalis auct. pt.). He зная в натуре ни одного из них, я был вынужден составить их характеристики и вышеприведенный родовой диагноз только по описаниям и рисункам Berlese и Michael’a.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ сГсГ
1 (2). Гнатосома необычно узкая, хелицеры суженные к основанию и образуют короткую «шею»; h е длинные; длина идиосомы около 290— 300 [1......................................1. М. corticalis (Mich.). „
2 (1). Гнатосома нормальной формы, хелицеры пе суженные к основанию; he короткие; длина идиосомы около 360—365 р........................
......................................2. М. berlesiana, nom. n.
99
1 (2). Общая длина тела около 750 р.............3. М. magna Berl.
2 (1). Общая длина тела но превышает 600 р.
3 (4). Стер дум сильно укороченный; ноги III и IV длиннее х/г ширины, идиосомы; общая длина тела около 450 р. 1. М. corticalis (Mich.).
14. MONIEZIELLA
МО,
4 (3). Стернум развит нормально; ноги III и IV короче ]/2 ширины идиосомы; общая длина тела 500—600 р. . . . 2. М. berleslnnn, нот. и,
1. Monieziella corticalis (Mich.) (фиг. 319—Г22, 358, 359).
Michael, 1885, Journ. R. Mier. Soo., ser. П, vol. V: 27, t. Ill, fig. 1—14; 1903: 66, t. XXVI i I (Hi?tiogaster').
d.Гомеоморфная фаза. Гнатосома очень узкая; хелицеры суженные к основанию, благодаря чему создается впечатление, что они образуют маленькую, вытянутую в длину «голову», отделенную от проподосомы короткой «шеей». Проподосома сильно суживающаяся кпереди. Края гистеросомы несколько выпуклые по всей длине. Не и 1 а длинные; sa е и р3 расположены у заднего края опистосомальной пластинки. Длина ног равна около 23 (I), 20 (II), 17 (III) и 15 (IV) % длины идиосомы. Общая длина тела, в среднем, 350 р.; длина идиосомы около 290—300 р., ее ширина около 90—120 р.. Гетероморфной фазы у d неизвестно.
Стернум сильно укороченный; иоги III и IV менее чем в 2 раза короче наибольшей ширины тела; длина пог составляет,,соответственно, около 17 (I, II и IV) и 16 (III) % длины идиосомы; генитальная щель расположена почти целиком в пространстве между эпимерами III. Общая длина тела, в среднем, 470 р.; длина идиосомы, соответственно, около 410 р., ее ширина 120 р..
Гипопус. Глаза очень крупные; эпимеры II короткие, далеко пе достигающие заднего края стернального щита, свободные. Присоски передней пары почти такого же размера, как вентральные; обе задние пары представлены очень маленькими присосками, расположенными в поперечный ряд. Парусные щетинки ног III и IV весьма длинные.
Распространение. Достоверно известен только в Англии (Уорвикгаайр и Дорсетшайр). Живет на отмерших стеблях тростника Phragmites communis Trin.
2. Monieziella berlesiana, nom. n: (фиг. 323—325, 360, 361).
mali Berlese, 1897: 107. —corticalis Berlese, 1890: LVI1, n° 7 (Histiogaster). —Non Histiogaster corticalis Michael (1885), nec Acarus malus Sehimer (1868).
Очень близок к предыдущему, но крупнее и отличается . еще следую,-щими признаками, d (гомеоморфная фаза): h короткие, гнатосома нормальной формы, хелицеры не суженные к основанию; длина идиосомы около-360—365 р., ширина 130 р.. ф. Стернум развит нормально; ноги короче; длина их составляет приблизительно 21—22 (I), 20 (II), 10—11 (III) и 12 (IV) % длины идиосомы; ноги III и IV, таким образом, в 2.5 раза короче наибольшей ширины тела. Генитальная щель лишь немного выдается Ингред за концы эпимер IV; длина идиосомы 450—525 р., ширина около 190 р.,
Гипопус. Глаза меньше; эпимеры II очень длинные; опнсыаал сложную кривую, они простираются назад до заднего края стернального щита., где соединяются с эпимеритами II; присоски передней лары много меньше
208
I. TYROGLYPHIDAE. 2. RBIZOGLYPHINAE
центральных и имеют почти такие же размеры, как задние 2 пары, расположенные здесь дугой.
Распространение. Достоверно известен только из Италии. Живет на трутовиках Polyp orus hirsutus.
К этому же роду, вероятно, относится и совершенно недостаточно они-
•санная:
3. Monieziella magna Bcrl.
Berlese, 1910, Redia, VI: 387.
Похожа на №. mali, по много крупнее. Половозрелая самка до 750
длины и 280 [л ширины. Найдена в Палермо (Сицилия).
1Б. Род THYBEOPHAGUS ROND.
Rondani, 1874, Boll. Soc Ent. Ital., VI: 67.
Тип рода: Acarus entomophagus Lab. 1852.
Гнатосома массивнее, чем у других родов трибы Rhizogtyphini. Идиосома узкая, вытянутая в длину, вальковатая. V е, sc i, h i, dx—d3 и 1 a отсутствуют; отсутствует также и одна из пар крестцовых (sa е); все имеющиеся щетинки представлены тонкими, гибкими, волосовидными микрохэтами (длиною в 18—50% длины идиосомы). Ноги короткие, довольно массивные; две задние пары далеко отодвинуты от передних; вооружение ног очень слабое, состоящее из тонких, волосовидных щетинок; т и 8 отсутствуют; е тонкая, палочковидная, расположенная впереди и несколько кнаружи от обонятельной палочки; наружная вершинная щетинка лапок I и II очень длинная, вентральных щетинок всего 2, обе волосовидные; строение средней вершинной щетинки лапок I различно у d и $; вентральные вершинные шипы в числе 2 (средний недоразвит) слабые, так же как и дорзальный. Коготки средней мощности; praetarsus крупный, хорошо развитый, одевающий большую половину коготка (как у Tyroglyphinae). Половой диморфизм резко выраженный.
сГ сГ представлены одной только гомеоморфной фазой. Задний конец идиосомы образует полукруглую, горизонтальную, довольно сильно хити-пизированную опистосомальную пластинку, расположенную на уровне вентральной поверхности гистеросомы; спинная поверхность последней сзади резко скошена, образуя крутой скат к находящейся внизу опистосомальной пластинке; вся эта скошенная поверхность плотно хитинизирована и окрашена в буроватый цвет; на спинной поверхности этой пластинки помещается только 1 пара щетинок — d4; sa е смещены на ее брюшную сторону; pj и р2 представлены крошечными микрохэтами, р3 волосовидные, длинные; средняя вершинная щетинка лапок I в виде короткой, притупленной на конце пластинки, обращенной вниз.
9. Задний конец тела не модифицирован, опистосомальная пластинка отсутствует; анальных щетинок 2 пары (1 аданальная и 1 постаналь-
15. THYREOPBAGUS
20»
пая); средняя вершинная щетинка лапок I и II щетинковйдная, загнутая кверху. ':
Личинка с недоразвитыми, пуговковидными грудными палочками. Развитие прямое, гипопальная стадия отсутствует.
Монотипический род.
,*1. Thyreophagus entomophagus (Lab.) (фиг.-201—203, 330, 334—336).
Laboulbfene, 1862, Ann. Soc. Entomol. Fr., Bulletin: 54 (deans); Laboulbfene et Robin, Ann. Soc. Entomol. Fr., (4): 317, t. X (Tyroglyphus); Berlese, 18.97: 107; 1898; XC, n° 1 (Monieziel'd); Michael, 1903: 55, t. XXVII (Histiogaster). —aleurophagus Sicher, 1894, in: Canestrini, Prospetto ‘d. Acarofauna Itai., VI: 815 (Histiogaster).
Вредоносность. Newstead & Duvall, 1919: 91. .
d. Идиосома в 2—2.3 раза длиннее своей ширины, с равномерно слабо выпуклыми боковыми краями, постепенно закругляющимися к основанию опистосомальной пластинки. Примерная длина щетинок спинной поверхности идиосомы (в процентах к ее длине): vi 15—16; see 35—40; he 24—28; Ip 18—20; sa i 23—26; рх помещаются-кнаружи от анальных присосок, р2 — непосредственно позади них. Длина ног составляет около 26— 28 (I), 24—25 (II), 22 (III) и 20 (IV) % длины идиосомы; лапки IV сильно укорочены (короче голеней). Длина 270—450 [i.
$. Идиосома с менее выпуклыми, более параллельными друг.другу боковыми краями и снетка приостреиным задним концом. Примерная длина крупных щетинок идиосомы (в процентах к ее длине): vi 12—13; sc е 28—30; h е 20; d3 10; 1 р 15; sa i 30—35; р а 20. Длина ног составляет около 23—25 (I и II), 19—21 (III) и 18—20 (IV) % длины идиосомы; лапки1 IV заметно длиннее голеней. Длийа 300—540 ;л.
Раскроет р а н е н и е. Франция, Италия, Англия, СССР; мне известен из Ленинграда, Москвы, Иванова и Барнаула.
Экология. Живет в старой муке, различных сухих лекарственных-веществах (спорынья, шпанская мушка и др.) и на сухих насекомых в коллекциях. Иногда вредит муке при ее продолжительном хранении. Известен только как синантропный вид.
SPECIES INCERTAE SEDIS t
* 1. Rhizoglyphus minor Dem.
Dementjev, 1913, Ztschr. Pflanzenkrankh., XI: 70, fig. 6.
Отличается от R. Caucasians Dem. (= echinopus F. et R.) меньшими размерами, светлой оранжево-красной, иногда отсутствующей окраской пог и хоботка (гнатосомы), узким, вытянутым в длину телом и цилиндрическим, закругленным на конце брюшком, и строением гипопуса. Из щетинок толп 4 направлены назад и 2 назад и в стороны, т. е. косо; все щетинки нелепые и малозаметные; 2 щетинки на эпистоме и 2 плечевых щетинки, кик у II. саи-, casicus. На верхней стороне 3-го членика I передних пог 2 миленьких щетинки неодинаковой длины, отходящие из одного места и соединяющиеся
14 Фауна СССР, Захваткин— 284
210
1. TYttOGLiPIHDAli. 2. JUllZOGLYPUINAti
таким образом, что очень трудно заметить, что их в действительности не 1, а 2. В теле яйценосной $ по наблюдалось больше одного зрелого или. полузрелого яйца... Странствующая личинка (гипопус) белого цвета, имеет только 2 щетинки па хоботке (гиитосоме). Нет щетинок на спинном щите, и он не образует выступа. Щиток (проподосомальный щит) оканчивается спереди округло, в остальном отот вид сходен с предыдущим (R. echinopus F. et R.).— Средняя длина 9 0.400 мм, 3 0.340 мм, странствующей личинки 0.200 мм. (По Дементьеву.)
Распространение. Кавказ (точное местонахождение по указано). Живот на корнях винограда, вместе с В. echinopus F. et R.
2. Rhizoglyphus trouessarti Berl.
Berlese, 1897: 110; Kramor, 1899: 143; Michael, 1903:144.
Ноги у гетероморфного-лишены шипов, за исключением 2 очень мелких на конце лапок. Очень мелкое существо (0.32 мм длиной), 4-ые ноги гетероморфного 3 не заходят за задний край брюшка й не заходят дальше назад, чем III пара ног. Лапки IV ног не длиннее голеней. 3 пары коротких, щетинок на заднем крае. (По Michael’io.)
Распространение. Описан из Франции.
3. Rhizoglyphus eutarsus Berl.
Berlese, 1921, Redia, XIV: 144—145.
Гетероморфный 3 довольно удлиненный, овальный, сзади закругленный и украшенный здесь несколько более длинными, чем у В. echinopus, щетинками, из которых по 2 задне-боковых, расположенных маргинально, длиннее других и почти равны ширине брюшка (гистеросомы); Плечевых, щетинок с каждой стороны по 2, из которых краевые (наружные) длиннее. Ноги длиннее и тоньше, чем у R. echinopus, что сразу бросается в глаза. Передние лапки в 4 раза длиннее своей ширины (по середине), длиною в 90 р, 15 р в диаметре. Отсутствует дорзальный шип впереди осязательного придатка, (обонятельной палочки). Снизу посередине лапки находятся 2 толстых, и крепких, довольно длинных шипа, сильно сближенных основаниями; нижпий шип, расположенный вблизи коготка, который хорошо развит у В. cchinopus, отсутствует у R. eutarsus. Основание penis короткое, коническое, стилет (дистальная часть) крайне длинный, на конце серповидно-изогнутый, втрое длиннее основания, 80 р длины. Копулятивные (анальные) присоски, почти как у R. nopus. Общая длина до 550 р, ширина 320 р.._ (По Berlese.)
Распространение. Италия (Палермо). Живет в почве.
4. Rhizoglyphus gcrmanlcus Berl.
Berlese, 1921, Redia, XIV: 145.
Гомеоморфный 3 овальный, сзади закругленный; 4 щетинки, расположенные в сильно дугообразно выгпутый кзади ряд, сильно приближенный.
SPECIES INCERTAE SED1S
211
к копулятивным (анальным) присоскам (рх и р2), имеют все одинаковые размеры, короткие, едва достигающие длины присосок. 2 вентральные щетинки между присосками и краем тела имеют такие же мелкие размеры. 2 задние 'вентральные щетинки, расположенные позади поперечного ряда (р3), небольшие; краевые и плечевые много длиннее, чем у II. echinopus, равные ширине брюшка (гистеросомы). Ноги тоньше и длиннее, чем у R. echinopus; передние лапки беднее вооружены, по все шипы, особенно пижпис (и на голенях), топкие п длинные. Лапки длиннее, чем у В. echinopus, свыше 3 раз длиннее своей ширины. Основание penis усечеппо-коническое, стилет не достигает длины основания, но острый, ножевидный. . Копулятивные присоски довольно похожи на таковые В. echinopus. Общая длина тела до 520 р., ширина 310 р.. (По Berlese.)
Распространение. Описан из Германии.
5. Rhizoglyphus globosus Berl.
Berlese, 1921, Redia, XIV: 145.
Гомеоморфный cl короткоовальиый, почти шаровидный, сзади закругленный и украшенный здесь многочисленными щетинками, из которых по 2 задне-боковых почти достигают ширины брюшка (гистеросомы), а но 3 с каждой стороны позади присосок, с которыми они сближены, короче; 2 вентральных, позади вышеупомянутых, у задне-боковых углов, небольших размеров. Ноги довольно короткие, но все же более удлиненные, чем у В. echinopus. Передние лапки приблизительно втрое длиннее своей ширины, вооруженные так же, как у В. echinopus, но дорзальный вершинный шип, расположенный над крупным серповидным коготком, почти равен коготку по размерам и сходен с ним по виду. Основание penis широко, коническое, стилет пестиковидный, выдвинутый из основания своей перед пей половиной, почти равный по длине основанию. Копулятивные (анальные) присоски своеобразного строения, с крупным внутренним диском, темным и радиально исчерченным. Общая6 7 длина тела до 380 [л, ширина 280 [г. 9 более удлиненноовальная (почти вдвое длиннее своей ширины), на задне-боковом крае брюшка, с каждой стороны, с 2 довольно длинными щетинками и такими же на боковых краях. Общая длина тела до 460 р., ширина 220 [г. (По Berlese.)
Распространение. Италия. В гниющей древесной трухе и в почве.
6 Rhizoglyphus occurrens Berl.
Berlese, 1921, Redia, XIV: 146.
Гетероморфный сГ удлиненноовальный, с явственными плечами, сзади закругленный и украшенный здесь с каждой стороны, у задно-бокопыч углов, 2 довольно длинными (почти равными ширине брюшка) щетинками, довольно сильно сближенными между собой. У заднего кран, aoHTpajiiiiio, имеются только мелкие щетинки, едва видимые при большом уполичоиип,
*
212
I. TYROGLYPHIDAB. 2. RHIZOGLYPllINAJi
из которых одна позади присосок, другая между присоской и наружным краем. Дорзо-маргинальпых щетинок по 2 с каждбй стороны, из которых -задняя сидит близ задне-бокового угла. Копулятивные (анальные) присоски довольно мелкие, округлые, с крайне мелким внутренним диском и хорошо хитинизированиыми краями. Основание penis широко усеченноконическое; стилет короткий, почти неотиковидпый, едва выдающийся из основания. .Задние йоги заходят за колец брюшка, средние вполне его достигают. Лапки передней пары короткие, одна в 2 раза длиннее своей ширины, вооруженные почти как у В. echinopus, вершинные шины у основания коготка снизу мелкие, в числе 4; нижний шип расположен чуть позади них. Ноги III пары сильно утолщенные, с дорзальным, довольно крупным шипом позади коготка и без зубца снизу у основания коготка, который здесь с 2 шипи-ками. Общая длина тела до 870 р, ширина 210 р.
$. Овальная, сзади угловатая, с каждой стороны с 2 довольно длинными щетинками. До 390 р длины, 260 р ширины (более обычны особи меньшего размера, длиною около 300 р). (По Berlese.)
Распространение. Италия. Во мху и в перегное.
7. Rhizoglyphus algidus Berl.
Berlese, 1921, Redia, XIV: 146.
Гетероморфный J удлиненноовальный, сзади прямо закругленный, украшенный довольно короткими щетинками, из которых плечевые и 2 задние почти равны 1/2 ширины брюшка. Между задними находятся 2 крайне мелкие щетинки, на заднем крае брюшка и на задне-боковых углах того же края, с каждой стороны, — по 2 коротких. Передние лапки короткие, едва в 2 раза длиннее своей ширины, вооруженные почти так же, как у, В. echinopus, но только с 1 нижней шпорой (вентральным шипом), расположенной субапикально. Ноги III пары едва в 2 раза толще ног IV, с таким же вооружением, как у В. echinopus, довольно длинные, почти достигающие до конца брюшка. Penis широкий, почти дисковидный, с 3-лопастным передним краем, без стилета.. Общая длина тела до 380 р, ширина 220 р..
Отличается от В. trouessarti несколько более крупными размерами и более удлиненной формой; более прямолинейным задним краем брюшка, украшенным более многочисленными и длинными щетинками; вооружением ланок; строением penis’a, и т. д. (По Berlese.)
Р а с п р о с т р и и о н и о. Италия. Найден во мху, в горах (на высоте 2600 м).
Фиг. 853—-367, Гипопусы различных Tyroglyphidae.
Фпг. 353. Acotyledon michaeli (Oude.), снизу (ио Michael’ю). Фиг. 354. A. alberti (A. Z.) сверху (ио Michael’ю). Фиг. 355. Caloglyphus berlesoi (Mich.) снизу (по Berlese). Фиг. 356. C.mycophagus (Megn.) снизу (по Megnln'y). Фиг. 357. С. mycophaf/us (M6gn.), гнатосома (по Megnin’y). Фиг., 358. Monieziella corticalis (Mich.) сверху (no Michacl’io). Фиг. 359. To же, снизу (по Michael’io). Фиг. 360. ЛГ. berlesiana, nom. п., снизу (по Berlese). Фиг. 361. То же, сверху (no Berlese). Фиг. 362. Rodionovia inopinata (Vtz.) сверху (no Vitzthum’y). Фиг. 363. То же, снизу (по Vitzthum’y). Фиг. 364. В. ino-pinata (Vtz.), гопень и лапка I сверху (по Vitzthum’y). Фиг. 365. Froriepia vimariensis Vtz. снизу (по Vitzthum’y). Фиг. 366. F. vimariensis Vtz., нога I (по Vitzthum’y). Фиг. 367. F. vimariensis
Vtz., нога II (no Vitzthum’y).
214 /. ТУ'НШПЛЧ'ШЬАЕ. 2. HHI7.0GLYI411N \ Е
16. Род LACKER BACERIA, gen. n.
Тип рода: Lackerbaueria cribratissima, sp. и.
Гипопус. Идиосома овальная, спинная поверхность ее умеренно выпуклая, очень сильно склеротизированная, в очень грубой и густой ямчатой скульптуре; усаживающие се щетинки относительно длинные, равные, в среднем, поперечнику голени I, тонкие, волосовидные. Гнатосома, как у Schulzea. Глаз нет. Псевдостпгмал иного органа я обнаружить не мог, Стернальпый щит хорошо очерченный с боков почти совершенно прямыми I
эпимеритами II и значительно слабее — сзади, где его прямая граница 1
обрисована только неутолщенцой складкой кожи; эпимеры II свободные, слабо дугообразные, далеко не достигающие заднего края стернального :
щита; коксальные поля III замкнутые, целиком обрамленные, как и лежащие позади них другие части вентрального щита, широкими краевыми ;
утолщениями, в пределах которых расположены обе пары коксальпых и крупные окологенитальные присоски; генитальный щит не обособленный. Присасывательный диск очень крупный, своеобразной формы, с 4 парами присосок (2 4-4 4-2); дополнительные присоски очень хорошо развиты. Ноги I и II длинные, стройные; вентральные щетинки вертлугов I короткие, вентральные щетинки бедер I и II и вертлугов I довольно длинные, гладкие, волосовидные; вептро-латеральные щетинки колен и голеней (I и II) тонкие, очень явственно перистые, как и дорзо-базальная щетинка колен, которая сильно смещена к внутреннему краю членика и резко дугообразно изогнута, как соответствующая щетинка у взрослых и нимф Glycyphagidae (например Gohieria)', вершина колен I с 1 (дорзальной) щетинкой; па лапках I присасывательная щетинка тонкая, с маленьким блюдцем; вершинные и предвершинные щетинки длинные, слабо ланцетовидно расширенные; ноги IV сильно укороченные; лапки III с 1, лапки IV с 2 очень длинными парусными щетинками.
Известен только 1 вид:
* 1. Lackerbaiicria cribratissima, sp. п. (фиг. 368, 369).-
Гипопус. Крупная форма. Окраска интенсивно ржаво-коричневая. Скульптура спинной поверхности состоит из различного размера круглых, глубоких ямок (па препарате щиты кажутся продырявленными), иногда сливающихся друг с другом; на переднем крае гистеросомального щита, вместо обычной поперечной штриховки, имеется мелкая пунктировка. . < Проподосомалынлй щит треугольный, со слабо выпуклыми передне-боко- । выми краями; длина его умещается в таковой гистеросомального щита немного более 2.3 раза. Гнатосома не выдается за вершину рострального выступа; аристы длинные, дугообразно расходящиеся, в стороны. Около- • генитальные присоски очень широко раздвинуты; присасывательный диск, по своей общей форме и строению, напоминает таковой некоторых Glycy-yhdgidae (роды Horstia, Neohorstia)-, в задней его части заметны3 сильно уплот- , :
ненных участка: один — между задними присосками и 2 — кнаружи от
-u,«.--ч-
16. LACKERBAlSERIA. 17. ROBLMSCA 21a
них; вентральные присоски не крупнее периферических, задние расположены эксцентрично в своей толстой, сильно хитинизированной рамке. Обонятельная палочка лапок I и II стройпая, с характерно перегнутым, приострениым концом; па лапках I и II по 4 ланцетовидно расширенных щетидки; на лапках III и IV их по 3 штуки; парусные щетинки лапок IV очень длинные. Длина 255—300 <л, ширина 170—210 jj..
Распространение. Известей только из окрестностей г. Иванова. Гипопусы найдены в очень большом количестве па осе Crabro fossorius L.
17. Род ROBINISCA, gen. п.
Тип рода: Tyroglyphus mycolichus Ouds. 1912.
Гипопусы. По многим признакам напоминают гипопусов рода Caloglyphus Beil. Идиосома правильной эллиптической формы, с гладкой (крайне мелкопористой), очень сильно выпуклой спинной.поверхностью. Все щетинки спинной поверхности представлены тонкими микрохэтами; расположение их не представляет каких-либо примечательных особенностей. Проподосомальный щпт спереди широко закругленный, без рострального выступа; v i крайне мелкие. Глаз нет. Псевдостигмальный орган крупный, толстый, S-образпо изогнутый. Гнатосома очень длинная и тонкая (не менее чем в 3 раза длиннее своей наибольшей ширины), нерасчлененная, значительно выдающаяся за широко закругленный передний край идиосомы; аристы слегка дугообразные, сходящиеся концами. Стернальный щит четко очерченный, с длинными боковыми краями; стернум далеко не доходит до заднего края стернального щита; эпимеры II изменчивой длины, то ’Свободные, то доходящие до заднего края стернального щита и тогда замыкающие коксальные поля II; коксальные поля' III замкнутые, почти соприкасающиеся вдоль средней линии тела; эпимеры IV смыкаются с продольным вентральным швом далеко позади переднего края вентрального щита; генитальный щит более или менее явственно обособленный; поверхность щнтов брюшной стороны тела равномерно хитинизированная, без особенно развитых краевых утолщений. Коксальные присоски хорошо развиты. Присасывательный диск крупный, близко придвинутый к концу тела, с 5 парами присосок (дополнительные угловые присоски хорошо развитые). Ноги I и II длинные пли средней длины, I целиком, II большей своей частью выступающие за края идиосомы; вентро-латеральные щетинки голеней н колен (I п II) превращены в уплощенные, пожевидные шипы, или же таковыми представлены только вентро-латеральные щетинки голеней, а на коленях они волосовидны; дорзальная щетинка тех же колеп чрезвычайно короткая, толстая; вершина их только с 1 щетинкой. Лапки I стройные; палочковидная сенсилла р расположена па одном уровне с глинной обонятельной палочкой или несколько более проксимально; сенсилла. Т отсутствует, о утолщенная в виде палочки; присасывательная щетинка •с очень крупным блюдцем, одна из вентральных щетинок шипообразпая,
216
I. TYROGLYPHIDAE. 2URHIZOGLYPHINAE
остальные вентральные и почти все вершинные щетинки несильно ланцетовидно расширенные; лапки IV с толстыми, короткими щетинками или шипами, без парусных микрохэт.
В род Robinisca я с уверенностью могу .поместить, помимо описываемых здесь В. mycolicha (Ouds.) и R. macrocnemis, sp. п.,'еще только бразильского Tyroglyphus fuscipes Ouds. J 924. Этот вид очень близок кВ. macrocnemis, от которого отличается значительно более крупными размерами (358— 380 р), гнатосомой, совершенно'спрятанной под передним краем идиосомы, и несколько иной конфигурацией последней; строение ног I и II и их вооружение, строение стернального и проподосомальпого щитов у R. fuscipes таково же, как у R. macrocnemis.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Голени I сильно гипертрофированные, почти в 2 раза длиннее колен I и больше чем в 1.5 раза длиннее своего диаметра; видимая часть проподосомальпого щита очень узкая, около 10 раз короче гпетеро-сомального; эпимеры II на значительное расстояние не доходят до-заднего края стернального щита; наружная латеральная щетинка колен I и II волосовидная; обонятельная палочка лапок I и II тонкая, цилиндрическая.......................* 1. В. macrocnemis, sp.в.
2 (1). Голени I нормального размера, короче колен. I и собственного диаметра; видимая часть проподосомальпого щита шире, только в 5— 5.3 раза короче гистеросомальпого щита; эпимеры II доходят до-заднего края стернального щита; наружные латеральные щетинки колен I и II шиповидные; обонятельная палочка лапок I и II толстая, булавовидная ............................ * 2. R. mycolicha (Ouds.).,
* 1. Robinisca macrocnemis, sp. в. (фиг. 177—179).
Г и п опус. Окраска довольно темная, буроватая. Идиосома широкоэллиптическая, очень высоко выпуклая; усаживающие ее щетинки чрезвычайно короткие. Проподосомальный щит очень короткий: длина его-видимой части составляет только около длины гистеросомальпого щита; sc выстроены в почти правильный поперечный ряд. Гнатосома в 3 раза длиннее своей максимальной ширины, с слегка выпуклыми боковыми краями, на конце но разделенная. Эпимеры II далеко пе доходят до заднего края стернального щита; эпимеры IV нс доходят до вентрального шва, простирающегося до заднего края нейтрального щита; вентральный и генитальный щиты и парастернальные склериты крупнопористые; продольная штриховка вентральных перегибов гистеросомальпого щита тоже резче обычного, и спереди, у плеч, переходит в ряды мелких бугорков. Голени I необычайно большие, почти в 2 раза длиннее колеи I и более чем в 1.5 раза длиннее своего максимального диаметра; голени II лишь немного длиннее колен, немного длиннее своего диаметра; лапки I и II очень строй-
17. ROBINISCA
217’
шло. Вентральные щетинки бедер очень мелкие на бедрах I, длинные на бедрах II; наружные латеральные щетинки колен I и II в виде тонких гибких щетинок, вершинная дорзальная щетинка колен I очень длинишц вентро-латеральные шипы голеней I и II с зазубренными краями. Сенсилла s очень длинная (почти равная по длине всей лапке), слабо дугообразно-изогнутая, постепенно утончающаяся к своему пуговковидио вздутому концу; шппообразная вентральная щетинка крупная, зазубренная; вершинные и предвершинные щетинки лапок I и II сильнее ланцетовидно-расширенные, чем у следующего вида. Па лапках IV дорзальная щетинка довольно длинная, вершинная дорзальная щетинка ланцетовидно расширена; ближайшая к основанию вентральная щетинка сильно зазубрена, остальные гладкие, довольно толстые, но не шиповидные. Длина 293 р, ширина 242 р..
Распространение. Вид описывается по 1 экз., найденному в Ивановской обл. (с. Болотове), в почве, густо перемешанной с отходами от-обмолота ржи, на колхозном току.
* 2. Robinisca mycolicha (Ouds.) (фиг. 175, 17G).
Oudemans, 1912, Entomol. Ber., Ill: 250 (Tyroglyphus). —wasmanni Oudemans, 1.918, Arch. Naturg., LXXIX, 10: 48, t.XVIII,fig. 17—18(Tyroglyphus).
Г и п о и у с. Окраска более бледная, светложелтоватая. Идиосома менее широкая, чем у В. macrocnemis, отношение её длины к максимальной ширине равно 2.7 : 2; спинная поверхность идиосомы не столь сильно выпуклая,, ее щетинки несколько длиннее. Видимая часть проподосомального щита, только в 5—5.3 раза короче гистеросомального; sc расположены вогнутой кпереди дугой; гнатосома в 4 раза длиннее своей ширины, с совершенно’ прямыми, параллельными краями, на конце расщепленная. Эпимеры II доходят до заднего края стернального щита, стернум заходит концом за. линию, проведенную через основания ацетабул II, но далеко пе достигает заднего края стернального щита; эпимеры IV доходят до продольного вентрального шва, простирающегося лишь до середины вентрального-щита; скульптура всех щитов брюшной поверхности одинаковая — крайне мелко пористая. Поры вентральных присосок присасывательного диска, разъединенные, каждая с самостоятельным ободком. Длина ног I составляет-около 45% общей длины идиосомы; голени I и II нормального размера,, короче соответствующих колен и собственного диаметра. Вентральные-щетинки бедер I и II короткие; наружные латеральные щетинки колен I л II в виде коротких, толстых шипов; вершинная щетинка колен I довольно-короткая; шипы голеней I и II гладкие; лапки I п II несколько короче и массивнее, чем у предыдущего вида; на лапках I сенсилла 7 сидит более проксимально, чем толстая, булавовидная обонятельная палочки; сенсилла е довольно короткая, прямая, цилиндрическая; стебелек прнеас.ы-натольпой щетинки очень толстый, в целом она грибовидная; нейтральная шипообразная щетинка короткая, вершинная и предвершинная щетинки.
218 I. TYROGLYPHIDAE. 2. RHIZOGLYPHINA£
лапок I и II очень слабо ланцетовидно расширенные. На лапках IV дорзальная щетинка короткая, вершинная дорзальная, как и большинство . вентральных, игловидная. Длина 165—231 р, ширина 130—180 р.
Распространение. Известен только из Голландии и СССР (окрестности Новочеркасска).
Экология. Экземпляр, послуживший Oudemans’y для описания вида, был найден в норе крота; единственный имеющийся у меня экземпляр собран в почве (поле пшеницы).
К тому же роду, невидимому, относится еще неизвестная мне в натуре:
3.? Robinisca tarsispina (Ouds.).
Oademans, 1912, Entomol. Her., Ill: 50 {Hypopus).
Гипопус. Очень близок к Hypopus spinitarsus Henn. Он отличается -от ранее известных видов следующими признаками: 1) передние края про-и мезосомы идут почти параллельно; просома, таким образом, в виде широкой полосы, а не треугольная; 2) нет теменных щетинок; 3) гипостом (гнато-сома) длинный и песет только 2 вершинных длинных волоска; 4) ноги пе только тоньше, но также и волоски, щетинки и обонятельные палочки длиннее. В гниющей соломе. Флоренция. (По Oudemans’y.)
Некоторые дополнения-к этому краткому диагнозу дает Vitzthuin в своем 'Описании Tyroglyphus тауаогшц. 1 Сравнивая этот вид с Т. tarsispinus Ouds., on пишет, что у последнего эпимеры I и II так же коротки (т. е. далеко не .достигающие заднего края стернального щита, свободные) и на гипостоме отсутствуют боковые щетинки. Этим исчерпывается все, что было опубликовано о Т. tarsispinus Ouds.
18. Род VIEDEBANTTIA OUDS.
Oiidemans, 1927, Entomol. Вег., VII: 485.
Тип рода: Viedebanttia schmitzi Ouds. 1927.
Совершенно недостаточно описанный род, известный только по гипо-шальной стадии и, невидимому, близкий к роду Robinisca. Родовой диагноз^ Oudemans’a сводится к следующему: Гнато-и идиосома приблизительно такого же строения, как у Zschachia necrophori Duj. 1849 (сем. Anoeti-dae)\ идиосома шире, как у Tyroglyphus. Снизу более или менее как у Tyroglyphus, по присасывательный диск дальше назад. Форма ног, как у Calo-..glyphus, по без ножовидных щетинок на ногах I и II.
Род Viedebanttia состоит только из 1 вида.
1. Viedebanttia schmttzi Ouds.
Oudemans, 1927, Entomol. Bor., VH: 485.
Окраска светложелтая, тогда как нимфы II Caloglyphus красновато-екоричневы. Гнатосома в 4 раза длиннее своей щирины. Проподосома очень
1 Arch, Naturg., LXXXVI ,10:5-1.
. 18. VlEDEHAiNTTIA. 1<J. TXOLPEAUIA 210
короткая, едва длиннее проксимального диаметра ланок I и II, имеет, таким образом, вид узкой поперечной полоски. Спинные волоски крайне мелкие и тонкие, трудно заметные; 2 явственных волоска на заднем крае. Иа ногах In II вертлуги и бедра более или менее одинаковой длины, тогда как лапки более или менее равны по длине голени -р колено. (По Oucle-jnans’y.)
Распространение. Голландия. Найден в поре крОта.
19. Род TROUPEAUIA, gen. и.
Тип рода: Tyroglyphus novus Ouds. 1907.
Гипопус. Идиосома эллиптическая, с умеренно и равномерно выпуклой, гладкой или пунктированной спинной поверхностью. Все щетинки идиосомы представлены очень мелкими п тонкими, трудно заметными микро-хэтамп. Гнатосома длиннее своей ширины, нерасчлеиенная, с параллельными боковыми краями; аристы прямые, расходящиеся под почти прямым углом. Глаз пет. Псевдостигмальный орган крупный, вполне явственный, обычной (щетинковидной) формы. Стернальный щит хорошо очерченный с боков, слабее — сзади; его боковые края, постепенно закругляясь, незаметно переходят в задний край, не образуя с пим хоть сколько-нибудь ощутимого угла; стернум не достигает середины щита; эпимеры II свободные, более или менее сильно дугообразно искривленные; концы их также далеко не достигают заднего края стернального щита; коксальные поля III замкнутые, смыкающиеся по средней линии тела на значительном протяжении; генитальный щит неясно обособленный от коксальных полей IV, длинный; поверхность этих щитов (как и стернального) равномерно, но не сильно склеротизирована. Коксальные присоски I, а иногда и II, заменены микрохэтами. Присасывательный диск округлой формы, близко придвинутый к заднему краю тела; дополнительные присоски неясные. Ноги I и II длинные, целиком выступающие за края тела; их ланки стройные, вентральные щетинки вертлугов и бедер I длинные; шипов па ногах нет — все щетинки, усаживающие ноги, тонкие, гибкие, более пли менее волосовидные ; вершина колен I только с 1 щетинкой; щетинка о лапок I в виде прямой, притупленной па конце палочки; вершинные и предвершинные щетинки длинные, тонкие, волосовидные (не расширенные листовидно).
К этому роду, вероятно, относится, помимо описываемых здесь 3 видов, еще и тропический Tyroglyphus novisimilis Vtzt. 1920, известный с Явы, Цейлона, из Индии и из Бразилии.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
.1 (2). Стернальный щит сильно трапециевидно вытянутый назад; эпимеры II S-образно искривленные, расходящиеся концами; спинные щиты гладкие; лапки III и IV каждая с 2 довольно длинными парусными
220
I. TYRGGLYPHIUAE. 2. RHIZOGLYPHINAE
щетинками; наружная вентро-латеральная щетинка голеней I и II отсутствует ...................................*1. Т. nova (Ouds.).
2 (1). Стернальный щит сзади широко закруглен; эпимеры II слабо дугообразные, сходящиеся концами; спинные щиты ямчато пунктированные; лапки III и IV без парусных щетинок; наружная вентро-латеральная щетинка голеней I и II хорошо развита. * 2. Т. crabronis, sp. в.
В таблицу не помещена недостаточно описанная Т. mycobora (Ouds.).
* 1. Troupeauia nova (Ouds.) (фиг. 37и, 37.1).
Oudemans, 1907, Abh. Naturwiiw. Ver. Bremen, XIX: 69, t. II, fig. 12—15 (Tyrogtyphus).
Гипопус. Окраска бледная, чуть желтоватая. Идиосома несколько более вытянутая в длину, чем у следующего вида; отношение ее длины к ширине составляет у моих экземпляров 1.6—1.7 : 1, по рисунку же Oudemans’a 1.6 : 1; вероятно, последний сделан с несколько расплющенного экземпляра, так как в тексте указана еще меньшая ширина, чем у моих особей (длина 232 р, при ширине всего в 100 р). Поверхность спинных щитов гладкая, усаживающие их щетинки мелкие, трудно заметные; v i очень короткие, sc расположены в виде трапеции. Проподосомальный щит крупный (равный 2/5 длины гистеросомального), спереди широко, но несколько угловато закругленный, с еле намеченным, почти не выдающимся ростральным выступом. Гнатосома с цельным, нерасщепленным дистальным краем; боковые щетинки расположены у середины ее длины, очень короткие. Стернальный щит сильно вытянутый назад в виде треугольника, с закругленной или почти прямо срезанной вершиной; его боковые края (эпимериты II) слабо вогнутые, ориентированные под острым углом друг к другу; стернум довольно длинный, эпимеры II S-образно искривленные, расходящиеся концами. Обе пары коксальпых присосок представлены очень мелкими микрохэтами (хорошо заметны лишь при пользовании иммер-зией); окологенитальные присоски раздвинуты менее широко, чем у Г. crabronis-, передние генитальные щупальцы далеко отодвинуты от задних; присасывательный диск небольшой; обе пары периферических присосок имеют почти одинаковые размерь!'; дополнительные присоски недифференцированные. Наружная вентро-латеральная щетинка голеней I и II отсутствует; длина обонятельной палочки лапок I и II составляет 1/2 общей длины лапки; она зиачительпо длиннее палочковидной сенсиллы е, которая слегка булавовидно утолщена; сенсилла 8 обычного размера, равная х/з обонятельной палочки. Лапки Ш и IV каждая с 2 парусными щетинками, длина которых но превосходит (а обычно значительно меньше) ширины идиосомы. Длина 170—282 |Л, ширина 100—150 р.
Фиг. 368—375. Гишшупы родоп Laclcerlaueria, Troupeauia и Schuteea.
Фиг. 368. Lackerbaiieria cribratinelrna> вр. П., идиосома сверху. Фиг. 369. L. cribratissima, ер. п.» общий вид снизу. Фиг. 370. Troupoauia nova (Oude.), идиосома сверху. Фиг. 371. Т. nova (Ouds.). общий вид снизу. Фиг. 373. Т. crabronis, ер. п., общий вид снизу. Фиг. 373. Schukea pamiren-sis, ер. n., идиосома сверху. Фиг. 37d. S. pamirensis, ер. n., общий вид снизу. Фиг. 375. S. cau-casica, ep. n., общий вид снизу
222 I; 1YR0GLYPU1DAE. 2. RUIZOGLYPIIINAE
Распространение. Известен пока только из Германии (окрестности Времена) и нескольких пунктов в окрестностях Москвы. Гипопусы были находимы на слонике Hylobius abietis L. и осе Crabro spinicollis Н. S.
*2. Troupeauia crahronis, sp. n. (фиг. 372).
Гипопус. Окраска такая ясс, как у Т. nova. Идиосома более широкая. Поверхность спинных щитов в очень густой ямчатой пунктировке. Цлина проподосомальпого щита составляет 1/3 длины гистеросомального; ростральный выступ сильно выдающийся. Гнатосома с вильчато расщепленным концом. Стернальный щит ио вытянутый в виде треугольника, сзади широко закругленный; его слегка выпуклые боковые края совершенно незаметно переходят в за,длин, образуя с ним непрерывную правильную дугу; стериум короткий, эпимеры II слабо дугообразные, сходящиеся концами. Волоска па месте коксальных присосок III я не мог ясно увидеть. Присасывательный диск крупнее, чем у предыдущего вида; задние присоски чуть крупнее боковых; доцолнительные присоски лучше оформлены. Наружная вентро-латеральная щетинка голеней I и II хорошо развита; длина, обонятельной палочки лапок I и II равна J/3 длины лапки; палочковидная сенсилла г одинаковой с ней длины, не утолщенная к концу; сенсилла о очень длинная, почти равная по длине обонятельной палочке, очень слабо утолщенная к концу. Парусных щетинок на лапках III и IV нет. Длина. 180 [х, ширина 118 [1.
Распространение. Известей пока только из одного пункта: Мытищи в окрестностях Москвы. Гипопусы, послужившие для описания вида, были найдены на нескольких экземплярах Crabro dives Lep., на. которых встречались по 1 —2 штуки. Надо думать, что и этот вид окажется обитателем гниющей древесины.
3. Troupeauia (?) my со bora (Ouds.).
Oudemans, 1912, Entomol. Вег., Ill: 188 (Tyroglyphus).
Мне в натуре неизвестен; видимо, близок к Т: crabronis и мог бы, с известной степенью вероятности, считаться идентичным с ним, если бы в его-описании не были'подчеркнуты необычно крупные размеры обонятельной палочки лапок I и II, чего никак нельзя сказать об этих органах у Т. crabronis, и скорее полукруглая форма проподосомальпого щита.
Известен только гипопус. Длина 155 |х, ширина 100 [л, по края тела, сильпо загнуты центрально, даже бока протеросомы спереди и в соседстве с ногами II загнуты вентрально. Этот вид очень близок к Т. mycophagus, от которого отличается следующими признаками. Ноги 1и11 тоньше (стройнее); лапки I и II дорзалыю с меньшим количеством волосков; их обонятельные палочки необычайно крупные; протеросома скорее полукруглая,, чем треугольная; жировые железы отстоят от протеросомы ла расстояние,, равное их собственной длило; па тазиках I нет присосок, по крайне мелкие волоски; на тазиках III присоски крайне мелкие; присасывательный диск.
If). TlififJPE\riA. 20. nOIHOyOVIA Ш
почти круглый (у T. myco'phagus почти четырехугольный); присоски па нем: 2 среднего размера, 2 крупные и 4 маленькие, расположенные вогнутой кпереди дугой. — В гниющих листьях. Гаагский лес. (По Oudemans’yA
20. Род RODIONOVIA, gen. 11.
Тип рода: Tyroglyphus inopinaius Vtz. 1924.
Гипопус. Идиосома продолговатая, в 1.7 раза длиннее своей максимальной ширины, приходящейся на передние углы гистеросомального-щита; спинная поверхность гладкая, умеренно выпуклая; все щетинки спинной поверхности представлены тонкими микрохэтами. Проподосо-мальный щит треугольный; боковые края гистеросомы почти прямые, задний— полого закругленный. Гнатосома, как у Troupeauia. Глаза и псевдо-стигмальный орган отсутствуют. Стернум короткий; коксальные поля II замкнутые, хорошо очерченные, почти смыкающиеся по средней линии тела f. сильно поперечные; строение вентрального щита такое же, каку Troupeauia.. Коксальные присоски нормального строения, мелкие. Присасывательный диск очень крупный, с обычными 4 парами присосок (дополнительные присоски не развиты). Ноги I и II довольно короткие, по почти целиком выступающие за края тела; вооружение 4 базальных члеников, как у рода-Troupeauia-, лапки умеренной длины, каждая с 3 листовидно-расширенными-щетинками; присасывательная щетинка лапок I длинная п тонкая, с небольшим овальным «блюдцем», напоминающим по своей форме цветок Calla,. лапок II — расширенная в очень крупную листовидную щетинку. Лапкж III и IV короткие, каждая с 4 листовидными щетинками.
К этому роду относится только 1 вид:
1. Rodioncvia inopinata (Vtz.) (фиг. 362—364).
Vitzthum, 1924, Arch. Naturg., XC, 10: 65, fig. 47—49 (Tyroglyphus).
Гипопус. Окраска беловатая. Проподосомальный щит с прямыми1 передне-боковыми краями, сходящимися кпереди под прямым углом; sc-намечают фигуру трапеции; sc i расположены значительно впереди sc е; длина проподосомального щита составляет чуть меньше 1/z длины гистеросомы. Гнатосома в 2 раза длиннее свей ширины, с параллельными боковыми краями и чуть расщепленной вершиной, целиком спрятанпая под; ростральным выступом (за вершину которого выдаются только аристы). Присасывательный диск поперечноовальный, очень крупный (диаметр превосходит наибольшей ширины тела); его присоски дальше, чем обычно, отодвинуты друг от друга; передние заметно мельче боковых И задних, расположенные значительно ближе к генитальным, чем к л рис,о скам центральной пары. Длина ног составляет, соответственно 30, Ш, .17 и 17% длины идиосомы; лапки I в 2.5, лапки II в 1.5 раза длиннее голени. Щетинки базальных члеников ног короткие; обонятельная палочка липок I и II крупная; дистальный конец ее образует массивную, широко закругленную на конце булаву. Длина 156 р, ширина — 92 р.
224 . I. TYROGLYPHIDAE. 2. RHIZOGLYPHINAE
Распространение. Известен только из Германии. Гипопус найден на слонике Cossonus linearis L., что позволяет предположить, что Л. inopinata является обитателем разлагающейся древесины.
21. Род (1ARSAULTIA OUDS.
Oudemans, 1916, Entomol. Вег., IV: 265.
Тип рода: Garsaullia tosludo Ouds. 1916.
Недостаточно описанный род, известный только по гипопусам. Родовой диагноз, данный Oudemans’ом, таков: главные признаки те же, как у гипопусов Tyroglyphus, 1 но присасывательный диск не имеет свободной перепончатой каймы и несет 10 присосок.
К этому роду относится только 1 вид.
1. Garsaultia testudo Ouds.
Oudemans, 1916, Entomol. Вег., IV: 265.
Гипопус, как у Tyroglyphus macgillavryi, с длинной проподосомой, -"прикрывающей, в виде защитного козырька или крыши, передние 4 ноги, и 10 присосками. Однако не непосредственно родствен ему, так как передние четыре .ноги массивные, с крепкими шипами, как у Tyroglyphus kramerii. Отличается от всех других гипопусов Tyroglyphidae присасывательным диском, который не имеет прозрачной, хотя и трудно заметной, свободной каймы, d своим хитинизированным краем непосредственно переходит в окружающие ого части. Длина 475 р., ширина 330 [л, высота 186 [л; широкоовальиый; в профиль выше, чем полушаровидный; кофейно-бурый. •Спинные волоски очень маленькие и тонкие. Спина гладкая. Генитальная щель длинная, глубоко вдается в присасывательную пластинку; внутренних генитальных щупальцев не видно. Десять присосок анальной пластинки расположены так: 4, 4, 2. Из передних 4 обе внутренние телескоповидные; из средних 4 две внутренние очень крупные; самые задние 2 меньше всех. Позади присасывательной пластинки — внутренняя, медиальная хитиновая палочка. (По Oudemans’y.)
Распространение. Найден в окрестностях Боппа (Германия) в гниющих листьях. :
22. Род SANOASSANIA OUDS
Oudemans, 1916, Entomol, Bor., IV: 265.
Тип рода: Sancassania chelono Ouds. 1916.
Так же как и предыдущий род, известен только по гипопальной стадии ш недостаточно полно описан. Оригинальный диагноз Oudemans’a сводится к следующему. По главным признакам напоминает гипопусов Tyroglyphus, во отличается от них,том, что между проподо- и гистеросомой находится
1 Род Tyroglyphus понимается здесь еще в широком смысле, в том объеме, который он имел . до 1923—1924 гг.
2/. GARSAULTIA. 22. SANCASSANIA. 23. SCHULZEA 22'i
линзообразный (чечевицеобразный) мягкий участок, резко отграничений как спереди, так и сзади.
Монотипический род.
1.’Sancassania chelone Ouds.
Oudemans, 1916, Entomol. Вег., IV: 266.
Гипопус. Так же как у Tyroglyphus macgillavryi, с длинной пропо-досомой, прикрывающей, в виде защитного козырька или крыши, 4 передние ноги. И все же не непосредственно родствен ему, так как передние 4 ноги массивны и торчат вперед перед щитом; имеется только 8 присосок; между проподо- и гистеросомой находится широкий серповидный участок поперечнополосатой, мягкой кожи. Спинные волоскп крайне короткие;, жировые железы далеко впереди; передние из 4 внутренних генитальных щупальцев конические. Размеры в описании не указаны. (По Oudemans’y.)
Распространение. Как и предыдущий вид,—найден в окрестностях Бонна, в гниющих листьях.
На мой взгляд, эта форма чрезвычайно близка, если нс идентична,.с гипопусом Caloglyphus polyphyllae. Наиболее характерный для нее признак— резко отграниченный серповидный участок поперечно штрихованной колеи па гистеросоме — также бросается в глаза и у названного вида Caloglyphus, но здесь он не мягкий, а имеет такую же плотную консистенцию, как вся остальная поверхность гистеросомального щита. Не может ли быть, что Oudemans счел этот участок мягкокожистым лишь, из-за покрывающей его поперечной штриховки, обычно (например у Cheyletidae и других ТготЫ-diiformes, многих Acaridiae и т. д.), связанной, как известно, с мягкой консистенцией кожи? Коническая форма генитальных щупальцев, характеризующая 8. chelone, также вполне явственна у Caloglyphus polyphyllae; к этому виду подходят и все другие приведенные в диагнозе 8. chelone признаки форма проподосомального щита, строение ног и т. д.).
23. Род SCHULZEA, gen. п.
Тип рода: Schulzea pamirensis, sp. n.
Гипопус. Идиосома широкоовальная, с умеренно выпуклой спинной поверхностью; спинные щиты гладкие, мелкопористые или пунктированные; все усаживающие их щетинки короткие, тонкие; расположение их не уклоняется от нормы. Гиатосома с явственно обособленными дистальными члениками. Глаз нет. Псевдостигмальный орган хорошо развитый. Коксаль-иыо поля II более или менее явственно замкнутые, широко раздвинутые; эпимеры II (боковые края стернального щита) сильно скошенные; сторпум далеко по достигает до уровня задних краев коксальных полей II; кокспль-1Н.!о поля III замкнутые, но с очень слабо намеченным внутренним контуром, расставленные почти так же широко, как коксальные поля II; гони-тпл1.Н1.|П щит пе обособленный; поверхность всех коксальных полей слабо
И. Фауна СССР, Захваткип—284
226
I. TYROGLYPHIDAE. 2. RBIZOGLYPHINAE
склеротизированная;. краевые, утолщения довольно хорошо развиты на вентральном щите и коксальпых полях I. Коксальные присоски хорошо-развиты, нормального строения; присасывательный диск крупный, несу-./ щий 4 пары присосок; присоски боковой пары расположены кнаружи и вне-, реди вентральных; дополнительные присоски довольно явственные. Вентральные щетинки вертлугов и бедер 1 и II очень длинные, волосовидные; вентро-латеральные щетинки колон и голеней и дорзо-базальная щетинка колен (I и II) в виде небольших жестковатых (по отнюдь не шипообразных), гладких щетинок; вершина колен I с 2 длинными, толстыми щетинками-(как у половозрелых Rhizoglyphinac и др.); присасывательная щетинка с небольшим «блюдцем», лапки IV с более или менее длинной «парусной» макро-хэтой.
. Сюда относятся 2 вида, известные т олько по гипопусам, собранным с одиночных пчел родов Megachile и Prosopis. Случайность нахождения их на этих переносчиках (оба вида были найдены каждый лишь на 1 экз. пчелы) делает вероятным предположение, что виды Schulzea слабо биологически-связаны с этими своими переносчиками и не являются постоянными обитателями' их гнезд.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Ноги массивные; лапки I короче, чем голень и колено I, взятые-вместе; гнатосома.короткая и массивная, целиком спрятанная под ростральным выступом; основной членик ее почти квадратный; парусная щетинка лапок IV короче половины ширины идиосомы . . ..
................................. . . *1. S. pamirensis, sp. и.
2 : (1). Ноги стройнее; лапки I значительно длиннее голени и колен I, вместе взятых; гнатосома длинная- ее дистальные членики (развилок) целиком выдаются за передний край рострального выступа, а основной более чем в 1.5 раза превосходит свою ширину у основания и еще суживается к вершине; парусная щетинка лапок IV почти равпа ширине идиосомы .....'........................ . *2. S. caucasica, sp.n..
*1. Schulzea pamirensis, sp. n. (фиг. 373—374).
Гипопус. Окраска интенсивно охрово-бурая. Спинная поверхность умеренно выпуклая, очень плотно хитипизированная, густопористая и, помимо того, покрытая частой пунктировкой из округлых неглубоких ямок. Проподосомальпый щит длиною в 3/г гистеросомального. Гнатосома короткая, целиком скрытая под вогнутой в виде камеростома нижней поверхностью рострального выступа; ее основной членик почти квадратный. Замкнутость коксальпых полей II не очень явственна, так как их контур, на значительном протяжении обрисован только тонкой, нехитинизирован-ной складкой кожи, а сами эпимеры II.далеко не достигают концами да загнутых внутрь концов эпимеритов II; внутренние контуры коксальных
23. SCHULZEA
227
нолей III и IV’ слабо намечены; краевые утолщения коксальпых полей относительно узкие. Коксальные присоски крупные; стернальные расположены у внутреннего края узких краевых утолщений коксальпых полей I, па более слабо хитшшзироваппой части этих полей, вентральные же присоски целиком окружены краевым хитиновым утолщением коксальпых полей III. Присасывательный диск крупнее и ближе» придвинут к заднему краю тела, чем у следующего вида. Ноги короче и массивнее,' лапки I короче, чем голепь и колено I, взятые вместе. Обопятельпая палочка лапок I и II довольно толстая (раз в 8—9 длиннее своего диаметра у основания); сенсилла 8 лапок I расположена почти на той же высоте, как обонятельная палочка, и. соответственно, много более проксимально, чем сенсилла г; парные вентральные и одна из вершинных (внутренняя) щетинок лапок I и II ланцетовидно расширенные. На лапках III и IV ланцетовидно расширены по 4 щетинки; одна из дорзальных щетинок лапок IV имеет довольно значительную длину, лишь немного уступающую-таковой всей ноги (IV). Длина 325—345 |i, Ширина 240—250 р..
Распространение. Описывается по материалу, собранному па Памире, без более точного указания местонахождения. Гипопусы были * найдены в довольно большом числе на тораксе пчелы Megachile grombccvslcii F. Mor.
* 2. Schulzea caucasica, sp. n. (фиг. 375).
Гипопус. Окраска бледножелтоватая. Отношение длины идиосомы к ее ширине 1.35:1; очертания ее на препарате более угловатые, чем у & pamirensis, что, очевидно, вызывается деформацией более сильно выпуклой, чем у последнего вида, спинной поверхности, которая слабее хитинизиро-вана; идиосома совершенно гладкая (крайне тонкопористая). Длина про-подосомапьиого щита составляет чуть больше ’/г длины гистеросомального щита; щетинки гистеросомы очень короткий; постапальпыо щетинки пс длиннее колена I. Гнатосома выдается всем развилком за конец рострального выступа; основной членик ее больше чем в 1.5 раза длиннее своей ширины у основания, несколько суживающийся к вершине.
Коксальные.поля II четко очерченные на всей окружности; внутренние контуры коксальпых полей III еле намечены, полей IV — совершенно неясные; краевые утолщения коксальпых полей широкие, особенно на полях I, где они занимают весь их наружный угол. Коксальные присоски несколько мельче, чем у S', pamirensis', стернальные присоски помещаются на территории краевых (угловых) утолщений, вентральные — у внутренней границы этих утолщений; присасывательный диск отделен от видного края тела относительно широким промежутком; он несколько меныно, чем у предыдущего вида; передняя пара периферических присосок несколько крупнее задней. Ноги длиннее и стройнее; лапки I значительно пропооходят длину голени и колена I, взятых вместе. Обонятельная палочки липок I и 11 чрезвычайно тонкая (раз в 20 длиннее своего диаметра у o('iiohhihiji);
*
228 I. TYROGLYPIIIDAE. 2.. RIHZOGLYPniNAE
сецсилла, § лапок I расположена значительно впереди основания обонятельной палочки, практически на одном уровне с сенсиллой е; вершинные и предвершинные щетинки лапок I и II волосовидные, как и все щетинки лапбк III и IV;, дорзальная щетинка лапок IV в.2 раза длиннее всей ноги IV. Размеры единственного экземпляра: длина 303 |л ширина 223 |i.
Распространение, . Известен ; пока, только из Грузии (Карт-вели). Гипопус найден на ближе по определенном виде рода Prosopis.
24. Род FHOBIEPIA VTZ.
Vitzthum, 1919,' Zobl. Anz., L: 189.
Тип рода: Froriepia vimariensis Vtz. 1919.
Гипопус. Легко отличается от всех остальных гипопусов сем. Tyroglyphidae необычайно длинными щетпнковидными коготками ног I и совершенно своеобразным строением коксо-стернального скелета. Идиосома имеет форму усеченного спереди овала; спинпая поверхность крайне тонкозернистая; щетинок на ней не удалось обнаружить; если таковые имеются, что следует предполагать, то они должны быть очень короткими • и тонкими. Гнатосома нерасчлененная, длиннее своей ширины. Глаз нет. Коксо-стернальный скелет крайне оригинального строения: сходящиеся под почти прямым утлом эпимеры II слиты концами с длинным стернумом, который в той же'точке слит с вентрумом и эпимерами III и тянется, не прерываясь, почти до генитальной щели; таким образом намечается фигура 6-лучевой звезды, от заднего, более длинного луча которой отходят, в свою очередь, слитые с ним эпимеры IV, .и коксальные поля. I, III и IV оказываются замкнутыми; эпимериты II отсутствуют, в соответствии с чем коксальные поля II и стернальный щит, .в целом, снаружи, не очерчены (и, следовательно, не обособлены от окружающих их полей). Коксальные присоски мелкие, нормального строения; генитальные присоски очень крупные; присасывательный диск с обычными 4 парами присосок (2 + 4 + 2). Ноги короткие и толстые, лишенные крупных шипов. Лапки очень короткие, все усаживающие их щетинки необычайно длинные, особенно на ногах I и II; коготки ног I почти такой же длины, как вся нога, щетинковидные, пе отличающиеся по своему строению от длинных щетинок лапки; па йогах II они несколько короче и резче искривлены, на ногах III и IV еще немного короче, но и здесь имеют все же совершенно исключительную длину и почти щетипковидпы; обонятельная палочка лапок I и II длинная, булавовидно вздутая па конце; сенсилла е лапок I почти такой же длины, как апикальная щетинка голени I; присасывательные щетинки лапок. I и II необычайно длинные; парусных щетинок на лапках III и IV.нет.
Известен, только 1 вид.
1. Froriepia vimnrionsls Vtz. (фиг. 365—367).
Vitzthum, 1919, Zool. Anz., L: .1.90, fig. 1—4.
Гипопус. Окраска беловатая. Идиосома в 1.5 раза длиннее своей максимальной ширины, приходящейся иа уровень коксальных присосок III.
Zf.'FRORIEPIA 229
Проподосомальный щит в виде чрезвычайно узкой полоски» (длиною по средней линии около 5 р, т. е. почтив 30 раз короче гистеросомального щита), окаймляющей спереди гистеросомальный щит; ростральный выступ слегка намеченный, слабо дугообразно выступающий вперед. Гнатосома длиной в 11 р, целиком спрятанпая.под передним краем идиосомы; ее конец слегка расщеплен; аристы очень, длинные (40 р). Присоски присасывательного диска все почти одинакового, размера (центральные лишь чуть крупнее периферических), передние способны далеко вытягиваться, телескоповидные, как у Forcellima. Ноги.;1 и II выдаются за передний край идиосомы частью голени и. лапкой; лапки I бутылковидные, с очень длинным (40 р), -щетинковидным, слабо изогнутым коготком и совершенно исключительного размера (49 р) присасывательной щетинкой, расширенной на конце нано* добие поршня; .кроме того, на лапке I сидят 2 очень длинные волосовидные щетинки и 2 короткие, почти шиповидные щетинки; обонятельная палочка почти достигающая концом вершины лапки, своеобразно приостренная. Лапки II сильно укороченные, их коготки имеют в длину 24 р и заметно S-образно изогнуты; присасывательная щетинка также очень длинна (36 р) и листовидно расширена на конце; обонятельная палочка далеко выдается за вершину лапки. Лапки III и IV также очень укорочены; их коготки имеют около 2.0 р в длину; одна из вершинных щетинок лапок III листовидно расширена на конце, все остальные щетинки волосовидные; па лапках IV, вероятно, также одна из щетинок расширена на конце, но это. не могло быть установлено с достоверностью. Длина 145 р, ширина 98 р...
Распространение. Известен пока только из окрестностей г. Вей» мара (Германия). Единственный экземпляр гипопуса этой своеобразной формы был найден на крыле, с? муравья Lasius fuliginosus Latr.
i И. Сем. 8APR0GLVPHIDAE
’ SaproglypMdae Ouds. 1924 + Winterschmidtiidde Ouds. 1924 Ensliniellidae Vtz. 1925 +
Czenspinskiidae Ouds. 1927 + Pontoppidaniidqe Ouds. 1925 4- Nanacaridae Ouds. 1923..
Характеристика семейства. Габитуально весьма напо-Н минают представителей сем. Tyroglyphidae. Идиосома всегда явственно раз-И делена на проподосому и гистеросому; подразделения последней на метапо-В досому и опистосому никогда не наблюдается. Кожа тонкая и нежная,
I перепончатой консистенции, в большинстве случаев совершенно гладкая
К (бесструктурная), иногда с очень тонкой продольной штриховкой; проподо-I сомальный щит то отсутствует (Saproglyphus, Nanacarus'p то хорошо развит I (Ensliniella, Шетга, Winterschmidtia} и тогда имеет такую же форму, КПК у Tyroglyphidae', у рода'Ensliniella его боковые края глубоко вырезаны перед серединой, а задний украшен ячеистой скульптурой, отсутствующе И у всех других родов; опистосомалЬного щита нет; обычно нет и хитиннаяропапных ? участков вокруг элементов коксо-стернального скелета, и только у 9 IVin-erschmidtia кожа несколько хитинизирована позади эпимер. Щ'1'1111111'
230
11. SAPROGLYPH1DAE
органы имеются у большинства видов, но слабо развиты. Все щетинки идио-сомы.тонкие и гладкие, волосовидные, чрезвычайно гибкие; длина их очень изменчива: v е отсутствуют; v i занимают такое же положение, как у 'Tyroglyphidae. и столь же константны; sc i всегда имеются, одинаковой длины с sc е или в 2—12 раз короче их; в большинстве случаев они сидят значительно позади линии, проведенной через основания sc е; псевдостигмальный орган простой, щетинковидный, иногда слегка перистый; плечевые щетинки в числе 3 пар (h i, h е и silt расположенные таким же образом, как у Tyroglyphidae, или смещенные дальше кнаружи. Taic что h i сидят маргинально, h е—субвентрально, a sh — совсем вентрально; положение дорзальных щетинок (d-—d2) очень изменчиво; боковых щетинок, так же как у Tyroglyphidae. 2 пары (1 а и 1 р), но их положение гораздо более изменчиво; крестцовых 2 пары — sa е и sa i (например у родов Ensliniella, Calvolia) или 1 пара (sae); в последнем случае (у Saproglyphus, Winterschmidtia) sa i замещены 4-й парой спин ых щетинок (d4), которые тогда расположены терминально. Жировые железы не имеют наружных выводных отверстий; анальные поры отсутствуют. Генитальные щупальпы хорошо развиты у обоих полов. Коксо-стернальный скелет развит, в общем, слабее, чем у Tyroglyphidae; эпимеры 1 соединены в V- или Y-образный стернум; эпимеры II-IV в большинстве случаев тонкие, с очень слабо развитыми аподемами; у рода Saproglyphus эпимеры II S-образно искривлены и снабжены овальными аподемами, а у сГсГ эпимеры III и IV своеобразно изогнуты и слегка раздвоены на конце; у рода Ensliniella модифицированы эпимеры IV, конец которых образует изящный завиток; эпимериты II слабо развиты и никогда не образуют самостоятельных склеритов, участвующих в определении гра-цицы между проподо- и гистеросомой (как'это имеет место у тироглифид). Длина и пропорции ног изменчивы в такой же степени, как у этих последних; общее строение их одинаково у обоих семейств и не уклоняется от типа. Praetarsi всегда хорошо развиты, более или менее явственно стебельчатые (суживающиеся к основанию); коготки очень мелкие, сидящие на дистальном конце praetarsus, с основанием, погруженным в его толщу, лишенным сочленовных склеритов и, соответственно, не: связанным с вершиной самих лапок; у рода Nanacarus коготки имеются только у нимф, а у половозрелой стадии отсутствуют. Хэтотаксия ног, в общем, близка к таковой тироглифид, ио несколько обеднена; все щетинки ног гладкие и тонкие, никогда не бывают шиповидиыми; вершинная щетинка голеней часто сильно отодвинута назад от дистального края членика; внутренняя вентральная щетинка голеней I иногда отсутствует (у cl cl Saproglyphus и, невидимому, у Winlerschmidlia). Вооружение лапок I сострит из стандартной для всей серин в целом обонятельной, палочки (а), расположенной у ее основания мелкой сенсиллы р, базальной палочки о (иногда отсутствую-, щей), 3 медиальных щетинок, из которых 2 помещаются’ вентрально и 1 (е) латерально. апикальной группы из ^тонких щетинок и I—3 вентральных- шипов ца самом концо;Лапки; у -рода; Winterschmidtia эти щйпы очень.
ХАРАКТЕРИСТИКА СЕМЕЙСТВА
s;it
велики, yCalvolian, особенно, у Ensliniella они сильно редуцированы и нрак-тически незаметны. Вторичные половые различия обычно невелики как по -объему затрагиваемых ими признаков, так и качественно; резче всего они у самивв рода Nanacarus.
с?. Анальные и тарзальные копулятивные присоски обычно отсутствуют. Генитальное отверстие расположено между основаниями ног IV или позади них, в соответствии с чем смещены назад и окружающие его щетипки; копулятивный аппарат имеет сложное строение и состоит из нескольких самостоятельных частей; анальная щель находится ближе к заднему краю тела, чем к генитальному отверстию (от которого, следовательно, отделена более или менее широким пространством); 2 пары анальных микрохэт, из которых передняя иногда помещается впереди анальной щели, а иногда, как и задняя, 'по ее сторонам; только I пара постанальных щетинок, имеющих обычно значительную длину. Вторичные половые отличия J d выражаются нередко также и в строении ног; так, у рода Saproglyphus лапки I и II несколько утолщены,- слегка сжаты с боков и снабжены снизу продольным килем; их вершинные вентральные шипы атрофированы; у родов Winlerschmidlia и Calvolia средний вершинный шип лапок I заменен перепончатой присоской; у рода Nanacarus ноги III и IV несколько утолщены и искривлены и вершинный шип их лапок увеличен. Кроме того, эпимеры IV у сГс? обычно имеют несколько другую форму, чем у 99, и часто соединены одна с другой тонкой поперечной линией. Диморфизма у. dd никогда не наблюдается.
9 Генитальное отверстие помещается в общем пространстве между эпимерами III и I V, между основаниями ног IV или даже значительно позади последних (Nanacarus)-, по своему строению, оно почти идентично с таковым тироглифид, с той только разницей, что внутренние края его клапанов слабее хитинизированы и более постепенно (часто от самого основания) расходятся кзади, не образуя угла на том месте, где начипается их расхо-. ждение. У многих родов имеется хорошо развитый эпигипий. Анальная щель всегда сдвинута к заднему краю идиосомы и нередко выдается за пего-концами своих клапанов. Обособленного копулятивного отверстия, невидимому, нет. Анальных щетинок всего 1 или 2 пары; в последнем случае; одна из них помещается впереди анальной щели, а вторая — по ее сторонам; постанальных щетинок 1 пара;. . • •
Нимфальные стадии известны лишь у очень немногих видов. В большинстве случаев -они сильно напоминают взрослых, отличаясь от них стандартными для всего надсемейства возрастными признаками; у рода, Nanacarus, впрочем, наблюдается любопытное отклонение от этого правила, заключающееся в том, что у нимф praetarsi вооружены хорошо ран-витыми коготками, отсутствующими у взрослой формы.: Личинам с хорошо развитыми грудными палочками; у формы с укороченными ас i, hl, <lh d2 и la эти последние иногда появляются только у взрослых (Мшшщт, по Oudemans’y). иногда же вполне развиты уже у личинок ни у одного из представителей семейств не наблюдалось ощутимых норе-
232
11. SAPROGLYPHIDAE
мещений щетинок в течение онтогенеза, аналогичных имеющим место в отношении ve у ряда Tyrophagini, и многих щетинок у большинства Glycy-’ phagidae. ,
Покоящихся гипопальных стадий пе образует, насколько известно, ни один вид. Расселительные гипопусы относятся к той же экологической группе — энтомохоров, что и гипопусы тироглифид, но признаки «типа сопротивления» и присасывательные приспособления развиты слабее, чем у них. Гнатосома сильно редуцирована; у некоторых родов она полностью-слита с идиосомой и представлена только парой арист (Ensliniella, Vidia), у других же (Calvolia) имеет вид прямоугольной пластинки, неподвижно-сращенной своим основанием с идиосомой и несущей дистально пару более или менее обособленных от псе апикальных члеников, на конце которых, сидят аристы. Идиосома обычно более вытянута в длину и сильнее суживается к параболически закругленному заднему концу, чем у Tyroglyphidae, спина менее выпукла, края слабо уплощены или совершенно закруглены и никогда не образуют периферического присасывательного кольца; про-подосомальный щит явственно отграничен от гистеросомы, но неподвижно-сращен с ней и целиком открыт. Спинные щиты покрывают всю или почти всю дорзальную поверхность идиосомы, но не загибаются на вентральную-сторону, не образуя, следовательно, эпиплевр; только у рода Calvolia • передние углы гистеросомального щита слегка загнуты на брюшную поверхность в виде коротких, быстро выклинивающихся кзади эпиплевр;. с другой стороны, у родов Ensliniella и Vidia задний спинной щит обнажает боковые края гистеросомы, одетые в таком случае мягкой, несколько склад-, чатой кожей; у большинства видов поверхность обоих щитов покрыта системой тонких вдавленных линий, более или менее анастомозирующих друг с другом и образующих сложный сетчатый орнамент, причем составляющие его угловатые, более или менее удлиненные ячеи обычно имеют на. проподосоме поперечное (в центре щита) И диагональное (по его периферию-направление, а на гистеросоме преимущественно продольное; у некоторых форм эти вдавленные линии образуют очень тесную продольную штриховку;, другие типы скульптуры (например ямчатая) в пределах семейства не встречаются, но нередки случаи, когда поверхность щитов вообще лишена-всякого орнамента и кажется (по крайней мере, на препаратах) совершенногладкой. Все щетинки спинной поверхности представлены крайне мелкими и топкими, трудно заметными микрохэтами; у рода Calvolia хорошо развит псевдостигмальлый орган, помещающийся в особом углублении (как у Оп-batei). У подавляющего большинства форм имеются хорошо развитые Парные глаза, помещающиеся на вершине всегда явственно намеченного-рострального выступа; они состоят из полушаровидной, кристально-про-зрачной линзы и находящейся под нею сильно пигментированной хитиновой ретины; у подрода Crabrovidia (род Vidia) глаза плохо развиты и представлены парой малорельефлых прозрачных выпуклостей, расположенных, дорзально у основания рострального выступа, и лишенных ретин; у под-'
ХАРАКТЕРИСТИКА СЕМЕЙСТВА й«!|
рода Vidia s. str. глаза совершенно отсутствуют. Стернальный щит болей или менее ясно очерчен, но довольно слабо хйтинизирован, как и вся вообще брюшная поверхность тела; строение стернума, эпимер и эпимеритов I Г, как у тироглифид; вентральный щит хорошо развит, слитый с генитальным в нерасчлененное целое; коксальные поля III всегда замкнуты и смыкаются по средней линии тела; эпимеры III слиты в круто выгнутую кпереди дугу, соединенную посередине с общим стволом эпимер IV, имеющих обычно форму перевернутого Y; коксальные поля IV не разделены по средней линии тела или разграничены более или менее развитым вентрумом. Коксальные и генитальные волоски более или менее сильно редуцированы, но никогда не превращены в присоски; присасывательный диск небольшой,, с 4 парами присосок; перепончатая рамка вокруг диска иногда свободная; в виде более или менее широкой, часто радиально штрихованной каймы. Генитальные шупальцы с конической дистальной частью, вытянутой в длинное тонкое острие..
Ноги I—III с хорошо развитыми амбулакрами, состоящими из стебельчатого, перепончатого praetarsus, достигающего часто большой длины, и очень маленького коготка, сидящего па конце praetarsus; лапки IV без амбулакр, с крупными (парусными) щетинками. Ноги I и II сродной длины или длинные, со стройными лапками; вооружение их проксимальных члеников отличается от такового у гипопусов Tyroglyphidae только тем, что составляющие его щетинки никогда не бывают утолщенными в виде шипов и что дорзально щетинка голеней обычно расположена посередине членика или даже в его проксимальной половине; вооружение лапок очень-изменчиво, но обычно состоит из меньшего числа органов чувств, чему Туго-glyphidae-, на лапках I и II имеются по 2—4 листовидно расширенных щетинки; Присасывательная щетийка отсутствует. Лапки III часто с 3—4-листовидными щетинками. Лапки IV иногда слиты с голенью в иерасчле* ненный tibiotarsus, вооружены 1—2 крупными парусными щетинками' и 2 более мелкими, без амбулакр; у рода Ensliniella они вооружены своеобразными пальцевидными и чешуевидными шипами. Размеры гипопусов-довольно однообразны : 176—270 [л.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (8). Sc 1 много короче sc е.
2 (3). Praetarsi лишены- коготков;, вершина лапок вентрально с 1 относительно крупным когтевидным, сильно отогнутым шипом; ноги: очень короткие (18—22% длины идиосомы), обе задние пары у сГ заметно утолщены; генитальное отверстие у обоих полов помещается; позади основания ног IV, непосредственно впереди анальной щели.. Очень мелкая форма, длиною не свыше 200 [л. . . . 6. Nanacarus Ouds..
3 (2). Praetarsi с явственными коготками; вершина лапок вентрально с 1—3- маленькими шипиками (иногда трудно заметными); ноги средней1
:234
П. SAPROGLYPHIDAE
длины или длинные (не короче 30—35% длины идиосомы), задние ноги у с? не утолщены; генитальное отверстие у 9 расположено между основаниями ног III и IV, ус? — между основаниями ног IV. Более крупные формы.
4 (5). с? с крупными, округлыми анальными присосками и 1 парой тарзальных копулятивных присосок, расположенных у самого основания лапок IV (по 1 на каждой); sc i, h i, dx, d2 и 1 а представлены микрохэтами (длиною около 10—12% длины идиосомы); sc е, h е, d3, d4 и 1р — волосовидными щетинками средней длины (30—50% длины идиосомы). Литоральная форма, живущая среди водорослей . . . ........................................... 1. Pontoppidaniii Ouds.
5 (4). d без анальных и тарзальных копулятивных присосок, с 2 парами анальных микрохэт; соотношение размеров щетинок иное. Сухопутные формы.
6 (7). На заднем конце тела 3 пары очень крупных щетинок — d4, 1 р> и sa е (длиною в 60—90% длины идиосомы), сидящих на конических бородавках; sc i в 8—10 раз короче sc е; эпимеры II резко искривленные, с очень широкими аподемами; ноги короче и массивнее; лапки I и II у <5 без присосок .....................2. Saproglyphus Berl.
7 (6). На заднем конце тела только 1 пара длинных щетинок — sa i (дли-, . ною в 35—55% длины идиосомы); sc i в 2.5—6 раз короче sc е; эпимеры II слабые, почти прямые, с узкими аподемами; ноги длинные и стройные; лапки I и II у d с крупной вентральной присоской . . . ............................................... 4. Calvolia Ouds.
8 (1). Обе цары sc приблизительно одинаковой длины.
9 (10)'. Ноги I и II очень массивные, много длиннее ног III и IV; лапки , очень короткие и толстые, их вершина вентрально с 3 крупными, .. изогнутыми шипами; sc значительно длиннее всех щетинок гистеросомы; хелицеры нормального строения; генитальное отверстие 9 расположено между основаниями ног IV, без эпигиния . .. . . . . ................. . ........... .... 3. Winterschmidtia Ouds.
10 (9). Все ноги стройные, I и II заметно короче ног III и IV; лапки длинные, тонкие, на конце без шипов; sc несколько короче большинства щетинок гистеросомы; хелицеры из широкого основания внезапно суживаются в тонкое остриё; генитальное отверстие 9 расположено в пространстве между основаниями ног III и IV, с. хорошо развитым эпигмпиом....................................... 5. Ensliniella Vtz.
Гипопусы
1 (2). Ноги IV сильно сдвинуты к заднему концу тела, очень толстые, _с чрезвычайно мощными вертлугами; их лапки на конце с крупным пальцевидным шипом и вторым, еще ббльшим, чешуевидным шипом, оттянутым на конце в тонкую ость, 2 крупными и 1 очень маленькой щетинками; гистеросома сзади с продольной: линейной аподемой;
1. PONTOPPIDANIA
235
ростральный выступ спереди с парой очень явственных, лолушаро-вндно выступающих глаз. На осах рода Odynerus . .5. Enslinic 1 In Vtz.
2 (1). Ноги IV расположены на большом расстоянии от заднего конца тела; вертлуги маленькие, лапки только с простыми волосовидными щетипками, без шипов; продольная аподема отсутствует. На различных других насекомых.
•3 (4). На вершине рострального выступа находится пара полушаровид-ных, сильно выступающих глаз, снабженных явственными ретинами; гнатосома хорошо развита............................. 4. Calvolia Ouds.
4 (3). Глаза отсутствуют или плохо дифференцированы, лишены ретин и расположены дорзально, у основания рострального выступа; гнатосома совершенно атрофирована и представлена только парой арист. . . . .......................................................7. Vidia Ouds.
1. Род PONTOPPIDANIA OUDS.
Oudemans, 1925, Entomol. Bor., VI: 302.
Тип рода: Tyroglyphus littoralis Halbert 1920.
Легко отличается от всех других родов наличием у ,d J тарзальных и анальных копулятивных присосок. Хелицеры нормального строения. Идиосома довольно широкая, у 9 почти грушевидная, тучная; проподосомаль-ный щит неявственный. Sc i много короче sc е; h е, d3, d4, 1 р и sa е приблизительно такой же длины, как sc е (30—40% длины идиосомы), тогда как h i, d4—d2 и 1 а даже уступают в размерах sc i (10—12% длины идиосомы); расположение щетинок больше напоминает таковое Tyroglyphidae, чем других родов Saproglyphidae. Строение коксо-стернального скелета не представляет каких-либо особенностей. Ноги довольно длинные и стройные; вентральные щетинки голеней умеренной длины; лапки стройные, на лапках I и II 8 и е волосовидные, такого же размера и строения,, как обе вентральные; вершинные щетинки лапок короткие; praetarsi довольно большие, коготок относительно крупный.
cf. Лапки I не модифицированы; ноги Щ и IV не утолщены. Лапки IV у основания с 1 копулятивной присоской каждая; генитальное отверстие .расположено позади основания ног IV; задняя пара анальных щетинок превращена в крупные круглые копулятивные присоски; имеются 3 пары коротких постанальных щетинок. .
9. Строение наружного полового аппарата не описано. — Неполовозрелые стадии неизвестны.
К этому роду относится только 1 вид.
1. Pontoppidania littoralis (Halb.) (фиг. 376—379).
Halbert, 1920, Proc. Irish Acad., XXXV, sect. B, n° 7: 136, t.XXII. fig, 20(Tyroglyphus)-, Oudemans,. 1925, Entomol. Вег., VI: 302 (Pontoppidania). . .
d. Много меньше самки, длиной около 360 р.; шириной 200 [л, наибольшая ширина приходится поперек передней части брюшка (гистеросомы )• зад
236
II. SAPROGLUPHIDAE
ний край явственно вырезан посередине; непосредственно над выемкой: находится маленький бугорок. Все волоски спинной поверхности, как у 9, но относительно много длиннее. Выделительные пузырьки (жировые железы) - очень’ крупные: Генитальные клапаны образуют полукруглый щит; имеются 2копулятивные присоски, очень напоминающие те же образования у Histiogaster entomophagus Lab. Ноги крепкие, характерного1 для рода строения, за исключенном последней пары; лапки последних, вместо того чтобы иметь 2 маленькие выпуклые присоски близ середины членика, имеют только 1 присоску, расположенную близко к основанию на верхней и внутренней поверхности.
9. Длина около G16 |л, ширина 418 р.. Животное, в целом, грушевидное. Покровы гладкие, прозрачные, выделительные пузырьки (жировые железы) представляются в виде крупных коричневых пятен. Головогрудь обычной формы, заметно уже брюшка, сильно суженная спереди, так что ее ростральная часть довольно длинна и узка, почти как у Calvolia heterосота (Mich.). Головогрудные волоски расположены в ряд; оба наружные очень длинные, внутренние очень короткие (длиной около 66 р.), ростральные волоски немного заходят за конец мандибул. Брюшко с довольно сильно выдающимися плечевыми углами, несколько суженное позади них, затем постепенно расширяющееся к заднему краю, который: выдается посередине в виде приостренного выступа. Невидимому, этот выступ не гомологичен трубчатой bursa copulatrix, наблюдаемой у рода Glycyphagus. На спине имеется 5 пар длинных волосков, из которых 3 расположены маргинально, и 4 пары относительно коротких волосков, расположенных на переднем крае или вблизи него. Эпимеральные поля и генитальное отверстие, почти как у Tyroglyphus sir о; у заднего края вентральной поверхности имеются 2 длинных волоска и несколько пар коротких. Ноги нормальные; 4-ые членики (голени) несут обычный длинный волосок, и крепкий изогнутый шип (обонятельная палочка) отходит от 5-го. членика (лапки) обеих передних пар. (По Halbert’y.)
Распространение. Известен пока только из Ирландии. 299 и С найдены во влажных гниющих водорослях у порта близ How th.
2. Род SAPROGLYPHUS BERL.
Berleso, 1890, LVH, n° 6.
Тип рода: Saproglyphus neglectus Berl. 1890.
Гнатосома нормального строения. Идиосома сильно вытянута в длину; . проподосомальпый щит исявствеяиый. Sc i представлены микрохэтами, в 8—12 раз короче sc о; hi, d1—<1а и 1 а также представлены микрохэтами;
Фиг. 876—384.
Фнг. 376. Pontoppidania littoralis (Mnlb.), (? сверху (по Halbert’y). Фиг. 377. Р. littoralis (Halb.), rf снизу (по Halbert’y). Фиг. 37U. р. littoralis (Halb.), лапка I сверху (по Halbert’y). Фиг. 379. Р. littoralis (Halb.), лапка IV d1 сверху (по Halbert’y). Фиг. 380. Ensliniellaparasitica Vtz., сГ сверху (по Vitzthum’y). Фиг. 381. То же, снизу (по Vitzthum’y). Фиг. 382. Е. parasitica Vtz., 2 сверху (по Vitzthum’y). Фиг. 383. То же, снизу (по Vitzthum’y). Фиг. 384. Е. parasitica Vtz., йога I сбоку (по Vitzthum’y).
238
II. SAPR0GLYPH1DAE
d4, ip, sae и p а очень длинные (50—90% длины идиосомы), расходятся-в стороны и образуют .такой же пышный вееровидный шлейф, как у Туго-phagini-, все эти крупные щетинки сидят на конических бородавках, придающих заднему копну тела весьма характерные угловатые очертания; щетинки всех дорзальных пар удалены друг от друга на приблизительно-одинаковое расстояние, образуя, таким образом, 2 почти параллельных, продольных ряда; da несколько ближе к <1чем к d3; d4 расположены терминально (на самом краю идиосомы); 1 а более удалены от d2, чем от d3;. 1 р почти па одиом уровне с d3, расположенные на самом краю тела, так: же как d4 и sa е; последние находятся на одинаковом расстоянии от 1 р. и d4; sa i отсутствуют. Коксо-стернальный скелет развит сильнее, чем у других родов; стернум длинный, Y-образный; эпимеры -II резко искривлены,, с очень широкими аподемами; эпимериты II почти незаметны. Ноги средней: длины, умеренно тонкие, мало отличающиеся друг от друга по своим размерам; вентральные щетинки голеней I и II крайне мелкие; вентральные-шипы конца лапок средней мощности, вершинные дорзальные щетинки: лапок короткие; 3 имеется; е крайне мелкая, палочковидная; praetarsi широкие, мясистые, не суживающиеся к основанию; коготки относительно' крупные.
<3. Лапки I и II короче и толще, чем у $, вентрально с тонким продольным килем и редуцированными вершинными шипами; ноги III и IV не утолщены; внутренние концы эпимер IV вильчато раздвоены, причем задние-ветви этого развилка соединены друг с другом тонкой поперечной линией; генитальное отверстие расположено между основаниями ног IV.
Лапки I и II не модифицированы, вентрально без продольного-киля и с 3 хорошо развитыми вершинными шипами; концы эпимер IV не раздвоены; внутренние концы эпимер III соединены друг с другом топкой поперечной линией; генитальное отверстие расположено в пространстве-между основаниями ног III и IV; эпигиний хорошо развит. Из неполовозрелых стадий известны только личинки; они очень похожи на взрослых,, но крупные щетинки гистеросомы заметно короче; грудные палочки длинные, стройные, булавовидно утолщающиеся к концу.
К роду Saproglyphus относится 1 европейский вид (8. neglectus Berl)., и 2 внепалеарктических (S. australis Ouds. 1924 и S.neolropicus Ouds. 1924).
1. Saproglyphus neglectus Berl. (фиг. 41—44, 389—393).
Berlese, 1890, LVII, n° 6.
Беловатый, с легким зеленоватым оттенком. Идиосома узкая, валькова-тая, в 2.1—2.5 раза длиннее своей ширины, с прямыми, параллельными друг другу боковыми краями и угловатым (из-за выступающих конических бородавок) задним краем; плечевые выпуклости явственные. Псевдостиг-мальный орган щетинковидпый, гладкий. Sc i расположены почти на одной поперечной линии с sc е; примерные размеры щетинок идиосомы (в процентах к ее длине): v i 12; see 50; sc i, h i, d1? d2 и da 4—5; d4 около 100;
2. SAPROGLYPHUS. 3. WINTERSCHMIDTLA Ш
1 a 6—7; 1 p 66—60; sa e 80; p a 50—55. Ноги I и II у d несколько короче, чем у 9, их размеры в процентах к длине идиосомы таковы у 9 36—38 ([), 32—34 (II), у d 32—33 (I), 30—31 (II), задние приблизительно одинаковой,, длины [32—34 (III), 38—40 (IV)% длины идиосомы]; щетинки бедер, колен и лапок средней длины; вентральные щетинки голеней чрезвычайно мелкие; у <5 одна из них — наружная — отсутствует; обонятельная палочка лапок I и II довольно толстая, почти прямая, закругленная на конце.. Длина d 220—305 р., ширина до 130 у; длина 9 250—310 р., ширина до 135 pi. Личинки сравнительно очень велики (до 160 X 72 у).
Распространение. Италия (Vigodarzere, близ Падуи), СССР: Калининская (Великолукский) и Одесская обл. (Нерубаевский лесопром-хоз). Berlese нашел колонию этого вида на гниющем трутовике (Polyporus hispidus); П. К. Чернышеву и мне S. neglectus попадался под отмершей корой дубовых пней.
3. Род WINTERSCHMIDTIA OUDS.
. Oudomans, 1923, Entomol. Вег., VI: 206.
Тип рода: Suidasia(?') hamadryas Vtz. 1923.
Хелицеры нормальные. Идиосома удлиненная, вальковатая; проиодо-сомальный щит вполне явственный, совершенно гладкий, слегка вырезанный по бокам. Обе пары sc приблизительно одинакового размера, заметно длиннее всех щетино!£ гистеросомы; последние — все однотипные, довольно короткие (20—25% длины идиосомы); d4 расставлены очень широко, так что они оказываются в близком соседстве chi; d2, наоборот, сильно приближены друг к другу (расстояние между ними составляет меньше 1/4 расстояния между. dx); d3 — почти на одинаковом расстоянии от боковых краев и средней линии тела; d4 расположены терминально; 1а — далеко-позади уровня d2 (ближе к d3); 1 р и sa е — на одной поперечной линии; sa i отсутствуют. Коксо-стернальный скелет средней мощности; стернум Y-образный, эпимеры очень слабо дугообразные, с узкими аподемами; эпимериты II плохо развиты. Ноги I и II средней длины, очень массивные (копательного типа), задние (III—IV) много короче и тоньше; вентральные щетинки голеней очень мелки (наружная, невидимому, отсутствует), дорзальная щетинка голеней III и IV сильно редуцирована; лапки толстые, укороченные, на конце с 3 большими, резко искривленными вентральными шипами (особенно крупными на передних ногах); средняя вершинная щетинка лапок очень длинная; 8 отсутствует; е волосовидная, смещенная па вентральную поверхность лапки (имеющую, таким образом, 3 вентральные щетинки). Praetaisi среднего размера, подушковидные, коготки крупные.
d. На лапках I средний вентральный шип заменен крупной прозрачной присоской; ноги III и IV не утолщены. Генитальное отверстие номоЩНОТОН между основаниями ног IV.
9. Средний вентральный шип лапок I хорошо развит. Генитальное отверстие расположено между основаниями ног IV; пет oiiiirniiiiji; uiiitJiliiliui
240
II. SAPROGLYPHIDAE
щель доходит до заднего края идиосомы.—Гипопус, личинка и протонимфа неизвестны. — Телебнимфа отличается от взрослой $ только значительно более короткими щетинками идиосомы.
До настоящего времени известен только 1 вид.
1. Winterschmidtia hainadryas (Vtz.) (фиг. 385—388).
Vitzthum, 1923, Arch. Nftturg., LXXXTX, 2: 1.5G, fig. 66-69 (Suidasia)-, Oudemans, 1923, Entomol. Вег., VI: 206; 1927, ibid., VI1: 240 (Winlmehmidtia).
Окраска (на препарате) бледная, но с заметным охристо-желтым оттенком. Идиосома вальковатая, в 2 раза длиннее своей наибольшей ширины, приходящейся па уровень явственно намеченных плеч; кожа у 9 гладкая, у с? на спинной стороне идиосомы, а также позади ног IV вентрально очень тонко продольно штрихованная; у 9 позади эпимер кожа несколько хити-низировапа. Псевдостигмальный орган утолщенный, в своей дистальной •части явственно перистый; щечные отростки короткие, конические, направлены вперед; sc i расположены заметно позади sc е; их основания соединены тонкой поперечной линией. Примерные размеры щетинок идиосомы в процейтах к ее длине (у 9): v i 12; sc е 38; sc i 36; h i и d-, 23; d2 20; d3 i d4 23; 1 а и 1 p 21; sa e 17; p a 26. Длина ног (у 9) равна приблизительно .28 (I и II) и 16 (III и IV) % длины идиосомы; задние далеко не достигают донцами до заднего края тела; обонятельная палочка лапок I и II тонкая, -цилиндрическая, слегка дугообразно изогнутая; из 3 вершинных щетинок этих же лапок 1 очень длинная, а 2 короче амбулакр. Длина сГ?; длина 9-•300—350 [1, ширина до 170 |х.
Распространение. Германия. Живет в ходах короеда Eccopto-gaster rugulosus Ratz. [на терносливе (Prunus insititia L.)]
4. Род CAEVOLIA OUDS.
Oudemans, 1911, Entomol. Ber., Ill: 187. —Czenspinskia Oudemans, 1927, ibid., VII: 246.
Тип рода: Calvolia hagensis Ouds. 1911 (гипопус).
Хелицеры нормального строения, с сильно выпуклым наружным контуром. Идиосома умеренной длины или довольно узкая; проподосомаль-пый щит у большинства видов неявственный. Sc i в 2.5—6 раз короче sc е; he много длиннее hi; все остальные щетинки гистеросомы, кроме h ей очень длинных sa i, короткие или средней длины, нередко несколько жестковатые; ра длинные, волосовидные; dx расположены значительно ближе друг к другу, чем к h i; da также сближены; d8n d4 расставлены несколько
Фиг. 385—399.
Фиг. 385. Winterschmidtia hamadryas (Vtz.), Q сверху (no Vitzthum’у). Фиг. 386. То же, снизу .(по Vitzthum’у). Фиг. 387. V/. hamadrvas (Vtz.), голень и лапка I 9 снизу (по Vitzthum’у). Фиг. 388. W. hamadryas (Vtz.), колено, голень и лапка I 9 снизу (по Vitzthum’y). Фиг. 389. Saprод typhus neglectus Berl., лапка I 9 сверху. Фиг. 390. То же, снизу. Фиг. 391. S. neglectus Berl., лапка I cf сверху. Фиг. 392. То же, снизу. Фиг. 393. S. neglectus Berl., клешня хелицер О сбоку. Фиг. 394. Calvolia heterocoma (Mich.), р снизу (по М1сЬаёГю). Фиг. 395. Nanacarus minutus (Ouds,.'), $ сверху (по Oudemans’у). Фиг. 39(1. То же, снизу (uo Oudemans’у). Фиг. 397. N. minutus (Ouds.), задний конец идиосомы с? снизу (ио Oudemans’у). Фиг. 398. N. minutus (Ouds.), голень и лапка f 9 сбоку (по Oudemans’y). Фиг. 399. N. minutus (Ouds.), нога IV О’ снизу (по Oudemans’У V
f
242
II. SAPROGIA’PHIDAE
шире; d4 помещаются дорзально, далеко от заднего края идиосомы;. 1 р> ближе к d4 чем к d3; sa е сильно удалены от 1р; sa i сидят терминально или несколько вентрально. Коксо-стсрпальпый скелет довольно слабый; стер-нум Y-образный, эпимеры с очень узкими аподемами; эпимериты II и IV явственны. Ноги довольно длинные, стройные; вентральные щетинки голеней хорошо развиты, верши иные щетинки ланок относительно короткие; 5 палочковидная; е такого же размера и строения, как обе вентральные щетинки; praetarsi, довольно крупные, коготок маленький.
<d. Анальные и тарзальные лопулппшпью присоски отсутствуют. Генитальное отверстие расположено между основаниями пог IV; эпимеры последних соединены топкой хитиновой перемычкой с кольцевидным Образованием, входящим в состав слоящого наружного полового аппарата. Лапки I и II значительно короче и массивнее, чем у q ; их вершина вентрально с крупной прозрачной присоской, вокруг которой располагаются вентральные щетинки, смещенные, таким образом, к самому концу лапки;, иоги III и IV не модифицированы, вершинное вооружение их лапок такое же, как у 9" '
9. Генитальное отверстие помещается в пространстве между основаниями ног III и IV; эпигиний имеется; эпимеры III свободные, нормального строения. Лапки I и II длинные и стройные; е и обе вентральные щетинки расположены в их среднем поясе; вершинное вооружение состоит из 1 медиального шипика и 2 очень коротких боковых щетинок.
Гипопус. Вершина рострального выступа несет пару полушаровид-лых, резко выступающих глаз с хрустально-прозрачными линзами и хорошо оформленными хитиновыми ретинами. Гнатосома хорошо развитая, состоящая из массивной основной части, неподвижно сращенной с идиосомой, и 2 более или менее обособленных от нее дистальных члеников, несущих хорошо развитые аристы. Структура спинных щитов очень разнообразна; гистеросомальный щит покрывает" всю спинную поверхность гистеросомы и спереди загибается на ее брюшную сторону, о всегда хорошо-развита и расположена более проксимально, чем обонятельная палочка; дистальное вооружение лапок I и II очень изменчиво. Ноги IV причленены к телу вентрально, на значительном расстоянии от заднего конца и боковых краев гистеросомы, так что за последние выдается только дистальная половина их лапок; вертлуги III и IV умеренных размеров, лапки без пальцевидных или каких-либо других шипов. Остальные признаки сильно изменчивы.
К роду Calvolia относятся 18 палеарктических видов, большинство которых (13) известно только по гипопусам. Последние довольноразнообразпы по своему строе, по, однако не настолько, чтобы нарушить естественность, создаваемой ими группы и вызвать необходимость ее дальнейшего расчленения. Я даже не счел возможным сохранить в виде самостоятельной группы род Csenspinskia Ouds. 1927 (тип рода Tyroglyphus heterocomus Mich. 1903)r так как гипопусы составляющих его видов не отличаются от таковых ти-
4. CALVOLIA
245
ничных Calvolia ни в одном существенном признаке (или характере их корреляции), а половозрелые формы последних до сих пор неизвестны.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
сс
1 (2). Задний край идиосомы прямоугольно закругленный, украшенный 6. более или менее полушаровидными бугорками; 1р длиною в 15— 17% длины идиосомы; эпимеры IV диагональные, образующие явственный тупой угол с хитиновой полоской, соединяющей их с генитальным отверстием; кожа в задней части гистеросомы штрихованная (особенно снизу ипо бокам); общая длина тела 323 (i*l.C.tuberculata,sp. п.
2 (1). Задний край идиосомы тупоугольно закруглен, без бугорков; 1 р длиною около 50% длины идиосомы; эпимеры IV поперечные, расположенные в том же направлении, как хитиновая соединительная линия; кожа совершенно гладкая; общая длина тела около 280 ...........................................* 2. С. heterocoma (Mich.).
99
1 (2). Кожа очень тонко поперечно штрихованная, особенно на брюшной стороне; одна из вершинных щетинок лапок I и II утолщена, палочковидна; средняя вентральная щетинка тех же лапок расположена ближе к основанию лапок, чем главная обонятельная палочка (а). Общая .. длина тела около 290 . ..............4. С. hebeclinii (Sieber).
2 (1). Кожа совершенно, гладкая или слегка штрихованная лишь в задней части гистеросомы; все вершинные щетинки лапок волосовидные; средняя вентральная щетинка лапок I и II расположена более дистально, чем главная обонятельная палочка. Общая длипа тела превышает 300 р-.
3 (4). Спинные щетинки довольно длинные (dx 14—15, da—d4 20—25% длины идиосомы), волосовидные, (очень тонкие и гибкие); 1а и 1р одинаковой длины с da—d8 и несколько короче d4; sc i в 2—2.5 раза . короче sc е. Более крупная форма (общая длина тела 450—480 ;л> '...................................... *5. С. kamenskii, sp. n.
4 (3). Спинные щетинки очень короткие (dx—d8 7—12, d4 9—14% длины идиосомы), немного жестковатые; Га и 1 р заметно длиннее всех d (вдвое длиннее dj.
5 (6). Задний край идиосомы с 2 мелкими полукруглыми бугорками; кожа в задней четверти гистеросомы слегка штрихованная; проподосомаль-ный щит явственный, е полукруглым задним контуром; sc i в 3.3— 3.6 раза короче sc е; 1 а заметно длиннее! р. *1. С. tubcrculata, нр. и.
(> (5). Задний край идиосомы без бугорков; кожа гладкая; соотношении размеров sc иные; I а не длиннее 1 р.
7 ^8). Sc i в 2—2.5 раза короче sc е; sa е заметно длинное боковых (I а и 1 р), размеры которых вдвое превышают таковые d1( <la и d8;
*
2Й
II. SAPBOGLYPHIDAE
h i около 2 раз короче h е; окраска живых особей желтоватаяили сцет-лооранжевая.........................* 2. С. heterocoma (Mich.).
8 (7). Sc i в 5 раз короче sc е; sa о заметно короче боковых; длина последних превышает таковую dx—d3 меньше, чем в 2 раза; окраска живых особей беловатая ........................*3. С. romanovae, sp. и.
Г и и опус ы
1 (2). Лапки J.V оканчиваются только 1 крупной (парусной) щетинкой
...........................................8. С. kneissli Krausse.
2. (1). Лапки IV оканчиваются 2 крупными (парусными) щетинками.
3 (4). Дистальные членики гнатосомы несут каждый по 2 длинных щетинки (аристы); в передних углах. гистеросомы находится по . овальному белому пятну, являющемуся, невидимому, глазом со слабо выпуклой .линзой; присасывательный диск с. очень, широким перепончатым г свободным краем . .......... . ./. .. ..... .18. С.,(?) zacheri Ouds.
4,(3). Дистальные членики гнатосомы несут каждый лишь по 1 длинной щетинке — аристе; в передних углах гистеросомы нет никаких образований, напоминающих глаза; присасывательный диск без свободной
перепончатой каймы.
5 (16). Вснтрум отсутствует; ноги IV чрезвычайно короткие, длиной в 10— < . 11% длины идиосомы; лапки IV обособлены от голеней, и тогда почти
полушаровидцы, или же слиты с ними в очень короткий, закруглен-
ный на конце tibibtarsns;. длина ног IV составляет не больше 18%
длины идиосомы, их лапки не длиннее бедер.
& (13). Спинные щиты покрыты явственной, густой продольной'Штриховкой или сетчатым рисунком с вытянутыми в продольном направлении ячеями; ретины обособлены друг от друга, дугообразные.
7 (8). Передние вентральные и внутренняя вершинная щетинки лапок - I и II чрезвычайно короткие, простые; вершинная щетинка голеней I й II очень длинная, далеко заходящая за конец амбулакр; аристы ' короткие, Доходящие кбнцами'только до основания голеней I; спинные щиты в густой продольной Штриховке ... 15. С. striata (Vtz.).
8 - (7). Передние вентральные и внутренняя вершинная щетинки-лапок I ; И II крупные, листовидно расширенные (ланцетовидные); вершинная щетинка голеней I и II короче, чуть заходящая за конец амбулакр.
9 (10). Идиосома правильной эллиптической формы, сзади очень широко (Полукругло) закругленная; спинные щиты с широким сетчатым рисунком; аристы короче, доходящие концами только до основания голеней I. На Ips acuminatus Eichh. .... * 17. С. elliptica, sp. n. 10’ (9). ИдиосоМа веретоповидная или' приблизительно яйцевидная, параболически суживающаяся кзадй; спинные щиты в густой продбль-
Фиг. 400—406. Гипопусы родов Ensliniella s Calvolia.
Фиг. 400. Ensliniella Kostylevi, Bp. tt.( сверху. Фиг. 401. To же, снизу. Фиг. 402. Е. parasitica
Vtz. снизу. Фиг. 403. Calvolia heteroepma (Mich.) сверху. Фиг. 404. С. elisabethae, sp. п., сверху.. Фиг. 405. То же, снизу. Фиг. 406. С. circumspectans Vtz. снизу (по Vitzthum’y).
246
IL SAPROGLYPHIDAE
ной штриховке, правильной на всей их поверхности или разреженной только вдоль средней линии тела и образующей здесь более или менее широкие, продолговатые ячеи; аристы длинные, доходящие до основания лапок I.
11 (12). Идиосома веретеповидпая, сильно и равномерно суживающаяся к приостренным переднему и заднему концам, в 1.7 раза длиннее своей ширины; породно-боковые края проподосомального щита равномерно слабо выпуклые ни всем своем протяжении, ростральный выступ необособлопиый; штриховка, спиппых щитов очень густая и правильная па всей их поверхности. Ila lilaslophagus. *14. С. fusiformis, sp.n.
12 (11). Идиосома приблизительно яйцевидная, в 1.45 раза длиннее своей ширины, кзади менее сильно суживающаяся; передне-боковые края проподосомы S-образные, образующие явственные прогибы позади глаз и обособляющие, таким образом, продолговатый ростральный выступ; штриховка спинных щитов разрежена вдоль средней линии (особенно на проподосомальном), образуя здесь широкую сеть с продолговатыми, замкнутыми ячеями. На Hylesinus. *16. C.nataliae, sp.n.
13 (6). Спинные щиты гладкие; ретины слиты передними концами в Х-об-разную фигуру.
14 (15). Praetarsi с очень длинным линейным стебельком; на лапках I— III по 4 крупных листовидных щетинки. На Ips stebbingi Stroh. . . ..........................................12. С. circumspectans Vtz.
15 (14). Praetarsi короткостебельчатые; на лапках I и II толькопо 2 ланцетовидных щетинки, па лапках III они совершенно отсуствуют . . ............................................. 13. С. ha gensis Ouds.
16 (5). Вентрум хорошо развитый; ноги IV менее укороченные, длиной в 15—20% длины идиосомы; их лапки слиты с голенями, образуя удлиненный, более или менее цилиндрический tibiotarsus, обычно слегка расширяющийся к своему усеченному концу, или же обособленные от голеней и тогда значительно длиннее своей ширины у основания; длина ног III составляет 30—33% длины идиосомы, редко меньше (у С. calliphorae nC.bulgaricay их лапки в 1.5—2 раза длиннее бедер.
17 (18). Ноги I и II очень длинные (55 и 45% длины идиосомы) и стройные; лапки I почти в 6 раз, голени I — в 2 раза длиннее своего диаметра у основания; идиосома сильно удлиненная, амфоровидная; вентральный щит узкий, в 1.7 раза длиннее своей ширины между точками прикрепления ног III, сзади с явственными краевыми утолщениями; обонятельная палочка лапок I и II короткая, отодвинутая от основания лапки на расстояние, превышающее ее длину. . * 9. С. thraca Vtz.
18 (17). Длина ног I не превышает 40, ног II — 36—38 % длины идиосомы; лапки I не более чем в 3.5 раза, голени I не-более чем в 1.7 раза длиннее своего диаметра у основания; идиосома другой формы, более широкая; вентральный щит шире (не более чем в 1.3 раза длин-
4. CAL VO LA
247
лее своей ширины), сзади не обрамленный, краевыми утолщениями; обонятельная палочка обычно длинная, придвинутая к основанию лапки.
19 (22). Длина проподосомального щита умещается в длине гистеросомального только 1.6 раза ноги III короткие, длиною в 16—22% длины идиосомы; лапки IV слиты с голенями в нерасчлененный tibiotarsus; щетинки голеней и лапок I и II и обонятельные палочки короткие; идиосома в 1.5—1.6 раза длиннее своей ширины.
20 (21). Передняя (наружная) ’ арусная щетинка ног IV сильно утолщенная, в 2 раза толще задней (внутренней); tibiotarsus IV вильчато раздвоен на конце; ретины слиты в округлое пигментированное пятно; задний конец тела правильно закруглен. *11. С. bulgarica Storkan.
21 (20). Передняя (наружная) парусная щетинка ног IV неутолщенная (тоньше задней); tibiotarsus IV на конце прямо усечен; ретины слиты в Х-образпую темную фигуру/ задний конец тела почти прямо усечен . .........................................10. С. calliphorae Vtz.
22 (19). Проподосомальный щит в 2 раза короче гистеросомального; длина ног III равняется 30—32% длины идиосомы; лапки IV обособлены от голеней; щетинки голеней и лапок I и II и обонятельные палочки длинные; идиосома широкая, в 1.25—1.4 раза длиннее своей ширины, сзади закругленная.
23 (24). Гйстеросомальный щит с явственным сетчатым рисунком; на центральной части щита часть 5- или 6-угольных ячей, составляющих этот орнамент, лишь немного длиннее своей ширины; лапки IV на конце прямо усеченные; обе парусные щетинки сидят терминально (на усеченной вершине лапки); вентральные щетинки бедер I и II чрезвычайно длинные и тонкие...................7. С. reticulata, sp.n.
24 (23). Орнамент на гистеросомальном щите обычно (на препаратах) неявственный, если же заметный, то все составляющие его ячеи значительно длиннее своей ширины; лапки IV со скошенным концом, так что одна из парусных щетинок (более короткая) сидит заметно ниже (более проксимально) другой; вентральные щетинки бедер I и II умеренной длины.
25 (26). Идиосома очень широкая, только в 1.25—1.3 раза длиннее своей ширины, сзади широко (полукругло) закругленная; эпимериты II крепкие, Хорошо заметные; расстояние между глазами равно поперечнику линз; praetarsi III с коротким стебельком и крупной, удлиненной пластинкой. Длина 170—180 |х. . . .*2. С. heterocoma (Mich.).
26 (25). Идиосома уже, в 1.4 раза длиннее своей ширины, с параболическим задним концом; эпимериты II совершенно неявственныо; расстояние между глазами в 2 раза больше поперечника линз; prnotni'Hi III с длинным линейным стебельком и короткой округлой пластинкой. Длина 200—220 [I.............................*6. С. oHsubethae, sp.n.
248
II. SAPEOGLYPHIDAE
* 1. Calvolia tuberculata, sp. и. (фиг. 407-^413).
Прижизненная окраска неизвестна. Кожа гладкая в передних 3/4 идиосомы, слегка штрихованная в ее задней четверти (особенно снизу и по бокам); у заднего края тела расположено несколько мелких полушаровид-ных бугорков. Проподосомальпый щит явственный, очень широкий, с полукруглым задним краем и 2 маленькими вырезками по бокам. Sc i в 3.3— 3.6 раза короче sc е; h е в 2—2.6 раза длиннее hi, длина которых едва превышает таковую d; sa о заметно короче I а и 1 р; последние превышают около 2 раз щетинки всех 4 пар <1, представленных короткими прямыми щетинками. Более крупные ив щетинок идиосомы сидят на небольших бугорках.
- d. Идиосома стройная, более чем в 2 раза длиннее своей максимальной; ширины, приходящейся па уровень основания ног III; ее боковые края слегка сходятся назад от этой точки до места прикрепления 1 р, затем конвергируют под прямым углом к выступающему в виде острия концу клапанов анальной щели; на заднем краю тела расположены 6 округлых бугорков, образующих группы по бокам анального выступа. L^p довольно короткие, не длиннее, чем у 9. Эпимеры IV имеют диагональное направление и образуют явственный тупой угол с тонкой хитиновой линией, соединяющей их с генитальным отверстием; передний край последнего расположен на одной поперечйой линии с задними краями ацетабул III, задний край — на одной линии с задними краями ацетабул IV. Длина 288—295 у, ширина 118—135 у; общая длина тела 323 у.
9. Задний край идиосомы с 2 мелкими бугорками. Все вершинные щетинки лапок волосовидные, неутолщенные; средняя вентральная ще-тийка лапок I и II расположена значительно более дистально, чем обонятельная палочка (а). Длина 386—400 у, ширина 164—185 у, общая длина, тела 440—450 у.
Гипопус и другие стадии развития неизвестны.
Распространение. Вид известен только из Караязского р-на-Грузинской ССР. Колония этого интересного вида была найдейа в початках кукурузы (в полевых условиях) в январе 193&г.; он находился здесь вместе-С Tyroglyphus farinae (L.), Tyrophagus tenuiclavus A. Z. и др.
.* 2. Calvolia heterocoma (Mich.) (фиг. 394, 403, 419, 420).
Michael, 1903:106, t. XXXIII (Tyroglyphus)-, Vitzthum, 1920, Arch. Naturg., LXXXVI.lOr 58 (Calvolia)-, Oudemans, 1927, Entomol. Вег., VII: 246 (Czenspinslcia).
/ Окраска живых особей свотложолтая или оранжевая,* 1 реже почти бесцветная; жировые желозы (точное — их содержимое) то красные, то ярко
1 «Окраска вариирует мояеду различными оттенками желтого и оранжевого цветов, невидимому, в связи с условиями обитания клеща и обычно употребляемой им пищей; те, которых я выводил в неволе, были окрашены много бледнее, чем находимые в природе, и те, которых я ловил, весной, были окрашены ярче находимых осенью. Животное иногда почти бесцветно, но обычно-его окраска светлооранжевая или желтая, более интенсивная, чем обычно у Tyroglyphi» (Michael,.
1. с.).
' 4. CALVOLIA 249
желтые. Кожа совершенно гладкая. Sc i около 2.3 раза короче не и и равны по длине hi; эти последние в 2 раза короче h е и почти вдвое дли в нов <1(, da и d3; sa е заметно длинее 1 а и 1 р, длина которых у Q приблизительно вдвое превышает таковую первых 3 пар d (d1—d3); все 4 пары (1 представлены короткими, слегка жестковатыми щетинками.. Приблизительные, размеры щетинок идиосомы (в процентах к ее длине) у 9 (по рисунку Michael ’.и): v i 12; sc е 36; sc i 15; h е 30; hi 15; dx и d2 7; d3 8; d4 9—10; 1 а и 1 p 15— 16; sa e 18—20; sa i 45—50; у <3 1 p очень длинные (размеры остальных щетинок неизвестны).
сГ. Идиосома около 1.7 раза длиннее своей максимальной ширины, приходящейся на ее заднюю четверть, далеко позади основания ног IV; ее боковые края слегка расходятся до точек прикрепления 1 р, затем сходятся под очень тупым углом к анальному выступу. L р очень длинные (равны приблизительно 7г длины идиосомы). Эпимеры IV имеют поперечное направление и составляют почти прямую линию с хитиновой полоской, соединяющей их с генитальным отверстием; последнее более смещено назад, чем у С. tuberculata; так что его передний край находится значительно позади (ниже) уровня задних краев ацетабул III, а задний па уровне конца бедер IV. Общая длина тела, в среднем, 280 у, ширина 130 у.
9. Все вершинные щетинки лапок волосовидные; средняя вентральная щетинка лапок I и II расположена много дистальнее, чем главпая обонятельная палочка.. Общая длина тела,' в среднем, 350 у, ширина 270 у.
Гипопус. Хорошо отличается от гипопусов других видов рода Calvolia своей очень широкой, полукругло закругленной сзади идиосомой, длина которой только в 1.25—1.3 раза превосходит ширину. Окраска бледножелтоватая. Поверхность спинных щитов покрыта неясным сетчатым орнаментом; составляющие его волнистые линии более или менее анастомозируют между собой, образуя удлиненные ячеи неправильной формы; все щ тинки спинной поверхности чрезвычайно короткие (заметные лишь под очень большим увеличением). Проподосомальный щит в 2.2—2.4 раза короче гистеросомы; передне-боковые края щита позади глаз чуть вогнутые, затем равномерно слабо выпуклые на всем протяжении, ростральный выступ мало выдающийся; sc намечают фигуру перевернутой трапеции; sc е находятся несколько позади и кнаружи от псевдостигмальных органов, sc i— далеко позади и кнутри от них, вблизи заднего края щита; расстояние между sc i равно промежутку, отделяющему каждую из них от sc е; sa i расположены заметно кнутри от р а. Пространство между глазами равно, диаметру линз; ретины сходятся кпереди под тупым углом и слиты друг с другом. Аристы умеренной длины, достигающие концами середины голеней I. Стернальный щит резко очерченный с боков крепкими, слегка дугообразными эпймеритами II, с очень слабо намеченным прямым задп и м кривы; вентральный щит широкий, с несколько трапециевидным передним краем, без краевых утолщений в задней части; эпимеры IV почти прямые, сходящиеся под прямым углом; их общий ствол спереди слипнется с эпимернмн
250
П. SAJPROGLYPHIDA.E
III; вентрум хорошо развитый, простирающийся почти до генитального отверстия. Присасывательный диск округлый; боковые присоски на одной линии с центральными, дополнительные несколько крупнее боковых; задний край хитиновой пластинки диска дугообразный; радиальной штриховки вокруг нее не заметно. Ноги I и II довольно длинные (38—40 и 35—37% длины идиосомы), средней толщины; лапки I в 3.5, голени I — в 1.5 раза длиннее своей ширины у основания; вентральные щетинки бедер и голеней довольно короткие; лапки слегка вздуты в основной половине, несколько суживаются к концу; обонятельная пилочка лапок I длиннее половины самой лапки, очень топкая, цилиндрическая, отстоящая па х/2 своей длины от основания лапки; вершинные и вентральные щетинки длинные, волосовидные; задняя вентральная щетинка близко придвинута к основанию лапки. Ноги Ш длиною в 28—30% длины идиосомы; их лапки в 1.5 раза длиннее бедер, вооруженные довольно длинными простыми щетинками; из которых 2. задние вентральные тоже придвинуты к основанию членика. Praetarsi I—III длинные, постепенно суживающиеся к основанию. Ноги IV относительно длинные (17% длины идиосомы); лапки явственно обособлены от голеней, в 2 раза длиннее своей ширины у основания, заметно расширяющиеся к своему косо срезанному концу; более короткая из двух парусных щетинок (задняя) сидит заметно ниже (более проксимально) другой. Длина 170—180 [х, ..ширина 130—140 |х.
Распространение. Англия, Голландия, СССР; у нас найден пока только на Дальнем Востоке.
Экология. Michael сбивал С. heterocoma с дубов и нашел ее в значительном количестве во мху, из которого было сделано летнее гнездо белки; он переносил добытых в этом гнезде особей на куски гриба (Boletus edulis L.), на которых они успешно развивались. Oudemans нашел одну самку под лишайниками на вязе (в январе), а другую на гелиотропе (Heliotropium corym-bosum R.et Pav.), в декабре. Имеющийся у меня гипопус был найден в шерсти белки (Sciurus vulgaris L.). Из всех этих данных можно сделать только тот вывод, что С. heterocomaMich.—преимущественно дендрофильная форма, ведущая отчасти открытый образ жизни и в какой-то степени биологически -связанная с белкой.
Гипопус с Дальнего Востока отличается от рис.- 4 и 5 таблицы Michael’n менее раздвинутыми глазами и наличием 4, а не 2 пар присосок на брюшном диске, но эти различия можно без особой натяжки отнести за счет несовершенства указанных рисунков, так как в тексте (стр. 102) Michael подчеркивает, что глаза расположены очень близко друг к другу и, следовательно, близко к средней линии (тела), а число присосок (8 + 2 более или менее развитых дополнительных) постоянно не только в роде Calvolia Duds., но и во всем сем. Saproglyphidae в целом.
Фиг, 407—413. Calvolia tuberculata, sp. n.
•Фиг. 407. С? сверху. Фиг. 408. То же, снизу. Фиг. 409. 9 снизу. Фиг. 410. Лапка I С? сверху. Фиг. 411. То же, снизу. Фиг. 412. Лапка I Q сверху. Фиг. 413. То же, снизу.
252
11. SAPROGLYPBIDAE
*3. Calvolia romanovae, sp, n. (фиг. 414, 416).
Q. Прижизненная окраска беловатая, с легким зеленоватым оттенком. Кожа совершенно гладкая. Sci в 4.5 — 5.5 раза короче see, длина которых составляет около 40—45% длины идиосомы; h е в 3 раза длиннее h i, длина которых равна или едва превышает таковую d2 и ds; 1 а и 1 р заметно длиннее всех пар d; sa е короче 1 а и 1 р; все 4 пары спинных (dx — d4), так же как и sc i, hi, sa е, la и 1 p, представлены короткими, несколько жестковатыми прямыми щетинками. Все верлпшлыо щетинки лапок волосовидные, неутолщенные; средняя нентрал иная щетинка лапок I и II расположена более дистально, чем главная обонятельная палочка. Длина 308—350 р, ширина 150—180 р; общая длина тела 355—380 р. — d и неполовозрелые стадии неизвестны.
Распространение. Известен пока только из Москвы. 299 этого вида были найдены в зерне (пшеница), хранившемся на складе, вместе с Tyroglyphus farinae (L.).
Этот вид очень близок к С. heterocoma, от которого отличается много более короткими sc i и sa е и некоторыми другими различиями в размерах щетинок идиосомы.
4. Calvolia heheclinii (Sicker) (фиг. 421).
Sicher, 1899, in: Canestrini, Prospetto d. Acarofauna Ital., VIII: 971, t. 99 (Glycyphagus). ‘ Судя по очень несовершенным рисункам и описанию автора, этот вид очень близок к предыдущему, имея, в частности, очень сходные размеры щетинок (sic о около 50%, sc i и все d 10—12% длины идиосомы); он отличается от С. romanovae и всех других видов рода Calvo На очень тонкой (но вполне явственной) штриховкой кожи, особенно заметной на брюшной поверхности тела, а также и тем, что одна из вершинных щетинок лапок I и II палочковидно утолщена, а на конце даже слегка булавовидно вздута, и что средняя вентральная щетинка тех же лапок расположена более проксимально, чем главная обонятельная палочка. Общая длина тела 9 около 290 р, ширина 145 р. d неизвестен.1
Яйцо овальное, вздутое у обоих полюсов, беловатого цвета, хорион крайне тонкий, иногда неправильно разорванный на брюшной стороне у развивающихся эмбрионов; размеры яиц 96 X 55. р. — Личинка с короткими, булавовидными грудными палочками. Длина 147 р, ширина 75 р. Гипопус неизвестен.
Распространение. Сицилия. Живет на Hebeclinium macrophyl-lum DC — тепличном растении, родом с Антильских островов, Гвианы, Бразилии, Леру и Новой Зеландии; держится обычно на нижней стороне
1 Sicher ошибочно принял за телеопимф, в чем легко убедиться из его рис, 1 d и 1 f.
Фиг, 414—418.
Фиг. 414. Calvolia romanovae, sp. n., Q снизу. Фиг. 415. To же, сверху. Фиг. 416. С. kamenskii, sp. n., 9 сверху (ногп не изображены).,Фиг. 417. С. kamenskii, sp. п., голень и лапка I. Фиг. 418.
С. kamenskii, sp. п.» колено, голень и лапка IV.
251
IT. SAPROGLYPHIDAE
листьев, реже на соцветиях, случайно па других частях растения; неполовозрелые стадии предпочитают подсыхающие листья. Медленная в своих движениях форма.
* б. Calvolia kamenskii, sp. и. (фиг. 416—418).
2. Легко отличается от. всех других известных видов рода Calvolia своими более крупными размерами и значительно более длинными, волосовидными спинными щетинками. Кожа совершенно гладкая. Sci в 2—2,3-раза короче sc е и приблизительно раины таковой hi; последние около 2.6 раза короче he, равны по длине dx и заметно (в 1.6 раза) короче d2 и d3; sa е явственно короче 1 а и 1 р, которые не превышают в размерах d2 и ds и несколько короче dx; все 4 пары спинных щетинок, а также sc i, h i, обе пары 1 и sa о длиннее, чем у других видов, очень тонки и гибки (волосовидны). Все вершинные щетинки лапок волосовидные, неутолщенные; средняя вентральная щетинка лапок I и II расположена более дистально, чем главная обонятельная палочка; г щетинковидная. Длина 380—440 |х, ширина 230—260 jx; общая длина тела 430—510 [х.
Распространение. Вид описывается по материалу из Башкирии (Стерлитамакский зерносовхоз). С. kamenskii была найдена в количестве-2 экз. ($$) в скоплениях растительных остатков на сетках комбайна.
* 6. Calvolia elisabethae, sp. п. (фиг. 404, 405, 443).
Известен только гипопус. Очепь похож па гипопуса С. heterocoma (Mich.), от которого отличается только следующими признаками: крупнее; идиосома заметно уже (отношение длины к ширине равняется 1.4 :1), скорее яйцевидная, с суживающимся, параболически очерченным задним концом; сетчатый орнамент спинных щитов составлен несколько более правильными, продолговато-шестиугольными ячеями; щетинки спинной поверхности длиннее, лучше заметные. Глаза широко раздвинутые. Стернальный щит обрисован сзади тупоугольной линией; эпимериты II очень слабые, представленные только тонкими прямыми линиями, очерчивающими стернальный щит с боков; задний край хитиновой пластинки присасывательного диска образует заметный выступ, отграниченный с боков пологими прогибами. Praetarsi I—III с длинным линейным стебельком и округлой дистальной пластинкой. Щетинки па погах значительно длиннее. Длина 200—220 [х, ширина 140—-156 [х.
Рае п р о о т р а и о и и о. Описывается по материалам из Старой Бухары. Найден па слепушонке (Jillobius talpinus Pall.). К этому же виду, вероятно, относится и плохо сохранившийся экземпляр с прометеевой мыши (Prometheomys shaposhnikovi Sat.) из Гудаура.
7. Calvolia reticulata, sp. n. (фиг. 428, 442).
Известен только гипопус. Очень похож на предыдущего, отличаясь от него только в следующем: поверхность спинных щитов с более густым, пра-
4. CALVOLIA
25»
пильным и четким сетчатым рисунком; на гистеросомалыюм щите многие-из 5- или 6-угольных ячей, составляющих этот орнамент, имеют приблизительно одинаковые ширину и длину; расстояние между глазами равно поперечнику линз; все щетинки ног I—III еще длиннее и тоньше. Praoinrui более короткие, почти не суживающиеся к основанию; лапки IV стройнее, почти цилиндрические, чуть заметно расширяющиеся к своей прямо усеченной вершине; обе парусные щетинки сидят терминально (на прямом дистальном крае лапки). Размеры: длина 235 р., ширина около 155 р.
Распространение. Швейцария (Sussillon в Альпах). Найден па пчеле Osmia (Anthocopa) dalmatina F. Mor.
8. Calvolia kneissli Krausse.
Krausse, 1919, Arch. Naturg., LXXXIII, 10:123, fig. 1—7
Гипопус. Мне в натуре неизвестен. Судя по несовершенным рисункам и очень поверхностному описанию Krausse, близок к С. reticulata, и другим видам типа С. heterocoma M;ch. по широкой форме тела, наличию ясного сетчатого рисуцка на спине и большой длине амбулакр, но отличается от всех других видов рода Calvolia тем, что лапки IV оканчиваются только 1 щетинкой (парусной), а не 2 более длинными и 2 короткими.
Распространение. Германия.
Экология. Найден в большом числе особей на короеде Ips laricis F.
* 9. Calvolia thraca Vtz. (фиг. 432, 447, 448).
Vitzthum, 1922, Zool. Jahrb., Syst., XLIV: 542, fig. T—U.
Гипопус. Легко отличается от других видов Своеобразной формой тела и очень длинными передними ногами. Окраска довольно интенсивная, буроватая; конец гнатосомы еще несколько темнее. Идиосома в 1.65—1.7 раза длиннее своей ширины, амфоровидная, резко суживающаяся к сильно, вытянутому, языковидно выступающему заднему концу; боковые края гистеросомы выпуклые до уровня основания ног IV, затем заметно вогнутые, чем и создается столь характерная для этого вида конфигурация тела. Поверхность спинных щитов покрыта продольной штриховкой, менее густой, чем у видов типа С. striata, и не столь явственной. Проподосомальный щит около 2.5 раза короче гистеросомы, с хорошо намеченными плечевыми выпуклостями и несколько волнистыми передне-боковыми краями, образующими явственные прогибы по обеим сторонам округлого рострального выступа; sc расположены слабо вогнутой кпереди дугой; расстояние между но I почти в 2 раза больше промежутка, отделяющего каждую из щетинок »то№ пары от see; ретины сливаются по средней линии в Х-образпую томную фигуру. Аристы очень длинные. Стернальный щит такого жо отроении, как у С. fusiformis, но менее вытянутый в длину. Эпимеры ill обринуlor почти правильный полукруг; их передний край отделен от нндного крип стернального щита заметным промежутком; эпимеры IV нормального ('трое
256
IL SAPROGLYPHIDAE
имя, вентрум длинный; боковые края коксальных полей IV в задней своей части оторочены узкими краевыми утолщениями. Присасывательный диск маленький; задний край ого хитиновой пластинки дугообразно выпуклый; вентральные присоски значительно крупнее боковых и задних, имеющих одинаковые размеры; передние очень мелкие; дополнительные крупнее боковых. .Ноги 1 и JI. очень длинные (55 и 45% длины идиосомы), стройные; лапки .1. и .11, в 2 раза длинное предыдущего членика (голени) и в 5.5 (лапки I) или 4 раза (ланки 11) длиннее своего диаметра у основания, в проксимальных а/в цилиндрические, затем ипозапно суживаются и дальше еще утончаются ic концу; обе вентральные щетинки голеней I и II и наружная щетинка колен длинные; обонятельная палочка очень короткая, прямая, несколько суживающаяся к концу, далеко отодвинутая от основания лапки; дорзальная вершинная щетинка не очень длинная, слегка расширенная на конце; наружная вершинная и обе передние вентральные щетинки короткие, простые, волосовидные; задняя вентральная и е длинные. Ноги III стройные, значительно длиннее, чем у других видов (около 30% длины идиосомы); их лапки длиннее голени и колена, взятых вместе, вооружены довольно длинными, простыми щетинками; листовидных щетинок па них также нет. Prae tarsi I—III очень длинные, с тонким, линейным стебельком и округлой или почти ромбической дистальной пластинкой. Длина ног IV составляет около 16% длины идиосомы; голень и лапка слиты в нерас-члененный,'цилиндрический tibiotarsus; резко скошенный па конце; наружная (передняя) паруспая щетинка много короче вершинной и прикреплена значительно более проксимально. Длина 185—200 р-, ширина 110—125 р.
Распространение. -Найден пока только- в Болгарии (окрестности Софии) и в СССР (Агрызский район Татарской АССР).
Экология. Экземпляр Vitzthum’a был обнаружен этим автором на ближе не определенной «маленькой металлически-зеленой мухе» \ экземпляр моей коллекции найден в навозе, в колхозном навозохранилище.
10. Calvolia calliphorae Vtz. (фиг. 422, 423).
Vitzthum, 1922, Zool. Jahrb., Syst., XLIV: 539, fig. P—Q.
Гипопус. Окраска светлоохристо-желтая; конец гнатосомы темно-коричневый, ноги бледножелтоватые. Идиосома в 1.6 раза длиннее своей максимальной ширины, с почти прямо усеченным задним краем и при-острешшм передним концом; боковые края гистеросомы очень слабо выпуклые, почти прямые. Поверхность спинных щитов гладкая. Проподосо-мальный щит только в 1.5 раза короче гистеросомы, с чуть намеченными плечевыми выпуклостями; ого передне-боковые края кпереди от этих выпуклостей почти строго прямолинейны, не образуют прогибов по сторонам рострального выступа; sc расположены сильно выгнутой вперед дугой, причем sc I находятся значительно впереди sc е, и расстояние между щетинками этой пары почти в 2 раза больше, чем между каждой из них и ве е; ретины слиты в темную Х-образную фигуру. Аристы короткие, еле захо
4. CALVOLIA . 257
дящие концами за основание голеней I. Стернальный щит шире своей длины, слабо очерченный с боков, с дугообразным задним краем; эпимеры Ш, образуют, как обычно, дугу; эпимеры IV сливаются по средней линии тела, -образуя фигуру перевернутого Y, но, в отличие от других видов, они не доходят на заметное расстояние до эпимер III (в связи с чем коксальные поля III остаются открытыми); вентрум короткий, но вполне явственный. Присасывательный диск вытянут в поперечном направлении; задний край его хитинизированной пластинки волнистый, с округлым срединным выступом, обособленным от дугообразных боковых краев легкими вырезками; боковые присоски почти такого же размера, как центральные, дополнительные присоски крупнее задних; задний край подушки присасывательного диска почти прямой, украшенный тонкой радиальной штриховкой, заметно выступающий за задний край гистеросомы. Ноги I и II средней длины (40 и 38% длины идиосомы), довольно массивные: лапки I и II около 1.6 раза длиннее голеней, тонкие, конусовидно расширяющиеся к основанию, вентрально с небольшим бугорком; вентральные щетинки голеней на рисунках Vitzthum’a не изображены, может быть, отсутствуют. Обонятельная палочка лапок I и II короткая, изогнутая, прикрепляющаяся вблизи основания лапки; дорзальная вершинная щетинка очень длинная, волосовидная; вентральные и наружная вершинная щетинки чрезвычайно короткие; пи одна из них не имеет листовидной формы. Ноги III умеренной длины (около 22 % длины идиосомы), стройные, с довольно длинной лапкой, значительно превосходящей по длине голень; дорзальная вершинная щетинка лапок III короткая, листовидных щетинок здесь также нет. Praetarsi ног I—III короткие, очень короткостебельчатые. Длина ног IV составляет около 16% Длины идиосомы; лапки слиты с голенью в нерасчлененный tibiotarsus, сильно расширенный к своему прямо усеченному копцу, несущему длинные парусные щетинки; из них наружная (передняя) лишь немного уступает в длине внутренней. Длина 208 pi, ширина 106 [х.
Распространение. Вид известен пока только из Германии (Веймар). Найден единственный раз Vitzthum’oM па синей мясной мухе (CalUphora vomitoria L.).
*1. Calvolia bulgarica Storkan (фиг. 424, 425).
Storkau, 1935, Vestn. ceskoslov. zool. spolecn., II: 92, 3 figg.
Гипопус. Близок к C. calliphosae Vtz.; легко отличается от всех других видов ретинами, слитыми в одно округлое оптическое пятно, и очень крупной, сильно утолщенной передней парусной щетинкой ног IV. Светло охристо-желтый. Идиосома в 1.5 раза длиннее своей ширины, приблизительно пятиугольная, с полукругло очерченным задним концом. Uoiuipx-иость спинных щитов тонкоморщинистая, что особенно заметно вдоль краев гистеросомы. Проподосомальпый щит в 1.5 раза короче гистеросомы, о почти равномерно выпуклыми на всем протяжении боковыми краями (па слегка приплюснутом препарате), параболически сходящимися кпереди; нс i соли-
17 Фауна СССР, Захваткин — 284
258
II. SAPROGT.YPHTDAE
жены и расположены па значительном расстоянии от заднего края щита; ретины слиты в округлое темно пигментированное поле. Аристы довольно. короткие. Стернальный щит явственно очерчен с боков слабо дугообразными эпиморптами П, без сколько-нибудь заметной задней границы; эпимеры Ш образуют'-'-’-образную дугу; эпимеры IV сливаются по средней липин тела в виде перевернутого Y; вептрум довольно длинный. Присасывательный диск, вытянут в поперечном направлении; задний край его хити-цизироваппоП H.ii.'ie.'i'iiiiitti и виде скобки (•—-); дополнительные присоски, одного размера, <1 центральными, периферические заметно мельче Ноги I и 11 довольно массивные п короткие (3(5 в 34% длины идиосомы); лапки почти в 2 разадлипнес.голеней, конусообразно расширяющиеся к основанию;, вентральные щетинки голеней короткие (на голенях I наружная щетинка отсутствует?). Обонятельная палочка лапок булавовидная, довольно короткая (особенно на лапках I), прикрепляющаяся вблизи основания лапки; наружная и внутренняя вентральные и наружная вершинная щетинки довольно длинные и крепкие, простые. Ноги III очень короткие (16% длины.9 идиосомы), с довольно длинной лапкой, значительно превосходящей подлине голень и бедро, без листовидных щетинок. Praetarsi I—III короткие, очень короткостебельчатые; коготки довольно крупные. Ноги IV длиною около 16% длины идиосомы; лапки слиты с голенью в нерасчлененный tibiotarsus, много превосходящий по длине бедро и сильно расширяющийся к своему вильчато расщепленному концу; обе парусные щетинки, сидящие-на его концах, очень длинны, причем передняя (наружная) много толще задней (у основания она в 2 раза толще последней). Длина 207 [л, ширина 140 |л.
Распространение. Описан из Болгарии: Юмрукчал в центральных Балканах. Недавно найден в овр. Смоленска на льняной., блошке (Aphtona euphorbiae) В. И. Волгиным.
12. Calvolia circumspectans Vtz. (фиг. 406).
Гипопус. Окраска охристо-желтая. Идиосома в 1.7 раза длйннее-своей максимальной ширины, обратно-яйцевидная, равномерно и довольно-сильно суживающаяся от заметно выступающих передних углов гистеросо-мального щита к параболически приостренному заднему концу и еще более резко — к переднему. Поверхность спинных щитов зеркально-гладкая. Проподосомальпый щит почти в 2 раза короче гистеросомального; sc образуют круто выгнутую вперед дугу; sc i отдалены друг от друга почти на. такое же расстояние, как каждая из них от sc е, ретины слиты друг с другом’ в д-образпую темную фигуру. Аристы длинные, достигающие основания лапок I. Стернальный щит шире своей длины, очень четко очерченный,, с дугообразно выпуклыми боковыми краями (эпимеритами II) и почти прямым, хитинизированным задним краем, с которым сливаются эпимеры III и намечающие фигуру перевернутого Y эпимеры IV; вентрум отсутствует. Присасывательный диск относительно крупный, сердцевидный; дополни
4. CAIFOZ/A
М!И>
тельные присоски слабо развитые. Ноги I и II умеренной длины (:к1 % длины идиосомы), довольно стройные; лапки почти в 2 раза длиппсо голнпоП, около 2.6 раза длиннее своего диаметра у основания, равномерно суживаю щиеся к концу Вентральные щетинки голеней на рисунках Vitzthnni’a иг изображены; обонятельная палочка тонкая, прямая, превосходящая 1/а длины ланки (на ногах I) или равная ей (па ногах II), расположенная у са-мого основания ланки; обе передние вентральные щетинки и обе вершинные— внутренняя и дорзальная — листовидно расширенные, длинные. Ноги III довольно короткие (18% длины идиосомы), не особенно толстые, с относительно длинной лапкой (почти равной по длине бедру), несущей дистально по 4 листовидных щетинки. Praetarsi I—III с длинным параллельно-крайним стебельком и овальной вершинной пластинкой. Ноги IV очень короткие (10% длины идиосомы), с полушаровидной лапкой, явственно обособленной от голени; вершинная парусная щетинка почти равна по длине идиосоме, внутренняя — на половину короче. Длина 175—181 р., ширина 105—111 |х.
Распространение. Известен только из Тибета. Гипопусы найдены на короеде Ips stebbingi Strohm.
13. Calvolia ha gensis Ouds.
Oudemans, 1911, Entomol. Ber., Ill: 187.
Гипопус. Мне в натуре неизвестен. Оригинальное описание Oude-mans’a в переводе таково: «Длина 185, ширина 100 р,. Протеросома длиннее, чем одна треть всего тела, 65 р., более или менее треугольная, совсем впереди снабженная 2 глазами (как у гипопуса Tyroglyphus heterocomus Michael). Ретины этих глаз каштановобурые. Ноги тонкие, но не настолько, как у Anoetus. Голень I с длинным, голень II с очень длинным осязательным волоском; лапка IV заканчивается 2 длинными щетинками («sleepharen»)». Недостаточность этого описания в известной мере восполняется Vitzthum’oM (1920), располагавшим, очевидно, неопубликованными рисунками самого Oudemans’a. По Vitzthum’y, С. ha gensis крайне близка к С. circumspectans, строение их точно совпадает почти во всех деталях, как и размеры. Существенное различие заключается в том, что у гипопуса С. hagensis амбулакры не так несоразмерно длинны и что они только на лапках I и II окружены листовидно расширенными волосками, которых только по 2, тогда как лапки III лишены этих образований. Лапки IV у голландского вида также менее богато вооружены, чем у центральноазиатского.
Распространение. Известен из Голландии (окрестности Гааги). Найден в гниющих листьях, в лесу.
* 14. Calvolia fusiformis, sp. п. (фиг. 429, 430, 445).
Гипопус. Окраска светлоохристо-желтая. Идиосома, in.iT.iiayтая в длину, веретеновидная, очень правильно и равномерно оужншнощаяся к одинаково приостренным переднему и заднему концам; со максимальная *
260
II. SAPROGLYPHIDkE
ширина (приходящаяся на середину ее длины) умещается в длине 1.8 раза: Поверхность спинных щитов покрыта тонкой густой продольной штриховкой; линии, составляющие эту штриховку, многократно ветвятся, но почти не образуют замкнутых ячей. ПрОподосомальный щит очень длинный, составляющий s/4 длины гистеросомалыюго. Sc выстроены в почти правильный поперечный ряд, причем промежуток между обеими sc i почти в 3 раза больше расстояния, отделяющего каждую из них от sc е; ретины дугообразные, обособленные друг от друга: прмсты очень длинные, достигающие концами середины лапок 1. Стернальный щит длиннее своей ширины, хорошо очерченный, со слабо выпуклыми боковыми краями, образующими с более резко выпуклым задним краем сильно закругленный, но все же довольно явственно намеченный угол; эпимеры III явственные. Присасывательный диск маленький, задний край его плотно хитинизированной пластинки образует явственный срединный выступ, обособленный от прямоугольных боковых углов дугообразными прогибами; дополнительные присоски мелкие, слабо развитые; радиальной штриховки вокруг присасывательного диска незаметно. Ноги I и II довольно длинные (41 и 37 % длины идиосомы) и массивные; лапки I и II приблизительно в 1.5 раза длиннее голеней, несколько суживающиеся к концу, начиная от середины. Наружная вентральная щетинка голеней I и II отсутствует; обонятельная палочка (лапок I и II) прямая, тонкая, расположена в ее проксимальной половине; дорзальная вершинная щетинка простая; обе передние вентральные и внутренняя вершинная щетинки листовидно расширены, узколанцетовидные. Ноги III йороткие (15% длины идиосомы), толстые, с короткой лапкой, приблизительно равной по длине голени. Praetarsi ног I—III длинные, постепенно , расширяющиеся от основания к дистальной трети и вновь равномерно суживающиеся к коготку. Ноги IV очень короткие (11% длины идиосомы), с почти полушаровидной лапкой, более или менее явственно обособленной от голени; вершинная парусная щетинка почти равна по длине всей идиосоме, передняя (наружная) короче ее половины. Длина 180 pi, ширина 105 [л.
Распространение. Описывается по 1 экз. из Клинского р-на Московской обл. Найден на короеде Blastophagus minor Ratz.
15. Calvolia striata (Vtz.) (фиг. 426, 427).
Vitzthum, 1.914, Zool. Anz., XLIV: 324, fig. (Vidia); 1922, Zool. Jahrb., Syst., XLIV: 539, fig. K-S (Calvolia).
Гипопус. Очень сходен с предыдущим. Идиосома менее веретеновидная, не так равномерно и менее сильно суживающаяся кзади; проиодо-
Фиг. 419—427.
Фиг. 419. Calvolia heterocoma (Mich.), 9 сверху (по Michael’to). Фиг. 420. То же, снизу (по М1-сЬаеГю). Фиг. 421. С. hebeclinii (Slchor), ланка I 2 (по Slcher’y). Фиг. 422. С. calliphorae Vtz., ципопус сверху (по Vitzthum’y). Фиг. 423. То же, снизу (по Vitzthum’y). Фиг. 424. С. bulgarica Stortan, гипопус сверху (по Stortiin’y). Фиг. 425. То же, снизу (по Storkan’y). Фиг. 426. С. striata (Vtz.), гипопус сверху (по Vitzthum’y). Фиг. 427. То же, снизу (по Vitzthum’y).
262
IL SAP.ROGLYPHIDAE
сомальный щит несколько короче, составляющий 3/8 всей длины идиосомы. Аристы много короче — их концы не заходят до основания голеней I. Задний край присасывательного диска полукруглый. Лапки I и II' длиннее и тоньше: длина лапок 1 превышает вдвое длину предыдущего чле- ' пика (голени); обонятельная палочка короче и, повидимому, массивнее; вершинная дорзальная щетинка, короче, не заходящая за конец коготка; вентральные и внутренняя норннтнпя щетинки чрезвычайно короткие, простые (пе листовидные). I l orn 111 еще короче, чем у С, fusiformis, с кольцевидными коленом и голенью; на. ногах IV .панки слиты с голенью в нерас-члененпое целое (tibiokirsus); парусные щетинки ланок IV короче —самая длинная из них по превышает а/4 длины идиосомы. Длина 168—182 щ ширина 91—101 |л.
Распространение. Описан из Западной Африки (Слоновый Берег) и впоследствии найден в Италии.
Экология. Гипопусы встречаются на мелких короедах Taphro-rhynchus sp. (в Италии) и Pityogenes lepidus Wichm. (в Африке). Весьма возможно, что Vitzthum ошибся, относя итальянские особи к своему африканскому виду.
* 16. Calvolia nataliae, sp. п. (фиг. 431, 446).
Гипопус. Крайне близок к С. fusiformis и к С. striata. Идиосома заметно шире (только в 1.45 раза длиннее своей ширины), скорее яйце- 1 видная; боковые края проподосомы образуют более длинный, чем у соседних видов, ростральный выступ (конфигурация проподосомы, в общем, напоминает таковую С. Лгаса)-, гистеросомальный щит с свободно выступающими передними углами, гораздо менее суживающийся кзади, чем у С. fusiformis. Штриховка спинных щитов менее правильная, сильно разреженная вдоль середины проподосомы, где она образует широкую сеть с вытянутыми в длину, неправильными ячеями; sc i более сближенные между собою, расстояние между ними только в 2 раза больше промежутка между каждой из них и sc е; аристы короче, доходящие концами только до основания голеней I. Присасывательный диск крупнее,, задний край его хитинизированной пластинки дугообразный. Вентральные щетинки бедер очень длинные; обонятельная палочка лапок I много короче, отстоящая от основания лапки на расстояние, превышающее ее длину. Praetarsi -уже, почти линейные, еле расширенные па конце. Длина 192 щ ширина 121 р..
Распространение. Описывается по материалу из окрестностей Москвы (с. Алексеевское.) Найден на короеде Hylesinus fraxini Pnz.
* 17. Calvolia elliptic», sp. 31. (фиг. 60, 61, 444).
Гипопус. Светлоохристо-желтый. Идиосома весьма правильной эллиптической формы, сзади очень широко (полукругло) закругленная, в 1.5 раза длиннее своей ширины. Поверхность спинных щитов с очень явственным сетчатым орнаментом, состоящим из широких, вытянутых
4. CALVOLIA
263
в продольном направлении ячей, частью открытых; проподосомальпый щит в 1.6—2 раза короче гистеросомы, с равномерно выпуклыми па. псом протяжении передне-боковыми краями и необособленным ростральным выступом; sc расположены сильно выгнутой кпереди дугой, расстояние между sc 1 в 2 раза больше, чем между каждой из них и sc е; ретины дугообразные, обособленные друг от друга. Аристы длинные, доходящие до основания лапок I. Стернальный щит короче своей ширины, очерченный сзади трапециевидной линией; эпимериты II неоформленные; вентральный щит довольно широкий, сзади очерченный явственными краевыми утолщениями; эпимеры IV в целом дугообразные, сходящиеся кпереди под очень острым углом и сливающиеся своим общим стволом с эпимерами III; веитрум отсутствует. Присасывательный диск относительно крупный, округлый; боковые присоски чуть позади центральных, дополнительные заметно крупнее боковых; хитинизированная пластинка диска состоит из 3 почти обособленных друг от друга бобовидных площадок; радиальной штриховки вокруг нее не заметно. Ноги I и II средней длины (34 и 32% длины идиосомы), стройные; лапки немного и довольно постепенно суживаются к вершине; вентральные щетинки бедер очень длинные; наружная вентральная щетинка голеней отсутствует; обонятельная палочка слегка дугообразная, слабо булавовидно утолщенная к концу, отстоящая от основания лапки на расстояние, не превышающее % ее длины; г слегка утолщенная на конце, 8 очень мелкая, придвинутая к а; обе передние вентральные и внутренняя вершинная щетинки ланцетовидно расширены, довольно крупные. Ноги III .длиною около 18% длины идиосомы; их лапки заметно короче бедер, несут по 4 ланцетовидных щетинки. Praetarsi I—III довольно длинные, постепенно расширяющиеся к своей овальной вершине. Ноги IV очень короткие (12% длины идиосомы), толстые; их лапки очень короткие, более или менее явственно обособленные от голеней, скошенные на конце, так что меньшая из двух парусных щетинок сидит заметно более проксимально, чем другая. Длина 188—190 у, ширина 118—120 |х.
Распространение. Известен пока только из окрестностей Москвы (Свиблово). Гипопусы найдены в числе 3 экз. на короеде Ips acumi-natus Eichh.(?).
18. Calvolia (?) zacheri Ouds.
Oudeman's, 1930, Entomol. Вег., VIII: 36.
Гипопус. Длина 190—200 р., ширина 125—135 jx. Эллиптической -формы; над тазиками I и II края очень мало вогнуты. Разделяющая линия (между проподосомой и гистеросомой) приблизительно на уровне % длины тела. Оба глаза — вблизи теменных щетинок. Последние расположены центрально и равны по длине бедру + колено I. Остальные волоски очень коротки и трудно заметны, кроме 2 на заднем крае, которые все же короче теменных волосков. Окраска желтовато-коричневая; передний край гистеросомы беловатый; расположенные приблизительно в середине длины жиро-
264
II. SAPRUGLYPHIDAE
вне железы темнокоричневые. В передних углах гистеросомы — овальное белое пятно, являющееся, повидимому, «глазом» со слабо выпуклой роговицей (линзой). С брюшной стороны нет ни присосок, ни волосков на тазиках (коксальных полях) Г и III. Никаких следов генитального отверстия и 4 внутренних генитальных щупальцев. Обе мелкие присоски, которые обычно находятся вблизи генитального отверстия, расположены здесь приблизительно посередине тазик,он IV. .Нет радиальной штриховки на очень широком перепончатом свободном крио присасывательного диска. Гнато-сома очевь короткая, нс видна сверху; каждое щупальце (дистальный членик, j’iia.TocoMi.i) с 2 длинными волосками. На всех ногах бедра и колени четко разделены, a practarsi 3-члеиистые. (По Oudcmans’y.)
Распространение. Описан из Германии (Берлин). Гипопусы найдены па заплесневевшем сыре.
5. Род ENSLINIELLA VTZ.
Vitzthum, 1925, Dtsch. Entomol. Ztschr., IV: 289
Тип рода: Ensliniella parasitica Vtz. 1926.
Хелицеры в основной части вздутые, кпереди внезапно суживающиеся в тонкое острие (представляющее собой необычайно утонченный неподвижный палец клешни). Идиосома умеренной длины; проподосомальный щит вполне явственный, с парой глубоких вырезок по бокам (перед серединой) и ячеистым орнаментом вдоль заднего края. Обе пары sc имеют одинаковую длину, несколько уступающую таковой большинства щетинок гистеросомы; последние все однотипные (длиною в 30—40% длины идиосомы), торчащие перпендикулярно к поверхности тела; dx сидят ближе друг к другу, чем: к h i; d2 расставлены значительно шире, чем dx; d3 очень широко раздвинуты (расположены гораздо ближе к боковым краям, чем к средней линии тела), и сильно смещены назад; так же широко раздвинуты и d4, расположенные' дорзально, на значительном расстоянии от заднего края тела; 1 а находятся приблизительно на одном уровне с d2; 1 р образуют с d3 поперечный ряд; обе пары 1 помещаются дорзально, на заметном расстоянии от краев идио-сомы; sa е ближе к d4, чем к 1 р; sai хорошо развиты и занимают терминальное положение (d) или же смещены на спинную сторону тела (9)* Коксо-стернальный скелет слабый; стернум V-образный, эпимеры II слегка дугообразные, с крайне узкими аподемами, III — сильно укорочены, IV — развиты сильнее, чем остальные, S-образно искривлены и крючковидно отогнуты концами кнаружи. Ноги средней длины, стройные; вентральные щетинки голеней длинные; лапки длинные, на конце без вентральных, шипов (или эти шипы настолько мелки, что практически незаметны); вершинные щетинки лапок короче самой лапки; 8 отсутствует, г волосовидная; praetarsi крупные, колокольчатые, с очень мелким коготком.
сГ. Лапки I и II пе модифицированы; ноги III и IV не утолщены; генитальное отверстие помещается между основаниями ног IV; анальная щель, отодвинута от заднего края тела на расстояние, превышающее ее длину..
5. ENSLINIELLA 2G5-
9. Генитальное отверстие расположено в пространстве между основаниями ног III и эпимерами IV, с хорошо развитым эпигинием; анальная щель несколько отодвинута от заднего края тела. —Нимфальные стадии неизвестны.
Гипопус. Тело удлиненно-яйцевидное, узкое. Спинные щиты с очепь неясным орнаментом, тонкопористые; проподосомальный щит в 2 раза короче гистеросомального; вершина рострального выступа занята парой сильно выпуклых, полушар обидных глаз, снабженных хорошо развитыми хитинизировапными ретинами; по краям гистеросомального щита остаются открытыми узкие участки мягкой, перепончатой кожи; от его заднего края тянется вперед линейная хитиновая аподема. Гнатосома совершенно редуцирована (представлена только аристами). Эпимериты II соединены с эпимерами III; вентральный щит сильно вытянут в’длину, сзади с очень рельефными краевыми утолщениями; вентрум хорошо развит. Присасывательный дйск расположен почти терминально, окруженный широкой прозрачной каймой, способной свободно расправляться и стягиваться; присоски также обладают способностью очень далеко выпячиваться; Ноги I и II длинные, стройные; лапки с 3 листовидными щетинками; е в виде длинной, толстой прямой щетинки; о отсутствует, но на ее месте иногда имеется округлая пора. Ноги III и IV очень короткие и толстые, с чрезвычайно мощными вертлугами; ноги IV сильно отодвинуты назад, к концу тела, и выдаются большей своею частью за его края; лапки III с 3 или 4 листовидными щетинками» иногда с крупным пальцевидным шипом; лапки IV крайне укорочены и вооружены 1 толстым пальцевидным шипом, 1 еще более крупным чешуевидным шипом, конец которого оттянут в тонкую ость, 2 крупными щетинками и 1 очень мелкой; самая крупная из этих щетинок (парусная) сидит на приподнятой овальной площадке.
К роду Ensliniella относятся 2 очень близких между собою вида, живущих в гнездах одиночных ос подрода Lionotus рода Odynerus (из этих видов один известен только по гипопусам).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
Гипопусы
1 (2). Лапки III значительно короче своих амбулакр, вооруженные таким же пальцевидным шипом, как лапки IV, и 4 листовидными щетинками; глаза почти соприкасающиеся; краевые утолщения вентрального щита дугообразные, сливающиеся с хитиновыми рамками, окружающими передние присоски; прозрачная кайма вокруг присасывательного диска очень широкая. На Odynerus (LionotusJ flwwtilu Sauss......................................*1. E. paniHltlBH Viz.
2 (1). Лапки III не уступают в длине своим амбулакрам, без пал ыщпидпого шипа и только с 3 листовидными щетинками; промежуток, между глазами почти равен диаметру линз; краевые утолщения нонтрпльного
266
II. SAPROGLYPHIDAE
щита угловатые, не соединенные с неполными хитиновыми рамками • передних присосок; прозрачная кайма вокруг присасывательного
диска узкая. На О dy nevus (Lionolus) rossii Lep................-
.........................................*2. E. kostylevi, sp. n.'
* 1. Ensliniella parasitica Viz. (фиг. 380—384, 402, 436—440).
Vitzthum, 1926, Disch. Entomol. 'Zincin'., 1V: 289.
Окраска чисто белая. 1’па.тосома., ноги и эпигиний (у 9) розоватые. Идиосома довольно тучная, в 1.7—1.8 раза длиннее своей максимальной ширины, приходящейся над основаниями ног IV; боковые края гистеросомы на всем протяжении сл"або выпуклые, плечевые углы слегка намечены; задний край проподосомального щита с поперечным рядом многоугольных ячей; sc i расположены немного позади sc е; sae значительно короче других щетинок гистеросомы (не считая стернальных, sh и анальных); обонятельная палочка лапок I и .II тонкая и длинная, дугообразно изогнутая, слегка утончающаяся к концу.
d. Щетинки идиосомы немного длиннее, чем у 9; Щ— d3 заметно короче плечевых и особенно длинных боковых; примерная длина щетинок (вычисленная по рис. 4 и б работы Vitzthum’a) в процентах к длине идиосомы: уi 18; sc е и sci 31; h е и hi 35; dx 29; d2 и d3 31; d4 35; 1 a 37; 1 p 40—41; sa e 16; sa i 31; p a 33. Bee 4 пары ног приблизительно одинаковой длины; 41 (I—II), 40 (III) и 43 (IV)% длины идиосомы. Длина 470 |л, ширина 260 pi. *
9. Щетинки идиосомы короче, размеры их более однообразны и, в процентах к ее длине, таковы: vi 14; see и sc i 27—29; he, hi, dls d2 и d4 .31; d3 25; 1 а и 1 p 29; sa e 19; sa i 26; p а около 30. Гетеротипия ног выражена резче — их длина составляет 35 (I), 34 (II), 37 (III) и 41 (IV) % длины идиосомы. Длина 610 р., ширина 8 0 р.
Гипопус. Глаза почти соприкасаются по средней линии тела. Ги-стеросомальный щит несколько уже, чем у следующего вида, сильнее суживающийся кзади; sc образуют почти прямой поперечный ряд; продольная хитиновая аподема простирается вперед дальше уровня d4. Аристы чрезвычайно короткие. Вентральный щит очень узкий, в 1.7—1.75 раза длиннее своей ширины между точками причленения ног III; вентрум короче; краевые утолщения задней части щита соединены в лирообразную фигуру и слиты с замкнутыми хитиновыми рамками передних присосок. Наружные присоски расположены заметно кпереди от центральных, дополнительные почти на одной линии с передними; задние присоски лежат в той же (вентральной) плоскости, как остальные. Длина ног I и II составляет, соответ-• ственно, 40 и 38% длины тола; вентральные щетинки бедер короче бедра и колена, взятых вместо'; па лапках по 3 довольно крупных ланцетовидных щетинки: 1 дорзальная и 2 вентральных, из которых наружная сдвинута на боковую поверхность лапки; 8 атрофирована, но на ее месте сохранилась округлая пора. Ноги III длиною около 18% длины тела; их лапки чрезвы-
5. ENSLINIELLA 267
чайпо короткие (много короче амбулакр III), вооруженные крупным iiihik-цевидным шипом и 4 ланцетовидными щетинками (2 дорзальными и у шчг тральными). Ноги IV очень короткие (13% длины тела);, трсуголыши Норн шинная площадка лапок широкая; пальцевидный шип почти цилиндрический, широко закругленный на конце; чешуевидный шип с короткой остью, в 1.5 раза длиннее пальцевидного; щетинка у его основания довольно длинная, тонкая; вершинная парусная щетинка почти в 3 раза длиннее ног IV и в 1.75 раза длиннее, чем субапикальная. Длина. 250—270 а, ширина 144—150 [X.
Распространение. Германия (долина Майна, окрестности Нго-ренберга), Армения (окрестности Эйлара), северо-западный Узбекистан (горы Ак-таш, близ Ташкента). Ареал хозяина охватывает, по Ю. А. Костылеву, Северную Африку, Западную Европу, горы Средней Европы, Крым, степи Сталинградской обл., восточное Закавказье, Среднюю Азию, южный Казахстан и Алтай.
Экология. Живет в гнездах осы Odynerus (Lionolus) floricdla Sauss. (0. delphinalis Gir.). Половозрелые Enshmella держатся на личинках этой осы (преимущественно на нижней поверхности первого грудного сегмента), питаясь, вероятно, их выделениями и не нанося им самим никакого заметного ущерба. Размножение клещей происходит перед самым окуклением хозяина; стадии личинки и протонимфы проводятся на куколке и к моменту вылета осы вся молодь Ensliniella успевает пройти стадию .протонифмы и превратиться в гипопусов, которые переползают на осу и прикрепляются к ней. Большинство гипопусов забирается в особое, характерное для Lionotus’o'B углубление под нависающим краем 1-го брюшного тер-гита, но некоторые иногда располагаются и на заднем скате алитрунка; на одной осе их часто усаживается до нескольких сот. Дальнейшая судьба гипопусов не прослежена, но весьма вероятно, что они сходят с осы в момент откладки ею яиц (с def Lionotus они, может быть, переходят на 99 при копуляции,'как это имеет место у триунгулипов жуков Meloidac и Slrcpsi-ptera) и таким образом заселяют новое гнездо, в котором протекает их дальнейшее развитие. Не выяснено также, имеет ли Ensliniella только одну годовую генерацию, подобно ее хозяину, или же в одном гнезде успевает развиться несколько поколений.
* 2. Ensliniella kostylevi, sp. п. (фиг. 400, 401, 441).
Известен только гипопус. Очень похож на гипопуса Е. parasitica. Глава более широко расставлены. Гистеросомальный щит немного шире и не так резко суживается кзади; продольная хитиновая аподема короткая. Аристы длинные, волосовидные. Стернум очень длинный, простирающийся пшшд почти до генитальной щели; краевые утолщения задней части щита имеют угловатые очертания, не соединены друг с другом по средней линии тела и не связаны с неполными хитиновыми рамками передних присосок, Пнруж лые присоски расположены почти на одной линии с центральными, депо,к-
268
II. SAPROGLWHDAE
нительные — значительно позади передних; задние ориентированы почти-прямо назад (при рассматривании сверху видны в профиль). Вентральные щетинки бедер I и II длиннее бедра и колена, взятых вместе. Сенсилла 8-совершенно отсутствует, а е немного тоньше. Ноги III несколько длиннее (21% длины идиосомы); ланки по столь укорочены и не уступают в длине амбулакрам; пальцевидны!! шин отсутствует, листовидных щетинок только по 3 (дорзальная и 2 вентральные). Ноги IV такого же размера, как у Е. parasitica: вершинная парус,пап л юти л км в 2 раза длиннее ног III и длиннее субапикалыюй. Длина 250—2Н0 [л, ширина 145—160 |л.
Распространение. Известен пока только из Белоруссии (Бело-веж, Vitzthum, 1925) и Крыма (Карадаг). Живет в гнездах Odynerus (Lionotus) rossii Lep. Ареал хозяина: Западная и Средняя Европа, Крым,. Закавказье.
6. Род NANACARUS OUDS.
Oudemans, 1903, Tijdschr., Entomol, XLVI: 8.—Froweinia Oudemans, 1903, Entomol. Ber. VI: 183 (нимфа); 1924, ibid.: 231.
Тип рода: Nanacarus minutus Cuds. 1903.
Крайне мелкие формы (длина 170—220 |i) Хелицеры нормального строения. Идиосома удлиненная, вальковатая; проподосомальный щит неявственный (отсутствует?). Sci в 4 раза короче sc е; все щетинки гистеросомы, кроме he и р а, очень короткие (6—10% длины идиосомы); d2 сближены между собой, d3 более раздвинуты, d4 ближе к краю идиосомы, чем к средней линии; sa i расставлены так же широко, как и d3. расположены терминально; 1 а сидят на уровне середины расстояния между d2 и d3; 1 р почти на одном уровне с d4: sae образуют с d4 прямой поперечный ряд; sa i очень короткие; р а много длиннее всех других щетинок идиосомы. Стернум V-образный; эпимеры слабые, слегка дугообразные, с совершенно незаметными аподемами; эпимериты II неявственны. Ноги очень короткие, но стройные;. IV пара их сильнее обычного смещена назад; дорзальные щетинки голеней I и II довольно короткие; на голенях III и IV они отсутствуют; средняя апикальная щетинка лапок I—III умеренной длины, на лапках IV — очень длинная; вершина ланок вентрально с 1 относительно крупным, сильно-отогнутым шипом. Praetarsi короткостебельчатые, лишенные коготка. Половой диморфизм резко выраженный.
сГ. Ноги III и IV заметно толще, чем у $, и невидимому, дугообразно-искривлены; вентральный вершинный шин их лапок особенно крупный, когтевидпый; генитальное отверстие помещается между основаниями ног IV, немного впереди anus.
9. Ноги III и IV пе утолщенные и не искривленные; вершинный шип -лапок не гипертрофирован; генитальное отверстие расположено позади эпимер IV, почти'у самого anus; эпигиний отсутствует.
Гипопус неизвестен. — Нимфы отличаются от половозрелых особей (помимо обычных возрастных признаков) наличием коготков на praetarsi и отсутствием sci, hi, d и 1. Монотипический род.
в. NANACARUS. 7. VIDIA
‘2(11)
*1. Nanacarus minutus (Ouds.) (фиг. 395—399).
Oudemans, 1902, Tiidschr. Ned. Dierk. Ver., (2), VII: 85, t. Ill, fig. 55—56 (Hypopus; нимфа); 1903, Tijdschr. Entomol., XLVI: 11, t. 2, fig. 23—28 (Nanacarus; J ,9).
Окраска беловатая или бледная. Идиосома вальковатая, около 2 раз длиннее своей ширины; боковые края гистеросомы почти прямые, параллельные друг другу, без плечевых выпуклостей; sc i расположены значительно позади sc е, короткие; примерные размеры щетинок идиосомы, в процентах к ее длине (у 9): vi 22; sc е 38; sc i 10; h e 17; d3.6: d3 8; d4 6; la 11; Ip 8; sai 5; sa e 6; p a 66—67. Длина пог (у 9) составляет приблизительно 20—22 (I и II) и 18—19% длины идиосомы; задние, несмотря на свою малую длину, заходят концами за задний край тела: обонятельная палочка лапок I и II короткая, сильно веретеновидно вздутая. Длина с? 172 fi, длина 9 220 |i, ширина 102 [i.
Распространение. Известен только из СССР (Смоленые), Германии и с о-ва Явы.
Экология. Одна нимфа была найдена на древесной пчеле Platyno-poda tenuiscapa Westw., другая — на летучей мыши Eptesicus serotinus Schreb. в окрестностях Времена; О' и 2 99 были обнаружены Oudemans’oM в пробирке с паразитами, собранными там же с землеройки Sorex araneus L. В. И. Волгиным N. minutus найден в зерне, в амбаре.
7. Род VIDIA OUDS.
Oudemans, 1905, Entomol. Вег., II: 21.
Тип рода: Vidia undulata Ouds. 1906.
Известны только гипопусы. Глаза совершенно отсутствуют или представлены парой слабо намеченных выпуклостей, расположенных дорзально у основания рострального выступа и лишенных хитиновых ретин. Гиато-сома совершенно редуцированная, представленная только парой щетинко-видных арист. Спинные щиты с явственным орнаментом, имеющим па про-подосоме поперечное и отчасти косое, а на гистеросоме — продольное направление; линии, составляющие этот орнамент, частично анастомозируют друг с другом, образуя отдельные узкие ячеи; иногда некоторые из этих ячей чепгуевидно приподняты. Гистеросомальный щит покрывает всю или почти всю спинную поверхность гистеросомы (по его сторонам иногда остается узкая каемка мягкой перепончатой кожи), но не загибается на ее брюшную сторону. Лапки I и II с 2—3 листовидно расширенными (ланцетовидными или серповидными) щетинками; о отсутствует; е более или менее утолщенная, часто палочковидная. Ноги IV причленены к телу нп значительном расстоянии от его заднего и боковых краев, за которые они выдаются только дистальной частью лапок; вертлуги III и IV небольшого размера. Лапки без пальцевидных или чешуевидных .шипов.
Этот род состоит из 4 палеарктических видов, биологически ciijiiuiи и ы х с обитающими в дереве перепончатокрылыми, и 1 ориентального (И, mata Ouds. 1909). найденного на мыши Rattus jordani Bly th ((Ннфннг, .Пни).
270
II. SAPROGLYPHIDAE
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦ А ВИДОВ
1 (4). Лапки I и II каждая с 3 ланцетовидными щетинками: 1 дорзальной и 2 вентральными; praetarsi 1 л II маленькие, нормального строения. На пчелах. (Подрод Vidia н. str.).
2 (3). Sai длиннее р а. На Trosopis conformis Forst..................
......................................... 4.. V. undulata Ouds.
3 (2). Sai короче pa, Па различных видах Megachile . .
.................................... . . . *3. V. lineata Ouds,.
4 (1). Лапки I и I I каждая с 3 серповидными щетинками, расположенными литералы io; practarsi I и II длинные, расширенные па конце в крупную блюдцевидную присоску. На осах рода Crabro. (Подрод Crabro-vidia, subg. n.).
б (6). Тело грушевидное, с резко суженным, языковидно выступающим задним концом; наружная серповидная щетинка лапок Пи II расположена значительно ближе к основанию дорзальной вершинной щетинки, чем к внутренней (вершинной) серповидной щетинке; обонятельная палочка цилиндрическая; tibiotarsus IV стройный, длиннее бедра IV....................................*1. V. gussakovskii, sp. n.
6,(5). Тело яйцевидное, постепенно суживающееся к заднему концу; наружная серповидная щетинка лапок I и II расположена значительно дальше от основания дорзальной вершинной щетинки, чем от внутренней (вершинной) серповидной; обонятельная палочка булавовидно вздутая на конце; tibiotarsus IV массивный, много короче бедра IV.
............. . . ........................., *2. V. popovi, sp. и.
' 1. Подрод Crabrovidia, subg. n.
Лапки I и II очень длинные и стройные, с 2 серповидными щетинками,, расположенными латеральном наружной (представляющей собой наружную из парных вентральных щетинок, смещенную на боковую поверхность-лапки) и 1 внутренней (внутренняя вершинная); обе эти щетинки сильно отогнуты от лапки и резко искривлены, очень напоминая аналогичные-щетинки взрослых Caloglyphus. Praetarsi I и II крупные, с сильно видоизмененной дистальной частью, образующей крупную блюдцеобразную присоску, способную отгибаться перпендикулярно в оси лапки. Аристы длинные,, крепкие. Ноги I и II длинные и стройные.
К этому подроду относятся 2 нидикольпых вида, биологически связанных с осами рода Crabro.
* 1. Vidla (Crabrovidia) gussakovskii, sp. n. (фиг. 433 , 449—451).
Гипопус. Окраска томная, каштаново-бурая. Тело грушевидное,, позади основания пог IV внезапно суживающееся в языковидный, несколько-
Фиг. 428—485. Гипопусы родов Calvolia и Vidia.
Фиг. 428. Calvolia reticulata, sp. П., сверху. Фиг. 429. С. fusiformis, sp. п., сверху. Фиг. 430.Тоже,. снизу., Фиг. 431. С. nataliae, sp. п., сверху. Фиг. 432. С. thraca Vtz. снизу. Фиг. 433. Vidia (Ста*-brovidia) gussakovskii, sp. п., сверху. Фиг. 434. V, (s. str.) lineata Ouds. сверху. Фиг. 435. To Hie. снизу.
272
II. SAPROGLYPHIDAE
уплощенный каудальный выступ, напоминающий аналогичное образование у Calvolia thraca Vtz. Проподосомальный щит в 2 раза короче гистеросомы; ростральный выступ несколько угловатый; у его основания дорзально расположена пара овальных, стекловидно прозрачных выпуклостей, слабо намеченных и не отграниченных от окружающих покровов; под ними заметно неправильное скопление томного пигмента, но нет оформленных ретин. Стернальный щит очерчен ('.вади н с боков непрерывной дугообразной линией, соприкасающийся посередине с эпимерами III; эпимеры IV слабо, но явственно дугообразно изогнутые, сливающиеся спереди по средней линии и достигающие до эпимер III. Присасывательный диск круглый; передние присоски много крупнее боковых и задних; дополнительные присоски очень крупные; хитиновые площадки овальные, широкие; задний край подушки присасывательного диска с явственной радиальной штриховкой. Ноги I и II длинные, стройные, составляющие, соответственно, около 47 и 40% длины тела; лапки очень длинные, равномерно утоньшающиеся от основания до точки прикрепления передних вентральных щетинок, затем резко суживающиеся и к концу вновь слегка расширяющиеся; длина их превышает свыше 4 раз диаметр у основания". Обонятельная палочка очень длинная, слегка утоньшающаяся к концу; дорзальная вершинная щетинка очень длинная, далеко отодвинутая от конца лапки; наружная серповидная щетинка расположена значительно ближе к ней, чем к основанию далеко вынесенной вперед внутренней (вершинной) серповидной щетинки, и на той же высоте, как впутренняя вентральная щетинка. Ноги III довольно длинные (22% длины тела). Лапки с узкими, почти прямыми ланцетовидными щетинками и очень длинным вершинным волоском; praetarsi III с длинным, параллельно-крайним стебельком и округлой дистальной пластинкой. Длина лапок IV составляет 17—18% длины тела; tibiotarsus стройный, длиннее бедра IV, с узкой, несколько вздутой на конце дистальной частью; наружная парусная щетинка отходит на значительном расстоянии от его вершины, очень длинная. Длина 196—265 |л, ширина 120—160 [т.
Расп р о странение. Описывается по материалам из Крыма (Карадаг) и нескольких пунктов Московской обл.
Гипопусы встречаются на различных видах рода Crabro: С. fossorius L., С. impressus Sm., С. vagus F. и С. solenius Ъ.
* 2. Vldln (Crubrovldhi) popovl, яр. в. (фиг.-452, 453).
Гипопус. Очень похож ла гипопуса V. gussakovskii, отличаясь от него следующими особенностями; тело правильной яйцевидной формы, не вытянутое сзади в языковидный выступ; в соответствии с меньшей длиной гистеросомы, длина лроиодосомального щита умещается в длине гистеросомы только 1.6 раза; глазные выпуклости очень неясные; задний край стернального щита, плохо обрисованный, отделенный значительным промежутком от эпимер 1,1.1; эпимеры IV не соединены с коротким линейным продольным склеритом, тянущимся назад от эпимер III; дополнитель-’
7. VIDIA
273
Ш.10 присоски чуть больше боковых и значительно уступают в рапиерах центральным. Ноги такого же строения, как у V. gussakovskii, по несколько короче; обонятельная палочка с булавовидно вздутым концом; наружная серповидная щетинка сидит много ближе к волосовидной дорзальной вершинной, чем к внутренней (вершинной) серповидной щетинке, и много впереди внутренней вентральной. Tibiotarsns IV очень короткий, значительно уступает в длине бедру IV; дистальный отдел его совсем короткий, постепенно суживающийся к концу. Длина 195—230 р, ширина 150—180 р.
Распространение. Описывается по материалу из Украины (Красноград, Полтавская обл.), Крыма (Карадаг), Абхазии (Синоп) и Узбекистана (окрестности Ташкента). Я нашел гипопусов этого вида на Crabro clypeatus L., С. alatius Pnz., С. wollmanni Kohl, и С. impressus Sm.; на последнем из названных видов встречался иногда совместно с гипопусами V. gussakovskii. '
2. Подрод Vidia в. str.
Лапки I и II короткие, массивные, с 3 широкими ланцетовидными щетинками, из которых две вентральные и одна дорзальная, расположенная рядом с волосовидной вершинной щетинкой; praetarsi I—III одинакового строения, маленькие, не модифицированные. Аристы довольно короткие. Положение dj и d2 нормальное (по крайней мере у V. lineata). Ноги I и II короткие, довольно массивные.
Сюда относятся также 2 вида, связанные с гнездящимися в древесине пчелами родов Prosopis и Megachile.
* 3. Vidia (s. str.) lineata Ouds. (фиг. 434, 435).
Oudemans, 1917, Entomol. Bcr., IV: 391.
Гипопус. Окраска буровато-(хитиново)-жолтая. Тело яйцевидное, очень изменчивое по своим пропорциям (отношение ого длины it ширине колеблется в пределах от 1 : 2 до 1.5 : 1). Проподосомальный щит в 1.5— 1.8 раза короче гистеросомы; sc выстроены в прямой поперечный ряд; ростральный выступ спереди широко закругленный. Глаз пет. Стернальный щит четко обрисован с боков прямыми эпимеритами II; задний край щита (точнее — коксальных полей II, так как посередине он прерван) еле намечен, отодвинут от эпимер III; эпимеры IV прямые, по отделенные от своего общего ствола; вентрум хорошо развит, длинный, простирающийся почти до генитальной щели. Очертания присасывательного диска несколько угловатые; передние присоски такого же размера, как задние и боковые; дополнительные присоски заметно крупнее; боковые хитиновые площадки очень крупные, дугообразно изогнутые, охватывающие основания наружных присосок и простирающиеся назад до уровня заднего края сродней площадки; эта последняя имеет почти прямоугольную форму. Ноги I и II довольно короткие (29 и 28% длины тела); лапки короткие, постепенно суживающиеся к вершине, около 2 раз длиннее своего диаметра у основания; обонятельная
18 Фаупа СССР, Захваткин — 284
214
II. SAPROGLYPIIIDAIi
палочка довольно массивная, прямая, слегка булавовидно утолщенная; дорзальная вершинная щетинка расположена на одном уровне с внутренней вентральной, на небольшом расстоянии от конца лапки; наружная вентральная щетинка расположена чуть ппже внутренней; задняя щетинка очень короткая; е в виде толстой, почти прямой щетинки, не утолщенная у основания. Ноги III довольно длинные (22% длины тела). Длина ног IV составляет около 18% длипы тела; tibiotarsus почти в 2 раза длиннее своего-диаметра у основания, с вильчато ришцсилеплым концом; обе парусные щетинки расположены терминально. Длина 210—266 jx, ширина 140— 180 |л.
Распространение. Вся Европа, Восточная, Северная и Средняя Азия. Крайние из известных мне пунктов нахождения V. lineata в СССР:; Парголово (Ленинград), Одесса, Ворошилов-Уссурийский, Кушка.
Экология. Живет в гнездах различных видов рода Megachile Latr.; Oudemans нашел гипопусов этого вида па М. pilicrus Р. Мог., мне они попадались на М. bdleina CklL, bombycina Rad., centunc'ularis L., cyprica CklL, dohrandti F. Mor., interrupla Spin., lagopoda L., maaki Rad., mandibularis F. Mor., maritima Ky., melanopyga Costa, nigriventris Schenck, octosignata Nyl., orientalis Mor., picicornis Mor., sericans sericans Fonsc., sericans gdlerbergi V. Popov и Osmia serrilabris F. Mor. и fusiformis Latr.
4. Vidia (s. str.) undulata Ouds.
Oudemans, 1905, Entomol. Вег., II:22,
Гипопус. Мне в натуре неизвестен. По Oudemans’y характеризуется следующими признаками: спина целиком покрыта 2 щитами. Просомаль-иый щит поперечнополосатый. Абдоминальный щит продольнополосатый. Полоски очень тонки и переходят одна в другую, так что кажутся более или менее'волнистыми. 4 просомальных волоска, 2 плечевых во-лоска, 4 пары субмедиальных волосков, 4 маргинальных волоска совсем сзади, Все эти волоски чрезвычайно мелкие, трудно заметные. Одна пара более длинных хвостовых волосков («sleepharen»). Лапка I и II дорзально с ланцетовидным волоском, вентрально с 2 такими же волосками. Лапка III как дорзально, так й вентрально с 2 ланцетовидными волосками. Лапка IV с вентральным коготком и оканчивается 4 волосками различной длины, .самый длинный из которых даже длиннее тела. Длина 210—250 у-, ширина 165—200 [л. Тело более или мепсе шестиугольное.
Р а с п р о с т р а и о н и о. Этот вид до сих пор известен только из Италии.
Фиг. 436—453. Детали строения гипопусов сем. Saproglyphidae.
Фиг. 436. Ensllniella parasitica Vtz., йога I сверху. Фиг. 437. Е. parasitica Vtz., голень и лапка снизу. Фиг. 438. Е. parasitica Vtz., ланка III сверху. Фиг. 439. То же, снизу. Фиг. 440. Е. parasitica Vtz., конец ноги IV спину. Фиг. 441. Е. kostylevi, sp. п., конец ноги IV снизу. Фиг. 442. Cal-volia reticulata, sp. n., tibiotarsus IV. Фиг. 443. C. elisabethae, sp. n., tibiotarsus IV. Фиг. 444. C. elliptica, sp. n., лапка I сверху. Фиг. 445. C. fusiformis, sp. n., лапка I сверху. Фиг. 446. С. nataliae, sp. n., лапка I сверху. Фиг. 447. C. thraca Vtz., нога I сверху. Фиг. 448. To же, снизу. Фиг. 449. Vidia (Crabrovidia) gussaliovshii, sp. n., нога I сверху. Фиг. 450. V. (Cr.) gussakonskii, sp. n., голень и лапка I снизу. Фиг. 451. V. (Cr.) gussakovskii, sp. n., нога IV снизу. Фиг. 452. V;
(Cr.) popovi, sp. n., лапка I снизу. Фиг. 453. V. (Cr.) popovi, sp. n., нога IV снизу.
246
III. G1ACY141AC,H)AE
Экология. Гипопусы были найдены на пчеле Prosopis conformis Forst. Мне не удалось просмотреть какие-либо материалы по Р. conformis, так как этот вид отсутствует в наших музеях; не удалось мне также найти V. undulata на других представителях рода Prosopis, которых я просмотрел очень много.
Различия между V. undulala и V. lincata неясны; приведенное выше описание вполне подходит и к V. linoaia. По Oudemans’y последний вид отличается от V. undulala только тем, что у пего наружные из двух нар волосков заднего конца тела (т. о. постоналиные) длиннее внутренних (sa i), нз чего следует, что у V. undulala имеют место обратные соотношения размеров этих волосков.
Ш. Сем. GLYCYPHAGIDAE
Glgeyphagidae (Berl.) Ouds. 1923 + Chortoglyphidae Ouds. 1923 + Carpoglyphidae Ouds-1923 -f- Lentungulidae Quds. 1923 + Ensliniellidae Vtz, 1931 (part.).
Характеристика, семейства. В противоположность обоим предыдущим семействам, характеризующимся морфологическим единством и большим габитуальным сходством составляющих их видов, представители сем. Glycyphagidae необычайно полиморфны. Идиосома у них, как правило, нерасчлененная; в тех немногих случаях, когда намечается ее подразделение на проподосому и гистеросому, разграничивающая их бороздка неглубока, не образует перетяжки между ними и более или менее явственна только па спипной стороне тела. Форма идиосомы необычайно, изменчива, но она почти всегда значительно шире, чем у Tyroglyphidae и Saproglyphidae, и часто уплощена (иногда очень сильно) в дорзо-вентральном направлении; у ряда видов имеется более или менее хорошо развитый ростральный выступ, частично или целиком прикрывающий сверху гнатосому и иногда отграниченный от остальной поверхности идиосомы заметным поперечным швом (многие Labidophorinae). Кожа в подавляющем большинстве случаев шероховатая, мелкобугорчатая, шиповатая, морщинистая или тонко штрихованная (Aeroglyphus), крайне редко гладкая, но тогда плотная, роговидной консистенции (Chortoglyphus, Fusacarus, 'Gohieria); у многих видов она более или менее сильно хитинизирована, окрашена в буроватые или красноватые топа и образует жесткий сплошной панцирь, иногда же — систему хитинизироваппых площадок, окруженных более мягкими участками. Про-подосомалышй щит хорошо развит лишь у очень немногих форм (роды Sennertia, Hericia, Ilyadcsia, A.8roglyphus)‘, у Glycyphagus s. str. он имеет вид узкого продольного склерита (теменной киль), аналогичного так называемому crista metopica многих Trombidiiformes; опистосомальный щит всегда отсутствует. Число, строение и расположение* щетинок спинной поверхности идиосомы крайне изменчивы. Нормой для семейства можно считать наличие 16—16 пар щетинок, распределяющихся по группам следующим образом: 2 пары теменных, 2 лопаточных, 2—3 плечевых, 4 спин-'
ХАРАКТЕРИСТИКА СЕМЕЙСТВА ill
пых, 3—б боковых и 2 крестцовых. Передние теменные (v а) имеются у всех видов, кроме некоторых Labidophorinae (Orycteroxenus; у Xenocaslor они сильно редуцированы), и, как правило, широко раздвинуты основаниями; задние теменные (v р) обычно расположены дорзально, далеко позади передних, но в некоторых случаях и латерально (как у Tyrophagus), на одном уровне с v a (Aeroglyphus, Chortoglyphus) или же в непосредственном соседстве с ними (Blomia); размеры их обычно не уступают таковым v а, а иногда даже превышают их; в редких случаях они отсутствуют. Наружные и внутренние лопаточные щетинки (sc е и sc i) образуют, как обычно, поперечный ряд; соотношения их размеров очень изменчивы. Плечевые щетинки обычно расположены латерально, передние (h а) — на значительном расстоянии от задних (Пр); у некоторых форм их 3 пары, в результате смещения кверху подплечевых (sb), располагающихся тогда между ha и hp. Положение спинных щетинок (dj— d4) очень изменчиво. Боковые (1х—13) расположены то маргинально, то дорзально; иногда 13 смещены на вентральную поверхность тела. Крестцовые (sa е и sa i) занимают обычно терминальное положение, но иногда и дорзальное (Ctenoglyphus и др.). Уклонения от этой схемы бывают как в сторону выпадения отдельных пар щетипок или даже целых их групп (например, у Orycteroxenus отсутствуют крестцовые), так и появления добавочных (5 пар боковых у ряда Ctenoglyphinae).
Строение коксо-стернального скелета исключительно разнообразно, но обычно сильно отличается от такового Tyroglyphidae. В такой же мере изменчиво и строение ног и вооружение, но характер наблюдаемой здесь изменчивости существенно отличается от таковой соседних семейств. Коготки всегда стебельчатые, т. е. сидящие на конце более или менее длинного перепончатого praetarsus, лишенные соединительных склеритов и, соответственно, не связанные с концом самих лапок. Характерными чертами хэтотаксии ног можно считать тенденцию к крупной перистости или гребенчатости щетинок колен и голеней, а иногда и других члеников ноги; случаев превращения этих щетинок в шипы в пределах семейства почти пе наблюдается. Генитальные щупальцы обычно имеются у обоих полов, редко только у о7 (Co-теtacarus) или только у $ (Sennertia); нормально они имеют кольцевидную форму и только у Carpoglyphus — пальцевидную; иногда они рудиментарны (Blomia thori) или отсутствуют.
С. Анальные и тарзальные копулятивные присоски отсутствуют у всех представителей семейства, кроме рода Chortoglyphus, у которого они имеют оригинальное строение, резко отличное от такового копулятивных присосок Tyroglyphidae. Положение генитального отверстия и строение копулятивного аппарата весьма разнообразны; у многих форм есть эпиапдрий; анальных микрохэт только 1 пара (в случае, если они превращены в присоски, как у Chortoglyphus, их нет совсем), расположенная впереди анальной щели; постанальных щетинок, как правило, 3 пары.
9. П сложение и строение генитального (яйцевыводного) отверстия очень изменчиво; обычно есть эпигиний, а у большинства Labidophorinae и хити
278
III. GLYCl'PHAGIDAE
новое циркумгепитальное кольцо; посередине генитальных клапанов обычно находится характерное полулунное вдавление. Копулятивное отверстие у большинства видов образует более или менее заметную цилиндрическую или коническую копулятивную трубку, достигающую иногда большой длины. Положение анальной гнели изменчиво; анальный комплекс щетинок состоит из двух групп: процнальной — представленный 2 парами микрохэт (рг а), образующих поперечный ряд впереди anus, и постанальной (р а) — складывающейся из У (у Aitroglyplius 4) пар более длинных щетинок, располагающихся треугольником по сторонам заднего конца анальной щели, по иногда и маргинально по заднему краю идиосомы (Ctenoglyphus).
Вторичные морфологические различия между полами исключительно разнообразны; наиболее велики эти различия у многих Labidophorinae и Ctenoglyphinac, у которых с? сГ теряют всякое сходство с 9$, но в других группах они подчас и незначительны. В разных подсемействах и родах эти отличия касаются самых различных признаков, начиная от размеров и общей, конфигурации идиосомы и кончая тонкими деталями вооружения пог. Диморфизма у dd никогда не наблюдается.
Нимфальные стадии иногда сильно отличаются от взрослых. Для личинок характерна редукция грудных палочек, лишенных вершинного купола и имеющих вид мясистых бугорков, очень коротких трубочек или кольцевидных образований. Зачаточные генитальные щупальцы протонимф ителео-цимф имеют, как правило, такую же кольцевидную форму, как у взрослых, по окружены особым овальным контуром, отсутствующим у последних (может быть, соответствует полулунным вдавлениям генитальных клапанов. 9);- у Chortoglyphus и, невидимому, у Blomia генитальные щупальцы развиты у нимф лучше, чем на половозрелой стадии, па которой они атрофированы. Строение гипопальных стадий настолько разнообразно, что трудно дать их общую характеристику, применимую ко всем представителям семейства. .
В принимаемом здесь объеме сем. Glycyphagidae соответствует б перечисленным выше «семействам» классификационной схемы Oudemans’a и Vitz-thum’a. К нему относятся около 70 палеарктических1 видов, образующих 26 родов, которые довольно естественно распределяются в 7 более или менее равноценных групп, трактуемых мною как подсемейства.
0П1ЧЖЕЛ11ТЕЛЫ1АЯ ТАБЛИЦА ПОДСЕМЕЙСТВ
сГсГ
1 (2). По сторонам анальной щели расположена пара крупных воронковидных копулятивных присосок; лапки IV с хорошо развитыми тарзальными копулятивными присосками; генитальное отверстие распо
1 Внепапеарктпческих видов известно сейчас около 20.
ТАБЛИЦА. ПОЛСЕМБЙСТБ
279
ложено между основаниями ног I, так что концы эпимер I широко • разобщены и стернум полностью отсутствует ... .-1. ChortoglyрhI пне.
:2 (1). Анальные и тарзальные копулятивные присоски отсутствуют; генитальное отверстие расположено между основаниями ног 11! ил и дальше назад; эпимеры I соединяются друг с другом по средней ли-- нии тела, образуя более или менее длинный стернум.
3 (4). Лапки I и II (а иногда и III и IV) сильно укорочены, оканчиваются очень крупным когтевидным шипом, у основания которого прикрепляется необычайно длинный, тонкий и гибкий praetarsus. Темно окрашенные морские формы, живущие среди водорослей на прибрежных камнях и сваях........................................6. Carpoglyphinae.
-4 (3). Лапки I и II не укорочены, без крупного когтевидного шипа; praetarsi нормального строения. Сухопутные формы.
А (6). Кожа сплошь покрыта очень тонкой и густой параллельной штриховкой; почти вся спинная поверхность идиосомы занята четко отграниченным полем, густо покрытым треугольными, шипиками (сквамои-дами); спинные щетинки длинные, равномерно убывают в размерах спереди назад (так что di длиннее прочих); vp расположены маргинально, на одной поперечной линии с v а. На древесных осах (Xylo-сора) й шмелях ...................................... 3. Aeroglyphinae.
•6 (б). Структура кожи иная; d3 и d4 длиннее d4 и d2, или же они все представлены микрохэтами или короткими шипиками; v р расположены далеко позади v а, если же на одной линии с ними, то они сидят дор-зально в непосредственном соседстве с v а (род Blomia). В имагиналь-ной стадии не встречаются на насекомых.
7 (10). Ноги III и IV более или менее резко модифицированы.
;8 (9). Ноги III много длиннее и тоньше ног IV, не искривленные; поги IV заметно вздуты и искривлены; гнатосома очень узкая; идиосома уплощенная, ромбической формы; коготки крупные, серповидные. Живут в вытекающем из деревьев соке ..................6. Carpoglyphinae.
-9 (8). Ноги III короче ног IV, одинакового с ними строения, более или менее резко искривленные и утолщенные, с косо срезанными на конце лапками; коготки очень мелкие; гнатосома довольно широкая, часто прикрытая сверху нависающим ростральным выступом; идиосома другой формы. Живут в гнездах млекопитающих и (род Gohieria Ouds.) в муке и других зернопродуктах.................б. Labidophorinae.
10 (7). Ноги III и IV нормального строения.
11 (12). Эпимеры II соединены концами с стернумом, образуя W-образную фигуру, эпимеры III и IV свободные; гнатосома очень узкая, сильно •подвижная; кожа гладкая; все щетинки спинной поверхпоедн идиосомы гладкие, короткие, длина их'составляет 4^—10% длины идиосомы (sc е до 17% длины идиосомы), только sa е и одна пи пир нот1-анальных очень длинные. .............. 0. Cni'|)iiu:ly|>lllHili'.
12 (11). Строение коксостернального скелета ипое; пропорции гпптош»м1.1
W
III. GLYCYPHAGIbAE
нормальные; кожа очень тонко точечно-бугорчатая; щетинки спинной поверхности идиосомы длинные, более или менее перистые (очень редко совсем короткие, шиповидные, ио тогда кожа спины густо, покрыта треугольными сквамоидами).
13 (14). Боковых щетинок & нар; все постанальные расположены маргинально, так что на спинной и боковых поверхностях идиосомы насчитывается в общей сложности 17—20 пар крупных щетинок; если их меньше, то они имеют листовидную форму; коксальпые ноля I—Ш уплотненные, (замкнутые, образующие 2 расходящихся назад ряда, тесно сближенных между собой коксальных щитов; penis обычно, очень длинный. Мелкие формы, живущие в сене и в ныли................
............................................ 4. Ctenoglypliinae.
14 (13). Боковых щетинок 3 пары, постанальные расположены на некотором расстоянии от заднего края идиосомы; на ее спинной и боковых поверхностях находится не больше 15 пар щетинок; строение коксостернального скелета иное.
15 (18). Лапки очень тонкие, шиловидные, коготки очень мелкие, заметные с трудом» все щетинки спинной и боковых поверхностей идиосомы длинные (иногда чрезвычайно длинные), густопушистые или тонкоперистые, гибкие. Преимущественно синантропные виды.
16 (17). V р расположены далеко позади v а; хелицеры нормального размера; идиосома эллипсоидальной формы, обычно с слегка вогнутыми посередине боковыми краями; Эпимериты II расположены диагонально, и соединены внутренними концами с эпимерами III; колена I дорзально с 2 вершинными сенсиллами..........................2. Glycyphaginae.
17 (16). V р расположены в непосредственном соседстве с v а, образуя с ними прямой или вогнутый поперечный ряд; хелицеры очень крупные и массивные; идиосома обратно-яйцевидной формы; эпимериты II свободные, расположенные поперечно; колена I дорзально только, с 1 утолщенной сенсиллой ........................1. Chortoglyphinae.
18 (15). Лапки менее тонкие, не шиловидные; коготки крупные; щетинки спинной поверхности идиосомы короткие или очень короткие (большую длину имеет только одна из пар гладких постанальных щетинок). Обитатели гнезд перепончатокрылых ... 7. Chaetodactylinae»
99
] (2). Кожа покрыта очень тонкой и густой параллельной штриховкой; почти вся спинная поверхность занята четко отграниченным полем» густо усаженным треугольными шипиками (сквамоидами); плечевых и боковых щетинок по 3 пары, постанальных — 4 пары; у р расположены маргинально, на одной поперечной линии с v а; размеры d убывают спереди назад. На Bornbus и Xylocopa ... 3. Aeroglyphinae.
2 (1). Структура кожи иная; постанальных щетинок 3 пары (число боковых и плечевых изменчиво); v р расположены иначе, если же маргинально;
ТАБМЩА. ПОДСЕМЕЙСТВ 28Г
и на одном уровне с v а, то кожа совершенно гладкая, сильно уилот-- ненная, и все щетинки идиосомы представлены микрохэтами; <1,( и <14
всегда крупнее dx и d2. В имагинальной стадии не встречаются ли насекомых.
3 (4). Периферические щетинки идиосомы опахаловидные, двусторонне-гребенчатые или листовидные; боковых обычно 5 пар, плечевые и постанальные щетинки расположены, так же как и опи, маргинально, образуя обычно вокруг тела красивую ажурную бахрому; кожа грубоморщинистая или усаженная высокими, расщепленными на конце бородавками. Живут преимущественно в сене ... 4. Ctenoglyphinae-
4 (3). Все щетинки идиосомы гладкие, пушистые или топкоперистые, узкие; боковых 3 пары, постанальные расположены вептрально, несколько отступя от заднего края тела; структура кожи иная.
5 (8). Все щетинки спинной и боковых поверхностей идиосомы длинные-(иногда чрезвычайно длинные), густопушистые или тонкоперистые, гибкие; идиосома пузыревидно-выпуклая; кожа очень нежная, бесцветная, крайне тонко точечнозернистая; лапки очень топкие, шило— видные; щетинки колен и голеней (кроме вершинных) густоисристые.
6 (7). V р расположены далеко позади v а; эпигиний имеется; генитальное отверстие продолговатое, почти целиком закрытое хорошо развитыми, генитальными клапанами; копулятивная трубка прямая, короткая; колена I дорзально с 2 вершинными сенсиллами...........................
...............................................2. Glycyphaginae..
7 (6). V р расположены,в непосредственном соседстве с v а, образуя с ними прямой или вогнутый поперечный ряд; эпигиний отсутствует; генитальное отверстие очень крупное, широко зияющее, с сильно редуцированными клапанами; копулятивная трубка длинная, очень узкая, дугообразно загнутая книзу; колена I дорзально только с 1 сенсиллой ...................................................1. Chortoglyphinae.
8 (5). Все щетинки спинной поверхности относительно короткие, гладкие или очень тонко и редкоперистые; в последнем случае они более илиь менее жесткие; форма идиосомы и строение кожи изменчивы, лапки-более массивные (не шиловидные).
9 (10). Лапки I и II сильно укорочены, оканчиваются очень крупным когтевидным йшпом, у основания которого прикрепляется необычайно-длинный, тонкий и гибкий praetarsus. Темно окрашенные морские формы, живущие среди водорослей на прибрежных камнях и сваях.
. .............................................6. Carpoglyphinae.
10 (9). Лапки I и II не укорочены, без крупного когтевидного шипа; ргпо-tarsi нормального строения. Сухопутные формы.
11 (16). Кожа совершенно гладкая, плотной роговидной icoiicncTotllllfli.
12 (16). Кожа бесцветная, прозрачная; ноги стройные, нормального строе ния.
13 (14). Эпимеры II свободные; эпимеры I слиты ио сродной линии толп.
:«82
III. GLYCYPHAGIDAE 1. CHORTOGLYPHINAE.
образуя широкую, сильно хитипизированную поперечную пластинку, задний край которой оттянут в очень короткий свободный стернум; эпигиний отсутствует; хелицеры очень крупные и массивные; все щетинки идиосомы представлены крайне тонкими, трудно заметными микрохэтами; щетинки ног гребенчатые . . . . 1. Chortoglyphinae. 14 (13). Стернум и эпимеры .11 слиты концами с массивным угловатым эпигинием; гнатосома очень узкая, весьма подвижная; sa е и одна из пар П0СТ111Н1ЛЫ1ЫХ щетинок очень длинные, волосовидные, осталь-1 ные — короткие, жестковатые; щетинки пог игольчатые.................
.............................................G. Carpoglyphinae.
15 (12). Кожа сильно пигментированная (буроватая); ноги I и II сильно : Сжаты с боков или украшены ребрами и килями........................
................. . . .............................5. Labidophorinae. 16 (11). Кожа зернистая, шиповатая или с какой-либо иной явственной . - скульптурой, различной консистенции.
17 (18). Гнатосома очень узкая; все щетинки идиосомы и ног гладкие, ' -жестковатые; эпигиний дугообразный, свободный. Живут в вытекающем из деревьев соке...............................6. Carpoglyphinae.
18 (17). Пропорции гнатосомы нормальные; эпигиний другой формы, обычно соединенный со стернумом. Живут в гнездах млекопитающих и перепончатокрылых. ;
19 (20). Коготки очень мелкие; длинных хвостовых щетинок нет. Обитатели гнезд насекомоядных и грызунов................5. Labidophorinae.
:20 (19). Коготки крупные или очень крупные, серповидные; sa е очень длинные, гладкие. Обитатели гнезд диких пчел ........................
...................................... 7. Chaetodactylinae.
1. Подсем. С h о г t о g 1 у р h i п а е
Гнатосома свободная. Хелицеры необычайно мощные и массивные, длиною около 30—35% длины идиосомы, с толстыми, круто закругленными на конце пальцами клешни и угловатым горизонтальным расширением у её основания. Идиосома обратно яйцевидная, сильно выпуклая, не разделенная на проподо- и гистеросому, без рострального выступа. Кожа бесцветная, гладкая и твердая или же перепончатой консистенции, крайне топко точечпобугорчатая. Щетинки спинной поверхности идиосомы то в виде очень тонких гладких микрохэт (Chortoglyphus), то длинные и густо •опушенные, сходные по строению с таковыми рода Glycyphagus (Blomia)-, в последнем случае (1а все же представлены гладкими микрохэтами; обе 'пары теменных щетинок (V а и V р) расположены у переднего края идиосомы, образуя поперечный ряд или трапециевидную фигуру; 3 пары плечевых щетинок (h a, h р и sli) и 3 пары боковых (1х—13); псевдостигмальный орган простой, щетинковидный или палочковидный, густо опушенный. Генитальные щупальцы отсутствуют или сильно редуцированы. Коксо7стерпальный ^скелет слабо развитый, эпимеры без пластинчатых аподем; эпимеры I слиты
ХАРАКТЕРИСТИКА ПОДСЕМЕЙСТВА
283
:fii"
•концами по средней линии тела, образуя фигуру в форме Y или скобки (^-), иногда свободные и тогда несколько модифицированные (у d Chorloglyphus); юпимериты II совершенно свободны, расположены в поперечном папраеле-иии или косо; эпимеры III и IV совершенно редуцированы, эпимериты IV отсутствуют. Ноги очень тонкие, «бегательного» типа, с очень длинными лапками; щетинки вертлугов и бедер очень тонкие, перистые, базальные щетинки колен и вентральные щетинки голеней гребенчатые (у рода Chor-toglyphus) или толстые и густоперистые, как у «волосатых» клещей (у рода Blomia); вершина колен I только1 с 1 органом чувств — цилиндрической или сильно утолщенной обонятельной палочкой (отсутствует расположенная рядом с ним щетинка, имеющаяся у всех других подсемейств); вершинные щетинки голеней I и II чрезвычайно длинные, голеней III и IV также длиннее обычного; обонятельная палочка лапок очень тонкая й длинная; Т, а иногда и р, отсутствуют; в медиальном поясе лапок I и II 4 щетинки <2. латеральные и 2 вентральные) перистые, гладкие или шипообразные; вершинные шипы лапок иногда сильно развиты. Prae tarsi удлиненные, хорошо развитые,, с мелким коготком. Вторичные половые различия крайне незначительны.
d. Строение копулятивного аппарата и связанного с ним комплекса структур совершенно различно у обоих относящихся сюда родов (см. родовые описания).
Q. Чрезвычайно крупное, широко зияющее генитальное отверстие расположено между основаниями ног III и IV; генитальные клапаны сильно .редуцированы; эпигиний отсутствует; у рода Blomia имеется очень длинная, дугообразно изогнутая копулятивная трубка, совершенно отсутствующая у Chortoglyphus; расположение щетинок анального комплекса также совершенно различно у обоих родов.
. Личинка (известна только у рода Chortoglyphus) с редуцированными трудными палочками; у прото-и телеонимф генитальные щупальцы развиты i лучше, чем у взрослых. Гипопальных стадий не образует, насколько известно, ни один из представителей данной группы.
К подсем. Chortoglyphinae я отношу 2 рода: Chortoglyphus Вег], с 2 видами и Blomia Ouds. с 3 или 4 видами. Объединение их в одну группу может показаться спорным из-за резких различий между ними в строении копу-' • дативного аппарата (имеющего типичное для семейства строение у рода Blomia и резко аберрантного, дополненного анальными и тарзальными L копулятивными присосками—у Chortoglyphus) и особенно бросающихся
। в глаза различий в размерах и строении, а отчасти и расположении щетинок
идиосомы; на самом же деле их систематическая близость очень велика, и этим комплексам признаков нельзя придавать большое таксономическое > значение, так как первые из них функционально связаны между собой 1 (и должны быть, следовательно, оценены как один признак), а инплогичппя .изменчивость в строении и размерах щетинок имеет место И ио многих других естественных группах Glycyphagidae, например у Labidophorinae.
284
Ш. GLYCYPHAGIbAE. 1. CHORTOGLYPHINAE
Представители нодсем. Chortoglyphinae известны почти исключительно* как синантропы, обитающие в зерне, муке и других растительных веществах. на складах и в амбарах и в пыли жилищ; все они — медленные в своих, движениях формы с малым биотическим потенциалом.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА родов
1 (2). Лее щетинки спиппоП поверхности идиосомы представлены гладкими,, топкими (подчас- трудно пмметлымп) микрохэтами; кожа совершенно гладкая, твердая (в соответствии с чем идиосома имеет более правильную и постоянную форму); щетинки голеней и колеи I и II гребенчатые, одна из вентральных щетинок лапок в виде крупного шипа; <5 с анальными и тарзальными копулятивными присосками и сильно-смещенным кпереди генитальным отверстием, помещающимся между раздвинутыми концами эпимер I. 9 без копулятивной трубки. . .. .......................................... 1. Chortoglyphus Вег!..
2 (1). Все щетинки спинной поверхности идиосомы (кроме d3) длинные,-довольно толстые, густо опушенные (некоторые из них иногда достигают длины всей идиосомы); кожа мягкая, перепончатой консистенции, очень мелко точечно-бугорчатая; щетинки голеней, колен и лапок довольно толстые, густоперистые; С без анальных тарзальных, копулятивных присосок, с генитальным отверстием, расположенным; между оспованиями ног III и IV; 9 с длинной, дугообразно изогнутой копулятивной трубкой...........................2. Blomia Ouds..
. 1. Род -CHORTOGbYPHUS BERL.
Berlese, 1884, XIV, n° 4; LXXXVIII, n° 9; Kramer, 1899: 137.
Тип рода: Tyroglyphus arcuatus Troupeau 1879.
Кожа довольно сильно уплотнена, твердой консистенции, совершенно» гладкая. Все щетинки идиосомы представлены очень тонкими (трудно заметными гладкими микрохэтами; v р длиннее всех остальных щетинок, спинной поверхности, явственно перистые, расположены латерально. по-сторонам гнатосомы; h а и h р находятся на одном уровне с d1? образуя, с ними дугообразный поперечный ряд; da расположены ближе к d3, чем. к dr Псевдостигмальный орган в виде тонкой перистой щетинки. Генитальные щупальцы отсутствуют. Базальные щетинки колен I и II и вентральные-щетшпш всех голепей гребенчатые; вершинная сенсилла колен I и II палочковидная; все щетинки лапок гладкие, одна из вентральных утолщена в виде-шипа; конец лапок с 2 довольно крупными, искривленными боковыми* шипами и 1 более мелким вентральным.
Фиг. 454—462.
<0иг. 454. Chortoglyphus arcuatus (Тг.), снизу. Фиг. 455. Ch. arcuatus (Тг.), хелицера с? сбоку' (с внутренней стороны). Фиг. 456. Ch. arcuatus (Тг.), 9 сверху. Фиг. 457. То же, снизу. Фиг. 458.. Ch. arcuatus (Тг.), нога I 9. Фиг. 459. Ch. arcuatus (Тг.), лапка IV сГ. Фиг. 460. Ch. arcuatus (Тг.> конец лапки I снизу. Фиг. 461. Blomia thorl A. Z., нога I 9. Фиг. 462. В. thori A. Z., хелицера..
286
III. GLYCYP11AGIDAE. 1. CHORTOGLYPHINAE
d. Генитальное отверстие расположено в пространстве между основаниями ног I и IT, в связи с чем эпимеры I свободны, широко раздвинуты концами и принимают участие в образовании генитальных клапанов; стер-пум, соответственно, отсутствует; апальпая щель расположена сейчас же позади основания пог IV, па значительном расстоянии от заднего конца тела; по ее сторонам помещается лара очень оригинально устроенных копулятивных присосок, состоящих из узкого трубкообразного, сильно хити-низировапиого основания и насаженной па пего широкой перепончатой воронки; рва отсутствуют;1 лапки IV несколько утолщены посередине и искривлены, снабжены парой цилиндрических копулятивных присосок.
9. Эпимеры I слиты по средней линии тела, образуя довольно широкую-поперечную хитинизироваиную пластинку, задний край которой оттянут в очень короткий стернум; задние края генитальных клапанов дугообразные, сходящиеся кпереди под тупым углом, сильно утолщенные и хитини-зированные; все рг а и р а представлены микрохэтами и расположены вдоль анальной щели, образуя по ее сторонам по продольному ряду (из 5 щетинок в каждом); копулятивная трубка совершенно отсутствует. — Личинки с крайне мелкими сосочковидными грудными палочками; у протонимф и телеонимф имеются зачаточные генитальные щупальцы.
Описаны только 2 вида: Ch. arcuatus (Troup.) и распространенный в Северной Америке Ch. gracilipes Banks.
*1. Chortoglyphus arcuatus (Troup.) (фиг. 56, 454—460).
Troupeau, 1879: 113, fig. 27, 29 (Tgroglyphus). —nudus Berlcse, 1884, XIV, n° 4; Canestrini, 1888a: 398, t. 34, fig. 4. —arcuatus Michael, 1903: t. I (Chortoglyphus').
При жизни полупрозрачный, с легким зеленоватым оттенком, сильно блестящий; ноги и гнатосома розоватые. Идиосома довольно правильной яйцевидной формы, в 1.4—1.5 раза длиннее своей максимальной ширины, приходящейся немного выше основания ног III. Все щетинки спинной поверхности идиосомы представлены очень тонкими микрохэтами, длиной в 3—5% длины идиосомы; vp заметно толще, длиною около 8%; щетинки брюшной поверхности длиннёе спинных. Ноги очень длинные и стройные; внутренняя базальная щетинка колен I и II гораздо длиннее наружной, красиво изогнута, крупногребенчатая; наружная щетипка прямая; вентральные щетинки голеней короткие; на йогах I они обе расположены лате-ральио па одной высоте, у самого конца членика; па ногах II наружная щетинка смещена на середину вентральной поверхности членика и сидит /много ниже внутренней.
- д'. Длина ног составляет около 50—54 (I), 47—50 (II и III) и 51—54 (IV) % длины идиосомы; лапки IV па 3/i короче двух предыдущих члеников ноги, взятых вместе, и только в 1.4 раза длиннее голеней IV; бедра и колени IV слегка искривлены; внутренняя вентральная щетинка голеней I длиннее
1 Имеются все основания считать, что именно они превращены в анальные копулятивные-, присоски.
1. CHOliTOGL YPHUS. 2. BLOMIA
28”'
наружной; обе вентральные щетинки лапок IV сидят почти на одном уровне,. Penis очень крупный, саблевидный; тарзальные копулятивные лрисишсл расположены на небольшом расстоянии друг от друга, в среднем отрезке лапок IV. Длина 270—330 р., ширина 150—220
9. Ноги I—III относительно несколько короче, чем у <5, ноги IV заметно длиннее, их размеры составляют около 46—47 (I), 45 (II и Ill) и 55—57 (IV)% длины идиосомы; лапки IV в 1.5 раза длиннее двух предыдущих члеников ноги, вместе взятых, и в 2.5 раза длиннее голеней IV; бедра и колени IV прямые; внутренняя вентральная щетинка голеней I короче наружной; вентральные щетинки ланок IV мельче, чем у tf, и сидят на большом расстоянии друг от друга. Длина 350—450 у, ширина 170—250 у.
Распространение. Франция, Италия, Англия, СССР; у нас найден пока только в Москве и ее ближайших окрестностях, в нескольких, пунктах Ивановской и Смоленской обл. и в Барнауле.
Экология. Преимущественно амбарный вид, живущий в муке, зерне, пшенице, ржи, овсе, семенах кормовых и огородных культур, а также в сене и старой соломе; иногда встречается и под открытым небом (в кучах старой соломы). Благодаря своей небольшой плодовитости и малой экологической валентности, Ch. arcuatus лишь очень редко приносит ощутимый вред заселяемым им продуктам. Биология его остается почти неизвестной.
2. Род BLOMIA OUDS,
Oudemang, 1928, Entomol. Вег., VII: 348; Захваткин, 1936, Бюлл. Моек. общ. иеп. прпр.,. XLV: 266—267 , 269—270.
Тип рода: Glycyphagus tjibodas Ouds. 1910.
Мелкие, нежные виды, напоминающие по своему габитусу протопимф рода Glycyphagus. Кожа очень тонкая, перепончатой консистенции, крайне мелко точечно-бугорчатая. D2 представлены топкими гладкими микрохэтами, все остальные щетинки спинной поверхности идиосомы, sli и р3 средней длины или длинные, густоперистые, похожие по своему строению на. таковые видов Glycyphagus-, соотношения их размеров также сходны с наблюдающимися в этом роде, а именно все щетинки, расположенные в передней половине идиосомы, значительно короче, чем сидящие ближе к ее заднему концу; sa i и sa е длиннее всех, достигая иногда 80—100 % длины идиосомы;, обе пары V сгруппированы близ середины переднего края идиосомы; 11 а помещаются на одном уровне с sc, h р заметно позади них; d2 расположены ближе к dj чем к d3. Псевдостигмальный орган в виде толстой пушистой палочки. Эпимеры I у обоих полов нормального строения, соединены между собой по средней линии тела, образуя короткий V-ortpntiiihiil стернум. Генитальные щупальцы отсутствуют или рудиментарны. IliillHAli* ные щетинки колен I и II, вентральные щетинки всех голонеП И нее |Це • тинки лапок (кроме вершинных и обонятельных) толстые, густонернстмо;
.288
III. GLYCYPHAGIDAE: 1. CHORTOGLYPHINAE
вершинная сенсилла колен I и II очень короткая и толстая; лапки на конце -с крайне мелкими, практически незаметными шипиками.
d. Генитальное отверстие расположено между основаниями ног IV, щрикрыто парой тесно смыкающихся по всей длине клапанов; анальная щель помещается непосредственно позади генитальной и простирается до заднего края тела; апалыше копулятивные присоски отсутствуют; рт а хорошо развиты; лапки IV не искривлены, лишены копулятивных при-.• сосок.
9. Задние края генитальных клапанов прямые, пеутолщенные, сходящиеся кпереди под острым углом; копулятивная трубка необыкновенно длинная, цилиндрическая, дугообразно загибающаяся концом книзу, .с плотными, склеротизированными стенками; рт а и р а расположены нормально. Неполовозрелые стадии неизвестны.
К роду Blomia Ouds. относятся 4 или 5 видов, известных пока только ,в качестве синантропов; все они очень редки, встречаются спорадически, -единичными особями или небольшими колониями, в домах и зернохранилищах.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
;1 (2). V расположены трапецией — внутренние заметно позади наружных; ha значительно короче hp; все щетинки идиосомы более длинные; внутренняя вентральная щетинка голеней I и II расположена у самого основания членика, много ниже наружной; задний край идиосомы у 9 с парой конических бугорков.................*3. В. kulagini A. Z.
(1) . V образуют прямой поперечный ряд; h а значительно длиннее h р; все щетинки идиосомы короче; вентральная щетинка голеней I и II расположена лишь немного позади середины членика, почти на том же уровне, как наружная; задний край идиосомы у 9 без бугорка . . .
............................................ *1. В. thori A. Z.
*1. Blomia thori A. Z. (фиг. 461, 462—466).
Захватила, 1936, Бюлл. Моск. общ. исп. прир., XLV: 267.
V образуют прямой поперечный ряд; все почти одинаковой длины; ;sc е заметно короче sc i; h а длиннее h р; щетинки идиосомы в общем короче, 'Чем у В. kulagini-, длина стернальных и других щетинок брюшной поверхности не уклоняется от нормы. Генитальные щупальцы рудиментарны, но .довольно хорошо заметны при большом увеличении. Перистые щетинки .ног довольно короткие и толстые; внутренняя вентральная щетинка голеней I и II расположена немного позади середины членика, почти на той же
Фиг. 463—471.
.-Фиг. 463. Blomia thori A. Z., 2 Сверху. Фиг. 464. То же, снизу. Фиг. 465. В. thori A. Z., псевдо-,-етигмальный орган. Фиг. 466. В. thori A. Z., голень и лапка IV 9- Фиг. 467. В. kulagini A. Z., 9 сверху. Фиг. 468. В. kulagini A. Z., сГ снизу. Фиг. 469. В. kulagini A. Z., нога I 9. Фиг. 476, ,В. kulagini A. Z., голень и лапка IV 9. Фиг. 471. В. kulagini, A. Z., псевдоетягмальныйорган.
290 Ш. GLYCYPHAGIDAE. 1. CIIORTOGLYPHINAE
высоте, как наружная; вентральная щетинка лапок IV много короче г/& длины лапки; вершинная сенсилла колен I и II в виде тупой выпуклой бляшки.
d1. Щетинки идиосомы длиннее, чем у 9; особенно длинны d4, на е и sa i, достигающие 70—fin % длины идиосомы; примерная длина других щетинок спинной поверхности (и процентах к длине идиосомы): v 20—23; sc е 24;. sc i 30—32; li a 38; h p 2d; d( 20; d2 4;d340;d460;lx и1234—36 ;13 около 60;. pru болен чем в зраза короче апальпой щели. Penis спереди широко закруглен. Длина 305 [л, ширина. 220 |л.
9. Все щетинки идиосомы короче, их примерная длина (в процентах к длине идиосомы): v 20—23; sc е 23; sc i 28; h а 25; 11 р 20; 16; d2 4;
d3 30; d448; Ij 16; 12и13 30; sa е nsa i около 50—55; внутренние pr а в 2 раза короче наружных, длина последних составляет меньше 1/2 длины анальной щели. Ширина генитального отверстия сзади приблизительно равна его длине (измеренной от заднего края до пе'реднегоюоединительного склерита); задний край тела без бугорков; копулятивная трубка сильно изогнута книзу, ее длина (в соответствующем раккурсе) значительно уступает длине, колен. Длина 320—375 у, ширина 245—280 у.
Распространение. Известен только из нескольких районов Смоленской обл., где был найден в семенах льна.
2. Blomia tjibodas (Ouds.).
Oudemans, 191.0, Entomol. Ber., 111:74; 1911,ibid.,Ill: 169; 1914, ibid., IV: 102—103 (Gly-cypliagus)-, 1928, ibid., VII: 348 (Blomia).
9. Длина (включая мандибулы) 362 у, ширина 168 у; почти совершенно эллиптическая. Мандибулы мощные. 4 теменных щетинки; посередине спины 2 маленьких, тонких волоска; 8 щетинок шлейфа, из которых две самые длинные имеют 285 у в длину. Псевдостигмальный волосок расположен дорзально над вертлугами I, выглядит приблизительно, как густопушистые щетинки колен I. На коленах I дорзо-дистально своеобразный, закругленный на вершине орган чувств. Вентро-проксимально на trochan-terofemur palpi по серповидному волоску равной длины с кальпами. Генитальное отверстие огромное, между 4 задними ногами. (По Oudemans’у.)
Распространение. Первоначально описан по 1 плохо сохранившейся 9 с Явы (Tjibodas), затем найден Oudemans’ом в Голландии (Arnhem), также в числе одной только 9; очень возможно, что эти экземпляры относятся па самом деле к 2 различным видам, но они описаны слишком поверхностно (упомянуты почти исключительно родовые признаки), чтобы можно было решить этот вопрос без детального изучения препаратов, автора.
Приведенное описание В. tjibodas, заимствованное из статьи 1914 г., было сделано с арнхэмской 9; как видно из этого описания, она отличается от 99 В. thori значительно меньшими размерами [общая длина последних (вместе с мандибулами) 410—465 у], большей длиной sa i и более длинной
2. НЮША
2<П
копулятивной трубкой, а от В. kulagini меньшими размерами и отсутствием конических выростов па заднем крае идиосомы.
*3. Blomia kulagini A. Z. (фиг. 467—471).
Захваткин, 1936, Бюлл. Моек. общ. исп. прпр., XLV: 270.
V расположены трапецией, внутренние заметно позади наружных; vi значительно длиннее, чем ve; sc 1 все приблизительно одинаковой длины (sc i чуть короче sc е); h а значительно короче й р; щетинки спинной поверхности длинные; стернальные, генитальные и рг а длиннее обычного, волосовидные. Генитальные щупальцы совершенно отсутствуют. Перистые щетинки ног длинные, тонкие; внутренняя вентральная щетинка голеней I и II помещается у самого основания членика, много ниже наружной; вентральная щетинка лапок IV почти равна длины лапки; вершинная сенсилла колен I и II в виде толстого конического шипа.
<5. Размеры щетинок спиппой поверхности идиосомы нс установлены (у обоих- имеющихся экземпляров они почти все отпали при заделке в препарат); рг а только в 2 раза короче анальной щели. Penis спереди вытянут в короткую трубку. Общая длина тела 305—325 р, длина идиосомы 235— 250 р, ширина 185—200 р.
$. Примерная длина щетинок идиосомы (в процентах к ее длине): v i 27; v е 21; sc е 33; sc i 31; 11 а 23—24; h р 30; d1 36; d2 4; d3 50; d4 80; Ij 50; 12 75; 13 80; sa e 75; sal 90; обе пары преанальных почти одинаковой длины, длиннее половины анальной щели. Ширина генитального отверстия сзади значительно уступает его длине; задний край идиосомы с парой довольно крупных конических выростов; копулятивная трубка длипою с колено ног I. Общая длина тела 400—415 р, длина идиосомы 315—330 р, шприца 235—245 р.
Распространение. Небольшая колония этого вида была, найдена в пшенице в одном из зернохранилищ г. Москвы.
2. Подсем. Glycyphaginae
Хелицеры умеренной величины, довольно изменчивые по своему строению. Идиосома сильно (пузыревидно) выпуклая, приблизительно эллиптической формы, нерасчлененная, без всяких следов поперечной бороздки и рострального выступа; ее боковые края обычно слегка вогнуты попади едва намеченных плечевых выпуклостей, задний край широко закругленный. Кожа бесцветная, нежной перепончатой консистенции, очень мелко точечно-бугорчатая. Щетинки спинной поверхности идиосомы чроипычпйно длинные, густо опушенные, у живых особей торчащие перпендикулярно к поверхности тела; v р расположены далеко позади v а; плочены» щетинок 2 пары, боковых 3 (1х—13); псевдостигмальный орган сильно нотниотыЙ;
$
292
HL GLYCYPUAGIDAE. 2. GLYCYPHAGhNAE
генитальные щупальцы вполне явственные, редко отсутствуют ($ Соте-tacarus}. Коксо-стернальный скелет слабо развитый, эпимеры без пластинчатых аподем; эпимеры I соединены ио средней линии тела, образуя—-образную фигуру; эпимеры II свободные, эпимериты II соединены внутренними концами с укороченными эпимерами III и IV, расположенные диагонально; эпимериты IV отсутствуют. Ноги очень тонкие, «бегательного» типа, с очепь длинными шиловидными лапками; большинство щетинок: ног густонсристые; исрпииш колен 1 с 2 органами чувств — обонятельной палочкой и расположенной рядом р кою щстипкой; обонятельная палочка лапок I и II средней длины, по бокам с 2 дополнительными сенсиллами; вершинные шипы лапок крайне мелкие, заметные с трудом. Praetarsi в виде узкой перепончатой лопастинки, с крайне мелким коготком. Вторичные половые различия обычно очень незначительны; если же они ясно выражены, то сказываются преимущественно в модификации некоторых щетинок ног I и II у <5.
С. Генитальное отверстие, овальное, небольшое, расположенное в пространстве между основаниями ног III и IV, обычно с небольшим эпиаидрием; penis маленький, трубчатый.
9. Генитальное отверстие помещается в пространстве между основаниями ног II и III, крупное, с хорошо развитыми клапанами, смыкающимися по средней линии на большом протяжении и небольшим, обычно полулунным эпигипием; копулятивная трубка цилиндрическая, довольно ши-' рокая.
Личинки с мелкими, но довольно явственными грудными палочками. Расселительных гипопусов не образует пи одна из относящихся сюда форм; покоящиеся гипопальные стадии имеются у ряда видов. ,
К этому подсемейству относится всего 2 рода, почти все представители которых являются синантропными, амбарными видами. Очень подвижные, быстрые в своих движениях формы.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). Самые крупные из щетинок идиосомы не больше чем в 2 раза превышают ее длину; v а тесно сближены между собой, v р широко расставлены основаниями; генитальные щупальцы мелкие, явственные у обоих полов; псовдостпгмальиый орган с 1 сильно ветвистым стволом ....................................... . 3. Glycyphagus Her.
2 (1). Самые крупные из щетинок идиосомы превышают ее длину в 3—3.5 раза, образуя ролочащпйся 'за пего «хвост», напоминающий хвост кометы; v а широко расставлены, v р тесно сближены основаниями; генитальные щупальцы у сГ довольно крупные, у 9 отсутствуют; псевдостигмальиый орган разделен до основания на 2 сильно ветвистых ствола...................................... 1. Cometacarus A. Z.
3. GLYCYPHAGUS
2II.T
3. Род GLYCYPHAGUS HERING
Hering, 1838, Nov. Act. Acad. Leop., XVIII, 2: 619; Caneatrini, 1888: 6; 1888a: 355; Berleso,. 1891:106; 1898, LXXXIX, n° 1; Kramer, 1839:144; Michael, 1901: 226 (partim). — Glycyphagus -|--f- Lepidoglyphus' Захватишь 1936, Бюлл. Моск. общ. псп. прир., XLV: 264.
Тип рода: Glycyphagus prunorum Hering 1838.
Хелицеры нормального строения, довольно короткие и массивные, режущие края обоих пальцев клешни сплошь сильно зубчатые. Перистые щетинки идиосомы цилиндрические, опушенные по всей окружности, очень-длинные, по все же не более чем в 1.5—2 раза длиннее тела. V а тесно сдвинуты, расположенные несколько отступя от переднего края идиосомы; v р-л sc широко расставлены; 2 пары плечевых (ha и sh); положение d 2 изменчиво; также изменчива и длина р а. Псевдостигмальный орган с 1 стволом. Наружные концы эпимеритов II свободные, направлены косо кнаружи и вперед, внутренние соединяются с эпимерами IV под прямым углом и не образуют крючковидного отростка. Генитальные щупальцы хорошо развиты у обоих полов. Лапки вооружены зазубренными щетинками средней длины или одеты пушистой чехловидном чешуей (представляющей собою модифицированную заднюю вентральную щетинку лапок); обонятельная палочка лапок I и II тонкая, цилиндрическая; вершинная щетинка голеней. III, примерно, такой же длины, как на голенях I и II.
б. Генитальное .отверстие расположено между основаниями пог III; есть хорошо развитый эпиандрий.
$. Генитальные клапаны хорошо развиты, смыкающиеся друг с другом на большом протяжеии по средней линии тела; эпигиний полулунный или полукруглый; передняя пара 'генитальных щетинок расположена позади эпигиния.
Род Glycyphagus насчитывает 9 видов, распространенных, невидимому, только в Палеарктике. Описанные из Соединенных Штатов «ГП.» robuslu» Banks 1906 и «GL» obesus Banks 1906 относятся не только к другим родам, но даже и к другим подсемействам (систематическое положение ориентального Gl. latipes Berl. 1912 и Gl. geniculatiis Vtz. 1919 рассматривается ниже).
В Номенклатуре и синонимике видов р. Glycyphagus до сих пор существовала большая путаница, вызванная трудностью хоть сколько-нибудь, достоверной аттрибуцип названий Schrank’a (1781) тому или иному из. различаемых теперь видов. Так, например, название «<?1. cadaverum» прима" няется современными авторами то для обозначения вида, именуемого здесь G. (Lepidoglyphus) destructor (Schrk.) Ouds., то для вида, впервые детально описанного Oudemans’oM под названием G. privatus и т. д. В ишугшНЦЙ работе я применяю эти названия, следуя их толкованию и отиты» Olltlih mans’a (1913 — «Glycyphagus groepeh» (*Entomol. Вег., TTI: JlliH).
S9A 111. GLYCYPllAGlllAH. 2. GLYCYPHAGINAE
ОИРВДИЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (10). Лапки одеты снизу и с боков плотно обхватывающим их чехлом, сплошь покрытым, очень мелкими волосками, вследствие чего они кажутся густо- и молкоомушенными; теменного киля нет; вентральная щетин КП. бедер Т гладкая, волосовидная. (Подрод Lepidoglyphus Л. Z.)
2 (5), Вала,лwiaii щотиика. колеи Ш превращена в крупную яйцевидную или ланцетную пушистую чешую; дистальные края голеней пластинчато расширены, образуя широки it раструб; пористые щетинки идиосомы и ног толстые, с более длинными бородками.
3 (4). Очень крупная форма: с? 450—560 р, 9 600—809 р; чешуевидная щетинка колен III продолговато-яйцевидная, около 2 раз длиннее своей максимальной ширины; волоски (хэтоиды), усаживающие ее края, почти такого же размера, как расположенные на ее поверхности волоски................................... *4. G. (L.) michaeli Ouds'.
4 (3). Более мелкий вид: <5 350—400 р; (9 неизвестна); чешуевидная щетинка колен III узколанцетовидная, в 6 раз длиннее своей максимальной ширины; ее краевые волоски образуют длинную бахрому . . ...................................... *5. G. (L.) burclianensis Ouds.
5 (2). Базальная щетинка колен III не расширена, такого же строения, как другие перистые щетинки ног; голени не образуют пластинчатых раструбов; перистые щетинки идиосомы тонкие.
6 (7). Перистые щетинки голеней и колен I и II очень широкие; вершинная щетинка колеи I чрезвычайно короткая, почти такой же длины, как расположенная рядом с нею обонятельная палочка; у О' обонятельная палочка колен II сильно увеличена в размерах и имеет вид крупного, массивного шипа, длиною с само колено; у 9 Длина генитального отверстия значительно превосходит расстояние между его задним краем и анальной щелью и задняя пара генитальных щетинок расположена на уровне ее заднего края ... *3. G. (L.) fustifer Ouds.
7 (6). Перистые щетинки голеней и колен I и II узкие; вершинная щетинка колен I в 4—6 раз длиннее расположенной рядом с нею обонятельной палочки; обонятельная палочка колен II у обоих полов нормального размера, очень маленькая; у 9 длина, генитального отверстия равна расстоянию между ее задним краем и анальной щелью; задняя пара генитальных щетинок расположена зпачительпо позади уровня зад-; него края генитального отверстия.
'8 (9). Очень мелкий вид: сГ 225 р; перистые щетинки идиосомы чрезвычайно .• длинные; d3, d4 и sa с в 2 раза, 18 в 1.5 раза длиннее идиосомы; у сГ рг а почти равны по длине анальной щели . 2. G. (L.) pilosus Ouds.
9 (8). Среднего размера вид: сГ 350—430 р, 9 380—560 р; перистые щетинки идиосомы много короче, самые крупные из них не более чем в 1.5 раза, длиннее идиосомы; у <5 рг а короче 1/2 длины анальной щели. . . .................................*1. G. (Ь.) destructor (Schrk.) Ouds.
3. GLYCYPHAGL8
295.
10 (1). Лапки лишены пушистого чехла, с несколькими довольно крупными зазубренными щетинками; имеется хитинизированный теменной киль; вентральная щетинка бедер I перистая. (Подрод Glycyphagus s. str.)
•11 (12). Va сидят на переднем конце линейного теменного киля, тянущегося назад до уровня sc i; d2 расположены нормально, много впереди d3 и на одной линии с 1т; обонятельная палочка колеи I и II короткая, нриостренная к концу ... *6. G. (s. str.) cadaverum (Schrk.) Ouds.
12 (11). V а-расположены посередине теменного киля, тянущегося назад только до уровня v р; d2 сильно смещены назад и помещаются между d3, на одной поперечной линии с ними или даже несколько позади нее, ы много дальше назад, чем 1Х; обонятельная палочка колен I и II тонкая, длинная, цилиндрическая.
13 (14). Вентральные щетинки голеней III и IV сидят в вырезе дистального края членика; вентральные щетинки лапок IV расположены на неодинаковом расстоянии друг от друга—обе проксимальные сильно сближены между собой и удалены от дистальной; теменной киль приострен к обоим концам; у <? лапки I и II резко искривлены, и наружная вентральная щетинка голеней I и II превращена в крупный, массивный треугольный гребень ......................*8. G. (s. str.) ornatus Kram.
14 (13). Вентральные щетинки голеней III и IV сидят на самом членике, заметно отступя от его края; вентральные щетинки лапок IV расположены на одинаковом расстоянии друг от друга; передняя половина теменного киля широкая, слабее хитинизирована, чем узкая задняя; лапки I и II у d не искривлены, обе вентральные щетинки голеней I и II нормального строения *7. G. (s. str.) domesticus(Deg.).
1. Подрод Lepidoglyphus A. Z.
Захваткин, 1936, Бюлл. Моск. общ. исп. прир., XLV: 264 , 269.
Теменной киль отсутствует. П2 всегда расположены много впереди da, на одной поперечной линии с 1Г Лапки одеты снизу и с боков плотно охватывающей их 1 чехлообразной чешуей (представляющей собою видоизмененную заднюю вентральную щетинку лапок), сплошь покрытой снаружи тонкими ресничками, вследствие чего лапки кажутся густо и мелко опушенными; остальные щетинки лапок сдвинуты к самому их концу, очень мелкие, гладкие; вентральная щетинка бедер I гладкая. — Личинки с очень мелкими бородайковидными, нехитинизированными грудными палочками. — Покоящиеся гипопусы с короткими, но вполне развитыми, расчлененными ногами, довольно заметным коксо-стернальаым скелетом и явственной генитальной щелью, с 2 парами округлых генитальных щупальцев; лапки I с удлиненными коготками и 2 крючковидно утолщенными на конце 1цотип-
1 На препаратах эта чешуя нередко отходит в сторону или даже совсем отшишипптоя, и лапки тогда представляются совершенно голыми (если не считать 3-4 краПпп мелких гладких голосков у самого их конца).
296
HI. GLYCYPHAGIDAE. 2. GLYCYPHAGINAE
ками (точнее — хэтоидами, так как они не артикулированы); на лапках II и III в такую же щетинку превращен их коготок. Стенки гипопальной камеры покрыты рельефным крупноячеистым сетчатым рисунком, почти совершенно прозрачные.
К этому подроду относятся б видов. — Glycyphagus geniculatus Vtz. 1919, распространенный в тропических частях Старого Света и весьма напоминающий представителей подрода Lepidoglyphus наличием чехловидной чешуи на лапках, относится, по всей вероятности, к особому роду, так как отличается от всех видов рода Glycyphagus рядом существенных хэто логических признаков, вздутостью бедер и колеи и т. д.; к G. geniculatus, повидимому, близок и совершенно недостаточно описанный яванский G. latipes Berl. 1912.
*1. Glycyphagus (Lepidoglyphus) destructor (Schrk.) Cuds. (фиг. 22,. 46—50, 52—66, 70—73, 75, 472—475).
Schrank, 1781, Enumer. Insect. Austr., n° 1057, t. 2, fig. H. (.Icarus) (sec. Oudemans, 1905). — spinipes C. L. Koch., 1841, Dtsch. Crust.,Myriap., Arachn.: 33, n° 1 (Acanis).— spinipes Fumouze et Robin, 1867, Journ. Anat. Physiol., V: 33, 41, t. 23; Berlese, 1884, XIV, n° 2; Canestrini, 1888a: 381, t. 27, fig. 1; Кйипег, 1899: 147 (Glycyphagus).—cadaverum Oudemans, 1903, Tijdschr. Ned. Dierk. Ver.,(2), VIII; 209, t.VIII, fig., 16—25, et t. IX, fig. 26—27(Glycyphagus).— destructor Oudemans, 1905, Entomol. Вег., I: 238; 1913, ibid., Ill: 328; Захваткпп, 1936, Бюлл. Моск. общ. исп. прир., XLV: 265.
Биология. Megnin, 1888, Ann. Soc. Entomol. Fr., I: 131.
V p расположены па приблизительно таком же расстоянии друг от друга, как sc i, так что основаниями этих 2 пар щетинок намечается фигура прямоугольника; пространство между v р и sc i, примерно, в 4 раза меньше расстояния между последними и в 1.5 раза меньше расстояния между va и. vp; базальные чашечки всех крупных щетинок небольшие, без выростов.. Перистые щетипки идиосомы относительно тонкие и гибкие, покрывающие-их реснички довольно коротки; длина самых крупных из щетинок идиосомы— d2, d4, J3 и sa е — не более чем в 1.6—1.4 раза превышает ее-длину, dx и ]2 обычно заметно уступают ей; рх почти в 2 раза короче р3;. р2 представлены гладкими микрохэтами. Псевдостигмальный орган с тонким стволом и многократно ветвящимися отростками. Эпимериты заметно-короче эпимер II. Голени и колени нормального строения, не образуют пластинчатых раструбов, перистые щетйнкн ног довольно тонкие; базальная щетинка колен III нормального строения (не чешуевидная); вентральные щетипки голеней III и IV сидят на порядочном расстоянии от дистального-края членика; вершинная щетинка колеи I в б—6 раз длиннее, чем. расположенная рядом с ней обонятельная палочка, но пе заходит концом за вершину колен.
cL Эпцандрий с коротким, слегка расширенным или раздвоенным па. конце отростком; гепптаЛьиые щетинки и рг а очень короткие (в 5—6 раз-короче sh); обонятельная палочка колен II не увеличена. Длина 340—430 р.
$. Длина генитального отверстия приблизительно равна расстоянию-между ним и анальной щелью; эппгиний полулунный; задняя пара гени-
3. GLYCYPHAGUS 297
тальпых щетинок помещается заметно позади края генитального отверстии. Длина 380—550 [л.
Покоящийся гипопус. Тело у живых особей овально!', в I ,з • 1.4 раза длиннее своей шпрппы, с тупоугольным передним краем и довольно явственной поперечной бороздкой, разделяющей его на проподосому и гистеросому; в препаратах шарообразное, без всяких следов разделения; вершина голеней I—III дорзалыю с небольшим коническим бугорком; базальная крючковидная щетинка лапок I едва длиннее апикальной, последняя в 2 раза длиннее коготка. Длина 220—260 р (в препаратах до 300 р).
Распространение. Вся Европа и палеарктическая Азия; может быть, заносится вместе с продуктами и в другие страны, по положительных данных об этом в литературе пет.
Экология. Один из самых распространенных и обычных видок амбарных клещей, живущий в запасах зёрна, круп, маслосемян, сухофруктов и других растительных материалов, а также в сепе и соломе; часто встречается также в коллекциях насекомых, ла сухих шкурках млекопитающих и других музейных объектах. В полевых условиях встречается значительно реже, менее обилен и заселяет преимущественно скопления субстратов, находящиеся вблизи жилья; известны отдельные случаи его нахождения в лесных и горных районах [например в Нарыпском районе Киргизской ССР, на дупеле Capella (Gallinago) solitaria Gr. ], но в основном, распространен в поле преимущественно в пределах земледельческой полосы. Экология G. destructor до сих пор никем пе изучалась; экспериментально-установлены только исключительно высокая жизнестойкость гипопусов-этого вида по отношению к отравляющим веществам (Смирнов и Полежаев,. 1936) и их долговечность (Moniez, 1889).
Характер и степень вредоносности G. destructor пе установлены; известно, что клещи могут выгрызать небольшие углубления на. поверхности зерен, но проникать в них (например в зародыш) они, видимо, не могут; высказано также предположение, что G. destructor существует в зерне,, главным образом, за счет пылевидной сорной примеси.
2. Glycyphagus (Lepidoglyphus) pilosus Cuds. (фиг. 513).
Oudemans, 1906, Entomol. Ber., 11:122.— setosus Oudemans, 1904 (non 0. L. Koch, 1841X-Tijdschr. Entomol., XLVII: 127, t. 8, fig. 5.1—53.
Очень близок к предыдущему, по гораздо мельче. Все щетинки идиосомы Много длиннее: d3, d4 и sa е в 2 раза, 13 в 1.5 раза длиннее идиосомы, dx и 1а. приблизительно равны ее длине; рг а и задняя пара генитальных щетинок только в 2 раза короче sh; р15 напротив, много короче, чем у G. destructor, едва больше рг а. Псевдостигмальный орган с утолщенным стволом, расщепленным дистально на 2 короткие, толстые ветви с короткими, слабо ветвистыми боковыми отростками. Стерпум и эпиапдрий отсутствуют; эпимеры П почти в 3 раза длиннее, чем эпимериты II. Строение и пооружю нпе ног,'Как у G. destructor. Длина 255 р, $ неизвестна.
598
III. GLYCYPtiAGIDAE. 2. GLYCYPilAGINAE
Распространение. Описан из Германии (Vegesack около 'Бремена). Единственный известный экземпляр этого вида был найден в муке вместе с Tyroglyphus farinac (L.).
*3. GlycyplnigUH (Lepidoglyphus) fustifer Ouds. (фиг. 480, 481).
Oudomans, 1.903, Entomol. Bor., 1:103; 1.904, Tijdschr. Entomol., XLVII: 129, t. 8, fig. 54—56.
V p расставлены так Жо широко, как sc i, так что их основаниями намечается фигура прямоугольника; пространство между v р и sc i в б раз меньше расстояния между последними и в 3—4 раза меньше расстояния между v р и v а; базальная чашечка d3 без отростка. Перистые щетинки идиосомы приблизительно такой же длины и толщины, как у G. destructor, d2 немного заходит концами за задний абрис тела; все 3 пары р а длинные, перистые. Эпимериты одинаковой длины с эпимерами II. Голени и колени нормального строения, не образующие пластинчатых раструбов; перистые щетинки голеней и колен I и II гораздо, шире', чем у G. destructor, базальная щетинка колен III представлена простой перистой щетинкой, не модифицирована; вентральные щетинки голеней III и IV сидят заметно отступя ют дистального края членика; вершинная щетинка колен I чрезвычайно короткая, едва длиннее расположенной рядом с нею мелкой обонятельной палочки.
сГ. Эпиандрий с коротким линейным отростком; генитальные щетинки и рг а короткие; обонятельная палочка колен II гипертрофирована, имеет вид массивного шипа, длиною с само колено. Длина 300—400 [л.
9- Генитальное отверстие очень крупное, значительно длиннее расстояния, отделяющего его от анальной щели; эпигиний полукруглый; задняя пара генитальных щетинок расположена на одной линии с задним краем генитального отверстия. Обонятельная палочка колен II нормального размера. Длина 415—180 [л. — Покоящийся гипопус неизвестен.
Распространение. Описан по 1 с? из окрестностей Бремена /Германия); в СССР найден пока только 1 раз в Ивановской обл. (Болотове).
Экология. Синантропный вид; первый раз.найден в жилом доме, второй — в колхозном амбаре, в семенной ржи, в довольно большом коли-честсе.
Возможно, что найденная у нас форма не идентична с бременской, так как Oudemans описывает и изображает гипертрофированную обонятельную палочку колен II сГ колбасовидной, т. е. почти цилиндрической, с закругленным концом, тогда как у наших экземпляров она слегка расширена в базальной четверти и постепенно суживается к своему немного приострен-
Фиг. 472—481.
Фиг. 472. Glycyphagus (Lepidoglyphus) destructor (Schrk.), нога II сУ сбоку. Фиг. 473. То же, с отошедшей центральной чешуей (чехлом). Фиг. 474. GI- (L.) destructor (Schrk.), нога III 9.
-Фиг. 475. Gl. (L.) destructor (Schrk.), генитальное отверстие d" • Фиг. 476. Gl. (L.) michaeli Ouds., генитальное отверстие о и окружающие его щетинки. Фиг. 477. GI. (Ь.) michaeli Ouds., нога III о. Фиг. 47В. GI. (L.) michaeli'Quds., голень и основание лапки IV снизу. Фиг. 479. Gl. (L.) burcha--nensis Ouds., колено III с ланцетовидной чешуей. Фиг. 480. GI. (Ь.) fustifer Ouds., нога II О'.
Фиг. 481. Gl. (L.) fustifer Ouds., генитальное отверстие о и окружающие его щетинки.
300
III. GLYCYPI1AGIDAE. 2. GLYCYPHAGINAE
ному концу; поскольку других уловимых различий между ними нет, я на счел возможным придать им значение самостоятельных форм (resp. подвидов).
*4. Glycyphagus (Lepidoglyplius) miehaeli Ouds. (фиг. 74, 76, 476—478).
Oudemans, 1903, Entomol. Bor.,Г: .1.03. —npinipes Michael, 1888, Journ. Linn. Soc. Lond.,. XX: 285, t. 16, fig. 8-15; .1901: 245-250, t. VII—VIII.
Самый крупный вид рода КЧ?д-2/2>/ш//пя. Vp расположены заметно ближе друг к другу, чем sc i, так что основаниями этих 2 пар щетинок намечается фигура трапеции; промежуток между V р и sc i в 3 раза меньше расстояния между последними; базальная чашечка d3 без отростка; перистые щетинки идиосомы толстые, усаживающие пх реснички длиннее, чем у видов группы destructor; длина самых крупных из щетинок идиосомы превосходит не более чем в 1.5 раза ее длину; d2 не доходят до ее заднего края; рг крупные, вдвое длиннее рх и в 6—8 раз длиннее очень мелких ра. Эпимериты длиннее эпимер II. Дистальные края голеней пластинчато расширены, образуя широкий раструб; аналогично, но в более слабой степени, расширены и концы колен; перистые щетинки пог очень широкие, толстые; базальная щетинка колен III превращена в очень крупную продолговато-яйцевидную чешую, длина которой только в 2 раза больше максимальной ширины; наружная поверхность этой чешуи густо покрыта тонкими ресничками, имеющими приблизительно одинаковую длину и на ее диске и по краям; вентральные щетинки голеней III и IV сидят в особом вырезе, образованном пластинчатыми краями членика; вершинная щетинка колец I крупная (далеко заходит за основание лапки), расположенная рядом с нею обонятельная палочка очепь маленькая, приостренная к концу,
d, Эпиандрий хорошо развит, отросток его линейный; генитальные-щетинки и рг а очень короткие; обонятельная палочка колен II не гипертрофирована. Длина 450—460 р..
9. Длина генитального отверстия в 11/2—2 раза меньше расстояния между ним и анальной щелью; его задний край расположен иа уровне передних концов эпимер III, т. е. много дальше кпереди, чем у других видов;, эпигиний крупный, полулунной формы; задняя пара генитальных щетинок находится далеко позади генитального отверстия. Длина 600—810 р.
Покоящийся гипопус с округлым, не расчлененным телом,, образующим спереди узкий параболический ростральный выступ; базальная; крючковидная щетинка лапок I (по рис. 17 табл. VIII Michael’я) вдвое-длиннее апикальной, которая имеет приблизительно такую же длину, как; коготок. Длина 240 {л. Гипопальпая капсула заметно суживающаяся кзади,, почти веретеновидпая; ее дверца сплошь покрыта сетчатым орнаментом..
Распространение. Достоверно известен только из Англии гт СССР; у нас найден в Москве и ее окрестностях, в Ленинграде, окрестностях' г. Киржача (Ивановской обл.), в Кировской обл. и'во многих пунктах Башкирии.
3. GLYCYPHAGUS ПО I
Экология. Живет в сене и соломе в старых стогах, на солопплах и в конюшнях и во влажном зерне (овес, пшеница) в амбарах и их подпольно; местами (например, в лесной полосе Башкирии и в окрестностях Ленинграда) очень распространен в зернохранилищах, иногда даже преобладая в них над другими амбарными видами Glycyphagus. Более стеновалентный и гигрофильный вид, нем G. destructor и G. domesticus, невидимому тяготеющий к более влажным районам; самый быстрый в своих движениях из амбарных клещей.
* б. Glycyphagus (Lepidoglyphus) burchanensis Ouds. (фиг. 479).
Oudemans, 1903, Entomol. Вег., 1:103; 1904, Tijdschr. Entomol., XLVII: 131, t. 9, fig. 57—60.
Близок к G. michaeli, по значительно меньше. Расстояние между v р и sc i почти в 4 раза меньше пространства между последними и в 2 раза меньше расстояния между va и vp; базальные чашечки всех перистых .щетинок гораздо крупнее, у d3 их задний край вытяпут в продолговатую хитиновую площадку; перистые щетинки идиосомы немного длиннее; концы •d2 заходят за задний край тела. Эпимериты в 2 раза короче эпимер II. Строение и вооружение. ног, как- у предыдущего вида, но раструбы голеней развиты слабее, а чешуевидная базальная щетинка колен III имеет узколанцетовидную форму, около 6 раз длиннее своей максимальной ширины и окаймлена более длинной бахромкой.
сГ. Эпиандрий довольно длинный, линейный, суженный к концу; рх и р2 •одинаковой длины, в 2 раза короче р3. Длина 350—400 [л. — § неизвестна.
Распространение. Описан с о-ва Боркум (Германия). У нас найден в Смоленске и в Житомире.
Экология. Попадается единичными экземплярами в зернохранилищах, в семепах льна и в пшенице.
2. Подрод Glycyphagus s. str.
Захваткин, 1936, Бюлл. Моск. общ. псп. прпр., XLV: 264, 268—269.
Идиосома спереди с хитинизированным продольным теменным килем. D2 •сильно смещены назад, образуя с d3 поперечный ряд, или расположены .нормально впереди последних. Лапки без чехлообразной чешуи, с несколькими довольно крупными зазубренными щетинками, из которых самая .длинная расположена близ середины их вентральной поверхности; вентральная щетинка бедер I пушистая. —- Личинки с относительно крупными, цилиндрическими, явственно хитинизированными грудными палочками. — Покоящиеся гппопусы в виде округлого, нерасчлепенного мешка, совершенно лишенные пог, коксо-стернальпого скелета и генитальной щели. Стенки гипопальной камеры пигментированы и покрыты очень густой волнистой гравировкой.
Подрод Glycyphagus s. str. состоит из 3 широко распространенных видов, различающихся между собой гораздо более резко, чем виды предыдущего
302
III. GIACYPHAGIDAE. 2. GLYCYPHAGIXAE
подрода; особенно любопытны глубокие различия в онтогенезе подродовых. признаков, наблюдающиеся у различных его видов. »
’ 1
*6. Glycyphagus (s. str.) cadavcruin (Schrank) Ouds. (фиг. 48G—488). >
Schrank, 1781, Enum. Ing. Austr., n° 1055 (/1 .earus); Oudemans, 1905, Entomol. Ber.,I: 238;.
1913, ibid., Ill: 328. —prunontm Tiering, 18118, Act. Acad. Leop., XVIII, 1: 619, t.45, fig. 16— 17.— privates Oudemans, 1903, Entomol. Ног., 1:103; 1904, Tijdschr. Ned. Dierk. Ver.,VIII: 232..
V а сидят па переднем конце длинного, линейного теменного киля, тянущегося назад до уровня но i;‘расстояние между последними и v р в 2—3 раза, меньше промежутка между v р и v а; (12 расположены, как у видов подрода Lepidoglyphus, т. е. много впереди d3, ла одной поперечной линии с 1х и на таком же расстоянии друг от друга, как от 1х. Перистые щетинки идиосомы довольно тонкие, гибкие; длина самых крупных из них равна длине идиосомы или немного превышает ее; рх более чем в 2 раза короче р2,. р3 немного длиннее, чем р2; все перистые. Голени не расширены в виде раструбов; перистые щетинки ног относительно тонкие; вентральные щетинки голеней III п IV отстоят на заметное расстояние от дистального края членика; вентральные щетинки лапок IV расположены на одинаковом расстоянии друг от друга; обонятельная палочка колен I и II короткая,, почти коническая; обонятельная палочка лапок II не утолщена.
<?. Эпиапдрий с очень коротким отростком; щетинки задней генитальной пары и рг а короткие; лапки I и II нормальные; перистые щетинки голеней I и II не модифицированы. Длина 280—360 ц.
Q. Генитальное отверстие очень крупное, простирающееся назад до заднего края ацетабул IV; расстояние, отделяющее его от анальной щели, заметно уступает его длине; эпигиний треугольный; задняя пара генитальных щетинок расположена на одной линии с задним краем генитального отверстия. Длина 375—450 р.. — Гипопус и другие неполовозрелые стадии неизвестны.
Распространение. Достоверно известен только из Голландии и СССР (Киржач, Ивановской обл.; Смоленск и ряд других пунктов Смоленской обл.; Калининская обл.; Башкирия).
Экология. Живет в домах и в складах, преимущественно в семенах кормовых трав (клевер) и льна, реже в зерне; в окрестностях г. Киржача найден также в поле на току, в кучах отходов от провеивания ржи.
* 7. Glycyphagus (s. str.) domesticus (Deg;) (фиг. 77, 482—485).
De Geer, 1778, M6m. hist. Ins., VH:87, n° 1, t. 5, fig. 1—11 (Acarus); Berlese, 1884, XIV,- ,
n° 3; Michael, 1901: 238, t. VI, VIII, fig. 3, 7—10, 12, 14, 16; Oudemans, 1904, Tijdschr. Ned..
Dierk. Ver., (2), VIII: 21.8, t. IX, fig. 28—34.—hippododos Hering, 1838, Act. Acad. Leop.,.
XVIII,.1: 607, t. 44,fig, 11 (Samples). —cursor Gervais, 1841, Ann. Sci. Nat., s4r. II, vol. XV: 18, t. 2, fig. 15; Fumouze ot Robin, 1867, Journ. Anat. Physiol., V: 569, 573; Troupeau,. 1879:110, t. 2, fig. 12—16, 18—19.
Биология. Michael, 1888, Journ. Linn. Soc. Lond., XX: 285; Hora, 1934, Ann. appl.. ;
Biol., XXI, n° 3:483—494.
V а сидят на особом расширении (area sensilligera) посередине теменного киля, на значительном расстоянии от переднего края идиосомы; перед
3. GLYCmiAdliS
няя половина киля (лейсащая кпереди от v а) широкая, слнЛон xilTIliiinui рована, чем более узкая задняя, идущая назад до уровня v р; piuMiToiiuiio между последними и sc i только в 1.5 раза меньше промежутка между обппмп sc i и менее чем в 1.5 раза меньше расстояния, отделяющего v а, от v р; d2 помещаются между d3, образуя с ними прямой поперечный ряд; расстояние-между обеими d2 почти в 2 раза меньше такового между каждой из пах п 1(, Перистые щетинки идиосомы довольно толстые, жестковатые; самые крупные из пих равны по длине идиосоме, редко немного длиннее ее; рг и р2 короткий, примерно одинаковой длины, в 2—2.5 раза короче р3. Голени нормального строения, не расширенные в виде раструбов; перистые щетинки ног довольно короткие и толстые; вентральные щетинки голеней III и IV сидят заметно отступя от дистального края членика; вентральные щетинки лапок IV расставлены на одинаковое расстояние друг от друга, обонятельная палочка колен I и II длинная, тонкая, цилиндрическая; обонятельная палочка лапок II значительно крупнее таковой лапок I.
с?. Эпиандрий спереди вытянут в квадратную площадку; задние генитальные щетинки и рг а длинные (почти равны по длине анальной щели); лапки и вентральные щетинки голеней I и II имеют нормальное строение. Длина 270—350 |г.
9. Генитальное отверстие меньше, чем у G. cadaverum, едва заходит за. уровень заднего края ацетабул III и короче промежутка, отделяющего его от анальной щели; эпигиний полулунный; задняя пара генитальных щетинок находится значительно позади заднего края генитальной щели. Копулятивная трубка очень широкая, цилиндрическая, перепончатой консистенции. Длина 350—680 ji.
У личинок теменной киль имеет такое же строение, как у взрослых, но слабее хитинизирован; d2 также занимают уже дефинитивное положение. — Гипопальная камера окрашена в бледный дымчато-буроватый цвет.
Распространение. Вся Европа и палеарктическая. Азия; найден также в Арктике (Земля Франца-Иосифа, Шпицберген, Гренландия) и на Огненной Земле, куда, вероятно, завозился с продуктами или. самими собиравшими его исследователями. Возможен завоз его и в другие страны..
Экология. Синантропный, преимущественно домовый вид, живущий на самых разнообразных сухих веществах растительного и животного происхождения и иногда размножающийся на них в таком огромном количестве, что приобретает серьезное хозяйственное значение. В амбарах и зернохранилищах встречается значительно реже, чем G. destructor, и ни играет видной роли в специально зерновом комплексе тироглифоидий. Был находим также в скирдах, стогах, ометах соломы и- т. п. Но данным Нога (1934), оптимальные для G. domesticus условия создаются при темпе ратуре около 23—25° и относительной влажности в 80—90%; при пляж ности в 60% клещи быстро гибнут, а при 70% развиваются слабо; при оптимальной температуре и влажности инкубационный период длится около 5, а постэмбриональное развитие — около 17 дней. Образование покоящихся
:to;
III. GLYCYPIIAGIDAL. 2. GLYCYPIIAGLYAH
гипопусов происходит независимо от внешних условий, и через эту стадию проходит около 40% всех особей; длительность гппопальной стадии колеблется от 5 дней до 6 месяцев.
=!'8. Glycyphagus (я. sir.) ornatus Кгатп. (фиг. 51, 489—492).
Kramw, I8HI„ Ztwhi'. ден. NulurwlHH,, LIV: '186; 1899:146; Berlesc, 1886, XXIX, n° 3; Cane-strini, 1888a: 1)85.
V а СИДЯТ на особом |).'ie!!!iipeiii!!i Посередине теменного КИЛЯ, ДОВОЛЬНО .далеко от переднего края идиосомы; обе половины киля одинаковой формы, суживающиеся к концу, сильнее хитиппзнрованы, чем у G. domesticus, задняя тянется пазад до уровня vp; расстояние между последними и sc i очень •немного уступает ширине промежутка между обеими sc i и расстоянию пиежду v а и v р. Da помещаются между d3, даже немного позади них, образуя с ними вогнутую кпереди дугу; промежуток между7 d2 в 2 раза меньше их расстояния от 1Г Перистые щетинки идиосомы толще п длиннее, чем у соседних видов, жестковатые; рх и р2 одинакового размера, довольно .длинные, вдвое короче р3; все перистые. Голени дистально слегка расширены, образуя неширокие раструбы; перистые.щетинки ног толстые,-сравнительно длинные; вентральные щетинки голеней III и IV сидят в глубоких вырезах дистальпого края членика (как у G. michaeli и G. hurchanensis); на лапках IV обе проксимальные щетинки сильно сближены между собой и .далеко отодвинуты от дистальной; обонятельные палочки колен I и II длинные, топкие, цилиндрические; обонятельная палочка лапок II значительно крупнее соответствующей сенсиллы лапок I.
с?. Эпиапдрий слабо развит, с коротким тупым отростком. Лапки I и II резко S-образно искривлены; наружная вентральная щетинка голеней I п II превращена в крупный треугольный гребень, несущий 8—10 (на голе-.пях I) или 4—5 (на голенях II) острых зубцов; внутренняя вентральная .щетинка тех же голеней укорочена. Длина 430—500 р.
9, Длина генитального отверстия значительно меньше расстояния .между ним и анальной щелью; его задний край находится на уровне эпп-.мер III; эпигиний небольшой, полулунный, с оттянутыми в стороны концами; -задняя пара генитальных щетинок расположена много позади генитального -отверстия. Вентральные щетинки голеней I и II не модифицированы, лапки прямые. Копулятивная трубка узкая, заметно суживающаяся к концу, -с утолщенными стенками. Длина 510—600 р.
Личинки без всяких следов теменного киля, с d2, расположенными, .как у G. cadaverum п видов подрода Lepidoglyphus (т. е. много впереди d3); все щетинки идиосомы, кроме d3, очень короткие. Зачаток теменного киля
Фиг. 482—489.
Фиг. 482. Glycyphagus (s. str.) domcalicus Deg., 2 сверху. Фиг. 483. Gl. (s. str.) domesiicus Deg., нога I O'. Фиг. 484. Gl. (s. str.) domesticue Deg., теменной киль. Фиг. 485. Gl. (s. str.) domesticus Deg., голень и лапка IV 9. Фиг. 488. Gl. (s. str.) cadaverum (Schrk.), О сверху. Фиг. 487. Gl. Xs. str.) cadaverum (Schrk.), теменной киль. Фиг. 488. Gl. (s. str.) cadaverum (Schrk.), голень н -лапка IV 9- Фиг. 489. Gl. (s. str.) ornatus Kram., теменной киль. Буквенные обозначения тс;ке, что к на фиг. 27.
20 Фауна СССР, Захвати ин—284
306
III. GLYCYPHAGIDAE. 2. GLYCYPHAGINAE
появляется только у протонимф, у которых он имеет вид узкой ланцетовидной полоски хитина, лежащей позади v а, и только у телеонимфы он получает свое дефинитивное строение; у протонимф d2 расположеныеще заметно-впереди d3, но ужо ближе к ним, чем у личинок. — Гипопус неизвестен.
Распростри и е и и о. Зарегистрирован в Германии, Англии, Голландии, Италии и СООР. У нас в Союзе найден в Москве и ее окрестностях, Ленинграде, Киеве и в Башкирии (Стерлитамакский район).
Эко л о г и я. Основным местообитанием итого вида являются поры грызунов и насекомоядных (крот, землеройки), в подстилке которых встречается иногда большими колониями; кроме того; G. ornaius живет в трухе сена в. сеновалах и конюшнях, а изредка встречается также в зернохранилищах, во влажном зерне (пшеница) ir маслосеменах; Poppe (1906) нахо-дил;>его ещё йа летучих-мышах, a Newstead и Duvall — в соре, счищенном в лошадей. / • '' .
4. Род COMETACARUS A. Z. ' '
- Захваткин, 1936, Бюлл. Моск. общ. исп. прир., XLV: .266 , 269. ' •
Тип рода: Cometacarus smirnovi, sp. n.
Характеризуется совершенно необычайной длиной перистых щетинок идиосомы, часть которых (d3, d4 и sa е) превосходит в 2.5—3 раза общую длину тела и образует волочащийся за ним «хвост», напоминающий хвост кометы. Хелицеры сильно вытянуты в длину, очень узкие; на обоих пальцах клешни вершинные зубцы отделены очень широким гладким пространством от коренного зубца (на неподвижном пальце) или соответствующей ему выемки (на подвижном). Перистые щетинки идиосомы уплощены дорзо-вентрально, опушены преимущественно по краям; v а широко раздвинуты, расположены на самом краю идиосомы; v р тесно сближены между собой; теменного киля нет; sc близко придвинуты друг к другу, образуя прямой поперечный ряд; 3 пары плечевых (h a, h р и sb); d2 сильно смещены назад, располагаясь между d3 и образуя с ними Поперечный ряд; р а довольно короткие, все, примерно, одинаковой длины, перистые. Псевдостигмальный орган от самого основания разделен на 2 ствола. Наружные концы эпимери-тов II елйты с эпимерами III; от точки стыка их внутренних концов с эпимерами IV отходит внутрь тонкий отросток, крючковидно загибающийся вперед. Лапки без пушистого чехла,, с 3—4 мелкими зазубренными щетинками (3 вентральными и 1 дорзальной на лапках I и II и только 3 вентральными на лапках III и IV); обонятельная палочка лапок I и II сильно веретеновидно вздутая; вершинная дорзальная щетинка голеней III короткая.
Фиг. 490—500.
Фиг. 490. Glycyphagus (s. str.) ornaius Kram., нога I д'. Фиг. 491. Gl. (s. str.) ornaius Kram., гребневидная щетинка голени II ci. Фиг. 492. Gl. (s. str.) ornatus Kram., голень и лапка IV 9. Фиг. 493. Cometacarus smirnovi, sp. n., O’ сверху. Фиг. 494. Cl. smirnovi, sp. П., хелицера $. Фиг. 495.
C. smirnovi, sp. n., cf снизу. Фиг. 400. C. smirnovi, sp. n., генитальный аппарат cC • Фиг. 497.
C. smirnovi, sp. n., нога I О'- Фиг. 498. C. smirnovi, sp. n., голень и лапка IV Q. Фиг. 499.
C. smirnovi, sp. n., то же. Фиг. 500. С. smirnovi, sp. п., генитальное отверстие О.
*
308 III. GLYCYPHAGIDAE. 2. GLYCYPHAGINAE
cf. Генитальное отверстие расположено между основаниями пог IV, с хорошо развитыми овальными щупальцами; эпиандрий отсутствует.
$. Гепитальные клапаны более или менее редуцированные, не смыкающиеся но средней линии тела.; генитальные щупальцы отсутствуют; эпи-гиний крышеппдпын; передняя пара генитальных щетинок расположена по бокам пингиння.
Сюда относятся а очень редких вида, ив которых 1 недостаточно известен и настоите,льпо требует вереоппсавня.
ОПРЕДЕЛИТЕЛЬНАЯ ТАВЛИЦА ВИДОВ
1 (2,).. Крайне мелкий вид: общая длина тела—<3 175 ц, $ 220 р; v р одинаковой-длины с sc i и на х/3 длиннее sc е; d2 в З раза короче d3; перистые щетинки ног очень крупные, наружная базальная щетинка колен I равна по длине лапке; сГ без опахаловидных пластинок на бедрах I и II .........................................2. С. oudemansi A. Z.
2 (1). Гораздо крупнее: общая длина тела— с? 400—420 р, $ 500—510 р; v р в 2 раза короче sc е и в 3 раза короче sc i; d3 в 2 раза короче d3; перистые щетинки ног умеренной длины, наружная базальная щетинка колен I вдвое короче лапки; вентральные щетинки бедер I и II у сГ превращены в крупные опахаловидные пластинки.........................
. . :....................................*1. С. smirnovi, sp. n.
С. setosus (0. L. Koch 1841) описан слишком поверхностно, чтобы его можно было включить, в таблицу.
*1. Cometacarus smirnovi, sp. н. (фиг. 493—500).
Захваткин, 1936, Бюлл. Моск. общ. исп. прпр., XLV: 265 (nom. nudum).
Довольно крупный вид. V р в 2 раза короче sc е и почти в 3 раза короче sc i; hi в 1.5 раза короче d2; последние только в 2 раза короче самых крупных щетинок идиосомы — d4, и едва больше чем в 1.5 раза уступают в размерах d3; d4 не более чем в 2.5 раза длиннее тела (включая гнатосому). Ноги I и II сравнительно немного мощнее задних; нижний дистальный угол колен I и II оттянут в остроугольный, резко выступающий зубец; перистые щетинки ног относительно короткие и тонкие; обонятельная палочка лапок I и II с закругленным концом.
(5. Генитальное отверстие расположено между основаниями ног IV, снабжено парой крупных овальных генитальных щупальцев. Penis очень тонкий, слабо S-образно изогнутый; эпиандрий отсутствует. Вентральные щетинки бедер I и II превращены в крупные опахаловидные пластинки, продольно сложенные пополам, с очень глубоко надрезанными, гребневиднозубчатыми краями; сильно модифицированы и вентральные щетинки голеней II, из которых наружпая имеет форму гребня с 5—6 тонкими отростками, а очень маленькая внутренняя дланевидно рассечена на 4 игольчатые доли; вентральные щетинки голеней I укорочены, но не модифицированы. Длина идиосомы 320—340 ц, общая длина тела 400—420 ц.
4. COMETACARUS
ЗОН
9. Генитальное отверстие относительно небольшое, целиком, умни,потен в пространстве между основаниями ног III и IV; эпигиний маленький, в соот ветствии с чем сидящие по его сторонам передние генитальные щетинки мало удалены друг от друга; вентральные щетинки всех голеней и колен нормальные. Длина идиосомы 400—430 р., общая длина тела около 500—510 р..
Распространение. Найден пока лишь 1 раз, в г. Иванове. Небольшая колония этого замечательного вида была обнаружена в семенах пастернака, в складе.
.2. Cometacarus oudemansi A. Z. (фиг. 514—517).
Захваткин, 1936, Билл. Моек. общ. исп. прпр., XLV: 265.— seiosus Oudemans, 1907, Abb. naturwiss. Ver. Bremen, XIX: 60, t. II, fig. 6—11 (non G. L. Koch, 1841) (Glyeyphagus).
Крайне мелкий вид, много уступающий в размерах всем другим представителям семейства, за исключением только карликовых СсГ рода Orycte-roxenus. V р одинаковой длины с sc i и на 1/3 длиннее sc е; h i едва короче, чем d2, эти последние в 3 раза короче d4 и почти в 2.5 раза короче d3; d4 в 3 раза длиннее тела (с гнатосомой). Ноги I и II значительно массивнее очень тонких ног III и IV; нижний дистальный угол колен I и II закругленный, совершенно не выступающий; перистые щетинки ног очень крупные, наружная базальная щетинка колен I и II равна по длине лапкам; обонятельная палочка лапок I и II с тонко заостренным концом.
сГ. В работе Oudemans’а сказано только, чтоб1 отличается от детально описываемой и изображаемой им 9 лишь гениталиями; отсюда можно сделать вывод, что вентральные щетинки голеней и бедер I и II у него не модифицированы. Общая длина тела 175 р..
9. Генитальное отверстие очень крупное, значительно выдающееся кпереди за концы эпимер III; расстояние между его задним концом и анальной щелью в 2.5 раза меньше его длины; эпигиний крупный, сидящие по его сторонам щетинки так же широко раздвинуты, как средние и задние генитальные щетинки; все перистые щетинки ног нормальные. Длина идиосомы около 165 pi, общая длина тела 200 рь — Неполовозрелые стадии неизвестны.
Распространение. Германия (окрестности Бремена). Местообитание не указано.
3. Cometacarus setosus (Koch) (фиг. 518).
0. L. Koch, 1841, Dtsch. Crust., Myriap., Arachn., XXXIII, no 3 (.learn).
Совершенно недостаточно известный вид, отличающийся от обоих предыдущих длинными, булавовидно, утолщающимися к концу базальными перистыми щетинками колен I и II. Размеры тела не указаны. Германия, в старом сене.
310
III. GLYCYPHAGIDAE. 8. AEROGLYPHINAE
3. Подсем. Aeroglyphinae
Хелицеры умеренной величины, довольно массивные, напоминающие по своей конфигурации таковые Chorioglyphinae. Идиосома широко-эллиптическая, уплощенная, без поперечной бороздки и рострального выступа. Кожа покрыта очень тонкой и густой поперечной штриховкой, отсутствующей только в области ясно намоченного проподосомального щита; спинная поверхность тела, усажена, кроме того, многочисленными треугольными шипиками. Щетинки спинной поверхности идиосомы умеренной длины (около 20—50% длины идиосомы), однотипные, несколько уплощенные, густо- и тонкопушистые, особенно по краям; sae и sai длиннее и тоньше других; vp расположены латерально, на одной поперечной линии ста, значительно уступают им в длине; плечевых и боковых щетинок по 3 пары, постанальных у сГ 2 пары, у g — 4; расположение их не уклоняется от нормы. Псевдостигмальный орган отсутствует. Генитальные, щупальцы мелкие, но вполне явственные. Коксо-стернальный скелет развит довольно слабо; стернум и эпимеры II—IV короткие, свободные; концы эпимеритов II соединены с эпимерами III. Ноги умеренной длины, довольно стройные; лапки чуть искривлены; щетинки колен и голеней слегка перистые; вершина колен I с 2 сенсиллами: маленькой обонятельной палочкой и расположенной рядом с нею длинной жесткой щетинкой; вершинные щетинки голеней II и III очень короткие. Praetarsi небольшие, с относительно крупными серповидными коготками. Вторичные половые различия очень слабо выражены.
сГ. Генитальное отверстие расположено между основаниями ног III и IV; penis очень маленький; эпиандрий отсутствует.
$. Генитальное отверстие помещается в пространстве между основаниями ног III и IV, довольно крупное, без эпигиния и циркумгенитального кольца. •— Неполовозрелые стадии неизвестны.
... К этому подсемейству относится только 1 род АёгодЪуplius, gen. n.,1 .биологически связанный с перепончатокрылыми; в отличие от других сим-фильных форм, Aeroglyphus расселяется в половозрелой стадии, переносимой хозяевами.
5. Род AEROGLYPHU S, gen. п.
Тип рода: Glycyphagus peregrinans Berlesc 1892.
Довольно крупные формы. Почти вся спинная поверхность идиосомы усажена мелкими треугольными сквамоидами, которые несколько увели-
1 Возможно, что сюда же следует отнести и североамериканского Glycyphagus rdbustus Banks 1906, но он недостаточно описан, чтобй это можно было утверждать.
Фиг. 501—507.
Фиг. 501. ASroglyphus peregrinans (Borl.), сверху. Фиг. 502. То же, снизу. Фиг. 503. A. peregrinans (Berl.), Q снизу. Фиг. 504. A. peregrinans (Berl.), нога I сверху. Фиг. 505. A. peregrinans (ВегЬ), нога III снизу. Фиг. 500. A. peregrinans (Berl.), нога IV сбоку. Фиг. 507. Ctenoglyphus plwmiger (С. L. Koch), нога I о".
512
111. GLYCYPHAGIDAE. 4. CTEAOGLYPHIAAE
чиваются в размерах и сгущаются по периферии занятой ими площади, придавая ей очень четкие границы; в передней части идиосомы кожа заметно уплотнена, образуя нроподосомальный щит с 2 невысокими гладкими продольными килями, которые простираются назад до уровця sc и затем дугообразно расходятся в стороны и сходят на-нет. Н а и h р помещаются дор-зально, sh— латеральио; размеры d и 1 постепенно убывают спереди назад, так что dj и lj длиннее других щетинок этих групп.
Мопотилический род.
*1. Aeroglyphus peregrinans (Berl.) (фиг. 501—506).
Berlese, 1892, LXV, n° 1; Kramer; 1899: 147 (Glycyphagus).
Идиосома в 1.25—1.3 раза длиннее своей ширины, с едва намеченными плечами и равномерно выпуклыми задними и боковым краями. Окраска бледная, с легким зеленоватым оттенком; ноги непигментированные. V а более чем в 1.5 раза длиннее три приблизительно равны по длине sc i; sc е немного длиннее sc i, образуют с ними прямой поперечный ряд. Примерная длина щетинок идиосомы, в процентах к ее длине: v а 26—30; v р 18—20; sc е 33—42; sc i 28—30; h а 33—37; sh 26—28; h p 38—41; d4 50—54; d2 48—50; d3 43—48; d4 37—40; Д 44—48; 12 38—45; 13 33—35; sa e 60—66; sai 54—60. Длина d 350—370 p, ширина 280—300 p; длина 9 до 415 p, ширина до 315 p.
Распространение. Описан из Италии; мне известен из Московской обл. (с. Лужки на Оке).
Экология. dd и 99 встречаются небольшими группами на Xylo-сора violacea L., X. valga К., а иногда также на шмелях (Bombus lapidarius L.).
4. Подсем. Ctenoglyphinae
Очень мелкие формы (180—300 р). Строение хелицер не уклоняется от нормы. Идиосома широкая, уплощенная дорзо-вентрально, с более или менее явственной разделяющей поперечной бороздкой или без таковой. Кожа тонкощагреневая, резкоморщинистая или усаженная крупными бородавками, более или менее сильно пигментированная, плотная. Щетинки спинной и боковых поверхностей идиосомы крупноперистые, опахаловидные (крупно двусторонне-гребенчатые) или листовидные; v р расположены! дорзалыго, позади v а (иногда отсутствуют); плечевых щетинок обычно. 3 пары, боковых — б пар; все они расположены маргинально, так же как и постанальные, и образуют в своей совокупности пышную бахрому вокруг тела; крестцовые иногда, смещены на спинную сторону идиосомы, всегда относительно короткие; исевдостигмальный орган отсутствует. Генитальные щупальцы мелкие, но вполне явственные. Коксо-стернальный скелет хорошо, развит, отличный по своему строению у d и $. Ноги довольно массивные, умеренной длины или длинные (у d Ctenoglyphus); щетинки ног гладкие,
ТАБЛИЦА РОЛОВ 313
тонкие; дорзальные вершинные щетинки голеней I и II олень длинные, вентральные иногда отсутствуют; вершина колен с 2 сенсиллами. Priieluini. небольшие; коготки среднего размера. Морфологические различия между <Г и ф часто очень велики и сказываются в размерах тела (сШ часто имеют карликовые размеры), форме щетинок идиосомы, строении коксо-стернального скелета, длине ног и т. д.
<5. Penis трубчатый, обычно очень длинный, лежащий в свободном пространстве между коксальными полями I—III, иногда достигающий концом до гнатосомы; эниандрий отсутствует.
9. Генитальное отверстие широкое, расположенное в пространстве между основаниями ног III и IV, с хорошо развитым эпигинием.
Нимфы характеризуются менее сложным строением щетинок. -—Гипопусов не образует, насколько известно, ни один вид; расселение осуществляется половозрелой формой, отличающейся высокой парусностью и, соответственно, легко разносимой ветром. Почти все члены подсем. Ctenoglyphinae живут в сене или пылевидных растительных остатках. L
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
1 (2). По периферии идиосомы расположены всего 9 пар асимметричных: листовидных щетинок с резкозубчатым наружным краем; v, sc и d клиновидные, с прямо усеченным зубчатым концом.....................
.....................................8. Nycteriglyphus, gen. в.
2 (1). По периферии идиосомы расположены 11—12 пар широкоперистых, опахаловидных или двусторонне-симметричных листовидных щетинок; v, sc и d такой же формы, как периферические щетинки, или тоньше ; их и покрыты густым мелким опушением.
3 (4). С С по общему виду и строению щетинок мало отличаются от 99 * у обоих полов кожа спины тонкошагреневая, щетинки спинной поверхности узкие, уплощенные, тонкоперистые по краям; v, sc и шире-других....................................6. Diamesoglyphus, gen. n...
4 (3). cTcf резко отличаются от 99 по общему виду и строению щетинок: идиосомы; кожа спины у них тонкозернистая или почти гладкая,-щетинки спинной поверхности длинные, относительно узкие, идиосома -, довольно выпуклая, с округлыми очертаниями; у 99 идиосома сильное уплощена, почти пятиугольная, кожа спины грубоморщинистая или. усажена высокими цилиндрическими бородавками с расщепленным концом, щетинки спинной поверхности (по крайней мере периферические) очень широкие, опахаловидные или листовидные...............
.........................................7. Ctenoglyphus Beil..
1 К этому же подсемейству, невидимому, мояяю отнести и Glycyphagu» (f)ennaeams) fuli-g-ineus Berlese, 1921 (Италия).
314
III. GLYCYPBAGIOAE. 4. CTENOGLYPBINAE
6. Род DIAMESOGLYPHUS, gen. n.
Тип рода: Glycyphagus intcrmedius Canestrini 1888.
Мелкая форма с совершенно не выраженным половым диморфизмом. Идиосома широко-эллиптическая, с слабо намеченными плечевыми выпуклостями и широко закругленным задним краем, без дорзальной поперечной бороздки. Кожа очень тонко шагреневая. Щетинки спинной поверхности идиосомы однотипные, уилошонпыо, но довольно изменчивые по своей ширине, топко- и густоперистыо по краям; v а помещаются дорзально, на некотором расстоянии от передних углов идиосомы, такого же строения, как vp; все плечевые и боковые расположены строго латерально; d3 расставлены так же широко, как d2; sa е и sa i сидят терминально; р а перистые, расположенные терминально. Все коксальные поля открыты.
сГ. Генитальное отверстие находится между основаниями ног IV; penis вредней длины, конически расширяющийся к основанию.
Q. Генитальное отверстие помещается в пространстве между основаниями ног III и IV, широкое, с явственным круговым контуром и слабо развитым эпигинием; копулятивная трубка редуцирована. —Нимфальные стадии неизвестны.
К роду Diamesoglyyhus относится 1 еще недостаточно известный вид, •описанный из Италии.
1. Diamesoglyphus intermedins (Сап.) (фнг. 527).
Canestrini, 1888 : 20; 1888а: 386, t. 26, fig. 2; Kramer, 1899: 147 (Glycyphiagus).
Отношение длины идиосомы к ее ширине равно приблизительно 1.4— 1.5 : 1. Ширина щетинок спинной поверхности постепенно убывает.кзади; v, sc и h а шире всех других; v а вдвое короче v р, имеющих одинаковую .длину с sc 1; see заметно короче sc i и приблизительно равны по длине •h a, h р и sh; длина боковых щетинок возрастает кзади до 14; все d очень .длинные; примерные размеры щетинок, в процентах к длине идиосомы у 9 (порисунку Canestrini): v а25; v р и sc i 60; sc и h 48—55; d1? d2nd488; ч!380; 14 и 12 60—63; I3 и15 75; 14 88—90; sa e 57—60; sa i около 80; p a 50—55. .Ноги длинные и стройные; наружная вентральная щетинка голеней I и II .•отсутствует. Общая длина тела 9 300 р., ширина 220 [л; размеры с? в ониса-лии но указаны.
Распространение. Италия.. Живет в сепе.
7. Род CTENOGLYPHES BERL.
Berlese, 1884, XIV, n° 1. —Glycyborus Oudemans, 1903, Tijdschr. Entomol., XLVI: 8.
Тип рода: Acarus plumigcr Koch 1835.
Половой диморфизм исключительно резок; общими обоим полам родо-щыми признаками можно считать только наличие более или менее ясной ^поперечной бороздки на спинной поверхности идиосомы, редукцию вен
6. DIAMESOGLYPHUS. 7. CTEA'OGLIPIIUS 313
тральных щетинок голеней I и II и вершинной дорзальной щстинки |ч»ли-ней IV и малые размеры коготка.
<5. Гораздо консервативнее в своих признаках, чем 9, которой он си,цып» уступает в размерах (длина не превышает 150—190 |л). Идиосома умеренно выпуклая, яйцевидная или почти грушевидная, заметно расширяющаяся кзади, с совершенно сглаженными плечами и полого закругленным задним краем. Кожа очень тонкозернистая или почти гладкая, часто с отдельными неправильными пятнами, покрытыми более явственной скульптурой. Вооружение спинной и боковых поверхностей идиосомы состоит из 16—19 пар длинных перистых, двусторонне-гребенчатых или частью листовидных щетинок; v а всегда двусторонне-гребенчатые; vp имеются у всех видов; 3 пары й, из которых передняя расположёна на пронодосоме; dx и d2 иногда редуцированы (или даже отсутствуют).; очень широко расставленные, длиннее всех других щетинок идиосомы (обычно равны длине идиосомы или даже превышают ее), всегда узкие, тонко- и густоперистые по краям; позади d4, на спинной поверхности, помещаются еще 2 пары щетинок, невидимому, гомологичные крестцовым—sa е и sa i, но занимающие совершенно необычное для последних положение; боковых щетинок 5 пар, постанальных О—3 пары; в соответствии с выпуклой формой идиосомы, ее периферические щетинки имеют очень разнообразное, но преимущественно дорзальное положение. Все коксальные поля (или, по крайней мере, поля первых з пар ног) замкнутые, с заметно уплотненной поверхностью, т. е. образуют хорошо оформленные коксальные щиты; они разъединены по средней линии расширяющимся кзади пространством (центральным полем), но щиты каждой •стороны тела очень тесно соприкасаются друг с другом. Генитальное отверстие очень длинное, занимает практически все свободное пространство между коксальными щитами; penis чрезвычайно длинный, саблевидный. Ноги очень длинные (достигающие 60—65% длины идиосомы), относительно толстые; вершинная дорзальная щетинка голеней I сильно гипертрофирована.
9. Значительно крупнее <5 (длина 250—300 р.). Идиосома сильно уплощенная в дорзо-вентральном направлении, с несколько пластинчатыми краями, приблизительно пятиугольная; гистеросома с резко выступающими плечами, боковые края прямые или слегка вогнутые (иногда неровные); задний обычно более или менее глубоко вогнут посередине. Кожа очень неровная, на спинной поверхности грубоморщинистая или усаженная крупными, сильно рельефными бугорками, снизу тонкозернистая. Вооружение спинной поверхности идиосомы состоит из 17—21 пар более коротких, чем у сГ, двусторонне-гребенчатых, перистых или листовидных щетинок, причем наблюдающееся колебание их числа может происходить :щ счет выпадения v р и sc I и крестцовых; 3 пары h, 5 пар 1,3 пары iioctuiiiijii.-ных, все расположены строго маргинально по окружности идиосомы; dt иногда атрофированы (у С. palmifer редуцированы также da 11 <ln); у части видов d3, а иногда и вообще все d и имеющиеся в таком случае на о и на i,
316
Ш. ЫУСУРНШВАЕ. 4. CTENOGLYPHINAE
представлены узкими тонко- и густоперистыми щетинками. Эпимеры: I соединяются по средней липни тела, образуя короткий стернум, все остальные свободные; эиимернты 11 короткие, соединенные с эпимерами III. Крупное, удлиненное генитальное отверстие расположено в пространстве-между основаниями пог ill и IV, иногда простираясь назад несколько дальше- 1 уровня задних крахи! ацетаиул ,111; эпигииий хорошо развит; копулятивная трубка длинная, цилиндрическая или конически расширяющаяся к основанию; анальная щ«лг. близко придвинута к. заднему краю идиосомы. Ноги умеренной длины, стройные; вершинная дорзальная щетинка голеней I развита слабее, чем ус?.
У личинок все щетинки спинной поверхности простые, слабо гребенчатые;: сложность их строения и формы приобретается лишь в последующих стадиях (иногда только в стадии телеонимфы); с другой стороны, у видов, лишенных в половозрелом состоянии v р (С. palmifer), они хорошо развиты, у нимф.
К этому роду относятся 4 вида, известных только из Европы.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РИДОВ
<3(3
1 (4). Все щетинки спинной поверхности более или менее однотипные,, двусторонне-гребенчатые или длинноперистые; d1 и d2 хорошо развиты.
2 (3). Все щетинки спинной поверхности, кроме d3,. двусторонне-гребенчатые, с 10—20 парами крепких, сильно отогнутых боковых отростков;. d3 довольно длинно- и топкоперистые; dx вдвое короче d2 . . . . .. .............................................*2. С. canestrinii (Arm.)..
3 (2). Все щетинки спинной поверхности длинно- и довольно тонкоперистые,, с очень многочисленными, слабо отогнутыми боковыми отростками;. d-L едва короче d2........................... *1. С. plumiger (Koch)..
4 (1). V а двусторонне-гребенчатые; v р, обе пары sc, - все h и 2 передние-пары боковых (1Х и 12) листовидные, узколанцетовидной формы; d3. и последние 3 пары 1 (13—15) тонкие, густо- и тонкоперистые по краям; d4, sa е и sa 1 листовидные, гораздо шире всех других щетинок спинной, поверхности идиосомы; dx и d2 редуцированы или отсутствуют . . .. ........................................... З.С. palmifer (F. et R.).
99
1 (4). V, sc и вес периферические щетинки идиосомы двусторонне-гребенчатые; кожа спинной поверхности усажена сильно рельефными бугорками, вершина которых обычно раздвоена или радиально расщеплена.
Фиг. 508—512.
Фиг. -508. Ctenoglyphus plumiger (С. L. Koch), сГ сверху. Фиг. 509. То же, снизу. Фиг. 510. С. plumiger (С. L. Koch), 9 сверху. Фиг. 511. То же, снизу. Фиг. 512. С. canestrinii (Arm.),. 9 сверху.
318
Ш. GLYCYPHAGIDAE. 7. CTENOGLYPHINAE
2 (3). Все дорзальные (dx—d4), sa e и sa i узкие, тонко- и длинноперистые по краям; боковые отростки двусторонне-гребенчатых щетинок прямые, отогнутые от ствола иод острым углом. . . *1. С. plumiger (Koch).
3 (2). Только d3 представлены узкими тонко- и длинноперистыми щетинками; dp d2 и d4 двусторонне-гребенчатые, точно такого же строения,, как периферически с щетинки идиосомы; боковые отростки последних отогнуты от ствола, иод прямым углом и загибаются концами в апикальном направлении..........................*2. С. canestrinii (Ann.).
4 (1). V, вс и все периферические щетинки идиосомы в виде широких листо-
. , видных чешуй приблизительно одинаковой, симметричной формы, ' шйроколанцетовидиые или эллиптические, слегка приострепные на конце; цельнокрайние, с 1 срединной продольной жилкой, от которой отводят в стороны многочисленные боковые' Ответвления; кожа Асцидной поверхности идиосомы резко морщинистая.
5 (6)f Pi и d2 представлены очень мелкими, слегка гребенчатыми щетинками;: листовидные щетинки длиннее, равные 20—36% длины идиосомы'. . . .Г. ..............................3. С. palmifer (F. et R.).
6 (5). Dj и d2 представлены такими же листовидными чешуями; как периферические щетинки идиосомы; последние короче, длиною в 15—20%.
длины идиосомы ........... 4. Ct. multifoliatus, nom. и.
*1. Ctenoglyphus plumiger (C_. L. Koch) (фиг. 507—511).
0. L. Koch, 1835, Dtsch. Crust., Myriap., Arachn., V, n° 15; Gervais, 1844, in Walckenaer; Hist. Nat. Ins. Apt., Ill: 263 (Acanis); Fumouze’et Robin, 1868, Journ. Anat. Physiol., V:67, t. VII; Canestrini, 1888:21; 1888a:388, t. XXVIII; Berlese, 1884, XIV, n° 1; Kramer,,1899:146; Michael, 1901:250, t. IX, XI, fig. 1—7 (Glycyphagus).
cJ. Идиосома заметно расширяющаяся кзади, с широко закругленным задним краем. Кожа местами гладкая, местами тонкозернистая. Все щетинки спинной поверхности идиосомы очень тонко и густо двусторонне-гребенчатые (или длинноперистые), длинные, но в общем очень изменчивые по своим, размерам; d3 по своему вооружению почти не отличаются от остальных,, но заметно длиннее их. Вершинная щетинка голеней I чрезвычайно длинная. Длина около 200 jx, ширина 120 |г.
9. Идиосома в 1.4—1.45 раза длиннее своей ширины, с широко закругленным задним краем. Кожа спинной поверхности усажена цилиндрическими-бородавками, более или менее расщепленными на конце в виде розетки. V р хорошо развиты; обе пары sc, все плечевые, боковые и постанальные.
Фиг. 513—526.
Фиг. 513. Glycyphagus pilosus Ouds., псевдостигмальный орган rf (по Oudemarts’y). Фиг. 514. Cometacarus oudemansi л. Z., 9 сверху (по Oudemans’y). Фиг. 515. То же, снизу (цо Oudemans’y). Фиг. 516. С. oudemansi Л. Z., хелицера (в профиль) (по Oudemans’y). Фиг. 517. С. oudemansi-A. Z., основание лапки! (по Oudemans’y). Фиг. 518. С. setosus (С. L. Koch) сверху (по С. L. Koch’у). Фиг. 519. Ctenoglyphus palmifer (F. et R.), o’сверху (no Michael’io). Фиг. 520. С. palmifer (F. etR.), 9 сверху (по Michael’io). Фиг. 521. С. palmifer (F. et R.), одна ив периферических щетинок идиосомы 9 (по Michael’io). Фиг. 522. То же, o’ (по Michael’io). Фиг. 523. Nycteriglyphus pteropho-rus (Berl.), 9 сверху (no Berlese). Фиг. 524. То же, снизу (по Berlese). Фиг. 525. N. pteropho-rus (Berl.), одна пз периферических щетинок 9 (по Berlese). Фиг. 526. N. pterophorus (Berl.),. нога I (по Berlese).
320
111. GLI’CYPHAGIDAE. 4. CTENOGLYPHINAE
крупные, двусторонне-гребенчатые (опахаловидные), с прямыми отростками, .-отклоненными иод острым углом к стволу; все спинные и обе пары sa узкие, ггибкие (лентовидные), густо- и топкоопушенные по краям; длина гребеи- чатых щетинок колеблется и пределах 20—50% длины идиосомы, dj равны, Примерно, 20, (1а 45, <1я 65, (14 50, sa е 40—45, sai 15% длины идиосомы. Длина 280=310 |Л, ширина 180—220 |Л.
Р а с п р о с т р а. и о и и о. Описан из Германии, затем найден во Фран-„цпи, в Англии и в СССР; мне известен из Ленинграда, окрестностей Москвы и Иванова.
Экология. Живет в сенной трухе па сеновалах и в конюшнях; ^Michael находил этот вид также в старых пчелиных сотах.
*2. Ctenoglyphus canestrini! (Апл.) (фиг. 512).
Armanelli, 1887, Acari del fieno della provincia di Padova: 7; Canestrini, 1888: 21, t. I; -1888a: 396, t. XXIX, fig. 1; Berlese, 1891, LVIII, n° 3; -Kramer, 1899:146; Michael, 1901: 255, s-i. X—XI, fig. 8—23 (Glycyphagus).
cJ. Форма тела й строение кожи, как у С. plumiger. Все щетинки спинной ^поверхности идиосомы, кроме d3, крепкие, двусторонне-гребенчатые, такого же строения, как гребенчатые периферические щетинки $ С. plumiger, длиною в 40—60 % длины идиосомы; d8 лентовидные, гибкие, тонкопушистые, /Почти равны по длине идиосоме. Вершинная щетинка голеней I’ умеренной длины. Длина около 200 р., ширина 130 р-.
9. Идиосома шире, чем у $ предыдущего вида, почти правильной пяти-• угольной формы, только на длиннее своей ширины; задний край посередине несколько вогнутый. Строение кожи, как у $ С. plumiger. V р отсутствуют; все щетинки спинной поверхности идиосомы, кроме d3, опахаловидные (двусторонне-гребенчатые), с очень длинными отростками, отклоненными от ствола под прямым углом и загибающимися концами по направлению к дистальному концу щетинки; d3 лентовидные, гибкие, густоопушен-ные-по краям; длина опахаловидных щетинок колеблется в пределах 18— .-40 % длины идиосомы, d3 почти равны ее длине; длина dr составляет около 25, %2 45% длины идиосомы. Длина 300—320 |х, ширина 240—250 |х.
Рас пр о с т р а н е п и е. Известен из Италии, Англии и СССР (окрестности Москвы, Астрахань). В остатках сена на сеновалах и в конюшнях, ..нередко вместе с предыдущим.
3. Ctenoglyphus palmifer (Fum. et Rob.) (фиг. 519—522).
Fnmouzo et Robin, 1868, Journ. Anat. Physiol.,V:69, t. 8—11; Michael, 1901:260, t.XII— yXIII (Glycyphagus').
cf. Боковые края идиосомы на значительном протяжении почти параллельны друг другу, задний край широко закругленный. Кожа равномерно тонкозернистая. V.a двусторонне-гребенчатые; vp, обе пары sc, все h и z2 передние пары боковых узколанцетовидные (по существу они также двухсторонне-гребенчаты, по между их отростками натянута тонкая, прозрачная перепонка,, оставляющая свободными только самые кончики этих от
7. CTENOGLYPHUS. 8. NYCTEItIGL YI4IUN Ul* I
ростков, выступающие в виде тонких зубчиков за. крал ЛИОТОНИДНОЙ IW|W» понки); d1 и d2 отсутствуют (или сильно редуцированы?); и 1я= 1ц уй1Н1Г| (лентовидные), густо- и тонкопушистые; d4 и обе пары нц листонндн'-Ш; широколанцетовидной формы, несколько искривленные, образующий крщ сивый букет в задней трети гистеросомы. Примерная длина щетинок идиосомы, в процентах к ее длине: v а 25; v р 50; sc е 33; sc i 48—50; li а и all 50—55; h р и 1х 00; 12 около 70; 13 80; 14 60; 15 35; d3110; d4 и обе пары на *’ около 50. Внутренний край голеней I вытянут в короткое острие, их дор зальная щетинка чрезвычайно длинная. Длина около 190 р., ширина 120 р„
$. Боковые края гистеросомы неровные, неправильно фестончатые, задний край посередине с узкой и глубокой вырезкой. Кожа грубоморщинистая. V р отсутствуют; все остальные щетинки, кроме dx и d2, листовидные, широколанцетовидной или почти эллиптической формы; 15 уже других, узколанцетовидные; dx и d2 очень короткие, палочковидные, с несколькими игольчатыми отростками, d3 и d4 листовидные. Размеры периферических листовидных щетинок колеблются в пределах 23—38, dx около 4, d2 5, d3 23—25, d4 10% длины идиосомы. Длина около'260 р, ширина 170 р.
Распространение. Известен пока из Франции, Великобритании, Италии и Германии. Живет в пыли погребов и в трухе сена в конюшнях.
4. Ctenoglyphus multifoliatus, пот. п.
palmifer Canestrnii, 1888а: 390, t. XXVI, fig. 1 (Glijcyphcujiis) (non Fumouze et Robin, 1868).
9. Отличается от $ C. palmifer(F. etR.) более закругленными и ровными контурами идиосомы, более короткими листовидными щетинками (длина которых не превышает 16—20% длины идиосомы), имеющими, невидимому, более однообразную эллиптическую форму, и тем, что dx и da представлены такими же эллиптическими листовидными чешуями. Общая длина тела около 300 р, ширина 180 р. — Совершенно недостаточно описанный и оставшийся неизображенным сГ отличается от такового С. palmifer наличием только одной (а не 4) пары тонких перистых щетинок на спинной поверхности идиосомы. Общая длина тела 200 р, ширина 140 р.
Распространение. Италия (окрестности Брешии). Найден в сене.
8. Род NYCTERI.GLYPHUS, gen. п.
Тип рода: Glycyphagus pterophorus Berlese 1892.
Идиосома приблизительно шестиугольная, с выступающим передним краем, почти прямыми и параллельными друг другу боковыми краями, сильно выдающимися (почти прямоугольными) задними углами и очень своеобразным, резко обособленным каудальным выступом, имеющим форму усеченного конуса; дорзальная поперечная бороздка вполне япотпоппц,
1 Гомологизация отдельных пар щетинок спорна; недостоверно, п члотююти, отошиотпле-пне щетинок заднего края идиосомы с зае и sai.
21 Фауна СССР, Захваткин—284
322
III. GLYCYPBAGIDAE. 4. CTEN0GLYPI1INAE
Вся поверхность кожи очень густо поперечпоморщинистая, складчатая. Вооружение спинной поверхности идиосомы состоит всего только из 13 пар щетинок; все эти щетинки, кроме sa е, представлены крупными прозрачными листовидными чешуями; sa е длинные и тонкие, слегка перистые. Постапальпых щетинок, невидимому, нет (или они крайне мелки?). Ноги короткие, довольно массивные; вершинная дорзальная щетинка голеней IV одинакового размера с таковой голеней Ш; коготки крупные (приблизительно равны но длине перепончатому pructarsua). — d1 пе описан.
$. Эпимеры I образуют V-образлую фигуру, вершина которой соединена с дугообразной линией, связывающей внутренние концы эпимер Пг благодаря чему коксальпые поля .1 совершенно замкнуты; эпимериты II необыкновенно длинные, тянущиеся от наружных углов ацетабул II до заднего края генитального отверстия, огибая изнутри основание ног III; эпимеры III и IV не развиты. Генитальное отверстие расположено между основаниями ног III, широкояйцевидное, с явственным круговым контуром и небольшим эпигипием; копулятивная трубка очень короткая, расположенная на вершине каудального выступа. Анальная щель очень сильно отодвинута от заднего края идиосомы.
Род устанавливается для одного оригинального вида, неизвестного мне в натуре и, к сожалению, недостаточно полно описанного.
1. Nycteriglyphus pterophorus (Berl.) (фнг. 523—526).
Berlese, 1892, LVIII, n° 6; Kramer, 1899: 146 (Glycyphagus).
Очень мелкий, бледно окрашенный вид. Отношение длины идиосомы к ее ширине (у $) равно 1.45 : 1. V а, обе пары sc и все 3 пары d обратно-клиновидные, с прямо усеченным зубчатым дистальным краем; sc значительно шире, чем v а и d, и заметно длиннее их, достигая 20—22% длины идиосомы (длина v а составляет около 16, dx—d3 13—14% длины идиосомы); d помещаются на одинаковом расстоянии друг от друга, образуя 2 параллельных продольных ряда; й а и h р, —13 и sai расположены строго-латерально по окружности идиосомы, все приблизительно одинаковой формы, асимметричные, скошенно-яйцевидные, с глубоко вырезанно-зубчатым наружным краем, образующим 5—7 острых зубцов, из которых апикальный крупнее других, и продольными жилками; боковые края всех этих чешуй пушистые; размеры их колеблются в пределах 18—25% длины идиосомы; длина sa о составляет около длины идиосомы. Общая длина тела: сГ до 200 р., § до 250 р..
Распространение. Известен только из Италии (Падуя). Несколько экземпляров этого вида были найдены на летучей мыши Nyctalus noctula L., в сообществе с хищным клещом Leiognathus arcuatus.
Фиг. 527-539.
Фиг. 527. Diamesoglyphus inlermedius (Can.), 9 сверху (по Canestrini). Фиг. 528—539. Xenoryetes krameri (Mich.) (no Michael’io). Фиг. 528. сГ сверху. Фиг. 529. То же, снизу. Фиг. 530. Нога I о". Фиг. 531. Нога II сь Фиг. 532. Нога III О’- Фиг. 533. 9 сверху. Фиг. 534. Тоже, снизу. Фиг. 535. Нога I 9. Фиг. 536. Базальная щетинка колена I. 9- Фиг. 537. Телеонимфа сверху.
Фиг. 538. Гипопус сверху. Фиг. 539. То же, снизу.
324
JJL GLYCYPUAGIDAE. 5. LABIDOPHORINAE
SPECIES INCERTAE SEDIS
1. Glycyphagus (hiirmneariis) iuligineus Berl.
Borleso, 1,921, Rndia, XIV: MR.
Овальный, серопптыЛ, цвета, разведенной сажи, украшенный очень короткими (у o’ едва заметными) 1цотинкн.мн; вся кожа гравирована грубыми штрихами, эпнстомальпые щетинки (v а.) листовидные, пушистые. Эпимеры 3-й и 4-й нар соединены вместо концами; у о удлипенно-шпателевидные. У 9 поги по толще, чем у <5; боковые щетинки тела, (позади лог IV) чрезвычайно короткие [наполовину короче, чем у G. fuscus Ouds. (fiohieria fusca Ouds.)], изогнутые, снаружи коротко зазубренные. Penis длинный, стилетовидный, тонкий и крайне острый (равный длине лапок 4-й пары, т. е. 60 |х длиной), направленный вперед, прикрепленный едва позади тазиков IV, придвинутый основанием к крупному, широкоовальному анальному отверстию. — о сзади прирстрена; щетинки тела слегка больше, чем у J, но не превышающие 30 ;х в длину, изогнутые, зазубренные снаружи. Генитальное отверстие (расположенное между тазиками III и IV) окружено трапециевидной фигурой (сзади слегка выпуклой назад, спереди прямой). Длина d до 350 [х, ширина 210 р; длина 9 до 380 |х, ширина 280 [х.
Распространение. Флоренция, в трухе сена. (По Berlese.)
Загадочная форма, могущая с одинаковым успехом быть относимой и к подсем. Labidophorinae. Помещая ее здесь, я руководствовался следующими из приводимых Berlese признаков: структурой кожи, листовидной формой v а, характерным для некоторых Ctehoglyphinae строением и особенно размерами penis, и местообитанием вида.
5. Подсем. Labidophorinae
Гнатосома в большинстве случаев прикрыта сверху нависающим в виде капюшона ростральным выступом, иногда даже погруженная в более или менее глубокое углубление на его вентральной поверхности (так называемый камеростом); хелицеры небольшие, относительно слабые. Идиосома нерасчлененная, очень разнообразной формы; ростральный выступ обычно хорошо развит, нередко отделенный от остальной идиосомы более или менее явственной поперечной бороздкой. Кожа в большинстве случаев сильно хитинизировапная, обычно окрашенная в буроватый или красноватый цвет, гладкая, зернистая или шиповатая, иногда с вдавленным сетчатым орнаментом. Щетинки спинной поверхности идиосомы короткие, нередко представленные микрохэтами или короткими шипиками, гладкие или слабо перистые; v а иногда редуцированы, vp нередко совершенно отсутствуют; число и расположение других щетинок также .очень изменчиво. У большинства видов, кнутри от плечевых щетинок, расположен поровидный орган; пара аналогичных образований имеется также в области жировых желез. Генитальные щупальцы обычно имеются и иногда достигают огромных раз-
ХАРАКТЕРИСТИКА ПОДСЕМЕЙСТВА
меров. Коксо-стернальный скелет сильно развитый, крайне разнообрапного строения. Ноги умеренной длины (ходильного типа) или короткие, ишн'Д.ч сильно модифицированные, с разнообразными пластинчатыми разращениями, ребрами ит. п.; строение усаживающих их щетинок очень разнообразно, по они никогда пе бывают густоперистыми; вершина колен I с 2 органами чувств — палочковидной сенсиллой и более длинным волоском; вершинные щетинки голеней I и II умеренной длины, III и IV — укорочен ные; конец лапок с 2 хорошо развитыми вершинными шипами. Praetarsi удлиненные, хорошо развитые, с очень мелким коготком.
Вторичные различия между полами, как правило, чрезвычайно резки и охватывают очень большое число признаков: общую конфигурацию тела, хэтотаксию, строение коксо-стернального скелета, строение и вооружение ног, а нередко и ряд других; крайним примером этих различий может служить род Orycteroxenus, dd которого абсолютно непохожи на 99, вообще более консервативных в своих признаках.
d. Ноги III и IV в большинстве случаев модифицированы. Положение генитального отверстия изменчиво; эпиандрий обычно хорошо развит; penis небольшой, ножевидный.
9. Генитальное отверстие крупное, занимающее обычно все пространство между основаниями ног II, III и IV, с хорошо развитыми парными клапанами и иногда добавочным непарным клапаном, замыкающим генитальное отверстие сзади; эпигиний хорошо развит и обычно соединен с элементами коксо-стернального скелета и с широким овальным хитиновым кольцом, окружающим генитальное отверстие (циркумгенитальное кольцо); такое же кольцо (циркуманальное) иногда окружает и анальную щель; копулятивная трубка обычно невелика. — Нимфальные стадии у многих родов резко отличаются от половозрелой.
РасселиТельные гипопусы (фиг. 64—65) имеются у многих видов. Они все относятся к терохорному типу и характеризуются следующими признаками. Гнатосома совершенно атрофирована, представлена только короткими арис-тами, которых обычно 2 пары. Тело бледно окрашенное, не очень сильно хитинизированное, уплощенное дорзовентрально. Проподосома треугольная, всегда явственно отделена от гистеросомы поперечным швом, но неподвижно сращена с нею; задняя часть гистеросомы обычно обособлена,, образуя покатую, более или менее четко отграниченную спереди опистосомаль-пую площадку. Спинная поверхность совершенно гладкая; у некоторых видов, впрочем, на опистосомальной площадке находятся небольшие поля, -покрытые неясным сетчатым рисунком, а вдоль боковых краев тела тянутся тонкие складки менее плотной в этих местах кожи. Все щетинки спинной поверхности представлены топкими, очень трудно заметными микрохэтами; у ряда форм, невидимому, многие из щетинок отсутствуют. Псевдостиг-мальныйюрган и глаза отсутствуют. На нижней поверхности тела пот четко очерченных щитов, и граница между проподо- и гистеросомой неясна. Коксостернальный: скелет умеренно развит; стернум и эпимеры 11 всегда свобод-
326
HI. GLYCYPIIAGIDAE. 5. LABIDOPHORINAE
пые; эпимеры III иногда соединены с эпимерами IV, замыкая, таким образом, широко раздвинутые коксальиые поля III; эпимсриты II обычно хорошо развиты; у некоторых видов па брюшной поверхности тела имеется медиальный продольный желобок, края которого поддерживаются отгибающимися вдоль лих концами эпимер II--IV if эпимеритов IV. Коксальпые щетинки отсутствуют. 'Генитальное отверстие крупное, с крупными дисковидными щупальцами, Присасывательный диск отсутствует и заменен своеобразным прикрепительным аппаратом, служащим для схватывания и удержания волос млекопитающих, являющихся переносчиками гипопусов. Этот аппарат состоит из 2 широких подвижных клапанов (способных отгибаться и захлопываться, как дверцы), внутренняя поверхность которых украшена системой более или менее рельефных поперечных ребер и 2 пар расположенных под ними зажимов — передней и задпей; вентральная поверхность зажимов также покрыта поперечными или диагональными ребрышками, обеспечивающими прочное удержание волоса, зажатого между ними и наружными клапанами. Пропорции ног изменчивы, но в большинстве случаев ноги довольно массивны; ноги I—III с хорошо развитым изогнутым коготком, без перепончатого praetarsus. Ноги IV обычно с редуцированным коготком и иногда с 1 длинной парусной щетинкой. Вооружение ног состоит из обычного набора щетинок, довольно изменчивых по своему строению; вершинные щетинки голеней у большинства форм палочковидные, более или менее сильно укороченные; концы лапок с простыми, листовидно расширенными или пуговковидно утолщенными на конце щетинками. Размеры расселительпых гипопусов колеблются в пределах 170—430 р.. Покоящихся гипопальных стадий нет.
К подсем. Labidophorinae я отношу 14 видов, распределяющихся по 8 родам; все его представители, за исключением рода Gohieria Ouds. 1939, — строго нидикольные формы, биологически связаны с млекопитающими (грызунами и насекомоядными), в гнездах которых они развиваются; многие из них используют своих хозяев и в качестве переносчиков расселительпых гипопусов. Из внепалеарктических видов, известных до настоящего времени, к этому подсемейству может быть отнесен только описанный из США Glycyphagus obesus Bnks. 1906, являющийся, невидимому, представителем особого рода.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ del
1 (2). Края идиосомы с глубокими надрезами, образующие 10 пар округлых лопастей в виде фестонов, часть которых несет на конце крепкие игольчатые щетинки .....................14. Talpacarus, gen. п.
2 (1). Идиосома цельнокрайняя, без краевых лопастей или. фестонов.
3 (4). Идиосома веретеновидная, сильно суживающаяся к обоим концам, очень сильно выпуклая (в поперечном сечении почти круглая); кожа совершенно гладкая, окрашенная в коричневый цвет; see волосо-
ТАБЛИЦА 1'ЩОН 3'47
видные, все остальные щетинки идиосомы в индо очень тонких, гибких микрохэт; ноги очень короткие, передние очень «плющены « боков, пластинчатые..............,...........16. Fhmkhii'hh Mhili,
4 (3). Идиосома другой формы, более или менее уплощенная в дорио-нон-тральном направлении; строение щетинок и пог иное.
5 (6). Крупные, бледно окрашенные формы; щетинки спинной поверхности идиосомы относительно длинные, более или менее перистые; кожа мягкая, топкозернистая; лапки III и IV с цельнокрайними листовидными продольными килями, остальные членики пог без пластинчатых разращений..............................10. Labidophorus Kram.
6 (5). Мелкие, темно окрашенные виды; щетинки спинной поверхности идиосомы короткие или очень короткие, гладкие, иногда шиповидные; кожа плотная, сильно хитинизированная, гладкая или грубозернистая; вооружение ног иное.
7 (8). Крайне мелкая форма (длина 170 р.); кожа спинной поверхности идиосомы густо усажена крупными полушаровидными бугорками; все щетинки спинной поверхности представлены короткими коническими шипами; коксо-стернальный скелет слабо развитый; ноги III и IV целиком скрыты под телом и направлены вперед..........................
. .............................. 13. Orycteroxenus, gen. n.
8 (7). Более крупные формы (длина 260—400 р.); кожа гладкая или довольно грубозернистая, очень плотная; щетинки спинной поверхности жестковатые (но не шиповидные), вполне явственные, или же представленные крайне мелкими, трудно заметными микрохэтами; коксо-стернальный скелет сильно развит; положение ног III и IV нормальное.
$ (10). Кожа гладкая; щетинки спинной поверхности длиною около 10— 15% длины идиосомы — жестковатые; ростральный выступ без продольных килей; базальные членики ног с тупыми продольными ребрами и пластинчато расширенным дистальным краем, без листовидных зубчатых килей. Амбарный вид, живущий в муке и других зер-нопродуктах......................................11. Gohieria Ouds.
10 (9). Кожа грубопористая и зернистая; ростральный выступ с парой неровных продольных килей; все щетинки идиосомы представлены крайне мелкими, трудно заметными микрохэтами; базальные членики пог I и II с зубчатыми пластинками, лапки с цельнокрайними продольными килями. В норах крота и грызунов..........................
.......................................12. Xenoryctes, gen. n.
99 ,
1 (4). Уплощенные боковые края идиосомы с глубокими надрезами, образующие ряд фестончатых или неправильно очерченных лопастей, на. нор-шинах которых сидят крепкие, болееилименее искривленные щетинки.
2 (3). Стернум и эпимеры II свободные; эпимеры III и IVсоединены попарно концами; генитальное отверстие не окружено хитиновым кольцом;
328
111. GLYCYPHAG1DAE. 5. LABIDOPHORINAE
все 4 пары спинных щетипок (dx—d4) хорошо развиты.............. ,
..........................................14. Talpacarus, gen. n.
3 (2). Стернум отсутствует; эпимеры I, II и III соединены концами с хитиновым кольцом, окружающим генитальное отверстие; эпимеры IV свободные, короткие; имеются только 2 пары спинных щетинок (d2 и ds), придвинутых к средней линии тела и тесно сближенных между собой. ...........................................13. Orycteroxenus, gen. n,
4 (1). Идиосома цельнокрайняя, без краевых лопастей или фестонов.
5 (6). Идиосома веретеновидпая (сильно суживающаяся к обоим концам), очень сильно выпуклая (в поперечном сечении почти круглая); кожа совершенно гладкая, окрашенная в коричневый цвет; все щетинки идиосомы, кроме вс е, представлены очень тонкими, гибкими микрохэ-тами, sc с волосовидные; ноги I и II пластинчатые, сильно сжатые с боков ..................................... 15. Fusacarus Mich.
(i (5). Идиосома другой формы, уплощенная в дорзо-вентральном направлении; строение кожи, щетинок и ног иное.
7 (8). Стернум и эпимеры II свободные; эпимеры III и IV соединены концами попарно; эпигиний обособлен от частей коксо-стернального-скелета, бобовидный; кожа с тонким сетчатым рисунком, беловатая; щетинки спинной поверхности гладкие, довольно длинные; ноги I и II укорочены. Крупная форма, живущая на бобре........................
...........................................9. Xenocastor, gen. n.
8 (7). Стернум соединен концом с эпигинием или с циркумгенитальным кольцом, или отсутствует, и тогда эпимеры I непосредственно соединены с эпигинием; структура кожи и щетинок иная; ноги I и П не укорочены.
9 (12). Кожа очень плотная, темнокоричневая; ростральный выступ хорошо развит, закрывающий сверху большую часть гнатосомы; щетинки спинной поверхности идиосомы очень короткие (иногда заметные лишь с большим трудом), гладкие; генитальное отверстие окружене хитиновым кольцом. Мелкие формы (300—450 ;л).
10 (И). Щетинки спинной поверхности идиосомы короткие, но вполне явственные; кожа гладкая, ростральный выступ без продольных килей; стернум отсутствует, эпимеры I непосредственно соединены с эпигинием; базальные членики пог ребристые, колени и голени с пластинчато расширенным дистальным краем. Амбарная форма, живущая в муке и других зерпопродуктах ... 11. Gohieria Ouds.
11 (10). Щетинки спинной поверхности идиосомы сильно редуцированы, едва заметные; кожа грубозернистая и пористая; ростральный выступ с 2 неровными продольными килями; стернум хорошо развит, соединен концом с эпигииисм; базальные членики ног цилиндрические, голени дорзально с более или менее высоким пластинчатым килем. В гнездах кротов и грызунов............................ . 12. Xenoryctes, gen. n.
12 (9). Кожа мягкая, бледно окрашенная, тонкозернистая; ростральный
9. XE.XOCASTOH
выступ отсутствует; щетинки спинной поверхности идинОбММ О‘ГНн сительпо длинные, более или менее перистые, jpipicyMri'iiiiTiiliblHHi'' кольца нет. Крупные формы (600—1000 |л) . 10. LabldophoruH К mill
Гипопусы
1 (2). Тело узкое, в 1.6—1.9 раза длиннее своей ширины; ноги I и II длин пые и стройные, вооруженные перистыми или гребенчатыми щетинками, с более или менее S-образио искривленными (в профиль) лапками; ноги IV очень короткие, оканчивающиеся длинной парусной-щетинкой, без коготка...........................16. Myacarus, gen. n.
2 (1). Тело более широкое; ноги I и II довольно короткие и массивные, вооруженные простыми или шиповидно утолщенными щетинками, с неискривленными, более или менее коническими лапками; ноги-III и IV немного короче передних.
3 (4). Коксальные поля III замкнутые; лапки IV без парусных щетинок;, задние зажимы шпателевидные, стройные, отделенные большим пространством от овальных, радиально-ребристых передних; опистосо-мальная площадка не обособлена................12. Xenoryctes, gen. n..
4 (3). Коксальные поля III открытые; если же замкнутые, то лапки IV оканчиваются длинной парусной щетинкой (Е. sciurinus Koch); задние-зажимы более или менее треугольные, близко придвинутые к передним и налегающие на них своими передними краями; опистосомальнаж площадка спереди четко отграничена .... 10. Ladidophorus Кг am,.
9. Род XEXOCASTOE, gen. л.
Тип рода: Xenocastor fedjushini, sp. n.
9. Гнатосома очень маленькая, целиком скрытая в углублении под., ростральным выступом. Идиосома широко-эллиптическая, заметно уплощена в дорзо-вентральном направлении, с крупным, необособлеииым ростральным выступом, округло выступающими плечевыми выпуклостями и чуть вогнутым посередине, широко закругленным задним краем; ее боковые края также заметно вогнуты близ середины, но не образуют лопастей. Кожа очень толстая (до 30 у.), по не склеротизированная, бесцветная, покрытая тонким сетчатым узором. V а сильно редуцированы, представлены крайне мелкими, толстыми палочками; v р отсутствуют; все остальные щетинки спинной поверхности идиосомы относительно длинные и топкие,* слегка жестковатые, гладкие; они все направлены назад и прижиты к ни» верхности тела. Жировые железы маленькие, но вполне явственные. Ацоти* булы всех 4 пар пог окружены очень мощными кольцевыми xhtiiiiiiiiiihiuimiu стернум Y-образный, свободный, отделенный широким Jipoiwpiitfli'miiM W необычайно маленького, бобовидного эпигиния; эпимеры Л оашЛодиым» III и IV — соединены друг с другом своими внутренними коицнми; ннимп»
9330
III. GLYCYPHAGIDAE. 5. LABIDOPHORINAE
риты II представлены короткими отростками, отходящими от середины эпимер III. Генитальное отверстие помещается в пространстве между основаниями ног III и IV, лишено хитиновой рамки и непарного заднего клапана;-генитальные щупал ьцы исключительно крупные; копулятивное отверстие помещается дорзалыю (значительно отступя от заднего края тела), не вытянуто в трубку; анальная щель и окружающие ее щетинкп далеко отодвинуты от заднего края идиосомы; цнркумальпого кольца нет. Ноги I и II короткие, слегка сжатые, с боков, ноги III и IVдлинные, стройные; поверхность члеников гладкая, без каких иы то ни было ребер и крыловидных килей; вооружение голеней и ланок состоит преимущественно из гладких, более пли менее искривленных шипообразпыхщетинок.— сГ неизвестен. — Гипопусов, насколько известно, не образует.
Известен только 1 вид, биологически связанный с бобром.
*1. Xenocastor fedjushini, sp. п. (фиг. 21, 540—543, 567).
$. Очень крупная, беловатая форма; гнатосома, ноги и окружающие их основания хитинизированные участки красновато-бурые. Идиосома в 1.4 раза длиннее своей наибольшей ширины. Размеры щетннок, в процентах к длине идиосомы: v а 1; sc е 17; sc i 14; h а и sh 15—16; hp 28; dx 14; d2 6; d3 35; d4 38; 14 12; 12 28; 13 21; sa e 55; sai 9. D2 расположены значительно позади lx, почти на одной линии с d3; анальные щетинки глад-:кие, волосовидные, р3 очень длинные. Генитальные щупальцы эллиптиче-•-ские, соприкасающиеся своими краями, необычайно крупные (передние 72 X 46 pi, задние 61 X 41 р.). Ноги I и II несколько сжатые с боков, длиною в 25 (I) и 27 (II) % длины идиосомы. Лапки около 3 раз длиннее своего диаметра у основания и чуть больше чем в 1.5 раза длиннее голеней, конически суживающиеся к концу, вентрально с 2 крупными шипами, сидящими .друг за другом, с очень короткой цилиндрической обонятельной палочкой и крупными притупленными боковыми вершинными шипами; длина praetarsi I составляет около 3/4 длины лапки; вентральные щетинки голеней шиловидные, сближенные между собой; колени дорзально с бугровидно выступающим основанием, разделенной почти до основания на 3—4 ветви базальной и очень короткой волосовидной наружной щетинкой. Ноги III немного . длиннее передних (28.5% длины идиосомы), но значительно стройнее; длина .лапок почти в 5.5 раза превосходит их диаметр у основания и более чем в 2 раза длину голени; лапки вооружены 3 крупными вентральными шипами и 1 вершинным, па конце с 3 топкими щетинками; вентральная щетинка голеней в виде длиицого прямого шипа. Ноги IV очень длинные (равные 40% длины идиосомы) и стройные; их лапки в 6 раз длиннее своего диаметра у основания, почти в 3 раза длиннее голеней и в 4.5 раза превышают длину
Фиг. 540—545.
Фиг. 540—543. Xenocastor fedjushini, sp. n.. 9- Фиг. 540. Общий вид сверху. Фиг. 541. То же, ..снизу. Фиг. 542. Лапка II сверху. Фиг. 543. Нога I сверху. Фиг. 544. Labidophoriis desmanae, sp. n.. 9, общий вид сверху, фиг. 545. То же, снизу.
332
ПГ. GLVCVPHAGIDAE. a. LABIPOPHORINAE
praetarsus; вооруженно пог IV сходно с таковым ног III, но вентральные шипы лапок длиннее, расположены более дистально и более сближены между' собой, образуя своего рода гребспь. Длина 1084 |л, ширина 760 |л.
Р а с и р о с т ране и и о. Найден в Белорусской ССР (Белорусский Гос. заповедник на. р. Березине).
9 к о л о г и я. .Живет на бобре. (Castor fiber vistulanus Matsch.). 2экз. этого оригиналыiого вида были найдены проф. А. В. Федюшиным весною-1928 г. в шерсти бобра; они «держались в самых густоволосых частях тела (на животе) л были обнаружены па. вычесанных пуховых волосах, после длительного вычесывания густым гребнем».1 Один из этих 2 экз. (§) сохранился в коллекции ЗИН и был любезно передан мне для изучения проф. В. В. Редикорцевым; судьба второго неизвестна.
Образ жизни X. fedjushini остается во многом неясным; не выяснено,, например, является ли он обитателем гнездовой подстилки хозяина, как все другие Lahidophorinae, или же настоящей эпизоической формой, подобно знаменитому Platypsyllus; последнее мне лично кажется вероятным вследствие наличия ясно выраженных эпизоических черт в организации Хепо-castor (уплощеннйя, обтекаемая форма тела, редукция v а и v р, прижатью к телу щетинки, направленные все назад, укорочение передних ног, шипы и гребни на ногах и т. д.) и отсутствия у него гипопуса. Обращает на себя внимание также и то, что X. fedjushini—живородящая форма; в $, исследованной мной, находятся 2 вполне развитых эмбриона (личинки); как известно,, случаи факультативного живорождения нередки у нормально яйцекладущих Tyroglyphoidea (Tyrophagus viviparus Ouds., различные виды Caloglyphusr Glycyphagus destructor Schrk.), по в данном случае можно думать, что оно является правилом.
10. Род LABIDOPHORUS KRAM.
Kramer, 1877, Arch. Naturg., XLIII, 1: 149. — Dermaearus Haller, 1880, Ztschr. wiss. Zool., XXXIV: 268.
Тип рода: Labidophorus talpae Kram. 1877 (гипопус).
Гнатосома среднего размера, свободная, частично видимая сверху. Идиосома более или менее сильно уплощена дорзо-вентрально, без сколько-нибудь ясного рострального выступа и фестончатых лопастей. Кожа довольно тонкая, мягкая, бесцветная или слабо пигментированная, тонкозернистая (шагреневая); v а хорошо развиты; v р часто редуцированы; плечевых щетинок 3 пары, боковых — 4 пары (1х—]4); d2 расположены много впереди d3; все щетинки спинной поверхности идиосомы умеренной длины (13—30%. длины идиосомы), жесткие, прямые, более или менее явственно перистые. Жировые железы хорошо развиты. Ноги длинные, стройные, не модифици-р ованные; поверхность беде р, колен и голеней ровная (без закраин, ребер или килей),бедра I и II не бокаловидны; их вооружение состоит преимущественно
1 А. В. Федюшин. Речной бобр. Изд. Главпушнины НКВТ, М., 1936, стр. 342.
10. LABIDOPHORUS
333
!из довольно тонких, гибких, слегка перистых щетинок; вентральная щетинка голеней IV хорошо развита.
сГ.1 Идиосома с параллельными сторонами и широко закругленным задним краем; v р и d2 хорошо развиты. Стернум, эпимеры и эпимериты II свободные (последние очень коротки); эпимеры III и IV слиты своими внутренними концами, дугообразно отогнутыми назад; эпимериты IV очень короткие; поверхность коксальных полей не уплотнена. Генитальное отверстие .расположено позади основания ног IV; penis (в состоянии покоя) обращен вершиной вперед; анальная щель длинная, ее клапапы немного выдаются .за край тела. Ноги I и II не модифицированы, без пластинчатых разращений; ноги III и IV массивнее передних, их лапки с цельнокрайними пластинчатыми килями; praetarsi гораздо крупнее, чем у ?, на задних ногах много крупнее, чем на передних.
9. Идиосома расширена кзади, с более или менее вогнутым задним краем, иногда почти сердцевидная; v р, а иногда и d2 редуцированы. Эпимеры I непосредственно слиты с крупным полулунным эпигииием, II — свободные, III и IV — соединены концами или свободные; коксальиые поля снаружи •с широкими хитиновыми утолщениями; генитальное отверстие расположено и пространстве между основаниями ног II и III, не окружено хитиновым кольцом; непарный задний клапан отсутствует; генитальные щупальцы среднего размера, несколько раздвинутые; копулятивная трубка отсутствует •(копулятивное отверстие помещается дорзально на небольшой выпуклости); анальная щель отделена некоторым расстоянием от заднего края идиосомы и не окружена хитиновым кольцом. Ноги без украшающих выростов, ргае-darsi не увеличены.
Личинки неизвестны. У прото; и телеонимф v р и d2 хорошо развиты, ле уступающие в размерах многим другим щетинкам спинной поверхности; •строение последних не отличается от дефинитивного или же они еще тоньше, волосовидны; зернистость кожи немного более крупная, но мало рельеф-лая; лопасти заднего края идиосомы слегка намечены.
Гипопус. Форма тела и его пропорции очень изменчивы, но оно всегда значительно шире, чем у Myacarus. Коксо-стернальный скелет хорошо .развит; эпимериты II всегда явственны, строение остальных его элементов у разных видов различно; очень изменчивы также строение прикрепительного аппарата и связанные с ним структуры заднего конца тела, но зажимы у всех видов сближены между собой и в покое более или менее сильно налегают друг на друга. Ноги массивные, лапки I—III с крупным коготком и 3—4 едва уплощенными, линейными или же совершенно не расширенными, шуговковидно вздутыми на конце щетинками; вершинная щетинка голеней у большинства видов палочковидная, все остальные щетинки пог гладкие •(иногда шиповидные); лапки IV с редуцированным коготком, без листовидно .расширенных щетинок.
1 Известен только у одного из видов.
334
Ш. GLYCYPHAGIDAE. 5. LABIDOPHORINAE
Я рассматриваю род Dermacarus Haller 1880 как синоним «гипопального» рода Labidophorus Кгаш. 1877 на том основании, что между взрослыми: D. sciurinus и таковым L. desmanae (вида, по строению своего гипопуса близкого к L. talpae и, несомненно, относящегося к одному с ним роду} пет различий родового масштаба. В состав расширенного таким образом рода Labidophorus входят 5 палеарктических видов, биологически связанных как с грызунами, так и с насекомоядными; из этих 5 видов з известны пока только по гипопусам.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА, ЕНДОВ
99
1 (2). Щетинки спинной поверхности явственно перистые; кожа почти? бесцветная, равномерно тонкобугорчатая; d2 редуцированы; h а находятся почти на одной линии с sc; эпимеры III и IV слиты своими внутренними концами. На белках и в их гнездах. * 1. L. sciurinus (Koch)..
2 (1). Щетинки спинной поверхности почти гладкие, с очень немногочисленными, редко сидящими короткими ресничками; кожа окрашена в бледный буроватый цвет, в задней части спины с группой крупных, плоских овальных бляшек; d2 хорошо развиты; 11 а помещаются много позади sc; эпимеры III и -IV свободные. На выхухоли и в ее гнездах . . .
................................. *2. L. desmanae, sp. n-
Гипопусы
1 (2). Коксальпые поля III замкнутые — эпимеры III сливаются концами с эпимерами IV; лапки IV заканчиваются 1 очень крупной (парусной} щетинкой, 2 более короткими щетинками и несколькими крепкими шипами; вентральные щетинки вертлугов I и II очень длинные, гибкие, волосовидные; длина до 350 р.. На белке. * 1. L. sciurinus (Koch).
2 (1). Коксальные поля III открытые — эпимеры III и IV свободные;.
. концы лапок IV без крупных (парусных) щетинок; вентральные щетинки вертлугов I и II короткие; длина не свыше 280 р. На других млекопитающих.
3 (4). Наружные вентральные щетинки голеней и колен I и II представлены массивными коническими шинами; проподосома сильно вытянута в длину, ес боковые края сходятся кпереди под острым, закругленным па вершине углом; вершинная щетинка голеней I и II длинная, немодифицированная; задние углы тела закругленные;, зажимы небольшого размера, с немногочисленными, сильно рельефными поперечными ребрышками. На дальневосточном кроте (M'ogera robusta Nchr.).....................5. L. mogerae, sp. n.
4 (3). Вентральные щетинки голеней и колен I и II тонкие, гибкие или в виде стройных шипов; проподосома короче, ее боковые края сходятся кпереди под тупым углом; вершинная щетинка голеней I и II короткая,.
10. LABIDOPHORUS ' 336
палочковидная, с закругленным концом; задние углы тела резко-выступающие; зажимы крупные, с многочисленными, тесно расположенными поперечными ребрышками.
5 (6). Тело удлиненное, приблизительно в 1.5 раза длиннее своей ширины; его задние углы выдаются в виде тупых цилиндрических рожков, слегка загнутых книзу; лапки IV на конце с 4 очень крупными, пальце-' видно расходящимися коническими шипами, 3 средней длины топкими щетинками и 2 маленькими шпповидпымп щетинками (нс считая редуцированного коготка); лапки I и II с 3 узколапцстовидпыми, расширенными щетинками. На выхухоли (Desmana moschala L.) . . ........................................... *2. L. desmanae, sp. в.
6 (5). Тело широкое, округлое или почти ромбическое, в 1.2—1.3 раза длиннее своей ширины; задние углы тела образуют перпендикулярно-отогнутую к его поверхности, резко выступающую па боках каемку; конец лапок IV без шипов, с 4 довольно длинными простыми и 1 короткой щетинками; щетинки лапок I и II простые или едва расширенные на конце.
7 (8). Длина 220—260 р.; аристы длинные; задние зажимы умеренной длины, закругленные на конце, с 10—12 поперечными ребрышками; тело ши--рокоовальное или почти круглое, более постепенно суживающееся, кзади. На кроте (Talpa europaea L.).............*3. L. talpae Кгаш.
8 (7). Длина 190—210 у; аристы очень короткие; задние зажимы чрезвычайно длинные, приблизительно с 15—20 поперечными ребрышками; тело скорее ромбическое, быстро суживающееся кзади. На различных видах землероек, но иногда также на кроте и разнообразный мышевидных грызунах.................................*4. L. soricis Ouds.
*1. Labidophorus sciurinus (C. L. Koch) (фиг. 64, 65, 577—579,584—586).
C. L. Koch, 1841, n° 7 (Dermaleichusy, 1842, Ubers. d. Arachn. Syst., III: 121, t. XIII, fig. 69; Canestrini, 1888a: 377 (Homopus): Haller, 1880, Ztschr. wiss. Zool., XXXIV: 268, t. IX, fig. 4—16; Kramer, 1899:150; Oudemans, 1910, Entomol. Ber., Ill: 23; 1915, ibid., IV: 199 (Per— macaws'); Michael, 1901: 283, t. XVIII—XIX; Berlese, 1897 : 106 (Glycyphagus); Oudemans, 1897, Tijdschr. Entomol., XL : 252 (Labidophorus).
Вледнолимонно-желтый, иногда с легким зеленоватым оттенком; гнато— сома, ноги и коксо-стернальный скелет красновато-бурые?Кожа почти бесцветная, спина в однообразной мелкобугорчатой скульптуре, только у переднего края идиосомы гладкая и здесь более сильно хитинизированпая. Щетинки спинной поверхности явственно перистые; псевдостигмальный орган перистый, напоминает по своей форме перистые щетинки видов Ctenoglyphus.
сГ. Идиосома в 2 раза длиннее своей ширины, с почти параллельными боковыми краями и широко закругленным задним концом; зернистость кожи на ее нижней поверхности гораздо тоньше, чем на спинной; v а гладкие (?); v р и d2 хорошо развиты, перистые; sc е гладкие, в 2.5 раза длиннее перистых sc i; ha расположены далеко позади sc; примерная длина щетинок идиосомы (в процентах к ее длине): va и vp ,10; sci 12—13;.
:33S III. GLYCYPHAGIDAE. 5. LABIDOPHOBINAE
sc e 32; ha 20; h p 27; dx 22; d2 7; d3 и d4 25—27; lx 13; ]2 и 13 35—38; 14 и sa e 22; sa i 10. Ноги пропорционально длиннее и значительно толще, чем у $, равные приблизительно 48 (I), 45 (11), 42 (111) и 50 (IV) % длины идиосомы; лапки III и IV снаружи с крупным, листовидным прозрачным килем, члепики ног I и II без выростов; praetarsi III и IV «сильно увеличенные, гораздо крупнее таковых I и II, значительно более крупных, чем у ф. Длина 370—655 р..
9. Форма идиосомы и ее пропорции меняются в зависимости от степени •ос растяжения яйцами; у молодых особей опа примерно в 1.4 раза длиннее «своей ширины и имеет почти параллельные друг другу боковые края и едва вогнутый посередине, широко закругленный задний край; у более старых, «сильно раздутых яйцами 99 идиосома только в 1.2 раза длиннее своей максимальной ширины (приходящейся теперь далеко позади ее середины) :и имеет почти сердцевидную форму. Скульптура брюшной поверхности такая же, как на спине; vp и d2 представлены микрохэтами; обе пары sc ..явственно перистые, sc е немного длиннее sc i; ha почти па одной поперечной линии с sc; ра расположены треугольником, р2 — значительно кнаружи от остальных; примерная длина щетинок идиосомы (в процентах к ее ..длине) у сильно раздутой 9: vp 19; sc i 15; see и ha 20; hp 24; dx .15; d3 25; d4 20; lx 13; 12 26; 13 20; 14 22. Генитальное отверстие очень .крупное, простирающееся назад до уровня верхнего края ацетабул IV; «боковые углы энигиния сильно оттянуты; эпимеры III и IV слиты своими, .внутренними концами, замыкая, таким образом, коксальные ноля III; утолщения хитина вокруг эпимер I и II очень широкие. Ноги гораздо тоньше, чем у сГ, особенно задние; примериая длина их составляет около 44 (I и II), -43 (III), и 60 (IV) % длины идиосомы; лапки без килей, praetarsi все одинакового (нормального) размера. Длина 460—920 |л.
У нимф щетинки спинной поверхности идиосомы имеют такое же строение, как у взрослых.
Гипопус. Вледножелтоватый, с легким оранжевым оттенком, особенно на боках; ноги и прикрепительный аппарат буровато-красные. Тело .довольно широкое, в 1.3—1.4 раза длиннее своей ширины, с тупоугольным передним краем, выпуклыми на всем протяжении, правильно закругляю-• .щимися кзади боковыми краями и полого вогнутым задним краем; углы последнего тупо закруглены; проподосомальный щит немного более чем в 2 раза короче гистеросомы; оиистосомальная площадка не обособлена. .Аристы очень короткие, образуют прямой поперечный ряд; наружные еще короче внутренних, палочковидные. Эпимеры I (=ветви переднего развилка стернума) в 2 раза короче самого стернума; концы эпимеритов II отделены .значительным промежутком от эпимер III; последние слиты концами с эпимерами IV, образуя вместе с ними сплошное хитиновое кольцо, замыкаю-.щее коксальные поля III; от нижней части этого кольца отходит внутрь .короткий отросток, связанный с очень тонкой S-образной линией, отграничивающей область генитального отверстия от коксальных полей IV; утол-
10. LABWOPHORVS 337
щепной каймы вокруг последних нет. Генитальное отверстие очень крупное; очень велики и. генитальные щупальцы, диаметр которых равен длине колеи III. Клапаны прикрепительного аппарата крупные, с при остренными, угловато выступающими задними углами, заметно выдающимися за задний край тела; внутренняя поверхность клапанов снабжена многочисленными, сильно хитинизированными поперечными ребрышками; передние зажимы грушевидной формы, задние — шпателевидные, все с 6 толстыми, редко расставленными ребрышками. Ноги I и II массивнее и короче, чем у других видов; длина их составляет примерно 30 (I) и 28 (II) % длины тела;щетинки вертлугов очень длинные, волосовидные, наружные щетинки колен и вентральные щетинки голеней относительно короткие, простые, гибкие; вершинная дорзальная щетинка голеней с притупленным концом, палочковидная, едва короче голени; лапки много короче голеней, с 3 слабо ланцетовидно расширенными щетинками; обонятельная палочка стройная, расположенная посередине лапки; слабо развита. Длина ног III равна приблизительно 23% длины тела; они довольно стройны, их лапки равны по длипе голеням и украшены 3 узколанцетовидными щетинками; вентральные щетинки голеней и колен тонкие, гибкие, вершинная дорзальная щетинка голеней игольчатая. Коготки I—III довольно длинные, стройные, слабо изогнутые. Ноги IV такой же длины, как ноги III; лапки короче голеней, вооруженные дистально 1 очень крупной парусной щетинкой (превосходящей 1/2 длины тела), 1 вдвое более короткой, 3 довольно крупными, стройными шипами и 4 мелкими щетинками; коготок отсутствует; щетинки базальных члеников такого же строения, как на ногах III. Длина 250—350 р. 1
. Распространение. Голландия, Италия, Германия, СССР. У нас найден пока только в Московской обл.
Экология. Живет в гнездах белок (Sciurus vulgaris L.), на которых гипопусы этого вида встречаются нередко в большом количестве. Пищей нимф и взрослых служат микроскопические древесные .водоросли (Proton соссасеае и Palmellaceae), невидимому соскабливаемые ими с веток, используемых белкой для постройки гнезда.
*2. Labidophorus desma пае, sp. п. (фиг. 544—550).
£. Идиосома окрашена (в препарате) в бледный буроватый цвет гнато-сома, ноги и коксо-стернальный скелет темнокоричневые. У сильно раздутой яйцами особи идиосома имеет широкояйцевидную форму, ее задний край едва вогнут. Кожа покрыта такой же мелкозернистой (шагреневой) скульптурой, как у L. sciurinus, но на задней четверти спипы (вдоль средней лилии) имеется группа крупных, плоских овальных бляшек; такие же, но несколько меньшего размера бляшки имеются в центре коксальных полей; копулятивная трубка усажена мелкими, очень рейьефными хитиновыми зернышками, Щетинки спинной поверхности почти гладкие, с редко сидящими короткими
1 Британские экземпляры L. sciurinus, описанные Michael’ем (1901), памотпо мельче германских и встречающихся в СССР; может быть, они представляют собой особую расу этого вида.
22 Фауна СССР, Захваткин—284
338
Ш. GLYCYPHAGIDAE. о. LABIDOPHORINAE
ресничками; v р представлены очень мелкими микрохэтами; d2 хорошо развиты, мало уступают в размерах lx; h а находятся далеке» позади sc; р выстроены в косой продольный ряд; длина щетинок мало отличается от таковой; у L. sciurinus-, у единственной Q, имеющейся в моем распоряжении, многие из них обломаны, так что я могу пригести размеры лишь следующих из них (в процентах к длине идиосомы): v р 2; sc е 20; h а 17; й р 20; d213;. Ij 15; 14 и sa е 24; sai 15. Псевдостигмальный орган расщеплен почти, до самого основания на 2 неправильно перистые ветви. Генитальное отверстие простирается назад только до уровня внутренних концов эпиме-ритов II; эпимеры III и IV и эпимериты III свободные, коксальные поля III довольно широко открыты. Ноги немного короче, чем у L. seiurinus, составляют соответственно 40 (I), 38—39 (II и III) и 46 (IV) % длины идиосомы; вентральные щетинки голеней и колен III и голеней IV в виде игольчатых шипов. Длина 688 pi.
Гипопус. Бледножелтоватый; ноги и прикрепительный аппарат-едва темнее тела. Тело в 1.4—1.5 раза длиннее своей ширины, с тупоугольным передним краем и сильно вогнутым — задним; боковые края слабо-выпуклы и почти параллельны друг другу на всем пространстве от плеч, до резко выступающих передних углов опистосомальной площадки, затем, явственно прогнуты и сходятся под почти прямым углом к задним углам, тела; эти последние сильно хитинизированы и выдаются в виде тупых, цилиндрических рожков, слегка загнутых книзу; sa i сидят на конце этих, рожков. Проподосомальный щит почти в 2 раза короче гистеросомы; описто-сомальная площадка очень четко обособленная, с полукруглым передним краем. Аристы короткие, расположенные прямоугольником. Стернум хорошо--развит; концы эпимеритов II почти касаются эпимер III; последние свободны и отделены очень широким промежутком от эпимер IV, которые, сливаясь с краевыми утолщениями коксальных полей IV, образуют замыкающую их рамку. Генитальное отверстие и щупальцы небольшие. Клапаны прикрепительного аппарата с тупыми задними углами, совершенно не выдающимися за края тела; ребристость их внутренней поверхности развита, слабо; передние зажимы овальные, косые, с приблизительно 10 поперечными ребрышками; задние имеют клинообразную форму и снабжены 14—15-такими ребрышками. Ноги I и II довольно короткие; щетинки вертлугов короткие; вентральные щетинки голеней и базальная щетинка колен, утолщены, почти шиповидные; вершинная дорзальная щетинка голеней, палочковидная, притупленная па конце, равна по длине голени. Лапки, заметно длиннее голеней, с 3 листовидно уплощенными, узколанцетными.
Фиг. 546—558. Гипопусы рода Labidophorus.
Фиг. 546. Labidophorus desmanae, ер. п., вид снизу. Фиг. 547. Тоже, вид сверху. Фиг. 548. L. desmanae, ер. п.> голень и лапка I сверху. Фиг. 549. L. desmanae, ер. п., конец лапки I снизу. Фиг. 550. £. desmanae, sp. п., лапка IV сверху. Фиг. 551. L. talpae Кгаш., общий вид снизу. Фиг. 558. То же, общий вид сверху. Фиг. 553. £. talpae Kram., левые зажимы прикрепительного-аппарата. Фиг. 554. £. soricis Ouds., очертания тела сверху. Фиг. 555. £. soricis Ouds., правые важимы прикрепительного аппарата. Фиг. 556. £. mogerae, sp. п., общий вид снизу. Фиг. 557».
£. mogerae, ер. п., очертания тела сверху. Фиг. 558. £. mogerae, sp. п., лапка IV сверху.
340
Ш. GLYCYPHAGIDAE. 5. LABIDOPLIORINAE
(почти линейными) щетинками. Ноги III равны по длине ногам II, довольно стройные; лапки с 3 такими яге листовидными щетинками, как лапки I и II; вентральные щетинки голеней и колен в виде прямых игольчатых шипов; дорзальная щетинка голеней короткая, слегка булавовидно утолщена., Ноги IV чуть длиннее ног HI; ланки почти вдвое длиннее голеней, заметно расширены к концу, несущему 4 очень крупных, пальцевидно растопыренных конических шипа, редуцированный коготок, 3 умеренно длинных тонких щетинки и 2 маленькие шнновндпыи щетинки. Длина 250—270 [а, ширина 170—1W [х.
Распространение. Вероятно, совпадает с таковым выхухоли (вид описан по материалу, собранному с безэтикетных рсобей этого животного).
Экология. Живет в гнездах выхухоли (Desmana mosdhata L.); весь материал, послуживший для описания L. desmanae, добыт фильтрацией осадка из банок с спиртовым материалом по выхухоли: 9 и 2 гипопуса — проф. В. В. Редикорцевым, протонимфа и 2 гипопуса — Е. М. Булановой.
*3. Labidophorus talpae Kram. (фиг. 551—553).
Kramer, 1877, Arch. Naturg., XLIII, 1:249, t. XVI. fig. 1—3; Oudemans, 1915, Entombl. Вег., IV: 198(Labidophorus)-, Berlese, 1897: 106; Kramer, 1899 ; 145 (Glycyphagus).
Известен только по гипопусам. Окраска бледножелтоватая, ноги и-прикрепительный аппарат не темнее тела. Тело в 12—1.3 раза длиннее своей ширины, широкояйцевидное, с тупоугольным передним краем, сильно выпуклыми боковыми и полого вогнутым задним краем, отороченным резко отогнутой пластинчатой каймой. Проподосомальный щит немного менее чем в 2 раза короче гистеросомы, в 2 раза короче своей ширины у основания; опистосомальная пластинка ясно очерченная, с сглаженными боковыми углами; лежащая впереди нее часть гистеросомы менее чем в 2 раза длиннее ее. Аристы выстроены в поперечный ряд, очень длинные, волосовидные. Стерпум хорошо развит; концы эпимеритов II соприкасаются с свободными эпимерами III; эпимеры и эпимериты IV слиты своими внутренними концами, так что коксальные поля IV замкнуты. Клапаны прикрепительного аппарата с отвернутыми задними углами, почти не вогнутыми спереди внутренними краями и тонкоребристой внутренней поверхностью; передние зажимы имеют форму усеченного сзади овала, крупные, приблизительно с 12 тонкими поперечными ребрышками; задние зажимы клиновидные, широкие, с закругленным концом, вооруженные 9—11 более редко расставленными ребрышками. Генитальное отверстие небольшое, с умеренно крупными щупальцами. Ноги I и II довольно короткие и массивные, равные приблизительно 33 (I) и 30 (II) % длины тела; вентральные щетинки голеней и боковые щетинки колен длинные, тонкие;, вершинная щетинка голеней очень короткая (почти в 2 раза короче голеней), палочковидная, притупленная на конце; лапки заметно короче голеней, с 4 длинными, пуговковидно утолщенными на конце щетинками; обонятельная палочка короткая, до-
10. LABIDOPHORUS 341
вольно массивная, расположенная в проксимальной трети лапки. Ноги III едва короче ног II, более стройные; голени без зубца, с длинной, гибкой вентральной и относительно крупной, булавовидно утолщенной к копну вершинной щетинками; лапки едва длиннее голеней, с б крупными простыми щетинками, снизу с 1 ланцетовидным шипом. Ноги IV заметно короче пог 1! I; вентральная щетинка голеней короткая, булавовидная; лапки заметно длиннее голеней, массивные, с 4 крупными щетинками (из которых одна равна длине всей ноги), 2 более мелкими щетинками и совершенно редуцированным коготком. Длина 226—260 р, ширина 175—260 |х.
Распространение. Известен из Германии, Голландии 'и ряда пунктов СССР (Московская, Ленинградская и Смоленская обл.).
Экология. Встречается на кроте (Talpa europaea L.); найден также на черной крысе (Rattus rattus L.) и на соне (Muscardinus avellanarius L.).
* 4. Labidophorus soricis Ouds. (фиг. 654, 655).
Oudemans, 1915, Entomol. Ber., IV:198.
Гипопус. Морфологически очень близок к L. talpae Kram. Значительно мельче; тело совершенно не пигментированное, более ромбической формы; /гистеросома заметно короче (только в 1.55—1.6 раза длиннее проподосомы) и сильнее суживается кзади; ее боковые края, слабо выпуклые в области плеч, становятся почти прямыми дальше назад; в пространстве между более ясно намеченными передними углами опистосомальной площадки и задним краем тела они более или менее явственно вогнуты; отвернутая пластинчатая кайма имеет более острые очертания. Аристы очень короткие. Концы эпимер I обычно соприкасаются на значительном протяжении по средней линии тела, но не образуют монолитного стернума. Внутренние края клапанов прикрепительного аппарата спереди с выемкой, передние зажимы меньшего размера, чем у Предыдущего вида; задние гораздо длиннее, очень узкие, вооруженные приблизительно 20 тонкими поперечными ребрышками. Ноги длиннее, чем у L. lalpae, особенно задние; длина их составляет приблизительно 38(1), 35(11), 36(111) и 38 (IV) % длины тела; вооружение ног, как у L. talpae. Длина 170—210 р., ширина 130—160 р.
Распространение. Голландия, Германия, СССР; мне известен из ряда пунктов Московской и Орловской обл., Абхазии и окрестностей Якутска.
Экология. Встречается преимущественно на различных видах землероек (Sorex araneus L., >S. minutus L., Neomys fodiens L.), но также и на кроте и многих видах грызунов (ArvicolaampliibiusL., Microtus oecunomvs Pall., Evotomys glareolus Schr., Micromys minutus L.).
6.< Labidophorus mogcrae, sp. и. (фиг. 556—558).
Известен только гипопус. Весь бледноохристый; тело удлиненное, в 1.6—1.75 раза длиннее своей ширины, значительно более ушсоо, чем у других видов; передне-боковые края проподосомы сходятся кпереди подострым
342
III. GLYCYPHAGIDAE. 5. LABIDOPHORINAE
углом, притупленным на конце; боковые края гистеросомы почти параллельны друг другу на большом протяжении, затем слегка закругляются и незаметно переходят в ее чуть вогнутый посередине задний край; задние углы тела сильно сглажены. Провод осомальный щит в 1.65 раза короче гистеросомы, у основания в 1.5 раза шире своей длины; опистосомальная площадка нелепо очерченная, с еле намеченными боковыми углами, Аристы короткие, расположенные поперечным рядом. Стернум нормальный; вдоль середины брюшной поверхности тела тянется широкий жолоб, боковые стенки которого поддерживаются отогнутыми назад концами эпимер II и III и эпимеритов II, концами эпимер IV й сильно развитыми, принявшими продольное направление эпимеритами IV; коксальные поля III и IV полуоткрыты. Генитальное отверстие сравнительно небольшое, с трудно заметными щупальцами. Зажимы клапанов прикрепительного аппарата мельче, чем -у других видов рода Labidophorus, каждый с 5 широко расставленными, сильно рельефными поперечными ребрышками; передни^ имеют форму прямоугольника с сглаженными углами, задние скорее треугольны. Ноги I и II довольно короткие, достаточно массивные; наружная вентральная щетинка голеней и наружная щетинка колен превращены в мощные, слегка зазубренные шипы; вершинная щетинка голеней длинная, гибкая, с едва пуговковидно утолщенным концом; лапки стройные, чуть длиннее голеней', с 4 длинными, слегка крючковидно загнутыми на конце щетинками; обонятельная палочка прямая, тонкая, расположенная в проксимальной четверти ланки; (3 шиновидная. ElQrH III очень стройные; голени дистально с коническим отростком и относительно длинной, немодифицированной дорзальной щетинкой; ланки с 5 довольно крупными, уплощенными (почти мечевидными) щетинками и редуцированным коготком. Ноги IV одинаковой длины с ногами III, но много массивнее (особенно толсты вертлуги и бедра); голени с таким же выростом, как голени III, с совершенно редуцированной вершинной щетинкой; лапки короче голеней, дорзально с толстым, искривленным шипом, на конце с редуцированным коготком и 5 расширенными у основания щетинками, из которых одна равна длине всей ноги. Длина. 255—270 |л, ширина 150—180 р..
Распространение. Описывается по материалам из Кореи (окрестности Сеула). Гипопусы найдены на дальневосточном кроте Модега robusta Nehr.
11. Род GOHIERIA. OUDS.
Oudemans, 1939, Zool. Am., OXXVII:185.—Ferminia Oudemans, 1928, Entomol. Ber., VII:347; Буланова, 1937, Бюлл. Моск. общ. исп. прир,, XLVI:304 (пот. ргаеосс.).
Тип рода: Glycyphagus fuscus Ouds. 1902.
Половой диморфизм выражен относительно слабо. Гнатосома среднего размера, почти целиком закрытая сверху ростральным выступом; идиосома у обоих полов эллиптическая (у уже, чем у 9), с параболическим (не очень явственным, вследствие своей нежной консистенции), необособленным ростральным выступом и широко закругленным задним краем, без
//. GOHIERIA
343
•фестончатых лопастей; пластинчато расширенные края ацетабул I—III несколько выдаются с боков, придавая им зубчатые очертания. Кожа сильно хитинизированная, гладкая .(тонкопористая), окрашенная в темный буроватый цвет; на коксальных полях она толще и покрыта более грубой пористой скульптурой; по окружности идиосомы, на самом ее краю, проходит очень узкая полоска перепончатой кожи, отделяющая спинной щит от брюшного, хорошо видимая при рассматривании животного в профиль. V а и v р хороша ;развиты; плечевых щетинок и боковых по 3 пары; d3 помещаются много впереди d3; все щетинки спинной поверхности идиосомы однотипны — .короткие, гладкие, довольно гибкие; на передней половине идиосомы они нормально направлены вперед, на задней — в стороны и назад; р а тонко-перистые. Ноги длинные, передние у обоих полов довольно массивные;-, бедра, колени и голени всех ног резко угловатые, как бы граненые, с многочисленными тупыми, продольными ребрами; на ногах I и II, кроме того, •с пластинчато расширенными, в виде раструбов, дистальными краями; бедра I и II бокаловидные, сильно суженные к основанию, с поперечным 1килем, отделяющим суженную проксимальную часть от расширенной дистальной (у § скульптура ног еще более рельефиа, чем yd); вооружение ног состоит из густоперистых и гребенчатых щетинок, вентральная щетинка .голеней IV хорошо развита.
d. Все коксальные поля целиком очень сильно хитинизированы и образуют сплошной стернальный панцырь, напоминающий таковой многих Sydracarina} не уплотнено только свободное пространство в центре тела, остающееся между внутренними концами эпимер; стернум хорошо развит, свободный; эпимеры II, III и IV соединены своими внутренними концами; эпимериты IV отсутствуют. Генитальное отверстие расположено позади основания ног IV; penis в состоянии покоя обращен вершиной назад; анальная щель длинная, с немного выдающимися за задний край тела клапанами. .Ноги без каких-либо специальных для сГ выростов, praetarsi не увеличены. Ноги III и IV заметно утолщены и ис..ривлены, с видоизмененными лапками.
9. Эпимеры I целиком слиты с эпигинием, образуя массивную поперечную пластинку впереди генитального отверстия; эпимеры II—IV свободные, но соприкасаются концами с овальным хитиновым кольцом, окружающим генитальное отверстие; последнее очень велико, занимает все пространство между основаниями ног II, III и IV; половина поверхности коксальных полей I и II и вся поверхность полей III и IV сильно хитинизирована; .генитальные щуиальцы очень мелки; копулятивная трубка отсутствует; анальная щель доходит до заднего края тела и даже выдается за псе концами своих клапанов; имеются хорошо развитые генитальные трахеи.
Нимфальные стадии с гораздо более нежными, непигмептированными жожными покровами; у личинок кожа равномерно мелкобугорчатая, щетинки идиосомы слегка перисты, имеются очень мелкие, сосочковидные грудные палочки; у прото- и телеонимфы кожа имеет структуру, напоминающую
ш
III. GLYCYPHAGIDAE. 5. LAB1D0PH0RINAE
так называемую крокодиловую кожу, щетинки спинной поверхности не-> . сколько уплощены, гладкие, лопасти заднего края идиосомы вполне явственны. — Гипопальной стадии, по всей вероятности, не -имеет.
Монотипический род.
1. Goliicria fusca (Grids.) (фиг. 559—566).
Oudemans, 1902, Entomol. Вег., 1:21; 1903, Tijdschr. Entomol., XLVT:15,t. II, fig. ol—32, et t. Ill, fig, 33—37 (GlycyiiJuiywy, 1928:347; Eyjiuuoua, 1937, Вюлл. Моск. общ. исп. прир., XLVI:304 (Eerminia). :
Биология. Буланова, 1937.
Одноцветно розовато-коричневый. Длина щетинок спинной поверхности обоих полов практически одинакова и колеблется в пределах 5—16% длины идиосомы; v a, sc i, dn d2 и sa i короче всех других (равные 5—6% длины идиосомы); sc е в 1.6—2 раза длиннее sc i, приблизительно одинакового размера с h a, h р, d3 и d4 (8—11% длины идиосомы), длиннее других 12—14 и sa е, достигающие 12—16 % длины идиосомы. Вентральные щетинки бедер I гладкие, бедер II и IV — тонкоперистые; базальные щетиньи колен I и II гребенчато-пильчатые, внутренняя — изящно изогнутая, наружная—почти прямая; вентральные щетинки голеней и средняя вентральная щетинка лапок I и II сравнительно короткие и толстые, гребенчато-перистые. У <5 ноги заметно длиннее и массивнее, чем у 9; их примерная длина составляет около 52(1), 50(11), 40—41(III) и 49—50(IV)% длины идиосомы; лапки III и IV едва длиннее голеней; длина ног 9 состав^ ляет примерно 42—43(1), 35—37 (II и III) и 41—42 (IV) % длины идиосомы; лапки III почти в 2 раза, лапки IV в 2—2.3 раза длиннее соответствующих голеней. Длина d 300—320 ширина 180—200 ;л; длина 9 386— 412 р., ширина 240.—260 р..
Распространение. Голландия, Франция, Англия, СССР. У нас найден в Москве, в нескольких пунктах Ивановской обл., на Кавказа (Моздок, Ваку), в Средней Азии (Ташкент, Ферганская долина: Пахталык-Куль) и в Западной Сибири (Барнаул).
Экология. Синантропный вид, живущий преимущественно в муке,, но найденный также в пшенице, рисе и жмыхах; в полевых условиях до сих .пор не обнаружен. В южных районах СССР (в Средней Азии, Закавказье) иногда встречается в массе и наносит ощутимый вред заражаемым им «продуктам; в других районах попадается лишь спорадически.
12. Род XENOKYCTES, gen. п.
Тип рода: Glycyphagus krameri M'chael 1886.
Половой диморфизм очень резкий. Идиосома довольно сильно уплощена дорзо-вентрально, с хорошо развитым, параболическим ростральным высту-
Фиг. 559—566. Gohima fvsev (Ouds.) (по Булановой). 1
Фаг. 659. cf снизу; Фиг. 560. Q сверху. Фиг. 561. То же, снизу. Фиг. 563. Нога I 9 сверху.. Фиг. 563. То же, снизу. Фиг. 564. Йога IV 9. Фиг. 565. Нога III о"- Фиг. 566. Нога IV о"-
346
III. GLYCYPHAGIDA& 5. LABIDOPHORINAE
-пом, без фестончатых лопастей. Кожа очень толстая, твердая, темноокра-шенная, грубозернистая, крупнопористая; ростральный выступ с парой S-образно изогнутых продольных килей; по окружности идиосомы проходит такая же линия ослабленного хитина, как у Gohieria, хорошо видимая сбоку. V а хорошо развиты, утолщены в виде рожков; v р отсутствуют; все остальные щетинки идиосомы представлены крайне мелкими, гладкими, толстыми микрохэтами, большинство которых трудно заметно (заметнее других периферические — ha, 12—la, sa i). Ноги довольно длинные, относительно стройные, с разнообразными пластинчатыми выростами; вершинная щетинка голеней сидит па трубчатом конце особого килевидного возвышения; щетинки бедер, колен и голеней (кроме вершинной) перистые, гребенчатые или так или иначе модифицированные; вентральная* щетинка голеней IV отсутствует.
<5. Идиосома эллиптической формы с почти параллельными боковыми краями и широко закругленным задним краем. Стернум, эпимеры II и IV и эпимериты II соединены концами с массивным хитиновым полукольцом, расположенным в центре тела, несколько впереди генитального отверстия; эпимеры III сильно редуцированы или отсутствуют; эпимериты IV хорошо развиты, свободные. Генитальное отверстие помещается между основаниями ног IV; penis в состоянии покоя направлен вперед; анальная щель очень маленькая, отставленная от заднего края идиосомы, окруженная тонким хитиновым кольцом. Ноги I и II сильно утолщены, с листовидными продольными килями на лапках (I и II) и на голенях (I) и сильно зубчатыми вентральными пластинками на голенях (I и II) и бедрах (I); голени I и II без вентральных щетинок. Ноги II] и IV мало искривленные; вентральные щетинки голеней и колен III модифицированы; лапки IV вентрально с таким же пластинчатым килем, как лапки I; praetarsi не увеличены.
9. Идиосома расширена кзади, с более или менее глубоко выемчатым задним краем, почти сердцевидная. Стернум хорошо развитый, длинный, -сливающийся концом с эпигинием или с особым хитиновым полукольцом, расположенным впереди генитального отверстия; концы эпимер II и III, а иногда и IV, также соединены с эпигинием или упомянутым полукольцом, которые, в свою очередь, связаны с массивным циркумгенитальным кольцом; анальная щель окружена массивной хитиновой рамкой. Ноги стройнее, чем у б, с более или менее развитыми килями на дорзальной стороне голеней; пластинчатые образования на вентральной поверхности голеней и колен отсутствуют; щетинки пог III не модифицированы, голени IV без вентральной щетинки.
Гипопус. Тело яйцевидное или овальное, с заметно выпуклой спин-ной поверхностью; обособленной опистосомальной площадки нет. Коксо-стернальный скелет слабый; эпимеры III соединены концами с эпимерами IV, явственно замыкая, таким образом, коксальные поля III. Передние -зажимы прикрепительного аппарата округлые, с немногими более или менее радиальными ребрышками; задние далеко отодвинуты от передних, узкие,
12. XENORYCTES
347
с 5—7 поперечными ребрышками. Ноги довольно длинные и стройные, вершинная щетинка голеней I и II длинная, нормального строения; лапки длинные, прямые, цилиндрические, с довольно изменчивым дистальным вооружением; лапки IV без парусной щетинки, с хорошо развитым коготком или без коготка.
Род Xenoryctes состоит из 2 видов, известных только из Европы; оба вида биологически связаны с насекомоядными.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
99
1 (2). Концы стернума и эпимер II и III соединены с тонкой хитиновой дугой, расположенной впереди циркумгенитального кольца и, в свою очередь, соединенной с ним своими задними концами; эпимеры IV свободные; голени I дорзально со слабо развитым продольным килем, вентрально только с 1 гладкой щетинкой . . 1. X. krameri (M.ch.).
2 (1). Концы стернума и эпимер II, III и IV непосредственно соединены с очень мощным полулунным эпигинием, слитым с циркумгениталь-ным кольцом; голени I дорзально с очень высоким секировидным килем, вентрально с 1 гладкой и 1 гребенчатой щетинками . . . .............................................*2. X. heptneri, sp. п.
Гипопусы
1 (2). Ростральный выступ очень узкий, сильно выдающийся вперед; концы клапанов прикрепительного аппарата выдаются сзади в виде острых ’ рожков, задний край тела между ними глубоко вогнут; ноги III и IV немного короче передних, лапки IV без.коготка, дистально с 4 длинными щетинками............................... 1. X. krameri (М ch.),
;2 (1). Ростральный выступ широко закруглен, совершенно не выдающийся;
концы клапанов прикрепительного аппарата не видны сверху; задний край тела едва вогнут; ноги III и IV много короче передних, лапки IV с хорошо развитым коготком и только с 1 длинной щетинкой . . . . ....................................*2. X. heptneri, sp. в.
, 1. Xenoryctes krameri (Mich.) (фиг. 628—539).
Michael, 1886, Joum. R. Mier. Soc., ser. II, vol. VI:377, t. X; 1901:275, t. XVI-XVII, 'fig. 5—11 (Glycyphagus). — talpae Canestrini, 1888:18, t. II, fig. 12—14; 1888a:379, t. XXXV, -fig. 3 (Homopus)', Berlese, 1897:106; Kramer, 1899:145 (Glycyphagus).
d. Темнокрасновато-коричневый. Идиосома в 2 раза длинное своей максимальной ширины, приходящейся над основаниями ног III, у живых особей с равномерно слабо выпуклым диском и слегка приподнятыми краями; длина рострального выступа составляет 1/8 общей длины идиосомы; его боковые края более параллельны,.чем у 9, дорзальные кили слабое расходятся .кзади. Кожа тонкозернистая, на спинной поверхности обычно с несколь
348
HI. GLYCYPHAGIDAE. 5. LABIPOPIIOP.LNAE
кими неправильными рельефными рубцами. Ноги гораздо массивнее,' чем у. 9; колена I сильно утолщены, много длиннее голеней, вентрально без-пластинчатых образований, с редуцированными (или совершенно отсутствующими'-) дорзальными щетинками; голени I вентрально с прозрачной' веерообразной пластинкой, расщепленной на 7—9 тонких расходящихся острий, дорзалыю с длинным трубчатым выростом, несущим вершинную-щетинку; ланки I дорзалыю с листовидным прозрачным килем, очень высоким в своей проксимальной части и постепенно сходящим на-нет к конечной трети лапки, вентрально с таким же, по более равномерно выгнутым килем, расположенным несколько дистальнее. Вооружение ног II сходно-с таковым пог I, отличаясь в том, что на вентральной поверхности колен помещаются 2 вееровидных зубчатых пластинки, из которых проксимальная меньше, имеет полукруглую форму и снабжена у основания центральным утолщением, а дистальная длиннее и напоминает по своей форме секиру; голени II дорзально с пластинчатым килем, листовидные кили лапок ниже, более равномерной ширины; внутренняя базальная щетинка колен хорошо-развита, гребенчатая. Ноги III лишены листовидных разращений; вентральная щетинка колен булавовидно вздута, шиповатая, вентральная щетинка голеней очень длинно гребенчатая, с 4—5 широко расставленными отростками. Лапки IV вентрально с невысоким пластинчатым килем. Длина около-250 Ji, ширина 130 р..
9. Немного светлее с?. Идиосома около 1.6 раза длиннее своей максимальной ширины, приходящейся значительно позади основания ног IV; дорзальная поверхность с более сложным рельефом, чем у d1. Кожа более грубозернистая; длина рострального выступа составляет около 1/а общей длины идиосомы, его боковые края и дорзальные кили сильнее расходятся Кзади Концы стернума и эпимер II и III соединены с тонким хитиновым полукольцом, находящимся впереди циркумгенитального кольца, с которым, в свою очередь, соединены концы этого полукольца; эпимеры IV свободные; циркумгенитальное кольцо поперечноэллиптическое, заметно шире своей длины. Ноги значительно стройнее, чем у d, длиною около 37 (I), 34—35 (II и III) и 43 (IV) % длины идиосомы; колени I и II заметно расширены к концу, несколько вздутые, невидимому без боковых килей; дорзальный и боковые кили голеней слабо развиты; базальные щетинки колен I и II одинаковой формы, толстые и довольно короткие резко гребенчатые, в дистальной половине с более крупными и редко расставленными отрост-<камй, чем в проксимальной; голени I и II вентрально с 1 (наружной) гладкой, щетинкой; щетинки лапок довольно длинные. Длина около 350 р., ширина 240 |i.
Телеонимфа. Беловатая, полупрозрачная. Проподосома довольно-явственно отделена от гистеросомы поперечной бороздкой, украшенная 16 небольшими, широко разделенными друг от друга гладкими площадками, расположенными обычно следующим образом: немного позади v а — 1 более крупная, непарная округлая площадка; позади нее — 2 более или менеетре-
/2. XENORYCTES ' 349 .
угольные, тоже довольно крупные площадки; дальше назад — поперечный ряд из 5меньшего размера площадок, из которых обе наружные значительно крупнее 3 внутренних; совсем сзади, вдоль поперечной бороздки,— ряд из 8 площадок, из которых крайняя наружная'пара крупнее других, следующая затем значительно меньше и обе вентральные очень мелки; аналогичные площадки гистеросомы имеют небольшие размеры и неправильные очертания и разделены широкими участками мягкой складчатой формы. V а неправильной формы, притупленные па конце; sa е хорошо развиты, одинакового размера с sa i, но меньше, чем 13.
Гипопус. Желтовато-белый; ноги и прикрепительный аппарат красноватые. Тело широкоовальное, с очень узким и сильно выдающимся ростральным выступом и глубоко вогнутым задним краем, за контур которого выдаются в виде острых рожков концы клапанов прикрепительного аппарата. Передние зажимы довольно крупные, с 5 резко расходящимися радиально ребрышками; задние зажимы линейные, не суженные к основанию, •с 7 поперечными ребрышками. Ноги очень стройные, особенно задние; вершинные щетинки лапок длинные; лапки IV без коготка, с 2 короткими и толстыми, загнутыми концами кверху щетинками и 4 длинными щетинками. Длина около 250 |i.
Распространение. Англия, Италия. Живет в норах крота {.Talpa europaea L.).
*2. Xenoryctes heptneri, sp. п. (фиг. 580—583, 593—595).
Icrameri Oudemans, 1915, Entomol. Вег., IV: 210 (Dermacarus) (non Michael).
9. Красновато-бурая. Идиосома в 1.4 раза длиннее своей максимальной ширины. Концы стернума и эпимер II, III и IV соединены непосредственно с очень мощным, неправильно очерченным эпигинием, обрамляющим спереди циркумгенитальную щель. Ноги очень стройные, длиною около 35 (I), 30—32 (II и III) и 40 (IV) % длины идиосомы; колени I и II слабо расширяются к концу, не вздутые, украшенные, так'же как и голени всех ног, очень явственными, тупыми продольными килями; все голени дорзально с вертикальным листовидным килем, со свободным трубчатым дистальным концом, несущим их вершинную щетинку; на голенях I этот киль необычайно высок и имеет форму секиры, на голенях II он значительно ниже, на задних Совсем низкий; голени II с 2 вентральными щетинками: гладкой и тонкой — наружной, и толстой, гребенчатой—внутренней. Длина 330—340 р., ширина 250 |i. — d неизвестен. , i
Телеонимфа. Отличается от соответствующей стадии X. krameri (на которую габитуально очень похожа) следующими признаками: почти вся спинная поверхность пропод'осомы занята 1 круцным гладким щитом, позади которого помещаются лишь 2 тесно придвинутых к нему ряда, из 4 площадок каждый; передний ряд составлен 2 овальными наружными й 2 крупными, сильно поперечными внутренними площадками; внутренняя лара площадок 2-го ряда Имеет такую же форму. Площадки гистеросомы
350
HI, GTWYPHAGIDAE. -5. LABTDOPHORINAE
крупнее, чем у X. krameri, более правильной округлой или овальной формы и разделены более узкими перепончатыми промежутками. V а правильной широколаицстовидной формы, sa е одинакового размера с 1 (13), sa v отсутствуют.
Гипопус. Беловатый, спепигментиррванными конечностями и желтоватым прикрепительным аппаратом. Тело более правильной, чем у гипопуса X. krameri, яйцевидной формы, с широко закругленным, совершенно-пе выдающимся ростральным выступом и едва вогнутым, закругленным задним краем. Клапаны прикрепительного аппарата с тупыми концами, не выдающимися за задний край тела; передние зажимы небольшие, со слабо дивергирующими ребрышками, задние — шпателевидные, сильно сужи-. вающиеся к основанию, с 5—6 поперечными ребрышками. Ноги, особеннозадние, значительно короче, чем у X. krameri-, лапки I—III с 3 более круп-' ными и 2 мелкими листовидными щетинками и 2 короткими простыми (из. которых наружная пуговковидно вздутая на конце); лапки IV с крупным, столь же хорошо развитым, как на передних ногах, коготком, дистально с 2 небольшими ланцетовидными щетинками и 1 довольно длинной простой, щетинкой. Длина 205—235 р,, ширина 130—155 |л.
Распространение. Известен пока только из Голландии и Московской обл.: дер. Шульгине, Кунцевского р-на, и в Можайском р-не.
Экология. Самки, нимфы и гипопусы найдены мною летом 1936 г. в норе ближе не определенной мыши, вместе с Glycyphagus ornatus Kram.,. Fusacarus laminipes Mich, и гипопусами Myacarus arvicolae (Duj.);. 1 экз. гипопуса добыт фильтрованием спирта из банки с. кротами.
13. Род ORYCTEROXENUS, gen. п.
Тип рода: Glycyphagus dispar Michael 1886.
Половой диморфизм настолько резок, что между d’d' и 99 не сохраняется практически никакого сходства.
<5. Карликовых размеров (общая длина тела около 170 |л), по общему виду несколько напоминает гипопусов. Идиосома щитовидной формы, с необычайно резко выступающими плечами, почти параллельными сторонами и широко закругленным задним краем; ростральный выступ гладкий, сильно хитинизированный, с тонким поперечным швом перед серединой, закрывающий всю гнатосому. Кожа всей остальной поверхности идиосомы, густо усажена полушаровидпыми бугорками, менее плотная, слабо пигмен-
Фиг. 567—583.
Фиг. 567. Xenocastor fedjushini, sp. П., личинка. Фиг. 568. Myacarus hypudaei (Koch), гипопус, очертания тела сверху. Фиг. 560. М. hypudaei (Koch), гипопус, общий вид снизу. Фиг. 570. М. hypudaei (Koch), гипопус, нога I сверху. Фиг. 571. М. hypudaei (Koch), гипопус, лапка I в профиль. Фиг. 572. М. arvicolae (Duj.), гипопус, очертания тела сверху. Фиг. 573. М. arvicolae (Duj.), гипопус, общий вид снизу. Фиг. 574. М. arvicolae (Duj.), гипопус, нога I снизу. Фиг. 575— 576. М. arvicolae (Duj.), гипопус, изменчивость вооружения лапок IV. Фиг. 577. Labidophorus-sciurinus (С. L. Koch), гипопус, голень и лапка I сверху. Фиг. 578. L. sciurinus (С. L. Koch), гипопус, лапка IV снизу. Фиг. 579. То же, сверху. Фиг. 580. Xenoryctes heptneri, sp. п., гипопус, общий вид снизу. Фиг. 581. X. heptneri, sp. п., гипопус, нога I снизу. Фиг. 582. X. heptneri, sp. п., гипопус, голень и лапка I сверху. Фиг. 583. X. heptneri, sp. п., гипопус, лапка IV.
352 7Я, GJ.YCYPHAGJDAE. 5. LABIDOPfTORINAE
тированная. Щетинки идиосомы представлены очень короткими толстыми шипами. Эпимеры I и II слабо развиты, соединены концами друг с другом и с наружными концами подковообразного эпиандрия; эпимеры III и IV и эпимериты IV совершенно отсутствуют. Генитальное отверстие помещается немного позади основания ног IT; penis короткий, в состоянии покоя обращен вершиной назад; анальная щель очень маленькая, окруженная тонким хитиновым кольцом, далеко отставленная от заднего края тела. Ноги I и II расположены нормально, довольно толстые, с бокаловидными бедрами, лишенные пластинчатых разращений; ноги III и IV немного стройнее, с длинными, бокаловидно суженными к основанию коленными члениками; они расположены почти на одном уровне, образуя дугообразный попереч-ный ряд (основания ног III смещены кнаружи, ног IV — вперед и внутрь), направлены вперед и целиком скрыты под идиосомой. 1
9. Много крупнее d; габитуально сходна с 9 Talpacarus. Идиосома несколько уплощена дорзо-вентрально, расширенная кзади, с очень резко обособленным ростральным выступом, гладким, угловато вогнутым посередине задним краем и сильно уплощенными, неправильно вырезанными боковыми краями. Кожа рострального выступа сильно'хитинизирована, гладкая, на остальной поверхности идиосомы покрытая шиповато-бугорчатой скульптурой, сходной с таковой Talpacarus. V а отсутствуют; v р хорошо развиты, прямые; sc е, обе пары плечевых и все 3 пары боковых расположены маргинально, на вершинах краевых лопастей идиосомы, крепкие, игольчатые, более или менее резко искривленные; dlt d4 и обе пары sa отсутствуют (или сильно редуцированы?), так что на диске идиосомы имеются только 2 пары щетинок, сильно придвинутые друг к другу и к средней линии тела. Концы эпимер I и II и эпимеритов II соединены с массивным хитиновым кольцом, окружающим генитальное отверстие; эпцмеры III и IV очень короткие, свободные. Копулятивная трубка хорошо развита, коническая; анальная щель сильно удалена от заднего края тела и окружена толстым хитиновым кольцом. Ноги длинные, тонкие, лишенные каких бы то ни было килей и выростов; колени и бедра I и II несколько бокаловидны; вентральная щетинка голеней IV отсутствует.
Нимфальные стадии не описаны. — Гипопальная стадия отсутствует. Известен только 1 вид.
1. Orycteroxcnus dispar (Mich.) (фиг. 687—590).
Michael, 1886, Joarn. Linn. Soc. bond., XIX: 280, t. XXXV, fig. 6—17;|Berlese, 1897:106; Kramer, 1899:145; Michael, 1901: 271, t. XV (Glycyphagus).
d. Тусклого светлосерого цвета, иногда с более или менее ясно выраженным красноватым оттенком. Идиосома в 1.3 раза длиннее своей ширины; все щетинки ее спинной поверхности имеют приблизительно одинаковую длину (3—5% длины идиосомы), шиповидные. Примерная длина ног состав
1 Судя по положению ног III и IV и их строению, можно думать, что они совершенно утратили свою локомоторную функцию и служат только для обхватывания § при копуляции.
13. ORYCTEROXENUS. 14. TALPACARUS
35?
ляет около. 40 (I и II) и 36 (IV) % длины идиосомы; голени I и II укороченные, в 1.5 раза короче вздутых колен и почти в 2.5 раза короче лапок; голени III и IV цилиндрические, почти равные по длине коленным членикам и лишь немного короче лапок; вентральные вершинное шипы лапок III сближены между собой, на лапках IV разделенные довольно широким расстоянием; praetarsi короче и шире, чем у 9. Длина около 170 |л, ширина 130 [л.
9. Прижизненная окраска красновато-коричневая. Идиосома в 1.4 раза длиннее своей максимальной ширины, приходящейся далеко позади ее середины; у живых особей уплощенные боковые поля отделены глубоким вдавле-нием от слегка выпуклой дистальной части спинной поверхности; вдоль середины последней тянется продольная выпуклость, спереди закругленцая и далеко не доходящая до основания рострального выступа, сзади расширяющаяся и занимающая весь задний край идиосомы (приобретающий, таким образом, большую рельефность). Vp длиною около 13% длины идиосомы, прямые, направленные вперед, так же как чуть более длинные sc i; sc е едва длиннее sc i, почти прямые, направлены горизонтально в стороны и немного назад; h а имеют в длину около 20% длины идиосомы, резко S-образно искривлены и направлены в стороны и назад; такое же направление имеют и дугообразно изогнутые h р, 1Ъ 12 и 13, размеры которых почти одинаковы (20—25% длины идиосомы). Длина ног составляет примерно 48 (I и II), 43 (III) и 52 (IV) % длищ>1 идиосомы; голени I и II немного короче колен; лапки I и III около 2 раз, IV — в 3.5 раза длиннее соответствующих голеней. Длина около 350 р., ширина около 240
Распространение. Известен только из, Англии. По Michael’ro, обычен в норах крота, свежих и обитаемых кротом в данное время; на самих кротах никогда не встречается.
14. Род TAIPACABUS, gen. п.
Тип рода: Glycyphagus platygaster Michael 1886.
Морфологические различия между полами не очень велики.
Идиосома несколько уплощена в дорзо-вентральном направлении, широко яйцевидная (9) или почти круглая (d), с очень резко обособленным ростральным выступом и уплощенными, неправильно фестончато вырезанными боковыми краями. Кожа рострального выступа сильно хитинизирована, гладкая, на остальной поверхности идиосомы — умеренной плотности, слабо пигментированная, покрыта мелкими коническими шипиками. V а хорошо развиты, слегка искривленные; vp длиннее v а; плечевых и боковых щетинок по 3 пары; все щетинкп спинной поверхности гладкие, игольчатые, довольно длинные; sc е, h a, h р, —13 и sa е (а у d еще и sa i) расположены совердпенно маргинально, на вершине краевых лопастей идиосомы; d2 не смещены назад. Ноги очень длинные и тонкие, I и II одинакового строения у обоих полов,-лишенные килей и пластинчатых выростов, вооруженные простыми или слабо зазубренными щетинками.
23 Фауна СССР, Захваткин—284 .
35i
III. GLYCYPHAGIDAE 5. LABIDOPHORlNAE
<5. Задний край идцосомы образует 4 округлые лопасти, более круп--ные, чем лопасти ‘боковых краев. Стернум свободный; концы эпимер II, III и IV соединены поперечной хитиновой перемычкой. Генитальное отверстие расположено в пространстве между основаниями ног III и IV; penis-в состоянии покоя направлен вершиной вперед; анальная щель отставлена от заднего края тела, окружена хитиновым кольцом. Ноги III и IV слабо искривлены.
$. Задний драп идиосомы цельный, прямой. Стсрпум свободный, короче, чем у сГ; энимсры II свободны, Ш и 1V соединены своими внутренними концами. Генитальное отверстие помещается между основаниями ног II и III, без хитинового кольца и непарного заднего клапана; эпйгиний крупный, стрельчатой формы, приостренный спереди; позади генитального отверстия находится более тонкий дугообразный склерит, также обращенный выпуклостью вперед; копулятивная трубка крупная, коническая; анальная щель длинная, почти доходящая до заднего край тела.
У нимф задний край идиосомы образует такие же выдающиеся (но болей узкие)-Допасти, как у <5, лопасти боковых краев разделены слабее; щетинки, такого же строения, как у взрослых, но короче; у личинок зернистость-, кожи, повидимому, несколько крупнее, чем у других стадий. — Гипопусов-, насколько известно, не образуют..
Монотипический род.
1. Talpacarus platygaster (Mich.) (фиг. 591, 592).
Michael, 1880, Journ. Linn. Soo. bond., XIX:276, t. XXXIV—XXXV, fig. 1—5; Beriese, 1897:106; Kramer, 1899:145; Michael, 1901:265, t. XIV, XVII» fig. 1—4 (Gtyeyphagus).
<3. Темнее $, розоватого цвета; ростральный выступ, гнатосома и ноги розовато-бурые. Идиосома (не считая рострального выступа) почти круглая, кожа с брлее грубой скульптурой, чем у $. Все щетинки спинной поверхности почти прямые; vp равны по длине sc i и почти вдвое короче ha; sa i, повидимому, отсутствуют; d4 расположены маргинально на -вершине медиальных выступов заднего края; примерная длина щетинок '(в процентах к длине идиосомы): vp 17; see 24; sc i 17—18; ha 30; hp 25; dj 20—21; d2 25; d3 45; d4 17; lx и 12 23—24; 13 16; sa e 25. Ноги -очень, длинные: 62 (I), 60 (II и III) и около 70 (IV) % длины идиосомы; III и IV — почти на краю тела, так что их вертлуги частично видны сверху. Длина: около 540 р., ширина около 440 р..
$. Кремово-желтая или пергаментного Двета, 'с розовыми боковыми краями, ростральным выступом и ногами. Идиосома широкояйцевидная, с прямо усеченным задним краем, постепенно расширяющаяся кзади до уровня 1Х, затем суживающаяся до точек прикрепления sa е; уплощенные
Фиг. 584—592 (по Michael’ю).
Фиг. 684. Labidopkorus salurinus (С. L. Koch), 9 сверху. Фиг. 585. То же, снизу. Фиг. 686». L. sciurinus (С. L. Koch), Cf сверху. Фиг. 587. Orycteroxenus dispar (Mich.), сГ сверху. Фиг. 588. То же, снизу. Фиг. 589. О. dispar (Mich.), 9 снизу- Фиг..590. То же, сверху. Фиг. 591» -Talpa-, carus platygaster (Mich.), 9 сверху. Фиг. 592. Т. platygaster (Mich.), o'сверху.
336
Ill. GLYCYPHAGIDAE. 5. LABIDOPHORINA G
боковые края отграничены изнутри продольным вдавлением, дискальная часть равномерно слабо выпуклая; очертания лопастей боковых краев более угловаты, чем у сГ, неявственно S-образно искривлены, ha, sc i и некоторые из периферических щетинок слегка изогнуты; v р в 1.5 раза длиннее sc i и немного Длинное h a; d4 вполне явственны; sa i расположены дорзально на некотором расстоянии от заднего края идиосомы; примерная длина щетинок (и процентах к длине идиосомы): va .8—9; vp и see 25; sc i 17; ha 23; hp 19; dx и <12 20—30; r'l3 45; 12; lx и 12 24—25; 1,
20—21; sa e 17; sa i около 20. Ноги много короче и тоньше, чем у с?, равные приблизительно 40 (I), 35—36 (II) и 60 (IV) % длины идиосомы; ноги III и IV прикреплены более медиально, их вертлуги и бедра целиком прикрыты краями идиосомы. Длина около 750 ;л, ширина 660 |л.
Распространение. Англия, Голландия, Германия. Живет в норах кротов.
15. Род FUSACARUS MI0H.
Michael, 1903:7.
Тип рода: Fusacarus laminipes Michael 1903.
Тело веретеновидное, очень сильно выпуклое (в поперечном сечении почти круглое), с узко закругленным передним концом и сильно заостренным задним; ростральный выступ совершенно не отграничен от идиосомы собственноу целиком закрывающий гнатосому; последняя помещается в глубоком камеростоме. Кожа твердая, сильно пигментированная, совершенно гладкая. Все щетинки спинной поверхности, кроме sc е, представлены очень тонкими и гибкими микрохэтами; see длинные, волосовидные; v а и vp хорошо развиты; постанальных щетинок у обоих полов только 1 пара. -Стернум отсутствует; эпимеры I слиты в дугообразную перемычку, окай--мляющую сзади камеростом; анальная щель расположена почти терминально, расщепляя приостренный задний конец идиосомы; генитальные щупальцы отсутствуют. Жировые железы не развиты. Ноги короткие; передние чрезвычайно сильно сжаты с боков, расширенные в дорзо-вент-ральном направлении, с крыловидным килем на вентральной поверхности бедер; задние менее сплющенные, с бокаловидными (сильно суживающимися ч к основанию) коленами; вооружение ног состоит из простых тонких щетинок. ^Вторичные половые различия невелики и сказываются, главным образом, в строении коксо-стернального скелета (помимо общих всем представителям группы различий в строении ног III и IV).
cf. Идиосома несколько уже; вентральная поверхность довольно равно-:мерно хитинизирована; эпимеры II дугообразные, свободные; эпимеры III и IV соединены друг с другом и за точкой их соединения продолжены внутрь :и затем изгибаются под прямым углом кпереди. Генитальное отверстие помещается в пространство между основаниями ног III и IV.
9. Вентральная поверхность подосомы имеет мозаичный вид, вследствие -того, что кожа местами сильно уплотнена, образуя широкие щитовидные .площадки, разделенные узкими перепончатыми полосками; одна из них
14. TALPACARUS. 15. FUSACARUS. 16. A1YACARUS
357
расположена непосредственно позади гнатосомы и простирается назад до-полукольцевидной перепончатой линии, окаймляющей спереди область, генитального отверстия; другие располагаются между этой линией и эпимерами II и III. Генитальное отверстие расположено между основаниями ног II и III, окруженное хитиновым кольцом, с очень крупным треугольным непарным задним клапаном и относительно узкими боковыми.
Нимфы отличаются от половозрелой стадии бледной и мягкой кожей и пеуплощепными ногами. — Гипопальной стадии нет.
Монотипический род.
*1. Fusacarus laminipes Mich. (фиг. 596—601).
Michael, 1903:9, t. XXI.
Более или менее темного красновато-бурого цвета, при жизни полупрозрачный, сильно блестящий. Идиосома у живых особей в 1.6 (9)—2.2 (d) раза, длиннее своей максимальной ширины, приходящейся между основаниями ног II 'и III. V а заметно длиннее v р; sc i около 10 раз короче see, достигающих 25% длины идиосомы; he и vp длиною около 10%; размеры всех остальных щетинок колеблются в пределах 2—6% длины, идиосомы.
сГ. Идиосома уже, чем у 9, сильнее приострена сзади, с более узким ростральным выступом. Ноги короче и еще шире, чем у 9, равные приблизительно 27 (I и II) и 25 (III и IV) % длины идиосомы; вентральная пластинка бедер I шире, вершинные щетинки голеней длиннее. Длина. 280—300 у, ширина 120—130 |л.
9- Идиосома заметно шире, менее сильно приостренная сзади, с менее-узким ростральным выступом. Ноги несколько длиннее, равные приблизительно 32 (I и II) и 26—28 (III и IV) % длины идиосомы; бедра I с менее развитой вентральной пластинкой. Длина 300—320 ;д, ширина 170—180 р.
Распространение. Описан из Англии; мне известен из окрестностей Москвы (Шульгино, Кунцевского р-на) и Иркутска. Живет в гнездах крота и землероек; экземпляр из Иркутска был найден в подполье зернохранилища.
16. Род MYACARUS, пот. п.
HomopuS 0. L. Koch, 1841, Dtsch. Crust., Jfyriap., Arachn., 39, n° 24 (дот. praeocc. in: Du-m6ril et Bibron, 1835, Reptilia).
Тип рода: Homopus hypudaei C. L. Koch 1841.
Известны только гипопусы. Тело узкое, более вытянутое в длину, чем у гипопусов других' родов; задний край уплощенный, очень узкий, полого вогнутый посередине, без закраин или иных разращений; опистосомальная площадка обрисована только с боков продольными или косыми опистосомальными килями, у основания с неясным сетчатым рисунком. Стернум, эпимеры и эпимериты II хитинизированы слабее^ чем обычно; эпимеры III свободные, серповидно изогнуты, очень четко
'358
HI. GLYCYPHAGIDAE. 5. LABIDOPHORINAE
очерченные. -Клапаны прикрепительного аппарата вытянуты в длину; задние зажимы расположены далеко позади передних, шпателевидной формы, с немногочисленными поперечными ребрами; передние — очень мелкие, поперсчноовалтшые. Ноги I и II длинные (35—40% длины идиосомы), очень стройные; .папки очень стройные, несколько сжатые с боков и в профиль S-образпо искри пленные, дистально с 3 короткими, по широкими листовидными щетинками и 1 длинной, пуговковидно вздутой на конце щетинкой; ближайшая к основа,пню вентральная щетинка лапок, вентральные щотипки голеней и обе базальные щетинки колен явственно перистые или гребенчатые; вершинная дорзальная щетинка голеней довольно короткая, притупленная на конце. Ноги 111 и IV очень короткие <15—20% длины идиосомы) и стройные; лапки III дистально с хорошо развитым коготком и 4 листовидными щетинками; лапки IV узкие, цилиндрические (в 2—4 раза длиннее своего диаметра), оканчивающиеся очень длинной парусной щетинкой, без коготка; остальные щетинки ног III и IV короткие, гладкие, более или менее игольчатые.
'Известны 2 вида, до сих пор смешивавшихся между собой.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Проподосомальный щит более чем в 1.5 раза шире (у основания) своей длины, спереди прямоугольный; эпимеры III узкие; лапки I и II значительно короче соответствующих голени и колена, вместе взятых; внутренняя вентральная щетинка голеней I и II расположена много выше наружной................... *1. М. arvicolae (Duj.).
2 (1). Проподосомальный щит не более чем в 1.3 раза шире своей длины, спереди вытянутый в очень острый угол; эпимеры III обычно очень широкие; лапки I и II длиннее, равные голени и колену, взятым вместе; обе вентральные щетинки голеней I и II расположены на одной высоте .............................................*2. М. hypudaei (Koch.).
*1. Myacarus arvicolae (Duj.) (фиг. 572—576).
Dujardin, 1849, Ann. Sci. Nat. Zool., s6r. 3, vol. XII:249, 264, t. II, fig. 15—16 (Hypo-Oudemans, 1900, Tijdschr. Entomol., XLVI:126, t. 6, fig. 39—40 (J)ermacarus).—hypudaei Oudemans, 1915, Entomol. Ber., IV:199 (Dermacants).
Г и п опус. Тело в 1.6—1.7 раза длинпее своей ширины; проподосома образует спереди прямой угол; ширина ее у основания более чем в 1.5 раза превышает длину; опистосомальные кили сходятся кпереди, заметно вогнутые. Плевральные поля с четкой дугообразной линией, идущей от наружного края ацетабул III к, основанию ног II, огибая снаружи эпимериты II; эпимеры III узкие. Длина клапанов прикрепительного аппарата составляет
Фиг. 593—601.
Фиг. 693. Xenoryctes heptneri, sp. n., Q снизу, фиг. 594. X. heptneri, sp. n., нога I (колено, голень и панна) 9 сбоку. Фиг. 595. X. heptneri, sp. п., тепеонимфа сверху (гнатосома и ноги не изображены). Фиг. 596. Fiisdcarus laminipes Mlcll., <Д сверху. Фиг. 597. То же, снизу, фиг, 598. F. lami-siipes Mich., нога I сГ сбоку. Фиг. 599. F. laminipes Mich., лапка IV cf снизу. Фиг. 600. F. laminipes Mich., 9 снизу. Фиг. 601. F. laminipes Mich., папка IV 9 снизу. .
t
360 HI, GLYCYPHAGIDAE. 5. LAIHDOPHOHINAE
Vs длины тела; их задние концы отделены от слабо вогнутого заднего края тела очень узким пространством; поперечные ребрышки на внутренней поверхности клапанов слабые, малозаметные; передние зажимы поперечноовальной формы, с 4 поперечными ребрышками, задние — шпателевидные, с 6—7 ребрышками. Лапки I значительно короче голени и колена, вместе взятых, в профиль слегка S-образно искривленные; обонятельная палочка прямая, слегка булавовидно вздутая на конце, расположенная значительнониже (более проксимально) середины ланки; р в виде довольно крупного-конического шипа; задняя вентральная щетинка помещается несколько выше, чем обонятельная палочка, но далеко не достает своим концом до вершины лапки, толстая, зазубренная; вентральные щетинки голеней и базальные щетинки колен I и II сравнительно короткие, тонко зазубренные; внутренняя вентральная щетинка голеней сидит много выше наружной. Лапки III довольно стройные, едва короче лапок IV; вентральная щетинка голеней III шиповидная. Длина 275—400 у, ширина 155—250 [л.
Распространение. Франция, Германия, Голландия, СССР. Мне известен из ряда пунктов Московской обл., северо-западного Казахстана (окрестности Кустаная), Киргизии и с Шантарских островов.
Экология. Встречается на полевках Microtus (Pytimys') subter-raneus Selys (Dujardin, 1849), M. arvalis L. и Evotomys glaredlus Schr. (Oudemans, 1915), водяной крысе Arvicola amphibius L. (Oudemans, 1. с.), хомяке (Cricetus cricetus fuscidorsis Arg.), хомячке (Cricetulus migratoriusPaAl.), степной пеструшке (Lagurus lagurus Pall.), бурундуке (Eutamias asiaticus orientals Bonn., о-в Шантар), кроте и обыкновенной землеройке (Sorex araneus L.).
*2. Myacarus hypudaei (C. L. Koch) (фиг. 568—571).
C. L. Koch, 1841, Dtsch. Crust., Myriap., Arachn., 39, n° 24 (Homopus).
Гипопус. Обычно крупнее, чем предыдущий; тело еще более вытянуто в длину, в 1.8—1.9 раза длиннее своей ширины; особенно удлинены опистосома и проподосома; последняя образует спереди очень острый угол и у основания только в 1.3 раза шире своей длины; опистосомальные кили почти параллельны друг другу, заметно дугообразно выпуклые. Плевральные дуги обычно отсутствуют; эпимеры III сильно расширены. Клапаны прикрепительного аппарата крупнее (их длина почти равна 1/i длины тела)-и отделены более широким промежутком от глубже вогнутого заднего киля тела; поперечные ребрышки па внутренней поверхности клапанов более рельефны и сильнее хитипизированы; передние зажимы косые, их ребрышки имеют диагональное направление; задние зажимы с 4—5 поперечными ребрышками. Лапки I длиннее (приблизительно равны длине голени и колена, вместе взятых) и резче искривлены (в профиль); обонятельная; палочка и окружающие ее щетинки расположены более дистально, ближе-к середине лапки; р очень мелкая, трудно заметная; задняя (проксимальная) вентральная щетинка длиннее, сильнее гребенчатая; вентральные-
/в. MY АСА RUS
36f
щетинки голеней и базальные щетинки колен I п II также длиннее и более явственно гребенчаты, чем у М. arvicolae’, внутренняя вентральная щетинка голеней сидит на одной высоте с наружной, ниже середины членика; лапки III короче и толще, вентральная щетинка голеней III не шиловидная. Длина 365—430 ;л, ширина 205—225 [Л.
Распространение. Германия, СССР. Известен мне из нескольких пунктов Московской обл. и из Приамурья.
Экология. Гипопусы встречаются на полевках (Microtus arval/is L. и Evotomys glareolus Schr.), кроте и землеройках (Crocidura lasiura Thos.).
Закрепляя Коховское название за этим видом, я руководствуюсь рисуп-ком этого автора, на котором очень ясно подчеркнуты большая длина и. узость проподосомы и ног.
6. Подсем. Carpoglyphinae
Гнатосома в большинстве случаев очень стройная, весьма подвижная;., хелицеры узкие, с очень тонкими, слабо вооруженными пальцами клешни. Идиосома эллипсоидальная, сильно выпуклая, или почти ромбической формы и тогда уплощенная дорзо-вентрально; у некоторых форм — с слегка намеченным подразделением на проподосому и гистеросому, без рострального выступа. Кожа довольно плотная, гладкая или шиповатая, иногда, с мозаикой из сильно хитинизированных округлых площадок (dd1 Hericia hericia F. et R.); проподосомальный щит обычно имеется. Все щетинки спинной поверхности идиосомы гладкие, жестковатые, короткие или умеренной длины; sa е и иногда одна из пар постанальных щетинок длинные, играющие роль «хвостовых» щетинок; число пар щетинок спинной и боковых поверхностей изменчиво, но не выше 15; их расположение более или менее спутано и в той или иной мере уклоняется от обычных норм. Псевдостигмальпый орган отсутствует. Строение коксо-стернального скелета несколько разнится у jh 9. Ноги умеренной длины (ходильного типа), довольно стройные или более или менее массивные, нередко-в той или иной мере модифицированные; все усаживающие их щетинки, гладкие; вершина колен с 2 крепкими щетинками.' Praetarsi крупные, иногда длинностебельчатые, на конце с относительно крупным коготком.
<5. Стернум очень длинный, слитый концом с эпиандрием, с наружными углами которого слиты также концы эпимер II, образуя в целом W-образ- ную фигуру. Генитальное отверстие помещается между основаниями ног IV . или дальше назад, обычно окруженное тонкой хитиновой рамкой; ноги 1 и II (у Carpoglyphus) или III и IV (Hericia, некоторые Hyadesia) более пли менее резко модифицированы.
$. Стернум и эпимеры II соединены концами с^ипшием или почти, касаются его; эпигиний подковообразный или дугообразный, очень большой, но тонкий; края генитальных клапанов более пли мопсе уплотнены.
362
HI. GLYCYPIIAGIEXE. 6. CARPOGLYPHINAE
Нимфальные стадии плохо известны. У Hericia и, может быть, у Carpoglyphus имеются рассолитслыгые гипопальные стадии.
В это подсемейство я объединяю 3 рода: 1) Carpoglyphus Rob. (составляю-, щий у Oudemans’a особое сом. Carpoglyphidae, к которому отнесен еще род .Ferminia Ouds.); 2) Hericia Сап., и 3) Hyadesia Megn. [также рассматривавшийся как продета!и 1тель особого семейства Lentungulidae(Mich.) Ouds.]; возможно, что в него войдет и малайская Fusohericia Vtz. Все относящиеся сюда виды — амфибнотичоскне формы, живущие преимущественно в жидких и полужидких средах.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОП
П (2). Лапки I и II сильно укороченные, заканчивающиеся очень крупным когтевидным шипом, у основания которого прикрепляется необычайно длинный, тонкий и гибкий praetarsus, несущий дистально небольшой крючковидный коготок; praetarsi III и IV короткие или совсем редуцированные, с более крупным, чем на передних ногах, коготком; идиосома дорзально с изогнутой поперечной бороздкой, намечающей ее подразделение на проподосому и гистеросому. Литоральные формы, живущие среди водорослей . . 17. Hyadesia Megn.
:2 (1). Все лапки нормального строения, не когтевидные, с лопастевидным praetarsus и крупным коготком; идиосома без всяких следов поперечной бороздки. Сухопутные формы, живущие по большей части' в жидких или полужидких средах.
^3 (4). Кожа гладкая; все щетинки идиосомы, кроме sa е и р3, очень короткие; генитальные щупальцы хорошо развиты; ноги III и IV одинакового строения у обоих полов; у $ эпимеры I и II соединены с эпи-гинием ........................18. Carpoglyphus Rob.
-4 (3). Кожа шероховатая, обычно усаженная многочисленными тонкими шипами; щетинки идиосомы более длинные и толстые; генитальные щупальцы рудиментарные; ноги III у d сильно удлинены, ноги IV резко вздуты и искривлены; эпимеры I и II у 9 свободные .... г................................................ . . . 19. Hericia Сап.
17. Род HYADESIA MEGN.
Mtfgnin, 1889, Miss. Scient. Gap. Horn, VI, 3, Protozoaires: 51; Andr6, 1931, Bull. Mus. Paris *(2), 111:496, fig. 1—6. —Lenlungula Michael, 1893, Proc. Zool. Soc. London: 262; Berlese, 1897:103; Lohmann, 1894, Wise. Moorcsuntors., I, 1:89; Kramer, 1899:136; Michael, 1901:192.
Тип рода: Hyadesia uncinifcr Megn. 1889.
Гнатосома довольйо массивная. Хелицеры нормального строения. Идиосома темно окрашенная,1 очень сильно выпуклая, с более или менее .явственной поперечйой бороздкой, намечающей ее расчленение на пропо-
1 Темная окраска идиосомы обусловлена цветом содержимого кишечника (и, может быть, резервных веществ), а не пигментацией кутикулы, которая прозрачна и почти бесцветна.
/7. HYADESIA
363
досому и гистеросому, сильно выступающими плечами и обычно слегка вогнутым посередине задним краем. Кожа гладкая или местами топко-.морщинистая, или топко пунктированная; проподосомальный щит более или менее явственный, вытянутый в поперечном направлении (иногда подковообразный); глаза отсутствуют. Щетинки спинной поверхности идиосомы разнообразной длины: sc е, h р, и sa е, а иногда также a, sa i и 13, длинные, остальные более или менее короткие; dx сильно приближены к плечевым; d2 расположены кнутри от dx, на одной поперечной линии с ними. Жировые железы отсутствуют. Генитальных щупальцев нет. Ноги I и 11 сильно сжатые с боков, широкие, с укороченной лапкой, заканчивающейся очень крупным когтевидным образованием (неподвижно сращенным с лапкой) и 1 небольшим шипом; у основания когтя прикрепляется необыкновенно длинный, тонкий и гибкий praetarsus, расширенный дистально в небольшую площадку, несущую тонкий крючковидный коготок; одна из вентральных щетинок голеней превращена в массивный шип, все остальные щетинки ног тонкие, более или менее гибкие. Ноги III и IV более стройные, цилиндрические, обычно с хорошо развитыми лапками, несущими .дистально нормальный по размерам или редуцированный praetarsus и более крупный, чем на передних ногах, коготок (у d Н. fusca лапки III и IV сильно укорочены, напоминая по своему строению лапки I и II). Вторичные половые различия обычно очень резки.
<5. Идиосома значительно стройнее, чем у $, и более резко суживается жзади; плечевые выпуклости более сглажены. Эпимеры III и IV всегда хорошо развиты, обособленные друг от друга. Основания пог IV сильно сдвинуты кпереди и помещаются между основаниями ног III, почти на одной поперечной линии с ними; на вентральной поверхности лапок I, III и IV находится округлая перепончатая присоска; у некоторых, видов лапки III и IV укорочены и сходны по своему строению с лапками передних пог. Генитальное отверстие расположено немного позади основания пог IV и окружено овальным хитиновым кольцом.
. Крупнее и толще d, с более резко выступающими плечевыми выпуклостями. Стернум и эпимеры II свободные. Эпимеры III и IV соединены друг с другом или неявственны. Ноги IV прикрепляются непосредственно позади основания ног III; лапки-без присосок; задние имеют всегда нормальное строение и много длиннее голеней. Генитальное отверстие располагается между основаниями ног III и IV или дальше назад, с хитинизи-рованными клапанами, снабженными более или менее явственным овальным вдавлением; эпигиний дугообразный или отсутствует.
О нимфальных стадиях известно только, что они очень похожи на взрослых. Гипопальная стадия отсутствует.
Все виды рода Hyadesia — литоральные формы, ведущие полу иодный образ жизни на скоплениях зеленых водорослей по берегам моря. Год имеет, повидимому, космополитическое распространение и состоит из G видов: Н. uncinifer Megn. (Огненная Земля), Н. algivorans Mich, (побережье
364
III. GLYCYPHAGIDAE. в. CARPOGLYPHINAE
Атлантического океана в Корнуэльсе), Н. fusca Lohm. (берега Северного' и Балтийского морей), Н. kerguelensis Lohm. (Кергуэленские острова), Н. curassavienvis Viets (Антильские острова) и Н. chelopus Trouess. (Индийский океан, без более точного указания местонахождения).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). V а очень длинные, мало уступающие в размерах sc е и h р; все d и 1 очень короткие, приблизительно одинакового размера; шиповидная вентральная щетинка голеней I и П в 4—5 раз короче диаметра голе-' ни, коническая; лапки III и IV у d сильно укорочены; генитальное отверстие <5 маленькое, отделенное большим расстоянием от заднего конца стернума; $ без эпигиния, с хорошо развитыми эпимерами III и IV...............................-........: . . . l.H. fusca (Lohm.).
2 (1). V а более чем в 2 раза короче sc е и h р; d и 1 довольно длинные (-13— 30% длины идиосомы), d2 заметно длиннее dx, 13 почти в 1.5 раза длиннее d2; шиповидная вентральная щетинка голеней I и II немного* короче диаметра голени, цилиндрическая; лапки III и IV у <5 не укорочены; генитальное отверстие <3 очень крупное, почти соприкасающееся с концом стернума; $ с крупным дуговидным эпигинием. и неявственными эпимерами III и IV. . . . 2. Н. algivorans (Mich.)..
1. Hyadesia fusca (Lohm.) (фиг. 24, 618—623).
Lohmann, 1896, Wiss. Meeresunters., 1:85, t. IV; Kramer, 1899:136 (Lentungula).
Идиосома темнокоричневая; проподосомальный щит, гнатосома, ноги и коксо-стернальный скелет красновато-бурые. Кожа местами очень тонкоморщинистая, местами гладкая; поверхность проподосомального щита в мелкой ячеистой пунктировке. V а очень длинные (30—50% длины идиосомы), мало уступающие в размерах sc е и h р; sc i, h а и h р, все d и —1^ короткие, все примерно одинакового размера (5—7% длины идиосомы); sa i вдвое короче весьма длинных sa е (равных приблизительно 1/2 длины идиосомы); d2 расставлены значительно шире, чем d3. Шиловидная вентральная щетинка голеней I и II коническая, в 4—5 раз меньше диаметра голени.. Половой диморфизм выражен резче, чем у следующего вида.
<5. Эпимериты II соединены наружными концами с ацетабулами II,. а внутренними концами с эпимерами III; последние простираются в медиальном направлении до конца эпимер IV, идя параллельно им на значительном протяжелии. Ноги III и IV заметно укорочены (длиною около' 25—28% длины идиосомы) в результате сильной редукции лапок, которые-в 3 раза короче голеней и вооружены на конце 2 изогнутыми шипами, напоминающими шипы лапок I и II, но далеко уступающими им в размерах;.
Фиг. 602—608. Carpoglyphug lactis (L.).
Фиг. 60S. о сверху. Фиг. 603. То же, сниву. Фиг. 604. Нога I (часть колена, голень и папка), сбоку. Фиг. 605. Лапка I снизу. Фиг. 606. Лапка III снизу. Фиг. 607. с? снизу. Фиг. 608. То же,. . папка I снизу.
605
366
III. GLYCYPHAGIDAE. в. CARPOGLYPIIINAE
praetarsi Ш и IV также сходны по строению с таковыми ног I и II, но много короче. Хитиновое кольцо, окружающее генитальное отверстие, очень' невелико, отделено большим пространством от конца стернума. Длина, около 400 |л.
Заметно крупнее d. Зиммеры III и IV хорошо развиты, образуют совместно с эпимсритамн II диагональную хитиновую полоску с 2 отростками (наружные части эпимер), идущими к основанию ног III и IV. Лапки 111 и IV заметно длинное голеней, постепенно утончающиеся к концу, лишенные шипов; их praetarsi хорошо развиты, значительно толще таковых I и II. Дугообразный энигиний отсутствует; передний конец генитального отверстия находится на одной поперечной линии с передними краями ацетабул IV. Длина около 500 |л.
Распространение. Германия: о-в Гельголанд, побережье Северного и Балтийского морей, Ирландия. Живет крупными, обычно узко локализованными колониями среди скоплений зеленых водорослей на камнях, сваях и т. п. у поверхности воды.
2. Hyadesia algivorans (Mich.) (фиг. 615—617, 624—627).
Michael, 1893. Proc. Zool. Soc. London: 262, t. XVIII; 1901:196, t. I; Kramer, 1899:136 (Len-tungula).
Идиосома оливково-бурая, обычно с бледножелтым мраморным рисунком; кожа бледножелтоватая, гнатосома, ноги и коксо-стернальный скелет-розовато-желтые. Проподосомальный щит неявственный; поверхность кожи совершенно гладкая. Va короткие (13—15% длины идиосомы), в 3 раза, короче see и Ь р, достигающих 45—50% длины идиосомы; sc i, h а и dt длиною около 14—15% длины идиосомы; d2 почти в 1.5 раза длиннее dx; длина d3 и 12 составляет 17—18, 1х 12, 13 около 27, sa е 30, sa i около 40%. длины идиосомы; sa i, таким образом, значительно длиннее sa е и размеры d и 1 очень различны; d2 гораздо более сближены между собой, чем очень широко расставленные d3. Шиповидная вентральная щетинка голеней I и II цилиндрическая, с приостренным концом, немного короче диаметра голени; лапки III и IV у обоих полов заметно длиннее голени, постепенно суживающиеся к концу, лишенные шипов; praetarsi III и IV очень короткие, одинакового строения у d и £. Длина d и Q 380 [±; вторичные половые различия, соответственно, невелики.
d. Эпимериты II не соединены ли с ацетабулами II, пи с йпимерами III;. концы последних едва достигают точки перегиба эпимер IV. Длина ног III составляет около 35, пог IV 40—42% длины идиосомы. Хитиновое кольцо, которым окружено генитальное отверстие, очень большое, почти соприкасающееся спереди с концом стернума.
9. Эпимеры III н IV пеявственны. Энигиний хорошо развит в виде широкого хитинового полукольца; передний конец генитального отверстия находится значительно впереди ацетабул III.
Распространение. Известен только из Англии: западное побережье Корнуэльса (мыс Land’s End). Найден единственный раз Michael’емс
11. HYADESIA. 18. CARPOGLYPHUS
367'
среди скоплений зеленых водорослей (Cladophora fracta) на прибрежных, скалах, в месте, где пресная вода небольшого потока стекала по поверхности, гранита в полосе прибоя.
18. Род CARPOGLYPHUS ROB.
Robin, 1869, Journ. Anat. Physiol., VI:197; Berlese, 1898, XCI, n° 13; Kramer, 1899:148;. Michael, 1903:42. —Phycdbius Canestrini, 1888a:355. ”
Тип рода: Acarus lactis L. 1758.
Гнатосома очень узкая, необыкновенно подвижная; хелицеры с очень топкой ножницеобразной клешней. Идиосома слегка уплощенная, продолговатоовальная, с полого закругленным или почти прямым задним краем и сглаженными, мало-выступающими плечами, одинаковой формы, у обоих полов; проподосома не отделена от гистеросомы. Кожа гладкая; проподосомальиый щит неявственный; ростральный выступ спереди с парой округлых глаз, лишенных пигментированных ретин. Все щетинки спинной поверхности идиосомы однотипные, короткие; sc е немного длиннее других; sa е очень длинные, расположенные терминально; очень длинны: также р3. Жировые железы хорошо развиты, смещенные к заднему краю идиосомы. Генитальные щупальцы хорошо развиты у обоих полов, усеченно-яйцевидные. Эпимеры III и IV свободные, широко расставленные концами. Ноги довольно стройные, немодифицированные; ноги IV прикрепляются значительно позади ног III; щетинки ног короткие, жестковатые; вершинные дорзальные щетинки всех голеней длинные. Praetarsi крупные, лопастевидные, одинакового строения на всех 4 парах ног; коготки крупные. Вторичные половые различия довольно велики по объему затрагиваемых ими признаков, но не столь резки, как у других родов.
сГ. Ноги I и II заметно толще, чем у $, с расширенными praetarsi и несколько отличным вершинным вооружением лапок. Генитальное отверстие расположено между основаниями ног III и IV, лишено хитинового кольца.
$. Ноги I и II стройные; их praetarsi такого же размера, как па ногах. III и IV. Стернум и эпимеры II слиты концами с массивным угловатым эпигинием; генитальное отверстие помещается в пространстве между основаниями ног II и III; его клапаны не хитинизированы.
У нимфальных стадий глаза относительно крупнее и лучше заметны,, чем у взрослых; личинки без грудных палочек. Гипопальная стадия, описана у 1 из видов.
К роду Carpoglyphus относятся 2 вида: вероятно космополитический С. lactis (L.) и С. alienus Banks 1906, найденный на Яве, в человеческой моче, и считающийся эндопаразитом. Недостаточно описанный С. vagabun-dus Berl. 1910 относится, по всей вероятности, к подсем. Chaetodar.lylinac.
*1. Carpoglyphus lactis (L.) (фиг. '20, 602—608).
Linne, 1758, Syst, Nat., ed-. X:1024, sp. 16 (Acarus). —dysenterian Linin'', 1758, 1. e., sp. Ifr (Aca/Ms). — •passularum Robin, 1869, Journ. Anat. Physiol., VI:197; Borleso, .1897:105; Kramer,.
..368
III. GLYCYPHAGIDAE. 6. CARPOGLYPHINAE
1899:148 (Carpoglyphus).—anonym,us Haller, 1882, Jahrcsb. Ver. Wiirttemb., XXXVIII:297 AGlycyphagus)-, Berlese, 1884, XIV, n° 10; Nalepa, 1885, Sitzber. Akad. Wiss. Wien, XCII, Abt. 1:1 ATrichodaclylus); Oanestrini, 1888:22; 1889a:392, t. XXXV (Phyeobiusy; Michael, .1903:44, t. XXV— XXVI, fig. 8—21 (Carpoglyphus)^
Полупрозрачный, почти бесцветный; ноги, гнатосома и коксостерналь-. ный скелет розоватые; прижизненная окраска сильновариирует в зависимости от цвета кормового субстрата. Каждый из пальцев клешни хелицер с 3 сближенными между собой зубцами, отделенными большим пространством от конца на.нг>ца. Размеры всех щетинок спинной поверхности идиосомы (кроме очень крупных на о) колеблются в незначительных пределах: 8—10% длины идиосомы; sc i несколько длиннее других (13—17% длины идиосомы), 13 и sai очень короткие (4—7%); длина sa c достигает 100, р3 70—80% длины идиосомы; v а сильно смещены назад, расположены значительно ближе к sc е, чем к переднему краю рострального выступа; очень -сильно сдвинуты.назад и sc i, сидящие заметно позади поперечной линии, проведенной через основания ha; h р на одном уровне с dp отделенные -большим расстоянием от h а, чем от 1Х; 13 расположены вентрально.
d1. Sc е почти в 2 раза длиннее sc i. Ноги I и II значительно короче (и толще) ног III; их лапки толстые, в 2—2.3 раза длиннее своего-диаметра у основания, с редуцированным средним вершинным шипиком, массивным наружным и только 1 расширенным шипом на внутренней поверхности. Длина 240—400 р..
9. Sc е в 1.5 раза длиннее sc i. Ноги I и II едва короче и толще ног III; ?их лапки стройные, в 3 раза длиннее своего диаметра у основания, вентрально с 3 одинаково развитыми, тонкими апикальными шипиками и 4-м, дополнительным, на внутренней поверхности членика. Длина 250—450 р..
Распространение. Вся'Европа и Северная Америка; вероятно, ^завозится с продуктами и в другие части света. У нас в Союзе найден пока в Москве, Ленинграде, Казани и Армавире.
Экология. Преимущественно синантропный вид, живущий на чрезвычайно разнообразных субстратах, как твердых, так и жидких, но почти : исключительно на тех из них, в которых содержатся молочная, уксусная .или янтарная кислоты. Встречается часто на сухих фруктах, старых медовых сотах, прокисшем варенье и других фруктовых изделиях, гниющей ..муке, старом сыре, гниющем картофеле, прокисшем молоке, гнилом дереве, кислом тесте, фруктовых напитках, пиве и вине. Размножаясь иногда в огромных количествах в этих субстратах, С. lactis может иметь ощутимое -экономическое и медицинское значение (имеются указания на то, что .им вызываются острые желудочно-кишечные заболевания). Несмотря на это, его биология никем детально нс изучалась.
Фиг. 609-617.
Фиг. 609. Hericia hericia (Rob.), о” сверху (по Michael’»). Фиг. 610. То же, снизу (по Michael’»). Фиг. 611. Н. hericia (Rob.), 2 сверху (по Michael’»). Фиг. 612. То же, снизу (по Michael’»). Фиг. 613. Я. hericia,(Rob.), гипопус сверху (noMichaer»).Фиг. 614. То же, снизу (по Michael’»). Фиг. 615. Hyadesia algivorans (Mlch.)> 9 сверху (по Michael’»). Фиг. 616. То же, снизу (по Michael’»). Фиг. 617. Я. algivorans (Mich.), cf снизу (по Michael’»).
24 Фауна СССР, Захватив—2S4
370
III. GLYCYPHAGIDAE. в. CARPOGLYPHINAE
19. Род HERICIA CAN.
Canestrini, 1888:6; 1888a :35G; Berlese, 1898, LXXXVII, n° 12; Kramer, 1899:143; Michael,. 1903:29.
Тип рода: Glycyphayus hericius Rob. 1868.
Гнатосома очень стройная и подвижная; клешня хелицер тонкая, пене в такой мере, как у рода Carpoglyphus. Идиосома уплощенная дорзо-вептральпо, приблизительно ромбической формы, без дорзальной поперечной бороздки. Ножа обычно сильно шероховатая, иногда густо усаженная шиновидными кутикулярными выростами; проподосомальный щит хорошо, развит, вытянутый в длину; глаза отсутствуют. Щетинки спинной поверхности идиосомы крепкие, довольно длинные; положение многих из них (sc, dx, h) и их размеры различны у с? и 9. Генитальные щупальцы сильно редуцированы. Жировые железы, повидимому, отсутствуют. Ноги длинные, резко отличающиеся по своему строению у с? и 9; лапки I и II не модифицированы; большая часть щетинок ног превращена в шипы. Ргае-tarsi всех 4 пар ног приблизительно одинакового строения, небольшие, с крупным коготком. Вторичные половые различия чрезвычайно резки.
с?. Идиосома шире в плечах и более уплощена, чем у 9, и резче суживается кзади; скульптура кожи грубее. Sa е и р3 очень длинные, da близко, придвинуты к плечевым и расположены на одной поперечной линии с d3.. Эпимеры III и IV соединены своими внутренними концами, эпимериты IV хорошо развиты. Ноги I и II более или менее сильно утолщены; ноги III длиннее и тоньше других, занимают латеральное положение и прикрепляются к телу па одной поперечной линии с смещенными кпереди основаниями ног IV; последние короче других, заметно утолщены и вооружены более тонкими и гибкими щетинками; вентральных присосок на лапках нет. Генитальное отверстие расположено позади основания ног IV, не окружено хитиновым кольцом.
9. Sae и р3 короче, dx расположены па значительном расстоянии от плечевых и много впереди d2. Стернум и эпимеры II—IV свободные, эпимериты IV не развиты. Ноги тонкие, длинные, все сходные по своим пропорциям; ноги IV расположены так же маргинально, как ноги III, на некотором расстоянии позади них. Генитальное отверстие помещается в пространстве между основаниями пог II и ПД с хитинизированными клапанами и крупным подковообразным эпигинием.
Фиг. 618-637.
Фиг. 618. Ilyadesia fusca (Lohm.), 9 сверху (по Lohmann’у). Фиг. 619. то же, снизу (по Lohmann’у). Фиг. 626. II. fusca (Lohm.), сГ снизу (по Lohmann’у). Фиг. 621. Н. fusca (Lohm.), конец, гопени и лапка IV 9 (по Lohmann’y). Фиг. 622. Н. fusca (Lohm.), дистальный конец колена, голень и лапка IV с( (по Lohmann’y). Фиг. 623. Н. fusca (Lohm.), проподосомальный щит (noLohmann’у). Фиг. 624. Я. alpicorans(Mich.), нога I (по Michael’»). Фиг. 625. II. algivoranstMiciii.), лапка I (по Michael’ro). Фиг. 626. II. algivorans (Mich.), penis (по Michael'»). Фиг. 627. Н. algivorans (Mich.), хелицера (no Michael’io). Фиг. 628. Hericia hericia (Rob.), нога IV сГ (no Michael’»). Фиг. 629. Я. hericia (Rob.), нога I rf (no Michael’»). Фиг. 630. H. hericia (Rob.), конец лапки. И амбулакра сГ (по Michael’»). Фиг. 631. Н. hericia (Rob.), нога II o’ (по Michael’»). Фиг. 632. Н. hericia(Rob.), нога IV 9 (по Michael’»). Фиг. 633. Н. hericia (Rob.), хелицера (по Michael’»). Фиг. 634. Н. hericia (Rob.), penia, окружающие его структуры (генитальные щупальцы, щетинки и пр.) (по Michael’»). Фиг. 685. я. hericia (Rob.), генитальное отверстие 9 с эпигинием (по Michael’»). Фиг. 636. Н. georgei Mich., нога II с? (по Michael’»). Фиг. 637. Н. georgei Mich., нога III СГ (по Michael’»).
372
III. GLYCYPHAGIDAE. 6. CARPOGLYPUINAE
Личинки характеризуются своей темной окраской, гладкой кожей и сильным развитием одной из вершинных щетинок лапок III.
, Гипопусы имеются у всех видов. Спинная поверхность идиосомы у них • целиком покрыта обоими дорзальными щитами, имеющими тонкозернистую структуру. Сторпум и эпимеры II—IV свободные. Очертания присасывательного диска неясны; передние присоски маленькие, способные очень сильпо выпячиваться; центральные присоски довольно велики, обычного строения; наружные и задние присоски очень мелкие, расположенные па крупном дугообразном склерите, лежащем параллельно заднему краю тела. Ноги длинные и стройные (особенно передние), все с хорошо развитыми овальными praetarsi и небольшими коготками; лапки III и IV с довольно длинными парусными щетинками.
2 европейских вида, из которых 1 очень плохо известен и требует пере-описания. Индийская Неricia greeni Ouds, 1916 относится, как убедительно показал Vitzthum (1924), к роду Sennertia (Chaetodactylinae).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Кожа густо покрыта довольно длинными шиповидными кутикулярными выростами; de? много крупнее 99, сильнее пигментированы и снабжены на дорзальной поверхности идиосомы многочисленными округлыми, вдавленными, сильно хитинизированными площадками; лапки II d вентрально с 2 крупными шипами, расположенными рядом, па одной высоте. Общая длина тела: d около 520—600 р, 9 500 р. .
. . . .................................. . 1. Н. hericia (Rob.).
2 (1). Кожа без шиповидных хэтоидов; dd мельче 99, одинакового с ними (беловатого) цвета, без хитинизированных площадок на спине; лапки II вентрально с 3 менее крупными шипами, расположенными друг за другом вдоль лапки. Общая длина тела: d около 460 р, 9 750 р. . . .
. . ...........................................2. Н. georgei Mich.
1. И. hericia (Rob.) (фиг. 609—614, 628—635).
Robin, 1868, Journ. Anat. Phvsiol., V:603 (Glycyphagus). —robinii Oanestrini, 1888:22; Ber-1080, 1897, LXXX, n° 5; Michael,’ 1903:31; t. XXIli—XXIV. — hericia Kramer, 1899:144 (He-ricia).
Прижизненная окраска светложелтовато-бурая, гпатосома и ноги с красноватым оттенком. Колса слегка зернистая, усаженная довольно длинными шиловидными хэтоидами; .поверхность проподосомального щита покрыта неглубокими вдавлепиями, напоминающими соты.
d. На спинной поверхности идиосомы симметрично расположены 8 пар округлых и овальных хитиновых цлощадок, густо пронизанных тонкими порами; у живых особей эти пластинки заметно вдавлены. V a, sc е и sc i, sh и d4 приблизительно одинаковой длины (19—23% длины идиосомы); dx короче других, d2, d8 и 12 примерно равны по длине (26—28%); h а, h р и 1Х длиною в 35—45, 13 до 60% длины идиосомы; sa е равны по длине
19. HERICIA
373
идиосоме, расположенные на сильно выступающих бугорках. Примерная длина ног составляет около 57 (I), 62 (II), 70 (III) и 54 (IV) % длины идиосомы; концы лапок (особенно лапок III) сильно скошены, с далеко выдвинутым вперед нижним краем, поддерживающем снизу короткий мешковидный praetarsus и основание коготка, с крупным дорзальным шипом; лапки II вентрально с 2 ножеридными шипами, расположенными рядом, на одном уровне; ноги IV дугообразно искривлены, вооружены слабее, чем другие пары. Длина, в среднем, 520 р, ширина 350 р.
9. Спинная поверхность идиосомы без хитинизированных площадок, h р и d2—d4 значительно длиннее, чем у <5, 13 и sa е много короче; примерная длина щетинок (в процентах к длине идиосомы): v а 20; see 15; sc i 18; h а 50; h p 35; sh 30; dx 18; d2 40; d3 48; d4 42; lx 42; 12 15; 13 40; sa e около 50. Длина ног составляет приблизительно 48(1), 50 (II), 53 (III) и 60 (IV) % длины идиосомы, они значительно стройнее, чем у б, и слабее вооружены: Длина около 500 р, ширина 330 р.
Гипопус. Тускложелтоватого цвета, при жизни с неправильным пятнистым и сетчатым буроватым рисунком. Тело широкоэллиптическое (в 1.3 раза длиннее своей ширины), спереди с узким, сильно выдающимся ростральным выступом, сзади широко закругленное. Длина около 170 р, ширина 130 р.
Распространение. Достоверно известен только из Франции и Англии. Живет на стволах вязов и дубов, в соке, вытекающем из щелей и поранений на них, или под сильно пропитанными им, отстающими участками коры.
Описание и рисунки Berlese отличаются во многих деталях от данных Robin’a и Michael’я, так что не исключена возможность, что Н. hericia окажется сборным видом; не имея материала по Hericia, я не могу сейчас решить этот вопрос. Приведенное здесь описание составлено по Michacl’io.
2. Hericia georgei Mich. (фиг. 636, 637).
Michael, 1903:38, t. XXXIX, fig. 5—12.,
Сомнительный, совершенно недостаточно описанный вид. Отличается от предыдущего своей беловатой окраской, кожей, лишенной шиповидных хэтоидов, отсутствием дорзального вершинного шипа на лапках I с? и наличием у d 3 вентральных шипов на лапках II, расположенных друг за другом вдоль лапки; идиосома имеет, повидимому, более правильную овальную форму и у <5 шире, чем у Н. Hericia Rob.; кроме того значительно мельче 9. Общая длина <5 около 460 |л, 9 750 |л.
Гипопус отличается от гипопуса только что названного вида более широко закругленным передним концом тела.
Распространение. Англия. Найден всего лишь раз, под отстающей корой тополя, поврежденного древоточцем Cossus cossun Jj.; встречался в огромном количестве на сильно увлажненных участках ствола,, пропитанных вытекавшим из него соком.
III. GLYCYPHAGIDAE. 7. CHAETODACTYLINAE
7. Подсем. Chaetodactylinae
Идиосома широкая (в 1.1—1.5 раза длиннее своей ширины), несколько . уплощенная, приблизительно шестиугольной формы, у некоторых форм с слегка намеченным подразделением на проподо- и гистеросому, без рострального выступа. Кожа мягкая, бесцветная, перепончатой консистенции, топкобугорчатая или покрытая треугольными кутикулярными выростами (сквамондамп), иногда с штриховкой, состоящей из поперечных рядов очень мелких бугорков (род Chactbdac.tylusy, у ряда форм имеется более или менее явственный проподосомальпый щит, нередко образующий по бокам пару хитйнизированных полос, тянущихся до основания ног II и напоминающих так называемые перитремы представителей подотряда Parasitiform.es. Щетинки спинной поверхности идиосомы перистые или зазубренные,. обычно довольно короткие; у некоторых видов почти все дискальные (sc, Щ—d4 и иногда и Щ—12) более или менее редуцированы; vp часто отсутствуют; плечевых и боковых по 3 пары; sa е обычно длиннее всех других щетинок тела и расположены терминально; псевдостигмальный орган простой, щетинковидный, или отсутствует. Строение коксо-стернального скелета изменчиво, но эпимеры I и.II никогда не соединены концами. Ноги стройные, с умеренно длинными и тонкими и иногда слегка искрив-ленными лапками; щетинки голеней й колен более или менее перистые или зазубренные; вершина колен I с 2 органами чувств — небольшой палочковидной сенсиллой и расположенной рядом с нею простой щетинкой; вершинные щетинки голеней I и II довольно короткие, голеней III и IV--- совершенно редуцированные; вооружение лапок состоит из нескольких гладких щетинок. Амбулакры состоят из небольшого перепончатого praetarsus, несущего дистально довольно крупный (у некоторых видов рода Sennertia очень,крупный) серповидный коготок. Вторичные половые различия очень не велики и совершенно не затрагивают вооружения ног.
2. Генитальное отверстие обычно очень крупное, с хорошо развитыми, слегка расходящимися кзади клапанами; в большинстве случаев оно сильно сдвинуто кпереди; энигиний отсутствует, но у многих видов родов 8еп-nerta и Horstia эпимеры I соединены хитиновой перемычкой, непосредственно примыкающей спереди к генитальному отверстию; генитальные щупальцы очень мелкие или отсутствуют; у некоторых видов есть слегка выступающая копулятивная трубка. '
Нимфальпые стадии плохо известны.
Расселительпые гипопусы имеются у всех без исключения видов (большинство их известно только по этой стадии) и относятся к энтомохорному типу. Гнатосома у них сильно редуцирована, представлена парой бугорков, несущих на конце недлинные аристы, или полностью слита с идиосомой. Ферма последней очень разнообразна; у многих форм она сильно уплощена, у других имеет более или менее выпуклую спинную поверхность. Разделение на проподосому и гистеросому неполное, намеченное только
ТАБЛИЦА РОДОВ 375
наличием 2 обособленных спинных щитов или же (у рода Sennerlia) совершенно не выраженное. Спинные щиты часто оставляют открытыми более или менее широкие участки мягкой, перепончатой кожи, имеющей в утих •случаях резко штрихованную структуру; поверхность щитов очень густопористая и украшена топким сетчатым рисунком такого же характера, как у гипопусов Saproglyphidae, или более или менее грубой ямчатой скульптурой. Щетинки спинной поверхности часто достигают большой длины и крепости. Нижняя поверхность тела покрыта мягкой, тонкой кожей. Строение коксо-стернального скелета разнообразно, но коксальные поля III и IV всегда разобщены широким промежутком; стернум нередко раздвоен на конце. Коксальные щетинки всегда имеются, по часто модифицированы. Присасывательный диск с 3. парами присосок; его пластинка обычно разделена на несколько самостоятельных склеритов. Ноги I—III массивные, часто занимающие совсем маргинальное положение (Sennerlia, Chaetodacty-lus). Ноги IV обычно короче других и в большинстве случаев оканчиваются 1—3 крупными (парусными) щетинками. Praetarsi узкие, цилиндрические, •более или менее складчатые или редуцированы; ноготки крупные, иногда огромных размеров и тогда спирально закрученные. Вооружение пог очень изменчиво, но все составляющие его щетинки всегда гладкие; на лапках некоторые из вершинных и вентральных щетинок, как правило, листовидно расширены. Покоящиеся гипопусы имеются у рода Chaelodaclylus (наряду с расселительными). ....................
Богатая видами группа; все ее представители биологически связаны с пчелами, в гнездах которых они обитают и которые сами служат переносчиками их гипопусам. Половозрелая стадия известна пока у очень немногих форм.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ
Взрослые
1(2). Проподосомальный щит отсутствует; идиосома в 1.4—1.5 раза длиннее своей ширины, с довольно явственной поперечной бороздкой, намечающей ее подразделение на проподосому и гистеросому; стернум хорошо развит у обоих полов; концы эпимер III и IV соединены тонкой хитиновой полоской, замыкающей, таким образом, коксальные поля III ...........................21. Chaetodactylus Bond,
2 (1). Проподосомальный щит хорошо развит, образует по бокам пиру
длинных хитинизированных полос, тянущихся до основания пог II и расширяющихся здесь в овальную площадку; идиосома очень широкая (в 1.1—1.3 раза длиннее своей ширины), без всяких олодоп поперечной бороздки;- у ,с? имеется V-образный стернум, у <t> эпимеры I непосредственно слиты с эпигинием; эпимеры III и IV (ШЛЫК) укорочены, свободные ................. 20. Sennei'ilu Ouds.
376
Ill. GLYCYPHAG1DAE. 7. CHAEIODACTYLLNAE
Гипопусы
1 (10). Ноги IV с хорошо развитым коготком; тело удлиненное (не менее чем в 1.5 раза длиннее своей ширины), более или менее яйцевидное; довольно сильно выпуклое; спинные щиты плотно хитинизированы, закрывают всю или почти всю дорзальную поверхность тела. По большей засти мелкие формы, нередко габитуально напоминающие гипопусов Raproylypliidac.
2 (5). Ланки IV без крупных (парусных) щетинок, но с 2—5 небольшими ланцетовидно расширенными щетинками; стернум простой, не раздвоенный на конце; коготки резко крючковидпо изогнуты, одинакового размера на всех ногах.
3 (4). Скульптура спинных щитов состоит из округлых или овальных» четко очерченных ямок, так что они кажутся продырявленными, как сито; присасывательный диск квадратный; задний край гистеросомы с более или менее глубокой выемкой. На шмелях................
.........................................25. Cerophagus Cuds.
4 (3). Скульптура спинных щитов состоит из тонких вдавленных линий, имеющих преимущественно продольное направление на гистеросо-мальном щите и поперечное и диагональное — на проподосоме; присасывательный диск округлый; задний край гистеросомы параболически закруглен ..........................26. Cerophagopsis, gen. n.
5 (2). Лапки IV с 1—3 очень крупными парусными щетинками, без листовидно расширенных щетинок, или же последние имеют форму узких, линейных лезвий; коготки нормального строения, почти прямые или винтообразно закрученные, и тогда коготки IV много меньше остальных; стернум на конце вильчато раздвоен.
6 (7). Коготки I—III очень крупные, винтообразно закрученные, окруженные у основания складчатыми, мясистыми ргаеtarsi; коготки IV много меньше остальных, но одинакового с ними строения; ноги Ги II очень мощные; лапки I—III с 3 крупными серповидными щетинками .....................................27. Sennertionyx, gen. n.
7 (6). Коготки нормального размера и строения, слабо дугообразные, все одинаковой величины; ргаеtarsi отсутствуют; ноги I и II обычного размера; лапки I—III с 4—5 крупными широколанцетовидными щетинками.
8 (9). Коксалыше и генитальные щетинки превращены в крупные, массивные яйцевидные образования, сидящие на короткой ножке; спинные щиты очень плотные, в грубой скульптуре из удлиненных, полулунных ямок; щетинки спинной поверхности очень длинные, уплощенные; лапки I и II с 5 ланцетовидными щетинками; на лапках IV 3 ножевидных, пластинчатых щетинки............24. Neohorstia, gen. iu
9 (8). Коксальные и генитальные щетинки редуцированные, представлены, маленькими кружками; спинные щиты умеренной плотности, с орна
20. SENNERTIA
377
ментом из тонких вдавленных линий; все щетинки спинной поверхности представлены (у палеарктических видов) микрохэтами, подчас очень трудно заметными; лапки I и II с 4 ланцетовидными щетинками; лапки IV вооружены только простыми щетинками . . 22. Horstia Ouds. 10 (1). Ноги IV без коготка.
11 (12). Коготки (I—III) простые, небольшого размера, слабо дугообразно^ изогнутые, сидячие; praetarsus отсутствует. Лапки I и 11с 3, лапки III с 5 ланцетовидными щетинками; лапки IV очень коротки, закруглены на конце; тело продолговатое, грушевидное или в форме вытянутого в длину шестиугольника, в 1.45—1.6 раза длиннее своей ширины. Мелкие формы (175—280 р)......................23. Tortonia Ouds.
12 (И). Коготки (I—III) чрезвычайно крупные, сильно винтообразно закрученные, сидящие на конце узкого, мясистого, цилиндрического' praetarsus; лапки I—III с 1—2 узкосерповидными щетинками или вовсе без расширенных щетинок; тело очень широкое, плоское, иногда почти круглое. По большей части крупные формы.
13 (14). Спинная поверхность с 2 хорошо развитыми щитами: треугольным — передним — и более или менее трапециевидным, расширяющимся кпереди — задним; базальная Щетинка колен I очень длинная. На. видахOsmia, Anthidium и Megachile .... 21. Chaetcdactylus Rond.
14 (13). Спинная поверхность только с 1 щитом — задним, более широким у заднего края тела и постепенно суживающимся кпереди; базальная I щетинка колен I короткая. На видах рода Xylocopa...................
.............................................20. Sennertia Ouds.
20. Род SENNERTIA OUDS. 1905
Oudemans, 1905, Entomol. Bor;, 11:21; Vitzthum, 1919, Arch, Naturg., LXXXV:31.
Тип рода: Acarus cerambycinus Scop. 1763.
Идиосома без дорзальной поперечной’бороздки, очень широкая (только в 1.15—1.3 раза длиннее своей ширины), пятиугольная, приблизительно одинаковой формы у обоих полов, с резко выступающими плечевыми выпуклостями и более или менее сильно вогнутым задним краем. Проподо-сомальный щит хорошо развит, образует по бокам длинные, плотно хитини-зированные полосы, тянущиеся до основания ног II и расширенные на конце в округлую или заостренную сзади площадку; эти полосы (псевдо-перитремы) в известной мере напоминают так называемые перитремы представителей подотряда. JParasitiformes. Щетинки спинной поверхности идиосомы довольно короткие, жесткие, более или менее густоперистые. Эпимеры III и IV сильно укорочены, свободные. Ноги длинные, стройные, передние иногда несколько уплощенные; вершинная щетинка голеней III и IV редуцирована. Praetarsi слабо развитые, узкие, короче коготков;, последние у многих видов очень велики и имеют серповидную форму.
3. Стернум V-образный, свободный; генитальное отверстие расположено в пространстве между основаниями ног II и III; penis очень крупный,
378
in. GLYCYPHAGIDAE. 7. CHAETODACTYLINAE
серповидный или саблевидный, у основания с 2 очень длинными палочковидными подпорками, расходящимися кзади под прямым или тупым углом; генитальные щупальцы отсутствуют; анальная щель придвинута к основанию penis.
9. Эпимеры I непосредственно сливаются с эпигипием, образующим более или менее широкую перемычку между их концами; генитальное отверстие расположено между основаниями пог II, на очень большом расстоянии от анальной щели; генитальные щупальцы имеются; копулятивной трубки пет; лапки IV иногда несколько искривлены.
Личинки неизвестны; покоящейся гинопалыгой стадии пе образует, насколько известно^ ни один из видов.
Р.ассе лите л ьный гипопус. Тело крупное (длина 220—515 у), сильно уплощенное, шестиугольное или почти круглое. Проподосомаль-ный щит совершенно отсутствует, гистеросомальный всегда много уже тела и оставляет открытой большую часть его спинной поверхности; кожа этих открытых участков имеет нежную перепончатую консистенцию и украшена резкой концентрической штриховкой, продолжающей аналогичный, но более слабый орнамент поверхности гистеросомального щита; иногда последний более или менее сильно редуцирован, и тогда почти вся спинная поверхность тела покрыта мягкой штрихованной кожей; у большинства видов, задний край гистеросомального щита завернут на брюпшую поверхность тела й здесь более или менее глубоко вырезан. Не менее 5 пар. щетинок спинной поверхности, а именно sc е, hi, he, 1 а и 1 р, представлены очень жесткими макрохэтами; степень развития- остальных щетинок изменчива, но обычно большая часть их редуцирована и представлена только округлыми порами; р а длинные, обычно расположенные, так же как и sa i, вентрально — па завернутом участке гистеросомального щита. Строение щетинок брюшной поверхности очень изменчиво. Гнатосома полностью слита с идиосомой; аристы очень короткие. Стернум простой; эпимеры II прямые или слабо дугообразные, свободные, широко расставленные концами; эпимеры III короткие, свободные или связанные с эпимеритами II; эпимеры IV сильно укорочены. Присасывательный диск относительно небольшой, лишенный перепончатой каймы, иногда окруженный неполной хитиновой рамкой. Ноги I—III очень мощные, узловатые, с огромными винтообразно скрученными коготками, сидящими на конце длинных мясистых praetarsi; последние образуют толстые поперечные складки и нередко несут более или менее развитый пальцевидный боковой отросток; лапки I—III короткие, обрывисто суживающиеся па конце, незаметно переходящем в praetarsus и несущем 3 длинных предвершинных щетинки, из которых крайние часто расширены дистально в узкую серповидную или ланцетовидную пластинку; дорзальное вооружение лапок I состоит из Обонятельной палочки, другой, такого же типа, сенсиллы, сидящей кпереди и кнаружи от нее, уже упомянутых 3 длинных щетинок (из которых средняя здесь прямая, палочковидная), расположенной еще более дистально на
20. SENNERTIA
379
ружной вершинной щетинки и 2 коротких вентральных щетинок. Ноги IV гораздо короче и тоньше остальных, оканчивающиеся очень мощной парусной щетинкой, обычно превосходящей длину тела, и 1—2 мелкими вентральными щетинками, без амбулакр.
Очень богатый видами род, распространенный преимущественно в тропиках и биологически связанный с древесными пчелами подсем. Xyloco-pinae; в пределах Палеарктики род Sennertia представлен 9 видами, пз которых 7 известны только по гипопусам.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
Взрослые
1 (2). Sc е и все 4 пары дорзальных щетинок (dx—d4) представлены короткими (4—6% длины идиосомы), толстыми микрохэтами; ha, hp и sa е очень толстые, дугообразно изогнутые, притупленные па конце; кожа спинной поверхности густо покрыта треугольными сквамоидами; коготки крупные, массивные. 4. S. cantabrica, sp. п.
,2 (1). Sc е и все d в виде довольно длинных (12—20% длины идиосомы), слегка перистых щетинок; такое же строение имеют и h а и 11 р; на о песколько толще, прямые; кожа спинной поверхности очень тонкозернистая; коготки маленькие .........*1. S. corambyclna (Scop.).
Гцпоцусы
1 (2). Sc i и d4 представлены макрохэтами, имеющими приблизительно такую же длину, как sc е (32—37% длины тела), так что-на спинной поверхности тела находится 7 пар однотипных-щетинок; d2 и d3 в виде довольно крепких щетинок, длиною в 6—15% длины тела. На видах группы Xylocopa olivieri Lep........ *7. S. zhelochovtsovl, sp. n.
2 (1). Sc i в 2—4 раза короче sc e или представлены крошечными микрохэтами; dx, d2 и d3 в виде чрезвычайно мелких микрохэт или совершенно редуцированные, имеющие вид мелких округлых пор; па спинной поверхности тела, таким образом, только 5 или 6 пар макрохэт. На других видах Xylocopa.
3 (6). Sc i длиною в 8—20 % длины тела, в 2—4 раза короче sc е, довольно крепкие, так что на спинной поверхности имеется 6 пар более или менее крупных щетинок.
4 1(5). Щетинки вентральной поверхности тела сильно утолщенные, шипо-видные; вентральные щетинки видоизменены в пару яйцевидных образований, сидящих на короткой ножке; спинные щетинки чрезвычайно мощные, саблевидные; sc i в 2 раза короче sc с, длиною, и I5--20% длины тела; гистеросомальный щит очень крушыП простирающийся вперед почти до sc i; одна из вентральных щедннок голеней
I и II и вентральная вершинная щетинка лапок в виде крепких конических шипов. На Xylcopa dissimilis Lor. . . (!. N. lifirrldu (Viz.).
380
III. GLYCYPnAGEDAE. 7. CnAETODACTYLINAE
5 (4). Все щетинки вентральной поверхности тела тонкие, волосовидные;, спинные макрохэты относительно тонкие и гибкие; sc i в 4 раза короче sc е, длиною в 8—10 % длины тела; гистеросомальный щит узкий, простирающийся вперед только до dx; все щетинки ног тонкие/ гибкие. Ila Xylocopa piclifrons Sm.............8. S. potanini, sp. n.
6 (3). Sc i представлены крошечными микрохэтами, так что крупных щетинок па спинной поверхности только 5 пар.
7 (8). Щетинки вентральной поверхности тела шиповидные; praetarsi I—Ш с сильно развитым, пальцевидным боковым отростком. На Xylocopa kalinovskii Rad.........................5. S. alfkeni (Ouds.)...
8 (7). Щетинки вентральной поверхности тела тонкие, волосовидные; praetarsi без выростов или с небольшим тупым отростком. На других видах Xylocopa.
9 (16). L р короче sc е или почти одинаковой с ними длины; гистеросомальный щит крупный, сильно хитинизированный и интенсивно окрашенный, простирающийся до dj или еще дальше вперед; наружные и задние присоски не очень мелкие, наружные расположены по сторонам центральных, значительно впереди задних.
10 (11). Очень крупная форма, длиною в 460—516 [г; гистеросомальный щит в 1.5 раза длиннее своей ширины у заднего края тела, равномерно суживающийся кпереди. На Xylocopa punctilabris Мог. и X. valga F.............................: . . . . *2. S. gargantua, sp. n.
11 (10). Более мелкие виды, не превышающие 340 у в длину; гистеросомальный щит в 1.75—2 раза длиннее своей ширины, с параллельными сторонами, спереди более или менее широко закругленный.
12 (13). Задний край гистеросомального щита прямо усеченный, совершенно не завернутый на брюшную сторону тела, в соответствии с чем sa i и р а расположены дорзально; присасывательный диск далеко отодвинутый от заднего края тела; вентральная щетинка лапок IV в 4 раза длиннее самой лапки; наружная вентральная щетинка лапок I и II неутолщенная, щетинковидная; 1 р в 1.5 раза короче sc е. На Xylocopa cantabrica Lep...................4. S. cantabrica, sp. n,
13 (12). Задний край гистеросомального щита закругленный, сильно завернутый на брюшную поверхность тела и снабженный здесь глубоким полукруглым вырезом, в который вдвинута задняя часть присасывательного диска; sa i и р а расположены вентрально; вентраль-
Фиг. 688—652.
Фиг. 638. Sennertia aerambycina (Scop.), Q сверху (no Berlese). Фиг. 639. S. aerambycina (Scop.)<-Cf снизу (no Berlese). Фиг. 640. S. aerambycina (Scop.), $ снизу (no Berlese). Фиг. 641. S.ceram-byaina (Scop.), конец лапки и амбулакра, снизу (по Berlese). Фиг. 642. То же, сбоку (по Berlese). Фиг. 643. S. aerambycina (Scop.), генитальное отверстие 9 (по Berlese). Фиг. 644. Cnaetodactylus osmiae(L. Duf.), О’сверху (по Michael’io). Фиг. 645. Ch. osmiae (I.. Duf.), 9 сверху (no Michael’io). Фиг. 646. Ch. osmiae (L. Duf.), нога I (no Michael 'ю). Фиг. 647. Ch. osmiae (L. Duf.) (no Michael 'to), нога IV. Фиг. 648. Ch. osmiae(L. Duf.), хелицера (в профиль) (no Michael'to). Фиг. 649. Ch.'osmiae (L. Duf.), структура кожи при сильном увеличении (по Michael’io). Фиг. 650. То же, при более слабом увеличении (по Michael’io). Фиг. 651. Horstia ornata (Ouds.), гипопус сверху (по Oude-mans’y). Фиг. 652. То же, снизу (по Oudemans’y).
382
III. GLYCYPHAGIDAE. 7. CHAETODACTYLINAE
ная щетинка лапок IV короне лапки или в 1.4—1.5 раза длиннее ее; 1 р чуть короче sc о.
14 (15). Гистеросомальнын щит в 1.75 раза длиннее своей ширины; 1 а., заметно короче sc о; наружная Субапикальная щетинка лапок I и II волосовидная, наружная вершинная щетинка резко искривленная, утолщенная, ложевндпая; вентральная щетинка лапок IV очень-маленькая, заметно короче лапки. Длина 270—330 |i. На видах группы Xylocopa violacca L.........................*1, S. cerambycina (Scop.).
15 (14). Глстеросомальпый щит уже, в 2 раза длиннее своей ширины;
1 а чуть длиннее sc е; наружная субапикальная щетинка лапок I и II ланцетовидно расширенная, наружная вершинная щетинка тонкая^ палочковидная; вентральная щетинка лапок IV заметно длиннее самой лапки. Длина 224—255 р. На Xylocopa dissimilis Lep. .....
....................................... . . . 3. S. dissimilis, sp. n.
16 (9). L p очень длинные, в 2 раза длиннее sc е; гистеросомальный щит-очень маленький, не достигающий даже уровня da, слабо хитинизи-рованный и почти не окрашенный, неясно обособленный от окружающей его мягкой кожи; наружные и задние присоски присасывательного диска очень мелкие, расположенные в поперечный ряд. На Xylocopa dissimilis Lep...........................9. S. on demansi, sp. n.
*1. Sennertia cerambycina (Scop.) (фиг. 638—643, 653—655).
Scopoli, 1763, Entomol. Carniol.: 386, n° 1054 (Pediculus). — xyloeopae Donnadieu, 1868: Ann. Sci. Nat., ser. V, vol. X: 69; Berlese, 1885, XVIII, n° 1 (Trichodaetylus)-, Canestrini, 1888a: 394, t. 36 (Tricho tarsus)-, Oudemans, 1905, Entomol. Ber., 11:21 (Sennertia).
Окраска бледнозеленоватая, гнатосома, псевдоперитремы и ноги интенсивно рыжие. Идиосома с сравнительно мало выступающими плечами и слабо вогнутым задним краем; кожа очень тонкозернистая; проподосомальный щит слабо хптинизированный; площадки псевдоперитрем сзади приостренные. Щетинки спинной поверхности средней длины, толстые, равномерно утончающиеся к концу, слегка перистые, все более или менее однотипные, за исключением 13, которые толще других, прямые, палочковидные, и sc i и h i, представленных микрохэтами; относительные размеры их у 9 (в процентах от длины идиосомы) приблизительно таковы: vi 9; v р 18; sc е 13; h а 20; h р 30; dx 20; da 22; d3 18; d4 16; 1418; la 15; 13 14; sai 55 и sa e 11. Ноги стройные, голени почти прямые; все щетинки ног довольно тонкие и гибкие, почти гладкие; вершинная щетинка голеней I и II короткая; вершила лапок снизу с пластинчатой треугольной шпорой; коготки относительно маленькие, стройные.
Фиг. 653—661. Гипопусы рода Sennertia.
Фиг. 653. Sennertiacerambycina (Scop.), гипопус сверху; буквенные обозначения см. в тексте, стр. 65. Фиг. 654. То же, конец ноги IV (и основание парусной щетинки) снизу. Фиг.' 655. S. cerambycina (Scop.), наружная субапикальная щетинка лапок I. Фиг. 656. S. gargantua, sp. п., очертания тепа гипопуса. Фиг. 657. S. gargantua, sp. п., конец ноги IV снизу. Фиг. 658. S. dissimilis, ер. п., папка I гипопуса. Фиг. 659. s. dissimilis, sp. п., наружная субапикальная щетинка лапки I. Фиг. 660.
S. dissimilis, sp. п., конец ноги IV снизу. Фиг. 661. S. zhelochovtsevi, sp. п., нога I сверху.
384
JU. GLYCYPIIAGIDAE. 7. CIIAETODACTYLINAE
сГ. Копулятивный аппарат помещается в пространстве между основаниями ног III и IV; penis серповидный; анальная щель очень близко придвинута к anus и отстоит па значительное расстояние от заднего края, тела.
9. Эпимеры I короткие, соединяющиеся под тупым углом с широким поперечным энигппием. Общая длина тела до 800
Гипопус. Тело в 1.1—1.2 раза длиннее своей ширины, округло-шсстиуголыюс. Гистеросомплы1ЫЙ щит крупный, закрывающий бблыпую часть спинной поверхности, почти в 1.5 раза шире своей длины, с параллельными сторонами, довольно широко закругленный спереди. Наспинной поверхности 5 пар крупных щетинок: sc е, h е и h i, 1 а и 1 р; sc е несколько длиннее прочих, достигая 29—30% длины тела, а остальные 4 пары имеют приблизительно одинаковые "размеры (23—26%); sc i расположены значительно позади sc е и пре; ставлены крошечными микрохэтами; такими же ликрохотами (или простыми порами) представлены все d; sa е очень тонкие, расположенные терминально; задний край гистеросомального щита довольно широко завернут на брюшную поверхность тела и здесь полукругло вырезан; р а расположены вентрально; все щетинки вентральной поверхности тела очень тонкие, волосовидные. Присасывательный диск, в целом, приблизительно яйцевидный, вдвинут своей задней половиной в соответствующий вырез, гистеросомального щита, лишен периферического хитинового кольца; наружные присоски средней величины, тесно придвинутые к центральным, расположенные по бокам и несколько кзади от них; •обе задние присоски помещаются на особом склерите полулунной формы. Ноги I и II довольно толстые; вентральные щетинки бедер и голеней, наружная щетинка колен и обе крайние предвершинные щетинки лапок тонкие, волосовидные; наружная вершинная щетинка лапок утолщенная, резко искривленная, ножевидная, вентральные вершинные щетинки очень мелкие и тонкие. Лапки IV слабо обособлены от голеней, заметно суживающиеся к концу; их вентральная щетинка очень мала, значительно короче самой лапки. Длина 270—380 р., ширина 240—300 р.. Остальные стадии развития неизвестны.
Распространение. Вся южная, средняя и восточная Европа, на север до 55° с. ш., Передняя и Средняя Азия до Тибета и Гашунского Гоби па восток и северного Ирана на юг.
Экология. Живет в гнездах различных видов Хуlocopa группы vio-lacea: X. violacea L., X. valga F. (включая subsp. pyrrhopyga Fr.), X.varen-tzovi F. Mor., X. nitidiventris Sm., X. przewalskii F. Mor.; гипопусы встречаются также на паразите ксилокоп Polyochrum repandum Sp., а иногда :(случайно?) на различных других перепончатокрылых: Osmia (Megalosmia') princeps F. Mor., 0. (Aceralosmia) macroglossa G-erst., Megachile argeniataJ?. н Anthidium christofi F. Mor.—Половозрелые особи S. ceramkycina были добыты до сих пор только 1 раз.
L
।
! 20. SENNERTIA ЯМК
*2: Sennertia gargantua, sp. n. (фиг. 656, 657).
Гипопус. Очень похож на гипопуса 8. cerambycina, но сразу же отличается своими огромными размерами. Гистеросомальный щит в 1.5 раза длиннее своей наибольшей ширины, приходящейся у заднего края тела, постепенно и довольно равномерно суживающийся кпереди (так что его края слегка выпуклы на всем своем протяжении и заметно конвергируют кпереди); sai утолщенные, шиповидные; вентральная щетинка ланок IV только в 1.5 раза короче tibiotarsus. Все остальные детали строения, как у 8. cerambycina. Длина 463—515 у, ширина 390—463 р.
Распространение. Узбекистан, Таджикистан (Кштут).
Экология. Гипопусы найдены на Xylocopa punctilabris F. Мог. и X. valga F.; на последнем виде они встречались совместно с гипопусами S. cerambycina. Судя по размерам гипопальной стадии, половозрелая форма этого вида должна также обладать исключительно крупными размерами.
3. Sennertia dissimilis, sp. н. (фиг. 658—660, 665).
Гипопус. Очень близок к 8. cerambycina, но несколько мельче и значительно уже. Тело в 1.23—1.3 раза длиннее своей ширины; гистеросомальный щит соответственно уже, чем у 8. cerambycina, в 2 раза длиннее своей ширины, с параллельными сторонами, довольно широко закругленный спереди; крупные щетинки тела немного короче (sc е 25, h е, 1 р и h 1 21—28, 1 а 26% длины тела); 1 а заметно длиннее 1 р и обеих плечевых и даже чуть больше sc е; наружные присоски сильнее смещены назад (относительно центральных) и несколько крупнее. Ноги более стройные; наружная субапикальная щетинка лапок I и II ланцетовидно расширена в своей дистальной половине; наружная вершинная щетинка тех же лапок прямая тонкая,,палочковидная; tibiotarsus IV почти в 2.5 раза длиннее своего диаметра и только в 1.25 раза длиннее щетинки, сидящей на его вентральной поверхности. Длина 245—255 pt., ширина 180—260 у.
Распространение. Япония. На Xylocopa dissimilis Lep., совместно с 8. horrida, S. oudemansi и Horstia helenae. На просмотренных мною экземплярах X. dissimilis из Ассама не встретилось ни одного гипопуса 8. dissimilis.
4. Sennertia cantabrica, sp. п. (фиг. 23, 662—664).
Телеонимфа. Идиосома с очень резко выступающими плечевыми выпуклостями и довольно сильно вогнутым задним краем; кожа спииноИ поверхности густо усажена треугольными сквамоидами; проподосомалюинП щит слабо хитинизированный, площадки псевдоперитрем сзади иакруг* ленные. Большинство щетинок спинной поверхности идиосомы — v I, обе пары sc, h i и все 4 пары d — представлены толстыми, почти iiiiiiioiiii/i,-ными микрохэтами; he, sh и sa е сильно утолщенные, ириту пленные па конце, дугообразные, с нижней стороны слегка зазубренные; на I прямые, палочковидные, явственно перистые; относительные размеры щетинок
25 Фауна СССР, Захваткип—284
386 III. GLYCYPHAGIDAE. 7. CHAETODACTYLINAE
(в процентах к длине идиосомы): v а 13; v р 4; sc е и sc i 6; h i 4; he 20;. sh25; dj—d44—5; lj 5; l2 20;sael2; sa i около 70. Ноги довольно толстые, голени слабо искривленные; базальная и наружная щетинки колен,I . и II и вентральная щетинка колен III широкие, густоперистые; вершинная щетинка голеней I и II довольно длинная; концы лапок снизу без пластинчатой шпоры; коготки очень крупные, массивные. Длина свежесли-нявшей особи 290 р., ширина около 260 р,.
Г и и о и у с. Сходен с гипопусами обоих предыдущих видов. Гистеро-сомальпыЙ щит узкий (в 2 раза длинное своей ширины), с параллельными краями, спереди параболически закругленный; he длинней всех других крупных щетинок тела (около 27 % его длипы), 1 р относительно очень короткие (15%); задний край гистеросомального щита прямой, совершенно не завернутый на брюшную поверхность тела', так что sa i и р а расположены дорзально; присасывательный диск сильно отодвинут вперед от заднего края тела, по своему строению почти не отличающийся от такового S. cerambycina. Обе предвершинные щетинки лапок I и II тонкие, волосовидные; наружная вершинная щетинка не утолщенная; базальная часть ргаеtarsi I—III тоньше, чем у S. cerambycina, без боковых выростов; tibiotarsus IV несколько суживающийся к концу, в 2 раза длиннее своего диаметра у основания; его вентральная щетинка длинная, в 2 раза длиннее-всего tibiotarsus. Длина 340 [i, ширина 285 р..
Распростра не н и е. Алжир (более точное местонахождение-неизвестно). Биологически связан с Xylocopa cantabrica Lep., на которой-и были найдены экзувии гипопусов и 1 свежеслипявшая телеонимфа этого вида.
5. Sennertia alfkeni (Ouds.) (фиг. 672—674).
Oudemans, 1901, Tijdschr. Entomol., XLIII:115, t. 5, fig. 18—20 (Tricholarsus).—japo*-nicus Оnlemana, 1901, ibid.: 146, t. 5, fig. 21 (Trichotarsus).
Гипопус. Интенсивно окрашенный вид. Гистеросомальный щит-такой же формы и приблизительно такого же размера, как у S. cerambycina; see, he, hi, la и Ip очень крупные, толстые, слегка притупленные на конце, сильно пигментированные; длина sc е равна 37—55, h 1 30—37, h е 22—27,1 а 33—47,1 р 40—47% длины тела; очень велики также и р а, достигающие 60—б5% длины тела и также заметно утолщенные; все остальные щетинки спинпой поверхности совершенно редуцированы, представленные мелкими поровидпыми образованиями; рай очень короткие sa i помещаются вентрально на слегка завернутом книзу участке спинного щита; щетинки брюшной поверхности тела (коксальные и генитальные)' сильно утолщены в основной половине, с концом, оттянутым в длинную-гибкую ость. Присасывательный диск небольшой, без хитиновой рамкп;. наружные присоски сильно смещены назад, образуя с задними дугообразный ряд, обращенный выпуклостью назад; задние присоски едва меньше-наружных, расположенных на отдельном полулунном склерите. Ноги,
L
20. SENNERTIA 387
I—Ш очень толстые; наружная базальная щетинка колен I и II сильна гипертрофирована, очень жесткая, темно окрашенная; наружная и внутренняя предвершинные щетинки лапок простые илп более или менее ланцетовидно расширены; наружная вершинная щетинка утолщенная, резко искривленная, вентральные вершинные щетинки слегка утолщены; ргае-tarsi с очень сильно развитым пальцевидным боковым отростком. Лапки IV явственно обособлены от голеней, едва суживающиеся к концу; вентральная щетинка очень длинная (равна по длине ноге IV). Длина 195—325 !, ширина 165—270 н.
Распространение. Япония, Корея, Китай. Гипопусы встречаются в большом количестве на Xylocopa kalinovskii Rad.
Обращает на себя внимание большая изменчивость в размерах, длине щетинок тела и степени листовидного расширения предвершинных щетинок ног I—III. Oudemans различал по последнему признаку 2 самостоятельных вида: S. alfkeni с волосовидными предвершинными щетинками ног и S. japonica, у которой они ланцетовидно расширены; исследование очень большого материала позволило установить, что в каждой популяции гипопусов можно встретить обе формы и все переходы между ними и что этот признак не связан с какими-либо другими различиями между ними.
6. Sennertia horrida (Vtz.) (фиг. 62—64).
Vitzthum, 1912, Ztschr. wiss. Insektenbiol., VIII:289 . (Trichotarsiis)-, 1919, Arch. Nature.,, LXXXV, 6:57, fig. 51—52 (Sennertia).
Гипопус. Интенсивно окрашенная, мощно вооруженная форма. Гистеросомальный щит крупнее, чем у других видов, заметно расширяющийся кпереди до уровня 1 а, спереди широко закругленный или несколько угловатый. Sc е, h е, h i, 1 а и 1 р очень крепкие, крупные, сильно пигментированные; sc i также представлены очень крепкими щетинками, по вдвое короче sc е; длина sc е составляет приблизительно 35—40, sc i 18—20, h i 40, h е 15—20,1 a 30—35,1 p 24—28 % длины тела; p а заметно утолщены, длиною до 55% длины тела; все Остальные щетинки спинной поверхности сильно редуцированы, представлены округлыми порами; щетинки брюшной поверхности тела сильно модифицированы: передние коксальные щетинки имеют такую же форму, как у S. alfkeni, sh и задние коксальнысг щетинки в виде толстых ножевидных шипов, а вентральные щетинки представлены яйцевидными, закругленными на конце пластинками, сидящими на короткой ножке; передние генитальные щетинки в виде тонких микрохэт. Присасывательный диск округлый, с почти замкнутой спереди хитиновой рамкой и парой линейных склеритов, идущих в диагональном, направлении от боковых присосок; боковые и задние присоски oiiin.iio-удалены от центральных, расположены вогнутой кпереди дугой и имеют вид массивных грушевидных образований. Ноги I и II массиппыо; uepiHtut-ная и боковая щетинки колен, наружная вентральная щетинка голеней, обонятельная палочка и расположенная рядом с пою сенсилла и наружная!
*
388 HI. GLYCYPHAGlbAE. 7. СНАЁТОЬШПШАЕ
й вентральная вершинная щетинки лапок превращены в более или менее толстые, острые шипы (массивнее других центральная вершинная щетинка лапок); наружная и внутренняя предвершинные щетинки лапок I и III крупные, расширенные на конце в широколанцетовидную пластинку. Лапки IV суживающиеся к концу, вентрально с очень длинной крепкой щетинкой (превосходящей длину ног IV). Длина 265—430 у., ширина. .205—340 р,.
Р а с и р о с т р а и с, и и с. Вид описан из Ватавии (Ява); мне известен •также из Японии и Ассама. Гипопусы встречаются в массе на Xylocopa ,dissimilis Lep.
*7. Sennertia zhelochovtsevi, sp. n. (фиг. 661, 666—668).
Гипопус. Тело широкое (1.15—1.2:1), бледно окрашенное. Гистеро-сомальный щит маленький, занимающий небольшую часть спинной поверхности, широкий в своей задней половине, затем быстро суживающийся кпереди и вдающийся в виде узкого языка между основаниями d2. На спинной поверхности тела 7 пар очень длинных, однотипных макрохэт: все, scц dj, la и 1р; длина всех этих щетинок почти одинакова и колеблется в пределах 32—50°/о длины тела; d2 длиною в 10—12, d3 около 6—7% длины тела, довольно крепкие; d4 и обе пары sa в виде более мелких и тонких, но вполне явственных микрохэт; все щетинки брюшной поверхности тела довольно длинные, волосовидные. Присасывательный диск, в целом, круглый; наружные присоски средней величины, расположенные по бокам центральных (на одной поперечной линии с ними), на небольших обособленных хитинизированных площадках; задние присоски такого же размера, как наружные, сидят на полукруглой, также обособленной площадке; обе эти пары присосок близко придвинуты к центральным. Ноги I и II •средней толщины; вентральные щетинки бедер и голеней, наружные щетинки колен и обе крайние субапикальные щетинки лапок чрезвычайно длинные, тонкие, волосовидные; наружная вершинная щетинка лапок почти прямая, слегка ланцетовидно расширенная. Praetarsi I—III без боковых отростков. Лапки IV немного менее чем в 1.5 раза длиннее своей ширины у основания, заметно суживающиеся к концу, с 2 очень мелкими вентральными щетинками. Длина 375—460 у., ширина 324—390 р,.
Распространение. Греция, Турция, Армения, среднеазиатские республики. Гипопусы встречаются па Xylocopa olivieri Lep. и близких к ней формах (X. ги/а F.). Экземпляры из Закавказья и Греции, в среднем, несколько мельче, чем бухарские, и спинные микрохэты у них обычно
Фиг. 662—668.
Фиг. 662. Sennertia cantabrica, sp. п., толеонимфа. Фиг. 663. S. cantabrica, sp. n., гипопус, очертания тела и гистеросомального щита сверху. Фиг. 664. S. cantabrica, sp. п., гипопус, дистальная часть ноги IV снизу. Фиг. 665. S. dissimilis, ер. п., гипопус, очертания тела и гистеросомального щита сверху. Фиг. 666. S. zhelochovtsevi, sp. и., гипопус, общий видео спинной стороны. Фиг. 667. JS. zhelochovtsevi, sp. п., гипопус, конец лапки I сбоку. Фиг. 666. S. zhelochovtsevi, sp. п., гипопус, конец ноги IV снизу.
665
390
III. GIACYPBAGIDAE. 7. CBAETODACT\LINAE
немного длиннее; возможно, что в дальнейшем их придется рассматривать -как особый подвид S. zhelochovtsevi.
8. Sennertia potanini, sp. n. (фиг. 669—671).
Гипопус. Тело очень широкое(1.1:1), бледно окрашенное. Гистеро- • сомальный щит узкий, языковидный, слегка расширяющийся у заднего конца тела. На спинной поверхности тела 5 пар крупных щетинок и 1 пара щетинок среднего размера (нс i); see заметно длиннее прочих (37—42% длины тола); sc i в 4 раза короче вс с; все щетинки брюшпой поверхности топкие, волосовидные. Присасывательный диск маленький, очерченный сзади и с боков несколько угловатым хитиновым полукольцом, па котором расположены очень мелкие наружные и боковые присоски. Ноги I и II относительно стройные; вентральные щетинки бедер и голеней и наружные щетинки колен тонкие, довольно короткие; обе крайние субапикальные щетинки лапок волосовидные; наружная вершинная щетинка утолщена и слабо искривлена, ножевидная. Praetarsi I—III без боковых отростков. Лапки IV в 1.5 раза длиннее своего диаметра у основания, заметно утончающиеся к концу, с длинной вентральной щетинкой (превосходящей в 3.5 раза длину лапки). Длина 370—415 р, ширина 350—366 р.
Распространение. Западный Китай: Сычуань, Ячжоу. Биологически связан с Xylocopa pictifrons Sm.
9. Sennertia oudemansi, sp.n. (фиг. 675—677).
Гипопус. Тело почти круглое (1.1:1). Гистеросомальный щит очень маленький, слабо хитинизировацный и почти не окрашенный, неясно обособленный от окружающей его мягкой кожи, параболической формы, простирающийся вперед только до d3; на-спинной поверхности тела 5 пар крупных щетинок, все они относительно очень тонкие; sc е и hi имеют одинаковые размеры; 1 а необычайно длинные, в 2 раза превосходящие 1 р и заметно превышающие половину длины тела (56%); 1 р короче всех других макрохэт, равные 27% длины тела; все остальные спинные щетинки в виде крайне мелких микрохэт или простых пор; все щетинки брюшной поверхности тела очень короткие и тонкие. Присасывательный диск очень маленький, без хитинового кольца; наружные и задние присоски очень мелкие, расположенные в слегка дугообразный поперечный ряд на одной узкой хитиновой площадке. Ноги Г и II довольно стройные; вентральные щетинки бедер и голеней и наружные Щетинки колен тонкие, очень короткие; наружная предвершинная щетинка ланцетовидно расширенная; наружная вершинная щетинка очень тонкая, волосовидная. Praetarsi I—III узкие, без боковых выростов. Лапки IV совершенно цилиндрические, в 3 раза длиннее •своего диаметра, с 2 крайне мелкими микрохэтами на вершине (по сторонам основания наружной щетинки), без щетинки на вентральной поверхности. Длина 303 р, ширина 467 р.
Распространение. Япония. На Xylocopa dissimilis Lep., вместе > с S. horn da и <S.. dissimilis.
20. SENNERTIA. 21. CHAETODACTYLUS
391
21. Род CHAETODACTYLUS ROND.
Roniani, 1.853, G-iorn. Agric. Industr. Ital., (3), V:12.—Trichodactulus L. Dufour, 1839, Anu. Set. Nat., s6r. II, vol. XI:276 (nom. praeoccnp.); Gervais, 1844,in: Walckenaer, Hist. Nat. Ins. .Apt., 111:286; Dmnadien, 1868, Ann. Sei. Nat., s6r. V, vol. X:69, —Triehotarsus Canestrini, 1888:7; 1888a:356; Berlese, 1897:105; Kramer, 1899:148; Oudemans, 1900, Tijdschr. Entomol., XVIII:115 (part.); 1905, Entomol. Ber., 11:21; 1911, ibid., 111:166; Vitzthum, 1919, Arch. Na-"turg., LXXXV, 5:31.
Тип рода: Trichodactylus osmiae Duf. 1839 (гипопус).
Идиосома более узкая, чем у соседних родов (в 1.4—1.5 раза длиннее своей ширины), приблизительно шестиугольная, с довольно явственными плечевыми выпуклостями и слегка вогнутым задним краем. Кожа в очень густой и тонкой поперечной штриховке, составленной рядами крайне мелких тупых бугорков. Щетинки спинной поверхности идиосомы средней длины (иногда некоторые из них редуцированы), слабо перистые или зазубренные; vp отсутствуют; sa гладкие, длиннее всех других щетинок тела. Эпимеры I соединяются под острым углом, образуя хорошо развитый стериум; эпимериты II неявственны (или отсутствуют?). Концы эпимер III и IV соединены тонкой хитиновой линией, замыкающей, таким образом, коксальные поля III; различия между с? и $ в строении коксо-стернального скелета невелики. Praetarsi довольно широкие, приблизительно равны по длине коготку.
с?. Стериум простой; генитальное отверстие расположено между основаниями ног IV или даже немного дальше назад; эпиандрий хорошо развит, в виде поперечной хитиновой дужки, утолщенной к обоим концам.
$. Задний конец стернума расширен в полулунную пластинку, при--остренные концы которой обращены назад;1 положение генитального отверстия не установлено.
Нимфы отличаются от взрослых более резким рельефом идиосомы, образующей по краям округлые лопасти, и невидимому, более однообразными размерами щетинок ее спинной поверхности. У обоих видов, биология которых более'или менее известна (Ch. osmiae Duf. и Ch. ludwiyi Troucss.), имеет место образование как расселительных, так и покоящихся гипопусов.
Рассе лите льный гипопус. Тело широкое, сильно уплощенное дорзо-вентрально, округленно-шестиугольное, обычно довольно крупное (до 450 р.). Спинные щиты уже тела, оставляют открытыми широкие периферические участки мягкой, явственно штрихованной кожи; пропо-досомальный щит треугольный, отделенный довольно широким промежутком от продолговато-трапециевидного, суживающегося кзади гистеросомального щита; задний край последнего завернут на брюшную сторону тела и образует здесь 2 или 3 более или менее выступающие вперед лопасти, разделенные глубокими вырезами; поверхность спинных щитов покрыта густой скульптурой из узких ланцетовидных ямок, имеющих поперечное .направление на проподосомальном щите и продольное — на гистеросо-
1 .Вероятна, представляет собой эпигииий.
392
III. GLYCYPHAGIDAE. 7. CHAETO DACTYLIN A E
мальном; промежутки между ямками густо- и тонкопористые. Щетинки-спинной поверхности частью (sc е, h. а и h р и иногда sc i, 1 а и 1 р) игольчатые, умеренной длины, частью короткие и тонкие; р а довольно короткие, расположенные вентрально па завернутом книзу участке гистеросомального щита. Строение щетинок брюшной поверхности изменчиво. Гнато-сома обычно представлена парой округлых или грушевидных лопастинок, несущих пару коротких арист. Стернум в большинстве случаев простой (только у Ch. claviiier вильчатый); строение дру их частей коксо-стерналь-пого скелета, в общем, как у Sennertia. Присасывательный диск маленький, без свободной перепончатой каймы. Ноги I—III очепь мощные, узловатые, с огромными винтообразными коготками, сидящими на конце длинных мясистых, складчатых praetarsi; лапки I—III короткие, обрывисто суживающиеся к концу, незаметно переходящему в praetarsus. Ноги IV обычно’ много слабее ног I—III, оканчивающиеся длинной парусной щетинкой и 2 более короткими, лишенные амбулакр. Дорзальное вооружение лапок I такое же, как у рода Sennertia.
К роду Chaetodaetylus относятся, помимо описываемых здесь 6 палеарктических видов, еще Ch. ludwigi Trouess. 1897 с Каролинских островов и; несколько еще не описанных неарктических видов.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
Гипопусы
1 (2). Стернальные щетинки превращены в массивные яйцевидные шипы, вентральные — сильно конически утолщенные; стернум на конце раздвоенный; ноги I и II сильно утолщенные; конец их лапок вен-
•* трально с очень крупным искривленным шипом, лапки IV с 3 очень, крупными парусными щетинками, Очень крупная форма: длина. 370—450 р,. На Osmia (Ceratosmia) tricornis Latr......... . .
.......................................4. Ch. claviger Ouds..
2 (1). Стернальные и вентральные щетинки очень тонкие, волосовидные;, стернум простой; ноги I и Пне утолщенные, конец их лапок без шипа; лапки IV с 1 очень крупной (парусной) и 2 гораздо более короткими;, и тонкими щетинками (иногда последние представлены микрохэтами).. Длина тела не превышает 375 р,.
3 (8). Наружные присоски расположены по бокам центральных (образуя, с ними поперечный ряд), много впереди задних.
4 (5). Sc i крепкие, имеющие такую же длину и толщину, как h i, и почти: не уступающие в размерах sc е, расположенные на одной поперечной;
Фиг. 669—677.
Фиг. 6в9. Sennertia potanini, sp. ц., гипопус, общий вид со спинной стороны. Фиг. 670. S. potanini, sp. п., гипопус, задний край тела и присасывательный диск (снизу). Фиг. 671. S. potanini, sp. н., гипопус, конец ноги IV снизу. Фиг. 672. S. alfkeni (Ouds.), гипопус, общий виц сверху. Фиг. 673. То же, вид снизу. Фиг. 674. S. alfkeni (Ouds.), гипопус, колено, голень и лайка! сбоку. Фиг. 675. S. oudemansi, sp. п., гипопус, общий вид сверху. Фиг. 676. То же, вид снизу. Фиг. 677.
S. oudemansi, sp. п., гипопус, конец ноги IV (лапка + дистальная часть голени).
:=^1Р тг-
394
Ш. GLYCYPHAGIDAE. 7. CHAETODACTYLINAE
линии с обеими парами плечевых (h i и h е); d3, sa е и sa i очень жест* кие, игольчатые; внутренняя вершинная щетинка лапок IV в 3 раза длиннее всей ноги IV и наружной вершинной щетинки. На Chelo-stoma florisomnis L..........................*3. Ch. birulai, sp. n.
5 (4). Sc i много короче и тоньше, чем h и sc е, расположенные значительно впереди поперочноЯ линии, проведенной через основания плечевых (h); d3, sae и sa i топкие, гибкие; внутренняя вершинная щетинка лапок IV по более чем в 2 раза длиннее ноги IV и наружной вершинной щетинки. На видах Osmia.
6 (7). Внутренняя вершинная щетинка лапок IV заметно короче ног IV и чуть длиннее наружной; sc е короче (10—11% длины тела), только в 1.5 раза длиннее sci; внутренняя субапикальная щетинка лапок I—III и наружная лапок III очень слабо ланцетовидно расширенные ...............................................*1. Ch. osmiae (Duf.).
7 (6). Внутренняя вершинная щетинка лапок IV в 2 раза длиннее ног IV и наружной вершинной щетинки; sc е длиннее (около 20% длины ' тела), в 3—4 раза длиннее sc i; внутренняя субапикальная щетинка лапок I—III и наружная щетинка лапок III сильно ланцетовидно расширенные. . .................................. .*2. Ch. reaumuri(Ouds.).
-8 (3). Наружные присоски расположены позади центральных, -на одной поперечной линии с задними. -
D (10). Длина тела около 350 р; спинные щиты светлокоричневые, с продольной ямчатой скульптурой;'присасывательный диск длиннее своей ширины. На Anthidium sticticum F.................5. Ch. anthidii (Ouds.).
10 (9). Длина тела 215—310 p; спинные щиты не пигментированные, с неправильным, нежным, сетчатым рисунком; присасывательный диск немного шире своей длины. На некоторых видах Megachile и на Pseudovespa germanica L..........................*6. Ch. dementievi, sp. n.
*1. Chaeto dactylus osmiae (Duf.) (фиг. 78, 79, 644—650, 678).
L. Dufour, 1839, Ann. Sci„ Nat., s6r. II, vol. XI:276, t. VIII, fig. 3; Donnadieu, 1868, ibid., зёг. V, vol. X:69 (Triihodactyhts)-, Canestrini, 1888a:394; Berlese, 1897:106; Kranier, 1899:149; Michael, 1903:17, t. XXII, XXXIX, fig. 13—15 (Trichotamis).
Биология. Trouessart, 1904, 0. R. Soc. Biol., LVI:241, 355; Popovici-Baznoseanu, 1913, Arch, zool., LII, Notes et Revue:33.
Прижизненная окраска очень бледножелтоватая, причем желтый оттенок более заметен в передней и на боковых частях идиосомы; гнатосома и ноги светлокрасноватые. Идиосома в 1.4—1.5 раза длиннее своей ширины. V а расположены вблизи переднего края идиосомы, d2 и d3 сильно редуцированы (или отсутствуют?); примерные размеры щетинок, в процентах к длине идиосомы (у $): v а и sc i 24—25; sc е 22; h а и sh 26—28; h р 33; dx 30; d4 22; lx 33—35; 12 22; 13 15—17; sa e около 100%. Щетинки ног гладкие. Дорзальная вершинная щетинка лапок длинная, особенно на ногах III и IV. Когбтки умеренной величины. Общая длина тела с? около -450 р, ширина 270 р; длина $ около 500 р, ширина 320 р.
21. CBAETODACTYLUS
395
Pповелительный гипопус. Спинные щиты плотные, сильно пигментированные, с типичной для рода ямчатой и пористой скульптурой; передние углы гистеросомального щита обычно оттянуты в виде узких треугольных выступов. Щетинки спинной поверхности довольно короткие; sc е, hi и h е жестковатые, длиннее и толще других; sc е в 1.5 раза длиннее sc i, помещающихся немного впереди линии, проведенной через основания h е; последние приблизительно равны по длине sc е (10—11% длины идиосомы) и заметно длиннее h i (8—9%); все остальные щетинки спинной поверхности длиной в 3—5% длины идиосомы, довольно тоийие и гибкие; щетинки брюшной поверхности тела тонкие, волосовидные. Коксо-стернальный скелет умеренно развит; стернум простой. Вырез завернутого на брюшную сторону участка гистеросомального щита с хорошо развитым медиальным выступом; боковые присоски расположены на одной попе-* речной линии с центральными; задние присоски такого же размера, как боковые, расположенные на овальной хитиновой площадке, отделенной широким промежутком от боковых склеритов (несущих боковые присоски), передние концы которых оттянуты в серповидный вырост. Ноги I—III умеренной толщины, вооруженные длинными, гибкими щетинками; дорзальная вершинная щетинка колен короче голени, субапикальныо щетинки лапок не расширены на конце; наружная щетипка колен Ш очень длинная. Ноги IV тонкие, значительно короче остальных, оканчивающиеся 1 очень длинной парусной и 2 короткими щетинками, из которых внутренняя заметно короче наружной, цо длине значительно уступающей таковой ноги IV. Длина 280—375 у, ширина 220—290 р..
Покоящийся гипопус (по Trouessart’y). Широкоэллип-«оидальной формы, по общему виду очень напоминает покоящихся гипопусов рода Glycyphagus. Ноги I и II конические, явственно сегментированные, ноги III и IV представлены кольцевидными образованиями, напоминающими присоски. Нет никаких следов присасывательного диска и генитальных присосок.
Распространение. Франция, Италия, Англия, Румыния, СССР. У нас пока найден только в окрестностях Сарепты и в Грузии (Тбилиси).
Экология. Живет в гнездах Osmia (Ceratosmia') rufa L., О. (С.) tricornis Latr. и 0. kohli Ducke, питаясь запасами пыльцы; Michael’ю удалось вывести взрослых из нимф, кормя их старым пчелиным воском.
*2. Chaetodactylus reaumuri (Ouds.) (фиг. 679).
Qudemana, 1906, Tijdschr. Entomol,, XLVIII: XXXI (Trichotarsus).
Гипопус. По общему виду и размерам очень напоминает гипопуса Ch. osmiae. See значительно длиннее, достигающие 18—20% длины тела; sc i в 3—4 раза короче sc е, очень тонкие; h равны 15—17% длины тела; 1аи1р несколько толще, чем у Ch. osmiae-, все d представлены тонкими микрохэтами, длиною в 4—5%. Задние и боковые присоски мельче (их диаметр в 2 раза меньше внутреннего диаметра хитиновой рамки вен-1
396
III. GLYCYPHAGIDAE. 7. CHAETODACTYLINAE
тральных присосок), задние сидят па 2 раздельных, тесно сближенных друг с другом хитиновых площадках; направленные вперед отростки боковых склеритов присасывательного диска длиннее и тоньше. Внутренняя суба-пикальпая щетипка лапок 1 и 11 и обе крайние субапикальные щетинки лапок III явственно ланцетовидно расширенные; наружная щетинка колен III короткая; внутренняя вершинная щетинка лапок IV длинная, в 2 раза длиннее пог IV и наружной вершинной щетинки. Длина 275— 375 ц, ширина 210—315 [>.. Другие стадии развития неизвестны.
Распространение. Известен из Германии и из Крыма (Артек) и Грузии (Тбилиси). Гипопусы встречаются па Osmia рапгегг F. Мог. и О. leucogastra F. Мог.
*3. Chaetodactylus birulai, sp. n. (фиг. 680).
Гипопус. Габитуально близок к обоим предыдущим. Почти все щетинки спинной поверхности однотипные — жесткие и игольчатые; sc е длиною в 18—20%; sc i чуть короче sc е (16%), но такой же толщины; dr и d2 относительно крупные (12—14%), равные по длине и крепости 1 а. и 1 р; d8 и d4 в виде тонких микрохэт; sa е довольно длинные (10—11%), толстые, искривленные, как и sa i. Расположение й относительные размеры присосок такие же, как у Ch. osmiae, но хитинизированные площадки, па. которых сидят задние присоски, не обособлены от боковых полуколец,, несущих наружные присоски. Ноги I и II неутолщенные; вершинные щетинки голеней короткие; наружная субапикальная щетинка лапок I и II и обе крайние субапикальные щетинки лапок III дистально очень слабо ланцетовидно расширены; внутренняя вершинная щетинка лапок IV в 3 раза длиннее наружной щетинки и всей ноги IV. Длина 290—335 р,, ширина. 230—265 |а. Прочие стадии развития неизвестны.
Распространение. Найден пока только в Кировской обл.г Лавы.
Экология. Гипопусы собраны в довольно большом числе на одном экземпляре Chelostoma florisomnis L. При осмотре серии этого вида в коллекции Зоологич. Института Акад. Наук СССР мне не попалось больше, ни одного гипопуса Ch. birulai.
4. Chaetodactylus claviger Ouds. (фиг. 681).
Oudemans, 1928, Entomol. Bor., VII:328.
Гипопус. Легко отличается от других видов своими крупными размерами и своеобразным, мощным вооружением брюшной поверхности тела и ног. Спинные щиты плотные, довольно сильно пигментированные, с типичной для рода ямчатой и пористой скульптурой; передние углы гисте-росомального щита закругленные. Щетинки спинной поверхности тела длин-
Фиг. 678—682.
Фиг. 678. Chaetodactylus osmiae (L. Duf.), гипопус сверху. Фиг. 679. Ch. rdaumuri (Ouds.), гипопус сверху. Фиг. 680. Ch. birulai, ер. п., гипопус сверху. Фиг. 681. Ch. claviger Ouds., гипопус снизу. Фиг. 682. Ch. dementjevi, sp. п., гипопус сверху.
398 III. GLYCYPBAGIDAE. 7. CBAETODACTYLINAE
нее, чем у других видов, жесткие, игольчатые; sc е и sc i одинакового размера (около 21—22% длины тела); sc i помещаются значительно впереди: линии, проведеппой через основания h; последние немного длиннее sc (23—24%); dx относительно очень длинные (16—17%); 1 а, I р и d2 примерно-одинаковой длипы (около 12—13%); d3, sa е и sa i заметно тоньше и короче всех других щетинок спинной поверхности; щетинки брюшной поверхности тела сильно модифицированы: стернальные превращены в крупные, массивные, яйцевидные образования, сидящие па короткой ножке, вентральные сильно конически утолщены к основанию, шиповидные; sh и р а тоже значительно толще, чем у других видов. Коксо-стернальпый скелет очень, мощный; стернум вильчато раздвоенный на конце; также раздвоены и концы эпимер II, причем короткие внутренние ветви их развилка почти соприкасаются с вершинами развилка стернума, а длинные наружные дугообразно отогнуты назад и в стороны; эпимериты II, эпимеры III и ГУ и краевые утолщения коксальных полей гораздо более мощные, чем у остальных, представителей рода Chaetodactylus. Вырез завернутого на брюшную поверхность края гистеросомального щита почти прямоугольный, без медиального выступа; задние и боковые присоски очень мелкие, выстроенные в почти прямой поперечный ряд позади центральных, сидящие все на одной и той же нерасчлененной хитиновой площадке своеобразной формы. Ноги I и. II чрезвычайно толстые, усаживающие их щетинки крепче, чем у других видов; вентральные щетинки бедер короткие, дорзальная вершинная щетинка; колен длиною с соответствующую голень; наружная и внутренняя предвершинные щетинки лапок практически пе расширенные; вершина лапок с очень., крупным, слегка искривленным шипом; наружная щетинка колен III очень, длинная. Ноги IV развиты гораздо сильнее, чем у прочих видов этого рода, не уступающие в длине ногам III (без их амбулакр), достаточно массивные,, оканчивающиеся 3 очень крупными щетинками, из которых внутренняя менее чем в 1.5 раза короче средней и лишь немного длиннее наружной. Длина 370—450 ширина 290—350 ;г. Прочие стадии развития неизвестны.
Распространение. Италия. Гипопусы встречаются только на. Osmia (Geratosmia) tricornis Latr., иногда совместно с таковыми Ch. osmiae.
5. Chaetodactylus anthidii (Ouds.).
Oudemans, 1911, Entomol. Ber., 111:165 (Triehotarsus).
Гипопус.. Мне в натуре неизвестен. Близок к Ch. osmiae и Ch., reaumuri. Отличается от обоих, между прочим, следующими признаками: лапки I и II несут только по 1 крайне трудно заметному ланцетовидному волоску; светлокоричпевый задний спинной щит усажен (так же как: у названных видов) тысячами чечевицеобразных ямок. Присоски на. присасывательной пластинке расположены иначе: впереди 2 маленьких,, затем 2 крупных, за ними 4 маленьких (у названных видов 2, 4, 2); кроме-того пластинка длиннее своей ширины. Длина около 350 fi. (По Oudemans’у.)
21. CHAETODACTYLUS. 22. HORSTIA
399:
Распространение. Тунис. Ha Anthidium sticticum F. На просмотренных мною экземплярах этой пчелы из Французского Марокко и Испании не было ни одного гипопуса Ch. anthidii.
*6. Chaetodactylus dementjevi, sp. n. (фиг. 682).
Гипопус. "Мелкая форма. Окраска очень бледная, спинные щпты нежные, почти не пигментированные, с тонким рисунком, составленным из более или менее волнистых вдавленных линий, местами соединяющихся друг с другом и образующих неправильные петли. Щетинки спинной поверхности довольно короткие и тонкие; sc е длиною в 15—16% длины идиосомы, в 3—4 раза длиннее sc i, представленные тонкими макрохэтами; обе пары h одинакового размера (14—16% длины тела); 1 а и I р заметно короче-плечевых (10—13%); все остальные щетинки в виде тонких, гибких микрохэт, длиною около 3—4% длины тела; щетинки брюшной поверхности тела очень, тонкие, волосовидные. Коксо-стернальный скелет слабо развитый; стернум простой, край завернутой вниз части гистеросомального щита почти прямой; задние и боковые присоски очень мелкие, расположенные в прямой-поперечный ряд позади центральных. Ноги I и II умеренной толщины; почти все усаживающие их щетинки очень тонкие и гибкие; вентральные щетинки бедер длинные; обе крайние субапикальпые щетинки лапок I—III нерасширенные, волосовидные; ноги IV короткие, очень стройные, оканчивающиеся 1 крупной (парусной) щетинкой и 2 микрохэтами, сидящими по бокам у ее основания. Длина 215—310 ширина 170—275 ;л. Прочие стадии развития неизвестны.
Распространение. Известен мне из окрестностей Ленинграда (Парголово), Углича, Усть-Каменогорска и с гор Ак-таш вблизи Ташкента. Гипопусы встречаются преимущественно на Megachile bombycina Rad. и М. ligniseca К1., но 1 Экз. был найден также па осе Pseudovespa germanica L. (Ак-таш).
22. Род HORSTIA OUDS.
Oudemana, 1905, Entomol. Ber., 11:21.
Тип рода: Trichotarsus ornatus Ouds. 1900.
Половозрелая стадия известна только у 1 из видов—у новогвинейской Н. trifilis Сап. 1897 —и описана очень поверхностно; она отличается от видов рода Sennertia, на которых, невидимому, довольно похожа, отсутствием генитальных щупальцев у $ и большей длиной женского генитального отверстия.
Гипопус. Идиосома продолговатая (в 1.4—1-.6 раза длиннее своей ширины), приблизительно яйцевидной формы, довольно сильно выпуклая; спинные щиты покрывают почти всю ее дорзальную поверхность, по не загибаются на вентральную; они довольно плотно хитинизировапы и украшены тонким рисунком из частично анастомозирующих вдавленных линий. Щетинки спинной поверхности представлены крайне мелкими, трудно
400 III. GLYCYPHAGIDAE. 7. CBAETODACTYLINAE
заметными микрохэтами (только у описанной из Венецуэлы Н. pulcherrima Vtz. 1912 они хорошо развиты и некоторые из них даже очень длинны). Гнатосома слегка обособлена от идиосомы, в виде подушковидной выпуклости. Стернум па конце внльчато раздвоен; концы его развилка часто слиты с эпимерами П и замыкают, таким образом, коксальные поля II; -коксальные поля III и IV обычно замкнуты благодаря слиянию концов эпимер II с эпиморитами II и эпимер III с эпимерами IV, и в таких случаях они раз'ьсдииепы ио средней линии тела обрамленным ими нродолговатым вентральным иолом. У ряда видов все эпимеры свободны, и коксальные поля, таким образом, открытые. Коксальные и генитальные щетинки редуцированные, представленные маленькими кружками. Присасывательный диск в большинстве случаев большой, с очень широкой прозрачной каймой, сильно выступающей за край идиосомы, и 3 парами крупных присосок. Ноги очень короткие, массивные, почти конические, все с очень короткими, почти прямыми коготками; praetarsi отсутствуют; щетинки колен I и II короткие; лапки I— III ие длиннее голеней, с 4 крупными листовидными щетинками; дорзальное вооружение лапок I состоит только из обонятельной палочки, второй, расположенной рядом с нею, палочковидной сенсиллы и 1 вершинной щетинки; лапки IV чрезвычайно короткие, заканчивающиеся 2 крупными и 3 более мелкими простыми щетинками.
К роду Horstia Ouds. относятся 5 видов, распространенных преимущественно в тропиках. Виды Horstia с свободными эпимерами (Н. helenae .Ouds., Н. glabra Vtz.) неправильно помещаются некоторыми авторами в род Tortonia Ouds.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
Гипопусы
1 (2). Крупная форма: длина 334 р., ширина 210 р.; все коксальные поля замкнутые, благодаря тому, что стернум сливается концами своего развидка с эпимерами II, последние, в свою очередь, — с эпи-меритами II, эпимеры III — с эпимерами IV, а эти последние — друг с другом, образуя короткий вентрум. На Xylocopa circum/oolans Sm. ...............................................1. Н. ornata (Ouds.).
;2 (1). Мелкая форма: длина 165—180 р.; все коксальные поля открытые: эпимеры II—IV, эпимериты II и стернум свободные. На Xylocopa dissimilisLep. и Platynopoda tenuiscapa Westw. 2. H. helenae (Ouds.).
1. Horstia ornata (Ouds.) (фиг. 651, 652).
Oademang, 1900, Tijdgehr. Entomol., XLIII:117, t. 6, fig. 22—23 (Trichotarsus)', 1905 Entomol. Ber., 11:21. (Horstia).
Крупная форма. Идиосома в 1.6 раза длиннее своей ширины, равномерно суживающаяся кзади, начиная от плеч; боковые края гистеросомы на большом протяжении почти прямые; v а довольно длинные; р а длиною в 20—
22. HORSTIA. 23. TORTONIA
401
22% длины идиосомы. Все коксальные поля замкнуты; стернум слипается концами своего развилка с эпимерами II, последние, в свою очередь, — •с-эпимеритами II, эпимеры III —с эпимерами IV, а эти последние — друг с другом, образуя короткий вентрум. Наружные присоски брюшного диска, много меньше центральных и расположены значительно впереди них; задние заметно крупнее наружных (но все же порядочно уступают в размерах центральным), раздвинуты гораздо шире, чем центральные, и отделены от них расстоянием, равным их диаметру. Листовидные щетппкп лапок I и II весьма широки; некоторые из них имеют довольно явственную среднюю жилку. Длина 334 [у ширина 210 [л.
Распространение. Япония (Кобе). На Xylocopa circumvo-lans Sm.
2. Horstia helenae (Ouds.) (фиг. 702, 703).
Oudemans, 1902, Tijdschr. Entomol., XLV:144, t. 12, fig. 44—45 (Trichotarsus); 1905> Entomol. Ber., 11:21 (Horstia)-, 1912, ibid., 111:166 (Tortonia).
Мелкая форма. Идиосома довольно правильной яйцевидной формы в 1.4—1.5 раза длиннее своей ширины; боковые края гистеросомы выпуклые на всем своем протяжении; v а и р а очень короткие. Все коксальные поля открытые: эпимеры II—IV, эпимериты II и стернум свободные, не соединены концами фуг с другом; вентрум отсутствует. Наружные присоски брюшного диска одинакового размера с центральными, расположенные почти на одном уровне с ними; задние несколько меньше наружных, отделены друг от друга приблизительно таким же расстоянием, как центральные, и близко придвинуты к ним. Листовидные щетинки лапок заметно уже. Длина 165—210 |л, ширина 110 ft.
Распространение. Япония, Индия, Ява. На Platynopoda tenuiscapa Westw. и Xylocopa dissimilis Lep.
23. Род TORTONIA OUDS.
Oudemans, 1911, Entomol. Ber., Ill : 166.
Тип рода: Trichotarsus intermedins Ouds. 1901.
Известны только гипопусы. Тело продолговатое, грушевидное или в форме вытянутого в длину шестиугольника, сильно выпуклое (у Т. intermedia) или уплощенное; спинные щиты то покрывают всю верхнюю поверхность тела, то более или менее сильно редуцированы и обнажают широкие участки мягкой, резко штрихованной кожи; длина щетинок спинной поверхности изменчива, но по крайней мере sc е представлены макрохэтами. Органы зрения отсутствуют. Гнатосома совершенно йе обособлена от идиосомы, представленная одними аристами. Стернум простой, свободный, так же как и эпимеры II—IV; последние мало искривлены и отделены большим пространством друг от друга; все коксальные поля, таким образом, широко открыты. Коксальные и генитальные щетинки в виде топких микрохэт.
26 Фауна СССР, Захватит!—284
502
III. GLYCYPHAGIDAE. 7. CHAETODACTYLINAE
Присасывательный диск небольшого размера, с несколько выступающей за края тела перепончатой каймой; дополнительные присоски иногда хорошо* развиты. Ноги средней длины или довольно короткие, I—III с маленьким, едва изогнутым коготком, отсутствующим на ногах IV; щетинки колен I и II короткие; лапки 1 и 1.1. с 3 крупными листовидными щетинками,, тонкой прямой обонятельной палочкой, 1 крупной дорзальной щетинкой,, палочковидной вершинной щетинкой над основанием коготка и 1 простой вентральной щетинкой; лапки III с Г> листовидными щетинками или вовсе без них; лапки IV пссут па конце очень длинную парусную щетинку, 2 более короткие и 1 или 2 совсем короткие щетинки.
Род Tortonia известен только из пределов Палеарктики; он состоит из 4 видов (из которых один, Т. fluctuata, очень недостаточно описан),, естественно распадающихся на 2 группы, которым я провизорно придаю значение подродов.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ
1 (2). Идиосома грушевидная, сильно пузыревидно вздутая; спинные щиты одевают всю ее дорзальную поверхность, покрыты резким сетчатым рисунком, местами приобретающим характер рельефных чешуй;. лапки I и II длинные, стройные, коготки относительно длинные и тонкие; sc выстроены в поперечный ряд; псевдостигмальный орган хорошо развитый. На видах Osmia и Stelis. (Подрод Tortonia s. str.). .................................... *1. Т. (s. str.) intermedia (Ouds.).
2 (1). Идиосома продолговато-шестиугольная, уплощенная дорзо-веитраль-но; спинные щиты покрывают только часть спинной поверхности (периферические части ее имеют мягкие перепончатые покровы) г с слабым продольным рисунком; лапки I и II короткие, массивные; коготки короткие и толстые; sc i расположены заметно впереди sc е;. псевдостигмальный орган отсутствует. На видах рода Anthidium. (Подрод Donnadieuia, subg. н.)
3 (4). Sc i, hi, dx—d2 и обе пары 1 (1 а и I p) в виде довольно длинных (13—28% длины идиосомы) мечевидных щетинок; sc е чрезвычайнодлинные (около 45% длины идиосомы), заходящие концами за основания 1 а; спинные щиты более узкие, покрывают небольшую часть, дорзальной поверхности идиосомы; задняя вершинная щетинка лапок IV короче ноги. На Anthidium septemdentatum Latr...............
....................................4. Т. (D). dogaressa, sp. n,
4 (3). Sc i, hi, dx—d3, la и Ip представлены очень мелкими, трудно заметными микрохэтами; sc е умеренной длины (20—25% длины идиосомы), доходящие концами только до основания d2; спинные щиты шире, покрывают несколько большую часть дорзальной поверхности идиосомы; задняя вершинная щетинка лапок IV больше чем вдвое-длиннее ноги IV. На Anthidium sticticum F.........................
...............................3. Т. (D.) smitsvanburgsti Ouds.
23. TORTONIA 403
1. Подрод Tortonia s. str.
Тело более или менее грушевидной формы, сильно вздутое, целиком покрытое сверху обоими спинными щитами; поверхность последних с резким сетчатым орнаментом, местами приобретающим характер рельефных чешуй; sc выстроены в поперечный ряд; есть псевдостигмальный орган; дополнительные присоски хорошо развиты; ноги стройные, с длинными лапками и тонкими коготками; число листовидных щетинок па лапках изменчиво.
*1. Tortonia (s. str.) intermedia (Ouds.) (фиг. 683—686).
Oudemans, 1902, Tijdsehr. Ned. Dierk. Ver., VII :308, t. X, fig. 49—60 (Triehotarsus); 191.1, Entomol. Ber., 111:166 (Tortonia).
Желтовато-(хитиново-)бурый. Тело в 1.45—1.6 раза длиннее своей ширины, грушевидное, сильно суживающееся к своему несколько уплощенному заднему концу; проподосомальный щит в 2.2—2.4 раза короче гистеросомального. Sc е крепкие, прямые, длиною около 20% длины тела; h е сходны с ними по строению, но значительно короче (12—14%); все остальные щетинки спинной поверхности тонкие, волосовидные, длиною в 4—8% длины тела. Длина ног составляет приблизительно 40—42 (I), 38 (II), 30—32 (III) и 16—18 (IV) % длины тела; лапки I и II более чем'в 3 раза длиннее своего диаметра у основания, с 3 крупными, но довольно узкими ланцетовидными щетинками; на лапках III находятся 2 такие же крупные листовидные щетинки и 3 гораздо меньших. Длина 195—220 р., ширина 120—155 [1.
Распространение. Описан из Германии;' мне известен еще из Швейцарии (кантон Vaud) и следующих пунктов СССР: Лужского р-на Ленинградской обл., Сибири (бассейн р. Нелькан) и Грузии (Боржоми).
Экология. Гипопусы встречаются на различных видах Osmia [О. leeana Kby., О. solskyi F. Mor., 0. nigriventris (Zett.) Nyl.] и на паразитирующих в их гнездах видах Stelis (например S. phaeoptera Latr.).
2. Tortonia (s. str.) fluctuata Ouds.
Oudemans, 1917, Entomol. Ber., IV: 391.
Гипопус. Мне в натуре неизвестен. Длина 186р., ширина 122р.. Отличается от Т. intermedia Ouds., между прочим, следующими признаками: ноги I и II длиннее; на заднем конце имеется более темный киль; лапки I и II имеют только по 1 ланцетовидному волоску, лапки III — пи одного; присасывательная пластинка маленькая, несет 10 присосок и не выдается за задний край. На Bombus proteus Gerst. var. tricolor Alfk. Германия (Времен). (По Oudemans’y.)
2. Подрод Donnadieuia, subg. n.
Тело имеет форму вытянутого в длину шестиугольника, уплощенное; спинные щиты покрывают только центральную часть спинной поверхности
404 111. (ILYCYPllAfilbAE. CHAETODACTYLINAE
тела; оставляя открытыми nii-ij>0K.uc участки мягкой, перепончатой, грубо штрихованной кожи; орнамент щитов нерезкий, обычного типа; sc расположены в виде трапеции; лсевдостигмальный орган отсутствует; дополнительные присоски слабо развиты; ноги-короткие и толстые, с короткими лапками и толстыми, почти коническими коготками; лапки 1 п II с 3, лапки III с Г» широкими листовидными щетинками.
2 вида, биологи чески связанных с, видами рода Anthidiiim.
3. Tortonia (honnadleula) smiIsvniilnirgsti Ouds. (фиг. 686, 704, 705).
Гипопус. Тело бледно окрашенное, в 1.6—1.7 раза длиннее своей Ширины; спинные щиты занимают довольно большуи? площадь дорзальной поверхности, слегка пигментированные. Sc е длиною в 2'3—25%'длины тела, гибкие, слегка лентовидно уплощенные,' доходящие концами- до основания (%; h е имеют в длину окОлоТб—16% длины тела; все остальные щетинки спинной поверхности представлены тонкими микрохэтами. Длина йбг составляет приблизительно 30—31 (I и II), 23 (III) й 16—18 (IV) % длины тела; лапки I и 11 не более чем в 2 раза' длиннее своего диаметра у основания; их волосовидная вентральная щетинка расположена на одинаковом расстоянии от основания и конца членика; на лапках IV задняя (3-я) крупная щетинка значительно длиннее всей ноги. Длина 230—280 р. ширина 140—175 [л.
Распространение. Описан из Туниса; мне известен еще из Алжира и Испании. Гипопусы встречаются исключительно на Anthidium sticticum F. -
; 4. Tortonia (Donnadieuia) dogairessa, sp, п. (фиг. 687).
Г и п опус. Близок к предыдущему. Спинные щиты занимают еще Мёйьшую площадь, оставляя открытыми более широкие участки перепон-'чатой, резко штрихованной кожи. Sc е-чрезвычайно длинные (около 45% длины тела), заходящие концами за основания la; he длиною около 20—22%; sc i, hi, dx—d3, la и 1р довольно длинные (13—18% длины тела), несколько лентовидно уплощенные, как и see и he; dx и обе пары sa представлены микрохэтами. Вентральные щетинки лапок I и •II придвинуты к основанию л апки; 3-я (задняя) крупная щетинка лапок TV короче ноги и заметно утолщена. Длина 175—220 р, ширина 120—145 р. Р а с п р о с т р а ц о и и с. Северная Италия: окрестности Венеции. Биологически связан с пчелой Anthidium seplemdentatum Latr., на которой его гипопусы найдены в очепР большом количестве.
Фиг. 683—696. Гипопусы родов Tortonia, Neoliorstia и Cerophagtis.
Фиг. 683. Tortonia intermedia (Ouds.), общий вид сверху. Фиг. 684. Т. intermedia (Ouds.), лапка I снизу. Фиг. 685. T. intermedia (Ouds.), лапка III снизу. Фиг. 686. Т. smitsvanburgsti Ouds., общий вид снизу. Фиг. 687. Т. dogaressa, sp. п., общий вид сверху. Фиг. 688. Neohorstia mamillata, sp. n., общий вид сверху. Фиг. 689. То же, общий вид снизу. Фиг. 690. jV. mamillata, sp. п., нога I сверху. Фиг. 691. N. mamillata, sp. n., голень и лапка I снизу. Фиг. 692. N. mamillata, sp. n., лапка IV сверху. Фиг. 693. Cerophagus granulatus (Duj.), общий вид сверху. Фиг. 694. То же, общий вид енцзу. Фиг. 695. С. granulatus (Duj.), лапка I снизу. Фиг. .696. С. granulatus (Duj..), конец лапки IV.
406 III. GLYCYPHAGIDAE. 7. CHAEIODACTYLINAE
24. Род NEOHORSTIA, gen. n.
Тип рода: Neohorstia mamillata, sp. n.
Известен только по гипопусам. Тело яйцевидное, довольно сильно выпуклое; спинные щиты покрывают всю его дорзальную поверхность, очень плотные, в грубой ямчатон скульптуре, состоящей пз глубоких лунок и штрихов, ориентированных более, пли менее параллельно краям тела; промежутки между ними тонкоиористые; на вершине рострального выступа помещается орган зрения, представленный слитыми в одно целое, сильно пигментированными хитиновыми ретинами. Щетинки спинной поверхности очень длинные; самые крупные из пих (sc, hi, dx—d3, la) несколько лентовидно уплощены. Гнатосома развита лучше, чем у других родов подсем. Glwetodactylinae, и имеет вид невысокой пластинки, расщепленной по средней линии и несущей дистально пару четко обособленных цилиндрических члеников, заканчивающихся аристами. Стернум на конце раздвоён в форме буквы Т; эпимеры II резко искривлены и почти соприкасаются концами с крепкими эпимеритами II, практически замыкая коксальные поля II; таким же образом искривлены и эпимеры III, также почти наглухо замыкающие соответствующие коксальные поля; концы эпимер IV отогнуты назад и тянутся, постепенно сходясь кзади, почти до генитальной щели; краевые утолщения всех коксальных полей очень широкие. Коксальные и генитальные щетинки превращены в массивные, короткостебельчатые яйцевидные образования и расположены на территории краевых утолщений коксальных щитов. Присасывательный диск крупный, без свободной перепончатой каймы, с 4 парами присосок. Ноги умеренной длины, довольно массивные, все с одинаково развитыми, небольшими, слегка крючковидно загнутыми на конце сидячими коготками, лишенными мясистого praetarsus; щетинки колен I и II средней длины; лапки I и II с 5 крупными листовидными щетинками, III — с 4 более узкими ножевидными щетинками, которые имеются также в числе 3 па лапках IV; лапки III с довольно длинной парусной щетинкой; лапки IV равны по длине 2 предыдущим членикам ноги, вместе взятым, снабженные 2 длинными парусными щетинками.
Известен 1 вид.
1. Neohorstia mamillata, sp. п. (фиг. 688—692).
Гипопус. Беловатый, с очень слабым охристо-желтым оттенком, более явственным в задней трети спины. Тело в 1.4—1.5 раза длиннее своей ширины, с полого закругленным, слегка притупленным и уплощенным задним концом. Проподосомальный щит немного более чем в 2 раза короче гистеросомы; sc очень длинные, гибкие, лентовидно уплощенные; длина see составляет около 30% длины тела и в 1.5 раза превосходит длину sci; hi, dx—d3 и 1 а длиною около 17—20% длины тела, такого же строения, как sc; h е, d4, 1 р и sa представлены топкими волосовид-
24. NEOHORSTIA. 25. CEROPHAGUS
407
яыми щетинками. Хитинизированная пластинка присасывательного диска образует 3 раздельные овальные площадки. Лапки короткие и толстые, I и II только в 1.5 раза длиннее своего диаметра у основания, лапки II I ,и IV едва длиннее его; обонятельная палочка лапок I и II прямая, постепенно суживающаяся к концу. Длина 200—262 р., ширина 144—180 р..
Распространение. Греция (Аттика), Иран (Хаджи-абад). Гипопусы найдены на Anthidium pubescens F. Мог. (Иран) п A. laticeps F. Мог. (Греция).
25. Род CEROPHAGUS OUDS.
Oudemans, 1904, Tijdsehr. Entomol., XLVI:12.
Тип рода: Hypopus granulatus Duj. 1849.
Известен только по гипопусам. Тело широкояйцевидное, довольно «сильно выпуклое, с сердцевидно вогнутым задним краем; спинные щиты покрывают всю его дорзальную поверхность и сзади слегка загибаются на брюшную, очень плотные, с чрезвычайно резкой однообразной скульптурой, состоящей из округлых и овальных, четко очерченных ямок, пролизывающих всю толщу хитина (так что они кажутся продырявленными подобно ситу); промежутки между ямками тонкопористые. Щетинки спинной стороны тела представлены мелкими, очень трудно заметными микро-хэтами. Органы зрения отсутствуют. Гнатосома слабо обособленная, в виде пары низких кольцевидных члеников, несущих на конце крепкие аристы. «Стернум простой, свободный, как и все эпимеры; все коксальные поля -открытые, разобщены по средней линии тела очень широким центральным полем; эпимериты II соединены с эпимерами 'III, несущими небольшой, направленный кпереди отросток. Коксальные и генитальные щетинки представлены тонкими микрохэтами. Присасывательный диск небольшой, квадратной формы, значительно удаленный от заднего края тела; обе задние присоски расположены на обособленной полукруглой хитиновой площадке; аподема присасывательного диска имеет продольное направление. Ноги все одинаково развиты, довольно стройные, с крупными, резко крючковидно изогнутыми коготками; praetarsi толстые, вентрально с зубовидным отростком; лапки I и II с 5, остальные с 4 сравнительно крупными листовидными щетинками; лапки III и IV без парусных щетинок, вооружение лапок I складывается из 13 щетинок.
Монотипический род.
1. Cerophagus granulatus (Duj.) (фиг. 693—696).
Dujardin, 1849, Ann. Sci. Nat., ZooL, (3), XIL253, 255 (Hypopus).—lomborum OiidciuiMM, 1902, Entomol. Ber., 1:20 (Glycyphagus); 1904, Tijdsehr. Entomol., XLVI: 14, t. 2, fig, 29—30, --.gracilis Vitzthum, 1912, Ztschr. wiss. Insektenbiol.. VIII:292, fig. 23 (Ceroplutgus).
Гипопус. Белого цвета, задний край тела слегка буроватый. Голо в 1.5 раза длиннее своей ширины; проподосомальный щит треугольный, только на х/3 короче гистеросомы, с почти прямыми боковыми краями, сходящимися кпереди под острым, закругленным па вершине углом; Йоко-
408
Ill. GLYCYPBAGIDAE. 7. CBAETODACEYLINAE
вые края гистеросомы на значительном протяжении почти параллельны друг другу, задний край, в целом, широко закруглен, ио образует посередине более или менее глубокую выемку. Псевдостигмальный орган хорошо-развит. Боковые присоски расположены значительно позади крупных, центральных, имеют одинаковые размеры с задними и образуют с ними слегка вогнутый кпереди поперечный ряд. Длина всех 4 пар йог почти одинакова, составляя около 32 (I и 11), 30—31 (III) и 29—30 (IV) % длины, тела; очень сходны они и но толщине и пропорциям отдельных члеников; лапки в 2.3—2.6 раза длиннее своего диаметра у основания, слегка суживающиеся к коицу; обонятельная палочка лапок I и II длинная, почти прямая, едва утолщенная к концу. Длина 195—220 р, ширина 130—150 р.
Распространение. Франция, Голландия, Германия.
Экология. По Dujardin’у гипопусы С. granulatus очень обычны на Bombus lapidarius L.; Oudemans и Vitzthum находили их на В. ter-restris L. Мне не удалось найти их ни на том, ни на другом из этих видов, шмелей, а только на В. argillaceus (Scop.) Schmdkn.
26. Род CEROPHAGOPSIS, gen. n.
Inn рода: Cerophagopsis slcorikovi, sp. n.
Известны только гипопусы. Тело продолговатояйцевидное, равномерно-суживающееся к параболически закругленному заднему концу; спинные щиты покрывают практически всю верхнюю поверхность тела, умеренно плотпые и украшенные типичным для мелких обитающих на пчелах гипопусов орнаментом из тонких вдавленных линий, имеющих поперечное и диагональное направление на проподосомальном щите и преимущественно-продольное —нагистеросомальном; все щетинки спинной поверхности тела представлены однотипными тонкими микрохэтами. Органов зрения нет.' Гнатосома в виде пары тесно сближенных друг с другом бугорков, несущих дистально довольно длинные аристы. Коксо-стернальный скелет, как у Sennertionyx, но стернум простой (не вильчатый) и краевые утолщения коксальных полей не прерваны посередине и имеют волнистые очертания. Коксальные и генитальные щетинки в виде тонких микрохэт. Присасывательный диск небольшой, лишенный свободной перепончатой каймы;, аподема присасывательного диска отсутствует. Ноги довольно длинные, III и IV несколько короче передних, по почти такой же толщины, все с довольно крупными, резко изогнутыми, слегка закрученными по спирали коготками; лапки I—Ш с 3, IV — с 2 серповидными щетинками, III и IV — без парусных щетинок.
Известен только 1 вид.
Фиг. 697—705. Гипопусы родов Horstia, Sennertionyx и Cerophagopsis.
Фиг. 697. Sennertionyx manicati (Giard), общий вид сверху. Фиг. 698. То же, снизу. Фиг. 699..
S. manicati (Giard), лапка I сверху. Фиг. 700. Cerophagopsis skorikovi, ер. п., общий вид снизу. Фиг. 701. С. skorikovi, ер. и., лапка I сверху. Фиг. 703. Horstia helenae Oude., общий вид снизу.
Фиг. 703. Н. helenae Oude., лапка I сверху. Фиг. 701. Tortonia smitsrianburgsti Oude., вид: сверху. Фиг, 705. Т. smitsvanburgsti Oude., лапка I сверху.
<410
HI. GLYCYPHAGIDAE. 7. CUAETODACTYLINAE
1. Cerophagopsis skorikovi, sp. и. (фиг. 700, 701).
Гипопус. Бледноохристо-жслтоватый. Тело в 1.4—1.5 раза длиннее своей ширины; пролодосома пойти в 2.5 раза короче гистеросомы. Присасывательный диск, округлый; боковые присоски расположены на одной поперечной линии с л,ентр;|льнымп, которым они сильно уступают в размерах; задние присоски так,ого же размера, как боковые; хитини-зироватпгая пластинка, диска, образует спади 3 раздельных площадки с несколько угловатыми очертаниями. Передние ноги значительно длиннее, но едва толще задних; лапки 1 nil в 2 раза,, III и IV — в 1.5 раза длиннее -своего диаметра у основания; обонятельная палочка лапок I и II слегка изогнутая, заметно вздутая на конце. Длина 220—285 [л, ширина 150— 185 [г.
Распространение. Япония (Киото). Гипопусы найдены в боль-лпом числе па Megachile doderleini Fr.
27. Род SENXERTIONYX, gen. и.
Тип рода: Trichotarsus manicati Giard 1900.
Известны только гипопусы. Тело продолговатояйцевидное, умеренно ;выпуклое; спинные щиты покрывают почти всю его дорзальную поверхность, оставляя открытыми лишь узкие полоски слабо штрихованной мягкой кожи; поверхность спинных щитов украшена неясным орнаментом такого же типа, как у большинства Saproglyphidae-, все щетинки спинной поверхности однотипные, в виде тонких гладких микрохэт. Орган зрения отсутствует. Гнатосома полностью слита с идиосомой, представленная • только короткими аристами. Стернум вильчато раздвоен на конце, свободный; эпимеры II очень мощные, резко искривленные, почти соприкасающиеся концами с короткими, направленными вперед отростками, отходящими от эпимер III; эпимериты II слиты концами с эпимерами III, концы последних — с эпимерами IV; эти последние также отогнуты назад и тянутся, постепенно сходясь кзади, почти до генитальной щели; таким образом все коксальные поля, кроме I, замкнуты и обрамляют продолговатое центральное поле, открытое только спереди; краевые утолщения коксаль-ных полей явственны, но довольно узки и прерваны посередине. Стернальные и коксальные щетинки представлены тонкими микрохэтами. При--сасывательпый диск средней величины, со слегка выступающим за края тела перепопчатым краем; на нем расположены, как обычно, 4 пары присосок. Ноги I и III очень мощные, с крупными, винтообразно закрученными ..коготками, сидящими па конце конического мясистого praetarsus; ноги IV много короче и тоньше остальных, с маленьким крючковидным когот-.ком; лапки I—III с 3 крупными серповидными щетинками, лапки IV •без листовидно расширенных щетинок, по с очень длинной парусной щетинкой и несколькими более короткими.
К роду Sennertionyx относится только 1 вид.
26. GE RO P BAGOPSIS. 27. SENNERTIONYX 411
1. Sennertionyx manicati (Giard) (фиг. 697—699).
Giard, 1900, Bull. Soc. Entomol. Er.: 375 (TrichotaTSus); Oudenrans, 1905, Entomol. Ber., ’11:21; Vitzthum, 1919, Arch. Naturg., LXXXV, 5:30 (Borstia).
Гипопус,. Светлобуровато-желтый. Тело приблизительно в 1.5 раза ..длиннее своей ширины, с параболически закругленным задним концом. Лроподосома почти в 2.5 раза короче гистеросомы; sc намечают фигуру .вытянутого в поперечном направлении прямоугольника. Боковые присоски почти одинакового размера с центральными, расположенные «заметно впереди них, образуя с ними вогнутую кпереди дугу; хитннпзированнаяпластинка присасывательного диска разделена на 3 самостоятельные овальные площадки. Длина ног в процентах к длине тела: 40 (I), 37—38 (II), 30 (III) и 20—23 (IV); лапки I и II немного менее чем в 2 раза длиннее своего диаметра у основания лапки, III и IV едва длиннее его. Длина 210—260 ;х, ширина 135—170 у..
Распространение. Вид описан из Франции; мне известен еще из Андалузии, нескольких пунктов Италии, о-ва Корфу и следующих местностей в СССР:окрестности Одессы, Грузия (Боржоми), Армения, Ко-петдаг (Кельте-чинар), северо-западный Казахстан (Воровское лесное хоз-во).
Экология. S. manicati биологически связан с видами рода Anthidium', его гипопусы найдены на A. manicatum L., A. florentinum F., А. diadema F., A. oblongatum Latr. и A. cingulalum Latr.
ВЁ8Ш1Ё
Liste des abreviations adoptees
Tarses IV des cfd des Tynglyphidae
ab — distance comprise entre la base de Particle et le bord proximal de la premiere ventouse copulatrice tarsale.
cd — distance entre les deux ventouses copulatrices tarsales.
ef —distance entre le bord distal de la second e ventouse copula trice tarsale et I’extremite de 1’article.
Soies de Tidiosome
an — soies anales. d — soies dorsales.
•dx — premiere paire de soies dorsales. d. — seconde paire de soies dorsales.
~d3 — troisieme paire de soies dorsales.
— quatrieme paire de soies dorsales. h — soies humtales.
he, — soies humtales exttaeures.
hi — soies humtales inttaeures.
.sh —soies subhum^rales.
I — soies lattales.
la — soies lattales antcrieures.
Ip — soies lattales posttaeures.
Z, — premiere paire de soies lattales I, — seconde paire de soies lattales l3 — troisieme paire de soies lattales Д — quatrieme paire de soies lattales 1- — cinquifeme paire de spies lattales pa — soies postanales.
seulement chez les Gtycyphagidae.
412
TYROGLYPHIDAE
I chez les males des Tyroglyphidae et les deux I sexes des Glyeyphagidae.
24 — premiere paire de soies postanales
24 — seconde paire de soies postanales p3 — troisieme paire de soies postanales pro,—soies preanales, sa — soies postfirieures.
sa e —soies posterieures extiiricures.
sa v— soies postfrienri’S intfrieures.
sc — soies scapulaires.
see —soies scapulaires cxtdricures. sei —soies scapulaires int/irieures.
v — soies vorticales,
va—soies vorticities antdrieures 1 , , я
vp -soies vertieales posterieures ) chcz lcs
vi - soies verticales'intdrieures | chez les ^oglyp^dae. et les SaproglypMae.
Sensilles des tarses I et II
a — batonnet «olfactif» principal.
fi — bMonnet «olfactif» accessoire.
7—sensille proximale du groupe basal. .
о — sensille distale du groupe basal.
s — sensille dbrsale du groupe intermediaire.
I. Famille TYROGLYPHIDAE
TABLEAU DES SOUS-FAMILLES
Adultes ....
1 (2).' Soies verticales exterieures (v e) disposees lateralement, pres des angles anterieurs de 1’ecusson propodosomal, longues, finement plumeuses, ou. bien, un peu plus en arriere (а x/4 environ de la distance entre les soies verticales interieures et les soies scapulaires) et alors representees par des microchetes; soies de la surface dorsale de 1’idiosome d’ordinaire longues et fines, finement et rarement ciliees; toutes les soies des pattes fines et flexibles, souvent barbelees; ongles assez petits, pretarscs bien • developpes, revetant la plus grande moitie de 1’ongle,. .........
. ................ . . . .................... 1. Sous-fam. Tyroglyphinae.
2 (1). Soies verticales exterieures (a e) representees par de minuscules microchetes lisses, disposees dorsalement pres du milieu des bords lateraux de 1’ecusson propodosomal, ou absentee; soies de la surface dorsale de 1’idiosome lisses, souvent rigides et renflees a la base, d’ordinaire assez courtes ou tres courtes; sc i, hi, dy, d2 (et quelquefois aussi ds et Id) souvent absentee; soies des pattes plus ou moins aciculees ou spini-formes; ongles robustes (surtout ccux des pattes anterieures), pretarses-peu developpds, revfitant rion quo la base de 1’ongle . . . ..........................
..............................................2. Sous-fam. Rhizoglyphinae.
TABLEAU GtiNtiiUQUE DES HYPOPES
1 (2). Ongles extrtoiement longs et fins; aux pattes I ils sont de la longueur de .toute la patte et ne different presque pas par leur structure et leur dimension des grandes soies des tarses; les soies adhesives des tarses I
TABLEAU DES GENRES
413
et II sont aussi d’une longueur excessive; epimeres II relies des bouts au sternum.........................................24. Froriepia Vtz.
Une seule espece, F. vimariensis Vtz. (Allemagnc). :2 (1). Ongles de dimension et structure normales, toujours bien plus courts que les tarses; la longueur des soies adhesives ne surpassc pas la moitie de celle des pattes I et II; epimeres non relies des bouts au sternum.
3 (4). Sur les bords lateraux du propodosome (au dcssus de la base des pattes I) est situee une paire de gros yeux hemispheriques . .........
................................................14. Monieziella Berl.
4 (3). Yeux absents, ou situes au sommet de la saillie restrain et denues de cornees convexes.
5' (36). Disque adhesif bien developpe, plus ou moins fortement chitinise, muni de 4 ou 5 paires de ventouses parfaitement developpees.
6 (35). Disque adhesif arrondi ou bien transversalement elliptique, pas plus long que large; ventouses exterieures toujours situees en avant des po-sterieures; soies adhesives des tarses I et II elargies aux bouts.
7 (8). Soies de 1’ecusson propodosomal (sc) tres longues, capilliformes, ayant 15—20% de la longueur totale de 1’idiosome; tarses IV avec une tres longue soie (macrochete) surpassant la moitie de la longueur.du corps, et une deuxieme plus courte...........................1. Tyroglyphus Latr.
:8 (7). Sc courtes, ne depassant pas 5—8% de la longueur de 1’idiosome.
9 (10). Gnathosome en forme de trapeze, sensiblement plus court que sa largeur basale; champs coxaux III femes, largement ecartes; ecusson genital non sspare de 1’ecusson ventral. Sur les bourdons et dans leur nids...................................................3. Kuzinia, gen. n..
10 (9). Gnathosome de forme differente, sensiblement plus long que large a sa base; dans les cas ou les champs coxaux III sont tres ecartes et fer-mes, Г eCusson genital est separe du ventral par une suture transversale tres visible.
11 (34). Sommet des genuaux I muni d’une seule soie.
12 (13). JEpimeres III et IV libres, n’atteignant pas la ligue mediane du corps: ecussons sternal et ventral non separes 1’un de 1’autre et des champs coxaux, formant avec eux une large surface monolithe; saillie rostrale, portant une paire d’yeux tres rapproches et representes par un amas de pigment fonce, avec des cornees faiblement differenciees, ou prives totalement de ces dernieres..........................12. Histiogaster Berl.
13 (12). Epimeres III joints ou fort rapproches des bouts, dessinant ainsi le bord anterieur de Гecusson ventral; champs coxaux III generaloment femes. Yeux absents.
14 (27). Les deux paires de ventouses coxales bien developpees, norinales.
J.5 (16). Tarses IV munis de 2 longues macrochetes, dont 1’uno egnle presqtie le corps en longueur; ecussons dorsaux converts d’uno (lense et gros-siere sculpture faveolee. Ventouses coxales disposees sur In surl’nce do tres forts epaississements coxaux; soie extero-ventrale des tibins I et II
414
TYROGIA’PHIDA.E
ainsi que I’extero-basale des genuaux (des memes pattes) longues, flexibles, plnmeuses.........................16. Lacker baiieria, gen. n„
16 (15). Tarses IV denufe de longues macrochetes; surface des ecussons dor-saux lisse (finement poreuse) ou pointillee plus finement et rare merit; epaississements des bords des champs coxaux pen visibles ou absents: soie basale des genuaux I et II lisse, cowte et robuste,
17 (18). Disque adhesif petit, dloignG du bord posterieur du corps d’une-distance siirpassant de beaiiconp salongueur; bords exterieurs des champs coxaux I et II longs, rectilignes; tarses III et IV longs et grSles, formant ordinairement angle droit avec le reste de la patte (dans les preparations)» .......................................................8. Acotyledon Ouds.
18 (17). Disque adhesif plus grand; 1’espace entre son bord posterieur et I’extremite posterieure du corps sensiblement plus petit que sa longueur; bords exterieurs des champs . coxaux III ordinairement-courts, plus ou moins convexes; tarses III et IV plus courts et plus robustes.
19 (20). Champs coxaux. II se joignant immediatement derriere le court, sternum de fagon a ce qu’il ne reste qu’un tres etroit interstice longitudinal entre leurs bords interieurs; diametre transversal du disque adhesif pres de 1.5 fois moindre que la largeur de 1’idiosome; soies adhesives des tarses II representees par un tres large appendice foliace. Especa tres petite. Longueur 155 p........................20. Rodionovia, gen. n.
Une seule espece, R. inopinata (Vtz.) (Allemagne).
20 (19). Champs coxaux II ne se rejoignant pas derriere le sternum, separes d’un large intervalle, souvent ouverts; diametre transversal du disque-adhesif ne depassant pas la moitie de la largeur de 1’idiosome. Longueur 200 p au moins.
21 (22). Sternum et epimeres II atteignant le bord posterieur de 1’ecusson. sternal, separant ainsi et fermant ]es champs coxaux I et II; ventouses exterieures placees fort en avant des centrales ... 6. Forcellinia Ouds.
22 (21). Sternum n’atteignant pas le bord posterieur de 1’ecusson ventral, laissant ainsi les champs coxaux I plus ou moins largement ouverts; ventouses exterieures du disque adhesif disposees a pen pres au тёте niveau que les ventouses centrales.
23 (24). Gnathosome long et etroit, й bords parallMes, non bifurque au bout;-bord anterieur de l’6cusson propodosomal arqud, parallele au bord ante-rieur de I’hystCosome et ne formant pas de saillie rostrale; v i absents; pattes I enticement visibles d’en haut..............17. Robinisca, gen. n.
24 (23). Gnathosome do forme differente; dans le cas ou il est etroit et allonge, les branches de sa fourche forment des petits articles independants, cylindriques; 6cusson propodosomal a peu pres triangulaire, formant une saillie rostrale distincte; dans les cas ou son bord anterieur est arque, 1’ecusson prend une forme demicirculaire, s’avangant en s forme de visiere et recouvrant presque enticement les pattes I et II;.
TABLEAU DES GENRES
25
26
27
28
29
30
-31
32
33
34
35
36
«i bien developpees, mais quelquefois refoules sur la surface ventrale-de 1’ecusson.
(26). Sensille г des tarses I et II et la sole apicale dorsale des genuaux I epaissies en bdtonnet, obtuses ou meme un peu renflees au bout; saillie -rostrale densement ponctuee......................10. Rhizoglyphus Clap..
Une seule espece cosmopolite, R: echinopus (F. et R.).
(25). Sensille г des pattes I et II ainsi que la soie dorso-apicale des genuaux I ayant Г aspect de poils plus on moins rigides, s’amincissant vers le bdut; saillie rostrale non ponctuee......................9. Caloglyphus Berl.
(14). Ventouses coxales anterieures totalement reduites ou remplacees par-de menus poils; les posterieures normalement developpees, ou bion aussi remplacees par des soies, ou completement reduites.
(31). Ventouses coxales posterieures bien developpees, de structure normale.
(30). Ventouses coxales anterieures remplacees par des poils; idiosome tres large, presque rond; ecusson propodosomal tres court...............
................................................8. Acotyledon Ouds.
(29). Ventouses coxales anterieures completement reduites; idiosome-allonge, 1.5 fois plus long que large; longueur de 1’ecusson propodosomal egale au quart de celle de I’hysterosome .... 13. Schwiebea Ouds.
(28). Ventouses coxales posterieures remplacees par des poils ou completement reduites.
(33). Les deux paires de ventouses remplacees par d’assez longs poils rigides; champs coxaux III et IV assez largement ecartes; gnathosome-avec des articles distaux distinctement separes . . 9. Caloglyphus Berl.
(32). Les deux paires de ventouses coxales remplacees par des micro-chetes peu distinctes ou les posterieures reduites; champs coxaux III et IV joints sur la ligne mediane du corps; gnathosome non articule, seulement legerement bifurque au bout...........19. Tropeauia, gen. n..
(11). Sommet des genuaux I muni de 2 soies dorsales: champs coxaux II et III cl os, largement ecartes; tons ayant des epaississements lateraux. distincts; ventouses exterieures du disque adhesif disposees sensiblement en avant les centrales.............................23. Schulzea, gen. n.
(6). Disque adhesif fort allonge, etroit, linguiforme; ventouses posterieures et laterales situees au тёте niveau, tres petites; soies adhesives des tarses I et II capilliformes, non elargics au bout............
............................................... 8. Acotyledon Ouds.
(5). Disque adhesif completement reduit. de la тёте consistance molle que les champs environnants, arme rien que de 2 paires de vontoiisoH fortement reduites, peu distinctes.................8. Acotyledon Ouds.
Les genres Viedebanttia Ouds., Sancassania Ouds. et GarsuuUia Ouds. insuffisamment decrits, n’ont pas pu Stre places dans ce tableau.
<16
TYROGLYPH IDAS
1, Sous-fam. Tyroglyphinae
TABLEAU DES GENRES
A d u 11 e s
.1 (2). Surface supcricuro dos articles des pattes I et II oriiee de carenes bi- furquees ou cruciformos; toutee les soies de la surface dorsale epaisscs, courtes, densemont pilmiscs; o' denue de ventouses copulatrices anales et tarsalcs, avec 2 paires de microchetes anales; orifice anal de la 9 fort £loign6 du bout posterieur du corps..........7. Scatoglyphus Bert.
Une seule espece, Sc. polytrematus Berl. (Italie).
.2 (1). Articles des pattes anterieures lisses, sans carenes ramifiees; soies du corps d’une autre structure, d muni de ventouses anales et tarsales bien developpees, ayant une seule paire de microchetes anales; fente anale de la 9 tres rapprochee du bord posterieur du corps.
-3(4). Dimorphisme sexuel tres prononce: chez le d les pattes I sont plus ou moins fusiformes, Jeiir femur est arme en dessous d’une forte epine conique; chez la 9 les pattes ont une structure normale; soie apicale interieure des genuaux tres longue, 6—10 fois plus longue que 1’exte-rieure, qui a la forme d’un batonnet arrondi au bout; extremite des tarses avec une seule petite epine ventrale; famulus absent (1. Tribu Tyroglyphini).......................................1. Tyroglyphus Latr.
4 (3). Caracteres sexuels secondaires peu accuses: pattes I normalement constitutes dans les deux sexes; soie apicale interieure des genuaux I pas plus de 3 fois plus longue que I’exterieure, qui d’ordinaire est aussi setiforme; extremite des tarses avec 3 ou 5 petites epines ventrales et souvent Une dorsale; famulus present (2. Tribu Tyrophagini).
-5 (8). Extremite des tarses avec une assez forte epine conique dorsalement et 5 epines vcntrales.
45 (7). V e tres longues (pres de 3/5 de la longueur des v г), finement et den-sement ciliees, disposees lateralement un peu derriere le niveau de la base des « i; elles flanquent de sorte le gnathosome, sont distinctement arquees, et leurs extremites sont dirigees obliquement en dedans et en has; organe pseudostigmal lanceole, tres plumeux; sensille г situee au meme niveau que le batonnet olfactif; chez le cf, les ventouses copulatrices tarsales sont disposees A peu pres a la meme distance de la base et de l’extr£mite du tarse IV.......................4. Tyrolichus Ouds.
"7 (6). V e repr£scnt6cs par des microchetes (4—6 fois plus courtes que les v г), lisses oufaiblement barbel£es, situ£es considerablemen t plus en arriere que le niveau transversal des v i\ organe pseudostigmal setiforme; г si-tuee'plus distalemcnt que le bdtonnet olfactif; ventouses copulatrices tarsales du d situees toutes les deux dans la moitie basale du tarse IV.
............................................ 6. Forcellinia Ouds.
-8 (5). Extremite des tarses avec 3 epines ventrales; ve disposees a peu pres comme dans le genre Tyrolichus, longues et ciliees.
TYROGLYPHUS, ALEUROGLYPIJUS 417
9 (10). L a tres longues, representees .„par des macrochetes; dz situees presque sur la meme ligne transversale que dj, extremement longues; tarses I et II ornes d’une сагёпе longitudinale forte, mais obtuse.................................................3. Kuzinia, gen. a.
Une seule espece, K. laevis (Duj.) (= Hypopus laevis = Tyroglyphus fucorum Ouds. =2’. lundbecki Drag.), habitant les nids des bourdons.
10 (9). L a tres courtes, de la тёте longueur que df, d2 situees beau-coup plus en arriere que d^, tarses I et II sans сагёпе longitudinale.
11 (12). Pattes relativement courtes et robustes; cheliceres tres fortes; ve droites ou faiblement arquees; sensille e situee au meme niveau que le bdtonnet olfactif.............................2. Aleuroglyphus, gen. n.
12 (11). Pattes relativement longues et greles; cheliceres de dimensions ordinaires; ve longues, fortement incurvees en has et au dedans; sensilie e des tarses I situee bien au deld du batonnet olfactif. . . .....................................................5. Tyrophagus Ouds.
1. TRIBU TYROGLYPHINI
TYROGLYPHUS (LATR.) OUDS.
1 (2). Sc i 2 ou 3 fois plus courtes que les sc e; dlt dz et Z a representees par des microchetes, toutes les autres soies de la surface dorsale de I’hyste-rosome tres longues, distinctement ciliees, formant une «traine» touf-fue, semblable a celle des Tyrophagus-, organe pseudostigmal long, seti-forme, faiblement cilie; tarses I du d avec une bosse au milieu de la surface ventrale. — URSS (Kazan, Ukraine). 2. T. tyrophagoides, sp. n.
2 (1). Sc i de la meme longueur que sc e; toutes les soies de l’hyst£rosome sauf h e, sa i et 1’une des paires postanales, tres courtes, presque lisses; le bord posterieur du corps n’est ainsi orne que de 2 paires de soies assez longues; organe pseudostigmal court, herisse de nombreux oils, elargi h la base; tarses I du d sans bosse sur sa surface inferieure. — Espece cosmopolite . . . ...............................1. T. farinae (L.).
4 2. TRIBU TYROPHAGINI
ALEUROGLYPHUS, gen. n.
1 (2). Sc i 2—2.5 fois plus courtes que sc e; h i, toutes les d et les 2 paires de Z (Z a et I p) sont representees par des soies tres courtes, finement cilices; h e, sa e, sa i et 1 ou 2 paires des postanales sont les soulos soies de I’hysterosome qui ont une longueur assez grande; elles foment on leur ensemble (h e exceptees) une maigre et courte «traine»; microcytes anales du d dispos£es sur la convexite meme des ventouses anales. — France. Hollande, URSS............................1. A. ovntus (Tv.).
27 Фауна СССР, Захватили—284
418 TYROGLYPHIDAE J
2 (1). Les deux paires de sc de longueur egale; la plupart des soies de l’hyst4’ "
rosome (h e, Ji i, d3, diy IP et les 2 paires de sa et 1 paire des postanales) 4
tres longues (mesurant 40—100% de la longueur de 1’idiosome), for- .. )
mant en leur ensemble une belle «traine» touffue etalee en eventail; '
microchetes anales du <5 fort eloignees des ventouses anales. — Sicile, ; -
URSS (Armdnie).........................2, A. Siculus (Fum. et Rob.).
TYROLIOIIUS OUDS.
1 (2). L a repr6sentdes par de longues soies, ayant Л peu pres les memes dimensions que les humerales; la surface dorsale de I’hystSrosome ne porte-ainsi qu’une seule paire de microchetes (dx). — Hollande, Angleterre, Allemagne, Italie, URSS (regions de Smolensk, deMoscou, d’lvanovo, j d’Odessa, Siberie orientale)..........................1. T. casei (Ouds.). f
2 (1). L a re presentees par des microchetes, de la тёте longue ur que dv — I
Allemagne..........................................2. T. lini (Ouds.). I
TYROPHAGUS OUDS.
1 (4). A 2.5—4 fois plus longues que dx; pattes relativement courtes, moins.
greles; tarses T pas plus longs que le tibia et le genual, pris ensemble; ;
ventouses copulatrices tarsales du cf disposees presqu’a la тёте di- '
stance de la base et de I’extremite des tarses IV.
2 (3). de 2.5 a 3 fois plus longues que d^, penis petit, fortement recourbe en S; organe pseudostigmal lanceole avec de longs appendices fortement inclines dans sa moitie basale elargie; tubercule coni que do la surface interieure des cheliceres situe beaucoup plus proximalement que la soie mandibulaire, allonge en fine et longue pointe; 6pine la-tero-coxale fusiforme; famulus ovoide. Espece considerablement plus \ petite. — URSS (Leningrad, Moscou, Novotcherkassk, Armavir, Azer- j
baidjan, Asie Centrale, Extreme Orient)............1. T. noxius A. Z. j
3 (2). D2 3.5-4 fois plus longues que dx; penis tres grand, гесоигЬё eh arc; ) organe pseudostigmal й tige etroite avec des appendices plus rares sur les 2/3 de sa longueur; le tubercule conique des cheliceres moins pointu, •’ plus rapproche de la soie mandibulaire; soie maxiilaire et famulus-faiblement epaissis. De taille plus grande; longueur: <5 450—540 p., 9 550—700 [л. — URSS (Kostroma, Caucase, Barnaoul).........................
.............................................2. T. perniciosus, sp. n.
4(1). Д de la meme longueur que dx ou bien un peu (pas plus de 1.6 fois) I plus longues; pattes longues et greles; tarses I souvent plus longs que le tibia et le genual, pris ensemble; ventouses copulatrices disposees d’une fagon variable.
5 (6). Pattes tres longues, et greles; tarses I presque 1.5 fois, tarses IV 1.45—
1 .6 fois plus longs que les tibias et genuaux respectifs, pris ensemble; soie maxiilaire assez longue, non epaissie; les deux soies ventrales des.
TYROLICHUS, TWIOPHAGUS 419
tarses IV disposees dans le tiers distal du tarse; idiosome relativement etroit, presque 2 fois plus long que large. — Angleterre, France, Italie ...........................................6. T. longior (Gcrv.).
6 (5). Pattes moins longues; tarses I pas plus de 1.15, tarses IV de 1.3 fois aussi longs que les tibias et genuaux respectifs, pris ensemble; soie maxiilaire courte, epaissie; soies ventrales des tarses IV placees pits du milieu du tarse; idiosome plus large.
7 (8). Z>2 1.3—1.6 fois plus longues que <2X; tarses I 1.04—1.16 fois, tarses IV 1.11—1.3 fois plus longs que les tibias et genuaux des pattes respec-tives, pris ensemble; penis fin, cylindrique, se retrecissant graduelle-ment vers le bout tres fin, tronque droit, tres semblable par sa forme au goulot d’une cafetiere; les deux ventouses copulatrices tarsales si-tuees dans la moitie proximale du tarse; batonnet olfactif des tarses I et II tres gr§le, long, sensiblement retreci vers le bout. — URSS (regions de Leningrad, de Kalinine, d’lvanovo, Caucase du Nord, Kam-tchatka)...................................... 5. T. tenuiclavus, sp. n.
8 (7). Д egalant dx en longueur; tarses I pas plus longs que le tibia ct lo genual pris ensemble; tarses IV pas plus de 1.08(cf)—1.2(9) fois plus longs que le tibia et le genual IV; penis de forme diffCrentc; ventouses tarsales copulatrices variablement disposees; batonnet olfactif des tarses I et II plus court et plus gros, sAlargissant vers le bout.
9 (14). Penis 2 fois coude, s’£largissant en entonnoir vers le bout oblique-ment tronque, robuste; tige de 1’organe pseudostigmal non elargie; famulus des tarses I non renfle. — Hollande, Allemagne, URSS (regions de Leningrad, de Moscou, d’lvanovo et d’Odessa, Eachkirie, Caucase du Nord, Siberie occidentale)...............4. T. humeresus Ouds.
10 (11). Courbe distale du penis faible; tarses IV plus courts, n’ayant quo 0.93—1.07 (O') ou 0.98—1.10 (9) de la longueur du tibia, et du gdnual pris ensemble; disposition des ventouses copulatrices tarsales tits variable; trongon ef egale 0.75—1.75 du trongon ab; ce dernier 1.5—3 fois plus long que 1’espace entre les ventouses; vari^td de petite taille: <5 288—370 p, Q 350—455 p. — URSS (Bachkirie, Caucase septentrional) ................................4c. T. humeresus tesquorum, ssp. n.
11 (10). Courbe distale du penis plus prononcee; tarses IV plus longs, ayant 1.03—1.08 (cf) ou 1.03—1.23 (9) de la longueur du tibia et du genual IV pris ensemble; trongon ef 1.5—2.5 fois plus long que ab.
12 (13). De plus grande taille (cf 470—766 p; 9 470—770 p); trongon (if des tarses IV 1.45—2.5 fois plus grand que le trongon ab; co dernier plus grand que 1’espace entre les ventouses (cd) 1.25—2.8 foia, =-Hollande, Allemagne, URSS. . . 4a. T. humerosus humeroHiis Ondii.
13(12). De plus petite taille; cf 280—380 p; 9 380—475 p.; trongon 0/2 - 2.2 fois plus grand que ab; ce dernier surpasse la distance entre leu ventouses 1.25—1.35 fois.—URSS (region d’Odcssn)...................... .
............................. 4b. T. humerosus Silvester, мир. nov.
Ш TYROGLYPHIDAE
14 (9). Penis court, recourbd en arc oil droit, a epaisses parois, arrohdi au bout; tige de 1’organo pseudostigmal lanceolee; famulus des tarses I ovoide, epaissi. Espfcce de grande taille. Longueur: d 515—520 p,; 9 —inconnue. — URSS (Saratov).....................3. T. molitor, sp. n.
POltOMXTNTA OUDS.
Л (I u 1 tea
1 (2). Z>x, d2 et I a repr&cntdcs par do fines microchetcs lisses de meme longueur (4—7% de la longueur de 1’idiosome), toutes les autres soies de la surface dorsale. de 1’idiosome tres longues (40—60% de la longueur de ce dernier), de structure normale —capilliformes, graduellement re-trdcies vers le bout, tres peu et finement ciliees; sc i notablement plus longues que sc e; penis extraordinairement long et fin. Facies des especes du groupe Tyrophagus humerosus Ouds. — Angleterre, partie europeenne de 1’URSS........................1. F. fungivora (Ouds.).
2 (1). Dr aplaties, elargies au bout, densement plumeuses lateralement, ayant pres de 15% de longueur de 1’idiosome, d2 et la presque 2 fois plus longues que dx; tout le reste des soies de la surface dorsale de moy-enne longueur, aplaties, s’elargissant vers le sommet, se terminant par une lamelie transparente ovale ou lanceolee, et tres plumeuses lateralement, ou bien plus longues et faiblement plumeuses avec une lamelie apicale tr&s (Stroite; toutes les deux paires des sc de meme longueur; penis de dimensions normales.
3 (4). Toutes les soies de la surface dorsale de 1’idiosome aplaties, elargies vers le bout, qui forme une lamelle ovale ou largement 1апсёо1ёе, relativement courte; chez le d px et p2 sont presque de la meme longueur. Habite exclusivement les fourmillieres. — Hollande, BohSme, URSS (regions de Moscouet de Kalinine).....................•...........
...........................................4. F. wasmanni (Mon.).
4 (3). spatuliformes, densement plumeuses; tout le reste des soies fines, assez faiblement et finement ciliees, se terminant par une fine lamelle lanceolee; leur dimensions tres variables (de 13 a 45% de la longueur de 1’idiosome); chez le d les soies p2 sont presque 2 fois plus longues que px. Habitent la couche de feuilles mortes des forets. — URSS (Moscou).......................'..................2. F. diamesa, sp. n.
H у p о p e s
1 (2). Gnathosome d6passant le bord anterieur de 1’idiosome de toute sa moitie distale, long (pres de 3 fois plus long que large), a bords paralleles; ecusson propodosomal a saillie rostrale petite, mats tres distincte, portant a son bout 2 petites soies («i) et ne recouvrant pas en visiere les bases des pattes I et II; coloration brune-rose. — Hollande,
FORCELLINIA Ш
Bohfeme, URSS (regions de Moscou et de Kalinine)....................
. ......................................... 4. F. wasmanni (Мои,).
2 (1). Gnathosome court (pas plus de 2 fois plus long que large a sa base), entierement сасЬё, ainsi que les articles proximaux des pattes I et II, sous 1’ecusson propodosomal, qui les recouvre comine une visiere; le bord anterieur de ce dernier presque droit (h peine arque), sans la moindre trace de saillie rostrale; vi absentes; coloration ocre-brune.
3 (4). Espece de petite taille (pres de 260 u.); champs coxaux II se retrecis-sant un peu en arriere de fagon a ce que leurs bo rds exterieurs ont une direction oblique; bord posterieur de 1’ecusson ventral ondule; ventouses peripheriques du disque adhesif un peu plus petites que les centrales.— Angleterre, partie europeenne de 1’URSS .......................
............................................1. F. fungi vora (Ouds.).
4 (3). Espece de tres grande taille (460 p); champs coxaux II s’elargissant distinctement vers Г arriere de fagon a ce que leurs bords exterieurs sont presque rigoureusement paralleles; bord posterieur de I’ecusson ventral arque; ventouses peripheriques:du disque adhesif beaucoup plus petites que les centrales. — Allemagne. 3. F. gigantonympha (Vtz.).
2. Sous-fam. R, h i z о g 1 у p h i n a о TABLEAU DES GEE RES
O’U
1 (4). V e toujours distinctes, representees par des microchetes; sc i, hi, sh, dt, d2 et I a bien dёveloppёes; soie apicale des tibias IV de structure normale; le batonnet olfactif n’est pas precede d’une epine (e se-tiforme, зйиёе considerablement devant lui, mais plus vers ГпТё-rieur); le bout posterieur du corps, non modifU, sans lamelle opistho-somale. (1. Tribu Acotyledonini).
2 (3). Les soies exterieure ventrale et interieure apicale des tarses I et II, quelquefois тёте des tarses III et TV, falquees, divergentes, dlargies au bout en une lamelle transparente; soies ventrales des tibias spini-formes; toutes les deux ventouses tarsales se trouvent dans la partie distale du tarse (IV). Especes de grande taille (600—1200 p) . . . . ....................................................9. Caloglyphus Berl.
3 (2). Soie exterieure ventrale ainsi que toutes les soies apicales des tarses I et II simples, cap ill if or mes, non elargies au bout (rien que chez les def de A. batsylevi la soie apicale exterieure est elargie en forme de lamelle ovale); ventouses copulatrices tarsales disposees presque dans la partie mediane du tarse IV. Especes assez petites (350—GOO p) , . ..........................’.........................8. Acotyledon Ou<Ih.
4 (1). V e absentes; sc i, h i, dr, d2 (et souvent aussi d3,1 a et sa i) .In plupnrt du temps absentes; dans les cas ou elles existent, ellcs sont горгёноп1ёен par des microchetes, et immediatement devant le bfltonnot olfactif st!
422
TYilOGLYPHIDAE
trouve une grosse 6pine (homologue de e); soie apicale des tibias IV transformee en Spine conique; bout posterieur du corps la plupart du temps fortement niodifiS, formant une lamelle horizontale chitinisee. (lamelle opisthosomalc). (2. Tribu Rhizoglyphini).
6 (6). Pattes IV fortement rcnflScs; bout posterieur du corps non modifiS, ' .n’ayant pas de lamelle opisthosomale...............12. Heteroglyphus Foa.
Une seule cspfcco insuffisainmont connue, H. vitium Foa (Italie).
6 (5). Pattes IV non renflSes; dans les can oft «lies sont un peu epaissies (chez Histiogasler), il cxiste une lamelle opisthosomale bien developpee portant 4 lobes transparents en forme d’£ventail.
7 (8). Lamelle opisthosomale absente, bout posterieur du corps non modifie; Pi et p2 longues; sc i, hi, et d2 representees par des microchetes; 2 paires de sa (sa e et sat); ventouses copulatrices anales n’ayant pas de cercle chitinise; soies ventrales exterieure et interieure ainsi que la soie apicale interieure des tarses I et II un peu elargies au bout, folia-cees................................................10. Rhizoglyphus Clap.
Une seule espece cosmopolite, R. echinopus (F. et R.).
8 (7). Lamelle opisthosomale bien developpee; pr ainsi que souvent aussi p2 representees pax des microchetes, ou bien p2 et p3 absentes; 1 paire de sa (sa г); ventouses anales entourees d’un cercle fortement chitinise; soies ventrales et apicales des tarses non elargies.
9 (10). Lamelle opisthosomale se retrecissant fortement vers la base, ayant un rebord festonne, portant 4 lobes hyalins disposes en eventail; pattes assez longues et greles; pattes IV sensiblement plus longues et plus robustes que les pattes III; ventouses anales entierement chitinisees, tres convenes, poils anaux lanceoles; une seule paire de soies postanales ................................................12. Histiogaster Berl.
10 (9). Lamelle opisthosomale a peu pres demi-circulaire, s’elargissant vers la base, ayant les bords unis, sans aucun appendice; pattes courtes, ordinairement robustes; pattes III un peu plus courtes et plus minces que les pattes IV, ou bien de memes dimensions; quelquefois les pattes III sont fort epaisses et modifiees (chez les dU heteromorphes); ventouses anales chitinisees rien qu’en cercle, moderement convexes, soies anales non aplaties; 3 paires de soies postanales.
11 (12). Une epine conique tres robuste se trouve immediatement devant le bdtonnet olfactif; soies ventrales exterieures des tibias et des genuaux I et II ainsi que 2 de celles des tarses representees par des epines plus ou moins coniques; pretarse totalement absent; lamelle opisthosomale situee au meme niveau que la surface dorsale de I’hysterosome, la continuant immediatement; la surface inferieure de Cette lamelle est oblique de fagon a ce que les ventouses anales, situees au-dessus, sont dirigees enarriere (vues de profit); tfU representes rien que par la phase hete-romorphe, ou bien cette phase est de beaucoup plus nombreuse que Thomeomorphe.................... . ...................13. Schwiebea Ouds.
RHIZOGLYPHINAE. TABLEAU DES GENRES
423
22 (11). Point d’epine (levant le bdtonnet olfactif; toutes les soies des tibias et des genuaux, ainsi que 2 des soies ventrales des tarses minces et flexibles plus ou moins capilliformes; pretarse bien developpe; cfd1 represents rien que par la phase homebmorphe.
13 (14). Vi souvent absentee; la bien developpees, assez longues; p2 longues, capilliformes; sc e et h e plus courtes (de 12 a 20% de la longueur de 1’idiosome); pretarse represents par une ou deux lamelles demicirculaires verticales, recouvrant des cotes la base de 1’ongle; structure de la lamelle opisthosomale comme chez Schwiebea ..... ....................................................14. Monieziella Berl.
14 (13). V i toujours bien developpes; la absentes; p2 representees par des microchetes minuscules; see et h e longues, (de 33—40% de la longueur de 1’idiosome); pretarse ayant Г aspect d’un lobe membraneux, recouvrant la plus grande partie de 1’ongle (comme chez les Tyrogly-phinaey, lamelle opisthosomale situee au meme niveau que la surface ventrale du corps; surface dorsale de 1’hysterosome abruptement de-clive par derriere, formant une surface oblique au-dessus de la lamelle (vue de profil)..................................16. Thyreophagus Rond.
Une seule espece, Th. entomophagus (Lab.). — Angleterre, France, Italie, URSS (Leningrad, Moscou, Ivanovo, Barnaoul).
. 99 '
1(4). Ve sont representees par des microchetes tres distinctes; sei, h i, <4 et d2 bien developpees; point d’epine devant le batonnet olfactif; (e setiforme, situee considerablement par devant et plus a 1’interieur que a); fente anale d’ordinaire situee au milieu de 1’opisthosome, a une grande distance du bout posterieur du corps; dans les cas memes oh elle est plus refoulee en arriere, la distance entre son extremite poste-rieure et le bord posterieur du corps n’est pas plus grande que la moitie de 1’espace entre son bord anterieur et la fente genitale; de 4 h 6 paires de microchetes anales (rarement 3 paires). (1. Tribu Acoty-ledonini).
2 (3). Soie exterieure ventrale ainsi que la soie interieure apicale des tarses I et II et souvent aussi des tarses III et IV falquees, robustes, divergeant en arc et elargies distalement en une lamelle transparente plus ou ,moins large; soies ventrales des tibias ayant 1’aspect de fortes epines coniques; pretarse moderement developpe, formant un etui a la base de 1’ongle. Especes de grande taille, quelquefois тёте gigantesques . . ................................................. . . .9. Caloglyphus Berl.
(2). Toutes les soies apicales et la ventrale exterieure des tarses simples, (capilliformes, non elargies au bout; soies ventrales des tibias non spi-miformes; pretarse fort developpe en forme de coussinct, recouvrant toute la moitie basale de 1’ongle. Formes de taille plus reduite ..................................................8. Acotyledon Ouds.
Ш
TYROGLYRIHDAE
4(1). Уе absentee; sei, hi, et d2 absentee egalement; dans les cas ou elles existent, elles sont representees par des microchetes et immedia-tement devant le bhtonnet olfactif se trouve une grossc epine conique (genre Rhizoglyphus)-, fente anale disposee plus en arriere, pres du bord posterieur du corps; quelqucfois mfrne ses lames se projcttcnt un pen au del& de cclui-ci; rien qu’uno paire de microchetes anales, ou bien celles-ci sont Lotalemeiit absentes. (2. Tribu Rhisoglyphini).
5 (6). Sei, hi, <1г et d2 sont. repUsontZos par des microchetes; organe pseudostigmal bien ddveloppe, s6tiformc; pattes robustes, armes de nom-breuses et fortes Spines. Forme de grande taille, tabs obfese..............
............................................ 10. Rhizoglyphus Clap.
Une seule espece cosmopolite, R. echinopus (F. et R.).
6 (5). Sei, hi, 1’organe pseudostigmal, et d2 absentee, ainsi que parfois. aussi d3 et la. Especes de plus petite taille (280—630 p.).
7 (10). Une Spine conique (e) se trouve immSdiatement devant le bhtonnet i olfactif; rien qu’une paire de soies sacrales; microchetes anales absentee; pretarse fortement reduit ou tout a fait absent.
8 (9). Sc e, he, d et sa e trSs longues (de 30 a 65% de la longueur de 1’idio-some); d3 de 15 a 40% de la longueur de 1’idiosome, pattes assez longues et grSles; poils ventraux exterieurs et intSrieurs des tarses I et II longs, capilliformes; sommet des gSnuaux I avec 2 poils dorsaux; Spi-meres III et IV non reliSs entre eux...............12. Histiogaster Berl..
9 (8). See, he, d et sa e courtes (de 7 a 25% de longueur de 1’idiosome); d3 et I a absentee ou bien reprSsentSes par des microchetes; pattes tres courtes (de 14 a 23% de longueur de 1’idiosome), robustes; soie ventrale-interieure des tarses I et II spiniforme; sommet des genuaux I avee une soie dorsale; lorsqu’il у en a 2, les epimeres III et IV sont reliespar leur bouts......................................13. Schwiebea Ouds.
10 (7). Point d’epine devant le batonnet olfactif; 2 paires de soies sacrales-(sa e et sa i)- une paire de microchetes anales; pretarse bien deve-loppe; idiosome tres allonge (de 2 a 4 fois plus long que large), cy-lindrique.
11 (12). V e souvent absentee; I a bien developpees; toutes les soies de 1’idiosome courtes, assez rigides; pretarse represents par 1 ou 2 lamelle.. verticales demi-circulaires, recouvrant des deux cotes la partie basale de I’ongle; idiosome extrfemement etroit, de 2 a 3.4 fois plus long que large..............’................................14. Monieziella Berl.
12 (11). V e bien developpees; I a absentee; toutes les soies de 1’idiosome tres longues et minces, capilliformes; pihtarse ayant 1’aspect d’un coussinet charnu, recouvrant la plus grande partie de I’ongle (de la. meme structure, que chez les Tyroglyphinae)-, idiosome de 2 a 2.2 fois-plus long que large ............................ 15. Thyreophagus Rond..
Une seule espece, Th. entomophagus (Lab.).,— Angleterre,.
< . France, Italic, URSS (Leningrad, Moscou, Ivanovo, Barnaoul).
ACOTYLEDON 423
ACOTYLEDON OUDS.
Ud
1 (2). Soies postanales disposees en 2 rangees longitudinales; I’intervalle entre' Pi et p2 est egal, de sorte qu’elles sont disposees on cane; distance entre I’extremite posterieure de la fente anale et le bord posterieur du corps depassant plus de 2 fois sa longueur; propodosome et gnathosome tres etroits. — Angleterre.............................5. A. michaeli (Ouds,).
2 (1). Position des soies postanales differente; jq toujours considerabl'e-ment plus ecartees que p2; 1’espace entre I’extremite* posterieure de la fente anale et le bord posterieur du corps ne depasse pas plus de 1.5 fois ' sa longueur; proportions du gnathosome et du propodosome normales.
3 (8). Sc ide 5 a 10 fois plus courtes que sc e; intervalle entre sc i depassant plus de 2 fois celui qui existe entre les poils de cette paire et sc e; organe-pseudostigmal setiforme, presque tout-a-fait lisse; poils ventraux des tarses non spiniformes; batonnet olfactif des tarses I et II claviforme.
4 (5). Sc i 10 fois plus courtes que sc e; elles sont separees d’une distance 6 fois plus grande que celle qui les separe de sc e; toutes les soies de 1’hysterosome extrfemement courtes; longueur de lie ne depassant pas 16, dA, I p et sa i — 5—7 % de la longueur de 1’idiosome; tarses let II moins de 2 fois plus longs que leur diametre. — URSS (Moscou, Kostroma) . ....................................1. A. rhizoglyphoides (A. Z.).
5 (4). Sc i de 5 a 6.5 fois plus Courtes que sc e; distance entre elles pas plus de 4—4.3 fois plus large que 1’espace qui les separe de sc e; quelques-unes des soies de 1’hysterosome assez longues: he egalant 20—30, dt, Ip et sa i 15—18% de la longueur de 1.’idiosome; pattes plus longues et plus greles; la longueur des tarses I et II depasse plus de 3 fois leur diametre au milieu.
6 (7). P2 situes au meme niveau que plt formant avec ces dcrniercs une rangee transversale; he plus longues, ayant jusqu’A 30% de la longueur de 1’idiosome. Pattes greles. — Italie...................4. A. agilis (Can.).
7 (6). P2 disposees bien plus en arriere que px, a peu pres au meme niveau que p3, formant avec celles-ci unerangee un peu arquee; he plus courtes, ayant pres de 20% de la longueur de 1’idiosome; pattes plus robustes. — URSS' (Leningrad, Moscou, Ivanovo, Bachkirie, Siberie occi-dentale).................................. : . . 2. A. sokolovi, sp. n.
8 (3). Sc г de 2 a 3 fois plus courtes que sc e; 1’interstice entre sc i no dc-passe pas plus de 1.5 fois la distance entre les soies de cette paire et sc e; organe pseudostigmal plumeux; bdtonnet olfactif des tarses I et II ne formant pas de massue bien delimitee.
9 (10). Idiosome fort alonge, presque cylindrique; sa longueur dtfpasse pres-que 2.5 fois sa largeur maximum; poils ventraux des tarses .1 e.L II non spiniformes. — Italie, URSS (region d’Odessa) . . 8. A. krinncii (Herl.).
10 (9). Idiosome pas plus de 1.9 fois plus long que large; poils median et ext<5ro-ventral des tarses I et II transformes en (Spines robustes, coniques.
326
TYROGLYPHIDAE
11 (12). Soies de 1’idiosome (surtout les plus courtes d’entre elles) droites, rigides, presque aciculees; p2 situees au meme niveau que p3. — Angle-terre...............................................7. A. alberti (A. Z.).
-12 (11). Soies de 1’idiosome plus ou moins fines, flexibles, non aciculees. .13 (14). Soies de 1’idiosome plus longues, toutes tres fines et flexibles; chez les exemplaires lioiiidomorphes, la longueur ded3et Ip est de pres de 22, I a do 1G a 17% do la longueur de 1’idiosome, p2 situees au тёте niveau quo 2?3, 3 fois plus longs que organe pseudostigmal epaissi, largement-lance.ole, portant quelques appendices piliformes sur sa surface dorsale; les individus heteromorphes ont 1’une dos soies apicales des tarses I et II elargie au bout, formant une lamelle transparente, ovale. — URSS: Vorochilovsk (Causase du Nord) .... 10. A. batsylevi, sp. n.
14 (13). Soies de 1’idiosome plus courtes; longueur de d3 et I p ne depasse pas de 16 a 17, celle de I a 9% de la longueur de 1’idiosome; quelques-unes d’entre elles — dlt d2, I a et quelquefois aussi d3, d^, I p et hi — obtuses, paraissant tronquees a faible grossissement; a un fort gros-sissement, on voit que leur bout est aplati en spatule arrondie; p2 si--tues considerablement devant p3, pas plus de 2 fois plus longues que рг; organe pseudostigmal plumeux rien qu’.aux bords; phase heteromorphe inconnue (sans doute absente). — URSS (Bachkirie, region de Kali-nine) .........................................6. A. redikorzevi (A. Z.).
15 (16). Soies de 1’idiosome plus courtes; longueur de sc i pres de 10, dx et d2 5.5—6.-5, Z a 14—15, sa e 14, sa i 25—27% de longueur de 1’idiosome, organe pseudostigmal plus large et plus aplati. — URSS (Bachkirie) ...........................6a. A. redikorzevi redikorzevi (A. Z.).
•16 (15). Soies de 1’idiosome plus longues; longueur de sc г de 14 a 15, dx et d2 de 9 a 10, I p pres de 17, sa e 23—24, sa i jusqu’a 32% de la longueur de 1’idiosome; organe pseudostigmal presque cylindrique. — URSS (region de Kalinine)....................6b. A. redikorzevi borealis, ssp. n.
99
1 (8). Sc i de 6 a 10 fois plus courtes que sc e; espace entre les deux soies de cette paire depassant plus de 2 fois 1’intervalle entre chacune d’elles et sc г; poils ventraux des tarses I et II non spiniformes; organe pseu-'dostigmal sfdiforme.
2 (3). Sc i plus de 10 fois plus courtes que sc e; distance entre elles 6 fois plus grande qu’entre chacune d’elles et see; toutes les soies de I’hystero-some tres courtes; longueur de d3 et d4 de 5 a 6, I p et sa i pres de 7, h e pas plus de 15% de la longueur de 1’idiosome; pattes courtes, tres ro-bustes; tarses I et II plus de 2 fois, tarses IV pres de 3 fois plus longs que leur diametre moyen; distance entre I’extremite posterieure de la fente anale et le bord posterieur du corps 2.5 fois moindre, que I’es-pace qui la separe de 1’orifice genital. — URSS (Moscou, Kostroma). ........................................... 1. A. rhizoglyphoides (A. Z.).
ACOTYLEDON 427
•;• = • е" -"'’-IF P|f |||||S|?cgV*11'Fjpt’Ei'4!Rf4
3 (2). Sc i de 6 a 7 fois plus courtes que sc e; 1’intervalle entre elles no d6-passe pas plus de 4—4.3 fois 1’espace entre elles et sc e; quelques-uncff des soies de I’hysterosome considerablement plus longues; pattes plus longues et plus gr61es; tarses I et II pas moins de 2 fois plus longs quo leur diametre au milieu; espace entre I’extremite posterieure de la fente anale et le bord posterieur du corps depassant la moitie de celui qui separe sonextremite anterieure de 1’orifice genital.
4 (5). Z>2 3 fois plus longs que egalant en longueur la moitie de d3; cos derniers ont pres de 25—27% de la longueur de 1’idiosome; espace entre les fentes genitale et anale a peu pres egal a la distance entre 1’extre-mite posterieure de cette derniere et le bord posterieur du corps; soies postanales situees fort en arriere de la fente anale .......................
...............................................5. A. michaeli (Ouds.).
3 (4). D2, egalant en longueur <4 et 5 fois plus courtes que д,я, ou de la тёте longueur que ces dernieres lorsque elles ont 1’aspect de microchetes; espace entre les fentes genitale et anale 1.7 fois plus grand que la distance entre I’extremite posterieure de celle-la et le bord posterieur du corps; soies postanales placees un peu en arriere de I’extremite de la fente anale.
6 (7). D3 ont 1’aspect de microchetes, de тёте que dx et d2; longueur de h e et di pres de 30% de la longueur de 1’idiosome.........................
..................................................4. A. agilis (Can.).
7 (6). Ds 5 fois plus longues que d2, egalant a peu pres 15% de la longueur de 1’idiosome; et sc i sont a peu pres de la тёте longueur (17—18%), he est de 20% de la longueur de 1’idiosome. — URSS (Leningrad, Moscou, Ivanovo, Bachkirie, Siberie occidentale)...........................
................................................2. A. sokolovl, sp.n.
8 (1). Sc г de 2 a 3 fois plus courtes que sc e; 1’espace entro les 2 soies do cotte paire depasse la distance qui les separe de sc e pas plus qu’ 1.5 fois; soies ventrales des tarses I et II ordinairement spiniformes; organe ' pseudostigmal plumeux.
; ’9 (10). Fente anale tr£s rapprochee du bord posterieur du corps; idoisome
2.1—2.2 fois plus long que large; soies ventrales des tarses I et II fines,
' aciculees; Ip 3 fois plus longues que la.—Italie, URSS (region
d’Odessa).......................................8. A. krameri (Berl.),
10 (9). Fente anale tres eloignSe du bord posterieur du corps. Idiosomo ран plus de 1.9 fois plus long que sa largeur maximum, I p pas plus do ,1 ,B
! fois plus longues que I a; soies ventrales des tarses I et II spiaifornioH.
| 21 (12). Soies de 1’idiosome droites, aciculees; ds courtes, do Ju тёшо loil-
! gueur, que d2; d3 egalant celles de d4 (12—15% do la Iongiioill' do
1’idiosome). — Angleterre.........................7. Л. и I bortl (A. Z,).
12 (11). Soies de 1’idiosome flexibles, non aciculdos.
13 (14). Soies de 1’idiosome longues, tres fines, сарШЯогтон; loligliour do et d4 Egalant de 24 h 25, I p et sa i pr£s de 21,—23, sa <i —do 17 Л 18%
428
TYROGLYPHIDAE
de la longueur de 1 ’idiosome; sc i un peu plus de 2 fois plus courtes que sc e; organe pseudostigmal aplati, large, lanceole, ayant quelques appendices spiniformes sur sa surface dorsale; les soies ventrales mediane-et interieure des tarses I et II ayant 1’aspect de longues et minces epines ........................................ 10. A. batsylevi, sp. n..
14 (13). Soies de I’idiosoinc plus courtes: longueur de ds et d4 ne dopasse quo de 15 й 1G, I p et sa a do 10 h 12, sa i 18% de la longueur de 1’idio-some; sc i 3 fois plus courtes quo sc c; une partie des soies de' la surface dorsale — dj, da, la et quelqucfois mGme ds, d4, I p et h i— obtuses aux bouts; organe pseudostigmal plus etroit, plumeux rien qu’aux bords; soies medianes et interieure de tarses I et II, ayant 1’aspect de courtes, epines robustes. — URSS (Bachkirie, region de Kalinine)...................
..........................................6. A. redikorzevi (A. Z.)..
16 (16). Soies de la surface dorsale de 1’idiosome plus courtes; la longueur de v i represents de lb a 12, sc e de 25 a 27, h e pres de 20, da 7 a 7.5, d$ de 12 a 13, la de 7.5 a 8% de la longueur de 1’idiosome; organe pseudostigmal plus large et plus lamelliforme. — URSS (Bachkirie) . . .. . . . .............................. 6a. A. redikorzevi redikorzevi A. Z.
16 (15). Soies de la surface dorsale de 1’idiosome plus longues; la longueur de v i atteint de 13.5 a 14, sc e de 30 a 33, h в pres de 25, % 10,5 a
11, ds de 15 a 20, Id—11% de la longueur de 1’idiosome; organe pseudostigmal plus etroit, presque cylindrique. URSS (region de Kali-nine) .......................... 6b. A. redikorzevi borealis, ssp. m
Hypo pes
1 (8). Bousson propodosomal a peu pres triangulaire, a bords obtus, non aplatis, n’occupant pas moins d’un quart la surface dorsale de 1’idiosome; plastron ventral de consistance membraneuse, presque atro-phie et portant alors de 1 a 2 paires de ventouses rudimentaires, ou bien fort allonge,etroit,linguiforme, avec 4pairesde ventouses plus ou moins rudimentaires; champs coxaux I et II faiblement chitinises, ne formant, pas d’ecusson sternal bien limite; la region de 1’ecusson genital n’est separee ni des champs coxaux IV, ni du plastron ventral.
2 (5). Plastron ventral avec 4 paires de ventouses (situees dans 1’ordre sui-vant: 2, 2, 4), dtroit, linguiforme, fortement allonge; venteuses des-paires antGrieure et centrale assez grandes, mais mal dessinees, les-posterieures toutes de la тёте dimension, petites et plus saillantes;. ventouses coxalcs plus ou moins distinctes................................
3 (4). Saillie rostrale en pointe aigue; taille plus forte. — URSS (Moscou,. Leningrad, Ivanovo, Bachkirie, Sibdrie occidentale).......................
...............................................2. A. sokolovi, sp. n.
4 (3) Saillie rostrale obtusement arrondie; taille plus faible. — URSS (Moscou, Kostroma, Jitomir).....................1. A. rhizoglyphoides (A. Z.).
ACOTYLEDON
429
5 (2). Plastron ventral tout-a-fait rudimentaire, ayant 2 paires de ventouses plus ou moins discernibles; ventouses coxales absentee.
6 (7). Fente ano-genitale tres longue, flanquee derriere le milieu d’une paire de ventouses assez bien dhveloppees; la seconde paire, tout-a-fait rudimentaire, se trouve un peu plus en arriere de la premibre et n’est reprh-sentbe que par leurs pores doubles, situees tres pres 1’une de 1’autre; epimerites II a peine distincts; le sternum est loin d’atteindre le bout des epimeres II. — URSS (Kiev, Irkoutsk) . . 3. A. paradoxa (Ouds.) 7 (6). Fente genitale courte, non reliee a la fente anale; les deux paires de ventouses sont disposees considerablement plus prbs du bord posterieur du corps, celles de la premibre paire bien plus ecartees que les poste-rieures, mais de la meme forme et de meme dimension; epimerites II assez accentues; sternum long, s’etendant presqu’au meme niveau que les epimeres II. — Angleterre...................6. A. michaeli (Ouds.).
8 (1). Ecusson propodosomal de forme differente, a bords aplatis (foliaces) recouvrant la base ou la plus grande partie des pattes I et II; sa longueur ne depasse pas Vs partie de celle de 1’idiosome; plastron ventral de structure normale, fortement chitinise, arrondi ou bien transversale-ment elliptique, ayant 4 paires de ventouses bien devolopphes; champs coxaux I et II egalement fort chitinises, formant un hcusson sternal ri-gide trbs bien dessine; champs coxaux III ouverts; dans le cas contraire ils sont tres hearths; ecusson genital distinctement delimite, separe de I’ecusson ventral et du plastron adhesif.
9 (10). Soies peripheriques de 1’hysterosome (surtout hi, he, sa i et pa) robustes, spiniformes. — Angleterre . ................7. A. alberti (A. Z.).
10 (9). Soies phripheriques de 1’hysterosome fines, flexibles, capilliformes. 11 (14). Bord anterieur de 1’hcusson propodosomal presque droit, faiblement ondule, a lobe median faiblement arque et echancrurcs lathrales peine marquhes; saillie rostrale dhplache sur la surface ventrale de I’ecusson, totalement recouverte par son bord, quelquefois absente; gnathosome a bords paralleles ou bien faiblement rhtrhei vers le bout.
12 (13). Espece robuste (380 p.) de couleur foncee; ecusson propodosomal extrhmement court, 36 fois plus court que 1’hysterosome; ecusson ventral considerablement plus long que le sternal, traverse dans toute sa longueur par une suture ventrale; champs coxaux plus quA moitid ouverts; pattes I et II courtes et robustes, plus qu’a moitih cachhos par I’ecusson propodosomal; soies exterieures des tibias et des ghiiiinux I et II robustes, en lame de coute.au. — URSS (Bachkirie)...................
.................................................11. A. volglnl, up. II.
13 (12). Petite espece faiblement coloree; I’ecusson propodosoinnI H fois seulement plus court que 1’hysthrosome; ecusson vonl.rnl boniicoiip plus court que le sternal, n’ayant pas de suture vontraIo; clininps coxaux II fermes, tres ecartes;pattes I et II longues et fines; pnlles I (Ibpassiint
430 TYROGLYPHIDAE
de beaucoup I’ecusson propodosomal (й commence! par le haut des femurs); les pattes II le depassant й commence! par I’extremite des genuaux; Spines exterieures des tibias et genuaux I et II minces, me-nues. — URSS (region d’Odessa)............ 12. A. tshernyshevi, sp. m
14(11). Bord anterieur de I’ecusson propodosomal profondement echancre en arc des deux cot£s do la saillie rostrale; celle-ci proeminente, de forme triangulaire, огпбо sur son sommet de soies bien developpees; les soies ventro-laterales des tibias et dos genuaux I et II elargies, cultriformes; gnathosome piriforme, pas plus de 2 fois plus long que large, proto n-dement fendu au bout; champs coxaux III ouverts, epimeres IV libres. 16(16). Saillie rostrale assez large, plus courte que les lobes lateraux de
I’ecusson propodosomal qui forment angle droit et ne sont arrondis qu’au sommet; 1’espace entre le bord posterieur du plastron ventral et le bord posterieur de 1’idiosome est de la longueur de I’ecusson sternal.— Italie, URSS (region d’Odessa)............8. A. krameri (Berl.).
16 (15). Saillie rostrale plus longue que les lobes lateraux de I’ecusson propodosomal, qui sont fortement arrondis presqu’en demi-cercle; cette saillie a la forme d’un appendice etroit tres saillant; espace entre le bord posterieur du plastron ventral et celui de 1’idiosome est plus petit que la longueur de I’ecusson sternal. — URSS (Bachkirie, region de Kalinine).......................................6. A. redikorzevi (A. Z.).
CALOGLYPHUS BERL.
UU
1 (6). JU heteromorphes.
2 (3). Organe pseudostigmal bien developpe, setiforme, egal en longueur au diametre des tibias I; peau lisse; p2 beaucoup plus longues que pn capilliformes.—Allemagne, URSS (Moscou, Leningrad, Ivanovo,. Jaroslavl, Barnaoul, Caucase du Nord, Bachkirie).......................
............................................1. C. rodionovi A. Z.
3 (2). Organe pseudostigmal ayant la forme d’un batonnet minuscule (5— 8 fi), visible seulement й un fort grossissement.
4 (5). Peau de 1’hysterosome distinctement rugueuse, organe pseudostigmal sensiblement epaissi vers la base, presque conique; p2 beaucoup pluslongues que pi, capilliformes; ventouses anales de dimensions normales.— Mandzhurie (Kharbine)......................3. C. mandzhur, sp. n.
5 (4). Peau absolument lisse; organe pseudostigmal cylindrique; p2 tres-courtes, ventouses anales tres petites. — Allemagne, URSS (Kiev) . . , ..............................................4. C. spinitarsus (Herm.).
6 (1). UU homeomorphes.
7 (12). Ventouses anales elliptiques, fortement chitinisees, obliques; leurs-bords posterieurs situes sensiblement plus bas que I’extremite de la fente anale; la longueur de cette derniere depasse mediocrement celle;
CALOGLYPBUS 431
des ventouses; soies de 1’idiosome fortes, ties rigides; especes de petite taille, n’ayant pas plus de 730 p. de longueur.
8 (9). Tarses IV munis de deux fortes soies falquees, d2 et d3 a peu pres de la meme longueur, sensiblement plus courtes que Zpet plus de 2 fois plus courtes que d4. — France, Hollande, Allemagne, URSS (environs de Moscou)...............................................6. C. moniezi A. Z.
9 (8). Tarses IV avec une seule soie falquee; d3 beaucoup plus longues que d2 et I p.
10 (11). Tons les tarses sont munis de fortes soies falquees; partie distale du penis grSle, droite; sc e, h e et Z p ne depassant pas 40—60, sa e 15%. de la longueur de 1’idiosome.— URSS: Transcaucasie (Batoumi) . . . ...................................................5. C. polyphyПае, sp. n.
11 (10). Tarses III et IV depourvus de fortes soies falquees; partie distale du penis recourbee, formant angle avec le socle, ce qui le fait paraitre obtus; longueur de sc e, 1г e et Z p 60—65, sa c 25% de la longueur de 1’idiosome. — Italie..............'................7. C. berltsei (Mich.).
12 (7). Ventouses anales arrondies ou ovoides, leurs bords posterieurs situ£s sensiblement devant I’extremite de la fente anale, cette derniere 2—3 fois plus longue que les ventouses; soies de 1’idiosome presque toujours assez fines et flexibles. Dimensions variables, souvent trbs fortes (pres de 1100 p).
13 (16). P2 plusieurs fois plus longues que sdtiformes, depassant de beaucoup le bord posterieur de 1’idiosome; soies de 1’hysterosome plus, longues.
14 (15). Peau de 1’hysterosome finement rugueuse; organe pseudostigmal ayant 1’aspect d’un ties petit batonnet (6—8 p.) conique; bord posterieur de I’ecusson propodosomal echancrb; 1’espace entre les p2 est 2 fois moindre que celui qui s epare les px; soies de 1’idiosome plus longues.— Mandzhurie (Kharbine).......................3. C. mandzhur, sp. n.
15 (14). Peau absolument lisse; organe pseudostigmal bien ddveloppe,. setiforme, egalant presqu’en longueur le diametre du tarse I; bord posterieur de I’ecusson propodosomal non echancre, droit ou faible-ment convexe; intervalle entre les p2 rien que 2—2.3 fois plus etroit qu’entre les px\ soies de 1’idiosome plus courtes. — Allemagne, URSS (Moscou, Leningrad, Jaroslavl, Barnaoul, Caucase du Nord, Bachkirie) ..............................................1. C. rodicnovi A. Z.
16 (13). P2 de memes dimensions que px, courtes; poils de I’hysterosomo plus courts.
17 (18). Batonnet olfactif des tarses I et II mince, cylindrique; idiosome fortement renfte, 1.5 fois plus long que sa largeur maximum; hypostome faiblement retreci anterieurement a des bords lateraux coiir-bes en S. — URSS (environs de Moscou) . . . 8. C. sphacroglister Л. Z.
18 (17). Bdtonnet olfactif des tarses I et II massif, fortement dpaissi au bout; idiosome assez svelte, presque 2 fois plus long que sa largeur maxi-
43'2 TYROGLYPHIDAE
mum, gnathosome tres etroit; hypostome tres retreci anterieurement, ses bords lateraux presque droits. — France........................
....................................... 9. C. mycophagus (Megn.).
99
1 (2). Bdtonnct olfactif des tarses I et II epais, fortement renfle en massue; gnathosome trfcs dtroit; hypostome fortement retreci anterieurement, a bords lateraux presque droits. — France...............................
..........................................9. C. mycophagus (M6gn.).
2 (1), Bdtonnet olfactif des tarses I et II mince, cylindrique; proportions du gnathosome normales, hypostome faiblement retreci par devant, a bords plus ou moins fortement sinues.
5 (4). Lamelles des soies falquees des tarses tres larges. Organe pseudostigmal absent......................................5. C. polyphyllae, sp. n.
4 (3). Lamelles des soies falquees des tarses plus etroites.lanceolees; organe pseudostigmal present (quoique souvent minuscule). '
5 (6). Organe pseudostigmal bien developpd, setiforme, egalant en longueur le diametre du tarse I; sa e tres courtes, de 8 a 9 fois plus courtes que sa i; Ip de 2 a 3.6 fois plus longues que la-,5 paires de microchetes anales, dont les 4 posterieures sont situees a une distance a peu ргёэ egale etforment une rangee arquee de chaque cdte de la fente anale. — Allemagne, URSS (Moscou, Leningrad, Jaroslavl, Barnaoul, Caucase-du Nord, Bachkirie)...............................1. C. rodionovi A. Z.
•6 (6). Organe pseudostigmal tres petit (4—8 p.), en forme de bdtonnet (visible seulement a de forts grossissements); de 3 a 6 paires de microchetes anales disposees d’une manure differente.
7 (8). Rien que 3 paires de microchetes anales; lamelles des soies falquees des tarses tres etroites. — Allemagne, URSS (Kiev)-.....................
................. .......................4. C. spin!tarsus(Herm.).
8 (7).4 a 6 paires de microchetes anales;lamelles des soies falquees des tarses plus larges.
® (10). Pattes tres greles; la longueur de d3, d4 et Ip egale 16 a 21% de la longueur totale de 1’idiosome; 4 paires de microchetes anales, dont deux sont situees pres de I’extremite anterieure. de la fente anale, une h mi-longueur et une paire au milieu de la distance entre la paire mediane et l’extr6mit<5 postOrieure de Г anus; palpes genitaux, atrium et bords de la f ente anale d’un roux ferruginoux. — URSS (environs de Moscou) ............................................. 8. C. sphaerogaster A. Z.
10 (9). Pattes robustes; lalongueur de ds, di et Ip ne depasse pas 8—11% de la longueur de 1’idiosome; 6 paires de microchetes anales, dont 2 paires sont situees a cote de I’extremite anterieure de la fente anale, une au milieu et trois autres sont groupees pres de son extremite poste-rieure; palpes genitaux, atrium et bords de la fente anale non pig-mentes.—Mandzhurie (Kharbine) ....... 3. C. mandzhur, sp. n.
CALOGLYPHUS
Ш
Hypopes
1(2). Ventouses coxales remplacees par des soies; articles apicaux'du gnathosome largement ecartes, longs (egalant en longueur son article basal, qui est саггё), subdivises en 2 articles: champs coxaux III et IV separes par un large intervalle longitudinal. — France...............
........................................9. C. mycophagus (Megn.).
2 (1). Ventouses coxales de structure normale; articles distaux du gnathosome rapproches, courts (toujours bien plus courts que 1’article basal), non subdivises ou bien meme peu distincts; champs coxaux III et IV separes par un etroit espace longitudinal.
3 (4). Pattes I et II tres courtes et epaisses, coniques, presque totalement cachees sous 1’ecusson propodosomal qui a la forme d’un croissant; son bord semicirculaire est a peine depasse par les bouts des tarses I et les soies foliacees des tarses II; v i sont situees sur la surface ventrale de 1’ecusson propodosomal a une assez grande distance de son bord ante-rieur. Еэрёсе de grande taille (400—420 p) qu’on rencontre sur 1’Os-moderma eremila Scop. — Allemagne, URSS (Kiev).......................
........................................4. C. spinitarsus (Herm.).
4 (3). Pattes I et II depassant les bords de I’bcusson propodosomal de trois articles au moins, plus greles; ecusson propodosomal plus ou moins triangulaire, a angle rostral saillant; vi sont situees au sommet de cet angle.
5 (12). Gnathosome totalement сасЬё sous la saillie rostrale de 1’ecusson propodosomal, au delA. duquel ressort seulement la partie distale des aristes.
6 (7). Ecusson propodosomal un peu plus court que la тоШё de I’hystero-some; gnathosome ovoidal; les branches de sa fourche courtes, non separees de 1’article basal; soie ventrale interieure des tibias II beau-coup plus grande que 1’exterieure, epimeres II convergents en angle obtus; ecusson sternal limite posterieurement par une ligne arquee. Petite espbce (200—236 p) blanchStre, se rencontrant en profusion sous les elytres de Geotrupes stercorosus Forst. —URSS, France, Allemagne ........................................ 10. C. geotruporum, sp. n.
7 (6). Ecusson propodosomal pas plus long que le х/4 de 1’hysterosoma!; branches de la fourche du gnathosome plus longues, visiblement separees de son article basal; epine ventrale interieure des tibias II plus courte que 1’exterieure; epimeres IV se rejoignant en angle aigu; ecusson sternal limit6 posterieurement par une ligne en forme de trapbze. Especes plus grandes.
8 (9). Ecusson propodosomal egale a x/6 dfi 1’ecusson hysterosoma!; pattes I et II courtes, tres robustes, depassant le.bord anterieur de 1’idiosome rien que des genuaux, tibias et tarses; ces derniers tr£s courts, considerablement plus courts que le genual et le tibia pris ensemble et rien
28 Фауна СССР, Захватит—284
434 TYROGLYPHIDAE
que 1.2—2 fois plus longs que leur diametre basal; epines ventrales exte-rieures des tibias I et II de la тёше longueur que les tarses; bords ante-rb-lateraux de I’dcusson propodosomal regulierement, quoique faiblement convexes sur toutc lour 6tendue; saillie rostrale a peine esquissee. Longueur 350—390 [i. Sur le Polyphylla fullo L.— URSS : Transcaucasie-(Batoumi)...................................5. C. polypliyllae, sp. n.
9 (8). Bousson propodosomal do 4.5 a 5 f ois plus court que celui de I’hystero-somc; pattes I et II plus greles; tarses dgalant on longueur ou depassant les genuaux et tibias respectifs, pris ensemble et de 3 a 4 fois plus longs-que leur diametre basal; epines exterieures ventrales des tibias I et II beaucoup plus courtes que les tarses; bords anterieurs de I’ecusson propodosomal distinctement sinues des deux cbtes de la saillie rostrale, qui est bien dessinee.
10 (11). Pattes II, ne depassant les bords du propodosome que de la moitie’ distale du genual, du tibia et du tarse; epines ventrales interieures des tibias I et II beaucoup plus fines que les exterieures; bords lateraux. du gnathosome droits, paralleles; saillie rostrale moins proeminente; la. longueur de I’ecusson ventral ne depasse pas sa largeur maximum; lai largeur du disque adhesif n’est que 3 fois nioindre que la largeur maximum du corps. Longueur 225—390 p. Commun sous les elytres des hannetons (Melolontha). — France, Hollande, Allemagne, URSS (environs de Moscou)......................................6. C. moniezi A. Z.
11 (10). Pattes II depassant le bord du propodosome de la moitie distale-des femurs, des genuaux, des tibias et des tarses; epines ventrales interieures des tibias I et II presque aussi robustes que les exterieures; bords-lateraux du gnathosome convexes, saillie rostrale tres proeminente; longueur de I’ecusson ventral depassant considerablement sa largeur maximum; la largeur du disque adhesif tient 4 fois dans celle du corps. Longueur 400—440 p. Hotes transporteurs inconnus. — Mandzhurie (Kharbine).........................................3. C. mandzhur, sp. n.
12 (5). Gnathosome, depassant le sommet de la saillie rostrale de toute la partie apicale de sa fourche ou тёте de toute sa moitie 'distale.
13 (14). Les branches distales du gnathosome ne sont pas separees de Particle basal; gnathosome piriforme, allonge, depassant le bord anterieur du. propodosome de toute sa moitie distale; bords lateraux sternal fortement recourbes en S; ventouses posterieures du disque adhesif un. peu plus petites que les anterieures; soies ventrales des tibias I et II fines, non spiniformes. Sous les Elytres cPOryctes nasicornis L. et-de PolyphyUa fullo L. — Italie . ... ... 7. 0. berlesei (Mich.).
14 (13). Branches distales du gnathosome formant des articles independants;, article basal a bords paralleles; bords lateraux de I’ecusson sternal presque droits; ventouses posterieures du disque adhesif beaucoup plus petites que les anterieures; soies ventrales des tibias I et II spiniformes.
-16 (16). Gnathosome depassant le sommet de la saillie rostrale d’un quart.
CALOGLYPHUS, HISTIOG ASTER
43^
ou d’un tiers de son article basal et de toute sa fotirche; son article basal de 3 & 3.5 fois plus long que large; Ecusson propodosomal de 6 h 8 fois plus court que Lhysterosome; soies interieures ventrales des tibiaslot II un peu plus courtes que les exterieures; soie exterieure ventrale des. genuaux II spiniforme; soies postanales plus courtes que les tibias II. — Allemagne, URSS (Moscou, Leningrad, Jaroslavl, Barnaoul, Caucase-du Nord, Bachkirie)..........’............ . . 1. С. ro dionovi A. Z.
16 (15). Gnathosome depassant le sommet de la saillie rostrale rien que du bout de ses articles distaux; article basal 2 fois plus long que large. Ecusson propodosomal 4 fois plus court que 1’hysterosome; soies interieures ventrales des tibias I et II de 2 a 3 fois plus courtes que les exterieures; soie exterieure ventrale des genuaux II pili-forme; soies postanales longues, 2 fois plus longues que les tibias II. — URSS (region d’Odessa)..........................2.* C. tshernyshevi, sp. m
HISTIOGASTER BERL.
dd
1 (2). La longueur dela lamelle opisthosomale egale sa largeur basale et n’est qu’1.5 fois plus petite que sa largeur maximum; lames hyalines de I’eventail sensiblement plus longues que larges; penis relativement petit; les pattes anterieures separees par un grand espace des paires posterieures (M. carpio Berl. 1883, non Kramer 1881). — Italic . . .
...................... . . ...............3. H. silenus, nom. n.
2 (1). Lamelle opisthosomale pres de 2 fois plus courte que large; lames hyalines deTeventail pas plus longues que larges; penis tres grand; espace entre les pattes anterieures et posterieures mediocre.
3 (4). Idiosome largement oval, 1.5—1.65 fois plus long (en comptant la lamelle opisthosomale) que large;, lames hyalines de I’eventail tout-a-fait separees; les festons divises par des echancrures assez pro-fondes. — Allemagne, URSS (Caucase, Abrau-Dursau)......................
............................................. 1. H. bacchus, sp. n.
4 (3). Idiosome allonge, plus de 2 fois plus long que large; lames hyalines de I’eventail confluentes a leur base, formant une bordure ininter -rompue a 4 lobes autour de la lamelle opisthosomale; les festons de cette derniere se touchent.de leurs bases. — Allemagne................
.............................................2. H. carpio (Kram.)..
Hypopes
1 (2). Gnathosome 1.5 fois plus long que large a sa base, en forme, de trapeze allonge; epimerites II n’atteignant pas de beaucoup les acetabules III;-leur bout donne naissance a unfih appendice ondule, dirige en avant; p a tres longues (pres de 25% de la longueur de 1’idiosome).. Lattes III
♦
436 TYROGLYPHIDAE
'et IV ayant 28—30% de longueur de 1’idiosome, avec de tres longues macrochetes; ventouses anterieures beaucoup plus grandes que les laterals et les posterieures. — Allemagne, URSS (Caucase, Abrau-Dur-sau)............................................ 1. H. bacchus, sp. n.
2 (1). Gnathosome 2—3.6 fois plus long que large, h cotes paralleles; epi-merites II rejoignant dos bouts le bord exterieur des acetabules III; epimeres III sans appcndice onduld; pa tr^scourtes(pas plus de8%de la longueur do 1’idiosomc); pattes III ct IV ayant 22—24% de la longueur de 1’idiosome;'macrochbtcs dos tarses III et IV courtes, ventouses anterieures de тёте dimension que les later ales, ou plus petites.
3 (4). Soies ventro-laterales des tibias et des genuaux I et II fines, non trans-formees en.epines; la soie dorso-apicale des genuaux tresgrande, voisine par ses dimensions de celle des tibias; bouts des epimeres II recour-bes exterieurement; ventouses coxales assez grandes; yeux disposes dans 1’axe longitudinal du corps, leurs bords sont par alleles sur toute leur etendue. —URSS (Vladivostok)...................4. H. Silippovi, sp. n.
4 (3). Soies ventro-laterales des tibias et des genuaux I et II ayant 1’aspect de courtes et robustes epines; soie dorso-apicale des genuaux petite; bouts des epimeres II convergeant, non recourbes й. I’exterieur, ventouses coxales tres petites; yeux disposes obliquement, presque relies des bouts anterieurs et divergeant largement en arriere.— Brout-Vernet (France)......................................5. H. ocellatus (Vtz.).
SCHWIEBEA OUDS.
1 (2). Sommet des genuaux I avec 2 soies; epines ventrales des tibias et des genuaux III fines, peu developpees (chez les exemplaires heteromor-phes); tarses I et II tres courts; p3 disposees pres du bord posterieur de la lamelie opisthosomale, auтёте niveau que les microchetes postanales (p2). (Sous-genre Megniniettia Jacot)...............................
........................................... 6. S. (M.) rossica, sp. n.
2 (1). Sommet des genuaux I avec une seule soie tres longue; les exemplaires heteromorphes ont les Spines des tibias et les genuaux III tres grosses, й bouts arrondis; tarses I et II plus longs; p3 situees sur les bords lateraux de la -surface ventrale de la lamelie opisthosomale — beaucoup au devant du niveau de la paire posterieure des microchetes anales (p2). (Sous-genre Schwiebea s. str.).
3 (4). D4 situees dans la partie centrale de la lamelie opisthosomale; d3 et la representes par des microchetes; sa e 3 fois plus courtes que p3 et plus de 2 fois plus courtes que Z p. — Allemagne...........................
............................................ 2. S. (s. str.) ipidis Vtz.
4 (3). situees tout au bord de la lamelie opisthosomale; d3 et I a absentee; sa e un peu plus longues que p3 et plus de 2 fois plus longues que Ip. — URSS (region d’Odessa). . . 1. S. (s. str.) tshernyshevi, sp. n.
SCHWIEBEA,. MONIEZIELLA И7
$9
1 (2), Epimeres III relies par les bouts interieurs aux epimeres IV; hohijiioI; du genuaux I avec 2 soies dorsales; soie exterieure ventrale des libins 1 et II absente; idiosome fortement allonge, de 2.2 & 2.4 fois plus long que large; bords lateraux de I’hysterosome droits, paralleles sur une grande etendue.Longueur 280—360 p. (Sous-genre Megniniettia Jacot). . ..............................................6. S. (M.) rossifca, sp. n.
2 (1). Epimeres II et IV libres, non.relies les uns aux autres; sommet des genuaux.I n’ayant qu’une soie dorsale; toutes les deux soies ventrales des tibias. I et II bien developpees; idiosome moins allonge, de 1.Б5 a 2 fois plus long que large; bord lateraux de I’hysterosome plus ou moins convexes. (Sous-genre Schwiebea s. str.).
3 (4). Largeur de 1’idiosome egale de 1.66 й 1.7 de sa longueur; I’extremite du corps etroitement arrondie,peau£paisse,faiblement coloree, d’un ton brundtre; ecusson propodosomal indistinct; soies de I’hysterosome tres fines, flexibles, capilliformes; pattes II et IV a peine plus courtes et plus minces que les deux paires anterieures; batonnet olfactif des tarses I et II droit, s’epaississant un peu vers le bout. Longueur 296— 300 p.—URSS (Moscou, region de Kalinine)..............................
...................................5. S. (s. str.) pachyderma, sp. n.
4 (3). La longueur maximum de 1’idiosome a un peu moins du double de sa largeur; partie posterieure du corps largement arrondie; peau de con-sistance normale, incolore; soies de I’hysterosome plus fortes et plus rigides; pattes III et IV beaucoup plus courtes et plus fines que les pattes I et II; batonnet olfactif des tarses I et II fortement recourbe, avec le bout renfle en bouton.
5 (8). L a et ds representees par des microchbtes bien distinctes.
6 (7). Longueur 690—630 p; ecusson propodosomal distinct; soies de 1’idiosome plus longues. — Allemagne. . . . 2. S. (s. str.) ipidis Vtz.
7 (6). Longueur 365—460 p; ecusson propodosomal indistinct; soies de 1’idiosome plus courtes. — Yougoslavie................................
.....................................3. S. (s. str.) cavernicola Vtz.
8 (5). La et ds absentee. — URSS (region d’Odessa).....................
...........:........................ 1. S. (s. str.) tshernyshevi, sp. n.
MOMEZIH.I.A BERL.
def
1 (2). Gnathosome excessivement btroit, cheliceres encore plus retrecies vers la base, formant un «сои» assez court; he longues; longueur de 1’idio-some pres de 290—300 p. — Angleterre . . . .1. M. corticalis (Mich,).
2 (1). Gnathosome de forme normale, cheliceres non retrecies vers la base; Jiecourtes; longueur de 1’idiosome pres de 350—365 p(IH. wwZt’Berl.).— Italie ..................................: ..2. M. berlesiana, nom. n.
Д38
TYR0GLTPH1DAE
99
•1 (2). Longueur generale du corps pres de 780 p.. — Italie...........
...............................................3. M. magna Berl.
2 (1). Longueur generalo du corps ne depass ant pas 600 p..
3 (.4). Sternum fort raceourci, pattes III et IV plus longues que la mi-lar-geur de 1’idiosome. Longueur generate du corps pres de 450 p.. — Angle-terre.........................................1. M. cortipalis (Mich.).
4 (3). Sternum d6vclopp6 noriualemcnt; pattes III et IV plus courtes que la mi-largeur de 1’idiosome, longueur gdndrale du corps 500—600 p. — Italie....................................... 2. M. berlcsiana, nom. n.
LACKERBAUERIA, gen. n.
Hypo pe. Espece de grande taille. Coloration intense, d’un brun ferru-gineux. Sculpture de la surface dorsale composee de foveoles rondes de differentes dimensions (dans les preparations les Ecussons paraissant cribles de trous), quelquefois confluantes entre elles; le bord anterieur de I’hysterosome est garni d’une ponctuation tres fine aulieu de stries trans vers ales. Ecusson propodosomal triangulaire avec des bords antero-lateraux f aiblement c onvexes; sa 1 ongueur est a peu pr6s 2.3 fois moindre que celle de 1’ecusson hysterosomal. Le gnathosome ne depasse pas le sommet de la saillie rostrale; aristes longues, arquees, fortement diver-gentes. Ventouses genitales tres largement ecartdes; disque adhesif rappelant par sa forme et sa structure celui de quelques-uns des Glycy-phagidae (Horstia, Neohorstia)- on distingue dans sa partie posterieure 3 ilotsfort compacts entre les ventouses posterieures et au dehors d’elles; ventouses centrales pas plus grosses que les peripheriques; les poste-rieures disposees excentriquement dans leur gros cadre fortement chi-tinise. Batonnet olfactif des tarses I et II grele, avec son bout aiguise singulierement recourbe; tarses I et II muni de 4 soies elargies, 1апсёо-lees; tarses III et IV n’en ayant que trois; macrochetes des tarses IV tres longues. Longueur 235—300 p., largeur 170—310 p. — URSS (environs d’Ivanovo); trouve sur Crabro fossorius L...............
.........................................L. cribratissima, sp. n.
ROBINISCA, gen. n.
II у P 0 p 0 S
1 (2). Tibias I hypertrophies, presque 2 fois plus longs que les genuaux I et plus de 1.5 fois plus longs que leur diametre; partie visible de Гecusson propodosomal tres etroite — plus de 10 fois plus courte que I’hysterosome; epimeres II n’atteignant pas le bord posterieur . de 1’ecusson sternal; soie extero-laterale des genuaux I et II capil-liforme; batonnet olfactif des tarses I et II fin, cylindrique. — URSS (region d’lvanovo)............................1. R.. macrocnemis, sp. n.
LACKERBAttERIA, ROBINISCA, TROUPEAI/IA, SCIHILgKA . 439
2 <(1). Tibias I de dimensions norrnales, plus courts que les gdimaux I ot quo leur pro pre diametre; part ie visible de 1’ecusson propodosomal rien que 5.3—5 fois plus courte que 1’ecussonhysterosomal; les Spimfcros II atteignent le bord posterieur de 1’ecusson sternal; soies ext6ro-!utc-rales des genuaux I et II spiniformes, batonnet olfactif des tarses I et II epais, en forme de massue. Hollande, URSS (environs de Novotchcr-kassk) . ...................................2. R. mycolicha (Ouds.).
4
'* TBOUPEAUIA, gen. n.
I*;..
Hypopes
'I 1 (2). Ecusson sternal fortement etendu posterieurement en forme de tra-| peze; epimeres II recourbes en forme de S, divergeant des bouts; ecus-
4 rsons dorsaux lisses; tarses III et IV ayantchacun 2 macrochetes assez
longues; soie exterieure latero-ventrale des tibias I et II absente.—
Allemagne, URSS (environs de Moscou). ..... 1. T. nova (Ouds.).
2 (1). Ecusson sternal largement arrondi posterieurement; epimeres III ' faiblement arquees, convergeant des bouts; ecussons dorsaux cribles
de fossettes; tarses III et IV sans macrochetes; soie exterieure ventro-laterale des tibias I et II bien developpee. — URSS (environs de Moscou) ......................................2. T. crahrohis, sp. n.
SCHULZEA, gen. n.
Hypopes
1 (2). Pattes robustes.; tarses I plus courts que le tibia et le genual I pris ensemble; gnathosome court et robuste, completement cache sous la saillie rostrale; son article basal est presque carre; macrochetes des tarses IV plus courtes que la moitie de la largeur de 1’idiosome. Espece plus grande. — Pamir.......................... 1. S. pamirensis, sp. n.
2 (1). Pattes plus grSles; tarses I considerablement plus longs que le tibia et le genua,! I pris ensemble; gnathosome long, ses articles distaux (fourche) dbbordant de toute leur, longueur; Particle basal depassant ' plus de 1.5 fois sa largeur, se retrecissaht graduellement vers le som- met; macrochete des tarses IV egale a la largeur de 1’idiosome.
Espece plus petite. — URSS: Georgie (Kartveli).................
......................................... 2. S. caucasica, sp. n.
Г II. Famine SAPROGLYPHIDAE
<•'’ TABLEAU DES GENRES
'*• A d u 11 e s ‘
1 (8). Sc i bien plus courtes que sc c.
2 (3). Pretarses prives d’ongles; sommet des tarses munis ventralement d’une grande epine fortement recourbee en forme de griffe; pattes tres
440
SAP ROG LT PH IDAS
’ courtes (18—22% de longueur de 1’idiosome), les deux paires posterieures du d1 sensiblement epaissies et arquees; orifice genital place derriere la base des pattes IV, immediatement devant la fente anale. Espece tres petite, ne depassant pas de 220jji. . . 6. Nanacarus Ouds.
Une seule espbce, N. minutus (Ouds.). —Java, Allemagne, URSS (Smolensk).
3 (2). Pretarsos muni d Angles distincts; partie apicale des tarses, muiiie vcntralcment de 1- 3 petites (Spines (parfois peu distinctes); pattes de moyenne longueur ou bien assez longues (pas plus courtes de 30—36%. de la longueur de 1’idiosome); pattes posterieures du O' non epaissies; orifice genital situe chez la 9 entre les bases des pattes III et IV; chez le U il se trouve entre les bases des pattes IV. Especes plus grandes.
4 (5). cf avec de grandes ventouses copulatrices anales et tarsales; ces der-nieres, situees ala base meme des tarses IV(par une surchaque tarse); sc i, In, i, dj,, d2 et la representees par des microchetes, ayant pres de 10—12% de la longueur de 1’idiosome; sc e,he, I p, d3 et d4 de moyenne longueur (30—60% de la longueur de 1’idiosome). Espece littorale, habitant parmis les algues........................1. Pontoppidania Ouds.
Une seule espece, P. littoralis (Halb.). — Irlande.
5 (4). d,. denue de ventouses anales et tarsales, ayant 2 paires de microchetes anales; proportion des soies diff^rente. Espbces terrestres.
6 (7). Trois paires de tres grandes soies sur la partie posterieure du corps (atteignant de 60—90% de la longueur de 1’idiosome), plantees sur des verrues coniques; sc i de 8 h 10 fois plus courtes que sc e; epimeres III fortement recourbes, avec de trbs larges apodemes; idiosome etroit, fort allonge; pattes plus courtes et plus robustes .. 2. SaproglVphus Berl.
Une seule espece europeenne, S. neglectus Berl. — Italic, URSS (regions de Kalinine et d’Odessa).
7 (6). Rien qu’une paire de longues soies sur la partie posteiieure du corps; sc i 2.5—6 fois plus courtes que sc e; epimeres III faibles, presque-droits, avec des apodemes etroites; idiosome plus court et plus large; pattes longues et giAles . . <..........................4. Calvolia Ouds.
8 (1). Toutes les deux paires de sc a peu pres de la тёте longueur.
9 (10). Pattes I et II robustes, bien plus longues que les pattes III et IV; tarses trbs courts et epais; leur partie apicale gamie ventralement de-3 grandes epines recourbees; see notablement plus longues que toutes-les soies de I’hystfrosome; ch61ici:res de structure normale; orificegenital de la 9 situ6 entre les bases des pattes IV, denue d’epigyne. . .
............................; ....... 3. Win ter schmidtia Ouds.
Une seule езрёсе, W. hamadryas (Vtz.). — Allemagne.
10 (9). Toutes les pattes greles, I et II visiblement plus courtes que les pattes III et IV; tarses longs, minces, sans epines au bout; sc quelque pern plus courtes que la plupart des soies de 1’hysterosome; cheliceres larges h la base et se retrecissant subitement en pointe aigue; orifice geni-
ENSLINIELLA, CALVOLIA 441
tai de la 9 situe dans 1’espace coinpris entre les biisos (1пн pnlh's Ш et IV, avec un epigyne bien developpe..........5. ЕпнНпЬ'Пи Viz.
H у p 0 p e s
1 (2). Pattes IV fortement rapprochees des bords posterieurs du corps, trC*H fortes, a trochanters extremement developpes et tarses armes au bo 111 d’une forte epine digitiforme, d’une seconde epine en forme d’ecaillo a sommet effile, de 2 fortes soies et d’une tres petite; hysterosome avec une apodeme longitudinale posterieure; saillie rostrale muni d’une paire d’yeux hemispheriques fortement proeminents. Sur les guepes du genre Odynerus..................................0. Ensliniella Vtz.
2 (1). Pattes IV disposees a une distance considerable de I’extremite posterieure du corps; trochanters petits; tarses sans epines, n’ayant que de simples poils setiformes; apodfcme longitudinale absente. Sur divers-autres insectes.
3 (4). Sur le sommet de la saillie rostrale se trouve une paire d’yeux hemispheriques fort proeminents et munis .de retines distinctes; gnathosome bien developpe......................................4. Calvolia Ouds.
4 (3). Yeux absents ou peu developpds, prives de retines et situes dorsale-ment h la base de la saillie rostrale; gnathosome absolument atrophie, represente rien que par une paire d’aristes............7. Vidia Ouds.
ENSLINIELLA VTZ.
Ну p 0 p e s
1 (2). Tarses III considerablement plus courts que leurs ambulacres, armes d’une epine" digitiforme, semblable a celle des tarses IV et de 4 soies foliacees; yeux presque contigus; epaississements marginaux de 1’ecus-son ventral arques, relies au cadres chitinises qui entourent les ventouses anterieures; rebord transparent, entourant le disque adhesif, tres-large. Sur Odynerus (Lionotus) floricola Sauss. — Allemagne, URSS-(Armenie, Ouzbekistan)......................... . . 1. E. parasitica Vtz.
2 (1). Tarses III de la тёте longueur que leurs ambulacres, sans epine-digitiforme, avec 3 soies foliacees seulement; intervalle entre les yeux egal au diametre des cornees; epaississements du bord de 1’ecus-son ventral anguleux, non relies aux cadres chitinises des ventouses-anterieures; rebord transparent autour ’du disque adhesif tres etroit. Sur Odynerus (Lionotus') rossii Lep. — URSS: Bielorussie, Crimee — (Karadagh)....................................2. E. kostylevi, sp. n.
CALVOLIA OUDS.
A d u 11 e s dd’ - .
1 (2). Bord posterieur de 1’idiosome saillant en angle droit, orne de 6 tuber-cules plus ou moins: hemispheriques; Ip de la longueur de 15—17%,
-142 SAPRQGLYPHIDAE
de celle de 1’idiosome; epimeres IV diagonaux, formant un angle obtus avec la ligno chitineuse qui les relie a 1’orifice genital; peau striee dans la partio posterieure de I’hysterosome, surtout au dessous et lateralement. Longueur totale du corps 323 y. — URSS: Georgie (Karajazy) , , ,............................. 1. C. tuberculata, sp, n.
:2 (1). Bord posterieur de I’idiosome formant un angle tres obtus, sans tuber-cules; Ip longues, de Г><)% a peu pres de la longueur de I’idiosome; epimeres [V transverse iix, disposes dans la direction de la ligne chiti-ncuso connective; peau absolument lisse. Longueur totale du corps pres de 280 p. —Angleterre, Hollande, URSS (Extreme Orient) .... ..................................................2. 0. heterocoma (Mich.).
9?
TL (2). Peau tres finement et legerement striee, surtout sur la surface abdomi-nale.; 1’une des soies apicales des tarses I et II epaissie en forme de ba-tohnet; soie ventrale mediane des memes tarses situde plus pres de la base des tarses que le bdtonnet olfactif. Longueur totale du corps pres de 290 у..— Italie................................4. C. hebeclinii (Sieber).
:2 (1). Peau absolument lisse ou legerement striee rien que dans la partie posterieure de I’hysterosome; toutes les soies apicales des tarses seti-formes; soie ventrale mediane des pattes I et II situee plus distalement que le batonnet olfactif. Longueur totale du corps depassant 300 p.
3 (4). Soies dorsales longues (dj atteignant 14—15, d2 et d3 20—25% de la longueur de I’idiosome), capilliformes (tres fines et flexibles); la et Ip de la тёте longueur que d.2—ds et quelque peu plus courtes que d4; sc i 2—2.5 fois plus courtes que sc e. Espece plus grande; longueur totale du corps 450—430 p. — URSS: Bachkirie (Sterlitamak) ..............................................5. C. kamenskii, sp. n.
4 (3).. Soies dorsales tres courtes (<ZX—d3 ne depassant pas de 7 a 12, d4 9— 14% de la longueur de I’idiosome), un peu rigides; I a et I p visiblement plus longues que toutes les d (2 fois plus longues que d2).
-o (6). Bbrd posterieur du corps muni de 2 petits tubercules hemispheriques; peau legerement striee dans le quart posterieur de I’hysterosome; ecusson propodosomal distinct, a contour posterieur arrondi; sc i 3.3—3.6 fois plus courtes que sc c; la visiblement plus longues que Z p. — URSS: Georgie (Karajazy)................................1. 0. tuberculata, sp. n.
6 (5). Bord posterieur do I’idiosome sans tubercules; peau lisse, proportions des sc Riff erentes; Z a pas plus longues que Z p.
7-(8). Sci 2—2.5 fois plus courtes que see; sa e visiblement plus longues que les laterales (I a et Z p), dont les dimensions depassent 2 fois celles de cZi, d2 et ds-, h i pres de 2 fois plus courtes que h e; coloration des exem-plaires vivants jaunatre ou bien orange clair. — Angleterre, Hollande, URSS (Extreme Orient).............................2. C. heterocoma (Mich.).
8 (7). Sc i 5 fois plus courtes que.se e; la longueur des laterales depasse celle
CALVOLIA
443
de dx—ds moins de 2 fois; sa e plus courtes que la et Ip; coloration des exemplaires vivants blanchatre. — URSS (Moscou)..............
. . ......................................3. C. romanovae, sp. n.
Hypopes
1 (2). Tarses IV se terminant par une forte macrochete. — Allemagne. . . .................................................8. C. kneissli Krause.
2 (1) Tarses IV tcrmincs par 2 fortes macrochetes.
3 (4). Articles distaux du gnathosome munis chacun de 2 soies (aristes); une tache ovale blanche dans chacun des angles anterieurs de 1’hys-. terosome, reprbsentant sans doute un oeil avec une lentille faiblement convexe; disque adhesif muni d’un bord libre tres large, membraneux.— Allemagne................... ....................18. C. (?)zacheri Ouds.
4 (3). Articles distaux du gnathosome portant chacun une seule longue soie (ariste); point de taches rappelant des yeux dans les angles de 1’hyste-. г о some; disque adhesif sans bordure membraneuse libre.
•5 (16). Ventrum absent; pattes IV extremement courtes, egalant A peine 10—11% de la longueur de I’idiosome; tarses IV separes des tibias, et alors hbmispheriques ou soudes avec eux et formant alors un tibiotar-sus court, arrondi au bout; longueur des tarses II ne depassant pas 18% de la longueur de I’idiosome; leurs tarses pas plus lohgs que les femurs.
€'(13). Ecussons dorsaux distinctement stries longitudinalement ou reticules dans la meme direction; retines separees, arquees.
7 (8). Soies ventrales des tarses I et II extremement courtes, simples; soies apicales des tibias I et II trbs longues, depassant de beaucoup le bout des ambulacres; aristes courtes, n’atteignant de leurs bouts que la base des tibias I; ecussons dorsaux stries longitudinalement. т Afrique occidentale, Italie......................... 16. C. striata (Vtz.).
•8 (7). Soies ventrales et intero-apicale des tarses I et II de fortes dimensions, foliacees, de forme lanceolee; soies apicales des tibias I et II plus 'courtes, depassant a peine le bout des ambulacres.
-9 (10). Idiosome de forme regulierement elliptique, tres largement arrondi en arriere; ecussons dorsaux a large dess in reticule; aristes plus courtes, n’atteignant de leurs bouts que la base des tibias I. Sur Ips acuminatus Eich.— URSS (Moscou)............................ 17. C. elliptica,sp. n.
1.0 .(9). Idiosome fusiforme ou presqu’ovoide, se retrecissant paraboliquement vers le bout posterieur; ecussons dorsaux densement stries longitudinalement d’une faqon regulibre sur toute leur surface ou a ornemen-tation plus espace le long de la ligne mediane du corps et formant ici des mailles plus ou moins larges, longitudinales; aristes plus longues, atteignant la base des tarses I.
11 (12). Idiosome fusiforme, se retrecissant fortement et regulierement vers ; les extremites anterieure et posterieure, 1.7 fois plus long que large;
444
SAPROGLYPHIDAE
bords antero-lateraux de 1’ecusson propodosomal regulierement et faib-. loment convexes sur toute leur etendue; saillie rostrale a peine marquee; ecussons dorsaux densement et regulierement.striees sur toute leur surface. Sur Blaslopbagus.— URSS (region de Moscou)................
................................ ............14. C. fusiformis, sp. n.
12 (11). Idiosome presqu’ovoido, 1.46 fois plus long que large, se retrecissant moins fort vers son extremite postcricurc; bords antero-lateraux 'du propodosomo sinueux, formant des concavitfe distinctes derriere les yeux et limitant ainsi la longue saillic rostrale; Ecussons dorsaux plus rarement strife le long de la ligne mddiane (surtout sur I’dcusson propodosomal), formant ici un large reseau a mailles oblongues fermees. Sur Hylesinus. —URSS (Moscou).............................16. C. nataliae,sp. n.
13 (6). Ecussons dorsaux lisses; retines reliees par leurs bouts anterieurs en forme de X.
14 (15). Pretarses a longue tige lineaire; tarses I—III avec 4 grandes soies-lanceolees. Sur Ips stebbingi Stroh. — Thibet...............................
.............................................12. C. circumspectans Vtz.
15 (14). Pretarses courtement pedonculfe, tarses I et II avec 2 soies lanceolees, tarses III totalement denufe de telles soies. — Hollande ....
. ......................................... , .13. C, hagensis Ouds.
16 (5). Ventrum bien developpe; pattes IV moins raccourcies, atteignant 15—20% de la longueur de 1’idiosome, leurs tarses soudfe aux tibias et formant un tibiotarsus allonge plus ou moins cylindrique, s’elargissant generalement vers son extremite qui est tronque, ou bien separe des tibias et alors beaucoup plus long que large; longueur des pattes III atteignant 30—33% de celle de 1’idiosome, rarement moindre (chez C. calliphorae et C. bulgarica pres de 22%); leurs tarses 1.5 fois plus longs que les femurs.
17 (18). Pattes I et II fort longues (35—45%de la longueur de 1’idiosome}et greles; tarses I presque 6 fois, tibias 2 fois plus longs que leur diametre basal; idiosome fortement allonge, ayant la forme d’une amphore; ecusson ventral etroit, 1.7 fois plus long que la largeur entre les points d’attache des pattes III, orne d’epaississements distincts aux bords poste-rieurs; batonnet olfactif des tarses I et II.court, eloigne de la base du. tarse a une distance depassant sa longueur. — Bui gar ie, URSS (Taf-tarie).....................................................9. C. thraca Vtz.
18 (17). Longueur des pattes I ne depassant pas 40%, celle des pattes II 35—38% de la longueur de 1’idiosome; tarses I pas plus de 3.5 fois, tibias pas plus de 1.7 fois plus longs que leur diametre basal; idiosome de forme differente, plus large; ecusson ventral pas plus de 1.3 fois plus, long que large, non encadre postdrieurement d’epaississements margi-naux; batonnet olfactif generalement long, rapproche de la base du tarse.
19 (22). Ecusson propodosomal seulement 1.5 fois plus court que I’hyste-rosome; pattes III courtes, n’ayant pas plus de 16—22% de la longueur
CALVOLIA, VIDIA
445
de 1’idiosome; tarses IV sondes aux tibias en un tibiotarsus; soies des tibias et des tarses I et II ainsi que les bdtonnets olfactifs courts; idiosome 1.Б—1.6 fois plus long que large, 1
20 (21). Macrochete anterieure des pattes IV fortement epaissie, 2 fois plus forte que la posterieure (interieure); tibiotarsus IV bifurque au bout; retines soudeesenune tache ronde, pigmentee; bout posterieur ducorps regulierement arrondi.—Bulgarie, URSS. ... 11. C. bulgarica storkan.
.21 (20). Macrochete anterieure des pattes IV non epaissie (plus mince que la posterieure); tibiotarsus IV tronque droit au bout; retines soudees, en une sorte de chevron fonce; bout posterieur du corps tronque presque droit. — Allemagne................................10. C. calliphorae Vtz.
22 (19). ficusson propodosomal 2 fois plus court que I’hysterosome; longueur des pattes III atteignant 30—32% de la longueur de 1’idiosome; tarses IV separes des tibias; soies des tibias et des tarses,let II ainsi que les bd-tonnets olfactifs, trfes longues; idiosome large, 1.Й6—1.4 fois plus long que large, arrondi posterieurement.
23 (24). Ecusson hysterosomal orne d’un dessin a reseau bien distinct; une partie des mailles penta- et hexagonales de la partie centrale de cette ornementation sont a peine plus longues que larges; tarses IV tronques au bout; ses deux macrochetes disposees terminalement (au bout tronque du tarse); soies ventrales des tibias I et II extremement longues et fines. — Suisse (Sussillon)....................7. C. reticulata, sp. n.
24 (23). Ornementation de 1’ecusson hysterosomal generalement indistincte (dans les preparations); dans le cas contraire on voit que les mailles sont considerablement plus longues que larges; tarses IV a bout oblique; 1’une des macrochetes (la plus courte) est plantee distinctement plus bas que 1’autre; soies ventrales des tibias I et II de longueur moyenne.
26 (26). Idiosome tres large, rien que 1.2Б—1.3 fois plus long que large, largement arrondi posterieurement; epimerites II forts, tres distincts; distance entre les yeux egale audiametre des cornees; pretarses III avec une tige courte et une grande lamelle allongee. Espece plus petite. Longueur 170—180 p.. — Angleterre, Hollande, URSS(Extr6me Orient) . .
..........................................2. C. heterocoma (Mich.).
26 (25). Idiosome plus Gtroit, 1.4 fois plus long que large, й bout posterieur parabolique; epimerites II peu distincts; distance entre les yeux 2 fois plus grande que le’diametre des cornees;. pretarses III avec un long pedoncule lineaire et une lamelle arrondie. Longueur 200—220 р..— URSS: Caucase, Bouchara..........................6. C. elisbethae, sp. n.
VIDIA OUDS.
Hypopes
1 (4). Tarses I et II ayant chacun 3 soies lanceolees: 1 dorsale et 2 ventrales; pretarses I et II petits, de structure normale. Sur les abeilles. (Sous-genre Vidia s. str.).
Й6
GLYCYPHAGIDAE
2 (3). Sa г plus longues que p a. Sur Prosopis conformis Forst. — Italie . . . ...............................................4. V.(s. str.) undulata Ouds.
3 (2). Sa i plus courtes que p a. Sur beaucoup d’especes du genre Megachile. — Europe, Asie.................................... 3. V. (s. str.) Jlneata Ouds.
4 (1). Tarses I et 11 ayant chacun 2 soies falquees, disposees lateralement; pretarses I et II longs, cJargis au bout, formant une ventouse en forme de cuillibre. Sur les guepes du genre Crabro. (Sous-genre Grabmidia, subg. n.).
6 (6). Corps piriforme d extremity postdrieurc fortement retrecie; soie falquee-exterieure des tarses I et II situde beaucoup plus pres do la base de la soie dorso-apicale que.de celle, de la seconde soie falqwSe; batonnet olfactif cylindrique; tibiotarsus IV grble, plus long que les femurs-IV. — URSS (region de Moscou, Crimee).......................................
................. . . .................1. V. (C.). gussakovskii, sp. n.
6 (6). Corps ovoide, se retrecissant graduellement vers I’extremiteposterieure;. soie falquee des tarses I et II situee sensiblement plus loin de la base . de la soie dorso-apicale que celle de la seconde soie falquee; batonnet.. olfactif renfle au bout en massue; tibiotarses IV robustes, beaucoup plus courts que le femur IV. — URSS: Ukraine (Krasnograd), Crimee-(Karadagh), Abkhasie, Ouzbekistan (Tachkent)................................
............................................ 2. V. (C.) popovi, sp. m.
III. Famine GLYCYPHAGIDAE
TABLEAU DES SOUS-FAMJLLES
1 (2). Fente analeflanquee d’une paire de ventouses copulatrices, ayant la. forme d’entonnoirs; tarses IV avec 2 paires de ventouses tarsales; ori-, fice genital situe entre les bases des pattes I, de faqon a ce que les epi-meres I sont largement disjoints et le sternum manque tbtalement. ................................................... 1. Chortoglyphinae.
2 (1). Aucune trace de ventouses copulatrices anales ou tarsales; orifice-genital situe plus en arriere; epimeres I se reliant par leur extremites et, formant un sternum plus ou moins long.
3 (4). Tarses I et II. (et quelquefois aussi III et IV )4res courts, se terminant par une forte epine en forme de griffe, a la base de laquelle prend nais-sance le pretarse extremement long, fin et flexible. Especes marines a coloration foncee....................................6. Carpoglyphinae.
4 (3). Tarses I et II de longueur normale, sans grosse epine terminale en forme de griffe; pretarses de structure normale. Especes terrestres.
6 (6). Peau tres finement et densement striee; presque toute la surface dorsale-de 1’idiosome est occupee par un champ distinctement limite convert, de petites epines (squamoides) triangulaires; soies dorsales longues,
TABLEAU DES- SOUS-FAMILLES MT
se raccomcissant graduellement en arrive, de fa?on a ce que sont beaucoup plus longues que les suivantes; v p disposees lateralement, au. meme niveau transversal que v a, Sur les abeilles du genre Xylocopa et les bourdons...................................3. Aeroglyphinae.
6 (6). Structure de la peau differente; d3 et dt plus longues que d, et d2 ou bien elles sont toutes representees par des microchetes ou de courtes epines; vp situees beaucoup plus enarriere que v a ou, dans le cas qu’elles. sont au тёте niveau que ces dernieres (genre Blomia), tres rappro-chees de v a. Les adultes n’habitent jamais sur des insectes vivants.
7 (10). Pattes III et IV plus ou moins fortement modifiees.
8 (9). Pattes III beaucoup plus longues et grGles que les1 pattes IV, non.cour-bees; gnathosome* tres etroit, idiosome aplati, rhomboidal; angles assez.. robustes, falques. Habitent dans le sue suintant du tronc des arbres.
.................• • •_.......................6. Carpoglyphinae.
9 (8). Pattes III plus courtes que les pattes IV, mais de la meme forme, plus ou moins fortement courbees et aplaties, avec des tarses obliquement tronques a I’extremite; ongles tres petits; gnathosome assez large, souvent cache par une saillie rostrale bien develbppee; forme de 1’idiosome* differente. Habitent dans les nids des mammiferes (rongeurs" et insec-tivores) et dans les greniers.........................Б. Labidophorinae.
10 (7). Structure des pattes III et IV normale.
11 (12). Epimeres II relies par leurs extremites au bout du sternum, formant * avec lui une figure enforme de W; epimeres III et IV libres; gnathosome tres fin et mobile; peau lisse; toutes les soies de 1’idiosome lisses, tres courtes, excepte sa i et 1’une des paires postanales qui sont representes. par de longues macrochetes flexibles..................6. Carpoglyphinae..
12 (11). Structure du squelette сохо-sternal differente; proportions du gnathosome normales; peau tres finement granulee, quelquefois settlement ridee; soies de la surface dorsale de 1’idiosome plus longues, plus ou. moins plumeuses, rarement tres courtes et alors spiniformes.
13 (14). Б paires de soies laterales; toutes les postanales sont disposees sur le bord meme de 1’idiosome, de sorte que sa surface dorsale porte de 17 a 20 paires de fortes soies; si il у en a moins, elles sont large-ment foliacees; champs coxaux sclerotises, clos, formant deux ran'gees obliques (divergentes en arrive) d’ecussons coxaux tres etroitement rapproches les uns des autres; penis ordinairement tres long. Especes de* trds petite taille, vivant dans le foin et la poussiere des caves........
................................................ 4. Ctenoglyphinae.
14 (13). 3 paires de soies lat6rales; les postanales sont situees a une distance notable du bord de 1’idiosome qui porte sur ses surfaces dorsales et laterales tout au plus 15 paires desoies; conformation du squelettecoxo-sternal differente.
15 (18). Tarses extremement fins, subules; ongles tres petits, a peine distincts; toutes les soies des surfaces dorsales et laterales de 1’idiosome (except^
•448
GLYCYPBAGIDAE
quelquefois d2) tres longues, densement pileuses ou plumeuses, flexibles. Especes principalement synanthropes.
16 (17). V p situees bien cn arriere des va; gnathosome de dimensions normales; idiosome ellipsoidal, h bords lateraux ordinairement un peu sinues; epimerites IT obliques, relies par leur extremite interieure aux epimeres III; extremity des gdnuaux I avec 2 sensilles dorsales . . . ......................................................2. Glycyphaginae.
17 (16). V p situees tout pres des « a, formant avec elles une rangee transversale droite ou anguleusc; gnathosome trbs fort; idiosome de forme ovoi’dale; epimerites II libres, transversaux; extrdmiWs des genuaux I avec une seule sensille dorsale......................1. Chortoglyphinae.
18 (16). Tarses moins greles, nonsubules; ongles de grande dimension, soies des surfaces dorsales et latdrales de I’idiosome assez courtes ou тёте tres courtes (une seule paire d’entre elles—:les sa e—sont longues, mais toujours lisses), rarement ciliees ou presque lisses. Habitants des nids de diff erents Hymenopteres.....................7. Chaetodactylinae.
99
1 (2). Peau tres finement et densement striee; presque toute la surface dor-sale de I’idiosome est occupee par un champ distinctement limite convert de petits accidents cuticulaires (squamoides); soies dorsales assez longues, se raccourcissant graduellement en arriere, de faijon a ce que sont beaucoup plus longues que les suivantes; v p disposees lateralement, au тёте niveau transversal que va; 4 paires de soies. postanales; sur les abeilles du genre Xylocopa et les bourdons. . . 3. Abroglyphinae.
.2 (1). Structure de la peau diff&ente; d3 et plus longues que dj. et ou bien toutes de longueur egale; position des v p differente; si elles sont dispo sees marginalement au meme niveau transversal que v a, la peau est absolument lisse et rigide et toutes les soies de la surface dorsale ont 1’aspect de fines microchetes. Les adultes ne se trouvent jamais sur des insectes vivants.
5 (4). Soies peripheriques- de I’idiosome largement plumeuses, bipectinees ou foliacees, au nombre de 13—21 paires, car les humerales et les post-‘ anales occupent une position strictement marginale, et il existe 5 paires de soies laterales (s’il у en a moins, elles sont foliacees); leur ensemble forme d’habitude une beliefrange entourant I’idiosome; peau grossiere-ment ridee ou ornee de hauts tubercules cylindriques a sommet divise, rarement chagrinee. Habitent principalement dans le foin et la poussieip des caves......................................... 4. Ctenoglyphinae. -
4 (3). Toutes les soies de I’idiosome ont une autre forme; 3 paires de soies laterales; les postanales sont disposees ventralement, a une certaine distance du bord posterieur de I’idiosome; structure de la peau diffe-rente.
© (8). Toutes les soies des surfaces dorsale et laterales de I’idiosome longues
TABLEAU DES SOUS-FAMILLES
449
(souvent extremement longues) et flexibles, densement pileuses ou finement plumeuses; idiosome fortement bombe; peau tres delicate, jjicolore, tres finement granule; tarses tres longs et fins, subuliformes; soies des genuaux et des tibias (1’apicale exceptee) densdment. plumcuscs.
6 (7). V p situees bien en arriere des v a; orifice genital oblong, presque en--tierement reconvert par- des James gdnitales bien developpees, muni d’un epigyne; tube copulateur court et droit; genuaux I avec 2 sensilles apicales ....................................... . . . . 2. Glycyphaginae.
7 (6). V p situees tout pres des va, formant avec elles une rangee transversale droite ou anguleuse; gnathosome tres fortement developpe; orifice genital tres grand, largement ouvert (grace a la reduction des lames genitales), sans epigyne; tube copulateur long, tres fin, fortement arque; genuaux I avec une seule sensille apicale ............1. Chortoglyphinae.
8 (5). Toutes les soies de la surface dorsale de I’idiosome relativement courtes, lisses ou finement et rarement ciliees; dans ce dernier cas elles sont plus ou moins rigides; forme de I’idiosome et structure de la peau variables, tarses plus robustes (non subuliformes). .
9 (10). Tarses I et II tres courts, se terminant en une tres forte griffe, a la base de iaquelle prend naissance le pretarse extremement long, fin et flexible. Especes marines a coloration sombre . . . . 6. Carpoglyphinae.
10 (9). Tarses I et II non abreges, sans forte griffe, pretarse de structure normale.
11 (16). Peau absolument lisse, rigide, de consistance plus ou moins cornee.
12 (1Б). Peau incolore, hyaline; pattes greles, de structure normale.
13 (14). Epimeres II libres; ceux de la premiere paire relies entre eux et formant une lamelie transversale fortement chitiniseo dont le bord posterieur se prolonge en arriere en un sternum libre trfcs court; pas d’epigyne; cheliceres tres robustes; toutes les soies de I’idiosome representees par* de tres fines microchetes difficiles a voir; soies des pattes pectinees . . ......................................................1. Chortoglyphinae.
14 (13). Sternum et epimeres II'relies par leur bouts aun fort epigyne angu-leux; gnathosome tres etroit et mobile; sa i et 1’une des paires postanales tres longues, capilliformes, toutes les autres soies de I’idiosome courtes, rigides, obtuses au bout; soies des pattes lisses. ... 6. Carpoglyphinae.
15 (12). Peau fortementpigmentee, brundtre; pattes I et II fortement compri-mees lateralement ou ornees de carenes longitudinales....................
............................................ . 5. Lahidophorinae.
16 (11). Peau granulee, epineuse on avec quelque autre sculpture distincte, de consistance variable.
17 (18). Gnathosome tres etroit; toutes les soies de I’idiosome lisse, rigides; epigyne arque, libre. Habitent dans le sue, suintant du tronc des ar-bres .................................................6. Carpoglyphinae.
18 (17). Gnathosome robuste; epigyne ayant une autre forme, ordinairement
29 Фауна СССР, Захватили—284
4аО
GLYCYPHAGJDA Е
relie au sternum. Habitent dans les nids des mammiferes et des insectes Hymenopthres..
19 (20). Ongles tres pc tits; aucune des soies de I’extremite postericure du corps ne depasse considerablement les dimensions des autrcs soies. Habitent dans les nids et les terriers des Rongeurs et des Insectivores . . .................................................. 5. Labidophorinae.
20 (19). Ongles robustes, souvent tres grands et ayant alors la forme d’une faucille; i tres longues, lisses, depassant de beaucoup la longueur de toutes les autrcs soies du corps. Habitants des nids do differentes abeil-les ................................................7. Chaeto dactylinae.
1. Sous-fam. Ch о г t о gl у p hi пае
’ TABLEAU DES GENRES
I (2). Toutes les soies de la surface dorsale de 1’idiosome extremement courtes, lisses, a peine visibles; peau tout-a-fait lisse, rigide; soies des tibias et des genuaux I et II pectinees, Tune des ventrales des tarses spiniforme; U arme de ventouses copulatrices anales et tarsales, a orifice genital fort deplace en avant et situe entre les bouts ecartes des epimeres I; 9 sans tube copulateur...................................l.Chortoglyphus Berl.
Urie espece palearctique, .C. arcuatus (Tr.). — Angleterre, France, Italie, URSS (Moscou, regions d’lvanovo et de Smolensk, Barnaoul).
2 (1). Toutes les soies de la surface dorsale de 1’idiosome (excepte d2) longues, assez fortes, densement plumeuses; peau de consistence membraneuse, tres finement granulee; soies des tarses, tibias et genuauxepaisses, densement plumeuses; <5 sans ventouses copulatrices, h orifice genital dispose entre les bases des pattes IlletIV; $ avec un tube copulateur tres long et fin..............................................2. Blomia Ouds.
BLOMIA OUDS.
1 (2). Soies verticales disposees en trapeze; palpes genitaux absents dans les deux sexes; soie ventrale interieure des tibias I et II situee pres de la base de 1’article, bien plus proximalement que 1’exterieure; chez la 9 le bord posterieur de 1’idiosome est arme de 2 tubercules coniques tres-distincts;.tube copulateur tres long. —URSS (Moscou) ....................
............................................. 3. B. kulagini A. Z.
2 (1). Soies verticales disposees en une rangee transversale presque droite; palpes genitaux presents, mais peu distincts; soie ventrale interieure des tibias I et II situee au milieu de 1’article, a peine plus bas que I’exte-rieure; 9 sans tubercules au bord posterieur de 1’idiosome, a tube copulateur plus court et plus arque. —URSS (region de Smolensk)', . . .
1. B. thori A. Z.
BLOMfA, GLYCYPHAGUS Ш-
2. Sous-fam. Glycyphaglnae
TABLEAU DES GENRES
1 (2). Les plus longues des soies de 1’idiosome ne depassent pas plus de 2 fois sa longueur; va tres rapprocliees, se touchant des bases, v p separees-par un ospace tres large; palpes genitaux petits, distincts* dans les deux sexes; organe pseudostigmal compose d’un seal tronc ramifie . . . ...............................................3. Glycyphagus Iler.
2 (1). Les plus longues des soies do 1’idiosome depassent sa longueur de 3 a З.Б fois et forment en leur ensemble une tres longue traine, ressemblant a une queue de comete; v a separee entre eux par un large espace, v p se touchant presque des bases; palpes genitaux du d asscz grands, absents, chez la 2; organe pseudostigmal divise en 2 troncs fortement ramifies .
...........................................4. Cometacarus A. Z.
GLYCYPHAGUS HER.
1 (10). Tarses revetus d’un etui ou ecaille entierement convert de petits-cils, ce qui les fait paraitre densement pileux; crista.metopica absente; soies ventrales des femurs I lisses, setiformes. (Sous-genre Lepidogly-phus A. Z.).
2 (Б). Soie basale des genuaux transformee en une grande ecaille ovale ou lanceolee, densement pileuse; bords distaux des tibias elargis en lamelle, formant une large genouillere; soies plumeuses de 1’idiosome et des-pattes plus fortes, plus longuement ciliees.
‘3 (4). Espece de tres grande taille: d 450—550р., 9 600—800 p; ecaille des-genuaux III de forme ovale, pres de 2 fois plus longue que sa largeur maximum; chetoi'des garnissant ses bords a peu pres de la meme dimension que ceux qui couvrent sa surface. — Angleterre, URSS (Leningrad,. Moscou, region d’lvanovo, Bachkirie) . . . 4. G. (L.) michaeli Ouds..
4 (3). Espece plus petite: <5 350—400 p. (9 inconnue); ecaille des genuaux III etroite, lanceolee, 6 fois plus longue que sa largeur maximum; les soies-qui la bordent foment une longue frange. — Allemagne, URSS (Smolensk....................................Б. G. (L.) burchanensis Ouds..
Б (2). Soie basale des genuaux III non elargie, ayant la тёте structure qim les autres sows plumeuses des pattes; tibias ne formant point.de genouil-leres lamelleuses; soies plumeuses de 1’idiosome fines.
6 (7). Soies plumeuses des tibias et des genuaux I et II tres larges; soie apicale des genuaux I tres courte, a peu pres de la meme longueur que le bdtonnet olfactif disposd a cote d’elle; chez les c?d le batonnet olfactif des genuaux II est fortement hypertrophie et presente 1’aspect d’une robuste1 epine, de la longueur du genual; chez la 9 la longueur de la fente genitale surpasse de beaucoup la distance entre son bord posterieur et la fente anale; paire posterieurc dos soies genitales placee a nivlau.
<52
GLYCYPIIAGIDAE
de son bord posterieur. — Allemagne, URSS (region d’lvanovo). . , .
.............................................3. G. (L.) fustifer Ouds. *
7 (6). Soies plumeuses des tibias et des genuaux I et II etroites; soie apicale des genuaux I do 4 a 5 fois plus longue que le batonnet olfactif plante pres d’elle; bAtonncts olfactifs des genuaux II de dimension normale, tris petits chez les deux sexes; longueur do lafente genitale de.la $ 6gale a la distance entre son bord posterieur ct la fente anale; paire posterieure des soies genitalcs plantce sensiblement plus on arriere que le bord posterieur- de la fente g6nitalo.
8 (9). Espece tres petite :U 225 y; soies plumeuses de 1’idiosome extremement longues; ds, et sa e 2 fois, Z3 1.5 fois plus longues que 1’idiosome; chez les cTd1 les pra egalent presque en longueur la fente anale. — Allemagne ......................................... .2. G. (L.) pilosus Ouds.
S (8). Espece de grandeur moyenne: d1 380—430р., 9 380—550 [л; les plus longues des soies plumeuses de 1’idiosome ne depassent pas plus de 1.5 fois la longueur de cette derniere; pra chez les <5(5 plus courtes que la moiti£ de la fente anale. — Europe, Asie palearctique.....................
................................. 1. G. (L.) destructor (Schrk.) Ouds.
10 (1). Tarses denues d’etui pileux, mais avec quelques soies barbelees assez grandes; crista metopica presente; soies ventrales des tibias I plumeuses. (Sous-genre Glycyphagus s. str.)
11 (12). V a plantees sur I’extremite anterieure de la crista metopica, qui s’etend en arribre jusqu’au niveau des sc; d2 placees normalement— bien au devant de d3 et au meme niveau que lx; batonnet olfactif des genuaux I et II court, s’attenuant vers le bout. — Hollande, URSS (regions d’lvanovo, de Smolensk et de Kalinine, Bachkirie)................
.............................6. G. (s. str.) cadaverum (Schrk.) Ouds.
12 (11). V a situes au milieu de la crista metopica, qui ne s’etend en arriere que jusqu’au niveau des v p-, d2 sont deplacees en arriere et se trouvent entre les d3, au тёте niveau qu’elles et beaucoup plus en arriere quelx; batonnet olfactif des genuaux I et II fin, long, cylindrique.
13 (14). Soies ventrales des tibias III et IV implantees dans 1’echancrure du bord distal de 1’article; soies ventrales des tarses IV disposees a une distance inegale 1’une de Г autre — les deux proximales tres j-approchees entre elles et 61oignees en тёте temps de la distale; les deux extremites de la crista metopica sont pointues; chez le d1, les tarses I et II sont fortement recourbees et la soie intero-ventralo des tibias I et II est transformee enunrobuste peigne triangulaire.,— Angleterre, Hollande, Italie, Allemagne, URSS (Mos'cou, Leningrad, Kiev, Bachkirie) . . .............................................8. G.(s. str.) ornatus Kram.
14 (13). Soies ventrales des tibias III et IV implantees sur 1’article тёте, eloignees de son bord; soies ventrales des tarses IV disposees a une distance egale; moitie anterieure de la crista metopica large, plus faible-
. • ment chitinisee que la posterieure qui est etroite; tarses I et II chez le <5 •
GLYCYPHAGUS, COMETACARUS, AEROGLYPHUS Ш
non recourbes, toutes les deux soies ventrales des tibias I et II de structure normale. — Europe et partie palearctique de 1’Asie, Arctiqti& (Terre de Frangois Joseph, Spitzberg, Greenland), Terre de Feu. . . . ............................ 9. G. (s. str.) domesticus (Deg,),.
COMETACARUS A. Z.
1 (2). Espece de tres petite taille: longueur totalc du c? 175, do la 9 200p; v p de la meme longueur quo sc i et d’un tiers plus longues que sc e; d9 3 fois plus courtes que d3; soies plumeuses des pattes tres longues, la basale exterieure des genuaux I aussi longue que le tarse; d sans lamelies-en forme d’eventail au femurs I et II. — Allemagne...............
....................................... 2. C. oudemansi A. Z.
2 (1). Taille beaucoup plus grande: longueur totale dud 400—420 p, de la 9 500—510 |л; v p 2 fois plus courtes que les sc e et 3 fois plus courtes que sei; d2 1.5 fois plus courtes que ds; soies plumeuses des pattes assez courtes, la basale exterieure des genuaux I ne ddpassant pas la moitie de la longueur’ du tarse; soies ventrales des fbmurs I et II transformees-chez le d en elegantes lamelles ayant a peu pres la forme d’un even-tail.— URSS (Ivanovo)......................... 1. C. smirnovi, sp. n..
3. Sous-fam. Aeroglyphinae
AEROGLYPHUS, gen. n.
Type du genre: Glycyphagus peregrinans Berl. 1892.
Taille assez grande(350—415 p). Presque toute la surface dorsale de 1’idiosome est occupee par un champ nettement limite convert de petits accidents cuticulaires ayant la forme d’epines triangulaires; sur la partie anterieure de 1’idiosome la peau est distinctement chitinisee et forme un ecusson. propodosomal orne de 2 carenes longitudinales atteignant en arriere le niveau des sc, ou elles se recourbent en dehors et disparaissent. Dimensions des soies dorsales et laterales baissant graduellement en arriere de fagon que et Zx sont plus longues que les suivantes. Differe biolo-giquement des autres Glycyphagidae par le fait que la propagation de 1’espece s’effectue par phoresie d I’etat adulte et non aux stades hypopi-aux. Une seule espece, trouvee sur les Xylocopa et les bourdons (Bomhus lapidarius L.) en Italie et en URSS (region de Moscou)...........
....................... . ...............A. peregrinans (Berl.).
4. Sous-fam. Cteno glyphinae
TABLEAU DES GENRES
A d u 11 e s
1 (2). Pourtour de Fidiosome avec seulement 7 paires de soies, dont 6 sont foliacees, asymetriques, a bord exterieur fortement dente, et 1 tres
43i
GLYCYPHXGlDMi
longue, simple; v, sc et d cuneiformes, tronquees au bout..............
..............................................8. Nycteriglyphus, gen. n.
Une seule espece, M. plerophorus (Berl.). — Italie.
2 (1). Pourtour de ridiosonie orne de 11—12 paires de soies largement plumeuses, palmees ou loliacces, mais toujours symetriques; v, sc et d de la тёте forme quo soies pthipheriques ou plus fines qu’elles et dense- 4
me nt pileuses (cilices'). i-
3 (4). Los dd1 different peu des 99 par leur aspect general et la conformation 1
des soies de I’idiosome; chez les deux sexes la peau de la surface dorsale i est finement chagrineo; les soies sont etroites, aplaties, largement et I’
finement plumees; v et sc plus large que les autres....................
............................. 6. Diamesoglyphus, gen. n.
Une seule espece, D. intermedins (Can.)..— Italie.
4 (3). cf cf tout-a-fait diff erents des $9 par leur aspect general et la conformation des soies de I’idiosome; la peau de la surface dorsale est chez eux finement chagrinee ou presque lisse, les soies sont longues, relative-ment etroites, I’idiosome distinctement bombe, й contours arrondis;
•chez les 99 I’idiosome est tres aplati, presque pentagonal, la peau de la surface dorsale est grossierement ridee ou couverte de hauls tuber- j
cules a sommet divise, les soies de la surface dorsale (ou tout au moins J
les soies peripheriques) tres larges, palmees ou foliacees.............
..............................................7. Ctenoglyphus Berl. j
• CTENOGLYPHUS BERL.
rfd
1 (4). Toutes les soies de la surface dorsale ayant a peu pres la meme structure — bipectinees ou plumeuses; d^ et d2 bien developpees.
2 (3). Toutes les soies de la surface dorsale de I’idiosome, excepte d3, bipectinees, portant de 10 a 20 paires de barbes robustes; d3 finement plumeuses; d2 deux fois plus courtes que dx. — Angleterre, Italie, URSS (Moscou, Astrakhan)................................... . . 2. C. canestrinii (Arm.).
3 (2). Toutes les soies de la surface dorsale finement plumeuses, portant de fins et tres nombreux cils; d2 a peine plus courtes que dx. — Angleterre, France, Allemange, URSS (Leningrad, Moscou, Ivanovo) . . . . .................................................1. G. plumiger (Koch).
4 (1). V a bipectinees; v p, les deux paires de sc, h a, h p et sh et les deux premieres paires de laterales — foliacees, etroitement lanceolees; t?3 et ls—к fines, densement et finement plumeuses; du sa e et sa i foliacees, beaucoup plus larges que toutes les autres soies de I’idiosome; ^et d2 absentee.— Angleterre, France, Allemagne, Italie.........................
................................3. C. palmifer (F. et R.).
CTENOGLYPHUS
455
?9 '
1 (4). V, sc et toutes les soies peri phen ques de I’idiosome bipectinees; peau de la surface dorsale couverte de haute tubercules cylindriques a sommet lobe.
2 (3). Toutes les soies dorsales (dx—dt), 'sa e ci sa i etroites, finement plumeuses; barbes des soies bipectinees droites, forment un angle aigu avec 1’axe de la soie. — Angleterre, France, Allemagne, URSS (Leningrad, Moscou, Ivanovo) . . . .............................1. C. plumiger (Koch).
3 (2). Rien que d3 sont etroites et finement plumecs; dr, d2 et d4 bipectinees, de la тёте forme que les soies peripheriques de I’idiosome; les barbes de ces dernieres sont fortement recourbees vers I’extremite de la soie et foment avec son axe un angle droit. — Angleterre, Italie, URSS (Moscou, Astrakhan)...............................2. C. canestrinii (Arm.).
4 (1). V, sc et toutes les soies peripheriques de I’idiosome representees par de larges ecailles foliacees; peau de la surface dorsale ridee, sans tubercules cylindriques.
5 (6). Drei d2 ayant 1’aspect de tres petites soies legerement pectindes;soies foliacees plus longues, mesurant 20—36% de la longueur de I’idiosome. — Angleterre, France, Allemagne, Italie .... 3. C. palmifer (F. et R.).
6 (5). et d2 foliacees, de la meme forme que les soies peripheriques; ces dernieres plus courtes, mesurant 15 — 20% de la longueur de I’idiosome. (Gl. palmifer Canestrini 1883, non Fumouze et Robin 1868). — Italie.........................................4. C. multifoliatus, nom. n.
5. Sous-fam. Lahido pho rimae
TABLEAU DES GENRES
cfcf
1 (2). Bords de I’idiosome profondement incises, formant 10 paires de lobes arrondis, dontplusieurs portent a leur sommet de fortes soies aciculees . . ............................................... 14. Talpacarus, gen. n.
Une seule espece, T. platygaster (Mich.). — Angleterre, Hollande.
2 (1). Bords de I’idiosome sans incisions ni lobes.
5 (4). Idiosome fusiforme, fortement retreci vers les deux bouts, tres convexe, peau absolument lisse, rigide, d’un brun fonce; sc e capilliformes, toutes les autres soies de I’idiosome representees par de fines et flexibles microchetes; pattes tres courtes, les anterieures fortement aplaties. late-ralement............................... . ...... . 15. Fusacarus Mich.
Une seule espece, F. laminipes Mich. — Angleterre, URSS (environs de Moscou et d’lrkoutsk).
4 (3). Idiosome de forme differente, plus ou moins aplati dans la direction dors'o-ventrale; conformation des pattes et des soies differente.
•5 (6). Espece de grande taille, a coloration pale; soies de la surface dorsale
О
GLYCYPHAGIDAE
de 1’idiosome assez longues, distinctement ciliees; peau molle, finement granulee; tarses III et IV avec des carenes foliacees longitudinales, les. autres articles des pattes sans excroissances lamellilormes............
............................................10. Labidophorus Kram.
6 (Б). Espece do petite taille, a coloration obscure; soies de la surface dorsale de I’idiosomo courtes ou tr«>s courtes, lisses, quelquefois spiniformes; peau epaisso, fortement chitinis6e, lisso ou grossierement granulee; armament dos pattes different.
7 (8). Espbcc do trbs petite taille (longueur 170 p.); peau. de la surface dorsale densement couverte de gros granules h(5misph(5riques; toutes ses soies ayant 1’aspect de courtes epines coniques; squelettc сохо-sternal faiblement developpb; pattes III et IV entierement cachees sous le corps, dirigees en avant................................13. Orycteroxenus, gen. n.
Une seule espece, 0. dispar (Mich.) — Angleterre.
8 (7). Taille plus grande (250—400jx); peaulisse ou grossierement granulee;. soies de la surface dorsale du corps rigides, mais non spiniformes, tres. distinctes ou bien representees par de fines microchetes a peine visibles; squelette сохо-sternal bien developpe; pattes III et IV normalement developpees.
9 (10). Peau lisse; soies de sa surface dorsale mesurant 10—1Б % de la longueur de 1’idiosome, assez rigides;saillie rostrale sans carenes longitudinales; articles basilaires des pattes ornes de carenes longitudinales basses et-obtuses, sans expansions foliacees et dentelees. Espece synanthroper habitant dans la farina at autres produits alimentaires.....................
................................................. 11. Gohieria Ouds.
Une seule espece, G. fusca (Ouds.) — Angleterre, France, Hollande, URSS (Moscou, region d’lvanovo, Caucase, Barnaoul, Asie centrale).
10 (9). Peau grossierement granulee et poreuse; saillie rostrale avec 2 carenes, longitudinales fortement granulees: toutes les soies de 1’idiosome ont 1’aspect de fines microchetes peu distinctes; articles basilaires des pattes I et II armes d’expansions foliaces a bord libre dentele, tarses avec de hautes carenes foliacees a bord entier. Espece nidicole, habitant, dans les terriers des taupes et des rongeurs . . . 12. Xenoryctes, gen. n.
99
1 (4). Bords lateraux de 1’idiosome profondement incises, formant des lobes, irreguliers, ou en forme de festons, portant au gommet de fortes soies-plus ou moins recourbees.
2 (3). Sternum et epimeres II libres; epimeres III relies par les bouts aux epimeres IV; orifice genital non entoure d’un cercle chitineux; toutes. les 4 paires de soies dorsales (dx—d4) bien developpees......................
.............................................14. Talpacarus, gen. n.
Une seule espece, T. platygaster (Mich.). — Angleterre, Hollande..
LABIDOPIIORINAE. TABLEAU DES GENRES 45T
3 (2). Sternum absent; epimeres I, II et III relies a un large cercle chitineux, entourant Г orifice genital; epimeres J V courts, libres; rien que 2 paires-de soies dorsales (d2 et d8) qui sont fortement rapprochees entre elles et. disposees presque sur la ligne mediane du corps.............................
............................................13. Orycteroxenu«, gen. n._ Une seule espece, 0. dispar (Mich.) — Angleterre..
4 (1). Bords lateraux de 1’idiosome entiers, sans lobes ni festons.
6 (6). Idiosome fusiforme (fortement attenue vers les deux bouts), tres con-vexe; peauabsolumentlisse, d’un brunfonce; toutes les soies de 1’idiosome-
r excepte see, ayant 1'aspect de tres fines et flexibles microchetes; see-
assez longues, capilliformes; pattes I et II tres fortement comprimees-lateralement, presque lameileuses..................16. Fusacarus Mich.
Une seule espece, F. laminipes Mich. Angleterre, URSS-(environs de Moscou et d’Irkoutsk).
6 (5). Idiosome de forme differente, aplati dans le sens dorso-ventral; conformation des pattes et des soies differente.
7 (8). Sternum et epimeres II libres, epimeres III relies a celles de la IV paire; epigyne reniforme, separe des elements du squelette coxo-sternal;. peau blanchdtre, molle, ornee d’un fin dessin reticule; soies do la surface .dorsale lisses, assez longues, pressees centre le corps; pattes I et IL raccourcies. Espece de grandes dimensions, habitant sur le castor . . . ...................................................9. Xenocastor, gen. n.
8 (7). Sternum relie par le bout a I’epigyne ou au cercle circumgenital, oik absent; dans le dernier cas les epimeres I sont directement reliees a. I’epigyne; conformation des soies differente; pattes I et II non raccourcies.
9 (12). Peau tres rigidc, brunatre; saillie rostrale bien developpee, cachanL hr plus grande partie du gnathosome; soies delasurface dorsale de 1’idio-some courtes ou tres courtes (quelquefois a peine visibles), lisses; orifice genital entoure d’un cercle chitineux. Especes de taille mediocre (300— 450 p.).
10 (11). Soies de la surface dorsale courtes, mais tres distinctes; peau lisse; saillie rostrale sans carenes longitudinales; sternum absent — les epimeres I sont relies directement a I’epigyne; articles basilaires des pattes carenes; bords distaux des genuaux et des tibias elargis en lamelle. Espece synanthrope, habitant dans la farine et autres produits alimen-taires..................................,................11. Gohieria Ouds..
Une seule espece, G. fusca (Ouds.) — Angleterre, France, Hollande, URSS (Moscou, region d’lvanovo, Caucase, Barnaoul, Asie Centrale).
11 (10). Soies de la surface dorsale de 1’idiosome extremement courtes, a. peine visibles; peau grossierement granulee et poreuse; saillie rostrale-avec 2 carenes longitudinales fortement granulees; sternum bien de-veloppe, relie par son extremite avec I’epigyne; articles basilaires des-
458
GLYCYPHAGIDAE
pattes cylindriques, tibias avec une haute carene dorsale foliacee. Especes nidicotes, habitant dans les terriers des taupes et des rongeurs. . . . . . ..........................................12. Xenoryctes, gen. n.
12 (9). Peau molle, de coloration pftle, tres finement granulee; saillie rostrale absente; soies ch; la surface dorsale de 1’idiosome relativement longues, plus ou moins cilices; especes do grande taille (600—1000 p.) . ... . ...................................................10. Labidophorus Kram.
.11 у p о p e s
1 (2). Corps etroit, 1.6—1.9'fois plus long que large; pattes I et II longues et greles, armees de soies plume uses ou pectinees, leurs tarses distincte-ment recourbes en S (dans un aspect lateral); pattes IV tres courtes et greles, se terminant par une tres longue macrochete, denuees d’ongle. ...................................................... 16. Myacarus, gen. n. .2 (1). Corps plus large; pattes I et II plus courtes et plus robustes, armees
de soies simples ou spiniformes; tarses I et II non recourbes en S, plus ou moins coniques; pattes III et IV mediocrement plus courtes que les anterieures.
3 (4). Champs coxaux III fermes; tarses IV sans macrochetes apicales; etaux posterieurs spatuliformes, fins, separes par un large espace des ante-rieurs, qui ont une forme ovale et sont armes de carenes disposees radia-lement; aire opisthosomale nonlimitee..............12. Xenoryctes, gen. n.
-4 (3). Champs coxauxIII ouverts; s’ils sont fermes, les tarses IV se terminent en une longue macrochete; etaux posterieurs a peu pres triangulaires, rapproches des anterieurs et les recouvrant partiellement de leur bord anterieur; aire opisthosomale anterieurement bien limitee....................
. . . ................................. 10.'Labidophorus Kram.
XENOCASTOR, gen. n.
?. Espece de tres grande taille, blanchatre; gnathosome, pattes et espaces chitinises entourant leurs bases, d’un rouge brunatre. Idiosome 1.4 fois plus long que large. Dimensions des soies, exprimees en% de la longueur de 1’idiosome; va 1; see 17; set 14; ha et sh 1Б a 16; Zip 28;
14; d2 6; d3 ЗБ; 38; Z4 12; Z2 28; Z3 21; sa e ББ; sa i 9;' d2 placees sensiblement en arriere de Z4 a peu pres au meme niveau que d3; soies anales lisses, capilliformes. Palpes genitaux contigus, exclusivement grands (les anterieurs 72 X 45 y, les posterieurs 61 X 41 p.). Pattes I et II quelque peu aplaties lateralement, ayant 25 (I) et 27 (II) % de longueur de 1’idiosome. Tarses pres de 3 fois plus longs que leur diametre basal et un peu plus de 1.Б fois plus longs que les tibias, coniquement retrecis vers le bout, avec 2 grandes epines ventrales plantees 1’une derriere 1’autre, un court batonnet olfactif cylindrique et de gran-1 des soies apicales"; longueur des pretarses I est egale aux 3/4 de la longueur du tarse; soies ventrales des tibias spiniformes; genuaux renfles en
XENOCASTOR, LABIDOPHORUS
459
bosse a la base dorsalement, soie basale divisee on 3—4 branches. Pattes III un-. peu plus longues que les anterieures (28.5 de longueur de 1’idiosome), mais sensiblement plus greles; la longueur des tarses surpasse 5.5foisleur diametre basal ot plus de 4 fois la longueur des tibias; tarses armes de 3 grandes epines ventraleset d’une apicale, avec3finessoies au bout; soies ventrales des tibias ayant 1’aspoct do longues epines droites. Pattes IV tres longues (40% do la longueur do 1’idiosome) et greles; leurs tarses 6 fois plus longs que lours diametre basal, presque 3 fois plus longs que les tibias et 4 fois plus longs que les pretarses; armement des pattes IV pareil a celui des pattes III, mais les soies ventrales des tarses plus longues et plus rapprochees, disposees plus distalement et formant ainsi une sorte de poigne. Longueur 1084 p., largeur 750 p.. — Trouve dans la Bielorussie, sur la Beresina. Habite sur le Castor (Castor fiber vistulanus Matschie)...........X. fedjushini, sp. n.
LABIDOPHORUS KRAM.
A d u 11 e s
99
' 1- (2). Soies de la surface dorsale distinctement plume uses; peau presqu’inco-lore, finement granulee; d2 tres reduites; h a a peu pres au meme niveau que see: epimeres III et IV relies par leur bouts interieurs. Sur les ecu-reuils et dans leurs nids. — Hollande, Allemagne, Italie, USSR (region de Moscou). . ....................................1.1. sciurinus (Koch).
' , 2 (1). Soies de la surface dorsale presque lisses, avec de courts oils tres rares;
; peau teintee de couleur brunatre-pdle, dans la partie dorsale posterieure
j avec un groupe de gros boutons ovales aplatis; bien developpees;
t h a placees bien en arriere de sc; epimeres HI et IV libres. Sur le Desman
* et dans ses terriers........................2: 1. desmanae, sp. n.
h s
L Hypopos
1 (2). Champs coxaux III clos (epimeres III reliees des bouts avec les epi-Г meres IV), tarses IV termines par une tres grande macrochete et quelques
epines plus courtes et fermes; soies ventrales des femurs I et II tres I longues, flexibles, capilliformes. Longueur 350 p.. Sur I’Ecureuil. —
!' Hollande, Allemagne, Italie, URSS (region de Moscou).............
....................................... 1. L. sciurinus. {Koch).
2 (1). Champs coxaux III ouverts — epimeres III et IV libres; extremite des tarses sans macrochetes, soies ventrales des femurs I et II courtes;
| longueur ne depassant pas 280 p.. Sur d’autres mammiferes.
|. 3 (4). Soies ventrales- exterieures des tibias et genuaux I et II representees
par de robustes epines coniques. Propodosome fort allonge; ses bords lateraux se relient par devant en angle obtus arrondi au sommet; soie ' apicale des tibias I et II longue, non modifies; angles posterieurs du
460
GLYCYPffAGIDAE
corps arrondis; «rapes» de petites dimensions avec de peu nombreuses-carenes transversales. Sur la taupe d’Extreme Orient (Mogera robusta Nehr.). — Koree: (Seoul)........................5. L. mogerae,sp. n.
4 (3). Soies ventrales des tibias et genuaux I et II fines, flexibles ou ayant 1’aspect de sveltes epincs; propodosome plus court, ses bords se re-joignent par devant en angle obtus; soie apicale des tibias I et II courte, a bout arrondi; angles postdrieurs du corps fort saillants; «rdpes» de-grandes dimensions avec de nombreuses carenes transversales.
6 (6). Corps allonge, a. peu pres 1.5 fois plus long quo large; ses angles poste-rieurs saillants en forme de petites comes cylindriques, recourbees en. bas; tarses IV avec de fortes epines coniques, 3 fines soies moyennes et 2 petites soies spiniformes (sans compter I’ongle reduit); tarses I et II avec 3 soies elargieg, lanc^olees. Sur le desman (Desmana moschata L} .....................................................2. L. desmanae, sp. n.
6 (5). Corps large, arrondi ou presque rliomboidal, 1.2—1.3 fois plus long que large; angles posterieure formant un rebord recourbe perpendicu-lairement a sa surface; bout des tarses IV sans epines, avec 4 soies assez. longues et une courte; soies des tarses I et II simples ou a peine e'largies au bout.
7 (8). De taille plus grande: longueur 220—250 p.; aristes longues; «rapes»-posterieures de longueur moyenne avec une dizaine de carenes transversales; corps arrondi, se retrecissant graduellement vers son extremite. Sur les taupes (Talpa europea L.). — Hollande, Allemagne, URSS (regions de Moscou, de Leningrad et de Smolensk). . 3. L. talpae Kram.
8 . (7). De plus petite taille: longueur 190—220 p; aristes tres courtes; «rapes» posterieures extremement longues, avec 16—20 petites carenes transversales; corps plutot rhombo'ldal, se retrecissant subitement vers le bout, posterieur. Sur differentes especes de musaraignes et parfois aussi sur la taupe et differents rongeurs. — Hollande, Allemagne, URSS (region de Moscou, Abkhasie, environs de Jakoutsk) .... 4. L. soricis Ouds.,
XENOKYCTES, gen. n.
99
1 (2). Bouts du sternum et des epimeres II et III relies a un fin arc cliitineuxr situd devant.le cercle circumgenital, lequel est a son tour relie avec coder ni er; epimeres IV libres; tibias I avec une carene dorsale peu deve-• loppee et une seule soie ventrale lisse. — Angleterre, Italic ......
...............................................1. X. krameri(Mich.)..
2 (1). Bouts du sternum et des epimeres II, III et IV directement relies & 1’epigyne tres robuste, confluant avec le cercle circumgenital; tibias-I avec une tres haute carene dorsale et deux soies ventrales, dont 1’une-lisse et la 2-me pectiniforme. — Hollande, URSS (Moscou)...................
...............................................2. X. heptneri, sp. n.
XEXORYCTES, MYACARUS
461
II у p оp e s
1 (2). Saillie rostrale tres ekoite, fort proemineute; les bouts des lames de Гар-pareil de fixation ressortent en arriere eh forme de petites cornes pointucs; bord posterieur do I’idiosome fortement echancre; pattes III et IV unpeu ; plus courtes Que les anterieures; tarses IV denues d’ongle, avec 4 longues
" soies distales. — Angleterre, Italie......1. X. krameri (Mich.),
ii 2 (1). Saillie rostrale arrondie, peu proeminente; extremity des lames de
[i 1’appareil de fixation obtuses, non visibles d’en haut; bord posterieur
de I’idiosome presque droit; pattes III et IV sensiblement plus courtes que les anterieures, tarses IV avec un ongle biendeveloppe. — Hollande, URSS (Moscou)................................2. X. heptneri, sp. n.
MYACABUS, nom. n.
Homopus Koch 1841, nom. praeoccup* к
Hypopes '
1 (2). Ecusson propodosomal plus de 1.5 fois plus large (a sa base) que long, a bord anterieur rectangulaire; Epimeres III Stroits; tarses I et III sensiblement plus courts que le genual et le tibia correspondantepris ensemble; soie ventrale interieure des tibias I et II plantee beaucoup plus haut que I’exterieure. —.France, Hollande, Allemagne, URSS (region de Moscou, 'nord-ouest du Kazakhstan, Kirghizie, lies de Chantar) . . . ...............................................1. M. arvicolae (Duj.).
2 (1). Ecusson propodosomal pas plus de 1.3 fois plus large (a la base) que long, formant par devant un angle aigu; epimeres III tres larges; tarses I et II plus longs, egalant en longueur les deux articles precedents des pattes pris ensemble; les deux soies ventrales des tibias I et II placees a la meme hauteur. — Allemagne, URSS (regions de Moscou et du fleuve
j Amour),. .................................2. M. hypudaei (Koch.).
6. Sous-fam. Carpoglyphinae
1 1 (2). Tarses I et II tres courts, se terminant en une forte griffe a la base
de laquelle prend naissance le pretarse extraordinairement long, fin et flexible, portant a son extremite un ongle relativement petit; idiosome avec un sillon transversal assez distinct.Especeslittorales, habitant parmi les algues . .........................17. Hyadesia Megn.
2 (1). Tous les tarses de structure normale, sans forte griffe terminale; pre-tames larges, de conformation normal; idiosome denue de toutes traces de sillon transversal. Especes terrestres, quoique habitant souvent les matieres liquides ou semiliquides.
462
GLYCYPHXG1DXE
3 (4). Peaulisse, toutes ]es soies de la surface dorsale de 1’idiosome, exccpte sa e, courtes, arroudies au bout; palpes genitaux bien developpes; pattes-111 et IV de conformation normale dans les deux sexes; chez la 9, les epimeres I et II sont relies a I’epigyne...........18. Carpoglyphus Rob.
Une seule, espece palearctique, C. lactis (L.). — Europe, Ame-rique du Norcl.
4 (3). Peau scabrciise, souvent couvertc do nombreuses petites epines cuti-culaircs; soies do 1’idiosomc plus longues et plus fortes; palpes genitaux rudimentaires; chez le d, les pattes III sont fortement allongees, celles-de la IV paire renfleeset incurvees; chez la 9, les epimeres I et II sont libres.................................................... 19. Hericia Can.
Ч
HfADESIA MEGN.
1 (2). Tarses III et IV du d pres de 3 fois plus courts que les tibias correspon-dants; v a tres longues, a peine plus courtes que sc e et h p; toutes les-d et I tres courtes, a peu pres de la meme longueur; epine ventrale des tibias I et II 4—5 fois plus courte que le diametre du tibias, conique;. orifice genital du d separe par un large espace de I’extremite du sternum; 9 sans epigyne. — Allemagne, Irlande. ..... 1. H. fusca (Lohm.).
2 (1). Tarses III et IV dans les deux sexes plus longs que les tibias c.orrespon-dants; v a plus de 2 fois plus courtes que sc e et h p; soies dorsales et laterales assez longues (13—30% de la longueur do 1’idiosome); d2 sensi-blement plus longues que dx, l3 presque 1.6 fois plus longues que d2, epine ventrale des tibias I et II mediocrement plus courte que le diametre du tibia; orifice genital du d tres large, touchant presque 1’extre-mite du sternum; 9 avec un grand epigyne arque. — Angleterre . . . .
........................................ 2. H. algivorans (Mich.).
HERICIA CAN.
1 (2). Peau densement couverte d’epines cuticulaires assez longues; dd plus grands que les 99, plus fortement pigmentes, a surface dorsale ornee t
de nombreux ilots chitineux un peu enfonces; tarses II du d avec 2 fortes epines ventrales placees A, la meme hauteur, 1’une pres de 1’autre. Longueur totale du d 520—600 p, de la 9 pres de 500 p. — Angleterre, France..............................................1. H. hericia (Rob.).
2 (1). Peausans epines cuticulaires; dd plus petits.que les 99, de la memo couleur qu’elles (blanchatres), sans ildts chitineux a la surface dorsale du corps; tarses II du d avec 3 epines moins fortes, disposees le long du tarse, 1’une derriere 1’autre. Longueur totale du d pres de 460 p, de la 9 750 p — Angleterre......................................2. H. georgei Mich.
СИ A ETODA C TYLLNA E. TABLEAU DES GE Al RES Ш
7. Sous-fam. Chaetodactylinae
TABLEAU DES GENRES'
A d U 11 e S
1 (2). Pas d’ecusson propodosomal; idiosome allonge 1.4—1.5 fois plus long que large, avec un sillon transversal assez distinct; sternum bien devc-loppe dans les deux sexes; extremites des epimeres III et IV relies entre elles par une fine ligne chitineusc, de sortc quo les champs coxaux III sont formes...........................21. Chaetodactylus Rond.
2 (1). ficusson propodosomal bien developpe, formant deux bandes laterales chitinisees qui atteignent la base des pattes II et se terminent ici ев. une lamelle plus ou moins ovale; idiosome tres large (1.1—1.3 fois seulement plus long que large), sans aucune trace de sillon transversal, d avec unsternum enforme de V; chez la 9 , les epimeres Isont directement relies a I’epigyne; epimeres III et IV tres courts, libres..
...........................................20. Sennertia Ouds.
Hypopes
1 (10). Pattes IV avec un ongle bien developpe; corps allonge (pas moins de 1.5 fois plus long que large), plus ou moins ovoi'de, assez convexe; ecussons dorsaux recouvrant toute ou presque toute la surface dorsale du corps. Especes de taille mediocre, souvent ressemblantes aux hypopes des Saproglyphidae.
2 (5). Tarses IV sans macrochetes, mais avec 2 ou 3 petites soies lanceolees;. sternum simple (non bifurque); ongles fortement recourbes en crochet,, a peu pres de memes dimensions a toutes les pattes.
3 (4). Sculpture des ecussons dorsaux tres grossiere, composee de fossettes ovales ou rondes a contours tres nets, ce qui fait paraitre les ecussons cribles de trous; disque adhesif carre; bord posterieur de l’hyst6rosome profondement sinue........................... 25. Cerophagus Ouds.
Une seule espece, C. granulatus (Duj.) — France, Hollande, Allemagne. "
4 (3). Sculpture des ecussons dorsaux tres delicate, composee de fines lignes, enfoncees, ayant une direction longitudinale sur I’hysterosome et oblique et transversale — sur le propodosome; disque adhesif arrondi; bord posterieur du corps parabolique......26. Cerophagopsis, gen. 11.
5 (2). Tarses IV avec 1—3 fortes macrochetes, sans soies foliacees ou celles-ci ont la forme de lames etroites, presque lineaires; ongles faiblement. arques ou contournes en spirale— dans ce dernier cas les ongles des pattesIV sont beaucoup plus petits que les autres; sternum a extremitc bifurquee.
6 (7). Ongles des pattes I—III tres robustes, contournes en spirale, plantes sur un pretarse charnu bien developpe; ongles des pattes IV beaucoup
464 GLYCYPHAGIDAE
plus petits que les autres, mais ayant la meme forme; pattes I et III tres robustes, beaucoup plus fortes que les posterieures (IV); tarses I— III ornes de 3 fortes soies falquees; soies de la surface dorsale extiAme-ment courtes et fines . ...................27. Sennertionyx, gen. n.
Uno scale espdcc, S. manicati (Giard)-—France, Andalousie, Italic, tie, do Corfoti; URSS (Odessa, Armenie, Kopet-dagh, nord-oacst du Kazakhstan).
"7 (6). Tons les ongles do la memo dimension, faiblement arques, sans pretarse charnu; pattes IV ayant a peu prte los mOmes dimensions que celles des autres paires; tarses I—III ornes de 4—5 soies largoment lanceolees.
•S (9). Soies de la surface dorsale represents sculement par de tres fines microchetes; sculpture des ecussons dorsaux indistincte, du meme type que chez Cerophagopsis-, tarses I—III avec 4 soies lanceolees, ceux de la IV paire ornes rien que de soies simples................22. Horstia Ouds.
(8). Soies dp la surface dorsale tres longues, un peu aplaties; sculpture des ecussons dorsaux composee de nombreuse fossettes allongees et plus ou moins vermiculees; tarses I—III avec 6 soies lanceolees; tarses IV ornes d’etroites soies foliacees................24. Neohorstia, gen. n.
10 (1). Pattes IV sans ongle.
11 (12). Ongles (I—III) simples, assez petits, faiblement arques, sessiles; pretarse absent; corps allonge, piriforme ou en hexagone allonge, 1.45—1.6 fois plus long que large. Especes de petite taille...............
................................................23. Tortonia Ouds.
.12 (11). Ongles (I—III) enormes, fortement contourn<5s en spirale, situe au bout d’un pretarse charnu plus ou moins long; corps tres large, aplati, souvent presque rond. Espece generalement de forte taille.
A3 (14). Deux ecussons dorsaux: le propodosomal, qui est de forme triangu-laire, et I’hysterosomal — en forme de trapeze renverse; soie basale des genuaux I tres longue. Sur les abeilles des genres Osmia. Anthidium et Megachile...................................21. Chaetodactylus Rond.
.14 (13). Un seul ecusson dorsal — I’hysterosomal; il atteint sa plus grande largeur pres du bord posterieur du corps et se rdtrecit plus ou moins graduellement par devant; soie basale des genuaux I courte. Sur diffe-rentes especes du genre Xylocopa.....................20. Sennertia Ouds.
SENNERTIA OUDS.
A d u 11 e s
A-(2). Sc et les 4 paires de soies dorsales (dt—d4) sont representees par de courtes et robustes microchetes; h a, h p, et sa i tres epaisses, ayant 1 ’aspect de fortes tiges recourbees en arc, obtuses au bout; peau de la surface dorsale densement couverte de squamoides triangulaires; ongles tres grands, robustes. — AJgerie........................4. S. cantabrica, sp. n.
SENNERTIA
465
2 (1). See et tons les d ayant 1’aspect d’assez longues soies (mesurant 12—20% de la longueur de 1’idiosome) un peu plumeuses; ha et hp ayant la meme structure; sa e un peu plus epaisses, droites; peau de la surface dorsale tres finement granulee; ongles petits. — Europe meridionals, centrale et orientale, partie palearctique de 1’Asie. . . ................................................1. S. cerambycina (Scop.).
H у p о p e s
1 (2). Sc i et representees par des macrochetes, ayant approximativement la longueur de sc e(32—37 % de la longueur du corps), de fa^on a ce que la surface dorsale du corps est munie de 7 paires de soies semblables; di&t, d2ont 1’aspect d’assezlongues etfortes soies, longues de 6—16% de la longueur du corps. Sur les especes du groupe Xylocopa olivieri Lep. — Grece, Turquie, Armenie, Asie Centrale. . . 7. S. zhelochovtsevi, sp. n.
2 (1). Sc i 2—4 fois plus courtes que sc e ou elles sont representees par des microchetes minuscules; <4, d2, et d3 ont 1’aspect de microchetes minuscules ou complfctement reduites, ayant la forme de petits pores arrondis; la surface dorsale n’est munie que de 5—6 paires de macrochetes. Sur d’autres especes de Xylocopa.
3 (6). Sc i longues de 8 a 20% de la longueur du corps, 2—4 fois plus courtes que sc e, assez fortes, ce qui fait que la surface dorsale porte 6 paires de soies plus ou moins longues.
4 (5). Soies de la surface ventrale du corps fortement epaissies, spiniformes; les ventrales transformees en une paire d’organes ovoides, fixes (й, une courte tige; soies dorsales extrSmement robustes; sc i 2 fois plus courtes que sc e, ayant de 15—20% de la longueur du corps; ecusson hystero-somal tres grand, atteignant presque sc i par devant; 1’une des soies ventrales des tibias I et II et la soie ventro-apicale des tarses ont la forme de robustes epines coniques. Sur Xylocopa dissimilishep. — Japon, Assam, Java............................................6. S. horrida (Vtz.).
5 (4). Toutes les soies de la surface ventrale du corps capilliformes; microchetes dorsales relativement fines et flexibles; sc i 4 fois plus courtes que sc e, longues de 8—10% de longueur du corps; ecusson hysterosoma! etroit, n’atteignant par devant que dx; toutes les soies des pattes fines, flexibles. Sur Xylocopa pictifrons Sm. — Sze-tchouan.................
............. . . ................................8. S. potanini, sp. n.
6 (3). Sc i representees par des microchetes minuscules, la surface dorsale ne. porte que 5 paires de fortes soies.
7 (8). Soies de la surface ventrale du corps spiniformes; pretarses I—III avec un appendice digitiforme fortement developpe. Sur Xylocopa kalinovskii Rad. (circumvolans Sm.). — Chine, Coree, Japon . .5. S.alfkeni (Ouds.).
8 (7). Soies de la surface ventrale du corps fines, capilliformes; pretarses sans excroissance ou avec un petit appendice obtus. Sur d’autres especes de Xylocopa.
30 Фауна СССР, Захваткин—284
466
GLYCYPHAGIDAE
9 (16). L p plus courtes que sc e ou presque la meme longueur; ecusson hyste-rosomal grand, fortement chitinise et intensement colore, s’etendant jusqu’h dj ou encore plus en avant; ventouses -exterieures et posterieures pas tres petites; les exterieures situees lateralement pres des centrales, considerablement eloignees des posterieures.
10 . (11). Espece ti-fcs grande, longue de 460—615 p; ecusson hysterosomal 1.6 fois plus long quo sa largeur h la base, se retrecissant reguli&ement vers rextremitd antCricurc. Sur Xylocopa punctilabrisF. Mor. etX. valga-G-erst. — Tadzhikistan, Turquie...................2. S. gargantua, sp. n.
11-(10). Especes plus petites, ne depassant pas 340 p. en longueur; ecusson hysterosomal 1.75—2 fois plus long que large, h bords paralleles, plus ou moins arrondi anterieurement.
12 (13). Bord posterieur de I’ecusson hysterosomal coupe droit, nonrecourbe-sur le c6te ventral du corps, ce qui fait que les sa i et p a sont disposees dorsalement; disque adhesif eloigne du bord posterieur du corps; soie ventrale des tarses IV 4 fois plus longue que les tarses eux memes,. I’exterieure ventrale des tarses I et II non epaissie, setiforme; I p 1.5 fois plus courte que see. Sur Xylocopa cantabrica hep. — Algerie......
.............................. 4. S.cantabrica, sp. n.
13 (12). Bord posterieur de I’ecusson hysterosomal arrondi, fortement recourbe sur la surface ventrale du corps et muni ici d’une profonde echancrure, dans laquelle entre la partie posterieure du-disque adhesif ; sa i et p a situees ventralement; soie ventrale des tarses IV plus courte que le-tarse ou 1.4—1.5 fois plus longue; I p h peine plus courte que sc e.
14 (15). Ecusson hysterosomal 1.75 fois plus long que large; la distincte-ment plus courtes que see; soie exterieure subapicale des tarses I et 1Г capilliforme, I’exterieure apicale fortement recourbee, epaissie em forme de couteau; la soie ventrale des tarses IVtres petite, distinctement plus courte que le tarse. Espece plus grande. Longueur 270 p, largeur 230 p.. Sur les especes du groupe Xylocopa violacea Lep.— Europe-meridionale,“ centrale et orientale, partie palearctique de 1’Asie. . . ...............-........................... . 1. S. cerambycina (Scop.).
16 (14). Ecusson hysterosomal plus etrojt, 2 fois‘-plus long que large; la a peine plus longues que sc e; 4a soie exterieure subapicale des tarses. I et Illanceolee, elargie, I’exterieure apicale fine, en forme' de batonnet;, soie ventrale des tarses IV sensiblement plus longue que le tarse lui. тёте. Taille plus petite; longueur 224—226 p.. Sur Xylocopa dissimilis Lep.—Japon......................................... . 3. S; dissimilis, sp. n.
16 (9). L p tres longues — 2 fois plus longues que sc e; ecusson hysterosomal tres petit, n’atteignant pas тёте le niveau de d2, faiblement chitinise-et presque incolore, indistinctement separe de la peau molle qui 1’en-toure; ventouses exterieures-et posterieures du disque adhesif tres petites,. disposees en rangee trans versale. Sur Xylocopa dissimilis Lep. — Japon.............................................9. S.oudemansi, sp. n.
SENNEIMIA, CHAETODACTYLUS 567
CHAETODACTYLUS ROND.
Hypopes
1 (2). Soies sternales transformees en robustes epines ovoides, les ventrales coniques, fortement epaissies; sternum bifurque au bout; pattes I et II tres robustes; le bout de leurs tarses a sur la surface ventrale une grosse epine recourbee; tarses IV avec 3 soies tres longues. Espece de forte taille. Longueur 370—450 [i. Sur Osmia (Ceratosmia) tricornis Latr. — Italie......................................4. Ch. claviger Ouds.
2 (1). Soies sternales et ventrales tres fines, capilliformes; sternum simple; pattes I et II non epaissies, boiit de leurs tarses depourvu d’epine; tarses IV avec une tres grosse macrochete et 2 soies beaucoup plus courtes et plus fines (quelquefois celles-ci sont representees par des microchetes). La longueur du corps ne'depasse pas 375 [л.
3 (8). Ventouses exterieures disposees lateralement pres des centrales (formant avec elles une rangee transversale), bien au devant des posterieures.
4 .(5). Sc i robustes, ayant la meme longueur et epaisseur que see et h p, disposees au тёте niveau que les 2 paires burner ales (ha et he); ds, sa e et sa i tres rigides, aciculees; soie intero-apicale des tarses IV 3 fois plus longue que toute la patte IV etque la soie extero-ventrale. Sur Chelostoma florisomnis L. — URSS (region de Kirov: Lavy) . . . ................................................ 3. Ch. birulai, sp. n.
5 (4). Sc i bien plus courtes et plus fines que h et sc e, situees sensiblement devant la ligne reliant les humerales (h); d3, sa e et sa i fines, flexibles; soie intero-apicale des tarses IV pas plus de 2 fois plus longue que la patte IV et que la soie extero-apicale. Sur les especes du genre Osmia.
6 (7). Soie intero-apicale des tarses IV sensiblement plus courte que les pat-' tes IV et un petit peu plus longue que I’exterieure; sc e plus courtes (10—11 % de la longueur du corps), rien que 1.5 fois plus longues que sc i; soie interieure subapicale des tarses I—III, ainsi que I’exterieure des pattes III tres faiblement elargie (etroitement lanceolee). — Angleterre, France, Italie, Roumanie, URSS (environs de Sarepta, et de Tbilisi) .......................................... 1. Ch. osmiae (L. Duf.).
7 (6). Soie intero-apicale des tarses IV 2 fois plus longue que les pattes IV .et 1’extero-apicale; sc e plus longues (pres de 20% de la longueur du? corps), de 3 a 4 fois plus longues que sc i; la soie interieure subapicale des tarses I—III et I’exterieure des tarses III fortement elargies, lan-ceolees. — Allemagne, URSS (Crimee, Georgie).............................
.......................................... 2. Ch. reauinuri (Ouds.).
8 (3). Ventouses exterieures disposees derriere les centrales, au meme niveau que les posterieures.
9 (10). Espece plus grande; longueur pres de 350 p.; ecussons dorsaux d’un
568
GLYCYPHAGIDAE
brun clair avec une sculpture poreuse et de nombreuses fossettes longitudinales; disque adhesif plus long que large. Sur Anthidium sticticum F. — Tunisie.............................5. Ch. anthidii (Ouds.).
10 (9). Espece plus petite; longueur du corps 215—310 [л; ecussons dorsaux non pigmentes, avec un delicat dessin reticule; disque adhesif un peu plus large que long. Sur quelques especes de Megachile et sur les guepes Pseudovespa germanica L. — URSS (Leningrad, Ouglitch, Oustj-Kame-nogorsk, Ak-tasch pr&s do Taschkent)........6. Ch. demcntjcvi, sp. n.
HORSTIA OUDS.
Hypopes
1 (2). Espece de grande taille (334 y);tous les champs coxaux clos; sternum relie par les extremites de sa fourche aux epimeres II, ceux-ci, a leur tour, aux epimerites II, les epimeres III — aux epimeres IV et ces der-niers confluents en un court ventrum. Sur Xylocopa circumvolans Sm. — Japon......................................1. H. ornata (Ouds.).
2 (1). Petite espece (165—180 p.); tous les champs coxaux ouverts; epimeres II—IV, epimerites II et sternum libres. Sur Xylocopa dissimilis Lep. et Platynopoda tenuiscapa, Westw. — Japon, Inde, Java................
........................................ 2. H. helenae (Ouds.).
TORTONIA OUDS.]
1 (2). Idiosome piriforme, fortement renfle; les ecussons dorsaux recouvrent toute la surface dorsale, converts d’un dessin reticule prononce qui prend parfois 1’aspect d’ecailles en haut relief; tarses I et II longs, greles, ongles relativement longs et fins; sc formant une rangee transversale; organe pseudostigmal bien developpe. Sur les especes des genres Osmia et Stelis. (Sous-genre Tortonia s. str.). — Allemagne, Suisse, URSS (region de Leningrad, Siberie, Georgie).........................
.........................................1. T. intermedia Ouds.
2 (1). Idiosome allonge, hexagonal, aplati dans le sens dorso-ventral; ecussons dorsaux ne couvrant qu’une partie de la surface dorsale (les parties peripheriques sont de consistance membraneuse, molies), a faible dessin longitudinal; tarses I et II courts, epais; ongles courts et gros; sc i disposees considerablement plus en avant que sc e; organe pseudostigmal absent. Sur les especes du genre Anthidium. (Sous-genre Donnadieuia, subg. n.).
3 (4). Sc i, h i, d2, d3et les deux paires de l2 (I a et I p) ayant 1’aspect d’assez longues soies (13—28% de la longueur de 1’idiosome) aplaties; see extre-mement longues (pres de 45% de la longueur de 1’idiosome), depassant de leur bouts la base de I a-, Ecussons dorsaux plus etroits, couvrant une petite partie de la surface dorsale de 1’idiosome; soie apicale posterieure
HORSTIA, TORTONIA, NEOHORSTIA, CEROPHAGOPSIS MS
des tarses IV plus courte que la patte. Sur Anthidium septemdentatnm-Latr. — Italie (Venise) ......................4. T. dogaressa, sp. n„
4 (3). Sc i, hi, d2, ds et I p representees par de minuscules microchetes A peine visibles; see de longueur moyenne (20—25% de la longueur de? 1’idiosome), atteignant a peine de leurs bouts la base de d2- ecussons dorsaux plus larges, couvrant une partie un peu plus grande de la surface-dorsale de 1’idiosome; soie postero-apicale des tarses IV 2 fois plus longues que la patte IV. Sur Anthidium sticticum F. — Espagne, Al-gerie, Tunisie...............................3. T. smitsvanburgsti Ouds»
NEOHORSTIA, gfen. n.
H у p o p e. Blanchdtre, faiblement tinte d’oere jaunatre, cette teinte plus prononcee dans la partie posterieure du dos. Corps 1.4—1.5 fois plus, long que large, h bord posterieur obtusement arrondi. Ecusson propodosomal un peu plus de deux fois plus court que I’hysterosome; sc tres longues, flexibles, distinctement aplaties; la longueur de sc i atteint pres de 30% de la longueur du corps et depasse la longueur de sc e; h ir —d3 et la ayant de 17 a 30% de la longueur du corps, de structure pareille a celle des sc i; le reste des poils represents par de fines soies capilliformes. Lamelle chitinisee du disque adhesif formant trois ilots ovales separes. Tarses courts, I et II rien que 1.5 fois, III et IV — a peine plus longs que leur diametre basal; bdtonnet olfactif des tarses I et II droit, se retrecissant graduellement yers le bout. Longueur 200—262 largeur 144—150 — Grece (Attique), Iran. Trouvesur Anthidium puhes-
cens F. Mor. (Iran) et A. laticeps F. Mor. (Grece). . N. mamillata, sp. n.
CEROPHAGOPSIS, gen. n.
H у p o p e. D’un jaune ocre pdle. Corps 1.4—1.5 fois plus long que large; propodosome 2.5 fois plus courte que I’hysterosome. Disque adhesif arrondi; ventouses laterales disposees au тёше niveau que les centrales, qui sont de dimensions beaucoup plus grandes; ventouses posterieu-res de la тёте grandeur queles laterales; lalamellechitinisee du disque forme en arriere trois il6ts separes, a contours uh peu anguleux. Pattes anterieures sensiblement plus longues, mais a peine plus grosses que les1 posterieures; tarses I et II 1.8—2 fois, III et IV —1.5 fois plus longs que leur diametre basal; batonnet olfactif des tarses I et II un peu re-courbe et renfle au bout. Longueur 280—285 p., largeur 150—175 p.— Japon (Koyto). Trouve en grand nombre sur Megachile doderleini Fr. ..................................................C. skorikovi, sp. n.
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ 1
Achropodophorus Rosas Costa 127,
Acotyledon Ouds. 80, 81, 82, 123, 125, 127, 4f4, 415, 421, 423, 425
Aeotyledonini 123, 125, 126, 421, 423
Aeroglyphinae 279, 280, 310, 447, 448, 453
Aijroglyphus gen. n. 310, 453
-africanus Ouds., Tyroglyphus 86
agilis (Can.), Acotyledon 129, 132, 140, 143, 425, 427
agilis Mich., Rhizoglyphus 140
alberti (A. Z.), Acotyledon 130, 133, 134, 146, 213, 426, 427, 429
Aleurobius Can. 85
Aleuroglyphus gen. n. 85, 92, 417
aleurophagus Sicher, Histiogaster 209 alikeni (Ouds.), Sennertia 380, 386, 393, 465
algidus Berl., Rhizoglyphus 68, 212
algivorans Mich., Hyadesia 364, 366, 369, 371, 462
•americanus Banks, Tyroglyphus 92, 106
anonymus Hall., Glycyphagus 368
anthidii (Ouds.), Chaetodactylus 394, 398, 468
arcuatus (Tr.), Chortoglyphus 27, 285, 286, 450
arcuatus Tr., Tyroglyphus 284
armipes Banks, Tyroglyphus 154 arvicolae (Duj.), Myacarus 351, 358, 461
australis Ouds., Saproglyphus 238
bacchus sp. n., Histiogaster 13, 131, 188, 190, 193, 195, 435, 436
batsylevi sp. n., Acotyledon 130, 133, 139, 148, 426, 428
berlesei (Mich.), Caloglyphus 155, 158, 170, 177, 185, 213, 431, 434
jberlesei Mich., Tyroglyphus 153
berlesiana nom. n., Monieziella 185, 206, 207, 213, 431, 434
birulai sp. n., Chaetodactylus 394, 396, 397, 467
Blomia Ouds., 284, 287, 450
borealis subsp. n., Acotyledon redikorzevi 130, 133, 146, 426, 428
brasiliensis Ouds., Caloglyphus 154
breviceps Banks, Tyroglyphus 92
brevipes Banks, Monieziella 198
bulgarica Storkan, Calvolia 247, 257, 261, 445
bitlleri WoL, Tyrophagus 103
burchanensis Ouds., Glycyphagus 294, 299, 301, 451
cadaverum Ouds., Glycyphagus 296
cadaverum (Schr.), Glycyphagus 293, 295, 302, 305, 452
calliphorae Vtz., Calvolia 247, 256, 261, 445
Caloglyphidae 68, 83, 180
Caloglyphus Berl., 81, 82, 123, 125, 153,
415, 421, 423, 430
Calvolia Ouds., 234, 235, 240, 440, 441
canestrinii (Arm.), Ctenoglyphus 316, 317, 318, 320, 454, 455
cantabrica sp. n., Sennertia 13, 379, 380, 385, 389, 464, 466
carpio (Kram.), Histiogaster 188, 192, 435
carpio Zacher, Histiogaster 190
Garpoglyphidae 276
Carpoglyphinae 279, 281, 282, 361, 446,
447, 449, 461
Carpoglyphus Rob. 362, 367, 462
casei Ouds., Tyroglyphus 96
casei (Ouds.), Tyrolichus 3, 21, 71, 75, 97, 417
castellanii Hirst, Tyrophagus 106
caucasica sp. n., Schulzea 221, 226, 227, 439
cavernicola Vtz., Schwiebea 200, 203, 437
cerambycina (Scop.), Sennertia 379, 381, 382, 383, 465, 466
cerambycinus Scop., Acarus 377
Ceroglyphus Vtz. 92
Cerophagopsis gen. n. 376, 408, 463, 469
Cerophagus Ouds. 376, 407, 463
Chaetodactylinae 68, 280, 282, 374, 448, 450, 463
Chaetodactylus Rond. 375, 377, 391, 463, 464, 467
chelone Ouds., Sancassania 224, 225
Chortoglyphidae 276
1 Названия подсемейств, семейств и.высших групп выделены жирным шрифтом, прочие названия — прямым шрифтом, причем роды и подроды начинаются с заглавной буквы, синонимы и прежние родовые названия для видов — курсивом. Жирными цифрами обозначены страницы, где даны описания, курсивными цифрами — страницы, где помещены рисунки.
УКАЗАТЕЛЬ
471
Chortoglyphinae 279, 280, 281, 282, 446, 448, 449, 45'0 л.
Chortoglyphus Berl. 284, 450
circumspectans Vtz., Calvolia 245, 246,
258, 444
-claviger Ouds., Chaetodactylus 392, 396, 397, 467
cocciphilus Banks, Tyroglyphus 92
-Coepophagus Megn. 180
Cometacarus A. Z. 292, 306, 451,-453
corticalis Berl., Histiogaster 207
corticalis (Mich.), Monieziella 185,206,
207, 213, 437, 438
•Cosinoglyphus Ouds. 127
crabronis sp. n., Troupeauia 220, 221, 222, 439
Crabrovidia subg. n. 270, 446
cribratissima sp. n., Lackerbaiieria 214, 221, 438
Ctenoglyphinao 280, 281, 312, 447, 448, 453
Ctenoglyphus Berl. 313, 314,- 454
cursor Gerv., Glycyphagus 302
Czenspinskia Ouds. 240, 242
Czenspinskiidae 229
dampfi Ouds., Acotyledon 129
dementievi sp. n., Chaetodactylus 394,
397, 399, 468
Dermacarus Hall. 332, 334
desmanae sp. n., Labidophorus 331, 334, 335, 337, 339, 459, 460
-destructor (Schr.), Glycyphagus 13, 25,
27, 33, 293, 294, 296, 299, 332, 452
diamesa sp. n., Forcellinia 111, 113, 116, 420
Diamesoglyphus gen. n. 313, 314, 454
-dimidiatus Herm., Tyrophagus 99, 100
dispar Mich., Glycyphagus 350
dispar (Mich.), Orycteroxenus 352, 355, 456, 457
-dissimilis sp. n., Sennertia 382, 383, 385, 389, 466
dogaressa sp. n., Tortonia 402, 404, 405, 469
domesticus (Deg.), Glycyphagus 33, 295, 302, 305, 453
Donnadieuia subg. n. 402, 403, 468
dujardini Clap.., Hypopus 182
dysenteriae L., Acarus 367
JSberhardia Ouds. 127, 141
Ebertia Ouds. 83, 92
Ebertiidae 83
echinopus (Fum. et. Rob.), Rhizoglyphus
71, 131, 181, 182, 189, 195, 415, 422, 424
echinopus Fum. et Rob., Tyroglyphus 180
echinopus Vtz., Rhizoglyphus 95
elisabethae sp. n., Calvolia 245, 267, 254, 275, 445
elliptica sp. n., Calvolia 29, 244, 262, 275
Ensliniella Vtz., 234, 235, 264, 441
Ensliniellidae 229, 276
entomophagus Lab., Acarus 208
entomophagus (Lab.), Thyreophagusldl, 189, 193, 209, 423, 424
eurynympha (Ouds.), Acotyledon 153 eutarsus Berl., Rhizoglyphus 68, 210
farinae L., Acarus 85
farinae (L.), Tyroglyphus 3, 15, 21, 29
33, 71, 75, 86, 89, 97, 417
/arris Ouds:, Aleurobius 86
farris Ouds., Tyroglyphus 90
fedjushini sp. n., Xenocastor 13, 329,
330, 331, 351, 459
Ferminia Ouds. 342
feytaudi (Ouds.), Acotyledon 148
filippovi sp. n., Histiogaster 190, 194, 195, 436
fluctuata Ouds., Tortonia 402, 403
Forcellinia Ouds. 81, 84, 110, 414, 416, 420
Forcelliniidae 68, 83, 91
Froriepia Vtz. 79, 228, 413
Froweinia Ouds. 268
fucorum Ouds., Tyroglyphus 95, 417
fuligineus Berl., Glycyphagus 313, 324
fungivora (Ouds.), Forcellinia 5, 71, 75, 83, 111, 113, 114, 115, 420, 421
Fusacarus Mich. 327, 328, 356, 455, 457
fusca (Ouds.), Gohieria 344, 345, 456, 457
fusca (Lohm.), Hyadesia 13, 364, 371, 462
fuscipes Ouds., Tyroglyphus 216
fusiformis sp. n., Calvolia 246, 259, 271,
275, 666
Fusohericia Vtz. 362
fustifer Ouds., Glycyphagus 294, 298, 299, 452
gargantua sp. n., Sennertia 380, 383, 385, 466
Garsaultia Ouds. 180, 224, 415
geniculatus Vtz., Glycyphagus 293
georgei Mich., Hericia 371, 372, 373, 462 geotruporum sp. n., Caloglyphus 157, 165, 178, 1'81, 433
germanicus Berl., Rhizoglyphus 68, 210,
gigantonympha (Vtz.), Forcellinia 114, 116, 119, 421
glabra Vtz., Horstia 400
globosus Berl., Rhizoglyphus 211
Glycyborus Ouds. 314
Glycyphagidae 67, 68, 276, 446
Glycyphagidae 276
Glycyphaginae 280, 281, 291, 448, 449, 451,
Glycyphagus Her. 292, 293, 451
Gohieria Ouds. 327, 328, 342, 456, 457
granulatus (Duj.), Cerophagus 405, 407, 463
granulatus Duj., Hypopus 407
greeni Ouds., Hericia 372
gussakovskii sp. n., Vidia 270, 271, 275, 446
hagensis Ouds., Calvolia 240, 246, 259, 444
hamadryas Vtz., Suidasia 239
hamadryas (Vtz.), Winterschmidtia 240, 241
hebecklinii (Sicher), Calvolia 243, 252, 261, 442
helenae Ouds., Horstia 400, 401, 409, 468
heptneri sp. n., Xenoryctes 347, 349, 351, 359, 460, 461
Hericia Can. 68, 362, 370, 462
kT:i
УКАЗАТЕЛЬ
hericia (Rob.), Hericia 389, 371, 372, 462
hericius Rob., Glycyphagus 370
heterocoma (Mich.), Calvolia 211, 243, 244, 245, tvi, 248, 261, 442, 445
heterocomus Mich., Tyroglyphus 242
Heteroglyphus Foa 123, 186, 422
he teromorphus Felt, Tyroglyphus 154
hippododos Her., Sarcoptes 302
Histiogaster Berl. 80, 124, 126, 187, 413, 422, 424
Homopus Koch 357
horrida (Vtz.), Sennertia 31, 379, 387, 465
Horstia Ouds. 377, 399, 464, 468
humerosus (Ouds.), Tyrophagus 71, 99, 100, 102, 106, 419'
humerosus (Ouds.), Tyrophagus humerosus 102, 105, 107, 419
hyacinlhi Boisd., Acarus, 182
hyacinthi (Boisd.), Rhizoglyphus 182
Hyadesia Megn., 362, 461, 462
Hypopus Ouds. 180
hypudaei Koch., Homopus 357
hypudaei (Koch), Myacarus 351, 358, 360, ’ 461
hypudaei Ouds., Dermacarus 358
infestans Berl., Tyrophagus 100, 109
inopinata (Vtz.), Rodionovia 213, 223, 414
inopinatus .Vtz., Tyroglyphus 223
intermedia (Ouds.), Tortonia 402, 403, 405, 468
intermedius (Can.), Diamesoglyphus 314, 323, 454
intermedius Ouds., Trichotarsus 401
ipidis Vtz., Schwiebea 185, 199, 200, 202, 436, 437
rtalica Ouds., Schwiebea 198
laponicus Ouds., Trichotarsus 386
kamenskii sp. n., Calvolia 243, 253, 254, 442
kneissli Kr., Calvolia 244, 255, 443
kostylevi sp. n., Ensliniella 245, 266, 267, 275, 441
krameri (Berl.), Acotyledon 130, 133, 135, 143, 147, 151, 425, 427, 430
krameri (Ouds.), Dermacarus 349
krameri (Mich. , Xenoryctes 323, 344, 345, 460, 461
kulagini A. Z., Blomia 288, 289, 291, 450
Kuzinia gen. n. 79, 85, 94, 413, 417
Labidophorinae 68, 279, 282, 324, 447, 449, 450, 455
Labidophorus Kram. 327, 329, 332, 456, 458 459
Lackerbaiieria gen. n. 80, 214, 414, 438
lactis (L.), Carpoglyphus 13, 365, 367,462
laevis Duj., Hypopus 94, 417
laevis (Duj.), Kuzinia 71, 75, 95, 101, 111, 417
laminipes Mich., Fusacarus 356. 357, 359, 455, 457
latipes Berl., Glycyphagus 293
leefmansi Ouds., Acotyledon 129
Lentungula Mich. 362
Lentungulidae 276, 362
Lepidoglyphus A. Z. 293, 294, 295, 45! lineata Ouds., Vidia 270, 271, 273, 446 lini (Ouds.), Tyrolichus 97, 98, 418 lintneri Osb., Tyroglyphus 92, 113 littoralis (Halb. j, Pontoppidania 235, 237,.
. 440
littoralis Halb., Tyroglyphus 235 longior (Gerv.), Tyrophagus 99, 100, 102' 109, 419
ludwigi Troucss., Chaetodactylus 391,392' lundbecki Trag., Tyroglyphus 95, 417
macrocnemis sp. n., Robinisca 115, 216, 438
magna Berl., Monieziella .206, 208, 438 mali Berl., Monieziella 206, 207 malus Schim., Acarus 207
mamillata sp. n., Neohorstia 405, 406, 469
mandzhur sp. n., Caloglyphus 21, 154, 155, 156, 158, 161, 163, 165, 171, 177,. 181, 430, 431, 432, 434
manicati (Giard), Sennertionyx 409, 411, 464
manicati Giard, Trichotarsus 410 megnini Haller, Tyroglyphus 182 Megninietta Jacot 198, 204
michaeli (Ouds.), Acotyledon 129, 132,.
134, 140, 143, 213, 425, 427, 429 michaeli Ouds., Glycyphagus 33, 294, 299,.
300, 451
minor Dem., Rhizoglyphus 68, 209 minutus (Ouds.), Nanacarus 241, 268, 269, 440
mogerae sp. n., Labidophorus 334, 339r 341, 460
molitor sp. n., Tyrophagus 103, 105, 106.. 420
moniezi A. Z., Caloglyphus 154, 158, 172,. 175, 181, 431, 434
Monieziella Berl. 79, 124, 126, 206, 413,. 423, 424, 437
multifoliatus nom. n., Ctenoglyphus 318„ 321, 455
Myacarus gen. n. 329, 357, 458, 461
Mycetoglyphus Ouds. 110
mycobora (Ouds.), Troupeauia 222 mycolicha (Ouds.), Robinisca 115, 217,.
439
mycolichus Ouds., Tyroglyhus 215 mycophagus (Megn.), Caloglyphus 155,, 156, 176, 177, 185, 213, 432, 433
mycophagus Berl., Tyroglyphus 170 mycophagus Moniez, Tyroglyphus 172 mycophagus Schulze, Tyroglyphus 159 Myrmoglyphus Vtz. 127
Nanacaridae 229
Nanacarus Ouds., 233, 268, 440 nataliae sp. n., Calvolia 246, 262, 271, 275, 444
neglectus Berl., Saproglyphus 23, 236, 238, 241, 440
Neohorstia gen. n. 376, 406, 464, 469’
yHA3A'lJi.lb
in
neo tropicus Ouds. , Saproglyphus 238
nova (Ouds.), Troupeauia 220, 212, 439
novisimilis Vtz., Tyroglyphus 219
novus Ouds., Tyroglyphus 219 «
noxius A. Z., Tyrophagus 21, 100, 101,
103, 105, 418
nudus Berl., Chortoglyphus 286
Nycteriglyphus gen. n. 313, 321, 454
obesus Banks, Glycyphagus 293, 326
occurens Berl., Rhizoglyphus 68, 211 ocellatus (Vtz.), Histiogaster 190, 196, 436
ornata (Ouds.), Horstia 381, 400, 468
ornatus Kram., Glycyphagus 27, 295, 304, 305, 307, 452
ornatus Ouds., Trichotarsus 399 Orvcteroxenus gen. n. 327, 328, 350, 456, 457
osmiae £L. Duf.), Chaetodactylus 33, 381, 394, 397, 467
osmiae L. Duf., Trichodactylus 391 '
oudemansi A. Z., Acotyledon 129
oudemansi A. Z., Cometacarus 308, 309, 319, 397, 453
oudemansi sp. n., Sennertia 382, 390, 393, 466
ovatus (Tr.), Aleuroglyphus 13, 21, 71, 75, 83, 92, 101, 417
ovatus Tr., Tyroglyphus 92
pachyderma sp. n., Schwiebea 198, 199, 200, 204, 437
palmifer (Fum. et Rob.), Ctenoglyphus 316, 318, 319, 320, 454, 455
palmifer Can., Glycyphagus 321
pamirensis sp. n„ Schulzea 221, 225, 226, 439
paradoxa Ouds., Acotyledon 127, 134, 138, 139, 151, 429
parasitica Vtz., Ensliniella 237, 245, 264, 265, 266, 275, 441
passularum Rob., Carpoglyphus 367
peregrinans (Berl.), A6roglyphus 311, 312, 453
peregrinans Berl., Glycyphagus 310 perniciosus sp. n., Tyrophagus 71, 100,
104, 105, 418
Phycobius Can. 367
pilosus Ouds., Glycyphagus 294, 297, 319, 452
platygaster Mich., Glycyphagus 353
platygaster (Mich.), Talpacarus 354, 355, 455, 456
plumiger Koch, Acarus 314
plumiger (Koch.), Ctenoglyphus 311, 316, 317, 318, 454, 455
polyphyllae sp. n., Caloglyphus 154, 155,
157, 168, 171, 175, 177, 181, 431, 432, 434
polytrematus Berl., Scatoglyphus 118, 120,
121, 416
Pontoppidania Ouds. 234, 235, 440
Pontoppidaniidae 229
popovi sp. n., Vidia 270, 272, 275, 446
potanini sp. n., Sennertia 380, 390, 393, 465
Povelsenia Ouds. 92
primtus Ouds., Glycyphagus 293, 302 prunorum. Her., .Glycyphagus 293, 302 pteroplmrus Berl., Glycyphagus 321 pterophorus (Berl.), Nycteriglyphus 319, 322, 454
pulcherrima Vtz., Horstia 400 putrescentiae (Schr.) Ouds., Tyroglyphus 99, 100, 106
reaumuri (Ouds.), Chaetodactylus 394, 395, 397, 567
redikorzevi (A. Z.), Acotyledon 130, 131, 133, 135, 139, 142, 151, 426, 428,_ 430 redikorzevi (A. Z.). Acotyledon redikorzevi 130, 133, 145, 426, 428
reticulata sp. n., Calvolia 247, 254, 271, 275, 445
Rhizoglyphidae 68, 180
Rhizoglyphinae 79, 121, 412, 421
Rhizoglyphini 123, 125, 179, 422, 424 rhizoglyphoides (A. Z.), Acotyledon 129, 132, 134, 135, 139, 151, 425, 426, 428
Rhizoglyphus Clap. 81, 123, 125, 180, 415, 424
robinii Can., Hericia 372
robinii Clap., Rhizoglyphus 182
Robinisca gen. n. 81, 215, 414, 438 robustus Banks, Glycyphagus 293, 310. rodionovi A. Z., Caloglyphus 131, 154, 155, 156, 158, 159, 161, 165, 171, 177, 181, 430, 431, 432, 435
Rodionovia gen. n. 80, 223, 414
romanovae sp. n., Calvolia 244, 252, 253, 443
rossica sp. n., Schwiebea 131, 198, 199, ' 200, 205, 436, 437
Sancassania Ouds., 180, 224, 415
Saproglyphidae 67, 68, 229, 349
Saproglyphus Berl. 234, 236, 440
Scatoglyphus Berl. 84, 118, 416
schmitzi Ouds., Viedebanttia 218
Schulzea gen. n. 82, 225, 415, 439 Schwiebea Ouds. 81, 124, 126, 197, 415,.
423, 424, 436
sciurinus (Koch.), Labidophorus 31, 334, 335, 351, 355, 459
Sennertia Ouds. 375, 377, 463, 464
Sennertionyx gen. n. 376, 410, 464
setosus (Koch.), Cometacarus 308, 309, 319
setosus Ouds., Glycyphagus 297, 309 siculus (Fum. et Rob.), Aleuroglyphus 92, 94, 101, 418
silenus nom. n., Histiogaster 187, 188, . 192, 435
Silvester subsp. n., Tyrophagus humerosus 102, 108, 419
siro Rob., Aleurobius 86
siro Can., Tyroglyphus 97
sironiformis Fum. et Rob., Acotyledon 1291 skorikovi sp. n., Cerophagopsis 408, 409, 410, 469
smirnovi. sp. n., Cometacarus 306, 307, 308, 453
УКАЗАТЕЛЬ
•smitsvanburgsti Ouds., Tortonia 402, 404, 405, 409, 469
sokolovi sp. n., Acotyledon 129, 132, 137, 139, 525, 427, 428
;soricis Ouds., Labidophorus 335, 339, 341, 460
spelaeus Berl., Tyroglyphus 154
.sphaerogaster A. Z., Caloglyphus 155, 156, 171, 173, 177, 431, 432
spinipes Koch, Acarus 296
spinitarsus (Herm.), Caloglyphus 154, 156, 157, 166, 175, 181, 430, 432, 433
spinitarsus Can., Rhizoglyphus 182
squamata Ouds., Vidia 269
striata (Vtz.), Calvolia 244, 260, 261, 443
talpa Ouds., Schwiebea 197, 198, 203
talpa Vtz., Schwiebea 202
Talpacarus gen. n. 326, 328, 353, 455, 456
talpae Can., Homopus 347
talpae Kram., Labidophorus 332, 335, 339, 340, 460
tarsispina (Ouds.), Robinisca 218
tenuiclavus sp. n., Tyrophagus 71, 75, 102, 105, 109, 419
tesquorum subsp. n., Tyrophagus humero-sus 102, 105, 108, 419
testudo Ouds., Garsaultia 117, 224 thori A. Z. Blomia 285, 288, 289, 450 thraca Vtz., Calvolia 246, 255, 271, 275, 555
Thyreophagus Rond. 125, 126, 208, 423
tjibodas Ouds., Glycyphagus 287
tjibodas (Ouds.), Blomia 290
Tortonia Ouds. 377, 401, 464, 468
Trichodactylus L. Duf. 391
Trichotarsus Can. 391
trifilis Can., Horstia 399
trouessarti Berl., Rhizoglyphus 68, 210
Troupeauia gen. n. 82, 219, 415, 439 tshernyshevi sp. n., Acotyledon 119, 135, 151, 152, 430
tshernyshevi sp. n., Caloglyphus 159, 163, 165, 181, 435
tshernyshevi sp. n., Schwiebea 189, 195, 199," 200, 201, 436, 437
tuberculata sp. n., Calvolia 243, 248, 250, 442
Tyroglyphidae 67, 68, 412
Tyroglyphidae 68, 83
Tyroglyphinae 78, 82, 412, 416
Tyroglyphini 84, 85, 416, 417
Tyroglyphopsis Vtz. 187, 188
Tyroglyphus Latr. 79, 84, 85, 413, 416, 417
Tyrolichus Ouds. 84, 96, 416, 418
Tyrophagidae 68, 83, 91
Tyrophagini 84, 91, 416, 417
tyrophagoides sp. n., Tyroglyphus 86, 89, 90, 417
Tyrophagus Ouds. 85, 99, 417, 418
ulmi Jacot, Monieziella 198
uncinifer Megn., Hyadesia 362, 363 undulata Ouds., Vidia 269, 270, 274, 446 unguiculata Vtz., Tyroglyphus 154
Vidia Ouds. 235, 269, 441, 445
Viedebanttia Ouds. 218
vimariensis Vtz., Froriepia 213, 228, 413 vitium Foa, Heteroglyphus 186, 422 viviparus Ouds., Tyrophagus 99, 100, 332 volgini sp. n., Acotyledon 119, 135, 150, 429
wasmanni (Mon.), Forcellinia 111, 113, 115, 117, 420, .421
wasmanni Mon., Tyroglyphus 110 wasmanni Ouds., Tyroglyphus 217 weeversi Ouds., Caloglyphus 154 Winterschmidtia Ouds. 234,' 239, 440 W interschmidtiidae 229
Xenocastor gen. n. 328, 329, 457, 458
Xenoryctes gen. n. 327, 328, 329, 344, 456, 458, 460
xylocopae Donn., Trichodactylus 382 xylophaga Banks, Monieziella 198
zacheri Ouds., Calvolia 244, 263, 443 zhelochovtsevi sp. n., Sennertia 379, 383, 388, 389, 465
Ob ДЕРЖАНИЕ
Предисловие.................................................................... V
«Систематический указатель видов..............................................VII
’Общая часть................................................................... 1
Внешнее строение половозрелой стадии.................................. 11
Краткий очерк внутреннего строения...................................... 16
История развития; строение неполовозрелых стадий........................ 19
Положение группы в системе Acaridiae и ее классификация................. 30
Географическое распространение.......................................... 36
Экология................................................................ 36
Хозяйственное значение................................................. 51
Меры борьбы........................................................... 57
Сбор и консервировка клещей............................................ 63
•Основная литература.......................................................... 65
‘Специальная часть............................................................ 67
Определительная таблица семейств.................................. • §7
Сем. Tyroglyphidae ..................................................... 68
Сем. Saproglyphidae.................................................... 229
Сем. Glycyptiagidae ................................................... 276
^Резюме (Resume) ........................................................... 411
Алфавитный указатель латинских названий..................................... 470
ФАУНА СССР
(новая серия)
ПАУКООБРАЗНЫЕ (ARACHNOIDEA)
Том I. Вып. 1. Скорпионы (Seorpiones); вып. 2. Ложноскорпионы (Pseudo-seorpiones); вып. 3; Сольпуги (Solifuga); вып. 4. Сенокосцы (Р halting idae) •
Том II—III. Пауки (Araneina).
Том IV. Клещи (Acari). Вып. I. Введение; вып. 2. Ixodoidea; вып. 3—5. Gamasides; вып. 6. Uropodina.
Том V. Клещи (Trombidiiformis). Вып. 1—3. Geacarina; вып. 4. Водные клещи (Hydracarina}, ч. 1; вып. 5. Водные клещи, ч. 2 (Halacaridae}; вып. 6—7. Trombidiidae; вып. 8—10. Tetrapodii.
Том VI. Клещи (Sarcoptiformes}. Вып. 1. Tyroglyphoidea; вып. 2. Anoetidae, Canestrinidae; вып. 3. Acaridae (Sarcoptidae), Psoroptidae и др.; вып. 4—5 Analgesidae; вып. 6—8. Oribatidae.
Подчеркнуты одной чертой — выпуски, подготовленные к печати; подчеркнуты двумя чертами — напечатанные.
ФАУНА СССР
(новая серия)
В ы ш л и и з п е ч а т и: '
Млекопитающие т. III, вып. 4 — Тушканчики (Б. С. Виноградов), 1937, цена 10 руб.; т. III, вып. 2 — Мыши и крысы (А. И. Аргиропуло), 1940, цена 10 руб. 50 коп.
Птицы т. I, вып. 2 — Основы орнитогеографического деления Палеарктики (Б. К. Штегман), 1938, цена 15 руб.; т. I, вып. 5 — Дневные хищники (Б. К. Штегман), 1937, цена 17 руб. .
Рыбы т. VI, вып. 1 — Triglidae (А. Н. Световидов) 1936, цена 1 р. 25 к.
Насекомые двукрылые т. III, вып. 2 — Москиты (П. П. Перфильев), 1937, цена 7 р.; т. III, вып. 5 — Кровососущие комары Culicinae (А, А. Штакель-берг), 1937, цена 12 р.; т/ VI, вып. 7 — Мошки Simulidae (И. А. Рубцов), 1940, цена 20 руб.; т. VII, вып. 2 — Слепни (Н. Г. Олсуфьев), 1937, цена'17 р.; т. IX, вып. 2—Жужжала, ч. 1 (С. Я. Парамонов), 1940; '
т. XIX, вып. 1 — Сем. Sarcophagidae, ч. 1 (Б. Б. Родендорф), 1937, цена 20 р.
Насекомые перепончатокрылые т. II, вып. 1 — Пилильщики Tenthredinodea, ч. 1 (В. В. Гуссаковский), 1935, цена 22 р. 75 к.; т. V, вып. 2 — Сем. Braconidae, Ч. 1 (Н. А. Теленга), 1936, цена 20 р. 75 к.; т. V, вып. 3 — Сем. Braconidae, ч. 2 (Н. А. Теленга), 1940.
Насекомые чешуекрылые или бабочки т. XIII, вып. 3 — CoBKii-Mgro«iziae'(n. В. Кожанчиков), 1937, цена 34 р. 50 к.
Насекомые жесткокрылые или жуки т. V, вып. 3 — Жуки-карапузики, ч. 1 (А. И. Рейхардт), 1940; т. XXI — Дровосеки, ч. 1 (Н. Й. Плавплыци- . ков), 1936, цена 22 р.; т. XXVI, вып. 1 — Листоеды Galerucinae (Д. А. Оглоблин), 1935, цена 17 р.; т. XXII.— Дровосеки, ч. 2 (Н. И. Плавпль- : щиков), 1940, цена 40 р. '
Насекомые кожпстокрылые (Г. Я. Бей-Биенко) 1936, цена 14 р. ’ / ;
Паукообразные т. I, вып. 3— Сольпуги (А. А. Бялыницкпй-Бируля), 1938, цена 11р.; т. V, вып. 4 — Водные клещи Нуdracarina, ч. 1 (II. И. Соколов), 1940, цена 25 руб.; т. VI, вып. 1 — Тироглифоидные клещи : (А. А. Захваткпн), 1940.
Ракообразные т. VII, вып.-?3— Равноногие Дальневосточных морей (К Ф. Гурьянова), 1936, цена 15 р. 50<к.; т. X, вып. 3 — Апотига (В. В. Мака-. ров), 1938, цена 21 р.
Моллюски т. III, вып. 1 — Сем. Unionidae (В. И. /Надин), 1938, цена 11 р.
Губки т. II, вып. 2 — Пресноводные губки (П. Д. Резвой), 1936, цена 6 р. 60 к.
Находятся в печати:
Птицы т. I, вып. 4 — Пластинчатоклювые (А. Я. Тугаринов).
Рыбы т. VII, вып. 4 — Трескообразные (А. Н. Световидов).
Насекомые жесткокрылые пли жуки т. X, вып. 3 — Щелкуны Eldteridae, ч. 2 (Л. А. Денисова).
Подготовлены к печати:
Млекопитающие т. I, вып. 2 — Олени (К. К. Флеров).
Птицы т. I, вып. 3 — Определитель отрядов и семейств, гагарообразные, буре-вестнпковые (Е. В. Козлова), аистообразные (А. Я. Тугаринов).
Насекомые двукрылые т. XX, вып. 6 — Оводы (И. А. Рубцов).
Насекомые перепончатокрылые т. II, вып. 2 — Пилильщики, ч. 2 (В. В. Гус-саковскпй); т. XII, вып. 1 — Муравьи, ч. 1 (К. В. Арнольди, В. А. Караваев); т. XIII, вып. 1 — Осы Vespidae (Ю. А. Костылев).
Насекомые жесткокрылые или жуки т. XIII, вып. 3 — Златки, ч. 1 (А. А. Рихтер); т. XIV, вып. 2 — Лжекороеды Bostrychidae (В. Н. Старк); т. XXIII, вып. 1 — Дровосеки, ч. 3 (Н. Н. Плавильщиков); т. XXVII, вып. 3 — Короеды (В. Н. Старк).
Насекомые сосущие т. VI, вып. 3 — Тли. Триба Eriosomini, (А. К. Мордвилко); т. VII — Щитовки и червецы (И. С. Борхсениус).
Насекомые прямокрылые т. I, вып. 2 — Саранчевые, ч. 1 (Л. Л. Мищенко).
Насекомые таракановые (Г. Я. Бей-Биенко).
Паукообразные т. V, вып. 5 — Водные клещи, ч. 2 (Й. II. Соколов).
Ракообразные т. II, вып.1 —Ракушковые (Ostracoda) континентальных водоемов (3. С. Бронштейн); т. III, вып. 3 — Cyclopoida пресных вод (В. М. Рылов); т. III, вып. 4 — Harpacticoida пресных вод (Е. В. Боруцкий); т. V — Паразитические Copepoda (А. П. Маркевич); т. VI, вып. 1 — Усоногие Girripedia thoracica (Н. И. Тарасов); т. VII, вып. 5. Пресноводные Isopoda (Я. А. Бирштейн); т. VIII, вып. 3 — Amphipoda Нуре-riidea (А. Л. Бенинг).
ТИПО-ЛЙТОГРАФЙЯ ИЗД ВА АН СССР
ОКОНЧАТЕЛЬНАЯ ПРОВЕРКА
№ 2
При обнаружении дефекта в книге Заводоуправление просит прислать данную книгу с приложением этого Талона по адресу:
Ленинград, 34. В. О., 9 линия, 12.
Заводоуправление