





































































































































































































































































































































Author: Богословская Л.С.
Tags: млекопитающие териология (маммалиология) морской сборник морские животные
ISBN: 5-86443-079-Х
Year: 2003




Text
НАСЛЕДИЕ БЕРИНГИИ
Л.С. БОГОСЛОВСКАЯ
КИТЫ ЧУКОТКИ
ПОСОБИЕ ДЛЯ МОРСКИХ охотников
165 170 175 180 175 170 165
Киты - одни из самых удивительных обитателей Земли. Миллионы лет назад они заселили все океаны, имевшие тогда другие очертания, потом пережили несколько ледниковых периодов. Люди появились, освоили морски побережья и познакомились с китами совсем недавно. На нашей планете существуют несколько мест, где киты тысячелетиями помогают жить людям и во многом определяют облик их древних культур.
Одно из таких мест - Чукотка.
Посвящается капитану китобойного судна <3вездныи> Л.М. Вотрогову и морским охотникам Чукотки
Российский научно-исследовательский институт культурного и природного наследия имени Д.С. Лихачёва
Фонд «Молодежная экологическая инициатива*
Совет по морским млекопитающим
KG БОГОСЛОВСКАЯ
КИТЫ ЧУКОТКИ
ПОСОБИЕ АЛЯ МОРСКИХ охотников
Наследие Берингии
Выпуск 1
Москва-Провидения-Анадырь 2003
ББК 28.693.369
Б 74
Богословская Л.С.
Киты Чукотки. Пособие для морских охотников.
М.: Институт Наследия, 2003. 324 с.; илл. ISBN 5-86443-079-Х
Пособие содержит биологические характеристики китов и дельфинов, обитающих в водах Чукотки, историю организации и результаты 25-летнего мониторинга китов силами коренных жителей Чукотки. Предназначено для морских охотников, школьников старших классов, студентов, а также для широкого круга читателей.
Рецензент:
доктор биологических наук, профессор В.А. Земский
Издание книги осуществлено на средства Департамента промышленной и сельскохозяйственной политики Правительства Чукотского автономного округа и Управления природных ресурсов Министерства природных ресурсов Российской Федерации по Чукотскому автономному округу.
ISBN 5-86443-079-Х
© Богословская Л.С., 2003
© Институт Наследия, 2003
Благодарность
Мне приятно выразить свою искреннюю признательность тем, кто на протяжении многих лет бескорыстно содействовал нашей экспедиции в изучении морских животных и древней морской зверобойной культуры Берингии. Я счастлива, что таких людей было много; не все они, к сожалению, здравствуют сегодня, но все будут всегда жить в моей памяти.
На Чукотке многие годы нам помогали Л.И. Айнана, Т.Ю. Ачиргина и другие члены «Общества эскимосов», Ю.М. Бабаев, начальник Провиденской гидробазы Министерства морского флота СССР, В. А. Беломестнов, капитан порта Провидения, А.Г. Вольфсон, заведующий Чукотским стационаром Института биологических проблем Севера ДВО АН СССР, А. Соколовская, И. и Т. Загребины, сотрудники Провиденского краеведческого музея, А.В. Комаров, начальник Управления сельского хозяйства Магаданского облисполкома, Д.М. Кажаев, директор совхоза «Заря коммунизма», А. и Н. Кузменовы, сотрудники того же совхоза, В. П. Мамонтов, командир экипажа вертолета МИ-8, А.И. Теущаков, радист совхоза «Ударник».
Успехи экспедиционной работы во многом были обеспечены доброжелательным отношением со стороны штаба Кенигсбергского ордена Красной Звезды пограничного отряда. Особая признательность Н.А. Пуршневу, начальнику погранкоменда-туры Чукотского района, В.А. Коренькову, начальнику погранзастав пос. Сиреники и острова Ратманова, В.М. Шкроботу, командиру экипажа вертолета МИ-8.
Я многим обязана зоологам, члену-корреспонденту РАН А.В. Яблокову и профессору В.Д. Ильичеву, которые в свое время поддержали мое стремление изучать животных Чукотки, а также профессору В. А. Земскому и кандидату биологических наук И.В. Смеловой за многолетнее сотрудничество в этой области.
Постоянную помощь во время работы над книгой оказывало мне руководство Российского института культурного и природного наследия. Н.П. Лакутина и Е.С. Богословская с любовью и тщательностью подготовили макет издания.
Л. Богословская
СОДЕРЖАНИЕ
ПОЧЕМУ НАПИСАНА ЭТА КНИГА.....................................8
1. КАПИТАН Л.М. ВОТРОГОВ ........._..........12
2 ИСТОРИЯ СОДРУЖЕСТВА УЧЕНЫХ И МОРСКИХ ОХОТНИКОВ ЧУКОТКИ ..................21
3. КАК УСТРОЕНО МОРЕ .........................47
4. О КИТАХ И ДЕЛЬФИНАХ .......................57
5. УСАТЫЕ КИТЫ...............................83
Сем. серых китов..........................92
Серый кит..............................92
Сем. гладких китов.......................136
Гренландский, или полярный, кит.......137
Южный (японский) гладкий кит..........172
Сем. полосатиковых.......................179
Синий, или голубой, кит...............180
Финвал, или сельдяной кит.............189
Сейвал, или сайдяной кит..............201
Малый, или остромордый, полосатик.....207
Горбач, или горбатый кит..............213
6. ЗУБАТЫЕ КИТЫ ............................231
Сем. дельфиновых.........................238
Косатка...............................238
Сем. морских свиней......................245
Обыкновенная морская свинья...........245
Белокрылая морская свинья.............249
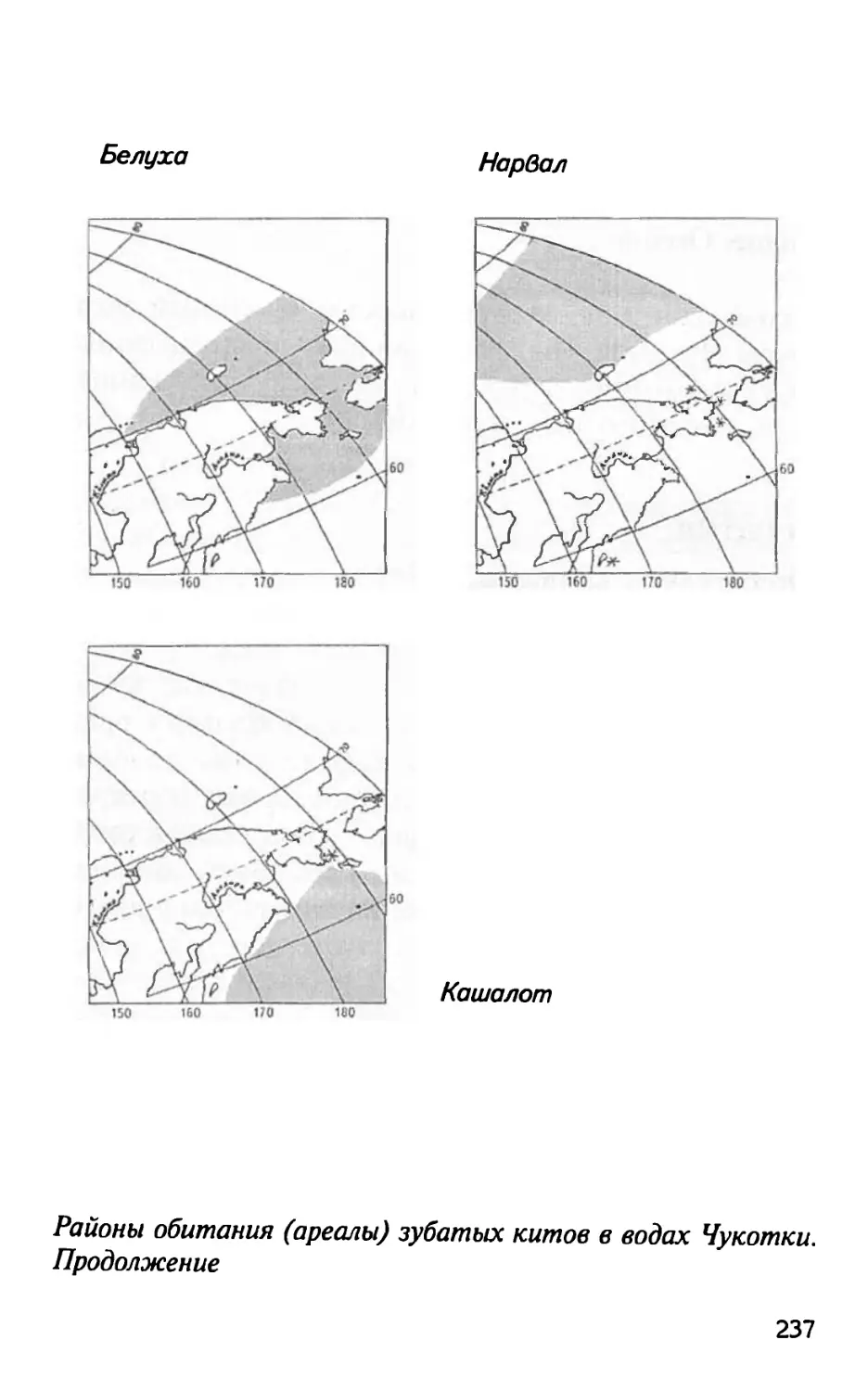
Сем. нарваловых........................253
Белуха..............................253
Нарвал..............................261
Сем. клюворыловых......................267
Северный плавун.....................267
Сем. кашалотовых.......................271
Кашалот.............................271
7. НАБЛЮДАЙТЕ ЗА КИТАМИ! .................283
НЕКОТОРЫЕ НАУЧНЫЕ ТЕРМИНЫ И ПОНЯТИЯ.................................295
НАУЧНО-ПОПУЛЯРНАЯ ЛИТЕРАТУРА ДЛЯ ДОПОЛНИТЕЛЬНОГО ЧТЕНИЯ ...............309
ОСНОВНЫЕ ЛИТЕРАТУРНЫЕ ИСТОЧНИКИ ..........310
Список авторов иллюстраций................315
Summary...................................320
Contents..................................322
Почему написана эта книга
Во второй половине XX в. на Чукотке работало много научных экспедиций из Москвы, Ленинграда, Магадана, Новосибирска и других академических центров нашей страны. С 1977 г. по 1992 г. там проводила полевые исследования и Чукотская зоологическая экспедиция Института эволюционной морфологии и экологии животных им. А.Н. Северцова Академии Наук СССР, руководить которой довелось автору этой книги.
Изучая морских млекопитающих и птиц в регионе Берингова пролива, мы, зоологи, познакомились со своими коллегами из Института этнографии и антропологии АН СССР, М.А. Членовым и И.И. Крупником. Родилась идея соместных этно-биологических морских экспедиций вдоль побережья Чукотского полуострова с целью изучения морской зверобойной культуры Берингии и ее биоресурсных основ. Было принято очень важное решение, повлиявшее на характер всех наших последующих работ -в экспедициях должны на равных правах участвовать коренные жители, те, кто создал и на протяжении тысячелетий развивал уникальный культурный комплекс Арктики.
Эту идею словом и делом поддержали капитан китобойного судна «Звездный» Л.М. Вотрогов, капитан порта Провидения В.А. Беломестнов и начальник Управления сельского хозяйства Магаданского облисполкома А.В. Комаров. Все вместе они помогли нам осуществить целый ряд длительных морских маршрутов на маломерных традиционных промысловых судах охотников Берингова пролива, кожаной байдаре (ее использовали зоологи) и деревянном вельботе (его предпочли этнографы). В течение 1980-х гг. нами было пройдено в общей сложности около 5000 км, сделан целый ряд зоологических и этнографических открытий, но, главное, был создан единый коллектив исследователей, для которых научные и традиционные знания имели одинаковую ценность.
Во время экспедиций каждый вечер мы с охотниками обсуждали все, увиденное за день, и, конечно, прежде всего поведение морских зверей и птиц. Охотников очень интересовало, откуда приходят весной и куда уходят на зиму серые киты, почему летом у восточного побережья Чукотки совсем мало гренландских китов, что такое миграции животных и еще многое-многое другое из жизни зверей и птиц. Мы с удовольствием отвечали на эти вопросы и сами расспрашивали охотников. Они рассказывали об известных нам животных много нового, такого, о чем невозможно прочитать ни в одном научном труде.
Опытнейшие охотники поселка Сиреники просили написать книгу о китах и ластоногих, куда бы вошли и традиционные знания — их беспокоило постепенное угасание традиций морского промысла, прежде всего исчезновение у молодых зверобоев уважительного отношения к зверю как к равному
себе. Мы обсуждали такую книгу с Андреем Анка-линым, Николаем Гальгаугье, Петром Тыпыхкаком и другими охотниками и, конечно же, с капитаном Л.М. Вотроговым, самым уважаемым человеком для морских охотников Чукотки и всех нас. Я обещала охотникам и капитану Вотрогову, что книга будет написана вместе с ними. К сожалению, этого не случилось — и капитан, и охотники, наши спутники по экспедициям, ушли из жизни один за другим.
Мы продолжали работать на Чукотке, все более и более ощущая себя хранителями полученных традиционных знаний о людях, море и животных Берингии. Эти знания нужно было передать молодым поколениям, вырастающим во времена разрыва культурных традиций, когда в 1990-х гг. практически не стало охотников-эскимосов старшего поколения и сильно сократился круг опытных зверобоев из береговых чукчей. Мой друг и коллега, этнограф Игорь Крупник, выполнил свой долг, написав замечательный труд «Пусть говорят наши старики», где собраны рассказы азиатских эскимосов, родившихся в начале XX в.
В определенной степени книга-пособие «Киты Чукотки» — тоже выполнение долга перед моими спутниками и друзьями. Однако она написана не только по материалам наших экспедиций, но и на основе великолепных работ М.М. Слепцова, Б.А. Зен-ковича, А. Г. Томилина, отечественных зоологов середины XX в., изучавших китов Чукотки. Надо отметить, что эти ученые с большим уважением относились к знаниям коренных жителей и приводили в своих работах мнения охотников из разных поселков. Нам посчастливилось в течение многих лет сотрудничать с профессором А. Г. Томилиным, который помог ре-
шить несколько трудных задач, например, определить видовую принадлежность черепов молодых китов, которых издавна добывали жители Мечигменс-кого залива.
М.М. Слепцов, Б.А. Зенкович и А.Г. Томилин были учеными и одновременно популяризаторами научных знаний о китах и дельфинах. Они выпустили в свет увлекательные книги о морских животных, которые давно стали библиографической редкостью, и потому я с удовольствием привожу в пособии большие цитаты из работ этих исследователей.
Несколько слов об иллюстрациях. Наверное, не стоит объяснять, как трудно не только рисовать, но и фотографировать китов и дельфинов, поэтому хорошие изображения китов пришлось искать в самых разных научных и популярных изданиях. Основная часть рисунков принадлежит известному художнику-анималисту Н.Н. Кондакову; он был современником трех ученых и иллюстратором практически всех их научных изданий. По точности и красоте никто до сих пор не смог превзойти рисунки морских зверей, сделанные Н.Н. Кондаковым.
Очень надеюсь, что эта небольшая книга, посвященная памяти всех, с кем я много лет вместе ходила в море и изучала китов, не пройдет мимо внимания молодых морских охотников Чукотки и побудит их внести свой вклад в науку о китах так же, как это сделали их далекие предки, деды и отцы.
Капитан китобойного судна «Звездный» Леонард Максимович Вотрогов. Чукотка, 1979 г.
1
Капитан ЛМ Вотрогов
Леонард Максимович Вотрогов родился и вырос во Владивостоке. Восемнадцати лет, после окончания техникума он поступил на работу в Дальневосточное управление китобойных флотилий (позже - Дальмо-репродукт) Министерства рыбного хозяйства СССР. В китобойном флоте он прослужил всю свою жизнь, в 1958 г. стал капитаном дальнего плавания, получил высшее образование.
С 1969 г. и до самой смерти Л.М. Вотрогов оставался бессменным капитаном китобойного судна «Звездный», четырехмоторного дизельэлекгрохода водоизмещением в 4000 т. Как и все китобойцы того времени, «Звездный» был боевым кораблем, минным тральщиком со снятым вооружением, слегка переделанным для охоты на китов. На Дальнем Востоке он стал единственным китобойцем, который каждый промысловый сезон уходил в автономное плавание, хотя формально, на бумаге, судно около десяти лет приписывалось к китобойной флотилии.
Один из самых опытных капитанов-китобоев вел свой «Звездный» из Владивостока далеко на север, туда, где соединяются Тихий и Ледовитый океаны,
Китобойное судно «Звездный». 1980 г.
в регион Берингова пролива. Там «Звездный» добывал серых китов для коренных жителей Чукотки. Так было и в 1982 г. Безнадежно бальной капитан Вотрогов отстоял на мостике свой последний сезон и только потом лег в больницу, откуда уже не вышел. Его хоронили жители Провидения, охотники из Сиреников и Чаплине, команды всех судов, стоявших в порту. На могиле Л.М. Вотрогова стоит красивая надгробная доска с изображением альбатроса, символа души моряка.
Морские охотники Чукотки ласково звали капитана Вотрогова «Максимыч», а мы, биологи, этнографы, врачи, художники, — словом, все, кто знал этого незаурядного человека, — «Капитан». Так мы и называем его до сих пор.
Для Капитана киты были не только добычей, но и прекрасными, во многом загадочными творениями природы. Прирожденный натуралист, Леонард Максимович много лет вел дневники наблюдений за китами и другими морскими животными. Он бескорыстно помогал множеству научных экспедиций и щедро делился своими знаниями с учеными. Буквально за два года до смерти им было опубликовано 15 научных работ, большинство из них на английском языке.
Капитан Вотрогов значительно дополнил современные знания о сером и гренландском китах. Он впервые составил карту районов концентрации серых китов у побережья Чукотки и детально проследил пути их сезонных перемещений в западной части региона Берингова пролива. Именно он из года в год настойчиво сообщал, что гренландские киты сохранились у берегов Чукотки в количестве, достаточном для возобновления ограниченного аборигенного промысла. В 1970-х гг. он первым отметил появление в водах Чукотки финвалов и горбачей,
которые, казалось, навсегда были истреблены безжалостным государственным китобойным промыслом.
Мне посчастливилось шесть сезонов изучать китов вместе с этим замечательным человеком. Леонард Максимович прекрасно знал внешний вид и особенности поведения всех китов, мог совершенно точно предсказать время и место встречи со своими любимыми серыми китами. Его интересовало буквально все — киты, моржи, тюлени, птицы, а главное — жизнь и традиции коренных народов Чукотки. Капитан как профессионал высоко оценил мореходные качества байдары, древней охотничьей лодки эскимосов и чукчей, и посоветовал мне использовать для длительных морских зоологических маршрутов именно байдару, а не вельбот Мы последовали совету Капитана и ни разу об этом не пожалели.
Л.М. Вотрогов сделал для Чукотки очень много. Он не только ежегодно обеспечивал национальные поселки мясом и жиром китов, но старался, чтобы люди не забывали истории своего края, сохраняли культуру и обычаи предков и гордились ими. В этих начинаниях его поддерживали известные деятели Чукотки, этнограф В.В. Леонтьев и врач А.Г. Вольфсон.
Капитан был смелым и независимым человеком. Когда в Польше начались антикоммунистические волнения, он уговорил всю команду «Звездного» стать коллективным членом Общества польско-советской дружбы. Взял отпуск и специально приехал для этого в Москву. Потом из Польши пришло очень теплое письмо, а родная парторганизация вынесла ему строгий выговор.
Не дождавшись разрешения властей Чукотки, Л.М. Вотрогов по собственной инициативе вместе с
Памятный знак (гарпун) в честь китобойной флотилии «Алеут». Установлен в 1970 г. в бухте Гильмымыл экипажем китобойного судна «Звездный» и жителями села Новое Чаплино
командой «Звездного» и коренными жителями поставил в Провиденском районе памятные знаки серому киту на полуострове Матлю, недалеко от села Новое Чаплине, и китобойной флотилии «Алеут» в бухте Гильмымыл пролива Сенявина. Возле села Янракыннот был воздвигнут памятник адмиралу Макарову, считавшему побережье Ледовитого океана «фасадом России»; в ясную погоду судовой колокол, венчающий памятник, издалека ввден проходящим кораблям. В 1980 г. команда «Звездного» вместе с нашей экспедицией установила памятник жителям Янракыннота, утонувшим в 1979 г. Китобоец спас тогда единственного оставшегося в живых пассажира вельбота.
Памятник адмиралу С. О. Макарову (1848-1904). Поставлен в 1974 г. возле села Янракыннот экипажем китобойного судна «Звездный», сельсоветом и жителями этого этого села
Будучи большим другом коренных жителей, Капитан обратил самое серьезное внимание на обшир
ные и глубокие традиционные биологические знания морских охотников, и понял, какое значение они могут иметь для науки. Во время кратких стоянок на рейде для сдачи китов Леонард Максимович приглашал в гости на чай старейших жителей поселка и записывал на магнитофон их рассказы о морских животных, просил вести дневники наблюдений, объяснял, что и как нужно заносить в них. Все считали за честь быть приглашенными в каюту Капитана.
В конце 1970-х гг. Л.М. Вотрогов вместе с нашей экспедицией приступил к созданию сети охотников-наблюдателей за морскими животными. Когда «Звездный» уходил на зиму во Владивосток, Капитан переписывался со многими наблюдателями, даже с теми, кого ни разу не видел, — жители Биллингса, Шмидта, Певека, островов Айон и Врангеля регулярно сообщали ему о китах, белых медведях, моржах и тюленях. Активную помощь оказывали также экипажи вертолетов Про-виденского погранотряда и штурманы судов торгового флота, доставлявших грузы в поселки Чукотки.
Весной 1981 г. на основе собственных данных, результатов нашей экспедиции и сведений, полученных от жителей Чукотки, Капитан подал в Главрыбвод Минрыбхоза СССР обширную записку о необходимости организации в регионе Берингова пролива системы морских заказников и заповедников для сохранения уникальной фауны морских млекопитающих; особый раздел был посвящен охране акваторий, где паслись самки и детеныши серого кита. Ответа он так и не дождался.
... В начале ноября 1978 г. был удивительно теплый день. Свежий ветер гнал легкие облака, сквозь которые по воде бежали солнечные блики. Успешно закончив промысловый сезон, «Звездный» возвращал
ся во Владивосток. Возле Сиреников пять бригад морских охотников вышли в море на байдарах проститься с китобойцем до следующей весны. «Звездный» встал на рейд, и Капитан пригласил всех на прощальный чай. Потом каждая бригада получила в подарок бухты троса, фленширные ножи для разделки китов и к ним рукоятки из бамбука, фал, брезент, робы и многое другое, столь необходимое охотникам. Такова была традиция, за все во Владивостоке платил Капитан. Последние рукопожатия, и байдары отходят от «Звездного». Ревут моторы, и за кормой появляется пенная ходовая волна. С байдар раздается залп из карабинов, гарпунная пушка китобойца отвечает десятью холостыми выстрелами. Капитан стоит на мостике и, прищурившись, смотрит на тройку серых китов, вдруг появившихся по левому борту. Он улыбается — охота на них уже закончена ...
Бронзовая доска на могиле капитана Л.М. Вотрогова. Авторы:
Д. и Н. Жилинские, И. Голицин, Д. Шаховской
2
История содружества ученых и морских охотников Чукотки
Промысел китов — традиционное занятие коренных жителей Восточной Чукотки, эскимосов и береговых чукчей. В течение двух с половиной тысячелетий они успешно добывают этих исполинов моря с промысловых кожаных лодок-байдар при помощи сложных поворотных гарпунов. Об удачных охотах предков современных охотников свидетельствуют огромные массы костей китов на местах древних поселений, найденные археологами многочисленные орудия китобойного промысла, а также наскальные изображения (петроглифы), записанные фольклорные тексты, рассказы и воспоминания стариков.
За долгую историю охоты на китов местные жители изучили особенности их поведения и пути передвижения. Недаром все крупные поселения Восточной Чукотки основаны в местах близкого подхода китов к берегу во время ежегодных весенних и осенних миграций. Большинство морских охотников умеют не только хорошо наблюдать, но тонко и кра-
Раздел написан совместно с И.И. Крупником.
сочно описывать повадки животных. Во многих семьях эскимосов и береговых чукчей профессия морского охотника передается от отца к сыну, а вместе с ней и весь опыт, накопленный поколениями предков.
Именно поэтому предлагаемая книга основана не только на данных специалистов, в том числе автора и его коллег, но и на сведениях, полученных от жителей Чукотки, главным образом, потомственных морских зверобоев. Многолетнее содружество ученых и коренных жителей, речь о котором пойдет ниже, оказалось плодотворным: в 1997—1998 гг. совместными усилиями была восстановлена традиционная охота на гренландских китов, запрещенная Правительством СССР в 1976 г.
ИНИЦИАТИВА КАПИТАНА ВОТРОГОВА
Начало сотрудничеству специалистов и морских зверобоев в деле изучения китов положил один человек — капитан китобойного судна «Звездный» Леонард Максимович Вотрогов. С 1969 г. его судно, обычный советский китобоец, каждое лето вело промысел в районе Берингова пролива и добывало серых китов «для нужд коренных жителей Чукотки». Капитан Вотрогов был совершенно особенным человеком, продолжателем традиции старых китобойных капитанов, которые внесли огромный вклад в научное познание морских животных. В этой традиции, идущей от англичанина Вильяма Скорсби (W. Scoresby) и американца Чарльза Скэммона (Ch. Scammon), российскому капитану Л.М. Вотро-гову принадлежит достойное место.
В течение всей своей работы на Чукотке Л.М. Вотрогов самостоятельно вел постоянные научные наблюдения за китами и другими морскими млекопитающими, а также регулярно опрашивал охотников в тех поселках, куда его китобоец доставлял убитых китов. В ответ охотники стали писать ему письма и сообщать о своих наблюдениях за китами, особенно в то время, когда китобоец находился далеко на юге, на своей основной базе во Владивостоке. В поселки арктического побережья Чукотки, не входившие в зону планового промысла, капитан Вотрогов рассылал личные письма со специальными вопросами и тоже получал ответы от местных жителей. Так были собраны важные сведения о состоянии льдов и датах появления гренландских китов и других морских млекопитающих вдоль побережья от Анадырского залива до Чаунской губы включительно.
Помимо этого Вотрогов опросил штурманов около 500 судов Морского флота и Гидрографической службы СССР, а также старых китобойных капитанов во Владивостоке о распределении разных видов китов вдоль всего тихоокеанского побережья России. Капитан был первым, кто предложил местным охотникам самим вести систематические долгосрочные наблюдения за морскими животными и записывать эти данные. Охотник Андрей Анкалин из пос. Сиреники по инициативе Вотрогова начал в 1977-1978 гг. наблюдать за весенней миграцией гренландских китов со своего охотничьего участка в бухте Анытыкук к востоку от Сиреников. Данные наблюдений А. Анкалина были опубликованы в 1982 г. на английском языке в Трудах Международной китобойной комиссии. Они впервые позволили поставить под сомнение результаты определения численности
Андрей Анкалин (1941-1994). Сиреники, 1980 г.
чукотско-беринговоморского стада гренландских китов, полученные американскими специалистами в мае 1977-1978 гг. в районе мыса Барроу. Эти данные в то время считались самыми достоверными.
СОЗДАНИЕ СЕТИ ОХОТНИКОВ-НАБЛЮДАТЕЛЕЙ
В 1979 г. уже несколько охотников вели подобные наблюдения за китами, ластоногими и морскими птицами в разных точках побережья Восточной Чукотки (Энмелен, Сиреники, Новое Чаплино, Янракыннот, Лорино, Уэлен, Инчоун). Свои дневники наблюдений они передавали капитану Вотрогову и начальнику Чукотской зоологической экспедиции Л.Богословской, которые сами также изучали морских животных и продолжали опрашивать жителей прибрежных поселков от Уэлькаля на юге до Ванкарема на севере. Вопросы касались не только морских животных, но и способов промысла современных совхозных бригад зверобоев и охотников предыдущих поколений.
В 1975-1979 гг. этнограф И. Крупник вел собственные записи и интервьюировал старейшин, родившихся в первые десятилетия XX в., о традиционных поселках, годовом хозяйственном цикле, прежних методах охоты и размерах морского промысла в каждом поселке, а также об отношении коренных жителей к разным видам животных.
Помимо опросов Л. Богословская и И. Крупник много раз выходили в море на охоту вместе с коренными жителями и совершали с ними многочисленные рейсы на байдаре и вельботе в пределах береговой линии совхозов. И. Крупник работал с эски
мосскими бригадами поселков Сиреники и Новое Чаплино, Л. Богословская — с чукотскими зверобоями пос. Янракыннот.
Осенью 1979 г. Л. Вотрогов и Л. Богословская встретились с И. Крупником, обсудили полученные к этому времени данные, прежде всего по гренландскому киту, и договорились работать вместе друг с другом и зверобоями Чукотки.
Начальным этапом таких работ стал предложенный И. Крупником морской маршрут на двух промысловых кожаных байдарах Сиреники-Кынлирак-Якук-Синрак-Агманырак-Кургу (октябрь 1979 г.), в результате которого появилось первое этнобиологичес-кое описание пяти старых поселков. Охотники детально объясняли, почему именно здесь, а не в других местах селились их предки, рассказывали о ледовой обстановке в этих местах, в какие сроки велась охота на гренландского кита, кто и когда в каждом поселке убил последнего гренландского кита и т.п. Л. Богословская и И. Крупник определяли по остаткам скелетов и черепов в разрушенных землянках-нынлю, какие виды китов и ластоногих добывались жителями поселков и каковы были масштабы промысла. После завершения маршрута и рассказа о его результатах всему коллективу морских охотников пос. Сиреники пятеро участников этой короткой экспедиции выразили готовность вести дневники наблюдений за морскими млекопитающими.
К 1980 г. накопилось уже достаточно материалов по разным вопросам традиционного морского зверобойного промысла, прежде всего по численности и распределению гренландских китов в водах Чукотки. Это позволило Л. Вотрогову, Л. Богословской и И. Крупнику сделать несколько докладов на науч
но-практической конференции «Комплексное экономическое и социальное развитие Магаданской области в ближайшей и долгосрочной перспективе», которая проходила 8-10 апреля 1980 г. в Магадане. Важнейшим стал совместный доклад «О возможности возобновления местного промысла гренландского кита на Чукотке». В нем на основе собственных данных авторов и наблюдений охотников впервые был поставлен вопрос о том, что реальная численность гренландских китов чукотско-беринговоморского стада выше, чем рассчитанная американскими специалистами, и, соответственно, вполне допустимо ведение ограниченного промысла этого вида в количестве 5-7 голов в год.
Сразу после конференции Л. Вотрогов и Л. Богословская прилетели в Сиреники, где провели наблюдения за массовой весенней миграцией гренландских китов, которые шли на север вместе с недавно родившимися детенышами. Данные ученых и коренных жителей по распределению и численности гренландских китов в водах Чукотки были изложены в обширной хорошо документированной статье, опубликованной в Трудах Международной китобойной комиссии (1982 г.). На основании собственных наблюдений авторов и, главное, документированных сообщений охотников Чукотки и Аляски в этой статье доказывалось, что весной гренландские киты мигрируют в двух направлениях: на восток — к мысу Барроу и на запад — к проливу Лонга и Чаунской губе. Но, к сожалению, американские специалисты отказывались верить кому-либо, кроме самих себя, и продолжали считать, что весной все гренландские киты идут только к мысу Барроу. Однако время показало, что правы были советские ученые и корен-
Николай Гальгаугье (1935-1995). Сиреники, 1980 г.
ные жители Чукотки, и теперь никто в этом уже не сомневается и не оспаривает данные, собранные охотниками за последующие два десятилетия.
ЭКСПЕДИЦИИ И МОРСКИЕ МАРШРУТЫ
В 1979-1992 гт. основой совместных работ с коренными жителями стали ежегодные полевые сезоны Чукотской зоологической экспедиции Института эволюционной морфологии и экологии животных имени А.Н. Север-цова АН СССР (руководитель — Л.С. Богословская). Их дополняли морские и сухопутные маршруты, проводимые совместно с этнографами из Института этнографии им. Н.Н. Миклухо-Маклая АН СССР (руководитель — М.А. Членов). Решающую роль в осуществлении этих маршрутов сыграли команды охотников из поселков Сиреники и Новое Чаплино. Биологи по совету Л.М. Вотрогова выбрали промысловую байда-ру-аньяпик, кожаную плоскодонную лодку длиной около Юм, которая к тому времени сохранилась на восточном побережье Чукотки только в пос. Сиреники. Этнографы отдали предпочтение деревянному вельботу чаплинских охотников.
1981 г. В июле-августе состоялась большая совместная экспедиция зоологов, этнографов и морских охотников, в которой участвовали два экспедиционных отряда.
Зоологический отряд:
Л.С. Богословская - научный руководитель;
А.С. Голубцов — ихтиолог;
Б.М. Звонов — орнитолог;
А. Анкалин - капитан байдары, рулевой;
Н. Гальгаугье - моторист;
В. Миенков — впередсмотрящий;
Этнографический отряд:
М.А. Членов — научный руководитель;
И.И. Крупник — этнограф;
С.А. Богословский — художник-фотограф;
Е. Паулин — капитан вельбота, рулевой;
Н. Панагиргин — моторист;
Д. Анагисяк — впередсмотрящий;
Ю. Ятта — матрос;
Г. Каяк — матрос
В. Тагитуткак- почетный член команды, старейшина общины чаплинских эскимосов.
Предполагалось обследовать побережье Восточной Чукотки между поселками Сиреники и Энурмино, однако неожиданный выход льдов из Чукотского моря в Берингов пролив блокировал экспедицию в заливе Лаврентия и заставил два отряда разделиться. Этнографы на вездеходе добрались до Уэлена и после отхода льдов продолжили работу с морскими охотниками этого поселка. Байдара зоологов вместе с командой чаплинского вельбота повернула на юг и, обгоняя льды, повторно прошла маршрут для того, чтобы внести уточнения в подсчеты колоний морских птиц.
Экспедиция оказалась очень успешной. Прежде всего было налажено рабочее взаимодействие с охотниками-членами команд байдары и вельбота. Они щедро делились знаниями о древних поселках и их обитателях, характере китобойного промысла в каждой общине, о распределении вдоль побережья гренландских и серых китов и особенностях их поведения. Охотники помогли нам провести опросы старейших
Члены экспедиции 1981 г. в пос. Лорино.
Слева направо: И. Крупник, Н. Гальгаугье, Н. Панагиргин, Е. Паулин, А. Анкалин,-М. Членов, Ю. Ятта, В. Миенков, Д. Анагисяк, В. Тагитуткак (перед ним стоит мальчик из пос. w Лорино), Б. Звонов, Л. Богословская, А. Голубцов. Внизу: С. Богословский и Г. Каяк
жителей в поселках Янракыннот, Лорино, Лаврентия, Уэлен, которые либо сами участвовали в традиционном китобойном промысле, либо хорошо помнили о нем.
Главным достижением экспедиции стало комплексное обследование и картирование древних и старых поселков Мечигменского залива, особенно древнего селения Масик (по-чукотски Мечигмен), что позволило описать новый для науки вариант традиционной китобойной культуры, основанный на промысле детенышей серых китов. Помимо этого на карту были положены районы прибрежной концентрации и пути сезонных миграций китов и ластоногих, залежки и лежбища последних, а также колонии морских птиц (птичьи базары).
1985 г. Трое зоологов (Л. Богословская, Б. Звонов, Н. Конюхов) и трое морских охотников (В. Миен-
Члены экспедиции 1985 г. Слева направо:
Л. Богословская, Т. Панаугъе, С. Нанухтак, Б. Звонов. Сидят: Н. Конюхов и В. Миенков
$
Тимофей Панаугье (1939-1995). Сиреники, 1980 г.
ков, С. Нанухтак, Т. Панаугье) на одиночной байдаре прошли вдоль побережья Чукотского полуострова на север, от пос. Сиреники до пос. Энурмино, и на запад, от пос. Сиреники до бухты Преображения. На всем протяжении были описаны и картированы древние и старые поселки, места концентрации китов и ластоногих, колонии морских птиц.
Замечательные результаты этого весьма протяженного и сложного маршрута были достигнуты благодаря великолепной и слаженной работе охотников.
1987 г. Целью экспедиции стал остров Ратмано-ва. После долгих переговоров со Штабом Погранвойск КГБ СССР удалось получить разрешение на обход острова Ратманова группой из шести человек на одиночной байдаре. На этот раз капитаном байдары шел П. Тыпыхкак, старейшина морских охотников пос. Сиреники, мотористом — В. Каваквыр-
гын, впередсмотрящим — К. Вашурин. Зоологические работы проводили Л. Богословская, Н. Конюхов, А. Паско. Маршрут проходил в тяжелейших погодных условиях — непрерывные дожди, шторма и туманы. Тем не менее вся работа была выполнена: на острове Ратманова описаны и картированы остатки трех поселков и грандиозное поле священных камней-Дагиров, многочисленные колонии морских птиц; изучено распределение серых китов в западной (советской) части акватории Берингова пролива. На материковом берегу, в бухте Поутен, был детально обследован и картирован большой старый поселок, жители которого, как и древние обитатели Мечиг-менского залива, специализировались на добыче детенышей серых китов.
Помимо описанных выше трех экспедиций в течение 1980-1990-х годов совместно с коренными жителями было проведено множество небольших морских и сухопутных маршрутов, позволивших составить детальное этнобиологическое описание побережья Восточной Чукотки и восстановить традиционную схему расселения.
МЕЖДУНАРОДНЫЕ ПРОГРАММЫ И ПРОЕКТЫ
С 1992 г. на созданной по инициативе капитана Вотрогов сети подготовленных наблюдателей начала базироваться российско-американская программа по традиционному природопользованию коренных жителей Восточной Чукотки, в которой особое внимание уделялось гренландскому киту. С американской стороны в ней участвовали сотрудники Департамента по управлению ресурсами диких животых Феде
рального округа Аляски «Самоуправление Северного склона» (Department of Wildlife Management, North Slope Borough) под руководством T. Алберта. С российской стороны в Программе сначала работали только члены национального товарищества «Наукан» под руководством его председателя М. Зеленского и научного консультанта В. Мельникова. В 1994 г. Программа была расширена, и в ней приняли участие охотники Провиденского района, объединенные Обществом эскимосов «Юпик» под руководством его председателя Людмилы Айнана и научных консультантов Н. Мымрина и Л. Богословской. Следует особо отметить большую работу по обучению наблюдателей, которую постоянно проводил Н. Мымрин.
Продолжением этих научно-практических исследований стала новая Программа, основанная на сотрудничестве коренных жителей со Службой Национальных парков США (U.S. National Park Service) в области изучения традиционного природопользования с учетом динамики морских промысловых ресурсов.
Кроме двух общих программ были выполнены еще три специализированные проекта. В 1995-1996 гг. Л. Богословская провела опрос коренных жителей Провиденского, Чукотского и Иультинского районов в рамках проекта Циркумполярной конференции инуитов (ICC) «Значение традиционного китобойного промысла в современной жизни эскимосов и чукчей». В те же годы Н. Мымрин вместе с опытными охотниками участвовал в российско-американском проекте «Традиционные экологические знания о белухе», а в 1998-1999 гг. выполнил инициативный проект «Традиционные экологические знания сибирских эскимосов о гренландском
ките», профинансированный Фондом Дж. и К. Ма-картуров.
ИНТЕГРАЦИЯ НАУЧНЫХ И ТРАДИЦИОННЫХ ЗНАНИЙ
Л. Вотрогов, Л. Богословская, И. Крупник, Н. Мым-рин были далеко не первыми специалистами, кто опрашивал население Восточной Чукотки об особенностях биологии морских животных. Указания на важность традиционных знаний местных жителей имеются в работах зоологов и этнографов 1930-1940-х годов — А.Г. Томилина, П.Н. Никулина, Б.А. Зенковича, В.А. Арсеньева, Е.С. Рубцовой и др. Однако работы на Восточной Чукотке в последние 25 лет принесли совершенно иное понимание ценности традиционных знаний и роли самих охотников в долговременных научных исследованиях. В результате многих лет совместной работы сложились и закрепились новые отношения ученых и коренных жителей, представляющих «академические» и традиционные знания.
Во-первых, стало возможным с полной уверенностью говорить о равноценности традиционных и научных знаний в плане их системности, глубины и точности. Вдобавок к этому, традиционные знания о природе всегда очень персональны, они несут на себе отпечаток личности не только самого охотника-информатора, но и тех, кто в свое время пере-дал ему эти знания. Кроме того, они окрашены совершенно особым отношением к животным: не как к объектам научного познания, а как к равным себе, продолжению своей личности и собственного мира.
Во-вторых, непрерывные наблюдения за животными (мониторинг), которые для ученых являются частью
научной работы, для коренных жителей служат залогом успешной охоты, а, значит, жизни. Каждый год откладывает в памяти охотника свои особенности погоды, поведения зверей, их распределения в море и бухтах. По всем этим признакам настоящий охотник складывает промысловую картину года, которая поможет принять правильное решение, если условия в той или иной степени повторятся. Охотнику надо не только хорошо чувствовать погоду, море и берег; он должен разбираться в повадках морских зверей, уметь предугадать их поведение. Отсюда и происходит удивительная системность знаний коренных жителей, которая связывает ветры, течения, льды и животных в единое целое. При этом, в отличие от узкоспециализированных научных программ, традиционные знания не имеют начала и конца — это вечный процесс. Мониторинг одних и тех же мест (участков экосистем) ведется непрерывно многими поколениями, знания накапливаются, постоянно подкрепляют друг друга и передаются в доступной форме следующим поколениям.
В-третьих, включение местных жителей как полноправных партнеров в систему научных исследований позволяет с новых позиций подойти к изучению и сохранению биологического разнообразия и биопродуктивности экосистем Арктики. Специалисты постоянно получают от охотников новые данные о сроках и путях сезонных миграций, распределении, колебаниях численности и особенностях поведения китов, что позволяет разрабатывать конкретные меры по их сохранению. Кроме того, интеграция традиционных и научных знаний о китах, основанная на многолетних непрерывных наблюдениях, может быть методически крайне полезна для организации подобных программ по другим редким и особо охраняемым видам.
Мониторинг морских млекопитающих в регионе Берингова пролива, проводимый в течение 25 лет силами охотников, — явление уникальное по обилию и ценности собранных материалов. Он позволил выявить изменения в экосистемах, связанные как с циклами естественных динамических процессов, так и с последствиями антропогенных воздействий разного происхождения. Опыт совместной работы создает новые основы для углубления концепции устойчивого развития, сохранения биологического и культурного разнообразия полярных регионов.
О НАБЛЮДАТЕЛЯХ
В заключение этого раздела один из его авторов (Л. Богословская) считает своим приятным долгом привести имена наблюдателей, чьи рассказы она в свое время записала или получила их в виде дневников и письменных сообщений во время выполнения программ и проектов, перечисленных выше. Безусловно, круг наблюдателей шире, и многие специалисты, вероятно, имели счастье работать с другими, не менее сведущими, коренными жителями.
Автор приносит свои извинения тем охотникам, имена которых не были записаны в свое время или в написание чьих фамилий вкрались ошибки из-за незнания чукотского и эскимосского языков.
Энмелен
Гыргольнаут Геннадий
Обнаружил группировку гренландских китов, остающуюся на лето у побережья пос. Энмелен, собрал ценнейший материал по сезонному распределе-
Владимир Акка (1922-1990) и Виктор Татыга (1943).
Сире ники, 1980 г.
Николай Каваугье (1935-1995). Сиреники, 1980 г.
нию и численности других китов и ластоногих. Его дневники могут служить образцом точности и широты охвата наблюдаемых явлений (помимо китов и ластоногих он отмечал даты прилета птиц, появления растений и т.д.).
Нунлигран
Кычи Николай
Сумел разобраться в сложном характере сезонных миграций китов и ластоногих в акватории родного поселка. Его данные подкрепили наблюдения Г. Гыр-гольнаута и охотников пос. Сиреники по направлению весенней миграции гренландских китов. Собрал ценный материал по миграциям белухи и серого кита.
Сиреники
Акка Владимир; Анкалин Андрей; Асыколян Николай; Вашурин Константин; Гальгаугье Николай; Исаков Олег; Кавакдыргын Степан; Каваквыргын Владимир; Канихин Анатолий; Коой; Кукильгин Андрей; Миенков Виктор; Панаугье Тимофей; Нанухтак Сергей; Прохоров Николай; Татыга Виктор; Тыпыхкак Петр; Ухтыкак Алексей.
Все вместе охотники сформировали основу современных знаний о гренландских китах в водах Чукотки. Они впервые сообщили о размножении китов в Сиреникской полынье, описали особую разновидность (морфу) гренландского кита, такышкак, выявили разницу весенних миграционных потоков у поселков Сиреники и Имтук, показали, что у берегов Чукотки весной киты идут намного дольше, чем у побережья Аляски. Эти данные помогли уяснить целый ряд особенностей биологии гренландского кита и поставить вопрос о существовании внутренней структуры чукотско-берингоморского стада. Кроме того, охотники составили хорошие описания мигра-
Коой (1914-1985). Сиреники, 1979 г.
ций белух и поведения косаток, в частности их охоты на серых и гренландских китов.
Новое Чаплино
Паулин Евгений; Рентин Сергей; Люнеут Павел; Панагиргин Николай; Тагитугкак Владимир; Ятта Юрий.
Рассказали о многочисленных наблюдениях за гренландскими, серыми и горбатыми китами в проливе Сенявина, районе мыса Чаплина, возле лагуны Кивак. Описали зимнее поведение белух, способы и приемы охоты на них. В. Тагитуткак сообщил о редких появлениях гладкого японского кита и кашалота в проливе Чирикова (между мысом Чаплина и островом Св. Лаврентия).
Янракыннот
Аксау Андрей; Ашкамакин Альберт; Виренеут Юрий (остров Аракамчечен); Кавранто Константин; Кутылин Леонид; Кымыечкын Константин; Омрырультат Николай; Этгувье Анатолий.
Степан Каваквыргын (1930-1987), Николай Асыколян (1927 -1989), Андрей Кукилъгин (1923-1985). Сиреники, 1984 г.
Ранав (1915 - ?). Масик-Мечигмен, 1981 г.
Детально описали сезонное распределение серых и гренландских китов, белух, малого полосатика и косаток в проливе Сенявина и южной части Мечиг-менского залива; сообщили важные сведения о состоянии льдов в пределах их охотничьей акватории. Рассказали о способе охоты на белух со льда и, со слов стариков, о единичных появлениях зимой нарвалов. Альберт Ашкамакин обладает незаурядными способностями натуралиста; в течение многих лет он в одиночку наблюдает китов и ластоногих у лагуны Гетлянген, где ему удалось обнаружить летнюю группировку гренландских китов.
Лорино
Аянто Михаил; Ранав
Детально описали распределение серых китов в акватории Мечигменского залива и традиционную охоту на их детенышей.
Уэлен
Армаиргин Роман; Еореле Василий; Рильхин; Тир-китегин; Тнанкергав Александр; Тулюкак; Элюч Владимир.
Сообщили о регулярном появлении гренландских китов, идущих весной на запад, в сторону Инчоуна, сезонных миграциях белух, поведении серых китов и косаток. Материалы от В. Еореле я получала по почте и, к сожалению, никогда не встречалась с самим наблюдателем.
Инчоун
Агранаут Алексей; Вуквутагин Яков; Гувалин Виктор; Пинеуги Петр; Ритлю Михаил (старый пос. Исэн); Тер-куквун Федор; Тототто Юрий; Чайвун Валерий.
Жители Инчоуна постоянно сообщали о весенней миграции гренландских китов в западном направлении. Их точные рассказы и описания, наряду
Федор Теркуквун (1944). Инчоун, 1980 г.
с материалами охотников пос. Сиреники, явились основанием для сомнений в достоверности американских данных о численности всего чукотско-бе-ринговоморского стада гренландских китов. Пинеу-ги рассказал, что в молодости видел двух нарвалов, а его наставник в морском деле Меето встречал их среди льдов несколько раз за свою жизнь.
Энурмино
Кеутегин Михаил.
Провел цикл детальнейших наблюдений и подсчетов гренландских китов во время сезонных миграций. Его данные подтвердили ранее записанные сообщения охотников Уэлена и Инчоуна о весеннем ходе гренландских китов на запад. С Кеутегиным, как и с Еореле, мы с Н. Мымриным списались по почте в начале 1990-х годов, а лично встретились много позже, на Аляске.
Нешкан
Имелеу; Калянто Николай; Лейвитеу; Ныпевги; Ээло.
Сообщили, что гренландские киты в некоторые годы проходят близко от поселка во время весенней и осенней миграций. В непогоду они часто следуют за белухами. Считают, что поселок расположен не слишком удачно для наблюдений за китами и поэтому лучше следить за этими животными из других мест.
Особой благодарности заслуживает помощь, которую оказывали автору и ее коллегам Людмила Айнана в Провиденском районе и Татьяна Ачиргина в Анадыре.
3
Как устроено море
В этом разделе много научных терминов, но совсем не обязательно запоминать их все, хорошо, если в памяти останутся хотя бы три слова — бентос, планктон и нектон — и такое совершенно необходимое биологическое понятие как первичная продукция.
Океаны и моря нашей планеты все вместе образуют Мировой океан, в котором существуют разные зоны жизни, населенные разными сообществами животных и растений. Система зон жизни, или по-научному, экологическая зональность морей и океанов, отражает смену условий существования организмов от поверхности в глубину и от прибрежья к открытым частям морских пространств. Начнем с того, что Мировой океан в целом делится на дно (бенталь) и толщу воды над ним (пелагиалъ).
ДНО И ДОННЫЕ ОРГАНИЗМЫ (бенталь и бентос)
Для населенного организмами дна морей и океанов принято название бенталь, в которой выделяют следующие вертикальные зоны:
1) супралитораль — зона заплеска и штормовых выбросов;
2) литораль — приливно-отливная зона;
3) сублиторалъ — зона шельфа, или материковой отмели (до 200-500 м глубины);
4) батиаль — зона континентального склона, или «свал глубин» (от 200-500 до 3000 м);
5) абиссаль — зона океанического ложа (3000-6000 м глубины);
6) ультраабиссаль — зона наибольших океанических глубин, так называемых впадин.
Бентос — это совокупность донных и придонных организмов; его подразделяют на растительный (фитобентос — морские травы, микроскопические и крупные водоросли) и животный (зообентос), включающий три больших группы животных, которые:
Ж обитают в грунте дна — некоторые иглокожие, многощетинковые черви, двустворчатые моллюски, ракообразные и др.;
* передвигаются по поверхности дна или прикрепляются ко дну — морские звезды, кораллы, губки, актинии, многощетинковые черви, большинство иглокожих, многие ракообразные, в том числе крабы, и др.;
* плавают вблизи дна и лишь периодически опускаются на дно — креветки, некоторые голотурии, множество придонных рыб и др.
По горизонтали для бентоса различают прибрежные, богатые жизнью области и бедные жизнью области в удаленных от берегов центральных районах океанов, а также переходные между ними зоны.
Общая масса (биомасса) бентоса в Мировом океане оценивается в 10-12 млрд. т. Ее величина убывает с глубиной: в прибрежных мелководных районах биомасса составляет до 5-10 кг/м2, а в бедных жизнью центральных океанических глубинах — всего лишь 0,01 г/м2 и менее. На долю расположенных вблизи материков мелководных районов, которые занимают менее 8% площади дна Мирового океана, приходится около 60% биомассы всего океанического бентоса. Именно к этой, богатой бентосом, части Мирового океана принадлежат моря Берингово, Чукотское и Восточно-Сибирское, омывающие берега Чукотки. А вот на долю участков дна, лежащих глубже 3000 м (так называемая абиссаль) и занимающих 3/4 дна Мирового океана приходится всего 10% биомассы бентоса.
ТОЛЩА ВОД И ЕЕ ОРГАНИЗМЫ (пелагиаль, планктон и нектон)
Вся толща вод, от поверхности до дна, называется пелагиаль, а растения и животные, населяющие ее, имеют общее название — пелагические организмы. В пелагиали вертикальная экологическая зональность выражена менее четко из-за перемещения вод и вертикальных миграций пелагических организмов на сотни, а порой и тысячи метров по глубине.
Тем не менее в пелагиали выделяют такие зоны: 1) эпипелагиаль — 0-200 м глубины;
2) мезопелагиалъ — 200-1000 м глубины;
3) глубоководная зона — глубже 1000 м.
Пелагические организмы делятся на две группы — планктон и нектон.
Планктон (по-гречески «блуждающий») — это совокупность мелких растений и животных, которые не могут самостоятельно передвигаться на большие расстояния, как это делают рыбы и киты, — их переносят с места на место морские течения. В состав планктона входят водоросли (фитопланктон), животные (зоопланктон) и бактерии (бактериопланктон). С глубиной количество и разнообразие планктона быстро убывают, однако зоопланктон и бактериопланктон населяют всю толщу вод до максимальных глубин.
Фитопланктон — основа пищи всех животных, обитающих в Мировом океане — это микроскопические растения, диатомовые водоросли, динофлагелляты и некоторые другие. С помощью солнечной энергии и хлорофилла (особое вещество, придающее растениям зеленый цвет) они преобразуют растворенные в воде неорганические вещества в органические вещества своего собственного организма, или, говоря по-научному, создают первичную продукцию моря. Хотя растения эти чрезвычайно малы, но число их столь велико, что, в конечном счете, они кормят весь океан. (На суше первичную продукцию точно так же создают наземные растения — травы, кустарники и деревья.)
Зоопланктон представлен множеством видов самых разных животных, среди них крохотные простейшие, состоящие всего из одной клетки, кишечно
полостные, крылоногие моллюски, оболочники, ракообразные, икра и мальки рыб, личинки многих беспозвоночных, в том числе и донных. Наибольшее видовое разнообразие зоопланктона — в тропических водах, а наибольшее его количество (масса) — в холодных водах. Размеры планктонных животных колеблются от тысячных долей метра до нескольких метров (гребневик «венерин пояс» — 1,5 м, медуза цианея диаметром 2 м и т.п.). У многих представителей зоопланктона выработались приспособления, облегчающие парение в воде.
Биомасса фитопланктона составляет от нескольких миллиграмм до нескольких грамм на кубометр воды; зоопланктона — от десятков миллиграмм до одного грамма и более на кубометр. Если говорить в целом о Мировом океане, то биомасса фитопланктона невелика по сравнению с биомассой всего зоопланктона — 1,5 и свыше 20 млрд, т, соответственно. Однако из-за быстрого размножения микроскопических водорослей их продукция в Мировом океане составляет около 550 млрд, т, а это почти в 10 раз больше общей продукции всего животного населения океана.
Нектон (по-гречески «плавающий», «плывущий») — это все активно плавающие пелагические животные, которые способны противостоять течениям и преодолевать значительные расстояния. Массовые представители нектона — рыбы, кальмары, морские змеи и китообразные. К нектону относятся и те животные, кто размножается на суше, а питается в воде, например, ластоногие, водяные черепахи, пингвины, чистиковые птицы, ловящие добычу в толще воды, и др. Нектон наиболее разнообразен и многочислен в верхних горизонтах вод в связи с обилием там пищевых ресурсов.
Океанический нектон — эволюционно молодое сообщество животных: современные океанические рыбы сформировались в палеогене (примерно от 66 до 25 млн. лет назад), головоногие моллюски (кальмары, осьминоги, каракатицы) и морские млекопитающие — в неогене (примерно от 25 до 1,8 млн. лет назад). Многие исследователи считают, что киты старше других морских зверей, поскольку остатки животных, очень похожих на современных нам морских исполинов, известны с середины палеогена.
ПИЩЕВЫЕ (трофические) ЦЕПИ МИРОВОГО ОКЕАНА
Итак, диатомовые водоросли и динофлагелляты съедаются мельчайшими животными, которые входят в состав зоопланктона, а их в свою очередь поедают более крупные животные. Вот таким образом пищевая (трофическая) цепь в конце концов протягивается от крохотной диатомеи через зоопланктон к гигантским кальмарам, а ими питаются самые большие зубатые киты, кашалоты. Трофическая цепь, идущая к гренландскому киту, заметно короче — он питается непосредственно зоопланктоном. А самую короткую пищевую цепь из всех морских млекопитающих имеют сирены (ламантины и дюгонь) — они едят водоросли, то есть непосредственно первичную продукцию моря.
Многочисленные виды мелких ракообразных, входящие в состав зоопланктона, сами кормятся диатомовыми водорослями и, подобно усатым китам, отфильтровывают свою добычу через усики-щеточки. Близкие к креветкам более крупные ракообразные, так называемые шримсовидные, служат
пищей для многих рыб, птиц и морских зверей. Одна из групп шримсовидных представлена самыми большими планктонными рачками, известными как эвфаузииды. От момента вылупления эвфаузиид из яиц до времени достижения ими полной зрелости проходит несколько лет, и тогда эти рачки собираются вместе в несметных количествах, обеспечивая китов богатой пищей. Массовые приповерхностные скопления зоопланктона, в которых основную часть составляют именно эвфаузииды, называются криль, что по-голландски означает «малыш, крошка, мелочь». Криль — это условное название, на самом деле животного с таким именем нет. В Антарктике криль состоит практически только из эвфаузиид.
В Арктике скопления зоопланктона представлены и множеством других животных, в том числе крохотными крылоногими моллюсками, или «морскими бабочками», которые сами питаются еще меньшими планктонными организмами. Иногда «морские бабочки» скапливаются в таком изобилии, что море на целые мили меняет свой цвет. Среди крылоногих моллюсков известен клион, маленькое слизнеобразное существо, обходящееся без всякой раковины. В высоких широтах, по окраинам Арктики огромные стаи клиона долго служили основной пищей когда-то многочисленных гренландских китов.
Можно привести еще много примеров пищевых связей между разнообразными растительными и животными организмами моря. Если на суше всякое мясо в конечном счете — трава, то в море всякая рыба, киты и все остальные животные в конечном счете — диатомовые водоросли, составляющие главную часть фитопланктона. Морские звери являются последним звеном в сложных пищевых (трофичес
ких) цепях, начало которых — морские травы, водоросли и фитопланктон. Все вместе эти растения осуществляют фотосинтез, без которого ни в море, ни на суше не было бы жизни.
Пищевые, или трофические, цепи служат единой цели — созданию и перераспределению вещества и энергии на огромных пространствах Мирового океана. Начало же этому процессу дают энергия солнца, хлорофилл диатомовых водорослей и определенные неорганические вещества, растворенные в морской воде. Именно они создают первичную продукцию моря.
МОЖЕТ ЛИ МОРЕ ЖИТЬ БЕЗ КИТОВ?
— А почему бы и нет, — наверное, скажут многие. — Что делают в море все эти киты, моржи, тюлени и морские котики? Едят и едят себе что ни попадется, да и нас, людей, здорово объедают. Ведь сколько рыбы нужно одному финвалу!
Рыбы финвалу действительно нужно много, и человек привык воспринимать морских зверей как своих весьма прожорливых пищевых конкурентов. Он не признает за ними никакой очевидной пользы, кроме той, что дает охота на них. Но так ли это на самом деле? Ведь прежде, когда в Мировом океане обитали огромные стада морских млекопитающих, почему-то рыбы, крабов и креветок было великое множество.
Давно замечено — у морских млекопитающих посезонно меняются пищевые объекты и места кормления, кроме того, животные разного возраста питаются разными обитателями моря. Нередко киты,
моржи, тюлени без всяких видимых причин бросают богатые кормовые угодья и возвращаются в них много времени спустя. Хотя еда — первая и постоянная потребность всех животных, но не только она определяет жизнь исполинов моря, их тысячекилометровые сезонные миграции и во многом загадочное поведение.
В настоящее время на Земле существуют две группы млекопитающих, которые полностью порвали связь с сушей, но сохранили, как и наземные позвоночные животные, легочный тип дыхания и теплокровность. Это китообразные — резко отклонившаяся от основного ствола млекопитающих группа морских и пресноводных зверей, которая объединяет усатых и зубатых китов. Вторая группа — сирены (ламантины и дюгонь). К сиренам принадлежала знаменитая морская корова Командорских островов, истребленная русскими промышленниками практически сразу после того, как ее открыла экспедиция Беринга. Другие группы водных млекопитающих в большей или меньшей степени сохранили связь с сушей, например, ластоногие, к которым относятся моржи и тюлени, они выходят на берег для отдыха или размножения.
Мы только начинаем улавливать, каким образом морские млекопитающие влияют на существование более простых организмов, в частности своей добычи. Постепенно к нам приходит понимание того, что странствия морских зверей — это не их «личное дело», а перенос и распределение вещества и энергии в Мировом океане, что без китов, дельфинов, сирен и ластоногих порвутся не видимые нам, но крепкие нити, которыми природа связала воедино всех обитателей моря.
Наконец, все более и более проясняется роль крупных морских животных в сохранении или изменении морских ландшафтов. Самое яркое и убедительное подтверждение этому — способ питания калифорнийского серого кита. Всасывыя донные организмы вместе с грунтом, он буквально спасает песчаное дно бассейна Чирикова от илистых выносов могучих рек Аляски, Юкона и Кускоквима, и тем самым сохраняет среду обитания для своих любимых пищевых объектов, рачков-амфипод, предпочитающих жить в песке. Серый кит делает это уже многие тысячелетия, со времен последнего ледникового периода, когда образовался и этот песок, и сам бассейн Чирикова вместе с примыкающим к нему на севере Беринговым проливом.
4
О китах и делыринах
Китообразные — одни из древнейших млекопитающих нашей планеты. Они распространены по всему Мировому океану, их можно встретить среди паковых льдов Арктики, в умеренных и тропических водах, у кромки ледяных полей Антарктики.
ИСТОРИЯ И СИСТЕМАТИКА КИТОВ
Поисками и изучением остатков древних животных, в том числе китообразных, занимается палеозоология. Она является частью обширной биологической науки палеонтологии, которая исследует животных и растения прошлых времен по их ископаемым остаткам и следам жизнедеятельности.
Ископаемых остатков китообразных известно относительно немного. Изучение их показало, что в очень далекие времена, более 35 млн. лет тому назад, на Земле существовали животные, имеющие все отличительные признаки китообразных: длинный позвоночник, типичный «китовый» череп, развитые передние конечности и почти исчезнувшие задние. Это были древние киты (Archaeoceti), вымершие около 25 миллионов лет тому назад. А ныне существующие
группы усатых китов (Mystacoceti) и зубатых китов (Odontoceti) начали формироваться не менее 20 миллионов лет тому назад.
Среди специалистов нет единого мнения о наземных предках китообразных. Ряд ученых высказываются в пользу их происхождения от древних копытных, с которыми они имеют общие черты строения (сложный желудок, двурогая матка, диффузная плацента). Анализ числа хромосом и изучение реакции преципитации (осаждения) белков крови говорят в пользу этой гипотезы.
Однако другие исследователи на основании сходства в строении скелета и зубной системы древних китообразных с креодонтами (предками современных хищных животных) полагают, что китообразные произошли от этой группы. Было высказано еще одно мнение — родоначальниками китообразных могут быть насекомоядные мелового периода, гораздо более древние животные, чем креодонты.
Заслуживает внимания гипотеза о двойственном происхождении китообразных. По мнению ее сторонников, принципиальные различия в строении усатых и зубатых китов очень велики и говорят об их независимом происхождении от двух предковых групп. А общее сходство объясняется тем, что историческое развитие (эволюция) и зубатых, и усатых китов в водной среде привело к появлению сходных признаков в связи со сходным образом жизни.
Так или иначе, но живущие ныне киты — потомки несчетных поколений, прошедших огромный путь развития в течение многих миллионов лет.
Теперь перейдем к систематике современных китообразных, но сначала несколько слов о самой науке. Систематика — это биологическая наука, которая опи-
сьгвает и обозначает все существующие и вымершие животные и растительные организмы, а также распределяет (классифицирует) их по группировкам разного уровня (ранга). В систематике каждый уровень, или ранг, вплоть до вида и подвида, имеет свое научное название на латинском языке (латыни), которое обязательно для ученых в любой стране мира. Это сделано для того, чтобы специалисты всех стран, на каких бы языках они сами ни говорили и писали, всегда могли понять друг друга и не спутать, например, один вид с другим из-за неточного перевода. Конечно, помимо латинского в каждом языке существуют и свои, национальные, названия, но чаще всего они есть только для «своих» животных и растений. В этой книге первые три научные латинские названия были приведены выше, в скобках, после русских названий вымерших и ныне живущих китообразных.
Итак, отряд современных китообразных (Cetacea) разделяется на два подотряда. Подотряд усатых китов (Mystacoceti) включает в себя 3 семейства: гладких китов, серых китов и полосатиковых; все вместе они объединяют 10 видов. Подотряд зубатых китов (Odontoceti) состоит из 5 или 7 семейств (по разным авторам), которые объединяют 71-77 видов. Надо сказать, что систематика китообразных, особенно зубатых китов, разработана еще недостаточно, и многие специалисты выделяют разное количество семейств, родов и видов.
Вид — это основная структурная единица в системе живых организмов. В отличие от латинских названий высших рангов, состоящих из одного слова, названия видов на латинском языке всегда содержат два слова: первое — название рода, к которому принадлежит данный вид, второе — его инди
видуальное «имя». Обычно после «имени» стоит фамилия ученого, придумавшего его, а через запятую — год, когда это имя было введено в научную литературу. Вот, например, описание систематического положения хорошо всем известной косатки на русском и латинском языках:
Класс Млекопитающие Отряд Китообразные Подотряд Зубатые киты Семейство Дельфиновые Род Косатки Вид Косатка обыкновенная
Klassis Mammalia Oido Cetacea Subordo Odontoceti Familia Delphinidae Genus Orcinus Species Orcinus orca
Linnaeus, 1758)
Здесь на каждом уровне, или ранге, подчеркнуты русские и соответствующие им по значению латинские названия. Последний, видовой, уровень говорит о том, что видовое «имя» orca дал косатке в 1758 г. Карл Линней (Linnaeus), великий шведский натура-лист XVIII века.
В этой книге для каждого кита или дельфина будут приведены только три последние латинские названия: семейство, род, вид. Если к одному и тому же семейству или роду принадлежат несколько видов, то русское и латинское названия такого семейства или рода приводятся только перед описанием первого по порядку вида.
КАК УСТРОЕНЫ КИТЫ И ДЕЛЬФИНЫ
Китообразные внешне очень похожи на рыб, но отличаются от них теплокровностью, легочным (а не
жаберным) типом дыхания, внутриутробным развитием детенышей и выкармливанием их молоком, а также многими другими чертами строения и функционирования различных органов.
Форма тела
Киты и дельфины прекрасно приспособлены к существованию в воде, об этом свидетельствует устройство каждой части их тела. Известно, что сопротивление воды при передвижении в ней в 800 раз больше, чем в воздухе. Чтобы снизить затраты энергии на его преодоление китообразные приобрели си-гаро- или торпедообразную форму тела, утратили ушные раковины и волосяной покров, которые имели наземные предки, и в результате их гладкое тело скользит в воде, вызывая чрезвычайно малое трение. Кроме того, подвижная и упругая кожа быстроходных китов и дельфинов, особым образом изменяя свою поверхность, способна гасить завихрения (турбулентность) воды, заметно снижающие скорость животных. Все это вместе помогает китам испытывать при движении минимальное сопротивление воды.
Некоторые киты все же сохранили на голове немного волос — это утолщенные осязательные волоски-вибриссы, или «усы», которые есть у всех наземных млекопитающих и ластоногих, особенно заметны вибриссы у моржей и тюленей. Функцию же полностью утраченного волосяного покрова — сохранение тепла в организме, постоянно находящемся в холодной воде, осуществляет у китообразных толстый подкожный жировой слой.
У быстрых и подвижных усатых китов-полосатиков поверхность горла, груди и живота до области пупка прорезана снаружи большим числом продоль-
Форма тела китообразных:
А — полосатик; Б — гренландский кит; В — дельфин;
Г — белуха; Д — кашалот.
Справа показаны поперечные сечения тела и головы
ных борозд длиной от одной до двух третей общей длины туловища кита. Борозды глубиной около 6 см делят кожу на ровные полосы, потому и появилось название этих китов - «полосатики». Когда кит питается, борозды растягиваются, заметно увеличивая емкость его пасти и брюшной полости.
У настоящих, или гладких, китов борозд на горле нет, но емкость их ковшеобразной пасти увеличивается иначе. Линия верхней челюсти у них сильно выгнута кверху, и потому нижняя челюсть отстоит далеко от верхней даже тогда, когда пасть сомкнута. Промежуток между верхней и нижней челюстями закрывается двумя половинами огромной нижней губы, они круто поднимаются вверх наподобие двух арок или луков. Недаром англичане так и назвали гренландского кита — «лукоголовый» (Bowhead).
Серый кит представляет собой промежуточную форму между полосатиками и настоящими китами: у него 2-4 короткие борозды на горле, а верхняя челюсть не имеет отчетливо выраженного сводчатого строения. Но для того, чтобы собирать корм со дна так, как это делает серый кит, совсем не нужно увеличивать объем пасти.
Подкожный жировой слой
Эта часть тела китообразных состоит из жировой ткани, в которой клетки, заполненные маслянистым веществом, связаны между собой волокнистой соединительной тканью. Благодаря этому подкожный жир — не мягкое, желеобразное вещество, а плотное и твердое. Толщина жирового слоя колеблется от 2 сантиметров у морской свиньи до 50 и более сантиметров у гладких китов. Жировой покров служит
изоляционным слоем, сохраняющим тепло в организме, а также запасом пищи на период голодовок китов во время миграций.
Но иногда даже киты испытывают потребность избавиться от «теплой одежды». У сухопутных млекопитающих излишки тепла при быстром движении выводятся из организма за счет учащенного дыхания или потения. Кит, находясь в глубинах моря, не может ни потеть, ни учащенно дышать. Но его жировой слой пронизан кровеносными сосудами, подводящими кровь непосредственно к поверхности тела. Когда под жировым слоем образуются излишки тепла, сосуды автоматически расширяются и регулируют температуру тела. Изменение притока крови к плавникам также играет существенную роль в терморегуляции китообразных.
Скелет
Скелет китов изменился очень сильно. Он заметно удлинился, важные перестройки произошли в конструкции передней (лицевой) части черепа, она стала сплющенной, низкой и заостренно-широкой, как волнорез. У настоящих китов глаза сдвинулись назад и расположились по бокам головы, поэтому поля зрения не пересекаются. Лицевая часть черепа значительно вытянулась вперед, так что верхняя и нижняя челюсти образовали длинное рыло (рост-рум). Носовые кости резко уменьшились, ноздри переместились с лицевой части черепа на мозговую и расположились на темени, то есть на самой верхней точке головы всех китообразных, кроме кашалота. При поднятии животного из воды первыми появляются на поверхности его ноздри, или дыхала, два — у усатых китов, одно — у зубатых китов.
Контуры тела и скелеты китов:
А — гренландский кит (1 — китовый ус;
2 — остатки таза;
Б — полосатик;
В — дельфин;
Г — кашалот (отдельно — нижняя челюсть кашалота)
В результате всех передвижений получилось своеобразное налегание одних костей черепа на другие, а из-за этого изменилась форма мозговой коробки: у всех китообразных она короткая, но зато широкая и высокая, и потому мозг часто имеет шаровидную форму, особенно у зубатых китов. Следует особо отметить, что череп усатых китов остался симметричным, а у зубатых китов он резко асимметричен, однако причины появления асимметрии до конца не выяснены.
Передние конечности всех китообразных превратились в грудные плавники, но сохранили свой скелет с небольшими изменениями.
Как и другие млекопитающие, киты имеют семь шейных позвонков, но они тонки, напоминают пластины, которые у многих видов срастаются в единую массу для поддержания огромной головы. Грудных позвонков в скелете усатых китов (полосатиков) насчитывается 12-16, у кашалотов — 11-12.
Скелет грудных плавников китообразных (по рентгенограммам):
А — гренландский кит; Б — финвал;
В — синий кит; Г — дельфин
Поясничных позвонков у усатых — 10-16, у кашалотов — 8-10. Хвостовых позвонков значительно больше, число их колеблется у усатых от 21 до 28, у кашалотов — 23-24.
Крестцовый отдел позвоночника полностью отсутствует, ведь он обеспечивает крепление задних конечностей, а они у китов исчезли. Сохранились лишь рудиментарные остатки тазовых костей, не соединенные с позвоночником и скрытые в брюшной мускулатуре. Некоторые виды китов имеют еще и пару небольших костей, представляющих собой остатки тазобедренной кости. Несмотря на свою незначительную величину, тазовые кости у китов все же не лишены функций: к ним прикрепляются некоторые мышцы органов размножения.
Органы движения и скорость
Главный орган движения всех китообразных — хвостовой отдел туловища. Плывущий кит посылает свое тело вперед мощными ударами хвоста, который на конце разделяется на две большие горизонтальные лопасти. Эти лопасти и спинной плавник — он имеется не у всех китов — состоят главным образом из кожи и плотной фиброзной ткани; внутри них нет костной основы, как у рыб, чьи плавники и хвост опираются на твердые плавниковые лучи. Поэтому специалисты называют хвостовые лопасти и спинной плавник китов бесскелетными образованиями.
Передние конечности китов, превратившиеся в плавники, используются главным образом для поддержания равновесия и для изменения направления движения.
Киты могут развивать очень большую скорость. Так, например, 25-метровый синий кит в течение
двух часов может плыть со скоростью свыше 50 км/час. И если учесть, что плывущий кит сравнительно медленно поднимает и опускает свой хвост, то эффективность тех усилий, которые он затрачивает при плавании, весьма высока. Размах лопастей его хвоста очень большой, и с каждым их ударом огромный объем воды отбрасывается назад, а тело движется вперед.
Детальный анализ энергетических затрат при скоростном движении китообразных выявил удивительный факт. Оказалось, что киты и дельфины плавают намного быстрее, чем это можно было ожидать от силовых возможностей их мускулатуры, — она не намного превосходит в этом отношении мускулатуру наземных млекопитающих, включая человека. Так, у мелких китообразных (морские свиньи и дельфины) скорость значительно больше, чем могло бы дать то количество энергии, которое они в состоянии затратить. Например, дельфин весом около 130 кг тратит 14 лошадиных сил, чтобы плыть под водой со скоростью 45-50 км/час. Этот показатель превышает более чем в шесть раз мускульное усилие, которое может «выжать из себя» самый тренированный атлет.
Тот парадокс, что дельфин плавает быстрее, чем это позволяет сила мышц, объясняется тем, что поверхность его тела, особым образом изменяя свою конфигурацию, гасит вихревые потоки, возникающие при движении. Это свойство кожи китообразных с успехом используется в конструкции обшивок самых современных подводных лодок.
Механизмы дыхания и погружения
У китообразных имеется целый ряд особенностей строения и функционирования внутренних органов, 68
Схема строения верхних дыхательных путей зубатых китов. Показано, каким образом они отделены от пищевода
которые все вместе обеспечивают продолжительную задержку дыхания без каких-либо вредных последствий для организма. Прежде всего дыхательная система надежно изолируется от воды снаружи и от пищи изнутри.
Киты погружаются в море очень глубоко, гораздо глубже, чем до сих пор может погрузиться человек. Как же им удается избежать кессонной болезни, от которой так страдают водолазы? Ответ очень прост — они избавлены от риска ею заболеть. Разница между китом и человеком при погружении состоит в том, что водолаз все время дышит сжатым воздухом, поступающим извне, так что азот может растворяться в его крови до полного насыщения, а кит уносит с собой в глубину лишь то количество воздуха, которое может поместиться при вдохе в его дыхательных путях и легких. Вот поэтому у него в крови и тканях содержится не слишком много азота. Следует добавить, что в отличие от человека у китов дыхательный центр обладает пониженной чувствительностью к накоплению в организме углекислоты.
Для китообразных характерно особое строение ряда кровеносных сосудов (артерий и вен), которое встречается и у некоторых наземных млекопитающих. В определенных местах крупные сосуды разделяются на множество переплетенных между собой ответвлений, которые сообщаются друг с другом, образуя густые сети. Они так и называются — чудесные сети.
Артериальные чудесные сети находятся на внутренней стороне позвоночного столба, в области шеи и грудной клетки, между ребрами, в основании головного мозга, вокруг спинного мозга. Венозные чудесные сети располагаются в основании черепа и области живота. Функция и деятельность чудесных сетей до сих пор не поняты до конца. Наличие чудесной сети, например, обеспечивает нормальное кровоснабжение головного мозга, наиболее чувствительного к дефициту кислорода. При этом мозг снабжается кровью не непосредственно из сонной артерии, а исключительно через такую чудесную сеть.
При длительных погружениях биение сердца китообразных резко замедляется: у дельфинов — от ста десяти ударов в минуту до пятидесяти и даже менее, у белухи пульс падает с тридцати до шестнадцати в минуту. Начинают действовать определенные механизмы, которые задерживают движение крови во всех кровеносных сосудах, кроме сосудов мозга и некоторых других органов. Однако к концу длительного погружения недостаток кислорода в тканях становится ощутимым, накапливается как бы «кислородный долг». Он возмещается, когда кит поднимается на поверхность и пускает фонтаны, обновляя запас воздуха в легких на 80-90% (для сравнения — чело
век способен обновить запас воздуха в легких только на 18-20%).
Итак, киты перед погружением набирают полные легкие воздуха, и их кровь предельно насыщается кислородом. Но это еще не все. Мышечная ткань, или, как говорят, красное мясо кита, интенсивного темно-красного цвета, потому что в ней содержится большое количество миоглобина — вещества, близкого по химическому составу и свойствам к гемоглобину — кислородному транспортеру крови. Перед погружением животного миоглобин также полностью насыщается кислородом, и таким образом «кислородный долг» в организме отодвигается на сравнительно длительное время.
Усатые киты обычно не погружаются глубже, чем на 50-100 л/, поскольку наибольшие скопления того зоопланктона, которым они питаются, как правило, находятся на глубине 10-20 м. Однако в случае необходимости — если, например, кит испуган — он в состоянии уйти на глубину 300-450 м. Когда киты-полосатики питаются, они обычно ныряют на 10-15 минут, а затем на 5-10 минут поднимаются на поверхность, чтобы подышать. Но они могут оставаться под водой и до 40 минут.
Гладкие киты и горбачи способны продержаться под водой еще меньше и уходят поэтому на меньшую глубину. А вот кашалот может не подниматься на поверхность от 30 минут до одного часа. Бутылконос, по некоторым предположениям, держится под водой до двух часов. Эти зубатые киты способны уходить под воду на значительно большую глубину, чем усатые киты. Рекордная глубина погружения кашалота была установлена, когда однажды был найден погибший зверь, который запутался
в подводном телеграфном кабеле на глубине около 1000 метров у тихоокеанского побережья Южной Америки. Напомним, что для обыкновенного человека предел задержки дыхания — примерно одна минута, и только хорошо тренированные искатели жемчуга могут пробыть под водой две-две с половиной минуты.
Способы питания
Киты поглощают огромное количество пищи и делают это двумя способами.
Усатые киты заглатывают свою пищу, в основном зоопланктон, «оптом», как травоядные животные на суше, которые собирают траву не по одному стебельку, а сразу целыми пучками. Усатые киты не имеют зубов, но снабжены совершенно особым механизмом, который позволяет им хватать свой корм огромными порциями. Это — цедильный аппарат из пластин китового уса. На верхней челюсти, там, где у других млекопитающих находятся зубы, у усатых китов имеется две, а то и три сотни плоских пластин, поставленных наискосок по краям нёба и свисающих с него. Пластины твердые, но гибкие, волокни
Расположение пластин китового уса в ротовой полости.
1 — межчелюстная кость;
2 — верхняя челюсть;
3 — нёбо;
4 — пластины китового уса;
5 — бахрома китового уса;
6 — нижняя челюсть;
7 — язык
Голова косатки
стые по структуре, внутренний их край расщеплен в виде длинной тонкой бахромы. Бахромчатые края соседних пластин находят один на другой и частично переплетаются, так что в пасти кита внутренние боковые стенки выглядят как огромные мохнатые занавески. Эти «занавески» служат фильтром для отцеживания планктона от воды.
Питание зубатых китов — совсем иное. Их корм состоит из более крупных животных, которых они заглатывают преимущественно поодиночке. Все зубатые киты, независимо от их размеров, — от огромного кашалота до обычной морской свиньи — питаются кальмарами и рыбой, но основная их пища — кальмары, которые в огромных количествах плавают в океане на самых различных глубинах. Хотя они очень подвижны и быстры, зубатые киты и дельфины ловят их во множестве, в основном небольших, длиной 30-60 см, а более крупные экземпляры становятся добычей кашалотов.
Большинство дельфинов имеет острые и, как правило, весьма многочисленные зубы. Обычно считают, что наличие множества зубов — следствие приспособления к питанию такими активными животными, как рыбы и кальмары. Но вот у представителей семейства клюворылов (Ziphiidae), которые тоже едят кальмаров, очень мало зубов. Каким образом эти киты захватывают свою столь проворную добычу, до сих пор неясно. В то же время некоторые из пресноводных дельфинов, например гангский дельфин, имеют множество острых, как иглы, зубов, которые служат не для схватывания быстро движущейся добычи, а для выбирания червей и лангустов из донного ила.
Размножение и развитие
Размножение китообразных — сложный период их жизни, занимающий много времени. Беременность длится от 10 до 16-17 месяцев, роды происходят под водой, плод выходит хвостом вперед, а пуповина рвется сразу у самого брюха, где она истончается еще в утробе матери. Единственный, хорошо развитый детеныш рождается очень крупным (от 1/6 до 1/4 длины тела матери). Двойни крайне редки - все зародыши, кроме одного, обычно рассасываются. Как только детеныш родится, мать начинает осторожно подталкивать его к поверхности воды, и китенок делает свой первый вдох автоматически, едва его дыхала (или дыхало) окажутся на воздухе.
Молочное выкармливание занимает от 4 месяцев (мелкие дельфины) до 13 месяцев (кашалот). Млечные железы у самки кита находятся под слоем подкожного жира. Два соска помещаются в кожных складках по обеим сторонам половой щели и высо-
Роды у дельфина-афалины
вываются наружу лишь на время кормления. Китенку необходимо дышать через небольшие интервалы, поэтому кормление должно занимать немного времени.
Когда малыш прикасается к материнскому соску, ему не приходится делать сосательных движений, так как молоко благодаря сокращениям специальных мускулов, окружающих млечные железы, само выжимается и льется ему прямо в рот. Содержание жира в молоке китов достигает 54%, и потому китенок растет в период молочного кормления необыкновенно быстро, у крупных видов малыши прибавляют в сутки до 100 кг.
Детеныш сразу после рождения должен уметь плавать и следовать за матерью. В первые месяцы жизни детеныши плывут с ней бок о бок, над ее спиной или у нее под головой — так они затрачивают меньше усилий Самки очень заботятся о своем потомстве и у большинства видов никогда не оставляют малыша в опасности.
Органы чувств и поведение
Из пяти органов чувств у зубатых китов осталось только четыре — они лишены обоняния, у них отсутствуют и обонятельный эпителий, и первичные обонятельные центры головного мозга (обонятельные луковицы). Полное исчезновение одного из органов чувств — случай исключительный в классе млекопитающих! Усатые киты все-таки сохранили остатки обонятельных луковиц, которые, правда, ничтожно малы. Причиной этого удивительного явления следует считать образ жизни китов. В воде их носовые проходы, где расположен обонятельный эпителий, плотно закрыты, а запахи атмосферы, когда киты делают краткие серии выдохов-вдохов, особого значения не имеют, ведь вся жизнь этих зверей проходит под водой.
Глаза китов приспособлены к тому, чтобы видеть в воде и на воздухе. Косатки, дельфины, кашалоты, полосатики и серые киты «выставляются» из воды, чтобы осмотреться. Особенно характерно это для косаток, которые в таком положении могут долго патрулировать лежбища, ожидая схода моржей в воду. У крупных китов глаз весит до 1 кг, у мелких дельфинов он примерно равен глазу собаки. Глаза китообразных отличаются от глаз сухопутных млекопитающих весьма толстой склерой, состоящей из очень жесткой волокнистой ткани. Склера — это внешний покров глаза, у человека он называется глазным белком. Возможно, массивность склеры объясняется тем, что она заменяет отсутствующую в черепе кита костную глазную впадину, которая защищает глазное яблоко.
Но даже самые лучшие глаза не могут далеко видеть под водой, как бы она ни была прозрачна.
Если водная среда не позволяет далеко видеть, то зато она позволяет хорошо слышать: вода проводит звук лучше, чем воздух. Поэтому слуховая система — самая главная из систем органов чувств китообразных, хотя еще совсем недавно исследователи думали, что слух у китов слабый, и только открытие способности зубатых китов к эхолокации опровергло это мнение. Именно необычайно чувствительная эхолокационная система (по-английски сонар) дает зубатым китам и дельфинам возможность хорошо ориентироваться даже в турбулентных потоках воды и в темноте на больших глубинах.
В эхолокационную систему зубатых китообразных входят:
— структуры среднего и внутреннего уха (киты не имеют наружного уха, то есть ушной раковины);
— многочисленные очень сложные центры головного мозга, содержащие миллионы нервных клеток;
— органы, производящие (генерирующие) ультразвуки. Это, по-видимому, подчерепные воздушные мешки и воздухоносные пути. Следует помнить, что все зубатые китообразные имеют только одну ноздрю (дыхало). Такое уникальное явление среди всех позвоночных животных пока не получило полного научного объяснения;
— лобная жировая подушка, или лобная линза, лобный выступ, — особое скопление жира над верхней челюстью. У кашалотов лобный жир называется спермацетом и обладает исключительными ранозаживляющими свойствами. Полагают, что лобная линза помогает фокусировать ультразвуковые импульсы в тонкий луч, напоминающий луч лазера. Им животное сканирует окружающую среду и распознает, где полыньи, берег или добыча.
Ориентация зубатых китов в паковых льдах с помощью эхолокатора (сонара):
А — нарвал в вертикальном положении изучает маленькую полынью в паковом льду;
Б — нарвал, лежа на спине, обследует нижнюю поверхность пакового льда и полынью
Итак, ультразвуки для локации испускают сами животные, причем некоторые зубатые киты способны производить сигналы частотой свыше 100 кГц. Воспринимают же они гораздо больший спектр частот — от низких звуков (инфразвуков) до ультразвуков частотой в 150-200 кГц.
Благодаря эхолокации зубатые киты обладают изумительной способностью ориентации в плохо освещенном пространстве и даже в полной темноте. Они обнаруживают и опознают различные предметы подводного мира на основании картины звуков, отражаемых ими. Если у птиц и многих дневных млекопитающих, включая человека, мир «видимый», то у зубатых китообразных — мир «слышимый». Кроме них истинными эхолокационными способностями наделены еще летучие мыши, активность которых приходится на сумерки и ночное время. Однако эхолокация в энергетическом плане весьма затратный механизм, особенно в воздушной среде, поэтому маленькие по величине летучие мыши активно используют дополнительные источники информации — прекрасно развитую обонятельную систему и глаза, весьма чувствительные к рассеянному ночному свету.
Усатые киты не обладают способностью к эхолокации. Возможности их слуховой системы лежат в зоне обычных звуков и инфразвуков, то есть в области средних и низких частот. Например, звуки серых китов, записанные Л. Вотроговым и автором этой книги в Мечигменском заливе, занимали спектр низких частот в пределах 400-450 Гц. Зато дальность действия слуховой системы усатых китов поразительна - сотни, а порой и тысячи километров! Объясняется это тем, что усатые киты умеют использовать для дальней связи друг с другом особые свойства так
называемых подводных акустических каналов, открытых Л.М. Бреховских. По этим каналам низкие звуки, не угасая, распространяются на огромные расстояния.
И зубатые, и усатые киты обладают хорошо развитой вестибулярной системой (системой равновесия), которая позволяет им легко передвигаться в трехмерном пространстве. Киты чувствуют себя в воде, словно птицы в воздухе. Они без труда движутся в любых направлениях, даже вниз головой. Эта же система помогает распознавать малейшую вибрацию, исходящую от живых объектов или возникающую во время опасных природных явлений — подводных и прибрежных тектонических процессов, море-и землетрясений, извержений подводных вулканов, цунами и т.п.
Итак, особенности органов чувств значительно отличают китов от всех наземных животных, но их поведение вполне сравнимо с поведением таких интеллектуальных групп наземных высших позвоночных, как слоны, обезьяны, попугаи и врановые птицы. Уже несколько десятков лет ученые исследуют поведение зубатых китов в океанариумах. Дельфины (в первую очередь афалина), косатка и белуха стали постоянными объектами изучения. Сотрудникам американского океанариума «Морской мир» удалось даже 7 месяцев наблюдать за ростом и развитием детеныша серого кита, самочки Джиджи, которую вернули в море, когда она подросла. Историю Джиджи описали в своей книге писательница Э. Корр и знаменитый исследователь китообразных У. Эванс, один из авторов этого удивительного проекта.
Наблюдения и эксперименты в океанариумах позволили узнать очень многое о поведении китов и
г
Примеры группового поведения зубатых китов (самцы нарвалов):
А — синхронный вертикальный выход из воды пары нарвалов; Б — два нарвала скрестили свои бивни в воздухе над спиной третьего, который выставил из воды только спину; В — три нарвала выставили головы из воды и скрестили в воздухе свои бивни; Г — два нарвала скрестили свои бивни под водой
возможностях их органов чувств. Однако жизнь в неволе сильно меняет многие черты поведения всех китообразных, содержащихся в океанариумах. Так, косатка, грозный и неумолимый хищник, становится в океанариуме миролюбивой по отношению к человеку и другим морским зверям, на которых она охотится в море. Именно поэтому наблюдения за китами в естественных условиях представляют очень большую ценность, особенно за редкими и малоизученными видами.
В заключение можно сказать, что всем китообразным свойственны сложные формы социального поведения. При этом киты, обитающие группами в прибрежных водах, (многие дельфины, серый кит и горбач) имеют более сложное поведение, чем виды, держащиеся в открытых водах одиночно или небольшими группами (синий кит, финвал, сейвал). Особенности поведения будут рассмотрены при описании каждого из пятнадцати видов китов, обитающих в водах Чукотки.
5
Усатые киты
(Mystacoceti)
Подотряд усатых китов состоит из трех семейств (Familia): серые киты, гладкие киты и полосатико-вые; семейства в свою очередь делятся на роды (Genus), а те — на виды.
В водах Чукотки обитают следующие виды уса
тых китов:
Русское название
Серый кит Гренландский кит Южный (японский) кит Синий кит Финвал Сейвал
Малый полосатик Горбач
Английское название
Gray (Whale)
Bowhead (Whale)
Научное (латинское) название
Eschrichtius robustus Balaena mysticetus
Right (Whale)
Blue (Whale)
Fin (Whale)
Sei (Whale) Minke (Whale) Humpback (Whale)
Eubalaena glacialis Balaenoptera musculus Balaenoptera physalus Balaenoptera borealis Balaenoptera acutorostrata Megaptera novaeangliae
Питание зоопланктоном стало, вероятно, главной причиной развития гигантизма среди усатых
китов — таких больших по размерам и массе животных, как синий кит, нет и никогда не было на Земле. Оно же сильно отразилось и в строении этих китов. Самые существенные изменения произошли в черепе: исчезли зубы и вместо них появились роговые пластины с бахромой, которые образуют идеальный цедильный аппарат.
Снабженные цедильным аппаратом усатые киты не могут выбирать те или иные виды пищи по своему желанию. Когда кит находит большое скопление рачков, он, раскрыв рот, врезается в их массу, и рачки попадают в его воронкообразную пасть в огромном количестве. Потом он закрывает пасть и, подняв язык, выталкивает из нее воду сквозь фильтр, образованный китовым усом. Бахромчатая завеса задерживает планктон, а вода вытекает из пасти через щели между пластинами. Огромный язык не только вытесняет из пасти воду, но продвигает пищу к глотке и в момент ее заглатывания прикрывает преддверие глотки, не пропуская туда воду.
Встречные токи воды оказывают на раскрытую пасть столь большое давление, что при слабо разви-
тие шнее строение усатого кита (горбач)
Работа цедильного аппарата усатых китов
той жевательной мускулатуре усатые киты (кроме серого кита) не в состоянии закрыть пасть - чтобы сомкнуть челюсти они должны повернуться на бок или лечь на спину. Верхняя челюсть, несущая роговые пластины, испытывает более значительное сопротивление воды, чем нижняя, и поэтому особым образом крепится к голове.
Увеличение полости рта усатых китов для того, чтобы можно было захватить как можно больше пищи, происходит двумя путями. Гладкие киты (гренландский и южный) становятся «головастиками» — голова занимает у них до 1/3 длины тела. Полосатики (синий кит, финвал, сейвал, малый полосатик и горбач) увеличивают объем своей пасти за счет растяжения горло-брюшных складок (полос). Чем больше корма в пасти, тем сильнее растягиваются складки! Среди усатых китов только серый кит является настоящим бентофагом (питающийся бентосом). Такой способ питания освобождает серого кита от необходимости иметь крупную голову или многочисленные складки-полосы. Голова у него относительно небольшая, а на горле всего от 2 до 4 коротких складок-полос.
Способ дыхания усатых китов также связан с особенностями питания зоопланктоном, который вылавливается при непродолжительном нырянии и сравнительно медленном движении животных. Парное дыхало оказывается над водой всего от 0,5 до 1,5 секунд, за это время киты успевают выдохнуть от 7000 до 14000 л воздуха и столько же снова набрать в легкие.
Из всех органов чувств усатых китов наиболее дальнодействующим является слух. Издавая серии низкочастотных звуков (инфразвуков), животные
общаются между собой на больших расстояниях. Это возможно благодаря сверхпроводимости низких звуков, которая возникает при определенных условиях в отдельных пластах морской воды. Такие пласты были открыты советским ученым-акустиком Л.М. Бреховских с соавторами и получили название «каналы Бреховских». По ним усатые киты могут передавать сообщения друг другу на десятки, сотни, а порой, и тысячи километров. По этим же каналам к животным приходят низкочастотные «сообщения» от многих природных явлений — надвигающегося шторма, приближающихся паковых льдов, земле- и моретрясений, извержений подводных вулканов и т.п. Низкочастотная компонента в шумах моторов позволяет усатым китам заранее узнавать о приближении судов.
Усатые киты неплохо видят в воде и на воздухе, но, как полагают специалисты, они близоруки и способны различать объекты на небольших расстояниях. Обоняние усатых китов развито слабо, судя по относительной величине обонятельных нервов и первичных обонятельных центров головного мозга. Органы осязания представлены вибриссами, которые располагаются на морде (роструме), включая верхние и нижние челюсти. Волосяная сумка каждой вибриссы снабжена весьма чувствительными нервными окончаниями, способными воспринимать малейшие внешние раздражения. Вибриссы помогают китам обнаруживать даже небольшие скопления зоопланктона на глубине, куда едва проникает свет, или в ночное время. Вкусовые сосочки языка редуцированы (см. редукция), что многие исследователи связывают со способом питания и захвата кормовых объектов.
Слабое развитие сплетений кровеносных сосудов, так называемых чудесных сетей, в грудной и брюшной полостях связано с непродолжительностью ныряния усатых китов. При этом, видимо, не нужно сильно насыщать кислородом кровь, как зубатым китам, которые могут длительное время находиться под водой.
Все усатые киты рождают крупных и быстро растущих детенышей, которых длительное время выкармливают молоком, содержащим до 54% жира.
Вибриссы усатых китов (сверху вниз): южного кита, финвала, серого кита
Роды происходят в теплых и умеренных водах; исключение составляет гренландский кит, производящий на свет потомство в холодных водах Арктики и Субарктики.
синий кит
Относительные размеры усатых китов, обитающих в водах Чукотки
Серый кит
Гренландский кит
Японский гладкий кит
Синий кит
Районы обитания (ареалы) усатых китов в водах Чукотки
Финвал
Сейвал
Гэрбач
Малый полосатик
Районы обитания (ареалы) усатых китов в водах Чукотки (продолжение)
Сем. серых китов (Fam. Eschrichtiidae) Род серых китов (EschrichUus)
Серый кит
(Eschrichtius robustus, Lilljeborg, 1861;
Eschrichtius gibbosus, Erxleben, 1777)
Серый кит — единственный представитель своего семейства — один из самых древних усатых китов, но это не мешает ему быть среди сородичей одним из самых приспособленных к различным морским экосистемам, от полярных морей до субтропических вод. Специалисты знают, что почти каждый вид китообразных в своем роде уникален и при ближайшем знакомстве поражает наблюдателя особыми, лишь ему свойственными, чертами биологии. В этом плане первое место среди усатых китов принадлежит серому киту, соперничать с ним может, наверное, только горбач.
ОБЩАЯ ХАРАКТЕРИСТИКА
Серого кита называют «живым ископаемым», потому что он сохранил целый набор признаков далеких пред-
ков: десятки чувствительных волосков-вибрисс на морде, подвижные голова и шейный отдел позвоночника, в котором позвонки не срастаются, крупные носовые кости черепа, увеличенные тазовые кости и многое другое. Из всех китов именно этот кит наиболее тесно связан с берегом — он размножается в теплых мелководных лагунах и кормится на небольших глубинах, захватывая ртом донный грунт со всеми живущими в нем и на нем организмами.
Серый кит уступает гладким и полосатым китам в величине и красоте. Его длина редко превышает 14 jw, причем самки в среднем на 0,3-1 м больше самцов. Вес взрослых китов составляет от 20 до 35 т. Тело, менее стройное, чем у полосатиков, окрашено в серый с буроватым оттенком цвет с многочисленными неясными светлыми пятнами различной формы. Кожа обычно покрыта бесчисленными колониями эктопаразитов, которые представлены усоногими раками в твердых раковинах-капсулах и китовыми вшами. Вместо изящного спинного плавника-фина природа наградила серого кита шишковатым хвостовым гребнем; он начинается с крупного шишковидного утолщения, а за ним по хвостовому стеблю располагаются от 6 до 14 более мелких бугров. Кожные складки на брюшной стороне отсутствуют, однако в области горла проходят 2, реже 3-4 глубокие борозды, достигающие в длину около 2 м.
Из всех усатых китов только у этого кита ротовая щель делит голову почти на две равные половины — верхнюю и нижнюю. Полость рта низкая; толстые светло-желтые пластины китового уса с грубой бахромой высотой всего 20-40 см. В каждой половине верхней челюсти у самцов и самок насчитывается 130-180 таких пластин, расположенных реже, чем у других усатых китов.
Серый кит, добытый и доставленный в поселок Сиреники китобойцем «Звездный»
Спина серого кита, вынырнувшего за порцией воздуха. Хорошо видны шишки хвостового гребня и открытые для вдоха парные дыхательные отверстия
Фонтан серого кита внизу раздвоенный, наверху — кучевидный; высота фонтана — 1,5-3 м.
В общем, красавцем серого кита никак не назовешь, да и обычная скорость его передвижения невелика, всего 7-9 км в час, и, хотя плавает он чуть быстрее гладких китов, но не может состязаться со стремительными полосатиками. Правда, напуганные серые киты способны развивать скорость до 18 км в час.
ГДЕ ЖИЛИ И ЖИВУТ СЕРЫЕ КИТЫ
Сравнительно недавно (несколько веков назад) серые киты обитали в Северном полушарии у побережий Тихого и Атлантического океанов. В Атлантике,
на восточном берегу Северной Америки, в Чесапикском заливе, найдены самые старые костные остатки американских атлантических серых китов, их возраст около 10 тыс. лет. А самые молодые, из Саутгемптона, что рядом с Нью-Йорком, отнесены к периоду ранней колонизации Америки, т.е. к XVI в. В Европе кости серых китов обнаружены в Исландии, Швеции, Голландии и на берегах пролива Ла-Манш. Полагают, что европейское атлантическое стадо как таковое исчезло в эпоху викингов, но небольшое количество серых китов, возможно, продержалось до XVII—XVIII вв. Трудно сказать, составляли ли атлантические серые киты Америки и Европы единое стадо или это были две самостоятельные популяции.
В Тихом океане до наших дней сохранились два стада. Первое, охотско-корейское, размножается в теплых мелководных заливах Южной Кореи и Китая и проводит лето в водах Приморья, Сахалина и Охотского моря. По последним данным, это стадо насчитывает сегодня около двухсот особей. Оно было почти истреблено массовым промыслом, начавшимся в 1899 г., и специалисты полагали, что охотско-корейские киты вот-вот исчезнут. Однако опытные китобойные капитаны Владивостока постоянно отмечали присутствие серых китов в заливе Петра Великого. Биологи поначалу не верили этим сообщениям, но потом сами начали встречать этих китов у берегов Сахалина, Курильских островов и в Охотском море.
Не так давно серые киты обитали и в прибрежных водах Японии, но жители этого островного государства на протяжении веков ловили их сетями в мелководных бухтах, а в наше время добывали с современных китобойных судов, пока не истребили полностью.
С 1980-х годов японские специалисты начали сообщать о появлении единичных серых китов у берегов своей страны. До сих пор точно не определено, были ли японские киты частью охотско-корейского стада или представляли самостоятельную популяцию.
История еще одного тихоокеанского стада, чукотско-калифорнийского, также полна драматизма, но, к счастью, не столь трагична. Неумеренный, сначала американский, а затем советский, коммерческий промысел XIX—XX вв. поставил его на грань исчезновения, но принятый в 1946 г. полный запрет на коммерческий промысел этого вида спас стадо. В настоящее время калифорнийские серые киты восстановили свою численность, которая, по разным оценкам, составляет от 15 до 26 тыс. особей. Они вновь стали обычными животными в водах Северной Америки и Северо-восточной Азии, и охота на них разрешена сейчас только коренным жителям Чукотки и Аляски. Вот об этих китах и пойдет речь дальше.
ВЕЛИКИЕ ПУТЕШЕСТВЕННИКИ
Медлительные и неизящные серые киты — великие путешественники. Дважды в год чукотско-калифорнийское стадо отправляется в миграционный путь, который по своей протяженности вполне сравним с перелетами птиц. В конце декабря киты приходят с севера в субтропические лагуны Калифорнии, удобные для спаривания и рождения детенышей, а в конце февраля-начале марта вместе с недавно родившимися китятами начинают обратный путь на север, стремясь к богатым мелководным пастбищам Берингова и Чукотского морей.
Миграционный путь чукотско-калифорнийского стада серых китов из субтропических лагун полуострова Калифорния к северным пастбищам Берингова и Чукотского морей.
1 — лагуна Ойо де Либре (Скэммона);
2 — лагуна Сан Игнасио;
3 — лагуна Магдалены.
Странствуя между кормовыми полями, расположенными в арктических и субарктических водах, и
зимними районами размножения в субтропических водах, калифорнийские серые киты преодолевают ежегодно свыше 18 тыс. км, расстояние, намного превышающее тот путь, который проходит за это же время любой другой усатый кит. Американские специалисты Д. Райс и А. Вулмен, занимавшиеся жизненным циклом серого кита, особо подчеркивают, что такой огромный миграционный путь, покрывающий по широте 50° (!), подвергает животных действию постоянно изменяющихся факторов внешней среды, в особенности сильных колебаний температуры и фотопериода.
Весенняя миграция серых китов очень растянута во времени и пространстве. Покинув теплые калифорнийские воды, киты регулярно проходят через некоторые проливы Алеутских островов, например, через пролив Унимак, затем большая часть стада продолжает движение на север вдоль побережья Северной 'Америки. В регион Берингова пролива киты приходят по мере взлома и разрежения льдов. Однако первые мигранты появляются у нас раньше в постоянных прибрежных полыньях и в районах с подвижным ледовым режимом. Они используют ветви мощного Поперечного течения, направленного от Северной Америки к берегам Северо-восточной Азии. Так, в конце апреля-начале мая серые киты одновременно идут в две акватории, разделенные обширным ледовым пространством: к побережью Корякии, между мысами Хатырка и Наварин, и к юго-восточной оконечности Чукотского полуострова, в Сиреник-скую полынью и бассейн Чирикова.
Возможно, есть еще один, южный, миграционный путь — вдоль Алеутской гряды и Командорских островов. Нельзя исключить и рассеянную миг
рацию серых китов напрямую через Тихий океан. Л.М. Вотрогов пять раз встречал их в открытом океане между 35° и 50° северной широты, в местах, где глубины превышают 2000 м.
Во время осенней миграции отход китов на юг начинается с середины октября и заканчивается во время льдообразования. Мне не раз доводилось видеть этих животных, спокойно пасущихся возле бухты Провидения в середине декабря, а охотники села Сиреники наблюдали единичных серых китов в Си-реникской полынье даже в январе-марте. Примечательно, что первыми приходят и первыми уходят беременные самки, а самцы могут задерживаться на кормовых полях, пока позволяет ледовая обстановка.
РАСПРЕДЕЛЕНИЕ НА МЕСТАХ НАГУЛА
Излюбленные места летне-осеннего обитания многих китов, среди которых преобладают беременные и кормящие самки с молодняком, расположены в регионе Берингова пролива — одном из самых высокопродуктивных районов Мирового океана. При этом наиболее ценными пастбищами являются его холодноводные участки, омывающие берега России. Именно сюда и направляется основная масса китов, а в более теплых водах Аляски кормится значительно меньше животных, которые обычно не идут дальше мыса Барроу, разделяющего моря Чукотское и Бофорта.
В водах России основная масса китов собирается в двух областях: южная (Корякское побережье) — от мыса Олюторский до мыса Барыкова и лагуны Тым-на в Беринговом море, северная (Чукотский полу-
Карта районов концентрации серых китов (1-22) у Корякского побережья и Чукотского полуострова. Составлена Л.М. Вотроговым в 1979 г. Указаны национальные поселки, население которых традиционно занимается китобойным промыслом
остров) — от залива Креста в Беринговом море до мыса Сердце-Камень и лагуны Нешкан в Чукотском море. Раньше думали, что серые киты в Чукотском море не встречаются западнее лагун Нутаугье и Тэн-кэргын-пылъгын (мыс Шмидта) и севернее 69 параллели, хотя и были отдельные указания, что они могут по разводьям среди дрейфующих льдов достигать о. Врангеля.
Однако не так давно российские исследователи обнаружили серых китов в прибрежных водах пролива Лонга и восточной части Восточно-Сибирского моря, от мыса Якан до мыса Биллингса. Кроме того, в Чукотском море были выявлены большие скопле
ния серых китов, от 400 до 2 тысяч голов, как возле берега, так и на удалении около 200 км от него. Полагают, что в результате роста численности чукотско-калифорнийского стада произошло расширение области его распространения на запад, вдоль арктического побережья России. Есть достоверные сведения о том, что в малоледные годы серые киты могут достигать даже устья реки Колымы.
Серые киты распределяются на мелководье, как правило, придерживаясь пятидесятиметровых глубин и редко удаляясь от берега далее 35-50 км. В прибрежной полосе они выбирают места с поверхностной соленостью воды ниже 30%,с ветровым перемешиванием, проникающим до дна, и с хорошим насыщением воды кислородом, но даже и в таких местах они сосредотачиваются только там, где концентрация донных организмов (бентоса), особенно мелких ракообразных, достаточно высока.
Для южной области и основной части северной области распространения серых китов Л.М. Вотро-говым в 1979 г. впервые была составлена карта районов концентрации этих животных на полях нагула.
Корякское побережье
Первые советские исследователи китообразных, Б.А. Зенкович и А.Г. Томилин, специально отмечали, что в Беринговом море летуют животные всех возрастов, как самки, так и самцы, но существует возрастное разделение стада — молодые киты скапливаются главным образом у Корякского побережья, от бухты Глубокая до мыса Наварин, а взрослые самки и самцы пасутся севернее, в водах Чукотского полуострова. Вот как описывает свое первое посещение лагун Корякского побережья Б.А. Зенкович.
«За мысом Наварина идем близко от берега, но не слишком — на картах береговая черта пунктиром показана, а это значит - глубины не обследованы... Перед нами открывается довольно широкая лагуна, идущая в юго-западном направлении и окруженная отвесными скалами с массой птиц. Птицы везде: и на берегу, и на скалах, и ... на спинах китов! И не только на спинах, но и на боках, так как вот два, нет три, еще больше! лежат на мели и птицы разгуливают по этим тушам. А в лагуне десятки фонтанов.
Бот тихо покачивается на малой волне, и нас подносит к мелякам, на которых застряли киты. Воды там метр-полтора, но много камней. Моросит мелкий дождь... Киты лежат неподвижно, но дышат и регулярно пускают фонтаны, изредка шевелятся — видно, не очень удобно на камнях. А чайки разгуливают по ним, расклевывают раковины паразитов и ловко хватают крупных «китовых вшей»... Совсем рядом с нами всплывают туши серых китов, они совершенно спокойны и не обращают на нас внимания. Но сколько здесь небольших китов, прямо китят, а не китов. И почти не видно крупных китов, какой-то детский сад.
Вот совсем рядом с нами высунулся чуть не весь (именно высунулся, а не выпрыгнул!) небольшой кит. Мне показалось, что он совершенно свободен от раковин паразитирующих усоногих раков. Не в пресной ли воде дело? Ведь многие морские организмы совсем не терпят пресной воды... Я проверил свои предположения, выяснилось, что все китовые паразиты — и коронулы1, и китовые вши погибали через полминуты-минуту после того, как я опускал
1 так называют паразитирующих на коже серых китов усоногих раков-криптолепасов
их в пресную воду. А в бухточках и лагунах вода полуопресненная, а часто и совсем пресная, так что киты, действительно, избавлялись от мучающих их паразитов...
Начинался прилив, и сидевшие на мели киты постепенно освобождались от своего плена. Мы видели, что они нисколько не пострадали, так как тут же начали нырять и питаться. А некоторые киты застывали надолго неподвижно и очень редко пускали фонтаны. Мне казалось, что они спали. Осторожно, на веслах, подошли к одному киту, ткнули его багром, и он никак на это не реагировал.
Удивительно только, что в эти лагуны редко заходили большие серые киты. Из добытых нами в этом районе серых китов старых было только 15 процентов. И все они были чистенькие, совершенно без паразитов. Действительно, санаторий для китов — обилие пищи, избавление от паразитов и всяких ран и болячек. А тишина какая...»2.
В 1979 г. капитан Вотрогов и я наблюдали сходную картину чуть южнее мыса Наварин, в устье реки Пикка. Около 40 серых китов среднего размера стояли бок о бок в маленькой опресненной лагуне. Был довольно сильный туман, шел дождь. Чайки сновали по спинам китов, которые, шумно дыша, пускали частые фонтаны и держались головами к устью реки. 23-26 августа 1983 г. при сплошном вертолетном обследовании лагун, бухт и заливов от мыса Африка на восточном берегу полуострова Камчатка до лагуны реки Катанваам в Анадырском заливе я насчитала свыше 400 серых китов; многие из них во
2 Б.А. Зенкович. «Вокруг света за китами». М. 1954 г., стр. 192-195
время отлива оставались на отмелях, совершенно не боясь обсыхания. Киты были разной величины, но в основном средних размеров.
Чукотский полуостров
Наблюдения последних лет и обработка результатов аборигенного промысла серых китов показали, что, в настоящее время у берегов Чукотского полуострова собираются преимущественно самки и молодые животные; последние занимают в основном прибрежное мелководье, а крупные киты держатся мористее, до глубин 50-60 м.
Наши совместные с капитаном Вотроговым наблюдения показали, что распределение животных по районам концентрации меняется не только из года в год, но и по месяцам в течение одного сезона, поскольку киты непрерывно перемещаются между полями нагула. Границы районов лучше определяются с моря, а не с берега - у береговых наблюдателей может возникнуть впечатление непрерывного потока китов, идущих вдоль побережья. А в море эти границы легче увидеть по разнице в поведении «проходных» и кормящихся китов.
У побережья Чукотского полуострова Л.М. Вотрогов нашел районы, где постоянно встречаются мелкие (молодые) киты: у лагуны Аччен и устья реки Курупка (районы 5 и 6), возле мыса Кыгынин острова Аракамчечен (район 9), в Мечигменском заливе, включая прилегающую к нему часть Мечигмен-ской губы (район 10), а также севернее мыса Нуня-мо — возле мысов Чини, Серый, Литке, Верблюжий (район 14). Биологи из Института проблем экологии и эволюции (Москва) определили радиоуглеродным методом возраст черепов маленьких серых китов в
древнем селении Масик на южной косе, отделяющей Мечигменскую губу от Мечигменского залива. Оказалось, что самки китов приводили и продолжают приводить сюда детенышей на протяжении последних полутора-двух тысяч лет.
КАК ПИТАЮТСЯ СЕРЫЕ КИТЫ
Среди усатых китов серый — единственный бентофаг, что означает «питающийся донными организмами». Поэтому он очень привязан к мелководьям, где
Голова серого кита.
Обратить внимание на то, что более узкая верхняя челюсть «вкладывается» в более широкую нижнюю челюсть. Видны пластины китового уса и колонии рачков-криптолепасов на голове и нижней губе кита
Череп серого кита. Хорошо видно «вложение» верхних челюстей в нижние
бентосные сообщества богаче, а корм добывать легче. В отличие от других морских млекопитающих серый кит кормится самым необычным способом: он поворачивается на бок так, чтобы рот был парал-* лелен морскому дну, и, втянув свой огромный (свыше тонны весом) язык, всасывает одной стороной
рта все, что находится на дне. Любопытно, что среди китов, как и среди людей, есть «правши» и «левши»: одни предпочитают питаться, лежа на правом боку, другие — на левом, что хорошо видно по степени стертости китового уса с каждой стороны пасти.
Наибольший объем пищи, который кит засасывает таким способом за один раз, составляет около 340 л. Естественно, что в рот попадает все, что находится на дне, — водоросли, песок, ил, галька. Порой из желудка добытого кита мы вычерпывали до 3-4 ведер грунта. Наполнив рот, кит выталкивает языком воду, а «сухой остаток» проглатывает. Суточное потребление корма взрослым серым китом, по данным А.Г. Томилина, составляет 380-2500 кг, по данным других исследователей, — 1100-1200 кг.
Список кормовых объектов серого кита включает около 60 видов мелких донных рачков-амфипод и 80-90 видов других беспозвоночных, а также водоросли и значительное количество песка, ила и гальки, которые присутствуют в желудках китов почти всегда. У побережья Чукотки основу питания китов составляют именно рачки-амфиподы, причем самыми распространенными объектами питания являются 18 видов из этой группы. Интересно, что в пределах районов, где держится основная масса серых китов, скопления этих ракообразных, видимо, образуют «кормовые пятна», чередующиеся с иными сообществами донных организмов, которыми питаются другие морские животные. Так, в районе, прилегающем к моржовому лежбищу острова Аракамче-чен, серые киты и моржи пасутся в непосредственной близости, нисколько не мешая друг другу.
Анализ проб питания китов, добытых китобойцем «Звездный», не показал возрастной и половой разницы по объектам питания, но выявил четкое различие по основным видам амфипод между южными (от острова Коса Мээчкин до бухты Провидения в Беринговом море) и северными (от мыса Нунямо в Беринговом море до мыса Сердце-Камень в Чукотском море) районами питания. Было установлено, что акватория между бухтой Провидения и мысом Нунямо характеризуется наиболее разнообразным составом донных беспозвоночных и киты там держатся, пока не подойдут тяжелые зимние льды.
В предыдущем разделе уже говорилось о том, что серые киты, питаясь, непрерывно перемещаются между кормовыми районами и задерживаются в каждом лишь на несколько дней.
«ПАХАРИ» МОРСКОГО ДНА
В конце 1970-х годов американский геолог К. Нельсон осматривал дно северо-восточной части Берингова моря в районе бассейна Чирикова. Он должен был оценить геологические факторы, потенциально опасные для установки в открытом море нефтяных вышек и трубопроводов. Нельсон и его помощники изучали дно с помощью ультразвукового локатора бокового обзора. Для записи рельефа судно буксирует под водой аппарат, который посылает в направлении дна расходящийся пучок звуковых импульсов и принимает отраженные сигналы. Принятые сигналы передаются на борт судна, где по ним самописец рисует сонограмму — картинку, напоминающую аэрофотоснимок поверхности земли.
Рассматривая такие сонограммы, Нельсон и его коллеги столкнулись с удивительным явлением — помимо ожидаемых опасностей на дне мелководных участков были видны ямы и борозды, которые никак нельзя было отнести за счет известных геологических процессов. И ученые сделали смелое предположение — это результат деятельности крупных морских млекопитающих, серых китов и моржей, которые в большом количестве приходят сюда кормиться.
К началу 1980-х годов американские специалисты убедились, что бассейн Чирикова действительно является в Беринговом море главным районом питания серых китов и моржей. Они попытались доказать, что ямы и борозды на сонограммах Нельсона сделаны этими животными во время кормления. Ученые решили определить количество пищи, которое дает бассейн Чирикова за год, а также выяснить, как влияет пищевая активность морских млекопитающих на состав верхнего слоя морского дна и на пищевые (трофические) цепи в северо-восточной части Берингова моря.
Сонограммы и пробы грунта показали, что дно бассейна Чирикова покрыто двухметровой толщей песка, сформировавшегося в конце последнего ледникового периода (12—10 тысячелетий назад). Когда лед растаял, уровень моря повысился, и оно затопило часть суши. В этом древнем осадочном слое среди роющих донных животных преобладают рачки-амфиподы. В песке они легко строят трубчатые ходы, которые пропитывают слизью; благодаря этому песок скрепляется в твердые пласты, не размываемые водой. Северный и западный края бассейна Чирикова выстланы в основном гравием и песком ледникового происхождения, а восточный край — залив Нор-110
тона — смесью ила и глины толщиной в несколько метров, поскольку здесь накапливаются выносы Юкона, великой реки Аляски.
Для района длиной около 4500 км геологи составили точную карту всех ям и борозд, предположительно созданных животными, и затем сопоставили сонограммы с прямыми наблюдениями аквалангистов за способами питания китов и моржей. Все предположения подтвердились.
Исследователи измерили более тысячи таких ям и сравнили свои данные с размерами пасти серого кита. Оказалось, что ширина раскрытого рта и длина одной стороны закрытого рта примерно равны и составляют 2,1м. Ямы же в среднем имели 2,5 м в длину, 1,5 м в ширину и 10 см в глубину. Ясно, что такую яму может сделать кит, когда он держит голову параллельно дну и всасывает обитателей дна одной стороной своей пасти. Иногда ямы сгруппированы очень характерно, их расположение соответствует последовательным актам заглатывания пищи китами, которые лежат на боку и не всплывают наверх за порцией воздуха. Ямы сосредоточены внутри района площадью 22 тыс. кв. км, главным образом в центре бассейна Чирикова и у южной оконечности острова Святого Лаврентия. В пределах самого бассейна эти ямы занимали 18% поверхности дна.
Наконец-то биологи получили уникальную возможность узнать, сколько пищи съедают серые киты на одном из важнейших кормовых полей. Суммарная площадь обнаруженных ям составляла 1200 кв. км из 22000 кв. км обследованной площади бассейна Чирикова, где каждый квадратный километр дна производит в год 171 т бентоса. Потребность в пище всего
чукотско-калифорнийского стада американские исследователи оценили примерно в 3168 млн. т (16 тыс. китов и 180 дней летне-осеннего сезона питания). Бассейн Чирикова дает китам 250 тыс. т, или 6,5—15% требуемой пищи, хотя площадь его составляет всего 2% от общей площади северных пастбищ этих животных. Все это наводит на размышления об особой экологической роли серых китов и моржей в регионе Берингова пролива.
Американские специалисты подсчитали, что за один сезон серые киты «перелопачивают» около 172 млн. т осадочного материала, а это почти в три раза больше количества илисто-глинистых осадков, выносимых рекой Юкон (60 млн. т в год). По самым осторожным оценкам, 200 000 моржей перекапывают еще примерно 100 млн. т грунта. Выходит, что вместе эти животные — основные борцы против заиления бассейна Чирикова выносами Юкона. В первую очередь именно благодаря серым китам песчаное дно бассейна Чирикова не заносится илом и глиной и остается прекрасным местом для обитания амфипод, любимой их пищи.
Серые киты не только поддерживают песок бассейна «в чистоте», но и сами обеспечивают себя кормом на следующие сезоны. Кит засевает молодью рачков взрытое им же дно — взрослые амфиподы удерживаются китовым усом в полости рта, а мелкие проскакивают сквозь его жесткую бахрому и падают обратно на дно. Киты и сеют амфипод, и подкармливают их. Вместе с поднятым осадком в воду попадают ранее захороненные под ним органические вещества, которые стимулируют рост планктона — основной пищи амфипод. Рачки-амфиподы являются пионерской (начальной, первой) группой беспозво
ночных, осваивающих геологические структуры дна. Они бурно размножаются и быстро укрепляют песок нарушенных участков дна своими трубчатыми ходами. Через сезон киты «снимают» урожай, снова «готовят» поле и «засевают» его.
Постоянное перепахивание дна кормящимися китами и моржами — непременное условие устойчивости морских экосистем региона Берингова пролива. Люди же могут грубо нарушить экологическое равновесие — ведь песчаный слой на морском дне очень тонок и современные геологические процессы не восстанавливают его; это может сделать только новый ледниковый период.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ
Инстинкт размножения заставляет серых китов покидать богатые пищей, но холодные арктические воды. Не питаясь, а живя за счет накопленных в Арктике запасов жира, киты проводят полтора-два зимних месяца в Мексике, у берегов полуострова Калифорния — именно там имеются подходящие условия для родов. Ведь китенок появляется на свет без подкожного слоя жира, и ему нужна очень соленая и очень теплая вода, чтобы не утонуть и не замерзнуть. Берег полуострова Калифорния оторочен голыми серыми дюнами, изрезан лабиринтом проливов и проходов. Прозвище «пустынные киты» пристало к серым китам за пристрастие именно к таким уединенным уголкам среди безжизненных песков.
Еще не так давно серые киты размножались и севернее, в удобных бухтах американского штата
Калифорния — Сан-Франциско и Сан-Диего. Но там выросли огромные города с большими современными портами, где вода столь грязна, а судов столь много, что об удобстве для тихих китов не может быть и речи. Для них остались только немногие лагуны мексиканской Калифорнии, которые правительство Мексики охраняет с похвальной строгостью.
Лагуны
Экспедиция известного морского натуралиста Ж.-И. Кусто, специально изучала в калифорнийских лагунах, прежде всего в лагуне Скэммона, образ жизни серого кита. По поведению животных было видно, что они прекрасно знают этот район. Много лет приходя сюда, они помнили «потайной ход», ведущий в залив, а приплывшие первый раз следовали за старшими, более опытными. По наблюдениям Кусто, будущие матери ищут уединения там, где повышенная соленость воды обеспечивает большую плавучесть детенышей. Правда, многие китихи рожают и вблизи пролива.
По данным американского зоолога Т. Уокера, самка серого кита рожает единственного детеныша на мелководье, лежа на спине, и тотчас выталкивает новорожденного на поверхность, чтобы он мог сделать свой первый вдох. Тело маленького китенка мягкое, словно резиновая губка, плавать он еще не может и, даже если работает хвостом, все равно не двигается с места. Да и на воде сам детеныш не держится — его удельный вес пока еще слишком велик. Подкожный жировой слой почти полностью отсутствует, а недостаточно развитая грудная клетка не обеспечивает нужное количество воздуху в легких 114
Семи-восьмимесячный эмбрион серого кита, самка длиной 247 см. На голове и нижней челюсти видны закладки вибрисс; пластины китового уса еще отсутствуют
для плавучести детеныша. Поэтому мать должна неусыпно смотреть за новорожденным; малыш крутится в воде, словно бочка, он лежит то на спине, то на боку, но мать все время поворачивает его животом вниз и следит, чтобы голова была на воздухе.
Длина новорожденного серого кита 3,5-5,5 л/, в среднем 4,5 м при весе 650 кг. Китенок растет очень быстро на питательном материнском молоке, жирность которого достигает 53%. По расчетам А.Г. Томилина, в среднем детеныш потребляет в сутки 36,6 кг молока, а за весь период молочного кормления около 5500 кг. Месяца через три, достигнув 6-7 м в длину, он будет достаточно крепок, чтобы приступить к первому в своей жизни походу на север. Конечно, если доживет до той поры.
Цикл размножения
Цикл размножения серых китов можно представить следующим образом. В стаде есть непрерывно размножающиеся самки, те, у кого сразу же за родами одного детеныша наступает новая беременность. Считают, что таких самок в стаде не менее половины, но кроме них есть и другие, у которых между рождением детеныша и новым зачатием проходит один год и более. Несомненно, за счет этих особей период спаривания может быть несколько растянут. Так, прямые наблюдения ряда исследователей и морских охотников Чукотки говорят о возможности спаривания серых китов в августе-сентябре в Беринговом море.
По-видимому, весь цикл размножения серых китов стабилизируется наличием подходящих условий для родов лишь в 1,5-2 зимних месяца в мелководных лагунах Калифорнии. Беременности, которые завершаются родами не в этот сравнительно короткий период, должны оканчиваться гибелью детеныша из-за неподходящих условий для родов и исключением самки из очередного цикла размножения.
Важно отметить, что часть лактирующих (кормящих) самок могут быть одновременно и беременными. Точно учесть их количество трудно, поскольку существует запрет на добычу самок с детенышами и такая добыча во время аборигенного промысла в водах России происходит лишь случайно. Однако за время наблюдений на Чукотке в 1965-1978 годы этих самок было встречено более 20%, что говорит о сравнительно широком распространении среди китов явления одновременной лактации и беременности.
Самцы серых китов в массе становятся половозрелыми при длине 11,5 м, самки — при длине
12,0 м. Есть основания полагать, что с возрастом у самок промежутки между беременностями увеличиваются. Спаривание китов длится с декабря по март, то есть самки спариваются вскоре после родов. Длительность беременности около года. Как и у всех больших животных, самки серых китов рожают одного детеныша.
История лагуны Скэммона
Для серых китов окаймленные манграми безлюдные бухты (или лагуны) Скэммона, Сан Игнасио и Магдалены в мексиканской Калифорнии — особый, чудом сохранившийся мирок, который едва не исчез в середине XIX века. Вот история трагических событий, случившихся в бухте Ойо де Либре, после которых она получила свое второе название — лагуна Скэммона.
Огромные просторы этой лагуны открываются сразу за узким и извилистым проливом, который вдается в глубь полуострова миль на тридцать. Уголок этот укрыт так хорошо, что самые ретивые китобои не знали о нем вплоть до середины XIX в.
Только в 1852 г. пролив был найден капитаном Ч. Скэммоном на бриге «Мери Хелен». Издали увидев фонтаны, он решил войти в лагуну, чтобы выяснить их происхождение, и обнаружил там огромные скопления китих с новорожденными китятами. Девять лет гарпунеры Скэммона сотнями били китов и наполняли тысячи бочек китовым жиром. Естественно, они строго хранили тайну своей сокровищницы, никто, кроме них, не знал о проливе, ведущем в самую благодатную бухту Калифорнийского полуострова, ныне известную как лагуна Скэммона. На десятый год конкуренты, снедаемые завистью
и полные решимости урвать свою долю добычи, наладили слежку за удачливым капитаном. В конце концов они проследили, куда водит свой бриг Скэм-мон, и тогда началась настоящая бойня. Как записал сам капитан в своем дневнике, «в сезон 1858— 1859 г.г. китоловами кишели не только лагуны и заливы — сплошная череда китобойных судов протянулась вдоль всего побережья: от Сан-Диего на юг до мыса Сан-Лукас».
В погоне за добычей китобои использовали материнский инстинкт самок — они обычно открывали огонь по детенышам, и тотчас матери шли в атаку на вельботы, а гарпунерам оставалось только бить их в упор. Но, несмотря на избиение, киты продолжали ежегодно приходить в лагуны Калифорнии, у них не было выбора, ведь только там китихи могли родить и выкормить детенышей.
Промысел серых китов достиг такого размаха, что к началу XX в. вид был на грани полного истребления. Его спасли международные соглашения, но почти полвека понадобилось, чтобы чукотско-калифорнийское стадо серых китов восстановилось после бойни, учиненной коммерческими китобойными судами.
Ныне серым китам угрожают множество опасностей, связанных с сильной индустриализацией юго-западного побережья Северной Америки. По мнению некоторых специалистов, серый кит — самый древний из сохранившихся до наших дней китов, настоящее живое ископаемое — обречен на вымирание, если будут затронуты последние из трех-четырех калифорнийских лагун, где он может размножаться. И это последнее убежище серых китов. В отличие от бухт американской Калифорнии здесь воды еще соне
храняют первозданную чистоту, если не считать нескольких заболоченных участков.
ПОВЕДЕНИЕ СЕРЫХ КИТОВ
Самые умные усатые киты
Многие специалисты и китобои считают серого кита самым умным из всех больших китов. Он умеет уходить от преследования, резко меняя курс, выныривая там же, где нырнул, плывя зигзагами или заходя на мелководье. При преследовании серые киты увеличивают свою обычную скорость до 18 км в час. Взрослые киты и детеныши не боятся льда. Они могут держаться в полыньях среди паковых льдов, мелкобитых сильно дрейфующих ледяных полей и у кромки полярных льдов. Но иногда такое поведение заканчивается трагически — киты попадают в ледяной плен и погибают. Так случилось в октябре
Один из трех серых китов в ледовом плену у мыса Барроу. Октябрь 1988 г.
1988 г. у мыса Барроу. Три кита оказались запертыми в небольшой полынье и, несмотря на все попытки людей спасти их, один за другим погибли.
Серые киты в водах Чукотки мирно уживаются с другими животными. На местах нагула они могут кооперироваться с горбачами и моржами. Например, возле крупных моржовых лежбищ на острове Аракамчечен, косе Руддера и острове Ратманова киты и моржи питаются в непосредственной близости друг от друга, и наблюдатели не отмечали при этом агрессивного поведения ни с той, ни с другой стороны. А среди многих сотен черепов серых китов в старом поселке Масик нами обнаружено несколько десятков черепов молодых горбачей. Это свидетельствует о том, что молодняк двух видов китов когда-то пасся вместе.
По отношению к людям серые киты ведут себя так же дружелюбно. В лагунах Калифорнии туристы гладят новорожденных и подросших малышей, играют с ними, и матери-китихи не препятствуют такому общению. Три кита, затертые льдами у мыса Барроу, тоже позволяли людям гладить себя. Порой серые киты наблюдают за людьми в лодке, аккуратно поднявшись над водой буквально в 1,5-2 м от борта. Именно так сделал один кит средних размеров, когда мы вместе с охотниками из села Сиреники проходили на байдаре мыс Кригуйгун. Около 20 мин. кит сопровождал байдару, выставившись из воды и двигаясь вместе с ней. К нам он был повернут боком, и его маленький глаз внимательно разглядывал все, что было в байдаре, в том числе и нас. Все это время нам казалось, что мы стоим на месте и смотрим друг на друга. Потом кит опустился под воду так же тихо, как вышел из нее.
Серые киты спокойно относятся к судам, от которых уходят лишь при настойчивом преследовании. С лежа-120
Различные позы серого кита.
Вверху: слева — серый кит наблюдает за происходящим на поверхности воды и берегу; справа — серый кит, как и другие усатые киты, любит выпрыгивать из воды и падать на спину. Внизу: серый кит погружается в воду. Часто именно в этот момент он издает звуковые сигналы для связи с сородичами
щего в дрейфе китобойца «Звездный» наблюдали, как пасущиеся животные дрейфовали по течению с той же скоростью, что и судно, причем не было заметно, что киты обращают на него особое внимание — они держались от китобойца на расстоянии 300 м. Если дистанция между китами и каким-либо судном становится меньше, то животные только отходят в сторону, но остаются на том же месте, а при большем расстоянии они вообще не обращают на судно внимания.
Группировки и социальное поведение
У берегов Чукотки серые киты обычно держатся небольшими группами по две-четыре особи, либо в одиночку и лишь иногда образуют значительные скопления. Одиночные животные (часто беременные самки) составляют до 44-65% от общего числа встреченных китов.
Почему же так много одиночек? В поисках ответа на этот вопрос Л.М. Вотрогов и я решили послушать, не переговариваются ли они между собой. Преодолев ряд технических трудностей, главная из которых — избежать посторонних шумов, мы убедились, что животные действительно «говорят». Пока киты держались тесной группой, бок о бок, нам не удавалось услышать их сигналы. Как только они
Одиночный серый кит выходит на поверхность для вдоха. Обратить внимание на характерную форму фонтана и белые пятна, оставшиеся от раковин эктопаразитов, рачков-криптолепасов
расходились на 800—1200 м и более, гидрофоны регистрировали редкие и короткие низкочастотные сигналы — киты «окрикивали» друг друга. Но не каждый кит отвечает голосом или поведением на звуковые сигналы сородичей, хотя, вероятно, знает, где они находятся и что делают.
Прекрасное подтверждение тому, что часть одиночек не теряют связи между собой, мне удалось получить во время работы на Корякском побережье в июле 1983 г. Целый день я наблюдала с берега за десятью китами, пасущимися поодиночке. Они рассеялись очень широко, и фонтаны самых дальних с трудом различались в бинокль. Наблюдатель с проходящего мимо судна безусловно оценил бы их как одиночных животных. Но во время длительного наблюдения было хорошо видно, что при всех поворотах и ныряниях они придерживаются общего направления на север. К вечеру киты сблизились, образовали три парные группы, одну из четырех особей и общим стадом быстро ушли к северу.
Интересно, что небольшие группы кормящихся или передвигающихся серых китов соблюдают свои собственные ритмы дыхания. При питании на глубинах 25-40 м киты выходят на поверхность для небольшой серии вдохов-выдохов, занимающей 5-15 секунд, через 3-4 мин. синхронно или с небольшим опережением друг друга. Удалось заметить, что порядок выхода на поверхность строго закреплен за каждым животным. Так, наблюдая за согласованным выходом одной пятерки китов, можно было уверенно предсказать, что сначала вынырнет «второй» кит, за ним появится «третий», потом одновременно-выйдут «первый» и «четвертый» и в заключение — «пятый», самый крупный. Какое-то время они побу
дут на цоверхности все вместе, а затем в том же порядке уйдут на глубину.
Несколько раз мы видели, как одиночный кит, присоединяясь к группе из трех-шести животных, сбивает ритм дыхания всей группы (киты начинают погружаться на более длительный срок) и уводит с собой напарника. Оставшиеся животные восстанавливают свой ритм, а новая пара вырабатывает свой собственный.
Несомненно, длительное соблюдение единого ритма дыхания, синхронного или с опережением, отражает согласованность поведения животных, их взаимную привязанность друг к другу. По мнению некоторых биологов, синхронное дыхание, точно в такт с партнером, — высший знак полного единения у китообразных. По-видимому, слишком большая сложность поддержания единого ритма дыхания в многочисленных группах является одной из главных причин их кратковременного существования. Группы свыше трех животных обычно непрочны и при преследовании распадаются.
Среди групп пары из взрослых китов наиболее привязаны друг к другу — во время кормежки и на переходах они всегда держатся вместе, часто оказывают партнеру помощь, доходящую иногда до самопожертвования. Следует специально подчеркнуть, что взаимная привязанность самок бывает столь же сильной, как самца к самке. Летом 1979 г., во время охоты китобойца «Звездный», один кит из пары подошел к загарпуненному и пытался поддержать его у поверхности, подводя под него свои хвост и голову. Было решено, что это самец, но оба крупных (длиной около 13 jw) животных при вскрытии оказались беременными самками.
Половой состав групп весьма разнообразен: встречаются пары из самца и самки, двух самок или двух самцов, в тройках бывают две самки и самец или одни самцы. Определить возраст животных в группах в большинстве случаев можно лишь по их размерам. В результате многочисленных наблюдений, проведенных вместе с охотниками Янракыннота, у нас сложилось впечатление, что в июле-августе начинается разделение групп по возрасту: китята отходят от матерей и собираются вместе в наиболее мелководных и богатых кормом районах Чукотки и Корякии.
Материнское поведение
На взаимоотношениях матерей и детенышей серых китов следует остановиться особо. Как полагает Томилин, питаться молоком китята прекращают, когда вырастают до 8, 5 м. Самки сильно привязаны к детенышам, и в прошлом китобои часто пользовались этим; так, например, поступал капитан Скэм-мон. Известны случаи нападения самок на вельботы охотников, ранивших детенышей. Об одном из них мне рассказывали старики Уэлена. Обычно у китов привязанность матери и детеныша обоюдная, но Б.А. Зенкович на основании своих наблюдений высказал мнение об односторонней привязанности самки серого кита к своему детенышу. Он видел уходы сосунков от убитых маток.
В начале августа 1981 г. члены совместной биолого-этнографической экспедиции АН СССР наблюдали у побережья старого поселка Масик трех маленьких серых китов. Они держались целый день на одном месте, то подходя вплотную друг к другу, то расходясь на 50-100 м. Дважды в сутки, примерно в 9 и 20 часов, к ним часа на два подходила пара
крупных китов, которые меняли свой ритм дыхания, подлаживаясь под малышей (выходы на поверхность через 15-45 сек. вместо 1,5-2 мин.). Очевидно, это матери приходили покормить своих китят.
На побережье Корякин в 1983 г. мне довелось наблюдать, как самка завела своего слабого малыша в очень удобную мелководную бухточку и устроила его на отмели, чтобы дыхало было лишь слегка прикрыто водой. Сама она ушла в море и навещала детеныша для кормления два-три раза в сутки. Обычно детеныш подплывает под мать и сосет ее. В этом же случае самка укладывалась боком на мелководье, а детеныш немного сползал с отмели и сосал ее непрерывно 30-60 секунд, затем делал перерыв на 15-20 секунд и снова начинал сосать. Кормление продолжалось около 10 минут. Затем мать начинала возиться с малышом, толкала его головой, заставляла плавать рядом в более глубоких местах, при этом китенок пристраивался у нее над спиной. Все это продолжалось около получаса, потом опять следовало 7-10-минутное кормление, после которого мать и детеныш на час-полтора замирали друг возле друга, выставив дыхала. Наконец, китенок высасывал за 4-6 минут последнюю порцию молока, и мать, немного повозившись с ним на глубине, выталкивала его на отмель и уходила в море. Китенок в ожидании матери спокойно переносил свое обсыхание во время отлива.
Однажды в бухточку зашли три взрослых кита, и один из них согнал малыша с отмели. Тот около трех часов плавал на глубоком месте, но бухту не покинул. Возвратившаяся для кормления самка моментально оценила ситуацию и нанесла обидчику, самому крупному киту, два сильных удара в верхнюю
челюсть, затем подлезла под него, подняв заднюю половину его тела на воздух. Кит без сопротивления подчинился разгневанной матери и, немного покружив по бухте вместе с партнерами, уплыл прочь.
Другие формы поведения
В водах Чукотки питание является, безусловно, главным, но не единственным фактором, определяющим поведение серых китов в летне-осенний период. В частности, возникающие внезапно для наблюдателя скопления китов не всегда, по нашему мнению, связаны с пищей. Например, в районах 4, 12, 14 (см. карту Л.М. Вотрогова) на небольшой площади собираются иногда 200-400 и более китов. В районе 4 такие скопления держатся до 2 суток, в 12 и 14 районах — 3-4 суток. Именно в этих случаях наблюдалось довольно сложное и непонятное групповое поведение китов. Они образуют временные группы, которые быстро распадаются, одиночные киты отбивают себе пару от тройки или четверки сородичей; животные часто переворачиваются на спину, выпрыгивают из воды, ложатся на бок и показывают грудные плавники. В это время они явно не питаются, а заняты взаимоотношениями друг с другом и более половины времени проводят на поверхности. Возможно, такое поведение связано с ложным или настоящим гоном, как полагают А.Г. Томилин и многие морские охотники Чукотки.
Подобные внезапные скопления животных, которые покидают хорошие кормовые угодья и в массе сосредотачиваются на небольших, но чем-то привлекательных участках, известны для многих млекопитающих и птиц. Часто животные склонны периодически менять кормовые участки. Без видимых причин
они могут покидать какой-либо район, хотя пищи в нем не убавилось, Некоторое время они посещают другой кормовой участок, а затем вновь возвращаются на старое место. Наши наблюдения позволяют утверждать, что серые киты ведут себя таким же образом.
Многие исследователи подробно описали специальные подходы серых китов, в опресненные мелководные лагуны или к устьям рек и катанье этих животных в береговой полосе для того, чтобы убить и снять с себя кожных паразитов. Уже рассказывалось, что киты готовы даже обсыхать на отмелых местах и подставлять свои спины и бока прожорливым чайкам, обирающим с них паразитов. По наблюдениям эскимосов Наукана, серый кит способен сойти с мели, если рядом есть глубокое место. Возможность изогнуться дугой, упершись мордой и хвостом в грунт, обеспечивают ему свободные (несрас-тающиеся) шейные позвонки.
Заключая настоящий раздел, можно сказать, что поведение серых китов характеризуется значительной пластичностью и дружелюбным отношением ко всем живым существам, если они ничем не угрожают детенышам. Это позволяет серым китам успешно использовать все многообразие прибрежных акваторий от субтропиков до полярных областей и вступать в контакт с самыми разными обитателями моря — другими китами, моржами, тюленями, птицами и даже людьми.
ПАРАЗИТЫ, КОНКУРЕНТЫ, ВРАГИ
Место серого кита в морских экосистемах региона Берингова пролива определяется его связями с пи
щевыми организмами, паразитами, комменсалами, врагами, а также его уникальным способом кормления, который сохраняет структуру осадочных пород морского дна.
Каждый серый кит — настоящий остров со своим населением, которое обитает и в нем, и на нем, перемещаясь вместе с хозяином на многие тысячи километров. Практически все серые киты в той или иной степени покрыты эктопаразитами. Тысячи усоногих рачков-криптолепасов врастают в кожу кита своими раковинами, закрывающимися особыми двойными крышечками. Удобно устроившись, рачки питаются вместе с китом, подбирая за ним и самостоятельно вылавливая очень мелких животных (зоопланктон). Больше всего они обычно поражают голову, плавники и хвост, но порой встречаются киты, сплошь покрытые раковинами. Издали такие страдальцы выглядят ослепительно белыми, как легендарный кит Моби-Дик американского писателя Мелвилла.
Вторая группа эктопаразитов — три вида китовых вшей-циамид, которые относятся к амфиподам — любимой пище китов. Если усоногие рачки-крипто-лспасы считаются комменсалами, т.е. животными, живущими на других животных или вместе с ними, но питающимися собственной добычей, то циами-ды — настоящие паразиты. Огромными массами они облепляют кита и «грызут» его кожу. Плотное плоское тело и лапки с цепкими крючьями позволяют им намертво прикрепляться к коже серых китов и, питаясь ею, путешествовать на своих жертвах.
Оба сорта кожных паразитов сильно досаждают китам, порой делая их жизнь просто невыносимой. Но киты нашли способ избавления от них, о котором рассказывалось в других разделах: они сначала
убивают своих маленьких врагов в опресненной воде лагун и устьев рек, а затем чешутся о скалы, катаются по песку и гальке в прибойной полосе и снимают с себя по крайней мере китовых вшей. Раковины умерших усоногих рачков сидят слишком глубоко, но постепенно кожа серого кита сама отторгает их, а на местах прикрепления остаются белые непигментированные пятна.
Помимо эктопаразитов серые киты возят внутри себя множество эндопаразитов, представленных паразитическими червями-гельминтами. Большинство из них — те же, что у моржа и морского зайца, которые заражаются ими, как и киты, во время питания донными и придонными организмами.
Эпизодические нахлебники (комменсалы) серых китов — многие морские птицы, чайки, чистиковые, трубконосые. Около 14 видов птиц активно используют пищу, невольно доставляемую им китами. Донные организмы застревают между пластинами усов и в концевой бахроме, и киты выносят их с собою на поверхность во время дыхательных актов. Птицы извлекают добычу и из водоворотов на месте ныряния китов — перемещения китов в толще воды во время питания, их резкие подъемы к поверхности тревожат мелких обитателей моря, которые на какое-то время становятся легкой добычей птиц. Интересно наблюдать, как птицы садятся на воду в самый момент погружения кита, не дожидаясь, пока он уйдет на глубину, или пристально следят за передвижениями китов под водой, видят, где они должны вынырнуть, и спешат туда. Большинство птиц прилетает к пасущимся китам с расположенных неподалеку птичьих базаров, а во второй половине августа появляются у берегов Чукотки колоссальные 130
кочующие стаи тонкоклювых буревестников, гнездящихся в Южном полушарии на островах возле Австралии. Этот вид очень часто кормится с разными китами, за что и прозван китобоями «финвальей птицей».
По-видимому, у серого кита нет никаких естественных врагов, кроме косаток, которые часто преследуют это достаточно тихоходное животное и при нападении прежде всего хватают его за концы грудных и хвостовых плавников, стараются открыть ему пасть и откусить язык. Киты спасаются от косаток, уходя на мелководье, часто даже за прибойную полосу. По данным охотников-наблюдателей, за год на побережье Чукотского полуострова между поселками Энмелен на юге и Нешкан на севере можно обнаружить от 8-10 до 15-20 трупов серых китов, забитых косатками.
ЧИСЛЕННОСТЬ
Сравнение всех известных на сегодняшний день данных по численности серых китов в первой половине XX века убеждает в том, что к моменту запрета коммерческого судового промысла, то есть к 1946 году, численность чукотско-калифорнийского стада не была столь низкой (всего несколько сотен животных), как считали многие зарубежные исследователи.
Известный морской биолог М.М. Слепцов полагал, что к 1946 г. в северо-западной части Берингова моря и в Чукотском море распределялось не менее 1500 серых китов, а к началу 60-х годов поголовье животных увеличилось вдвое и достигло 2500-3000 голов. В 1970-х годах Л.М. Вотрогов ежегодно отме
чал с борта китобойного судна «Звездный» за один китобойный сезон (конец июня-начало ноября) около 2200 серых китов, но, безусловно, часть из них встречалась на пути «Звездного» не один раз. Следует также сказать, что многие кормовые поля серых китов вообще не посещались специалистами или обследовались достаточно бегло и поверхностно. Поэтому можно думать, что в свое время серых китов, как и моржей, просто подсчитывали недостаточно полно.
В связи с этим увеличение количества животных не столь значительно, а темпы прироста всего стада в 1940-60-е годы не столь высоки, как это предполагалось некоторыми биологами вопреки закономерностям размножения крупных млекопитающих. В 1980 году Научный комитет Международной китобойной комиссии (МКК) сообщил, что на 1979/80 годы численность чукотско-калифорнийского стада серого кита составляет 15647 голов и, следовательно, ежегодный прирост с 1967/68 годов примерно равен обычным 2,5%.
Проводимые в течение ряда последних лет подсчеты на местах размножения китов в лагунах Калифорнии дали основания полагать, что общая численность чукотско-калифорнийских китов достигла 26000 особей и сейчас даже начался спад, поскольку количество китов, как предполагают специалисты, уже превысило экологическую емкость мест их обитания.
СТАТУС
В наше время серые киты, принадлежащие к двум уцелевшим стадам (популяциям), охотско-корейскому и чукотско-калифорнийскому, являются эндеми
ками Северной Пацифики. Они занесены в Красную книгу Международного союза охраны природы (МСОП) и Красную книгу России, которая является нормативно-правовым документом нашей страны. Охотско-корейская популяция отнесена к первой категории (находящаяся под угрозой исчезновения); чукотско-калифорнийская — к пятой категории (восстановленная и восстанавливающаяся).
ОХРАНА
Положение чукотско-калифорнийской популяции серых китов в настоящее время можно оценивать как благополучное. Однако, ее существованию сейчас реально угрожают множество опасностей, связанных с сильной индустриализацией побережья Северной Пацифики: факторы беспокойства на местах размножения, резко возросшее загрязнение прибрежных акваторий, лов крабов и рыбы на местах миграций и летнеосеннего нагула.
С 1974 г. китобойное судно «Звездный» ежегодно добывало одного-двух китов, несущих на хвостовом стебле и лопастях обрывки синтетических тросов от рыболовных снастей или глубокие, до 25 см, следы от такой намотки. Судя по состоянию ран, киты носили на себе обрывки снастей не менее трех-пяти месяцев. Следует подчеркнуть, что все киты, добытые с обрывками тросов, были взрослыми крупными животными, сумевшими порвать нейлоновые снасти, тогда как сосунки и молодые особи далеко не всегда могут сделать это. Кроме того, с конца 1980-х годов появились больные киты, мясо которых имеет неприятный запах и не пригодно для питания коренных жителей.
На долгом миграционном пути многие серые киты сталкиваются с рыболовными снастями и тросами крабовых ловушек. На снимке — намотка 15 м троса толщиной 3,5 см на хвосте взрослого серого кита, добытого китобойцем «Звездный» в 1980 г.
Особенно страшной угрозой для серых китов является разработка нефтегазовых месторождений на шельфе. Весь мир знает о трагедии охотско-корейских китов, пасущихся у побережья острова Сахалин, — места их летнего нагула совпали с местами установки плавучих буровых платформ. Самый древний из сохранившихся до наших дней китов обречен на вымирание, если будет развиваться добыча нефти в тех местах, где расположены его кормовые поля и проходят пути сезонных миграций.
В настоящее время Мексика строго охраняет последние несколько лагун, где размножаются серые киты чукотско-калифорнийского стада. В России на полях нагула серых китов пока не установлено никакого природоохранного статуса.
Начавшееся интенсивное хозяйственное освоение Аляски и Северо-Востока России, в том числе поиски и добыча полезных ископаемых на шельфе и береговой полосе, угрожают существованию очень хрупких и трудно восстановимых сообществ морских организмов, в которых серый кит играет весьма заметную роль.
За свою долгую историю жизни в течение нескольких геологических эпох серые киты пережили ледниковые периоды, смогли устоять в тяжелые времена коммерческого промысла, но теперь они снова в опасности!
Семейство гладких китов
(Familia Balaenidae)
За исключением рода карликовых гладких китов, семейство объединяет крупных животных, причем самки больше самцов. Тело укороченное, толстое, очень неуклюжее. Голова большая (от 1/4 до 1/3 длины тела), ротовая полость громадная, высокая. Дыхательные отверстия расставлены широко, и фонтан двухструйный. Линия ротовой щели сбоку сложно изогнута, нижняя губа очень высокая. Спинного плавника нет, грудные плавники короткие широкие, лопатообразные. Полос-складок нет, кожа гладкая по всему телу. Пластины уса эластичные, узкие и очень длинные, бахрома пластин тонкая, мягкая, длинная. Правый и левый ряды пластин в передней части верхней челюсти не соединяются. Киты тихоходные, медлительные, неповоротливые. Населяют теплые, умеренные и холодные воды обоих полушарий. Во внутренние моря не заходят.
В семействе 3 рода (каждый с одним видом): южных гладких китов, гренландских китов и карликовых гладких китов. В отечественных водах обитают представители южных гладких и гренландских китов.
Род гренландских китов (Genus Balaena)
В роде один вид — гренландский кит. Он занимает совершенно особое место в традиционной морской зверобойной культуре эскимосов и чукчей, которые добывают это животное в течение двух с половиной
тысячелетий. Охота на гренландского кита — вершина промысловой деятельности каждой береговой общины, каждого морского охотника. Все крупные селения в регионе Берингова пролива основаны в местах близкого подхода китов к побережью во время ежегодных весенних и осенних миграций. Кости гренландских китов присутствуют в руинах всех береговых селений, ведь без них невозможно было бы построить полупод-земные жилища, нынлю. О культовом значении гренландского кита в жизни эскимосов убедительно свидетельствуют многочисленные ритуальные сооружения из черепов и челюстей гигантских животных, а также сложные и длительные обряды встречи и проводов каждого добытого кита.
Гренландский кит
(Balaena mysticetus Linnaeus, 1758)
Гренландские киты обитают в холодной зоне северного полушария, где придерживаются кромки арктических льдов. Они совершают сезонные миграции, сроки которых в значительной степени зависят от положения паковых льдов и характера их дрейфа. Кроме арктических вод гренландские киты на-
селяют также Охотское море, но в наше время небольшое по численности охотоморское стадо полностью изолировано от китов, живущих в полярных морях. В бассейне Ледовитого океана существуют три достаточно обособленные стада (популяции).
Западногренландское стадо населяет восточную часть Канадского Арктического архипелага, включая Гудзонов и Баффинов заливы, проливы Ланкастер и Девиса, по нему киты поднимаются к северу почти до 80° с.ш. Зимуют киты этого стада к югу от проливов Девиса и Гудзонова. Воды западной части Канадского Арктического архипелага отделяют западногренландских китов от чукотско-беринговоморского стада.
Шпицбергенское стадо обитает у восточных берегов Гренландии, в водах Исландии, у островов Ян-Майен и Медвежий, у архипелага Шпицберген и к северу от него, в северной и западной частях Баренцева моря. Во времена большой численности стада киты появлялись даже в западной части Карского моря. Полагают, что киты шпицбергенского стада зимуют восточнее Южной Гренландии.
Чукотско-беринговоморское стадо распространено в северной и восточной (почти до Бристольского залива) частях Берингова моря, включая Анадырский залив и Берингов пролив, затем в Чукотском море и в море Бофорта до залива Амундсена на востоке. Северная граница распространения определяется расположением кромки паковых льдов и прилежащей системой стационарных полыней и ледовых трещин.
Весной стадо делится на две части. Одна часть идет вдоль берегов Аляски в восточную часть Чукотского моря, море Бофорта и к заливу Амундсена. Другая часть направляется вдоль берегов Чукотского
полуострова в Чукотское море, достигает островов Врангеля и Геральд, а в благоприятные годы проникает в восточную и даже среднюю части Восточно-Сибирского моря. Начиная с августа, по мере наступления паковых льдов, киты постепенно отходят к югу, на свои обычные места зимовки.
Изолированность трех арктических популяций гренландского кита достаточно условна, о чем еще полвека назад писал М.М. Слепцов:
«Мы уверены, что в отдельные годы, когда арктические льды оттесняются к северу, стада могут смешиваться. Об этом можно судить по добыче в Беринговом проливе гренландского кита, в теле которого оказался гарпун голландских китобоев, упустивших его во время охоты у Шпицбергена. Ввиду слабой изученности биологии данного вида, вопрос об отдельных стадах, особенностях их весенних и осенних миграций из арктических вод в субарктические нужно считать открытым.»1
За последние 20 лет в восточной части Карского моря, море Лаптевых и западной части Восточно-Сибирского моря обнаружены небольшие группировки гренландских китов, которые как бы соединяют шпицбергенское и чукотско-беринговоморское стада.
Далее будут изложены материалы по биологии китов чукотско-беринговоморского стада. За долгую историю аборигенного промысла гренландских китов коренные жители Чукотки хорошо изучили особенности их поведения и пути передвижения. Именно поэтому предлагаемый очерк основан не только на собственных данных автора, его коллег и литератур
1 Слепцов М.М. Китообразные дальневосточных морей. Владивосток, 1952, стр. 122.
ных источниках, но и на традиционных знаниях морских охотников Чукотки.
ОБЩАЯ ХАРАКТЕРИСТИКА
Самый крупный кит семейства, длина взрослого животного 16-18 л/, максимальная — 21 м. Общая масса до 100 т и более. Даже при такой значительной длине гренландский кит кажется толстым и неуклюжим из-за большой величины поперечного сечения тела.
Внешний вид гренландского кита на рисунке из книги Ч. Скэммона (1874 г.). Художник верно изобразил характерные особенности этого животного: огромную голову, дугообразный изгиб нижней губы, хорошо выраженный шейный перехват и хвостовой «горб»
Скелет гренландского кита на рисунке из книги Ч. Скэммо -на (1874 г.). Правильно отражены пропорции животного, размеры и форма верхней и нижней челюстей, особенности скелета передней конечности, превратившейся в грудной плавник, положение и величина остатков тазовых костей
Очень крупная голова (до 1/3 длины тела) отделена от туловища хорошо выраженным шейным перехватом, который является характерной особенностью этого вида усатых китов. На голове и нижней губе несколько десятков осязательных волосков, но роговых наростов нет. Ротовая полость огромная, нижняя губа высокая, ее верхняя кромка имеет форму почти полукруга. Пластины уса эластичные, узкие и очень длинные, их максимальная высота — 4,5 м. Бахрома пластин мягкая, тонкая и необычайно длинная (до 60 см). Пластины и бахрома черные у взрослых животных и серые у молодых. С каждой стороны верхней челюсти располагается в среднем по 400 пластин.
Спинного плавника нет, но у большинства китов примерно в области перехода хвостового стебля в
Голова гренландского кита, вынырнувшего для выдоха-вдоха. Хорошо видны парные дыхательные отверстия (дыхала), расположенные под острым углом друг к другу, и шейный перехват
Схема строения хвостовой части новорожденного гренландского кита длиной 4,5 м.
h — хвостовой «горб»; f — поперечное сечение хвостового плавника; п — выемка между хвостовыми плавниками. Все размеры даны в сантиметрах
линию спины имеется заметное утолщение (по-английски hump, то есть «горб»); оно хорошо выражено даже у эмбрионов длиной менее 15 см. На закругленных вершинах широких грудных плавников нет заостренных кончиков. Толщина подкожного жирового слоя наибольшая среди всех китов, в условном месте она нередко превышает 50 см.
Окраска всего тела обычно одинаковая, черная, но у некоторых животных в области горла может быть светлое пятно неопределенной формы.
К настоящему времени накоплены данные, позволяющие говорить о полиморфизме гренландских китов чукотско-беринговоморского стада. Еще в 1874 г. знаменитый китобойный капитан Ч. Скэммон писал, что помимо типичных гренландских китов многие китобои сообщали о трех вариантах внешнего вида
этих животных: great polar 'whale (большой полярный кит), arctic ice whale (кит арктических льдов) и banchback (горбатый).
Эскимосы Чукотки и Аляски также неоднократно рассказывали ученым о «других» гренландских китах: инутук, инутувук и такышкак. Специалисты США, исследовав в 1980 г. несколько особей инутук морфологическими и генетико-биохимическими методами, пришли к заключению, что инутук — морфа (вариант внешнего вида) гренландского кита, которая появляется среди молодых самок. Вариация инутувук пока не изучена, но, возможно, это взрослые самки инутук.
Вариант такышкак хорошо известен старшему поколению эскимосов в поселках Сиреники и Новое Чаплино. Именно такой кит был добыт 27 мая 1998 г. бригадой семьи Тыпыхкак из поселка Сиреники. Такышкак в отличие от обычных гренландских китов (аг "вы пик) имеет каплевидную форму — он заметно короче и толще известных науке представителей этого вида. Встречается, как правило, весной, во время последней волны миграции (конец мая-начало июня).
РАСПРОСТРАНЕНИЕ И МИГРАЦИИ
Наиболее полные сведения о распределении и сезонных миграциях гренландских китов в водах Чукотки были получены коренными жителями и специалистами в период совместной работы по двум программам:
1. Программа Чукотской зоологической экспедиции Института эволюционной морфологии и экологии
животных им. А.Н. Северцова Академии Наук СССР (1977-1991);
2. Российско-американская программа Территории Самоуправления Северного Склона Аляски (1992-1996). Карты и таблицы, сделанные на основе материалов охотников-наблюдателей, впервые дают детальное представление о распределении, численности и направлении движения гренландских китов у побережья Восточной Чукотки в течение всего года.
Летние районы обитания
Во время американского коммерческого китобойного промысла считалось, что июль-сентябрь — лучшее время охоты на гренландских китов в северной части Берингова и южной части Чукотского морей, а также в море Бофорта вплоть до острова Банкс. Промысловые карты, составленные американскими исследователями по судовым журналам прошлых лет, показывают, что заметное число животных в недавнем прошлом постоянно оставалось на лето в водах Чукотки, от Анадырского залива в Беринговом море до западной части Чукотского моря включительно.
Именно эти киты были почти поголовно истреблены коммерческими судами во второй половине XIX века, но все же и до наших дней сохранились немногочисленные особи, проводящие лето возле берегов Чукотки. Как правило, это одиночки или группы из двух-трех китов. Летом 1970-1975 гг. капитан Л.М. Вотрогов отмечал таких животных в разных частях Анадырского залива и Берингова пролива. О них сообщали нам в 1977-1987 гг. местные жители, много лет наблюдавшие гренландских китов вдоль побережья Чукотки, от залива Крес-144
Самка гренландского кита, добытая китобойным судном «Звездный» 27 июня 1971 г. £ и доставленная в поселок Энмелен. Длина кита более 14 м, рассчитанный вес — 90 т
Миграционные пути гренландских, китов в водах Чукотского полуострова. Летне-осенний период. А-киты, летующие у Чукотского полуострова; В—киты, летующие в Чукотском и Восточно- Сибирском морях; С—киты, летующие в водах Аляски и Канады
та до мыса Биллингса. В архивах Л.М. Вотрогова сохранились уникальные снимки гренландских китов, сделанные в июле-августе 1971 и 1974 годов.
Одиночных китов и группы животных ежегодно наблюдают охотники и местные жители в июне, июле, августе у побережья Чукотского полуострова, между поселками Энмелен и Нешкан. Более того, выявлены две устойчивые локальные группировки гренландских китов, проводящих лето у поселка Энмелен (наблюдатель Г. Гыргольнаут) и в районе лагуны Гетлянген (наблюдатель А. Ашкамакин).
Осенняя миграция
По наблюдениям охотников и местных жителей, самая западная точка появления гренландских китов в конце лета — прибрежные воды о. Айон в Чаун-ской губе Восточно-Сибирского моря. Китов здесь видят в августе-сентябре, но нерегулярно. К поселку Биллингс в западной части пролива Лонга киты в это же время подходят значительно чаще, объединяясь в группы до 10 голов. Жители поселка неоднократно наблюдали, как киты «играют», выбрасываясь из воды. Пока еще слишком мало данных, чтобы обсуждать статус китов, появляющихся у о-ва Айон и пос. Биллингс, — мигрирующие или летующие это животные.
Жители поселка Шмидта в восточной части пролива Лонга китов видят регулярно в конце августа-сентябре. Сначала подходят одиночки, затем группы до 60 и более животных, которые обычно появляются вместе с образованием молодого льда или со старыми дрейфующими льдами. В октябре гренландские киты отмечены в акватории между островами Врангеля и Геральда; вполне возможно, что эти киты
приходят со стороны Аляски, как об этом не раз писали российские и американские специалисты.
Однако хорошо выраженный ход китов на восток, к Берингову проливу, начинается в сентябре-октябре у поселка Ванкарем, где постоянно видят их крупные скопления. Далее к востоку, вдоль побережья от мыса Сердце-Камень до поселка Уэлен, гренландские киты мигрируют во второй половине октября и особенно в первой половине ноября; на этом участке их движение в сторону Берингова пролива выражено наиболее четко.
В те же сроки (вторая половина октября—первая половина ноября) массовый ход гренландских китов наблюдается и в более южных поселках, от мыса Дежнева до залива Креста. Здесь осенняя миграция происходит в более сжатые сроки, чем весенняя. Она мало зависит от силы ветра, снегопадов и температуры, но тесно связана с подходом льдов.
Очень хорошо осенняя миграция прослеживается в поселке Сиреники, мимо которого наибольшее количество животных идет в первой половине ноября. Обычно в это время из-за сильных штормов охотники редко выходят в море и наблюдают китов с берега.
Интересно, что осенью, пока не подошел лед, киты охотно посещают район пролива Сенявина, заходя даже в его узкие ответвления, разделяющие острова. Охотник из поселка Новое Чаплино Е. Паулин в ноябре 1976 г. видел, как в проливе Сенявина, южнее острова Кынкай 6-7 гренландских китов взламывали спинами молодой лед. Сюда же, в бухту Пенкигней, в ноябре приходят кормиться до 200-300 китов (наблюдения охотников из Янракыннота К. Кавранто, К. Кымыечкина, Л. Кутылина).
В зависимости от ледовых условий и других, пока не известных, факторов пик осенней миграции может сдвигаться на месяц. Это хорошо видно по таблицам учета китов в 1994 и 1995 годах. Если в 1994 г. основное количество китов мигрировало в сентябре-октябре, то в 1995 г. пик миграции пришелся на ноябрь-декабрь. В ноябре 1995 г. на определенных участках побережья наблюдатели отмечали очень много кормящихся китов: пос. Нешкан — 272 кита, мыс Нетген — 247, пос. Энурмино — 588, бухта Пенкигней — 276-307 китов.
Таким образом, у азиатских берегов Чукотского моря осенняя миграция гренландских китов начинается очень рано, с конца августа-начала сентября. Важно отметить, что миграция протекает достаточно синхронно (одновременно) вдоль всего Чукотского полуострова. Так, первая миграционная волна возникает сразу в нескольких точках побережья на фоне продолжающих летовку китов. В некоторые годы множество китов одновременно появляются у поселков Сиреники и Уэлен. Например, в 1977, 1980, 1985, 1987 гг. основной ход китов был с 4 по 12 ноября в Уэлене, и всю первую половину ноября — в Си-рениках, причем особенно много гренландских китов возле обоих поселков проходило 7 и 10 ноября. Скорость миграции гренландских китов такова, что за этот период только небольшая часть «уэленских» китов могла достигнуть акватории поселка Сиреники. Затем в конце ноября к Сиреникам обычно подходит вторая миграционная волна — это уже могут быть «уэленские» киты.
Синхронизация миграционных процессов в разных видовых частях ареала описана и у других животных, например, моржей.
g 1994 г. ДАННЫЕ О ВСТРЕЧАХ ГРЕНЛАНДСКИХ КИТОВ
В ВОДАХ ЧУКОТСКОГО ПОЛУОСТРОВА (наблюдения с берега и лодок)
№ Месяц Место III IV V VI VII VIII IX X XI XII всего
1 Уэлька ль 0
2 Энмелен 5 59 44 2 2 2 114
3 Нунлигран 10 8 26 44
4 Сиреники 3 14 >80 34 3 2 9 >145
5 Мыс Чукотский 8 13 5 26
6 Чечен-Сиволкут 1 3 4
7 Янракыннот 2 2 10 14
8 Гетлянген 3 1 4
9 Лорино 15 1 3 19
10 Аккани 3 19 13 35
11 Нутспснмен 8 8
12*13 Пинакулъ-Нунямо 2 2
14 Уэлен 2 17 1 43 63
15 Инчоун 7 95 298 400
16-17 Энурмино-Неттен 7 382 13 402
18 Нешкан 30 61 91
всего 3 >113 148 59 50 565 389 19 0 >1381
1995 г. ДАННЫЕ О ВСТРЕЧАХ ГРЕНЛАНДСКИХ КИТОВ В ВОДАХ ЧУКОТСКОГО ПОЛУОСТРОВА (наблюдения с берега и лодок)
№ сеян Место 1 11 Ш IV V VI vii Vni IX X XI ХИ сего
i Уэлькаль
2' Энмслсн 7 7 26 14 2 ' 6 8 *70
3 Нуплигран Т 1 25 3 5 1 39 13 89
4 Сирсники 4 3 5 29 1 1 80 33 156
Аван 31 41 72
6 Чечен - Сиволкут J 1
7 Яяракыннот id 265-294 35-67 330-371
S Гстляш-ен 2 $ 6 11-13 31-33
5 Лорино К я
га Аккани 8 1 48 29 52 1 139
н Нутенепмен 1 1
П Пинакуль 1 23 24
Я Нунямо 3 3 4 10
14 Умел "Т“ 3 1 73 171-181 249-259
15 Иичоун 1 9 3 52 181-261 246-266
iZ Энурмино 4 15 60 53-61 588 28 748-756
П Неттсн 3 13 74 247 68 405
15 Нешкан 10 8 114 33-34 172 437-438
всего 2 5 ЗЯ 18 5Й 41 43 82 312 126-135 1751-1782 550-592 3016-3098
Зимние районы обитания
Зимой распределение гренландских китов у побережья Чукотки тесно связано с положением кромки паковых льдов, образованием в некоторых участках мощного припая и, главное, системой стационарных Польшей и ледовых трещин, позволяющей китам выживать в тяжелых зимних условиях. В этом плане особенно важна крупнейшая Сиреникская полынья, которая дает возможность китам, а также другим морским млекопитающим и птицам обитать у южного берега Чукотского полуострова даже в очень суровые зимы с 90—100% ледовым покрытием моря. Наблюдатели из поселков, расположенных у Сиреникской полыньи (Энмелен, Нунлигран, Сиреники), отмечают, что гренландские киты устойчиво держатся в полынье с октября-ноября вплоть до июня, то есть до окончания последней волны весенней миграции, а в начале весны часть из них рожают здесь детенышей.
Вдоль восточного побережья Чукотского полуострова постоянные полыньи и разводья располагаются в основном у поворотных мысов, например, у мысов Лысая Голова, Чаплина, Кригуйгун, Нунямо, Дежнева. Очень важна декабрьская полынья в районе бухты Пенкигней, куда поздней осенью-зимой сотни китов приходят питаться. Кроме того, большие стационарные полыньи существуют у северного и южного берегов острова Св. Лаврентия (США), а морское пространство в горле Берингова пролива, между островом Ратмано-ва и районом мыса Дежнева, отличается очень динамичным ледовым режимом с постоянно возникающими и закрывающимися разводьями чистой воды, полыньями и ледовыми трещинами.
Хатырско-Наваринский район продолжает оставаться важным местом зимовки основной части чу-
Миграционные пути гренландских китов в водах Чукотского полуострова. Зимне-весенний период. А - киты, летующие у Чукотского полуострова; В—киты, летующие в Чукотском и Восточно-Сибирском морях; С— киты, летующие в водах Аляски и Канады
котско-беринговоморского стада. Отмеченные нами при аэровизуальном наблюдении в ноябре 1990 г. и в декабре-январе 1994-1995 гг. несколько крупных групп китов численностью свыше 90 голов двигались именно в этот район вдоль условной линии, соединяющей мысы Чаплина и Наварин. Две группы китов, 54 и 11 животных, замечены практически в районе мыса Наварин. Наблюдатели в старом поселке Аван неоднократно видели объединения в 20-40 китов, также шедшие на юг, в сторону Хатырско-Наваринского района.
Весенняя миграция
Многие авторы отмечали постоянство гренландских китов как в отношении районов обитания, так сроков и путей весенней миграции. Известный исследователь Восточной Чукотки К. Богданович писал, что, несмотря на урон, нанесенный этому киту американским коммерческим промыслом, «он придерживается до сих пор мыса Чаплина при своем весеннем продвижении на север.»1
Много позже, в 1993 г., американские исследователи сообщили, что на севере Аляски первое появление китов всегда наблюдается в марте, затем следуют три волны весенней миграции, причем они дифференцированы по половозрастному составу и очень постоянны по срокам, несмотря на то, что фронт льда весьма меняется из года в год.
Первая волна: конец апреля-начало мая; детеныши.
1 К. Богданович Очерки Чукотского полуострова. СПб., 1901, стр. 20.
Вторая волна: середина мая; животные разного возраста и пола.
Третья волна: конец мая-середина июня; самки с новорожденными детенышами и крупные самцы.
У побережья Чукотского полуострова весной также наблюдаются три миграционные волны, первая из них начинается в конце марта, то есть на месяц раньше первой волны на Аляске, а последняя заканчивается примерно на месяц позже, чем третья волна на Аляске. Направление и степень выраженности этих трех волн различны в разных точках побережья Чукотки, и ярче всего эти различия выявляются в районе Сиреникской полыньи:
поселок Энмелен — весенний ход китов направлен в основном юг;
поселок Нунлигран — в марте-июне киты разделяются на две части и мигрируют в двух противоположных направлениях: одни идут на запад, в сторону Энмелена, другие — на восток, к поселку Сиреники;
поселок Сиреники — с конца марта-начала апреля все киты идут вдоль побережья на юго-восток, к мысу Чукотскому, самой южной точке Чукотского полуострова.
Во время весенней миграции наибольшие скопления китов отмечены в районе старого поселка и одноименной лагуны Имтук. Есть основания полагать, что в акватории от Имтук и до мыса Чаплина киты из Сиреникской полыньи встречаются с китами, зимующими в других частях Берингова моря, в том числе в Хатырско-Наваринском районе. Хотя Сиреники и Имтук разделяет всего 8 километров, но это небольшое расстояние является границей между двумя районами — акваторией, где идет миграция
«сиреникских» китов, и акваторией, в которой происходит соединение «сиреникских» китов с китами из других зимних группировок. На это указывают постоянно фиксируемые охотниками различия в численности мигрирующих китов у поселка Сиреники и возле лагуны Имтук.
Например, 25 мая 1978 г. возле поселка Сиреники отмечен только 1 кит, но в тот же день А. Ан-калин записал, что «у лагуны Имтук китов такое множество, что охотники осторожно обходили их на байдарах, чтобы случайно не наехать.»1 25—29 мая 1994 г. возле поселка Сиреники двумя опытными наблюдателями не отмечено ни одного кита, а у лагуны Имтук Т. Панаугье наблюдал две большие группы гренландских китов, которые «были настолько многочисленны, что их невозможно было сосчитать. Киты двигались в сторону бухты Провидения»2 Многие старые охотники из поселка Сиреники неоднократно рассказывали мне о том, что когда киты не шли мимо их поселка, они ехали в Имтук и дальше, к бухте Провидения, где всегда в конце мая-начале июня были большие киты.
Для своего дальнейшего продвижения к Берингову проливу киты используют систему стационарных полыней и ледовых трещин, а также разводья чистой воды, постоянно возникающие среди многолетних льдов. Следуя вдоль сплошного берегового припая, который закрывает Мечигменский залив, поток китов разделяется на две части.
1 Богословская Л.С., Вотрогов Л.М., Крупник И.И. Гренландский кит в водах Чукотки... В кн. Морские млекопитающие М.: 1984, с. 198—199.
2 Полевой дневник Т. Панаугье за май 1994 г.
Одна часть направляется на восток, к берегам Аляски. Вторая часть идет между мысом Дежнева, где полвека назад был крупнейший китобойный поселок Наукан, и островом Ратманова, затем поворачивает на запад и продвигается в Чукотском море к проливу Лонга. Киты движутся вдоль припайных льдов северного побережья Чукотского полуострова, используя обширную заприпайную Чукотскую полынью, которая тянется от Уэлена до Колючинской губы. Не исключено, что на этом пути некоторые группы и одиночные животные отделяются и уходят к берегам Аляски по дугообразным ледовым трещинам, появляющимся весной в Чукотском море. В июле киты также продолжают двигаться на запад, но уже по чистой воде.
О западном миграционном пути гренландских китов в Чукотском море впервые в научной литературе упоминает В.К. Арсеньев в 1980 г. О нем же Л.М. Вотрогову и мне в 1978 г сообщили охотники поселков Уэлен, Инчоун, Энурмино, которые видят этих китов с припая практически ежегодно, особенно в конце мая-июне. Со слов охотников, этот путь был описан нами в 1982 и 1984 годах, а затем подтвержден прямыми наблюдениями с берега и судов В.В. Мельниковым и А.В. Бобковым в 1991-1993 годах, а также аэровизуальным обследованием морей Российской Арктики, выполненным мной в мае-июне 1993 г. и в последующие годы.
Первоначально существование западного пути вызывало большие сомнения американских специалистов, но хорошо документированные наблюдения морских охотников, которые велись непрерывно в течение 20 лет, положили конец этим сомнениям.
Сопоставление сроков весенней миграции гренландских китов у берегов Чукотки и Аляски позволило заключить, что на азиатской стороне основной поток китов начинает движение к северу на месяц раньше, а подходит к бассейну Ледовитого океана приблизительно на месяц позже, чем у побережья Америки.
ГДЕ ДОБЫВАЛИ КИТОВ ДРЕВНИЕ ЖИТЕЛИ ЧУКОТКИ
Важные сведения о распределении гренландских китов у берегов Восточной Чукотки в течение последних двух с половиной тысяч лет дало изучение древних и старых китобойных поселков, где при строительстве полуподземных жилищ обязательно использовались челюсти этих китов — в арктических тундрах не было лучшего материала для опорных балок кровли. Из черепов и челюстей гренландских китов построены многочисленные культовые сооружения. Крупнейшее из них, так называемая «Китовая аллея», находится на острове Ыттыгран.
Первое специальное зоологическое обследование руин древних поселений и культовых сооружений на побережье Чукотского полуострова было проведено в 1979-1981 гг. И. И. Крупником и мной в рамках многолетнего совместного проекта этнографов и зоологов по морской зверобойной культуре региона Берингова пролива. В 1985, 1987 и 1988 гг. зоологи, работая самостоятельно, завершили обследование
Нижняя челюсть гренландского кита, поставленная в качестве одного из священных столбов «Китовой аллеи» — знаменитого памятника эскимосской культуры XV-XVII веков. Рядом стоит фотограф Чукотской зоологической экспедиции Н. Перов (рост 184 см). Остров Ыттыгран, Чукотка, 1978 г.
поверхностно лежащих костных остатков в древних сооружениях на побережье от бухты Руддера до поселка Энурмино, включая острова пролива Сенявина и остров Ратманова.
Гренландский кит был главным объектом китобойного промысла коренных жителей южного и юго-восточного побережий Чукотского полуострова, от Энмелена до мыса Чаплина (древнее селение Уна-зик) и поселков пролива Йергин включительно. Кости в развалинах поселка Аван свидетельствуют, что жители добывали здесь, в основном, крупных взрослых животных. Напротив, заброшенное селение Сингак (к западу от поселка Сиреники) выстроено из черепов очень маленьких гренландских китов, возможно, новорожденных.
Далее к северу, по берегам Мечигменского залива, гренландских китов добывали в гораздо меньших количествах, а на первое место выходил молодняк серого кита. Это, безусловно, связано с существованием мощных припайных льдов, которые закрывают до конца июня-начала июля практически весь залив, препятствуя тем самым промыслу гренландского кита в период его весенней миграции.
Множество костей гренландских китов было обнаружено в остатках поселков Яндогай и Нунямо, стоявших на входных мысах залива Лаврентия, а также в более северных селениях: Наукане, расположенном в самом горле Берингова пролива, и на побережье Чукотского моря, от Уэлена до Нешкана.
К западу от мыса Сердце-Камень кости гренландских китов встречаются в развалинах селений вплоть до мыса Шмидта, а, судя по петроглифам в долине реки Пегтымель, древние жители Чукотки промышляли гренландского кита до мыса Биллингса включительно.
Все поселки, круглогодично добывавшие в прошлом большое количество гренландских китов, располагались либо на поворотных мысах (Чаплино, Яндогай, Нунямо, Энурмино), либо у стационарных полыней и ледовых трещин или в местах с подвижным ледовым покровом (Сиреники, Аван, Наукан, Уэлен, Инчоун и др.). Поселки, основанные в местах, где зимой образуется береговой припай, могли охотиться на гренландских китов только летом или во время их осенней миграции (поселки Мечигмен-ского залива).
В заключение этого раздела следует отметить, что, гренландские киты обитают у берегов Восточной Чукотки круглый год, образуя значительные скопления во время весенней и осенней миграций. Необходимо подчеркнуть, что весенняя миграция вдоль восточного побережья Чукотки начинается на месяц раньше и продолжается на месяц дольше, чем у берегов Аляски, так что многие «чукотские» киты попадают в бассейн Ледовитого океана лишь к концу лета. Вероятно, это связано с большей холодноводностью и, следовательно, более высокой биопродуктивностъю вод Чукотки по сравнению с водами Аляски, что позволяет гренландским китам активно кормиться во время весенней миграции. Для осенней миграции характерна синхронность ее протекания в различных точках побережья.
Можно также полагать, что в прибрежных водах Чукотки пути весенней и осенней миграций совпадают, но неясно, одни и те же группы китов движутся по ним весной и осенью или в разное время года эти пути используют разные части стада.
ПИТАНИЕ
Основу питания гренландских китов составляют мелкие планктонные ракообразные (главным образом калянусы) длиной 3-4 мм, образующие в арктических морях громадные скопления в верхних горизонтах моря. Порой киты как бы погружены в «питательный бульон», состоящий из зоопланктона, и странствуют вместе с ним. В некоторых местах ареала китов большое значение в питании имеет клион, один из мелких крылоногих моллюсков. В желудках гренландских китов находили и несколько других видов планктонных ракообразных, которые, однако, не имеют для них существенного значения.
Специализированное устройство цедильного аппарата (длинные эластичные пластины уса, длинная мягкая волосовидная бахрома) обеспечивает этим гигантским животным, как и их ближайшим родственникам, южным гладким китам, возможность добывать крошечных планктонных ракообразных и моллюсков в количествах, необходимых для столь огромного организма.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ
Гренландский кит — единственный вид усатых китов, приносящий потомство в холодных водах. Период родов и спаривания определен лишь приблизительно — это февраль-май, когда киты находятся в южной части ареала, но и в это время они обитают в холодной прикромочной зоне. Длина новорожденного около 4,5 м. Еще в утробе матери у него образуется толстый (до 15 см) слой подкожного сала. Это предохраняет
ЛОПАТКА
РОДНИЧОК
ХВОСТОВОЙ ТОРБ-
ГЛАЗ
РОСТРУМ
ПУПОВИНА
ЖЕЛТОЧНЫЙ МЕШОК
ФАЛАНГИ
НИЖНЯЯ
ЧЕЛЮСТЬ
Строение эмбриона гренландского кита длиной 13,4 см. Даже на столь раннем периоде внутриутробного развития хорошо виден хвостовой «горб»
детеныша от гибели при родах в холодной воде. Продолжительность молочного кормления детеныша не установлена. Ко времени перехода на самостоятельное питание длина китенка достигает 7 м, а к годовалому возрасту — 9 м.
Возраст, при котором наступает половая и физическая зрелость китов, неизвестен, зато в последние годы появились данные американских специалистов о длительном, до 100 лет и более, сроке жизни гренландских китов. Эти сведения — результат изучения нескольких случаев находок старинных гарпунов второй половины XIX века в китах, добытых эскимосами Чукотки и Аляски в середине-конце XX века. Один такой гарпун был найден в теле гренландского кита охотниками поселка Сиреники в 1952 г.; об этом рассказал И.И. Крупнику Н. Галь-гаугье.
ПОВЕДЕНИЕ
Гренландские киты могут встречаться как среди сплоченных льдов, так и на открытой воде. Обитание в ледовой зоне — их обычный образ жизни; эти киты способны пересечь район плотных дрейфующих льдов протяженностью в сотни километров, используя лишь незначительные полыньи и разводья. Так, гренландские киты порой отмечаются жителями поселков Нунлингран и Сиреники в феврале-марте среди льдов 100-процентной сомкнутости. Охотники поселка Янракыннот сообщают, что подход китов поздней осенью в район бухты Пенкигней часто совпадает с появлением в проливе Сенявина сплошного молодого льда, но животные его не боятся. Они поднимают и взламывают своими спинами лед толщиной до 30 см, оставляя очень характерные образования, которые охотники называют «домики». Все же и гренландским китам иногда суждено попадать в ледовый плен, и, если не произойдет подвижек льдов, они могут погибнуть. Нередко киты оказываются на долгое время «запертыми» в полыньях.
Возле поселка Сиреники охотники в последние 15-20 лет постоянно видят весной гренландских китов разной величины, вплоть до очень маленьких, «в два моржа длиной». В мае к востоку от этого поселка, в местечке Ангытыкук, охотники много лет наблюдают игры гренландских китов на мелководье, всего в 150-200 метрах от берега. Животные выпрыгивают из воды, крутятся на месте, показывая грудные плавники. А. Анкалин, впервые сообщивший об этом, слышал также особые звуки типа низкочастотных свистов, сопровождавшие такие игры.
Гренландские киты вместе с белухами в полыньях возле мыса Дежнева. За животными наблюдал и зарисовал их распределение В.М. Шкробот, командир экипажа вертолета МИ-8, он сообщил, что киты смогли уйти из полыней только 4 января 1987 г.
Летом гренландские киты скоплений не образуют, обычно они наблюдаются поодиночке, реже мелкими группами (до 5 голов). Во время пиков сезонных миграций (май, ноябрь) животные идут большими группами, спокойно, не погружаясь в воду и не торопясь. Наблюдатели сравнивают их с караваном барж. Такие крупные скопления китов являются, очевидно, временными объединениями животных, движущихся в одном направлении. В это время киты подходят близко к берегу, не боясь шума людей и механизмов. За такими группами следуют одиночные мигранты, которые, как бы догоняя ушедших, движутся быстро, часто заныривают (сообщения П. Тыпыхкак и А. Канихина).
Гренландские киты ныряют на короткое время и находятся под водой не более 10 минут, поскольку питаются только в верхних горизонтах моря. После погружения они держатся у поверхности моря примерно до 3 минут, часто выныривают и выпускают характерные для гладких китов двухструйные фонтаны, высота которых не превышает 5-6 м. Интересной особенностью поведения этих китов является так называемое стояние. Кит может около суток держаться на одном и том же месте непосредственно у поверхности воды, время от времени слегка погружаясь и вновь всплывая. Некоторые охотники считают, что так животные отдыхают и даже спят.
Гренландские киты тихоходны, при спокойном плавании их скорость до 7-8 км/час, максимальная скорость, вероятно, не превышает 15-17 км/час.
СТРУКТУРА ЧУКОТСКО-БЕРИНГОВОМОРСКОГО СТАДА
Вопрос о структуре чукотско-беринговоморского стада детально никогда не рассматривался. Однако материалы наблюдений Общества эскимосов Чукотки за 1994-1996 годы, Национального товарищества «На-укан» за 1992-1996 годы, мои собственные данные, а также ряд литературных источников позволяют с достаточной долей осторожности подойти к решению этого вопроса на основании следующих фактов.
1. Установлены два пути весенней миграции гренландских китов из района Берингова пролива:
а) на восток, вдоль побережья Аляски. Этот путь хорошо известен, описан в литературе и используется для подсчета китов на мысе Барроу;
б) путь на запад, вдоль берегов Чукотки, в направлении пролива Лонга. Киты идут по системе стационарных полыней, ледовых трещин, в разводьях среди сплоченных льдов. Этот путь с давних времен известен охотникам Берингова пролива, но специалисты узнали о нем сравнительно недавно.
2. Установлено, что часть китов остается на лето в водах Чукотского полуострова. Помимо одиночных китов и групп, из года в год наблюдаемых в разных точках побережья, выявлены две небольшие локальные группировки в районе села Энмелен и у лагуны Гетлянген.
3. Установлено присутствие гренландских китов весной, летом и осенью в проливе Лонга, Чукотском, Восточно-Сибирском морях и море Бофорта.
По моему мнению, эти данные свидетельствуют о том, что чукотско-беринговоморское стадо состоит, по меньшей мере, из трех группировок китов:
— группировка А, летующая у юго-восточного и восточного побережий Чукотского полуострова (Анадырский залив, бассейн Чирикова, Мечигменский залив, Берингов пролив);
— группировка В, летующая у северного побережья Чукотки (западная часть Чукотского моря, Восточно-Сибирское море);
— группировка С, летующая у берегов Аляски и Канады (восточная часть Чукотского моря, море Бофорта).
Киты из этих группировок встречаются вместе во время сезонных миграций и в местах зимовок. Все три группировки образуют чукотско-беринговоморское стадо гренландских китов, подобно тому, как три репродуктивные группировки моржей, в Бристольском заливе, Хатырско-Наваринском регионе и Сиреникской полынье, образуют вместе стадо тихоокеанского моржа.
Вопрос о степени генетического обмена между этими группировками, между стадом в целом и другими крупными объединениями китов (например, в морях центрального сектора Российской Арктики) остается пока открытым.
ПАРАЗИТЫ, КОНКУРЕНТЫ, ВРАГИ
Эктопаразиты гренландского кита такие же, как у японского гладкого кита. Конкурентов у китов, принадлежащих к трем арктическим стадам, нет — эти киты нечасто встречаются с другими видами планк-тоноядных китов, которые стремятся избегать льдов. Гренландские киты, особенно новорожденные и молодые, иногда подвергаются нападениям косаток, но, в отличие от серых китов, они не являются 168
объектами регулярной охоты этих хищников. Главная опасность для существования вида в целом — современная промышленная деятельность человека, прежде всего разработка нефтяных и газоконденсатных месторождений в тех участках шельфа, где обитают киты.
ЧИСЛЕННОСТЬ
Когда-то гренландские киты были довольно многочисленны во всех частях своего ареала, но в результате многолетнего нерегулируемого коммерческого промысла все четыре стада их оказались доведенными до катастрофического состояния. Теперь, под влиянием многолетнего запрета коммерческой добычи, численность этого вида постепенно увеличивается.
Представление о современной численности гренландских китов чрезвычайно приближенное. Еще недавно специалисты полагали, что в наиболее крупном чукотско-беринговоморском стаде насчитывается от 1500 до 3000 китов, несколько меньше численность западногренландского стада, а в самых маленьких стадах, шпицбергенском и охотоморском, сохранилось всего лишь по нескольку десятков голов. Расчеты специалистов дали численность гренландских китов в Северной Пацифике до начала их коммерческого промысла в 12-20 тыс. голов. Затем она резко снизилась, но благодаря длительному запрету промысла чукотско-беринговоморское стадо насчитывает сейчас около 8000 голов. Возникает вопрос — какова доля «чукотских» китов в этом стаде?
— Группировка А явно немногочисленна. Анализ встреч с животными этой группировки в 1970-
2000 гг. позволяет думать, что в настоящее время в Анадырском заливе, бассейне Чирикова, Мечигмен-ском заливе и Беринговом проливе проводят лето не более 60-100 китов.
— Группировка В объединяет гораздо больше гренландских китов. Так, еще в 1981 г. известный специалист А.А. Берзин считал, что количество китов, летующих у арктических берегов Чукотки может достигать 1000-1500 особей. Результаты береговых подсчетов, выполненных коренными жителями, не опровергают мнение А.А. Берзина.
Данные специалистов и береговые подсчеты охотников показали, что на фоне круглогодичного присутствия гренландских китов в водах Чукотки выявляются два пика численности: май-июнь (весенняя миграция) и сентябрь-декабрь (осенне-зимняя миграция). Особенно много китов проходит у побережья Чукотского полуострова осенью. Это могут быть животные всех трех группировок, включая группировку С, которая представлена животными, летующими в водах Аляски и, возможно, Канадской Арктики и проводящими зиму в полыньях острова Св. Лаврентия и Хатырско-Наваринском районе. Все имеющиеся на сегодня данные позволяют полагать, что во время миграций в водах Чукотки присутствуют не менее 3000-5000 животных.
СТАТУС
Коммерческий промысел гренландского кита был запрещен в 1935 г. Вид занесен в Приложение I СИТЕС (Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения) и в Красную книгу России, 170
категория статуса редкости — 1 (находящийся под угрозой исчезновения).
ОХРАНА
В России не предпринято никаких специальных мер по охране гренландских китов чукотско-беринговоморского стада. Одной из главных задач, связанных с его сохранением, является разработка и осуществление программы по максимально полному количественному учету всех группировок гренландских китов в водах Чукотки. Поскольку уже показано, что весенняя миграция на юго-востоке Чукотского полуострова протекает в те же сроки, что и у мыса Барроу, следует проводить подсчеты китов в нескольких точках Чукотского полуострова по единой методике и синхронно с подсчетами на мысе Барроу, а также и на месяц позже.
Род южных гладких китов
(Genus Eubalaena)
В роде один вид — южный гладкий кит.
Южный (японский) гладкий кит (Eubalaena glacialis Muller, 1776)
На основании главным образом географической изоляции, но при отсутствии резких морфологических различий вид делят на три подвида:
бискайский южный гладкий кит — обитатель Северной Атлантики;
японский южный гладкий кит, обитает в северной половине Тихого океана; этого кита часто азывают просто японским китом или южным китом;
австралийский южный гладкий кит населяет воды южного полушария.
Нередко эти три подвида называют просто бискайский, японский и австралийский киты, а вид, к которому они принадлежат, — южный кит, или гладкий кит.
ОБЩАЯ ХАРАКТЕРИСТИКА
Японский кит — наиболее крупный из подвидов, по своей величине не уступает гренландскому киту. Средняя длина тела 14—16 jw, максимальная до 18 и даже до 20-21 м. Общая масса от 50 до 100 тп. Тело короткое и толстое. Голова большая, до 1/4 длины тела. На голове и нижней челюсти сидят более 200 осязательных волосков-вибрисс. На передней части рыла расположен большой роговой нарост, называемый шляпкой, по бокам головы имеется два продольных ряда некрупных наростов, а еще два ряда расположены по бокам нижней челюсти. Нижняя губа высокая, по верхней кромке сложно изогнутая, имеет ряд фестончатых выростов. На верхней челюсти с каждой стороны расположено до 300, иногда более, пластин китового уса; оба ряда пластин не соединены между собой, между ними есть промежуток. Пластины узкие и длинные, высотой до 2,5 м; тонкая, шелковистая бахрома пластин образу-
Вынырнувший на поверхность южный гладкий кит. Видны роговые наросты на голове и дугообразный изгиб нижней губы
Скелет и контуры тела гладкого кита
ет очень густой фильтр. Пластины и бахрома темные, почти черные, у взрослых животных и серые у детенышей. Спинного плавника нет. Широкие грудные плавники имеют на закругленной вершине заостренные кончики. Подкожный жировой слой очень мощный, толщиной до 50 см. в условном месте.
Окраска тела, как правило, однотонная, обычно темная или почти черная, на брюшной стороне тела нередки небольшие белые отметины. Во времена коммерческого промысла изредка добывались киты с пестрым брюхом, белым брюхом и боками и даже полностью пестрые животные.
РАСПРОСТРАНЕНИЕ И МИГРАЦИИ
Три подвида южного гладкого кита населяют теплые и умеренные зоны обоих полушарий, в южном полушарии киты заходят в воды Антарктики до ледовой зоны.
В американской части Северной Пацифики японские киты зимуют в водах Южной Калифорнии и Мексики, а жир нагуливают в районе Алеутской гряды и восточной части Берингова моря. На азиатской
стороне японский кит зимует на уровне 20-40° с.ш., в водах Тайваньского пролива, островов Бонин и прилегающих районов, На лето киты идут на север до Охотского моря, юго-восточного побережья Камчатки, Командорских островов и южной части Берингова моря возле побережья Корякии. Севернее 60° с. ш. этот кит поднимается очень редко. Однако есть три сообщения о присутствии японских китов у юго-восточного побережья Чукотского полуострова:
1969 г. — одиночного крупного кита заметил в августе между островами Аракамчечен и Ыттыгран (пролив Йергын) охотник из пос. Н.Чаплино В. Та-гитуткак, который сообщил, что в детстве слышал о подходах таких китов в район мыс Чаплина — пролив Сенявина.
1971 г. — одиночный крупный кит в июле обнаружен возле старого поселка Имтук. Внешний вид кита точно описали охотники А. Анкалин и Т. Па-наугье из пос. Сиреники;
1975 г. — одиночного небольшого кита наблюдали в августе у материкового побережья пролива Сенявина охотники пос. Янракыннот (опознан К. Кавранто по наросту на голове и извилистому краю нижней губы);
Южный гладкий кит совершает регулярные сезонные миграции, при этом не только каждый подвид, но и его отдельные популяции движутся по своим миграционным путям и в различные сроки. В принципе, все киты весной перемещаются с мест зимовки из теплых районов ареала в умеренные и холодные воды на лето, а осенью возвращаются на места зимовки и размножения.
ПИТАНИЕ
Устройство цедильного аппарата свидетельствует о том, что южный гладкий кит кормится мелкими планктонными ракообразными. Основными объектами питания в северном полушарии являются различные виды рачков-калянусов. Иногда в желудках китов находили моллюсков, которые, вероятно, захватывались китом случайно, но никогда не обнаруживали рыб. Питаются гладкие киты в верхних горизонтах моря и потому в поисках пищи ныряют неглубоко, на 15-18 м, могут заходить и в небольшие проливы глубиной всего лишь 12 м.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ
Роды и спаривание у гладких китов северного полушария происходят, вероятно, в декабре-феврале в южных частях ареала каждого подвида. Беременность длится около года. Длина новорожденного 4,5-6 м, за период молочного питания, продолжающегося 6-7 месяцев, его длина увеличивается до 10-11 м. Достигнув таких размеров, детеныш переходит к самостоятельному питанию. Предполагают, что половая зрелость наступает в возрасте 10 лет. К этому времени самцы имеют длину в среднем 14,5-15,5, самки — 15-16 м. Время наступления физической зрелости не установлено.
ПОВЕДЕНИЕ
Летом японские киты держатся в большинстве случаев поодиночке, реже — парами и совсем редко — тройками. Только в местах концентрации объектов
питания могут образовывать небольшие скопления до нескольких десятков голов. Все наблюдатели отмечают большую привязанность самки к детенышу, она не покидает даже мертвого китенка. Ныряют эти киты не более чем на 10 минут, между погружениями находятся у самой поверхности, выныривая и давая фонтаны через каждые 30 сек. Фонтан ясно выраженный, двухструйный, пушистый, высотой около 5 м. Часто кит ныряет вертикально и тогда выставляет из воды громадные лопасти хвостового плавника. Наблюдались игры, когда животное выставляется из воды высоко или даже почти выпрыгивает и затем с шумом падает на воду. В спокойном состоянии японский кит плывет со скоростью 7 км/час, а напуганный — до 14 км/ час.
ПАРАЗИТЫ, КОНКУРЕНТЫ, ВРАГИ
Японский кит страдает от китовых вшей, которые особо досаждают ему в области роговых наростов. Тело кита покрывают многочисленные усоногие рачки-коронулы, чьи раковины глубоко внедряются в его кожу. Эндопаразиты изучены недостаточно хорошо, но фауна гельминтов такая же, как у других тихоходных китообразных в этих водах.
Конкурентом японского кита мог бы быть гренландский кит, но, за исключением Охотского моря, их ареалы отделены друг от друга географически (пространственно).
Главным врагом японского кита является человек, превративший его из обычного морского зверя Северной Пацифики в редкое животное нашей планеты.
ЧИСЛЕННОСТЬ
В результате интенсивного промысла в конце прошлого и в начале текущего столетия численность всех подвидов южного гладкого кита катастрофически сократилась. Полагают, что до начала коммерческого промысла в северной части Тихого океана насчитывалось 10000 японских китов, в наше время их осталось всего 500-600 голов. Именно поэтому сведения о встречах японских китов представляют большой интерес.
СТАТУС
Коммерческий промысел японского кита был запрещен в 1935 г. Этот подвид южного гладкого кита занесен в Приложение I СИТЕС (Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения) и в Красную книгу России, категория статуса редкости — 1 (находящийся под угрозой исчезновения).
ОХРАНА
В России не предпринято никаких специальных мер по охране японского гладкого кита, кроме внесения его в Красную книгу. Очень важны сообщения о встречах этого кита в водах Чукотки севернее мыса Наварии, то есть за пределами его ареала.
Семейство полосатиковых (Familia Balaenopteridae)
Семейство объединяет преимущественно крупных китов, исключение составляет малый полосатик. Тело у настоящих полосатиков удлиненное, стройное, относительно тонкое, у горбача — укороченное, толстое. Голова сравнительно небольшая. Дыхательные отверстия расположены близко по отношению друг к другу, фонтан одноструйный. Ротовая щель почти прямая, нижняя губа низкая. Спинной плавник хорошо развит. Грудные плавники у настоящих полосатиков ланцетовидные, несколько удлиненные, у горбача — очень длинные (до 4 л*). На горле и брюхе много продольных полос-складок, тянущихся до области пупка. Пластины уса грубые, короткие и широкие, бахрома пластин толстая и грубая, за исключением сейвала и кита Брай-да. Правый и левый ряды пластин на конце верхней челюсти соединены специальными стержневидными роговыми образованиями. Населяют воды всех океанов и открытых морей в обоих полушариях от Арктики до Антарктики. В семействе два рода — настоящих полосатиков и горбачей.
Род настоящих полосатиков (Genus Balaenoptera)
Род состоит из 5 видов, из них в водах Чукотки 4 вида: синий, или голубой, кит, финвал, сейвал, малый полосатик. Опытные морские охотники легко различают виды полосатиков по фонтанам и по характерным Для каждого вида движениям у поверхности воды.
Синий, или голубой, кит
(Balaenoptera muscidiis Linnaeus, 1758)
Вид делят на 3 подвида, которые, однако, признаны не всеми специалистами:
— северный синий кит — средний среди трех подвидов по размерам тела; он обитает в северной половине Атлантического и Тихого океанов;
— южный синий кит — самый крупный подвид, обитает в водах южного полушария во всех трех океанах, широко распространен в Антарктике;
— карликовый синий кит иногда признается самостоятельным видом. Этот наиболее мелкий из трех подвидов (длина на 2-3 м меньше, чем у двух других) обитает преимущественно в Индийском океане.
Авторитетный советский специалист М.М. Слепцов считал сомнительным выделение трех подвидов исключительно по результатам промысла. Такой подход, по его мнению, не учитывает возрастные особенности группировок синих китов в различных промысловых районах. Кроме того, на телах синих китов северного полушария неоднократно находили эктопаразитов из Антарктики, а это может означать только одно — северный и южный подвиды синего кита географически (пространственно) не изолированы друг от друга.
ОБЩАЯ ХАРАКТЕРИСТИКА
Синий кит — самое крупное животное из всех когда-либо обитавших на Земном шаре. Средняя длина тела 23-24 м, общая масса 70-80 т. Макси-
Скелет синего кита, вписанный в контур его тела
мальной из числа измеренных китов была самка, добытая в Антарктике, ее длина равнялась 33,3 м, масса, вероятно, превышала 150 т, то есть она весила больше, чем 50 африканских слонов. В северном полушарии наибольшим оказался синий кит, зашедший в Панамский канал 23 января 1922 г. Такой же исполин, весивший 135 т, был добыт у Алеутских островов в 1964 г. Самки синих китов в среднем крупнее самцов.
Длинное, стройное тело этих китов окрашено в темно-серый цвет с голубоватым оттенком и покрыто светло-серыми пятнами и мраморным узором. Пятен в задней половине тела и на брюхе больше, чем в передней половине и на спине; голова имеет наиболее темную окраску. Синие киты в холодных водах обрастают диатомовыми водорослями, благодаря чему приобретают зеленоватый оттенок; прежде китобои так и называли их «зелеными» китами.
Голова синего кита небольшая относительно размеров тела, сверху широкая, с сильно выдающимися
в стороны краями. На конце морды несколько десятков осязательных волосков. Пластины уса широкие, высотой до 130 см, почти треугольной формы. На каждой стороне верхней челюсти находится в среднем 360 пластин. Как уже говорилось, спереди правый и левый ряды пластин соединены между собой роговыми стерженьками. Бахрома китового уса толстая, грубая. Окраска пластин и бахромы смолянисто-черная, а нёбо между пластинами черное.
В области основания грудных плавников расположено от 70 до 120 (в среднем около 80) полос-складок, которые тянутся до области пупка. Спинной плавник по сравнению с громадным телом маленький, высотой всего около 30 см, сдвинут далеко назад и находится позади вертикали анального отверстия. Грудные плавники относительно узкие и длинные, концы их заострены. Толщина подкожного жирового слоя в условном месте (на боку тела животного, по его средней линии на вертикали анального отверстия) от 7 до 15 см.
РАСПРОСТРАНЕНИЕ И МИГРАЦИИ
Синий кит населяет практически все открытые моря и океаны, но не заходит в такие «закрытые» моря, как Средиземное, Балтийское, Красное. Раньше в летнее время эти киты были обычны в Беринговом море, встречались в Анадырском заливе и Беринговом проливе, заходили в Чукотское море. Вот что писал в 1952 г. М.М. Слепцов:
«В районах китобойного промысла в дальневосточных водах голубые киты встречаются ежегодно, но в небольшом количестве. Они появляются с ап
реля-мая и остаются до октября. Обычно эти киты встречаются группами по 2-3 особи в каждой или одиночками; их видели у побережья Камчатки — в районе Авачинской губы, бух. Моржовой, в Олю-торском заливе. Севернее указанных районов, видимо, проникают редко. Томилин и Зенкович указывают, что во время работы китобойной флотилии «Алеут» в Чукотском море голубые киты не встречались, но по опросным данным известно, что чукчи прекрасно знают голубого кита. Во время разведки китов в Чукотском море на китобойце «Пурга» в начале сентября мы видели несколько пар этих китов в районе м. Сердце-Камень. Капитаны транспортных судов, курсирующих в Чукотском море, утверждают, что голубые киты достигают о. Врангеля, где их видели среди разреженных льдов и в крупных полыньях. Все это говорит о том, что в наших водах голубые киты бывают ежегодно, но в ограниченном количестве. ... За 17 лет промысла флотилия «Алеут» добыла всего около 50 голубых китов, что свидетельствует о ничтожных запасах этого вида в северной половине Тихого океана.»1
В результате интенсивного коммерческого промысла синие киты в водах Чукотки теперь единичны. Так, за тринадцать лет своей работы на Чукотке капитан Л.М. Вотрогов отметил их всего три раза: 1973 г. — в 50 км от мыса Наварин; 1977 г.-в 20 км к северу от пос. Уэлен; 1979 г. — в 35 км к северу от мыса Сердце-Камень. А дольше, чем он, никто из китобоев или ученых в этом регионе не работал.
1 Слепцов М.М. Китообразные дальневосточных морей. Известия ТИНРО. 1952. Т.38. Стр. 120-121
Синие киты совершают регулярные дальние миграции. Схематично их можно охарактеризовать как перемещения из районов зимовки и размножения, расположенных в теплых зонах, в воды высокой биологической продуктивности летом, то есть в умереннохолодные и холодные воды северного и южного полушарий. Дальность миграционных путей составляет тысячи километров. В северной половине Тихого океана эти киты зимуют в прибрежных водах Японии, включая Бонинские острова, Кореи, Калифорнии, а также в других южных районах. Известны случаи встречи с голубыми китами в северной части Японского моря, у Курильских островов.
ПИТАНИЕ
Строение пластин и бахромы китового уса свидетельствует о том, что основными объектами питания синих китов являются крупные планктонные беспозвоночные, в первую очередь ракообразные. Вместе с ними в желудках синих китов встречаются и мелкие ракообразные (копеподы), некоторые виды рыб и головоногих моллюсков, которые, однако, являются второстепенными объектами питания. Количество пищи, поедаемой этими огромными животными, очень велико. В наполненном желудке синего кита может содержаться 1,2-2 т ракообразных.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ
Периоды родов и спаривания растянуты, но основная масса самок в северном полушарии рожает в декабре и спаривается в январе, а в южном полуша
рии — соответственно в июне и июле. Темп размножения медленный, самки приносят одного детеныша в 2-3 года.
Беременность длится 11-12 мес. Длина новорожденных достигает 7-8 л/, масса около 2 т. Молочное питание детенышей — 6-7 месяцев, за это время их длина увеличивается до 16 м и более, а масса до 20-23 /и, причем самки уже в это время бывают крупнее самцов. За сутки детеныш прибавляет в весе 60-100 кг и вырастает на 4 см.
Половая зрелость у синих китов наступает в возрасте 4-5 лет, у самок при длине тела в среднем около 24, а у самцов — около 23 м. Рост этих китов прекращается по достижении ими физической зрелости (полное окостенение позвоночника) — в возрасте 10-11 лет. Максимальный возраст добытых китобоями самок синего кита определен в 40-45 лет.
ПОВЕДЕНИЕ
Обычно синие киты не скопляются в стада, а держатся группами по 2-3 особи, порой даже поодиночке. Только в местах концентрации кормового планктона они могут образовывать довольно крупные скопления, внутри которых, как правило, сохраняются характерные для них мелкие группки. По сравнению с другими видами китообразных синий кит обладает наиболее мощным фонтаном, в тихую погоду он напоминает узкий конус, обращенный вершиной вниз, его высота составляет от 6-9, до 12 м.
Питаясь в верхних горизонтах моря (100—200 jw), синий кит в поисках корма держится под водой не более 10-12 минут. Но после длительного (от 5 до
50 минут) и глубокого ныряния следует серия из 6-15 коротких выходов на поверхность и небольших погружений. Каждый выход длится всего 6-7 секунд, а неглубокое погружение — 15-40 секунд, во время него кит успевает проплыть 40-50 м под самой поверхностью воды. Наиболее высокими выходами из воды в такой серии бывают первый (после подъема из глубины) и последний (перед погружением в глубину).
В первом случае кит, слабо изгибая тело, показывает сначала макушку головы с дыхалами, потом широкую спину, спинной плавник и, наконец, хвостовой стебель. Во втором случае он круто изгибается, наклоняясь вниз, и высоко выставляет верхнюю кромку согнутого дугой хвостового стебля. Поэтому спинной плавник показывается, когда голова и передняя часть спины уже успевают скрыться под водой, и занимает наивысшую точку в тот момент, когда «дуга» хвостового стебля максимально поднимается из воды. Потом «дуга» становится все ниже, и зверь скрывается, не показав лопастей хвостового плавника.
Пасущийся синий кит движется со скоростью 11-15 км/час, часто и резко меняя под водой направление движения, а напуганный и уходящий от погони зверь, наоборот, идет по прямой линии, развивая скорость до 33-40 км/час, выныривая и давая небольшие фонтаны через каждые 30-40 секунд. Но с такой скоростью он может идти не более 10-15 минут.
Этапы погружения синего кита
ПАРАЗИТЫ, КОНКУРЕНТЫ, ВРАГИ
На коже синих китов живут эктопаразиты и комменсалы из класса ракообразных, в том числе усоногие рачки, ракушки которых своими основаниями погружаются в кожу зверей. В ротовой полости на
Эктопаразиты на спинном плавнике синего кита
китовом усе найдены веслоногие рачки-баленфилу-сы и круглый червь одонтобиус. В северном полушарии синие киты заражены этими паразитами меньше, чем в южном. В холодноводных районах нагула кожа синих китов, как и всех других полосатиков, обрастает зеленой пленкой из диатомовых водорослей, которая исчезает в умеренных и теплых водах. Такие «зеленые» киты бывают жирнее еще не обросших, которые пришли в районы нагула позднее. Максимальное количество «зеленых» китов встречается в конце сезона нагула.
Возможно, конкурентами синих китов до начала эры коммерческого китобойного промысла были другие животные, поедающие большие количества зоопланктона и рыбы, но теперь об этом трудно судить. Единственным конкурентом и одновременно врагом этих китов сейчас является только человек.
ЧИСЛЕННОСТЬ
Количество всех синих китов до начала интенсивного коммерческого китобойного промысла определено с помощью математических расчетов в 200 тыс. голов. Поскольку при переработке этих китов получали значительно большее количество продукции, чем при переработке других видов, они были самыми важными объектами охоты для китобоев всех стран. Это привело к катастрофическому сокращению запасов синих китов, и в 1971 г. их численность в северной части Тихого океана составляла только 1760 особей, а в целом по всему Мировому океану она уже не превышала 5-10 тыс. голов.
СТАТУС
В 1965 г. Международная китобойная комиссия приняла решение о запрете промысла синих китов в Антарктике, а в 1966 г. — и в остальных промысловых районах. С 1967 г. этот вид строго охраняется по всему ареалу государствами-членами МКК и внесен в Приложение 1 СИТЕС (Конвенции «О международной торгов-ле видами дикой фауны и флоры, находящимися под угрозой исчезновения»). Вид внесен в Красную книгу России, категория статуса редкости — 1 (находящийся под угрозой исчезновения).
ОХРАНА
В России не предпринято никаких специальных мер по охране синего кита, кроме внесения этого вида в Красную книгу.
Финвал, или сельдяной кит
(Balaenoptera physalus Linnaeus, 1758)
Морские охотники Чукотки, особенно старшее поколение, хорошо знают этого кита и умеют ^правильно определить его даже в те мгновения, когда финвал показывается из воды для вдоха.
Этот вид иногда делят на два подвида:
северный финвал — более мелкая форма, обитает в северном полушарии (в Тихом и Атлантическом океанах и прилежащих водах Северного Ледовитого океана);
южный финвал крупнее северного, обитает в южном полушарии, где населяет воды всех трех океанов, а также моря Антарктики и Субантарктики.
ОБЩАЯ ХАРАКТЕРИСТИКА
По своим размерам финвал уступает лишь синему киту. Средняя длина самок из северного полушария 19,3, самцов — 18,3 м, в южном полушарии — соответственно, 21 и 20 м. Максимальные размеры самки из Антарктики 27,7, самца — 27,8 м. Общая масса финвала составляет около 50 т. В связи с интенсивным промыслом этих китов их средние размеры за последние десятилетия снизились.
Тело финвала удлиненное, относительно тонкое и стройное, его толщина в 6-6,5 раз меньше длины. Спинной плавник сравнительно высокий (до 50 см), расположен сразу за вертикалью анального отверстия. Грудные плавники удлиненные и узкие, ширина их в 4-5 раз меньше длины. Среднее количество горло-брюшных складок в области основания грудных плавников 70-90. Толщина жирового слоя в условном месте от 7 до 12-15 см.
Голова небольшая, не более 1/4 длины тела, она заметно более узкая, чем у синего кита; нижняя челюсть длиннее верхней. На переднем конце морды расположены 40-50 осязательных волосков. Пластины уса широкие и короткие, длиной не более 90 см\ бахрома грубая, но несколько тоньше, чем у синего кита. В каждом ряду в среднем по 355-365 пластин. Цвет китового уса характерен для вида: большинство пластин однотонные, серо-голубые, но есть одна удивительная особенность, свойственная только финвалу, — 20-30% передних пластин правого ряда имеют белую окраску. Ротовая полость и нёбо между рядами китового уса белые или розоватые.
Окраска тела двухтонная: спинная сторона шиферно-серая, на боках по мере продвижения к брюху цвет постепенно светлеет и на уровне грудных плавников без резкой границы переходит в белый.
Вся горловая и брюшная части тела белые, внутренняя сторона грудных и нижняя поверхность хвостовых лопастей плавников тоже белые. Характерный видовой признак — асимметричная окраска головы: с правой стороны четвертая часть нижней челюсти и кончик верхней челюсти (там, где расположены белые пластины уса) — белые или почти
белые, но по мере удаления от конца морды белая окраска темнеет и переходит в серую. Левая нижняя челюсть на всем протяжении имеет такой же темный цвет, как голова и бока тела.
РАСПРОСТРАНЕНИЕ И МИГРАЦИИ
Финвал — космополит, в открытых водах морей и океанов он встречается от Арктики до Антарктики. Как и синий кит, финвал переносит большие колебания температуры воды и проникает в высокие широты обоих полушарий вплоть до ледяных полей. Населяет и сравнительно закрытые моря (Красное, Средиземное), иногда появляется даже в Балтийском море и Горле Белого моря. В обоих полушариях финвалы совершают регулярные сезонные миграции из умеренных и теплых вод в холодные.
В северной половине Тихого океана финвал распространен от острова Тайвань, островов Бонин и побережья полуострова Калифорния на юге до Берингова пролива и Чукотского моря на севере. На Дальнем Востоке он чаще встречается в Беринговом и Чукотском морях, меньше — в Охотском и Японском. В Северной Атлантике наблюдается повсеместно; отмечен во многих местах Канадского Арктического архипелага.
В южной половине Тихого океана известен вдоль всего побережья Южной Америки (почти от экватора) до Австралии и Новой Зеландии. В южной части Атлантического океана обитает у берегов южно-американского и африканского материков. Широко распространен в Индийском океане, водах Антарктики и Субантарктики.
В пределах своего обширного ареала финвалы держатся достаточно обособленными популяциями. В северной половине Тихого океана существуют по меньшей мере два стада — азиатское и североамериканское. Первое стадо мигрирует весной от Желтого моря и южных вод Японии до северных вод Охотского моря, Камчатки и Чукотского полуострова, а осенью в обратном направлении. Второе стадо весной движется на север от Нижней Калифорнии до побережья Британской Колумбии и Аляски, затем вдоль Алеутской гряды до Камчатки, Корякского побережья, Анадырского залива, Берингова пролива и Чукотского моря. Таким образом, у берегов Чукотки на полях нагула кормятся оба стада, но не установлено, держатся ли они при этом отдельно или могут смешиваться.
ПИТАНИЕ
Основу пищи составляют беспозвоночные животные, главным образом ракообразные, и рыбы. Число кормовых видов довольно велико, при этом видовой состав объектов питания изменяется в зависимости от района обитания, а также сезона года даже в одном и том же районе.
В северной половине Тихого океана главными объектами питания финвалов являются планктонные ракообразные (несколько видов эуфаузиид и веслоногих раков), затем головоногие моллюски (кальмары) и, в меньшей степени, рыбы. В желудках финвалов находили терпугов, сайру, минтая, анчоусов, по-донему, сардин, мойву. Плотно наполненный желудок финвала содержит до 1000 кг пищи. Наибольшее
количество кальмаров в желудке одного кита достигало 2800 шт. (560 кг), сайры — 4640 (550 кг), сельди — 2550 (610 кг). Вот как описывает Б.А. Зенкович охоту финвалов за сельдью в Беринговом море у мыса Олюторский.
«Огромные стаи сельди шли на нерест к берегам, и среди сельди находилось до тридцати финвалов, деятельно охотившихся за нею. Оказывается, и наши финвалы не прочь отказаться от планктонной диеты и после мелких рачков перейти на рыбий стол. ... Громадные туши, ясно видимые в прозрачной воде, поворачивались на бок и с широко открытой пастью бросались на густые косяки сельди, заглатывая ее десятками. А камчатская сельдь — не маленькая приморская сслсдка, она чуть меньше горбуши. Тут же недалеко вертелось несколько косаток. Я читал много рассказов о нападении косаток на китов и ужасе, который будто бы испытывают все киты при приближении косаток. А здесь косатки тоже питались сельдью, и киты не обращали на них внимания.»1
РАЗМНОЖЕНИЕ И РАЗВИТИЕ
Роды и спаривание могут происходить практически в любое время года, но большая часть самок приносит потомство в относительно сжатые сроки. Длительность беременности равна приблизительно году (11-11,5 месяцев), следовательно, роды и спаривание происходят в одни и те же месяцы. В северной части Тихого океана большинство самок рожают детенышей и спариваются с декабря по февраль с
1 Зенкович Б.А. Вокруг света за китами. М.: Географгиз, 1954. С. 140
пиком в январе, в южном полушарии — в июне-августе.
Как правило, самки приносят одного детеныша один раз в два года, т. е. цикл размножения — двухлетний. Средняя длина новорожденного в северном полушарии около 6 м, масса до 2 т. Молочное питание длится 6-7 мес., за этот срок длина тела детеныша увеличивается до 12-13 м, а масса — до 10-11 т.
Форма тела новорожденных и взрослых даже в деталях одинакова. Видимо, детеныши быстроходных полосатиков с момента рождения наделены гидродинамическими свойствами своих родителей. Длина годовалых финвалов в северном полушарии около 14 м. Половой зрелости финвалы северного полушария достигают в возрасте 6-8 лет при длине тела около 18 л/, физической зрелости — в возрасте 22-25 лет (самки — при длине тела около 20 м, самцы — около 19 м).
Предельный возраст жизни финвалов не определен, в Антарктике добывали меченых финвалов, носивших в теле метку более 30 лет.
ПОВЕДЕНИЕ
В северной части Тихого океана финвалы обычно встречаются одиночками, парами или небольшими группами. Одиночными чаще оказываются половозрелые киты, пары более чем в 50% случаев состоят из самца и самки. Реже парами держатся два самца или две самки. На хороших кормовых участках финвалы образуют относительно крупные скопления. Группировки финвалов имеют разный состав. В одних случаях группа состоит из крупных половозре-
Характерный вид спины и спинного плавника в момент погружения финвала
лых самцов или самок, в других — из неполовозрелых китов, в третьих — из молодых половозрелых, но не участвующих в размножении животных. Характерно образование объединений одноразмерных китов, причем в них иногда входят киты разных видов: финвалы с мелкими синими китами или финвалы с крупными сейвалами. Даже в тех случаях, когда образуются скопления в сотни китов, последние всегда состоят из множества групп, которые трудно разъединяются при преследовании.
На полях нагула финвалы держатся на чистой воде, за пределами кромки льда. Оказавшись в полынье, финвал, как сейвал или малый полосатик, кладет кончик морды на край льдины и, опираясь на нее, то выставляет дыхало на воздух, то погружает его на несколько минут в воду. Под поверхностью воды финвалы могут издавать стоны, напоминающие иногда звуки флейты от низкого до высокого тона; этот сигнал звучит около секунды и повторяется до 30 секунд.
Продолжительность пребывания финвалов под водой обычно не превышает 20-30 мин. Питающийся
кит плывет с открытой пастью на боку на глубине до 100 м, затем поворачивается на спину и закрывает пасть, после чего фильтрует большой объем воды, захваченный им при этом. Во время кормления финвалы ныряют на 5-8 минут, редко на больший срок, а потом выныривают 5-8 раз подряд с интервалом в 10-30 секунд, выпуская каждый раз фонтан высотой 5-6, иногда до 9 м. При промежуточных погружениях тело кита почти прямое, но, уходя на глубину, финвал очень круто изгибает хвостовой стебель, выставляя его из воды в виде полукруга. Обычная скорость хода китов не более 9-10 км/час. При дальних переходах киты идут со скоростью 22-24 км/час, испуганный кит делает рывок и плывет со скоростью 33-37 км/час.
Б.А. Зенкович оставил одно из лучших описаний процесса дыхания и особенностей поведения финвала.
«Фонтан сельдяного кита напоминает довольно стройную колонну, расширяющуюся в верхней части. Первый наиболее мощный фонтан достигает 8-10 м высоты и процесс выпускания фонтана — выдоха испорченного воздуха — длится 4-5 секунд. При этом слышен характерный и довольно сильный звук, напоминающий звук выпускаемого под давлением пара.
Во время жировки, когда финвалы усиленно питаются, в спокойном состоянии сельдяной кит выбрасывает от трех до шести фонтанов (чрезвычайно редко больше шести) с интервалами в 10-15 сек. Мы сотни раз проверяли с секундомером в руках эти интервалы, после чего и вывели эти средние цифры. У поверхности воды, во время «фонтанирования»-дыхания, киты эти находятся до двух минут. Затем они снова ныряют в глубину, оставаясь там от 3-х до 6 минут.
Перед нырянием на сравнительно длительное время финвал делает последний продолжительный вдох, набирая запас воздуха, Когда финвал ныряет на длительное время, он показывает голову, всю спину со спинным плавником и часть хвостового стебля, при этом тело его сильно изгибается, выпячивая спину, выгибая хвостовой стебель, и исчезает он, как будто бы вращаясь. Но он никогда не показывает лопастей своего хвоста.
На поверхность же финвал поднимается очень отлого, показывая все свое огромное туловище почти в одной плоскости; фонтан появляется еще ранее того, как покажется голова, так как он открывает дыхала еще под водой, а потом уже появляется верхняя часть головы с открытыми дыхалами и передняя часть спины. Как только финвал выпустит первый фонтан и вдохнет запас свежего воздуха, он медленно двигается вблизи поверхности воды и все его движения очень хорошо видны...
Ночью финвалы проводят почти все время у поверхности воды, куда из глубины поднимаются мириады планктонных ракообразных и крылоногих моллюсков. Днем эти рачки и моллюски почти всегда находятся в сумраке глубины, они избегают яркого света, но ночью совершают «суточные миграции» вверх, как говорят гидробиологи. Впрочем, когда небо мрачно, а в наших водах это обычное явление, то эти рачки так и остаются у поверхности, окрашивая воду в буроватокрасные тона. Тогда и финвалы не ныряют, а ходят почти у поверхности и питаются ими. Конечно, и фонтаны тогда совсем другой конфигурации и силы.
Сельдяные киты, играя, никогда не выпрыгивают из воды, они очень спокойно и даже монотонно ведут себя. Но при этом никто и никогда не мог угадать, в каком месте они появятся после длитель
ного ныряния. Но плавают под водой они быстро, так как за те минуты, что они находятся под водой, они проплывают иногда более трех миль.
Что же касается глубины, на которую ныряют обычно эти киты, то она невелика и не доходит даже и до 100 метров. Мы часто проверяли это, сверяя улов нашей планктонной сетки с содержимым желудков финвалов, добытых в тех же местах. Основная масса планктонных рачков находилась на глубине в 50 м, следовательно, киту и не нужно было нырять глубже. Осенью эти рачки опускаются на значительные глубины, и мы убедились, что в таких местах у китов либо пустые желудки, либо же они питались рыбой.
Инстинкт материнства и товарищества у сельдяных китов-финвалов развит слабо. Если ранена или убита кормящая самка, то сосунок убегает и держится вдалеке. Если ранен детеныш, матка к нему не приближается и уходит. Не отличаются они и привязанностью самца к самке и наоборот.
Особой памятливостью эти киты также не обладают. Некоторых китов задевали гарпуном по нескольку раз, и все же через известное время они вели себя по-прежнему, ничего не опасаясь.
Спят эти киты крепко. Мы иногда подходили к такому спящему киту и легко его убивали.»1
ПАРАЗИТЫ, КОНКУРЕНТЫ, ВРАГИ
Наружные и внутренние паразиты финвалов примерно такие же, как у синего кита. В печени и кишечни
1 Зенкович Б.А. Вокруг света за китами. М.: Географгиз, 1954. С. 149-151.
ке отмечены паразитические черви-гельминты (трематоды, нематоды, акантоцефалы). Некоторые из паразитов свойственны совершенно определенным частям Мирового океана. Соответственно, по их присутствию на коже или во внутренних органах кита можно уверенно сказать, какие районы он посещал. Так, в Беринговом море Б.А. Зенкович регулярно находил на финвалах паразитическое ракообразное, пэнеллу антарктическую. Этот эктопаразит «метит» китов тогда, когда они приходят в воды Антарктики. Пэнелла глубоко внедряется в кожу и подкожную жировую клетчатку, а снаружи выглядит как веточка с метелочкой и длинными волосками-жгутиками на свободном конце. Как и синие киты, финвалы обрастают диатомовыми водорослями, которые образуют на их коже коричневатую пленку.
Возле китов нередко промышляют птицы, чайки и трубконосые. Один из видов трубконосых, тонкоклювый буревестник, прозван китобоями «финва-льей птицей» — он держится возле китов, кормясь вместе с ними.
На молодых финвалов изредка нападают косатки, но их главным врагом в последние полтора века был и остается человек: раньше он охотился на них, теперь загрязняет море, что отрицательно сказывается на кормовой базе и среде обитания финвалов.
ЧИСЛЕННОСТЬ
Финвал был наиболее многочисленным видом усатых китов, однако в результате коммерческого промысла величина его популяций значительно сократилась. За период 1868-1990 гг. во всех морях и океанах было добыто 878530 этих китов. Популяции
этого вида в южном полушарии, особенно в водах Антарктики, более многочисленны, чем в северном полушарии.
В первой половине XX в большие стада финвалов встречались в Беринговом проливе и Чукотском море, более мелкие — у берегов Камчатки и Командорских островов. С 1932 г. по 1990 г. китобойные флотилии СССР выбили здесь 17928 финвалов? В настоящее время принято считать, что оптимальный уровень численности финвалов в северной части Тихого океана — около 27000, но реальное количество этих китов сейчас не превышает 10-15 тыс. животных: у азиатского побережья 5-7 тыс., остальные киты — в американских водах. Возле берегов Чукотки этот кит продолжает оставаться редким, хотя порой встречаются группировки из 40 животных.
СТАТУС
В 1976-77 гг. коммерческий промысел финвала был запрещен по решению Международной китобойной комиссии. Вид был внесен в Приложение I СИТЕС (Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения). Северный подвид финвала внесен в Красную книгу России, категория статуса редкости — 2 (сокращающийся в численности).
ОХРАНА
В России не предпринято никаких специальных мер по охране финвала, кроме внесения его северного подвида в Красную книгу.
Сейвал, или сайдяной кит
(Balaenoptera borealis Lesson, 1828)
Сейвал — редкий кит в водах Чукотки, особенно их мелководной северной части. С большей вероятностью его можно встретить в южных районах Берингова моря, вплоть до мыса Наварин, поскольку он придерживается континентального свала глубин.
ОБЩАЯ ХАРАКТЕРИСТИКА
Средняя длина самок сейвала из северного полушария 13,5—14, самцов — 13-13,5 м, из Антарктики — соответственно, 15,5 и 14,6 м; максимальная длина северных сейвалов до 18 м, южных — до 19 м.
По сравнению с финвалом сейвал менее вытянут, тело его относительно толще, грудные плавники короче и уже (длина плавника менее чем в 4 раза превышает его ширину). А вот спинной плавник и абсолютно и относительно высокий, до 85 см; он сдвинут вперед, к началу задней трети тела сейвала. Горло-брюшные полосы-складки короче, чем у других видов, они заканчиваются, не достигая области пупка; их количество у основания грудных плавников колеблется от 40 до 60. Толщина жирового слоя в условном месте невелика, всего 4-7 см.
Небольшая голова составляет приблизительно 1/4 длины тела. При этом у молодых она меньше, чем у старых, и у самок меньше, чем у самцов. Нижняя челюсть немного длиннее верхней. Число осязательных волосков на морде обычно не превышает 70. Нёбо узкое, белое или розовое. Пластины уса жесткие, короткие, высота наибольших достигает 80 см. Они, как правило, имеют однородную темно-серую окраску разных оттенков, бахрома всегда несколько светлее самих пластин. В отличие от других видов полосатиков у сейвала волосовидная бахрома, длинная, тонкая, извитая и очень мягкая, образует частый фильтр. В среднем с каждой стороны верхней челюсти располагается около 350 пластин, из них несколько десятков передних — белые.
Окраска спинной части тела в большинстве случаев темно-серая, порой почти черная, боков — серая с явным голубоватым оттенком, а на брюхе — непостоянная, от серой до частично белой но весь низ никогда не бывает белым, как у финвала. Иногда в передней части брюшной поверхности имеется белое пятно, в задней части туловища, на боках — светлосерые пятна. Спинной плавник и лопасти хвоста сверху того же цвета, что и спина, а грудные плавники сверху такие же, как бока тела. Снизу хвостовые лопасти и грудные плавники обычно серые.
РАСПРОСТРАНЕНИЕ И МИГРАЦИИ
Сейвал распространен так же широко, как и финвал, но более теплолюбив, чем другие полосатики, и далеко в холодные воды не проникает. Поэтому в
Арктике и Антарктике он обычно избегает льдов, появляясь там позже крупных полосатиков. Этот кит предпочитает открытые воды морей и океанов, особенно окраины континентальных ступеней.
В северной половине Тихого океана распространен от острова Тайвань, прибрежных вод южной Японии, островов Бонин и Мексики до северной части Берингова моря. Известны случаи проникновения в Берингов пролив и прилежащие воды Чукотского моря. В Японском, Охотском и Беринговом морях, в водах Калифорнии довольно малочислен, наиболее обычен у восточного побережья Японии, в водах Курильских островов и у побережья Британской Колумбии.
Сезонные миграции обычно носят менее регулярный характер, чем у других видов усатых китов, хотя схема миграций такая же. В северной части Тихого океана киты весной покидают места зимовки (воды островов Бонин, полуострова Калифорния, Мексики) и уходят на север к Курильским и Командорским островам, к Камчатке и в другие места Берингова моря, а осенью возвращаются обратно. В Северной Атлантике миграции сейвалов не так ясно выражены из-за мощного теплого течения Гольфстрим. Сезонные миграции в южном полушарии изучены слабо.
ПИТАНИЕ
Строение цедильного аппарата (густая волосовидная бахрома) указывает на питание сейвала преимущественно мелкими планктонными беспозвоночными. В северной части Тихого океана в пище этих китов
преобладают копеподы, некоторое значение имеют и другие животные — кальмары, крупные планктонные ракообразные и даже рыбы. В желудках китов находили анчоусов, сайру, морского окуня, макрель, мойву, минтая и некоторые другие виды рыб. Наполненный желудок сейвала в среднем содержит до 200 кг пищи. Максимально наполненный желудок сейвала, добытого у Курильских островов, содержал 3100 кальмаров общим весом 600 кг.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ
Беременность у сейвалов длится почти 12 мес., поэтому роды и спаривание происходят приблизительно в одно и то же время. В северном полушарии наибольшее количество самок приносит потомство и спаривается в январе-феврале, меньше — в декабре и марте. В южном полушарии пик родов и спаривания наблюдается в июле и августе. Цикл размножения этих китов, по-видимому, двухлетний, и темп пополнения популяции довольно медленный. Средняя длина новорожденного сейвала южного полушария 4,5-5 л/, у сейвалов северного полушария — несколько меньше. Молочное питание длится 5-6 мес., за этот период длина китенка увеличивается до 8-9 м. Возраст достижения половой зрелости не установлен; длина тела достигших половой зрелости самок из северной части Тихого океана колеблется от 12,5 до 14 м, самцов — от 12 до 13 м.
Физическая зрелость наступает в возрасте около 10 лет. Продолжительность жизни не установлена.
ПОВЕДЕНИЕ
Крупных концентраций сейвалы не образуют, держатся преимущественно одиночками или парами, но в местах с изобильной пищей образуют группы в несколько десятков голов. Как наиболее теплолюбивый кит сейвал в северном полушарии далеко в холодные воды не заходит, но в Антарктике не только достигает ледовой зоны, но и встречается среди льдов.
Сейвал, в отличие от финвала, погружается отлого, слабо изгибаясь; поэтому кромка его хвостового стебля либо совсем не показывается из воды, либо выставляется низко, не образуя полукруга. Форма фонтана — обычная для полосатиков, но высота конуса не превышает 5 м. Количество фонтанов между погружениями нерегулярное, от 1 до 5, с паузами между ними от 4 до 22 секунд. Погружение продолжается от полминуты до 12 минут. Зверь плавает обычно неглубоко, и тогда на поверхности видны водовороты, вызванные работой хвостовых лопастей.
Китобои считают сейвалов самыми быстроходными китами, наибольшая скорость передвижения преследуемого кита в первые моменты может достигать 40-50 км/час. Однако пасущиеся сейвалы движутся со скоростью 5 км/ час.
Специалисты полагают, что паразиты, конкуренты и враги сейвала те же, что у финвала.
ЧИСЛЕННОСТЬ
Общая численность сейвалов невысока. Самые мелкие популяции населяют Северную Атлантику,
где на громадной акватории добывалось всего лишь около 200 голов в год. В северной половине Тихого океана численность сейвалов выше, ориентировочно ее исходное значение (до коммерческого китобойного промысла) определено в 42-47 тыс., современное — 8,6-14 тыс. голов. Более многочисленны популяции этого вида в южном полушарии. Предполагается, что величина популяций антарктических сейвалов равна 50-70 тыс. голов.
СТАТУС
Коммерческий промысел сейвала был запрещен Международной китобойной комиссией в 1979 г. Этот вид внесен в Приложение 1 СИТЕС (Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения). Сейвал северного полушария, обитающий в водах России, внесен в Красную книгу России, категория статуса редкости — 3 (редкий).
ОХРАНА
В России не предпринято никаких специальных мер по охране сейвала, кроме внесения его в Красную книгу. Очень важны сообщения о встречах с этим китом в водах Чукотки, где проходит северная граница его ареала в Тихом океане.
Малый, или остромордый, полосатик;
кит Минке
(Balaenoptera acutorostrata Laeepede, 1804)
Обычный, но немногочисленный вид у берегов Чукотки. Чаще всего малого полосатика можно наблюдать в проливе Йергын между островами Аракамче-чен и Ыттыгран. Все морские охотники Чукотки хорошо знают этого кита.
Как и другие киты рода полосатиков, малые полосатики разделяются на две крупные группы, населяющие Мировой океан:
северный малый полосатик — более мелкий представитель вида, обитает в водах морей и океанов северного полушария, имеет хорошо заметный отличительный признак, белую поперечную полосу на грудных плавниках;
южный малый полосатик — более крупный кит, населяет воды южного полушария, в том числе все моря Антарктики. Внешние отличия южных малых полосатиков от северных очень четкие: они крупнее, не имеют белого пояска на грудных плавниках, у них белые усовые пластины с темным наружным краем и более толстой бахромой, на морде большее число осязательных волосков-вибрисс.
ОБЩАЯ ХАРАКТЕРИСТИКА
Самый мелкий представитель семейства полосатиков, длина взрослого кита 7-10 м, масса 7-9 т. Тело кита Минке толще, короче и не такое стройное, как у других видов рода. Спинной плавник высокий (до 35 см), с сильно загнутой назад вершиной, расположен впереди вертикали анального отверстия. Грудные плавники узкие, ланцетовидные. Горло-брюшных полос около 60.
Голова небольшая, нижняя челюсть с сильно заостренным концом и длиннее верхней челюсти. На морде 30-40 осязательных волосков, причем у китов в северном полушарии число их меньше, чем в южном. А. Г. Томилин связывает это явление с большей рыбоядностью северных полосатиков и планктоноядно-стью южных китов. Высота пластин уса не более 30 см, бахрома короткая, жесткая. Пластины и бахрома имеют белую или желтовато-белую окраску. Ежегодно пластины отрастают на 4-4,5 см и на столько же снашиваются с вершин. В каждом ряду по 270-330 пластин.
Спинная часть тела имеет темно-серую окраску, бока светлее спины, брюшная поверхность белая. Переходы в окраске нерезкие. Только у малых полосатиков северного полушария имеется характерный признак — белая поперечная полоса на темносерых грудных плавниках.
РАСПРОСТРАНЕНИЕ И МИГРАЦИИ
Широко распространен в умеренных и холодных зонах Мирового океана, в теплых, особенно тропических, водах редок. Повсеместно обитает в Антарктике, где
постоянно проникает в зону дрейфующих льдов. Здесь малые полосатики собираются в стада до сотни голов и далеко заходят в ледяные поля, используя проходы, сделанные айсбергами. Случается, что льды отрезают целые группы этих китов, вынуждая их зимовать в полыньях. В северной половине Тихого океана населяет Восточно-Китайское, Желтое, Японское, Охотское и Берингово моря, тихоокеанские воды Японии, воды Курильских и Алеутских островов, прибрежные воды США и Канады. У берегов Чукотки идет на север до ледовой зоны Чукотского моря. По сравнению с крупными полосатиками этот кит держится ближе к берегам, часто заходит в бухты и заливы.
Малым полосатикам свойственны сезонные миграции с мест зимовки на места летнего нагула, однако пути и сроки их изучены недостаточно.
ПИТАНИЕ
В отличие от других видов рода малые полосатики питаются преимущественно рыбой, особенно в северном полушарии, и в меньшей степени — планктонными ракообразными и головоногими моллюсками. Соотношение отдельных видов животных в пищевом рационе китов в разных местах ареала различно, причем видовой состав объектов питания значительно меняется по сезонам года. В водах Японии малые полосатики поедают главным образом ракообразных, тогда как у Курильских островов — рыб. Из ракообразных в желудках китов находили копепод и более крупных эуфаузиид (несколько видов). Очень разнообразен набор видов рыб: обнаружены сельдь, скумб
рия, навага, треска, минтай, мойва, сайра, терпуг, морские окуни и некоторые другие. Масса содержимого наполненного желудка достигает 200 кг.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ
Период размножения малых полосатиков определяется весьма приближенно. Полагают, что в северном полушарии большинство самок приносит потомство с ноября по март, а спаривание происходит с февраля до мая. Продолжительность беременности составляет приблизительно 10-11 мес. Длина новорожденного 2,1-2,7 м. Молочное питание продолжается 5-6 месяцев, за этот период длина китенка увеличивается до 5 м. В двухлетнем возрасте, когда длина самцов достигает 7,5, самок — 8 jw, наступает половая зрелость и рост китов замедляется. Физической зрелости малые полосатики достигают при длине тела 8,5-9 л*, возраст китов в этот период не установлен.
ПОВЕДЕНИЕ
В летние месяцы, в период нагула киты обычно держатся очень маленькими группами, парами и даже одиночками, но в местах изобилия пищи нередко образуют скопления в десятки и даже сотни голов.
Киты Минке уходят под воду на 10-15 мин. и, питаясь, выныривают через каждые 3-4 мин., находясь у поверхности моря не более 2 минут. При этом кит выставляется из воды лишь на 2-3 секунды, из которых сам дыхательный акт занимает всего 1,2 секунды. При выдохе кита на расстоянии до 50 м ощущается специфический запах. Фонтан малого по
лосатика небольшой, всего до 2 м, быстрый, в виде опрокинутого конуса и плохо различим на расстоянии. При нырянии малый полосатик изгибает хвостовой стебель меньше, чем финвал, и движется медленнее крупных полосатиков. Его скорость достигает 26-33 км/час. Иногда, следуя за судами, этот кит целиком выпрыгивает из воды; изредка можно видеть на берегу обсохшего малого полосатика.
ПАРАЗИТЫ, КОНКУРЕНТЫ, ВРАГИ
Эктопаразиты заражают этот вид намного реже, чем других полосатиков, и не вызывают на его коже появления светлых пятен. А.Г. Томилин полагает, что это связано с меньшим размахом миграций по направлению к экватору, а следовательно, и с меньшими возможностями нападения эктопаразитов.
В водах Чукотки конкурентов у малого полосатика нет — немногочисленные киты пасутся в местах, куда редко заходят другие полосатики. Косатки, видимо, не нападают на этих достаточно скоростных китов, во всяком случае, морские охотники Чукотки пока ни разу не сообщали об этом.
Как и для всех других китообразных, главным врагом является человек, источник шума и загрязнения акваторий, где обитают эти небольшие киты.
ЧИСЛЕННОСТЬ
Общая численность малых полосатиков в Мировом океане предположительно определена в 350 тыс. Вид, по-видимому, немногочислен, так как в пределах
огромного ареала большие скопления этих китов отмечаются довольно редко, преимущественно в Антарктике, там численность их приближенно оценена в 180 тыс. голов. В северной части Тихого океана обитает 25-30 тыс. малых полосатиков.
СТАТУС
Коммерческий промысел малого полосатика был запрещен Международной китобойной комиссией в 1986 г., однако Норвегия и Япония считают охоту на этого кита традиционным видом деятельности своих народов и настаивают на ее возобновлении. Вид внесен в Приложение 1 СИТЕС (Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения), но он не внесен в Красную книгу России.
ОХРАНА
В России не предпринято никаких специальных мер по охране малого полосатика.
Род горбатых китов
(Genus Megaptera)
В роде один вид — горбатый кит, или горбач.
Горбатый кит, или горбач
(Megaptera novaeangliae Borowski, 1781)
В настоящее время горбач восстанавливает свою численность у берегов Чукотки. Многие морские охотники легко узнают этого кита по ряду характерных особенностей его внешнего вида и поведения.
Выделяют два подвида горбатого кита: северный горбач — более мелкая форма, обитает в океанах и морях северного полушария, включая полярные воды Арктики;
южный горбач — населяет воды всех океанов и морей южного полушария, включая полярные моря Антарктики.
Все биологические ритмы северного и южного подвидов горбача (спаривание, роды, молочное выкармливание детенышей и т.п.) сдвинуты приблизительно на 6 месяцев.
ОБЩАЯ ХАРАКТЕРИСТИКА
Средняя длина горбачей из северной части Тихого океана около 12, из Антарктики — 12,5 м. Максимальная длина самки северного полушария 15,9, из Антарктики — 17,4 м. Самки на 1-1,5 м крупнее самцов. Средний вес горбатого кита около 40 т. Соотношение между длиной и толщиной туловища иное, чем у настоящих полосатиков. Тело у горбачей сравнительно короткое и толстое, особенно в передней части, поэтому они кажутся неуклюжими. Полосы-складки на горле и брюхе широкие, тянутся до области пупка, их насчитывается всего от 12—15 до 30—35. Спинной плавник широкий, сравнительно
Горбач, выпрыгивающий из воды. Большая уплощенная голова кита покрыта наростами-шишками; грудные плавники необычайно длинные
низкий (менее 30 см), имеет характерную ступенчатую форму спереди и выемку сзади, расположен несколько впереди вертикали анального отверстия. Замечательны грудные плавники горбача — их длина доходит до 1 /3 длины туловища и у крупных китов достигает 4 м. Они изогнуты по форме нижней части тела, их передние края извилисты и несут до 10 крупных бугров-наростов. Верхняя поверхность плавников темная, нижняя — светлая. Именно из-за таких огромных плавников горбача когда-то называли «длинноруким китом». Подкожный жировой слой мощный, порой его толщина в условном месте бывает более 15 см.
Крупная плоская и притупленная голова занимает до трети всей длины животного. Нижняя челюсть намного шире и несколько длиннее верхней. Сверху на морде расположено от 3 до 5 рядов бородавчатых кожных наростов-шишек, средний ряд насчитывает 5-8 наростов, а боковые — по 5-15. Такие же наросты в количестве 10-15 распределены вдоль каждой половины нижней челюсти. По форме и размерам наросты похожи на половинку апельсина, и каждый из них имеет по одному осязательному волоску, или вибриссе. На переднем конце нижней челюсти находится большой, до 30 см в диаметре, кожный нарост неправильной формы. Общее количество наростов на голове горбача — от 40 до 70. На верхней и нижней челюстях расположены около 50 вибрисс, которые «сидят» главным образом на этих наростах. В ротовой полости на передней части неба проходят две продольные борозды. Грубые и широкие пластины китового уса относительно высоки — до 1 м; в каждой половине верхней челюсти их насчитывается от 270 до 400. Они пепельно-черного цвета с жесткой бурой бахромой.
Группа из трех горбачей. Хорошо виден «ступенчатый» плавник одного ныряющего кита и хвостовые лопасти («бабочка») другого зверя, уходящего на глубину
Спинная поверхность тела и бока обычно имеют однотонную темную (почти черную) окраску, брюшная поверхность и хвостовые лопасти различную: черную, пеструю с разным сочетанием цветов и даже почти белую. Б.А. Зенкович сообщал, что окраска горбачей в одном и том же стаде (группе) варьирует от белой и розовой до черной на брюхе, и только спина и голова у всех черные. Тем не менее многие специалисты считают, что особенности окраски могут служить надежным признаком для установления родственных связей между китами. Различают также несколько типов окраски китов из северной части Тихого океана и Антарктики, характерные для отдельных популяций.
Горбачи часто ныряют, показывая хвостовые лопасти, или «бабочку», окраска которой индивидуальна и по ней можно узнать «в лицо» каждого кита.
Для китов, обитающих возле Гавайских островов, в 1979 г. был даже составлен альбом персональных фотографий их хвостовых лопастей.
РАСПРЕДЕЛЕНИЕ И МИГРАЦИИ
Горбачи обитают на огромной акватории Мирового океана, от тропических вод до ледовых полей Арктики и Антарктики, заходят они также и в «закрытые» моря, Средиземное и Балтийское. В северной половине Тихого океана они встречаются от тропиков до северной части Берингова моря, Берингова пролива и прилегающих вод Чукотского моря включительно.
Эти киты — типичные мигрирующие животные. Они совершают регулярные перемещения с мест зимовки в теплых водах к районам летнего нагула, которые расположены в холодных водах обоих полушарий. В пределах своего обширного ареала горбатые киты образуют несколько более или менее обособленных популяций, причем каждая из них имеет свои пути и сроки миграций в пределах общей схемы сезонных передвижений. По наблюдениям Б.А. Зенковича, горбачи северного полушария совершают свои сезонные миграции в непосредственной близости от берегов. Вот почему на них и можно было охотиться даже древним народам с самым примитивным вооружением, в том числе устанавливать сети в узких проходах, бухтах и лагунах.
В тихоокеанских водах России обитают две популяции горбачей. Азиатская популяция зимует у островов Хонсю, Бонин, Марианские и Маршалловы, а весной мигрирует к Курилам, Камчатке, Командорским островам и Корякскому побережью.
Северо-западная популяция проводит зиму в теплых водах американской Калифорнии и Мексики, весной движется к Берингову проливу и южной части Чукотского моря, достигая кромки полярных льдов в проливе Лонга и акватории острова Врангеля.
Итак, в воды Чукотки приходят киты из обеих популяций, однако остается неизвестным, смешиваются ли они между собой на местах летне-осеннего нагула.
ПИТАНИЕ
Видовой состав пищи горбачей гораздо разнообразнее, чем у других полосатиков, ведь они придерживаются прибрежных вод, где набор объектов пита
Группа из трех горбачей во время питания. Характерный для этих китов вид спины и спинного плавника в момент погружения
ния намного шире и более значителен в количественном отношении. Так, горбачи поедают не только пелагических, но и придонных животных. Основными кормовыми объектами являются ракообразные, как относительно крупные (эуфаузииды), так и мелкие виды (калянусы), а также рыбы; меньшее значение имеют головоногие моллюски. Из рыб в желудках этих китов находили мойву, терпугов, сельдь, минтая, треску, песчанку, морских окуней, некоторых лососевых и др. В разных частях ареала объекты питания горбатых китов различны, причем наблюдается их смена по сезонам года. В полном желудке кита может быть до 800 крупных сельдей. Полагают, что суточное потребление пищи антарктического горбача может превышать 3 т.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ
Роды и спаривание у горбатых китов совершаются приблизительно в одни и те же сроки, поскольку беременность длится около года, но эти сроки сильно растянуты. В северном полушарии они происходят приблизительно с декабря по май-июнь, в южном — с августа по ноябрь. Цикл размножения горбачей более короткий, чем у других видов усатых китов, так как большая часть самок приносит детенышей ежегодно. В связи с этим восстановление их численности происходит быстрее, чем у других видов.
Длина новорожденного около 5 м. Молочное питание продолжается 5—6 месяцев, и за этот период длина детеныша увеличивается вдвое, но с переходом на самостоятельное питание рост замедляется. Маленький детеныш постоянно держится над спиной
Группа из двух взрослых горбачей и детеныша, снятая под водой. Китенок располагается непосредственно над спиной матери; такая поза характерна для детенышей большинства видов китов
матери, которая при движении как бы несет своего малыша в слое воды, перемещающемся вместе с нею.
К моменту достижения половой зрелости (в возрасте 4—5 лет) длина самцов из северного полушария составляет около 11, из южного — около 13 ти, самок — соответственно, 12 и 13,5 м. Физическая зрелость наступает приблизительно в возрасте 10 лет при различной длине тела. Продолжительность жизни горбачей не определена.
ПОВЕДЕНИЕ
Крупных скоплений горбатые киты не образуют, чаще всего держатся парами или тройками. Состав пар различный, иногда это два взрослых кита разного пола,
иногда два самца или две самки, но в летние месяцы, как правило, это самка с детенышем. В местах концентрации объектов питания такие мелкие группки порой образуют нечто вроде стада.
По наблюдениям китобоев, горбачи отличаются от других китов-полосатиков гораздо большим умом и сообразительностью, однако непугливость и тихо-ходность этих китов сделали их легким объектом охоты.
«Горбачи, пожалуй, самые беспечные среди всех китов. Плавают они очень часто вблизи берегов, входят в бухты и лагуны, на подход судна внимания не обращают, — впрочем, и кого им бояться? Косатка разве нападет, да и то это бывает чрезвычайно редко; если и случается, что, не находя другой добычи, голодные косатки и набросятся на горбача, то и он сможет ответить им мощными ударами своих «рук» и хвоста. Вообще же горбачи косаток не боятся и их не избегают. Мы не раз были свидетелями весьма безразличного отношения горбачей к приближению косаток, хотя в косяке горбачей были кормящие матки с сосунками. Косатки тоже не обнаруживали агрессивных намерений. ...
Очень многие из осмотренных мною горбачей имели повреждения на грудных плавниках и иногда на животе. В нескольких случаях можно было заметить следы, похожие на укус акулы. Но помня о привычках горбатых китов проходить через узкие проливы, входить в мелкие бухты и лагуны, допускаю, что эти повреждения получены ими от соприкосновения с подводными камнями.»1
1 Зенкович Б.А. Вокруг света за китами. М.: Географгиз, 1954. С. 178-179.
Горбатые киты обладают сильно развитым инстинктом взаимной привязанности, но если самец никогда не покидает раненой или загарпуненной самки, то она неизменно спасается бегством, когда ходивший с ней в паре самец оказывается загарпуненным. Однако самки горбачей очень привязаны к детенышам и всегда защищают их. Записано много случаев трогательного стремления матери спасти раненого малыша. Когда это не удавалось сделать, самка в ярости нападала на вельбот и даже небольшой китобойный катер, стремясь уничтожить врага.
У горбачей не только мать стремится спасти своего детеныша — взрослые горбачи оказывают друг другу помощь, доходящую до самопожертвования. Во времена расцвета китобойного промысла собрано большое количество сообщений о том, что к раненому киту бесстрашно подходили товарищи и пытались, крепко прижавшись к нему с двух сторон, поддержать его на поверхности, чтобы он мог дышать. Опытные китобои сообщали, что некоторые киты могли предугадать действия охотящихся за ними людей и легко уходили от преследования.
Горбач — единственный кит, который специально прыгает в косяках сельди, оглушая ее своими длинными плавниками, чтобы потом полакомиться ею. Более того, он кружит вокруг косяка сельди, уплотняя рыбу до предела, прежде чем схватить ее своей огромной пастью. Также поступают эти киты и с крилем (так называют большие скопления разных видов мелких ракообразных красновато-буроватого цвета). Обнаружив скопление криля, горбачи начинают выпускать мощные потоки мелких пузырьков воздуха и ходить вокруг криля по кругу, все время сужая его. Таким образом киты создают сильное
круговое течение, захватывающее этих рачков и собирающее его в центре плотной массой, которая еще более уплотняется и отделяется от окружающего пространства благодаря действию воздушно-пузырьковой завесы. Как только это происходит, горбачи ложатся на бок и устремляются к центру образовавшейся воронки, заглатывая каждый огромное количество рачков. Утверждают, что другие киты этого не делают, а довольствуются тем, что находят на своем пути.
Время пребывания горбача под водой не превышает 10-15 минут. Между двумя погружениями кит держится у поверхности моря, регулярно через короткие промежутки времени выныривает, выбрасывая серию фонтанов. Фонтан горбача по высоте (около 5 м) мало уступает среднему фонтану финвала, но значительно шире в начале колонны и как бы кучевидней. Эти киты довольно тихоходны, их максимальная скорость, вероятно, не превышает 28 км/час.
Иногда горбачи играют — громадные киты выпрыгивают из воды, полностью отрываясь от ее поверхности, размахивают в воздухе длинными грудными плавниками и с громким всплеском падают на бок. При нырянии на относительно долгий срок
Двойное сальто горбача при выпрыгивании из воды
горбач опускается почти вертикально и очень часто обнажает лопасти хвостового плавника — показывает «бабочку». По ее характерному виду и окраске и по столь же характерной «сгорбленности» тела во время ныряния горбачей легко отличить от других видов китообразных.
В водах Чукотки горбатые киты нередко входят в состав группировок столь же тихоходных серых китов и достаточно долгое время пасутся и передвигаются вместе с ними, прилаживаясь к их ритмам дыхания.
Горбач, выпрыгивающий из воды. Видны полосы на брюхе, белая окраска внутренней поверхности правого плавника и темная — наружней поверхности левого плавника, скопления эктопаразитов (обрастания) нижней части головы и правого плавника
Гигантская «бабочка» горбача. Хорошо видны зазубренные края хвостовых лопастей
Среди всех усатых китов только горбачи способны издавать очень сложные сочетания звуков, напоминающие пение. Поют они обычно в теплых водах, на зимовках, при этом каждая популяция обладает собственным типом песни. Записано несколько дисков с песнями горбатых китов, производящих на слушателей впечатление безысходной грусти.
ПАРАЗИТЫ, КОНКУРЕНТЫ, ВРАГИ
Горбачи больше, чем все остальные киты, страдают от эктопаразитов. Они покрыты огромным числом китовых вшей и обрастают множеством раковин усоногих рачков, коронул. На этих раковинах в свою очередь селятся «морские уточки», еще один вид из того же отряда усоногих ракообразных, которые свешиваются с раковин коронул, как бахрома. Обычно китовых вшей и рачков-паразитов на ките столь много, что зверь одет ими, как панцирем; особенно большое их количество располагается на голове, горле и у полового отверстия.
Один из мучителей горбатого кита — китовая вошь. Этот паразит намертво прикрепляется к коже кита тремя парами ног с крючьями
Два вида эктопаразитов горбатого кита.
На раковине усоногого рачка, глубоко внедрившейся в кожу кита, поселился другой паразит — усоногое ракообразное «морская уточка»
Б.А. Зенкович определил, что один кит может носить на себе до 500 кг этих маленьких врагов, которые не только доставляют ему физические мучения, но, безусловно, уменьшают скорость его передвижения. Из эндопаразитов горбатых китов отмечены нематоды и акантоцефалы.
Множество морских птиц сопровождают кормящихся горбачей, ловко пользуясь оглушенной и обездвиженной добычей, будь то рыба или ракообразные. Серьезной конкуренции этим китам ни морские птицы, ни другие виды морских млекопитающих не составляют.
Выше говорилось, что горбачи могут питаться придонными организмами; нередко они делают это вместе с серыми китами. В таких случаях можно говорить о возможной конкуренции этих двух видов.
То же самое происходит, когда огромные скопления планктона привлекают одновременно южного гладкого кита, настоящих полосатиков и горбача. Такой случай мне довелось наблюдать лишь однажды, севернее мыса Олюторский, — пара финвалов, один гладкий кит, три малых полосатика и две пары горбачей с разных сторон зашли в большое красноватое «облако» планктона и начали поедать его, словно мягкий пирог. Однако все киты вели себя спокойно, и никакой конкуренции за пищу между ними я не заметила. Первым ушел гладкий кит, за ним — малые полосатики, а финвалы и горбачи продолжали трудиться и после захода солнца.
Единственным врагом горбатых китов является человек, который значительно сократил их численность во всех районах Мирового океана.
Внешний вид горбача на рисунке из книги Ч. Скэммона (1874 г.). Художник тщательно изобразил два вида рачков-паразитов, сидящих на коже кита
ЧИСЛЕННОСТЬ
Численность горбатых китов неодинакова в различных частях ареала. Наиболее многочисленны они были в Антарктике, где общая величина популяций, возможно, до коммерческого промысла была близка к 100 тыс. голов. Однако в результате промысла к середине 60-х годов количество горбачей снизилось там примерно до 3000 голов, и с 1964 г. промысел их в южном полушарии был запрещен. В Северной Атлантике популяции горбачей всегда были малочисленны, и теперь этот зверь здесь редок. Первоначальная численность горбатых китов северной части Тихого океана, по расчетам, составляла 8,5-10 тыс.; в результате промысла к концу 1950-х годов она сократилась до 5000 голов. Современную численность северного тихоокеанского горбача специалисты определяют в 1,5 тыс. голов.
СТАТУС
В 1963 г. Международная китобойная комиссия наложила запрет на добычу горбачей Северной Пацифи-ки, в последующие годы промысел этого вида был запрещен и в других частях его ареала. Вид внесен в Приложение 1 Конвенции «О международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения» (СИТЕС). Северный горбатый кит внесен в Красную книгу России, категория статуса редкости — 1 (находящийся под угрозой исчезновения).
Бесстрашные серебристые чайки выхватывают рыбу прямо из пасти горбатого кита
ОХРАНА
В России не принято никаких специальных мер по охране северного горбача, кроме внесения его в Красную книгу.
6
Зубатые киты
( Odontoceti)
В водах Чукотки обитают представители пяти семейств зубатых китообразных:
Русское название
Белуха
Нарвал Косатка Обыкновенная морская свинья Белокрылая морская свинья Кашалот
Северный плавун
Английское название
Belukha (White Whale) Narwhal
Killer Whale (Orca)
Harbor Porpoise
Dall’s Porpoise
Sperm Whale
Baird’s Beaked Whale (Giant Bottelenose)
Научное (латинское) название
Delphinapterus leucas
Monodon monoceros Orcinus orca
Phocoena phocoena
Phocoienoides dalli Physeter catodon (macrocephalus) Berardius bairdi
Зубатые китообразные представлены большим количеством видов и освоили значительно большие водные пространства, чем усатые киты. Некоторые из них стали превосходными глубоководными ныряльщиками (кашалот, клюворылы), другие смогли
Внешнее строение зубатого кита (дельфин)
приспособиться к жизни в пресных водах крупных тропических рек и озер (семейство речных дельфинов). Все зубатые китообразные — хищники, питающиеся в основном головоногими моллюсками и рыбой. Только косатка, единственная среди всех китов, нередко охотится и на теплокровных животных — других морских млекопитающих (серых китов, настоящих и ушастых тюленей, моржей), а также морских птиц, когда те сидят на воде.
Объем ротовой полости зубатых китов сравнительно невелик, язык короткий, зубы разнообразны по величине, форме и расположению; у некоторых видов зубной аппарат развит неполностью (клюворылые). Зубатые киты схватывают зубами и всасывают достаточно крупных одиночных животных.
Зубы некоторых зубатых китообразных
Скелет зубатых китов отличается от скелета усатых китов большей плотностью, меньшей пористостью и меньшим содержанием жира в костях (за исключением кашалота). Эти отличия прямо связаны с более высокой скоростью движения зубатых китов, охотников на активную и нередко весьма скоростную добычу, с величиной и силой их мускулатуры и увеличением удельного веса тела. Больший вес тела усиливает инерционный эффект, что позволяет экономить мышечную энергию и запас кислорода при погружении. Укорочение и слияние шейных позвонков сближает голову с туловищем, благодаря чему передние отделы спинных мышц прикрепляются в затылочной области черепа. Все это прочно фиксирует череп и устраняет вибрацию головы во время быстрого движения и молниеносных атак на добычу.
Зубатые киты имеют не два наружных дыхательных отверстия (две ноздри), как усатые киты и все другие млекопитающие, а только одно дыхало (одну ноздрю), которое находится на темени, за лобной (жировой) подушкой. Лишь у кашалота дыхало сдвинуто вперед, расположено с левой стороны и открывается под углом в 45°.
Верхние дыхательные пути зубатых китов (носовой канал с воздушными мешками). А — нарвал; Б — кашалот. Черепа китов заштрихованы, жировые подушки над ними выделены точками
Серьезные изменения происходят в строении черепа, который у всех зубатых китов асимметричен. Это уникальное явление до конца не понято, но полагают, что асимметрия черепа связана с особенностями строения верхних дыхательных путей и способами генерации (производства), испускания и приема эхолокационных ультразвуковых сигналов. Направленному излучению этих сигналом помогает лобный выступ (жировая подушка), которой нет у усатых китов.
Зубатые киты обладают самой сложной среди всех животных слуховой системой. Не так давно ученые открыли ее удивительные возможности. Киты могут одновременно испускать два типа звуковых сигналов: один тип сигналов обеспечивает связь с сородичами, другой служит для эхолокационного изучения ближнего пространства и поиска в нем добычи. Принимая эхо (отражение) от собственных ультразвуковых сигналов, зубатые киты безошибочно определяют, на кого они охотятся, как велико расстояние до берега, скалистый он или песчаный и многие другие характеристики окружающей среды. Но все это возможно
Косатка, хватающая одиночную добычу, слева — детеныша нерпы, справа — рыбу
только на близком расстоянии, ведь коротковолновые звуки быстро гаснут в воде. А во время шторма работа эхолокатора зубатых китов крайне затруднена — низкочастотные шумы волн поглощают ультразвуковые сигналы. Предполагают, что массовые обсыхания зубатых китов и их выбрасывания на берег в той или иной степени связаны с нарушением работы эхолокатора этих животных.
Зрение зубатых китов и дельфинов, по крайней мере, у обследованных видов, развито неплохо, с его помощью они уверенно ориентируются и в воде, и на воздухе, но только на небольших расстояниях. Осязание сильно редуцировано у большинства видов, но у некоторых речных дельфинов, охотящихся в мутной воде, вибриссы не только сохранились на морде, но появились даже на теле этих животных. А вот обоняние у представителей этого подотряда китообразных исчезло полностью, в этом плане зубатые китообразные — уникальное явление среди всех позвоночных животных, от самых примитивных до человека включительно.
Таким образом, целый ряд важнейших характеристик строения зубатых китов, их способов ориентации в пространстве и питания отличают эту группу от усатых китов. Кроме того, большинство видов зубатых китообразных, прежде всего дельфины, принадлежат к животным с высоким интеллектом, таким, как обезьяны, слоны, врановые птицы, попугаи. Дельфины способны решать весьма сложные задачи, хорошо дрессируются в неволе, у них чрезвычайно развито исследовательское поведение. Сложная организация поведения и удивительные возможности слуховой системы выделяют зубатых китов из всего ряда высших позвоночных животных, к которым принадлежат млекопитающие и птицы.
Обыкновенная морская свинья
Косатка
Белокрылая морская Северный плавун
свинья
Районы обитания (ареалы) зубатых китов в водах Чукотки
Белуха
Нарвал
Кашалот
Районы обитания (ареалы) зубатых китов в водах Чукотки. Продолжение
Семейство дельфиновых, или настоящих дельфинов (Familia Delfinidae)
Род косаток
(Genus Orcinus)
В роде один вид, косатка, хорошо известный охотникам Чукотки. По традиционным представлениям морских охотников Чукотки, косатка — священный зверь, которого нельзя убивать.
Косатка
(Orcinus orca Linnaeus, 1758)
ОБЩАЯ ХАРАКТЕРИСТИКА
Косатка — хищный дельфин, самый крупный представитель семейства с ясно выраженным половым диморфизмом. Длина тела самцов может достигать 10 л/, самок — до 8 м; средняя длина самцов около 6,5 jw, самок — около 6 м. Тело более мощное, относительно более короткое и толстое, чем у других
Череп косатки. Хорошо видно, каким страшным оружием являются зубы этого дельфина
дельфинов. Голова крупная с округлой передней частью, клюва нет. Грудные плавники широкие и короткие, округлой формы. Спинной плавник необычайно высок (видовая особенность) — у самцов до 1,5 jw, у самок гораздо ниже. Такое различие — самый заметный признак полового диморфизма у этого вида. Зубы массивные, покрытые эмалью и очень прочные; они сжаты и несколько загнуты спереди назад для того, чтобы легче было схватить и удержать крупных и сильных животных. На обеих челюстях 40-48 зубов.
Окраска своеобразная, характерная для вида. Спина и бока черные, горло и брюхо белые. В задней части туловища белая окраска двумя языками поднимается по бокам кверху, иногда достигая спины. Два белых пятна расположены перед глазами и над ними. У самцов белое или беловатое пятно позади спинного плавника. Отмечена значительная географическая изменчивость окраски, связанная с существованием внутри вида многих популяций и устойчивых родственных групп, так называемых подов.
РАСПРОСТРАНЕНИЕ И МИГРАЦИИ
Населяет почти весь Мировой океан, от кромки льдов Арктики до кромки льдов Антарктики. Встречается как вблизи берегов, так и над большими глубинами. В арктических морях распространение этого вида прерывистое: косатка не отмечена в море Лаптевых и в водах центральной части Канадского Арктического архипелага.
Косаток наблюдают на большей части ареала в течение почти всего года. Однако некоторые сезонные перемещения они, безусловно, совершают, так как численность их в отдельных районах в разные периоды года колеблется. Очень хорошо это заметно в холодных водах обоих полушарий.
Вдоль побережья Чукотского полуострова, от залива Креста в Беринговом море до поселка Ванка-рем в Чукотском море насчитывается 7-9 локальных групп косаток по 6-10 животных в каждой группе.
Косатка выпрыгивает из воды
Наибольшая активность косаток отмечена в июле-сентябре; в ноябре, с появлением тяжелых льдов, косатки отходят к югу и возвращаются обратно в начале-середине мая, вместе с серыми китами.
ПИТАНИЕ
Единственный вид китообразных, который наряду с другими объектами питания поедает и теплокровных животных. В желудках косаток находили остатки морских птиц, тюленей, морских львов, моржей, дельфинов, малых полосатиков и сейвалов. В последние два десятилетия появилось много сообщений о нападении косаток на китов и зарегистрированы многочисленные следы мощных зубов косаток на телах и плавниках многих видов китов, прежде всего серого кита и горбача. Но все же основу питания этих хищных дельфинов составляют рыбы и головоногие моллюски (преимущественно кальмары). Количество видов рыб, поедаемых косаткой, довольно велико (лососевые, сельдь и другие сельдевые, треска и мойва, палтусы и скаты, различные виды мелких акул, тунцы и другие рыбы).
В огромных скоплениях рыб косатки могут питаться вместе с другими морскими зверями (сивучами, морскими котиками, китами и дельфинами), не нападая на них. У берегов Чукотки объектами охоты косаток чаще всего становятся небольшие серые киты и моржи. Нередко хищники терпеливо дежурят возле моржовых лежбищ, выбирая наиболее легкую добычу. У серых китов косатки прежде всего вырывают язык, а тушу кита могут оставить нетронутой. Дважды, в декабре 1981 г. и ноябре 1983 г., охотники пос. Сиреники наблюдали нападение косаток на гренланд
ских китов средней величины. В обоих случаях атака длилась более 4 часов; в 1981 г. кит был убит, а в 1983 г. косатки бросили гренландского кита и переключились на проходившую мимо небольшую группу моржей.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ
Роды и спаривание происходят в летние месяцы. Продолжительность беременности косаток точно не установлена, но большинство специалистов считают, что она длится 16 месяцев. Детеныш рождается длиной 2,5—2,7 м, к годовалому возрасту он вырастает до 3,5 м. Возраст, при котором у самцов и самок наступает половая зрелость, не определен.
ПОВЕДЕНИЕ
Косатки — животные с высоким уровнем интеллекта, сильно выраженным стремлением к коллективности и развитым игровым поведением. Как
У косаток даже малыши движутся в строю вместе с взрослыми животными
в открытом море, так и вблизи берегов они встречаются обычно локальными группами (подами), основой которых являются родственные связи между зверями. Иногда косатки образуют скопления в сотни голов. Небольшие группы чаще всего передвигаются как бы строем, что легко наблюдать по выступающим над поверхностью моря громадным спинным плавникам.
На тюленей, дельфинов и китов косатки охотятся стаей, причем их охотничье поведение поразительно напоминает поведение стаи волков. Вот что писала об этом Е.С. Рубцова: «Эскимосы считали, что зимой косатка превращается в волка, а летом снова надевает шкуру косатки. Эскимосы полагали, что волк и косатка одно и то же, что это, собственно, человек в другом одеянии».1
Эти звери могут заходить в крупные реки и оставаться в пресной воде до 15 суток. Во время поисков пищи (исключая моменты охоты) скорость движения косаток 12-14 км/час, максимальная скорость хода составляет до 55 км/час. Обычно косатки уходят под воду не более чем на 2-5 мин. Высота фонтана не превышает 2 м, и он плохо заметен.
Косатки хорошо приживаются в океанариумах, быстро привыкают к человеку, легко обучаются обычным трюкам дельфиньего цирка. Удивительно, но в неволе они не проявляют никаких хищных инстинктов по отношению к людям и дельфинам других видов, которые живут вместе с ними в бассейне.
У косаток хорошо развито водное и воздушное зрение. Во льдах они нередко выставляются из воды
1 Е.С. Рубцова. Материалы по языку и фольклору эскимосов. 1954. Стр. 156.
и, найдя подходящую добычу (нерпу или маленького моржа), стремятся перевернуть льдину или, выбросившись на нее, столкнуть свою жертву в воду. Однако главным для косаток является слух, прежде всего прекрасно развитая система эхолокации. В океанариумах проводят специальные опыты по изучению сигналов, которые эти дельфины используют во время охоты, для ориентации в пространстве и связи со своими сородичами.
ПАРАЗИТЫ, КОНКУРЕНТЫ, ВРАГИ
Эктопаразиты у косатки неизвестны; эндопаразиты представлены 8 видами гельминтов.
Косатка не имеет конкурентов и врагов, кроме человека.
ЧИСЛЕННОСТЬ
Общая численность всех популяций этого вида не определена даже приблизительно. Для Берингова моря расчетная численность косаток находится в пределах 900 голов.
СТАТУС И ОХРАНА
Косатка — обычный и многочисленный вид, который в настоящее время не нуждается в охране.
Сем. Морских свиней (Familia Phocoenidae)
Семейство появилось в отряде китообразных недавно — прежде был род морских свиней, входивший в семейство дельфиновых. Современные специалисты сочли различия дельфинов и морских свиней существенными и повысили уровень этого рода на одну ступень, сделав его семейством. В семействе четыре вида, два из них встречаются в водах России. Морские свиньи — небольшие звери, более тяжелого и грубого телосложения, чем дельфины, с короткой головой без «клюва».
Род морских свиней (Genus Phocoena)
Обыкновенная морская свинья
(Phocoena phocoena Linnaeus, 1758)
Вид делят на три достоверных подвида по районам их обитания:
североатлантическая обыкновенная морская свинья — наиболее крупная из трех подвидов;
черноморская обыкновенная морская свинья — средняя по своим размерам;
тихоокеанская обыкновенная морская свинья —
самая мелкая из трех подвидов.
ОБЩАЯ ХАРАКТЕРИСТИКА
Один из самых мелких дельфинов, средняя длина 1,5-1,6 м, максимальная — не более 2 м. Масса 55-75 кг. Три очень крупные самки из северной части Тихого океана длиной 178,5 см, 173 см и 168 см весили соответственно 75,2 кг, 70,3 кг и 73,4 кг.
Тело морской свиньи укороченное и утолщенное, голова небольшая, лобовое утолщение (жировая подушка) развито слабо, клюва нет. Спинной плавник низкий, укороченные грудные плавники округлой формы. Окраска спинной стороны тела темная, иногда почти черная, брюшной — светлая. Граница раздела цветов на боках, как правило, неровная,
РАСПРОСТРАНЕНИЕ И МИГРАЦИИ
Обитает только в северном полушарии, где населяет умеренные и холодные воды. В северной половине Тихого океана обыкновенная морская свинья распространена широко, но избегает открытого моря, придерживаясь прибрежных мелководий. В азиатских водах обитает в Чукотском море, Беринговом проливе и Беринговом море, вдоль берегов Камчатки, Командорских и Курильских островов, в Охотском и Японском морях, Японии и Кореи. В восточной части встречается от мыса Барроу (Аляска) в Чукотском море, вдоль побережий США и Канады до полуострова Калифорния и Мексики.
Миграции не изучены, хотя некоторые сезонные перемещения, особенно заметные в северных частях ареала, безусловно, имеют место. Можно с уверенностью говорить, что в северных частях Тихого океана и Чукотском море морские свиньи проводят только летние месяцы, а на зиму уходят на юг.
ПИТАНИЕ
Материалы по питанию этого подвида свиньи крайне скудны. М.М. Слепцов и А.Г. Томилин сообщали, что он питается рыбой (треска, мойва, навага, сайка,сельдь, шэд, акула, угольная рыба, сиг), креветками и головоногими моллюсками.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ
Данные по биологии размножения имеются лишь для черноморского подвида. Здесь приведены только те факты, которые, безусловно, являются общими для всех трех подвидов. Беременность длится 9-10 мес. Длина новорожденных около 75 см, масса от 2,8 до 7,9 кг. Молочное питание продолжается до 4 месяцев или несколько дольше. Молоко может содержать почти 46% жира. Самки приносят детенышей, очевидно, ежегодно, т. е. темп пополнения стада относительно быстрый. Время спаривания и родов неизвестно, продолжительность жизни этих дельфинов не установлена.
ПОВЕДЕНИЕ
Обитает главным образом в прибрежных мелководных районах. Животное стадное, но чаще встречается мелкими группами. Зверь сравнительно тихоходный; время ныряния — до 6 минут, при погружении характерно сильное изгибание тела. Из воды обычно не выпрыгивает.
ПАРАЗИТЫ, КОНКУРЕНТЫ, ВРАГИ
По количеству видов эндопаразитов морские свиньи занимают одно из первых мест среди всех китообраз
ных — у них зарегистрировано 18 видов гельминтов, которые найдены почти во всех внутренних органах.
Конкурентами морской свиньи за рыбу могут быть некоторые ластоногие.
Наиболее опасным врагом этого вида у берегов Чукотки являются косатки.
ЧИСЛЕННОСТЬ
Тихоокеанская обыкновенная морская свинья занимает обширный ареал, однако конкретные данные об общей численности подвида отсутствуют. У берегов Чукотки этот зверь явно немногочислен — о встречах с ним морские охотники сообщают крайне редко.
СТАТУС
Вид внесен в Приложение II СИТЕС (Конвенция о международной торговле дикими видами фауны и флоры, находящимися под угрозой исчезновения). Тихоокеанская обыкновенная морская свинья внесена в Красную книгу России, категория статуса редкости — 4 (неопределенные по статусу).
ОХРАНА
В России не предпринято никаких специальных мер по охране тихоокеанской обыкновенной морской свиньи, кроме внесения ее в Красную книгу. Сообщения о встречах с этим зверем в водах Чукотки представляют большой интерес.
Род белокрылых морских свиней
(Genus Phocoenoides)
В роде один вид — белокрылая морская свинья. По рассказам старейших охотников пос. Сиреники, когда-то этот зверь использовался в ритуальной охоте, имитируя собой гренландского кита. Такая охота проводилась, если долгое время не было настоящих гренландских китов.
Белокрылая морская свинья
(Phocoenoides dalli True, 1885)
Вид обитает в северной части Тихого океана, имеет две хорошо выраженные формы: северную и южную. У северной формы расположенные на брюшной стороне и боках белые пятна далеко не достигают уровня грудных плавников. У южной формы белое пятно на боках заходит вперед за основания грудных плавников и почти достигает области глаз. На брюхе оно оканчивается у основания грудных плавников.
ОБЩАЯ ХАРАКТЕРИСТИКА
Мелкое китообразное, но крупнее обыкновеной морской свиньи; длина тела около 1,8 м, максималь-
ные размеры немногим более 2 ж, общая масса 135-220 кг. Тело укороченное, утолщенное и мощное; голова небольшая, клюва нет. Нижняя челюсть немного длиннее верхней. Спинной плавник имеет форму почти равнобедренного треугольника с отогнутой назад заостренной вершиной. Грудные плавники широкие, округлые, темного цвета. Окраска всей верхней поверхности тела темная, почти черная, большая часть брюшной поверхности белая, на боках два больших продольных белых пятна. Спинной плавник частично (иногда полностью) белый.
РАСПРОСТРАНЕНИЕ И МИГРАЦИИ
Этот вид обитает только на западе и востоке северной половины Тихого океана; распространен от Японии и побережья полуострова Калифорния до Берингова пролива и южных районов Чукотского моря. В водах России встречается в Японском и частично Охотском морях, у Курильских островов, восточного побережья Камчатки, в Анадырском заливе, Беринговом проливе и прилегающих водах Чукотского моря.
Сезонные миграции четко выражены в северной части ареала, которую белокрылые морские свиньи покидают на зиму и вновь заселяют после исчезновения льда весной.
ПИТАНИЕ
Главными объектами питания являются стайные рыбы и головоногие моллюски. В желудках этих
зверей находили светящихся рыб, скумбрию, хека, ставриду, сельдь, несколько видов кальмаров. Значение отдельных видов животных в питании белокрылой морской свиньи в разных местах ареала различно.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ
Размножение происходит в летние месяцы, роды, вероятно, в июле-августе, спаривание — в августе-сентябре. Беременность длится 11-12 месяцев. Длина новорожденного около 90 см. В период молочного питания детеныши растут очень быстро, а с переходом на самостоятельное питание темп роста замедляется. Самки достигают половой зрелости не ранее чем в 2-летнем возрасте, самцы — в 3-летнем и даже позже.
ПОВЕДЕНИЕ
Животные стадные, но обычно держатся мелкими группами, реже образуют скопления в 100 голов и более. Обитают как в прибрежных, так и в открытых водах. Относятся к высокоскоростным видам; часто выбрасываются из воды, совершая в воздухе длинные прыжки. Нередко сопровождают корабли, играя и легко обгоняя даже быстроходные суда. Ныряют на 2-4 минуты, глубина погружения не известна. При выходе на поверхность часто оставляют за собой «петушиный хвост» — широкий след в воздухе из водяных брызг.
ПАРАЗИТЫ, КОНКУРЕНТЫ, ВРАГИ
Эндопаразиты практически не изучены. О конкурентных отношениях белокрылой морской свиньи с другими видами китообразных и рыбоядными ластоногими ничего не известно. Косатки не представляют опасности для этого быстроходного зверя.
ЧИСЛЕННОСТЬ
При относительно небольшом ареале вид сравнительно многочисленный, хотя даже приближенного цифрового определения общего количества этих животных не сделано.
СТАТУС
Эндемик Северной Пацифики. Вид внесен в Приложение II СИТЕС (Конвенция о международной торговле дикими видами фауны и флоры, находящимися под угрозой исчезновения.
ОХРАНА
В России не предпринято никаких специальных мер по охране белокрылой морской свиньи.
Семейство нарваловых (Familia Monodontidae)
Семейство нарваловых объединяет китообразных средних размеров (длина 3-6 м, общая масса в среднем 500, максимальная — до 1000 кг). Тело короче и толще, чем у дельфинов; Голова небольшая, округлая, с большой жировой подушкой; клюва нет. Спинной плавник отсутствует. Грудные плавники широкие, округлой формы. Отмечается ясно выраженная возрастная изменчивость окраски. Животные стадные, образуют группы различной численности.
Обитают в холодных и частично умеренных водах северного полушария.
Семейство состоит из двух родов — рода белух и рода нарвалов, оба представлены в водах России.
Род белух
(Genus Delphinapterus)
В роде один вид — белуха. Это животное хорошо известно морским охотникам Чукотки, которые правильно описывают многие особенности его биологии и поведения.
Белуха
(Delphinapterus leucas Pallas, 1776)
Белухи, обитающие в морях России, образуют три стада (популяции):
беломорская белуха — длина тела в среднем 3,12 м, обитает в Белом и частично в Баренцевом морях;
карская белуха — длина тела около 3,9 м, населяет Баренцево, Карское моря и море Лаптевых;
дальневосточная белуха — наиболее крупная, средняя длина тела 4,25 м\ распространена в северной части Японского моря, Охотском, Беринговом и Чукотском морях.
ОБЩАЯ ХАРАКТЕРИСТИКА
Максимальная длина тела самцов до 6 м, самок — до 5 м. Тело умеренно удлиненное; хорошо выражена шея. Голова сравнительно небольшая, округлая, с сильно развитой и очень подвижной жировой подушкой. Простые колышковидные зубы (общее число до 52) расположены на обеих челюстях и посажены редко. Грудные плавники на концах изогнуты, причем у старых особей это выражено резче.
Помимо размеров тела самцы и самки имеют и другие различия. У самцов корпус относительно длиннее, хвостовые плавники крупнее, число зубов больше и лобный выступ (жировая подушка) развит сильнее, чем у самок.
Окраска белух однотонная, изменяющаяся с возрастом. Детеныши темного цвета, который постепенно светлеет, проходя через цветовые стадии (серую и голубую). Взрослые животные имеют чисто белую окраску, иногда с небольшим желтоватым оттенком, хвостовые лопасти с темным краем.
РАСПРОСТРАНЕНИЕ И МИГРАЦИИ
Белухи населяют северные части Тихого и Атлантического океанов, а также моря Северного Ледовитого океана почти кругополярно. Дальневосточная белуха обитает на севере Японского моря (проливы Татарский и Лаперуза), в Охотском, Беринговом и Чукотском морях. «Чукотская» белуха спускается к югу до района мыса Наварин, а на север проникает до 75° с.ш., а в некоторых местах и еще дальше. В Восточно-Сибирское море белуха только заходит.
Белухам свойственны сезонные миграции, которые хорошо прослеживаются в водах Чукотки. Весной (апрель-май) белухи покидают Берингово море и уходят в бассейн Ледовитого океана, распределяясь там на большом пространстве, от Восточно-Сибирского моря до моря Бофорта. Меньшая часть этих зверей остается летовать в Анадырском заливе, питаясь лососевыми во время хода рыбы на нерест в крупные реки, впадающие в залив, особенно, в реке Анадырь. Отдельные звери странствуют по всему Анадырскому заливу. Обычно каждое лето несколько белух попадают в рыбачьи сети у поселков Энмелен и Нунлигран, где больше всего проходной рыбы.
Осенью (октябрь-ноябрь) белуха идет с севера вместе с другими морскими млекопитающими, в основном гренландскими китами и моржами. Её зимнее распределение у побережья Чукотки зависит от скоплений рыбы, прежде всего сайки. Охотники хорошо знают и предвидят, где пойдет белуха и в каких местах она будет держаться.
ПИТАНИЕ
На всем обширном ареале главными объектами питания служат различные рыбы, причем поедаются преимущественно виды, образующие скопления, а число их довольно велико. Однако наибольшее значение имеют лососевые (кета и горбуша), сельдь, навага, мойва, сайка, сиговые рыбы. Меньшую роль играют некоторые виды ракообразных, ими питаются главным образом молодые звери, а также некоторые головоногие и другие моллюски. Считают, что молодые белухи, переходящие с молочного питания на самостоятельное, начинают поедать ракообразных, затем переходят на питание мелкими рыбами и, лишь достигнув определенного возраста, могут ловить таких крупных рыб, как дальневосточные лососевые.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ
Период родов и спаривания растянут на 5-6 месяцев, но для большей части самок эти сроки более сжаты, причем различны в зависимости от географических районов ареала. Вероятно, пик спаривания приходится на май-июнь. Беременность длится 11-12 месяцев. Длина новорожденных детенышей 1,5-1,8 м. В период молочного выкармливания, которое длится 8-10 месяцев, они растут быстро. У десятимесячных детенышей прорезываются зубы, и они переходят на питание ракообразными. Длина детеныша в возрасте 1 года достигает почти 3 м.
По мере роста белух изменяется и их окраска. Детеныши темного окраса; в первой цветовой груп
пе, названной «серые», животные имеют сине-серую окраску и все они неполовозрелые. Следующая груп-па — «голубые» — является переходной от группы «серых» к группе белых. «Голубые» представлены как неполовозрелыми, так и уже созревшими зверями. Последняя группа — «белые» — включает только взрослых половозрелых животных обоего пола, причем у более молодых животных этой группы окраска чисто белая, у старых — с желтоватым оттенком. Половой зрелости самки достигают в возрасте 4-6, самцы — 6-9 лет. Продолжительность жизни не установлена.
ПОВЕДЕНИЕ
Типичное стадное животное, образующее группы различной численности. Одиночные звери встречаются редко. В погоне за проходными рыбами, на-
Вынырнувшая из воды белуха, осматривается и издает звуки. Жировая подушка (лобный выступ) сдвинута вперед
пример, лососевыми, порой образуют громадные быстро идущие косяки. Вершину такого косяка обычно составляют взрослые самцы, в середине и в конце косяка идут самки (в том числе самки, сопровождаемые детенышами) и неполовозрелые животные. Мелкие группы, как правило, состоят из животных разного пола и возраста.
Белухи обитают преимущественно в зоне прибрежных мелководий, опускаясь в поисках пищи на глубину 10 м и более. Под водой держатся около 5 минут, но могут оставаться и до 15 минут. При спокойном передвижении идут со скоростью около 9-10 км/час, максимальная скорость передвижения, вероятно, не превышает 18-20 км/час. В летние месяцы, питаясь проходными рыбами, белухи подолгу находятся в опресненной воде возле устьев рек, более того, в погоне за рыбой нередко заходят в крупные реки, поднимаясь по ним иногда на расстояние до 1000 км.
Льдов белуха не избегает, в некоторых случаях даже зимует в больших полыньях, хотя порой попадает в ледовый плен. Может проходить сотни километров через районы сплоченных льдов, стремясь достичь чистой воды. Полагают, что предвидеть подвижки льдов и прокладывать себе путь под ледяными полями этим животным помогают их незаурядные способности к эхолокации. Охотники отмечают также, что во время штормовой погоды белух не видно; возможно, мощные инфразвуковые шумы шторма снижают их навигационные возможности, особенно вблизи берегов.
Помимо эхолокационных сигналов белухи издают разнообразные звуки, которые служат средством общения между животными.
Исследователи и морские охотники отмечают необыкновенную осторожность белух, их чуткость ко всем звукам в воде и на воздухе, а также перемещениям по льду. Прежде на белуху охотились у полыней, поджидая, когда зверь вынырнет для вдоха. Охотники одевали белую маскировочную одежду и буквально замирали на месте — животные могли осторожно и незаметно вынырнуть поблизости, разглядеть человека и уйти от опасного места. Однако в погоне за рыбой белуха может потерять осторожность и подойти под выстрел.
ПАРАЗИТЫ, КОНКУРЕНТЫ, ВРАГИ
У белухи найдены 13 видов эндопаразитов-гельминтов. Наружные паразиты не известны. Пищевыми конкурентами являются некоторые ластоногие, прежде всего ларга. К врагам белух можно отнести косатку и белого медведя, но большого ущерба эти звери белухам не приносят.
ЧИСЛЕННОСТЬ
Весьма приближенно численность белух во всех стадах (популяциях) мира определена в 150-200 тыс. голов. Точное количество белух, обитающих в водах Чукотки, никогда не определялось, но, по некоторым расчетам, их не более 10-15 тыс. Однако в период миграций их число может возрастать за счет особей, приходящих с американской стороны Берингова и Чукотского морей.
СТАТУС
Дальневосточная белуха, как и другие, внесена в Приложение II СИТЕС (Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения).
ОХРАНА
В России не предпринято никаких специальных мер по охране дальневосточной белухи.
Несколько лет назад международные и российские экологические организации выразили резкий протест по поводу коммерческой добычи белухи на Дальнем Востоке, и под давлением общественности Правительство России запретило промысел белух в торговых целях.
Род нарвалов
(Genus Monodon)
В роде один вид — нарвал, крайне редкий и малоизвестный морской зверь у берегов Чукотки.
Нарвал, или единорог
(Monodon monoceros Linnaeus, 1758)
ОБЩАЯ ХАРАКТЕРИСТИКА
Животное средних размеров, максимальная длина самцов немногим больше 6 м, самок — около 4,5 м. Масса до 1,6 т. Тело несколько укороченное, без спинного плавника, но у многих животных вдоль слегка выпуклой спины тянется невысокая кожная складка до 5 см высотой и 75 см длиной. Шейный перехват выражен слабее, чем у белухи. Голова небольшая с сильно развитой жировой подушкой, придающей ей округлую, даже шарообразную форму. Рот маленький, расположен снизу. Верхняя губа выступает несколько вперед над нижней.
Зубная система весьма своеобразна, именно благодаря ей самцы нарвала имеют совершенно уникальный облик. На нижней челюсти зубов нет, на верхней — всего два (по одному с каждой стороны). У самок они не прорезываются, а у самцов правый зуб, как правило, также остается скрытым в десне, а левый превращен в громадный бивень, винтообразно изогнутый по всей длине и направленный горизонтально вперед. Его длина может достигать 300 см. У некоторых особей иногда развивается и правый бивень, но он всегда короче левого.
Асимметрия черепа имеет необычный характер в связи с присутствием на левой стороне бивня: челюстная и межчелюстная кости более развиты с левой стороны, а мозговой отдел черепа сильнее развит на правой стороне.
Окраска изменяется с возрастом. Молодые животные имеют однотонную темную окраску, взрослые —
Череп нарвала. Слева — нормальное развитие левого зуба в бивень, хорошо заметна асимметрия черепа. Справа — редкий случай развития обоих верхних зубов в бивни
I
Бивень самца нарвала, добытого 26 июля 1982 г.
в Пойнт-Инлет на о. Баффинова Земля (Канада)
светлую, желтовато-серую. По верхней части туловища беспорядочно разбросаны небольшие темные пятна неправильной формы, которые могут сливаться.
РАСПРОСТРАНЕНИЕ И МИГРАЦИИ
Нарвал обитает только в холодных водах Арктического бассейна, где распространен по арктическим морям и всему Ледовитому океану, однако чаще встречается в североамериканском секторе Арктики и водах Гренландии. Известны отдельные дальние заходы нарвалов к югу. На северо-востоке России нарвалы или их остатки отмечены у мыса Сердце-Камень, в Беринговом проливе, в районе острова Аракамчечен и даже на острове Беринга (Командорские острова).
Нарвалы совершают регулярные миграции, сроки и дальность которых определяются движением кромки арктических льдов. В летние месяцы звери держатся в районе 80-85° с. ш., на зиму вместе со льдами спускаются к югу. Районы и летнего, и зимнего обитания зависят от гидрометеорологических и ледовых условий года.
ПИТАНИЕ
Главной пищей нарвалов являются головоногие моллюски, на втором месте рыбы. В желудках нарвалов найдены остатки нескольких видов кальмаров, а также лососевых рыб, сельди, трески, скатов, палтусов и камбал.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ
Новорожденный детеныш нарвала.
Видны остатки пуповины
Роды и спаривание могут проходить в течение большей части года. Продолжительность беременности не установлена. Рождается обычно один детеныш длиной около 1,5 м; двойни крайне редки.
ПОВЕДЕНИЕ
Как правило, держатся небольшими группами, реже — одиночками, вдали от берегов, к которым подходят только в поисках пищи. Группы могут состоять из животных разного пола и возраста, только из самцов или только из самок с детенышами. Порой
Самцы нарвалов с помощью бивней выясняют, кто сильней
группа неподвижно лежит на поверхности моря, и тогда бивни всех самцов направлены вверх. Пребывание во льдах для нарвалов является обычным.
ПАРАЗИТЫ, КОНКУРЕНТЫ, ВРАГИ
На коже нарвала найдено два вида китовых вшей, а во внутренних органах — три вида гельминтов.
Реальных конкурентов у нарвалов нет. Врагами могут быть белые медведи, а для молодняка — полярные акулы.
ЧИСЛЕННОСТЬ
В пределах всего ареала вид очень малочисленный, почему может быть даже отнесен к группе редких животных. Есть немало указаний, что в прошлом встречались стада нарвалов в тысячи голов, но в последние несколько десятилетий таких крупных скоплений никто уже не наблюдал.
СТАТУС
Нарвал внесен в Приложение II СИТЕС (Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения). Внесен в Красную книгу России, категория статуса редкости — 3 (редкий).
ОХРАНА
В России не предпринято никаких специальных мер по охране нарвала.
Семейство клюворылов (Familia Zi phiidae )
В семейство входят средние и крупные зубатые киты длиной от 5-6 до 10-12 м. Распространены в теплых и холодных водах обоих полушарий. В семействе 5 родов, из них в морях России представлены 4 рода — плавуны, бутылконосы, клюворылы и ремнезубы.
Род плавунов
(Genus Berardius)
Крупнейшие в семействе киты с хорошо выраженным клювом. В роде два вида: южный плавун и северный плавун, который встречается в водах Чукотки.
Северный плавун, или берардиус Бэрда
(Berardius bairdi Stejneger, 1883)
ОБЩАЯ ХАРАКТЕРИСТИКА
Крупный кит; самки в среднем длиннее самцов: длина тела самок до 12,5 jw, самцов — до 11 м. Масса северного плавуна 8-10 т. Небольшая голова с характерным сферическим «лбом», полулунное дыхало обращено рожками назад, явно выраженный
клюв удлинен и уплощен. Верхняя челюсть в области клюва чуть уже и короче нижней. Зубов две пары, обе располагаются в передней части нижней челюсти (у самок чаще одна передняя пара). На верхней челюсти зубов нет. У молодых китов зубы колышкообразные, у старых — сдавленные с боков, треугольной формы при рассматривании их сбоку. Массивные зубы первой пары (шириной до 85 мм) так сдвинуты вперед, что при закрытой пасти торчат снаружи. На горле две небольшие, но глубокие продольные борозды, расходящиеся в стороны задними концами. Спинной плавник сравнительно низкий, но длинный. Грудные плавники короткие и широкие.
Окраска спинной поверхности тела и боков у взрослых китов бурая, часто темно-бурая, брюшной поверхности — несколько светлее. Иногда имеются белые отметины на «лбу», у пупка и в других местах. Молодые животные окрашены светлее, чем старые.
РАСПРОСТРАНЕНИЕ И МИГРАЦИИ
Эндемик северной половины Тихого океана, где широко распространен и у западного, и у восточного побережий. В водах Азии встречается от южной оконечности Японских островов до Анадырского залива в Беринговом море (около 62° с.ш.), населяя Японское и Охотское моря, тихоокеанские воды у Японских, Курильских и Командорских островов и Южной Камчатки, а также западную часть Берингова моря. В водах Северной Америки обитает вдоль всего побережья, от полуострова Калифорния до
залива Аляска, Алеутских островов и восточной части Берингова моря. Сезонные миграции из умеренных вод в холодные и обратно выражены четко, но пути их не прослежены.
ПИТАНИЕ
Основными объектами питания являются головоногие моллюски (несколько видов), меньшее значение имеют рыбы, в основном придонные виды (скаты и их яйца, морские ерши, тресковые), а также крабы и крупные ракообразные.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ
Роды и спаривание происходят в зимние месяцы во время пребывания китов на местах зимовки. У большей части самок роды проходят в декабре, хотя период родов растянут. Длина новорожденных около 4,5jw. Продолжительность беременности около 10 месяцев. Длительность молочного кормления не установлена. Пик спаривания отмечен в феврале. Половая зрелость у самцов и самок наступает приблизительно в возрасте 3 лет или немного позже.
ПОВЕДЕНИЕ
Северные плавуны предпочитают держаться над глубинами и к берегам подходят редко. Животные стадные, как правило, образуют группы от 2-10 до 40 голов, реже кормятся поодиночке. Нередко молодые киты формируют отдельные группы, а взрослые сам
цы объединяются вместе с самками и детенышами. Иногда животные собираются в группы смешанного состава. Киты дружно ныряют на 8-20 минут (очень редко до часу и больше). На поверхности бывают 3-4 минуты, выпуская 10-15 быстрых фонтанов высотой до 1,5 м.
ПАРАЗИТЫ, КОНКУРЕНТЫ, ВРАГИ
Северный плавун относится к слабо изученным видам, в связи с чем достоверные сведения по данному разделу отсутствуют.
ЧИСЛЕННОСТЬ
Плавунов можно отнести к видам со сравнительно небольшой численностью, хотя количество этих животных не определено даже приближенно.
СТАТУС
Малочисленный эндемичный вид Северной Паци-фики, запрещен к промыслу Международной китобойной комиссией; внесен в Приложение II СИТЕС (Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения).
ОХРАНА
Никаких специальных мер по охране северного плавуна в России не предпринято.
Семейство кашалотов (Familia Physeteridae)
В семействе 2 рода (с одним видом в каждом): кашалот (Physeter), самый крупный представитель зубатых китов длиной до 20 м, и карликовый кашалот, или когия (Kogia), длиной немногим более 3 м. Ареал семейства охватывает все океаны, включая Ледовитый. В водах России обычен кашалот; возможно, у Южных Курил встречается когия.
Род кашалотов
(Genus Physeter)
В роде один вид — кашалот, который делят на северный и южный подвиды:
северный кашалот — более мелкий подвид северного полушария. Обитает в водах до Анадырского залива, отдельные звери проникают к северу до мыса Чаплина.
южный кашалот по размерам крупнее северного. Распространен до тропиков до кромки льдов Антарктики.
Кашалот
(Physeter catodon, Linnaeus, 1758)
ОБЩАЯ ХАРАКТЕРИСТИКА
Киты с резко выраженным половым диморфизмом. Самцы — самые крупные животные в подотряде, средний размер 15-16 л/, максимальный — до 18-20 м, масса до 40 т и более. Самки — животные среднего размера, длина тела 11—12 jw, максимальная — около 16 м. У самцов тело значительно утолщенное в переднем отделе. У самок тело удлиненное и относительно менее утолщенное. Кожа на теле имеет морщинистый вид.
Огромная, тупо срезанная спереди и сдавленная с боков голова определяет характерный облик кашалота. У самцов она крупнее и занимает до 1/3 длины туловища; голова самок сравнительно небольшая (не более 1/4 длины тела), спереди довольно округлая Череп резко асимметричен, в его лицевой части образуется ложе для жировой подушки, заполненной спермацетом, которого бывает до 6 т. Жировая подушка выступает резко вперед за кончик узкой нижней челюсти, и вся ротовая щель оказывается снизу головы.
Кожа на теле кашалота имеет характерный морщинистый вид
Дыхательное отверстие у кашалотов, в отличие от дыхательного отверстия всех других китообразных, смещено на конец морды, где открывается только с левой ее стороны, имеет форму латинской буквы S и вытянуто по оси тела. Правый носовой канал, не открываясь наружу, связан спереди и сзади с левым носовым каналом обширными воздушными мешками.
Полость рта покрыта шершавым эпителием, мешающим выскальзыванию добычи. На узкой нижней челюсти расположено 18—30 пар зубов без эмали, однотипных, похожих на колышки; крупнейшие зубы весят почти по 1,6 кг. Широкая верхняя челюсть обычно лишена зубов, которые просто не прорезываются, оставаясь скрытыми в деснах. Строение рта (отсутствие верхних зубов, длинная нижняя челюсть и углубление снизу головы, в которое входят зубы) позволяет кашалотам активно всасывать добычу.
Зуб кашалота. Слева — внешний вид, справа — продольный разрез
Спинной плавник в виде горба, позади которого располагается еще несколько небольших бугров. Грудные плавники широкие и короткие, тупо закругленные.
Окраска тела темная, довольно однотонная, с мелкой пятнистостью, варьирует от серо-коричневой до почти черной. Верхняя часть тела всегда темнее нижней; на брюхе обычно расположено белое пятно различной величины и формы.
РАСПРЕДЕЛЕНИЕ И МИГРАЦИИ
В Тихом океане кашалоты обитают в Охотском и Беринговом морях, где достигают северной части Берингова моря (мыс Наварин — остров Святого Лаврентия, 62°-63в с. ш.); порой отдельные самцы подходят к мысу Чаплина (сообщение В. Тагитуткак и других старейших охотников Чаплинской общины). Отсюда северная граница распространения этого вида проходит по полосе резкого уменьшения глубин к островам Прибылова. В Берингов пролив, как правило, эти киты не заходят.
Самцы распространены на большей площади, чем самки, и мигрируют дальше их. В водах России самцы кашалота чаще всего встречаются около Курильской гряды, в южных частях Охотского моря, у Командорских островов и Камчатки.
Самки живут в гаремах, размножаются в тропиках и редко выходят за пределы субтропической зоны.
Кашалотам свойственны сезонные миграции, система которых сложна. Сроки и пути миграций в отдельных районах громадного ареала различны.
В схеме эти миграции представляют собой перемещения с мест зимовки в теплых водах в умеренные и частично холодные районы с высокой биологической продуктивностью весной и обратные миграции к местам размножения осенью.
Миграции различных групп китов различаются по дальности. Все самки и неполовозрелые киты, за редким исключением, не проходят севернее (или южнее) границы умеренных вод и в холодные воды не заходят. На Дальнем Востоке такой примерной границей служит гряда Алеутских островов (52°-54° с. ш.). Большая же часть половозрелых самцов, отделяясь от основного стада, мигрирует в холодные воды обоих полушарий, оставаясь там в течение летних месяцев. Осенью все самцы возвращаются в теплые воды, где проводят зимние месяцы вместе со стадом.
ПИТАНИЕ
В питании кашалотов во всех частях ареала преобладают головоногие моллюски: главным образом кальмары и, в меньшей степени, осьминоги. В большинстве случаев кашалоты питаются некрупными кальмарами, длиной не более 100 см, но иногда в их желудках обнаруживают громадных головоногих длиной 9-10 м и массой около 200 кг. Чудовищные кальмары, защищаясь от кашалота, оставляют на его голове следы своих присосок в виде вдавленных кружочков диаметром до 20 см. Известно около 40 видов головоногих моллюсков, составляющих более 90% от общей массы пищи, съедаемой этими китами.
Второй группой пищевых объектов кашалота являются рыбы (до 50 видов), но значение их невели
ко — они составляют около 5% пищи, съедаемой этими китами. Поскольку основная часть кормовых животных представлена глубоководными видами, можно считать, что эти киты в большинстве случаев питаются на глубинах около 500 м или даже глубже.
Наряду с обычной пищей кашалоты иногда заглатывают крабов, моллюсков, губок, водоросли, камни. Такие камни устойчивы к кислотам, не разрушаются желудочным соком и, видимо, используются, как жернова, для механического перетирания пищи. В прямой кишке самцов иногда обнаруживают куски пахучего и очень ценного продукта — амбры, используемой в парфюмерном производстве. У одного кашалота в Антарктике был найден кусок амбры массой в 420 кг.
РАЗМНОЖЕНИЕ И РАЗВИТИЕ
Кашалоты — полигамные животные, образующие гаремы, в каждом обычно 10-15 самок, сосунки и крупный самец. Если гаремы объединяются в стадо, то при нем держатся несколько самцов. У кашалотов, обитающих в северном и южном полушариях, сроки всех биологических ритмов различаются приблизительно на 6 месяцев. Роды у кашалотов могут происходить в любое время года, но в северном полушарии большинство самок приносят потомство в июле-сентябре. Вслед за родами наступает период спаривания. Беременность длится 16-18 месяцев. К моменту рождения длина эмбриона достигает 4 jw, а масса до 1000 кг. Длина годовалого кита до 6 м, а трехгодовалого — до 8 м. Период молочного выкармливания детенышей длится почти 2 года, поэтому цикл размножения кашалотов
составляет около 3-х лет, а темп пополнения стада очень медленный.
Самки кашалотов достигают половой зрелости в возрасте около 4 лет при 8-10 слоях дентина в зубах и средней длине тела 8,9-9,3 м. Самцы — в возрасте 5 лет, когда в зубах бывает 9-10 слоев дентина, а длина тела составляет 9,5-10 м. Физически зрелыми дальневосточные самки становятся в 15-17 лет, имея длину тела 11 м, самцы — в 23-25 лет, при длине 15,9 м.
Возраст самых старых самок определен в 30-35 лет, самцов — 27-32 года. На основании количества слоев дентина в зубах полагают, что кашалоты живут не более 45-50 лет.
ПОВЕДЕНИЕ
Животное стадное, в районах размножения кашалоты держатся группами примерно по 15 голов, представляющими, как правило, гаремы с одним взрослым самцом. Остальные взрослые самцы, не участвующие в размножении, плавают поодиночке. Наблюдения последних десятилетий дают основания полагать, что гаремы имеют сложную структуру и в них входят киты, состоящие в родстве, и что кроме старого крупного самца в размножении могут участвовать и другие, иногда более мелкие, самцы. Нередко много гаремов собираются в относительно небольшом районе и образуют скопление, или стадо.
В местах летнего обитания самок и неполовозрелых китов формирование групп имеет свою специфику. В большинстве случаев группы состоят из одноразмерных китов, причем в группы молодых входят пре
имущественно одновозрастные звери, а в группы старых — разновозрастные. Это обеспечивает одинаковые возможности и условия питания на определенных глубинах для всех китов, входящих в ту или иную группу.
Среди всех морских животных кашалот — один из самых лучших ныряльщиков. В поисках пищи киты опускаются на глубину до 1000 м и более. Зарегистрировано более 20 случаев обнаружения кашалотов, запутавшихся в подводных кабелях, как правило, на глубине около 500 м, в нескольких случаях отмечена глубина свыше 1000 м, а однажды запутавшийся в кабеле кашалот был обнаружен на глубине 2200 м.
Кормящийся кашалот ныряет до 50 минут, но может оставаться под водой и до полутора часов. При этом он уходит круто вниз, выставляя над водой хвостовые лопасти. После длительного ныряния
Два варианта запутывания кашалотов в подводных телеграфных кабелях. Вверху — кашалот, запутавшийся в 1940 г. у берегов Перу на глубине 830 м. Внизу — кашалот, запутавшийся в 1931 г. у берегов Эквадора на глубине около 980 м.
Последовательные стадии погружения кашалота на большую глубину
он лежит на поверхности воды, почти не двигаясь, по 10-15 минут и дышит, пуская фонтаны 60-70 раз с интервалами в 20-30 секунд. Так как дыхало у кашалотов расположено на левом конце морды, фонтан направлен не вертикально вверх, как у всех других китов, а влево вперед и вверх от конца морды под углом 450°. Он имеет как бы шарообразную форму, высота его около 3 м. Крупные (старые) кашалоты дают больше фонтанов и ныряют дольше, чем мелкие (молодые). Если кит, ныряя, не показывает хвост, он погружается мелко. Самки чаще возвращаются на поверхность, чем взрослые самцы. Под водой все кашалоты великолепно ориентируются с помощью эхолокации, издавая три типа звуков: короткие и частые щелканья, стонущие скрипы и частый треск.
Последовательные положения кашалота, выскакивающего из воды
Вертикальная и горизонтальная позы кашалота
Кашалот тихоходен. Скорость пасущегося кита 5-6, ходового — 9-13, преследуемого или раненого — 16-30 of/час. Ходовые киты, в отличие от пасущихся, ныряют неглубоко, настойчиво придерживаясь одного направления. Возбужденные кашалоты целиком выскакивают из воды, падая обратно с оглушительным всплеском, громко хлопают по воде хвостовыми лопастями, либо поднимают ими тучу брызг. Иногда «столбом» выставляют голову из воды или подолгу лежат на боку.
Известно много случаев обсыхания кашалотов на берегу. Наряду с одиночными китами находили большие группы по 20-30 китов. Часто группы обсохших кашалотов состоят только из самцов. Обсыхая на берегу, кашалоты издают сильный рев.
ПАРАЗИТЫ, КОНКУРЕНТЫ, ВРАГИ
Из многочисленных эндопаразитов опасен для кашалота гигантский (длиной до 8,5 м) круглый червь плацентонема, живущий в плаценте самок. Из эктопаразитов на зубах животного поселяется усоногий рак — конходерма ауритум, а на коже — веслоногий рачок-пенелла с гроздьями усоногих рачков другого вида, конходерма виргатум.
Конкурировать с кашалотом может северный плавун, также питающийся кальмарами, однако о взаимоотношении этих двух видов на кормовых полях ничего не известно. Естественных врагов, кроме поедаемых гигантских кальмаров, кашалот не имеет, а вот человек представляет для него, как и для других морских млекопитающих, постоянную угрозу: снача-ло он резко сократил численность кашалотов, а теперь загрязняет моря, в которых обитают эти киты.
ЧИСЛЕННОСТЬ
Кашалот обычен на всем своем громадном ареале. Общая численность составляет несколько сотен тысяч животных, что позволяет считать его одним из наиболее многочисленных видов китов, несмотря на беспощадный коммерческий промысел всех популяций кашалота в течении ста лет. В настоящее время в северной части Тихого океана насчитывается 167 тыс. самцов и 181 тыс. самок.
СТАТУС
Широко распространенный многочисленный вид. Коммерческая добыча запрещена в 1979 г. Внесен в Приложение II СИТЕС (Конвенция о международной торговле видами дикой фауны и флоры, находящимися под угрозой исчезновения).
ОХРАНА
Никаких специальных мер по охране кашалота в России не предпринято.
7
Наблюдайте за китами!
Много есть замечательных зверей, но киты лучше всех. Когда они идут в море мимо байдары, такие великие и спокойные, сразу понимаешь свое место на Земле и внутри тебя тепло становится... На китов глядеть очень интересно, но, главное, полезно и необходимо, чтобы полноценным человеком и охотником стать. Киты дают нам спокойствие и красоту жизни.
Николай Гальгаугъе, поселок Сиреники
Именно эти слова открывают уникальную книгу об аборигенном китобойном промысле и его значении в жизни эскимосов — «Inuit, Whaling, and Sustainability». Ее написали в 1998 г. специалисты Канады, Гренландии, России и США вместе с коренными жителями этих стран. То, что сказал о китах Николай Галъ-гаугье, было признано самым ярким и образным из всех высказываний эскимосов.
Действительно, смотреть на китов очень интересно, но и очень трудно. Трудно потому, что киты и дельфины лишь на мгновения показываются из воды и угадать место и время их следующего выхода на поверхность крайне сложно. Интересно потому, что китообразные, во многом устроенные не так, как
другие млекопитающие, очень умны и обладают наряду со слонами, обезьянами, некоторыми ластоногими и врановыми птицами наиболее развитым интеллектом среди всех животных.
В мировой литературе накопилось очень много сведений о необычных, поражающих воображение поступках морских гигантов. При этом наблюдатели отмечают особую сложность поведения дельфинов, а также серого кита и горбача, которые обитают в наполненных жизнью и весьма изменчивых прибрежных водах. Многие сообщения морских охотников Чукотки об усатых китах стали настоящей сенсацией для науки и послужили основой для интеграции традиционных и научных знаний по биологии морских млекопитающих. Они существенно расширили наши представления о характере миграционного, пищевого и социального поведения китов.
В 1980-х гг. выходя в море вместе с охотниками, я убедилась, что они прекрасно различают виды китов, быстро определяют размеры, а, следовательно, и возраст животных, хорошо подсчитывают их количество, если оно не превышает 15-20 особей. Однако уже в то время далеко не все молодые зверобои обладали знанием китов, умением и терпением наблюдать их «просто так». Вот что написал мне в 1982 г. Анкалин, брат Николая Гальгаугье, о разнице в отношении к китам старшего и младшего поколений охотников: «В море все время думаю о гренландском ките, надеюсь увидеть, услышать дыхание. Бью моржа, лахтака, а думаю о нем. Это для нас самый великий зверь. Наверное, в Африке — слон, а у нас — гренландский кит...
Теперь молодежь мало видит, да и не любят они просто так, для себя, на море смотреть. Когда
в Анитикок охочусь на песца, всегда смотрю как играют гренландцы. Сам становлюсь китом, верчусь в воде вместе с ними. Ты смеяться будешь — один раз с берега свалился — забыл, что человек, и пошел к ним».
Вспоминаю, как Виктор Миенков рассказывал о нападении косаток на гренландского кита. Это было в конце декабря у входа в бухту Провидения. Стоял сильный мороз, но охотники три часа наблюдали с вельбота битву китов и пошли домой, как только косатки бросили гренландца, не сумев одолеть его, — израненный кит забился возле мыса Столетия в узкую щель между торчащими из воды камнями и стал недоступен для хищников.
Кроме чисто познавательного и научного интереса есть еще и практическая необходимость в наблюдениях и количественных учетах китов — ведь нужно знать, в каком состоянии находится, например, та часть стада серых китов, которая является основой китобойного промысла коренных жителей Чукотки.
После непродолжительного подъема численности всего чукотско-калифорнийского стада (последний учет в 1997/98 гг. — 26.635 особей) резко упала рождаемость этих животных: в 1997 г. в лагунах Мексиканской Калифорнии был подсчитан 1.431 детеныш; 1998 г. — 1.388; 1999 г. - 427; 2000 г. - 279; 2001 г. -100 детенышей. Эти цифры говорят о том, что следует ожидать значительного снижения общего количества серых китов, в том числе и у берегов Чукотки. При подсчетах обязательно следует учитывать не только число живых зверей, но и их трупы, выброшенные на берег, а также китов с обрывками троса на хвостовом стебле и еще так называемых «вонючих» китов, мясо которых не пригодно для питания людей и ездовых собак.
Я могу дать лишь несколько советов тем, кто хочет внимательно приглядеться к китам и увидеть их не как свою добычу, а как равных человеку обитателей нашей планеты.
Сначала необходимо научиться быстро определять, какой вид кита вы встретили. Если в этот момент рядом с вами будет опытный охотник, то он укажет на характерные признаки, по которым отличают, например, серого кита от горбача. Но можно это освоить и самому. Нужны только желание, время и терпение, а помочь могут рисунки и фотографии разных китообразных, а также схемы фонтанов и силуэтов этих зверей над поверхностью воды. В конце главы приведена одна из лучших схем такого рода, ее составил в 1952 г. М.М. Слепцов.
Когда вы сумеете уверенно определять виды, можно попробовать узнавать китов «в лицо», то есть индивидуально распознавать животных по каким-либо приметным деталям внешнего облика, главным образом по особенностям окраски. Об этом стоит рассказать подробнее.
За тысячелетия взаимоотношений людей и китов накопилось много данных о животных, приходящих в одно и то же место в течение длительного времени, иногда десятков лет. В основном это разные виды дельфинов и косатка. Австралийский зоолог Дакин описал совместные действия китобоев и косаток при лодочном промысле горбачей в заливе Туфолд на юго-восточном берегу Австралии. Косатку Старый Том — самого активного помощника — китобои непрерывно наблюдали в этом заливе почти 50 лет. Он погиб здесь же в 1930 г., вероятно от старости. В качестве признания заслуг Старого Тома в китобойном промысле его смонтированный 286
скелет был выставлен на главной улице городка Иден.
В 1980-х годах охотники-эскимосы пос. Сиреники очень почтительно относились к косаткам и никогда даже не стреляли в их сторону. Они считали, что косатки приходят помочь в охоте на моржей и угощали их табачком как своих друзей -крошили в воду сигареты. Одна из косаток, большой самец, имела приметную, чуть загнутую влево, вершину огромного спинного плавника. А. Анкалин и Н. Гальгаугье в 1981 г. сказали, что между собой называют этого кита Рыглыналг’и (Вызывыющий любопытство у других), видят его уже 8 лет и всегда примерно в одном и том же месте, возле старого поселка Синг’ак. Мне довелось несколько раз встретить столь заметного зверя и однажды долго наблюдать за его поведением. Последнее сообщение о Рыглыналг’и Николай Гальгаугье прислал незадолго до своей гибели в сентябре 1995 г.
Современные исследователи китов не прошли мимо всех этих историй и попробовали использовать достижения науки и новейшую фототехнику для индивидуального опознания животных. Оказалось, что разные виды китов обладают разными признаками, которые наиболее успешно можно использовать для узнавания животных. Так, горбачей легче всего различать по окраске хвостовых лопастей, или «бабочки». Она строго индивидуальна и никогда не повторяется. Для одной из популяций горбачей был даже составлен и опубликован альбом фотографий хвостовых лопастей; с его помощью ученые узнавали отдельных китов и следили за их поведением и передвижениями. Л.М. Вотрогову и мне удавалось несколько раз опознавать серых китов, имевших де-
фекты хвостовых лопастей (вероятно, куски хвоста были оторваны косатками) или бросающиеся в глаза особенности расположения белых пятен от раковин усоногих рачков-коронул.
У косаток установлены групповые (популяционные, семейные) и индивидуальные отличия в окраске, главным образом это проявляется в величине и расположении белых пятен. Столь же изменчива черно-белая окраска тела и спинного плавника белокрылой морской свиньи. А у кашалота обнаружено, что уникальные особенности окраски могут наследоваться. Конечно, нужен большой опыт и «хороший глаз» для того, чтобы узнавать китов по внешнему виду, но стоит попробовать заняться этим, — когда видишь «знакомого» кита, испытываешь радость, как от встречи со старым и любимым другом.
Весьма полезно и для себя и для науки вести дневник наблюдений за животными, в котором при каждой встрече с китами необходимо записать следующие данные (в скобках приведены примеры таких записей):
1. Дата и время наблюдения (06 июля 2002, 20 час);
2. Место наблюдения (мыс Кыгынин острова Ара-камчечен);
3. Погодные и ледовые условия (волнение моря 3 балла, северный ветер (С-, или N-ветер) до 8-10 м/сек, слабый дождь);
4. Виды наблюдаемых китов, их количество, группировки (2+3 серых кита + 2 горбача);
5. Удаленность китов от берега, направление их движения (800-1000 м, медленно движутся на северо-восток (С-В, или NO);
6. Особенности поведения (все три группы китов держатся близко друг к другу, расстояния между ними 50-100 м\ часто выходят на поверхность, погружаются, не показывая хвостовых лопастей);
7. Примечания (здесь можно записать какие-либо дополнительные данные или наблюдения за другими животными, например, за птицами, которые нередко сопровождают кормящихся китов).
По этой схеме, оформленной в виде таблицы, вот уже 25 лет ведут наблюдения за китами морские охотники Чукотки. Полученные ими материалы не только обогатили науку новыми сведениями по биологии китов, но помогли восстановить квоту на добычу гренландского кита и сохранить высокий уровень квоты на промысел серого кита.
Старшие поколения зверобоев внесли свой неоценимый вклад в познание китов и дельфинов. Дело теперь за молодыми охотниками, которые, я верю, смогут достойно продолжить традиции своих отцов, дедов и прадедов. Словами одного из них, Тимофея Панаугье, я завершаю эту книгу:
— Мы не хотим плохого китам, мы хотим, чтобы они были всегда.
Голубой кит
Серый кит
Гренландский
Южный кит
Белуха
Фонтаны и силуэты китов над водой
Индивидуальные особенности окраски хвостовых лопастей, или «бабочки», горбачей в водах Гавайских островов
Варианты окраски боковой поверхности белокрылой
морской свиньи
Варианты формы и окраски спинного плавника белокрылой морской свиньи
Случай полного совпадения деталей уникальной окраски самки кашалота (А), ее эмбриона (Б) и другой взрослой самки (В), добытых в одной и той же группе
Некоторые научные термины и понятия
Статьи, объясняющие термины и понятия, расположены в алфавитном порядке; в них курсивом выделены термины, для которых существуют самостоятельные статьи, или отсылки к таким статьям.
Абиссаль — см. Бенталь.
Амбра — воскоподобное вещество, образующееся в кишечнике некоторых кашалотов; содержит остатки клювов кальмаров, которыми питаются эти киты. Куски амбры могут достигать веса 420 кг. Это очень дорогое вещество используется в парфюмерии для закрепления запахов духов.
Ареал — часть земной поверхности (наземной и/или водной), в пределах которой распространен и проходит полный цикл своего развития вид, род, семейство, отряд и т.д. каких-либо живых организмов. Ареалы многих видов животных и растений претерпевают изменения под влиянием деятельности человека. Например, китобойный промысел и промышленное освоение побережья Северной Пацифики сократили ареалы серого кита и горбача.
Бенталь — населенное организмами дно океанов и морей, в котором выделяют следующие зоны:
1) супралитораль — зона заплеска и штормовых выбросов;
2) литораль — приливно-отливная зона;
3) сублитораль — зона шельфа, или материковой отмели (до 200-500 м глубины);
4) батиаль — зона континентального склона, или свал глубин (от 200-500 м до 3 о/);
5) абиссаль — зона океанического ложа (3-6 км глубины);
6) ультраабиссаль — зона наибольших океанических глубин, или так называемых океанических впадин.
Бентос — совокупность организмов, обитающих в бентали. в лрунте и на грунте дна или лишь периодически опускающихся на дно. Бентос разделяется на растительный (фитобентос) и животный (зообентос). Суммарная биомасса бентоса в Мировом океане оценивается в 10-12 млрд, т, причем ее величина убывает с глубиной. Так, 60% всей биомассы приходится на долю мелководий (литорали и сублиторали глубиной до 200 м), которые занимают менее 8% площади дна Мирового океана, а на долю абиссали. занимающей 75% дна Мирового океана, — менее 10%.
Бентофаг — животное, питающееся бентосом (серый кит, морж и др.).
Биомасса — суммарная (общая) масса всех особей одного вида, группы видов или сообщества организмов, которую выражают обычно в единицах массы сухого или сырого вещества, отнесенных к единицам площади или объема (кг/га; кг/м2; кг/мЗ; г/м2; г/мЗ). Вся биомасса нашей биосферы составляет 1800 млрд, т сухого вещества, из которого около 90% приходится на долю зеленых растений и лишь 10% — на все остальные организмы Земли. Для морских экосистем и крупных внутренних водоемов характерна малая биомасса растений. Так, биомасса животных Мирового океана составляет около 6 млрд. т, что в 20 раз больше общей биомассы водных растительных организмов (300 млн. т). Определение биомасс используют для изучения продуктивности каких-либо групп организмов, отдельных частей экосистем и биосферы в
целом, а также при прогнозировании последствий хозяйственной деятельности человека.
Биосфера — оболочка Земли, состав, структура и энергетика которой определяются совместной деятельностью всех живых организмов, как ныне существующих, так и существовавших в прошлые геологические эпохи. Биосфера охватывает часть атмосферы Земли до высоты озонового слоя (20-25 км), часть верхней геологической оболочки Земли (литосферы), прежде всего кору выветривания, и всю гидросферу (океаны, моря, реки, озера). Нижняя граница биосферы опускается в среднем на 2-3 км вглубь суши и на 1-2 км ниже дна океана. Целостное учение о биосфере разработал русский ученый-геохимик В.И. Вернадский, который ввел в науку понятие о живом веществе.
Согласно Вернадскому, биосфера — область жизни, включающая наряду с живыми организмами и среду их обитания, а живое вещество — это «совокупность всех живых организмов, в данный момент существующих, численно выраженное в элементарном химическом составе, в весе, в энергии. Оно связано с окружающей средой биогенным током атомов: своим дыханием, питанием и размножением».
Живое вещество распределено в биосфере крайне неравномерно. Его максимум приходится на приповерхностные участки суши (особенно велика биомасса тропических лесов) и гидросферы.
Основной источник происхождения и развития биосферы — солнечная энергия, с ее помощью зеленые растения и некоторые микроорганизмы создают в процессе фотосинтеза органическое вещество, которое затем обеспечивает пищей и энергией все остальные организмы Земли. Это органическое вещество называется первичной продукцией.
Биота — исторически сложившаяся совокупность живых организмов, объединенных общей областью распространения. В состав биоты входят виды, которые могут и не иметь экологических связей друг с другом (например, рыба-бычок и лемминг в фауне Чукотского полуострова).
Вид — основная структурная единица в системе живых организмов. В науке до сих пор не существует строгого общепринятого понятия «вид». Обычно «вид» понимается как совокупность популяций особей, которые населяют определенный ареал, обладают наборами общих структурных и функциональных признаков и способов взаимодействия с окружающей средой и друг с другом, а также могут скрещиваться между собой с образованием плодовитого потомства.
Генотип — генетическая (наследственная) конституция организма, или совокупность всех его наследственных задатков.
Генофонд — совокупность генов, которые имеются у особей данной популяции, группы популяций или вида. Генофонд создается в результате полового процесса, который обеспечивает генетическую целостность популяции благодаря постоянному обмену генами (наследственным материалом) между организмами. Таким образом формируется единый генофонд популяции, куда в каждом поколении особями разного генотипа вносится больший или меньший вклад наследственного материала. Важнейшая особенность единого генофонда (популяции, группы популяций, вида) — его глубокая неоднородность (дифференцированность) и способность изменяться под действием различных факторов.
Детрит — мелкие органические частицы (остатки растений и разложившихся трупов животных вместе с содер
жащимися в них бактериями), осевшие на дно или взвешенные в толще воды. Детрит играет важную роль в круговороте органического вещества. Существует детритная трофическая цепь, есть животные-детритофаги, то есть питающиеся частицами детрита.
Диатомовые водоросли — мелкие и микроскопические водоросли размером от 4 до 2000 микрометров. Эти одноклеточные одиночные или колониальные растительные организмы имеют твердую двустворчатую кремнеземную оболочку-панцирь, содержат хлорофилл и фукоксантин, придающий им бурый цвет. Диатомовые водоросли являются важнейшими продуцентами (производителями) органического вещества: им принадлежит около 25% мировой первичной продукции, создаваемой растениями нашей планеты с помощью фотосинтеза. Диатомовые водоросли широко распространены в морях и океанах, континентальных (наземных) водоемах, почве. Они образуют плотный коричневый налет на нижней стороне льдов Антарктики, а также на поверхности многих морских организмов, в том числе на синих китах, отчего эти животные вопреки своему названию выглядят коричневыми. Диатомовые водоросли — древние организмы Земли, они известны с юрского периода (от 190-195 до 130-140 млн. лет тому назад).
Зообентос — часть бентоса, представленная животными организмами, которые делят на четыре группы:
1) обитающие в толще грунта (инфауна) — многие многощетинковые черви, двустворчатые моллюски, некоторые иглокожие и др.;
2) передвигающиеся по поверхности грунта (онфау-на) — различные ракообразные, большинство иглокожих, некоторые многощетинковые черви и моллюски;
3) прикрепляющиеся ко дну и расположенному на нем субстрату (эпифауна) — губки, актинии, различные
кораллы, гидроиды, мшанки, некоторые двустворчатые моллюски, морские желуди и др.;
4) плавающие вблизи дна и лишь периодически опускающиеся на дно (нектобентос) — креветки, ми-зиды, некоторые голотурии, придонные рыбы и др.
Зоопланктон — часть планктона, представленная самыми разнообразными морскими животными величиной от нескольких микрометров до 1,5-2 м (гребневик «венерин пояс», медуза цианея и др.). В зоопланктон входят многочисленные простейшие, кишечнополостные, оболочники, различные ракообразные, крылоногие моллюски, икра и личинки рыб, личинки многих беспозвоночных, в том числе донных. Многие из них совершают регулярные вертикальные миграции с амплитудой в сотни метров, а иногда свыше километра, что способствует перенесению пищевых ресурсов в глубины морей и океанов. Значительная часть планктонных животных питается фитопланктоном. В свою очередь зоопланктон — основной корм многих морских млекопитающих и птиц, для некоторых животных он вообще является единственным кормом (гладкие киты и др.). Часто планктонные животные собираются в огромных количествах, например, эвфаузиевы рачки (эвфаузииды), скопления которых называют «криль». Биомасса зоопланктона составляет от десятков миллиграмм до 1 г на кубический метр воды.
Комменсал — животное-нахлебник, вступающее в особые (комменсальные) отношения с животным-хозяином. Основой таких отношений могут быть общее пространство, передвижение, пища и т.п. Используя особенности образа жизни или строения хозяина, комменсал извлекает из этого одностороннюю пользу. Комменсалами китов являются усоногие ракообразные, врастающие своими раковинами в их кожу, подбирающие за ними остатки пищи и путешествующие на китах.
Криль — см. Зоопланктон.
Литораль — см. Бенталь.
Нектон — совокупность активно плавающих пелагических животных, способных противостоять течениям и самостоятельно преодолевать большие расстояния. Массовые и постоянные представители нектона — рыбы, кальмары, морские змеи, китообразные, сирены. К нектону относят также размножающихся на суше, но питающихся в воде животных (водяные черепахи, пингвины и другие морские птицы, калан, ластоногие и т.п.). Некгонные животные имеют обтекаемую форму тела и хорошо развитые органы движения. Нектон наиболее разнообразен и многочислен в верхних слоях пелагиали в связи с обилием там пищевых ресурсов, прежде всего зоопланктона. Океанический нектон — эволюционно молодая группа животных.
Паразитизм — форма взаимоотношений двух различных организмов, когда один вид (паразит) использует другой вид (хозяина) в качестве среды обитания (среда 1-го порядка) или источника пищи (среда 2-го порядка). Паразитизм известен на всех уровнях организации живого, начиная с вирусов и бактерий и кончая высшими растениями и животными. Пути проникновения паразитов в организм хозяина различны; многие паразитирующие виды имеют помимо основного хозяина еще нескольких промежуточных хозяев. Различают два типа паразитов: эктопаразиты (наружные паразиты) обитают на поверхности тела хозяина, эндопаразиты (внутренние паразиты) живут во внутренних полостях, органах, тканях и клетках хозяина. Географическое распространение паразитов тесно связано с распространением их хозяев и с особенностями среды обитания. Часто ареалы паразитов и их хозяев совпадают, но иногда определенные паразиты встречаются только в небольшой части ареала хозя
ина. Такие случаи известны для рыб, некоторых видов китообразных и ластоногих
Пелагиаль — толща воды морей и океанов от дна до поверхности. Основные зоны: эпипелагиаль (0-200 м глубины), мезопелагиаль (200-1000 м глубины) и глубоководная зона, которую делят на три подзоны. Вертикальная экологическая зональность выражена менее четко, чем в бентали вследствие постоянного перемещения вод (ветры, течения и пр.) и вертикальных миграций пелагических организмов, которые охватывают сотни, иногда тысячи метров по глубине.
Пелагические организмы — растительные, бактериальные и животные организмы, обитающие в толще пелагиали и образующие две большие группы: планктон и нектон.
Первичная продукция — создается наземными зелеными растениями, водорослями и некоторыми бактериями. Все они используют для построения своего организма СО2 (углекислый газ) в качестве единственного или главного источника углерода, обладают системой ферментов для усвоения СО2 с помощью энергии солнечного света, то есть осуществляют фотосинтез, и способны самостоятельно строить все компоненты клеток, из которых состоят. Именно эти растения и бактерии являются продуцентами (производителями) первичного органического вещества, или первичной продукции, в биосфере — они поставляют в биосферу около 162 млрд, т в год. Наземные растения производят 2/3 этого количества.
Первичную продукцию водных экосистем создает главным образом фитопланктон.
В высокопродуктивных водах, несмотря на малую биомассу фитопланктона, но благодаря его быстрому росту и размножению, первичная продукция может достигать в энергетическом выражении столь же высоких величин,
как и в наземных экосистемах. Однако биомасса самого фитопланктона остается низкой из-за того, что его поедает зоопланктон. Высокопродуктивные воды составляют небольшую долю от общей площади Мирового океана, и потому первичная продукция морских экосистем в целом намного меньше первичной продукции экосистем суши.
Планктон — совокупность организмов, которые населяют толщу воды пресноводных водоемов, морей и океанов и не способны противостоять переносу их течениями. В состав планктона входят: фитопланктон, населяющий поверхностные воды при достаточной для фотосинтеза освещенности (в морях до 50-100 м глубины), бакгериопланктон и зоопланктон, распространенные в пелагиали от поверхностных слоев до максимальных глубин. У многих планктонных организмов существуют приспособления, облегчающие парение в воде. С глубиной количество и видовое разнообразие планктона быстро убывают. Наибольшее видовое разнообразие планктона — в тропических водах.
Половая зрелость - возраст животного, когда оно становится способным к размножению.
Популяция — совокупность (группа) особей одного вида, которые обладают общим генофондом и занимают определенную общую территорию. Контакты и связи (в том числе половые) между особями внутри одной популяции происходят чаще, чем между особями разных популяций. Внутри популяции можно выделить более мелкие подразделения (семьи, поды, демы, парцеллы и др.). Любая популяция характеризуется общей численностью особей, их плотностью (число особей на единицу площади) и особенностями пространственного распределения, а также упорядоченностью структуры. Структура популяции проявляется в определенном количественном соотноше
нии особей разного возраста, пола, размера, разных генотипов и т.п. Соответственно различают возрастную, половую, размерную, генетическую и др. структуры популяции.
У большинства видов животных и растений численность популяций более или менее постоянна, но у некоторых видов она регулярно сильно меняется (полевки, лемминги, некоторые насекомые). Обычно такие вспышки численности сопровождаются миграциями животных. Колебания численности популяций получили название «волны жизни». Они могут быть (1) периодическими, например, сезонными, генетически обусловленными, и (2) апериодическими, несезонными, в этом случае обусловленными воздействием различных факторов окружающей среды. Часто «волны жизни» сопровождаются пульсирующими изменениями территориальных границ популяции. Считают, что «волны жизни», или популяционные волны, — один из четырех основных факторов эволюции, три другие — мутации, изоляция и естественный отбор.
Редукция (органа или структуры) — недоразвитие или полное исчезновение у живых организмов какого-нибудь органа (структуры), нормально развитого у предков или на ранних стадиях индивидуального развития. В таких случаях говорят о редуцированном органе (структуре).
Северная Пацифика — северная часть Тихого океана, который по-английски называется Pacific Ocean.
Сонар — см. эхолокация
Спермацет — жировосковое вещество, заполняющее два громадных спермацетовых мешка в верхней части головы кашалота. Функциональное значение спермацета не вполне ясно, полагают, что он играет определенную роль
в проведении эхолокационных сигналов кашалота. Спермацет обладает ранозаживляющим и омолаживающим свойствами, употребляется в парфюмерии как основа кремов и мазей. От взрослых китов удается получить 1-5,7 т спермацета.
СуБЛИТОраль — см. Бенталь.
Толщина жира в условном месте — толщина подкожного жирового слоя кита в точке на пересечении средней линии бока кита с вертикалью, проходящей через анальное отверстие. Во времена коммерческого китобойного промысла по этому показателю определялась упитанность кита и, соответственно, предполагаемый выход товарного жира.
Трофическая цепь (пищевая цепь, цепь питания) — взаимоотношения между организмами через пищу, благодаря которым в экосистемах происходит трансформация вещества и энергии.
Физическая зрелость — возраст животного, когда прекращается рост его тела. У китов этот период наступает при полном окостенении позвоночника.
ФитоБентос — часть бентоса, представленная растительными организмами. Морской мелководный фитобентос составляют водоросли и морские травы, а также скопления донных диатомовых водорослей. В глубинах на дне обитают только низшие грибы и бактерии.
Фитопланктон — совокупность водных растительных организмов, в основном производящих органическое вещество, за счет которого существуют все остальные живые существа, то есть В морях фитопланктон представлен главным образом диатомовыми водорослями и
некоторыми другими группами микроскопических по своим размерам растений, а также рядом бактерий, роль которых в составе планктона очень значительна — они перерабатывают детрит и таким образом включают его вновь в трофические цепи. Фитопланктон населяет поверхностные воды (до 50-100 м глубиной), куда проникает солнечный свет, необходимый для процесса фотосинтеза, поэтому в умеренных и холодных водах проявляется сезонность в его развитии Его биомасса составляет от нескольких миллиграмм до нескольких грамм на кубический метр воды, а суммарная масса — всего 1,5 млрд, т — в 13 раз меньше, чем суммарная масса зоопланктона (20 млрд. т). Однако из-за быстрого размножения микроскопических водорослей продукция фитопланктона в Мировом океане составляет около 550 млрд, т — почти в 10 раз больше суммарной продукции всего животного населения океана. Обилие фито-планктона в различных участках морей зависит от содержания в поверхностном слое пелагиали биогенных веществ (фосфаты, соединения азота и пр.). Фитопланктон является пищей мелких животных, которыми питаются более крупные, в связи с этим в районах развития фитопланктона обильны зоопланктон и нектон. Фитопланктон — начальное звено большинства трофических цепей Мирового океана.
Фотосинтез — процесс создания органических веществ (первичной продукции} с помощью энергии света в клетках высших растений, водорослей и некоторых бактерий, которые содержат особые светочувствительные пигменты (хлорофиллы и некоторые другие). Фотосинтез — единственный процесс на Земле, ведущий к увеличению свободной энергии биосферы за счет внешнего источника (Солнца) и обеспечивающий существование растений, всех животных и человека. Ежегодно в результате фотосинтеза на нашей планете образуется 150 млрд, т
органического вещества и выделяется около 200 млрд, т свободного кислорода.
Экологическая зональность морей и океанов — существование в Мировом океане различных зон жизни, населенных разными организмами и их сообществами. Главные зоны — бенталь и пелагиаль.
Экологическая зональность отражает смену условий существования живых существ от поверхности в глубину и от прибрежья к открытым частям морей.
Экосистема (экологическая система) — совокупность совместно обитающих организмов и условий их существования, которые связаны между собой системой закономерных отношений различной сложности. Понятие «экосистема» употребляется очень широко и его можно применять к объектам различной величины и сложности, например, экосистема Берингова моря и экосистема Анадырского лимана; экосистема тундр Арктики и экосистема тундры острова Врангеля и т.д.
Эктопаразиты — см. Паразитизм.
Эндемики (эндемы) — виды, роды, семейства и т.д. растений и животных, ограниченные в своем распространении определенной, чаще всего небольшой, географической областью. Возникновение и развитие эндемизма связано с географической (территориальной) изоляцией, климатическими условиями и некоторыми биотическими факторами. Больше всего эндемиков в географических изолятах (на океанических островах, в горных системах и т.п.).
Эндопаразиты — см. Паразитизм.
Эхолокация — способность некоторых животных излучать и воспринимать отраженные, как правило, высокочас
тотные звуки (ультразвуки) с целью ориентации в пространстве, обнаружения добычи и определения ее свойств. Кроме того, эхолокационные сигналы служат для связи животных друг с другом. Эхолокация развита у летучих мышей, зубатых китов, некоторых мелких млекопитающих и птиц. Летучие мыши и дельфины производят ультразвуковые сигналы частотой до 150-180 кГц, длительностью от 0,2 до 4-5 мсек. Система органов и нервных центров животного, обеспечивающая эхолокацию, получила у специалистов название сонара от англ, sound navigation and randing («звуковое наведение и определение расстояния» — так назывался технический эхолокатор, применявшийся для обнаружения подводных объектов).
Научно-популярная литература для дополнительного чтения
Артюхин Ю.Б., Бурканов В.Н. Морские птицы и млекопитающие Дальнего Востока России. М.: Аст.1999. 214 с.
Атлас морских млекопитающих СССР (ред. В.А. Земский). М.: Пищевая промышленность. 1980. 184 с.6
Богословская Л.С. Охотники Берингова пролива, ж. Вокруг света. 1989. № 6, с. 36-42.
Богословская Л.С. Серый кит. ж. Природа. 1996. № 12, с. 46-60.
Дозье Т. Удивительный мир диких животных. Киты и другие морские млекопитающие. М.: Мир, 1980. 129 с. 7
Земский В.А. Киты - гиганты океана. М.: Пищевая промышленность. 1971. 124 с.
Зенкович БА. Вокруг света за китами. М.: Географгиз, 1954. 408 с.
Кит (ред. К.К. Чапский). Л.: Гидрометеоиздат. 1973. 160 с. 7
Корр Э., Эванс У. Кит в океанариуме. М.: Знание, 1988. 128 с.
Крупник И.И. Пусть говорят наши старики. Рассказы азиатских эскимосов-юпик. М.: Институт Наследия, 2000. 528 с.
Кусто Ж.-И., Диоле Ф. Могучий властелин морей. М.: Мысль, 1977. 188 с.
Моуэт Ф. Кит на заклание. Л.: Гидрометеоиздат, 1979. 168 с.
Мымрин Н.И. Традиционные экологические знания сибирских эскимосов о гренландском ките. Провидения, 1998-1999. Препринт. 88 с.
Слепцов М.М. Гиганты океанов. Владивосток, 1948. 95 с. 6
Томилин А.Г. История слепого кашалота. М.: Наука, 1965. 192 с.*
Томилин А.Г. Отряд китообразных. В кн.: Жизнь животных. М.: Просвещение, 1971. Т. 6. С. 251-300.
Томилин А.Г. Снова в воду. М.: Знание, 1977. 144 с. + 8 вкл.
Томилин А.Г. В мире китов и дельфинов. М.: Знание, издание 2-ое, 1980. 224 с.
Хантингтон Т.П., Мымрин Н.И. Традиционные экологические знания о белухе. Анкоридж, ICC, 1996. Препринт. ПО с.+ 13 карт.
Основные литературные источники
Белькович В.М. Ориентация дельфинов. Механизмы и модели. М., Изд-во НЦССХ РАМН, 2001.240 с.
Берзин А.А. Кашалот. М.: Пищевая промышленность, 1983. 368 с.
Берзин А.А. Современное состояние популяций гренландского кита // Природа. 1981. № 6. С. 81-83.
Биологический энциклопедический словарь (гл. ред. Гиляров М.С.). М.: Советская энциклопедия, 1986. 832 с.
Блохин С.А. Распределение, численность и поведение серых китов (Eschrichtius robustus) американской и азиатской популяций в районах их летнего распределения у берегов Дальнего Востока // Экологические исследования морских млекопитающих Дальнего Востока. Изв. ТНИРЦ, 1996. Т. 121. С. 36-53.
Богословская Л.С. Структура чукотско-беринговоморского стада гренландского кита // Морские млекопитающие Голарктики. М.: Межвед. ихтиол, ком. и др., 2002. С. 39-41.
Богословская Л. С. Распределение и численность калифорнийского серого кита в водах России // Морские млекопитающие Голарктики. М.: Межвед. ихтиол, ком. и др., 2002. С. 41-42.
Богословская Л. С., Вотрогов Л.М., Крупник И.И. Гренландский кит в водах Чукотки: История и современное состояние популяции // Морские млекопитающие. М.: Наука, 1984. С. 191-212.
Динесман Л.Г., Киселева Н.К, Савинецкий А.Б., Хасанов Б.Ф. Вековая динамика прибрежных экосистем северо-востока Чукотки. М.: Аргус, 1996. 190 с.
Здор Э.В. Современное состояние аборигенного китобойного промысла на Чукотке // Морские млекопитающие Голарктики. М.: Межвед. ихтиол, ком. и др., 2002. С. 116-118.
Зенкович Б.А. Еще о сером калифорнийском ките // Вестник ДВФ АН СССР. 1937. Вып. 23, с. 91-103.
Ильяшенко В.Ю. Таксономический и правовой статус наземных позвоночных животных России. М.: Экоцентр МГУ, 2001. 150 с.
Клейненберг С.Е., Яблоков А.В., Белькович В.М., Тарасевич Н.М. Белуха. М.: Наука, 1964. 455 с.
Кочнев АЛ Гибель китообразных (CETACEA) в Чукотском море и проливе Лонга: видовой состав, распределение и причины гибели // Зоол. ж. 1998. Т. 77. № 5. С. 601-605.
Мельников В.В., Бобков А.В. Миграции гренландских китов в Чукотском море // Биология моря. 1993. № 3. С. 60-67.
Млекопитающие Советского Союза. Ластоногие и зубатые киты (гл. ред. В.Г. Гептнер). М.: Высшая школа, 1976. 719 с.
Никулин П. Г. О распределении китообразных в морях, омывающих Чукотский полуостров // Изв. ТИНРО. 1947. Вып. 22. С. 255-257.
Рубцова Е.С. Материалы по языку и фольклору эскимосов (Чаплинский диалект). Часть 1. М.-Л.: Изд-во АН
СССР, 1954. 556 с.
Слепцов М.М. Китообразные дальневосточных морей // Изв. ТИНРО. 1952. Вып. 38. 165 с.
Слепцов М.М. О колебании численности китов в Чукотском море в разные годы // Тр. ИМЖ АН СССР. 1961. Вып. 34. С. 54-64.
Соколов В.Е. Систематика млекопитающих. М.: Высшая школа, 1979. 528 с.
Томилин А.Г. Киты Дальнего Востока // Ученые записки Моск. Университета. 1937. Сер. биол. Вып. 12. С. 119-167.
Томилин А.Г. Китообразные фауны морей СССР. М.: Изд-во АН СССР, 1962. 211 с.
Эванс В., Яблоков А.В. Изменчивость окраски китообразных. М.: Наука, 1983. 136 с.
Alaska Whales and Whaling. Alaska Geographic (Anchorage). 1978. V. 5. #4. 144 pp.
Bogoslovskaya, L.S. 1986. On the Social Behaviour of Gray Whales off Chukotka and Koryaka. In: Rep. Int. Whal. Commn. (Special Issue 8). Pp. 243-251.
Bogoslovskaya, L.S. 1993. List of villages of Chukotka Peninsula. In: Beringian Lett. (Anchorage, NPS). Ser. 2. Iss. 2. Pp.2-10.
Bogoslovskaya, L.S., Votrogov, L.M. and Krupnik, 1.1. 1982. The Bowhead Whales of Chukotka: Migrations and Aboriginal Whaling. In: Rep. Int. Commn. 32. Pp.391-399.
Bogoslovskaya, L.S., Votrogov, L.M. and Semenova, T.N. 1982. Distribution and Feeding of Gray Whales off Chukotka in the Summer and Autumn of 1980. In: Rep. Int. Whal. Commn. 32. Pp.385-389.
Darling, J.D. 1988. Whales. An Era of Discovery. In: Nat. Geogr. V. 174. # 6. Pp. 872-909.
Field Guide to the Gray Whale (The Oceanic Society). Seattle, Sasquatch Books, 1989. 51 pp.
Fraker, M.A. 1984. Balaena mysticetus: Whales, oil, and whaling in the Arctic. Anchorage, Sohio Alaska Petr. Comp, and BP Alaska Expl. Inc. 64 pp.
Freeman, M., Bogoslovskaya, L.S, Caulfield, R., Krupnik, LI., and Stevenson, M. 1998. Inuit, Whaling, and Sustainability. Walnut Creek, AltaMira Press. 208 pp.
George, J.C., Bada, J., Zeh, J., Scott, L., Brown, S.E., O'Hara, T., and Suydam, R. 1999. Age and Growth Estimates of Bowhead Whales (Balaena mysticetus) via Aspartic Acid Racemization. In: Can. J. Zool. 77. Pp. 571-580.
Gordon, D.G., and Baldridge, A. 1991. Gray Whales. Monterey, Monterey Bay Aquarium, 61 pp.
Hoyt, E. 1984 (1950). The Whale Watcher’s Handboook. Toronto, Madison Press Books. 208 pp.
In Pursuit of Agvig (Compiler Harrit, R.K.). Anchorage, UAA, 2001. 96 pp.
Krupnik, L, and Bogoslovskaya, L. 1999. Old Records, New Stories. Ecosystem Variability and Subsistence Hunting in the Bering Strait Area // Arctic Research U.S. V. 13. Pp. 15-24.
Krupnik, 1.1., Bogoslovskaya, L.S., and Votrogov, L.M. 1983. Gray Whaling off the Chukotka Peninsula: Past and Present Status. In: Rep. Int. Whal. Commn. 33. 557-562 pp.
Pilleri, G. 1983. Auf Baffinland. Zur Erforschung des Narvals (Monodon monoceros). Croydon, Verlag des Himanatomischen Institutes (Berne). 80 S. + 22 Tafel.
Pryor, К. Lads Before the Wind. N.-Y.,San Francisko, London, Harper f. Row, 1975.
Rice, D.W., and Wolman, A.A. 1971. The Life History and Ecology of the Gray Whales (Eschrichtius robustus). Am. Soc. Mamml. Spec. Publ. # 3. 142 pp.
Scammon, Ch.M. 1968 (1874). The Marine Mammals of the Northwestern Coast of North America. N.-Y., Dover Publ. Inc. 319 + V pp.
Slijper, EJ. 1965. A Hundred Years of Modern Whaling. Amsterdam, NCINP. 43 pp.
The Bowhead Whale (Eds. Burns, J.J., Montague, J.J., and C.J. Clowles). 1993. Soc. Marine Mamm. Special Publ. 2. Lawrence, Allen Press Inc. 787 pp.
The Gray Whale Eschrichtius robustus (Eds. Jones, M.L., Swartz, S.L., and Leatherwood, St.). Orlando-San Diego- etc., Academic Press, 1984. 600 pp.
Votrogov, L.M., and Bogoslovskaya, L.S. 1980. Gray Whales off Chukotka Peninsula. In: Rep. Int. Whal. Commn. 30. Pp. 435-437.
Votrogov, L.M., and Bogoslovskaya, L.S. 1986. A Note on Gray Whales off Kamchatka, the Kuril Islands and Peter the Great Bay. In: Rep. Int. Whal. Commn. 36. Pp. 281-282.
Wexo, J.B. 1987. Whales. Zoobooks. San Diego. Frye&Smith. 18 pp.
Wynne, K., and Folkens, P. 1992. Guide to Marine Mammals of Alaska. Fairbanks, UAF. 75 pp.
Yablokov, A.V., and Bogoslovskaja, L.S. 1984. A Review of Russian Research on the Biology and Commercial Whaling of the Gray Whale. In: The Gray Whale Eschrichtius robustus (Eds. Jones, M.L., Swartz, S.L., and Leatherwood, St.).Orlando-San Diego- etc., Academic Press. Pp. 465-485.
СПИСОК АВТОРОВ ИЛЛЮСТРАЦИЙ
В списке указана страница, на которой находится иллюстрация, затем ее автор, год, когда она была сделана или опубликована, и источник, откуда она взята. Рисунки Н. Кондакова 1971 г. являются иллюстрациями известной работы А. Г. Томилина Отряд китообразных в книге Жизнь животных, 1971, т. 6.
АЧЗЭ — архив Чукотской зоологической экспедиции.
РИСУНКИ И ФОТОГРАФИИ
Стр.
1. КАПИТАН ВОТРОГОВ
12 фото П. Блузмы, 1978. АЧЗЭ
14 фото С. Богословского, 1980. АЧЗЭ
18 фото П. Блузмы, 1978. АЧЗЭ
20 фото Н. Конюхова, 1985. АЧЗЭ
2. ИСТОРИЯ СОДРУЖЕСТВА УЧЕНЫХ И МОРСКИХ ОХОТНИКОВ ЧУКОТКИ
24 фото С. Богословского, 1980. АЧЗЭ
28 фото С. Богословского, 1980. АЧЗЭ
31 фото С. Блохина, 1981. АЧЗЭ
32 фото Б. Звонова, 1985. АЧЗЭ
33 фото С. Богословского, 1980. АЧЗЭ
39 фото С. Богословского, 1980. А ЧЗЭ
40 фото С. Богословского, 1980. АЧЗЭ
42 фото И. Крупника, 1975. Собств. автора
43 фото Н. Конюхова, 1984. АЧЗЭ
44 фото С. Богословского, 1981. АЧЗЭ
45 фото С. Богословского, 1980. АЧЗЭ
3. КАК УСТРОЕНО МОРЕ
47 рис. Н. Кондакова, 1956. Зенкович Б.А. Вокруг света за китами
4. О КИТАХ И ДЕЛЬФИНАХ
62 рис. Н. Кондакова, 1971. Жизнь животных, т. 6
65 рис. Н. Кондакова, 1971
66 рис. Н. Кондакова, 1971
69 рис. Н. Кондакова, 1971
72 рис. Н. Кондакова, 1971. По Hentschel (1937).
Slijper, Е. 1965. A Hundred Years ...
73 рис. Н. Кондакова, 1971
75 рис. Н. Кондакова, 1971
78 рис. G. Pilleri, 1983. Auf Baffinland. Zur Erforschung des Narvals ...
81 рис. H. Silverman, 1979. Pilleri, G. 1983. Auf Baffinland ...
5. УСАТЫЕ КИТЫ
84 рис. P. Folkens, 1992. Wynne, K. Guide to Marine Mammals of Alaska
85 рис. В. Hoopes, 1987. Wexo, J.B. Whales. Zoobooks
88 рис. H. Кондакова, 1971
89 рис. H. Кондакова, 1971
92 рис. Е. Богословской, 2002. Собств. автора
94 фото Н. Перова, 1979. АЧЗЭ
95 фото А. Лемберга, 1978. АЧЗЭ
106 фото Н. Перова, 1979. АЧЗЭ
107 фото Н. Перова, 1979. АЧЗЭ
115 фото П. Блузмы, 1978. АЧЗЭ
119 фото J. Smith, 1988. Post card
121 рис. Р. Folkens, 1983. Field Guide to the Gray Whale, 1989
122 фото А. Лемберга, 1978. АЧЗЭ
134 фото Л. Вотрогова, 1977. АЧЗЭ
137 рис. Н. Кондакова, 1971
140 рис. Ch. Scammon, 1874 (1968). Marine Mammals ...
141 фото T. Walker, 1995. Smith, D., Walker, T.
Alaska's Mammals
142 рис. F. Durham, 1980. Marine Fish. Review, V. 42, it 9-10
145 фото Л. Вотрогова, 1971. АЧЗЭ
158 прорисовка петроглифа. Диков Н.Н. 1971.
Наскальные загадки древней Чукотки
159 фото А. Лемберга, 1979. АЧЗЭ
163 рис. F. Durham, 1980. Marine Fish. Review, И 42, It 9-10
165 рис. В. Шкробота, 1986. АЧЗЭ
172 рис. Н. Кондакова, 1971
173 фото G. Joyce, 1984. Marine Fish. Review, V. 46, # 4
174 рис. H. Кондакова, 1980. Атлас морских
млекопитающих
180 рис. Н. Кондакова, 1956. Зенкович Б.А. Вокруг света за китами
181 рис. Н. Кондакова, 1971
186 рис. Н. Кондакова, 1971
187 фото F. Nicklin, 1988. Nat. Geograph., И. 174, it 6
189 рис. Н. Кондакова, 1952. Слепцов М.М.
Китообразные дальневосточных морей
195 фото S. Mizcoch, 1984. Marine Fish. Review, V. 46, it 4
201 рис. H. Кондакова, 1952. Слепцов М.М.
Китообразные дальневосточных морей
207 рис. Н. Кондакова, 1952. Слепцов М.М.
Китообразные дальневосточных морей
213 рис. Н. Кондакова, 1971
214 фото неизв. автора. Интернет
216 фото К. Kleist, 2001. Indig. Affairs, it 4
218 фото К. Kleist, 2001. Indig. Affairs, it 4
220 фото неизв. автора. Интернет
223 рис. Н. Кондакова, 1971
224 фото R. Sanford. Роу/ card
225 фото неизв. автора. Интернет
226 рис. Н. Кондакова, 1971
227 рис. Н. Кондакова, 1971
228 рис. Ch. Scammon, 1874 (1968). Marine Mammals ...
230 фото J. Gibbs, 1984. Hoyt, E. The Whale Watcher's Handbook
6. ЗУБАТЫЕ КИТЫ
232 рис. P. Folkens, 1992. Wynne, K. Guide to Marine
Mammals of Alaska
232 рис. H. Кондакова, 1971
233 рис. H. Кондакова, 1971
234 рис. L. Foster, 1988. Nat. Geograph., V. 174, # 6
238 рис. H. Кондакова, 1971
239 рис. Н. Кондакова, 1971
240 фото неизв. автора. Интернет
242 фото J. Foott. Post card
245 рис. Н. Кондакова, 1971
249 рис. Н. Кондакова, 1971
253 рис. Н. Кондакова, 1971
257 фото неизв. автора. Интернет
261 рис. Н. Кондакова, 1971
262 рис. Н. Кондакова, 1971
263 фото G. Pilleri, 1983. Auf Baffinland. Zur Erforschung des Narvals ...
265 рис. H. Кондакова, 1971
265 фото F. Nicklin, 1988. Nat. Geograph., V. 174, it 6
267 рис. H. Кондакова, 1971
271 рис. H. Кондакова, 1971
272 фото G. Joyce, 1984. Marine Fish. Review, V. 46, # 4
273 рис. H. Кондакова, 1971
278 рис. В. Heezen, 1957. Берзин А.А. 1971. Кашалот
279 рис. D. Gaskin, 1964. Берзин А.А. 1971.Кашалот
280 рис. D. Gaskin, 1964. Берзин А.А. 1971. Кашалот
281 рис. D. Gaskin, 1964. Берзин А.А. 1971. Кашалот
7. НАБЛЮДАЙТЕ ЗА КИТАМИ!
290 рис. Н. Кондакова, 1952. Слепцов М.М. Китообразные дальневосточных морей
291 рис. S. Katona et al., 1979. Эванс В., Яблоков А. 1983. Изменчивость окраски ...
292 рис. В. Эванса, А. Яблокова, 1983. Изменчивость окраски ...
293 рис. В. Эванса, А. Яблокова, 1983. Изменчивость окраски ...
294 рис. Г. Вейнгера, 1983. Эванс В., Яблоков А. Изменчивость окраски ...
На 4-ой странице обложки воспроизведен фрагмент рисунка из альбома Beringia Conservation Program.
National Audubon Society, 1991.
КАРТЫ
90-91
98
101
146
153
236-237
Л. Богословская, наст, издание
В. Prochnow, 1983. Field Guide to the Gray
Whale. 1989
Л. Вотрогов, 1979. АЧЗЭ
Л. Богословская, 1995. Наст, издание
Л. Богословская, 1995. Наст, издание
Л. Богословская, наст, издание
SUMMARY
The book «The Whales of Chukotka» has been written for young hunters of Chukotka, Asian Eskimos and maritime Chukchi as a text-book on biology of whales dwelling in the water of the part of Russia concerned. The objective of the book is conservation and passing on to younger generations of traditional knowledge on whales and its integration with scientific knowledge. The accuracy and authenticity of traditional knowledge on marine mammals of Chukotka have been noted by the distinguished Russian biologists M.M. Sleptsov, A.G. Tomilin, B.A. Zenkovich and later L.M. Votrogov, captain of the whaler Zvyozdnyi, who harvested gray whales for the subsistence of indigenous people of Eastern Chukotka in 1969-1982.
L.M. Votrogov, initiated a network of whale watchers from the Aioyn Island in Chaunskaya Inlet of Eastem-Siberian Sea to the Mainopylgin village in the Bering Sea. The watchers monitored baleen whales, primarily, the gray whale and the bowhead whale. A substantial portion of the data was obtained under the guidance of the present author in 1977-1991 by members of the Chukotka Zoological Expedition of the Institute of Evolutionary Morphology and Ecology of Animals, USSR Academy of Sciences (Severtsov Institute) — both scientists and hunters. In addition, zoologists and Eskimos of the Sireniki village in 1981, 1985, 1987 in a traditional skin boat, or umiak navigated along the coast of the Chukchi Peninsula. As a result, some new information was obtained on the abundance, dates and migration routes of bowhead and gray whales in Chukotka waters, and also the first data on the recovery of the populations of the humpback whale and fin whale.
The book is a popular account of the properties of biology of baleen and toothed whales, providing information about 15 species of whales of Chukotka. The last chapter advises on watching whales and recording observations. The book is supplied with a dictionary of scientific terms.
CONTENTS
WHY HAS THIS BOOK BEEN WRITTEN? ....................8
1. CAPTIAN L.M. VOTROGOV ..........................12
2. HISTORY OF COLLABORATION OF SCIENTISTS AND MARINE HUNTERS OF CHUKOTKA ........................21
3. THE STRUCTURE OF THE SEA.......................47
4. ON WHALES AND DOLPHINS ........................57
5. BALEEN WHALES .................................83
Family of gray whales ..........................92
Gray whale....................................92
Family of right whales ........................136
Bowhead whale, or polar whale ...............137
Northern right whale ........................172
Family of finback whales ......................179
Blue whale ..................................180
Finwhales ...................................189
Sei whale....................................201
Minke whale..................................207
Humpback whale...............................213
6. TOOTHED WHALES................................231
Family of dolphins ............................238
Killer whale.................................238
Family of porpoises............................245
Common porpoise .............................245
Dall’s porpoise .............................249
Family of narwhales ...........................253
White whale .................................253
Narwhale ....................................261
Family of beaked whales ..................267
Baird’s whale ...........................267
Family of Sperm whales ...................271
Sperm whale .............................271
7. WATCH WHALES! ............................283
SOME SCIENTIFIC TERMS AND CONCEPTS ..........295
POPULAR SCIENTIFIC LITERAURE FOR SUPPLEMENTAL READING.....................309
REFERENCES...................................310
LIST OF FIGURES..............................315
SUMMARY......................................320
Научное-популярное издание
Людмила Сергеевна Богословская
КИТЫ ЧУКОТКИ.
Пособие для морских охотников
Рецензент: доктор биологических наук, профессор В.А. Земский
Утверждено к печати Редакционно-издательским советом Российского научно-исследовательского института культурного и природного наследия имени Д.С. Лихачёва
Лицензия ЛР №020730 от 3 марта 1998 г.
Дизайн обложки Е.С. Богословской Компьютерная вёрстка Н.П. Лаку тиной Компьютерная обработка иллюстраций Е.С. Богословской, Н.П. Лакутиной
Подписано в печать 4.08.03
Формат 60 х 90 1/16. Гарнитура TimesET Бумага офсетная. Печать офсетная.
Усл. псч. л. 20,3 Тираж 600 экз. Зак. № 318.
Российский научно-исследовательский институт культурного и природного наследия имени Д.С. Лихачёва
129366, Москва, ул. Космонавтов, 2
Отпечатано в типографии ИПП «Гриф и К» Тула, ул. Октябрьская. 81а
ЛР 065454 от 13.10.97, тел. (0872) 49-76-96
Богословская Людмила Сергеевна закончила биолого-почвенный факультет Московского государственного университета им. М.В. Ломоносова, доктор биологических наук, руководитель центра традиционной культуры природопользования Российского института культурного и природного наследия им. Д.С. Лихачева.
Главные научные интересы - традиционные культуры коренных народов и старожилов Севера, этноэкология, биология морских зверей и птиц. Особым направлением деятельности является интеграция научных и традиционных знаний в области природопользования и экологии природных комплексов Севера.
Более 30 лет Л.С.Богословская работает в Арктике, от Шпицбергена и Новой Земли до региона Берингова пролива включительно.
В 1977-1991 годах она возглавляла Чукотскую зоологическую экспедицию Института эволюционной морфологии и экологии животных им. А.Н. Северцова Академии Наук СССР.
Под ее руководством и при непосредственном участии впервые нанесены на карту миграционные пути и места концентрации китов, залежки и лежбища ластоногих, колонии морских птиц, а также древние селения на побережье Восточной Чукотки между бухтой Руддера и Колючинской губой. В течение всех лет работы постоянными членами Чукотской экспедиции были морские охотники из поселков Сиреники и Янракыннот.
Как эксперт Государственной Думы федерального Собрания РФ Богословская многие годы участвует в разработке законопроектов, имеющих отношение к природе и коренному населению Севера.
Автор 150 научных публикаций, в том числе пяти монографий.
В течение последнего миллиона лет Азия и Северная Америка неоднократно соединялись обширной сушей, получившей название Берингия. Во время ледникового периода, 18 тыс. лет назад, сухопутный мост между материками имел ширину более 10ОО км, уровень моря был ниже современного на 90 м. Берингов пролив, соединивший Тихий и Ледовитый океаны, появился всего 14-12 тыс. лет назад.
Берингия оказала мощное воздействие на формирование растительного и животного мира Земли и на историю людей, впервые перешедших по ней из Азии в Америку. Все природные и культурные явления этой части планеты можно считать Наследием Берингии.
Что делали киты, когда ещё не было пролива? Ушли все к югу от Берингии? Или часть их всё время оставалась в арктических водах? Наука пока не знает ответа на эти вопросы.