






























































































































































































































































































































Author: Тиходеев О.Н.
Tags: психология учебники и учебные пособия по психологию молекулярная биология
ISBN: 978-5-7695-8051-2
Year: 2011




Text
Высшее профессиональное образование
Учебник
О. НЛиходеев
ОСНОВЫ
ПСИХОГЕНЕТИКИ
ПСИХОЛОГИЯ
ACADEM А
УЖК 159.9(075.8)
BSK 88я73
Т462
Рецензенты:
щоктор биологических наук, академик РАН, заведующий кафедрой генетики
и селекции Санкт-Петербургского государственного университета
С. Г. Инге-Вечтомов,
днюктор психологических наук, доцент, заведующий кафедрой медицинской
психологии и психофизиологии Санкт-Петербургского государственного
университета О. Ю. Щелкова
Ъгходеев О. Н.
Т4в<>2 Основы психогенетики : учебник для студ. учреждений
высш. проф. образования / О. Н.Тиходеев. — М.: Издатель-
ский центр «Академия», 2011. — 320 с.
ISBN 978-5-7695-8051-2
Учебник создан в соответствии с Федеральным государственным
образовательным стандартом по направлению подготовки 030300 «Пси-
хология» (квалификация «бакалавр»).
Рассмотрены концептуальные положения молекулярной биологии,
важнейшие признаки формирования индивидуальных признаков живо-
го организма, молекулярные процессы, лежащие в основе функциони-
рования нервной системы человека, его психики, а также методы со-
временной психогенетики и их возможности. Приведены данные о
влиянии конкретных генов на психологические признаки животных и
человека и молекулярных механизмах такого влияния. Освещены вопро-
сы истории возникновения психогенетики, перспективы ее развития,
современного представления об эволюции человеческой психики и
этических аспектах психогенетики.
Для студентов учреждений высшего профессионального образова-
ния.
УДК 159.9(075.8)
ББК 88я73
Оригинал-макет данного издания является собственностью
Издательского центра * Академия*, и его воспроизведение любым способом
без согласия правообладателя запрещается
ISVTN 978-5-7695-8051-2
© Тиходеев О. Н., 2011
© Образовательно-издательский центр «Академия», 2011
© Оформление. Издательский центр «Академия», 2011
ПРЕДИСЛОВИЕ
Вопрос о влиянии генов на человеческую психику обсуждает-
ся психологами и генетиками уже более ста лет. Но только совсем
недавно стало возможным перейти от косвенных доводов к вы-
явлению генов, непосредственно участвующих в контроле психо-
логических признаков. Это связано с бурным развитием молеку-
лярной генетики и разработкой специальных методов генетиче-
ского анализа, применимых к такому сложному объекту, как
человек.
Основная задача данного учебника — ознакомить студентов-
психологов на доступном для них уровне с идеологией и важней-
шими методами современной психогенетики, с ее достижениями
и ближайшими перспективами. Особое внимание уделено психо-
логическим признакам, для которых уже установлены конкретные
гены. Это эпизодическая память, пластичность мышления, вни-
мательность, склонность к самоубийству, отношение к пище,
а также предрасположенность к алкоголизму. Влияние конкретных
генов на перечисленные признаки продемонстрировано множе-
ством независимых исследований и убедительно подтверждено
мета-анализом.
Необходимо отметить, что термин «психогенетика» главным
образом характерен для отечественной литературы. Зарубежным
аналогом является «генетика поведения». Мы будем использовать
термин «психогенетика» ко всем аспектам генетики поведения
человека за исключением отчетливо выраженных психических
аномалий, традиционно являющихся объектами психиатрии.
В основу настоящего учебника положен курс «Психогенетика»,
читаемый автором в течение ряда лет на факультете психологии
Санкт-Петербургского университета. Автор выражает глубокую
признательность Н. И. Абрамсон, М. С. Березанцевой, Е. В. Голубко-
вой, Е. В. Даеву, J1. А. Мамон, П. А. Шапчицу, О. В. Щербаковой за
ценные рекомендации, высказанные ими при подготовке руко-
писи к изданию.
Глава 1
ОСНОВНЫЕ ПОЛОЖЕНИЯ МОЛЕКУЛЯРНОЙ
БИОЛОГИИ
1.1. Место биологии среди естественных
наук
Приступая к краткому знакомству с современной биологией,
необходимо начать с концептуального вопроса о том, каково ее
место среди естественных наук. К этим наукам относят астроно-
мию, геологию, физическую географию, химию, биологию и
физику (с учетом их многочисленных разделов). Астрономия за-
нимается всесторонним исследованием космоса, геология изуча-
ет недра земного шара, физическая география — поверхность
нашей планеты. Химия исследует строение и свойства веществ.
Биология изучает жизнь во всех ее проявлениях, а физика — все-
общие, фундаментальные законы мироздания.
Опираясь на это общепринятое описание наук, нетрудно
прийти к принципиально важному выводу. Если физика изучает
всеобщие законы природы, это значит, что любая другая есте-
ственная наука имеет дело с теми же самыми законами, но с
учетом специфики конкретных исследуемых явлений. Соответ-
ственно все естественные науки можно рассматривать в качестве
частных физических дисциплин (табл. 1.1). Астрономия, по сути,
является физикой космоса, геология — физикой недр земного
шара, физическая география — физикой поверхности Земли.
Химия представляет собой физику веществ на атомарно-молеку-
лярном уровне их организации. И, наконец, биология — это
физика живого.
Насколько оправданы такого рода представления? Попробуем
выяснить это на примере биологии. В 1944 г. выдающийся австрий-
ский физик Эрвин Шредингер опубликовал свою книгу «Что такое
жизнь?*. В этой книге он обратил внимание на то, что целый ряд
явлений живой природы был в то время абсолютно необъясним с
позиции физики. Поэтому, предполагал Шредингер, в результате
дальнейшего развития биологии будут открыты принципиально но-
вые физические законы, выполняющиеся только в живой природе.
4
Таблица 1.1. Естественные науки как частные физические дисциплины
Наука Объект изучения Примеры исследуемых явлений Основные причины явлений
Астрономия Космос Смена лунных фаз Движение Луны по орбите вокруг Земли; падение солнечных лучей на поверхность Луны
Формирование марсианских кратеров Взаимодействие поверхности Марса с ме- теоритами
Развитие Солнечной системы Постепенное формирование и уплотнение разнообразных скоплений в гигантском об- лаке космического газа и пыли
Старение звезд Истощение запасов водорода в недрах звезд
Геология Недра Земли Извержение вулканов Образование разрывов в земной коре; вы- брос расплавленных компонентов мантии на земную поверхность
Формирование осадочных пород Разрушение горных пород; смывание об- разующихся частиц в водоемы и осаждение на дне
Движение материковых плит Постепенное перемещение расплавленных пород в толще мантии
Формирование месторождений нефти и природного газа Образование мощных отложений из остан- ков растений; преобразование этих остан- ков под действием высоких температуры и давления
Окончание табл. 1.1
Наука Объект изучения Примеры исследуемых явлений Основные причины явлений
Физическая география Поверхность Земли Разнообразие осадков Варьирование температуры и влажности воздуха в различных участках атмосферы
Сезонная смена ветров Сезонные изменения температуры на определенных участках земной поверхности
Формирование речной долины Размывание земной коры потоками воды; изменение русла в результате вращения Земли; отложение осадков на берегах при разливах
Развитие горной системы Деформация земной коры при движении ма- териковых плит; разрушение образовавших- ся складок водными потоками, ветром и др.
Разнообразие климатических поясов Неравномерное распределение тепла, по- ступающего на земную поверхность с солнечными лучами
Химия Строение и свой- ства веществ Разнообразие химических эле- ментов Варьирование количества протонов в ядрах атомов
Перенос электронов при окислительно-восстановительных реакциях Стабилизация электронных оболочек у взаимодействующих атомов
Кислотно-щелочные свойства веществ Характер диссоциации молекул при взаимо- действии с растворителем
Ускорение химической реакции в присутствии катализаторов Способность катализатора взаимодейство- вать с участниками реакции с образованием высокоактивных промежуточных соединений
Биология Жизнь Питание организма Поглощение необходимых молекул из окру- жающей среды и их преобразование в собственные молекулы организма
Дыхание организма Окисление части собственных органических веществ с выделением энергии, используе- мой на нужды организма
Развитие многоклеточного орга- низма из одноклеточного заро- дыша Деление клеток зародыша; неравномерное распределение регуляторных молекул по разным клеткам; задействование разных генов в различных клетках
Подвижность яшерицы Использование энергии молекул АТФ для перемещения белковых нитей в мышечных клетках
Разворачивание листьев на побеге картофеля под действием света Восприятие рецепторными молекулами квантов света; активация молекул, обеспе- чивающих развитие листьев
С тех пор биология совершила гигантский скачок, что связано
с переносом исследований на молекулярный уровень. Уже раз-
гаданы многие тайны живой природы, однако это пока не при-
вело ни к каким существенным изменениям в области физики.
Действительно, практически для каждого живого существа вы-
полняются все без исключения физические законы: закон сохра-
нения вещества и энергии, законы электро- и термодинамики,
законы механики, оптики и многие другие. Более того, даже в
сложнейших биологических закономерностях все отчетливее про-
слеживаются конкретные молекулярные механизмы, имеющие
под собой сугубо физическую основу. Наконец, организмы со-
стоят из таких же атомов, что и неживая природа. Варьирует
только соотношение между разными атомами (табл. 1.2).
Таблица 1.2. Относительное содержание химических элемен-
тов в живой и неживой природе
Химический элемент Содержание, % по массе
во Вселенной в земной коре в организме человека
Водород (Н) 87 0,14 10
Гелий (Не) 12 < 0,000001 < 0,000001
Углерод (С) 0,03 0,03 18
Азот (N) 0,008 0,0001 3
Кислород (О) 0,06 46 65
Натрий (Na) 0,0001 2,5 0,1
Магний (Mg) 0,0003 2,3 0,03
Алюминий (А1) 0,0002 8 0,00009
Кремний (Si) 0,003 27 0,001
Фосфор (Р) 0,00003 0,1 1,2
Сера(S) 0,002 0,05 0,2
Хлор (О) 0,000008 0,02 0,2
Калий (К) 0,000007 2,2 0,2
Кальций (Са) 0,0001 4,1 1.5
Железо (Fe) 0,002 5 0,006
8
Таким образом, биологию с современной точки зрения вполне
правомочно рассматривать как физику живого. В соответствии с
этим мы вправе ожидать, что любые биологические явления, будь
то развитие, подвижность, размножение или поведение, воз-
никают в результате обмена веществ и энергии, т.е. очень
сложных, но сугубо материальных процессов.
1.2. Молекулярная специфика живого
Одной из ключевых особенностей живой природы является
сложнейший химический состав. Так, даже в примитивной бак-
териальной клетке содержатся десятки тысяч различных соедине-
ний, а в организме взрослого человека — многие миллионы.
Подавляющее большинство из них относится к органическим
соединениям.
Молекула любого органического вещества содержит в своем
составе атомы углерода, причем, как правило, в довольно боль-
шом количестве. Эти атомы ковалентно связаны между собой,
образуя достаточно прочные углеродные цепочки. Разнообразие
таких цепочек поразительно велико. Во-первых, они очень силь-
но варьируют подлине. Во-вторых, они могут быть разными по
своему строению, а именно — линейными, разветвленными или
кольцевыми. В-третьих, связи между соседними атомами угле-
рода бывают не только одинарными, но и двойными или даже
тройными. В результате всего лишь из нескольких атомов угле-
рода можно «построить» большое разнообразие подобных цепо-
чек.
Кроме того, в состав органических молекул входят атомы и
некоторых других химических элементов. Чаще всего это атомы
водорода, кислорода, азота, фосфора или серы. Они ковалентно
присоединены к углеродным цепочкам, образуя друг с другом
различные сочетания. При этом одна и та же углеродная цепочка
служит «скелетом» для множества разных соединений, что при-
водит к огромному разнообразию органических веществ.
Как известно, в процессе обмена веществ и энергии органи-
ческие соединения вступают в сложнейшие взаимодействия.
В результате формируются разнообразные молекулярные ком-
плексы, обеспечивающие необходимые для организма химиче-
ские реакции. Нужно отметить, что каждый подобный комплекс
содержит лишь строго определенные молекулы, причем в кон-
кретном соотношении между собой. Что же позволяет молекулам
правильно распознавать друг друга среди тысяч и тысяч различных
соединений?
9
Рис. 1.1. Формирование молекулярного комплекса с учетом формы молекул
(а) и их зарядов (б)
Прежде всего, взаимодействующие органические молекулы
точно соответствуют друг другу по размеру и форме (рис. 1.1, а).
Однако в составе одного и того же организма обычно содержится
много различных соединений, молекулы которых близки и по
форме, и по размеру. Таким образом, чтобы молекулы распозна-
вали друг друга, должно соблюдаться еще одно важное условие,
а именно — взаимное соответствие их зарядов (рис. 1.1, б). В этом
случае возникает электростатическое притяжение, позволяющее
молекулам сформировать необходимый комплекс.
Подобные заряды обусловлены тем, что некоторые атомы,
входящие в состав органической молекулы, существенно разли-
чаются своей электроотрицательностью, т.е. способностью
притягивать электроны (табл. 1.3). В результате образуются кова-
лентные полярные связи, за счет которых молекула и приобрета-
ет определенные заряды. Такие связи обеспечиваются группами
разных атомов, ковалентно присоединенных к углеродному ске-
лету. Чаще всего это следующие группы: —ОН, —СО, —СООН,
—NH2, —SH и —Н2РО4. Их названия и свойства представлены в
табл. 1.4.
Таблица 1.3. Типы химических связей
Тип связи Основные особенности Примеры*
Ковалентная неполярная Образуется между атомами, имею- щими одинаковую электроотрица- тельность. Электронная плотность не смещается ни к одному из взаимо- 2 О О hi II | z О п Q ? Г) I 1 11
Окончание табл. 1.3
Тип связи Основные особенности Примеры*
действующих атомов. Дополнитель- ных зарядов на них не возникает. Достаточно прочная связь
Ковалентная полярная Образуется между атомами, имеющи- ми разную электроотрицательность. Электронная плотность частично смещается к одному из атомов, в результате чего возникают слабые электростатические заряды. Доста- точно прочная связь Т Q Z X х й< I II । J I । J U О Z z О
Ионная Образуется между атомами, резко отличающимися своей электроотри- цательностью. Один или несколько электронов полностью переходят к более электроотрицательному атому, в результате чего возникают сильные заряды. Непрочная связь, в растворе легко диссоциирует с образованием ионов н+сг К+ОН- Na*Cl- Са2+СО32
Водородная Образуется между атомами, несущи- ми слабые противоположные по зна- ку электростатические заряды. Очень слабая связь. Легко разрушается С* = о^ • •
* Знаками <6+» и «6-» показаны слабые положительные и отрицательные за-
ряды, возникающие в результате частичного смещения электронной плотности к
более электроотрицательному атому. Знаками «+» и «-» показаны сильные заря-
ды, возникающие в результате полного смещения электрона к одному из атомов.
Именно от разнообразия заряженных групп, от их количества
и взаимного расположения на углеродном скелете зависят свой-
ства конкретной органической молекулы. При этом обычно вы-
полняется следующее правило: чем больше различных заряжен-
ных групп у молекулы, тем сложнее ее химические свойства.
В свою очередь, от этого зависят и биологические функции,
выполняемые данными молекулами в организме.
Необходимо особо подчеркнуть, что в живой природе встречают-
ся далеко не все органические вещества. Для удобства их принято
называть биоорганическими соединениями. Именно этими соеди-
нениями обусловлено все разнообразие явлений живой природы.
11
Таблица 1.4. Характеристика наиболее распространенных заряженных групп атомов
Название группы Химическая формула группы Распределение зарядов* Результат для молекулы
Гидроксильная -ОН -О^Н4* Молекула приобретает два слабых электростатических за- ряда, противоположных друг другу по своим знакам. На поверхности молекулы оказывается слабый положи- тельный заряд
Карбонильная -<° 5- —с^° Молекула приобретает два слабых электростатических заряда, противоположных друг другу по своим знакам. На поверхности молекулы оказывается слабый отрицатель- ный заряд
Карбоксильная -<° он I л о от Связь О—Н в карбоксильной группе является ионной. Такие связи легко разрушаются в водной среде. В ре- зультате молекула, содержащая карбоксильную группу, обычно приобретает сильный отрицательный заряд
Амино —N н тд 5+ 6-/Н —N—Н+ чн8+ Атом азота в составе аминогруппы способен присоеди- нять протон из водной среды. В результате молекула, содержащая аминогруппу, обычно приобретает сильный положительный заряд
Тио -SH -S6 н^ Молекула приобретает два слабых электростатических заряда, противоположных друг другу по своим знакам. На поверхности молекулы оказывается слабый положитель- ный заряд
Фосфатная О II —о—Р—он 1 он о —о—р—О’ 1 О" Связи О —Н в фосфатной группе являются ионными. Такие связи легко разрушаются в водной среде. В резуль- тате молекула, содержащая фосфатную группу, обычно приобретает сильный отрицательный заряд
См. пояснения к табл. 1.3.
1.3. Важнейшие классы биоорганических
соединений
1.3.1. Углеводы
Знакомство с наиболее важными биоорганическими соедине-
ниями мы начнем с класса углеводов. Эти соединения получили
свое название за то, что состоят из многочисленных звеньев
Н —С —ОН, ковалентно соединенных друг с другом в различные
цепочки. В результате углеводы, как правило, соответствуют об-
щей химической формуле С„(Н2О),„.
Все углеводы благодаря своим многочисленным группам —ОН
довольно охотно контактируют с водой, т.е. являются гидрофиль-
ными веществами, а многие из них прекрасно в ней раство-
римы. Такие хорошо растворимые в воде углеводы обычно об-
ладают приятным сладким вкусом и называются сахарами.
В природе широко распространены два типа сахаров: это моно-
сахариды и дисахариды (табл. 1.5).
Молекула любого моносахарида представляет собой одну срав-
нительно небольшую цепочку, образованную несколькими зве-
ньями Н —С —ОН. Количество этих звеньев, в зависимости от
конкретного соединения, обычно варьирует в пределах от трех до
семи. Кроме того, молекулы многих моносахаридов способны
замыкаться в кольцо через один из своих атомов кислорода. При-
мером такого циклического моносахарида является хорошо из-
вестная вам глюкоза С6Н)2О6.
Дисахариды имеют более сложное строение. Каждый из них
образован двумя моносахаридами, соединенными друг с другом
через атом кислорода. Как правило, в составе подобной молекулы
содержатся разные типы моносахаридов. Например, молекула
сахарозы, т.е. обычного пищевого сахара, включает в себя глю-
козу и фруктозу.
Существуют и еще более сложные углеводы. Это сложно орга-
низованные полимерные молекулы, состоящие из многочислен-
ных остатков моносахаридов. Такие углеводы называют поли-
сахаридами, к их числу относятся крахмал, гликоген, целлю-
лоза и хитин. Из-за очень крупного размера своих молекул эти
соединения уже практически нерастворимы в воде и не обладают
сладким вкусом.
Функции углеводов. Несмотря на свое многообразие, все
углеводы имеют достаточно сходный план строения. Поэтому их
химические свойства и биологические функции тоже не могут
быть очень разнообразными. Наиболее важная функция углево-
дов — энергетическая. Дело в том, что подавляющее большин-
14
Таблица 1.5. Важнейшие природные моно- и дисахариды
Тип сахара Название Химическая формула Распространение
Моносахариды Глюкоза С6н12о6 Практически у всех живых существ
Фруктоза С6н12о6* В сочных плодах растений
Галактоза С6н12о* В клетках млечных желез у млекопитающих
Дисахариды Сахароза (тростнико- вый сахар) С|2Н22О|| (остаток глюко- зы, соединенный с остатком фрук- тозы) Практически у всех наземных растений
Лактоза (мо- лочный сахар) С|2Н220ц (остаток глюко- зы, соединенный с остатком галак- тозы) В молоке млеко- питающих
Мальтоза (солодовый сахар) С12Н220ц (два соединенных друг с другом остатка глюкозы) Практически у всех животных и грибов
* Молекулы глюкозы, фруктозы и галактозы имеют одинаковую химическую
формулу, но при этом отличаются взаимным расположением отдельных атомов.
Такие молекулы называют изомерами. К изомерам относятся и все перечислен-
ные дисахариды.
ство живых существ получает необходимую для их жизнедеятель-
ности энергию в основном за счет окисления глюкозы. Неудиви-
тельно, что именно глюкоза является самым распространенным
в природе углеводом.
С энергетической функцией углеводов тесно связана и их за-
пасная роль. Действительно, у многих организмов запас пита-
тельных веществ представлен преимущественно полисахаридами.
У растений эту функцию обычно выполняет крахмал, иногда со-
ставляющий до 90% сухого веса (например, в зернах риса или в
клубнях картофеля). Между тем у подавляющего большинства
15
животных и грибов основным запасным полисахаридом является
гликоген. Молекулы каждого из этих углеводов содержат многие
тысячи звеньев глюкозы и при необходимости могут быть под-
вергнуты расщеплению. В результате высвобождается необходи-
мое количество глюкозы, которая затем используется на нужды
организма.
Еще одна важная функция углеводов — строительная. Для
примера достаточно вспомнить о клеточных стенках: их основу
составляют именно полисахариды, у растений — целлюлоза, а у
грибов — хитин.
По ходу дальнейшего изложения материала мы познакомимся
и с некоторыми другими углеводами.
1.3.2. Жиры
В отличие от углеводов все жиры весьма неохотно контакти-
руют с водой (т.е. являются сильно гидрофобными веществами).
Это связано с тем, что любая молекула жира содержит три длин-
ных углеводородных «хвоста», не имеющих существенных элек-
тростатических зарядов и потому избегающих взаимодействия с
водой (рис. 1.2, а). Обычно в составе одной молекулы жира при-
сутствуют разные углеводородные «хвосты». Они отличаются друг
от друга своими размерами, а также количеством и расположени-
ем двойных ковалентных связей С=С. Впрочем, несмотря на
указанные различия, все жиры устроены достаточно однообразно,
а потому способны выполнять лишь ограниченный круг биоло-
гических функций.
Функции жиров. Самая важная из этих функций — запасная.
Действительно, у многих организмов основной запас питательных
веществ образован именно жирами. Например, маслянистые пло-
ды и семена некоторых растений (оливкового дерева, облепихи и
подсолнечника) или жировые отложения у млекопитающих.
Вторая функция жиров — энергетическая. Дело в том, что
различные жиры, как и глюкоза, тоже могут подвергаться окис-
лению, в результате чего высвобождается необходимая энергия.
Хорошо известно, что жиры обладают низкой теплопроводно-
стью. Поэтому у теплокровных животных (млекопитающих и
птиц) жиры выполняют и термоизолирующую роль. Неудиви-
тельно, что отложения жира в основном расположены не внутри
организма, а непосредственно под кожей. Этот слой должен быть
особенно толстым у животных, постоянно подвергающихся риску
переохлаждения (у китов, тюленей, пингвинов, белых медведей
и др.). В частности, у синего кита этот слой достигает толщины
в I м.
16
н оннннннннннннннн
I II I I I I I I I I I I I I I I I
н-с-о-с-с-с-с-с-с-с-с-с-с-с-с-с-с-с-с-н
I I I I I I I I I I I I I I I
ннннннннннннннн
оннннннннннннннннн
II I I I I I I I I I I I I I I I I I
н-с-о-с-с-с-с-с-с-с-с-с-с-с-с-с-с-с-с-с-с-н
I I I I I I I I I I I I I I I I I
ннннннннннннннннн
оннннннннннннннннн
II I I I I I I I I I I I I I I I I I
н-с-о-с-с-с-с-с-с-с-с-с=с-с-с-с-с-с-с-с-с-н
I I I I I I I I I I I I I I I I
н ннннннн нннннннн
Углеводородные «хвосты»
Гидрофобные
«хвосты»
Гидрофильная
«головка»
б
Рис. 1.2. Жиры: обший план строения молекулы — углеводородные «хво-
сты» (а), схематичное изображение фосфолипида (6), двойная фосфолипид-
ная пленка, являющаяся основой биологических мембран (в)
Очень важную биологическую функцию выполняют родствен-
ные жирам фосфолипиды. Они образуют основу клеточных
мембран. Вместо одного из трех углеводородных «хвостов» моле-
кула фосфолипида содержит сложный радикал с заряженной
группой — Н2РО4. Благодаря наличию сильных электростатических
зарядов эта группа способна охотно контактировать с водой. Та-
ким образом, в молекуле фосфолипида можно выделить два раз-
ных по свойствам участка: гидрофильную «головку» и сильно
гидрофобные «хвосты» (рис. 1.2, б). Поэтому в водной среде (на-
пример, в цитоплазме клетки) молекулы фосфолипидов распола-
гаются так, чтобы их гидрофильные «головки» контактировали с
водой, а гидрофобные «хвосты» были обращены друг к другу.
В результате происходит формирование различных структур, в
том числе и двухслойных фосфолипидных мембран (рис. 1.2, в).
Итак, и углеводы, и жиры являются важными биооргани-
ческими соединениями. В основном они выполняют запасную
и энергетическую функции, а в ряде случаев — и некоторые
17
другие. Тем нс менее из-за однообразия своего химического
строения ни углеводы, ни жиры не способны обеспечить все
остальные необходимые для жизни функции.
1.3.3. Белки
Данны» класс соединений заслуживает особого внимания. Это
связано с тем, что именно белки считаются наиболее сложными
природными веществами: они не только имеют полимерное
строение, но и состоят из множества разных мономеров
Мономерами белков являются различные аминокислоты. Так
называют соединения, отвечающие химической формуле NH2—
CHR — СООН (рис. 1.3, а). Нетрудно заметить, что в составе
любой аминокислоты к одному и тому же атому углерода присое-
динены две заряженные группы: — Ь1Н2и —СООН. Первая обла-
дает сильным положительным зарядом, вторая — напротив, силь-
ным отрицательным. Эта особенность строения позволяет амино-
кислотам принимать участие в сложных химических реакциях,
в том числе — в образовании длинных полимеров (рис. 1.3, б).
Наряду с двумя перечисленными выше группами каждая ами-
нокислота содержит определенный радикал R. Структура данно-
го радикала может быть различной, от одного единственного
атома водорода и до сложно организованных многоатомных груп-
пировок. В принципе, за счет варьирования этого радикала воз-
н R о
\ I '/
N-C-C
\ т
N-C-C
н' А 4 он
N-C-C
Рис. 1.3. Аминокислоты: обшее строение (я) и принцип формирования по-
лимера (б). Связи между соседними аминокислотами называют пептидными.
Поэтому в качестве синонима слова «белок* очень часто неполную! слово
«полипептид»
18
можно огромное разнообразие аминокислот. Однако все извест-
ные живые существа используют для построения своих белков
универсальный «набор» из 20 вариантов аминокислот (табл. 1.6).
Необходимо особо подчеркнуть, что их радикалы различаются не
только по форме и размеру, но и по заряду. Он может либо от-
сутствовать, либо присутствовать, быть слабым или сильным,
положительным или отрицательным.
Таблица 1.6. Универсальный набор аминокислот
Аминокислота Ее обозначение
Русскоязычное Международ- ное Однобуквен- ное
Аланин Ала Ala A
Аргинин Apr Arg R
Аспарагин Асн Asn N
Аспарагиновая кислота Асп Asp D
Валин Вал Vai V
Гистидин Гис His H
Глицин Гли Gly G
Глутамин Глн Gin Q
Глутаминовая кислота Глу Glu E
Изолейцин Иле lie I
Лейцин Лей Leu L
Лизин Лиз Lys К
Метионин Мет Met M
Пролин Про Pro P
Серин Сер Ser S
Тирозин Тир Туг Y
Треонин Тре Thr T
Триптофан Трп Trp W
Фенилаланин Фен Phe F
Цистеин Цис Cys C
19
Вы уже знаете, что свойства и функции биоорганической мо-
лекулы определяются нюансами ее химического строения. Осо-
бенно важную роль играют заряды. Их количество, разнообразие
и расположение зависят от того, в какой последовательности
соединены друг с другом различные аминокислоты. Поэтому
свойства и функции конкретного белка определяются его
последовательностью аминокислот.
Сколько же разных последовательностей можно составить на
основе универсального аминокислотного набора? Очевидно, что
это зависит от их размера. Если каждая последовательность со-
держит по 2 аминокислоты, можно получить 400 разных вариан-
тов (202). Для последовательностей, содержащих по 3 аминокис-
лоты, количество вариантов будет в 20 раз больше (8 000, т.е. 203).
Между тем среднестатистическая молекула белка состоит из не-
скольких сотен аминокислотных остатков. Таким образом, в от-
личие от жиров и углеводов белки обладают огромным струк-
турным разнообразием. Соответственно биологические
функции белков тоже поразительно разнообразны.
Функции белков. Важнейшая из функций белков — фермен-
тативная. Многие необходимые организму химические реакции
не могут осуществляться «сами по себе», а если и протекают, то
с крайне низкой скоростью. Эту проблему решают особые бел-
ки — ферменты. Они работают в качестве биологических ката-
лизаторов, ускоряя необходимые реакции в миллионы раз. Каж-
дую реакцию обычно обслуживает особый фермент. Для примера
достаточно вспомнить о пищеварительных ферментах: одни обес-
печивают расщепление только углеводов, другие — только белков,
а третьи — только жиров.
Очень важна и рецепторная функция белков. Любому, даже
примитивному одноклеточному организму необходимо своевре-
менно ощущать разнообразные изменения, происходящие в
окружающей среде и в нем самом. Это обеспечивается особыми
белками — рецепторами, каждый из которых воспринимает
определенные воздействия.
Многие белки выполняют в организме регуляторную функ-
цию. Проиллюстрируем это на очень простом примере. Допустим,
что некий фермент (обозначим его Ф) способен проявлять свою
активность только в том случае, если он взаимодействует с моле-
кулой белка Р. Это значит, что белок Р регулирует активность
фермента Ф.
Еще одна важная функция белков — строительная. Многие
белки являются строительным материалом для организма. В част-
ности, к таким белкам относится кератин — основной компонент
верхнего слоя кожи и его разнообразных производных (волос, ногтей,
копыт, перьев, игл, роговых чешуй и др.) у позвоночных животных.
20
Некоторые белки выполняют двигательную функцию. Имен-
но они обеспечивают сокращение мышц, движение жгутиков,
изменение формы клеток и др.
Белки, для которых характерна защитная функция, позволя-
ют организму бороться с различными инфекциями. У позвоноч-
ных животных к таким белкам относятся антитела, вырабаты-
ваемые организмом при иммунном ответе.
Известны белки, выполняющие транспортную функцию, т.е.
отвечающие за перенос в организме определенных молекул.
В частности, у многих животных белок гемоглобин обеспечивает
транспортировку кислорода по кровеносной системе.
Безусловно, данный перечень можно продолжить. Однако уже
не вызывает сомнения тот факт, что подавляющее большинство
необходимых для жизни функций обеспечивается в организ-
мах именно белками.
1.3.4. Пространственная организация молекул
белка
Говоря об особенностях строения белковых молекул, принято
различать четыре уровня структур. Если обсуждают первичную
структуру белка, речь идет о его непосредственной аминокислот-
ной последовательности. Следующие уровни касаются укладки
молекулы.
В составе одной и той же молекулы белка содержатся сотни, а
то и тысячи заряженных атомов. Между некоторыми из них воз-
никает электростатическое отталкивание, а между некоторыми,
напротив, электростатическое притяжение. Например, атом кис-
лорода в группе С=О и атом азота в группе N — Н несут на себе
слабые противоположные заряды и потому способны притяги-
ваться друг к другу. В результате между ними образуется водо-
родная связь (см. подразд. 1.2, табл. 1.3). Эти связи значительно
слабее ковалентных. Тем не менее если они достаточно много-
численны, определенные участки молекулы могут скручиваться в
так называемые вторичные структуры.
В белках различают два типа вторичных структур: а-спирали
и P-листы (рис. 1.4). При этом в составе одной и той же молеку-
лы могут присутствовать разные вторичные структуры. Для при-
мера обратимся к лизоциму — небольшому белку, выпол-
няющему в организме важную защитную функцию: он разруша-
ет оболочки различных бактериальных клеток. В молекуле
лизоцима можно выделить 13 участков: 5 а-спиралей, 1 p-лист и
7 нескрученных. В результате дополнительных взаимодействий
между этими участками молекула лизоцима приобретает третич-
21
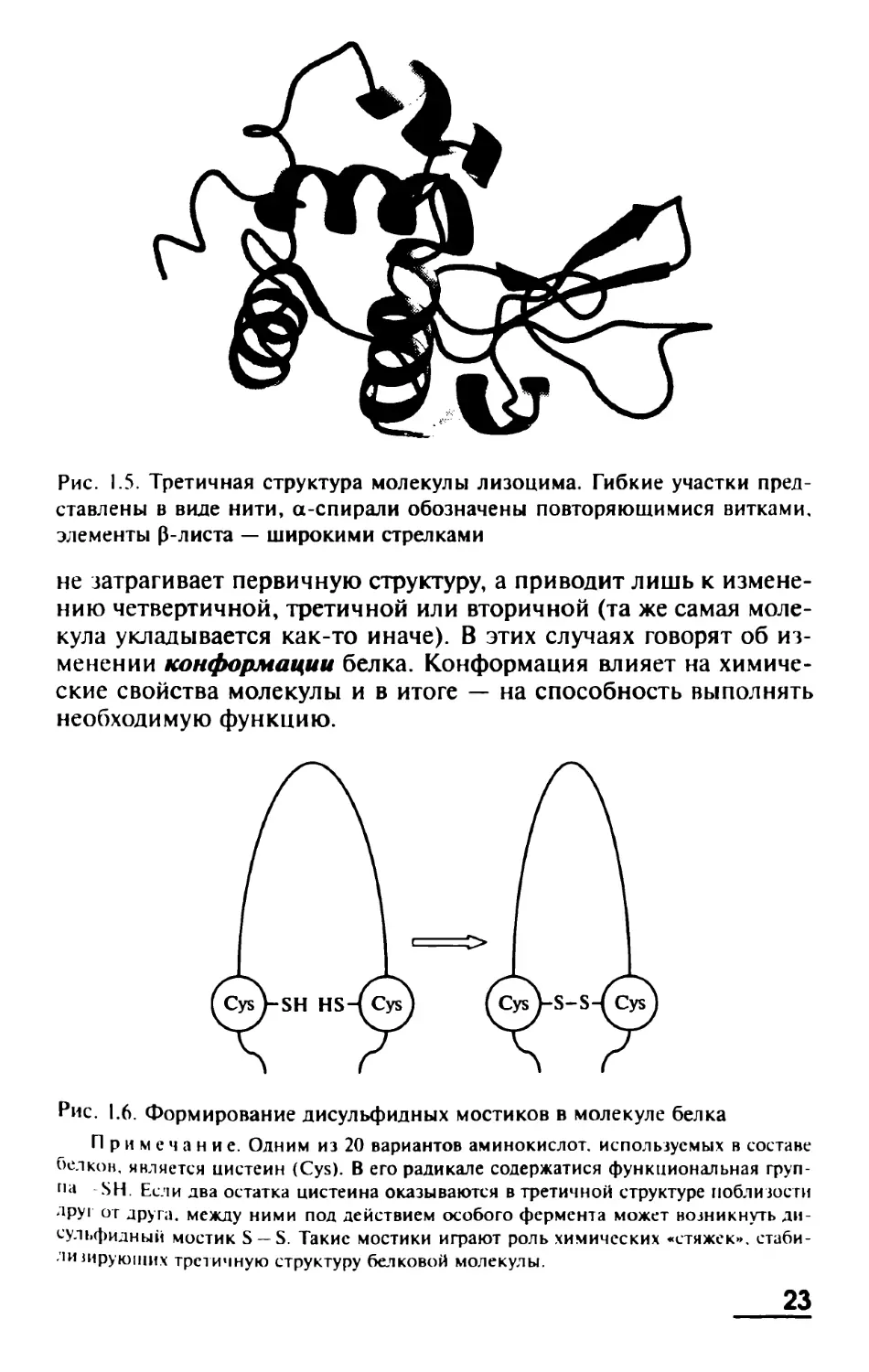
Рис. 1.4. Вторичные структуры белка: а-спираль (а) и p-лист (б)
ную структуру, т. е. некую сложно организованную трехмерную
укладку (рис. 1.5).
Что же именно приводит к образованию третичной структуры?
Во-первых, участки, богатые гидрофобными радикалами, стре-
мятся как можно теснее приблизиться друг к другу, чтобы умень-
шить площадь контакта с молекулами воды. Во-вторых, многие
заряженные группы атомов не участвуют в процессе формирова-
ния вторичных структур. Такие группы способны вступать во
взаимодействие друг с другом и обеспечивать дополнительное
скручивание молекулы белка. В-третьих, между определенными
остатками аминокислот могут возникнуть прочные дисульфидные
мостики (рис. 1.6). Эти мостики служат своеобразными химиче-
скими «стяжками», закрепляющими необходимую трехмерную
укладку молекулы.
Наконец, говоря о четвертичной структуре белка, имеют
в виду различного рода молекулярные комплексы. Классическим
примером является гемоглобин, состоящий из двух молекул
а-глобина, двух молекул р-глобина, гема и иона железа.
Под действием некоторых внешних или внутренних факторов
может происходить изменение структуры белка. Очень часто оно
22
Рис. 1.5. Третичная структура молекулы лизоцима. Гибкие участки пред-
ставлены в виде нити, а-спирали обозначены повторяющимися витками,
элементы 0-листа — широкими стрелками
не затрагивает первичную структуру, а приводит лишь к измене-
нию четвертичной, третичной или вторичной (та же самая моле-
кула укладывается как-то иначе). В этих случаях говорят об из-
менении конформации белка. Конформация влияет на химиче-
ские свойства молекулы и в итоге — на способность выполнять
необходимую функцию.
Рис. 1.6. Формирование дисульфидных мостиков в молекуле белка
Примечание. Одним из 20 вариантов аминокислот, используемых в составе
белков, является цистеин (Cys). В его радикале содержатися функциональная груп-
па SH. Если два остатка цистеина оказываются в третичной структуре поблизости
друг от друга, между ними под действием особого фермента может возникнуть ди-
сульфидный мостик S —S. Такие мостики играют роль химических «стяжек», стаби-
лизирующих третичную структуру белковой молекулы.
23
Таким образом, функции конкретной молекулы белка за-
висят не только от ее аминокислотной последовательности,
но и от ее конформации в данный момент. Давайте рассмот-
рим несколько наглядных примеров.
В обычном состоянии белок куриного яйца является почти
прозрачным и полужидким. Но при высокой температуре он бе-
леет и становится плотным, что связано с необратимым измене-
нием его конформации. В результате сильно нагретый яичный
белок уже не может выполнять свои биологические функции, дру-
гими словами, оказывается необратимо инактивированным. Сход-
ным образом ведут себя и многие иные белки: достаточно вспом-
нить о жареном мясе или вареной рыбе. Необходимо отметить, что
в организме человека необратимая инактивация некоторых важных
белков начинается уже при температуре 42...43°C. Вот почему
температуру около 42 °C считают для человека смертельно опас-
ной.
На конформацию белка влияет и концентрация ионов. В этом
нетрудно убедиться, если сырой яичный белок посыпать крупны-
ми кристаллами поваренной соли. Можно увидеть, что вокруг
упавших кристаллов постепенно возникнет тонкое мутноватое
кольцо. Это за счет высокой концентрации ионов Na+ и СГ из-
менилась конформация молекул яичного белка.
В отличие от высокой температуры даже сильное изменение
концентрации ионов обычно не приводит к необратимой инак-
тивации белков. Действительно, восстановив первоначальную
концентрацию ионов, можно вернуть белку его исходную кон-
формацию, а значит, и утраченную активность.
Таким образом, изменяя в своих клетках концентрацию
некоторых ионов, организм способен тонко регулировать
биологическую активность собственных белков.
1.3.5. Нуклеиновые кислоты
Биосинтез любого белка — очень сложный процесс. В част-
ности, для него необходима информация о том, в какой последо-
вательности должны быть соединены различные аминокислоты.
Такую информацию называют наследственной, или генетической.
Она содержится в особых биоорганических молекулах, относя-
щихся к классу нуклеиновых кислот. Без этих молекул биосин-
тез белка принципиально невозможен.
Подобно белкам, любые нуклеиновые кислоты тоже являются
сложными полимерными соединениями. Их мономерами служат
различные нуклеотиды, в состав которых входят три обязательных
компонента: это определенный циклический моносахарид, груп-
па —Н2РО4 и азотистое основание (рис. 1.7).
Первые два из перечисленных компонентов обеспечивают
соединение нуклеотидов в полимерные цепочки. При этом в ка-
честве циклического моносахарида может использоваться либо
рибоза, либо дезоксирибоза. Соответственно различают
два типа нуклеиновых кислот: рибонуклеиновые и дезоксирибо-
нуклеиновые кислоты (сокращенно — РНК и ДНК).
Третий компонент нуклеотида (азотистое основание) предна-
значен непосредственно для записи генетической информации и
в этом плане может рассматриваться как своеобразная «буква».
По аналогии, всю генетическую информацию организма можно
представить в качестве огромной «базы данных», записанной с
помощью различных азотистых оснований.
Как видно на рис. 1.8, а, используемые в нуклеиновых кисло-
тах азотистые основания (их набор совершенно одинаков у всех
живых существ) отличаются друг от друга строением углеродного
скелета, а главное — расположением слабых электростатических
зарядов. В результате строго определенные азотистые основания
способны притягиваться и образовывать так называемые компле-
ментарные пары (рис. 1.8, б). Именно способность к формиро-
ванию комплементарных пар позволяет нуклеиновым кислотам
выполнять свои функции.
он он он н
Рибоза Дезоксирибоза
Рис. 1.7. Строение нуклеотида
25
Гуанин
Тимин
Цитозин
СН
I
Аденин
С_
। Тимин
О
б
Рис. 1.8. Азотистые основания, характерные для ДНК: их строение (а) и
комплементарные пары (б).
Условные обозначения: - — водородная связь; -> — связь, соединяющая
азотистое основание с остатком дезоксирибозы
26
Рис. 1.9. Принцип удвоения ДНК
Примечание. Специальные белки расплетают двухцепочечную молекулу ДНК,
после чего начинается синтез комплементарных цепочек. Старые цепи показаны на
рис. 1.9 светлыми, новые — затемнены. Азотистые основания аденин, гуанин, тимин
и цитозин обозначены латинскими буквами соответственно A, G, Т и С.
Таблица 1.7. Характерные особенности ДНК и РНК
Тип нуклеино- вой кислоты Общий план строения Используемый в нуклеотидах моносахарид Используемые в нуклеотидах азотистые основания Важнейшие биоло- гические функции
ДНК Как правило, двухцепочеч- ные молекулы дезокси- рибоза Аденин (А), Гуанин (G), Цитозин (С), Тимин(Т) Хранение генети- ческой информа- ции
РНК Как правило, одноцепочеч- ные молекулы рибоза Аденин (А), Гуанин (G), Цитозин (С), Урацил (U) Участие в процес- сах реализации генетической информации
27
Убедимся в этом на примере молекулы ДНК. Она состоит из
двух полимерных цепочек, комплементарных друг другу по всей
своей длине (рис. 1.9). Однако взаимное притяжение между эти-
ми цепочками обеспечивается только за счет водородных связей.
Как известно, такие связи очень непрочны. Они при определен-
ных условиях легко разрываются, в результате чего исходная
молекула ДНК может распасться на две самостоятельные ну-
клеотидные цепочки. В свою очередь, каждая из них способна
стать основой для биосинтеза нового поколения комплементар-
ных цепей. В итоге возникают две идентичные молекулы, явля-
ющиеся точными копиями исходной. Таким образом, благодаря
своему химическому строению молекулы ДНК не только хра-
нят генетическую информацию, но и обеспечивают возмож-
ность ее точного копирования. Без этого размножение орга-
низмов было бы невозможным.
Характерные особенности строения молекул РНК приведены
в табл. 1.7. С некоторыми важными функциями этих молекул вы
познакомитесь в следующих подразделах.
1.4. Процесс реализации генетической
информации
В любом организме синтезируются тысячи разных белков.
Необходимая для этого генетическая информация представлена
в молекулах ДНК в виде множества генов. Однако мы еще не
выяснили, что такое «ген». А ведь именно это понятие будет
центральным во всех вопросах, рассматриваемых в данном учеб-
нике.
Понятие «ген» является одним из наиболее важных, но вместе с
тем и наиболее сложных понятий в биологии. Поэтому мы не даем
его строгое определение, а ограничиваемся весьма упрощенной
формулировкой, впрочем, достаточной для восприятия дальнейше-
го материала. Мы будем считать, что ген — это участок молекулы
ДНК, содержащий генетическую информацию о неком белке.
Каким же образом информация, содержащаяся в конкретном
гене, определяет аминокислотную последовательность соответ-
ствующего белка? Для этого в организмах осуществляется специ-
альный процесс, получивший название реализация генетической
информации. Он состоит из нескольких этапов. С их деталями
мы будем знакомиться в следующих подразделах, а пока лишь
кратко остановимся на главных моментах.
На первом этапе (его называют транскрипцией) происходит
считывание информации, хранящейся в гене. Сначала между
28
двумя цепочками ДНК возникает локальный разрыв водородных
связей. В этом месте комплементарные цепочки отходят друг от
друга, а одна из них используется для синтеза молекулы РНК.
Данный синтез тоже основан на принципе комплементарности
(рис. 1.10). В соответствии с этим в образующейся молекуле РНК
содержится та же информация, что и непосредственно в 1ене.
Именно в виде такой одноцепочечной молекулы (ее называют
информационной РНК} генетическая информация пригодна к
дальнейшему использованию.
Следующий этап, получивший название трансляция, проте-
кает на особых молекулярных комплексах — рибосомах. Они
сканируют молекулу информационной РНК (иРНК) и в строгом
соответствии с ее нуклеотидной последовательностью соединяют
различные аминокислоты в единую цепочку. В результате проис-
ходит биосинтез определенного белка.
Таким образом, молекула иРНК является посредником, пере-
дающим генетическую информацию от гена на рибосомы.
Итак, в процессе реализации генетической информации уча-
ствуют три типа биоорганических полимеров: это молекулы ДНК,
РНК и белков Их биосинтез осуществляется по матричному
Цепь ДНК,
Рис. 1.10- Принцип транскрипции
Примечание. Процесс транскрипции основан на принципе комплемент ар-
ности Единственное существенное отличие от удвоения ДНК заключается в том, что
в составе рибонуклеотидов (нуклеотидов, используемых в качестве мономеров РНК)
вместо тимина присутствуетуряция (сокращенно — U). Обратите внимание: молеку-
ла иРНК своей нуклеотидной последовательностью очень похижа на некодирующую
Цепочку ДНК Поэтому последовггельность нуклеотицсч» каждого гена принято За-
писывать по ею некодирующей цепи
29
принципу. Это значит, что последовательность мономеров в одном
полимере служит образцом (или матрицей) при синтезе другого.
Действительно, нуклеотидная последовательность конкретного
гена используется в качестве матрицы для молекулы и PH К.
В свою очередь, нуклеотидная последовательность данной моле-
кулы представляет собой матрицу для соответствующего белка.
В конечном итоге соблюдается общая закономерность: ами-
нокислотная последовательность конкретного белка опреде-
ляется нуклеотидной последовательностью соответству-
ющего гена, поэтому все без исключения особенности орга-
низма, в формирование которых вовлечены молекулы белков,
неизбежно находятся под генетическим контролем.
1.5. Молекулярные основы транскрипции
От общих принципов реализации генетической информации
перейдем к конкретным механизмам этого процесса. И для на-
чала остановимся на первом этапе — транскрипции.
Как известно, подавляющее большинство химических реакций,
необходимых организму для поддержания своей жизнедеятельности,
осуществляется не самопроизвольно, а под действием ферментов.
Это правило полностью справедливо и в отношении транскрипции:
она возможна только в присутствии специального фермента, по-
лучившего название РНК-полимераза. Именно за счет активности
данного фермента происходит считывание информации с различных
генов и биосинтез соответствующих им молекул РНК.
Чтобы приступить к транскрипции определенного гена, РНК-
полимераза должна к нему присоединиться. Для этого в гене
имеется особый участок (промотор), нуклеотидная последова-
тельность которого специфически узнается РНК-полимеразой и
служит ей в качестве своеобразной «посадочной площадки». Если
структуру промотора существенно изменить, ген не будет рас-
познан РНК-полимеразой, и содержащаяся в нем информация
останется несчитанной.
Связавшись с промотором, РНК-полимераза начинает посте-
пенно перемещаться по молекуле ДНК к той части гена, которая
содержит наследственную информацию. Направление этого пере-
мещения задано промотором (если перевернуть промотор на 180°,
РНК-полимераза «пойдет» в противоположную сторону). Таким
образом, промотор является молекулярным сигналом, опреде-
ляющим начало и направление транскрипции гена.
По мере своего продвижения по гену РНК-полимераза раз-
рывает водородные связи между двумя комплементарными це-
30
Промотр
Кодирующая область гена
- - - I I I I I Г~П VI I I I I I I I I 'I I I I I I I I I I I I I I I I - - -
Область посадки Кодирующая цепочка
РНК-пол имеразы
Точка начала
транскрипции
а
Ген X Ген Y *]
--Till I I I I I I I I I I I I I I I I I I I I I I П I I I I I I I I I I I I I I I I I I I I ! I I I I 1 I П I ! И I I I I I I I I I I I I I I I I I Jl ip I I I ГГр-
--|| -------------------------------------------------------------------
б
Рис. 1.11. Роль промотора в выборе начала (а) и направления (б) транс-
крипции
Примечания. I. РНК-полимераза распознает не весь промотор, а только впол-
не определенную нуклеотидную последовательность, находящуюся лишь в одной из
двух цепей ДНК. Именно этим обусловлен выбор кодирующей цепочки. Направление
транскрипции тоже задано промотором: перемещение РНК-полимеразы по цепи ДНК
может происходить лишь строго однонаправленно («слева направо*, если судить по
нижней цепи на рис. I.II, а).
2. Для разных генов в одной и той же молекуле ДНК кодирующие цепочки могут
быть разными (рис. I.I1, б).
почками ДНК. При этом нуклеотидная последовательность одной
из цепочек используется как матрица для синтеза молекулы РНК.
Выбор используемой цепочки далеко не случаен. Дело в том, что
в составе практически каждого гена лишь одна из цепочек несет
генетическую информацию, а вторая выполняет сугубо вспомо-
гательную функцию: участвует в процессе удвоения молекулы
ДНК. Именно «нужная» цепочка и используется для транскрип-
ции (рис. 1.11).
Пройдя через всю кодирующую область гена, РНК-полимераза
достигает специального участка, который является еще одним
важным молекулярным сигналом. На этом участке (его называют
терминатором) фермент прекращает синтезировать РНК и от-
деляется от гена. Тем самым транскрипция гена оказывается за-
вершенной. В дальнейшем освободившаяся РНК-полимераза
способна вступать во взаимодействие с другими промоторами,
обеспечивая транскрипцию множества разных генов.
Итак, в гене имеются специальные участки, служащие
сигналами для начала и окончания транскрипции — соот-
ветственно промотор и терминатор. Между ними нахо-
дится кодирующая область, которая непосредственно со-
держит генетическую информацию.
31
1.6. Молекулярные основы трансляции
В ходе трансляции — второго этапа реализации генетической
информации — считанная с гена молекула и PH К используется
в качестве матрицы для синтеза белка. Этот процесс обеспечива-
ют специальные ферменты, организованные в сложные комплек-
сы — рибосомы.
Рибосома способна распознавать нуклеотидные последователь-
ности, характерные для одного из концов молекулы и PH К. Ис-
пользуя эти последовательности как «посадочную площадку»,
рибосома присоединяется к молекуле и PH К, а затем начинает
перемещаться к противоположному концу. Достигнув участка,
который содержит генетическую информацию, рибосома при-
ступает к синтезу соответствующего белка.
В белках используются 20 вариантов аминокислот. Между тем
в состав любой молекулы иРНК входит всего 4 варианта нуклео-
Таблица 1.8. Генетический код
Положение нуклеотида в триплете и PH К
1-с 2-е 3-e
и c A G
и UUUXph. ucu UAU\ ~ UGU \ U
UUC J Phe ucc 1 _ UAC / Tyr UGC J Cys c
UUA\ UUG J Leu UCA [ Ser UCG J UAA'i ~ UAG } *СТОП- UGA «Стоп» UGG Trp A G
С CULM CCU CAU\ „. CGU'T U
cue f ecc 1 D CAC J H,s CGC .
CUA [ Leu CCA fPro CAA \ Q| CGA [ Arg y^
CUG J CCG J CAG J ° CGG J G
у^ AUU''j ACU AAU\ . AGU 1 Ser U
AUC f lie ACC л Thr AAC f A^n AGC J c
AUA J АСА [ " AAA 1 Lvs AGA \ Arg A
AUG Met ACG J AAGJ Ly^ AGG J G
G GUU"| GCU 'I GAU\ . GGU "I U
GUC Lval GCC L Ala GAC J Asp GGC 1 c
GUA 1 GCA [ GAATn„ GGA |Gly A
GUG J GCG J GAG J Glu GGG J G
* Триплетам (JAA, UAG и UGA не соответствует ни одна из аминокислот.
Перечисленные триплеты обозначают окончание трансляции.
32
Аминокис по гы.
Рибосома
Рис. 1.12. Принцип трансляции:
а — к шум первым триппстам иРНК подошпи два посредника с соответствующи-
ми аминокислотами (этими посредниками являются особые РНК); б — рибосома
соединила две аминокислоты (между ними образовалась пептидная связь) и пере-
несла их на посредник 2; л — рибосома выбросила использованный посредник I и
переместилась по и РНК на один триплет. К третьему триплету подошел очередной
посредни» с соответствхюшен аминокислотой; г — рибосома перенесла две первые
аминокислоты на третью, затем снова переместится на один триплет, присоединит
очередной посредник и т.д.
тидов. При этом совершенно очевидно, что один нуклеотид не
может кодировать одну аминокислоту. То же самое справедливо
и для двух соседних нуклеотидов (возможно лишь 16 вариантов
подобных пар). В результате соблюдается следующая обшая за-
кономерность: одна аминокислота в молекуле белка кодиру-
ется тремя соседними нуклеотидами в и PH К. Подобные
тройки нуклеотидов называют триплетами Существует 64 вари-
анта триплетов, что с избытком достаточно для кодирования
W аминокислот (табл. 1.8).
33
Соседние триплеты не перекрываются друг с другом. В связи
с этим кодирующую область молекулы иРНК можно представить
в виде определенной последовательности триплетов. Рибосома
постепенно сканирует данную последовательность и в соответ-
ствии с ней синтезирует надлежащий белок. Пройдя через всю
кодирующую область иРНК, рибосома достигает специального
молекулярного сигнала, обозначающего точку прекращения
трансляции. Это может быть любой из трех бессмысленных три-
плетов: UAA, UAG или UGA. Здесь рибосома отделяется от иРНК,
после чего, оказавшись в свободном состоянии, готова приступить
к трансляции новой матрицы.
Особого внимания заслуживает тот факт, что триплеты не
способны к комплементарному взаимодействию с аминокисло-
тами, поэтому центральную роль в биосинтезе белка играют
специальные молекулы-посредники. В каждой из них наиболее
важными компонентами являются определенный триплет ну-
клеотидов и соответствующая данному триплету аминокислота.
С помощью своего триплета молекула-посредник распознает
комплементарный триплет в составе иРНК и вступает с ним в
строго специфичное кратковременное взаимодействие. В резуль-
тате присоединенная к посреднику аминокислота оказывается
именно в нужном положении по отношению к последователь-
ности триплетов в иРНК. Аналогично действуют и другие вари-
анты посредников, обеспечивая необходимую «расстановку»
аминокислот. Рибосома отслеживает правильность этой расста-
новки и ковалентно соединяет аминокислоты в единую цепочку
(рис. 1.12). Тем самым достигается строгое соответствие
между последовательностью триплетов в молекуле иРНК
и аминокислотной последовательностью синтезируемого
белка.
1.7. Как формируются признаки организма
Каждый из многочисленных белков, синтезируемых в организ-
ме, предназначен для выполнения строго определенных функций.
В свою очередь, от того, как выполняются данные функции, за-
висит соответствующий признак организма. Для наглядности
обратимся к следующему примеру.
Как известно, люди различаются по цвету волос. Этот при-
знак (имеется в виду лишь естественный цвет) обусловлен специ-
альными пигментами — меланинами, характерными для подав-
ляющего большинства позвоночных животных. Существуют две
основные разновидности меланина: эумеланин — ярко-черного
или коричневого цвета и феумеланин — ярко-красного или
34
желтого цвета. Именно от соотношения между этими двумя
пигментами и зависит окраска волос конкретного человека.
Откуда же появляются меланины в человеческом организме?
Они синтезируются в результате химических реакций, протека-
ющих в особых клетках кожного покрова. Эти реакции не могут
идти самопроизвольно: для каждой необходимо присутствие
определенного фермента. И если какой-то из ферментов неакти-
вен, организм не способен осуществлять соответствующую реак-
цию, а потому биосинтез меланинов оказывается нарушенным.
В конечном итоге это отражается и на цвете волос. Таким обра-
зом, цвет волос конкретного человека непосредственно зависит
от функционирования ферментов, обеспечивающих синтез эу- и
феумеланина (рис. 1.13).
Все ферменты с химической точки зрения являются белками,
а каждый белок кодируется определенным геном. В итоге совер-
шенно очевидно, что признак «цвет волос* находится под
контролем ряда генов, кодирующих ферменты биосинтеза
меланинов. Однако можно ли ожидать, что и в основе любого
другого признака организма тоже лежит генетический контроль?
На первый взгляд, это кажется довольно сомнительным, особен-
но в случае разнообразных психологических характеристик. Дей-
ствительно, правомочно ли связывать с конкретными генами
Тирозиназа
Рис. 1.13. Схема биосинтеза эу- и феумеланина
Примечание. Биосинтез эу- и феумеланина начинается с одной и той же хи-
мической реакции, которую обеспечивает фермент тирозиназа. В дальнейшем пути
биосинтеза меланинов расходятся. Если у человека нарушен самый первый этап, оба
варианта меланинов будут отсутствовать, и как следствие волосы окажутся практиче-
ски белыми. Если первый этап биосинтеза протекает нормально, но в дальнейшем
работает только один из путей, волосы будут либо черными, либо ярко-рыжими. Если
же все этапы осуществляются нормально, волосы будут неопределенного темного
Цвета (подобных людей принято называть шатенами). Важную роль играет и интен-
сивность каждого этапа. Так, у людей с недостаточно активной тирозиназой синтези-
руется небольшое количество обоих пигментов, и в конечном итоге волосы получа-
ются русыми.
35
настолько тонкие и сложные характеристики человека, как лжи-
вость, талант, чувство меры и многие другие? Попробуем кратко
ответить на этот вопрос.
Начнем с того, что каждая из перечисленных выше характери-
стик хотя бы косвенно связана с функционированием нервной
системы. В свою очередь, данная система состоит из нейронов,
осуществляющих проведение нервных импульсов по организму.
Эти импульсы обусловлены изменениями мембранного потен-
циала, т.е. изменениями концентрации некоторых ионов (в пер-
вую очередь — Na\ К? и Са2+) по разные стороны от поверхност-
ной мембраны нейрона. Одним из важнейших механизмов таких
изменений является избирательный транспорт ионов через мем-
брану. Его обеспечивают специальные белки — разнообразные
ионные насосы и каналы. Но ведь каждый белок кодируется со-
ответствующим геном! Следовательно, даже тончайшие психоло-
гические особенности должны в какой-то степени зависеть от
генотипа.
Впрочем, было бы наивным предполагать, что признаки орга-
низма обусловлены исключительно генами. Важную роль играют
и факторы окружающей среды. Дело в том, что очень многие
из этих факторов могут оказывать прямые или косвенные воз-
действия на процесс реализации генетической информации.
С механизмами подобных воздействий мы познакомимся позже,
а пока ограничимся следующим примером. Известно, что скорость
химических реакций существенным образом зависит от темпера-
туры. Поэтому температура окружающей среды неизбежно влия-
Рис. 1.14. Процессы, лежащие в основе формирования признаков
36
ет на реализацию генетической информации, и как следствие — на
очень многие признаки организма.
Итак, в основе формирования признаков организма лежит
процесс реализации генетической информации с учетом раз-
личных воздействий окружающей среды (рис. 1.14). Забегая
вперед, добавим, что еще одним фактором, оказывающим замет-
ное влияние на формирование признаков, является стадия раз-
вития (возраст) организма.
1.8. Дифференциальная экспрессия генов
У любого организма, даже у мельчайшей бактерии, генетиче-
ская информация представлена тысячами генов. Причем эти гены
используются не одновременно, а в строгом соответствии с про-
граммой развития организма. В результате «работа* каждого
конкретного гена приурочена к совершенно определенным
этапам жизнедеятельности. Это явление называют диффе-
ренциальной экспрессией генов. Оно характерно для всех живых
существ, в особенности — для сложно устроенных многоклеточ-
ных организмов, где разные клетки предназначены для выполне-
ния разных функций.
Продемонстрируем данное явление на следующем примере.
Представим, что один из генов многоклеточного организма (в
дальнейшем мы будем называть его геном Z) кодирует белок,
который необходим для формирования жгутиков. При этом жгу-
тики формируются далеко не у всех, а только у некоторых, особым
образом специализированных клеток. Именно в них и должен
синтезироваться соответствующий белок, для остальных же кле-
ток он совершенно бесполезен. В связи с этим экспрессия гена Z
идет дифференциально: в клетках, формирующих жгутики, она
осуществляется, однако во всех остальных она подавлена.
Каковы же механизмы дифференциальной экспрессии генов?
Оказывается, в клетках имеются специальные молекулы, регули-
рующие процесс реализации генетической информации. Суще-
ствуют различные варианты подобной регуляции. С некоторыми
из них мы познакомимся в дальнейшем, а пока остановимся на
самом распространенном варианте: на регуляции, которая за-
трагивает стадию транскрипции.
Известно, что центральную роль в процессе транскрипции
играет фермент РНК-полимераза. Но он «не умеет» самостоя-
тельно распознавать, какие именно гены должны быть транскри-
бированы. Эту функцию выполняют особые регуляторные белки,
способные связываться с промоторами строго определенных генов
37
и либо усиливать активность РНК-полимеразы, либо, напротив,
значительно ее подавлять. Такие белки получили название транс-
крипционные факторы (рис. 1.15).
В составе одного и того же многоклеточного организма функ-
ционируют сотни различных транскрипционных факторов, при-
чем их работа четко скоординирована. Для наглядности вновь
обратимся к гену Z. Допустим, что его дифференциальную экс-
прессию обеспечивает транскрипционный фактор ТФ„, который
работает, подавляя активность РНК-полимеразы. Очевидно, что
в клетках, способных к формированию жгутиков, этого транс-
крипционного фактора быть не должно. Все остальные клетки,
напротив, должны его содержать. Таким образом, дифференци-
альная экспрессия гена Zобусловлена неравномерным распреде-
лением молекул ТФЛ по разным клеткам одного и того же орга-
низма.
Но тогда возникает совершенно естественный вопрос: почему
же рассматриваемый нами транскрипционный фактор, в свою
очередь, распределен по организму неравномерно? Наиболее
логично следующее объяснение: экспрессия гена, кодирующего
ТФЯ, происходит в организме тоже дифференциально под кон-
тролем какого-то другого транскрипционного фактора (назовем
его ТФЯ_|). Казалось бы, получился классический замкнутый круг.
Ведь если и дальше придерживаться этой же логики, ее пришлось
бы продолжать до бесконечности, привлекая все новые и новые
транскрипционные факторы, каждый из которых распределен по
организму неравномерно. И все же подобная логика верна.
Дело в том, что на всем протяжении развития организма, на-
чиная со стадии одноклеточного зародыша (зиготы) и заканчивая
PH К-полимераза
T ранскрипционный
фактор
Промотор Кодирующая область гена
Рис. 1.15. Принцип действия транскрипционных факторов
Примечание. Существует большое разнообразие транскрипционных факторов.
Некоторые из них, присоединившись к конкретному промотору (для упрощения
представлена только кодирующая цепь), подавляют активность РНК-полимеразы и
тем самым ослабляют транскрипцию данного гена. Некоторые, напротив, стимули-
руют PH К-пол и меразу, в этом случае транскрипция гена будет усилена. Таким об-
разом, интенсивность транскрипции конкретного гена зависит от того, какие транс-
крипционные факторы находятся на его промоторе в данный момент.
38
Неравномерное Дифференциальная Неравномерное
распределение ТФ| - транскрипция гена,---► распределение ТФ2---- • • • *
по зиготе кодирующего ТФ2 по организму
Дифференциальная Неравномерное
_ —. транскрипция гена, • распределение ТФ„ *
кодирующего ТФЛ по организму
К______________-___>
Дифференциальная
транскрипция гена Z
Рис. 1.16. Каскадный принцип дифференциальной транскрипции генов
Примечание. Фигурными скобками выделены однотипные этапы каскада. ТФ|
и ТФ, — транскрипционные факторы соответственно первого и второго «эшелонов».
Прочие пояснения — в тексте.
неизбежными процессами старения, транскрипционные факторы
действуют по каскадному принципу. Так, на ранних этапах раз-
вития зародыша функционируют транскрипционные факторы
«первого эшелона». Будучи неравномерно распределенными по
зиготе, они обеспечивают дифференциальную экспрессию генов,
кодирующих транскрипционные факторы «второго эшелона», а
те, в свою очередь, — третьего и т. д., вплоть до рассматриваемо-
го нами ТФ„ (рис. 1.16).
Остается уточнить всего лишь одну деталь: почему транскрип-
ционные факторы «первого эшелона» сами распределены по
зиготе неравномерно? Как правило, это закладывается материн-
ским организмом еще на ранних этапах созревания яйцеклетки.
В итоге каскады дифференциальной экспрессии генов законо-
мерно передаются из поколения в поколение.
1.9. Молекулярные основы формирования
биологических ответов
Каждый организм подвергается многочисленным воздействи-
ям. Эти воздействия можно подразделить на две большие группы:
витальные — непосредственно влияющие на жизнеспособность
(например, смертельная травма, причиненная хищником), и сиг-
нальные — несущие информацию о внешней среде. Очевидно,
что на многие сигналы необходимо реагировать, причем возни-
кающие реакции должны быть адекватными. Каковы же механиз-
мы формирования подобных реакций?
Различают два типа сигналов: химические и физические.
К первым относят разнообразные молекулы и ионы, ко вторым —
механические воздействия (в частности, звуки), кванты света,
тепловые излучения и др. При этом на каждый вариант получен-
ных сигналов организм вырабатывает определенный биологиче-
ский ответ.
39
Физический
сигнал I
Химический
сигнал V
Мембрана
Рецептор 1
Рецептор 2
б
Рис. 1.17. Принципы функционирования рецепторов
Примечание. Рецепторы обычно расположены на внешней мембране. При
наличии химического сигнала (например, конкретного типа молекул) соответствующий
рецептор вступает во взаимодействие с ним и вследствие этого изменяет свою кон-
формацию. При наличии физического сигнала (например, квантов света) соответ-
ствующий рецептор поглощает его энергию, что опять-таки приводит к изменению
конформации рецептора. Каждый рецептор строго специфичен, т.е. способен рас-
познавать лишь определенные сигналы.
Общую схему формирования биологического ответа можно
представить в виде трех последовательных этапов. На первом
этапе происходит восприятие сигнала. Эту функцию выполняет
конкретный рецепторный белок, имеющий высокую специфич-
ность к данному сигналу, а потому способный распознавать его
среди многих других. Обнаружив сигнал, рецептор изменяет кон-
формацию и тем самым извещает организм о наличии воздействия
(рис. 1.17).
Рецепторы — это своеобразные «сенсорные устройства», без
которых биологические ответы были бы невозможны. Между тем
непосредственными исполнителями биологических ответов яв-
ляются совсем другие структуры организма, иногда находящиеся
от рецепторов на значительном удалении. Поэтому воспринятый
сигнал необходимо преобразовать и передать в преобразованном
виде на соответствующие структуры. Данный этап называют
трансдукцией сигнала. Как правило, его осуществляют специ-
альные белки, а также различные вспомогательные молекулы и
ионы.
Белки, принимающие участие в трансдукции сигнала (для
удобства мы будем называть их белкам и-п осредниками),
не должны функционировать, пока сигнал не поступил. Это воз-
можно в одной из следующих ситуаций:
• либо белки-посредники изначально отсутствуют и синтези-
руются только в результате восприятия сигнала;
• либо белки-посредники изначально присутствуют, однако
находятся в неактивном состоянии и активируются только в ре-
зультате восприятия сигнала.
40
Нетрудно заметить, что первая схема весьма напоминает диф-
ференциальную экспрессию генов. И действительно, в ней часто
используется тот же механизм, что и в дифференциальной экс-
прессии, а именно — каскадная регуляция на стадии транскрип-
ции. Отличие состоит лишь в том, что данные каскады запускают-
ся не с самого начала индивидуального развития, а с момента из-
менения конформации определенных рецепторов (рис. 1.18, а).
Между тем активация уже существующих белков происходит,
как правило, посредством их фосфорилирования (т.е. при-
соединения к ним фосфатных групп). Напомним, что фосфатная
группа сильно заряжена. Поэтому ее присоединение к молекуле
белка приводит к заметному изменению его конформации, в ре-
зультате чего белок и становится активным. Столь важные про-
цессы не могут идти самопроизвольно. Они обеспечиваются
Рецептор изменяет Запуск Запуск
Сигнал —- конформацию —*• транскрипции гена, —- биосинтеза —► • * • —-
и становится T<Dt кодирующего ТФ2 ТФ2
Запуск Запуск
транскрипции гена, —- биосинтеза —►
кодирующего ТФЛ ТФ„
ч______________ "______________)
Запуск
транскрипции
гена,
кодирующего
белок У
Запуск
—* биосинтеза —*
белкаY
Сигнал —*•
Рецептор изменяет
конформацию
и становится ПК]
Фосфорили-
рование ПК2
Активация
ПК2
Фосфорили- Активация _ рование ПК„ ПКЯ < J Фосфорили- Активация _ * рование —* белка W —Ответ белкаW оелка w
б
Рис. 1.18. Трансдукция сигнала на основе транскрипционных (а) и протеин-
киназных (б) каскадов
Примечание. Фигурными скобками выделены однотипные этапы каскада. ТФ|
и ТФ, — транскрипционные факторы первого и второго «эшелонов» соответственно.
ПК, и ПК2 — протеинкиназы первого и второго «эшелонов» соответственно. Белки
У и W — ключевые белки, обеспечивающие реализацию неких биологических ответов.
Существуют многочисленные варианты представленных схем. Во-первых, молекуляр-
ный каскад может ветвиться, приводя к реализации сразу нескольких биологических
ответов. Во-вторых, разные каскады могут сходиться, обеспечивая один и тот же ответ
на разные сигналы. В-третьих, в одном и том же молекулярном каскаде могут при-
сутствовать и транскрипционные, и протеинкиназные этапы. И наконец, далеко не
все рецепторные белки сами становятся ТФ( или ПК,: между рецептором и первым
Участником каскада может быть задействована специальная система посредников
(подробнее см. подразд. 2.6).
41
особыми ферментами (п роте и н к и н аза м и), имеющими вы-
сокую степень специфичности и зачастую тоже работающими по
каскадному принципу (рис. 1.18, б).
По какой бы схеме ни протекала трансдукция сигнала, в ко-
нечном итоге она достигает необходимых структур и приводит к
запуску третьего, последнего этапа, т.е. к непосредственной реа-
лизации биологического ответа. При этом некоторые ответы
настолько существенны, что способны заметно отразиться на при-
знаках организма. Например, под действием интенсивного солнеч-
ного излучения кожа человека становится более пигментированной,
и европеец может преобразиться в «смуглого мулата».
Следует подчеркнуть, что при разных механизмах трансдукции
скорость формирования ответов весьма неодинакова. В частности,
процессы каскадной активации белков могут осуществлять-
ся в организме очень быстро (иногда всего лишь за тысячные
доли секунды) и поэтому обусловливают быстрые биологи-
ческие ответы. Между тем каскады с использованием
транскрипционных факторов требуют достаточно дли-
тельных промежутков времени (нескольких минут, часов
или даже дней) и в итоге приводят к медленным биологиче-
ским ответам.
Разные клетки одного и того же организма являются по отноше-
нию друг к другу окружающей средой. Поэтому не следует делать
особых различий между ответами на внешние и внутренние сигналы:
и те, и другие основаны на сходных молекулярных механизмах.
1.10. Молекулярные основы возникновения
и действия мутаций
Знакомясь с особенностями строения нуклеиновых кислот, мы
выяснили, что молекулы ДНК способны к копированию. Теперь
остановимся на этой способности более подробно.
По аналогии со многими другими химическими реакциями,
необходимыми организму для поддержания своей жизнедеятель-
ности, копирование ДНК не может идти самопроизвольно. Его
обеспечивает целый ряд ферментов, важнейшим из которых явля-
ется ДНК-полимераза. Именно за счет активности данного фер-
мента происходит подбор необходимых комплементарных нуклео-
тидов и их соединение в цепочку по матричному принципу.
В любом, даже очень точно отлаженном процессе неизбежно
возникают хотя бы единичные ошибки. Это правило выполняет-
ся и в случае копирования ДНК. Действительно, осуществляя
подбор комплементарных нуклеотидов, ДНК-пол и мераза время
42
от времени ошибается с частотой приблизительно 10"5. В столь
важном процессе, как копирование генетической информации,
даже такая частота ошибок чрезвычайно опасна, поэтому суще-
ствуют специальные молекулярные механизмы, обеспечивающие
выявление и исправление возникающих ошибок. Однако и здесь
абсолютная точность недостижима. В результате некоторые ошиб-
ки ДНК-пол и меразы все же остаются необнаруженными и неис-
правленными. Их частота составляет примерно 10_,° (1 ошибка
на 1О10 нуклеотидов).
ДНК, в которой сохранилась неисправленная ошибка, тоже
способна участвовать в процессе копирования. При этом воз-
никшие копии оказываются неодинаковыми. Первая, синтезиро-
ванная на основе «правильной» цепочки, полностью идентична
исходной молекуле ДНК, а вторая, полученная на основе цепоч-
ки с ошибкой, несет соответствующее изменение нуклеотидной
последовательности. Но теперь изменение затрагивает уже не
одну, а обе комплементарные цепочки в молекуле ДНК, и потому
должно сохраняться при дальнейшем копировании. Подобные
изменения принято называть мутациями (рис. 1.19).
Некоторые мутации относительно безвредны для организма,
поскольку возникают не в генах, а в участках между ними. Но
другие мутации приводят к весьма ощутимым последствиям.
В частности, если мутация возникнет в промоторе и повредит
«посадочную площадку» для РНК-полимеразы, транскрипция
Рис. 1.19. Принцип возникновения мутаций:
а — исходная молекула ДНК (буквами А, Т, G и С обозначены нуклеотиды с со-
ответствующими азотистыми основаниями); б — молекула ДНК, возникшая при
копировании исходной молекулы (в качестве матрицы использована «верхняя» цепь).
В результате ошибки ДНК-полимеразы в «нижнюю» цепь был вставлен неверный
нуклеотид. Данная ошибка осталась незамеченной и сохранилась до следующего ко-
пирования ДНК; в — молекула ДНК, возникшая при копировании неисправленной
молекулы (в качестве матрицы использована цепь с ошибкой). В 8-м положении этой
молекулы вместо исходной комплементарной пары А —Т находится комплементарная
пара G — С, способная стабильно воспроизводиться в последующих поколениях
данного гена может быть нарушена. Если же будет затронута ко-
дирующая область, возможны изменения в структуре соответ-
ствующего белка. Его функции при этом тоже могут измениться.
Но окажутся ли они улучшенными или ухудшенными, или у бел-
ка возникнет новая функция? Чтобы ответить на этот вопрос,
воспользуемся аналогией.
Представим, что в электрической схеме телевизора случайным
образом изменили несколько контактов. Безусловно, существует
определенная вероятность того, что новая схема окажется лучше
по сравнению с исходной. Однако такая вероятность крайне мала.
Скорее всего, измененная схема телевизора будет работать не-
много хуже, нежели исходная, а возможно, и вовсе окажется ис-
порченной. И уж тем более трудно представить, чтобы это из-
менение привело к возникновению какой-либо новой функции,
например, превратило телевизор в стиральную машину. Эта ло-
гика полностью применима и в отношении мутаций. Если в
каком-то из генов возникает мутация, существенно влия-
ющая на функцию кодируемого белка, данный белок обычно
оказывается дефектным.
1.11. Аллели гена
В результате неизбежно возникающих разнообразных му-
таций один и тот же ген может встречаться в разных
вариантах, имеющих определенные различия в нуклеотидной
последовательности. Такие варианты гена называют аллелями.
Теоретически возможно огромное их количество (табл. 1.9). На
самом же деле, по каждому конкретному гену обычно встречает-
ся «всего лишь» несколько сотен аллелей. Однако и этого доста-
точно, чтобы в пределах вида наблюдалось значительное разно-
образие по соответствующему признаку. Обратимся к одному из
примеров подобного разнообразия.
Таблица 1.9. Теоретически возможное разнообразие аллелей
гена, состоящего из 10 тыс. нуклеотидов
Тип изменений Количество возможных вариантов
Для одного положения Для всего гена
Замена одного нуклеотида 3 JI0*
Замена двух нуклеотидов — |3-104]2 = 9-10*
Замена трех нуклеотидов — [3104]’ = 31012
44
Окончание табл. 1.9
Тип изменений Количество возможных вариантов
Для одного положения Для всего гена
Отсутствие одного нуклеотида 1 104
Отсутствие двух нуклеотидов — |104]2= 10»
Отсутствие трех нуклеотидов — |ют = го12
Вставка лишнего нуклеотида 1 I04
Вставка двух нуклеотидов — 1ЮТ= io»
Вставка трех нуклеотидов — [ 104]3 = го12
Вы уже знаете, что синтез эу- и феумеланина может идти в
организме человека только в присутствии активного фермента
тирозиназы (см. рис. 1.13). Этот фермент кодируется геном TYR,
по которому существуют тысячи разных аллелей. Одна из них
содержит «правильную» нуклеотидную последовательность —
именно ту, что нужна для образования активного фермента.
Данная аллель (ее называют нормальной) обусловливает высокий
уровень биосинтеза меланинов и как следствие — сильную пиг-
ментацию волос. Между тем для мутантных аллелей этого же гена
характерны различные изменения нуклеотидной последователь-
ности, приводящие к заметному снижению активности тирози-
назы. Если активность тирозиназы отсутствует полностью, во-
лосы получаются практически бесцветными — белыми. Но если
фермент сохраняет частичную активность, небольшое количество
меланинов все же синтезируется, и окраска волос варьирует от
слабо-желтой до золотисто-серой или даже светло-коричневой.
Таким образом, разные аллели гена TYR обуславливают разную
степень пигментации волос.
У организмов, возникших в результате полового размножения,
многие гены представлены не в одном, а в двух экземплярах.
В этом случае возможны различные сочетания аллелей. Если ор-
ганизм несет две одинаковые аллели, его называют гомозиготой,
а если две разные — гетерозиготой. Отметим, что у гетерозигот-
ного организма могут одновременно присутствовать аллели, рез-
ко отличающиеся друг от друга по своим проявлениям. В качестве
удобного примера представим человека, обладающего двумя ал-
лелями гена TYR, одна из которых нормальная, а другая — му-
тантная. При этом будем считать, что мутантная аллель соответ-
ствует ферменту с полным отсутствием активности и в отличие
от нормальной аллели гена не способна придавать волосам даже
45
Первый этап биосинтеза меланинов
осуществляется с интенсивностью 100 %
Функция выполняется Функция выполняется
Молекулы
активной [
тирозиназы
Молекулы
активной
тирозиназы
| Трансляция |
иРНК--------------- -----------------иРНК
| Транскрипция |
Нормальная аллель
Нормальная аллель
Первый этап биосинтеза меланинов
осуществляется с интенсивностью 50 %
Функция выполняется Функция не выполняется
t t
Молекулы ч Молекулы
активной I ~l I У ~1 дефектной
тирозиназы тирозиназы
| Трансляция |
иРНК--------------- ---------X------иРНК
| Транскрипция |
Нормальная аллель Мутантная аллель
Первый этап биосинтеза меланинов
не осуществляется
Функция не выполняется Функция не выполняется
t I
Молекулы ч ч Молекулы
дефектной i ~| i ~1 дефектной
тирозиназы тирозиназы
| Трансляция |
иРНК---------------------------------иРНК
| Транскрипция |
Мутантная аллель Мутантная аллель
в
46
Рис. 1.20. Процесс реализации генетической информации у гомозигот по нор-
мальной аллели (а), гетерозигот (б) и гомозигот по мутантной аллели (в) гена
TYR
Примечание. В гетерозиготном организме одновременно присутствуют и ак-
тивные, и дефектные молекулы тирозиназы. Первый этап биосинтеза меланинов
осуществляется, но его интенсивность приблизительно в 2 раза ниже по сравнению с
организмами, гомозиготными по нормальной аллели. В данном случае двукратное
снижение активности фермента никак не отражается на степени проявления при-
знака (50%-ной актйвности тирозиназы вполне достаточно для синтеза необходимо-
го количества допахинона — общего предшественника эу- и феумеланина). Такая
ситуация характерна только для ферментов, имеющих ярко выраженную избыточную
активность. Если же фермент не обладает избыточной активностью, степень прояв-
ления признака у гетерозигот может быть слабее, чем у гомозигот по нормальной
аллели. Косым крестом обозначена мутация в гене TYR и обусловленные ею наруше-
ния в иРНК и белке.
слабую пигментацию. Так какими же будут волосы у данного
человека?
Чтобы выяснить это, обратимся к рис. 1.20. Как вы видите,
в клетках рассматриваемой нами гетерозиготы происходит экс-
прессия обеих имеющихся аллелей. В результате образуются два
варианта тирозиназы: активная (она соответствует нормальной
аллели) и дефектная (ее кодирует мутантная аллель). Активный
фермент обеспечивает биосинтез меланинов, дефектный — никак
не участвует в этом процессе, но и не может препятствовать ему.
В итоге волосы будут пигментированными.
Итак, у этого гетерозиготного организма проявляется только
одна из присутствующих аллелей, а вторая оказывается как бы
замаскированной. Сходная ситуация характерна для большинства
гетерозигот, о каком бы конкретном гене мы ни говорили. Соот-
ветственно, принято различать два типа аллелей: доми-
нантные (т.е. способные проявляться в гетерозиготе) и рецес-
сивные (не проявляющиеся у гетерозигот).
Обратите внимание, в приведенном примере доминантной
оказалась нормальная аллель гена TYR, В этом прослеживается
глубокий биологический смысл. Действительно, выполнение не-
кой молекулярной функции, скорее всего, должно доминировать
над невыполнением. Таким образом, нормальные аллели обыч-
но доминантны, в то время как мутантные — рецессивны.
1.12. Генотип и фенотип
По каждому гену существует хотя бы несколько аллелей. В ре-
зультате в пределах любого биологического вида, будь то лев,
инфузория-туфелька или бледная поганка, неизбежно наблюда-
47
ется огромное разнообразие генотипов — конкретных комбина-
ций наследственных задатков. Это правило полностью справед-
ливо и для человека.
В генетике разработана специальная система символов, по-
зволяющая описывать любые возможные генотипы. Каждый ген
обозначают строго определенным сочетанием, составленным из
букв латинского алфавита иногда с добавлением одной или не-
скольких цифр. При этом обязательно указывают конкретные
аллели. Если в работе анализируют всего две аллели, одна из ко-
торых доминантна, а вторая рецессивна, доминантную обычно
пишут заглавными буквами (например, К, STM или GL3), а ре-
цессивную — строчными (у, stm, gl3). Если в анализ вовлекают
много аллелей, принято использовать более сложную символику.
В этом случае вводят различные дополнительные индексы, припи-
сывая их к обозначению гена через дефис (lys2-87, lep-\G133y AdhJ-
Fm335)y надстрочно (Л1, Notch?2) или подстрочно (Ph 56).
Иногда исследователей интересует не ген целиком, а только стро-
го определенный его участок, по которому наблюдается естественное
разнообразие. Такие участки принято называть полиморфными
сайтами. Их количество в гене может быть достаточно велико. На-
пример, в уже знакомом нам гене TYR обнаружено около 1 000 по-
лиморфных сайтов. Подавляющее большинство из них являются
однонуклеотидными. Каждый сайт в зависимости от своего располо-
жения в гене и характера варьирования получает специальное обо-
значение. Примеры обозначений представлены в табл. 1.10.
Генотип (с учетом воздействий окружающей среды) обуслов-
ливает всю совокупность признаков организма, включая любые
особенности его строения, физиологии, развития, поведения и
др. Эту совокупность называют фенотипом. Важными элемен-
тами фенотипа конкретного человека являются и все его психо-
логические особенности.
Фенотип организма охватывает тысячи признаков, генотип пред-
ставлен тысячами разных генов. При этом обычно между генами и
признаками организма существуют достаточно сложные взаимоот-
ношения. Продемонстрируем их на следующем примере.
Меланины являются компонентами не только волос, но и не-
которых других структур человеческого организма, в частности,
кожи и радужных оболочек глаз. Соответственно полная неспособ-
ность синтезировать меланины, возникающая в результате мутаций
по гену TYR, неизбежно влияет на каждую из этих структур. Волосы
человека получаются практически белыми, его кожный покров —
очень бледным и легко обгорающим, а радужные оболочки глаз —
розовато-сиреневыми. Таких людей называют альбиносами.
Как мы видим, ген TYR обладает множественным проявлени-
ем, т.е. одновременно контролирует несколько признаков. По-
48
Таблица 1.10. Примеры однонуклеотидных полиморфных сайтов в гене TYR
Полиморфный сайт Локали- зация* Характер полимор- физма Полиморфизм на уровне иРНК Полиморфизм на уровне белка
С - 248Т -248 Нуклеотидная за- мена С/Т — —
Met 1 Vai 1 Нуклеотидная за- мена A/G 1-й триплет AUG/GUG** 1 -е аминокислотное положение метионин/валин
MetlThr 2 Нуклеотидная за- мена Т/С 1-й триплет AUG/ACG 1 -е аминокислотное положение метионин/треонин
Leu9Pro 26 Нуклеотидная за- мена Т/С 9-й триплет CUG/CCG 9-е аминокислотное положение лейцин/пролин
Тгр39Х 116 Нуклеотидная за- мена G/A 39-й триплет UGG/UAG*** Нормальный белок/укороченный белок
223delG 223 Н ал ичие/отсутствие G/- 75-й триплет GAU/AU**** Нормальный белок/белок с из- мененной последовательностью аминокислот, начиная с 75-го положения
* Номер нуклеотида по отношению к началу кодирующей области гена. Знак «-» показывает, что нуклеотид находится в про-
моторе. т.е. еще до начала кодирующей области.
** Жирным шрифтом выделен полиморфный сайт.
*** UAG является «стоп»-триплетом, на нем происходит остановка синтеза белка. В обозначении полиморфного сайта это от-
мечено знаком «X».
**** Отсутствие нуклеотида приводит к «сдвигу рамки считывания»: рибосома неправильно читает все последующие триплеты.
добные ситуации встречаются достаточно часто. Это связано с
тем, что продукт определенного гена может использоваться
в разных биологических процессах и в итоге оказывать влия-
ние на разные признаки. В то же время практически каждый
признак организма контролируется не одним, а как минимум
несколькими генами. Для наглядности вновь обратимся к цвету
волос. За него отвечает не только тирозиназа, но и не менее де-
сятка других белков. Именно от них зависит, в каком соотноше-
нии будут синтезироваться эу- и феумеланин, а значит, какого
цвета окажутся волосы: черными, рыжими, каштановыми, пе-
пельными и др. Совершенно естественно, что все соответству-
ющие гены тоже участвуют в контроле цвета волос.
Если достаточно сложный генетический контроль характерен
даже для такого простейшего признака, что же говорить о тонких
психологических чертах. Поэтому крайне наивны представления
о том, что существует «ген агрессивности», «ген алкоголизма»,
«ген ослабленного внимания» и др. Безусловно, все эти признаки
контролируются генетически. Однако в основе формирования
любого из них лежит совместное действие множества генов, сре-
ди которых трудно выделить какой-либо главный. Каждый в от-
дельности лишь вносит определенный вклад в ту или иную пси-
хологическую особенность человека. Мы убедимся в этом в по-
следующих разделах учебника.
СПИСОК ЛИТЕРАТУРЫ
1. Беркинблит М. Б., Глаголев С. М., Фуралев В. А. Общая биология.
Т. I. - М. : МИРОС, 1999.
2. Инге-Вечтомов С. Г. Генетика с основами селекции. — СПб.: НЛ,
2010.
3. Корочкин Л. И. Введение в генетику развития. — М. : Наука, 1999.
4. Шредингер Э. Что такое жизнь с точки зрения физики? — М.: Гос.
изд-во иностр, лит-ры, 1947.
5. ReesJ. L. Genetics of hair and skin color // Annual Review in Genetics. —
2003. - V. 37. - P. 67-90.
КОНТРОЛЬНЫЕ ВОПРОСЫ
1. Охарактеризуйте важнейшие классы биоорганических соединений.
2. В чем состоит процесс реализации генетической информации?
Опишите его важнейшие этапы.
3. Опишите механизм дифференциальной экспрессии генов.
4. Сравните молекулярные механизмы быстрых и медленных био-
логических ответов. В чем они сходны, чем различаются?
5. Почему нормальная аллель гена TYR доминирует над мутант-
ной?
Глава 2
МОЛЕКУЛЯРНЫЕ ОСНОВЫ
ФУНКЦИОНИРОВАНИЯ НЕРВНОЙ СИСТЕМЫ
2.1. Основное назначение нервной системы
Закономерности, рассмотренные ранее, одинаково справедли-
вы для всех без исключения организмов. Тем не менее на базе
таких универсальных закономерностей существует огромное раз-
нообразие частных вариаций, обуславливающих отличительные
особенности разных видов, разных органов, тканей, клеток и др.
Определенные особенности есть и у нервной системы. Именно
им посвящена настоящая глава.
Мы не будем подробно рассматривать нервную систему: это
чрезвычайно сложная и объемная задача, далеко выходящая за
рамки данного учебника. Остановимся только на некоторых ха-
рактерных чертах, наиболее важных для дальнейшего изложения
материала. И начнем мы с общего назначения нервной системы.
Поведение любого живого существа складывается из множества
взаимосвязанных биологических ответов на ту или иную совокуп-
ность полученных сигналов. При этом соотношение между мед-
ленными и быстрыми ответами у разных организмов крайне неоди-
наково. В частности, у всех бактерий, растений и грибов заметно
преобладают медленные биологические ответы. Дело в том, что
характерной особенностью перечисленных организмов является
наличие прочных клеточных стенок.
Такие структуры обеспечивают клеткам надежную защиту, од-
нако значительно ограничивают их подвижность. В соответствии
с этим и бактерии, и растения, и грибы выработали относительно
пассивную стратегию поведения: они приспособились пережидать
неблагоприятные условия, что не требует широкого использования
быстрых ответов.
Животные в отличие от бактерий, растений и грибов не фор-
мируют каких бы то ни было клеточных стенок, зато компенси-
руют этот «недостаток» незамедлительными реакциями на изме-
нения внешней среды. Совершенно естественно, что у животных
51
(в особенности многоклеточных) быстрые ответы используются
очень широко. Более того, для обслуживания этих ответов возник
целый ряд высоко специализированных структур:
• различные органы чувств, каждый из которых предназначен
для восприятия определенного рода сигналов;
• нервная система, преобразующая сигналы в нервные им-
пульсы и быстро передающая их к исполнительным клеткам;
• мышцы, способные обеспечивать подвижность организма в
ответ на совокупность полученных нервных импульсов. Как пра-
вило, мышцы находятся в тесной взаимосвязи с теми или иными
опорными структурами организма, одновременно выполняющи-
ми функцию механической защиты (например, с хитиновым
панцирем у членистоногих или же с внутренним скелетом у по-
звоночных). Но в отличие от клеточных стенок такие структуры
не ограничивают подвижность каждой конкретной клетки и тем
самым не мешают активному поведению организма.
По-видимому, на ранних этапах своей эволюции нервная си-
стема имела сугубо экологическую направленность, т.е. обслу-
живала реакции организма на внешние сигналы. Но затем по мере
возникновения все новых органов появилась необходимость тон-
ко координировать их работу и увязывать ее с ответами на внеш-
ние воздействия. Поэтому нервная система в процессе эволюции
приобрела и вторую направленность — координирующую. Даль-
нейшее развитие этих координирующих функций привело к
формированию сложных аналитических способностей, а в конеч-
ном итоге — к возникновению мышления.
Таким образом, основное назначение нервной системы —
обеспечивать формирование быстрых биологических ответов
за счет передачи нервных импульсов на исполнительные
клетки.
2.2. Типы строения нервной системы
Как известно, главными элементами нервной системы,
непосредственно обеспечивающими проведение нервных им-
пульсов, являются высоко специализированные клетки —
нейроны (рис. 2.1). Впервые возникнув еще у древних кишечно-
полостных, эти клетки мало изменялись в процессе эволюции.
Зато многократно происходили существенные изменения, затра-
гивающие общую организацию нервной системы, а именно —
характер взаимного расположения нейронов, их количество,
распределение контактов между ними и др. В конечном итоге на
основе примерно одинаковых клеток сформировались различные
типы нервной системы.
52
Рис. 2.J. Схема строения нейрона
Примечание. Даже в пределах одного и того же организма можно обнаружить
огромное разнообразие нейронов. Но общий план их строения достаточно сходен.
Обычно нейрон имеет компактное тело (здесь находится ядро и большая часть цито-
плазмы) и множество сильно разветвленных нитевидных отростков. Эти отростки
служат для проведения нервных импульсов. У самых примитивных животных (кишечно-
полостных) все отростки одного нейрона приблизительно равноценны и могут про-
водить импульсы в обоих направлениях. У более высокоразвитых животных ситуация
иная: каждый отросток проводит импульсы однонаправлено. В этом случае принято
различать два типа отростков: дендриты, несущие импульсы к телу нейрона (обычно
такие отростки достаточно многочисленны), и аксоны, проводящие импульсы от тела
нейрона (как правило, аксон представлен в единственном числе).
Наиболее примитивна диффузная нервная система. В этом
случае нейроны расположены диффузно (поодиночке), образуя
своими отростками плотную сеть (рис. 2.2, а). Если в одном из
нейронов возникает возбуждение, оно равномерно распростра-
няется во всех направлениях и в итоге обычно охватывает весь
организм. В результате возможны лишь относительно простые
реакции (например, проталкивание пищи в ротовое отверстие
или же сокращение тела в ответ на травму). На данном этапе раз-
вития нервной системы ее координирующие функции прослежи-
ваются слабо.
Узловая нервная система значительно сложнее: нейроны
расположены не диффузно, а в виде скоплений, которые называ-
ют ганглиями, или нервными узлами (рис. 2.2, б). Каждый узел
иннервирует определенную группу органов. В соответствии с этим
организм получает возможность дифференцированно управлять
различными участками тела, что обеспечивает широкое разноо-
бразие поведенческих реакций. В то же время, поскольку узлы
взаимосвязаны друг с другом, организм способен координировать
свои реакции: некоторые — усиливать, а некоторые — наоборот,
подавлять. Ведущую роль в такой координирующей деятельности
53
Гидра
Головной
нервный
узел
Нервный
тяж
Нервы
Головной
нервный
узел
Цепочка
нервных
узлов
Насекомое
Планария Дождевой
3
3
Рис. 2.2. Типы строения нервной системы:
а — диффузная нервная система (характерна для кишечнополостных); б — узловая
нервная система (характерна для червей, моллюсков и членистоногих); в — трубчатая
нервная система (характерна для позвоночных)
Примечание. Если в организме имеются скопления нейронов (например,
нервная трубка у позвоночных животных или нервные узлы у червей, членистоногих
и моллюсков), эти скопления называют центральной нервной системой. Ее главная
задача — координировать поведенческие реакции. От скоплений нейронов отходят
многочисленные нервы, образованные длинными нейронными отростками. Нервы
составляют периферическую нервную систему, она служит для проведения импульсов
по всему организму.
обычно играют головные (окологлоточные) узлы, непосредствен-
но получающие информацию от органов чувств.
Особого развития узловая нервная система достигает у голо-
воногих моллюсков и насекомых. Для них характерны крупные
нервные центры, возникшие в результате слияния отдельных
узлов. Эти центры обладают мощными координирующими спо-
собностями, позволяющими вырабатывать сложные варианты
поведения с ярко выраженными элементами высшей нервной
деятельности. В частности, хорошо известно, что медоносные
пчелы легко запоминают дорогу от улья к пище и с помощью
«танцев» передают эти сведения своим соплеменницам.
У животных, имеющих трубчатую нервную систему, пода-
вляющее большинство нейронов «собрано» вместе в виде длин-
ного полого тяжа (нервной трубки). Эта трубка проходит прак-
тически через все туловище, обеспечивая интегральное управле-
ние всем организмом. Как правило, в ней можно вычленить два
отдела (рис. 2.2, в):
• собственно туловищный участок (спинной мозг), контро-
лирующий сравнительно простые поведенческие реакции напо-
добие болевого, зрачкового или сосательного рефлексов;
• расширенный передний участок (головной мозг). Именно
здесь происходит анализ воспринятых сигналов, здесь координи-
руются различные биологические ответы, здесь осуществляются
процессы высшей нервной деятельности.
По мере эволюционного развития трубчатой нервной системы
относительные размеры головного мозга постепенно увеличива-
лись, что способствовало значительному усложнению поведенче-
ских реакций. Вершиной этого направления являются птицы и
млекопитающие (табл. 2.1). Неслучайно наиболее сложные вари-
анты поведения характерны именно для представителей данных
классов.
Таблица 2.1. Относительные размеры головного мозга у раз-
ных позвоночных (по Jerison, 1973 с изменениями)
Класс Объект Энцефализация*
Костные рыбы Угорь 0,002
Карась 0,005
Пресмыкающиеся Аллигатор 0,004
Варан 0,013
Ящерица амейва 0,027
Птицы Страус 0,019
55
Окончание табл. 2. /
Класс Объект Эн цефализация*
Колибри 0,087
Ворона 0,223
Млекопитающие Кенгуру 0,067
Крот 0,054
Кролик 0,054
Крыса 0,063
Кошка 0,099
Слон 0,164
Волк 0,234
Дельфин 0,464
Горилла 0,247
Шимпанзе 0,261
Австралопитек 0,374
Человек разумный 0,787
* Энцефализацию (Е) рассчитывают по формуле
р _ Мгм .
Е-^Г'
где А/гм — масса головного мозга,; А/т — масса тела, г.
Необходимость использования степени 2/з продиктована тем, что размеры го-
ловного мозга растут с увеличением тела нелинейно.
2.3. Синапсы
Многочисленные нейроны одного и того же организма
связаны друг с другом в единую «информационную сеть».
Связь между ними осуществляется с помощью синапсов, т. е.
особого рода межклеточных контактов, специально предназна-
ченных для передачи нервных импульсов (рис. 2.3, а).
Принципы синаптической передачи весьма консервативны:
они не претерпели почти никаких изменений на всем протяжении
эволюции нервной системы. В этом плане, абстрагируясь от мно-
гих второстепенных нюансов, можно подразделить все синапсы
56
натри группы: элекгрпческие, химические и смешанные (электро-
химические).
Главнои отличительной особенностью электрических синап-
сов является очень узкая синаптическая щель, не создающая для
нервных импульсов значитетьных препятствий. При этом мембра-
ны обеих контактирующих клегок обычно пронизаны многочис-
ленными сквозными перемычками — коннексонами (рис. 2.3, б).
В резульгате импульсы передаются практически без задержки и,
Направление
нервного
Рис. 2.3. Строение синапсов:
а — три компонента синапса (пресинап! ический участок, т.е. нервное окончание,
по которому нервные импульсы подаются на синапс; синаптическая щель, т.е. узким
межклеточный просвет, разделяющий мембраны двух контактирующих клеток и
постсинаптичсскии участок — это может быть либо какой-то другой нейрон, либо
исполнительная клетка — мышечная или железистая), б — строение электрического
синапса; в — строение и принцип функционирования химического синапса (пузырьки,
содержащие нейромедиатор, формируются в тедс нейрона и затем транспонируются
к синапсу по аксону)
57
как правило, одинаково успешно в обоих направлениях. Такие
синапсы очень надежны и высоко эффективны, однако почти не
поддаются регуляции и используются в основном с одной един-
ственной целью: обеспечить синхронное возбуждение группы
нейронов для мощного и строго одновременного сокращения
мышц.
Химические синапсы значительно медленнее электрических.
В этом случае синаптическая щель во много раз шире и никаких
сквозных перемычек между клетками нет. Импульсы передаются
с помощью молекул-посредников, получивших название нейро-
медиаторы, или нейротрансмиттеры (рис. 2.3, в). Известно
несколько сотен подобных соединений. К важнейшим из них от-
носятся глицин, глутаминовая и у-аминомасляная кислоты, до-
фамин, серотонин, норадреналин, ацетилхолин, а также разно-
образные нейропептиды (например, эндорфины). Обычно в
пределах одного и того же организма используется довольно ши-
рокий спектр нейромедиаторов (в частности, у человека — не
менее 150). Но при этом каждый конкретный химический синапс
«обслуживается» вполне определенным нейромедиатором. Соот-
ветственно, различают глутаматэргические синапсы, дофаминэр-
гические, холинэргические и др.
Несмотря на некоторую задержку в проведении импульсов,
химические синапсы обладают ярко выраженными достоинства-
ми. Во-первых, они функционируют строго однонаправленно и
тем самым препятствуют хаотичному распространению импуль-
сов. Во-вторых, они подвержены сложной многоуровневой регу-
ляции, что очень важно для координирующей деятельности
нервной системы. Неудивительно, что в нервной системе высших
позвоночных безоговорочно преобладают именно химические
синапсы (у человека их доля превышает 95 %).
Смешанные синапсы являются своеобразными «химерами»,
совмещающими оба рассмотренных выше принципа. Благодаря
взаимодействию между химическими и электрическими компо-
нентами такие синапсы тоже подвержены сложной регуляции.
Но, к сожалению, пока они изучены крайне слабо. Поэтому мы
только упоминаем об их существовании, не вдаваясь в какие-либо
конкретные функциональные характеристики.
Некоторые нейроны формируют лишь единичные синапсы, но
некоторые — десятки, сотни и даже тысячи. В результате инфор-
мация от одного возбужденного нейрона может непосредственно
передаваться на множество других. Это явление, называемое ди-
вергенцией нервных путей, позволяет организму в ответ на
определенный сигнал согласованно задействовать различные свои
структуры. Вместе с тем наблюдается и конвергенция нервных
путей (передача информации от разных нейронов на один и тот
58
же), позволяющая тонко координировать ответные реакции на
множество одновременно воспринимаемых организмом сигна-
лов.
Судя по современным оценкам, головной мозг человека со-
стоит приблизительно из 100 млрд нейронов. И каждый образует
примерно по тысяче синапсов. В результате формируется слож-
нейшая нейронная сеть, обладающая огромными координиру-
ющими и аналитическими способностями. По-видимому, имен-
но эти феноменальные способности и лежат в основе всех про-
явлений человеческой психики.
2.4. Функциональная специализация
нейронов
На ранних этапах эволюции нервной системы все нейроны в
организме были практически равноценными. Однако вскоре на-
чалась их функциональная специализация, зачатки которой про-
слеживаются даже у кишечнополостных. Для примера достаточно
обратиться к пресноводной гидре. У этого животного с прими-
тивной нервной системой обнаружено два типа нейронов: чув-
ствительные и моторные. Первые воспринимают сигналы и
преобразуют их в импульсы, а вторые передают импульсы на
мышечные клетки (рис. 2.4, а).
У животных с узловой или трубчатой нервной системой эта
специализация заходит значительно дальше. Возникает большое
разнообразие чувствительных нейронов, отличающихся друг от
друга по спектру используемых рецепторов, а значит, по спектру
воспринимаемых ими сигналов (табл. 2.2). Заметно специализи-
рованными становятся и моторные нейроны. Действительно, они
уже не связаны в единую сеть и могут функционировать доста-
точно автономно друг от друга, что позволяет дифференцирован-
но управлять различными органами.
Иногда чувствительный и соответствующий ему моторный
нейроны напрямую контактируют друг с другом своими отрост-
ками. В результате образуется простейшая рефлекторная дуга,
обеспечивающая автоматическую реакцию на данный сигнал (рис.
2.4, б). Однако обычно ситуация значительно сложнее: в состав
рефлекторной дуги наряду с перечисленными входит и некоторое
количество дополнительных нейронов. Их называют вставочны-
ми, или ассоциативными. Своими многочисленными отростка-
ми они контактируют с другими отделами центральной нервной
системы и тем самым способствуют координации различных от-
ветов (рис. 2.4, в). Прекрасным примером может служить реф-
59
Неспециализированный
нейрон
Чувствительные нейроны
Спинной мозг
Чувствительные
нейрона
б
Отростки,
идущие
в головной
мозг
Отростки,
идущие
из головного
мозга
Тела
Белое Тело
вещество моторного
нейрона
Серое
вещество
вставочн
нейронов
Тело чувствительного
нейрона
Чувствительные
окончания
Аксон
моторного
нейрона
Исполнительные
клетки
Дендрит
чувствительного
нейрона
Рис. 2.4. Передача нервных импульсов на исполнительные клетки:
а - кишечнополостные (даже у таких примитивных животных, как гидра, большин-
ство нейронов специализировано на чувствительные и моторные. Нервный импульс
исходно формируется чувствительным нейроном и затем через моторные поступает
на мышечные клетки — двусторонними стрелками показана способность моторных
нейронов передавать нервные импульсы в обоих направлениях. Лишь некоторые ней-
роны не имеют специализации: они сами воспринимают сигнал, формируют нервные
импульсы и сами доводят их непосредственно до мышечных клеток); б — простейшая
рефлекторная дуга у позвоночных (стрелками показаны направления нервного импуль
са) ; в — классическая рефлекторная дуга у позвоночных (пояснения — в тексте)
лекторная дуга, используемая в случае чесательного рефлекса у
собаки (в ответ на раздражение небольшого участка кожи воз-
никают строго согласованные сокращения мышц, причем не
одной или двух, а нескольких десятков).
Вставочные нейроны вовлечены и в аналитическую деятель-
ность. Дело в том, что подавляющее большинство нейронов го-
ловного мозга относится не к чувствительным или моторным, а
именно к вставочным. Они способны сохранять полученную ин-
формацию, накапливать ее в течение жизнедеятельности организ-
ма, сопоставлять со вновь поступающей и др. Это позволяет ор-
ганизму выстраивать свое поведение с учетом ранее приобретен-
ного индивидуального опыта.
Для примера давайте представим, что некому рыболову по-
счастливилось выудить крупных размеров рыбу. Снимая ее с
крючка, он укалывается о плавник, что приводит к задействова-
нию соответствующей рефлекторной дуги. Если бы вставочных
нейронов не существовало, человек автоматически отдернул бы
уколотую руку и тем самым избавился от источника болевых
ощущений. Но в действительности аналитические отделы голов-
ного мозга мгновенно просчитывают весь спектр возможных
последствий и в итоге «приходят к выводу», что в данной ситуации
намного важнее не избавиться от боли, а удержать трофей. Поэ-
тому к моторному нейрону рефлекторной дуги посылаются нерв-
ные импульсы, препятствующие его возбуждению, несмотря на
наличие ярко выраженного болевого воздействия.
Итак, в сложно организованной нервной системе, незави-
симо от того, узловая она или трубчатая, наблюдается
отчетливая функциональная специализация нейронов', чув-
ствительные нейроны обеспечивают восприятие сигналов и
их дальнейшее преобразование в нервные импульсы, мотор-
ные — передают эти импульсы на исполнительные клетки,
а вставочные — выполняют координирующие и аналитиче-
ские функции.
61
Таблица 2.2. Основные типы чувствительных нейронов у человека
Тип нейронов Локализация чувстви- тельных окончаний Соответствующие рецепторы
Название Число типов Принцип функционирования
Палочки Сетчатка глаза Родопсин (черно- белое зрение) 1 Рецепторы, связанные с G-бел- ками. Вторичный мессенджер- цГМФ*, его содержание снижается за счет стимулирования фосфо- диэстеразы
Колбочки Фотопси ны (цветное зрение) 3
Обонятельные Слизистая носовой полости Обонятельные рецепторы Более 300 Рецепторы, связанные с G-бел- ками. Вторичный мессенджер- цАМФ, его содержание возрастает за счет стимулирования аденилат- циклазы
Вкусовые** Слизистая языка Рецепторы слад- кого 2 Рецепторы, связанные с G-бел- ками. Системы вторичных мессенджеров исследованы недостаточно под- робно
Рецепторы горь- кого Около 30
Рецепторы юма- ми*** 2
Рецепторы кис- лого Неизвестно По-видимому, протонные каналы
Рецепторы со- леного Неизвестно По-видимому, натриевые каналы
Тактильные Кожа Механорецептор- ные каналы Не менее 2 Ионные каналы с двумя высту- пающими антеннами. Внутренняя антенна прочно заякорена в цито- плазме. Механическое воздействие на внешнюю антенну приводит к открыванию канала
Проприоцептивные Мышцы и сухожи- лия
Бароцептивные Стенки внутренних органов
Слуховые* ** Улитка внутреннего уха
Вестибулярные** Вестибулярный аппарат
Термочувствитель- ные Преимущественно кожа и слизистая ротовой полости Терморецепторы Около 10 Температуро-зависимые ионные каналы, протеинкиназы и др.
Ноцицептивные (чувствительные к боли) Различные ткани Ноцицепторы Не менее 10 Ионные каналы, чувствительные к температуре, осмотическому дав- лению и некоторым агрессивным веществам
* Цикло-гуанозинмонофосфат. Наряду с цАМФ используется многими нейронами в качестве вторичного мессенджера (см.
подразд. 2.6).
** Указанные нейроны воспринимают сигналы не напрямую, а через клетки-посредники. Такие посредники (рецептирующие
клетки) способны самостоятельно распознавать соответствующий сигнал, а затем с помощью выделяемого нейромедиатора «ин-
формировать» об этом чувствительный нейрон.
*** Вкус глутамата кальция.
2.5. Молекулярные основы нервного импульса
От общих задач и принципов организации нервной системы
перейдем к молекулярным механизмам ее функционирования.
К сожалению, многие из них пока еще не выяснены. Мы не зна-
ем, как именно сохраняется индивидуальный опыт, как организм
прогнозирует свои дальнейшие действия, как происходит рас-
познавание образов и др. В то же время не вызывает сомнений,
что эти механизмы тесно связаны с формированием и проведе-
нием нервных импульсов. Давайте же кратко познакомимся с
данными процессами.
Начнем с того, что импульсы формируются очень быстро: в не-
которых случаях — за тысячные доли секунды с момента вос-
приятия нейроном соответствующего сигнала. Это значит, что все
необходимые для импульса белки уже синтезированы и только
«ожидают» своей активации. И действительно, все без исключе-
ния нервные импульсы от каких бы чувствительных нейро-
нов они ни исходили и каким бы исполнительным клеткам
ни предназначались^ осуществляются по принципу каскадной
активации белков.
Упрощенную схему подобных каскадов мы рассматривали
в подразд. 1.9 (см. рис. 1.18, б). Эта схема удобна для понимания
сути вопроса, но требует целого ряда серьезных уточнений. Дело
в том, что различные белки активируются по-разному: некото-
рые — за счет фосфорилирования, как на указанной схеме, не-
которые — под действием специальных молекул-посредников,
а некоторые — в зависимости от концентрации определенных
ионов. Соответственно разные этапы одного и того же каскада
зачастую обслуживаются разными молекулярными механизмами.
Это правило полностью применимо и к нервному импульсу. При-
чем большинство участвующих в импульсе белков активиру-
ется за счет изменений мембранного потенциала.
Известно, что каждая клетка, в том числе и нейрон, содержит
многочисленные органические и неорганические ионы. Они сво-
бодно диффундируют по всей цитоплазме, однако не могут само-
стоятельно проходить сквозь мембраны (для заряженных частиц
двойной фосфолипидный слой непроницаем). Поэтому в мем-
бранах имеются особые белки, служащие для ионов своеобраз-
ными «пропускными пунктами». Такие белки в зависимости от
характера их функционирования принято подразделять на две
большие группы: ионные насосы и ионные каналы. Первые
транспортируют ионы против разницы концентраций, что требу-
ет от клетки значительных энергетических затрат. Вторые — по
разнице концентраций, без затраты энергии (рис. 2.5).
64
Рис. 2.5. Схема функционирования ионных насосов (а) и ионных каналов (б)
Примечание. Ионные насосы и каналы высоко избирательны. В каждом из
них имеется специальный проход, способный пропускать только строго определенные
ионы (на рисунке обозначены серыми кружками). Насосы и каналы функционируют
не постоянно. Изменяя свою конформацию, они могут переходить из «открытого»
состояния в «закрытое» и наоборот. На этом основана регуляция транспорта ионов.
К числу наиболее важных представителей первой группы от-
носится натрий/калиевый насос. Используя энергию молекул
АТФ, он выкачивает Na+ из цитоплазмы во внеклеточное про-
странство, одновременно транспортируя К+ в обратном направ-
лении. При этом строго выдерживается следующее соотношение:
на 3 удаляемых из клетки иона натрия приходится 2 поглощаемых
иона калия. Таким образом, при каждом срабатывании данного
насоса цитоплазма теряет один положительный заряд, а внекле-
точное пространство, напротив, его приобретает. И в результате
на мембране формируется разность потенциалов.
В этом процессе принимают участие и кальциевые насосы.
Они эффективно выкачивают Са2+ из цитоплазмы, транспортируя
его, во-первых, во внеклеточное пространство, а во-вторых, в
многочисленные полости эндоплазматической сети, которые
служат главными накопителями данных ионов. Очевидно, что в
ходе такого перераспределения ионов кальция цитоплазма до-
полнительно лишается положительных зарядов, что увеличивает
разность потенциалов на наружной мембране.
Эта разность зависит от типа и состояния клетки. В частности,
у большинства нейронов в состоянии покоя ее значение равно
приблизительно -70 мВ («-» показывает, что внутренняя сторона
мембраны заряжена отрицательно). В этих условиях все ионные
каналы, как правило, закрыты, что способствует стабильному
поддержанию мембранного потенциала. Однако с приходом сиг-
нала ситуация изменяется. Дело в том, что сигнал запускает ка-
скадные процессы, приводящие к массовому открытию ионных
65
каналов и как следствие — к деполяризации мембраны нейрона
(рис. 2.6).
Обычно восприятие каждого конкретного сигнала происходит
на относительно небольшом участке нейрона (например, в преде-
лах специализированного чувствительного окончания или же в
области синапса с предыдущим нейроном). Поэтому возникшее
изменение мембранного потенциала имеет поначалу сугубо локаль-
ный характер. Тем не менее даже такой локальной деполяризации,
если она достигает определенного порога, вполне достаточно, что-
бы вызвать открытие ионных каналов на соседних участках этой
же мембраны. В конечном итоге возникает волна кратковременной
деполяризации, быстро распространяющаяся, по всей мембране
нейрона. Именно эта волна и является нервным импульсом.
Цитоплазма
а
каналы
приводящие к открытию
ближайших каналов
б
Рис. 2.6. Деполяризация мембраны под действием сигнала:
а — наружная мембрана нейрона в состоянии покоя (ионные каналы закрыты);
б — с приходом сигнала открываются особые ионные каналы, расположенные в
непосредственной близости от рецептора (открытые каналы отмечены серым цве-
том). В результате происходит частичная деполяризация мембраны, т.е. небольшое
снижение мембранного потенциала; в — при снижении мембранного потенциала до
определенного уровня открываются многочисленные потенциалзависимые каналы
(электрический потенциал не только снижается до нуля, но и кратковременно меняет
свой знак на противоположный — калиевые каналы открываются позже натриевых,
и когда концентрация ионов Na4 почти уравнялась ионы К4 еще удерживаются на
внутренней стороне мембраны, придавая ей положительный заряд. Небольшая за-
держка в открывании калиевых каналов показана в виде пунктирных стрелок); г — все
ионные каналы закрываются (они смогут открыться лишь после того, как ионные
насосы полностью восстановят мембранный потенциал, т.е. нейрон вернется в со-
стояние покоя и будет готов к восприятию нового сигнала)
66
Если рассматривать импульс более подробно, в его «судьбе»
можно выделить три основные стадии:
• формирование импульса;
• его проведение по нейрону;
• передачу через синапс на следующую клетку.
Давайте остановимся на каждой из перечисленных стадий и
познакомимся с их ключевыми молекулярными механизмами.
2.6. Молекулярные механизмы
формирования нервного импульса
Исходной причиной, лежащей в основе формирования им-
пульса, является восприятие нейроном определенного сигнала.
Эту функцию выполняют молекулы соответствующего рецептора,
расположенные, как правило, на наружной мембране нейрона.
Обнаружив сигнал, рецепторы изменяют свою конформацию и
тем самым воздействуют на соседние ионные каналы, заставляя
их перейти из закрытого состояния в открытое.
Механизмы такого воздействия могут быть разными. В не-
которых случаях (например,, в чувствительных нейронах^
обеспечивающих восприятие тепловых или механических
сигналов) сами рецепторы одновременно являются и ионны-
ми каналами, поэтому активация данных рецепторных
белков напрямую приводит к значительной деполяризации
мембраны и в результате — практически к мгновенному
формированию импульса. Такие рецепторы принято назы-
вать ионотропными. Но чаще всего между рецепторами и
ионными каналами функционируют достаточно сложные
каскады посредников.
Наиболее распространенные варианты подобных каскадов
схематично представлены на рис. 2.7. Обратите внимание: не-
смотря на многочисленные нюансы, эти каскады организованы
по сходному молекулярному принципу.
И действительно, в них участвуют следующие «наборы» ком-
понентов:
• рецепторы, насквозь пронизывающие наружную мембрану;
• G-белки, расположенные на внутренней стороне мембраны и
активируемые в результате изменения конформации рецепторов;
• фермент, активность которого регулируется (7-белками.
В одном варианте каскада это аденилатциклаза, в другом — фос-
фолипаза С или D;
• вторичные мессенджеры, т. е. небольшие регуляторные
молекулы, служащие внутриклеточными переносчиками инфор-
67
мации о сигнале. К ним относится циклический аденозинмоно-
фосфат (сокращенно — цАМФ), синтез которого обеспечивает-
ся аденилатциклазой, а также инозитолтрифосфат и диацилгли-
церол, образуемые за счет активности фосфолипазы. Нужно
отметить, что появление в клетке инозитолтрифосфата приводит
к выбросу Са2+ из эндоплазматической сети, и эти ионы, быстро
диффундируя по всей цитоплазме, тоже становятся важными
вторичными мессенджерами;
• белки, активируемые под действием вторичных мессенд-
жеров. Это особые ионные каналы, обеспечивающие начальную
стадию деполяризации мембраны (см. рис. 2.6). Кроме того, это
Сигнал 1 । Сигнал 2
Рис. 2.7. Системы молекулярных посредников, обеспечивающих передачу
информации от рецепторов на ионные каналы и протеинкиназы:
а — аденилатциклазная система (I и 2 — регуляторная и каталитическая субъедини-
цы протеинкиназы А. Появление цАМФ в цитоплазме должно быть импульсным,
поэтому излишки данного вторичного мессенджера уничтожаются специальным фер-
ментом — фосфодиэстеразой); б — фосфолипазная система (3 и 4 — регуляторная и
каталитическая субъединицы Са3*-зависимой протеинкиназы; 5 и 6 — регуляторная и
каталитическая субъединицы протеинкиназы С. Фосфолипаза синтезирует инозитол-
трифосфат и диацил глицерол из молекулярных компонентов наружной мембраны); <7,
и G2 — G-белки, обслуживающие рецепторы соответственно I и 2 (шрифтом выделены
вторичные мессенджеры; пунктирными стрелками показаны молекулярные каскады,
запускаемые соответствующими протеинкиназами. Прочие пояснения — в тексте)
68
определенные прогеинкиназы, активно участвующие в регуляции
последующих процессов за счет фосфорилирования различных
мембранных белков.
Активация одной единственной молекулы рецептора приводит
к едва ощутимому изменению потенциала. Между тем для того,
чтобы открылись натриевые каналы (а без этого импульс прин-
ципиально не может сформироваться), необходима довольно
существенная деполяризация мембраны: как правило, не менее
чем на 20 мВ. И если воспринятый сигнал недостаточно интен-
сивен, т.е. активирует слишком мало молекул рецептора, воз-
никшее изменение не достигает необходимого порога и быстро
компенсируется различными ионными насосами. В результате
мембрана возвращается к первоначальному состоянию, и выра-
ботки нервного импульса не происходит.
Казалось бы, данный принцип функционирования нейронов
(он получил название «все или ничего») в корне противоречит
тому общеизвестному факту, что организм не только распознает
наличие сигналов, но и способен оценивать их интенсивность.
Однако в действительности никакого противоречия нет. Взаимо-
действие между сигналом и молекулами рецептора, как и любые
другие молекулярные взаимодействия, строго подчиняется стати-
стическим (т.е. вероятностным) закономерностям. Это значит,
что при действии сигнала фиксированной силы существует
определенная вероятность формирования импульса. Чем мощ-
нее сигнал, тем эта вероятность выше. Соответственно, чем
выше интенсивность воспринимаемого сигнала, тем чаще ней-
рон вырабатывает нервные импульсы. Именно это и служит
оценкой интенсивности сигнала.
2.7. Молекулярные механизмы проведения
нервного импульса
Напомним, что процессы формирования нервного импульса
затрагивают сравнительно небольшой участок мембраны.
На этом участке разность потенциалов стремительно
снижается и кратковременно меняет свою полярность на
противоположную (обычно с - 70 до +40мВ). В результате
возникают локальные электрические токи, приводящие к
достаточно существенной деполяризации мембраны и на
соседних, еще не возбужденных ее участках. В свою очередь,
это обеспечивает немедленную активацию находящихся там
потенциалзависимых ионных каналов и как следствие при-
водит к расширению области возбуждения. Сходным образом
69
возбуждение передается и на следующие участки и в итоге быстро
распространяется по всей мембране. Причем, независимо от
пройденного по мембране расстояния, амплитуда изменения по-
тенциала не угасает, составляя примерно все те же ПО мВ.
Молекулярные основы проведения нервного импульса очень
близки у всех многоклеточных животных, начиная от примитив-
ных кишечнополостных и заканчивая млекопитающими. То же
самое касается и нейронов с различной специализацией. Но ско-
рость проведения импульсов может варьировать. В частности, чем
больше диаметр отростка нейрона, тем ниже сопротивление ци-
топлазмы в данном отростке, а значит, тем выше скорость про-
ведения импульса. Для примера: тончайшие окончания чувстви-
тельных нейронов, обеспечивающих восприятие болевых сигналов
у млекопитающих, проводят импульсы со скоростью всего лишь
1 м/с. Между тем для гигантских аксонов кальмаров и членисто-
ногих эта скорость в среднем приблизительно на два порядка
выше.
Характерной особенностью многих нейронов у позвоночных
животных является наличие так называемой миелиновой обо-
лочки, окружающей аксоны и некоторые длинные дендриты.
Обладая очень высоким электрическим сопротивлением, эта
оболочка выполняет функцию своеобразной изоляции наподобие
резиновой или пластиковой оплетки на проводах. Впрочем,
миелиновая оболочка далеко не сплошная. В ней периодически
встречаются небольшие разрывы, сквозь которые мембрана
контактирует с внеклеточным пространством. Именно здесь за-
мыкаются локальные электрические токи, образующиеся в про-
цессе проведения нервного импульса. В результате импульс
распространяется по мембране скачкообразно: он быстро пере-
скакивает от одного разрыва к другому, минуя участки, непо-
средственно окруженные миелиновой оболочкой. Такой меха-
низм проведения нервного импульса наиболее экономичен с
точки зрения расходования ионов и обеспечивает наивысшую
скорость распространения возбуждения (в некоторых случаях до
120 м/с).
Известно, что скорость химических реакций зависит от тем-
пературы. Это правило полностью применимо и к молекулярным
процессам, обеспечивающим формирование и проведение нерв-
ного импульса. Соответственно у теплокровных животных (птиц
и млекопитающих) импульс распространяется со значительно
большей скоростью, нежели в аналогичных нейронах у холодно-
кровных. Неудивительно, что нервная система млекопитающих
и птиц способна функционировать с очень высокой эффектив-
ностью и быстро обрабатывать огромные объемы информа-
ции.
70
2.8. Молекулярные механизмы
синаптической передачи нервного
импульса
Дойдя по мембране нейрона до области синапса, нервный
импульс может быть передан на следующую клетку. Самые общие
представления об этом процессе мы получили в гл. 1. Но теперь
необходимо существенно их детализировать. При этом мы огра-
ничимся только химическими синапсами как наиболее широко
распространенными и подробно изученными.
Напомним, что импульс передается через химический си-
напс с помощью молекул определенного нейромедиатора. Эти
молекулы содержатся в небольших синаптических пузырьках,
сосредоточенных в расширенной области пресинаптического
окончания.
Мембрана, окружающая каждый подобный пузырек, имеет в
своем составе особый белковый комплекс, чувствительный к из-
менению внутриклеточной концентрации ионов Са2+. Пока кон-
центрация кальция в цитоплазме низка (это значит, что нейрон
находится в состоянии покоя), такие белковые комплексы оста-
ются неактивными и не могут вступать во взаимодействие с на-
ружной мембраной. Однако при возбуждении нейрона ситуация
изменяется. Благодаря кратковременному открытию ионных ка-
налов цитоплазма обогащается кальцием из межклеточного про-
странства, а также из многочисленных полостей эндоплазмати-
ческой сети. Упомянутые белковые комплексы изменяют конфор-
мацию, активируются и приобретают к мембране высокое
сродство. В результате пузырьки сливаются с наружной мембра-
ной, а нейромедиатор «выбрасывается» в синаптическую щель.
Оказавшись в синаптической щели, молекулы нейромедиато-
ра диффундируют к противоположной (постсинаптической)
мембране и вступают в специфическое взаимодействие с ее ре-
цепторами. Дальнейшие события зависят от множества факто-
ров:
• от конкретного варианта используемого нейромедиатора;
• интенсивности его выделения в синаптическую щель;
• типа рецептора;
• специализации постсинаптической клетки;
• ее физиологического состояния и ряда других.
В частности, показано, что в некоторых холинэргических си-
напсах нейромедиатор способствует возбуждению постсинапти-
ческой клетки, а в некоторых, напротив, — производит тормозя-
щий эффект. Подробнее о синапсах, выполняющих функцию
торможения, вы сможете узнать из подразд. 2.9.
71
Отметим, что действие подавляющего большинства нейроме-
диаторов ограничено довольно коротким промежутком времени
(обычно всего лишь несколькими миллисекундами). Дело в том,
что молекулы нейромедиатора в синаптической щели, как прави-
ло, подвергаются мощной ферментативной деградации, а также
импортируются обратно в пресинаптическое окончание. Это
позволяет решить две важные задачи: во-первых, «очистить» си-
напс для следующего импульса, а во-вторых, экономно исполь-
зовать молекулы нейромедиатора с возможностью их повторного
выброса в синаптическую щель. Добавим, что активированные
молекулы рецепторных белков, в свою очередь, тоже подвергают-
ся быстрой деградации и вскоре заменяются новыми, еще не
активированными.
Хотя синаптические щели совсем невелики, суммарное вре-
мя, необходимое для диффузии молекул нейромедиаторов,
является одним из наиболее существенных факторов, ограни-
чивающих скорость распространения импульсов по нервной
системе. Таким образом, для быстрого срабатывания рефлек-
торной дуги число преодолеваемых синапсов должно быть ми-
нимальным. Именно поэтому нейроны обладают длинными
отростками: каждый отросток способен функционировать зна-
чительно быстрее, нежели сходная по длине цепочка из не-
скольких нейронов.
Каждый из перечисленных этапов функционирования
синапса (выброс молекул нейромедиатора в синаптическую
щель, их диффузия., деградация, импорт в пресинаптическое
окончание, взаимодействие с соответствующими рецеп-
торами и т.п.) обычно подвержен сложной многоуровневой
регуляции. Это явление обеспечивается различными молеку-
лярными механизмами, и в частности, — влиянием широкого
спектра нейрогормонов. Таким образом, в зависимости от воз-
раста, физиологического состояния и всей совокупности вос-
принятых ранее сигналов организм способен существенно по-
разному реагировать даже в ответ на абсолютно одинаковые
воздействия.
2.9. Молекулярные механизмы
синаптических взаимодействий
Наряду с передачей информации от нейрона к нейрону (и в
конечном итоге — на соответствующие исполнительные клетки)
многие синапсы выполняют еще одну важную задачу: осущест-
вляют необходимую обработку передаваемой информации. Эта
72
способность достигается разнообразными синаптическими взаимо-
действиями. Познакомимся с наиболее распространенными их
вариантами.
Начнем с того, что однократного срабатывания химического
синапса для возбуждения постсинаптической клетки, как прави-
ло, недостаточно (мембрана деполяризуется всего лишь на не-
сколько милливольт). Однако подобное изменение мембранного
потенциала обычно компенсируется не сразу, а с некоторым за-
позданием. И если синапс задействован с высокой частотой,
эффекты отдельных импульсов суммируются друг с другом, и в
результате потенциал достигает порогового уровня. Это явление
называют временной суммацией импульсов. Оно позволяет ре-
гулировать передачу возбуждения в зависимости от частоты ак-
тивации данного синапса (рис. 2.8, а).
Теперь обратимся к рисунку 2.8, б. На нем схематично пока-
зано несколько синапсов, независимо передающих информацию
на один и тот же нейрон. Каждый вызывает сравнительно не-
большую деполяризацию и потому самостоятельно не приводит
к возбуждающему эффекту. Но если нескольких синапсов сраба-
тывает синхронно, суммарное изменение потенциала может быть
таким, что рассматриваемый нами нейрон становится возбужден-
ным. В этом случае говорят о пространственной суммации им-
пульсов.
Существуют и еще более сложные механизмы взаимодействия.
Дело в том, что мембрана одного и того же нейрона зачастую со-
держит различные рецепторные белки. Некоторые из них, рас-
познав соответствующий нейромедиатор, обеспечивают частич-
ную деполяризацию постсинаптической мембраны и тем самым
увеличивают вероятность возбуждения клетки. В то же время
активация других рецепторных белков производит на клетку
Постсинаптическая мембрана
а б
Рис. 2.8. Основные типы синаптических взаимодействий:
а - временная суммация импульсов; б — пространственная суммация импульсов; в —
взаимодействие возбуждающих и тормозящих импульсов (первые показаны стрелкой,
вторые — знаком «1»)
73
диаметрально противоположный эффект: мембрана нейрона под-
вергается гиперполяризации, что существенно понижает вероят-
ность его возбуждения.
Соответственно в зависимости от типа используемых рецеп-
торов все синапсы принято подразделять на две большие группы:
к первой относят возбуждающие, ко второй — тормозные. При
этом один и тот же постсинаптический нейрон, как правило, со-
вместно иннервируется и теми, и другими (рис. 2.8, в). Их коли-
чество может исчисляться сотнями и даже тысячами. В результа-
те вероятность возбуждения постсинаптического нейрона опреде-
ляется тем, в каком конкретном соотношении задействованы
синапсы первого и второго рода.
Благодаря разнообразным вариантам синаптических взаимо-
действий организм способен «отсекать» несущественные сигналы,
координировать параллельно реализуемые биологические ответы,
выбирать оптимальные программы дальнейшего поведения, уста-
навливать их иерархию и др. (для примера достаточно вспомнить
ситуацию с рыболовом, рассмотренную нами в подразд. 22.4).
По-видимому, перечисленные механизмы играют важную роль и
во всем разнообразии проявлений человеческой психики.
СПИСОК ЛИТЕРАТУРЫ
1. Воронов Н.В., Климова Н.М., Менджерицкий А.М. Анатомия цен-
тральной нервной системы. — М. : Аспект Пресс, 2005.
2. Жуков Д. А. Биология поведения: гуморальные механизмы. — СПб. :
Речь, 2007.
3. Корочкин Л. И., Михайлов А. Т. Введение в нейрогенетику. — М. :
Наука, 2000.
4. Николлс Дж. Г., Мартин А. Р., Валлас Б. Дж., Фукс П.А.<Ут нейрона
к мозгу. - М.: УРСС, 2003.
5. Шулъговский В. В. Основы нейрофизиологии. — М. : Аспект Пресс,
2000.
6. Clapham D.E. TRP channels as cellular sensors // Nature. — 2003. —
V. 426. - P. 517-524.
7. Frings S. Chemoelectrical signal transduction in olfactory sensory neurons
of air-breathing vertebrates — Cellular & Molecular Life Sciences. — 2001. — V.
58. - P. 510-519.
8. Gillespie P.G. and Walker R.G. Molecular basis of mechanosensory
transduction — Nature. — 2001. — V. 413. — P. 194 — 202.
9. Hardie R. C. Phototransduction: Shedding light on translocation // Current
Biology. - 2003. - V. 13. - P. 775-777.
10. Jerison H.J. Evolution of the Brain and Intelligence. — New York:
Academic Press, 1973.
11. Lee Y., Lee C.-H. and Oh U. Painful Channels in Sensory Neurons //
Molecules & Cells. - 2005. - V. 20. - P. 315-324.
КОНТРОЛЬНЫЕ ВОПРОСЫ
1. Каковы молекулярные механизмы формирования и проведения
нервного импульса?
2. Опишите принципы функционирования химических и электри-
ческих синапсов.
3. Что такое вторичные мессенджеры? Опишите принципы их функ-
ционирования.
4. Охарактеризуйте разнообразие чувствительных нейронов.
5. Как функционирует классическая рефлекторная дуга?
Глава 3
ОСНОВНЫЕ МЕТОДЫ
СОВРЕМЕННОЙ ПСИХОГЕНЕТИКИ
3.1. Выявление элементарных
психологических характеристик
Любое исследование в области психогенетики предполагает
работу с конкретными психологическими характеристиками,
которые должны быть, по возможности, максимально простыми.
Это требование обусловлено несколькими причинами.
Первая связана с закономерностями молекулярной генетики.
Каждому гену соответствует конкретный белок, выполняющий в
организме строго определенные функции. В свою очередь, за счет
выполнения данных функций происходит формирование опреде-
ленного элементарного признака. Продемонстрируем это на
примере гена TYR. Известно, что он контролирует несколько
признаков, включая цвет глаз, цвет кожи и цвет волос. Однако
ни один из них не относится к элементарным, поскольку «слага-
ется» из множества более простых (интенсивности синтеза эуме-
ланина, интенсивности синтеза феумеланина, их распределения
по клеткам и т.д.). Элементарным же признаком в случае гена
TYR является способность к синтезу предшественников мелани-
нов. Подобные признаки нужно искать и в человеческой психике,
чтобы в дальнейшем исследовать их связь с конкретными гена-
ми.
Вторая причина имеет общенаучные корни. Человеческая
психика настолько сложна и многогранна, что пытаться познать
ее целиком практически невозможно. В подобных случаях при-
нято использовать универсальное правило: чтобы решить очень
сложную научную проблему, необходимо «разбить» ее на множе-
ство частей. Действительно, каждая из них значительно проще, а
потому существенно легче поддается исследованию. И чем более
дробным окажется это разбиение, тем глубже мы сможем «про-
никнуть» в изучаемую проблему. Этот подход называют научным
анализом. Однако, разбивая исходную проблему на части, мы
получаем лишь массу отрывочных, разрозненных сведений. По-
76
этому после детального и всестороннего анализа необходимо
приступить ко второму этапу работы: собрать все сведения в еди-
ную непротиворечивую картину. Данный подход называют на-
учным синтезом. Таким образом, поиск элементарных пси-
хологических характеристик представляет собой страте-
гически важную задачу, без решения которой последующий
синтез принципиально невозможен.
В течение первой половины XX в. психология уже занималась
подобным дроблением. Возникло целое направление — струк-
турализм. расчленявшее человеческую психику на множество
составляющих и именно в этом видевшее путь к познанию пси-
хики. Однако к заметным успехам оно не привело. На смену
структурализму пришли разнообразные концепции, напротив,
рассматривающие психику как нечто целое, и эти концепции
продолжают доминировать и теперь. Такая ситуация вовсе не
удивительна. На первых этапах развития каждой науки (а пси-
хология, по сути, — совсем молодая дисциплина, во многих
случаях еще не умеющая четко сформулировать, что именно
является предметом ее изучения) исходную проблему разбивают
довольно случайно. Соответственно, первые циклы анализа —
синтеза обычно приводят к одному и тому же результату: осо-
знанию, что исходное разбиение было сделано неверно. Но это
уже огромный шаг вперед! Постепенно, опираясь на накоплен-
ный опыт, приводящий к отрицательным результатам, ученые
приближаются к адекватным вариантам анализа, и рано или
поздно эти варианты будут найдены. Таким образом, поиск
элементарных психологических характеристик в современном
понимании смысла этой задачи продолжает представлять огром-
ную научную ценность. В дальнейшем мы увидим множество
конкретных примеров.
Психологическую характеристику можно считать элементар-
ной:
• если она достаточно стабильна, т.е. статистически воспро-
изводится в сходных условиях на протяжении всей жизни чело-
века или хотя бы в определенные возрастные периоды. При этом
особого внимания заслуживает тот факт, что подавляющее боль-
шинство психологических характеристик являются вероятност-
ными (например, глупый человек отличается от умного не тем,
что он не совершает умных поступков, а лишь с достоверно мень-
шей их частотой). Только в редких случаях характеристику оце-
нивают по одному единственному событию;
• относительно автономна от других психологических харак-
теристик;
• неделима на более простые составляющие. В этом плане
совершенно очевидно, что многие характеристики, широко ис-
77
Таблица 3.1. Примеры дробных характеристик темперамента
у грудных детей*
Характеристика Способ оценки
Активность Ребенок предоставлен сам себе. Оценивают число спонтанных движений, совершаемых им в течение определенного времени
Порог реактивности Оценивают минимальную интенсивность внешнего сигнала, способную вызвать у ребен- ка ответную реакцию (вздрагивание, поворот головы и др.)
Настойчивость Перед ребенком кладут любимую игрушку, но так, чтобы он не мог до нее дотянуться. Оце- нивают, как долго он будет пытаться достать ее самостоятельно (без плача и переключения внимания на что-либо другое)
Сосредоточенность Ребенку дают его любимую игрушку. Пока он играет, оценивают интенсивность внешнего сигнала, способную отвлечь внимание ребенка
Л юбопытство/насто- роженность Ребенку показывают не знакомый ему объект и оценивают первую реакцию: приближение или удаление
Настроение Средняя продолжительность здорового, сыто- го, сухого бодрствования без плача
* Ребенок должен быть здоровым, сытым, сухим и бодрствующим.
пользуемые в современных психологических исследованиях (ин-
теллект, креативность, экстраверсия, нейротизм и др.), никак не
могут претендовать на роль элементарных. Все они слишком
сложны и многокомпонентны, а значит, должны быть «раздро-
блены» на более простые.
В табл. 3.1 представлено несколько характеристик, более или
менее отвечающих перечисленным требованиям. Нетрудно за-
метить, что каждая из них несет достаточно существенную физио-
логическую окраску. Но это не является поводом для сомнений.
Ведь чем ближе мы подходим к пониманию конкретных процес-
сов, лежащих в основе определенной психологической характе-
ристики, тем отчетливей и ярче становится ее «физиологичность».
Поэтому в целях корректности следовало бы говорить об элемен-
тарных психофизиологических характеристиках.
78
3.2. Близнецовый анализ
Обнаружив некую элементарную психологическую характери-
стику, можно приступать к изучению ее генетического контроля.
Однако необходимо учитывать, что признаки организма обуслов-
лены не только генотипом, но и средой. Поэтому в первую оче-
редь нужно определить, насколько обнаруженная психиоло-
гическая характеристика зависит от генотипа, а насколь-
ко — от воздействий окружающей среды. Эту задачу успешно
решают с использованием близнецов.
Близнецы возникают у человека сравнительно редко. Дело в
том, что у женщины в каждом менструальном цикле созревает,
как правило, только по одной яйцеклетке. Впрочем, возможны и
некоторые сбои. Иногда (вероятно, в связи с гормональными на-
рушениями) происходит одновременное созревание нескольких
яйцеклеток. В результате на свет появляются дизиготные близ-
нецы, генетически различающиеся точно в такой же степени, как
и обычные родные братья или сестры (рис. 3.1, а). Кроме того, из
одной оплодотворенной яйцеклетки по каким-то еще не совсем
понятным причинам могут сформироваться несколько полноцен-
Оплодотворение
Зигота 1
Зигота 2
деление зиготы
Развитие зародышей
Дизиготные близнецы
(DZ)
Монозиготные близнецы
(MZ)
Рис. 3.1. Схема возникновения дизиготных (а) и монозиготных (6) близне-
цов
79
ных зародышей. Это приводит к рождению монозиготных близ-
нецов, идентичных друг другу своими генотипами (рис. 3.1, б).
Разработано несколько вариантов близнецового анализа. Са-
мый распространенный базируется на следующих соображениях.
Как правило, с момента своего появления на свет близнецы на-
ходятся примерно в одинаковых условиях: получают воспитание
от одних и тех же родителей, становятся свидетелями одних и тех
же событий, играют в одни и те же игры и др. Соответственно, у
монозиготных близнецов одинаковы не только генотипы, но и
среда. Дизиготные тоже находятся в одинаковой tpene, но отли-
чаются друг от друга своими генотипами. Таким образом, срав-
нивая моно- и дизиготных близнецов, можно выявить влияние
генотипа на исследуемый признак.
Таблица 3.2. Принцип расчета конкордантности на примере
чувства юмора
Номер пары* Близнецы** *** Тип пары Номер пары* Близнецы** Тип пары
1 2 1 2
1 +*♦* + К 9 + + К
2 + — Д 10 + — д
3 — — К 11 + + к
4 — + д 12 + + к
5 + + к 13 — — к
6 — 4- д 14 + — д
7 + — д 15 + + к
8 — — к
Общее число конкордантных пар = 9
Конкордантность (доля конкордантных пар) = 9/15 = 60 %
* Для получения достаточно надежных результатов количество как монози-
готных, так и дизиготных пар должно быть не менее 50.
** Одну таблицу заполняют для монозиготных, другую — для дизиготных
близнецов.
*** В таблице использованы следующие обозначения: «+» наличие чувства
юмора у конкретного близнеца; *—» отсутствие чувства юмора у конкретного
близнеца; «К* — конкордантная пара (пара близнецов, сходных поданному при-
знаку); «Д» — дискордантная пара (пара близнецов, различающихся поданному
признаку).
80
Чтобы результаты сравнения были убедительными, необходи-
ма достаточно большая выборка близнецов: не менее сотни пар,
а желательно и более. При этом следует обращать особое внима-
ние на точность диагностики каждой анализируемой пары. Раз-
нополые близнецы, несомненно, являются дизиготными, с одно-
полыми же ситуация значительно сложнее. Дело в том, что даже
очевидное внешнее сходство не может служить гарантией их
монозиготности. Поэтому точная диагностика однополых близ-
нецов возможна только с использованием молекулярных марке-
ров, таких как группа крови, резус-фактор, //L/1-антиген и целый
ряд других.
Разделив близнецов на моно- и дизиготные, приступают к не-
посредственному анализу данной характеристики. В том случае,
если она является дискретной, рассчитывают конкордантностъ
(табл. 3.2) отдельно для моно-, отдельно для дизиготных близне-
цов и сравнивают эти показатели друг с другом. В итоге возмож-
на одна из следующих ситуаций:
• конкордантность монозиготных близнецов (Кмз) достоверно
больше, чем дизиготных (Кдз). Это значит, что изучаемая психо-
логическая характеристика существенным образом зависит от
генотипа;
• Кмз не имеет достоверных отличий от Кдз. Из этого следует,
что роль генотипа весьма незначительна, т.е. характеристика за-
висит в основном от среды;
• Кмз достоверно меньше, чем Кдз. Такой результат противо-
речит здравому смыслу и свидетельствует о грубых ошибках в
проведенной работе.
Если же изучаемая характеристика является количественной,
высчитать ее кон корда нтность не представляется возможным.
В этом случае сравнивают коэффициенты внутрипарной кор-
реляции (гмз и гдз, соответственно; формула расчета данных
коэффициентов приведена в приложении 1).
Чтобы полученные результаты были более наглядными, можно
рассчитать коэффициент наследуемости характеристики, т.е.
степень ее зависимости от генотипа:
А2=^мг2да или А2 = *мз-ЛДз
1-гдз 100-/ГДз
где Кмз и /^дз выражены в процентах. Значения этого коэффици-
ента для некоторых признаков приведены в табл. 3.3.
Долгое время было принято считать, что близнецовый анализ
является важнейшим методом психогенетики. Однако такие пред-
ставления явно устарели. Очевидно, что сам по себе близнецовый
анализ не способен ответить на главные вопросы: какие гены
81
Таблица 3.3. Вклад генотипа в формирование некоторых
психологических признаков (по Loehlin, 1992; Равич-Шербо и др.,
1999; с дополнениями)
Психологическая характеристика Вклад генотипа* Психологическая характеристика Вклад генотипа*
IQ у детей** 0,2 Поиск новизны 0,45
IQ у подростков** 0,5 Добросовестность 0,38
1Q у взрослых** 0,81 Откровенность 0,45
Общий фактор ин- теллекта g 0,52 Способность к со- гласию 0,35
Математические способности 0,38 Радикализм 0,65
Пространственные представления 0,46 Консервативность мышления 0,54
Освоение навыков чтения вслух 0,7 Авторитарность 0,62
Беглость речи 0,3 Раздражител ьность 0,37
Дивергентное мыш- ление 0,22 Жестокость у муж- чин 0,55
Вербальная понят- ливость 0,5 Агрессивное анти- общественное по- ведение у детей 0,66
Логичность 0,48 Склонность к тяж- ким преступлениям 0,5
Точность 0,46 Предрасположен- ность к самоубий- ству 0,35
Успешность в освоении языков 0,46 Склонность к об- жорству 0,45
Успешность в осво- ении гуманитарных дисциплин 0,48 Склонность к опи- атной зависимости 0,25
Успешность в осво- ении естественных дисциплин 0,3 Предрасположен- ность к алкоголизму 0,55
82
Окончание табл. 3.3
Психологическая характеристика Вклад генотипа* Психологическая характеристика Вклад генотипа*
Внимание 0,75 Предрасположен- ность к курению у женщин 0,45
Экстраверсия 0,39 Предрасположен- ность к курению у мужчин 0,6
Нейротизм 0,31 Интенсивность курения 0,75
Альтруизм 0,35 Гомосексуализм 0,4
Импульсивность 0,38 Аутизм 0.61
* Усредненные данные по многим независимым исследованиям.
* * Возрастные изменения вклада генотипа и среды характерны для очень мно-
гих психологических признаков. В частности, интеллектуальная деятельность ре-
бенка в основном зависит от факторов внешней среды: внимания к нему, интен-
сивности речевого общения, использования развивающих методик и др. Однако
по мере того, как ребенок взрослеет, все более значимую роль играет генотип.
И в конечном итоге интеллект взрослого человека главным образом определяется
именно наследственными задатками, а воспитание и образование играют второ-
степенную роль.
контролируют данную характеристику, сколько их и что именно
они кодируют. Таким образом, в современных психогенетических
исследованиях близнецовый анализ рассматривают как предва-
рительный этап, позволяющий выявить наиболее перспективные
характеристики (те, что находятся под существенным генетиче-
ским контролем). Однако для непосредственного изучения этого
контроля нужен совершенно иной методический арсенал. В част-
ности, различные молекулярно-генетические подходы.
3.3. Молекулярно-генетические методы
Методы молекулярной генетики настолько разнообразны и
вместе с тем настолько технически сложны, что требуют высоко-
го уровня специальной подготовки. Поэтому даже поверхностное
их описание никак не входит в задачи данного учебника. Отметим
лишь, что молекулярно-генетические методы позволяют
83
найти в ДНК конкретный ген, определить его точную ну-
клеотидную последовательность, выяснить функции коди-
руемого белка и многое другое. В результате открываются
поистине огромные возможности.
Для примера представим, что некая исследовательская груп-
па занимается всесторонним изучением музыкального слуха.
С помощью близнецового анализа она продемонстрировала вы-
сокую зависимость этой характеристики от генотипа и теперь
приступает к поиску соответствующих генов. Такая задача может
быть решена различными путями. Мы остановимся только на
одном.
Сначала необходимо сформировать две контрастные выборки:
одну — из людей с абсолютным музыкальным слухом, другую —
из тех, кому «медведь на ухо наступил». При этом желательно,
чтобы все остальные факторы (возраст, образование, социальное
положение и др.) были максимально выровнены в обеих выборках.
У каждого испытуемого берут образец ДНК и подвергают его
детальному молекулярно-генетическому анализу. К сожалению,
сразу по всей совокупности генов, а их у человека приблизитель-
но 25 тыс., провести подобный анализ весьма затруднительно.
Намного реальнее работать с конкретными генами. Но какие же
именно выбрать для этого анализа?
Практически все гены человека уже идентифицированы. Для
многих выяснены функции соответствующих белков, а для не-
которых — даже особенности дифференциальной экспрессии.
В частности, обнаружено несколько тысяч генов, экспрессиру-
ющихся преимущественно в нейронах головного мозга. Вполне
очевидно, что именно такие гены относятся к числу наиболее
вероятных кандидатов на участие в генетическом контроле музы-
кального слуха. Сделать конкретный выбор из этих генов можно
на основании неких дополнительных соображений, а при их от-
сутствии — более или менее случайно.
Допустим, что выбор исследователей пал на ген Z, в котором
известно 250 полиморфных сайтов. Анализировать каждый из них
весьма трудоемко. Для удобства выбирают один или несколько
сайтов, затрагивающих наиболее важные участки гена, и проводят
анализ только по этим сайтам. Выясняют, какие именно их алле-
ли присутствуют в генотипе каждого конкретного испытуемого.
Затем подсчитывают, какое соотношение разных аллелей харак-
терно для первой выборки, какое — для второй, и статистически
сравнивают друг с другом полученные соотношения. Если раз-
личия окажутся недостоверными, значит, ген Z не играет суще-
ственной роли в процессах, ведущих к формированию музыкаль-
ного слуха. Если же соотношения будут достоверно различаться,
значит, ген Z принимает заметное участие в генетическом кон-
84
Выборка 1 Выборка 2
(люди с абсолютным слухом) («медведь на ухо наступил»)
Испытуемый 1 А ' Испытуемый 1 1 1 1 1 I 1 99 1 1 1 1 J
Испытуемый 2 А А Испытуемый 2 1 1 1 1 1 1 1 1 1 1 1 1
Испытуемый 3 G Испытуемый 3 1 1 1 1 1 1 =7 9 1 1 1 1
Испытуемый 4 G Испытуемый 4 — — -q--_
А ► PiA: p|G — — — к PjA : P2g
Испытуемый 5 А Испытуемый 5 1 1 1 1 1 1 9? 1 1 1 1
Испытуемый 6 А А Испытуемый 6 1 1 1 1 1 1 г? 1 1 1 1
Испытуемый 7 А А Испытуемый 7 1 1 1 1 1 1 9=7 1 1 1 1 к
Рис. 3.2. Схема поиска генетических ассоциаций между признаком «музы-
кальный слух» и неким геном Z
Примечание. Предположим, <<70 группа исследователей пытается выяснить,
влияет ли некий ген Z на музыкальный слух. Для анализа выбран один полиморфный
сайт, затрагивающий функционально важный участок гена и представляющий собой
нуклеотидную замену А/G (напомним, что нуклеотидную последовательность гена
принято записывать по некодирующей цепи ДНК, что показано на рис. 1.10). По
данному полиморфному сайту возможны три генотипа: АА (гомозиготы по аллели >4),
GG (гомозиготы по аллели (7), а также AG или GA (гетерозиготы). Установив генотип
каждого конкретного испытуемого, рассчитывают частоты аллелей А и G и сравни-
вают их соотношение в анализируемых выборках. р>А и P/G — частоты аллелей А и
G в выборке I, р2А и р/7 — частоты аллелей А и G в выборке 2. Прочие пояснения —
в тексте.
троле рассматриваемой нами характеристики. Этот подход на-
зывают поиском генетических ассоциаций (рис. 3.2).
Предположим, что исследовательской группе очень повезло, и
«нужный» ген был выявлен с первой же попытки. Однако это не
означает завершения работы. В самом деле, наивно считать, что
музыкальный слух находится под контролем одного единствен-
ного гена. Поэтому приведенную выше экспериментальную схему
придется повторять и с другими генами-кандидатами. Безусловно,
такая работа весьма трудоемка, но зато ученые будут постепенно
приближаться к выявлению всей совокупности искомых генов.
Подобная схема исследования может быть использована по
отношению к любой психологической характеристике человека.
Причем это только самые первые этапы. В дальнейшем для каж-
дого гена потребуется выяснить конкретные молекулярные функ-
ции соответствующего белка, степень их нарушения в результате
85
различных мутаций, влияние внешних факторов на рассматри-
ваемые функции, изменение экспрессии с возрастом и др. Более
того, необходимо детально исследовать все многообразие взаимо-
действий между этими генами. И в результате шаг за шагом станут
прорисовываться молекулярные механизмы изучаемой психоло-
гической характеристики.
3.4. Мета-анализ
Любое молекулярно-генетическое исследование на человеке
охватывает сравнительно небольшое количество испытуемых. Как
правило, несколько десятков, в лучшем случае — сотен. Это свя-
зано, во-первых, со сложностью молекулярных методик, во-
вторых, с жесткими критериями отбора испытуемых и, в-третьих,
с ограниченными возможностями самих экспериментаторов.
Между тем при использовании выборок небольшого объема
очень остро встает проблема разнообразных случайностей 9
способных существенно отразиться на результатах. Неуди-
вительно, что выводы, полученные разными авторами при вы-
полнении, казалось бы, практически одинаковых исследований,
нередко бывают диаметрально противоположными.
В подобных ситуациях возникает закономерный вопрос: какие
из полученных результатов соответствуют действительности? Эту
задачу решают с помощью мета-анализа. Сначала выявляют все
публикации поданной теме. Затем критически анализируют каж-
дую из них с точки зрения грамотности подходов к отбору ис-
пытуемых, адекватности использованных методов и др. «Отсеи-
вают» все работы, вызывающие сомнение. В итоге остаются пу-
бликации, каждая из которых строго соответствует всем
предъявленным требованиям и, по-видимому, содержит заслужи-
вающие доверия результаты.
После этого тщательно сравнивают оставшиеся работы на
предмет независимости использованного в них материала. Если
какие-то исследования дублируют друг друга (выполнены на одних
и тех же выборках испытуемых), их тоже отсеивают, оставляя лишь
наиболее обширное. В конечном итоге из всей совокупности рас-
сматриваемых публикаций отбирают лишь самые качественные
и неперекрывающиеся.
Затем объединяют данные всех этих работ. Суммарные вы-
борки во много раз больше по сравнению с исходными и обычно
насчитывают по несколько тысяч испытуемых, а в некоторых
случаях — даже десятки тысяч. На этом уровне различные случай-
ности уже не столь существенны. Поэтому статистическая обра-
86
ботка объединенных выборок, как правило, дает достаточно на-
дежные выводы.
Итак, единичное молекулярно-генетическое исследование
(а тем более выполненное на выборках небольшого объема)
само по себе не представляет значительного интересакакие
бы именно результаты не были получены. Необходимо как
минимум несколько независимых подтверждений, а в случае
противоречивых данных — мета-анализ.
Проиллюстрируем эту необходимость конкретным примером.
К настоящему времени выполнены сотни работ по поиску генов,
контролирующих отдельные свойства темперамента. Пока к ощу-
тимому успеху они не привели. Наиболее обнадеживающие дан-
ные были получены в отношении признака «поиск новизны».
Десятки независимых исследований свидетельствуют о том, что
этот признак контролируется геном DRD4 (подробнее о нем см.
подразд. 5.6). Казалось бы, можно говорить о доказанном факте.
Однако полученные результаты крайне неоднородны. В разных
работах одна и та же аллель «ведет себя» по-разному: в некоторых
исследованиях ассоциирует с низкими показателями, а в некото-
рых, напротив, с высокими показателями признака. Наконец, в
десятках работ ассоциация не выявлена. Три независимо выпол-
ненных мета-анализа тоже не смогли обнаружить наличие ассо-
циации. Таким образом, следует признать, что ген DRD4 не
оказывает существенного влияния на поиск новизны. Вероятно,
основная проблема заключается в том, что данный признак слиш-
ком далек от элементарного и потому для генетического анализа
малопригоден.
3.5. Использование животных в качестве
модельных объектов
Казалось бы, главным объектом психогенетических исследо-
ваний должен являться не кто-либо, а сам человек. Однако такое
впечатление довольно обманчиво. Дело в том, что любые экспе-
рименты, проводимые на человеке, неизбежно сопряжены с се-
рьезными методическими ограничениями. Невозможно целена-
правленно конструировать необходимые генотипы, получать
различных мутантов по интересующим нас признакам, изменять
особенности дифференциальной экспрессии генов в клетках го-
ловного мозга и т. п. Таким образом, человек неудобен как объект
для исследований.
Несравненно более широкие методические возможности от-
крываются в случае использования лабораторных животных.
87
Именно они на протяжении последних десятилетий стали основ-
ными модельными объектами психогенетики. Однако насколь-
ко адекватен подобный подход? Правомочно ли с помощью ре-
зультатов, полученных на животных, пытаться объяснить меха-
низмы человеческой психики?
Судя по имеющимся данным, вполне правомочно. Сегодня уже
не вызывает сомнения тот факт, что в основе функционирования
нервной системы человека и любого другого сложного многокле-
точного животного, будь то улитка, муха, мышь или шимпанзе,
лежат одни и те же молекулярные механизмы. Различия сводятся
только к определенным нюансам. Это позволяет переносить по-
лученные результаты с модельных объектов на человека и наобо-
рот. Например, если у какого-то модельного объекта выявлен ген,
контролирующий уровень спонтанной агрессии, весьма вероятно,
что родственный ген имеется и у человека. Более того, предпо-
лагаемый человеческий ген тоже должен быть вовлечен в контроль
агрессивности. В дальнейшем мы познакомимся с множеством
подобных примеров.
Излюбленными модельными объектами современной психо-
генетики служат небольшие, удобные в разведении животные,
а именно крысы, мыши и мухи-дрозофилы. При этом использу-
ют не взятые из природы экземпляры, а специально созданные
инбредные линии. Так называют группы организмов, полученные
путем близкородственного скрещивания на протяжении как ми-
нимум нескольких десятков поколений. В результате все пред-
ставители одной и той линии имеют строго определенный гено-
тип. Напротив, у разных линий генотип различен.
Обычно исследование проводят по следующей схеме. Выбрав
для анализа несколько инбредных линий, сравнивают их по
изучаемому поведенческому признаку (разумеется, сравнение
проводят на одновозрастных организмах в максимально выров-
ненных условиях среды). Выявив линии, четко отличающиеся
друг от друга, скрещивают их и анализируют полученное по-
томство в первом и втором поколениях гибридов. Особенно
пристального внимания заслуживают скрещивания, в которых
первое поколение гибридов фенотипически однородно, а во
втором наблюдается расщепление 3: /. Этот факт свидетельству-
ет о том, что обнаруженное различие контролируется всего одним
единственным геном (является моногенным\ рис. 3.3). В даль-
нейшем с помощью молекулярно-генетических методов при-
ступают к непосредственному поиску данного гена, выяснению
особенностей его дифференциальной экспрессии, изучению
функций соответствующего белка и др. Наконец, полученные
результаты переносят на человека и проверяют всем арсеналом
доступных методов.
88
Нормальные
мыши
Гибриды
2-го поколения
4^1
Я^э rFs? 7^?
Нормальные Нормальные
мыши мыши
Нормальные Вальсирующие
мыши мыши
Суммарное
соотношение
3 нормальные
(доминантные)
1 вазьсирующая
(рецессивная)
J
Рис. 3.3. Пример моноген ною наследования поведенческого признака
Примечание Hci норне линии мышей (и к называют «вальсирующими*) от-
шчаются от нормы быстрым движением по кругу. Ьще в начале XX в. было показано,
что эта особенность поведения наследуется монотонно в соответствии с первым и
вторым законами Менделя (первый закон — единообразие 1ибриюв Fh второй — рас-
щепление 3 доминантных : I рецессивный в Р2). И действительно, впоследствии вы-
яснилось, что вальсирующие мыши являются мутантами по гену Cdh23. Мутантная
аллель отмечена звездочкой.
Существуют и более эффективные подходы. В частности, благо-
даря достижениям генной инженерии стало возможным получать
мышей и крыс, у которых целенаправленно разрушен конкретный
ген. Работая с такимиюрганизмами, нетрудно установить, за что же
именно «отвечает» ссчлгветствующии белок. Этот подход называют
нокаутированием генов. В перспективе он позволит выявить все
гены, контролирующие высшую нервную деятельность у крыс и
мышей. Очевидно, что подобные сведения будут очень ценны для
понимания молекулярных механизмов человеческой психики.
Итак. основными моде /ьными объекта ъ и психогенетики
являются крысы, мыши и дрозофилы. Результаты, получен-
ные на > мих модельных объектах, могут быть успешно
перенесены и на человека (конечно, с учетом определенных
корректив, отражающих специфику его молекулярной орга-
JB9
низации). Однако чем ближе модельный объект к человеку, тем
менее существенными оказываются такие коррективы. Поэтому
в последнее время в психогенетических исследованиях все чаще
используют различные виды обезьян. Это самые перспективные
объекты психогенетики.
СПИСОК ЛИТЕРАТУРЫ
1. Александров А. А. Психогенетика. — СПб. : Питер, 2004.
2. Баранов В. С., Баранова Е.В., Иващенко Т.Э. Геном человека как
научная основа предиктивной медицины : в кн.: В. И. Иванов, Л.Л. Киселев
Геномика — медицине. — М. : Академкнига, 2005. — С. 361 —379.
3. Голимбет В. Е. Геномика в психологии и психиатрии // Молекулярная
биология. - 2004. - Т. 38. - № I. - С. 165- 170.
4. Канаев И. И. Близнецы и генетика. — Л. : Наука, 1968.
5. Малых С. Б., Егорова М.С.. Мешкова Т.А. Психогенетика. T.I. —
СПб. : Питер, 2008.
6. Равич-Щербо И. В., Марютина Т.М., Григоренко E.JJ. Психогенети-
ка. — М. : Аспект-Пресс, 2003.
7. Тихомирова М.М. Генетический анализ. — Л. : Изд-во ЛГУ, 1990.
8. Boomsma D., Busjahn A. and Peltonen L. Classical twin studies and beyond.
Nature Reviews. Genetics. — 2002. — V. 3. — P. 872 — 882.
9. de Bakker P. I. W. et al. Efficiency and power in genetic association studies.
Nature Genetics. - 2005. - V. 37. - P. 1217- 1223.
10. Khusnutdinoua E. K. et al. Use of DNA fingerprints for human population
genetic studies. Molecular and General Genetics. — 1995. — V. 247. — P. 488—493.
II. Norman S.-L.T. Tutorial in biostatistics. Meta-analysis: formulating,
evaluating, combining, and reporting. Statistics in Medicine. — 1999. — V. 18. — P.
321-359.
12. Schinka J. A., Letsch E.A. and Crawford F. C. DRD4 and novelty seeking:
results of meta-analyses. Am J Med Genet. — 2002. — V. 114. — P. 643 — 648.
13. Sutton A. J. et al. Methods for Meta-analysis in Medical Research. London:
John Wiley, 2000.
КОНТРОЛЬНЫЕ ВОПРОСЫ
1. В чем состоит смысл поиска элементарных психологических при-
знаков?
2. Как проводят близнецовый анализ в психогенетике?
3. В чем заключается поиск генетических ассоциаций?
4. Какую роль в психогенетике играет мета-анализ?
5. Для чего в современной психогенетике используют модельные жи-
вотные объекты?
Глава 4
ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ ОБУЧАЕМОСТИ
И ПАМЯТИ
4.1. Выявление мутантов по обучаемости
и памяти у дрозофилы
Одной из основных задач современной психогенетики являет-
ся изучение молекулярно-генетических механизмов, непосред-
ственно связанных с формированием когнитивных способно-
стей.
Некоторые из этих способностей (обучаемость и память)
характерны для всех без исключения позвоночных животных и
даже для многих беспозвоночных, например насекомых. Поэтому
в генетических исследованиях обучаемости и памяти нередко ис-
пользуют различные виды беспозвоночных, и в первую очередь —
плодовую муху-дрозофилу.
Существует большое разнообразие экспериментальных
подходов, позволяющих оценивать обучаемость и память
дрозофилы. Однако их базовый принцип, как правило, сходен',
у мух вырабатывают определенный условный рефлекс и
отслеживают^ с какой эффективностью он формируется
и насколько стабильно сохраняется с течением времени.
С этой целью обычно используют рефлекс избегания, а в каче-
стве условных сигналов — различные запахи. Данный подход
называют обонятельным ассоциативным обучением. Для при-
мера познакомимся с одним из его вариантов, наиболее часто
встречающихся в современных исследованиях:
I) в специальную небольшую камеру с электропроводящими
стенками помещают около 100 одновозрастных дрозофил, при-
надлежащих одной и той же инбредной линии, а значит, имеющих
строго одинаковый генотип;
2) в течение I мин на них воздействуют определенным запахом
(запах № I), одновременно подвергая серии электрических раз-
рядов;
3) продувают камеру чистым воздухом, и мухи отдыхают в
течение нескольких минут;
91
4) после этого на дрозофил воздействуют запахом № 2. Данное
воздействие тоже продолжается 1 мин, однако не сопровождается
электрическими разрядами;
5) опять продувают камеру чистым воздухом и вновь позволя-
ют мухам отдохнуть;
6) несколько раз повторяют весь цикл обучения. В результате
у мух вырабатывается условный рефлекс, обеспечивающий рас-
познавание «опасного» и «безопасного» запахов;
7) приступают к проверке результатов обучения. Для этого
переносят дрозофил в новую камеру, в одном из ответвлений
которой присутствует запах № 1, а в противоположном — запах
№ 2. Подсчитывают, сколько прошедших обучение мух выбирает
потенциально опасное ответвление камеры (обозначим это число
как Л|), сколько — безопасное ответвление (л2), и на их основе
вычисляют индекс обучаемости:
Д=^—\
N
где — общее число используемых дрозофил. Необходимость
такого расчета обусловлена тем, что в безопасном ответвлении
камеры могут находиться не только обученные мухи, но и необу-
чившиеся (выбравшие данное ответвление абсолютно случайно).
К сожалению, точное их число определить невозможно, и его
принимают приблизительно равным
Рис. 4.1. Динамика угасания памяти у дрозофилы при разных методах обо-
нятельного обучения:
/ — между циклами обучения были 15-минугные интервалы. Возникающая при этом
память способна сохраняться в течение 7 суток и даже дальше; 2 — циклы обучения
следовали непосредственно друг за другом. Возникающая при этом память значительно
слабее и полностью угасает примерно на четвертые сутки
Примечание. В качестве эталона нормы у дрозофилы принято использовать
инбредную линию Canton-S. После десяти циклов обонятельного обучения эта линия
демонстрирует высокий индекс обучаемости (Л - 90%). Однако в зависимости от
нюансов используемой методики динамика угасания памяти сильно варьирует.
92
8) в дальнейшем через фиксированные промежутки времени
проводят повторные измерения индекса обучаемости и тем самым
отслеживают динамику угасания памяти.
С помощью таких относительно несложных подходов можно
анализировать десятки инбредных линий, сравнивая их друг с
другом по когнитивным способностям. При этом необходимо
учитывать следующее обстоятельство. В зависимости от выбора
конкретной методики обучения, количества циклов, интервалов
между ними и т.п. показатели обучаемости и памяти могут силь-
но варьировать даже у одной и той же инбредной линии (рис. 4.1).
Таким образом, чтобы сравнение было адекватным и позволило
отличить мутантные линии от нормальных, нужно строго при-
держиваться определенного экспериментального подхода.
4.2. Разнообразие мутантов по обучаемости
и памяти у дрозофилы
В общей сложности к настоящему времени у дрозофилы вы-
явлено несколько десятков интересующих нас мутантов. У многих
одновременно нарушены и память, и обучаемость, но у некото-
рых — только одна из перечисленных характеристик. Именно на
таких мутантах мы и остановимся.
У мутантов по генам latheo и linotte значение Л сразу по окон-
чании процесса обучения значительно ниже по сравнению с
нормальными мухами, но динамика угасания памяти практически
неизменна (рис. 4.2, а). Не нарушено у них и восприятие исполь-
зуемых сигналов. Эти факты свидетельствуют о том, что пере-
численные гены в основном контролируют не процесс хранения
информации, а интенсивность ее исходного «фиксирования в
организме» (в данном случае именно это понимают под термином
«обучаемость»).
Теперь обратимся к мутантам с нарушенной памятью. Среди
них наблюдается весьма существенное разнообразие. Так, для
мутантов по генам dunce и rutabaga характерно очень резкое
угасание памяти в первые же 30 мин по окончании обучения
(рис. 4.2, б). Между тем у некоторых мутантов по гену СгеЬ2
нарушение памяти становится заметным не сразу, а только по
прошествии нескольких часов (рис. 4.2, в). Судя по этим данным,
ген СгеЬ2 «отвечает» у дрозофилы непосредственно за долго-
временную память, а гены dunce и rutabaga — за кратковре-
менную.
Как выяснилось, мухам присуща и средневременная память,
контролируемая генами amnesiac и DC0. Таким образом, процесс
93
Рис. 4.2. Результаты обонятельного обучения (10 циклов с 15-минутными
интервалами) некоторых мутантов дрозофилы по сравнению с нормой:
а — мутант по обучаемости (динамика угасания памяти соответствует норме, но
исходное значение Л заметно снижено); б — мутанты по кратковременной памяти.
В течение 30 мин по окончании обучения память угасает значительно быстрее, чем в
норме. Затем наблюдается небольшое увеличение Л — организм пытается сформиро-
вать средневременную память, но эта попытка оказывается малоуспешной); в — му-
тант по долговременной памяти (в течение нескольких часов по окончании обучения
память угасает практически так же, как в норме. Затем нарушение памяти становится
очевидным. Через четверо суток память утрачивается полностью)
формирования памяти у дрозофилы можно представить в виде
трех последовательных этапов. На первом этапе формируется
кратковременная память, на втором (приблизительно через
30 мин) — средневременная и на третьем (примерно через 8 ч) —
долговременная, способная сохраняться неделю и даже больше.
Кратковременная и средневременная память весьма нестабиль-
ны: они легко разрушаются различными анестезирующими воз-
действиями, например, при использовании непродолжительного
холодового шока. В этом случае процесс не доходит до третьего
этапа, и как следствие долговременная память уже не вырабаты-
вается. Но если она сформировалась еще до воздействия, анесте-
зия не оказывает на нее никакого эффекта.
94
Для того чтобы выработать у дрозофилы долговременную па-
мять, необходимо соблюсти и некоторые дополнительные условия.
В частности, требуются многочисленные циклы обучения с до-
статочно длительными периодами отдыха между ними: как пра-
вило, 10 циклов с 15-минутными интервалами. Если же циклы
следуют непосредственно друг за другом, формируется консоли-
дированная, но менее устойчивая память. Она не является ослаб-
ленным вариантом долговременной, поскольку практически не
зависит от гена СгеЬ2 и не может быть заблокирована с помощью
анестезирующих воздействий. Эта память (ее называют устой-
чивой к анестезии) полностью отсутствует у мутантов по гену
radish. Остальные виды памяти у данных мутантов не затронуты.
Таким образом, судя по результатам генетического ана-
лиза , для дрозофилы характерно четыре вида памяти: крат-
ковременная, средневременная, долговременная и устойчивая
к анестезии. Первые три вида памяти формируются по-
следовательно, а четвертый — относительно независимо
от них (рис. 4.3).
Консолидация
памяти
б
Рис. 4.3. Схема формирования различных видов памяти у дрозофилы:
° — предполагаемые пути формирования различных видов памяти; б — современные
представления о динамике формирования и угасания различных видов памяти: О —
общая динамика угасания памяти, К — кратковременная память, С — средне временная
память. Д — долговременная память, У — память, устойчивая к анестезии
95
4.3. Функции генов, контролирующих
обучаемость и память у дрозофилы
Как известно, современные методы молекулярной генетики
позволяют найти в ДНК определенный ген, выяснить особен-
ности его дифференциальной экспрессии и даже конкретные
функции кодируемого белка. В частности, эти задачи успешно
решены и для целого ряда интересующих нас генов.
Некоторые из них (например, latheo и linotte) в основном экс-
прессируются на личиночной стадии развития, обеспечивая про-
цесс формирования нервной системы. Соответственно у дрозо-
фил, мутантных по этим генам, наблюдается частичная недораз-
витость «головного мозга», что, по-видимому, и приводит к
заметному снижению обучаемости.
Гены dunce, rutabaga, DCOh Creb2 экспрессируются преиму-
щественно не в личинке, а во взрослой дрозофиле и не оказыва-
ют существенного влияния на развитие организма. Это значит,
что они контролируют какие-то процессы, непосредственно свя-
занные с обработкой и хранением информации. Для начала по-
знакомимся с функциями гена rutabaga. Его «продуктом» явля-
ется фермент аденилатциклаза. Как вы помните, мы уже встре-
чались с этим ферментом: он обеспечивает синтез циклического
аденозин монофосфата — одного из наиболее важных вторичных
мессенджеров (см. подразд. 2.6, рис. 2.7). Таким образом, мухи,
мутантные по гену rutabaga, отличаются от нормальных пони-
женным содержанием цАМФ.
Ген dunce тоже принимает непосредственное участие в кон-
троле метаболизма циклического аденозинмонофосфата, однако
играет диаметрально противоположную роль. Этот ген кодирует
одну из разновидностей фосфодиэстераз — ферментов, которые
в отличие от аденилатциклазы не синтезируют цАМФ, а, наобо-
рот, его разрушают. В результате у мух, мутантных по данному
гену, содержание цАМФ значительно выше нормы.
Какое же отношение имеет цАМФ к обучаемости и памяти?
На этот вопрос мы попробуем ответить в следующих параграфах,
а пока ограничимся лишь самыми общими соображениями. Как
известно, вторичные мессенджеры (и в частности, цАМФ) явля-
ются промежуточными звеньями в молекулярных каскадах, ис-
пользуемых для передачи информации о поступивших сигналах.
Поэтому нет ничего удивительного в том, что недостаток или из-
быток данного химического соединения существенно отражается
на когнитивных способностях организма.
Теперь обратимся к гену DCO. Он кодирует каталитическую
субъединицу протеинкиназы А. Этот фермент участвует в том
96
же каскаде, что и цАМФ. Более того, как видно на рис. 2.7 (см.
подразд. 2.6) именно цАМФ регулирует активность данной про-
теинкиназы. Таким образом, ген DCO контролируют следующее
после цАМФ молекулярное звено в механизме передачи инфор-
мации о воспринятом сигнале.
Продуктом гена СгеЬ2 является белок, который, хотя и не по-
казан на рис. 2.7, тоже участвует в рассматриваемом сигнальном
каскаде. Этот белок относится к числу транскрипционных фак-
торов. В обычных условиях он остается практически неактивным,
однако, будучи фосфорилированным протеинкиназой А, вклю-
чается в регуляцию экспрессии целого ряда генов и тем самым
запускает дальнейшие этапы каскада.
Итак, в контроле обучаемости и памяти у дрозофилы при-
нимают участие как минимум две группы генов*, первая
«отвечает* за развитие нервной системы, а вторая — за
передачу воспринятой нейронами информации.
4.4. Локализация молекулярных процессов,
обеспечивающих обонятельную
обучаемость и память у дрозофилы
Все перечисленные ранее мутанты дрозофилы (а также и мно-
гие другие) были выявлены в тестах с использованием обонятель-
ного обучения. В этих тестах на мух воздействовали определен-
ными запахами в комбинации с достаточно сильными электри-
ческими разрядами и отслеживали формирование условного
рефлекса избегания. Способность дрозофилы вырабатывать по-
добные рефлексы предполагает конвергенцию соответствующих
нейронных путей. Давайте же кратко познакомимся с этими пу-
тями и попробуем выяснить, где именно они конвергируют. Нач-
нем с путей передачи информации о запахах.
Органами обоняния дрозофилы являются антенны. На их по-
верхности расположены многочисленные чувствительные нейро-
ны, несущие на мембранах специальные обонятельные рецепто-
ры. Существует около 60 вариантов подобных рецепторов, и
каждый из них кодируется отдельным геном. При этом в каждом
конкретном обонятельном нейроне экспрессируется, как прави-
ло, только один из соответствующих генов, а значит, присутству-
ет только один вариант рецепторов.
Принцип распознавания запахов достаточно сложен. Дело в
том, что одно и то же пахучее соединение обеспечивает активацию
нескольких вариантов рецепторов. В свою очередь, один и тот же
обонятельный рецептор способен взаимодействовать с разными
97
пахучими соединениями, имеющими друг с другом значительное
структурное сходство. Тем не менее даже для очень похожих за-
пахов совокупности активируемых рецепторов не совпадают. По-
видимому, именно это позволяет дрозофиле тонко различать
огромное разнообразие запахов.
Импульсы, возникающие в возбужденных обонятельных ней-
ронах, передаются в антеннальные доли головного мозга. Каждая
состоит из 43 «клубочков» (гломерул). При этом строго соблюда-
ется следующая закономерность: нейроны, несущие идентичные
обонятельные рецепторы, контактируют не со всеми, а только с
определенными гломерулами (зачастую с одной единственной в
каждой из долей). Таким образом, в зависимости от действующе-
го на дрозофилу запаха активируются разные участки головного
мозга. Этот принцип называют пространственным кодировани-
ем запахов.
В дальнейшем полученная дрозофилой обонятельная инфор-
мация поступает в высшие отделы головного мозга, и в частности,
в так называемые грибовидные тела. )десь тоже используется
принцип пространственного кодирования, однако конкретные
закономерности пока не установлены. И, наконец, происходит
возбуждение моторных нейронов, непосредственно обеспечива-
ющих ответные реакции организма.
Пути передачи информации об электрических разрядах, к со-
жалению, продолжают оставаться практически невыясненными.
Удалось обнаружить лишь одно из промежуточных звеньев: это
пара симметрично расположенных ДПМ-нейронов, образующих
многочисленные контакты с грибовидными телами (рис. 4.4).
Таким образом, грибовидные тела получают информацию не
только о запахах, но и разрядах электрического тока. Этот факт
Грибовидное гело
доля
головного мозга
Обонятельный нерв
Рис. 4.4. Элементы ценгральнои нервной системы дрозофилы, принима-
ющие непосрецс! венное участие в обонятельном обучении (показана правая
половина головною мозга)
98
позволяет предположить, что ключевые события, обеспечивающие
способность дрозофилы обучаться и запоминать (по крайней
мере, в случае обонятельного обучения), локализованы преиму-
щественно в данном отделе мозга. И действительно, именно в
нейронах грибовидных тел зафиксирован наиболее высокий
уровень экспрессии многих генов, участвующих в контроле
обучаемости и памяти.
4.5. Молекулярные основы обонятельной
памяти у дрозофилы
Обонятельная память вырабатывается у дрозофилы не сразу
же с началом обучения, а только по прошествии как минимум
нескольких минут. Это значит, что в основе рассматриваемого
нами процесса наряду с проведением импульсов лежат и другие,
существенно более медленные молекулярные механизмы. К их
числу, по-видимому, относятся некоторые каскады, обеспечиваю-
щие внутриклеточную передачу полученной информации.
Как мы выяснили, процесс формирования обонятельной па-
мяти локализован у дрозофилы преимущественно в грибовидных
телах. Нейроны, расположенные в этих отделах мозга, получают
информацию по двум независимым путям: во-первых, через обо-
нятельные доли (о воспринятых запахах), а во-вторых, через
ДПМ-нейроны (об электрических разрядах). Предполагают, что
непосредственным результатом такой конвергенции является
одновременный запуск нескольких каскадов, взаимодействие
которых и приводит к формированию памяти.
Что именно происходит в нейронах грибовидных тел при по-
лучении обонятельной информации, до сих пор неизвестно. Зато
достаточно подробно исследован каскад, запускаемый при по-
ступлении информации об электрических разрядах. В нем уча-
ствует не менее десятка различных компонентов, а судя по кос-
венным данным, значительно больше. Этот каскад начинается с
восприятия нейромедиатора, выделяемого синаптическими окон-
чаниями ДПМ-нейронов. Изменяется конформация соответ-
ствующих рецепторных белков, что приводит к активации моле-
кул аденилатциклазы и как следствие — к резкому усилению
биосинтеза цАМФ.
Являясь одним из важнейших вторичных мессенджеров, цАМФ
очень быстро диффундирует по цитоплазме нейрона и вызывает
активацию следующих компонентов каскада, в частности — про-
теинкиназы А. Данный фермент, в свою очередь, фосфорилирует
различные белки, непосредственно или косвенно вовлеченные в
99
формирование памяти. К их числу относится и транскрипцион-
ный фактор СгеЬ2. Он запускает экспрессию целого ряда генов,
контролирующих возникновение новых отростков нейрона, а в
итоге — образование новых межнейронных контактов.
Оказалось, что разные нарушения этого каскада по-разному
отражаются на процессе формирования памяти: чем раньше на-
рушение, тем сильнее наблюдаемые последствия.
В самом деле, для мух, мутантных по гену rutabaga (напомним,
этот ген кодирует аденилатциклазу), характерно резкое ослабление
обонятельной памяти в первые же 30 мин по окончании обучения.
В результате такие мутанты не могут сформировать ни долговре-
менную, ни средневременную, ни кратковременную память.
Между тем у дрозофил, мутантных по гену СгеЬ2, нарушение
памяти проявляется значительно позже. В этом случае заметно
ослаблена лишь долговременная память, а кратковременная и
средневременная практически неизменны.
Наконец, у мутантов по гену DC0, кодирующему одну из
субъединиц протеинкиназы А, наблюдаемые изменения носят
промежуточный характер.
Безусловно, не следует думать, что кратковременная память
возникает непосредственно в результате биосинтеза цАМФ,
средневременная — в результате активации протеинкиназы А,
а долговременная — при задействовании транскрипционного
фактора СгеЬ2. Эти молекулы являются лишь важными проме-
жуточными звеньями в сложных каскадах, обеспечивающих фор-
мирование обонятельной памяти. Конкретные «носители» памя-
ти пока не установлены. Тем не менее напрашиваются вполне
определенные выводы, а именно:
• у дрозофилы формирование долговременной обонятель-
ной памяти связано с возникновением новых межнейронных
контактов, т.е. со структурными изменениями головного
мозга. Неудивительно, что этот вид памяти достаточно
стабилен и практически нечувствителен к различным внеш-
ним воздействиям;
• формирование кратковременной и средневременной па-
мяти обусловлено более быстрыми молекулярными процес-
сами, для запуска которых необходим биосинтез цАМФ.
По-видимому, конечным результатом подобных процессов
является образование неких белковых комплексов, легко раз-
рушаемых различными внешними воздействиями. Именно
поэтому кратковременная и средневременная память не-
стабильны.
Сходные выводы получены и на других беспозвоночных жи-
вотных: пчелах и морских улитках Aplysia californica. А теперь
обратимся к результатам исследований на грызунах.
100
4.6. Генетический контроль обучаемости
и памяти у крыс и мышей
Грызуны, по сравнению с дрозофилой и другими беспозвоноч-
ными, имеют значительно более развитую нервную систему и
вследствие этого способны к намного более сложному поведению.
В частности, они демонстрируют хорошие результаты не только
в различных вариантах ассоциативного обучения (обонятельно-
го, зрительного, слухового, тактильного и др.), но и в тестах,
требующих элементарных логических операций.
Кратко рассмотрим один из подобных тестов. На крыс воз-
действовали определенным световым сигналом, после которого
следовал либо звуковой сигнал, либо кратковременный доступ к
источнику пищи. В результате у животных сформировались устой-
чивые связи «свет -> звук», «свет -> пища», а также «звук -> пища».
Последняя заслуживает особенного пристального внимания, ведь
она не вытекает непосредственно из схемы обучения. По-види-
мому, данная связь возникла потому, что и «пища», и «звук» рас-
сматривались крысами как равноценные последствия одного и
того же сигнала. Дополнительные исследования подтвердили та-
кое предположение. Более того, оказалось, что подопытные кры-
сы отличали сигналы, возникавшие «сами по себе», от сигналов,
которые порождались действиями соседа (например, при его на-
жатии лапой на педаль).
Безусловно, у животных с такими когнитивными способностя-
ми количество генов, участвующих в контроле обучаемости и
памяти, должно быть значительно выше, чем у дрозофил. И дей-
ствительно, к настоящему времени у крыс и мышей обнаружено
уже около 80 интересующих нас генов. Их функции непосред-
ственно связаны с молекулярными каскадами, обеспечивающими
процессы формирования нервного импульса, а также внутрикле-
точную передачу воспринятой информации. В частности, среди
соответствующих генных продуктов широко представлены:
• различные рецепторные белки;
• ферменты, участвующие в метаболизме вторичных мессенд-
жеров;
• каталитические и регуляторные субъединицы протеинки-
наз;
* ионные каналы;
• транскрипционные факторы и др.
Необходимо особо отметить, что спектры белков, контроли-
рующих обучаемость и память у грызунов и дрозофилы, суще-
ственным образом перекрываются (табл. 4.1). Этот факт
свидетельствует о том, что у разных животных, незави-
101
Таблица 4.1. Гены, контролирующие обучаемость и память
у грызунов, гомологичные* соответствующим генам дрозофилы
Ген Соответствующий белок Молекулярные функции
Adcyi Аденилатци клазы Биосинтез цАМФ в нейронах го- ловного мозга
Adcy8
Pde2a Фосфодиэстеразы Расщепление цАМФ
Pde4d
Pde5a
Prkar2b Регуляторная субъеди- ница протеинкиназы А Регуляция каталитической субъ- единицы протеинкиназы А
Crebl Транскрипционный фактор, фосфорили- руемый протеинкина- зой А Стимуляция формирования и ро- ста нейронных отростков
Prkcb Протеинкиназы С Фосфорилирование определенных белков при повышении концен- трации диацил глицерола
Prkcz
Camk2A Кальций/ кальмодулин- зависимые протеин- киназы А Фосфорилирование определенных белков при повышении концен- трации Са2+ в цитоплазме
Camkkl
Dbh Дофамин Р- гидроксилаза Расщепление дофамина
Dnml Динамин Формирование пресинаптических пузырьков
Fyn Протеинкиназа Src- типа Регуляция клеточных делений
Gabra5 а-субъединица рецеп- торов ГАМК Регуляция активности нейронов посредством ГАМК
Gria 1 Ионотропные рецеп- торы глутаминовой кислоты Регуляция активности нейронов посредством глутаминовой кис- лоты
Grin 1
Grin2A
Grin 2 В
Itga3 Инте грины а Регуляция формирования межкле- точных контактов
Itga5
Itga8
102
Окончание табл. 4.1
Ген Соответствующий белок Молекулярные функции
Кспа 1 Потенциалзависимый К* канал Формирование нервного импульса
Nfl Нейрофибромин Регуляция молекулярных каскадов с участием протеинкиназы ras
Notch 1 Мембранный рецеп- тор, обладающий свойствами транс- крипционного фак- тора Регуляция экспрессии определен- ных генов на этапе транскрипции
Rps6kb 1 Rps6kb2 Субъединицы проте- инкиназы S6 Регуляция молекулярных каска- дов, в том числе с участием транс- крипционного фактора CREB1
Yivhae Yivhag Ywhaz Белки, активирующие ти рози н гидрокс илазу Регуляция молекулярных каскадов с участием нейротрофинов
* Гомологичные гены кодируют сходные белки, что свидетельствует о высокой
степени их эволюционного родства и близости выполняемых ими молекулярных
функций.
симо от того, позвоночные они или беспозвоночные, в осно-
ве когнитивных способностей лежат принципиально сходные
молекулярные механизмы. Такие же механизмы, по-видимому,
используются и у человека.
4.7. Изучение генетического контроля
обучаемости у человека. Основные подходы
Очевидно, что люди значительно отличаются друг от друга по
способности усваивать новую для них информацию. В этих раз-
личиях большую роль играет наследственность. Однако конкрет-
ные данные чрезвычайно скудны. Это связано с трудностями
генетического анализа человека: у людей невозможно получать
когнитивных мутантов, скрещивать их, конструировать необхо-
димые генотипы, измерять интенсивность экспрессии конкрет-
ного гена в различных участках головного мозга и др. Кроме того,
обучаемость — очень сложная характеристика (особенно у такого
103
объекта, как человек) и поэтому совершенно непригодна для
генетических исследований, пока не будет «раздроблена» на более
простые. Именно на стадии поиска таких характеристик и на-
ходится современная генетика обучаемости человека.
Как известно, в человеческом обществе имеются структуры,
специально предназначенные для обучения людей. Одной из
подобных структур является школа. В принципе, ее главные за-
дачи весьма немногочисленны: научить человека читать, писать
и считать (мы говорим о самом распространенном, заурядном
случае). Соответственно, сугубо с формальной точки зрения,
обучаемость индивида можно оценивать по этим трем критери-
ям.
Разные люди различным образом справляются с перечислен-
ными задачами, многие — достаточно успешно (в зависимости от
усердия). Но у некоторых людей обнаруживаются систематические
проблемы, полностью снять которые обычно не удается даже при
использовании специальных компенсаторных методик. В связи с
этим выделяют три следующие психологические особен-
ности.
• Дислексия — нарушение формирования навыков чтения.
В результате чтение сопровождается нечетким произношением,
перестановками отдельных букв или целых слогов, изменением
окончаний, частыми запинками и т.п.
* Дисграфия — нарушение формирования навыков письма.
В этом случае наблюдаются многочисленные грамматические
ошибки: пропуски, замены и перестановки букв или слогов, раз-
рывы в словах или слитное их написание.
• Дискалькулия — нарушение формирования навыков сче-
та. В результате заметно затруднены операции с числами: чело-
век допускает многочисленные арифметические ошибки, с
большим трудом осваивает таблицу умножения, плохо понима-
ет разрядную структуру чисел (разбиение на единицы, десятки,
сотни и т.д.).
Исследования, посвященные генетическим основам дис-
калькулии, находятся пока на сугубо предварительной ста-
дии: известны лишь единичные работы по близнецовому
анализу. Они свидетельствуют о довольно существенной роли
генотипа, однако не получили никакого конкретного продол-
жения. Возможно, в ближайшей перспективе ситуация изменит-
ся благодаря масштабному научно-исследовательскому проекту,
официально объявленному в 2002 г. Национальным институтом
здоровья ребенка и развития человека совместно с Департаментом
образования (США). Этот проект направлен на выявление и
всестороннее изучение генетических, культурных, этнических,
социально-экономических, возрастных, физиологических, эмо-
104
циональных и прочих факторов, оказывающих влияние на фор-
мирование математических способностей, в том числе и на спо-
собность производить арифметические действия. Впрочем, уже
сегодня не вызывает сомнений, что разные формы дискалькулии
имеют разную этиологию и поэтому могут контролироваться
разными генами.
Дисграфия, к сожалению, исследована еще в меньшей
степени. В литературе отсутствуют какие бы то ни было
сведения даже по близнецовому анализу этой характери-
стики. Тем не менее, судя по некоторым косвенным данным
(например, по частотам встречаемости дисграфии среди род-
ственников), роль генотипа в ее формировании достаточно ве-
лика.
Наибольшие успехи в генетике обучаемости человека связаны
с исследованиями генетического контроля дислексии. На резуль-
татах этих исследований остановимся более подробно.
4.8. Современное состояние проблемы
генетического контроля дислексии
Дислексия — довольно распространенная когнитивная анома-
лия. Ее частота независимо от национальности, культурных тра-
диций, образования и социально-экономического статуса людей
обычно варьирует в пределах от 5 до 10 %. Нов некоторых семьях
она встречается во много раз чаще, что свидетельствует о высокой
наследуемости этой аномалии. И действительно, судя по резуль-
татам близнецового анализа, дислексия в основном обусловлена
именно генотипом. Его вклад оценивают приблизительно в
70%.
К сожалению, пока мы не можем с уверенностью утверж-
дать^ какие именно гены участвуют в контроле дислексии.
Однако не вызывает сомнений, что таких генов много. Дело
в том, что к настоящему времени в геноме человека уже
обнаружено, по меньшей мере, семь районов, имеющих не-
посредственное отношение к рассматриваемому признаку.
К их числу относятся районы: 1р36-р34 (1-я хромосома), 2р16-
р/5 (2-я хромосома), 3pl2-ql3 (3-я хромосома), 6р22 (6-я хромо-
сома), 15q21 (15-я хромосома), 18р1 /.2(18-я хромосома) и Xq27.3
(Х-хромосома).
Каждый из этих районов достаточно велик и содержит как
минимум десятки различных генов. Осталось выяснить, какие же
именно контролируют дислексию. Учитывая эффективность со-
временных молекулярно-генетических методов, эта задача может
105
быть решена в ближайшие годы. Некоторые «гены-кандидаты»
уже известны.
Так, в районе 3pl2-ql3 основное внимание сосредоточено на
гене ROBO1. Этот ген кодирует мембранный рецепторный белок,
облегчающий взаимное распознавание аксонов и дендритов и тем
самым способствующий формированию межнейронных контак-
тов. Как выяснилось, сходный белок имеется и у дрозофилы. Его
дефектность влияет на развитие некоторых аксонов, что отража-
ется на строении головного мозга насекомого. Этот факт заслу-
живает особенно пристального внимания. Дело в том, что у людей
с тяжелыми формами дислексии обнаружено значительное недо-
развитие белого вещества (различного рода нейронных отростков)
в затылочно-височных областях больших полушарий. Вероятно,
одной из причин подобных нарушений являются мутации в гене
ROBO1.
Данный ген представлен у человека множеством аллелей.
В настоящее время проводят анализ их нуклеотидных последо-
вательностей, чтобы выяснить, какие именно изменения в гене
коррелируют с теми или иными симптомами дислексии. Воз-
можно, это некие мутации в промоторной области, приводящие
к ослаблению экспрессии гена ROBOL Но четкого эксперимен-
тального подтверждения пока не получено. Наконец, нельзя
исключать и возможность того, что дислексия контролируется
какими-то другими генами из числа локализованных в районе
3pl2-ql3.
Наиболее вероятными кандидатами в районе 6р22 считаются
гены DCDC2 и KIAA0319. Хотя их конкретные функции еще не
установлены, показано, что оба гена играют важную роль в фор-
мировании различных отделов головного мозга, и в частности тех,
которые активируются непосредственно при чтении. Кроме того,
для каждого из упомянутых генов обнаружена связь между не-
которыми мутациями и наличием дислексии. Однако полученные
результаты пока неоднозначны и требуют тщательной проверки
в дальнейших исследованиях.
Основным кандидатом в районе 15q21 является ген с еще не-
известными биологическими функциями, получивший обозначе-
ние DYX1C1. Его экспрессия происходит в различных тканях
организма, включая определенные нейроны головного мозга.
О том, что рассматриваемый ген контролирует дислексию, сви-
детельствует достаточно большое количество фактов, но эти
факты пока не укладываются в четкую закономерность. В част-
ности, одни и те же мутантные аллели в некоторых выборках
тесно коррелируют с дислексией, а в некоторых, напротив, —
с нормальной способностью к чтению. Чем обусловлено такое
противоречие, пока непонятно.
106
4.9. Генетический контроль дислексии.
Ближайшие перспективы
Одним из условий, определяющих успешность генетического
анализа, является четкая диагностика исследуемых признаков.
Между тем оказалось, что многие симптомы дислексии могут
быть следствием широкого спектра других аномалий, таких как
дефекты зрения, слуха или артикуляции, умственная отсталость,
низкий интеллект и т.д. Поэтому в настоящее время под терми-
ном «дислексия» понимают лишь строго определенные наруше-
ния чтения: только те, что формируются на фоне нормального
интеллекта и не связаны с дефектами сенсорики, артикуляции
или психики.
Но даже при этом симптомы дислексии весьма разнообразны.
К ним относят:
• различные нарушения фонематической грамотности.
Например, у дислексиков возникают значительные трудности при
попытках подразделять слова на отдельные фонемы, сравнивать
их друг с другом и др.;
• проблемы, связанные с распознаванием прочитываемых
слов. Дислексики плохо понимают смысл прочитанного, не от-
личают друг от друга близкие по написанию слова или, напротив,
по-разному читают одно и то же слово;
• затруднения при быстром перечислении однотипных объ-
ектов. букв, цифр, цветов, предметов и др. Из-за этого чтение
медленное, с многочисленными запинками;
• проблемы, непосредственно связанные с проговариванием
слов. Особые трудности вызывают необычные сочетания букв,
например, при использовании произносимых, но бессмысленных
конструкций.
Обычно все перечисленные симптомы сопутствуют друг другу.
Нов некоторых случаях это правило может не выполняться. Так,
оказалось что нарушения фонематической грамотности слабо
коррелируют с проблемами в проговаривании слов. В связи с этим
многие исследователи высказывают мнение, что дислексия не яв-
ляется элементарной психологической характеристикой, а сла-
гается как минимум из нескольких более простых.
В настоящее время проводится поиск подобных характеристик.
С этой целью разрабатывают новые диагностические методики
для количественной оценки отдельных аспектов дислексии.
В дальнейшем предполагается использовать такие методики для
сравнения симптомов дислексии у разных людей, в особенно-
сти — у мутантов по разным генам-кандидатам. Вероятнее всего,
это позволит расчленить дислексию на несколько относительно
107
самостоятельных когнитивных нарушений, каждое со своим ав-
тономным генетическим контролем.
Таким образом, ближайшие перспективы в изучении дис-
лексии связаны:
• с тщательным исследованием ее симптомов и анализом их
степени независимости друг от друга. Это позволит существенно
приблизиться к пониманию того, из каких элементарных харак-
теристик слагается дислексия;
• с поиском генов, контролирующих указанные элементарные
характеристики. Уже сейчас очевидно, что этих генов не менее
семи (по количеству соответствующих районов в геноме челове-
ка);
• с уточнением роли наиболее вероятных генов-кандидатов
(ROBO1, DCDC2, К1АА0319и DYX1C1).
4.10. Какие именно процессы нарушены
при дислексии?
Как известно, современная наука не располагает методиками,
позволяющими напрямую оценивать обучаемость организма.
Поэтому любые оценки результатов обучения являются косвен-
ными и обычно основаны на исследовании памяти (см. подразд.
4.2, рис. 4.2). Таким образом, анализируя способность человека
читать, мы лишь формально проводим оценку его обучаемости.
На самом же деле мы исследуем нечто иное, а именно — способ-
ность использовать полученные знания для быстрого пере-
вода зрительной информации в звуковую.
Познакомимся с основными этапами этого процесса на при-
мере прочитывания некого конкретного слова. Сначала вос-
принятая сетчаткой зрительная информация передается по
специальной системе проводящих путей и в итоге проецирует-
ся на первичную зрительную кору. Затем начинается обработ-
ка воспринятой информации. В первую очередь происходит
распознавание последовательности букв. В этом процессе за-
действованы высшие зрительные области и прилегающие к
ним участки ассоциативной коры. На следующем этапе, про-
текающем в зоне Вернике, распознанная последовательность
букв осмысливается как слово. Соответствующая информация
поступает в зону Брока, где для этого слова подыскивается
звуковой эквивалент. Наконец, активируются области двига-
тельной коры, непосредственно управляющие мускулатурой
речевого аппарата, и человек произносит определенное соче-
тание звуков.
108
Очевидно, что первый и последний из перечисленных этапов
имеют отчетливо выраженную физиологическую окраску. Такие
этапы безоговорочно находятся под генетическим контролем. Но
промежуточные отличаются существенно большей «психологич-
ностью»: они непосредственно связаны с процессом обработки
информации, распознаванием образов, их осмыслением и т.п.
Распространяется ли генетический контроль и на эти этапы?
Или, быть может, они контролируются некими механизмами,
имеющими принципиально иную, негенетическую природу?
Напомним, что в соответствии с современными представле-
ниями о дислексии эта особенность не связана с нарушениями
сенсорных систем, а также систем, обеспечивающих способность
человека к артикуляции. Таким образом, у людей, страдающих
классической дислексией, нарушены именно процессы обра-
ботки информации, А поскольку дислексия контролируется
преимущественно генотипом, нет сомнений, что и эти
«сугубо психологические* процессы тоже имеют молекулярно-
генетическую основу.
Мы отмечали, что люди с тяжелыми формами дислексии от-
личаются от нормы недоразвитием белого вещества в затылочно-
височных областях больших полушарий. Этот факт хорошо со-
гласуется с тем обстоятельством, что большинство перечисленных
выше участков коры (зрительные и примыкающие к ним ассо-
циативные области, зона Вернике, зона Брока и целый ряд других)
расположено либо в затылочной и височной долях, либо в непо-
средственной близости от них. Различные нарушения взаимодей-
ствий между этими участками, обусловленные, например, недо-
статочным количеством синапсов или функциональными дефек-
тами отдельных нейронов, вероятно, и являются основными
причинами дислексии. Обнаружение конкретных генов, контро-
лирующих дислексию, позволит приблизиться к пониманию таких
взаимодействий, и как следствие — к выявлению ключевых мо-
лекулярных механизмов, обеспечивающих распознавание образов
и осмысление информации.
4.11. Родь гена HD в контроле пластичности
мышления человека
Обучаемость человека зависит от множества факторов, в том
числе и от степени пластичности мышления. Очевидно, что люди
различаются по этому показателю: некоторые мгновенно улавли-
вают новые идеи, а некоторые, наоборот, воспринимают их с
явным трудом. Причины подобных различий еще не установлены.
109
Однако постепенно накапливается все больше данных, свидетель-
ствующих о весьма заметной роли генотипа. Например, оказалось,
что пластичность мышления человека резко падает при развитии
генетически обусловленного заболевания, известного как хорея
Гентингтона.
Эта болезнь (ее средняя встречаемость — одно заболевание на
10 тыс. человек) обычно проявляется у человека лишь в зрелом
возрасте, не ранее 35 лет. Сначала возникают небольшие когни-
тивные проблемы, как правило, практически незаметные для
окружающих. Затем происходит прогрессирующее нарушение
моторики: человек совершает резкие неконтролируемые движе-
ния, испытывает затруднения с речью и при глотании. В дальней-
шем симптомы заболевания постепенно усиливаются, затрагива-
ют различные психические и физиологические функции и, на-
конец, через 10— 15 лет приводят к смерти. Впрочем, бывают и
исключения из этого правила. Так, легкие формы хореи Гентинг-
тона зачастую проявляются только в глубокой старости и про-
грессируют значительно медленнее, в течение десятилетий. На-
против, наиболее тяжелые формы заболевания начинают разви-
ваться еще с 20-летнего возраста и приводят к летальному исходу
за 3—4 года.
Начало развития и степень тяжести хореи Гентингтона зависят
в основном от одного единственного гена, получившего обозна-
чение HD (qt Huntington disease). В этом гене обнаружен весьма
примечательный участок, образованный многократно повторяю-
щимися триплетами CAG. Их число может варьировать от 10 до
115. Из-за этого в соответствующем участке кодируемого белка
может присутствовать разное количество остатков глутамина
(глутамин — аминокислота, кодируемая триплетом CAG).
Белки, содержащие длинные полиглутаминовые тракты (т.е.
множество расположенных друг за другом остатков глутамина),
представляют для клетки достаточно серьезную угрозу. Их молеку-
лы взаимодействуют друг с другом с образованием амилоидов —
крупных аномально организованных белковых комплексов, способ-
ных постепенно увеличиваться в своих размерах за счет присоеди-
нения все новых и новых молекул. Такие комплексы, по-видимому,
оказываются токсичными для клетки и в итоге значительно уско-
ряют ее отмирание. Именно это происходит при развитии хореи
Гентингтона: нейроны, локализованные в области полосатого тела,
накапливают множество амилоидов и затем погибают. Со временем
количество погибших нейронов увеличивается, что приводит к по-
степенному усилению симптомов данного заболевания.
Чем длиннее тракт, тем быстрее формируются амилоиды, тем
раньше развивается и тяжелее протекает болезнь. Соответственно
выделяют четыре группы аллелей гена HD:
110
• аллели, несущие не более 26 повторов CAG. Они не приво-
дят к развитию хореи Гентингтона;
• аллели, содержащие от 27 до 35 повторов CAG. Облада-
тели таких аллелей относятся к группе риска. Каждый их них
предрасположен к хорее Гентингтона, но вероятность развития
заболевания сравнительно невелика: не более 10 % при 35 повто-
рах. Если болезнь проявляется, то ближе к старости и при этом
обычно прогрессирует невысокими темпами;
• аллели, несущие от 36 до 40 повторов CAG. Эти аллели
вызывают заболевание с высокой вероятностью. Но все же не-
которые обладатели таких аллелей имеют шанс провести всю
жизнь без признаков болезни;
• аллели с более чем 40 повторами CAG. В этих случаях
болезнь оказывается практически неизбежной. Если же число
повторов превышает 60, возникает особенно тяжелая форма за-
болевания, начинающая стремительно прогрессировать с 20-
летнего возраста.
Судя по результатам анализа историй болезни, первыми
когнитивными нарушениями при хорее Гентингтона (для
всех окружающих человек еще абсолютно здоров) оказыва-
ются нежелание прислушиваться к чужому мнению и
резкое снижение восприимчивости к новым идеям. Это не
значит, что каждый консервативно мыслящий человек нахо-
дится на ранней стадии хореи Гентингтона. Однако в основе
консервативного склада мышления могут лежать какие-то
молекулярно-генетические нюансы, имитирующие начало раз-
вития данной болезни.
4.12. Моделирование симптомов хореи
Гентингтона у животных
Консервативность, возникшая в результате хореи Гентингтона,
отчетливо проявляется при решении разнообразных задач, и в
частности — в тестах на распознавание «правильного объекта».
В этих тестах человеку предлагают пары изображений, отлича-
ющихся друг от друга одновременно по нескольким параметрам,
один из которых является значимым, а остальные — нет (рис. 4.5, а).
После каждого правильного выбора следует подкрепление. Люди,
находящиеся на ранней стадии заболевания, достаточно быстро
вычленяют значимый параметр, после чего способны к практи-
чески безошибочному выбору. Если использовать новые пары
изображений, для которых значимым остается все тот же параметр
(рис. 4.5, б), испытуемые успешно справляются и с этой задачей.
111
Неверно Неверно Верно
а
Верно Верно Верно
б
Неверно Верно Неверно
в
Рис. 4.5. Пример задачи на распознавание «правильного объекта». Испытуемый
заранее предупрежден, что положение объекта на листе значения не имеет:
а — методом проб и ошибок испытуемый выясняет, что «правильными объектами»
являются белые фигуры вне зависимости от их формы и размера (выбранные объекты
отмечены стрелкой): б — вычленив значимый параметр (белый цвет), испытуемый
легко выбирает «правильные объекты» даже при использовании новых вариантов изо-
бражений; в — в процессе тестирования значимый параметр был изменен (теперь это
не белый цвет, а маленький размер), и старая тактика приводит к многочисленным
ошибкам. Необходимо заново вычленить значимый параметр. Для человека на ранней
стадии хореи Гентингтона эта задача достаточно трудна: он продолжает придерживать-
ся старых «правил игры», хотя они со всей очевидностью уже не работают
Но если произвести замену значимого параметра (рис. 4.5, в),
у испытуемых возникают весьма серьезные затруднения даже при
использовании знакомых пар изображений. Иными словами,
люди, страдающие хореей Гентингтона, с трудом переключаются
при изменении «правил игры».
В сходных тестах экспериментально доказано, что и другие
млекопитающие (мыши, крысы и различные виды обезьян) тоже
способны к вычленению значимого параметра. Более того, для
каждого из перечисленных видов продемонстрирована достаточ-
но высокая степень переключаемое™ при замене значимого па-
раметра по ходу исследования. Однако под действием определен-
ных химических препаратов, блокирующих функционирование
112
ГАМК-эргических синапсов (синапсов, в которых в качестве
нейромедиатора используется у-аминомасляная кислота)^ спо-
собность организма переключаться существенно падает. Таким
образом, степень когнитивной пластичности млекопитающих во
многом зависит от передачи информации через данные синапсы.
Этот вывод вполне справедлив и в отношении человека. Дело в
том, что нейроны, отмирающие при хорее Гентингтона, исполь-
зуют в качестве нейромедиатора именно ГАМК.
Как видим, у разных млекопитающих, включая и человека,
нарушение функционирования конкретного типа синапсов при-
водит к достаточно сходным когнитивным эффектам. Не менее
показательны и результаты генетических исследований. Установ-
лено, что в геномах практически всех млекопитающих присут-
ствуют гены, гомологичные (т.е. родственные) гену HD. В свою
очередь, в каждом из этих генов имеется участок, образованный
многократно повторяющимися триплетами CAG. Но в целом
такие участки сравнительно невелики и, по-видимому, недоста-
точны для формирования амилоидных комплексов.
С помощью методов молекулярной генетики и генной инже-
нерии у крыс и мышей произвели замену их собственных гомо-
логов на аллели с более чем сотней повторов CAG (таки£ аллели
были синтезированы искусственно). В результате у животных на-
блюдались нейродегенеративные процессы, характерные для
людей, страдающих хореей Гентингтона. Эти процессы сопрово-
ждались появлением следующих симптомов:
• значительным снижением когнитивной пластичности при
замене значимого параметра в вышеописанных тестах;
• нарушениями общей обучаемости и памяти;
• в некоторых случаях — нарушением координации движений.
Нетрудно заметить, что все перечисленные нами симптомы впол-
не соответствуют различным проявлениям хореи Гентингтона.
Таким образом, сходные изменения в гомологах гена HD
приводят к сходным когнитивным эффектам у крыс, мышей
и человека. Этот факт свидетельствует о том, что кры-
сы и мыши действительно являются адекватными модель-
ными объектами для изучения генетического контроля
когнитивной пластичности.
4.13. Основные подходы к изучению
генетического контроля памяти у человека
На примере дрозофилы мы убедились, что даже беспозвоноч-
ным свойственно как минимум несколько видов памяти. Человек
113
в этом плане, конечно, значительно сложнее. И действительно,
используя различные варианты классификации, можно выделить
десятки видов человеческой памяти, в том числе:
• в зависимости от способа восприятия информации — зри-
тельную, слуховую, обонятельную, вкусовую и осязательную*
а также висцеральную (основанную на внутренних ощущениях);
• по степени стабильности — сенсорную, кратковременную
и долговременную. В близком, но несколько ином варианте клас-
сификации — сенсорную, первичную, вторичную и третичную
(табл. 4.2);
* с точки зрения специфики вывода хранящейся информа-
ции — моторную, эмоциональную, образную и словесно-логиче-
скую (вербальную);
• в зависимости от характера использования полученных зна-
ний — процедурную (способность непосредственно их реализо-
вать) и декларативную (способность описать их словесно-
логически). В свою очередь, декларативную память можно подраз-
делить:
на семантическую, оперирующую сугубо абстрактными зна-
ниями (наподобие «Волга впадает в Каспийское море» без какой
бы то ни было связи с географической картой);
эпизодическую, оперирующую знаниями о конкретных ситуа-
циях (об особенностях и взаимном расположении различных
объектов, о последовательности событий, их продолжительности
и др.);
• по механизму формирования — произвольную (человек спе-
циально старался что-то запомнить) и непроизвольную (возник-
шую как бы случайно).
Увы, подавляющее большинство перечисленных видов памяти
очень слабо исследовано даже близнецовым анализом. Дело в том,
что до недавнего времени основное внимание было приковано к
интегральным показателям когнитивных способностей, таким как
IQ, или общий фактор интеллекта &. Как выяснилось, подобные
показатели в значительной степени (не менее чем на 50 %) зависят
от генотипа и при этом достаточно тесно коррелируют друг с дру-
гом. В результате исследователи не видели особой необходимости
в более дробном генетическом анализе когнитивных способно-
стей.
Сегодня не вызывает сомнений, что эти способности слагают-
ся из множества относительно самостоятельных характеристик,
количество которых, по-видимому, исчисляется десятками. К по-
добным характеристикам относят и различные виды памяти.
Показано, что некоторые из них независимы друг от друга (на-
пример, эпизодическая память утрачивается быстрее по сравне-
нию с семантической или процедурной). Соответственно воз-
114
Таблица 4.2. Основные характеристики сенсорной, первичной, вторичной и третичной памяти человека
Характеристика Виды памяти
Сенсорная Первичная* Вторичная Третичная
Емкость Малая Мала Очень велика Очень велика
Длительность Доли секунды От нескольких секунд до не- скольких минут От нескольких минут до нескольких лет Не ограничена
Ввод информации Автоматически в про- цессе восприятия Посредством осо- знания восприня- той информации В результате повто- рений** В результате мно- гократных частых повторений**
Доступ к информа- ции Ограничен скоростью восприятия Очень быстрый Медленный Очень быстрый
Механизм забыва- ния Быстрое угасание и «стирание» Старая информа- ция вытесняется новой Подавление доступа По-видимому, не происходит
Чувствительность к стрессовым воз- действиям Высокая Высокая Сравнительно низкая Низкая
* Первичная память соответствует кратковременной, вторичная и третичная память составляют долговременную память.
** В некоторых случаях при наличии феноменальной памяти или при длительном поиске соответствующей информации (ор-
ганизм заранее подготовлен к ее восприятию) возможно запоминание с одного единственного раза. Кроме того, одиночное, но
яркое событие многократно «прокручивается* в сознании индивида, что равноценно повторению данного события.
никает все больший и больший интерес к поиску генов, контро-
лирующих конкретные виды памяти.
Казалось бы, эта задача сравнительно проста. У крыс обнару-
жено более 80 генов, принимающих участие в контроле обучае-
мости и памяти. Гомологи всех этих генов присутствуют и у че-
ловека. Логично предположить, что как минимум некоторые из
них тоже оказывают существенное влияние на память. Осталось
провести молекулярно-генетический анализ и выявить связь кон-
кретных видов памяти с конкретными гомологами.
К сожалению, реальная ситуация значительно сложнее. Во-
первых, приходится работать только с теми аллелями, которые
естественным образом встречаются у людей. Это резко ограни-
чивает эффективность генетического анализа. Во-вторых, не вы-
зывает сомнений, что каждый вид памяти совместно контроли-
руется множеством разных генов. Поэтому эффект мутации в
исследуемом гене может быть «смазан» или полностью нивели-
рован мутациями в других. В-третьих, необходимо учитывать, что
память человека зависит и от многих факторов внешней среды.
А ведь их невозможно выровнять для всех испытуемых. Таким
образом, нет ничего удивительного в том, что пока мы имеем
лишь крайне отрывочные сведения о влиянии конкретных генов
на память человека.
4.14. Влияние гена АРОЕ на память человека
У человека известны десятки наследственных патологий, со-
провождающихся теми или иными расстройствами памяти. Уда-
лось обнаружить и некоторые соответствующие гены. Очень
многие из них (табл. 4.3) являются гомологами генов, принима-
ющих участие в контроле обучаемости и памяти у крыс и мышей.
Этот факт свидетельствует о том, что у разных млекопитающих
молекулярные механизмы памяти очень близки.
Таблица 4.3. Генетически обусловленные заболевания челове-
ка, сопровождающиеся нарушениями памяти
Заболевание Основные симптомы Ключевые гены
Болезнь Альцгеймера Прогрессирующее ослабление памяти, постепенно переходя- щее в различные физиологиче- ские проблемы АРР*
PSEN1
PSEN2
АРОЕ
116
Продолжение табл. 4.3
Заболевание Основные симптомы Ключевые гены
Синдром Коффина- Лоури Задержка умственного разви- тия, многочисленные аномалии скелета RPS6KA3**
Синдром Рубинштейна-Тайби Задержка умственного разви- тия, аномалии черепа, пробле- мы при дыхании и глотании CREBBP**
Синдром ломкой Х-хромосомы Удлиненный череп, оттопырен- ные уши, пониженный мышеч- ный тонус, низкий интеллект FMR1
Хорея Гентингтона Снижение пластичности мыш- ления, проблемы с координа- цией движения HD
Болезнь Паркинсона Повышенное напряжение мышц, тремор конечностей PARK2
Болезнь Готе Аномалии развития печени и селезенки, дефекты скелета, судороги, слабоумие GBA
Метахроматическая лейкодистрофия Слепота, судороги, паралич, слабоумие ARSA
Болезнь Александера Судороги, задержка умственно- го развития GFAP
Болезнь Наито- Оянаги Судороги, атаксия, слабоумие ATN1
Нейрофиброматоз Множественные пигментные пятна и опухоли на коже, судо- роги, сниженная обучаемость и память NF1
Синдром Юнертана Хождение на четвереньках, судороги, слабоумие VLDLR
Синдром Ретта Недоразвитость головного мозга, стереотипные движения, аутизм, слабоумие МеСР2
117
Окончание табл. 4.3
Заболевание Основные симптомы Ключевые гены
Синдром Ангельмана Эпилепсия, беспричинное ве- селье, искаженные черты лица, слабоумие UBE3A
* Полужирным шрифтом выделены гены, гомологи которых принимают уча-
стие в контроле обучаемости и памяти у крыс и мышей.
** Гены, участвующие в контроле аденилатциклазной системы.
Подавляющее большинство подобного рода заболеваний при-
водит далеко не только к ослаблению памяти, но и ко многим
другим психическим и физиологическим аномалиям. Причем от-
четливо выраженные нарушения памяти характерны обычно лишь
для поздних этапов болезни. В связи с этим возникает вполне
обоснованный вопрос: можно ли считать такие наследственные
патологии примерами влияния генотипа непосредственно на
память? Ответ, вероятнее всего, должен быть отрицательным.
Ярким исключением является болезнь Альцгеймера: наруше-
ние памяти — ее ключевой симптом. Это заболевание, как и хорея
Гентингтона, обусловлено массовым отмиранием нейронов го-
ловного мозга в результате постепенного накопления в них ами-
лоидов. Но состав амилоидных комплексов совершенно иной,
в случае болезни Альцгеймера они образуются из фрагментов
белка, кодируемого геном АРР. Функции данного белка пока не-
известны. Неясны и причины, вызывающие его амилоидизацию
(в этом белке нет длинных полиглутаминовых трактов, характер-
ных для продукта гена HD, см. подразд. 4.11).
У разных пациентов болезнь протекает по-разному, но общая
картина примерно одна и та же. Первые симптомы (ослабление
кратковременной памяти) обычно принимаются окружающими
за старческую рассеянность или последствия каких-либо стрес-
совых ситуаций. В дальнейшем начинает страдать и долговремен-
ная память, могут наблюдаться повышенная раздражительность
и агрессивность, резкие перемены настроения, нарушения речи.
Когнитивные проблемы дополняются сугубо физиологическими.
Смерть наступает в среднем через 7 лет после появления пер-
вых симптомов заболевания. К сожалению, лечить болезнь Альц-
геймера пока невозможно.
Среди людей, не достигших 60-летнего возраста, данная пато-
логия встречается достаточно редко (приблизительно 40 случаев
на 100 тыс. чел.). Среди людей старше 60 лет вероятность воз-
118
Таблица 4.4. Генетические факторы, повышающие вероятность развития болезни Альцгеймера
Ген Соответствующий белок Его молекулярные функции Мутации, ассоциирую- щие с болезнью Альцгей- мера Их эффект
АРР Предшественник ами- лоидов Неизвестны Около 30 Повышают вероятность ранних форм болезни
PSEN1 Преселенин 1 Более 150
PSEN2 Преселенин 2 Около 10
АРОЕ Аполиполипротеин Е Транспорт стеринов и жиросодержащих соединений е4 (эпсилон 4) В гомозиготном состоя- нии повышает вероят- ность поздних форм болезни*
У европеоидов и монголоидов; для представителей других рас результаты неоднозначны.
никновения заболевания неуклонно растет. Так, у людей от 60 до
75 лет доля больных составляет уже около I %, от 76 до 85 лет —
20 %, а от 86 лет и выше — 40 %. Обнаружены и некоторые другие
факторы риска. К их числу относятся излишнее содержание хо-
лестерина, ожирение, диабет, повышенное давление крови.
Между тем интенсивная и регулярная интеллектуальная деятель-
ность достоверно снижает вероятность болезни Альцгеймера.
По-видимому, это связано с тем, что интеллектуальная деятель-
ность стимулирует холинэргические нейроны мозга, замедляя их
амилоидизацию и отмирание (при болезни Альцгеймера они от-
мирают в первую очередь).
Немаловажную роль играет и генотип. Судя по результатам
нескольких близнецовых исследований, ее оценивают приблизи-
тельно в 50...70 %. К настоящему времени выявлено четыре гена,
мутации которых увеличивают риск заболевания (табл. 4.4). Мы
сконцен грируем внимание на гене АРОЕ.
Одна из дефектных аллелей данного гена (она получила обо-
значение с4) повышает вероятность развития поздней формы
болезни. Особенно четкий эффект наблюдается у гомозигот: они
страдают болезнью Альцгеймера в 8 раз чаще по сравнению с
людьми, не имеющими этой аллели. Тем не менее о строгой пред-
определенности говорить не приходится. Многие гомозиготы по
рассматриваемой нами аллели не проявляют никаких симптомов
болезни Альцгеймера, даже дожив до очень глубокой старости.
Десятки независимых исследований свидетельствуют о
том, что здоровые люди, гомозиготные по аллели е4, от-
личаются от нормы ослабленной эпизодической памятью.
Этот вывод подтвержден и результатами мета-анализа.
Таким образом, ген АРОЕ оказывает влияние не только на веро-
ятность развития болезни Альцгеймера, но и непосредственно на
уровень эпизодической памяти.
4.15. Влияние гена BDNF на память человека
Обратимся к генам, не связанным с ярко выраженными пато-
логиями. В геномах подавляющего большинства позвоночных
животных присутствует ген, получивший обозначение BDNF.
Судя по результатам исследований на крысах и мышах, этот ген
(его функции мы рассмотрим в следующем параграфе) участвует
в контроле пространственной памяти организма. Естественно,
напрашивается предположение, что и в случае человека он вы-
полняет примерно такую же биологическую функцию, а именно —
вовлечен в контроль эпизодической памяти.
120
В данном гене известно около 600 полиморфных сайтов. Сре-
ди них наиболее подробно исследован один — нуклеотидная за-
мена G196A. Эта замена отражается на структуре кодируемого
белка: в 66-м положении аминокислотной последовательности
вместо валина присутствует метионин. В результате соответству-
ющие аллели гена BDNF получили обозначение 66иа1 и ббтег
Комбинируясь друг с другом, они дают три генотипа: 66val/66val
(гомозиготы по нормальной аллели), 66metl66met (гомозиготы
по мутантной аллели) и 66иа1/66те1 (гетерозиготы). Каждый из
генотипов встречается достаточно часто, что облегчает проведение
необходимых генетических исследований.
Существует большое разнообразие экспериментальных под-
ходов, позволяющих оценивать эпизодическую память человека.
Например, испытуемому предлагают небольшой рассказ, описы-
вающий ситуацию из 50 взаимосвязанных элементов, и через
некоторое время (допустим, через 30 мин) просят как можно
точнее ее восстановить. Результаты учитываются в баллах по спе-
циальной шкале и служат количественной оценкой данного вида
памяти.
Оказалось, что люди, гомозиготные по аллели 66met, де-
монстрируют заметно ослабленную эпизодическую память
по сравнению с представителями двух остальных генотипов.
Этот вывод получен в шести независимых исследованиях. К со-
жалению, мета-анализ пока не проведен. Тем не менее с высокой
вероятностью мы можем ожидать, что достоверность обнаружен-
ной ассоциации будет подтверждена.
В завершение необходимо отметить следующее обстоятельство.
У людей, страдающих ярко выраженными симптомами шизоф-
рении, показатели эпизодической памяти одинаково низки вне
зависимости от генотипа по гену BDNF. Возможны различные
объяснения такой ситуации. Во-первых, вполне вероятно, что
развитие шизофрении приводит у человека к определенным функ-
циональным нарушениям, напоминающим результаты мутации в
рассматриваемом гене. Во-вторых, шизофрения относится к на-
следственным заболеваниям. Поэтому нельзя исключить, что
какие-то мутации, вызывающие у человека высокую предрасполо-
женность к шизофрении, сами оказывают существенное влияние
на память и потому нивелируют разнообразие по гену BDNF.
4.16. Молекулярные функции гена BDNF
Проведенные исследования показали, что ген BDNFкодирует
регуляторный белок из группы нейротрофинов. С белками по-
121
добного рода мы еще не встречались. Поэтому кратко рассмотрим
их главные особенности.
Нейротрофины оказывают сильное регуляторное влияние на
все основные аспекты жизнедеятельности нейронов:
• стимулируют их дифференцировку;
• поддерживают жизнеспособность;
• обеспечивают направленный рост аксонов и дендритов;
• способствуют формированию новых межнейронных контак-
тов;
• участвуют в регуляции активности синапсов.
В настоящее время известно несколько типов нейротрофинов.
Каждый синтезируется определенными клетками организма, вы-
деляется в окружающее их околоклеточное пространство и воз-
действует на мембранные рецепторы ближайших нейронов. При
этом конкретные результаты такого воздействия зависят от стадии
развития нервной системы, от сочетания различных нейротрофи-
нов и их концентрации, от типа нейрона, его состояния и т. д. Для
нас наиболее интересен последний эффект, а именно — регуляция
активности синаптических контактов. На нем мы и сосредоточим
свое внимание.
Оказавшись в области синапса, молекулы нейротрофинов
активируют особые рецепторы на каждой из мембран. В резуль-
тате осуществляется запуск каскадных процессов, обеспечива-
ющих, во-первых, усиленный выброс нейромедиатора, а во-вторых,
повышенную чувствительность постсинаптического окончания.
Соответственно, проводимость синапса резко возрастает и может
поддерживаться на этом же уровне до нескольких дней. Судя по
имеющимся данным, такая регуляция играет важную роль в функ-
ционировании нервной системы, и в частности — в процессах,
ведущих к формированию памяти.
Теперь перейдем непосредственно к гену BDNF. Его экспрессия
наблюдается во многих тканях организма, но главным образом —
в нейронах головного мозга. Именно с этим и связано обозначение
данного гена: оно происходит от brain derived neurotrophic factor
(нейротрофический фактор головного мозга).
Особенно интенсивная экспрессия гена BDNFхарактерна для
нейронов, расположенных в области гиппокампа. Между тем в
многочисленных исследованиях экспериментально доказано, что
именно в гиппокампе происходят ключевые события, лежащие в
основе эпизодической памяти человека и ее ближайших аналогов
у других млекопитающих (например, пространственной памяти
у крыс и мышей). Таким образом, логично предположить^ что
пространственная память контролируется у человека не
только геном BDNF^ но и другими генами, обеспечивающими
функционирование нейронов гиппокампа.
122
Уже получены и косвенные подтверждения этой гипотезы. По-
казано, что многие нейроны, расположенные в гиппокампе, ис-
пользуют в качестве нейромедиатора глутаминовую кислоту.
При этом искусственная инактивация глутаматэргических синап-
сов приводит к значительному ослаблению эпизодической памя-
ти. По-видимому, сходное проявление должно быть характерно
и для мутантов с нарушенной структурой соответствующих ре-
цепторов.
4.17. Основные итоги изучения генетического
контроля памяти
Несмотря на крайнюю отрывочность имеющихся данных,
можно сделать несколько вполне определенных выводов.
Во-первых, не только у человека, но даже у дрозофилы
обнаружено как минимум несколько типов памяти, резко
отличающихся друг от друга по степени стабильности.
У человека это сенсорная, кратковременная и долговременная
память, а у дрозофилы — кратковременная, средневременная и
долговременная. Наивно думать, что они абсолютно тождествен-
ны друг другу. Тем не менее отчетливо прослеживаются и общие
моменты. В частности, и у дрозофилы, и у человека одним из
главных условий формирования долговременной памяти являет-
ся многократное повторение «нового материала», причем пред-
почтительно с достаточно длительными перерывами. В этом
плане крылатая фраза «повторение — мать учения» отражает ка-
кие-то фундаментальные молекулярные закономерности, одина-
ково справедливые и для насекомых, и для человека.
Во-вторых, экспериментально доказано, что кратковременная
память (независимо от того, у какого объекта она сформирована)
легко разрушается различными стрессовыми воздействиями: хо-
лодом, электрическим шоком и т.д. Но долговременная практи-
чески нечувствительна к подобным воздействиям. Это значит, что
в процессе поэтапного формирования памяти происходит пере-
ключение с одних механизмов на другие. Сначала возникают
сугубо функциональные изменения (запуск молекулярных каска-
дов, коррекция мембранного потенциала, регуляция активности
синапсов и т.д.).
Такие изменения нестабильны и легко обратимы. Но в даль-
нейшем начинаются существенные структурные перестройки:
возрастает количество синапсов между некоторыми нейронами,
стимулируется образование новых аксонов и дендритов и их на-
правленный рост в определенные области мозга. Тем самым
123
формируются новые межнейронные контакты, потенциально
способные сохраняться на протяжении всей жизни. В результате
память консолидируется, т.е. становится стабильной. Эта схема
экспериментально подтверждена для нескольких объектов, вклю-
чая дрозофилу, улитку Aplysia, крыс и мышей. У человека прямых
подтверждений пока не получено из-за отсутствия четких мутан-
тов по долговременной памяти. Но косвенные данные (см. под-
разд. 4.14, табл. 4.3) достаточно хорошо согласуются с предложен-
ной схемой. Таким образом, весьма вероятно, что консолидация
памяти происходит у разных объектов по единому принципу.
В-третьих, даже у насекомых, не говоря о млекопитающих,
память находится под контролем множества генов: как минимум
нескольких десятков, а возможно, и сотен. При этом соответ-
ствующие продукты весьма разнообразны, начиная с мембранных
рецепторов и протеинкиназ и заканчивая различного рода транс-
крипционными факторами (см. подразд. 4.6, табл. 4.1).
Таким образом, формирование памяти — многоэтапный
процесс, обусловленный не только проведением нервных им-
пульсов, но и значительно более медленными молекулярными
механизмами.
В-четвертых, судя по результатам сравнительных исследований,
у разных объектов память обслуживают сходные группы
генов. Для примера достаточно обратиться к гену BDNF. Как мы
выяснили, у человека он контролирует эпизодическую память, а у
крыс и мышей — ее ближайший аналог, пространственную. Более
того, у каждого из перечисленных объектов этот ген преимуще-
ственно экспрессируется в нейронах гиппокампа и обеспечивает
сходную регуляцию активности синапсов. Нет сомнений, что по
мере дальнейшего развития психогенетики количество подобных
примеров будет быстро расти, подтверждая универсальность ме-
ханизмов формирования памяти.
4.18. Перспективы генетики обучаемости
и памяти человека
Какие же гены относятся к наиболее вероятным кандидатам
на участие в контроле обучаемости и памяти у человека? Прежде
всего, это гомологи многочисленных генов, контролирующих
обучаемость и память у модельных объектов: дрозофилы, крыс,
мышей и некоторых других. Но и на этом список кандидатов не
исчерпывается.
К настоящему времени накоплен обширный фактический
материал о влиянии различных соединений на память человека.
124
Например, как уже было отмечено, введение в гиппокамп анта-
гонистов глутаминовой кислоты (т.е. веществ, подавляющих
активность глутаматэргических синапсов) приводит к значитель-
ному ослаблению эпизодической памяти. Сходный эффект вы-
зывают и некоторые антагонисты дофамина при введении в
зрительные центры среднего мозга. Таким образом, вполне веро-
ятно, что многие гены, вовлеченные в контроль функционирова-
ния соответствующих синапсов, оказывают на эпизодическую
память хотя бы косвенное влияние. Аналогичные прогнозы воз-
можны и для других видов памяти.
Нет сомнений, что в основе способности обучаться и запо-
минать лежит не только проведение импульсов по определенным
нейронам, но также и различного рода молекулярные каскады,
обеспечивающие внутриклеточную передачу информации о сиг-
налах. Например, молекулярные каскады с использованием
цАМФ и последующей активацией транскрипционного фактора
CREB абсолютно необходимы для формирования долговременной
памяти и у беспозвоночных животных, и у позвоночных. По-
видимому, они столь же важны и в случае человека. Поэтому все
гены, обслуживающие подобные каскады, тоже должны рассма-
триваться в качестве кандидатов.
Итак, суммарное количество генов-кандидатов на участие
в контроле обучаемости и памяти у человека исчисляется^
по меньшей мере^ несколькими сотнями. Однако это вовсе не
означает, что для каждого из них будет доказано непосредственное
влияние на память и обучаемость. Дело в том, что один и тот же
ген может «работать» в достаточно широком спектре клеток. На-
пример, интенсивная экспрессия гена NR1 (его продуктом явля-
ется один из двух типов субъединиц, входящих в состав рецепто-
ров глутаминовой кислоты) обнаружена у человека не только в
области гиппокампа, но и в миндалине, мозжечке, лобных долях
больших полушарий и некоторых других отделах головного мозга.
Таким образом, в организме, мутантном по гену NR1, одновре-
менно должны быть нарушены многие процессы. В свою очередь,
это приведет к многочисленным эффектам, затрагивающим раз-
личные стороны жизнедеятельности организма. А возможно, и к
резкому снижению его жизнеспособности. Соответственно, му-
тации по целому ряду генов, вовлеченных в контроль обучаемости
и памяти человека, могут характеризоваться настолько разно-
образными проявлениями, что нарушения когнитивных способ-
ностей останутся без внимания.
Кроме того, влияние некоторых генов может оказаться слабым
и трудно различимым. Особенно на фоне неконтролируемых
воздействий среды. В этом плане весьма перспективны исследо-
вания на детях. Ведь ребенок (в особенности, если речь идет о
125
дошкольнике), как правило, еще не испытывает той лавины воз-
действий, с которой приходится сталкиваться взрослому челове-
ку и которая значительно влияет на когнитивные способности.
Поэтому показатели обучаемости и памяти ребенка должны на-
много теснее коррелировать с его наследственными задатками,
нежели аналогичные показатели взрослого человека. И действи-
тельно, судя по результатам близнецового анализа, у детей в
возрасте от трех до семи лет память практически полностью за-
висит от генотипа.
Безусловно, необходимо учитывать, что психика ребенка су-
щественным образом отличается от психики взрослого. В связи
с этим далеко не все аспекты памяти могут быть в полном объе-
ме исследованы у детей. Но по некоторым аспектам это вполне
возможно. Таким образом, мы вправе ожидать, что многие све-
дения о генетическом контроле обучаемости и памяти у человека
будут со временем получены именно на детях.
СПИСОК ЛИТЕРАТУРЫ
1. Величковский Б.М. Когнитивная наука: Основы психологии познания.
В 2 т. — М. : Издательский центр «Академия», 2010.
2. Голимбет В. Е. и др. Поиск мутации в гене, кодирующем белок пред-
шественник Р-амилоида, у больных деменциями альцгеймеровского типа //
Журнал клиники неврологии и психиатрии им. С. С. Корсакова. — 1996. —
Т. 96. — № 1.-С. 75-78.
3. Малых С. Б., Егорова М. С., Мешкова Т.А. Основы психогенетики. —
М. : Эпидавр, 1998.
4. Нуркова В. В. Память. Общая психология. В 7 т. / под ред. Б. С. Братуся.
Т. 3 — М. : Издательский центр «Академия», 2006.
5. Равич-Щербо И.В.^ Марютина Т.М., Григоренко Е.Л. Психогенети-
ка. — М. : Аспект-Пресс, 2003.
6. Савватеева Е.В. Генетический контроль систем вторичных посред-
ников и их роль в обучении // Успехи современной генетики. — Т. 17. —
1991. - С. 33-99.
7. Alarcon М. et al. A twin study of mathematics disability // J. Learning
Disabilities. - 1997. - V. 30. - P. 617-623.
8. Blaisdell A. P. et al. Causal reasoning in rats // Science. — 2006. — V. 311. —
P. 1020-1022.
9. Brandt J. et al. Trinucleotide repeat length and clinical progression in
Huntington's disease // Neurology. — 1996. — V. 46. — P. 527 — 531.
10. Caylak E. A review of association and linkage studies for genetical analyses
of learning disorders //American J. Medical Genetics. Part B. — 2007. — V. 144B. —
P. 923-943.
11. Deutsch G. K. et al. Children’s reading performance is correlated with white
matter structure measured by diffusion tensor imaging // Cortex. — 2005. — V. 41. —
P. 354-363.
126
12. Dunning J. and During M.J. Molecular mechanisms of learning and
memory // Expert Reviews in Molecular Medicine. — 2003. — V. 5. — P. I —11.
13. Ertekin-Taner N. Genetics of Alzheimer’s disease: A centennial review //
Neurol Clin. — 2007.
14. FisherS. E. and Francks C. Genes, cognition and dyslexia: learning to read
the genome // TRENDS in Cognitive Sciences. — 2006. — V. 10. — P. 250 — 257.
15. Kamyshev N. G. et al. Novel memory mutants in Drosophila: Behavioral
characteristics of the mutant nemyP153 // BMC Neuroscience. — 2002. — V. 3:9.
16. Koppel J. and Goldberg T. The genetics of episodic memory // Cognitive
Neuropsychiatry. — 2009. — V. 14. — P. 356—376.
17. McGrath L. M. et al. Breakthroughs in the search for dyslexia candidate
genes // TRENDS in Molecular Medicine. — 2006. — V. 12. — P. 334 — 341.
18. McGuire S. E. et al. Thirty years of olfactory learning and memory research
in Drosophila melanogaster // Progress in Neurobiology. — 2005. — V. 76. — P.
328 - 347.
19. Plomin R. and Walker S. O. Genetics and educational psychology // British
J. Educational Psychology. — 2003. — V. 73. — P. 3—14.
20. Schott B.H. et al. The dopaminergic midbrain participates in human
episodic memory formation: evidence from genetic imaging // J. Neuroscience. —
2006. - V. 26. - P. 1407- 1417.
21. Small B.J. et al. Apolipoprotein E and cognitive performance: a meta-
analysis // Psychol. Aging. — 2004. — V. 19. — P. 592—600.
22. Willemsen R. et al. The fragile X syndrome: from molecular genetics to
neurobiology // Mental Retardation & Developmental Disabilities Research
Reviews. - 2004. - V. 10. - P. 60-67.
КОНТРОЛЬНЫЕ ВОПРОСЫ
1. Как у дрозофилы выявляют мутантов по обучаемости и памяти?
2. Какие молекулярные дефекты привадят к нарушению памяти у
дрозофилы?
3. Охарактеризуйте современное состояние и перспективы изучения
генетического контроля дислексии.
4. Опишите влияние гена HD на пластичность мышления.
5. Что известно о генах, контролирующих память у человека?
Глава 5
ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ ГИПЕРАКТИВНОСТИ
И ОСЛАБЛЕННОГО ВНИМАНИЯ
5.1. Синдром гиперактивности
и ослабленного внимания
Успеваемость школьника зависит от множества факторов.
В том числе и от способности сконцентрироваться на определен-
ной задаче. Практически в каждом классе есть хотя бы один ре-
бенок с нормальными или даже высокими показателями интел-
лекта, но при этом испытывающий серьезные затруднения в
учебе из-за своей чрезмерной невнимательности и непоседливо-
сти.
В соответствии с современной классификацией психических
расстройств речь идет о синдроме гиперактивности и ослаблен-
ного внимания. Это самая распространенная поведенческая ано-
малия у детей. Она наблюдаются приблизительно у 5 % школьни-
ков, а по некоторым оценкам — даже у I0 %.
Проявления данного синдрома весьма разнообразны. В част-
ности, к ним относят следующие.
Дефицит внимания*.
• ребенок не в состоянии следовать инструкциям (особенно
если нужно учитывать какие-то нюансы);
• неспособен надолго сконцентрироваться на конкретной за-
даче;
• регулярно допускает ошибки по невнимательности;
• не слушает то, что говорят непосредственно ему;
• часто теряет вещи;
• легко отвлекается;
• не может самостоятельно организовать работу или игру;
• регулярно забывает, что необходимо сделать.
Гиперактивность*.
• ребенок почти все время находится в движении (вертится,
суетится, двигает руками и ногами);
• излишне подвижен, когда такое поведение недопустимо;
• не может играть в спокойные («тихие») игры;
128
• вскакивает с места;
• чрезмерно много говорит.
Разработаны соответствующие опросники для родителей и учи-
телей. Диагноз ставят в том случае, если у ребенка регулярно на-
блюдают не менее 6 симптомов. В зависимости от того, какие из
них преобладают, различают 3 основные формы этого расстройства:
во-первых, ослабленное внимание совместно с гиперактивно-
стью во-вторых, ослабленное внимание без гиперактивности,
в-третьих, гиперактивность без ослабленного внимания.
Синдром начинает проявляться еще в раннем детстве, но ста-
новится особенно выраженным к 6 —8 годам. Долгое время этот
факт объясняли социальными причинами, а именно — вступле-
нием ребенка в школьную жизнь (новые требования, новое окру-
жение и т.д.). Однако в недавних исследованиях было показано,
что даже у пациентов, никогда не ходивших в школу, в этом воз-
расте наблюдаются практически те же изменения. Таким образом,
усиление синдрома к 6 —8 годам обусловлено скорее возрастны-
ми, а не социальными причинами. В дальнейшем выраженность
синдрома постепенно снижается, и некоторые проявления могут
бесследно исчезнуть. Тем не менее приблизительно у двух третей
пациентов некоторые симптомы сохраняются до подросткового
возраста, а примерно у половины продолжают наблюдаться и в
зрелом возрасте.
В последнее время все больше исследователей склоняется к
мнению, что в основе синдрома гиперактивности и ослаблен-
ного внимания лежит как минимум несколько разных пато-
логий. И в перспективе он должен быть «разъят» на не-
сколько составляющих, каждую из которых необходимо
рассматривать по отдельности.
5.2. Генетический анализ гиперактивности
и ослабленного внимания. Основные
подходы
Судя по результатам семейного и близнецового анализа, роль
генотипа в формировании синдрома очень велика. Ее оценивают
приблизительно в 60...90 %. Соответственно, возникает вопрос о
«генах-кандидатах».
Многочисленные клинические исследования свидетельствуют
о том, что проявления синдрома можно ослабить определенными
медикаментами. С этой целью обычно используют метилфени-
дат. Он легко преодолевает гемо-энцефалический барьер, про-
никает в различные отделы головного мозга и значительно уси-
129
ливает активность дофаминовой системы (с конкретным меха-
низмом мы познакомимся в подразд. 5.4). Кокаин, амфетамины
и другие препараты, стимулирующие эту же систему, тоже оказы-
вают на пациентов благотворное действие. Таким образом, основ-
ная причина возникновения синдрома тесно связана с недоста-
точностью дофаминовой системы регуляции.
В этой системе центральную роль играет дофамин — один из
важнейших нейрогормонов и нейромедиаторов. Нейроны, ис-
пользующие данный нейромедиатор, в основном сосредоточены
в миндалине, гипоталамусе, вентральной покрышке среднего
мозга и компактной части черной субстанции ствола. Они иннер-
вируют лобные доли больших полушарий, срединное возвышение
гипофиза, прилежащее ядро и некоторые другие области голов-
ного мозга.
Принято считать, что гл а в н а я функция дофамина—
запуск программ, приводящих организм в состояние удоволь-
ствия. В самом деле, в исследованиях на человеке и других мле-
копитающих убедительно показано, что многие дофаминэргиче-
ские синапсы активируются в ответ на различные позитивные
сигналы (например, при утолении голода или оргазме), либо
нейтральные, но достаточно тесно ассоциированные с позитив-
ными. Напротив, при наличии сигналов негативного характера
(вызванных болью, голодом, страхом и т.д.) наблюдается резкая
инактивация этих синапсов. Соответственно «правильные» дей-
ствия получают подкрепление и впоследствии могут охотно и
многократно воспроизводиться, а «неправильные» приводят к
возникновению неприятных эмоций, и в дальнейшем организм
старается их избегать.
Кроме того, дофаминэргические синапсы участвуют в регуля-
ции сна, различных мотиваций, настроения, памяти, внимания
и моторной активности. Две последние функции представляют
особый интерес, поскольку имеют самое непосредственное от-
ношение к синдрому. Наконец, существенное ослабление био-
синтеза дофамина часто приводит к развитию у человека болезни
Паркинсона (неконтролируемым сокращениям некоторых мышц).
Все эти данные хорошо согласуются с представлениями о том, что
основной причиной рассматриваемого нами синдрома является
недостаточная активность дофаминовой системы.
Итак, к числу наиболее вероятных кандидатов на участие
в контроле гиперактивности и ослабленного внимания от-
носятся гены, «обслуживающие* дофаминовую систему
регуляции. В настоящее время проводят исследования 10 генов:
TH, DDC, SNAP25, DAT1, DRD1, DRD2, DRD3, DRD4, DRD5
и СОМТ. Для нескольких из них получены вполне надежные
подтверждения.
130
5.3. Роль генов, контролирующих биосинтез
и секрецию дофамина
Биосинтез дофамина осуществляется в два этапа, первый из
которых обеспечивается тирозин гидроксилазой, а второй — де-
карбоксилазой ароматических аминокислот (рис. 5.1). Соот-
ветствующие гены получили обозначения TH и DDC. К сожале-
нию, в анализе гиперактивности и ослабленного внимания эти
гены используются явно недостаточно: в общей сложности из-
вестно всего лишь несколько работ. И ни в одной не обнаружено
четкой ассоциации с синдромом. Однако из этого не следует, что
перечисленные гены не принимают в контроле синдрома ника-
кого участия.
Дело в том, что каждый из них высоко полиморфен, а иссле-
довано к настоящему времени только по две аллели. Таким об-
разом, для того чтобы сделать окончательные выводы, необходи-
мо вовлечь в анализ и другие аллели.
Синтезированный дофамин поступает в синаптические пу-
зырьки (см. подразд. 2.3, рис. 2.3). В их мембранах содержатся
сложные белковые комплексы, способные при возбуждении ней-
рона изменять конформацию и вступать во взаимодействие с
белками пресинаптической мембраны. В результате такого взаи-
модействия пузырьки раскрываются и выбрасывают накопленный
дофамин в синаптическую щель.
Белки, принимающие участие в этом процессе, не являются
непосредственными компонентами дофаминовой системы. Они
работают во всех вариантах химических синапсов независимо от
типа используемого нейромедиатора и, строго говоря, должны
рассматриваться отдельно. Тем не менее, учитывая их «место»
Дигидрокси-
Тирозин фенилаланин
Рис. 5.1. Биосинтез дофамина
131
в дофаминовой регуляции, мы для удобства включили их имен-
но в этот параграф.
Показано, что выброс нейромедиатора в синаптическую щель
обеспечивается взаимодействием как минимум десятка белков.
Центральную роль среди них шрают продукты трех генов: VAMP1,
STX1B и SNAP25. Последний представляет для нас особый ин-
терес. Это связано с результатами исследований на модельных
животных. У мышей получена инбредная линия Coloboma с от-
сутствием одной из двух копий гомологичного гена. Количество
соответствующего белка у таких животных приблизительно в два
раза меньше по сравнению с нормой, что приводит к недостаточ-
но эффективному выбросу дофамина и как следствие — к повы-
шенному уровню моторной активности. Нужно отметить, что этот
эффект снимается амфетаминами, т.е. вызван нарушениями
именно дофаминовой системы.
Результаты исследований на мышах позволяют предположить,
что нечто подобное должно наблюдаться и у человека. И действи-
тельно, одна из аллелей гена SNAP25 (нуклеотидная замена 239G)
ассоциирует с синдромом гиперактивности и ослабленного вни-
мания. Наличие ассоциации подтверждено в нескольких иссле-
дованиях, однако для окончательного вывода их недостаточно.
Таким образом, в настоящее время нет четких данных о
влиянии генов, контролирующих биосинтез и секрецию до-
фамина, на внимание и моторную активность у человека.
Но поскольку у мышей это влияние экспериментально доказано
(по крайней мере, для гомолога гена SNAP25), мы вправе ожи-
дать, что у человека оно тоже будет обнаружено.
5.4. Роль гена DAT1, кодирующего
транспортер дофамина
Как известно, действие подавляющего большинства нейроме-
диаторов ограничено очень короткими промежутками времени.
Этот эффект обусловлен несколькими молекулярными механиз-
мами. В частности тем, что выделенные нейроном молекулы
нейромедиатора остаются в синаптической щели совсем нена-
долго (обычно на сотые или даже тысячные доли секунды), после
чего подвергаются реимпорту в пресинаптическое окончание.
Подобный процесс происходит и в дофаминэргических синапсах.
Его обеспечивает мембранный белок — транспортер дофамина,
являющийся продуктом гена DAT1. Показано, что некоторые ме-
дикаменты (например, метилфенидат, о котором мы упоминали в
подрадд. 5.2) специфически подавляют активность данного белка.
132
В результате дофамин накапливается в синаптических щелях, более
эффективно воздействует на соответствующие рецепторы и вызы-
вает формирование более выраженных биологических ответов.
Именно с этим связана способность метилфенидата ослаблять сим-
птомы гиперактивности и дефицита внимания.
Если подавление функций транспортера дофамина очень ярко
отражается на внимании и моторной активности, логично пред-
положить, что сходные поведенческие эффекты должны возникать
и вследствие мутаций в гене DAT1. Справедливость такого пред-
положения доказана для мышей, а в последние годы — и непо-
средственно для человека.
Человеческий ген DAT1 высоко полиморфен. В нем выявлено
несколько участков варьирующего размера, а также около I 000
нуклеотидных замен. В генетический анализ гиперактивности и
ослабленного внимания вовлечено пока менее 10 полиморфных
сайтов, причем в большинстве исследований используют лишь
один — 40-нуклеотидные повторы в конце транскрибируемой
области. Эти повторы (они встречаются в количестве от трех до
тринадцати) способны существенно влиять на экспрессию гена.
В самом деле, аллели, содержащие 10 повторов, экспрессируются
значительно интенсивнее по сравнению с остальными и приводят
к избыточному содержанию транспортера дофамина, а в итоге —
к недостаточно эффективной дофаминовой регуляции.
Судя по результатам многочисленных независимых исследова-
ний, такие аллели ассоциируют с гиперактивностью и ослаблен-
ным вниманием. В ряде работ обнаружить эту связь не удалось.
И все же она подтверждается мета-анализом.
Таким образом, аллели гена DAT1 с десятью экземплярами
40-нуклеотидных повторов, хотя и слабоу но все же стати-
стически достоверно повышают предрасположенность к
гиперактивности и дефициту внимания.
В последнее время в литературе появились свидетельства того,
что с синдромом ассоциируют и некоторые другие полиморфные
сайты, например количество 30-нуклеотидных повторов в ин-
троне 8. Однако этот вывод пока остается предварительным.
5.5. Роль гена СОМТ, контролирующего
деградацию дофамина
Показано, что в некоторых дофаминэргических синапсах со-
держание транспортера дофамина сравнительно невелико. В этих
случаях удаление нейромедиатора из синаптических щелей обе-
спечивается преимущественно катехол-О-метилтрансферазой.
Данный фермент обнаружен в различных тканях и органах. Его
133
задача — быстро разрушать излишки катехоламинов (т.е. до-
фамина, адреналина и норадреналина) и тем самым регулировать
концентрацию перечисленных соединений.
У многих млекопитающих, и в частности — у человека, суще-
ствует две формы катехол-О-метил трансферазы. Одна локализо-
вана на мембранах, вторая — в цитоплазме, и обе являются про-
дуктами гена СОМТ (рис. 5.2). Для нас наиболее интересна
мембранная форма. Именно она «отвечает» за деградацию до-
фамина, выделяемого в синаптические щели соответствующими
нейронами.
Судя по тому, что катехол-0-метилтрансфераза одновременно
используется в нескольких регуляторных системах, ее дефектность
Л-
Э-1
Э-1
,,.... Мембранная форма
катехол-О-метилтрансферазы
I Трансляция
------------------------Зрелая иРНК|
Вырезание интронов,| сшивание экзонов
------------------------——————— — Незрелая иРНК|
И-1 |-И-2 И-3 И-4 И-5
---Ген СОМТ
Э-6
Э-2 Э-3 Э-4 Э-5
а
Цитоплазматическая форма ____
- метилтрансферазы
| Трансляция
- ---Зрелая иРНК-2
Вырезание интронов, | сшивание экзонов
——- । ----------— Незрелая иРНК2
И-1 И-2 И-3 И-4 f И-5
------------------1-------—----------- - Ген СОМТ
Э-2 Э-3 Э-4 Э-5 Э-6
б
Рис. 5.2. Механизм образования мембранной (а) и цитоплазматической (б)
форм катехол-О-метилтрансферазы
Примечание. Ген СОМТ содержит два альтернативных промотора. В связи с
этим, его транскрипция может начинаться с двух разных точек (отмечены черными
треугольниками). В том случае, если задействован «левый» промотор, охватывается
вся кодирующая область гена с образованием мембранной формы катехол-О-
метилтрансферазы. Если же транскрипция начинается с «правого» промотора, воз-
никает укороченная форма данного фермента. Она не способна заякориваться в
мембране и потому функционирует к толще цитоплазмы. Как и многие другие гены
растений и животных, ген СОМТ содержит экзоны и интроны (отмечены буквами
«Э» и «И» соответственно). Интроны не несут информации о колируемом белке, они
вырезаются из молекулы иРНК и быстро деградируют. Оставшиеся участки (экзоны)
сшиваются друг с другом, образуя зрелую молекулу и РНК, которая и служит основой
для синтеза белка.
134
должна отражаться на многих признаках. И действительно, по-
давление активности этого фермента вызывает у мышей:
• усиление исследовательского поведения;
• ослабление обучаемости и некоторых видов памяти;
• эффекты, напоминающие действие антидепрессантов;
• расстройство пищеварительной системы.
Однако непосредственного влияния на моторную активность
нив одном из проведенных исследований обнаружить не удалось.
Добавим, что у мышей, нокаутированных по гену СОМТ, т.е.
вовсе не имеющих катехол-О-метилтрансферазы, тоже не наблю-
дается существенных изменений подвижности. В чем причины
такой ситуации, пока непонятно. Вполне возможно, что в синап-
сах, контролирующих моторную активность, основным механиз-
мом очистки является реимпорт, а деградация играет сугубо
второстепенную роль и потому почти не влияет на их функцио-
нирование.
По-видимому, нечто подобное характерно и для человека.
Во-первых, одна из мутаций гена СОМТ (она получила обо-
значение 158Met) приводит к значительному снижению актив-
ности фермента. Тем не менее люди, гомозиготные по этой му-
тации, не проявляют симптомов гиперактивности или ослаблен-
ного внимания.
Во-вторых, у человека описана хромосомная аномалия, при-
водящая к отсутствию одной из двух копий данного гена. Люди,
имеющие такую хромосомную аномалию, отличаются от нормы
многочисленными тяжелыми патологиями, в частности — повы-
шенной агрессивностью, навязчивыми идеями, склонностью к
совершению различных антисоциальных поступков, а иногда и
отдельными симптомами шизофрении. Но внимание и моторная
активность практически не затронуты.
Итак, полученные данные свидетельствуют о том, что ген СОМТ9
хотя и обслуживает дофаминовую систему, не участвует в
контроле гиперактивности и ослабленного внимания.
5.6. Роль генов, кодирующих рецепторы
дофамина
Несмотря на кратковременное пребывание в синаптической
щели, многие молекулы дофамина успевают ее пересечь и вступить
во взаимодействие с соответствующими рецепторными белками.
Это приводит к передаче импульса на следующий нейрон.
У человека известно 5 типов рецепторов дофамина'. DI, D2,
D3. D4 и £>5, каждый из которых кодируется отдельным геном
135
Таблица 5.1. Основные характеристики рецепторов дофамина у человека
Тип рецептора Принцип функционирования Результат активации Основная локализация Контролируемые процессы Ген
D1 Рецепторы, связанные с G-бед кам и. Вторичный мессенд- жер — цАМФ, его содержание возрастает за счет стимулирова- ния аденилатциклазы Постсинаптиче- ское возбуждение Головной мозг, почки, стенки кровеносных со- судов Состояние удоволь- ствия, кровообра- щение, мочеотде- ление DRD1
D2 Рецепторы, связанные с G-белками. Вторичный мессенд- жер — цАМФ, его содержание снижается за счет стимулирова- ния фосфодиэстеразы Постсинаптиче- ское торможение Головной мозг, почки,стенки кровеносных со- судов Состояние удоволь- ствия, кровообра- щение, мочеотде- ление DRD2
D3 Головной мозг (преимуществен- но полосатое тело и прилежащее ядро) Состояние удоволь- ствия, иммунитет DRD3
D4 Головной мозг, сердце, стенки кровеносных со- судов Моторная актив- ность, внимание, кровообращение DRD4
D5 Рецепторы, связанные с G-бел- ками. Вторичный мессенджер — цАМФ, его содержание возрас- тает за счет стимулирования аденилатциклазы Постсинаптиче- ское возбуждение Неизвестна Моторная актив- ность, внимание DRD5
(табл. 5.1). Наиболее подробно исследован ген DRD4. Его продукт
присутствует в центральной нервной системе, а также в широком
спектре периферических органов: кровеносных сосудах, сердце,
почках и некоторых других. Но основная масса молекул данного
рецептора сосредоточена в лобных долях коры больших полуша-
рий, т.е. именно в тех областях головного мозга, что «отвечают»
за когнитивные способности, эмоции и внимание.
Ген DRD4 относится к высоко полиморфным: в нем обнару-
жено более 150 нуклеотидных замен и множество участков, ва-
рьирующих по своему размеру. Таким образом, с учетом комби-
нирования различных мутаций этот ген существует в виде не-
скольких тысяч аллелей.
К настоящему времени в анализ вовлечено около десятка сайтов.
Но особенно тщательно исследуется только один — 48-нуклеотидные
повторы в экзоне 3. Их количество может варьировать от двух до
одиннадцати. Из-за этого соответствующий участок рецептора D4
тоже значительно варьирует по своей протяженности, что влияет на
его взаимодействие с (7-белками и в результате — на запуск даль-
нейших молекулярных каскадов. Это влияние наиболее заметно при
сравнении аллелей с четырьмя и семью экземплярами повторов
(такие аллели принято обозначать 4R и 7R). Если первые обуслав-
ливают нормальное функционирование рецептора, то вторые сни-
жают его эффективность в несколько раз.
Многочисленные независимые исследования свидетель-
ствуют о том, что аллели 7R ассоциируют с гиперактив-
ностью и ослабленным вниманием. В некоторых работах не
обнаружено такой ассоциации. Но она убедительно подтвержда-
ется мета-анализом.
Для генов DRD1, DRD2 и DRD3 не показано связи с рассма-
триваемым нами синдромом. Более перспективная ситуация с
геном DRD5. Дело в том, что в промоторной области данного гена,
приблизительно за 18 тыс. пар нуклеотидов (т. п. н.) до старта транс-
крипции, обнаружена высоко полиморфная нуклеотидная после-
довательность, отдельные варианты которой ассоциируют с синд-
ромом. Но пока этот вывод носит предварительный характер.
5.7. Роль генов, контролирующих
норадреналиновую систему
Дофаминовая система тесно связана с норадреналиновой. Во-
первых, хорошо известно, что именно дофамин является непосред-
ственным предшественником норадреналина, а во-вторых, в обе-
их системах есть общие компоненты. С одним из них мы кратко
познакомились в предыдущих параграфах: речь идет о ферменте
137
катехол-О-метилтрансферазе, обеспечивающем деградацию и до-
фамина, и норадреналина. Добавим, что некоторые рецепторы
(например, D4) активируются молекулами и того, и другого нейро-
медиатора. В связи с этим различные нарушения норадреналиновой
системы могут заметно влиять на дофаминовую и наоборот.
Процесс преобразования дофамина в норадреналин осущест-
вляется ферментом дофамин р-гидроксилазой. Соответству-
ющий ген получил обозначение DBH. В нем выявлено более 450
нуклеотидных замен и несколько участков, варьирующих по сво-
ей протяженности. Из трех полиморфных сайтов, вовлеченных в
анализ, лишь один (нуклеотидная замена А2 в интроне 5) ассо-
циирует с синдромом гиперактивности и ослабленного внимания.
Этот вывод подтвержден двумя независимыми исследованиями,
но в пяти ассоциацию обнаружить не удалось. Для мета-анализа
имеющихся данных пока недостаточно. Таким образом, вопрос
об участии гена DBH в контроле интересующих нас признаков
остается открытым.
Норадренэргические нейроны весьма многочисленны. Они
сконцентрированы в некоторых ядрах ствола головного мозга
(преимущественно в районе так называемого голубого пятна) и
иннервируют обширные области центральной нервной системы, в
том числе и лобные доли коры больших полушарий. Важную роль
в регуляции работы соответствующих синапсов играет мембранный
белок транспортер норадреналина, являющийся продуктом гена
NET1 и обеспечивающий реимпорт нейромедиатора в пресинап-
тические окончания. Показано, что подавление функций данного
белка при помощи некоторых три-циклических антидепрессан-
тов смягчает симптомы гиперактивности и ослабленного внима-
ния. В связи с этим логично предположить, что ген NET1 тоже
участвует в контроле рассматриваемых нами признаков.
В настоящее время известны результаты всего двух работ, и ни
в одной не удалось обнаружить четкой ассоциации. Однако не-
обходимо учитывать, что в обоих исследованиях в общей слож-
ности проанализировано 4 полиморфных сайта, а в гене NET1
их выявлено около 600. Они тоже должны быть вовлечены в ге-
нетический анализ.
Принято считать, что многие норадренэргические нейроны на-
ходятся под непосредственным контролем адренэргических. Поэ-
тому гены, обслуживающие функционирование адреналиновой
системы, тоже относятся к числу потенциальных кандидатов.
Действительно, судя по результатам восьми исследований,
нуклеотидная замена —1291G в премоторной области гена
Adra2A (он кодирует а.2А-рецепторы адреналина) ассоцииру-
ет с некоторыми симптомами ослабленного внимания.
И лишь в одной работе обнаружить эту связь не удалось. К со-
138
жалению, мета-анализ пока не проведен. Однако с высокой ве-
роятностью можно ожидать, что статистическая значимость ас-
социации будет доказана.
5.8. Синдром нечувствительности
к тироксину
Ослабленное внимание и повышенная моторная активность
могут быть связаны и с некоторыми другими молекулярными де-
фектами. Например, с недостаточной чувствительностью организ-
ма к тироксину — одному из важнейших гормонов щитовидной
железы. Данный гормон влияет на множество разных процессов:
• во-первых, участвует в регуляции развития и роста (хрони-
ческая нехватка тироксина в детском возрасте приводит к недо-
развитости скелета и головного мозга);
• во-вторых, стимулирует общий обмен веществ;
• в-третьих, повышает эффективность адреналиновой системы;
• в-четвертых, влияет на множество «частных» характеристик,
таких как сердечный ритм, перистальтика кишечника, скорость
поглощения кислорода, температура тела, иммунитет, кроветво-
рение, возбудимость и моторная активность.
Тироксин вырабатывается фолликулами щитовидной железы,
затем поступает в кровь, разносится по организму и оказывает
регуляторное воздействие на все типы клеток, в том числе и на
нейроны центральной нервной системы. Это воздействие суще-
ственно отличается по своему механизму от эффектов, возника-
ющих при выбросе нейромедиаторов. Дело в том, что рецепторы,
обеспечивающие восприятие тироксина, расположены не на внеш-
них мембранах, а в клеточных ядрах и, по сути, являются своео-
бразными транскрипционными факторами, активность которых
регулируется данным гормоном. Если такие рецепторы не связаны
с тироксином, они «садятся» на премоторные области определен-
ных генов и, как правило, резко подавляют их транскрипцию. Но
в присутствии тироксина конформация рецепторов изменяется.
В результате они утрачивают способность присоединяться к про-
моторам, и транскрипция соответствующих генов значительно
усиливается.
Таким образом, в основе регуляторных эффектов тирок-
сина лежит изменение экспрессии определенных генов.
Доставка тироксина из внеклеточного пространства в ядро
осуществляется при помощи специальных транспортных белков.
В этом процессе обычно участвуют и некоторые ферменты. Они
преобразуют тироксин в трийодтиронин — соединение с точно
139
такими же гормональными свойствами, но значительно более
активное по сравнению с тироксином.
Оба гормона воздействуют на одни и те же рецепторы. У че-
ловека известно два типа подобных белков, получившие обозна-
чения TRa и 77?р. И те, и другие присутствуют практически во
всех типах клеток, однако в существенно разных количественных
соотношениях (p-рецепторы преобладают в клетках печени, а а-ре-
цепторы — в нейронах тловного мозга).
Известно, что интенсивность работы щитовидной железы в
значительной степени зависит от содержания тиротропина. Этот
гормон выделяется гипофизом при недостатке тироксина и ока-
зывает на щитовидную железу стимулирующее действие. Тем
самым обеспечивается принцип обратной связи, и уровень ти-
роксина в организме возвращается к норме.
Центральную роль в оценке гипофизом содержания ти-
роксина, по-видимому, играют ft-рецепторы: их дефект-
ность воспринимается как резкая нехватка гормона. В ре-
зультате вырабатывается избыточное количество тиро-
тропина , что приводит к чрезмерному стимулированию
щитовидной железы вплоть до значительного увеличения ее
размеров. Содержание тироксина в организме заметно по-
вышается, вызывая многочисленные физиологические и пси-
хологические расстройства, в том числе учащение сердечно-
го ритма, эмоциональные проблемы, низкий интеллект,
гиперактивность и ослабленное внимание.
Ни один из перечисленных симптомов не является обязатель-
ным. Так, излишне высокая моторная активность характерна при-
близительно для 60% больных, низкие показатели интеллекта —
примерно для 40 %, а увеличенная щитовидная железа — для 80 %.
ДлН того чтобы четко диагностировать данное заболевание (его
называют синдромом нечувствительности к тироксину), недо-
статочно учитывать только внешние его проявления. Необходимо
применять специальные молекулярные методики, позволяющие
непосредственно оценивать чувствительность рецепторов.
5.9. Генетический анализ синдрома
нечувствительности к тироксину
Роль генотипа в формировании синдрома очень велика. Судя
гюрезультатам семейного и близнецового анализа, она составля-
ет не менее 80%. Один из соответствующих генов уже установ-
лен- это ген, кодирующий ^-рецепторы тироксина. В данном
геце (он получил обозначение Thrb) обнаружено более 4 тыс.
му-^ций. Подавляющее большинство локализовано в интронных
141
участках и никак не связано с рассматриваемым нами синдромом,
но некоторые очень тесно с ним ассоциируют. Таких мутаций
известно уже около 50. Они затрагивают строго определенные
области гена (а именно — экзоны с седьмого по десятый) и при-
водят к значительным дефектам рецепторного белка.
P-Рецептор в норме способен выполнять две функции: во-
первых, «садиться» на промоторы конкретных генов, а во-вторых,
специфически распознавать молекулы тироксина. Соответствен-
но, в рецепторе имеется два разных участка: один для связывания
с промоторами, второй — с гормоном. Мутации, тесно ассоции-
рующие с данным заболеванием, вызывают существенные нару-
шения второго участка, но при этом никак не влияют на функцию
первого. В результате, несмотря на достаточное количество ти-
роксина, p-рецепторы слабо взаимодействуют с его молекулами
и сохраняют конформацию, необходимую для связывания с про-
моторами, как если бы содержание гормона было бы низким.
Разные клетки по-разному реагируют на эту ситуацию. В част-
ности, в поперечно-полосатой мышечной ткани существенно
ослабляется синтез АТФ. Остеобласты (клетки, формирующие
костную ткань) хуже справляются со своими биологическими
функциями, из-за чего развитие скелета может задерживаться.
Между тем определенные нейроны в составе гипофиза выраба-
тывают избыточное количество тиротропина и в итоге чрезмерно
стимулируют щитовидную железу. В свою очередь, это приводит
к избытку тироксина. Именно он, по-видимому, является главной
причиной, вызывающей у людей с дефектными р-рецепторами
целый ряд симптомов гиперактивности и ослабленного внимания.
В пользу такого предположения свидетельствует тот факт, что
различные доброкачественные опухали щитовидной железы, при-
водящие к повышенному содержанию тироксина в организме,
тоже вызывают чрезмерную суетливость и невнимательность.
Клинические исследования показывают, что интересующие
нас мутации присутствуют как минимум у 90% людей, страда-
ющих синдромом нечувствительности к тироксину. При этом даже
одной мутантной аллели вполне достаточно для развития симпто-
мов данного заболевания. Но особенно ярко оно проявляется у
гомозигот. В этом случае возникают тяжелые формы болезни, при
которых отчетливо выражены все ее симптомы, включая и осла-
бленное внимание, и гиперактивность.
К настоящему времени ни у одного из исследованных больных
не обнаружено мутантных аллелей гена Thra (этот ген кодирует
^.-рецепторы тироксина). По-видимому, подобные аллели ока-
зываются летальными и достаточно быстро отсеиваются есте-
ственным отбором. Этот вывод недавно подтвержден в экспери-
ментах на мышах. Таким образом, можно предположить, что
141
а-рецепторы играют более важную роль в жизнедеятельности
организма, нежели продукты гена Thrb.
5.10. Моделирование синдрома
нечувствительности к тироксину у мышей
Тироксиновая система характерна для всех позвоночных. При
этом ее основные молекулярные компоненты, впервые возник-
шие у животных наподобие ланцетника, не претерпели суще-
ственных изменений в ходе эволюции. В частности, а- и р-ре-
цепторы тироксина обнаружены не только у человека и прочих
млекопитающих, но и у птиц, пресмыкающихся, земноводных
и даже у рыб. В свою очередь, p-рецепторы разных позвоночных
(например, человека и лосося или мыши и лягушки) отличают-
ся друг от друга лишь несколькими аминокислотными заменами,
что открывает широкие возможности для сравнительных иссле-
дований.
Кратко остановимся на одной из подобных работ. У пациента,
страдающего синдромом нечувствительности к тироксину, взяли
образец ДНК и из этого образца клонировали (выделили) мутант-
ную аллель гена Thrb. Соответствующая мутация затрагивала 10-й
экзон и вызывала полную невосприимчивость рецептора к гор-
мону. Затем с помощью методов генной инженерии клонирован-
ную аллель внедрили в геном мышей. В результате животные
имели три копии гена: две собственные (нормальные) и одну
человеческую (мутантную).
Рецепторы, кодируемые собственными аллелями гена, адек-
ватно реагировали на присутствие молекул тироксина и успешно
выполняли свою регуляторную функцию. Между тем продукты
мутантной человеческой аллели были полностью лишены способ-
ности взаимодействовать с гормоном. Эти рецепторы, несмотря
на достаточное содержание тироксина, сохраняли конформацию,
необходимую для связывания с промоторами, и продолжали по-
давлять экспрессию определенных генов (вели себя практически
так же, как у исходного больного).
Полученные трансгенные мыши отличались от нормы:
• значительно увеличенным содержанием тиротропина и ти-
роксина;
• меньшей массой;
• несколько замедленным развитием скелета;
• сниженными показателями обучаемости и некоторых видов
памяти;
• существенно повышенной моторной активностью.
142
Все перечисленные физиологические и поведенческие нарушения
являются типичными симптомами нечувствительности к тироксину,
и каждое было отчетливо выражено у исходного пациента. Таким
образом, мутантная аллель человеческого гена Thrb вызывает
у мышей примерно те же эффекты, что и у человека.
Этот факт еще раз убедительно свидетельствует о том, что ре-
зультаты, полученные в исследованиях на модельных животных,
правомочно переносить на человека и наоборот.
5.11. Перспективы генетики гиперактивности
и ослабленного внимания
Итак, судя по результатам проведенных исследований, в гене-
тическом контроле гиперактивности и ослабленного внимания
принимают участие как минимум три гена: DAT1, DRD4 и Thrb.
Первый и второй гены обслуживают дофаминовую систему,
третий — тироксиновую систему регуляции. Логично предпо-
ложить, что и некоторые другие гены, имеющие непосредственное
отношение к перечисленным системам, тоже оказывают влияние
на рассматриваемые признаки.
В одном из предыдущих параграфов мы уже отмечали, что по-
давление функций транспортера норадреналина при помощи
различных три-циклических антидепрессантов значительно смяг-
чает проявления данного синдрома. Таким образом, гены, обслу-
живающие норадреналиновую систему, тоже представляют
безусловный интерес. В первую очередь это касается гена NET1,
продуктом которого и является упомянутый белок.
Многие люди, страдающие гиперактивностью и дефицитом
внимания, отличаются от нормы диспропорцией головного мозга:
лобные доли существенно уменьшены, а затылочные — увеличены,
поэтому гены, контролирующие развитие лобных долей (напомним,
что именно эти отделы мозга отвечают у человека за способность
концентрировать внимание), тоже относятся к числу вероятных
кандидатов. В результате суммарное число интересующих нас генов
может исчисляться десятками и даже сотнями.
Изучение синдрома гиперактивности и ослабленного внимания
значительно затрудняется нечеткостью его диагностики. Как из-
вестно, отдельные проявления невнимательности и непоседливо-
сти можно обнаружить практически у любого ребенка. Поэтому
диагноз ставят только в том случае, если у пациента в течение
длительного времени (в наиболее типичной ситуации — не менее
полугода) регулярно наблюдают несколько разных симптомов.
В соответствии с современными представлениями достаточно
143
шести. Но если присутствует меньшее количество симптомов,
такого ребенка обычно считают здоровым. Нет сомнений, что
подобная диагностика не очень надежна.
К сожалению, и сами симптомы не отличаются четкостью.
В частности, неспособность ребенка следовать инструкциям, до-
жидаться своей очереди или полностью выслушивать вопросы
зачастую ошибочно воспринимается как невоспитанность,
а ослабленное внимание медлительного, вялого ребенка нелегко
отличить от отсутствия надлежащей мотивации. В результате по-
лучаемые оценки весьма субъективны. Неудивительно, что даже
высоко квалифицированные специалисты не всегда одинаково
оценивают конкретного индивида.
Два основных проявления данного синдрома (ослабленное
внимание и повышенная моторная активность) могут встречать-
ся в разных сочетаниях. В связи с этим все больше исследователей
склоняется к мнению, что каждое из перечисленных проявлений
относительно автономно и, возможно, имеет «свой собственный»
генетический контроль. Соответственно их нужно анализировать
строго по отдельности. И действительно, одна из мутаций в гене
Adra2a (нуклеотидная замена — 1291G) ассоциирует с дефицитом
внимания, но не с гиперактивностью.
Таким образом, следует ожидать, что синдром нечувстви-
тельности к тироксину будет в перспективе «разъят* на
несколько составляющих, каждая с относительно независи-
мым генетическим контролем. И чем более дробными ока-
жутся эти составляющие, тем адекватнее будут резуль-
таты генетического анализа.
СПИСОК ЛИТЕРАТУРЫ
I. Белоусова Е.Д., Никанорова М.Ю. Синдром дефицита внимания/
гиперактивности // Российский вестник перинатологии и педиатрии. —
2000. — № 3. — С. 39-42.
2. Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition,
Text Revision. — Washington. DC: American Psychiatric Association, 2000.
3. Faraone S. V. and Khan S.A. Candidate gene studies of attention-deficit/
hyperactivity disorder // J. Clinical Psychiatry. — 2006. — V. 67. — Suppl. 8. —
S. 13-20.
4. Girault J. and Greengard P. The neurobiology of dopamine signaling //
Archives Neurology. — 2004. — V. 61. — P. 641 —644.
5. Giros B. et a!. Hyperlocomotion and indifference to cocaine and amphe-
tamine in mice lacking the dopamine transporter // Nature. — 1996. — V. 379. —
P. 606-612.
6. Gizer /. /?., Ficks C. and Waldman I. D. Candidate gene studies of ADHD:
a meta-analytic review // Hum Genet. — 2009. — V. 126. — P. 51 —90.
144
7. Lasky-Su J. et al. Evidence for an association of the dopamine d5 receptor
gene on age at onset of attention deficit hyperactivity disorder // Annuals Human
Genetics. - 2007. - V. 71. - P. 648-659.
8. Levy F.y Hay D.A., McStephen M., Wood C. and Waldman /. Attention-
deficit hyperactivity disorder. A category or a continuum? Genetic analysis of a
large-scale twin study // J. American Academy Child Adolescent Psychiatry. —
1997. - V. 36. - P. 737 - 744.
9. McDonald M.P.., Wong R., Goldstein G., Weintraub B.y Cheng S.-y. and
Crawley J. N. Hyperactivity and learning deficits in transgenic mice bearing a
human mutant thyroid hormone bl receptor gene // Learning & Memory. —
1998. - V. 5. - P. 289-301.
10. Ono S. et al. Homozygosity for a dominant negative thyroid hormone
receptor gene responsible for generalized resistance to thyroid hormone //J. Clinical
Endocrinology & Metabolism. — 1991. — V. 73. — P. 990 — 994.
II. Polanczyk G. et al. Association of the adrenergic alpha2A receptor gene
with methylphenidate improvement of inattentive symptoms in children and
adolescents with attention-deficit/hyperactivity disorder // Archives General
Psychiatry. - 2007. - V. 64. - P. 218-224.
12. Refetoff S.y Weiss R.E. and Usala S.J. The syndromes of resistance to
thyroid hormone // Endocrine Reviews. — 1993. — V. 14. — P. 348 — 399.
13. Stevenson J. Evidence for a genetic etiology in hyperactivity in children //
Behavior Genetics. — 1992. — V. 22. — P. 337 — 344.
14. Volkow N. D. et al. Is methylphenidate like cocaine? Studies on their
pharmacokinetics and distribution in the human brain // Archives General
Psychiatry. - 1995. - V. 52. - P. 456-463.
15. Wang E. et al. The genetic architecture of selection at the human dopamine
receptor D4 (DRD4) gene locus // American J. Human Genetics. — 2004. — V.
74. - P. 931-944.
16. Wilson M. C. Coloboma mouse mutant as an animal model of hyperkinesis
and attention deficit hyperactivity disorder // Neuroscience & Biobehavioral
Reviews. - 2000. - V. 24. - P. 51-57.
КОНТРОЛЬНЫЕ ВОПРОСЫ
I. Какие факты свидетельствуют о роли дофаминовой системы в регу-
ляции моторной активности и внимания?
2. Перечислите важнейшие компоненты дофаминовой системы.
3. Опишите механизм влияния гена DAT1 на моторную активность
и внимание.
4. Опишите роль генов, кодирующих рецепторы дофамина, в контроле
моторной активности и внимания.
5. К каким эффектам приводят мутации в гене Thrb?
Глава 6
ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ АГРЕССИВНОСТИ
6.1. Биологический смысл агрессивного
поведения
В обиходе, сугубо с эмоциональной точки зрения, термин
«агрессия» имеет отчетливо негативную окраску: этим термином,
как правило, принято обозначать поведение, непосредственно
или косвенно направленное на причинение вреда. Тем не менее
подобное поведение играет важную роль в адаптивных реакциях
большинства высокоразвитых животных.
Каждому организму необходимы определенные ресурсы. Для
животного это вода, источники пищи, территория обитания, по-
ловой партнер и т.д. При этом многие ресурсы строго лимитиро-
ваны и недостаточны для поддержания жизнедеятельности всех
претендентов. В результате неизбежно возникает жесткая конку-
ренция. Особенно жесткой она оказывается среди организмов,
проявляющих максимально сходные потребности в ресурсах, т.е.
среди представителей одного и того же вида. Это не значит, что
все взаимоотношения в пределах вида имеют исключительно не-
гативный, конкурентный характер. Встречаются и другие взаимо-
отношения, вплоть до альтруистических. Однако чтобы выжить в
окружении многочисленных соперников, организму приходится
отстаивать свои личные интересы и время от времени прибегать
к откровенно агрессивному поведению.
Агрессивное поведение характерно для многих животных,
включая большинство позвоночных и даже беспозвоночных. Для
примера обратимся к, казалось бы, безобидной дрозофиле. Ее
самцы «сражаются» друг с другом за доступ к пище. Победители
получают преимущество не только в питании, но и в спаривании,
поскольку пища приманивает самок. Среди самок тоже проис-
ходят аналогичные «бои». И в результате наибольший успех в
питании и размножении, как правило, сопутствует тем самкам и
самцам, которые чаще других побеждают своих конкурентов.
Организм, совершенно не способный к агрессивному поведе-
нию, имеет мало шансов на выживание и оставление потомства.
146
Но чрезмерно высокая агрессивность тоже невыгодна. Во-первых,
у такого организма велика вероятность получить в многочислен-
ных столкновениях серьезную травму. Во-вторых, неизбежным
результатом чрезмерной агрессивности является нерациональное
расходование времени и сил. Вместо того, чтобы переключиться
на другие формы поведения, более биологически значимые в
сложившейся ситуации, организм продолжает демонстрировать
высокую агрессию и в итоге не успевает справляться с остальны-
ми задачами.
Этот вывод экспериментально подтвержден для целого ряда
объектов. В частности, у дрозофилы получены инбредные линии,
агрессивность которых повышена в несколько раз. Оказалось, что
самцы этих линий, по сравнению с нормой, отличаются досто-
верно пониженной успешностью спаривания.
Таким образом, только адекватный уровень агрессии способ-
ствует выживанию особи и оптимальному размножению.
6.2. Разнообразие форм агрессии
у животных
Задачи агрессивного поведения весьма разнообразны. Они
варьируют в зависимости от видовой принадлежности организма,
возраста, пола, функционального состояния и прочих особен-
ностей, а также от конкретной ситуации, в которой он находится.
Различают множество форм агрессии, среди которых наиболее
распространенными являются следующие.
• Межсамцовая агрессия, обусловленная конкуренцией за
самку или за лидирующее положение в определенной группе (как
правило, обе задачи тесно взаимосвязаны). Подобное поведение
встречается очень широко: достаточно вспомнить о петушиных
боях, тетеревиных «турнирах», брачных поединках оленей и др.
Но не следует забывать, что для некоторых видов характерна
вполне аналогичная агрессия между самками. Один из ярчайших
примеров — пятнистые гиены. У них доминируют самки, а не
самцы. И если в одном помете оказываются «сестры», они с рож-
дения проявляют друг к другу такую агрессию, что вскоре одна
убивает всех остальных.
* Агрессия, связанная с защитой своей территории. Для
примера обратимся к соответствующему поведению тигров. Эти
хищники ведут одиночный образ жизни (не считая сравнительно
коротких периодов спаривания) и ревностно защищают свою
охотничью территорию от посягательств любого соперника неза-
висимо от пола.
147
• Материнская агрессия, обеспечивающая защиту потом-
ства. Подобное поведение характерно для многих животных.
Вместе с тем необходимо учитывать, что у целого ряда видов (в
частности, у страусов нанду и некоторых рыб) забота о потомстве
«возложена» исключительно на самцов. В этих случаях следует
говорить об отцовской агрессии.
• Агрессия, обусловленная страхом перед некой угрозой.
В частности, заяц, спасающийся от нападения волка, в безвы-
ходной ситуации может сам перейти в атаку и попытаться укусить
врага своими мощными зубами.
• Агрессия, вызванная различного рода препятствиями,
не дающими организму достичь желаемого результата. Например,
обезьяна, пытающаяся расколоть орех, способна перейти в весь-
ма агрессивное состояние, если ей никак не удается справиться
с задачей.
• Агрессия хищника, направленная на добычу. Нужно отме-
тить, что данная форма агрессивного поведения существенно
отличается своими проявлениями от всех предыдущих. И дей-
ствительно, кошка, подкрадывающаяся к воробью или к мыши,
не шипит, не выгибает спину, не оскаливает зубы, не выпускает
когти, не поднимает шерсть и т.д. В связи с этим некоторые ис-
следователи высказывают мнение, что действия хищника по от-
ношению к добыче являются не агрессией, а всего лишь специфи-
ческими приемами добывания пищи. Тем не менее многочислен-
ные факты свидетельствуют об обратном (подробнее об этом — см.
далее).
Судя по результатам исследований на модельных объектах,
перечисленные формы агрессии относительно независимы, и каж-
дая в значительной степени контролируется генотипом.
Таким образом, даже у животных, не говоря о человеке,
агрессивность не является элементарным поведенческим
признаком. Она слагается из множества относительно ав-
тономных характеристик, каждую из которых необходимо
анализировать по отдельности.
Познакомимся с несколькими примерами подобного анализа.
6.3. Генетический анализ агрессивности
хищника у крыс и мышей
Первым условием, необходимым для проведения генетическо-
го анализа, является подбор контрастно различающихся форм.
Применительно к рассматриваемому признаку это означает, что
среди представителей одного и того же вида должны существовать
148
как «хищные*, так и «нехищные* особи. Но в действительности
подобное требование трудно выполнимо. Ведь хищник, не агрес-
сивный по отношению к потенциальной добыче, не сможет себя
прокормить и погибнет от голода еще на достаточно ранних эта-
пах развития.
Решением проблемы оказались факультативные хищни-
ки — виды, для которых хищничество не является обязательным.
К животным подобного рода относятся и мыши. Они в основном
используют растительную пищу, но при случае могут поедать и
некоторых беспозвоночных. В частности, экспериментально по-
казано, что «дикие» мыши охотно нападают на сверчков и род-
ственных насекомых и убивают их одним или несколькими
укусами в голову. Эта способность характерна и для многих
инбредных линий. Однако у некоторых линий она полностью от-
сутствует. Таким образом, в зависимости от действий по отноше-
нию к сверчкам, мышей подразделяют на убивающих и неуби-
вающих.
При скрещивании убивающих самок с неубивающими самца-
ми все гибриды первого поколения оказываются убивающими.
Из этого следует, что высокая агрессивность доминирует над
низкой.
Однако теоретически возможно и другое объяснение. Можно
представить, что данная форма афессивного поведения переда-
ется потомкам не «через гены*, а за счет воспитания. При этом
ведущая роль отводится самкам, поскольку именно они вына-
шивают и выкармливают детенышей.
Такую гипотезу не представляет труда экспериментально про-
верить: нужно скрестить неубивающих самок с убивающими
самцами и исследовать поведение потомков по отношению к
сверчкам. Если оно формируется за счет воспитания, первое по-
коление гибридов должно быть неубивающим. Но в действитель-
ности опять получаются убивающие потомки. Это значит, что
изучаемое поведение контролируется генотипом.
К сожалению, анализ второго поколения гибридов не приводит
к каким бы то ни было конкретным результатам из-за сложности
и недостаточной четкости наблюдаемого расщепления. Поэтому
вопрос о количестве соответствующих генов, об их локализации
и молекулярных функциях остается открытым. Тем не менее
можно сделать несколько важных выводов.
Во-первых, судя по результатам многочисленных исследований,
отношение к сверчкам не зависит от других форм агрессии,
а значит, имеет «свой собственный» генетический контроль.
Во-вторых, необходимо учитывать, что убивающие линии
поддерживаются в лабораторных условиях десятками поколе-
ний, не вступая со сверчками ни в какие непосредственные
149
контакты. Из этого следует, что данная форма агрессии мышей
основана на генетически запрограммированном распознавании
образов, а именно — на распознавании образов потенциальной
добычи. Аналогичные выводы получены и в экспериментах на
крысах (их агрессию обычно тестируют по отношению к мы-
шам).
В подразд. 6.2 было отмечено, что некоторые исследователи
считают поведение хищника не формой агрессии, а специфиче-
скими приемами, необходимыми для добывания пищи. Между
тем хорошо известно, что многие хищники убивают потенциаль-
ную добычу, даже будучи сытыми. Сходным образом ведут себя
убивающие мыши и крысы. Напротив, животные, относящиеся
к неубиваюшим линиям, не становятся хищниками даже при
длительном голодании. Эти факты свидетельствуют о том, что
поведение хищника не имеет строгой «привязки» к добыванию
пищи и может рассматриваться в качестве особой формы агрес-
сии.
6.4. Генетический анализ межсамцовой
агрессивности у мышей
Как известно, мыши относятся к социальным животным. Они
образуют крупные семейные группировки (демы) со сложной
многоуровневой системой иерархических отношений. Важным
фактором в установлении и поддержании такой иерархии явля-
ется межсамцовая агрессия в пределах дема.
Агрессивное поведение самцов по отношению друг к другу
выражается у мышей в особых угрожающих позах, в отталкивании
соперника, в нанесении ему укусов (преимущественно в спину,
бока и основание хвоста), в ударах ногами и, наконец, в пресле-
довании побежденного противника. Эти действия можно коли-
чественно учитывать в эксперименте и тем самым оценивать
агрессивность конкретного самца.
Обычно используют метод попарного ссаживания. Двух сам-
цов помещают в одну и ту же клетку и в течение строго опреде-
ленного промежутка времени (10—15 мин) учитывают агрессив-
ные действия каждого из них. Усредняя результаты нескольких
аналогичных повторностей, получают количественную оценку
межсамцовой агрессивности для каждой интересующей исследо-
вателя ин бред ной линии.
Этот метод удобен и прост, но не лишен недостатков. Если
один из самцов проигрывает схватку, он переходит преимуще-
ственно к защитному или подчиненному поведению и уже почти
150
не атакует победившего соперника. Соответственно, оценка его
агрессивности оказывается заниженной. Более адекватна моди-
фикация данного метода, основанная на использовании так на-
зываемого стандартного тестера. В этом случае исследуют
агрессивность лишь одного из самцов, а второй выступает в ка-
честве «провокатора агрессии», причем не способного оказать
существенного отпора. Таким провокатором обычно является
кастрированный самец, имеющий строго определенный (стан-
дартный) генотип. Подобная особь не воспринимается нормаль-
ным самцом как соперник, поэтому, для того чтобы вызвать
межсамцовую агрессию, на провокатора предварительно наносят
капли мочи, полученной от нормальных самцов исследуемого
Таблица 6.1. Анализ материнского влияния на межсамцовую
агрессию у мышей (по Roubertoux and Carlier, 1988)
Исследуемая линия Доля атакующих самцов (%)* Количество атак** Длительность первой атаки (сек)
Н 28 ±8 1,5 ± 0,5 330 ±9
Н, воспитанные самками N 20 ± Ю 1,4 ± 1,2 340 ± 10
Н, рожденные гибридными самками*** 23 ±9 1,9 ± 1,0 320 ± 20
N 80 ± 8 6,5 ± 1,2 150 ± 25
N, воспитанные самками Н 9I ±6 6,3 ±0,9 153 ± 25
N, рожденные гибридными самками**** 80 ±9 8,8 ± 1,3 123 ± 30
* Доля самцов, совершающих хотя бы I атаку в течение 6 мин после подсажи-
вания к тестерному самцу.
** Среднее количество атак, совершаемых самцами в течение 6 минут после
подсаживания к тестерному самцу (первая атака обычно значительно продолжи-
тельнее последующих).
*** Самкам, полученным от скрещивания HxN, пересаживали яичники от
самок Н. Затем этих самок скрещивали с самцами Н. В результате получали мы-
шей, имеющих такой же генотип, как и у линии Н, но рожденных от матерей
гибридного генотипа.
**** Самкам, полученным от скрещивания HxN, пересаживали яичники от
самок N. Затем этих самок скрещивали с самцами N. В результате получали мы-
шей, имеющих такой же генотип, как и у линии N, но рожденных от матерей
гибридного генотипа.
151
генотипа (у мышей именно в моче содержится информация о
возрасте, поле и функциональном состоянии организма).
Многочисленные исследования показали, что у разных инбред-
ных линий уровень межсамцовой агрессивности значительно
варьирует. Такие различия, судя по результатам генетического
анализа, преимущественно обусловлены не родительским вос-
питанием, а генотипом. Этот вывод убедительно подтвержден в
специальных экспериментах с использованием «приемных роди-
телей» или пересадкой яичников (табл. 6.1).
Селекция на повышенную или пониженную межсамцовую
агрессивность практически не влияет на прочие формы агрессии.
Этот факт свидетельствует о том, что рассматриваемая характе-
ристика находится под относительно автономным генетическим
контролем. Более того, отдельные ее составляющие (длительность
латентного периода перед вступлением в драку, количество сты-
чек, а также суммарная их продолжительность) тоже наследуются
относительно независимо друг от друга.
Таким образом, в генетическом контроле межсамцовой
агрессивности, несомненно, принимает участие множество
генов. Некоторые уже обнаружены и подробно изучены. Кратко
рассмотрим наиболее важные результаты.
6.5. Молекулярные основы межсамцовой
агрессивности у мышей
Судя по данным, представленным в табл. 6.2, интенсивность
межсамцовой агрессии находится под контролем нескольких тес-
но взаимодействующих молекулярных систем. Одна из них свя-
зана с восприятием обонятельной информации. Дело в том, что
у мышей и многих других млекопитающих основным языком
общения являются запахи. Соответственно, ключевую информа-
цию о своих соплеменниках каждая мышь получает именно через
органы обоняния (точнее, через так называемый вомероназаль-
ный орган — особо чувствительную зону слизистой носа, специ-
ально предназначенную для восприятия индивидуальных запахов).
И действительно, у мышей с дефектами вомероназальных
нейронов уровень межсамцовой агрессивности значительно
снижен.
Вторая система обеспечивает половую дифференцировку и
регулирует широкий спектр поведенческих программ, непосред-
ственно или косвенно связанных с половым размножением.
Важнейшими компонентами этой молекулярной системы явля-
ются мужские и женские половые гормоны. Известно, что их
152
153
Таблица 6.2. Гоны, контролирующие межсамцовую агрессивность у мышей
Система Ген Соответствующий белок Молекулярные функции
Обонятельная Gai2 G-белок Передача информации от вомероназальных рецепторов на следующие компоненты сигналь- ного каскада
Тгр2 Ионный канал Формирование нервных импульсов вомерона- зальными нейронами
Гормональная Аг Рецептор андрогенов Регуляция активности нейронов посредством андрогенов
Esra а-Рецептор эстрогенов Регуляция активности нейронов посредством эстрогенов
Esrb 0-Рецептор эстрогенов
Серотониновая Tph2 Триптофан гидроксилаза 2 Синтез ключевого предшественника серотони- на в нейронах головного мозга
Маоа Моноамин оксидаза А Деградация серотонина
Slc6a4 Транспортер серотонина Реимпорт серотонина из синапса в пресинапти- ческие окончания
5-HtlaR 1А-Рецептор серотонина Регуляция активности нейронов посредством серотонина
5-HtlbR IB-Рецептор серотонина
Прочие Co mt Катехол-О-метилтрансфераза Деградация дофамина, адреналина и норадре- налина
Окончание табл. 6.2
Система Ген Соответствующий белок Молекулярные функции
aCamkll Са+2-зависимая протеинкиназа Регуляция молекулярных каскадов, активируе- мых фосфолипазой
nNos Синтетаза оксида азота Регуляция активности нейронов посредством оксида азота
Avprlb Рецептор вазопрессина Регуляция активности нейронов посредством вазопрессина
Rgs2 Регулятор активности G-белка Регуляция формирования нервных импульсов в нейронах гиппокампа
Hrhl Рецептор гистамина Регуляция активности нейронов посредством гистамина
Ncaml Белок, участвующий в адгезии нейронов Регуляция формирования межнейронных кон- тактов
116 Интерлейкин Регуляция функционирования иммунной систе- мы
соотношение контролируется генотипом. При этом ведущая роль
отводится генам, локализованным в половых хромосомах X и К
У самок (для них характерны две %-хромосомы) преобладают
женские половые гормоны — эстрогены, а у самцов (XY) — муж-
ские, т.е. андрогены. И чем выше уровень андрогенов у данно-
го самца, тем отчетливее выражена его межсамцовая
агрессивность.
Различного рода нарушения этой системы тоже существенно
влияют на рассматриваемый признак. Например, у самцов, ли-
шенных половых желез, уровень андрогенов снижается практи-
чески до нуля. В результате такие самцы не проявляют агрессии
ни по отношению к нормальным, ни друг к другу. Сходным об-
разом ведут себя и мутанты по гену Аг. В этом случае уровень
андрогенов сохраняется в норме, но нарушены функции соот-
ветствующих рецепторных белков. Между тем при отсутствии
некоторых рецепторов эстрогенов (например, у мутантов с но-
каутированным геном Esrb) интенсивность межсамцовой агрессии
значительно возрастает. По-видимому, организм воспринимает
такую ситуацию как дополнительный сдвиг гормонального балан-
са в сторону андрогенов.
Важную роль в рассматриваемой форме агрессивного поведе-
ния играет и серотониновая система. Ее ключевым компонентом
является серотонин — один из широко используемых нейроме-
диаторов. Нейроны, выделяющие этот нейромедиатор, обнаруже-
ны в гиппокампе, гипоталамусе, базальных ядрах, лобных долях
коры больших полушарий и некоторых других отделах головного
мозга. В зависимости от своей конкретной локализации они ре-
гулируют разные аспекты жизнедеятельности организма: сон,
температуру тела, чувство голода, настроение, половое поведение
и агрессивность.
У многих млекопитающих прослеживается четкая закономер-
ность: чем ниже активность серотониновой системы, тем
выше наблюдаемый уровень межсамцовой агрессии. В связи
с этим принято считать, что серотониновая система оказывает на
межсамцовую агрессию тормозящий эффект. Впрочем, чрезмерно
высокое содержание серотонина (например, при отсутствии мо-
ноаминокс идазы — ключевого фермента, разрушающего данный
нейромедиатор) тоже приводит к усилению межсамцовой агрес-
сии.
В чем причина такого эффекта, пока непонятно. Вполне воз-
можно, что серотонинэргические синапсы нормально функцио-
нируют только при эффективной очистке. По-видимому, в кон-
троле рассматриваемого нами признака участвуют и некоторые
Другие молекулярные системы. Но пока подтвердить это экспе-
риментально не удалось.
155
6.6. Генетический анализ агрессивности
животных по отношению к человеку
Исследования генетического контроля данной формы агрессии
предпринимались неоднократно на разных модельных объектах.
Но основные результаты получены в многолетних экспериментах,
начатых около 50 лет назад в Институте цитологии и генетики
(Новосибирск) под руководством академика Дмитрия Констан-
тиновича Беляева.
Исходным толчком для проведения этих уникальных экс-
периментов послужила вполне конкретная прикладная задача.
Дело в том, что в 1950-е гг. одним из объектов пушного зверо-
водства в нашей стране являлась серебристо-черная лисица.
Однако даже при длительном разведении на зверофермах эти
животные оставались пугливыми по отношению к человеку,
что негативно отражалось на их плодовитости и качестве меха.
Более того, при непосредственном контакте с человеком они
проявляли довольно высокую степень агрессии и при укусе
могли нанести значительные повреждения. Поэтому перед ге-
нетиками была поставлена задача: осуществить селекцию
серебристо-черных лисиц на пониженный уровень агрессии по
отношению к человеку. Такую селекцию называют одомашни-
ванием, или доместикацией.
Работу проводили по следующей схеме. У животных, содержа-
щихся по одиночке в небольших вольерах, оценивали реакцию
на приближение незнакомого человека. При этом использовали
специальную балльную шкалу, отражающую интенсивность ис-
следуемой формы агрессии. В результате было выявлено четыре
типа особей:
• высоко агрессивные (при попытке человека зайти в вольеру
они рычали, щелкали зубами, бросались на него, старались уку-
сить протянутую палку и т.п.);
• злобно-трусливые (они проявляли и страх, и агрессию);
• трусливые (основная реакция — ярко выраженный страх,
жалобное повизгивание);
• реагирующие на человека относительно спокойно.
Из нескольких десятков исследованных «полудиких» лисиц
были отобраны особи с наименьшей степенью агрессии. Их скре-
стили друг с другом, а затем из полученного потомства опять
отобрали наименее агрессивных животных. В итоге уже через не-
сколько поколений селекции средние показатели агрессивности
значительно снизились: лисицы спокойно реагировали на незна-
комого человека, а некоторые позволяли себя гладить и даже брать
на руки. Наконец, приблизительно через десять поколений от-
156
бора были получены настолько дружелюбные к человеку лисицы,
что их поведение мало отличалось от поведения собак.
Параллельно гинетики осуществляли отбор лисиц и в противо-
положном направлении. Эта селекция тоже оказалась весьма
продуктивной.
Таким образом, на основе исходных «полудиких» лисиц были
получены контрастно различающиеся инбредные линии: одна —
ручная, а вторая — повышенно агрессивная к человеку. Как по-
казали дальнейшие исследования двух этих линий, различия
между ними имеют генетическую природу и, по-видимому, за-
трагивают небольшое количество генов. К числу вероятных кан-
дидатов на данную роль относятся гены, контролирующие функ-
ционирование серотониновой системы. И действительно, уста-
новлено, что активность триптофангидроксилазы (ключевого
фермента биосинтеза серотонина) у одомашненных лисиц
значительно выше по сравнению с агрессивными. Любопытно,
что одомашненные лисицы отличались от агрессивных и по цело-
му ряду сугубо морфологических признаков: шерсть стала более
тусклой и существенно погрубела, а хвосты завились «крючком»,
как у некоторых собак. Таким образом, гены, участвующие в
контроле агрессивности, оказывают определенное влияние и на
внешние признаки.
Отметим, что оба рассмотренных варианта селекции серибрис-
то-черной лисицы не отразились на уровне материнской и меж-
самцовой агрессии.
Аналогичные результаты были получены и в исследованиях на
крысах (в этом случае оценивали реакцию на протянутую перчат-
ку).
Перечисленные факты еще раз свидетельствует о том, что раз-
личные формы агрессии независимы друг от друга и находятся
под относительно автономным генетическим контролем.
6.7. Основные итоги изучения генетического
контроля агрессивности животных
На примере мышей и серебристо-черных лисиц мы познако-
мились с некоторыми формами агрессивного поведения и убеди-
лись, что каждая из них контролируется генотипом. Безусловно,
существенную роль играет и среда (табл. 6.3). Однако обычно она
лишь «накладывается» на генетические различия, способствуя
или препятствуя их проявлению.
Разные формы агрессивного поведения во многом похожи. Это
прослеживается не только в их сходном биологическом предна-
157
Таблица 6.3. Факторы внешней среды, влияющие на интенсив-
ность межсамцовой агрессии у мышей
Факторы внешней среды Оказываемый эффект Молекулярный механизм воздействия
Генотип соперника* Варьирует в зависи- мости от генотипа каждого из самцов Восприятие специфи- ческого запаха со- перника
Количество сопер- ников При возрастании ко- личества соперников агрессивность повы- шается Восприятие специфи- ческих запахов каж- дого из соперников
Успех в предыдущих схватках Повышение агрессив- ности Неизвестен
Неудача в предыду- щих схватках Ослабление агрессив- ности Неизвестен
Наличие самок Варьирует в зависи- мости от генотипа всех участников Восприятие женских феромонов
Территория, на которой происходит схватка На «своей» террито- рии агрессивность значительно возрас- тает, в то время как на «чужой», наоборот, значительно падает Восприятие пахучих меток территории
П редварител ьная изоляция в течение нескольких недель Резкое и, по-ви- димому, патологи- ческое повышение агрессивности** Неизвестен
* Для каждого организма все остальные живые существа являются факторами
окружающей среды.
** Подвергнутые изоляции самцы при попадании на общую территорию на-
падают на самок, наносят друг другу укусы в жизненно важные органы, атакуют
неживые объекты и даже крыс.
значении (причинять другим организмам какой-либо вред), но и
в конкретных путях решения соответствующих задач. И действи-
тельно, особи одного и того же вида обычно пользуются пример-
но одинаковыми «техническими приемами» во всех вариантах
конфликтных ситуаций.
158
В частности, собака, обороняющая своих щенков, лает, рычит,
поднимает шерсть и оскаливает зубы практически так же, как
если бы она отстаивала свою территорию, любимую игрушку,
лидерство, пищу и т.д. Из этого следует, что разные формы агрес-
сивного поведения имеют достаточно сходную физиологическую
основу и, вероятно, контролируются близкими молекулярными
системами.
Одной из них является система серотониновой регуляции.
Многочисленные исследования показывают, что уровень серото-
нина влияет на все без исключения виды агрессии. Но тогда воз-
никает вполне закономерный вопрос: почему же различные
формы агрессивного поведения проявляют значительную степень
автономности друг от друга? По-видимому, это обусловлено не-
сколькими причинами.
Во-первых, необходимо учитывать, что агрессивное поведение
контролируется не только серотониновой, но и другими система-
ми, и в частности — мужскими и женскими половыми гормонами.
Известно, что их соотношение играет важную роль в регуляции
материнской, межсамцовой и территориальной агрессии, однако
практически не влияет на агрессию хищника. При этом материн-
ская агрессия усиливается эстрогенами, в то время как межсам-
цовая и территориальная — андрогенами. И в результате различ-
ные формы агрессивного поведения оказываются в некоторой
степени независимыми друг от друга.
Во-вторых, даже сама серотониновая система влияет на раз-
ные формы агрессии неодинаково. Этот факт убедительно про-
демонстрирован в исследованиях на мышах с искусственно по-
вышенным содержанием серотонина. Как выяснилось, у особей,
подвергнутых такому воздействию, некоторые формы агрессии
(например, межсамцовая) ослаблены по сравнению с нормой
довольно существенно, а некоторые (агрессия хищника) — от-
носительно слабо.
В-третьих, «стандартная программа» агрессивного поведения,
несомненно, управляется множеством пусковых механизмов.
Каждый задействуется в определенном спектре ситуаций и, по-
видимому, имеет свой собственный генетический контроль.
И если один из механизмов оказывается дефектным, это ослабля-
ет определенную форму агрессии, в то время как все остальные
соответствуют норме.
Таким образом, в зависимости от конкретного генотипа
организм может быть агрессивным в одной ситуации, но не
способным выполнить те же самые действия в другой.
Поэтому, описывая поведенческие признаки организма, не-
корректно говорить о его агрессивности в целом, каждую
Форму агрессии нужно оценивать по отдельности.
159
6.8. Разнообразие форм агрессии
у человека
Человек, подобно другим высокоразвитым животным, тоже
способен к различным проявлениям агрессии. Возникает зако-
номерный вопрос: как их классифицировать? С одной стороны,
очевидно, что в человеческом обществе можно встретить все
формы агрессии, характерные для животных: межсамцовую, ма-
теринскую, отцовскую, территориальную и т.д. С другой стороны,
агрессивные действия любого человека можно рассматривать с
сугубо социальной точки зрения. В результате предложены десят-
ки различных классификаций, что затрудняет сравнение данных,
полученных разными авторами. Поэтому в последнее время пред-
принимаются попытки критически осмыслить существующие
классификации агрессии и разработать на их основе более или
менее унифицированные. Постепенно растет понимание того,
что все разнообразие проявлений человеческой агрессии мож-
но описывать четырьмя параметрами.
Первый параметр — причина возникновения агрессии.
Эти причины весьма разнообразны и, как показывает анализ, не
имеют отчетливого деления на социальные и биологические (под-
робнее см. подразд. 6.18 и 9.12.1). Соответственно, подавляющее
большинство современных исследователей предпочитает не вда-
ваться в тонкие подробности и для удобства рассматривает всего
две формы агрессии:
• защитную, или реактивную, агрессию (она возникает в
ответ на провоцирующие внешние воздействия);
• инструментальную, или проактивную, агрессию (ее непо-
средственным инициатором является сам человек).
Второй параметр — способ проявления агрессии. В этом
плане обычно выделяют три формы агрессии:
• физическую агрессию (удары, толчки, нанесение различных
повреждений);
• вербальную агрессию (грубая речь, оскорбления, угрозы);
• ментальную, или скрытую, агрессию (любые агрессивные
действия, совершаемые мысленно).
Третий параметр — объект, подвергаемый агрессии. Со-
ответственно выделяют:
• агрессию по отношению к предметам'.
• агрессию по отношению к животным. Нанесение вреда
«неодушевленным» живым объектам (например, сбивание ногой
мухоморов в лесу) приравнивают обычно к действиям из преды-
дущего пункта;
• агрессию по отношению к самому себе’.
160
• агрессию по отношению к другим людям. При этом раз-
личают прямую и опосредованную агрессию (первая направлена
непосредственно на конкретного человека, а вторая — на раз-
личные связанные с ним объекты).
Четвертый параметр — интенсивность агрессии. Дан-
ный параметр можно оценивать в разных ситуациях и поэтому
его можно расчленить его на несколько более дробных:
• враждебность, или общий негативизм (отношение чело-
века к окружающему миру при отсутствии каких бы то ни было
провоцирующих воздействий);
• раздражительность (необходимый уровень провоциру-
ющих воздействий для перехода человека от обычного поведения
к агрессивному);
• злобность или жестокость (степень выраженности спро-
воцированной агрессии).
К сожалению, приведенная классификация весьма несовер-
шенна. Дело в том, что она не учитывает, какими именно при-
чинами (а значит, какими конкретными пусковыми механизма-
ми) обусловлены те или иные агрессивные действия. Соответ-
ственно в одну и ту же группу попадают различные, хотя и
близкие по своему проявлению формы агрессии. Тем не менее
несмотря на указанный недостаток, такая классификация имеет
рациональное зерно: она направлена на поиск элементарных
психологических характеристик, из которых слагается агрессив-
ное поведение человека. И чем более дробными окажутся эти
характеристики, тем адекватнее они для использования в гене-
тических исследованиях.
6.9. Методы оценки агрессивного поведения
человека
Изучение генетического контроля агрессивности человека было
начато около 80 лет тому назад (первая публикация, посвященная
этому вопросу, вышла в 1929 г.). Поначалу использовали различ-
ные косвенные оценки, в основном — наличие судимостей или
прочих взысканий, вызванных грубыми нарушениями обществен-
ного порядка. Оказалось, что склонность человека к таким право-
нарушениям в значительной степени (на 40...60 %) зависит от
генотипа. Эта зависимость особенно высока для криминальных
поступков.
Однако со временем выяснилось, что подобные оценки не
являются адекватными показателями агрессивности человека.
Во-первых, нередко происходят судебные ошибки, поэтому само
161
по себе наличие судимости не означает, что данный человек пред-
расположен к агрессии. Во-вторых, различные формы криминаль-
ного поведения обладают достаточно высокой степенью автоном-
ности (с одним из примеров можно будет познакомиться в под-
разд. 6.17). Соответственно, необходимы другие методические
подходы, позволяющие оценивать агрессивность более точно.
К таким подходам относится использование специализиро-
ванных опросников. Испытуемому предлагают ответить на ряд
вопросов, непосредственно или косвенно связанных с его агрес-
сивностью, и тем самым получают оценку исследуемого призна-
ка. Одним из классических считается опросник Басса—Дарки,
построенный по принципу «да —нет» и позволяющий оценивать
как «общую агрессивность», так и некоторые ее составляющие.
Этот опросник исходно содержал 66 позиций, но впоследствии
был преобразован в более краткие версии, получившие в совре-
менных исследованиях широкое распространение. Особенно
популярна версия Басса — Перри. Она содержит 29 позиций,
объединенных в четыре блока: физическая агрессия, вербальная
агрессия, враждебность и злобность. При этом используется пя-
тибалльная система оценок: «да», «скорее да, чем нет», «затруд-
няюсь ответить», «скорее нет, чем да» и «нет».
Главное и несомненное достоинство подобных опросников —
возможность работать с практически неограниченными выбор-
ками испытуемых и очень быстро оценивать различные параме-
тры агрессивности. При этом экспериментально показано, что
при повторных опросах наблюдается достаточно высокая вос-
производимость результатов. Однако необходимо учитывать, что
такие оценки, хотя и стабильны, но отражают лишь собственное
мнение испытуемых, а потому неизбежно носят субъективный
характер. Ведь даже в том случае, если испытуемый предельно
откровенен, его мнение о себе не во всем соответствует действи-
тельности.
Наиболее объективную оценку агрессивности человека обе-
спечивают непосредственные наблюдения за испытуемыми.
Некоторые исследования основаны на длительных наблюдениях
продолжительностью от нескольких дней до нескольких месяцев,
в ходе которых учитывают весь спектр агрессивных проявлений.
Безусловно, такие исследования весьма трудоемки и не могут
охватывать очень крупные выборки испытуемых. Поэтому раз-
работан целый ряд экспериментальных подходов, позволяющих
быстро оценивать агрессивность индивида. Например, ребенку
дают определенную куклу и учитывают, сколько раз и как сильно
он ее ударит. В результате исследователь получает экспресс-оценку
физической агрессивности ребенка по отношению к предмету.
Близкие подходы разработаны и для взрослых.
162
6.10. Основные подходы к генетическому
анализу предрасположенности
к самоубийству
Самоубийство — это крайняя форма агрессивного поведения,
направленного индивидом непосредственно на себя самого. По
оценкам Всемирной организации здравоохранения (ВОЗ), еже-
годно в мире совершается до I млн самоубийств, причем их чис-
ло с каждым годом неуклонно возрастает.
Подобная агрессия известна не только у человека. Случаи на-
несения себе тяжелых физических повреждений, иногда приво-
дящих даже к летальному исходу, описаны и у ряда других мле-
копитающих: крыс, мышей, макак, горилл и шимпанзе. Такие
травмы являются результатом патологической агрессивности,
спровоцированной либо аномалиями нервной системы, либо
введением некоторых фармакологических препаратов, либо раз-
личного рода стрессовыми воздействиями: болью, страхом, дли-
тельной изоляцией и т.д. Поэтому вполне вероятно, что в основе
самоубийств тоже лежат определенные физиологические откло-
нения, непосредственно связанные с патологическим повышени-
ем агрессивности.
Как известно, подавляющее большинство попыток самоубий-
ства совершается людьми в состоянии тяжелой депрессии, вы-
званной различного рода стрессовыми ситуациями. В то же время,
судя по результатам близнецового анализа, вероятность суицида
на 30...60 % зависит от генотипа. Эти данные свидетельствуют о
том, что причиной самоубийств, скорее всего, является взаимо-
действие двух факторов:
• во-первых, генетически контролируемой предрасположен-
ности к суициду;
• во-вторых, условий, способствующих проявлению генети-
ческой предрасположенности к суициду, т.е. стрессовых ситуаций,
тяжелых переживаний и др.
Обычно исследование проводят по следующей схеме. В каче-
стве испытуемых используют либо самоубийц (а точнее — их
посмертно взятые образцы ДНК), либо людей, совершавших по-
пытки суицида. При этом, осуществляя отбор конкретных ис-
пытуемых, стараются обеспечить их максимально возможную
однородность по прочим факторам, способным спровоцировать
самоубийство. Например, отбирают исключительно из числа ши-
зофреников или людей, страдающих тяжелыми формами депрес-
сии. В качестве контроля используют больных с такими же диаг-
нозами, но никогда не совершавших попыток самоубийства, или
же обычных психически здоровых людей.
163
Напомним, что агрессивное поведение у крыс и мышей в зна-
чительной степени определяется системой серотониновой регу-
ляции. То же самое характерно для макак, серебристо-черных
лисиц, некоторых птиц, рыб и ракообразных. Напрашивается
естественное предположение, что и у человека эта система тоже
вовлечена в контроль за агрессивностью. И действительно, судя
по результатам клинических исследований, низкое содержание
5-гидроксиинозитолуксусной кислоты (основного производного
серотонина в спинномозговой жидкости) коррелирует со склон-
ностью к агрессии, и в частности — к самоубийству.
Таким образом, к числу наиболее вероятных кандидатов на
участие в генетическом контроле предрасположенности к
суициду относятся гены, обеспечивающие функционирование
серотониновой системы.
Безусловно, количество этих генов достаточно велико. В на-
стоящее время основное внимание сосредоточено на семи (ТРН1,
ТРН2, SLC6A4, HTR1A, HTR1B, HTR2A и МАОА\ контроли-
рующих ключевые этапы биосинтеза серотонина, его реимпорта
в клетку, рецепции и деградации. Кратко рассмотрим важнейшие
результаты проведенных исследований.
6.11. Роль генов, контролирующих синтез
серотонина
Серотонин синтезируется у всех млекопитающих из триптофа-
на, одного из 20 вариантов аминокислот. Данный процесс состо-
ит из двух последовательных этапов (рис. 6.1). Первый этап обе-
спечивается триптофангидроксилазой — ферментом, имеющим
высокую субстратную специфичность и участвующим исключи-
тельно в процессе биосинтеза серотонина. Второй обслуживается
декарбоксилазой ароматических аминокислот. Этот фермент об-
ладает широкой субстратной специфичностью и принимает уча-
стие во многих молекулярных процессах, затрагивающих произ-
водные триптофана, фенилаланина и тирозина.
Показано, что первый этап является лимитирующим: именно
от него зависит количество серотонина, синтезируемого в орга-
низме того или иного человека. Поэтому в генетических исследо-
ваниях предрасположенности к самоубийству основное внимание
уделяют именно триптофан гидроксилазе. Данный фермент пред-
ставлен у человека двумя изоформами, значительно различающи-
мися по своей тканевой специфичности. Одна изоформа (ее ко-
дирует ген ТРИ/) функционирует преимущественно в стенках
двенадцатиперстной кишки, а также в печени, сердце, легких,
164
почках и надпочечниках. Она обнаружена и в некоторых отделах
головного мозга, однако в значительно меньших концентрациях.
Между тем вторая изоформа триптофан гидроксилазы (ее коди-
рует ген ТРН2) характерна исключительно для нейронов голов-
ного мозга.
Ген ТРН1 существует в виде множества аллелей, отличающих-
ся друг от друга заменами отдельных нуклеотидов. Такие замены
обнаружены примерно в 200 положениях, преимущественно в
промоторной области или в интронах. В исследованиях, посвя-
щенных генетике суицидного поведения, к настоящему времени
использовано 11 из этих положений.
Первые полученные результаты свидетельствовали о том, что
замены 218А и 779С (обе локализованы в пределах 7-го интрона)
тесно ассоциируют со склонностью к суицидному поведению.
Этот вывод подтвержден и в некоторых последующих работах.
Однако в подавляющем большинстве проведенных исследований
обнаружить искомую ассоциацию не удалось.
Не вносит окончательной ясности и мета-анализ. Так, анализ
17 независимых исследований не обнаружил ассоциации ни с
одной из перечисленных замен. Между тем в трех других, более
строгих аналитических работах показана слабая но все же
статистически значимая связь между склонностью к суи-
циду и заменой 218А. Данная нуклеотидная замена встре-
чается у самоубийц несколько чаще, чем в контрольных
группах. Однако обнаруженное различие сравнительно невелико
и статистически подтверждается только для очень крупных вы-
борок. Таким образом, ни одна из исследованных нуклеотидных
Триптофан НО /=\ L Триптофан- / \ гидроксилаза ' nh2 н НО Декарбоксилаза у ароматических аминокислот / Рис. 6.I. Биосинтез серотонина Г идрокситриптофан но но / NH2 н =\ NH2 N H Серотонин
165
замен не оказывает на склонность к самоубийству ярко выражен-
ного влияния.
Ген ТРН2 обнаружен у человека сравнительно недавно. Для него
характерна высокая степень полиморфизма: к настоящему време-
ни известно более 900 положений, по которым встречаются за-
мены отдельных нуклеотидов. В генетический анализ предрасполо-
женности человека к самоубийству вовлечено приблизительно
30 сайтов. Судя по имеющимся данным, суицидное поведение
ассоциирует с заменами 519Т, 2058G, 40237G и 59665С. Од-
нако пока не получено независимых подтверждений, этот вывод
необходимо рассматривать только как предварительный.
6.12. Роль гена SLC6A4, кодирующего
транспортер серотонина
Серотонин является одним из важнейших нейромедиаторов.
Соответствующие синапсы обнаружены в центральной нервной
системе (преимущественно в миндалине, ядрах шва, гиппокампе,
гипоталамусе, полосатом теле и лобных долях коры больших по-
лушарий), пищеварительном тракте и некоторых других перифе-
рических органах. При этом в зависимости от своей конкретной
локализации они регулируют различные аспекты жизнедеятель-
ности организма: сон, чувство голода, жажду, температуру тела,
половое поведение, общее настроение и беспокойство. Два по-
следних аспекта тесно связаны со склонностью к самоубийству.
Но, к сожалению, пока неизвестно, какие именно синапсы непо-
средственно вовлечены в контроль суицидного поведения.
Молекулы серотонина, попавшие в синаптическую щель, под-
вергаются быстрому реимпорту в пресинаптическое окончание.
Этот процесс обеспечивается специальным мембранным белком,
получившим название «транспортер серотонина* и явля-
ющимся продуктом гена SLC6A4. В данном гене обнаружено
более 500 нуклеотидных замен, а также два участка, варьирующих
по своей протяженности:
первый — 44-нуклеотидная последовательность в промото-
ре, приблизительно за I т. п.н. до старта транскрипции. Эта по-
следовательность может присутствовать или отсутствовать, при-
водя к существованию двух вариантов промотора: длинного и
короткого (соответственно L и 5);
второй — / 7-нуклеотидные повторы в интроне 2, их может
быть 9, 10 или 12.
В генетическом анализе предрасположенности к суицидному
поведению к настоящему времени используется следующий по-
лиморфизм:
166
• варианты L и 5-промотора. Показано, что 5-промотор менее
эффективен: он обусловливает пониженный уровень транскрип-
ции гена. В результате из-за нехватки транспортера серотонина
соответствующие синапсы должны очищаться недостаточно.
В свою очередь, это, казалось бы, должно привести к существен-
ному повышению активности серотониновой системы (по анало-
гии с дофаминовой — см. подразд. 5.4). Но в действительности
наблюдается прямо противоположная картина: 5-промотор сни-
жает активность серотониновой системы. В чем причина такой
ситуации, пока непонятно;
• нуклеотидный полиморфизм по положению -179. Он инте-
ресен тем, что замена -179G нарушает важный участок промотор-
ной области и тоже приводит к ослаблению транскрипции гена;
• число повторов в интроне 2. Показано, что аллели, содер-
жащие 12 повторов, транскрибируются с большей эффективно-
стью, чем остальные.
Наиболее изучен полиморфизм размеров премоторной обла-
сти. При этом отчетливо просматривается следующая закономер-
ность: S-промотор повышает вероятность «жестких» са-
моубийств. Речь идет о суициде с использованием различных
видов оружия, самосожжении, повешении, прыжках с высо-
ты и т.д. Между тем для относительно «легких» форм
ухода из жизни (вскрытие вен или прием смертельных доз
снотворного) ни в одном из исследований достоверной связи
не обнаружено.
Таким образом, склонность к «жесткому» или «легкому»
суициду нужно рассматривать как две самостоятельные
поведенческие характеристики. Этот вывод убедительно под-
твержден и мета-анализом (табл. 6.4).
Если все виды самоубийства рассматривать вместе, обнаружить
влияние 5-промотора обычно не удается. Об этом же свидетель-
ствуют и результаты мета-анализа при обобщении данных 18 не-
зависимых работ. Однако в двух других обширных мета-анализах
наличие слабой ассоциации статистически подтверждено. Воз-
можно, такая ассоциация действительно существует, но крайне
слаба и значима лишь в многотысячных выборках.
В чем же различие между «жесткими» и «легкими» формами са-
моубийства? В первом случае человек желает погибнуть немедленно,
во втором — планирует умереть в ближайшем будущем и знает, что
смерть его может быть предотвращена в результате своевременного
медицинского вмешательства. Таким образом, люди, прибегающие
к «легкому» суициду, настроены на уход из жизни менее решитель-
но по сравнению с людьми, использующими «жесткие» формы.
5-промотор тесно ассоциирует не только с «жесткими», но и с
некоторыми другими формами суицидного поведения. В част-
167
168
Таблица 6.4. Мета-анализ ассоциации между S-промотором гена SLC6A4 и различными формами суи
цидного поведения (по U and Не, 2007)
Анализируемые выборки* Результат анализа
СП Контроль
Частота варианта S-промотора в анализируемых выборках
Пациенты, прибегавшие к «жестким» формам СП Норма S-промотор слабо ассоциирует с «жесткими» формами СП (р = 0,0396)
Пациенты, прибегавшие к «легким» формам СП Норма Ассоциации не обнаружено (р = 0,7558)
Пациенты, прибегавшие к «жестким» формам СП Пациенты, не имеющие суицидно- го опыта S-промотор ассоциирует с «жесткими» формами СП (р = 0,0009)
Пациенты, прибегавшие к «легким» формам СП Пациенты, не имеющие суицидно- го опыта Ассоциации не обнаружено (р = 0,4901)
Частота генотипов SS + SL в анализируемых выборках
Пациенты, прибегавшие к «жестким» формам СП Норма + Пациенты, не имеющие суицидного опыта SS + LS ассоциирует с «жесткими» формами СП (р < 0,0001)**
Пациенты, прибегавшие к «легким» формам СП Норма + Пациенты, не имеющие суицидного опыта Ассоциации не обнаружено (р = 0,7897)
* В таблице использованы следующие обозначения и понятия:
СП — суицидное поведение; пациенты — люди с различными поведенческими отклонениями (алкоголики, наркоманы, ши-
зофреники и др ), либо имеющие суицидный опыт, либо не имеющие его; норма — психически полноценные люди, не имеющие
суицидного опыта.
** Предрасположенность человека к «жестким» формам самоубийства тесно ассоциирует с наличием в его генотипе хотя бы
одного экземпляра гена SLC6A4 с короткой формой промотора. Данный вариант промотора проявляется как доминантный.
ности, показана его связь с повторными попытками, а также с
попытками, приводящими к тяжелым последствиям. Таким об-
разом, суицидное поведение на фоне S’-промотора реализуется
намного более интенсивно, нежели на фоне полноразмерного
L-варианта.
Судя по результатам четырех независимых работ, нуклеотидный
полиморфизм в положении -179 не оказывает существенного
влияния на склонность к самоубийству. Ситуация с повторами в
интроне 2 не столь однозначна. По некоторым данным, аллели с
10 повторами достаточно тесно ассоциируют со склонностью к
суициду. Но в большинстве исследований такой ассоциации не
обнаружено. Не выявляется она и при использовании мета-анализа.
Поэтому в настоящее время принято считать, что этот полимор-
физм не связан с предрасположенностью к самоубийству.
6.13. Роль гена MAQA, контролирующего
деградацию серотонина
Значительная часть реимпотрированного из синапса серото-
нина подвергается в пресинаптическом окончании быстрой де-
градации. Этот процесс совместно обеспечивается несколькими
ферментами, важнейшим из которых является моноаминоксида-
за. Данный фермент участвует в деградации не только серотони-
на, но и целого ряда других биологических аминов, в том числе —
дофамина, адреналина и норадреналина. Поэтому он способен
играть очень важную роль в регуляции активности многих синап-
тических контактов и тем самым влиять на различные формы
поведения.
Моноамин оксидаза представлена у человека двумя высоко
гомологичными изоформами (А и В), являющимися продуктами
генов МАОА и МАОВ. Обе функционируют в нейронах головно-
го мозга, преимущественно в лобных долях коры больших по-
лушарий, гиппокампе, миндалине, гипоталамусе и некоторых
других. Однако, несмотря на сходство обеих изоформ, основное
внимание пока уделяют только одной из них, а именно — моно-
аминоксидазе А.
В соответствующем гене обнаружено более 800 нуклеотидных за-
мен, а также три участка, варьирующих по своей протяженности:
• первый — 30-нуклеотидные повторы в промоторе, при-
близительно за 1.2 т. п.н. до начала транскрипции. Они встреча-
ются в трех, трех с половиной, четырех или пяти экземплярах;
• второй — 23-нуклеотидные повторы в интроне 7;
• повторы (СА)„ в интроне 2.
169
К настоящему времени в генетический анализ предрасполо-
женности к суициду вовлечено четыре полиморфных сайта в гене
Л/ЛСМ:
• 30-нуклеотидные повторы в промоторной области. Известно,
что аллели с тремя с половиной или четырьмя повторами транс-
крибируются в несколько раз интенсивнее по сравнению с осталь-
ными и обуславливают повышенное содержание моноаминокси-
дазы А (а значит, более эффективную деградацию серотонина,
что в принципе должно приводить к усилению агрессивности);
• повторы (СА)Л в интроне 2;
• нуклеотидные замены Т/G в положении 941 (экзон 8). По-
казано, что наличие гуанина в данном положении приводит к
существенному усилению активности фермента;
• нуклеотидные замены С/Т в положении 1460 (экзон 14). Эти
замены тоже влияют на активность фермента: в присутствии ти-
мина она повышается в несколько раз.
Судя по результатам большинства проведенных исследова-
ний^ полиморфизм по этим сайтам не связан со склонностью
к самоубийству. Нов двух работах ассоциация все же обнаруже-
на. В одной — с предрасположенностью к «жестким» формам
самоубийства (ассоциируют три с половиной и четыре копии по-
второв в промоторе), в другой — со склонностью к суициду толь-
ко у мужчин (ассоциирует нуклеотидная замена 1460Т). Поэтому
вполне вероятно, что остальные исследования не смогли обнару-
жить влияния гена МАОА по причине недостаточно дробного
анализа признака.
6.14. Роль генов, кодирующих рецепторы
серотонина
У человека известно 14 типов рецепторов серотонина. Неко-
торые из них расположены на постсинаптических мембранах и
обеспечивают восприятие передаваемых нервных импульсов, а
некоторые, напротив, имеют пресинаптическую локализацию и
регулируют количество выбрасываемого нейромедиатора. При
этом в зависимости от конкретного типа рецепторов их активация
может способствовать либо возбуждению, либо, наоборот, тор-
можению следующей клетки.
Каждый из четырнадцати упомянутых типов рецепторов коди-
руется в организме человека отдельным геном (табл. 6.5). В ис-
следования, посвященные предрасположенности к суицидному
поведению, пока вовлечено только 9 их этих генов: HTR1A,
HTR1B, HTRlDa, HTR1E, HTR1F, HTR2A, HTR2C, HTR5A и
170
Таблица 6.5. Основные характеристики рецепторов серотонина у человека
Тип рецептора Принцип функционирования Результат активации Основная локализация Контролируемые процессы Ген
1А Рецепторы, связанные с G-белками. Вторичный мес- сенджер — цАМФ. Его содержа- ние снижается за счет стимули- рования фосфодиэстеразы Постсинаптиче- ское торможение цнс Сон, насыщение, термо- регуляция, беспокойство HTR1A
1В Пресинаптиче- ское торможение Давление крови HTR1B
ID-а Моторная активность, давление крови HTRlDa
ID-P HTRlDb
1Е Неизвестен Неизвестны HTR1E
1F HTR1F
2А Рецепторы, связанные с G-белками. Вторичные мес- сенджеры — инозитол-3-фосфат и диацил глицерол. Их содержа- ние возрастает за счет стимули- рования фосфолипазы Постсинаптиче- ское возбужде- ние Обучение, давление кро- ви, сокращения гладких мышц HTR2A
2В Неизвестен Желудок Сокращения желудка HTR2B
2С ЦНС Выделение спинномоз- говой жидкости HTR2C
3 Серотонин-зависимый Na+ и К‘ канал. Активация рецептора деполяризует мембрану ЦНС, пне Беспокойство, рвота HTR3
172
Окончание табл. 6.5
Тип рецептора Принцип функционирования Результат активации Основная локализация Контролируемые процессы Ген
4 Рецептор, связанный с G-белками. Вторичный мес- сенджер — цАМФ. Его содержа- ние возрастает за счет стимули- рования аденилатциклазы Постсинаптиче- ское возбужде- ние цнс, сердце, пищева- рительный тракт Сокращения пищевари- тельного тракта HTR4
5А Рецептор, связанный с G-белками. Вторичный мес- сенджер — цАМФ. Его содержа- ние снижается за счет стимули- рования фосфодиэстеразы Неизвестен ЦНС Неизвестны HTR5A
6 Рецептор, связанный с G-белками. Вторичный мес- сенджер — цАМФ. Его содержа- ние возрастает за счет стимули- рования аденилатциклазы HTR6
7 Рецептор, связанный с G-белками. Вторичный мес- сенджер — цАМФ. Его содержа- ние возрастает за счет стимули- рования аденилатциклазы ЦНС, пищевари- тельный тракт HTR7
HTR6. Причем для большинства имеются лишь единичные ра-
боты с использованием всего одной или двух нуклеотидных замен.
Это касается генов HTRlDa, HTR1E, HTR1F, HTR2C, HTR5A
и HTR6. Ассоциации с предрасположенностью к суициду для них
не обнаружено.
Несколько более подробно исследованы гены HTR1A, HTRIB
и HTR2A. В первом проанализированы три нуклеотидные заме-
ны, одна из которых {-1019Г} достаточно тесно ассоциирует со
склонностью к самоубийству. Однако пока не получено незави-
симых подтверждений, эти данные необходимо рассматривать в
качестве предварительных. В гене HTR1В тоже исследованы три
замены, но ни одна не оказывает влияния на суицидное поведе-
ние. Наибольшее внимание уделяется гену HTR2A'. в нем проана-
лизировано семь нуклеотидных замен. Для двух из них, располо-
женных в промоторе (-1420Т и rs631 /), продемонстрирована
слабая ассоциация со склонностью к самоубийству, но эти данные
пока не получили убедительного подтверждения. Более отчетливая
ассоциация была исходно продемонстрирована для замены 102С
в экзоне L Однако в дальнейших исследованиях она не подтвер-
дилась. Не удалось обнаружить ее и с помощью мета-анализа.
Таким образом, в настоящее время нет четких данных о
влиянии генов, кодирующих рецепторы серотонина, на склон-
ность человека к суицидному поведению.
6.15. Роль генов, контролирующих другие
молекулярные системы
Серотонин выполняет в организме множество функций. Одна
из важнейших — усиление активности дофаминовой системы.
Поэтому в генетический анализ предрасположенности к само-
убийству безоговорочно необходимо вовлекать и соответствующие
гены. Но, к сожалению, пока они остаются без должного внима-
ния. Ярким примером служит ситуация с геном TH. Его продук-
том является фермент тирозингидроксилаза — ключевой участ-
ник биосинтеза дофамина и норадреналина. Однако известны
лишь две экспериментальные работы, анализирующие влияние
этого гена на суицидное поведение. В одной показано, что у шве-
дов, страдающих психическими расстройствами, аллель КЗ тесно
ассоциируете попытками самоубийства. В другой продемонстри-
ровано, что у носителей этой же аллели существенно снижено
содержание З-метокси-4-гидроксифенилгликоля — основного
производного норадреналина. Но этих данных пока недостаточно
для четкого вывода.
173
Генам, кодирующим различные рецепторы дофамина, тоже
посвящены всего лишь единичные исследования. Изучение по-
лиморфизма гена DRD4 не обнаружило связи между количеством
повторов в экзоне 3 и вероятностью совершения попыток само-
убийства. Отрицательный результат получен и для гена £>/?Р2при
анализе выпадения цитозина в положении -141. В то же время
нуклеотидная замена в интроне 8 ассоциирует у алкоголиков с
множественными попытками суицида.
Важную роль в регуляции активности дофаминовой системы
играет катехол-О-метилтрансфераза — ключевой фермент,
обеспечивающий деградацию дофамина, а также адреналина и
норадреналина. Этот фермент является продуктом гена СОМ7\
в котором известно около 300 сайтов полиморфизма. Но основное
внимание уделяется только одному — Vall58Met в экзоне 4. По-
казано, что замена 158Met приводит к существенному снижению
активности фермента. При этом приблизительно в половине про-
веденных исследований продемонстрирована ее ассоциация со
склонностью к самоубийству, причем не с любыми, а преимуще-
ственно с «жесткими» формами. Наличие связи убедительно под-
тверждается и мета-анализом.
Дофаминовая система тесно связана с адреналиновой. В связи
с этим в генетический анализ предрасположенности к суициду
был вовлечен и ген ADRA2A, кодирующий а2А рецептор адрена-
лина. Для трех полиморфных сайтов в премоторной области
(G-1800T, С-1291Г и G-261A) не удалось обнаружить связи со
склонностью к самоубийству. Вместе с тем описана редкая ну-
клеотидная замена, характерная только для нескольких само-
убийц. Случайность ли это, пока остается неясным.
Данные по другим молекулярным системам мы не рассматри-
ваем, они еще носят сугубо предварительный характер.
6.16. Основные итоги изучения генетического
контроля предрасположенности к суициду
Предрасположенность человека к суицидному поведению
находится под контролем как минимум двух генов, имеющих
отношение к серотониновой и дофаминовой системам. Речь
идет о генах:
SLC6A4, продуктом которого является транспортер серотони-
на (ассоциирует S’-вариант промотора);
СОМТ, кодирующем катехол-О-метилтрансферазу (ассоции-
рует нуклеотидная замена 158Met).
При этом особого внимания заслуживает тот факт, что 5-про-
мотор гена SLC6A4 ассоциирует лишь с некоторыми формами
174
суицидного поведения, а именно: с «жесткими» способами са-
моубийства, с попытками, приводящими к тяжелым медицинским
последствиям, а также с повторными (множественными) попыт-
ками суицида.
В то же время в отношении «легких» форм ухода из жизни
обнаружить ассоциацию с 5-про мотором, как правило, не удает-
ся. Это значит, что «жесткие» и «легкие» формы самоубийства
обладают достаточно высокой степенью автономности и пред-
ставляют собой вполне самостоятельные поведенческие характе-
ристики. Сходный вывод напрашивается и в случае гена СОМТ.
Действительно, нуклеотидная замена 158Met ассоциирует преиму-
щественно с «жесткими» формами самоубийства.
Судя по имеющимся данным, в суицидном поведении можно
вычленить и некоторые другие автономные характеристики. На-
пример, одиночные и множественные попытки самоубийства,
а также попытки с летальным эффектом и без него (т.е. реальные
самоубийства и их различные «имитации»).
Возможно, основной причиной отрицательного результата для
прочих исследованных к настоящему времени генов является от-
сутствие дробного подхода к признаку. В этом плане весьма при-
мечательно, что в гене ТРН2опт из замен (19918G) достаточно
тесно ассоциирует с реальными самоубийствами, однако практи-
чески не связана с попытками суицида. Кроме того, аллели гена
МАОА с четырьмя или тремя с половиной повторами в промото-
ре ассоциируют только с «жесткими» формами самоубийства.
Таким образом, вполне вероятно, что при дробном подходе (т.е.
учете степени реализации суицидных намерений, конкретных
способов, использованных для сведения счетов с жизнью, а так-
же причин, спровоцировавших человека на самоубийство) роль
прочих генов тоже окажется статистически значимой.
В заключение необходимо отметить еще один нюанс. Человек,
неоднократно совершавший попытки самоубийства, несомненно,
имеет ярко выраженные суицидные наклонности. Но если само-
убийство реализовано с первой же попытки, без каких бы то ни
было предварительных намеков на суицид, о наличии соответ-
ствующих наклонностей у данного человека приходится судить по
единственному событию. Специфика исследуемого признака здесь
такова, что оценить его степень стабильности уже невозможно. Тем
не менее и с этим признаком успешно работают. Дело в том, что
каждый конкретный случай самоубийства официально подтверж-
дается службами правопорядка на основе необходимых судебно-
медицинских экспертиз.
Таким образом, из всех проявлений человеческой агрессии
именно суицид фиксируется наиболее четко и пока наиболее адек-
ватен для генетического анализа.
175
6.17. Генетический анализ несуицидных
форм агрессии
Анализ агрессивного поведения, не связанного с суицидом, пред-
ставляет с методической точки зрения очень сложную задачу. Во-
первых, это связано с тем, что по сравнению с самоубийствами такая
агрессия значительно труднее поддается учету. Во-вторых, ее про-
явления намного разнообразней. И в-третьих, пока не существует
единых подходов, позволяющих четко классифицировать эти про-
явления. Неудивительно, что результаты, полученные в данной
области, крайне отрывочны и пока во многом противоречивы.
Принято считать, что мужчины агрессивнее женщин. Это мне-
ние, прочно укоренившееся в общественном сознании, в основ-
ном опирается на данные криминальной статистики. И действи-
тельно, из всей совокупности тяжких преступлений около 90%
совершается мужчинами. В то же время по остальным, более
легким проявлениям агрессии, женщины обычно не только не
уступают мужчинам, но по некоторым показателям могут их даже
превосходить.
Однако вернемся к тому, что насилия и убийства совершают-
ся мужчинами во много раз чаще, чем женщинами. Причины
такой ситуации до конца не ясны. Тем не менее, судя по резуль-
татам близнецового анализа, это различие в значительной степе-
ни контролируется генотипом и обусловлено разными наборами
половых хромосом (XX — у женщин, XY — у мужчин). Иными
словами, наличие у человека К-хромосомы существенно повы-
шает его предрасположенность к насилию и убийству.
У человека описано множество хромосомных аномалий.
В частности, у некоторых мужчин (примерно у одного из I 000)
имеется не один, а два экземпляра Y-хромосомы. Казалось бы,
совершенно логично, что такие индивиды должны отличаться от
нормальных мужчин избыточной агрессивностью и относиться к
числу потенциальных насильников и убийц. Исследования, вы-
полненные в нескольких американских тюрьмах, показали, что
лишняя копия У-хромосомы встречается у осужденных прибли-
зительно в Ю раз чаще, чем среди обычного мужского населения.
Сходный вывод был сделан и для пациентов психиатрических
клиник. В результате гипотеза о высокой агрессивности мужчин,
имеющих лишнюю копию К-хромосомы, получила в научных
кругах широкое признание и даже вошла в ряд учебников по
психологии. Более того, в Соединенных Штатах Америки обсуж-
дали вопрос о поголовном тестировании детей на предмет вы-
явления индивидов с набором хромосом XYY и их изоляции от
общества в раннем возрасте.
176
Между тем в проведенных исследованиях был существенный
недостаток. Авторы не использовали в своей работе ни соответ-
ствующих опросников, ни непосредственных наблюдений за ис-
пытуемыми. Все выводы опирались лишь на косвенные оценки
агрессивности. И как только данная оплошность была устранена,
стало ясно, что наличие дополнительной У-хромосомы практи-
чески не влияет на степень агрессивности человека. К тому же
большинство заключенных с набором хромосом XYY отбывали
наказание не за убийства, а за жульничество и воровство.
В результате дальнейших исследований было установлено, что
мужчины с двумя экземплярами Y-хромосомы отличаются
от нормы значительно повышенной импульсивностью, а по-
тому легко вовлекаются в различные аферы. Второй харак-
терной особенностью этих мужчин является несколько
сниженный уровень интеллекта. По-видимому, именно со-
четание двух этих факторов и приводит к повышенной ве-
роятности оказаться в тюрьме или в психиатрической
лечебнице (человек совершает какие-то сомнительные дей-
ствия, не особо задумываясь об ответственности и воз-
можных последствиях). Тем более, что мужчины с лишней
У-хромосомой обычно имеют очень высокий рост и оттого при-
влекают повышенное внимание окружающих.
В последние годы появились экспериментальные работы по
поиску связи между склонностью к насилию и конкретными ге-
нами. Показано, что S’-промотор гена SLC6A4 ассоциирует с
совершением особо тяжких преступлений, с повторными насиль-
ственными преступлениями у алкоголиков, а также с проявле-
ниями насилия у психических больных.
Менее однозначная ситуация с геном СОМТ. Несколько не-
зависимых исследований свидетельствует о том, что нуклеотидная
замена 158Met ассоциирует со склонностью к насилию, особен-
но у мужчин, но остальные исследования не подтверждают этой
закономерности.
Для гена МАО А тоже получены противоречивые результаты.
В одной из голландских семей обнаружена мутация, приводящая
к полной дефектности соответствующего фермента. Мужчины,
несущие данную мутацию, отличаются от нормы приступами
агрессивного поведения, особенно при наличии внешних прово-
цирующих факторов, но эти данные плохо согласуются со сле-
дующим фактом. У человека известна небольшая хромосомная
аномалия, при которой отсутствуют оба гена моноамин оксидазы.
Такие люди страдают целым рядом тяжелых патологий (например,
быстро прогрессирующей потерей зрения), но по уровню агрессии
вполне соответствуют норме. В нескольких независимых иссле-
дованиях было показано, что слабо транскрибируемые аллели гена
177
Таблица 6.6. Гены, контролирующие несуицидную агрессию у человека (предварительные данные)
Молекулярная система Ген Соответствующий белок Молекулярные функции Контролируемые про- явления агрессии
Серотонино- вая ТРНГ Триптофан гидро- ксилаза Синтез ключевого предшественника серотонина в соматических органах (кишечнике, печени, сердце и т.п.) Общий негативизм Раздражительность Жестокость
ТРН2 Триптофан гидро- ксилаза Синтез ключевого предшественника серотонина в нейронах головного мозга Раздражительность
SLC6A4 Транспортер серото- нина Реимпорт серотонина из синаптиче- ской щели Общий негативизм Раздражительность Жестокость Физическая агрес- сия
МАОА Моноамин оксидаза А Деградация серотонина Раздражительность Жестокость Физическая агрес- сия
HTR2A Рецептор серотонина Восприятие импульсов, передаваемых через серотонинэргические синапсы Жестокость
HTR3 Серотонин- зависимый Na* и К* канал Восприятие импульсов, передаваемых через серотонинэргические синапсы Раадражител ьность Вербальная агрес- сия
Дофаминовая DRD4 Рецептор дофамина Восприятие импульсов, передаваемых через дофаминэргические синапсы Жестокость
сомт Катехол-О- метил трансфераза Деградация дофамина, адреналина и норадреналина Общий негативизм Раздражител ьность Жестокость Физическая агрес- сия
Адреналино- вая ADRA2A Рецептор адреналина Восприятие импульсов, передаваемых через адренэргические синапсы Раздражительность Вербальная агрессия
ADRB2 Рецептор адреналина Восприятие импульсов, передаваемых через адреналинэргические синапсы Жестокость
Шрифтом выделены гены, гомологи которых участвуют в контроле агрессивного поведения у крыс и мышей.
МАОА ассоциируют со склонностью к воспроизведению «цикла
насилия» (человек, подвергавшийся насилию в раннем детстве,
впоследствии сам становится источником жестокости). Но по-
следующие работы не подтвердили этого факта.
Изучаются и другие аспекты несуицидной агрессии (табл. 6.6).
Однако пока не получено четких подтверждений, все эти данные
следует рассматривать в качестве предварительных.
6.18. Влияние социальных факторов
на агрессивное поведение
6.18.1. Роль материнской заботы у макак
Агрессивное поведение конкретного животного или человека в
значительной степени зависит от факторов окружающей среды.
Особого внимания заслуживают социальные факторы, обусловлен-
ные различного рода взаимодействиями с другими представителями
того же вида. В частности, применительно к межсамцовой агрессии
у мышей такими факторами являются «своя» или «чужая» террито-
рия, наличие самок, число соперников и многое другое (см. подразд.
6.7, табл. 6.3). Возникает закономерный вопрос: какие механизмы
лежат в основе социального контроля агрессивного поведения?
Этой проблеме пока посвящены лишь единичные исследова-
ния. Одно из них выполнено на макаках резусах (Масаса mulatto).
Данный вид очень близок человеку с генетической точки зрения
и потому представляет собой удобный модельный объект для
изучения молекулярных механизмов психической деятельности.
К сожалению, у макак еще не созданы инбредные линии. По
этой причине в исследованиях приходится использовать живот-
ных, имеющих друг с другом многочисленные генетические раз-
личия. Такие исследования проводят по той же схеме, что и в
случае человека, т.е. посредством сравнения контрастных выборок
(см. подразд. 3.3).
Установлено, что около Ю % макак в ответ на достаточно уме-
ренные стрессовые воздействия демонстрируют неадекватно вы-
сокий уровень агрессии, вплоть до нанесения повреждений самим
себе. Эта поведенческая аномалия тесно ассоциирует с хрониче-
ски низкой активностью серотониновой системы. В свою очередь,
в основе дефектности серотониновой системы лежит взаимодей-
ствие генетических и социальных факторов.
У макак имеется гомолог гена SLC6A4, очень близкий челове-
ческому не только по нуклеотидной последовательности, но и по
варьированию размеров промоторной области. Как и в ситуации
с человеческим геном SLC6A4, эта область может быть длинной
180
(L) или короткой (5), причем первый вариант существенно эф-
фективнее второго. Напрашивается естественное предположение,
что и в случае макак 5-промотор способствует повышенному
уровню агрессии. Проведенные исследования подтвердили это
предположение. В самом деле, животные, гомозиготные по 5-про-
мотору, отличаются от прочих более выраженным агрессивным
поведением. Но у разных особей оно проявляется по-разному: у
одних — сравнительно слабо, а у других — очень ярко. Как вы-
яснилось, основная причина подобных различий тесно связана с
социальными факторами на ранних этапах жизни.
Исследования на макаках обычно проводят в специальных
питомниках. При этом некоторых детенышей отнимают от мате-
рей и искусственно вскармливают в отсутствие контактов со
взрослыми особями. В дальнейшем, по достижении детенышами
определенного возраста, их переводят в своеобразные «детские
сады» (группы из нескольких искусственно вскормленных свер-
стников), а затем, еще через некоторое время — в общую группу.
Макаки, прошедшие через такие «детские сады», отличаются от
тех, что воспитаны собственными матерями, заметно большей
обособленностью, сниженной эмоциональностью, существенно
менее сложным социальным поведением, а иногда и чрезмерно
высоким уровнем агрессии. Причем аномальная агрессивность,
как правило, наблюдается лишь у животных, гомозиготных по
5-варианту промотора. Таким образом, макаки в зависимости от
своего генотипа по-разному реагируют на отсутствие материнской
заботы: гомозиготы 55 оказываются гиперагрессивными,
в то время как прочие особи — вполне адекватными.
Особого внимания заслуживает следующее обстоятельство.
У макак, гомозиготных по 5-варианту промотора и воспитанных
в условиях отсутствия материнской заботы, возни кает хроническое
изменение метаболизма серотонина. В результате содержание
серотонина заметно снижается, что и приводит к резкому усиле-
нию агрессивного поведения. Между тем у особей генотипа LL
или LS таких изменений в серотониновой системе не происходит,
и в итоге уровень агрессии остается нормальным. Это значит, что
рассмотренные социальные факторы, а именно наличие или
отсутствие материнской заботы, взаимодействуют с ге-
нотипом на уровне молекулярных механизмов. Соответствен-
но, возникает ряд принципиальных вопросов:
• какие ферменты изменяют активность при отсутствии мате-
ринской заботы;
• на каком из этапов реализации генетической информации
происходит изменение активности этих ферментов;
• какие молекулярные системы позволяют детенышу ощущать
наличие или отсутствие материнской заботы;
181
• какие пути используются для передачи воспринятой инфор-
мации;
• почему у особей генотипа LL или LSотсутствие материнской
заботы практически не отражается на эффективности серотони-
новой системы?
В результате можно с полным основанием говорить о зарож-
дении нового направления в психогенетике — изучение моле-
кулярных механизмов действия социальных факторов.
6.18.2. Роль социально-экономического статуса
у человека
Если у макак, имеющих определенный генотип, серотониновая
система чувствительна к социальным факторам, не выполняется
ли та же закономерность и в случае человека? Это предположение
удалось экспериментально проверить. Исследование проводили на
выборке здоровых добровольцев, для каждого из которых был
установлен его генотип (речь идет о размерах промоторной области
гена SLC6A4). В качестве социального фактора в данной работе
использовали социально-экономический статус индивида. Его
оценку осуществляли на основе двух следующих параметров:
• суммарного времени, затраченного индивидом на свое обу-
чение в школе, колледже, вузе и т.д.;
• годового дохода индивида.
Каждый параметр оценивали в баллах по специальной шкале,
после чего выводили их усредненное значение. Разумеется, по-
добный подход далек от совершенства. Но в первом приближении
и он оказался приемлемым. Он позволил разделить испытуемых
на две группы: с условно высоким и условно низким социально-
экономическим статусом.
У каждого испытуемого тестировали активность серотониновой
системы. Это возможно с помощью нескольких стандартных ме-
тодик.
Об одной из них мы кратко упоминали ранее: спинномозговую
жидкость конкретного индивида исследуют на содержание основ-
ного производного серотонина (5-гидроксиинозитолуксусной
кислоты). Чем выше содержание этого химического соединения,
тем выше суммарная активность серотониновой системы.
Другая методика основана на использовании фенфлурами-
на — препарата, стимулирующего мощный выброс серотонина и
в то же время резко подавляющего его реимпорт. В результате
серотонин накапливается в синаптических щелях, что приводит
к значительному усилению надлежащих ответов, и в частности, —
к повышению концентрации пролактина в крови (пролактин
182
относится к числу нейрогормонов, выделяемых гипофизом при
активации серотонинэргических синапсов). Следовательно, из-
меряя концентрацию пролактина в крови при введении опреде-
ленного количества фенфлурамина, можно оценивать суммарную
активность серотониновой системы. Эта методика и была ис-
пользована в данном эксперименте.
Оказалось, что людям, гомозиготным по £-промотору, свой-
ственна одинаковая активность серотониновой системы вне за-
висимости от их социально-экономического статуса. Совсем иная
закономерность наблюдается у генотипа 55: активность при вы-
соком статусе соответствует норме, но при низком существенно
падает (приблизительно на треть). Сходная ситуация обнаружена
и у гетерозигот, однако падение активности у них не столь за-
метно. Напомним, что пониженная активность серотониновой
системы тесно связана с чрезмерно высокой степенью агрессии.
Таким образом, низкий социально-экономический статус
является фактором, способным провоцировать излишнюю
агрессивность, но не у всех людей, а только у обладателей
определенных генотипов’, гомозигот SS и гетерозигот LS.
При этом мы вновь убеждаемся, что социальные факторы взаи-
модействуют с генотипом на уровне молекулярных механизмов,
а значит, не имеют принципиальных отличий от биологических.
6.19. Перспективы генетики агрессивного
поведения человека
Как известно, генетический анализ может дать результаты
только в том случае, если признак очерчен достаточно четко.
Между тем в исследованиях агрессивного поведения человека
большинство используемых признаков крайне расплывчато и в
значительной степени носит субъективный характер. Это связано
с множественностью проявлений человеческой агрессии и от-
сутствием общепринятых подходов к их классификации. Соот-
ветственно необходимо разрабатывать такие подходы. И чем
более дробной окажется полученная классификация, тем адек-
ватнее она для использования в генетических исследованиях.
В этом плане показательна ситуация с суицидным поведением.
Судя по имеющимся данным, оно неоднородно и варьирует у
разных индивидов по нескольким параметрам (способу реализа-
ции, количеству попыток и степени их серьезности). Если рас-
сматривать каждый из параметров строго по отдельности, роль
генов SLC6A4 и СОМТпрослеживается отчетливо. В противном
же случае генетический анализ безрезультатен.
183
Исследования на модельных объектах свидетельствуют о том,
что некоторые внешне похожие формы агрессии обслуживаются
разными «пусковыми механизмами». В самом деле, в зависимости
от своего конкретного генотипа организм может быть агрессив-
ным в одной ситуации, но не способным к выполнению таких же
действий в другой (ярким примером являются серебристо-черные
лисицы, сохраняющие межсамцовую агрессию, но дружелюбные
к человеку). Вполне вероятно, что нечто подобное встречается и
у людей. Поэтому при анализе агрессивного поведения человека
очень важно учитывать причины возникновения агрессии. В част-
ности, непосредственным поводом для вскрытия вен у одних
индивидов становятся угрызения совести, у других — крах карье-
ры, у третьих — любовная неудача. Следовало бы провести их
генетическое сравнение друг с другом, а также с людьми, имев-
шими в жизни похожие проблемы, однако не совершавшими
попыток самоубийства. В принципе, такими путями можно вы-
явить гены, контролирующие именно «пусковые механизмы»
агрессии.
Большого внимания заслуживает поиск молекулярных меха-
низмов, посредством которых агрессия регулируется социальны-
ми факторами. Пока эта область исследований очень молода. Тем
не менее полученные результаты настолько принципиальны, что
позволяют объяснить целый ряд серьезных противоречий. Обще-
признано, что криминальное поведение многих преступников
обусловлено тяжелыми обстоятельствами детства и юности. В то
же время известно огромное число примеров, где трудное детство
не помешало нормальному развитию личности, и в итоге из ре-
бенка сформировался прекрасный человек. Причина различий,
по-видимому, в разных генотипах. Действительно, судя по резуль-
татам исследований на макаках, отсутствие материнской заботы
усиливает агрессивность особей, гомозиготных по 5-промотору
гена SLC6A4, но не влияет на гомозигот LL и гетерозигот. Учи-
тывая генетическое сходство макаки и человека, логично предпо-
ложить, что такая же или близкая закономерность должна вы-
полняться и в случае людей. Тем более, что низкий социально-
экономический статус (а именно он, как правило, соответствует
трудному детству) снижает активность серотониновой системы
только у обладателей генотипов 55 или LS.
Таким образом, ближайшие перспективы в генетике агрессив-
ного поведения человека связаны с развитием следующих на-
правлений:
• вычленение элементарных проявлений человеческой агрес-
сии;
• поиск генов, контролирующих эти элементарные проявле-
ния;
184
• изучение генетического контроля «пусковых механизмов»,
регулирующих уровень агрессии в разных ситуациях;
• исследование молекулярных механизмов, с помощью которых
агрессивное поведение контролируется социальными факторами.
СПИСОК ЛИТЕРАТУРЫ
I. БеляевД. К. Генетические аспекты доместикации животных // Проблемы
доместикации животных и растений. — М. : Наука, 1972. — С. 39—45.
2. Куликов А. В., Попова Н.К. Формы агрессивного поведения и их ге-
нетическая детерминация // Исследования по генетике. — Т. 17. — 1991. —
С. 131-151.
3. Куликов А. В., Жанаева Е.Ю.У Попова Н.К. Изменение активности
триптофан гидроксилазы в мозге серебристо-черных лисиц и серых крыс-
пасюков в ходе селекции по поведению // Генетика. — 1989. — Т. 25. —
С. 346-349.
4. Никулина Э.М.Ч Попова Н.К. Генетический анализ агрессивности
хищника у мышей // Генетика. — 1983. — Т. 19. — № 7. — С. 1105— 1110.
5. Трут Л.Н. Генетика и феногенетика доместикационного повеления.
В: Вопросы общей генетики (под ред. Ю.П. Алтухова). — М.: Наука, 1981. —
С.323-332.
6. Bethea C.L. et al. Anxious behavior and fenfluramine-induced prolactin
secretion in young rhesus macaques with different alleles of the serotonin reuptake
transporter polymorphism (5HTTLPR) // Behavior Genetics. — 2004. —
Vol. 34. - P. 295-307.
7. Bjorkquist K. Sex differences in physical, verbal, and indirect aggression: A
review of recent research // Sex Roles. — 1994. — V. 30. — P. 177— 188.
8. Bondy B., Buettner A. and Zill P. Genetics of suicide // Molecular
Psychiatry. — 2006. — V. 11. — P. 336 — 351.
9. Buss A. F. and Durkee A. An inventory for assessing different kinds of hostility
11 J. Consulting Psychology. — 1957. — V. 21. — P. 343 — 349.
10. Buss A. F. and Perry M. The aggression questionnaire // J. Personality &
Social Psychology. — 1992. — V. 63. — P. 452 — 459.
II. Caspi A. et al. Role of genotype in the cycle of violence in maltreated
children // Science. — 2002. — V. 297. — P. 851 —854.
12. Dierick H.A. and Greenspan R.J. Molecular analysis of flies selected for
aggressive behavior // Nature Genetics. — 2006. — V. 38. — P. 1023— 1031.
13. Hallikainen T. et al. Association between low activity serotonin transporter
promoter genotype and early onset alcoholism with habitual impulsive violent
behavior// Molecular Psychiatry. — 1999. — V. 4. — P. 385 — 388.
14. Kia-Keating В. M., Glatt S.J. and Tsuang M. T. Meta-analyses suggest
association between COMT, but not HTR1B, alleles, and suicidal behavior //
American J. Medical Genetics. B. Neuropsychiatric Genetics. — 2007. —
V. 144B. - P. 1048-1053.
15. Li D. and He L. Meta-analysis supports association between serotonin
transporter (5-HTT) and suicidal behavior // Molecular Psychiatry. — 2007. —
V. 12. - P. 47 - 54.
185
16. Liao D. L. et al. Possible association between serotonin transporter promoter
region polymorphism and extremely violent crime in Chinese males //
Neuropsychobiology. — 2004. — V. 50. — P. 284 — 287.
17. Manuck S. B. et al. Socio-economic status covaries with central nervous
system serotonergic responsivity as a function of allelic variation in the serotonin
transporter gene-linked polymorphic region // Psychoneuroendocrinology. —
2004. - V. 29. - P. 651-668.
18. Moyer K.E. Kinds of aggression and their physiological basis //
Communications in Behavioral Biology. — 1968. — V. 2A. — P. 65 — 87.
19. Popoua N. K. From genes to aggressive behavior: the role of serotonergic
system // Bioessays. — 2006. — V. 28. — P. 495 — 503.
20. Roubertoux P.L. and Carlier M. Differences between CBA/H and NZB
mice on intermale aggression. II. Maternal effects// Behavior Genetics. — 1988. —
V. 18. - P. 175-184.
21. Rujescu D., Thalmeier A., Moller H.-J., Bronisch T. and Giegling I.
Molecular genetic findings in suicidal behavior What is beyond the serotonergic
system? // Archives of Suicide Research. — 2007. — V. II. — P. 17 — 40.
22. Statham D.J. et at. Suicidal behaviour: An epidemiological and genetic
study // Psychological Medicine. — 1998. — V. 28. — P. 839 — 855.
23. Suomi S.J. Aggression and social behaviour in rhesus monkeys // Novartis
Found Symposium. 2005. — V.268. — P.216 — 222.
24. Theilgaard A. A psychological study of the personalities of XYY — and
XXY-men Ц Acta Psychuatrica Scandinavica. Supplements. — 1984. — V. 315. —
P. 1-133.
25. Vitaro F., Brendgen M. and Barker E. D. Subtypes of aggressive behaviors:
A developmental perspective // International J. Behavioral Development. —
2006. -V. 30. - P. 12-19.
26. Weinshenker N.J. and Siegel A. Bimodal classification of aggression:
affective defense and predatory attack // Aggression & Violent Behavior. — 2002. —
V. 7. - P. 237 - 250.
27. Young S. E. et al. Clinical validation of possible links between MAO-A
genotype and maltreatment in the risk for adolescent conduct disorder // American
J. Psychiatry. - 2006. - V. 163. - P. 1 -7.
КОНТРОЛЬНЫЕ ВОПРОСЫ
1. Опишите биологическую роль и разнообразие видов агрессии
у животных.
2. Какие гены контролируют межсамцовую агрессию у мышей?
3. Опишите методы оценки агрессивного поведения человека.
4. Что известно о генетическом контроле суицидного поведения?
5. Каковы поведенческие особенности мужчин с набором хромосом
XYY?
Глава 7
ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ
ПРЕДРАСПОЛОЖЕННОСТИ К ПЕРЕЕДАНИЮ
7.1. Актуальность проблемы переедания
В развитых странах, и в первую очередь — в США, значитель-
ная часть населения страдает от переедания. Это связано как
минимум с несколькими причинами. Во-первых, с достаточно
высоким уровнем жизни при относительно низкой стоимости
продуктов питания. Во-вторых, с преимущественно сидячим об-
разом жизни. В-третьих, с развитой сетью быстрого питания,
предлагающей чрезмерно обильную и калорийную пищу. В-чет-
вертых, с огромным количеством соответствующей рекламы.
И наконец, с широким распространением пищевых продуктов,
специально направленных на разжигание аппетита. К таким про-
дуктам, в частности, относятся чипсы и кола: после сухих и со-
леных чипсов хочется пить, а после шипучей колы — что-нибудь
съесть. В итоге возникает своеобразная «цепная реакция», по-
буждающая человека к систематическому перееданию.
Непосредственным результатом переедания является ожирение.
Напомним, что жир — это важное запасное вещество. При острой
необходимости (например, при длительном голодании) оно исполь-
зуется в качестве «внутреннего источника пищи» и позволяет орга-
низму пережить неблагоприятное время. У взрослых людей нор-
мальной комплекции жир составляет I5...2O% общей массы тела.
К настоящему времени разработан целый ряд специальных
методик, напрямую оценивающих количество жира в организме.
Но чаще применяют различные косвенные оценки. Наиболее
распространенной является индекс массы тела. Его рассчиты-
вают как отношение массы тела к квадрату роста (m/h2), где
масса выражена в килограммах, а рост — в метрах. В норме зна-
чение этого индекса находится в пределах от 18,5 до 24,9. При
более высоких значениях (от 25 до 29,9) принято говорить о на-
личии избыточной массы. Если же индекс достигает 30 и более,
это считают свидетельством явного ожирения (табл. 7.1).
187
Таблица 7.1. Оценка комплекции с помощью индекса массы
тела
Значение индекса массы тела* * Диагноз
< 18,5 Недостаток массы
18,5...24,9 Норма
25 ...29,9 Избыточная масса
30... 34,9 1 степень ожирения
35... 39,9 II степень ожирения
>40 П1 степень ожирения
* Для человека в возрасте 25 — 30 лет.
Ожирение характерно приблизительно лля ’/з американцев и
с каждым годом становится все более и более массовым. Ситуация
усугубляется тем, что у граждан США настолько гипертрофиро-
вано чувство личной свободы, что многие ничуть не смущаются
своих размеров даже при наличии ярко выраженных симптомов
ожирения. Однако определенные проблемы все же возникают.
Чрезмерная тучность негативно отражается на подвижности и
здоровье, часто приводит к существенным затруднениям в транс-
порте, требует нестандартной одежды и т.д. С такими проблема-
ми можно бороться традиционными методами: регулярными
физическими нагрузками и строгой диетой. Не каждый человек
обладает достаточной силой воли, чтобы стойко выдерживать не-
обходимые длительные самоограничения. Поэтому возникло не-
сколько альтернативных подходов, позволяющих бороться с пе-
рееданием без особых усилий. В том числе:
• использование низкокалорийных продуктов питания (чело-
век не ограничивает себя в приеме пищи, но не страдает от ее
избытка);
• применение лекарственных препаратов, подавляющих ап-
петит (человек не испытывает позывов к приему пищи и обхо-
дится существенно меньшим ее количеством).
Разумеется, для нас интересен последний подход. Первые си-
стематические исследования в этом направлении были начаты
еще в середине прошлого столетия. Оказалось, что у всех мле-
копитающих, и в частности у человека, имеется высоко
консервативная молекулярная система, обеспечивающая
регуляцию количества потребляемой пищи.
Рассмотрим ее основные компоненты.
188
7.2. Общая схема регуляции чувства голода
Потребность в пище — одна из фундаментальных особенностей
живого. Питаясь, организм получает «строительный материал»,
а также энергию, необходимую для поддержания своей жизне-
деятельности. В природных условиях пищевые ресурсы всегда
ограничены и, как правило, являются объектами жесткой конку-
ренции. Потому у многих животных (особенно примитивных)
знакомое нам чувство насыщения практически отсутствует. Такие
животные поглощают столько пищи, сколько сумеют.
Пищевое поведение млекопитающих значительно сложнее.
Оно меняется в зависимости от количества и качества пищи, за-
трат энергии, объема накопленных жировых запасов, конкретной
окружающей обстановки и прочих факторов. Центральную роль
в подобной регуляции играет гипоталамус, и в первую очередь —
его аркуатное ядро, в котором имеется два типа конкурирующих
нейронов: возбуждение одних приводит к усилению чувства го-
лода, а возбуждение других — к диаметрально противоположному
эффекту.
В свою очередь, степень активности этих нейронов зависит от
целого ряда молекулярных сигналов, непрерывно поступающих
от разных типов тканей и отражающих текущее состояние энер-
гетического баланса. К таким сигналам относятся небольшие
регуляторные белки грелин, инсулин, лептин и пептид YY. Все
они выделяются в кровь, разносятся по организму, успешно
преодолевают гемо-энцефалический барьер и воздействуют на
конкретные нейроны аркуатного ядра (рис. 7.1).
Для начала рассмотрим биологические функции грелина. Этот
белок вырабатывается определенными клетками желудка в ответ
на недостаточную загруженность пищеварительной системы.
Попав в аркуатное ядро, молекулы грелина препятствуют воз-
буждению нейронов, ослабляющих чувство голода. Конкуриру-
ющие нейроны оказываются более активными, и организм ощу-
щает ярко выраженное желание поесть. После еды биосинтез
грелина резко ослабляется, и вскоре чувство голода тоже начина-
ет затухать.
Инсулин синтезируется клетками поджелудочной железы при
поступлении в кровь значительного количества глюкозы (обычно
это связано с усвоением переваренной пищи). Напомним, что
главная биологическая функция глюкозы — служить организму
эффективным источником энергии. Поэтому достаточно высокое
содержание инсулина является молекулярным сигналом, свиде-
тельствующим о том, что организм не испытывает проблем с
источниками энергии в ближайшее время и не нуждается в до-
189
Рис. 7.1. Важнейшие молекулярные сигналы, регулирующие активность
нейронов аркуатного ядра
Примечание. Воздействия, приводящие к возбуждению нейронов, показаны
стрелками. Тормозящие воздействия показаны знаком «1». Нейромедиаторы пред-
ставлены в виде кружков; Y— нейропептид; Y, А — агути-подобный белок; П — про-
опиомеланокортин; ГАМК — у-аминомасляная кислота. Прочие пояснения —
в тексте.
полнительном питании. Соответственно под действием инсулина
чувство голода ослабляется.
Пептид YY выделяется клетками толстого кишечника в ответ
на принятие значительного количества пищи. Таким образом,
данный белок, подобно инсулину, сигнализирует об отсутствии
проблем с питательными веществами и тоже способствует ослаб-
лению чувства голода.
Избыток полученных организмом питательных веществ от-
кладывается про запас преимущественно в виде жира. Этот жир
накапливается в крупных овальных клетках, специально предна-
значенных для выполнения данной функции и потому получив-
ших название «жировые». Подобные клетки присутствуют во
многих внутренних органах, но особенно многочисленны в под-
кожной жировой клетчатке, которая служит главным хранилищем
жировых запасов.
Каждая жировая клетка вырабатывает лептин. При этом от-
четливо просматривается следующая закономерность: чем больше
накоплено жира в конкретной клетке, тем большее количество
лептина она выделяет. Таким образом, высокое содержание леп-
190
Таблица 7.2. Циклическое влияние молекулярных сигналов на нейроны дуговидного ядра гипоталамуса
Исходная ситуация Ее последствия Результат
физиологические молекулярные* на уровне нейронов дуговидного ядра общий
Организм голоден 1. Принятие пиши 1. Начинается синтез пеп- тида YY 1. Торможение нейронов, усили- вающих чувство голода Организм сыт
2. Переваривание пищи в желудке, а затем в кишечнике 2. Ослабляется синтез грелина 2. Повышение активности нейро- нов, ослабляющих чувство голода
3. Всасывание пита- тельных веществ из кишечника в кровь 3. Клетки поджелудочной железы начинают синтези- ровать инсулин 3. Повышение активности нейро- нов, ослабляющих чувство голода
Организм сыт 1. Организм до- статочно долго не принимает пишу 1. Ослабляется синтез пеп- тида YY I. Повышение активности нейро- нов, усиливающих чувство голода Организм голоден
2. Количество пиши в желудке и кишеч- нике сокращается 2. Усиливается синтез грелина 2. Торможение нейронов, ослаб- ляющих чувство голода
3. Всасывание пита- тельных веществ не происходит 3. Падение концентрации глюкозы в плазме крови. Прекращают синтез инсу- лина 3. Понижается активность нейро- нов, ослабляющих чувство голода
* Обычно за время между двумя приемами пищи запасы жира не успевают претерпеть существенных изменений, поэтому
лептиновая регуляция является долгосрочной и в данной таблице не отражена.
тина в организме свидетельствует о значительных запасах пита-
тельных веществ, не требующих немедленного пополнения извне.
Неудивительно, что и этот белок подавляет чувства голода.
Для организма^ хронически испытывающего недостаток
пищич характерна острая нехватка глюкозы и жира. В та-
ком организме вырабатывается мало инсулина и лептина.
Невелико и содержание пептида YY. Напротив^ уровень гре-
лина довольно высок. Все эти факторы способствуют ярко
выраженному чувству голода. При избыточном питании
возникает обратная ситуация, из-за которой чувство голо-
да практически отсутствует. И наконец, при более или
менее адекватном питании чувство голода иногда затухает,
а иногда обостряется в результате циклических воздействий
на аркуатное ядро (табл. 7.2).
7.3. Генетический контроль чувства голода
у мышей
Мыши, подобно человеку, практически всеядны. Они потреб-
ляют не только растительную, но и животную пищу, включая
разнообразные продукты с нашего стола. Тем не менее при рабо-
те с мышами в лабораторных условиях обычно используют специ-
ально подобранные комбинированные корма, имеющие опти-
мальный состав и оптимальную калорийность. В результате жи-
вотные получают стандартное питание. При этом в зависимости
от цели конкретного исследования количество предоставляемого
корма может быть различным.
Если подопытные мыши не ограничены в пище, они способны
самостоятельно регулировать свой рацион. В норме, с учетом
специфики лабораторного содержания (небольшие размеры клетки,
комфортная температура, отсутствие необходимости в поиске пищи
и т.п.) он составляет приблизительно 3...4 г в сутки. Между тем у
мутантов по целому ряду генов регуляция чувства голода существен-
но нарушена, что приводит к значительным изменениям этого по-
казателя. К настоящему времени известно более 60 подобных генов.
И для многих уже установлены их конкретные функции.
Помимо четырех перечисленных выше гормонов (грелина,
лептина, инсулина и пептида YY) в регуляции чувства голода за-
действованы и другие молекулы:
• во-первых, рецепторы упомянутых выше гормонов. Такие
рецепторы характерны для многих типов клеток, и в частности,
непосредственно для нейронов аркуатного ядра. Напомним, что
именно это ядро гипоталамуса оценивает текущее состояние
192
энергетического баланса и вырабатывает нервные импульсы,
регулирующие чувство голода;
• во-вторых, небольшие белки, используемые в качестве ней-
ромедиаторов для передачи импульсов от нейронов аркуатного
ядра на следующие за ними вставочные и исполнительные ней-
роны (см. рис. 7.1);
• в-третьих, рецепторы, воспринимающие эти нейромедиа-
торы;
• и, наконец, десятки других регуляторных белков, влияющих
на активность нейронов головного мозга, метаболизм жиров и
некоторые иные процессы.
Логично предположить, что эти же молекулярные системы
участвуют в регуляции чувства голода и у человека.
7.4. Основные подходы к генетическому
анализу предрасположенности
к перееданию у человека
Нет сомнений, что склонность человека к ярко выраженному
перееданию может быть связана с различного рода «негенетиче-
скими» причинами: примером родителей, собственными при-
вычками, перенесенными болезнями, отсутствием других инте-
ресов и т.д. Вместе с тем в целом ряде семейных и близнецовых
исследований продемонстрирована высокая наследуемость данной
характеристики (роль генотипа варьирует от 30 до 60%). Соот-
ветственно встает вопрос о поиске генов, принимающих участие
в контроле предрасположенности к перееданию.
Эта задача с методической точки зрения весьма непроста. Для
всех испытуемых нужно создать одинаковые условия со строгим
учетом количества потребляемой пищи, причем эта пища тоже
должна быть совершенно одинаковой. В действительности ис-
пользуют значительно менее строгие подходы: склонность чело-
века к перееданию оценивают не напрямую, а по косвенным
признакам. Например, по стилю питания. В этом случае ис-
пытуемым предлагают ответить на вопросы, так или иначе свя-
занные с потреблением пищи. На основе полученных ответов
принято выделять:
• нормальное питание. Испытуемый потребляет столько
пищи, сколько ему необходимо, не прибегая к каким-либо диетам
и очистительным процедурам;
• ограниченное питание. Испытуемый постоянно ограничи-
вает себя в приеме пищи, поскольку считает, что в противном
случае он будет переедать;
193
• ограниченное питание, чередующееся с перееданием.
Иногда испытуемый придерживается строгой диеты, но в осталь-
ное время он ярко выражен но переедает;
• систематическое переедание. Испытуемый ест существен-
но больше, чем ему необходимо. Об этой склонности могут сви-
детельствовать и другие характеристики:
привычка обильно есть в вечернее время;
частые «перекусывания» между регулярными приемами пищи;
неспособность устоять перед «чем-нибудь вкусненьким»;
привычка есть за телевизором, в кинотеатре и т.д.
Такие опросы становятся все более и более популярными.
Однако отметим, что они выявляют лишь субъективное мнение
испытуемых о самих себе, а это мнение далеко не всегда соот-
ветствует реальности. Например, человек может думать, что он
переедает, хотя в действительности его суточный рацион вполне
адекватен. Нередко встречаются и прямо противоположные си-
туации. Поэтому получаемые результаты не слишком надежны.
Еще одним косвенным признаком, свидетельствующим о пе-
реедании, служит наличие у человека ярко выраженного ожире-
ния. Но и здесь не приходится говорить о строгой корреляции.
Иногда ожирение вовсе не связано с перееданием, а обусловлено
определенными дефектами обмена веществ. В частности, у мышей
получен целый ряд мутантов, для которых характерно нормальное
потребление пищи, но при этом наблюдаются тяжелые формы
ожирения. Подобные ситуации встречаются и у человека. В свя-
зи с этим при анализе людей, страдающих ожирением, необходи-
мо учитывать не только их индекс массы тела, но и хотя бы при-
близительные количества потребляемой пищи.
Наконец, у человека известны патологические состояния,
непосредственно связанные с нарушениями пищевого поведе-
ния'.
• булимия — неспособность контролировать себя в приеме
пищи, что в конечном итоге приводит к хроническому перееда-
нию, лишь иногда прерываемому попытками «очистить» орга-
низм;
• анорексия — резко негативное отношение к пище, обуслов-
ленное навязчивым страхом набрать лишний вес. При этом чув-
ство голода может быть очень сильным, но оно преодолевается
опасением «испортить фигуру». Стараясь избавиться от мнимых
излишков веса, больные злоупотребляют слабительными или
мочегонными средствами, искусственно вызывают рвоту и др.
Принято различать две формы этой патологии:
• ограничительную — человек потребляет мало пищи, но счи-
тает, что и такое количество слишком велико и потому регулярно
«очищает» свой организм;
194
• очистительную — не выдержав голода, человек объедается,
после чего приступает к длительному «очищению» организма,
которое вновь заканчивается обжорством и т.д.
Использование соответствующих пациентов в генетическом
анализе представляется одним из наиболее перспективных под-
ходов. Однако и здесь, к сожалению, возможны серьезные про-
блемы.
Симптомы обоих заболеваний довольно расплывчаты, к тому
же нередко встречаются промежуточные формы (чередование
приступов обжорства и «очищения» организма). Тем не менее уже
обнаружено несколько генов, контролирующих склонность чело-
века к хроническому перееданию. Кратко остановимся на основ-
ных полученных результатах.
7.5. Роль генов, контролирующих грелиновую
систему регуляции
О грел и не мы уже упоминали выше, когда рассматривали об-
щие принципы регуляции чувства голода. Теперь необходимо
познакомиться с ним более подробно. Начнем с того, что гре-
лин — это очень короткий белок, состоящий всего из 28 амино-
кислот. Но продукт соответствующего гена значительно крупнее.
Данный продукт (его называют препрогрелином) исходно состоит
из 117 аминокислот, а затем укорачивается за счет отщепления
нескольких фрагментов. Подобный процесс характерен для мно-
гих секреторных белков и играет немаловажную роль в регуляции
их активности.
Гредин сигнализирует нейронам аркуатного ядра о недостаточ-
но высокой загруженности пищеварительной системы и тем са-
мым приводит к усилению чувства голода. Как выяснилось, это
не единственная функция грелина. Но мы сконцентрируем вни-
мание исключительно на ней, поскольку она наиболее тесно
связана с пищевым поведением.
Способность молекул грелина обострять чувство голода под-
тверждена не только в экспериментах на модельных животных,
но также в клинических исследованиях непосредственно на че-
ловеке.
Действительно, введение грелина нормальным испытуемым
приводит к значительному увеличению их рациона. То же самое
наблюдается и у больных с ослабленным чувством голода. Логич-
но предположить, что пониженное содержание грелина должно
вызывать в организме диаметрально противоположный эффект.
Однако у мышей, нокаутированных по соответствующему гену
195
(этот ген получил обозначение GHRL}, количество потребляемо-
го корма почти не изменяется.
Причины такого противоречия пока неизвестны. Одно из
возможных объяснений заключается в том, что существуют и
некие другие молекулярные системы, независимо сигнализи-
рующие гипоталамусу о недостатке питания и способные под-
держивать адекватное чувство голода даже при полном отсут-
ствии грел и на.
Если предложенная гипотеза соответствует действительности,
мы вправе ожидать, что различного рода мутации, усиливающие
экспрессию человеческого гена GHRL, должны ассоциировать со
склонностью к перееданию. Между тем мутации, ослабляющие
экспрессию гена, не должны существенно влиять на рассматри-
ваемый признак.
Результаты исследований, выполненных к настоящему време-
ни, в основном хорошо согласуются с этой гипотезой. В частно-
сти, нуклеотидная замена 3056С, приводящая к повышенному
содержанию грелина в плазме крови, ассоциирует с высокими
значениями индекса массы тела, а также со склонностью к
чередованию обжорства и «очищения*. В то же время нуклео-
тидная замена 346А, вызывающая значительное снижение уровня
грелина, практически никак не влияет на отношение к пище. Эти
выводы основаны на нескольких независимых исследованиях, но
для надежности требуют дополнительных экспериментальных
подтвержде н и й.
Достигнув аркуатного ядра, молекулы грелина вступают во
взаимодействие с определенными рецепторными белками, яв-
ляющимися продуктами гена GHSR. Мыши, нокаутированные
по этому гену, нечувствительны к грел и ну. Но количество потре-
бляемой ими пищи соответствует норме, что согласуется с гипо-
тезой о наличии альтернативных систем, сигнализирующих ней-
ронам гипоталамуса о недостатке питания.
Представляется вполне вероятным, что и в случае человека
лаже полное нарушение функций рецептора грелина не должно
существенно отражаться на отношении к пище. Между тем мута-
ции, повышающие активность рецептора, должны ассоциировать
с чрезмерным чувством голода и склонностью к перееданию. По-
добная связь продемонстрирована для нескольких нуклеотидных
замен, но пока не получила подтверждения в независимых ис-
следованиях и не может рассматриваться в качестве установлен-
ного факта.
Таким образом, имеющиеся данные свидетельствуют о том,
что склонность человека к перееданию может быть связана
с мутациями, повышающими активность грелиновой систе-
мы (например, с заменой 3056С в гене GHRL}.
196
7.6. Роль генов, контролирующих систему
пептида YY
В организме, ожидающем очередного принятия пиши, содержа-
ние грелина в плазме крови постепенно растет и достигает макси-
мального уровня непосредственно перед едой. В этот момент ор-
ганизм испытывает наивысшее чувство голода. Но уже через не-
сколько минут ситуация изменяется. Во-первых, происходит
подавление биосинтеза грелина. А во-вторых, начинается выработ-
ка молекулярных сигналов, под действием которых чувство голода
быстро затухает. Один из подобных сигналов — пептид YY.
Данный пептид является продуктом гена PYY. Показано, что
мыши, нокаутированные по этому гену, отличаются от нормы по-
вышенным потреблением пищи и в конечном итоге — тяжелыми
формами ожирения. Но если вводить нокаутированным животным
пептид YY, их суточный рацион и комплекция нормализуются.
У человека прослеживается примерно такая же закономерность:
при регулярном введении данного пептида количество потребляе-
мой пищи значительно сокращается. Таким образом, весьма веро-
ятно, что некоторые мутации, затрагивающие структуру человече-
ского гена PYY, тоже способны влиять на исследуемый признак.
Эта гипотеза недавно получила серьезное подтверждение.
Известно, что многие индейцы из племени Пима (основное
коренное население штата Аризона, США) страдают чрезвычай-
но тяжелыми формами ожирения, наследуемость которых очень
высока. Для анализа было выбрано около 400 испытуемых с ин-
дексами массы тела не менее 45. Как показало сравнение этих
людей с их нормальными соплеменниками, столь тяжелая форма
ожирения тесно ассоциирует с двумя нуклеотидными заменами в
гене PYY: -1746Т и -1653С. Обе затрагивают промоторную об-
ласть гена и, по-видимому, резко ослабляют его экспрессию.
Любопытно, что каждая из перечисленных нуклеотидных замен
ассоциирует со склонностью к ожирению только у мужчин. Этот
факт еще не получил однозначного объяснения. Наиболее вероят-
ным представляется следующий механизм. Известно, что экспрес-
сия гена PYYнаходится под контролем мужских половых гормонов
и потому осуществляется у мужчин и женщин по-разному. Соот-
ветственно одна и та же аллель интересующего нас гена может
иметь неодинаковое проявление у женщин и мужчин.
Установлено, что в аркуатном ядре гипоталамуса пептид YY
воздействует на рецепторы Y2. Они характерны для многих от-
делов нервной системы, но в основном локализованы именно в
гипоталамусе. Эти рецепторы кодируются геном NPY2R. Судя по
результатам исследований на модельных животных, его отсутствие
197
приводит к повышенному потреблению пищи и как следствие
к резкому увеличению массы тела. Поэтому мы вправе ожидать,
что некоторые мутации, нарушающие структуру человеческого
гена NPY2R, должны вызывать приблизительно такой же фено-
типический эффект. В данном гене известно около 100 нуклео-
тидных замен. Но в анализ пока вовлечено чуть более десяти. Для
двух из них (-4448А и -4179Т) продемонстрирована тесная ас-
социация с тяжелым ожирением. При этом использованные вы-
борки достаточно велики, что позволяет надеяться на адекват-
ность полученных результатов.
Безусловно, наличие у человека ярко выраженного ожирения
не является доказательством того, что он переедает. Возможны и
другие причины излишнего веса. Например, определенные де-
фекты обмена веществ. Поэтому пока мы не можем с уверен-
ностью утверждать, что гены, контролирующие систему
пептида YY, непосредственно влияют у человека на склон-
ность к перееданию. Однако с учетом результатов исследо-
ваний на мышах такое влияние представляется весьма и
весьма вероятным.
7.7. Роль генов, контролирующих
инсулиновую и лептиновую системы
Чувство голода подавляется не только пептидом YY. Сходное
действие оказывают и некоторые другие белки, синтезируемые
организмом в ответ на обильное питание. К их числу относятся
лептин и инсулин.
Молекулы инсулина информируют аркуатное ядро об избы-
точном содержании глюкозы в плазме крови, что обычно наблю-
дается непосредственно после еды. В результате вырабатываются
импульсы, подавляющие чувство голода. Соответственно недо-
статочная активность инсулиновой системы должна провоциро-
вать организм на избыточное потребление пиши. И действитель-
но, мыши, нокаутированные по гену Insr (его продуктом являет-
ся рецептор инсулина), отличаются от нормы перееданием и
склонностью к ожирению.
У человека ситуация далеко не столь однозначна. Дело в том,
что хроническая недостаточность инсулиновой системы сопрово-
ждается развитием диабета — тяжелой болезни с множествен-
ными нарушениями обмена жиров и углеводов. Люди, страда-
ющие диабетом, как правило, полны. Но связана ли их полнота
с дефектами метаболизма или со склонностью к систематическо-
му перееданию, остается неясным. Поэтому говорить о влиянии
198
Таблица 7.4. Мутации, инактивирующие лептиновую систему человека
Ген Соответствующий белок Мутация Молекулярные последствия Распространение Особенности проявле- ния
LEP Лептин 105Tip Нарушен участок моле- кулы лептина, отвечаю- щий за ее способность секретироваться в кровь. В результате лептин не покидает жировые клетки и подвергается быстрой деградации Обнаружена у 17 представителей одной и той же семьи (из Турции) Гетерозиготы обыч- но не отличаются от нормы или имеют сравнительно легкую форму ожирения. Индекс массы тела от 23 до 35. Гомозиготы стра- дают тяжелейшими формами ожирения по причине чрезмер- но высокого чувства голода. Индекс массы тела обычно не менее 50
AG133 Образуются укороченные молекулы лептина, полно- стью лишенные своих биологических функций Обнаружена у 15 представителей трех семей (пакистанцы)
LEPR Рецептор лептина 1А в 16-м интроне Нарушается сплайсинг 16-го интрона. В результа- те образуется аномальный вариант рецептора, не способный активировать соответствующие G-белки Обнаружена у 7 пред- ставителей одной и той же семьи (алжир- цы)
661г
инсулиновой системы на потребление пищи у человека пока
преждевременно.
Теперь обратимся к лептиновой системе. Напомним: она сиг-
нализирует аркуатному ядру о состоянии имеющихся в организме
жировых отложений. Если содержание лептина чрезмерно велико
(что свидетельствует об излишнем количестве накопленного жира),
чувство голода оказывается в значительной степени подавленным.
Напротив, при нехватке лептина оно гипертрофировано.
ЭТа схема подтверждена не только экспериментами на мышах.
У человека описано несколько очень редких мутаций, затраги-
вающих кодирующую область гена LEP и приводящих к полной
дефектности лептина. Показано, что люди, гомозиготные по
этим мутациям, отличаются патологически высоким чувством
голода и как следствие страдают тяжелейшими формами ожире-
ния (табл. 7.4). Но если таким пациентам в течение длительного
времени регулярно вводить необходимое количество лептина, они
снижают потребление пищи и нормализуют комплекцию.
Лептин воздействует на рецепторы, кодируемые геном LEPR.
Их дефектность тоже приводит к неутолимому чувству голода.
Соответствующие мутанты получены у крыс и мышей, а также
описаны непосредственно у человека (см. табл. 7.4). В этих слу-
чаях организм практически нечувствителен к лептину, а потому
дополнительное введение данного белка не оказывает влияния на
комплекцию и отношение к пище.
Таким образом, участие генов LEP и LEPR в контроле пред-
расположенности к перееданию не вызывает сомнений.
7.8. Роль генов, контролирующих систему
проопиомелонокортино
Получив информацию о состоянии энергетического баланса
(см. табл. 7.3), нейроны аркуатного ядра ее преобразуют и в виде
импульсов посылают в другие участки гипоталамуса. В этом про-
цессе задействовано множество разных белков. Но мы сконцен-
трируем внимание только на тех, что изучены к настоящему вре-
мени наиболее подробно. К их числу относится проопиоме-
ланокортин — крупный белок (приблизительно 240 аминокислот),
расщепляемый на несколько относительно коротких фрагментов,
каждый из которых очень важен для нервной системы.
Нейроны, входящие в состав аркуатного ядра, различают-
ся по своему влиянию на чувство голода: одни усиливают его,
другие — наоборот, ослабляют. Установлено, что молекулы про-
опиомеланокортина (а точнее, одного из продуктов его расще-
200
Таблица 7.5. Основные характеристики рецепторов а-меланоцитстимулирующего гормона у человека
Тип рецептора Принцип функционирования Результат активации Основная локализация Регулируемые признаки Соответствующий ген
MCI Рецепторы, связан- ные с G-белками. Вторичный мессенд- жер — цАМФ (его содержание при ак- тивации рецептора повышается за счет стимулирования аденилатциклазы) Постсинаптиче- ское возбуждение Определенные клетки кожи Пигментация кожи MC1R
МСЗ Головной мозг, преимущественно гипоталамус Эффективность усвоения пищи MC3R
МС4 Чувство голода MC4R
МС5 Определенные клетки кожи Активность желез наружной секреции MC5R
пления — а-меланоцитстимулирующего гормона) используются
нейронами только второго типа. Таким образом, данный белок
подавляет чувство голода. В самом деле, при его искусственном
введении в гипоталамус количество потребляемой пищи резко
снижается. Напротив, у мышей, нокаутированных по гену РОМС
(именно он кодирует проопиомеланокортин), наблюдается чрез-
мерное чувство голода и сильное ожирение.
Мы вправе ожидать, что дефектность человеческого гомолога
тоже должна приводить к хроническому перееданию. Но пока
убедительных подтверждений получить не удалось. Скорее всего,
это связано с тем, что к настоящему времени в генетический
анализ вовлечено лишь несколько мутаций.
Молекулы а-меланоцитстимулирующего гормона распознают-
ся у человека четырьмя вариантами рецепторов: MCI,
МСЗ, МС4 и МС5. Каждый является продуктом отдельного гена
и характерен для строго определенного типа тканей (табл. 7.5).
Судя по результатам исследований на крысах и мышах, рецепто-
ры MCI, МСЗ и МС5 не имеют непосредственного отношения
к регуляции чувства голода и потому не представляют для нас
особого интереса. Гораздо большего внимания заслуживают ре-
цепторы МС4. Показано, что мыши, лишенные соответствующе-
го гена (он получил обозначение MC4R), страдают ярко выражен-
ным перееданием и как следствие — ожирением. Сходный эффект
наблюдается и при использовании химических препаратов, вы-
зывающих специфическую инактивацию данных рецепторов.
В связи с этим логично предположить, что и в случае человека
нарушение функций рецепторов МС4 должно приводить к чрез-
мерно обильному потреблению пищи. И действительно, в чело-
веческом гене MC4R обнаружено более 30 различных мута-
ции, обусловливающих аномально высокое чувство голода, а
в итоге — хроническое переедание с тяжелым ожирением.
Особенно ярко оно проявляется у гомозигот. Около 4 % всех
случаев ожирения связано с мутациями именно этого гена.
7.9. Роль гена AGRP, кодирующего
агути-подобный белок
Вернемся к рис. 7.1 (см. подразд. 7.2). Как видите, нейроны,
обеспечивающие ослабление чувства голода, возбуждаются под
действием молекул лептина и инсулина. В организме, питающем-
ся более или менее адекватно, содержание обоих гормонов до-
статочно высоко: наличие первого обусловлено накопленными
жировыми запасами, а второго — регулярными поступлениями
202
глюкозы в плазму крови. Казалось бы, упомянутые нейроны ар-
куатного ядра должны находиться в возбужденном состоянии
практически непрерывно и тем самым удерживать чувство голода
на низком уровне. Но в действительности наблюдается намного
более сложная ситуация.
Даже при высоком содержании лептина и инсулина чувство
голода время от времени способно возрастать. Причина — по-
степенное опустошение пищеварительной системы. В результате
происходит усиление биосинтеза грелина и каких-то других, пока
не известных сигнальных молекул, вызывающих массовое воз-
буждение конкурирующих нейронов.
Возникшие импульсы передаются в другие районы гипотала-
муса. В этом процессе участвует несколько разных нейромедиа-
торов, в том числе небольшой агути-подобный белок. Он тоже
воздействует на рецепторы МС4У но в отличие от а-меланоцит-
стимул и рующе го гормона обеспечивает значительное снижение
их активности. Таким образом, молекулы агути-подобного белка
затрудняют передачу нервных импульсов, подавляющих чувство
голода.
Данный белок кодируется геном AGRP. Мыши и крысы с по-
вышенной экспрессией этого гена (т.е. с избыточным содержа-
нием агути-подобного белка и как следствие — с малой активно-
стью рецепторов МС4) отличаются хроническим перееданием и
сильным ожирением. Напротив, у особей, дефектных по гену
AGRPy потребление пиши по сравнению с нормой существенно
снижено, что приводит к нехватке жира и недостаточному весу.
Вполне вероятно, что мутации гомологичного гена у человека
должны влиять на отношение к пище приблизительно так же.
В человеческом гене AGRP к настоящему времени обнару-
жено около 30 нуклеотидных замен. Среди них основное вни-
мание уделяется одной'. 67Thr в экзоне 3. Судя по результа-
там пяти независимых исследований, эта замена ассоцииру-
ет с пониженным индексом массы тела, относительно
небольшим содержанием жира в организме и даже некото-
рыми симптомами анорексии. И лишь в одной работе получен
отрицательный результат. К сожалению, мета-анализ пока
не проведен. Однако с высокой вероятностью можно ожи-
дать, что он подтвердит наличие искомой ассоциации.
7.10. Роль генов, контролирующих систему
нейропептида Y
Нейроны аркуатного ядра, усиливающие чувство голода, пере-
дают свои импульсы одновременно по нескольким направлениям
203
204
Таблица 7.6. Основные характеристики рецепторов нейропептида У у человека
Тип рецептора Принцип функциони- рования Локализация в си- напсах Результат активации Специфичность* Соответствующий ген
Y1 Рецепторы, связан- ные с G-белками. Вторичные мессенд- жеры: цАМФ и (или) ионы Са2*. Содержание первого при активации ре- цептора снижается, содержание вто- рого — напротив, возрастает Преимущественно пресинаптические мембраны Пресинаптиче- ское торможение; постсинаптическое возбуждение У> УУ» БПЖ NPY1R
Y2 Пресинаптические мембраны Пресинаптическое торможение УУ> У» БПЖ NPY2R
Y3 Неизвестна Неизвестен У» УУ> БПЖ Неизвестен**
Y4 Неизвестна Неизвестен БПЖ » УУ > У NPY4R
Y5 Преимущественно пресинаптические мембраны Пресинаптиче- ское торможение; постсинаптическое возбуждение У> УУ> БПЖ NPY5R
* В таблице использованы следующие обозначения: Y— нейропептид Y; УУ— пептид УУ; БПЖ — белок поджелудочной желе-
зы (выделяется клетками поджелудочной железы при заполнении пищеварительной системы, подавляет чувство голода).
* * Ген, кодирующий рецепторы Y3, еще не клонирован.
и при этом используют три разных нейромедиатора. Во-первых,
уже знакомый нам агути-подобный белок. Во-вторых, у-амино-
масляную кислоту (ее молекулы обеспечивают торможение кон-
курирующих нейронов; см. рис. 7.1). И в-третьих, так называемый
нейропептид Y.
Речь идет о небольшом белке (36 аминокислот), широко ис-
пользуемом в качестве нейромедиатора как в центральной, так и
в периферической нервной системе. Его молекулы в зависимости
от их локализации регулируют различные аспекты жизнедеятель-
ности организма, в том числе сердечную деятельность, давление
крови, настроение, суточную активность, температуру тела, ин-
тенсивность обмена веществ и потребление пищи.
При искусственном введении нейропептида Y в гипоталамус
наблюдается резкое усиление чувства голода. Этот факт убеди-
тельно продемонстрирован на многих позвоночных: млекопита-
ющих, птицах, рептилиях и даже рыбах. Логично предположить,
что отсутствие данного белка должно вызывать в организме диа-
метрально противоположный эффект, т.е. способствовать ощу-
щению сытости и сокращению рациона. Однако у животных,
лишенных соответствующего гена (этот ген получил обозначение
NPY), количество поглощаемой пищи практически в норме. Ве-
роятно, существуют какие-то «обходные пути», позволяющие
компенсировать нехватку нейропептида Y.
В человеческом гене ^РКосновное внимание уделяется толь-
ко одной из выявленных мутаций — нуклеотидной замене Рго7
в экзоне 1. По некоторым данным, она не связана с индексом
массы тела, а согласно другим — слабо ассоциирует с ею повы-
шенными значениями. Таким образом, вопрос об участии гена
NPY в контроле предрасположенности к перееданию оста-
ется открытым.
У человека известно как минимум пять типов рецепторов,
способных распознавать молекулы нейропептида Y (табл. 7.6). Эги
рецепторы имеют в основном пресинаптическую локализацию,
регулируя количество выбрасываемого нейромедиатора. Но некото-
рые встречаются и на постсинаптических мембранах, обеспечивая
восприятие передаваемых нервных импульсов. Судя по результатам
исследований на модельных животных, в нейронах, получающих
информацию от аркуатного ядра, особенно многочисленны рецеп-
торы Y1 и Y5. Их активация приводит к усиленному потреблению
пищи. Тем не менее мыши, нокаутированные по соответствующим
генам, не отличаются от нормы своим среднесуточным рационом,
зато страдают достаточно выраженными формами ожирения. В чем
причины такого эффекта, пока непонятно.
Ситуация дополнительно осложняется еще и тем, что эти ре-
цепторы могут взаимодействовать и с другими сигналами, непо-
205
средственно участвующими в регуляции пищевого поведения (см.
табл. 7.6). В результате очень трудно определить, какие именно
сигналы наиболее значимы для каждого типа упомянутых рецеп-
торов. Особенно с учетом их широкой локализации в организме.
Для генов, кодирующих рецепторы нейропептида К пока не
выявлено отчетливой связи со склонностью к перееданию. Но это
не значит, что подобной связи вовсе не существует. Наиболее
перспективный кандидат — ген NPY2R, поскольку некоторые
мутации в его промоторной области ассоциируют с тяжелыми
формами ожирения (см. подразд. 7.6).
7.11. Роль генов, обслуживающих
иные молекулярные системы
Помимо рассмотренных выше молекулярных систем, в регу-
ляции чувства голода задействованы и многие другие. К сожале-
нию, мы не сможем подробно остановиться на каждой и ограни-
чимся кратким упоминанием только о тех, для которых получены
достаточно надежные данные.
Прежде всего, необходимо отметить серотониновую систему.
Известно, что многие нейроны в составе гипоталамуса иннерви-
руются при помощи серотонинэргических синапсов. Соответ-
ственно, мы вправе ожидать, что некоторые мутации, нарушаю-
щие нормальное функционирование этих синапсов, тоже заметно
отражаются на отношении к пище. И действительно, S-промотор
гена SLC6A4 (см. подразд. 6.12) демонстрирует отчетливую
ассоциацию с анорексией. Этот вывод основан на пяти не-
зависимых работах и к тому же статистически подтверж-
дается мета-анализом.
Серотониновая система регуляции тесно связана с дофамино-
вой. Таким образом, гены, контролирующие дофаминовую си-
стему, тоже заслуживают самого пристального внимания. Три
независимых исследования свидетельствуют о том, что замена
158Уа! в гене СОМТ (см. подразд. 5.5) хотя и слабо, но все же
повышает вероятность анорексии. Однако с окончательным вы-
водом торопиться не следует, поскольку использованные выбор-
ки не очень велики, а мета-анализ еще не проведен.
Постепенно становится все более и более очевидным, что су-
щественную роль в процессе регуляции чувства голода играют
различного рода нейротрофины, и в частности — нейротрофиче-
ский фактор головного мозга (мы познакомились с ним в подразд.
4.16). Судя по результатам восьми независимых исследований ч
нуклеотидная замена 6 6Met в гене BDNfF тесно ассоциирует
206
с различными формами анорексии. Механизм этой связи пока
остается неясным. Но некоторые детали известны уже сегодня.
Установлено, что продукт аллели 66Met секретируется значитель-
но хуже по сравнению с нормальным. Соответственно у людей,
несущих данную аллель, молекулы нейротрофического фактора
головного мозга выделяются в межклеточное пространство в не-
достаточном количестве и, по-видимому, выполняют свои функ-
ции не в полной мере.
Любопытно, что низкое содержание указанного нейротрофина
характерно не только для пациентов, страдающих анорексией, но
также для людей с ярко выраженной склонностью к перееданию.
Более того, в трех независимо выполненных работах обнаружена
связь между мутацией 66Met и булимией. Получается, что один
и тот же молекулярный дефект (недостаток нейротрофического
фактора головного мозга) в некоторых случаях приводит к от-
торжению пищи, а в некоторых, наоборот, способствует хрони-
ческому обжорству. Одно из вероятных объяснений этого проти-
воречия будет рассмотрено в следующем параграфе.
Имеются данные и по некоторым иным молекулярным систе-
мам. Однако все они требуют дополнительных подтверждений и
пока и не могут рассматриваться в качестве окончательных.
7.12. Генетический анализ
предрасположенности человека
к перееданию. Основные итоги
и ближайшие перспективы
Итак, многочисленные исследования свидетельствуют о том,
что отношение человека к пише находится под контролем как
минимум пяти генов: LEP, LEPR, MC4R, SLC6A4 и BDNF. Де-
фектность трех первых приводит к гипертрофированному чувству
голода и в итоге — к тяжелым формам хронического переедания
(особенно в случае гомозиготности по соответствующим мутаци-
ям). Короткий вариант промотора гена SLC6A4. напротив, спо-
собствует проявлениям анорексии. И наконец, замена 66Met в
гене BDNFассоциирует как с анорексией, так и с булимией.
Последний факт, казалось бы, противоречит здравому смыслу.
Разве может одна и та же нуклеотидная замена вызывать у разных
людей противоположные эффекты? Давайте попробуем разобрать-
ся в этом вопросе. В переводе с латыни «анорексия» означает «от-
сутствие чувства голода». Однако, как показывают детальные на-
блюдения за пациентами, в действительности ситуация значитель-
207
но более сложная. Подавляющее большинство пациентов, страда-
ющих анорексией, постоянно испытывает голод, причем ярко
выраженный. Но он преодолевается навязчивым страхом потол-
стеть. В отдельные моменты чувство голода настолько обострено,
что пациент теряет способность себя контролировать и в резуль-
тате безостановочно объедается какое-то время. Затем наступает
тяжелое длительное раскаяние: пациент старается «очистить» свой
организм и вновь ограничивает себя в потреблении пищи.
Перечисленные симптомы во многом напоминают булимию.
В данном случае тоже наблюдаются периоды «очищения», но их
продолжительность сильно уступает периодам обжорства. Таким
образом, вполне вероятно, что обе патологии (или хотя бы не-
которые их разновидности, связанные с чередованием обжорства
и последующего «очищения») формируются приблизительно на
одной и той же основе. Этой основой может служить излишнее
чувство голода. Если человек не способен ему противостоять,
скорее всего, возникнет одна из форм булимии. Но если человек
по каким бы то ни было причинам (болезнь, воспитание, соб-
ственные убеждения и др.) воспринимает свой голод как нечто
негативно-постыдное и способен длительное время его преодо-
левать, в конечном итоге может развиться анорексия.
Судя по результатам исследований на модельных животных,
одной из причин, обусловливающих повышенное чувство голода,
является недостаток продукта гена BDNF. Напомним, что имен-
но к такому молекулярному эффекту приводит нуклеотидная
замена 66Met. И в результате складывается достаточно стройная
картина. По-видимому, рассматриваемая нами нуклеотидная за-
мена вызывает в организме существенное повышение чувства
голода, способное у одних индивидов спровоцировать булимию,
а у других, наоборот, привести к развитию анорексии.
Таким образом, склонность человека к хроническому перееда-
нию формируется в результате различных молекулярных де-
фектов, непосредственно или косвенно усиливающих чувство
голода. В частности, это могут быть дефекты лептиновой
системы, проопиомеланокортиновой и некоторых других.
Генетические основы анорексии значительно сложнее.
В этом случае теоретически возможны мутации трех ти-
пов:
• во-первых, значительно ослабляющие чувство голода (чело-
век почти не испытывает потребности в пище и даже уверен, что
он регулярно переедает). К подобного рода мутациям, по-
видимому, относится короткий промотор гена SLC6A4;
• во-вторых, оказывающие диаметрально противоположное
действие, при условии, что оно воспринимается крайне негатив-
но. Речь идет о замене 66Met в гене BDNF\
208
• в-третьих, никак не влияющие на чувство голода, но приво-
дящие к паническому страху набрать лишний вес. Примеры по-
добных мутаций пока неизвестны.
Как видим, необходимо различать два следующих параметра:
степень выраженности чувства голода и отношение к нему. К со-
жалению, оценивать их по отдельности очень трудно. Дело в том,
что результаты, полученные при помощи опросов, отражают ис-
ключительно субъективное мнение испытуемых, а оно в этих
случаях, как правило, далеко от реальности. До сих пор не раз-
работаны и строго экспериментальные подходы. Наиболее пер-
спективными представляются долговременные исследования,
в ходе которых каждого конкретного испытуемого начинали бы
оценивать еще с младенческого возраста, когда чувство голода
выражено максимально отчетливо. В дальнейшем, по мере того,
как испытуемый взрослеет, стало бы ясно и его отношение к
своему чувству голода. Такой подход позволит намного точнее
диагностировать различные типы пищевых патологий человека,
что будет способствовать более четкому генетическому анализу.
СПИСОК ЛИТЕРАТУРЫ
I. Коркина М. В., Цивилько М.А., Марилов В. В. Нервная анорексия. —
М. : Медицина, 1986.
2. Мазаева Н.А., Осипова А. А. Нервная анорексия — проблема, далекая
от решения // Журнал неврологии и психиатрии им. С.С. Корсакова. —
2007. - Т. 107. - № 10. - С. 85-94.
3. Циркин С.Ю.' Гладышев О. А., Бабин А. Г. Нервная булимия: критерии
и типология // Соц. и клинич. психиатрия. — 2000. — Т. 10. — № 1. —
С. 68-72.
4. Ando Т. et al. Variations in the preproghrelin gene correlate with higher body
mass index, fat mass, and body dissatisfaction in young Japanese women //
American J Clinical Nutrition. — 2007. — V. 86. — P. 25 — 32.
5. Argyrvpoulos G. et al. The agouti-related protein and body fatness in humans //
international J. Obesity. — 2003. — V. 27. — P. 276 — 280.
6. Beck B. Neuropeptide Y in normal eating and in genetic and dietary-induced
obesity /1 Philosophical Transactions Royal Society, B. — 2006. — V. 361. —
P. 1159-1185.
7. Bulik С. M., Sullivan P. E, Wade T. D. and Kendler K.S. Twin studies of
eating disorders: a review 11 International J Eating Disorders. — 2000. — V. 27. —
P. I - 20.
8. Chaudhri O. et al. Gastrointestinal hormones regulating appetite // Philo-
sophical Transactions Royal Society. B. — 2006. — V. 361. — P. 1187— 1209.
9. Clement K. et al. A mutation in the human leptin receptor gene causes obesity
and pituitary dysfunction // Nature. — 1998. — V. 329. — P. 398 — 401.
10. Gorwood P.„ Kipman A. and Foulon C. The human genetics of anorexia
nervosa // European J. Pharmacology. — 2003. — V. 480. — P. 163— 170.
209
11. Hashimoto К. et al. Role of brain-derived neurotrophic factor in eating
disorders: Recent findings and its pathophysiological implications // Progress Neuro-
Psychopharmacology & Biological Psychiatry. — 2005. — V. 29. — P. 499 — 504.
12. Houseknecht K. L. et al. The biology of leptin: A review // J. Animal
Sciences. - 1998. - V. 76. - P. 1405- 1420.
13. Ma L. et al. Variations in peptide YY and Y2 receptor genes are associated
with severe obesity in Pima Indian men // Diabetes. 2— 005. — V.— 54. —
P. 1598-1602.
14. MacKenzie R. G. Obesity-associated mutations in the human melanocortin-4
receptor gene // Peptides. — 2006. — V. 27. — P. 395 — 403.
15. Naslund E. and Hellstrom P. M. Appetite signaling: From gut peptides and
enteric nerves to brain // Physiology & Behavior. 2007. V.92. P.256 — 262.
16. O’Rahilly S., Farooqi I.S., Yeo G.S.H. and Challis B.G. Minireview:
Human obesity —lessons from monogenic disorders // Endocrinology. — 2003. —
V. 144. — P. 3757-3764.
17. Perusse L. et al. The human obesity gene map: The 2004 update // Obesity
Research. - 2005. - V. 13. - P. 381 -490.
18. Siddiq A. et al. Single nucleotide polymorphisms in the neuropeptide Y2
receptor (NPY2R) gene and association with severe obesity in French white subjects //
Diabetologia. - 2007. - V. 50. - P. 574-584.
19. Slof-Op't Landt M. C. et al. Eating disorders: from twin studies to candidate
genes and beyond // Twin Research & Human Genetics. — 2005. — V. 8. —
P. 467-482.
20. Strobel A. et al. A leptin missense mutation associated with hypogonadism
and morbid obesity // Nature Genetics. — 1998. — V. 18. — P. 213 — 215.
21. Williamson D.A. et al. Microanalysis of eating behavior of three leptin de-
ficient adults treated with leptin therapy // Appetite. — 2005. — V. 45. — P. 75 — 80.
КОНТРОЛЬНЫЕ ВОПРОСЫ
I. Как оценивают отношение к пище у мышей и человека?
2. Опишите молекулярные механизмы регуляции чувства голода.
3. К каким эффектам приводят мутации по генам LEP и LEPR?
4. Каков механизм влияния гена MC4R на чувство голода?
5. Опишите роль генов SLC6A4 и BDNFn контроле отношения к пище
у мышей и человека.
Глава 8
ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ ЗАВИСИМОГО
ПОВЕДЕНИЯ
8.1. Что такое зависимость?
Под зависимостью в психологии понимают чрезмерное увле-
чение, систематически выходящее за рамки самоконтроля. Такие
поведенческие аномалии могут проявляться:
• в готовности проводить много времени за своим увлечением;
• неспособности самостоятельно контролировать это время;
• постепенном возрастании расходов на свое увлечение в
ущерб другим, даже более важным жизненным целям;
• стремлении использовать свое увлечение как универсальное
средство для ухода от всех возникающих проблем;
• неспособности достигать удовольствия иными путями;
• непринятии критики по отношению к подобному образу
жизни;
• смирении с потерей репутации, работы, друзей или семьи;
• в особо тяжелых случаях — в разрушении личности.
Предметом зависимости может быть все что угодно: никотин,
алкоголь, наркотики, определенные виды пищи, деньги, работа,
секс, экстремальные ситуации, конкретные люди, домашние жи-
вотные, физические нагрузки, музыка, книги, покупки, коллек-
ционирование и др. Крайние формы фанатизма (спортивного,
религиозного, политического, культурного, научного и др.) тоже,
по сути, являются ярко выраженными зависимостями. В последнее
время стремительное развитие электронных технологий привело к
появлению целого ряда новых зависимостей: от компьютерных игр,
мобильной связи, общения в Сети, плееров, телесериалов и др.
По оценкам ВОЗ, как минимум каждый третий человек стра-
дает хотя бы одной из перечисленных аномалий (табл. 8.I). Воз-
можно, такая оценка еще и занижена. Дело в том, что некоторые
аспекты зависимого поведения часто воспринимаются как осо-
бенности характера или стиля жизни и не всегда попадают в поле
зрения специалистов.
211
Таблица 8.1. Распространенность некоторых видов зависимо*
го поведения, %*
Страна Вид зависимости
Никотиновая Алкогольная Наркоти- ческая Игровая
г 2 d 9 -I...3 -5...10
Австрия 48,1 46,5, 5,2 1,8
Аргентина 28,9 22,6 6,7 1,7
Бельгия 30 25 9,5 3,6
Бразилия 21,4 12,7 17,1 5,7
Великобритания 28 24 7,5 2,1
Германия 37,1 30,5 6 1,5
Египет 59,3 2,7 0,4 0
Канада 18,8 15,9 14 4,5
Китай 57,4 3,5 6,6 0,2
Польша 38 25,6 23,3 4,1
Россия 61,3 15 18 5
США 19,8 15,4 10,8 4,8
Турция 49,4 17,6 1,7 0,7
Украина 62 17 18,5 4,5
Финляндия 27 20 6,5 1,5
Франция 30 21,2 13,3 4,1
Южная Корея 52,8 5,8 6,9 1,7
ЮАР 27,1 8,2 27,6 9,9
* Доля от населения старше 15 лет (сведения за период 2005 — 2008 гг.).
Безусловно, вероятность развития конкретной зависимо-
сти в значительной степени определяется факторами внеш-
ней среды', воспитанием, культурными традициями, уровнем
образования, кругом общения, социальным статусом, про-
212
фессиональными навыками. Однако довольно существенна и
роль генотипа. В этом плане наиболее изучены три вида за-
висимостей: наркотическая, алкогольная, а также никоти-
новая. Именно на них мы и сосредоточим свое внимание.
Нередко названные три зависимости сопутствуют друг другу.
Тем не менее каждая из них относительно автономна и, по-
видимому, имеет «свой собственный* генетический контроль,
поэтому мы будем рассматривтаь их по отдельности.
8.2. Разнообразие наркотиков
Для начала определимся с понятием «наркотики*. Оно не
имеет однозначного толкования. При использовании в самом ши-
роком, обыденном смысле это понятие означает практически
любое воздействие, способное со временем превратиться в предмет
зависимости. Подобная трактовка встречается достаточно часто,
например, в утверждении «власть — сильнейший наркотик», но с
научной точки зрения, конечно, не выдерживает критики.
В более узком смысле к наркотикам относят вещества, не
входящие в число необходимых для поддержания жизнедея-
тельности, но оказывающие заметное влияние на человече-
скую психику.
Формально под это определение вполне подпадают и героин,
и этиловый спирт, и клеевые растворители, и никотин, и гашиш,
и даже запах любимых цветов, если он существенным образом
повышает настроение. Поэтому приходится уточнять, что понятие
«наркотики» охватывает исключительно те психоактивные соеди-
нения, которые законодательно выведены из свободного оборота
из-за опасности использования в немедицинских целях.
К настоящему времени известно более 100 подобных веществ.
Они различаются своим строением, механизмами действия, а,
самое главное — характером вызываемых эффектов (табл. 8.2).
В частности, некоторые наркотики, например ЛСД, вызывают у
человека исключительно психологическую зависимость, т. е.
желание вновь испытать определенные эмоции без какой бы то
ни было физиологической потребности в них. Между тем героин
и многие другие наркотики приводят к серьезным изменениям в
обмене веществ и в итоге становятся необходимыми его компо-
нентами. В результате возникает тяжелая физическая зависи-
мость: человек не способен самостоятельно отказаться от нарко-
тика, а в его отсутствие испытывает мучительную «ломку».
Еще раз отметим, что некое вещество считают наркотиком,
если оно внесено в соответствующие законодательные акты, за-
213
Таблица 8.2. Важнейшие классы наркотиков
Класс наркотиков Представители Основные эффекты Тип зависимости
Опиаты* Морфин, кодеин, героин, метадон Эйфория, обезболивание, общее успокоение Психологическая и физическая
Психостимуляторы Кокаин, первитин, амфетамин Бодрость, усиление внима- ния, общее возбуждение
Каннабиноиды Гашиш, марихуана, тетра- гид ракан наби нол Обезболивание, обострение чувств, изменение сознания, в небольших количествах — легкая дисфория Преимущественно психологическая
Седативные средства Кетамин, пентобарбитал, триазолам Общее успокоение, подавле- ние страха и депрессии
Галлюциногены ЛСД, мескалин, псилоцибин Различного рода галлюцина- ции
Чужеродные для организма химические вещества, действующие наподобие опиума.
□решающие или строго ограничивающие его оборот. Таким об-
разом, этиловый спирт и никотин, хотя и приводят к развитию
очень тяжелых зависимостей, не могут рассматриваться в качестве
наркотических средств.
Существует и еще более узкое определение наркотиков. Оно
вытекает из исходного смысла понятия «наркоз» — одурманива-
ние, утоление боли, приведение в оцепенение. В этом смысле к
наркотикам относят различные опиаты, т.е. опиум и ряд других
химических веществ, вызывающих обезболивание, эйфорию и
общее успокоение. Опиатная зависимость считается самой тяже-
лой. Она очень быстро перерастает из психологической в физи-
ческую и в дальнейшем требует постепенного увеличения дозы,
что ведет к разрушению личности и скорой смерти. Поэтому из
множества видов наркотической зависимости мы подробно рас-
смотрим именно опиатную.
8.3. Молекулярные основы опиатной
зависимости
Иногда человеку приходится перенапрягаться. В таких ситуа-
циях «включаются» специальные молекулярные системы, позво-
ляющие организму преодолевать усталость и боль. В частности,
резко усиливается синтез эндорфинов.
Эндорфинами называют небольшие регуляторные белки,
выполняющие функцию внутренней анестезии (табл. 8.3). Эти
белки вырабатываются определенными группами нейронов,
проникают во все отделы центральной нервной системы и обе-
спечивают активацию нескольких типов рецепторов. В результате
осуществляется запуск молекулярных каскадов, приводящих к
подавлению боли, усталости и беспокойства (другими словами, у
человека «открывается второе дыхание»). К усилению биосинтеза
эндорфинов приводит и секс. В этом случае помимо подавления
негативных эмоций эндорфины вызывают ярко выраженное удо-
вольствие — эйфорию (рис. 8.1).
Оказалось, что рецепторы эндорфинов могут быть активиро-
ваны и под действием некоторых чужеродных для организма ве-
ществ. При введении подобных веществ (их называют опиатами)
ослабляются все виды боли, исчезает волнение, проходит уста-
лость, наступает состояние полного счастья. Эти эффекты на-
столько приятны и настолько сильны, что очень быстро, всего
лишь за несколько приемов наркотика, вызывают развитие тяже-
лой психологической зависимости, самостоятельно преодолеть
которую человек не способен.
215
Таблица 8.3. Важнейшие типы эндорфинов
Тип эндорфинов Основные эффекты Строение Соответству- ющие гены
р-эндорфины Ослабление боли, успокое- ние, эйфория Небольшие белки приблизительно из 30 ами- нокислот, образующиеся при расщеплении проопиомеланокортина РОМС
Динорфины Ослабление боли, дисфо- рия* Небольшие белки приблизительно из 20 ами- нокислот, образующиеся при расщеплении проди норфи на PDYN
Энкефалины Ослабление боли Короткие белки из 5 аминокислот, образую- щиеся при расщеплении проэнкефалина, про- опиомеланокортина и продинорфи на PENK, РОМС, PDYN
* Высокие концентрации динорфинов вызывают симптомы отчетливого неудовольствия (дисфории).
Рис. 8.1. Важнейшие молекулярные системы, регулирующие состояние удо-
вольствия у млекопитающих
Примечание. Центральную роль в регуляции состояния удовольствия играют
дофаминэргические нейроны миндалины, компактной части черной субстанции
ствола, а также вентральной покрышки среднего мозга. Чем больше дофамина вы-
деляется этими нейронами, тем острее и ярче приятные ощущения организма. В свою
очередь, степень активности данных нейронов регулируется целым рядом молекуляр-
ных систем, главным образом — серотониновой и эндорфиновой. И та, и другая
повышают активность дофам ин эрги ческих нейронов. Соответственно, при соверше-
нии каких-либо действий, сопровождающихся выбросом серотонина или эндорфинов
(например, во время еды или занятия сексом), организм испытывает приятные ощу-
щения, эйфорию. Воздействия, приводящие к возбуждению нейронов, показаны
стрелками. Тормозящие воздействия показаны знаком «±». Нейромедиаторы пред-
ставлены в виде кружков; Д — дофамин; С — серотонин; ГАМК — у-аминомасляная
кислота. Прочие пояснения — в тексте.
Вскоре возникшая зависимость перерастает в физическую.
Дело в том, что в организме, регулярно принимающем опиаты,
происходят значительные изменения обмена веществ. В первую
очередь резко ослабляется синтез эндорфинов. Тем самым орга-
низм утрачивает природную способность самостоятельно подав-
лять усталость, волнение и боль, что приводит к развитию абсти-
нентного синдрома, т.е. к мучительным «ломкам» в отсутствие
наркотика. Это ведет к приему все новых и новых доз, а в итоге —
к дальнейшему усилению опиатной зависимости.
Со временем происходит еще одно важное изменение: посте-
пенно снижается чувствительность организма к наркотику. В ре-
зультате при использовании некой «стандартной» дозы человек
уже не может испытывать прежнего удовольствия. Это побуждает
217
его увеличивать дозу или переходить к еще более сильному нар-
котическому средству.
Наряду с перечисленными основными эффектами опиаты вы-
зывают и множество побочных, в том числе:
• расширение кровеносных сосудов в толще кожи, вызыва-
ющее неестественный румянец на шее и щеках;
• повышенную потливость;
• значительное ослабление иммунной системы;
• потерю концентрации;
• подавление кашля;
• ослабленное дыхание.
По мере того как человек увеличивает дозы наркотика, по-
степенно усиливаются и их побочные эффекты. В конечном
итоге очередная доза оказывается летальной. Наиболее распро-
страненная причина — остановка дыхания. Таким образом,
смерть наркомана от передозировки не является результа-
том его случайной ошибки. Как правило, это абсолютно
закономерный финал.
8.4. Моделирование опиатной зависимости
у мышей
Эндорфиновая система характерна для всех позвоночных. Этот
факт позволяет моделировать опиатную зависимость у различных
лабораторных животных, например у мышей.
Обычно используют следующую экспериментальную схему.
Животное помещают в достаточно просторную клетку, состоящую
из нескольких соединенных друг с другом отсеков. В тот момент,
когда мышь находится в определенном отсеке, ей вводят фикси-
рованное количество опиатного наркотика. Через некоторое
время повторяют эту же процедуру, причем в том же отсеке, что
и первоначально.
В результате подопытное животное быстро запоминает, в какой
части клетки оно получало наркотик, и в дальнейшем преимуще-
ственно находится именно там.
Подобное поведение свидетельствует о возникшей зависимо-
сти, т.е. желании повторно испытать приятные эмоции. И дей-
ствительно, если животному предложить две поилки, одна из
которых содержит обычную воду, а вторая — такую же воду, но с
добавкой наркотика, регулярным предпочтением будет пользо-
ваться вторая поилка.
В многочисленных исследованиях показано, что введение
опиатов вызывает у мышей подавление беспокойства и боли. При
этом организм очень быстро «привыкает» к наркотику и посте-
218
пенно становится все менее чувствительным к нему. Соответ-
ственно, для того чтобы достичь определенного эффекта, напри-
мер блокировать стандартное болевое воздействие, необходимо
постепенно повышать вводимую дозу. В результате усиливаются
разнообразные косвенные эффекты, такие как чрезмерное на-
пряжение кишечной мускулатуры, подавление иммунной системы
и ослабление дыхания. Со временем это может привести к леталь-
ному исходу.
У мышей, регулярно получающих опиатный наркотик, резко
понижена выработка собственных эндорфинов. И если впослед-
ствии прекратить введение опиата, наблюдается широкий спектр
негативных реакций: слезотечение, рвота, понос, отсутствие ап-
петита, нарушения сна, высокая раздражительность, депрессивное
состояние, болевые симптомы, тремор конечностей и даже судо-
роги. Это очень похоже на классический абстинентный синдром,
возникающий у опиатного наркомана в отсутствие наркотика.
Таким образом, мыши — прекрасный модельный объект для
изучения механизмов формирования опиатной зависимости.
Необходимо особо отметить, что у нормальных мышей эта за-
висимость формируется быстро, за 3 — 4 приема. Но некоторые
линии заметно отличаются от нормы. Данный факт указывает на
то, что отношение к опиатам в значительной степени опре-
деляется генотипом организма. И некоторые соответ-
ствующие гены уже обнаружены. В первую очередь, это
гены, кодирующие рецепторы эндорфинов.
8.5. Генетический контроль отношения
к опиатом у мышей. Роль генов, кодирующих
рецепторы эндорфинов
У мышей обнаружено три типа рецепторов эндорфинов: ц,
к и 5 (табл. 8.4). Первые принято считать наиболее важными. Если
подавить их активность специфическими ингибиторами, чувстви-
тельность организма к опиатам резко снижается. Аналогичные
результаты получены и в генетических исследованиях.
Показано, что мыши, нокаутированные по гену Оргт и тем
самым полностью лишенные \i-рецепторов, почти не испытыва-
ют обезболивания при введении опиатов и демонстрируют слабую
зависимость от этих наркотиков.
Понижена чувствительность и к некоторым другим психоак-
тивным соединениям, в том числе — к никотину, этиловому
спирту и тетрагидроканнабинолу (табл. 8.5). Таким образом,
р-рецепторы принимают участие в формировании как минимум
219
Таблица 8.4. Основные характеристики рецепторов эндорфинов
Тип рецептора Принцип функционирования Локализация в синапсах Результат активации Распознаваемые эндорфины Регулируемые признаки Ген
Ц (мю) Рецепторы, свя- занные с G-бел- ками. Вторичный мес- сенджер — цАМФ. Его содержание снижается за счет стимулирования фосфодиэстеразы В основном на преси- наптических мембранах ГАМК- эргических синапсов Главным об- разом преси- наптическое торможение Преимуществен- но р-эндорфины и энкефалины Чувствитель- ность к боли, настроение, волнение и др. Оргт*
к(кап- па) Преимуществен- но динорфины Чувствитель- ность к боли, настроение Оргк
б (дель- та) Преимуществен- но энкефалины Чувствитель- ность к боли, настроение Oprd
* Аббревиатура Орг (от opioid receptor) связана с исторически сложившимся термином «опиоиды». Этим термином, возник-
шим еше до понятия «эндорфины», обозначают собственные молекулы организма, способные вызывать эффекты наподобие
опиатов.
Таблица 8.5 Особенности мышей, нокаутированных по генам рецепторов эндорфинов
Ген Реакция на психоактивные соединения Спонтанное поведение
Морфин Алкоголь Никотин Тетрагидроканнабинол Чувствительность к боли Подвижность Беспокойство
Оргт Резко ослаблена Ослаблена Ослаб- лена Ослаблена Резко повышена Понижена* Понижено
Оргк Ослаблена Слегка ослаблена — Слегка ослаблена Повышена Не изме- нена Не изменено
Oprd Ослаблена Усилена — Не изменена Повышена Повышена Повышено
* Только по отношению к висцеральной боли.
нескольких видов зависимости, включая опиатную, никотиновую,
алкогольную и каннабиноидную.
У животных, нокаутированных по гену Оргк, чувствительность
к каждому из перечисленных выше соединений тоже заметно
снижена по сравнению с нормой. Но степень снижения относи-
тельно невелика. Этот факт свидетельствует о том, что к-рецепторы
тоже участвуют в формировании упомянутых зависимостей, но
играют не главную, а второстепенную роль.
Ситуация с Ъ-рецепторами более сложная. Дело в том, что
нокаутирование гена Oprd приводит к значительному ослаблению
чувствительности к опиатам, но практически не влияет на отно-
шение к тетрагидраканнабинолу и, наконец, достоверно усили-
вает алкогольную зависимость. Причины такого эффекта пока
неясны. По-видимому, 6-рецепторы существенно отличаются от
рассмотренных выше ц- и к-рецепторов и даже выполняют ча-
стично противоположные функции. Этот вывод хорошо согласу-
ется с тем обстоятельством, что мыши, нокаутированные по гену
Оргт, проявляют пониженный уровень активности и беспокой-
ства, в то время как мыши, дефектные по гену Oprd, напротив,
чрезмерно встревожены и гиперподвижны (табл. 8.5).
Таким образом, степень выраженности опиатной зависи-
мости находится у мышей под сложным генетическим
контролем, в котором главную роль играет ген Оргт. Этот
же ген влияет и на некоторые другие характеристики,
в том числе на отношение к алкоголю и каннабиноидам,
а также на чувствительность к боли, подвижность и бес-
покойство.
8.6. Генетический анализ отношения
к опиатам у человека
Результаты близнецовых исследований свидетельствуют о том,
что развитие опиатной зависимости у конкретного человека в
основном определяется факторами внешней среды, а именно —
длительностью приема опиатных наркотиков. Вместе с тем от-
четливо прослеживается и роль генотипа: ее оценивают равной в
20...30%.
Возникает закономерный вопрос: какие же механизмы могут
лежать в основе такого генетического контроля? Во-первых,
вполне вероятно, что некие мутации вызывают хроническое
состояние неудовольствия (дисфорий) и тем самым прово-
цируют человека на поиск средств, способных хотя бы крат-
ковременно улучшать настроение. Этими средствами могут
222
оказаться и опиатные наркотики. Во-вторых, судя по результа-
там исследований на мышах, разные генотипы имеют разную
чувствительность к опиатам. Таким образом, мы вправе ожи-
дать, что у некоторых людей даже в случае довольно продол-
жительного приема наркотика возникает сравнительно слабая
форма зависимости.
В первой ситуации наиболее вероятными кандидатами явля-
ются гены, участвующие в контроле дофаминовой системы (см.
подразд. 5.3 —5.6), а во второй — гены, обслуживающие эндор-
ф и новую систему. И те, и другие вовлечены в анализ интересую-
щего нас признака.
Для начала обратимся к генам эндорфиновой системы. Эта
система у всех млекопитающих очень консервативна. В частности,
ее основные компоненты у человека практически те же, что и у
мышей. Единственное различие касается спектра рецепторов:
если у мышей он представлен тремя вариантами (ц, 5 и к), то
человеку свойственен еще и четвертый — а. К сожалению, его
функции пока недостаточно ясны. Однако экспериментально по-
казано, что и в случае человека центральная роль в формирование
зависимости от опиатов тоже принадлежит рецепторам ц-типа.
Поэтому логично предположить, что соответствующий ген (он
получил обозначение OPRM1) заметно влияет на отношение к
опиатным наркотикам.
В этом гене известно около 2,5 тыс. полиморфных сайтов.
В генетический анализ предрасположенности к опиатной зави-
симости к настоящему времени вовлечено лишь около 20, среди
которых подробно исследован только один — нуклеотидная за-
мена А118G. Этой замене посвящено более 30 независимых работ.
В нескольких обнаружено наличие слабой ассоциации, но осталь-
ные исследования ее не подтверждают. Об отсутствии связи сви-
детельствует и мета-анализ.
Имеются данные и о других генах эндорфиновой системы
(табл. 8.6). Однако пока не получено независимых подтверждений,
все они носят сугубо предварительный характер.
Роль генов дофаминовой системы изучена еще меньше: каж-
дому из них посвящены лишь единичные работы с использова-
нием одного или двух полиморфных сайтов. Ассоциация с опи-
атной зависимостью, как правило, отсутствует (см. табл. 8.6).
И все же некоторые результаты заслуживают внимания. В част-
ности, показано, что нехватка определенного нуклеотида в про-
моторной области гена DRD2 (эту мутацию обозначают -141delC)
ассоциирует с героиновой зависимостью только у наркоманов,
потребляющих наркотик в виде ингаляций. Если же героин ис-
пользуют в виде инъекций, возникающая зависимость не связана
с данным полиморфизмом.
223
224
Таблица 8.6. Поиск ассоциации между генами эндорфиновой и дофаминовой систем и опиатной за-
висимостью (предварительные данные)
Молекулярная система Ген Соответствующий белок Полиморфные сайты Результат исследования
Всего Исследовано
Эндорфиновая РОМС Про-он иомел анокортин. Расщепляется с образовани- ем p-эндорфинов и энкефа- линов -90 7 Некоторые гаплотипы* ассоциируют с опиатной за- висимостью
PDYN Про-динорфин. Расщепля- ется с образованием динор- финов и энкефалинов - 30 1 Ассоциации не обнаружено
PENK Про-энкефалин. Расщепля- ется с образованием энке- фалинов - 100 7 Некоторые гаплотипы ассо- циируют с опиатной зависи- мостью
OPRM1 ц-ренептор эндорфинов - 2 500 - 30 • Ассоциации не обнаружено
0PRD1 5-рецептор эндорфинов - 600 - 10 1 гаплотип тесно ассо- циирует с зависимостью от героина
OPRK1 к-рецептор эндорфинов - 330 - 20 1 сайт ассоциирует с зависи- мостью от героина
MDR1 Транспортер метадона** - 2 100 3 1 сайт ассоциирует с чув- ствительностью к метадону
Дофаминовая DAT1 Транспортер дофамина - 1000 1 Ассоциации не обнаружено
DRD2 D2 рецептор дофамина -600 - 10 1 сайт и некоторые гаплоти- пы ассоциируют с зависимо- стью от героина
DRD3 D3 рецептор дофамина - 500 2 Ассоциации не обнаружено
DRD4 D4 рецептор дофамина - 150 3 1 сайт слабо ассоциирует с зависимостью от героина
DRD5 D5 рецептор дофамина - 80 1 Ассоциации не обнаружено
СОМТ Катехол-О- метилтрансфераза - 500 3 Ассоциации не обнаружено
* Гаплотипами называют различные сочетания аллелей по нескольким полиморфным сайтам.
• • Метадон относится к числу синтетических опиатов. Он оказывает на человека мощное анестезирующее действие, но при
этом вызывает сравнительно слабую зависимость и потому применяется при лечении зависимости от героина.
225
Возможно, такая ситуация объясняется тем, что при разных
способах введения опиатного наркотика получаемые эффекты
несколько различны. В самом деле, зависимость, вызванная ин-
галяцией героина, развивается медленнее и легче поддается лече-
нию по сравнению с зависимостью при инъекциях того же нар-
котика. Соответственно в генетическом анализе рассматриваемо-
го признака необходимо учитывать каждый из перечисленных
аспектов.
Итак, в настоящее время нельзя утверждать, что гены,
контролирующие дофаминовую или эндорфиновую системы,
влияют на вероятность развития опиатной зависимости.
Однако вполне вероятно, что при дробном подходе (учете
способа потребления конкретного наркотика, длительности
приема, эффективности лечения и др.) роль этих генов ока-
жется статистически значимой.
8.7. Молекулярные основы алкогольной
зависимости
У всех млекопитающих, и в частности — у человека, в основе
процесса формирования положительных эмоций лежит возбуж-
дение определенных дофаминэргических нейронов. Чем чаще они
возбуждаются, тем сильнее удовольствие. В свою очередь, степень
активности этих нейронов негативно регулируется у-амино-
масляной кислотой. Ее молекулы резко подавляют выброс до-
фамина и тем самым затрудняют запуск программ удовольствия
(см. подразд. 8.3, рис. 8.1).
Некоторые соединения препятствуют действию ГАМК. К их
числу относится и этиловый спирт (этанол). Являясь побочным
продуктом обмена веществ, он постоянно присутствует в орга-
низме в небольших концентрациях, обычно не превышающих
0,01 %. В таких концентрациях этанол абсолютно безвреден. Но
если его содержание существенно возрастает (например, в резуль-
тате употребления алкогольных напитков), это может заметно
отразиться на рецепторах ГАМК.
Что именно с ними происходит, пока непонятно. По некото-
рым данным, молекулы этилового спирта вызывают частичное
«разжижение» клеточных мембран, что косвенно влияет на
функции рассматриваемых рецепторов. Однако нельзя исклю-
чить и прямого воздействия. Как бы то ни было, излишнее со-
держание этилового спирта приводит к снижению чувстви-
тельности рецепторов ГАМК и в итоге — к повышенной
активности дофаминовой системы, человек расслабляется,
226
становится благодушным, испытывает эмоциональное
раскрепощение и др.
В дальнейшем возможны и некоторые другие эффекты, в том
числе достаточно неприятные для организма: усталость, депрес-
сия, головные боли, сухость во рту, тошнота, а в особо тяжелых
случаях — рвота (о причинах их возникновения в следующем
подразделе). Но даже с учетом таких негативных последствий
многие люди регулярно используют алкогольные напитки для
быстрого «снятия стресса» и достижения эйфории. Эта привычка
легко перерастает в алкогольную зависимость.
На ранних этапах развития данной зависимости человек ис-
пытывает к спиртному неосознанное влечение и утрачивает спо-
собность контролировать количество выпитого. Все чаще воз-
никают тяжелые формы опьянения. Устойчивость организма к
алкоголю постепенно растет, что ведет к потреблению все боль-
ших и больших доз. Со временем происходит изменение обмена
веществ, и в итоге зависимость от спиртного становится физиче-
ской. Человек уже практически не может обходиться без выпивки.
При отсутствии алкоголя возникает болезненное состояние (де-
прессия, повышенная раздражительность, головные боли, уча-
щенное сердцебиение, тошнота, дрожание рук, бессонница,
озноб, потеря аппетита и др.), побуждающее к принятию очеред-
ной порции спиртного. В результате пьянство принимает запой-
ный характер или становится ежедневным, т.е. постоянным.
Алкоголь влияет не только на рецепторы ГАМК. Многочислен-
ные клинические исследования свидетельствуют о том, что высокие
концентрации этанола токсичны для организма и постепенно при-
водят к нарушению всех его функций. Особенно страдают печень
и нервная система. Поэтому типичными «спутниками» хрониче-
ского алкоголизма являются цирроз печени и острые психозы
(в том числе, так называемая «белая горячка*).
8.8. Молекулярные механизмы утилизации
этилового спирта
Поскольку этанол относится к токсичным соединениям, его
избыток представляет для организма серьезную опасность. Если
концентрация алкоголя в крови человека превышает нормальные
показатели приблизительно на порядок, велика вероятность раз-
вития острого отравления, а если на два — неизбежен смертель-
ный исход. В связи с этим в организме существуют молекулярные
системы, позволяющие утилизировать излишки этилового
спирта. Важнейшая из них представлена на рис. 8.2.
227
Алкоголь-
Н3С—СН2 дегидрогеназа
ОН
Этиловый
спирт
z°
Н3С-С
ХН
Уксусный
альдегид
Алыегид-
дегидрогеназа
✓°
н3с-с
хон
Уксусная
кислота
Рис. 8.2. Молекулярный механизм утилизации этилового спирта
Эта система преобразует этанол з уксусную кислоту, которая
в соответствующих концентрациях абсолютно безвредна. Однако
непосредственным предшественником данного соединения явля-
ется высоко токсичный уксусный альдегид. Даже совсем неболь-
шие его концентрации могут вызвать общую.слабость, сердечную
аритмию, головокружение, сухость во рту, головные боли, тош-
ноту и множество других неприятных эффектов. В обычных
условиях (при отсутствии излишков этанола) количество уксус-
ного альдегида в организме ничтожно. Но после принятия значи-
тельной дозы спиртного оно постепенно растет и через некоторое
время способно весьма негативно отразиться на жизнедеятель-
ности. Таким образом, высокое содержание этилового спирта
поначалу вызывает у человека состояние эйфории (это связано
со снижением чувствительности рецепторов ГАМК), но затем
стимулирует синтез уксусного альдегида, что в итоге приводит к
классическим симптомам похмелья. Особенно неприятны ощуще-
ния на следующее утро, когда излишки этилового спирта уже. из-
расходованы, а концентрация уксусного альдегида еще высока.
Ферменты, непосредственно участвующие в утилизации эта-
нола (алкогольдегидрогеназа и альдегиддегидрогеназа), обна-
ружены в широком спектре тканей и органов, но основная масса
сосредоточена в клетках печени. Дело в том, что печень выпол-
няет барьерную функцию — обезвреживает различного рода
токсичные вещества. В частности, она утилизирует и излишки
этанола. Соответственно наивысшие концентрации уксусного
альдегида наблюдаются у пьющего человека именно в печени, что
ведет к тяжелым поражениям данного органа.
Несмотря на способность этанола вызывать эйфорию, разные
люди по-разному относятся к алкогольным напиткам. Это связа-
но со степенью активности упомянутых ферментов. В наиболее
распространенном случае и тот, и другой обладают достаточно
высоким уровнем активности, обусловливая стандартную реакцию
на избыток алкоголя: сначала эйфория, а через некоторое время —
похмелье. Однако встречаются и менее стандартные реакции.
Если активность обоих ферментов очень высока, этанол так
быстро преобразуется в уксусную кислоту, что человек почти не
испытывает алкогольного опьянения даже от весьма значительных
доз спиртного. Подобные люди нередко бравируют своими воз-
228
можностями, еще больше увеличивают дозу и в конечном итоге
попадают в тяжелейшую зависимость от алкоголя.
Совершенно иная картина возникает в том случае, если актив-
ность первого фермента (ал ко гольде гидрогеназы) значительно
выше, нежели второго (альдегиддегидрогеназы). Этанол эффек-
тивно преобразуется в уксусный альдегид, однако тот не успевает
перерабатываться в уксусную кислоту и постепенно накаплива-
ется в организме все в большем количестве. В результате, еще не
успев ощутить эйфорию, человек оказывается в состоянии, близ-
ком к похмелью. Такие люди, как правило, испытывают отвра-
щение к спиртному или же относятся к нему с большой осторож-
ностью.
Наконец, возможен еще один вариант. Если алкоголь дегидро-
геназа недостаточно активен, процесс утилизации этанола прак-
тически не идет. Алкоголь сохраняется в организме значительно
дольше и тем самым надолго инактивирует рецепторы ГАМК.
В этом случае даже небольшие количества спиртного вызывают
у человека ярко выраженное алкогольное опьянение, после кото-
рого обычно не бывает симптомов похмелья. Человек очень бы-
стро пьянеет и охотно спивается.
Итак, отношение человека к алкогольным напиткам за-
висит от степени активности алкогольдегидрогеназы и
альдегиддегидрогеназы — двух ключевых ферментов, обе-
спечивающих утилизацию этилового спирта. В связи с этим
логично предположить, что соответствующие гены должны
оказывать заметное влияние на склонность к алкоголизму.
8.9. Генетический контроль
предрасположенности к алкоголизму. Роль
генов, кодирующих алкогольдегидрогеназу 2
У человека известно семь типов алкогольдегидрогеназы
(табл. 8.7). В принципе, все они выполняют сходную функцию
(окисляют различные спирты до альдегидов), но при этом каждый
имеет определенные особенности, в том числе и по степени срод-
ства к этиловому спирту. Например, ферменты III и IV типа
эффективно воздействуют на молекулы крупных спиртов, но
почти не проявляют активности в отношении этанола. Их влияние
на алкогольную зависимость маловероятно.
В этом плане среди всех разновидностей алкогольде гидрогена-
зы наибольший интерес представляют ферменты / типа'. 1а, 10
и 1у. Именно они играют главную роль в окислении этанола. Осо-
бого внимания заслуживают 0- и у-варианты. В отличие от алко-
229
Таблица 8.7. Типы алкогольдегидрогеназы у человека
Тип фермента Молекулярные функции Основная локализация Роль в утилизации этилового спирта Соответ- ствующий ген
1а Окисление раз- личных спиртов до альдегидов* Печень (у детей) Ведущая (—70 %) ADH1 (ADH1A)
Ф Печень (у взрослых) ADH2 (ADH1B)
h Печень (у взрослых) ADH3 (ADH1C)
II Печень, роговица Второстепенная ADH4
III Печень, желудок По-видимому, отсутствует ADH5
IV Желудок, пищевод ADH7
V Печень — ADH6
* К спиртам относят разнообразные органические соединения, несущие в
своем составе гидроксильную группу, к альдегидам — соединения, несущие кар-
бонильную группу (см. подразд. 1.2, табл. 1.4).
гольдегидрогеназы 1а, характерной преимущественно для ранних
этапов развития, эти ферменты активно работают во взрослом
организме, который, как правило, и сталкивается с излишками
алкоголя. Таким образом, мы вправе ожидать, что соответству-
ющие гены непосредственно вовлечены в контроль рассматри-
ваемого признака.
В гене, кодирующем алкогольдегидрогеназу 10, обнаружено
около 200 полиморфных сайтов. Наиболее подробно исследована
нуклеотидная замена, в результате которой в 47-м положении
фермента вместо аргинина присутствует гистидин. Люди, име-
ющие хотя бы одну такую аллель (ее обозначают ADH2*2)> ста-
новятся алкоголиками как минимум в 2 раза реже, чем люди,
гомозиготные по отсутствию данной аллели.
Этот вывод убедительно подтвержден и мета-анализом, осо-
бенно в отношении представителей монголоидной расы: китай-
цев, корейцев, японцев, тайцев и др.
230
Как показывают биохимические исследования, упомянутая
аллель многократно повышает активность ал ко гольде гидрогеназы.
Излишки этанола утилизируются значительно быстрее и, по-
видимому, меньше воздействуют на нервную систему. К тому же
повышенная активность данного фермента должна способство-
вать усиленному синтезу уксусного альдегида и тем самым сти-
мулировать негативные реакции на алкоголь. Эти эффекты со-
вместно препятствуют алкогольной зависимости.
Роль гена, кодирующего ал ко гольде гид роген азу 1у, пока оста-
ется в значительной степени неясной. В нескольких работах по-
казано, что одна из его аллелей, а именно — ADH3*2 тоже за-
метно снижает вероятность алкоголизма. Однако необходимо
учитывать, что эта аллель тесно сцеплена, т.е. обычно наследует-
ся совместно с аллелью ADH2*2, которая существенным образом
препятствует алкоголизму. Возможно, именно она и обусловли-
вает эффекты, причисляемые аллели ADH3*2.
8.10. Роль гена, кодирующего
альдегиддегидрогеназу 2
Как следует из анализа данных табл. 8.8, в организме человека
функционирует 10 типов альдегиддегидрогеназы. Они тоже
отличаются друг от друга по многим параметрам. В процессе
утилизации излишков этилового спирта (а точнее, на втором
этапе данного процесса — в окислении уксусного альдегида до
уксусной кислоты) центральную роль играет альдегиддегидроге-
наза 2. В том случае, если этот фермент недостаточно активен,
алкоголь вызывает многочисленные неприятные эффекты в ре-
зультате накопления в организме уксусного альдегида. Таким
образом, мы вправе ожидать, что некоторые мутации, приводящие
к дефектности альдегидде гидрогеназы 2, снижают вероятность
развития алкогольной зависимости.
Проведенные исследования подтверждают это предположение.
Известно, что у многих представителей монголоидной расы на-
блюдается резко негативная реакция на алкоголь. Даже от срав-
нительно небольшого количества спиртного человек начинает
испытывать общую слабость, учащение пульса, головную боль и
приступы тошноты. Лицо покрывается неестественно ярким ру-
мянцем. Заметно ослабевает способность сохранять равновесие,
нередко возникают бредовые состояния. Если же порция алкого-
ля была велика, возможны и значительно более неприятные эф-
фекты вплоть до тяжелых судорог и потери сознания.
Оказалось, что негативные реакции на спиртные напитки
очень тесно ассоциируют с одной из нуклеотидных замен,
231
Таблица 8.8. Типы альдегиддегидрогеназы у человека
Тип фермента Молекулярные функции Основная локализация Роль в утилиза- ции этилового спирта Соответ- ствующий ген
1 Окисление раз- личных альдеги- дов до органиче- ских кислот* Печень, желудок Второстепен- ная ALDH1
2 Печень, желудок Ведущая Ь 90%) ALDH2
3 Желудок Незначитель- ная или отсут- ствует ALDH3
4 Печень, почки ALDH4
5 Семенники, печень ALDH5
6 Слюнные железы, желудок ALDH6
7 Почки, легкие ALDH7
8 Околоушная область ALDH8
9 Печень, почки, мышцы ALDH9
10 Печень, сердце, мышцы ALDH10
* К органическим кислотам относят разнообразные соединения, несущие в
своем составе карбоксильную группу (см. табл. 1.4).
обнаруженных в гене альдегиддегидрогеназы 2. Эта аллель
(ее обозначают ALDH2 *2) приводит к практически полной
дефектности фермента, что значительно ослабляет ути-
лизацию уксусного альдегида. Соответственно люди, имею-
щие данную аллель, не испытывают положительных эмоций
при принятии спиртного и как следствие редко страдают
алкогольной зависимостью. Особенно ярко это выражено у
гомозигот. Среди них алкоголики встречаются исключитель-
но редко' до сих пор известно всего лишь три примера.
Нетрудно заметить, что аллель ALDH2*2 проявляет себя в ге-
терозиготе как доминантная. Казалось бы, это противоречит гене-
тической логике, в соответствии с которой (см. подразд. 1.11) до-
232
Таблица 8.9. Разнообразие тетрамеров альдегиддегидрогена-
зы 2 у гетерозигот по аллели ALDH2'2
Типы тетрамеров* Их соотношение** Ферментативная активность Общее соотношение тетрамеров
НЛ I 4- I 15 активные неактивные
НЛ 4 —
НЛ 6 —
Н|Дз 4 —
НЛ I —
* Н-нормальные молекулы фермента, Д-дефектные молекулы фермента.
♦* При случайном комбинировании нормальных и дефектных молекул.
минировать должна не мутантная, а нормальная аллель. Однако
никакого противоречия здесь нет. Дело в том, что молекулы рас-
сматриваемого нами фермента не способны выполнять свою
функцию по одиночке: они «работают» только в составе тетра-
мерных комплексов, т.е. объединившись друг с другом по четыре.
Причем взаимодействующие молекулы должны быть нормальны-
ми. Если же хотя бы одна из них дефектна, активность всего те-
трамера резко снижается. Именно это и происходит у гетерозигот:
в результате случайного комбинирования разных молекул пода-
вляющее большинство тетрамеров оказывается неактивным (табл.
8.9). И в итоге мутантная аллель доминирует над нормальной.
Добавим, что среди негроидов и европеоидов аллель ALDH2+2
встречается крайне редко. Поэтому о ее влиянии на склонность
к алкоголизму можно судить только по результатам исследований
на людях монголоидной расы.
8.11. Роль генов, кодирующих рецепторы
у-ОМИНОМОСЛЯНОЙ кислоты
Эйфория, возникающая под действием алкогольных напитков,
обусловлена снижением чувствительности рецепторов ГАМК (см.
подразд. 8.3). Таким образом, гены, кодирующие эти рецепторы,
тоже относятся к числу вероятных кандидатов на участие в гене-
тическом контроле предрасположенности к алкоголизму.
У человека известно два типа рецепторов ГАМК: А и В (табл.
8.10). Несмотря на значительные структурные и функциональные
233
Таблица 8.10. Разнообразие рецепторов ГАМК у человека
Тип рецепторов Принцип функционирования Локализация в синапсах Типы субъединиц Соответ- ствующие гены
А ГАМК-зависимые хлорные каналы* Главным образом, на постсинаптиче- ских мембранах al GABRA1
a2 GABRA2
a3 GABRA3
а4 GABRA4
а5 GABRA5
аб GABRA6
01 GABRB1
02 GABRB2
рз GABRB3
?! GABRG1
Г2 GABRG2
уЗ GABRG3
5 GABRD
с GABRE
тс GABRP
0 GABRQ
pl** GABRR1
р2 GABRR2
рЗ GABRR3
В Рецепторы, связан- ные с G-белками*** Преимущественно на пресинаптиче- ских мембранах BI GABBR1
В2 GABBR2
* Канады состоят из пяти субъединиц. Самое распространенное сочетание —
две субъединицы al, две р2 и одна у2. Однако встречается и множество других
сочетаний.
•• Рецепторы, состоящие из р-субъединиц, иногда выделяют в особый тип — С.
*** Димеры из двух субъединиц (ВI и В2). Вторичный мессенджер —цАМФ,
его содержание при активации рецептора снижается за счет подавления функции
аденилатциклазы.
234
Таблица 8.11. Поиск ассоциации между генами, кодирующи-
ми рецепторы ГАМК, и алкогольной зависимостью (предвари-
тельные данные)
Ген Соответствующая субъединица рецептора Число полиморф- ных сайтов Результат исследования
Всего Исследо- вано
GABRA1 al -500 - 20 5 сайтов ассоциируют с алкогольной зависимостью
GABRA2 a2 -I 400 -60 Более 30 сайтов ассоцииру- ют с алкогольной зависи- мостью*
GABRA4 a4 -700 6 Ассоциации не обнаружено
GABRA5 a5 -700 4 Ассоциации не обнаружено
GABRA6 a6 - 10 6 2 сайта ассоциируют с алкогольной зависимостью
GABRB1 Pl -3 800 7 1 сайт ассоциирует с алко- гольной зависимостью
GABRB2 P2 -2 200 1 Ассоциации не обнаружено
GABRB3 ₽3 -60 6 1 сайт ассоциирует с тяжелыми формами алко- голизма
GABRG1 yl -900 6 Некоторые гаплотипы ассоциируют с алкогольной зависимостью
GABRG2 -800 2 Ассоциации не обнаружено
GABRG3 -5 500 - 10 7 сайтов слабо ассоциируют с алкогольной зависимостью
♦ Для одного из них (а именно /4596(7) наличие ассоциации подтверждено не-
зависимыми исследованиями.
различия, и те, и другие выполняют одну и ту же задачу: регули-
руют активность нейронов путем торможения. Это самые много-
численные тормозные рецепторы в организме. Они характерны
для всех отделов центральной нервной системы и влияют на все
235
без исключения аспекты жизнедеятельности, в том числе и на
процесс формирования положительных эмоций.
Для нас наиболее интересны рецепторы A-типа. Предпола-
гают, что именно они играют главную роль в возникновении
эйфорического состояния под действием алкоголя. Эти рецепто-
ры являются ГАМ К-зависимыми хлорными каналами. Их акти-
вация приводит к тому, что ионы СГ, в обычных условиях удаляе-
мые из клетки наружу, перемещаются по разнице концентраций в
обратном направлении и тем самым дополнительно увеличивают
мембранный потенциал (внутренняя сторона мембраны заряжена
отрицательно, и этот заряд лишь усиливается с переносом ионов
СГ). Соответственно происходит торможение данного нейрона.
Существует большое разнообразие рецепторов А -типа. Дело в
том, что они представляют собой пентамеры, т.е. комплексы,
состоящие из пяти субъединиц. Причем это могут быть различные
субъединицы (их известно 19 вариантов; табл. 8.10). В результате
формируются десятки разных сочетаний, каждое со своими кон-
кретными функциональными особенностями.
Среди генов, кодирующих субъединицы рецепторов ГАМК,
к настоящему времени наиболее исследован ген GABRA2 (его
продуктом является субъединица а2). Показано, что неко-
торые из его нуклеотидных замен достаточно тесно ассо-
циируют с зависимостью от алкоголя. Особенно присталь-
ного внимания заслуживает замена, получившая обозначение
rs279858‘. ее связь с алкоголизмом продемонстрирована в семи
исследованиях. К сожалению, мета-анализ еще не проведен.
Тем не менее с высокой вероятностью можно ожидать, что
он подтвердит наличие искомой ассоциации.
Данные по генам, кодирующим другие субъединицы, для удоб-
ства приведены в табл. 8.11. Пока они носят сугубо предваритель-
ный характер.
8.12. Роль генов, контролирующих
дофаминовую, серотониновую
и эндорфиновую системы
Напомним: непосредственной причиной состояния эйфории,
возникающего в результате употребления алкогольных напитков,
является возбуждение определенных дофаминэргических нейро-
нов. Соответственно гены, обслуживающие дофаминовую систе-
му (см. подразд. 5.3 и 5.4), тоже представляют весьма существен-
ный интерес.
236
Для начала обратимся к гену DRD2. Несколько десятков работ
свидетельствует о том, что аллель, получившая обозначение АЦ
повышает вероятность возникновения алкогольной зависи-
мости. Этот вывод подтвержден и результатами мета-
анализа. Но механизм обнаруженного эффекта пока непо-
нятен. Более того, как показали недавние исследования,
мутация затрагивает не сам DRD2, а расположенный с ним
по соседству ген ANKK1, функции которого еще не установ-
лены. Таким образом, вопрос об участии гена DRD2 в контроле
предрасположенности к алкоголизму остается открытым. Данные
о других генах дофаминовой системы (они сведены в табл. 8.12)
в большинстве своем тоже не могут считаться окончательными.
Как известно, дофаминовая система тесно связана с серото-
ниновой (повышение активности последней стимулирует первую).
Поэтому гены, участвующие в контроле серотониновой системы,
также рассматриваются в качестве вероятных кандидатов.
Многочисленные независимые исследования позволяют ут-
верждать, что короткий вариант промотора гена SLC6A4 (см.
подразд. 6.12) достоверно повышает вероятность возникно-
вения алкоголизма. Об этом же свидетельствуют и резуль-
таты мета-анализа. Однако обнаруженный эффект сравни-
тельно слаб и, по-видимому, играет лишь некую второстепенную
роль. Среди прочих генов, обслуживающих серотониновую си-
стему (табл. 8.12), наиболее перспективным является ген МАОА.
В его промоторе имеются 30-нуклеотидные повторы, количество
которых варьирует от трех до пяти. Судя по результатам четырех
независимых работ, аллели с тремя экземплярами данных повто-
ров ассоциируют с тревожно-депрессивными формами алкого-
лизма. Но если исследуют так называемых «тихих алкоголиков»,
обнаружить влияние гена МАОА обычно не удается. Этот факт
позволяет предположить, что разные формы алкоголизма нахо-
дятся под относительно автономным генетическим контролем, и
потому их нужно анализировать строго по отдельности.
Еще раз вернемся к рис. 8.1 (см. подразд. 8.3). Как вы видите,
активность дофаминэргических нейронов, непосредственно обе-
спечивающих запуск программ удовольствия, регулируется не
только при помощи серотонина и ГАМК. В этом процессе при-
нимают участие и некоторые эндорфины. Неудивительно, что
гены, обслуживающие эндорфиновую систему, тоже вовлечены в
анализ предрасположенности к алкоголизму.
Напомним: важнейшими компонентами данной системы яв-
ляются p-эндорфины, энкефалины и динорфины, а также три
типа рецепторов (ц, 6 и к; см. подразд. 8.3 и 8.6). В генетических
исследованиях, посвященных алкогольной зависимости, основное
внимание приковано к нуклеотидной замене А —> G в 118 по-
237
Таблица 8.12. Поиск ассоциации между генами, контролирующими дофаминовую, серотониновую
оо и эндорфиновую системы, и алкогольной зависимостью (предварительные данные)
Молекулярная система Ген Соответствующий белок Количество поли- морфных сайтов Результат исследования
Всего Исследо- вано
Дофаминовая TH Тирозингидроксилаза ^120 - 10 2 сайта ассоциируют с алкоголь- ной зависимостью
DATI Транспортер дофамина - 1000 2 1 сайт ассоциирует с алкоголь- ной зависимостью
DRD1 D1 рецептор дофамина -50 2 1 сайт ассоциирует с тяжелыми формами алкоголизма
DRD2 D2 рецептор дофамина -600 - 20 3 сайта слабо ассоциируют с алкогольной зависимостью*
DRD3 D3 рецептор дофамина - 500 1 Ассоциации не обнаружено**
DRD4 D4 рецептор дофамина - 150 2 Ассоциации не обнаружено**
COMT Катехол-0-метил трансфераза - 500 5 1 сайт ассоциирует с количе- ством употребляемого алкоголя
DBH Дофамин-р-гидроксилаза -450 2 1 сайт ассоциирует с алкоголь- ной зависимостью
Серотониновая TPH1 Триптофангидроксилаза - 200 2 Ассоциации не обнаружено
239
ТРН2 Триптофан гидроксилаза *900 1 Ассоциации не обнаружено
SLC6A4 Транспортер серотонина * 500 2 S-промотор ассоциирует с алко- гольной зависимостью***
МАОА Моноаминоксидаза А - 850 2 1 сайт ассоциирует с тревожно- депрессивным алкоголизмом
МАОВ Моноаминоксидаза В * 500 1 Ассоциации не обнаружено
HTR1A 1А рецептор серотонина - 40 1 Ассоциации не обнаружено
HTR1B 1В рецептор серотонина *40 * 10 2 сайта ассоциируют с алкоголь- ной зависимостью
HTR2A 2А рецептор серотонина * 700 2 1 сайт ассоциирует с алкоголь- ной зависимостью
HTR2C 2С рецептор серотонина *2 500 1 Ассоциации не обнаружено
Эндорфиновая РОМС Проопиомеланокортин (предшественник 0-эндор- финов и энкефалинов) *90 7 2 сайта слабо ассоциируют с алкогольной зависимостью
PEN К Проэнкефалин (предше- ственник энкефалинов) * 100 7 Ассоциации не обнаружено
PDYN Продинорфин (предше- ственник динорфинов и энкефалинов) *2 500 * 20 Около 10 сайтов ассоциируют с алкогольной зависимостью
Окончание табл. 8.12
Молекулярная система Ген Соответствующий белок Количество поли- морфных сайтов Результат исследования
Всего Исследо- вано
OPRM1 ц-рецептор эндорфинов -450 - 20 1 сайт ассоциирует с восприим- чивостью к лечению алкоголизма налтрексоном* ****
OPRD1 5-рецептор эндорфинов - 600 -20 Несколько гаплотипов ассоции- рует с алкогольной зависимостью
OPRK1 к-рецептор эндорфинов - 330 - 15 1 сайт ассоциирует с алкоголь- ной зависимостью
• Без учета аллели А1, поскольку мутация локализована вне гена DRD2.
** Вывод основан на нескольких независимых исследованиях.
*** Подтверждено многочисленными исследованиями и мета-анализом.
**** Подтверждено четырьмя независимыми исследованиями.
ложен и и гена OPRM 1. Это единственная известная к настояще-
му времени мутация, влияющая на функции ц-рецепторов у че-
ловека (она значительно повышает их способность взаимодей-
ствовать с эндорфинами).
Судя по результатам нескольких десятков исследований, упо-
мянутая мутация не влияет на склонность к алкоголизму. Однако
постепенно накапливаются свидетельства того, что люди, несущие
аллель 118G, более восприимчивы к терапевтическому дей-
ствию налтрексона (медикамента, широко используемого
при лечении алкоголизма), чем гомозиготы по аллели 118А.
Этот вывод подтвержден четырьмя независимыми работами. Для
прочих генов, контролирующих эндорфиновую систему, четкой
связи с интересующим нас признаком пока не выявлено.
8.13. Основные результаты и ближайшие
перспективы генетического анализа
предрасположенности к алкоголизму
Итак, в настоящее время не вызывает сомнений, что
склонность человека к алкоголизму зависит от генотипа.
В этом контроле принимает участие как минимум пять генов:
• ADH2, кодирующий алкогольде гидрогеназу 2;
• ALDH2, кодирующий альдегиддегидрогеназу 2;
• GABRA2, продуктом которого является а2-субъединица
рецепторов ГАМК;
• ANKK1, функции которого пока неустановлены;
• SLC6A4, кодирующий транспортер серотонина.
Степень влияния этих генов на отношение к алкоголю неоди-
накова. Наиболее значимую роль играет ген ALDH2. В самом
деле, гомозиготы по мутации ALDH2*2 проявляют ярко выра-
женное негативное отношение к алкоголю независимо от своего
генотипа по всем прочим генам. Соответственно в генетическом
анализе рассматриваемого признака необходимо одновременно
учитывать несколько генов, а иначе влияние одних может быть
скрыто другими.
Неудивительно, что в течение достаточно продолжительного
времени, несмотря на высокую наследуемость интересующего нас
признака (роль генотипа оценивают в 50 — 60%), обнаружить
ассоциацию с конкретными генами не удавалось. Поэтому еще в
конце XX в. широко бытовало мнение, что склонность к алкого-
лизму определяется исключительно факторами внешней среды:
примером родителей, социальным окружением и др. Но теперь
подобные взгляды безоговорочно устарели.
241
Количество генов, контролирующих предрасположенность к
алкоголизму, вероятнее всего, не ограничено перечисленными пя-
тью. Этот вывод основан на результатах исследований на животных.
Многочисленные эксперименты показывают, что мыши и крысы
охотно потребляют разбавленный этиловый спирт и, как правило,
даже предпочитают его чистой воде. В результате может сформиро-
ваться алкогольная зависимость. Такие животные:
• преимущественно находятся в той части клетки, где располо-
жен источник этилового спирта;
• при наличии регулярного доступа к этому источнику по-
степенно становятся все менее чувствительными к алкоголю и как
следствие увеличивают потребляемую ими дозу;
• а в отсутствие алкоголя страдают повышенным беспокойством,
чрезмерным уровнем агрессии и судорожными припадками.
Если в клетку с подопытным животным поместить две поилки,
в одной из которых находится чистая вода, а другой — вода с до-
бавлением этилового спирта, и предоставить неограниченный
доступ к обеим поилкам, можно измерить количество свободно
потребляемого алкоголя. Этот параметр зависит от множества
разных факторов, в том числе от пола и возраста данного орга-
низма, его физиологического состояния, калорийности пищи,
применения определенных медикаментов и др.
Немаловажную роль играет и генотип. Показано, что многие
инбредные линии мышей достоверно отличаются друг от друга
по интересующему нас признаку. В этом плане их можно подраз-
делить на три большие группы: норма, «чрезмерно пьющие» и
наоборот. Аналогичные линии описаны и у крыс.
Некоторые из соответствующих генов уже установлены: к на-
стоящему времени их известно более 70. Если же учитывать не
только свободное потребление, но и другие реакции организма
на алкоголь (различные аспекты чувствительности, динамику
привыкания, степень выраженности абстинентного синдрома и
др.), количество выявленных генов приближается к 100. Вполне
вероятно, что некоторые их гомологи контролируют склонность
к алкоголизму и у человека.
8.14. Молекулярные основы никотиновой
зависимости
Многочисленные клинические исследования свидетельствуют
о том, что в процессах, обеспечивающих запуск программ удо-
вольствия, заметную роль играет и ацетилхолин. Данное соеди-
нение относится к важнейшим нейромедиаторам. Оно использу-
242
ется в различных отделах нервной системы и регулирует огромный
спектр биологических функций, в том числе — мотивацию, память
и положительные эмоции, т.е. функции, имеющие непосредствен-
ное отношение к зависимостям.
Механизмы влияния ацетилхолина на положительные эмоции
довольно сложны, и потому мы не будем их подробно рассма-
тривать. Отметим лишь, что в вентральной покрышке среднего
мозга (ключевом отделе нервной системы, ответственном за удо-
вольствие) ацетилхолин вызывает возбуждение дофаминэргиче-
ских нейронов. В результате значительно усиливается выброс
дофамина, и как следствие возникает состояние легкой эйфо-
рии.
Подобно любому другому нейромедиатору, ацетилхолин воз-
действует на строго определенные рецепторы. Оказалось, что эти
рецепторы могут быть активированы и с помощью некоторых
чужеродных для организма веществ, например содержащегося в
табаке алкалоида никотина. При введении никотина в организм
человека (наиболее распространенный способ — курение та-
бака) дофаминовая система начинает функционировать
интенсивнее, вызывая целый ряд ярко выраженных положи-
тельных эмоций. На этой основе очень быстро формируется
психологическая зависимость. Организм постепенно привыкает
к воздействию никотина и становится все менее и менее чувстви-
тельным к нему. В результате приходится курить все чаще и
чаще.
Это приводит к значительным изменениям обмена веществ, и
в итоге психологическая зависимость перерастает в физическую.
Организм уже не может обходиться без никотина. В его отсутствие
человек:
• с трудом концентрирует внимание;
• практически постоянно пребывает в подавленном настрое-
нии;
• испытывает сильное беспокойство;
• становится раздражительным;
• страдает бессонницей, головными болями и др.
Обычно перечисленные эффекты настолько сильны, что око-
ло 95% интенсивных курильщиков не способны самостоя-
тельно избавиться от этой зависимости. И даже при ис-
пользовании современных методов лечения положительный
результат наблюдается сравнительно редко, не более чем в
1О...15% случаев. Таким образом, никотиновая зависимость
очень устойчива.
По оценкам ВОЗ, это самая распространенная из всех извест-
ных зависимостей: она характерна приблизительно для 1,5 млрд
человек. При этом необходимо учитывать, что интенсивное ку-
243
рение приводит к тяжелым заболеваниям дыхательной системы,
и в частности — к высокой вероятности рака легких. Показано,
что около половины заядлых курильщиков умирает от проблем,
непосредственно связанных с курением, а около четверти не до-
живает даже до 50 лет.
8.15. Основные подходы к генетическому
анализу предрасположенности
к никотиновой зависимости
Если человек никогда не потреблял никотина (не курил, не
пользовался нюхательным табаком и др.), никотиновая зависи-
мость, конечно, ему не грозит. Поэтому, говоря о причинах ее
возникновения, в первую очередь обращают внимание на внеш-
ние факторы:
• пробовал ли данный человек когда-нибудь курить;
• если да, то с какой интенсивностью и как долго он курит;
• как относятся к курению его друзья и ближайшие родствен-
ники;
• страдает ли он другими зависимостями, например алкоголь-
ной (курение зачастую сопутствует алкоголизму), и др.
Между тем хорошо известно, что некоторые люди не
страдают ярко выраженной зависимостью от никотина
даже при довольно продолжительном опыте курения, а не-
которые становятся зависимыми практически сразу. Эти
факты свидетельствуют о существенной роли генотипа.
И действительно, судя по результатам близнецовых иссле-
дований, она составляет приблизительно 45% у женщин и
около 60% у мужчин. Для отдельных аспектов (например,
для интенсивности курения) роль генотипа еще более значи-
ма — до 80%. Соответственно возникает вопрос о генах-кан-
дидатах.
Обычно исследование проводят по следующей схеме. Испы-
туемые заполняют специально разработанные опросники, по-
зволяющие оценить наличие опыта курения, его продолжитель-
ность, интенсивность, вызываемые никотином эмоции, способ-
ность самостоятельно бросить курить и др. На основе этих данных
создают контрастные выборки, различающиеся по тем или иным
аспектам курения (табл. 8.13), и затем проводят их генетический
анализ.
Обратите внимание: в подобного рода исследованиях некор-
ректно сравнивать всех курящих со всеми некурящими. Дело в
том, что обе эти выборки неоднородны. Некоторые курящие не
244
Таблица 8.13. Примеры выборок для изучения аспектов куре-
ния
Исследуемый аспект Экспериментальная выборка Контрольная выборка
Желание попро- бовать покурить* Испытуемые, хотя бы однажды про- бовавшие покурить (независимо от того, стали они куриль- щиками или нет) Испытуемые, ни разу не пробовавшие курить
Наличие нико- тиновой зависи- мости Испытуемые, стра- дающие ярко выра- женной никотино- вой зависимостью Испытуемые, имеющие определенный опыт куре- ния (например, выкурив- шие не менее 100 сигарет), но при этом никогда не страдавшие никотиновой зависимостью
Среднее коли- чество сигарет, выкуриваемых в сутки** Интенсивные ку- рильщики (более 25 сигарет в сутки) Неинтенсивные курильщи- ки (менее 5 сигарет в сутки при регулярном курении)
* Этот признак не имеет непосредственного отношения к курению. Он кос-
венно отражает другую психологическую характеристику — желание испытать
что-то новое, «поиск новизны».
** Данный параметр отражает чувствительность организма к никотину: чем
она ниже, тем больше сигарет приходится выкуривать.
испытывают потребности в никотине и «балуются табачком» лишь
за компанию или по привычке. Их, конечно, не следует объеди-
нять с заядлыми курильщиками.
Некурящие тоже представляют собой гетерогенную группу.
Многие из них хотя бы однажды пробовали курить, но в дальней-
шем смогли легко обходиться без никотина. Такие люди, по-
видимому, не имеют предрасположенности к курению. Между
тем, о людях, которые не курили ни разу (они тоже формально
относятся к числу некурящих), ничего определенного сказать пока
невозможно, поскольку провоцирующего воздействия они не ис-
пытывали. Вполне вероятно, что некоторые из них — потенци-
альные курильщики.
Таким образом, в генетическом анализе предрасположен-
ности к курению необходимо очень четко вычленять иссле-
дуемую характеристику, иначе полученные результаты
будут неадекватными.
8.16. Генетический контроль
предрасположенности к никотиновой
зависимости
Наибольшее количество работ к настоящему времени посвя-
щено влиянию на никотиновую зависимость гена ANKK1 (на-
помним, он участвует в контроле предрасположенности к алко-
голизму, а алкогольная и никотиновая зависимости, как правило,
сопутствуют друг другу). В нескольких работах показано, что ал-
лель А1 ассоциирует с отдельными аспектами никотиновой за-
висимости. Но в большинстве исследований ассоциация не об-
наружена. Мета-анализ тоже не дает однозначного результата:
в зависимости от способа обработки полученных данных он то
подтверждает наличие ассоциации, то опровергает. Таким обра-
зом, вопрос об участии гена ANKK1 в контроле предрасположен-
ности к курению пока не решен.
В последние годы все более пристальное внимание уделяется
генам, кодирующим рецепторы ацетилхолина. Ведь именно на
эти рецепторы и воздействует никотин. Поэтому познакомимся
с ними немного подробнее. Существует большое разнообразие
данных рецепторов (табл. 8.14). В принципе, все они имеют срод-
ство с никотином, однако у некоторых оно относительно невели-
ко. В связи с этим выделяют два типа рецепторов ацетилхолина:
• никотиновые (имеющие высокое сродство с никотином);
• мускариновые (сравнительно слабо чувствительные к нему,
но зато обладающие высоким сродством с мускарином — ядови-
тым веществом из мухоморов и бледных поганок).
Для нас наиболее интересны никотиновые рецепторы, именно
они участвуют в регуляции положительных эмоций. Эти рецеп-
торы являются сложными молекулярными комплексами, состоя-
щими из пяти одинаковых или разных субъединиц (наподобие
/4-рецепторов ГАМК; см. подразд. 8.11). У человека известно
17 типов таких субъединиц. Они образуют друг с другом десятки
различных сочетаний, каждое со своими характерными функ-
циональными особенностями.
В анализ вовлечено 16 соответствующих генов. Поскольку ис-
следования были начаты сравнительно недавно, однозначных ре-
зультатов пока получить не удалось. Однако предварительные
данные весьма обнадеживающие (табл. 8.15). В частности, на вы-
борке из 15 тыс. испытуемых (такие выборки не всегда проводятся
встречаются и в мета-анализах) показано, что некоторые аллели
гена CHRNA3 тесно ассоциируют с высокой интенсивностью
курения. И даже несмотря на отсутствие независимых подтверж-
дений, этот вывод представляется достаточно надежным.
246
Таблица 8.14. Разнообразие рецепторов ацетилхолина у чело-
века
Тип рецепторов Принцип функционирования Локализация в нервной системе Типы субъ- единиц Соответству- ющие гены
Никотиновые Ацетил холин- зависимые калий- натриевые каналы* Как в центральной, так и в перифе- рической нервной системе а! CHRNA1
а2 CHRNA2
аЗ CHRNA3
а4 CHRNA4
а5 CHRNA5
аб GABRA6
а7 CHRNA7
а8 CHRNA8
а9 CHRNA9
аЮ CHRNA 10
PI CHRNB1
₽2 CHRNB2
РЗ CHRNB3
Y CHRNG
5 CHRND
Е CHRNE
Мускариновые Рецепторы, связан- ные с G-белками** Преимущественно в периферической нервной системе Ml CHRM1
М2 CHRM2
М3 CHRM3
М4 CHRM4
М5 CHRM5
Мб CHRM6
* Состоят из пяти субъединиц. Самые распространенные сочетания: в цен-
тральной нервной системе — (а4)2(02)3 или (а7)5; в ганглиях — (аЗ)2(04)3; в мыш-
цах — (al )2|315е. Однако встречаются и многие другие.
** Вторичные мессенджеры: либо инозитолтрифосфат (его содержание воз-
растает за счет усиления функции фосфолипазы С), либо цАМФ (его содержание
снижается за счет подавления функции аденилатциклазы).
247
Таблица 8.15. Поиск ассоциации между генами рецепторов
ацетилхолина и никотиновой зависимостью (предварительные
данные)
Ген Соответствую- щая субъедини- ца рецептора Количество поли- морфных сайтов Результат исследования
Всего Иссле- довано
CHRNA/ а! -250 3 Ассоциации не обнаружено
CHRNA2 а2 - 300 - 20 Ассоциации не обнаружено
CHRNA3 аЗ -500 -20 5 сайтов ассоциируют с никотиновой зависимостью; 3 сайта — с интенсивностью курения
CHRNA4 а4 - 500 - 10 4 сайта ассоциируют с нико- тиновой зависимостью; 3 сайта — с интенсивностью курения
CHRNA5 а5 - 500 - 20 6 сайтов ассоциируют с никотиновой зависимостью
CHRNA6 аб - 250 4 1 сайт ассоциирует с никоти- новой зависимостью
CHRNA 7 а7 -1600 - 10 Ассоциации не обнаружено
CHRNA9 а9 - 350 - 10 Ассоциации не обнаружено
CHRNA 10 аЮ - 100 3 Ассоциации не обнаружено
CHRNB1 PI - 150 - 10 1 сайт ассоциирует с наличи- ем опыта курения
CHRNB2 ₽2 - 150 4 1 сайт ассоциирует с наличи- ем опыта курения
CHRNB3 РЗ -500 - 10 5 сайтов ассоциируют с нико- тиновой зависимостью
CHRNB4 Р4 - 200 - 15 1 сайт ассоциирует с никоти- новой зависимостью; 2 сайта — с наличием опыта курения
CHRNG Y - 150 6 4 сайта ассоциируют с нико- тиновой зависимостью
248
Окончание табл. 8.15
Ген Соответствую- щая субъедини- ца рецептора Количество поли- морфных сайтов Результат исследования
Всего Иссле- довано
CHRND 5 ~ 100 3 2 сайта ассоциируют с нико- тиновой зависимостью
CHRNE Е - 100 3 Ассоциации не обнаружено
Таким образом, с высокой вероятностью можно ожидать, что
гены, кодирующие никотиновые рецепторы ацетилхолина, и в
первую очередь — ген CHRNA3, оказывают существенное влия-
ние на склонность к курению.
8.17. Основные итоги и ближайшие
перспективы генетического контроля
зависимого поведения
Многочисленные исследования свидетельствуют о том, что
каждая из трех рассмотренных нами зависимостей пристрастия к
курению табака и алкоголю обусловлена не только факторами
внешней среды. Весьма немаловажную роль играет и генотип. Он
особенно значим для алкогольной и никотиновой зависимостей
(его вклад в этих случаях — не менее 50 %).
Очевидно, что все варианты зависимого поведения тесно свя-
заны с процессами запуска положительных эмоций. Поэтому
логично предположить, что разные зависимости имеют достаточ-
но сходную физиологическую основу и контролируются пример-
но одними и теми же генами. В данном предположении, несо-
мненно, есть доля истины. Как показано в исследованиях на
мышах, ген OPRM (он кодирует ц-рецепторы эндорфинов — см.
подразд. 8.5) одновременно влияет на чувствительность и к
опиатам, и к этанолу, и к никотину, и к тетрагидраканнабинолу.
Вполне вероятно, что сходная ситуация характерна и для челове-
ка. Но убедительных подтверждений пока получить не удалось.
Тем не менее, сравнивая различные зависимости друг с другом
(например, опиатную, никотиновую и алкогольную), нетрудно
заметить, что каждая из них относительно автономна. Начнем с
249
того, что никотин, опиаты и этиловый спирт воздействуют в ор-
ганизме на разные молекулярные мишени. В результате их эф-
фекты, хотя и близки, но все же неодинаковы (так, эйфория,
возникающая под действием никотина, во много раз слабее, чем
при использовании опиатов и, как правило, не сопровождается
мощной анестезией). Неодинакова и скорость привыкания к
этим соединениям: если опиатная зависимость формируется бы-
стро, всего лишь за несколько приемов определенного наркотика,
то алкогольная и никотиновая зависимости формируются обычно
значительно медленнее. Наконец, обнаружено как минимум пять
генов (ADH, ALDH, GABRA2, SLC6A4 и ANKK1Y контролирую-
щих вероятность развития алкоголизма. Но на другие зависимо-
сти эти гены, по-видимому, не влияют.
Некоторые аспекты одной и той же зависимости также нахо-
дятся под разным генетическим контролем. Проиллюстрируем
это на примере алкоголизма. Судя по имеющимся данным (см.
подразд. 8.12), наличие трех экземпляров 30-нуклеотидных по-
второв в промоторной области гена МАОА ассоциирует с
тревожно-депрессивными формами алкоголизма, однако не об-
наруживает связи с его «тихими» формами. Кроме того, мутация
/ 18G в гене OPRM/ не влияет на вероятность возникновения
алкогольной зависимости, но повышает эффективность ее лече-
ния с помощью налтрексона.
Таким образом, и здесь выполняется общее правило: чем силь-
нее «дробят* исследуемую психологическую характеристику,
тем успешнее оказывается ее генетический анализ. Именно
с этим подходом связаны ближайшие перспективы в изучении
генетического контроля зависимого поведения. Необходимо:
• каждую зависимость исследовать по отдельности;
• вычленять различные аспекты одной и той же зависимости,
проявляющие относительную автономность друг от друга;
• строго по отдельности анализировать каждый аспект.
СПИСОК ЛИТЕРАТУРЫ
1. Анализ полиморфизма генов дофаминергической системы у больных
алкоголизмом, якутов и чукчей по этнической принадлежности // Медицин-
ская генетика. — 2008. — Т. 7. — № 4. — С. 1—9.
2. Анохина И. П. и др. Современные проблемы генетики зависимости от
психоактивных веществ // Наркология. — 2004. — № 6. — С. 71—77.
3. Сравнительный анализ вариантов полиморфизма генов дофаминовых
рецепторов DRD2 и DRD4 у больных с зависимостью от разных видов
ПАВ If Наркология. 2007. - Т. 4. - № 4. - С. 31 - 38.
4. Сравнительная характеристика основных групп психоактивных веществ.
Перечень наркотических средств, психотропных веществ и их прекурсоров,
250
подлежащих контролю в Российской Федерации. Списки сильнодействующих
и ядовитых веществ // Наркология. — 2002. — № 5. — С. 8 — 15.
5. Arias A. et al. Association of an Asn40Asp (A118G) polymorphism in the
ц-opioid receptor gene with substance dependence: A meta-analysis // Drug &
Alcohol Dependence. — 2006. — V. 83. — P. 262 — 268.
6. Berrettini H< et al. ot-5/а-З nicotinic receptor subunit alleles increase risk
for heavy smoking 11 Molecular Psychiatry. — 2008. — V. 13. — P. 368 — 373.
13. Borinskaya S. et al. Distribution of alcoholism protecting alcohol
dehydrogenase ADHlB*47His allele in Eurasia // Amer. J. Hum. Genet. —м
2009. - V. 84. - P. 89-94.
7. Bressan R.A. and Crippa J. A. The role of dopamine in reward and pleasure
behaviour. Review of data from preclinical research // Acta Psychiatr Scand
Suppl. - 2005. - № 427. - P. 14-21.
8. Crabbe J.C., Phillips T.J., Harris R.A., Arends M.A. and Koob G.F.
Alcohol-related genes: contributions from studies with genetically engineered
mice//Addiction Biology. — 2006. — V. II. — P. 195 — 269.
9. Feinn R. et al. Meta-analysis of the association of a functional serotonin
transporter promoter polymorphism with alcohol dependence // American J
Medical Genetics. Part B. - 2005. - V. I33B. - P. 79-84.
10. Gaueriaux C. and Kieffer B. L. Opioid receptor genes inactivated in mice:
The highlights // Neuropeptides. — 2002. — V. 36. — P. 62 — 71.
11. Kohnke M. D. Approach to the genetics of alcoholism: A review based on
pathophysiology. Biochem Pharmacol. — 2008. — V. 75. — P. 160— 177.
12. Lobo I. A, Harris R.A. GABAa receptors and alcohol // Pharmacology
Biochemistry and Behavior. — 2008. — V. 90. — P. 90—94
13. Luczak S.E. et al. Meta-analyses of ALDH2 and ADH1B with alcohol
dependence in Asians // Psychological Bulletin. — 2006. — V. 132. — P. 607—621.
14. Munaf M.R. et al. Association of the DRD2 gene TaqlA polymorphism
and alcoholism: a meta-analysis of case-control studies and evidence of publication
bias // Mol Psychiatry. — 2007. — V. 12. — P. 454 — 461.
15. Olsen R. W. et al. GABAa receptor subtypes: the «one glass of wine»
receptors // Alcohol. —2007. —V. 41. —P. 201 —209.
КОНТРОЛЬНЫЕ ВОПРОСЫ
I. Что называют зависимостью? Как она формируется?
2. Опишите молекулярные механизмы действия опиатных наркоти-
ков.
3. К каким эффектам приводят мутации по генам рецепторов эндор-
финов у мышей?
4. Какие гены участвуют к контроле предрасположенности человека к
алкоголизму? Опишите механизм этого контроля.
5. Как проводят генетический анализ предрасположенности человека
к курению?
Глава 9
ИСТОРИЯ РАЗВИТИЯ И ПЕРСПЕКТИВЫ
ПСИХОГЕНЕТИКИ
9.1. Истоки психогенетики
Психогенетика имеет достаточно глубокие корни. О том, что
люди различаются по умственным способностям, справедливости,
смелости, тщеславию, аккуратности, наглости и целому ряду других
психологических черт, было хорошо известно еще в Древнем мире.
Иногда эти знания находили практическое применение. В частно-
сти, в Древнем Китае и Вавилоне существовали жесткие испытания
для будущих чиновников. Александр Македонский, набирая себе
личную охрану, проверял у каждого кандидата реакцию на испуг:
если человек бледнел, его отвергали, если же краснел, его считали
пригодным (предполагалось, что такие люди в критической ситуа-
ции будут действовать намного решительнее и быстрее других).
О причинах психологических различий задумывались мало.
Главную роль обычно отводили неодинаковому воспитанию. По-
казательны мнения Аристотеля и Теофраста: они полагали, что
характер человека является отражением накопленного индивиду-
ального опыта и с возрастом может существенным образом из-
меняться.
Возникали новые теории. Согласно одним учениям, характер
человека определялся его именем, другим — расположением звезд
в момент рождения, третьим — пищевыми пристрастиями и др.
Несмотря на огромное разнообразие подобных теорий, все они
имели принципиально важное сходство: главным фактором в
формировании характера человека считалась среда. Этот взгляд
доминировал на протяжении многих столетий и сохраняется в
общественном сознании и сегодня.
Первые научные опровержения данного взгляда были получе-
ны лишь в середине XIX в. Их представил английский исследо-
ватель Френсис Гальтон. Это был широко образованный одарен-
ный человек, увлекавшийся многими разделами естествознания,
в том числе географией, метеорологией и антропологией. Прово-
252
дя статистический анализ интеллектуальных способностей, он
заметил, что яркая одаренность встречается редко (в среднем
приблизительно 1 на 4 тыс. чел.) и, по-видимому, представляет
собой некое отклонение от нормы. Между тем, как показало
изучение 300 родословных, у гениального человека родственники
тоже часто оказываются выдающимися людьми. И чем ближе
родство, тем выше вероятность таланта. Будучи убежденным сто-
ронником теории эволюции Гальтон предположил, что уровень
интеллектуальных способностей в основном определяется не вос-
питанием, а наследственными задатками.
Гальтона интересовали не только интеллектуальные способ-
ности, он исследовал десятки различных характеристик. Опираясь
на созданный им близнецовый метод, он утверждал, что многие
психологические признаки человека наследуются в той же
степени, как и физические.
Несомненные заслуги Гальтона заключаются в следующем:
• он впервые исследовал большие выборки людей, что позво-
лило получать статистически надежные данные;
• разработал многочисленные тесты и экспериментальные
подходы для оценки различных физических и психических
свойств. Некоторые успешно применяются до сих пор. С иссле-
дований Гальтона началась экспериментальная психология',
• стремясь к максимальной точности своих результатов, ши-
роко использовал различного рода измерения;
• подвергал полученные данные статистическому анализу, что
потребовало создания специального математического аппарата.
Эту деятельность продолжили его ученики и последователи,
в частности — Карл Пирсон и Джон Холдейн, заложившие осно-
ву современной математической статистики;
• разработал близнецовый метод, с помощью которого можно
сравнивать влияние наследственности и внешней среды на про-
цесс формирования того или иного признака.
9.2. Развитие статистических методов
В конце, а тем более в середине XIX в., когда Гальтон уже за-
нимался наследованием гениальности, наука под названием «ге-
нетика» еще не возникла. Существовали лишь самые общие
представления о наследственности, вытекавшие из дарвиновской
теории естественного отбора, поэтому о генах и их роли в фор-
мировании психики не могло быть и речи. Идеология исследова-
телей была совершенно иной.
Все признаки в зависимости от особенностей их варьирования
можно подразделить на качественные и количественные. Для
253
первых характерны достаточно четкие градации (например, на-
личие или отсутствие шерсти у кошек). Вторые отличаются не-
прерывными вариационными рядами, где вычленить какие-либо
градации практически невозможно (типичный пример — IQ; рис.
9.1). Очевидно, что с точки зрения генетического анализа наи-
более удобны качественные признаки. Говоря современным язы-
ком, они ближе к элементарным, а потому изучать их наследова-
ние сравнительно просто.
Работать с количественными признаками намного сложнее.
Дело в том, что наличие непрерывного вариационного ряда, осо-
бенно если он близок нормальному распределению, свидетель-
ствует о влиянии множества разных факторов, роль каждого из
которых в отдельности невелика. И как следствие, вычленять эти
факторы крайне трудно.
Признаки, непосредственно связанные с поведением человека,
за редкими исключениями являются именно количественными и
потому чрезвычайно сложны для генетического анализа. Эти труд-
ности вполне осознавал уже сам Гальтон. Он считал, что бессмыс-
ленно вычленять наследственные факторы, принимающие участие
в контроле определенного признака. Свою задачу он видел лишь
в детальном описании признаков, в поиске статистических за-
кономерностей их варьирования, в оценке суммарного вклада
наследственности и среды.
Этой же стратегии придерживались и последователи Гальтона.
Их работы носили статистический характер и сводились, по сути,
к трем основным направлениям. Во-первых, к анализу варьиро-
вания признаков в популяциях (в эту задачу входит построение
вариационных рядов, вычисление средних значений, дисперсий
и др.). Во-вторых, к оценке влияния среды и наследственности.
И наконец, к поиску корреляций между разными признаками.
Рис. 9.1. Вариационный ряд 1Q у взрослых людей
254
Наиболее продуктивным оказалось последнее направление.
В результате был создан ряд статистических методов, в том
числе корреляционный, факторный и регрессионный анализы,
получившие широкое распространение не только в психологии,
но и во многих других научных областях.
Постепенно росло количество накопленных данных. Косвенно
они подтверждали гипотезу о том, что многие, а возможно и все
психологические признаки в значительной степени обусловлены
наследственными факторами. Но до строгих доказательств было
еще далеко.
9.3. Возникновение классической
психогенетики
Первая публикация с описанием законов наследования (Грегор
Мендель, «Опыты над растительными гибридами») вышла в свет
в 1865 г. Однако современники не оценили этого открытия. Оно
получило широкое признание значительно позже — лишь в 1900 г.,
когда выводы Менделя нашли подтверждение в трех независимых
исследованиях. Поэтому именно 1900 г. считают официальной
датой возникновения генетики.
Новая наука развивалась стремительными темпами. Законы
Менделя оказались применимы не только к растениям (гороху,
кукурузе, пшенице и многим другим), но и к различным животным,
включая млекопитающих. Стало ясно, что генетические законо-
мерности универсальны, поэтому основное внимание было со-
средоточено на нескольких наиболее удобных модельных объектах,
среди которых самой «популярной» оказалась дрозофила: она име-
ет сравнительно короткий жизненный цикл, высоко плодовита,
легко поддерживается в лабораторных условиях. В результате уже в
середине 1920-х гг. выдающийся американский генетик Томас Хант
Морган на основе исследований, выполненных с дрозофилой,
сформулировал хромосомную теорию наследственности.
Эти успехи не могли не отразиться на психогенетике, однако
их влияние было неоднозначным. С одной стороны, развитие
генетической идеологии привело к пониманию того, что в близ-
нецовом анализе при выявлении моно- и дизиготных близнецов
недостаточно опираться на степень их внешнего сходства. Для
надежности необходимо учитывать и биохимические признаки:
группу крови, резус-фактор и др.
С другой стороны, между генетиками и биометристами (это
название прочно закрепилось за последователями Гальтона в
связи с их пристрастием к измерениям и статистическим методам)
255
постепенно возникло отчетливое противостояние. Генетики в
основном работали с качественными признаками, в то время как
биометристы — преимущественно с количественными. В этом
случае даже в потомстве от контрастных родителей не наблюда-
ется ничего похожего на менделевские закономерности. В итоге
биометристы твердо уверовали в то, что законы дискретного
наследования, изучаемые генетикой, к психологическим при-
знакам человека неприменимы.
Отголоски этого противостояния ощущаются и теперь. В част-
ности, во многих учебниках по психогенетике ее цель до сих пор
формулируется следующим образом: сравнение роли наследствен-
ности и внешней среды в формировании психологических при-
знаков человека. Точно так же, как это делал еще сам Гальтон!
И это несмотря на то, что в последние годы психогенетика всту-
пила в принципиально новый этап: началось выявление и иссле-
дование конкретных генов, непосредственно участвующих в
контроле психологических черт. Однако традиционные взгляды
очень устойчивы, поэтому нет ничего удивительного в том, что
современная молекулярно-генетическая идеология, оперирующая
дискретными единицами (молекулами и генами), неохотно вос-
принимается классической психогенетикой.
9.4. Методы классической психогенетики.
Использование близнецового анализа
для оценки наследуемости
Основная задача классической психогенетики заключалась в
сравнении роли наследственности и среды в формировании того
или иного психологического свойства. Эту задачу можно решать
различными способами, и в первую очередь — с помощью близ-
нецового метода. Напомним, его впервые использовал Гальтон.
Однако в то время (в конце XIX в.) еще не было четкого понима-
ния биологических различий между монозиготными и дизигот-
ными близнецами. Поэтому исследования не отличались особой
корректностью. Только в 1924 г. Герман Сименс сформулировал
концепцию близнецового анализа в том виде, в котором она су-
ществует сегодня. Во-первых, он разработал надежные методы,
позволяющие четко идентифицировать тип близнецов. До этого
диагностика основывалась на интуиции. Во-вторых, он включил
в анализ дизиготных близнецов (изначально, как правило, огра-
ничивались лишь монозиготными).
Наиболее популярная версия близнецового анализа была рас-
смотрена в подразд. 3.2. Остановимся на ней подробнее.
256
Фенотип близнецов зависит от следующих факторов:
• генотипа (он совпадает у монозиготных и заведомо раз-
личается у дизиготных близнецов);
• внешней среды, которую принято подразделять на так на-
зываемую общую среду (одинаковые условия для обоих предста-
вителей пары: например, степень финансовой обеспеченности
семьи, психологические особенности родителей, общие интересы)
и индивидуальную среду (любые неодинаковые условия: разный
круг знакомых, личные увлечения и др.).
Каждый из перечисленных факторов существует и оказывает
определенное влияние на фенотип близнецов, однако непосред-
ственные измерения в этом случае невозможны. Приходится
прибегать к сугубо косвенным оценкам. Для этого обычно ис-
пользуют следующие формулы (поскольку психологические при-
знаки являются количественными, расчеты ведут по коэффици-
ентам внутрипарной корреляции):
• влияние генотипа (коэффициент наследуемости):
^2 __ гмз ~ гдз.
1 -гдз
• влияние общей среды:
Es - гмз ~ Л2;
• влияние индивидуальной среды:
^NS ~ । “ ГМЗ-
Получаемые при этом оценки весьма условны. Они отражают
лишь среднестатистическую ситуацию без учета множества ню-
ансов и тонкостей, специфичных для каждой конкретной пары
близнецов, и могут значительно варьировать в разных исследова-
ниях (табл. 9.1). Таким образом, эти оценки заслуживают доверия
Таблица 9.1. Роль генотипа, общей и индивидуальной среды
в формировании одной и той же психологической характери-
стики по результатам четырех независимых исследований
(по Loehlin, 1992; с дополнениями)
Признак Исследование N h2 Es Ens
MZ DZ
Экстраверсия Финское 117 135 0,23 0,1 0,67
Шведское 4 987 7 790 0,38 0,13 0,49
150 204 0,51 0,03 0,46
Миннесотское 2I7 114 0,55 0,08 0,37
257
Окончание табл. 9.1
Признак Исследование N h2 Es Ens
MZ DZ
Нейротизм Финское 117 135 0,24 0,08 0,68
Ш ведекое 4 987 7 790 0,35 0,15 0,5
151 204 0,22 0,19 0,59
Миннесотское 217 114 0,22 0,33 0,46
* MZ — монозиготные близнецы, DZ — дизиготные близнецы.
только при наличии многих независимых работ, более или менее
совпадающих по своим результатам. Важнее другое: если по дан-
ным проведенных исследований внутрипарная корреляция
монозиготных близнецов достоверно превышает внутрипар-
ную корреляцию дизиготныхч можно говорить о существен-
ной роли генотипа и приступать к использованию моле-
кулярно-генетических методов.
9.5. Варианты близнецового метода оценки
наследуемости
Помимо классической схемы близнецового анализа существу-
ют и некоторые другие его варианты. Кратко рассмотрим их
главные отличительные особенности.
• Лонгитюдные (т.е. долговременные) близнецовые ис-
следования. Этот метод основан на длительных, многолетних
наблюдениях за одними и теми же парами близнецов. В допол-
нение к классической версии близнецового анализа он позволя-
ет оценивать стабильность психологических признаков, отсле-
живать их возрастную динамику и др.
• Метод контрольного близнеца. Исследование проводят на
монозиготных близнецах. Одного представителя от каждой ана-
лизируемой пары подвергают определенному воздействию (на-
пример, обучению), а второго используют в качестве контроля.
С помощью данного метода можно установить, как влияет кон-
кретное воздействие на исследуемый признак.
• Метод разлученных близнецов. Некоторые близнецы прак-
тически с самого рождения разлучены друг с другом и воспиты-
ваются в разных условиях.
258
Причины таких ситуаций, как правило, трагические: нежелание
воспитывать собственных детей, лишение родительских прав или
гибель родителей. Но для науки подобные случаи крайне инте-
ресны. Действительно, если два монозиготных близнеца нахо-
дятся с раннего детства в разных условиях, все психологические
различия между ними должны быть обусловлены исключительно
внешней средой. В свою очередь, это наглядно свидетельствует
о том, как соотносятся роли генотипа и среды в формировании
того или иного психологического признака. Это самый надежный
из всех близнецовых подходов. Однако описанная ситуация на-
столько редка, что возможности данного метода весьма ограни-
чены.
• Метод частично разлученных близнецов. Многие со-
вместно выросшие близнецы рано или поздно обзаводятся соб-
ственными семьями, разъезжаются по разным городам и даже
странам и в итоге оказываются в достаточно разных условиях.
В некоторых случаях (при длительном проживании врозь) воз-
никает ситуация, напоминающая разлученных близнецов. Но при
этом необходимо учитывать, что их «разлучение» произошло не
с самого детства, а значительно позже. Соответственно, используя
подобного рода близнецов, можно выяснить, влияет ли длитель-
ность проживания врозь на ту или иную психологическую харак-
теристику.
• Сравнение близнецов с одиночнорожденными людьми.
Близнецы у человека — достаточно редкое явление. В связи с этим
возникает вполне закономерный вопрос: можно ли использовать
результаты близнецового анализа применительно ко всей сово-
купности людей, подавляющее большинство из которых — оди-
ночнорожденные? Другими словами, нет ли каких-то существен-
ных различий между людьми, рожденными совместно и пооди-
ночке? Как показывает целый ряд исследований, различия есть.
Близнецы значительно чаще оказываются недоношенными, что
приводит к некоторому снижению интеллектуальных способно-
стей (среднее значение IQ у близнецов на 7 баллов ниже средне-
популяционного). Имеются определенные отличия и в постна-
тальном развитии. Близнецы, находясь в постоянном контакте
друг с другом, образуют вполне самодостаточную социальную
группу и зачастую вырабатывают свой собственный автономный
язык. При этом в развитии полноценной человеческой речи они
могут существенно отставать от обычных детей. Впрочем, со вре-
менем эти отличия ослабевают, и у взрослых людей, как правило,
практически незаметны.
• Метод близнецовых семей. Он представляет собой соче-
тание близнецового метода с семейным и будет рассмотрен в
подразд. 9.6.
259
Каждый из перечисленных вариантов близнецового анализа
позволяет решать определенные научные задачи, тесно связанные
с наследуемостью психологических свойств человека. Но ни один
не способен ответить на главные вопросы: какие гены контроли-
руют конкретный психологический признак и каковы молекуляр-
ные основы его формирования? Эти задачи решаются только
молекулярными методами.
9.6. Методы классической психогенетики.
Семейный анализ
Некоторые сведения о характере наследования признака мож-
но получить с помощью семейного метода. Если исследуемый
признак является качественным, составляют родословную каж-
дой анализируемой семьи (таких родословных должно быть до-
статочно много) и прослеживают его передачу из поколения в
поколение, пытаясь выяснить:
• доминантен или рецессивен признак (пояснения на рис. 9.2,
а,
• проявляется ли он и у женщин, и у мужчин;
• от кого он передается потомкам — от отца или матери, или
же в равной степени от обоих родителей.
В свою очередь, это позволяет сделать вывод о том, не связан
ли данный признак с половыми хромосомами, а если связан, то
с какой конкретно — X или Y (см. рис. 9.2, в).
Если же нас интересует количественный признак, весь анализ
приходится строить на косвенных данных, а именно — на коэф-
фициентах фенотипической корреляции. Этот подход основан
на следующих соображениях. Разные члены одной и той же семьи
имеют друге другом разное генетическое сходство. У монозиготных
близнецов оно стопроцентное, у детей по отношению к их родите-
лям — в 2 раза меньше, у внуков по отношению к их бабушкам и
дедушкам — еще в 2 раза меньше и т.д. (табл. 9.2). Поэтому если
в формировании некого признака достаточно существенную роль
играет генотип, корреляция должна соответствовать степени род-
ства.
И действительно, судя по результатам многочисленных иссле-
дований, чем выше генетическое сходство между родственниками,
тем больше их коэффициент фенотипической корреляции (для
примера см. табл. 9.3). Это косвенно свидетельствует о наличии
генетического контроля. Однако ничего конкретнее сказать не-
возможно, поскольку не оценивается вклад окружающей среды,
которая тоже способна воздействовать на фенотип.
260
a
в
Рис. 9.2. Генетический анализ родословных
Условн ые обозначения: О — женщина; □ — мужчина; черным цветом
отмечены индивиды с отклонением от нормы.
Примечания. 1. Пример наследования рецессивной аномалии (рис. 9.2, а).
О ее рецессивности свидетельствует тот факт, что аномалия проявляется не
в каждом поколении. Генотипы индивидов 2, 6, 7, 9 и 10 сомнений не вы-
зывают. Генотипы индивидов 8 и 11 на основе представленной схемы уста-
новить невозможно. Каждый из них может быть либо гетерозиготой, либо
гомозиготой по доминантной (нормальной) аллели. Индивид I, скорее всего,
является доминантной гомозиготой, поскольку все его 4 ребенка фенотипи-
чески нормальны. Тем не менее он может быть и гетерозиготой.
2. Пример наследования доминантной аномалии (рис. 9.2, б}. У каждого
индивида с аномальным проявлением признака хотя бы один из родителей
имеет такой же фенотип (аномалия проявляется в каждом поколении).
3. Пример наследования рецессивной аномалии, обусловленной мутацией
в Х-хромосоме (рис. 9.2, в). В данной родословной аномалия встречается
только у мужчин. При этом она не связана с У-хромосомой (мужчины 5 и 6
получили ее от деда по материнской линии). Таким образом, аномалия обу-
словлена рецессивной мутацией, затрагивающей один из генов Х-хромосомы.
В женском организме имеется две %-хромосомы (одна — от матери, другая —
от отца), в мужском — только одна, доставшаяся от матери (от отца мужчина
получает У-хромосому). Соответственно, если мать гетерозиготна, кому-то из
ее сыновей достанется нормальная, а кому-то — «мутантная» Х-хромосома
(показана серым цветом).
261
Таблица 9.2. Генетическое сходство между разными род-
ственниками
Степень родства Примеры Генетическое сходство, %
I Монозиготные близнецы Дизиготные близнецы Сибсы* Родитель — ребенок 100 В среднем около 50 В среднем около 50 50
2 Дедушка или бабушка — внук или внучка Дядя или тетя — племянник или племянница Двоюродные братья и сестры Полусибсы** В среднем около 25
3 Двоюродные дедушка или бабушка — двоюродные внук или внучка Троюродные братья и сестры В среднем около 12,5
* Разные одиночнорожденные дети от одной и той же родительской пары.
•• Дети одного и того же человека от разных браков.
Таблица 9.3. Связь внутрипарной корреляции по общему фак-
тору интеллекта д со степенью генетического родства
(по Bouchard and McGue, 1981; Егорова, 1988)
Степень родства Генетиче- ское сход- ство, % г N
Один и тот же человек, тестированный дважды 100 0,87 456
Монозиготные близнецы 100 0,86 4 672
Дизиготные близнецы - 50 0,6 5 546
Сибсы - 50 0,47 26 473
Ребенок и один из его родителей 50 0,42 8 433
Приемный ребенок и один из его био- логических родителей 50 0,22 814
Приемный ребенок и один из его усыно- вителей 0 0,19 1397
262
Таким образом, данные, полученные семейным анализом,
не являются строгими доказательствами наследуемости
признака (разумеется, речь не идет о признаках качественных).
Иногда семейный анализ совмещают с близнецовым, в этом
случае говорят о методе близнецовых семей. Обратите внимание:
дети монозиготных близнецов с точки зрения их генетического
сходства соответствуют полусибсам (детям от разных браков одно-
го человека), но воспитываются в заведомо разных условиях
среды. Это позволяет оценить роль среды и генотипа. Кроме того,
с помощью данного метода можно изучать материнские и от-
цовские эффекты. Если среди детей монозиготных сестер на-
блюдается более высокая фенотипическая корреляция, чем среди
детей монозиготных братьев, признак преимущественно насле-
дуется от материнского организма. В обратной ситуации преоб-
ладает отцовский эффект.
9.7. Методы классической психогенетики.
Анализ формирования приемных детей
Логика данного метода очень проста. У ребенка, взятого на
воспитание в другую семью, нет генетического сходства с при-
емными родителями, но зато он находится с ними в общих усло-
виях. В то же время со своими непосредственными биологиче-
скими родителями он имеет 50%-ное генетическое сходство, но
развивается в совершенно иных условиях среды. Таким образом,
сравнивая выборку усыновленных детей с обоими вариантами
родителей (приемными и биологическими), можно выяснить,
какую роль в формировании признака играет окружающая среда,
а какую — генотип.
Наличие высокой корреляции с приемными родителями
свидетельствует о решающей роли внешних условий. Обрат-
ная ситуация — о значительном вкладе генотипа. Однако
эти выводы надежны только в том случае, если дети были усы-
новлены еще в раннем возрасте (в идеале — в первые же месяцы
после рождения) и с тех пор со своими биологическими родите-
лями не общались.
Данный метод можно использовать в нескольких вариантах.
В частности, особого внимания заслуживают семьи, где наряду с
приемным ребенком есть и свой собственный. Такие дети нахо-
дятся в одной и той же среде, но имеют только случайное гене-
тическое сходство. Поэтому степень фенотипической корреляции
между ними служит четкой оценкой влияния внешних условий.
Впрочем, возможности метода не безграничны. Во-первых,
очень невелико количество детей, усыновленных практически
263
сразу после рождения. Во-вторых, имеются вполне обоснованные
опасения, что матери, решившие отказаться от собственных детей,
в большинстве своем страдают различного рода патологиями
(алкоголизмом, психическими расстройствами, наркоманией
и др ), а потому их дети не являются адекватной моделью для
изучения психологических признаков в обществе в целом. На-
конец, для того чтобы взять приемного ребенка, необходимо со-
ответствовать достаточно жестким требованиям, а значит, при-
емные родители далеко не случайны. В результате возникают
сомнения в адекватности метода.
Перечисленные выше проблемы можно обойти, но для этого
требуется тщательное планирование эксперимента. Яркий при-
мер — колорадское исследование приемных детей, проводимое
на протяжении более чем трех десятилетий в Институте генетики
поведения университета Колорадо (США). Оказалось, что можно
найти социальную группу, в которой психические здоровые, не-
деградировавшие матери готовы в определенной ситуации отка-
заться от ребенка. Эту группу составляют молодые незамужние
женщины, воспитанные в традициях строгого католицизма. Если
у них возникает нежелательная беременность, они не могут в силу
своих религиозных убеждений прибегнуть к аборту или оставить
ребенка себе и в итоге обычно соглашаются на усыновление.
Колорадский проект охватывает к настоящему времени около
250 усыновленных детей, а также их биологических и приемных
родителей. Каждый ребенок общался со своей биологической
матерью не более одной недели после рождения, был усыновлен
в течение первого месяца и в дальнейшем уже никогда с ней не
контактировал. Для надежности в анализ вовлечены и обычные
семьи, они выступают в качестве контрольной группы. Исследо-
вание носит люнгитюдный характер, для некоторых семей оно
длится уже 20 лет.
Многие специалисты считают, что рассмотренный метод —
самый строгий из методов классической психогенетики: он по-
зволяет максимально четко развести друг от друга средовое и
генетическое влияние на признак. Но если тайна усыновления
охраняется законом (такая ситуация характерна и для России),
использовать данный метод практически невозможно.
9.8. Основные достижения классической
психогенетики
С момента становления классической психогенетики прошло
уже более 80 лез. За это время она накопила огромный материал
264
и ответила на множество стоявших перед ней вопросов. Кратко
перечислим ее основные успехи.
Во-первых, разработан большой методический арсенал,
позволяющий оценивать генетическое и средовое влияние.
Во-вторых, показано, что самые разные признаки, имею-
щие непосредственное отношение к поведению человека, в зна-
чительной степени зависят от генотипа, в среднем, примерно на
40...45 % (см. подразд. 3.2, табл. 3.3).
В-третьих, отслежена возрастная динамика признаков.
Оказалось, что их чувствительность к внешней среде неодинако-
ва на разных этапах индивидуального развития.
И наконец, проведен анализ средовых воздействий. Как вы-
яснилось, влияние общей и индивидуальной среды для разных
психологических признаков сильно варьирует, но чаще всего роль
последней более значима (табл. 9.4).
Основное внимание уделяли когнитивным способностям: 1Q
и общему фактору интеллекта£. Большинство публикаций по-
свящалось именно им. Следующее место занимали экстраверсия
и нейротизм. Остальные характеристики исследовались менее
интенсивно.
Но самая серьезная проблема заключалась не в этом. Пока не
наступила эпоха молекулярной биологии количество четко иден-
тифицированных генов человека, т.е. генов, для которых установ-
лен характер наследования и место, занимаемое на той или иной
хромосоме, было еще совсем невелико. И ни один из генов не
имел непосредственного отношения к поведению. Таким образом,
классический этап развития психогенетики можно смело назвать
«психогенетикой без генов».
Таблица 9.4. Соотношение вкладов общей (£$) и индивидуаль-
ной (Fmj) среды в формирование некоторых психологических
признаков (обобщенные данные по Loehlin, 1992; с дополне-
ниями)
Признак Л2 Es Е V.s
Общий фактор интеллекта g 0,52 0,33 0,15
Экстраверсия 0,39 0,02 0,59
Нейротизм 0,3! 0,07 0,62
Альтруизм 0,35 0,1! 0,54
Импульсивность 0,38 0,07 0,55
Поиск новизны 0,45 0,06 0,49
Антиобщественное поведение у детей 0,4! 0,16 0,43
265
Большинство экспериментов строилось по следующей схеме.
Проведя анализ определенного психологического признака, уче-
ные пытались интерпретировать полученные данные с помощью
различного рода математических моделей. В эти модели заклады-
вали разное количество генов, разные типы взаимодействия
между ними, разные средовые эффекты и др. И если результаты,
предсказанные какой-то моделью, достаточно хорошо совпадали
с реальными данными, задачу генетического анализа считали вы-
полненной. На самом же деле это были теоретические построения
с совершенно абстрактными «наследственными факторами», не
имеющими подсобой никакой реальной основы. Неудивительно,
что в первых учебниках по психогенетике ни одного конкретного
гена не упомянуто.
Ситуация стала существенным образом изменяться только во
второй половине 1980-х гг. Стремительное развитие молекулярно-
генетических методов позволило выявить многие тысячи генов,
в том числе обеспечивающих функционирование нервной систе-
мы. Были разработаны и получили широкое применение методы
поиска генетических ассоциаций. И наконец, в 1990 г. было вы-
полнено первое молекулярно-генетическое исследование, посвя-
щенное психологическому признаку (а именно — алкоголизму).
Психогенетика вступила в современный этап, базирующийся на
молекулярно-генетической идеологии.
9.9. Современное состояние развития
психогенетики
Несмотря на свой юный возраст, молекулярная психогенети-
ка уже смогла добиться ощутимых успехов. Перечислим лишь
факты, подтвержденные мета-анализом.
• Ген HD (конкретные функции пока неизвестны) контроли-
рует уровень пластичности мышления. Аллели с повышенным
количеством повторов CAG вызывают развитие хореи Гентинг-
тона, первые симптомы которой — нарастающая консерватив-
ность и нежелание прислушиваться к мнению других людей.
• Эпизодическая память зависит от гена А РОЕ, кодирующего
аполипопротеин Е. У здоровых людей гомозиготность по аллели
е4 способствует ослаблению данного вида памяти.
• В генетическом контроле внимания и моторной активности
принимают участие как минимум следующие гены:
DAT1, его продуктом является транспортер дофамина. Аллели
с десятью 40-нуклеотидными повторами ослабляют внимание и
повышают моторную активность;
266
DRD4, кодирующий четвертый тип рецепторов дофамина.
Наличие семи повторов в экзоне 3 тесно ассоциирует с гиперак-
тивностью и дефицитом внимания;
THRB, кодирующий p-рецепторы тироксина. Дефектные ал-
лели (их известно около 50) приводят к широкому спектру фено-
типических нарушений, в том числе — к ослабленному вниманию
и чрезмерной активности.
• Предрасположенность к «жестким» формам самоубийства
находится под контролем не менее чем двух генов:
SLC6A4, продуктом которого является транспортер серотони-
на. Склонность к жесткому суициду ассоциирует с 5-промото-
ром;
СО МТ, кодирующем катехол-О-метилтрансферазу. У людей с
нуклеотидной заменой 158Met вероятность «жесткого» самоубий-
ства достоверно повышена.
• В генетическом контроле отношения человека к пище при-
нимают участие, по меньшей мере, четыре гена:
LEP, продуктом которого является лептин. Гомозиготность по
дефектным аллелям данного гена приводит к практически неуто-
лимому чувству голода и в конечном итоге — к тяжелейшим
формам ожирения;
LEPR, кодирующий рецептор лептина. Ситуация такая же, как
в случае с геном LEP;
MC4R, его продукт — четвертый тип рецепторов меланокор-
тина. Дефектные аллели (их известно около 30) повышают чувство
голода и способствуют хроническому ожирению;
SLC6A4 (см. выше). S-промотор повышает вероятность раз-
вития анорексии.
• Склонность к алкогольной зависимости находится под кон-
тролем как минимум четырех генов. К ним относятся:
ADH2, продуктом которого является алкогольдегидрогеназа 2.
Наличие аллели ADH2+2 значительно снижает вероятность раз-
вития алкоголизма;
ALDH2, кодирующий альдегиддегидрогеназу 2. Аллель
ALDH2+2 вызывает в организме негативное отношение к спирт-
ному и тем самым препятствует развитию алкогольной зависимо-
сти;
ANKK1, функции которого пока неизвестны. Аллель А! по-
вышает вероятность алкоголизма;
SLC6A4 (см. выше). 5-промотор тоже способствует алкоголь-
ной зависимости.
Таким образом, к началу 2010 г, обнаружено, по меньшей
мере, 13 генов, непосредственно участвующих в контроле
психологических признаков. Еще для семи (SNAP25, DRD5,
ADRA2A4 AGRP4 OPRM14 BDNFи GABRA2) получены много-
267
численные независимые подтверждения, но мета-анализ по
ним пока не проведен, И хотя с окончательными выводами то-
ропиться не следует, вполне вероятно, что уже в ближайшее вре-
мя роль каждого из этих генов будет доказана.
9.10. О чем свидетельствуют данные
психогенетики
Подведем основные итоги изложенного.
Во-первых, подавляющее большинство психологических
признаков хотя бы в некоторой степени зависит от гено-
типа. Для одних роль наследственности очень и очень существен-
на, для других, наоборот, сравнительно невелика. Однако важны
не конкретные числовые значения, а сам факт наличия генети-
ческого контроля, открывающий огромные перспективы для
дальнейших исследований.
Во-вторых, для целого ряда психологических свойств обна-
ружена четкая ассоциация с конкретными генами. К этим
свойствам относятся внимательность, эпизодическая память,
пластичность мышления, предрасположенность к «жестким»
формам самоубийства, отношение к пище, склонность к алкого-
лизму и поиск новизны. На подходе и некоторые другие характе-
ристики, в том числе успешность в освоении навыков чтения,
а также склонность к курению табака.
В-третьих, один и тот же психологический признак кон-
тролируется совместно как минимум несколькими генами.
Этот вывод основан не только на общих соображениях (сложность
молекулярной организации нервной системы, близость вариаци-
онных рядов к нормальному распределению), но и на данных
непосредственно генетического анализа. Для примера обратимся
к контролю отношения к пище. В нем участвует, по меньшей мере,
четыре гена: LEP, LEPR, MC4R и SLC6A4. При этом их вклад
неодинаков. Постепенно выявляются молекулярные системы,
контролирующие ту или иную психологическую характеристику.
В частности, центральную роль в регуляции внимания играют две
системы: тироксиновая (ген THRB) и дофаминовая (гены DAT1
и DRD4).
В-четвертых, влияние одного и того же гена может рас-
пространяться на разные психологические признаки. В этом
плане показателен ген SLC6A4. Он влияет как минимум на три
различные характеристики: предрасположенность к «жестким»
формам самоубийства, отношение к пище и склонность к алко-
голизму. Причины такой ситуации, по-видимому, в следующем.
268
Данный ген контролирует активность серотониновой системы.
Известно, что серотонинэргические синапсы присутствуют во
многих отделах головного мозга и регулируют множество разных
аспектов жизнедеятельности, в том числе агрессивность, чувство
голода и положительные эмоции. Неудивительно, что каждый из
трех перечисленных признаков ассоциирует с геном SLC6A4.
В-пятых, у модельных животных и человека сходные гены
отвечают за сходные поведенческие признаки. Мы убедились
в этом на множестве конкретных примеров. Особенно показа-
тельна ситуация с геном HD: излишнее количество повторов CAG
приводит и у человека, и у мышей к поражению одной и той же
области мозга (ГАМК-эргических нейронов полосатого тела) и
вызывает значительное снижение когнитивной пластичности.
Таким образом, данные, полученные на модельных животных,
вполне применимы к человеку и наоборот.
В-шестых, генетический анализ оказывается эффектив-
ным только при четкой диагностике исследуемых признаков,
поэтому, работая с интегральными психологическими характери-
стиками, в большинстве своем крайне размытыми и неконкрет-
ными, обнаружить соответствующие гены обычно не удается.
Необходимо использовать более дробные характеристики. При-
мер — генетический анализ предрасположенности к самоубийству.
5-промотор гена SLC6A4 и мутация 158Met в гене СОМТ ассо-
циируют с «жесткими» формами суицидного поведения, но не
связаны с «легкими» способами ухода из жизни. И если к само-
убийствам подходят недифференцированно, влияние перечислен-
ных генов, как правило, незаметно. Таким образом, стратегиче-
ская задача современной психогенетики — вычленять элементар-
ные психологические характеристики и проводить генетический
анализ каждой в отдельности.
9.11. Перспективы дальнейшего развития
психогенетики
Рассмотрев современное состояние психогенетики, попытаем-
ся в общих чертах представить ее будущее. Для этого обратимся
к генетике развития, тоже возникшей из недр молекулярной
биологии и стремительно прогрессирующей в последние десяти-
летия. Эта наука сформировалась раньше современной психоге-
нетики и потому успела пройти более длительный путь. Именно
он и будет служить для нас ориентиром.
Для многих живых существ, как растений, так и животных,
подробные описания всех стадий индивидуального развития были
269
сделаны еще к середине XIX в. Но причины развития продолжа-
ли оставаться загадкой. Их объясняли стремлением организма к
самоусовершенствованию, таинственными «внутренними силами»
и прочей абстракцией. И лишь со временем стали накапливаться
конкретные факты.
Сначала ученые обратили Внимание на то, что развитие во
многом зависит от внешних условий. В этом плане прекрасным
примером являются растения: При полном отсутствии света они
сильно вытягиваются, не распускают зачатки листьев и не цветут.
Внешняя среда влияет и на развитие животных. Так, у крокодилов
и некоторых других пресмыкающихся пол определяется темпера-
турой инкубации яиц.
Следующий шаг к пониманию механизмов развития был сделан
уже в начале XX в. Оказалось, что их можно регулировать хими-
ческими соединениями. Более того, подобные «химические регу-
ляторы» были найдены непосредственно в самих живых суще-
ствах. И постепенно, шаг за шагом возникло понимание, что в
основе развития любого организма лежат какие-то процессы об-
мена веществ.
На следующем этапе у некоторых модельных объектов (глав-
ным образом мышей, дрозофцЛЬ1 и кукурузы) было выявлено
множество мутантов с нарушециями развития. Их анализ по-
казал, что любая структура организма формируется под четким
контролем со стороны генотипа. Поначалу создавалось впечат-
ление, что ситуация проста: каждая структура контролируется
одним-единственным геном. Однако такое представление было
ошибочным.
Таблица 9.5. Число генов, контролирующих развитие опреде-
ленных органов
Объект Орган Число соответствующих генов
Дрозофила Нога Около 170
Крыло Около 220
Глаз Около 300
Арабидопсис* Цветок Не менее 90
Горох
Азотфиксирующий клубенек Не менее 50
• Небольшое цветковое растение (блцзкиЯ родственник редиса и капусты),
широко используемое как модельный объект генетики развития
270
По мере получения все новых и новых мутантов становилось
очевидным, что развитие каждой структуры контролируется до-
статочно большим количеством генов: настолько большим, что
счет идет на десятки (табл. 9.5). Эти гены находятся в тесном
функциональном взаимодействии, обеспечивая четко скоорди-
нированную генетическую программу, основанную на принци-
пах каскадной регуляции. Таким образом, развитие каждой кон-
кретной структуры контролируется не отдельными генами, а целой
программой. Добавим, что некоторые программы взаимодейству-
Таблица 9.6. Сопоставление генетики развития и психогене-
тики
Этапы становления Генетика развития Психогенетика
Научный анализ !• Развитие организма зави- сит от внешних условий: температуры, наличия света и т.п. Поведение человека зависит от внешних усло- вий: воспитания, непо- средственного окружения и др.
2* Развитие можно регу- лировать химическими соединениями,напри- мер, различными гормо- нальными препаратами. Аналогичные соединения имеются и в самих орга- низмах Поведение можно модифицировать хими- ческими соединениями: алкоголем, наркотиками, специальными медика- ментами. Аналогичные соединения имеются и в самих организмах
3 Существуют гены, кон- тролирующие процессы развития Существуют гены, кон- тролирующие поведение человека
Научный синтез 4 Существуют сложные генетические программы развития, регулируемые определенными фактора- ми внешней среды Еще не наступил
5 Разные генетические программы взаимосвяза- ны, обеспечивая согла- сованное развитие всего организма Еще не наступил
• Этапы, предшествовавшие возникновению данных наук.
271
ют друг с другом. В результате развитие разных структур организ-
ма происходит не строго независимо, а согласованно.
Психогенетика идет примерно по тому же пути. Но пока ее
достижения значительно скромнее: она находится на стадии вы-
явления отдельных генов (табл. 9.6). Соответственно мы вправе
ожидать, что в ближайшей перспективе число выявленных генов
будет расти, а затем начнется этап научного синтеза. Он позволит
вычленять ключевые генетические программы, контролирующие
формирование психологических признаков человека, и суще-
ственно приблизиться к пониманию человеческой психики во
всем разнообразии ее индивидуальных проявлений.
9.12. Наиболее перспективные направления
психогенетики
9.12.1. Молекулярные механизмы действия
социальных факторов
Основная задача современной психогенетики — поиск генов,
участвующих в контроле психологических признаков. Однако не
следует забывать, что на все эти признаки оказывает значительное
влияние и окружающая среда. И естественно, встает вопрос о
механизмах влияния.
Если бы мы говорили не о человеке или хотя бы не о психоло-
гических характеристиках, подобный вопрос не вызвал бы особых
проблем. В настоящее время имеется много литературных источ-
ников, посвященных молекулярным механизмам внешних воздей-
ствий. В их основе лежат различные каскадные процессы, начи-
нающиеся с активации того или иного рецептора и заканчиваю-
щиеся строго определенным биологическим ответом (см. подразд.
1.13 и 2.6). Но справедлива ли данная схема для нашего случая?
Мы достаточно хорошо представляем молекулярные каскады,
запускаемые с помощью химических и физических сигналов.
Между тем в процессах формирования психологических призна-
ков главную роль отводят различным социальным факторам.
Механизмы их действия должны быть совершенно иными.
Принято считать, что это безоговорочно так. Но если вдумать-
ся, возникает все больше и больше сомнений. Во-первых, необ-
ходимо отметить, что понятие «социальное» не означает чего-то,
присущего только человеку. В самом деле, сложно организованные
социальные отношения характерны для многих млекопитающих
и даже для насекомых (классическим примером являются пчелы
и муравьи). При этом в основе подобного рода отношений всегда
272
лежат или химические, или физические сигналы. Достаточно
вспомнить о роли феромонов у мышей.
Во-вторых, постепенно накапливаются свидетельства того, что
под действием некоторых «сугубо социальных» факторов проис-
ходят вполне определенные молекулярные изменения. В част-
ности, у самок зебровой амадины (тропического родственника
российского домового воробья) при прослушивании брачной
песни, издаваемой самцом, изменяется экспрессия генов в ней-
ронах головного мозга. Существуют две разновидности этой
песни. Одна предназначена непосредственно для самки, вторая
не имеет конкретного адресата. И у самок в зависимости от типа
услышанной песни экспрессия генов изменяется по-разному.
Еще один яркий пример описан у макак, гомозиготных по
5-промотору гена SLC6A4. Как мы видели (см. подразд. 6.18),
отсутствие материнской заботы у таких обезьян приводит к сни-
жению активности серотониновой системы и в конечном итоге,
как правило, к гиперагрессивности. Конкретный механизм пока
остается неясным, но, скорее всего, здесь тоже затронута экс-
прессия генов.
Вряд ли у самок амадины имеются рецепторы, напрямую вос-
принимающие различные типы песни, а у макак — рецепторы на
отсутствие материнской заботы. Тем не менее перечисленные
факторы все же распознаются и приводят к вполне определенным
молекулярным последствиям. В связи с этим должны существо-
вать молекулярные системы, позволяющие косвенно оценивать
социальные воздействия и запускать в организме надлежащие
биологические ответы. Нет сомнений, что поиск подобного рода
систем — одна из интереснейших задач психогенетики будущего.
9.12.2. Изучение механизмов генотип-средовых
отношений
Говоря о влиянии генотипа и внешней среды на процесс фор-
мирования психологических признаков человека, наивно думать,
что эти влияния просто суммируются. Между ними бывают зна-
чительно более сложные отношения. В частности, одно и то же
внешнее воздействие может вызвать у разных генотипов разные
эффекты. К числу наиболее широко известных примеров отно-
сится влияние обогащенной и обедненной среды на обучаемость
«умных» и «глупых» инбредных крыс. Обогащенная среда повы-
шает обучаемость «глупых», однако на успехи «умных» практиче-
ски не влияет. Обедненная среда приводит к обратному эффекту:
почти не оказывает влияния на обучаемость «глупых», но суще-
ственным образом снижает способности «умных».
273
Подобные ситуации встречаются достаточно часто, их назы-
вают взаимодействием генотипа и среды. Пока не наступила
эпоха молекулярной биологии, это явление оставалось загадочным
и необъяснимым. Многочисленные научные дискуссии по дан-
ному вопросу не имели возможности опираться на конкретные
механизмы и все больше и больше сводились к абстрактным рас-
суждениям.
Первые проблески появились в конце 1970-х гг. У дрозофилы
был выявлен целый ряд мутантов, память которых зависит от
внешних условий. Так, у мутанта Notch'52 при низкой температу-
ре 18 °C формируется вполне нормальная долговременная память,
а при повышенной температуре (30 °C) — резко ослабленная.
Оказалось, что мутантный белок недостаточно стабилен и утра-
чивает свою активность при повышенной температуре. Соответ-
ственно долговременная память не может сформироваться. Между
тем активность нормального варианта белка при таком же повы-
шении температуры почти не меняется, обеспечивая стандартный
уровень формирования памяти.
Стало ясно, что внешняя среда взаимодействует с гено-
типом на уровне протекающих в организме молекулярных
процессов. Этот вывод в настоящее время не вызывает сомнений:
его справедливость доказана для сотен различных признаков у
разных модельных объектов, включая и человека. В итоге сфор-
мировалось целое научное направление, получившее название
экологическая генетика (генетика взаимодействий организмов
с внешней средой). Появились примеры и из области человече-
ской психики.
Кратко остановимся только на одном из них.
Существуют различные способы улучшения настроения. К их
числу относится прием алкогольных напитков. В результате про-
исходит активация дофаминовой системы, человек ощущает при-
лив положительных эмоций, становится более общительным и
раскрепощенным. Однако у некоторых людей (в основном мон-
голоидов) алкогольные напитки вызывают противоположный
эффект: слабость, учащение пульса, приступы тошноты, сильную
головную боль, неестественный румянец. Речь едет о гомозиготах
по аллели ALDH2*2. Они не способны утилизировать уксусный
альдегид и даже от совсем небольшого количества спиртного мо-
гут оказаться в состоянии алкогольного отравления (см. подразд.
8.9 и 8.10).
Детальное изучение подобного рода примеров представляет
для психогенетики огромный интерес. Оно позволит значительно
приблизиться к пониманию механизмов, лежащих в основе ин-
дивидуальных психологических различий. Нет сомнений, что эти
механизмы окажутся молекулярными.
274
9.13. Важнейшие методологические
направления современной психогенетики
Завершая рассмотрение истории и перспектив психогенетики,
необходимо еще раз напомнить, что любая наука развивается по
одной и той же универсальной схеме. На первом этапе использу-
ют научный анализ', разбивают изучаемую проблему на более
простые и кропотливо исследуют каждую из них по отдельности.
В дальнейшем по мере накопления фактического материала на-
чинают сводить его в единую картину, т.е. переходят от анализа
к научному синтезу (см. подразд. 3.1).
Несмотря на более чем столетнюю свою историю, психогене-
тика, по сути, — совсем молодая наука. Она находится еще на
стадии научного анализа: выявляет элементарные психологиче-
ские характеристики, ведет поиск генов, участвующих в их фор-
мировании. Таким образом, ос н о в н у ю задачу современ-
ной психогенетики можно выразить словами: «дробить^
дробить и дробить». Перечислим наиболее важные методоло-
гические направления, позволяющие обеспечить выполнение
данной задачи.
• Максимально дробный подход к исследуемым признакам
при объективной и четкой диагностике каждого из них
(переход от опросников к непосредственным эмпирическим
оценкам). Мы касались этих аспектов уже неоднократно и по-
тому не будем дополнительно на них останавливаться.
• Дифференциальный анализ признака у женщин и муж-
чин. Как известно, мутации, локализованные в %-хромосоме,
проявляется у мужчин отчетливей, у женщин (мужчина имеет одну
Х-хромосому, а женщина — две). Кроме того, женские и мужские
половые гормоны оказывают на нервную систему неодинаковое
воздействие. В связи с этим у особей мужского и женского пола
формирование одной и той же психологической характеристики
может осуществляться с некими специфическими нюансами. На-
пример, мужчины заметно отличаются от женщин по общему
вкладу генотипа в предрасположенность к курению. Генетический
анализ подобного рода признаков необходимо проводить отдель-
но для каждого пола.
• Дифференциальный анализ признака для разных возрас-
тов. Многие анатомические и физиологические признаки инди-
вида (пропорции тела, мышечная масса, число зубов, упругость
кожи, острота зрения и др.) закономерно изменяются в процессе
развития организма. То же самое свойственно и некоторым чер-
там поведения. В частности, у маленьких детей интеллектуальные
способности в основном определяются факторами внешней сре-
275
ды, а у взрослого человека, напротив, — его генотипом (см. под-
разд. 3.2). Успешность генетического анализа в таких ситуациях
в значительной степени зависит от возраста испытуемых.
• Совместный анализ по нескольким генам-кандидатам.
В генетическом контроле любой психологической характеристи-
ки принимает участие как минимум несколько генов. При этом
их вклад может быть весьма неодинаковым (например, вероят-
ность развития алкоголизма зависит от гена ALDH2 намного
сильнее, чем от генов ANKK1 и SLC6A4). Соответственно, влия-
ние сравнительно «слабого» гена трудно выявить без учета гете-
рогенности по «более сильным».
Накопив большое количество фактического материала по ге-
нетическому контролю различных психологических признаков,
психогенетика со временем приступит и к научному синтезу. Но
для этого ей нужно пройти еще длительный путь.
9.14. История развития психогенетики
в России
Вопросы, связанные с наследованием психологических свойств,
поднимались в отечественной литературе уже давно. Так, еще в
1865 г. вышли в свет очерки Василия Марковича Флоринского
«Усовершенствование и вырождение человеческого рода». Автор
высказывал мнение, что от качеств супругов непосредственно
зависят качества их потомков, и обсуждал, какие именно комби-
нации родителей способствуют физическому и нравственному
здоровью детей.
Выдающийся педагог и антрополог Константин Дмитриевич
Ушинский, признавая сам факт наследуемости характера, связы-
вал его исключительно с наследованием привычек. Попытка экс-
периментально проверить эту гипотезу была предпринята в 1923 г.
Нобелевским лауреатом Иваном Петровичем Павловым: он изу-
чал, наследуются ли условные рефлексы у собак. Первоначально
Павлов считал, что наследование есть, но затем убедился в несо-
стоятельности этого вывода.
Первые систематические исследования по психогенетике были
начаты в России только в 1930-х гг. Они проводились в Медико-
биологическом институте (Москва), который в 1935 г. был пере-
именован в Медико-генетический институт. В нем изучали на-
следственные основы различных заболеваний, моторных функ-
ций, внимания, памяти и интеллекта. Руководил исследованиями
Соломон Григорьевич Левит, психологическое направление воз-
главлял Александр Романович Лурия, вопросами математической
276
обработки близнецового анализа занимался Михаил Васильевич
Игнатьев.
Близнецовый анализ был одним из важнейших направлений.
Уже в 1934 г. он охватывал около 700 пар близнецов, а в 1936 г. —
в 2 раза больше. При институте функционировали особые ясли
и детсад, в которых близнецы могли находиться круглосуточно
под постоянным наблюдением психологов и врачей. Успешно
использовали метод контрольного близнеца, планировали вести
пожизненные лонгитюдные исследования.
К сожалению, дальнейшая судьба института трагична. Наступила
печально известная эпоха Лысенко, генетика подверглась гонениям
и полному запрету. В 1937 г. институт был закрыт. Еще через год
арестовали С. Г. Левита, и впоследствии он был расстрелян по лож-
ному обвинению. Генетические исследования в СССР были пре-
кращены, в том числе и работы по генетике поведения человека.
Лысенковщина закончилась к началу 1960-х гг. Но даже со
снятием всех официальных запретов отечественная генетика не
могла возродиться в одночасье: в стране почти не осталось ква-
лифицированных кадров, отсутствовала необходимая для иссле-
дований материальная база. На возрождение генетики ушло еще
несколько лет.
Исследования в области генетики поведения человека возоб-
новились в 1972 г. на базе НИИ общей и педагогической психо-
логии (ныне — Психологический институт Российской академии
образования). Эти исследования были направлены на проверку
гипотезы о врожденности индивидуальных свойств нервной си-
стемы. В качестве основных индикаторных характеристик ис-
пользовали биоэлектрическую активность мозга и различного рода
вызванные потенциалы (зрительные, слуховые, моторные и др.).
Все исследования носили сугубо статистический характер и вы-
полнялись в русле классической психогенетики. Поэтому о кон-
кретных генах речи не шло.
В 1980-е гг. проблематика постепенно сместилась в сторону
анализа генотип-средовых отношений (подробно об этих отно-
шениях было рассмотрено в подразд. 9.14). Оказалось, что одна
и та же психологическая характеристика демонстрирует разную
чувствительность к внешней среде в зависимости от возраста ис-
пытуемых, их функционального состояния и даже от семантиче-
ской структуры используемого стимула. В результате возникла
достаточно продолжительная дискуссия о специфике психологи-
ческих признаков человека.
Перестройка и последовавший за ней распад СССР не способ-
ствовали развитию отечественной науки. Разумеется, это сказа-
лось и на психогенетике, особенно с учетом того, что в 1990-е гг.
она вступила в свой принципиально новый этап, базирующийся
277
на молекулярно-генетической идеологии и требующий высоко
технологичных дорогостоящих методов.
И все же, молекулярная психогенетика есть и в России. Ис-
следования проводятся в Научном центре психического здоровья
РАМН (Москва), Институте биохимии и генетики Уфимского
научного центра РАН, НИИ медицинской генетики Томского
научного центра РАН, в Институте общей генетики (Москва),
Национальном научном центре наркологии Росздрава (Москва),
в НИИ физико-химической медицины Росздрава (Москва), НИИ
экологии человека и гигиены окружающей среды РАМН (Мо-
сква), НИИ гигиены и охраны здоровья детей и подростков (Мо-
сква), Научном центре здоровья детей (Москва), а также в ряде
других научных учреждений.
Хочется верить, что уже в ближайшей перспективе российская
молекулярная психогенетика получит мощное развитие и займет
в мировой науке достойное место.
СПИСОК ЛИТЕРАТУРЫ
1. Анохин А. П. Генетика, мозг и психика человека: тенденции и перспек-
тивы исследований. — М. : Изд-во ИПАН, 1988.
2. Баранов В. С. и др. Геном человека и гены «предрасположенности».
Введение в предиктивную медицину. — СПб. : Интермедика, 2000.
3. Боринская С. А., Гасемианродсари Ф., Кальина Н.Р., Соколова М.В.,
Янковский Н.К. Полиморфизм гена алкогольдегидрогеназы ADHIB в вос-
точнославянских и ираноязычных популяциях // Генетика. — 2005. — Т. 41. —
№ II. - С. 1563-1566.
4. Булаева К. Б. Генетические основы психофизиологии человека. — М.:
Наука, 1991.
5. Галыпон Ф. Наследственность таланта. Законы и последствия. — М.:
Мысль, 1996.
6. Голимбет В. Е. и др. Связь генов дофаминергической системы с экс-
траверсией и поиском новизны И Журнал высшей нервной деятельности. —
2006. - Т. 56. - №4. - С. 457-463.
7. Голимбет В.Е., Алфимова М.В.У Митюшина Н.Г. Полиморфные
варианты гена рецептора серотонина (5HTR2A) и особенности личности //
Молекулярная биология. — 2004. — Т. 38. — № 3. — С. 404 — 412.
8. Гумерова О. В. и др. Анализ ассоциаций генов, регулирующих работу
серотонинэргической нейромедиаторной системы, с уровнем интеллектуаль-
ного развития человека // Генетика. — 2007. — Т. 4. — № 6. — С. 835 — 840.
9. Егорова М.С., Зырянова Н.М. Влияние генотипа на соотношение
показателей интеллекта и когнитивного поля. — Генетика. — 1997. — Т. 33. —
№ 1. - С. ПО- 115.
10. Егорова М. С., Зырянова Н. М., Пьянкова С.Д. Возрастные изменения
генотип-средовых соотношений в показателях интеллекта // Вопросы пси-
хологии. — 1993. — С. 106— 108.
278
11. Ильин Е.П. Психология индивидуальных различий. — СПб. : Питер,
2011.
12. Корочкин Л. И. Введение в генетику развития. — М. : Наука, 1999.
13. Левит С. Г. Некоторые итоги и перспективы близнецовых исследо-
ваний И Труды медико-биологического института. — 1934. — Т. 3. —
С. 5-17.
14. Малых С. Б. Количественные методы в генетике поведения. В кн.
Генетика поведения: количественный анализ психологических и психофи-
зиологических признаков в онтогенезе. — М. : Sociologos, 1995. —
С. 4-96.
15. Малых С. Б. Исследования генетической детерминации ЭЭГ челове-
ка И Вопросы психологии. — 1997. — № 6. — С. 109— 128.
16. Марютина Т.М. Роль Генотипа и изменчивости вызванных потен-
циалов человека на разных этапах онтогенеза // Высшая нервная деятель-
ность. - 1993. - Т. 53. - № 3. - С. 636-644.
17. Небылицын В.Д. Темперамент. Психофизиологические исследования
индивидуальных различий. — М. : Наука, 1976.
18. Bouchard T.J. Jr and McGue M. Familial studies of intelligence :
a review 11 Science. — 1981. — V. 212. — P. 1055— 1059.
19. Cooper R. and Zubeck J. Effects of enriched and restricted early
environments on the learning ability of bright and dull rats // Canadian J.
Psychology. - 1958. - V. 12. - P. 159- 164.
20. Filipenko M. L. et al. Repeated experience of social defeats increases
serotonin transporter and monoamine oxidase A mRNA levels in raphe nuclei of
male mice // Neurosci Lett. — 2002. — V. 321. — P. 25—28.
21. Gaysina D. et al. The serotonin transporter gene: polymorphism and
haplotype analysis in Russian suicide attempters // Neuropsychobiology. — 2006. —
V. 54. - P. 70-74.
22. Ge X. et al. Notch signaling in Drosophila long-term memory formation //
Proc. Natl. Acad. Sci. USA. - 2004. - V. 101. - P. 10172-10176.
23. Loehlin J.C. Genes and Environment in Personality Development //
Thousand Oaks, CA, US: Sage Publications, Inc. — 1992. — 145 p.
24. Plomin R. Genetics and Experience. The Interplay Between Nature and
Nurture // Thousand Oaks: Sage Publications, Inc. — 1994. — 208 p.
25. Teramitsu /. and White S.A. FoxP2 regulation during undirected singing
in adult songbirds // J Neurosci. — 2006. — V. 26. — P. 7390 —7394
КОНТРОЛЬНЫЕ ВОПРОСЫ
1. Опишите основные этапы становления психогенетики.
2. Какие варианты близнецового анализа используют в психогене-
тике?
3. Каковы возможности семейного анализа в психогенетике?
4. Опишите примеры взаимодействия генотипа и среды.
5. Каковы ближайшие перспективы развития психогенетики?
Глава 10
КТО ЖЕ ТАКОЙ ЧЕЛОВЕК?
10.1. Молекулярное родство человека
с другими млекопитающими
Говоря о механизмах формирования психологических призна-
ков, мы в конечном итоге приходим к неизбежному вопросу: что
же именно отличает человека от прочих животных? Обратимся к
систематике — специальной биологической науке, изучающей
степень родства между разными видами.
Все детали строения и физиологии современного человека
(официальное название вида — «человек разумный») позволяют
безоговорочно отнести его к царству животных, типу хордо-
вых. классу млекопитающих, отряду приматов. К этому от-
ряду принадлежит около 300 видов. Среди них наиболее значи-
тельным сходством с человеком обладают шимпанзе, гориллы
и орангутаны, составляющие группу человекообразных обе-
зьян.
Пока не наступила эпоха молекулярной биологии, было при-
нято считать, что люди и человекообразные обезьяны представ-
ляют два близких, но все-таки разных семейства. Однако в
дальнейшем выяснилось, что это не так. Стремительное развитие
молекулярно-генетических методов привело к возникновению
нового направления систематики, в основу которого положен
анализ ДНК. Данное направление называют ееносистемати-
кой. Оно показало, что люди и человекообразные обезьяны на-
ходятся в более близком эволюционном родстве, и теперь их
объединяют в одно семейство — гоминиды (табл. Ю.1). Среди
всех современных представителей этого семейства (человек
разумный, шимпанзе, карликовый шимпанзе, горилла и оран-
гутан) нашими ближайшими родичами являются шимпанзе.
Геномы шимпанзе и человека отличаются друг от друга в 3 раза
меньше, чем геномы мыши и крысы (табл. I0.2). Более того,
каждый третий белок шимпанзе имеет точно такую же амино-
кислотную последовательность, как и его человеческий гомолог.
280
Судя по результатам молекулярно-генетического анализа, пред-
ки человека и шимпанзе разошлись в эволюции около 6 млн лет
назад (рис. 10.1).
Таблица 10.1. Современные представления о положении
человека в системе живого
Таксономический ранг Название группы Основные особенности
Царство Животные (Anima На) Эукариотическое строение клеток (наличие многочислен- ных внутриклеточных отсеков, каждый из которых выполняет определенные функции) Гетеротрофное питание (соб- ственные вещества синтезиру- ются из чужих органических) Отсутствие клеточных стенок (высокая подвижность) Многоклеточность с четко вы- раженными тканями и органами (разделение функций между разными типами клеток)
Тип Хордовые (Chordata) Внутренний скелет (надежная опора, не ограничивающая под- вижности организма) Нервная система представлена нервной трубкой (интегральное управление всем организмом)
Класс Млекопитающие (Mammalia) Выкармливание детенышей молоком (необходимая дете- нышам пища производится непосредственно материнским организмом) Теплокровность (интенсивный обмен веществ, независящий от температуры окружающей среды) Дифференцированные зубы (способность откусывать и тща- тельно пережевывать пишу) Достаточно развитые боль- шие полушария (способность к сложному приспособительно- му повелению)
281
Окончание табл. 10.1
Таксономический ранг Название группы Основные особенности
Отряд Приматы (Primates) Глазницы направлены вперед (объемное зрение, хорошая ори- ентация в пространстве) Пятипалые конечности хвата- тельного типа (способность к сложным манипуляциям) Особо чувствительные участки кожи на ладонях и подои! вах (способность к тонкому осяза- нию)
Семейство Гоминиды (Hominidae) Дополнительное развитие больших полушарий и связанное с этим изменение пропорций черепа Отсутствие хвоста Способность хотя бы к непро- должительному прямохождению
Род Человек (Ното) Резкое увеличение больших полушарий Прямохождение Противопоставленность большого пальца руки четырем остальным Способность изготавливать орудия
Вид Человек разум- ный (Ното sapiens) Разум (?) Речь (?)
Таблица 10.2. Сравнение геномов* некоторых млекопитающих
(по данным 2010 г.)
Вид Размер генома Число выявленных генов Степень сходства геномов
Человек разумный - 3,2 10’п.н. - 22 600 -97%
Шимпанзе ~ 3,1 10’п.н. - 14 000**
282
Окончание табл. JO. 2
Вид Размер генома Число выявленных генов Степень сходства геномов
Серая крыса -2,8 Ю’п.н. - 21 000 -90%
Домовая мышь -2,9 Ю’п.н. - 22000
* Под геномом понимают все нуклеотидные последовательности в составе
одного гаплоидного набора хромосом.
** Процесс выявления генов шимпанзе еще не закончен.
Australopithecus
у boisei
2-1
z
z
z
z" Australopithecus
--------robustus
2-1,5
Homo
neanderthalensis
,41,35—0,03
Общий предок,
с шимпанзе
Australopithecus Australopithecus Homo
— qfarensis --------- <
Homo Homo
africanus-----habilis—erectus---
3-2 2,2-1,6 1,4-0,6 “P™
Ьвсгрллоакпюи
Человек
б
Рис. 10.1. Эволюция высших приматов:
а — эволюционное древо современных высших приматов. Многочисленные группы
дриопитеков, не оставившие современных потомков, показаны в виде одной пунктир-
ной линии. Числами обозначено приблизительный период, миллионов лет назад, рас-
хождения предков человека с предками других высших приматов; б — предполагаемый
путь эволюционного становления человека после расхождения с предками шимпанзе
по представлениям второй половины XX в. (в результате обнаружения новых чело-
векоподобных ископаемых форм эта схема теперь считается устаревшей. Реальные
эволюционные связи в ней пока неясны. Числами обозначен приблизительный период
существования, в миллионах лет назад, каждой из представленных форм)
283
К настоящему времени получены подробные данные о геномах
не только человека и шимпанзе, но и макаки, свиньи, крупного
рогатого скота, крысы, мыши, собаки, лошади, утконоса, курицы
и некоторых видов рыб. Оказалось, что многие гены и соот-
ветствующие белки высоко консервативны практически у
всех позвоночных. Для примера обратимся к нейропептиду F:
у человека, курицы^ аллигатора^ лягушки и угря он варьирует
всего лишь по нескольким аминокислотам (табл. 10.3). Таким
образом, можно не сомневаться, что в каждом из нас работают
приблизительно те же молекулярные механизмы, что и у осталь-
ных млекопитающих, в особенности обезьян.
Таблица 10.3. Сравнение аминокислотной последовательно-
сти нейропептида Y у разных позвоночных
Класс Объект Число замен при сравнении с человеком
Млекопитающие Шимпанзе 0*
Макака резус 0
Зеленая мартышка 0
Серая крыса 0
Домовая мышь 0
Кролик 0
Африканский слон 0
Лошадь 1
Собака 0
Кошка 0
Крупный рогатый скот 1
Птицы Курица 1
Зебровая амадина 1
Пресмыкающиеся Миссисипский аллигатор 0
Земноводные Прудовая лягушка 0
284
Окончание табл. 10.3
Класс Объект Число замен при сравнении с человеком
Костные рыбы Карп 3
Полосатый дан ио 3
Треска 5
Дальневосточный лосось 5
Обыкновенный угорь 7
Хряшевые рыбы Скат торпедо 2
Зимний скат 5
* Размер нейропептида У — 36 аминокислот.
10.2. Какие признаки видоспецифичны
для человека разумного?
Несмотря на достаточно тесное родство с обезьянами, совре-
менный человек существенно от них отличается. Для него харак-
терны измененные пропорции черепа (значительно более крупный
мозговой отдел и, напротив, резко уменьшенные челюсти), пря-
мохождение, отсутствие сплошного волосяного покрова, противо-
поставленность большого пальца руки четырем остальным, на-
личие подбородка, разум, членораздельная речь. Казалось бы, все
перечисленные нами признаки должны считаться для Ното
sapiens видоспецифичными. Однако реальная ситуация намного
сложнее.
Во-первых, большинство упомянутых выше особенностей воз-
никло еще у предков человека разумного. Так, прямохождение
появилось у австралопитеков — древнейших представителей
человеческой ветви приматов. Заметное увеличение размеров
головного мозга и связанное с этим изменение пропорций чере-
па происходило в эволюции человека неоднократно (рис. 10.2).
Противопоставленность большого пальца руки имела место
у Homo habilis и Homo erectus. Приблизительно такой же, как
у нас, волосяной покров был характерен для неандертальцев,
а возможно, и для более древних представителей рода Ното.
Во-вторых, все предполагаемые предки человека известны нам
только в форме ископаемых останков. Их возраст исчисляется
285
Шимпанзе
Australopithecus Australopithecus
afarensis africanus
Homo erectus
Homo sapiens
Рис. 10.2. Постепенное увеличение размеров мозга у высших приматов (аб-
солютные размеры черепов не отражены)
сотнями тысяч лет. В подобных случаях очень трудно использовать
геносистематику и, как правило, приходится ограничиваться
анатомическими признаками. Соответственно остается множество
нерешенных вопросов. Например, сколько видов относится к роду
Ното! Многие современные исследователи считают, что три:
Homo habilis, Homo erectus и Homo sapiens. Но другие ученые
придерживаются иной точки зрения и выделяют более десятка
самостоятельных видов, имеющих друг с другом определенные
анатомические различия (табл. 10.4).
Весьма показательна ситуация с неандертальцами. В настоящее
время существует три точки зрения: неандертальцы являлись
вполне самостоятельным видом, они были одной из разновид-
ностей Homo erectus и представляли собой раннюю форму Ното
sapiens. Анатомические данные не дают однозначного ответа.
Возможно, он будет получен в ближайшем будущем в результате
выполнения проекта «Геном неандертальца». Этот проект был
начат в 2006 г. на основе хорошо сохранившихся останков, возраст
которых «всего» 38 тыс. лет.
Для других ископаемых представителей рода Ното такие ге-
нетические исследования пока невозможны. В результате до сих
пор не существует единого мнения о том, кто относится,
а кто не относится к человеку разумному, и каковы отли-
чительные признаки данного вида.
Чаще всего отличительными признаками Homo sapiens счита-
ют способность к речи и наличие разума. Кратко остановимся
на каждой из этих особенностей.
286
Таблица 10.4. Виды, выделяемые в составе рода Ното
Вил Возраст остан- ков, млн лет Места обнаружения Рост*, м Масса*, кг Объем мозга, см3 Количество находок Год первого описания
Н. habilis 2,2-1,6 Африка 1...1,5 33-55 -660 Множество 1960
Н. rudolfensis 1,9 Кения — — — 1 1972
Н. georgicus 1,8 Грузия — — -600 Несколько 1999
Н. ergaster 1,9-1,4 Африка - 1,9 — 700...850 Множество 1975
Н. erecrus 1,4-0,2 Африка, Европа, Азия - 1,8 -60 850...1 100 Множество 1891
Н. antecessor 1,2-0,8 Испания - 1,75 -90 - 1000 2 1997
Н. cepranensis 0,9-0,8 Италия — — - 1000 1 1994
Н. heidelbergensis 0,6-0,35 Африка, Европа, Азия ' 1,8 - 60 1 100...1400 Множество 1908
Н. rhodesiensis 0,3-0,12 Замбия — — - 1300 Несколько 1921
Н. neanderthalensis 0,35-0,03 Европа, Азия -1,6 55-70 1200... 1900 Множество 1829
Н. sapiens 0,2 — настоя- щее время Все части света 1,4...1,9 50-100 1000...1850 Множество —
Н. floresiensis 0,1-0,012 Индонезия - 1 - 25 -400 7 2003
Для взрослых особей.
287
10.3. Генетический анализ способности
к устной речи. Роль гена FOXP2
Возникновение речи — одно из важнейших событий на всем
эволюционном пути становления человека. Оно позволило обме-
ниваться опытом, согласовывать действия, совместно анализировать
и обобщать полученные знания, накапливать их и передавать из
поколения в поколение. В результате мы можем выстраивать свое
поведение, опираясь далеко не только на собственный опыт.
Как известно, новорожденные дети говорить не умеют. Для
того чтобы у ребенка сформировалась полноценная речь, он дол-
жен пройти через длительный процесс обучения, успешность
которого зависит от множества факторов. В частности, развитие
речи сильно затруднено при отсутствии слуха или низком уровне
интеллекта. Необходимо отметить еще один важный нюанс: если
ребенок до 3-4-летнего возраста не имел речевого общения с
другими людьми, последующее обучение к успеху уже не приво-
дит. Таким образом, запуск процессов формирования речи воз-
можен лишь в первые годы жизни ребенка.
Даже при самом благоприятном стечении обстоятельств (нор-
мальный слух, высокий интеллект, своевременное обучение) не-
которые люди страдают дефектами речи. Это может выражаться
в различных нарушениях артикуляции, интонации, громкости,
плавности, связности, темпа и ряда других речевых характеристик
(табл. I0.5). Чаще всего подобного рода дефекты возникают в
результате физических или психических травм. Вместе с тем опре-
деленную роль играет и генотип.
Первые конкретные шаги в изучении этой роли были сделаны
в 1990 г. В поле зрения генетиков попала большая семья, в кото-
рой около половины всех ее членов несмотря на нормальных слух
и средний интеллект изъяснялись примитивными фразами, край-
не невнятно, с многочисленными нарушениями элементарных
правил грамматики. Причем перечисленные проблемы наследу-
ются моногенно, т.е. обусловлены одним единственным геном.
Через несколько лет этот ген удалось идентифицировать. Оказа-
лось, что он кодирует транскрипционный фактор, регулирующий
развитие широкого спектра органов, в том числе — кишечника,
легких и головного мозга. Данный ген получил обозначение
FOXP2. У каждого из плохо говорящих членов семьи одна из двух
копий гена является мутантной. Это приводит к недостаточному
развитию базальных ядер, зоны Брока и некоторых других райо-
нов мозга, принимающих участие в иннервации речевого аппа-
рата. Соответственно люди с мутацией в гене FOXP2 плохо
владеют мускулатурой нижней части лица и потому не способны
288
Таблица 10.5. Разнообразие дефектов речи у человека
Тип нарушений Проявление
Ал алия Полное отсутствие или недоразвитость речи в ре- зультате локальных поражений головного мозга на ранних этапах индивидуального развития
Диел алия Замена одних звуков другими (картавость, шепе- лявость и т. п.) Пропуски отдельных звуков
Дизартрия Невнятная речь («каша во рту»)
Брад ил алия Чрезмерно замедленная речь
Тахилалия Чрезмерно ускоренная речь
Заикание Судорожные нарушения темпа и ритма речи
Логоклония Многократное повторение отдельных слогов слова
Олигофазия Бедная речь с малым запасом слов
Парафазия Неправильный порядок слов в предложении
Акатофазия Замена слов сходными по звучанию, но не подхо- дящими по смыслу
Шизофазия Бессмысленный набор слов, облеченный в грам- матически правильно построенные предложения
Логорея Неудержимая быстрая и многословная речь
Афазия Полная или частичная утрата речи в результате локальных поражений головного мозга
к четкой артикуляции. Испытывая серьезные трудности с выго-
вариванием слов, такие люди уже не обращают внимания на
грамматику и стараются использовать самые короткие предложе-
ния, зачастую прибегая к всевозможным поясняющим жестам.
Белок FOXP2 поразительно консервативен. При крупном
размере (более 700 аминокислот) он различается у ящерицы,
попугая, свиньи и шимпанзе всего лишь полутора десятками
аминокислотных замен. Среди приматов его разнообразие совсем
невелико. У павиана, макаки, гиббона, шимпанзе и гориллы этот
белок практически одинаков. Казалось бы, он должен быть таким
же и у человека.
Однако реальная ситуация совершенно иная. Человек, не-
смотря на близкое родство с обезьянами, отличается от них
структурой белка FOXP2 в большей степени, чем обезьяны от
289
Таблица 10.6. Число аминокислотных замен в белке FOXP2 у разных позвоночных при сравнении
с шимпанзе (по Goodman et al., 2005; Li et al., 2007; Benton and Donoghue, 2007; с дополнениями)
Класс Отряд Объект Число замен* Приблизительное время эволюционного расхождения предков, млн лет
Земноводные Бесхвостые Шпорцевая лягушка 29 340
Пресмыкающиеся Чешуйчатые Ящерица анолис 8 320
Черепахи Красноухая черепаха 5 320
Птицы Попугаеобразные Волнистый попугайчик 5 320
Воробьи нообразн ые Зебровая амадина 4 320
Млекопитающие Насекомоядные Азиатская водяная землеройка 3 100
Обыкновенная бурозубка 2 100
Парнокоп ытные Свинья 3 100
Коза 1 100
Хищные Свиной барсук 3 100
Кошка 1 100
Непарнокопытные Осел 1 100
Хоботные Африканский слон 1 100
Неполнозубые Девятипоясный броненосец 0 100
Грызуны Серая крыса 2 85
Домовая мышь 1 85
Зайцеобразные Кролик 1 85
Приматы Серый мышиный лемур 1 70
Павиан анубис 0 28
Макака резус 0 28
Белорукий гиббон 0 14
Орангутан 1 10
Горилла 0 8
Человек разумный 2 6
Карликовый шимпанзе 0 < 1
* Размер белка FOXP2 — около 715 аминокислот. У разных видов он незначительно варьирует за счет изменения длины моно-
тонных участков, состоящих исключительно из остатков глутамина. На функциях белка это не отражается.
броненосца или слона (представителей достаточно древних от-
рядов животных; табл. 10.6).
Обнаруженные различия, скорее всего, неслучайны. Они по-
зволили человеку, по сравнению с другими приматами, значи-
тельно тоньше управлять мускулатурой лица и тем самым из-
давать очень сложные акустические сигналы, без чего возник-
новение речи было бы невозможным.
Способность использовать сложные акустические сигналы
характерна и для некоторых других млекопитающих. В этом
плане особенно интересны летучие мыши. Они ориентируются
в пространстве с помощью эхолокации, что требует, с одной
стороны, тончайшего слуха, а с другой — соответствующего
развития голосового аппарата. Установлено, что разные виды
летучих мышей резко различаются по спектру издаваемых звуков.
Возникает закономерный вопрос: не связано ли это с изменения-
ми структуры интересующего нас белка? И действительно, его
разнообразие у летучих мышей намного выше, чем у всех осталь-
ных млекопитающих. Так, у бесхвостого листоноса и ночницы
Рикетта он различается двадцатью четырьмя аминокислотными
заменами (гораздо сильнее, чем у ящерицы и шимпанзе).
Интереснейшие данные получены при исследовании мышей.
У этого объекта с помощью генной инженерии заменили его ген
FOXP2 человеческим гомологом. Разумеется, подопытные жи-
вотные не начали говорить. Но тембр издаваемых звуков у них
изменился, подтверждая влияние гена на голосовой аппарат.
Люди, несущие мутацию в гене FOXP2, отличаются от
нормы не только невнятностью речи. Они испытывают
определенные трудности при письме и плохо понимают слож-
ные лингвистические конструкции, например, сложноподчи-
ненные предложения. Таким образам, некоторые когнитивные
аспекты речи тоже находятся под контролем данного гена.
10.4. Только ли человек способен
пользоваться речью?
Еще недавно, каких-нибудь 40 лет назад, ответом на вопрос,
вынесенный в заголовок, было твердое «да». Считалось, что глав-
ную роль в возникновении речи сыграли социальные факторы, и
в частности — труд. Биологические факторы практически полно-
стью исключались. В связи с этим любые, даже самые робкие идеи
о наличии каких-то зачатков речи у животных казались абсолют-
но нелепыми и антинаучными. Способность говорить признава-
ли исключительно за человеком.
292
Первые обоснованные сомнения в справедливости этого взгля-
да были высказаны только в конце 1960-х гг. Американские ис-
следователи Алан и Беатрис Гарднеры попытались обучить
10-месячную самку шимпанзе элементам амслена — упрощен-
ного языка глухонемых. За три года обезьяна освоила 130 знаков,
научилась складывать из них короткие фразы и вступала с людь-
ми в полноценные двухсторонние отношения: комментировала
чужие и собственные действия, отвечала на вопросы, сама их за-
давала, поправляла себя при ошибках, шутила, ругалась. Она
четко различала знаки «я», «ты», «мой», «твой» и правильно рас-
ставляла их в зависимости от смысла фразы. Например, обраща-
ясь к человеку с какой-либо просьбой, она в 90 % случаев стави-
ла знак «ты» перед знаком «мне» («ты дать мне»), а предлагая
предмет, — наоборот («я дать тебе»).
Поначалу отношение к этим фактам было крайне скептиче-
ским. Однако по мере выполнения аналогичных исследований
стало ясно, что выводы Гарднеров — не мистификация. Более
того, способность осваивать амслен была экспериментально до-
казана и для горилл.
Кратко суммируем наиболее важные результаты. В многочис-
ленных работах показано, что гориллы и шимпанзе (если не все,
то хотя бы некоторые из них) способны:
• осваивать элементы жестового языка, включая имена людей,
названия предметов, глаголы, характеристики размера, цвета и
вкуса, местоимения («я», «ты», «мой», «твой», «этот», «тот»),
оценки («хорошо», «плохо», «неправильно», «жаль»), указания
места («вверх», «вниз», «отсюда», «внутрь»), обозначения эмо-
циональных состояний («смешно», «страшно», «больно»), числи-
тельные в пределах 10, сравнительные наречия («скорее», «еще»),
отрицания («нет»), этикетные междометия («пожалуйста», «спа-
сибо») общим объемом до 150 знаков;
• использовать эти знаки в соответствии с правилами синтак-
сиса;
• обобщать, применяя усвоенные знаки к незнакомым объ-
ектам этой же категории;
• создавать новые осмысленные комбинации знаков;
• употреблять некоторые знаки в переносном смысле (напри-
мер, «грязный» в значении «плохой»);
• общаться с помощью знаков с другими обезьянами;
• комментировать собственные действия себе самому.
Обезьяны успешно осваивают и другие языки. Они могут об-
щаться с человеком посредством компьютера, нажимая на кла-
виши с абстрактными графическими символами, каждый из ко-
торых соответствует определенному слову. Результаты примерно
такие же, как и с амсленом. Наконец, некоторые человекообраз-
293
ные обезьяны, находящиеся с самого рождения среди людей,
спонтанно, т.е. в отсутствие целенаправленных поощрений, на-
чинают понимать устную человеческую речь. Рекордсмен — кар-
ликовый шимпанзе по имени Канзи. Он освоил лексикон из
нескольких сотен слов и хорошо понимает разнообразные их
комбинации с учетом особенностей синтаксиса английского язы-
ка. В частности, он различает такие фразы, как «налей кока-колу
в лимонад» и «налей лимонад в кока-колу» или «вынеси морков-
ку на улицу» и «выйди на улицу и найди там морковку».
Получив 600 неповторяющихся устных заданий в ходе специ-
ально разработанного многодневного тестирования, Канзи спра-
вился с 81 % из них. Между тем успехи двухлетней девочки Али
(дочери одного из участвовавших в исследовании сотрудников) были
заметно скромнее — 64%. Таким образом, по степени понимания
устной речи обезьяна ничуть не уступала двухлетнему ребенку.
Даже у самых смышленых горилл и шимпанзе способность
использовать знаки в процессе общения весьма далека от уровня
взрослого человека. Поэтому некоторые ученые продолжают счи-
тать, что обезьяны только имитируют человеческую речь, заучивая
те или иные комбинации действий. Ситуация осложняется тем,
что в современной лингвистике существуют различные толкова-
ния понятия «речь», от очень широких («передача информации с
помощью знаков») до очень узких, касаться которых мы не станем.
Общение обезьян посредством компьютера или жестов под не-
которые толкования подпадает, а под некоторые — нет.
Вопрос о том, какая формулировка наиболее адекватна, ре-
шать, безусловно, не нам, а профессиональным лингвистам. Тем
не менее вполне очевидно, что некоторые способности, еще
недавно приписываемые только человеку (например, способ-
ность использовать правила синтаксиса), в зачаточной
форме присутствуют и у обезьян. В этом плане различия
между человеком и обезьянами, по-видимому, носят не ка-
чественный, а количественный характер.
10.5. Только ли человек способен
к разумному поведению?
Несмотря на широкое использование в научной литературе,
понятие «разум» пока не имеет четкого определения. Разные ис-
следователи трактуют его по-разному. Предлагаемые при этом
формулировки настолько расплывчаты, что оперировать ими, увы,
практически невозможно (табл. Ю.7).
Для того чтобы ответить на поставленный в заголовке вопрос,
необходимо обратиться к элементарным проявлениям разума и
294
выяснить, встречаются ли они среди животных. К таким прояв-
лениям разума обычно относят:
• способность узнавать себя в зеркале, телевизоре и др. Этой
способностью обладают и шимпанзе, и гориллы. Некоторые даже
узнают свои фотографии и откладывают их в ту же стопку, что и
человеческие, причисляя себя не к животным, а именно к людям;
• способность изготовлять и использовать простейшие
орудия. Например, шимпанзе, гориллы и орангутаны могут до-
ставать удаленные от них предметы (пищу, любимую игрушку и
др.) с помощью различного рода подручных средств, в том числе
стыкуя друг с другом короткие палки. Карликовый шимпанзе
Канзи, наблюдая за человеком, научился изготовлять простейшие
каменные ножи и использовать их для разрезания веревок. Более
того, он придумал «собственную технологию»: не бить камнем по
камню, как ему показали, а бросать их на что-нибудь твердое,
пока не расколются;
• элементы логического мышления. Одной из важнейших
операций логического вывода является так называемое транзи-
ентное заключение (если А связано с В, а В — с С, из этого
следует, что А связано с С). К такой операции способны не толь-
ко обезьяны, но и многие другие млекопитающие, включая крыс
(см. подразд. 4.6);
• способность находить решение путем инсайта (мыслен-
ной оценки результатов возможных действий с получением «го-
тового» решения без предварительных проб). Это свойство харак-
терно для всех человекообразных обезьян. Например, при наличии
Таблица 10.7. Примеры определений понятия «разум»
Определение Источник
Дискурсивная способность души, которая продвигается от известного к неизвестному и выводит одно из другого в четкой и правильной последовательности суждений Локк, 1988. - С. 21
Всеобщий и спокойный аффект, который ис- ходит из всестороннего и осуществляемого издали рассмотрения объекта и побуждает волю, не возбуждая какой-либо чувственной эмоции Юм, 1996. - С. 171
Способность, дающая нам принципы априорно- го знания Кант, 1994. - С. 44
Наивысшая ступень мышления Гегель, 1972. - С. 23
Способность понимания и осмысления БЭС, 2000. - С. 993.
295
нескольких разных палок обезьяна использует не любую, а имен-
но ту, что подходит по длине, толщине, прочности и др. Шим-
панзе успешно справляются с различными задачами наподобие
компьютерных игр «выйди из лабиринта». Это значит, что они
способны анализировать ситуацию, планировать действия с уче-
том промежуточных результатов, выбирать оптимальный план из
нескольких возможных, следовать ему и корректировать по мере
необходимости;
• способность к обобщению. Обезьяны, обученные тому или
иному языку, могут спонтанно переносить знакомые им знаки на
другие объекты этого же класса. В частности, самка шимпанзе по
имени Уошо изначально использовала знак «открыть» для ком-
натных дверей, но в дальнейшем самостоятельно стала его при-
менять к холодильнику, буфету, различного рода контейнерам,
коробкам, бутылкам и даже к водопроводному крану;
• передача приобретенных навыков из поколения в поко-
ление. Общаясь со своими более взрослыми сородичами, молодые
обезьяны постепенно перенимают их навыки. Например, самец
шимпанзе по имени Лулис научился «говорить на амслене» не-
посредственно у Уошо. За 5 лет он освоил около 50 знаков.
В основном обучение шло путем подражания и не требовало ак-
тивных вмешательств со стороны Уошо. Но иногда такие вмеша-
тельства все же происходили. Однажды, получив тарелку с овся-
ной кашей, Уошо показала Лулису знак «пища» (сложила пальцы
щепоткой и поднесла их ко рту), а затем проделала то же самое с
его собственными пальцами. В результате Лулис быстро освоил
этот знак. Подобного рода ситуации не единичны и, по сути,
являются зачатками культурной преемственности.
Итак, основные элементы разумного поведения наблюда-
ются в зачаточной форме и у высших обезьян. Безусловно, не
все исследователи с этим согласны. Кто-то продолжает использо-
вать классическую аргументацию «этого не может быть, потому
что этого не может быть никогда». Однако постепенно накапли-
вается все больше фактов, убедительно свидетельствующих
о том, что психика человека отличается от психики обезьян
не качественно, а количественно.
10.6. Современные представления
об эволюции человека
Около 20 млн лет назад существовала особая группа обезьян —
дриопитеки. Они отличались отсутствием хвоста, широкой груд-
ной клеткой, высокой подвижностью запястий и плечевого пояса,
296
небольшими челюстями и крупным головным мозгом. Дриопи-
теки вели полудревесный образ жизни и использовали при пере-
движении все четыре конечности. Широко расселившись по
Африке, Европе и Азии, они дали начало нескольким эволюци-
онным ветвям (см. рис. 10.3), одна из которых привела к возник-
новению Homo sapiens.
Как именно это происходило, мы, конечно, не знаем, но общая
картина уже начинает просматриваться. Основные события раз-
ворачивались в Восточной Африке. В результате очередного гло-
бального изменения климата площадь тропических лесов значи-
тельно сократилась, а площадь саванн, напротив, существенно
возросла. В связи с этим возникли новые формы дриопитеков —
перешедшие к наземному обитанию в условиях саванн.
В саванне главную опасность для любого примата представляют
крупные кошачьи (леопарды и львы), активные преимущественно
в ночное время суток. Чтобы снизить вероятность столкновения с
этими хищниками, дриопитекам пришлось освоить дневной образ
жизни. Но днем в африканской саванне очень жарко. Дриопитеки
подверглись жесткому естественному отбору в пользу особей,
успешно справлявшихся с угрозой перегрева. Этот отбор привел,
во-первых, к прямохождению (узкое, вертикально расположенное
тело в наименьшей степени нагревается солнечными лучами), а во-
вторых, к утрате сплошного волосяного покрова и значительному
увеличению количества потовых желез (резко повысилась эффек-
тивность терморегуляции).
Прямохождение не только уменьшило опасность перегрева, но
и открыло широкие возможности для использования рук. Даль-
нейшие этапы эволюции к современному человеку были связаны
с увеличением размера головного мозга, усилением когнитивных
способностей и развитием речи (не имея ни острых когтей, ни
крупных клыков, ни какой бы то ни было другой надежной за-
щиты от смертельно опасных хищников и конкурентов, наши
предки «пошли по пути» повышения интеллекта).
За каждым из перечисленных нами эволюционных шагов
стояли случайно возникшие генетические изменения,
оказавшиеся полезными для данного направления эволюции и
потому подхваченные естественным отбором. Кратко рассмотрим
наиболее вероятные из них.
• Переход предков человека к прямохождению. Некоторые
особи человекообразных обезьян могут передвигаться, не опира-
ясь на передние конечности. В этом плане особенно интересен
шимпанзе Оливер: он ходит практически так же, как человек.
Причины его прямохождения пока неизвестны. Вероятнее всего,
они связаны с некими мутациями, повышающими устойчивость
вертикального положения тела. У человека известны мутации с
297
обратным эффектом. Например, для мутантов по гену VLDLR
(этот ген участвует в развитии головного мозга) характерны тя-
желые психические и физические аномалии, в том числе — не-
способность полностью выпрямить колени и удерживать тело в
вертикальном положении. Такие люди могут передвигаться толь-
ко на четвереньках, напоминая тем самым наших далеких предков.
Поэтому мы вправе предположить, что ген VLDLR сыграл в эво-
люции прямохождения заметную роль.
• Утрата сплошного волосяного покрова. У многих млеко-
питающих (кошек, собак, лошадей, морских свинок, тушканчиков,
крыс, хомяков и др.) известны мутации, приводящие к отсутствию
шерсти. Более того, у целого ряда грызунов этот признак закре-
пился в качестве видоспецифичного. Что-то подобное произошло
и в эволюции человека. Однако волосяной покров у человека был
утрачен не полностью. Он сохранился хотя бы на некоторых участ-
ках тела (в основном на обращенной непосредственно к солнцу
голове) и в редких случаях может восстанавливаться.
• Значительное увеличение размера головного мозга. Пред-
полагают, что главными «виновниками» этого процесса являлись
гены MCPHL ASPM. CDK5RAP2, CENPJ и STIL (табл. 10.8).
Люди, мутантные по любому из перечисленных генов, отличают-
ся резко уменьшенным объемом мозга при нормальных размерах
лицевого отдела черепа. Особенно сильно затронуты большие
Таблица 10.8. Гены, контролирующие размер головного мозга
у человека
Ген Кодируемый белок Молекулярные функции Результат мутации
МСРН1 Микро- цефалии 1 Поддержание нормальной структуры ДНК Окружность череп- ной коробки как минимум в 2 раза меньше по сравне- нию с нормой. Резко уменьшен объем го- ловного мозга, осо- бенно его больших полушарий. Других аномалий развития мозга нет. Размеры лицевой части че- репа соответствуют норме
ASPM Микро- цефалии 5 Регуляция процесса расхождения хромосом
CENPJ Белок J, ассо- циированный с центромерами
ST1L Микро- цефалии 7 Регуляция кле- точных делений
CDK5RAP2 Регуляторная субъединица циклинзави- симой протеин- киназы 5 Регуляция диф- ференцировки нейронов
298
полушария (иногда они почти такие же, как у шимпанзе). Доба-
вим, что данная патология обычно соПровождаеТся тяжелыми
формами задержки умственного развитНя> но не ведет к каким.то
другим психическим отклонениям. Поэтому вполне вероятно, что
указанные гены имеют непосредственнее отношение и к следую-
щему генетическому изменению.
• Резкое усиление когнитивных способностей. У всех совре-
менных человекообразных обезьян обнаРужены зачаточные формы
разумного поведения. Вряд ли у каждого ВИда они возникли неза-
висимо. Скорее всего, определенные зачЦтки разума имедись еще у
ранних гоминид, а затем по мере эволюционного становления че-
ловека постепенно расширялись. углуб.;(ялись и совершенствова-
лись. В этом процессе могли быть задействованы многие гены,
контролирующие развитие и функционийование нервной системы,
в том числе и ответственные за размер г<)ловного МОзга
Возникновение устной речи. Достаточно высокий уровень
когнитивных способностей, позволяющий ОСВОить элементарные
знаковые системы, характерен для всех Человекообразных обезьян.
Например, им доступны простейшие хитовые языки. Обезьяны,
с раннего возраста воспитываемые людьми, постепенно привы-
кают к устной человеческой речи и мОгут понимать ее не хуже
двухлетнего ребенка. Однако примитивное строение голосового
аппарата не дает им возможности тонко артикулировать, а значит,
самим использовать устную речь. Такац возможность появилась
у предков человека в результате «оптимизации» целого ряда генов
и в первую очередь - гена FOXP2 (сц. подразд. 10.3). Вполне
человеческая версия данного гена былп характерна еще для не-
андертальцев. Поэтому нельзя исключЦть, что и неандертальцы
уже имели хотя бы примитивную устную речь
Освоение человеком новых возможней шло постепенно. Оно
слагалось из множества отдельных «маленьких открытий», совер-
шаемых разными индивидами по ходу их жизни, а затем обобщест-
вляемых с помощью подражания или Печи. Накопленный опыт
передавался из поколения в поколение и тем самым закладывал
основы культурных традиций. Предполагают, что когнитивные воз-
можности Homo sapiens к настоящему времени реализованы еще
не полностью и в будущем могут быть з)|ачигельно расширены.
10.7. Что отличает человека от высших
обезьян?
Еще недавно в научных кругах не вызывало сомнений, что
главными отличиями Homo sapiens от црочих приматов являют-
299
ся его уникальные психические способности и в первую очередь —
наличие разума и речи. Этот взгляд принимали безоговорочно,
как аксиому. Однако его очевидность оказалась обманчивой. Дело
в том, что сами понятия «разум» и «речь» не имели (и сейчас не
имеют) четкого определения. Их рассматривали в качестве неких
человеческих свойств. В результате получался классический замк-
нутый круг: понятие «человек» объясняли через разум и речь,
а понятия «речь» и «разум» — через человека.
Попытки уйти от предвзятого отношения к проблеме появи-
лись только в конце XX в. Оказалось, что все составляющие
разума и речи, будь то логическое мышление, самосознание,
способность к планированию с учетом промежуточных результа-
тов, способность следовать элементарным правилам синтаксиса,
способность к обобщению или культурная преемственность, в за-
чаточной форме присущи и высшим обезьянам. Поэтому ни речь,
ни разум не могут считаться ключевыми отличиями человека от
прочих приматов.
В чем же тогда состоит специфика Homo sapiens? Однознач-
ного ответа пока, к сожалению, нет. Тем не менее становится все
более и более очевидным, что различия между человеком и дру-
гими приматами имеют не качественный, а лишь количественный
характер. Впервые этот взгляд был высказан Чарльзом Дарвином
и с тех пор получил многочисленные косвенные подтверждения.
В частности, десятки исследований свидетельствуют о том, что
психика взрослой человекообразной обезьяны вполне соответ-
ствует уровню двухлетнего ребенка. А если обезьяна с рождения
воспитывалась людьми, ее достижения могут быть существенно
выше.
Достаточно высокий уровень когнитивных способностей был
свойственен, по-видимому, еще дриопитекам. Современные шим-
панзе, гориллы и орангутаны ушли от него сравнительно недале-
ко, поскольку сохранили предковый образ жизни. Между тем
представители человеческой ветви эволюции были вынуждены
осваивать новую среду обитания, что потребовало от них весьма
значительных изменений как в строении организма, так и в по-
ведении.
В результате в геноме закрепился целый ряд мутаций, обеспе-
чивших дальнейшее увеличение головного мозга, резкое повы-
шение уровня когнитивных способностей, усложнение голосово-
го аппарата и т. п. Вероятно, именно эта совокупность мутаций и
является главной особенностью Homo sapiens.
Геномы человека разумного и шимпанзе совпадают прибли-
зительно на 97 %. Причем подавляющее большинство обнару-
женных различий не затрагивает функционально важные области
генов. В связи с этим возникает вполне закономерный вопрос:
300
неужели процесс эволюционного становления человека был
основан на столь «незначительных» генетических изменениях?
Как известно, даже одна единственная мутация, если она за-
трагивает важный ген, способна привести к весьма ощутимым
последствиям, а если подобных мутаций накапливается несколь-
ко, их совместные эффекты могут быть очень серьезными. Для
примера достаточно обратиться к домашним собакам. Все они
относятся к одному и тому же виду и имеют высокую степень
генетического сходства — около 99,5 %. Тем не менее для них
характерно огромное разнообразие как по внешнему облику, так
и по особенностям поведения (табл. 10.9).
Отметим, что это разнообразие было получено всего за каких-
нибудь 20 тыс. лет — крохотный период времени с точки зрения
эволюции.
Таблица 10.9. Особенности поведения некоторых пород до-
машних собак (по Фогл. 2004)
Породы Особенности поведения
Возбуди- мость* Дрессируе- мость Агрессив- ность**
Английский бульдог, бобтейл, бладхаунд, бассет Очень низкая Низкая Очень низкая
Самоед, сенбернар, боксер, далматин, чау-чау Низкая Низкая Высокая
Лабрадор-ретривер, венгерская легавая, ньюфаундленд, колли Низкая Высокая Низкая
Немецкая овчарка, доберман, ротвейлер Очень низкая Очень высокая Очень высокая
Коккер-спаниель, пекинес, гончая, ирландский сеттер Высокая Низкая Средняя
Шелти, пудели, спрингер- спаниель Высокая Очень высокая Средняя
Чихуахуа, фокстерьер, такса, скотч-терьер Высокая Средняя Очень высокая
* Общая активность, склонность кусать детей, неумеренный лай, потреб-
ность в любви.
** Способность охранять территорию, сторожевой лай, агрессивность к дру-
гим собакам.
301
С момента расхождения предков шимпанзе и человека прошло
уже около 6 млн лет. Неудивительно, что степень различий меж-
ду этими видами млекопитающих в конечном итоге оказалась
настолько существенной, но кардинальных различий, по-види-
мому, так и не возникло.
10.8. «Белые пятна»
Успехи современной биологии трудно переоценить, однако
впереди еще много нерешенных вопросов, имеющих непосред-
ственное отношение к человеческой психике. Кратко остановим-
ся на наиболее важных.
До сих пор неизвестны механизмы формирования памяти.
Мы знаем, что данный процесс идет поэтапно: сначала возника-
ют нестабильные виды памяти, и лишь затем при соблюдении
целого ряда условий сохраненная информация может консолиди-
роваться.
На первом этапе одну из центральных ролей играют молеку-
лярные каскады с участием цАМФ, на втором образуются новые
межнейронные контакты (см. подразд. 4.5 и 4.17). Но как именно
осуществляется запись и хранение информации и что служит ее
носителем, пока непонятно. То же самое касается и механизмов
реализации памяти.
Еще более сложный вопрос — что такое мысль. На протяжении
долгого времени было принято считать, что мысль по своей при-
роде нематериальна и потому нс может быть предметом биологи-
ческих исследований. Причина подобного взгляда вполне очевид-
на: до сих пор не существует ни одного прибора, позволяющего
«прочитать» хотя бы простейшую мысль. Но это не значит, что
такие приборы создать невозможно. И если когда-нибудь их,
действительно, сконструируют, отношение к проблеме придется
в корне менять.
Все более весомую роль в современной науке играют стреми-
тельно развивающиеся компьютерные технологии. Они показы-
вают, что некие аналоги мыслительных процессов (пускай и весь-
ма примитивные по сравнению с человеческими) могут быть
созданы на сугубо материальной основе. Кроме того, разработаны
специальные устройства, с помощью которых, не подключаясь к
конкретному компьютеру, можно выяснить, над чем он «думает»
в данный момент. По сути, это очень похоже на чтение мыслей.
Таким образом, по крайней мере для компьютерной аналогии
доказать материальность мысли уже удалось.
Многие люди считают, что человеческое мышление обуслов-
лено действием некой «психической энергии». Такие взгляды
302
особенно популярны среди индивидов, выдающих себя за раз-
личного рода целителей, экстрасенсов, магов, колдунов, ворожей
и др. Подавляющее большинство из них — типичные шарлатаны.
Но если психическая энергия все же существует и через некоторое
время будет обнаружена, не вызовет ли это крушения современ-
ной науки?
«Энергия» — одно из важнейших научных понятий. Оно озна-
чает способность совершать какую-либо работу, например: пере-
мещать тела, воздействовать на заряды, разрывать химические
связи, изменять температуру. Причины такой способности могут
быть разными. Соответственно выделяют различные виды энер-
гии, в том числе механическуюу химическую, электромагнит-
ную, тепловую и ядерную (табл. 10.10). Каждый из них при со-
блюдении определенных условий может переходить в другие (табл.
10.11), но суммарное количество энергии в изолированной систе-
ме при любых ее преобразованиях остается неизменным. Это
значит, что энергия не может возникать ниоткуда или, напротив,
бесследно исчезать.
Таблица 10.10. Некоторые виды энергии
Вил энергии Его проявления
в неживых объектах в живых существах
Механическая Движение планет, сход снежной лавины, порывы ветра, морской прибой Полет птицы, сужение зрачка, кровообраще- ние. вибрация голосовых связок
Химическая Горение торфа, коррозия металла, взрыв метана в угольном пласте Фотосинтез, пищеваре- ние, репликация ДНК, транскрипция
Электромаг- нитная Молния, магнитное поле Земли, солнечный свет Разряды электрического ската, нервный импульс, свечение глубоководных рыб
Тепловая Тепловое излучение Солнца, броуновское движение частиц Теплокровность мле- копитающих и птиц, диффузия нейромедиато- ра через синаптическую щель
Ядерная Радиоактивность урана и стронция, термоядерные реакции на Солнце Неизвестны
303
Таблица 10.11. Примеры преобразования энергии
Вид преобразования Его примеры
в неживых объектах в организме человека
Химическая -> механическая Взрыв метана Изменение конформации рецептора под действием химического сигнала
Химическая -> электромагнитная Работа щелочной батарейки Формирование потенциала покоя на мем- бране нейрона
Химическая -> тепловая Горение угля Использование энергии АТФ для поддержа- ния температуры тела
Электромагнитная -> механическая Работа электромотора Изменение конформации потенциалзависи- мых ионных каналов
Электромагнитная —► химическая Зарядка аккумулятора от сети Усиление биосинтеза меланинов в коже под действием солнечного света
Электромагнитная -► тепловая Работа микроволновой печи Неизбежные потери тепла при проведении нервных импульсов
Механическая -> химическая Активация химической грелки при ее деформации Изменение химических свойств молекулы белка при изменении ее конформации
Механическая —► электромагнитная Генерирование электрического тока на гидроэлектростанции Формирование нервного импульса при активации механорецепторов
Механическая —► тепловая Падение метеорита Согревание организма при частых сокраще- ниях мышц
Тепловая —> химическая Самовозгорание торфа в жар- кую погоду Ускорение обмена веществ при повышении температуры
Тепловая -> электромагнитная Генерирование электрического тока на теплоэлектростанции Формирование нервного импульса при активации терморецепторов
Тепловая —► механическая Извержение вулкана Изменение конформации терморецепторов при повышении температуры
§
Данный закон (закон сохранения энергии) был сформулирован
еще в середине XIX в. В то время ученые знали и умели измерять
лишь две разновидности энергии: тепловую и механическую.
Поэтому закон распространялся только на них. Однако в даль-
нейшем выяснилось, что в некоторых случаях, например под
действием электромагнитного поля или в процессе химической
реакции, закон сохранения энергии не выполняется. Причем при
одних и тех же условиях эксперимента наблюдается строго опре-
деленное нарушение закона. Стало ясно, что кроме тепловой и
механической энергии существуют и некие другие ее разновид-
ности. Именно это позволило ученым постепенно обнаружить
другие известные ныне виды энергии и подробно исследовать
закономерности их преобразования.
Если психическая энергия действительно существует (этого
мнения придерживался Б. Г. Ананьев), рано или поздно ее ждет
такая же судьба. Будут созданы специальные приборы для ее из-
мерения, выявлены количественные закономерности ее преоб-
разования в тепловую, химическую и электромагнитную энергию
и наоборот, будут найдены ключевые молекулярные системы
организма, принимающие непосредственное участие в этих пре-
образованиях, и выявлены гены, контролирующие данные систе-
мы.
Таким образом, если психическую энергию обнаружат (при
желании ее могут называть и по-другому: «биополем», «аурой»,
«духом», «душой» и т.п.), это не внесет никаких существенных
изменений в суть представлений о материальности мыслитель-
ных процессов и о роли генетической информации в их фор-
мировании. Но пока говорить о наличии психической энергии
преждевременно.
СПИСОК ЛИТЕРАТУРЫ
I. Ананьев Б. Г. Человек как предмет познания. — СПб. : Питер, 2001.
2. Выготский Л. С. Мышление и речь. — М. : Лабиринт, 1999.
3. Зорина ЗА., Смирнова А. А. О чем рассказали «говорящие» обезьяны:
Способны ли высшие животные оперировать символами? — М. : Языки
славянских культур, 2006.
4. Ладыгина-Котс Н.Н. Предпосылки человеческого мышления. — М.:
Наука, 1965.
5. Ламберт Д. Доисторический человек. Кембриджский путеводитель. —
Л. : Недра, 1991.
6. Леонтьев А. Н. Проблемы развития психики. — М. : Наука, 1981.
7. Мэрион Д. Б. Физика и физический мир. — М. : Мир, 1975.
8. Панов Е.Н. Знаки, символы, языки. — М. : Товарищество научных
изд-в КМ К, 2005.
306
9. Резникова Ж. И. Современные подходы к изучению языкового поведе-
ния животных : в кн.: Разумное поведение и язык. Вып. I / сост. А. Д. Кошелев.
Т. В. Черниговская. — М. : Языки славянских культур, 2008. — С. 293 — 336.
10. Фогл Б. О чем думает собака. — М. : Астрель, 2004.
II. Черниговская Т.В. Что делает нас людьми: почему непременно ре-
курсивные правила? (взгляд лингвиста и биолога) : в кн.: Разумное поведе-
ние и язык. Вып. 1 / сост. А.Д. Кошелев, Т. В. Черниговская. — М. : Языки
славянских культур, 2008. — С. 395 — 412.
12. Brudno М. et al. Automated whole-genome multiple alignment of rat,
mouse, and human // Genome Research. — 2004. — V. 14. — P. 685 — 692.
13. Charles A. and Williams M.D. Macrocephaly syndromes // R.C. P. U.
Newsletters. — 2008. — V. 20. — No. 1/ — P. 1—4.
14. Ely J. J., Leland M., Martino M., Swett W. and Moore C.M. Technical
note: chromosomal and mtDNA analysis of Oliver // Am. J. Phys. Anthropol. —
1998. - V. 105. - P. 395-403
15. Enard W. et al. A humanized version of Foxp2 affects cortico-basal ganglia
circuits in mice 11 Cell. — 2009. — V. 137. — P. 961 —971.
16. Gardner R.A. and Gardner В. T. Teaching sign language to a chimpanzee //
Science. - 1969. - V. 165. - P. 664-672.
17. Kaindl A. M. et al. Many roads lead to primary autosomal recessive
microcephaly. Prog Neurobiol. — 2010. — V. 90. — P. 363 — 383.
18. Li G., Wang J., Rossiter S.J., Jones G.t Zhang S. Accelerated FoxP2
evolution in echolocating bats // PLoS ONE. — 2007. — V. 2. — Issue 9. —
e900. - P. 1-10. - doi: 10.1371.
19. Ozcelik T. et al. Mutations in the very low-density lipoprotein receptor
VLDLR cause cerebellar hypoplasia and quadrupedal locomotion in humans //
PNAS. - 2008. - V. 105. - P. 4232-4236.
20. Savage-Rumbaugh E.S., Shanker S. and Taylor T.J. Apes, Language and
the Human Mind // N. Y.: Wiley and Sons. Inc.. — 1994. — 299 p.
КОНТРОЛЬНЫЕ ВОПРОСЫ
1. Что отличает человека разумного от других высших приматов?
2. Какие факты свидетельствуют о роли гена FOXP2 в возникновении
устной речи?
3. Что такое в современном понимании разум?
4. Каковы современные представления о происхождении человека
разумного?
5. Если удастся обнаружить психическую энергию, приведет ли это к
крушению современной науки? Почему?
Глава 11
ЭТИЧЕСКИЕ АСПЕКТЫ ПСИХОГЕНЕТИКИ
11.1. Психогенетика и евгеника
Френсис Гальтон являлся родоначальником не только психо-
генетики. Он создал несколько новых научных направлений, одно
из которых получило название «евгеника*. Эта наука, по мнению
Гальтона, должна была способствовать улучшению наследствен-
ных качеств человеческой породы.
Аналогичные идеи неоднократно высказывались и раньше.
Еще Платон настаивал на государственном регулировании браков
и советовал, как подбирать идеальных супругов для получения
физически и нравственно выдающихся потомков. В Древней
Спарте селекцию людей осуществляли на практике, убивая боль-
ных и недостаточно развитых младенцев.
Ф. Гальтон придерживался более гуманной позиции. Он считал,
что нужна широкая просветительская работа, разъясняющая
основные положения эволюционной теории и возможности их
применения непосредственно к человеку. Человек должен сам
управлять своей эволюцией. Эта концепция вполне соответство-
вала духу времени и быстро нашла поддержку в развитых странах.
В частности, Совет английского евгенического общества пред-
лагал для улучшения расы следующие меры:
• тщательный подбор супругов по определенным критериям;
• контроль зачатий в семьях с низкими и посредственными
способностями;
• ограничение иммиграции для генетически неполноценных
людей;
• стерилизацию и сегрегацию безнадежно дефектных индиви-
дов.
В ряде стран евгенические идеи применялись на практике,
особенно активно в США. Так, в 1907 г. в штате Индиана был
принят первый закон о принудительной стерилизации. За этим
примером последовали еще 29 штатов. В большинстве из них до-
308
статочным показанием для принудительной стерилизации счита-
лись криминальные наклонности, хронический алкоголизм,
проституция, олигофрения и различные психозы. В 1924 г. Кон-
гресс США ввел резкие ограничения на иммиграцию евреев и
цыган, а также выходцев из стран Восточной и Южной Европы.
Впоследствии (с 1933 г.) аналогичные законы действовали и в
фашистской Германии. Представителей «неполноценных рас» не
только стерилизовали, но и использовали в изуверских медицин-
ских экспериментах, подвергали массовому истреблению в конц-
лагерях. В связи с этим, начиная с середины 1930-х г., евгениче-
ские идеи утратили исходную привлекательность и с тех пор
воспринимаются обществом крайне негативно.
Несмотря на свои очень близкие исторические корни, психо-
генетика пошла по совершенно иному пути: никогда не стреми-
лась усовершенствовать человеческую психику, не разделяла
людей на полноценных и неполноценных, не пыталась ограничить
размножение кого бы то ни было. Поэтому многочисленные пре-
тензии , предъявляемые к евгенике, никакого отношения к
психогенетике не имеют.
11.2. Этические аспекты современной
психогенетики
Существуют профессии, связанные с риском для окружающих.
Например, водитель автобуса или трамвая отвечает не только за
собственную жизнь, но и за жизнь других участников движения
(пассажиров, пешеходов, водителей, кондукторов и др.). Человек,
желающий получить такую профессию, должен соответствовать
достаточно жестким требованиям, в том числе и по ряду психо-
логических аспектов. Человек должен быть внимательным, с хо-
рошей реакцией, не терять самоконтроля в сложных стрессовых
ситуациях, не иметь наркотической или алкогольной зависимо-
сти.
Работодатель стремится к максимальному снижению рисков.
В связи с этим подробная информация о генотипе работника
представляет для него достаточно большой интерес. Но здесь воз-
никает серьезная правовая проблема. Генотип, если мы говорим
о психологических признаках, не является гарантией строго
определенного поведения. Он свидетельствует лишь о некой
врожденной предрасположенности. Поэтому отсев претенден-
тов по генетическим основаниям считается одним из ва-
риантов дискриминации и в некоторых странах юридически
запрещен.
309
Здесь отчетливо видны противоречия между обществом и ин-
дивидом. Люди, регулярно пользующиеся общественным транс-
портом, с точки зрения своей безопасности заинтересованы в
том, чтобы водители не имели нежелательных генетических пред-
расположенностей. Для индивида, пытающегося получить работу
водителя, напротив, важна презумпция его невиновности', пока
он здоров, он ничем не хуже остальных. Либеральные нормы от-
дают предпочтение индивиду. Таким образом, пока не появились
клинические симптомы, человек имеет право претендовать на
любую профессию даже при наличии нежелательной генетической
предрасположенности. Но это, естественно, приводит к опреде-
ленному риску.
Информация о генотипе интересна не только работодателям.
Конкуренты, желтая пресса и страховые компании с удовольстви-
ем использовали бы эти сведения в собственных целях. Поэтому
любые данные о генотипе индивида являются врачебной
тайной и огласке не подлежат. В некоторых развитых странах,
особенно в США, разглашение подобной информации уголовно
преследуется. Соответственно при опубликовании полученных
результатов сообщают лишь самые общие сведения об испытуе-
мых (возраст, пол, этническую принадлежность и др.) без каких
бы то ни было имен и адресов.
Еще одна проблема связана с результатами исследований. При
анализе генотипа родителей и их потомков могут выявиться дети,
зачатые вне брака. К сожалению, подобные случаи — вовсе не
редкость. По имеющимся оценкам, в США и Западной Европе в
среднем около 4% отцов воспитывают не своих биологических
детей, будучи абсолютно уверенными, что это их собственные.
И если исследование выявило внебрачного ребенка, полученная
информация должна сохраняться в тайне.
При анализе ряда психологических признаков наподобие ал-
когольной зависимости или анорексии широко используют па-
циентов соответствующих клиник. Но если характеристика не
имеет «медицинского оттенка», приходится набирать испытуемых
среди добровольцев. В этом случае возникает серьезная методи-
ческая проблема: соответствуют ли добровольцы поставленной
задаче? Допустим, группа исследователей заинтересовалась во-
просом, существует ли генетический контроль успеха в политике,
и обратилась к лидерам крупнейших мировых государств с пред-
ложением сдать образцы их ДНК. Трудно поверить, что эта прось-
ба будет удовлетворена. Безусловно, найдется достаточно много
добровольцев, но они не будут входить в мировую элиту и потому
окажутся непригодными в качестве испытуемых. Таким образом,
этот чрезвычайно интересный вопрос, вероятнее всего, никогда
не получит ответа.
310
11.3. Не принижает ли человека
молекулярная психогенетика?
Некоторые люди, узнав о существовании психогенетики, ис-
кренне удивляются, зачем она нужна. Для чего человеку вторгать-
ся в «святая святых», погружаться в сокровенные тайны челове-
ческой психики, выяснять тончайшие механизмы ее формирова-
ния? Это может привести к весьма нежелательным последствиям.
Во-первых, к возможности манипулировать чужим сознанием.
Во-вторых, к низведению человека до «обычного животного» и
тем самым — к разрушению морально-этических устоев.
Попытаемся кратко прокомментировать эти опасения. Безу-
словно, зная механизмы человеческой психики, можно научиться
манипулировать другими людьми. Однако не следует забывать, что
такие манипуляции получили широкое распространение в чело-
веческом обществе еще задолго до возникновения современной
психогенетики. Гипноз, наркотические средства, политические
технологии, различные виды рекламы и пиара способны оказы-
вать на психику значительное воздействие и активно использу-
ются в самых разных целях. Проблема манипуляции человеческим
сознанием непосредственного отношения к психогенетике не
имеет.
Теперь обратимся к другой проблеме. Для многих людей, осо-
бенно гуманитарного склада, присущи этика и мораль, лишь они
удерживают человека от «природного варварства». Если человек
поймет, что он «просто зверь», он не будет испытывать потреб-
ности ни в этике, ни в морали и в конечном итоге опустится до
скотского состояния.
Казалось бы, это опасение вполне обосновано. Однако даже
такие возвышенные качества, как любовь, гуманность, ответ-
ственность и самопожертвование, несомненно, имеют глубокие
биологические корни и характерны далеко не только для чело-
века (табл. ПЛ). Вместе с тем человеку необходимы и противо-
положные свойства, без которых невозможно отстаивать свои
жизненные интересы. Именно по этой причине в человеческой
натуре одновременно присутствуют как «зло», так и «добро».
Варьирует лишь соотношение между ними. А оно в значительной
мере определяется генотипом: большинство характеристик, от-
ражающих данное соотношение, имеют коэффициент наследуе-
мости не ниже 0,4 (см. подразд. 3.2, табл. 3.3). У кого-то преоб-
ладает «зло», у кого-то — «добро», но от степени биологической
грамотности это не зависит. Например, многие тысячи совре-
менных ученых, понимая, что они, по сути, молекулярные ма-
шины, не прекращают совершать хорошие, добрые поступки.
311
Таблица 11.1. Биологическое значение некоторых «человеческих» качеств (по Докинз, 1993; Дольник,
1994; Эфроимсон, 1995)
Качество Его проявление у человека Аналог у других животных Биологический смысл
Любовь Глубокая привязанность одного индивида к дру- гому, сопровождающаяся обычно ярко выражен- ным сексуальным влече- нием Для волков характерны прочные мо- ногамные отношения. Самец и самка, как правило, остаются друг с другом пожизненно Стабильные взаимовыгодные отношения между «супруга- ми» повышают выживаемость каждого из них и способствуют более эффективному уходу за потомством
Гуманность Способность проявлять жалость к побежденному Проигравший схватку волк в знак своего подчинения подставляет по- бедителю шею. Победитель получает возможность добить побежденного, но за редкими исключениями не пользу- ется ею Победитель экономит силы и время. Кроме того, добивая побежденного соперника, он рисковал бы получить опас- ные ранения (оказавшись в безвыходной ситуации, даже слабый соперник способен от страха проявить запредельную агрессию)
Самопо- жертвова- ние Способность пожертво- вать жизнью ради другого человека (как правило, родственника, соплемен- ника или единоверца) У многих видов муравьев имеются солдаты — бесплодные самки, обеспе- чивающие защиту всей семьи. Бес- страшно атакуя любого «нарушителя спокойствия», эти особи, как правило, оказывается смертниками Пожертвовав собой, организм повышает вероятность выжи- вания своих родственников. Тем самым он способствует воспроизведению хотя и не собственной, но все же очень близкой генетической инфор- мации
Ответствен- ность Способность выполнить взятые на себя обязатель- ства лаже в том случае, если они обременитель- ны Для павианов характерна четкая ие- рархия самцов. Лидер имеет неогра- ниченный доступ к самкам и пресе- кает попытки спаривания со стороны остальных. Подчиненные самцы иногда «договариваются» о взаимопо- мощи. Один отвлекает лидера на себя, а второй в это время спаривается с какой-то из самок. Через некоторое время они меняются своими ролями. Казалось бы, это невыгодно первому самцу. Однако если он не выполнит своих «обязательств», обманутый напарник не станет с ним больше со- трудничать Выполняя свои «обязатель- ства», организм демонстрирует, что он является надежным партнером и тем самым создает основу для нового сотрудни- чества
В то же время многие тысячи религиозных фанатиков, искренне
веря в свою «одухотворенность», с легкостью готовы пойти на
чудовищные зверства исключительно ради господства своей
идеологии.
Бояться современной психогенетики не стоит. Человек лишь
пытается познать самого себя, и ничего опасного или постыдно-
го в этом нет. Очень близкую задачу решает художественная ли-
тература: она тоже исследует «глубины человеческой души». Как
известно, получаемая картина — не самая приятная. Тем не менее
мы восхищаемся Достоевским и Гоголем, Шекспиром и Пушки-
ным, Мольером и Гете, Булгаковым и Эзопом, Салтыковым-
Щедриным и Сервантесом, Селинджером и Чеховым, гениально
показавшими всю сложность этих глубин.
Литература только описывает человеческую сущность, а совре-
менная паука начинает ее объяснять. Постепенно становится все
более и более понятным, почему «аппетит приходит во время еды»,
«сколько волка не корми, он все в лес смотрит», «повторение —
мать учения» и др. Все более и более подкрепляется народная
мудрость, что бесполезно пытаться «переделывать» близкого чело-
века: его нужно принимать таким, каков он есть, а иначе, скорее
всего, вы его потеряете. Следовательно, данные, полученные мо-
лекулярной психогенетикой, уже сегодня имеют определенное
прикладное значение, и со временем это значение будет все более
и более возрастать.
11.4. Самые распространенные
заблуждения по поводу психогенетики
Психогенетика настолько сложна, что едва ли доступна для
непрофессионала. В результате возникает большое число заблуж-
дений. Кратко прокомментируем несколько самых распростра-
ненных.
• История знает достаточно много примеров, когда под
влиянием каких-либо внешних обстоятельств характер че-
ловека существенным образом изменялся. Этот факт не-
редко считают «доказательством* того, что психические
особенности индивида от генов не зависят. Однако такая
интерпретация в корне неверна. Необходимо учитывать, что экс-
прессия многих генов, в том числе и обеспечивающих работу
нервной системы, в значительной степени регулируется гормона-
ми, а их содержание в конкретном организме непостоянно. Оно
меняется, во-первых, в процессе индивидуального развития (клас-
сические примеры — половое созревание и климакс), а во-вторых,
314
в различного рода стрессовых ситуациях. И действительно, по-
давляющее большинство изменений характера приурочено к
переходному возрасту, периоду беременности, родам, климаксу,
сильным эмоциональным потрясениям, т.е. к резким перестрой-
кам гормональной системы организма, неизбежно влияющим на
экспрессию многих генов.
• Из школьного курса биологии известно, что поведением
человека управляют нервные импульсы. Эти импульсы передают-
ся быстро, за доли секунды. Генетическая информация считыва-
ется гораздо медленнее. Соответственно, многие люди уверены
в том, что гены просто «не успевают* влиять на поведение,
а значит, оно не должно зависеть от генов. Этот вывод осно-
ван на двух принципиальных ошибках. Во-первых, не учитыва-
ется, что главные молекулярные компоненты, обеспечивающие
формирование и передачу нервных импульсов (рецепторы,
(7-белки, различные протеинкиназы, мембранные насосы, каналы
и др.), сами являются продуктами определенных генов и в зависи-
мости от генотипа работают по-разному. Во-вторых, в основе
функционирования нервной системы лежат далеко не только
нервные импульсы, но и значительно более медленные молеку-
лярные процессы, в том числе и изменение экспрессии некоторых
генов. Для примера достаточно вспомнить о рецепторах тирок-
сина, представляющих собой транскрипционные факторы (см.
подразд. 5.9 и 5.10).
• Нередко встречается и прямо противоположное мнение.
Соглашаясь, что поведение человека зависит от генов, мно-
гие считают, что эта зависимость полная, и каждый по-
ступок заранее предрешен генотипом. Причина — в неверном
понимании слова «зависимость». Обыватель, как правило, не
знает, что в современной науке это слово означает «наличие ста-
тистической связи» и отражает вероятность осуществления
определенных событий. Более того, он хочет получать от науки
только четкие и строго однозначные ответы, а потому к вероят-
ностным закономерностям относится с недоверием.
• Опираясь на безграмотные сообщения в желтой прессе,
многие люди твердо уверены в том, что существуют гены
алкоголизма, самоубийства и т.д. Однако в действительности
ничего подобного нет. Во-первых, любая характеристика, осо-
бенно психологическая, контролируется не одним единственным,
а множеством генов. Во-вторых, эти гены «отвечают» не за кон-
кретные действия, а за некие аспекты работы нервной системы
и лишь косвенно связаны с той или иной характеристикой, т.е.
имеются гены, влияющие на склонность к алкоголизму, склон-
ность к курению, склонность к самоубийству и др., но ни один
из генов не предназначен непосредственно для этого.
315
• Очень часто неверно истолковывают термин «насле-
дуемость*. Например, говоря о том, что наследуемость дислек-
сии составляет приблизительно 70%, многие люди считают, что
данное нарушение передается потомкам именно с такой вероят-
ностью. На самом же деле коэффициент наследуемости показы-
вает, насколько фенотипическое разнообразие по некоему при-
знаку определяется разнообразием людей по их генотипам. По-
нятие «наследуемость» не применимо к конкретным индивидам
с их конкретными генотипами и условиями внешней среды. Оно
используется только по отношению к признаку в целом.
Для того чтобы успешно преодолевать подобные заблуждения,
необходимо вести широкую просветительскую работу, до-
ступно., но грамотно отражающую современную психогене-
тику , ее задачи, методы и уже имеющиеся достижения.
Иначе в общественном сознании будут сформированы совершен-
но искаженные представления об этой науке, что может негатив-
но отразиться на перспективах ее развития.
СПИСОК ЛИТЕРАТУРЫ
1. Голимбет В.Е., Трубников В. И. Этические проблемы молекулярно-
генетической диагностики в психиатрии // Социальная и клиническая
психиатрия. — 19%. — Т. 6. — № 1. — С. 109— 112.
2. Докинз Р. Эгоистичный ген. — М. : Мир, 1993.
3. Дольник В. Непослушное дитя биосферы. — М. : Педагогика-Пресс,
1994.
4. Иванов В. И., Ижевская В.Л. Геномика и этика : в кн. : В. И. Иванов,
JI. JI. Киселев. Геномика — медицине. — М.: Академкнига, 2005. — С. 349 — 360.
5. Эфроимсон В. П. Генетика этики и эстетики. — СПб. : Талисман, 1995.
6. Lucassen A., Parker М. Revealing false paternity: some ethical consi-
derations // Lancet. — 2001. — V. 357. — P. 1033— 1035.
КОНТРОЛЬНЫЕ ВОПРОСЫ
1. Что такое евгеника? Как она возникла?
2. Как относится современное общество к евгенике? Почему?
3. Опишите этические аспекты современной психогенетики.
4. Принижает ли человека молекулярная психогенетика? Почему?
5. Опишите самые распространенные заблуждения по поводу психо-
генетики.
ОГЛАВЛЕНИЕ
Предисловие............................................................3
Глава 1. Основные положения молекулярной биологии......................4
1.1. Место биологии среди естественных наук............................4
1.2. Молекулярная специфика живого.....................................9
1.3. Важнейшие классы биоорганических соединений......................14
1.3.1. Углеводы....................................................14
1.3.2. Жиры........................................................16
1.3.3. Белки.......................................................18
1.3.4. Пространственная организация молекул белка..................21
1.3.5. Нуклеиновые кислоты.........................................24
1.4. Процесс реализации генетической информации.......................28
1.5. Молекулярные основы транскрипции.................................30
1.6. Молекулярные основы трансляции...................................32
1.7. Как формируются признаки организма...............................34
1.8. Дифференциальная экспрессия генов................................37
1.9. Молекулярные основы формирования биологических ответов...........39
1.10. Молекулярные основы возникновения и действия мутаций............42
1.11. Аллели гена.....................................................44
1.12. Генотип и фенотип...............................................47
Глава 2. Молекулярные основы функционирования нервной
системы...............................................................51
2.1. Основное назначение нервной системы..............................51
2.2. Типы строения нервной системы....................................52
2.3. Синапсы..........................................................56
2.4. Функциональная специализация нейронов............................59
2.5. Молекулярные основы нервного импульса............................64
2.6. Молекулярные механизмы формирования нервного импульса............67
2.7. Молекулярные механизмы проведения нервного импульса..............69
2.8. Молекулярные механизмы синаптической передачи нервного
импульса..............................................................71
2.9. Молекулярные механизмы синаптических взаимодействий..............72
Глава 3. Основные методы современной психогенетики....................76
3.1. Выявление элементарных психологических характеристик.............76
3.2. Близнецовый анализ...............................................79
3.3. Молекулярно-генетические методы..................................83
3.4. Мета-анализ......................................................86
3.5. Использование животных в качестве модельных объектов.............87
Глава 4. Генетический контроль обучаемости и памяти...................91
4.1. Выявление мутантов по обучаемости и памяти у дрозофилы...........91
4.2. Разнообразие мутантов по обучаемости и памяти у дрозофилы........93
4.3. Функции генов, контролирующих обучаемость и память
У дрозофилы...........................................................96
4.4. Локализация молекулярных процессов, обеспечивающих
обонятельную обучаемость и память у дрозофилы.........................97
4.5. Молекулярные основы обонятельной памяти у дрозофилы..............99
4.6. Генетический контроль обучаемости и памяти у крыс и мышей........101
4.7. Изучение генетического контроля обучаемости у человека.
Основные подходы......................................................ЮЗ
317
4.Х. Современное состояние проблемы генетического контроля
дислексии..............................................................105
4.9. Генетический контроль дислексии. Ближайшие перспективы............107
4.10. Какие именно процессы нарушены при дислексии?....................108
4.11. Роль гена HD в контроле пластичности мышления человека...........109
4.12. Моделирование симптомов хореи Гентингтона у животных.............Ill
4.13. Основные подходы к изучению генетического контроля памяти
у человека.............................................................113
4.14. Влияние гена АРОЕ на память человека.............................116
4.15. Влияние гена BDNFна память человека..............................120
4.16. Молекулярные функции гена BDNF...................................121
4.17. Основные итоги изучения генетического контроля памяти............123
4.18. Перспективы генетики обучаемостии памяти человека................124
Глава 5. Генетический контроль гиперактивности и ослабленного
внимания...........................................*...................128
5.1. Синдром гиперактивности и ослабленного внимания...................128
5.2. Генетический анализ гиперактивности и ослабленного
внимания. Основные подходы.............................................129
5.3. Роль генов, контролирующих биосинтез и секрецию дофамина..........131
5.4. Роль гена DAT1, кодирующего транспортер дофамина..................132
5.5. Роль гена СОМТ, контролирующего деградацию дофамина...............133
5.6. Роль генов, кодирующих рецепторы дофамина.........................135
5.7. Роль генов, контролирующих норадреналиновую систему.............. 137
5.8. Синдром нечувствительности к тироксину............................139
5.9. Генетический анализ синдрома нечувствительности к тироксину.......140
5.10. Моделирование синдрома нечувствительности к тироксину
у мышей................................................................142
5.11. Перспективы генетики гиперактивности и ослабленного
внимания...............................................................143
Н1ава 6. Генетический контроль агрессивности...........................146
6.1. Биологический смысл агрессивного поведения........................146
6.2. Разнообразие форм агрессии у животных.............................147
6.3. Генетический анализ агрессивности хищника у крыс и мышей..........148
6.4. Генетический анализ межсамцовой агрессивности у мышей.............150
6.5. Молекулярные основы межсамцовой агрессивности у мышей.............152
6.6. Генетический анализ агрессивности животных по отношению
к человеку.............................................................156
6.7. Основные итоги изучения генетического контроля
агрессивности животных.................................................157
6.8. Разнообразие форм агрессии у человека.............................160
6.9. Методы оценки агрессивного поведения человека.....................161
6.10. Основные подходы к генетическому анализу
предрасположенности к самоубийству.....................................163
6.11. Роль генов, контролирующих синтез серотонина.....................164
6.12. Роль гена SLC6A4, кодирующего транспортер серотонина.............166
6.13. Роль гена МАОА, контролирующего деградацию серотонина............169
6.14. Роль генов, кодирующих рецепторы серотонина......................170
6.15. Роль генов, контролирующих другие молекулярные системы...........173
6.16 .Основные итоги изучения генетического контроля предрасположенности
к суициду...............................................................174
6.17. Генетический анализ нссуицидных форм агрессии....................176
6.18. Влияние социальных факторов на агрессивное поведение
6.18.1. Роль материнской заботы у макак............................180
6.18.2. Роль социально-экономического статуса у человека...........182
6.19. Перспективы генетики агрессивного повеления человека.............183
318
Слава 7. Генетический контроль предрасположенности
к перееданию..........................................................187
7.1. Актуальность проблемы переедания.................................187
7.2. Общая схема регуляции чувства голода.............................189
7.3. Генетический контроль чувства голода у мышей.....................192
7.4. Основные подходы к генетическому анализу предрасположенности
к перееданию у человека...............................................193
7.5. Роль генов, контролирующих грелиновую систему регуляции..........195
7.6. Роль генов, контролирующих систему пептида YY....................197
7.7. Роль генов, контролирующих инсулиновую и лептиновую системы......198
7.8. Роль генов, контролирующих систему проопиомеланокортина..........200
7.9. Роль гена AGRP, кодирующего агути-подобный белок.................202
7.10. Роль генов, контролирующих систему нейропептида Y...............203
7.11. Роль генов, обслуживающих иные молекулярные системы.............206
7.12. Генетический анализ предрасположенности человека
к перееданию. Основные итоги и ближайшие перспективы..................207
Глава 8. Генетический контроль зависимого поведения...................211
8.1. Что такое зависимость?...........................................211
8.2. Разнообразие наркотиков..........................................213
8.3. Молекулярные основы опиатной зависимости.........................215
8.4. Моделирование опиатной зависимости у мышей.......................218
8.5. Генетический контроль отношения к опиатам у мышей. Роль генов,
кодирующих рецепторы эндорфинов........................................219
8.6. Генетический анализ отношения к опиатам у человека...............222
8.7. Молекулярные основы алкогольной зависимости.......................226
8.8. Молекулярные механизмы утилизации этилового спирта...............227
8.9. Генетический контроль предрасположенности к алкоголизму. Роль генов,
кодирующих алкогольдегидрогеназу 2.....................................229
8.10. Роль гена, кодирующего альдегиддегидрогеназу 2...................231
8.11. Роль генов, кодирующих рецепторы у-аминомасляной кислоты........233
8.12. Роль генов, контролирующих дофаминовую, серотониновую
и эндорфиновую системы................................................236
8.13. Основные результаты и ближайшие перспективы генетического
анализа предрасположенности к алкоголизму.............................241
8.14. Молекулярные основы никотиновой зависимости.....................242
8.15. Основные подходы к генетическому анализу
предрасположенности к никотиновой зависимости.........................244
8.16. Генетический контроль предрасположенности к никотиновой
зависимости...........................................................246
8.17. Основные итоги и ближайшие перспективы генетического
контроля зависимого поведения.........................................249
Слава 9. История развития и перспективы психогенетики.................252
9.1. Истоки психогенетики.............................................252
9.2. Развитие статистических методов..................................253
9.3. Возникновение классической психогенетики.........................255
9.4. Методы классической психогенетики. Использование
близнецового анализа для оценки наследуемости.........................256
9.5. Варианты близнецового метода оценки наследуемости................258
9.6. Методы классической психогенетики. Семейный анализ...............260
9.7. Методы классической психогенетики. Анализ формирования
приемных детей........................................................263
9.8. Основные достижения классической психогенетики...................264
9.9. Современное состояние развития психогенетики.....................266
9.10. О чем свидетельствуют данные психогенетики......................268
9.11. Перспективы дальнейшею развития психогснетики...................269
319
9.12. Наиболее перспективные направления психогенетики................272
9.12.1. Молекулярные механизмы действия социальных факторов......272
9.12.2. Изучение механизмов генотип средовых отношений...........273
9.13. Важнейшие методологические направления современной
психогенетики.........................................................275
9.14. История развития психогенетики в России.........................276
Глава 10. Кто же такой человек?.......................................280
10.1. Молекулярное родство человека с другими млекопитающими..........280
10.2. Какие признаки видоспецифичны для человека разумного?...........285
10.3. Генетический анализ способности к устной речи. Роль гена FOXP2..288
10.4. Только ли человек способен пользоваться речью?..................292
10.5. Только ли человек способен к разумному поведению?...............294
10.6. Современные представления об эволюции человека..................296
10.7. Что отличает человека от высших обезьян?........................299
10.8. «Белые пятна»...................................................302
Глава 11. Этические аспекты психогенетики.............................308
11.1. Психогенетика и евгеника........................................308
11.2. Этические аспекты современной психогенетики.....................309
11.3. Не принижает ли человека молекулярная психогенетика?............311
11.4. Самые распространенные заблуждения по поводу психогенетики......314
Учебное издание
Тйходеев Олег Николаевич
Основы психогенетики
Учебник
Редактор Н. В. Шувалова
Технический редактор О. Н. Крайнова
Компьютерная верстка: С. Ф. Фёдорова
Корректор Н. В. Савельева, Т. Н. Чеснокова
Изд. № 101115883. Подписано в печать 18.07.2011. Формат 60 x 90/16.
Гарнитура «Ньютон*. Бумага офс. № I. Печать офсетная. Уел. печ. л. 20,0.
Тираж I 500 экз. Заказ № 5677
ООО Издательский центр «Академия*, www.academia-moscow.ru
125252, Москва, ул. Зорге, д. 15, корп. I, пом. 266.
Адрес для корреспонденции: 129085, Москва, пр-т Мира, 101 В. стр. I, а/я 48.
Тел./факс: (495)648-0507, 616-0029.
Санитарно-эпидемиологическое заключение № РОСС RU. AE5I. Н 14964 от 21.12.2010.
Отпечатано с электронных носителей издательства.
ОАО «Тверской полиграфический комбинат*. 170024, г. Тверь, пр-т Ленина, 5.
Телефон: (4822) 44-52-03, 44-50-34. Телефон/факс (4822) 44-42-15
Home page — www.tvcrpk.ru. Электронная почта (E-mail) — sales@tverpk.ru