









































































































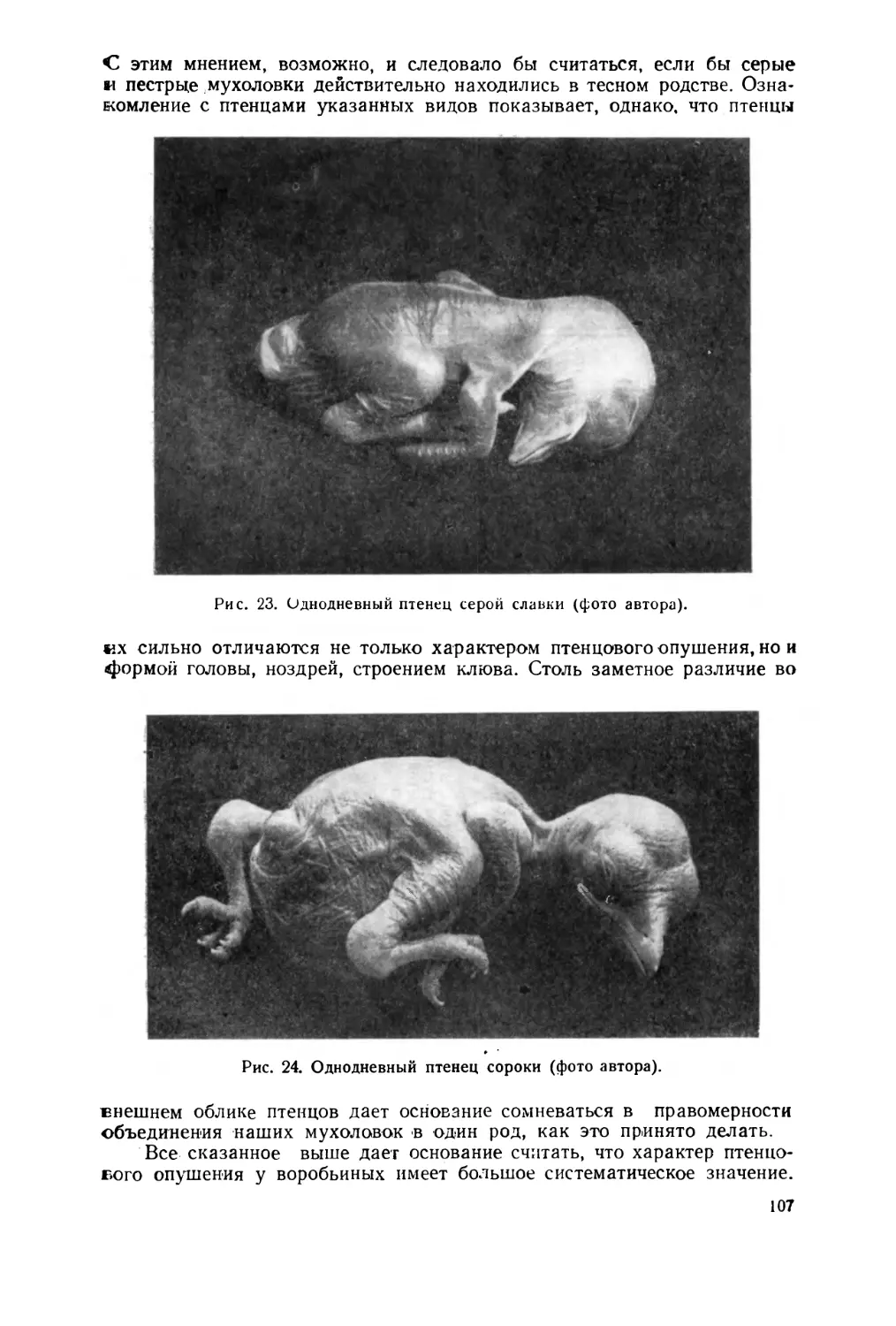



















































































































































































Text
zoomet.ru
А С. МАЛЬЧЕВСКИЙ
H ЕЗДОВАЯ жизнь ПЕВЧИХ " птиц
ЛЕНИНГРАДСКИЙ ОРДЕНА ЛЕНИНА ГОСУДАРСТВЕННЫЙ
УНИВЕРСИТЕТ имени А. А. ЖДАНОВА
А. С. МАЛЬЧЕВСКИЙ
ГНЕЗДОВАЯ ЖИЗНЬ
ПЕВЧИХ ПТИЦ
РАЗМНОЖЕНИЕ И ПОСТЭМБРИОНАЛЬНОЕ РАЗВИТИЕ ЛЕСНЫХ ВОРОБЬИНЫХ ПТИЦ ЕВРОПЕЙСКОЙ ЧАСТИ СССР
ИЗДАТЕЛЬСТВО ЛЕНИНГРАДСКОГО УНИВЕРСИТЕТА 19 5 9
Печатается по постановлению Редакционно-издательского совета Ленинградского университета
Книга представляет собою оригинальное исследование и одновременно критическую сводку данных по биологии размножения и постэмбриональному развитию наиболее многочисленных птиц — лесных воробьиных. Впервые в литературе последовательно разбираются вопросы, касающиеся всего цикла их размножения, начиная с пения и гнездострое-ния и кончая воспитанием птенцов.
Монография состоит из двух частей: первая — посвящена общим вопросам, связанным с гнездовым периодом, вторая— состоит из сорока семи очерков о жизни различных видов певчих птиц. Книга иллюстрирована фотографиями, автора.
Издание рассчитано на специалистов-орнитологов, преподавателей и студентов вузов, работников лесного хозяйства, а также на любителей природы, интересующихся широким кругом вопросов биологии.
Ответственный редактор проф. Г. А. Новиков
ВВЕДЕНИЕ
Воробьиные (Passeriformes)—самый многочисленный и широко-распространенный отряд птиц. Представители его встречаются во всех географических зонах СССР, но чаще всего в пределах лесной и лесостепной полосы. Здесь сконцентрирована богатая фауна воробьиных, составляющих основу птичьего населения наших лесов, садов и парков. Экологической характеристике гнездового периода этих птиц и посвящена настоящая работа.
Поскольку именно среди лесных воробьиных птиц приспособления, обеспечивающие успешное воспитание потомства, достигли наибольшей сложности и разнообразия, изучение их гнездовой жизни может дать особенно ценный материал по ряду вопросов биологии размножения птиц, представляющих обще биологический интерес. В частности, для понимания процесса эволюции птиц большое значение имеет выявление возможных путей формирования их биологических признаков, проявляющихся в период размножения (характер песни, тип гнездовой постройки, размеры кладки и т. п.), а также установление степени пластичности гнездовых инстинктов и биологического значения различных форм поведения. Особый интерес представляет слабо освещенный в литературе вопрос об адаптивном и систематическом значении птенцовых признаков воробьиных. Недостаточно хорошо изучен и процесс постэмбрионального развития многочисленных представителей данной группы птиц. Тем не менее накопление сведений о птенцах, особенно в их раннем возрасте, должно иметь определенное значение для дальнейшего развития представлений о родственных взаимоотношениях между видами, родами и семействами отряда воробьиных, в целом еще слабо разработанного в систематическом отношении.
Изучение биологии размножения лесных воробьиных тесно связано также с рядом практических вопросов. Подавляющее большинство воробьиных нашей фауны—полезные насекомоядные птицы, играющие важную роль в охране деревьев и кустарников от вредных насекомых. С целью использования этих птиц для борьбы с вредителями разрабатываются мероприятия по охране, привлечению и переселению птиц. Естественно, что успешное осуществление этих мероприятий невозможно без всестороннего знания экологии птиц и в первую очередь— условий их гнездования.
3
Вследствие^ очень большого разнообразия представителей группа лесных воробьиных, в сущности, еще плохо изучена в экологическом отношении. Приступая к специальному изучению биологии размножения воробьиных автор преследовал цель в какой-то мере восполнить этот пробел. В процессе полевой работы обращалось внимание на различные стороны жизнедеятельности птиц, но прежде всего на такие, которые казались наименее изученными, а именно: на степень изменчивости гнездовых инстинктов и степень 'растянутости сроков размножения у членов одной популяции, на выяснение судьбы кладок и выводков, а также на возрастные признаки, характер питания и первоначальный способ передвижения птенцов. При изложении материала автор стремился разбирать явления в той очередности, в которой они следуют одно за другим в общем цикле размножения.
Наиболее трудоемкой частью работы оказался сбор данных по питанию птенцов, а также выяснению судьбы отдельных кладок и выводков. Естественно, что без помощников эти сведения не могли быть собраны одним автором в необходимом количестве. Основную помощь в этой работе автору оказывали: С. И. Божко, Е. Р. Гагинская, И. А. Нейфельдт, Н. П. Овчинникова, О. П. Подлевских, И. В. Покровская, И. В. Прокофьева, Ю. К. Эйгелис, а также Т. Б. Ардамацкая, Г. И. Баранова, Д. С. Бекжанова, С. В. Васильев, Т. Н. Геракова, В. А. Зар-хидзе, Г. А. Носков, Р. Л. Потапов, П. П. Стрелков и О. М. Татаринова. Большое количество оригинальных неопубликованных данных по гнездовой жизни воробьиных сообщили автору Н. П. Кадочников и Г. А. Новиков.
В основу исследования была положена методика прижизненного наблюдения за отдельными особями птиц, их гнездами, кладками и птенцами. Прижизненная методика исследования требовала проведения стационарных работ, позволяющих ежедневно и в течение длительного отрезка времени следить за одними и теми же птицами и их гнездами. Местами постоянных наблюдений были ближайшие окрестности Ленинграда (1946—1948), район ст. Кузнечное Ленинградской области (1950 и 1954), учлесхоз «Лес на Ворскле» Белгородской области (1949) и Савальский лесхоз Воронежской области (1951—1953).
При подготовке рукописи автор получал ценные советы и указания от профессоров П. В. Терентьева, Г. А. Новикова, А. И. Иванова, Ь. К. Штегмана, Л. А. Портенко, старшего научного сотрудника Зоологического института АН СССР К. А. Юдина. Всем указанным выше лицам автор выражает свою глубокую признательность за помощь и дружеское участие в его работе.
Часть первая ГНЕЗДОВОЙ ПЕРИОД
В ЖИЗНИ ВОРОБЬИНЫХ ПТИЦ
ГЛАВА I
ЯВЛЕНИЕ ТОКОВАНИЯ, ЕГО ФОРМЫ, СРОКИ И БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ
Несмотря на то, что по вопросу весеннего токования птиц накопилась большая, в основном, правда, иностранная литература, это явление пока еще нельзя считать хорошо изученным. Различные формы проявления тока у птиц недостаточно хорошо известны нам даже с фактической стороны.
Исходя из этого, а также учитывая, что в нашей литературе явление токования применительно к труппе лесных воробьиных птиц специальному разбору не подвергалось, автор счел уместным на основании собственных наблюдений, а также литературных материалов дать краткий анализ данного явления. Токование птиц — проблема разносторонняя. Ее можно обсуждать с позиций самых различных направлений биологической науки. Не претендуя на исчерпывающее освещение проблемы, в настоящей главе автор сознательно ограничил себя лишь некоторым кругом вопросов, избранных им с точки зрения интереса в основном полевого орнитолога.
ФОРМЫ ПРОЯВЛЕНИЯ ТОКА
Весенние токовые явления у воробьиных могут выражаться в воздушных играх, позах ухаживания и, наконец, в различных звуках, издаваемых птицами во время тока.
Воздушные игры. Воздушные игры, или токовые полеты, характерны в основном для воробьиных—обитателей опушек, вырубок или кустарниковых зарослей, т. е. более или менее открытых мест. Они наблюдаются у лесного жаворонка, лесного конька, а также ястребиной и серой славок, варакушки. Поют на лету самцы чечеток и нередко коноплянок. Известны своими весенними воздушными играми обыкновенные скворцы и некоторые врановые, особенно те, которые значительную часть своей жизни проводят в открытых стациях (ворон, серая ворона, грач и галка).
Для настоящих лесных воробьиных воздушные игры во время токования в целом не характерны. Однако и среди этих птиц есть виды, у которых токование нередко сопровождается полетами. Чаще всего это наблюдается опять-таки у птиц, живущих в более разреженных лесах, либо у птиц, имеющих обыкновение держаться весной высоко, на вершинах деревьев. Последние вообще используются как место то
1
кования очень многими воробьиными. На верхушках деревьев очень часто поют дрозды (певчий, белобровик, рябинник и деряба), зеленушка, чиж, чечевица, зяблик, клест-еловик, обыкновенная овсянка, лесной конек, пеночка-теньковка, обыкновенная горихвостка, поползень и мн. др. Даже держащаяся в нижних ярусах леса зарянка в часы наиболее интенсивного пения, в сумерках, садится на вершины деревьев. Наиболее скрытные из наших лесных воробьиных — крапивник и лесная завирушка, которых в обычное время бывает очень трудно заметить, — весной, во время токования, занимают места повыше. Одним словом, поющая и токующая птица всегда стремится занять место повиднее и всем своим поведением как бы заявить о своем присутствии. Средства, которые при этом пускаются в ход, обычно зависят от экологической обстановки. Обитатели более или менее открытых мест обращают на себя внимание не только голосом, но и своеобразными полетами, которые делают даже весьма скромно окрашенного лесного конька в это время заметным на большом расстоянии. Лесные же виды используют в основном свои голосовые средства; если они и совершают токовые полеты, то на опушках, в редколесье или над лесом, где они еще могут быть заметны издали.
Особи, принадлежащие к одному виду, но живущие в различных экологических условиях, могут вести себя во время тока по-разному. Индивидуальные отличия в поведении во время тока можно наблюдать, например, у зябликов, лесных коньков и пеночек-трещоток. У этих видов одни особи, поселившиеся на опушке или в разреженном лесу,, часто поют на лету, совершая характерные планирующие (зяблик) или полукруговые (пеночка-трещотка) перелеты от дерева к дереву; другие, живущие среди густых зарослей, почти никогда не поют на лету. В то же время нельзя не указать на виды (например, чечевица или садовая овсянка), самцы которых совсем не совершают токовых полетов, даже в случае обитания в совершенно открытом ландшафте. Подобных примеров, однако, немного, но они указывают на специфические черты поведения видов, которые могут не укладываться в общую схему.
Позы «ухаживания». Характер поведения самцов певчих птиц в момент «ухаживания» их за самкой изучен еще недостаточно хорошо, а для многих видов остается совсем неизвестным. Это объясняется тем, что подобные интимные стороны жизни птиц очень трудно наблюдать и тем более фотографировать. Дело в том, что энергичное «ухаживание» самца, во время которого он принимает типичные для вида позы, бывает, как правило, очень непродолжительным, повторяется не часто и протекает обычно в скрытых от глаза наблюдателя местах. Самцы многих видов лесных певчих птиц нередко «ухаживают»' за самочками на земле, среди травянистой и кустарниковой растительности.
Позы «ухаживания» обычно принимаются непосредственно' перед спариванием или же при попытке к спариванию. Поведение «ухаживающих» самцов поражает необычностью движений и характером издаваемых при этом звуков. Автор, например, был очень поражен своеобразием поз самца первогодка чечевицы в момент ухаживания. Сидя на поваленном стволе березы и находясь всего в нескольких сантиметрах от самки, самец запрокидывал голову назад и до того сильно вытягивал вверх шею, что на ней даже становились заметными аптерии. Хвост тоже был поднят вертикально кверху, а дрожащие крылья полностью расправлены и немного приподняты. В такой позе самец медленно и очень плавно кружился около самки. По временам
8
он особенно энергично трепетал крыльями, раскрывал клюв и начинал издавать щебечущий тихий звук, медленно и 'плавно покачивая из стороны в сторону сильно вытянутой шеей. В момент наибольшего возбуждения помимо щебечущего звука он в пол-голоса издавал характерную видовую песню. «Ухаживание» происходило всего в нескольких метрах от гнезда, с которого только что слетела самка, отложившая в него свое первое яйцо. Продолжалось оно около минуты.
Надо сказать, что большинство певчих птиц во время «ухаживания» издает тихие щебечущие звуки или поет видовую песнь «под сурдинку». В это время многие птицы сильно вытягивают кверху шею и запрокидывают назад голову. Помимо чечевицы такая поза наблюдалась у сорокопута-жулана и зяблика, а также у зеленушки (Walker, 1949), варакушки (Natorp, 1928) и некоторых других птиц. Гораздо реже «ухаживание» сопровождается громкими звуками; самец сорокопута-жулана, например, в этот момент издает громкое верещание, напоминающее скрипение немазанной коляски; «ухаживание» крапивника сопровождается особой трелью, еще более звонкой, чем его обычная песня (Nelder, 1948).
В момент «ухаживания» у самцов многих наших певчих птиц приходят в движение также и перья: то они плотно прижимаются к телу, благодаря чему птица становится тонкой и как бы приглаженной, то на 'определенных участках тела они вдруг поднимаются, и птица приобретает уже совершенно иной облик. Характер движений и принимаемых при этом поз может быть самый различный. У обыкновенной горихвостки бросаются в глаза прежде всего резкие движения из стороны в сторону ее распущенного ярко окрашенного хвоста. Подобные же движения хвоста характерны для «ухаживающего» снегиря. Щегол резко раскачивается всем телом и поворачивается то одним, то другим боком; серая мухоловка вся вытягивается вперед и забавно взмахивает поочередно крыльями; самец белой трясогузки кланяется, распуская хвост и одно крыло, и «чертит» вокруг самки наподобие петуха и т. п. Одним словом, для каждого вида характерен определенный набор движений, свой «ритуал», совершаемый самцом перед самкой. При этом все поведение самца указывает на то, что он находится в состоянии крайнего возбуждения. Например, обычно флегматичный сорокопут-жулан становится совершенно непохожим на себя. В это время он чрезвычайно подвижен, одна поза быстро сменяется другой: то он выпятит вперед грудь и запрокинет голову назад, то вдруг резко присядет, а потом снова весь вытянется до предела. Эти странные ужимки и поклоны сопровождаются постоянным подергиванием хвоста, иногда трепетанием крыльев и громким скрипучим верещаньем.
Со времен Дарвина (1871) в орнитологической литературе накопилось большое количество наблюдений, отмечающих, что самцы птиц во время «ухаживания» за самками стараются выставить напоказ наиболее ярко окрашенные участки своего оперения. Существуют также указания на то, что в это время самцы (производят именно такие движения, которые делают наиболее заметной разницу в окраске определенных участков оперения птицы, благодаря чему, очевидно, достигается наибольший эффект возбуждающего действия самца на нервную систему самки. Отмечают, например, что резкие смены вертикального и горизонтального положения тела характерны для птиц, имеющих различно окрашенные участки на нижней стороне тела (Morris, 1954). Этим же принципом объясняются быстрые повороты тела ухаживающего за самкой щегла, как бы показывающего желтые участки своих полураспущенных крыльев.
9»
Не отрицая возможной-справедливости указанного выше предположения, нельзя также не согласиться и с тем, что значение этой яркой окраски в процессе размножения, в частности певчих птиц, не должно слишком преувеличиваться. В этом отношении вполне справедливо замечание Гексли (Huxley, 1938), обращающего внимание на то, что стимуляция полового возбуждения самки, помимо демонстрации специальных «украшений» и ярких окрасок самцов во время токования, может достигаться принятием причудливых поз. Действительно, типичные позы «ухаживания» самцы певчих птиц принимают, находясь обычно на очень близком расстоянии от самки, чаще всего в нескольких сантиметрах от нее. На таком расстоянии на нервную систему самки в первую очередь должны действовать именно позы, выражающие крайнее возбуждение, нежели окраска оперения.
Сказанное, на наш взгляд, хорошо подтверждается наблюдениями за размножением видов, самцы которых приобретают яркую окраску оперения лишь к концу второго или даже третьего года жизни, но вполне успешно размножаются уже в конце первого года. Подобное явление наблюдается, например, у ряда вьюрковых (клест, чечевица, щур и др.). Судя потому, что у обыкновенной чечевицы среди птиц, участвующих в размножении, ежегодно встречается очень большой процент молодых самцов, не имеющих, как известно, в окраске оперения никаких признаков, отличающих их от скромно окрашенных самок, можно предположить, что эти самцы-первогодки с таким же успехом участвуют в размножении, как и ярко окрашенные старые самцы.
Чтобы закончить разбор явления весеннего «ухаживания» самцов за самками, необходимо остановиться еще на особом способе передачи корма от самца к самке. У английских орнитологов эта церемония обозначается специальным термином «courtship-feeding», чем подчеркивается то, что она рассматривается как один из моментов общего «ритуала» брачного поведения птиц. Получая от самца корм, самка подает голос и принимает определенную позу. Как позой, так и голосом в этот момент она напоминает птенца слетка, просящего есть. Описываемое явление чаще всего наблюдается в период насиживания кладки. Оно отмечено даже у пеночек (Мау, 1947), у которых самцы, как правило, регулярно не кормят самок в период насиживания. Как форма брачного «ритуала» эта процедура в целом для воробьиных все же менее характерна, чем, например, для голубей или попугаев. Среди воробьиных она чаще всего наблюдается у вьюрковых и врановых. Если бы подобное поведение птиц не наблюдалось до периода откладки яиц и насиживания, а также не отмечалось бы у птиц, для которых выкармливание самки самцом вообще не характерно (пеночки), можно было бы подозревать, что эта церемония не имеет ничего общего с процедурой «ухаживания», а является чисто пищевым актом.
Звуки, издаваемые во время тока. Весной и летом, как известно, поют самые различные птицы: куриные, голуби, кулики, кукушки и др., но ни в одной группе пение не достигл о такого разнообразия и силы, как в группе воробьиных. Недаром большинство представителей этого отряда получило название певчих птиц. Однако все ли звуки, издаваемые птицами в брачную пору, можно назвать пением и для всех ли наших певчих птиц характерно весеннее пение? Можно ли, например, называть песней весеннее цыканье самца серой мухоловки, тихое нерегулярное скрипение снегиря или, наконец, -громкий брачный посвист поползня, похожий больше на призывный крик?
Для, того чтобы ответить на поставленные вопросы, необходимо сначала определить, что следует понимать под весенним пением птиц.
ю
Существующие б литературе определения этого понятия недостаточно точны. Например, определение пения как звукового выражения избыточной энергии, накопившейся в организме птицы, данное Л. Б. Бёме (1952, стр. 8—9), слишком общо. Оно страдает тем недостатком, что не отличает пения от других типов брачных звуков, издаваемых птицами, а также не отражает видовой специфики и биологического смысла явления. Нельзя полностью согласиться и с формулировкой, данной Никольсоном (Nicholson, 1927, стр. 39), который вслед за Говардом (Howard, 1920) полагает, что биологический смысл весенней песни самца заключается в охране гнездовой территории. С Никольсоном можно согласиться лишь в той части формулировки, где он отмечает, что настоящее пение обязательно должно представлять собою регулярную комбинацию следующих друг за другом резких или хотя бы одинаковых звуков. Поэтому, например, весенний посвист поползня, воспроизводимый с достаточной регулярностью, можно считать его весенней песней.
Пение, с нашей точки зрения, есть приспособление, облегчающее, встречу полов. Именно поэтому настоящая весенняя песня имеет демонстративный характер, т. е. издается громко и обычно с заметного места, и прежде всего этой чертой она отличается от других звуков, издаваемых птицами в пору размножения—от так называемой «подпесни» и призывов к спариванию во время «ухаживания» самца за самкой. О последних мы уже говорили. Чаще всего они издаются вполголоса и нерегулярно. Точно так же и «подпеоня» («sub-song»), как на это указывал Никольсон, отличается от настоящей песни меньшей силой и определенностью звуков. Обычно она бывает слышна лишь с небольшого расстояния. Например, набор тихих свистовых и щебечущих звуков («подпесня»), издаваемых иволгой, можно услышать с расстояния всего нескольких десятков метров, в то время как ее громкий флейтовый свист (настоящая песня) иногда бывает слышен более чем за километр. То же самое можно сказать и про негромкое скрипение («подпесня») и сбегающую гамму громких свистовых звуков (настоящая песня) дрозда-белобровика. Описывая пение пестрого дрозда, К. А. Юдин (1952) отмечает, что оно состоит из меланхолического громкого свиста, слышимого более чем за 1 км, и щебетания, которое удается слышать только на очень близком расстоянии. Количество подобных примеров может быть умножено. Кстати заметим, что в нашей орнитологической литературе за последнее в<ремя появились указания на то, что будто бы тихое щебетание иволги надо понимать как настоящее пение, а громкий флейтовый свист—как вид призыва (Козлов, 1950; Корелов, 1954). Надо полагать, что за настоящее пение иволги надо все-таки принимать ее громкий флейтовый свист.
Иногда «подпесня» воспроизводится не только самцами, но и самками и даже неполовозрелыми особями. Ее принято считать более примитивной формой пения, и с этим нельзя не согласиться, ибо молодые птицы по существу начинают петь с «подпесни». Очень часто «подпесня» входит в состав настоящей песни. Если же она воспроизводится отдельно, то в большинстве случаев бывает слышна нерегулярно. Чаще, чем настоящая песня, она звучит осенью и зимой. Таким образом, «подпесня» не имеет демонстративного значения и меньше связана с определенным сезоном, полом и возрастом, чем настоящая песня. Иногда, правда, бывает трудно решить, имеем мы дело с настоящей песней или с «подпесней». Весеннее скрипение дубоноса, снегиря, сойки или сорокопута-жулана обычно принято назы
11
вать пением, хотя по существу здесь мы имеем дело с «подпесней». Итак, настоящее весеннее пение птиц — это регулярное повторение-комбинации громких и специфичных для вида звуков, используемых самцами для привлечения самок.
Как, однако, следует понимать в таком случае песню лесной малиновки (Hippolais icterina), скворца, болотной и садовой камышовок и некоторых других птиц, широко известных своим пересмешни-чеством? Можно ли считать их подражательное пение настоящей видовой песней, если звуки, издаваемые ими, в основном заимствованы у других певчих птиц? Поскольку «пересмешники» воспроизводят звуки, отличающиеся большой силой и регулярностью, и в их песне все-таки бывают слышны звуки, типичные только для своего вида, и даже заимствованные фразы воспроизводятся в характерном для своего вида темпе и ритме, с полным основанием можно считать, что они имеют настоящую видовую весеннюю песнь. Орнитолог легко распознает, например, подражательную песню лесной малиновки или садовой камышовки.
Интересно, что во время подражательного пения обыкновенный скворец поет обычно вполголоса, свои же видовые звуки воспроизводятся им с гораздо большей силой. Это бывает особенно заметно во* время пролета самок. Завидя пролетающих самок, самцы обычно совсем прекращают подражательное пение и начинают издавать громкие, пронзительные, свойственные только скворцам звуки, сопровождая их взмахами крыльев. Все поведение самца в это время указывает на стремление обратить на себя внимание летящих мимо самок. Таким образом, в данном случае самостоятельные видовые звуки, воспроизводимые скворцами, имеют значение настоящей песни, а его* подражательное булькание, шипение и посвистывание должны быть скорее отнесены к категории «подпесни». Правда, здесь так же, как во многих других случаях, разницу между песней и «подпесней» иногда бывает трудно уловить. Сказанное не противоречит необходимости разделения звуков, издаваемых птицами в пору размножения, на несколько категорий. Наоборот, этот пример лишний раз убеждает нас в том, что у многих птиц настоящая песня в процессе эволюции могла выработаться на основе «подпесни».
Почему некоторые певчие птицы не имеют весенней видовой песни? По отношению к врановым этот вопрос задавался уже давно. Еще Дарвин обратил внимание на несоответствие между грубыми и однообразными звуками, издаваемыми воронами, воронами и сороками, и их тонко дифференцированной нижней гортанью, которая по* сложности устройства мало чем отличается от голосового аппарата хорошо поющих певчих птиц. В дальнейшем из наблюдений как в неволе, так и в природе стало известно, что врановые птицы, кажущиеся столь грубоголосыми, способны издавать и обычно воспроизводят в природе весьма разнообразные по своему диапазону звуки. Тем более для нас должно быть интересно, почему врановые птицы не имеют демонстративной, т. е. громкой, регулярной песни.
В свое время Витчелл (Witchell, 1896), придававший большое* значение окружающей обстановке в формировании весенней песни у птиц, предполагал, что ворон, ворона, грач и сорока перестали петь в -связи с переходом из лесной среды в открытые стации, а также в связи с увеличением размеров их тела. Можно также предположить, что объясняется это скрытным образом жизни, который ведут в гнездовой период наши врановые, а также не имеющие регулярной весенней песни сорокопуты, и наполовину хищническим способом питания;
12
«большинства представителей этих групп. Хищнику, охотящемуся в районе своего гнезда, конечно, не выгодно оглашать окрестности громкой песней и тем самым выдавать место своего постоянного прерывания и охоты. Это предположение, однако, не объясняет причину отсутствия настоящей регулярной песни у некоторых птиц, в частности у ремеза, хохлатой и длиннохвостой синиц.
Нельзя согласиться с Никольсоном, а также Говардом, считающими, что птицы, не воспроизводящие в период размножения регулярной и громкой песни, не нуждаются в таковой по причине отсутствия у них постоянных гнездовых участков, на которых они принуждены были бы собирать корм для своего потомства. Однако про хохлатую и длиннохвостую синиц, ремеза, всех наших сорокопутов, а также лесных врановых птиц никак нельзя сказать, что они собирают корм для своих птенцов в местах, удаленных от гнезда. Во всяком случае эти птицы гораздо более «территориальны» в гнездовой период, нежели очень многие, громко поющие весной: скворец, иволга, большинство вьюрковых, а также дроздов рода Turdus. Таким образом, так называемая «теория гнездовой территории» не может объяснить причину отсутствия у ряда певчих птиц громкой и регулярной весенней песни.
Среди лесных певчих птиц непоющих видов сравнительно немного. Тем не менее одинакового объяснения причин малозаметного поведения самцов этих видов в период размножения, по нашему мнению, не может быть дано: очень уж различны представители этой группы птиц по своей экологии и истории развития. В некоторых случаях причиной тому, может быть, действительно послужил переход к частично хищному образу жизни; в других — это произошло в связи с возникновением громких брачных криков, биологически заменяющих весеннее пение; наконец, у ряда видов громкая весенняя песня не выработалась, возможно потому, что образование пар у них происходит еще при жизни в стаях ранней весной, зимой или даже, может быть, осенью.
ПРОДОЛЖИТЕЛЬНОСТЬ И СТЕПЕНЬ РЕГУЛЯРНОСТИ ПЕНИЯ
Весеннее пение певчих птиц в той или иной мере связано с их размножением. Как правило, оно указывает на готовность поющей птицы к спариванию. Выяснение сроков весеннего пения птиц важно для установления степени растянутости периода размножения популяции, особенно в тех случаях, когда мы не располагаем достаточным количеством находок гнезд.
Регулярное пение самцов может продолжаться в течение всего гнездового периода и даже после вылета птенцов из гнезда. Иногда юно прекращается в середине цикла размножения или еще раньше. € целью сравнения общей продолжительности сроков регулярного пения самцов у разных видов воробьиных автор на основании наблюдений в окрестностях Ленинграда в 1946—1954 гг. составил график пения наших лесных певчих птиц (рис. 1). Из графика видно, что продолжительность регулярного пения самцов у певчих птиц весьма различна. У ряда видов — зеленушка, щегол, чиж, коноплянка, зяблик, обыкновенная овсянка, большая синица, пухляк, желтоголовый королек, дрозды (белобровик и певчий), зарянка, крапивник — она равна 3—3,5, даже 4 мес. У других — скворец, обыкновенная чечевица, поползень, пищуха, -славки (завирушка, Черноголовка и садовая), пеночка-пересмешка, лесной и обыкновенный сверчки, садовая камы
13
шовка, соловей, обыкновенная горихвостка и, по всей вероятности, дрозд-деряба — период активного и регулярного пения продолжается всего лишь 1 —1,5 мес. или немного дольше. Третью, промежуточную, группу составляют птицы, самцы которых активно поют примерно 2—2,5 мес. Сюда относятся: иволга, лесной конек, лазоревка, мухоловка-пеструшка, пеночки — желтобровка, весничка и теньковка, серая славка.
Рис. 1. Сроки и продолжительность весеннего и осенне-зимнего пения лесных певчих птиц в окрестностях Ленинграда.
Сплошная линия — регулярное пение, пунктир — нерегулярное, точки — осеннее.
Столь существенные различия в продолжительности активного пения самцов прежде всего следует объяснить наблюдающимися различиями в сроках размножения: чем сильнее растянуты сроки размножения вида, тем, как правило, продолжительнее бывает токование самцов. В свою очередь сильно растянутые сроки размножения зависят от ряда причин и прежде всего от количества выводков в се
14
зоне. Большая продолжительность регулярного пения самцов наблюдается прежде всего у зимующих либо рано к нам прилетающих птиц. Такими являются перечисленные выше виды, характеризующиеся 3—4-месячным периодом пения самцов; почти все они в условиях Ленинградской области могут дважды в лето выводить птенцов.
В то же время среди зимующих у нас видов имеются и такие, например поползень и пищуха, у которых период токования чрезвычайно укорочен: в Ленинградской области их пение прекращается уже в конце апреля — начале мая*. Столь ранний спад регулярного пения объясняется тем, что в данном случае мы имеем дело с птицами, которые гнездятся очень рано и размножаются только лишь один раз в сезон. Скворец, выводящий птенцов в средних и северных широтах только один раз в лето, тоже имеет короткий период регулярного пения, несмотря на свой ранний прилет. Вполне понятны также причины непродолжительности активного пения у большинства тех видов, у которых он исчисляется 1 —1,5 мес.: это почти все поздно прилетающие птицы с моноциклическим типом размножения. Непонятными для нас остаются только причины раннего прекращения токования дрозда-дерябы и славки-завирушки — видов, у которых размножение растянуто очень сильно.
Относительно третьей группы птиц, у которых регулярное пение самцов длится 2—2,5 мес., можно сказать, что ее составляют в основном моноциклические виды. Значительную протяженность токования самцов у них следует, по всей вероятности, объяснять сильной растянутостью размножения, возникающей вследствие неодновременности начала гнездования отдельных пар или частых случаев разорения гнезд.
Несколько необычно долго для моноциклического вида тянется период активного пения у иволги. В средних широтах он продолжается в течение всего гнездового периода, вплоть до начала августа. После ’же вылета птенцов из гнезд голоса самцов начинают звучать даже чаще обычного. В конце гнездового периода весьма активно поют также самцы пеночки-веснички. Лэк (Lack, 1953), отмечая эту особенность для зарянки, считает, что для тех видов, у которых песня не наследуется, позднее пение самца имеет серьезное значение для обучения пению молодых.
С подобным объяснением запоздалого пения самцов согласиться нельзя уже по одному тому, что возрастание активности пения самцов наблюдается и у видов с наследуемой песней, например у козодоя (Мальчевский и Нейфельдт, 1954). Нельзя также отрицать и того, что у очень многих птиц, у которых молодые усваивают видовую песнь главным образом путем подражания старым (скворец, горихвостка, соловей, дрозд-белобровик и др.), никакого подъема активности пения самцов к концу гнездового периода не наблюдается, а наоборот, она резко падает. Так, например, у обыкновенной горихвостки после вылупления птенцов количество песен уменьшается с 239 до 25 в час (Смогоржевский, 1950); соловей практически прекращает петь в Воронежской области уже к 20 июня, когда в большинстве гнезд еще есть яйца.
В случаях с иволгой и пеночкой-весничкой активное пение самцов в самом конце гнездового периода, когда приступать к размножению уже поздно, следует расценивать, очевидно, как явление, сходное с осенним пением птиц. Иволга дает нам пример птицы, у которой нет разрыва между весенне-летним (гнездовым) и осенним пением. У боль
15
шинства других видов, у которых известно осеннее пение (см. рис. 1), наблюдается период затишья, приходящийся на конец гнездования. Он может быть весьма коротким либо, наоборот, весьма продолжительным. У пеночки-веснички, например, период между окончанием регулярного гнездового пения самцов (середина июля) и началом их осеннего пения, которое в условиях Ленинградской области бывает в первых числах августа, равен примерно двум неделям, а у обыкновенного скворца он составляет около двух месяцев.
Нерегулярное пение самцов (отмечено на рис. 1 пунктирной линией) бывает в начале и в конце размножения. Весеннее нерегулярное пение самцов оседлых видов объясняется непостоянством климатических условий поздней зимой и ранней весной, когда птицы поют лишь в ясную или теплую погоду, а также неодновременностью полового созревания отдельных особей популяции. У перелетных видов начало регулярного весенного пения определяется по существу сроками и степенью растянутости прилета, сильно варьирующими в разные годы. Нерегулярность пения самцов в конце гнездового периода объясняется, с одной стороны, неодновременностью, а с другой — постепенностью прекращения их половой активности. Холостые самцы, .а также самцы, бросившие или потерявшие свою самку, как известно, сохраняют способность петь дольше, чем птицы, гнездившиеся успешно.
В южных широтах птицы в общем поют в течение более продолжительного времени, хотя и прекращают петь, как правило, несколько раньше, чем на севере. Удлинение периода регулярного пения самцов здесь происходит в связи с более ранним их прилетом и, очевидно, за счет более продолжительного гнездового периода. Сравнение дат наиболее позднего пения птиц в учлесхозе «Лес на Ворскле» (лесостепная зона) и в окрестностях Ленинграда, несмотря на элемент случайности, который всегда имеет место при определении крайних сроков пения птиц, показывает тем не менее, что многие виды лесных певчих птиц в лесной зоне кончают петь несколько позднее, чем в лесостепной полосе (табл. 1).
Таблица 1
Даты наиболее позднего пения некоторых певчих птиц в лесной и лесостепной зонах
Название вида Дата наиболее позднего пения
в окрестностях Ленинграда (данные автора) в учлесхозе „Лес на Ворскле* (данные Новикова)
Иволга 12 VIII « VIII
Зеленушка 8 VIII 26 VII
Зяблик 27 VII 25 VII
Овсянка обыкновенная 6 VIII 20 VII
Лесной конек 22 VII 10 VII
Пеночка-пересмешка 23 VII 19 VII
Пеночка трещотка 23 VII 8 VIII
Славка-черноголовка 29 VII 20 VII
Славка садовая 6 VIII 24 VII
Славка серая 23 VII 28 VII
Дрозд певчий 22 VII 15 VII
Зарянка 2 VIII 25 VII
Соловей восточный 5 VII 27 VI
46
МЕСТНЫЕ НАПЕВЫ И ГЕОГРАФИЧЕСКАЯ ИЗМЕНЧИВОСТЬ ПЕНИЯ
Факт географической изменчивости пения певчих птиц известен с очень давних времен. Любителям птиц уже давно было известно, что соловьи в разных местах поют неодинаково. Отдельные местные напевы были настолько определенными, что опытный птицелов по характеру песни мог сказать, где была поймана птица. В прежние времена на московском рынке различали курских, киевских, черниговских, бердичевских, польских и других соловьев. Позднее были отмечены и описаны географические вариации напевов у ряда видов: певчего дрозда, зарянки, варакушки, славки-черноголовки, у зяблика, обыкновенной овсянки и др. В настоящее время орнитология располагает уже значительным количеством фактов, указывающих на то, что явление географической, изменчивости пения распространено среди птиц очень широко и свойственно почти всем представителям отряда воробьиных. Известно, что степень этой изменчивости не у всех видов одинакова. Примером птиц со слабо варьирующей песней могут служить многие камышовки-сверчки, пеночки и некоторые другие птицы. Наоборот, пение дроздов, зарянок, варакушек, соловьев может видоизменяться весьма значительно.
В нашей литературе особенно много уделено внимания различиям, которые были обнаружены в пении зябликов. Так, Н. И. Дергунов (1925), изучавший зябликов в Ленинградской и Московской областях, а также в зоопарке Аскания-Нова и на Кавказе, установил у них большие изменения характера конечных звуков и темпа пения в различных районах. А. Н. Промптовым (1930) были обнаружены и статистически обработаны различия в напевах московских и уральских (по долине р. Уфы) зябликов. Пение уфимских птиц оказалось в большинстве случаев короче и примитивнее московских. Центрально-русские зяблики вообще оказались поющими наиболее звонко и задорно.
У громадного большинства видов песнь варьирует все же не так сильно, чтобы нельзя было узнать птицу по ее голосу, г^ому известна песнь зяблика или обыкновенной овсянки, тот без труда распознает этих птиц повсюду, ибо даже в весьма отдаленных один от другого районах они поют в общем все-таки сходно. Однако у некоторых видов местная вариация напева бывает настолько непохожей на обычную видовую песню, что даже специалист-орнитолог, слыша ее впервые, становится втупик и думает,. что он имеет дело с какой-нибудь особой, до сих пор ему неизвестной птицей. Столь резкая географическая изменчивость пения характерна, по-видимому, для очень немногих видов. Среди европейских лесных певчих птиц в этом отношении чрезвычайно интересен дрозд-белобровик, у которого на территории Ленинградской области автору настоящей работы удалось обнаружить исключительное разнообразие местных напевов.
У данного вида дрозда, как известно, песнь состоит из двух частей. Первая часть—демонстрационная, слышимая на далеком расстоянии, состоит из короткого, но очень звучного свистового напева. Вторую же часть, которую скорее следует назвать «подпесней», составляет довольно неопределенное скрипение, исполняемое как бы вполголоса и слышимое обычно недалеко. Продолжительность этого скрипения бывает весьма различной, но характер его более или менее одинаковый у всех белобровиков. Первая же, свистовая часть песни, по которой обычно издали узнаёшь этот вид дрозда, варьирует чрезвычайно сильно. Пишущий эти строки ежегодно, начиная с 1946— 1947 гг., отмечал различия в пении дроздов, гнездящихся в следую-
2 А. С. Мальчевский
17
Таблица 2"
Вариации напевов дрозда-белобровика в окрестностях Ленинграда
Вариации | Запись напева Характер звуков Район
1-я свистовая часть (продолжительность 1,5—2 сек.) 2-я скрипучая (продолжительность неопределенная)
1 Скрипение Короткий восходящий свист, затем нисходящая неторопливая хроматическая гамма чистых звуков, напоминающих тонкий человеческий свист Парки Ст. Петергофа и г. Ломоносова
2 ТИ^ ^РРИ ТЮ 9 Обычно три чистых неторопливых высвиста, средний из которых издается со значительным понижением Парк Мариен-бург—Гатчино
3 /ТИ ить Чтич ₽и 9 Первый звук короткий и слабый; далее идут три чистых понижающихся неторопливых звука, напоминающих тонкий человеческий свист Охтинский лесопарк
4 Хт X 'т₽ю 9 Ряд быстро понижающихся дребезжащих высвистов; один из наиболее широкораспространенных напевов Парк Лесотехнической академии и др. пригороды Ленинграда
5 X. % ИТю 9 Нежная, равномерно сбегающая гамма торопливых звуков Ст. Лаврики окраина Павловского парка,, район поселка Саблино
б 9 Равномерно восходящая хроматическая гамма торопливых дребезжащих звуков Ст. Кузнечное (Приозерский район)
7 ™х J » Протяжный, сперва понижающийся, а потом повышающийся дребезжащий посвист Ст. Кузнечно (Приозерский район)
18
щих местах: а) парк Лесотехнической академии им. С. М. Кирова, расположенный в черте г. Ленинграда; б) парк Биологического института Ленинградского университета в Ст. Петергофе; в) Гатчинский парк; г) Павловский парк; д) Охтинский лесопарк; е) северо-восточная часть Карельского перешейка близ ст. Кузнечное (Приозерский район Ленинградской области), характеризующаяся скалисто-лесным ландшафтом. Было установлено, что в каждом из этих мест дрозды-белобровики поют на свой лад, причем вполне определенно. Используя буквенно-графический метод записи пения птиц приблизительно в том виде, как его применил в свое время Л., А. Портенко (1937), автор схематически записал эти напевы и свел их в таблицу (табл. 2).
Птицы, гнездящиеся в парке Лесотехнической академии, имеют наиболее типичный для ленинградских дроздов-белобровиков напев, состоящий из ряда отдельных дребезжащих, постепенно понижающихся высвистов (вариация 4). У большинства дроздов, населяющих окраины Павловского парка, а также окрестности поселка Саблино, эти высвисты сливаются в непрерывную, равномерно сбегающую торопливую гамму (вариация 5)* Немногосложно и неторопливо поют и охтенские дрозды (вариация 3). Песнь гатчинских белобровиков отличается краткостью свистовой ее части, состоящей обычно из трех неторопливых высвистов, средний из которых издается со значительным понижением (вариация 2). Дрозды, гнездящиеся всего в двух десятках километров от Гатчины, в парке Ст. Петергофа, имеют, как этр на первый взгляд ни странно, совершенно иной напев, обнаруженный нами еще только у дроздов из парка в г. Ломоносове. Здесь белобровики первоначально издают короткое «фюить», за которым следует нисходящая, хроматическая гамма, состоящая из очень чистых звуков, сходных со свистом человека у верхней границы его диапазона (вариация 1). Наконец, в Приозерском районе Ленинградской области (ст. Кузнечное) дрозды-белобровики имеют две не похожие одна на другую вариации, каждая из которых очень сильно отличается от напевов птиц, населяющих ближайшие пригороды Ленинграда. Большинство птиц высвистывает здесь торопливую восходящую дребезжащую хроматическую гамму, звучащую весьма мелодично (вариация 6). Некоторая же часть особей издает протяжный, сперва понижающийся, а потом повышающийся дребезжащий посвист (вариация 7). Последняя вариация совершенно не похожа на обычную песнь ленинградских белобровиков. Небезынтересно, что в самые последние годы в районе ст. Кузнечное все меньше и меньше попадались нам птицы, поющие этим напевом. Дрозды-белобровики, гнездящиеся южнее Приозерска, в районе ст. Сосново, воспроизводят уже не восходящую гамму свистовых звуков, как на севере Карельского перешейка, а более обычную сбегающую гамму.
Наблюдения за пением одних и тех же самцов, проведенные в течение более или менее продолжительного отрезка времени, показали, что некоторые птицы могут последовательно воспроизводить два сильно уклоняющихся варианта песни. В тех случаях, когда разные вариации следовали непосредственно одна за другой, пение дрозда-белобровика производило впечатление не одноколенного в его свистовой части, как это типично для вида, а уже двух- или даже многоколейного. Во всех случаях, правда, белобровики отдавали явное предпочтение какому-нибудь одному напеву, обычно характерному для данной местности. Так, в районе ст. Кузнечное автором было установлено, что отдельные самцы белобровика исполняют сначала вариант 6 (несколько раз подряд), а затем один или два раза воспроизводят вариацию 7. В Павловском парке мы неоднократно отмечали одних и тех же дроздов, которые на-о* 19
свистывали вариации 5 и 3 или 5 и 1. Здесь же 9 мая 1954 г. мы наблюдали одного самца, последовательно воспроизводившего 5 и 6 вариации напевов.
Способность одних и тех же самцов воспроизводить различные модификации видовой песни уже отмечалась у зябликов (Kayser, 1912; Промптов, 1930), а также у обыкновенных овсянок (Промптов, 1957). Неодинаковое 'исполнение песни одним и тем же самцом у этих видов неоднократно слышал и автор. У зябликов и овсинок это выражается, однако, в гораздо менее яркой форме, чем у дроздов-белобровиков.
В связи с изложенными-выше фактами возникает целый ряд вопросов: какова причина возникновения местных напевов у птиц? Каким образом местные вариации напевов могут сохраняться из года в год? Каким образом и как объяснить то, что отдельные птицы могут усвоить одновременно несколько напевов? Насколько, наконец, постоянны местные напевы при столь узкой их локализации, как это наблюдается, например, у дрозда-белобровика в окрестностях Ленинграда? Ниже мы попытаемся последовательно ответить на поставленные вопросы.
а) Каковы причины возникновения местных напевов у певчих птиц? Вызываются ли они внутренними причинами, являясь, например, результатом неодинакового строения голосового аппарата или они зависят от внешних факторов, действующих по-разному на птиц, обитающих в разных местах?
Нет никакого сомнения в том, что объяснение причин бросающейся в глаза географической изменчивости пения у птиц, способных к имитации звуков, надо искать во внешней «звуковой среде», которая действительно может по-разному влиять на птиц, обитающих в местах, удаленных одно от другого. Непричастность же анатомических особенностей голосового аппарата к тому типу географической изменчивости пения птиц, о котором только что говорилось, доказывается наблюдающимися у дроздов-белобровиков фактами последовательного воспроизведения одной и той же птицей двух совершенно различных напевов. Известно также, что у птиц, для которых характерно явление голосовой мимикрии, особи одного и того же вида с однотипным строением голосового аппарата могут петь как совершенно одинаково, так и совершенно различно. Все известное нам о пересмешничестве певчих птиц (Промптов, 1944) и их большой восприимчивости к звукам окружающей среды позволяет предполагать, что общий хор птичьих голосов, который редко бывает одинаковым в разных удаленных друг от друга местах, является основной причиной, вызывающей вариации видовой песни. Данная мысль была высказана еще И. К. Шамовым (1910), писавшим, что разнообразие и звучность соловьиного пения в известной мере зависит от полноты хора птичьих голосов той местности, где гнездится соловей.
Идея влияния окружающих звуков на видовые напевы развивалась Витчеллом (1896), а также Фрилингом (Frieling, 1937), который стремился показать, что звуки, издаваемые птицами, обитающими в сходных ландшафтах, имеют между собой много общего. Оба эти автора несколько переоценивают роль звуков ландшафта в эволюции птичьего пения и, наоборот, недооценивают наследственной стороны явления, значение которой несомненно. Тем не менее формирующая роль «звуковой среды» в эволюции песни как видового признака птиц несомненна. Иначе, нам кажется, становится невозможным понять причины существования географической и индивидуальной изменчивости пения большинства птиц.
Очень возможно, что отсутствие разнообразных звуков, которые могли бы слышать вокруг себя некоторые попавшие на Галапагосские
20
острова предки ныне живущих там дарвиновых вьюрков (Geospiza, Camarhynchus и др.), и обусловило то, что все виды этих вьюрков, как это видно из сообщения Лэка (1947а), поют удивительно однообразно, примитивно* и сходно друг с другом. Эти птицы, эволюция которых длительное время протекала в обстановке полной изоляции от других певчих птиц, наоборот, могли даже по причине малого разнообразия слышимых ими звуков упростить характер своего первоначального пения, как это, очевидно, и произошло с ними, а также с галапагосским пересмешником Neosomimus. Лэк указывает на то, что бедность и примитивность птичьих напевов, по всей вероятности, представляет собою общую тенденцию, характеризующую орнитофауну изолированных островов. Если подобная тенденция у птиц, населяющих океанические острова, действительно существует, ее, по нашему мнению, надо объяснять скорее всего однообразием «звуковой среды».
В том, что звуки, которые птица слышит вокруг себя, стимулирующе действует на ее пение, можно легко убедиться, наблюдая за птицами, содержащимися в неволе. Известно, например, что певчие птицы часто начинают петь, когда слышат человеческий голос, свист или игру на музыкальном инструменте. Особенно восприимчивы они к шуршащим и свистовым звукам.
Решающее значение звуков окружающей среды в формировании напевов у птиц-имитаторов не вызывает сомнений. У некоторых «пересмешников» эта закономерность проявляется в наиболее чистом виде. Во многих же случаях влияние окружающей среды выступает, видимо, в-завуалированной форме: птица лишь приближает характер воспроизводимых ею звуков к тем, которые она постоянно слышит около себя, но не копирует их. Благодаря этому у птиц, склонных к подражанию, очевидно, и могут возникать различные местные уклонения от видового напева,.
Гораздо сложнее решается вопрос о причинах географической изменчивости птиц, у которых не установлена способность к подражанию* звукам. На территории СССР таким видом, например, является пеноч-ка-теньковка, различные подвиды которой поют хотя и сходно, но неодинаково. Так, по наблюдениям Л. М. Шульпина (1928), на Алтае (подвид tristis) пение этой пеночки отличается большим разнообразием звуков и более 'быстрым темпом исполнения, отчего песня кажется более слитной. Поскольку в данном случае мы сталкиваемся с различием в напевах подвидов, т. е. в физиологическом отношении неодинаковых птиц,, которые, кстати, раньше считались даже за самостоятельные виды, по-видимому, есть основание говорить о том, что и различия в пении обусловливаются здесь более глубокими, наследственными причинами. Хотя для того чтобы быть окончательно уверенным в этом, нужны, конечно,, дополнительные эксперименты.
С другой стороны, географическую изменчивость наследственно закрепленной песни можно рассматривать как явление того же порядка,, что и изменчивость позывов, характер которых, как это известно (Wit-chell, 1896; Heinroth, 1924; Промптов и Лукина, 1945), у птиц передается по наследству. Тем не менее в природе можно наблюдать как географическую, так и индивидуальную изменчивость позывов, издаваемых птицами. Известно, например, что у разных подвидов одного и того же вида сходные по своему биологическому значению позывы имеют разные оттенки.
Поскольку врожденные позывы подвержены у .птиц географической изменчивости, нас не должны удивлять отмеченные выше факты изменяемости песни строго наследуемого типа. Объяснить причину геогра
21
фической изменчивости подобного рода автор в настоящее время не берется. Можно только констатировать, что врожденные звуки (позывы, песни некоторых видов), издаваемые птицами, варьируют гораздо слабее, нежели пение большинства видов певчих птиц, у которых, как уже отмечалось, напев формируется в значительной мере условнорефлекторным путем.
б) Каким образом могут сохраняться из года в год местные напевы у птиц при столь узкой их локализации, как, например, в описа-санном выше случае с дроздом-белобровиком на территории Ленинградской области?
На основании существования у певчих птиц местных «диалектов» пения еще в начале текущего столетия Луканус (Lucanus, 1907) сформулировал положение о регулярном возврате дикой перелетной птицы па старые места гнездования. Это замечательное обобщение, как известно, вскоре подтвердилось большим количеством документальных фактов, полученных путем кольцевания. Действительно, факт существования достаточно стойких и столь узколокализованных географических вариаций местных напевов, какие, например, были обнаружены автором на территории Ленинградской области у дроздов-белобровиков, нельзя понять, если не признавать того, что из года в год определенные группы птиц, характеризующиеся определенным пением, со своих мест зимовок возвращаются для размножения в один и тот же район, даже в один и тот же парк. То, что известно нам о способности молодых птиц перенимать и точно копировать пение окружающих их взрослых птиц, помогает понять процесс передачи местного напева последующим поколениям.
Сразу же, однако, возникает вопрос: имеем ли мы дело в каждом месте, где существует своя вариация пения, с определенной группой особей и именно с их потомками или данный местный напев в равной степени могут усваивать молодые птицы-первогодки из соседних областей, которые впервые осели на гнездовье в местах, удаленных от тех, где они сами появились на свет? Иными словами, можно ли вкладывать генетический смысл в понятие популяции, когда речь идет, например, <о петергофских, гатчинских, охтинских и других группах дроздов-белобровиков, столь отчетливо различающихся характером напева? Некоторые авторы на этот вопрос отвечают положительно. Фрилинг (1937), например, считает, что различие в птичьих «диалектах» может в дальнейшем привести к изоляции популяции и образованию нового вида. На такой же точке зрения стоит и Майр (Мауг, 1944), отмечающий, что значение таких негенетических изменений, как местные «диалекты» птичьего голоса, в образовании генетических рас несомненно, ибо они содействуют изоляции. Представления о наличии у птиц весьма стойких местных популяций или стай, проводящих совместно большую часть года и пополняющихся главным образом за счет возвращения молодых птиц на места своего рождения, придерживается также Ю. А. Исаков (Исаков, 1949; Исаков и др., 1953).
Некоторые данные, однако, заставляют сильно сомневаться в существовании у ленинградских дроздов-белобровиков отдельных мелких популяций в генетическом смысле слова. Судя по тому, как неодновременно прилетают эти дрозды в места гнездования и как неодновременно они приступают к размножению и, окончив гнездование, покидают наши парки, можно заключить, что вряд ли, например, петергофских белобровиков можно рассматривать как стойкую местную популяцию, как стаю, члены которой проводят совместно большую часть жизни.
22
Знание локальных напевов позволило провести наблюдения за сроками прилета местных дроздов-белобровиков в пригородные парки г. Ленинграда. Эти наблюдения показали, что первые одиночные особи с местным напевом появляются обычно во второй декаде апреля—15 апреля (1950, 1952), 18 апреля (1954). В дальнейшем идет постепенное увеличение количества птиц, насвистывающих напев, характерный для того или иного парка. Позднее прилетающими бывают преимущественно молодые птицы-первогодки (Тугаринов, 1950; Weigold, 1926). При весьма растянутой картине прилета на места гнездования и при более позднем прилете молодых птиц вполне естественно предположить, что птицы-первогодки часто оседают на гнездовья не строго в тех местах, где они появились на свет. В таких случаях молодые птицы должны, очевидно, усваивать напев той местности, где они впервые гнездятся. И действительно, самцы-первогодки, судя по многочисленным наблюдениям, научаются петь, как окружающие их взрослые птицы, уже на местах гнездования в конце первого года жизни.
Старые птицы, если они случайно собьются с пути или примкнут во время зимовки или перелета к птицам других местностей, даже гнездясь не на своей «родине», обычно сохраняют напев, усвоенный ими в первую весну размножения. Этим, видимо, можно объяснить тот факт, что среди основной массы птиц данной местности, поющих чрезвычайно сходно, иногда попадаются особи, воспроизводящие напевы, характерные для других местностей.
Например, в конце мая 1954 г. в парке Ст. Петергофа автор отметил 10—12 самцов дрозда-белобровика, насвистывавших типичный для данного места вариант 1, и одного дрозда, который упорно пел вариант 4, т. е. так, как поют дрозды в парке Лесотехнической академии; 25 мая 1954 г. на протяжении 6-километрового пути из дер. Пудость в Гатчино автор насчитал 8 дроздов, певших вариант 2, и одного — насвистывавшего вариацию видовой песни, близкую «к 4; 14 июня 1950 г. в Гатчинском парке мы насчитали около 15 птиц, воспроизводивших вариацию 2, и 2 — певших вариацию 5. В районе ст. Кузнечное в период с 10 по 15 июня 1954 г. было отмечено 27 самцов белобровиков с типичным для той местности напевом 6 и 3 дрозда, певших совершенно необычно для данного района, и т. п. Таким образом, судя по характеру пения, приблизительно 1/10 часть местного населения дроздов составляют старые птицы, по какой-то причине покинувшие место своего прежнего гнездования. По отношению к основному «ядру» старых птиц данной местности они, естественно, составляют более значительную часть.
Среди молодых дроздов-первогодков процент птиц, гнездящихся не на своей «родине», по всей вероятности, еще более высок, ибо известно, что так называемый «веер разлета» молодых птиц значительно больше, нежели у старых птиц. Поэтому вряд ли правы те, кто полагает, что образование местных узколокализованных «диалектов» у птиц может привести к изоляции и образованию генетически обособленных рас.
в) Каким же образом получается, что отдельные птицы могут усваивать одновременно несколько напевов?
В этих случаях мы, по всей вероятности, имеем дело с молодыми птицами-первогодками, которые в период формирования своего индивидуального напева могут усвоить два варианта, если постоянно будут их слышать. Надо полагать, что такой усложненный тип песни птицы мо-гут сохранять и в течение последующих лет и «передавать» его впоследствии другим птицам.
23
В каких же случаях молодая птица может одновременно слышать два различных напева? Думается, что одиночные старые особи, залетевшие в необычные для них места, но сохранившие своеобразие напева «родной» местности, о которых только что шла речь, не могут сильно повлиять на пение окружающих их молодых птиц, поскольку последние будут отдавать, видимо, предпочтение типичному для данной территории варианту, действующему на них с большей частотой и регулярностью. На границе же районов, населенных птицами, поющими различные варианты видовой песни, усвоение двойного напева или вырабатывание среднего промежуточного варианта вполне вероятно. Такая же возможность возникает и при случающихся иногда в природе массовых переселениях птиц из одной местности, где они постоянно гнездились, в другую; в результате может получиться смешение двух групп птиц, характеризующихся различными вариантами видовой песни. Таким образом, в ходе рассуждений мы подошли к ответу на последний из поставленных ранее вопросов.
г) Насколько постоянны местные напевы при столь узкой их локализации, как это наблюдается у дрозда-белобровика в окрестностях Ленинграда?
Как уже отмечалось, на севере Карельского перешейка, в районе ст. Кузнечное, в последнее время стало встречаться меньше дроздов^ высвистывающих вариацию 7. С 1954 г. по нашим наблюдениям в Гатчинском парке помимо птиц, поющих вариант 3, появились также дрозды с напевом, который автор раньше здесь не слыхал. Песня этих птиц не имеет характера сбегающей или восходящей гаммы, как это типично для большинства дроздов-белобровиков Ленинградской области, а представляет собою ряд дребезжащих однотонных звуков «ти-трютрю-трютрютрю», среди которых лишь первый свист отличается более высоким тоном.
В плане ведущеюся обсуждения чрезвычайно интересное наблюдение мы находим в работе Д. Н. Кайгородова (1909). В 1903 г. в парке Лесного института (ныне парк Лесотехнической академии), а также в Гатчино, Петергофе и других местах Кайгородов отметил внезапное появление большого количества дроздов-белобровиков, отличавшихся особым напевом, отличным от напева этих дроздов, гнездившихся или встречавшихся на весеннем пролете в этих местах. Как-сообщает Кайгородов, прежде громкая часть песни у дроздов-белобровиков состояла здесь лишь из 3—4 отрывистых высвистов, У дроздов же нового происхождения эта часть песни воспроизводилась сверху вниз красивой хроматической гаммой из 7—9 полутонов. Этот в то время новый для данной местности напев сохранился до настоящего времени во многих пригородах Ленинграда. Он ближе всего подходит к описанным выше вариациям 4 и 5. Более древний, судя по описанию Кайгородова, для окрестностей Ленинграда напев (вариации 2 и 3) представлен сейчас в Гатчинском парке и Охтинском лесничестве.
Есть все основания предполагать, что своеобразное пение, наблюдающееся сейчас у белобровиков, гнездящихся в парке Ст. Петергофа (вариации 1), имеет сравнительно недавнее происхождение. К сожалению, в подтверждение этого автор может сослаться лишь на устное сообщение А. Н. Промптова, который в 1947 г. отметил, что до Великой Отечественной войны эти дрозды не обращали на себя внимания особой оригинальностью пения. История Петергофского парка насчитывает около 25Q лет. За это время дрозды-белобровики, населявшие парк^ могли, видимо, несколько раз изменить характер своего местного напева. И сейчас, направляясь весной в парк Ст. Петергофа, автор этих
24
строк не бывает полностью уверен в том, что он услышит здесь точно такой же напев дроздов, каким он знал его раньше.
При столь большой географической изменчивости пения, какая наблюдается в настоящее время у дроздов-белобровиков Ленинградской области, случаи быстрого изменения местных напевов вполне вероятны и сравнительно легко объяснимы. Территориальная близость отдельных «популяций» этих дроздов с резко отличающимися вариациями видовой песни приводит к частым случаям смешения птиц с разными напевами на местах гнездования. Близкое соседство неодинаково поющих птиц обусловливает влияние одного напева на другой. Со временем это может привести к образованию нового варианта песни у преобладающего большинства птиц, населяющих данную местность.
БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ПЕНИЯ ПТИЦ
Существующие в литературе высказывания по поводу биологического значения пения птиц можно свести к следующим точкам зрения
а) Пение певчих птиц есть одна из форм проявления полового отбора: самцы птиц поют для того, чтобы стать возможно заметнее для самок, которые выбирают себе наиболее активного самца. Как известно, эта точка зрения была высказана Дарвином (1871). Дарвиновскому принципу следовали многие орнитологи конца прошлого и начала нынешнего столетия. Однако еще Альтум (Altum, 1868) и позднее Лука-нус (1907) рассматривали пение самцов не только как средство привлечения самок, но и как средство запугивания соперника и определения границ гнездовой территории. В свое время дарвиновская теория полового отбора была подвергнута резкой критике и большинством ученых оставлена. Выступавший в защиту данной теории Гексли (1938) в понимании биологического смысла пения самцов певчих птиц придерживался тем не менее теории гнездовой территории, которая, с нашей точки зрения, противоречит дарвиновскому пониманию биологического значения пения птиц.
б) Пение певчих птиц является в первую очередь средством отпугивания особей того же вида в целях максимального сохранения кормовых запасов вблизи гнезда, что необходимо для успешного воспитания жизненно стойкого потомства. По мнению создателя данной теории Говарда (1920, 1929), вся гнездовая жизнь большинства птиц подчинена стремлению сохранить за собой определенный участок местности, на котором птица могла бы прокормить себя и свое потомство. Ранний прилет самцов на места гнездования, их интенсивное весеннее пение а драки — все это, по мнению Говарда и его последователей, есть различные формы проявления борьбы за гнездовую территорию. Громко и регулярно поющий самец как бы извещает остальных самцов того же вида о том, что данная территория занята. В связи с таким поведением самцов каждую весну происходит быстрое и равномерное распределение птиц по угодьям. Наиболее активно поющие самцы становятся обладателями лучших кормовых участков, и поэтому они с большим успехом участвуют в размножении. Более слабые и менее энергичные особи бывают принуждены селиться в менее благоприятных условиях, а иногда, как пишет Говард, по причине занятости всех(!) подходящих для гнездования мест другими самцами и вовсе не гнездятся. Таким образом, по мнению Говарда, пение самцов полезно для вида, так как оно предотвращает вредные последствия перенаселения, которое иначе могло бы постоянно возникать. Видовой песне как средству привлечения самки Говард придавал подчиненное значение.
25
Теория гнездовой территории сильно повлияла на взгляды орнитологов. Даже пересмешничеству стали придавать территориальное значение (Hingston, 1933; Промптов, 1944, и др.). Многие орнитологи усмотрели зависимость между звучностью пения птиц и размерами их гнездовых участков (Nicholson, 1927, и др.). Некоторые исследователи, полагая, что каждый вид может быть охарактеризован определенной площадью гнездовой территории, одной из задач полевой орнитологии -стали считать определение размеров гнездовых участков. В большинстве случаев такое исследование сводилось по существу к количественному учету птиц какого-либо вида, гнездящихся на территории, и последующему арифметическому вычислению площади, занимаемой одной парой, или к определению среднего расстояния между гнездами, половина величины которого принималась за радиус гнездовой территории вида (Лукашкин, 1936).
Подобного рода изыскания могут представлять интерес, однако, лишь с точки зрения количественного учета птиц. Размеры же гнездовых участков таким способом, по нашему мнению, определять нельзя, ибо в природе, как правило, мы не встречаем предельных плотностей населения певчих птиц. Вообще надо сказать, что само понятие гнездового участка настолько неопределенно, что выражать его конкретными цифрами почти бессмысленно. Как правило, мы бываем не в состоянии учесть все чрезвычайно многообразные и постоянно меняющиеся условия, могущие повлиять на расстояние места добывания корма птицы от ее гнезда.
С другой стороны, нельзя отрицать того, что у многих видов отдельные пары стремятся селиться на известном расстоянии друг от друга и что определение этих расстояний бывает важно для практики привлечения, например некоторых птиц дуплогнездников. Факт гнездования многих птиц на известном расстоянии друг от друга не должен, •однако, обязательно означать того, что эти птицы имеют строго определенные кормовые участки и тем более что песня самца есть своего рода заявка на участок. Наблюдения показывают, что даже так называемые «территориальные» птицы в период выкармливания птенцов часто летают за кормом в обильные пищей места, значительно удаленные от их гнезд, где пищу они собирают бок о бок с птицами из других гнезд, не проявляя при этом никакой враждебности друг к другу. Точно так же и поблизости от своих гнезд певчие птицы почти никогда не вступают в конкурентные взаимоотношения из-за пищи для своих птенцов. На это указывали уже зоологи, критиковавшие теорию кормовых участков Говарда (Смогоржевский, 1950; Lack, 1953, и Др.). Отдельные же стычки между самцами следует, видимо, расценивать как результат присущей для них в период размножения враждебности друг к другу, как столкновение из-за самок, а не из-за кормового участка.
Против теории Говарда можно привести еще целый ряд аргументов. Во-первых, многочисленные наблюдения говорят, что у большинства певчих птиц активность пения самцов резко снижается к моменту появления птенцов, т. е. как раз к тому времени, когда потребность в корме резко возрастает; если бы пение действительно являлось средством отпугивания особей того же вида в целях максимального сохранения запасов корма вблизи гнезда, то оно, казалось бы, наоборот, к этому времени должно было усилиться. Во-вторых, известно, что после разорения гнезда птицы приступают к строительству нового гнезда очень часто за пределами своего первоначального участка; если бы в природе так называемые гнездовые территории действительно тесно
25
примыкали друг к другу и так ожесточенно охранялись, как полагал это Говард, то птицы не могли бы так легко менять своего гнездования при вторичной постройке гнезда. В-третьих, целый ряд видов не имеет весенней песни, хотя и собирает корм обычно в непосредственной близости от гнезд. Наконец, в-четвертых, наблюдающиеся случаи выкармливания выводка тремя, а иногда и четырьмя взрослыми птицами вряд ли могли иметь место, если бы птицы вели постоянную и непримиримую борьбу за территорию; случаи подобного рода наблюдались у длиннохвостых синиц (Makatsch, 1951), у лазоревок (Орлов, 1947), у обыкновеной каменки (Sutton and Parmelee, 1954), у чечевиц (наблюдение автора). Количество подобного рода доводов мы могли бы увеличить.
в) Видовая песня — это сигнал угрозы, служащий средством отпугивания других особей того же вида, главным образом самцов, в целях образования и поддержания на определенный срок известного постоянства пары, что необходимо для успешного воспитания потомства. Точно так же, как и позы угрозы, громкая песня самца, как полагает Лэк (1953), предохраняет птиц от излишних столкновений, могущих оказаться вредными для вида. Пению же самцов как средству привлечения самки он так же, как и Говард, придает подчиненное значение.
Полностью соглашаясь с Лэком в том, что гнездование так называемых «территориальных» птиц на известном расстоянии пары от пары объясняется не стремлением сохранить за собой кормовой участок, а природной враждебностью самцов этих видов друг к другу в период размножения, мы, тем не менее, никак не можем согласиться с тем, что пение у певчих птиц имеет в первую очередь отпугивающее значение и выполняет то же назначение, что и угрожающие позы. Нельзя согласиться также и с тем, что пение самца относится исключительно к категории так называемого «территориального» поведения, как почему-то полагает Лэк, следующий в этом отношении за Говардом. Известно, например, что очень многие птицы активно поют на пролете. Это касается не только колониальных или полуколониальных птиц, но и типичных «территориальных» птиц, т. е. таких, отдельные пары которых гнездятся, как правило, на известном расстоянии друг от друга. В Ленинградской области очень интенсивно, например, поют пролетные пеночки-теньковки, зеленые пеночки, камышовки-сверчки, соловьи и многие другие птицы. По наблюдениям Гроте (Grote, 1930), пролетные соловьи начинают петь, находясь еще в Африке и южной Аравии. Наблюдения также показывают, что самцы многих птиц активно поют не только в районе своего гнезда, но и на весьма значительном расстоянии от него. Это бывает, например, у иволог, черных дроздов, зеленушек, коноплянок и многих других птиц, у которых такое поведение никак уже нельзя признать «территориальным».
Далее, многие исследователи отмечали, что активность пения самцов резко падает после появления на гнездовом участке самки и, наоборот, возрастает или бывает повышенной при внезапной потере самки или при отсутствии ее (Montagu, 1802; Дарвин, 1871; Buxton, 1950; Смогоржевский, 1950; Lack, 1953; Птушенко, 1954). Автору это явление приходилось наблюдать весной 1954 г. в районе ст. Кузнечное. Вблизи одного дома появился самец серой мухоловки, который с 10 по 15 июня постоянно находился на вершине радиомачты, громко и пронзительно цыкал, взмахивал крыльями и т. п., одним словом, вел себя •очень заметно. Затем прилетела самочка, и наступил период «ухаживания», в течение которого самец уже редко показывался на своем наблюдательном пункте. В это время птицы постоянно держались вместе.
27
В гнезде, построенном в стене сарая, 18 июня появилось первое яйцо. Когда утром 22 июня самка откладывала последнее яйцо, ее поймала кошка. После гибели самки поведение самца резко изменилось и стало вновь таким же, как и в первое время после прилета: он снова стал садиться на вершины деревьев и радиомачту, начал усиленно цыкать,, одним словом, всячески заявлять о своем присутствии. Такое поведение продолжалось несколько дней, после чего птица куда-то исчезла.
Если бы песня действительно являлась средством отпугивания соперников и предупреждала схватки между ними, то, казалось бы, с появлением на гнездовом участке самки активность пения самца не должна была бы снижаться, так как именно в этот период самцы должны больше всего охранять свою самку. В своих работах Говард (1920„ 1929) и полагал, что если бы песня предназначалась в первую очередь для самки, то пение с ее появлением должно было бы не ослабевать,, а усиливаться. На самом же деле, как это он отмечал и сам, после появления самки у самцов начинается как бы вторая фаза тока: они. усиленно «ухаживают» за самочкой. Далее идет спаривание, совпадающее обычно с периодом гнездостроения и откладки яиц. С началом насиживания кладки пение самцов возобновляется, и, по мнению Говарда, это-то как раз и указывает на то, что песня сама по себе не имеет прямого отношения к самке, а воспроизводится самцом главным образом в целях охраны территории. По нашему мнению, активность пения самцов с появлением самки затухает вследствие того, что средство, которое пускалось в ход для привлечения самки (громкое и регулярное пение, слышимое на значительное расстояние), достигло своей цели. Далее начинается наиболее эмоциональная фаза тока — период «ухаживания», часто сопровождаемая пением, но уже не громким, а обычно издаваемым вполголоса.
Возобновление активности пения самцов после того, как самка начала насиживать кладку, объясняется, на наш взгляд, физиологическим состоянием самца, половая активность которого продолжается и который громким и регулярным пением снова стремится привлечь к себе самок. Этот второй подъем активности пения наблюдается далеко не у всех видов птиц. У поползня и обыкновенной пищухи, например, в период насиживания кладки самцы обычно уже не поют. У ряда видов активное пение самцов продолжается до вылупления птенцов, у некоторых же — до конца гнездового периода. Биологический смысл растянутости периода пения самцов, с нашей точки зрения, огромный. Благодаря позднему пению самцов самка, потерявшая кладку или своего партнера, имеет возможность вновь встретиться с готовым к размножению самцом >и приступить к повторному гнездованию.
За последнее время в литературе все чаще и чаще стали встречаться описания случаев наблюдавшейся у певчих птиц бигамии (Gosnell, 1949; Chappel, 1949; Лукина, 1951; Haartman, 1949). Последний из названных авторов установил, что некоторые из окольцованных им самцов мухоловки-пеструшки первоначально спаривались с одной самкой, а затем с другой, прилетевшей позднее или потерявшей своего самца. В 1954 г. на территории Савальского лесхоза один из самцов горихвостки принимал участие в выкармливании двух выводков, расположенных друг от друга на расстоянии 150 м (устное сообщение Н. П. Кадочникова). По всей вероятности, этот самец являлся отцом птенцов обоих выводков. Макач (1951), ссылаясь на работу Руитера (Ruiter, 1941), упоминает о многих случаях, когда один и тот же самец, обыкновенной горихвостки воспитывал птенцов в двух различных гнездах. По всей вероятности, случаи последовательного спаривания самца
28
с несколькими самками у певчих птиц не так уже редки. Повторное же спаривание самок с те'м же самым или другим партнером после гибели первой, а иногда и второй кладки имеет место очень часто; судя по количеству разоряющихся гнезд, оно бывает примерно у 40% самок, причем некоторые из них повторно спариваются не один раз вследствие неоднократного разорения их гнезд.
Естественно, что самка, потерявшая первую кладку, скорее всего будет вторично спариваться с тем же самым самцом. Из этого, однако, не следует, что самцы в период насиживания кладки продолжают интенсивно петь только ради этого возможного случая. Они будут спариваться с любой другой готовой к размножению самкой, не имеющей самца, что, видимо, нередко и случается в природе и что, безусловно, не может не иметь значения для успехалразмножения вида. Во всяком случае наблюдения Е. В. Лукиной, Л. Хаартмана и других орнитологов, отмечавших факты бигамии у певчих птиц, показывают, что оба выводка птенцов, имевших общего отца, успешно заканчивают свое развитие. При этом большое значение имеет, очевидно, неодновремен-ность сроков откладки яиц и насиживания у разных самок.
Таким образом, биологический смысл растянутого периода токования самцов следует связывать с растянутым периодом размножения и целесообразностью имеющих место в природе повторных спариваний.
Исходя из всего вышеизложенного, нам кажется, что можно следующим образом сформулировать биологический смысл весеннего пения птиц.
г) Весеннее пение птиц есть приспособление, облегчающее встречу полов. Так, примерно, представляли себе значение пения птиц еще Montagu (1802), Дарвин (1871) и за ними другие орнитологи, считавшие, что основное значение песни заключается в том, что оно служит призывом самца к самке и что у птиц самки ищут самцов, а не наоборот. Нельзя, видимо, даже говорить о второстепенном предостерегающем значении песни, которая все же признается большинством орнитологов. Вряд ли также пение самца, после того как данная пара уже образовалась, служит в дальнейшем для более прочного закрепления моногамии и предотвращения возможных случаев спаривания птиц из разных пар (Смогоржевский, 1950). Среди птиц мы находим много ви-дов-моногамов (снегирь, сорокопут-жулан, длиннохвостая синица, вра-новые и др.), у которых сезонное постоянство пар поддерживается и без регулярного и громкого пения самцов, вообще для них не характерного, а среди птиц-полигамов встречается немалое количество видов (фазан, перепел, коростель <и др.), самцы которых чрезвычайно активно токуют в течение весьма длительного времени и на совершенно определенных участках. Значение моногамии в процессе размножения певчих птиц несомненно, но пение самцов, как нам кажется, с этим явлением не связано.
Во всех случаях активного пения и токования самцов на строго определенных местах, будь это в начале сезона размножения или в конце его, у моногамных видов или у полигамов создаются наиболее благоприятные условия для встречи самца и свободной самки. В этом, по нашему мнению, и заключается биологический смысл громкой и регулярной весенней песни и фиксированного местонахождения токующих самцов у большинства птиц. Это наглядно выявляется на примере поведения очень редких в той или иной местности видов певчих птиц, но регулярно там гнездящихся. К таковым на территории Ленинградской области можно отнести, например, зеленую пеночку, гнезда которой здесь находил А. Н. Промптов (1949). Самцы этого вида, как известно,
29
в период размножения ведут себя весьма активно. Песня у них очень звонкая, издается часто и регулярно и, как правило, в одном и том же* месте в течение длительного времени. При чрезвычайно малой численности этого вида в Ленинградской области (1—3 встречи за сезон при почти ежедневном экскурсировании!) вряд ли могли бы самки зеленой пеночки, прилетающие позднее самцов, найти себе пару и вывести потомство, если самцы не обладали бы столь звонкой песней и не имели бы своего постоянного местопребывания. То же самое можно было бы сказать о садовой камышовке, соловье, обыкновенном сверчке и о некоторых других птицах, численность которых во многих районах Ленинградской области ничтожна. В данном случае просто нелепо говорить об отпугивающем значении песни и о кормовой территории.
Строгая приуроченность настоящего пения к определенному полу также указывает на значение весеннего пения птиц прежде всего для встречи полов и никак не вяжется с идеей предостерегающего значения пения.
В заключение следует указать еще на две малораспространенные среди зоологов точки зрения по вопросу о биологическом значении птиц.
д) Пение птиц следует трактовать как вынужденное явление, обеспечивающее вентиляцию абдоминальных воздушных мешков и регулирование температуры семенников в сторону их охлаждения, что необходимо для нормального хода сперматогенеза у птиц. По мнению Г. А. Васильева (1947, 1948), пение птиц и принятие ими различных,, подчас причудливых поз «ухаживания» должны трактоваться как «безобидные упражнения» самцов, регулирующие внутреннюю температуру их семенников. Таким образом, согласно этой теории, самцы птиц, токуют и поют весной главным образом для самих себя и отнюдь не для особей другого пола.
С данной точкой зрения трудно согласиться. Громкие, регулярно повторяющиеся и специфичные для вида звуки, издаваемые в период размножения одним из полов, наблюдаются не только у птиц, но и у многих млекопитающих, амфибий и даже насекомых, т. е. у таких групп животных, у которых никак нельзя говорить о необходимости охлаждения семенников для нормального хода сперматогенеза. Полагаем, что биологический смысл этих звуков во всех случаях один и тот же — они обеспечивают встречу самца и самки.
Вообще трудно себе представить, чтобы строгая видовая специфичность издаваемых звуков и принимаемых птицами поз «ухаживания» возникла бы из-за подобной необходимости. Для этого, по нашему мнению, совсем необязательно пускать в ход голосовые средства, ибо циркуляция воздуха в абдоминальных воздушных мешках, если уже она действительно так обязательна, с большим успехом может быть достигнута другими способами, не требующими регулярного воспроизведения строго определенной комбинации звуков, называемых нами пением птиц. Если стоять на точке зрения Васильева, то становится совершенно непонятно, почему самцы резко снижают активность своего пения с момента появления самки, и наоборот, начинают снова петь интенсивнее, когда они теряют самку. Непонятно также и то, зачем во время пения самец стремится занять наиболее заметную позицию и почему позы «ухаживания» он принимает главным образом в непосредственной близости от самки. То обстоятельство, что у некоторых птиц (большая синица, пухляк, пищуха, королек, крапивник и др.) пение самцов начинается задолго до созревания половых желез, не должно нас смущать, ибо оно нисколько не противоречит взгляду, рассматри
30
вающему пение как приспособление, облегчающее встречу полов. Наблюдения показывают, что образование пар у этих птиц часто происходит задолго до того, как оба партнера станут готовыми к размножению.
е) Пение птиц есть выражение избыточной энергии, накопившейся в организме птицы (Бёме, 1952). Такое понимание птичьего пения чаще всего встречается в кругах любителей птиц, считающих, что ломать особенно голову над тем, почему поют птицы и какой биологический смысл имеет видовое пение у птиц, не стоит. Данная точка зрения не приемлема уже по одному тому, что она вообще снимает вопрос о биологическом смысле весеннего пения птиц. Наблюдения показывают, что зяблик в период своего наибольшего весеннего возбуждения воспроизводит видовую песню до 412 раз в час (Heinroth, 1938), обыкновенная горихвостка — 342 раза (Смогоржевский, 1950), лесной конек — до 418 раз (Гептнер, 1958), а зеленая пеночка — до 478 раз (наблюдения автора). Вряд ли просто от избытка энергии самцы этих, а также и других птиц столь регулярно, часто и громко воспроизводят свой строго специфичный для каждого вида напев.
Итак, весеннее пение птиц мы понимаем как видовой биологический признак, выработавшийся в процессе эволюции как приспособление, обеспечивающее или облегчающее встречу полов.
ГЛАВА II
ГНЕЗДОВАНИЕ ЛЕСНЫХ ВОРОБЬИНЫХ
ГНЕЗДОСТРОЕНИЕ
Весной, в 'пору размножения, строят гнезда самые различные птицы. Однако ни в одной из групп этого класса инстинкт гнездостроения не /Гостит такого разнообразия, полноты и совершенства, как у воробьиных. Большинство неворобьиных имеет примитивные гнезда; к тому же среди преобладающего числа отрядов этих птиц мы находим целые группы видов, вовсе не строящих гнезд. Среди воробьиных, напротив, нет таких представителей: при нормальных условиях все они строят гнезда. Большинство же сооружает довольно искусные постройки; особенно характерно это для вадов, живущих в лесу. Таким образом, лесные воробьиные могут дать наибольшее количество материала, необходимого для анализа рассматриваемого явления и обсуждения вопросов, связанных с эволюцией инстинкта гнездостроения.
Изучение инстинкта гнездостроения воробьиных важно и с практической точки зрения. Успешное привлечение этой полезной группы птиц в лесные посадки, сады и парки возможно лишь при всестороннем знании условий их гнездования. В этом отношении более или менее благополучно обстоит дело лишь с некоторыми птицами из группы дуплогнездников, обобщение материалов но гнездованию которых в естественных условиях позволило успешно разработать эффективные меры их привлечения в искусственные гнездовья (Благосклонов, 1957). Методы же привлечения полезных открыто гнездящихся воробьиных до сих пор основываются лишь на общих положениях и не могут считаться удовлетворительно разработанными, что в первую очередь объясняется недостаточностью сведений о гнездовании птиц в различных условиях.
Материалы, на основании которых составлена настоящая глава, собраны нами в основном в Ленинградской области (1946—1954) и на территории Савальского лесхоза (1951—1953). В этих районах автор вместе со своими помощниками нашел 1738 гнезд 47 видов лесных воробьиных птиц. Из них 925 гнезд (31 вид) было найдено в Савальском лесу и 813 гнезд (39 видов) —на территории Ленинградской области. Суммарные данные о их расположении приведены в соответствующих таблицах (см. табл. 3—13). Более подробные сведения по гнездованию отдельных видов изложены во второй части этой книги.1
1 Автор располагает также данными по распределению птичьих гнезд в заповеднике «Лес на Ворскле» (Белгородская область). Сообщение об этих наблюдениях опубликовано ранее (Мальчевский, Покровская, Овчинникова и Геракова. 1954).
32
Среди различных сторон жизнедеятельности птиц в период размножения выбор подходящего места для помещения гнезда является весьма ответственным актом, от которого нередко зависит успех выведения потомства. Наблюдения за поведением птиц в период, непосредственно предшествующий гнездостроению, показывают, что, прежде чем приступить к постройке гнезда, птицы некоторое время осваиваются с окружающей обстановкой. В этот период большинство воробьиных ведет себя скрытно и осторожно. Нередко, начав строить гнездо в одном месте, птица бросает его и начинает постройку в другом. Такое поведение птиц указывает на наличие у них известной избирательности по отношению к месту будущего гнезда. Какие же причины заставляют птицу остановить свой выбор на определенном месте? Для того чтобы ответить на этот вопрос, надо прежде всего знать, как распределены птичьи гнезда среди лесной растительности.
Места (расположения гнезд. В пределах лесной и лесостепной зон Европейской части СССР основными местами, на которых лесные воробьиные предпочитают располагать свои гнезда, являются взрослые деревья, подлесок, подрост, пни, хворост, поверхность земли, а также сооружения человека.
а) Взрослые деревья. Взрослый древостой является местом гнездования многих массовых видов воробьиных. В лесах и парках Ленинградской области, например, из 813 найденных гнезд (39 видов) 347 гнезд (27 видов) было на взрослых деревьях (табл. 3). Примерно такую же роль играет древостой и в Савальском лесу (269 находок из 925, табл. 4). В старых высокоствольных лесостепных дубравах, например в «Лесу на Ворскле», где имеется большое количество старых дуплистых дубов и лип, значение взрослого древостоя для гнездования птиц становится еще более заметным.
Среди воробьиных, связанных гнездованием со взрослыми деревьями, надо указать прежде всего на многочисленных птиц, поселяющихся в дуплах: галку, скворца, полевого воробья, поползня и пищуху, различных синиц, горихвостку и некоторых др. Гнездование в дуплах части воробьиных следует, видимо, расценивать прежде всего как приспособление, направленное к избежанию вредного действия хищников. Эта же причина заставляет иногда поселяться в дуплах птиц, гнездящихся обычно открыто (серая мухоловка, сойка, певчий дрозд, белобровик). Некоторые представители воробьиных находятся как бы на «полпути» к дуплогнездничеству (зарянка).
К группе птиц, гнездящихся открыто на взрослых деревьях, принадлежат большинство врановых, а также иволга, чиж, щегол, зяблик, длиннохвостая синица, мухоловки — серая и малая, дрозды — деряба, рябинник и частично белобровик, черный и певчий и некоторые др. Кроме того, на взрослых деревьях нередко гнездятся птицы, обычно располагающие свои постройки на кустах или молодой древесной поросли. Таковыми являются: зеленушка, снегирь, дубонос, лесная малиновка, крапивник и изредка даже сорокопут-жулан и славки — садовая и Черноголовка. Многие из этих птиц, поселяясь на крупных деревьях, часто используют мутовки стволовой поросли, весьма характерные для лиственных пород, а также нижние боковые ветви хвойных деревьев.
Преобладающее большинство птиц способно использовать для помещения своего гнезда самые различные древесные породы (табл. 3— 4). В то же время в каждом районе можно выделить группу пород деревьев, наиболее охотно заселяемых птицами. В различных районах эти породы могут быть разными и необязательно относящимися к числу преобладающих в данной местности. Как правило, однако, они бы-
3 А. С. Мальчевский
33
Таблица 3
G5 .1*
Распределение гнезд воробьиных по породам взрослых деревьев в Ленинградской области
Название типа Количество найденных гнезд
на хвойных деревьях на лиственных деревьях
ч о сосна лиственница береза ольха серая липа клен остролистный \о черемуха ильмовые рябина осина ивы боярышник тополь всего
Ворона серая 5 8 1 — — 1 1 — — — — — — — 1 17
Сорока 1 5 — 6
Скворец — — 1 5 — 4 — 2 — — — — — — — 12 о
Иволга — — — — «— —- 1 2 — — — — — — — О
Зеленушка 6 — 6
Чиж 1 1 2
Снегирь 2 2
Зяблик 15 5 1 11 10 3 5 2 3 3 1 — — 3 — 62
Клест-еловик 1 1 2
Поползень • — — — 1 — 2 — — — — — — — — — 3
Синица большая — — — 2 — 3 3 1 ’— — — 2 — — 1 12
Лазоревка — — — 1 — — — 1 — — — — — — — 2
Пухляк — — — — — — —— — —— — — 1 — — — 1
Синица длиннохвостая — — — 1 — — — — — — — — — — — 1
Сорокопут-жулан 1 — — — 1 — 1 — — — — — — — — 3
Мухоловка серая 7 7 1 8 2 5 — — — 2 2 2 — — — 36
Мухоловка-пеструшка — 1 — 2 1 2 — — — 1 1 5 — — — 13
Мухоловка малая — — — 1 — — — — — — — — — — — 1
Малиновка лесная 2 — 1 2 — — 1 — — — — — — — — 6
Славка садовая — — — — — — 1 — — — — — — — — 1
Дрозд-рябинник 14 12 2 13 7 1 3 1 — 4 2 — 3 — — 62
Дрозд певчий 6 18 — 3 — 1 — — 1 — — — — — — 29
Дрозд-белобровик 11 4 1 2 15 4 2 1 8 1 1 1 1 — — 52
Дрозд черный — — — — 1 — — — — — — — — — — 1
Горихвостка обыкновенная .... — — — 1 — 1 — 3 — — 3 — — — — 8
Зарянка — — — 1 — 1 — — — — — — — — — 2
Крапивник — — — — 1 — — — — — 1 — — — — 2
Итого, . . . , 7? | 62 ) 1 8 | 54 38 | 28 | 18 I 1 13 12 11 1 " 11 4 3 2 1 347
Таблица 4
Расположение гнезд воробьиных на взрослых деревьях в Савальском лесу
Количество найденных гнезд
на хвойных деревьях
на лиственных деревьях
Название вида
х
о
1
о
М о
X
3 ю X
и
s
х х
05 X
о х
и
о
х
3
х
1
1
2
Сорока ..............
Сойка ...............
Скворец..............
Иволга...............
Дубонос .............
Зеленушка...........
Щегол...............
Зяблик ..............
Синица большая . . . Лазоревка ...........
Сорокопут-жулан . . . Мухоловка серая . . . Малиновка лесная . . . Дрозд-деряба.........
Дрозд-рябинник .... Дрозд певчий ........
Дрозд черный.........
Горихвостка обыкновенная .................
7
10
24
2 14
3
3
16
23
2
8
10
Итого. .
79
11
2
5 62 51 29
П 19
5
14
5
5
2
48
10
2
3
61
2
2
20
13
32
10
1 269
5
2
9
1
1
2
2
2
1
3
3
9
1
4
2
4
2
8
2
6
1
1
1
2
1
1
1
2
I
1
4
6
2
1
1
1
1
1
1
1
1
2
1
1
5
4
И
3
2
8
1
7
3
7
3
3 1
2
вают общими для очень многих видов. Это указывает на то, что известная избирательность у птиц существует, iho что обусловливается она не частными, а какими-то общими требованиями, сходными для многих птиц. Даже у таких, казалось бы,'узко специализированных в отношении характера гнездования видов, как клест и чиж, не существует строгой привязанности к определенной породе дерева. В Ленинградской области свои гнезда они располагают как на елях, так и на соснах, т. е. на породах, имеющих совершенно различную структуру ветвей.
Иволга и лесная малиновка, обычно зачисляемые в группу птицг гнездование которых тесно связано с лиственными породами деревьев,, на самом деле охотно селятся также и на хвойных: лесная малиновка, часто помещает гнездо на елях, иволга — на соснах. То же можно сказать о щегле. Среди певчих птиц нашей фауны, кажется, лишь один желтоголовый королек пока что может служить примером вида, гнездящегося лишь на одной породе—на елях (Palmgren, 1932).
Большинство мелких воробьиных, связанных гнездованием с древостоем, при выборе места для помещения гнезда предъявляет к растительности сходные требования. Важно, чтобы ветви или изъяны в стволе дерева являлись достаточно надежной опорой, могущей обеспечить устойчивое положение гнезда, а листва — скрыть кладку, птенцов и самих птиц от глаз хищников. На севере этим требованиям лучше всего отвечает ель, а в южных лесах — широколиственные породы: дуб,, вяз и отчасти липа, способные образовывать мутовки стволовой поросли 3* 35
на сравнительно небольшой высоте, столь охотно занимаемые мелкими птицами. Наименее 'благоприятные условия для строительства гнезд создают кроны ясеня и осины.
В одних случаях птица находит удобную для укрепления гнезда и хорошо укрытую мутовку сразу, в других — после ряда неудачных попыток построить гнездо в мало подходящем месте. Об этом свидетельствуют часто встречающиеся в природе неоконченные постройки, брошенные птицами, а также прямые наблюдения за поведением птиц. В процессе наблюдения за строительством гнезда серой вороны автору удалось установить, что птица, прежде чем окончательно построить гнездо, три раза меняла место «закладки» фундамента своего сооружения. Первоначально ворона таскала сучья в основание развилки двух крупных ветвей дерева. Вскоре, однако, весь принесенный ею материал вследствие весьма неустойчивого его положения упал на землю. Это произошло в присутствии самой птицы. Следующую ветку она принесла, уже на соседнее дерево, где облюбовала место столь же -неудобное, как и первое, ибо уже после четвертого ее прилета принесенные сучья опять посыпались вниз. В третий раз ворона начала постройку в месте, более удобном, и на этот раз закончила ее успешно.
б) Подлесок и подрост. Когда говорят о гнездовании птиц в нижних ярусах леса, очень часто не различают подроста и подлеска, в; связи с чем роль каждого из них для гнездования птиц остается почти невыясненной. Анализ собранного материала позволяет говорить о том, что значение кустарников в гнездовой жизни птиц обычно преувеличивается, а роль подроста и кустообразной поросли древесных пород, наоборот, недооценивается. Дело в том, что многие породы кустарников (сирень, боярышник, калина, бересклеты, желтая акация, крушина, орешник) в гнездовой период мало привлекательны для птиц. При естественном произрастании ветви их сильно вытягиваются и не образуют мутовок, пригодных для укрепления гнезда. Они становятся удобными для гнездования птиц лишь после специальной их подрезки или механического повреждения ветвей.
Как уже отмечалось в литературе (Доппельмаир, 1939), в природе иногда имеет место и естественная подстрижка кустарников — обгрызание верхушечных побегов скотом и дикими копытными. На Карельском перешейке нам неоднократно приходилось находить, например, гнезда садовых славок, помещенные на значительном удалении от земли (до 3,5 м) в правильных мутовках прибрежного ивняка, образовавшихся в результате обгрызания кустов лосями. Эти мутовки, представляющие собою три расходящиеся под углом побега, очень удобны для укрепления гнезд славки.
При естественном произрастании лишь немногие кустарники привлекают на гнездовье разнообразную фауну птиц. К таковым следует прежде всего отнести спирею рябцнолистную, а также красную бузину. Введение последней в качестве подлеска в сосновые культуры Саваль-ского лесхоза чрезвычайно обогатило местный лес орнитофауной. Здесь в 1951—1953 гг. в кустах бузины нами было найдено 146 гнезд 17 различных видов воробьиных и только 36 гнезд на всех остальных породах кустарников (бересклет бородавчатый, желтая акация, жимолость, малина и др.), достаточно многочисленных в Савальской лесной даче (табл. 5). Особенно часто в кустах бузины нам приходилось находить гнезда зеленушки и сорокопута-жулана. Поселялись здесь также сорока, сойка, дубонос, зяблик, различные виды славок, лесная малиновка, черный и певчий дрозды. Привлекательность бузины как места гнездования для большого количества видов птиц объясняется тем, что
36
Таблица 5
Распределение гнезд воробьиных по породам кустарников и травянистых растений в Савальском лесу
Количество найденных гнезд
Название вида
Сорока ............
Сойка .............
Дубонос ...........
Зеленушка..........
Зяблик ............
Овсянка обыкновенная Сорокопут-жулан . . Мухоловка серая . . Малиновка лесная . . Славка ястребиная . . Славка-черноголовка Славка серая .... Славка-завирушка . .
Славка садовая . . . Дрозд-рябинник . . . Дрозд певчий .... Дрозд черный ....
1
6
5
26
3
1
43
1
5
1
7
1
4
4
17
21
11-1
1
6
5
26
3
1
46 I
5
1
16 3
1
25 4
17 21
Итого
146 12 6 6 4 2 1 1 1 1 1 1 182
этот кустарник удобен не только для .птиц, укрепляющих свои гнезда обычно среди тонких веточек, но и для птиц, требующих для помещения своих сравнительно 'Крупных построек более надежной опоры. Гнезда, ^расположенные среди ветвей бузины, вследствие ее густой листвы обычно бывают хорошо скрыты от глаз хищников, что имеет немаловажное значение для мелких птиц.
Спирея рябинолистная, встречающаяся в большом количестве в наших парках и садах, более всего привлекательна для славок — садовой и Черноголовки. Однако на ветвях этого кустарника нам приходилось находить также гнезда сорокопута-жулана, чечевицы, садовой камышовки, лесной малиновки, славки-завирушки и дрозда-белобровика (табл. 6).
Бузина, спирея рябинблистная, отчасти татарская жимолость, барбарис и шиповник хорошо заселяются птицами, но они, как правило, не имеют широкого распространения в наших лесах, и поэтому роль их вообще невелика. Савальский лес — это единственное место, где нам пришлось наблюдать концентрацию птичьих гнезд в кустарниках. Если бы, однако, в этом лесу не было красной бузины, искусственно введенной и распространенной самими же птицами, то роль кустарников в гнездовании птиц была бы и здесь совсем мало заметной.
Подрост, особенно кустообразная поросль его, часто встречающаяся в лесу, имеет для гнездования мелких воробьиных птиц обычно большее значение, нежели подлесок (кустарники). Это было установлено нами в «Лесу на Ворсиле» (Мальчевский, Покровская, Овчинникова, Геракова, 1954). В Ленинградской области роль подроста и кустообразной древесной поросли оказалась еще более заметной (табл. 7).
37
Таблица 6
Распределение гнезд воробьиных по породам кустарников и травянистых растений в Ленинградской области
Название вида Количество найденных гнезд
спирея рябинолистная бузина акация желтая боярышник смородина малина сирень крушина шиповник жимолости барбарис жасмин хмель папоротник орешник всего
Коноплянка *. 1 1
Чечевица . 2 — 3 1 6
Сорокопут-жулан . . . 3 — — 3 1 — 2 1 1 1 — — — — — 12
Камышовка садовая . . 1 1 1 3
Малиновка лесная . . . 1 1 2
Славка-черноголовка 4 1 1 1 — 7
Славка серая — 1 — — 1 1 — — — — — 1 — — — 4
Славка-завирушка . . . 2 2
Славка садовая . . . 8 4 2 1 1 — 16
Дрозд-белобровик . . . 1 2 3 1 — — 2 1 1 1 12
Крапивник — — 2 — — — — — — — — — — — — 2
Итого. . 22 10 9 4 4 4 4 2 2 1 1 1 1 1 1 67
Таблица 7
Количество гнезд воробьиных, найденных в подлеске и в подросте
Район наблюдения Количество найденных гнезд
в подросте в подлеске
Учлесхоз „Лес на Ворскле* .... 64 31
Ленинградская область 202 67
Итого 266 98
Породы подроста, избираемые птицами для помещения гнезда, более определенны, чем породы взрослых деревьев. В лесостепной полосе птицы чаще всего используют поросль и /молодые деревца ильмовых п дуба. В Савальском лесу на маленьких вязах гнездятся даже серые мухоловки. Здесь в 1951—1953 гг. на вязовом и дубовом подросте нами было найдено 14 Жилых гнезд этого вида (табл. 9).
В лесной зоне очень многие виды, гнездящиеся в нижних ярусах леса, отдают явное предпочтение еловому подросту (табл. 8). Особенно охотно заселяют его зеленушка, коноплянка, снегирь, чечевица, сорокопут-жулан, славки — Черноголовка н садовая, певчий дрозд и белобровик. Интересно, что среди перечисленных видов мы встречаем тех же птиц, которые в Савальском лесхозе гнездились преимущественно в кустах бузины (зеленушка, сорокопут-жулан, певчий дрозд). Этот-пример наглядно демонстрирует справедливость мысли, высказанной Н. А. Северцовым еще в 1855 г. (изд. 1950 г., стр. 91), указывавшего на то, что выбор породы для постройки гнезда «зависит от местных обстоятельств и вместе с ними изменяется».
38
Таблица 8
Распределение гнезд воробьиных на молодых деревцах (до 3 м) и кустообразной поросли древесных пород (Ленинградская область)
^Название вида
Количество найденных гнезд
на хвойных породах
на лиственных породах
Зеленушка.........
Коноплянка . . . . Снегирь ..........
Чечевица .........
Зяблик............
<орокопут-жулан . . Мухоловка серая . . Малиновка лесная . Славка-черноголовка Славка-завирушка . Славка садовая . . . Дрозд-рябинник . . .Дрозд певчий . . . Дрозд-белобровик . Крапивник .........
2 4 3 8 4
9
2 12
1
7
7
21
25
3
3
1
~2
4
13
4
2 9
3
10 11 23
1 9
30 1
22 8
36 33
4
Итого. .
108
202
1
в) Старые пни и кучи хвороста. Имеющиеся в лесу кучи хвороста, .а также старые пни используются для помещения гнезда целым рядом .видов .воробьиных. В северных лесах, где обычно бывает много гнилых, трухлявых обломков стволов и 'пней, последние, как известно, являются излюбленным местом гнездования зарянки, серой мухоловки, дрозда-•белобровмка, а также пухляка и хохлатой синицы. В кучах хвороста располагают свои гнезда крапивник, лесная завирушка, черный дрозд, ^сорокопут-жулан, пеночка-теньковка. На низких пнях можно найти даже гнезда певчего дрозда и рябинника.
В старых, выгнивших, полуоткрытых пнях нередко на уровне земли гнездятся иногда горихвостка, черный дрозд и даже серая мухоловка. В Савальском лесу в одном из низких березовых пней с прогнившей сердцевиной поселилась большая синица, расположившая свое гнездо на дне вертикального хода.
• г) Сооружения человека. Помимо галки, полевого воробья, скворца, большой синицы, серой мухоловки и обыкновенной горихвостки, гнездование которых в постройках человека — явление обычное, среди группы лесных воробьиных можно указать еще на несколько видов, использующих иногда сооружения человека. Это — дрозд-белобровик, певчий дрозд и рябинник, а также крапивник. Гнезда последнего вида мы находили между балок в старых землянках и блиндажах. Дрозды чаще всего поселяются в старых, заброшенных домах или сараях, расположенных неподалеку от леса. Такой способ гнездования особенно ^характерен для птиц, населяющих Финляндию (Siivonen, 1939) и Ка-
39
Табл и'ц а &
Распределение гнезд воробьиных на молодых деревцах (до 3 м) и кустооб* разной поросли древесных пород в Савальском лесу
Название вида
Количество найденных гнезд
на хврйных породах
на лиственных породах
Сорока ............
Иволга.............
Дубонос ...........
Зеленушка..........
Щегол..............
Зяблик ............
Сорокопут-жулан . . Мухоловка-серая . . Пецочкатеньковка . Малиновка лесная . . Славка-черноголовка Славка серая . . . . Славка-завирушка . . Славка садовая . . . Дрозд-рябинник . . . Дрозд черный . . . .
Итого.. 3 7 17
1 2
1 1
1
1
3
2
1
4
б
14
2
2
3
4
12
28
1
3
87
рельский перешеек (наблюдения автора). Поселяясь в постройках^ дрозды обычно помещают гнездо на верхних балках снаружи или внутри дома или сарая, почти всегда под крышей. На Карельском перешейке в районе ст. Кузнечное (Приозерский район) подобным, образом гнездятся 3 вида дроздов — рябинник, белобровик и певчий.
Иногда на различных сооружениях человека гнездятся и такие-птицы, которых даже трудно заподозрить в этом. Известны, например, находки гнезд зяблика на балках строений (Kalitsch, 1927), длиннохвостой синицы на плетне (Гавриленко, 1929) и сойки в углу стропил на балке пожарной вышки, сооруженной среди леса (устное сообщение Н. П. Кадочникова).
Таким образом, использование человеческих сооружений в целях гнездования — явление достаточно широко распространенное среди лесных воробьиных. Многочисленные находки гнезд в различных постройках, а также на грибах трутовиках, наростах и т. п., свидетельствуют о том, что для помещения гнезд птицы часто нуждаются не столько в. самих деревьях и кустарниках, сколько в тех механических и защитных условиях, которые создает им растительность (рис. 2, 3). .
д) Поверхность земли. Для известной части воробьиных в период размножения наиболее надежным убежищем оказалась поверхность, земли. Наземное гнездование характерно, например, для обыкновенной и садовой овсянок, лесного конька, лесного жаворонка, лугового чекана, варакушки, восточного соловья и почти для всех пеночек. За; исключением пеночек, перечисленные виды не могут быть названы типичными лесными птицами. Они гнездятся обычно на опушках, выруб-
40
ис. 2. Гнездо дрозда-рябинника на грибе-трутовике (ст. Кузнечное» май 1956 г. Фото автора).
ис. 3. Гнездо дрозда-рябинника под крышей сарая (ст. Кузнечное, июль 1954 г. Фото В. А. Попова).
жах или среди кустарников вне леса. Их ближайшие родственники яв-.ляются представителями полей, лугов и степей. Сами они, видимо, недавно заселили лесные биотопы. Поселяясь иногда среди настоящего леса, эти птицы сохраняют свой первоначальный наземный характер гнездования, единственно В'озможный ib открытых стациях;- Труднее понять причину наземного гнездования пеночек, которые, судя по всем повадкам, являются настоящими лесными птицами.
Можно предположить, что пеночки некогда помещали .свои гнезда среди ветвей деревьев или кустарников, как это делает большинство настоящих лесных воробьиных. У * некоторых видов пеночек способность помещать гнездо над поверхностью земли сохранилась и сейчас (пе-ночка-теньковка, зеленая пеночка и некоторые др.). Однако и у этих видов наблюдается тенденция располагать свою постройку у самой •земли или на земле. Очевидно, поверхность почвы является для пеночек более надежным убежищем, нежели ветви деревьев и кустарников. Вряд ли данное приспособление выработалось у них в связи с необходимостью избежать неблагоприятные климатические условия, вообще редко являющиеся у лесных воробьиных причиной гибели кладки или птенцов. Больше оснований полагать, что наземное гнездование явилось следствием частого разорения их гнезд пернатыми врагами.
Случаи наземного расположения гнезда известны также для многих видов дроздов и славок, горихво&гки, сорокопута-жулана. Серая славка, например, свою постройку так же часто помещает на земле, дсак и над землей. Часто гнездится на земле и дрозд-белобровик: из 143 гнезд этого вида, найденных нами в Ленинградской области, 20 (т. е. около 15%) располагалось прямо на земле, среди кустиков черники, мха, злаков и другой растительности. Высота расположения гнезд у этого вида вообще очень незначительная: более половины их (82 из 143) помещалось не выше 1 >м от земли.
Высота расположения гнезд. Приведенный выше пример с дроздом-белобровиком является лишь частным случаем более общей закономерности: расположение гнезд на незначительном удалении от земли характерно для большинства мелких открыто гнездящихся лесных воробьиных. Наоборот, крупные виды воробьиных (ворон, грач, серая ворона) гнездятся, как правило, на значительной высоте (табл. 10, И).
Среди мелких певчих птиц нашей фауны на значительном удалении от земли часто гнездятся клест-еловик, иволга, дрозды — рябинник и деряба; однако и для этих птиц высокое расположение гнезда не является правилом. Для дрозда-рябинника, например, большая высота гнездования характерна в основном на территориях, густо населенных, где этих птиц часто тревожат. В местах же, удаленных от человеческих поселений, они нередко гнездятся на небольших елочках, на л высоте 1 —1,5 м от земли. У ’дуплогнездников высота гнездования варьирует обычно сильнее и в среднем бывает более значительной, чем у птиц, строящих открытые гнезда.
Низкое расположение гнезд мелких воробьиных под лесным пологом или на нижних ветвях деревьев отчасти объясняется тем, что здесь, в нижних «этажах» лесной растительности, птицы находят более благоприятные микроклиматические условия и больше мест, удобных для укрепления гнезда.1 Громадное значение имеет и то, что низко распо-
1 Это особенно справедливо для северной тайги, где приуроченность гнезд к нижним ярусам леса связана преимущественно со своеобразной архитектоникой деревьев и более благоприятными внизу микроклиматическими условиями (примеч. ред.).
42
Таблица 10
Высота расположения гнезд воробьиных на территории Ленинградской области
Количество найденных гнезд
на высоте (в м)
на поверхности земли
Название вида
6-8
8-Ю 10-1515-20
до 1 1-2 2-3 3-4 4—6
Ворона серая — — — — — 1 — 1 11 5
Сорока — — — — 5 — — — ——
Скворец — 1 4 2 7 8 3 1 — —
Иволга . 1 1 — — —— — 1 —
Зеленушка —. — 2 1 1 4 ' — — — —
Чиж Л — - — — 2 — — —
Коноплянка 10 —
Снегирь — 3 2 — — — — — —
Чечевица — 11 4 1 — — — — — —
Зяблик — 24 24 12 7 5 1 — —
Клест-еловик — — — 1 1 — —
Овсянка обыкновенная . 5
Поползень __ 1 2 — — —
Синица большая .... — 3 4 3 4 2 1 — —
Лазоревка — — 1 — 1 — — — —
Пухляк — 1 1 — — — — — —
Синица длиннохвостая . — — — 1 — — — —
Сорокопут-жулан . . . — 23 10 5 — — — — — —
Мухоловка серая . . . 1 11 17 19 6 6 — — —. —
Мухоловка-пеструшка . — — 10 2 3 4 — — — —
Мухоловка малая . . — — — — — — 1 — —
Леночка-весничка . . . 31
Пеночка-трещотка . . 14
Конек лесной 13
Камышовка садовая . . — 3
Малиновка лесная . . . — 1 9 2 2 2 1 — — —
Славка-черноголовка — 26 10 1 — — — — — —
Славка серая 4 4
Славка-завирушка . . . — 3 — — — — — — — —
Славка садовая .... 3 30 5 2 1 1 —. — — —
.Дрозд-рябинник .... — 1 14 26 13 15 1 6 5 —
Дрозд певчий 1 20 28 15 2 2 1 1 — —
Дрозд-белобровик . . . 20 62 45 13 3 — — — — —
Дрозд черный — 1
Чекан луговой .... 6
Горихвостка обыкновенная 1 4 6 1
Соловей восточный . . 2
Зарянка — 3 1 — — — — — — —
-Крапивник — 7 3 — — — — — — —
♦ Итого. . 1 100 222 199 126 61 56 15 12 17 5
-ложенное гнездо менее заметно для глаз пернатых врагов, которых у мелких воробьиных птиц очень много.
Престон и Норрис (Preston and Norris, 1947) на примере птиц Западной Пенсильвании показали, что количество птичьих гнезд в лесу закономерно убывает по направлению к вершинам деревьев. Думается, однако, что указанные авторы не смогли бы установить подобной закономерности, если бы мелкие воробьиные птицы, гнездящиеся в нижних «этажах» леса, не составляли бы преобладающего большинства лесного пернатого населения. Вряд ли поэтому правы Престон и Норрис,
43
Таблица 1Г
Высота расположения гнезд воробьиных на территории Савальского лесхоза
Название вида Количество найденных гнезд
на поверхности земли на высоте (в м)
до 1 1-2 2-3 3—4 4-6 6-8 8-10 10—15 15-20
Сорока 3 4 3 3
Сойка — — И 9 5 1 — 1 — —
Скворец — — 2 — 11 16 26 3 2 — -
Иволга — — — 1 4 5 1 2 2 —
Дубонос — — 8 3 — — 2 — — —
Зеленушка — — 24 7 2 — — — —
Щегол — — 1 2 — — — — — —
Зяблик — — 13 16 7 11 5 2 1 —
Овсянка обыкновенная . 25 — 1 — — — — —р —
Овсянка садовая 38
Синица большая .... — 3 3 15 28 10 — — — —
Лазоревка — — 2 — 2 2 — — — —
Сорокопут-жулан . . . — 27 25 3 1 — — — — —
Мухоловка серая . . . 2 20 30 14 4 7 5 1 1 —
Мухоловка белошейка . — — — — 10 10 8 6 — —
Пеночка-теньковка . . 2 3 — — — — — — —
Пеночка-трещотка . . . 23
Конек лесной 60 — —
Малиновка лесная . . . — 2 5 — — — 2 — — —
Славка-ястребиная . . . — 1 — — — — — — — —
Славка-черноголовка — И 4 4 — — — — — —
Славка серая 1 7 — — — — — — — —
Славка-завирушка . . . — 13 — — — — — — — —
Славка садовая .... 2 50 3 — — — — — — —
Дрозд-деряба — — 1 — — — 1 — — —
Дрозд-рябинник .... — — 7 4 2 6 2 3 2 —
Дрозд певчий 1 6 19 7 2 1 — — 1 —
Дрозд черный Горихвостка обыкновен- 10 20 25 7 3 2 2 — — —
ная 11 5 6 12 12 — 1 1 — —
Соловей восточный . . 27
Зарянка — 1 —
Итого. . 202 169 193 108 96 74 55 19 9 —
объяснявшие данную закономерность будто бы существующим у всех птиц стремлением три отсутствии отпугивающего влияния человека и домашних животных гнездиться на земле. Такое объяснение совершенно игнорирует наличие исторически сложившихся связей между лесной растительностью и птицами, выражающихся в многообразных (Приспособлениях к гнездованию на ветвях деревьев.
Итак, для успеха размножения птиц прежде всего важно, чтобы гнездо было расположено в недоступном или малозаметном месте. Большинство мелких птиц прячет свои гнезда невысоко над землей — среди нижних ветвей деревьев, в древесной поросли и кустарниках, отдавая при этом предпочтение породам, которые помимо удобных мутовок имеют достаточно густую крупную листву или хвою. Именно такие породы деревьев и кустарников следует в первую очередь вводить в. лесные культуры и использовать при организации «птичьих городков».
Хорошо защищенная и удобная для укрепления гнезда мутовка,, расположенная на высоте от 0,5 до 3 м, всегда будет привлекать вни
44
мание птиц. Стремиться создавать возможно более густые мутовки путем ежегодной подрезки кустарников и молодых деревьев на одном уровне не следует: слишком густая «щетка» ветвей часто только мешает гнездованию. На тех участках, где проводятся 'мероприятия по привлечению птиц, очень важно также помимо взрослых деревьев и кустарников иметь поросль лиственных пород, молодые елочки и кусты можжевельника. Помимо того, что сама эта поросль может служить местом гнездования древесных и кустарниковых птиц, она будет создавать также необходимое укрытие для птиц, располагающих свои гнезда на земле.
О ПЛАСТИЧНОСТИ ГНЕЗДОВЫХ инстинктов
У большинства воробьиных характер расположения гнезд не бывает строго определенным. У дрозда-белобровика, например, гнезда могут быть на земле, в кустах, на пнях, на взрослых деревьях, в постройках человека; обыкновенная горихвостка может гнездиться почти открыто на земле, в выгнивших пнях или изъянах стен каменных построек и в то же время поселяться в глубоких дуплах и скворечниках; зарянка помещает свои гнезда в дуплах, но часто гнездится и открыто между корней деревьев или у оснований гнилых пней. Серая мухоловка использует для устройства гнезда изъяны стволов, пни, карнизы зданий, ветви крупных деревьев, но в некоторых случаях располагает свою постройку на молодых деревцах, кустах и даже на земле и т. п.
Степень внутривидового разнообразия -гнездостроения у разных видов различна. Существуют так называемые «эв^и-нидные» виды с разнообразным характером гнездования и «стенонидные» виды, положение гнезда у которых более определенно (Формозов, 1934). Деление птиц на указанные группы до известной степени условно, ибо так называемые «стенонидные» виды также нередко обнаруживают способность видоизменять свои гнездостроительные инстинкты и располагать гнезда в совершенно необычных местах. Например, типичный дуплогнездник — большая синица — при наличии благоприятных кормовых условий может гнездиться в молодых лесопосадках, где нет естественных дупел, поселяясь в закрытых сверху сорочьих гнездах. В отдель-. ных случаях этот вид может переходить даже к открытому гнездованию, располагая свое гнездо на основе старого гнезда другого вида. Обыкновенная овсянка, гнездящаяся на земле, в отдельных случаях помещает свое гнездо на кустах; сойка, гнездящаяся обычно на деревьях, вьет гнезда также в кустах, а иногда устраивается на бревнах сооружений и даже поселяется в дуплах (Dereck, 1953; Tutt, 1953); полевой воробей, являясь дуплогнездником или обитателем человеческих построек, в отдельных случаях, как это характерно и для домового воробья, сооружает шарообразное гнездо среди ветвей деревьев (Ка-litsch, 1927).
Об изменчивости инстинкта гнездостроения свидетельствуют также данные анализа строительного материала гнезд воробьиных, на что указывал еще А. Н. Промптов (1946). Известно, например, что многие лесные птицы, поселяясь вблизи человеческого жилья, могут вить гнезда из ваты, бумаги, бинтов, тряпок, веревок, пряжи, ниток, опилок и даже обрезков тонкого целлофана. Собирается, по-видимому, все, что в той или иной степени подходит по своим механическим свойствам.
Птицы, однако, неодинаково относятся к такого рода искусственному материалу. Некоторые используют его весьма охотно, другие — в значительно меньшей степени, наконец третьи почти никогда не со-
45
бирают его, несмотря на то, что очень часто селятся в таких местах,, где он имеется в изобилии. Способность использовать для постройки гнезда вату, бумагу, тряпки и т. п. была отмечена у зяблика, зеленушки, серой славки (Промптов, 1946), серой мухоловки, лесной малиновки, сорокопута-жулана (Мальчевский, 1950), мухоловки-пеструшки (Благосклонов, 1957), белой трясогузки (Лукина, 1953), а также у иволги, елавки-черноголовки, чижа, конорлянки, полевого воробья (наблюдения автора). Наоборот, в гнездах певчего дрозда, дрозда-белобровика, соловья, пеночки-трещотки, лесного конька, обыкновенной, овсянки и других птиц подобных материалов, кажется, еще никто не находил. Таким образом, по отношению к строительному материалу, так же как и по месту расположения гнезда, воробьиных можно груба разделить на эври- и стенонидных. В природе имеют место и индивидуальные вариации формы, величины и конструкции самих построек,,, о чем речь у нас будет несколько позднее.
Учет различных отклонений от видового «стереотипа» гнездостроения дает материал для теории эволюции этого важнейшего биологического признака птиц. На значение подобного рода исследований в-нашей литературе впервые было обращено внимание А. Н. Промпто-вым (1945, 1946). Обсуждение вопроса было позднее продолжено Е. В. Лукиной (1953). Автор настоящей работы также обращал внимание на степень пластичности гнездостроительных инстинктов. Помимо некоторых фактов, опубликованных ранее (Мальчевский, 1950; Мальчевский и др., 1954), удалось собрать новые материалы, часть которых сведена в табл. 12, 13. Цель этих таблиц — показать возможную степень изменчивости инстинкта гнездования у некоторых лесных воробьиных в пределах одного района.
Ознакомление с таблицами показывает, что особи одного вида, живущие бок о бок в сходных условиях, например среди сосновых насаждений Савальской дачи, могут, тем не менее, располагать свои гнезда по-разному. В нескольких десятках шагов от гнезда обыкновенной горихвостки, помещающегося в дупле дерева, можно найти другое гнездо этого же вида, но на земле, под валяющейся сосновой веткой. В одном и том же лесном квартале черные и певчие дрозды, сойкиг зеленушки, зяблики и многие другие птицы могут гнездиться как на-крупных деревьях, так и в кустах бузины и т. п.
Интересно, что в зависимости от места расположения гнезда* обычно меняется форма и конструкция самой постройки. Например, серые мухоловки, гнездящиеся в полудуплах, вьют, как известно,,, довольно небрежные гнезда, имеющие различную толщину стенок, уплощенную форму и неглубокий лоток. Совсем иной вид имеет гнезда этой птицы, расположенное среди тонких ветвей молодого деревца, например на вязовом подросте.. Такие гнезда необычны по своей форме, конструкции и даже строительному материалу. Они скорее походят на постройку зяблика, нежели на обычные гнезда серой мухоловки, так как имеют высокие и плотные боковые стенки равной толщины, глубокий лоток, а снаружи убраны коконами наездника.
Весьма упрощенное строение имеет иногда гнездо дрозд а-бел о б ровика на земле. Одно из гнезд этого вида, найденное автором на севере Карельского перешейка, состояло лишь из небольшого количества сухой травы, выстилавшей дно ямки, в которой оно помещалось. Самога каркаса гнезда, изготовляющегося дроздами обычно из земли, не было. Боковыми стенками ему служили, с одной стороны, камень, с другой — плотный мох. Таким образом, в связи с особым местом расположения гнезда у данной особи выпал целый ряд звеньев инстинкта, и весь акг
46
Таблица 12й
Вариации расположения гнезд лесных воробьиных на территории Савальского лесхоза
Количество найденных гнезд
закрытое гнездование открытое гнездование
Название вида уплах дере-в нилых пнях, tax хвороста и ъ местах [остройках овека и скво- никах взрослых евьях одросте и корой поросли кустах и на вах пнях и кучах роста а* о СО >> а CU о а о о земле о
о f, j — С Ч tr О О CU со С И О со СО CU о со са о ч о СО о
а а Й » h са tr си Д *=С и а а н а х са ег а а
Сорока — — 11 1 1 13
Сойка + — — 19 — 6 — 2 — 27'
Скворец 5 — 55 — — — — — 60'
Иволга — — — 14 1 — — — — 15
Дубонос — — — 5 3 5 — — — 13
Зеленушка — — — 5 2 26 — — — 33
Щегол — — — 2 1 — — — — 3
Зяблик . — — — 48 4 3 — — — 55
Овсянка обыкновенная . — — — — — 1 — — 25 26
Овсянка садовая . — — — — — — — 38 38
Синица большая . . . 9 1 48 1 — — — — — 59
Лазоревка 2 — 4 — — — — — — б
Сорокопут-жулан . . . — — — 3 6 46 1 — — 53
Мухоловка серая . . . 1 — — 60 14 1 5 1 2 84
Мухоловка-белошейка . — — 34 — — — — — — 34
Пеночка-теньковка — — — — 2 — 1 — 2 5
Пеночйа-трещотка . . — — — — — — — — 23 23
Конек лесной — — — — — — — — 60 60
Малиновка лесная . . . — — — 2 2 5 — — — 9
Славка ястребиная . . — — — — — 1 — — — 1
Славка-черноголовка — — — 3 16 — — — 19
Славка серая . ... — — — — 4 3 — — 1 8
Славка-завирушка . . . — — — — 12 1 — — — 13
Славка садовая . . — — — — 28 25 — — 2 55
Дрозд-деряба — — — 2 — — — — — 2
Дрозд-рябинник .... — — — 20 1 4 1 — — 26
Дрозд певчий .... — — — 18 — 17 1 — 1 37
Дрозд черный — — — 32 3 21 3 — 10 69
Горихвостка обыкновенная 10 12 22 4 48
Соловей восточный . . — — — — — — — — 27 27
Зарянка — — — — ~ ! — 1 — — 1
Итого. . 27 13 163 242 87 182 13 3 195 925
Примечание. Гнезд соек, расположенных в дуплах, автор не находил; они были обнаружены Н. П. Кадочниковым и Ю. К. Эйгелисом в 1954 и 1955 гг.
гнездостроения свелся к последней стадии — к приготовлению внутренней выстилки лотка.
Неодинаковую конструкцию имеют гнезда славок-черноголовок, расположенные в вертикальной мутовке и в горизонтальней развилке ветви. В первом случае гнездо имеет обычную чашеобразную форму, во втором — вид гамачка, напоминающего в миниатюре гнездо иволги
47
Таблица 13
Вариации расположения гнезд лесных воробьиных в Ленинградской области
Количество найденных гнезд
закрытое гнездование открытое гнездование
Название вида X 6 са СО X я
О) г=С к Я Я X о я Я X Я Я я
X X □Я о S * ч н х я со Я СО <d
СО 3 П, Я Я ” Q о. « СО я СО X н >> Я я
Я ч Я о са я о о я Н X О СО Я о I-1 о я S о о
О Я е 3 5 (са с О X СО е S 8 ч О)
я я са и s1 а Я Си са я н я я Я я
Ворона серая — — 17 17
Сорока — — — 6 — — — — — 6
Скворец 12 — 14 — — — — — — 26
Иволга — — — 3 — — — — — 3
Зеленушка — — — 6 2 — — — — 8
Чиж — — — 2 — — — — — 2
Коноплянка — — — 9 1 — — — 10
Снегирь — — — 2 3 — — — — 5
Чечевица — — — — 10 6 — — — 16
Зяблик — — — 62 11 — — — — 73
Клест-еловик — — — 2 — — — — — 2
Овсянка обыкновенная . 3 5 5
Поползень — — — — — — — — 3
Синица большая .... 12 — 5 — — — — — — 17
Лазоревка 2 1 2
Пухляк 1 — — — — — — — 2
Синица длиннохвостая . — — — 1 — — — — — 1
Сорокопут-жулан . . . — — — 3 23 12 — — — 38
Мухоловка серая . . . — — — 36 1 — 4 18 1 60
Мухоловка-пеструшка . 13 — 6 1 — — — — — 19
Мухоловка малая . . . — — — — — — — — 1
Пеночка-весничка . . . 31 31
Пеночка-трещотка . . . — — — — — — — — 14 14
Конек лесной 13 13
Камышовка садовая . . — — — — — 3 — — — 3
Малиновка лесная . . . — — — 6 9 2 — — — 17
Славка-черноголовка . . — — — — 30 7 — — — 37
Славка серая — —- — — — 4 — — 4 8
Славка-завирушка . . . — — — — 1 2 — — — 3
Славка садовая .... — — — 1 62 22 16 — — 3 42
Дрозд-рябинник .... — — — 8 — 2 9 — 81
Дрозд певчий — — — 29 36 — 1 3 1 70
Дрозд-белобровик . . . — — — 52 33 12 8 18 20 143
Дрозд черный — — — 1 — — — — — 1
Чекан луговой .... Горихвостка обыкновен- — — — — — — — 6 6 12
ная 7 — 3 1 — — — 1 —
Соловей восточный . . — — — — — — — — 2 2
Зарянка . 2 о — — — — — — — 4
Крапивник — — — 2 4 2 — 2 — 10
Итого . . . 52 3 28 295 202 67 15 51 100 813
(Мальчевский и др., 1954). Известно, что в зависимости от местоположения гнезда и сроков размножения сильно варьируют также по ха-
48
рактеру укрепления и форме гнезда-ремеза (Штегман, 1951; Портенко, 1955).
Очень изменчив характер гнезд и у обыкновенной горихвостки. Они могут иметь различную форму и конструкцию. Помещенные, например, среди сучьев хвороста, они бывают вполне оформленными, с ясно выраженными лотком и боковыми стенками. Постройки, расположенные на земле или <в дупле, более примитивные. Особенно простые гнезда у птиц, поселяющихся в узких дуплах, имеющих небольшую поверхность дна. В отдельных случаях горихвостка совсем даже не вьет гнезда, а откладывает яйца прямо на древесную труху, как это делают вертишейки и дятлы.
Выпадение гнездостроительного акта из репродуктивного цикла наблюдается иногда и у некоторых других воробьиных — дуплогнездников, обычно строящих гнезда, например у пухляка (Шнитников, 1913) и галки (Makatsch, 1950; Эйгелис, 1958). Это бывает, когда птице приходится самой выгребать ямку среди трухи на дне дупла или выдалбливать помещение в гнилой древесине. По всей вероятности, аналогичная причина привела в процессе эволюции к полному исчезновению инстинкта гнездостроения у дятлов, которые Стали выдалбливать себе дупла, соответствующие размерам их тела, в связи с чем строительство гнезда оказалось излишним.
Всё приведенные выше факты противоречат той точке зрения, которая рассматривает гнездостроение как акт, от начала до конца врожденный, в который «отдельные особи не могут внести чего-либо своего; нового» (Heinroth, 1938). Если бы это было действительно так, то эволюция Инстинкта гнездостроения застыла бы на месте. В этом отношении мы полностью согласны с мнением А. Н. Промптова (1946) и Е. В. Лукиной (1953), уже обращавших внимание на несоответствие подобной точки зрения с фактами, наблюдаемыми в природе, и с результатами лабораторных экспериментов, указывающих на значительную изменчивость инстинктивных гнездостроитёльных реакций у птиц.
Как же, однако, следует понимать эту изменчивость инстинкта гнездостроения? Имеем ли мы здесь дело с наследственно закрепленными внутривидовыми типами гнездования, или наблюдаемое в природе разнообразие характера Гнездования есть лишь проявление экологической пластичности особей?
Индивидуальные отличия в характере гнездования следует, невидимому, объяснять в основном экологической пластичностью особей, способностью птиц активно отыскивать в зависимости от сложившейся обстановки наиболее благоприятные для их жизни места. Генетически закрепленная экологическая дифференциация вида, как правило, возникает при наличии географической изоляции (см. ниже). Для воробьиных, имеющих большое количество врагов, постоянно разоряющих их гнезда, гораздо важнее обладать широкой экологической пластичностью гнездостроительных инстинктов, нежели быть «узкими специалистами», приспособленными к строго определенным условиям среды, которые, кстати сказать, постоянно изменяются. Отбор у этих птиц идет, видимо, в направлении формирования пластичности гнездострои-тельного инстинкта, а не его консервативности.
Наблюдения за последовательным гнездованием одних и тех же особей показывают, что, несмотря на разорение их гнезд, птицы обычно находят такие места, где им в конце концов удается вывести потомство. При повторном гнездовании они обнаруживают способность к избирательности и помещают гнезда в более безопасном месте, которое у особей одного и того же вида может быть различным в зависи-
4 А. С. Мальчевский
49
мости от сложившейся обстановки. Изменение же места расположение гнезда неизбежно влечет за собой, как это было показано выше, изменение конструкции самой постройки. В зависимости от того или иного места гнездования гнездостроительный акт может усложняться или^ наоборот, упрощаться, а иногда и полностью выпадать из общего цикла размножения. В отдельных географических районах благодаря специфике местных условий подобные отклонения от нормы могут принимать массовый характер. Таким путем возникает имеющая место природе географическая изменчивость гнездостроения. При достаточно* длительной географической изоляции эти особенности могут оказаться^ наследственно закрепленными.
ЯВЛЕНИЕ ГНЕЗДОВОГО КОНСЕРВАТИЗМА
Вопрос о так называемом «гнездовом консерватизме» птиц — одитг из наиболее животрепещущих вопросов современной орнитологии. От правильного его решения зависит правильность наших представлений, о природе популяций птиц. Различия в понимании данного явления по существу приводят к различиям во взглядах на ход эволюционного процесса у птиц.
Существующие в литературе высказывания по вопросу о гнездовое консерватизме птиц противоречивы. В большинстве случаев мнение исследователей сводятся к признанию существования у птиц строгой привязанности к 'местам своего рождения и старым (местам гнездования.
На основании изучения биологии отдельных видов многие орнитологи пришли к заключению, что не только старые птицы из года в го^ сохраняют за собой раз уже занятый ими гнездовой участок, но и молодые птицы — первогодки гнездятся преимущественно в непосредственной близости от того места, где они вывелись. Одним словом, было* признано, что правило регулярного возврата на родину в одинаковой мере применимо к старым и молодым птицам. Имеющиеся в литературе указания на нахождение закольцованных птиц, гнездящихся вдали от мест их рождения, стали расценивать как исключение, и особей*, не возвратившихся на родину, считать в основном погибшими. По невозвращению птиц стали судить о возрастном составе популяции и проценте смертности молодняка. При этом «родину», понимали достаточно узко, как территорию лесного массива, лесопарка, долины и т.п. т е. такие участки местности, на которых исследователь мог рассчитывать на более или менее достоверный учет возвращения окольцованных птиц. Таким образом возникли сильно укоренившиеся среди орнитологов представления о чрезвычайно высоком проценте гибели птиц на первом году их жизни. «Нельзя забывать,—пишут Ю. А. Исаков*. В. М. Поливанов и Н. Н. Титаева (1953),—что в естественных условиях он (отход молодых на первом году жизни, — А. М.) достигает 90—95%». Поэтому, по мнению сторонников теории строгого гнездового консерватизма, нет ничего удивительного, что процент возврата птиц, окольцованных в гнездах птенцами, во всех случаях массового' кольцевания получается очень низким.
Биологический смысл местных «микропопуляций», по мнению некоторых орнитологов, заключается в том, что такие популяции лучше приспособлены к конкретным местным условиям существования, ибо рефлексы и навыки, облегчающие птицам существование в определенной экологической обстановке, легче всего создаются на первом году их жизни (Исаков, 1954). По мнению Исакова, местная популяция иа 50
года в год населяет определенную территорию и пополняется главным образом за счет своих 'потомков. Таким образом, в понятие «элементарной» или «микропопуляции» вкладывается определенный генетический. смысл.
'С другой стороны, орнитологическая наука накопила уже немало фактов, указывающих на существование у птиц явления расселения и постоянного и широкого генетического обмена между группами особей, населяющих соседние районы. Эти данные, заставляющие с осторожностью относиться к только что изложенной точке зрения, можно систематизировать следующим образом.
а) Наблюдения, отмечающие сравнительно быстрое расширение ареалов некоторыми видами птиц. Среди певчих птиц нашей фауны отмечено расширение ареалов, например у зеленой пеночки, дубровника, домашнего воробья, горихвостки-чернушки, желчной овсянки, черноголовой овсянки, синехвостки, канареечного вьюрка, майны» дрозда-рябинника и у некоторых других видов. Надо сказать, что сами по себе факты исторического расселения видов в целом не противоречат концепции гнездового консерватизма даже в узком смысле этого слова. Сторонники данной точки зрения признают расселение видов и по-разному его объясняют.
Однако приводимые объяснения применимы далеко не ко всем случаям. Так, например, причину расселений видов иногда видят в переуплотнении населения вида внутри ареала, главным образом на его границах. На это можно ответить, что история орнитологии знает примеры, когда расселение шло при чрезвычайно низкой плотности населения расселяющегося вида (зеленая пеночка, дубровник, синехвостка— в пределах Восточной Европы). Речь, однако, здесь должна идти о фактах быстрого расселения, какое, например, имело1 место у домашнего воробья (Судиловская, 1952), майны (Рустамов, 1946; Дементьев, 1953), зеленой пеночки и некоторых других птиц. Вряд ли виды «строго консервативные» могут быстро изменять границу своего ареала. Нельзя не согласиться со справедливыми высказываниями В. В. Станчинского (1927), Н. А. Гладкова (1949), а также Г. П. Дементьева и Е. П. Спангенберга (1949) относительно того, что, несмотря на существующие сезонные перемещения птиц, их способность ориентироваться в пространстве обусловливает известное постоянство их географического размещения, приводящее к образованию географических форм. Несомненно, что оставшиеся в живых как старые, так и молодые птицы, гнездившиеся или появившиеся на свет, например в Ленинградской области, на следующий год в преобладающем большинстве своем снова возвратятся в эту же область для размножения. Из этого, однако, не следует, что мы должны соглашаться с теорией «микропопуляций».
Известное постоянство ареалов или чрезвычайная медленность завоевания большинством видов новых для них территорий еще не доказывает строгой консервативности мест гнездования у птиц. Прежде всего виды птиц могут сильно отличаться друг от друга по степени своей «консервативности». На это указывает хотя бы то, что границы ареалов разных видов изменяются с разной скоростью. Говорить о гнездовом консерватизме птиц вообще, как это часто делается, нельзя, необходимо различать консерватизм вида и консерватизм особей. Несомненно, что одно связано с другим и одно из другого вытекает» но несомненно и то, что ареал всегда более постоянен, чем места обитания особей. Доказательством данного положения служат следующие наблюдения.
51
4*
б) Наблюдения, отмечающие изменения птичьего населения, происходящие в пределах ареала вида. Сюда прежде всего можно отнести многочисленные «работы, констатирующие сравнительно быстрое заселение птицами полезащитных лесных полос, водоемов «и других биотопов, пригодных для гнездования птиц, возникающих в связи с хозяйственной деятельностью человека. Изменения в составе орнитофауны юго-востока Европейской части СССР, отмечаемые Н. П. Дубининым (1953), скорее всего следует отнести к данной группе фактов, а не к предыдущей, как полагает сам автор, ибо в большинстве случаев имеют место все-таки изменения, происшедшие внутри прежних ареалов видов. Тем не менее нельзя не согласиться с Дубининым (1955) в том, что для объяснения сравнительно быстрых изменений орнитофауны, происходящих, например, под влиянием полезащитного лесоразведения, недостаточно говорить только лишь об одном гнездовом консерватизме. Наряду с консерватизмом у птиц существует известная склонность к «дисперсии» и изменению мест гнездования*
в) Наблюдения, отмечающие быстрое увеличение численности птиц в связи с вывешиванием в лесу искусственных гнездовий. Сюда следует отнести серию работ, посвященных успешным опытам привлечения дуплогнездников в искусственные гнездовья. Эти опыты показывают, что если данное место вообще является благоприятным для жизни птиц, то проведение мероприятий по привлечению может значительно увеличить здесь их численность, причем за весьма короткий срок и сразу на большой территории. Это особенно характерно для чистых сосновых насаждений, обычно бедных птичьим населением в гнездовое время. Столь быстрая реакция целого ряда видов (мухоловка-пеструшка, большая синица, обыкновенный скворец, полевой воробей, обыкновенная горихвостка и даже гоголь) на вывешивание искусственных гнездовий говорит не в пользу очень строгого гнездового консерватизма всех особей; и вряд ли будет правильным считать, Ч1о наблюдающееся при этом увеличение численности птиц происходит в основном за счет молодняка данного же «(микрорайона». Во всяком случае работы орнитологов по мухоловке-пеструшке (Haartman, 1949; Creutz, 1955; Михельсон и Чаун, 1957; Чаун, 1958), а также результаты кольцевания птенцов (ряда видов на территории Савальского лесхоза (табл. 14) не подтверждают последнее. С другой стороны, тот, кто признает это, должен, следовательно, признать рассредоточение молодняка и в тех Случаях, когда привлечение не проводится.
Существует, правда, мнение (Лихачев, 1955), объясняющее чрезвычайно низкий процент гнездования первогодков мухоловки-пеструшки в местах своего рождения тем, что основная масса этих птиц в природных условиях из-за нехватки мест гнездования совсем не гнездится в конце первого года жизни, а включается в размножение лишь в последующие годы по мере освобождения дупел или искусственных гнездовий в результате естественной смертности старых птиц. Не отрицая возможности такого положения, мы, тем не менее, считаем, что им одним нельзя объяснить отмечаемый обычно чрезвычайно низкий процент возвращения птиц на места своего рождения. Дело в том, что количество возвратившихся молодых птиц, как правило, не превышает 5%, а чаще бывает ниже, даже в тех (случаях, когда удается зарегистрировать всех возвратившихся птиц в течение ряда лет подряд.
г) Наблюдения, отмечающие значительные изменения численности или нерегулярность гнездования некоторых птиц на 'определенном участке местности. В этом отношении могут представлять интерес работы, освещающие изменение фауны какой-либо территории на протяжении
52
более или менее длительного отрезка времени. Для такого рода исследований наиболее удобны некоторые городские и пригородные парки, территории которых достаточно велики для заселения разнообразной орнитофауной и в то же время достаточно ограничены и изолированы, что дает возможность проводить регулярные учеты всего птичьего населения.
С этой целью в 1947—1955 гг. автором был взят под наблюдение парк Лесотехнической академии, который ранее являлся местом систематических наблюдений Д. Н. Кайгородова (1886, 1896, 1909) и впоследствии В. В. Строкова (1938). Таким образом автору представилась возможность провести сравнение видового состава и количества населявших этот парк птиц на протяжении довольно значительного отрезка времени (Мальчевский, 1954).
Здесь отметим лишь, что качественные и количественные изменения орнитофауны, наблюдавшиеся на территории парка Лесотехнической академии в конце прошлого и начале нынешнего столетий, во многих случаях происходили вследствие перемещения птичьего населения: птицы либо покидали парк, либо внезапно заселяли территорию парка, прилетая, очевидно, из соседних районов. Особенно нерегулярно гнездились в парке зеленушка, щегол, чиж, крапивник, скворец и дрозд-рябинник. На протяжении рассматриваемого периода эти птицы несколько раз исчезали из парка и затем появлялись вновь. Д. Н. Кай-городов (1909) отмечает, что гнездящиеся дрозды-белобровики впервые появились в парке в 1901 г. и после этого уже стали гнездиться здесь каждый год. Численность данного вида особенно резко возросла в 1903 г., что произошло, как считает Кайгородов, за счет вселения в окрестности Ленинграда каких-то стай белобровиков, залетевших из другой местности, о чем можно было судить по особой вариации их напева, ранее здесь не отмечавшейся. А. Г. Гржибовский (1911) также отметил внезапное население большим количеством дроздов-белобровиков окрестностей Смоленска. В 1910 г. он был поражен количеством гнездящихся белобровиков, превосходивших по численности всех других дроздов, так как до этого в течение многих лет подряд он не находил здесь гнезд этого дрозда.
Нерегулярность летнего пребывания наблюдается также у другого вида дрозда—дерябы. Например, в районе ст. Кузнечное (Ленинградская область) в период с 1947 по 1956 г. дрозд-деряба гнездился только в 1956 г., когда он появился здесь как-то внезапно' и сразу в большом количестве.
Приведенные выше примеры позволяют говорить о том, что в природе иногда происходят заметные перемещения птичьего населения. Очевидно, что, помимо клестов, свиристелей и розовых скворцов, известных непостоянством мест своего гнездования, среди европейской фауны птйц существуют и другие виды, места гнездования которых могут изменяться из года в год или через ряд лет в зависимости от степени обилия их основных кормов или от каких-либо других причин. Из певчих птиц к таковым, по нашему мнению, могут быть отнесены многие представители вьюрковых (зеленушка, щегол, коноплянка, чиж), дрозды (деряба, рябинник, отчасти белобровик), кра|пивник, обыкновенная каменка и некоторые др.
д) Данные, указывающие на существование у птиц значительного «веера разлета» молодых птиц-первогодков и известного рассеивания старых птиц. Выше уже говорилось о том, что во всех случаях, когда производилось массовое кольцевание гнездовых птенцов певчих птиц» процент их возврата на места рождения оказывался чрезвычайно
53
низким. Различные исследователи по-разному расценивают эти результаты. Некоторые, как уже говорилось, объясняют это в основном колоссальной смертностью молодых птиц, другие придают большое значение также существующей в природе закономерности расселения молодых птиц. Так, например, А. Н. Промптов и Е. В. Лукина (1937), проводившие многолетнее кольцевание больших синиц, отметили исчезновение молодых птиц из того района, где они появились на свет, и поэтому предположили существование у этого вида значительного «разлета» молодняка. Г. Н. Лихачев (1957) также сообщает, что ни одна из более чем 1400 окольцеванных им молодых синиц не была обнаружена в последующие годы в районе кольцевания.
Это не означает, конечно, что молодые синицы никогда не возвращаются на места своего рождения. Из окольцованных в 1951—1953 гг. 273 птенцов большой синицы на территории Савальского лесхоза «в последующие годы все же один был обнаружен поблизости от места своего рождения. Вряд ли, однако, местная популяция больших синиц могла бы существовать долгое время, если бы она пополнялась столь ничтожным количеством молодых птиц. Из существующих в литературе данных по кольцеванию птенцов этого вида нам известна работа Л. В. Шапошникова (1949), отмечающая более значительную оседлость молодых птиц: из 11 окольцованных птенцов 2 продолжали держаться в районе кольцевания не только осенью и зимой, но и весной, летом и осенью следующего года. Возможно, что в данном случае более значительная оседлость молодых птиц была вызвана подкормкой их в осенне-зимнее время (птицы отлавливались на кормушках).
Результаты кольцевания скворцов в Литве (Иванаускас, 1953) тоже показали, что -в противоположность старым скворцам молодые птицы обычно не возвращаются на место своего рождения, а гнездятся в других колониях. Это же оказалось характерным и для такого оседлого вида, как домовый воробей (Summers-Smith, 1954).
Весьма обстоятельная работа по кольцеванию мухоловок-пеструшек была проделана в Финляндии Хаартманом (1949). Его исследование показало, что очень многие птицы-первогодки оседают в смежных и более отдаленных районах. В то же время вновь появлявшиеся на участке наблюдений неокольцованные пеструшки были в основном однолетками, прилетевшими из других мест. Среди старых птиц самцы оказались более консервативными. Точно так же и Михельсон (1958) отмечает, что вновь вывешенные искусственные гнездовья заселяются преимущественно молодыми мухоловками-пеструшками, значительная часть которых не возвращается на места выведения. Закономерное возвращение старых птиц и значительное рассеивание молодежи, подмеченные у большой синицы, скворца и мухоловки-пеструшки, свойственны, вероятно, рчень многим нашим лесным певчим птицам.
На территории Савальского лесхоза в 1951—1953 гг. автор настоящей работы совместно с Н. П. Кадочниковым проводил кольцевание птенцов певчих птиц. Взрослые птицы кольцевались лишь при случае. Всего было закольцовано 8 взрослых птиц (5 горихвосток и 3 большие синицы) и 1229 гнездовых птенцов и птенцов слетков 29 различных видов певчих птиц (табл. 14). Старые птицы кольцевались двумя кольцами в различной комбинации, молодые — одним кольцом. В .течение трех последующих лет (с 1952 по 1954 г.) все птицы, гнезда которых мы находили, а также и другие встречавшиеся взрослые птицы внимательно обследовались. Всего было просмотрено 1588 птиц, и только у 9 были обнаружены кольца! Почти все они были отловлены в западню и вновь -выпущены. Весьма примечательно, что 4 из этих 9 оказа-‘
54
Таблица 14
Результаты кольцевания певчих птиц в Савальском лесхозе
Название вида Окольцованные птицы в 1951—1953 гг. Гнездившиеся птицы в 1952—1954 гг.
взрослыми птенцами общее количество обследованных количество окольцованных взрослыми количество окольцованных птенцами
Сорока — 18 50 — —
Сойка — 47 31 1*
Скворец — 88 176 — —
Иволга — 6 32 — —
Дубонос — 20 6 — —
Зеленушка — 23 47 — —
Зяблик — 28 75 — —
Овсянка обыкновенная . — 28 58 — —
Овсянка садовая .... — 36 54 — —
Конек лесной — 87 102 — —
.Лазоревка — 31 24 — —
.Большая синица .... 3 273 190 1 1
•Сорокопут-жулан . . . г 49 88 — —
Мухоловка серая . . . Мухоловка-белошейка . — 63 88 — —
— 16 42 — —
Пеночка-трещотка . . . — 75 47 — 1
Пеночка-теньковка . . — 4 6 — —
Малиновка-лесная . . . — 13 10 —
Славка ястребиная . . . — 3 2. — —
Славка-завирушка . . . — 9 20 —— —
Славка-черноголовка — 7 36 — —
Славка садовая .... — 24 80 — —
Дрозд-деряба — 4 2 — —
.Дрозд-рябинник .... — 59 44 — —
.Дрозд певчий — 15 39 — —
.Дрозд черный — 89 92 — —
Соловей восточный . . — 52 27 —- —
Зарянка (Горихвостка обыкновен- — 7 2 — —
ная 5 55 118 3 , 2**
Итого ... 8 1229 1588 4 5
^лись птицами, которых мы кольцевали взрослыми. Таким образом, половина окольцованных нами взрослых птиц возвратилась на старые места гнездования. В то же время из 1229 птиц, помеченных в птенцовом возрасте, в последующие годы нам удалось обнаружить всего лишь 5 особей. Отметим попутно, что из 5 горихвосток, окольцованных взрослыми, 3 птицы (2 самца и 1 самка) были обнаружены на следующий год на том же участке леса, на расстоянии 200—500 м от места их прошлогоднего гнездования. По сообщению Н. П. Кадочникова, продолжившего наши наблюдения в 1954—1956 гг., эти горихвостки по возвращении на «родину» гнездились уже в новом составе пар. Из них 2 птицы, составляющие в 1953 г. «брачную» пару, возвращались в один и тот же лесной квартал вплоть до 1956 г., т. е. гнездились здесь 4 года подряд. Спаривались они, однако, уже с какими-то другими птицами.
* Факт гнездования установлен не был.
*♦ Одна из них не была отловлена, вследствие чего история ее кольца осталась {невыясненной.
55
Полагаем, что при столь низком проценте возвращающихся на места своего рождения молодых численность птиц в Савальском лесхозе не могла бы возрастать, как это наблюдалось, но даже и оставаться на прежнем уровне (табл. 15). По крайней мере в случае с большой синицей, горихвосткой и мухоловкой-белошейкой, почти все гнезда которых были на учете, можно уверенно сказать, что увеличение численности этих птиц следует объяснять притоком их со стороны, а отнюдь не успешным пополнением «рядов» за счет возвратившегося потомства местных птиц. Оставшиеся в живых молодые местного происхождения в основном, очевидно, оседали на гнездовье в смежных или в более отдаленных районах.
Таблица 15
Количество гнезд горихвосток, мухоловок-белошеек, больших синиц, найденных в 1951—1954 гг. в Савальском лесхозе
Название вида 1951 г. 1952 г. 1953 г. 1954 г.
Горихвостка .обыкновенная . . . 10 18 20 ?
Мухоловка-белошейка 0 3 9 10
Синица большая 5 12 26 59
У преобладающего большинства видов воробьиных «микропопуляция» в основном пополняется, видимо, не за счет собственной молодежи, а за счет молодежи, появившейся на свет в смежных районах. Если это действительно так, то взрослые птицы—второгодки, третье-годки и т. д., составляющие основное «ядро постоянных туземцев», представляют собою сборище особей, произошедших из разных районов. Если даже допустить, что молодые птицы-первогодки на 50.% состоят из птиц, появившихся на свет здесь же, чего у большинства лесных певчих птиц, по-видимому, никогда не бывает, то и в данном случае нельзя будет говорить о генетической обособленности «микропопуляций». Поэтому вряд ли в природе имеет место лишь частичное смешение так называемых «микропопуляций», позволяющих все же, как это предполагает В. М. Поливанов (1954, 1957а), говорить об их обособленности. По нашему мнению, в громадном большинстве случаев у лесных певчих птиц постоянно происходит очень сильный обмен особями, приводящий к полному генетическому смешению популяций, населяющих смежные микрорайоны;
Таким образом, «родиной» птицы в узком смысле этого слова следовало бы считать не тот микрорайон, где она появилась на свет, а тот участок местности, на котором она впервые гнездилась, ибо* лишь после первого года размножения можно говорить о более или менее строгом ее гнездовом консерватизме. «Родина» в широком смысле у старых и молодых птиц, как правило, конечно одна, но понимать ее нужно значительно шире, чем территория лесного массива, долины и т. п.- В данном отношении нельзя не согласиться с А. Н. Промпто-вым (1936), полагавшим, что у большинства лесных певчих птиц, гнездовой биотоп которых занимает большие площади, свободно скрещивающиеся популяции относительно велики и что лишь пространственный фактор может создать некоторую генетическую неоднородность у групп особей, обитающих в более удаленных точках ареала.
56
Нельзя отрицать того, что внутри географических форм могут существовать отдельные популяции, имеющие различную историю, отличающиеся путями пролета, местами зимовок и т. п. Это, однако, не означает, что каждую географическую форму надо обязательно дробить на бесчисленное количество маленьких локальных популяций. Во всяком случае автор не может согласиться с Майром (1944) в том, что описанные Сиком (Sick, 1939) в окрестностях Штутгарта популяции зябликов, отличающиеся «местным диалектом» так называемого «дождевого сигнала», следует рассматривать как генетически обособленные группы особей и что такого рода преграда, как участок железнодорожных построек, могла помешать гибридизации двух популяций зябликов,, отдаленных друг от друга всего на 500 м. Подобная точка зрения может возникнуть только у того, кто глубоко убежден в существовании строгого гнездового консерватизма как у старых, так и у молодых птиц.
Какое же принципиальное значение имеет то, признаем ли мы факт существования «микропопуляций» с вкладыванием в этот термин генетического смысла или мы признаем существование у певчих птиц значительно более крупных популяций?
Признание строгого гнездового консерватизма и существования у птиц генетически обособленных «микропопуляций» может легко привести, с одной стороны, к переоценке значения экологической изоляции в процессе видообразования у птиц и, с другой — к недооценке роли экологической пластичности особей в эволюции птиц.
. Дело в том, что после того, как возвращение дикой перелетной птицы на старые места гнездования и места своего рождения стало считаться правилом, в орнитологии все большее значение начали придавать так называемой «экологической изоляции» как одному из основных факторов эволюции, тфиводящему к образованию внутривидовых биологических групп. Стали, например, считать, что при гнездовании одной части особей в одном биотопе, а другой части в другом биотопе, благодаря существованию у птиц строгого гнездового консерватизма и постоянному стремлению из поколения в поколение гнездиться в одном и то же месте, местная популяция легко может распасться на две практически не скрещивающиеся группы птиц. Обособленность же групп с течением времени может привести к тому, что они будут отличаться по характеру питания, типу гнездостроения, особенностям напева и т. п. Одним словом, было признано, что таким путем возникают биологические группы птиц или «экотипы», которые могут дать начало новым видам.
Так, подмосковные зяблики были разбиты А. Н. Промптовым (1934а) на две биологические группы, отличающиеся стацией, и характером гнездования; а у зябликов окрестностей Штутгарта Майр, основываясь на данных Сика, допускает возможность существования генетически обособленных микропопуляций, отличающихся характером голоса. В. М. Поливанов (1954,1957а), объясняя причину обособленности и привязанности членов каждой микропопуляции к определенному гнездовому району приспособленностью к кормовым и гнездовым условиям района их обитания, приводит пример со скворцами, которые в центральной усадьбе Дарвинского заповедника выкармливают птенцов преимущественно гусеницами, а. на затопленных островах — пауками. В данном случае микропопуляция понимается, очевидно, в очень узком смысле слова.
Количество описанных в литературе примеров, приводившихся исследователями для подтверждения факта существования в природе
57
индивидуальной, групповой и микропопуляционной специализации, может быть значительно увеличено. Отметим только, что сейчас мы говорим лишь о тех случаях, когда речь шла не о проявлении географической изменчивости биологических признаков и не об экологической дифференциации встречающихся на одной территории двух географических форм, а о внутрипопуляционном разнообразии «стереотипа поведения» в пределах одного географического района, которое в силу действия правила строгого гнездового консерватизма якобы может привести к изоляции и образованию микропопуляций, отличающихся генетически закрепленными биологическими особенностями.
Так, например, у обыкновенных горихвосток, гнездящихся в сосновых насаждениях часто на земле (под упавшими ветками), а также в скворечниках или дуплах, следуя примеру сторонников данной точки зрения, можно было бы различать две экологические группы птиц и предполагать в них возможное начало образования внутривидовых биологических групп. Однако наблюдения за окольцованными птицами показали, что в данном случае имеет место лишь проявление большой экологической пластичности этих птиц, а не индивидуальная и тем более не перенимаемая молодыми птицами от старых групповая специализация к условиям гнездования.
В подтверждение чрезвычайного непостоянства условий, при которых могут гнездиться одни и те же птицы и их потомки, можно привести факты из практики нашей работы с горихвостками на территории Савальского лесхоза.1 Упомянутые выше наблюдения Хаартмана (1949) за окольцованными мухоловками-пеструшками также показали, что одна и та же птица в течение ряда лет подряд может гнездиться в совершенно различных биотопах. Вряд ли в подобных случаях можно говорить о специализации. Многие примеры индивидуального отклонения от видового стереотипа поведения птиц, по всей видимости, следует объяснять как проявление экологической пластичности, как способность птиц активно отыскивать в зависимости от тех или иных причин наиболее благоприятные для их жизни места. К числу таких примеров можно отнести случаи дуплогнездничества сойки, гнездование галок в зданиях и одновременно и в непосредственной близости в дуплах деревьев, гнездование домовых воробьев в щелях строений, а в конце .лета, при нехватке свободных мест, — в кронах деревьев, гнездование дроздов-белобровников на земле и на деревьях и т. п.
С другой стороны, нельзя отрицать и значения закрепления удачного опыта гнездования в определенных условиях путем возвращения на старые места отдельных птиц. Факты многолетнего гнездования окольцованной птицы в совершенно сходных условиях (иногда буквально в том же самом месте) неоднократно описывались в литературе. Нельзя отрицать и того, что в природе существует не только индивидуальная, но и групповая специализация, обусловливаемая различиями экологических условий отдельных «микротерриторий». Тем не менее подобного рода специализация обычно не препятствует свободному скрещиванию потомков различно специализированных особей, если только не имеет места значительная территориальная разобщенность (географические формы) или встреча на одной территории двух популяций, географических форм или близких видов, исторически возникших в разных местах. Одним словом, мы не отрицаем понятия «микро-популяции», если только не вкладывать в него генетического смысла.
1 Мальчевский А. С. 1957. Явление гнездового консерватизма у воробьиных птиц. Вестник ЛГУ, № 9.
'58
Биологическое значение свободного скрещивания между особями их потомками, населяющими смежные «микротерритории», осуществляющегося /ПО причине постоянного и значительного рассеивания молодых птиц и нередко наблюдающегося изменения места гнездования старых птиц, помимо препятствования вредному влиянию инбридинга (Дементьев и Спангенберг, 1949) заключается, видимо, также в быстром распространении возникающих у отдельных особей полезных наследственных отклонений, которые скоро разносятся по значительной территории и становятся достоянием многих особей.
Итак, не отрицая фактов существования индивидуальной и групповой специализации в питании и характере гнездования (биотоп, месторасположение гнезда и т. п.) внутри популяций, мы, тем не менее, считаем, что у преобладающего большинства воробьиных птиц подобного рода специализация не может привести к длительной изоляции и образованию генетически обособленных «микропопуляций» или «экотипов» по -причине имеющего место в природе постоянного и интенсивного взаимопроникновения и скрещивания особей и их потомков, населяющих смежные микрорайоны и в случае экологически пластичных видов -различные биотопы. Вот почему нам кажется, что сторонники теории строгого гнездового консерватизма часто переоценивают значение экологической изоляции в процессе видообразования.
Сказанное вовсе не означает того, что следует вообще отрицать значение экологической изоляции (особенно в условиях горного ландшафта) и возможности существования в одной географической точке двух или, может быть, даже большего количества биологических рас или географических форм одного вида. За исключением, однако, биологических рас птиц — гнездовых паразитов, эволюция которых идет особым путем, в орнитологической литературе пока что нет убедительных примеров изолированного сосуществования таких биологических групп птиц, которые являлись бы автохтонами одного географического района. Даже у птиц, живущих на океанических островах, явление адаптивной радиации в большинстве случаев объясняют географической изоляцией (существование на разных островах в различных условиях или неодновременное проникновение одного или разных видов с континента) и последующей встречей экологически разошедшихся форм или видов (Майр, 1944; Лэк, 1947а). Известные в литературе примеры изолированного сосуществования близких форм или видов у континентальных птиц, как это, например, показано в работах Б. К. Штегмана (1930), Г. П. Дементьева (1936) и И. А. Долгушина (1948), также трактуются как встреча на одной территории двух расселившихся •форм, образовавшихся в разных географических условиях и не скрещивающихся между собой по причине экологической дифференциации, -обусловливающей возможность дальнейшего расхождения сосуществующих форм.
ГЛАВА III
РАЗМЕРЫ, СРОКИ И СУДЬБА КЛАДОК
КОЛИЧЕСТВО яиц В КЛАДКЕ
Количество яиц, откладываемых воробьиными нашей фауны, хорошо известно орнитологам. Это, однако, не означает, что вопрос больше не заслуживает внимания. Для большинства видов певчих птиц еще мало накоплено данных по индивидуальной, сезонной и географической изменчивости размеров кладки. Хотя по этим вопросам в литературе уже имеются обобщающие высказывания (Дементьев, 1946; Lack, 19476, 1948, 1949, 1954; Рустамов, 19546, 1955, и др.), они остаются еще освещенными недостаточно полно. Для дальнейшей разработки вопроса об изменчивости величины кладки у птиц особенно важны, как нам кажется, детальные наблюдения в природе за отдельными птицами и судьбой отдельных кладок.
Путем ежедневных наблюдений за гнездами птиц автор стремился собрать возможно большее количество таких данных. Неоконченные кладки не учитывались. Кладка принималась за полную лишь в тех случаях, когда была уверенность, что птица отложила в гнездо последнее яйцо. Повторные (дополнительные) кладки, даже с сильно сокращенным числом яиц, считались за полные. В гнезде, в котором были уже птенцы, размер кладки определялся по количеству птенцов. Результаты этих наблюдений сведены в табл. 16.
У большинства лесных видов певчих птиц среднее количество яиц. в кдадке в общем сходно и варьирует от 3 до 6. Сойка и обыкновенная горихвостка могут отложить в одно гнездо до 9 яиц. Наиболее высоким показателем плодовитости отличаются синицы. Среди них наиболее замечательна большая синица, кладка которой иногда состоит из 15—16 и даже 17 яиц (Glasewald, 1928). Наименьшее количество яиц среди лесных воробьиных нашей фауны откладывает иволга, у которой кладка, обычно состоит из 3—4 яиц.
Причины, которые определяют видовые особенности размеров кладки, могут быть различными. У одних видов количество потомства, как уго полагает Лэк (1954), может определяться пищевым фактором и_ возможностью выкормить лишь определенное количество птенцов, у других — количеством яиц, которое способна покрыть птица своим телом, у третьих — размерами -и положением гнезда и т. п. Можно предположить, в частности, что ограниченное количество яиц в кладке у иволги в какой-то степени связано с особым расположением ее гнезда на кон-60
Таблица 16
Количество яиц в кладках некоторых воробьиных (по наблюдениям автора и его помощников)
Название вида Количество найденных кладок Количество гнезд, содержащих кладку яиц (в шт.) Среднее количество яиц в кладке
2 3 4 5 6 7 8 9 10 И 12 13—16
Ворона серая .... 5 2 3 — — — — — — — 4,6
Сорока 18 — — 2 2 7 — — — — — — 6,0
Сойка 19 — — 5 2 4 5 1 2 — — — — 6,0
Скворец обыкновенный 17 2 8 7 4,3
Иволга 14 — 8 5 1 3,4
Дубонос 9 — 1 4 4 4,3
Зеленушка 32 — 1 6 22 3 — 4,8
Щегол 4 4,0
Чиж 2 — 2 ?
КЫГопляйка . ♦ . . > 12 — — 1 7 3] 1 — — — — — — 5,3
Снегирь 5 — — 2 1 2 5,0
Чечевица 12 — 1 4 7 4,5
Зяблик 48 — 6 14 26 2 4,5
Овсянка обыкновенная 39 1 5 15 14 4 — 4,4
Овсянка садовая . . 33 — 2 11 15 5 4,7
Конек лесной . . . 72 — 5 20 37 10 4,7
Синица большая . . 36 — —— — 1 — — 6 2 9 9 5 4 10,5
Лазоревка 5 — — — — — — 3 1 —ь — 1 — 9,0
Синица длиннохвостая 1 1 ?
Сорокопут-жулан . . 63 — 5 13 30 14 1 — — — — — — 4,9
Мухоловка серая . . 79 — 6 35 36 2 4,4
Мухоловка-белошей-ка 10 __ 1 2 4 3 — 4,9
Мухоловка-пеструшка 10 1 2 2 5 — 5,1
Пеночка-теньковка . 5 — — 2 2 1 4,8
Пеночка-весничка 24 — — 2 2 10 10 — — — — — — 6,2
Пеночка-трещотка . 38 — 1 5 12 12 7 1 — — -— — — 5,6
Малиновка лесная . 20 — 4 10 5 1 4,2
Славка-черноголовка 73 1 6 25 33 8 4,6
Славка серая .... 13 — 2 5 5 1 4,4
Славка-завирушка . 16 — — 5 9 2 4,8
Славка садовая . . . 74 1 9 20 44 4,5
Славка ястребиная . 6 — 1 5 — — — — — — — — 4,8
Дрозд-рябинник . . 50 — 5 8 21 14 2 — — — — — — 5,0
Дрозд-деряба .... 3 — — 3 -— — — — — — — — — 4,0
Дрозд певчий .... 64 — 4 18 38 4 4,7
Дрозд-белобровик . 89 — 3 12 54 20 5,0
Дрозд черный . . . 47 — 1 15 22 8 1 — — — — — — 4,8
Чекан луговой . . . 6 — — — 3 3 — — — — —— — — 5,5
Горихвостка обыкновенная 43 1 3 6 17 10 5 1 — — — — 6,2
Соловей восточный . 33 — 3 8 21 1 4,6
Зарянка 5 — — — 4 — 1 — — — — — — 5,4
Итого. . . 1154 3 85 293 511 166 45 16 6 10 9 6 4
61
цах ветвей деревьев. При сравнительно небольших его размерах птенцы^ если бы их было, например, 5—6, легко могли бы вывалиться из гнезда,, постоянно раскачиваемого ветром. Таким образом, более высокая плодовитость иволги вряд ли была бы полезна для вида.
Почему, однако, у воробьиных птиц обычно наблюдаются значительные вариации в количестве яиц, откладываемых особями одного* вида? Приведенные в табл. 16 материалы демонстрируют это достаточно наглядно. У сойки, например, в пределах весьма ограниченной территории Савальского лесхоза количество яиц в отдельных кладках варьирует от 4 до 9. То же можно сказать и относительно кладки обыкновенной горихвостки и многих других птиц.
Каковы причины столь сильного варьирования числа яиц в кладке-у особей одного вида? Зависит ли это в первую очередь от различий в наследственной природе отдельных особей, или индивидуальные отличия в размерах кладки определяются главным образом физиологически; ми причинами?
Некоторые исследователи, например В. Карпов и В. Паровщиковг (1941), полагают, что у мухоловки-пеструшки число яиц, откладывае-г мых одной самкой, остается неизменным, даже если самка бросает первую кладку и приступает ко второй. Наоборот, в работе Крейца (Creutz^ 1955) мы находим указание на то, что в случае дополнительных кладок мухоловка обычно откладывает меньшее количество яиц. Лэк (1948^ 1949, 1954), который посвятил рассматриваемому вопросу целую сериюь работ, считает, что вариации в величине кладки одной и той же особи,, зависящие главным образом от кормовых условий, места и времени размножения, бывают все же не столь значительными, как различия в величине кладки у разных особей. Таким образом, основной причиной существующих различий в числе яиц, откладываемых особями одного вида, является, по Лэку, наследственная природа организма, неодинаковая у разных особей. Естественный отбор закрепляет плодовитость, наиболее выгодную для данных условий. Это положение Лэк кладет в основу при объяснении существующей в природе внутривидовой географической, индивидуальной и сезонной изменчивости размеров кладки. По мнению других исследователей (Kendeigh, 1941), уменьшение количества откладываемых птицами яиц во второй половине сезона размножения является результатом непосредственного действия света в связи с изменением продолжительности дневного освещения.
Нет сомнения в том, что степень плодовитости отдельных членов популяции может быть неодинаковой. Эти различия могут обусловливаться не только генетическими, но и возрастными особенностями организма. Известно, например, что самки, размножающиеся впервые*, откладывают яиц меньше, чем в последующие годы (Nice, 1937; Лихачев, 1955 а, б, и др.). Нельзя не согласиться также и с тем, что продолжительность дневного освещения в какой-то степени влияет на числом откладываемых птицей яиц, ибо стимулирующее действие светового1 фактора на половые железы птиц общеизвестно и доказано экспериментально. Не могут не сказываться на количестве яиц в кладке также и годовые, сезонные и местные отличия в условиях питания.
Однако никак нельзя согласиться с тем, что генетические причины и непосредственное действие абиотических факторов среды могут обусловить столь широкую амплитуду колебания числа яиц в кладках у особей одного вида, гнездящихся на сравнительно ограниченном пространстве при сходных климатических и кормовых условиях, как это, например, имеет место у сойки или горихвостки в Савальском лесхозе или у дрозда-белобровика в пригородах Ленинграда (см. табл. 17). Основ-62
ной причиной резких колебаний числа откладываемых птицами яиц, обусловливающих так называемую «индивидуальную изменчивость плодовитости» отдельных особей, является, по нашему мнению, частое разорение гнезд хищниками, приводящее к возникновению нередко нескольких дополнительных кладок, содержащих, как правило, уменьшенное количество яиц.
Этот мощный фактор должен быть положен в основу объяснения причины сезонных отличий в размерах кладок, находимых в гнездах, строящихся в разные сроки. Таким образом, наряду с индивидуальной изменчивостью плодовитости отдельных особей, которая в целом не так уже, видимо, велика, в природе всегда имеют место более существенные различия в величине кладки одних и тех же особей, вынужденно приступающих к строительству гнезда и откладке яиц иногда несколько раз в течение одного сезона размножения.
На территории Ленинградской области в период с 1946 по 1954 г. нами было найдено 89 полных кладок дрозда-белобровика. Количество* яиц в обнаруженных кладках варьировало от 3 до 6. Среднее количество яиц оказалось равным примерно 5. Сравнение сроков появления яиц в отдельных гнездах показало, что кладки, содержавшие 3 или 4 яйца, во всех случаях были закончены после 1 июня. Ни одна из майских кладок не содержала сокращенного (до 3 или 4) количества яиц. В то же время в мае было найдено наибольшее количество гнезд с 5 и. 6 яйцами. Поэтому среднее количество яиц в майских кладках оказалось выше, чем в кладках, отложенных в июне или в июле (табл. 17).
Таблица 17'
Количество яиц в кладках дрозда-белобровика по месяцам (Ленинградская область)
Время откладки яиц Количество гнезд, содержащих кладку яиц (в шт.) Среднее количество яиц в кладке
3 4 5 б
Май 37 12 5.2
Июнь ...... 3 10 16 8 4,8
Июль — 2 1 — 4,3
Итого. . 3 12 54 20 5,0
Таким образом, в июне не наблюдается общего равномерного снижения количества откладываемых дроздами яиц, которое позволило бы расценивать это явление как результат закрепленного отбором специального наследственного приспособления птиц к кормовым и климатическим условиям, характерным именно для июня. В этом месяце наряду с нормальными кладками, состоящими из 5 или 6 яиц, появляется значительное количество кладок с резко уменьшенным числом яиц, за счет которых и снижается средний показатель.
Итак, уменьшение откладываемых птицами яиц к концу сезона следует рассматривать в основном не как приспособление, а как вынужденное явление. Приспособительной в данном случае является способность большинства воробьиных неоднократно восстанавливать утерянную кладку и начинать цикл размножения сызнова. Птица, гнездо которой оказалось разоренным, отнюдь не «выбывает из строя», как член популяции, не давший в этом году приплода, а вскоре приступает к постройке нового гнезда и повторной откладке яиц, количество которых по причи
63^
не нарастающего истощения организма самки, бывает, как правило, меньше, чем при первой попытке размножения.
У некоторых птиц возможна дополнительная кладка даже после гибели птенцов. Так, в Савальсксм лесхозе наблюдался случай повторной откладки яиц горихвосткой уже через 4 дня после гибели ее 8—9-дневных птенцов. Во второй раз самка отложила на 1 яйцо меньше, чем в первый раз. Штейн (Stein, 1927) описывает случай повторного гнездования пухляка после гибели птенцов первого выводка в возрасте около 12 дней. В первой кладке у этой синицы было 10 яиц. Вторая, дополнительная, кладка состояла из 7 яиц. Подобные случаи наблюдались также у большой синицы.
ПРИЧИНЫ РАСТЯНУТОСТИ ПЕРИОДА ОТКЛАДКИ ЯИЦ
При определении степени растянутости периода размножения отдельных видов в основу наблюдений был положен метод, предложенный
Название вида
Сорока ..................
Сойка ...................
Скворец обыкновенный . . Иволга "обыкновенная . . . Дубонос обыкновенный . . Зеленушка................
Зяблик ..................
Овсянка обыкновенная . . . Овсянка садовая .........
Конек лесной ............
Синица большая...........
Сорокопут-жулан .........
Мухоловка серая..........
Мухоловка-белошейка . . . Пеночка-трещотка.........
Пересмешка...............
Славка-черноголовка .... Славка серая ............
Славка-завирушка.........
Славка садовая ..........
Дрозд-рябинник...........
Дрозд певчий.............
Дрозд черный.............
Горихвостка обыкновенная . Соловей восточный . .
Рис. 4. Степень растянутости периода возможных сроков откладки яиц у некоторых воробьиных в Ленинградской области (по наблюдениям в 1946—1956 гг.).
в свое время еще В. Л. Бианки (1917—1918). Отыскивалось возможно большее количество птичьих гнезд, и путем регулярных наблюдений за каждым гнездом выяснялись точные сроки размножения отдельных пар птиц. Срок размножения считался выясненным лишь в тех случаях, когда удавалось установить время появления в гнезде яиц, день вылупления или точный возраст птенцов. По этим данным можно было судить
64
о времени появления первого яйца в найденных нами гнездах. Насиженные кладки, дальнейшее наблюдение за которыми провести не удалось, в расчет не принимались. Результаты таких наблюдений изложены в видовых очерках (см. часть вторую). Здесь мы приводим лишь обобщенные данные, которые для наглядности изображаем в виде графиков {рис. 4, 5). В этих графиках сплошная линия против каждого вида соответствует продолжительности сроков начала откладки яиц, установленной для района наблюдения. Вопросительный знак ставился в тех случаях, когда, с нашей точки зрения, возможны более ранние или более поздние сроки размножения, нежели те, которые удалось определить.
Название вида
Ворона серая ............
Скворец обыкновенный Зеленушка................
Чиж......................
Коноплянка ..............
Снегирь .................
Чечевица ................
Зяблик ..................
Овсянка обыкновенная . . . Конек лесной.............
Синица большая...........
Сорокопут-жулан .........
Мухоловка серая..........
Мухоловка-пеструшка . . . Пеночка-весничка ........
Пеночка-трещотка.........
Пересмешка...............
Славка-черноголовка . . . . Славка-завирушка.........
Славка садовая ..........
Дрозд-рябинник...........
Дрозд певчий.............
Дрозд-белобровик.........
Чекан луговой ...........
Горихвостка обыкновенная . Зарянка .................
Возможные сроки начала откладки яиц
апрель май
июнь июль август
Рис. 5. Степень растянутости возможных сроков откладки яиц у некоторых воробьиных в Савальском лесу (по наблюдениям в 1951—1953 гг.).
Анализ данных показывает, что для всех видов воробьиных, охваченных исследованием, характерна сильная растянутость сроков размножения. Значительная протяженность периода откладки яиц наблюдалась каждый год и во всех пунктах, где проводилась стационарная работа. У некоторых видов период откладки яиц продолжался около 3 недель (серая ворона, иволга), у других — 4—6 недель (скворец, чечевица, садовая овсянка, сорокопут-жулан и Др.); наконец, у третьих — 7—10 и даже более недель. Явление растянутости сроков размножения сильнее всего выражено у зеленушки, чижа, коноплянки, обыкновенной овсянки, большой синицы, черного и певчего дроздов, дрозда-белобровика и зарянки. В гнездах этих птиц первые отложенные яйца могут быть обнаружены с конца апреля до начала и даже середины июля.
5 А. С. Мальчевский
65
Какие же причины вызывают столь сильную растянутость сроков, откладки яиц у воробьиных? Факторы, обусловливающие явление растянутости периода размножения популяций, могут быть различными. Основные из них, по нашему мнению, следующие.
а) Частые случаи разорения гнезд. Об этой особенности гнездовой жизни птиц нам уже приходилось говорить. Укажем еще раз, что в случае разорения гнезда или оставления кладки птицы, как правило, спустя некоторое время начинают размножаться повторно. У многих особей таких «попыток» бывает несколько. Дополнительная кладка иногда возникает даже после гибели довольно крупных птенцов. В этих случаях разница в сроках откладки яиц при первом и последующем гнездованиях бывает особенно значительной.
Естественно, что разорение гнезд сильнее всего влияет на ход размножения тех видов птиц, гнезда которых располагаются открыто^ Дуплогнездники страдают от хищников меньше, в связи с чем вынужденные повторные кладки встречаются у них реже. Этим и объясняется то, что у моноциклических видов птиц-дуплогнездников (скворец, мухоловки— пеструшка и белошейка) период откладки яиц обычно бывает менее продолжительный, чем у птиц, гнездящихся открыто.
б) Растянутость прилета птиц в гнездовую область. Известно, что-особи одного вида прилетают в район гнездования далеко не одновременно. Явление прилета растягивается иногда почти на целый месяц. Например, первые местные дрозды-белобровики в 1954 г. были отмечены автором под Ленинградом 18 апреля, в то же время 16 мая наблюдались небольшие стайки этих дроздов, еще не приступивших к размножению. Наиболее же ранняя кладка белобровика под Ленинградом в 1954 г. была начата 3 мая. По наблюдениям Крейца (1955), прилет и оседание местных мухоловок-пеструшек в одном из парков Германии растянулся с 12 апреля по 10 мая.
Известно также, что пролет молодых птиц-первогодков идет, как. правило, позднее (Weigold, 1926). В гнездовую область они попадают нередко тогда, когда старые птицы уже насиживают кладки. Постепенностью включения особей в репродуктивный цикл в основном и объясняется значительная растянутость сроков откладки яиц в первой половине периода размножения (табл. 18 и 19). Неодновременность начала гнездования особенно характерна для рано прилетающих птиц. В рав-
Таблица 18
Начало кладки (по декадам) некоторых лесных воробьиных на территории Ленинградской области в 1946—1954 гг.
Название вида Количество гнезд, в которых кладка началась
в апреле| в мае 1 S । июне I в июле
ш | I п III 1 I 1 11 111 I I 1 Ч
Зяблик 3 10 10 3 2 1
Сорокопут-жулан . . . — — 10 14 6 2 — — —
Мухоловка серая . . . — — — 6 21 11 2 1 —
Пеночка-весничка . . . — — 1 8 11 4 2 2 —
Пеночка-трещотка . . . — — 9 2 2 — 1 —
Малиновка лесная . . . — — — — 6 5 2 — —
Славка-черноголовка — — 2 9 10 4 3 1 —
Славка садовая .... — — — 1 16 9 7 1 —
Дрозд-рябинник .... 4 13 16 10 8 5 1 — —
Дрозд певчий — 5 11 8 12 7 3 3 —
Дрозд-белобровик . . . 3 27 24 19 15 10 13 1 1
66
Таблица 19
Начало кладки (по декадам) некоторых лесных воробьиных на территории Савальского лесхоза в 1951 — 1953 гг.
Название вида Количество гнезд, в которых кладка началась
вапреле| в мае в июне | в июле
III 1 I 1 = И J in 1 1 11 1 Ш 1 1 L .
Зяблик 1 6 б 5 7 2 1
Овсянка обыкновенная . — 4 5 9 4 4 4 — —
Овсянка садовая .... — 1 7 18 5 4 2 — —
Конек лесной —• 5 9 12 11 11 4 2 —
Синица большая .... 2 7 5 3 5 7 7 — —
Сорокопут-жулан . . . — — — 12 8 б 5 3 —
Мухоловка-серая . . . — — — 10 24 15 9 2 —
Пеночка-трещотка . . . — — 10 10 2 — 1 1 —
Славка садовая .... — —. — 12 . 15 7 6 2 —
Дрозд певчий — 1 4 4 2 2 5 — —
Дрозд черный Горихвостка обыкновен- 1 2 10 8 6 6 6 3 2
ная — 2 13 12 8 8 2 1 —
Соловей восточный . . — — 9 11 5 5 — — —
ной степени это типично и для оседлых видов. У многих из них «пик» размножения наступает спустя 2—3 недели после появления первых кладок. Наоборот, птицы, прилетающие к нам поздно (сорокопут-жулан, славки, соловей), приступают к размножению более дружно.
в) Позднее весеннее похолодание. Как известно, весенняя погода чрезвычайно изменчива. Вначале весна нередко бывает дружной и теплой. Птицы, прилетевшие первыми, оказываясь в благоприятных для размножения условиях, очень скоро приступают к откладке яиц. Затем погода резко меняется, и наступают длительные холода, которые задерживают откладку яиц у птиц, прилетающих в гнездовую область позднее. Таким, например, в Ленинградской области был 1954 г. В этом году в окрестностях Ленинграда в середине мая можно было видеть кочующих, не приступивших еще к размножению, дроздов-белобровиков, зябликов, скворцов и в то же время в первой декаде мая находить гнезда этих же видов с полными кладками. В менее резкой форме такая картина наблюдается под Ленинградом почти каждый год. Сильная растянутость сроков размножения, обусловленная непостоянством весенней погоды, отмечалась и в других местах. Например, в Тульских засеках в связи с резкими сменами погодных условий весной 1950 и 1951 гг. наблюдалась неодновременность начальных сроков размножения у мухоловки-пеструшки (Лихачев, 1953). В теплые дни количество пеструшек, приступающих к откладке яиц, резко повышалось и, наоборот, сокращалось в период холодов и дождей.
г) Нехватка мест, пригодных для гнездования. Нехватка мест, пригодных для гнездования, отражается на сроках размножения, в основном птиц-дуплогнездников. На то, что в природе нередко действительно имеет место недостаток свободных дупел, указывают прежде всего многочисленные факты, подтверждающие наличие межвидовой конкуренции у дуплогнездников из-за места гнездования, а также случаи вынужденного заселения дуплогнездниками старых построек врановых птиц (большая синица), гнезд хищников (полевой воробей, иногда горихвостка), куч хвороста (горихвостка) и т. п. мест.
Отношение птиц к искусственным гнездовьям — дуплянкам и скворечникам, также указывает на то, что природные ресурсы естественных
5*
67
дупел часто не могут полностью удовлетворить потребностей популяции. Например, случаи позднего гнездования скворцов в тех же самых дуплах или скворечниках, в которых воспитание потомства уже закончилось в северных и средних широтах, можно трактовать, как вынужденное запоздалое размножейие птиц, которым до момента массового вылета скворчат свободного дупла или скворечника найти не удалось.
д) Наличие нормальных вторых кладок, В большинстве случаев нельзя еще с полной достоверностью ответить на вопрос, имеет ли тот или иной вид птицы одну кладку в году или несколько. Количество точных данных, полученных путем наблюдения за окольцованными птицами, невелико и касается в основном лишь небольшого числа дуплогнездников. У преобладающего большинства воробьиных нашей фауны о числе выводков судят по косвенным показателям, главным образом по степени растянутости сроков откладки яиц. Случай нахождения поздней свежей кладки часто считается доказательством наличия у вида двух или даже трех кладок в году.
Рис. 6. Даты появления первого яйца у некоторых воробьиных в Савальском лесу.
и — синица большая, б — дрозд черный, в — сорокопут-жулан, г — горихвостка обыкновенная.
Правильно решить вопрос о количестве циклов размножения очень трудно прежде всего из-за большого числа дополнительных кладок, постоянно возникающих и постепенно увеличивающихся в числе от весны к лету. Эти вынужденные кладки сильно запутывают общую картину размножения популяции, которую было бы нетрудно выявить статистическим путем, если бы гнезда птиц не разорялись так часто. У дуплогнездников, кладки которых гибнут сравнительно редко, наличие или отсутствие второго цикла размножения в некоторых случаях удается установить довольно легко даже при ограниченном количестве материала. У большой синицы, например, на территории Савальского лесхоза совершенно отчетливо выражены два «пика» интенсивности размножения — в первой половине мая и во второй половине июня (рис. 6), соответствующие массовому появлению первых и вторых кладок. В этом отношении наши данные соответствуют выводам В. И. Таращука (1953) и Г. Нл Лихачева (1954), установившим двойной цикл размножения у большой синицы на Украине и в Тульских засеках. Приходится сомневаться лишь в справедливости заключения В. И. Таращука о существовании у этой синицы тройного цикла размножения.
Решение вопроса, является ли вид моно- или полициклическим, чрезвычайно осложняется также наличием индивидуальных отклонений и нередко влиянием климатических условий года. Популяция в целом не бывает полициклической. Среди птиц, обычно гнездящихся дважды в сезон (например, большая синица), всегда имеется часть особей, кото-68
рая при нормальных условиях не приступает к откладке яиц во второй раз. В то же время среди однократно размножающихся птиц иногда встречаются представители, выводящие потомство дважды.
На основании сопоставления степени растянутости периода откладки яиц у различных видов как в Ленинградской области, так и в Са-вальском лесничестве автор пришел к заключению, что по характеру размножения охваченные исследованием виды могут быть разделены на четыре группы.
Первая группа. Виды, у которых большинство особей имеет две нормальные кладки в сезон. Для представителей этой группы птиц характерна значительная протяженность периода откладки яиц (свыше 2 мес.) и ясно выраженный двойной «пик» интенсивности размножения. Среди лесных воробьиных нашей фауны к таковым следует отнести большую синицу, полевого воробья, коноплянку, зеленушку, чижа и щегла (рис. 6, большая синица).
Рис. 7. Даты появления первого яйца у некоторых воробьиных в Ленинградской области.
а — серая мухоловка, б — славка-черноголовка, в — дрозд-белобровик.
Вторая группа. Виды, у которых лишь часть особей имеет две кладки, большинство же успевает вывести потомство один раз. Второе гнездование после успешного воспитания первого выводка наблюдается, как правило, лишь у тех членов популяции, которые в данный сезон раньше других приступили к размножению. Период возможных сроков откладки яиц у этих видов растянут также более, чем на 2 мес. Кривая интенсивности размножения у них далеко не всегда имеет явно выраженный двухвершинный характер. Вторая (меньшая) вершина кривой (рис. 7, дрозд-белобровик), соответствующая времени появления новых свежих кладок птиц, приступающих к размножению снова, часто сглаживается из-за большого количества запоздалых дополнительных кладок, максимум которых бывает как раз во второй половине сезона (рис. 6, черный дрозд).
Преобладающее большинство видов, принадлежащих к данной группе, прилетает в гнездовую область рано и начинает откладку яиц в конце апреля — начале мая. Таковыми являются: дрозд-белобровик, черный и певчий дрозды, зарянка, обыкновенная овсянка, лесной конек и, по всей вероятности, зяблик, крапивник и дрозд-рябинник.
69
Третья группа. Моноциклические виды, у которых лишь очень немногие особи выводят потомство дважды в лето. Среди лесных воробьиных нашей фауны к этой группе можно отнести, например, обыкновенную горихвостку и мухоловку-пеструшку. Первая из них, видимо, больше склонна к полициклии, нежели вторая. На возможность последовательного воспитания двух выводков одной и той же самкой горихвостки указывают Е. В. Лукина (1951) и Н. П. Кадочников (1957). В достоверности этих сведений не может быть сомнений, ибо в обоих случаях исследователи имело дело с окольцованными птицами.
Случаи вторичного гнездования у мухоловки-пеструшки очень редки. В Германии, например, за 17-летний период работы с большим количеством птиц этого вида, помеченных цветными кольцами, ни у одной из самок не удалось установить двойного цикла размножения (Creutz, 1955). Н. Г. Лихачеву (1955) в течение многих лет работы в Тульских засеках пришлось наблюдать такой случай только один раз.
Четвертая группа. Виды, у которых до сих пор случаев вторичного размножения после успешного воспитания первого выводка не наблюдалось. У птиц, гнезда которых часто разоряются, период возможных сроков откладки яиц иногда растягивается почти на 2 мес. (сорока, сойка). Наоборот, у тех птиц, кладки которых гибнут сравнительно редко, откладка яиц продолжается всего 3—4 недели (ворона). Кривая интенсивности размножения у представителей данной группы птиц имеет явно выраженный одновершинный характер (рис. 7, серая мухоловка).
Из числа изученных видов представителями последней группы являются ворона, сорока, сойка, иволга, чечевица, садовая овсянка, сорокопут-жулан, мухоловка серая, пеночка-весничка, пеночка-трещотка, лесная малиновка, славка-черноголовка, садовая славка, восточный соловей, луговой чекан и, по всей вероятности, обыкновенный скворец и славка-завирушка.
Таким образом, сильную степень растянутости периода размножения можно объяснить наличием нормальных вторых кладок только у некоторых видов птиц. В большинстве же случаев влияние оказывают другие рассмотренные выше причины.
СУДЬБА КЛАДОК
Много ли гибнет в природе яиц и гнездовых птенцов воробьиных и каковы основные причины отхода?
В отечественной литературе уже накопилось немало различных сообщений, указывающих на то, что весенние холода, затяжные дожди и ливни, град и паводки нередко приводят к гибели кладок и птенцов (Аверин, 1909; Ефремов, 1941; Барабаш-Никифоров и Павловский, 1948; Данилов, 1949; Лихачев, 1953, и Др.). Еще чаще приходится находить указания на отрицательную роль хищников, разоряющих большое количество птичьих гнезд. В местах, примыкающих к населенным пунктам, гнезда певчих птиц, кроме того, растаптываются скотом. В некоторых районах птичьи яйца до сих пор являются объектом своеобразного «мальчишеского промысла» или коллекционирования, на пагубные последствия которого в свое время уже обращал внимание В. В. Стаи-чинский (1927). За исключением, однако, немногих видов, в литературе накоплено еще мало сведений об отходе яиц и смертности гнездовых птенцов воробьиных, основанных на выяснении судьбы отдельных кладок. Между тем лишь таким путем можно получить материал, необходимый для суждения о том, какие из действующих причин смертности эмбрионов и гнездовых птенцов являются основными, а какие — второ
70
степенными. Такие сведения необходимы для правильного понимания путей формирования биологических признаков птиц, в частности для понимания процесса формирования поведения воробьиных птиц в гнездовой период.
Всего в период с 1946 по 1954 г. автору совместно с помощниками удалось проследить судьбу 1266 кладок 34 видов лесных воробьиных. Из этого количества 775 кладок — на территории Савальского лесхоза, 95 — в заповеднике «Лес на Ворскле» и 396 — на территории Ленинградской области. Таким образом, в пределах лесостепной полосы была выяснена 1история 870 гнезд, а в лесной зоне — 396 гнезд.
Результаты наблюдений по каждой из зон сведены в табл. 20 и 21. Эти наблюдения показали, что отход яиц и гнездовых птенцов лесных
Таблица 20
Выживаемость потомства в гнездах воробьиных в лесной зоне (Ленинградская область)
Название вида Количество кладок под наблюдением Общее количество яиц под наблюдением Количество вылупившихся птенцов Количество птенцов, покинувших гнездо
абсолютное в % к первоначальному количеству яиц
Ворона серая .... 5 23 21 17 74
Сорока 2 14 11 11 78
Зеленушка 5 22 18 17 77
Коноплянка .... 6 34 22 22 65
Зяблик 25 115 95 ,61 53
Снегирь 5 25 8 8 32
Чечевица Овсянка обыкновен- 9 41 17 9 22
ная 3 16 15 11 68
Конек лесной .... 9 43 37 30 69
Сорокопут-жулан . . 30 146 82 76 52
Мухоловка серая . . Мухоловка-пеструш- 29 132 95 91 69
ка 10 51 27 22 43
Пеночка-трещотка . 12 63 63 49 77
Пеночка-весничка 24 148 111 99 67
Малиновка лесная . 13 55 47 39 70
Славка-черноголовка 24 109 74 59 54
Славка серая .... 4 18 11 4 24
Славка садовая . . . 28 129 77 56 43
Дрозд-рябинник . . 33 186 134 112 60
Дрозд певчий .... 36 165 124 107 65
Дрозд-белобровик . Горихвостка обыкно- 77 384 266 210 55
венная 7 41 39 33 80
Итого ... 396 1960 1394 1143 58
воробьиных птиц значителен как в наших северных лесах, так и в южных. Если суммировать данные по всем изучавшимся видам, то количество вылетающих из гнезд птенцов по отношению к первоначальному количеству отложенных яиц составляет примерно 60%. Следовательно, около 40% откладываемых птицами яиц не превращаются в конечном счете в птенцов-слетков. Лишь у некоторых дуплогнездников (скворец и большая синица) процент отхода яиц и птенцов оказался значительно
71
Таблица 2Т
Выживаемость потомства в гнездах воробьиных в лесостепной зоне (Савальский лесхоз и „Лес на Ворскле")
Название вида Количество кладок, судьба которых известна Общее количество яиц под наблюдением Количество вылупившихся птенцов Количество птенцов, покинувших гнездо
абсолютное в % к первоначаль- ному количеству яиц>
Сорока 8 43 35 28 65
Сойка 16 96 61 39 40
Скворец 63 280 261 249 89
Иволга * 14 49 44 35 71
Дубонос 10 41 37 30 73
Зеленушка 22 104 68 53 65
Щегол * 4 20 20 20 100
Коноплянка * . . . . 5 25 25 20 80
Зяблик 23 101 88 79 78
Овсянка обыкновенная * 30 138 98 64 46
Овсянка садовая . . 33 155 101 79 51
Конек лесной * . . . 63 297 223 153 50
Синица большая . . 148 1416 1228 1073 76
Сорокопут-жулан . . 33 162 106 87 54
Мухоловка серая . . 50 217 152 112 52
Мухоловка-белошей-ка 34 178 90 89 50
Пеночка-трещотка * 26 149 121 88 59
Пеночка-теньковка . 5 24 23 14 58
Малиновка лесная . 7 28 27 27 96
Славка-черноголов-ка * 49 224 179 157 70
Славка серая .... 9 39 19 10 26
Славка-завирушка * . 16 77 56 44 57
Славка-садовая * . . 46 200 117 87 43
Дрозд-рябинник . . 12 64 45 36 54
Дрозд певчий .... 28 133 81 68 51
Дрозд черный * . . . 47 228 184 140 61
Горихвостка обыкновенная 36 225 143 130 58
Соловей восточный * 33 152 120 91 60
Итого . . . 870 4865 3752 3102 64
Примечание. Виды, отмеченные звездочкой, изучались на территории Са-вальского лесхоза и в заповеднике „Лес на Ворскле", для них приводятся суммарные-данные. Материалы по остальным видам собраны только в Савальском лесхозе.
ниже среднего. Сходные показатели естественного отсева до вылета птенцов из гнезда у воробьиных птиц получены также европейскими орнитологами (Лэк, 1954).
Каковы же основные причины большого отхода яиц и гнездовых птенцов воробьиных?
Причины отхода яиц. Наблюдения показывают, что причины отхода яиц воробьиных птиц могут быть самыми различными. Основные из них следующие.
а) Расхищение кладок различными врагами птиц. Деятельность-хищников, в широком смысле этого слова, является главной причиной„
72
обусловливающей высокий процент отхода яиц воробьиных птиц. Так,, например, из находившихся под наблюдением в лесной и лесостепной полосе 6825 яиц различных видов воробьиных вылупилось 5146 птенцов. Таким образом, 1679 яиц не дали в конечном счете птенцов. Из этого количества 1072 яйца (около 64% всего отхода) было расхищено различными врагами птиц.
Большое количество яиц расхищалось не только из гнезд, расположенных на земле, но также и из гнезд, находящихся в местах, малодоступных для наземных хищников. Это указывало на то, что у воробьиных немало пернатых врагов.
б) Оставление кладок самими птицами. Известно, что птицы, будучи сильно потревоженными у гнезда, иногдгт бросают его. Чаще всего это бывает, если птицу напугать в период постройки гнезда или откладки яиц. Во время насиживания кладки привязанность птицы к гнезду выражена значительно сильнее, но и в это время она нередко бросает гнездо и приступает к постройке нового, которое располагает обычно в более укромном месте. В случае сильного испуга птица может бросить гнездо даже с птенцами или в период их вылупления. Так иногда поступают, например, большие синицы, пойманные на гнезде и тут же выпущенные. Подчас птицы бывают принуждены бросить кладку из-за муравьев, поселившихся в толще гнездовой подстилки. Подобные случаи наблюдались у пухляка (Basecke, 1927; Осмоловская и Формозов, 1950), у сорокопута-жулана (Мальчевский, 1950), серой мухоловки (устное сообщение Г. А. Новикова). В целом эта «статья расхода» птичьих яиц оказалась незначительной, сильно уступающей отходу яиц по причине разорения гнезд хищниками, и составила 19% от общего числа яиц, не давших в конечном счете птенцов. Оставление полных кладок автору чаще всего приходилось наблюдать у чечевицы, снегиря, сорокопута-жулана, пеночки-веснички и садовой славки.
Оставление кладки, видимо, следует рассматривать как одну нз-приспособительных форм поведения птиц. Наблюдения показывают, что-хищник, например сойка или горностай, обнаруживший гнездо и утащивший из него яйцо или птенца, обычно посещает его повторно и утаскивает все, что находится в нем. Подобной особенностью поведения некоторых хищников и объясняются довольно частые в природе случаи поочередного исчезновения яиц и птенцов из гнезд. Способность воробьиных строить новое гнездо и возобновлять кладку дает им возможность уйти от врага в более безопасное место. Некоторые особи, делающие это после первого же посещения хищником их гнезда, часто оказываются в более выгодном положении, нежели те, которые упорно продолжают насиживать кладку до тех пор, пока из их гнезда не будет утащено последнее яйцо. Многое зависит также и от сроков размножения особи, определяющих ее физиологическое состояние и способность организма к дальнейшей откладке яиц. Весной и в начале лета птицы чаще бросают свои кладки, нежели в конце сезона размножения, когда многие из них, гнездясь в течение данного сезона уже не в первый раз, оказываются неспособными возобновить кладку.
в) Наличие яиц, из которых птенцы по той или иной причине не вылупляются. Как правило, такие яйца не выбрасываются птицами, а остаются лежать в гнезде. Их нередко приходится находить под птенцами, уже готовыми к вылету. Изучение этих яиц показывает, что причины невылупления птенцов могут быть разными. Чаще всего приходится иметь дело с так называемыми болтунами, т. е. неоплодотворен-ными яйцами. Несколько реже яйца, оставшиеся лежать в гнезде, содержат мертвых недоразвившихся зародышей (это обычно яйца, помятые
73
«самой птицей) или птенцов, погибших в яйце непосредственно перед актом вылупления. Последних иногда называют «задохликами». Это название надо признать удачным, поскольку оно в целом правильно отражает причину гибели эмбриона, который действительно погибает от недостатка кислорода. На это указывает, в частности, то, что клюв у такого зародыша обычно бывает открытым. Далеко не во всех случаях гибель зародыша наступает по причине нехватки кислорода в воздушной камере, как это обычно принято считать (Рольник, 1946). Иногда невы-лупление происходит из-за неправильного положения эмбриона. В нормальных случаях голова зародыша обращена к тупому концу яйца, где, как известно, находится воздушная камера, в которую и попадает клюз птенца в критический для него период перехода от аллантоисного дыхания к легочному. Бывают, однако, такие случаи, когда птенец перед вылуплением занимает обратное положение, при котором голова его обращена к острому концу яйца.
Болтуны бывают далеко не в каждой кладке, если же они встречаются, то чаще всего по одному, реже по два на гнездо. Однажды в кладке дрозда-рябинника автор обнаружил 3 болтуна, а у большой синицы 5 таких яиц. В отдельных случаях даже вся кладка может состоять из неоплодотворенных яиц. Так, на территории Савальского лесхоза известны случаи, когда у мухоловки-белошейки кладки целиком состояли из болтунов (устное сообщение Н. П. Кадочникова). Всего из
Т а б л и ц а 22
Отход яиц в гнездах воробьиных в лесной зоне (Ленинградская область)
Название вида Количество яиц под наблюдением Причина отхода Вылупилось птенцов
брошено погибло из-за разорения гнезда болтуны и .задохлики"
Ворона серая 23 2 21
Сорока 14 — 3 — 11
Зеленушка 22 4 — — 18
Коноплянка 34 6 5 1 22
Зяблик 115 — 14 6 95
Снегирь 25 17 — — 8
Чечевица 41 10 14 — 17
Овсянка обыкновенная 16 — — 1 15
Конек лесной 43 — 4 2 37
Сорокопут-жулан 146 26 33 5 82
Мухоловка серая 132 5 27 5 95
Мухоловка-пеструшка 51 — 23 1 27
Пеночка-трещотка 63 —— — — 63
Пеночка-весничка 148 13 20 4 111
Малиновка лесная 55 3 4 1 47
Славка-черноголовка 109 5 28 2 74
Славка серая 18 — 4 3 11
Славка садовая 129 16 35 1 77
Дрозд-рябинник 186 3 39 10 134
Дрозд певчий 165 9 28 4 124
Дрозд-белобровик 384 26 79 13 266
Горихвостка обыкновенная . ... 41 — 2 39
Итого 1960 | 143 360 63 1394
74
178 яиц белошейки оказалось 26 неоплодотворенных, т. е. примерно 14%. Интересно, что относительно высокий процент болтунов отмечался л'акже для мухоловки-пеструшки: 6% (Тауриньш, 1951), 8,6—13% (Лихачев, 1953) и даже 25% (Карпов и Паровщиков, 1941). Надо сказать, что далеко не всякое яйцо, в котором не происходит развитие зародыша, можно считать болтуном. У большой синицы, например, часть яиц иногда теряется в гнездовой подстилке, благодаря чему не инкубируется. Вообще говоря, у птиц, строящих открытые гнезда, болтуны и «задохлики» встречаются, как правило, реже, чем у дуплогнездников. В целом по всем изучавшимся нами видам количество яиц, из которых по той или иной причине птенцы ;не вылупились, составило около 4% от первоначально отложенных и 17% от общего отхода яиц.
Гибели кладок воробьиных птиц из-за понижения температуры воздуха, дождей и т. п. неблагоприятных воздействий климата автору наблюдать не приходилось. В пределах средних широт подобного рода случаи вообще, видимо, редки.
Таблица 23
Отход яиц в гнездах воробьиных в лесостепной зоне (Савальский лесхоз и .Лес на Ворскле*)
Название вида Количество яиц под наблюдением Причина отхода Вылупилось птенцов
брошено погибло из-за разорения гнезда болтуны и .задохлики*
•Сорока 43 7 1 35
Сойка 96 7 25 3 61
Скворец 280 5 3 11 261
Иволга 49 — 3 2 44
Дубонос . • .4 41 2 — 2 37
Зеленушка 104 — 34 2 68
Щегол 20 — — — 20
Коноплянка 25 — — 25
Зяблик • 101 9 4 88
Овсянка обыкновенная 138 5 29 6 98
Овсянка садовая 155 — 49 5 101
Конек лесной 297 23 40 11 223
Синица большая 1416 23 73 92 1228
Сорокопут-жулан 162 27 26 3 106
Мухоловка серая 217 5 48 12 152
Мухоловка-белошейка 178 24 38 26 90
Пеночка-трещотка 149 — 26 2 121
Пеночка-теньковка 24 — 1 23
Малиновка лесная 28 — 1 27
Славка-черноголовка 224 4 36 5 179
Славка серая 39 — 20 — 19
Славка-завирушка 77 — 16 5 56
Славка садовая 200 20 49 14 117
Дрозд-рябинник 64 6 11 2 45
Дрозд певчий 133 5 47 — 81
Дрозд черный 228 20 20 4 184
Горихвостка обыкновенная 225 5 73* 4 143
Соловей восточный 152 — 30 2 120
Итого 4865 181 712 220 3752
Примечание. Звездочкой отмечены данные, которые включают и гибель 1яиц из-за кукушки.
75
Более полные данные по отходу яиц у отдельных видов птиц приведены нами в табл. 22 и 23, а также в очерках, помещенных в части второй .настоящей работы. Здесь отметим лишь, что относительно „ очень-высокий процент отхода яиц (около 32%) в гнездах обыкновенной горихвостки на территории Савальского лесхоза (табл. 21) объясняется тем,, что яйца этого вида уничтожались не только хищниками, но и кукушкой, паразитирующей в данном районе исключительно на горихвостке.
Причины гибели гнездовых птенцов. Причины гибели гнездовых птенцов воробьиных чрезвычайно разнообразны, а общий процент отхода весьма значителен (табл. 24 и 25). Иногда птенцы погибают самым необычным образом. На. рис. 8, например, изображен желудок и пищевод птенца обыкновенной овсянки, до отказа набитые конским волосом из подстилки гнезда. Птенец, каким-то образом наглотавшийся волоса, запутался в нем и погиб от истощения, так как не мог уже заглатывать пищу (рис. 9). Случаи подобного рода, конечно, редки.
Таблица 24-
Отход птенцов в гнездах певчих птиц в лесной зоне (Ленинградская область)
Название вида Количество птенцов под наблюдением Причина отхода Количество птенцов-слетков
разорение гнезда иные причины
Ворона серая 21 4 17
Сорока П — — 11
Зеленушка 18 — 1 17
Коноплянка . . . . • 22 — — 22
Зяблик 95 19 15 61
Снегирь 8 — — 8
Чечевица 17 8 — 9
Овсянка обыкновенная 15 4 11
Конек лесной . . . . .... 37 5 2 30
Сорокопут-жулан 82 6 — 76’
Мухоловка серая 95 — 4 91
Мухоловка-пеструшка 27 5 — 22
Пеночка-весничка 111 11 1 99
Пеночка-трещотка 63 14 49-
Малиновка лесная 47 8 39
Славка-черноголовка 74 9 6 59
Славка серая 11 7 — 4-
Славка садовая 77 18 3 56-
Дрозд-рябинник 134 22 — 112
Дрозд певчий 124 13 4 107
Дрозд-белобровик 266 52 4 21<>
Горихвостка обыкновенная 39 6 — 33
Итого ' 1394 207 44 1143
Иногда наблюдается гибель всего выводка или части птенцов из-за «неряшливой» постройки гнезда и плохого прикрепления его к ветвям. Такие гнезда, особенно после дождливой погоды, скоро начинают крениться на тот бок, на который чаще всего садится птица, и иногда опрокидываются, а птенцы вываливаются (Мальчевский, 1948). Известны также случаи гибели отдельных птенцов и целых выводков от питающихся кровью личинок паразитических мух Protocalliphora. В Англии, например, личинки этих мух были найдены в гнездах 22 видов воробьи-
76
пых (Owen, 1954). Оуэн указывает, что они способны умертвить даже крупных птенцов сороки. В одном из гнезд большой синицы он нашел 70 личинок этих мух, погубивших всех птенцов, когда им было уже 12 дней от роду.
Таблица 25
Отход птенцов в гнездах воробьиных в лесостепной зоне (Савальский лесхоз и «Лес на Ворскле*)
Название вида Количество птенцов под наблюдением Причина отхода Количество птенцов-слетков
разорение гнезда иные причины
Сорока 35 7 28
Сойка • 61 22 — 39
Скворец 261 3 9 249
Иволга 44 7 2 35
Дубонос 37 7 — 30
Зеленушка 68 15 — 53
Щегол 20 — — 20
Коноплянка 25 — 5 20
Зяблик 88 9 — 79
Овсянка обыкновенная 98 27 7 64
Овсянка садовая 101 14 8 79
Конек лесной 223 65 5 153
Синица большая 1228 40 115 1073
Сорокопут-жулан 106 19 — 87
Мухоловка серая 152 31 9 112
Мухоловка-белошейка 90 — 1 89
Пеночка-теньковка 23 9 — 14
Пеночка-трещотка 121 31 2 88
Малиновка лесная 27 — — 27
Славка-черноголовка 179 22 — 157
Славка серая 19 9 — 10
Славка-завирушка . • 56 12 — 44
Славка садовая 117 26 4 87
Дрозд-рябинник 45 9 — 36
.Дрозд певчий 81 9 4 68
Дрозд черный 184 31 13 140
Горихвостка обыкновенная .... 143 6 7 130
Соловей восточный 120 25 4 91
Итого 3752 455 1 195 | 3102
Гибель отдельных птенцов иногда наступает из-за неравномерного распределения пищи между членами выводка. Маленькие и слабые птенцы, вылупившиеся последними, оттесняются и затаптываются более крупными и сильными. Это чаще всего случается у птиц, воспитывающих сразу -по многу птенцов. Например, в гнездах большой синицы нередко приходится находить высохших плоских птенцов, погибших в различном возрасте (рис. 10); количество их может составлять свыше половины общего отхода потомства (табл. 25). В естественных дуплах, которые обычно бывают более тесными, чем искусственные гнездовья, отход птенцов, очевидно, еще значительнее. В одном дупле у большой синицы автор находил до 4 погибших птенцов. Подобное же явление нам приходилось наблюдать у лазоревки, длиннохвостой синицы, пеночек, -.а также у зеленушек, дрозда-белобровика, сорокопута-жулана и некото-
77
Рис. 8. Желудок птенца обыкновенной овсянки, переполненный конским волосом из подстилки гнезда.
рых др. Многие птицы выбрасывают погибших птенцов из гнезда, так что для установления факта гибели слабых птенцов необходимы ежедневные наблюдения.
В плохую погоду птицы, выкармливающие птенцов насекомыми,, прилетают кормить их реже обычного. Иногда это приводит к гибели части выводка. Такие случаи автор наблюдал, например, у обыкновенной овсянки в период затянувшихся дождей и резкого снижения температуры воздуха. Во всех случаях погибшие птенцы были самыми маленькими. Судя по указаниям, имеющимся в литературе, затянувшиеся холода и дожди, а также сильная засуха могут повлечь за собой даже массовую гибель птенцов насекомоядных птиц целыми выводками. Эти указания относятся главным образом к птенцам мухоловки-пеструшки (Кнорре, 1947; Ефремов, 1941; Осмоловская и Формозов, 1950; Лихачев, 1953; Тима, 1954).
Массовая гибель птенцов воробьиных по причине неблагоприятного действия климатических факторов и бескормицы бывает, однако, далеко не каждый год. Во всяком случаев периоде 1946 по 1954 г. ни у одного из видов воробьиных, за гнездами которых проводилось регулярное наблюдение, нами не было отмечено массовой гибели птенцов. Несмотря на большую чувствительность птенцов к дождю и непосредственному воздействию солнечных лучей, погибают от этого они сравнительно редко. Отрицательное влияние климата на птенцов воробьиных часто, по-видимому, преувеличивается, что происходит от недооценки значения поведения взрослых птиц, очень чутко реагирующих на всякое изменение среды в сторону, неблагоприятную для жизни :и развития их птенцов, и защищающих их своим телом в случае необходимости.
Наши наблюдения позволяют говорить о том, что не только прямое, но и косвенное воздействие климата не уничтожает такого количества птенцов, как многочисленные враги воробьиных птиц. Во всех пунктах, где велись систематические наблюдения за птичьими гнездами, деятельность хищных и врановых птиц, а также различных млекопитающих была основной и наиболее постоянно действующей причиной отхода гнездовых птенцов,
В литературе часто приходится находить указание-
на то, что гнезда разоряются главным образом человеком. При этом имеются в виду подростки, которые иногда специально отправляются на поиски птичьих гнезд. И хотя подобное безобразие действительно еще имеет место, было бы неправильным считать, что основная масса гнезд гибнет из-за деятельности человека. Даже в густо населенных районах и пригородах в этом наиболее повинны.вороны, кошки и нередко собаки.
В местах, удаленных от населенных пунктов, процент разоряемых птичьих гнезд также бывает высоким. Например, в безлюдных районах севера Ленинградской области гнезда воробьиных разоряются почти так же часто, как и в пригородах Ленинграда или в Савальском лесхозе. Если в населенных местах главной причиной гибели птенцов воробьиных являются вороны, кошки и частично человек, то в девственных лесах на смену им приходят сойка, сарыч, перепелятник, тетеревятник, лисица белка и др.
78
Рис. 9. Птенец обыкновенной овсянки, наглотавшийся конского волоса из подстилки гнезда (Савальский лесхоз, июль 1953 г. Фото автора).
Рис. 10. Мумия птенца большой синицы, затоптанного собратьями (Савальский лесхоз, июль 1953 г. Фото автора).
От разорения гнезд гибнет птенцов почти в 3 раза больше, нежели -от всех других причин, вместе взятых. Лишь у некоторых дуплогнездников (большая синица, скворец) птенцы, как мы видели выше, чаще погибают от скученности и недоедания, нежели от хищников.
ВРАГИ ВОРОБЬИНЫХ В ГНЕЗДОВОЙ ПЕРИОД
Многочисленных врагов лесных воробьиных в гнездовой период гру-<5о можно разбить на следующие 3 категории: а) хищники, в широком -смысле слова, питающиеся воробьиными птицами, их яйцами и птенцами; б) конкуренты по месту гнездования, нередко выбрасывающие или расклевывающие кладки, и в) различные паразиты, вызывающие гибель птенцов или заставляющие взрослых птиц иногда бросать кладки. На разборе последней группы мы, однако, останавливаться не будем, так как паразиты птичьих гнезд — тема специального исследования.
а) Первая группа врагов наиболее опасна для воробьиных в гнездовой период. К ней следует отнести хищных и врановых птиц, сорокопутов, хищных млекопитающих, некоторых грызунов, а также змей. Хищные птицы помимо птенцов уничтожают и взрослых птиц, отчего нередко гибнут кладки и гнездовые птенцы. Специальные работы, посвященные изучению питания хищных птиц и их птенцов (Жарков и Теплое, 1932; Птушенко, 1936; Якшис, 1952; Измайлов, 1940, и др.), показывают, что лесные воробьиные и их птенцы, хотя и не являются основным видом корма хищников, тем не менее неизменно присутствуют в их пищевом рационе.
Из врановых птиц основными врагами мелких лесных воробьиных следует признать серую ворону и сойку, чаще других разоряющих гнезда. Многочисленные наблюдения указывают на то, что ворона и сойка способны отыскивать и разорять гнезда различных певчих птиц, расположенные как на кустах и деревьях, так и на земле и даже в дуплах (Басил ьчук, 1915; Christoleit, 1927; Мау, 1947; Осмоловская и Формозов, 1950; Попов и др., 1954; Goodwin, 1953; Кадочников и Эйгелис, 1954). Серая ворона, в большинстве мест более многочисленная, все же наще беспокоит мелких птиц и разоряет их гнезда, нежели сойка. Ее деятельность особенно сильно ощущается в пригородных районах, где она наряду с бродячими кошками, по нашему мнению, должна быть признана основным врагом певчих птиц.
Сорока, которая в гнездовой период собирает корм преимущественно среди мелколесья или на открытых местах, не так часто разоряет гнезда лесных певчих птиц, как ворона или сойка. Однако и она при случае ворует яйца и птенцов и притаскивает их сорочатам. После вылета птенцов из гнезд сороки нередко переселяются в леса, где кочуют целыми выводками в поисках корма. В этот период они становятся более опасными для певчих птиц. Там, где держится выводок сорок, постоянно бывают слышны жалобные, тревожные крики различных птиц, не успевших еще вывести птенцов.
Несмотря на то, что сорокопут-жулан питается в основном насекомыми, он тем не менее с полным правом может быть назван врагом певчих птиц. В Ленинградской области зарегистрированы случаи нападения жулана на птенцов чечевицы, лазоревки, варакушки, пеночки-веснички. Ширман (Schiermann, 1927) видел, как этот «хищник» уносил в клюве только что вылупившегося птенчика пеночки-теньковки. Н. В. Шарлемань (1915) отмечает факт похищения сорокопутом птенца зяблика и т. п.
Из хищных млекопитающих гнезда певчих птиц чаще всего разоряют мелкие куньи (ласка и горностай), лисица и бродячие кошки. Дея-
-80
тельность последних весьма ощутительна в местах, прилегающих к населенным пунктам. По наблюдениям К. Н. Благосклонова (1950а), на участке Болыпевской биологической станции в некоторые годы около трети' всех вылетевших из гнезд птенцов уничтожается кошками. Горностай и ласка становятся особенно опасными для мелких птиц в годы низкой численности мышевидных грызунов. Автор этих строк однажды лаже поймал горностая, который спал в гнезде полевого воробья на трупе своей жертвы. Гнездо помещалось в дупле старого вяза на высоте 2 м от земли. Оно было все переворошено, яйца все расколоты, а у самки воробья была отъедена голова. В заповеднике «Лес на Ворскле» удалось наблюдать также, как горностай, взобравшись по стволу молодого дуба, проник в гнездо сорокопута-жулана, проделал в дне его отверстие, умертвил всех находившихся в гнезде птенцов и затем стал по очереди их куда-то утаскивать
Гнезда, расположенные на земле или на небольшой высоте, нередко разоряются лисицей, барсуком, волком, енотовидной собакой и другими хищниками. Разорителями гнезд иногда являются также некоторые грызуны (белка, сони) и насекомоядные (еж). Об этом свидетельствуют указания целого ряда авторов (Лихачев, 1953; Птушенко, 1949; Благо-склонов, 1950а; Формозов, 1952; Семенов, 19566).
. Наконец, следует указать на змей, заползающих иногда в птичьи гнезда и пожирающих кладки и птенцов. В этом отношении очень интересны наблюдения Т. Б. Ардамацкой (устное сообщение), сообщающей о случаях разорения гнезд некоторых птиц-дуплогнездников четырехпо-лосым полозом в Черноморском заповеднике. В северных районах не-редкр разоряет гнезда мелких птиц, в частности пёночек-весничйк, лес-ная гадюка (устное сообщение Т. А. Воропановой и Ю. Г. Галеева).
б) Вторую группу врагов воробьиных (конкуренты по месту гнездования) составляют в основном различные виды птиц-дуплогнездников, которые, будучи крупнее и энергичнее многих мелких певчих птиц, изгоняют их из занятых ими дупел, при этом нередко выкидывают и расклевывают их кладки, к таковым следует отнести прежде всего полевого и городского воробьев, отрицательная роль которых отмечалась в литературе неоднократно (Васильчук, 1915; Дергунов, 1928; Мальчевский, 1947; Благосклонов, 1950а; Гаращук, 1953 и Др.). Аналогичные указания имеются и в отношении вертишейки (Благосклонов, 1957).
Иногда причиной гибели кладки является большой пестрый дятел, залезающий в дупла, занятые мелкими птицами, с целью ночевки, а также похищения недавно вылупившихся птенцов. Подобная «хищническая» деятельность большого пестрого, дятла отмечалась в Германии (Schnurre, 1937; Pfeifer, 1952; Werner, 1954.), на южном Урале (Кири-ков, 1952), в Латвии (устное сообщение К. А. Вилкса), в Воронежской области (устное сообщение Н. П. Кадочникова), а также в Ленинградской области (наблюдения автора). Зарегистрированы случаи разорения гнезд мухоловки-пеструшки и белошейки, горихвостки, большой синицы, зарянки, крапивника, зяблика и некоторых других мелких воробьиных.
Мы перечислили здесь далеко не полный состав врагов и конкурентов лесных воробьиных, тем не менее и приведенных примеров 'Достаточно для того, чтобы представить, насколько полна опасностями жизнь певчих птиц в гнездовой период, и понять, почему так часто гибнут кладки и исчезают из гнезд птенцы,
Так как при регистрации фактов гибели яиц или птенцов далеко ие всегда возможно определить вид хищника, разорившего гнездо, количественная характеристика значения отдельных видов хищников при таком
6 А. С. Мальчевский
81
Позвоночные, вызывающие у лесных певчцх птиц „сигнал* тревоги (по наблюдениям в Ленинградский области)
Таблица 26
Виды птиц, подававших .сигнал* тревоги Число тревог причины которых Число тревог причинами которых являлись
выяснялись выяс- 1 немы 1 сарыч 2.5 * 01'4) X CG Я, пустельга дерб-ник ушастая сова kedao ЧХ1ЧЗ -ВЭН болыщ пест-фЫЙ дятел об К 1 1 сорока сойка сорокопут-4 жулан воро- 1 бей полевой большая синица лисица горностай белка кошка
Скворец 3 3 1 2
Иволга 3 3 1 1 —- — — 1 — - — —— — — — — — — —
Зеленушка 3 3 — — — 1 — • — — 1 — — — — — — — — 1
Щегол 2 2 — — 4 — ) — — — —— — 1 — — — —
Чиж 2 2 1 — 1 — —в — — — — ——- —— — —
Чечевица 2 2 — 1 — 1
Зяблик 30 25 1 5 — — — 3 — 5 3 6 — т— — — — 1 1
Овсянка обыкновенная 1 1 1
Синица большая 2 2 — 1 1
Пухляк 1 1 —« — • — — -г- — — — -г- — 1 — — — — — —
Синица длиннохвостая 1 1 — 1
Сорокопут-жулан 3 3 -т— 1 — — — — — 1 1 — — — — — — —
Мухоловка серая 6 5 — 1 — — —— — — — — 1 1 — — — — — 2
Мухоловка-пеструшка 14 14 — 2 — — 1 1 2 — — 2 2 2 1 — — — 1
Пеночка-весничка 8 7 1 1 — — 1 —- 1 “Г* 2 — —— 1 —— — —
Пеночка-трещотка 8 8 1 2 — —5 1 — —> 3 1 — — — — ' —- —
Малиновка лесная 2 2 — 1 1
Славка-черноголовка 2 2 1 — 1
Славка садовая 8 7 1 — — — —— —— — —• —г- 2 — — — — 4
Славка серая 2 2 2
Славка ястребиная 1 1 1
Дрозд-рябинник 27 27 1 1 — — — 2 — 22 — — 1 — — — — — —
Дрозд певчий 10 2 1 — —— —— — — — — — — — — 1 —— —
Дрозд-белобровик 16 9 —- 1 1 — — 2 — 3 — — — — — — — — 2
Горихвостка 8 7 1 • — —- i — — 1 — — — — — — 4
Соловей восточный 2 2. 1 — — — — —— — — — — — 1
Зарянка 8 2 — 1 — — — — — — — — — — » 1 — 1 —
Итого 175 145 11 21 1 3 1 12 2 35 5 14 15 2 1 2 1 2 17
Позвоночные, вызывающие у лесных певчих птиц „сигнал" тревоги (по наблюдениям в Савальском лесхозе)
Таблица 27
Виды птиц, подавших .сигнал" тревоги Число тревог, причины которых Число тревог, причинами которых являлись
выяснялись выяснены коршун сарыч перепелятник чеглок 1 пустельга 1 ушастая сова больш. пестр. дятел кукушка ворона сорока сойка сорокопут-жулан 1 волк лисица горностай кошка уж обыкновенный
Скворец 8 8 - 3 3 — 2 —
Иволга 26 25 3 9 2 2 — —— 2 2 4 1 — — — — — —
Зеленушка 1 1 — 1
Щегол 3 3 2 1 — — —— — — — —. — — — -— — — —- —
Зяблик 41 36 4 9 — — —. . 2 — — 1 2 18 — — — — — —
Овсянка обыкновенная 3 2 2
Овсянка садовая 3 2 1 1
Конек лесной 23 21 3 И — 1 1 — — — — 5 — —- — —— — —
Синица большая 5 5 — — —— 1 — — — — 4 — — — — — —
Сорокопут-жулан 1 1 — 1
Уухоловка серая 38 34 2 9 1 — — 1 1 2 — — 16 2 — — — — —
Мухоловка-белошейка 2 2 — — — — — — 1 — — — 1 — — — — — —
Пеночка-трещотка 17 17 5 б — — — 2 — 1 — — 3 — 1 — — — —
Малиновка лесная 12 11 — 4 — — — — — 1 — — 5 — 1 — — — —
Славка-черноголовка 3 3 — 1 1 1
Славка садовая 14 14 — 5 — — 1 1 — 3 — — 1 1 1 — — — 1
Славка-завирушка 1 1 1
Дрозд-деряба 2 2 — 1
Дрозд-рябинник 13 12 — 1 11
Дрозд-черный 16 13 — 4 1 2 — 1 — — — — 3 — — 1 1 — —
Горихвостка 15 14 1 3 1 — — 1 —- 1 — 4 1 1 1 — — —
Соловей восточный 20 18 1 1 — — 3 — — 4 3 5 — -— — 1 —• —
Итого , . 267 245 22 : 66 5 4 2 12 3 10 23 12 70 4 4 2 2 2 2
методе исследования невозможна. Для того чтобы хоть сколько-нибудь восполнить этот недостаток, автор применил косвенный учет врагов певчих птиц. В процессе полевых исследований эта методика получила название «хождения на птичьи переполохи». Она заключалась в возможно более скором внимательном обследовании участка, где раздались беспокойные крики птиц. Такие наблюдения требуют большой осторожности и терпения, ибо далеко не всегда птичий враг позволяет выявить себя сразу. Иногда причина тревожного поведения птиц так и остается неустановленной.
В Савальском лесхозе певчих птиц чаще всего беспокоили сойка, сарыч, ворона и коршун. Сарыча и коршуна мы, как правило, обнаруживали вблизи птенцов дроздов, овсянок, соловьев, уже оставивших гнездо. Хищники обычно неподвижно сидел и в кроне дерева и выслеживали прыгающих по земле слетков. Сойка и ворона попадались нам преимущественно у гнезд. В Ленинградской области наиболее обычными посетителями гнездовых участков оказались ворона и ястреб-перепелятник; несколько реже беспокоили птиц сойка, сорокопут-жулан, серая неясыть и сарыч. Из млекопитающих чаще всего попадались на глаза бродячие кошки.
Анализируя данные наших учетов в целом, можно сказать, что в гнездовой период хищные и врановые птицы значительно чаще беспокоят мелких лесных воробьиных, чем млекопитающие (табл. 26 и 27).
О ПОВЕДЕНИИ ВОРОБЬИНЫХ У ГНЕЗД И ПТЕНЦОВ
Большое количество врагов, окружающее мелких лесных воробьиных, частые случаи разорения их гнезд, опасность, которая буквально на каждом шагу подстерегает их совершенно беспомощных птенцов, — все это привело к возникновению соответствующих-приспособлений, направленных на защиту потомства или как-то компенсирующих частые случаи потери кладок. К числу последних следует прежде всего отнести способность воробьиных легко восстанавливать утерянную кладку и начинать цикл размножения сызнова. Об этой особенности репродуктивного цикла воробьиных мы говорили выше. Сообщалось также о наблюдающейся у мелких птиц тенденции располагать свои гнезда в недоступных или мало заметных для хищников местах. Но, кроме этого, у воробьиных птиц возникали различные приспособительные формы поведения в случае непосредственной опасности, угрожающей гнезду или выводку.
Многие млекопитающие, будучи потревоженными у нор, гнезд или логовищ, перетаскивают своих детенышей в более безопасные места, у птиц же, за исключением поганок, некоторых уток (Книпперт, 1956) и куликов, подобный способ уберегать свое потомство не выработался. Факт переноса птенцов ястребом-тетеревятником, приведенный в книге Макача (1951), является для птенцовых птиц пока что единственным и нуждается в проверке. На появление врага вблизи гнезда или выводка воробьиные реагируют по-иному. Можно различать следующие формы их поведения, имеющие, с нашей точки зрения, приспособительное значение.
а) Оставление кладки, переселение в другое, более безопасное место, постройка! нового гнезда и повторная откладка яиц. Об этой форме поведения птиц мы уже говорили.
б) Отвод врага в сторону от гнезда или птенца. Здесь следует прежде всего указать на удивительную способность многих воробьиных имитировать движения больной или раненой птицы. Некоторые особи
84
садятся на выдающиеся ветви кустов и оттуда демонстративно падают на землю, затем, сгорбившись, отбегают в сторону, распустив крылья и хвост. В том случае, если враг не обращает внимания на птицу и продолжает находиться вблизи гнезда, этот «прием» иногда повторяется несколько раз подряд. Наоборот, если за такой птицей побежать, на что и рассчитано ее поведение, она, после того как отведет преследователя на некоторое расстояние в сторону от гнезда, внезапно «выздоравливает» и улетает. Биологический смысл подобного поведения очевиден. Результат действия птицы иногда бывает поразительным. Даже человек, знающий, что птица «притворяется», часто невольно бросается ее преследовать.
Имитирование раненой птицы — явление довольно широко распространенное среди воробьиных. Автор наблюдал его у 16 видов (табл. 28). В большинстве случаев, однако, подобное поведение было отмечено лишь у очень немногих особей того или другого вида. Только для представителей рода Sylvia оно оказалось обычным. Явление «отвода» мы наблюдали у большинства встреченных нами особей славок-черноголо-вок, завирушек и ястребиных, садовая и особенно серая славки «притворялись» больными значительно реже.
Интересно, что большинство птиц, гнездящихся на земле (овсянки, пеночки, луговой чекан, восточный соловей и лесной конек), для которых данное явление, казалось бы, должно быть наиболее характерным, сравнительно редко имитирует раненых птиц. Не наблюдали мы его также у многих птиц, гнездящихся в кустах невысоко над землей (коноплянка, чечевица, сорокопут-жулан).
Для некоторых видов птиц характерен другой прием отвода: птица близко подлетает к врагу, садится на видное место и издает беспокойный позыв, но при первой попытке хищника приблизиться к ней быстро отлетает в сторону. Этот прием очень типичен, например, для обыкновенной горихвостки. Иногда его применяют также пеночки, серая мухоловка, лесная малиновка и другие птицы. Подобный способ отвлечения внимания врага также эффективен, как и имитация раненой птицы. Автору неоднократно приходилось наблюдать, как горихвостки, применявшие описанный выше прием, совершенно сбивали с толку кошек и отводили их в сторону от птенцов, только что покинувших гнездо.
в) Отпугивание врага. Приемы отпугивания, которые применяют воробьиные, опять-таки могут быть разными. Некоторые птицы с силой налетают на врага и ударяют его клювом, лапами или всем телом. Этот способ защиты потомства почти всегда применяют, например, дрозды-рябинники по отношению к своему основному врагу — вороне. Таким же образом дрозды-рябинники, нередко и белобровики, находясь в состоянии сильного возбуждения, иногда нападают даже на человека, близко подошедшего к их гнезду или птенцам. Активное нападение на врага характерно также для иволги и врановых птиц, в частности для серой вороны и сойки.
Агрессивный способ защиты своего потомства применяют иногда и совсем мелкие виды воробьиных: сорокопут-жулан, серая мухоловка, лесная малиновка, обыкновенная горихвостка. Например, один из сорокопутов-жуланов, гнездившихся в 1947 г. в парке Лесотехнической академии, был замечателен тем, что каждый раз, когда кто-нибудь близко подходил к его гнезду, он с налета пикировал и наносил довольно сильный удар в голову. Сходным поведением отличалась и одна самка обыкновенной горихвостки, гнездившаяся в этом же парке. Чтобы отогнать от своих гнезд сойку, серые мухоловки и малиновки всегда норовят ее ударить.
85
Таблица 28
Характер поведения некоторых лесных воробьиных при опасности, угрожающей потомству
Название вида Приемы, отводящие врага в сторону от гнезда Приемы, отпугивающие врага Скрытное поведение
имитация раненой птицы крик вблизи с последующим отлетом в сторону активное нападение отпугивание голосом и движениями крик в стороне молчаливое и скрытное поведение
Ворона серая иногда иногда иногда иногда
Сорока — — л обычно 9
Сойка — — иногда - иногда обычно
Скворец — — обычно иногда
Иволга — — 9 9 л
Дубонос — — — обычно W
Щегол иногда иногда — — м ш
Зеленушка п п — — V
Коноплянка — • — — V
Снегирь — — — — »
Чечевица — иногда — — л
Клест-еловик — — — —
Зяблик иногда обычно — — иногда
Овсянка обыкновенная . иногда — — обычно
Овсянка садовая .... » обычно — — иногда
Конек лесной п иногда — — обычно
Синица большая .... — — обычно и
(на гнезде)
Синица длиннохвостая . — — — — 9 я
Сорокопут-жулан . . . — обычно иногда иногда иногда м
Мухоловка серая . . . оч. редко V 9 9 » •
Мухоловка-белошейка . — — — — обычно V
Мухоловка-пеструшка . — — иногда иногда иногда
Пеночка-весничка . . . оч. редко обычно — — иногда м
Пеночка-трещотка . . . иногда V — — » 9
Малиновка лесная . . . — W иногда иногда л л
Славка ястребиная . . . обычно часто — —
Славка-черноголовка . . п иногда оч. редко оч. редко V
Славка садовая .... часто часто — — часто
Славка-завирушка . . . обычно иногда — — часто
Славка серая иногда — — обычно иногда
Дрозд-деряба — — иногда обычно иногда
Дрозд-рябинник .... — — часто •
Дрозд черный оч. редко иногда — оч. редко обычно
Дрозд-белобровик . . . п п иногда обычно иногда я
Дрозд певчий — 9 оч. редко оч. редко обычно я
Чекан луговой —’ 9 — — я
Горихвостка —— обычно иногда иногда иногда я
Соловей восточный . . иногда иногда — — обычно -
Зарянка — 9 — — 9
Завирушка лесная . . . — — — — — обычно
Крапивник — иногда — — обычно иногда
Чаще, однако, птицы, пикируя на врага, взмывают над самой головой и как раз в это время внезапно издают резкий, неприятный крик, как бы желая испугать неприятеля. Так часто делают иволги и дрозды — белобровик, деряба и рябинник.1 Устрашающие позы и угрожающее ши
1 Интересно отметить, что поведение остальных видов наших дроздов — черного и певчего — совсем иное: они почти никогда не отпугивают человека от своего гнезда, а обычно держатся в стороне.
86
пение характерно, например, для большой синицы, которая внезапностью и своеобразностью издаваемого звука может напугать даже человека, ^заглядывающего в дупло*
Наконец, некоторые виды птиц, атакуя неприятеля, применяют совершенно своеобразный прием «бомбардировки» своими жидкими экскрементами. Среди воробьиных птиц этот способ защиты характерен для дроздов-рябинников и иногда для ворон.
г) Скрытное поведение. Большой скрытностью поведения у своих гнезд отличаются многие птицы. Чрезвычайно, например, трудно увидеть у гнезда лесную завирушку, сойку, кукшу, кедровку, снегиря. Не любят показываться на глаза также черный и певчий дрозды, некоторые синицы и мухоловки. В период насиживания кладки мало заметны у гнезд сорокопут-жулан, зарянка, восточный соловей. Кроме того, среди птиц, держащихся обычно открыто, часто встречаются очень осторожные особи. Наблюдения за такими птицами, показывают, что они не индифферентно относятся, например, к человеку, появившемуся в районе их гнезда. Они, как правило, бывают при этом возбуждены, иногда подают «сигнал» тревоги, но тем не менее продолжают держаться в стороне. Подобное поведение чаще всего приходится наблюдать у птиц, гнездящихся в глухом лесу, в отдалении от человеческого жилья.
У многих птиц осторожность является исходной формой поведения, в то время как доверчивость — вторичной, приобретенной птицами благодаря постоянному контакту с людьми. Бывает, однако, и так, что доверчивая птица после сильного испуга становится очень осторожной. Таким образом, причина скрытного поведения 1птиц у гнезд часто заключается в их инстинкте самосохранения. В то же время такое поведение может оказаться полезным и для потомства. Дело в том, что, беспокоясь у гнезда, птицы иногда сами выдают место его расположения: чем ближе к гнезду враг, тем сильнее тревожится птица, и наоборот. Опытный зверь, который уже неоднократно разорял птичьи гнезда под «аккомпанемент» интенсивного крика птиц, может в дальнейшем начать руководствоваться этим криком при разыскивании гнезда или птенцов. Наблюдения показывают, что громкий беспокойный крик, издаваемый птицами у гнезда или выводка, действительно привлекает внимание различных хищников. Особенно часто прилетают к «месту происшествия» вороны и сойки, а оказавшись поблизости от гнезда, они легко его находят и разоряют.
Из табл. 28 можно видеть, что особи одного и того же вида в случае опасности, угрожающей потомству, могут вести себя по-разному. Остается не совсем ясным, каким образом следует объяснять столь сильную индивидуальную изменчивость поведения. Почему, например, большинство зябликов, зеленушек, лесных коньков или садовых овсянок даже при очень сильном возбуждении не имитируют раненую птицу, в то время как некоторые особи этих видов делают это?
Если бы все представители одного вида в случае опасности вели себя совершенно однотипно, как это и считает Хейнрот (1938), то тогда, может быть, и следовало бы рассматривать подобное «притворство» птиц как явление «от начала до конца врожденное». Однотипности поведения птиц, однако, не существует. Имеет место прежде всего различная степень возбуждаемости птиц, которую, очевидно, надо объяснять различным жизненным опытом отдельных особей. Следуя мнению А. Н. Промп-това и Е. В. Лукиной (1940), можно предположить, что те особи, гнезда которых уже разорялись, ведут себя более активно. В то же время нельзя не учитывать также способности птиц в зависимости от сложившейся обстановки быстро изменять характер своего поведения и переходить от
87
одного «приема» защиты потомства к другому. Автору не раз приходилось наблюдать, как славки-черноголовки, первоначально реагируя на появление человека у гнезда обычным для вида способом, имитируя раненую птицу, вдруг переставали «притворяться» и переходили к активному нападению. Физиологическая природа такого поведения остается* не вполне понятной. Можно только сказать, что воробьиные птицы в течение своей индивидуальной жизни способны не только вырабатывать свои приемы защиты потомства от врагов, но и изменять их в зависимости от сложившейся обстановки.
ГЛАВА IV
ВЫЛУПЛЕНИЕ И ПТЕНЦОВЫЕ ПРИЗНАКИ
Птенцы воробьиных, особенно в их раннем возрасте, незаслуженно мало привлекали внимание орнитологов. Поэтому в наших определителях и сводках почти полностью отсутствуют точные сведения о птенцовых признаках разбираемой группы птиц. Весьма ограничены также работы, посвященные характеристике процесса вылупления воробьиных. Между тем этот процесс, в течение которого совершается превращение эмбриона в птенца, не может быть оставлен без внимания.
ВЫЛУПЛЕНИЕ ПТЕНЦОВ
Процесс вылупления птенца из яйца до сих пор изучался главным образом у домашних птиц, преимущественно у кур и уток. Работ, основанных на наблюдениях за дикими видами, очень немного (Heinroth О. u. М., 1924—1933; Steinmetz, 1932; Makatsch, 1951; Промптов, 1956). Основными объектами наших наблюдений были различные виды воробьиных: певчий и черный дрозды, сорокопут-жулан, серая мухоловка,, большая синица и различные виды славок. Для сравнения изучалось также вылупление и у неворобьиных птиц — у обыкновенной кукушки, погоныша и некоторых др. Собранный материал дает возможность подвергнуть обсуждению лишь некоторые вопросы, связанные с процессом вылупления.
Характер движений птенца при вылуплении. Перед вылуплением птенец занимает в яйце положение, типичное для созревшего птичьего эмбриона. Спина его сильно изогнута и своим изгибом обращена к тупому концу яйца. Голова подвернута под правое крыло-и . почти целиком скрыта под ним и под правой ногой. Клюв, вооруженный яйцевым «зубом», выдается из-под правого крыла примерно на уровне 7з яйца от вершины тупого конца и направлен к скорлупе под. острым углом или даже почти параллельно ей. В связи с этим небезынтересно заметить, что яйцевой «зуб» — главное орудие, при помощи которого птенец разбивает скорлупу, почти всегда сидит на известном' расстоянии от конца клюва.
При вылуплении птенец время от времени пытается разогнуть спину, поднять голову и оттолкнуться ногами от скорлупы нижней (острой) части яйца. В результате этих попыток выпрямиться клюв птенца приобретает движение в направлении снизу вверх. При каждом движении
8»’
головы яйцевой «зуб» касается скорлупы яйца и царапает ее внутреннюю стенку. В некоторых местах он проламывает ее. Снаружи эти места имеют вид характерных звездочек или бугорков, которые, кстати, иногда бывает легче обнаружить на ощупь, чем на глаз, особенно на яйцах с темной окраской скорлупы.
Поскольку птенец в своем стремлении освободиться от скорлупы производит довольно сильные и резкие движения, его положение внутри яйца несколько изменяется. В связи с этим меняется соответственно и положение клюва по отношению к скорлупе яйца, что приводит к проламыванию скорлупы в разных местах. Очень часто, когда птенец поворачивается в яйце в одном направлении, не изменяя оси вращения, эти проломы располагаются строго по одной линии на расстоянии 2—3 мм один от другого.
Иногда, однако, птенец поворачивается в яйце, изменяя ось вращения. В этих случаях места пролома располагаются весьма неопределенно. В результате намечается не линия, а целая область нарушения целостности скорлупы. В месте наибольшего разрушения мелкие кусочки скорлупы обычно отваливаются. Возникает довольно значительное отверстие, через которое бывают видны клюв, голова, а также правые передняя и задняя конечности птенца. Такой характер вылупления автор наблюдал, например, у погоныша, а также у черного дрозда и кукушки. Таким образом, указанные отличия зависят, по-видимому, от индивидуальных особенностей положения зародыша и не могут считаться характерными для каких-либо определенных систематических групп птиц.
Освобождение птенца от скорлупы начинается после того, как целостность скорлупы бывает нарушена примерно на V4 или Уз окружности яйца. В результате нескольких резких движений головы, а также крыльев и ног, после 'ряда попыток птенцу, наконец, удается выпрямиться и тем самым наполовину раскрыть скорлупу яйца. Через образовавшуюся широкую щель обычно появляются передние конечности, которые теперь уже полностью освобождены. Затем птенец, упираясь головой в верхнюю (тупую) часть скорлупы яйца, а ногами в нижнюю (заостренную) ее часть, резким движением окончательно разламывает скорлупу. В местах разлома край скорлупы бывает обычно неправильной формы. Участок, прорезанный яйцевым «зубом», наоборот, ломается обычно по окружности и имеет лишь небольшие зубчики. Последние, как правило, бывают загнуты внутрь. Это обстоятельство ни в коей мере не может являться доказательством участия самки в вылуплении птенца, как это предполагали В. А. Вагнер (1900) и вслед за ним Т. П. Шеварева и Е. Т. Бровкина (1954). Мы полностью согласны с мнением А. Н. Промптова (1956), считавшего, что загнутость зубчатых краев внутрь вызывается наличием пленки, подстилающей скорлупу и стягивающей ее края. Автору настоящей статьи ни разу не приходилось наблюдать, чтобы у воробьиных взрослые птицы каким-либо образом помогали птенцу освобождаться от скорлупы. Весь процесс вылупления совершается исключительно благодаря усилиям самого птенца.
Как бы «зевающие» движения челюстей, которые А. Н. Промптов наблюдал лишь во время вылупления, когда птенец уже почти освобождался от скорлупы, могут, видимо, начинаться значительно раньше. Во всяком случае в яйце, частично вскрытом за сутки до нормального вылупления, птенцы черного и певчего дроздов, по нашим наблюдениям, уже совершали подобные движения челюстями. Возможно, что эта двигательная реакция связана с установлением нормального дыхания* Од-нако наблюдения за птенцами самых различных видов показывают, что акт дыхания совершается при закрытом рте птенца как до окончательного 40
освобождения от скорлупы, так и позднее. Скорее можно предположить, что в данном случае мы имеем дело с зачаточной формой пищевой реакции, которая к моменту вылупления птенца обычно бывает уже сформирована. Неоднократно, например, приходилось наблюдать активную пищевую реакцию, сопровождавшуюся вытягиванием шеи и открытием рта у птенцов некоторых воробьиных, еще полностью не освободившихся от скорлупы. Это явление особенно характерно для тех видов, у которых птенцы с первого же дня после вылупления отличаются большой активностью, например у серой мухоловки и всех видов славок. Естественно, что все это может происходить при оптимальных температурных условиях, необходимых для нормальной активности птенца. В природных условиях это удается наблюдать в тех случаях, если в период вылупления птенцов самка бывает спугнута с гнезда.
Только что высказанное предположение по поводу значения зевательных движений эмбриона будет более убедительным, если учесть последние работы по питанию эмбрионов кур (Рагозина, 1954; Шейнис, 1954). Этим авторам удалось показать, что после поступления белка в полость амниона происходит активное заглатывание эмбрионом амниотической жидкости, содержащей белок, и усвоение этого белка в кишечном тракте. В связи со сказанным небезынтересно заметить, что на внутренних стенках скорлупы яйца, у его острого конца, к которой обращено анальное отверстие птенца, нам неоднократно приходилось обнаруживать беловатые сгустки. Очень возможно, что эти сгустки представляют собой не что иное, как экскремент эмбриона.
Характер звуков при вылуплении. Птенцы воробьиных, будучи еще в яйце, редко подают голос. Это бывает лишь в случаях сильно затянувшегося вылупления. Как правило, птенцы начинают подавать голос (правда, едва слышимый писк) только после окончательного освобождения от скорлупы. Писк птенца при вылуплении более характерен для выводковых птиц, а из птенцовых — для дневных хищников. Цыпленок может подавать голос за два дня до вылупления, и по этому признаку можно судить о том, что он уже попал клювом в воздушную камеру (Howard, 1952). При наклюнутом яйце голос птенца у выводковых бывает иногда довольно громким. Например, писк вылупляющегося погоныша может быть услышан на расстоянии нескольких метров.
Изучение звуков, издаваемых птенцом при вылуплении, привело к установлению довольно любопытного факта, который до последнего времени оставался неизвестным. Наблюдение за процессом вылупления целого ряда птиц, принадлежащих к различным систематическим группам, позволило выявить особый, чрезвычайно характерный при вылуплении род звука — слабое, но вполне отчетливое и регулярное пощелкивание. Этот звук был известен многим и ранее, но его принимали за постукивание яйцевого «зуба» о скорлупу, полагая, что так называемое «стучание яйца» есть не что иное, как первая фаза вылупления, в течение которой птенец регулярными ударами клюва разбивает скорлупу. Следует, однако, отметить, что треск скорлупы, соскабливаемой или разламываемой яйцевым «зубом», бывает слышен нерегулярно и лишь в те моменты, когда птенец предпринимает попытку выпрямиться и освободиться от скорлупы. В этих случаях яйцевой «зуб» приходит в соприкосновение со скорлупой яйца, в результате чего и возникает неясное и нерезкое трещание — звук совсем иного рода, нежели описанное выше регулярное пощелкивание.
Как показали наблюдения, этот щелкающий звук возникает в связи с легочным дыханием птенца. Он совпадает с выдохом и вызывается,
91
возможно, расхождением еще влажных и не вполне сформировавшихся дыхательных путей. По числу звуков можно судить о количестве сделанных птенцом вдохов и выдохов. Для доказательства того, что данная категория звука не связана с разбиванием скорлупы яйцевым «зубом», автор проделал следующий эксперимент. Во время вылупления птенца' был удален весь участок скорлупы, примыкающей к клюву. Несмотря на то, что клюв птенца свободно выдавался наружу, регулярное пощелкивание продолжалось. В дальнейшем было установлено, что регулярный щелкающий звук птенцы воробьиных издают в течение нескольких дней и после вылупления. Например, птенцы большой синицы весьма отчетливо «щелкают» в возрасте 4—5 дней. У таких птенцов установить связь данного звука с дыханием не представляется затруднительным.
Пощелкивание, связанное с дыханием птенца, воспроизводится с большой регулярностью. Частота его, как показали наблюдения, резко колеблется в зависимости от температуры среды, в которой находится птенец или яйцо с зародышем. При охлаждении яйца «стучание» постепенно затухает. Оно даже может совсем прекратиться и на довольно долгое время. Это, однако, не означает того, что зародыш погиб. При. пониженных температурах он, по-видимому, может довольно долго не пользоваться легкими, а довольствоваться аллантоисным дыханием. При повышении температуры тела у эмбриона ускоряется обмен веществ, что, естественно, влечет за собой учащение дыхания. При температуре, близкой к температуре тела самки (яйцо помещалось на грелке -с температурой воды около 40° С), число дыханий у птенца черного дрозда, находившегося в яйце с уже проломленной скорлупой, (за несколько часов до вылупления), по наблюдениям автора, варьировало от 27 до 32 раз в 1 мин.
Установить точный срок начала функционирования легких у эмбриона в полевых условиях довольно трудно. Судя по «стучанию» яйца у мелких видов воробьиных птенцы начинают в какой-то мере пользоваться легочным дыханием примерно за сутки до вылупления. Однако» при неблагоприятных условиях процесс вылупления может затягиваться, в связи с чем продолжительность периода легочного дыхания внутри яйца становится более значительной.
Длительность вылупления. Если в последние дни инкубации самку кто-либо беспокоит и ей часто приходится покидать гнездо,, происходит периодическое охлаждение яиц, что замедляет и даже временно приостанавливает развитие эмбриона. Это в свою очередь неизбежно отражается на скорости процесса вылупления. Иногда вылупление затягивается на несколько дней, что часто наблюдается у дневных хищников, чаек и других птиц с толстой скорлупой яиц. У воробьиных выход птенца из яйца наступает обычно через несколько часов после проклевывания скорлупы, но и у этих птиц процесс вылупления может растянуться более чем на сутки. Временным охлаждением кладки в период вылупления, очевидно, и следует объяснить в основном значительную разницу в продолжительности инкубационного периода у особей одного вида — явление, часто наблюдающееся в природе.
Наиболее интенсивно процесс вылупления протекает, видимо, У кукушки, птенцы которой иногда освобождаются от скорлупы менее, чем через 10 мин. после первого ее проклева (Промптов, 1956). Однако даже у этого вида вылупление протекает по существу в значительно более растянутые сроки, ибо, с нашей точки зрения, за начало процесса вылупления надо принимать не первый проклев скорлупы, а момент, когда птенец яйцевым «зубом» разрывает подстилающую скорлупу пленку, что происходит намного раньше. Этот момент знаменуется началом
92
-«стучания» яйца, т. е. началом смены аллантоисного дыхания на легочное.
В целом вылупление у птиц продолжается, по нашему мнению, значительно дольше, нежели это принято считать. Процесс смены аллантоисного дыхания на легочное также представляется довольно постепенным. В известный период жизни, предшествующий вылуплению, эмбрион, ло-видимому, обладает как бы двойным дыханием, которое функционально может заменяться одно другим. Двойное дыхание имеет для раз^ вивающегося эмбриона, видимо, очень большое значение. Его можно рассматривать как приспособление, обеспечивающее жизнедеятельность эмбриона в период вылупления при разных температурных условиях. Как уже отмечалось, в природе нередки случаи охлаждения вылупляющихся эмбрионов. В эти периоды дыхание зародыша осуществляется, по-видимому, лишь при помощи аллантоиса. Напротив, при постоянном обогреве его самкой основную функцию дыхания начинают выполнять легкие. Иначе трудно понять, каким Образом вылупляющийся птенец, уже начавший дышать легкими, не погибает при довольно длительных охлаждениях и отсутствии явных признаков, указывающих на наличие у него легочного дыхания. С другой стороны, более интенсивное легочное дыхание внутри яйца при все возрастающей потребности вылупляющегося птенца в кислороде заставляет его разбить скорлупу яйца, что птенец и делает на определенной стадии развития.
ПТЕНЦОВЫЕ ПРИЗНАКИ ВОРОБЬИНЫХ
В процессе работы автором были составлены описания птенцов воробьиных, найденных им в лесной й лесостепной ронах^Евдфйеиской части СССР. Кроме Того, были испоадз^ваны спиртовые коллекции -Зоологического-института АНЛЗССР, а также птенцы некоторых видов, любезно’ предоставленные К* А. Воробьевым, Н. П. Кадочниковым, <). М. Татариновой и Ю. К. Эйгелисом. В отдельных случаях, когда собственного материала оказывалось недостаточно, внешний облик птенца устанавливался по литературным источникам, главным образом по справочнику Визерби и др. (Witherby and oth., 1952).
По большей части изучались новорожденные или однодневные .птенцы, описание которых производилось с возможно большей подробностью. Обращалось внимание не только на характер и степень опушен-ности тела, окраску ротовой полости и наружных краев клюва, но и на цвет кожи, когтей, яйцевого «зуба» и форму ноздрей — признаки, которые могут иметь иногда диагностическое значение. Характер распределения пуха на теле птенца описывался по схеме, предложенной в свое время Тайсхорстом (Ticehurst, 1908).
Видовые характеристики птенцов изученных видов приведены в специальной части настоящей работы, в данном же разделе автор преследует цель дать сравнительную характеристику основных птенцовых признаков воробьиных и обсудить материал с точки зрения их адаптивного и систематического значения.
Яйцево й «з у б». Яйцевой «зуб», или, как его иногда называют, «яйцевая мозоль», представляет собою, как известно, затвердевшее роговое утолщение, которое располагается у конца надклювья птенца (рис. II). Помимо этого образования у птенцов некоторых птиц, например дятлов, -имеется еще второй яйцевой «зуб», располагающийся на конце подклювья.
Известно, что яйцевой «зуб» характерен не только для птиц, но и для ^многих рептилий, а также яйцекладущих млекопитающих. Он представ
93
ляет собою приспособление для разбивания скорлупы яйца в момент вылупления зародыша. Функционирует он лишь в сравнительно очень короткий период жизни (от момента первого проклева скорлупы до освобождения птенца от 'нее), но тем не менее у птенцов воробьиных сохраняется обычно в течение 12—14 дней.
У сравнительно медленно развивающихся и долго находящихся в гнезде птенцов дневных хищников, цапель и некоторых других яйцевой «зуб» сохраняется еще дольше. По наблюдениям Макача (1951), у птенца серой цапли он бывает заметен и в 45-дневном возрасте.
У выводковых, которые с первого же дня начинают пользоваться своим клювом при разыскивании корма, яйцевой «зуб» исчезает значительно раньше, нежели у птенцовых. Например, у цыпленка, а также у птенца большого кроншнепа (Noll, 1924) он отпадает часто уже в первые сутки после вылупления.
Рис. 11. Однодневный птенец иволги. На конце надклювья виден белый яйцевой язубв (фото автора).
Таким образом, длительность сохранности яйцевого «зуба» зависит от типа развития птенца и того образа жизни, который он начинает вести сразу после вылупления.
У воробьиных скорость исчезновения яйцевого «зуба» зависит от характера расположения гнезда и степени активности птенцов. Птенцы, воспитывающиеся в дуплах, как правило, теряют яйцевой «зуб» значительно раньше, нежели птенцы открыто гнездящихся видов. Находясь в большой тесноте и отличаясь, к тому же, большой активностью, они постоянно касаются клювом стенок дупла, в связи с чем яйцевой «зуб» отваливается у них на 5—6 день. Среди открыто -гнездящихся птиц относительно скоро (на 6—8 день) пропадает яйцевой «зуб» у птенцов многих вьюрковых (зеленушка, коноплянка, чечевица), что, видимо, надо связывать с относительно очень быстрым ростом их рамфотеки. Чем быстрее растет клюв, тем скорее должен отлететь нерастущий яйцевой «зуб». Обычно он отпадает вместе с тонкой роговой пластинкой, отслаивающейся от рамфотеки, по мере-роста клюва. Вообще же исчезновение яйцевого «зуба» очень часто зависит от совершенно случайных механических причин, в связи с чем разница в продолжительности существования яйцевого «зуба» может быть весьма значительной даже у птенцов одного выводка.
94
Окраска кожи, когтей и клюва. Характер окраски кожиг клюва и когтей у птенцов воробьиных в общем довольно однообразен,, хотя в отдельных случаях он бывает весьма специфичным и настолько определенным, что может иногда служить хорошим диагностическим признаком.
До того, как птенец начинает покрываться пеньками контурных перьев, окраска его кожи бывает в большинстве случаев розовато-красной или желтовато-оранжевой. У птенцов птиц типичных дуплогнездников она обычно менее яркая, чем у птенцов, развивающихся в открытых гнездах, хотя и среди последних встречаются представители с бледной окраской кожи, например птенцы сорокопута-жулана. Интенсивно окрашенную кожу имеют птенцы некоторых врановых и вьюрковых. Для последних очень характерны мясо-красные тона, а для птенцов врановых — грязно-серые (ворона), ярко-оранжевые (сорока) или даже оливковозеленые (сойка). Во всех случаях интенсивность Окраски кожи усиЛи-
вается с возрастом птенца, но лишь в течение (нескольких первых дней его Жизни. Окраска кожи может быть различной у представителей даже одного рода. Например, среди различных славок птенца серой славки легко отличить по характерному свинцовому или скорее слабому фиолетовому оттенку его кожи. Птенцы пеночки-веснички отличаются особенно бросающейся в глаза желтизной кожи и т. п.
Первоначальный цвет когтей у птенцов преобладающего большинства воробьиных светлый. Когти начинают темнеть лишь на 4—6 день жизни птенца.
Рис. 12. Характер рисунка на клюве однодневных птенцов садовой славки (а) и славки-черно-головки (б).
Есть, однако, виды, у которых когТи с первого же дня бывают темными (зяблик, щур) или желтыми (пеночка-весничка).
Клюв первоначально бывает светло-серого, рогового или воскового цвета, иногда с розовым (иволга) или желтым (пеночка-весничка, чиж) оттенком. Окраска его может быть однообразно светлой либо иметь характерный темный рисунок в виде вершинной или предвершинной темной полосы, темных отметин вокруг ноздрей и т. п. По этому рисунку, например, можно распознать очень сходных птенцов близких видов славок (рис. 12).
Окраска ротовой полости. Орнитологи уже неоднократно высказывались по поводу биологического значения этого признака у птенцов -воробьиных (Pycraft, 1907; Ingram, 1907; Heinroth О. u. М.„ 1924—1933; Makatsch, 1951; Промптов, 1956, и др.).
Общий смысл этих высказываний примерно следующий: яркая раскраска ротовой полости служит своеобразным раздражителем, постоянно побуждающим родителей приносить птенцам пищу и облегчающим взрослой птице определять местонахождение глотки птенца, особенно если гнездо находится в темном месте. В справедливости подобного взгляда убеждает нас прежде всего то, что яркая окраска рта встречается у птенцов именно тех птиц, которые выкармливают свое потомство особым способом, вкладывая корм глубоко в раскрытый рот птенца. В свою очередь птенец у таких птиц способен широко раскрыть свою пасть и долго держать ее в таком положении. В 'немецкой литературе эту особенность птенцов принято называть особым термином «sperren». Помимо воробьиных эта черта свойственна, кажется, только удодам
95
и кукушкам, птенцы которых тоже имеют яркую раскраску ротовой полости.
Характер окраски ротовой полости и краев клюва у птенцов, воспитывающихся в закрытых и открытых гнездах, бывает обычно разным.
Известно, что птенцы дупло-
гнездников, например большой синицы или скворца, отличаются наличием широкой и светлой складки, опоясывающей раскрытую ротовую полость. Эта белая оторочка, бросающаяся в глаза, если смотреть сверху на сидящих в темном дупле птенцов, несомненно помогает птице быстро ориентироваться при выкармливании птенцов. Гистологическое изучение боковых выростов клюва у птенцов скворца, проведенное Вак* кернагелем (Wackernagel, 1954), подтверждает сцравед-
Рис. 13. Птенец лесной малиновки „просит 'есть*. На корне -языка заметны два черных пятна (Савальский лесхоз, июль 1951 г. Фото автора).
ливость такой точки зрения.
У птенцов, воспитывающихся в открытых гнездах, оторочка клюва выражена
обычно значительно менее ярко. В то же время среди этой группы птиц можно наблюдать иные приспособления, не встречающиеся у птиц-дуплогнездников. Например, у
камышовок, лесной малиновки (рис. 13) и славок (рис. 14), а также у лесной завирушки на языке имеются темные пятна. У славок эти пятна часто имеют неясные очертания и не столь интенсивно окрашены, как, -например, у камышовок.
Окраска самой ротовой полости у птенцов наших лесных воробьиных чаще всего бывает желтой или желтовато-оранжевой (скворцы, дрозды, горихвостки, соловьи, чеканы, пеночки, камышовки, сорокопуты, синицы, славки и др.), а также темно-розовой или мясо-красной (врановые, иволги, вьюр-
Рис. 14. Птенец садовой славки „просит есть'. На языке видны темные, неясно очерченные пятна (Савальский лесхоз, июль 1953 г. Фото автора).
ковые, славки). окраска оранжевой (коньки) или разноцветной, как, например, у дубоноса или снегиря.
Обычно представители одного рода, а иногда и ряда близких родов
овсянки, некоторые Реже внутренняя рта бывает ярко-
имеют в общем сходную окраску ротовой полости. Известное исключение представляют славки, птенцы которых по раскраске рта могут быть раз-
«6
биты на две группы: малиново-красноротых (наши лесные виды — садовая и Черноголовка) • и желторотых (славки более открытых стаций— ястребиная, серая и завирушка).
Поскольку окраска ротовой полости — признак весьма стойкий, и у птенцов родственных форм, как правило, бывает выражен весьма сходно, его, наряду с другими признаками, необходимо учитывать при установлении родственных взаимоотношений между видами. Например, пеночки, судя по совокупности некоторых птенцовых признаков, отличающихся от других славковых (язык без черных пятен, опушенные птенцы), занимают, видимо, несколько особое положение в семействе славковых.
Яркая окраска ротовой полости, являясь типичным птенцовым признаком, тускнеет после определенного возраста. Тайсхорст (1908) отмечает, что у птенца лесной завирушки часть пятен во рту исчезает уже на 4—5 день. У птенцов садовых славок темные пятна на корне языка, по нашим наблюдениям, начинают тускнеть примерно в 7—8-дневном
Рис. 15. Форма клюва у птенцов чечетки (а), чижа (б), щегла (в), зяблика (г), полевого воробья (д) и обыкновенной овсянки (е).
возрасте, когда птенцы находятся еще в гнезде. В большинстве случаев, однако, яркая окраска рта сохраняется в течение всего гнездового периода и некоторое время после него. Уже прилично летающих 3-недель-ных птенцов лесной малиновки или садовой камышовки можно всегда узнать по сохранившимся на корне языка двум темным точкам.
Боковые выросты клюва. Боковые выросты клюва или, как их часто называют, «клювные валики» — тоже типичный птенцовый признак, исчезающий по мере перехода птенца на самостоятельное питание. В той или иной степени он выражен у птенцов всех наших певчих птиц, а из неворобьиных очень характерен для птенцов удода и дятлов. Эти выросты обычно окрашены в светлые тона и бывают ярко-белыми, бледно-желтыми или бледно-розовыми. Иногда они могут достигать весьма значительной величины, что характерно, например, для птенцов птиц-дуплогнездников (Ingram, 1920; Понанин, 1949). Средн наших птиц этот признак особенно ярко выражен у птенцов обыкновенного скворца. Довольно крупные боковые складки клюва имеют также птенцы мухоловок, синиц и воробьев. Среди птиц, воспитывающихся в открытых гнездах, баковые выросты клюва хорошо заметны у птенцов ворона, вороны серой, грача, сойки, овсянок и многих дроздовых: черного и певчего дроздов, соловья и др.
Интересные отличия наблюдаются в размерах боковой складки клюва у птенцов вьюрковых — у чижа, щегла и чечетки она очень маленькая и представлена только в углах рта, а у птенца зяблика, наоборот, она широкая и большая (рис. 15). Бросается в глаза также существенная разница в ширине разреза ротовой полости: у первых трех видов разрез рта узкий, в то время как у птенца зяблика он широкий. Указанные выше морфологические особенности птенцов следует, видимо,
7 А. С. Мальчевский
97
объяснять особенностью их питания: птенцы щегла, чижа и чечетки пре* имущественно растительноядны, а птенцы зяблика исключительно насекомоядны. Птенцы родственных групп — воробьев и овсянок, выкармливающиеся главным образом насекомыми, тоже имеют широкий разрез? ротовой полости и довольно значительную складку в углах рта (рис. 15). Таким образом, птенцы некоторых птиц, принадлежащих во взрослом состоянии к группе зерноядных (воробьи, овсянки, некоторые вьюрки),, имеют ряд признаков,’ характерных для птенцов насекомоядных птиц.
Насекомые с их длинными членистыми ногами, усиками, крыльями представляют собою весьма объемистый корм, трудный для захвата маленьким птом.-К тому же они подвижны. Иногда'к гнезду птица при-
Рис. 16. Птенец обыкновенного скворца „просит есть* (фото автора).
носит в клюве целый пучок живых насекомых, который удивительно ловко и быстро распределяет между птенцами. Быстрая и экономная передача объемистого, подвижного и цепляющегося корма, каким являются насекомые, может успешно осуществляться не только благодаря ловкости взрослой птццы, но и благодаря каким-то приспособлениям, свойственным птенцам. К числу таких приспособлений и следует отнести боковые выросты клюва и широкоротость птенцов насекомоядных птиц. Птенцы насекомоядных птиц, воспитывающиеся в тесных дуплах или закрытых гнездах, особенно поражают вместительностью* своей ротовой полости (рис. 16). Колоссальные размеры ротовой воронки достигаются здесь главным образом за счет боковых выростов клюва, которые, например, у птенца скворца, увеличивают ширину ротовой воронки примерно в 2 раза. При сравнении птенцов скворца и щегла в момент выпрашивания корма особенно заметна узкоротость последнего (рис. 17).
Таким образом, боковые выросты клюва следует в первую очередь расценивать как приспособление, возникшее у птенцов большинства воробьиных в связи с их насекомоядностью. Величина клювных валиков
98
изменяется по мере роста птенца (Portman, 1938). У новорожденных и однодневных птенцов размеры их бывают незначительными. В возрасте 4—5 дней они обычно достигают своей максимальной величины. Например, у новорожденного птенца скворца ширина складки равна 2,5 мм, а 5-дневного — 5 мм (рис. 18).
Рис. 17. Птенец щегла с максимально широко раскрытым клювом (фото автора).
Прогнатизм нижней челюсти. Явление нижнечелюстного прогнатизма, выражающееся в увеличении размеров нижней челюсти
Рис. 18. Форма клюва у птенцов обыкновенного скворца: новорожденный (а), 2-дневный {б) и 5-дневный (в).
птенца, известно для дятлов, удодов и некоторых других птиц-дуплогнездников. Исследователи, касавшиеся данного явления (Ingram, 1920; Heinroth, 1924—1933; Красовский, 1936; Познании, 1938, 1949 и др.), сходятся в следующей оценке его биологического значения: благодаря
7*
99
удлиненной и расширенной нижней челюсти в условиях дупла облегчается процесс захвата пищи, принесенной родителями.
Для птенцов воробьиных птиц нижнечелюстной прогнатизм в общем не характерен, однако нельзя считать, что он совсем отсутствует у птенцов воробьиных. Этой особенностью, например, отличаются птенцы обыкновенного скворца, у которых нижняя челюсть значительно шире и длиннее верхней. Таким образом, связь разбираемого явления с условиями гнездования заметна и на примере воробьиных птиц.
Иногда, однако, полагают, что гнездование в дуплах должно обязательно приводить к увеличению размеров нижней челюсти птенцов и что все случаи отсутствия прогнатизма нижней челюсти у птенцов дуплогнездников надо объяснять недавним переходом этих птиц к гнездованию в дуплах. У птенцов большинства воробьиных птиц, живущих в дуплах, отсутствие прогнатизма модкет быть действительно объяснено сравнительно недавним возникновением Дуплогнездничества. Тем не менее в ряде случаев такое объяснение Нельзя признать удачным. Причина возникновения нижнечелюстного прогнатизма кроется, видимо, не только в условиях гнездования, но и в характере корма и способе его передачи птенцам. Например, отсутствие прогнатизма нижней челюсти у птенцов клинтуха, а также других голубей, гнездящихся в закрытых местах, вряд ли можно объяснить только лишь тем, что голуби являются недавними дуплогнездниками. Наоборот, странно даже было бы ожидать увеличения нижней челюсти у голубей, выкармливающих птенцов пищей, отрыгиваемой из зоба. При таком способе кормления удлиненная нижняя челюсть могла бы только помешать нормальному выкармливанию птенцов.
Пяточная мозоль. Известно, что этот признак вообще характерен для живущих в дуплах дятлов, туканов и бородастиков (Шуль-пин, 1940). О наличии его у представителей отряда воробьиных обычно не упоминают. Между тем следует отметить, что птенцы многих даже открытогнездящихся видов лесных воробьиных имеют утолщение на задней стороне интертарзального сустава. Объясняется это, очевидно, тем, что в первые дни жизни птенца, пока он не в состоянии еще по настоящему опираться на пальцы ног, этот сустав служит главной точкой опоры во время приема пищи, когда его тело принимает вертикальное положение (рис. 19). Среди европейских видов воробьиных, утолщение на задней стороне интертарзального сустава заметно у птенцов полевого воробья, большой синицы, мухоловки-белошейки, но лучше всего оно выражено у птенцов обыкновенного скворца, у которого имеет вид небольшой мозоли.
У птенцов воробьиных пяточная мозоль, однако, никогда не достигает столь сильного развития, как это наблюдается у птенцов дятловых птиц. Согласно представлениям Пайкрафта (1907), а также Хейнрота (1938), пяточная мозоль у птенцов дятлов предохраняет их ноги от стирания, которое будто бы может иметь место в дупле, лишенном подстилки. Судя же по тому, что у птенцов воробьиных, несмотря на наличие гнездовой подстилки, иногда все же наблюдается образование пяточной мозоли, мы полагаем, что и у птенцов дятлов основное назначение этого образования сводится к обеспечению необходимой устойчивости положения в тот момент, когда они тянутся за кормом.
Птенцовое опушение. Птенцы воробьиных, как известно, появляются на свет либо совершенно голыми, либо одетыми в эмбриональный пух, количество которого у разных видов сильно варьирует. Естественно, что у исследователей, занимавшихся изучением птенцового опушения воробьиных, должен был возникнуть вопрос, как следует
100
объяснять наблюдающиеся у птенцов резкие отличия в степени их опу-шенности. Многие авторы считают, что в данном отношении решающее значение имеют условия гнездования: у птиц, строящих закрытые гнезда или гнездящихся в закрытых местах ,птенцы вылупляются голыми или слабо опушенными, птенцы же видов, гнездящихся открыто, бывают опушены сильнее. Эта точка зрения, придающая птенцовому опушению воробьиных значение серьезного утепляющего фактора, была первоначально высказана Инграмом (Ingram, 1920), а затем Л. П. Познаниным
Рис. 19. Птенец мухоловки-белошейки, выпрашивающий корм. 'Интертарзальное сочленение является местом главной опоры (фото автора).
(1949), Р. В. Никитиной (1954), М. Н. Денисовой (1958а) и др. В настоящее время она пользуется довольно широким признанием. Не разделяя ее, автор для проверки фактического положения дела сопоставил характер гнездования и внешнего облика птенцов у 132 видов воробьиных из 176 известных для Европы. О птенцах 44 видов не удалось собрать нужных данных.
Результаты проведенного исследования показали, что так называемое «правило» закономерной редукции птенцового пуха в связи с закрытым гнездованием в группе воробьиных имеет гораздо больше исключений, нежели случаев, подтверждающих его. Среди открыто гнездящихся воробьиных птиц, встречающихся в Европе, облик птенцов которых в настоящее время известен, насчитывается 25 видов птиц, имеющих голых
101
птенцов (Garrulus, Bombycitla, Panurus, Lanius, Sylvia, Cisticola, Hip-polais, Acrocephalus, Erythropygia). Наоборот, среди всех воробьиных Европы, гнездящихся в дуплах, а также в закрытых или полузакрытых гнездах, мы находим всего 6 видов (Pica, Passer, Remiz, Aegithalos), птенцы у которых вылупляются голыми (табл. 29).
Таблица 29
Распределение видов воробьиных птиц Европы, имеющих голых птенцов, по типам гнездования
Тип гнездования Число видов, облик птенцов которых известен Число видов, имеющих голых птенцов
п в Н
Закрытый 29 5 18,5
Полузакрытый 16 1 6,3
Открытый 87 25 28,8
Итого 132 31 23
Таким образом, если судить по современным условиям гнездования, у во<робьи;ных нельзя установить правила закономерной редукции птенцового «пуха в связи с закрытым гнездованием. Ссылки некоторых зоологов (Познанин, 1949; Денисова, 1958а) на якобы иной тип гнездования многих воробьиных в прошлом, при попытках как-то объяснить многочисленные исключения из этого «правила», нельзя признать удачными. Считать, например, что обыкновенный скворец лишь сравнительно недавно перешел на новый способ гнездования в дуплах и что именно поэтому птенцы у него сейчас сравнительно сильно опушены, — нет достаточно веских оснований, потому что по другим птенцовым признакам (прогнатизм нижней челюсти, наличие пяточной мозоли, очень широкая боковая складка клюва, замедленный тип развития) скворца, наоборот, следует признать одним из наиболее древних дуплогнездников средн воробьиных. Нельзя также объяснять отсутствие птенцового опушения у части славковых птиц и сорокопутов их южным происхождением: среди птиц южного происхождения немало видов, имеющих значительно опушенных птенцов (иволги, скворцы и др.), а среди северных птиц есть такие, у которых птенцы бывают совсем голыми (свиристели).
Можно привести много примеров существования резких отличий в < степени опушенности птенцов у воробьиных, несмотря на однотипность в расположении их гнезд, и, наоборот, случаев сходства птенцов у видов, гнездящихся в разных условиях. Птенцы славок, камышовок, лесной малиновки, выращиваемые в открытых гнездах, совсем лишены эмбрионального пуха, а у пеночек, у которых гнезда закрытого типа, птенцы имеют на теле пух. Птенцы обыкновенной каменки значительно сильнее опушены, нежели птенцы лугового чекана, между тем каменка гнездится в более закрытых местах, чем чекан. Среди гнездящихся в закрытых местах или строящих закрытые гнезда ткачиковых мы находим совсем голых птенцов у домового и полевого воробьев и довольно значительно опушенных у каменного воробья. Гнездящаяся в дуплах или щелях построек галка и строящий открытые гнезда грач имеют в общем сходно опушенных птенцов. В то же время у кедровки, серой вороны и сойки, устраивающих в одинаковой степени открытые гнезда, птенцы разные:
102
у сойки они совершенно голые, а у серой вороны (рис. 20) и у кедровки «опушенные.
Сроки размножения тоже не играют роли в определении степени спушенности птенцов. Иначе трудно было бы объяснить отсутствие специальных приспособлений в строении и количестве пуха на теле птенцов клеста, гнездящегося в различное время года, даже зимой. Степень юпушенности у них примерно такая же, как у других вьюрковых, размножающихся в теплое время года, например у щура или чечевицы.
Одной из чрезвычайно характерных черт гнездового поведения самок, а иногда и самцов воробьиных является постоянное обогревание птенцов в первые дни после их вылупления (Kendeigh, 1952). В этот период птица постоянно находится на гнезде, редко и на очень короткий срок оставляя птенцов. Воробьиные почти беспрерывно обогревают птенцов в первые дни их жизни независимо от того, гнездятся ли эти виды в закрытых или открытых местах и имеют ли голых или опушенных
Рис. 20. Однодневный птенец серой вороны (фото автора).
птенцов. Иногда, правда, вследствие крайней осторожности птиц трудно «бывает наблюдать насиживание птенцов, так как часто птицы при приближении человека незаметным образом слетают с гнезда. Однако недавнее присутствие самки на гнезде обычно удается установить по температуре тела птенцов, которые, несмотря на свою холоднокровность, всегда в таких случаях бывают горячими.
Высокая температура тела, сообщаемая птенцам самкой, постоянно находящейся в гнезде, им жизненно необходима. В противном случае птенцы, будучи в этом возрасте пойкилотермными, не могли бы быстро усваивать пищу и расти. Если же птица по какой-либо причине перестает постоянно находиться в гнезде и обогревать птенцов, они скор© начинают коченеть независимо от того, опушены они или нет. У преобладающего же числа видов воробьиных птенцы опушены настолько слабо, что приходится вообще сомневаться в том, может ли эмбриональный пух иметь для них сколько-нибудь серьезное обогревающее значение.
Современные приборы, которыми пользуются в экспериментальной экологии, несомненно, могут уловить некоторые отличия в скорости падения температуры тела у опушенных и неопушенных птенцов, после того как обогревавшая маленьких птенцов птица спугнута с гнезда.
103
Однако вряд ли эти различия имеют существенное значение для жизни птенцов. При неблагоприятных температурных условиях, равно как и во время дождя, птицы всегда садятся на гнездо с маленькими птенцами» а если и слетают, то на очень непродолжительное время. В данном случае, может быть, скорее следует говорить о значении моногамии, у воробьиных, об известной стойкости их птенцов, способных переносить довольно длительные понижения температуры тела, нежели придавать серьезное значение эмбриональному пуху. К тому же надо иметь в виду» что поскольку в постэмбриональный период пух у воробьиных уже не растет, утепляющее значение его резко падает с первого же дня жизни птенца по мере увеличения его тела.
Нельзя, конечно, отрицать, что в некоторых случаях эмбриональное опушение птенцов может оказаться в каком-нибудь отношении полезным. Например, у степных жаворонков оно может иметь маскирующее значение. Довольно густой пух у птенцов дубоноса, иволги или скворца иногда, возможно, играет роль некоторого утеплителя. Это, однако, не означает того, что он имеет решающее значение для их нормального развития й что птенцовый пух воробьиных вообще следует рассматривать каю специальное приспособление к открытому гнездованию.
Иногда полагают, что у лесных воробьиных, гнездящихся низко и в-затененных местах, птенцы имеют более темное опушение» нежёлй у видов, устраивающих свои гнезда высоко на деревьях и на осветленных учайках (Ingram, 1920). Считают, например, что белая окраска пуха на Теле птенцов щегла или дубоноса объясняется тем, что гнезда у этих видов располагаются В осветленных местах. Такое объяснение Мало убедительно, так как можно привести много других примеров, даже в пределах той жё группы вьюрковых птйц, когда у видов, располагающих ёвои гнёзда в местах, не менеё осветлённых, окраска птёнцбвогй пуха темйая: чиж, клест, чечетка, чечевица. Нельзя также предполагать, что светлая окраска пуха у птенцов дубоноса и щегла связана ё более южной областью распрЬстранёийя этих видов, ибо гнездящийся в северных шихтах topoK тоже ймеет белый пух.
Окраска эмбрионального пуха у птенцов воробьиных вообще довольно разнообразна. Она может быть белой или почти белой (у дубоноса, щегла, юрка), серой (у скворца, лугового чекана, зяблика, зеленушки и мн. др.), темной или даже почти черной (обыкновенная? горихвостка, восточный соловей, зарянка и др.), охристой или золотисторыжей (дрозд-рябинник, белобровик и певчий дрозд) и, наконец, светло-желтой (иволга, пеночка-весничка). Чаще всего первичный покров бывает окрашен в серый цвет. Приведенные примеры показывают/что окраска пуха может быть одинаковой у птенцов, воспитывающихся в-совершенно различных условиях.
В строении эмбрионального пуха у воробьиных, как правило,, не удается установить существенных отличий. И хотя у близких видов» пух иногда бывает различно окрашен, а у представителей разных родов; он может довольно сильно отличаться не только по цвету, но и по длине' и толщине бородок, в целом, однако, пушинки, взятые от самых различных представителей отряда воробьиных, очень схожи. Приспособительная дифференциация окраски пуха, как это характерно для выводковых,, на теле птенцов воробьиных не наблюдается. Различия в размерах и числе бородок следует, видимо, объяснять различной степенью деградации отдельных пушинок. Количество бородок второго порядка весьма непостоянно даже в пушинках вполне нормальных размеров. Число ихг у воробьиных варьирует в пределах от 6 до 14, но чаще всего их бывает 8—10.
104
При сравнении птенцов воробьиных с птенцами других групп птиц» у которых птенцовое опушение, без сомнения, возникло и существует как утепляющее приспособление, обнаруживаются, значительные отличия в развитии и характере распределения пуха на теле птенпов. Например, у утиных, куликов, чаек, дневных хищников, сов и других пух более-или менее равномерно покрывает все тело птенца, чего у певчих птиц не бывает. Однако и этот наряд, несмотря на первоначальную его значительную густоту, вследствие быстрого роста птенца начинает постепенно-редеть и терять значение. Естественно, что птенцы даже этих птиц в-возрасте уже нескольких дней могли бы страдать от холода, если бы в их покровах не происходили бы последующие приспособительные изменения: появление дополнительного «второго» пуха, вырастающего в промежутках между «первым» (дневные хищники), возникновение непосредственно за первой генерацией второй генерации пуха (трубконосые» пингвины) или, наконец, очень скорое вырастание на теле птенца в промежутках между птенцовым пухом дефинитивного пуха (утиные) и т. п.
Рис. 21. Схема расположения пуховых птерилий нателе птенцов воробьиных (по Тайсхорсту, 1908).
) — надглазничная, 2 — глазная, 3 — затылочная, 4 — плечевая, 5 - локтевая, 6 — спинная, 7 — бедренная, 8 — брюшная, 9 — голенная птеридин.
Подобных приспособлений в строении и развитии покровов у птенцов воробьиных на ранних стадиях их развития мы не встречаем. Единственным серьезным средством защиты птенцов от охлаждения до тех: пор, пока они- не начнут покрываться перьями, остается обогрев их телом взрослой птицы.
Проведенное сравнение степени опушенности и характера распределения пуха на теле птенцов одного и того же вида, но взятых из разных районов, не обнаружило заметной географической изменчивости данного-явления. Например, присутствие или отсутствие пуха на брюшной стороне у лесного конька и зяблика было обнаружено у птенцов, добытых в различных областях. В то же время подобные вариации наблюдаются иногда даже у птенцов одного выводка. Сильно варьирует и общее количество пушинок на теле отдельных птенцов.
Наличие сильной индивидуальной изменчивости в опушении птенцов позволяет предположить, что в данном случае мы имеем дело с признаком, претерпевающим большие изменения. Так бывает с признаками, либо начинающими приобретать какое-нибудь значение, либог наоборот, теряющими значение для организма и существующими в силу исторических причин. Второе предположение по отношению к опушению птенцов воробьиных птиц, по-видимому, вернее.
105*
На прилагаемом рисунке (рис. 21) отмечены все пуховые птерилии, которые могут быть на теле птенцов воробьиных: глазные, надглазничные, затылочная, спинная, плечевые, локтевые, бедренные, голенные и брюшные. Помимо них у многих видов, имеющих даже неопушенных птенцов, на местах будущих маховых и рулевых перьев можно обнаружить присутствие маленьких, видоизмененных, щетинкообразных «пушинок», которые (предшествуют (появлению перьевых пеньков. Эти пушинки имеют сокращенное количество бородок первого порядка и сильно редуцированные бородки второго порядка. У птенцов сорокопугатжулана • сходные образования были обнаружены и на брюшной стороне тела виде двойного ряда очень мелких, едва заметных для простого глаза
Рис. 22. Однодневный птенец сорокопута-жулана. На брюшной стороне заметен рудиментарный пух (фото автора).
.-светлых пушинок (рис. 22). Наличие этого удивительно стойкого и в то ^ке время не имеющего, видимо, никакого значения в жизни птенца признака дает основание рассматривать его как рудимент птенцового опушения, каким обладали, вероятно, предки этого вида.
С составом и характером распределения пуховых птерилий по телу птенцов некоторых воробьиных можно ознакомиться по табл. 30. В наиболее полном виде эти птерилии представлены у птенцов ряда вьюрковых, а также у лесного конька и иволги; почти полный набор птерилий имеют птенцы скворца, овсянок, некоторых врановых. Однако птенцы целого ряда воробьиных почти или совершенно лишены пуха: все славки (рис. 23), камышовки (Acrocephalus), малиновки (Hippolais), воробьи, ремез, сойка, сорока, (рис. 24), сорокопуты и некоторые др.
Ознакомление с таблицей, кроме того, показывает, что у видов, принадлежащих к одному роду, птенцы, как правило, имеют очень сходное расположение птерилий и сходную степень опушенности. Известное исключение из правила представляют по-разному опушенные птенцы мухоловок: серой мухоловки, с одной стороны, и белошейки и пеструшки — с другой. Различие в степени их опушенности обычно объясняют -имеющими место различиями в характере гнездования этих видов.
406
этим мнением, возможно, и следовало бы считаться, если бы серые и пестрые мухоловки действительно находились в тесном родстве. Ознакомление с птенцами указанных видов показывает, однако, что птенцы
Рис. 23. Однодневный птенец серой славки (фото автора).
«х сильно отличаются не только характером птенцового опушения, но и формой головы, ноздрей, строением клюва. Столь заметное различие во
Рис. 24. Однодневный птенец сороки (фото автора).
внешнем облике птенцов дает основание сомневаться в правомерности объединения наших мухоловок в один род, как это принято делать.
Все сказанное выше дает основание считать, что характер птенцового опушения у воробьиных имеет большое систематическое значение.
107
OD Характер опушения птенцов некоторых воробьиных Таблица 30
Птерилии
Название вида Окраска и густота опушения Максимальная длина о 3 X S “ п 3 о . 3 2 аг X СО Я <Ь я* О) X о ГУ X а
пуха (в мм) СП со аг S О) (V Ч 3 £ £ о 55 Ч £ S 3 2 о о 5
ч Я X со О сп Ч с С CQ ИМ ч и \о X 2 я \О х
Ворон Темно-серый, не густой 13 + + + 4 4 4 — ±
Ворона серая То же 14 — + + + 4 4 4 — 4-
Грач Темно-серый, редкий 9 — — — + 4 4 4 — —
Галка То же 10 — —— — + 4 4- 4- — —
Сорока Пух отсутствует — — — — — — — — — —
Сойка То же — — — — — — — —
Скворец обыкновенный Светло-серый, густой 18 — + 4- 4 4- -f- 4 4 U _1_
Иволга обыкновенная Светло-желтый, густой 10 + + 4 + 4 4 ± 4
Дубонос обыкновенный Белый, густой 12 — + + 4 4 4 4 4- 4
Зеленушка Грязно-серый, густой 9 — + + 4 4- 4 4 4- ‘ 4-
Щегол Беловатый, густой 8 ± + + 4 4 4 4 4- 4-
Чиж Темный, довольно густой 8 + + + 4 4 4 4 4- ’Г
Чечетка Пепельно-серый, густой 9 4- + + 4 —1- 4 4- ; 4- 4-
Коноплянка Темно-серый, густой ? — + + 4 4- 4 4 4- 4-
Снегирь То же 12 — + + 4 4 4 4 4- 4-
Чечевица То же 11 ± + + 4 4 4 4 4- 4-
Щур Дымчатый, густой 14 ± + + 4 4 4 4- 4- 4-
Зяблик Грязно-серый, густой 10 4- + + 4 4 4 + 4- ±
Юрок Белый, густой ? 4- + -j- 4 4 4 4- 4 4-
Клест-еловик Темно-серый, густой 11 + + 4- 4 4 4 4- 4- 4-
Овсянка обыкновенная Бурый, густой 12 — + + 4 4 4 4- 4-
Овсянка садовая Серый, густой 13 .— + + 4 4 4 4- 4- 4-
Подорожник лапландский Довольно густой 12 4- + + 4 4 4 4- 4" —
Жаворонок черный То же 20 — + + 4- 4- 4 4- — —
Воробей полевой Пух отсутствует — — — —- — — — — — —
Воробей домовой То же — — —- —- — — — — — —
Воробей каменный Довольно густой 11 — 4“ + 4- 4- — — — —
Конек лесной Темно-серый 15 4- 4- 4* 4 4- 4- 4- 4- ±
Лазоревка Светло-серый, редкий 10 — + 4 — 4* — — — —
Синица большая Дымчатый, редкий 12 — + + 4- 4-
Продолжение табл.
Название вида Окраска и густота опушения Максимальная длина пуха (в мм) Птерилии
глазные надглазничные затылочные спинная плечевые локте- вые 1 бедренные голен-ные брюш-1 ные
Пухляк Дымчатый, редкий 10 — + 4* + + — — —
Ремез Пух отсутствует — — — — •— — — — —
Сорокопут-жулан Светлый, едва заметный 1 — — — — — — — — 4~
Мухоловка серая Дымчатый, не густой 7 + + + 4- + 4- + — —
Мухоловка-пеструшка Серый, редкий 9 ± + + 4- + — — — —
Мухоловка-белошейка Светло-серый, редкий 8 ± 4- + 4- + — — — —
Пеночка-теньковка Светлый, редкий 8 — + + — + — — — —
Пеночка-весничка Очень светлый, редкий 9 — + + — + — — — —
Пеночка-трещотка Светлый, редкий 12 — + + — 4- — — — —
Славка ястребиная Пух отсутствует — — — — — — — — — —
Славка садовая То же — — — — — — — — — —
Славка-завирушка То же — — — — — — — — — —
Славка серая Го же — — — — — — — — — —
Славка-черноголовка То же — — — — — — — — — —
Малиновка лесная То же — — — — — — — — — —
Камышовка-барсучок То же — — — — — — — — — —
Дрозд черный Буровато-серый, редкий 17 — + + 4- + + — — —
Дрозд-белобровик Светло-охристый, редкий 13 — + + + + 4- — — —
Дрозд певчий Рыжеватый, редкий 13 — 4- + 4- + 4- — — ——
Дрозд-рябинник То же 17 ± 4- + 4- + + — — —
Горихвостка обыкновенная Почти черный, не густой 10 + + 4- + + — — — —
Соловей восточный Почти черный, довольно густой . . 14 — + 4- 4- + — — — —
Варакушка Темный, довольно густой 10 + 4- 4- 4- + — — — —
Зарянка Почти черный, не густой 15 + + 4- 4- + — — — —
Каменка обыкновенная Довольно густой ? — + 4- + + —— 4' — —
Чекан луговой Буровато-серый, редкий 14 — + 4- 4- + — — — —
Крапивник Редкий 8 — + 4- + —— — — 1 1
2 Примечание. Знак ± означает, что на данном участке тела йух бывает не всегда.
Наряду с другими птенцовыми признаками оно может служить хорошим показателем степени родства между видами. Это важное положение,, на которое в свое время уже обращалось внимание (Ticehurst, 1926) „ оказалось незаслуженно забытым в связи с существующим стремлением объяснить характер и степень опушенности птенцов воробьиных только лишь условиями гнездования.
Итак, разобранные нами птенцовые признаки воробьиных можно разделить на две категории: а) признаки адаптивного характера, основное назначение которых сводится к облегчению процесса передачи корма от родителя к птенцу (окраска ротовой полости, боковые выросты" клюва и др.), и б) признаки, не имеющие явного адаптивного значения,., существующие, видимо, в силу исторических причин (птенцовое опушение) .
Большинство птенцовых признаков имеет серьезное систематическое-значение. В своей совокупности они могут служить критерием родовой принадлежности вида.
ГЛАВА V
ПИТАНИЕ ВОРОБЬИНЫХ В ГНЕЗДОВОЙ ПЕРИОД
Изучение питания лесных воробьиных в гнездовой (период производилось путем перевязывания пищевода у птенцов — по методике, уже-ранее нами описанной (Мальчевский и Кадочников, 1953). Часть мате-
Рис. 25. Длиннохвостая синица с пучком веснянок в клюве. Задача исследователя — перехватить пучок насекомых в момент передачи его птенцам (ст. Кузнечное, июнь 1954 г. Фото автора).
риалов была собрана путем отбирания принесенного птенцам корма непосредственно из клюва взрослой птицы после предварительного ее приручения (рис. 25, 26). Кроме того, были использованы некоторые данные исследования желудков, собранные автором еще в довоенные годы. Исследованием было охвачено 39 видов певчих птиц. Количество подопытных птенцов равнялось 892. Из их пищеводов при помощи пе-
11k
«ревязок было извлечено 4000 пищевых комков. Общее количество насекомых и других животных, обнаруженных в этих пищевых комках, составило около 11000 экз.
Рис. 26. Длиннохвостая синица прилетела кормить своих птенцов, посаженных ваклетку (ст. Кузнечное, июнь 1964 г. Фото автора).
О КОРМОВОЙ СПЕЦИАЛИЗАЦИИ ВОРОБЬИНЫХ
Одним из вопросов, встающих перед исследователем, занимающимся изучением питания птиц как взрослых, так и «птенцов, является вопрос о видовой специфике кормового рациона. Существует ли у воробьиных в гнездовой период избирательность в питании или они приносят своим птенцам и поедают сами насекомых и других животных пропорционально их встречаемости?
Этот вопрос обсуждался в литературе неоднократно. Некоторые исследователи полагали, что для большинства насекомоядных птиц характерен строго определенный набор насекомых, составляющих основу питания вида и определяющих положение его в биоценозе. Сторонники подобной точки зрения были убеждены, что исследователь, детально изучивший питание отдельных видов, в состоянии отразить характер взаимоотношений «между птицами и «насекомыми путем построения определенных схем (Фридолин, 1936, и др.)* Дальнейшее накопление материала по питанию насекомоядных птиц показало, однако, что подобные схемы, как правило, не могут отразить действительное положение вещей, ибо для преобладающего большинства птиц характерна известная многоядность и способность быстро переключаться с одного вида корма на другой в зависимости от динамики популяций насекомых и той обстановки, в которой живет птица (Формозов, 1950).
С другой стороны, данные, говорящие о высокой степени пластичности поведения большинства насекомоядных птиц, послужили основанием для признания частью орнитологов противоположной точки
112
зрения, выражающейся в идее примерно пропорциональной поедаемо-сти насекомых, выдвинутой в свое время Мак-Ати (Mc-Atee, 1932). Этот взгляд был подвергнут критике С. С. Шварцем (1950) и Н. П. Дубининым (1955).
Не ставя своей целью доказать или опровергнуть принцип пропорциональности поедания насекомых, автор настоящей работы тем не менее считает, что при существующих несовершенных методах учета численности энтомофауны, а также из-за неточности методики изучения питания птиц, особенно если учесть легкую переключаемость птиц с одного корма на другой, решение данного вопроса путем сопоставления частот встречаемости насекомых в желудках птиц и в природе, особенно в лесу, вряд ли вообще возможно. Известно, например, что в желудках птиц орнитологи обнаруживали таких насекомых, которых
Таблица 31
Встречаемость различных животных кормов в пище птенцов певчих птиц
Название вида Количество экземпляров животных в исследованных порциях корма
беспозвоночных позвоночных
всего насекомые пауки черви моллюски многонож-ки грызуны яйца птиц птенцы гады рыбы
Ворона серая 2325 2050 39 10 74 5 10 18 6 3 ПО
Сорока 1044 921 35 21 15 8 — 4 6 34 —
Сойка 942 709 201 — 15 — — 5 12 — —
Скворец 234 220 6 4 — 4 — — — — —
Иволга 25 20 3 — 2 — — — — — —
Дубонос • 168 167 1 —
Зеленушка 37 37 —
Щегол 6 6 —
Чечевица 6 6 —
Шур 79 70 8 — 1 — — — — — —
Клест-еловик 6 6 —
Зяблик 23 18 5 —•
Овсянка обыкновенная .... 186 171 14 — 1 — — — — — —
Овсянка садовая 405 400 5
Конек лесной 180 150 26 — 3 1 — — — — —
Синица большая 933 666 262 — 5 — — — — — —
Лазоревка 15 9 6 —
Синица длиннохвостая .... 896 880 15 — 1 — — — — — —
Сорокопут-жулан 154 124 12 — 7 — 3 — 8 — —
Мухоловка серая 77 70 3 — 4 — — — — — —
Мухоловка-белошейка .... 410 252 142 — 14 2 — — — — —
Мухоловка-пеструшка .... 17 14 2 — 1 — — — — — —
Пеночка-теньковка 87 74 11 — 2 —— — — — — —
Пеночка-весничка 203 177 19 — 7 — — — — — —
Пеночка-трещотка 67 23 38 — 6 — — — — — —
Малиновка лесная 46 39 7 —
Славка-черноголовка 178 90 79 — 9 — — — — — —
Славка садовая 210 144 46 — 20 — — — — — —
Дрозд-деряба 12 9 1 — — 2 — — — —
Дрозд певчий 251 186 5 35 24 — — — — 1 —
Дрозд-рябинник 40 19 — 20 1 — — — — — —
Дрозд-белобровик 201 130 9 39 23 — — — — — —
Дрозд черный *10 376 8 18 5 3 — — — — —
Горихвостка обыкновенная . . 1153 706 393 — 48 6 — — — — —
Соловей восточный 241 206 29 — 6 — — — — — —
3 А. С. Мальчевский
113
энтомологи, несмотря на специальные (поиски, в данной местности не находили. В то же время на основании простой глазомерной оценки, иногда нетрудно убедиться в том, что насекомоядные птицы избегают употреблять в (пищу некоторых насекомых — шпанских мушек, гусениц кольчатого шелкопряда, клопов-солдатиков, различных жалящих перепончатокрылых.
В тех же случаях, когда мы имеем дело со съедобными и легко доступными насекомыми, то чем многочисленнее вид насекомого, тем,, естественно, больше у него шансов стать жертвой птицы. Поэтому в желудках многих птиц отдельные группы беспозвоночных иногда, и находятся приблизительно ;в тех же количественных отношениях, в каких они встречаются в той местности, где кормятся эти птицы.
Если бы, однако, все виды птиц ловили бы насекомых только-, лишь пропорционально их встречаемости, то характер питания различных видов птиц и их птенцов, гнездящихся в одном биотопе и собирающих корм в сходных условиях, был бы примерно одинаковым. Так, однако, не бывает или бывает очень редко. Если мы, например, срав-
Степень поедаемости различных групп
Количество
Название вида бабочки жуки мухи и комары прямокрылые муравьи прочие перепончатокрылые
Ворона серая 452 788 154 1 570 2
Сорока 175 416 14 38 242 12
Сойка 639 25 17 14 3 5
Скворец 79 88 41 — 6 2
Иволга 14 4 1 — । —
Дубонос 140 9 1 — — 17
Зеленушка 29 8 — — — —
Щегол —. 6 — — — —
Чечевица 3 3 — — —
Щур 13 15 15 — —
Зяблик 9 — 8 —
Овсянка обыкновенная . . 80 7 37 44 2
Овсянка садовая 245 74 32 46
Конек лесной 41 4 62 6 1 i
Синица большая 605 1 — : 60
Лазоревка 9 — — — — (.
Синица длиннохвостая . . 61 8 4 — — —
Сорокопут-жулан 11 79 2 8 — 22
Мухоловка серая 10 1 54 — — 3 3
Мухоловка-белошейка . . 128 44 21 — — 2
Мухоловка-пеструшка . . 4 — 6 — — 4
Пеночка-теньковка .... 33 1 9 —- —
Пеночка-весничка 46 6 96 — 4
Пеночка-трещотка .... 13 — 8 — — —
Малиновка лесная .... 8 1 25 2 —
Славка-черноголовка . . . 74 8 — — — 1
Славка садовая 111 3 13 — — 2
Дрозд-деряба 3 4 2 — — —
Дрозд певчий 96 48 42 — — —
Дрозд-рябинник 4 14 1 — — —
Дрозд-белобровик .... 24 20 7 — 76 3
Дрозд черный 123 80 31 21 106 1
Горихвостка обыкновенная 339 54 54 69 54 26
Соловей восточный .... 52 74 7 2 1 2
114
ним набор кормов птенцов большой синицы и обыкновенной горихвостки (табл. 31—32) —птиц, очень часто гнездящихся бок о бок в совершенно однотипных условиях, то наряду с некоторыми общими чертами их питания (ярко выраженная насекомоядность и поедание значительного» количества пауков) нетрудно уловить и существенные различия. Большая синица в условиях сосновых насаждений Савальского лесхоза приносит птенцам почти исключительно бабочек или их гусениц и личинок некоторых перепончатокрылых (пилильщиков); гнездящиеся же рядом горихвостки выкармливают птенцов бабочками, жуками, мухами, саранчовыми, муравьями, клопами и другими насекомыми, добываемыми на различных фазах их развития. В данном случае существенные отличия в питании указанных видов обусловлены в первую очередь различиями в способах кормодобывания. Горихвостки схватывают насекомых на лету, подбирают их на земле или в траве, ловят в листве, на ветвях или стволах деревьев или кустарников; большая синица собирает насекомых в гнездовой период почти исключительно
Таблица 32 насекомых птенцами певчих птиц
обнаруженных экземпляров
клопы стрекозы цикады S ь сетчатокрылые верблюдки веснянки ручейники прочие неопреде- . ленные о ь-<и» CQ
7 .76 2050
13 — — — — — — 1 10 921
2 4 — — — — — 709
2 2 — — — — — — — 220
— — 1 — — — — — — 29
— — — — — — — — — 167
— — — —— — — — — — 37
— — — —i — — — — — 6
— — — — — — — — — 6
— — — 27 — — — — — 79
— — — — — — — — 1 18
— —— — — __ — — 1 171
1 — — — — 1 — — 1 409
1 — — 30 — 4 — — i 150
— — — — — — — — — 669
— —— — 9
— 2 14 13 — — 778 — 880
— 1 — — — — — — 1 124
— — — __ — 1 1 70
— — — 14 2 28 — 4 9 252
—— —— — — — ——1 — 14
— — 2 29 — — — — — 74
— 3 12 10 — — — — — 177
— 1 — — — —— —— — 1 23
2 1 — — — — — — 39*
— — — — 4 3 90
*— 4 — — 3 — — — 8 144
— — — —— — — 9
—— — — — — — — — — 186
— — — — — — — — — 19
__ — — — — — 130
10 — — — — 1 — 1 2 376
15 — — 14 26 — 26 29 706
3 — 2 3 — 53 — — Т 206
115
на деревьях. В связи с этим набор ее кормов и оказывается менее разнообразным.
Таким образом, круг животных, которых ловит птица, определяется в первую очередь способом ее охоты. Одни птицы отыскивают главным образом малоподвижную добычу (гусениц, пауков), добывая их преимущественно на деревьях (синицы), другие охотятся в основном на земле, где находят больше всего жуков и их личинок, червей, муравьев, а также наземных моллюсков (дрозды); третьи ловят насекомых главным образом в воздухе (серая мухоловка), в связи с чем в их рационе -преобладают летающие формы — мухи и бабочки; наконец, четвертые, как, например, горихвостка, мухоловка-белошейка, сорокопут-жулан и другие, способны схватывать как очень подвижную, так и малоподвижную добычу, будучи в состоянии ловить насекомых в воздухе, на деревьях, в траве и на земле.
Многие вьюрковые (чечевица, щегол, зеленушка), будучи в основном приспособленными к добыванию растительной нищи, очень ограничены в выборе насекомых и поедают, как правило, лишь малоподвижных гусениц, личинок цветочных мух и жучков, которых иногда находят в тех местах, где кормятся семенами или зеленью. Щегол, например, кормящийся обычно на соцветиях травянистых растений, наряду с семенами и завязями иногда поедает живущих в соцветиях гусениц бабочек и личинок мух. Такими образом, межвидовые отличия в питании неизбежны уже по одному тому, что для каждого вида характерен свой способ охоты и кормления, свои присущие виду повадки.
В определении видовой специфики питания воробьиных большое значение имеют также величина птицы и характер ее «вооружения», обусловливающие определенный круг жертв, с которыми данная птица может справиться. Например, небольшие размеры гусениц большинства наших бабочек позволяют справляться с ними самым мелким птицам; в то же время они достаточны для того, чтобы привлечь внимание даже сравнительно крупных птиц. Поэтому число врагов у мелких гусениц бабочек среди пернатых особенно велико, то же можно сказать и относительно мелких видов жуков. Жучки Harpalus были найдены нами в желудках 13 видов насекомоядных птиц полезащитных полос Заволжья. Самые разнообразные по величине птицы (от серой славки до грача) поедали их в довольно значительном количестве.
С майским жуком может справиться уже далеко не всякая птица. Несмотря на массовое размножение этих жуков в 1952 г., на территории Савальского лесхоза ими совершенно не питались горихвостки, лесные коньки, славки, овсянки и соловьи, не говоря уже о более мелких пеночках. В то же время гнездцвшиёся на тех же лесных участках, подчас совсем рядом черные и певчие дрозды, а также сойки приносили этих жуков своим птенцам довольно часто. Иволга и сорокопут-жулан также оказались способными поедать майских жуков.
Наконец, очень крупных жуков, например жука-оленя, умертвить может только сильная птица с достаточно мощным клювом, в связи с чем преобладающее большинство воробьиных птиц не употребляет его в пищу, несмотря на его съедобность. В учлесхозе «Лес на Ворскле» автор неоднократно наблюдал, как поползень, обладающий, как известно, довольно сильным клювом, безуспешно нападал на самца жука-оленя средних размеров. Каждый раз при приближении поползня жук принимал угрожающую позу, выставлял навстречу свои «рога»-жвалы и отпугивал птицу. В Савальском лесхозе жук-олень — обычная жертва серой вороны. Остатки этого жука были обнаружены также
Ю. К. Эйгелисом (1957) в погадках ворона. Также только лишь у врановых птиц (серая ворона, сорока и сойка) были обнаружены крупные жуки-красотелы (Calasoma) и оленьки (Dorcus parallelopipedus).
Интересно, с другой стороны, * что очень мелкие насекомые, например кровососущие комары, как сообщает Г. А. Новиков (1956), несмотря на их 'многочисленность, также очень редко поедаются птицами по причине их малой значимости. Из воробьиных птиц кровососущих комаров чаще всего поедают пеночка-весничка, а также, по наблюдениям В. С. Андриевской и С. И. Божко (устное сообщение), обыкновенная пищуха.
Итак, степень поедаемости насекомых зависит не только от их массовости, но и от степени съедобности, размеров, способа передвижения, приметности и всего образа жизни насекомого, равно как и характер питания насекомоядной птицы определяется в первую очередь ее видовыми особенностями: способом охоты, величиной и т. п. Если иметь в виду какой-либо определенный вид птицы, например сойку, то по поводу ее питания в гнездовой период вполне законно сказать, что питается она в основном наиболее массовыми и для нее наиболее доступными видами беспозвоночных, как это и было показано Н. П. Кадочниковым и Ю. К. Эйгелисом (1954). Это совсем не должно означать того, что сойка на территории своего охотничьего участка должна поедать всех насекомых в количестве, пропорциональном их встречаемости. Например, муравьев, весьма многочисленных в тех местах, где охотится в Савальском лесхозе сойка, она почти не употребляет в пищу. В связи с большой доступностью последних offa, возможно, и стала бы потреблять их ,в большом количестве в том случае, если бы ее излюбленные в гнездовой период корма — гусеницы и куколки шелкопрядов и других бабочек, а также пауки — по какой-либо причине вдруг иссякли бы. Это было бы, однако, для нее вынужденным питанием, подобным известным примерам, когда клесты-еловики при неурожае или отсутствии семян хвойных деревьев переходили на питание ягодами и семенами травянистых растений (Новиков, 1952) или личинками тлей, которых выклевывали из галлов на листьях деревьев (Мальчевский, 1947).
О СТЕПЕНИ МНОГОЯДНОСТИ ПТЕНЦОВ
Известно, что некоторые дятлы выкармливают своих птенцов в течение всего гнездового периода почти исключительно муравьями. Следовательно, муравьи и их куколки по калорийности и возможности добыть их в достаточном количестве могут являться подходящим кормом для воспитания потомства. Изучение питания птенцов, например, вертишейки, проводившееся автором совместно с Е. Р. Гагинской и И. В. Прокофьевой на территории Ленинградской (район ст. Кузнечное), Воронежской (Савальский лесхоз) и Белогородской (учлесхоз «Лес на Ворскле») областей, показало, что в различных частях ареала вертишейка выкармливает своих многочисленных (до 14) птенцов почти исключительно муравьями, их личинками и куколками. Подобный же корм получают и птенцы вертишеек, гнездящихся в Дарвинском заповеднике (Титаева и Поливанов, 1953), в Латвии (Тима, 1958), в Центрально-черноземном заповеднике (Хватова, 1956), а также других местах. Одним словом, принцип выборочности в питании у этого вида выступает чрезвычайно наглядно.
Можно ли, однако, среди европейских видов певчих птиц найти пример подобной стенофагии птенцов? Среди насекомоядных певчих
117
птиц, по-видимому, нет таких, которые выкармливали бы своих птенцов какой-либо одной, строго определенной группой насекомых. Исключая клестов, которые, как известно, выкармливаются семенами хвойных деревьев, для птенцов наших певчих птиц вообще не характерна узкая специализация. Все они в той или иной степени многоядны.
Птенцы многих видов наряду с животной пищей способны усваивать также и растительную. К этой группе относится большинство представителей европейских видов врановых и вьюрковых птиц, скворец, иволга, воробьи, а также некоторые овсянки, славки, дрозды и горихвостки (таблх 33). По наблюдениям И. В. Прокофьевой (1958) в Лужском районе Ленинградской области, выкармливает своих птенцов растительной пищей (семена ели) довольно часто также лесная завирушка.
Среди врановых птиц растительную пищу чаще всего получают птенцы кедровки, выкармливаемые, по наблюдениям Сванберга (Swanberg, 1951), а также А. А. Меженного (устное сообщение) и Н. Ф. Реймерса (1957), в основном кедровыми орехами, запасаемыми взрослыми птицами еще с осени. Семена различных сельскохозяйственных растений часто приносят своим птенцам серая ворона, а также прач и галка. Наоборот, птенцы сойки и сороки получают растительный корм чрезвычайно редко и крайне нерегулярно.
Из вьюрковых птиц наиболее растительноядны птенцы у клестов, щегла, коноплянки, зеленушки, чечевицы, а наиболее насекомоядны — у зяблика. Птенцы перечисленных выше зерноядных птиц наряду с основным растительным кормом очень часто получают некоторое количество и животного корма. Даже птенцы клеста-еловика иногда получают животный корм в виде зимующих в шишках насекомых. Так, в «зобу» одного птенца клеста, добытого под Ленинградом 2 марта 1955 г., автор обнаружил несколько личинок каких-то мелких мух.
Преимущественно насекомоядными являются птенцы дубоноса, питающиеся в основном гусеницами и личинками и лишь изредка потребляющие растительную пищу. Чечетки выкармливают своих птенцов в основном растительной пищей, щуры — смешанной, а вьюрки главным образом насекомыми, хотя изредка приносят семена и зелень {наблюдения О. Татариновой и В. Зимина). Изучение питания снегиря, проведенное С. И. Божко на территории Ленинградской области, показало, что птенцы этого вида получают в основном растительный корм. Животная же пища (пауки, мелкие гусеницы и т. п.) была обнаружена лишь в виде незначительной примеси к семенам.
По характеру строения ротового аппарата птенцы чижа ближе всего стоят к группе растительноядных. В литературе, однако, имеются сведения о значительной степени насекомоядности птенцов этого вида (Покровская, 1956). По данным И. В. Покровской (1953) зеленушки в условиях лесостепной дубравы «Лес на Ворскле» ранние выводки выкармливают почти исключительно насекомыми, а поздние — семенами. Это, видимо, не следует считать правилом, ибо очень часто даже ранние выводки зеленушка выкармливает семенами. В конце мая и в июне птенцы зеленушек, гнездившихся в Савальском лесхозе, получали семена главным образом вяза — древесной породы, отличающейся, как известно, ранним созреванием семян. .
Птенцы многих вьюрковых птиц (коноплянка, зеленушка, щегол) способны с первого дня своего существования одинаково хорошо усваивать как растительные, так и животные корма. Таким образом, у весьма специализированных зерноядных птиц птенцы потенциально многоядны. Это указывает на относительность их пищевой специализации,
X 18
Таблица 33
Степень животноядности и растительноядности птенцов певчих птиц
Название вида Общее количество исследованных порций корма Количество порций, состоявших из корма:
только животного смешанного только растительного
только из беспозвоночных смешанного только из позвоночных
Ворона серая 129 61 31 18 18 1
•Сорока 150 120 23 6 1 —
Сойка 404 390 8 5 1 —
Скворец обыкновенный . . 131 114 — — — 17
Иволга обыкновенная . . . 25 25 — — — —
. Дубонос обыкновенный . . 50 42 — — 5 3
Зеленушка 128 — — — 20 108
Щегол 34 2 — —. 4 28
Чиж 3 — — — 3
Коноплянка 30 — — — 30
•Снегирь 68 — — — 42 26
Чечевица 22 — — — 3 19
Щур 16 — — —- 16 —
I Клест-еловик 3 — — — 1 2
Зяблик 14 13 -— — 1 —
Овсянка обыкновенная . . 68 68 — — — —
•Овсянка садовая 192 190 — — 12 —
Конек лесной 61 61 —— — —
Синица большая 742 742 — — — —
Лазоревка 13 13 — — — —
'Синица длиннохвостая . . 104 104 — — — —
"Сорокопут-жулан 129 118 — И — —
Мухоловка серая 37 37 — — —— —
Мухоловка-белошейка . . 210 210 — — — —
.Мухоловка-пеструшка . . 9 9 —— — — —
Л1еночка-теньковка .... 31 31 — — — —
Пеночка-весничка .... 107 107 — —. — —
Пеночка-трещотка .... 42 42 — — —
МалиноВка лесная .... 16 16 — — —
•Сла^ка-черноголовка . . . 140 140 — — — —
Славка садовая 158 157 — — — 1
Дрозд-рябинник 27 27 — — — —
.ДрОзд певчий 119 118 — 1 — —
Дрозд-белобровик .... 89 81 — — 7 1
. Дрозд-деряба 12 12 — — — —
Дрозд черный 180 180 — — — ——
^орцхвостка обыкновенная 400 393 — — 7 —-
Соловей восточный .... 64 64 — — — —
Примечание. За .порцию” корма здесь принимается 1 комок пищи, извлеченный яз пищевода птенца, содержимое одного желудка или зоба, а в случае с со-ьрйкопутом-жуланом—также остатки пищи, найденные в гнезде или поблизости от гнезда. ' •
не закрывающей у современных видов зерноядных вьюрков возникновения новых приспособлений даже в направлении насекомоядности, как это, например, и имело -место в процессе эволюции некоторых raJ ла лагосских вьюрков.
Помимо некоторых врановых и вьюрковых птиц пища растительного происхождения довольно часто 'поглощается птенцами воробьев.
11$
Гораздо реже и обычно лишь при позднем гнездовании получают растительную пищу птенцы обыкновенного скворца. В лесостепной дубраве «Лес на Ворскле» автор, наблюдая за поздними выводками скворцов,, неоднократно отмечал, как со времени созревания вишен (конец июня — начало июля) родители начинали приносить их птенцам. По мере созревания ягод ими иногда выкармливаются птенцы славок,, дроздов и горихвостки. Для преобладающего большинства видов воробьиных нашей фауны растительный корм в гнездовой период не является, однако, главным, а лишь второстепенным или случайным. Только у некоторых зерноядных вьюрковых, а из врановых у кедровки,, он составляет основу питания .птенцов.
Другой формой проявления многоядности птенцов воробьиных, следует признать склонность к плотоядности, наблюдающуюся у ряда видов данной группы. Это прежде всего относится к некоторым врано-вым птицам и сорокопутам, которые довольно часто кормят птенцов-мясом позвоночных. Как исключительно редкое явление, мелких позвоночных получают от своих родителей птенцы дроздов. Например,. Ю. К. Эйгелис извлек однажды из пищевода гнездового птенца певчего* дрозда мелкий экземпляр прыткой ящерицы, а И. В. Покровская1 (1956) установила случаи поедания птенцами дрозда-рябинника голо-, вастиков травяной лягушки. Наиболее разнообразных позвоночных приносят своим птенцам ворон и серая ворона. По наблюдениям Н. П. Кадочникова, изучавшего питание птенцов серой вороны в Са-вальском лесхозе в 1952 г., оказалось, что птенцы этого вида получают здесь грызунов, яйца и птенцов мелких певчих птиц, лягушек, ящериц и особенно много мелких рыб (табл. 31).
По данным Н. В. Смирновой (устное сообщение), в Аскании-Нова самых различных позвоночных приносят своим птенцам также грачи. В желудках грачат, взятых из гнезд в середине мая 1954 г., Н. В. Смирнова обнаружила значительное количество грызунов и ящериц, а также-яйца и птенцов птиц, земноводных (жабы) и даже рыбу. Интересно,, что видовой состав грызунов, принесенных грачами своим птенцам^ оказался довольно разнообразным: в желудках грачат Н. В. Смирнова* обнаружила остатки малого суслика, полевой и домовой мыши, серого хомячка, степной пеструшки, общественной и обыкновенной полевок.
Сорока и сойка чаще всего нападают на гнезда мелких насекомоядных птиц. Их яйца и птенцов они нередко приносят своим птенцам. Сорока иногда отваживается нападать на птенцов даже сравнительно крупных птиц — сойки и даже пустельги и кобчика. В гнездовой период, у сороки и мелких соколов, гнездящихся в ее гнездах, отношения иногда напоминают описанные Г. Н. Лихачевым (1951) взаимоотношения1 ворона и балобана. В степных и полупустынных районах птенцы сорок» из позвоночных чаще всего получают мелких рептилий. По наблюдениям автора в системе Богдинских полезащитных полос, ящурки (Еге-mias arguta) в желудках гнездовых птенцов сороки по объему занимали примерно 50% всего их содержимого.
Склонность сорокопута-жулана к хищничеству общеизвестна. В северных частях ареала он, однако, нападает на позвоночных чаще, нежели на юге. Так, несмотря, например, на равное количество обследованных гнезд, из 11 известных нам случаев нападения жулана на позвоночных, о которых мы судили по находкам жертв, наколотых в. районе гнезда на сучки, 9 было обнаружено в лесной зоне и лишь 2 в лесостепной. Существует мнение (Лэк, 1954) о том, что сорокопут-жулан легко добывает пищу в хорошую, но не в сырую погоду и кладовая служит резервом на дождливые дни. По всей вероятности, воз
120
растание хищничества к северу следует объяснить тем, что на северег где чаще бывают дожди и холода, жулан не всегда может найти достаточное количество насекомых. В холодные и дождливые годы склонность жулана к хищничеству, по-видимому, также возрастает.
Как видно из табл. 31, среди изучавшихся птиц нет ни одного вида, который выкармливал бы своих птенцов исключительно насекомыми, хотя в большинстве случаев последние и составляют основу рациона. Для птенцов всех исследованных нами видов птиц оказалось характерным поедание весьма разнообразных отрядов насекомых и различных классов наземных беспозвоночных. В последнее время стало известно (Покровская и Герд, 1955), что некоторые 'виды лесных певчих птиц при соответствующих условиях используют для выкармливания птенцов даже представителей водной фауны.
Таким образом, наряду с известными видовыми особенностями питания определенная степень многоядности является характерной чертой гнездовых птенцов почти всех воробьиных нашей фауны.
О СПЕЦИФИКЕ ПТЕНЦОВОГО ПИТАНИЯ
Подбирают ли птицы для своих птенцов специальный корм или они выкармливают их примерно той же пищей, которую они потребляют в этот же период сами?
С одной стороны, несомненно, что в целом характер питания птенцов соответствует летнему питанию взрослых птиц. Так, например, щегол, коноплянка, чечевица и некоторые другие вьюрковые птицы, будучи сами в основном растительноядными, выкармливают птенцов главным образом растительной пищей; всеядная ворона приносит своим птенцам не только различных животных, но и корм растительного происхождения; питающиеся иногда ягодами скворцы, дрозды, славки и горихвостки по мере созревания ягод начинают приносить их и своим птенцам; известные потребители дождевых червей — дрозды чаще, нежели какие-либо другие птицы, приносят их своим птенцам и т. д. Одним 'Словом, остается совершенно очевидным, что приспособления, определяющие характер пищи взрослых птиц, определяют и характер питания их птенцов.
С другой стороны, не лишено оснований весьма распространенное мнение о том, что большинство птиц кормит потомство более нежной пищей, чем та, которой питаются сами. Предпринятое в 1939 г. в системе Тимашевских лесных полос (левобережная часть Куйбышевской области) изучение питания взрослых полевых воробьев и их птенцов показало, что взрослые птицы в период выкармливания птенцов потребляют значительно больше растительного корма, чем их птенцы (табл. 34).
Таблица 34
Питание взрослых полевых воробьев и их птенцов в гнездовой период
Возраст Количество исследованных желудков Характер пищи (в %) от всего содержимого желудков
семена с.-х растений насекомые
Взрослые птицы ! | ‘ г 125 81 19
Гнездовые птицы 36 29 71
12 Г
Во второй половине гнездового периода, т. е. когда созревают се-тмена сельскохозяйственных растений, последние начинают играть более заметную роль в питании как взрослых воробьев, так и их птенцов. Однако в связи с тем, что на ранних стадиях развития (птенцы воробья, как правило, не получают растительной пищи, последняя в целом составляет значительно меньшую часть их рациона, нежели у взрослых птиц. Существенные различия в питании взрослых птиц и птенцов были установлены А. Л. Келейниковым (1953), а также Б. К. Штегманом (1956) у среднеазиатских и казахстанских воробьев.
Аналогичная картина выявилась и при сравнении летнего питания взрослых садовых овсянок и их птенцов. Оказалось, что семена сельскохозяйственных растений и сорняков наряду с насекомыми неизменно присутствуют в желудках взрослых овсянок в течение всего гнездового периода (рис. 27)„ в то время как в составе пищевого рациона птенцов они отмечаются в исключительно редких случаях
Рис. 27. Летнее питание садовой овсянки (на основании анализа содержимого 34 желудков взрослых птиц, добытых в Заволжье в 1939 г.).
5С°10 — насекомые, 12°/0 — семена диких растений, 38°10 — семена с.-х. растений.
Наблюдения И. А. Нейфельдт и Н. В. .Смирновой, проводившиеся летом 1952 и 1953 гг. на территории Савальского лесхоза, показали» что взрослые садовые и обыкновенные овсянки охотно поедали овес, а зяблик — просо, рассыпанные вблизи их гнезд. Особенно охотно птицы подбирали семена в холодную и пасмурную погоду; однако своим^ птенцам эти птицы семян не давали, продолжая выкармливать их на-> секомыми. В то же время более зерноядная зеленушка не только сам# поедала рассыпанные семена подсолнуха, но и приносила их в гнездо своим птенцам.
С целью сравнения характера питания взрослых насекомоядных птиц и их птенцов однажды (28 июня 1954 г. на Карельском перешейке) нами был убит самец дрозда-белобровика у гнезда, в котором находились 7-дневные птенцы. Вскрытие его желудка показало, что по-’ следний содержит многочисленные остатки муравьев и ‘их куколок. Пища же птенцов, извлеченная в этот же день из их пищеводов, а также из желудка одного птенца, состояла в основном из дождевых червей и совсем не содержала муравьев.
- В противоположность, дятловым, воробьиные, видимо, сравнитель-' но редко прйносят; муравьев своим птенцам. Среди 38 видов птиц, пи-; такие, которых нами изучалось, муравьи в большинстве случаев в* зесьма ограниченном количестве были обнаружены в пище птенцов 122
только 10 видов (табл. 32). Наоборот, в списках животных кормов, характеризующих летнее питание взрослых птиц, они фигурируют очень часто (Таращук, 1953; Новиков, 1956, и др.). Высказанное предположение требует, однако, специальной проверки. При выяснении вопроса надо учитывать способность птиц изменять характер питания в зависимости от времени и места гнездования, времени суток и даже погоды и поэтому особо строго соблюдать принцип единства места и времени исследования. Правильнее всего сравнивать питание членов одной семьи птенцов и их родителей.
Существенной особенностью питания гнездовых птенцов певчих птиц является относительно высокий процент потребления мелких и сочных кормов, как-то: гусениц бабочек и пауков. Почти все виды .лесных насекомоядных птиц, приносят этот корм своим птенцам в значительном количестве (табл. 31 и 35). По данным А. А. Сафронова (1954), проводившего изучение питания птенцов и взрослых больших синиц и мухоловок-пеструшек в Мордовском заповеднике, оказалось, что степень поедания птицами пауков в гнездовой период, судя по ана--лизу содержимого желудков птенцов, выше, чем в пред- и послегнездо-вой периоды. По наблюдениям С. М. Семенова (1956а), мухоловка-пеструшка кормит своих птенцов гусеницами и пауками главным образом в первую половину их гнездовой жизни.
Быстрый рост и развитие птенцов невозможны без достаточного количества влаги. До сих пор, однако, никому еще не удавалось наблюдать, чтобы мелкие певчие птицы приносили своим птенцам воду. Можно только предполагать, что подобные действия свойственны некоторым нашим вьюрковым птицам, выкармливающим птенцов главным образом семенами. Взрослые птицы имеют возможность летать на водопой, посещать лужи, пить росу или дождевые капли. Птенцы же получают необходимую для них влагу вместе с сочными гусеницами и пауками. По наблюдениям А. Н. Промптова и Е. В. Лукиной (1938), -большие синицы иногда даже просто выдавливают полужидкое содержимое пауков и гусениц в раскрытые глотки птенцов. Подобное же действие характерно и для мухоловки-пеструшки при выкармливании однодневных птенцов (Милованова, 1956).
Большинство видов птиц, питание которых нами специально изучалось, гораздо чаще приносили своим птенцам бабочек, чем жуков, ,а гусениц бабочек чаще, чем бабочек имаго. Для преобладающего количества видов оказалось характерным также поедание большого количества мух и крупных комаров (Tipula) —добычи, как известно, тоже достаточно нежной и легко перевариваемой. Особенно характерным поедание двукрылых оказалось для птенцов лесного конька, серой мухоловки, пеночки-веснички, лесной малиновки. У ряда видов существенное значение в питании птенцов имеют личинки жуков (певчий и чер-‘ ный дрозды, обыкновенная горихвостка, серая ворона, мухоловка-бе-л ошейка) и пилильщиков (большая синица, дубонос). Личинки мухи прямокрылых поедаются птенцами лесйых певчих птиц сравнительно редко (табл. 35). Жуки (имаго) преобладали в питании птенцов только тех видов, крупные размеры и строение клюва которых позволяли им отчленять голову и освобождать насекомое от твердого хитинового панциря (вороца, сорока, сброКопут-жулан). ;
Крупным (гусеницам и личинкам птицы обычно тоже отрывают* головы. При выкармливании маленьких'птенцов в возрасте нескольких дней птицы обычно подбирают мелких насекомых и сильно деформи-1 ;руют даже гусениц и паукбв. Эта черта характерна, по-видимому, длн ^преобладающего большинства певчих птиц. По нашим наблюдений^
т
Степень поедаемости птенцами певчих птиц
Название вида Количество обнаружен
бабочки жуки
имаго гусеницы и куколки кладки имаго личинки и куколки
Ворона серая Сорока Сойка Скворец обыкновенный Иволга обыкновенная Дубонос обыкновенный Зеленушка Щегол Клест-еловик Чечевица Щур Зяблик Овсянка обыкновенная Овсянка садовая Конек лесной Синица большая Лазоревка Синица длиннохвостая Сорокопут-жулан Мухоловка серая Мухоловка-белошейка Мухоловка-пеструшка Пеночка-теньковка Пеночка-весничка Пеночка-трещотка Малиновка лесная Славка-черноголовка Славка садовая Дрозд-деряба Дрозд певчий Дрозд-рябинник Дрозд-белобровик Дрозд черный Горихвостка обыкновенная Соловей восточный > 4 8 52 5 3 1 25 189 12 278 1 13 7 7 94 1 10 6 1 3 11 17 । 33 38 248 18 448 158 587 74 И 140 29 3 13 8 55 56 29 326 8 48 4 3 34 3 23 40 12 5 63 94 2 63 4 24 85 91 34 1 1 1 1 1 1 1 1 1 1 1 1 1 1 - 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 709 414 25 88 4 1 8 1 6 74 3 8 79 1 10 1 6 1 8 3 2 25 14 20 41 33 67 79 2 8 6 3 14 1 1 34 2 23 39 21 7
различные виды дроздов разрывают на части дождевых червей, прежде чем дать их своим маловозрастным птенцам, а сойка рвет на куски гусениц и куколки шелкопрядов; дубонос предварительно разжевывает гусениц, а славки подбирают мелких гусениц и паучков с тем,, чтобы птенцы могли их проглотить.
Нужно, однако, сказать, что даже однодневные птенцы мелких певчих птиц способны проглатывать относительно крупную добычу. Так, например, однодневные птенчики садовой и серой славок заглатывают гусениц пядениц до 17 мм длиной, а 3-дневный птенец славки-Черноголовки— гусениц совок до 30 мм в длину; 4-дневный птенец черного-' дрозда способен проглотить крупную личинку бронзовки, а также майского жука; 10-дневные птенцы сойки свободно заглатывают очень крупных гусениц соснового шелкопряда длиной до 70 мм.
124
Таблица 35
насекомых различных возрастных фаз
ных экземпляров______________________
мухи и комары прямокрылые муравьи прочие перепончатокрылые прочие насекомые
имаго личинки имаго личинки имаго личинки имаго личинки имаго личинки
2 152 1 3 567 2 7 76
14 — 16 22 69 173 12 — 14 10
17 .— 14 — 3 — 5 — 6 —
41 — — — 1 5 2 — 4 —
1 — — — — — — — 1 —
г 1 — — — — — — 17 —। —
— — — — — — — — — —
— — — — — — — — — —
— 6 — — — — — — — —
— — — — — — — — — —
15 — — — — — — — 27 —
8 — —— __ — — — — 1 —
37 — 43 1 2 — — 1 1 —
32 46 — — — — — 3 —
62 — 6 — 1 — — — 36 —
1 — — — — — — 60 — —
.— — — — — — — — — —
4 — 807 —
2 — 6 2 — — 22 — 2 —
54 — — — — — 2 1 2 —
11 10 — — — — — 2 53 4
6 — — — — 4 — —_ —
9 — — — — 31 —
96 — — — — — 4 — 25 —
8 — — — — — 2 —
24 1 — 2 — — — — 2 1
— — — 1 — 7 —
5 8 — — — — 2 — 15 —
2 — — — — — — — — —
42 — — — — — — — — —
1 — — — — — — —
3 4 — — 45 31 2 1 — —
23 8 15 6 106 — — 1 14 —
54 — 25 44 54 — 17 9 40 70
7 — 2 — 1 — 2 — 66 2
В том случае, если птенцам приносятся мохнатые гусеницы, например соснового шелкопряда (сойка, иволга, черный и певчий дрозды, восточный соловей и др.), птицы (предварительнообрабатывают их так,
что колючая гусеница превращается во влажную, покрытую слизью пилюлю. Для этого птицам, очевидно, приходится частично раздавли-
вать ее и выдерживать некоторое время в пищеводе.
Одним словом, наблюдения показывают, что птицы часто подбирают специальный корм для маленьких птенцов либо совершают определенные действия, обеспечивающие более легкое проглатывание насекомого (рис. 28). Надо полагать, что специальная обработка пищи, деформирование и подбор мелких насекомых имеют большое значение при выкармливании птенцов в слепом возрасте, когда они еще слабые jh не могут проглатывать крупных насекомых.
125
У самих птенцов избирательность по отношению к корму выражена, по-видимому, очень слабо. Будучи голодными, они готовы заглатывать почти все, что им положат в рот.
Рис. 28. Майские жуки, извлеченные из пищеводов 4-дневных птенцов черного дрозда. Прежде чем отдать жуков птенцам, взрослые птицы оторвали у них надкрылья и головы (фото автора).
МИНЕРАЛЬНЫЕ КОРМА И ГАСТРОЛИТЫ
Известно, что минеральные соли, а в особенности соли кальция играют в жизни птиц большую роль. Известно также, что в период, размножения птиц значение минеральных солей возрастает (Дементьев,. 1947). В этот .период соли кальция необходимы, например, для образования твердой скорлупы, для нормального построения скелета развивающегося птенца. При нарушении кальциевого обмена или вследствие-недостатка кальция в природе, откуда птицы могли бы его достать,, возникают различные аномалии в строении или развитии птенцов. Иногда птица откладывает мягкие яйца, лишенные твердой известковой оболочки. Однако факты, которые бы указывали на нарушение кальциевого обмена, встречаются у диких видов птиц крайне редко. Следовательно, птицы где-то находят необходимые для них минеральные вещества.
Известно, что у птиц, так же, как и у некоторых млекопитающих, существуют специальные места, куда они летают клевать землю, богатую минеральными солями. М. Н. Корелов (1953), описавший это явление. назвал такие места «птичьими солонцами». Скопление птиц на подобного рода «солонцах» можно наблюдать, однако, далеко не всюду. Например, многие лесные виды птиц, собирающие корм вблизи гнезда, в течение всего периода размножения иногда пребывают на сравнительно ограниченном участке заболоченного леса, где трудно
126
предполагать существование солонцов. Где же, спрашивается; такие4 птицы находят необходимые для нормального развития птенцов минеральные корма?
Прежде всего обращает на себя внимание то, что почти все изучавшиеся нами лесные певчие птицы приносили своим птенцам наземных моллюсков (табл. 31). Во всех случаях мелкие моллюски скармливались целиком в неизмененном виде. Когда же птицы, например дрозды или иволги, приносили крупных моллюсков, то их раковины хотя и разламывались, но всегда обязательно также отдавались птенцу. Подобного рода факты позволяют предполагать, что наземные моллюски с их известковыми панцирями используются певчими птицами при выкармливании птенцов не только в качестве мясной пйщи, но и: как минеральный корм. На значение известкового' панциря моллюсков-для удовлетворения потребности в солях кальция по отношению к взрослым птицам указывает в своей работе А. Кеве (Keve, 1952—1955).
Более или менее закономерным оказалось отсутствие моллюсков, в пище птенцов ряда вьюрков — щегла, зеленушки, чечевицы и некоторых других птиц, которые в поисках корма отлетают от гнезда далеко. Весьма вероятно, что они посещают в это время «птичьи солонцы», где и добывают необходимый для птенцов минеральный корм. В условиях культурного ландшафта, особенно вблизи человеческих поселений, многие птицы находят скорлупу куриных яиц, которую и притаскивают своим птенцам.
Как уже отмечалось нами (Мальчевский и Кадочников, 1953), применение методики временного перетягивания пищеводов у гнездовых птенцов помогло установить одну малоизвестную до сего- времени деталь, связанную с их питанием. Оказалось, что многие насекомоядные птицы (табл. 36) приносят своим птенцам специальные порции «корма», состоящие исключительно из земли. Комочки земли приносятся:
Таблица 36
Список видов птиц Савальского лесхоза, у которых отмечен факт приноса комочков земли птенцам
Название вида
Количество извлечений из пищевода
всего содержащих комочки земли
Иволга...........................
Овсянка обыкновенная ............
Конек лесной ....................
Синица большая ..................
Мухоловка-белошейка..............
Пеночка-трещотка.................
Славка садовая ..................
Дрозд черный.....................
Горихвостка обыкновенная ........
Соловей восточный................
25 65 61
742 200
45 158 180 400
64
1
2
2
43
11
2
1
1
23
1
птенцам не очень часто и преимущественно в первую половину гнездовой жизни птенцов. Из пищевода птенца садовой славки автор извлекал их уже на второй день после вылупления.
Иногда птицы дают птенцам также довольно крупные камешки.
В желудках 2-дневных птенцов обыкновенной овсянки автор находил камешки размером до 3,5 мм. Совершенно очевидно, что столь отно-
127
сительно крупные камни да еще в значительном количестве не могли быть принесены птенцу случайно. Песок и гравий мы находили также в желудках птенцов коноплянки, чечевицы, горихвостки, Дрозда-белобровика. Основное значение песка и камешков, по-видимому, механическое. Они выполняют роль гастролитов, помогая еще слабому желудку птенца перетирать корм, который иногда бывает довольно грубым (жуки, коконы, семена).
ПРОДОЛЖИТЕЛЬНОСТЬ ВЫКАРМЛИВАНИЯ ПТЕНЦОВ
В каком возрасте птенцы начинают и прекращают получать корм •от своих родителей?
В литературе существует утверждение, будто бы некоторые птицы, в частности певчие, начинают кормить своих птенцов лишь на 3-й день их жизни. Так, например, В. М. Модестов (1937), наблюдавший гнездование певчего дрозда, пришел к заключению, что родители совсем не кормят своих птенцов в течение первых 2 дней их жизни. Это мнение укрепилось в литературе (Гладков, 1954, стр. 431), тем не менее мы сочли необходимым подвергнуть данное положение специальной проверке. Вопрос выяснялся путем перевязки шеи недавно или только что вылупившихся птенцов. Оказалось, что не только птенцы певчего дрозда, но и всех других изученных нами видов птиц (черного дрозда и дрозда-белобровика, обыкновенной горихвостки, большой синицы, садовой славки, сойки, зеленушки, коноплянки, щегла) получали корм в 1-й же день их постэмбриональной жизни. У коноплянки и щегла нам удавалось отмечать семена и зелень в «зобу» у еще не вполне обсохших птенцов. Е. Р. Гагинская, перевязавшая шею птенца дрозда-белобровика в момент его вылупления, отметила, что свою первую пищу (кусок земляного червя) этот птенец получил через 2 час. 15 мин. В подобных же экспериментах Н. П. Кадочникова и Ю. К. Эйгелиса (1954) установлено, что птенцы сойки получали корм уже через 3 час. после их вылупления.
Итак, певчие птицы обычно начинают кормить своих птенцов очень скоро после их появления на свет, часто даже тогда, когда в гнезде еще не окончилось вылупление всех птенцов.
Гораздо труднее ответить на вопрос: когда птенцы становятся вполне самостоятельными и перестают получать корм от своих родителей? Дело в том, что после того, как птенцы покинут гнездо, за ними в большинстве случаев становится чрезвычайно трудно наблюдать. Отчасти причиной тому бывает скрытный образ жизни птиц, который они начинают вести, как только их птенцы выскочат из гнезда, отчасти — чрезвычайная непоседливость выводков. Наблюдения показывают, что выводки певчих птиц часто начинают перемещаться тогда, когда птенцы еще не умеют летать, а лишь прыгают по земле (дрозды, соловьи, зарянки, славки и др.). Вследствие такой подвижности птиц наблюдатель в громадном большинстве случаев теряет из вида выводок специально помеченных им птенцов уже на 3—4-й день после вылета их из гнезда. Среди певчих птиц известным исключением в этом отношении является сорокопут-жулан, чрезвычайно крикливые птенцы которого очень долго держатся в районе гнезда. Вследствие этого наблюдения за выводками сорокопутов-жуланов не представляют трудности; и это, пожалуй, единственный вид, по которому автору удалось собрать более или менее точный материал, имеющий отношение к поставленному вопросу. По нашим наблюдениям, птенцы сорокопута-жулана становятся самостоятельными примерно в возрасте 38—40 дней. Оставляют 128
они гнездо на 13-й день и затем около 3, 5 недель (24—25 дней) продолжают держаться неподалеку от гнезда. Так, например, в парке Лесотехнической академии выводок жуланов, покинувший- гнездо 8 июля (1947), оставался на том же участке до 3 августа. В течение всего этого времени родители активно кормили птенцов, несмотря на то, что уже в возрасте около 20 дней они могли хорошо летать. В конце периода выкармливания птенцы жулана стали почти не отличимы от взрослых птиц.
При воспитании в неволе птенцы воробьиных, видимо, раньше научаются самостоятельно брать корм, чем в природе. Так, например, воспитывавшийся в неволе Ю. К. Эйгелисом гнездовой птенец обыкновенной горихвостки, вылупившийся 4 июля, уже 19 июля, т. е. в возрасте 15 дней, перешел на самостоятельное поедание корма, находившегося в клетке. В естественных условиях на 16-й день птенцы горихвостки обычно только лишь покидают гнездо, и нам никогда не приходилось наблюдать, чтобы в этом возрасте они могли самостоятельно схватывать корм. Весьма интересно, что упомянутый выше птенец горихвостки через 5 дней после того, как он сам научился подбирать корм, т. е. в возрасте всего лишь 19 дней, стал выкармливать муравьиными куколками подсаженных к нему 14-дневных птенцов-слетков того же вида! Вряд ли подобный случай можно наблюдать в природе, где птенцы выкармливаются родителями даже в месячном возрасте.
Известно, например, что птенцы воробьиных продолжают выпрашивать (и получать) корм от взрослых птиц в том возрасте, когда, казалось бы, сами уже вполне способны добывать себе пропитание. Птенец уже достиг размера взрослой птицы, хорошо летает, хвост у него почти полностью отрос, и тем не менее он продолжает летать за родителями и «клянчить» у них корм. Подобное явление характерно для очень многих видов воробьиных.
Длительная привязанность молодой птицы к своим родителям имеет, видимо, большое значение, так как именно за это время она должна пройти «основы жизненного воспитания», заключающегося в постоянном подражании поведению родителей.
О КОЛИЧЕСТВЕ ПИЩИ, ПОЕДАЕМОЙ ПТЕНЦАМИ
О количестве поедаемой гнездовыми птенцами пищи обычно судят по числу кормовых прилетов взрослых птиц к гнезду, а также среднему количеству насекомых, приносимых птицей своим птенцам за один раз. Выяснению характера суточной ритмики активности птиц у гнезд было уделено большое внимание. В результате в орнитологической литературе накопилось достаточно большое количество материала, позволяющее судить о частоте кормления птенцов у многих видов (Промптов, 1940; Kendeigh, 1952; Haartman, 1954; Королькова, 1954; Зверев, 1954; Тима, 1954; Шеварева и Бровкина, 1954, и др.).
Было установлено, что каждому виду птиц свойствен свой ритм активности. В то же время наблюдения показали, что количество кормовых прилетов сильно варьирует в зависимости от возраста и количества птенцов, а также от характера корма, погоды и расстояния, на которое приходится летать птице в поисках корма. В экспериментальных условиях число кормлений может быть увеличено в несколько десятков раз (Благосклонов, 1956). Все это указывает на то, что степень интенсивности кормления птенцов — понятие весьма относительное, а цифровое выражение его — величина непостоянная. Тем не ме-
9 А. С. Мальчевский
129
нее по степени интенсивности выкармливания птенцов различные виды воробьиных могут отличаться очень сильно. Например, длиннохвостая синица прилетает к гнезду (примерно в 10 раз чаще, чем зеленушка, и в 20 раз чаще, чем сойка (табл. 37). По данному (признаку лесные воробьиные могут быть разбиты на птиц, (прилетающих кормить своих птенцов очень часто (обычно несколько сот раз в сутки), и на птиц, прилетающих к гнезду сравнительно редко (обычно несколько десятков раз).
Таблица 37
Частота кормовых прилетов у некоторых певчих птиц
Дата наблюдения Название вида Продолжительность наблюдений (в час.) Общее число кормовых прилетов Среднее число прилетов (в 1 час.) зличество енцов гнезде Возраст птенцов (в днях) Место наблюдения
с п
9/VII 1951 г. Сойка 16 20 1,2 5 14 Балашовская об-
ласть
31/VII 1957 г. Зеленушка . . . 12 24 2 5 10 Ленинградская область
16/VI 1954 г. Длиннохвостая си-
ница 8,5 191 22,5 9 12-13 Там же
Примечание. Данные по сойке получены автором совместно с Н. П. Кадочниковым, а сведения по длиннохвостой синице-при участии Е. Р. Гагинской.
К первой группе птиц, прилетающих к гнезду весьма часто, относится преобладающее большинство мелких певчих птиц, выкармливающих птенцов исключительно или преимущественно мелкими беспозвоночными. Вторую группу составляют крупные воробьиные (врановые) и та часть небольших по размеру птиц, которые выкармливают птенцов /преимущественно семенами (клесты, зеленушка, щегол, коноплянка и некоторые другие вьюрки). Представители последней группы летают за кормом обычно далеко, набирают его сразу помногу и с каждым прилетом кормят всех или большинство птенцов, находящихся в-гнезде. У этих птиц наблюдается развитие специальных приспособлений, дающих возможность транспортировать относительно крупные комки пищи. У многих врановых эту функцию в гнездовой период выполняет горловой, или подъязычный, мешок. Горловые мешки, служащие для транспортировки пищи птенцам, были обнаружены также1 у горных вьюрков рода Leucosticte (Miller, 1941), а также у щура (French, 1954).
Сой^а, многие вьюрковые, а также воробьи, приносящие своим птенцам сразу крупные порции корма, переносят пищу в сильно растяжимом пищеводе. У этих птиц пищевод функционально уже мало чем отличается от настоящего зоба. Сойка, например, за один прилет способна принести 5 крупных куколок соснового шелкопряда, которых она набирает в ротовую полость и пищевод так же, как это она делает с желудями в осеннее время. Весьма значительное количество пищи за один раз приносят к гнезду самцы вьюрковых птиц, выкармливающие самку и птенцов семенами. Особенно это характерно для клестов, птенцы которых в связи с суровыми условиями развития должны полу
130
чать усиленный паек. По наблюдению автора, проведенному 2 марта 1955 г. в окрестностях Ленинграда, в «зобу» одного из трех только что накормленных птенцов клеста-еловика помещалось 158 еловых семян (рис. 29). Очевидно, самец клеста, накормивший сразу трех птенцов, за один прилет принес примерно в три раза больше, т. е. около 450 семян.
По иному пути пошло развитие приспособлений у мелких птиц, выкармливающих своих птенцов насекомыми. Как правило, они летают недалеко от своего гнезда и вынуждены часто подлетать к нему скормом, так как много принести за один раз не могут. Вследствие этого с каждым прилетом они кормят обычно одного птенца. Мелкие порции пищи очень часто полностью не удовлетворяют птенца, и он сразу же
Рис. 29. Птенец клеста-еловика с наполненным „зобом*. Сильно растяжимый пищевод птенца способен вместить значительное количество- корма (ст. Комарове, февраль 1955 г. Фото автора).
снова начинает просить есть. Иногда один наиболее активный и занявший наиболее выгодную позицию птенец несколько раз подряд получает корм.
В том случае, когда насекомоядная птица находит скопление мелких беспозвоночных, ей, конечно, выгодно сразу набрать в клюв их как можно больше. Наблюдения показывают, что многие насекомоядные птицы способны таскать в клюве целые пучки насекомых, которые они с удивительной ловкостью, не роняя и очень быстро могу г распределять между птенцами. Это особенно характерно для птиц, собирающих мелких насекомых, — длиннохвостой синицы, пищухи и др. Наоборот, птицы, собирающие более крупных беспозвоночных — дождевых червей, слизняков, гусениц, личинок, крупных жуков, кобылок, кузнечиков и т. п. (дрозды, иволга, сорокопут-жулан, синицы), — прилетают к гнезду с 1 или 2 экз. в клюве. Следует, однако, сказать, что и дрозды и синицы, в том случае, когда они нападают на скопления .мелких насекомых, способны набирать их в клюв помногу. Эта черта характерна, по-видимому, для всех насекомоядных птиц, и ее надо рассматривать как приспособление, до некоторой степени заменяющее
9*
131
насекомоядным птицам отсутствующие у них «зоб» или подъязычный мешок.
Пучки корма, приносимого птенцам, которые автору удавалось отнимать у длиннохвостой синицы, состояли обычно из 10—15 экз. насекомых. Всего было проанализировано 104 таких пучка. Один из них, содержавший 24 экз., состоял из 17 экз. веснянок, 6 тлей и 1 паука. Однако в отдельных случаях, когда эта синица приносила бабочку или крупную гусеницу, она прилетала к гнезду, держа в клюве единственное насекомое. Таким образом, количество экземпляров беспозвоночных, приносимое за один раз, сильно варьирует и зависит главным образом от того, какой корм собирает птица.
Применяя методику перевязок, которая дает возможность отбирать от птенцов весь корм, приносимый родителями, автор совместно с Н. П. Кадочниковым попытался установить вес суточной порции корма, съедаемого птенцами сойки. Для этого было установлено дежурство у гнезда в течение всего периода активности птиц. Работа проводилась в Савальском лесхозе 9 июля 1951 г. у гнезда сойки, в котором находилось 5 двухнедельных птенцов. За прилетом взрослых птиц следили из-за укрытия, 'в стороне от гнезда. Поскольку сойки прилетают кормить своих птенцов редко (см. табл. 37), мы успевали не только отбирать принесенный корм от всех птенцов, но и накормить последних во время отсутствия родителей. Таким образом, наше вмешательство в жизнь птиц не должно было оказывать заметного влияния на частоту кормовых прилетов. Птенцы сойки прекрасно переносили 16-часовую перевязку'пищевода с короткими перерывами на время их кормления. За весь период суточной активности, начавшийся в 4 час. утра и кончившийся^ 20 час., сойки подлетали к гнезду 20 раз и принесли птенцам 63 очень крупных куколки соснового шелкопряда, 9 крупных пау-ков-крестбвиков, 3 зеленых кузнечика и 1 бражника. Общий вес корма, принесенного птенцам, равнялся 170 г. Следовательно, на каждого птенца пришлось в среднем по 34 г пищи. Средний вес 14-дневного птенца соййи равняется около 117—118 г. Таким образом, суточная норма одного птенца в данном случае оказалась равной примерно Уз (34,7%) его собственного веса. Существующее убеждение — будто бы гнездовые птенцы певчих птиц потребляют колоссальное количество пищи, чуть ли не равное их собственному весу, — по всей вероятности, неправильно и, по-видимому, справедливо лишь по отношению к птенцам-слеткам, отличающимся большой прожорливостью.
Сильно преувеличено также представление относительно неспособности гнездовых птенцов переносить голодовку. Иногда полагают, будто маленькие птенчики певчих птиц легко могут погибнуть, если не получат корма в течение нескольких часов. Наблюдения, однако, показывают, что птенцы многих певчих птиц, во всяком случае в первый период их гнездовой жизни, способны голодать довольно долго (Карпов и Парювщиков, 1941; Благосклонов, 1956). Для этого необходимо только, чтобы птенцы не переохлаждались. Автор держал своих подопытных птенцов в помещении, где температура воздуха колебалась от 26 до 35°. В таких условиях 2—3-дневные птенцы черного дрозда, сойки, скворца и обыкновенной горихвостки свободно переносили голодовку в течение 24 час. Двухдневный птенец садовой славки, взятый из гнезда в 8 час. утра, до 20 час. не получал пищи. За это время он не только не погиб, но стал даже более активным. В ответ на любое тактильное раздражение он сразу же принимал характерную для птенцов славок позу пищевого рефлекса — вытягивал до предела шею, занимал вертикальное положение и тряс головой. Несмотря на 12-часо-
132
вую голодовку, он был в состоянии сохранять такую позу, требующую, вероятно, от него большого напряжения в течение нескольких минут подряд. Об истощении и упадке сил у этого птенца, очевидно, не могло быть и речи. После эксперимента птенец был помещен в гнездо другой пары садовых славок, которая успешно воспитала его наравне со своими птенцами.
Способность маленьких воробьиных птиц переносить многочасовую голодовку нередко спасает их от гибели в случаях затянувшейся ненастной погоды, препятствующей насекомоядным птицам собирать корм и вынуждающей их основное время тратить на обогревание птенцов.
ГЛАВА VI
ПОСТЭМБРИОНАЛЬНОЕ РАЗВИТИЕ ВОРОБЬИНЫХ
Существующие в отечественной литературе данные по развитию воробьиных птиц скудны. В иностранных работах разбираемый вопрос также освещен недостаточно полно. Замечательные исследования Хейнрот (Heinroth О. u. М., 1924—1933), несмотря на обилие фактического материала, не затрагивают многих сторон, наблюдение которых возможно лишь в природной обстановке.
Большинство исследователей, занимавшихся изучением развития птенцов воробьиных, главное внимание обращало на постэмбриональное изменение веса птенцов или темпы роста отдельных частей их тела. Работ, которые бы характеризовали развитие птенцов по разным показателям, очень мало. Между тем накопление ‘подобного рода сведений важно не только для выяснения степени влияния условий гнездования на темпы развития птенцов, но и для правильного понимания истории отдельных видов. Особый интерес представляют сведения по наиболее ранним, а также наиболее поздним стадиям постэмбриональ-ного развития птенцов. Первые из них могут служить известным критерием для определения степени родства между родами и семействами, вторые — между видами. Сбор таких данных по возможно большему количеству видов даст возможность конкретизировать общие положения о развитии птенцов воробьиных, разработке которых мы обязаны трудам А. Портмана (1938, 1954), Г. А. Васильева (1938, 1940), А. Н. Промптова (1946, 1948, 1956) «и др.
В процессе изучения развития птенцов певчих птиц автор стремился для каждого из изучаемых (видов определить время открытия слуховых проходов и глаз, начала развертывания опахал маховых и рулевых перьев, длительность пребывания птенцов в гнезде, начало активного полета. Кроме того, обращалось внимание на возрастные изменения голоса, а также на способы передвижения птенцов в первые дни после оставления гнезда. Всего удалось собрать данные по развитию 32 видов лесных воробьиных; эти наблюдения сведены в табл. 38. Более подробно о развитии птенцов каждого вида сообщается в специальной части настоящей работы. В данной же главе автор хотел бы поделиться некоторыми общими соображениями, возникшими у него в процессе изучения постэмбрионального развития воробьиных.
О весовых показателях возраста птенцов. Весовыми показателями роста птенцов воробьиных интересовался целый 134
ряд исследователей (Heinroth О. u. М., 1924—1933; Portman, 1938; Банников, 1939; Познании, 1946, 1948; Lack and Silva, 1949; Lees, 1949; Подлевских, 1952; Иванаускас, 1954; Денисова, 19586, и др.). Их трудами были установлены основные закономерности роста птенцов данной группы птиц. У самых разнообразных видов воробьиных характер роста оказался однотипным. Первоначально вес птенца увеличивается очень быстро и, достигнув скоро определенного максимума, становится более или менее постоянным или даже несколько снижается .перед вылетом птенцов из гнезда.
Чрезвычайно большая определенность роста птенцов воробьиных птиц позволяет использовать весовые показатели при определении примерного возраста птенца. При выработке соответствующей шкалы необходимо, однако, учитывать, что в раннем возрасте у нормально развивающихся птенцов возможны значительные колебания веса в зависимости от степени наполнения желудка и кишечника. По данным Т. Л. Иванаускаса, в вечерние часы вес -содержимого желудка 3-дневного птенца скворца составляет до 25% веса его тела. В связи с большой наполненностью желудков вечером утренний вес птенцов нередко бывает меньше вечернего веса предшествующего дня. Вес маленьких птенцов может также существенно измениться после акта дефекации.
К сказанному следует добавить, что в первые дни после вылупления нередко наблюдаются индивидуальные вариации веса у птенцов «одного выводка, что является следствием различных размеров яиц. Разница в весе птенцов обычно скоро сглаживается -и лишь в отдельных случаях, наоборот, усиливается. У насекомоядных птиц причиной этому иногда бывают затянувшееся похолодание или продолжительные дожди, сокращающие птицам возможность добывания корма. В результате нехватки пищи последняя распределяется между птенцами неравномерно: более сильные и энергичные перехватывают ее у более слабых и вялых. Птенец, получающий меньше корма, начинает постепенно отставать в росте и иногда даже гибнет. Такую картину автору приходилось наблюдать в гнездах обыкновенной овсянки, большой синицы, черного дрозда, сорокопута-жулана, серой мухоловки и некоторых других птиц.
Большая разница в размерах и весе птенцов одного выводка гораздо чаще, однако, бывает по причине неодновременности вылупления птенцов, что в свою очередь является следствием начала насижи вания до окончания откладки яиц. Известно, что разновозрастность птенцов одного выводка для воробьиных в целом не характерна. Принято считать, что среди воробьиных это явление наблюдается лишь у врановых и клестов, у которых самки, как правило, приступают к насиживанию до откладки последнего яйца. Однако это положение нуждается в уточнении, так как явление растянутости вылупления птенцов из яиц одной кладки было обнаружено и у других видов воробьиных. •Особенно характерным оно оказалось для щегла, зеленушки, коноплянки, большой синицы, дрозда-рябинника, черного дрозда и белобровика, у которых вылупление птенцов в отдельных случаях растягивается на 3—4 дня.
Причины преждевременного насиживания кладки, приводящего к неодновременному вылуплению птенцов у певчих птиц, еще недостаточно хорошо выяснены. Изучая гнездование дроздов-белобровиков в Ленинграде, автор (Мальчевский, 1948) обратил внимание на то, что преждевременное насиживание кладки чаще всего приходится наблюдать при запоздалых сроках размножения. Тогда же было установ
135
лено, что это характерно в основном для дроздов, которые гнездятся повторно, после разорения гнезда с первой кладкой. В дальнейшем эта-особенность поведения птиц наблюдалась нами также и у других видов певчих птиц. У мухоловки-пеструшки значительная растянутость-вылупления птенцов при повторном размножении отмечается также Крейцем (1955).
Как уже отмечалось, неодновременность вылупления часто влечет за собой нарушение нормального роста птенцов, вылупившихся последними. В таких случаях очень трудно бывает определить возраст птенца по его весу. Другие показатели развития птенца, рассматриваемые нами ниже, более стойки и в меньшей степени, нежели вес, зависят от количества потребляемой птенцами пищи.
Сроки прозревания и открытия слуховых • лрохо-дов. Как известно, птенцы воробьиных птиц появляются на свет слепыми и с закрытыми слуховыми проходами. Последнее обстоятельство не означает, однако, что они глухи. Способность птенцов реагировать на звуки в первые дни жизни была обнаружена в свое время Кульманом (Kuhlmann, 1909), а также Г. А. Васильевым (1938). А. Н. Промптов (1956), правда, считает, что 1—2-суточные птенцы воробьиных реагируют на звуки в результате тактильной, а не слуховой рецепции. По его мнению, роль тонкого тактильного анализатора играет эмбриональное опушение, благодаря которому птенец может воспринимать через кожу даже звуковую волну. Наблюдения, однако, показывают, что неопушенные птенцы, например, садовой славки не менее чувствительны к звуковым раздражениям, чем опушенные птенцы серой мухоловки, Вряд ли птенцовое опушение следует рассматривать как специальное приспособление, способствующее возбуждению у птенцов пищевой реакции. По всей видимости, звуки воспринимаются все-таки через стенки черепа.
Естественно, что при закрытых слуховых проходах возможности анализатора птенца ограничены. Тихих звуков однодневные птенчики вообще не воспринимают, но если сила звука достаточно велика, то на звуки различного диапазона—от тонкого свиста до грубого голоса — •птенцы отвечают одинаковой пищевой реакцией. Пищевая реакция появляется на всякий звуковой раздражитель даже у подросших птенцов. Например, 2-недельные птенцы сойки, имеющие не только открытые слуховые проходы, но и уже полностью открытые глаза, приподымаются на ногах, вытягивают шеи и открывают рты в ответ на любой звук, даже на такой, который они никогда не слышат в природе, например на щелкание фотоаппарата. Сходное поведение наблюдали мы и у птенцов других видов.
Незадолго до оставления гнезда в поведении птенцов наступает переломный момент: они начинают пугаться незнакомых им звуков. К этому времени у них вырабатывается дифференцировка пищевого рефлекса на голос родителей.
Как сообщает Портман (1938), передняя (височная) и задняя (затылочная) стенки слухового прохода птенца в момент вылупления плотно прижаты одна к другой. Благодаря этому кожа здесь несколько отстает и собирается в складки. В дальнейшем стенки слухового прохода расходятся, и начинает образовываться наружное слуховое отверстие. Сначало оно имеет вид небольшой дырочки, которую бывает даже трудно отыскать в складках кожи. По истечении же нескольких дней эта дырочка превращается в широкое отверстие. Способность птенца к различению звуков к этому времени, естественно^ возрастает.
136
Рис. 30. Однодневный птенец щегла „просит есть". В момент выпрашивания корма может приоткрываться глазная щель (фото автора).
Рис. 31. 4-дневный птенец лесного конька в позе пищевого рефлекса (фото автора).
Поскольку образование наружного слухового отверстия связано с ростом и формированием костей черепа птенца, следовало ожидать .большой определенности этого процесса. Специальное обследование птенцов различных7 видов подтвердило это предположение. Сроки открытия слуховых проходов оказались столь же определенными, как и сроки открытия глаз или начала развертывания опахал маховых перьев.
Выяснилось, что ушные отверстия появляются в возрасте от 3 до ..Б—6 дней. У всех дроздовых, славок и пеночек, а также у лесного конька и садовой овсянки они открываются -в конце третьего дня жизни, т. е. раньше, нежели приоткрываются глаза. У птенцов серой мухоловки, сорокопута-жулана и зяблика, у которых слуховой проход открывается в возрасте 4 дней, нами было установлено совпадение во времени образования слухового прохода и приоткрывания глаз. У птенцов некоторых вьюрковых (щегол, зеленушка) ушные отверстия появляются позднее, чем открываются глаза, — лишь на 5—6 день. Причина столь позднего образования слухового прохода у этих видов для нас пока что неясна; возможно, это зависит от особой конфигурации черепной коробки.
Переходя к рассмотрению процесса прозревания птенцов певчих птиц, необходимо прежде всего сказать несколько слов о терминологии, ts орнитологической литературе нередко употребляется выражение «глаза прорезаются». Это выражение к птицам, видимо, вообще, неприменимо, ибо прорезания гл аз у птенцов не происходит. Специальное изучение процесса развития глаз (Slonaker, 1925; Portman, 1938) показало, что веки глаз заканчивают свое формирование уже ко дню вылупления. У новорожденного птенца края век плотно примыкают друг к другу, но не срощены. В связи с этим лучше употреблять выражение приоткрывание, а не прорезание глаз.
Согласно наблюдениям Слонакера и Портмана, в первые дни постэмбриональной жизни мускулатура век совершенно не действует, благодаря чему веки продолжают оставаться плотно сомкнутыми в течение первых 5—6 суток. Эти выводы, полученные на основе изучения развития птенцов домового воробья и обыкновенного скворца, справедливы, видимо, лишь по отношению к птенцам, воспитывающимся в закрытых гнездах, и распространять их на всю группу воробьиных нельзя. У некоторых открыто гнездящихся птиц мускулатура, управляющая движением век, в какой-то мере действует уже с первого дня жизни птенца. Автору, например, удалось наблюдать расхождение век у однодневного щегла во время пищевого рефлекса (рис. 30). С возрастом степень расхождения век в момент выпрашивания корма становится все более и более значительной. У большинства видов птенцы, достигшие 4—5-дневного возраста, имеют уже хорошо заметные глазные щели (рис. 31). У дуплогнездников же, действительно, веки способны приоткрываться несколько позже. Стадия развития глаза, характерная для 4—5-дневных птенцов дрозда или славки, у скворца и большой синицы достигается лишь в возрасте 7—8 дней.
Первое время даже уже прозревшие птенцы, выпрашивая корм, тянутся только кверху и не обращают внимания на источник зрительного раздражения. Распознавание окружающих предметов начинается, видимо, лишь за несколько дней до вылета из гнезда. До этого поведение птенцов бывает одинаковым как при виде подошедшего к гнезду человека, так и при виде подлетевшей к гнезду птицы — ее родителя. В обоих случаях птенец просит есть, вытягивая шею и открывая рот. Перелом в поведении наступает довольно неожиданно. Еще сегодня „доверчивые птенцы — на следующий же день вдруг становятся необы-
•138
Рис. 32. Гнездовые птенцы сойки в возрасте 15—16 дней выпрашивают корм даже у человека (Савальский лесхоз, июнь 1952 г. Фото автора).
Рис. 33. Те же птенцы через сутки, когда наметился перелом в их поведении-При виде человека они принимают позу страха или угрозы и издают громкий крик (фото автора).
чайно пугливыми. При виде человека они прижимаются ко дну гнезда,, принимают угрожающие позы или ударяются в бегство. Подобного рода изменение в поведении птенцов у разных видов воробьиных наступает в различном возрасте: у славок обычно на 7—8-й, у дроздов — на 9—10-й, у иволги — на 12—13-й, а у птенцов сойки—лишь на 16— 17-й день жизни (рис. 32, 33).
Резкие изменения поведения гнездовых птенцов происходят, видимо, в связи с тем, что к этому времени достигает определенного уровня развития центральная нервная система.
Следует обратить внимание еще на одну особенность поведения, птенцов воробьиных, связанную с работой зрительного рецептора. Она заключается в частом стремлении гнездовых птенцов смыкать веки. Эту особенность можно наблюдать даже в первые дни послегнездовой жизни. Смыкание век необязательно надо связывать с испугом или сонным состоянием птенца. Часто можно наблюдать бодрствующих птенцов-слетков, которые подолгу сидят на ветвях с закрытыми глазами и в то же время регулярно воспроизводят видовой птенцовый ‘ позыв, по которому родители время от времени находят их и кормят. Каждый раз, когда прилетает с кормом взрослая птица, птенец открывает глаза, трепыхает крыльями и кричит, но после получения корма снова закрывает глаза (рис. 34).
Таким образом, несмотря на ранние сроки открытия глаз, последние сравнительно мало используются птенцами в течение почти всего периода их гнездовой жизни. Ведущую роль во взаимоотношениях птенца с родителями долгое время играет, по-видимому, орган слуха.
Возрастные изменения голоса. Способен ли новорожденный птенец использовать свой голосовой аппарат и воспроизводить звуки? Относительно выводковых, а также некоторых птенцовых (дневные хищники) птиц известно, что их птенцы могут издавать писк, находясь еще в яйце. Птенцы воробьиных в этом отношении еще недостаточно хорошо изучены. Имеющиеся наблюдения очень скудны и противоречивы. Саттон и Пармели (1954), наблюдавшие гнездование обыкновенной каменки, отмечают, что новорожденные птенцы этого вида безголосы даже в момент действия пищевого рефлекса и едва слышный для человеческого уха звук они начинают издавать лишь с 5-дневного возраста. С другой стороны, Крейц (1955) сообщает о том, что однодневные птенцы мухоловки-пеструшки в ответ на звуковые и тактильные раздражения поднимают кверху голову, открывают рот и издают тихий «цыкающий» звук. По нашим наблюдениям, новорожденные птенцы многих видов воробьиных при выпрашивании корма способны воспроизводить тихий отрывистый писк, который, однако, можно услышать лишь на очег*-> близком расстоянии. Это приходилось наблюдать у птенцов славок, пеночек, синиц, полевого воробья. Птенцы дуплогнездников с самого начала отличаются более сильным голосом, нежели птенцы, развивающиеся в открытых гнездах; голос 3-дневного птенца большой синицы хорошо слышен с расстояния 10 м. Разница в силе голоса особенно заметна позднее, когда птенцы станут оперяться. К этому времени птенцы многих дуплогнездников начинают систематически подавать голос.
По-иному ведут себя птенцы, воспитывающиеся в открытых и легко доступных для хищников гнездах. Их крикливое поведение могло бы повлечь за собою весьма нежелательные последствия: хищник мог бы легко обнаружить гнездо по крику птенцов и уничтожить весь выводок. Очевидно, в связи с этим птенцы мелких певчих птиц, гнездящихся открыто, сидят в гнездах, как правило, тихо и голоса не подают. Про-
140
иеос выкармливания у них также протекает без особого шума. Лишь в период, непосредственно 'предшествующий вылету из гнезда, они начинают выдавать свое присутствие громким голосом, издаваемым в момент прилета родителей. Особенно это характерно для некоторых видов врановых птиц, гнездящихся высоко или в дуплах. Очень громко, грубыми голосами кричат птенцы ворона, грача, серой вороны, галки. Наоборот, у сойки и сороки, гнездящихся в нижних ярусах леса, выкармливание птенцов, пока последние находятся еще в гнездах, протекает обычно в обстановке почти полной тишины.
В день оставления гнезда, а в некоторых случаях накануне этого дня птенцы с особенной силой и частотой начинают издавать особый птенцовый позыв, имеющий призывное значение. Этот позыв
Рис. 34. Птенец серой мухоловки, только что покинувший гнездо (Зеленогорск, июль 1955 г. Фото автора).
по характеру звука обычно сильно отличается от крика, который издают птенцы при виде подлетевшего с кормом родителя. Последний бывает громче и воспроизводится лишь в течение короткого времени, пока птенец выпрашивает корм. Призывный же крик воспроизводится регулярно и при совсем ином, сравнительно спокойном, физиологическом •состоянии. Издавая его, птенец либо неподвижно сидит на ветке, либо передвигается в каком-нибудь направлении. Этот позыв является основным средством связи между членами выводка и родителями. Можно определенно сказать, что в первые дни послегнездовой жизни птенцов «кормит» голос. Разве могли бы родители найти и накормить всех своих не умеющих летать птенцов, сидящих в разных местах, если последние не давали бы о себе знать?
Кроме того, голос птенцов способствует сохранению целостности выводка. Покинув гнездо, птенцы обычно не сидят на одном месте, а начинают перемещаться в каком-нибудь направлении. Передвигаясь среди густой травы, часто не видя друг друга, птенцы тем не менее продолжают держаться вместе, выводком. Иногда птенец впервые
141
издает призывный крик сразу после того, как очутится вне гнезда-В данном случае акт вылета как бы побуждает птенца воспроизводить соответствующий позыв.
Призывный крик птенцов следует рассматривать как типичный птенцовый признак, ибо функционирует он временно, лишь в определенном возрасте. По мере дальнейшего развития птенца и перехода его на самостоятельное питание этот признак .постепенно превращается в видовой призывный крик взрослой птицы. Несмотря на постепенное изменение воспроизводимого звука, характер первоначального (птенцового) и окончательного (видового) позывов различен.
В отдельных случаях птенцовый позыв строго специфичен для вида. Чрезвычайно характерно, например, кричат птенцы садовой славки, позыв которых звучит как короткий и высокий выкрик «жюй», чередующийся с низким и более часто воспроизводимым криком «век-век». Из неворобьиных птиц данный признак очень ярко выражен у птенцов вертишейки, высокие голоса которых звенят, как тоненькие колокольчики. Обычно птенцовые позывы близких видов бывают сходными, трудно различимыми для человеческого уха. Очень сходно, например, кричат птенцы-слетки дроздов —певчего и белобровика, позывы которых представляют собою короткий визгливый выкрик. По прошествии же некоторого времени у дрозда-белобровика этот позыв преобразуется в тонкое и протяжное «цсси», а у певчего дрозда — в короткое «цк». Весьма похожие позывы имеют птенцы мухоловок, многих синиц, некоторых вьюрков. В то же время у представителей одного семейства птенцовые позывы могут быть совершенно разными. Например, призывный чирикающий крик птенцов зяблика гораздо более похож па соответствующий позыв молодого воробья, чем на позывы птенцов большинства вьюрковых птиц.
У некоторых видов певчих птиц нет резких отличий между позывом, имеющим призывное значение, и криком, издаваемым в момент выпрашивания корма. Последний вид голосовой реакции у них является по существу лишь более громкой и учащенной вариацией позыва первого типа.
В заключение .нашего короткого очерка о голосе птенцов певчих птиц укажем на характерный для них третий вид голосовой реакции. Это резкий крик, издаваемый ими в момент сильного испуга. Этим криком часто сопровождается бегство чем-либо напуганных птенцов из гнезда. Он издается только тогда, когда птенца, кто-либо преследует или схватывает. По характеру воспроизводимого звука данная голосовая реакция мало чем отличается от крика взрослой птицы в момент ее сильного возбуждения. Так, например, птенец-слеток иволги, будучи внезапно схваченным, кричит так же, как кричат взрослые иволги, когда они, очень сильно беспокоясь у гнезда, вьются над своим врагом. Пойманный слеток дрозда-белобровика или рябинника верещит точно так же, как кричит, например, подстреленный взрослый дрозд.. Крик чем-либо напуганных птенцов сойки напоминает соответствующий крик взрослых птиц этого вида и т. п. Одним словом, этот вид голосовой оеакции не может быть причислен к разряду птенцовых, ибо-физиологическое состояние и характер звука, выражающий это состояние, у птенца и взрослой птицы сходны.
Точно так же не .может быть причислен к разряду птенцовых голосовых реакций и четвертый тип позыва, который обычно начинает воспроизводиться птицей еще в птенцовом возрасте. Имеются в виду звуки, выражающие состояние беспокойства. Сюда следует отнести «коканье» дроздят, «тиканье» каменок, «чеканье» сорокопутов, «щелканьем
142
славок и камышовок и т. п. голоса молодых птиц, по характеру не отличимые от соответствующих позывов взрослых птиц и впервые воспроизводимые птенцами, когда они умеют уже летать, но еще продолжают просить корм у своих родителей.
Итак, у певчих птиц следует различать две птенцовые голосовые реакции: птенцовый призывный крик и крик при выпрашивании корма. Первая из указанных реакций, преобразующаяся постепенно в соответствующий видовой позыв взрослой 'птицы, исчезает с переходом птенца во взрослое состояние. Вторая реакция иногда проявляется вновь. Это бывает у самок в пору размножения, когда их выкармливают самцы, что особенно ярко выражено у некоторых врановых и вьюрковых птиц.
О развитии перьев крыла и хвоста. Возраст, в котором у птенцов воробьиных начинают раскрываться трубочки растущих маховых, весьма определенен. Задержка в росте перьев и запаздывание сроков развертывания опахал наступает лишь в редких случаях сильного голодания. При нормальном же ходе развития птенцов сколько-нибудь значительных индивидуальных отклонений в данном явлении установить не удается. В то же время почти всегда наблюдается некоторая растянутость процесса, .выражающаяся в неодновременном начале развертывания опахал отдельных перьев крыла.
Если учитывать не только маховые, но и кроющие перья, то начало9 появления кисточек перьев растягивается обычно на 1,5—2 суток. Раньше других появляются опахала кроющих, затем второстепенных и, наконец, .первостепенных маховых. Пеньки крайних первостепенных. раскрываются обычно последними. Такая .последовательность развертывания опахал перьев крыла наблюдалась нами у птенцов пеночки-веснички, обыкновенной горихвостки, лесного конька, серой мухоловки, сорокопута-жулана и ряда других птиц. Подобная же закономерность была отмечена С. И. Божко (1954) для дубоноса и зеленушки. Иногда, однако, в последовательности появления кисточек перьев крыла трудно усмотреть какой-либо определенный порядок. Например, у славок отдельные пеньки второстепенных и первостепенных маховых начинают раскрываться примерно в одно время; лишь кисточки перьев кроющих крыла у них, как и у большинства птиц, появляются несколько раньше. В связи с относительно очень быстрым темпом роста перьев на крыльях процесс (развертывания опахал у птенцов славок вообще протекает очень дружно.
Видовые различия сроков появления кисточек маховых перьев у воробьиных в целом незначительны. У преобладающего большинства открыто гнездящихся видов мелких воробьиных пеньки маховых перьев начинают раскрываться в возрасте 6—8 дней. Наиболее ранние сроки характерны для всех видов славок, пеночек, а также для серой мухоловки, зяблика, дубоноса и овсянок (табл. 38 и рис. 35). Относительно очень рано, если учесть их сравнительно крупные размеры, появляются кисточки опахал маховых перьев у дроздов — в возрасте 7 дней. Примерно в этом же возрасте раскрываются пеньки маховых у лесного1 конька. У зеленушки, щегла, сорокопута-жулана, а также у восточного соловья это происходит лишь в возрасте 8 дней. Таким образом, по состоянию перового покрова у мелких воробьиных разница в сроках развития не превышает двух суток.
Отмеченное уже в литературе (Промптов, 1957) несколько замедленное развитие птенцов вьюрковых птиц характерно не для всех представителей данного семейства. В частности, птенцы зяблика и обыкновенного дубоноса развиваются примерно так же быстро, как и птенцы серой мухоловки. Эти данные наряду с упоминавшимися в главе IV
143>
Таблица 38
Показатели развития птенцов некоторых воробьиных
Название вида Возраст в днях (не считая дня вылупления)
открываются слуховые проходы приоткрываются глаза раскрываются трубочки маховых 1 раскрываются трубочки рулевых происходит оставление гнезда (нор- мальное) птенец способен к активному полету
Ворона серая ? ? ? ? 30-32 33—34
Сорока 5 6—7 10-11 7 24—25 ?
•Сойка 5 5-6 9 И 18-20 23-24
Скворец 4-5 7-8 10-11 ? 20-22 19-20
Иволга 4—5 6-7 8 ? 14—15 18- 19
.Дубонос '--4 4-5 6-7 9 12-13 14-15
Щегол л -<> 4-5 1 8 ? ? ?
.Зеленушка 5—6 4—5 8 8-9 13-14 ?
Коноплянка ! ? 4-5 7 ? 13—14 14-15
.Зяблик i 4 4 6—7 8 12—13 14-15
Воробей полевой 1 ? 6-7 8 ? 15-16 14-15
Овсянка обыкновенная . . . i ? 4 6-7 7 9-10 7
Овсянка садовая 3 4 6-7 ? 8-9 14-15
Конек лесной 1 з 4 7 7—8 9-11 14—15
Синица большая ! 4 7 8-9 8-9 18—19 17-18
Сорокопут-жулан 4 4 8 9 12—13 17-18
Мухоловка серая 4 4 6 7-8 12—13 13—14
Пеночка-весничка 3 4—5 6-7 6 11—12 13—14
Пеночка-трешотка 3 4-5 6-7 6—7 11—12 13-14
Пеночка-теньковка ? 4—5 6—7 6-7 12-13 13-14
Малиновка лесная ? 4-5 6-7 7-8 11 — 13 13-14
Славка-черноголовка 3 4-5 6 7 9-11 ?
Славка садовая 3 4-5 6 8 9-11 13-14
Славка-завирушка 1 ? 4-5 6 7 9—11 7
Дрозд-рябинник .i 4—5 7 7 12-13 16—17
Дрозд-черный 3 4—5 7 8 12—13 16-17
Дрозд-белобровик 3 4—5 7 8 11 — 12 1Ь-17
Дрозд певчий 3 4—5 7 8 12-13 16—17
Горихвостка обыкновенная . . 3 -4 5-6 8 9 15-16 15-16
Соловей восточный 4-5 8 9 10-11 15-16
указывают на то, что семейство вьюрковых генетически весьма неоднородно.
У крупных представителей фауны певчих птиц, например у врановых, кисточки маховых перьев 'появляются позднее, чем у мелких (табл. 38). Тем не менее темпы роста пера у крупных видов воробьиных значительно более быстрые, нежели у мелких. Например, у сойки ежедневный прирост 9-го махового пера равен, по нашим наблюдениям, 6 мм (среднее за 19 дней), в то время как у серой мухоловки всего лишь 3,4 мм (среднее за 12 дней). В то же время у сойки формирование маховых перьев идет значительно быстрее, чем у равной ей по величине сизоворонки. За 19-дневный период развития птенцов 9-е маховое перо сойки на 30 мм обгоняет в росте соответствующее перо птенца сизоворонки. Разница же в размерах образующихся к этому времени опахал еще более существенная: опахала маховых перьев птенца сойки примерно в 2 раза больше (рис. 36). Столь значительные различия в темпах развития перового покрова у птенцов упомянутых выше видов птиц нельзя объяснять только лишь различиями в условиях гнездования, ибо сойка и сизоворонка — представители разных отрядов. В целом развитие перового покрова у воробьиных идет более
444
быстрыми темпами, нежели у представителей других отрядов с птенцовым типом развития.
Для того чтобы передние конечности птенца превратились в крылья, необходимо прежде всего образование сплошной несущей поверхности крыла. Этот процесс связан с образованием опахал не только маховых,
но
и кроющих перьев.
а
б
в
А
7 дней.
8 дней.
Пока опахала последних не достигают оснований опахал маховых, птенец летать не может по той причине, что его крыло еще проницаемо для воздуха. Простран-ства^ не покрытые перьями, дольше всего сохраняются в области нер вскрывшихся оснований первостепенных маховых (рис. 37), но «зарастание» их идет очень быстрыми темпами: крыло, которое еще вчера имело «дыры»,— сегодня оказывается уже сплошным. Быстрые темпы формирования несущей поверхности крыла достигаются встречным ходом увеличения поверхности опахал маховых (за счет освобождения оснований от охватывающих их чехлов) и кроющих перьев кры-
Рис. 36 Внутреннее первостепенное маховое 19-дневного птенца сойки (а) и равного ему по возрасту птенца сизоворонки (б).
Рис. 35. Размеры опахала внутреннего первостепенного махового у птенцов некоторых воробьиных в различном возрасте.
а — серая мухоловка, б — лесной конек, в — черный дрозд.
ла. В ускорении этого процесса известную роль играют и сами птенцы, которые незадолго перед -вылетом из гнезда начинают как бы усиленно «чиститься» (эти движения иногда принимают за очищение птенцов от паразитов). Они постоянно теребят клювом нераскрывшиеся части маховых, помогая тем самым их освобождению от рогового чехла.
Кроющие крылья у птенцов певчих птиц начинают и заканчивают свое развитие раньше, чем окончательно сформируются опахала махо-
10 А. С. Мальчевский
145
вых перьев. Среди последних раньше других прекращают расти внутренние второстепенные и внутренние первостепенные маховые. Наружные же первостепенные перья и иногда крайние второстепенные растут" дольше всего. Вначале эти перья особенно заметно отстают в развитии.
Благодаря различным темпам роста и определенной последовательности образования опахал перьев на различных участках крыла достигается постепенное накопление качеств, свойственных птичьему крылу, именно в той последовательности, в какой это необходимо длж развивающегося птенца по мере перехода его к самостоятельному образу жизни. Для птенца, недавно оставившего гнездо, в целях самосохранения очень важно как можно скорее приобрести способность хоть, как-нибудь летать. Наблюдения показывают, что птенцы большинства наших певчих птиц становятся летными уже тогда, когда их маховые
Рис. 37. Крыло 2-недельного птенца певчего дрозда (внутренние кроющие крыла выдернуты).
еще примерно на !/з находятся в трубочках. Тем не менее перья, слагающие крыло, уже в этом возрасте образуют сплошную поверхность^ достаточную для выполнения функции примитивного полета (см. рис. 37). Это достигается главным образом благодаря быстрому росту кроющих крыла, опахала которых очень скоро начинают полностьк> прикрывать пространства между нервскрывшимися основаниями маховых перьев.
Крыло птенца-слетка, только что научившегося летать, в некоторых отношениях напоминает крыло первоптиц: вследствие недоразвитости крайних первостепенных маховых оно туповершинно, имеет ограниченную несущую поверхность и отдельные маховые перья его еще слабо дифференцированы. Первоначально крылья выполняют весьма примитивную функцию: они служат молодой птице для коротких перелетов,, перепархиваний с ветки на ветку или взлета с земли. Птенец, впервые научившийся передвигаться по воздуху, летит прямолинейно, очень тяжело, с предельной весовой нагрузкой. Вследствие этого посадка его очень неуверенная. Дальность полета крайне невелика. В первый день она обычно не превышает нескольких десятков метров. Тем не менее даже такие крылья имеют колоссальное значение для выживаемости птенцов, ибо, несмотря на свою несовершенность, они все же дают основное — возможность использовать воздушную среду при встрече с наземными врагами.
146
Совершенствование летных качеств 'птенца идет очень быстрыми темпами. Через 4—5 дней после приобретения способности с трудом перемещаться по воздуху птенец уже прилично летает, без труда набирает высоту, может делать повороты, приземляться и уверенно садиться на ветки. В этом возрасте он уже сам начинает гоняться за родителями, выпрашивая у них корм, а не сидит, как прежде, на месте в расчете на то, что взрослые птицы найдут и накормят его. Такое быстрое совершенствование в полете является не столько результатом тренировки молодой птицы, сколько изменением аэродинамических свойств ее крыла. Дело в том, что в связи с продолжающимся ростом маховых перьев неуклонно увеличивается общая площадь несущей поверхности
Рис. 38. Крыло птенца лесного конька в день оставления гнезда (а) и примерно месяц спустя (б).
крыла. Поскольку вес тела молодой птицы в этот период жизни существенно уже не меняется, начинает быстро уменьшаться весовая нагрузка на крыло; это и дает возможность птенцу-слетку совершать все более и более сложные движения в воздухе.
С прекращением роста самых длинных первостепенных маховых, а иногда и крайних внутренних второстепенных, заканчивается формирование несущей поверхности, и к этому времени крыло приобретает типичную для вида форму. У мелких лесных певчих птиц, например у лесного конька, это наступает примерно в полуторамесячном возрасте (рис. 38), приблизительно через месяц после первого полета.
Вследствие того, что рулевые перья начинают развиваться позднее и растут более медленными темпами, чем маховые, хвост у птенцов певчих птиц долгое время продолжает оставаться относительно очень коротким. У большинства лесных воробьиных кисточки перьев на хвосте появляются, как правило, на 1—2 дня позднее, чем на крыле, и лишь у некоторых видов (пеночки, синицы, овсянки) это происходит примерно в одно время. Тем не менее даже в этих случаях рулевые
10*
147
перья сильно отстают в росте от маховых. Когда опахала маховых перьев развернутся уже больше чем на половину и позволяют птенцу уже перепархивать с ветки на ветку, размеры хвостовых перьев у птенцов воробьиных бывают, как правило, весьма незначительными (рис. 39). Отставание роста рулевых перьев особенно характерно для тех видов птиц, у которых птенцы первое время после оставления гнезда ведут наземный образ жизни. Это типично, например, для многих
дроздовых.
В запаздывании сроков развития рулевых перьев можно усмотреть определенный биологический смысл. Раннее отрастание хвоста в тот период, пока птенцы находятся еще в гнезде и постоянно касаются гузкой дна и стенок гнезда, неизбежно привело бы к его искривлению и преждевременному износу. Это и случается у птиц, у которых птенцы долго сидят в гнезде, цапримёр у синиц. Кроме того, для птенцов многих видов, воспитывающихся в тесных гнездах, большой хвост мог бы оказаться даже вредным, противоречащим требов аниям необх о ди м о й э ко но м ии места. Дело в том, что у большинства певчих птиц дефицит «жилплощади» в гнезде начинает ощущаться уже за несколько дней до вылета птенцов. Например, полный выводок птенцов дрозда-белобровика (6—7 птенцов) может разместиться в гнезде лишь при условии определенного порядка расположения отдельных его членов. Есте-Рис. 39. Внутренние первостепенны е ственно, что при отросших хвостах маховые (слева) и крайние рулевые птенцы .не МОгли бы сидеть в гнезде (справа) перья 12-дневного птенц а _ м
обыкновенной горихвостки (а) и зяб - столь компактно, как это обычно име-лика (б). ет место в период, предшествующий
их вылету.
У большинства наших открыто гнездящихся птиц заметное отрастание хвоста наступает примерно :в возрасте 17—25 дней, т. е. через 5—10 дней после оставления гнезда, в тот период, когда птенец перестает сидеть на месте, а начинает сам гоняться за взрослыми птицами.
Зависимость скорости развития птенцов от условий гнездования. В какой мере место расположения гнезда сказывается на скорости развития птенцов у певчих птиц?
Обычно принято считать, что в связи с наземным расположением гнезда развитие птенцов соответствующих видов идет значительно быстрее, чем у птиц, гнездящихся на деревьях. По отношению К воробьиным это положение справедливо лишь отчасти. По многим показателям птенцы видов, гнездящихся на земле, развиваются не быстрее, а в некоторых случаях даже медленнее, чем птенцы видов, располагающих свои гнезда на деревьях. Это прежде всего видно по срокам появления кисточек опахал маховых перьев. Например, у овсянок опахала маховых перьев начинают развертываться не раньше, чем у птенцов серой мухоловки, зяблика или дубоноса, а у птенцов лесного конька или восточного соловья даже позднее. Темпы развития птенцов наземно гнездящихся птиц существенно не отличаются от большинства других воробьиных также и по срокам открытия слуховых проходов и глаз (табл. 38).
Представление об ускоренном развитии птиц, устраивающих гнезда на земле, справедливо главным образом в отношении сроков пре-
148
бывания птенцов в гнезде, которые у данной экологической группы действительно по большей части сокращенные. Эта черта особенно характерна для птенцов садовой овсянки, которые очень часто покидают гнездо в возрасте 8—9 дней с едва раскрывшимися опахалами маховых. Очень рано выскакивают из гнезд птенцы обыкновенной овсянки и, по наблюдениям А. Н. Промптова (19346, 1956), овсянки-дубровника. По всей видимости, все овсянки отличаются ранней способностью к самостоятельному передвижению. Для птенцов других воробьиных, гнездящихся на земле, — коньков, соловьев, жаворонков — также характерны ранние сроки оставления гнезд, чего нельзя, однако,
сказать относительно пеночек, которые нормально выпрыгивают из гнезд в значительно более зрелом состоянии лишь через 11—13 дней после вылупления, т. е. примерно в том же возрасте, в котором обычно выскакивают значительно более крупные и редко гнездящиеся на земле дрозды. Следовательно, наземное расположение гнезда не во всех случаях влечет за собою даже раннее его оставление.
Более тесная зависимость между темпами развития и условиями гнездования существует у птиц, птенцы которых воспитываются в дуплах. Сроки появления кисточек опахал маховых перьев у них в целом поздние (табл. 38). Особенно это характерно для большой синицы и скворца. В связи с более медленным
Рис. 40. Крыло 2-недельного птенца большой синг-цы (а) и 13-дневного пгенца серой мухоловки (б).
ростом маховых, а также кроющих крыла у этих птиц позднее происходит формирование несущей поверхности крыла, благодаря чему они позже становятся способными к полету. В то время как у птенце в большинства открыто гнездящихся мелких видов певчих птиц образование сплошной несущей поверхности крыла наступает уже в возрасте 13—14 дней, крыло птенца большой синицы в этом возрасте имеет еще большие «дыры»: опахала кроющих перьев не дорастают еще до оснований опахал маховых и не прикрывают своими концами нерас-крывшиеся их части (рис. 40).
Как уже отмечалось выше, птенцы у дуплогнездников развиваются более медленными темпами также .и по другим показателям.
Оставление гнезда и первоначальный способ передвижения птенцов. Продолжительность пребывания птенцов в гнезде у воробьиных варьирует чрезвычайно сильно (табл. 38). Птенцы врановых сидят в гнезде от 18—20 (сойка) до 40
149
дней (ворон). У мелких птиц птенцы пребывают в гнезде 8—11 дней (овсянки, славки, коньки), 11 —14 (дрозды, вьюрки), 15—16 (воробьи, горихвостки), 18—19 (большая синица) или 20—22 дня (скворец) :и т. п.
Поздние сроки вылета птенцов из дупел нельзя объяснять только лишь медленными темпами развития птиц-дуплогнездников. Как правило, птенцы этой группы птиц покидают гнезда в относительно более зрелом возрасте. Таким образом, судить о скорости развития птенцов
Рис. 41. Недавно покинувший гнездо птенец иволги в момент опасности (Савальский лесхоз, июль 1953 г. Фото автора).
по продолжительности пребывания их в гнезде нельзя. Например, скорость развития птенцов обыкновенной горихвостки в случае наземного расположения гнезда и гнездования в дупле одинаковая; тем не менее, как показывают наблюдения, в дупле птенцы этого вида сидят 15—16 дней, а в наземном гнезде всего 13 дней. Восточный соловей и горихвостка примерно в одном возрасте приобретают способность к полету, но птенцы соловья, развиваясь в открытом наземном гнезде, оставляют его обычно на 5—6 дней раньше птенцов горихвостки.
Возраст, в котором птенец покидает гнездо, зависит также и от различных случайностей, побуждающих иногда птенцов выскакивать из гнезда на 2—3 дня раньше нормального срока. Это относится главным образом к птицам, гнездящимся открыто. Подобное поведение птенцов следует
расценивать как приспособление, направленное к обеспечению выживаемости потомства. Поскольку специальных
средств защиты или отпугивания у птенцов воробьиных нет, единственным средством избежать опасность является бегство и последующее затаивание. Поза притаившегося птенца-слетка весьма характерна и сходна у разных видов. В момент опасности птенец весь вытягивается, принимает вертикальное положение, закидывает голову назад (рис. 41) и становится неподвижным. Такая поза часто принимается птенцом в ответ на «сигнал тревоги» взрослых птиц.
Ранняя способность в случае необходимости покидать гнездо и уходить в сторону от него—черта, присущая птенцам почти всех воробьиных, но среди наших птиц она особенно характерна для птенцов различных видов славок и овсянок, оставляющих иногда гнездо уже в недельном возрасте. Наблюдения за птенцами, рано выскочившими из гнезда, показывают, что они способны существовать вне гнезда, не возвращаясь больше в него. Описанные в литературе (Southern, 1949а)
150
олучаи обратного возвращения птенцов в гнездо у лесных воробьиных ^чрезвычайно редки. Гораздо чаще птенцам приходится существовать вне гнезда, и, несмотря на то, что в момент оставления гнезда очень часто они еще плохо оперены и нуждаются в обогреве, птенцы обычно не погибают. Сразу же после оставления гнезда, как только минует опасность, у птенцов возникает стремление вновь соединиться. Благодаря способности к самостоятельному передвижению, а также развившемуся к этому времени голосу, они вскоре находят друг друга и, тесно прижавшись один к другому, образуют общий комок, в котором иногда бывает даже трудно определить количество составляющих его птенцов (рис. 42). Для птенцов, у которых еще окончательно не уста-
Рис. 42. Птенцы длиннохвостой синицы (4 птенца) в день оставления гнезда (ст. Кузнечное, июль 1954 г Фото автора).
повилась терморегуляция, но которые уже находятся вне гнезда, взаимный обогрев друг друга чрезвычайно важен, ибо обогревание птенцов телом родителя после момента оставления гнезда в природе никем еще, кажется, не наблюдалось.
Потребность быть вместе и сидеть тесной группой, так, как это было в гнезде, возникает у птенцов-слетков и после нормальных сроков оставления гнезда. Но при преждевременном выскакивании птенцов это явление имеет, видимо, особое значение. Иначе не могли бы нормально продолжать развиваться живущие «на воле» полуоперившиеся птенцы славок, коньков, овсянок, рано выскакивающие из гнезд. В том, что такие птенцы выживают и нередко дорастают до летного состояния, убеждают нас факты, полученные в процессе наблюдения за окольцованными особями. Например, на территории Савальского лесхоза окольцованные птенцы славки-черноголовки выскочили из гнезда 22 июня на 8-й день после вылупления. Через 2 дня (24 июня) автор нашел весь выводок в 30 м от гнезда.
Акт оставления гнезда нельзя расценивать как результат стремления птенцов к рассредоточению. Он обусловливается, видимо, в первую очередь нарастающей потребностью птенцов к передвижению и пере
151
мене места. Чем раньше птенцы приобретут способность к самостоятельному -передвижению, тем выгоднее для вида, ибо, повторяем,, основным средством спасения от хищников птенцов мелких певчих птиц в этом возрасте является движение, приводящее к изменению* места их пребывания.
Способы передвижения у птенцов разных видов могут быть различными. Главные из них следующие: прыганье или беганье по земле» перепархивание с ветки на ветку и, наконец, полет. Птенцы, например, славок или зяблика могут комбинировать первый и второй способы передвижения, птенцы синиц — второй и третий и т. п. Для большинства видов» характерен, однако, какой-нибудь основной способ первоначального передвижения. Различать его, нам кажется, необходимо, особенно при решении вопросов, связанных с историей видов.
Перемещение по земле как первоначальный способ движения, характерно для птенцов овсянок, коньков, соловья, зарянки, всех дроздов. Передвигаясь по земле прыжками (большинство) или семенящими шагами (коньки), не умеющие еще летать птенцы в состоянии тем не менее очень быстро перемещаться из одного места в другое. Поймать такого птенца бывает очень трудно. Длина прыжка нелетного птенца черного дрозда равна примерно 20 см. Ноги у птенцов рассматриваемой группы птиц отличаются относительно сильно развитой мускулатурой, а также большой длиной цевки и высоким «подъемом» ступни (рис. 43). Возможность раннего использования задних конечностей в целях передвижения и обусловливает то, что птенцы данной группы птиц, как правило, очень скоро покидают гнезда.
У некоторых видов птиц птенцы по выходе из гнезда передвигаются путем коротких перепархиваний с ветки на ветку. К этой группе от-представителей наших лесных певчих птиц:
иволга, сорокопуты, пеночки, славки, камышовки, большинство вьюрковых и др. Птенцы почти всех перечисленных видов птиц могут также передвигаться и по земле—прыжками. Однако этот способ передвижения нельзя признать у них основным, ибо как только птенцу представляется возможность взобраться на ветви кустов или деревьев, он стремится это сделать. Кроме того, при передвижении по земле птенцы этих птиц, как правило, пользуются не одними задними конечностями» а пускают в ход также и крылья, благодаря чему сохраняется тот же характер движения путем коротких перепархиваний.
Лапы птенцов таких птиц отличаются большой цепкостью, способностью уже в раннем возрасте удерживать тело птенчика на тонких ветках. Особенной цепкостью обладают задние конечности птенцов» иволги. Птенцы этого вида оставляют гнездо неспособными к полету и долгое время обитают высоко в кронах деревьев на тонких качающихся ветвях. Наземный образ жизни совершенно не характерен как для молодых, так и для взрослых иволог. Это исключительно древесные птицы, отличающиеся способностью прыгать и лазать по ветвям» начиная с раннего возраста. Подобный образ жизни выработал у этих.
Рис. 43. Нога и коготь птенцов-слетков певчего дрозда (а) и иволги (б).
носится большинство
152
птиц лапу особого типа, удобную для обхватывания ветвей; приспособительные черты строения ноги птенца иволги особенно наглядно' выступают при сравнении ее с лапой птенца, ведущего наземный образ, жизни, например певчего дрозда (см. рис. 43). Ноги птенца-слетка иволги отличаются прежде всего сильно укороченными толстыми цевками, плоскими (без «подъема») и расширенными основаниями ступней и пальцев, а также очень острыми и круто загнутыми когтями. Если посадить птенца-слетка иволги на палец, то человек, держащий, птенца, испытывает довольно резкую боль, настолько сильно птенец впивается своими острыми когтями. Только что выбравшиеся из гнезда,, не умеющие еще летать птенцы иволги способны к удивительным «акробатическим» движениям. Падающему птенцу иволги достаточно-зацепиться одним коготком за тонкую веточку, чтобы из положения «вниз головой» вскарабкаться кверху и принять нормальную позу.
У других видов рассматриваемой группы способность птенцов уверенно и крепко держаться на тонких веточках достигается за счет образования на нижней поверхности тонких и гибких пальцев сильно увеличенных мягких шероховатых «подушечек», помогающих птенцу крепко зажимать ветки различной толщины.
Птицы, у которых птенцы сразу по выходе из гнезда могут летать, среди лесных воробьиных не многочисленны. К данной группе могут быть прежде всего отнесены виды, гнездящиеся в дуплах. В связи с более длительным периодом пребывания птенцов в гнезде их крылья, несмотря даже на замедленные темпы развития, успевают превратиться в орган полета. Однако исходный способ передвижения и у птенцов птиц-дуплогнездников может быть различным. Например, для птенцов синиц это будет перепархивание с ветки на ветку, а у слетков горихвостки—прыганье по земле. Указанные типы движений наиболее характерны для птенцов упомянутых видов еще до оставления гнезда и приобретения способности к полету. По поводу птенцов мухоловок можно сказать, что в нелетном состоянии они в силу своих относительно маленьких и слабо развитых задних конечностей совсем плохо передвигаются по земле и по ветвям деревьев. Основным способом передвижения птенцов этих птиц с самого начала является полет. Этим, очевидно, и следует объяснять относительно быстрые темпы развития маховых и кроющих перьев крыла, обусловливающих раннюю способность к полету птенцов серой мухоловки.
Приобретение способности к полету. Возраст, в котором птенцы начинают летать, определялся путем установления дня их вылупления и последующего ежедневного наблюдения за окольцованными птенцами в течение возможно более длительного периода времени. За начало полета принимался момент приобретения молодой птицей способности летать, набирая высоту. Неполнота наших наблюдений объясняется трудностью проведения наблюдений за птенцами в лослегнездовой период. Эти трудности возникают главным образом по причине обычного непостоянства мест обитания выводков и тех внезапных перемещений, которые начинают совершать птицы вместе со* своими птенцами иногда сразу же после оставления гнезда. В связи с этим судить о возрасте, в котором птенцы того или иного вида начинают летать, а также о скорости, с которой они удаляются от места своего рождения после того, как покинут гнездо, приходилось по небольшому количеству повторных встреч окольцованных нами птенцов.
В каком возрасте начинают летать певчие птицы? Прежде всего это зависит от их размеров. Крупные виды, отличающиеся более длительным периодом роста тела и маховых перьев, начинают летать
153-
позднее мелких. Птенцы серой вороны впервые становятся летными в возрасте 33—34 дней, сойки — в 23—24, дрозда-рябинника — в 16—17, а лесной малиновки — в 13—14 дней. Преобладающее большинство мелких видов певчих птиц нашей фауны начинает летать примерно в 2-недельном возрасте. Имеются и известные исключения. Сравнительно поздно «поднимаются на крыло» птенцы иволги (18—19 дней), скворца (19—20), большой синицы (17), сорокопута-жулана (17—18), обыкновенной горихвостки и восточного соловья (15—16 дней).
Помимо размеров птицы и условий гнездования возраст, в котором птенцы научаются летать, зависит от видовой специфики развития птенцов. Например, многие птицы, птенцы которых по выходе из гнезда могут быстро передвигаться^ по земле при помощи своих сильных ног, как это характерно для большинства представителей дроздовых, приобретают способность к полету несколькими днями позднее большинства остальных лесных певчих птиц.
Начало перемещения выводков. Как скоро после оставления гнезда начинается переселение молодых птиц и можно ли связывать начало перемещения выводков певчих птиц со временем приобретения способности птенцов к полету?
На этот вопрос одинакового ответа для всех видов воробьиных не может быть. Известно, что многие птицы изменяют место своего пребывания и кормежки почти сразу же после того, как их птенцы покинут гнездо. К таковым относятся многие виды дуплогнездников, птенцы которых оставляют гнездо, уже умея хорошо летать. Кольцевание птенцов скворца, проводившееся в Литве (Иванаускас, 1953), показало, что почти сразу же после вылета из гнезд молодые скворцы начинают перелет. Наблюдались даже случаи, когда во время перелета некоторые скворцы еще кормили своих птенцов. По нашим наблюдениям, направленные перемещения выводков у чернолобого сорокопута начинаются также до прекращения выкармливания молодых. Птенцы большой синицы, по наблюдениям А. Н. Промптова и Е. В. Лукиной (1938), уже через час-два после вылета из дупла откочевывают за пределы ближайших окрестностей гнезда. По наблюдениям Г. Н. Лихачева (19556), почти сразу же после вылета птенцов покидают «гнездовую территорию» также и мухоловки-пеструшки. У данного вида, как и у скворцов, начало послегнездовых перемещений выводков по существу является началом откочевки из гнездовой области. В связи с ранним отлетом вскоре после вылета птенцов из гнезд эти птицы вообще исчезают из района гнездования. По наблюдениям Н. А. Гладкова (1953), примерно так же ведет себя и садовая овсянка.
Видимо, многие виды лесных певчих птиц совершают перекочевки •сразу же после того, как их птенцы научатся летать. Это, однако, не всегда сразу бросается в глаза: из-за неодновременности сроков размножения отдельных пар, а также в связи с появлением новых для данного места выводков птиц, прилетающих с соседних территорий, создается впечатление известного постоянства обитания птиц в после-•гнездовой период на территории того или иного микрорайона.
Наблюдения за окольцованными молодыми птицами показали, что у некоторых видов смена места обитания выводка часто начинается уже и тогда, когда птенцы даже летать еще не умеют. Пишущему эти строки приходилось неоднократно наблюдать, как не умеющие еще летать птенцы садовых славок и славок-завирушек пересекали лесные поляны, двигаясь всем выводком в каком-нибудь определенном направлении. Перемещения выводков нелетных птенцов приходилось отмечать также у лесных малиновок, восточных соловьев, черных и пев-
154
’чих дроздов, дроздов-белобровиков и ряда других 1птиц. Однако в некоторых случаях птенцы этих же самых видов, покинув гнездо, продолжали держаться поблизости от него в течение многих дней. Так, например, окольцованный автором на территории Савальского лесхоза 28 июня 1951 г. 12-дневный птенец певчего дрозда через 8 дней (6 июля) был обнаружен в 150 м от места кольцевания. В другом случае птенцы славки-завирушки через 2 недели после вылета из гнезда были встречены всего лишь в 300 м от него. Подобные наблюдения могут быть приведены для выводков обыкновенной горихвостки, восточного соловья и других птиц.
Таким образом, у целого ряда певчих птиц нашей фауны возраст, в котором птенцы начинают удаляться от гнезда и совершать перекочевки, весьма неопределенен. Птенцы могут либо сразу же по выходе из гнезда, не умея еще летать, переместиться в другой участок леса, либо, наоборот, остаться в районе гнезда даже после того, как - научатся летать. Причины, которые побуждают птенцов этих птиц в раннем возрасте уходить из района своего гнезда, могут быть разными. Иногда перемещение выводка происходит вследствие того, что птиц кто-нибудь сильно потревожил; в некоторых случаях птицы просто уходят в места, более богатые кормом; в других — переселение бывает вызвано поисками более надежных убежищ и т. п. Например, если гнездо горихвосток располагается в (сосновом лесу, лишенном подседа, го птенцы по выходе из гнезда сразу начинают перемещаться в поисках участков леса с подседом. Поскольку горихвостки в послегнездо-вой период обычно кормятся на земле, их стремление уйти в такие места, где в лесу есть подлесок и подрост, вполне понятно: здесь они лучше скрыты от глаз хищников, нежели в чистых сосняках.
Среди наших лесных воробьиных встречаются, наконец, такие ©иды, которые в противоположность большинству птиц чрезвычайно долго держатся в районе гнезда после вылета птенцов. К таковым прежде всего следует отнести сорокопута-жулана. Наблюдения за окольцованными птенцами этого вида ..показали, что выводки сорокопута-жулана после того, как птенцы покинут гнездо, обычно в течение 2—3 недель, а иногда и дольше держатся вблизи гнезда. Эта особенность жуланов объясняется, очевидно, тем, что птенцы данного вида очень поздно приобретают самостоятельность. Таким «образом, сорокопут-жулан не только характером питания и способом охоты, но и некоторыми чертами своего постэмбрионального развития напоминает дневных хищных птиц. Эта особенность гнездовой жизни среди воробьиных характерна еще для врановых птиц, в частности для сороки и серой вороны, птенцы которых после вылета обычно тоже долго держатся в районе гнезда. Так, например, окольцованный автором 1июля 1951 г. птенец сороки, только что оставивший гнездо, через месяц (3 августа) был обнаружен в этом же районе.
В заключение приведем таблицу признаков для определения примерного возраста птенцов певчих птиц, составленную автором на основании изучения процесса развития наиболее часто встречающихся у нас лесных видов. Эта таблица является результатом попытки дальнейшего усовершенствования соответствующих определителей, составленных в свое время В. Л. Бианки (1913) и А. Н. Промптовым (1957).
В процессе работы автор стремился выработать шкалу возрастных ’Признаков, пригодную для определения возраста птенцов всех представителей певчих птиц нашей фауны. Это, однако, не удалось по причине ^сильных отклонений в темпах развития отдельных групп (врано-
155
вые, скворцы, синицы). Поэтому при определении возраста птенцов надо помнить, что таблица 'Применима лишь к мелким (от пеночки до дрозда) и -открыто гнездящимся лесным видам воробьиных нашей фауны.
Таблица
Примерный возраст в днях (не считая дня вылупления) Внешний облик Характер поведения
1-2 Либо совсем голые (славки, камышовки, сорокопуты, пересмешник), либо местами покрыты пухом (вьюрки, овсянки, коньки, мухоловки, пеночки, дрозды, чеканы, соловьи); трубочки будущих маховых и рулевых перьев не обозначились (хотя мелкие зубчики и могут быть), ушные отверстия закрыты; веки сомкнуты (но не срощены) В ответ на сотрясение гнезда,, прикосновение, громкий звук и т. п. тянут кверху uieip и открывают рот
3—5 Тело местами покрыто пеньками контурных перьев или их зачатками; трубочки маховых и рулевых перьев обозначились или уже отросли, но на концах еще не раскрылись; ушные отверстия, как правило, открыты; глаза в виде узких щелей Тело местами покрыто наполовину раскрывшимися пеньками контурных перьев; трубочки маховых перьев раскрылись и несут на своих вершинах кисточки или небольшие опахала; глаза открыты, но полностью еще не округлились В ответ на сотрясение гнезда,, прикосновение, звук и т. п. приподымаются, вытягивают шею и открывают рот
6-8 При осмотре гнезда часто закрывают глаза и прижимаются, в некоторых случаях (славки) даже выскакивают из гнезда
9—10 Тело оперено не полностью (брюшко еще голое); маховые развернулись меньше чем наполовину (длина раскрывшейся части меньше половины длины всего пера); опахала кроющих крыла не достигают опахал маховых При осмотре гнезда прижимаются, принимают оборонительную-позу или выскакивают из гнезда^ летать еще не могут
11-12 Оперены почти полностью; маховые развернулись наполовину и даже больше (длина раскрывшейся части равна половине длины пера или больше половины) При осмотре гнезда обычно выпрыгивают из него; хорошо прыгают по‘земле и перепархивают с ветки на ветку; летать еще не могут; при попытке летать садятся обычно на землю; птенцы овсянок, коньков, славок, соловьев в этом возрасте живут уже вне гнезда
13—16 Маховые развернулись почти полностью; опахала кроющих крыла налегают на опахала маховых; хвост сильно укорочен; у птенцов, появляющихся на свет в пуху, последний обычно сохраняется еще на голове и спине Могут летать, но высоту набирают с трудом; находятся обычна вйе гнезда
17—25 Вполне оперившиеся, лишь хвост несколько укорочен Летают хорошо, без труда набирая высоту, посадка уверенная;, гоняются следом за родителями^ выпрашивая у них корм
Часть вторая
МАТЕРИАЛЫ
ПО БИОЛОГИИ РАЗМНОЖЕНИЯ И ПОСТЭМБРИОНАЛЬНОМУ РАЗВИТИЮ ОТДЕЛЬНЫХ ВИДОВ
ВОРОНА СЕРАЯ
(CORVUS CORONE CORN IX L.)
Токование. В Ленинградской области разгар весенних «игр»' ворон приходится на середину марта—начало апреля. Однако образование пар и занятие гнездовых участков часто происходит значительно-раньше. Вероятнее всего, что весной особенно интенсивно токуют самцы, не имеющие пары. Птицы, живущие в парах зимой, находят друг друга,., наверное, еще осенью. Может быть, в этом и надо усматривать биологический смысл осенних воздушных «игр» у данного вида, которые обычно бывают в конце сентября — октябре.
Расположение гнезда. В пригородах Ленинграда автору чаще всего приходилось находить вороньи гнезда на соснах (8 находок. из 17), реже на елях (5 находок) или лиственницах, а в центральных районах города, где хвойных деревьев мало,— на высоких тополях, кленах, липах. В заповеднике «Лес на Ворскле» серая ворона обычно селится на дубах (14 находок из 15) на большой высоте. Судя по литературным данным (Асписов, 1932; Попов и др., 1954), в поймах рек этот вид часто гнездится довольно низко, иногда даже в кустах на высоте 2—3 м.
Сроки размножения. Сроки размножения серой вороны варьируют значительно меньше, нежели у большинства других воробьиных. Объясняется это, очевидно, во-первых, тем, что гнезда этих птиц, разоряются не часто, благодаря чему повторное гнездование у них — сравнительно редкое явление, и, во-вторых — отсутствием у этого вида нормальных вторых кладок. Наибольшая разница в сроках размножения отдельных пар вопон в окрестностях г. Ленинграда в 1954 г. равнялась 15 дням (табл. 40).
Максимальная растянутость сроков размножения в разные годы достигала здесь 20 дней. Наиболее ранний для Ленинградской области вылет птенцов из гнезда был отмечен нами 27—28 мая (1954), а наиболее-поздний— 16 июня (1947). Сравнение этих данных с соответствующими данными, приводимыми для других широт, показывает, что в. окрестностях Ленинграда ворона размножается относительно очень рано. Так,. по наблюдениям А. И. Иванова (1952), на юго-западном побережье Каспия строительство гнезд у этого вида начинается лишь в последней декаде марта; по указаниям С. К. Даля (1954), начало кладки серой вороны в Армянской ССР приходится на конец апреля — начало мая,
159 •
Таблица 40
Сроки размножения отдельных пар серых ворон в окрестностях Ленинграда
№ гнезда Год наблюдения Откладка яиц Вылупление птенцов Оставление гнезда птенцами
1 1947 Не прослежена 10-12 V Не прослежено
2 1947 а* М Не прослежено 15 VI
3 1947 12—14V 16 VI
4 1950 • » Не прослежено 4 VI
5 1954 3-7IV. 24-26 IV 27-28 V
6 1954 Не прослежена 5- 8V Не прослежено
7 1954 - 6- 9V 9-10 VI
8 1954 9 9 8-11 V Не прослежено
а вылет птенцов из гнезд происходит в начале июля. По данным А. П. Чмутовой (1953), самый ранний срок начала кладки серой вороны под Москвой — 7 апреля, а в районе Краснодара — даже 10 апреля. В Ленинградской же области кладка иногда может начаться уже в первых числах апреля. В самом Ленинграде нередко можно наблюдать, как уже < конца декабря—начала января вороны парами прилетают на ночевку в одно и то же место, и здесь же в дальнейшем у этой пары обычно появляется гнездо. Птицы либо строят гнездо на основе одного из старых, либо вьют его заново.
Весной 1954 г. представилась возможность проследить жизнь одной пары в течение всего гнездового периода. Первая ветка для этого гнезда была принесена воронами 15 марта. Первоначально постройка гнезда шла очень медленно, а в холодные дни, сопровождавшиеся снегопадом, даже совсем прекращалась. Однако каждый раз, как наступало потепление, птицы начинали вить гнездо активнее. Период строительства продолжался до 1 апреля. Начиная с 4 апреля самку можно было уже постоянно видеть сидящей на гнезде. Первый птенец вылупился утром 24 апреля, а последний (5-й) — вечером 26 апреля. Судя по дате вылупления птенцов, откладка яиц в гнезде началась 3 апреля.
Размеры и судьба кладок. Наблюдения по этому вопросу проводились лишь в Ленинградской области. Всего под постоянным наблюдением находилось 23 яйца (5 гнезд), из них вылупился 21 пте-. нец, 2 яйца оказались болтунами. Одно гнездо с 4 птенцами было разорено людьми. Таким образом, благополучно закончило развитие и вылетело из гнезд 17 птенцов, что составило в среднем 3—4 птенца на 1 гнездо.
Поведение взрослых птиц у гнезда. Во время насиживания кладки, а также в течение длительного последующего периода обогревания птенцов, когда самка редко слетает с гнезда, ее кормит самец. Иногда кормление происходит на гнезде, а иногда поблизости, на ветвях дерева. Во время приема пищи самка ведет себя, как птенец-слеток: трепыхает крыльями и громко каркает.
«Насиживание» птенцов — явление широко распространенное среди воробьиных. У серой вороны в связи с ранними сроками размножения оно имеет особенно большое значение для птенцов. В одном из известных автору случаях птенцы вылупились 24—26 апреля. Когда им было всего 2—4 дня от роду температура воздуха упала до —1°, выпал снег, который пролежал целые сутки. Все это время самка неотлучно находилась на гнезде, благодаря чему на состоянии птенцов такая погода никак не отразилась.
160
Поведение взрослых птиц при обнаружении человека у гнезда бывает различным. Некоторые вороны ведут себя весьма агрессивно: налетают, принимают угрожающие позы, иногда опрыскивают жидкими экскрементами, — точно так, как это делают дрозды-рябинники у своих гнезд. У других птиц наблюдается иной способ защиты своих птенцов: они начинают кругами летать над гнездом и громко каркать. На их крик обычно прилетают другие вороны, после чего птицы действуют уже сообща. По отношению к пернатому хищнику, например к тетеревятнику, такой способ защиты, видимо, более эффективен, нежели первый.
Развитие птенцов. Однодневный птенец серой вороны покрыт темно-серым пухом, довольно густым и длинным на спине (до 14 мм), плечах и бедрах и коротким на голове, особенно на затылке (около 4 мм). Пух .расположен на надглазничных (часть пушинок сидит на верхнем крае глазного яблока), затылочном, спинном (от лопаток до хвоста), плечевых, локтевых, бедренных и брюшном участках тела. На последнем пух очень редкий и короткий (2—3 мм). На местах будущих маховых и рулевых — редкие темные щетинистые пушины. Клюв бледно-розовый в основании и серый в вершинной половине. Ноздри мелкие и круглые. Яйцевой «зуб» и когти светлые. Кожа грязно-розового цвета. Ротовая полость мясо-красная или интенсивно розовая. Язык, как и у птенцов других врановых,-относительно короткий. Складка в углах рта бледно-мясного цвета, очень широкая.
Облик птенца начинает меняться с первых же дней постэмбрионального развития: боковая складка клюва увеличивается еще больше, изменяется форма ноздрей, цвет кожи, когтей и т. д. У 5-дневного вороненка ноздри становятся овальными, ширина боковой складки достигает 5,5 мм, когти становятся серыми, а кожа темнеет. Яркая окраска ротовой полости перед вылетом из гнезда тускнеет, хотя у молодых ворон примесь розового цвета во рту сохраняется еще и в октябре. В дальнейшем, по данным А. П. Чмутовой (1948), ротовая полость становится белой, а затем со второго года жизни начинает темнеть.
В гнезде, за которым велось систематическое наблюдение, птенцы находились 32 дня, считая со дня появления на свет первого птенца. В этом отношении наши наблюдения не расходятся с большинством литературных данных (Niethammer, 1937; Рустамов, 1954а), но, по А. П. Чмутовой (1955), вылет воронят из гнезда происходит на 28-й день. Первые дни после оставления гнезда птенцы обычно сидят около гнезда. Дня через 2—3 они уже могут перелетать с дерева на дерево.
Питание гнездовых птенцов. По питанию птенцов серой вороны автор располагает лишь материалами Н. П. Кадочникова, собранными на территории Савальского лесхоза весной 1952 г. Анализ содержимого 126 пищевых комков, принесенных воронами своим птенцам, и 3 желудков показал, что птенцы серой вороны так же, как и взрослые птицы этого вида, всеядны. Основу питания составляют, однако, различные мелкие беспозвоночные, в первую очередь насекомые. Второе местр в пищевом рационе воронят занимает мясо позвоночных. Растительный корм (семена пшеницы, подсолнуха, гречихи и других с.-х. растений) поедается ими сравнительно редко. Из 126 исследованных пищевых комков 61 комок состоял исключительно из беспозвоночных, 49 — из позвоночных или беспозвоночных животных вместе с позвоночными и только в 19 комках, помимо корма животного происхождения* был обнаружен также и растительный корм.
Животные, которых серые вороны добывают для своих птенцов, могут быть самой различной величины — от личинок муравьев, мелких жучков Harpalus, мелких гусениц зимней пяденицы или дубовой листо
11 А. С. Мальчевский
161
вертки до самых крупных у нас жуков-оленей, красотелов, прытких ящериц, птенцов дроздов, а также мышей, полевок и даже сусликов. Средин беспозвоночных, приносимых серыми воронами своим птенцам, преобладали жуки имаго, а среди позвоночных — рыба. Гусеницы бабочек*, а также личинки муравьев и мух хотя и поедаются в довольно значительном количестве, тем не менее по причине их мелких размеров существенного значения в пищевом рационе не имеют. Из жуков птенцы чаще всего получают крупные виды — майских жуков и их личинок, красо-телов (Calosoma inquisitor и С. sycophanta), различных навозников^ (Geotrupes stercorosus, Copris lumarisи др.), оленьков (Dorcus parallelo-pipedus), бронзовок и их личинок (Cetonia aurata) и др.; из бабочек — мелких гусениц пядениц и листоверток (Operophtera brumata,, Tortrix viridana) и др.
Очень часто ворона таскает яйца и птенцов дроздов-рябинников*, крупные гнезда которых заметны и доступны для нее. По сообщению В. А. Попова и др. (1954), в Татарии серая ворона разоряет более 50% гнезд этих дроздов. Контратаки последних нередко оказываются безуспешными. Приходилось также наблюдать, как серая ворона разоряла гнезда зяблика, серой мухоловки, дрозда-белобровика, горихвостки it других певчих птиц. Серая ворона может вытащить даже птенца скворца из скворечника^ Общеизвестен, наконец, вред, приносимый вороной? охотничье-промысловому хозяйству (Формозов, 1940; Теплов и Туров*. 1956), а также заповедникам, в частности Астраханскому (Дюнин*. 1936; Дубинин, 1940), где ворона питается за счет гнездовых колоний? цапель, караваек, а также гусей, уток, лысух и других ценных охотничьих птиц. Таким образом, в гнездовой период серая ворона является-врагом самых различных птиц; однако, повторяем, в лесу и в парках: чаще всего она беспокоит все же дроздов-рябинников.
СОРОКА (PICA PICA L.)
Ра сположение гнезда. Как в Ленинградской области, так: и на территории Савальского лесхоза гнезда сороки чаще всего мы находили на молодых соснах (12 находок из 19), а в более южных районах — в крупных колючих кустарниках. В Богдинских лесных полосах сорока гнездится, например, преимущественно на лохе. Как сосна, так: и лох по своей архитектонической структуре удобны для укрепления тяжелых, вымазанных внутри землей и глиной сорочьих гнезд. Последние обычно очень прочно* сидят на ветвях, и снять их бывает довольно трудно. Высота расположения найденных гнезд варьировала от 1,5 до 6 м. В некоторых местах, где сороку часто тревожат, она помещает свои постройки и на более значительной высоте. В литературе имеется указание на то, что сорока иногда проникает далеко в область безлесных болот, где гнездится в тростниках (Gerd, 1927). *
Сроки размножения. В наиболее ранних гнездах, найденных в Ленинградской области, откладка яиц происходила в начале последней, декады апреля, в наиболее поздних — в середине мая (табл. 41).
Таким образом, разница в сроках размножения отдельных пар сорок один и тот же 1954 г. равнялась 23 дням.
В Савальском лесхозе сроки размножения сороки были более растянуты. Здесь в наиболее ранних гнездах откладка яиц происходила в конце апреля, а вылет птенцов — в конце I декады июня (примерно тогда же, когда оставляли гнезда ранние выводки сорок в Ленинградской области). Наряду с этим свежие кладки сорок мы находили здесь
162
'Г а б л и ц a 41
Сроки размножения отдельных пар сорок в окрестностях Ленинграда
Год наблюдения Откладка яиц Вылупление птенцов Оставление гнезда птенцами
1954 22—29 IV 16-17 V 9 VI
1955 30IV-6 V Не прослежено Не прослежено
1955 1— 7 V -
1954 9—15 V 2-3 VI 27—2s VI
1955 13—19 V 5-6 VI Не прослежена
1954 15—19 V 5-6 VI 30 VI—1 VII
в I и II декадах июня. Неоднократно также в конце июля в лесу нам попадались еще не умеющие летать сорочата. Наиболее позднее гнездо со свежей кладкой было найдено 28 июня 1951 г. Столь значительную растянутость сроков размножения можно объяснить только лишь частыми случаями разорения гнезд сороки.
Размеры и судьба кладок. Количество яиц в 18 кладках сороки было следующим: в 2 — по 4 яйца, в 2 — по 5, в 7 — по 7. Из: находившихся под наблюдением на территории Савальского лесхоза 43 яиц сороки (8 гнезд) вылупилось 35 птенцов: 1 яйцо оказалось болтуном и 7 погибло при разорении одного из гнезд. Затем было разорено еще 1 гнездо с 7 птенцами. Таким образом, благополучно закончили развитие 28 сорочат, что составило в среднем 3—4 птенца на 1 гнездо.
Количество вылетающих из гнезд сорочат сильно варьирует (от 2 до 7), что объясняется прежде всего частым разорением гнезд, особенно в местах, населенных человеком. После разорения гнезда сороки приступают к повторному размножению. Если первые кладки, которые в средних широтах бывают в конце апреля—начале мая, содержат 6—7 яиц, то поздние (июньские) обычно 4—5.
Поведение взрослых птиц у гнезда. При подходе человека к гнезду сороки обычно поднимаются высоко в воздух или садятся поодаль на вершины высоких деревьев, откуда и ведут наблюдение, издавая беспокойное трещание. На человека они не нападают, даже если видят, что их птенец пойман.
Птенцы. Новорожденный птенец сороки совершенно голый (см. рис. 24). Кожа светло-оранжевая с розоватым оттенком. Клюв восковой с потемнением в области яйцевого зуба. Ноздри первоначально круглые, затем овальные. Яйцевой «зуб» белый, когти светлые. Роговая полость мясного цвета с оранжевым оттенком; складка в углах рта широкая, бледно-розовая. Однодневный птенец вялый, малоподвижный. Опираться на передние конечности и привставать во время кормления начинает лишь на 2—3-й день.
Ушные отверстия становятся заметными через 5 дней после вылупления, глаза приоткрываются в возрасте 6—7 дней, а трубочки маховых перьев раскрываются на 10—11-й день жизни птенца. Таким образом, если судить по перечисленным выше признакам, развитие сорочат по сравнению с мелкими видами воробьиных идет с запозданием всего на 2—3 дня. Позднее, в связи с более длительным периодом роста крупных перьев, разница в скорости развития начинает ощущаться сильнее; в среднем сорочата сидят в гнезде в 2 раза дольше, чем птенцы большинства мелких воробьиных птиц. В одном из гнезд птенцы находились
11*
163
23 дня, в других — 25 дней. По наблюдениям Ю. К. Эйгелиса (устное сообщение), в некоторых случаях птенцы сороки остаются в гнезде до 28-дневного возраста. Оставление гнезда происходит до приобретения способности к полету. Первое время сорочата сидят на крыше гнезда или поблизости на ветвях. При преследовании они, прыгая с ветки на ветку, стремятся забраться выше; попадая на землю, они могут легко стать жертвой наземных хищников. В Савальском лесхозе, например, молодыми сороками питаются лисицы, у нор которых постоянно валяются остатки уже вполне оперившихся птенцов.
Выводок сорочат обычно очень долго продолжает держаться в районе гнезда. Например, окольцованные автором 6 июля 1951 г. в Савальском лесхозе только что оставившие гнездо птенцы 3 августа были обнаружены в 500 м от гнезда. Аналогичные наблюдения были сделаны нами в районе Зеленогорска (Ленинградская область), где оставивший гнездо 30 июня 1954 г. выводок сорок продолжал держаться почти в том же самом месте вплоть до 3 августа.
Питание гнездовых птенцов. Питание птенцов сороки изучалось в 1940 г. в полезащитных лесных полосах полупустынной зоны в районе ст. Богдо, а также на территории Савальского лесхоза (1951—1953).1 Всего было исследовано 19 желудков и 130 порций корма, извлеченных из пищеводов птенцов. Анализ показал, что пища птенцов сороки менее разнообразна, нежели пища птенцов вороны. В ней почти полностью отсутствуют растительные корма; только в одном случае в желудке была обнаружена ягода шелковицы. Менее разнообразен и состав позвоночных, поедаемых птенцами. Не имея привычки посещать в гнездовой период берега водоемов, сорока не носит своим птенцам рыбу, как это делает ворона. Из позвоночных сорочата чаще всего получают ящериц, реже яйца и птенцов мелких птиц. Так, например, одним из основных кормов птенцов сороки в полупустынных районах южного Заволжья является разноцветная ящурка (Eremias arguta) (Мальчевский, 1947). Ящерицы оказались характерным кормом сорок в первой половине лета также и по наблюдениям Б. А. Голова и В. И. Осмоловской (1955), проводивших исследование на трассе лесных полос между Камышиным и Сталинградом. Весьма характерным кормом гнездовых птенцов сороки в степном Центрально-черноземном государственном заповеднике им. В. В. Алехина оказалась веретенница (Anguis fragilis) (Елисеева и Хватова, 1957). По данным И. В. Покровской (1956), птенцы сороки иногда получают ящериц и в Ленинградской области.
Проводившийся автором специальный учет «птичьих переполохов» показал, что сорока значительно реже беспокоит лесных птиц у их гнезд, чем ворона или сойка. Она собирает корм преимущественно среди мелколесья или на открытых местах, где в большем количестве, чем птиц, находит ящериц. Среди многочисленных данных по питанию сорок, собранных Б. А. Головым и В. И. Осмоловской, остатки птиц были обнаружены в ничтожном количестве (в 4 желудках из 275 изученных). В Савальском лесхозе птенцы и яйца птиц приносились сорочатам все же несколько чаще (в 10 из 130 порций корма).
Из беспозвоночных сорока чаще всего собирает жуков, затем гусениц или коконы бабочек. Других насекомых, а также пауков, червей, моллюсков и многоножек она ловит для своих птенцов сравнительно редко. В районе Богдо среди жуков, поедавшихся птенцами сороки,
1 Помимо автора в сборе материалов принимали участие Н. П. Кадочников, Г. И. Баранова и И. В. Прокофьева.
164
преобладали хлебные жуки (Anisoplia segetum), в Савальском лесхозе в 1952 г. — майские жуки. Кроме того, сорока часто ловит жужелиц. (Carabus, Harpalus), навозников (Scarabeus, Copris и др.) и различных долгоносиков (Psalidium, Bothynoderes и др.). В наибольшем количестве приносились птенцам гусеницы или коконы соснового, кольчатого и непарного шелкопрядов (Савальский лесхоз), а также гусеницы бабочки Dicranura vinula (Богдо).
В пище сорочат мы обнаруживали фаланг (Galeodes sp.), шпанских мушек (Litta vesicatoria), гусениц кольчатого шелкопряда (Malacasoma neustria), а также жалящих перепончатокрылых (Apidae); чаще, чем птенцы других птиц, сорочата получают муравьев и их личинок.
СОЙКА
(GARRULUS GLANDARIUS L.)
Токование. Так называемое весеннее «пение» сойки — ее негромкое подражательное скрипение — воспроизводится птицей крайне нерегулярно. В Ленинградской области «песню» сойки можно услышать в апреле — начале мая. Осеннее же «пение» автор слышал всего один раз — 25 августа. Весной сойки иногда издают особые короткие и резкие звуки, совершенно непохожие на их обычные крики. При этом они гоняются друг за другом и бывают сильно возбуждены. Судя по поведению птиц, этот своеобразный позыв имеет, видимо, брачное значение. Весеннее возбуждение соек длится, однако, недолго. С началом строительства гнезда и откладки яиц они становятся, наоборот, чрезвычайно скрытными и молчаливыми.
Расположение гнезда. Из 27 гнезд сойки, найденных на территории Савальского лесхоза в 1951—1953 гг., большинство помещалось на соснах (10 находок) и на кустах бузины (6). Гнезда этого вида были обнаружены также на дубе (4), яблоне (2), березе, вязе и липе. 2 гнезда сойки, принадлежавшие одной и той же птице, были обнаружены Н. П. Кадочниковым (195-7) в углу стропил на балке пожарной, вышки, построенной среди леса.
Большинство (20) гнезд располагалось на высоте от 1,5 до 2 м, остальные помещались в 3”-7 м от земли. Приуроченность гнезд к подлеску и нижним частям крон деревьев в Савальском лесхозе следует, по-видимому, объяснять наличием там большого количества опасных для яиц ц птенцов соек пернатых хищников. В отдельных случаях сойка, гнездится здесь даже в дуплах деревьев (устное сообщение Ю. К. Эйге-лиса). Случаи гнездования соек в дуплах известны также в Англии (Tutt, 1953; Dereck, 1953).
Сроки размножения. Период размножения у сойки растянут очень сильно. В Савальском лесхозе разница в сроках откладки яиц в отдельных гнездах в течение одного сезона 1953 г. составляла примерно 1,5 мес.; в наиболее раннем из найденных в этом году гнезде первое яйцо появилось 3 мая, а в наиболее позднем — 17 июня (табл. 42).
В 1952 г. Ю. К. Эйгелисом было найдено гнездо, в котором, судя по возрасту находившихся в нем птенцов, вылупление происходило 16—18 мая. Следовательно, откладка яиц в этом гнезде началась в последних числах апреля.
Размеры и судьба кладок. Количество яиц в известных автору кладках сойки было следующее: в 5 — по 4 яйца, в 2 — по 5, в 4 — по 6, в 5 — по 7, в 1—8 и в 2 — по 9. Отход яиц и птенцов в гнездах оказался весьма значительным. В Савальском лесхозе под постоянным наблюдением находилось 16 гнезд. Из 96 находившихся в этих
165
Таблица 42
Сроки размножения отдельных пар соек на территории Савальского лесхоза в 1953 г.
Откладка яиц Вылупление птенцов Оставление гнезд птенцами
3-11 V Не прослежено Гнездо было разорено
19-23 V 7— 8 VI 27-28 VI
30 V—4 VI 22—23 VI 12 VII
5— 9 VI : 5—26 VI 15 VII
' 7 — 12 VI 28-29 VI Гнездо было разорено
13-17 VI 2— 3 VII 21 VII
17-23 VI 8- 9 VII Гнездо было разорено
гнездах яиц вылупился 61 птенец: 25 яиц было растащено хищниками и ребятами, одна кладка из 7 яиц была брошена сойкой и 3 яйца оказались болтунами. Благополучно вылетело из гнезд только 39 птенцов,
Рис. 44. Сойка на гнезде в момент вылупления птенцов. Открытый рот указывает на сильный испуг (Савальский лесхоз, июнь 1952 г. Фото автора).
остальные погибли от различных хищников, разоривших гнезда. Таким образом, всего лишь 40% яиц превратилось в конечном счете в летных птенцов. Количество вылетающих из гнезд птенцов в наблюдавшихся нами случаях варьировало от 3 до 7. Основная масса яиц и птенцов гибнет от хищников, часто разоряющих гнезда соек. В ряде случаев было отмечено, что птенцов сойки утаскивали сороки. Одно гнездо было разорено сарычом.
Поведение взрослых птиц у гнезда. В гнездовой период сойки ведут себя обычно очень скрытно, в связи с чем на глаза попадаются редко (рис. 44). Заслышав, однако, громкий крик встревожен-
166
пых птенцов, они обычно сразу же прилетают к месту происшествия. Поведение их при этом часто бывает агрессивным. Они. отваживаются нападать даже на человека.
Птенцы. Однодневный птенец совершенно голый, с желтовато-ро--зовым цветом кожи, в возрасте 4—5 дней приобретающий желтовато-зе--леный налет. Клюв розоватый с потемнением на конце. Ноздри мелкие, круглые, позднее с заметным сужением спереди. Яйцевой «зуб» светлый; когти светло-серые. Ротовая полость, первоначально розовато-мясного цвета, к недельному возрасту становится почти малиновой. Складка в углах рта белая со слабым розовым налетом и довольно широкая (у недельного птенца до 3,5 мм). Язык относительно очень короткий и широкий.
На 1-й день после вылупления птенцы сойки, как,впрочем, и птенцы других врановых птиц, бывают очень вялыми, открывают рот, по большей части не поднимая головы. На 2-й день птенец уже в состоянии .держать голову и опираться на передние конечности. На 5-й день, когда у соечат начинают открываться слуховые проходы, а на крыле появляются уже заметные трубочки 'маховых, активность птенцов возрастает: при выпрашивании пищи они принимают вертикальную позу и подолгу держат сильно вытянутую кверху шею. Движения их, однако, продолжают оставаться вялыми и медлительными. Глаза приоткрываются на 6-й день. Пеньки маховых перьев раскрываются в возрасте 9 дней, и рулевых — 2 днями позднее. К этому времени птенцы становятся более активными: при выпрашивании корма высоко поднимаются на ногах, широко раскрывают рот и сильно вытягивают шею, мерно и очень долго покачивая ею из стороны в сторону. При этом вплоть до вылета из гнезда птенцы не издают громких криков, а лишь слабо попискивают. "Таким образом, кормление птенцов сойки протекает обычно в обстановке почти полной тишины, что имеет, видимо, немаловажное значение для успешного их воспитания.
В возрасте 11—12 дней птенцы сойки уже многим напоминают взрослых птиц: уже хорошо заметен голубой цвет на перьях крыла, становится белым надхвостье и т. п. 2-недельные птенцы все еще остаются доверчивыми. В возрасте 16—17 дней поведение их резко меняется: увидя человека, они прижимаются к дну гнезда, сильно втягивают в себя шеи, приоткрывают рот (см. рис. 33), при этом они иногда закрывают глаза и запрокидывают голову назад (поза страха). При первом прикосновении к ним они, как по команде, хором начинают, кричать очень громко, Протяжно и неприятно.
Птенцы сойки сидят в гнезде обычно 19—20 дней, но, будучи сильно обеспокоенными, могут выскочить из гнезда на 1—2 дня раньше. Гнездо покидается, когда птенцы не умеют еще летать. Крыло в этом возрасте еще не может служить для полета, так как слагающие его перья не образуют сплошной несущей поверхности. Но лапы их к этому времени становятся очень крепкими и цепкими, благодаря чему птенцы .могут держаться даже на тонких веточках. Первое время соечата сидят на ветвях поблизости от гнезда, а затем, перепархивая с ветки на ветку, выводок начинает перемещаться. В Савальском лесхозе излюбленным местом пребывания птенцов-слетков являются густые сосновые жердняки. На ночь птенцы одного выводка соединяются. Один из выводков сойки, окольцованный нами 7 июля 1952 г., когда птенцам было 19—20 дней от роду, 11 июля вечером был обнаружен в гнезде сороки в 50 м от своего гнезда. Эти птенцы, возраст которых равнялся 23—24 дням, Могли уже перелетать от дерева к дереву, не теряя высоты. Полет их, однако, был еще неуверенным и напряженным.
167
Питание гнездовых птенцов. Н. П. Кадочников и Ю. К-Эйгелис (1954) детально исследовали питание птенцов сойки в условиях Савальского лесхоза, в связи с чем мы здесь подробно не будем останавливаться на этом вопросе. Укажем лишь, что птенцы сойки так же,, как и птенцы сороки, в основном жцвотноядны, растительную пищу получают чрезвычайно редко. Из позвоночных животных родители при* носят главным образом яйца и птенцов мелких лесных птиц. В лесостепных борах, где количество гнездящихся соек весьма значительно, они доставляют много беспокойства целому ряду мелких певчих птиц и в-первую очередь зябликам и серым мухоловкам, располагающим свои гнезда открыто на деревьях^ Особенно часто гнезда разоряются сойками в конце июня и в июле, когда в большом количестве появляются их выводки, кочующие в поисках корма.
Из беспозвоночных в условиях Савальского лесхоза птенцы чаще всего получают гусениц и куколок различных бабочек (преимущественно шелкопрядов) и крупных (весом до 1 г) пауков. В противоположность сороке и вороне жуки, а также муравьи поедаются редко. В незначительном количестве приносятся птенцам также мухи (главным образом ктыри), прямокрылые, клопы и стрекозы. В пище птенцов сойки были отмечены также наземные моллюски.
Таким образом, из всех врановых, питание птенцов которых мы рассмотрели, сойка является наиболее специализированным видом, охотящимся главным образом в кронах деревьев и собирающим малоподвижную добычу.
СКВОРЕЦ. ОБЫКНОВЕННЫЙ (STURNUS VULGARIS L.)
Токование. Период регулярного пения у скворцов длится недолго. В Ленинградской области в зависимости от сроков прилета пение начинается либо с конца марта, либо с начала апреля, достигает наибольшей интенсивности во второй половине апреля — начале мая и уже в двадцатых числах мая почти полностью прекращается. Период наибольшей интенсивности пения совпадает с массовым пролетом и прилетом самок, а момент затухания совпадает с вылуплением птенцов. В конце мая—начале июня поют только отдельные, видимо, холостые или потерявшие пару самцы. Такое запоздалое пение отдельных самцов приводит иногда к образованию пар с настолько поздним сроком размножения, что часто даже возникает вопрос, не имеем ли мы дело со* второй кладкой.
Расположение гнезда. В Ленинградской области помимо-скворечников и естественных дупел данный вид нередко поселяется в изъянах стен каменных построек. Таким образом, в отношении степени пластичности гнездового инстинкта северные популяции, очевидно, мало* отличаются от скворцов степных районов европейского юга и северного* Казахстана, где, как известно, колонии скворцов иногда размещаются в трещинах и пустотах различных построек, оврагов и т. п. местах.
Сроки размножения. В условиях Ленинградской области двойного цикла размножения у обыкновенного скворца, видимо, не бывает. На это прежде всего указывает отсутствие второго подъема интенсивности пения самцов. Наличие вторых кладок у скворцов не подтверждается также результатами наблюдений за сроками размножения этих птиц. Например, массовый вылет скворчат из гнезд в окрестностях Ленинграда происходит в зависимости от хода весны между 2 и 20 июня (табл. 43). Сроки массового вылета в обычные годы являются по суще*
168
Таблица 43
Сроки массового вылета скворчат из гнезд в окрестностях Ленинграда
Год наблюдения Даты массового вылета Характер весны
1946 7-9 VI Долгое таяние снегов вследствие многоснежной
зимы; температурные условия без особых аномалий.
1950 3— 5 VI Дружная и довольно теплая
1952 2— 3VI m * *
1954 9—12 VI Очень затяжная ’ и холодная, особенно в 1955 г.,
1955 16—20 VI с резким падением температуры в конце апреля, мае-
и начале июня
Примечание. В 1955 г. часть скворцов, видимо, не гнездилась совсем; в конце мая—начале июня наблюдались небольшие группы старых негнездившихся. птиц.
ству и сроками наиболее1раннего вылета скворчат. Вовремя массового1 вылета скворчат из гнезд шочти всегда можно наблюдать, что некоторые скворцы продолжают кормить еще своих птенцов, а иногда даже насиживать кладку. Наиболее поздний вылет птенцов был зарегистрирован автором 2 июля (1954), спустя примерно 20—22 дня после того, как основная масса скворцов покинула свои гнезда. Подобная разница крайних дат размножения отдельных пар в Ленинградской области не дает основания предполагать у скворцов наличия двух кладок в сезон.
Гораздо труднее решить вопрос о возможности вторых кладок у скворцов в средних широтах, где размножение еще более растянутое и где разница в крайних сроках размножения отдельных пар еще значительнее (табл. 44). Основываясь на поздних (июньских) находках све-
Таблица 44
Сроки вылета птенцов скворцов из гнезд на территории Савальского лесхоза
Год наблюдения Наиболее ранние даты Массовый вылет Наиболее поздние даты
1951 30V 30—31 V 8 VII
1952 30V 30 V-l VI 7 VII
1953 31 V 3—4 VI 8 VII
жих кладок скворцов, а также на случаях повторного заселения этими, птицами искусственных гнездовий или естественных дупел, многие авторы (Сомов, 1897; Шнитников, 1913; Schilling, 1927; Аверин, 1941; Та-ращук, 1953; Спангенберг, 1954, и др.) предполагают наличие двух нормальных кладок у скворца. Е. П. Спангенберг и И. М. Олигер (1949) приходят к заключению, что лишь некоторая часть скворцов, гнездящихся в Дарвинском заповеднике, выводит птенцов 2 раза в лето. Однако работавшему здесь позднее с закольцованными скворцами В. М. Поливанову (19576) этого установить не удалось. По данным Шнейдера (Schneider, 1927, 1952), в Германии скворцы гнездятся нормально один
169»’
раз в лето, но в отдельные годы около 50% самок дают вторые кладки часто в том же скворечнике. Такая же картина наблюдается в некоторых местах и на юге нашей страны, например в районе Черноморского заповедника (устное сообщение Т. Б. Ардамацкой). В табл. 44 приводятся результаты наблюдений за сроками вылета молодых скворцов из гнезд на территории Савальского лесхоза в 1951—1953 гг. Эти данные, как нам кажется, не позволяют говорить о двойном цикле размножения скворцов в условиях Воронежской области.
Позднее гнездование части особей иногда объясняют нехваткой подходящих мест для размещения гнезд. Со справедливостью этого утверждения нельзя не согласиться. Эта нехватка особенно должна ощущаться в южных и средних широтах, где в силу прекрасных кормовых условий концентрируется много скворцов, а угодья, пригодные для гнездования этих птиц, территориально ограничены. Н. П. Кадочников сообщил нам, что в 1952 г. на территории Савальского лесхоза часть скворечников была вывешена в середине и в конце мая, т. е. в период, непосредственно предшествовавший вылету скворчат из гнезд. Несмотря на это, нашлись птицы, которые заняли эти гнездовья, построили гнезда и отложили яйца. Таким образом, случаи позднего размножения известной части скворцов в Савальском лесхозе скорее всего следует объяснять недостатком подходящих мест для гнездования.
Поведение взрослых птиц у гнезда. В день вылета скворчат из гнезд взрослые птицы ведут себя очень беспокойно. Они беспрестанно кричат, вьются над скворечником или дуплом, присаживаются к летку и вновь отлетают к птенцам, уже покинувшим гнездо. Птенцов в это время они почти не кормят. Птицы подлетают к скворечнику, держа пищу в клюве, но не отдают ее птенцам, а только показывают и вновь улетают. Эти действия взрослых птиц трудно расценить иначе, как «выманивание» птенцов из дупла или скворечника.
В первые дни после массового вылета птенцов, что бывает обычно в начале июня, около опустевших дупел и скворечников часто можно наблюдать появление взрослых птиц, которые ведут себя очень возбужденно: заглядывают в отверстие, иногда поют и таскают внутрь гнездовой материал. Вскоре, однако, большинство этих птиц исчезает, но некоторые из них остаются и приступают к размножению.
Описанное выше поведение скворцов, наблюдающееся в начале июня, внешне очень 1напоми.нает явление осеннего (августовского или сентябрьского посещения ими мест гнездования, неоднократно отмечавшееся в литературе (Гавриленко, 1929; Иванаускас, 1953, и Др.). Осеннее повышение активности скворцов никогда, однако, не приводит к образованию пар и откладке яиц. Его, очевидно, следует расценивать как явление, сходное с осенним токованием птиц, в то время как июньское возбуждение — как поведение, связанное с их запоздалым размножением.
Птенцы. Птенец покрыт довольно густым светло-серым пухом, длина которого варьирует от 12 до 18 мм (на спине). Пух расположен на надглазничных, затылочных, спинном, плечевых, локтевых, бедренных участках тела; едва заметный пух имеется также на брюшной стороне и голени. Кожа бледная, слабо желтоватая. Клюв плоский, светло-желтый, с незначительным потемнением на конце и сильно вздутый в области ноздрей, имеющих овальную форму. Ротовая полость ярко-желтая. Складка в углах рта светло-желтая, очень широкая, особенно на нижней челюсти, которая несколько выдается вперед. Размеры боковой складки клюва увеличиваются с возрастом от 2,5 (новорожденный птенец) до 5 мм (5-дневный). Поданным Портмана (1938), максимальные
170
(размеры боковых выростов клюва наблюдаются у птенца скворца в возрасте 7—8 дней. Позднее эта складка начинает постепенно уменьшаться. Из приспособлений к закрытому гнездованию у птенцов скворца следует отметить еще наличие пяточной мозоли и чрезвычайно большую активность птенцов, начиная с первого же дня их жизни.
Слуховые проходы открываются раньше, чем глаза, но разница в сроках развития этих органов у скворца более значительная, чем у открыто гнездящихся видов. 4-дневный скворчонок имеет уже открытые слуховые проходы, глаза же у него начинают приоткрываться лишь в возрасте 7—8 дней. Еще сильнее отстает развитие пера. Трубочки маховых перьев раскрываются только в 10—11-дневном возрасте. Оставление же гнезда у скворца происходит на 21-й день, т. е. на 8—9 дней позднее, чем, например, у равных ему по величине дроздов. Правда, оставляя гнездо, птенцы скворца уже в состоянии хорошо летать, в то время как дроздята приобретают способность к полету спустя несколько дней.
Сразу же после оставления гнезд выводки начинают перемещаться, откочевывая в поля и речные поймы. Эта смена стаций особенно бросается в глаза в годы с дружными сроками размножения скворцов. .Местность, сегодня изобилующая птицами, на следующий день совершенно пустеет.
В результате столь стремительной откочевки скворцов некоторые отставшие в развитии птенцы бывают брошены своими родителями в гнезде.‘Пытаясь присоединиться к выводку, они выскакивают из гнезда, но, будучи не в состоянии лететь, падают на землю, где становятся жертвой голода или хищников.
В целом выводки у скворцов дружные. Это обусловливается тем, что скворцы редко приступают к насиживанию яиц до окончания кладки. Поэтому вылупление птенцов у них, как правило, происходит в сжатые сроки и растягивается не больше, чем на сутки. Только при запоздалом гнездовании скворцов нам приходилось наблюдать более значительную разницу в сроках вылупления птенцов одного выводка (до2 суток).
Питание гнездовых птенцов. По питанию обыкновенного скворца в гнездовой период в настоящее время в литературе накоплен довольно значительный материал, указывающий на способность скворцов в большом количестве уничтожать насекомых—массовых вредителей не только сельского, но и лесного хозяйства. Имеющиеся по ^этому вопросу литературные данные достаточно полно сведены В. И. Осмоловской и А. Н. Формозовым (1950). Подробно касаться данного вопроса здесь мы не будем. Укажем лишь, что из массовых вредителей леса скворцы, гнездившиеся в Савальском лесхозе в' 1953 г., в большом количестве приносили своим птенцам майских жуков, гусениц зимней пяденицы, и дубовой листовертки. По количеству экземпляров эти вредители составляли примерно половину всех беспозвоночных, извлеченных из пищеводов птенцов. В довольно значительном количестве приносились птенцам также крупные комары (Tipula). Остальные насекомые, а также пауки и многоножки поедались птенцами в ограниченном количестве. Таким образом, несмотря на весьма разнообразный состав беспозвоночных, приносимых скворцами своим птенцам, основу питания последних все же составляли массовые виды вредителей леса. Дождевые черви в южных районах поедаются, видимо, редко. На севере же, например в Ленинградской области, скворцы приносят их птенцам до-ч.цольно часто, особенно в прохладную и дождливую погоду. Излюбленным кормом скворцов, как известно, являются саранчовые, однако в свя-ДЗиГс раннцми сроками гнездования птенцы скворца могут получать их j дшшь в; случае запоздалых кладок.
171
Кормовой рацион скворчат позднего вывода помимо присутствия в нем прямокрылых иногда отличается еще и наличием известного количества растительной пищи, в частности ягод. В конце июня — начале-июля 1949 г. скворцы, гнездившиеся в заповедной дубраве «Лес на Вор-скле», приносили своим птенцам ягоды вишни. На земле под некоторыми гнездами мы находили и косточки вишен, попавшие сюда вместе с экскрементами птенцов. Интересно, что в некоторых косточках, поврежденных вишневым долгоносиком (Rhynochites auratus), нами были найдены его живые личинки. Это указывает на то, что ягоды вишни в желудке птенцов долго не задерживаются.
ИВОЛГА ОБЫКНОВЕННАЯ
(ORIOLUS ORIOLUS L.)
Токование. Самцы иволги поют в течение почти'всего периода пребывания их в гнездовой области. В окрестностях Ленинграда наиболее ранняя дата весеннего пения иволги отмечена 15 мая (1949), а наиболее поздняя— 12 августа (1946). По данным Г. А. Новикова (устное сообщение), в «Лесу на Ворскле» самое раннее пение этого вида наблюдалось 4 мая (1941), а наиболее позднее — 8 августа (1946). В конце-июля и в начале августа поют лишь немногие птицы и то преимущественно по утрам.
Интересно, что интенсивность пения самцов не понижается по мере-вылупления и роста птенцов в гнездах, как это характерно для большинства певчих птиц. В период вылета птенцов из гнезд пение этих птиц раздается иногда даже чаще обычного. На территории Савальского лесхоза такой подъем интенсивности пения ежегодно наблюдался нами в. последней декаде июля.
«Сигналом отпугивания», предупреждающим о занятости гнездового участка, пение иволги, как это иногда полагают, служить не может. Несмотря на то, что иволги гнездятся всегда на известном расстоянии одна пара от другой, они тем не менее не имеют определенных гнездовых (кормовых) участков. Быстрый сильный полет позволяет им удаляться от гнезда на весьма значительные расстояния. В сосняках Савальского лесхоза в разгар гнездового периода автору неоднократно-приходилось отмечать концентрацию иволог (до 10 поющих самцов на на 1 га) на участках леса, изобилующих гусеницами соснового шелкопряда. Видно было, как птицы прилетали сюда из разных мест. Во время кормежки самцы продолжали громко петь.
Расположение гнезда. Хвойные и лиственные породы используются иволгой для гнездования одинаково часто. На это в свое-время обращал внимание В. Н. Шнитников (1913). В борах, где иволга бывает иногда весьма многочисленной, она селится преимущественно на соснах, в дубравах — на дубах, кленах и липах, в садах и парках — на соснах, кленах, липах, осинах, тополях. На соснах иволга гнездится, как правило, высоко, в верхних частях крон, не на лиственных породах гнездо ее располагается иногда низко. Одно из гнезд, найденных в-«Лесу на Ворскле», было помещено на нижней ветке молодого клена-, всего лишь в 1,5 м от земли.
Сроки размножения. Сроки размножения у иволги растянуты значительно меньше, чем у большинства других лесных воробьиных нашей фауны. В Савальском лесхозе максимальная разница в датах откладки яиц по 12 гнездам за 3 года наблюдений (1951—1953)-равнялась примерно 20 дням. В наиболее раннем гнезде яйца появились-в самых последних числах мая, а в наиболее позднем — в конце II де-172
кады июня (табл. 45). Сравнительно дружные сроки размножения являются, очевидно, следствием дружных и сжатых сроков прилета.
Таблица 45
Сроки размножения отдельных пар мволог на территория Савальского лесхоза в 1951—1953 гг.
Год наблюдения Откладка яиц Вылупление птенцов Оставление гнезда птенцами
1952 29-31 V 13—14 VI 28 VI
1953 31 V-2 VI 16-17 VI Не прослежено
1953 2— 4 VI 19-20 VI л* m
1952 3- 6 VI 20-21 VI
1953 5- 7 VI Не прослежено 9 Я
1951 5- 8 VI 22-23 VI 7 VII
1951 6— 9 VI 22-23 VI Не прослежено
1953 7- 9 VI 22—23 VI 6— 7 VII
1953 10—12 VI 25—26 VI 9—10 VII
1953 13-16 VI 30 VI—1 VII 14—15 VII
1951 17-19 VI 3 VII 17 VII
1951 17-19 VI 3—4 VII 18 VII
Размеры и судьба кладок. У автора имеются данные по «истории 14 гнезд иволги, найденных в лесостепной зоне. В этих гнездах находилось 49 яиц, из которых вылупилось 44 птенца. Отход яиц был незначительным: 2 яйца оказались болтунами и 3 яйца (1 кладка) пропало вместе с гнездом во время лесохозяйственных работ в лесу. Два выводка (7 гнездовых птенцов) погибло от пернатых хищников. Общеё количество птенцов, покинувших гнезда, равнялось 35. Размеры выводка в момент оставления гнезда варьировали от 2 (1 гнездо) до 4 (3 гнезда) птенцов. В остальных 7 случаях выводок состоял из 3 птенцов. Первоначальное количество яиц в кладках чаще всего равнялось 3 (8 случаев) или 4 (5 случаев). Только 1 кладка состояла из 5 яиц.
Малое по сравнению с другими певчими птицами количество яиц .в кладке у иволги, 'возможно, следует объяснять особенностями расположения гнезда, помещающегося, как известно, на концах тонких ветвей деревьев. При небольших размерах и постоянно качающемся открытом гнезде вряд ли возможно успешное воспитание большого потомства.
Поведение взрослых птиц у гнезда. При появлении человека некоторые птицы проявляют большую осторожность и не подлетают даже тогда, когда птенцы, взятые в руки, испуганно кричат. Другие, наоборот, активно защищают не только птенцов, но и гнездо с яйцами. В таком случае птица молча пикирует на врага, взмывает над самой его головой, издавая в это время резкий, неприятный крик. Поведение как бы рассчитано на то, чтобы испугать неприятеля.
Птенцы. Птенец покрыт светло-желтым пухом, довольно коротким и тонким, но густым. Пух представлен на глазных, надглазничных, затылочной, спинной, плечевых, локтевых, брюшных и иногда на голен-ных птерилиях. Цвет кожи светло-оранжевый, клюв восковой. Ноздри продолговатые, яйцевой «зуб» и когти белые. Ротовая полость розовая, с возрастом становится более яркой. У новорожденного птенца обращают на себя внимание относительно крупные-передние и задние конечности. Кисть равна предплечью. Короткая и толстая цевка, очень широкая подошва лап, а также чрезвычайно острые и круто загнутые
173
когти указывают на наличие приспособлений к древесному образу жизни-уже в птенцовом возрасте. Ширина подошвы ногу птенца-слетка иволги: равна 6,5 мм.
По сравнению с птенцами, например, дроздов развитие иволги идет несколько более медленными темпами, запаздывая примерно на сутки. Слуховые проходы открываются в конце 4-го — начале 5-го дня. Глаза приоткрываются через 6—7 дней после вылупления, а пеньки маховых перьев начинают развертываться лишь в возрасте 8 дней. На 11—12-й день жизни птенцы иволги еще очень доверчивы.
Способ выпрашивания корма напоминает таковой у птенцов славок: они сильно вытягивают шею и производят частые колебательные движения головой. При этом птенцы издают очень слабый писк. Непосредственно перед оставлением гнезда и долгое время после этого голос птенцов при выпрашивании корма бывает громким, представляя собою резкие выкрики «ки-ки-ки». Этот звук они издают также и прииспуге* Будучи же внезапно схваченными, птенцы иволги кричат «кошкой», как., кричат взрослые птицы при очень сильном беспокойстве.
На 12—13-й день поведение птенцов резко изменяется: увидя человека, они прижимаются ко дну гнезда и иногда выскакивают из него. Нормальное оставление гнезда наступает на 14—15-й день. Поскольку в этом возрасте птенцы еще не могут летать, особое значение имеет для. них цепкость лап, так как в это время они обитают высоко в кронах на* тонких качающихся ветвях. Способность к полету приобретается примерно на 19—20-й день их жизни. Так, в заповеднике «Лес на Ворскле» птенцы иволги, покинувшие гнездо 27 июня, через 5 дней могли уже, правда с большим трудом, перепархивать с дерева на дерево.
Питание гнездовых птенцов. Все отобранные от птенцов--насекомые и другие беспозвоночные отличались крупными размерами. Среди них отметим чаще всего встречавшихся крупных (до 5 см в длину) гусениц соснового шелкопряда, очень крупных гусениц липового бражника, цикад (Cicadetta), пауков-крестовиков (до- 1,2 г весом) и, наконец, майских хрущей, дававшихся птенцам без голов и с отломанными надкрыльями. Моллюски, принесенные птенцам, также отличались-крупными размерами; хотя раковины их и разламывались птицами, тем* не менее они тоже отдавались птенцам. Мохнатые гусеницы соснового шелкопряда отдавались птенцам в «обработанном» виде — покрытыми слизью и с оторванной головой, благодаря чему колючие гусеницы превращались во влажные мягкие комки и легко могли проглатываться' птенцами.
В желудках птенцов, добытых 7 мюля 1939 г. в Тимашевских полосах Куйбышевской области, тоже были обнаружены преимущественно' крупные гусеницы бабочек: ильмового ногохвоста, лунки серебристой, липового бражника и др., а также цикады (Cicadetta montana). Таким* образом, характерной чертой питания птенцов иволги является заглатывание крупных экземпляров беспозвоночных. Иногда птенцы получают растительный корм, в частности ягоды вишни (Осмоловская и Формозов, 1950).
Так же, как и многие другие насекомоядные птицы, иволга изредка1 приносит своим птенцам комочки земли, камешки, а иногда и кусочки: стекла.
ДУБОНОС ОБЫКНОВЕННЫЙ (COCCOTHR AUSTE S СОССОTHRAUSТЕS L.)
Расположение гнезда. На территории Савалъского лесхоза-из 13 находок гнезд дубоноса 5 было в кустах бузины, 5 — на взрослых.
174
и молодых дубах, 3 — на грушах и небольших деревцах вяза. Высота’ расположения гнезд варьировала в пределах от 1,5 до 8 м.
Сроки размножения. Наблюдения С. И. Божко (1954), а также наблюдения автора, проводившиеся в Савальском лесхозе в 1951—1953 гг., показали, что период размножения у дубоноса растянут довольно сильно. Разница сроков появления яиц в разных гнездах в течение одного сезона оказалась равной 36 дням. Так, в 1951 г. некоторые пары приступили к откладке яиц в начале II декады мая, в то время как в наиболее поздних гнездах кладка началась только лишь в конце II декады июня. В 1952 г. 16 июля автор встретил выводок не умеющих еще летать птенцов дубоноса, только что оставивших гнездо. Судя по возрасту птенцов, они вышли из яиц, отложенных в самом конце II — начале III декады июня.
Размеры и судьба кладок. Под постоянным наблюдением находилось 10 гнезд, в которых первоначально было 41 яйцо. Количество1 яиц в кладках было следующим: в 4 кладках по 5, в 4 — по 4, в 2 — по 3 и в 1 — 2 яйца. 2 яйца (неполная кладка) были брошены после того, как гнездом завладел сорокопут-жулан, подправивший постройку и начавший в нем откладку своих яиц; в 2 яйцах оказались мертвые эмбрионы. Таким образом, вылупилось 37 птенцов, 30 из них благополучно закончили развитие, а 7 были кем-то утащены из гнезд. Есть основания предполагать, что воровством птенцов занимается гнездящийся в сходных стациях сорокопут-жулан.
Поведение взрослых птиц у гнезда. В гнездовой период взрослые птицы обычно очень скрытны и осторожны. Их трудно-увидеть даже у гнезда с птенцами. Свое беспокойство они выражают резким двойным или тройным цыканьем или коротким взвизгиванием, при этом, однако, держатся в стороне и близко не подлетают. Только* однажды автору удалось наблюдать «потерявшего самообладание» дубоноса, который с громким цыканьем летал у самой земли возле человека, поймавшего его птенца. По наблюдению. Г. А. Новикова, а также Н. П. Овчинниковой, в «Лесу на Ворскле» некоторые насиживающие яйца самки подпускали к гнезду, не слетая с него, на 1—1,5 м.
Птенцы. Однодневные птенцы дубоноса покрыты густым пухом белого цвета. Первое время они имеют вид белых пуховых комочков. Пух расположен на спине, на плечах, предплечье, на бедрах, голени (очень немного) и брюшной стороне. Воскового цвета клюв очень массивный, резко вздутый в области круглых ноздрей. Яйцевой «зуб» белый, когти светлые. Окраска ротовой полости разноцветная: малиновокрасная со светлыми, голубоватыми и фиолетовыми участками.
Однодневные птенцы дубоноса довольно вялые. Однако на 2-й день-активность их значительно возрастает, и они становятся способными, приподниматься на передних конечностях и запрокидывать голову назад. При выпрашивании корма они медленно приподнимаются и долго держат рот открытым. Медлительность движений — черта, характерная вообще для многих вьюрковых в первый период жизни их в гнезде.
Развитие дубоноса идет несколько быстрее, чем некоторых других вьюрковых. Это прежде всего видно по раннему открытию слуховых проходов и быстрому развитию маховых перьев. В то время, как у щегла, зеленушки и некоторых других вьюрковых слуховые проходы открываются только к концу 5-го дня жизни, у птенцов дубоноса это происходит на 4-й день. В этом же возрасте намечаются пеньки маховых перьев. Глазные щели приоткрываются на 5-й день, st трубочки растущих маховых открываются уже в возрасте 6—7 дней. Рулевые перья отрастают
175
позднее и начинают развертываться только в 9-дневном возрасте. На 9—10-й день птенцы становятся очень пугливыми: при виде человека «они иногда выскакивают из гнезда. Нормальное оставление гнезда происходит на 12—13-й день. В этом возрасте птенцы уже перепархивают, но летать по-настоящему еще не могут: летят вниз по наклонной и садятся обычно на землю. Способность к полету приобретается, однако, очень скоро. 2-недельный птенец может уже некоторое время лететь, не теряя высоты.
Питание гнездовых птенцов. Мощный челюстной аппарат птенцов, казалось бы, должен указывать на то, что пища их должна быть преимущественно растительной. Однако изучение питания показало, что дубонос в гнездовой период в основном — насекомоядная Г1тица. Основным кормом птенцов дубоноса являются гусеницы бабочек и отчасти .личинки пилильщиков и жуков.
В 1951 и 1953 гг. от птенцов дубоноса в Савальском лесхозе было отобрано 168 экз. беспозвоночных животных. Среди них обнаружены гусеницы пядениц (64), гусеницы листоверток (54),. гусеницы совок (7), гусеницы прочих бабочек (15), личинки березового пилильщика (17), личинки и куколки жуков (8), майские жуки (1), мухи (1) и пауки (1). За один прилет дубонос приносит сразу несколько насекомых и кормит нескольких птенцов. Прежде чем отдать птенцам насекомых, он почти всегда их предварительно деформирует, разжевывая клювом. Пища растительного происхождения хотя и приносится птенцам иногда даже в 1-й день их жизни, тем не менее играет для них второстепенную роль.
ЗЕЛЕНУШКА
(CHLORIS CHLORIS L.)
Токование. Регулярное пение продолжается всю весну и все лето, с середины первой недели апреля вплоть до конца I декады августа. Поют зеленушки и позднее, но уже нерегулярно, по-осеннему. Наиболее позднее осеннее пение отмечено нами в Ленинградской области 5 октября (1954).
В южных районах изредка попадаются самцы, поющие совершенно необычно. Резко уклоняющийся от нормы вариант песни автору пришлось слышать в 1949 г. в заповеднике «Лес на Ворскле». Птица через определенные промежутки времени регулярно повторяла звук «виззирр» и никогда не издавала характерных трелей. Самец с аберрантной песней держался всегда на одних и тех же деревьях и пел в течение большей части июня. В начале июля он замолчал, но с 10 июля снова запел с прежней энергией и на том же самом месте. Подобные же вариации песни были отмечены впоследствии и у других зеленушек, гнездившихся в заповеднике. Двойной период активности пения одних й тех же особей зеленушек, по всей вероятности, следует связывать с характерным для этого вида двойным циклом размножения. Падение активности пения совпадает, очевидно, с* периодом спаривания, которое бывает во время постройки гнезда и откладывания яиц. В течение же всего периода насиживания кладки и выкармливания птенцов зеленушки поют весьма активно.
Расположение гнезда. В Ленинградской области зеленушка располагает свои гнезда почти исключительно на елях как на молодых, так и на взрослых. По данным В. Н. Шнитникова (1913), в Минской области этот вид также гнездится преимущественно на елках* В Са-вальском лесхозе излюбленным местом гнездования этого вида оказались кусты бузины, растущие среди соснового леса или на вырубках
176
(26 находок из 33), а в заповеднике «Лес на Ворскле» — тополь, дуб и липа; образующиеся на них мутовки стволовой поросли хорошо удерживают гнездо и делают его мало заметным. Таким образом, в разных частях гнездового ареала зеленушка тяготеет к различным породам. Приуроченность к тем или иным деревьям или кустарникам зависит в первую очередь от экологической обстановки района. В местах, часто посещаемых людьми, бродячими кошками и собаками (например, в парках пригородной зоны Ленинграда), зеленушки гнездятся, как правило, на высоте 4—6 м, в районах более глухих — значительно ниже, иногда в 1,5 м от земли на маленьких елочках, чаще всего на окраине лесного массива.
Сроки размножения. Как в лесной, так и -в лесостепной зонах период размножения у зеленушки растянут очень сильно. В Ленинградской области свежие, ненасиженные кладки этого вида можно находить начиная с конца последней декады апреля и до начала III декады июля. В наиболее раннем из известных автору гнезд зеленушки, найденном под Ленинградом, откладка яиц происходила в период с 27 апреля но 2 мая (1950). В 1954 г. 28 мая нами был пойман птенец, только что покинувший гнездо. Судя по возрасту (13—14 дней), он вылупился из яйца, отложенного тоже в самом конце апреля — начале мая. Наряду с очень ранними (майскими) выводками мы неоднократно находили гнезда, в которых откладка яиц происходила в конце I — начале II декады июля. В наиболее позднем из найденных гнезд .первое яйцо было отложено 15 июля, а последнее — 20 июля (1946). Птенцы из этого гнезда вылетели 13—14 августа. Таким образом, максимальная разница в сроках появления кладок зеленушки под Ленинградом оказалась равной 79 дням.
Сильная растянутость сроков размножения наблюдается и у зеленушек, гнездящихся в лесостепной зоне. Например, в Савальском лесхозе за 3 года наблюдений были установлены следующие даты появления первого яйца: 10, 12, 17, 20, 25, 27, 29, 30 мая, 2, 5, И, 23 июня и 4 июля. В заповеднике «Лес на Ворскле» в 1949 г. в некоторых гнездах зеленушек мы находили яйца еще 25 июля. Здесь же в 1950 г. И. В. Покровская наблюдала вылупление птенцов зеленушки 8 августа, а вылет птенцов — 20 августа.
Зеленушка, без сомнения, имеет 2 цикла размножения в сезон. На это указывают не только очень сильно растянутые сроки ее размножения, но и наблюдения за активностью пения отдельных самцов. Известным доказательством того, что поздние (июльские) кладки у зеленушек часто бывают не дополнительными, а вторыми, является, по нашему мнению, то, что они, как правило, содержат нормальное для вида количество яиц.
Размеры и судьба кладок. В Савальском лесхозе под наблюдением находилось 22 гнезда. Общее количество яиц в этих гнездах равнялось 104. Преобладающее большинство кладок (16) состояло из 5 яиц. В 3 гнездах было по 4 яйца и в 2 — по 6. 34 яйца погибло в результате разорения 7 гнезд и 2 яйца оказались болтунами, вылупилось всего 68 птенцов. Впоследствии было разорено еще 3 гнезда с 15 птенцами. Таким образом, общее количество вылетевших птенцов равнялось 53, что составило примерно 50% от первоначального количества отложенных яиц. В заповеднике «Лес на Ворскле» под! наблюдением находилось 6 гнезд с 29 яйцами. Из этих гнезд вылетело 24 птенца. В окрестностях Ленинграда из 22 яиц (5 гнезд) в конечном счете получилось 17 слетков. Количество птенцов, вылетевших из гнезда, у зеленушки варьировало от 3 до 6.
12 А. С. Мальчевский
177
При поздних сроках размножения, когда птицы начинают насиживать яйца до окончания кладки и когда в гнездах наблюдается большая разновозрастность птенцов, вылупившиеся последними иногда погибают. Однако в большинстве случаев, несмотря на очень сильную растянутость, вылупления и большое количество птенцов, они все выживают.
Какие хищники разоряют гнезда зеленушек, установить не удалось. Можно было лишь предполагать, что поочередное исчезновение птенцов-зеленушки из гнезд, неоднократно отмечавшееся в Савальском лесхозе, следует связывать с «воровскими» налетами весьма многочисленного здесь и гнездящегося в сходных стациях сорокопута-жулана.
Пов едение взрослых птиц у гнезда. Иногда считают,, что большая загрязненность гнезда зеленушки является следствием того, что взрослые птицы этого вида не уносят помета птенцов, как это делают другие воробьиные (Михеев, 1957). Это, однако, не подтвердилось нашими наблюдениями. Оказалось, что и самец и самка зеленушкц регулярно и до последнего дня выносят белые комочки экскрементов-Вблизи одного гнезда, найденного в парке Лесотехнической академии 31 июля 1948 г., в котором находились уже почти готовые к вылету птенцы, автор в течение 7 час. следил в бинокль за процессом выкармливания птенцов. За это время птицы накормили птенцов 15 раз (10 раз. самка и 5 самец) и унесли 12 комочков экскрементов (8 раз самка и 4 самец). Подлетевшая к гнезду птица всегда кормила сразу несколько птенцов, и каждый раз посте этого почти все они начинали поворачиваться и испражняться на край гнезда. Уносила же птица только один комочек экскремента. Таким образом, загрязненность гнезда зеленушек следует объяснить не отсутствием соответствующего инстинкта у взрослых птиц, а их редкими прилетами к гнезду и резким несоответствием: между количеством помета, уносимого и поступающего на край гнезда. Это несоответствие особенно значительно в конце гнездового периода жизни птенцов.
У гнезда с птенцами зеленушки обычно ведут себя довольно спокойно. Несколько раз, однако, автору приходилось наблюдать у самок явление «отвода»: птица, слетая с гнезда, снижалась к земле и летела медленным трепещущим полетом.
Птенцы. Птенец покрыт довольно густым, но не длинным пухом грязно-серого цвета. Пух расположен на надглазничных, затылочном^ спинном, плечевых, локтевых, а также на бедренных, брюшных и голен-ных участках тела, где он- очень редок. Ротовая полость мясо-красного цвета. Складка в углах рта маленькая. Разрез рта небольшой. Клюв, массивный, с темной предвершинной перевязью. Ноздри круглые. Яйцевой «зуб» £ерый, когти светлые. Пищевод сильно растяжим и функционирует, как зоб.
Развитие птенца зеленушки можно охарактеризовать следующим образом: глаза открываются сравнительно рано — на 4—5-й день, в то же время открытие слуховых проходов происходит в конце 5-го — начале 6-го дня жизни птенца, т. е. на целых 2 дня позднее, чем у большинства мелких воробьиных. Кисточки на концах пеньков растущих маховых начинают появляться в возрасте 8 дней. Оставление гнезда обычно происходит на 13—14-й день. В этом возрасте птенцы по-настоящему еще не умеют летать: выскакивая из гнезда, они летят вниз по наклонной и обычно садятся вд землю. При сильном испуге птенцы могут выскочить из гнезда и значительно раньше, в 10—11-дневном возрасте.
В гнездах зеленушек нередко приходится наблюдать большую разницу в размерах между птенцами одного выводка. Один птенец иногда бывает в 2 раза больше другого. Как правило, это зависит от неодно-
178
временности вылупления птенцов и бывает в тех случаях, когда самка начинает насиживать яйца до окончания кладки. Подобные явления нам приходилось наблюдать главным образом при позднем размножении птиц. Так, например, в гнезде, в котором первое яйцо было отложено 15 июля 1946 г., а последнее--20 июля, самка приступила к насиживанию 17 июля после откладки 3-го яйца (пригороды Ленинграда). В результате вылупление птенцов растянулось на 3 дня; 7 августа вес птенцов в этом гнезде равнялся 9,5; 14; 16; 16,5 и 18 г. В другом гнезде, найденном в Савальском лесхозе, вылупление птенцов растянулось на 4 дня и происходило с 17 по 20 июля (1953). Различия в величине птенцов в таких случаях бывают особенно заметными в первые дни после вылупления. Перед вылетом из гнезда размеры и вес птенцов выравниваются, и возрастные различия между птенцами заключаются в степени развитости перового покрова.
Перед вылетом из гнезда подвижность птенцов заметно возрастает. Они часто садятся на край гнезда или на соседние ветви, затем снова впрыгивают в гнездо. Голос птенцов к моменту оставления гнезда становится громким, слышимым за несколько десятков метров. Звуки, воспроизводимые ими, бывают в основном двух родов: очень'высокий дребезжащий, издаваемый птенцами при выпрашивании корма, и позыв «тюй-тюй», имеющий призывное значение. Этот звук издается птенцами особенно часто и регулярно первые дни после оставления гнезда.
Питание гнездовых птенцов. Всего на территории Ленинградской и Воронежской областей было исследовано 128 порций корма, извлеченных из пищеводов. Основная масса этих порций (108) состояла исключительно из растительного корма и лишь 20 — из смешанной растительной и животной пищи. В конце мая, в июне и начале июля 1952—1953 гг. зеленушки, гнездившиеся на территории Савальского лесхоза, приносили своим птенцам преимущественно семена вяза и различную зелень. По наблюдениям И. В. Покровской (1953), зеленушка часто приносит своим птенцам также семена петушьего проса (Echinochloa).
Из насекомых как в Савальском лесхозе, так и в заповедной дубраве «Лес на Ворскле» птенцы чаще всего получали мелких гусениц пядениц и листоверток, жучков-долгоносиков и божьих коровок. Таким образом, так же как и многие другие зерноядные вьюрковые, зеленушка весьма ограничена в выборе насекомых и поедает, как правило, мелких гусениц и мало подвижных жучков, которых находит в местах, где кормится семенами или зеленью.
• ЩЕГОЛ
(CARDUELIS CARDUELIS L.)
Токование. У щегла, как и у многих других наших вьюрковых птиц, регулярное пение продолжается в • течение • нескольких Месяцев. В Ленинградской области активное пение щеглов можно слышать с конца марта до середины августа. Столь продолжительный период пения следует связывать с двойным циклом размножения этого вида, что, по всей вероятности, имеет место не только в лесостепной, но и в лесной зоне.
Рас п*о ложение гнезда. Найденные 7 гнезд располагались на дубах, соснах, вязе, остролистном клене и яблоне. Даже это весьма ограниченное количество данных указывает на разнообразие древесных пород, используемых щеглом для гнездования. Высота расположения гнезд варьировала от 1,5 до 8 м. По наблюдениям Г. А. Новикова в «Ле-
12*
179
су на Ворскле», щеглы нередко располагают гнезда на высоте 10 м от земли.
Сроки размножения. В заповеднике «Лес на Ворскле» в 1949 г. вылет птенцов из известных нам гнезд произошел 24 июля и 15 июля. Вместе с тем 9 июня этого же года мы наблюдали уже хорошо летавших птенцов-слетков, которых родители еще продолжали выкармливать. Еще большая растянутость сроков размножения наблюдалась (1954) в окрестностях Ленинграда. Птенцы-слетки примерно одинакового возраста были отмечены нами 23 июня и 31 августа. В конце августа в парке Лесотехнической академии автору приходилось неоднократно наблюдать выводки молодых щеглов, выпрашивающих корм у взрослых птиц. По всей видимости, как в северных, так и в средних широтах щеглы выводят птенцов дважды в лето.
В некоторых случаях воспитание второго выводка протекает, видимо, в местах, удаленных от тех, где они гнездились в первый раз. Так, в 1952 г. в Савальском лесу щеглы появились как-то внезапно и сразу в большом количестве лишь в конце июня. Они держались либо парочками, либо небольшими стайками, в которых наряду со старыми птицами были ставшие уже вполне самостоятельными молодые. 7 июля 1952 г. нами был добыт молодой щегол-сеголеток, который по своим размерам уже ничем не отличался от взрослых птиц. В то же время все поведение старых птиц указывало на то, что они приступили к размножению: самцы активно пели и сопровождали самок, таскавших гнездовой материал. Подобная же картина повторялась и в 1953 г. В двух известных нам в этом году гнездах щегла первые яйца появились 3 и 6 июля.
Размеры и судьба кладок. Все найденные кладки состояли из 5 яиц. Из всех находившихся под наблюдением 20 яиц вылупились птенцы, которые благополучно закончили свое развитие.
Поведение взрослых птиц у гнезда. Самки систематически обогревали птенцов в течение первых 5—6 дней после вылупления. Инстинкт обогревания (насиживания) у некоторых птиц был настолько сильно выражен, что они садились даже на пустое гнездо и обогревали его в течение всего времени, пока производилось обследование и взвешивание птенцов. При этом птицы были очень доверчивыми и позволяли чуть ли не касаться их рукой. Одна самка, слетая с птенцов, несколько раз отводила от гнезда, имитируя раненую птицу: она падала комком на землю и билась внизу, как подстреленная.
Птенцы. Птенец покрыт светло-серым, довольно густым (исключая брюшную сторону и голень) коротким пухом, который расположен на глазных (не всегда), надглазничных, затылочной, спинном, плечевых, локтевых, бедренных, голенных и брюшных участках тела. Кожа желтовато-оранжевая. Клюв воскового цвета с темной вершиной. Ноздри круглые. .Яйцевоцч«зуб» серый, очень маленький, когти светлые. Ротовая полость мясного цвета. Складка в углах рта белая, но небольшая. Разрез ротовой полости неширокий. Сильно растяжимый пищевод — «зоб» способен вместить значительное количество корма.
Питание гнездовых птенцов. Было исследовано 34 комка пищи, извлеченных из пищеводов щеглят в Савальском лесхозе в период с 18 по 24 июля. Анализ этих данных показал, что птенцы щегла почти исключительно растительноядны. С первых же часов своего существования они получают от родителей незрелые семена и завязи цветов главным образом травянистых растений. Кормясь на травах, щеглы находят живущих в них личинок мух и гусениц бабочек, которых изредка приносят птенцам. В комках пищи у птенцов щегла были обнаружены, 180
в частности, безногие гусеницы Metzneria lapella, живущие в головках репейника. Из 34 обследованных «зобов» щеглят в 28 случаях корм был исключительно растительного происхождения, в 4 — пища состояла главным образом из семян и незначительного количества (1—2) мелких личинок цветочных мух и гусениц бабочек Gelichiidae, и в 2 случаях в «зобах» птенцов были найдены только гусеницы или личинки.
чиж
(SPINUS. SPINUS L.)
В Ленинградской области весеннее оживление у чижей иногда начинается уже с конца марта. Регулярное пение самцов продолжается со Ц декады апреля до начала августа. В августе активно поют лишь немногие птицы. Осеннее пение можно слышать вплоть до конца октября.
Из 2 гнезд, найденных нами на территории Ленинградской области, одно было обнаружено автором в районе ст. Елизаветино в мелком заболоченном елово-сосновом лесу. Располагалось оно почти у самой вершины невысокой елки, вблизи ствола, на высоте 6,5 м. Наружные стенки гнезда была построены из мха, внутренняя же выстилка — из золотисто-медных стебельков кукушкиного ’льна. Другое гнездо чижа бьрю найдено недалеко от ст. Кузнечное (Приозерский район). Оно помещалось на высоте 8 м на конце боковой ветви крупной сосны. Лишайники, обильно покрывавшие ветви дерева, почти целиком, скрывали гнездо, так что заметить его даже на близком расстоянии было очень трудно. Боковые стенки гнезда состояли в основном из лишайников, основание — из ваты, а лоток выстлан конским волосом.
Наблюдения за сроками размножения чижей дают основание предполагать у этого вида наличие двух нормальных кладок. Так, в окрестностях Ленинграда можно наблюдать молодых чижей, хорошо летающих, но еще выпрашивающих корм, с первой половины июня до конца августа. Разница в сроках размножения, отдельных пар птиц может быть свыше 2 мес. В одном из гнезд,. найденном 15 июня 1954 г., птенцы, судя по их возрасту, вылупились 10—11 июня, в то время как в другом гнезде, обнаруженном 25 августа 1947 г., вылупление птенцов должно было произойти 15—16 августа. Таким образом, откладка яиц в последнем гнезде происходила примерно на 65-г-бб дней позднее, чем в первом.
В обоих гнездах чижей, найденных нами на территории Ленинградской области, было по 3 птенца.
Птенец чижа покрыт довольно коротким темным пухом средней густоты, расположенным на глазных (всего несколько пушинок), надглазничных, затылочных, плечевых, локтевых, бедренных, голенных и брюшных участках тела. Клюв высокий, резко суживающийся к концу, одноцветно-желтый. Ноздри крупные, круглые. Яйцевой «зуб» и когти светлые. Цвет кожи грязно-розовый. Ротовая полость мясо-красная. Имеется «зоб» — сильно растяжимый пищевод, наполняющийся кормом после заглатывания пищи. В гнезде, найденном 15 июня 1954 г., птенцы, которым было примерно 4 дня от роду, имели уже открытые глазные щели; в то же время ушные отверстия их были почти полностью закрыты. При-' нимая позу пищевого рефлекса, птенцы издавали едва слышимый писк. Незадолго перед вылетом из гнезда птенцы поражали своей доверчивостью и в то же время очень большой подвижностью и способностью уже в возрасте 10—12 дней уверенно лазать и прыгать по еловым веткам. При выпрашивании корма они громко и подолгу кричали звонкими высокими голосами. 25 августа (1947) пищеводы всех птенцов были туго набиты семенами какого-то растения.
181
КОНОПЛЯНКА
(ACANTHIS CANNABINA L.)
Токование. В Ленинградской области период активного пения самцов захватывает почти 5 мес. Продолжается оно с апреля до начала последней декады августа. В начале апреля и в августе пение бывает нерегулярным, так как в это время поют далеко не все самцы. Столь длительный период токования коноплянок следует связывать с очень сильно растянутым периодом размножения этих птиц.
Расположение гнезда. В Ленинградской области коноплянка так же, как и зеленушка, явно предпочитает селиться на хвойных породах: 8 из 10 найденных гнезд было расположено не выше 1 м среди ветвей небольших елочек или сосен, 1 — на можжевельнике и 1 — на
Рис. 45. Самка коноплянки на гнезде („Лес на Ворскле*, июль 1957 г. Фото автора).
смородине. Излюбленными стациями являются здесь лесные опушки, сады, защитные насаждения вблизи железнодорожных путей. На территории учлесхоза «Лес на Ворскле» в 1.949 г. большинство гнезд было устроено в кустах татарской жимолости (6 находок из 9), 2 гнезда располагались на ветвях сирени и 1 — в кусте барбариса. Максимальная высота гнездования достигала здесь 2 м.
Сроки размножения. В Ленинградской области период возможных сроков откладки яиц растянут почти на 3 мес. Так, в наиболее раннем гнезде 1-е яйцо появилось 28 апреля (1950), а в наиболее позднем — 26 июля (1953).
Как известно, коноплянки гнездятся небольшими колониями, поселяясь в количестве 3—5 пар на каком-либо ограниченном участке местности. В 1949 г. автор проследил жизнь нескольких пар коноплянок, гнездившихся на усадьбе учлесхоза «Лес на Ворскле» (рис. 45). Здесь было найдено 9 гнезд. 2 из них были обнаружены уже после вылета птенцов. В остальных случаях нам удалось установить сроки откладки яиц, вылупления или вылета птенцов (табл. 46).
182
Таблица 46
Сроки размножения отдельных пар коноплянок из одной колонии („Лес на Ворскле", 1949 г.)
№ гнезда Откладка первого яйца Вылупление птенцов Вылет птенцов из гнезда
1* 5V Не прослежено 6 VI
2# 19V 19 VI
3 6 VI 22—24 VI 5 VII
4 10 VI 27-29VI 9 VII
5 2 VII 19—21 VII Птенцы погибли в гнезде
в возрасте 2—3 дней
6 6 VII 24—25 VII Не прослежено
7 10 VII 27—29 VII
Примечание. Звездочкой отмечены гнезда, даты откладки яиц в которых <5ыли определены по возрасту птенцов.
В данном случае растянутые сроки откладки яиц зависели как от неодновременности созревания половых желез у отдельных членов колонии, так и от того, что мы имели здесь случаи второго гнездования после успешного воспитания первого выводка. На это, в частности, указывало также и то, что при наличии на территории усадьбы заповедника 9 жилых гнезд коноплянок здесь держалось не более 5 пар этих птиц.
Размеры и судьба кладок. В окрестностях Ленинграда в *6 известных нам гнездах коноплянки, история которых была прослежена до конца, количество яиц было следующим: 7, 6, 6, 5, 5 и 5. Из этих 34 яиц вылупилось только 22 птенца: 1 гнездо с 5 яйцами было разорено, видимо, людьми, 1 кладка из 6 яиц была брошена птицами и 1 яйцо •оказалось болтуном. Все 22 вылупившихся птенца благополучно закончили свое развитие и вылетели из гнезд. Число птенцов-слетков в выводках у коноплянок (по наблюдениям за 11 гнездами) варьировало от 4 до 7.
Интересно, что в гнезде коноплянки, найденном в Ленинградской юбласти, в котором выводок состоял из 7 птенцов, кладка началась поздно — в начале июля. Таким образом, при поздних сроках размножения у коноплянок не всегда бывает уменьшенное количество яиц в кладке. В данном случае получилось даже наоборот: количество яиц оказа-.лось максимальным и, кстати, превышающим число яиц, известное у нас до сих пор (Бёме, 1954, стр. 206).
Птенцы. У коноплянки нередко приходится наблюдать неодно- ’ временность вылупления птенцов. В одном из гнезд вылупление 6 птенцов растянулось на 3 дня — с 23 по 25 июня. Птенцы выходит из яйца одетыми в темно-серый пух, довольно густой, расположенный на надглазничных, затылочной, спинной, плечевых, локтевых, бедренных, го-ленных и брюшных птерилиях. Окраска ротовой полости мясо-красная. Складка в углах рта небольшая. Клюв высокий, резко сужйвающийся к концу. Ноздри, как и у птенцов других вьюрковых, круглые.
Из гнезд птенцы вылетают на 12—14-й день жизни, а в случае сильного испуга даже на 11-й день. Способными к активному полету они становятся уже в 2-недельном возрасте. Например, птенец, появившийся на свет 26 июня, через 14 дней (10 июля) мог уже прилично перелетать на расстояние 25—30 м.
183
Питание гнездовых птенцов. Во всех обследованных нами «зобиках» птенцов коноплянок в возрасте от 1 до 12 дней как в лесной, так и в лесостепной зонах мы обнаружили лишь растительную (пищу, состоящую, как правило, из незрелых семян различных травянистых растений. В районах учлесхоза «Лес на Ворскле» пищеводы птенцов коноплянок чаще всего были наполнены семенами редьки (Ra-phanus), а также размельченными семенами подсолнуха. В «зобу» птенцов, найденных 28 июля в Ленинградской области, были обнаружены в основном семена короставника полевого (Knautia arvensis) с незначительной примесью семян белой мари (Chenopodium album). Изучение птенцов коноплянки, проведенное И. В. Покровской (1956) на территории Ленинградской области, показало, что помимо растительной пищи птенцы этого вида получают также значительное количество насекомых.
СНЕГИРЬ
(PYRRHULA PYRRHULA L.)
Токование. В Ленинградской области «скрипение» снегирей чаще всего бывает слышно ранней весной — с конца февраля и до середины апреля. В это время птицы держатся здесь в большом количестве. Наиболее поздняя дата весеннего «пения» снегиря была отмечена нами 17 мая 1953 г. в саду Ленинградского университета. Пели явно пролетные, хотя и державшиеся уже парами птицы. Во время гнездования снегири совсем не поют.
Расположение гнезда. Все известные автору 6 гнезд снегиря, найденных в пригородах Ленинграда, располагались на больших (2 гнезда) или, наоборот, на очень мелких елках на высоте от 1,5 до 3 м от земли. Одно из этих гнезд помещалось на конце нижней ветки крупной ели. На предпочтение, отдаваемое снегирем еловым деревьям,, указывает также А. М. Каминский (1916). В литературе известны, однако, случаи гнездования снегиря и на других породах — на можжевельнике, сосне и даже на березе (Бёме, 1954).
Сроки размножения. О сроках размножения снегирей в Ленинградской области автор имеет возможность судить лишь по 6 гнездам этого вида, найденным в-окрестностях Ленинграда. Из табл. 47 видно, что в большинстве случаев откладка яиц у этого вида в Ленинградской области начинается, видимо, в конце мая — первой половине июня. Однако иногда снегири приступают к размножению и в июле.
Таблица 47
Сроки размножения отдельных пар снегирей в окрестностях Ленинграда
Год наблюдения Количество яиц в кладке Откладка первого яйца Вылупление птенцов Оставление гнезда птенцами
1949 6 25 V Птицы бросили кладку
1949 5 26 V То же —
1953* 4 31 V 17—18 VI 2 VII
1949 Ь 10 VI Птицы бросили кладку —
1953* 4 13 VI 30 VI— 1 VII 13 VII
1955 5 4 VII 19 VII —
Примечание. Звездочкой отмечены случаи, в которых даты откладки первого яйца были определены по возрасту птенцов.
184
Поведение взрослых птиц у гнезда. В период гнездования снегири ведут очень незаметный образ жизни. Увидеть их у гнезда удается редко. Во время беспокойства они держатся почти всегда в стороне, изредка издавая свой обычный позыв «фю». Осторожность птиц выражается также в том, что многие особи очень легко бросают гнезда даже с насиженной кладкой. Однако и среди снегирей попадаются -отдельные птицы, отличающиеся чрезвычайной доверчивостью. В 1955 г. в парке Ст. Петродворца гнездилась самка снегиря, которая не слетела с гнезда даже тогда, когда из-под нее,брали птенцов (сообщение С. И. Божко).
Птенцы. Птенец опушен. Пух довольно густой и длинный темносерого цвета. Расположен на голове (над глазами и на затылке), на спине, на плечах и предплечье, на бедрах, голени и брюшной стороне. Ротовая полость мясо-красного цвета с фиолетовым оттенком на отдельных участках. Клюв массивный, рогового цвета с потемнением к концу. Яйцевой «зуб» темный. Кожа мясного цвета.
Питание гнездовых птенцов., У всех 4 птенцов снегиря, найденных автором 26 июня 1953 г. в парке Ст. Петродворца, «зобики» были .наполнены какими-то крупными зелеными семенами. Более подробные сведения по питанию птенцов этого вида в пригородах Ленинграда были получены С. И. Божко (устное сообщение). Снегири, за которыми она наблюдала, приносили птенцам главным образом семена различных растений (в частности, черники). Паучки и гусеницы бабочек хотя и были обнаружены в 42 порциях корма из 68, составляли тем не менее незначительную часть рациона, ибо были представлены лишь единичными экземплярами.
По наблюдениям Г. А. Новикова (1952), на Кольском полуострове взрослые птицы в гнездовое время также питаются почти исключительно семенами.
ЧЕЧЕВИЦА ОБЫКНОВЕННАЯ (ERYTHRINA ERYTHRINA PALL.)
Токование. Результаты наших наблюдений за прилетом чечевиц в окрестности Ленинграда несколько отличаются от данных Д. Н. Кай-городова (1922), указывавшего для этих мест 21 мая как наиболее раннюю дату прилета чечевиц за последнее тридцатилетие. С 1946 по 1955 г. чечевицы неоднократно прилетали к нам раньше указанного срока. Так, например, первая песня чечевицы была отмечена 16 мая (1949), 19 мая (1954) и 20 мая (1948).
Период регулярного пения чечевицы в Ленинградской области продолжается около 1,5 мес. — с последних чисел мая до конца I декады июля. Позднее интенсивность пения резко падает, хотя отдельные «амцы и поют вплоть до конца июля. В 1946 г. пение чечевицы слышали . до 30 июля. Эта дата является для Ленинградской области, конечно, исключительно поздней. В большинстве случаев самцы чечевицы совсем прекращают петь уже во II декаде июля.
Расположение гнезда. В Ленинградской области большинство гнезд чечевицы было найдено на молодых елочках (8 гнезд из 16), а также в подрезанных кустах желтой акации (3 находки), расту-Щихпо краям аллей пригородных парков. Некоторые гнезда помещались на лиственном подросте (дуб, ольха) и в кустах спиреи, барбариса. Таким образом, породы деревьев и кустарников, используемых чечевицей для размещения гнезд, довольно разнообразны. Высота гнездования, как правило, очень небольшая, чаще всего 0,5—1 м.
185
Сроки размножения. В наиболее позднем гнезде чечевицы, найденном под Ленинградом, откладка яиц, судя по дате вылупления птенцов, должна была начаться 25 июня (1947), а самая ранняя дата ►откладки первого яйца была установлена 28 мая (1949). Таким образом, период откладки яиц растянут у чечевицы примерно на 1 мес. (табл. 48).
Из таблицы видно, что сроки размножения и количество яиц в кладках у чечевиц в различные годы были неодинаковыми: в 1949 г. в большинстве гнезд первые яйца были отложены в период с 28 мая по 3 июня, а количество яиц во всех найденных нами кладках равнялось 5. В 1954 г. первые яйца появились позднее (3 июня— 13 июля), и кладки в большинстве случаев состояли из 4 яиц.
Эти различия следует, по нашему мнению, объяснять неблагоприятными метеорологическими условиями весны 1954 г., которая была очень затяжной и холодной и сопровождалась резкими падениями температуры в конце мая и начале июня.
Размеры и судьба кладок. Размножение чечевиц не имело успеха в большинстве из 12 известных нам гнезд. Из табл. 48 видно, что в 9 гнездах, судьба которых была нами прослежена до конца, находилось 41 яйцо, из них вылупилось всего 17, а вылетело из гнезда только 9 птенцов, что составило всего лишь около 22% от первоначального количества отложенных яиц.
Таблица 48
Сроки размножения отдельных пар чечевиц в окрестностях Ленинграда
Год наблюдения Количество отложенных яиц Дата откладки первого яйца Вылупление птенцов Вылет птенцов
1949 5 28 V Гнездо разорено до вылупления птенцов —
1949 5 30 V Гнездо брошено до вылупления птенцов —
1949* 5 1 VI 17—18 VI Птенцы погибли до вылета
1949 5 3VI 19 VI 28 VI
1946 5 3 VI Гнездо разорено до вылупления —
1954* 4 3VI 19-20 VI Не прослежено
1954 4 5 VI Гнездо разорено до вылупления птенцов —
1954* 5 7 VI 23-24 VI Не прослежено
1954 4 . 12 VI 28 VI Не прослежено
1954* 4 13 VI 30 VI-1 VII 9 VII
1950 5 15 VI Гнездо брошено перед вылуплением птенцов —
1947* 3 25 VI 9—10 VII Птенцы погибли в возрасте 3—4 дней
Примечание. Звездочкой отмечены случаи, в которых дата откладки первого яйца была определена по дате вылупления птенцов.
Столь высокий процент гибели яиц и гнездовых птенцов чечевицы зависит в первую очередь от очень частого разорения их гнезд, а также от того, что некоторые птицы очень пугливы и, будучи потревоженными, легко бросают гнезда даже с сильно насиженными кладками.
Поведение взрослых птиц у гнезда. У гнезда чечевицы. ведут себя по-разному. Наблюдается резкая индивидуальная измен
186
чивость: есть очень осторожные особи, которые слетают с гнезда незаметно и близко к наблюдателю не подлетают, даже если он берет в руки птенцов. Эти птицы легко бросают даже насиженные кладки. Встречаются, однако, и такие, которые во время насиживания или обогревания птенцов не слетают с гнезда даже в том случае, если до них дотрагиваются, а лишь принимают угрожающую позу, шипят и клюются.
Птенцы. Пух, покрывающий тело птенца, темно-серый, густой, но не очень длинный. Расположен на глазных, надглазничных, затылочном, спинном, плечевых, Локтевых, бедренных, а также голенных и •брюшных участках тела (на последних пух очень редкий и короткий). Клюв рогового цвета с сильным. потемнением в вершинной части, несколько вздутый в области круглых ноздрей. Яйцевой зуб темный, когти желтоватые. Кожа грязно-розового цвета. Складка в углах рта небольшая. Пищевод способен сильно растягиваться и постоянно бывает набит незрелыми семенами различных растений. .
В тех случаях, когда птенцы чем-либо потревожены, они покидают гнездо очень рано — на 9—10-й день после вылупления. Нормальное оставление гнезда происходит в возрасте 13—14 дней. К этому времени перья крыла уже образуют сплошную несущую поверхность, благодаря чему птенец может перепархивать.
Питание гнездовых птен ц о в. Почти все обследованные автором «зобики» птенцов чечевицы из гнезд, найденных им в окрестностях Ленинграда, были наполнены семенами растений. Только в 3 случаях из 22 в пищеводах птенцов помимо семян, составляющих основу пищевого комка, были также мелкие гусеницы и жучки, которых птицы, очевидно, подобрали там, где они кормились семенами. В разгар периода выкармливания птенцов чаще всего нам приходилось встречать взрослых чечевиц, собиравших семена мятлика (Роа annua).
КЛЕСТ-ЕЛОВИК (LOXIA CURVIROSTRA L.)
За десятилетний период наблюдений с 1946 по 1956 г. на территории Ленинградской области клесты-еловики .в большом количестве появлялись в 1945—1946, 1948—1949, 1952—1953 и 1954—1955 гг. «Клестовые годы» являлись следствием обильного урожая семян ели, наблюдавшегося осенью 1945, 1948, 1952 и 1954 гг. Судя по сообщениям, появлявшимся в литературе (Воробьев, 1955; Бубнов, 1956), а также по устным сообщениям отдельных лиц, в указанные годы клесты-еловики были многочисленны в Московской, Новгородской, Ярославской, Костромской и других областях Европейской части СССР.
Появление клестов-еловиков в Ленинградской области каждый раз сопровождалось их размножением, которое, судя по поведению птиц, состоянию их половых желез, а также по находкам отдельных гнезд у большинства пар происходило в период с конца января по март включительно. Помимо обычного времени токования клестов (ноябрь— февраль) активное пение у этого вида под Ленинградом отмечалось в марте, апреле и мае 1946 г., в июне и Июле 1948 г. и в октябре 1954 г. В период с марта по октябрь пение клестов было единичным.
О степени растянутости периода размножения клестов в пределах одного и того же года мы можем судить лишь по известным нам находкам двух гнезд клеста-еловика, обнаруженных в 1955 г. на Карельском перешейке. Одно гнездо было найдено Р. Л. Потаповым, другое — Л. Н. Поповым. Первое гнездо располагалось у вершины крупной сосны, несколько отступя от главного ствола, на высоте около 15 м. 2 марта
187
в гнезде было 3 птенца в возрасте 5—6 дней. Примерно такого же возраста были птенцы, найденные Л. Н. Поповым 29 марта в гнезде, располагавшемся на елке на высоте примерно 8 м. Таким образом, разница в сроках вылупления птенцов в этих гнездах была равна примерно 1 мес.
Судя по литературным данным, разница в сроках размножения отдельных птиц в пределах одного и того же года может быть значительнобольшей. Так, по сообщению Г. П. Дементьева (1941) и Д. В. Тернов-ского (1954), зимой 1940—1941 гг. размножение клестов-еловиков под Москвой началось в декабре и продолжалось в январе и феврале. Кроме того, под Звенигородом в том же году кладка клест а-ел овика была обнаружена в июле. На способность клестов при наличии определенных условий размножаться не только зимой и ранней весной, но и летом указывал в свое время также Рейникайнен (Reinikainen, 1937).
В ельниках Волосовсксго района Ленинградской области 20 октября 1954 г. автором был обнаружен выводок клеста-еловика, состоявший из 4 уже хорошо летавших молодых птенцов, которые с характерным,, очень похожим на. позыв молодых зеленушек криком летали за старой самкой клеста. Судя по возрасту птенцов, размножение клестов в данном случае должно было протекать в сентябре. На возможность сентябрьского размножения клестов-еловиков на Южном Урале указывает С. В. Кириков (1952), отмечающий, что эта вторая волна размножения клестов бывает там в годы совпадения урожая семян сосны и лиственницы. Закономерность, .отмеченная С. В. Кириковым для сосноволиственничных лесов Южного Урала, в известной мере может проявляться и в окрестностях Ленинграда. Дело в том, что как в черте самого города, так и в пригородной зоне существует немало парков с лиственничными насаждениями, которые в годы урожая семян могут прокормить некоторое количество клестов в течение всего лета вплоть до нового урожая семян ели.
Есть основания предположить, что на сроках пребывания и размножения клестов-еловиков может сказаться не только обилие урожая семян ели и лиственницы, но и обилие галлов вязовой тли, представляющих источник питания в период, наиболее бедный для клестов кормами. В литературе (Козлова, 1930; Мальчевский, 1947) имеются указания на способность клестов расклевывать галлы на тополях. По всей вероятности, галлообразующие насекомые при массовом их размножении вообще могут играть немаловажную роль в жизни клестов.
Птенец клеста опушен довольно сильно. Пух темно-серого цвета, довольно густой, но не очень длинный. Максимальная длина пушин на спине и на затылке достигает 11 мм. На других участках тела пух еще короче. Он расположен на глазных, надглазничных, затылочной, спинной, плечевых, локтевых, бедренных птерилиях, а также на брюшных и голенных участках. Ноздри круглые, клюв темно-серый, широкий в основании и несколько горбатый. Яйцевой «зуб» светлый, края рта светло-желтые. Ротовая полость мясо-красного цвета. Когти серые. Кожа сверху темно-серая, снизу мясного цвета. Разрез рта довольно широкий,, и складка в углах рта достигает заметного развития, несмотря на преимущественную растительноядность птенцов. Пищевод способен растягиваться весьма сильно и вмещать очень много корма. Птенцы клестов, которые, как это хорошо известно, выкармливаются в зимних условиях семенами хвойных деревьев, не являются, как это на первый взгляд ня странно, исключительно растительноядными. В «зобу» одного птенца, взятого из гнезда 2 февраля 1955 г., помимо многочисленных семян ели автор обнаружил также 6 небольших личинок какой-то мухи. Возможно, что присутствие в кормовом рационе птенцов клеста белка животного
188
происхождения в виде зимующих в шишках личинок насекомых даже необходимо для нормального развития птенцов.
ЗЯБЛИК (FRINGILLA COELEBS L.)
Токование. За истекшее десятилетие (1946—1955) в окрестностях Ленинграда самая ранняя первая песня зяблика была отмечена 1 апреля (1949), а наиболее поздняя—16 апреля (1955). Последний год отличался небывало холодной, многоснежной и затяжной весной. Обычно же регулярное пение зябликов под Ленинградом начинается в конце I—начале II декады апреля и продолжается до начала II декады июля. Нерегулярное пение отдельных самцов продолжается вплоть до конца июля. В 1954 г. в районе ст. Кузнечное (Приозерский район Ленинградской области) автор слышал песню одного зяблика 27 июля. Наиболее поздняя дата «рюмящего» позыва самца была отмечена 22 августа (1955). Основная масса зябликов прекращает «рюмить» уже в конце июля.
Расположение гнезда. Породы деревьев, на которых зяблик вьет гнезда, весьма разнообразны. В лесной и лесостепной зонах автору приходилось находить постройки этого вида на следующих породах: ель, сосна, лиственница, можжевельник, береза, различные ильмовые, дуб, липа, клен, тополь, ива, рябина, серая ольха, черемуха, боярышник и бузина. В северных широтах (Ленинградская область) гнезда зяблика чаще всего бывают расположены на елках (19 находок из 73),-березах (13) и ольхах (11), реже на соснах (5), черемухе (3), клене (5) и др. В лесостепной полосе зяблик гнездится преимущественно на дубе, вязе, березе, клене. В чистых сосновых посадках зяблик, как известно, не многочислен. Однако в тех местах, где в сосновом лесу встречается хотя бы незначительное количество лиственных деревьев, эта птица становится весьма обычной. Например, в искусственных борах Савальского лесхоза, где зяблика довольно много, он гнездится главным образом на вязах, березах и дубах, растущих по краям сосновых массивов. Здесь же нами были зарегистрированы случаи гнездования зяблика в подлеске, на кустах бузины.
Большинство найденных нами гнезд зяблика располагалось низко, в 1,5—3 м от земли (77 из 128 известных гнезд). На высоте от 3 до 6 м было найдено 37 гнезд и от 6 до 12 м —всего 14.
Характерная для зяблика черта — инкрустирование гнезд светлыми лишайниками и Пленками березы в случае расположения их на деревьях со светлой корой (береза, ольха), — безусловно,'имеет определенное биологическое значение; такие гнезда заметить, действительно, очень трудно. В тех же случаях, когда зяблик помещает свое светлое гнездо на деревьях с темной корой, что бывает не так уже редко, всякий смысл отмеченной особенности теряется. Факты подобного рода указывают на то, что действия птиц не всегда бывают целесообразны.
Сроки размножения. О сроках размножения зяблика в Ленинградской области мы можем судить по 29 гнездам, найденным в период с 1946^ по 1954 г. В наиболее раннем из гнезд первое яйцо появилось 1 мая", в наиболее позднем — 24 июня. Таким образом, период откладки яиц у зяблика в Ленинградской области оказался растянутым почти на 2 мес. В наибольшем количестве известных нам гнезд начало кладки падало на II и III декады мая (табл. 49).
Июньские кладки, по всей' вероятности, являются дополнительными, возникшими вынужденно после разорения гнезда. Неодновременное же
189
Таблица 49 Сроки (по декадам) начала откладки яиц у зяблика в Ленинградской и Воронежской областях
Район наблюдений Количество гнезд, в которых откладка яиц началась
в мае в июне в июле
' 1 II III 1 1 II III I II
Ленинградская область . . . Воронежская область .... 3 1 10 6 10 6 3 5 2 7 1 2 1 —-
гнездование зябликов в мае следует, видимо, объяснять растянутостью сроков прилета и различной степенью готовности к размножению отдельных особей. Возможно, что отдельные птицы в условиях Ленинградской области и успевают вывести потомство дважды в сезон. Однако такие случаи являются скорее исключением, нежели правилом.
На территории Савальского лесхоза было найдено 28 гнезд, в которых удалось установить время появления яиц. Откладка яиц происходила с начала I декады мая вплоть до середины I декады июля. Наряду^ с этим здесь отмечено большое количество свежих кладок, появившихся во II декаде июня; отдельные же особи продолжали откладывать яйца еще в конце июня—начале «юля. 4 июля (1953) автор наблюдал спаривание птиц, а 18 июля нашел гнездо, в котором самка еще насиживала кладку. Таким образом, общий спад волны размножения в Воронежской области наметился значительно позднее, чем в Ленинградской. Поздние сроки размножения у большого количества зябликов, обитающих в лесостепной зоне, нельзя объяснить только лишь повторным их гнездованием после разорения первых гнезд, так как очень многие кладки из найденных нами в июне содержали по 5 яиц, т. е. не были уменьшенными. Трудно себе представить также, чтобы среди зябликов имелись особи, которые впервые приступали бы к размножению во II декаде июня. Таким образом, для того чтобы объяснить причину появления большого* количества свежих кладок во II декаде июня, остается допустить наличие двойного цикла размножения у известной части зябликов, гнездящихся в лесостепной зоне. Такая точка зрения, как известно, существовала уже давно (Сомов, 1897; Мензбир, 1895).
Размер ы и судьба кладок. На территории Ленинградской: области история кладки и выводка была прослежена лишь в 25 случаях. В большинстве гнезд (в 16 из 25) кладка состояла из 5 яиц. в 8 гнездах — из 4 и в 1 — из 3. Ни в одном из гнезд не было более 5 яиц. Общее количество яиц во всех 25 кладках составляло 115, из них вылупилось 95 птенцов. Таким образом, отход яиц был сравнительно небольшой: 14 яиц погибло при разорении 3 гнезд и 6 яиц оказались неоплодо-творенными. Впоследствии было разорено еще 5 гнезд с 19 птенцами.. Кроме того, 15 птенцов погибло в гнездах по неизвестной для нас причине: птенцы постепенно теряли активность, становились холодными и* погибали. Обычно это случалось со всем выводком. В таких случаях гибель выводка в гнезде надо, видимо, связать с гибелью самки. Всего из-115 яиц в конечном счете остался 61 птенец; это составило в среднем1 2,4 птенца на .1 гнездо. Число птенцов в тнездах-в момент оставления их варьировало от 2 до 5.
На территории Савальского лесхоза удалось проследить историю-23 кладок. Общее количество яиц во всех этих гнездах составляло 10Е
190
яйцо. Количество яиц в отдельных кладках было следующее: в 2 — по 6,. в 10 — по 5, в 6 — по 4, в 5 — по 3 яйца. Вылупилось 88 птенцов, так как 9 яиц погибло при разорении 2 гнезд, видимо вороной или сойкой,, и 4 яйца оказались болтунами или содержали погибшие эмбрионы. Впоследствии было разорено еще 2 гнезда с 9 птенцами. Таким образом, общее количество птенцов, вылетевших из 23 гнезд, составило 79, т. е. в среднем на 1 гнездо пришлось 3—4 птенца.
Основными врагами зяблика в гнездовой период являются хищные и врановые птицы. Особенно часто тревожат зябликов у гнезд сойка, ворона, сорока, ястреб-перепелятник и сарыч.
Поведение взрослых птиц у гнезда. При появлении человека вблизи гнезда зяблики ведут себя неодинаково. Некоторые удивительно спокойно сидят в стороне и выдают свое присутствие лишь обычным «сигналом тревоги». Другие, наоборот, возбуждаются настолько сильно, что даже «притворяются» ранеными. Явление «отвода» у зябликов мы наблюдали неоднократно даже в период насиживания кладки. Чаще всего это было в случае низкого расположения гнезда. Сходное поведение самки зяблика отмечает Сазерн (Southern, 19496).
Самки, беспокоясь у гнезда, только «пинькают», самцы же помимо «пиньканья» издают рюмящий позыв (немецкий «Regenruf»). Кроме того, в возбужденном состоянии зяблик изредка воспроизводит тонкий протяжный свист, также, видимо, имеющий значение «сигнала тревоги». На севере Карельского перешейка наблюдается индивидуальная изменчивость воспроизведения рюмящего позыва. Здесь помимо птиц, регулярно издающих во время беспокойства у гнезда типичный дребезжащий позыв («трю» или «рюм»), были обнаружены особи, у которых этот же позыв звучал, как «фюить», почти неотличимо от позыва горихвостки. Доказательством того, что этот позыв является вариацией обыкновенного рюмения зяблика, могут служить факты последовательного воспроизведения одной и той же птицей обоих вариантов позыва. Интересно, что вариации рюмящего позыва зяблика, описанные в Германии и Дании, были по существу те же, что и в нашем случае. Это «riid» и «Rotschwanz-huit» (Sick, 1939) в Германии, «ryd» и «uid» в Дании (Poulsen, 1951).
По поводу биологического смысла «рюмящего» позыва в орнитологической литературе нет единства взглядов. Большинство полагает, что зяблики «рюмят» перед дождем (Кайгородов, 1923; Першаков, 1932; Бёме, 1954, и др.). Некоторые авторы (Heider, 1954; Peitzmeier, 1955)r специально посвятившие свои работы выяснению смысла данного позыва зяблика, придают ему двоякое значение. Они рассматривают его, с одной стороны, как звук, издаваемый в связи с определенными метеорологическими условиями, и, с другой — как тревожный крик при появлении вблизи гнезда сойки, кошки и других врагов.
Если бы зяблики действительно «рюмили» перед дождем, то вряд ли бы этот позыв мог быть тесно связан с полом и периодом размножения. Кроме того, если бы «рюмение» вызывалось определенными метеорологическими условиями, то при наступлении соответствующих условий начинать «рюмить», казалось бы, должны были все зяблики или во всяком случае большинство. На самом же деле «рюмение» зябликов никогда не бывает массовым явлением. Далее, «рюмение» обычно бывает связано с другим позывом — «пиньканием», который уже, бесспорно, является «сигналом» тревоги. Это должно указывать на сходство физиологического состояния птицы в момент последовательного воспроизведения обоих позывов. Одним словом, все говорит за то, что «рюмение» зяблика следует рассматривать лишь как воспроизводимый в период
191
размножения позыв, выражающий тревожное состояние птицы. Специальные наблюдения за беспокоящимися у гнезда самцами зябликов показали, что в зависимости от степени возбуждения зяблик «рюмит» в различном темпе. Часто он «рюмит» и одновременно «пинькает». Наблюдения также показывают, что зяблик может активно «рюмить» как перед плохой погодой, так и перед хорошей. В теплых, сухих районах нашей страны, например в Воронежской области, где дожди выпадают всего несколько раз в лето, зяблики «рюмят» столь же часто, как и в условиях влажного и дождливого лета Ленинградской области. Таким •образом, «рюмение» перед дождем, по нашему мнению, надо расценивать как случайное совпадение.
С целью выявления причин, вызывающих у зябликов «рюмящий» позыв, автор неоднократно подходил К «рюмящим» зябликам и внимательно обследовал место, где находилась птица. В 61 случае из 71 автору удалось обнаружить причину возбужденного состояния зяблика. Оказалось, что тревожили зябликов, как правило, хищные или врановые птицы. Суммированные результаты нашего обследования, проводившегося на территории Ленинградской области и Савальского лесхоза Воронежской области, приведены ниже:
Число случаев Чисто случаев
Коршун . . 4 Ворона . . . . 6
Сарыч . . 10 Сорока . . . . 5
Перепелятник . . . 5 Сойка . . . . 24
Ушастая сова . . . . . 2 Белка . . . . 1
Неясыть . . 3 Кошка . . . . 1
Надо сказать, что далеко не всегда причину беспокойства зяблика удавалось выяснить сразу. Первоначально создавалось впечатление, будто птица «рюмит» без всякой к тому причины. Однако после длительных поисков мы нередко все-таки выпугивали притаившуюся ворону, сойку, белку или какое-нибудь другое животное, беспокоившее зяблика. Полагаем, что те случаи (10 из 71), когда причина «рюмения» не была нами выяснена, следует объяснять в основном тем, что нам просто не удавалось выследить «хищника». Надо также иметь в виду, что иногда возбужденное состояние зяблика угасает очень медленно, и он продолжает «рюмить» еще долго после того, как опасность, угрожавшая гнезду или птенцам, миновала. В этих случаях очень легко счесть, что птица «рюмит» сама по себе.
Птенцы. Птенец покрыт грязно-серым пухом, густым, довольно длинным. Пух расположен на глазных, надглазничных, затылочном, спинном, плечевых, локтевых, бедренных, голенных и брюшных (на последних не всегда) участках тела (рис. 46). Кожа грязного желтоваторозового цвета. Клюв сверху серый, ноздри круглые, яйцевой «зуб» светлый, очень мелкий. Когти темные. Ротовая полость малиново-красная; складка в углах рта довольно широкая. Разрез рта широкий. По это^у признаку птенцы зяблика мало чем отличаются от птенцов большинства насекомоядных птиц, что согласуется с характером их питания.
В первые дни после вылупления птенцы зяблика мало активны. Даже в 3—4-дневном возрасте они неспособны к резким движениям. При выпрашивании корма они обычно тянут кверху только шею, не откидываясь назад, как это делают птенцы многих видов. Малая подвижность птенцов зяблика особенно бросается в глаза при сравнении их с птенцами, например, славок, которые уже в 3-дневном возрасте могут рывком принять вертикальную позу и долго сохранять ее, сидя на гузке.
192
Рис. 46. Однодневный птенец зяблика (фото автора).
Рис. 47. Птенец зяблика в момент оставления гнезда. На голове видны остатки эмбрионального пуха (Савальский лесхоз, июль 1952 г. Фото автора).
Глаза и ушные проходы у птенцов зяблика начинают приоткрываться в возрасте 4 дней; трубочки первостепенных маховых перьев раскрываются в возрасте 7 дней, а рулевых — днем позднее. Кисточки второстепенных маховых и кроющих крыла появляются в конце 6-го дня жизни. На И—10-й и даже 9-й день своей жизни птенцы при испуге уже-могут выскочить из гнезда. Нормальное же оставление гнезда происходит обычно в возрасте 12—13. реже 14 дней (рис. 47). Как правило,, оставляя гнездо, птенцы по-настоящему еще не умеют летать. Вылетая: из гнезд, они очень часто попадают на землю, где и принуждены бывают существовать 1—2 дня, шока не научатся летать. Слетки зяблика,, сидящие в траве, — явление столь же обычное, как, например, и прыгающие по земле птенцы дроздов. Только некоторые птенцы, оставляющие гнездо в возрасте 14 дней, могут уже перелетать с дерева на дерево..
Призывный крик птенцов-слетков по характеру очень напоминает чириканье молодых воробьев. Обычно он начинает воспроизводиться незадолго до вылета птенцов из гнезда.
Питание гнездовых птенцов. Из пищеводов птенцов, зябликов удалось извлечь всего лишь 23 экз. беспозвоночных животных, и 1 семечко растения. Среди насекомых преобладали гусеницы бабочек (пяденицы), а также крупные мухи (Asilidae и др.). Кроме того, зяблики приносили своим птенцам уховерток и пауков. В Ленинградской области взрослые зяблики в гнездовой период (май—начало июня) охотно поедают семена ольхи, которые остаются лежать на земле после-того, как сойдет снег. Особенно большое значение этот корм имеет для них в периоды длительных весенних холодов, когда активность насекомых почти совсем прекращается. И. В. Прокофьева (устное сообщение) установила, что семена ольхи и ели зяблики нередко приносят также и своим птенцам. В целом, однако, среди европейских видов вьюрковых: птиц птенцы зяблика отличаются наибольшей степенью насекомоядное™.
ВОРОБЕЙ ПОЛЕВОЙ (PASSER MONTANUS L.)
Сроки размножения. Гнездование полевого воробья изучалось-в Заволжье, в системе Тимашевских лесных полос, в 1939 г. Большинство полевых воробьев приступает здесь к размножению сравнительнопоздно. Наиболее ранние кладки этого вида были найдены нами лишь, в середине мая. Однако, судя по тому, что в начале июня появляются первые птенцы-слетки, отдельные особи начинают, видимо, гнездиться уже в самом начале мая. В то же время в 20-х числах мая здесь можно встретить не разбившихся еще на пары, явно негнездовых воробьев. Период откладки яиц продолжается все лето, вплоть до начала августа. В середине августа во многих гнездах были еще птенцы. В табл. 50 приведены результаты периодических осмотров гнезд полевого воробья^ проведенных в период с 19 мая по 15 августа 1939 г.
Таким образом, вылет птенцов полевого воробья из гнезд происходит в течение всего -июня, июля и значительной части августа. Столь-сильную растянутость периода размножения следует объяснять тремя причинами: а) разными сроками готовности к размножению отдельных особей; б) наличием запоздалых дополнительных кладок, возникающих в случае гибели первых; в) характерным для многих особей двойным циклом размножения. По всей вероятности, первая из указанных причин часто является следствием двух последующих: птенцы, по той или иной причине появившиеся на свет в конце июля или начале августа, на сле-
194
Таблица 50
Сроки размножения полевого воробья в системе Тимашевских лесных полос
Дата наблюдения Обнаружены гнезда Примечание
строящиеся с яйцами с птенцами
19 V 18 1 — Повсюду заметны воробьи,, летающие с гнездовым материалом
• 27 V 7 11 —
7 VI 14 18 9 Замечены первые птенцы,, покинувшие гнезда; постройка, гнезд продолжается
16 VI 11 13 11 Молодые птицы держатся, стайками по 20—30 шт.; постройка гнезд продолжается
21 VII 2 <13 22 На полях заметны стаи в.
150—200 особей; отдельные воробьи строят еще гнезда
15 VIII — — 10 На полях держатся тысячные стаи воробьев
дующий год приступают к размножению, видимо, позднее птенцов, вылетевших из гнезд в начале июня. Старые же птицы (по второму, третьему и т. д. году) начинают откладку яиц, по всей вероятности, еще-раньше.
Птенцы. Птенец совершенно голый, цвет кожи розоватый. Клюв короткий и массивный, воскового цвета. Яйцевой «зуб» одноцветный с клювом, когти белые. Ротовая полость желтовато-розовая. Складка в углах рта белая, довольно широкая. Пяточная мозоль небольшая. Прогнатизм нижней челюсти не выражен. Разрез ротовой полости широкий. Пища не задерживается в пищеводе, а сразу проходит в желудок, как это характерно для птенцов насекомоядных птиц. Лишь у подросших: птенцов, когда пища их становится преимущественно растительной, пищевод начинает выполнять функцию зоба. С первого же дня своего существования птенцы полевого воробья отличаются чрезвычайно большой активностью. В возрасте 2—3 дней они уже способны совершать направленные передвижения, энергично работая всеми четырьмя конечностями.
Судя по литературным данным (Келейников, 1953), глаза у птенцов полевого воробья открываются на 6—7-й день, трубочки перьев начинают раскрываться на 8-й день, а из гнезда они вылетают в возрасте 15—16 дней. К. Н. Благосклонов (19506) сообщает, что в этом возрасте птенцы полевого воробья уже хорошо летают. По всей вероятности, способность к полету приобретается за день-два до вылета из гнезда. Таким образом, воробьята развиваются лишь немногим медленнее птенцов открыто гнездящихся видов. Сильнее всего запаздывает развитие глаз,, что вообще характерно для птенцов, воспитывающихся в дуплах или закрытых гнездах.
Питание гнездовых птенцов. Анализ содержимого 36 желудков птенцов, добытых в 1939 г. (Куйбышевская область), показал^ что в первой половине лета полевые воробьи выкармливают своих птен-13* 195
цов исключительно насекомыми. В этот период года взрослые воробьж также предпочитают, питаться животной пищей. Позднее, когда пшеница достигает стадии молочной зрелости, полевые воробьи переключаются в основном на растительный корм, который приносят также и птенцам. По А. Л. Келейникову (1953), растительный корм птенцы начинают получать, однако, не раньше 4-дневного возраста. В целом в гнездовой период птенцы полевого воробья более насекомоядны, чем взрослые птицы. Основу их питания составляют вредители полеводства. В желудках птенцов полевого воробья чаще всего встречались листогрызы (Ну-pocassida, Cassida и др.), долгоносики (Tanimecus, Bothynoderus), клопы (Aelia, Eurygaster), а также саранчовые (Chorthippus и др.); из семян сельскохозяйственных культур ими поедались семена пшеницы и проса. По наблюдениям Г. А. Новикова (устное сообщение) в «Лесу на Ворскле», полевые воробьи в первой половине лета поедают большое количество семян ильмовых пород, после того как они опадут на землю.
ОВСЯНКА ОБЫКНОВЕННАЯ (EMBERIZA CITRINELLA L.) •
Токование. В Ленинградской области пение обыкновенной .овсянки можно услышать уже с середины марта. В это время поют лишь немногие самцы, и песня их звучит крайне нерегулярно. Регулярное пение начинается с конца I декады апреля и продолжается до последней декады июля. Отдельные самцы иногда поют еще и в начале августа. Так, в 1946 г. 6 августа в районе ст. Елизаветино Ленинградской области автор наблюдал поющего самца. Рядом находилась самка, таскавшая корм птенцам, сидевшим ib гнезде. Таким образом, в данном случае пение -самца, несмотря на его чрезвычайно поздние .сроки, указывало на гнездование птиц.
Расположение гнезда. В лесостепной зоне обыкновенная овсянка в гнездовой период ведет себя как настоящий лесной вид. Здесь она гнездится, как правило, внутри лесных массивов. Обычные же для нее места — опушки, вырубки и выгоны — в лесостепи заняты садовой овсянкой. Подобная картина распределения гнездовых стаций между двумя указанными видами наблюдалась нами как в районе заповедника «Лес на Ворскле», так и .на территории Савальского лесхоза.
Почти все найденные нами гнезда обыкновенной овсянки располагались на земле, под защитой кустов, древесной поросли или среди высокой травы. -В местах, лишенных лесного полога и травянистой растительности, этот вид, как правило, не гнездится. Иногда обыкновенная овсянка помещает свое гнездо над землей. Одно такое гнездо было найдено на территории ,Савальского лесхоза, другое — в учлесхозе «Лес на Ворскле». Помещались они в кустах бузины и сирени на высоте 1 и 1,8 м от земли. Случаи гнездования обыкновенной овсянки на кустах отмечали также Н. А. Зарудный (1910) и В. Н. Шнитников (1913).
Сроки размножения. В Ленинградской области свежие кладки обыкновенной овсянки можно находить начиная с первых .чисел мая. Птенцов, только что оставивших гнездо в возрасте 9—10 дней, автор неоднократно ловил в самом начале июня (наиболее ранняя дата—1 июля 1948 г.), в то же время 6 августа (1946) автор наблюдал обыкновенных овсянок, выкармливавших насекомыми птенцов, находившихся еще в гнезде. Таким образом, в лесной зоне на широте
196
Ленинграда откладка яиц у обыкновенных овсянок возможна вплоть до -второй декады июля. Разница же между наиболее ранними и поздними датами появления яиц .в отдельных гнездах может быть равной почти 2,5 мес. Очень -сильная растянутость сроков размножения позволяет подозревать существование двойного цикла гнездования у отдельных птиц.
У овсянок, гнездящихся в лесостепной зоне, несмотря на значительно большее количество собранных данных, нам не приходилось находить свежих июльских кладок. Здесь наиболее поздние сроки появления яиц в гнездах 'были отмечены лишь в последней декаде июня. Самый ранний срок откладки яиц, который удалось установить,— 5 ,мая. Таким образом, весь период размножения у савальской популяции оказался примерно на 15 дней короче, нежели у овсянок, гнездящихся под Ленинградом. Сроки размножения обыкновенной овсянки в Савальском лесхозе изучались в ,1951 —1953 гг. Ниже приводятся данные о количестве гнезд, в которых откладка яиц началась:
В I декаде мая..........4
Во II декаде мая..........5
В III декаде мая..........9
,В I декаде июня..........4
Во II декаде июня..........4
В III декаде июня..........4
Размеры и судьба к л а д о к. В пределах лесостепной зоны под наблюдением находилось 36 гнезд обыкновенной овсянки. Количество яиц в этих гнездах было следующим: в 4 — по 2, в 5 — по 5, в 15 — по 4, в 12 — по 5, в 3—по 6 яиц.
История кладки и выводка была прослежена до конца в 30 гнездах, в которых первоначально было 138 яиц. Из них вылупилось 98 птенцов: 29 яиц (6 кладок) погибло в результате разорения гнезд, 1 кладка (5 яиц) была брошена птицами в период насиживания-и 6 яиц оказались болтунами. Отход птенцов также был значительным: из 98 птенцов выжило и покинуло гнездо 64. Основной причиной высокой смертности птенцов явились хищники как наземныё, так и воздушные. Среди последних чаще других разоряют гнезда обыкновенных овсянок, видимо, ворона и сойка. Из наземных хищников в похищении птенцов овсянок больше всего -следует подозревать ласку или горностая.
Поведение взрослых птиц у гнезда. Несмотря на наземное расположение гнезда обыкновенная овсянка обычно не «отводит» от гнезда. Самка, сидящая на гнезде, при приближении к ней с шумом вылетает да-под самых ног наблюдателя либо незаметно отбегает в сторону и затем уже взлетает. После этого в большинстве случаев овсянки сидят в стороне, изредка воспроизводя позывы, выражающие ,их возбуждение. Эти позывы могут быть двух родов: птица тонко и протяжно свистит или издает короткий, отрывистый, «цыкающий» звук.
Птенцы. Птенец покрыт буровато-серым, довольно длинным и густым пухом, который расположен на надглазничных, затылочных, спинном, бедренных и (очень жидкий и короткий) на брюшных и го-лен.ных участках тела. Кожа розовато-оранжевая. Клюв сравнительно слабый, серый, с темной предвершинной полосой. Ноздри продолговатые, Когти светлые, яйцевой «зуб» белый. Ротовая полость мясо-красного цвета, сходного с окраской рта вьюрковых. Боковая складка клюва довольно широкая, бледно-желтая. Разрез рта тоже широкий. Пища, принесенная родителями, не задерживается в пищеводе, как это характерно для птенцов большинства вьюрковых птиц, а сразу проходит в желудок.
197
Принято считать, что в связи с наземным расположением гнезд рост и развитие птенцов овсянок идет значительно быстрее, чем у птиц, располагающих гнезда на деревьях. Наблюдения, однако, показывают, что по многим показателям 'скорость развития птенцов обыкновенной овсянки (Примерно такая же, как, например, у птенцов зяблика. В обоих случаях глаза становятся «приоткрытыми в возрасте 4 дней, а трубочки маховых перьев раскрываются в 6—7-дневном возрасте. Из перьев крыла раньше других начинают раскрываться трубочки кроющих и .второстепенных маховых. Ранняя способность ’птенцов к самостоятельному передвижению и быстрые темпы развития контурного пера — характерные особенности развития овсянок, которые можно связывать с наземным гнездованием. Развертывание пеньков контурного лера у обыкновенной овсянки начинается уже в возрасте 4 дней.
Птенцы оставляют гнездо обычно уже в возрасте 9—10 дней. В отдельных случаях птенцы покидают гнездо даже в возрасте 7 дней, с еще едва-едва раскрывшимися пеньками перьев.
Способность к полету приобретается спустя несколько дней после оставления гнезда. Наземный период жизни у птенцов овсянок продолжается дольше, чем у птенцов большинства других видов воробьиных. Они предпочитают держаться на земле даже тогда, когда станут уже вполне летными. Даже взрослые овсянки имеют обыкновение в летнюю пору ночевать на земле, забившись в траву где-нибудь на вырубке или на опушке. Эти особенности поведения обыкновенных овсянок наряду с некоторыми другими их биологическими .признаками (наземное расположение гнезда, тяготение к вырубкам и т. п.) указывают на исторические связи данного вида с открытым ландшафтом.
Питание гнездовых птенцов. Материалы по питанию птенцов обыкновенной овсянки, собранные на территории Савальского лесхоза, частично уже опубликованы (Нейфельдт, 1956). Здесь укажем лишь, что среди 186 экз. насекомых и других беспозвоночных, извлеченных из пищеводов птенцов этого вида, чаще всего .встречались гусеницы бабочек (совок, пядениц, листоверток, шелкопрядов), мелкие прямокрылые (кобылки) и двукрылые (комары-долгоножки, ктыри). Нередко птенцы обыкновенной овсянки получали от родителей также бабочек имаго (кольчатый шелкопряд, различные совки) и пауков. Другие группы беспозвоночных (жуки, перепончатокрылые, клопы и т. п.) ими почти не поедались.
Растительный корм в пищевом рационе птенцов обнаружен не был. В то же время взрослые овсянки в пору размножения поедают иногда семена травянистых растений; последнее обычно наблюдается в пасмурную, холодную погоду, когда снижается активность насекомых, и птицам становится трудно их раздобывать.
Маленьким птенцам иногда приносятся комочки земли и довольно крупные камешки, играющие, очевидно, роль гастролитов.
ОВСЯНКА САДОВАЯ
(EMBERIZA HORTULANA L.)
Токование. Песни отдельных самцов несколько отличаются по тону, а также по количеству воспроизводимых звуков. Большинство птиц поет неторопливую 4-сложную песню. Однако отдельные самцы слагают свою песню из 5 и даже 6 звуков. Время, затрачиваемое на воспроизведение разных вариантов, одинаковое. Поэтому 6-сложные напевы звучат более поспешно, чем обычные 4-сложные.
198
Расположение гнезда. Почти все найденные нами гнезда садовой овсянки располагались под защитой небольших кустиков древесной -поросли или травы. В Тимашевских лесных полосах одно из гнезд было обнаружено под листом железа. На территории Савальского лесхоза некоторая часть садовых овсянок гнездится в глубине лесного массива, среди разреженных сосняков.
Сроки размножения. Сроки размножения садовой овсянки на территории Савальского лесхоза изучались в 1951—1953 гг. Самая ранняя кладка началась 8 мая (1951). Преобладающее большинство гнезд со свежими яйцами было найдено в конце II, а также в III декаде мая. В июне размножались уже немногие особи. Кладки, появившиеся в июне, чаще всего состояли из 4, реже—из 5 или 3 яиц, тогда 5<ак майские кладки, как правило, содержали 5, иногда 6 или 4 яйца. Гнезд с 3 яйцами в мае месяце нами найдено не было. Сокращенное количество яиц в июньских кладках позволяет предполагать, что в -большинстве своем они были дополнительными, возникшими после гибели первых. Предполагать наличие двух нормальных кладок у садовой овсянки нет оснований.
Ниже приводятся данные о количестве гнезд, в которых началась откладка яиц:
В I декаде мая . . «... 1
Во II декаде мая...........7
В III декаде мая..........18
В I декаде июня..........5
Во II декаде июня..........4
В III декаде июня..........2
Размеры и судьба кладок. Всего мы располагаем данными по истории 33 кладок этих птиц. Общее количество яиц в них составляло 155. Из них вылупился только 101 птенец: 49 яиц (10 кладок) пропало при разорении гнезд, и 5 яиц оказались болтунами. Отход гнездовых птенцов был не столь значительным: из 101 птенца выжило 79. Таким образом, показатель выживаемости потомства у садовой овсянки оказался равным 2,4 птенца на 1 гнездо. Количество птенцов в выводке в момент оставления гнезда варьировало от 3 до 6. Большая часть пропавших яиц и птенцов была, видимо, растащена хищниками. 3 одном случае кладка была раздавлена коровой, в другом — яйца погибли в связи с распашкой участка, на котором гнездились птицы. Один раз автор был свидетелем гибели гнездовых птенцов садовой овсянки от перегрева.
Пов едение взрослых птиц у гнезда. Во время беспокойства у гнезда садовые овсянки в целом ведут себя более возбужденно, нежели обыкновенные. Они менее пугливы, держатся более заметно и чаще подают голос. При виде человека вблизи гнезда или птенцов лишь некоторые особи «притворяются» ранеными.
Систематическое обогревание птенцов самкой продолжается лишь первые 3—4 дня. Позднее самка присутствует на гнезде нерегулярно, защищая птенцов от сильного дождя или яркого солнца. Автор наблюдал садовую овсянку, которая, расправив крылья, стояла над 4— -5-дневными птенцами, защищая их от палящих солнечных лучей, падавших как раз на гнездо. В этот день температура воздуха у земли на солнце доходила до 4-53° С. Все поведение птицы указывало на то, нто ее организм испытывает большое напряжение; овсянка широко -раскрывала рот и очень тяжело дышала. Находившиеся под ней птенцы тоже сидели с открытыми ртами. Спугнутая с гнезда взрослая птица долгое время не возвращалась. Наблюдения за ней показали, •что она, продолжая тяжело дышать, сидела в тени. Во время ее от
199
сутствия все птенцы погибли от перегрева. Поза у всех была однотипная — они лежали на боку, вытянув шеи и приоткрыв рты.
Чувствительность птенцов к воздействию прямых солнечных лучей, видимо, чрезвычайно высока. По нашим наблюдениям, 1—2-дневный птенец садовой овсянки, помещенный на открытое солнце, когда? температура приземных слоев воздуха равна 50—53° С, почти сразу же-резко учащает дыхание, через 2—3 мин. начинает судорожно открывать рот, а через 5—10 мин. после начала эксперимента совсем прекращает дыхание и гибнет. Тем не менее птенцы садовой овсянки сравнительно редко гибнут от перегрева, так как гнезда этого вида располагаются под кустиками травы, крупными листьями и другими предметами, образующими тень. Большое значение имеет также и поведение взрослых птиц, защищающих птенцов своим телом в том случае, если солнечные лучи все же падают на гнездо. Начиная примерно* с недельного возраста, птенцы могут уже сами избежать неблагоприятного воздействия солнечных лучей: будучи в этом возрасте способными к самостоятельному передвижению, они в случае, если их начинает беспокоить солнце, выбираются из гнезда и прячутся под листья лопуха и другие укрытия.
Птенцы. Птенец покрыт светлым желтовато-серым, густым л длинным пухом, расположенным на надглазничных, затылочном, спинном, плечевых, локтевых, бедренных, голенных и брюшных участках: тела. В отличие от обыкновенной овсянки пух на брюшке довольно-густой. Кожа мясного цвета с оранжевым оттенком. Клюв сверху бурый, с более темной вершиной и резко вздутый в области ноздрей. Ноздри овальные. Яйцевой «зуб» темный, когти светлые. Ротовая полость мясо-розовая.
Развитие птенцов садовой овсянки протекает примерно так жег как и у птенцов предыдущего вида. Очень рано, в конце 3-го дня жизни, открываются слуховые проходы; глаза бывают приоткрыты уже на* 4-й день, а опахала растущих маховых начинают развертываться по* истечении 7 дней. Оставление гнезда происходит, как правило, в возрасте 8—9, иногда даже 7 дней. Способность к активному полету приобретается примерно на 14—15-й день. Целую неделю после оставления гнезда птенцы садовой овсянки ведут наземный образ жизни и не уходят далеко от гнезда. Только с приобретением способности к полету начинаются более значительные перекочевки. Окольцованные-автором 8-дневные птенцы садовой овсянки, покинувшие гнездо* 13 июня (1953), через 5 дней (18 июня), т. е. в возрасте 13 дней, были; обнаружены в 50 м от гнезда. Птенцы еще не могли летать по-настоящему. Через 2 дня они уже были способны перелетать с места на место.
Питание гнездовых птенцов. Исследование проводилось, на территории Савальского лесхоза. Всего из пищеводов птенцов этого* вида было извлечено 190 порций корма, содержавших 405 экз. различных беспозвоночных. Из них больше половины (245) составляли бабочки и их гусеницы. Из бабочек чаще всего поедалась сосновая пяденица, особенно в 1954 г. — в год массового ее размножения. На втором’ месте по количеству экземпляров (74) стояли жуки, главным образом июньский и садовый хрущики. Были обнаружены также саранчовые-(46) и различные двукрылые (32): ктыри, комары-долгоножки и др. Июньские хрущи отдавались птенцам с оторванными надкрыльями и. головогрудью. Саранчовые в более или менее заметном количестве-стали попадаться в пище птенцов лишь с июля месяца. Пауки поедались птенцами сравнительно редко (всего 5 экз.). Наземные моллюск»
200
совсем не были обнаружены в пищеводах птенцов. Растительный корм был зарегистрирован только в 2 случаях. Взрослые же садовые овсянки хотя и в незначительном количестве, но регулярно поедают семена растений в течение всего весенне-летнего сезона (см. рис. 27).
КОНЕК ЛЕСНОЙ
(ANTHUS TRIVIAL IS L.)
Токование. Наиболее ранняя дата первой песни лесного конька, слышанная нами в окрестностях Ленинграда, — 27 апреля (1950); наиболее позднее пение отмечено 22 июля (1947); регулярное пение бывает слышно с конца I декады мая до конца II декады июля, т. е. продолжается свыше 2 мес. В конце июня — первой половине июля в-Ленинградской области наблюдается заметное повышение активности пения самцов, которое продолжается около 2 недель и затем резко падает. В Воронежской области активность пения лесных коньков также заметно повышается к концу июня—. началу июля. В поисках самок самцы лесных коньков меняют места своего постоянного пребывания иногда даже в середине лета. Например, на территории парка Лесотехнической академии 29 июня 1947 г. внезапно появился активно поющий самец, который обосновался на участке, где лесные, коньки не наблюдались в течение всей предшествующей части сезона.
Расположение гнезда. В Ленинградской области лесной конек гнездится обычно на опушках и вырубках, а в лесостепной зоне, как правило, — в глубине лесных массивов. На территории Са-вальского леса, в заповеднике «Лес на Ворскле», а также в Куйбышевской области (район Тимашева) преобладающее большинство гнезд этого вида было найдено нами среди спелого древостоя. Располагались они, как правило, под защитой кустов, травы или лежащих на земле ветвей. Одно из найденных автором гнезд помещалось под корнями срубленного вяза, на 6 см ниже уровня лесной подстилки.
Сроки размножения. На территории Савальского лесхоза в 1951—1953 гг. период возможных сроков откладки яиц у лесного конька оказался растянутым примерно на 2 мес. В наиболее ранних гнездах яйца появились в середине I декады мая (6—7), а в наиболее поздних — в первых числах июля. Заметного «пика» размножения выявить не удалось: начиная со II декады мая и до конца II декады июня можно было находить большое количество гнезд с совершенно свежими кладками. Спад интенсивности размножения наметился только лишь в конце июня.
Ниже приводятся данные о количестве гнезд, в которых откладка яиц началась:
В I декаде мая..........5
Во II декаде мая..........9
В III декаде мая.........12
В I декаде июня..........11
Во II декаде июня..........11
В III декаде июня............4
В I декаде июля..............2
По всей вероятности, часть коньков, успешно воспитавшая свое потомство в мае, приступает к размножению вторично в июне. На вероятность двойного цикла размножения у части особей косвенным образом указывает также двойной период повышенной активности пения самцов.
В Ленинградской области, несмотря на более поздние начальные сроки размножения, некоторые птицы, по-видимому, тоже успевают вывести потомство 2 раза в лето. Здесь, как и в лесостепной полосе,
201
сроки появления свежих кладок варьируют в пределах более, чем 2 мес. (табл. 51).
Таблица 51
Сроки размножения отдельных пар лесных коньков в Ленинградской области.
Год наблюдений Откладка первого яйца Вылупление птенцов Оставление гнезда птенцами
1950 9 V 27 V 6 VI
1946 10 V 27 V Не прослежено
1946 14 V 31 V 10 VI
1954 16 V 3 VI 14 VI
1954 26 V 12 VI 23 VI
1950 14 VI 1 VII Не прослежено
1950 16 VI 2 VII 11 VII
1950 20 VI 6-7 VII Гнездо разорено
1950 20 VI 6 VII Не прослежено
1946 15 VII Не прослежено То же
Размеры и судьба кладок. В лесостепной зоне под наблюдением находилось 63 гнезда, история которых была прослежена до конца. Количество яиц в отдельных кладках варьировало от 3 до 6. Наибольшее количество гнезд содержало 5 яиц (32 находки), затем 4(17), 6(9) и, наконец, 3 яйца (5 находок). Под наблюдением находилось 297 яиц, из них вылупилось 223 птенца. Отход яиц составил 73 яйца: 40 погибло при разорении 9 кладок хищниками, 22 яйца (5 кладок) было брошено птицами во время насиживания, и И яиц оказались болтунами или содержали мертвые эмбрионы. Из 223 птенцов погибло 75. Таким образом, из 297 яиц в конечном счете получилось 153 птенца, что в среднем составило 2—3 птенца на 1 гнездо. По отношению к первоначальному количеству яиц общий отход яиц и птенцов был равен 50%. Основная причина высокого процента отхода — частое разорение гнезд различными хищниками. Среди последних следует указать на ласку, горностая, бродячих кошек и других наземных хищников, а также на сойку, ворону и сарыча. Взрослые птицы во время насиживания кладки и обогревания птенцов сравнительно редко попадают в Лапы хищников: из 72 самок, за которыми мы наблюдали, только одна была схвачена каким-то наземным хищником.
Поведение взрослых птиц у гнезда. При появлении человека вблизи гнезда лесные коньки ведут себя обычно очень беспокойно. Держатся они почти всегда на почтительном расстоянии в кронах деревьев, беспрестанно издавая свой короткий «цыкающий» позыв. Этот позыв, кажущийся тихим, человеческое ухо может воспринимать на расстоянии 300 м. При сильном волнении помимо «цыкающего» позыва лесные коньки иногда воспроизводят высокую непродолжительную трель, очень напоминающую крик молодых вертишеек. Этот крик тоже имеет значение «сигнала тревоги». В присутствии человека лесные коньки почти никогда не опускаются к гнезду, даже в период наиболее интенсивного кормления птенцов. Поэтому выследить гнездо, руководствуясь поведением птицы, как это легко сделать, например, у пеночек, 'обычно не удается.
Явление «отвода» для лесных коньков не характерно. Только несколько раз нам приходилось наблюдать самок, «отводящих» от гнезда.
Птенцы. Пух, покрывающий тело птенца, темно-серый, длинный и густой, особенно на голове; расположен на глазных, надглазничных, 202
затылочной, спинной, плечевых, локтевых, бедренных, голенных и брюшных птерилиях (рис. 48). На последних пух очень редкий и бывает не всегда. Кожа грязно-розового цвета. Клюв воскового цвета, с заметным (потемнением на конце, вздутый в области, лежащей кнутри от ноздрей. Ноздри 'продолговатые. Яйцевой «зуб» темный, когти светлые. Ротовая полость ярко-оранжевая, складка в углах рта светло-желтая.
Процесс развития птенцов протекает очень однотипно. Во всех наблюдавшихся нами случаях (около 50 птенцов) ушные отверстия открывались в -конце 3-го дня жизни, а глаза—несколько позднее,
Рис. 48. Полуторадневный птенец лесного конька (фото автора).
в 4-дневном возрасте. Однодневные птенцы лесного конька выпрашивают корм молча или издавая слабый, едва различимый писк. Голос их становится слышимым на расстоянии 10—15 м только на 7—8-й день, т. е. примерно за 2—3 дня до оставления гнезда.
Несмотря на наземное расположение гнезда, рост и развитие перового покрова идет не быстрее, чем у большинства мелких воробьиных, гнездящихся на деревьях. Кисточки маховых перьев появляются лишь в возрасте 7 дней. Первыми раскрываются пеньки второ-степенных и внутренних первостепенных, а также кроющих крыла. Кисточки контурного пера на спине и груди появляются в конце 6-го дня жизни птенца. Оставление гнезда происходит обычно в 9—11-дневном 'возрасте. При сильном испуге птенцы иногда выскакивают и на 8-й день. В этом возрасте они уже прилично передвигаются по земле. При этом они, как взрослые коньки, поочередно переступают ногами.
Способность к полету приобретается спустя несколько дней после оставления гнезда (рис. 49). Все это время птенцы держатся на земле, ловко скрываясь в траве.
203
Питание гнездовых птенцов.1 Чаще всего лесные коньки -приносили птенцам двукрылых, главным образом настоящих мухг затем гусениц и бабочек, тлей и пауков. Жуки и клопы поедались крайне редко, а прямокрылые (бескрылые кобылки) —лишь при позднем, гнездовании — в конце июня — начале июля. Изредка птенцы получали, мелких наземных моллюсков, которых птенцы проглатывали вместе с раковинками. Один раз была принесена многоножка — кивсяк.
Таким образом, лесные коньки выкармливают птенцов главным, образом мелкими беспозвоночными: мухами, пауками, тлями, мелкими
Рис. 49. Птенец лесного конька, недавно научившийся летать. Ему немногим больше 2 недель (Савальский лесхоз. Июнь 1952 г. Фото автора).
бабочками и их гусеницами. Несколько раз было отмечено, правда,» и поедание сравнительно крупных насекомых — мух семейства Asilidae и бабочек кольчатого шелкопряда.
Подлетая к гнезду, лесной конек каждый раз -кормит обычно только одного птенца, который иногда получает корм несколько раз подряд, пока не насытится.
СИНИЦА БОЛЬШАЯ
(PARUS MAJOR L.)
Токование. Среди птиц, не обнаруживающих склонности к пе-ресмешничеству, трудно найти другой вид, у которого бы столь резко была выражена индивидуальная изменчивость весенней песни. Отдельные самцы этого вида отличаются не только по тембру и ритму исполнения песни, но и по количеству издаваемых звуков. Встречаются певцы, напевающие трех-, двух- и даже односложные колена. В последнем
1 В изучении питания птенцов лесного конька принимала участие И. А. Ней-фельдт, материалы которой уже опубликованы (Нейфельдт, 1956).
204
«случае песня большой синицы походит на напев пухляка -и звучит, как ритмичное повторение одного звука, воспроизводимого в одном и том же тоне. В том случае, когда синица высвистывает двух- или трехсложный напев, он может быть однотонным или двутонным (с повышением или понижением на последнем слоге). Птицы, поющие трехсложные и одновременно трехтонные песни, встречаются очень редкол Как правило, один и тот же самец может исполнять несколько вариантов песни, хотя всегда бывает ведущая, наиболее часто воспроизводимая вариация. Сколько-нибудь заметной географической изменчивости пения у нашего подвида большой синицы автору наблюдать не приходилось.
Пение большой синицы можно услышать почти в любое время года, котя настоящее громкое весеннее пение этого вида звучит только лишь в конце зимы, весной и летом. Осеннее пение отличается меньшей силой звука и поспешностью исполнения. Регулярное пение больших синиц в окрестностях Ленинграда продолжается около 4 мес. и бывает слышно с середины марта до II декады июля. В конце апреля — первой половине мая, а также во второй половине июня — начале июля большие синицы поют особенно активно. Два периода повышенной активности пения самцов обусловлены двойным циклом размножения синиц. Второй период повышенной активности самцов особенно ярко выражен в лесостепной зоне.
Расположение гнезда. Характер расположения гнезд большой синицы говорит о пластичности ее гнездовых инстинктов. Она заселяет расположенные на различной высоте дупла с разнообразной величиной входного отверстия, а также гнездится в гнилых пнях, щелях и изъянах зданий. Интересный факт сообщает Б. К. Штегмап (1951): одна пара синиц гнездилась несколько лет подряд в выхлопной трубе старого мотора.
При наличии благоприятных кормовых условий большая синица "может выводить потомство также и в молодых лесопосадках, где нет естественных дупел, поселяясь в других закрытых убежищах, в частности в сорочьих гнездах. В отдельных случаях этот вид может переводить даже к совершенно открытому гнездованию. Например, на участке молодых лесопосадок Савальского лесхоза в 1951 г. автор нашел гнездо с птенцами большой синицы, которое располагалось на ветвях невысокой елки, на высоте около 2 м. Оно состояло лишь из толстой войлочной подстилки и небольшого количества мха; основой ему служило старое гнездо сойки. О втором «случае открытого гнездования большой синицы сообщил автору С. М. Марин, нашедший в полезащитных полосах Каменной степи свежую кладку этой синицы в старом сорочьем гнезде, лишенном крыши.
Другой особенностью гнездового поведения рассматриваемого вида является способность совмещать процесс откладки яиц с гнездострое-тшем. Обычно, не закончив еще строительства, птица приступает уже к откладке яиц. Автору неоднократно приходилось наблюдать синиц, таскавших гнездовой материал (пучки шерсти) в скворечник, в котором были уже яйца. В результате еще не оконченная кладка оказывалась покрытой слоем шерсти. На следующий день в скворечнике мы находили новое отложенное яйцо, а сверху опять появлялся войлок. Подобное поведение большой синицы наблюдается как при первом, так и при втором цикле размножения, за исключением тех случаев, когда синица вторично гнездится в том же гнезде.
Обычно принято считать, что синица в период кладки специально прикрывает яйца шерстью с тем, чтобы предохранить их от возможного охлаждения во время ее отсутствия (Лихачев, 1954). Однако синица
205
не прикрывает яйца шерстью после начала насиживания, когда развитие эмбриона уже началось и когда, казалось бы, яйца больше нуждаются -в предохранении. По нашему мнению, рассматриваемая особенность поведения большой синицы возникает в результате совмещения периода откладки яиц и гнездостроения. Судя по указанию Штейна (Stein, 1927), другие виды синиц (лазоревка, пухляк) также покрывают яйца гнездовым материалом, продолжая таскать его в тот период,, когда откладка яиц еще не окончена.
Совмещение двух этапов репродуктивного цикла у большой синицы имеет определенный биологический смысл. В связи с большими размерами кладки и длительным периодом воспитания потомства продолжительность цикла размножения синицы примерно на 2 недели больше, чем, например, у славок, пеночек, дроздов и других воробьиных. В то же время она -обычно успевает вывести 2 семьи в лето. Сокращение: периода размножения даже на несколько дней имеет большое значение. Это и достигается путем совмещения во времени различных этапов гнездовой жизни, которые у других птиц обычно закономерно сменяют друг друга. Наблюдения показывают, что большая синица способна совмещать процесс откладки яиц не только с гнездостроением^ но даже и с воспитанием птенцов. Так, например, М. А. Воинственский. (1949), Г. Н. Лихачев (1954) и другие отмечают, что эта синица при переходе ко второму циклу размножения первые яйцй откладывает часто тогда, когда в гнезде находятся еще птенцы первого выводка.
Укажем, наконец, на. способность большой синицы совмещать процесс выкармливания и обогревания птенцов с гнездостроением. Это-было установлено экспериментальным путем. У одной пары синицг у которой недавно вылупились птенцы, 2 июля 1953 г. было отобрано все гнездо, а птенчики были положены на дно скворечника на слой. сухих листьев. 10 июля в этом скворечнике под птенцами мы обнаружили слой войлока, принесенный взрослыми птицами, сухие же листья сказались выброшенными наружу. Впоследствии этот эксперимент был. повторен автором 2 раза. В одном случае синицы выбросили из дуплянки сухие листья и притащили вместо них зеленый мох, а в другом — окружили 2—4-дневных птенцов кольцом строительного материала, состоявшим из конского волоса, корешков и небольшого количества^ шерсти. Поведение птиц в данном случае было весьма целесообразным: окружавший птенцов валик не позволял им расползаться в стороны,, а заставлял сидеть компактной кучкой, без чего не мог осуществляться процесс их обогревания.
Сроки размножения. Период возможных сроков откладки, яиц у большой синицы примерно 2,5 мес. — с начала последней декады апреля и до конца июня. Максимальная разница в сроках откладки яиц в разные гнезда в пределах одного и того же года оказалась равной 66 дням. Судя по датам нахождения свежих кладок, большая синица в Савальском лесхозе имеет два цикла размножения. Это подтверждается и наблюдениями за отдельными окольцованными самками. В этом отношении большая синица в Савальском лесхозе ведет себя так же, как в Латвии (Тауриньш и Тима, 1953), в Тульских засеках и Приокско-Террасном заповеднике (Лихачев, 1954), на Украине (Таращук, 1953) и в других местах, где было установлено, что этот: вид синицы имеет обычно 2 кладки в сезон.
Сроки размножения отдельных пар как при первом, так и при втором гнездовании далеко не всегда одинаковы. Обычно наблюдаются две сильно растянутые волны появления свежих кладок со слабо выраженными пиками в начале мая и во второй половине июня.
206
Некоторые синицы, по всей вероятности, впервые приступают к размножению лишь в конце мая— начале июня (молодые птицы из-прошлогодних вторых выводков). Они, видимо, размножаются только-1 раз в сезон. На это указывает то, что во второй .половине июня среди кочующих по лесу стай молодняка всегда присутствует некоторое количество взрослых -особей. Старые птицы, начинающие откладку яиц. в конце апреля — начале мая, вторично гнездятся во II и III декаде июня.
В Ленинградской области были установлены следующие начальные сроки откладки яиц в гнездах большой синицы: 2 мая, 12 маяг 1 июня (дважды), 22 июня и 23 июня. Эти даты не являются, конечно, предельными. Вполне вероятны случаи как более раннего, так и более позднего размножения. Период возможных сроков откладки яиц у большой синицы растянут здесь тоже примерно на 2 мес. Часть больших сйниц успевает вывести потомство дважды в лето и на широте Ленинграда. Наблюдения за . отдельными окольцованными синицами,, проводившиеся Е. В. Лукиной (1951), показали, что самка, отложившая кладку в первой половине мая, может вторично загнездиться уже в начале III декады июня.
Размеры и судьба кладок. На территории Савальского-лесхоза в 1952—1955 гг. под наблюдением 1 находилось 148 гнезд, в которых первоначально было 1416 яиц (в среднем 9,5 на 1 гнездо). Из-них вылупилось 1228 птенцов (в среднем 8,6 на 1 гнездо). Общий отход составил 188 яиц: 23 яйца (5 неполных кладок) было брошено синицами, 73 яйца (12 кладок) погибло в результате разорения гнезд большим пестрым дятлом и серой вороной и, наконец, 92 яйца оказалось болтунами. Относительно очень высокий процент болтунов (около 6,5) следует объяснять тем, что в результате совмещения периода откладки яиц и гнездостроения, о чем говорилось выше, отдельные яйца оказываются затерянными в гнездовой подстилке и изолированными от источника тепла.
Отход птенцов в гнездах был сравнительно небольшим: из 1228 птенцов погибло 155. Остальные 1073 благополучно закончили развитие и вылетели из гнезд.
Основной причиной смертности гнездовых птенцов является отход отдельных наиболее слабых членов выводка. По этой причине в одном гнезде может погибнуть сразу несколько (до 5) птенцов, чаще всего это случается с птенцами, вылупившимися последними. Подобная участь постигла 92 птенцов из 155 погибших. Остальные 63 птенца погибли в результате разорения гнезда или по другим неизвестным для нас причинам.
Поведение взрослых птиц у гнезда. Некоторые особи, если их сильно испугать, довольно легко бросают не только свежие, но и сильно насиженные кладки. При сильном испуге, например после отлова в скворечнике, большие синицы бросают даже птенцов. Известно также, что синица нередко бросает кладку из-за скопления в гнезде блох. По наблюдениям Г. Н. Лихачева (1954), это чаще всего случается, когда синица после первого вывода птенцов второй раз гнездится в том же дупле или скворечнике. Подобные же случаи наблюдались нами на территории Савальского лесхоза, -где синицы из-за блох иногда бросали кладку непосредственно перед вылуплением птенцов.
1 Помимо автора в наблюдениях участвовали Н. П. Кадочников и студенты: С. В. Васильев, Ю. К. Эйгелис и Д. С. Бекжанова.
207
Птенцы. Пух, покрывающий тело птенца, дымчато-серый, довольно длинный, но редкий, особенно на спине и плечах; расположен на надглазничных, затылочной, плечевой и спинной птерилиях (рис. 50). Цвет кожи розовато-оранжевый. Клюв светлый с предвершинной темной полосой. Ноздри круглые. Складка в углах рта ярко-белая и широкая. Яйцевой «зуб» светлый, когти темные. Ротовая полость желтая. Будучи вынутыми из дупла, птенцы в своем стремлении забраться в темное место уже в 3-дневном возрасте показывают способность к направленному передвижению, что совершенно не характерно для слепых еще птенцов открыто гнездящихся видов воробьиных. Во время передвижения птенец большой синицы вытягивает шею и начинает уси-
Рис. 50. Однодневный птенец большой синицы (фото автора).
ленно работать всеми четырьмя конечностями: резко отталкивается задними, приподымаясь на передних. На тактильное или звуковое раздражение он моментально реагирует резким выбросом головы кверху и настолько быстрым выпрямлением туловища, что нередко опрокидывается на спину. Голос птенцы начинают подавать с первого же дня их жизни.
Большую подвижность птенцов следует связывать с закрытым гнездованием и большим количеством птенцов в выводке. Птенец слабый, малоразвитый легко может оказаться затоптанным своими чрезвычайно энергичными собратьями. Без повышенной активности птенцов у большой синицы вряд ли могло осуществиться равномерное выкармливание столь многочисленного выводка в условиях тесного и узкого дупла.
Время открытия глаз и начала развертывания опахал маховых перьев по сравнению с большинством мелких воробьиных, строящих открытые гнезда, запаздывает у большой синицы нд 1—2 суток. Наоборот, ушные отверстия открываются примерно тогда же, когда и у остальных видов, — в возрасте 4 дней. Глаза приоткрываются в воз-
208
расте 7 дней, и только на 9-й день появляются кисточки маковых перьев. Кисточки рулевых появляются почти одновременно с маховыми. Лететь, набирая высоту, птенец большой синицы может в возрасте 17 дней; оставление же гнезда происходит обычно, когда птенцам исполнится 18 или 19 дней. Как это уже упоминалось в литературе (Промптов и Лукина, 1938), перед вылетом из дупла птенцы начинают особенно часто воспроизводить позывы, имеющие призывное значение, звучащие как «цизмзи-цизизизи». После оставления гнезда этот позыв они воспроизводят беспрестанно. Биологическое значение егоогромно:,он служит основным связующим звеном между выводком, постоянно меняющим место своего пребывания, и родителями, продолжающими выкармливать птенцов. У большой синицы нередко бывает значительная разница в величине птенцов в выводке—результат разновозрастное™, возникающей в »авязи с началом насиживания ^иц до окончания кладки. Чаще всего вылупление растягивается на 2 суток. Нередко, однако, птенцы вылупляются в течение 3, а иногда и А суток. Так, например, в одном из гнезд, найденных в Ленинградской области (1954), в котором кладка состояла из 11 яиц, вылупление птенцов началось утром 22, а кончилось во второй половине дня 24 июня. В другом гнезде, найденном в Савальском лесхозе (1953), вылупление птенцов происходило в течение 5 дней и продолжалось с 28 июня по 9 июля. В результате птенец, вылупившийся последним, был в 2 с лишним раза легче своего старшего собрата.
Питание гнездовых птенцов. Всего в Савальском лесхозе от птенцов было отобрано 742 порции корма, которые содержали 933 экз. беспозвоночных. За один прилет синица обычно кормила одного или двух птенцов. Изредка пища отдавалась сразу трем птенцам. Как правило, с каждым прилетом синицы приносили своим птенцам 1—2, реже — 3 экз. беспозвоночных.
Основу питания птенцов составляют гусеницы и куколки бабочек (326 экз. из 933), бабочки-имаго (278) и пауки (262). В более или менее значительном количестве птенцы получают также личинок пилильщиков-ткачей (60), главным образом березового пилильщика (Cimbex). Остальные насекомые и другие беспозвоночные, за исключением единичных экземпляров наземных моллюсков и мух, совсем не встречаются. Среди гусениц бабочек преобладают пяденицы, шелкопряды и листовертки. Во взрослом состоянии чаще всего поедаются бабочки сосновой пяденицы и кольчатого шелкопряда. Таким образом, среди насекомых, уничтожаемых большими синицами в гнездовой период, количественно преобладают опасные вредители лесного хозяйства.
Предпочтение, которое отдают большие синицы во время выкармливания птенцов бабочкам (имаго, гусеницам и куколкам) и паукам, оказалось чертой, характерной для птиц, живущих в самых различных географических районах — в Ленинградской области (Промптов и Лукина, 1938), на территории Мордовского заповедника (Сафронов, 1954) и в заповеднике Аскания-Нова (сообщение Н. В. Смирновой). Судя по материалам, собранным на территории Савальского лесхоза, эти группы беспозвоночных являются главной пищей птенцов как первых, так и вторых выводков в течение всего периода их нахождения в гнезде. Питание птенцов ранних возрастов отличается лишь тем, что в первые дни они получают обычно мелкие экземпляры насекомых. А. Н. Промптовым и Е. В. Лукиной было установлено, что синицы иногда выдавливают в глотки маленьких птенцов жидкое содержимое гусениц и пауков. Однако даже в возрасте 1—2 дней пте.нцы часто получают целых бабочек, гусениц и пауков. Комочки земли большие
14 а. С. Мальчевский
209
синицы приносят птенцам, видимо, чаще, нежели другие насекомоядные птицы: из 742 порций корма 43 состояли из земли*.
Результаты к о л ь ц ев а н и я. Из числа окольцованных автором совместно с Н. П. Кадочниковым на территории Савальского лесхоза в 1951—1953 гг. 3 взрослых и 273 молодых синиц в последующие-годы в районе кольцевания было обнаружено всего 2 синицы, причем одна из них оказалась из числа 3 птиц, окольцованных во взрослом состоянии. Эта синица (самка) на следующий год гнездилась дважды и оба раза успешно выводила птенцов. Первое гнездо было устроено* на расстоянии всего лишь 80 м от места прошлогоднего гнездования^ а второе — на расстоянии 300 м.
Молодая синица-первогодок (самец) была окольцована в птенцо-дом возрасте наряду -с 15 другими птенцами этого же выводка. На следующий год окольцованная птица была обнаружена гнездящейся примерно в 200 м от места ее рождения. О 'случае гнездования большой, синицы-первогодка вблизи от того места, где она появилась на свет„ сообщил нам также Г. А. Носков. Птенец, окольцованный им в 1953 г. в районе ст. Мельничный Ручей Ленинградской области, на следующий год был обнаружен гнездившимся в 250 >м от гнезда его родителей. Подобные же факты (2 случая) были описаны Л. В. Шапошниковым (1949). Однако случаи гнездования молодых синиц вблизи от гнезд; родителей довольно редки. Более типичным является расселение молодняка. Об этом свидетельствуют факты чрезвычайно низкого процента возвращения молодых птиц, а также результаты изучения миграций большой синицы (Лихачев, 1957).
ЛАЗОРЕВКА (PARUS COERULEUS L.)
Токование. Наиболее ранняя песня лазоревки была отмечена нами в окрестностях Ленинграда 15 марта (1955), самая поздняя — 22 июля (1953). Осеннее пение бывает слышно в августе, сентябре и. октябре. Регулярное весеннее пение начинается со второй половины апреля и продолжается до начала июля. Во второй половине июня иногда наблюдается подъем активности пения самцов, что, очевидно^ следует связывать со вторым циклом размножения у известной части особей.
Расположение гнезда. Известные автору 4 гнезда лазоревки располагались в дуплах липы, дуба, березы и вяза на высоте 6, 2 и 1,5 м от земли. Судя по литературным данным, лазоревка менее' пластична при выборе места для гнезда, однако Б. К. Штегману (1951) пришлось однажды наблюдать -случай гнездования лазоревки в рукаве тулупа, вывешенного для проветривания.
Сроки размножения. Из 2 гнезд лазоревки, найденных в. окрестностях Ленинграда в 1947 г., вылет птенцов произошел 21 и 24 июня. В Савальском лесхозе из одного гнезда птенцы вылетели. 15 июня (1952), из другого — 5 июля (1951). В известных автору гнездах лазоревки кладка состояла из 8, 8, 9, 9, и 12 яиц.
Развитие птенцов. Новорожденный птенец лазоревки опушен несколько слабее, нежели большой синицы. Он имеет жидкий светло-серый пух, расположенный лишь на надглазничных, затылочной и плечевых птерилиях. Окраска ротовой полости желтая, складка в углах рта широкая, светлая. Уже с первого дня существования птенец проявляет большую активность; в этом возрасте он способен принимать
210
вертикальную позу и высоко «подымать голову -при выпрашивании корма.
Из гнезда, за которым проводили постоянное наблюдение, птенцы вылетели в возрасте 16 дней. Выводок очень 'скоро исчез из гнездового района. Эта особенность —очень скоро оставлять район.гнездования,— видимо, вообще характерна для синиц.
Питание гнездовых птенцов. Характер питания птенцов лазоревки чрезвычайно близок к таковому большой синицы. При выкармливании птенцов лазоревки отдают явное предпочтение паукам и бабочкам. По наблюдениям С. В. Васильева (устное сообщение), помимо бабочек и пауков, составлявших основу питания, птенцы лазоревки изредка получают также наземных моллюсков, мух и перепончатокрылых.
ПУХЛЯК
(PARUS ATRICAPILLUS L.)
Токование. Свою весеннюю песню пухляк начинает петь очень рано, еще зимой. В Ленинградской области мы отмечали пение пухляка уже в начале II декады января. В конце зимы эта синица поет, однако, крайне нерегулярно, преимущественно в -солнечные и теплые дни. Регулярное пение начинается примерно с середины — конца марта и продолжается до середины июня. Отдельные самцы поют иногда до конца июля. В середине лета пухляк часто воспроизводит свою песню во время сбора корма для птенцов. Помимо настоящей песни у пухляка существует еще особая замысловатого типа трель, которую можно услышать в различные сезоны года, но особенно отчетливо она издается в период ухаживания самца за самкой, часто непосредственно перед спариванием.
Расположение гнезда. Одно из найденных гнезд пухляка помещалось в трухлявом пне, другое — в гнилом стволе сломанной осины; входные отверстия располагались на высоте 1 и 1,5 м от земли. В одном из этих дупел совсем не было гнездовой выстилки — птенцы^ сидели на древесной трухе. Случаи подобного рода уже отмечалйс?> в литературе (Шнитников, 1913). В одном гнезде птенцы вылупились 5 июня (1955), из другого вылетели 21 июня .(1954). ;
Поведение взрослых птиц у гнезда. Как правило, пухляки ведут себя у гнезда очень доверчиво. Выкармливая птенцов, взрослые птицы безбоязненно влетают и вылетают из дупла иногда в 1 м от наблюдателя.
Развитие птенцов. Однодневный птенец пухляка покрыт редким, но довольно длинным пухом дымчатого цвета. Пух представлен на надглазничных, затылочной, плечевых и спинной птерилиях. Ротовая полость желтая, складка в углах рта широкая, светло-желтая. Ноздри круглые, яйцевой «зуб» белый, когти серые, кожа розоватооранжевая.
Птенцы пухляка очень активны с первого же дня своего существования. Уже вскоре после вылупления птенец может подолгу держать ♦ высоко поднятую голову с широко открытым ртом.
СИНИЦА ДЛИННОХВОСТАЯ (AEGITHALOS CAUDATUS L.)
Токование. Своеобразные щебечуще-булькающие звуки, издаваемые этими птицами в различные сезоны года, следует скорее всего
14*
211
отнести к категории «подпесни». Подобно хохлатой синице, ополовни-чек держится в брачное время тихо и скрытно. Самцы громкой и регулярной песни .не имеют.
Расположение гнезда. Единственное гнездо длиннохвостой синицы было найдено на Карельском перешейке (ст. Кузнечное) в сосново-березовом заболоченном лесу. Оно располагалось на высоте 6 м в развилке трех вертикальных суков березы. Наружные стенки гнезда были убраны теми же лишайниками, которые покрывали кору березы. Первоначально в нем было 10 птенцов, но один из них погиб за 2 дня до вылета. Птенцы покинули гнездо 16 -июня (1954). Весь .процесс
Рис. 51. Выводок птенцов длиннохвостой синицы (ст. Кузнечное, июнь 1954 г. Фото автора).
оставления гнезда продолжался 10—15 мин. При первом же полете все птенцы оказались способными лететь, не снижая высоты. Вскоре, однако, начался обратный процесс концентрации. Возникла усиленная перекличка, и птенцы стали слетаться в кучу. Стремление к соединению обусловлено, видимо, потребностью во взаимном обогреве, которую начинают ощущать птенцы вскоре после оставления гнезда (рис. 51).
Поведение взрослых птиц и птенцов. Прежде всего следует отметить чрезвычайную доверчивость и способность этих птиц к быстрому приручению. В течение нескольких дней работы нам удалось добиться того, что длиннохвостые синицы стали садиться к нам на руку и даже позволяли брать себя в руки (рис. 52). Вряд ли эта особенность поведения характерна для всех особей рассматриваемого вида. Очевидно, имеет место индивидуальная изменчивость поведения.
212
Каждый раз, когда на гнездовом участке появлялся ястреб-перепелятник, синицы издавали протяжный, тонкий, дребезжащий свист, напоминающий призывный крик свиристеля. При воспроизведении этого звука птенцы, выглядывавшие из гнезда, прятались внутрь. При более легком возбуждении синицы издавали свой характерный позыв «циррк-циррк», который они часто воспроизводят также во время осенне-зимних кочевок. Заслышав этот звук, птенцы обычно переставали подавать свои «позывные», но не прятались внутри гнезда.
При дефекации птенцы используют иногда небольшие щели и трещины в стенках гнезда, так как доступ к входному отверстию иногда
Рис. 52. Длиннохвостая синица после 4-дневного приручения (ст. Кузнечное, июнь 1954 г. Фото автора).
бывает затруднительным. В этих случаях получается впечатление, будто комочек экскремента проходит прямо через стенку гнезда. Подобные факты показывают, насколько сильно бывает у подросших птенцов стремление испражняться обязательно за пределы гнезда.
Из отверстия гнезда одновременно могут выглядывать 3 или 4 птенца, т. е. примерно только 7з выводка (рис. 53). Получает же пищу один, реже два или три птенца. Занятие выгодной позиции у входного отверстия происходит не без столкновений. Об этом можно судить по сильной возне и тем толчкам, которые испытывают стенки гнезда непосредственно перед появлением птенцов у входного отверстия. Естественно, что при большой тесноте и многочисленности потомства отставшие в развитии или ослабевшие птенцы могут легко погибнуть.
Птенцы длиннохвостой синицы очень походят на птенцов настоящих синиц желтым цветом ротовой полости и широкой белой складкой в углах рта. По этому признаку они резко отличаются от птенцов уса
213
той синицы, с которой длиннохвостую синицу в настоящее время сближают систематики.
Характером голоса и всем своим поведением птенцы длиннохвостой синицы тоже очень напоминают птенцов настоящих синиц. Призывный крик, издаваемый ими, звучит как 3—4 раза повторяющийся тонкий свист обычного синичьего типа: «сисисиси-сисиси». Сходный позыв воспроизводят и взрослые птицы. Постоянная перекличка между родителями, разыскивающими корм, и молодыми птицами устанавливается еще тогда, когда все птенцы находятся в гнезде и родители при отыскивании их руководствуются главным образом звуковыми восприятиями.
Выпиленную нами часть ствола с гнездом и птенцами мы помещали в разные места на расстоянии 25—30 м от прежнего места расположения гнезда. Во всех случаях взрослые птицы, прилетавшие кормить
Рис. 53. Птенцы длиннохвостой синицы выглядывают из гнезда в ожидании прилета своих родителей (ст. Кузнечное, июнь 1954 г. Фото автора).
птенцов, сразу же находили их и к гнезду, помещенному в новое место, подлетали без малейшей задержки, не обращая внимания .на изменение окружающей обстановки (рис. 54). При этом мы неоднократно имели возможность убедиться в совершенстве органа слуха птенцов. Находясь в закрытом гнезде, они начинали реагировать на позыв, издаваемый родителями тогда, когда мы этот позыв различать еще могли. О приближении родителей к гнезду мы судили первоначально по поведению птенцов, которые, заслышав голос взрослых птиц, начинали воспроизводить ответный позыв и наперебой стремились занять выгодную позицию у входного отверстия. Лишь спустя некоторое время после этого мы начинали слышать голоса подлетающих к гнезду взрослых синиц.
Питание гнездовых птенцов. Изучение питания птенцов длиннохвостой синицы проводилось автором совместно с Е. Р. Га-
214
тинской путем постепенного приручения взрослых птиц и отбирания жомков корма непосредственно из клюва птицы при помощи пинцета. Питание изучалось в период, непосредственно предшествующий вылету птенцов из гнезда. До того, как начать работу, мы отпилили кусок дерева, на котором помещалось гнездо, и спустили его на землю. Затем в течение 2 дней мы подсаживались к гнезду на все более и более близкое расстояние, постепенно приучая птиц не бояться близкого присутствия человека.
После того, как птенцы покинули гнездо, 7 из .них были пойманы и посажены в небольшую клетку, после чего снова оказалось возможным проводить наблюдения. Поведение одной из взрослых птиц не
Рис. 54. Длиннохвостая синица у гнезда с птенцами (ст. Кузнечное, июнь 1954 г. Фото автора).
изменилось— она продолжала с той же интенсивностью выкармливать «пленников». Вторая птица держалась в стороне и выкармливала двух оставшихся на свободе птенцов. В конце периода приручения доверчи-.вость длиннохвостой синицы дошла до того, что она стала кормить птенцов, которых мы держали в руках. Птица позволяла даже брать себя в руки. Мы отбирали из клюва птицы пучок насекомых, а затем отпускали ее. Через несколько 'минут она появлялась с новым пучком жорма, и вся процедура повторялась. Работая таким образом, мы отобрали от птицы весь корм, который они принесла птенцам за 4, 5 часа. За это время птица подлетала к птенцам 104 раза. В 104 порциях корма, которые она принесла своим птенцам, оказалось 896 экз. различных беспозвоночных животных, из них: веснянок — 778, гусениц бабочек— 48, бабочек (имаго)—13, жуков (слоники)—8, цикадок—14, •стрекоз (лютки)—2, комаров — 3, мух—1, пауков—15, моллюсков — 1.
215
Обилие веснянок в кормовом рационе птенцов в данном случае объяснялось близостью водоема. Несмотря на преобладание веснянок, излюбленным кормом длиннохвостой синицы были все-таки голые гусеницы бабочек* Об этом можно было судить по поведению прирученной нами длиннохвостой синицы, которая вела себя довольно спокойно, когда мы отнимали у нее веснянок, но реагировала очень бурно,. Когда из клюва ее вытаскивалась гусеница бабочки. Птица вступала в драку, пыталась выхватить гусеницу и активно бросалась подбирать ее, если она падала на землю, и т. п. Одним словом, по всему было» Видно, что голая гусеница являлась для нее более «лакомым куском», Нежели веснянки, жучки, комары и другие насекомые.
Количество насекомых, приносимых длиннохвостой синицей за; один раз, варьировало от 1 до 25.
КОРОЛЕК ЖЕЛТОГОЛОВЫЙ (REGULUS REGULUS L.)
По сообщениям Д, Н. Кайгородова (1923), а также Е. В. Лукиной: (1951), песню желтоголового королька в окрестностях Ленинграда при соответствующей погоде Можно услышать уже в конце февраля — начале марта. Регулярное пение, однако, начинается лишь с середины, апреля. Продолжается оно до начала июля. Отдельные самцы поют в течение всего июля. Во второй половине июня наблюдается некоторый подъем активности пения, что, по-видимому, связано с наличием у некоторых пар двойного цикла размножения. В 1954 г. на севере Карельского перешейка народном и том Же лесном участке автор в течение всего лёта наблюдал поющего кбролька и дважды с перерывом, в 1,5 -нёс. отмечал Лыводки; птенцов-слетков, выкармливаемых родителям:». Первый раз вывёдок, состоявший из 6 птёйЦов, был обнаружен 26 июня. Птенцы сидели на одной- из верхних ветвей крупной ели, тесно прнжавшйсц друг к другу. Родители беспрестанно подлетали и кормили Их. СаМёц во время отыскивания корма пел очень активно. На», этой же самой елке вторично мы наблюдали кормление птенцов в период с 17 по 24 августа. Весьма вероятно, что оба выводка принадлежали одной в той же родительской паре.
сорокопут-жулан (LANIUS COLLURIO L.)
Токование. Настоящей весенней песни сорокопут-жулан, видимо, не имеет. В период образования пар самцы издают громкие и резкие выкрики. При этом они обычно занимают видную позицию — вершину дерева илй куста, иногда эти звуки издаются ими на лету. Во» время подражательного щебетания, которое бывает крайне нерегулярным, жулан «поет» обычно вполголоса; издаваемые 'при этом звуки носят характер «подпесни». В Ленинградской области автор слышал, как сорокопут-жулан подражал -пению серой славки, лугового чекана it издавал звуки, напоминавшие отдельные строфы певчего дрозда. Е. Я. Ефимов (1915) сообщает, что жулан может копировать также-вяблика, соловья, пересмешника, болотную камышовку.
В момент ухаживания самец в непосредственной близости от самки принимает уморительнейшие позы, сопровождая их громким скрипучим верещанием. По окончании периода ухаживания и образования: пар жуланы становятся чрезвычайно молчаливыми и скрытными. Так.
216
продолжается обычно вплоть до вылета птенцов из гнезда, после чего птицы снова бывают заметными.
Расположение гнезда. Обычно гнездо прячется в кустах или среди древесной поросли. Породы деревьев или кустарников могут быть самыми различными. В Ленинградской области 38 гнезд сорокопута-жулана были найдены на 16 видах растений. Чаще всего, однако, постройки жулана располагаются здесь на небольших елочках, растущих на лесных опушках или вырубках. Кустарники используются реже. В Савальском лесу сорокопут-жулан отдает предпочтение кустам бузины (43 находки из 56), густая листва которой делает гнезда, расположенные на ее ветвях, малозаметными. Помимо бузины жулан гнездится здесь также на терне, калине, яблоне, черемухе, молодых вязах, дубах, кленах, липах.
Высота расположения гнезда редко бывает выше 2 М. Из 94 гнезд, найденных в Ленинградской области и на территории Савальского лесхоза, 50 располагалось на высоте до 1 м, 35 — на высоте 1—2 м, 8— 2—3 м и только 1 гнездо — в 3—4 м от земли. Постройки жулана, помещенные в большинстве выше 2 м, были найдены в городских парках, где гнезда жулана очень часто разоряются.
Строительный материал, используемый птицами, живущими по соседству с человеком, иногда бывает очень своеобразным. Автору приходилось находить гнезда жулана, наполовину состоявшие из ваты, бумаги, кусков пряжи, (бинтов, тряпок и т. п.
Сроки размножения. Сроки откладки яиц, несмотря на наличие у данного вида одной кладки в сезон, растянуты довольно значительно. Это оказалось особенно характерным для сорокопутов, гнездящихся в лесостепной полосе. В Савальском лесхозе в период с 1951 по 1953 г. нам удалось выяснить точные сроки откладки яиц в 34 гнездах. В наиболее раннем гнезде первое яйцо было отложено 21 мая/’ (1952), а в наиболее позднем — 8 июля (1952). Таким образом, в пределах одного лесного массива в течение одного сезона размножения разница в датах появления первого яйца в отдельных гнездах оказалась равной 48 дням. Максимальное количество свежих кладок было найдено в конце мая. Во многих гнездах откладка яиц началась в первых'числах июня. В дальнейшем в течение почти всего июня — начала июля в каждые 5 дней нам не приходилось находить более 3 свежих кладок (табл. 52).
В Ленинградской области, судя по 33 находкам свежих кладок, сроки размножения сорокопута-жулана несколько более поздние и не столь сильно растянутые, как в лесостепной полосе. Здесь наиболее ранний срок откладки первого яйца был отмечен 24 мая (1949) и наиболее поздний — 25 июня (1947). Пик размножения падает на последние числа мая — начало июня. Начиная же со второй половины II декады июня количество особей, приступающих к размножению, постепенно уменьшается. Таким образом, большинство особей откладывает яйца здесь примерно на 5 дней позднее, чем в лесостепной полосе (табл. 52). Растянутость периода размножения у жулана объясняется сильным разорением гнезд и частыми случаями возникновения дополнительных кладок.
Размеры и судьба кладок. История 33 гнезд была прослежена на территории Савальского лесхоза и 30 гнезд в окрестностях Ленинграда. В Савальском лесу количество яиц в полных кладках
1 В литературе (Кадочников, 1957) имеется указание на более ранние сроки размножения сорокопута-жулана в Савальском лесхозе.
217
Таблица 52
Сроки размножения сорокопута-жулана в окрестностях Ленинграда и в Савальском лесхозе
Количество гнезд, в которых откладка яиц началась
Район наблюдения 20-25 V 26—31 V 1-5 VI 6-10 VI 11-15 VI 16-20 VI 21-25 VI 26-30 VI 1 1-5 VII 1 1 6—10 VII
Ленинградская область 1 9 , 8 6 4 2 2
Воронежская область 7 5 5 3 3 3 3 2 2 1
жулана варьировало от 3 до 6 (то 3 яйца в 2 гнездах, по 4 — в 8, по J5 — в 14, по 6 — в 9), а в Ленинградской области — от 3 до 7 (по .3 яйца в 3 гнездах, по 4 — в 5, по 5 — в 16, по 6 — в 5 и по 7 в 1).
Количество птенцов-слетков в выводках варьировало от 2 до 6. Отход яиц и птенцов оказался значительным и равнялся примерно 46—48% по отношению к числу отложенных яиц. Из 162 яиц, найденных в Воронежской области, вылупилось 106 птенцов: 27 яиц (5 кладок) было брошено птицами, 26 — погибло при разорении гнезд хищниками, 3 яйца оказались болтунами. Из 106 птенцов 19 погибло при .разорении гнезд, остальные 87 благополучно покинули гнезда. В Ленинградской области под наблюдением было 146 яиц; из них вылупилось .82 птенца: 26 яиц (6 кладок) было брошено птицами, 33 — погибло при разорении гнезд, и 5 яиц оказались болтунами. В дальнейшем было .разорено еще 1 гнездо с 6 птенцами. Таким образом, всего вылетело из гнезд 76 птенцов.
Гнезда разоряются не только воздушными (ворона, сойка), но и наземными хищниками, в частности лаской и горностаем. Подобная деятельность мелких куньих особенно заметна в годы низкой численности мышевидных лесных грызунов, когда ласка и горностай частично переключаются на питание яйцами и птенцами мелких птиц, гнездящихся на земле, в кустах и на молодых деревцах (см. главу III), Поведение взрослых птиц у гнезда. Будучи потревожен у гнезда в период откладки или насиживания яиц, сорокопут-жулан очень часто бросает гнездо и строит его в новом месте. Приступая к повторному размножению, жулан обычно устраивается в более безопасном месте. В 1947 г. в парке Лесотехнической академии все 5 первоначально построенных гнезд этого вида были разорены или брошены. Тем не менее наблюдением было установлено (Мальчевский, 1950), что все 5 пар жуланов при повторном гнездовании благополучно вывели .потомство благодаря тому, что новые гнезда их располагались уже в более укромных местах.
На появление человека вблизи гнезда жуланы реагируют обычно по-разному. Когда в гнезде находятся яйца, то птицы редко приходят lb сильное возбуждение. После вылупления птенцов поведение жуланов резко меняется. При приближении наблюдателя к гнезду они почти всегда начинают выдавать свое присутствие резким «чеканьем». Первым замечает опасность, как правило, самец, который в районе гнезда имеет несколько «наблюдательных пунктов». Обычно он еще издали встречает идущего к гнезду человека. В некоторых случаях птицы ведут себя агрессивно и нападают. Например, в парке Лесотехнической академии один из гнездившихся там в 1947 г. жуланов отличался тем,
•J218
^что никому «не позволял» близко подходить к его гнезду с птенцами. Каждый раз, когда мимо кто-либо проходил, сорокопут молча «пикировал» и с налета наносил довольно сильный удар в голову клювом и .лапами. Он делал это так быстро и неожиданно, что случайно подошедший к гнезду человек невольно пугался и отходил в сторону. Очень похожее поведение жулана наблюдал в 1948 г. также Майре (Myres, 1949) в окрестностях Винчестера. Будучи достаточно активными, жуланы могут отогнать от*своего гнезда птиц значительно крупнее, чем они сами, например обыкновенную пустельгу. Наиболее агрессивные особи держатся обычно молчаливее остальных.
Пте н ц ы. Однодневный жулан -с первого взгляда кажется совершенно голым. При тщательном рассмотрении на его брюшной стороне можно заметить 2 ряда очень коротких светлых пушинок. Наличие едва заметного первичного пуха у птенцов сорокопута-жулана позволяет предположить, что в данном случае мы имеем дело с рудиментарным признаком.
Птенец жулана в «слепом возрасте» напоминает собою птенцов славок, но может легко быть отличен от последних по одноцветной окраске клюва и языка и круглой форме ноздрей. Яйцевой «зуб» и когти у него светлые. Кожа бледно-розовато-желтая. Ротовая полость желтая, язык не имеет темных отметин. Складка в углах рта светло-желтая. Зубец на надклювье, характерный для взрослых сорокопутов, начинает образовываться ко времени самостоятельной охоты молодых жуланов примерно в месячном возрасте. По данным П. П. Сушкина (1902), аналогичный признак у соколов является весьма древним и выражен еще у эмбриона. У сорокопутов, наоборот, зубец на надклювье, по-видимому, является сравнительно недавним приобретением.
В первые дни после вылупления птенцы жулана отличаются малой подвижностью. Выпрашивают корм они обычно лежа, не приподымаясь на передние конечности и не поднимая высоко головы. Голос (едва слышимый писк) подают сразу же по выходе из яйца. С 5-дневного возраста они становятся заметно активнее. Громкий визгливый писк, характерный для птенцов-слетков, издается ими примерно с 10— И-дневного возраста. Крикливость молодых птиц становится, однако, ’особенно заметной после того, как птенцы оставят гнездо и научатся летать.
У птенцов, за которыми автор вел ежедневные наблюдения, ушные «отверстия открылись в возрасте 4 дней. В это же время у них начали приоткрываться глаза и намечаться пеньки будущих перьев. У 6-дневных птенцов на спине впервые появились кисточки контурных перьев. Пеньки кроющих крыла раскрылись в возрасте 7 дней. Днем позднее появились кисточки маховых, и еще через день, т. е. у 9-дневных птенцов, начали раскрываться пеньки рулевых. В этом возрасте при виде человека птенцы уже прижимаются, а при сильном испуге могут даже выскочить из гнезда. Нормальное оставление гнезда происходит в возрасте 12—13 дней (рис. 55). Гнездо покидается обычно не одновременно, иногда вылет растягивается на 2—3 дня. К активному полету птенцы жулана становятся способными в 17—18-дневном возрасте. Например, окольцованные автором птенцы, вылупившиеся 18 и 19 июня (1952) и оставившие гнездо 1—2 июля, впервые стали летать без потери высоты только 6—7 июля. В связи с поздней самостоятельностью молодые жуланы продолжают оставаться на гнездовом участке и держаться выводком еще долгое время и после того, как научатся летать. Птенцы одндго выводка, за которым наблюдал автор, покинули гнездо •8 июля, но оставались на том же участке вплоть до 3 августа.
219
Питание гнездовых птенцов. Всего от птенцов сорокопута-жулана нами было отобрано 154 экз. различных животных. Из них 11 приходится на долю Позвоночных. В окрестностях Ленинграда, найдены наколотыми на сучки в районах гнезда: полевая мышь — 2 раза, рыжая полевка —1 раз, -гнездовой птенец чечевицы — 4 раза,, гнездовой птенец варакушки и слеток лазоревки — по 1 разу; в учлес-хозе «Лес на Ворскле» — слеток овсянки обыкновенной, а в Савальском лесхозе — мясо какого-то позвоночного. Во всех случаях у жертвы была оторвана голова.
Несмотря на примерно равное количество обследованных гнезд сорокопута-жулана в лесной и в лесостепной зонах (30 и 33), боль-
Рис. 55. Выводок сорокопутов-жуланов (фото автора).
шинство позвоночных, наколотых на сучки, было найдено в северных районах, в частности на территории Ленинградской области. Здесь по причине частых холодов и дождей сорокопут-жулан, по всей вероятности, не всегда может найти достаточное количество насекомых, в связи с чем и переходит чаще, чем на юге, на питание позвоночными. Проводившийся нами учет «птичьих переполохов» также показал, что в северных районах сорокопут-жулан гораздо чаще является причиной беспокойства мелких птиц, нежели в южных.
Среди -беспозвоночных животных первое место в пище птенцов •сорокопута-жулана занимают жуки, количество которых равнялось 7*> из 143 экз. беспозвоночных, извлеченных из пищеводов птенцов или найденных в подстилке гнезда. В Савальском лесхозе это были в основном жуки средних размеров (Anisoplia, Melalontha, Carabus); в. Ленинградской области чаще всего уничтожались различные жужелицы и навозники. Среди жуков, принесенных жуланами своим птенцам, -следует еще отметить усачей (Rhagium), могильщиков и мертвоедов. В гнездах сорокопута-жулана на дне лотка были обнаружены мертвыми дикие пчелы, шмели и осы.
220
Помимо жуков и перепончатокрылых сорокопуты приносили своим птенцам также гусениц и бабочек, преимущественно совок, среди которых были обнаружены, например, крупные гусеницы большой южной совки. Из прямокрылых они нападали на медведок и на кобылок, а при позднем гнездовании — на зеленых кузнечиков. В годы массового размножения саранчовых последние имеют первостепенное значение в питании птенцов. Так, например, было в жаркое и засушливое лето 1951 г. на территории Савальского лесхоза.
Пауки и наземные моллюски поедаются птенцами регулярно, но в сравнительно ограниченном количестве.
МУХОЛОВКА СЕРАЯ (MUSCICAPA STRIATA PALL.)
Токование. В период образования пар самцы серых мухоловок часто садятся на вершины деревьев и издают довольно определенный набор цыкающих звуков, слышимых благодаря их высокой частоте на значительном расстоянии. Эти звуки имеют определенный ритм и последовательность, благодаря чему их можно назвать песней. Ведуг себя при этом самцы весьма демонстративно, и совершенно очевидно, что издаваемые в это время самцом звуки предназначены для привлечения самки.
В моменты «ухаживания» за самочкой самец тоже усиленно цыкает и поскрипывает, но значительно менее определенно, нежели во время пения. Принимаемые им при этом позы весьма забавны: то он весь вытянется и подымет перья на голове, то, приняв горизонтальное положение и вытянув шею, вдруг поочередно или вместе начнет судорожно взмахивать крыльями.
Расположение гнезда. Серая мухоловка — один из наиболее пластичных видов в отношении инстинкта гнездостроения. Эта пластичность проявляется не только в помещении гнезд в самых разнообразных местах и на различной высоте, но и в способности в зависимости от места гнездования сооружать постройки, сильно отличающиеся по форме, конструкции и строительному материалу. При гнездовании в лесу серая мухоловка может располагать свои гнезда в основании толстых сучков деревьев, в различных изъянах стволов, за отставшей корой, на пнях и лесных изгородях, на кучах хвороста и в других местах. Постройки этой птицы автору приходилось находить в старых гнездах дроздов и зяблика. В редких случаях, когда серая мухоловка не находит мест, удобных для помещения гнезда, она поселяется даже прямо на земле. Автор нашел 3 таких гнезда в молодых сосновых посадках. Вблизи человеческих поселений мухоловка вьет гнезда на карнизах домов, на срубах, в изъянах стен, в стары* гнездах деревенских ласточек. Одно гнездо было найдено нами на обрезе рукоятки большой лопаты, приставленной к стене строящегося дома.
Более всего, однако, интересны гнезда серых мухоловок, укрепленные на сравнительно тонких ветвях подрастающих деревьев. Подобного рода гнездование мухоловок мы наблюдали на территории Савальского лесхоза, где из 86 найденных гнезд 14 были свиты на молодых вязах. Эти гнезда поражали необычайностью формы, конструкции и строительного материала. Они имели высокие и плотные боковые стенки одинаковой толщины, глубокий лоток и снаружи были убраны коконами наездников. Первоначально автор даже полагал, что серая мухоловка лишь достраивает старые гнезда зяблика. Это
221
предположение, однако, отпало после того, как была прослежена работа 'серой мухоловки от начала до конца -постройки.
Гнезда этого вида, расположенные на земле, были, наоборот,, чрезвычайно примитивны -и представляли собою очень рыхлое сооружение, сделанное из ограниченного количества строительного материала. Нет сомнения в том, что одна и та же птица способна вить, гнезда различной конструкции. Все зависит от того, где производится постройка.
Сроки размножения. Сроки откладки яиц удалось установить в 101 гнезде. Из этого количества 41 гнездо было найдено в-Ленинградской области и 60 — на территории Савальского лесхоза. Период возможных сроков размножения у мухоловок, гнездящихся в лесной и лесостепной зонах, в общем одинаковый и продолжается 40— 41 день. Так, например, в Ленинградской области наиболее ранняя дата откладки первого яйца была отмечена 26 мая (1950), а наиболее поздняя — 3 июля (1947). Почти такие же сроки появления первых яиц в самой ранней и самой поздней кладках были установлены нами и в Савальском лесхозе: 25 мая (1951)1 и 3 июля (1952). Наряду с этим было отмечено, что на севере период размножения проходит более дружно (табл. 53).
Таблица 53
Сроки размножения серой мухоловки в Ленинградской и Воронежской областях
Количество гнезд, в которых откладка яиц началась.
Район наблюдения 20—25 V 26-31 V 1-5 VI 6—10 VI 11-15 VI 16-20 VI 21-25 VI 26-30 VI |
Ленинградская область . . . 6 13 8 7 4 1 1
Воронежская область .... 1 9 12 12 9 6 4- 5
Значительную степень растянутости сроков размножения у этого вида следует объяснять неодновременностью включения отдельных особей в репродуктивный цикл, а также дополнительными кладками, возникающими после разорения гнезда. Поздние гнезда серых мухоловок обычно содержат 3 или 4 яйца, в то время как в начале периода размножения кладка, как правило, состоит из 5 яиц.
Размеры и судьба кладок. Прослежена история 79 гнезд,, из них 50 наблюдались в Воронежской области и 29 — в Ленинградской. Большинство гнезд в Ленинградской области содержало кладки, из 5 яиц, в то время как в Воронежской области нам чаще приходилось находить гнезда с 4 яйцами (табл. 54).
Количество вылетевших птенцов по отношению к первоначальному количеству отложенных яиц в Ленинградской области составляло 69%, в то время как в Воронежской области — 52%. Из 217 яиц в Савальском лесхозе вылупилось всего 152 птенца: 48 яиц (11 кладок) погибло при разорении гнезд различными хищниками, 12 — оказалось болтунами или «задохликами», и 5 яиц (одна кладка) было брошено птицами в период насиживания. Из 152 птенцов выжили и
1 По наблюдениям Н. П. Кадочникова (1957), в Савальском лесхозе серая мухоловка в отдельные годы (1954) может начать кладку уже 15 мая.
222
Таблица 54
Количество яиц в кладках серой мухоловки
Район наблюдения Количество гнезд, содержащих кладку яиц (в шт.) Среднее количество яиц на 1 гнездо
3 1 4_ 1 5 L_6 __
Ленинградская область .... 2 9 18 4,55
Воронежская область .... 4 26 18 2 * 4,36
покинули гнезда 112 птенцов: 31 птенец (8 выводков) -погиб от хищников, а 9 (2 выводка) — упав на землю вместе с гнездом.
Столь высокий процент отхода яиц и птенцов серых мухоловок на территории Савальского лесхоза следует объяснять обилием в этом лесу соек, являющихся основными врагами мухоловок в гнездовой период. Автор несколько раз наблюдал соек в то время, как они разоряли гнезда этих птиц. Кроме сойки, серых мухоловок беспокоят сарыч, коршун, перепелятник, ушастая сова, большой пестрый дятел и сорокопут-жулан. В отдельных случаях виновницей гибелй кладки или-выводка была сама -птица: небрежно построенное и плохо укрепленное гнездо постепенно кренилось на бок и падало на землю. Один раз автор обнаружил на гнезде мертвую самку, покрывавшую своим телом 5 насиженных яиц. Причина смерти птицы осталась неизвестной.
Поведение взрослых птиц у гнезда. В течение 3 первых дней после вылупления птенцов взрослая птица почти не сходит с гнезда. С 4-го дня систематическое обогревание птенцов прекращается. В случае резкого понижения температуры воздуха, дождя или. ярких солнечных лучей, падающих на гнездо, самка защищает своим телом даже уже совсем подросших птенцов. В последнем случае птица привстает на ноги и расправляет крылья, создавая таким образом значительную тень.
В поведении серых мухоловок вблизи гнезда наблюдаются индивидуальные отклонения. Некоторые птицы держатся всегда в стороне и ведут себя довольно спокойно, даже когда трогают их птенцов. Другие, наоборот, проявляют сильное беспокойство. Автору приходилось встречать птиц, которые с особым трещанием крутились над головой и, «пикируя», старались ударить своим телом наблюдателя, осматривавшего гнездо. Есть особи, которые -при виде человека, стоящего у их гнезда, летают вокруг особым трепещущимся полетом, как бы «притворяясь» неспособными к быстрым движениям. Более определенную имитацию раненой птицы отмечал у серых мухоловок Чэппл (Chappel, 1948). В целом для серых мухоловок характерен агрессивный способ защиты птенцов. На сойку мухоловки всегда нападают с большой стремительностью. Они «пикируют» и иногда даже сильно ударяют ее с налета своим телом. При этом серые мухоловки издают совершенно особое трещанье, являющееся, ^очевидно, их криком угрозы.
Помимо своего обычного позыва беспокойства серая мухоловка при появлении опасности у гнезда часто воспроизводит тонкий протяжный свист, который тоже имеет значение «сигнала тревоги». Первоначально автор полагал, что этот свист воспроизводится птицами лишь в случае появления ястреба или другого пернатого хищника, пролетающего мимо, ибо неоднократно отмечал совпадение этих явлений. В дальнейшем, однако, оказалось, что точно такой же свист птица из-
223
дает иногда и при -виде человека или собаки, подошедших близко к ее гнезду.
Развитие птенцов. Птенец покрыт довольно редким пухом дымчатого цвета. Голова опушена сильнее остальных частей тела. Пух расположен на глазных, надглазничных, затылочном участках тела, а также на спинной, плечевых, локтевых и бедренных птерилиях. На последних пух особенно жидкий. Кожа желтоватая. Клюв плоский, одноцветно-серый, слабо вздутый в области ноздрей. Яйцевой «зубэ бурый, когти светлые. Ноздри овальные. Окраска ротовой полости желтая.
Птенцы серой мухоловки становятся активными очень рано. Еще в процессе освобождения от скорлупы птенчик может высоко тянуть шею и подолгу держать поднятой голову с раскрытым клювом. В
Рис. 56. Однодневный птенец серой мухоловки, выпрашивая корм, привстает, опираясь на передние и задние конечности (фото автора).
конце первого дня жизни птенец, просящий есть, уже свободно приподымается на всех четырех конечностях (рис. 56). В возрасте 6— 7 дней птенцы уже пугаются вида людей — прижимаются, закрывая глаза. На 10-й день они иногда выбрасываются из гнезда, хотя в этом возрасте летать еще совсем не умеют и передвигаются по земле плохо.
Развитие птенцов идет быстрее, чем у многих других открыто гнездящихся птиц. Ушные отверстия начинают открываться тогда же, когда и глаза, —в возрасте 4 дней. У 5—6-дневных птенцов глаза уже почти полностью открыты. Опахала маховых перьев начинают развертываться в возрасте 6 дней. Первыми раскрываются пеньки второстепенных маховых, затем первостепенных. Несколько позднее (у 8-днев-ных птенцов) появляются кисточки рулевых.
Если птенцов никто не пугает, они обычно сидят в гнезде вплоть до приобретения способности к полету, которая у них наступает в возрасте 13—14 дней (рис. 57). Очень часто оставление гнезда происходит в возрасте 12 дней, когда птенец еще не умеет летать по-настоя-
224
тему. В этом возрасте птенец летит обычно по наклонной вниз. На землю молодые мухоловки падают, однако, редко. Обычно им все же удается сесть на ветку куста или дерева. Первое время после оставления гнезда 'выводок продолжает оставаться на том же участке. Окольцованные -в гнезде 1птенцы серых мухоловок неоднократно встречались ;нами неподалеку от того места, где они вывелись, спустя 5— 6 дней после их вылета. В некоторых -случаях, однако, выводок переселяется в другой район уже через 1—2 дня.
Место, где держится выводок, легко определить .по призывному крику птенцов, который они постоянно издают, разместившись повыше в кроне дерева. Этот позыв птенцы начинают регулярно воспро-
Рис. 57. Выводок птенцов серой мухоловки (фото автора).
изводить лишь в период, непосредственно предшествующий оставлению гнезда. Кажущееся негромким «цыканье» птенцов серых мухоловок можно отчетливо слышать на расстоянии (нескольких десятков метров.
Питание гнездовых птенцов. Всего из пищеводов 'птенцов серой мухоловки было извлечено 77 экз. беспозвоночных животных. Чаще всего птенцы поедали двукрылых (всего 54 экз.) —настоящих мух, реже слепней и ктырей. Таким образом, даже судя по питанию птенцов, этот вид вполне оправдывает данное ему название. Кроме того, птенцам приносились бабочки (шелкопряды), реже гусеницы (всего 10 экз.), а также характерные для пищевого рациона птенцов почти всех лесных певчих птиц пауки (3) и наземные 'моллюски (4). Один раз была принесена уховертка. В Кандалакше, по наблюдениям О. М. Татариновой, серая мухоловка помимо двукрылых и бабочек приносит птенцам шмелей, ручейников и жуков-мягкотелок.
По справедливому замечанию Э. Анчарова (Осмоловская и Формозов, 1950), серая мухоловка после дождя часто изменяет свой обыч-
15 А. С. Мальчевский
225
ьый способ «охоты и начинает собирать насекомых с земли. Так делает она, однако, значительно реже, чем белошейка и мухоловка-пеструшка. Основным способом охоты ее все-таки остается ловля насекомых в воздухе.
Как и многие другие дневные птицы, охотящиеся за летающим» насекомыми, серая мухоловка по вечерам долго продолжает оставаться активной. В Савальском лесхозе во второй половине июля серые мухоловки продолжают еще выкармливать своих птенцов в сумерках, между 19 час. 30 мин. и 20 час., в то время как большинство других дневных птиц уже устраивается на ночевку. На наших глазах серые мухоловки схватывали уже начавших к этому времени летать бабочек кольчатого шелкопряда и отдавали их птенцам.
МУХОЛОВКА-ПЕСТРУШКА (MUSCICAPA HYPOLEUCA PALL.)
7
Токование. Песця мухоловки-пеструшки подвержена значительной индивидуальной изменчивости. На севере Ленинградской области (Приозерский район) в 1954 г. ярко окрашенные самцы пеструшки насвистывали более продолжительные и разнообразные напевы, нежели самцы, одетые в скромный самочий наряд. В частности, правильная гаМма свистовых звуков, которую пеструшки часто воспроизводят в конце песни, была отмечена нами только у ярко окрашенных самцов. Интересно, что все местные (приозерские) мухоловки-пеструшки, для которых это колено,было характерно, воспроизводили не нисходящую, как это типично для мухоловок из центральных районов. Европейской части СССР (Благосклонов, 1954), а восходящую гамму чистых звуков.
Как известно, копировка звуков — явление вообще характерное для мухоловок-пеструшек. В Ленинградской области некоторые птицы включают в песню призывный крик клеста, другие — посвисты лесного конька и 'большой синицы, третьи — куски песни певчего дрозда или зарянки.
После вылупления птенцов активность пения самца резко падает и вскоре совсем прекращается. Наиболее поздние даты пения мухоловки-пеструшки для Ленинградской области — 2 июля (1954) и 13 июля (1955), наиболее ранние — 27 апреля (1951»), 2 мая (1949).
Расположение гнезда. В Ленинградской области мухоловка-пеструшка помимо скворечников гнездится в дуплах самых различных пород деревьев: березы, липы, рябины, вяза, ольхи, сосны. На Карельском перешейке чаще всего, однако, нам приходилось находить ее гнезда в дуплах старых осин, в том числе и выдолбленных дятлом. Высота расположения гнёзда, как правило, небольшая, чаще всего от 1 до 2 м, реже — на высоте 5—6 м.
Сроки размножения. В гнездах, найденных в Ленинградской области, откладка яиц началась 20 мая (1949), 24 мая (1947), 28 мая, 29 мая (1954), 1 июня (1950), 8 июня, 9 июня, 12 июня и 13 июня (1954). Таким образом, размножение пеструшек протекает в более сжатые сроки, чем у большинства других мелких лесных певчих птиц.
Размеры и судьба кладок. Под наблюдением находилось 51 яйцо (10 кладок), из них вылупилось только 27 птенцов. 23 яйца (4 кладки) погибло при разорении .гнезд, и 1 яйцо оказалось болтуном. В дальнейшем погибло еще 5 гнездовых птенцов. Таким образом, вылетело из гнезд всего 22 птенца. Большинство гнезд, за которыми мы
226
наблюдали, было разорено мальчишками. В одном случае гнездо пеструшек с 5 птенцами 'пострадало от большого пестрого дятла. О разорении гнезд мухоловок-пеструшек большим пестрым дятлом в Латвии сообщал нам также К. А. Вилкс.
Вдали от человеческих поселений, там, где «деятельность» разорителей гнезд не ощущается столь сильно, процент отхода яиц и птенцов, в гнездах мухоловок-пеструшек значительно ниже. Об этом свидетельствует обстоятельное исследование Г. Н. Лихачева (1953).
Поведение взрослых птиц у гнезда. Птицы, гнездящиеся в населенных пунктах, иногда бывают очень доверчивы. Наоборот, пеструшки, живущие вдали от человеческих селений, держатся у гнезда, как правило, очень осторожно. Особенной осторожностью отличаются старые, интенсивно окрашенные самцы, а также некоторые самки. Автор знал гнездо пеструшек, располагавшееся в низком дупле» у которого в период насиживания кладки практически невозможно было увидеть самку, настолько она была осторожна. При подходе к гнезду птица заблаговременно вылетала из дупла -и возвращалась к нему только тогда, когда наблюдатель уходил.
В случае очень сильного возбуждения мухоловки-пеструшки прибегают к активной защите своего гнезда и птенцов. Однажды автор наблюдал, как молодой самец пеструшки прогонял большого пестрого дятла, пытавшегося проникнуть в дупло, в котором помещалось гнездо. Каждый раз, как дятел заглядывал в дупло, пеструшка, издавая «боевой клич», стремительно «пикировала» и с силой ударяла дятла всем телом в спину. В результате дятел, не выдержав столь энергичного натиска, улетел.
Птенцы. Птенец мухоловки-пеструшки покрыт редким и недлинным серым пухом, который расположен на глазных (не всегда), на надглазничных, затылочном, спинном и плечевых участках тела. Клюв высокий, довольно короткий *и массивный, неширокий, несколько вздутый в области ноздрей. Ноздри круглые, а не продолговатые, как у серых мухоловок. Когти темные. Ротовая полость желтая с оранжевым оттенком. Складка в углах рта желтовато-белая, толстая и довольно широкая.
Как это вообще характерно для птиц, гнездящихся в дуплах, птенцы оставляют гнездо, уже будучи способными летать. Очутившись вне гнезда, птенец стремится сесть на ветку повыше, при этом предпочитает прятаться в кронах лиственных деревьев.
Питание гнездовых птенцов. По питанию птенцов мухоловки-пеструшки в литературе накопилось значительное количество данных. Все исследователи, изучавшие питание птенцов пеструшки (Кнорре, 1947; Осмоловская и Формозов, 1950; Сафронов, 1954; Haartman, 1954; Семенов, 1956а; Милованова, 1956; Поливанова, 1957, и др.), указывали, с одной стороны, на большое количество видов поедаемых насекомых‘и, с другой, — на разнообразие способов охоты пеструшки. Хаартман (1954)/^проводивший специальный количественный учет движений птиц во время охоты за насекомыми, отметил, что в Финляндии мухоловка-пеструшка чаще всего схватывает свою жертву на земле. Этот же исследователь обратил внимание на то, что в кормовом рационе птенцов пеструшки встречается летающих насекомых не больше, чем, например, в рационе птенцов пеночек. Оказалось, что в питании птенцов мухоловки-пеструшки большое значение имеют пауки, а при массовом размножении мелких чешуекрылых пеструшка может легко переключаться на питание бабочками и их гусеницами. Одним словом, имеющиеся в литературе сведения указывают на боль
15*
227
шое сходство характера питания птенцов пеструшки и белошейки (см. далее). В кормовом рационе птенцов обоих видов двукрылые, в частности мухи, имеют второстепенное значение. Таким образом, оба вида с меньшим правом могут быть названы мухоловками, чем, например, пеночка-весничка, лесной конек или лесная малиновка.
МУХОЛОВКА-БЕЛ ОШЕЙКА
(MUSCICAPA ALBICOLLIS ТЕММ.)
Расположение гнезда. В заповеднике «Лес на Ворскле» белошейка весьма обыкновенна и гнездится чаще всего в дуплах старых лип, расположенных на высоте от 1,5 до 3 м. На территории Савальского леса все найденные гнезда этого вида располагались в искусственных гнездовьях. Мухоловки селятся здесь как в дубняках, так и в смешанных и чистых сосновых насаждениях.
Сроки размножения. Период возможных сроков откладки яиц у мухоловки-белошейки растянут примерно на 1 мес. В наиболее раннем из гнезд первое яйцо было отложено 15 мая (1952), а в наиболее позднем — кладка появилась во II декаде июня (1953).
Размеры и судьба кладок. Материалы по истории гнезд мухоловки-белошейки предоставил в наше распоряжение Н. П. Кадочников, изучавший биологию размножения этого вида на территории Савальского лесхоза. Всего под его наблюдением находилось 34 гнезда, в которых находилось 178 яиц (в среднем 5,2 яйца на 1 гнездо). Из них вылупилось только 90 птенцов. Столь значительный отход яиц объясняется высоким процентом болтунов (26 яиц, около 14,5%), частыми случаями бросания кладок (24 яйца, 4 кладки) и разорения гнезд (38 яиц, 8 кладок).. По наблюдениям Н. П. Кадочникова, гнезда мухоловки-белошейки в Савальском лесхозе разоряются главным образом большим пестрым дятлом, который расширяет летки у скворечников, залезает внутрь их и расклевывает яйца. Гнезда белошеек разорялись также серой вороной и вертишейкой. Несколько кладок было брошено птицами после долгого насиживания яиц. ,Н. П. Кадочников сообщил нам, что одна самка сидела на яйцах 22 дня, после чего покинула гнездо. Оказалось, что яйца, на которых она сидела, совсем не содержали зародышей — они, очевидно, были болтунами. Таких кладок, в которых все яйца оказались впоследствии болтунами, было несколько. Смертность гнездовых птенцов белошеек была, наоборот, ничтожной: из 90 вылупившихся птенцов погиб только 1.
Поведение взрослых птиц у гнезда. У гнезд мухолов-ки^белошейки — довольно осторожные и скрытные птицы. При осмотре гнезда они чаще всего держатся в стороне, издавая свой монотонный позыв, выражающий беспокойство. Лишь очень немногие особи ведут себя доверчиво. Позывы беспокойства белошейки сходны с позывами малой мухоловки: помимо монотонного свиста, как и малая мухоловка, белошейка иногда потрескивает и быстро цыкает. Однако по большинству других позывов этот вид напоминает скорее пеструшку (Haartman u. Lohrl, 1950).
Птенцы. Птенец опушен примерно так же, как и птенец пеструшки. Светло-серый пух, еще более редкий и короткий, расположен на глазных (не всегда), надглазничных, затылочном, спинном и плечевых участках тела. Цвет кожи грязно-розовый. Клюв по форме напоминает таковой у птенцов пеструшек, светлый, с небольшим потемнением на конце. Яйцевой «зуб» и когти серые. Ноздри круглые или слег
228
ка овальные. Ротовая полость желтая. Складка в углах рта белая, толстая <и довольно широкая.
Птенцы чрезвычайно подвижные. Будучи вцнутыми из гнезда, они начинают быстро двигаться, работая всеми четырьмя конечностями, и стремятся куда-нибудь забиться. Птенцы, вылупившиеся утром 25 июня (1953), 1 июля, т. е. через 6 дней, имели уже открытые слуховые проходы и могли держать приоткрытыми глазные веки, но глаза у них были еще совершенно мутные. Кисточки маховых перьев и кроющих крыла у птенцов мухоловки-белошейки появляются на 8-й день жизни.
Оставляют гнездо птенцы в возрасте 16 дней, уже способными довольно хорошо летать. Оставив гнездо, они рассаживаются обычно на ветвях, высоко в кронах деревьев.
Окраска оперения птенцов-слетков белошейки не совсем одинаковая. Одни имеют выраженный светлый «ошейник», а также белый цвет на наружных опахалах и 'основаниях внутренних второстепенных маховых и на концах кроющих крыла. У других «ошейника» нет, основания внутренних второстепенных маховых темные, а белый цвет на концах кроющих крыла и наружных опахалах внутренних первостепенных заменен охристым. Указанные отличия следует скорее всего рассматривать как половой диморфизм, проявляющийся у данного вида еще тогда, когда птенцы находятся в гнезде.
Питание гнездовых птенцов. Судя по представленным в наше распоряжение материалам, собранным на территории Савальского лесхоза в 1952—1954 гг. Н. П. Кадочниковым и практикантами С. В. Васильевым и Д. С. Бекжановой, состав кормов птенцов мухоловки-белошейки разнообразнее, нежели таковой у птенцов серой мухоловки. Это объясняется более разнообразными способами ее охоты. Белошейка способна ловить насекомых в воздухе, собирать их с ветвей и листьев деревьев, а также с земли, что чаще всего она делает в пасмурную или дождливую погоду.
Среди беспозвоночных животных, извлеченных из пищеводов гнездовых птенцов (всего 410 экз), наиболее многочисленными оказались пауки (142 экз.), а также бабочки и их гусеницы (128 экз.). Обе эти группы кормов составили примерно 2/3 общего количества беспозвоночных, которых мухоловки-белошейки принесли своим птенцам. Кроме них, птенцы выкармливались жуками (44 экз.), мухами и комарами (21 экз..), верблюдками (28 экз.), тлями (14 экз.), ручейниками (4 экз.), мелкими моллюсками (14 экз.) и кивсяками (2 экз.). Среди бабочек преобладали пяденицы, различные виды совок и хохлаток. Среди жуков были обнаружены личинки «коровок», а также усачики и хрущики. Довольно часто белошейка приносит своим птенцам комочки земли. По данным Г. Е. Корольковой (1954), изучавшей питание белошейки в Теллермановском лесничестве, основу рациона птенцов составляют бабочки, пауки же не играют существенной роли. По всей вероятности, расхождение данных следует объяснять не только местными особенностями условий, но и различием в методике работы.
ПЕНОЧКА-ТЕНЬКОВКА
(PHYLLOSCOPUS COLLYBITUS V I Е I L L.)
Токование. В Ленинградской области песню пеночки-теньков-ки можно слышать начиная с конца апреля вплоть до поздней осени, когда поют уже пролетные птицы. Наиболее ранняя дата пения тень-ковки отмечена автором 28 апреля (1946) в Пашском районе. Регулярное пение начинается в конце I декады мая и продолжается до' послед
229
ней декады июля, хотя отдельные самцы поют по-весеннему до начала августа. Осеннее пение, которое для этого вида чрезвычайно характерно, бывает слышно с середины августа вплоть до конца октября. Оно не регулярно, и поющая птица не имеет постоянного места обитания, но по характеру и силе звуков оно не отличается от весенне-летнего.
Расположение гнезда. Постройки этого вида, найденные в Савальском лесу и в учлесхозе «Лес на Ворскле», частично помещались на земле, частично возвышались над поверхностью почвы, располагаясь на кучах хвороста, молодых деревцах и т. п.
Сроки размножения. Период размножения у пеночки-тень-ковки сильно растянут. В Савальском лесхозе в разных гнездах вылупление птенцов происходило 29 мая, 18 июня, 25 июня и 28 июня. В одном из гнезд самка насиживала еще кладку 12 июля (1952).
Поведение взрослых птиц у гнезда. Во всех случаях кормила птенцов только одна самка; самцы в это время пели где-то на вершинах деревьев. В большинстве самки очень доверчивы. При появлении человека у гнезда теньковки выражают свое беспокойство почти так же, как и веснички. Позывы, которые при этом издают птицы, у обоих видов очень похожи, но свист теньковки как бы двойной, несколько выше по тону и воспроизводится более поспешно.
Птенцы. Птенец пеночки-теньковки имеет довольно редкий, но относительно длинный пух светло-серого цвета, расположенный на голове (надглазничные и затылочный участки) и на плечах. 6-дневные птенцы пеночки-теньковки имеют совершенно открытые глаза и уже пугаются наблюдателя. В этом же возрасте у них появляются кисточки контурных перьев на груди и спине. Кисточки первостепенных маховых, а также рулевых перьев начинают показываться на 7-й день жизни птенца. Нормальное оставление гнезда—на 12—13-й день после вылупления. Только что оставившие гнездо птенцы теньковки способны уже совершать короткие перелеты. Выводок очень скоро начинает перемещаться. В одном случае окольцованные птенцы теньковки в первый же день вылета из гнезда были обнаружены в 60—70 м от гнезда. Птенцы держались на ветвях деревьев на высоте около 3 м от земли.
Питание гнездовых птенцов. Материалы по питанию птенцов пеночки-теньковки, собранные на территории Савальского лесхоза, опубликованы Нейфельдт (1956). Чаще всего пеночки приносили здесь своим птенцам гусениц бабочек, тлей, пауков и мух. В учлесхозе «Лес на Ворскле» в июле 1957 г. птенцы выкармливались в основном мухами сем. Larvivoridae.
ПЕНОЧКА-ВЕСНИЧКА (PHYLLOSCOPUS TROCHILUS L.)
Токование. В некоторые годы пение пеночек-весничек на территории Ленинградской области можно слышать уже в конце апреля. Регулярное же пение начинается обычно с конца I декады мая и продолжается примерно до середины июля. Отдельные самцы поют до последних чисел июля. Осенняя песнь звучит обычно в августе, реже — в начале сентября. Факта гнездования весничек автору не удалось установить ни в учлесхозе «Лес на Ворскле» (1949 и 1957 гг.), ни в Савальском лесхозе (1951 —1953), где в 1953 г. все же был отмечен экземпляр самца, который регулярно пел (с 7 июня по 6 августа) на одном и том же участке леса. Однако ни разу здесь не удавалось наблюдать одновременно двух пеночек и слышать их беспокойного крика, как это бывает характерно для гнездящихся птиц. Поэтому автор не •счел возможным включить пеночку-весничку в описок гнездящихся
230
птиц Савальского лесхоза. По всей вероятности, южная граница области нормального гнездования этого вида должна проходить несколько севернее Савальского лесхоза и учлесхоза «Лес на Ворскле».
Расположение гнезд а. Все гнезда пеночки-веснички, найденные автором в Ленинградской области, располагались на земле среди мха или прошлогодних сухих листьев под защитой маленького деревца или кустарника. Лоток гнезда обычно бывает выстлан перьями различных куриных, на что уже обращал внимание Г. А. Новиков (1948). На Карельском перешейке в гнездах, расположенных вдали от поселков, мы, как правило, обнаруживали перья глухаря (самца и самки), тетерева и рябчика. У домов пеночка чаще всего использует перья домашних кур. В некоторых гнездах наряду с куриными мы обнаруживали также перья скворца, дрозда-рябинника и серой вороны. Весничка может таскать перья в гнездо и тогда, когда в нем находятся уже птенцы. Если из такого гнезда вытащить часть перьев и разбросать их поблизости, то пеночка иногда собирает и затаскивает их обратно.
Сроки размножения. Период возможных сроков откладки яиц у пеночки-веснички на территории Ленинградской области продолжается примерно 50 дней. Так, первое яйцо в наиболее раннем гнезде появилось 15 мая (1949), а самая поздняя кладка началась 5 июля -(1954). Максимальное количество гнезд со свежей кладкой было найдено в последней декаде мая и I декаде июня. Начиная со II декады июня количество птиц, приступающих к размножению, резко снижается. В поздних гнездах мы находили, как правило, 4—5 яиц, в то время как у птиц, размножавшихся в нормальные сроки, кладка обычно состояла из 6—7 яиц. Это, по нашему мнению, указывает на то, что кладки, найденные в конце июня —начале июля, являются дополнительными, а не вторыми, как это иногда считают.
Ниже приведено количество гнезд, в которых кладка началась:
В I декаде мая..........—
Во II декаде мая..........1
В III декаде мая..........8
В I декаде июня........11
Во II декаде июня...........4
В III декаде июня...........2
В I декаде июля...........2
Во II декаде июля...........1
Таким образом, «пик размножения» веснички в Ленинградской области приходится на конец мая — I декаду июня. Второй вспышки размножения не наблюдается.
Размеры и судьба кладок. В Ленинградской области в период с 1946 по 1954 г. была прослежена судьба 24 кладок. Количество яиц в этих кладках было следующее: по 4 яйца — в 2 кладках, по 5—в 2, по 6 — в 10, по 7 яиц — в 10 гнездах. Среднее количество яиц на 1 гнездо оказалось, таким образом, равным 6.
Из 148 яиц, находившихся под наблюдением, вылупилось 111 птенцов: 13 яиц (2 кладки) было брошено -птицами в период насиживания, 20 яиц (3 кладки) погибло при разорении гнезд, и 4 яйца оказались «болтунами. В дальнейшем было разорено еще 2 гнезда с 11 птенцами. Кроме того, 1 птенец по неизвестной причине был выброшен взрослыми птицами из гнезда и погиб. Таким образом, вылетело из гнезд 99 птенцов, что составило 67% от первоначального количества яиц. Количество птенцов в выводке в момент оставления гнезда варьировало от 4 до 7.
Поведение взрослых птиц у г н ез д а. Поведение большинства взрослых птиц во время беспокойства у гнезда довольно однотипно: пеночки обычно держатся на нижних ветвях деревьев и, издавая растянутый тонкий свист, постоянно перелетают с места на место.
231
Автору никогда не приходилось наблюдать, чтобы пеночки-веснички: «отводили» от гнезда, хотя подобные наблюдения имеются. Некоторые особи проявляют удивительную доверчивость и спокойно кормят птенцов в непосредственной близости от наблюдателя. В начале гнездового периода веснички, наоборот, (пугливы и легко бросают гнезда с неоконченной или слабо насиженной кладкой.
Птенцы. Птенец имеет нежный светло-желтый пух, кажущийся иногда почти белым; он 'представлен на теле птенца отдельными пучками — над глазами, ма затылке и на плечах. У однодневных птенчиков, бросается в глаза желтый цвет, который окрашивает самые различные участки тела в значительно большей степени, нежели у других видов пеночек. Цвет кожи и ротовой полости оранжево-желтый, ноги, когти,, клюв и яйцевой «зуб» желтоватые, складка в углах рта лимонно-желтая. Желтоватая окраска лап и когтей, продолжающая оставаться яркой до 7—8-дневного возраста, в дальнейшем постепенно бледнеет и сереет, так что у птенца-слетка в возрасте 11 —12 дней желтый цвет остается только на нижней стороне пальцев.
Как и у других видов мелких воробьиных, развитие птенцов идет быстрыми темпами. Ушные отверстия бывают открыты в конце 3-го дня жизни, глаза приоткрываются на 4—5-й день. Пеньки маховых перьев растут чрезвычайно интенсивно в период с 3-го по 6-й день, и уже на 7-й день на их концах появляются кисточки опахал. Развертывание опахал кроющих крыла, а также рулевых начинается несколько раньше— в конце 6-го дня. 6-дневные птенцы пеночки-веснички располагаются в гнезде уже в определенном порядке: головы всех бывают обращены в направлении входного отверстия. В недельном возрасте они становятся пугливыми и при виде человека прижимаются ко дну гнезда. Будучи сильно потревоженными, они могут выскочить из гнезда уже на 10-й день после вылупления.
Испуганный птенец издает особый «скрипящий» крик, который очень сильно возбуждает остальных: птенцы, сидевшие до того спокойно, начинают поочередно выпрыгивать из гнезда.
Нормальное оставление гнезда наступает спустя И, а чаще—12 дней после вылупления. В этом возрасте птенчики пеночки-веснички бывают оперены уже почти полностью. Опахала кроющих заходят на основания опахал маховых примерно на 10 мм, в результате чего образуется сплошная несущ,ая поверхность крыла. Птенцы, только что оставившие гнездо, могут пролетать не более 10—15 м без снижения высоты. Поймать птенца, недавно выскочившего из гнезда, очень трудно. Быстро перепархивая с ветки на ветку, он забирается все выше и выше и вскоре становится недосягаем. Этот способ первоначального передвижения путем коротких, стремительных, быстро следующих один за* другим перелетов с ветки на ветку и стремление как можно скорее забраться повыше в кроны деревьев характерны для птенцов всех видов пеночек, а также для птенцов лесной малиновки.
Питание гнездовых птенцов. Питание птенцов пеночки-веснички изучалось автором совместно с Е. Р. Гагинской летом 1954 г. в районе ст. Кузнечное Ленинградской области. Кроме того, были использованы некоторые материалы, собранные на территории Кандалакшского заповедника О. М. Татариновой. Всего из пищеводов птенцов было отобрано 203 экз. насекомых и других беспозвоночных. Чаще всего птенцы веснички получают от своих родителей различных двукрылых, которые в целом составили около половины (96) всех беспозвоночных, полученных птенцами. Среди них преобладали хищные мухи (Rhagionidae, Empididae и Asilidae), а также слепни и комары-
232
долгоножки. Поедались, кроме того, иногда настоящие мухи, цветочные мухи, синие мухи Calliphora и комары-звонцы. На втором месте по количеству съеденных экземпляров (40) стояли гусеницы пядениц, совок, голубянок и других бабочек. Бабочки имаго приносились птенцам лишь изредка (6). В обоих районах исследования птенцы весничек получали мелких цикад (12), мелких стрекоз и тлей. Помимо насекомых из пищеводов птенцов весничек постоянно извлекались пауки-крестовики (19) и реже —наземные моллюски (7).
ПЕНОЧКА-ТРЕЩОТКА
(PHYLLOSCOPUS SIBILATOR В Е С Н S Т.)
Токование. Во время активного токования самцы пеночек-трещоток помимо своей обычной трели издают еще специфичный для вида «тюкающий» позыв, воспроизводящийся в более учащенном темпе, нежели во время беспокойства у гнезда. Как и песня, этот позыв имеет брачное значение. По нашим наблюдениям, особенно часто он воспроизводится самцом в период спаривания, постройки гнезда и откладки яиц.
В 1950 г. пеночка-трещотка начала петь в парке Лесотехнической академии 23 апреля. Однако регулярное пение пеночек под Ленинградом обычно начинается лишь в конце I декады мая. В конце II декады июля активность пения резко падает, но отдельные самцы поют и позднее. Последняя песня отмечена нами 20 июля (1946), 21 июля (1950), 23 июля (1954). Самцы поют активно в течение всего гнездового периода, вплоть до вылета птенцов из гнезда.
Сроки размножения. В Ленинградской области период возможных сроков откладки яиц у пеночки-трещотки продолжается с конца мая до начала июня (табл. 55). В наиболее раннем из гнезд первое яйцо появилось 20 мая (1948 и 1949 гг.), в наиболее позднем — 3 июля (1954). Максимальное количество свежих кладок было найдено в III декаде мая. В июне и в июле приступают к размножению лишь немногие особи, очевидно те, у которых по какой-либо причине пропала первая кладка.
В лесостепи, на территории Савальского лесхоза, период размножения у трещоток оказался еще более растянутым. Наиболее ранний вылет птенцов в Савальском лесу произошел 10 июня (1951). Судя по возрасту птенцов, откладка яиц началась в этом гнезде примерно 10 мая. В наиболее позднем из найденных гнезд первое яйцо появилось 1 июля (1953). В целом откладка яиц у этого вида пеночки идет довольно дружно и в лесостепной зоне. Основная масса птиц приступает к размножению во II и III декаде мая. Начиная уже с I декады июня количество особей, откладывающих яйца, резко сокращается. В конце июня и начале июля гнездятся, по-видимому, лишь птицы, потерявшие первые кладки. На это указывает уменьшенное количество яиц (4 и 5) в их гнездах. Возможность второго цикла размножения после успешного воспитания первого выводка мало вероятна.
Размеры и судьба кладок. Всего под наблюдением находилось 38 гнезд: 12 в лесной зоне и 26 в лесостепной. В лесной зоне 3, 6, 7 и 8 яиц, найдены в гнезде по одному разу и 5 яиц — 7 раз. В лесостепи по 4 яйца насчитывалось в 4, по 5—в 5, по 6—в И, по 7— в 6 гнездах. Кладки, содержавшие 6 и 7 яиц, в лесостепи встречались чаще, чем в лесной полосе. В связи с этим среднее количество яиц на 1 кладку здесь оказалось 5,73, против 5,25 в лесной зоне.
233
Таблица 55
Сроки размножения пеночки-трещотки (по декадам)
Район наблюдения Количество гнезд, в которых откладка яиц началась
в мае в июне в июле
II III 1 II Ill I
Ленинградская область . . 9 2 2 1
Воронежская область . . . 10 10 2 — 1 1
Однако отход яиц и птенцов в лесостепной полосе более значителен: из 149 яиц вылупился 121 птенец, 26 яиц (4 кладки) пропало при разорении гнезд, и 2 яйца оказались неоплодотворенными. Впоследствии было разорено еще 6 гнезд с 31 птенцом. Кроме того, 2 птенца в 2 гнездах погибли по неизвестной причине незадолго до вылета. Всего благополучно закончили развитие 88 птенцов. Показатель выживаемости оказался равным, таким образом, 3—4 птенца на 1 гнездо.
На территории Ленинградской области из 63 яиц вылупилось 63 птенца. Из них погибло 14 (2 выводка), остальные 49 птенцов благополучно оставили гнезда. В среднем на каждое гнездо пришлось по 4 птенца. Более низкий отход яиц и птенцов в северных широтах (лесная зона) следует объяснять, очевидно, меньшим количеством врагов, а также менее интенсивным выпасом скота в лесу.
Поведение взрослых птиц у гнезда. Самка, спугнутая с гнезда, в некоторых случаях «притворяется» раненой. Обычно же трещотки сидят на гнезде настолько плотно, что иногда позволяют даже поймать себя рукой. Из-за этого они нередко погибают под ногами пасущегося в лесу скота. Первые 4—-5 дней после вылупления птенцов самка очень много времени проводит в гнезде. В дальнейшем систематическое обогревание сменяется временным в случае холодной или дождливой погоды. Приходилось наблюдать трещоток на птенцах, которым было уже 9—10 дней. Подобное поведение указывает на то, что биологический смысл «крыши» гнезда у пеночек заключается, видимо, больше в защите потомства от глаз хищников, нежели от возможных неблагоприятных последствий непогоды.
Птенцы. Пух у птенцов светло-желтовато-серый, редкий, но довольно длинный, особенно на плечах. Расположен он, как и у всех пеночек, над глазами, на затылке и на плечах. Клюв короткий и широкий в основании. Ноздри продолговатые, с несколько приподнятыми краями. Яйцевой «зуб» и когти светлые. Ротовая полость желтая, язык без пятен*. Складка в углах рта относительно широкая, светло-желтая.
Пеньки будущих маховых обозначаются на 3-й день жизни птенца. Слуховые проходы открываются в 3-дневном возрасте; глаза приоткрываются на 4—5-й день. У 6-дневных птенцов глаза уже полностью открыты. В недельном возрасте птенцы начинают пугаться людей, прижимаются ко дну гнезда и начинают угрожающе «шипеть». Кисточки маховых перьев появляются у них в конце 6-го — начале 7-го дня жизни. Примерно в этом же возрасте появляются и кисточки на концах пеньков рулевых перьев. 10-дневные птенцы имеют уже хорошо развитые опахала рулевых и маховых. При сильном испуге они могут уже выскочить из гнезда. Нормально птенцы покидают гнездо в возрасте 12 дней. Покинув гнездо, птенец обычно по земле удаляется от него
234
на 25—30 м, затем, перепрыгивая с ветки на ветку, очень скоро оказывается на 3—5-метровой высоте. Здесь он успокаивается и начинает регулярно воспроизводить призывный крик. Этот птенцовый позыв напоминает звуки, .издаваемые самцом трещотки в начале песни.
О времени, когда выводки начинают совершать перекочевки, трудно сказать что-либо определенное. В одних случаях автор обнаружил окольцованных птенцов в районе гнезда спустя 7 дней после оставления гнезда, в других — выводок уже на следующий день перемещался на расстояние 100 м в сторону.
Питание гнездовых птенцов. Материалы по питанию <5ыли собраны в Савальском лесхозе в 1952 г. И. А. Нейфельдт (1956) и автором в 1953 г. Всего от птенцов этого вида было отобрано 23 экз. насекомых, 38 пауков и 6 моллюсков. Из насекомых поедались почти исключительно гусеницы бабочек, мухи или крупные комары.
Результаты кольцевания. В 1951 —1952 гг. на территории Савальского лесхоза было окольцовано 75 птенцов-слетков пеночек-трещоток, из них только один (самка) был обнаружен гнездящимся на следующий год. Гнездо было построено в 1,5 км от места рождения птицы.
ПЕНОЧКА ЗЕЛЕНАЯ (PHYLLOSCOPUS TROCHILOIDES SUND.)
Пригородные сады и парки, видимо, являются наиболее характерным местом обитания зеленых пеночек в Ленинградской области. Наибольшее количество встреч этого редкого у нас вида зарегистрировано в Павловском и Петергофском парках, а также в парке Лесотехнической академии имени С. М. Кирова. Найденные в Ленинградской области гнезда этого вида (Промптов, 1949) тоже были обнаружены в парке (пос. Колтуши). В конце мая — начале июня пение зеленой пеночки можно было слышать в 1952 и 1954 гг. в саду Ленинградского университета на Менделеевской линии. В 1955 г. на севере Карельского перешейка (Приозерский район) активное пение пролетного самца автор слышал 13 мая. Интересно, что эта чрезвычайно ранняя дата весеннего пролета совпала с необыкновенно холодной и затяжной весной.
Не более многочисленной эта пеночка оказалась и на территории Савальского лесхоза. В течение 3 лет (1951—1953) встречено всего 2 самца, оба в 1953 г., которые держались на строго определенных участках леса в продолжение почти всего июня. Судя по поведению птиц, это были холостые самцы. В разгар брачной поры самцы зеленой пеночки повторяют свою задорную песню чрезвычайно часто. 14 июня 1953 г. в Савальском лесхозе автор следил за одним самцом в продолжение 2 час. За эго время птица воспроизвела свою песню 956 раз.
КАМЫШОВКА САДОВАЯ (ACROCEPHALUS DUMETORUM BLYTH.)
Пение садовой камышовки в Ленинградской области можно слышать начиная с конца мая вплоть до конца II декады июля. Наиболее ранняя дата первой песни отмечена нами 28 мая в 1954 г. Регулярное пение продолжается с конца I декады июня до середины июля, т. е. примерно в течение месяца. Из звуков, заимствованных у других птиц, во время пения садовой камышовки бывают слышны обрывки песни певчего дрозда, беспокойное «пиньканье» зяблика, позывы серой слав
235
ки, сорокопута-жулана, чижа, большой* синицы, а также козодоя (по* зыв «уик»).
Наиболее северной точкой, где автору приходилось наблюдать активно поющих самцов садовых камышовок (10—16 июня 1954 г.), является ст. Кузнечное, лежащая в 150 км к северу от Ленинграда. Гнез-да садовой камышовки, найденные в окрестностях Ленинграда, были расположены в кустах бузины, смородины и таволги рябинолистной на высоте 0,5—0,9 м. В одном из гнезд первое яйцо появилось 2 июня, в. другом — 6 июня и, наконец, в третьем, судя по возрасту птенцов, откладка яиц началась 21—22 июня.
Птенцы у садовой камышовки появляются на свет совершенно голыми, ротовая полость у них окрашена в желтый цвет, а на корне языка имеются два очень ярких черных пятна. Эти пятна сохраняют интенсивность окраски и тогда, когда птенцы покидают гнездо. Молодых камышовок, только что оставивших гнездо, поймать трудно, хотя они и не умеют летать. Они быстро могут передвигаться по веткам кустарников, среди которых предпочитают держаться в послегнездовой период, жизни.
МАЛИНОВКА ЛЕСНАЯ, ИЛИ ПЕРЕСМЕШНИК (HIPPOLAIS ICTERINA VIEILL.)
Токование. В окрестностях Ленинграда в течение последнего десятилетия наиболее ранняя дата первой песни пересмешника отмечена нами 14 мая 1949 г., а наиболее поздняя — 31 мая 1951 г. Таким образом, период нерегулярного пения благодаря разнице в сроках прилета составляет более 2 недель. Регулярное пение продолжается до начала II декады июля, хотя отдельные самцы поют до середины последней декады этого месяца.
Как известно, пение лесной малиновки поражает разнообразием, и точностью копировки звуков, заимствованных у других птиц. Наблюдения показывают, что пересмешник заимствует в основном не пение других птиц, как это принято считать, а различные позывы, причем главным образом позывы, выражающие состояние беспокойства птиц. Эти наблюдения служат, как нам кажется, хорошим доводом против-мнения некоторых орнитологов, которые рассматривают пере-смешничество как «территориальный сигнал» расширенного значения. Лесные малиновки, гнездящиеся в местах, населенных различной орнитофауной, поют по-разному. В Ленинградской области они помимо позывов широко распространенных видов птиц имитируют голоса дрозда-белобровика (тревожный сигнал), чижа (позыв), клеста-еловика, кулика-черныша (тревожный крик), пеночки-веснички (позыв и обрывки песни). В Воронежской области часто бывают слышны гол-оса представителей лесостепи: золотистой щурки, черного дрозда (сигнал тревоги), иволги (пение и тревожный крик), пустельги и др.
Расположение гнезда. В окрестностях Ленинграда лесная малиновка помимо лиственных деревьев (береза, рябина, клен, вяз) гнездится нередко и на хвойных. Неоднократно приходилось находить гнезда на елях (4 находки из 17), причем не только на подросте, но и на крупных деревьях. В последнем случае гнезда располагались на конце лапы одной из нижних ветвей. Как уже отмечалось (Птушенко, 1954), лесная малиновка чаще гнездится на молодых деревцах, чем на взрослых. Из 17 гнезд этого вида, найденных нами в Ленинградской области, 9 располагалось в подросте, 6 — на крупных деревьях и только 2 — в кустарниках (спирея рябинолистная, бузина) .
236
В Савальском лесхозе большинство гнезд было обнаружено на Лузине (5 из 9). Высота расположения гнезд варьировала от 0,6 до 10 м.
Сроки размножения. Во всех гнездах пересмешника, найденных в Ленинградской области, откладка яиц происходила в июне. В наиболее раннем из этих гнезд первое яйцо появилось 1 июня (1950), в самом позднем — 24 июня (1946). В преобладающем количестве случаев размножение птиц начиналось в I и II декаде июня.
На территории Савальского лесхоза кладка началась 10 июня, 11 июня (1951), 18 июня, 21 июня (1952), 22 июня (1951), 23 июня (1953) и 7 июля (1952).
Размеры и судьба кладок. Количество яиц варьировало ют 3 до 6. Чаще всего встречались кладки, содержавшие 4 или 5 яиц. Количество яиц в отдельных гнездах было следующим: 3, 4, 4, 4, 4, 4, 5 (Савальский лесхоз) и 3, 3, 3, 4, 4, 4, 4, 4, 5, 5, 5, 5 и 6 (Ленинградская область). В Савальском лесу из 28 яиц вылупилось 27 птенцов (одно яйцо оказалось болтуном), и все они благополучно вылетели из гнезд. В Ленинградской области из 55 яиц вылупилось 47 птенцов: одна кладка (3 яйца)' была брошена, 4 яйца (одна кладка) была кем-то утащена, и 1 яйцо оказалось неоплодотворенным; вылетело из гнезд 39 птенцов (2 гнезда с 8 птенцами были разорены каким-то хищником). Таким образом, количество птенцов, покинувших гнездо, составило здесь 70% от количества первоначально отложенных яиц.
Поведение взрослых птиц у гнезда. Во время насиживания яиц или обогрева птенцов пересмешник сидит на гнезде очень плотно и иногда даже позволяет дотронуться до него рукой. Будучи же вспугнутым с гнезда, он, как правило, сильно возбуждается и часто переходит в наступление: принимает угрожающие позы, громко щелкает клювом, стремительно «пикирует» и ударяет врага своим маленьким тельцем. Пересмешников, «притворяющихся» ранеными, автору наблюдать не приходилось.
Птенцы. Птенцы лесной малиновки совершенно голые и так же, как камышовки рода Acrocephalus, имеют на основании языка два резко очерченных черных пятна. Ротовая полость у них ярко-желтая с •оранжевым оттенком.
В однодневном возрасте птенцы вялые. Активность заметно возрастает с 4—5-дневного возраста, когда у птенцов приоткрываются глаза и намечаются пеньки маховых и рулевых перьев. Развертывание опахал маховых начинается в конце 6-го — начале 7-го дня жизни, пеньки рулевых раскрываются на сутки позднее. К этому времени птенец почти весь покрывается кисточками растущих перьев, только на лбу и шее пеньки остаются неразвернувшимися вплоть до вылета птенцов из гнезда. Если птенцов не пугать, они оставляют гнездо на 12—13-й день. В случае же сильного испуга птенцы начинают выскакивать уже с 9— 10-дневного возраста. Способность к полету приобретается в возрасте 13—14 дней. Птенцы-слетки очень быстро перемещаются в кронах деревьев. Один еще нелетный птенец, на наших глазах выскочивший из гнезда, расположенного у самой земли, через 10 мин. оказался уже сидящим на ветке дерева на высоте 3 м и в 60 м от гнезда. На следующий день он уже был способен перелетать с дерева на дерево, не снижая высоты полета.
Питание гнездовых птенцов. Несмотря на ограниченность материалов, собранных в Савальском лесу, они все же позволяют говорить о предпочтении, которое отдают пересмешники при выкармливании птенцов двукрылым, главным образом мухам (24 экз. из 46). Довольно часто птенцы поедают бабочек и их гусениц (8), а также пау
237
ков (7). Жуки (божья коровка), прямокрылые (личинки саранчовых), клопы (личинки Pentatomidae) и мелкие стрекозы были принесены птенцам в количестве 1—2 экз. Среди чешуекрылых были обнаружены гусеницы совок, огневок и пядениц.
СЛАВКА ЯСТРЕБИНАЯ (SYLVIA NISORIA BECHST.)
Пение ястребиной славки очень напоминает песню садовой славки; от последней она отличается большим количеством скрипучих звуков и тем, что издается очень часто на лету. В пригородах Ленинграда наиболее раннее пение этого вида отмечено 22 мая (1954).
В гнезде, обнаруженном на территории Савальского лесхоза в. 1952 г., вылупление птенцов произошло 15 июня. В Ленинградской области, в окрестностях Зеленогорска, птиц, беспокоившихся около птенцов-слетков, автор наблюдал 13 июля (1955).
Во время беспокойства у гнезда или птенцов ястребиные славки возбуждаются, как правило, очень сильно. Обычно они очень близко-подлетают к наблюдателю, воспроизводят громкие «чекающие» или «трещащие» позывы, принимают позы больной или раненой птицы.
Птенцы у ястребинки появляются на свет, лишенные эмбрионального пуха. Ротовая полость у них желтая с оранжевым оттенком. На корне языка заметны 2 темных пятна. По окраске ротовой полости иг наружных краев клюва птенцы ястребиной славки ближе всего стоят к птенцам серой славки и завирушки.
СЛАВКА-ЧЕРНОГОЛОВКА (SYLVIA ATRICAPILLA L.)
Токование. В течение последнего десятилетия прилет славок-черноголовок в окрестности Ленинграда происходил обычно между 15 и 31 мая. В первые дни после прилета поют лишь отдельные самцы. Регулярное пение славок-черноголовок в окрестностях Ленинграда можно слышать начиная с последних чисел мая вплоть до начала II декады июля. Отдельные самцы в некоторые годы поют еще в течение последней декады июля: 22 и 29 июля (1954).
Расположение гнезда. Сравнение материалов по гнездованию славки-черноголовки, собранных в трех различных пунктах Европейской части СССР (окрестности Ленинграда. Савальский лес и заповедник «Лес на Ворскле»), показало, что в каждом из этих мест Черноголовка имеет излюбленные растения, на которых она предпочитает помещать сбои гнезда. В окрестностях Ленинграда таковыми оказались еловый подрост и поросль черемухи, в заповеднике «Лес на Ворскле» — поросль широколиственных древесных пород (дуба, вяза, липы, клена)., а в Савальском лесу — кусты бузины и отчасти бересклета бородавчатого. Помимо перечисленных выше пород деревьев и кустарников гнезда этой птицы мы находили также на желтой акации,, спирее рябинолистной, малине, можжевельнике, ольхе, березе, рябине, жимолости и некоторых др. Таким образом, экологической зависимости Черноголовки от какого-либо определенного вида растения в пределах ее ареала установить не удалось.
Все это подчеркивает значительную степень пластичности гнездовых инстинктов вида, следствием которой является его способность-применяться к различным условиям. На это же указывают и сравнительно редкие, правда, факты нахождения гнезд на ветвях крупных
238
деревьев, на высоте 3—4 м от земли. Такие гнезда приходилось находить в Минской области В. Н. Шнитникову (1913), а также Н. П. Овчинниковой в лесостепной дубраве «Лес на Ворскле» и Г. А. Новикову в Шиповом лесу Воронежской области.
Сроки размножения. В Ленинградской области период возможных сроков откладки яиц продолжается с конца II декады мая до* конца I декады июля. Здесь самая ранняя из известных нам кладок началась 17 мая (1950), а самая поздняя—10 июля. Установленные нами крайние сроки размножения славки-черноголовки в Ленинградской области почти точно соответствуют данным В. Л. Бианки (1917— 1918). Наиболее ранний срок появления первого яйца в гнезде славки-черноголовки В. Л. Бианки отметил в 1910 г. в парке Ст. Петергофа* 19 мая. Ровно через 40 лет в этом же самом парке автор нашел гнездо, в котором кладка началась 17 мая. В такой же степени сходными оказались и даты наиболее позднего гнездования.
Таблица 56*
Сроки размножения славки-черноголовки (по декадам)
Из табл. 56 видно, что «пик» интенсивности размножения у славок,, гнездящихся в Ленинградской области, приходится на конец мая — начало июня, а у птиц Белгородской области — на II и III декады мая. Сравнительно поздно начинается откладка у птиц, гнездящихся в восточных районах лесостепи (Воронежская область). На протяжении 3* лет работы в Савальском лесхозе нам не удалось найти ни одного гнезда, в котором степень насиженности яиц или возраст птенцов указывал бы на то, что откладка яиц началась до 25 мая. До 20 июня не наблюдали мы <и птенцов-слетков данного вида.
У Черноголовки, по нашему мнению, существует только одна кладка в году. На это указывает отсутствие второй «волны» размножения в конце лета. Свежие кладки этого вида, появляющиеся во второй половине июня — начале июля, по всей видимости, являются не вторыми,, как это иногда считают, а дополнительными.
Размеры и судьба кладок. Автор располагает данными по истории 73 гнезд: из них судьба 49 гнезд была прослежена в пределах лесостепи («Лес на Ворскле» и Савальский лес), а история 24 гнезд — в лесной зоне, главным образом в окрестностях Ленинграда.. Количество яиц в кладках варьировало от 2 до 6. Чаще всего встречались кладки, состоявшие из 5 яиц, несколько реже — из 4 (табл. 57).
Из 224 яиц, найденных в лесостепной зоне, вылупилось 179 птенцов: 36 яиц (8 кладок) погибло при разорении гнезд, 4 яйца (1 кладка) было брошено самой птицей, и 5 яиц оказались неоплодотворенными. Из 173 птенцов 157 благополучно закончили свое развитие и покинули гнезда, 22 птенца (5 выводков) было уничтожено различными хищни-
239
Таблица 57
Количество яиц в кладках славки-черноголовки
Зона Количество гнезд, содержащих кладку яиц (в шт.) Среднее количество яиц на 1 гнездо
2 3 4 5 6
Лесная зона 1 1 8 12 2 4,54
Лесостепная зона — 5 17 21 6 4,57
Рис. 58. Только что оставивший гнездо птенец славки-черноголовки („Лес на Ворскле", июль 1957 г.
Фото автора).
ками. По отношению к первоначальному количеству отложенных яиц количество вылетевших из гнезд птенцов в лесостепной зоне составило, таким образом, около 70%. В окрестностях Ленинграда показатель степени выживаемости потомства был более низким — 54%. Основной причиной отхода яиц и птенцов здесь также явились наземные хищники, в частности бродячие кошки.
Поведение взрослых птиц у гнезда. Самцы славок-черноголовок принимают деятельное участие как в насиживании яиц, так и в обогревании птенцов. Специальный учет, проведенный автором и Н. П. Овчинниковой, показал, что самцы почти так же часто сидят на яйцах и птенцах, как и самки. Из 120 наблюдений за гнездами в. 73 случаях на них сидели самки, в 47—самцы. Самцы славок-черноголовок, которых мы отлавливали в западню или убивали у гнезд, имели наседное пятно.
При появлении человека вблизи гнезда некоторые особи, главным образом птицы, гнездящиеся вдали от жилья человека, ведут себя удивительно осторожно. Однако в большинстве случаев Черноголовки бывают очень активны: подлетают близко и «притворяются» ранеными. Ярко выраженное явление «отвода» удалось установить у 40 из 72 наблюдавшихся особей; многие «отводили» от гнезда да
же тогда, когда в нем были ненаси-женные кладки. Имитация раненой птицы в одинаковой степени характерна как для самцов, так и для самок славок-черноголовок. Иногда сам-
цы ведут себя даже более активно. Автору пришлось однажды наблюдать самца Черноголовки, который 15 раз подряд повторял один и тот же прием «отвода»: бросался под ноги и, трепыхаясь, отползал в сторону. Птенцы. Птенец первоначально совершенно голый. Очень похож на птенца садовой славки, но отличается от последнего наружной расцветкой клюва, имеющего вокруг ноздрей резко очерченные темные участки. Яйцевой «зуб» серый, когти светлые. Ноздри продолговатые. Ротовая полость мясо-красная, на корне языка 2 неясных темных пятна. Кожа желтовато-розовая. Со 2-го дня жизни становятся отчетливо
240
заметными подкожные потемнения на местах‘будущих маховых и контурных перьев. В конце 3-го дня у птенцов открываются слуховые проходы, глаза приоткрываются на 4—5-й день. В возрасте 6 дней начинают появляться кисточки маховых перьев, а также кроющих крыла и контурных перьев спины. При сильном испуге птенец выскакивает из гнезда иногда в недельном возрасте. Оставление гнезда служит обычно стимулом для воспроизведения короткого мелодичного выкрика — позыва, имеющего призывное значение. После того, как птенцы покинут гнездо (рис. 58), этот -позыв издается птенцом регулярно.
Питание гнездовых птенцов. Материалы по питанию были собраны на территории Савальского лесхоза в основном в 1952 г. Всего удалось извлечь из пищеводов птенцов 178 экз. различных беспозвоночных. Среди них на первом месте оказались пауки (79), а на втором— гусеницы и куколки бабочек (63). Взрослых бабочек (11), а также жуков (8) и прочих насекомых (верблюдки, ручейники, перепончатокрылые— всего 8 экз.) славки-черноголовки приносили своим птенцам редко. Среди бабочек, уничтожавшихся славками, численно преобладали гусеницы и куколки листоверток, пядениц и хохлаток. Были обнаружены также гусеницы и бабочки совок и несколько экземпляров мохнатых гусениц шелкопрядов. Из жуков птенцам давались преимущественно мягкотелки. Помимо того, в пищеводах птенцов было обнаружено 9 экз. моллюсков. Таким образом, наиболее характерной пищей птенцов Черноголовки являются пауки и голые гусеницы бабочек.
СЛАВКА СЕРАЯ (SYLVIA COMMUNIS LATH.)
В Ленинградской области наиболее ранняя дата первой песни отмечена 11 мая (1954), а наиболее поздняя песнь — 23 июля (1954). Большинство птиц прекращает, однако, пение в середине июля.
Половина найденных гнезд (4 из 9) помещалась на земле, между стеблей крапивы или других трав; остальные располагались на высоте ют 0,1 до 0,8 м в кустах бузины, смородины, малины и жасмина. В Савальском лесхозе серая славка в большинстве случаев гнездилась на кустах (бересклет, полынь) или среди прикорневой поросли вяза, липы, вишни, осины; только одно из гнезд было расположено на земле.
В строительстве гнезда деятельное участие принимает самец. Иногда самец почти до конца выстраивает подряд несколько гнезд, затем появляется самка, которая быстро доканчивает постройку одного из гнезд и начинает откладывать в него яйца. Например, весной 1947 г. мы наблюдали в парке Лесотехнической академии самца серой славки, который в течение многих дней (с 2 по 20 июня) держался на одном и том же участке, беспрерывно напевая свою песню. За это время он выстроил по крайней мере 2 гнезда в 6 м одно от другого. Самка на этом участке впервые была замечена лишь 20 июня. Вечером этого дня она подправила одно из гнезд, выложила лежащую на дне его вату на край лотка и уже утром 21 июня снесла первое яйцо. Второе гнездо так и осталось без употребления. Активность пения самца после появления самки резко снизилась.
На территории Савальского лесхоза даты начала откладки яиц в разных гнездах отмечались 23 мая (1952), 30 мая (1951), 24 июня (1953), 27 июня (1951) и 6 июля (1953). В последнем случае самка отложила в гнезде Только 3 яйца.. Это была, вероятно, дополнительная кладка. По наблюдениям В. Л. Бианки (1917—1918), период возможных сроков откладки яиц у серой славки в Ленинградской области
16 А. С . Мальчевский 241
продолжается примерно 1 мес. — с конца II декады мая до начала III декады июня. В 4 гнездах, найденных здесь нами, откладка яиц началась 9 и 10 июня (1954), 12 июня (1950) и 21 июня (1947). Из 4 гнезд,, содержавших в общей сложности 18 яиц (4, 4, 5 и 5), только в одном* птенцы успешно закончили свое развитие. Остальные гнезда были разорены каким-то хищником. На территории Савальского лесхоза из 39* яиц (9 кладок: 3, 3, 4, 4, 4, 5, 5, 5 и 6 яиц) вылупилось 20 птенцов, и из них только 10 (3 выводка) нормально покинули гнезда, остальные погибли от хищников.
Птенец серой славки лишен пуха. Он имеет желтую окраску ротовой полости и в основании языка два хорошо заметных темных пятна,. Боковые выросты клюва бледно-желтые. Клюв воскового цвета с заметным потемнением на конце и сзади ноздрей. Яйцевой «зуб» серый, когти светлые. Ноздри продолговатые. Цвет кожи грязно-розовый с фиолетовым оттенком. Уже к концу первого дня у птенчика бывают заметны скрытые под кожей пеньки будущих маховых и плечевых перьев. Для7 птенцов серой славки так же, как и для других славок, типична ранняя способность к самостоятельному передвижению. Они оставляют гнездэ за несколько дней до приобретения способности к полету.
СЛАВКА-ЗАВИРУШКА (SYLVIA CURRUCA L.)
Токование. Наиболее ранняя первая песня славки-завирушки в течение последнего десятилетия отмечена нами в окрестностях Ленинграда 6 мая (1950), а наиболее поздняя песня — 9 июля (1955). Период, регулярного пения длится примерно с середины мая и до середины последней декады июня. Вообще это наиболее скрытная и молчаливая славка. У гнезда она поет редко даже в разгар брачного периода.
Расположение гнезда. По наблюдениям В. Л. Бианки (1917—1918), в Ленинградской области славка-завирушка располагает свои гнезда главным образом среди молодой поросли хвойных пород, (можжевельник, ель). В Савальском лесу также заметна тенденция помещать гнезда на хвойном подросте: из 13 известных нам гнезд 6 были укреплены на молодых елочках, 4 — на подросте вяза, 2 — на молодых: сосенках и 1 — на бузине. Высота расположения гнезда варьировала от 0,1 до 1 м.
Сроки размножения. В окрестностях Ленинграда в одном из гнезд первое яйцо появилось 23 мая (1947), в другом—1 июня (1948). В гнезде третьей пары насиживание кладки, состоявшей из 4 яиц, началось лишь 16—17 июля (1955). На территории Савальского лесхоза в наиболее раннем гнезде откладка яиц началась 25 мая (1952), а в самом позднем—25 июня (1951).
Размеры и судьба кладок. Наблюдения проводились только в лесостепной зоне — на территории Савальского лесхоза и в заповеднике «Лес на Ворскле». Под наблюдением находилось 16 кладок; в» 5 из них было по 4 яйца, в 9 — по 5 и в 2 — по 6. Из 77 яиц вылупилось 56 птенцов: 16 яиц (3 кладки) было утащено хищниками, и 5 яиц оказались неоплодотворен.ными или содержали замершие зародыши. Из^ 56 птенцов нормально закончили развитие 44, остальные погибли при последующем разорении гнезд. Количество благополучно вылетевших, птенцов составило таким образом 57% от первоначального количества яиц.
Поведение взрослых птиц у гнезда. Некоторые особи держатся у гнезда скрытно и осторожно даже в тот период, когда в их 242
гнездах находятся птенцы; другие, наоборот, имитируют раненую птицу уже в начале периода насиживания. Из 15 особей этого вида 6 птиц, держались у гнезда очень осторожно, а 9 особей при появлении человека вблизи гнезда применяли приемы «отвода». Одна птица, за которой автор шел следом, медленно ползла по земле в течение нескольких минут и отвела, таким образом, на 80 м. Некоторые особи садились, на выдающиеся ветки кустов и оттуда демонстративно падали на землю, затем, хромая, бежали в сторону, распустив хвост и волоча крылья.
Рис. 59. Славки-завирушки („Лес на Ворскле**, июль 1957 г. Фото автора). Вверху — птенцы и самка с добычей, внизу — самка и только что накормленный птенец.
Птенцы. Птенцы появляются на свет совершенно голыми. Ротовая полость у них желтого цвета, а наружные края клюва светлые с желтоватым оттенком. На корне языка два отчетливо выраженных тем.-ных пятна.
Ход развития птенцов примерно такой же, как и у других видов славок: глаза приоткрываются на 4—5-й день, пеньки маховых перьев раскрываются в возрасте 6 дней, а рулевых — в возрасте 7 дней. Из 4 гнезд, за которыми наблюдал автор, птенцы выскочили через 9, 10, 11
16*
243
vi И дней после вылупления. Первые дни после оставления гнезда птенцы скрываются в траве или лесном подседе. Они не могут еще летать, но по ветвям кустарников'передвигаются очень ловко (рис. 59). После того, как птенцы покинут гнездо, выводок иногда почти сразу начинает совершать перекочевку из одного участка леса в другой. Автор наблюдал, как не умеющие еще летать 3 птенца славки-завирушки пересекали лесную поляну, двигаясь в одном направлении на расстоянии 8— 10 м друг от друга. Птенцы то прыгали по земле, то перепархивали с сетки на ветку среди соснового подроста. При этом они все время перекликались, издавая сходные с позывом взрослых птиц «чекающие» звуки, а также типичное только для птенцов короткое повизгивание.
СЛАВКА САДОВАЯ
(SYLVIA BORIN В О D D.)
Токование. Наиболее раннее появление садовой славки в окрестностях Ленинграда было отмечено в 1954 г. 16 мая. Обычно же садовые славки прилетают сюда в последних числах мая. Их регулярное пение можно слышать примерно с 25—28 мая до середины июля. Во второй половине июля и начале августа поют лишь отдельные самцы. Наиболее позднее пение садовой славки было отмечено автором под Ленинградом 6 августа (1946) и 18 августа (1955). В августе славки поют крайне нерегулярно и обычно не в полную силу.
Расположение гнезда. Найденные нами гнезда садовой славки располагались на весьма разнообразных породах деревьев, кустарников и трав. В Ленинградской области они были обнаружены на 12 видах растений (по 42 находкам) и в Савальском лесу — на 16 (по 55 находкам). Древесная поросль и кустарники использовались примерно в равной степени. В пригородных садах и парках гнезда садовой славки мы находили чаще всего на кустах спирей, реже — на бузине, малине и на пневой поросли черемухи и липы, в местах, удаленных от жилья человека, — на маленьких елочках и можжевельнике.
На территории Савальского лесхоза гнезда этого вида нам попадались на кустах бересклета бородавчатого, бузины, малины, а также >на поросли дуба, вяза, березы и др. Какв Ленинградской, так и Воронежской областях были найдены гнезда, помещенные прямо на земле, между стеблей крапивы и других травянистых растений (5 случаев из 97). Высота расположения гнезд, как правило, не превышала 1 м (85 находок), 8 гнезд было найдено на высоте от 1 до 2 м, 2 — на высоте от 2 до 3 м, и, наконец, 2 постройки помещались в 4 и 5,5 м от земли. В последнем случае гнездо располагалось на тонких боковых ветках ^<лена. Оно было найдено в парке Лесотехнической академии 17 июля 1947 г. Кладка была еще слабо насиженной. Поздний срок размножения позволял предполагать, что птица поместила свое гнездо в столь необычном месте после ряда неудачных попыток гнездиться в кустах. На Карельском перешейке автор находил гнезда садовых славок, расположенные на значительном удалении от земли (до 3,5 м), в прибрежных зарослях ивняка, подстриженного лосем.
Сроки размножения. В Савальском лесхозе в самом раннем гнезде первое яйцо появилось 23 мая (1952), а в наиболее позднем—5 июля (1953). В 1952 г. самая поздняя кладка началась 4 июля. Таким образом, в течение одного года период откладки яиц у садовой славки продолжается 42 дня. Большинство птиц, однако, приступает -к гнездованию в конце мая — начале июня. Поздние (июльские) кладки состояли в одном случае из 2, а в другом —из 3 яиц. По всей вероятности,
244
они принадлежали птицам, уже не один раз терявшим кладки в этом сезоне.
В Ленинградской области основная масса свежих кладок была найдена в I декаде июня. Из 35 известных гнезд только в одном кладка началась в конце мая. Таким образом, большинство птиц размножается здесь на 5—10 дней позднее, чем в Воронежской области. В 1947 г. на Карельском перешейке 12 августа автор нашел гнездо садовой славки, в котором было 4 четырехдневных птенца. Случаи столь позднего размножения у садовой славки, конечно, очень редки. Обычно самые поздние кладки начинаются не позднее I декады июля (табл. 58).
Таблица 58 Сроки размножения садовой славки (по декадам)
Район наблюдения Количество гнезд, в которых кладка началась
в мае в июне в июле
III ' 1 II III I II III
Ленинградская область . . Воронежская область . . . 1 12 16 15 9 7 7 б 1 2 — 1
Сильная растянутость периода размножения садовой славки объясняется главным образом частыми случаями разорения гнезд этого вида, влекущими за собой появление запоздалых дополнительных кладок. Предполагать наличие у этой славки двойного цикла размножения нет достаточных оснований.
Размеры и судьба кладок. Количество яиц в период насиживания кладки варьировало от 2 до 5. Кладок из 6 яиц ни в одном из 74 случаев не наблюдалось (табл. 59).
Т аблица 59 Количество яиц в кладках садовой славки
Зона Количество гнезд, содержащих кладку яиц (в шт.) Среднее количество» яиц на 1 гнездо
2 3 4 5
Лесная 1 5 17 23 4,35
Лесостепная — 4 3 21 4,61
В лесостепной зоне из 200 яиц вылупилось 117 птенцов: 49 яиц. (12 кладок) погибло при разорении гнезд хищниками или во время лесохозяйственных работ, 20 яиц (5 кладок) было брошено в период насиживания и, наконец, 14 оказались болтунами либо с замершими зародышами. Из 117 птенцов в конечном счете выжило 87; 26 'птенцов (6 выводков) погибло от хищников и 4 — по неизвестной причине. По отношению к первоначальному количеству яиц птенцы, вылетевшие из гнезд, составили 43%.
Сходный процент выживаемости потомства был у садовых славок, гнездившихся в Ленинградской области: из 129 яиц (28 кладок) вылупилось 77 птенцов, и из них только 56 оставили гнезда. Остальные (35 яиц и 21 птенец) погибли в результате деятельности различных хищни
245
ков, 16 яиц (4 кладки) было брошено птицами, и 1 яйцо оказалось пеоплодотворенным.
Высокий процент отхода яиц у садовых славок объясняется частично тем, что птицы, будучи потревоженными у гнезда, сравнительно легко бросают свои кладки. Из 329 яиц садовые славки в период насиживания в общей сложности бросили 36’ яиц, что составило около 11%. Бросив кладку, славка очень скоро приступает к постройке нового гнезда, которое располагает обычно, в 15—20 м от прежнего. Иногда таких «попыток» бывает несколько; Кладки, содержащие сокращенное количество яиц, чаще всего встречаются в гнездах, строящихся во второй половине лета; очевидно, они принадлежат птицам, гнездившимся в первый раз неудачно.
Из врагов, которые беспокоят славок вблизи их гнезд, отметим ворону, сойку, сорокопута-жулана, обыкновенного ужа, а также бродячих кошек, которым иногда удается не только загубить птенцов, но и поймать взрослую птицу на гнезде. Из 74 самок, за которыми мы наблюдали, двух птиц постигла такая участь. Интересно, что после гибели самки мы в обоих случаях в этот же день отмечали резкий подъем активности пения самца.
Поведени ев з рослых птиц у гнезда. Явление «отвода» характерно для садовых славок в меньшей степени, чем для славок-черноголовок. Из. 38 особей, характер поведения которых нам был известен, имитация раненой птицы наблюдалась в 14 случаях.
Птенцы. Птенец первоначально совершенно голый с красновато-розовым цветом кожи. Клюв светлый, на конце имеет широкую темную предвершинную полосу. Потемнений в области ноздрей нет. Яйцевой «зуб» и когти светлые. Ноздри продолговатые. Окраска ротовой полости яркая, розовато-красного цвета. Складка в углах рта белая. На корне языка 2 темных пятна, бледнеющих к моменту вылета славок из гнезда.
Слуховые проходы открываются в конце 3-го дня, глаза приоткрываются на 4—5-й день. Кисточки контурных перьев на спине, затылке, боках и груди появляются в конце 5-го дня жизни; трубочки маховых перьев начинают развертываться в возрасте 6 дней; кисточки рулевых перьев появляются в возрасте 8 дней. С первого же дня птенцы садовой славки проявляют большую активность. Через несколько часов после вылупления они уже бодро тянут кверху шею и просят есть. В конце 1-го дня птенцы приподымаются, опираясь на передние конечности, а в возрасте 2—3 дней при выпрашивании пищи могут принимать вертикальную позу. При этом они чрезвычайно сильно вытягивают шею и трясут из стороны в сторону головой.
6-дневный птенец способен уже стоять, опираясь на пальцы ног (рис. 60), а в возрасте 7 дней может передвигаться прыжками по земле, что иногда ему и приходится делать при вынужденном оставлении гнезда. Чаще птенцы покидают гнездо через 9—10 дней после вылупления. В том случае, если их никто не тревожит, они иногда сидят в гнезде 11 дней. Птенцы, оставившие гнездо, не способны еще к полету. Тем не менее уже в 1-й день своей жизни вне гнезда птенцы иногда совершают небольшие перекочевки. Птенцовый призывный крик двоякого рода: низкий и чаще воспроизводимый звук «век-век» и короткий высокий выкрик «жюй». Услышав тревожный «сигнал» родителей, птенцы обычно настораживаются (рис. 61) и прекращают подавать тол ос.
Покинувшие гнездо птенцы садовых славок передвигаются путем короткого перепархивания с ветки на ветку. Иногда они соединяются
146
Рис. 60. 6-дневный птенец садовой славки стоит, опираясь на пальцы ног (фото автора).
Рис. 61. Птенцы садовой славки в возрасте 10 дней перед вылетом из гнезда; поза ориентировочной реакции (Савальский лесхоз, июль 1953 г. Фото автора).
вместе и сидят на высоте 2—3 м от земли, тесно прижавшись друг к: другу. В случае преследования они спрыгивают на землю и прячутся в траве. Успокоившись, они снова стремятся забраться повыше.
Питание гнездовых птенцов. Питание птенцов в основном изучалось на территории Савальского лесхоза. Всего удалось отобрать 210 экз. различных беспозвоночных, принесенных птенцам примерно за 160 прилетов. В большинстве случаев птицы приносили птенцам одно, но зато Довольно крупное насекомое. Среди 210 беспозвоночных оказалось 144 экз. насекомых, 46 пауков и 20 моллюсков. Из насекомых наибольшее значение имели бабочки, их куколки и гусеницы (.111). Преобладали гусеницы хохлаток, пядениц и совок, были обнаружены также куколки кольчатого, соснового и непарного шелкопрядов. Жуки, двукрылые, перепончатокрылые, стрекозы, сетчатокрылые существенного значения в питании птенцов, видимо, не имеют.
Птенцы садовой славки оказались до некоторой степени также и растительноядными: по мере созревания ягод последние начинают попадаться в пище птенцов.
ДРОЗД-ДЕРЯБА
(Т URDUS VISCIVORUS L.)
На территории Ленинградской области в довольно значительном количестве поющих самцов деряб мы встречали лишь в сосновых лесах Пашского района (28 апреля—13 мая 1946 г.) и окрестностей Зеленогорска (30 апреля — 20 мая 1954). Для окрестностей пос. Комарово (Карельский перешеек) наиболее раннее появление этих Дроздов-P. Л. Потапов отмечает 12 апреля (1949). Регулярное пение начинается в последней декаде апреля. Даты окончания пения нами не установлены, но, судя по всему, период активного пения у дрозда-дерябы непродолжительный.
В одном из гнезд дрозда-дерябы, найденном в Савальском лесхозе в 1953 г., откладка яиц началась в первых числах мая, в другом—первое яйцо появилось 26 мая (1952). Эти гнезда располагались на соснах на высоте 1,5 и 7 м от земли. Гнездо, найденное в Ленинградской области, также было расположено на сосне на высоте* 4 м. Птенцы в нем вылупились 25 мая (1956). Во всех гнездах кладка состояла из 4 яиц. Две из этих кладок не имели успеха: яйца и птенцы были уничтожены каким-то хищником. В третьем гнезде 4 птенца благополучно закончили свое развитие. Оставили гнездо они 2? июня (1952). Летать птенцы не могли, но по земле передвигались быстро. Через 2 дня, т. е. когда им было примерно 15 дней, они продолжали еще держаться на земле в 50—60 м от гнезда. К этому вре-мени полет их еще не установился, но они могли уже перемещаться по воздуху на 20—25 м, летя низко над землей. По всему было видно, что птенцы находятся накануне дня приобретения способности к активному помету.
ДРОЗД-РЯБИННИК
(Т URDUS PILARIS L,)
Токование. Можно ли считать, что у дрозда-рябинника имеется настоящая видовая весенняя песня? Громкое, разноголосое скрипение, издаваемое этими дроздами в период размножения, соответствует принятому нами (см. гл. I) определению настоящей видовой песииг оно специфично для вида, достаточно громко и регулярно, а главное —
248
в момент его воспроизведения птица ведет себя весьма демонстративно. Рябинник поет обычно на вершинах деревьев или на лету перед, посадкой. С другой стороны, мы должны признать, что песня дрозда-рябинника по своему характеру близка к «подпесне» белобровика. Это указывает на то, что между категориями «песня» и «подпесня» иногда может и не быть резкой границы, и в этом нет никакого противоречия, ибо настоящая песня, как мы уже говорили, может возникнуть на основе «подпесни».
Расположение гнезда. Породный состав деревьев, на которых рябинник располагает свои гнезда, весьма разнообразен. Постройки этого вида мы находили, например, на соснах, елях, лиственницах, березах, ольхах, кленах, ивах, вязах, рябинах, дубах, липах, а также на кустах бузины, что характерно, например, для дроздов, гнездящихся в Савальском лесхозе. В Ленинградской области рябинник чаще всего гнездится на елях, березах, соснах или ольхах. На Карельском перешейке излюбленным местом его гнездования являются также старые заброшенные строения, расположенные неподалеку от леса. В этих случаях гнездо обычно помещается на верхних балках, снаружи или внутри дома, под крышей (см. рис. 3).
Высота расположения найденных нами гнезд варьировала очень сильно — от 0,8 до 14 м. Поселяясь в многолюдных пригородных парках, рябинник, как правило, гнездится значительно выше, чем в местах, удаленных от жилья человека. В лесах Ленинградской области нам неоднократно приходилось находить гнезда этого вида на маленьких елочках в 1 — 1,5 м от земли.
Сроки размножения. Наибольшее количество данных па размножению дрозда-рябинника удалось собрать в Ленинградской области: здесь, в период с 1946 по 1954 г. было найдено 57 гнезд. В самом раннем гнезде кладка началась 28 апреля (1949), а в самом позднем — 23 июня (1954). В большинстве гнезд, однако, яйца появились в мае. В июне к размножению приступали лишь немногие особи, в основном, видимо, те, у которых по какой-либо причине пропали первые кладки. У преобладающего большинства птиц бывает только одна кладка в сезоне. На это указывает отсутствие второй волны появления свежих кладок в июне. Количество особей, приступающих к размножению, постепенно сокращается начиная с конца II декады мая (табл. 60). На территории Савальского лесхоза большинство птиц приступает к размножению также во II декаде мая.
Таблица1 ба
Сроки размножения дрозда-рябинника
Район наблюдения Количество гнезд, в которых кладка началась
в апреле в мае в июне
III I II III I II III
Ленинградская область . . 4 13 16 10 8 5 1
Воронежская область . . . — — 4 1 1 2 2
Случаи второго гнездования после успешного воспитания первого выводка в лесостепи более вероятны, чем в лесной зоне.
Количество яиц в кладках варьировало от 3 до 7 (табл. 61). Столь резкие колебания следует в первую очередь обьяснять нали-
249
Таблица 61
Количество яиц в кладках дрозда-рябинника
Район наблюдения Количество гнезд, содержащих кладку яиц (в шт.) Среднее количество яиц на 1 гнездо
3 4 5 6 4 7
Ленинградская область . . 5 5 18 9 1 4,9
Воронежская область . . . — 3 3 5 1 5,3
чием у этого вида значительного числа повторных уменьшенных кладок вследствие частого разорения гнезд.
Отход яиц и птенцов был довольно большим в обоих районах наблюдения. По отношению к первоначальному количеству отложенных яиц он оказался равным 40—44%. Например, из 186 яиц этого вида, находившихся под наблюдением в Ленинградской области, вылупилось 134 птенца: 39 яиц было растащено главным образом воронами, 3 яйца (1 кладка) было брошено дроздами в период насиживания, и 10 оказались неоплодотворенными. В гнездах рябинников нам приходилось отмечать до 3 неоплод отворенных яиц. Из вылупившихся 134 птенцов вылетело из гнезд 112; 22 птенца были утащены, по всей видимости, воронами. В Воронежской области «из 64 яиц вылупилось 45 птенцов, 36 из них успешно закончило свое развитие. Отход яиц и птенцов здесь так же, как и в Ленинградской области, был обусловлен главным образом хищнической деятельностью вороны.
Поведение взрослых птиц у гнезда. Обычно дрозды-рябинники очень бурно реагируют на появление человека вблизи их гнезда. Поведение отдельных особей при этом не бывает однотипным. Некоторые птицы выражают свое возбуждение беспрестанным трещанием, но сами держатся в стороне. Другие нападают на пришельца, «пикируют» на него, а иногда ударяют и обливают жидкими экскрементами. По отношению к своему основному врагу — серой вороне дрозды-рябинники почти всегда прибегают к этим приемам. Удары их, по всей видимости, бывают ощутительными, так как ворона всегда обращается в бегство, даже когда на нее нападает всего один дрозд.
Вороны все же очень часто разоряют гнезда дроздов. По наблюдениям В. А. Попова и др. (1954), в Татарии вороны почти полностью разоряют колонии дроздов при первом их гнездовании. Приступая к повторным кладкам, дрозды обычно не образуют уже колоний, а гнездятся отдельными парами. Упомянутые выше авторы сообщают, что при одиночном гнездовании гнезда дроздов разоряются значительно реже.
Сходная картина наблюдается также и в Ленинградской области. Распадение колоний происходит постепенно, по мере разорения отдельных гнезд. Пара, у которой пропала кладка или утащены птенцы, при постройке нового гнезда перемещается на несколько десятков, а иногда и на 100—200 м в сторону. Таким образом, постепенно возникает рассредоточенное гнездование и разнобой в сроках размножения. В литературе имеются указания на то, что в одной колонии рябинников может «быть до 3 и более десятков гнезд. На территории Ленинградской и Воронежской областей нам не приходилось находить таких скоплений дроздов. Наиболее крупные из колоний были образованы 6—8 парами.
Птенцы. Птенец покрыт не густым, но длинным пухом. Пух зо-^ютисто-рыжий, расположен на глазных (иногда), надглазничных, спин-:250
ной, плечевых и локтевых птерилиях. У однодневных птенцов клюв одноцветный, светлый. Ноздри продолговатые. Яйцевой «зуб» и когти светлые. Ротовая полость желтая, складка в углах рта желтовато-белая, довольно широкая.
Птенчики рябиннйка сравнительно мало подвижные, хотя уже в однодневном возрасте могут приподымать голову и приподыматься на передних конечностях. Несмотря на сравнительно крупные размеры этого .дрозда, птенцы развиваются очень быстро. Ушные отверстия открываются в конце 3-го дня жизни, а глаза—'на 4—5-й день. Кисточки маховых перьев появляются в 7-дневном возрасте. На 10—11-й день после вылупления птенцы в случае испуга могут уже выскочить из гнезда. ’Нормальное оставление гнезда происходит, однако, через 12—13 дней .после вылупления.
По нашим наблюдениям, способность к полету приобретается у птенцов спустя 3—4 дня после оставления гнезда, т. е. примерно в возрасте 16 дней. Птенцы у дрозда-рябинника далеко не всегда вылупляются дружно. Вылупление птенцов одного выводка растягивается иногда на 2 .и даже 3 дня. В одном из гнезд, найденном в Ленинградской области, вылупление 4 птенцов происходило 25, 26 и 27 июня (1954), следствием этого явилась большая разница в величине птенцов. Сходную картину -большой разновозрастности птенцов одного выводка автор наблюдал также у дроздов-рябинников, гнездившихся на территории Савальского лесхоза.
Большая растянутость вылупления птенцов дрозда-рябинника может являться также следствием начала насиживания до окончания откладки яиц, особенно характерного для позднего размножения (вторые или повторные кладки).
Питание гнездовых птенцов. Автор располагает всего 40 экз. беспозвоночных, отобранными от птенцов этого вида в Ленинградской области, где основу питания птенцов составили земляные черви (20). В пище птиц, гнездившихся в сосняках Савальского лесхоза, преобладали жуки (главным образом Melolontha). Кроме того, были обнаружены гусеницы пядениц и слепень (Ленинградская область), а также гусеницы соснового шелкопряда и 1 наземный моллюск (Са-вальский лесхоз).
Иногда дрозды-рябинники выкармливают своих птенцов частично растительным кормом. Так, В. И. Осмоловская и А. Н. Формозов (1950) отметили факт выкармливания птенцов ягодами малины, а И. В. Покровская (1956) —ягодами клюквы.
ДРОЗД ПЕВЧИЙ (Т URDUS ERICETORUM TURT.)
Токование. В окрестностях Ленинграда наиболее ранние даты первой песни певчего дрозда в течение последнего десятилетия были отмечены 15 апреля (1949) и 18 апреля (1954), наиболее позднее пение мы слышали 22 июля (1950). Регулярное пение продолжается с начала последней декады апреля до середины июля. В начале июня пение певчих дроздов под Ленинградом несколько ослабевает, но во второй половине июня — в начале июля снова становится активным. Второй подъем активности следует, по-видимому, связывать с наличием у некоторых пар второго цикла размножения.
Расположение гнезда. Характер распределения гнезд певчего дрозда по породам деревьев в Ленинградской области чрезвычайно определенен: как правило, птица помещает их на хвойных породах, при
251
чем главным образом на елках. Так, например, из 70 известных автору гнезд певчего дрозда 39 было найдено на елях (подрост и взрослые деревья) , 13 — на кустах можжевельника и 6 — на соснах. Остальные 12 гнезд помещались на березах (3 находки), черемухе, липе, на балках заброшенных домов (3 гнезда, найденные на Карельском перешейке) или просто на земле, в обочине канавы (1 находка). На территории Савальского лесхоза примерно половина гнезд (17 из 37) была обнаружена нами в кустах бузины. Из древесных пород, несмотря на преобладание сосны, чаще всего использовался, однако, дуб, особенно зимняя его-форма, способная на небольшой высоте образовывать мутовки стволовой поросли, охотно занимаемые певчим дроздом. В заповеднике «Лес на Ворскле» певчие дрозды также гнездятся главным образом на дубах (23 находки из 37).
Преобладающее число известных нам гнезд располагалось ниже 2 м. Очень часто, особенно /в лесной зоне, где певчий дрозд любит гнездиться на маленьких елочках и кустах можжевельника, его постройки приходилось находить в 0,5—1 м от земли. Высокое расположение гнезда для данного вада не характерной наблюдается в местах, часто посещаемых людьми, бродячими кошками и собаками (Быков, 1896). Из 144 гнезд, найденных нами в лесной и лесостепной зонах, высота расположения только в 20 случаях превышала 4 м. Гнезда, помещенные выше 6 м. составляют редкое исключение.
Как это было уже показано Сиивоненом (Siivonen, 1939), материал, из которого певчий дрозд свивает гнездо, может быть различным. Наружные стенки иногда бывают построены целиком из мха или лишайников, из сухих листьев или прошлогодней травы, наконец из тонких прутиков ели. Внутренняя обмазка обычно состоит из плотно утрамбованного тонкого слоя гнилой древесины, реже из мха, конского навоза и т. п. Гнезд, вымазанных изнутри глиной, илом или землей, как об этом; иногда пишут, автор не находил.
С р«о ки размножения. В Ленинградской области период возможных сроков откладки яиц у певчего дрозда продолжается с начала' мая до середины I декады июля (табл. 62). Самая ранняя из известных автору кладок началась 1 мая (1950). Большинство птиц откладывает яйца во II декаде мая. В I декаде июня количество птиц, приступающих, к размножению, опять несколько увеличивается, но затем снова сокращается и продолжает оставаться незначительным вплоть до начала» июля. Наиболее поздняя из найденных нами кладок, судя по дате вылупления птенцов, началась 6 июля (1950). Таким образом, в 1950 г. пе: риод откладки яиц продолжался 67 дней. Вторые кладки у певчего дрозда бывают, но они немногочисленны. Дважды в лето, по всей вероятности, гнездятся лишь птицы, приступившие к размножению в начале мая, так как их птенцы выходят из гнезд в конце того же" месяца, и в I декаде июня они могут приступить ко второй кладке. В Финляндии, по мнению Сиивонена, количество кладок у певчего дрозда в году также варьирует от одной до двух.
То же можно сказать и про певчих дроздов, гнездящихся в лесостепной зоне, например, в Савальском лесхозе, хотя здесь, судя по» имеющимся материалам, дрозды начинают размножаться несколько позднее, чем в Ленинградской области. Самая ранняя из известных нам кладок началась здесь 5 мая (1952). Максимальное же число гнезд со свежими кладками было найдено во II—III декаде мая и в начале III декады июня (табл. 62).
Размеры и судьба кладок. Была прослежена судьба 64 кладок, из них 36 в Ленинградской области и 28 в Савальском лесхозе и 252
Таблица 62
Сроки размножения певчего дрозда
Район наблюдения Количество гнезд, в которых кладки начались
в мае в июне в июле
I И Ill I II III I II
Ленинградская область . . . 5 11 8 12 7 3 3
Воронежская область .... 1 4 4 2 2 5 — —
учлесхозе «Лес на Ворскле». Во всех районах наблюдения преобладали кладки из 5 яиц (табл. 63).
Таблица 63
Количество яиц в кладках певчего дрозда
Место наблюдения Количество гнезд, содержащих кладку яиц (в шт.) Среднее количество яиц на 1 гнездо
3 4 5 6
Лесная зона 3 10 22 1 4,58
Лесостепная полоса 1 8 16 3 4,75
Отход яиц и птенцов в лесной зоне был 35°/0, а в лесостепной оказался более значительным и составил примерно 50 %. Из 133 яиц певчих дроздов, найденных в заповеднике «Лес на Ворскле» и Савальском лесхозе, вылупился 81 птенец: 47 яиц растащили хищники и 5 (1 кладка) было брошено птицей в период насиживания. Благополучно закончилось развитие 68 птенцов. В лесной зоне из 165 яиц вылупилось 124 птенца, и из них 107 вылетело из гнезд. Основная масса яиц и птенцов певчего дрозда гибнет, видимо, от ворон, соек, бродячих кошек и других хищников.
Поведение взрослых птиц у гнезда. При появлении человека вблизи гнезда певчий дрозд, как правило, незаметно слетает с него и скрывается с глаз. Наблюдения, однако, показывают, что он обычно продолжает держаться ib районе гнезда и следит за пришельцем, перелетая с дерева на дерево. При более сильном возбуждении птица беспокойно кричит. Лишь несколько раз автору приходилось наблюдать птиц, которые вели себя почти так же, как белобровики: угрожающе щелкали клювом и стремительно налетали на наблюдателя. Будучи сильно напуганным, певчий дрозд легко бросает гнездо даже с насиженной кладкой. Очень редко встречаются особи, отличающиеся большой доверчивостью, которые сидят на гнезде настолько плотно, что до них можно дотронуться рукой.
Птенцы. Птенец имеет буроватый или золотисто-рыжий пух, расположенный на надглазничных, затылочном, спинном, плечевых и локтевых участках тела. Ноздри овальные. Яйцевой «зуб» и когти светлые. Ротовая полость ярко-желтая, складка в углах рта бледно-желтая.
Наши наблюдения за развитием птенцов в целом соответствуют дан-/ ным В. М. Модестова (1937). В дополнение к сведениям, сообщенным В. М. Модестовым, укажем лишь на то, что ушные отверстия у птенцов этого дрозда открываются в конце 3-го дня жизни, кисточки маховых тперьев появляются в 7-дневном, а кисточки на концах пеньков. руле-
253
вых — в 8-дневном возрасте. 9-дневные. птенцы уже очень сильно пугаются вида человека. Поза «страха» весьма определенна: птенец прижимается ко дну гнезда, запрокидывает голову назад и открывает рот. При сильном испуге птенцы могут выскочить из гнезда уже в возрасте' 10 дней.
Нормальное оставление гнезда наступает через 12, реже— 13 дней: после вылупления. В этом отношении наши наблюдения несколько расходятся с данными В. М. Модестова, отмечавшего, что кормление птенцов певчего дрозда в гнезде продолжается 14—16 дней. 16-дневный птенец уже способен взлетать с земли и садиться на деревья, что не характерно для птенцов, только что оставивших гнездо. Первый период после-гнездовой жизни птенцы певчего дрозда проводят на земле, скрываясь в лесном подседе. Наземный образ жизни характерен для них и после того, как они научатся летать.
Питание гнездовых птенцов. Материалы по питанию собирались в Савальском лесхозе и на территории Ленинградской области. Всего от птенцов было отобрано 250 экз. различных, животных. Это были в основном насекомые (186), дождевые черви (35) и наземные моллюски (24). Среди последних поедались голые слизни очень крупных размеров. Весьма значительной величины (до 8 см в . длину) бывают и дождевые черви, заглатываемые птенцами. Иногда певчие* дрозды приносят своим птенцам даже мелких позвоночных. Так, однажды из пищевода птенца певчего дрозда был извлечен мелкий экземпляр прыткой ящерицы. В Савальском лесхозе птенцы певчего дрозда чаще всего получали гусениц бабочек хохлаток, пядениц, совок и листоверток. Среди жуков преобладали майские жуки, их личинки, личинки бронзовок и щелкунов. Пауки поедались сравнительно редко. В Ленинградской области певчие дрозды приносили своим птенцам главным образом дождевых червей и крупных голых слизней.
ДРОЗД-БЕЛОБРОВИК (Т URDUS MUSICUS L.)
Токование. Первая песня дрозда-белобровика в окрестностях Ленинграда, как правило, бывает слышна в период с 10 по 20 апреля: 10 апреля (1948), 15 апреля (1950 и 1952 гг.), 18 апреля (1954), 20 апреля (1946). Только в необычайно холодную и затяжную весну 1955 г. песню этого дрозда нам впервые удалось услышать лишь 1 мая. Регулярное пение в нормальные годы продолжается до середины июля, наиболее позднее отмечено нами 25 июля (1948). Осеннее пение для белобровиков не характерно, хотя автору приходилось слышать пение некоторых пролетных птиц в конце августа и в сентябре.
Первое время после прилета в пении отдельных дроздов нет однообразия. Однотипность пения наступает примерно лишь через месяц, после прилета первых местных птиц, дату появления которых нам удавалось определить по своеобразию их местного напева. Так, например» в 1954 г. первый из местных дроздов-белобровиков в парке Ст. Петергофа появился 18 апреля, а однообразие в пении птиц, гнездившихся на территории этого парка, установилось только во второй половине мая.
Знание местных напевов дроздов в некоторых случаях может помочь выяснению пролетных направлений популяций. Например, на весеннем пролете в ближайших окрестностях Ленинграда автору никогда не приходилось слышать птиц, высвистывающих вариацию песни, характерную для дроздов, гнездящихся на севере Карельского перешейка (район ст. Кузнечное), где, однако, мы неоднократно отмечали птиц, поющих 254
так, как поют белобровики южнее Ленинграда. Отсюда можно было заключить, что часть птиц, гнездящихся под Ленинградом, равно как и дрозды, гнездящиеся на севере Карельского перешейка, весной попадают сюда не с юга, как это следовало бы ожидать, а с северо-запада, из Финляндии.
Расположение гнезда. Старое название белобровика — «ольховый дрозд» до некоторой степени отражает характер его гнездования: ольха действительно является деревом, на котором белобровик располагает свои гнезда. В равной степени, однако, он может быть назван также и «еловым» или «земляным» дроздом, ибо гнезда его, помещенные на елках или просто на земле, среди травянистой растительности, явление столь же обычное. В некоторых садах и парках, где отсутствуют ель и ольха, белобровик обычно вьет гнезда на кустах акации, сирени, бузины. Постройки этого вида нам приходилось находить также на кустах спиреи, шиповника, боярышника, крушины, где его» с полным правом можно было бы назвать «кустарниковым» дроздом. Во многих местах белобровик гнездится на деревянных заборах, на сараях или в заброшенных домах. Таким образом, характер гнездования этого вида чрезвычайно разнообразен. Среди воробьиных нашей фауны это одна из наименее прихотливых птиц, способная применяться к самым различным условиям. Очень возможно, что наблюдавшееся за последнее время расширение гнездового ареала белобровика и заселение им новых стаций (города, поселки, сады и парки) оказалось возможным благодаря большой пластичности гнездовых инстинктов этого вида. Из 143 гнезд белобровика, найденных нами в Ленинградской области, на взрослых деревьях располагались 52 гнезда, на молодых деревцах (до 3 м) —33, на кустах— 12, на пнях и кучах хвороста — 8, в сооружениях человека— 18 и, наконец, на земле — 20 гнезд.
Сроки размножения. Период возможных сроков откладки яиц у дрозда-белобровика в Ленинградской области продолжается примерно в течение 2,5 мес. — с конца апреля до середины июля. В наиболее раннем из известных нам гнезд первое яйцо появилось 27 апреля (1952), а в наиболее позднем—13 июля (1947). Апрельские кладки наблюдались только в 1950 г., когда весна была ранняя и дружная. В другие годы первые яйца чаще всего появлялись 1 мая. В 1954 г.— в год с особенно поздней весной — самые ранние кладки начались 4 и 6 мая. Ниже приведено количество гнезд дрозда-белобровика, найденных нами в окрестностях Ленинграда, откладка яиц в которых началась:
В III декаде апреля......3 Во II декаде июня...........10
В I декаде мая..........27 В III декаде июня...........13
Во II декаде мая..........24 В I декаде июля............1
В III декаде мая..............19 Во II декаде июля............1
В I декаде июня..............15
Таким образом, основная масса птиц приступает к размножению в начале мая. Некоторый подъем интенсивности размножения во второй половине июня можно объяснить влиянием второго цикла размножения, который, по всей вероятности, имеет место у особей, приступающих к откладке яиц в начале мая. Вторые кладки, видимо, могут начаться уже в I и И декадах июня. Количество птиц, приступающих к откладке яиц во второй раз, варьирует в зависимости от климатических условий года. Например, в холодное затяжное лето 1954 г. это явление у дроздов-белобровиков почти не наблюдалось. Наоборот, в обычный по своим климатическим условиям 1949 г. было отмечено заметное увеличение числа свежих кладок, появившихся в III декаде июня (табл. 64). Всего в пределах Ленинградской области нами было найдено 89 полных
255
Таблица 64
Сроки размножения дрозда-белобровика в Ленинградской области по наблюдениям в 1949 и 1954 гг.
Год наблюдения Количество гнезд, в которых кладка началась
в мае в июне
I II Ш I II III
1949 6 7 5 2 2 7
1954 14 10 4 2 3 1
кладок данного вида. Количество яиц в них варьировало от 3 до 6. Преобладающее число кладок содержало 5 яиц (54). Кладки из 6 яиц' (20) и 4 яиц (12) встречались уже значительно реже; наконец, гнезда, в которых было 3 яйца, встречались совсем редко (3). Таким образом, кладки белобровика, состоящие из 3 или 4 яиц, в общей сложности •составляли всего лишь 17% (15 из 89).
Ни одна из майских кладок не содержала уменьшенного количества яиц. Все кладки из 3 или 4 яиц начались в июне или в июле (табл. 65).
Таблица 65
Сроки откладки яиц в гнезда с уменьшенными кладками
Количество яиц в кладке Даты окончания откладки яиц Место и год наблюдения
4* 1- 2VI Парк Лесотехнической академии, 1950
4* 1- 2 VI Район ст. Кузнечное, 1950
4* 2— 3 VI Парк ст. Петергофа, 1949
4* 3- 4 VI 1950
4* 11-12 VI Охтинское лесничество, 1946
4* 16-17 VI Парк ст. Петергофа, 1950
4* 19—20 VI 1949
3 24 VI Район ст. Кузнечное, 1954
4 24 VI Парк Лесотехнической академии, 1947
4 24 VI Парк ст. Петергофа, 1949
3 25 VI , „ 1949
4 26 VI „ . 1949
3* 29—30 VI Район ст. Кузнечное, 1950
4* 1—2 VI 1950
4 8 VII Парк ст. Петергофа, 1948
Примечание. Звездочкой отмечены случаи, когда даты откладки яиц определялись по срокам вылупления или возрастным признакам птенцов.
В то же время в мае было найдено наибольшее количество гнезд, содержавших кладки из 5 и 6 яиц.
Однако среди белобровиков иногда встречаются птицы, откладывающие максимальное для данного вида количество яиц в середине июня, а кладки в 5 яиц появляются иногда даже в середине июля. Все это дает основание предполагать наличие у белобровика запоздалых вторых кладок.
Отход яиц и птенцов довольно значительный. Из 384 яиц вылупилось 266 птенцов, из которых только 210 благополучно закончило свое
256
развитие. По отношению к первоначальному количеству отложенных яиц это составляет примерно 55%. Основная масса яиц и птенцов (79 и 52) была утащена различными хищниками, среди которых прежде всего следует указать на ворону. В районах, прилежащих к населенным пунктам, опасным врагом белобровика является также кошка. Автору несколько раз приходилось заставать бродячих кошек у гнезд белобровиков на «месте преступления». Несколько кладок (26 яиц) было брошено птицами IB период насиживания. Число неоплодотворенных яиц и замерших зародышей было в целом невелико—13 из 384.
Поведение взрослых птиц у гнезда. Во время беспокойства у гнезда белобровики ведут себя довольно однотипно: они обычно налетают на человека с характерным стрекотанием. Достаточно, однако, отойти от гнезда на 30—40 м, как дрозды успокаиваются и замолкают. Инбгда в ответ на крик пойманного птенца белобровики имитируют поведение раненой птицы. Это бывает, однако, очень редко. В целом для белобровиков характерен агрессивный способ защиты птенцов. При сильном возбуждении некоторые из них «отваживаются» нападать на человека. Однако среди белобровиков встречаются и такие особи, которые держатся у гнезда очень скрытно и осторожно, стараясь не попадаться на глаза наблюдателю. Свое возбуждение они выражают «коканьем» либо высоким, но тихим протяжным свистом.
Белобровики, сильно потревоженные в момент строительства гнезда, как правило, бросают его. Известны также случаи оставления гнезд в разгаре периода насиживания.
Птенцы. Однодневные птенцы опушены несколько слабее, чем у рябинника. Окраска пуха светло-охристая. Пух расположен на надглазничных, затылочном, спинном, плечевых и локтевых участках тела. Длина пуха на голове до 8 мм, а на спине — от 11 до 13 мм. Так же как у птенцов певчего дрозда и рябинника, количество, длина и интенсивность окраски пуха на теле птенцов белобровиков варьирует индивидуально. Одни птенцы бывают покрыты более коротким и темным пухом, другие — более длинным и светлым. Новорожденные птенцы этого вида имеют клюв одноцветно-светлый, ноздри овальные, яйцевой «зуб» и когти светлые. Кожа желтовато-розовая. Ротовая полость желтая с зеленым оттенком на нёбе. Складка по краям клюва белая и довольно широкая.
Первые 3—4 дня после вылупления птенцы мало активны. В это время они почти непрерывно обогреваются самкой, редко слетающей с гнезда. Заметное повышение активности наступает на 5—6-й день, т. е. ко времени прозревания птенцов. Ушные отверстия открываются несколько раньше — к концу 3-го дня. У 6-дневного птенца появляются уже кисточки контурных перьев, а также кроющих маховых. Развертывание опахал маховых начинается в 7-дневном возрасте. Пеньки рулевых перьев раскрываются на сутки позднее. В возрасте 8—9 дней птенцы уже пугаются вида человека и в случае очень сильного испуга могут даже выскочить из гнезда. В этом возрасте они уже бывают способны передвигаться по земле прыжками. Нормальное оставление гнезда происходит через 11 —12 дней после вылупления. Покинув гнездо, выводок нередко сразу же начинает совершать перекочевку в другой участок леса. В одном случае птенцы дрозда-белобровика, окольцованные в день оставления, гнезда в возрасте 12 дней, на следующий же день оказались в 100 м от гнезда. Эти птенцы очень хорошо передвигались по земле, но летать еще не могли. Через 3 дня выводок был встречен примерно в том же месте. Птенцы, которым было уже 16 дней от роду, могли подыматься с земли и садиться на деревья.
17 А. С. Мальчевский
257
Уже научившись летать, птенцы белобровика имеют тем не менее обыкновение держаться на земле.
При повторных и поздних кладках белобровики начинают насиживать яйца иногда за несколько дней до окончания откладки яиц. В этих случаях вылупление птенцов протекает неодновременно. Вылупившиеся позднее, будучи более слабыми и получая меньше корма, чем их более-энергичные «собратья», иногда совсем слабеют и гибнут. В одном и» гнезд, найденном в 1947 г. в окрестностях Ленинграда, вылупление-птенцов растянулось на трое суток: 6 июля вылупилось 2 птенца, 7 июля — 3-й, а 4-й появился лишь 8 июля утром. Этот птенец так и продолжал оставаться самым маленьким и слабым. Когда начались дождливые дни, родители стали прилетать к гнезду реже обычного. Меньше-всех получал пищи самый маленький птенец. 14 июля он был еще жив,, но сиделец самом низу, под другими птенцами, а 15 погиб и был выброшен родителями из гнезда. При вскрытии его желудок оказался совершенно пустым.
Сильную растянутость процесса вылупления птенцов при поздних: кладках у белобровика наблюдала также О. П. Подлевских (1952). В одном из гнезд вылупление птенцов, растянувшееся на 4 дня, происходило с 3 по 5 июля (1949). 6 июля вес птенцов в этом гнезде равнялся: 17,1; 16,2; 9,3 и 4,3 г. Птенец, вылупившийся последним, погиб* уже на 3-й день. Вес остальных птенцов вскоре выравнялся.
Питание гнездовых птенцов. Пища птенцов этого вида оказалась довольно разнообразной. Птенцы получают не только животный корм, но и растительный. О. М. Татаринова в Кандалакше зарегистрировала 7 случаев приноса птенцам ягод черники. По сообщению’ И. В. Покровской (1956), черника дается иногда птенцам белобровика и в Ленинградской области.
Животный корм, по данным анализа 80 пищевых комков, состоит из-насекомых (130), дождевых червей (39), моллюсков (23 слизня) и пауков (9). Дождевые черви были обнаружены в пище у птенцов главным образом в Ленинградской области; одним из основных кормов белобровиков в районе Кандалакши являлись слизни. Из насекомых белобровики таскали своим птенцам гусениц бабочек, жуков (усачей, щелкунов,, долгоносиков), а также двукрылых, пилильщиков, муравьев и других перепончатокрылых; в пищеводе одного птенца была обнаружена оса.
ДРОЗД ЧЕРНЫЙ (Т и R D и S М Е R U L A L.)
Токование. На территории Ленинградской области пение самцов черного дрозда мы отмечали в районе ст. Кузнечное и в парке-Ст. Петергофа.. На территории Савальского лесхоза регулярное пение черного дрозда продолжается до середины июля. Отдельные птицы поют до конца этого месяца.
Расположение гнезда. Из 69 гнезд черного дрозда, найденных в Савальском лесхозе, 32 располагались на взрослых деревьях (главным образом на дубах и вязах, реже — на соснах, яблонях, ивах^ черемухе, черной ольхе), 21 — в кустах бузины, 10 — прямо на земле,. 3 — на молодых деревцах (подрост клена и черемухи) и, наконец, 3 — на пнях и кучах хвороста. Таким образом, среди различных пород деревьев и кустарников бузина оказалась особенно привлекательной для' этого дрозда. В заповеднике «Лес на Ворскле» большинство гнезд этого-вида располагалось на дубах (24 находки из 40). Гнездо, найденное в Ленинградской области (район ст. Кузнечное) было помещено Hai высоте 0,6 м от земли в изгибе ствола ольхи, растущей на берегу озера.
258
Высота расположения гнезд незначительная. В Савальском лесу в большинстве известных нам случаев она не превышала 2 м (55 из 69). Выше 3 м было найдено всего 7 гнезд.
Сроки размножения. В Савальском лесу период размножения начинается в последних числах апреля—начале мая. Так, в 1951 — 1953 гг. количество гнезд, в которых началась откладка яиц, было:
В III декаде апреля . . . . 1
В I декаде мая.............2
Во II декаде мая.........10
В III декаде мая..........8
В I декаде июня............6
Во II декаде июня..........6
В III декаде июня..........6
В I декаде июля............3
Во II декаде июля..........2
Наиболее ранний выводок птенцов, только что покинувших гнездо, был обнаружен 21 мая (1951). Судя по возрасту птенцов, откладка яиц у данной пары началась в конце апреля. Большинство птиц начинает откладывать яйца во II декаде мая. В июле размножаются только отдельные экземпляры. Наиболее поздняя из известных нам кладок черного дрозда началась 11 июля (1953). В гнезде черного дрозда, обнаруженном автором на севере Карельского перешейка, первое яйцо появилось 29 апреля (1953). В этом же районе мы наблюдали птенцов черного дрозда, только что покинувших гнездо, 21 мая и 9 июня.
Размеры и судьба кладок. Прослежена история 47 кладок этого вида (40 гнезд в Савальском лесхозе и 7 в заповеднике «Лес на Ворскле»).
Число яиц в полных кладках этого дрозда варьировало от 3 до 7, а число птенцов, вылетевших из гнезда, от 1 до 6. Основной причиной отхода яиц и птенцов являлось разорение гнезд различными хищниками, которые уничтожили в общей сложности 20 яиц и 44 птенца. Кроме того, 20 яиц (4 кладки) были брошены птицами в период насиживания, из них 1 кладка была оставлена накануне дня вылупления птенцов. Всего* из 228 яиц вылупилось 184 птенца, из них 140 вылетело из гнезд. Общий отход яиц и птенцов по отношению к первоначальному количеству яиц составил, таким образом, 39%. Из врагов, беспокоивших черных дроздов у их гнезд, укажем на ворону, сойку, сарыча, черного коршуна, ушастую сову, горностая и лисицу.
Поведение взрослых птиц у гнезда. При появлении человека вблизи гнезда взрослые птицы обычно держатся в стороне, скрываясь в кронах деревьев. Иногда птицы даже не подают голоса. Значительно чаще, однако, бывает слышен их характерный беспокойный позыв. В некоторых случаях тревожный «сигнал» черного дрозда представляет собою тонкий протяжный свист, напоминающий по тону свист рябчика, издаваемый обычно в стороне от гнезда. Несколько раз нам приходилось наблюдать самцов и самок черного дрозда, которые, подлетев на очень близкое расстояние, начинали медленно прыгать в сторону от гнезда, пригнувшись к земле и полураспустив крылья. Явление «отвода» у черного дрозда отмечалось в иностранной литературе (Firth, 1947).
Птенцы. Пух у однодневного птенца буровато-серый, редкий, но длинный; распределен на надглазничных, затылочном, спинном, плечевых и локтевых участках тела. Клюв светлый, без темных отметин. Ноздри продолговатые. Яйцевой «зуб» и когти светлые. Кожа бледно-розовая. Ротовая полость желтая, складка в углах рта желтовато-белая. В первые дни после вылупления птенцы черного дрозда очень вялы; резкость движений приобретается ими лишь в возрасте 6—7 дней, когда на ощупь они становятся горячими.
17* 259
Развитие протекает примерно так же, как и у других видов дроздов. Ушные отверстия открываются на 3-й день, глаза — на 4—5-й. У 6-дневных птенцов глаза уже почти полностью открыты. В этом возрасте у них впервые появляются кисточки контурных перьев на груди. Развертывание опахал маховых перьев начинается в возрасте 7 дней, пеньки рулевых перьев раскрываются в конце 8-го дня жизни. Оставление гнезда происходит обычно через 12—13 дней после вылупления (рис. 62).
Первые дни своей послегнездовой жизни птенцы черного дрозда живут на земле. Начинают летать они в возрасте 16—17 дней. Окольцо-
Рис. 62. Птенцы черного дрозда, выпрашивающие корм; воспитаны в нево-ле (фото автора).
ванные автором 26 июня (1952) 13-дневные птенцы дрозда 28 июня были уже способны, поднявшись с земли, садиться на нижние ветки деревьев.
Вследствие того, что черный дрозд очень часто начинает насиживать еще в период откладки яиц, у этого вида нередки случаи большой растянутости вылупления птенцов одного выводка. Например, в гнезде, найденном в Савальском лесу 5 июня (1951), вылупление 7 птенцов происходило между 6 и 10 июня. И июня вес этих птенцов равнялся 42,1; 39,3; 30,4; 32,7; 23,9; 19,1 и 9,5 г. Самый маленький птенец из этого выводка погиб на 3-й день своей жизни и был выброшен птицами из гнезда; 6 остальных птенцов благополучно закончили свое развитие и покинули гнездо 18 и 19 июня. Накануне дня оставления гнезда (17 июня) вес птенцов выравнялся. В гнезде, найденном 24 мая (1951), вылупление 5 птенцов происходило 25—28 мая. В результате 28 мая птенец, вылупившийся первым, весил почти в 3 раза больше птенца, появившегося на свет последним (20,5 и 6,6 г.)- В гнезде, найденном
260
8 июля, насиживание началось после откладки второго яйца. В результате вылупление 4 птенцов растянулось на 3 дня и происходило 25—27 июля.
Количество примеров, указывающих на то, что растянутость периода вылупления птенцов в гнездах черного дрозда — явление довольно обычное, мы могли бы увеличить.
Питание гнездовых птенцов. Питание птенцов изучалось нами лишь на территории Савальского лесхоза. Из пищеводов птенцов было извлечено 410 экз. различных беспозвоночных. Оказалось, что пища птенцов черного дрозда в целом значительно разнообразнее таковой птенцов других видов дроздов. В пищевом рационе птенцов были зарегистрированы в довольно значительном количестве бабочки, их куколки и гусеницы (всего 123), жуки и их личинки (80), прямокрылые и их личинки (21), личинки мух и взрослые экземпляры различных двукрылых (31), клопы (10) и муравьи (106). В то же время оказалось, что черный дрозд реже, чем, например, певчий или дрозд-белобровик, приносит своим птенцам дождевых червей (19) и особенно наземных моллюсков, поедание которых, вопреки нашим ожиданиям, для него оказалось совсем не характерным (5 экз. из 410). Изредка птенцам давались пауки (8) и многоножки (3 экз.).
Среди насекомых на первом месте по количеству уничтоженных экземпляров стояли чешуекрылые, причем главным образом их гусеницы. Чаще птенцам приносились очень крупные гусеницы, например, лунки серебристой, соснового шелкопряда, соснового бражника, хохлатки. Иногда птенцы получали и взрослых бабочек; листоверток, шелкопрядов, совок. Из жуков чаще всего поедались майские жуки и их личинки, а также личинки бронзовок. Из прямокрылых зарегистрированы кобылки (Podisma pedestris) и прусс (Calliptamus italicus), а среди клопов — черепашка (Eurygaster austriacus).
Если черный дрозд приносит своим птенцам крупных насекомых, он подвергает их соответствующей обработке. У майских жуков отрываются головогрудь и надкрылья, у саранчи — голова и ноги, у крупных гусениц и личинок — голова и т. п.
ГОРИХВОСТКА ОБЫКНОВЕННАЯ (PHOENICURUS PHOENICURUS L.)
Токование. Самая ранняя песня в окрестностях Ленинграда отмечена 27 апреля (1951), а самая поздняя—19 июля (1955). Регулярное пение самцов продолжается, однако, всего лишь немногим больше месяца — с середины I декады мая и до конца II декады июня. Позднее поют лишь очень немногие птицы. В парке Лесотехнической академии автор однажды слышал пение горихвостки 19 августа. Этот единственный случай осеннего пения горихвостки был отмечен в 1955 г., отличавшемся необыкновенно теплым августом. В разгар брачного периода — в середине мая и начале июня — горихвостки поют в течение большей части суток, замолкая очень поздно и начиная петь с полуночи. В конце июня — начале июля активное пение самцов бывает лишь по утрам.
Расположение гнезда. Гнезда этого вида автор находил в самых различных местах: в скворечниках и дуплах деревьев, в каменных зданиях, в соломенных крышах сараев, в гнилых пнях у самой земли, в поленницах дров, в кучах хвороста, наконец прямо на земле, под прикрытием небольшой сухой веточки. На Украине гнезда этого вида находили высоко на деревьях, в толще гнезда сарыча (В. М. Зу-баровский, 1958). Таким образом, в различных случаях горихвостка
261
ведет себя как дуплогнездник и как свободно гнездящаяся птица. В зависимости от места расположения гнёзда имеют различную форму и конструкцию. Помещенные в куче хвороста, они, как правило, бывают вполне оформленными, с глубоким лотком и хорошо выраженными бортами. Постройки, расположенные на земле или в дупле, имеющем широкое дно, обычно более примитивны. В узком же дупле, имеющем небольшую поверхность дна, птицы устраивают очень простые гнезда, представляющие собой лишь ямку, выстланную незначительным количеством травинок, перьев и т. п. В отдельных случаях горихвостка совсем не вьет гнезда, а откладывает яйца прямо на древесную труху, как это делают вертишейки, дятлы ,и иногда пухляки. Одно такое «гнездо» было найдено автором в Савальском лесу. Оно располагалось в телеграфном столбе, в небольшой выемке, выдолбленной пестрым дятлом. Дно этой выемки как раз соответствовало размерам внутреннего лотка обычной постройки горихвостки. Всякий дополнительный материал в данном случае только лишь помешал бы птице откладывать и насиживать яйца, в связи с чем гнездо и не было выстроено.
Сроки размножения. На территории Савальского леса период возможных сроков откладки яиц у горихвостки продолжается около 2 мес. — с конца I декады мая до начала июля. В наиболее раннем из гнезд кладка началась 9 мая (1952 -и 1953 гг.), а в самом позднем— 2 июля (1952). Большинство птиц приступает к размножению во II и III декадах мая. В конце июня — начале июля гнездятся лишь отдельные, очень немногие птицы. Поздние кладки содержат обычно сильно уменьшенное количество яиц, что указывает на то, что они, как правило, являются не вторыми, а дополнительными.
Вторая нормальная кладка после успешного воспитания первого выводка у.обыкновенной горихвостки бывает очень редко. В Савальском лесхозе двойной цикл размножения удалось установить только однажды. Преобладающее большинство горихвосток воспитывало только один выводок.
Ниже мы указываем количество гнезд, найденных в 1951—1953 гг., в которых откладка яиц началась:
В III декаде апреля .... 1 В I декаде июня..........6
В I декаде мая...............2 Во II декаде июня..........6
Во II декаде мая..........10 В III декаде июня..........6
В III декаде мая ............8 В I декаде июля..........3
Во II декаде июля...........2
В Ленинградской области самая ранняя кладка началась 18 мая (1947), а наиболее поздняя — в самых последних числах июня, птенцы из этого гнезда вылетели лишь 30 июля (1955).
Размеры и судьба кладок. Количество яиц в найденных нами кладках обыкновенной горихвостки варьировало от 3 до 9. Кладка из 9 яиц была найдена 1 раз, из 8 яиц — 5 раз, из 7 — 8 раз, из 6— 14, из 5 — 5, из 4 — 2 и из 3 яиц — 1 раз. По устному сообщению Н. П. Кадочникова, в 1954 г. в Савальском лесхозе было найдено гнездо горихвостки, содержавшее 10 яиц. Кладки, содержавшие 3 или 4 яйца, относились к числу наиболее поздних. Так, в одном из гнезд, содержавших 1 яйца, вылупление птенцов происходило только лишь 17—18 июля, в другом, — где было 3 яйца, откладка происходила в 20-х числах июня. Было установлено, что это гнездо принадлежало птицам, которые до этого уже дважды приступали к размножению и оба раза неудачно.
В Савальском лесхозе из находившихся под наблюдением 225 яиц вылупилось 143 птенца: 33 яйца погибло при разорении гнезд, 5 яиц 262
<1 кладка) было брошено птицами, 4 яйца содержали замершие заро-дыши или оказались неоплодотворенными, и, наконец, 40 яиц погибло из-за кукушки, которая в Савальском лесхозе подкладывает яйца в доступные для нее гнезда обыкновенной горихвостки (рис. 63). Часть этих яиц была похищена из гнезд взрослыми кукушками при подбрасывании своего яйца, а другая часть — выброшена или задавлена кукушатами. Отход птенцов был значительно ниже. Погибло всего 13 (из 143; птенцов. Из них только 6 птенцов (1 выводок) было уничтожено каким-то хищником, остальные (7 птенцов из разных выводков) погибли в гнезде го неизвестной причине. Мертвых птенцов горихвостка часто выбрасывает из гнезда.
Рис. 63. Самка горихвостки кормит птенца кукушки (Савальский лесхоз, июль 1953 г. Фото автора).
На территории Ленинградской области была прослежена судьба только 7 гнезд горихвостки. Количество яиц в гнездах горихвосток, найденных в Ленинградской области, было следующее: 7, 7, 6, 6, 6, 6 и 4. Кладка, состоявшая из 4 яиц, была чрезвычайно поздняя (птенцы покинули гнездо 30 июля) и, видимо, дополнительная.
Поведение взрослых птиц у гнезда. Появление человека вблизи гнезда обычно вызывает довольно бурную реакцию со стороны его хозяев. Птицы подлетают на близкое расстояние, садятся на видные места, учащенно свистят и пощелкивают, приседают, трясут хвостом и т. п. При первой попытке приблизиться к ним они отлетают в сторону от гнезда. Подобный прием, примененный по отношению, напри-1иер, к кошке, не менее эффективен, нежели имитация движений большой или раненой птицы. Иногда горихвостки применяют и активный
263
способ защиты своего потомства. Мы наблюдали, как они стремительно налетали на большого пестрого дятла и сойку, появлявшихся в районе их гнезда. При очень сильном возбуждении некоторые птицы нападают даже на человека. В противоположность большинству некоторая часть горихвосток ведет себя чрезвычайно осторожно и молчаливо.
В некоторых случаях не удается видеть у гнезда самцов. Это бывает, однако, не всегда по причине большой осторожности птиц. Иногда самцы отсутствуют вообще, и выводок воспитывает одна самка. Чаще всего это случается при позднем размножении птиц. Например, в 1955 г. в парке Лесотехнической академии автор нашел гнездо, около которого в течение нескольких дней наблюдений держалась только одна самка. Птенцы из этого гнезда вылетели очень поздно — 30 июля. Данное явление иногда можно трактовать как результат бигамии, уже отмечавшейся у горихвосток (Ruiter, 1941), а также у мухоловок-пеструшек (Haartman, 1949). Самец, первоначально живущий с одной самкой,, через некоторое время спаривается с другой, прилетевшей в данную-местность позднее или потерявшей своего первоначального партнера.. В таких случаях основную заботу о потомстве самец проявляет по отношению к первому выводку. Вторая же самка принуждена бывает воспитывать выводок одна. Один из окольцованных автором в 1953 г. в Савальском лесу старых самцов горихвостки в 1954 г., по наблюдениям Н. П. Кадочникова, принимал участие в выкармливании двух выводков,, находившихся в разных гнездах на расстоянии 150 м один от другого. Есть основания полагать, что данный самец являлся отцом птенцов-обоих выводков.
Развитие птенцов. Птенец горихвостки покрыт сравнительно* густым, но Не очень длинным пухом темно-серого, почти черного цвета; расположен пух на глазных, надглазничных, затылочном, спинном и плечевых участках тела. Клюв у однодневного птенца одноцветно-светлый, вздутый в области мелких, чуть продолговатых ноздрей. Яйцевой: «зуб» и когти светлые. Цвет кожи желтовато-оранжевый. Ротовая полость желтая с оранжевым оттенком. Складка в углах рта белая со слабым желтоватым налетом.
Уже через несколько часов после вылупления птенец горихвостки Может высоко поднять голову и долго сохранять позу пищевого рефлекса. К концу первого дня жизни он уже способен приподыматься, опираясь на передние коне^йюсти (рис. 64). Ушные отверстия начинают' открываться лишь на 4-й день жизни, а глаза приоткрываются на 5— 6-й день; кисточки маховых перьев появляются через 8 дней после вылупления. Пеньки кроющих перьев крыла и хвоста начинают раскрываться на сутки раньше. Рулевые перья сильно отстают в развитии.. У 7-дневного птенца пеньки рулевых перьев почти незаметны. В этом возрасте видимость «хвоста» создают отрастающие кисточки верхних кроющих хвоста. Способность к активному полету, по нашим наблюдениям, устанавливается через 15—16 дней после вылупления. Оставление-гнезда происходит примерно в этом же возрасте. Из гнезд, расположенных на земле, птенцы выскакивают раньше, на 13-й день жизни. Птенцы, преждевременно оставившие гнездо, быстро и уверенно передвигаются прыжками по земле. Своими повадками они сходны с птенцами: зарянки и соловья. Последних они напоминают также своим позывом,, имеющим характер сухого, низкого, непродолжительного потрескивания^
Даже научившись хорошо летать, птенцы горихвостки часто держатся на земле. При этом они выбирают участки леса с подседом it густой травой, где они могли бы прятаться от глаз хищников. В тол*’ случае, если гнездо располагается в лесу, лишенном травы и подлеска,.
264
выводок сразу же начинает перекочевку в поисках таких участков леса, где были бы выражены нижние яруса растительности.
Питание гнездовых птенцов. Питание птенцов горихвостки изучалось лишь на территории Савальского лесхоза.1 От птенцов было отнято 1153 экз. беспозвоночных: 706 насекомых, 393 паука, 48 моллюсков и 6 многоножек. Из насекомых чаще других поедались чешуекрылые (бабочки и гусеницы), которые составляли больше половины (339) всех принесенных птенцами насекомых. По числу экземпляров бабочки уступали лишь паукам. Из чешуекрылых, особенно в 1954 г., наиболее часто поедались гусеницы и бабочки сосновой пяденицы и различных совок. Было отмечено также поедание гусениц листоверток
Рис. 64. Однодневный птенец обыкновенной горихвостки, выпрашивающий корм (фото автора).
и соснового шелкопряда. Жуки приносились птенцам значительно реже (всего 54). Из них следует отметить мелких хрущиков, личинок божьих коровок и щелкунов. Среди прямокрылых (69) чаще всего поедались бескрылые кобылки и личинки саранчовых. Несколько раз из пищеводов птенцов извлекались некрупные зеленые кузнечики с оторванными ногами и головой. Кроме того, горихвостки приносили птенцам различных двукрылых (главным образом мух), клопов, муравьев, личинок пилильщиков, верблюдок, ручейников и др. Во второй половине лета горихвостки иногда приносят птенцам ягоды бузины.
СОЛОВЕЙ ВОСТОЧНЫЙ
(LUSCINIA LUSCINIA L.)
Токование. В период наших наблюдений, за последние десять лет наиболее ранние даты песни соловья в окрестностях Ленинграда
1 Помимо автора в работе принимали участие Н. П. Кадочников, И. А. Ней-фельдт, С. В. Васильев и Д. С. Бекжанова.
265
'были следующие: 5 мая (1950), 8 мая (1954), 13 мая (1947), 14 мая (1948), 16 мая (1946). Последняя дата соответствует примерно началу регулярного пения соловьев под Ленинградом. В конце июня интенсивность пения соловья резко падает. Отдельные птицы поют до середины 1 декады июля. В годы с очень холодной весной и низкой майской и ..июньской температурой соловьи поют очень нерегулярно и в течение короткого отрезка времени.
В лесостепи пение соловьев прекращается обычно несколькими днями раньше, нежели в лесной зоне. В 1949 г. в заповеднике «Лес на Ворскле» последнее (очень вялое) пение соловья было отмечено нами .27 июня. На территории Савальской дачи пение соловьев в годы наблюдений (1951—1953) прекращалось в последних числах июня.
Расположение гнезда. Случаев гнездования восточного со-.ловья на кустах, как это иногда бывает (Шнитников, 1913; Петров, 1954), зарегистрировано нами не было. Все 37 гнезд этого вида, найденные в Савальском лесу, в учлесхозе «Лес на Ворскле» и в окрестностях .Ленинграда, располагались на земле, среди сухих листьев или в корнях различных трав.
Сроки размножения. В лесостепной полосе, на территории Савальского лесхоза, период сроков откладки яиц у восточного соловья «оказался растянутым примерно на 1 мес. В наиболее раннем из известных нам гнезд откладка яиц, судя по дате вылупления птенцов, началась примерно 16 мая (1953), а в наиболее позднем—14 июня (1951 и 1952 гг.). Большинство птиц приступает к откладке яиц во II и III декаде мая. В июне размножаются уже немногие птицы, очевидно те, которые по какой-либо причине потеряли свои первые кладки. Ниже приве-лено количество гнезд, найденных в 1951—1953 гг., в которых откладка яиц началась:
Во II декаде мая..........9 В I декаде июня.............5
В III декаде мая..........11 Во II декаде июня...........5
Из гнезд восточного соловья, найденных автором в пригородах .Ленинграда, птенцы выскочили в возрасте 9—11 дней, 19 июня и 26 июня (1947).
Размеры и судьба кладок. Наблюдения проводились в учлесхозе «Лес на Ворскле» и на территории Савальского лесхоза. Прослежена судьба 33 кладок. Количество яиц варьировало от 3 до 6. Преобладающее число кладок состояло из 5 яиц (21 находка). Гнезд с 4 яйцами было значительно меньше (8); (кладок из 3 яиц было найдено всего 3, а из 6 яиц — только 1. Все кладки, состоявшие из 3 яиц, появи-.лись во второй половине июня. По всей вероятности, они принадлежали птицам, приступившим к размножению в этом сезоне повторно.
Отход яиц и птенцов в гнездах был довольно значительным и составлял примерно 40%. Из 152 яиц вылупилось 120 птенцов, и из них только 91 птенец благополучно закончил развитие. Основная причина гибели яиц и птенцов — действие хищников, которые иногда вместе с кладкой или выводком уничтожали также и взрослую птицу. Подобная участь .постигла 3 самок из 33, за которыми мы следили. Количество неоплодо-творенных яиц было крайне незначительно: 2 из 152.
Поведение взрослых птиц у гнезда. Соловей почти никогда не подлетает к гнезду прямо, в связи с чем выследить гнездо, руководствуясь поведением взрослых птиц, очень трудно. Прежде чем ‘Опуститься к гнезду, соловей, прилетевший кормить своих птенцов, долго осматривается и несколько раз опускается на землю в разные тместа,'Как бы запутывая свои ,движения. На близкое расстояние соловей
Г266
(подлетает обычно лишь тогда, когда слышит крик пойманного птенца. Е этом случае некоторые птицы иногда даже имитируют движение раненой птицы, что в целом для соловьев не характерно, несмотря на наземное гнездование. Лишь изредка среди соловьев попадаются особи, отличающиеся большой доверчивостью.
От наземных хищников соловьи защищают свое потомство, примерно так же, как и горихвостки: садятся вблизи и назойливо кричат, перед самой мордой зверя, при первой же попытке быть схваченными, они стремительно бросаются в сторону. Автор неоднократно наблюдал подобное поведение по отношению к кошке.
Птенцы. Птенцы восточного соловья появляются на свет одетыми в очень темный, почти черный пух, который расположен на надглазничных, затылочном, спинном и плечевых участках тела. Клюв одноцветный, со слабым потемнением в области ноздрей. Яйцевой «зуб» темный, когти светлые. Ротовая полость желтовато-оранжевая, наружные края клюва белые с желтоватым оттенком.
У птенцов соловья сравнительно рано (на 6—7-й день) появляются кисточки контурного пера на груди, спине, боках и плечах. Пеньки кроющих крыла и хвоста раскрываются в 7-дневном возрасте. Первостепенные маховые начинают развертываться только через 8 дней после вылупления. Рулевые перья отстают в развитии еще сильнее. Опахала рулевых перьев начинают появляться лишь в возрасте 9 дней, когда общая длина пеньков равна всего 3—4 мм.
Птенцы соловья покидают гнездо обычно очень рано. Нормальное оставление гнезда происходит в возрасте 10—И дней. В этом возрасте они уже довольно хорошо оперены, хотя крылья и особенно хвост долго еще продолжают оставаться короткими. Окольцованные автором птенцы, встреченные через 4 дня после оставления гнезда примерно в возрасте 14—15 дней, летать еще не могли, но по земле они передвигались чрезвычайно стремительно, так что поймать их было очень трудно.
В Савальском лесхозе слетки особенно любят скрываться в густых зарослях крапивы, растущей куртинами вблизи ям и канав. Выводки обычно держатся рассредоточенно. Птенцы постоянно перемещаются из одного места в другое, все время перекликаясь короткими трещащими позывами. Кормление слетков происходит почти всегда на земле.
Питание гнездовых птенцов. Материалы по питанию птенцов были собраны на территории Савальского лесхоза. От птенцов было отобрано 239 экз. беспозвоночных: 204 насекомых, 29 пауков и <> моллюсков. Среди насекомых преобладали различные жуки (главным образом дровосеки и долгоносики), а также бабочки, куколки и гусеницы листоверток и шелкопрядов. Сравнительно часто поедались вер-блюдки. Остальные группы насекомых (двукрылые, клопы, цикады, тли, перепончатокрылые, прямокрылые) приносились птенцам в весьма ограниченном количестве.
ЗАРЯНКА (ERITHACUS RUBECULA L.)
Регулярное пение зарянок в окрестностях Ленинграда продолжается обычно с конца II декады апреля до середины июля. Отдельные самцы поют до начала августа. Осеннее пение зарянки в пределах Ленинградской области слышится редко и бывает крайне нерегулярным. Мы отмечали его лишь в августе и сентябре.
В хвойных лесах Ленинградской области автор находил открытые гнезда зарянок, расположенные почти у самой земли в углублениях ста
267
рых пней. В парках пригородной зоны Ленинграда этот вид часто поселяется в дуплах деревьев. По наблюдениям Г. А. Новикова (устное сообщение), в заповеднике «Лес на Ворскле» зарянка также является главным образом дуплогнездником.
На широте Ленинграда период сроков откладки яиц у зарянки растянут почти на 2,5 мес. Так, в одном из гнезд первое яйцо появилось 2 мая (1950), в другом — 31 мая (1949), в третьем гнезде птенцы вылупились лишь 2 августа (1954). Из гнезда, найденного на территории: Сдвальского лесхоза, птенцы выскочили 18 июня (1952).
Рис. 65. Птенец зарянки, только что оставивший гнездо (Павловский парк. Ленинградская область, июль 1955 г. Фото автора).
Новорожденный птенец зарянки похож на птенца горихвостки, только пух у него более длинный (на голове до 15 мм). Цвет пуха темно-бурый, почти черный. Он расположен на глазных, надглазничных, затылочной, спинной и плечевых птерилиях. Клюв желтовато-восковой, наружные края белые с желтоватым налетом. Ноздри продолговатые с приподнятыми внутренними краями. Яйцевой «зуб» серый, когти светло-желтые. Ротовая полость ярко-желтая. Цвет кожи желтоватооранжевый. Голос подается птенцом с первого же дня; он представляет собою слабое цыканье, напоминающее цыканье других птенцов воробьиных в этом же возрасте.
Молодые зарянки выскакивают из гнезда, не умея летать (рис. 65). Характер их передвижения очень напоминает таковой у птенцов соловья. Они очень уверенно держатся на ногах и быстрыми скачками передвигаются по земле, ловко скрываясь среди травы и кустов.. В том
268
•случае, если их преследуют, они начинают комбинировать прыжки с короткими перепархиваниями. Быстро переместившись на 15—20 м, птенец обычно забирается под кочку и там затаивается^ Характерный для взрослых птиц позыв — звонкое потрескиванье — молодые птенцы начинают воспроизводить вскоре после оставления гнезда.
КРАПИВНИК
(TROGLODYTES TROGLODYTES L.)
Д. Н. Кайгородов (1923) отмечает, что пение крапивника в окрестностях Ленинграда можно услышать уже в начале марта. Однако лишь с середины II декады апреля песня крапивника в Ленинградской области звучит более или менее регулярно. Период активного пения тянется вплоть до конца июля, продолжаясь, таким образом, около 3,5 мес. Один раз автор слышал пение двух крапивников 16 сентября (1946). Это было, однако, уже осеннее пение, которое в пределах Ленинградской области слышится довольно редко.
Из 10 совершенно свежих гнезд крапивника, найденных в Ленинградской области, только 2 оказались жилыми, содержавшими яйца. В этих 2 гнездах была богатая подстилка из перьев рябчика и тетерки и других птиц. На большое количество встречающихся в природе свеже-выстроенных, но пустых гнезд крапивника обратил внимание еще В. Н. Шнитников (1913), который указывал также на то, что далеко не во всех гнездах ему приходилось находить внутреннюю перьевую выстилку.
Как известно, у крапивников строит гнездо в основном самец. Последний обычно устраивает несколько сооружений, но лишь одно из них впоследствии избирается птицами для выведения потомства. Выбор производит самка. Она же, по всей вероятности, и натаскивает перья в облюбованное ею гнездо. Остальные постройки так и остаются недоде--ланными. Видимо, в связи с этим мы и находим лишь в некоторых гнездах крапивника внутреннюю перьевую выстилку..
Найденные гнезда располагались в различных местах. Большинство (6 находок) помещалось на небольших елочках и соснах или в подрезанных кустах желтой акации в 0,5—1,5 м qr зем^и. 2 гнезда были обнаружены между балок старых землянок, наконец в 2 случаях постройка укреплялась в густой мутовке стволовой поросли крупных деревьев на 2-метровой высоте. Некоторые гнезда были построены почти целиком из мха, другие же — из прошлогодних сухих листьев.
В двух жилых гнездах откладка яиц началась 22 июня и 6 июля (1950). Длительный период активного пения самцов, а также находки поздних свежих кладок позволяют предполагать наличие у части особей двойного цикла размножения.
Новорожденный птенец крапивника имеет темно-бурый пух, довольно густой и длинный (до 8 мм) только на надглазничных птерилиях. *На других участках тела (затылок и спина) пух редкий, особенно на •спине, где всего имеется несколько пушин.
Во время беспокойства у гнезда взрослые птицы держатся в стороне, скрываясь в зарослях подлеска. Свое возбуждение они выражают криком, сходным с тем, который издают и в другие сезоны года. '
ЛИТЕРАТУРА
Аверин В. Г. 1909. К орнитофауне Харьковской губернии. Тр. О-ва испыт. природы при Харьк. ун-те, т. XLIII.
Аверин В. Г. 1941. О мероприятиях по обогащению свекловичных полей полезной' дикой птицей. Зап. Харьк. с.-х. ин-та, юбилейный вып.
Асписов Д. И. 1932. Некоторые данные о хозяйственном значении серой вороны в условиях поймы низовий р. Камы. Работы Волжск.-Камск. зон. охотн. пром, биост., вып. 2. Казань.
Банников А. Г. 1939. Материалы к постэмбриональному изменению веса некоторых представителей Passeriformes. Зоол. журн., т. XVIII, вып. 1.
Барабаш-Никифоров И. И. и Н. К. Павловский. 1948. Фауна наземных, позвоночных Воронежского государственного заповедника. Тр. Воронежск. гос. запов., вып. 2.
Бёме Л. Б. 1952. Певчие птицы. Изд. Моск, об-ва испыт. природы.
Бёме Л. Б. 1954. Род щеглы. В кн.: «Птицы Советского Союза», т. 5. Изд. «Сов. наука», М.
Бианки В. Л. 1913. Наставления для собирания птиц, их гнезд и яиц. Программы и наставления для набл. и собир. коллекций по ест. истории, изд. 7-е. Изд.. СПб.-го о-ва естествоисп.
Бианки В. Л. 1917—1918. Опыт обработки наблюдений над гнездованием птиц Петроградской губернии. Ежегодн. Зоол. муз. АН, т. XXII.
Благосклонов К- Н. 1950а. Техника привлечения и охрана лесных птиц. В кн.: А. Н. Формозова, В. И. Осмоловской, К. Н. Благосклонова «Птицы и вредители-леса». Изд. Моск, о-ва испыт. природы.
Благосклонов К. Н. 19506. Биология и сельскохозяйственное значение полевого воробья в полезащитных насаждениях юго-востока. Зоол. журн., т. XXIX, вып. 3.
Благосклонов К- Н. 1954. Мухоловковые. В кн.? «Птицы Советского Союза»,, т. 6. Изд. «Сов. наука», М.
Благосклонов К. Н. 1956. Гнездовые рефлексы птиц при перевозке птенцов с кормилицами. В кн.: «Пути и методы использования птиц в борьбе с вредными насекомыми». Изд. Мин. с.-х., М.
Благосклонов К. Н. 1957. Охрана и привлечение птиц, полезных в сельском хозяйстве, изд. 4-е, испр. и дополн. Учпедгиз. М.
Божко С. И. 1954. Материалы по биологии размножения дубоноса и зеленушки, в условиях искусственных насаждений Савальской лесной дачи. Уч. зап. ЛГУ,.. № 181, зоол. позвоночных.
Бубнов М. А. 1956. Клест-еловик (Loxia curvirostra L.). Зоол. журн., т. XXXV^. вып. 2.
Быков А. М. 1896. Список и описание коллекции по биологии птиц Привислинского края. Коллекция зоол. каб. Варшавского ун-та, вып. 3.
Еагнер В. А. 1900. Городская ласточка (Delichon urbica), ее индустрия и жизнь как материал сравнительной психологии. Зап. Имп. АН, т. X, № 6.
Васильев Г. А. 1938. Дальнейшие материалы по изучению рефлексов у птенцов, птенцовых птиц и сравнение птенцов корвидов в отношении их нервной деятельности. Тезисы докладов на III совещании по физиологическим проблемам..
270
Васильев Г. А. 1940. Нервно-гуморальная реакция птенцового поведения. Тезисы докладов VII совещания по проблемам высшей нервной деятельности, посвященного памяти акад. И. П. Павлова.
Васильев Г. А. 1947. К вопросу о возможном терморегуляторном значении некоторых периодических явлений в жизни птиц. Изв. АН СССР, сер. биол., № L
Васильев Г. А. 1948 Новое в теории терморегуляторного значения брачного поведения самцов птиц. «Охрана природы», № 5.
Васильчук А. Н. 1915. Опыт привлечения птиц в искусственные гнездовья. Тр. по* лесному опытн. делу в России, вып. 55.
Воинственский М. А. 1949. Пищухи, поползни, синицы УССР. Изд. Киевск.. гос. ун-та.
Воинственский М. А. 1954. Синицевые. В кн.: «Птицы Советского Союза», т. 5'-Изд. «Сов. наука».
Волчанецкий И. Б. 1954. Славковые. В кн.: «Птицы Советского Союза», т. 6... Изд. «Сов. наука».
Воробьев К. А. 1955. Зимнее гнездование клестов. «Природа», № 6.
Гавриленко Н. И. 1929. Птицы Полтавщины. Полтава.
Г е п т н е р В. Г. 1958. Интенсивность пения некоторых птиц. Уч. зап. МГУ, вып. 1977-Орнитология.
Гладков Н. А. 1949. Полет, перелет и географическое распространение птиц. Извд АН СССР, сер. биол., № 1.
Гладков Н. А. 1953. Хозяйственное значение диких птиц и методы его определения. В кн.: «Перелеты птиц в Европейской части СССР». Изд. АН Латв. ССР^ Рига.
Гладков Н. А. 1954. Дроздовые. В кн.: «Птицы Советского Союза», т. 6. Изд.-«Сов. наука».
Голов Б. А. и В. И. Осмоловская. 1955. Биология и хозяйственное значение* Сороки в естественных и искусственных лесных насаждениях юго-востока Европейской части СССР. Тр. Ин-та географии АН СССР, т. LXVI. Материалы по биогеограф. СССР., вып. 2.
Гржибовский А. Г. 1911. О гнездовании белобрового дрозда (Turdus iliacus)i в окрестностях г. Смоленска. Орнитол. вест., № 1.
Даль С. К. 1954. Животный мир Армянской ССР, т. I. Позвоночные животные. Изд. АН Армянск. ССР, Ереван.
Данилов Н. Н. 1949. Влияние паводка на жизнь птиц в дельте Волги. Уч. зап.. ЛГУ, № 117, вып. I.
Дарвин Ч. 1871. Происхождение человека и половой отбор. СПб.
Дементьев Г. П. 1936. К вопросу о границах основных систематических категорий птиц. Зоол. журн., т. XV, вып. 1.
Дементьев Г П. 1940. Птицы. Руководство по зоологии, т. VI. Изд. АН СССР,. М.—Л.
Дементьев Г. П. 1941. Зимнее гнездование клестов в Московской области. «Сов. охотник», № 4.
Дементьев Г. П. 1946. Концепция целостности организма и некоторые задачи* систематики (материалы к проблеме «Животное и ландшафт»). Зоол. журн.. т. XXV, вып. 6.
Дементьев Г. П. 1947. Периодические явления и йх эволюция у наземных позвоночных. Бюлл. Моск, о-ва испыт. природы, отд. биол., т. II, вып. 5.
Дементьев Г. П. 1953. Новые материалы по распространению и биологии майны1 в Туркмении. Изв. АН Туркм. ССР, № 3.
Дементьев Г. П. и Е. П. Спангенберг. 1949. Некоторые экологические проблемы, связанные с заселением птицами полезащитных насаждений. Зоол. журн „ т. XXVIII, вып. 4.
Денисова М. Н. 1958а. О значении опушения у птенцовых птиц. Уч. зап. Моск, обл. пед. ин-та им. Н. К. Крупской, т. LXV. вып. 3.
Денисова М. Н. 19586. Особенности роста и развития птиц в связи с условиями гнездования. Уч. зап. Моск. обл. пед. ин-та им. Н. К. Крупской, т. LXV, вып. 3.
Дергунов Н. И. 1925. Биологические заметки: а) Песня зяблика; б) К заметкам' «Песня зяблика». Листки биост. юн. натур, им. Тимирязева, №№ 15, 19.
Дергунов Н. И. 1928. Дикая фауна Аскании. В кн.: «Степной заповедник Чапли-Аскания-Нова», Госиздат, М.
Динесман Н. И. 1940. Материалы к гнездовому периоду постэмбриональнэго роста некоторых видов птиц отряда воробьиных. Сб. студ. научн. работ МГУ, № 12.
Долгушин И. А. 1948. Об экологической дифференциации близких форм воробьев Ю.-В. Казахстана. «Охрана природы», № 5.
Доппельмаир Г. Г. 1939. Значение архитектоники деревьев и кустарников для гнездования птиц. «Природа», № 12.
27 Г
Дубинин В. Б. 1940. Некоторые наблюдения над биологией серой вороны в дельте Волги. «Природа»» № 6.
Дубинин Н. П. 1953. Птицы лесов нижней части долины р. Урала, ч. I, Тр. Ин-та леса АН СССР, т. XVIII.
Д у б и н и н Н. П. 1955. О проблемах изменения фауны птиц в лесах юго-восточной части СССР. Сообщения Ин-та леса АН СССР, вып. 4.
Дюнин А. Г. 1936. Бакланы в дельте Волги. Научн. тр. гос. заповедников, сер. 1, Астраханок, запов.. вып. 1.
Елисеева В. И. и Л. П. Хват о в а. 1957. Данные о питании некоторых птиц в Центрально-черноземном заповеднике. Тр. Центрально-черноземного запов. им. проф. В. В. Алехина, вып. 4.
Ефимов Е. Я. 1915. Птицы истоков Оки. Материалы к познанию природы Орловской губ., № 20.
Ефремов П. Г. 1941. К орнитофауне Марийской АССР. Уч. зап. Марийск. пед. ин-та, т. II, юбилейный вып.
Жарков И. В. и В. П. Теплое. 1932. Материалы по питанию хищных птиц Тат. республики. Уч. зап. Казанск. ун-та, кн. 7—8. вып. 2.
За рудный Н. А. 1910. Птицы Псковской губернии. Зап. АН, т. XXV, № 2.
Зверев М. Д. 1954. Изучение питания птиц непосредственными наблюдениями и с применением счетчиков. В кн.: «Привлечение и переселение полезных птиц в лесонасаждения». Изд. Мин. с. х., М.
Зуб а ронский В. М. 1958. Из наблюдений по биологии обыкновенной кукушки (Cuculus canorus L.). Зоол. журн., т. XXXVII, вып. 1.
Иванаускас Т. Л. 1953. Перелеты птиц по Латвийской ССР. В кн.: «Перелеты птиц в Европейской части СССР». Изд. АН Латв. ССР, Рига.
Иванаускас Т. Л. 1954. Исследование орнитофауны юго-восточной части Латвийской ССР. Тезисы докладов II Прибалт, орнитол. конференции по проблеме миграции птиц. Таллин.
Иванов А. И. 1940. Птицы Таджикистана. Изд. АН СССР, М.—Л.
Иванов А. И. 1952. Весенние наблюдения над птицами юго-западного побережья Каспия. Тр. Зоол. ин-та АН СССР, т. IX, вып. 4.
Иванова С. А. 1935. Изучение механизма действия света на семенники птиц. Биол. журн., т. IV, вып. 4.
Измайлов И. В. 1940, Фауна птиц и млекопитающих Хоперского государственного заповедника. Тр. Хоперск. гос. запов., вып. 1.
Исаков Ю. А. 1949. К вопросу об элементарных популяциях у птиц. Изв. АН СССР, сер. биол., вып. 2.
Исаков Ю. А. 1954. Биологические основы переселения* перелетных птиц. Сб. «Привлечение и переселение полезных птиц в лесонасаждения». М.
Исаков Ю. А., В. М. Поливанов, Н. Н. Титаева. 1953. Опыты переселения лесных насекомоядных птиц. В кн.: «Преобразование фауны позвоночных нашей страны», Изд. Моск, о- ва испыт. природы.
Кадочников Н. П. 1957. Птицы Савальского лесничества Воронежской области. Тр. Всесоюзного ин-та защиты растений, вып. 8.
Кадочников Н. П. и Ю. К. Эйгелис. 1954. Питание гнездовых птенцов сойки (Garrulus glandarius L.) в искусственных сосновых и дубовых насаждениях Савальского лесхоза Воронежской области. Зоол. журн., т. XXXIII, вып. 6.
Кайгородов Д. Н. 1886. Птицы парка Лесного института. Ежегодн. Лесного ин-та, т. I.
Кайгородов Д. Н. 1898. Орнитофауна парка Лесного института и изменения, происшедшие в ней за десятилетие. Ежегодн. Лесного ин-та, вып. 1.
Кайгородов Д. Н. 1909. Орнитофауна парка Лесного института и изменения, происшедшие в ней за десятилетие. Изв. Лесного ин-та, вып. 18.
Кайгородов Д. Н. 1922. Материалы по фенологии Петрограда. Изв. Геогр. ин-та, вып. 3.
Кайгородов Д. Н. 1923. Из царства пернатых. Госиздат, М.—Пг.
Каминский А. М. 1916. Наблюдения над редкими и малоизвестными птицами Московской губернии. Орнитол. вести., № 3.
Карпов В. и В. Паровщиков. 1941. Пеструшка или пестрая мухоловка. «Природа и соц. хоз.». Сб. VIII, ч. 2.
Келейников А. Л. 1953. Экология домового и полевого воробьев как массовых вредителей зерновых культур в южных районах СССР. Автореф. дисс. МГУ.
Кир и ко в С. В. 1952. Птицы и млекопитающие в условиях ландшафтов южной оконечности Урала. Изд. АН СССР. М.
Книпперт К. К. 1956. Материалы по размножению кряквы. Тр. Ин-та зоологии АН Казахск. ССР, т. VI, зоол.
Кнорре Е. П. 1947. Опыты привлечения в очаги сосновой пяденицы насекомоядных птиц как меры борьбы с этим вредителем. Научно-метод. зап. Гл. упр. по запов., вып. IX.
272
Козлов П. С. 1950. Птицы леса. Саратов.
Козлова Е. В. 1930. Птицы Юго-Западного Забайкалья, Северной Монголии и Центральной Гоби. Материалы Комисе, по исслед. Монгольской и Тувинской народных республик и Бурят-Монг. АССР, вып. 12.
Коре лов М. Н. 1953. Птичьи солонцы. Изв. АН Казахск. ССР, сер. биол., вып. 8.
Корелов М. Н. 1954. Иволговые. В кн.: «Птицы Советского Союза», т. 5. Изд. «Сов. наука».
Королькова Г. Е. 1954. Значение птиц в истреблении массовых вредных насекомых. Сообщения Ин-та леса АН СССР, вып. 2.
Красовский С. К. 1936. Морфология черепа дятлов в связи с вопросом их происхождения и эволюции. Изв. Научн. ин-та им. Лесгафта, т. XIX, вып. 2.
Лихачев Г. Н. 1951. Размножение и питание ворона в Тульских засеках. Бюлл. Моск, о-ва испыт. природы, отд. биол., т. VI, вып. 5.
Лихачев Г. Н. 1953. Наблюдения над размножением мухоловки-пеструшки в искусственных гнездовьях. Бюлл. ДАоск. о-ва испыт. природы, отд. биол., т. LVIII, вып. 2.
Лихачев Г. Н. 1954. О взаимоотношениях большой синицы и мухоловки-пеструшки при заселении искусственных гнездовий. В кн.: «Привлечение и переселение птиц в лесонасаждения». Изд. Мин. с. х., М.
Лихачев Г. Н. 1955а. Большая синица и ее связь с гнездовой территорией. Бюлл. Моск, о-ва испыт. природы, отд. биол., т. LX, вып. 4.
Лихачев Г. Н. 19556. Мухоловка-пеструшка и ее связь с гнездовой территорией. Тр. Бюро кольцевания, вып. 8.
Лихачев Г. Н. 1957. Оседлость и миграции большой синицы (Partis major). Тр. Бюро кольцевания, вып. 9. Изд. /Мин. с. х., М.
Лукашкин В. Ф. 1936. Опыт учета гнездящихся пар птиц и разделение между ними территории. Бюлл. Моск, о-ва испыт. природы, отд. биол., т. XLV, вып. 1.
Лукина Е. В. 1951. Птичий городок. Изд. Моск, о-ва испыт. природы.
Лукина Е. В. 1953. Изменчивость некоторых инстинктивных реакций у птиц. «Природа», Ко 7.
Мальчевский А. С. 1946. О залете клеста-еловика в южное Заволжье. «Природа», Ко 6.
Мальчевский А. С. 1947. Роль птиц в полезащитных полосах Заволжья. Вест. ЛГУ, сер. биол. наук, № 4.
Мальчевский А. С. 1948. Из жизни птиц в гнездовой период. «Естествознание в школе», № 3.
Мальчевский А. С. 1950. О гнездовании птиц в городских условиях. Тр. Лен. о-ва естественен., вып. 4.
Мальчевский А. С. 1954. Орнитофауна парка Лесотехнической академии им. С. М. Кирова (г. Ленинград) и ее изменения с 1880 по 1950 г. Уч. зап. ЛГУ, сер. биол., № 181, вып. 38.
Мальчевский А. С. и Н. П. Кадочников. 1953. Методика прижизненного изучения питания птенцов насекомоядных птиц. Зоол. журн., т. XXXII, вып. 2.
Мальчевский А. С. и И. А. Н е й ф е л ь д т. 1954. Материалы по биологии размножения и питанию обыкновенного козодоя. Уч. зап. ЛГУ, № 101, вып. 38, зоол. позвоночных.
Мальчевский А. С., И. В. П о к р о в с к а я, Н. П. Овчинникова, Т. Н. Г е-ракова. 1954. Об экологических закономерностях распределения птичьих гнезд в лесу. Уч. зап. ЛГУ, № 181, вып. 38.
Мензбир М. А. 1895. Птицы России, т. II, М.
Милованова Г. А. 1956. Материалы по питанию мухоловки-пеструшки и большой синицы. В кн.: «Пути и методы использования птиц в борьбе с вредными насекомыми». Изд. Гл. упр. по запов., М.
Михеев А. В. 1957. Определитель птичьих гнезд. Учпедгиз, М.
Михельсон Г. А. 1958. Биологические основы привлечения мухоловки-пеструшки. Привлечение полезных птиц-дуплогнездников в лесах Латвийской СОР. Изд. АН Латвийской ССР.
Михельсон Г. А. и М. Г. Чаун. 1957. Миграции мухоловки-пеструшки, ее привязанность к гнездовой территории и расселение по данным кольцевания в Латвийской ССР. Труды II Прибалт, орнитол. конференции. М.
Модестов В. М. 1937. К вопросу о гнездовом периоде певчего дрозда. Зоол. журн., т. XVI, вып. 4.
Нейфельдт И. А. 1956. Материалы по питанию гнездовых птенцов некоторых лесных насекомоядных птиц. Зоол. журн., т. XXXV, вып. 3.
Никитина Р. В. 1954. Адаптивные особенности пухового покрова воробьиных. Автореф. канд. дисс. Моск. обл. пед. ин-та им. Н. К. Крупской.
Новиков Г. А. 1948. Строительный материал гнезд птиц Кольского полуострова. Научн. бюлл. ЛГУ, № 19.
18 А. С. Мальчевский
273
Новиков Г. А. 1952. ^Материалы по питанию лесных птиц Кольского полуострова.. Тр. Зоол. ин-та АН СССР, т. IX, вып. 4.
Новиков Г. А. 1956. Еловые леса как среда обитания и роль в их жизни млекопитающих и птиц. В кн.: «Роль животных в жизни леса». Изд. МГУ, М.
Орлов П. II. 1947. К биологии некоторых птиц (по наблюдениям в Черкасской районе Киевской области). «Природа», № 8.
Осмолрвская В. И. и А. Н. Формозов. 1950. Очерки экологии некоторых полезных птиц леса. В кн. А. Н. Фо>рмозова, В. И. Осмоловской, К. Н. Благо> склонова. «Птицы и вредители леса». Изд. Моск, о-ва испыт. природы.
Першаков А. А. 1932. Птицы нагорных дубрав Чувашской республики. Уч. зап. Казанск. гос. ун-та, т. 92.
Петров В. С. 1954. К орнитофауне поймы Нижнего Днепра. Тр. Научно-иссл. ин-та* биол. Харьк. гос. ун-та, т. 20.
ПодлевскихО. П. 1952. О гнездовом периоде дрозда-белобровика. Уч. зап. ЛГУ,, сер. биол. наук, № 145, вып. 31.
Познании Л. П. 1938. Материалы по онтогенетическому развитию дятлов (Picidae). Зоол. журн., т. XVII, вып. 2.
Познании Л. П. 1946. Постэмбриональный рост некоторых лесных птиц в связи с особенностями их экологии. Изв. АН СССР, нов. сер., т. LIV, вып. 3.
Познании Л. П. 1948. Адаптивные особенности постэмбрионального роста некоторых птиц. Изв. АН СССР, сер. биоХ, № 3.
Познании Л. П. 1949. Экологическая морфология птиц, приспособленных к древесному образу жизни. Тр. Ин-та морфол. животн., т. III, вып. 2.
Покровская И. В. 1953. Материалы по экологии птиц, связанных гнездованием с древостоем. Уч. зап. Лен. гос. пед. ин-та им. А. И. Герцена, т. 91.
Покровская И. В. и С. Я. Герд. 1955. Использование лесными птицами водных; беспозвоночных при выкармливании птенцов. Уч. зап. Лен. гос. пед. ин-та им. А. И. Герцена, т. ПО.
Покровская И. В. 1956. Материалы по питанию гнездовых птенцов лесных птиц. Ленинградской области. Зоол. журн., т. XXXV, в. 1.
Поливанов В. М. 1954. Наблюдения над формированием и динамикой местных популяций у некоторых перелетных птиц. Сб. «Привлечение и переселение полезных птиц в лесонасаждения». М.
Поливанов В. М. 1957а. Местные популяции у птиц и степень их постоянства. Тр. Дарвинск. гос. запов., вып. 4. Вологда.
Поливанов В. М. 19576. Сезонное размещение и некоторые черты биологии скворца (Sturnus vulgaris L.) по данным кольцевания. Тр. Бюро кольцевания,, вып. IX. М.
Поливанова Н. Н. 1957. Питание птенцов некоторых видов полезных насекомоядных птиц в Дарвинском заповеднике. Тр. Дарвинск. гос. запов., вып. 4. Вологда.
Поликарпова Е. Ф. 1941. Внешние факторы и половой цикл птиц. Журн. общ., биол., Ко 1.
Попов В. А., Ю. К. Попов, Г. П. Приезжее, Т. М. Кулаева и др. 1954. Результаты изучения животного мира зоны затопления Куйбышевской ГЭС*. Тр. Казанск. фил. АН СССР, сер. биол. наук, вып. 3
Портенко Л. А. 1937. Фауна птиц Северного Урала. Изд. АН СССР, М.—Л.
Портенко Л. А. 1955. Обзор форм палеарктических ремезов (Remiz pendulinus L.>-и их таксономическая оценка. Тр. Зоол. ин-та АН СССР, т. XVIII.
Прокофьева И. В. 1958. О питании гнездовых птенцов некоторых видов лесных, птиц. Уч. зап. Лен. гос. пед. ин-та им. А. И. Герцена, т. 143, вып. 7.
Промптов А. Н. 1930. Географическая изменчивость песни зяблика (Fringilla сое-lebs L.) в связи с общими вопросами сезонных перелетов птиц. Зоол. журн.,. т. X, вып. 3.
Промптов А. Н. 1934а. Об экологических факторах изоляции у птиц. Зоол. журн.,. т. XIII, вып. 4.
Промптов А. Н. 19346. Биология дубровника в связи с вопросом о расселении, видов птиц. Зоол. журн., т. XIII, вып. 3.
Промптов А. Н. 1936. Эколого-генетические факторы эволюционной дивергенции* у птиц. Биол. журн., т. V, вып. 6.
Промптов А. Н. 1940. Изучение суточной активности птиц в гнездовой период.. Зоол. журн., т. XIX, вып. 1.
Промптов А. Н. 1944. Голосовая имитация воробьиных птиц как одно из специфических свойств их высшей нервной деятельности. ДАН СССР, т. XLV, № 6.
Промптов А. Н. 1945. Физиологический анализ инстинкта гнездостроения у птиц. Изв. АН СССР, отд. биол., № 1.
Промптов А. Н. 1946. Об условнорефлекторных компонентах в инстинктивной; деятельности птиц. Физиол. журн. СССР, т. XXXII, вып. 1.
274
Пр ом пт о в А. Н. 1947. Опыт классификации имитационных явлений на основе экспериментального изучения поведения птиц. Физиол. журн. СССР, т. XXXIII, № 5.
ПромптовА. Н. 1948. О некоторых закономерностях онтогенетического поведения птиц в связи с проблемой эволюции функций нервной системы. Журн. общ. биол., т. IX, № 2.
Промптов А. Н. 1949. Некоторые наблюдения над птицами в дни войны и блокады Ленинграда. «Охрана природы», № 6.
Промптов А. Н. 1956. Очерки по проблеме биологической адаптации поведения воробьиных птиц. Изд. АН СССР, М.—Л.
Промптов А. Н. 1957. Птицы в природе. Изд. 3-е, Учпедгиз, Л\
Промптов А. Н. и Е. В. Лукина. 1937. Изучение оседлости синиц (Paridae, Aves) методом кольцевания. Зоол. журн., т. XVI, вып. 4.
Промптов А. Н. и Е. В. Лукина. 1938. Опыты по изучению биологии и питания большой синицы (Parus major L.) в гнездовой период. Зоол. журн., т. XVII, вып. 5. .
Промптов А. Н. и Е. В. Лукина. 1940. О биологических взаимоотношениях кукушки и некоторых видов птиц, ее воспитателей. Бюлл. Моск, о-ва испыт. природы, отд. биол., т. XIX, вып. 5—6.
Промптов А. Н. и Е. В. Лукина. 1945. Условнорефлекторная дифференцировка позывов у воробьиных птиц и ее биологическое значение. ДАН СССР, т. XXXXVI, Кз 9.
Птушенко Е. С. 1936. К вопросу о пищевом режиме птенцов черного коршуна. Бюлл. Моск, о-ва испыт. природы, отд. биол., т. XV, вып. 1.
Птушенко Е. С. 1949. О заселении птицами полезащитных насаждений Сталинградской области. «Охрана природы», № 9.
Птушенко Е. С. 1954. Славковые. В кн.: «Птицы Советского Союза», т. 6. Изд. «Сов. наука», М.
Рагозина М. Н. 1954. Роль белковой оболочки яйца в развитии цыпленка. Тр. Ин-та морфол. животн. им. А. Н. Северцова, вып. 12.
Реймерс Н. Ф. 1957. Наблюдение за гнездом кедровок. «Природа», № 7.
Рольник В. В. 1946. Влияние изменения состава газов в воздушной камере яйца на развитие эмбриона и повышение выводимости цыплят. ДАН СССР, т. II, №9. Рустамов А. К. 1946. Расселение майны в бассейне Аму-Дарьи. «Природа», № 2. Рустамов А. К. 1954а. Вороновые. В кн.: «Птицы Советского Союза», т. 5. Изд. «Сов. наука», М.
Рустамов А. К. 19546. Птицы пустыни Кара-Кум. Ашхабад.
Рустамов А. К. 1955. К вопросу о понятии «жизненные формы» в экологии животных. Зоол. журн., т. XXXIV, вып. 4.
Сафронов А. А. 1954. Опыт изучения питания мухоловки-пеструшки и большой синицы. В кн.: «Привлечение и переселение полезных птиц в лесонасаждения». Изд. Мин. с. х., М.
С е в ер ц о в Н. А. 1855. Периодические явления в жизни зверей, птиц и гад Воронежской губернии. Изд. АН СССР, М., 1950.
Семенов С. М. 1956а. Материалы по питанию мухоловки-пеструшки в гнездовой период. В кн.: «Пути и методы использования птиц в борьбе с вредными насекомыми». Изд. Гл. упр. по запов., М.
Семенов С. М. 19566. Лесная соня как истребитель птиц в искусственных гнездовьях. В кн.: «Пути и методы использования птиц в борьбе с вредными насекомыми». Изд. Гл. упр. по запев., М.
Смогоржевский Л. А. 1950. К вопросу о значении гнездовой территории и пения у птиц. Тр. Зоол. муз. Киевск. ун-та, т. II, вып. 6.
Спангенберг Е. П. 1954. Скворцовые. В кн.: «Птицы Советского Союза», т. 5. Изд. «Сов. наука», М.
Спангенберг Е. П. и И. М. Олигер. 1949. Орнитологические исследования в Дарвинском заповеднике в 1946 и 1947 гг. Тр. Дарвинск. гос. запов., вып. 1.
Сомов Н. Н. 1897. Орнитологическая фауна Харьковской губернии. Харьков.
Станчинский В. В. 1927. Птицы Смоленской губернии. Научн. изв. Смоленск, гос. ун-та, т. IV.
Строков В. В. 1938. Орнитофауна парка Лесотехнической академии им. С. М. Кирова по наблюдениям в 1936—1937 гг. Сб. научно-исслед. работ студентов лесотехнических вузов.
Судиловская А. М. 1952. О расселении домашнего воробья. Бюлл. Моск, о-ва исслед. природы, отд. биол., т. VII, вып. 5.
Сушкин П. П. 1902. К морфологии скелета птиц. I. Сравнительная остеология дневных хищных птиц (Accipitres) и вопросы классификации. II. Сокола и их ближайшие родственники. Уч. зап. Моск, ун-та, отд. ест. истории, вып. XVII.
Гаращук В. И. 1953. Птицы полезащитных насаждений степной зоны УССР и возможность использования их для борьбы с вредителями. Изд. АН УССР, Киев.
Й8*
275
Тауриньш Э. Я. 1951. Материалы по экологии мухоловки-пеструшки в искусственных гнездовьях в сосновом бору в условиях Латвийской ССР. Тезисы докладов II эколог, конференции, вып. 3.
Тауриньш Э. Я. и Ч. Б. Т и м а. 1953. Наблюдения по биологии некоторых видов птиц, гнездящихся в искусственных гнездовьях в условиях Латвийской ССР. В кн.: «Перелеты птиц в Европейской части СССР».
Те плов В. П. и И. С. Туров. 1956. Значение серой вороны в пойменных охотничьих угодьях среднего течения р. Оки. Зоол. журн., т. XXXV, вып. 5.
Терновский Д. В. 1954. Зимнее гнездование клестов. Бюлл. Моск, о-ва испыт. природы, отд. биол., № LIX, вып. 1.
Тима Ч. Б. 1954. Роль дуплогнездников в борьбе с вредителями хвойных насаждений Латвийской ССР. Автореф канд. дисс. Рига.
Тима Ч. Б. 1958. Материалы по корму дуплогнездников в сосновых насаждениях Латвийской ССР. Привлечения полезных птиц-дуплогнездников в лесах Латвийской ССР. Изд. АН Латвийск. ССР.
Титаева Н. Н. и В. М. Поливанов. 1953. О методике изучения питания мелких насекомоядных птиц в гнездовой период. Бюлл. Моск, о-ва испыт. природы» отд. биол., т. LV1II, вып. 2.
Тугаринов А. Я. 1950. Происхождение миграций птиц Палеарктики. Памяаи ака.и П. П. Сушкина. Изд. АН СССР, М.
Формозов А. Н. 1934. Хищные птицы и грызуны. Зоол. журн., т. XIII, вып. 4.
Формозов А. Н. 1940. Практическое значение птиц. Руководство по зоологии» т. VI. Птицы. Изд. АН СССР, М.—Л.
Формозов А. Н. 1950. О некоторых чертах биологии птиц в связи с вопросами охраны от вредителей лесов и лесопосадок. В кн. А. Н. Формозова, В. И. Осмоловской, К. Н. Благосклснова. «Птицы и вредители леса». Изд. Моск, о-ва испыт. природы, М.
Формозов А. Н. 1952. Спутник следопыта. Изд. Моск, о-ва испыт. природы, М.
Фридолин В. Ю. 1936. Животно-растительное сообщество горной страны Хибин. Тр. Кольской базы им. С. М. Кирова АН СССР, вып. 3.
Хватова Л. Т. 1956. Питание большой синицы, полевого воробья и вертишейки. В кн.: «Пути и методы использования птиц в борьбе с вредными насекомыми». Изд. Гл. упр. по запов., М.
Чаун М. Г. 1958. Состав и динамика местных популяций мухоловки-пеструшки в искусственных гнездовьях. Привлечение полезных птиц-дуплогнездников в лесах Латвийской ССР. Изд. АН Латвийск. ССР.
Чмутова А. П. 1948. Возрастные признаки и группы серых ворон. ДАН СССР» нов. сер., т. XI, № 4.
Чмутова А. П. 1953. Особенности развития и размножения птиц (серая ворона) в различных географических зонах СССР. Бюлл. Моск, о-ва испыт. природы» отд. биол., т. LVIII, вып. 6.
Чмутова А. П. 1955. Постэмбриональное развитие серой вороны. Бюлл. Моск, о-ва испыт. природы, отд. биол., т. LX, вып. 4.
Ш а м о в И. К. 1910. Наши певчие птицы. Изд. 3-е, М.
Шапошников Л. В. 1949. Об оседлости большой синицы. «Охрана природы», № 7.
Шарлемань Н. В. 1915. Наблюдения над птицами хвойных лесов окрестностей г. Киева. «Птицеведение и птицеводство», вып. 2—3.
Шварц С. С. 1950. Эффективность криптической окраски. Тр. Лен. о-ва естество-исп., т. XX, вып. 4.
Шеварева Т. П. и Е. Т. Бровкина. 1954. Материалы к сравнительной экологии гнездования ласточек. Уч. зап. Моск. гор. пед. ин-та им. В. П. Потемкина» т. XXVIII, вып. 2.
Шейнис С. А. 1954. Питание куриного зародыша в последние дни инкубации. ДАН СССР, т. 96; № 5.
Шнитников В. Н. 1913. Птицы Минской губернии. Материалы к познанию фауны и флоры Российской империи, т. XII.
Ш те гм а н Б. К. 1951. В тростниках Прибалхашья. Алма-Ата.
Штегман Б. К. 1956. Воробьи в Казахстане, их вредная деятельность и борьба с ними. Тр. Республ. станции защиты раст. Казахск. филиала ВАСХНИЛ.
Шульпин Л. М. 1928. Заметки о птицах Алтая. Ежегодн. Зоол., муз. АН СССР» т. XXIX.
Шульпин Л. М. 1940. Орнитология. Изд. ЛГУ.
Эйгелис Ю. К. 1957. Материалы по питанию и хозяйственному значению ворона в условиях лесостепной дубравы «Лес на Ворскле». Вест. ЛГУ, № 9, сер. биол.» вып. 2.
Эйгелис Ю. К. 1958. Биология размножения галки в условиях лесостепной дубравы «Лес на Ворскле». Вестник ЛГУ, № 3, сер. библ., вып. 1.
Юдин К. А. 1952. Птицы Красноярского края. Тр. Зоол. ин-та АН СССР, т. IX,. вЫп. 4.
276
Я к ш и с Я. Я. 1952. к вопросу об изучении питания хищных птиц. Зоол. журн. т. XXX. вып. 6.
А 1 turn. 1868. Der Vogel und sein Leben.
Basecke K. 1927. Vernichten der Bruten durch Ameisen. Beitr. zur Fortpfl. biol. d. VOgel, Nr 3,
Buxton J. 1950. The Redstart.
Chappel В. M. A. 1948. .Injury-Feigning* of Spotted Flycatcher. British Birds, vol. XVI.
Chappel В. M. A. 1949. Presumed Bigamy of Rook. British Birds, vol. XLII, No 3. Chisholm A. H. 1937. The Problem of Vocal Mimicry. The Ibis, pp. 703—721.
Christoleit W. 1927. Ueber Nest und Gelege des Zwergfliegenfangers (Muscicapa p. parva Bachst.). Beitr. zur Fortpfl. biol. der Vdgel, Nr 5.
C re utz G. 1955. Der Trauerschnapper (Muscicapa hypoleuca). Eine Populationsstudie. J. f. Ornith., Bd. 96, Hf. 3.
Dereck G. 1953. Jays Nesting in Hollows in Trees. British Birds, vol. 46, No 3.
Firth F. M. 1947. .Injury-Feigning* of the Blackbird. British Birds, vol. XL, No 5.
French N. R. 1954. Notes on Breeding Activities and Gular Sacks in the Pine Grosbeak. Condor, 56.
Frieling H. 1937. Die Stimme der Landschaft. Munch, u. Berlin.
Gerd H. 1927. Dobrudschareise 1925. J. f. Ornith. (Makatsch, 1950).
Glasewald R. 1928. Brutergebnisse bei hohlenbriitenden VOgeln im Jahre 1928.
Beitr. zur Fortpfl. biol. der VOgel, Nr 6.
Goodwin D. 1953. The Reactions of Some Nesting Passerines Towards Live and Stuffed Jays. British Birds, vol. 46, No 6.
Gosnell H. T. 1949. Polygamy in Spotted Flycatcher. British Birds, vol. XLI. No 3.
Groebbels F. 1937. Der Vogel. II Band. Geschlecht-und Fortpflanzung.
Grote H. 1930. Wanderungen und Winterquartiere der palaarktischen Zugfogel in Africa. Mitt. Zool. Mus„ Berlin, B. 16, Nr 1.
Haartman L. 1949. Der Trauerfliegenschnapper. I. Ortstreue und Rassenbilding. Acta Zool. Fennica, 56.
Haartman L. 1954. Der Trauerfliegenschnapper. III. Die Nahrungsbiologie. Acta Zool. Fennica, 83.
Haartman L. u. H. Lohrl. 1950. Die Lautausserungen des Trauer-und Halsband-fliegenschnappers Muscicapa h. hypoleuca (Pall.) und M. a. albicollis (Temmink).* Orn. Fennica, Nr 4.
H e i d e r Dr. Richard. 1954. 1st der Regenruf des Buchfinken (Fr. coelebs) wetterbe-dingt? Ornith. Mitt., 6, Jahrgang, Hf. 9.
Heinroth O. 1924. Lautausserungen d. Vogel, J. f. Ornith., Bd. 72, Hf. 2.
Heinroth O. u. Frau Magdalen. 1924—1933. Die Vogel Mitteleuropas, Berlin.
Heinroth O. 1938. Aus dem Leben der Vogel (русск. перевод, ИЛ, M., 1947).
Kingston R. W. G. 1933. The Meaning of Animal Colour and Adornment (Chisholm, 1937).
Howard H. 1920. Territory in the Bird Life. London.
Howard H. 1929. An Introduction to the Study of Bird Behaviour, Cambridge.
Howard H. 1952. Lillie’s Development of the Chick.
Huxley J. S. 1938. The Present Standing of the Theory of Sexual Selection. Evolution, ed. by G. R. de Beer Oxford, pp. 11—42.
I ngram C. A. 1907, Tongue-marks in Joung Birds. Ibis, vol. I, p. 576.
Ingram C. A. 1920. Contribution to the Study of Nestling Birds. Ibis, vol. II.
Kalitsch L. 1927. Freistehendes Nest des Feldsperlings. Beitr. zur Fortpfl. biol. der Vogel, Nr 2.
Kayser. 1912. Finkenschlage und Finkenliebhaberei. Gef. Welt, B. 41.
Kendeigh S. Ch. 1941. Length of Day and Energy Requirements for Ganad Development and Egg-laying in Birds. Ecology, vol. 22, No 3.
Kendeigh S. Ch. 1952. Parental Care and Its Evolution in Birds. Ulin. Biological Monographs, vol. XXII.
Keve A., 1952—1955. A Madarak csigataplaleka, IV. Aquila, vol. 63—64, p. 69.
Kuhlmann F. 1909. Some Preliminary Observations on the Development of Instincts and Habits in Joung Birds. Psychol. Rev. (Psychol. Monogr.), vol. II, No 1.
Lack D. 1939. The Behaviour of the Robin. Proceed, of the Zool. Society of London, vol. 109.
Lack D. 1947a. Darvin’s Finches (русск. перевод, ИЛ, M., 1949).
Lack D. 1947b. The Significance of Clutchsize; Pts la. 2, Ibis, 89, Pt. 3, 90.
Lack D. 1948. Natural Selection and Family Size in the Starling. Evolution, vol.
II, No 2.
Lack D. 1949. Family Size in Certain Thrushes (Turdidae). Evolution, vol. Ill, No 1.
Lack D. 1953. The Life of the Robin. A Pelikan Book.
277
Lack D. 1954. The Natural Regulation of Animal Numbers, Oxford (русск. перевод. ИЛ, M., 1957).
Lack D. a E. T. Silva. 1949. The Weight of the Nestling Robins. Ibis, 91.
Lees J. 1949. Weights of Robins, Ibis, 91.
L u k a n u s. 1907. Lokale Gesangserscheinungen und Vogeldialekte, ihre Uhrsachen und Entstehung. Ornithol. Monatsber., Hf. 7—8.
Makatsch W. 1949. Der Vogel und sein Ei. Die neue Brehm-Bucherei. Leipzig.
Makatsch W. 1950. Der Vogel und sein Nest. Die ireue Brehm-Bucherei. Leipzig.
M’akatsch W. 1951. Der Vogel und seine Jungen. Die neue Brehm-Bucherei. Leipzig!.
May D. Y. 1947. Observations on the Territory Breeding Behaviour of the Willowwarbler. British Birds, vol. XI, No 1.
Mayr E. 1944. Sistematics and the Origin of Species from the Viewpoint of a Zoologis (русск. перевод, ИЛ, M., 1947).
M c-A t e e. 1932. Effectiveness in Nature of the So-called Protective Adaptations in the Animal Kingdom Chiefly as Illustrated by the Foodhabits of Nearctic Birds. Smithsonian misc. collection, vol. 85, No 7.
Miller A. H. 1941. The Buccal Food-carrying of the Rosy Finch. The Condor, vol. 43. Montagu G. 1802. Ornithological Dictionary Introduction (Lack, 1953).
Morris D. 1954. The Reproductive Behaviour of the Zebra Finch. (Poephila guttata) with Special Reference to Pseudofemale Behaviour and Displacement Activities. Behaviour, No 4.
My res M. T. 1949. Red-Backed Shrike Striking Human Intruders. British Birds, vol. XLII, N 9.
N atorp O. 1928. Blaukehlchens Stern. Beitr. zur Fortpfl. biol. der Vogel, Nr 4.
Nelder I. A. 1948. Display of Wren. British Birds, vol. XII, No 3.
Nice M. M. 1937. Territory and Mating with the Song-Sparrow. The Proceed, of the 8-th Intern. Ornith. Congress.
Nicholson E. M. 1927. How Birds Live.
Niethammer. 1937. Handbuch der deutschen Vogelkunde. Bd. 1. Passeres.
Noll H. 1924. Sumpfvogelleben. Wien (Makatsch, 1951).
Owen D. F. 1954. Protocalliphora in Birds Nests. British Birds, vol. XLVII, No 7.
Palmgren P. 1932. Zur Biologie von R. regulus und P. atricapillus borealis. Acta Zool. Fennica, 14.
Peitzmeier. I. 1955. Zur Deutung des „Regenrufes* des Buchfinken (Fringilla coe-lebs). J. f. Ornith. B. 96. Hf. 2.
Pfeifer S. 1952. Der Grope Buntspecht als Nestrauber. Vogelwelt, 73.
Port man A. 1938. Beitrage zur Kenntnis der postembrionalen Entwicklung der Vogel. Rev. Suisse de Zoologie, 45.
Portman A. 1955. Die Postembrionale Entwicklung der Vogel als Evolutionsprob-lem. Acta XI Congressus intern. Ornithologica. Basel. 1954,
Poulsen H. 1951. Inheritance and Learning in the Song of Chaffinch, Fringilla coe-lebs L. Proceed, of the X-th Intern. Ornithol. Congress. Uppsala.
Preston a. Norris. 1947. Nesting Heights of Breeding Birds. Ecology, 28(3).
Py craft W. 1907. Nestling Birds and Some of the Problems they Present. British Birds, vol. 1, No 4, 5 and 6.
Reinikainen A. 1937. The Irregular Migration of the Crossbill. Orn. Fennica, No 2.
Ruiter C. J. S. 4941. Waarnemingen omtrent de levenswijze van de Gekraagde Rood-staart Phoenicurus phoenicurus L. Ardea, 30 (Makatsch, 1951).
Schiermann G. 1927. Notizen fiir »Brutnachbarschaften> der Vogel. Beitr. zurFortpfl. biol. der Vogel, Nr 5.
Schilling Ph. 1927. Brutet Sturnus vulgaris (L.) zweimal? Ornithol. Monatschrift, S. 71.
Sqhneider W. 1927. Erfahrungen bei der Starenberingung. Mitt, des Vereines sa-chsischen Ornithologen, S. 72.
Schneider W. 1952. Beitrag zur Zebensgeschichte des Stars Sturnus v. vulgaris L. Beitr. Vogelkunde, B. 3.
Schnurre. 1937. Zum Vogelfang des Gropen Buntschpechts. Beitr. zur Fortpfl. der Vogel., Nr 13.
Sick H. 1939. Dialektbildung beim Regenruf des Buchfinken. J. f. Ornith., 87.
Siivonen L. 1939. Zur Okologie und Verbreitung der Singdrossel (Turdus ericeto-rum philomelos Brehm.). Annales Zoologica Societatis. Zool.-Bot. Fennica, 7, Nr 1.
SlonakerJ. K. 1925. The Development of the Eye and its Accessary Parts in the English Sparrow. J. of Morph. Physiol., 35 (Portman, 1938).
Summers-Smith D. 1954. Colonial Behaviour in the House Sparrow. British Birds, vol. XLVII, No 8.
278
Southern J. 1949a. Joung Birds Returning to the Nest. British Birds, vol. XLH, No 4.
Southern J. 1949b. „Injury-Feigning* of Chaffinch. British Birds, vol. XLII, No 8.
S t e g m a n B. 1930. Uber die Formen der palearctischen Rotrucken- und Rotschwanz-wurger und deren taxonomischem Wert. Ornith. Monatsber, 4.
Stein G. 1927. Weitere Mitteilungen zur Fortpflanzung der markischen Weidenmeisen.
Beitr. zur Fortpfl. biol. der Vogel, Nr 5.
Steinmetz H. 1932. Beobachtungen und Untersuchungen uber den Schltipfakt. f. Ornith.
Stresemann E. 1932—1937. Aves. Handbuch der Zoologie, Bd. VII, Hf. 2.
Sutton G. a. D. P a r m e 1 e e. 1954. Nesting of the Greenland Weather on Baffin Island. Condor, 56, No 5.
Swanberg. 1951. Food Storage, Territory and Song in the Thickbilled Nutcracker, Proceed. X-th Int. Ornith. Congr., 1950.
Ticehurst С. B. 1908. On the Down-Plumage and Mouth Coloration of Some Nesting Birds. British Birds, vol. II, No
Ticehurst С. B. 1926. On the Down-Plumages of Some Indian Birds. J. of the Bombay Natural Hist. Society. August.
Tutt H. R. 1953. Notes on the Nesting of a Pair of Jays inside a Hollow Tree. British Birds, vol. XLVI, No 3.
Wackernagel H. 1954. Der Schnabelwulst des Stars (Sturnus vulgaris). Ref. Suisse Zool., vol. 61, N 1.
Walker A. B. 1949. Unusual Display of Greenfinch. British Birds, vol. XLII, No 5.
Weigold H. 1926. Masse, Gewichte und Zug nach Alter und Geschlecht bei Helgo-lander Zugvogeln. Wiss. Meeresuntersuch. Abt. Helg. Neue Folge, Nr 15.
Werner Keif. 1954. Groper Buntschpecht pliindert Bruten von Freibrutern. OrnithoL Mitteilungen, 6 Jahrgang, Hf. 9.
W it ch ell Ch. 1896. The Evolution of Bird Song with Observations on the Influence of Heredity and Imitation. London.
W i t h e r b у H., F. C. R. J о u r d a i n, N. E. T i c e h u r s t a. B. W. T u с к e r. 1952» The Handbook of British Birds. London.
ОГЛАВЛЕНИЕ
Введение.....................................................• ... 3
ЧАСТЬ ПЕРВАЯ
ГНЕЗДОВОЙ ПЕРИОД В ЖИЗНИ ВОРОБЬИНЫХ ПТИЦ
Глава I. Явление токования, его формы, сроки и биологическое значение .............................................................. 7
Формы проявления тока............................................... —
Продолжительность и степень регулярности пения.................... 13
Местные напевы и географическая изменчивость пения................ 17
Биологическое значение пения птиц.................................. 25
Глава II. Гнездование лесных воробьиных................................ 32
Гнездостроение ....................................................... —
О пластичности гнездовых инстинктов.................................. 45
Явление гнездового консерватизма .................................... 50
Глава III. Размеры, сроки и судьба кладок................................. 60
Количество яиц в кладке.............................................. —
Причины растянутости периода откладки яиц........................... 64
Судьба кладок ...................................................... 70
Враги воробьиных в гнездовой период................................. 80
О поведении воробьиных у гнезд и птенцов............................ 84
Глава IV. Вылупление и птенцовые признаки................................ 89
Вылупление птенцов.................................................. —
Птенцовые признаки воробьиных....................................... 93
Глава V. Питание воробьиных в гнездовой период......................... 111
О кормовой специализации воробьиных.................................. 112
О степени многоядности птенцов ...................................... 117
О специфике птенцового питания....................................... 121
Минеральные корма и гастролиты............................• .... 126
Продолжительность периода выкармливания птенцов...................... 128
О количестве пищи, поедаемой птенцами................................ 129
Глава VI. Постэмбриональное развитие воробьиных.......................... 134
ЧАСТЬ ВТОРАЯ
МАТЕРИАЛЫ ПО БИОЛОГИИ РАЗМНОЖЕНИЯ И ПОСТЭМБРИОНАЛЬНОМУ РАЗВИТИЮ ОТДЕЛЬНЫХ ВИДОВ
Ворона серая (Corvus corone cornix L.)................................... 159
Сорока (Pica pica L.).................................................... 162
Сойка (Garrulus glandarius L.)........................................... 165
•80
Скворец обыкновенный (Sturnus vulgaris L.).........................• . 168
Иволга обыкновенная (Oriolus oriolus L.)............................... 172
Дубонос обыкновенный (Coccothraustes coccothraustes L.)................ 174
Зеленушка (Chloris chloris L.) . . . •................................. 176
Щегол (Carduelis carduelis L.) ........................................ 179
Чиж (Spinus spinus L.)................................................. 181
Коноплянка (Acanthis cannabina L.)..................................... 182
Снегирь (Pyrrhula pyrrhula L.) ........................................ 184
Чечевица обыкновенная (Erythrina erythrina Pall.)...................... 185
Клест-еловик (Loxia curvirostra L.) ................................... 187
Зяблик (Fringilla coelebs L.).......................................... 189
Воробей полевой (Passer montanus L.) .................................. 194
Овсянка обыкновенная (Emberiza citrinella L.) . .*..................... 196
Овсянка садовая (Emberiza hortulana L.)................................ 198
Конек лесной (Anthus trivialis L.)..................................... 201
Синица большая (Parus major L.)........................................ 204
Лазоревка (Parus coeruleus L.)......................................... 210
Пухляк (Parus atricapillus L.)......................................... 211
Синица длиннохвостая (Aegithalos caudatus L.)............................ —
Королек желтоголовый (Regulus regulus L.).............................. 216
Сорокопут-жулан (Lanius collurio L.)..................................... —
Мухоловка серая (Muscicapa striata Pall.).............................. 221
Мухоловка-пеструшка (Muscicapa hypoleuca Pall.)........................ 226
Мухоловка-белошейка (Muscicapa albicollis Temm.)....................... 228
Пеночка-теньковка (Phylloscopus collybitus Vieill.).................... 229
Пеночка-весничка (Phylloscopus trochilus L.)........................... 230
Пеночка-трещотка (Phylloscopus sibilator Bechst.)...................... 233
Пеночка зеленая (Phylloscopus trochiloides Sund.)...................... 235
Камышовка садовая (Acrocephalus dumetorum Blyth.)........................ —
Малиновка лесная, или пересмешник (Hippolais icterina Vieill.)......... 236
Славка ястребиная (Sylvia nisoria Bechst.)............................. 238
Славка-черноголовка (Sylvia atricapilla L.).............................. —
Славка серая (Sylvia communis Lath.)................................... 241
Славка завирушка (Sylvia curruca L.)................................... 242
Славка садовая (Sylvia borin Bodd.)..........•......................... 244
Дрозд-деряба (Turdus viscivorus L.) . . •.............................. 248
Дрозд-рябинник (Turdus pilaris L.) ................................... —
Дрозд певчий (Turdus ericetorum Turt.)................................. 251
Дрозд-белобровик (Turdus musicus L.)................................... 254
Дрозд черный (Turdus merula L.)........................................ 258
Горихвостка обыкновенная (Phoenicurus phoenicurus L.).................. 261
Соловей восточный (Luscinia luscinia L.)............................... 265
Зарянка (Erithacus rubecula L.)........................................ 267
Крапивник (Troglodytes troglodytes L.) . . . •......................... 269
Литература................................................................. 270
Мальчевский Алексей Сергеевич Гнездовая жизнь певчих птиц Редактор О. Л. Петровичева Техн, редактор С. Д. Водолагина Корректоры:
Р. И. Гольдина, А. А. Кузьмин
Сдано в набор 13 VIII 1958 г. М-09648. Подп. к печ. 16-XII-58 г.
Уч.-изд. л. 25,2. Печ. л. 24,1. Бум. л. 12,05. Форм. бум. 70ХЮ81/1б. Тираж 2150 экз. Заказ 970.
Типография ДОЛГУ. Ленинград, Университетская наб., 7/9.
ИСПРАВЛЕНИЯ И ОПЕЧАТКИ
Страница Строка Напечатано Следует читать
11 13 сверху резких разных
64 7—8 снизу в Ленинградской области в Савальском лесу
65 14 снизу в Савальском лесу в Ленинградской области
186 10 сверху июля июня
214 8 снизу могли не могли
278 11 снизу Zebensgeschichte Lebensgeschichte
Зак. 970